
再構築された挿入部位を含有する組換え改変ワクシニアアンカラ(MVA)ワクシニアウイルス
本発明は、ウイルスゲノムの遺伝子間領域(IGR)であってワクシニアウイルスゲノムの2つの隣り合った必須オープンリーディングフレーム(ORF)の間に位置するIGRの中へ、異種核酸配列を組み込むのに有用な、再構築部位を含有する組換え改変ワクシニアアンカラ(MVA)ウイルスであって、隣り合った必須ORFは該組換えMVAウイルスの構築に使用された親MVAウイルス中では隣り合っていないことを特徴とする組換えMVAウイルスに関し、かつ、ワクシニアウイルスのゲノムに異種DNAを挿入するのに有用な関連核酸構築物に関し、さらに、医薬品またはワクチンとして開示された該ウイルスの使用に関する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、MVAゲノム中への異種DNA配列の安定的組込みに有用な挿入部位に関する。より具体的には、本発明は、必須遺伝子および非必須遺伝子の組み合わせを包含する改変ワクシニアアンカラ(MVA)ウイルスゲノムの領域を再構築して、異種DNAが該ゲノム中への安定的な組込み状態を維持するようにする方法に関する。
【背景技術】
【0002】
ポックスウイルス・ファミリーを構成するウイルスは、数百種のタンパク質をコードしている大きな二本鎖DNAゲノムを有する(非特許文献1)。ポックスウイルスは、脊椎動物および昆虫の宿主域を踏まえて、コードポックスウイルス亜科(Chordopoxvirinae)およびエントモポックスウイルス亜科(Entomopoxvirinae)に分けられる。コードポックスウイルス亜科は8つの属すなわち:オルソポックスウイルス属(Orthopoxvirus)、パラポックスウイルス属(Parapoxvirus)、アビポックスウイルス属(Avipoxvirus)、カプリポックスウイルス属(Capripoxvirus)、レポリポックスウイルス属(Leporipoxvirus)、スイポックスウイルス属(Suipoxvirus)、モルシポックスウイルス属(Molluscipoxvirus)、およびヤタポックスウイルス属(Yatapoxvirus)で構成されている。オルソポックスウイルス属の原型株はワクシニアウイルスである。ワクシニアウイルス(VACV)は、無関係な病原体に対する防御免疫応答を引き起こすことが示された最初の組換えウイルスであり(非特許文献2;非特許文献3)、家畜用および野生生物用ワクチン剤のためのベクターとして使用されている(非特許文献4)。しかしながら、ヒトに使用するための組換えVACVの開発は安全性の問題により阻まれてきた。この理由から、免疫不全の動物においても模範的な安全プロファイルを備えた極めて弱毒化された天然痘ワクチンである改変VACVアンカラ(MVA)に、関心が集まっている(非特許文献5〜非特許文献7)。MVAのゲノム配列(非特許文献8)は、ほとんどの哺乳動物細胞において増殖することができず、かつ組換えワクチンベクターの優れた候補であることが知られており(非特許文献9ならびに非特許文献10)、ニワトリ胚線維芽細胞において570回を越えて継代され、その間に、親の野生型アンカラ株に対して6つの主な欠失が、宿主域の厳しい制限を伴って発生した(非特許文献11)。MVAは宿主域が厳しく制限されており、ほとんどの哺乳動物細胞において、ビリオンのアセンブリ阻止が原因でほとんどまたは全く繁殖しない(非特許文献9)。組換えMVA(rMVA)を用いた初期実験から実証されたその能力は、外来タンパク質を確実に発現すること(非特許文献9)、ならびに防御的な体液性免疫および細胞性免疫を誘導することであった(非特許文献10)。現在、種々様々な病原体の遺伝子を発現するrMVA候補ワクチンが、動物およびヒトの試験を受けている(非特許文献12)。
【0003】
候補のヒト免疫不全ウイルス(HIV)ワクチンおよびその他のワクチンを開発している間に、突然変異体rMVAが組織培養継代後に外来タンパク質を発現する能力を失うことが観察された(非特許文献13〜非特許文献15)。しかしながら、この不安定性は、個々のプラークが単離されて分析されなければ当初は察知されずにいる可能性がある。しかし、集団内において一度確立されると、非発現体は元のrMVAを圧倒して急速に増殖することができる。上記を考慮することは、rMVAの大規模なワクチンシードストックの製造にとって特に重要である。MVAにおけるクローン化遺伝子の不安定性は驚くべきことである、というのも、MVAはすでにニワトリ胚線維芽細胞(CEF)における数百回の継代を通じてその適応時の遺伝的変化を経ており、現在は完全に安定しているからである。実際、3つの独立したプラーク単離物(受託番号U94848、AY603355、およびDQ983236)について、アントワーヌらにより、同一の167,000bpのゲノム配列が報告されている(非特許文献16)。遺伝子挿入物の不安定性の原因はこれまで調査されていないが、組換えタンパク質の有害な影響が非発現性突然変異体の選択優位性において役割を果たすように思われる。したがって、パラインフルエンザウイルスおよび麻疹ウイルスの膜貫通型タンパク質の発現レベルを低減すること、ならびにHIV Envの細胞質尾部の一部を欠失させることにより、rMVAの安定性が改善される(非特許文献13、非特許文献14、非特許文献15)。しかしながら、発現の低減は免疫原性も低下させる可能性があり、したがって不適当な場合がある。(非特許文献17)。
【0004】
ワクチンとしてのrMVAの潜在的価値を考慮すれば、この有害な不安定性の問題を理解し、かつ安定な組換えMVAウイルスを構築する方法を開発することが重要である。さらに、安定性の問題についての理解は、他のDNA発現ベクターに当てはまる洞察を提供することも考えられる。本発明はそのような洞察を提供し、かつ安定な組換えMVAウイルスの構築という問題の解決策を提供する。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】D.M.クナイプ(Knipe)、P.M.ハウリー(Howley)、D.E.グリフィン(Griffin)、R.A.ラム(Lamb)、M.A.マーティン(Martin)、B.ロイズマン(Roizman)、およびS.E.シュトラウス(Straus)編、「フィールズ・ウイルス学(Fields Virology)」第5版、モス(Moss)、B.著、「ポックスウイルス科:ウイルスおよびその複製(Poxviridae: The Viruses and Their Replication)」、米国フィラデルフィア州フィラデルフィア、リッピンコット・ウィリアムズ・アンド・ウィルキンズ(Lippincott Williams and Wilkins)、2007年
【非特許文献2】モス(Moss)、B.、G.L.スミス(Smith)、J.L.ゲリア(Geria)、およびR.H.パーセル(Purcell)、「生きている組換えワクシニアウイルスはB型肝炎からチンパンジーを防御する(Live recombinant vaccinia virus protects chimpanzees against hepatitis B.)」、ネイチャー誌(Nature)、1984年、第311巻、p.67−69
【非特許文献3】パオレッティ(Paoletti)、E.、B.R.リピンスカス(Lipinskas)、C.サムソノルフ(Samsonolf)、S.R.マーサー(Mercer)、およびD.パニカリ(Panicali)、「遺伝子組換えポックスウイルスを使用する生ワクチンの構築;B型肝炎ウイルス表面抗原および単純ヘルペスウイルス糖タンパク質Dを発現するワクシニアウイルス組換え体の生物学的活性(Construction of live vaccines using genetically engineered poxviruses; biological activity of vaccinia virus recombinants expressing the hepatitis B virus surface antigen and the herpes simplex virus glycoprotein D)」、米国科学アカデミー紀要(Proc. Natl. Acad. Sci. USA)、1984年、第81巻、p.193−197
【非特許文献4】モス(Moss)、B.、「組換え遺伝子の発現、ワクチン接種、および安全性のための遺伝子組換えポックスウイルス(Genetically engineered poxviruses for recombinant gene expression. vaccination, and safety.)」、米国科学アカデミー紀要(Proc. Natl. Acad. Sci. USA)、1996年、第93巻、p.11341−11348
【非特許文献5】マイア(Mayr)、A.、V.ホッホスタイン‐ミンツェル(Hochstein−Mintzel)、およびH.スティックル(Stickl)、「弱毒化ワクシニアウイルス株MVAの継代経歴、特性、および適用性(Passage history, properties, and applicability of the attenuated vaccinia virus strain MVA.)」、インフェクション誌(Infection)、1975年、第3巻、p.6−14(独語)
【非特許文献6】スティックル(Stickl)、H.、V.ホッホスタイン‐ミンツェル(Hochstein−Mintzel)、A.マイア(Mayr)、H.C.フーバー(Huber)、H.シェーファー(Schafer)、およびA.ホルツナー(Holzner)、「天然痘のMVAワクチン接種:弱毒化された生ワクシニアウイルス株(MVA)の臨床試験(MVA vaccination against smallpox: clinical trial of an attenuated live vaccinia virus strain (MVA).)」、ドイチェ・メディツィニッシェ・ボーヘンシュリフト誌(Dtsch. Med. Wschr.)、1974年、第99巻、p.2386−2392(独語)
【非特許文献7】スティッテラー(Stittelaar)、K.J.、T.クイケン(Kuiken)、R.L.デ・スワルト(de Swart)、G.ファン・アメロンゲン(van Amerongen)、H.W.ウォス(Vos)、H.G.ニーステルス(Niesters)、P.ファン・スハルクウェイク(van Schalkwijk)、T.ファン・デル・クワスト(van der Kwast)、L.S.ワイアット(Wyatt)、B.モス(Moss)、およびA.D.オスターハウス(Osterhaus)、「免疫抑制マカクにおける改変ワクシニアウイルス・アンカラ(MVA)の安全性(Safety of modified vaccinia virus Ankara (MVA) in immune−suppressed macaques.)」、ワクチン誌(Vaccine)、2001年、第19巻、p.3700−3709
【非特許文献8】マイア(Mayr)、A.ら、ツェントラルブラット・フュール・バクテリオロギー(Zentralbl Bakteriol)、1978年、第167巻、p.375−390
【非特許文献9】サッター(Sutter)、G.およびモス(Moss)、B.、「非複製型ワクシニアベクターは組換え遺伝子を効率的に発現する(Nonreplicating vaccinia vector efficiently expresses recombinant genes.)」、米国科学アカデミー紀要(Proc. Natl. Acad. Sci. USA)、1992年、第89巻、p.10847−10851
【非特許文献10】サッター(Sutter)、G.、L.S.ワイアット(Wyatt)、P.L.フォーリー(Foley)、J.R.ベニンク(Bennink)、およびB.モス(Moss)「宿主域が制限され、かつ高度に弱毒化されたワクシニアウイルスMVA株に由来する組換えベクターはマウスにおいてインフルエンザウイルスに対する防御免疫を活性化する(A recombinant vector derived from the host range−restricted and highly attenuated MVA strain of vaccinia virus stimulates protective immunity in mice to influenza virus)」、ワクチン誌(Vaccine)、1994年、第12巻、p.1032−1040
【非特許文献11】マイヤー(Meyer)、H.ら、ジャーナル・オブ・ジェネラル・ビロロジー誌(J Gen Virol)、1991年、第72巻、p.1031−1038
【非特許文献12】ゴメス(Gomez)、C.E.、J.L.ナジェラ(Najera)、M.クルーパ(Krupa)、およびM.エステバン(Esteban)、「ポックスウイルスベクターMVAおよびNYVAC 感染症およびがんに対するワクチン接種のための遺伝子送達システム(The poxvirus vectors MVA and NYVAC a gene delivery systems for vaccination against infection diseases and cancer)」、カレント・ジーン・セラピー誌(Curr. Gene Ther.)、2008年、第8巻、p.97−120
【非特許文献13】スティッテラー(Stittelaar)、K.J.、L.S.ワイアット(Wyatt)、R.L デ・スワルト(de Swart)、H.W.ウォス(Vos)、J.グローエン(Groen)、G.ファン・アメロンゲン(van Amerongen)、R.S.ファン・ビンネンダイク(van Binnendijk)、S.ローゼンブラット(Rozenblatt)、B.モス(Moss)、およびA.オスターハウス(Osterhaus)「受動的獲得抗体の存在下における改変ワクシニアウイルスアンカラ系の麻疹ウイルスワクチンを接種されたマカクの防御免疫(Protective immunity in macaques vaccinated with a modified vaccinia virus Ankara−based measles virus vaccine in the presence of passively acquired antibodies)」、ジャーナル・オブ・ビロロジー誌(J. Virol)、2000年、第74巻、p.4236−4243
【非特許文献14】ワイアット(Wyatt)、L.S.、I.M.ベリアコフ(Belyakov)、P.L.アール(Earl)、J.A.ベルゾフスキー(Berzofsky)、およびB.モス(Moss)、「組換えMVAによって発現されたHIV Envの自然発生的切断に起因する、細胞表面発現、免疫原性および遺伝的安定性の増大(Enhanced cell surface expression, immunogenicity and genetic stability resulting from a spontaneous truncation of HIV Env expressed by a recombinant MVA)」ビロロジー誌(Virology)、2008年、第372巻、p.260−272
【非特許文献15】ワイアット(Wyatt)、L.S.、S.T.ショル(Shors)、B.R.マーフィー(Murphy)、およびB.モス(Moss)、「動物モデルにおけるパラインフルエンザウイルス3感染に対して有効な複製欠損型組換えワクシニアウイルスワクチンの開発(Development of a replication−deficient recombinant vaccinia virus vaccine effective against parainfluenza virus 3 infection in an animal model.)」、ワクチン誌(Vaccine)、1996年、第14巻、p.1451−1458
【非特許文献16】アントワーヌ(Antoine)、G.、F.シェイフリンガー(Scheiflinger)、F.ドルナー(Dorner)、およびF.G.フォークナー(Falkner)、訂正記事10「改変ワクシニアアンカラ(MVA)株の完全なゲノム配列:他のオルソポックスウイルスとの比較(The complete genomic sequence of the modified vaccinia Ankara (MVA) strain: comparison with other orthopoxviruses)」、ビロロジー誌(Virology)、2006年、第350巻、p.501−502、[1998年第244巻p.365の訂正]
【非特許文献17】ワイアット(Wyatt)、L.S.、P.L.アール(Earl)、J.フォークト(Vogt)、L.A.エラー(Eller)、D.チャンドラン(Chandran)、J.リウ(Liu)、H.L.ロビンソン(Robinson)、およびB.モス(Moss)、「組換え改変ワクシニアウイルスアンカラHIVワクチンの免疫原性とin vitro発現レベルとの関連(Correlation of immunogenicities and in vitro expression levels of recombinant modified vaccinia virus Ankara HIV vaccines)」、ワクチン誌(Vaccine)、2008年、第26巻、p.486−493
【発明の概要】
【0006】
本発明は、改変ワクシニアアンカラ(MVA)ウイルスのゲノムを、該ゲノムの領域を再構築することによってより安定させることができるという発見に関する。特に、本発明者らは、非必須遺伝子を含有するゲノム領域が遺伝学的に不安定であることを発見した。さらに、そのような領域は、非必須DNAを除去し、これらの領域の必須遺伝子を互いに隣り合わせることによってより安定させることが可能である。必須遺伝子の喪失は増殖上の不都合を有するウイルスを生じるので、そのようなウイルスはウイルス集団から急速に失われて、必須遺伝子および任意の介在DNAが維持されたウイルスの集団が生じる。
【0007】
本開示は、組換え改変ワクシニアアンカラ(MVA)ウイルスであって、MVAウイルスゲノムの2つの隣り合った必須オープンリーディングフレームの間に異種核酸配列を含むウイルスを提供する。必須ORFの選択は、該ORFが、本発明の組換えウイルスを構築するために使用される親MVAウイルスのゲノム中で隣り合っていないように行われる。すなわち、該必須ORFは少なくとも1つの非必須ORFによって隔てられている。しかしながら、組換え改変ワクシニアアンカラ(MVA)子孫ウイルスでは、該必須ORFは隣り合わせにされている。すなわち、必須ORFの間には介在する非必須ORFが存在しない。従って、必須ORFの間の領域は安定であり、ウイルス集団内で維持される。従って、この領域は、異種核酸配列の挿入のための新規かつ有用な部位を提供する。そのような異種核酸配列は、抗原のような、治療上有用なタンパク質をコードすることができる。
【0008】
本開示はさらに、本発明の組換え改変ワクシニアアンカラ(MVA)ウイルスを構築するために使用することができる核酸構築物も提供する。そのような構築物は、親MVAウイルス由来であって親ウイルスにおいては隣り合っていない必須ORFを含んでいる。しかしながら、開示される核酸構築物では、これらの必須ORFは互いに隣り合わせにされている。さらに、必須ORFの間に遺伝子間領域を含有する構築物が開示され、これらは異種核酸配列の挿入に使用することができる。
【0009】
最後に、本発明のウイルスを疾患の予防および治療のために使用する方法も開示される。
【図面の簡単な説明】
【0010】
【図1】pol遺伝子塩基配列の同一性に基づいたHIV‐1およびHIV‐2の系統関係を示す図。SIVcpzおよびSIVsmmは、それぞれチンパンジーおよびスーティーマンガベイから検出された、ヒトに近い霊長類のレンチウイルスである。
【図2】4つの異なるSIVcpz単離物を用いた、完全長pol遺伝子塩基配列に基づいたHIV‐1のグループM、NおよびOの系統関係を示す図。バーは遺伝的距離0.1(10%のヌクレオチド相違)を示し、アステリスクはenvの配列に基づいてグループNのHIV‐1単離物を位置付けている。
【図3】HIV‐1単離物の指向性および生物学的特性を示す図。
【図4】HIVにコードされたタンパク質を示す図。HIV遺伝子の位置、一次翻訳産物(場合によってはポリプロテイン)のサイズ、およびプロセシングされた成熟ウイルスタンパク質が示されている。
【図5】成熟HIV‐1ビリオンの概略図。
【図6】HIV‐1 Env糖タンパク質を直線的に表す図。矢印は、gp160が切断されてgp120およびgp41となる部位を示す。gp120では、斜線の付されたエリアは可変ドメイン(V1〜V5)を表わし、無地のボックスは保存配列(C1〜C5)を示している。gp41エクトドメインでは、いくつかのドメインが示されている、すなわち:N末端融合ペプチド、および2つのエクトドメインへリックス(NへリックスおよびCへリックス)である。膜貫通ドメインは黒色のボックスによって表わされている。gp41細胞質ドメインでは、Tyr‐X‐X‐Leu(YXXL)エンドサイトーシス・モチーフ(配列番号1)および2つの予測へリックスドメイン(ヘリックス‐1および‐2)が示されている。アミノ酸番号が示されている。
【図7】pLW‐73核酸構築物(配列番号2および3)を示す図。
【図8】導入用ベクターpLW‐73のヌクレオチド配列を示す図(上側の鎖、配列番号2;下側の鎖、配列番号3)。
【図9】ウガンダ人のクレードD Envタンパク質(単離物AO7412)をコードしているヌクレオチド配列を示す図(配列番号4)。
【図10】ウガンダ人のクレードD gagpolタンパク質(単離物AO3349)をコードしているコドン変更されたヌクレオチド配列を示す図(配列番号5)。
【図11】組換えMVAの生成および挿入遺伝子の安定性の分析について示す図。A)envおよびgagpolがそれぞれDel II部位およびDel III部位へ挿入されることを示す概略図。B)免疫染色による安定性の評価。
【図12】MVA/65A/G envにおけるenv突然変異の種類および頻度を示す図。
【図13】I8R/G1L IGRにEnv、Del IIIにGagPolが挿入されることを示す図。
【図14】安定性を増大するためのA/G構築物への改変を示す図。
【図15】プラーク継代後のEnvの発現について示す図。
【図16】個々のクローンのPCRおよびウエスタンブロット解析を示す図。
【図17】二重組換えMVAによるA/G envの発現を示す図。
【図18】ウガンダ人のHIV‐1単離物由来のenvおよびgagpolを発現する組換えウイルスを示す図。
【図19】MVA/UGD4a − 非染色envプラークの分析について示す図。
【図20】組換えMVAにおけるUGD env遺伝子の改変を示す図。
【図21】MVA/UGD4b − 非染色gagプラークの分析について示す図。*はGまたはC残基が4〜6個連続した箇所の位置である。
【図22】組換えMVAにおけるUGD gagpol遺伝子の改変を示す図。
【図23】UGDのenvおよびgagpolを発現する安定した組換えMVAの構築を示す図。
【図24】MVA/UGD4dによって誘発された細胞応答を示す図。
【図25】MVA/UGD4dによって誘発された抗体応答を示す図。
【図26】MVAウイルスゲノムのdel III部位を再構築する方法の概要を示す図。
【図27】pLW‐76核酸構築物(配列番号21および22)を示す図。
【図28】del III部位の再構築によるrMVAのシンシチウム表現型を示す図。
【図29】様々な組換えMVAウイルスにおける異種核酸の安定性の比較を示す図。
【図30】様々な組換えMVAウイルスにおけるUGD Envタンパク質発現レベルの比較を示す図。
【図31】導入用ベクターpLW‐76のヌクレオチド配列(配列番号21および22)を示す図。
【発明を実施するための形態】
【0011】
微生物の寄託
次の微生物は、ブダペスト条約の条項に従って記載の日時に米国バージニア州マナッサス所在のアメリカン・タイプ・カルチャー・コレクション(ATCC)に寄託済みである。
【0012】
【表1】
MVA 1974/NIHクローン1は、2003年3月27日、米国バージニア州20110‐2209、マナッサス、ユニバーシティ・ブールバード10801、アメリカン・タイプ・カルチャー・コレクション(ATCC)に、ATCC受託番号PTA‐5095として寄託された。本寄託は、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約およびその規定(ブダペスト条約)に従って行われた。これは、寄託日時から30年間にわたり該寄託物の生存培養物の維持を保証するものである。寄託物はブダペスト条約の条項の下でATCCより入手可能となり、また、関連する米国特許の発行または任意の米国もしくは外国特許出願の公開のいずれか早い方に際して公衆が該寄託物の培養物の子孫を恒久的かつ無制限に入手可能であることを保証し、かつ米国特許法第122条およびこれに関連する長官が定める規則(米国特許法施行規則第1.14条を含む)に従って権利を与えられることが米国特許商標庁長官によって決定された者が該子孫を入手可能であることを保証する、出願人とATCCとの間の合意に従うことになる。寄託された株が入手可能であるということは、任意の政府の権限の下で該政府の特許法に従って与えられた権利に違反して本発明を実行することが許されると解釈されるものではない。
【0013】
好ましい実施形態の詳細な説明
本発明についてさらに説明する前に、了解されるべきことは、本発明は記載される特定の実施形態に限定されるものではなく、それ自体が当然多様であってよいことである。さらに当然ながら、本発明の範囲は特許請求の範囲によってのみ限定されるので、本明細書中で使用される専門用語は特定の実施形態のみについて説明するためのものであって、限定するようには意図されていない。
【0014】
別途定義のないかぎり、本明細書中で使用される技術用語および科学用語は本発明の属する分野の当業者が一般に理解するのと同じ意味を有する。例えば、シングルトン(Singleton)Pおよびセインズベリー(Sainsbury)D.、「微生物学および分子生物学の辞典(Dictionary of Microbiology and Molecular Biology)」第3版、米国ニューヨーク州チチェスター、J.ワイリー・アンド・サンズ(J. Wiley and Sons)、2001年、ならびに、D.M.クナイプ(Knipe)、P.M.ハウリー(Howley)、D.E.グリフィン(Griffin)、R.A.ラム(Lamb)、M.A.マーティン(Martin)、B.ロイズマン(Roizman)、およびS.E.シュトラウス(Straus)編、「フィールズ・ウイルス学(Fields Virology)」第5版、米国フィラデルフィア州フィラデルフィア、リッピンコット・ウィリアムズ・アンド・ウィルキンズ(Lippincott Williams and Wilkins)、2007年)を参照のこと。本発明の実施または試験においては、本明細書に記載のものに類似または等価の任意の方法および材料も使用可能であるが、好適な方法および材料についてここで説明する。本明細書中で言及された出版物はすべて、該出版物が関連して引用された方法または材料のうち少なくともいずれかを開示かつ説明するために、参照により本願に組み込まれる。
【0015】
本発明によれば、単離されたタンパク質(または核酸分子)とは、その天然の環境から取り出されたタンパク質(または核酸分子)である。単離されたタンパク質(または核酸分子)は、例えば、その天然の供給源から得られてもよいし、組換えDNA技術を使用して生産されてもよいし、化学的に合成されてもよい。そのため、単離された、とは、タンパク質または核酸分子が精製された状態や程度を反映するものではない。
【0016】
留意せねばならないのは、本明細書および添付の特許請求の範囲において使用されるように、単数形の「1つの(a)」、「1つの(an)」、および「その(the)」は、文脈がそうでないことを明白に規定していないかぎり、複数の指示物を含むことである。さらに留意すべきは、特許請求の範囲はあらゆる任意選択の要素を除外して作成されうることである。そのため、この記述は、請求項の構成要件の記述に関する「専ら(solely)」、「〜のみ(only)」などのような排他的な用語の使用、または「否定的な(negative)」限定の使用について、先行記述としての役割を果たすことが意図されている。
【0017】
本明細書中で使用されるように、用語「1つの(a)」物または「1つの(an)」物は、1つ以上のその物を指すことが理解されよう。例えば、(1つの(a))核酸分子は1つ以上の核酸分子を指す。そのため、用語「1つの(a)」、「1つの(an)」、「1つ以上の(one or more)」および「少なくとも1つの(at least one)」は互換的に使用可能である。同様に、用語「〜を含む(comprising)」、「〜を含む、〜を備える(including)」および「〜を有する(having)」は互換的に使用可能である。
【0018】
移行語「〜を含む(comprising)」は、「〜を含む、〜を備える(including)」、「〜を含有する(containing)」、または「〜を特徴とする(characterized by)」と同義であり、包括的すなわちオープンエンドであって、記述されていないさらなる構成要件または方法ステップを除外しない。
【0019】
移行句「〜からなる(consisting of)」は、請求項において指定されていないあらゆる構成要件、ステップ、または成分を除外するが、通常付随する不純物のような、本発明とは無関係な追加の構成成分またはステップは除外しない。
【0020】
移行句「本質的に〜からなる(consisting essentially of)」は、請求項の範囲を、指定された材料またはステップ、およびクレームに記載された発明の基礎的かつ新規な特徴に実質的に影響を及ぼさないものに限定する。
【0021】
本明細書中で議論された出版物は、専ら本願の出願日以前の開示について提供される。本明細書は、本発明が先行発明に基づくそのような出版物に先行することは認められないという了解として解釈されるべきではない。さらに、提示された出版日は実際の出版日とは異なる可能性があり、独立に確認する必要がある場合もある。
【0022】
当然のことであるが、明確さのために別個の実施形態において記載されている本発明のある種の特徴が、単一の実施形態において組み合わせて提供されることも可能である。反対に、簡潔にするために単一の実施形態を背景として記載されている本発明の様々な特徴が、別々に、または任意の適切な小さな組み合わせ(サブコンビネーション)で提供されてもよい。実施形態の全ての組み合わせが、本発明によって明確に包含され、かつ、ありとあらゆる組み合わせが個々に明示的に開示されているかのように、本明細書中に開示される。さらに、小さな組み合わせも全て、本発明によって明確に包含され、かつ、ありとあらゆる小さな組み合わせが個々に明示的に本明細書中に開示されているかのように、本明細書中に開示される。
【0023】
全ゲノム塩基配列は、コードポックスウイルスの各々の属のウイルスのうち少なくとも1つについて、および2つのエントモポックスウイルスについて報告されている。ほぼ100個の遺伝子がすべてのコードポックスウイルスにおいて保存されており、これらの約半分がエントモポックスウイルスにも存在する。上記に基づいて、いくつかの一般化を行うことができる。すなわち、遺伝子は、大部分はオーバーラップしておらず、ゲノムの近いほうの端部に向かってブロック単位で存在する傾向があり、高度に保存されていて必須の複製機能に関係している場合は通常は中央領域に位置し、可変的であり宿主との相互作用に関係している場合は通常は端部領域に位置している。中央の遺伝子の配置構成は、すべてのコードポックスウイルスにおいて著しく類似している。ゲノム全体の塩基配列決定以前に始まり、その後ワクシニアウイルスのコペンハーゲン株の完全配列について使用された、ワクシニアウイルスの遺伝子またはORF(オープンリーディングフレーム)の命名のための慣例は、HindIII制限エンドヌクレアーゼによるDNAフラグメントの文字、次にフラグメント内のORF番号(左から右へ)、そして、ORFの方向に応じてLまたはRを使用することからなる。この規則にはHindIII Cフラグメントについて例外が作られた。すなわち、ORFについて、高度に可変的なゲノムの左端から始まるのを避けるため、右から番号付けが行われた。ポリペプチド名は、LまたはRが外される以外は遺伝子名に対応する。ほとんどの後続の完全長ポックスウイルスゲノム配列では、ORFはゲノムの一端から他端へ連続的に番号付けが行われた。しかしながら、文献上の連続性を提供するために古い文字表示が一般名として保持されてきた。ワクシニアウイルスのウエスタン・リザーブ(WR)株のORF番号は、この株が生化学および遺伝学研究の大多数について使用されてきたため、参考図書において共通して示されている。
【0024】
本発明の発明者らは、改変ワクシニアアンカラ(MVA)ウイルスのゲノム内に外来DNA配列の安定な挿入を行うための新たな部位、ならびに新たな部位を作出する方法を特定した。本発明は、安定な組換えMVAウイルスを構築する方法の特定を目的とした研究から生じた。従来観察されてきたように、MVAゲノムに挿入された異種DNA配列を含有している組換えMVAを得ることが可能である一方、これらの挿入物は多くの場合不安定であった。この不安定性の調査から、ORFとORFとの間の場所への、ウイルス増殖に必須でない異種DNA配列の挿入は、同じくウイルスによって欠失させられる恐れがある、という結論が生じた。したがって、安定な組換えMVAウイルスを構築するための改良された戦略の必要性が認識された。
【0025】
本明細書中で使用されるように、オープンリーディングフレーム(ORF)とは、タンパク質のアミノ酸をコードする一続きの連続したヌクレオチドを意味する。そのようなタンパク質はペプチド、ポリペプチドであってよく、単一アミノ酸より大きい任意の長さであってよい。当然のことながら、ORFは、アミノ酸をコードしない終止コドンも含むことができる。当業者には当然のことであるが、組換え事象により、一部のORFはその本来のコード能力の一部を失っており、よって機能を持たないタンパク質をコードしている。そのようなORFはORFフラグメントと呼ばれることがある。ORFは、コード領域の外側に位置する調節要素(例えばプロモーター、転写制御要素、エンハンサーなど)を含まない。それに対し、遺伝子は、ORF(終止コドンを含む)および該ORFの転写を調節することができる調節要素を指す。
【0026】
ORFは、隣り合っているとされる場合もあれば隣り合っていないとされる場合もある。本明細書中で使用されるように、2つのORFは、該ORFが同じ核酸分子中に存在し、かつこれらの最も近接した2つの端部が別のポックスウイルスORFによって隔てられていない場合は、隣り合っている。隣り合っていないORFとは、その最も近接した2つの端部が別のポックスウイルスORFによって隔てられているORFである。隣り合ったORFとは、一方のORFに属する末端コドンと他方のORFに属する末端コドンとの間に他のヌクレオチド配列がないことを意味する、連続したものであってもよい。末端コドンとは、ORFの最初のコドンまたは最後のコドン(終止コドンを含む)を意味する。末端コドンの一例は、ORFによってコードされるタンパク質の最初の5’アミノ酸をコードするコドンである。末端コドンの別の例は、ORFによってコードされるタンパク質の最後の3’アミノ酸をコードするコドンである。末端コドンのさらに別の例は、ORFの終止コドンである。
【0027】
隣り合ったORFが核酸配列によって隔てられている場合もある。そのような配列は遺伝子間領域(intergenic region)と呼ばれる。本明細書中で使用されるように、遺伝子間領域とは、隣り合ったORFの最も近接した末端コドンの間の核酸配列であって、ポックスウイルス転写制御要素以外に、ワクシニアウイルスに由来するヌクレオチド配列を含んでいない配列を意味する。IGR配列は隣り合ったORFの終止コドンの外側に位置し、よってこれらの隣り合ったORFによってコードされるタンパク質のいかなる部分もコードしていない。IGR配列はポックスウイルス転写制御要素を含有する場合がある。IGRは、ポックスウイルス以外の生物に由来する配列を含有する場合もある。好ましくは、IGRは、ポックスウイルス転写制御要素の一部ではないいかなるポックスウイルス配列も有していない。一実施形態では、IGRは少なくとも1つの異種核酸配列を含む。そのような配列は、IGRの中に本来存在するかまたは他の異種核酸配列を挿入する目的でIGRに導入済みの、制限酵素認識部位すなわち制限部位に挿入可能である。
【0028】
ORFのヌクレオチド配列はタンパク質をコードするが、2つのORFの間の遺伝子間領域(IGR)にはコード能力はない。したがってIGRは、いかなるウイルスタンパク質の生産にも影響せずに異種DNAを挿入することができる部位としての役割を果たす可能性がある。しかしながらIGRは、ウイルス遺伝子発現の転写制御に必須であるかまたは関与する、調節要素、結合部位、プロモーター配列またはエンハンサー配列のうち少なくともいずれかを含む場合がある。したがって、IGRは、ウイルスの生活環の調節管理に関与する可能性がある。それでも本発明者は、IGRが、MVAの典型的な特徴および遺伝子発現に影響を及ぼしたり変化させたりせずにMVAゲノムに異種核酸配列を安定的に挿入するために使用可能であることを見出した。この新しい挿入部位は、MVAのORFやコード配列が変更されないので、特に有用である。
【0029】
本発明についてさらに説明する前に、ポックスウイルスゲノムにおける遺伝子の配置について理解することは有用である。ORFのヌクレオチド配列は、規則的に開始コドンで始まり終止コドンで終わる。2つの隣り合ったORFの向きに依存して、これらのORFの間の領域であるIGRは、2つの隣り合ったORFの2つの終止コドンによって挟まれるか、2つの隣り合ったORFの2つの開始コドンによって挟まれるか、第1のORFの終止コドンおよび第2のORFの開始コドンによって挟まれるか、または、第1のORFの開始コドンおよび第2のORFの終止コドンによって挟まれる。
【0030】
従って、IGRへの外来DNA配列の挿入部位は、第1のORFの終止コドンの下流すなわち3’側となる場合がある。第2のORFとも呼ばれる隣りのORFが第1のORFと同じ向きを有する場合、第1のORFの終止コドンの下流のこの挿入部位は、第2のORFの開始コドンの上流すなわち5’側に位置する。
【0031】
第2のORFが第1のORFに対して逆の向きを有する場合、これは2つの隣り合ったORFの向きが互いに向かっていることを意味するが、この場合挿入部位は両ORFの終止コドンの下流に位置する。
【0032】
第3の別例として、2つの隣り合ったORFが逆方向に読まれるが該2つの隣り合ったORFの向きが互いに離れる向きである場合、これは2つのORFの開始コドンが互いに隣り合っていることを特徴とする配置と同義であるが、この場合外来DNAは両方の開始コドンに対して上流に挿入される。
【0033】
MVAゲノム中のORFは2つのコード方向で存在する。従って、mRNA合成活動は、左から右すなわち順方向、および、同様に、右から左(逆方向)に生じる。ORFを、ゲノムの様々なHindIII制限酵素切断フラグメントにおける向きおよび配置によって同定することは、ポックスウイルス学における慣例であり、ワクシニアウイルスの標準的な分類となっている。命名法については、様々なHindIIIフラグメントはそのサイズの大きい順に対応して降順の大文字により示される。ORFは各HindIIIフラグメント上で左から右に番号付けされ、ORFの向きは、大文字のL(右から左への転写を表わす)またはR(左から右への転写を表わす)によって示される。さらに、MVAゲノム構造についてはごく最近の刊行物があるが、該刊行物は、ゲノムの左端から右端へと単純にORFを番号付けし、その向きを大文字のLまたはRで示す、異なる命名法を使用している(アントワーヌ(Antoine)、G.ら、ビロロジー誌(Virology)、1998年、第244巻、p.365−396)。例として、古い命名法によるI8R ORFは、アントワーヌらによる069R ORFに相当する。
【0034】
HIV遺伝子を発現する改変ワクシニアウイルス・アンカラ(MVA)の組換え体をワクチン剤候補として作製する努力において、本発明者らは、不安定性の原因の1つは外来遺伝子および隣接するMVA配列の欠失に起因することを究明した。この問題を克服する試みにおいて、本発明者らは、組換えMVAにおいて生存可能な欠失が生じるのを防止するために、保存された遺伝子の間に外来遺伝子を挿入しようと計画した。そのような欠失を備えたウイルスは生育上有利であり、よってrMVAウイルス集団において異常増殖することになる。ワクシニアゲノム中の保存された遺伝子(これらの遺伝子はワクシニアウイルスの複製に必要と考えられ、したがって「必須遺伝子」である)の間に外来遺伝子を挿入すれば、必須遺伝子のいかなる欠失もウイルス複製を阻害し、したがって組換えMVAを抑えて異常増殖することはないと思われる。したがって、rMVA集団の安定な発現が維持される。組換え体を作製するために本発明者らが使用してきたMVAの株は、該発明者らによって米国疾病管理予防センター(CDC)に提供され、その後アカンビス(Acambis)により塩基配列決定がなされた(Genbank受託番号AY603355)。ババリアン・ノルディック(Bavarian Nordic)が同社の国際公開公報第03/097845号の基礎としてきたMVAの株は、アントワーヌ(Antoine)、G.ら、ビロロジー誌(Virology)、1998年、第244巻、p.365−396で塩基配列決定がなされたワクシニアウイルス株改変ワクシニア・アンカラ(Genbank受託番号U94848)である。(所与の遺伝子についての上記2つの配列における遺伝子番号が異なることに注意されたい。)
本発明者らは当初、ポックスウイルス科において保存された遺伝子およびコードポックスウイルス亜科(脊椎動物のポックスウイルス)において保存された遺伝子に着目していた(アプトン(Upton)、C.ら、ジャーナル・オブ・ビロロジー誌(Journal of Virology)、2003年、第77巻、p.7590−7600)。これらの遺伝子は、ワールドワイドウェブのpoxvirus.orgに見出されるポックスウイルス・バイオインフォマティクス・リソース・センター(Poxvirus Bioinformatics Resource Center)に掲載されたコペンハーゲン・ワクシニアウイルス(Genbank受託番号M35027)の命名法でリストされている。これらの遺伝子は、ポックスウイルス科における合計49個の保存遺伝子と、コードポックスウイルスにおいて保存された41個の追加の遺伝子とであり、合計90個の保存遺伝子である。これらの90個の保存遺伝子から、本発明者らは、保存遺伝子の対の間の遺伝子間部位をリストした。これらの遺伝子対を以下の表1に列挙する。(ババリアン・ノルディックの国際公開公報第03/097845号に含まれていない遺伝子には印が付されていることに注意されたい)。
【0035】
【表2】
これらの遺伝子の向きは様々であり、あるものは右に、あるものは左に向かって転写される。このことは、遺伝子間部位のうちのいくつかは、挿入ベクターの構築において保存されるべきであろうプロモーターを含有することを意味している。さらに、オーバーラップしている保存遺伝子については、ベクター構築の際に、繰り返し配列を最小限にするために代替コドンを使用して遺伝子が再構築されるべきであろう。
【0036】
本発明者らは、保存遺伝子の3’終止コドンが互いに近接した、該遺伝子の向きが「終端対終端」である保存遺伝子に注目した。これらの部位で使用される導入ベクターの構築は、終止コドンと終止コドンとの間のこの領域にはプロモーターは存在しないであろうという事実によって容易になる。終止コドンを隔てる遺伝子間ヌクレオチドが存在する場合、挿入ベクターの構築は簡単である。一方の遺伝子の終止コドンが他方の遺伝子の3’末端の内部にある場合、プラスミド導入ベクターの構築の際に、遺伝子は、繰り返し配列を最小限にするために代替コドンを使用して再構築されてもよいし、または、オーバーラップ部分の長さに応じて、オーバーラップしないように隣接部のPCRにおいて単純に調整されてもよい。表2は、「終端対終端」である保存遺伝子の向きの要件を満たす遺伝子間部位を示す。灰色で強調表示された遺伝子間部位にはオーバーラップしている終端がなく、したがって構築が最も簡単である。
【0037】
【表3】
このリストから、本発明者らは、終端がオーバーラップしていない6つの遺伝子間部位に注目した。実施例においては、これら6つのうち遺伝子間部位071‐072(I8R‐G1L)が、異種遺伝子を挿入する部位として選択された。この遺伝子間部位を使用した組換えMVAウイルスの構築、および得られたウイルスの特徴については、実施例1に、および参照により全体が本願に組み込まれる国際公開公報第2008/142479号A2に、記載されている。
【0038】
上述された保存遺伝子および対応する遺伝子間部位に加えて、本発明者らは異種核酸配列の挿入に有用な他の部位を発見した。例えば、任意の遺伝子であってその欠失(または不活性化)が力価の0.5log、0.75logまたは1log(10倍)の低減をもたらすことが実験的に実証された遺伝子は、「必須遺伝子」と考えることができるかもしれない。同様に、必須遺伝子とは、該当する遺伝子が欠失または不活性化されなかったウイルスと比較して力価の少なくとも50%、少なくとも75%、または少なくとも90%の低減をもたらす任意の遺伝子である。この遺伝子が別の必須遺伝子に隣り合っていれば、2つの遺伝子の間の遺伝子間部位は異種核酸配列の挿入に有用な部位になるであろう。これらのORFの1つ以上が、間に入っている異種核酸配列とともに欠失しても、ウイルスの増殖は妨げないと思われる一方、これらのORFを含有しているウイルスと比較して増殖率の低下がもたらされるであろう。したがって、時間とともに、1つ以上の必須ORFを失ったウイルスは徐々にウイルス集団全体の中でより小さい割合を占めるようになり、十分な時間を経ると、ウイルス集団から完全に消失するであろう。
【0039】
したがって、本発明の1つの実施形態は、組換え改変ワクシニアアンカラ(MVA)ウイルスであって、MVAウイルス由来の2つの隣り合った必須ORFの間に、または該ORFに挟まれた、異種核酸配列を含むウイルスである。一実施形態では、隣り合ったORFは遺伝子間領域(IGR)によって隔てられている。既述のように、IGRは異種核酸配列を含有することができる。したがって、一実施形態は、組換え改変ワクシニアアンカラ(MVA)ウイルスであって、MVAウイルス由来の2つの隣り合った必須ORFの間に、または該ORFに挟まれた、遺伝子間領域内に異種核酸配列を含むウイルスである。
【0040】
本明細書中で使用されるように、異種の、または外来の核酸配列とは、自然界において通常は本発明によって使用されるポックスウイルスに関して見出されることのない配列である。本発明のさらなる実施形態によれば、外来の核酸配列は少なくとも1つのコード配列を含む。該コード配列は、転写制御要素に、好ましくはポックスウイルスの転写制御要素に、作動可能に連結される。さらに、ポックスウイルスの転写制御要素と、例えば内部リボソーム導入部位との間の組み合わせも使用可能である。
【0041】
さらなる実施形態によれば、異種核酸配列は、1個または数個の転写制御要素に連結された2以上のコード配列を含むこともできる。好ましくは、コード配列は1つ以上のタンパク質をコードする。いくつかの実施形態では、該タンパク質は抗原であるか、または、抗原エピトープであって特に治療面から見て興味深い遺伝子のものを含む。
【0042】
本発明による、治療面から見て興味深い遺伝子とは、疾患を引き起こす病原性または感染性の微生物の遺伝子に由来するかまたは相同である遺伝子であってよい。従って、本発明に関しては、そのような治療面から見て興味深い遺伝子は、特異的免疫応答に影響を及ぼす、好ましくは特異的免疫応答を引き起こすために生物体の免疫系に提示され、その結果として、該生物体に、微生物による感染に対するワクチン接種すなわち予防的保護をもたらす。本発明のさらに好ましい実施形態では、治療面から見て興味深い遺伝子は、感染性ウイルス、例えば、限定するものではないが、デング熱ウイルス、B型肝炎ウイルスもしくはC型肝炎ウイルス、またはHIVのようなヒト免疫不全ウイルスの遺伝子から選択される。
【0043】
本発明の好ましい実施形態によれば、異種核酸配列はHIVに由来し、かつHIV envをコードするものであって、好ましくは該HIV env遺伝子は隣り合ったORFの間のIGRに挿入される。後天性免疫不全症候群(AIDS)の病原因子は、ヒト免疫不全ウイルス(HIV)と呼ばれる、レンチウイルス属の典型的な特徴を示すレトロウイルスであることが認識されている。ヒトレンチウイルスの系統関係を図1に示す。HIV‐2は、HIV‐1に対するよりも、野生のスーティーマンガベイから単離されたウイルスであるSIVsmmに対してより近縁な関係にある。現在、HIV‐2はヒトへのSIVsmmの人獣共通感染を表わすものと考えられている。捕獲チンパンジー由来の、SIVcpzと表された一連のレンチウイルス単離物は、HIV‐1の遺伝的近縁種である。
【0044】
最も初期のHIV‐1単離物の系統解析は欧州/北米およびアフリカ由来の試料に集中し、世界のこれら2つのエリアから別々のウイルス群が同定された。その後、HIV‐1の個別の遺伝学的サブタイプまたはクレードが定義されて3つのグループすなわち:M(メジャー);O(アウトライヤー);およびN(MでもOでもない)に分類された(図2)。全世界のウイルス単離物の95%以上が含まれる、HIV‐1のMグループは、全ウイルスゲノムの配列に基づけば、少なくとも8つの別々のクレード(A、B、C、D、F、G、HおよびJ)で構成される。HIV‐1のグループOに属するものは、カメルーン、ガボンおよび赤道ギニアで生活する個体から回収され、これらのゲノムは、グループMのウイルスとはヌクレオチド配列で50%未満の同一性を共有する。さらに最近発見されたグループNのHIV‐1株は、カメルーン人感染者において同定され、標準的な全ウイルスの酵素結合抗体免疫吸着アッセイ(ELISA)では血清学的に反応しないが、従来のウエスタンブロット解析では容易に検出可能である。
【0045】
HIV‐1の遺伝的多様性に関する最新の知見は、多様な地理的起源のグループMのウイルスに関する研究からもたらされている。過去十年間に集められたデータは、個々の感染者に存在するHIV‐1集団がヌクレオチド配列で6%〜10%異なる可能性があることを示している。1つのクレード内のHIV‐1単離物は、gagコード配列で15%およびgp120コード配列で最大30%のヌクレオチド距離を示す場合がある。クレード間の遺伝的多様性は、分析される遺伝子に依存して30%〜40%の範囲となりうる。
【0046】
HIV‐1のグループMサブタイプはすべてアフリカにおいて見出すことができる。クレードAのウイルスは遺伝学的に最も異なっており、流行の初期にはアフリカで最も一般的なHIV‐1サブタイプであった。1990年代半ばから後期の間にHIV‐1がアフリカ南部へ急速に広がるとともに、クレードCのウイルスが主要なサブタイプとなり、現在は世界中のHIV‐1感染の48%を占めている。クレードBのウイルスは最も集中的に研究されたHIV‐1サブタイプであり、欧州および北米において絶えず最も優勢な単離物である。
【0047】
高い遺伝子組換え率はレトロウイルスの特徴である。遺伝学的に相異なるウイルス株による同時感染は、HIV‐1のリスクにさらされた個体において成立しないであろうと当初は考えられていた。しかしながら、1995年までに、HIV‐1グループMの世界規模の多様性のうちかなりの割合が、クレード間のウイルス組換え体を含んでいることが明らかになった。現在では、HIV‐1組換え体は、多数のHIV‐1サブタイプが共存し、かつ流行しているHIV‐1株の10%超を占める可能性のある、アフリカ、南米および東南アジアのような地理的なエリアで見出されるであろうと認識されている。分子レベルでは、これらの組換えウイルスのゲノムは寄せ集めのモザイクに似て、並置された相異なるHIV‐1サブタイプのセグメントを備えており、このことはその世代に寄与する多数の交差事象を反映している。ほとんどのHIV‐1組換え体はアフリカで発生し、大多数は元々クレードAのウイルスに由来するセグメントを含有している。タイ国では、例えば、優勢な流行している株の組成は、クレードAのgag+pol遺伝子セグメントおよびクレードEのenv遺伝子からなる。タイ国のHIV‐1株のクレードEのenv遺伝子は、中央アフリカ共和国由来のウイルス単離物中に存在するクレードEのenvと近縁関係にあるので、最初の組換え事象はアフリカで生じ、その後子孫ウイルスがタイ国に伝わったと考えられる。興味深いことに、完全長のHIV‐1サブタイプE単離物(すなわちサブタイプEのgag、pol、およびenv遺伝子を備えたもの)は現在まで報告されていない。
【0048】
αおよびβケモカイン受容体が感受性CD4+細胞へのウイルスの融合および侵入の共受容体として機能するという発見は、HIV‐1のための改定分類体系をもたらした(図3)。ここで、HIV‐1 gp120およびCD4+共受容体のタンパク質が別々の細胞で発現される融合アッセイにおけるケモカイン受容体の利用に基づいて、単離物をグループ化することができる。図3に示されるように、CXCR4受容体を使用するHIV‐1単離物(ここではX4ウイルスと呼ぶ)は通常はT細胞株(TCL)指向性のシンシチウム誘導型(SI)の株である一方、専らCCR5受容体を利用するもの(R5ウイルス)は主にマクロファージ(M)指向性かつ非シンシチウム誘導型(NSI)である。患者単離物の大多数を含み、かつ一連の指向性表現型を示す二重指向性R5/X4株は、多くの場合SIである。
【0049】
複製能を有するすべてのレトロウイルスの場合のように、3つのHIV‐1一次翻訳産物(すべて構造タンパク質をコードしている)は最初にポリプロテイン前駆体として合成され、該前駆体はその後ウイルスまたは細胞のプロテアーゼによってプロセシングされて成熟粒子関連タンパク質となる(図4)。55kdのGag前駆体であるPr55Gagは、切断されてマトリックス(MA)、カプシド(CA)、ヌクレオカプシド(NC)およびp6タンパク質となる。160kdのGag‐PolポリプロテインであるPr160Gag‐Polの自己触媒反応により、プロテアーゼ(PR)、ヘテロダイマー性逆転写酵素(RT)、およびインテグラーゼ(IN)タンパク質が生じるが、細胞性酵素によるタンパク質分解は、グリコシル化160kd Env前駆体gp160をgp120表面(SU)およびgp41膜貫通(TM)分解産物に変換する。残る6つのHIV‐1にコードされたタンパク質(Vif、Vpr、Tat、Rev、Vpu、およびNef)は、スプライシングされたmRNAの一次翻訳産物である。
【0050】
Gag
HIVのGagタンパク質は、他のレトロウイルスのGagタンパク質と同様に、非感染性のウイルス様粒子の形成にとって必要かつ十分である。レトロウイルスのGagタンパク質は一般にポリプロテイン前駆体として合成され、HIV‐1のGag前駆体はそのみかけの分子量に基づいてPr55Gagと命名された。先に示したように、Pr55GagのmRNAはスプライシングされない9.2kb転写物(図4)であり、該転写物は細胞質中での発現にRevを必要とする。polのORFが存在すると、ウイルスのプロテアーゼ(PR)が細胞からの出芽時または出芽直後にPr55Gagを切断して、成熟Gagタンパク質p17(MA)、p24(CA)、p7(NC)、およびp6を生成する(図4を参照)。ビリオンでは、MAはウイルスエンベロープの脂質二重層のすぐ内側に局在化し、CAは粒子の中心において円錐形のコア構造の外側部分を形成し、NCはウイルスRNAゲノムとのリボ核タンパク質複合体の状態でコアの中に存在する(図5)。
【0051】
HIVのPr55Gag前駆体はその翻訳に続いてオリゴマー化して原形質膜に向かい、原形質膜ではEMで視認可能な十分なサイズおよび密度の粒子がアセンブリされる。Pr55Gagによるウイルス様粒子の形成は、Gag前駆体に沿った多重ドメインの間で生じる重要なGag‐Gag相互作用を伴う、自己アセンブリのプロセスである。ウイルス様粒子のアセンブリには、ゲノムRNA(ただし核酸の存在は不可欠のようである)、polにコードされた酵素、またはEnv糖タンパク質の関与を必要としないが、感染性ビリオンの産生には、ウイルスRNAゲノムのカプシド形成ならびにEnv糖タンパク質およびGag‐Polポリプロテイン前駆体Pr160Gag‐Polの取り込みを必要とする。
【0052】
Pol
gagの下流にはHIVゲノムの最も高度に保存された領域であるpol遺伝子が存在し、3つの酵素すなわちPR、RT、およびINをコードしている(図4を参照)。RTおよびINはそれぞれ、ウイルスRNAゲノムの二本鎖DNAコピーへの逆転写、およびウイルスDNAの宿主細胞染色体内への組込みに必要である。PRは成熟した感染性ビリオンの産生を調節することにより生活環後期において重大な役割を果たす。pol遺伝子産物は、Pr160Gag‐Polと呼ばれる160kdのGag‐Pol融合タンパク質の酵素的切断によって得られる。この融合タンパク質は、Pr55Gag翻訳時のリボソームのフレームシフトによって生じる(図4を参照)。他の多くのレトロウイルスにも利用されている、Gag‐Pol発現のフレームシフト機構は、polから生じるタンパク質が低レベル(Gagのおよそ5%〜10%)で発現されることを確実にする。Pr55Gagと同様に、Pr160Gag‐PolのN末端はミリスチル化されて原形質膜を対象として方向づけられる。
【0053】
プロテアーゼ
鳥類のレトロウイルスを用いて実施された初期のパルス追跡研究は、レトロウイルスのGagタンパク質が当初はポリプロテイン前駆体として合成され、切断されてより小さな産物を生成することを明白に示した。その後の研究から、プロセシング機能は細胞性酵素ではなくウイルス性酵素により提供されること、GagおよびGag‐Pol前駆体のタンパク質分解がウイルスの感染性にとって不可欠であることが実証された。レトロウイルスのPRの配列分析は、PRがペプシンおよびレニンのような細胞の「アスパラギン酸」プロテアーゼとの関連を有することを示した。これらの細胞性酵素と同様に、レトロウイルスのPRは、標的タンパク質中のペプチド結合の加水分解を触媒する水分子を配位するために活性部位の2つの並んだAsp残基を使用する。偽ダイマーとして機能する(活性部位を生成するために同一分子内の2つの折り畳み部分を使用する)細胞性アスパラギン酸プロテアーゼとは異なり、レトロウイルスのPRは真のダイマーとして機能する。HIV‐1 PRのX線結晶学的データは、2つのモノマーが、各モノマーのN末端およびC末端両方に由来する4本の逆平行βシートによって部分的に結びついていることを示している。基質結合部位は2つのモノマーの間に形成された裂溝内にある。細胞のホモログと同様に、HIV PRダイマーは、結合部位の上に張り出して裂溝内の基質を安定させる柔軟な「フラップ」を含有し、活性部位のAsp残基はダイマーの中心にある。興味深いことに、活性部位残基を囲む何らかの限定的なアミノ酸ホモロジーが観察されるが、レトロウイルスのPRの一次配列は非常に多様であり、それでもその構造は非常に類似している。
【0054】
逆転写酵素(Reverse Transcriptase)
定義によれば、レトロウイルスは感染過程の初期段階においてその一本鎖RNAゲノムを二本鎖DNAに変換する能力を有している。この反応を、その関連するRNaseH活性とともに触媒する酵素がRTである。レトロウイルスのRTは3つの酵素活性すなわち(a)RNA依存性のDNA合成(マイナス鎖DNA合成のため)、(b)RNaseH活性(DNA‐RNAハイブリッド中間体に存在するtRNAプライマーおよびゲノムRNAの分解のため)、ならびに(c)DNA依存性のDNA合成(第2鎖またはプラス鎖DNA合成のため)を有する。
【0055】
成熟したHIV‐1 RTホロ酵素は66kdサブユニットおよび51kdサブユニットのヘテロダイマーである。51kdサブユニット(p51)は、66kd(p66)サブユニットから、PRによるp66のC末端15kd RNaseHドメインのタンパク質分解による除去によって生じる(図4を参照)。HIV‐1 RTの結晶構造は、p66サブユニットおよびp51サブユニットの配向が大きく異なっている極めて非対称的なフォールディングを示している。p66サブユニットは右手として視覚化することが可能であり、手のひらの内部にポリメラーゼ活性部位を、かつ手のひら、指、および親指のサブドメインによって形成された鋳型結合性の深い裂溝を備えている。ポリメラーゼドメインは接続サブドメインによってRNaseHに連結される。手のひらに位置する活性部位は、近接した3つの重要なAsp残基(110、185および186)と、2つの配位されたMg2+イオンとを含有している。これらのAsp残基の突然変異はRTの重合活性を消失させる。3つの活性部位Asp残基の配向は、他のDNAポリメラーゼ(例えば大腸菌のDNA polIのクレノウ・フラグメント)で観察されるものと同様である。p51サブユニットは剛性のようであり、重合用の裂溝を形成せず、このサブユニットのAsp110、185および186は分子内部に埋まっている。プライマー‐鋳型二重鎖のおよそ18塩基対が、核酸結合性の裂溝内にあり、ポリメラーゼ活性部位からRNaseHドメインへと伸びている。
【0056】
RT‐プライマー‐鋳型‐dNTP構造物では、プライマーの3’末端にジデオキシヌクレオチドが存在することにより、新たに入って来るdNTPに対して反応開始する直前に捕捉された触媒複合体の視覚化が可能となる。先に得られた構造物との比較から、新たなdNTPに対するプライマーの3’‐OHの求核攻撃に先立って鋳型およびdNTPを捕捉するために、指サブドメインが閉じるモデルが示唆される。成長している鎖に新たなdNTPが付加された後は、指サブドメインがより大きく開いた形態をとることにより、ピロリン酸を放出し、かつRTが次のdNTPに結合するのを可能にすることが提唱されている。HIV‐1 RNaseHの構造もX線結晶構造解析によって決定されており、このドメインは大腸菌のRNaseHに類似の球状のフォールディングを示す。
【0057】
インテグラーゼ
レトロウイルスの複製の卓越した特徴は、逆転写に続いて宿主細胞の染色体内にウイルスゲノムのDNAコピーが挿入されることである。この組込まれたウイルスDNA(プロウイルス)は、ウイルスRNA合成のための鋳型としての役割を果たし、感染細胞が生存している間は宿主細胞ゲノムの一部として維持される。組込み能力が不完全なレトロウイルス突然変異体は一般に増殖性感染を確立できない。
【0058】
ウイルスDNAの組込みは、HIV‐1 Gag‐PolポリプロテインのC末端部分がPRにより切断されて生成される32kdタンパク質であるインテグラーゼ(図4を参照)によって触媒される。
【0059】
レトロウイルスのINタンパク質は、構造的かつ機能的に異なる3つのドメインすなわち、N末端のジンクフィンガー含有ドメイン、コアドメイン、および比較的非保存的なC末端ドメイン、から構成されている。その溶解度が低いことから、288アミノ酸のHIV‐1 INタンパク質全体を結晶化することは未だ可能ではない。しかしながら、3つのドメインすべての構造がX線結晶構造解析またはNMR法によって独立に解明されている。トリ肉腫ウイルスのINのコアドメインの結晶構造も決定済みである。N末端ドメイン(残基1〜55)の構造はNMR分光法によって解明されたが、該ドメインはアミノ酸His‐12、His‐16、Cys‐40、およびCys‐43によって配位された亜鉛を備えた4つのヘリックスから構成されている。このN末端ドメインの構造は、いわゆるヘリックス・ターン・ヘリックス・モチーフを含むヘリックス系DNA結合タンパク質を連想させるが、HIV‐1構造においては、このモチーフはダイマー形成に寄与している。当初は、難溶性であることがコアドメインの構造を解明する努力の妨げとなった。しかしながら、INの残基185におけるPheからLysへの変更がin vitroでの触媒活性を妨げることなく溶解度を大幅に高めることが観察された時、結晶構造解析の試みは成功した。HIV‐1 INのコアドメイン(IN残基50〜212)の各モノマーは、ヘリックスに挟まれた5本ストランドのβシートから構成されており、この構造は、RNaseHおよびバクテリオファージMuAトランスポサーゼを含む他のポリヌクレオチジルトランスフェラーゼに非常に類似している。高度に保存された3つの残基が、他のポリヌクレオチジルトランスフェラーゼの類似の位置に見出されており、HIV‐1 INではこれらはAsp64、Asp116およびGlu152、いわゆるD,D‐35‐Eモチーフである。これらの位置における突然変異はin vivoおよびin vitroいずれにおいてもHIV INの機能を妨げる。トリ肉腫ウイルスおよびHIV‐1のいずれのコアドメインの結晶構造においても上記の3つのアミノ酸が近接していることは、これらの残基が、組込みプロセスの中心にあるポリヌクレオチド転移反応の触媒作用において中心的な役割を果たすという仮説を支持している。C末端ドメインの構造はNMR法によって解明済みであるが、該ドメインはSrcホモロジー3(SH3)ドメインを連想させる5本ストランドのβバレルのフォールディングトポロジーをとる。最近になって、コアドメインおよびC末端ドメインの両方を包含しているSIVおよびラウス肉腫ウイルスのINタンパク質フラグメントのX線構造が解明された。
【0060】
Env
HIV Env糖タンパク質はウイルスの生活環において主要な役割を果たす。該タンパク質は、CD4受容体および共受容体と相互作用する決定基を含有し、またウイルスエンベロープの脂質二重層と宿主細胞原形質膜との間の融合反応を触媒する。さらに、HIV Env糖タンパク質は、診断薬およびワクチンのいずれの開発の観点からも重要な免疫応答を誘発するエピトープを含有している。
【0061】
HIV Env糖タンパク質は、単独にスプライシングされた4.3kbのVpu/Envバイシストロン性mRNAから合成され(図4を参照)、翻訳は粗面小胞体(ER)に結合したリボソームで行われる。160kdポリプロテイン前駆体(gp160)は、成熟したTM Env糖タンパク質gp41となるように予定されているドメインの疎水性の輸送停止シグナルによって細胞膜につなぎ留められる、内在性膜タンパク質である(図6)。gp160は翻訳時にグリコシル化され、ジスルフィド結合を形成し、ERにおいてオリゴマー化する。主なオリゴマー型はトライマーのようであるが、ダイマーおよびテトラマーも観察される。gp160はゴルジ体に輸送され、ゴルジ体では、他のレトロウイルスのエンベロープ前駆体タンパク質と同様に、細胞性酵素によりタンパク質分解によって切断されて、成熟したSU糖タンパク質gp120およびTM糖タンパク質gp41となる(図6を参照)。高度に保存されたLys/Arg‐X‐Lys/Arg‐Argモチーフ直後のレトロウイルスEnv前駆体の切断に関与する細胞性酵素は、フーリンまたはフーリン様プロテアーゼであるが、その他の酵素がgp160のプロセシングを触媒する場合もある。gp160の切断はEnv誘導型の融合活性およびウイルス感染性に必要である。gp160の切断に続いて、gp120およびgp41は、ゴルジ体から細胞表面へのEnv複合体の輸送に重要な非共有結合性会合を形成する。gp120‐gp41相互作用は非常に弱く、相当量のgp120が、Envを発現している細胞の表面から流される。
【0062】
HIV Env糖タンパク質複合体、特にSU(gp120)ドメイン)は、非常に強くグリコシル化されており、gp160の分子量のおよそ半分はオリゴ糖側鎖で構成されている。ER内のEnv合成部位から原形質膜までのEnvの輸送中に、側鎖の多くが複合糖類の付加によって修飾される。多数のオリゴ糖側鎖は、宿主の免疫認識からgp120の大部分を遮る糖の雲として想像できるようなものを形成する。図6に示されるように、gp120は、散在する保存された(C1〜C5)ドメインおよび可変(V1〜V5)ドメインを含有している。様々な単離物のgp120に存在するシステイン残基は高度に保存されており、大きなループの中の最初の4つの可変領域を連結するジスルフィド結合を形成する。
【0063】
ウイルスのEnv糖タンパク質の主要な機能は、ウイルスエンベロープの脂質二重層と宿主細胞の細胞膜との間の膜融合反応を促進することである。この膜融合事象はウイルスのコアが宿主細胞の細胞質内に入り込むのを可能にする。gp120およびgp41の両方の多くの領域が、Envを介した膜融合に直接または間接的に関与するとされてきた。オルトミクソウイルスのHA2ヘマグルチニンタンパク質およびパラミクソウイルスのFタンパク質に関する研究から、これらのタンパク質のN末端の高疎水性ドメインは、融合ペプチドと呼ばれ、膜融合に重大な役割を果たすことが示された。突然変異解析は、HIV‐1、HIV‐2およびSIVのTM糖タンパク質のN末端に相似ドメインがあることを実証した(図6を参照)。gp41のこの領域内の非疎水性置換はシンシチウム形成を大幅に低減または阻害し、その結果非感染性の子孫ビリオンを生じた。
【0064】
gp41融合ペプチドのC末端側は、膜融合において中心的な役割を果たす2つの両親媒性へリックスドメインである(図6を参照)。Leuジッパー様の7アミノ酸繰り返しモチーフを含有するN末端へリックス(N‐へリックスと呼ばれる)における突然変異は、感染性および膜融合活性を低下させ、これらの配列に由来するペプチドは培養物中で強力な抗ウイルス活性を示す。HIV‐1およびSIVのgp41のエクトドメインの構造、特に2つのへリックスモチーフは、近年の構造分析の中心となってきた。へリックスドメインを含有する融合タンパク質、N‐ヘリックスおよびC‐ヘリックスに由来するペプチドの混合物、または、SIV構造の場合には残基27〜149の完全なgp41エクトドメイン配列、のいずれかについて、X線結晶構造解析またはNMR分光法により構造が決定された。これらの研究からは、基本的に同様のトライマー構造であって2つのヘリックスドメインが逆平行方式でパッキングされて6ヘリックス束を生成している構造が得られた。N‐へリックスは該ヘリックス束の中心にコイルドコイルを形成するとともに、C‐へリックスは外側の疎水性の溝の中にパッキングされる。
【0065】
膜融合をもたらすステップでは、CD4の結合がEnvの立体配座変化を引き起こし、該変化が共受容体の結合を促進する。gp120/CD4/共受容体三重複合体の形成に続いて、gp41は、融合ペプチドが標的の脂質二重層内に挿入されるのを可能にする仮説上の立体配座をとる。gp41の6ヘリックス束(gp41のN‐ヘリックスとC‐ヘリックスとの間の逆平行相互作用を伴っている)の形成はウイルスおよび細胞の膜を結び付け、膜融合が生じる。
【0066】
さらに、本発明による治療面から見て興味深い遺伝子は、増殖性疾患、がんまたは代謝疾患に対して治療効果を有する疾患関連遺伝子も含む。例えば、がんに関する治療面から見て興味深い遺伝子とは、特異的な抗がん性免疫反応を誘導する能力を有するがん抗原であることが考えられる。
【0067】
本発明のさらなる実施形態によれば、異種核酸配列は少なくとも1つのマーカー遺伝子または選択遺伝子を含む。
選択遺伝子は、細胞に対して特定の耐性を形質導入することにより、ある種の淘汰法が可能となる。当業者は、ポックスウイルスの系において使用可能な様々な選択遺伝子に精通している。その中には、例えばネオマイシン耐性遺伝子(NPT)またはホスホリボシルトランスフェラーゼ遺伝子(gpt)がある。
【0068】
マーカー遺伝子は、形質導入細胞において発色反応を引き起こし、該反応は形質導入細胞を同定するために使用することができる。当業者は、ポックスウイルスの系において使用可能な様々なマーカー遺伝子に精通している。その中には、例えばβ‐ガラクトシダーゼ(β‐gal)、β‐グルコシダーゼ(β‐glu)、緑色蛍光タンパク質(EGFP)、または青色蛍光タンパク質をコードする遺伝子がある。
【0069】
本発明のさらに別の実施形態によれば、異種核酸配列は、ポックスウイルスの転写制御要素および/または異種核酸配列中のコード配列を、隣り合うORFの終止コドンおよび/または開始コドンから隔てる、スペーシング配列を含む。隣り合うORFの終止/開始コドンと異種核酸配列中の挿入コード配列との間のこのスペーサー配列は、挿入された異種核酸配列を、したがって得られる任意の組換えウイルスを、安定化するという長所を有する。スペーサー配列のサイズは、該配列がそれ自体のコード機能または制御機能を持たない限り、様々であってよい。
【0070】
さらなる実施形態によれば、ポックスウイルスの転写制御要素および/または異種核酸配列中のコード配列を隣り合うORFの終止コドンから隔てるスペーサー配列は、少なくとも1ヌクレオチドの長さである。
【0071】
本発明の別の実施形態によれば、ポックスウイルスの転写制御要素および/または異種核酸配列中のコード配列を隣り合うORFの開始コドンから隔てるスペーシング配列は、少なくとも30ヌクレオチドである。特に、典型的なワクシニアウイルスのプロモーター要素が開始コドンの上流に確認される場合、異種核酸配列の挿入は、該プロモーター要素を隣り合うORFの開始コドンから隔てることができない可能性がある。典型的なワクシニアプロモーター要素は、例えば後期プロモーターについては配列「TAAAT」(デービソン(Davison)およびモス(Moss)、ジャーナル・オブ・モレキュラー・バイオロジー誌(J. Mol. Biol.)、1989年、第210巻、p.771−784)および早期プロモーターについてはA/Tリッチドメインを精査することにより同定可能である。約30ヌクレオチドのスペーシング配列は、ORFの開始コドンの上流に位置するポックスウイルスのプロモーターが影響を受けないことを確実にする、好ましい距離である。加えて、さらに好ましい実施形態によれば、挿入された異種核酸配列と隣り合うORFの開始コドンとの間の距離は約50ヌクレオチド、より好ましくは100ヌクレオチドである。
【0072】
本発明のさらに好ましい実施形態によれば、スペーシング配列は、隣りのORFの転写を制御することができる追加のポックスウイルス転写制御要素を含む。
ここまで、本開示は親MVAウイルス中で隣り合っているORFを使用した組換えMVAウイルスに注目してきた。しかしながら、本発明はさらに、組換えウイルス、およびそのようなウイルスを作製する方法であって、異種核酸配列が隣り合った必須ORF技術の間に挿入され、挿入に使用されるORfは親MVAウイルス中で隣り合っていないものも含む。すなわち、ウイルスは、組換えMVAウイルスにおいて隣り合っているORFが親MVAウイルス中では1つ以上のポックスウイルスORF(介在ORF)によって隔てられているように、構築可能である。本明細書中で使用されるように、親MVAウイルスとは、子孫の組換えウイルスが構築される元のウイルスである。親MVAウイルスの一例は、MVA 1974/NIHクローン1である。親ウイルスを使用して、本明細書中に開示された技術を使用して組換えウイルスを構築し、構築プロセスの間に介在ORFが除去可能であるようにすることができる。ポックスウイルス分野の当業者には当然のことであるが、注意深く選択されたポックスウイルスORFを含む核酸分子を使用することにより、それら2つのORFの間のウイルスゲノムの一部を、相同組換えのプロセスを通じて欠失させることが可能である。例えば、非必須ORFを含有している1キロベースのゲノム領域によって2つの必須ORFが隔てられていることを前提として、その2つの必須ORFが、例えばプラスミド内で隣り合うようにクローニングされた核酸構築物を作製可能である。該核酸構築物がポックスウイルス感染細胞(例えば親MVAウイルス感染細胞)に導入されると、必須ORFは親ウイルスのウイルスゲノム中の対応するORFとの組換えを行うことになる。当業者には理解されるさらなる組換え事象を通して、1キロベースの領域はウイルスゲノムから切除され、その結果2つの必須ORFが隣り合うようになる。したがって、本発明の1つの実施形態は、MVAウイルスゲノム由来の2つの隣り合った必須ORFの間に位置する異種核酸配列を含む組換え改変ワクシニアアンカラ(MVA)ウイルスであって、親MVAウイルス中の対応する必須ORFの間に存在する非必須ORFを欠くことを特徴とする組換えMVAウイルスである。したがって、異種核酸配列は、親MVAウイルス中においては隣り合っていない必須ORFによって挟まれている。必須ORFは、非必須ORFによって隔てられた、MVAゲノム中に存在する必須ORF対から選択される。一実施形態では、必須ORFは、A50R(MVA163)、B1R(MVA167)、F10(MVA‐039)、F12(MVA042)、F13L(MVA043)、F15L(MVA045)、F17L(MVA047)、E4L(MVA051)、E6L(MVA053)、E8L(MVA055)、E10L(MVA057)、I1L(MVA062)、I3L(MVA064)、I5L(MVA066)、J1R(MVA085)、J3R(MVA087)、D7L(MVA104)、D9L(MVA106)、A24R(MVA135)、およびA28R(MVA139)からなる群から選択される。一実施形態では、2つの必須ORFは、A50R‐B1R(MVA163‐MVA167)、F10‐F12(MVA039‐MVA042)、F13L‐F15L(MVA043‐MVA045)、F15L‐F17L(MVA045‐MVA047)、E4L‐E6L(MVA051‐MVA053)、E6L‐E8L(MVA053‐MVA055)、E10L‐I1L(MVA057‐MVA062)、I3L‐I5L(MVA064‐MVA066)、J1R‐J3R(MVA085‐MVA087)、D7L‐D9L(MVA104‐MVA106)、およびA24R‐A28R(MVA135‐MVA139)からなる群の中の必須ORF対から選択される。一実施形態では、必須ORFはA50R(MVA163)およびB1R(MVA167)から選択される。一実施形態では、一方の必須ORFはA50R(MVA163)であり、他方の必須ORFはB1R(MVA167)である。
【0073】
先に議論されたように、細胞培養物中での大規模な継代の結果、MVAウイルスゲノムは、Del I、II、II、IV、VおよびVIと呼ばれる6つの主な欠失を含有している。歴史的には、およそ31,000ヌクレオチドの欠失であるDel III周辺の領域が異種核酸配列の挿入に使用されてきた。したがって、本発明の1つの実施形態では、組換えMVAウイルスの構築の際に欠失させられる非必須ORFは、野生型MVAウイルス中でDel III領域に隣接している。
【0074】
説明してきたように、組換えMVAウイルスは、2つの隣り合った必須ORFの間に、IGRおよび/または異種核酸配列のような追加の配列を含有することができる。そのような配列は本明細書中に記載されている。したがって、本発明の1つの実施形態は、MVAウイルスゲノム由来の2つの隣り合った必須ORFの間に位置する異種核酸配列を含む組換え改変ワクシニアアンカラ(MVA)ウイルスであって、親MVAウイルス中の対応する必須ORFの間に存在する非必須ORFを欠くこと、および異種核酸配列がIGRに挿入されていることを特徴とする組換えMVAウイルスである。本開示において別記されているように、異種核酸配列は、転写制御要素に制御されるコード配列を含有することができる。
【0075】
本発明者らは特定の必須ORFおよびその配列について開示してきたが、本発明は、開示されたORFの一部またはバリアントを使用する組換えMVAウイルスおよび該組換えMVAウイルスを作製する方法も含む。例えば、本発明はORF A50Rおよびその一部(配列番号11および配列番号14)、ならびにORF B1Rおよびその一部(配列番号16および配列番号19)を開示しているが、本発明は、これらの配列のバリアントを含む組換えMVAウイルスを、そのバリアントORFが対応する野生型ORFによってコードされるタンパク質とほぼ同じ機能を有するタンパク質をコードする限りにおいて、含む。2つのタンパク質は、それぞれのタンパク質を含むMVAウイルスが、同じ細胞株を使用して増殖させたとき互いに対して約10%、約20%、約30%または約40%以内の力価を生じる場合に、ほぼ同じ機能を有すると見なされる。したがって、本発明の1つの実施形態は、2つの隣り合うORFの間に位置する異種核酸配列を含む組換え改変ワクシニアアンカラ(MVA)ウイルスであって、該隣り合うORFはMVA由来の必須ORFと少なくとも90%、少なくとも95%、少なくとも97%または少なくとも99%の配列同一性のヌクレオチド配列を含むことを特徴とする。一実施形態では、隣り合うORFは、A50R(MVA163)、B1R(MVA167)、F10(MVA‐039)、F12(MVA042)、F13L(MVA043)、F15L(MVA045)、F17L(MVA047)、E4L(MVA051)、E6L(MVA053)、E8L(MVA055)、E10L(MVA057)、I1L(MVA062)、I3L(MVA064)、I5L(MVA066)、J1R(MVA085)、J3R(MVA087)、D7L(MVA104)、D9L(MVA106)、A24R(MVA135)、およびA28R(MVA139)からなる群から選択された必須ORFに対して少なくとも90%、少なくとも95%、少なくとも97%または少なくとも99%の同一性を有するヌクレオチド配列を含む。好ましい実施形態では、2つの隣り合うORfは同じ必須ORFに由来するものではない。一実施形態では、2つの隣り合うORFは、A50R‐B1R(MVA163‐MVA167)、F10‐F12(MVA039‐MVA042)、F13L‐F15L(MVA043‐MVA045)、F15L‐F17L(MVA045‐MVA047)、E4L‐E6L(MVA051‐MVA053)、E6L‐E8L(MVA053‐MVA055)、E10L‐I1L(MVA057‐MVA062)、I3L‐I5L(MVA064‐MVA066)、J1R‐J3R(MVA085‐MVA087)、D7L‐D9L(MVA104‐MVA106)、およびA24R‐A28R(MVA135‐MVA139)からなる群の必須ORF対と少なくとも90%、少なくとも95%、少なくとも97%または少なくとも99%の同一性を有するヌクレオチド配列を含む。一実施形態では、一方の隣り合うORFは配列番号A50R(MVA163)と少なくとも90%、少なくとも95%、少なくとも97%または少なくとも99%の配列同一性のヌクレオチド配列を含み、他方の隣り合うORFは、第2の必須ORFと少なくとも90%、少なくとも95%、少なくとも97%または少なくとも99%の配列同一性のヌクレオチド配列を含む。一実施形態では、一方の隣り合うORFは配列番号B1Rと少なくとも90%、少なくとも95%、少なくとも97%または少なくとも99%の配列同一性のヌクレオチド配列を含む。
【0076】
本発明はさらに、本発明の組換えウイルスを生産するのに有用な核酸構築物も開示する。本明細書中で使用されるように、核酸構築物は、MVAウイルス由来の少なくとも1つの必須ORFの少なくとも一部を含む組換え核酸分子である。該核酸構築物は、環境内、例えば限定するものではないが生物体内、組織内、または細胞培養物中の細胞への、有用な核酸配列の輸送を可能にする。本開示の核酸構築物はヒトの介在によって生産される。該核酸構築物は、DNA、RNAまたはそのバリアントであってよい。核酸分子は、直鎖DNA、DNAプラスミド、ウイルスベクター、またはその他のベクターであってよい。一実施形態では、核酸分子はDNAプラスミドであってよい。一実施形態では、核酸分子は、ウイルス成分、プラスミド成分、転写制御要素、ならびに核酸分子の送達および発現を可能にする当業者に周知の任意のその他の有用な要素を含むDNAプラスミドであってよい。組換え核酸分子の一般的な構築のための方法は良く知られている。例えば、サムブルック(Sambrook)ら、「分子クローニング:実験の手引き(Molecular Cloning: a Laboratory Manual)」第3版、コールド・スプリング・ハーバー・ラボラトリー・プレス(Cold Spring Harbor Laboratory Press)、2001年、およびオースベル(Ausubel)ら編、「分子生物学における最新プロトコール(Current Protocols in Molecular Biology)」、ジョン・ワイリー・アンド・サンズ(John Wiley and Sons)、1994年を参照されたい。
【0077】
本発明の1つの実施形態は、単離された核酸構築物であって、(a)改変ワクシニアアンカラ(MVA)ウイルスゲノム由来の第1の必須ORFに由来するかまたは該ORFに相同な第1の核酸配列と、(b)MVAウイルスゲノム由来の第2の必須ORFに由来するかまたは該ORFに相同な第2の核酸配列とを含み、第1および第2の必須MVAウイルスORFは、MVAウイルスゲノム中では少なくとも1つの非必須ORFによって隔てられていることと、第1および第2の核酸配列は単離された核酸構築物中では互いに隣り合っていることと、第1および第2の核酸配列は、それぞれ第1および第2の必須MVA ORF由来の少なくとも25個連続したヌクレオチドを含むことと、を特徴とする。そのような核酸構築物は、相同組換えのプロセスを通じて組換えMVAウイルスを構築するのに有用である。このプロセスを使用して、本発明の単離された核酸構築物は、親MVAウイルスにおいては隣り合っていない(すなわち他の非必須MVA ORFによって隔てられている)ORFが、子孫である組換えMVAウイルスにおいては隣り合わせにされる組換えMVAウイルスを構築するために使用可能である。これは、例えば、親MVAウイルス由来の隣り合っていないORFをプラスミドのような核酸分子中へクローニングし、かつその間にある非必須ORFのクローニングは伴わないことにより、行うことができる。こうして、隣り合っていないORFは核酸構築物中で隣り合わせにされる。説明してきたように、MVAウイルスゲノムの中にそのような核酸構築物を組み換えることにより、間に存在する親MVAウイルス由来の非必須ORFの欠失がもたらされ、その結果として、もとは隣り合っていないORFが隣り合っている子孫組換えMVAウイルスが生じることになる。したがって、好ましい実施形態では、第1および第2の核酸配列は、親MVAウイルスにおいては隣り合っていない第1および第2の必須MVA ORFにそれぞれ由来するかまたは該ORFと相同である。すなわち、第1および第2の必須ORFは、親MVAウイルスゲノム中では少なくとも1つの非必須ORFによって隔てられている。
【0078】
本明細書中で使用されるように、「〜に由来する」という語句は、その核酸配列が得られた起源の核酸(すなわちORF)を指す。したがって、この点では、核酸配列は起源となるORFの全部または一部と同一であってよい。しかしながら、核酸配列は起源となるORFとは配列が異なっていてもよい。したがって、MVA ORFに由来する核酸配列は、得られた核酸配列において元のORFの機能が維持される限り、MVA ORFの全部(または一部)と配列が同一であっても同一でなくてもよい。例えば、当分野では当然のことであるが、ポックスウイルスの近縁種の核酸分子は、そのような分子の配列が同一でなくても、組換えを起こすことができる。したがって、本発明の1つの実施形態では、第1および第2の核酸配列は、該核酸配列の由来する元の必須MVA ORFとの十分な配列同一性を有し、第1または第2の核酸配列を含む核酸分子と、そのような配列の由来する元の必須MVA ORFを含む核酸分子との間の相同組換えを可能にする。一実施形態では、第1および第2の核酸配列は、該核酸配列の由来する元の必須MVA ORFの少なくとも一部に対して、少なくとも75%、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも97%、または少なくとも99%同一である。一実施形態では、核酸配列は、該核酸配列の由来する元の必須MVA ORFの少なくとも一部と同一である。
【0079】
同様に当分野では当然のことであるが、小さなポリヌクレオチド分子でも相同組換えの過程に関与することができる。従って、本発明の核酸構築物中に存在する核酸配列は、該核酸構築物がMVAウイルスゲノム中への組換えを生じうるために必須MVA ORFの全配列を含む必要はない。実際、長さ20塩基相応の小さなポックスウイルスゲノムのフラグメントが、ウイルスゲノム中のそれぞれの配列との相同組換えに関与することが可能であることが示されている。したがって、本発明の1つの実施形態では、第1および第2の核酸配列は、必須MVA ORFからの25、30、35、40、45、50、100、150、200、250、または300ヌクレオチドを含むことができる。本発明の1つの実施形態は、単離された核酸構築物であって、(a)第1の必須MVA ORFからの少なくとも25個連続したヌクレオチドを含む第1の核酸配列と、(b)第2の必須MVA ORFからの少なくとも25個連続したヌクレオチドを含む第2の核酸配列とを含み、第1および第2の必須MVAウイルスORFは、MVAウイルスゲノム中では少なくとも1つの非必須ORFによって隔てられていること、ならびに第1および第2の核酸配列は単離された核酸構築物中では互いに隣り合っていることを特徴とする。一実施形態では、第1の核酸配列は、A50R(MVA163)、B1R(MVA167)、F10(MVA‐039)、F12(MVA042)、F13L(MVA043)、F15L(MVA045)、F17L(MVA047)、E4L(MVA051)、E6L(MVA053)、E8L(MVA055)、E10L(MVA057)、I1L(MVA062)、I3L(MVA064)、I5L(MVA066)、J1R(MVA085)、J3R(MVA087)、D7L(MVA104)、D9L(MVA106)、A24R(MVA135)、およびA28R(MVA139)からなる群から選択された必須ORFからの25個連続したヌクレオチドを含む。一実施形態では、第2の核酸配列は、A50R(MVA163)、B1R(MVA167)、F10(MVA‐039)、F12(MVA042)、F13L(MVA043)、F15L(MVA045)、F17L(MVA047)、E4L(MVA051)、E6L(MVA053)、E8L(MVA055)、E10L(MVA057)、I1L(MVA062)、I3L(MVA064)、I5L(MVA066)、J1R(MVA085)、J3R(MVA087)、D7L(MVA104)、D9L(MVA106)、A24R(MVA135)、およびA28R(MVA139)からなる群から選択された必須ORFからの25個連続したヌクレオチドを含む。一実施形態では、第1の核酸配列は、配列番号11または配列番号14からの少なくとも25個連続したヌクレオチドを含み、第2の核酸配列は、配列番号16または配列番号19からの少なくとも25個連続したヌクレオチドを含む。
【0080】
本発明の核酸構築物は、MVAウイルスのゲノムに異種核酸配列を送達するために使用される。したがって、一実施形態、本発明の核酸構築物は、第1および第2の核酸配列の間に異種核酸分子を含む。典型的な異種核酸配列は本開示において別記されている。本明細書中に開示されたいずれの異種核酸配列も本発明の核酸構築物に含めるのに適している。
【0081】
本発明の核酸構築物は親MVAウイルスのゲノムと組換えを行うことができるので、ウイルスゲノムに異種核酸配列を挿入するために使用可能である。したがって、本発明の1つの実施形態では、本発明の核酸コントラスト(nucleic acid contrast)は、第1および第2の核酸配列の間に遺伝子間領域を含有している。遺伝子間領域は、転写制御要素、制限部位および非ワクシニアのオープンリーディングフレームのようなものを含むことができる。したがって、遺伝子間領域は転写制御要素に制御される遺伝子を含む異種核酸配列を挿入するために使用可能である。核酸構築物がMVAウイルスゲノムとの組換えを行うと、異種核酸配列は、MVAウイルスゲノム中の、核酸構築物中において該核酸配列を挟んでいる2つの隣り合った必須ORFに対応する必須ORFの間に、挿入されることになる。得られるMVAウイルスは、MVAウイルスゲノム中に安定的に組み込まれた異種核酸配列を含有している組換えMVAウイルスとなる。
【0082】
一実施形態では、本発明の核酸構築物は、ウイルスゲノムの2つの隣り合ったORFの間に位置するIGR配列の完全長フラグメントまたは部分フラグメントを含む。好ましくは、核酸構築物は、前記IGRに由来する配列中に挿入された少なくとも1つのクローニング部位であって、対象とする異種DNA配列を挿入するための、および好ましくは前記異種DNA配列に作動可能に連結されたポックスウイルス転写制御要素を挿入するための、クローニング部位を含む。任意選択で、核酸構築物はレポーター遺伝子カセットおよび/または選択遺伝子カセットを含む。核酸構築物は、好ましくは、IGR配列の前記完全長フラグメントまたは部分フラグメントに隣接している2つの隣り合ったORFの配列も含む。
【0083】
ヌクレオチド配列を含まないいくつかのIGRが同定されている。これらの場合、プラスミドベクターは、IGRに隣接している配列のDNA配列(すなわち2つの隣り合ったORFのDNA配列)を含む。好ましくは、異種DNA配列の挿入用のクローニング部位がIGRに挿入される。IGRに隣接している配列のDNAは、MVAゲノムの対応するIGR中へ外来DNA配列を挿入するために使用される。そのようなプラスミドベクターは、異種DNA配列と、任意選択でレポーター遺伝子カセットおよび/または選択遺伝子カセットとを挿入するためのクローニング部位を含むIGR配列の完全長フラグメントまたは部分フラグメントをさらに含むことができる。
【0084】
本発明の一実施形態は、安定な組換え改変ワクシニアアンカラウイルスを作製する方法である。そのような方法は、本明細書中に開示された核酸構築物を使用する。したがって該方法は、第1に、MVAウイルスゲノムの2つの隣り合った必須オープンリーディングフレーム(ORF)の間にあるかまたは該ORFに挟まれた異種核酸配列を含む核酸構築物を得る工程を含み、該MVAウイルスは、親のMVAウイルス中の対応する2つの必須ORFの間に存在する非必須ORFまたはORFフラグメントを欠いている。例えば、適切な核酸構築物を得るために、必須MVA ORFの核酸配列が単離されてpBluescript(ストラタジーン(Stratagene))のような標準的なクローニングベクターにクローニングされ、該核酸配列がMVAゲノムに挿入されるべき異種DNAに隣接するようにされてもよい。その後、この構築物は、当業者に周知の方法(例えばトランスフェクション)を使用して細胞に導入可能である。該核酸構築物を含有している細胞はその後MVAウイルスに感染させられ、核酸構築物とMVAウイルスゲノムとの間の相同組換えを可能にするのに適した条件下で培養される。その後適切な時期に細胞が採取され、組換えMVAウイルスが単離される。得られたウイルスは安定な組換えMVAウイルスとなる。そのようなウイルスは誘導体ウイルスと呼ばれる場合もある。当然ながら、核酸構築物を細胞に導入するステップおよび該細胞を感染せしめるステップの順序は逆にされてもよいし、これら2つのステップが同時に行われてもよい。
【0085】
核酸構築物中の異種核酸配列をMVAゲノムに導入する一般的な方法および組換えMVAを得る方法は当業者にはよく知られており、さらに、J.サムブルック(Sambrook)、E.F.フリッシュ(Fritsch)およびT.マニアティス(Maniatis)、「分子クローニング:実験の手引き(Molecular Cloning: a Laboratory Manual)」第2版、コールド・スプリング・ハーバー・ラボラトリー・プレス(Cold Spring Harbor Laboratory Press)、1989年、ならびに「分子生物学における最新プロトコール(Current Protocols in Molecular Biology)」、ジョン・ワイリー・アンド・サンズ・インコーポレイテッド(John Wiley and Sons Inc.)、1998年、第16章第IV節「ワクシニアウイルスベクターを使用する哺乳動物細胞におけるタンパク質の発現(Expression of proteins in mammalian cells using vaccinia viral vectors)」、から導き出すことが可能である。
【0086】
本発明によるDNA配列は、本発明によるMVAまたはその誘導体、および本発明によるMVAに感染した細胞または個体を、同定または単離するために使用可能である。該DNA配列は、例えば、PCRプライマー、ハイブリダイゼーションプローブを生成するために使用され、またはアレイ技術において使用される。
【0087】
本発明による誘導体ウイルスなどの用語は、親ウイルスと同じ特長を示すがそのゲノムの1つ以上の部分において違いを示す子孫ウイルスを指す。用語「MVAの誘導体」は、MVAと同じ機能持性を有するウイルスについて述べている。例えば、MVA 1974/NIHクローン1の誘導体は、MVA 1974/NIHクローン1の特長を有する。MVA 1974/NIHクローン1またはその誘導体のこれらの持性のうちの1つは、その弱毒化および宿主域の厳格な制限である。
【0088】
本発明による組換えMVAは医薬品またはワクチンとして有用である。したがって、本発明の一実施形態は、本発明の組換えMVAウイルスを使用して個体を疾患から防御する方法である。
【0089】
本発明の組換えMVAウイルスは標的細胞内への外来コード配列の導入に使用することも可能であり、前記配列は標的細胞のゲノムに対して同種であっても異種であってもよい。標的細胞内への外来コード配列の導入は、タンパク質、ポリペプチド、ペプチド、抗原または抗原エピトープを生産するためにin vitroで行われてもよい。この方法は、本発明の組換えMVAを用いた宿主細胞の感染、適切な条件下での感染宿主細胞の培養、ならびに、前記宿主細胞によって生産されるポリペプチド、ペプチド、タンパク質、抗原、エピトープおよび/またはウイルスの単離および/または濃縮を含む。
【0090】
さらに、細胞内に1つ以上の同種配列または1つ以上の異種配列を導入するための方法は、in vitroおよびin vivoの治療法に適用可能である。in vitroの治療法については、本発明による組換えMVAに前もって(ex vivoで)感染させられた単離細胞が、免疫応答に影響を及ぼすため、好ましくは免疫応答を引き起こすために、生きている動物の身体に投与される。in vivoの治療法については、本発明による組換えポックスウイルスは、免疫応答に影響を及ぼすため、好ましくは免疫応答を引き起こすために、生きている動物の身体に直接投与される。この場合、接種部位の周囲の細胞、またウイルスが輸送されるのに介在した場所の、例えば血流の細胞も、in vivoで本発明の組換えMVAに直接感染する。感染後、これらの細胞は、外来コード配列によってコードされている治療遺伝子のタンパク質、ペプチドまたは抗原エピトープを合成し、続いて、それらまたはその一部を細胞表面上に提示する。免疫系の特殊化した細胞は、そのような異種のタンパク質、ペプチドまたはエピトープの提示を認識し、特異的免疫応答を開始する。
【0091】
MVAは増殖が高度に制限されており、従って高度に弱毒化されているので、ヒトを含む様々な哺乳動物(免疫力が低下した動物またはヒトを含む)の治療に有用である。本発明はさらに、ヒトを含む生きている動物の身体において免疫応答を引き起こすための医薬組成物およびワクチンを提供する。
【0092】
医薬組成物は一般に、1つ以上の、医薬として許容可能であるかまたは認可されている担体、添加剤、抗生物質、保存剤、アジュバント、希釈剤および/または安定化剤を含むことができる。そのような補助物質は、水、生理食塩水、グリセロール、エタノール、湿潤剤もしくは乳化剤、pH緩衝物質、または同様のものであってよい。適切な担体は、典型的には大型で徐々に代謝される分子、例えばタンパク質、多糖、ポリ乳酸、ポリグリコール酸、アミノ酸重合体、アミノ酸共重合体、脂質凝集体(lipid aggregate)などである。
【0093】
ワクチンの調製については、本発明の組換えポックスウイルスは生理学的に許容可能な形態に変換される。これは、天然痘に対するワクチン接種に使用されたポックスウイルス・ワクチン剤の調製における経験(スティックル(Stickl)、H.ら、ドイチェ・メディツィニッシェ・ボーヘンシュリフト誌(Dtsch Med Wochenschr.)、1974年、第99巻、p.2386−2392)に基づいて行うことができる。例えば、精製されたウイルスは、約10mM トリス、140mM NaCl(pH7.4)中で5×10E8 TCID50/mlの力価に製剤化されて−80℃に保存される。ワクチン注射剤の調製については、例えば、10E2−10E8のウイルス粒子が、アンプル(好ましくはガラス製アンプル)中で2%ペプトンおよび1%ヒトアルブミンの存在下にて100mlのリン酸緩衝生理食塩水(PBS)中で凍結乾燥される。別例として、ワクチン注射剤は、製剤中のウイルスの段階的な凍結乾燥によって製造されてもよい。この製剤は、追加の添加剤、例えばマンニトール、デキストラン、糖質、グリシン、ラクトースもしくはポリビニルピロリドン、または他の助剤、例えば酸化防止剤もしくは不活性ガス、安定化剤もしくはin vivo投与に適した組換えタンパク質(例えばヒト血清アルブミン)を含有することができる。その後、ガラス製アンプルは密閉され、4℃〜室温で数か月間保存可能である。しかしながら、必要がない限りは、アンプルは−20℃未満の温度で保存されることが好ましい。
【0094】
ワクチン接種または治療については、凍結乾燥品は0.1〜0.5mlの水性溶液、好ましくは生理食塩水またはトリス緩衝液に溶解されて、全身投与または局所投与、すなわち非経口投与、皮下投与、筋肉内投与、スタンプ式(by scarification)投与、または当業者に周知の任意の他の投与経路によって、投与されうる。投与方式、用量および投与回数は、周知のやり方で当業者によって最適化可能である。しかしながら、最も一般的には、患者は最初のワクチン接種注射の約1か月〜6週間後に、2回目の注射でワクチン接種される。
【0095】
本発明の一実施形態は、抗原に対する免疫応答を生成する方法である。そのような応答は、CD8+T細胞免疫応答または抗体応答であってよい。特に、本発明は、初回免疫組成物の投与によって誘発された免疫応答が追加免疫組成物の投与によって増強(ブースト)される「プライム・アンド・ブースト」免疫化法に関する。本発明は、有効な追加免疫が、改変ワクシニアアンカラ(MVA)ベクターを使用して、組換えMVA自体を含む様々な異なる種類の初回免疫組成物のうち任意のものを用いた初回免疫の後で達成されうる、という先の実験証明に基づいている。
【0096】
数多くの病原体に対する免疫応答の主要な防御成分は、細胞傷害性リンパ球(CTL)としても知られているCD8+型のTリンパ球によって仲介される。CD8+細胞の重要な機能はγインターフェロン(IFN)の分泌であり、これはCD8+T細胞免疫応答の尺度を提供する。免疫応答の別の成分は、病原体のタンパク質に対する抗体である。
【0097】
本発明は、先の実験が示すように、様々な異なる初回免疫組成物のうち任意のものを使用して抗原に対して初回免疫されたCD8+T細胞免疫応答を増強するため、さらには抗体反応を誘発するための、有効な手段であることが見出されたMVAを使用する。
【0098】
特に、先の実験研究は、本発明の先行物の使用により、HIV抗原を発現する組換えMVAウイルスが、DNAワクチンによって初回免疫されたCD8+T細胞免疫応答を増強し、さらには抗体応答を誘発することが可能となることを実証している。該MVAは、免疫化後にCD8+T細胞応答を誘発することが見出される場合がある。組換えMVAは、組換えMVAの1回以上の接種により増強される免疫応答を初回免疫することが示される可能性もある。
【0099】
プラスミドDNAで免疫化され、MVAで追加免疫された非ヒト霊長類は、生ウイルスを用いた粘膜内抗原投与に対して有効に防御された(アマラ(Amara)ら、サイエンス誌(Science)、2001年、第292巻、p.69−74)。好都合なことに、本発明者らは、初回免疫および追加免疫の両方に皮内免疫、筋肉内免疫または粘膜免疫を用いるワクチン接種法を使用することができ、これは、例えばヒトにおいてCD8+T細胞を誘導し、さらに抗体応答を誘発するのに適した一般的な免疫化法を構成すると考える。
【0100】
本発明の種々の態様および実施形態は、HIV抗原をコードする核酸の事前投与によって初回免疫された該抗原へのCD8+T細胞免疫応答を増強し、さらに抗体応答を誘発するために、該抗原をコードするMVAベクターを使用する。
【0101】
本発明の一般的な態様は、HIV抗原に対するCD8+T細胞免疫応答を増強し、さらに抗体応答を誘発するためのMVAベクターの使用を提供する。
本発明の1つの態様は、個体においてHIV抗原に対するCD8+T細胞免疫応答を増強し、さらに抗体応答を誘発する方法を提供し、該方法は、該抗原をコードする核酸を含むMVAベクターであって、該核酸が該核酸からの発現により個体において抗原を生産するための調節配列に作動可能に連結されているMVAベクターを、個体の体内に提供し、これによって事前に該個体において初回免疫された抗原へのCD8+T細胞免疫応答が増強されることを含む。
【0102】
HIV抗原に対する免疫応答は、プラスミドDNAを用いた免疫化によって、または感染性物質による感染によって初回免疫されうる。
本発明のさらなる態様は、個体においてHIV抗原に対するCD8+T細胞免疫応答を誘導し、さらには抗体応答を誘発する方法を提供し、該方法は、該個体に、抗原をコードする核酸を含む初回免疫組成物を投与する工程と、その後、該抗原をコードする核酸を含むMVAベクターであって、該核酸が該核酸からの発現により個体において抗原を生産するための調節配列に作動可能に連結されているMVAベクターを含む追加免疫組成物を投与する工程とを含む。
【0103】
さらなる態様は、開示されているように、HIV抗原に対するCD8+T細胞免疫応答を増強し、さらには抗体応答を誘発するために哺乳動物に投与するための医薬品の製造における、MVAベクターの使用を提供する。そのような医薬品は一般に、抗原をコードする核酸を含む初回免疫組成物の事前投与に続いて投与するためのものである。
【0104】
初回免疫組成物は、抗原をコードするDNAであって、そのようなDNAが好ましくは哺乳動物細胞内で複製することができない環状プラスミドの形態であるものを含むことができる。いかなる選択可能なマーカーも、臨床的に使用される抗生物質に対して耐性であるべきではなく、したがって、例えばカナマイシン耐性がアンピシリン耐性よりも好ましい。抗原の発現は、哺乳動物細胞において活性を有するプロモーター、例えばサイトメガロウイルス最初期(CMV IE)プロモーターによって、駆動されるべきである。
【0105】
本発明の様々な態様の特定の実施形態では、初回免疫組成物の投与に続いて、追加免疫組成物、または第1および第2の追加免疫組成物であって同一であるかまたは互いに異なる第1および第2の追加免疫組成物を用いて、追加免疫が行われる。さらに別の追加免疫組成物が、本発明から逸脱することなく使用される場合もある。一実施形態では、三重免疫化法は、DNA、次いで第1の追加免疫組成物としてアデノウイルス、次いで第2の追加免疫組成物としてMVA、次いで任意選択でさらなる(第3の)追加免疫組成物またはその後の同一もしくは異なるベクターのうち一方もしくは他方もしくは両方の追加免疫投与を使用する。別の選択肢は、DNA、次いでMVA、次いでアデノウイルス、次いで任意選択でその後の同一もしくは異なるベクターのうち一方もしくは他方もしくは両方の追加免疫投与である。
【0106】
それぞれの初回免疫組成物および追加免疫組成物においてコードされる抗原は(いかに多くの追加免疫組成物が使用されても)同一である必要はないが、少なくとも1つのCD8+T細胞エピトープを共有するべきである。抗原は完全抗原に相当してもよいし、そのフラグメントに相当してもよい。より効率的に、抗原中の不要なタンパク質配列および1つまたは複数のベクター中のコード配列を切り取って、ペプチドエピトープまたは人為的なエピトープ連続物が使用されてもよい。1つ以上の追加のエピトープ、例えば、Tヘルパー細胞によって認識されるエピトープ、特に様々なHLA型の個体において認識されるエピトープが含まれてもよい。
【0107】
組換えMVAウイルスによってコードされる本発明のHIV抗原には、少なくとも1つのCD8+T細胞エピトープとしてHIVのEnv、Gag、Pol、Vif、Vpr、Tat、Rev、Vpu、またはNefアミノ酸配列のアミノ酸配列によって誘発される免疫原活性を有するポリペプチドが挙げられる。このアミノ酸配列は、実質的に、既知のHIV EnvもしくはPolの少なくとも1つの10〜900アミノ酸フラグメントおよび/または共通配列;または、既知のHIV Gagの少なくとも1つの10〜450アミノ酸フラグメントおよび/または共通配列;または、既知のHIV Vif、Vpr、Tat、Rev、Vpu、もしくはNefの少なくとも1つの10〜100アミノ酸フラグメントおよび/または共通配列に相当する。
【0108】
完全長Env前駆体配列が本発明での使用に関して提示されているが、Envは任意選択で部分配列を欠失している。例えば、gp120表面型切断産物およびgp41膜貫通型切断産物の領域を欠失していてもよい。
【0109】
完全長Gag前駆体配列が本発明での使用に関して提示されているが、Gagは任意選択で部分配列を欠失している。例えば、マトリックスタンパク質(p17)の領域、カプシドタンパク質(p24)の領域、ヌクレオカプシドタンパク質(p7)の領域、およびp6(GagポリプロテインのC末端ペプチド)の領域を欠失していてもよい。
【0110】
完全長Pol前駆体配列が本発明での使用に関して提示されているが、Polは任意選択で部分配列を欠失している。例えば、プロテアーゼタンパク質(p10)の領域、逆転写酵素タンパク質(p66/p51)の領域、およびインテグラーゼタンパク質(p32)の領域を欠失していてもよい。
【0111】
そのようなHIV Env、Gag、またはPolは、既知のEnv、Gag、またはPolタンパク質のアミノ酸配列に対して少なくとも50%の全体的な同一性、例えば50〜99%の同一性、またはその中の任意の範囲もしくは値の同一性を有する一方で、HIVの少なくとも1つの株に対する免疫原性応答を誘発することができる。
【0112】
同一性(%)は、例えば、ウィスコンシン大学遺伝学コンピュータ・グループ(University of Wisconsin Genetics Computer Group)(UWGCG)から入手可能なGAPコンピュータプログラム、バージョン6.0を使用して配列情報を比較することにより、測定することができる。該GAPプログラムは、ニードルマン(Needleman)およびブンシュ(Wunsch)のアライメント方法(ジャーナル・オブ・モレキュラー・バイオロジー誌(J Mol Biol)、1970年、第48巻、p.443)を、スミス(Smith)およびウォーターマン(Waterman)によって修正されたようにして(アドバンシズ・イン・アプライド・マスマティックス誌(Adv Appl Math)、1981年、第2巻、p.482)、利用する。簡潔に述べると、GAPプログラムは、同一性を、アライメントされた同一のシンボル(すなわちヌクレオチドまたはアミノ酸)の数を2つの配列のうち短いほうのシンボルの総数で除算したもの、として定義する。GAPプログラム用の好ましい初期設定パラメータには、(1)ユニタリ比較行列(同一について1および非同一について0の値を含む)ならびにグリブスコフ(Gribskov)およびバージェス(Burgess)(ヌクレイック・アシッズ・リサーチ誌(Nucl Acids Res)、1986年、第14巻、p.6745)の重み付き比較行列、シュワルツ(Schwartz)およびデイホフ(Dayhoff)(編、「タンパク質の配列および構造の図解集(Atlas of Protein Sequence and Structure)」、米国ワシントンD.C.所在のナショナル・バイオメディカル・リサーチ・ファウンデーション(National Biomedical Research Foundation)、1979年、pp.353−358)の記述によるもの;(2)各ギャップについて3.0のペナルティ、および各ギャップの各シンボルについてさらに0.10のペナルティ;ならびに(3)エンドギャップについてはペナルティなし、が含まれる。
【0113】
好ましい実施形態では、本発明のEnvは少なくとも1つのHIVエンベロープタンパク質の1つのバリアント型である。好ましくは、Envはgp120と、gp41の膜貫通部およびエクトドメインとで構成され、gp41の細胞質ドメインの一部または全部を欠いている。
【0114】
既知のHIV配列は、民間およびGENBANKのような公的なHIV配列データベースから、または公開された編集物として、例えばマイヤーズ(Myers)ら編、「ヒト・レトロウイルスおよびAIDS、核酸およびアミノ酸配列の編集および分析(Human Retroviruses and AIDS, A Compilation and Analysis of Nucleic Acid and Amino Acid Sequences)」、第I巻および第II巻、米国ニューメキシコ州ロスアラモス、セオレティカル・バイオロジー・アンド・バイオフィジックス(Theoretical Biology and Biophysics)、1993年、もしくはワールドワイドウェブのhiv−web.lanl.gov/において、容易に入手可能である。
【0115】
本発明の組換えMVAウイルスで使用される核酸によってコードされる、新たなHIV Env、GagまたはPolを得るためのHIV Env、GagまたはPolの置換または挿入には、少なくとも1つのアミノ酸残基(例えば1〜25個のアミノ酸)の置換または挿入が挙げられる。別例として、少なくとも1つのアミノ酸(例えば1〜25個のアミノ酸)をHIV Env、GagまたはPolの配列から欠失させてもよい。好ましくは、そのような置換、挿入または欠失は、安全上の特性、発現レベル、免疫原性、およびMVAの高い複製速度との両立性に基づいて特定される。
【0116】
本発明のHIV Env、GagまたはPolにおけるアミノ酸配列の変異は、例えば、DNA中の突然変異によって作製することができる。そのようなHIV Env、GagまたはPolは、例えば、アミノ酸配列中の様々なアミノ酸残基をコードするヌクレオチドの欠失、挿入または置換を含んでいる。言うまでもなく、HIV Env、GagまたはPolをコードする核酸において作製されることになる突然変異は、配列をリーディングフレームから外してはならず、第2のmRNA構造物を生じる可能性のある補足的なドメインを作らないことが好ましいであろう。
【0117】
本発明のHIV Env、GagまたはPolをコードする核酸は、本明細書中に示された教示および手引きに基づいて、HIVのEnv、GagまたはPolをコードするDNAまたはRNA中のヌクレオチドの増幅または部位特異的変異誘発、およびその後の、HIV Env、GagまたはPolをコードするDNAまたはRNAを生産するための該コードDNAの合成または逆転写によって調製されることも可能である。
【0118】
本発明のHIV Env、GagまたはPolを発現する組換えMVAウイルスには、本明細書中に示された教示および手引きに基づいて、当業者であれば過度の実験作業を伴うことなく日常作業で得ることができる置換ヌクレオチドとして、HIV Env、GagまたはPolをコードする配列の有限な集合が含まれる。タンパク質の化学的性質および構造の詳細な説明については、シュルツ(Schulz)、G.E.ら、「タンパク質構造の原理(Principles of Protein Structure)」、米国ニューヨーク州ニューヨーク、シュプリンガー・フェアラーク(Springer−Verlag)、1978年、およびクライトン(Creighton)T.E.、「タンパク質:構造および分子特性(Proteins: Structure and Molecular Properties)」、米国カリフォルニア州サンフランシスコ、W.H.フリーマン社(W. H. Freeman and Co.)、1983年を参照されたい。コドン選択のような、ヌクレオチド配列置換の表示については、オースベル(Ausubel)ら編、「分子生物学における最新プロトコール(Current Protocols in Molecular Biology)」、米国ニューヨーク州ニューヨーク、グリーン・パブリシング・アソシエーツ(Greene Publishing Assoc.)、1994年、§§A.1.1〜A.1.24、およびサムブルック(Sambrook)、J.ら、「分子クローニング:実験の手引き(Molecular Cloning: a Laboratory Manual)」第2版、米国ニューヨーク州コールド・スプリング・ハーバー、コールド・スプリング・ハーバー・ラボラトリー・プレス(Cold Spring Harbor Laboratory Press)、1989年、付録CおよびD、を参照されたい。
【0119】
したがって、当業者は、本明細書中に示された教示および手引きがあれば、置換バリアント、欠失バリアントまたは挿入バリアントを含む代替HIV Env、Gag、またはPolを得るためには、HIVのenv、gag、またはpolのDNAまたはRNAの他の部位の他のアミノ酸残基をどのようにして置換するかが分かるであろう。
【0120】
MVAベクター内では、コードされた抗原を発現するための調節配列はプロモーターを含むことになる。「プロモーター」とは、ヌクレオチドの配列であって、該配列から、下流に(すなわち二本鎖DNAのセンス鎖上の3’方向に)作動可能に連結されたDNAの転写が開始されうるところの配列を意味する。「作動可能に連結され」とは、転写がプロモーターから開始されるように適切に配置かつ配向されて、同じ核酸分子の一部として連結されることを意味する。プロモーターに作動可能に連結されたDNAは、プロモーターの「転写開始調節下」にある。ターミネーターフラグメント、ポリアデニル化配列、マーカー遺伝子およびその他の配列を含む他の調節配列が、当業者の知識および慣行に従って適正に含められてもよい。例えば、D.M.クナイプ(Knipe)、P.M.ハウリー(Howley)編、「フィールズ・ウイルス学(Fields Virology)」の、モス(Moss)、B.著「ポックスウイルス科:ウイルスおよびその複製(Poxviridae: The Viruses and Their Replication)」、フィラデルフィア所在のリッピンコット・ウィリアムズ・アンド・ウィルキンズ(Lippincott Williams and Wilkins)、2001年、p.2849〜2883、を参照されたい。例えば核酸構築物の調製、突然変異誘発、塩基配列決定、細胞内へのDNAの導入および遺伝子発現における核酸の取扱、ならびにタンパク質の分析のための数多くの既知の技法およびプロトコールは、オースベル(Ausubel)ら編、「分子生物学における最新プロトコール(Current Protocols in Molecular Biology)」、ジョン・ワイリー・アンド・サンズ(John Wiley and Sons)、1998年に詳細に記載されている。
【0121】
本発明の態様および実施形態で使用されるプロモーターは、ポックスウイルスの発現系に適合しうるものであって、天然の配列、改変された配列および合成配列が含まれうる。
初回免疫組成物および追加免疫組成物のうち一方または両方は、顆粒球マクロファージコロニー刺激因子(GM‐CSF)またはこれをコードする核酸のようなアジュバントを含んでもよい。
【0122】
追加免疫組成物の投与は、一般に初回免疫組成物の投与の約1〜6か月後、好ましくは約1〜3か月後である。
好ましくは、初回免疫組成物、追加免疫組成物、または初回免疫組成物および追加免疫組成物両方の投与は、皮内免疫、筋肉内免疫または粘膜免疫である。
【0123】
MVAワクチン剤の投与は、ウイルスの懸濁液を注入するために注射針を使用することにより達成されてもよい。代替例は、ウイルス懸濁液を投与するための無針注射デバイス(例えばBiojector(登録商標)無針注射器を使用)、または、冷蔵を必要としない個別調製された単回投与量の製造をもたらす、再懸濁されたワクチン含有凍結乾燥粉末を使用することである。これは、アフリカの農村地域において必要とされるワクチンについては大きな利点となるであろう。
【0124】
MVAはヒトの免疫化において優れた安全上の実績を備えたウイルスである。組換えウイルスの生成を簡単に行うことが可能であり、大量に再現性よく製造することができる。したがって、組換えMVAウイルスの皮内、筋肉内、または粘膜投与は、CD8+T細胞応答によって制御可能なAIDSに対するヒトの予防的または治療的ワクチン接種に非常に適している。
【0125】
個体は、該抗原の送達および該抗原に対するCD8+T細胞免疫応答の生成が有益であるか、または治療上有益な効果を有するようなAIDSに罹患している個体であってよい。
【0126】
恐らくは、投与は、感染または症状発現の前にHIVまたはAIDSに対する免疫応答を生成するための予防的な目的を有することになる。
本発明に従って投与される成分は医薬組成物として製剤化されうる。これらの組成物は、薬学的に許容可能な賦形剤、担体、緩衝剤、安定化剤または当業者に良く知られた他の材料を含むことができる。そのような材料は無毒でなければならず、かつ有効成分の有効性を妨げてはならない。担体またはその他の材料の厳密な性質は、投与経路、例えば静脈内経路、皮内または皮下経路、経鼻、筋肉内経路、腹腔内経路に応じて変化しうる。
【0127】
言及したように、投与は好ましくは皮内投与、筋肉内投与、または粘膜投与である。
生理食塩水、デキストロースもしくはその他糖類の溶液またはグリコール、例えばエチレングリコール、プロピレングリコールもしくはポリエチレングリコールが含まれてもよい。
【0128】
静脈内、皮内、皮下、筋肉内、もしくは粘膜への注射、または罹患部位での注射については、有効成分は、発熱性物質を含まず適切なpH、等張性および安定性を有する非経口的に許容可能な水性溶液の形態となろう。当業者は、例えば、生食注射液、リンガー液、乳酸加リンガー液のような等張の媒体を使用して、適切な溶液を調製することができる。必要に応じて、保存剤、安定化剤、緩衝剤、酸化防止剤および/またはその他の添加剤が含まれていてもよい。
【0129】
徐放製剤が使用されてもよい。
MVA粒子の生産および任意選択のそのような粒子の組成物中への製剤化に続いて、該粒子は、個体、特にヒトまたはその他の霊長類に投与されうる。投与は、別の哺乳動物、例えばマウス、ラットもしくはハムスターのようなげっ歯動物、モルモット、ウサギ、ヒツジ、ヤギ、ブタ、ウマ、ウシ、ロバ、イヌまたはネコに対するものであってもよい。
【0130】
投与は、「予防的に有効な量」または「治療上有効な量」(場合に応じて予防は治療と考えることもできるが)であることが好ましく、これは個体に対する有益性を示すのに十分ということである。投与される実際の量、ならびに投与の速さおよび時間的投与条件は、治療対象者の性質および重症度に応じて変わることになろう。治療の処方、例えば投与量の決定などは、一般臨床医およびその他の医師、または獣医学的状況では獣医師の担当事項であり、典型的には、治療される病気、個々の患者の状態、送達部位、投与方法、および当業者に周知の他の要因を考慮する。上述の技法およびプロトコールの例は、オソル(Osol)、A.編、「レミングトンの製薬科学(Remington’s Pharmaceutical Sciences)」、第16版、1980年、に見出すことができる。
【0131】
1つの好ましい投与法では、DNAが注射1回あたり投与量300μg〜3mgで投与された後、MVAが注射1回あたり投与量106〜109個の感染性ウイルス粒子として投与される。
【0132】
組成物は単独で投与されてもよいし、他の治療と組み合わされて、治療すべき状態に応じて同時に、または順次、投与されてもよい。
ヒト以外の哺乳動物への送達は必ずしも治療目的である必要はなく、実験的状況で、例えば、対象とする抗原に対する免疫応答、例えばHIVまたはAIDSからの防御のメカニズムの研究に使用するためであってもよい。
【0133】
実施例
以下の実施例は、実施形態を作製および使用する方法の完全な開示および説明を当業者に提供するために提示されており、本発明者らが発明と見なすものの範囲を限定するようには意図されておらず、また下記の実験が全てであるかまたはその実験しか実施されないということを示すように意図されたものでもない。使用される数値(例えば量、温度など)に関する精度を保証する努力がなされてはいるが、何らかの実験上の誤差および偏差が考慮されるべきである。別途記載の無いかぎり、部は重量部であり、分子量は重量平均分子量であり、温度は摂氏度である。標準的な略語が使用される。
【0134】
実施例1
以下の実施例は、シャトルプラスミド、組換えMVA/HIV1臨床用ワクチン構築物、ならびに、外来遺伝子のコドン変更および2つのワクシニアウイルス必須遺伝子の間への外来遺伝子の挿入による、組換えMVAにおいて完全な外来遺伝子挿入物を維持するメカニズムを実証する。本開示は以下のメカニズムを提示する、すなわち:
・ MVAの複製にとって必須の2つのワクシニアウイルス遺伝子の間に完全な外来遺伝子を挿入することにより該外来遺伝子を維持するメカニズム。外来遺伝子の欠失はその組換えMVAに生育上の著しい優位性をもたらす可能性があり、継代が繰り返された際に完全な外来遺伝子を含有するMVAと競合する場合がある。しかしながら、外来遺伝子のほとんどの欠失は、隣接するワクシニアウイルスDNAのある程度の部分の喪失も含んでいる。そのワクシニアウイルスDNAが必須であれば、欠失を備えたウイルスは複製せず、完全な外来遺伝子を含有しているMVAと競合しないことになる。この方法論は、臨床試験で使用するためなど大規模に増幅させる必要のある組換えワクシニアウイルスの生産に有用であろう。また、
・ 組換えウイルスを繰り返し継代した後に容易に突然変異を生じる特異的「ホットスポット」を変質させることにより、外来遺伝子挿入物を安定させるメカニズム。この方法論は、臨床試験で使用するためなど大規模に増幅させる必要のある組換えウイルスの生産に有用である。
【0135】
そして、
・ 2つの必須ワクシニアウイルス遺伝子の間に外来遺伝子を挿入するために使用されるシャトルプラスミドpLW‐73、および
・ 上記2つのメカニズムの使用を具体化する材料である、組換えMVA/HIV‐1臨床用ワクチン構築物MVA/UGD4d
について説明する。
【0136】
安定な組換えMVAウイルスの生成
様々な地理的位置からのHIV‐1単離物由来のenvおよびgagpol遺伝子を発現する改変ワクシニアウイルスアンカラ(MVA)組換え体が構築された。外来遺伝子は、MVAの2つの部位、欠失II(Deletion II)および欠失III(Deletion III)に挿入された。組織培養物中で組換えMVAを繰り返し継代した後の上記遺伝子の安定性は変化しやすいことが判明した。本発明者らは、この不安定性が、外来遺伝子全体および一部の隣接DNAの欠失に起因するか、または外来遺伝子を発現しないため生育上有利である子孫ビリオンの増殖をもたらす特異的点突然変異に起因するかのいずれかであることを実証した。ここで本発明者らは、完全な外来遺伝子組換えMVAを保持する2つの新規な方法について述べる。第1に、本発明者らは、ゲノムの保存された中央領域の2つの必須ワクシニアウイルス遺伝子の間に外来遺伝子を挿入させる導入ベクターを構築した。この遺伝子挿入部位の使用は、必須ワクシニアウイルスDNAを含んだ大きな欠失部を含有するバリアントの増殖を防止する。加えて、このプラスミドは、組換えウイルスへのさらなる遺伝子の挿入に使用することができる。第2に、点突然変異を備えた単離物の分析から、翻訳中に早期終了を引き起こす単一塩基の挿入または欠失の傾向を備えたある種の「ホットスポット」が明らかとなった。本発明者らは、これらの部位におけるサイレント突然変異の生成が、挿入遺伝子の安定性をもたらすことを示した。
【0137】
I.新規な導入ベクターの構築および適用
新規な導入ベクターpLW‐73の構築
1. MVAゲノムの中央領域K7R‐A24Rについて、1)ポックスウイルス科またはコードポックスウイルス亜科において保存された遺伝子対、および2)遺伝子の3’末端どうしが近接するように逆方向を向いた遺伝子であって、その結果としてワクシニアプロモーターを妨害しないであろう挿入部位を提供する遺伝子、に関する検査が行われた。新しい挿入部位として選択された部位は、2つの必須遺伝子I8RおよびG1Lの間であった。
【0138】
2. 新しいベクターの左側隣接部は次の方法で構築された:プラスミドLAS‐1が、del III MVA隣接部、GFP、およびMVA隣接部のダイレクトリピートを取り出すために、制限酵素EcoRIおよびXhoIで切断された。この挿入物はAscIおよびSacIで切断されて、GFPフラグメントが単離された。I8R遺伝子末端の531塩基対(TAA終止コドンを含む)が、PCR産物の末端にEcoRIおよびAscI制限部位を備えるようにPCR増幅された。(TAA終止コドンを含むI8R遺伝子末端からの)229塩基対のダイレクトリピートのPCR増幅が、SacIおよびXhoI制限部位を含有するオリゴヌクレオチドを用いて行なわれた。全4片のDNAすなわち、1)EcoRI末端およびXhoI末端を備えたベクター骨格、2)EcoRI末端およびAscI末端を備えたI8R末端を含有する新しい左側隣接部、3)AcsI末端およびSacI末端を備えたGFP、ならびに4)SacI末端およびXhoI末端を備えたI8R隣接部のダイレクトリピート、が合わせてライゲーションされてプラスミドpLW‐72が作製された。
【0139】
3. 右側隣接部は以下のように作製された:pLW‐72が制限酵素PstIおよびHindIIIで切断され、該プラスミド中のMVAのdel III隣接部が放出された。G1L遺伝子末端の702塩基対が、両端にPstIおよびHindIII制限酵素部位を備えるようにPCR増幅され、pLW‐72ベクター中にライゲーションされてpLW‐73が作製された(図7)。pLW‐73の配列を図8に示す。
【0140】
4. pLW‐73の顕著な特色は次のとおりである:1)ベクターは、MVAゲノム中の必須遺伝子の間に外来遺伝子を挿入するために設計された。左側隣接部はI8R遺伝子の終端部からなり、右側隣接部はG1L遺伝子の終端部からなる。2)組換えウイルスの簡単な初期選択のためにGFP遺伝子が含まれている。3)GFPはI8R遺伝子のダイレクトリピートに隣接しており、組換えウイルスが繰り返し継代されるとGFPが失われることになるためGFPの一時的発現が可能となっている。国際公開公報第2004/087201号を参照すると、特色2および3は、MVA/HIV組換え体pLAS‐1およびpLAS‐2を作製するために使用された初期プラスミドにも含まれていた。
【0141】
pLW‐73の適用
1. HIV‐1のクレードBのADA単離物由来のenv遺伝子がpLW‐73にクローニングされ、組換えMVAウイルスが作製された。DNA塩基配列決定により、env遺伝子の位置および完全性が確認された。
【0142】
2. MVAの欠失II(Deletion II)部位にウガンダのクレードD(単離物AO7412)のenv遺伝子を発現する組換えMVAウイルス(図9)は、不安定であること、すなわち、培養物中の連続継代が繰り返された後に遺伝子が大部分のウイルス子孫から欠失することが判明した。その後、同じ遺伝子がpLW‐73にクローニングされ、組換えMVAウイルスが作製されて特徴解析された。env遺伝子挿入物は培養物中で連続継代が繰り返された(8×)後も安定していた、すなわち、挿入遺伝子または隣接するMVA領域の欠失は見出されなかった。加えて、遺伝子がこの部位に挿入された時は他の突然変異が発生しなかった。
【0143】
II.「ホットスポット」の点突然変異
点突然変異の分析
MVAの欠失III(Deletion III)部位にウガンダのクレードD(単離物AO3349)gagpol遺伝子を発現する組換えMVAウイルスは、不安定であることが判明した。主な遺伝子変化は、GまたはC残基が4〜6個連続した箇所における単一の点突然変異の生成であった(表3)。加えて、同様の点突然変異は、ケニアのクレードAのHIV‐1単離物およびタンザニアのクレードCの単離物由来のgagpol遺伝子を発現する同様の組換えウイルスからの非染色プラークに見出された。
【0144】
ホットスポットの突然変異誘発および組換えウイルス中の安定性の分析
部位特異的突然変異誘発を使用して、ウガンダのHIV‐1単離物由来のgag遺伝子の6つのそのような領域にサイレント突然変異を作製した。この変異遺伝子UGD4d gagpol orf(図10)はpLAS‐1にクローニングされ、不安定なウイルスの構築で行われたようにしてMVAの同じ欠失III(Deletion III)部位へ組み込まれた。培養物中で連続継代を繰り返した(8×)後、発現のないプラークは見出されなかった。継代8代目のウイルスストックのDNA塩基配列決定から、gagpol遺伝子の完全性が維持されていることが確認された。
【0145】
III. 二重組換え体の構築
MVA/UGD4dウイルス
MVA/UGD4dウイルス、すなわちウガンダのサブタイプD AO7412のエンベロープおよびAO3349のgagpolを発現する組換えウイルスは、次の方法で構築された:エンベロープおよびgagpolの遺伝子は、それぞれシャトルプラスミドpLW‐73およびpLAS‐1を利用した相同組換えによってMVA 1974/NIHクローン1に挿入された。MVA/UGD4dは、ニワトリ胚線維芽細胞における6回のプラーク精製によって単離され、その後増幅されて特徴解析された。
【0146】
概要
1. MVAゲノムの保存された中央領域の2つの必須遺伝子I8RおよびG1Lの間への外来遺伝子の組み込みを行うプラスミド導入ベクターが構築された。この部位の使用により、挿入遺伝子/MVA隣接部の欠失を有する変異ウイルスの選択が阻止されることが示された。
【0147】
2.GおよびC残基が連続した極めて変異しやすい箇所は部位特異的突然変異誘発によって変更され、コード配列中のサイレント突然変異が生成された。この変化は、遺伝子がMVAの欠失III(Deletion III)に挿入された場合に該遺伝子を安定させることが示された。
【0148】
3. 上記の2つの方法を利用して、ウガンダのクレードDのenvおよびgagpolの両方を安定して発現するUGD4d二重MVA組換え体が構築された。
実施例2
様々な単離物からのHIV‐1 env遺伝子およびgagpol遺伝子を発現する組換えMVAが作製された。組織培養物中で継代が繰り返された後の挿入遺伝子の安定性は変化しやすいことが判明した。ここで本発明者らは、(1)不安定性は、自然突然変異または挿入遺伝子の欠失と、発現のない突然変異の選択との組合せを表わすことを実証し、(2)不安定性を低減する新規な方法について述べる。
【0149】
概観
多数の異なる単離物からのenvおよびgagpolを発現する組換えMVAが構築された。各ウイルスは、臨床試験用ウイルスの製造に必要な大規模増幅を模倣するためにニワトリ胚線維芽細胞において繰り返し継代された。挿入物の安定性は、個々のプラークのenvおよびgagを免疫染色することによってモニタリングされた。いくつかの組換えウイルスについては、envおよび/またはgagの発現がウイルス集団のかなりの割合において急速に失われることが見出された。発現喪失のメカニズムを同定するために、個々のプラークが単離され、突然変異の性質が特徴解析された。いくつかの事例では、単一ヌクレオチドの付加または欠失によって突然変異する傾向を備えた特定のDNA塩基配列が同定された。そのような突然変異の生成は、予測される翻訳産物を変化させることなくコドンを変更することにより、回避することができた。他の事例では、発現の喪失は、隣接している非必須MVA遺伝子の中まで及ぶことの多い大規模な欠失によって引き起こされた。これが生じるのを防止するために、2つの必須MVA遺伝子の間に外来遺伝子を挿入させることを目指した新しいシャトルプラスミドが構築された。この部位への組込みは外来DNAの欠失を低減した。しかしながら、ある事例では、高レベルのHIV env発現に関連した毒性が非常に強いため、希少な突然変異体を選択してもなお不安定な集団が得られた。この場合、envの膜貫通ドメインを短縮化して初めて安定した組換えMVAの構築が可能となった。
【0150】
組換えMVAの生成および挿入遺伝子の安定性の分析
Env遺伝子およびgagpol遺伝子はMVAシャトルベクターにクローニングされた。発現および機能は一時的発現アッセイによって分析された。gagpolはMVA 1974/NIHクローン1に組み込まれた。組換えMVAは6〜8回プラーク精製され、その後ウイルスの増幅が行われた。Envは該MVA/gagpol単離物に組み込まれ、二重組換えMVA(図11A)は6〜8回プラーク精製され、増幅された。挿入物の安定性を評価するために、ウイルスは、大規模製造を模倣するために〜1pfu/細胞の感染多重度(m.o.i.)を使用してCEF細胞中で連続的に継代された。安定性は、モノクローナル抗体で免疫染色することにより測定された、envまたはgagを発現する細胞の割合(%)の測定により評価された(図11B)。
【0151】
組換えMVAの安定性
様々な地理的位置からのHIV‐1単離物由来の遺伝子を発現する組換えMVAが構築された。env遺伝子およびgagpol遺伝子は、それぞれMVAの欠失IIおよびIIIに挿入され、いずれも改変H5プロモーターの制御下におかれた。7つの組換えMVAのenv遺伝子およびgagpol遺伝子の安定性が表4に示されている。様々な程度の不安定性がこの7つのウイルスにおいて観察された。MVA/65A/Gでは、envの発現は急速に失われて、継代6代目までにはわずか25%のビリオンしかenvを発現しなかった。MVA/UGD4aでは、envおよびgagpol両方の発現は連続的なウイルス継代とともに徐々に失われた。第I相試験用の大量のウイルスの製造には少なくとも6〜7回の継代が必要なので、これら2つのウイルスは不適当であると考えられた。
【0152】
MVA/65A/Gの発現の分析
図12を参照すると、MVA/65A/GのP3およびP5から13個のプラークが無作為に採取され、T‐24 mAb(結合部位をaに示す)を用いた免疫染色、ウエスタンブロッティング、PCR、および配列決定により分析された。5種類のプラークが見出され、各種類について得られたこれらのプラークの数は図12の右側に示されている。プラークa、bおよびcは染色されたが、bおよびcは、塩基置換(終止コドンを生じるもの)(b)、ならびに、env遺伝子の端部およびMVA隣接部の一部の欠失(c)に起因する短縮型であった。染色されないプラークdおよびeは、Gが5個連続した箇所へのGの付加を原因とするフレームシフト(d)、ならびに、env遺伝子全体およびMVA隣接部の一部の大規模な欠失(e)から生じていた。したがって、塩基対付加、置換、および欠失はすべてMVA/65A/Gのenv遺伝子の不安定な発現に寄与していた。このA/G envは検討した中で最も不安定な例であり、安定性を増強しうる改変を研究するために採取された。
【0153】
安定性を増大させるためのA/G構築物の改変
1. 点突然変異を防止するために、GおよびCが4個および5個連続した箇所をサイレント突然変異によって除去することにより合成エンベロープが作製された。
【0154】
2. ベクターI8/G1すなわちpLW‐73は、遺伝子およびMVA隣接部の欠失が生存可能とならないように、必須遺伝子I8RおよびG1Lの間の挿入部位を用いて構築された。MVAのI8R遺伝子の端部(500bp)およびG1L遺伝子の端部(750bp)がPCRによって増幅され、外来遺伝子の発現を制御するワクシニアウイルス初期/後期mH5プロモーターを含有するベクターに挿入された。このI8/G1ベクターは、相同組換えによってMVAに外来遺伝子を挿入するために使用された(図13)。挿入遺伝子および該挿入遺伝子に隣接しているMVAの欠失は、必須遺伝子の部分が欠失することになるので生存可能とはならなくなる。したがって、これらの突然変異を備えたウイルスは、正常な生育上の利点を備えた集団において異常増殖することができなくなる。
【0155】
3. A/G gp140エンベロープは、gp41の膜貫通ドメインおよび細胞質尾部を欠失させることにより変異させられ、分泌タンパク質を生じた。
安定性を増大させるための試験的改変
図14に示されるように、envの改変および/または新しいベクターの使用により7つの単一組換えウイルスが作製された。改変がエンベロープの安定的発現を増強したかどうかを判定するために、各ウイルスの5つのプラークが単離されてCEF中で独立に継代された。継代されたプラークは、mAb T‐43(結合部位はenvの101〜125aaにマッピングされた)を用いた免疫染色、ウエスタンブロッティング、PCR、および配列決定により分析された。
【0156】
プラーク継代後のEnv発現
図15を参照すると、上記に列挙された7つの組換え体それぞれの5つの独立に継代されたプラーク単離物が、継代1、3、5および7代目において、mAb T‐43(gp120の101〜125a.a.の間に結合する)を用いた免疫染色によって特徴解析された。7種のウイルスのうち4つ(図15のa、b、c、e)は、5つの継代プラークそれぞれにおいて不安定なタンパク質発現を有し、(図15f)の2つのプラーク継代物も不安定なenv発現を有していた。これらは、MVAゲノムのdel II(図15c)および必須遺伝子部位(図15f)の両方に合成envを備えたウイルスを含んでいた。短縮された分泌型gp140としてのエンベロープを含有している組換えウイルスだけが、エンベロープを安定に発現し続けた(図15のdおよびg)。
【0157】
ウエスタンブロッティング、PCRおよび配列分析
選択されたプラーク継代物からクローンを採取し、ウエスタンブロッティング、PCRおよび配列分析によってタンパク質発現を分析した(図16)。ウエスタンブロット解析については、クレードAのエンベロープの先頭部および終端部にそれぞれ結合するT‐24およびT‐32が、部分的エンベロープのみが作製されたか完全長エンベロープが作製されたかを測定するために使用された。cと印付けされた対照ウイルスが各ブロットの右側にある。MVAの欠失IIにおいて作製された3つのウイルス(図16a、bおよびc)については、図16c(すなわちgp140クローン)にのみ、全クローンがウエスタン分析で検出可能なタンパク質を発現していた。このタンパク質(T‐32によって計測)は短縮されていなかった。エンベロープがベクターI8/G1によって必須遺伝子部位へ挿入されると(図16d、eおよびf)、この場合も、gp140エンベロープだけがすべてのクローンにおいて発現されており、短縮されていなかった。I8/G1ベクターの使用はenv配列の突然変異生成を防止しなかったが、del IIに挿入されたエンベロープで見られていた欠失は防止した。(I8/G1ベクター由来の試験されたすべてのクローンの陽性PCR産物、対してdel IIベクターを使用して試験されたクローンの陰性PCR産物に注目されたい。)
クレードA/G二重組換え体におけるEnvの発現
1つのenv分析を用いた先の結果に基づいて、gp140または合成gp160遺伝子とともにgagpolを発現する二重組換え体が作製され、env発現の安定性について試験された(図17)。先述のようにして各々から5つのプラークが単離され、env発現の安定性を分析するために7回継代された。継代7代目で、継代されたプラークはT‐43およびT‐32 mAbs(それぞれgp120およびgp41に結合する)両方を用いて免疫染色された。T‐43 mAbでは、合成エンベロープを発現する組換え体の5つのクローンのうちの1つが染色されないプラークのみで構成されていた。続いてこれらのプラークのT‐32染色は、別のプラークが短縮されたエンベロープを発現することを示した。gp140エンベロープを含有する二重組換え体からの継代プラークは全て、T‐43およびT‐32両方の免疫染色により、安定しているように見えた。力価も他の二重組換え体を用いるより2log高かった。したがって、安定してエンベロープを発現するクレードA/G二重組換え体は、gp140エンベロープを用いた場合にのみ作製可能であった。
【0158】
ウガンダのHIV‐1単離物由来のenvおよびgagpolを発現する組換えウイルス
ウガンダの単離物AO7412およびAO3349由来のHIV‐1 envおよびgagpol遺伝子を発現する組換えMVAウイルスは、図18に示されるようにして構築された。各々4〜6個の独立な単離物が連続的に継代され、両遺伝子が単独で発現されても組み合わせで発現されても不安定であることが分かった(表5)。対照的に、膜結合型gp160の代わりとしてgp140の発現は、連続継代後のenv遺伝子の安定性をもたらした(図18および表5)。
【0159】
MVA/UGD4a − 非染色envプラークの分析
不安定性のメカニズムを究明するために、24個の個々の非染色プラーク(Mab T‐43を使用)がMVA/UGD4aの継代6代目から単離され、増幅され、特徴解析された。2つの小さな欠失(1.2kbおよび0.3kb)がPCR増幅およびDNA塩基配列決定によって同定された(図19)。他の単離物はすべて、隣接するMVAにまで及ぶ非常に大きな欠失を含んでいた。これらの欠失に関するおよその切断点は、env遺伝子または隣接するMVA領域の内側からプライマー対を使用して同定された。
【0160】
組換えMVAにおけるUGD env遺伝子の改変
UGD env遺伝子の不安定性の問題を改善するために、新しいベクターI8/G1を使用して、AO7412 env遺伝子をMVAに挿入した。このベクターI8/G1は、2つの必須ワクシニアウイルス遺伝子I8およびG1の間に外来遺伝子を組み込み、かつ改変H5プロモーターを使用する(図20)。独立した4つのプラークが連続的に継代され、継代5代目でMab T‐43およびT‐32を用いて免疫染色することによりenv発現について分析された。すべての単離物において、遺伝子は安定していた(表6)。
【0161】
MVA/UGD4b − 非染色gagプラークの分析
gag遺伝子の不安定性のメカニズムを究明するために、8個の個々の非染色プラーク(Mab 183‐H12‐5C(NIAID AIDSリポジトリ)を使用)がMVA/UGD4bの継代6代目から採取され、増幅され、gagpol挿入物が配列決定された(表7)。7個の単離物には、564〜569位における単一のG残基の挿入または欠失が見出された。1つの単離物では、530〜534位の配列CCCCからC残基が欠失していた。更に、MVA/KEAおよびMVA/TZCの多数回継代ストック由来の非染色プラークから、同様の突然変異ホットスポット、すなわち564〜569位が明らかとなった。UGD AO7412 gagpol遺伝子の完全配列の調査から、22箇所の4個以上連続したGまたはC残基が実証された(図21)。
【0162】
組換えMVAにおけるUGD gagpol遺伝子の改変
gagpol遺伝子の不安定性のメカニズムが主として4〜6個の連続したGまたはC残基内の単一ヌクレオチドの挿入または欠失であったので、この遺伝子の安定性を改善する戦略は、そのような部位でサイレント突然変異を生成させることであった。したがって、p17およびp24 gagの6部位における部位特異的突然変異誘発(表3)が使用された。結果として得られた、MVAの同じ場所すなわち欠失III(Deletion III)に挿入されたコドン変更された(c.a.)遺伝子は、連続継代しても安定であることが証明された(図22および表8)。
【0163】
UGD envおよびgagpolを発現する安定な組換えMVAの構築
MVAのI8/G1座においてUGD env遺伝子、および欠失III(Deletion III)においてコドン変更されたgagpol遺伝子を発現する組換えウイルスが構築された(図23)。連続継代では、いずれの遺伝子の不安定性も示されなかった。更に、タンパク質発現のレベルおよびDNA塩基配列は継代を経ても不変であった(表9)。
【0164】
結論
env挿入物およびgagpol挿入物の不安定性は、点突然変異および欠失の生成ならびに非発現性MVA突然変異が生育上優位であることに起因する。不安定性は一般に、コドン変更、またはMVAゲノムの必須領域への挿入のうち少なくともいずれか一方によって低減することが可能である(MVA/UGD4d)。しかし、1つの事例ではenvを変更しなければならなかった(MVA/65A/G)。
【0165】
実施例3
BALB/cマウスにおけるMVA/UGD4dの免疫原性
マウス10匹の群が、それぞれ感染単位106または107のMVA/UGD4dを用いて腹腔内経路により免疫化された。マウス5匹の群が、それぞれ親のMVA‐1974を用いて同様に免疫化された。マウスは第0週および第3週で免疫化処置され、第0週、第3週および第5週で採血された。第5週で脾臓が採取された。
【0166】
細胞の応答は細胞内のサイトカイン染色によって新鮮な脾細胞で計測された。脾細胞は、下記すなわち:1)免疫優勢であるgagペプチド(AMQMLKETI(配列番号6))、2)envペプチド(DTEVHNVWATHACVP(配列番号7)およびQQQSNLLRAIEAQQH(配列番号8))、3)polペプチド(ELRQHLLRWGLTT(配列番号9)およびHGVYYDPSKDLIAE(配列番号10)の単一アミノ酸バリアントを備えた8つのペプチド)、ならびに4)MVAを用いて別々に刺激された。
【0167】
細胞は、CD4およびCD8の表面発現について染色され、次いでIFN‐γおよびIL2またはTNFのいずれかの細胞内発現について染色された。図24に示されるように、MVA/UGD4dはgagペプチド、polペプチドおよびMVAに対してCD8/IFN‐γ応答を誘発した。gagペプチドの応答は、IFN‐γと、IL2またはTNFのいずれかとの両方を発現する多機能性であった。さらに、CD4/IFN‐γ応答がenvペプチドの集合物に対して誘発された。
【0168】
体液性応答はELISAによって計測された(図25)。UGD envに対する強い応答が1回の免疫化処置の3週間後に実証され、第2の免疫化処置によって増強された。加えて、強いワクシニアウイルス応答が1回および2回の免疫化処置の後で誘発された。
【0169】
【表4】
【0170】
【表5】
【0171】
【表6】
【0172】
【表7】
【0173】
【表8】
【0174】
【表9】
【0175】
【表10】
実施例4
この実施例は、安定な組換えMVAウイルスを生成するためのさらなる挿入部位の使用について実証する。MVAウイルスゲノムのDel III領域はいくつかの非必須遺伝子および遺伝子フラグメントを含有しており、よって歴史的に異種核酸配列を挿入するために使用されてきた。したがって、MVAのdel III挿入部位周辺の隣接領域が、フラグメント化遺伝子または非必須遺伝子の存在について分析された。いくつかの細胞においてVACVの複製に重要であることが知られている遺伝子、すなわちA50R DNAリガーゼおよびB1Rキナーゼは、del III挿入部位からそれぞれ約1kbpおよび1.8kbpに位置していた。本発明者らは、Del III挿入部位に隣接する非必須遺伝子を除去すれば、より安定な挿入部位にすることができるであろうと推論した。この目的に向けて、A50R DNAリガーゼORFの3’末端部分(左)、ならびにB1R ORFの5’末端、およびプロモーター(右)を含む隣接配列を備えた核酸構築物(例えばシャトルベクター)が以下のように構築された。これにより、相同組換えが生じた時に上記2つの重要な遺伝子の間の非必須遺伝子の領域が有効に除去されるであろう。
【0176】
A. A50R/B1Rシャトルベクターの調製:
MVAゲノム中のdel III挿入部位周辺の隣接領域(bp数143552、アカンビス(Acambis)3000 Genbank AY603355))の分析から、いくつかの細胞においてVACVの複製に重要であることが知られている少なくとも2つの遺伝子が明らかとなった。具体的には、A50R DNAリガーゼ(ORF163;ACAM3000_MVA_163;配列番号11)およびB1Rキナーゼ(ORF167;ACAM3000_MVA_167;配列番号16)が、del III挿入部位からそれぞれ約1kbpおよび1.8kbpに位置していた。したがって、ORF163とORF167との間にある非必須遺伝子またはフラグメント化遺伝子が除去の標的とされた。具体的には、ORF164(A51R‐A55のフラグメント)、ORF165(A56Rプロモーターの一部を欠いている)、ORF166、およびフラグメント化したA57Rが除去の標的となった。これらの非必須遺伝子およびフラグメント化遺伝子の除去を行うために、核酸構築物(すなわちシャトルベクター)が、MVAゲノムのORF163とORF167との間に相同組換えによって組換えを生じ、その結果として介在配列を除去することができるように設計された。そのような組換えを達成すると、核酸構築物は、ACAM3000_MVA_163由来の1つの核酸配列(左側の隣接配列)と、ACAM3000_MVA_167由来の1つの核酸配列(右側の隣接配列)とを含むことになる。これらの配列は該核酸構築物中で隣り合うことになり、これは、該配列がいかなるポックスウイルスORFによっても隔てられていないことを意味する。より具体的には、左側隣接部はA50RリガーゼORFのC末端を含有することになり、右側隣接部はプロモーター領域およびB1R ORFのN末端を含有することになる。ベクターの設計は図26に示されている。
【0177】
シャトルベクターを構築するために、隣接部はそれぞれ別々に作出された。再構築Del IIIベクターの左側隣接部は最初に以下のようにして構築された。
緑色蛍光タンパク質(GFP)をコードする遺伝子およびダイレクトリピートと共に左側隣接部全体(I8R遺伝子の一部を含有する隣接部1)を切り出すために、プラスミドLW‐73(図7)がEcoRIおよびXhoIで消化された。その後、GFPを含有するフラグメントが制限酵素AcsIおよびSacIで消化されてGFP遺伝子が取り出された。
【0178】
ORF163のC末端部分を含んでいる左側隣接部を作出するために、プライマーLW470(配列番号23)およびLW471(配列番号24)を使用したポリメラーゼ連鎖反応(PCR)法によってMVAゲノムからDNAフラグメントが増幅された。PCR増幅は標準的な条件を使用して実施された。次に、ORF163の末端のダイレクトリピートが、プライマーLW‐472(配列番号25)およびLW‐473(配列番号26)を使用してMVAゲノムから増幅された。最後に、EcoRI部位およびXhoI部位を備えたベクター骨格、AcsI部位およびSacI部位を備えたGFP遺伝子、EcoRI部位およびAscI部位を含有するORF163フラグメント(左側隣接部)、ならびにSacI部位およびXhoI部位を含有するORF163のC末端領域由来のダイレクトリピートが合わせてライゲーションされて、中間プラスミド#2743が形成された。
【0179】
ORF167のN末端部分(そのプロモーター領域を含む)を含有している右側隣接部を作出するために、中間プラスミド#2743は制限酵素PstIおよびHindIIIで消化されて右側隣接部が放出された。次に、プライマーLW‐474(配列番号27)およびLW‐475(配列番号28)を使用してMVAゲノムからDNAフラグメントがPCR増幅された。このフラグメントは制限酵素PstIおよびHindIIIで消化され、該消化フラグメントは同様に消化されたシャトルベクター骨格にライゲーションされてLW‐676核酸構築物が作られた。(図27)
pLW‐76の顕著な特色は次のとおりである:
1) 該ベクターは、MVAゲノム中のA50R DNAリガーゼ遺伝子(ORF163)の端部と、プロモーターおよびB1Rキナーゼ遺伝子(ORF167)のN末端部分との間に外来遺伝子を挿入するために設計されている。左側隣接部はA50Rリガーゼ遺伝子の端部からなり、右側隣接部はプロモーターおよびB1Rキナーゼの先頭部からなる。
【0180】
2) 組換えウイルスの簡単な初期選択のためにGFP遺伝子が含まれている。
3) GFPはA50Rリガーゼ遺伝子のダイレクトリピートに隣接していることにより、組換えウイルスの継代が繰り返されるとGFPが失われるのでGFPの一時的発現を可能にする。特色2および3は、MVA/HIV組換え体を作製するために使用された初期のプラスミドpLAS‐1およびpLAS‐2にも含まれていた。
【0181】
その後、ウガンダのクレードDのヒト免疫不全ウイルス(HIV)(単離物AO7412)由来のenv遺伝子が新しいpLW‐76構築物にクローニングされた。このenvを含有する核酸構築物はその後細胞にトランスフェクションされ、該細胞をMVAウイルスに感染させることでHIV ENVタンパク質を発現する組換えMVAウイルスrMVA/UGDenv(delIIIrst)が生成された。次いでこのウイルスが特徴解析された。
【0182】
ニワトリ胚繊維芽細胞(CEF)において生育させると、rMVA/UGDenv(delIIIrst)による感染がシンシチウム型の細胞質効果(CPE)をもたらすことが観察された。これは、再構築されたdel III部位で生じた組換えの際の非必須A56ヘマグルチニン遺伝子の欠失に起因していた。正常なrMVAはフレイ(flay)フォーカス(図28A)を有していたが、rMVA/UGDenv(delIIrt)に感染すると、シンシチウム形成を示すフォーカスを示し、凝縮されたシンシチウムへと進行した。(図28B)。
【0183】
その後、rMVA/UGDenv(delIIIrst)は、挿入された異種核酸配列の安定性に関して特徴解析された。これは、CEF細胞中でウイルスを繰り返し継代すること、および、各世代について、発現されたHIV ENVタンパク質の存在を試験することにより行われた。ENVタンパク質の検出は、HIVエンベロープタンパク質に対するモノクローナル抗体を用いてウイルスのプラークをスクリーニングすることにより行われた。rMVA/UGDenv(delIIIrst)の安定性は、del II領域にenv遺伝子を含有しているウイルス、およびenv遺伝子が中央の保存領域へ挿入されたウイルスと比較された。この比較の結果は図29に示されている。発現されているENVタンパク質のレベルも、HIV ENVタンパク質に対するモノクローナル抗体を使用してウエスタンブロット法により計測された。
【0184】
図29は、MVA/UGDenv(del II)が、env内部に生じ、かつ隣接するMVAに及んでいる欠失が原因で、明らかに不安定であったことを示している。UGD envが、MVA/UGDenv(I8/G1)のように2つのVACV必須遺伝子の間に置かれると、生存可能な欠失が防止された。最後に、HIV env遺伝子のrMVA/UGDenv(del IIIrst)への組み入れは、少なくとも11回の継代を通じて安定であることが観察された。
【0185】
図30は、11種のウイルス構築物が同様の量のENVタンパク質を発現したことを示している。
したがって、これらの研究の結果は、MVAウイルスゲノムのdel III領域が、非必須遺伝子を除去してdel III部位を再構築することにより、より安定となったことを示唆している。
【0186】
本発明は明瞭さおよび理解を目的としてある程度詳細に説明されているが、当業者であれば、本発明の真の範囲から逸脱することなく形態および詳細の様々な変更を行なうことができることを認識するであろう。上記に引用されたすべての図面、表および付録、ならびに特許、出願および公開は、参照により本願に組込まれる。
【技術分野】
【0001】
本発明は、MVAゲノム中への異種DNA配列の安定的組込みに有用な挿入部位に関する。より具体的には、本発明は、必須遺伝子および非必須遺伝子の組み合わせを包含する改変ワクシニアアンカラ(MVA)ウイルスゲノムの領域を再構築して、異種DNAが該ゲノム中への安定的な組込み状態を維持するようにする方法に関する。
【背景技術】
【0002】
ポックスウイルス・ファミリーを構成するウイルスは、数百種のタンパク質をコードしている大きな二本鎖DNAゲノムを有する(非特許文献1)。ポックスウイルスは、脊椎動物および昆虫の宿主域を踏まえて、コードポックスウイルス亜科(Chordopoxvirinae)およびエントモポックスウイルス亜科(Entomopoxvirinae)に分けられる。コードポックスウイルス亜科は8つの属すなわち:オルソポックスウイルス属(Orthopoxvirus)、パラポックスウイルス属(Parapoxvirus)、アビポックスウイルス属(Avipoxvirus)、カプリポックスウイルス属(Capripoxvirus)、レポリポックスウイルス属(Leporipoxvirus)、スイポックスウイルス属(Suipoxvirus)、モルシポックスウイルス属(Molluscipoxvirus)、およびヤタポックスウイルス属(Yatapoxvirus)で構成されている。オルソポックスウイルス属の原型株はワクシニアウイルスである。ワクシニアウイルス(VACV)は、無関係な病原体に対する防御免疫応答を引き起こすことが示された最初の組換えウイルスであり(非特許文献2;非特許文献3)、家畜用および野生生物用ワクチン剤のためのベクターとして使用されている(非特許文献4)。しかしながら、ヒトに使用するための組換えVACVの開発は安全性の問題により阻まれてきた。この理由から、免疫不全の動物においても模範的な安全プロファイルを備えた極めて弱毒化された天然痘ワクチンである改変VACVアンカラ(MVA)に、関心が集まっている(非特許文献5〜非特許文献7)。MVAのゲノム配列(非特許文献8)は、ほとんどの哺乳動物細胞において増殖することができず、かつ組換えワクチンベクターの優れた候補であることが知られており(非特許文献9ならびに非特許文献10)、ニワトリ胚線維芽細胞において570回を越えて継代され、その間に、親の野生型アンカラ株に対して6つの主な欠失が、宿主域の厳しい制限を伴って発生した(非特許文献11)。MVAは宿主域が厳しく制限されており、ほとんどの哺乳動物細胞において、ビリオンのアセンブリ阻止が原因でほとんどまたは全く繁殖しない(非特許文献9)。組換えMVA(rMVA)を用いた初期実験から実証されたその能力は、外来タンパク質を確実に発現すること(非特許文献9)、ならびに防御的な体液性免疫および細胞性免疫を誘導することであった(非特許文献10)。現在、種々様々な病原体の遺伝子を発現するrMVA候補ワクチンが、動物およびヒトの試験を受けている(非特許文献12)。
【0003】
候補のヒト免疫不全ウイルス(HIV)ワクチンおよびその他のワクチンを開発している間に、突然変異体rMVAが組織培養継代後に外来タンパク質を発現する能力を失うことが観察された(非特許文献13〜非特許文献15)。しかしながら、この不安定性は、個々のプラークが単離されて分析されなければ当初は察知されずにいる可能性がある。しかし、集団内において一度確立されると、非発現体は元のrMVAを圧倒して急速に増殖することができる。上記を考慮することは、rMVAの大規模なワクチンシードストックの製造にとって特に重要である。MVAにおけるクローン化遺伝子の不安定性は驚くべきことである、というのも、MVAはすでにニワトリ胚線維芽細胞(CEF)における数百回の継代を通じてその適応時の遺伝的変化を経ており、現在は完全に安定しているからである。実際、3つの独立したプラーク単離物(受託番号U94848、AY603355、およびDQ983236)について、アントワーヌらにより、同一の167,000bpのゲノム配列が報告されている(非特許文献16)。遺伝子挿入物の不安定性の原因はこれまで調査されていないが、組換えタンパク質の有害な影響が非発現性突然変異体の選択優位性において役割を果たすように思われる。したがって、パラインフルエンザウイルスおよび麻疹ウイルスの膜貫通型タンパク質の発現レベルを低減すること、ならびにHIV Envの細胞質尾部の一部を欠失させることにより、rMVAの安定性が改善される(非特許文献13、非特許文献14、非特許文献15)。しかしながら、発現の低減は免疫原性も低下させる可能性があり、したがって不適当な場合がある。(非特許文献17)。
【0004】
ワクチンとしてのrMVAの潜在的価値を考慮すれば、この有害な不安定性の問題を理解し、かつ安定な組換えMVAウイルスを構築する方法を開発することが重要である。さらに、安定性の問題についての理解は、他のDNA発現ベクターに当てはまる洞察を提供することも考えられる。本発明はそのような洞察を提供し、かつ安定な組換えMVAウイルスの構築という問題の解決策を提供する。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】D.M.クナイプ(Knipe)、P.M.ハウリー(Howley)、D.E.グリフィン(Griffin)、R.A.ラム(Lamb)、M.A.マーティン(Martin)、B.ロイズマン(Roizman)、およびS.E.シュトラウス(Straus)編、「フィールズ・ウイルス学(Fields Virology)」第5版、モス(Moss)、B.著、「ポックスウイルス科:ウイルスおよびその複製(Poxviridae: The Viruses and Their Replication)」、米国フィラデルフィア州フィラデルフィア、リッピンコット・ウィリアムズ・アンド・ウィルキンズ(Lippincott Williams and Wilkins)、2007年
【非特許文献2】モス(Moss)、B.、G.L.スミス(Smith)、J.L.ゲリア(Geria)、およびR.H.パーセル(Purcell)、「生きている組換えワクシニアウイルスはB型肝炎からチンパンジーを防御する(Live recombinant vaccinia virus protects chimpanzees against hepatitis B.)」、ネイチャー誌(Nature)、1984年、第311巻、p.67−69
【非特許文献3】パオレッティ(Paoletti)、E.、B.R.リピンスカス(Lipinskas)、C.サムソノルフ(Samsonolf)、S.R.マーサー(Mercer)、およびD.パニカリ(Panicali)、「遺伝子組換えポックスウイルスを使用する生ワクチンの構築;B型肝炎ウイルス表面抗原および単純ヘルペスウイルス糖タンパク質Dを発現するワクシニアウイルス組換え体の生物学的活性(Construction of live vaccines using genetically engineered poxviruses; biological activity of vaccinia virus recombinants expressing the hepatitis B virus surface antigen and the herpes simplex virus glycoprotein D)」、米国科学アカデミー紀要(Proc. Natl. Acad. Sci. USA)、1984年、第81巻、p.193−197
【非特許文献4】モス(Moss)、B.、「組換え遺伝子の発現、ワクチン接種、および安全性のための遺伝子組換えポックスウイルス(Genetically engineered poxviruses for recombinant gene expression. vaccination, and safety.)」、米国科学アカデミー紀要(Proc. Natl. Acad. Sci. USA)、1996年、第93巻、p.11341−11348
【非特許文献5】マイア(Mayr)、A.、V.ホッホスタイン‐ミンツェル(Hochstein−Mintzel)、およびH.スティックル(Stickl)、「弱毒化ワクシニアウイルス株MVAの継代経歴、特性、および適用性(Passage history, properties, and applicability of the attenuated vaccinia virus strain MVA.)」、インフェクション誌(Infection)、1975年、第3巻、p.6−14(独語)
【非特許文献6】スティックル(Stickl)、H.、V.ホッホスタイン‐ミンツェル(Hochstein−Mintzel)、A.マイア(Mayr)、H.C.フーバー(Huber)、H.シェーファー(Schafer)、およびA.ホルツナー(Holzner)、「天然痘のMVAワクチン接種:弱毒化された生ワクシニアウイルス株(MVA)の臨床試験(MVA vaccination against smallpox: clinical trial of an attenuated live vaccinia virus strain (MVA).)」、ドイチェ・メディツィニッシェ・ボーヘンシュリフト誌(Dtsch. Med. Wschr.)、1974年、第99巻、p.2386−2392(独語)
【非特許文献7】スティッテラー(Stittelaar)、K.J.、T.クイケン(Kuiken)、R.L.デ・スワルト(de Swart)、G.ファン・アメロンゲン(van Amerongen)、H.W.ウォス(Vos)、H.G.ニーステルス(Niesters)、P.ファン・スハルクウェイク(van Schalkwijk)、T.ファン・デル・クワスト(van der Kwast)、L.S.ワイアット(Wyatt)、B.モス(Moss)、およびA.D.オスターハウス(Osterhaus)、「免疫抑制マカクにおける改変ワクシニアウイルス・アンカラ(MVA)の安全性(Safety of modified vaccinia virus Ankara (MVA) in immune−suppressed macaques.)」、ワクチン誌(Vaccine)、2001年、第19巻、p.3700−3709
【非特許文献8】マイア(Mayr)、A.ら、ツェントラルブラット・フュール・バクテリオロギー(Zentralbl Bakteriol)、1978年、第167巻、p.375−390
【非特許文献9】サッター(Sutter)、G.およびモス(Moss)、B.、「非複製型ワクシニアベクターは組換え遺伝子を効率的に発現する(Nonreplicating vaccinia vector efficiently expresses recombinant genes.)」、米国科学アカデミー紀要(Proc. Natl. Acad. Sci. USA)、1992年、第89巻、p.10847−10851
【非特許文献10】サッター(Sutter)、G.、L.S.ワイアット(Wyatt)、P.L.フォーリー(Foley)、J.R.ベニンク(Bennink)、およびB.モス(Moss)「宿主域が制限され、かつ高度に弱毒化されたワクシニアウイルスMVA株に由来する組換えベクターはマウスにおいてインフルエンザウイルスに対する防御免疫を活性化する(A recombinant vector derived from the host range−restricted and highly attenuated MVA strain of vaccinia virus stimulates protective immunity in mice to influenza virus)」、ワクチン誌(Vaccine)、1994年、第12巻、p.1032−1040
【非特許文献11】マイヤー(Meyer)、H.ら、ジャーナル・オブ・ジェネラル・ビロロジー誌(J Gen Virol)、1991年、第72巻、p.1031−1038
【非特許文献12】ゴメス(Gomez)、C.E.、J.L.ナジェラ(Najera)、M.クルーパ(Krupa)、およびM.エステバン(Esteban)、「ポックスウイルスベクターMVAおよびNYVAC 感染症およびがんに対するワクチン接種のための遺伝子送達システム(The poxvirus vectors MVA and NYVAC a gene delivery systems for vaccination against infection diseases and cancer)」、カレント・ジーン・セラピー誌(Curr. Gene Ther.)、2008年、第8巻、p.97−120
【非特許文献13】スティッテラー(Stittelaar)、K.J.、L.S.ワイアット(Wyatt)、R.L デ・スワルト(de Swart)、H.W.ウォス(Vos)、J.グローエン(Groen)、G.ファン・アメロンゲン(van Amerongen)、R.S.ファン・ビンネンダイク(van Binnendijk)、S.ローゼンブラット(Rozenblatt)、B.モス(Moss)、およびA.オスターハウス(Osterhaus)「受動的獲得抗体の存在下における改変ワクシニアウイルスアンカラ系の麻疹ウイルスワクチンを接種されたマカクの防御免疫(Protective immunity in macaques vaccinated with a modified vaccinia virus Ankara−based measles virus vaccine in the presence of passively acquired antibodies)」、ジャーナル・オブ・ビロロジー誌(J. Virol)、2000年、第74巻、p.4236−4243
【非特許文献14】ワイアット(Wyatt)、L.S.、I.M.ベリアコフ(Belyakov)、P.L.アール(Earl)、J.A.ベルゾフスキー(Berzofsky)、およびB.モス(Moss)、「組換えMVAによって発現されたHIV Envの自然発生的切断に起因する、細胞表面発現、免疫原性および遺伝的安定性の増大(Enhanced cell surface expression, immunogenicity and genetic stability resulting from a spontaneous truncation of HIV Env expressed by a recombinant MVA)」ビロロジー誌(Virology)、2008年、第372巻、p.260−272
【非特許文献15】ワイアット(Wyatt)、L.S.、S.T.ショル(Shors)、B.R.マーフィー(Murphy)、およびB.モス(Moss)、「動物モデルにおけるパラインフルエンザウイルス3感染に対して有効な複製欠損型組換えワクシニアウイルスワクチンの開発(Development of a replication−deficient recombinant vaccinia virus vaccine effective against parainfluenza virus 3 infection in an animal model.)」、ワクチン誌(Vaccine)、1996年、第14巻、p.1451−1458
【非特許文献16】アントワーヌ(Antoine)、G.、F.シェイフリンガー(Scheiflinger)、F.ドルナー(Dorner)、およびF.G.フォークナー(Falkner)、訂正記事10「改変ワクシニアアンカラ(MVA)株の完全なゲノム配列:他のオルソポックスウイルスとの比較(The complete genomic sequence of the modified vaccinia Ankara (MVA) strain: comparison with other orthopoxviruses)」、ビロロジー誌(Virology)、2006年、第350巻、p.501−502、[1998年第244巻p.365の訂正]
【非特許文献17】ワイアット(Wyatt)、L.S.、P.L.アール(Earl)、J.フォークト(Vogt)、L.A.エラー(Eller)、D.チャンドラン(Chandran)、J.リウ(Liu)、H.L.ロビンソン(Robinson)、およびB.モス(Moss)、「組換え改変ワクシニアウイルスアンカラHIVワクチンの免疫原性とin vitro発現レベルとの関連(Correlation of immunogenicities and in vitro expression levels of recombinant modified vaccinia virus Ankara HIV vaccines)」、ワクチン誌(Vaccine)、2008年、第26巻、p.486−493
【発明の概要】
【0006】
本発明は、改変ワクシニアアンカラ(MVA)ウイルスのゲノムを、該ゲノムの領域を再構築することによってより安定させることができるという発見に関する。特に、本発明者らは、非必須遺伝子を含有するゲノム領域が遺伝学的に不安定であることを発見した。さらに、そのような領域は、非必須DNAを除去し、これらの領域の必須遺伝子を互いに隣り合わせることによってより安定させることが可能である。必須遺伝子の喪失は増殖上の不都合を有するウイルスを生じるので、そのようなウイルスはウイルス集団から急速に失われて、必須遺伝子および任意の介在DNAが維持されたウイルスの集団が生じる。
【0007】
本開示は、組換え改変ワクシニアアンカラ(MVA)ウイルスであって、MVAウイルスゲノムの2つの隣り合った必須オープンリーディングフレームの間に異種核酸配列を含むウイルスを提供する。必須ORFの選択は、該ORFが、本発明の組換えウイルスを構築するために使用される親MVAウイルスのゲノム中で隣り合っていないように行われる。すなわち、該必須ORFは少なくとも1つの非必須ORFによって隔てられている。しかしながら、組換え改変ワクシニアアンカラ(MVA)子孫ウイルスでは、該必須ORFは隣り合わせにされている。すなわち、必須ORFの間には介在する非必須ORFが存在しない。従って、必須ORFの間の領域は安定であり、ウイルス集団内で維持される。従って、この領域は、異種核酸配列の挿入のための新規かつ有用な部位を提供する。そのような異種核酸配列は、抗原のような、治療上有用なタンパク質をコードすることができる。
【0008】
本開示はさらに、本発明の組換え改変ワクシニアアンカラ(MVA)ウイルスを構築するために使用することができる核酸構築物も提供する。そのような構築物は、親MVAウイルス由来であって親ウイルスにおいては隣り合っていない必須ORFを含んでいる。しかしながら、開示される核酸構築物では、これらの必須ORFは互いに隣り合わせにされている。さらに、必須ORFの間に遺伝子間領域を含有する構築物が開示され、これらは異種核酸配列の挿入に使用することができる。
【0009】
最後に、本発明のウイルスを疾患の予防および治療のために使用する方法も開示される。
【図面の簡単な説明】
【0010】
【図1】pol遺伝子塩基配列の同一性に基づいたHIV‐1およびHIV‐2の系統関係を示す図。SIVcpzおよびSIVsmmは、それぞれチンパンジーおよびスーティーマンガベイから検出された、ヒトに近い霊長類のレンチウイルスである。
【図2】4つの異なるSIVcpz単離物を用いた、完全長pol遺伝子塩基配列に基づいたHIV‐1のグループM、NおよびOの系統関係を示す図。バーは遺伝的距離0.1(10%のヌクレオチド相違)を示し、アステリスクはenvの配列に基づいてグループNのHIV‐1単離物を位置付けている。
【図3】HIV‐1単離物の指向性および生物学的特性を示す図。
【図4】HIVにコードされたタンパク質を示す図。HIV遺伝子の位置、一次翻訳産物(場合によってはポリプロテイン)のサイズ、およびプロセシングされた成熟ウイルスタンパク質が示されている。
【図5】成熟HIV‐1ビリオンの概略図。
【図6】HIV‐1 Env糖タンパク質を直線的に表す図。矢印は、gp160が切断されてgp120およびgp41となる部位を示す。gp120では、斜線の付されたエリアは可変ドメイン(V1〜V5)を表わし、無地のボックスは保存配列(C1〜C5)を示している。gp41エクトドメインでは、いくつかのドメインが示されている、すなわち:N末端融合ペプチド、および2つのエクトドメインへリックス(NへリックスおよびCへリックス)である。膜貫通ドメインは黒色のボックスによって表わされている。gp41細胞質ドメインでは、Tyr‐X‐X‐Leu(YXXL)エンドサイトーシス・モチーフ(配列番号1)および2つの予測へリックスドメイン(ヘリックス‐1および‐2)が示されている。アミノ酸番号が示されている。
【図7】pLW‐73核酸構築物(配列番号2および3)を示す図。
【図8】導入用ベクターpLW‐73のヌクレオチド配列を示す図(上側の鎖、配列番号2;下側の鎖、配列番号3)。
【図9】ウガンダ人のクレードD Envタンパク質(単離物AO7412)をコードしているヌクレオチド配列を示す図(配列番号4)。
【図10】ウガンダ人のクレードD gagpolタンパク質(単離物AO3349)をコードしているコドン変更されたヌクレオチド配列を示す図(配列番号5)。
【図11】組換えMVAの生成および挿入遺伝子の安定性の分析について示す図。A)envおよびgagpolがそれぞれDel II部位およびDel III部位へ挿入されることを示す概略図。B)免疫染色による安定性の評価。
【図12】MVA/65A/G envにおけるenv突然変異の種類および頻度を示す図。
【図13】I8R/G1L IGRにEnv、Del IIIにGagPolが挿入されることを示す図。
【図14】安定性を増大するためのA/G構築物への改変を示す図。
【図15】プラーク継代後のEnvの発現について示す図。
【図16】個々のクローンのPCRおよびウエスタンブロット解析を示す図。
【図17】二重組換えMVAによるA/G envの発現を示す図。
【図18】ウガンダ人のHIV‐1単離物由来のenvおよびgagpolを発現する組換えウイルスを示す図。
【図19】MVA/UGD4a − 非染色envプラークの分析について示す図。
【図20】組換えMVAにおけるUGD env遺伝子の改変を示す図。
【図21】MVA/UGD4b − 非染色gagプラークの分析について示す図。*はGまたはC残基が4〜6個連続した箇所の位置である。
【図22】組換えMVAにおけるUGD gagpol遺伝子の改変を示す図。
【図23】UGDのenvおよびgagpolを発現する安定した組換えMVAの構築を示す図。
【図24】MVA/UGD4dによって誘発された細胞応答を示す図。
【図25】MVA/UGD4dによって誘発された抗体応答を示す図。
【図26】MVAウイルスゲノムのdel III部位を再構築する方法の概要を示す図。
【図27】pLW‐76核酸構築物(配列番号21および22)を示す図。
【図28】del III部位の再構築によるrMVAのシンシチウム表現型を示す図。
【図29】様々な組換えMVAウイルスにおける異種核酸の安定性の比較を示す図。
【図30】様々な組換えMVAウイルスにおけるUGD Envタンパク質発現レベルの比較を示す図。
【図31】導入用ベクターpLW‐76のヌクレオチド配列(配列番号21および22)を示す図。
【発明を実施するための形態】
【0011】
微生物の寄託
次の微生物は、ブダペスト条約の条項に従って記載の日時に米国バージニア州マナッサス所在のアメリカン・タイプ・カルチャー・コレクション(ATCC)に寄託済みである。
【0012】
【表1】
MVA 1974/NIHクローン1は、2003年3月27日、米国バージニア州20110‐2209、マナッサス、ユニバーシティ・ブールバード10801、アメリカン・タイプ・カルチャー・コレクション(ATCC)に、ATCC受託番号PTA‐5095として寄託された。本寄託は、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約およびその規定(ブダペスト条約)に従って行われた。これは、寄託日時から30年間にわたり該寄託物の生存培養物の維持を保証するものである。寄託物はブダペスト条約の条項の下でATCCより入手可能となり、また、関連する米国特許の発行または任意の米国もしくは外国特許出願の公開のいずれか早い方に際して公衆が該寄託物の培養物の子孫を恒久的かつ無制限に入手可能であることを保証し、かつ米国特許法第122条およびこれに関連する長官が定める規則(米国特許法施行規則第1.14条を含む)に従って権利を与えられることが米国特許商標庁長官によって決定された者が該子孫を入手可能であることを保証する、出願人とATCCとの間の合意に従うことになる。寄託された株が入手可能であるということは、任意の政府の権限の下で該政府の特許法に従って与えられた権利に違反して本発明を実行することが許されると解釈されるものではない。
【0013】
好ましい実施形態の詳細な説明
本発明についてさらに説明する前に、了解されるべきことは、本発明は記載される特定の実施形態に限定されるものではなく、それ自体が当然多様であってよいことである。さらに当然ながら、本発明の範囲は特許請求の範囲によってのみ限定されるので、本明細書中で使用される専門用語は特定の実施形態のみについて説明するためのものであって、限定するようには意図されていない。
【0014】
別途定義のないかぎり、本明細書中で使用される技術用語および科学用語は本発明の属する分野の当業者が一般に理解するのと同じ意味を有する。例えば、シングルトン(Singleton)Pおよびセインズベリー(Sainsbury)D.、「微生物学および分子生物学の辞典(Dictionary of Microbiology and Molecular Biology)」第3版、米国ニューヨーク州チチェスター、J.ワイリー・アンド・サンズ(J. Wiley and Sons)、2001年、ならびに、D.M.クナイプ(Knipe)、P.M.ハウリー(Howley)、D.E.グリフィン(Griffin)、R.A.ラム(Lamb)、M.A.マーティン(Martin)、B.ロイズマン(Roizman)、およびS.E.シュトラウス(Straus)編、「フィールズ・ウイルス学(Fields Virology)」第5版、米国フィラデルフィア州フィラデルフィア、リッピンコット・ウィリアムズ・アンド・ウィルキンズ(Lippincott Williams and Wilkins)、2007年)を参照のこと。本発明の実施または試験においては、本明細書に記載のものに類似または等価の任意の方法および材料も使用可能であるが、好適な方法および材料についてここで説明する。本明細書中で言及された出版物はすべて、該出版物が関連して引用された方法または材料のうち少なくともいずれかを開示かつ説明するために、参照により本願に組み込まれる。
【0015】
本発明によれば、単離されたタンパク質(または核酸分子)とは、その天然の環境から取り出されたタンパク質(または核酸分子)である。単離されたタンパク質(または核酸分子)は、例えば、その天然の供給源から得られてもよいし、組換えDNA技術を使用して生産されてもよいし、化学的に合成されてもよい。そのため、単離された、とは、タンパク質または核酸分子が精製された状態や程度を反映するものではない。
【0016】
留意せねばならないのは、本明細書および添付の特許請求の範囲において使用されるように、単数形の「1つの(a)」、「1つの(an)」、および「その(the)」は、文脈がそうでないことを明白に規定していないかぎり、複数の指示物を含むことである。さらに留意すべきは、特許請求の範囲はあらゆる任意選択の要素を除外して作成されうることである。そのため、この記述は、請求項の構成要件の記述に関する「専ら(solely)」、「〜のみ(only)」などのような排他的な用語の使用、または「否定的な(negative)」限定の使用について、先行記述としての役割を果たすことが意図されている。
【0017】
本明細書中で使用されるように、用語「1つの(a)」物または「1つの(an)」物は、1つ以上のその物を指すことが理解されよう。例えば、(1つの(a))核酸分子は1つ以上の核酸分子を指す。そのため、用語「1つの(a)」、「1つの(an)」、「1つ以上の(one or more)」および「少なくとも1つの(at least one)」は互換的に使用可能である。同様に、用語「〜を含む(comprising)」、「〜を含む、〜を備える(including)」および「〜を有する(having)」は互換的に使用可能である。
【0018】
移行語「〜を含む(comprising)」は、「〜を含む、〜を備える(including)」、「〜を含有する(containing)」、または「〜を特徴とする(characterized by)」と同義であり、包括的すなわちオープンエンドであって、記述されていないさらなる構成要件または方法ステップを除外しない。
【0019】
移行句「〜からなる(consisting of)」は、請求項において指定されていないあらゆる構成要件、ステップ、または成分を除外するが、通常付随する不純物のような、本発明とは無関係な追加の構成成分またはステップは除外しない。
【0020】
移行句「本質的に〜からなる(consisting essentially of)」は、請求項の範囲を、指定された材料またはステップ、およびクレームに記載された発明の基礎的かつ新規な特徴に実質的に影響を及ぼさないものに限定する。
【0021】
本明細書中で議論された出版物は、専ら本願の出願日以前の開示について提供される。本明細書は、本発明が先行発明に基づくそのような出版物に先行することは認められないという了解として解釈されるべきではない。さらに、提示された出版日は実際の出版日とは異なる可能性があり、独立に確認する必要がある場合もある。
【0022】
当然のことであるが、明確さのために別個の実施形態において記載されている本発明のある種の特徴が、単一の実施形態において組み合わせて提供されることも可能である。反対に、簡潔にするために単一の実施形態を背景として記載されている本発明の様々な特徴が、別々に、または任意の適切な小さな組み合わせ(サブコンビネーション)で提供されてもよい。実施形態の全ての組み合わせが、本発明によって明確に包含され、かつ、ありとあらゆる組み合わせが個々に明示的に開示されているかのように、本明細書中に開示される。さらに、小さな組み合わせも全て、本発明によって明確に包含され、かつ、ありとあらゆる小さな組み合わせが個々に明示的に本明細書中に開示されているかのように、本明細書中に開示される。
【0023】
全ゲノム塩基配列は、コードポックスウイルスの各々の属のウイルスのうち少なくとも1つについて、および2つのエントモポックスウイルスについて報告されている。ほぼ100個の遺伝子がすべてのコードポックスウイルスにおいて保存されており、これらの約半分がエントモポックスウイルスにも存在する。上記に基づいて、いくつかの一般化を行うことができる。すなわち、遺伝子は、大部分はオーバーラップしておらず、ゲノムの近いほうの端部に向かってブロック単位で存在する傾向があり、高度に保存されていて必須の複製機能に関係している場合は通常は中央領域に位置し、可変的であり宿主との相互作用に関係している場合は通常は端部領域に位置している。中央の遺伝子の配置構成は、すべてのコードポックスウイルスにおいて著しく類似している。ゲノム全体の塩基配列決定以前に始まり、その後ワクシニアウイルスのコペンハーゲン株の完全配列について使用された、ワクシニアウイルスの遺伝子またはORF(オープンリーディングフレーム)の命名のための慣例は、HindIII制限エンドヌクレアーゼによるDNAフラグメントの文字、次にフラグメント内のORF番号(左から右へ)、そして、ORFの方向に応じてLまたはRを使用することからなる。この規則にはHindIII Cフラグメントについて例外が作られた。すなわち、ORFについて、高度に可変的なゲノムの左端から始まるのを避けるため、右から番号付けが行われた。ポリペプチド名は、LまたはRが外される以外は遺伝子名に対応する。ほとんどの後続の完全長ポックスウイルスゲノム配列では、ORFはゲノムの一端から他端へ連続的に番号付けが行われた。しかしながら、文献上の連続性を提供するために古い文字表示が一般名として保持されてきた。ワクシニアウイルスのウエスタン・リザーブ(WR)株のORF番号は、この株が生化学および遺伝学研究の大多数について使用されてきたため、参考図書において共通して示されている。
【0024】
本発明の発明者らは、改変ワクシニアアンカラ(MVA)ウイルスのゲノム内に外来DNA配列の安定な挿入を行うための新たな部位、ならびに新たな部位を作出する方法を特定した。本発明は、安定な組換えMVAウイルスを構築する方法の特定を目的とした研究から生じた。従来観察されてきたように、MVAゲノムに挿入された異種DNA配列を含有している組換えMVAを得ることが可能である一方、これらの挿入物は多くの場合不安定であった。この不安定性の調査から、ORFとORFとの間の場所への、ウイルス増殖に必須でない異種DNA配列の挿入は、同じくウイルスによって欠失させられる恐れがある、という結論が生じた。したがって、安定な組換えMVAウイルスを構築するための改良された戦略の必要性が認識された。
【0025】
本明細書中で使用されるように、オープンリーディングフレーム(ORF)とは、タンパク質のアミノ酸をコードする一続きの連続したヌクレオチドを意味する。そのようなタンパク質はペプチド、ポリペプチドであってよく、単一アミノ酸より大きい任意の長さであってよい。当然のことながら、ORFは、アミノ酸をコードしない終止コドンも含むことができる。当業者には当然のことであるが、組換え事象により、一部のORFはその本来のコード能力の一部を失っており、よって機能を持たないタンパク質をコードしている。そのようなORFはORFフラグメントと呼ばれることがある。ORFは、コード領域の外側に位置する調節要素(例えばプロモーター、転写制御要素、エンハンサーなど)を含まない。それに対し、遺伝子は、ORF(終止コドンを含む)および該ORFの転写を調節することができる調節要素を指す。
【0026】
ORFは、隣り合っているとされる場合もあれば隣り合っていないとされる場合もある。本明細書中で使用されるように、2つのORFは、該ORFが同じ核酸分子中に存在し、かつこれらの最も近接した2つの端部が別のポックスウイルスORFによって隔てられていない場合は、隣り合っている。隣り合っていないORFとは、その最も近接した2つの端部が別のポックスウイルスORFによって隔てられているORFである。隣り合ったORFとは、一方のORFに属する末端コドンと他方のORFに属する末端コドンとの間に他のヌクレオチド配列がないことを意味する、連続したものであってもよい。末端コドンとは、ORFの最初のコドンまたは最後のコドン(終止コドンを含む)を意味する。末端コドンの一例は、ORFによってコードされるタンパク質の最初の5’アミノ酸をコードするコドンである。末端コドンの別の例は、ORFによってコードされるタンパク質の最後の3’アミノ酸をコードするコドンである。末端コドンのさらに別の例は、ORFの終止コドンである。
【0027】
隣り合ったORFが核酸配列によって隔てられている場合もある。そのような配列は遺伝子間領域(intergenic region)と呼ばれる。本明細書中で使用されるように、遺伝子間領域とは、隣り合ったORFの最も近接した末端コドンの間の核酸配列であって、ポックスウイルス転写制御要素以外に、ワクシニアウイルスに由来するヌクレオチド配列を含んでいない配列を意味する。IGR配列は隣り合ったORFの終止コドンの外側に位置し、よってこれらの隣り合ったORFによってコードされるタンパク質のいかなる部分もコードしていない。IGR配列はポックスウイルス転写制御要素を含有する場合がある。IGRは、ポックスウイルス以外の生物に由来する配列を含有する場合もある。好ましくは、IGRは、ポックスウイルス転写制御要素の一部ではないいかなるポックスウイルス配列も有していない。一実施形態では、IGRは少なくとも1つの異種核酸配列を含む。そのような配列は、IGRの中に本来存在するかまたは他の異種核酸配列を挿入する目的でIGRに導入済みの、制限酵素認識部位すなわち制限部位に挿入可能である。
【0028】
ORFのヌクレオチド配列はタンパク質をコードするが、2つのORFの間の遺伝子間領域(IGR)にはコード能力はない。したがってIGRは、いかなるウイルスタンパク質の生産にも影響せずに異種DNAを挿入することができる部位としての役割を果たす可能性がある。しかしながらIGRは、ウイルス遺伝子発現の転写制御に必須であるかまたは関与する、調節要素、結合部位、プロモーター配列またはエンハンサー配列のうち少なくともいずれかを含む場合がある。したがって、IGRは、ウイルスの生活環の調節管理に関与する可能性がある。それでも本発明者は、IGRが、MVAの典型的な特徴および遺伝子発現に影響を及ぼしたり変化させたりせずにMVAゲノムに異種核酸配列を安定的に挿入するために使用可能であることを見出した。この新しい挿入部位は、MVAのORFやコード配列が変更されないので、特に有用である。
【0029】
本発明についてさらに説明する前に、ポックスウイルスゲノムにおける遺伝子の配置について理解することは有用である。ORFのヌクレオチド配列は、規則的に開始コドンで始まり終止コドンで終わる。2つの隣り合ったORFの向きに依存して、これらのORFの間の領域であるIGRは、2つの隣り合ったORFの2つの終止コドンによって挟まれるか、2つの隣り合ったORFの2つの開始コドンによって挟まれるか、第1のORFの終止コドンおよび第2のORFの開始コドンによって挟まれるか、または、第1のORFの開始コドンおよび第2のORFの終止コドンによって挟まれる。
【0030】
従って、IGRへの外来DNA配列の挿入部位は、第1のORFの終止コドンの下流すなわち3’側となる場合がある。第2のORFとも呼ばれる隣りのORFが第1のORFと同じ向きを有する場合、第1のORFの終止コドンの下流のこの挿入部位は、第2のORFの開始コドンの上流すなわち5’側に位置する。
【0031】
第2のORFが第1のORFに対して逆の向きを有する場合、これは2つの隣り合ったORFの向きが互いに向かっていることを意味するが、この場合挿入部位は両ORFの終止コドンの下流に位置する。
【0032】
第3の別例として、2つの隣り合ったORFが逆方向に読まれるが該2つの隣り合ったORFの向きが互いに離れる向きである場合、これは2つのORFの開始コドンが互いに隣り合っていることを特徴とする配置と同義であるが、この場合外来DNAは両方の開始コドンに対して上流に挿入される。
【0033】
MVAゲノム中のORFは2つのコード方向で存在する。従って、mRNA合成活動は、左から右すなわち順方向、および、同様に、右から左(逆方向)に生じる。ORFを、ゲノムの様々なHindIII制限酵素切断フラグメントにおける向きおよび配置によって同定することは、ポックスウイルス学における慣例であり、ワクシニアウイルスの標準的な分類となっている。命名法については、様々なHindIIIフラグメントはそのサイズの大きい順に対応して降順の大文字により示される。ORFは各HindIIIフラグメント上で左から右に番号付けされ、ORFの向きは、大文字のL(右から左への転写を表わす)またはR(左から右への転写を表わす)によって示される。さらに、MVAゲノム構造についてはごく最近の刊行物があるが、該刊行物は、ゲノムの左端から右端へと単純にORFを番号付けし、その向きを大文字のLまたはRで示す、異なる命名法を使用している(アントワーヌ(Antoine)、G.ら、ビロロジー誌(Virology)、1998年、第244巻、p.365−396)。例として、古い命名法によるI8R ORFは、アントワーヌらによる069R ORFに相当する。
【0034】
HIV遺伝子を発現する改変ワクシニアウイルス・アンカラ(MVA)の組換え体をワクチン剤候補として作製する努力において、本発明者らは、不安定性の原因の1つは外来遺伝子および隣接するMVA配列の欠失に起因することを究明した。この問題を克服する試みにおいて、本発明者らは、組換えMVAにおいて生存可能な欠失が生じるのを防止するために、保存された遺伝子の間に外来遺伝子を挿入しようと計画した。そのような欠失を備えたウイルスは生育上有利であり、よってrMVAウイルス集団において異常増殖することになる。ワクシニアゲノム中の保存された遺伝子(これらの遺伝子はワクシニアウイルスの複製に必要と考えられ、したがって「必須遺伝子」である)の間に外来遺伝子を挿入すれば、必須遺伝子のいかなる欠失もウイルス複製を阻害し、したがって組換えMVAを抑えて異常増殖することはないと思われる。したがって、rMVA集団の安定な発現が維持される。組換え体を作製するために本発明者らが使用してきたMVAの株は、該発明者らによって米国疾病管理予防センター(CDC)に提供され、その後アカンビス(Acambis)により塩基配列決定がなされた(Genbank受託番号AY603355)。ババリアン・ノルディック(Bavarian Nordic)が同社の国際公開公報第03/097845号の基礎としてきたMVAの株は、アントワーヌ(Antoine)、G.ら、ビロロジー誌(Virology)、1998年、第244巻、p.365−396で塩基配列決定がなされたワクシニアウイルス株改変ワクシニア・アンカラ(Genbank受託番号U94848)である。(所与の遺伝子についての上記2つの配列における遺伝子番号が異なることに注意されたい。)
本発明者らは当初、ポックスウイルス科において保存された遺伝子およびコードポックスウイルス亜科(脊椎動物のポックスウイルス)において保存された遺伝子に着目していた(アプトン(Upton)、C.ら、ジャーナル・オブ・ビロロジー誌(Journal of Virology)、2003年、第77巻、p.7590−7600)。これらの遺伝子は、ワールドワイドウェブのpoxvirus.orgに見出されるポックスウイルス・バイオインフォマティクス・リソース・センター(Poxvirus Bioinformatics Resource Center)に掲載されたコペンハーゲン・ワクシニアウイルス(Genbank受託番号M35027)の命名法でリストされている。これらの遺伝子は、ポックスウイルス科における合計49個の保存遺伝子と、コードポックスウイルスにおいて保存された41個の追加の遺伝子とであり、合計90個の保存遺伝子である。これらの90個の保存遺伝子から、本発明者らは、保存遺伝子の対の間の遺伝子間部位をリストした。これらの遺伝子対を以下の表1に列挙する。(ババリアン・ノルディックの国際公開公報第03/097845号に含まれていない遺伝子には印が付されていることに注意されたい)。
【0035】
【表2】
これらの遺伝子の向きは様々であり、あるものは右に、あるものは左に向かって転写される。このことは、遺伝子間部位のうちのいくつかは、挿入ベクターの構築において保存されるべきであろうプロモーターを含有することを意味している。さらに、オーバーラップしている保存遺伝子については、ベクター構築の際に、繰り返し配列を最小限にするために代替コドンを使用して遺伝子が再構築されるべきであろう。
【0036】
本発明者らは、保存遺伝子の3’終止コドンが互いに近接した、該遺伝子の向きが「終端対終端」である保存遺伝子に注目した。これらの部位で使用される導入ベクターの構築は、終止コドンと終止コドンとの間のこの領域にはプロモーターは存在しないであろうという事実によって容易になる。終止コドンを隔てる遺伝子間ヌクレオチドが存在する場合、挿入ベクターの構築は簡単である。一方の遺伝子の終止コドンが他方の遺伝子の3’末端の内部にある場合、プラスミド導入ベクターの構築の際に、遺伝子は、繰り返し配列を最小限にするために代替コドンを使用して再構築されてもよいし、または、オーバーラップ部分の長さに応じて、オーバーラップしないように隣接部のPCRにおいて単純に調整されてもよい。表2は、「終端対終端」である保存遺伝子の向きの要件を満たす遺伝子間部位を示す。灰色で強調表示された遺伝子間部位にはオーバーラップしている終端がなく、したがって構築が最も簡単である。
【0037】
【表3】
このリストから、本発明者らは、終端がオーバーラップしていない6つの遺伝子間部位に注目した。実施例においては、これら6つのうち遺伝子間部位071‐072(I8R‐G1L)が、異種遺伝子を挿入する部位として選択された。この遺伝子間部位を使用した組換えMVAウイルスの構築、および得られたウイルスの特徴については、実施例1に、および参照により全体が本願に組み込まれる国際公開公報第2008/142479号A2に、記載されている。
【0038】
上述された保存遺伝子および対応する遺伝子間部位に加えて、本発明者らは異種核酸配列の挿入に有用な他の部位を発見した。例えば、任意の遺伝子であってその欠失(または不活性化)が力価の0.5log、0.75logまたは1log(10倍)の低減をもたらすことが実験的に実証された遺伝子は、「必須遺伝子」と考えることができるかもしれない。同様に、必須遺伝子とは、該当する遺伝子が欠失または不活性化されなかったウイルスと比較して力価の少なくとも50%、少なくとも75%、または少なくとも90%の低減をもたらす任意の遺伝子である。この遺伝子が別の必須遺伝子に隣り合っていれば、2つの遺伝子の間の遺伝子間部位は異種核酸配列の挿入に有用な部位になるであろう。これらのORFの1つ以上が、間に入っている異種核酸配列とともに欠失しても、ウイルスの増殖は妨げないと思われる一方、これらのORFを含有しているウイルスと比較して増殖率の低下がもたらされるであろう。したがって、時間とともに、1つ以上の必須ORFを失ったウイルスは徐々にウイルス集団全体の中でより小さい割合を占めるようになり、十分な時間を経ると、ウイルス集団から完全に消失するであろう。
【0039】
したがって、本発明の1つの実施形態は、組換え改変ワクシニアアンカラ(MVA)ウイルスであって、MVAウイルス由来の2つの隣り合った必須ORFの間に、または該ORFに挟まれた、異種核酸配列を含むウイルスである。一実施形態では、隣り合ったORFは遺伝子間領域(IGR)によって隔てられている。既述のように、IGRは異種核酸配列を含有することができる。したがって、一実施形態は、組換え改変ワクシニアアンカラ(MVA)ウイルスであって、MVAウイルス由来の2つの隣り合った必須ORFの間に、または該ORFに挟まれた、遺伝子間領域内に異種核酸配列を含むウイルスである。
【0040】
本明細書中で使用されるように、異種の、または外来の核酸配列とは、自然界において通常は本発明によって使用されるポックスウイルスに関して見出されることのない配列である。本発明のさらなる実施形態によれば、外来の核酸配列は少なくとも1つのコード配列を含む。該コード配列は、転写制御要素に、好ましくはポックスウイルスの転写制御要素に、作動可能に連結される。さらに、ポックスウイルスの転写制御要素と、例えば内部リボソーム導入部位との間の組み合わせも使用可能である。
【0041】
さらなる実施形態によれば、異種核酸配列は、1個または数個の転写制御要素に連結された2以上のコード配列を含むこともできる。好ましくは、コード配列は1つ以上のタンパク質をコードする。いくつかの実施形態では、該タンパク質は抗原であるか、または、抗原エピトープであって特に治療面から見て興味深い遺伝子のものを含む。
【0042】
本発明による、治療面から見て興味深い遺伝子とは、疾患を引き起こす病原性または感染性の微生物の遺伝子に由来するかまたは相同である遺伝子であってよい。従って、本発明に関しては、そのような治療面から見て興味深い遺伝子は、特異的免疫応答に影響を及ぼす、好ましくは特異的免疫応答を引き起こすために生物体の免疫系に提示され、その結果として、該生物体に、微生物による感染に対するワクチン接種すなわち予防的保護をもたらす。本発明のさらに好ましい実施形態では、治療面から見て興味深い遺伝子は、感染性ウイルス、例えば、限定するものではないが、デング熱ウイルス、B型肝炎ウイルスもしくはC型肝炎ウイルス、またはHIVのようなヒト免疫不全ウイルスの遺伝子から選択される。
【0043】
本発明の好ましい実施形態によれば、異種核酸配列はHIVに由来し、かつHIV envをコードするものであって、好ましくは該HIV env遺伝子は隣り合ったORFの間のIGRに挿入される。後天性免疫不全症候群(AIDS)の病原因子は、ヒト免疫不全ウイルス(HIV)と呼ばれる、レンチウイルス属の典型的な特徴を示すレトロウイルスであることが認識されている。ヒトレンチウイルスの系統関係を図1に示す。HIV‐2は、HIV‐1に対するよりも、野生のスーティーマンガベイから単離されたウイルスであるSIVsmmに対してより近縁な関係にある。現在、HIV‐2はヒトへのSIVsmmの人獣共通感染を表わすものと考えられている。捕獲チンパンジー由来の、SIVcpzと表された一連のレンチウイルス単離物は、HIV‐1の遺伝的近縁種である。
【0044】
最も初期のHIV‐1単離物の系統解析は欧州/北米およびアフリカ由来の試料に集中し、世界のこれら2つのエリアから別々のウイルス群が同定された。その後、HIV‐1の個別の遺伝学的サブタイプまたはクレードが定義されて3つのグループすなわち:M(メジャー);O(アウトライヤー);およびN(MでもOでもない)に分類された(図2)。全世界のウイルス単離物の95%以上が含まれる、HIV‐1のMグループは、全ウイルスゲノムの配列に基づけば、少なくとも8つの別々のクレード(A、B、C、D、F、G、HおよびJ)で構成される。HIV‐1のグループOに属するものは、カメルーン、ガボンおよび赤道ギニアで生活する個体から回収され、これらのゲノムは、グループMのウイルスとはヌクレオチド配列で50%未満の同一性を共有する。さらに最近発見されたグループNのHIV‐1株は、カメルーン人感染者において同定され、標準的な全ウイルスの酵素結合抗体免疫吸着アッセイ(ELISA)では血清学的に反応しないが、従来のウエスタンブロット解析では容易に検出可能である。
【0045】
HIV‐1の遺伝的多様性に関する最新の知見は、多様な地理的起源のグループMのウイルスに関する研究からもたらされている。過去十年間に集められたデータは、個々の感染者に存在するHIV‐1集団がヌクレオチド配列で6%〜10%異なる可能性があることを示している。1つのクレード内のHIV‐1単離物は、gagコード配列で15%およびgp120コード配列で最大30%のヌクレオチド距離を示す場合がある。クレード間の遺伝的多様性は、分析される遺伝子に依存して30%〜40%の範囲となりうる。
【0046】
HIV‐1のグループMサブタイプはすべてアフリカにおいて見出すことができる。クレードAのウイルスは遺伝学的に最も異なっており、流行の初期にはアフリカで最も一般的なHIV‐1サブタイプであった。1990年代半ばから後期の間にHIV‐1がアフリカ南部へ急速に広がるとともに、クレードCのウイルスが主要なサブタイプとなり、現在は世界中のHIV‐1感染の48%を占めている。クレードBのウイルスは最も集中的に研究されたHIV‐1サブタイプであり、欧州および北米において絶えず最も優勢な単離物である。
【0047】
高い遺伝子組換え率はレトロウイルスの特徴である。遺伝学的に相異なるウイルス株による同時感染は、HIV‐1のリスクにさらされた個体において成立しないであろうと当初は考えられていた。しかしながら、1995年までに、HIV‐1グループMの世界規模の多様性のうちかなりの割合が、クレード間のウイルス組換え体を含んでいることが明らかになった。現在では、HIV‐1組換え体は、多数のHIV‐1サブタイプが共存し、かつ流行しているHIV‐1株の10%超を占める可能性のある、アフリカ、南米および東南アジアのような地理的なエリアで見出されるであろうと認識されている。分子レベルでは、これらの組換えウイルスのゲノムは寄せ集めのモザイクに似て、並置された相異なるHIV‐1サブタイプのセグメントを備えており、このことはその世代に寄与する多数の交差事象を反映している。ほとんどのHIV‐1組換え体はアフリカで発生し、大多数は元々クレードAのウイルスに由来するセグメントを含有している。タイ国では、例えば、優勢な流行している株の組成は、クレードAのgag+pol遺伝子セグメントおよびクレードEのenv遺伝子からなる。タイ国のHIV‐1株のクレードEのenv遺伝子は、中央アフリカ共和国由来のウイルス単離物中に存在するクレードEのenvと近縁関係にあるので、最初の組換え事象はアフリカで生じ、その後子孫ウイルスがタイ国に伝わったと考えられる。興味深いことに、完全長のHIV‐1サブタイプE単離物(すなわちサブタイプEのgag、pol、およびenv遺伝子を備えたもの)は現在まで報告されていない。
【0048】
αおよびβケモカイン受容体が感受性CD4+細胞へのウイルスの融合および侵入の共受容体として機能するという発見は、HIV‐1のための改定分類体系をもたらした(図3)。ここで、HIV‐1 gp120およびCD4+共受容体のタンパク質が別々の細胞で発現される融合アッセイにおけるケモカイン受容体の利用に基づいて、単離物をグループ化することができる。図3に示されるように、CXCR4受容体を使用するHIV‐1単離物(ここではX4ウイルスと呼ぶ)は通常はT細胞株(TCL)指向性のシンシチウム誘導型(SI)の株である一方、専らCCR5受容体を利用するもの(R5ウイルス)は主にマクロファージ(M)指向性かつ非シンシチウム誘導型(NSI)である。患者単離物の大多数を含み、かつ一連の指向性表現型を示す二重指向性R5/X4株は、多くの場合SIである。
【0049】
複製能を有するすべてのレトロウイルスの場合のように、3つのHIV‐1一次翻訳産物(すべて構造タンパク質をコードしている)は最初にポリプロテイン前駆体として合成され、該前駆体はその後ウイルスまたは細胞のプロテアーゼによってプロセシングされて成熟粒子関連タンパク質となる(図4)。55kdのGag前駆体であるPr55Gagは、切断されてマトリックス(MA)、カプシド(CA)、ヌクレオカプシド(NC)およびp6タンパク質となる。160kdのGag‐PolポリプロテインであるPr160Gag‐Polの自己触媒反応により、プロテアーゼ(PR)、ヘテロダイマー性逆転写酵素(RT)、およびインテグラーゼ(IN)タンパク質が生じるが、細胞性酵素によるタンパク質分解は、グリコシル化160kd Env前駆体gp160をgp120表面(SU)およびgp41膜貫通(TM)分解産物に変換する。残る6つのHIV‐1にコードされたタンパク質(Vif、Vpr、Tat、Rev、Vpu、およびNef)は、スプライシングされたmRNAの一次翻訳産物である。
【0050】
Gag
HIVのGagタンパク質は、他のレトロウイルスのGagタンパク質と同様に、非感染性のウイルス様粒子の形成にとって必要かつ十分である。レトロウイルスのGagタンパク質は一般にポリプロテイン前駆体として合成され、HIV‐1のGag前駆体はそのみかけの分子量に基づいてPr55Gagと命名された。先に示したように、Pr55GagのmRNAはスプライシングされない9.2kb転写物(図4)であり、該転写物は細胞質中での発現にRevを必要とする。polのORFが存在すると、ウイルスのプロテアーゼ(PR)が細胞からの出芽時または出芽直後にPr55Gagを切断して、成熟Gagタンパク質p17(MA)、p24(CA)、p7(NC)、およびp6を生成する(図4を参照)。ビリオンでは、MAはウイルスエンベロープの脂質二重層のすぐ内側に局在化し、CAは粒子の中心において円錐形のコア構造の外側部分を形成し、NCはウイルスRNAゲノムとのリボ核タンパク質複合体の状態でコアの中に存在する(図5)。
【0051】
HIVのPr55Gag前駆体はその翻訳に続いてオリゴマー化して原形質膜に向かい、原形質膜ではEMで視認可能な十分なサイズおよび密度の粒子がアセンブリされる。Pr55Gagによるウイルス様粒子の形成は、Gag前駆体に沿った多重ドメインの間で生じる重要なGag‐Gag相互作用を伴う、自己アセンブリのプロセスである。ウイルス様粒子のアセンブリには、ゲノムRNA(ただし核酸の存在は不可欠のようである)、polにコードされた酵素、またはEnv糖タンパク質の関与を必要としないが、感染性ビリオンの産生には、ウイルスRNAゲノムのカプシド形成ならびにEnv糖タンパク質およびGag‐Polポリプロテイン前駆体Pr160Gag‐Polの取り込みを必要とする。
【0052】
Pol
gagの下流にはHIVゲノムの最も高度に保存された領域であるpol遺伝子が存在し、3つの酵素すなわちPR、RT、およびINをコードしている(図4を参照)。RTおよびINはそれぞれ、ウイルスRNAゲノムの二本鎖DNAコピーへの逆転写、およびウイルスDNAの宿主細胞染色体内への組込みに必要である。PRは成熟した感染性ビリオンの産生を調節することにより生活環後期において重大な役割を果たす。pol遺伝子産物は、Pr160Gag‐Polと呼ばれる160kdのGag‐Pol融合タンパク質の酵素的切断によって得られる。この融合タンパク質は、Pr55Gag翻訳時のリボソームのフレームシフトによって生じる(図4を参照)。他の多くのレトロウイルスにも利用されている、Gag‐Pol発現のフレームシフト機構は、polから生じるタンパク質が低レベル(Gagのおよそ5%〜10%)で発現されることを確実にする。Pr55Gagと同様に、Pr160Gag‐PolのN末端はミリスチル化されて原形質膜を対象として方向づけられる。
【0053】
プロテアーゼ
鳥類のレトロウイルスを用いて実施された初期のパルス追跡研究は、レトロウイルスのGagタンパク質が当初はポリプロテイン前駆体として合成され、切断されてより小さな産物を生成することを明白に示した。その後の研究から、プロセシング機能は細胞性酵素ではなくウイルス性酵素により提供されること、GagおよびGag‐Pol前駆体のタンパク質分解がウイルスの感染性にとって不可欠であることが実証された。レトロウイルスのPRの配列分析は、PRがペプシンおよびレニンのような細胞の「アスパラギン酸」プロテアーゼとの関連を有することを示した。これらの細胞性酵素と同様に、レトロウイルスのPRは、標的タンパク質中のペプチド結合の加水分解を触媒する水分子を配位するために活性部位の2つの並んだAsp残基を使用する。偽ダイマーとして機能する(活性部位を生成するために同一分子内の2つの折り畳み部分を使用する)細胞性アスパラギン酸プロテアーゼとは異なり、レトロウイルスのPRは真のダイマーとして機能する。HIV‐1 PRのX線結晶学的データは、2つのモノマーが、各モノマーのN末端およびC末端両方に由来する4本の逆平行βシートによって部分的に結びついていることを示している。基質結合部位は2つのモノマーの間に形成された裂溝内にある。細胞のホモログと同様に、HIV PRダイマーは、結合部位の上に張り出して裂溝内の基質を安定させる柔軟な「フラップ」を含有し、活性部位のAsp残基はダイマーの中心にある。興味深いことに、活性部位残基を囲む何らかの限定的なアミノ酸ホモロジーが観察されるが、レトロウイルスのPRの一次配列は非常に多様であり、それでもその構造は非常に類似している。
【0054】
逆転写酵素(Reverse Transcriptase)
定義によれば、レトロウイルスは感染過程の初期段階においてその一本鎖RNAゲノムを二本鎖DNAに変換する能力を有している。この反応を、その関連するRNaseH活性とともに触媒する酵素がRTである。レトロウイルスのRTは3つの酵素活性すなわち(a)RNA依存性のDNA合成(マイナス鎖DNA合成のため)、(b)RNaseH活性(DNA‐RNAハイブリッド中間体に存在するtRNAプライマーおよびゲノムRNAの分解のため)、ならびに(c)DNA依存性のDNA合成(第2鎖またはプラス鎖DNA合成のため)を有する。
【0055】
成熟したHIV‐1 RTホロ酵素は66kdサブユニットおよび51kdサブユニットのヘテロダイマーである。51kdサブユニット(p51)は、66kd(p66)サブユニットから、PRによるp66のC末端15kd RNaseHドメインのタンパク質分解による除去によって生じる(図4を参照)。HIV‐1 RTの結晶構造は、p66サブユニットおよびp51サブユニットの配向が大きく異なっている極めて非対称的なフォールディングを示している。p66サブユニットは右手として視覚化することが可能であり、手のひらの内部にポリメラーゼ活性部位を、かつ手のひら、指、および親指のサブドメインによって形成された鋳型結合性の深い裂溝を備えている。ポリメラーゼドメインは接続サブドメインによってRNaseHに連結される。手のひらに位置する活性部位は、近接した3つの重要なAsp残基(110、185および186)と、2つの配位されたMg2+イオンとを含有している。これらのAsp残基の突然変異はRTの重合活性を消失させる。3つの活性部位Asp残基の配向は、他のDNAポリメラーゼ(例えば大腸菌のDNA polIのクレノウ・フラグメント)で観察されるものと同様である。p51サブユニットは剛性のようであり、重合用の裂溝を形成せず、このサブユニットのAsp110、185および186は分子内部に埋まっている。プライマー‐鋳型二重鎖のおよそ18塩基対が、核酸結合性の裂溝内にあり、ポリメラーゼ活性部位からRNaseHドメインへと伸びている。
【0056】
RT‐プライマー‐鋳型‐dNTP構造物では、プライマーの3’末端にジデオキシヌクレオチドが存在することにより、新たに入って来るdNTPに対して反応開始する直前に捕捉された触媒複合体の視覚化が可能となる。先に得られた構造物との比較から、新たなdNTPに対するプライマーの3’‐OHの求核攻撃に先立って鋳型およびdNTPを捕捉するために、指サブドメインが閉じるモデルが示唆される。成長している鎖に新たなdNTPが付加された後は、指サブドメインがより大きく開いた形態をとることにより、ピロリン酸を放出し、かつRTが次のdNTPに結合するのを可能にすることが提唱されている。HIV‐1 RNaseHの構造もX線結晶構造解析によって決定されており、このドメインは大腸菌のRNaseHに類似の球状のフォールディングを示す。
【0057】
インテグラーゼ
レトロウイルスの複製の卓越した特徴は、逆転写に続いて宿主細胞の染色体内にウイルスゲノムのDNAコピーが挿入されることである。この組込まれたウイルスDNA(プロウイルス)は、ウイルスRNA合成のための鋳型としての役割を果たし、感染細胞が生存している間は宿主細胞ゲノムの一部として維持される。組込み能力が不完全なレトロウイルス突然変異体は一般に増殖性感染を確立できない。
【0058】
ウイルスDNAの組込みは、HIV‐1 Gag‐PolポリプロテインのC末端部分がPRにより切断されて生成される32kdタンパク質であるインテグラーゼ(図4を参照)によって触媒される。
【0059】
レトロウイルスのINタンパク質は、構造的かつ機能的に異なる3つのドメインすなわち、N末端のジンクフィンガー含有ドメイン、コアドメイン、および比較的非保存的なC末端ドメイン、から構成されている。その溶解度が低いことから、288アミノ酸のHIV‐1 INタンパク質全体を結晶化することは未だ可能ではない。しかしながら、3つのドメインすべての構造がX線結晶構造解析またはNMR法によって独立に解明されている。トリ肉腫ウイルスのINのコアドメインの結晶構造も決定済みである。N末端ドメイン(残基1〜55)の構造はNMR分光法によって解明されたが、該ドメインはアミノ酸His‐12、His‐16、Cys‐40、およびCys‐43によって配位された亜鉛を備えた4つのヘリックスから構成されている。このN末端ドメインの構造は、いわゆるヘリックス・ターン・ヘリックス・モチーフを含むヘリックス系DNA結合タンパク質を連想させるが、HIV‐1構造においては、このモチーフはダイマー形成に寄与している。当初は、難溶性であることがコアドメインの構造を解明する努力の妨げとなった。しかしながら、INの残基185におけるPheからLysへの変更がin vitroでの触媒活性を妨げることなく溶解度を大幅に高めることが観察された時、結晶構造解析の試みは成功した。HIV‐1 INのコアドメイン(IN残基50〜212)の各モノマーは、ヘリックスに挟まれた5本ストランドのβシートから構成されており、この構造は、RNaseHおよびバクテリオファージMuAトランスポサーゼを含む他のポリヌクレオチジルトランスフェラーゼに非常に類似している。高度に保存された3つの残基が、他のポリヌクレオチジルトランスフェラーゼの類似の位置に見出されており、HIV‐1 INではこれらはAsp64、Asp116およびGlu152、いわゆるD,D‐35‐Eモチーフである。これらの位置における突然変異はin vivoおよびin vitroいずれにおいてもHIV INの機能を妨げる。トリ肉腫ウイルスおよびHIV‐1のいずれのコアドメインの結晶構造においても上記の3つのアミノ酸が近接していることは、これらの残基が、組込みプロセスの中心にあるポリヌクレオチド転移反応の触媒作用において中心的な役割を果たすという仮説を支持している。C末端ドメインの構造はNMR法によって解明済みであるが、該ドメインはSrcホモロジー3(SH3)ドメインを連想させる5本ストランドのβバレルのフォールディングトポロジーをとる。最近になって、コアドメインおよびC末端ドメインの両方を包含しているSIVおよびラウス肉腫ウイルスのINタンパク質フラグメントのX線構造が解明された。
【0060】
Env
HIV Env糖タンパク質はウイルスの生活環において主要な役割を果たす。該タンパク質は、CD4受容体および共受容体と相互作用する決定基を含有し、またウイルスエンベロープの脂質二重層と宿主細胞原形質膜との間の融合反応を触媒する。さらに、HIV Env糖タンパク質は、診断薬およびワクチンのいずれの開発の観点からも重要な免疫応答を誘発するエピトープを含有している。
【0061】
HIV Env糖タンパク質は、単独にスプライシングされた4.3kbのVpu/Envバイシストロン性mRNAから合成され(図4を参照)、翻訳は粗面小胞体(ER)に結合したリボソームで行われる。160kdポリプロテイン前駆体(gp160)は、成熟したTM Env糖タンパク質gp41となるように予定されているドメインの疎水性の輸送停止シグナルによって細胞膜につなぎ留められる、内在性膜タンパク質である(図6)。gp160は翻訳時にグリコシル化され、ジスルフィド結合を形成し、ERにおいてオリゴマー化する。主なオリゴマー型はトライマーのようであるが、ダイマーおよびテトラマーも観察される。gp160はゴルジ体に輸送され、ゴルジ体では、他のレトロウイルスのエンベロープ前駆体タンパク質と同様に、細胞性酵素によりタンパク質分解によって切断されて、成熟したSU糖タンパク質gp120およびTM糖タンパク質gp41となる(図6を参照)。高度に保存されたLys/Arg‐X‐Lys/Arg‐Argモチーフ直後のレトロウイルスEnv前駆体の切断に関与する細胞性酵素は、フーリンまたはフーリン様プロテアーゼであるが、その他の酵素がgp160のプロセシングを触媒する場合もある。gp160の切断はEnv誘導型の融合活性およびウイルス感染性に必要である。gp160の切断に続いて、gp120およびgp41は、ゴルジ体から細胞表面へのEnv複合体の輸送に重要な非共有結合性会合を形成する。gp120‐gp41相互作用は非常に弱く、相当量のgp120が、Envを発現している細胞の表面から流される。
【0062】
HIV Env糖タンパク質複合体、特にSU(gp120)ドメイン)は、非常に強くグリコシル化されており、gp160の分子量のおよそ半分はオリゴ糖側鎖で構成されている。ER内のEnv合成部位から原形質膜までのEnvの輸送中に、側鎖の多くが複合糖類の付加によって修飾される。多数のオリゴ糖側鎖は、宿主の免疫認識からgp120の大部分を遮る糖の雲として想像できるようなものを形成する。図6に示されるように、gp120は、散在する保存された(C1〜C5)ドメインおよび可変(V1〜V5)ドメインを含有している。様々な単離物のgp120に存在するシステイン残基は高度に保存されており、大きなループの中の最初の4つの可変領域を連結するジスルフィド結合を形成する。
【0063】
ウイルスのEnv糖タンパク質の主要な機能は、ウイルスエンベロープの脂質二重層と宿主細胞の細胞膜との間の膜融合反応を促進することである。この膜融合事象はウイルスのコアが宿主細胞の細胞質内に入り込むのを可能にする。gp120およびgp41の両方の多くの領域が、Envを介した膜融合に直接または間接的に関与するとされてきた。オルトミクソウイルスのHA2ヘマグルチニンタンパク質およびパラミクソウイルスのFタンパク質に関する研究から、これらのタンパク質のN末端の高疎水性ドメインは、融合ペプチドと呼ばれ、膜融合に重大な役割を果たすことが示された。突然変異解析は、HIV‐1、HIV‐2およびSIVのTM糖タンパク質のN末端に相似ドメインがあることを実証した(図6を参照)。gp41のこの領域内の非疎水性置換はシンシチウム形成を大幅に低減または阻害し、その結果非感染性の子孫ビリオンを生じた。
【0064】
gp41融合ペプチドのC末端側は、膜融合において中心的な役割を果たす2つの両親媒性へリックスドメインである(図6を参照)。Leuジッパー様の7アミノ酸繰り返しモチーフを含有するN末端へリックス(N‐へリックスと呼ばれる)における突然変異は、感染性および膜融合活性を低下させ、これらの配列に由来するペプチドは培養物中で強力な抗ウイルス活性を示す。HIV‐1およびSIVのgp41のエクトドメインの構造、特に2つのへリックスモチーフは、近年の構造分析の中心となってきた。へリックスドメインを含有する融合タンパク質、N‐ヘリックスおよびC‐ヘリックスに由来するペプチドの混合物、または、SIV構造の場合には残基27〜149の完全なgp41エクトドメイン配列、のいずれかについて、X線結晶構造解析またはNMR分光法により構造が決定された。これらの研究からは、基本的に同様のトライマー構造であって2つのヘリックスドメインが逆平行方式でパッキングされて6ヘリックス束を生成している構造が得られた。N‐へリックスは該ヘリックス束の中心にコイルドコイルを形成するとともに、C‐へリックスは外側の疎水性の溝の中にパッキングされる。
【0065】
膜融合をもたらすステップでは、CD4の結合がEnvの立体配座変化を引き起こし、該変化が共受容体の結合を促進する。gp120/CD4/共受容体三重複合体の形成に続いて、gp41は、融合ペプチドが標的の脂質二重層内に挿入されるのを可能にする仮説上の立体配座をとる。gp41の6ヘリックス束(gp41のN‐ヘリックスとC‐ヘリックスとの間の逆平行相互作用を伴っている)の形成はウイルスおよび細胞の膜を結び付け、膜融合が生じる。
【0066】
さらに、本発明による治療面から見て興味深い遺伝子は、増殖性疾患、がんまたは代謝疾患に対して治療効果を有する疾患関連遺伝子も含む。例えば、がんに関する治療面から見て興味深い遺伝子とは、特異的な抗がん性免疫反応を誘導する能力を有するがん抗原であることが考えられる。
【0067】
本発明のさらなる実施形態によれば、異種核酸配列は少なくとも1つのマーカー遺伝子または選択遺伝子を含む。
選択遺伝子は、細胞に対して特定の耐性を形質導入することにより、ある種の淘汰法が可能となる。当業者は、ポックスウイルスの系において使用可能な様々な選択遺伝子に精通している。その中には、例えばネオマイシン耐性遺伝子(NPT)またはホスホリボシルトランスフェラーゼ遺伝子(gpt)がある。
【0068】
マーカー遺伝子は、形質導入細胞において発色反応を引き起こし、該反応は形質導入細胞を同定するために使用することができる。当業者は、ポックスウイルスの系において使用可能な様々なマーカー遺伝子に精通している。その中には、例えばβ‐ガラクトシダーゼ(β‐gal)、β‐グルコシダーゼ(β‐glu)、緑色蛍光タンパク質(EGFP)、または青色蛍光タンパク質をコードする遺伝子がある。
【0069】
本発明のさらに別の実施形態によれば、異種核酸配列は、ポックスウイルスの転写制御要素および/または異種核酸配列中のコード配列を、隣り合うORFの終止コドンおよび/または開始コドンから隔てる、スペーシング配列を含む。隣り合うORFの終止/開始コドンと異種核酸配列中の挿入コード配列との間のこのスペーサー配列は、挿入された異種核酸配列を、したがって得られる任意の組換えウイルスを、安定化するという長所を有する。スペーサー配列のサイズは、該配列がそれ自体のコード機能または制御機能を持たない限り、様々であってよい。
【0070】
さらなる実施形態によれば、ポックスウイルスの転写制御要素および/または異種核酸配列中のコード配列を隣り合うORFの終止コドンから隔てるスペーサー配列は、少なくとも1ヌクレオチドの長さである。
【0071】
本発明の別の実施形態によれば、ポックスウイルスの転写制御要素および/または異種核酸配列中のコード配列を隣り合うORFの開始コドンから隔てるスペーシング配列は、少なくとも30ヌクレオチドである。特に、典型的なワクシニアウイルスのプロモーター要素が開始コドンの上流に確認される場合、異種核酸配列の挿入は、該プロモーター要素を隣り合うORFの開始コドンから隔てることができない可能性がある。典型的なワクシニアプロモーター要素は、例えば後期プロモーターについては配列「TAAAT」(デービソン(Davison)およびモス(Moss)、ジャーナル・オブ・モレキュラー・バイオロジー誌(J. Mol. Biol.)、1989年、第210巻、p.771−784)および早期プロモーターについてはA/Tリッチドメインを精査することにより同定可能である。約30ヌクレオチドのスペーシング配列は、ORFの開始コドンの上流に位置するポックスウイルスのプロモーターが影響を受けないことを確実にする、好ましい距離である。加えて、さらに好ましい実施形態によれば、挿入された異種核酸配列と隣り合うORFの開始コドンとの間の距離は約50ヌクレオチド、より好ましくは100ヌクレオチドである。
【0072】
本発明のさらに好ましい実施形態によれば、スペーシング配列は、隣りのORFの転写を制御することができる追加のポックスウイルス転写制御要素を含む。
ここまで、本開示は親MVAウイルス中で隣り合っているORFを使用した組換えMVAウイルスに注目してきた。しかしながら、本発明はさらに、組換えウイルス、およびそのようなウイルスを作製する方法であって、異種核酸配列が隣り合った必須ORF技術の間に挿入され、挿入に使用されるORfは親MVAウイルス中で隣り合っていないものも含む。すなわち、ウイルスは、組換えMVAウイルスにおいて隣り合っているORFが親MVAウイルス中では1つ以上のポックスウイルスORF(介在ORF)によって隔てられているように、構築可能である。本明細書中で使用されるように、親MVAウイルスとは、子孫の組換えウイルスが構築される元のウイルスである。親MVAウイルスの一例は、MVA 1974/NIHクローン1である。親ウイルスを使用して、本明細書中に開示された技術を使用して組換えウイルスを構築し、構築プロセスの間に介在ORFが除去可能であるようにすることができる。ポックスウイルス分野の当業者には当然のことであるが、注意深く選択されたポックスウイルスORFを含む核酸分子を使用することにより、それら2つのORFの間のウイルスゲノムの一部を、相同組換えのプロセスを通じて欠失させることが可能である。例えば、非必須ORFを含有している1キロベースのゲノム領域によって2つの必須ORFが隔てられていることを前提として、その2つの必須ORFが、例えばプラスミド内で隣り合うようにクローニングされた核酸構築物を作製可能である。該核酸構築物がポックスウイルス感染細胞(例えば親MVAウイルス感染細胞)に導入されると、必須ORFは親ウイルスのウイルスゲノム中の対応するORFとの組換えを行うことになる。当業者には理解されるさらなる組換え事象を通して、1キロベースの領域はウイルスゲノムから切除され、その結果2つの必須ORFが隣り合うようになる。したがって、本発明の1つの実施形態は、MVAウイルスゲノム由来の2つの隣り合った必須ORFの間に位置する異種核酸配列を含む組換え改変ワクシニアアンカラ(MVA)ウイルスであって、親MVAウイルス中の対応する必須ORFの間に存在する非必須ORFを欠くことを特徴とする組換えMVAウイルスである。したがって、異種核酸配列は、親MVAウイルス中においては隣り合っていない必須ORFによって挟まれている。必須ORFは、非必須ORFによって隔てられた、MVAゲノム中に存在する必須ORF対から選択される。一実施形態では、必須ORFは、A50R(MVA163)、B1R(MVA167)、F10(MVA‐039)、F12(MVA042)、F13L(MVA043)、F15L(MVA045)、F17L(MVA047)、E4L(MVA051)、E6L(MVA053)、E8L(MVA055)、E10L(MVA057)、I1L(MVA062)、I3L(MVA064)、I5L(MVA066)、J1R(MVA085)、J3R(MVA087)、D7L(MVA104)、D9L(MVA106)、A24R(MVA135)、およびA28R(MVA139)からなる群から選択される。一実施形態では、2つの必須ORFは、A50R‐B1R(MVA163‐MVA167)、F10‐F12(MVA039‐MVA042)、F13L‐F15L(MVA043‐MVA045)、F15L‐F17L(MVA045‐MVA047)、E4L‐E6L(MVA051‐MVA053)、E6L‐E8L(MVA053‐MVA055)、E10L‐I1L(MVA057‐MVA062)、I3L‐I5L(MVA064‐MVA066)、J1R‐J3R(MVA085‐MVA087)、D7L‐D9L(MVA104‐MVA106)、およびA24R‐A28R(MVA135‐MVA139)からなる群の中の必須ORF対から選択される。一実施形態では、必須ORFはA50R(MVA163)およびB1R(MVA167)から選択される。一実施形態では、一方の必須ORFはA50R(MVA163)であり、他方の必須ORFはB1R(MVA167)である。
【0073】
先に議論されたように、細胞培養物中での大規模な継代の結果、MVAウイルスゲノムは、Del I、II、II、IV、VおよびVIと呼ばれる6つの主な欠失を含有している。歴史的には、およそ31,000ヌクレオチドの欠失であるDel III周辺の領域が異種核酸配列の挿入に使用されてきた。したがって、本発明の1つの実施形態では、組換えMVAウイルスの構築の際に欠失させられる非必須ORFは、野生型MVAウイルス中でDel III領域に隣接している。
【0074】
説明してきたように、組換えMVAウイルスは、2つの隣り合った必須ORFの間に、IGRおよび/または異種核酸配列のような追加の配列を含有することができる。そのような配列は本明細書中に記載されている。したがって、本発明の1つの実施形態は、MVAウイルスゲノム由来の2つの隣り合った必須ORFの間に位置する異種核酸配列を含む組換え改変ワクシニアアンカラ(MVA)ウイルスであって、親MVAウイルス中の対応する必須ORFの間に存在する非必須ORFを欠くこと、および異種核酸配列がIGRに挿入されていることを特徴とする組換えMVAウイルスである。本開示において別記されているように、異種核酸配列は、転写制御要素に制御されるコード配列を含有することができる。
【0075】
本発明者らは特定の必須ORFおよびその配列について開示してきたが、本発明は、開示されたORFの一部またはバリアントを使用する組換えMVAウイルスおよび該組換えMVAウイルスを作製する方法も含む。例えば、本発明はORF A50Rおよびその一部(配列番号11および配列番号14)、ならびにORF B1Rおよびその一部(配列番号16および配列番号19)を開示しているが、本発明は、これらの配列のバリアントを含む組換えMVAウイルスを、そのバリアントORFが対応する野生型ORFによってコードされるタンパク質とほぼ同じ機能を有するタンパク質をコードする限りにおいて、含む。2つのタンパク質は、それぞれのタンパク質を含むMVAウイルスが、同じ細胞株を使用して増殖させたとき互いに対して約10%、約20%、約30%または約40%以内の力価を生じる場合に、ほぼ同じ機能を有すると見なされる。したがって、本発明の1つの実施形態は、2つの隣り合うORFの間に位置する異種核酸配列を含む組換え改変ワクシニアアンカラ(MVA)ウイルスであって、該隣り合うORFはMVA由来の必須ORFと少なくとも90%、少なくとも95%、少なくとも97%または少なくとも99%の配列同一性のヌクレオチド配列を含むことを特徴とする。一実施形態では、隣り合うORFは、A50R(MVA163)、B1R(MVA167)、F10(MVA‐039)、F12(MVA042)、F13L(MVA043)、F15L(MVA045)、F17L(MVA047)、E4L(MVA051)、E6L(MVA053)、E8L(MVA055)、E10L(MVA057)、I1L(MVA062)、I3L(MVA064)、I5L(MVA066)、J1R(MVA085)、J3R(MVA087)、D7L(MVA104)、D9L(MVA106)、A24R(MVA135)、およびA28R(MVA139)からなる群から選択された必須ORFに対して少なくとも90%、少なくとも95%、少なくとも97%または少なくとも99%の同一性を有するヌクレオチド配列を含む。好ましい実施形態では、2つの隣り合うORfは同じ必須ORFに由来するものではない。一実施形態では、2つの隣り合うORFは、A50R‐B1R(MVA163‐MVA167)、F10‐F12(MVA039‐MVA042)、F13L‐F15L(MVA043‐MVA045)、F15L‐F17L(MVA045‐MVA047)、E4L‐E6L(MVA051‐MVA053)、E6L‐E8L(MVA053‐MVA055)、E10L‐I1L(MVA057‐MVA062)、I3L‐I5L(MVA064‐MVA066)、J1R‐J3R(MVA085‐MVA087)、D7L‐D9L(MVA104‐MVA106)、およびA24R‐A28R(MVA135‐MVA139)からなる群の必須ORF対と少なくとも90%、少なくとも95%、少なくとも97%または少なくとも99%の同一性を有するヌクレオチド配列を含む。一実施形態では、一方の隣り合うORFは配列番号A50R(MVA163)と少なくとも90%、少なくとも95%、少なくとも97%または少なくとも99%の配列同一性のヌクレオチド配列を含み、他方の隣り合うORFは、第2の必須ORFと少なくとも90%、少なくとも95%、少なくとも97%または少なくとも99%の配列同一性のヌクレオチド配列を含む。一実施形態では、一方の隣り合うORFは配列番号B1Rと少なくとも90%、少なくとも95%、少なくとも97%または少なくとも99%の配列同一性のヌクレオチド配列を含む。
【0076】
本発明はさらに、本発明の組換えウイルスを生産するのに有用な核酸構築物も開示する。本明細書中で使用されるように、核酸構築物は、MVAウイルス由来の少なくとも1つの必須ORFの少なくとも一部を含む組換え核酸分子である。該核酸構築物は、環境内、例えば限定するものではないが生物体内、組織内、または細胞培養物中の細胞への、有用な核酸配列の輸送を可能にする。本開示の核酸構築物はヒトの介在によって生産される。該核酸構築物は、DNA、RNAまたはそのバリアントであってよい。核酸分子は、直鎖DNA、DNAプラスミド、ウイルスベクター、またはその他のベクターであってよい。一実施形態では、核酸分子はDNAプラスミドであってよい。一実施形態では、核酸分子は、ウイルス成分、プラスミド成分、転写制御要素、ならびに核酸分子の送達および発現を可能にする当業者に周知の任意のその他の有用な要素を含むDNAプラスミドであってよい。組換え核酸分子の一般的な構築のための方法は良く知られている。例えば、サムブルック(Sambrook)ら、「分子クローニング:実験の手引き(Molecular Cloning: a Laboratory Manual)」第3版、コールド・スプリング・ハーバー・ラボラトリー・プレス(Cold Spring Harbor Laboratory Press)、2001年、およびオースベル(Ausubel)ら編、「分子生物学における最新プロトコール(Current Protocols in Molecular Biology)」、ジョン・ワイリー・アンド・サンズ(John Wiley and Sons)、1994年を参照されたい。
【0077】
本発明の1つの実施形態は、単離された核酸構築物であって、(a)改変ワクシニアアンカラ(MVA)ウイルスゲノム由来の第1の必須ORFに由来するかまたは該ORFに相同な第1の核酸配列と、(b)MVAウイルスゲノム由来の第2の必須ORFに由来するかまたは該ORFに相同な第2の核酸配列とを含み、第1および第2の必須MVAウイルスORFは、MVAウイルスゲノム中では少なくとも1つの非必須ORFによって隔てられていることと、第1および第2の核酸配列は単離された核酸構築物中では互いに隣り合っていることと、第1および第2の核酸配列は、それぞれ第1および第2の必須MVA ORF由来の少なくとも25個連続したヌクレオチドを含むことと、を特徴とする。そのような核酸構築物は、相同組換えのプロセスを通じて組換えMVAウイルスを構築するのに有用である。このプロセスを使用して、本発明の単離された核酸構築物は、親MVAウイルスにおいては隣り合っていない(すなわち他の非必須MVA ORFによって隔てられている)ORFが、子孫である組換えMVAウイルスにおいては隣り合わせにされる組換えMVAウイルスを構築するために使用可能である。これは、例えば、親MVAウイルス由来の隣り合っていないORFをプラスミドのような核酸分子中へクローニングし、かつその間にある非必須ORFのクローニングは伴わないことにより、行うことができる。こうして、隣り合っていないORFは核酸構築物中で隣り合わせにされる。説明してきたように、MVAウイルスゲノムの中にそのような核酸構築物を組み換えることにより、間に存在する親MVAウイルス由来の非必須ORFの欠失がもたらされ、その結果として、もとは隣り合っていないORFが隣り合っている子孫組換えMVAウイルスが生じることになる。したがって、好ましい実施形態では、第1および第2の核酸配列は、親MVAウイルスにおいては隣り合っていない第1および第2の必須MVA ORFにそれぞれ由来するかまたは該ORFと相同である。すなわち、第1および第2の必須ORFは、親MVAウイルスゲノム中では少なくとも1つの非必須ORFによって隔てられている。
【0078】
本明細書中で使用されるように、「〜に由来する」という語句は、その核酸配列が得られた起源の核酸(すなわちORF)を指す。したがって、この点では、核酸配列は起源となるORFの全部または一部と同一であってよい。しかしながら、核酸配列は起源となるORFとは配列が異なっていてもよい。したがって、MVA ORFに由来する核酸配列は、得られた核酸配列において元のORFの機能が維持される限り、MVA ORFの全部(または一部)と配列が同一であっても同一でなくてもよい。例えば、当分野では当然のことであるが、ポックスウイルスの近縁種の核酸分子は、そのような分子の配列が同一でなくても、組換えを起こすことができる。したがって、本発明の1つの実施形態では、第1および第2の核酸配列は、該核酸配列の由来する元の必須MVA ORFとの十分な配列同一性を有し、第1または第2の核酸配列を含む核酸分子と、そのような配列の由来する元の必須MVA ORFを含む核酸分子との間の相同組換えを可能にする。一実施形態では、第1および第2の核酸配列は、該核酸配列の由来する元の必須MVA ORFの少なくとも一部に対して、少なくとも75%、少なくとも85%、少なくとも90%、少なくとも95%、少なくとも97%、または少なくとも99%同一である。一実施形態では、核酸配列は、該核酸配列の由来する元の必須MVA ORFの少なくとも一部と同一である。
【0079】
同様に当分野では当然のことであるが、小さなポリヌクレオチド分子でも相同組換えの過程に関与することができる。従って、本発明の核酸構築物中に存在する核酸配列は、該核酸構築物がMVAウイルスゲノム中への組換えを生じうるために必須MVA ORFの全配列を含む必要はない。実際、長さ20塩基相応の小さなポックスウイルスゲノムのフラグメントが、ウイルスゲノム中のそれぞれの配列との相同組換えに関与することが可能であることが示されている。したがって、本発明の1つの実施形態では、第1および第2の核酸配列は、必須MVA ORFからの25、30、35、40、45、50、100、150、200、250、または300ヌクレオチドを含むことができる。本発明の1つの実施形態は、単離された核酸構築物であって、(a)第1の必須MVA ORFからの少なくとも25個連続したヌクレオチドを含む第1の核酸配列と、(b)第2の必須MVA ORFからの少なくとも25個連続したヌクレオチドを含む第2の核酸配列とを含み、第1および第2の必須MVAウイルスORFは、MVAウイルスゲノム中では少なくとも1つの非必須ORFによって隔てられていること、ならびに第1および第2の核酸配列は単離された核酸構築物中では互いに隣り合っていることを特徴とする。一実施形態では、第1の核酸配列は、A50R(MVA163)、B1R(MVA167)、F10(MVA‐039)、F12(MVA042)、F13L(MVA043)、F15L(MVA045)、F17L(MVA047)、E4L(MVA051)、E6L(MVA053)、E8L(MVA055)、E10L(MVA057)、I1L(MVA062)、I3L(MVA064)、I5L(MVA066)、J1R(MVA085)、J3R(MVA087)、D7L(MVA104)、D9L(MVA106)、A24R(MVA135)、およびA28R(MVA139)からなる群から選択された必須ORFからの25個連続したヌクレオチドを含む。一実施形態では、第2の核酸配列は、A50R(MVA163)、B1R(MVA167)、F10(MVA‐039)、F12(MVA042)、F13L(MVA043)、F15L(MVA045)、F17L(MVA047)、E4L(MVA051)、E6L(MVA053)、E8L(MVA055)、E10L(MVA057)、I1L(MVA062)、I3L(MVA064)、I5L(MVA066)、J1R(MVA085)、J3R(MVA087)、D7L(MVA104)、D9L(MVA106)、A24R(MVA135)、およびA28R(MVA139)からなる群から選択された必須ORFからの25個連続したヌクレオチドを含む。一実施形態では、第1の核酸配列は、配列番号11または配列番号14からの少なくとも25個連続したヌクレオチドを含み、第2の核酸配列は、配列番号16または配列番号19からの少なくとも25個連続したヌクレオチドを含む。
【0080】
本発明の核酸構築物は、MVAウイルスのゲノムに異種核酸配列を送達するために使用される。したがって、一実施形態、本発明の核酸構築物は、第1および第2の核酸配列の間に異種核酸分子を含む。典型的な異種核酸配列は本開示において別記されている。本明細書中に開示されたいずれの異種核酸配列も本発明の核酸構築物に含めるのに適している。
【0081】
本発明の核酸構築物は親MVAウイルスのゲノムと組換えを行うことができるので、ウイルスゲノムに異種核酸配列を挿入するために使用可能である。したがって、本発明の1つの実施形態では、本発明の核酸コントラスト(nucleic acid contrast)は、第1および第2の核酸配列の間に遺伝子間領域を含有している。遺伝子間領域は、転写制御要素、制限部位および非ワクシニアのオープンリーディングフレームのようなものを含むことができる。したがって、遺伝子間領域は転写制御要素に制御される遺伝子を含む異種核酸配列を挿入するために使用可能である。核酸構築物がMVAウイルスゲノムとの組換えを行うと、異種核酸配列は、MVAウイルスゲノム中の、核酸構築物中において該核酸配列を挟んでいる2つの隣り合った必須ORFに対応する必須ORFの間に、挿入されることになる。得られるMVAウイルスは、MVAウイルスゲノム中に安定的に組み込まれた異種核酸配列を含有している組換えMVAウイルスとなる。
【0082】
一実施形態では、本発明の核酸構築物は、ウイルスゲノムの2つの隣り合ったORFの間に位置するIGR配列の完全長フラグメントまたは部分フラグメントを含む。好ましくは、核酸構築物は、前記IGRに由来する配列中に挿入された少なくとも1つのクローニング部位であって、対象とする異種DNA配列を挿入するための、および好ましくは前記異種DNA配列に作動可能に連結されたポックスウイルス転写制御要素を挿入するための、クローニング部位を含む。任意選択で、核酸構築物はレポーター遺伝子カセットおよび/または選択遺伝子カセットを含む。核酸構築物は、好ましくは、IGR配列の前記完全長フラグメントまたは部分フラグメントに隣接している2つの隣り合ったORFの配列も含む。
【0083】
ヌクレオチド配列を含まないいくつかのIGRが同定されている。これらの場合、プラスミドベクターは、IGRに隣接している配列のDNA配列(すなわち2つの隣り合ったORFのDNA配列)を含む。好ましくは、異種DNA配列の挿入用のクローニング部位がIGRに挿入される。IGRに隣接している配列のDNAは、MVAゲノムの対応するIGR中へ外来DNA配列を挿入するために使用される。そのようなプラスミドベクターは、異種DNA配列と、任意選択でレポーター遺伝子カセットおよび/または選択遺伝子カセットとを挿入するためのクローニング部位を含むIGR配列の完全長フラグメントまたは部分フラグメントをさらに含むことができる。
【0084】
本発明の一実施形態は、安定な組換え改変ワクシニアアンカラウイルスを作製する方法である。そのような方法は、本明細書中に開示された核酸構築物を使用する。したがって該方法は、第1に、MVAウイルスゲノムの2つの隣り合った必須オープンリーディングフレーム(ORF)の間にあるかまたは該ORFに挟まれた異種核酸配列を含む核酸構築物を得る工程を含み、該MVAウイルスは、親のMVAウイルス中の対応する2つの必須ORFの間に存在する非必須ORFまたはORFフラグメントを欠いている。例えば、適切な核酸構築物を得るために、必須MVA ORFの核酸配列が単離されてpBluescript(ストラタジーン(Stratagene))のような標準的なクローニングベクターにクローニングされ、該核酸配列がMVAゲノムに挿入されるべき異種DNAに隣接するようにされてもよい。その後、この構築物は、当業者に周知の方法(例えばトランスフェクション)を使用して細胞に導入可能である。該核酸構築物を含有している細胞はその後MVAウイルスに感染させられ、核酸構築物とMVAウイルスゲノムとの間の相同組換えを可能にするのに適した条件下で培養される。その後適切な時期に細胞が採取され、組換えMVAウイルスが単離される。得られたウイルスは安定な組換えMVAウイルスとなる。そのようなウイルスは誘導体ウイルスと呼ばれる場合もある。当然ながら、核酸構築物を細胞に導入するステップおよび該細胞を感染せしめるステップの順序は逆にされてもよいし、これら2つのステップが同時に行われてもよい。
【0085】
核酸構築物中の異種核酸配列をMVAゲノムに導入する一般的な方法および組換えMVAを得る方法は当業者にはよく知られており、さらに、J.サムブルック(Sambrook)、E.F.フリッシュ(Fritsch)およびT.マニアティス(Maniatis)、「分子クローニング:実験の手引き(Molecular Cloning: a Laboratory Manual)」第2版、コールド・スプリング・ハーバー・ラボラトリー・プレス(Cold Spring Harbor Laboratory Press)、1989年、ならびに「分子生物学における最新プロトコール(Current Protocols in Molecular Biology)」、ジョン・ワイリー・アンド・サンズ・インコーポレイテッド(John Wiley and Sons Inc.)、1998年、第16章第IV節「ワクシニアウイルスベクターを使用する哺乳動物細胞におけるタンパク質の発現(Expression of proteins in mammalian cells using vaccinia viral vectors)」、から導き出すことが可能である。
【0086】
本発明によるDNA配列は、本発明によるMVAまたはその誘導体、および本発明によるMVAに感染した細胞または個体を、同定または単離するために使用可能である。該DNA配列は、例えば、PCRプライマー、ハイブリダイゼーションプローブを生成するために使用され、またはアレイ技術において使用される。
【0087】
本発明による誘導体ウイルスなどの用語は、親ウイルスと同じ特長を示すがそのゲノムの1つ以上の部分において違いを示す子孫ウイルスを指す。用語「MVAの誘導体」は、MVAと同じ機能持性を有するウイルスについて述べている。例えば、MVA 1974/NIHクローン1の誘導体は、MVA 1974/NIHクローン1の特長を有する。MVA 1974/NIHクローン1またはその誘導体のこれらの持性のうちの1つは、その弱毒化および宿主域の厳格な制限である。
【0088】
本発明による組換えMVAは医薬品またはワクチンとして有用である。したがって、本発明の一実施形態は、本発明の組換えMVAウイルスを使用して個体を疾患から防御する方法である。
【0089】
本発明の組換えMVAウイルスは標的細胞内への外来コード配列の導入に使用することも可能であり、前記配列は標的細胞のゲノムに対して同種であっても異種であってもよい。標的細胞内への外来コード配列の導入は、タンパク質、ポリペプチド、ペプチド、抗原または抗原エピトープを生産するためにin vitroで行われてもよい。この方法は、本発明の組換えMVAを用いた宿主細胞の感染、適切な条件下での感染宿主細胞の培養、ならびに、前記宿主細胞によって生産されるポリペプチド、ペプチド、タンパク質、抗原、エピトープおよび/またはウイルスの単離および/または濃縮を含む。
【0090】
さらに、細胞内に1つ以上の同種配列または1つ以上の異種配列を導入するための方法は、in vitroおよびin vivoの治療法に適用可能である。in vitroの治療法については、本発明による組換えMVAに前もって(ex vivoで)感染させられた単離細胞が、免疫応答に影響を及ぼすため、好ましくは免疫応答を引き起こすために、生きている動物の身体に投与される。in vivoの治療法については、本発明による組換えポックスウイルスは、免疫応答に影響を及ぼすため、好ましくは免疫応答を引き起こすために、生きている動物の身体に直接投与される。この場合、接種部位の周囲の細胞、またウイルスが輸送されるのに介在した場所の、例えば血流の細胞も、in vivoで本発明の組換えMVAに直接感染する。感染後、これらの細胞は、外来コード配列によってコードされている治療遺伝子のタンパク質、ペプチドまたは抗原エピトープを合成し、続いて、それらまたはその一部を細胞表面上に提示する。免疫系の特殊化した細胞は、そのような異種のタンパク質、ペプチドまたはエピトープの提示を認識し、特異的免疫応答を開始する。
【0091】
MVAは増殖が高度に制限されており、従って高度に弱毒化されているので、ヒトを含む様々な哺乳動物(免疫力が低下した動物またはヒトを含む)の治療に有用である。本発明はさらに、ヒトを含む生きている動物の身体において免疫応答を引き起こすための医薬組成物およびワクチンを提供する。
【0092】
医薬組成物は一般に、1つ以上の、医薬として許容可能であるかまたは認可されている担体、添加剤、抗生物質、保存剤、アジュバント、希釈剤および/または安定化剤を含むことができる。そのような補助物質は、水、生理食塩水、グリセロール、エタノール、湿潤剤もしくは乳化剤、pH緩衝物質、または同様のものであってよい。適切な担体は、典型的には大型で徐々に代謝される分子、例えばタンパク質、多糖、ポリ乳酸、ポリグリコール酸、アミノ酸重合体、アミノ酸共重合体、脂質凝集体(lipid aggregate)などである。
【0093】
ワクチンの調製については、本発明の組換えポックスウイルスは生理学的に許容可能な形態に変換される。これは、天然痘に対するワクチン接種に使用されたポックスウイルス・ワクチン剤の調製における経験(スティックル(Stickl)、H.ら、ドイチェ・メディツィニッシェ・ボーヘンシュリフト誌(Dtsch Med Wochenschr.)、1974年、第99巻、p.2386−2392)に基づいて行うことができる。例えば、精製されたウイルスは、約10mM トリス、140mM NaCl(pH7.4)中で5×10E8 TCID50/mlの力価に製剤化されて−80℃に保存される。ワクチン注射剤の調製については、例えば、10E2−10E8のウイルス粒子が、アンプル(好ましくはガラス製アンプル)中で2%ペプトンおよび1%ヒトアルブミンの存在下にて100mlのリン酸緩衝生理食塩水(PBS)中で凍結乾燥される。別例として、ワクチン注射剤は、製剤中のウイルスの段階的な凍結乾燥によって製造されてもよい。この製剤は、追加の添加剤、例えばマンニトール、デキストラン、糖質、グリシン、ラクトースもしくはポリビニルピロリドン、または他の助剤、例えば酸化防止剤もしくは不活性ガス、安定化剤もしくはin vivo投与に適した組換えタンパク質(例えばヒト血清アルブミン)を含有することができる。その後、ガラス製アンプルは密閉され、4℃〜室温で数か月間保存可能である。しかしながら、必要がない限りは、アンプルは−20℃未満の温度で保存されることが好ましい。
【0094】
ワクチン接種または治療については、凍結乾燥品は0.1〜0.5mlの水性溶液、好ましくは生理食塩水またはトリス緩衝液に溶解されて、全身投与または局所投与、すなわち非経口投与、皮下投与、筋肉内投与、スタンプ式(by scarification)投与、または当業者に周知の任意の他の投与経路によって、投与されうる。投与方式、用量および投与回数は、周知のやり方で当業者によって最適化可能である。しかしながら、最も一般的には、患者は最初のワクチン接種注射の約1か月〜6週間後に、2回目の注射でワクチン接種される。
【0095】
本発明の一実施形態は、抗原に対する免疫応答を生成する方法である。そのような応答は、CD8+T細胞免疫応答または抗体応答であってよい。特に、本発明は、初回免疫組成物の投与によって誘発された免疫応答が追加免疫組成物の投与によって増強(ブースト)される「プライム・アンド・ブースト」免疫化法に関する。本発明は、有効な追加免疫が、改変ワクシニアアンカラ(MVA)ベクターを使用して、組換えMVA自体を含む様々な異なる種類の初回免疫組成物のうち任意のものを用いた初回免疫の後で達成されうる、という先の実験証明に基づいている。
【0096】
数多くの病原体に対する免疫応答の主要な防御成分は、細胞傷害性リンパ球(CTL)としても知られているCD8+型のTリンパ球によって仲介される。CD8+細胞の重要な機能はγインターフェロン(IFN)の分泌であり、これはCD8+T細胞免疫応答の尺度を提供する。免疫応答の別の成分は、病原体のタンパク質に対する抗体である。
【0097】
本発明は、先の実験が示すように、様々な異なる初回免疫組成物のうち任意のものを使用して抗原に対して初回免疫されたCD8+T細胞免疫応答を増強するため、さらには抗体反応を誘発するための、有効な手段であることが見出されたMVAを使用する。
【0098】
特に、先の実験研究は、本発明の先行物の使用により、HIV抗原を発現する組換えMVAウイルスが、DNAワクチンによって初回免疫されたCD8+T細胞免疫応答を増強し、さらには抗体応答を誘発することが可能となることを実証している。該MVAは、免疫化後にCD8+T細胞応答を誘発することが見出される場合がある。組換えMVAは、組換えMVAの1回以上の接種により増強される免疫応答を初回免疫することが示される可能性もある。
【0099】
プラスミドDNAで免疫化され、MVAで追加免疫された非ヒト霊長類は、生ウイルスを用いた粘膜内抗原投与に対して有効に防御された(アマラ(Amara)ら、サイエンス誌(Science)、2001年、第292巻、p.69−74)。好都合なことに、本発明者らは、初回免疫および追加免疫の両方に皮内免疫、筋肉内免疫または粘膜免疫を用いるワクチン接種法を使用することができ、これは、例えばヒトにおいてCD8+T細胞を誘導し、さらに抗体応答を誘発するのに適した一般的な免疫化法を構成すると考える。
【0100】
本発明の種々の態様および実施形態は、HIV抗原をコードする核酸の事前投与によって初回免疫された該抗原へのCD8+T細胞免疫応答を増強し、さらに抗体応答を誘発するために、該抗原をコードするMVAベクターを使用する。
【0101】
本発明の一般的な態様は、HIV抗原に対するCD8+T細胞免疫応答を増強し、さらに抗体応答を誘発するためのMVAベクターの使用を提供する。
本発明の1つの態様は、個体においてHIV抗原に対するCD8+T細胞免疫応答を増強し、さらに抗体応答を誘発する方法を提供し、該方法は、該抗原をコードする核酸を含むMVAベクターであって、該核酸が該核酸からの発現により個体において抗原を生産するための調節配列に作動可能に連結されているMVAベクターを、個体の体内に提供し、これによって事前に該個体において初回免疫された抗原へのCD8+T細胞免疫応答が増強されることを含む。
【0102】
HIV抗原に対する免疫応答は、プラスミドDNAを用いた免疫化によって、または感染性物質による感染によって初回免疫されうる。
本発明のさらなる態様は、個体においてHIV抗原に対するCD8+T細胞免疫応答を誘導し、さらには抗体応答を誘発する方法を提供し、該方法は、該個体に、抗原をコードする核酸を含む初回免疫組成物を投与する工程と、その後、該抗原をコードする核酸を含むMVAベクターであって、該核酸が該核酸からの発現により個体において抗原を生産するための調節配列に作動可能に連結されているMVAベクターを含む追加免疫組成物を投与する工程とを含む。
【0103】
さらなる態様は、開示されているように、HIV抗原に対するCD8+T細胞免疫応答を増強し、さらには抗体応答を誘発するために哺乳動物に投与するための医薬品の製造における、MVAベクターの使用を提供する。そのような医薬品は一般に、抗原をコードする核酸を含む初回免疫組成物の事前投与に続いて投与するためのものである。
【0104】
初回免疫組成物は、抗原をコードするDNAであって、そのようなDNAが好ましくは哺乳動物細胞内で複製することができない環状プラスミドの形態であるものを含むことができる。いかなる選択可能なマーカーも、臨床的に使用される抗生物質に対して耐性であるべきではなく、したがって、例えばカナマイシン耐性がアンピシリン耐性よりも好ましい。抗原の発現は、哺乳動物細胞において活性を有するプロモーター、例えばサイトメガロウイルス最初期(CMV IE)プロモーターによって、駆動されるべきである。
【0105】
本発明の様々な態様の特定の実施形態では、初回免疫組成物の投与に続いて、追加免疫組成物、または第1および第2の追加免疫組成物であって同一であるかまたは互いに異なる第1および第2の追加免疫組成物を用いて、追加免疫が行われる。さらに別の追加免疫組成物が、本発明から逸脱することなく使用される場合もある。一実施形態では、三重免疫化法は、DNA、次いで第1の追加免疫組成物としてアデノウイルス、次いで第2の追加免疫組成物としてMVA、次いで任意選択でさらなる(第3の)追加免疫組成物またはその後の同一もしくは異なるベクターのうち一方もしくは他方もしくは両方の追加免疫投与を使用する。別の選択肢は、DNA、次いでMVA、次いでアデノウイルス、次いで任意選択でその後の同一もしくは異なるベクターのうち一方もしくは他方もしくは両方の追加免疫投与である。
【0106】
それぞれの初回免疫組成物および追加免疫組成物においてコードされる抗原は(いかに多くの追加免疫組成物が使用されても)同一である必要はないが、少なくとも1つのCD8+T細胞エピトープを共有するべきである。抗原は完全抗原に相当してもよいし、そのフラグメントに相当してもよい。より効率的に、抗原中の不要なタンパク質配列および1つまたは複数のベクター中のコード配列を切り取って、ペプチドエピトープまたは人為的なエピトープ連続物が使用されてもよい。1つ以上の追加のエピトープ、例えば、Tヘルパー細胞によって認識されるエピトープ、特に様々なHLA型の個体において認識されるエピトープが含まれてもよい。
【0107】
組換えMVAウイルスによってコードされる本発明のHIV抗原には、少なくとも1つのCD8+T細胞エピトープとしてHIVのEnv、Gag、Pol、Vif、Vpr、Tat、Rev、Vpu、またはNefアミノ酸配列のアミノ酸配列によって誘発される免疫原活性を有するポリペプチドが挙げられる。このアミノ酸配列は、実質的に、既知のHIV EnvもしくはPolの少なくとも1つの10〜900アミノ酸フラグメントおよび/または共通配列;または、既知のHIV Gagの少なくとも1つの10〜450アミノ酸フラグメントおよび/または共通配列;または、既知のHIV Vif、Vpr、Tat、Rev、Vpu、もしくはNefの少なくとも1つの10〜100アミノ酸フラグメントおよび/または共通配列に相当する。
【0108】
完全長Env前駆体配列が本発明での使用に関して提示されているが、Envは任意選択で部分配列を欠失している。例えば、gp120表面型切断産物およびgp41膜貫通型切断産物の領域を欠失していてもよい。
【0109】
完全長Gag前駆体配列が本発明での使用に関して提示されているが、Gagは任意選択で部分配列を欠失している。例えば、マトリックスタンパク質(p17)の領域、カプシドタンパク質(p24)の領域、ヌクレオカプシドタンパク質(p7)の領域、およびp6(GagポリプロテインのC末端ペプチド)の領域を欠失していてもよい。
【0110】
完全長Pol前駆体配列が本発明での使用に関して提示されているが、Polは任意選択で部分配列を欠失している。例えば、プロテアーゼタンパク質(p10)の領域、逆転写酵素タンパク質(p66/p51)の領域、およびインテグラーゼタンパク質(p32)の領域を欠失していてもよい。
【0111】
そのようなHIV Env、Gag、またはPolは、既知のEnv、Gag、またはPolタンパク質のアミノ酸配列に対して少なくとも50%の全体的な同一性、例えば50〜99%の同一性、またはその中の任意の範囲もしくは値の同一性を有する一方で、HIVの少なくとも1つの株に対する免疫原性応答を誘発することができる。
【0112】
同一性(%)は、例えば、ウィスコンシン大学遺伝学コンピュータ・グループ(University of Wisconsin Genetics Computer Group)(UWGCG)から入手可能なGAPコンピュータプログラム、バージョン6.0を使用して配列情報を比較することにより、測定することができる。該GAPプログラムは、ニードルマン(Needleman)およびブンシュ(Wunsch)のアライメント方法(ジャーナル・オブ・モレキュラー・バイオロジー誌(J Mol Biol)、1970年、第48巻、p.443)を、スミス(Smith)およびウォーターマン(Waterman)によって修正されたようにして(アドバンシズ・イン・アプライド・マスマティックス誌(Adv Appl Math)、1981年、第2巻、p.482)、利用する。簡潔に述べると、GAPプログラムは、同一性を、アライメントされた同一のシンボル(すなわちヌクレオチドまたはアミノ酸)の数を2つの配列のうち短いほうのシンボルの総数で除算したもの、として定義する。GAPプログラム用の好ましい初期設定パラメータには、(1)ユニタリ比較行列(同一について1および非同一について0の値を含む)ならびにグリブスコフ(Gribskov)およびバージェス(Burgess)(ヌクレイック・アシッズ・リサーチ誌(Nucl Acids Res)、1986年、第14巻、p.6745)の重み付き比較行列、シュワルツ(Schwartz)およびデイホフ(Dayhoff)(編、「タンパク質の配列および構造の図解集(Atlas of Protein Sequence and Structure)」、米国ワシントンD.C.所在のナショナル・バイオメディカル・リサーチ・ファウンデーション(National Biomedical Research Foundation)、1979年、pp.353−358)の記述によるもの;(2)各ギャップについて3.0のペナルティ、および各ギャップの各シンボルについてさらに0.10のペナルティ;ならびに(3)エンドギャップについてはペナルティなし、が含まれる。
【0113】
好ましい実施形態では、本発明のEnvは少なくとも1つのHIVエンベロープタンパク質の1つのバリアント型である。好ましくは、Envはgp120と、gp41の膜貫通部およびエクトドメインとで構成され、gp41の細胞質ドメインの一部または全部を欠いている。
【0114】
既知のHIV配列は、民間およびGENBANKのような公的なHIV配列データベースから、または公開された編集物として、例えばマイヤーズ(Myers)ら編、「ヒト・レトロウイルスおよびAIDS、核酸およびアミノ酸配列の編集および分析(Human Retroviruses and AIDS, A Compilation and Analysis of Nucleic Acid and Amino Acid Sequences)」、第I巻および第II巻、米国ニューメキシコ州ロスアラモス、セオレティカル・バイオロジー・アンド・バイオフィジックス(Theoretical Biology and Biophysics)、1993年、もしくはワールドワイドウェブのhiv−web.lanl.gov/において、容易に入手可能である。
【0115】
本発明の組換えMVAウイルスで使用される核酸によってコードされる、新たなHIV Env、GagまたはPolを得るためのHIV Env、GagまたはPolの置換または挿入には、少なくとも1つのアミノ酸残基(例えば1〜25個のアミノ酸)の置換または挿入が挙げられる。別例として、少なくとも1つのアミノ酸(例えば1〜25個のアミノ酸)をHIV Env、GagまたはPolの配列から欠失させてもよい。好ましくは、そのような置換、挿入または欠失は、安全上の特性、発現レベル、免疫原性、およびMVAの高い複製速度との両立性に基づいて特定される。
【0116】
本発明のHIV Env、GagまたはPolにおけるアミノ酸配列の変異は、例えば、DNA中の突然変異によって作製することができる。そのようなHIV Env、GagまたはPolは、例えば、アミノ酸配列中の様々なアミノ酸残基をコードするヌクレオチドの欠失、挿入または置換を含んでいる。言うまでもなく、HIV Env、GagまたはPolをコードする核酸において作製されることになる突然変異は、配列をリーディングフレームから外してはならず、第2のmRNA構造物を生じる可能性のある補足的なドメインを作らないことが好ましいであろう。
【0117】
本発明のHIV Env、GagまたはPolをコードする核酸は、本明細書中に示された教示および手引きに基づいて、HIVのEnv、GagまたはPolをコードするDNAまたはRNA中のヌクレオチドの増幅または部位特異的変異誘発、およびその後の、HIV Env、GagまたはPolをコードするDNAまたはRNAを生産するための該コードDNAの合成または逆転写によって調製されることも可能である。
【0118】
本発明のHIV Env、GagまたはPolを発現する組換えMVAウイルスには、本明細書中に示された教示および手引きに基づいて、当業者であれば過度の実験作業を伴うことなく日常作業で得ることができる置換ヌクレオチドとして、HIV Env、GagまたはPolをコードする配列の有限な集合が含まれる。タンパク質の化学的性質および構造の詳細な説明については、シュルツ(Schulz)、G.E.ら、「タンパク質構造の原理(Principles of Protein Structure)」、米国ニューヨーク州ニューヨーク、シュプリンガー・フェアラーク(Springer−Verlag)、1978年、およびクライトン(Creighton)T.E.、「タンパク質:構造および分子特性(Proteins: Structure and Molecular Properties)」、米国カリフォルニア州サンフランシスコ、W.H.フリーマン社(W. H. Freeman and Co.)、1983年を参照されたい。コドン選択のような、ヌクレオチド配列置換の表示については、オースベル(Ausubel)ら編、「分子生物学における最新プロトコール(Current Protocols in Molecular Biology)」、米国ニューヨーク州ニューヨーク、グリーン・パブリシング・アソシエーツ(Greene Publishing Assoc.)、1994年、§§A.1.1〜A.1.24、およびサムブルック(Sambrook)、J.ら、「分子クローニング:実験の手引き(Molecular Cloning: a Laboratory Manual)」第2版、米国ニューヨーク州コールド・スプリング・ハーバー、コールド・スプリング・ハーバー・ラボラトリー・プレス(Cold Spring Harbor Laboratory Press)、1989年、付録CおよびD、を参照されたい。
【0119】
したがって、当業者は、本明細書中に示された教示および手引きがあれば、置換バリアント、欠失バリアントまたは挿入バリアントを含む代替HIV Env、Gag、またはPolを得るためには、HIVのenv、gag、またはpolのDNAまたはRNAの他の部位の他のアミノ酸残基をどのようにして置換するかが分かるであろう。
【0120】
MVAベクター内では、コードされた抗原を発現するための調節配列はプロモーターを含むことになる。「プロモーター」とは、ヌクレオチドの配列であって、該配列から、下流に(すなわち二本鎖DNAのセンス鎖上の3’方向に)作動可能に連結されたDNAの転写が開始されうるところの配列を意味する。「作動可能に連結され」とは、転写がプロモーターから開始されるように適切に配置かつ配向されて、同じ核酸分子の一部として連結されることを意味する。プロモーターに作動可能に連結されたDNAは、プロモーターの「転写開始調節下」にある。ターミネーターフラグメント、ポリアデニル化配列、マーカー遺伝子およびその他の配列を含む他の調節配列が、当業者の知識および慣行に従って適正に含められてもよい。例えば、D.M.クナイプ(Knipe)、P.M.ハウリー(Howley)編、「フィールズ・ウイルス学(Fields Virology)」の、モス(Moss)、B.著「ポックスウイルス科:ウイルスおよびその複製(Poxviridae: The Viruses and Their Replication)」、フィラデルフィア所在のリッピンコット・ウィリアムズ・アンド・ウィルキンズ(Lippincott Williams and Wilkins)、2001年、p.2849〜2883、を参照されたい。例えば核酸構築物の調製、突然変異誘発、塩基配列決定、細胞内へのDNAの導入および遺伝子発現における核酸の取扱、ならびにタンパク質の分析のための数多くの既知の技法およびプロトコールは、オースベル(Ausubel)ら編、「分子生物学における最新プロトコール(Current Protocols in Molecular Biology)」、ジョン・ワイリー・アンド・サンズ(John Wiley and Sons)、1998年に詳細に記載されている。
【0121】
本発明の態様および実施形態で使用されるプロモーターは、ポックスウイルスの発現系に適合しうるものであって、天然の配列、改変された配列および合成配列が含まれうる。
初回免疫組成物および追加免疫組成物のうち一方または両方は、顆粒球マクロファージコロニー刺激因子(GM‐CSF)またはこれをコードする核酸のようなアジュバントを含んでもよい。
【0122】
追加免疫組成物の投与は、一般に初回免疫組成物の投与の約1〜6か月後、好ましくは約1〜3か月後である。
好ましくは、初回免疫組成物、追加免疫組成物、または初回免疫組成物および追加免疫組成物両方の投与は、皮内免疫、筋肉内免疫または粘膜免疫である。
【0123】
MVAワクチン剤の投与は、ウイルスの懸濁液を注入するために注射針を使用することにより達成されてもよい。代替例は、ウイルス懸濁液を投与するための無針注射デバイス(例えばBiojector(登録商標)無針注射器を使用)、または、冷蔵を必要としない個別調製された単回投与量の製造をもたらす、再懸濁されたワクチン含有凍結乾燥粉末を使用することである。これは、アフリカの農村地域において必要とされるワクチンについては大きな利点となるであろう。
【0124】
MVAはヒトの免疫化において優れた安全上の実績を備えたウイルスである。組換えウイルスの生成を簡単に行うことが可能であり、大量に再現性よく製造することができる。したがって、組換えMVAウイルスの皮内、筋肉内、または粘膜投与は、CD8+T細胞応答によって制御可能なAIDSに対するヒトの予防的または治療的ワクチン接種に非常に適している。
【0125】
個体は、該抗原の送達および該抗原に対するCD8+T細胞免疫応答の生成が有益であるか、または治療上有益な効果を有するようなAIDSに罹患している個体であってよい。
【0126】
恐らくは、投与は、感染または症状発現の前にHIVまたはAIDSに対する免疫応答を生成するための予防的な目的を有することになる。
本発明に従って投与される成分は医薬組成物として製剤化されうる。これらの組成物は、薬学的に許容可能な賦形剤、担体、緩衝剤、安定化剤または当業者に良く知られた他の材料を含むことができる。そのような材料は無毒でなければならず、かつ有効成分の有効性を妨げてはならない。担体またはその他の材料の厳密な性質は、投与経路、例えば静脈内経路、皮内または皮下経路、経鼻、筋肉内経路、腹腔内経路に応じて変化しうる。
【0127】
言及したように、投与は好ましくは皮内投与、筋肉内投与、または粘膜投与である。
生理食塩水、デキストロースもしくはその他糖類の溶液またはグリコール、例えばエチレングリコール、プロピレングリコールもしくはポリエチレングリコールが含まれてもよい。
【0128】
静脈内、皮内、皮下、筋肉内、もしくは粘膜への注射、または罹患部位での注射については、有効成分は、発熱性物質を含まず適切なpH、等張性および安定性を有する非経口的に許容可能な水性溶液の形態となろう。当業者は、例えば、生食注射液、リンガー液、乳酸加リンガー液のような等張の媒体を使用して、適切な溶液を調製することができる。必要に応じて、保存剤、安定化剤、緩衝剤、酸化防止剤および/またはその他の添加剤が含まれていてもよい。
【0129】
徐放製剤が使用されてもよい。
MVA粒子の生産および任意選択のそのような粒子の組成物中への製剤化に続いて、該粒子は、個体、特にヒトまたはその他の霊長類に投与されうる。投与は、別の哺乳動物、例えばマウス、ラットもしくはハムスターのようなげっ歯動物、モルモット、ウサギ、ヒツジ、ヤギ、ブタ、ウマ、ウシ、ロバ、イヌまたはネコに対するものであってもよい。
【0130】
投与は、「予防的に有効な量」または「治療上有効な量」(場合に応じて予防は治療と考えることもできるが)であることが好ましく、これは個体に対する有益性を示すのに十分ということである。投与される実際の量、ならびに投与の速さおよび時間的投与条件は、治療対象者の性質および重症度に応じて変わることになろう。治療の処方、例えば投与量の決定などは、一般臨床医およびその他の医師、または獣医学的状況では獣医師の担当事項であり、典型的には、治療される病気、個々の患者の状態、送達部位、投与方法、および当業者に周知の他の要因を考慮する。上述の技法およびプロトコールの例は、オソル(Osol)、A.編、「レミングトンの製薬科学(Remington’s Pharmaceutical Sciences)」、第16版、1980年、に見出すことができる。
【0131】
1つの好ましい投与法では、DNAが注射1回あたり投与量300μg〜3mgで投与された後、MVAが注射1回あたり投与量106〜109個の感染性ウイルス粒子として投与される。
【0132】
組成物は単独で投与されてもよいし、他の治療と組み合わされて、治療すべき状態に応じて同時に、または順次、投与されてもよい。
ヒト以外の哺乳動物への送達は必ずしも治療目的である必要はなく、実験的状況で、例えば、対象とする抗原に対する免疫応答、例えばHIVまたはAIDSからの防御のメカニズムの研究に使用するためであってもよい。
【0133】
実施例
以下の実施例は、実施形態を作製および使用する方法の完全な開示および説明を当業者に提供するために提示されており、本発明者らが発明と見なすものの範囲を限定するようには意図されておらず、また下記の実験が全てであるかまたはその実験しか実施されないということを示すように意図されたものでもない。使用される数値(例えば量、温度など)に関する精度を保証する努力がなされてはいるが、何らかの実験上の誤差および偏差が考慮されるべきである。別途記載の無いかぎり、部は重量部であり、分子量は重量平均分子量であり、温度は摂氏度である。標準的な略語が使用される。
【0134】
実施例1
以下の実施例は、シャトルプラスミド、組換えMVA/HIV1臨床用ワクチン構築物、ならびに、外来遺伝子のコドン変更および2つのワクシニアウイルス必須遺伝子の間への外来遺伝子の挿入による、組換えMVAにおいて完全な外来遺伝子挿入物を維持するメカニズムを実証する。本開示は以下のメカニズムを提示する、すなわち:
・ MVAの複製にとって必須の2つのワクシニアウイルス遺伝子の間に完全な外来遺伝子を挿入することにより該外来遺伝子を維持するメカニズム。外来遺伝子の欠失はその組換えMVAに生育上の著しい優位性をもたらす可能性があり、継代が繰り返された際に完全な外来遺伝子を含有するMVAと競合する場合がある。しかしながら、外来遺伝子のほとんどの欠失は、隣接するワクシニアウイルスDNAのある程度の部分の喪失も含んでいる。そのワクシニアウイルスDNAが必須であれば、欠失を備えたウイルスは複製せず、完全な外来遺伝子を含有しているMVAと競合しないことになる。この方法論は、臨床試験で使用するためなど大規模に増幅させる必要のある組換えワクシニアウイルスの生産に有用であろう。また、
・ 組換えウイルスを繰り返し継代した後に容易に突然変異を生じる特異的「ホットスポット」を変質させることにより、外来遺伝子挿入物を安定させるメカニズム。この方法論は、臨床試験で使用するためなど大規模に増幅させる必要のある組換えウイルスの生産に有用である。
【0135】
そして、
・ 2つの必須ワクシニアウイルス遺伝子の間に外来遺伝子を挿入するために使用されるシャトルプラスミドpLW‐73、および
・ 上記2つのメカニズムの使用を具体化する材料である、組換えMVA/HIV‐1臨床用ワクチン構築物MVA/UGD4d
について説明する。
【0136】
安定な組換えMVAウイルスの生成
様々な地理的位置からのHIV‐1単離物由来のenvおよびgagpol遺伝子を発現する改変ワクシニアウイルスアンカラ(MVA)組換え体が構築された。外来遺伝子は、MVAの2つの部位、欠失II(Deletion II)および欠失III(Deletion III)に挿入された。組織培養物中で組換えMVAを繰り返し継代した後の上記遺伝子の安定性は変化しやすいことが判明した。本発明者らは、この不安定性が、外来遺伝子全体および一部の隣接DNAの欠失に起因するか、または外来遺伝子を発現しないため生育上有利である子孫ビリオンの増殖をもたらす特異的点突然変異に起因するかのいずれかであることを実証した。ここで本発明者らは、完全な外来遺伝子組換えMVAを保持する2つの新規な方法について述べる。第1に、本発明者らは、ゲノムの保存された中央領域の2つの必須ワクシニアウイルス遺伝子の間に外来遺伝子を挿入させる導入ベクターを構築した。この遺伝子挿入部位の使用は、必須ワクシニアウイルスDNAを含んだ大きな欠失部を含有するバリアントの増殖を防止する。加えて、このプラスミドは、組換えウイルスへのさらなる遺伝子の挿入に使用することができる。第2に、点突然変異を備えた単離物の分析から、翻訳中に早期終了を引き起こす単一塩基の挿入または欠失の傾向を備えたある種の「ホットスポット」が明らかとなった。本発明者らは、これらの部位におけるサイレント突然変異の生成が、挿入遺伝子の安定性をもたらすことを示した。
【0137】
I.新規な導入ベクターの構築および適用
新規な導入ベクターpLW‐73の構築
1. MVAゲノムの中央領域K7R‐A24Rについて、1)ポックスウイルス科またはコードポックスウイルス亜科において保存された遺伝子対、および2)遺伝子の3’末端どうしが近接するように逆方向を向いた遺伝子であって、その結果としてワクシニアプロモーターを妨害しないであろう挿入部位を提供する遺伝子、に関する検査が行われた。新しい挿入部位として選択された部位は、2つの必須遺伝子I8RおよびG1Lの間であった。
【0138】
2. 新しいベクターの左側隣接部は次の方法で構築された:プラスミドLAS‐1が、del III MVA隣接部、GFP、およびMVA隣接部のダイレクトリピートを取り出すために、制限酵素EcoRIおよびXhoIで切断された。この挿入物はAscIおよびSacIで切断されて、GFPフラグメントが単離された。I8R遺伝子末端の531塩基対(TAA終止コドンを含む)が、PCR産物の末端にEcoRIおよびAscI制限部位を備えるようにPCR増幅された。(TAA終止コドンを含むI8R遺伝子末端からの)229塩基対のダイレクトリピートのPCR増幅が、SacIおよびXhoI制限部位を含有するオリゴヌクレオチドを用いて行なわれた。全4片のDNAすなわち、1)EcoRI末端およびXhoI末端を備えたベクター骨格、2)EcoRI末端およびAscI末端を備えたI8R末端を含有する新しい左側隣接部、3)AcsI末端およびSacI末端を備えたGFP、ならびに4)SacI末端およびXhoI末端を備えたI8R隣接部のダイレクトリピート、が合わせてライゲーションされてプラスミドpLW‐72が作製された。
【0139】
3. 右側隣接部は以下のように作製された:pLW‐72が制限酵素PstIおよびHindIIIで切断され、該プラスミド中のMVAのdel III隣接部が放出された。G1L遺伝子末端の702塩基対が、両端にPstIおよびHindIII制限酵素部位を備えるようにPCR増幅され、pLW‐72ベクター中にライゲーションされてpLW‐73が作製された(図7)。pLW‐73の配列を図8に示す。
【0140】
4. pLW‐73の顕著な特色は次のとおりである:1)ベクターは、MVAゲノム中の必須遺伝子の間に外来遺伝子を挿入するために設計された。左側隣接部はI8R遺伝子の終端部からなり、右側隣接部はG1L遺伝子の終端部からなる。2)組換えウイルスの簡単な初期選択のためにGFP遺伝子が含まれている。3)GFPはI8R遺伝子のダイレクトリピートに隣接しており、組換えウイルスが繰り返し継代されるとGFPが失われることになるためGFPの一時的発現が可能となっている。国際公開公報第2004/087201号を参照すると、特色2および3は、MVA/HIV組換え体pLAS‐1およびpLAS‐2を作製するために使用された初期プラスミドにも含まれていた。
【0141】
pLW‐73の適用
1. HIV‐1のクレードBのADA単離物由来のenv遺伝子がpLW‐73にクローニングされ、組換えMVAウイルスが作製された。DNA塩基配列決定により、env遺伝子の位置および完全性が確認された。
【0142】
2. MVAの欠失II(Deletion II)部位にウガンダのクレードD(単離物AO7412)のenv遺伝子を発現する組換えMVAウイルス(図9)は、不安定であること、すなわち、培養物中の連続継代が繰り返された後に遺伝子が大部分のウイルス子孫から欠失することが判明した。その後、同じ遺伝子がpLW‐73にクローニングされ、組換えMVAウイルスが作製されて特徴解析された。env遺伝子挿入物は培養物中で連続継代が繰り返された(8×)後も安定していた、すなわち、挿入遺伝子または隣接するMVA領域の欠失は見出されなかった。加えて、遺伝子がこの部位に挿入された時は他の突然変異が発生しなかった。
【0143】
II.「ホットスポット」の点突然変異
点突然変異の分析
MVAの欠失III(Deletion III)部位にウガンダのクレードD(単離物AO3349)gagpol遺伝子を発現する組換えMVAウイルスは、不安定であることが判明した。主な遺伝子変化は、GまたはC残基が4〜6個連続した箇所における単一の点突然変異の生成であった(表3)。加えて、同様の点突然変異は、ケニアのクレードAのHIV‐1単離物およびタンザニアのクレードCの単離物由来のgagpol遺伝子を発現する同様の組換えウイルスからの非染色プラークに見出された。
【0144】
ホットスポットの突然変異誘発および組換えウイルス中の安定性の分析
部位特異的突然変異誘発を使用して、ウガンダのHIV‐1単離物由来のgag遺伝子の6つのそのような領域にサイレント突然変異を作製した。この変異遺伝子UGD4d gagpol orf(図10)はpLAS‐1にクローニングされ、不安定なウイルスの構築で行われたようにしてMVAの同じ欠失III(Deletion III)部位へ組み込まれた。培養物中で連続継代を繰り返した(8×)後、発現のないプラークは見出されなかった。継代8代目のウイルスストックのDNA塩基配列決定から、gagpol遺伝子の完全性が維持されていることが確認された。
【0145】
III. 二重組換え体の構築
MVA/UGD4dウイルス
MVA/UGD4dウイルス、すなわちウガンダのサブタイプD AO7412のエンベロープおよびAO3349のgagpolを発現する組換えウイルスは、次の方法で構築された:エンベロープおよびgagpolの遺伝子は、それぞれシャトルプラスミドpLW‐73およびpLAS‐1を利用した相同組換えによってMVA 1974/NIHクローン1に挿入された。MVA/UGD4dは、ニワトリ胚線維芽細胞における6回のプラーク精製によって単離され、その後増幅されて特徴解析された。
【0146】
概要
1. MVAゲノムの保存された中央領域の2つの必須遺伝子I8RおよびG1Lの間への外来遺伝子の組み込みを行うプラスミド導入ベクターが構築された。この部位の使用により、挿入遺伝子/MVA隣接部の欠失を有する変異ウイルスの選択が阻止されることが示された。
【0147】
2.GおよびC残基が連続した極めて変異しやすい箇所は部位特異的突然変異誘発によって変更され、コード配列中のサイレント突然変異が生成された。この変化は、遺伝子がMVAの欠失III(Deletion III)に挿入された場合に該遺伝子を安定させることが示された。
【0148】
3. 上記の2つの方法を利用して、ウガンダのクレードDのenvおよびgagpolの両方を安定して発現するUGD4d二重MVA組換え体が構築された。
実施例2
様々な単離物からのHIV‐1 env遺伝子およびgagpol遺伝子を発現する組換えMVAが作製された。組織培養物中で継代が繰り返された後の挿入遺伝子の安定性は変化しやすいことが判明した。ここで本発明者らは、(1)不安定性は、自然突然変異または挿入遺伝子の欠失と、発現のない突然変異の選択との組合せを表わすことを実証し、(2)不安定性を低減する新規な方法について述べる。
【0149】
概観
多数の異なる単離物からのenvおよびgagpolを発現する組換えMVAが構築された。各ウイルスは、臨床試験用ウイルスの製造に必要な大規模増幅を模倣するためにニワトリ胚線維芽細胞において繰り返し継代された。挿入物の安定性は、個々のプラークのenvおよびgagを免疫染色することによってモニタリングされた。いくつかの組換えウイルスについては、envおよび/またはgagの発現がウイルス集団のかなりの割合において急速に失われることが見出された。発現喪失のメカニズムを同定するために、個々のプラークが単離され、突然変異の性質が特徴解析された。いくつかの事例では、単一ヌクレオチドの付加または欠失によって突然変異する傾向を備えた特定のDNA塩基配列が同定された。そのような突然変異の生成は、予測される翻訳産物を変化させることなくコドンを変更することにより、回避することができた。他の事例では、発現の喪失は、隣接している非必須MVA遺伝子の中まで及ぶことの多い大規模な欠失によって引き起こされた。これが生じるのを防止するために、2つの必須MVA遺伝子の間に外来遺伝子を挿入させることを目指した新しいシャトルプラスミドが構築された。この部位への組込みは外来DNAの欠失を低減した。しかしながら、ある事例では、高レベルのHIV env発現に関連した毒性が非常に強いため、希少な突然変異体を選択してもなお不安定な集団が得られた。この場合、envの膜貫通ドメインを短縮化して初めて安定した組換えMVAの構築が可能となった。
【0150】
組換えMVAの生成および挿入遺伝子の安定性の分析
Env遺伝子およびgagpol遺伝子はMVAシャトルベクターにクローニングされた。発現および機能は一時的発現アッセイによって分析された。gagpolはMVA 1974/NIHクローン1に組み込まれた。組換えMVAは6〜8回プラーク精製され、その後ウイルスの増幅が行われた。Envは該MVA/gagpol単離物に組み込まれ、二重組換えMVA(図11A)は6〜8回プラーク精製され、増幅された。挿入物の安定性を評価するために、ウイルスは、大規模製造を模倣するために〜1pfu/細胞の感染多重度(m.o.i.)を使用してCEF細胞中で連続的に継代された。安定性は、モノクローナル抗体で免疫染色することにより測定された、envまたはgagを発現する細胞の割合(%)の測定により評価された(図11B)。
【0151】
組換えMVAの安定性
様々な地理的位置からのHIV‐1単離物由来の遺伝子を発現する組換えMVAが構築された。env遺伝子およびgagpol遺伝子は、それぞれMVAの欠失IIおよびIIIに挿入され、いずれも改変H5プロモーターの制御下におかれた。7つの組換えMVAのenv遺伝子およびgagpol遺伝子の安定性が表4に示されている。様々な程度の不安定性がこの7つのウイルスにおいて観察された。MVA/65A/Gでは、envの発現は急速に失われて、継代6代目までにはわずか25%のビリオンしかenvを発現しなかった。MVA/UGD4aでは、envおよびgagpol両方の発現は連続的なウイルス継代とともに徐々に失われた。第I相試験用の大量のウイルスの製造には少なくとも6〜7回の継代が必要なので、これら2つのウイルスは不適当であると考えられた。
【0152】
MVA/65A/Gの発現の分析
図12を参照すると、MVA/65A/GのP3およびP5から13個のプラークが無作為に採取され、T‐24 mAb(結合部位をaに示す)を用いた免疫染色、ウエスタンブロッティング、PCR、および配列決定により分析された。5種類のプラークが見出され、各種類について得られたこれらのプラークの数は図12の右側に示されている。プラークa、bおよびcは染色されたが、bおよびcは、塩基置換(終止コドンを生じるもの)(b)、ならびに、env遺伝子の端部およびMVA隣接部の一部の欠失(c)に起因する短縮型であった。染色されないプラークdおよびeは、Gが5個連続した箇所へのGの付加を原因とするフレームシフト(d)、ならびに、env遺伝子全体およびMVA隣接部の一部の大規模な欠失(e)から生じていた。したがって、塩基対付加、置換、および欠失はすべてMVA/65A/Gのenv遺伝子の不安定な発現に寄与していた。このA/G envは検討した中で最も不安定な例であり、安定性を増強しうる改変を研究するために採取された。
【0153】
安定性を増大させるためのA/G構築物の改変
1. 点突然変異を防止するために、GおよびCが4個および5個連続した箇所をサイレント突然変異によって除去することにより合成エンベロープが作製された。
【0154】
2. ベクターI8/G1すなわちpLW‐73は、遺伝子およびMVA隣接部の欠失が生存可能とならないように、必須遺伝子I8RおよびG1Lの間の挿入部位を用いて構築された。MVAのI8R遺伝子の端部(500bp)およびG1L遺伝子の端部(750bp)がPCRによって増幅され、外来遺伝子の発現を制御するワクシニアウイルス初期/後期mH5プロモーターを含有するベクターに挿入された。このI8/G1ベクターは、相同組換えによってMVAに外来遺伝子を挿入するために使用された(図13)。挿入遺伝子および該挿入遺伝子に隣接しているMVAの欠失は、必須遺伝子の部分が欠失することになるので生存可能とはならなくなる。したがって、これらの突然変異を備えたウイルスは、正常な生育上の利点を備えた集団において異常増殖することができなくなる。
【0155】
3. A/G gp140エンベロープは、gp41の膜貫通ドメインおよび細胞質尾部を欠失させることにより変異させられ、分泌タンパク質を生じた。
安定性を増大させるための試験的改変
図14に示されるように、envの改変および/または新しいベクターの使用により7つの単一組換えウイルスが作製された。改変がエンベロープの安定的発現を増強したかどうかを判定するために、各ウイルスの5つのプラークが単離されてCEF中で独立に継代された。継代されたプラークは、mAb T‐43(結合部位はenvの101〜125aaにマッピングされた)を用いた免疫染色、ウエスタンブロッティング、PCR、および配列決定により分析された。
【0156】
プラーク継代後のEnv発現
図15を参照すると、上記に列挙された7つの組換え体それぞれの5つの独立に継代されたプラーク単離物が、継代1、3、5および7代目において、mAb T‐43(gp120の101〜125a.a.の間に結合する)を用いた免疫染色によって特徴解析された。7種のウイルスのうち4つ(図15のa、b、c、e)は、5つの継代プラークそれぞれにおいて不安定なタンパク質発現を有し、(図15f)の2つのプラーク継代物も不安定なenv発現を有していた。これらは、MVAゲノムのdel II(図15c)および必須遺伝子部位(図15f)の両方に合成envを備えたウイルスを含んでいた。短縮された分泌型gp140としてのエンベロープを含有している組換えウイルスだけが、エンベロープを安定に発現し続けた(図15のdおよびg)。
【0157】
ウエスタンブロッティング、PCRおよび配列分析
選択されたプラーク継代物からクローンを採取し、ウエスタンブロッティング、PCRおよび配列分析によってタンパク質発現を分析した(図16)。ウエスタンブロット解析については、クレードAのエンベロープの先頭部および終端部にそれぞれ結合するT‐24およびT‐32が、部分的エンベロープのみが作製されたか完全長エンベロープが作製されたかを測定するために使用された。cと印付けされた対照ウイルスが各ブロットの右側にある。MVAの欠失IIにおいて作製された3つのウイルス(図16a、bおよびc)については、図16c(すなわちgp140クローン)にのみ、全クローンがウエスタン分析で検出可能なタンパク質を発現していた。このタンパク質(T‐32によって計測)は短縮されていなかった。エンベロープがベクターI8/G1によって必須遺伝子部位へ挿入されると(図16d、eおよびf)、この場合も、gp140エンベロープだけがすべてのクローンにおいて発現されており、短縮されていなかった。I8/G1ベクターの使用はenv配列の突然変異生成を防止しなかったが、del IIに挿入されたエンベロープで見られていた欠失は防止した。(I8/G1ベクター由来の試験されたすべてのクローンの陽性PCR産物、対してdel IIベクターを使用して試験されたクローンの陰性PCR産物に注目されたい。)
クレードA/G二重組換え体におけるEnvの発現
1つのenv分析を用いた先の結果に基づいて、gp140または合成gp160遺伝子とともにgagpolを発現する二重組換え体が作製され、env発現の安定性について試験された(図17)。先述のようにして各々から5つのプラークが単離され、env発現の安定性を分析するために7回継代された。継代7代目で、継代されたプラークはT‐43およびT‐32 mAbs(それぞれgp120およびgp41に結合する)両方を用いて免疫染色された。T‐43 mAbでは、合成エンベロープを発現する組換え体の5つのクローンのうちの1つが染色されないプラークのみで構成されていた。続いてこれらのプラークのT‐32染色は、別のプラークが短縮されたエンベロープを発現することを示した。gp140エンベロープを含有する二重組換え体からの継代プラークは全て、T‐43およびT‐32両方の免疫染色により、安定しているように見えた。力価も他の二重組換え体を用いるより2log高かった。したがって、安定してエンベロープを発現するクレードA/G二重組換え体は、gp140エンベロープを用いた場合にのみ作製可能であった。
【0158】
ウガンダのHIV‐1単離物由来のenvおよびgagpolを発現する組換えウイルス
ウガンダの単離物AO7412およびAO3349由来のHIV‐1 envおよびgagpol遺伝子を発現する組換えMVAウイルスは、図18に示されるようにして構築された。各々4〜6個の独立な単離物が連続的に継代され、両遺伝子が単独で発現されても組み合わせで発現されても不安定であることが分かった(表5)。対照的に、膜結合型gp160の代わりとしてgp140の発現は、連続継代後のenv遺伝子の安定性をもたらした(図18および表5)。
【0159】
MVA/UGD4a − 非染色envプラークの分析
不安定性のメカニズムを究明するために、24個の個々の非染色プラーク(Mab T‐43を使用)がMVA/UGD4aの継代6代目から単離され、増幅され、特徴解析された。2つの小さな欠失(1.2kbおよび0.3kb)がPCR増幅およびDNA塩基配列決定によって同定された(図19)。他の単離物はすべて、隣接するMVAにまで及ぶ非常に大きな欠失を含んでいた。これらの欠失に関するおよその切断点は、env遺伝子または隣接するMVA領域の内側からプライマー対を使用して同定された。
【0160】
組換えMVAにおけるUGD env遺伝子の改変
UGD env遺伝子の不安定性の問題を改善するために、新しいベクターI8/G1を使用して、AO7412 env遺伝子をMVAに挿入した。このベクターI8/G1は、2つの必須ワクシニアウイルス遺伝子I8およびG1の間に外来遺伝子を組み込み、かつ改変H5プロモーターを使用する(図20)。独立した4つのプラークが連続的に継代され、継代5代目でMab T‐43およびT‐32を用いて免疫染色することによりenv発現について分析された。すべての単離物において、遺伝子は安定していた(表6)。
【0161】
MVA/UGD4b − 非染色gagプラークの分析
gag遺伝子の不安定性のメカニズムを究明するために、8個の個々の非染色プラーク(Mab 183‐H12‐5C(NIAID AIDSリポジトリ)を使用)がMVA/UGD4bの継代6代目から採取され、増幅され、gagpol挿入物が配列決定された(表7)。7個の単離物には、564〜569位における単一のG残基の挿入または欠失が見出された。1つの単離物では、530〜534位の配列CCCCからC残基が欠失していた。更に、MVA/KEAおよびMVA/TZCの多数回継代ストック由来の非染色プラークから、同様の突然変異ホットスポット、すなわち564〜569位が明らかとなった。UGD AO7412 gagpol遺伝子の完全配列の調査から、22箇所の4個以上連続したGまたはC残基が実証された(図21)。
【0162】
組換えMVAにおけるUGD gagpol遺伝子の改変
gagpol遺伝子の不安定性のメカニズムが主として4〜6個の連続したGまたはC残基内の単一ヌクレオチドの挿入または欠失であったので、この遺伝子の安定性を改善する戦略は、そのような部位でサイレント突然変異を生成させることであった。したがって、p17およびp24 gagの6部位における部位特異的突然変異誘発(表3)が使用された。結果として得られた、MVAの同じ場所すなわち欠失III(Deletion III)に挿入されたコドン変更された(c.a.)遺伝子は、連続継代しても安定であることが証明された(図22および表8)。
【0163】
UGD envおよびgagpolを発現する安定な組換えMVAの構築
MVAのI8/G1座においてUGD env遺伝子、および欠失III(Deletion III)においてコドン変更されたgagpol遺伝子を発現する組換えウイルスが構築された(図23)。連続継代では、いずれの遺伝子の不安定性も示されなかった。更に、タンパク質発現のレベルおよびDNA塩基配列は継代を経ても不変であった(表9)。
【0164】
結論
env挿入物およびgagpol挿入物の不安定性は、点突然変異および欠失の生成ならびに非発現性MVA突然変異が生育上優位であることに起因する。不安定性は一般に、コドン変更、またはMVAゲノムの必須領域への挿入のうち少なくともいずれか一方によって低減することが可能である(MVA/UGD4d)。しかし、1つの事例ではenvを変更しなければならなかった(MVA/65A/G)。
【0165】
実施例3
BALB/cマウスにおけるMVA/UGD4dの免疫原性
マウス10匹の群が、それぞれ感染単位106または107のMVA/UGD4dを用いて腹腔内経路により免疫化された。マウス5匹の群が、それぞれ親のMVA‐1974を用いて同様に免疫化された。マウスは第0週および第3週で免疫化処置され、第0週、第3週および第5週で採血された。第5週で脾臓が採取された。
【0166】
細胞の応答は細胞内のサイトカイン染色によって新鮮な脾細胞で計測された。脾細胞は、下記すなわち:1)免疫優勢であるgagペプチド(AMQMLKETI(配列番号6))、2)envペプチド(DTEVHNVWATHACVP(配列番号7)およびQQQSNLLRAIEAQQH(配列番号8))、3)polペプチド(ELRQHLLRWGLTT(配列番号9)およびHGVYYDPSKDLIAE(配列番号10)の単一アミノ酸バリアントを備えた8つのペプチド)、ならびに4)MVAを用いて別々に刺激された。
【0167】
細胞は、CD4およびCD8の表面発現について染色され、次いでIFN‐γおよびIL2またはTNFのいずれかの細胞内発現について染色された。図24に示されるように、MVA/UGD4dはgagペプチド、polペプチドおよびMVAに対してCD8/IFN‐γ応答を誘発した。gagペプチドの応答は、IFN‐γと、IL2またはTNFのいずれかとの両方を発現する多機能性であった。さらに、CD4/IFN‐γ応答がenvペプチドの集合物に対して誘発された。
【0168】
体液性応答はELISAによって計測された(図25)。UGD envに対する強い応答が1回の免疫化処置の3週間後に実証され、第2の免疫化処置によって増強された。加えて、強いワクシニアウイルス応答が1回および2回の免疫化処置の後で誘発された。
【0169】
【表4】
【0170】
【表5】
【0171】
【表6】
【0172】
【表7】
【0173】
【表8】
【0174】
【表9】
【0175】
【表10】
実施例4
この実施例は、安定な組換えMVAウイルスを生成するためのさらなる挿入部位の使用について実証する。MVAウイルスゲノムのDel III領域はいくつかの非必須遺伝子および遺伝子フラグメントを含有しており、よって歴史的に異種核酸配列を挿入するために使用されてきた。したがって、MVAのdel III挿入部位周辺の隣接領域が、フラグメント化遺伝子または非必須遺伝子の存在について分析された。いくつかの細胞においてVACVの複製に重要であることが知られている遺伝子、すなわちA50R DNAリガーゼおよびB1Rキナーゼは、del III挿入部位からそれぞれ約1kbpおよび1.8kbpに位置していた。本発明者らは、Del III挿入部位に隣接する非必須遺伝子を除去すれば、より安定な挿入部位にすることができるであろうと推論した。この目的に向けて、A50R DNAリガーゼORFの3’末端部分(左)、ならびにB1R ORFの5’末端、およびプロモーター(右)を含む隣接配列を備えた核酸構築物(例えばシャトルベクター)が以下のように構築された。これにより、相同組換えが生じた時に上記2つの重要な遺伝子の間の非必須遺伝子の領域が有効に除去されるであろう。
【0176】
A. A50R/B1Rシャトルベクターの調製:
MVAゲノム中のdel III挿入部位周辺の隣接領域(bp数143552、アカンビス(Acambis)3000 Genbank AY603355))の分析から、いくつかの細胞においてVACVの複製に重要であることが知られている少なくとも2つの遺伝子が明らかとなった。具体的には、A50R DNAリガーゼ(ORF163;ACAM3000_MVA_163;配列番号11)およびB1Rキナーゼ(ORF167;ACAM3000_MVA_167;配列番号16)が、del III挿入部位からそれぞれ約1kbpおよび1.8kbpに位置していた。したがって、ORF163とORF167との間にある非必須遺伝子またはフラグメント化遺伝子が除去の標的とされた。具体的には、ORF164(A51R‐A55のフラグメント)、ORF165(A56Rプロモーターの一部を欠いている)、ORF166、およびフラグメント化したA57Rが除去の標的となった。これらの非必須遺伝子およびフラグメント化遺伝子の除去を行うために、核酸構築物(すなわちシャトルベクター)が、MVAゲノムのORF163とORF167との間に相同組換えによって組換えを生じ、その結果として介在配列を除去することができるように設計された。そのような組換えを達成すると、核酸構築物は、ACAM3000_MVA_163由来の1つの核酸配列(左側の隣接配列)と、ACAM3000_MVA_167由来の1つの核酸配列(右側の隣接配列)とを含むことになる。これらの配列は該核酸構築物中で隣り合うことになり、これは、該配列がいかなるポックスウイルスORFによっても隔てられていないことを意味する。より具体的には、左側隣接部はA50RリガーゼORFのC末端を含有することになり、右側隣接部はプロモーター領域およびB1R ORFのN末端を含有することになる。ベクターの設計は図26に示されている。
【0177】
シャトルベクターを構築するために、隣接部はそれぞれ別々に作出された。再構築Del IIIベクターの左側隣接部は最初に以下のようにして構築された。
緑色蛍光タンパク質(GFP)をコードする遺伝子およびダイレクトリピートと共に左側隣接部全体(I8R遺伝子の一部を含有する隣接部1)を切り出すために、プラスミドLW‐73(図7)がEcoRIおよびXhoIで消化された。その後、GFPを含有するフラグメントが制限酵素AcsIおよびSacIで消化されてGFP遺伝子が取り出された。
【0178】
ORF163のC末端部分を含んでいる左側隣接部を作出するために、プライマーLW470(配列番号23)およびLW471(配列番号24)を使用したポリメラーゼ連鎖反応(PCR)法によってMVAゲノムからDNAフラグメントが増幅された。PCR増幅は標準的な条件を使用して実施された。次に、ORF163の末端のダイレクトリピートが、プライマーLW‐472(配列番号25)およびLW‐473(配列番号26)を使用してMVAゲノムから増幅された。最後に、EcoRI部位およびXhoI部位を備えたベクター骨格、AcsI部位およびSacI部位を備えたGFP遺伝子、EcoRI部位およびAscI部位を含有するORF163フラグメント(左側隣接部)、ならびにSacI部位およびXhoI部位を含有するORF163のC末端領域由来のダイレクトリピートが合わせてライゲーションされて、中間プラスミド#2743が形成された。
【0179】
ORF167のN末端部分(そのプロモーター領域を含む)を含有している右側隣接部を作出するために、中間プラスミド#2743は制限酵素PstIおよびHindIIIで消化されて右側隣接部が放出された。次に、プライマーLW‐474(配列番号27)およびLW‐475(配列番号28)を使用してMVAゲノムからDNAフラグメントがPCR増幅された。このフラグメントは制限酵素PstIおよびHindIIIで消化され、該消化フラグメントは同様に消化されたシャトルベクター骨格にライゲーションされてLW‐676核酸構築物が作られた。(図27)
pLW‐76の顕著な特色は次のとおりである:
1) 該ベクターは、MVAゲノム中のA50R DNAリガーゼ遺伝子(ORF163)の端部と、プロモーターおよびB1Rキナーゼ遺伝子(ORF167)のN末端部分との間に外来遺伝子を挿入するために設計されている。左側隣接部はA50Rリガーゼ遺伝子の端部からなり、右側隣接部はプロモーターおよびB1Rキナーゼの先頭部からなる。
【0180】
2) 組換えウイルスの簡単な初期選択のためにGFP遺伝子が含まれている。
3) GFPはA50Rリガーゼ遺伝子のダイレクトリピートに隣接していることにより、組換えウイルスの継代が繰り返されるとGFPが失われるのでGFPの一時的発現を可能にする。特色2および3は、MVA/HIV組換え体を作製するために使用された初期のプラスミドpLAS‐1およびpLAS‐2にも含まれていた。
【0181】
その後、ウガンダのクレードDのヒト免疫不全ウイルス(HIV)(単離物AO7412)由来のenv遺伝子が新しいpLW‐76構築物にクローニングされた。このenvを含有する核酸構築物はその後細胞にトランスフェクションされ、該細胞をMVAウイルスに感染させることでHIV ENVタンパク質を発現する組換えMVAウイルスrMVA/UGDenv(delIIIrst)が生成された。次いでこのウイルスが特徴解析された。
【0182】
ニワトリ胚繊維芽細胞(CEF)において生育させると、rMVA/UGDenv(delIIIrst)による感染がシンシチウム型の細胞質効果(CPE)をもたらすことが観察された。これは、再構築されたdel III部位で生じた組換えの際の非必須A56ヘマグルチニン遺伝子の欠失に起因していた。正常なrMVAはフレイ(flay)フォーカス(図28A)を有していたが、rMVA/UGDenv(delIIrt)に感染すると、シンシチウム形成を示すフォーカスを示し、凝縮されたシンシチウムへと進行した。(図28B)。
【0183】
その後、rMVA/UGDenv(delIIIrst)は、挿入された異種核酸配列の安定性に関して特徴解析された。これは、CEF細胞中でウイルスを繰り返し継代すること、および、各世代について、発現されたHIV ENVタンパク質の存在を試験することにより行われた。ENVタンパク質の検出は、HIVエンベロープタンパク質に対するモノクローナル抗体を用いてウイルスのプラークをスクリーニングすることにより行われた。rMVA/UGDenv(delIIIrst)の安定性は、del II領域にenv遺伝子を含有しているウイルス、およびenv遺伝子が中央の保存領域へ挿入されたウイルスと比較された。この比較の結果は図29に示されている。発現されているENVタンパク質のレベルも、HIV ENVタンパク質に対するモノクローナル抗体を使用してウエスタンブロット法により計測された。
【0184】
図29は、MVA/UGDenv(del II)が、env内部に生じ、かつ隣接するMVAに及んでいる欠失が原因で、明らかに不安定であったことを示している。UGD envが、MVA/UGDenv(I8/G1)のように2つのVACV必須遺伝子の間に置かれると、生存可能な欠失が防止された。最後に、HIV env遺伝子のrMVA/UGDenv(del IIIrst)への組み入れは、少なくとも11回の継代を通じて安定であることが観察された。
【0185】
図30は、11種のウイルス構築物が同様の量のENVタンパク質を発現したことを示している。
したがって、これらの研究の結果は、MVAウイルスゲノムのdel III領域が、非必須遺伝子を除去してdel III部位を再構築することにより、より安定となったことを示唆している。
【0186】
本発明は明瞭さおよび理解を目的としてある程度詳細に説明されているが、当業者であれば、本発明の真の範囲から逸脱することなく形態および詳細の様々な変更を行なうことができることを認識するであろう。上記に引用されたすべての図面、表および付録、ならびに特許、出願および公開は、参照により本願に組込まれる。
【特許請求の範囲】
【請求項1】
組換え改変ワクシニアアンカラ(MVA)ウイルスであって、MVAウイルスゲノムの2つの隣り合った必須オープンリーディングフレーム(ORF)の間に異種核酸配列を含み、親MVAウイルス中の対応する2つの必須ORFの間に存在する非必須ORFを欠いていることを特徴とする、組換えMVAウイルス。
【請求項2】
異種核酸配列は、2つの隣り合った必須ORFの間にある遺伝子間領域(IGR)内にある、請求項1に記載の組換えMVAウイルス。
【請求項3】
非必須遺伝子はMVAウイルスのDel III挿入部位に隣接している、請求項1に記載の組換えMVAウイルス。
【請求項4】
隣り合った必須ORFのうち少なくとも一方は、配列番号11、配列番号14、配列番号16または配列番号19と少なくとも90%の同一性を有する、請求項1に記載の組換えMVAウイルス。
【請求項5】
隣り合った必須ORFのうち一方は配列番号11または配列番号14と少なくとも90%の同一性を有し、隣り合った必須ORFのうち他方は配列番号16または配列番号19と少なくとも90%の同一性を有する、請求項1に記載の組換えMVAウイルス。
【請求項6】
隣り合った必須ORFのうち一方は配列番号11または配列番号14を含み、隣り合った必須ORFのうち他方は配列番号16または配列番号19を含む、請求項1に記載の組換えMVAウイルス。
【請求項7】
CDC/アカンビス(Acambis)による命名法を用いると、隣り合った必須ORFのうち一方は163であり、隣り合った必須ORFのうち他方は167である、請求項1に記載の組換えMVAウイルス。
【請求項8】
異種核酸配列は、転写制御要素に転写制御される少なくとも1つのコード配列を含む、請求項1に記載の組換えMVAウイルス。
【請求項9】
コード配列は1つ以上のタンパク質をコードする、請求項8に記載の組換えMVAウイルス。
【請求項10】
転写制御要素はポックスウイルスの転写制御要素である、請求項8に記載の組換えMVAウイルス。
【請求項11】
コード配列はヒト免疫不全ウイルス(HIV)に由来する、請求項8に記載の組換えMVAウイルス。
【請求項12】
コード配列はHIVエンベロープタンパク質をコードする、請求項8に記載の組換えMVAウイルス。
【請求項13】
コード配列は配列番号4と少なくとも90%の同一性を有するヌクレオチド配列を含み、コードされているタンパク質が、配列番号4によってコードされるタンパク質に対する免疫応答の誘発能を有する、請求項8に記載の組換えMVAウイルス。
【請求項14】
コード配列は配列番号4を含む、請求項8に記載の組換えMVAウイルス。
【請求項15】
単離された核酸構築物であって、
(a)第1の必須MVAウイルスORF由来の少なくとも25個連続したヌクレオチドを含む第1の核酸配列と、
(b)第2の必須MVAウイルスORF由来の少なくとも25個連続したヌクレオチドを含む第2の核酸配列と
を含み、
第1および第2の必須MVAウイルスORFはMVAゲノム中で少なくとも1つの非必須ORFによって隔てられており、第1および第2の核酸配列は、単離された核酸構築物において互いに隣り合っていることを特徴とする、核酸構築物。
【請求項16】
a)第1の核酸配列は、第1の必須MVAウイルスORFの少なくとも100個連続したポリヌクレオチド領域と少なくとも75%の同一性を有する少なくとも100個連続したポリヌクレオチド領域を含み、かつ
(b)第2の核酸配列は、第2の必須MVAウイルスORFの少なくとも100個連続したポリヌクレオチド領域と少なくとも75%の同一性を有する少なくとも100個連続したポリヌクレオチド領域を含む、
請求項15に記載の核酸構築物。
【請求項17】
CDC/アカンビスによる命名法を用いると、第1のORFはACAM3000_MVA_163であり、第2のORFはACAM3000_MVA_167である、請求項15に記載の核酸構築物。
【請求項18】
第1および第2の核酸配列の隣り合った端部は、(a)遺伝子間領域および(b)制限酵素認識部位からなる群から選択された少なくとも1つのヌクレオチド配列を含む第3の核酸配列によって隔てられている、請求項15に記載の核酸構築物。
【請求項19】
第1および第2の核酸配列の隣り合った端部は、異種核酸配列を含む第3の核酸配列によって隔てられていることを特徴とする、請求項15に記載の核酸構築物。
【請求項20】
異種核酸配列は、転写制御要素に転写制御される少なくとも1つのコード配列を含む、請求項19に記載の核酸構築物。
【請求項21】
転写制御要素はポックスウイルスの転写制御要素である、請求項20に記載の核酸構築物。
【請求項22】
コード配列は1つ以上のタンパク質または低分子干渉RNAをコードする、請求項20に記載の核酸構築物。
【請求項23】
異種核酸配列はヒト免疫不全ウイルス(HIV)に由来する、請求項21に記載の核酸構築物。
【請求項24】
異種核酸配列はHIVエンベロープタンパク質をコードする、請求項21に記載の核酸構築物。
【請求項25】
pLW‐76の配列(配列番号21)を含む、請求項15に記載の核酸構築物。
【請求項26】
安定な組換え改変ワクシニアアンカラ(MVA)ウイルスを製造する方法であって、
(a)請求項15に記載の核酸構築物で細胞をトランスフェクションする工程と、
(b)トランスフェクションした細胞をMVAウイルスに感染させる工程と、
(c)核酸構築物とMVAウイルスゲノムとの間の相同組換えを可能とするのに適した条件下で感染細胞を培養する工程と、
(d)組換えMVAウイルスを単離する工程と
を含む方法。
【請求項27】
対象者において免疫応答を誘導する方法であって、請求項8に記載の組換えMVAウイルスを該対象者に投与する工程を含む方法。
【請求項28】
in vitroでタンパク質を生産する方法であって、
(a)宿主細胞を請求項8に記載の組換えMVAウイルスに感染させる工程と、
(b)感染宿主細胞を適切な条件下で培養する工程と、
(c)宿主細胞による生産物を単離する工程と
を含む方法。
【請求項1】
組換え改変ワクシニアアンカラ(MVA)ウイルスであって、MVAウイルスゲノムの2つの隣り合った必須オープンリーディングフレーム(ORF)の間に異種核酸配列を含み、親MVAウイルス中の対応する2つの必須ORFの間に存在する非必須ORFを欠いていることを特徴とする、組換えMVAウイルス。
【請求項2】
異種核酸配列は、2つの隣り合った必須ORFの間にある遺伝子間領域(IGR)内にある、請求項1に記載の組換えMVAウイルス。
【請求項3】
非必須遺伝子はMVAウイルスのDel III挿入部位に隣接している、請求項1に記載の組換えMVAウイルス。
【請求項4】
隣り合った必須ORFのうち少なくとも一方は、配列番号11、配列番号14、配列番号16または配列番号19と少なくとも90%の同一性を有する、請求項1に記載の組換えMVAウイルス。
【請求項5】
隣り合った必須ORFのうち一方は配列番号11または配列番号14と少なくとも90%の同一性を有し、隣り合った必須ORFのうち他方は配列番号16または配列番号19と少なくとも90%の同一性を有する、請求項1に記載の組換えMVAウイルス。
【請求項6】
隣り合った必須ORFのうち一方は配列番号11または配列番号14を含み、隣り合った必須ORFのうち他方は配列番号16または配列番号19を含む、請求項1に記載の組換えMVAウイルス。
【請求項7】
CDC/アカンビス(Acambis)による命名法を用いると、隣り合った必須ORFのうち一方は163であり、隣り合った必須ORFのうち他方は167である、請求項1に記載の組換えMVAウイルス。
【請求項8】
異種核酸配列は、転写制御要素に転写制御される少なくとも1つのコード配列を含む、請求項1に記載の組換えMVAウイルス。
【請求項9】
コード配列は1つ以上のタンパク質をコードする、請求項8に記載の組換えMVAウイルス。
【請求項10】
転写制御要素はポックスウイルスの転写制御要素である、請求項8に記載の組換えMVAウイルス。
【請求項11】
コード配列はヒト免疫不全ウイルス(HIV)に由来する、請求項8に記載の組換えMVAウイルス。
【請求項12】
コード配列はHIVエンベロープタンパク質をコードする、請求項8に記載の組換えMVAウイルス。
【請求項13】
コード配列は配列番号4と少なくとも90%の同一性を有するヌクレオチド配列を含み、コードされているタンパク質が、配列番号4によってコードされるタンパク質に対する免疫応答の誘発能を有する、請求項8に記載の組換えMVAウイルス。
【請求項14】
コード配列は配列番号4を含む、請求項8に記載の組換えMVAウイルス。
【請求項15】
単離された核酸構築物であって、
(a)第1の必須MVAウイルスORF由来の少なくとも25個連続したヌクレオチドを含む第1の核酸配列と、
(b)第2の必須MVAウイルスORF由来の少なくとも25個連続したヌクレオチドを含む第2の核酸配列と
を含み、
第1および第2の必須MVAウイルスORFはMVAゲノム中で少なくとも1つの非必須ORFによって隔てられており、第1および第2の核酸配列は、単離された核酸構築物において互いに隣り合っていることを特徴とする、核酸構築物。
【請求項16】
a)第1の核酸配列は、第1の必須MVAウイルスORFの少なくとも100個連続したポリヌクレオチド領域と少なくとも75%の同一性を有する少なくとも100個連続したポリヌクレオチド領域を含み、かつ
(b)第2の核酸配列は、第2の必須MVAウイルスORFの少なくとも100個連続したポリヌクレオチド領域と少なくとも75%の同一性を有する少なくとも100個連続したポリヌクレオチド領域を含む、
請求項15に記載の核酸構築物。
【請求項17】
CDC/アカンビスによる命名法を用いると、第1のORFはACAM3000_MVA_163であり、第2のORFはACAM3000_MVA_167である、請求項15に記載の核酸構築物。
【請求項18】
第1および第2の核酸配列の隣り合った端部は、(a)遺伝子間領域および(b)制限酵素認識部位からなる群から選択された少なくとも1つのヌクレオチド配列を含む第3の核酸配列によって隔てられている、請求項15に記載の核酸構築物。
【請求項19】
第1および第2の核酸配列の隣り合った端部は、異種核酸配列を含む第3の核酸配列によって隔てられていることを特徴とする、請求項15に記載の核酸構築物。
【請求項20】
異種核酸配列は、転写制御要素に転写制御される少なくとも1つのコード配列を含む、請求項19に記載の核酸構築物。
【請求項21】
転写制御要素はポックスウイルスの転写制御要素である、請求項20に記載の核酸構築物。
【請求項22】
コード配列は1つ以上のタンパク質または低分子干渉RNAをコードする、請求項20に記載の核酸構築物。
【請求項23】
異種核酸配列はヒト免疫不全ウイルス(HIV)に由来する、請求項21に記載の核酸構築物。
【請求項24】
異種核酸配列はHIVエンベロープタンパク質をコードする、請求項21に記載の核酸構築物。
【請求項25】
pLW‐76の配列(配列番号21)を含む、請求項15に記載の核酸構築物。
【請求項26】
安定な組換え改変ワクシニアアンカラ(MVA)ウイルスを製造する方法であって、
(a)請求項15に記載の核酸構築物で細胞をトランスフェクションする工程と、
(b)トランスフェクションした細胞をMVAウイルスに感染させる工程と、
(c)核酸構築物とMVAウイルスゲノムとの間の相同組換えを可能とするのに適した条件下で感染細胞を培養する工程と、
(d)組換えMVAウイルスを単離する工程と
を含む方法。
【請求項27】
対象者において免疫応答を誘導する方法であって、請求項8に記載の組換えMVAウイルスを該対象者に投与する工程を含む方法。
【請求項28】
in vitroでタンパク質を生産する方法であって、
(a)宿主細胞を請求項8に記載の組換えMVAウイルスに感染させる工程と、
(b)感染宿主細胞を適切な条件下で培養する工程と、
(c)宿主細胞による生産物を単離する工程と
を含む方法。
【図8】

【図8−1】

【図8−2】

【図8−3】

【図8−4】

【図8−5】

【図8−6】

【図8−7】

【図8−8】
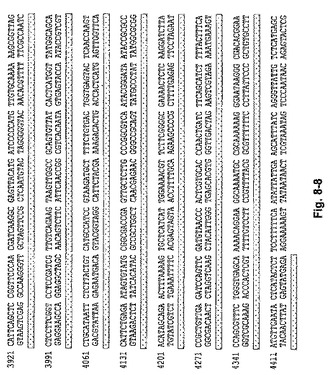
【図8−9】

【図9】

【図10】

【図10−1】

【図10−2】

【図18】
【図20】
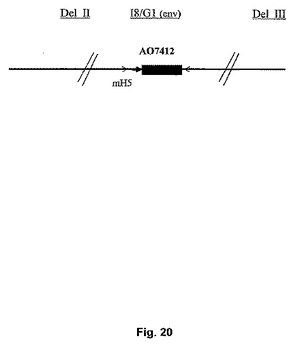
【図22】

【図23】
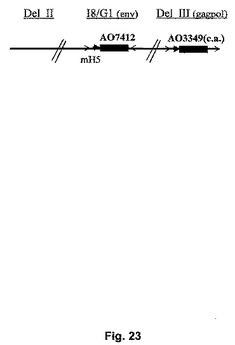
【図30】
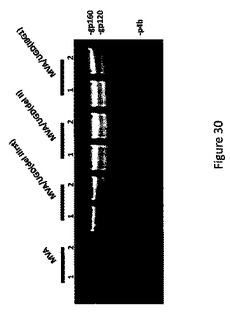
【図31−1】

【図31−2】

【図31−3】

【図31−4】

【図31−5】

【図31−6】

【図1】
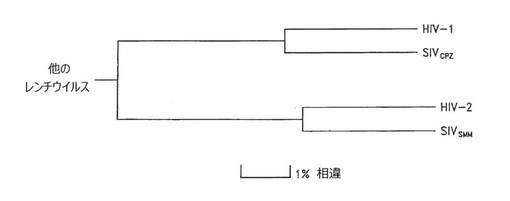
【図2】
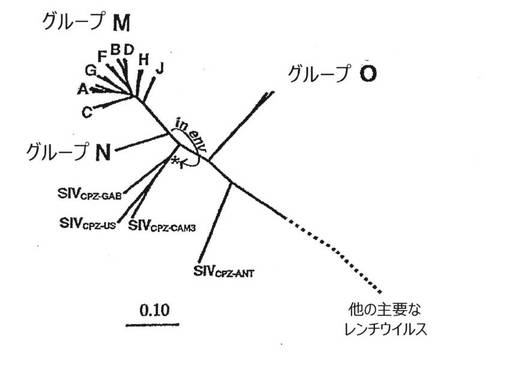
【図3】
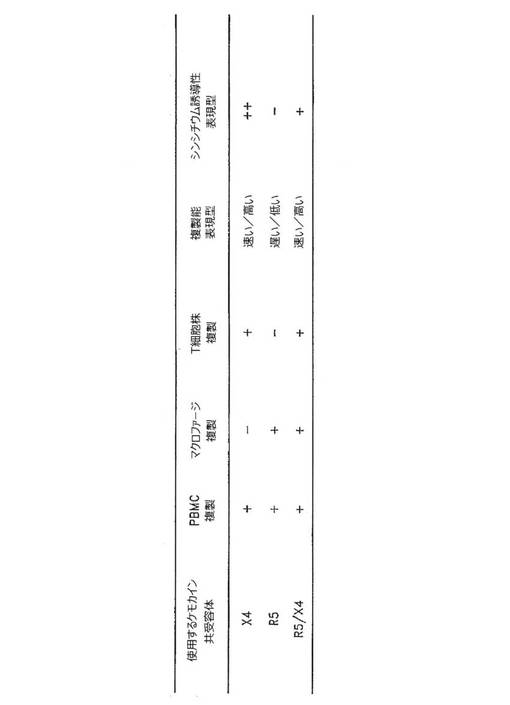
【図4】
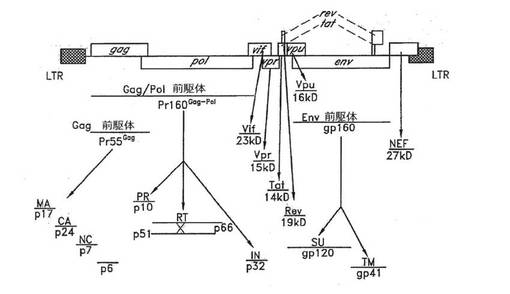
【図5】
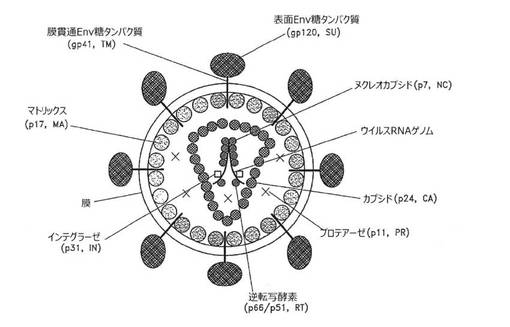
【図6】
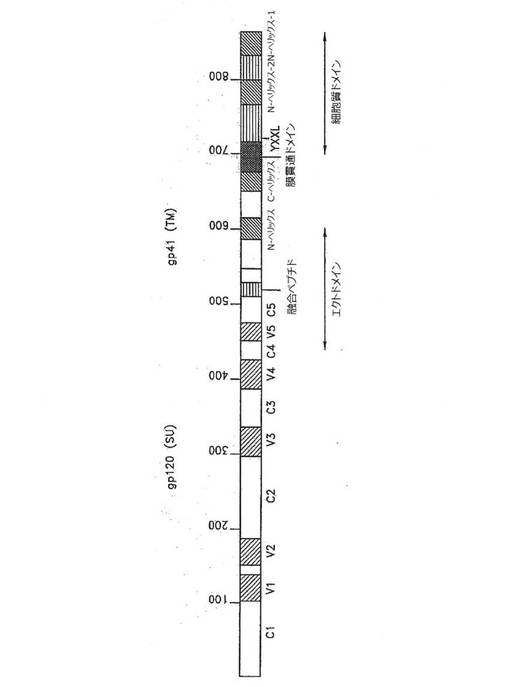
【図7】
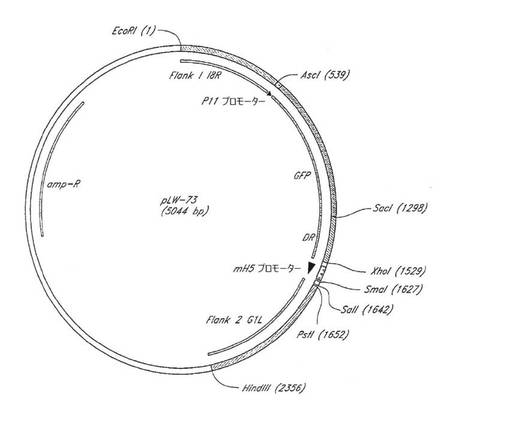
【図11】
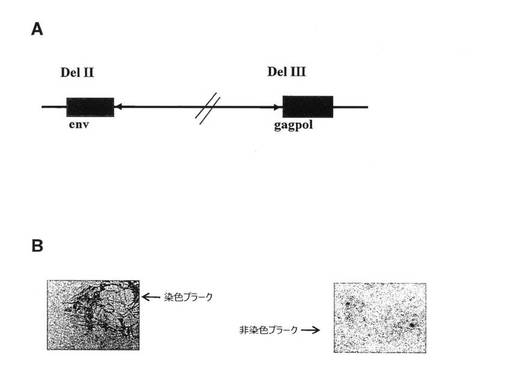
【図12】
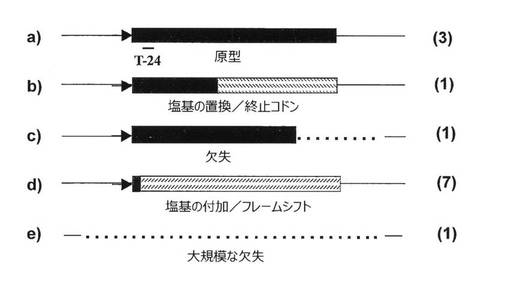
【図13】
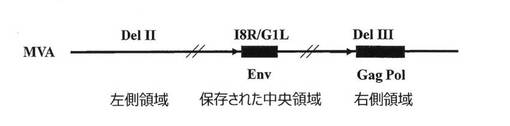
【図14】
【図15】
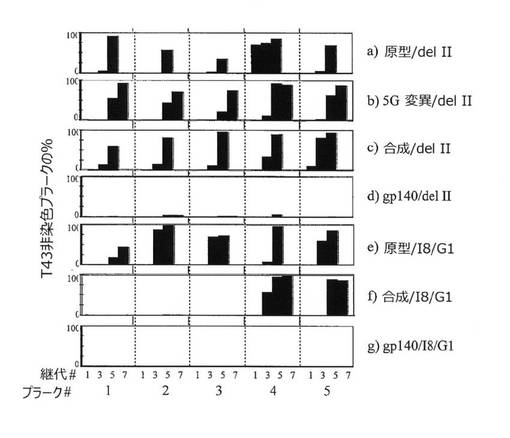
【図16】
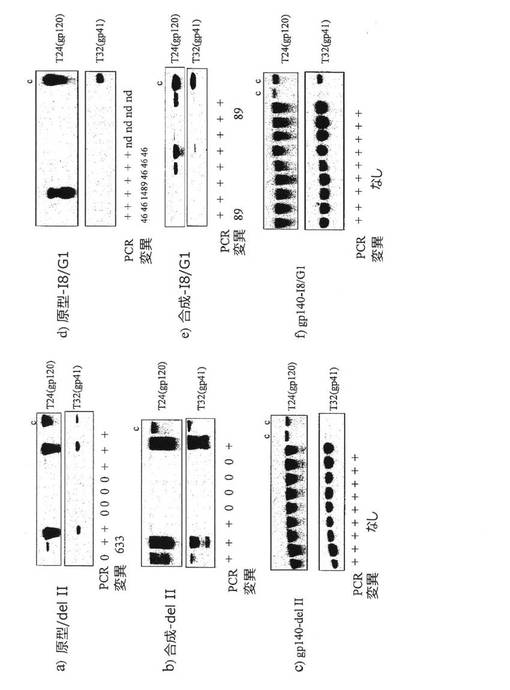
【図17】
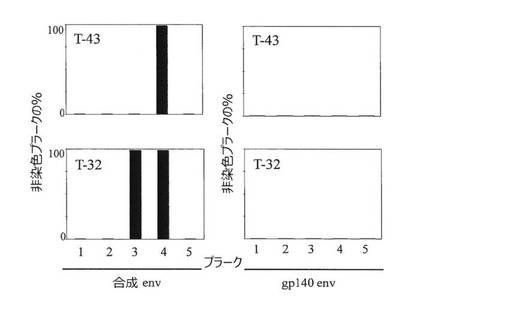
【図19】
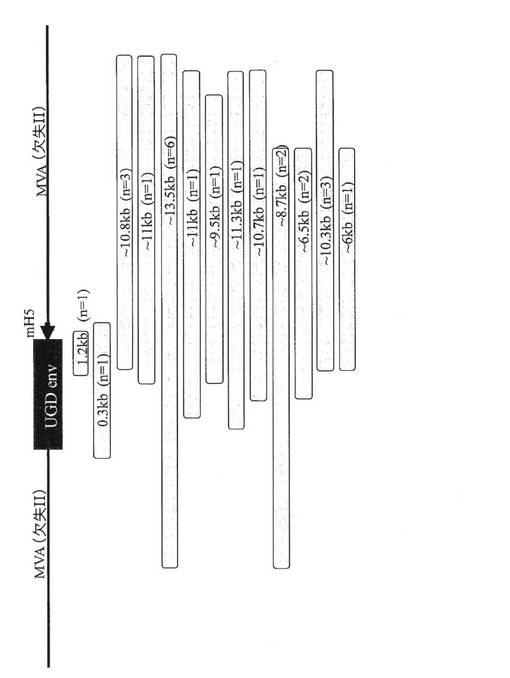
【図21】
【図24】

【図25】
【図26】
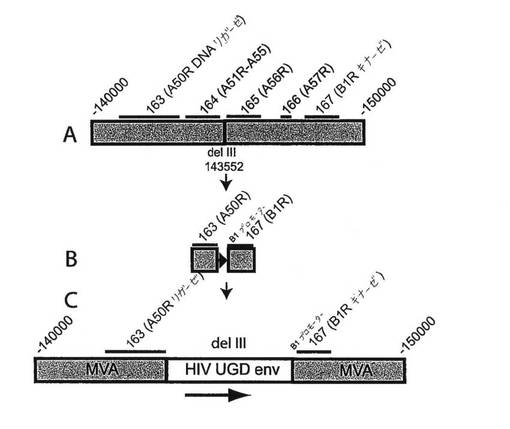
【図27】
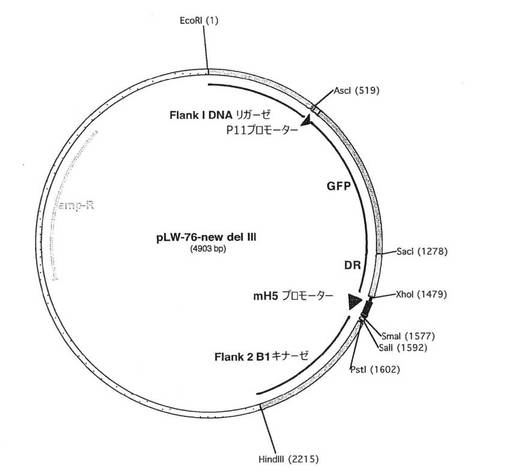
【図28】

【図29】
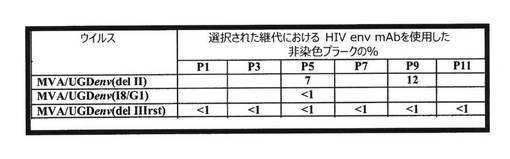
【図8−1】
【図8−2】
【図8−3】
【図8−4】
【図8−5】
【図8−6】
【図8−7】
【図8−8】
【図8−9】
【図9】
【図10】
【図10−1】
【図10−2】
【図18】
【図20】
【図22】
【図23】
【図30】
【図31−1】
【図31−2】
【図31−3】
【図31−4】
【図31−5】
【図31−6】
【図1】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図19】
【図21】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【公表番号】特表2013−507935(P2013−507935A)
【公表日】平成25年3月7日(2013.3.7)
【国際特許分類】
【出願番号】特願2012−534414(P2012−534414)
【出願日】平成22年10月15日(2010.10.15)
【国際出願番号】PCT/US2010/052929
【国際公開番号】WO2011/047324
【国際公開日】平成23年4月21日(2011.4.21)
【出願人】(309004585)ザ ユナイテッド ステイツ オブ アメリカ アズ リプリゼンテッド バイ ザ セクレタリー、デパートメント オブ ヘルス アンド ヒューマン サービシーズ (2)
【氏名又は名称原語表記】THE U.S.A.AS REPRESENTED BY THE SECRETARY,DEPARTMENT OF HEALTH AND HUMAN SERVICES
【Fターム(参考)】
【公表日】平成25年3月7日(2013.3.7)
【国際特許分類】
【出願日】平成22年10月15日(2010.10.15)
【国際出願番号】PCT/US2010/052929
【国際公開番号】WO2011/047324
【国際公開日】平成23年4月21日(2011.4.21)
【出願人】(309004585)ザ ユナイテッド ステイツ オブ アメリカ アズ リプリゼンテッド バイ ザ セクレタリー、デパートメント オブ ヘルス アンド ヒューマン サービシーズ (2)
【氏名又は名称原語表記】THE U.S.A.AS REPRESENTED BY THE SECRETARY,DEPARTMENT OF HEALTH AND HUMAN SERVICES
【Fターム(参考)】
[ Back to top ]