
分子アダプター
本発明は、試料中の分析物の検出に用いるための膜貫通タンパク質細孔に関する。細孔は、細孔と分析物との間の相互作用を助長する分子アダプターを含む。アダプターは、細孔を用いて分析物を検出することを可能にする向きで、細孔と共有結合している。

【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、確率的センシング(stochastic sensing)によって分析物を検出するために有用な膜貫通タンパク質細孔に関する。分子アダプターが、分析物の検出を可能する向きで、細孔と結合している。
【背景技術】
【0002】
発明の背景
確率的検出は、分析物分子と受容体との間の個々の結合イベントの観察に依拠する、センシングのためのアプローチである。確率的センサーは、ナノメートル寸法の単一の細孔を絶縁膜中に配置して、細孔を通しての電位駆動(voltage-driven)イオン輸送を分析物分子の存在下で測定することによって作り出すことができる。電流の揺らぎの発生頻度から、細孔の中に結合する分析物の濃度が明らかとなる。分析物の実体は、その特徴的な電流サイン(current signature)、とりわけ電流遮断の持続時間および程度を通じて明らかとなる(Braha, O., Walker, B., Cheley, S., Kasianowicz, J. J., Song, L., Gouaux, J. E., and Bayley, H. (1997) Chem.Biol. 4, 497-505(非特許文献1);およびBayley, H., and Cremer, P. S. (2001) Nature 413, 226-230(非特許文献2))。
【0003】
細菌性細孔形成毒素α-ヘモリシン(αHL)の工学的に作製した型のものは、多くのクラスの分子の確率的センシングのために用いられている(Bayley, H., and Cremer, P. S. (2001) Nature 413, 226-230(非特許文献2);Shin, S.-H., Luchian, T., Cheley, S., Braha, O., and Bayley, H. (2002) Angew.Chem.Int.Ed. 41, 3707-3709(非特許文献3);およびGuan, X., Gu, L.-Q., Cheley, S., Braha, O., and Bayley, H. (2005) ChemBioChem 6, 1875-1881(非特許文献4))。これらの研究の過程で、低分子有機分析物と結合するようにαHLを直接的に工学的に作製するという取り組みは負担が大きく、しかも成功例は稀であることが見いだされた(Guan, X., Gu, L.-Q., Cheley, S., Braha, O., and Bayley, H. (2005) ChemBioChem 6, 1875-1881(非特許文献4))。幸いなことに、非共有結合的に結合させた分子アダプター、とりわけシクロデキストリン(Gu, L.-Q., Braha, O., Conlan, S., Cheley, S., and Bayley, H. (1999) Nature 398, 686-690(非特許文献5))、さらには環状ペプチド(Sanchez-Quesada, J., Ghadiri, M. R., Bayley, H., and Braha, O. (2000) J.Am.Chem.Soc. 122, 11758-11766(非特許文献6))およびカボチャ型分子(cucurbituril)(Braha, O., Webb, J., Gu, L.-Q., Kim, K., and Bayley, H. (2005) ChemPhysChem 6, 889-892(非特許文献7))を利用する、1つの異なる戦略が発見された。シクロデキストリンはαHL細孔内に一過性に停留して、実質的ではあるが不完全なチャネル遮断を生じさせる。シクロデキストリンの疎水性内部の中に結合する有機分析物は、この遮断を増強して、分析物の検出を可能にする(Gu, L.-Q., Braha, O., Conlan, S., Cheley, S., and Bayley, H. (1999) Nature 398, 686-690(非特許文献5))。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Braha, O., Walker, B., Cheley, S., Kasianowicz, J. J., Song, L., Gouaux, J. E., and Bayley, H. (1997) Chem.Biol. 4, 497-505
【非特許文献2】Bayley, H., and Cremer, P. S. (2001) Nature 413, 226-230
【非特許文献3】Shin, S.-H., Luchian, T., Cheley, S., Braha, O., and Bayley, H. (2002) Angew.Chem.Int.Ed. 41, 3707-3709
【非特許文献4】Guan, X., Gu, L.-Q., Cheley, S., Braha, O., and Bayley, H. (2005) ChemBioChem 6, 1875-1881
【非特許文献5】Gu, L.-Q., Braha, O., Conlan, S., Cheley, S., and Bayley, H. (1999) Nature 398, 686-690
【非特許文献6】Sanchez-Quesada, J., Ghadiri, M. R., Bayley, H., and Braha, O. (2000) J.Am.Chem.Soc. 122, 11758-11766
【非特許文献7】Braha, O., Webb, J., Gu, L.-Q., Kim, K., and Bayley, H. (2005) ChemPhysChem 6, 889-892
【発明の概要】
【0005】
本発明者らは、驚いたことに、分子アダプターを、膜貫通タンパク質細孔と、ある特定の向きで共有結合させることができることを実証した。本発明者らはまた、驚いたことに、分子アダプターを、膜貫通細孔と、分析物が細孔を通して流れる電流に対してその分析物に特異的な様式で影響を及ぼすことを可能にする向きで共有結合させることができることも実証した。したがって、分子アダプターは、確率的センシングを介して分析物を検出するために細孔を用いることを可能にする向きで、細孔と共有結合させることができる。細孔は好ましくは、分子アダプターのその配向を助長するように修飾(modify)される。
【0006】
分子アダプターの固定的な性質は、分析物が細孔と相互作用する時には常に、特徴的な電流が細孔を通して流れることを意味する。その結果として、本発明の膜貫通タンパク質細孔は、確率的センシングのための、特にヌクレオチドの検出または核酸のシークエンシングのための有用なツールとなる。
【0007】
したがって、本発明は、試料中の分析物の検出に用いるための膜貫通タンパク質細孔であって、細孔と分析物との間の相互作用を助長する分子アダプターを含んでおり、そのアダプターが、分析物を細孔を用いて検出することを可能にする向きで細孔と共有結合している細孔を提供する。
【0008】
本発明はまた、以下も提供する:
‐本発明の膜貫通タンパク質細孔を作製する方法であって:
(a)膜貫通タンパク質細孔に、細孔と分析物との間の相互作用を助長する分子アダプターを共有結合させる段階;および
(b)該アダプターが、分析物を細孔を用いて検出することを可能にする向きで細孔と結合しているか否かを判定する段階、
を含む方法;
‐分析物の有無を判定する方法であって:
(a)分析物を、本発明の膜貫通タンパク質細孔と、分析物が細孔と相互作用するように接触させる段階;および
(b)相互作用の間に細孔を通過する電流を測定し、それによって分析物の有無を判定する段階、
を含む方法;
‐個々のヌクレオチドを同定する方法であって:
(a)ヌクレオチドを、本発明の膜貫通タンパク質細孔と、ヌクレオチドが細孔と相互作用するように接触させる段階;および
(b)相互作用の間に細孔を通過する電流を測定し、それによってヌクレオチドの実体を決定する段階、
を含む方法;
‐標的核酸配列のシークエンシングの方法であって:
(a)個々のヌクレオチドを、標的配列の一方の末端から、連続移動性(processive)エキソヌクレアーゼを用いて消化する段階;
(b)ヌクレオチドを、本発明の膜貫通タンパク質細孔と、ヌクレオチドがアダプターと相互作用するように接触させる段階;
(c)相互作用の間に細孔を通過する電流を測定し、それによってヌクレオチドの実体を決定する段階;および
(d)段階(a)から(c)までを核酸配列の同じ末端で繰り返し、それによって核酸の配列を決定する段階、
を含む方法;ならびに
‐本発明による膜貫通タンパク質細孔および連続移動性エキソヌクレアーゼを含む、核酸のシークエンシングのためのキット。
【図面の簡単な説明】
【0009】
【図1】pdbファイル(Montoya and Gouaux, unpublished)から生成された(M113F-RL2)7・βCDおよび(M113N-RL2)7・βCDのX線構造を示している。a)αHL細孔の側面図。アミノ酸108〜120および138〜150が黄色で強調表示されている。b)(M113F)7・βCD。αHLのアミノ酸108〜120は黄色で描かれており、βCDは青色のスティックで示されている。Phe-113およびLys-147の側鎖もスティック形態で示されている。βCDの第一級ヒドロキシルのO原子とPhe-113の最近接フェニル環の中心との間の距離は約6.5Åである(点線)。c)(M113N-RL2)7・βCD。αHLのアミノ酸108〜120は黄色で描かれており、βCDは青色のスティックで示されている。Asn-113、Lys-147およびThr-117の側鎖もスティック形態で示されている。βCDの第一級ヒドロキシルのO原子とThr-117の最近接O原子との間の距離は約6.0Åである(点線)。
【図2】βCD-PDPの化学構造およびスティック表示を示している。分子はChemDraw 3Dで描写し、続いてPyMolにてレンダリングした。
【図3】非共有結合複合体αHL(M113F-RL2)7・βCDおよび共有結合物αHL(M113F-RL2)6(M113C-D8RL2)1-βCDの特性の比較を示している。各実験は少なくとも6回繰り返した。a)突然変異型αHL細孔(M113F-RL2)7のアミノ酸108〜120のカートゥーン表示。Phe-113残基はスティック形態で表示されている。25mM Tris・HCl、pH 8.0、1M KCl中、+100mVでの(M113F-RL2)7細孔からの単一チャネル電流軌跡。b)(M113F-RL2)7・βCDのX線構造によって判明した、βCDが結合したαHL(M113F-RL2)7のカートゥーン表示。矢印は、βCDが細孔を自由に出入りして動くことを指し示している。この電流軌跡は、最初は占有されていなかった細孔へのβCDの結合、およびその後の解離を指し示している(平均τoff=316±62ms)。c)αHL(M113F-RL2)6(M113C-D8RL2)1のアミノ酸108〜120のカートゥーン表示。Phe-113残基はスティック形態で表示されており、Cys-113は褐色に彩色されて同じくスティックで表示されている。(c)の拡大図を図4に見ることができる。矢印は、βCD-PDPが細孔に入ると第一級ヒドロキシルがPhe-113の側鎖と相互作用して、Cys-113がβCD-PDPのピリジルジスルフィドと反応してジスルフィド結合を形成することを指し示している。電流軌跡は反応の間の電流振幅の変化を示している。電流遮断85%の過渡状態に印を付している(*)。d)ジチオトレイトール(DTT)による(M113F-RL2)6(M113C-D8RL2)1-βCDの切断。矢印は、DTTによるジスルフィド結合の切断後にシクロデキストリン誘導体が113位から離脱することを指し示している。電流軌跡は切断の間の電流振幅の変化を示している。電流遮断98%の過渡状態に印を付している(*)。e)(M113F-RL2)7・βCD(τ)およびαHL(M113F-RL2)6(M113C-D8RL2)1-βCD(■)に関する単一チャネルI-V曲線。条件:25mM Tris・HCl、pH 8.0、1M KCl。
【図4】図3cの拡大図を示している。αHL(M113F-RL2)6(M113C-D8RL2)1のアミノ酸108〜120のカートゥーン表示。Phe-113残基はスティック形態で表示され、Cys-113は褐色に彩色されて同じくスティックで表示されている。
【図5】非共有結合複合体αHL(M113N-RL2)7-βCDおよび共有結合物αHL(M113N-RL2)6(T117C-D8RL3)1-βCDの特性の比較を示している。各実験は少なくとも6回繰り返した。a)突然変異型αHL細孔(M113N-RL2)7のアミノ酸108〜120のカートゥーン表示。Asn-113残基はスティック形態で表示されている。25mM Tris・HCl、pH 8.0、1M KCl中、-100mVでの、(M113N-RL2)7細孔からの単一チャネル電流の軌跡。b)(M113N-RL2)7・βCDのX線構造によって判明した、βCDが結合したαHL(M113N-RL2)7のカートゥーン表示。矢印は、βCDが細孔を自由に出入りして動くことを指し示している。この電流軌跡は、最初は占有されていなかった細孔へのβCDの結合、およびその後の解離を指し示している(平均τoff=10.7±1.5s)。c)αHL(M113N-RL2)6(T117C-D8RL3)1のアミノ酸108〜120のカートゥーン表示。Asn-113残基はスティック形態で表示され、Cys-117は褐色に彩色されて同じくスティックで表示されている。図5cの拡大図を図6に見ることができる。矢印は、βCD-PDPが細孔に入ると第二級ヒドロキシルがAsn-113の側鎖と相互作用し、Cys-117がβCD-PDPのピリジルジスルフィドと反応してジスルフィド結合を形成することを指し示している。この電流軌跡は、Cys-117とβCD-PDPのピリジルジスルフィドとの間で反応が起こった時の電流振幅の変化を示している。電流遮断80%の過渡状態に印を付している(*)。d)DTTによる(M113N-RL2)6(T117C-D8RL3)1-βCDの切断。矢印は、ジスルフィド結合の切断後にシクロデキストリン誘導体が117位から離脱することを指し示している。電流軌跡は切断の間の電流振幅の変化を示している。電流遮断90%の過渡状態に印を付している(*)。e)(M113N-RL2)7・βCD(τ)およびαHL(M113N-RL2)6(T117C-D8RL3)1-βCD(■)に関する単一チャネルI-V曲線。条件:25mM Tris・HCl、pH 8.0、1M KCl。
【図6】図5cの拡大図を示している。αHL(M113N-RL2)6(T117C-D8RL3)1のアミノ酸108〜120のカートゥーン表示。Asn-113残基はスティック形態で表示され、Cys-117は褐色に彩色されて同じくスティックで表示されている。
【図7】共有結合したβCDを含むαHL細孔による2-アダマンタンアミンの検出を示している。a)25mM Tris・HCl、pH 8.0、1M KCl中、+100mVでの、単一の(M113F-RL2)6(M113C-D8RL2)1-βCD細孔からの電流軌跡。b)シスチャンバーにおける30μMの2-アダマンタンアミンの存在下での電流軌跡。c)トランスチャンバーにおける30μMの2-アダマンタンアミンの下での電流軌跡。d)25mM Tris・HCl、pH 8.0、1M KCl中、+100mVでの、単一の(M113N-RL2)6(T117C-D8RL3)1-βCD細孔からの電流軌跡。e)シスチャンバーにおける30μMの2-アダマンタンアミンの下での電流軌跡。f)トランスチャンバーにおける30μM 2-アダマンタンアミンの下での電流軌跡。
【図8】シス-リトコール酸と(M113F-RL2)6(M113C-D8RL2)1-βCDとの長寿命結合イベントを示している。a)分子アダプター(M113F-RL2)6(M113C-D8RL2)1-βCDが結合したαHL、およびシス-リトコール酸の構造。b)25mM Tris・HCl、pH 8.0、1M KCl中、-100mVでの、シス-リトコール酸(50μM)と(M113F-RL2)6(M113C-D8RL2)1-βCDとの特徴的な結合イベント。平均τoff=8.6±7.8s(イベントの数=60)。レベル1:(M113F-RL2)6(M113C-D8RL2)1-βCD;レベル2:(M113F-RL2)6(M113C-D8RL2)1-βCD・リトコール酸、遮断92%;レベル3:(M113F-RL2)6(M113C-D8RL2)1-βCD・リトコール酸、遮断98%。
【図9】(M113F-RL2)6(M113C-D8RL2)1-βCDおよび(M113F-RL2)7がいずれも50μMのβCDの存在下でシス-リトコール酸の非存在下で閉鎖を示し、それはシス-リトコール酸結合と混同される可能性があるもののイベントはより短いことを示している。a)図6と同じ条件下でのシス-リトコール酸の非存在下における(M113F-RL2)6(M113C-D8RL2)1-βCD。レベル1:(M113F-RL2)6(M113C-D8RL2)1-βCD;レベル2:細孔の自然発生的ゲート開閉。イベントの持続時間は30〜300msの間でさまざまであった。b)25mM Tris・HCl、pH 8.0、1M KCl中、-100mVでの、50μMのβCDの存在下における(M113F-RL2)7。τoff=0.68±0.12s。レベル1:(M113F-RL2)7;レベル2:(M113F-RL2)7・βCD;レベル3:細孔複合体(M113F-RL2)7・βCDの自然発生的ゲート開閉。
【図10】25mM Tris・HCl、pH 8.0、1M KCl中、+100mVでの、(M113F-RL2)6(M113C-D8RL2)1-βCDの単一チャネル電流の変動を温度とともに示している。プロット中に示されている値は、3つの異なる実験の平均値である。単一チャネル電流は温度に直線的に依存する。I(pA)=14.307+0.5402T(℃)。
【図11a】(M113F-RL2)6(M113C-D8RL2)1-βCDおよび(M113N)6(T117C-D8RL3)1-βCDのイオン選択性を示している。シス:200mM KCl;トランス:1000mM KCl(n=3)を伴う25mM Tris・HCl、pH 8.0中で行った記録に基づく、(M113F-RL2)6(M113C-D8RL2)1( )および(M113F-RL2)6(M113C-D8RL2)1-βCD(■)に関するI-V曲線。逆転電位(Vr)には矢印を付している。
【図11b】(M113F-RL2)6(M113C-D8RL2)1-βCDおよび(M113N)6(T117C-D8RL3)1-βCDのイオン選択性を示している。シス:1000mM KCl;トランス:200mM KCl(n=3)を伴う25mM Tris・HCl、pH 8.0中で行った記録に基づく、(M113F-RL2)6(M113C-D8RL2)1( )および(M113F-RL2)6(M113C-D8RL2)1-βCD(■)に関するI-V曲線。逆転電位(Vr)には矢印を付している。
【図12】αHL-D8RL3(オクタ-アスパラギン酸尾部(octa-aspartate tail)を含む)をコードする配列を示している。この配列はSEQ ID NO:18に示されている。αHL-D8RL3をコードする配列(SEQ ID NO:18)は、実施例で用いているT117C-D8RL3をコードするSEQ ID NO:15のヌクレオチド13〜918と、SEQ ID NO:15が残基361〜363にTGC(ACGではなく)を有する点を除いて同一である。
【0010】
配列表フリーテキスト
配列の説明
SEQ ID NO:1は、野生型α-ヘモリシンの1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:2は、野生型α-ヘモリシンの1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:3は、α-ヘモリシンM113H-RL2の1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:4は、α-ヘモリシンM113H-RL2の1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:5は、α-ヘモリシンM113K-RL2の1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:6は、α-ヘモリシンM113K-RL2の1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:7は、α-ヘモリシンM113R-RL2の1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:8は、α-ヘモリシンM113R-RL2の1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:9は、実施例で用いているα-ヘモリシンM113F-RL2の1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:10は、実施例で用いているα-ヘモリシンM113F-RL2の1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:11は、実施例で用いているα-ヘモリシンM113N-RL2の1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:12は、実施例で用いているα-ヘモリシンM113N-RL2の1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:13は、実施例で用いているα-ヘモリシンM113C-D8RL2の1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:14は、実施例で用いているα-ヘモリシンM113C-D8RL2の1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:15は、実施例で用いているα-ヘモリシンT117C-D8RL3の1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:16は、実施例で用いているα-ヘモリシンT117C-D8RL3の1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:17は、λエキソヌクレアーゼのアミノ酸配列を示している。この配列は、集合して三量体となる3つの同一なサブユニットの1つである。
SEQ ID NO:18は、αHL-D8RL3(図12中の配列)をコードする配列を示している。
【発明を実施するための形態】
【0011】
発明の詳細な説明
開示される生成物および方法の種々の適用物は、当技術分野における具体的な要求に合わせて適応させうることが理解されるべきである。また、本明細書で用いる専門用語は特定の態様を説明することのみを目的としており、限定的であることは意図していないことも理解されるべきである。
【0012】
さらに、本明細書および添付の特許請求の範囲で用いる場合、単数形の「1つの(a)」、「1つの(an)」および「その(the)」は、その文脈で明らかに別の指示がなされない限り、複数のものに関する言及も含む。したがって、例えば、「1つの分析物(an analyte)」に対する言及は「複数の分析物(analytes)」を含み、「1つの膜貫通タンパク質細孔」に対する言及は2つまたはそれ以上のそのような細孔を含み、「1つの分子アダプター」に対する言及は2つまたはそれ以上のそのようなアダプターを含むなどである。
【0013】
本明細書で引用したすべての刊行物、特許および特許出願は、上記または下記のいずれであるかを問わず、その全体が参照により本明細書に組み入れられる。
【0014】
膜貫通タンパク質細孔
本発明は、分析物の検出に用いるための膜貫通タンパク質細孔に関する。膜貫通タンパク質細孔は、分析物との相互作用を助長する分子アダプターを含む。アダプターは、細孔と特定の向きで共有結合している。アダプターは、分析物と細孔との間の相互作用の間に、分析物が、細孔を通して流れる電流に対して、その分析物に特異的な様式で影響を及ぼすことを可能にするような向きにある。アダプターはしたがって、確率的センシングを介して分析物を検出するために細孔を用いることを可能にする向きで、細孔と共有結合している。
【0015】
本発明の膜貫通タンパク質細孔は、確率的センシングのための有用なツールである。分子アダプターの固定的な性質は、細孔から得られる信号が細孔のバレル内またはチャネル内の分析物の存在に完全に依存し、細孔からのアダプターの解離によっては影響されないことを意味する。換言すれば、アダプターの固定的な性質は、分析物が細孔と相互作用する時には常に、特徴的な電流が細孔を通して流れることを意味する。これはその結果として:(1)分析物の濃度が低い場合などのような、稀な相互作用イベントの検出;(2)分析物濃度が高い場合などのような、頻度の高い相互作用イベントの検出;(3)アダプターに対する分析物の親和性が高い場合などのような、長寿命(long-lived)相互作用イベントの検出;および(4)アダプターに対する分析物の親和性を測定しうるようにする分析物の滞留時間の測定、を可能にする、より高感度な系をもたらす。最も重要なこととして、信号に中断が全く存在しないことは、細孔の中に入ったすべての分析物が検出されることを意味する。そのような効率的な検出系は、核酸のシークエンシングを容易にする。これについて以下にさらに考察する。
【0016】
本発明の膜貫通タンパク質細孔にはほかにも利点がある。実施例の項に記載しているように、本発明の膜貫通タンパク質細孔は、最高55℃まで、または最高100℃までというような高温で機能することができる。これは、細孔が分析物を極限条件下で検出するために適していることを意味する。
【0017】
本発明の分子アダプターの固定的な性質は、それが細孔と無関係に分析物と自由に相互作用することはないことを意味する。これは干渉を減少させるともに、分析物の濃度のより正確な測定を得ることができることを意味する。
【0018】
実施例の項に記載しているように、細孔に対する分子アダプターの結合はそのイオン選択性を永続的に変化させる。このことは特定のイオン選択性を有する膜貫通タンパク質細孔の作製を可能にする。
【0019】
さらになお、本発明の分子アダプターの固定的な性質は細孔およびアダプターを一緒に保管することを可能にし、その結果、直ちに使用できる(ready-to-use)センサーの製造を可能にする。
【0020】
膜貫通タンパク質細孔
膜貫通タンパク質細孔とは、イオンが印加電圧に沿って膜の一方の側からもう一方に流れることを許容するポリペプチドのことである。細孔は好ましくは、分析物が印加電圧に沿って膜の一方の側からもう一方に流れることを許容する。
【0021】
細孔は、典型的にはオリゴマーである。細孔は好ましくは、6個、7個または8個のサブユニットといった数個の繰り返しサブユニットで構成される。細孔はより好ましくは七量体である。細孔は典型的には、イオンが流れることができるバレルまたはチャネルを含む。細孔のサブユニットは典型的には中心軸を取り囲み、ストランドを膜貫通性のβバレルもしくはチャネル、または膜貫通性のα-ヘリックス束もしくはチャネルへと導くのに寄与する。
【0022】
細孔のバレルまたはチャネルは典型的には、分析物との相互作用を助長するアミノ酸を含む。これらのアミノ酸は好ましくは、バレルまたはチャネルの狭窄部(constriction)の付近に位置する。ヌクレオチドまたは核酸の検出に用いるための細孔は、典型的には、アルギニン、リジンまたはヒスチジンといった、1つまたは複数の正に荷電したアミノ酸を含む。これらのアミノ酸は典型的には、ヌクレオチド中のリン酸基と相互作用することによって、またはヌクレオチド中の塩基とのπ-カチオン相互作用によって、細孔とヌクレオチドとの間の相互作用を助長する。ヌクレオチドまたは核酸の検出に用いるための細孔は、好ましくは、バレルまたはチャネルの狭窄部の付近に位置する、アルギニン、リジンまたはヒスチジンといった正に荷電したアミノ酸の環を有する。正に荷電した各アミノ酸は、典型的には、細孔サブユニットのそれぞれによって提供される。
【0023】
本発明に従って用いるための細孔は、β-バレル細孔またはα-ヘリックス束細孔でありうる。β-バレル細孔は、β-シートから形成されたバレルまたはチャネルを含む。適したβ-バレル細孔には、α-ヘモリシンおよびロイコシジンといったβ-毒素、ならびに恥垢菌(Mycobacterium smegmatis)ポーリンA(MspA)、外膜ポーリンF(OmpF)、外膜ポーリンG(OmpG)、外膜ホスホリパーゼA、およびナイセリア菌(Neisseria)自己輸送体リポタンパク質(NalP)といった細菌の外膜タンパク質/ポーリンが非限定的に含まれる。α-ヘリックス束細孔は、α-ヘリックスから形成されたバレルまたはチャネルを含む。適したα-ヘリックス束細孔には、内膜タンパク質およびα外膜タンパク質、例えばWZAなどが非限定的に含まれる。
【0024】
本発明に用いるために最も好ましい細孔は、α-ヘモリシンまたはその変異体である。α-ヘモリシン細孔は7個の同一なサブユニットで形成される(すなわち、それは七量体である)。α-ヘモリシンの1つのサブユニットの配列はSEQ ID NO:2に示されている。変異体は、7個のサブユニットのうち1つまたは複数がSEQ ID NO:2のものとは違いのあるアミノ酸配列を有し、かつ細孔活性を保っている七量体細孔である。変異体は、以下に考察するような、アダプターの共有結合またはアダプターの配向を助長する修飾(modification)を含んでもよい。
【0025】
変異体α-ヘモリシン中のサブユニットの1個、2個、3個、4個、5個、6個または7個が、SEQ ID NO:2のものとは違いのあるアミノ酸配列を有してもよい。変異体細孔の中の7個のサブユニットすべては同一であってもよいが典型的には異なり、これは、サブユニットの1つまたは複数が、以下に考察するようにアダプターの共有結合またはアダプターの配向を助長するように修飾されている場合には特にそうである。
【0026】
ヌクレオチドまたは核酸の検出に用いるためのα-ヘモリシンの好ましい変異体は、バレルまたはチャネルの狭窄部の付近に位置する、アルギニン、リジンまたはヒスチジンといった1つまたは複数の正に荷電したアミノ酸を有する。細孔は好ましくは、バレルまたはチャネルの狭窄部の付近に位置する、アルギニン、リジンまたはヒスチジンなどの4個、5個、6個または好ましくは7個の正に荷電したアミノ酸の環を有する。環内の各アミノ酸は、典型的には、変異体サブユニットのそれぞれによって提供される。適した変異体は、正に荷電したアミノ酸を各サブユニットの113位に含む。ヌクレオチドまたは核酸の検出に用いるための細孔は、好ましくは、SEQ ID NO:4に示されているような7個のサブユニットを含むα-ヘモリシン(M113K-RL2)7、または好ましくは、SEQ ID NO:6に示されているような7個のサブユニットを含むα-ヘモリシン(M113H-RL2)7、または最も好ましくは、SEQ ID NO:8に示されているような7個のサブユニットを含むα-ヘモリシン(M113R-RL2)7である。
【0027】
α-ヘモリシンの他の好ましい変異体は、バレルまたはチャネルの狭窄部の付近に位置する、アスパラギンなどの1つもしくは複数の非荷電アミノ酸、またはフェニルアラニンなどの1つもしくは複数の芳香族アミノ酸を有する。これには、β-CD-PDPと結合してそれを共有結合反応のために配向させること、および共有結合後のβ-CD-PDPの向きを維持することという目的がある。細孔は好ましくは、バレルまたはチャネルの狭窄部の付近に位置する、4個、5個、6個または好ましくは7個の非荷電アミノ酸または芳香族アミノ酸の環を有する。環内の各アミノ酸は、典型的には、変異体サブユニットのそれぞれによって提供される。適した変異体は、非荷電アミノ酸または芳香族アミノ酸を各サブユニットの113位に含む。本発明に従って用いるための細孔は、好ましくはα-ヘモリシン(M113F-RL2)6(M113C-D8RL2)1である。
【0028】
変異体は、生物体によって、例えばブドウ球菌(Staphylococcus bacterium)によって発現される、天然に存在する変異体であってよい。また、変異体には、組換え技術によって作製された非天然の変異体も含まれる。SEQ ID NO:2のアミノ酸配列の全長にわたって、変異体のサブユニットは好ましくは、その配列に対して、アミノ酸同一性に基づいて少なくとも50%相同であると考えられる。より好ましくは、サブユニットポリペプチドは、SEQ ID NO:2のアミノ酸配列に対して、配列全体にわたって、アミノ酸同一性に基づいて少なくとも55%、少なくとも60%、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、より好ましくは少なくとも95%、97%または99%相同であってよい。200個またはそれ以上、例えば230個、250個、270個または280個またはそれ以上の連続したアミノ酸にわたって、少なくとも80%の、例えば少なくとも85%、90%または95%のアミノ酸同一性が存在しうる(「厳密な相同性(hard homology)」)。
【0029】
アミノ酸置換を、例えば最大で1個、2個、3個、4個、5個、10個、20個または30個までの置換を、SEQ ID NO:2のアミノ酸配列に対して施すことができる。保存的置換は、例えば、以下の表1に従って施すことができる。
【0030】
(表1)保存的置換
第2列の中の同じブロック内の、好ましくは第3列の中の同じ行内のアミノ酸は、互いに置換することができる。
【0031】
SEQ ID NO:2のアミノ酸配列の1つまたは複数のアミノ酸残基を、代替的または追加的に、削除することができる。最大で1個、2個、3個、4個、5個、10個、20個もしくは30個またはそれ以上の残基を削除することができる。
【0032】
変異体は、SEQ ID NO:2の断片でできたサブユニットを含んでもよい。そのような断片は細孔形成活性を保っている。断片は、少なくとも50、100、200または250アミノ酸長でありうる。そのような断片は、キメラ性細孔を作製するために用いることができる。断片は好ましくは、SEQ ID NO:2の細孔形成ドメインを含む。
【0033】
変異体には、SEQ ID NO:2の断片または部分を含むキメラ性タンパク質細孔が含まれる。キメラ性タンパク質細孔は、それぞれがSEQ ID NO:2の断片または部分を含むサブユニットから形成される。キメラ性タンパク質細孔の細孔部分またはチャネル部分は、典型的には、SEQ ID NO:2の断片または部分によって形成される。
【0034】
1つまたは複数のアミノ酸を、代替的または追加的に、上記のポリペプチドに付加することができる。伸張は、SEQ ID NO:2のアミノ酸配列、またはそのポリペプチド変異体もしくは断片のN末端またはC末端に与えることができる。伸張は、極めて短くてもよく、例えば1〜10アミノ酸長であってもよい。または、伸張がより長く、例えば、最大で50または100アミノ酸であってもよい。担体タンパク質を、本発明によるアミノ酸配列と融合させてもよい。
【0035】
相同性を決定するためには、当技術分野における標準的な方法を用いることができる。例えば、UWGCG Packageは、相同性を計算するために、例えばそのデフォルト設定で用いることのできる、BESTFITプログラムを備えている(Devereux et al (1984) Nucleic Acids Research 12, p387-395)。PILEUPアルゴリズムおよびBLASTアルゴリズムは、例えば、Altschul S. F. (1993) J Mol Evol 36:290-300;Altschul, S.F et al (1990) J Mol Biol 215:403-10に記載されているように、相同性を計算するため、または配列を整列させるため(等価な残基または対応する配列を(典型的にはそれらのデフォルト設定で)同定することなど)に用いることができる。
【0036】
BLAST分析を行うためのソフトウエアは、National Center for Biotechnology Information(http://www.ncbi.nlm.nih.gov/)から公に入手可能である。このアルゴリズムでは、データベース配列中の同じ長さのワードとアラインメントを行った場合に何らかの正値の閾値スコアTと一致するか、またはそれを満たす、長さWの短いワードをクエリー配列中に同定することにより、高スコア配列ペア(high scoring sequence pair)(HSP)をまず同定する。Tは近隣ワードスコア閾値(neighbourhood word score threshold)と呼ばれる(Altschulら、前記)。これらの初期の近隣ワードでのヒットは、それらを含むHSPを見いだすための検索を開始するシードとなる。ワードヒットは、累積アラインメントスコアが増加する限り、各配列の両方向に対して延長される。各方向へのワードヒットの延長は以下の場合に停止する:累積アラインメントスコアが最大達成値に比べて量X低下した場合:1つもしくは複数の負スコアの残基アラインメントの蓄積のために累積スコアがゼロもしくはそれ未満になった場合;または配列のいずれかの末端に達した場合。BLASTアルゴリズムのパラメーターであるW、TおよびXは、アラインメントの感度および速度を決定する。BLASTプログラムはデフォルトとしてワード長(W)11、BLOSUM62スコアリングマトリックス(Henikoff and Henikoff (1992), Proc. Natl. Acad. Sci. USA 89: 10915-10919を参照)アラインメント(B)50、期待値(E)10、M=5、N=4および両ストランドの比較を用いる。
【0037】
BLASTアルゴリズムは、2つの配列間の類似性に関する統計分析も行う;例えば、Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90: 5873-5787を参照。BLASTアルゴリズムによって得られる類似性の指標の1つは最小合計確率(smallest sum probability)(P(N))であり、これは2つのアミノ酸配列間の一致が偶然に起こる確率の指標を与える。例えば、ある配列は、第1の配列と第2の配列との比較による最小合計確率が約1未満、好ましくは約0.1未満、より好ましくは約0.01未満、最も好ましくは約0.001未満の場合に、もう1つの配列と類似しているとみなされる。
【0038】
膜貫通タンパク質細孔は、例えば、それらの同定もしくは精製を助けるためのヒスチジン残基もしくはアスパラギン酸残基の付加によって、またはそのポリペプチドがそのような配列を天然には含まない場合に細胞からのそれらの分泌を促進するためのシグナル配列を付加することによって、修飾することができる。
【0039】
細孔は、顕在化標識(revealing label)で標識することができる。顕在化標識は、細孔を検出することを可能にする任意の適した標識であってよい。適した標識には、蛍光分子、放射性同位体、例えば125I、35Sなど、酵素、抗体、抗原、ポリヌクレオチド、およびビオチンなどのリガンドが非限定的に含まれる。
【0040】
細孔は、黄色ブドウ球菌(Staphylococcus aureus)などの細孔産生性の生物体から単離すること、または合成的に、もしくは組換え手段によって作製することができる。例えば、細孔をインビトロの翻訳および転写によって合成することができる。細孔のアミノ酸配列は、非天然のアミノ酸を含むように、または化合物の安定性を高めるように修飾することができる。細孔を合成的手段によって作製する場合には、そのようなアミノ酸を作製中に導入することができる。また、細孔を、合成的または組換え的な作製の後に改変することもできる。
【0041】
また、細孔をD-アミノ酸を用いて作製することもできる。これは、そのようなタンパク質またはペプチドを作製することに関して当技術分野で慣習的である。
【0042】
上記および下記において詳細に考察しているように、細孔は、アダプターの結合および配向を助長するための、1つまたは複数の特異的な修飾を含むことができる。細孔はまた、他の非特異的な修飾も、それらがアダプターの結合および配向と干渉しない限りは含んでよい。さまざまな非特異的な側鎖修飾が当技術分野で公知であり、細孔の側鎖に対して施すことができる。そのような修飾には、例えば、アルデヒドとの反応に続いてNaBH4による還元を行うことによるアミノ酸の還元的アルキル化、アセトイミド酸メチルによるアミド化、または無水酢酸によるアシル化が含まれる。
【0043】
組換え型の膜貫通細孔は、当技術分野で公知の標準的な方法を用いて作製することができる。細孔または細孔サブユニットをコードする核酸配列は、当技術分野における標準的な方法を用いて、単離して複製させることができる。細孔または細孔サブユニットをコードする核酸配列を、当技術分野において標準的な手法を用いて、細菌宿主細胞において発現させることもできる。細孔または細孔サブユニットは、組換え発現ベクターからのポリペプチドのインサイチュー発現によって細胞内で産生させることができる。発現ベクターは任意で、ポリペプチドの発現を制御するための誘導性プロモーターを保有する。
【0044】
細孔または細孔サブユニットをコードする核酸配列は、当技術分野における標準的な方法を用いて、単離して複製させることができる。染色体DNAを、細孔産生性の生物体、例えば黄色ブドウ球菌などから抽出する。細孔または細孔サブユニットをコードする遺伝子を、特異的プライマーを伴うPCRを用いて増幅する。増幅された配列を、続いて、クローニングベクターなどの複製可能な組換えベクター中に組み入れる。このベクターは、適合性のある宿主細胞内で核酸を複製させるために用いることができる。このように、細孔または細孔サブユニットをコードする核酸配列は、細孔または細孔サブユニットをコードするポリヌクレオチドを複製可能なベクター中に導入し、ベクターを適合性のある宿主細胞内に導入して、ベクターの複製をもたらす条件下で宿主細胞を増殖させることによって作ることができる。ベクターは宿主細胞から回収することができる。細孔または細孔サブユニットをコードするポリヌクレオチドのクローニングのために適した宿主細胞は当技術分野で公知であり、以下にさらに詳細に説明する。
【0045】
細孔または細孔サブユニットをコードする核酸配列は、適した発現ベクター中にクローニングすることができる。発現ベクター中で、細孔または細孔サブユニットをコードする核酸配列は、典型的には、宿主細胞によるコード配列の発現をもたらすことのできる制御配列と機能的に連結される。そのような発現ベクターは、細孔または細孔サブユニットを発現させるために用いることができる。
【0046】
「機能的に連結された」という用語は、記載された成分が、それらの意図した様式でそれらが機能することを可能にする関係で並列されていることを指す。コード配列と「機能的に連結された」制御配列は、制御配列と適合性のある条件下でコード配列の発現が達成されるような様式で連結されている。同じまたは異なる、細孔または細孔サブユニットの遺伝子の複数のコピーを、ベクター中に導入することができる。
【0047】
続いて、発現ベクターを適した宿主細胞内に導入することができる。このため、1つの種類のサブユニットのみを含む単量体性の細孔またはオリゴマー性の細孔の場合には、本発明の方法は、細孔または細孔サブユニットをコードする核酸配列を発現ベクター中に導入して、ベクターを適合性のある細菌宿主細胞内に導入して、細孔または細孔サブユニットをコードする核酸配列の発現をもたらす条件下で宿主細胞を増殖させることによって作製された細胞に対して実施することができる。組換え的に発現された細孔サブユニットは、宿主細胞膜の中で細孔へと自己集合すると考えられる。または、このようにして作製された組換え細孔を宿主細胞から単離して、別の膜に挿入することもできる。少なくとも2種類の異なるサブユニットを含むオリゴマー性の細孔の場合には、異なるサブユニットを上記のように異なる宿主細胞内で別々に発現させ、宿主細胞から取り出して、別の膜、例えばウサギ細胞膜などの中で細孔へと集合させることができる。
【0048】
ベクターは例えば、複製起点、任意で前記核酸配列の発現のためのプロモーター、および任意でプロモーターの調節要素(regulator)を備えた、プラスミドベクター、ウイルスベクターまたはファージベクターであってよい。ベクターは、1つまたは複数の選択マーカー遺伝子、例えばテトラサイクリン耐性遺伝子を含むことができる。プロモーターおよび他の発現調節シグナルは、それに対して発現ベクターを設計しようとする宿主細胞と適合性があるように選択することができる。T7、trc、lac、araまたはλLプロモーターが典型的には用いられる。
【0049】
宿主細胞は典型的には、細孔または細孔サブユニットを高レベルで発現する。細孔または細孔サブユニットをコードする核酸配列によって形質転換される宿主細胞は、その細胞を形質転換するために用いる発現ベクターと適合性があるように選択される。宿主細胞は典型的には細菌であり、好ましくは大腸菌(Escherichia coli)である。λ DE3溶原を有する任意の細胞、例えばC41(DE3)、BL21(DE3)、JM109(DE3)、B834(DE3)、TUNER、OrigamiおよびOrigami Bは、T7プロモーターを含むベクターを発現することができる。
【0050】
細孔または細孔サブユニットは、細孔産生性の生物体から、または上記の組換え発現後の、任意のタンパク質液体クロマトグラフィーによる精製を経て、大規模に生産することができる。典型的なタンパク質液体クロマトグラフィーシステムには、FPLC、AKTAシステム、Bio-Cadシステム、Bio-Rad BioLogicシステムおよびGilson HPLCシステムが含まれる。続いて、天然のまたは組換え的に作製された細孔または細孔サブユニットを、本発明による使用のために天然のまたは人工的な膜の中に挿入することができる。
【0051】
細孔または細孔サブユニットは、当技術分野で公知の任意の方法を用いて膜の中に挿入することができる。例えば、国際出願第PCT/GB2006/001057号(WO 2006/100484として公開)に記載された「ピック・アンド・プレース(pick and place)」法を用いて、細孔または細孔サブユニットを膜の中に導入することができる。
【0052】
分子アダプター
膜貫通細孔は、細孔と分析物との間の相互作用を助長する分子アダプターを含む。アダプターの存在は、細孔および分析物のホスト-ゲスト化学作用(host-guest chemistry)を改善する。ホスト-ゲスト化学作用の原理は当技術分野において周知である。アダプターは細孔の物理的または化学的な特性に影響を及ぼして、分析物とのその相互作用を改善する。アダプターは典型的には、細孔のバレルもしくはチャネルの電荷を改変して、または分析物と特異的に相互作用もしくは結合して、それによって細孔とのその相互作用を助長する。
【0053】
好ましくは、アダプターの少なくとも一部は、細孔のバレルまたはチャネルの内側にある。より好ましくは、アダプターのすべてが細孔のバレルまたはチャネルの内側にある。
【0054】
アダプターは、分析物と細孔との間の相互作用を媒介する。分析物は好ましくは、アダプターを介して、またはアダプターとともに細孔と可逆的に結合する。分析物は、最も好ましくは、それが膜を横切って細孔を通過する際に、アダプターを介して、またはアダプターとともに細孔と可逆的に結合する。分析物はまた、それが細孔膜を横切って細孔を通過する際に、アダプターを介して、またはアダプターとともに細孔のバレルまたはチャネルと可逆的に結合することもできる。アダプターは好ましくは、バレルまたはチャネルを、それが分析物と相互作用しうるように狭窄させる。アダプターはより好ましくは、バレルまたはチャネルを狭窄させ、細孔を通してのイオンの流れを制限する。細孔を通してのイオンの流れは、アダプターの存在が、細孔を通しての、および膜を横切ってのイオンの動きと干渉するならば制限される。細孔を通してのイオンの流れは、分析物の検出のために必要である。上記で考察したように、分析物と細孔との間の相互作用は、その分析物に特異的な様式で、細孔を通して流れるイオン電流に影響を及ぼす。アダプターは典型的には環状である。アダプターは好ましくは、細孔と同じ対称性を有する。細孔が七量体である(例えば、中心軸の周りに、14本のストランドを1つの膜貫通βバレルに提供する7個のサブユニットを有する)場合には、7回対称性(seven-fold symmetry)を有するアダプターが典型的には用いられる。同様に、細孔が六量体(例えば、中心軸の周りに、12本のストランドを1つの膜貫通βバレルに提供する6個のサブユニットを有するか、または12本のストランドのβバレルである)場合には、6回対称性を有するアダプターが典型的には用いられる。
【0055】
アダプターは典型的には、分析物と相互作用する、例えば結合する。アダプターと分析物との間の相互作用、例えば結合などは、分析物が細孔と相互作用する尤度を高める。アダプターは典型的には、ホスト-ゲスト化学作用を介して分析物と相互作用する。アダプターの一部分が、典型的には分析物と相互作用することができる。その部分は、分析物と相互作用することのできる1つまたは複数の化学基を含む。その1つまたは複数の化学基は、好ましくは、疎水性相互作用、水素結合、ファンデルワールス力、π-カチオン相互作用および/または静電力といった非共有結合相互作用によって分析物と相互作用する。分析物と相互作用することのできる1つまたは複数の化学基は、好ましくは、アミノ基および/またはヒドロキシル基を含む。アミノ基またはヒドロキシル基は、第一級、第二級または第三級炭素原子と結合していてよい。
【0056】
分析物と相互作用することのできるアダプターの部分は、好ましくは、π-カチオン相互作用を介して分析物と相互作用することのできる芳香環を含む。分析物と相互作用することのできるアダプターの部分は、より好ましくは、アミノ基またはヒドロキシル基の環、例えば、6個、7個または8個のアミノ基またはヒドロキシル基の環を含む。その部分は最も好ましくは、7個のアミノ基またはヒドロキシル基の環を含む。プロトン化アミノ基の環は、分析物中の負に荷電した基、例えばヌクレオチド中のリン酸基などと相互作用することができる。アミノ基の環は、細孔のバレルまたはチャネルの狭窄部において、正に荷電したアミノ酸の環との組み合わせで、分析物と相互作用することができる。ヒドロキシル基の環は、水素結合を介して分析物と相互作用することができる。ヒドロキシル基の環は、細孔のバレルまたはチャネルの狭窄部において、非荷電アミノ酸の環との組み合わせで、分析物と相互作用することができる。
【0057】
アダプターは、最も好ましくは、(1)バレルまたはチャネルを狭窄させ、それによって細孔を通してのイオンの流れを制限する、かつ(2)分析物と相互作用するか、または結合する。
【0058】
以下により詳細に考察するように、細孔のバレルまたはチャネルの中でのアダプターの正しい配向は、アダプターと細孔との間のホスト-ゲスト化学作用によって助長される。アダプターは好ましくは、細孔内の1つまたは複数のアミノ酸と相互作用することのできる、1つまたは複数の化学基を含む。アダプターはより好ましくは、非共有結合性の相互作用、例えば疎水性相互作用、水素結合、ファンデルワールス力、π-カチオン相互作用および/または静電力をなどを介して、細孔内の1つまたは複数のアミノ酸と相互作用することのできる、1つまたは複数の化学基を含む。細孔内の1つまたは複数のアミノ酸と相互作用することのできる化学基は、典型的にはヒドロキシルである。ヒドロキシル基は、第一級、第二級または第三級炭素原子と結合していてよい。ヒドロキシル基は、細孔内の、好ましくは細孔のバレル内またはチャネル内の非荷電アミノ酸と水素結合を形成することができる。以下により詳細に考察するように、アダプター内の1つまたは複数の化学基と、細孔内の1つまたは複数のアミノ酸との相互作用は、アダプターを正しい向きに保たせるために用いることができる。細孔内の1つまたは複数のアミノ酸と相互作用してアダプターを正しい向きに保たせる、アダプター内の1つまたは複数の化学基は、分析物と相互作用することのできる1つもしくは複数の化学基と同じでもよく、または分析物と相互作用することのできる1つもしくは複数の化学基と異なってもよい。
【0059】
細孔と分析物との間の相互作用を助長する、任意のアダプターを用いることができる。適したアダプターには、シクロデキストリン、環状ペプチドおよびカボチャ型分子が非限定的に含まれる。アダプターは好ましくは、ヘプタキス-6-アミノ-β-シクロデキストリン(am7-β-CD)または6-モノデオキシ-6-モノアミノ-β-シクロデキストリン(am1β-CD)である。以下の表2は、細孔およびアダプターの好ましい組み合わせを示している。
【0060】
(表2)細孔およびアダプターの適した組み合わせ
【0061】
好ましくは、アダプターは、ペプチド、ポリペプチド、タンパク質、核酸またはポリヌクレオチドなどのような、反復性サブユニットでできた線状ポリマーではない。より好ましくは、アダプターはプロテインキナーゼインヒビターペプチドではない。
【0062】
共有結合
アダプターは細孔と共有結合している。アダプターは、当技術分野で公知の任意の方法を用いて細孔と共有結合させることができる。アダプターは、細孔と直接結合させることができる。アダプターは好ましくは、二官能性架橋剤(bifunctional crosslinker)を用いて細孔と結合される。適した架橋剤は当技術分野において周知である。好ましい架橋剤はサクシニミジル3-(2-ピリジルジチオ)プロピオネート(SPDP)である。典型的には、アダプターは、アダプター/架橋剤複合体を細孔と共有結合させる前に二官能性架橋剤と共有結合させるが、二官能性架橋剤/細孔複合体をアダプターと結合させる前に二官能性架橋剤を細孔と共有結合させることも可能である。
【0063】
共有結合の部位は、アダプターが分析物と細孔との相互作用を助長し、それによって分析物の検出を可能にするように選択される。共有結合の部位は、分析物が、細孔を通して流れる電流に対して、その分析物に特異的な様式で影響を及ぼすように選択される。
【0064】
好ましくは、アダプターは、細孔のバレルまたはチャネルと共有結合される。アダプターは、アダプターが分析物と細孔との相互作用を助長し、それによって分析物の検出を可能にする限り、バレルまたはチャネル内の任意の部位に共有結合させることができる。好ましくは、アダプターは、アダプターとバレルまたはチャネルとの相互作用が起こる部位の近くにある、バレルまたはチャネル内のアミノ酸と共有結合される。アダプターを、アダプターとバレルまたはチャネルとの間の相互作用が起こる部位とは隔たっている、バレルまたはチャネル内のアミノ酸と共有結合させる場合には、アダプターが、アダプターとバレルまたはチャネルとの間の相互作用が起こるバレルまたはチャネル内の部位に到達しうるように、適した長さの二官能性架橋剤を用いるとよい。アダプターは、細孔と接触する際にそれが可逆的に相互作用するバレルまたはチャネル内の部位と結合していなければならないわけではない。
【0065】
細孔は好ましくは、分子アダプターと二官能性架橋剤との共有結合を助長するように修飾される。細孔のバレルまたはチャネルは、より好ましくは、分子アダプターと二官能性架橋剤との共有結合を助長するように修飾される。細孔は、当技術分野で公知の任意の方法を用いて修飾することができる。上記で考察した変異体の任意のものを、分子アダプターまたは二官能性架橋剤の共有結合を助長するために用いることができる。
【0066】
修飾は典型的には、アダプターと二官能性架橋剤との共有結合を助長するための、細孔内への、好ましくは細孔のバレルまたはチャネル内への、1つまたは複数の、例えば1個、2個、3個、4個、5個、6個または7個などのアミノ酸の導入を伴う。1つまたは複数のアミノ酸を、細孔の同じまたは異なるサブユニット中に導入することができる。共有結合を形成することのできる任意のアミノ酸、例えばシステインなどを導入することができる。アミノ酸は天然または非天然のいずれでもよい。1つまたは複数のアミノ酸は、好ましくは置換によって導入される。以下の表3は、特定の化学基を介したアダプターまたは二官能性架橋剤の共有結合を助長するために細孔内に導入することのできるアミノ酸の種類の例を示している。
【0067】
(表3)アダプターの共有結合を助長するための細孔の修飾
【0068】
好ましい態様においては、SEQ ID NO:14および16に示されているサブユニットの1つを用いて、七量体性細孔内にシステイン残基を導入する。
【0069】
アダプターの向き
アダプターは、分析物を細孔を用いて検出することを可能にする向きで細孔と共有結合される。アダプターは、分析物が、細孔を通して流れる電流に対して、その分析物に特異的な様式で影響を及ぼすような向きに置かれる。アダプターは、それが細孔および分析物のホスト-ゲスト化学作用を改善するような向きに置かれる。アダプターは、それが細孔の物理的または化学的な特性に影響を及ぼして、分析物とのその相互作用を改善するような向きに置かれる。アダプターが分析物と特異的に相互作用すること、または結合することができる場合には、アダプターは、それが分析物と特異的に相互作用すること、または結合することができるような向きに置かれる。後者の態様において、分析物と相互作用するアダプターの部分は、好ましくは、分析物がそこを通って入る細孔の末端に向けて置かれる。より好ましくは、分析物と相互作用することのできる、アダプター内の1つまたは複数の化学基は、分析物がそこを通って入る細孔の末端に向けて置かれる。基は好ましくはアミノ基またはヒドロキシル基である。分析物がそこを通って入る細孔の末端は、シス末端またはトランス末端であってよい。末端は好ましくはトランス末端である。
【0070】
アダプターの向きは、アダプターと細孔との間の共有結合および/またはホスト-ゲスト化学作用によって決定される。共有結合は、アダプターが正しい向きに置かれるように設計することができる。例えば、アダプターが細孔と共有結合する部位(例えば、アミノ酸)を、アダプターが正しい向きに置かれるように設計することもでき、および/または、二官能性架橋剤を、アダプターが正しい向きに置かれるように用いることもできる。
【0071】
好ましくは、細孔は、アダプターの配向を助長するように修飾される。細孔の修飾は、アダプターと細孔との間のホスト-ゲスト化学作用を改善し、それによってアダプターを正しい向きに保たせることができる。より好ましくは、細孔のバレルまたはチャネルは、アダプターの配向を助長するように修飾される。細孔のバレルまたはチャネルは、アダプターの配向を助長する任意の部位で修飾することができる。好ましくは、細孔のバレルまたはチャネルは、共有結合されたアダプターと細孔との間でホスト-ゲスト化学作用が起こる部位で修飾される。最も好ましくは、共有結合されたアダプターと非共有結合性の相互作用を介して相互作用することができる細孔のバレルまたはチャネル内の1つまたは複数のアミノ酸は、アダプターの配向を助長するように修飾される。修飾されるアミノ酸は、細孔と分析物との相互作用を助長する、上記で考察したアミノ酸の1つまたは複数であってよい。細孔は、当技術分野で公知の任意の方法を用いて修飾することができる。上記で考察した変異体の任意のものを、分子アダプターの配向を助長するために用いることができる。
【0072】
修飾は典型的には、アダプターの配向を助長するための、細孔内への、または好ましくはバレルまたはチャネル内への、1つまたは複数の、例えば1個、2個、3個、4個、5個、6個または7個などのアミノ酸の導入を伴う。1つまたは複数のアミノ酸を、同じまたは異なるサブユニット中に導入することができる。1つまたは複数のアミノ酸は、好ましくは、アダプターを正しい向きに保たせるアミノ酸である。1つまたは複数のアミノ酸は、より好ましくは、疎水性相互作用、水素結合、ファンデルワールス力、π-カチオン相互作用および/または静電力といった非共有結合性の相互作用を介してアダプターを正しい向きに保たせるアミノ酸である。1つまたは複数のアミノ酸は、天然または非天然のいずれでもよい。1つまたは複数のアミノ酸は、好ましくは非荷電アミノ酸である。非荷電アミノ酸はアスパラギンのように極性であってもよく、またはフェニルアラニンのように非極性であってもよい。細孔内の非荷電アミノ酸は、アダプター内のヒドロキシル基と水素結合を介して相互作用することによって、アダプターを方向づけることができる。1つまたは複数のアミノ酸は、好ましくは置換によって導入される。以下の表4は、種々の化学基を含むアダプターを正しく方向づけるために細孔内に導入することのできるアミノ酸の種類を示している。
【0073】
(表4)アダプターを配向させるための細孔の修飾
【0074】
好ましい態様において、七量体性細孔は、第一級炭素原子と結合したヒドロキシル基を含むアダプター、例えば6-モノデオキシ-6-モノアミノ-β-シクロデキストリン(am1βCD)などを細孔のシス側の方に配向させることのできる、6個のフェニルアラニン残基を含む。この七量体性細孔は、SEQ ID NO:10に示されたサブユニット6個を含むことができる。
【0075】
別の好ましい態様において、七量体性細孔は、第二級ヒドロキシル基を含むアダプター、例えば6-モノデオキシ-6-モノアミノ-β-シクロデキストリン(am1βCD)などを細孔のトランス側の方に配向させることのできる、6個のアスパラギン残基を含む。この七量体性細孔は、SEQ ID NO:12に示されたサブユニット6個を含むことができる。
【0076】
最も好ましくは、細孔は、アダプターの共有結合を助長するように、および配向を助長するように修飾される。そのような態様において、共有結合の部位と、アダプターの配向を助長するように修飾される細孔の部位との間の空間的関係は、アダプターが正しい向きに確実に保たれるように設計される。修飾は、アダプターが正しい向きに保たれる限り、細孔内の、好ましくはバレルまたはチャネル内の任意の場所に施すことができる。好ましくは、アダプターの共有結合を助長する修飾、および配向を助長する修飾は、細孔内で空間的に近接している。より好ましくは、アダプターの共有結合を助長する修飾、および配向を助長する修飾は、細孔のバレルまたはチャネル内の同一のアミノ酸の環内に作製される。環内の1つのアミノ酸をアダプターの共有結合を助長するように修飾し、一方、環内の他のアミノ酸のうち1つまたは複数、例えば1個、2個、3個、4個、5個または6個などをアダプターの配向を助長するように修飾することができる。共有結合を助長する修飾がアダプターの配向を助長する修飾から隔たっている場合には、アダプターが、アダプターの配向を助長する修飾の近くに配置され、それによって正しく配向されるように、適切な長さの二官能性架橋剤を用いるとよい。上記で考察した修飾の任意のものを用いることができる。
【0077】
最も好ましい態様においては、SEQ ID NO:10に示されているサブユニット6個およびSEQ ID NO:14に示されているサブユニット1個を用いて、またはSEQ ID NO:12に示されているサブユニット6個およびSEQ ID NO:16に示されているサブユニット1個を用いて、七量体性細孔を作製する。
【0078】
本発明の膜貫通タンパク質細孔を作製する方法
本発明はまた、本発明の膜貫通タンパク質細孔を作製する方法にも関する。本方法は、膜貫通タンパク質細孔と、細孔と分析物との間の相互作用を助長する分子アダプターとを共有結合させる段階を含む。アダプターは、当技術分野で公知の任意の方法を用いて、細孔と共有結合させることができる。
【0079】
上記で考察した細孔、アダプターおよび二官能性架橋剤の任意のものを、本方法に用いることができる。共有結合の部位は、上記で考察したように選択される。
【0080】
細孔は好ましくは、アダプターの共有結合を助長し、および/またはアダプターの配向を助長するように修飾される。これについては以上により詳細に考察している。
【0081】
本方法はまた、アダプターが、分析物を細孔を用いて検出することを可能にする向きで細孔と結合しているか否かを判定する段階も含む。これは、膜貫通タンパク質細孔を、分析物の有無を判定するために用いうるか否かを判定することを伴う。これは、以下により詳細に説明するようにして行うことができる。分析物の有無を判定することができるならば、アダプターは正しい向きにあり、本発明の膜貫通タンパク質細孔が作製されたことになる。分析物の有無を判定することができないならば、アダプターは正しくない向きにある可能性が高く、本発明の膜貫通タンパク質細孔は作製されなかったことになる。
【0082】
分析物を検出する方法
本発明はまた、分析物の有無を判定する方法にも関する。本方法は、分析物を、本発明の膜貫通タンパク質細孔と、分析物が細孔と相互作用するように接触させる段階、および相互作用の間に細孔を通過する電流を測定し、それによって分析物の有無を判定する段階を含む。本発明の膜貫通タンパク質細孔の任意のものを用いることができる。分析物を検出するために本発明の膜貫通タンパク質細孔を用いることに伴う利点については上記で考察している。
【0083】
電流がその分析物に特異的な様式で細孔を通って流れるならば(すなわち、その分析物に伴ってみられる特徴的な電流が細孔を通って流れるのが検出されたならば)、その分析物は存在する。電流がその分析物に特異的な様式で細孔を通って流れないならば、その分析物は存在しない。
【0084】
本発明はこのため、分析物の確率的センシングを伴う。本発明は、構造の類似した分析物を、それらが膜貫通タンパク質細孔を通過する電流に対して及ぼす影響の違いに基づいて識別するために用いることができる。本発明はまた、試料中の特定の分析物の濃度を測定するために用いることもできる。
【0085】
また、本発明を、バルクセンシング用途に多数のまたは数千個もの本発明の細孔を用いるセンサーに用いることもできる。
【0086】
本方法は、本発明の膜貫通タンパク質細孔を膜の中に挿入する任意の適した膜/細孔系を用いて実施することができる。本方法は典型的には、(i)本発明の膜貫通タンパク質細孔を含む人工膜、(ii)本発明の膜貫通タンパク質細孔を含む、単離された天然に存在する膜、または(iii)本発明に従って修飾された膜貫通タンパク質細孔を発現する細胞、を用いて実施される。本方法は好ましくは人工膜を用いて実施される。膜は、本発明の膜貫通タンパク質細孔に加えて、他の膜貫通タンパク質および/または膜内タンパク質、さらには他の分子も含むことができる。
【0087】
本発明の方法は、典型的にはインビトロで実施される。
【0088】
膜
膜はイオンおよび分析物の流れに対する障壁を形成する。膜は好ましくは脂質二重層である。本発明による使用のために適した脂質二重層は、当技術分野で公知の方法を用いて作製することができる。例えば、脂質二重層膜は、Montal and Mueller (1972)の方法を用いて形成させることができる。本発明の方法は、リン脂質、糖脂質、コレステロールおよびそれらの混合物を非限定的に含む任意の膜脂質から形成された脂質二重層を用いて実施することができる。脂質二重層は好ましくは、1,2-ジフィタノイル-sn-グリセロ-3-ホスホコリンから形成される。
【0089】
脂質二重層などの膜の中に細孔を挿入するための方法は、当技術分野で公知である。例えば、細孔を、脂質二重層を含む溶液中に精製形態で懸濁させ、それが脂質二重層へ拡散して、脂質二重層との結合および機能的状態への集合によって挿入されるようにすることができる。または、細孔を、M. A. Holden, H. Bayley. J. Am. Chem. Soc. 2005, 127, 6502-6503に記載された方法を用いて膜の中に直接挿入することもできる。
【0090】
細孔と分析物との間の相互作用
分析物は、膜のいずれかの側で細孔と接触させることができる。分析物は、膜のいずれかの側で細孔に導入することができる。分析物は、分析物が細孔を通って膜の他方の側へと通過することを可能にする膜の側と接触させられる。例えば、分析物は、その本来の環境においてイオンまたは低分子、例えば分析物などが細孔のバレルまたはチャネル内に入ることを可能にする細孔の末端と、イオンまたは低分子が細孔を通過しうるように、接触させられる。そのような場合、分析物は、それが膜を横切って細孔のバレルまたはチャネルを通過する際に、細孔および/またはアダプターと相互作用する。または、分析物を、分析物がアダプターを介して、またはアダプターとともに細孔と相互作用し、細孔から解離して、膜の同じ側に残ることを可能にする膜の側と接触させることもできる。本発明は、アダプターの向きが固定されている細孔を提供する。その結果として、分析物は好ましくは、アダプターが配向している方の細孔の末端と接触する。最も好ましくは、分析物は、分析物と相互作用するアダプターの部分が方向づけられている細孔の末端と接触させられる。
【0091】
分析物は、任意の様式かつ任意の部位で、細孔と相互作用することができる。上記で考察したように、分析物は好ましくは、アダプターを介して、またはアダプターとともに細孔と可逆的に結合する。分析物は最も好ましくは、それが膜を横切って細孔を通過する際に、アダプターを介して、またはアダプターとともに細孔と可逆的に結合する。分析物はまた、それが膜を横切って細孔を通過する際に、アダプターを介して、またはアダプターとともに細孔のバレルまたはチャネルと可逆的に結合することもできる。
【0092】
分析物と細孔との間の相互作用の間に、分析物は、細孔を通して流れる電流に対して、その分析物に特異的な様式で影響を及ぼす。例えば、特定の分析物は、特定の平均期間にわたって特定の程度で、細孔を通して流れる電流を減少させると考えられる。換言すれば、細孔を通して流れる電流は、特定の分析物に関して特徴的である。対照実験を、細孔を通して流れる電流に対して特定の分析物が及ぼす効果を決定するために実施することができる。続いて、試料中の特定の分析物を同定するため、または試料中に特定の分析物が存在するか否かを判定するために、被験試料に対して本発明の方法を実施したことによる結果を、そのような対照実験から得られたものと比較することができる。細孔を通して流れる電流が特定の分析物を指し示す様式で影響を受ける頻度を、試料中のその分析物の濃度を決定するために用いることができる。
【0093】
装置
本方法は、膜貫通タンパク質細孔が膜の中に挿入されている膜/細孔系を調べるために適した任意の装置を用いて実施することができる。本方法は、確率学的センシングのために適した任意の装置を用いて実施することができる。例えば、装置は、水溶液を含むチャンバーおよびチャンバーを2つの区域に分ける障壁を含む。障壁は、細孔を含む膜が形成されている開口部を有する。分析物は、分析物をチャンバー内に導入することによって細孔と接触させることができる。分析物は、チャンバーの2つの区域のいずれかの中に導入することができる。
【0094】
本発明の方法は、分析物との相互作用の間に、細孔を通過する電流を測定することを伴う。したがって、装置はまた、電位を加えて膜および細孔を横切る電気信号を測定することのできる電気回路も含む。本方法は、パッチクランプ(patch clmp)または電圧固定(voltage clamp)を用いて実施されうる。本方法は、好ましくは、電圧固定の使用を伴う。実施例は、電圧固定法を実施するための1つのやり方を開示している。
【0095】
分析物
分析物は、試料中の任意の物質でありうる。適した分析物には、金属イオン、無機塩、重合体、例えば重合酸または塩基など、色素、脱色剤、医薬品、診断用薬、娯楽薬(recreational drug)、爆発物および環境汚染物質が非限定的に含まれる。
【0096】
分析物は、細胞から分泌される分析物であってもよい。または、分析物は、本発明が実施可能となる前に分析物を細胞から抽出しなければならないような、細胞の内側に存在する分析物であってもよい。
【0097】
分析物は好ましくは、アミノ酸、ペプチド、ポリペプチドまたはタンパク質である。アミノ酸、ペプチド、ポリペプチドまたはタンパク質は天然または非天然のいずれでもありうる。ポリペプチドまたはタンパク質は、その内部に合成アミノ酸または修飾アミノ酸を含みうる。アミノ酸に対するさまざまな異なる種類の修飾が当技術分野で公知である。本発明の目的には、分析物を、当技術分野において入手可能な任意の方法によって修飾しうることが理解されるべきである。
【0098】
タンパク質は、酵素、抗体、ホルモン、増殖因子または増殖調節タンパク質、例えばサイトカインなどでありうる。サイトカインは、インターロイキン、好ましくはIFN-1、IL-2、IL-4、IL-5、IL-6、IL-10、IL-12もしくはIL-13、インターフェロン、好ましくはIL-γ、または他のサイトカイン、例えばTNF-αなどから選択することができる。タンパク質は、細菌タンパク質、真菌タンパク質、ウイルスタンパク質または寄生生物由来タンパク質であってよい。それを細孔と接触させる前に、タンパク質をアンフォールディングさせてポリペプチド鎖を形成させ、それによってそれが細孔のバレルまたはチャネルの中に入り、アダプターを介して、またはアダプターとともに細孔と相互作用することを可能にすることもできる。
【0099】
ヌクレオチドおよび核酸の検出については、以下にさらに詳細に考察している。
【0100】
分析物は任意の適した試料中に存在する。本発明は典型的には、分析物を含むことが分かっているか、または分析物を含むことが疑われる試料に対して実施される。本発明を、その実体が不明である1つまたは複数の分析物を含む試料に対して実施することもできる。または、本発明を、試料中に存在することが分かっているかまたは予想される、1つまたは複数の分析物の実体を確認するために、試料に対して実施することもできる。
【0101】
試料は生物試料であってよい。本発明は、任意の生物体または微生物から得られた、または抽出された試料に対して、インビトロで実施することができる。生物体または微生物は、典型的には原核生物または真核生物であり、典型的には、植物界、動物界、菌界、モネラ界および原生生物界という5つの界の1つに属する。本発明は、任意のウイルスから得られた、または抽出された試料に対してインビトロで実施することができる。試料は好ましくは流体試料である。試料は典型的には患者の体液を含む。試料は、尿、リンパ、唾液、粘液または羊水でありうるが、好ましくは血液、血漿または血清である。典型的には、試料はヒトが起源であるが、またはそれは、ウマ、ウシ、ヒツジもしくはブタなどの商業的な飼育動物など別の哺乳動物由来であってもよく、またはネコもしくはイヌなどのペットであってもよい。
【0102】
試料は非生物試料であってもよい。非生物試料は好ましくは流体試料である。非生物試料の例には、手術液、水、例えば飲用水、海水または河川水など、および臨床検査のための試薬が含まれる。
【0103】
試料は典型的には、アッセイされる前に、例えば遠心分離によって、または望ましくない分子もしくは細胞、例えば赤血球などを濾別する膜を通過させることによって処理される。試料は、採取次第直ちに測定してもよい。また、試料は典型的には、アッセイの前に、好ましくは-70℃未満で保存してもよい。
【0104】
条件
本発明の方法は、分析物との相互作用の間に細孔を通過する電流を測定することを伴う。膜貫通タンパク質細孔を通るイオン電流を測定するために適した条件は当技術分野で公知であり、実施例に開示されている。本方法は、膜および細孔をまたいで印加される電圧を用いて実施される。用いられる電圧は、典型的には-250mV〜+250mVである。用いられる電圧は、好ましくは、-200mV、-150mV、-100mV、-50mV、-20mVおよび0mVから選択される下限、ならびに+10mV、+20mV、+50mV、+100mV、+150mVおよび+200mVから独立に選択される上限を有する範囲にある。
【0105】
本方法は、任意のアルカリ金属塩化物塩の存在下で実施される。上記で考察した例示的な装置において、塩はチャンバー内の水溶液中に存在する。塩化カリウム(KCl)、塩化ナトリウム(NaCl)または塩化セシウム(CsCl)が、典型的には用いられる。KClが好ましい。塩濃度は、典型的には0.1〜2.5M、0.3〜1.9M、0.5〜1.8M、0.7〜1.7M、0.9〜1.6M、または1M〜1.4Mである。塩濃度は好ましくは約1Mである。
【0106】
本方法は、典型的には、緩衝液の存在下で実施される。上記で考察した例示的な装置において、緩衝液はチャンバー内の水溶液中に存在する。任意の緩衝液を本発明の方法に用いることができる。1つの適した緩衝液は、Tris-HCl緩衝液である。本方法は、典型的には4.0〜10.0、4.5〜9.5、5.0〜9.0、5.5〜8.8、6.0〜8.7、または7.0〜8.8、または7.5〜8.5のpHで実施される。用いられるpHは好ましくは約8.0である。
【0107】
本方法は典型的には、0℃〜100℃、15℃〜95℃、16℃〜90℃、17℃〜85℃、18℃〜80℃、19℃〜70℃、または20℃〜60℃で実施される。本方法は好ましくは室温で実施される。
【0108】
個々のヌクレオチドを同定する方法
本発明はまた、個々のヌクレオチドを同定する方法にも関する。本方法は、ヌクレオチドを、本発明の膜貫通タンパク質細孔と、ヌクレオチドが細孔と相互作用するように接触させる段階;および相互作用の間に細孔を通過する電流を測定し、それによってヌクレオチドの実体を決定する段階を含む。本発明はこのため、個々のヌクレオチドの確率的センシングを伴う。本発明は、構造の類似したヌクレオチドを、それらが膜貫通タンパク質細孔を通過する電流に対して及ぼす影響の違いに基づいて識別するために用いることができる。個々のヌクレオチドを、それらが細孔と相互作用する時のそれらの電流振幅から単分子レベルで同定することができる。本発明はまた、特定のヌクレオチドが試料中に存在するか否かを判定するために用いることもできる。本発明はまた、試料中の特定のヌクレオチドの濃度を測定するために用いることもできる。
【0109】
個々のヌクレオチド
本発明による個々のヌクレオチドとは、単一のヌクレオチドのことである。個々のヌクレオチドとは、別のポリヌクレオチドとヌクレオチド結合によって結合していないもののことである。ヌクレオチド結合は、ヌクレオチドのリン酸基の1つが、別のヌクレオチドの糖基(sugar group)と結合したものを含む。個々のヌクレオチドは、典型的には、少なくとも5、少なくとも10、少なくとも20、少なくとも50、少なくとも100、少なくとも200、少なくとも500、少なくとも1000、または少なくとも5000ヌクレオチドの別のポリヌクレオチド配列とヌクレオチド結合によって結合していないものである。例えば、個々のヌクレオチドは、DNAストランドまたはRNAストランドなどの標的ポリヌクレオチド配列から消化される。
【0110】
本発明の方法は、任意のヌクレオチドを同定するために用いうる。ヌクレオチドは天然または人工的のいずれであってもよい。ヌクレオチドは、典型的には、核酸塩基、糖、および少なくとも1つのリン酸基を含む。核酸塩基は、典型的には複素環式である。適した核酸塩基には、プリンおよびピリミジン、より具体的にはアデニン、グアニン、チミン、ウラシルおよびシトシンが含まれる。糖は、典型的には五炭糖である。適した糖には、リボースおよびデオキシリボースが非限定的に含まれる。ヌクレオチドは、典型的には、リボヌクレオチドまたはデオキシリボヌクレオチドである。ヌクレオチドは、典型的には、モノホスフェート、ジホスフェートまたはトリホスフェートを含む。
【0111】
適したヌクレオチドには、以下のものが非限定的に含まれる:アデノシン一リン酸(AMP)、アデノシン二リン酸(ADP)、アデノシン三リン酸(ATP)、グアノシン一リン酸(GMP)、グアノシン二リン酸(GDP)、グアノシン三リン酸(GTP)、チミジン一リン酸(TMP)、チミジン二リン酸(TDP)、チミジン三リン酸(TTP)、ウリジン一リン酸(UMP)、ウリジン二リン酸(UDP)、ウリジン三リン酸(UTP)、シチジン一リン酸(CMP)、シチジン二リン酸(CDP)、シチジン三リン酸(CTP)、サイクリックアデノシン一リン酸(cAMP)、サイクリックグアノシン一リン酸(cGMP)、デオキシアデノシン一リン酸(dAMP)、デオキシアデノシン二リン酸(dADP)、デオキシアデノシン三リン酸(dATP)、デオキシグアノシン一リン酸(dGMP)、デオキシグアノシン二リン酸(dGDP)、デオキシグアノシン三リン酸(dGTP)、デオキシチミジン一リン酸(dTMP)、デオキシチミジン二リン酸(dTDP)、デオキシチミジン三リン酸(dTTP)、デオキシウリジン一リン酸(dUMP)、デオキシウリジン二リン酸(dUDP)、デオキシウリジン三リン酸(dUTP)、デオキシシチジン一リン酸(dCMP)、デオキシシチジン二リン酸(dCDP)およびデオキシシチジン三リン酸(dCTP)。ヌクレオチドは、好ましくはAMP、TMP、GMP、UMP、dAMP、dTMP、dGMPまたはdCMPである。
【0112】
ヌクレオチドを、リボ核酸(RNA)またはデオキシリボ核酸などの核酸配列の消化によって導くこともできる。単一の核酸配列由来の個々のヌクレオチドを、その核酸の全体または一部のシークエンシングのために、連続的な様式で細孔と接触させることができる。本発明の第2の態様に従った核酸のシークエンシングについて、以下により詳細に考察する。
【0113】
ヌクレオチドは、核酸配列の消化によって導かれる場合のように、典型的には非修飾である。または、ヌクレオチドを修飾すること、または損傷させることもできる。ヌクレオチドは典型的にはメチル化される。ヌクレオチドを顕在化標識で標識することもできる。顕在化標識は、ヌクレオチドが検出されることを可能にする任意の適した標識であってよい。適した標識には、蛍光分子、放射性同位体、例えば125I、35Sなど、およびビオチンなどのリンカーが含まれる。
【0114】
ヌクレオチドは、典型的には、任意の適した生物試料中に存在する。適した生物試料については上記で考察している。
【0115】
本方法の実施
分析物の検出にかかわる、特に用いうる細孔、膜、装置および条件にかかわる上記の考察のすべては、この方法に対して等しく適用される。
【0116】
条件に関しては、用いられる電圧は、好ましくは、デオキシリボヌクレオチド5'一リン酸、例えばdAMP、dTMP、dGMPおよびdCMPなどについては約+130mVであり、リボヌクレオチド5'一リン酸、例えばAMP、TMP、GMPおよびUMPなどについては+110mVである。本方法は好ましくは、デオキシリボヌクレオチド5'一リン酸、例えばdAMP、dTMP、dGMPおよびdCMPなどについてはpH 8.0、1M KClにて+130mVで実施され、リボヌクレオチド5'一リン酸、例えばAMP、TMP、GMPおよびUMPなどについてはpH 8.0、1M KClにて+110mVで実施される。
【0117】
核酸のシークエンシングの方法
本発明はまた、標的核酸配列のシークエンシングの方法にも関する。本方法は、(a)個々のヌクレオチドを、標的配列の一方の末端から、連続移動性エキソヌクレアーゼを用いて消化する段階;(b)ヌクレオチドを、本発明の膜貫通タンパク質細孔と、ヌクレオチドが細孔と相互作用するように接触させる段階;(c)相互作用の間に細孔を通過する電流を測定し、それによってヌクレオチドの実体を決定する段階;および(d)段階(a)から(c)までを核酸配列の同じ末端で繰り返し、それによって核酸の配列を決定する段階、を含む。それ故に、本方法は、核酸のシークエンシングのための、連続的な様式での、核酸配列の各々の単一ヌクレオチドの確率的センシングを伴う。
【0118】
本発明の膜貫通タンパク質細孔は、この方法のために特に適している。核酸の有効なシークエンシングのためには、核酸中のすべてのヌクレオチドが連続的な様式で確実に同定されるようにすることが重要である。本発明の膜貫通タンパク質細孔におけるアダプターの固定的な性質は、各々の連続的なヌクレオチドが細孔と相互作用する時には常に、特徴的な電流が細孔を通して流れることを意味する。
【0119】
核酸の全体または一部のみを、この方法を用いてシークエンシングすることができる。核酸は任意の長さでありうる。例えば、核酸は、少なくとも10、少なくとも50、少なくとも100、少なくとも150、少なくとも200、少なくとも250、少なくとも300、少なくとも400、または少なくとも500ヌクレオチド長でありうる。核酸は天然または人工的のいずれであってもよい。例えば、本方法は、製造されたオリゴヌクレオチドの配列を検証するために用いることができる。本方法は典型的にはインビトロで実施される。
【0120】
この方法の段階(b)および(c)は、上記で考察したヌクレオチドを同定する方法において実施される段階と概ね同一である。分析物の検出にかかわる、特に用いうる細孔、膜、装置および条件にかかわる上記の考察のすべては、この方法に対して等しく適用される。核酸は典型的には、上記で考察したような任意の生物試料中に存在する。
【0121】
連続移動性エキソヌクレアーゼ
標的核酸のシークエンシングの方法は、核酸を連続移動性エキソヌクレアーゼと接触させて、個々のヌクレオチドを核酸の一方の末端から遊離させることを伴う。連続移動性エキソヌクレアーゼとは、典型的には、核酸の一方の末端に取り付き、配列を消化してその末端から1回に1個ずつのヌクレオチドを分離させる酵素のことである。連続移動性エキソヌクレアーゼは、核酸を5'から3'の方向に、または3'から5'の方向に消化することができる。連続移動性エキソヌクレアーゼが結合する核酸の末端は、典型的には、用いる酵素の選択を通じて、および/または当技術分野で公知の方法を用いて決定される。核酸配列の特定の末端に対する連続移動性エキソヌクレアーゼの結合を防止または助長するためには、典型的には、核酸配列のいずれかの末端でヒドロキシル基またはキャップ構造を用いるとよい。
【0122】
任意の連続移動性エキソヌクレアーゼ酵素を、本方法に用いることができる。本方法における使用のために好ましい酵素は、λエキソヌクレアーゼである。λエキソヌクレアーゼの1つのサブユニットの配列は、SEQ ID NO:17に示されている。3つの同一のサブユニットが相互作用して、三量体性エキソヌクレアーゼを形成する。λエキソヌクレアーゼの変異体は、SEQ ID NO:17のそれとは違いのあるアミノ酸配列を有し、かつ連続移動性エキソヌクレアーゼ活性を保っているポリペプチドサブユニットから形成された酵素である。これらの変異体は、上記のSEQ ID NO:2の変異体に関して上記で考察したのと同じ様式で、かつ同じ程度で、SEQ ID NO:17とは違っていてよい。変異体は、好ましくは、核酸と結合して核酸を消化する原因となるドメイン(触媒ドメイン)を含む。変異体は、好ましくは、野生型酵素と比較して、必要に応じて上昇または低下した酵素活性速度、および/またはより高い塩耐性を有する。連続移動性エキソヌクレアーゼは、膜貫通タンパク質細孔の作製に関して上記で考察した方法の任意のものを用いて作製することができる。
【0123】
標的核酸配列のシークエンシングの方法は、核酸配列を、連続移動性エキソヌクレアーゼと、上記で考察したようにヌクレオチドが各々の個々のヌクレオチドの同定を可能にする速度で核酸の末端から消化されるように、接触させることを含む。これを行うための方法は当技術分野において周知である。例えば、ポリペプチドの末端から単一のアミノ酸を連続的に消化して、それらを高速液体クロマトグラフィー(HPLC)を用いて同定しうるようにするために、エドマン分解が用いられる。類似の方法を本発明に用いることもできる。
【0124】
連続移動性エキソヌクレアーゼは、好ましくは、膜貫通タンパク質細孔と共有結合される。連続移動性エキソヌクレアーゼを細孔と共有結合させるための方法は、当技術分野において周知である。
【0125】
連続移動性エキソヌクレアーゼが機能しなければならない速度は、典型的には、野生型連続移動性エキソヌクレアーゼの至適速度よりも遅い。第2の態様の方法における連続移動性エキソヌクレアーゼの適した活性速度には、1秒当たり0.5〜1000ヌクレオチド、1秒当たり0.6〜500ヌクレオチド、1秒当たり0.7〜200ヌクレオチド、1秒当たり0.8〜100ヌクレオチド、1秒当たり0.9〜50ヌクレオチド、または1秒当たり1〜20もしくは10ヌクレオチドの消化が含まれる。速度は、好ましくは1秒当たり1、10、100、500または1000ヌクレオチドである。連続移動性エキソヌクレアーゼの適した活性速度は、さまざまなやり方で達成することができる。例えば、至適活性速度が低下している変異型連続移動性エキソヌクレアーゼを本発明に従って用いることができる。
【0126】
連続移動性エキソヌクレアーゼの活性は、典型的にはpH依存性であり、pHが低下するにつれてそれらの活性が低下する。それ故に、第2の態様の方法は、典型的には、7.5〜8.0、または7.7〜8.0というpHで実施される。用いられるpHは、好ましくは約8.0である。
【0127】
連続移動性エキソヌクレアーゼの活性速度は、典型的には、塩濃度が上昇するにつれて低下する。しかし、極めて高い塩濃度は、典型的には、酵素の活性に対して有害な影響を及ぼす。酵素の速度を制限するもう1つのやり方は、活性に有害な影響を及ぼすことなしに酵素の活性速度を低下させる塩濃度で、本方法を実施することである。例えば、本方法を0.5〜1Mの塩濃度で実施することができる。塩濃度は好ましくは約1Mである。
【0128】
キット
本発明はまた、標的核酸配列のシークエンシングの方法を実施するために用いうるキットにも関する。したがって、本キットは、核酸配列のシークエンシングのために適している。キットは、本発明の膜貫通細孔および連続移動性エキソヌクレアーゼを含む。
【0129】
キットは、上述した方法の態様の任意のものを実施することを可能にする1つまたは複数の他の試薬または器具をさらに含みうる。そのような試薬または器具には、以下の1つまたは複数が含まれる:適した緩衝液(水溶液)、対象から試料を得るための手段(例えば、注射針を含む容器または器具)、核酸配列を増幅するための手段、上記で定義した膜、または電圧固定もしくはパッチクランプの装置。試薬は、キット中に、流体試料が試薬を再懸濁させるような乾燥状態で存在しうる。キットはまた、任意で、キットを本発明の方法に用いることを可能にする説明書、または本方法を使用しうる患者に関する詳細を含みうる。キットは任意で、ヌクレオチドを含みうる。
【0130】
以下の実施例は、本発明を例示的に説明している。
【0131】
実施例
この実施例において、本発明者らは、β-シクロデキストリン(β-CD)をαHL細孔に対して、アダプターを共有結合のために配向させ、かつ反応後の細孔内腔の中でそれらを安定化するための113位の突然変異を用いて共有結合させた。このアプローチには2つの重要な成果があった。第1に、アダプターはナノポア(nanopore)から解離することができない。第2に、細孔のトランス入口(trans entrance)に面した第一級ヒドロキシルまたは第二級ヒドロキシルのいずれかを用いて、シクロデキストリンの向きを制御することができる。
【0132】
1.材料および方法
1.1 αHL細孔
α-ヘモリシン突然変異型細孔(M113F-RL2)7(WTバックグラウンド;SEQ ID NO:10)、(M113N-RL2)7(WTバックグラウンド;SEQ ID NO:12)、(M113F-RL2)6(M113C-D8RL2)1(SEQ ID NO:10および14)、(M113N)6(T117C-D8RL3)1(SEQ ID NO:12および16)を、以前に記載した通りに発現させ、集合させて精製した。
【0133】
RL2は、膜貫通βバレルをコードする配列のカセット突然変異誘発を可能とするように考案された半合成遺伝子の産物である(Cheley, S., Braha, O., Lu, X., Conlan, S., and Bayley, H. (1999) Protein Sci. 8, 1257-1267)。これは、6個のサイレント制限部位、およびコードされるポリペプチド配列中に5個の改変アミノ酸を含む(K8A、V124L、G130S、N139QおよびI142L)。D8RL2は、オクタ-アスパラギン酸尾部(octa-aspartate tail)を有するRL2である。
【0134】
RL3は、アミノ酸レベルではWT αHLポリペプチドと同一であるが、この遺伝子は、ループをコードする領域内に、カセット突然変異誘発の一助となる6個のサイレント突然変異を含む(S. Cheley, unpublished)。D8RL3は、オクタ-アスパラギン酸尾部を有するRL3である。図12は、pT7-RL3-D8の完全ポリヌクレオチド配列を示している。
【0135】
1.2 化学物質
試薬は以下の通りに入手した:1,2-ジフィタノイル-sn-グリセロ-3-ホスホコリン(Avanti Polar Lipids)、ペンタン(JT Baker)、ヘキサデカン(99+%、Sigma-Aldrich)、塩酸6-モノデオキシ-6-モノアミノ-β-シクロデキストリン(99%、Cyclolabs Budapest, Hungary)、Trizma塩基(99.9%、Sigma-Aldrich)、濃HCl(分析用試薬等級、Fisher Scientific)、塩化カリウム(99%、Sigma-Aldrich)、2-アダマンタンアミン・HCl(99%、Aldrich)、サクシニミジル3-(2-ピリジルジチオ)プロピオネート(SPDP、95%、Fluka)、トリエチルアミン(99.5%、Fluka)。
【0136】
1.3 6-モノデオキシ-6-[3-(2-ピリジルジチオ)プロピオニル]モノアミノ-β-シクロデキストリン(βCD-PDP)の合成
塩酸6-モノデオキシ-6-モノアミノ-β-シクロデキストリン(am1βCD)(11.7mg、0.01mmol)およびサクシニミジル3-(2-ピリジルジチオ)プロピオネート(SPDP、6.24mg、0.02mmol)を、5mLのMeOH/H2O中に溶解させた(1:1)。トリエチルアミン(100μL)を5分間かけて添加し、この混合物を一晩撹拌した。所望の産物は白色沈殿物として出現し、これを濾別して、冷水およびアセトンで連続して洗浄した。分析用試料は、DMSOおよび水からの再結晶化によって得られた。
【0137】
1.4 単一チャネル電流の記録
1,2-ジフィタノイル-sn-グリセロ-3-ホスホコリン(Avanti Polar Lipids)の二重層を、平面状二重層チャンバーをシスおよびトランスという2つの区画に区分するポリカーボネートフィルム(20μm厚、Goodfellow, Malvern, PAによる)中の直径100〜150μmの開口部全体にわたって形成させた。いずれの区画も緩衝液1mLを含んでいた。工学的に作製したタンパク質細孔をシス区画に添加し、それを接地させた。βCD-PDPをトランス区画に添加し、それを増幅器のヘッドステージ(head-stage)に接続した。
【0138】
別に指示する場合を除き、実験はすべて、25mM Tris・HCl、1M KCl、pH 8.0中、22.5±2℃にて±100mVで実施した。タンパク質およびβCD-PDPの新たな凍結アリコートを毎日用いた。電流はパッチクランプ増幅器(Axopatch 200B;Axon instruments, Foster City, CA)で記録し、内蔵の四極Besselフィルターにより10kHzでローパスフィルター処理を行った上で、Digiデータ 1200 A/Dコンバーター(Axon instruments)を装備したコンピュータによって20kHzでサンプリングを行った。
【0139】
1.5 データ分析
電流軌跡は、pClamp 9.0ソフトウエア(Axon Instruments)を用いて分析した。イベントはEvent Detection機能を用いて検出し、それを用いて振幅および滞留時間のヒストグラムを作成した。Origin(Microcal, Northampton, MA)を、曲線フィッティングおよびグラフ提示のために用いた。
【0140】
1.6 分子モデル
Michelle MontoyaおよびEric Gouauxによって提供された七量体性α-ヘモリシン細孔(M113F-RL2)7および(M113N-RL2)7のpdbファイルを、PyMOL(DeLano Scientific, San Carlos, CA)に表示した。βCD-PDP構造も、構築ツールを用いることによってPyMOLに表示した。
【0141】
2.結果
2.1 リンカーの設計および工学的に作製したαHL細孔のβCD-PDPによる誘導体化
(M113F-RL2)7・βCDおよび(M113N)7・βCDのX線構造(図1、Montoya and Gouaux, unpublished)から、αHL細孔のβバレル内の非共有結合的に結合したβCDの位置が判明している。(M113F-RL2)7・βCDでは、7個のPhe-113のフェニル環がβCDの7個の第一級6-ヒドロキシル基との疎水性相互作用に関与し、その結果、第一級ヒドロキシルがシス口部(cis mouth)の方を向く、細孔内でのβCDの特異的な配向をもたらす(図1b)。対照的に、(M113N-RL2)7・βCDでは、7個のAsn-113のアミド基およびLys-147のε-アミノ基がそれぞれ、βCDの7個の2-第二級ヒドロキシル基および7個の3-第二級ヒドロキシル基の水素結合距離の内部にある。この向きでは、βCDの第一級ヒドロキシル基は細孔のトランス入口の方を向く(図1c)。
【0142】
分子モデリングにより、本発明者らは、αHLバレルの内腔内の、βCDを結合させるのに適した位置を同定した。(M113F-RL2)7・βCDでは、Phe-113のフェニル環の中心とβCDの第一級ヒドロキシル基の最近接O原子との間の距離は6.5±0.5Åである(7つの位置の平均)(図1b)。(M113N-RL2)7・βCDでは、βCDの第一級ヒドロキシル基のO原子とThr-117の最近接O原子との間の距離は平均6.0±0.5Åである(図1c)。本発明者らは、6-モノデオキシ-6-モノアミノ-β-シクロデキストリン(am1βCD、これはβCDの単一の第一級ヒドロキシル基がアミノ基によって置換されている)を、位置-113または位置-117にあるシステイン残基と連結させるために、二官能性架橋剤サクシニミジル3-(2-ピリジルジチオ)プロピオネート(SPDP)を選んだ。本発明者らはまず、am1βCDをSPDPとカップリングさせてβCD-PDPを形成させた(図2)。βCD-PDPでは、リンカーの末端にあるピリジルジスルフィドを、αHL細孔内のシステイン残基の遊離チオールによって攻撃してジスルフィド結合を形成させることができる。リンカーの長さは、βCD-PDPをPyMolの中で構築することによって測定した;伸長コンフォメーションでは、am1βCDのアミド結合内のN原子と、ピリジルS原子との間の距離はおよそ7.0Åである。したがって、このリンカーは、am1βCDを、(M113F-RL2)7に見られる向きにあるβCDを用いて位置-113のシステインと連結させるため、または(M113N-RL2)7に見られる向きにあるβCDを用いて位置-117のシステインと連結させるために必要とされるよりも幾分長い。
【0143】
これらのモデリング研究に基づき、位置-113に6個のフェニルアラニン残基を有し、第7の位置に単一のシステインを有する、ヘテロ七量体性αHL細孔(M113F-RL2)6(M113C-D8RL2)1を工学的に作製した。M113Cサブユニット中のシステイン残基は、シクロデキストリンの第一級ヒドロキシルがM113Fサブユニットの6個のPhe残基の近隣に保たれるような様式で、二重層のトランス側から提示されるβCD-PDPのピリジルジスルフィドと反応しうるように設計した。反応後に、βCDは永続的な分子アダプターとしてαHL細孔の内側に係留されると考えられる。もう1つの構築物において、本発明者らは、工学的に作製した細孔(M113N-RL2)6(T117C-D8RL3)1内の117位にシステイン残基を配置した。この細孔は、M113Nサブユニット内の残基Asn-113およびLys-147と水素結合を形成した第二級ヒドロキシルの相互作用の安定化を維持しながらβCD-PDPと反応するように設計した。
【0144】
本発明者らは、PyMOLの中で、所望の共有結合複合体(M113F-RL2)6(M113C-D8RL2)1-βCDおよび(M113N-RL2)6(T117C-D8RL3)1-βCDを描き出した。(M113F-RL2)6(M113C-D8RL2)1-βCDにおいて、am1βCDのアミドN原子と、113位のシステインのS原子との間の距離は5.6Å(図3cおよび4)である。(M113N-RL2)6(T117C-D8RL3)1-βCDにおいて、am1βCDのアミドN原子と、117位のシステインのSとの間の距離は6.2Åである(図5cおよび6)。これらの値は非共有結合複合体のX線構造から求められた対応する測定値に近く(上記参照)、このことはリンカーの長さの選択が正しいことを裏づけている。
【0145】
2.2 (M113F-RL2)6(M113C-D8RL2)1-βCDの電気特性決定
単一の(M113F-RL2)6(M113C-D8RL2)1細孔をシスチャンバーから脂質二重層内に導入した場合には、本発明者らが測定したイオン電流は98±5pAであった(+100mV、25mM Tris・HCl、pH 8.0、1M KCl、n=6)。細孔との反応を観察するために、βCD-PDPをトランスチャンバーに添加して、電位を+100mVに保った。全電流の70%および95%に相当する2種類の過渡的遮断イベントが観察された。対照的に、βCDが(M113F-RL2)7と相互作用する時には、70%の電流遮断しか観察されない。このため、その第一級ヒドロキシルがPhe-113側鎖と相互作用し、リンカーがシス側に向けて突出している細孔内にβCD-PDPが結合する時には、70%の遮断が生じる可能性が非常に高い。95%の遮断は、ピリジル環がシクロデキストリンの中空部の内側に停留している細孔にβCD-PDPが入り、それによって電流の流れを低下させる時に起こる可能性がある。最終的に、5分間から1時間までの範囲にわたる期間の後には、電流レベルは70%遮断の状態に永続的に固定される。このイベントの前には、48±7msにわたって続く85%遮断がみられた(n=5)(図3cおよび4)。70%遮断状態から非修飾細孔のそれへの電流の回復は、24時間という全記録期間中に全く観察されなかった(16回の実験を通じて)。これに比して、(M113F-RL2)7と非共有的に結合したβCDの平均τoffは、同じ条件下で316±62msである。この遮断状態は、位置113でのジスルフィド結合を通じて共有結合したβCDを有するαHL細孔を表していると推定される(図3cおよび4)。この考え方に沿う形で、ジスルフィド結合はシスチャンバーへの2mM DTTの添加によって切断された。切断の間に、寿命が150±12ms(n=5)で電流遮断98%の過渡的な中間状態が観察された(図3d)。(M113F-RL2)6(M113C-D8RL2)1-βCDにおける70%遮断は、±250mVの範囲内で印加電位を上昇させたり低下させたりした後も維持された。αHL細孔内でのβCDなどの中性分子の結合は電圧依存性であり、それは電気浸透の影響によることを指摘しておきたい(Gu, L.-Q., Cheley, S., and Bayley, H. (2003) Proc.Natl. Acad. Sci. USA 100, 15498-15503)。したがって、共有結合は、非共有結合複合体が短寿命である電位でβCD解離を防止する。
【0146】
電流-電圧(I-V)曲線を、(M113F-RL2)6(M113C-D8RL2)1-βCDおよび非共有結合複合体(M113F)7・βCDに関して計測した。I-V曲線は+150〜-60mVの間では有意な違いがなかった。しかし、印加電位が-60mV未満の場合には、(M113F-RL2)6(M113C-D8RL2)1-βCDに関するI-V曲線の方が(M113F-RL2)7・βCDのそれよりも整流的であった(図3e)。
【0147】
2.3 (M113N-RL2)6(T117C-D8RL3)1の電気特性決定
単一の(M113N-RL2)6(T117C-D8RL3)1タンパク質細孔をシスチャンバーから脂質二重層内に導入した場合には、本発明者らが測定したイオン電流は-80±5pAであった(-100mV、25mM Tris・HCl、pH 8.0、1M KCl、n=6)。βCD-PDPをトランス区画に添加した後には、(M113N-RL2)7・βCDで観察された65%遮断に近い、70%の電流遮断という1種類の過渡的遮断イベントのみが観察された。1分間から20分間までの範囲にわたる期間の後には、寿命が5±2ms(n=5)で電流遮断80%という中間状態が観察され、その後には永続的な70%電流遮断が続いた(図4および5c)。(M113F-RL2)6(M113C-D8RL2)1-βCDの場合と同じように、この70%遮断は合計24時間の間には回復しなかったが(16回の実験を通じて)、一方、(M113N-RL2)7・βCDに関する平均τoffは同じ条件下で10.7±1.5sに過ぎなかった。この場合も、印加電位を±250mVの範囲内で上げ下げしても細孔の遮断解消は生じなかった。したがって、この状態は、ジスルフィド結合を通じて117位に共有結合したβCDを有するαHL細孔を表すと推定される(図4および5c)。この場合も、ジスルフィド結合はシスチャンバーへの2mM DTTの添加後に切断された(図5d)。切断の間に、電流遮断80%で寿命が300±20ms(n=5)という過渡的中間状態が観察された(図5d)。
【0148】
電流-電圧(I-V)曲線を、付加生成物(M113N-RL2)6(T117C-D8RL3)1-βCDに関して計測した。この複合体のI-V曲線は±100mVの範囲内では(M113N-RL2)7・βCDのそれに類似しており、高度の正電位および負電位では幾分逸脱する(図5e)。
【0149】
2.4 βCDを逆向きに連結させる試み
共有結合したβCDに関して提唱した向きについてさらに調べ、確認するために、本発明者らは、X線構造で立証されているように、βCD-PDPとタンパク質との間の共有結合形成が、113突然変異が通常支えると考えられるシクロデキストリンの向きとは反対の向きをいずれの場合にも生じさせる、ヘテロ七量体(M113N-RL2)6(M113C-D8RL3)1および(M113F-RL2)6(T117C-D8RL2)1を調製した(図1)。
【0150】
(M113N-RL2)6(M113C-D8RL2)1の場合には、βCD-PDPとCys-113との共有結合は、各30分ずつの10回の試行後には起こらなかった。対照的に、ヘテロ七量体(M113F-RL2)6(T117C-D8RL3)1は、永続的な80%の電流遮断によって示されたように、βCD-PDPと反応した。前述した2つの場合と同じように、結合したβCDは±250mVの電位の上げ下げに耐え、シスチャンバーへの2mM DTTの添加によって離脱させることができた(提示せず)。推定される共有結合物である(M113F-RL2)6(T117C-D8RL3)1-βCDについて、25mM Tris・HCl、pH 8.0、1M KCl中、±100mVでの電気記録によって検討した(n=3)。単一チャネル電流(24.0pA)は(M113N-RL2)6(T117C-D8RL3)1-βCDのそれ(40.0pA)よりも小さく、正電位および陰電位の両方で頻回な完全遮断を呈した(データ非提示)。加えて、(M113F-RL2)6(T117C-D8RL3)1-βCDの脇を通過する電流は、(M113N-RL2)6(T117C-D8RL3)1-βCDで観察されたものよりもはるかにノイズが多く、このために、それを有機分析物の確率的センシングのために用いることはできない(下記参照)。
【0151】
2.5 共有結合したβCDアダプターを有する工学的に作製したαHL細孔を用いた分析物検出
新たに作り出した構築物をバイオセンサー検出素子として評価するために、本発明者らはモデル分析物である2-アダマンタンアミン(2-AdNH2)の確率的センシングを行った(12,15)。上記の通りに、インサイチューで、βCDアダプターを(M113F-RL2)6(M113C-D8RL2)1または(M113N-RL2)6(T117C-D8RL3)1と結合させた。続いて、2-アダマンタンアミン・HClをシスチャンバーまたはトランスチャンバーのいずれかに添加し、過渡的な電流の準安定状態(substate)を観察した(図7)。これらのイベントは、工学的に作製したαHL細孔内での個々の2-AdNH2・βCD複合体の形成および解離を表す。平均滞留時間(τoff)は、2-AdNH2を非共有結合複合体(M113F-RL2)7・βCDおよび(M113N-RL2)7・βCDに添加した場合に観察されたものと同程度であった(以下の表5)。τoffのわずかな違いはおそらくリンカーによる干渉によって生じたものと考えられる。
【0152】
(表5)βCDおよびαHLの共有結合性および非共有結合性の付加物による、2-アダマンタンアミン(2-AdNH2)の確率的センシング
条件:25mM Tris・HCl、pH 8.0、1M KCl、+100mVで、30μMの2-AdNH2・HCl(シスまたはトランス)を伴う。個々の実験の数は括弧内に示されている。
【0153】
2.6 共有結合したアダプターを有するαHL細孔を用いた長寿命結合イベントの検討
共有結合させたアダプターを用いることで、非共有結合的に結合したアダプターを用いて検討することがより困難な、長寿命イベントおよび稀なイベントという2種類のイベントの動態を研究することが可能である。シス-リトコール酸はβCDに関して知られている最も強固な結合分子である(Yang, Z., and Breslow, R. (1997) Tetrahedron Lett. 38, 6171-6172)。このため、本発明者らはこの分子を選び、長時間にわたる結合イベントを研究するために新たに構築した細孔の試験を行った。(M113F-RL2)6(M113C-D8RL2)1-βCDを選択した理由は、分析物の非存在下でのバックグラウンドシグナルのノイズが少ないためである(例えば、図7参照)。シス-リトコール酸(50μM、図8a)を25mM Tris・HCl、pH 8.0、1M KCl中、+100mVのシスチャンバーに導入したところ(n=3)、驚いたことに、全くシグナルが観察されなかった。これはおそらく、βCDをバレル壁に結合させるリンカーによる(M113F-RL2)6(M113C-D8RL2)1-βCDのシス側に対する結合の立体障害に起因すると考えられる。シス-リトコール酸(50μM)を-100mVのトランスチャンバーに導入したところ(n=3)、本発明者らは多くの極めて長寿命のイベントを観察した。これらのイベントは98%の電流遮断を示し、各イベントの最初の200±100ms、およびイベントの最後の150±100msの間は92%の遮断であった(図8b、図9)。イベントの平均τoffは8.6±7.8sであり(イベントの数=60;範囲0.5s〜80s)、これは同じ条件下の(M113F-RL2)7・βCD複合体におけるβCDに関するτoff(0.68±0.12s)よりもかなり長寿命である、すなわち、βCDは、リトコール酸結合イベント中には非共有結合性の(M113F-RL2)7・βCDから最も高い頻度で解離すると考えられる。これは、長寿命の結合イベントを検討することに関しての、非共有結合複合体を上回る、共有結合させたアダプターの利点を示している。
【0154】
2.7 高温での、共有結合したアダプターを有するαHL細孔
本発明者らは、CDの滞留時間が減少する高温では、共有結合したCDを有するαHL細孔が、誘導体化されていない細孔よりも優れた働きをする可能性があると推測した(Kang, X., Gu, L.-Q., Cheley, S., and Bayley, H. (2005) Angew. Chem. Int. Ed. Engl., 1495-1499)。最高55℃までの(M113F)6(M113C-D8RL2)1-βCDの単一チャネル電流軌跡を得た。単一チャネル電流は温度に直線的に依存した(図10)。25mM Tris・HCl、pH 8.0、1M KCl中、+100mVでI(pA)=14.3+0.54T(℃)であり、これは非共有結合的に結合したβCDを有するαHL細孔で以前に観察されたものと同様の温度依存性である(Kang, X., Gu, L.-Q., Cheley, S., and Bayley, H. (2005) Angew. Chem. Int. Ed. Engl., 1495-1499)。これらの実験中に、(M113F-RL2)6(M113C-D8RL2)1-βCD細孔からのβCDの解離は観察されなかった。例えば、(M113F-RL2)6(M113C-D8RL2)1-βCDの脇を通過する電流は、55℃での5分間の測定中に一定に保たれたが、一方、(M113F-RL2)6(M113C-D8RL2)1と結合したβCDに関するτoffは同じ条件下で56±3.8msに過ぎなかった。これらのデータは、共有結合したCDを有するαHL細孔が、極限条件下の水溶液中での確率的センシングを可能とすることを示している。
【0155】
2.8 共有結合したアダプターを有するαHL細孔におけるイオン選択性の永続的な改変
αHL細孔内に停留した非共有結合性の分子アダプターは、イオン輸送に関する荷電選択性を劇的に改変することができる(Gu, L.-Q., Dalla Serra, M., Vincent, J. B., Vigh, G., Cheley, S., Braha, O., and Bayley, H. (2000) Proc.Natl.Acad.Sci. USA 97, 3959-3964)。例えば、pH 7.5では、WT-αHLの荷電選択性(PK+/PCl-、KCl:シス200mM、トランス1000mM)は、βCDが内腔に停留するようになった後には0.55±0.02から0.25±0.01に変化し、すなわち細孔はよりアニオン選択的になる(Gu, L.-Q., Dalla Serra, M., Vincent, J. B., Vigh, G., Cheley, S., Braha, 0., and Bayley, H. (2000) Proc.Natl.Acad.Sct. USA 97, 3959-3964)。このため、本発明者らは、βバレルの内側に共有結合したβCDは、αHL細孔のイオン選択性を永続的に改変すると予想した。本発明者らは、(M113F-RL2)6(M113C-D8RL2)1および(M113F-RL2)6(M113C-D8RL2)1-βCDを被験事例として採用し、シス/トランスおよびトランス/シスKCl勾配の両方において記録した単一チャネル電流に関してI-V曲線を作成した(図11)。非誘導体化細孔および誘導体化細孔の両方の、測定されたコンダクタンス値(g)および逆転電位(Vr)、ならびにGoldman-Hodgkin-Katz式から計算された荷電選択性(PK+/PCl-)(Hille, B. (2001) Ion channels of excitable membranes, 3rd edition,, Sinauer, Sunderland, MA, USA)を、表6(下記)に示している。
【0156】
(表6)(M113F-RL2)6(M113C-D8RL2)1および(M113F-RL2)6(M113C-D8RL2)1-βCDの逆転電位(Vr)、計算された荷電選択性(PK+/PCl-)およびコンダクタンス値(g)
各項目に関して、3回の別々の実験を行った。逆転電位(Vr)は、記述した条件下での平均値(n=3)である。荷電選択性(PK+/PCl-、n=3)およびコンダクタンス値(n=3)は、平均±SDとして引用している。PK+/PCl-は、活性および実験上のVr値(20)を用いることにより、Goldman-Hodgkin-Katz式を用いて計算した。
* 25mM Tris・HCl、pH 8.0。塩濃度(KCl)はmM単位で提示されている。
† いずれのチャンバーにおいても、+100mV、1M KCl、25mM Tris・HCl、pH 8.0
【0157】
共有結合させた分子アダプターはαHL細孔の荷電選択性を実際に改変し、この効果は永続的であった。この場合も、細孔はよりアニオン選択性となり、PK+/PCl-について観察された変化は、非共有結合的に結合したβCDで認められたものと同程度である(Gu, L.-Q., Dalla Serra, M., Vincent, J. B., Vigh, G., Cheley, S., Braha, 0., and Bayley, H. (2000) Proc.Natl.Acad.Sct. USA 97, 3959-3964)。この特徴は、イオン浸透の基礎研究およびバイオテクノロジーにおける応用のいずれについても、膜チャネルのデノボ設計において有用なはずであり、修飾された特性を有する細孔のさまざまな組み立てには大きな有用性があると考えられる(Hwang, W. L., Holden, M. A., White, S., and Bayley, H. (2007) submitted for publication;およびHolden, M. A., Needham, D., and Bayley, H. (2007) J Am Chem Soc, in press)。
【0158】
3.結論
確率的検出のための有機分析物の結合を可能にするためのαHL細孔の工学的操作はごく稀にしか成功していないが(Guan, X., Gu, L.-Q., Cheley, S., Braha, O., and Bayley, H. (2005) ChemBioChem 6, 1875-1881)、非共有結合性の分子アダプターは、絶え間なく起こる細孔との会合および解離によって課せられるある種の制約にもかかわらず、この点に関して有用であることが立証されている。後者の問題を改善するために、本発明者らは今回、βCDアダプターをαHL細孔の内側に共有結合させた。さらに、βCDの向きに関して制御して、すなわち、βCDの第一級ヒドロキシルを細孔のトランス入口またはシス入口のいずれかに面するようにさせて、結合を行わせた。共有結合は検出に欠落がないことを意味し、βCDの向きに対する制御は、ある種の分析物が2つの入口のうち一方のみを通ってβCD中空部と結合する、またはそれらが入る側に応じて異なる様式で結合するという可能性が高いことから重要である(Kang, X. F., Cheley, S., Guan, X., and Bayley, H. (2006) J Am Chem Soc 128(33),10684-10685)。共有結合させたβCDを有するαHL細孔の確率的センシングにおける有用性を例示するために、本発明者らは本明細書で、細孔を用いて、βCD内部での滞留時間が長寿命である分析物を検出しうることを示している。この場合には、βCDは通常、分析物がβCDから解離する前に細孔から解離して、信号の情報量を劣化させると考えられる(Braha, O., Webb, J., Gu, L.-Q., Kim, K., and Bayley, H. (2005) ChemPhysChem 6, 889-892)。絶え間なく起こるCDの会合および解離の影響は高温で悪化するが(Kang, X., Gu, L.-Q., Cheley, S., and Bayley, H. (2005) Angew.Chem.Int.Ed.Engl., 1495-1499)、本明細書で実証しているように共有結合はこの問題も改善する。本発明者らはまた、共有結合させたβCDを有するαHL細孔がイオン選択性を永続的に改変し、それが例えば、ナノバッテリー(nanobattery)を構築するために有用な可能性があることも示している(Hwang, W. L., Holden, M. A., White, S., and Bayley, H. (2007) submitted for publication;およびHolden, M. A., Needham, D., and Bayley, H. (2007) J Am Chem Soc, in press)。
【0159】
また、単分子DNAシークエンシング(Bayley, H. (2006) Curr Opin Chem Biol 10, 628-637)も、共有結合したβCDを含むタンパク質ナノポアの使用によって恩典を受けると考えられる。1つのアプローチでは、デオキシリボヌクレオシド5'-一リン酸を、連続移動性エキソヌクレアーゼによって個々のDNAストランドから遊離させる(Jett, J. H., Keller, R. A., Martin, J. C., Marrone, B. L., Moyzis, R. K., Ratliff, R. L., Seitzinger, N. K., Shera, E. B., and Stewart, C. C. (1989) J.Biomol.Struct.Dynam. 7, 301-309)。元々明示されたものでは、完全に置換されて転写されたDNAストランドから蛍光性塩基類似体が遊離されるが、それは遂行するのが極めて困難なことが判明している。しかし、本発明者らは最近、非共有結合性のαHL(M113R-RL2)7・am7βCD複合体(式中、am7βCDはヘプタキス-(6-デオキシ-6-アミノ)-β-シクロデキストリンである)を用いて、4種類のデオキシリボヌクレオシド5'-一リン酸を識別しうることを示した(Astier, Y., Braha, O., and Bayley, H. (2006) J Am Chem Soc 128(5), 1705-1710)。これは一歩前進ではあるが、シークエンシングを成功させるためには、エキソヌクレアーゼによって遊離させられるすべての塩基を検出素子によって捕捉して同定しなければならない。このため、分子アダプターを有するαHL細孔を用いるのであれば、共有結合させたアダプターが、シークエンシングにおける欠落を回避する助けになると考えられる。
【0160】
以下の表7〜16は、配列情報を示している。
【0161】
(表7)SEQ ID NO:1および2‐黄色ブドウ球菌由来の野生型α-ヘモリシン
SEQ ID NO:2はSEQ ID NO:1の残基1〜879によってコードされる
【0162】
(表8)SEQ ID NO:3および4‐α-ヘモリシンM113H-RL2
SEQ ID NO:4はSEQ ID NO:3の残基39〜920によってコードされる
n=a、c、gまたはt
場所5にあるXはLeuまたはPheを表す
場所7にあるXはTyr、Trp、Cys、Ser、LeuまたはPheを表す
【0163】
(表9)SEQ ID NO:5および6‐α-ヘモリシンM113K-RL2
SEQ ID NO:6はSEQ ID NO:5の残基39〜920によってコードされる
n=a、c、gまたはt
場所5にあるXはLeuまたはPheを表す
場所7にあるXはTyr、Trp、Cys、Ser、LeuまたはPheを表す
【0164】
(表10)SEQ ID NO:7および8‐α-ヘモリシンM113R-RL2
SEQ ID NO:8はSEQ ID NO:7の残基39〜920によってコードされる
n=a、c、gまたはt
場所5にあるXはLeuまたはPheを表す
場所7にあるXはTyr、Trp、Cys、Ser、LeuまたはPheを表す
【0165】
(表11)SEQ ID NO:9および10‐α-ヘモリシンM113F-RL2
SEQ ID NO:10はSEQ ID NO:9の残基39〜920によってコードされる
n=a、c、gまたはt
場所5にあるXはLeuまたはPheを表す
場所7にあるXはTyr、Trp、Cys、Ser、LeuまたはPheを表す
【0166】
(表12)SEQ ID NO:11および12‐α-ヘモリシンM113N-RL2
SEQ ID NO:12はSEQ ID NO:11の残基39〜920によってコードされる
n=a、c、gまたはt
場所5にあるXはLeuまたはPheを表す
場所7にあるXはTyr、Trp、Cys、Ser、LeuまたはPheを表す
【0167】
(表13)SEQ ID NO:13および14‐α-ヘモリシンM113C-D8RL2
SEQ ID NO:14はSEQ ID NO:13の残基39〜944によってコードされる
n=a、c、gまたはt
場所5にあるXはLeuまたはPheを表す
場所7にあるXはTyr、Trp、Cys、Ser、LeuまたはPheを表す
【0168】
(表14)SEQ ID NO:15および16‐α-ヘモリシンT117C-D8RL3
SEQ ID NO:16はSEQ ID NO:15の残基13〜918によってコードされる
【0169】
(表15)SEQ ID NO:17‐λエキソヌクレアーゼ
【0170】
(表16)SEQ ID NO:18‐α-ヘモリシン-D8RL3
【技術分野】
【0001】
発明の分野
本発明は、確率的センシング(stochastic sensing)によって分析物を検出するために有用な膜貫通タンパク質細孔に関する。分子アダプターが、分析物の検出を可能する向きで、細孔と結合している。
【背景技術】
【0002】
発明の背景
確率的検出は、分析物分子と受容体との間の個々の結合イベントの観察に依拠する、センシングのためのアプローチである。確率的センサーは、ナノメートル寸法の単一の細孔を絶縁膜中に配置して、細孔を通しての電位駆動(voltage-driven)イオン輸送を分析物分子の存在下で測定することによって作り出すことができる。電流の揺らぎの発生頻度から、細孔の中に結合する分析物の濃度が明らかとなる。分析物の実体は、その特徴的な電流サイン(current signature)、とりわけ電流遮断の持続時間および程度を通じて明らかとなる(Braha, O., Walker, B., Cheley, S., Kasianowicz, J. J., Song, L., Gouaux, J. E., and Bayley, H. (1997) Chem.Biol. 4, 497-505(非特許文献1);およびBayley, H., and Cremer, P. S. (2001) Nature 413, 226-230(非特許文献2))。
【0003】
細菌性細孔形成毒素α-ヘモリシン(αHL)の工学的に作製した型のものは、多くのクラスの分子の確率的センシングのために用いられている(Bayley, H., and Cremer, P. S. (2001) Nature 413, 226-230(非特許文献2);Shin, S.-H., Luchian, T., Cheley, S., Braha, O., and Bayley, H. (2002) Angew.Chem.Int.Ed. 41, 3707-3709(非特許文献3);およびGuan, X., Gu, L.-Q., Cheley, S., Braha, O., and Bayley, H. (2005) ChemBioChem 6, 1875-1881(非特許文献4))。これらの研究の過程で、低分子有機分析物と結合するようにαHLを直接的に工学的に作製するという取り組みは負担が大きく、しかも成功例は稀であることが見いだされた(Guan, X., Gu, L.-Q., Cheley, S., Braha, O., and Bayley, H. (2005) ChemBioChem 6, 1875-1881(非特許文献4))。幸いなことに、非共有結合的に結合させた分子アダプター、とりわけシクロデキストリン(Gu, L.-Q., Braha, O., Conlan, S., Cheley, S., and Bayley, H. (1999) Nature 398, 686-690(非特許文献5))、さらには環状ペプチド(Sanchez-Quesada, J., Ghadiri, M. R., Bayley, H., and Braha, O. (2000) J.Am.Chem.Soc. 122, 11758-11766(非特許文献6))およびカボチャ型分子(cucurbituril)(Braha, O., Webb, J., Gu, L.-Q., Kim, K., and Bayley, H. (2005) ChemPhysChem 6, 889-892(非特許文献7))を利用する、1つの異なる戦略が発見された。シクロデキストリンはαHL細孔内に一過性に停留して、実質的ではあるが不完全なチャネル遮断を生じさせる。シクロデキストリンの疎水性内部の中に結合する有機分析物は、この遮断を増強して、分析物の検出を可能にする(Gu, L.-Q., Braha, O., Conlan, S., Cheley, S., and Bayley, H. (1999) Nature 398, 686-690(非特許文献5))。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Braha, O., Walker, B., Cheley, S., Kasianowicz, J. J., Song, L., Gouaux, J. E., and Bayley, H. (1997) Chem.Biol. 4, 497-505
【非特許文献2】Bayley, H., and Cremer, P. S. (2001) Nature 413, 226-230
【非特許文献3】Shin, S.-H., Luchian, T., Cheley, S., Braha, O., and Bayley, H. (2002) Angew.Chem.Int.Ed. 41, 3707-3709
【非特許文献4】Guan, X., Gu, L.-Q., Cheley, S., Braha, O., and Bayley, H. (2005) ChemBioChem 6, 1875-1881
【非特許文献5】Gu, L.-Q., Braha, O., Conlan, S., Cheley, S., and Bayley, H. (1999) Nature 398, 686-690
【非特許文献6】Sanchez-Quesada, J., Ghadiri, M. R., Bayley, H., and Braha, O. (2000) J.Am.Chem.Soc. 122, 11758-11766
【非特許文献7】Braha, O., Webb, J., Gu, L.-Q., Kim, K., and Bayley, H. (2005) ChemPhysChem 6, 889-892
【発明の概要】
【0005】
本発明者らは、驚いたことに、分子アダプターを、膜貫通タンパク質細孔と、ある特定の向きで共有結合させることができることを実証した。本発明者らはまた、驚いたことに、分子アダプターを、膜貫通細孔と、分析物が細孔を通して流れる電流に対してその分析物に特異的な様式で影響を及ぼすことを可能にする向きで共有結合させることができることも実証した。したがって、分子アダプターは、確率的センシングを介して分析物を検出するために細孔を用いることを可能にする向きで、細孔と共有結合させることができる。細孔は好ましくは、分子アダプターのその配向を助長するように修飾(modify)される。
【0006】
分子アダプターの固定的な性質は、分析物が細孔と相互作用する時には常に、特徴的な電流が細孔を通して流れることを意味する。その結果として、本発明の膜貫通タンパク質細孔は、確率的センシングのための、特にヌクレオチドの検出または核酸のシークエンシングのための有用なツールとなる。
【0007】
したがって、本発明は、試料中の分析物の検出に用いるための膜貫通タンパク質細孔であって、細孔と分析物との間の相互作用を助長する分子アダプターを含んでおり、そのアダプターが、分析物を細孔を用いて検出することを可能にする向きで細孔と共有結合している細孔を提供する。
【0008】
本発明はまた、以下も提供する:
‐本発明の膜貫通タンパク質細孔を作製する方法であって:
(a)膜貫通タンパク質細孔に、細孔と分析物との間の相互作用を助長する分子アダプターを共有結合させる段階;および
(b)該アダプターが、分析物を細孔を用いて検出することを可能にする向きで細孔と結合しているか否かを判定する段階、
を含む方法;
‐分析物の有無を判定する方法であって:
(a)分析物を、本発明の膜貫通タンパク質細孔と、分析物が細孔と相互作用するように接触させる段階;および
(b)相互作用の間に細孔を通過する電流を測定し、それによって分析物の有無を判定する段階、
を含む方法;
‐個々のヌクレオチドを同定する方法であって:
(a)ヌクレオチドを、本発明の膜貫通タンパク質細孔と、ヌクレオチドが細孔と相互作用するように接触させる段階;および
(b)相互作用の間に細孔を通過する電流を測定し、それによってヌクレオチドの実体を決定する段階、
を含む方法;
‐標的核酸配列のシークエンシングの方法であって:
(a)個々のヌクレオチドを、標的配列の一方の末端から、連続移動性(processive)エキソヌクレアーゼを用いて消化する段階;
(b)ヌクレオチドを、本発明の膜貫通タンパク質細孔と、ヌクレオチドがアダプターと相互作用するように接触させる段階;
(c)相互作用の間に細孔を通過する電流を測定し、それによってヌクレオチドの実体を決定する段階;および
(d)段階(a)から(c)までを核酸配列の同じ末端で繰り返し、それによって核酸の配列を決定する段階、
を含む方法;ならびに
‐本発明による膜貫通タンパク質細孔および連続移動性エキソヌクレアーゼを含む、核酸のシークエンシングのためのキット。
【図面の簡単な説明】
【0009】
【図1】pdbファイル(Montoya and Gouaux, unpublished)から生成された(M113F-RL2)7・βCDおよび(M113N-RL2)7・βCDのX線構造を示している。a)αHL細孔の側面図。アミノ酸108〜120および138〜150が黄色で強調表示されている。b)(M113F)7・βCD。αHLのアミノ酸108〜120は黄色で描かれており、βCDは青色のスティックで示されている。Phe-113およびLys-147の側鎖もスティック形態で示されている。βCDの第一級ヒドロキシルのO原子とPhe-113の最近接フェニル環の中心との間の距離は約6.5Åである(点線)。c)(M113N-RL2)7・βCD。αHLのアミノ酸108〜120は黄色で描かれており、βCDは青色のスティックで示されている。Asn-113、Lys-147およびThr-117の側鎖もスティック形態で示されている。βCDの第一級ヒドロキシルのO原子とThr-117の最近接O原子との間の距離は約6.0Åである(点線)。
【図2】βCD-PDPの化学構造およびスティック表示を示している。分子はChemDraw 3Dで描写し、続いてPyMolにてレンダリングした。
【図3】非共有結合複合体αHL(M113F-RL2)7・βCDおよび共有結合物αHL(M113F-RL2)6(M113C-D8RL2)1-βCDの特性の比較を示している。各実験は少なくとも6回繰り返した。a)突然変異型αHL細孔(M113F-RL2)7のアミノ酸108〜120のカートゥーン表示。Phe-113残基はスティック形態で表示されている。25mM Tris・HCl、pH 8.0、1M KCl中、+100mVでの(M113F-RL2)7細孔からの単一チャネル電流軌跡。b)(M113F-RL2)7・βCDのX線構造によって判明した、βCDが結合したαHL(M113F-RL2)7のカートゥーン表示。矢印は、βCDが細孔を自由に出入りして動くことを指し示している。この電流軌跡は、最初は占有されていなかった細孔へのβCDの結合、およびその後の解離を指し示している(平均τoff=316±62ms)。c)αHL(M113F-RL2)6(M113C-D8RL2)1のアミノ酸108〜120のカートゥーン表示。Phe-113残基はスティック形態で表示されており、Cys-113は褐色に彩色されて同じくスティックで表示されている。(c)の拡大図を図4に見ることができる。矢印は、βCD-PDPが細孔に入ると第一級ヒドロキシルがPhe-113の側鎖と相互作用して、Cys-113がβCD-PDPのピリジルジスルフィドと反応してジスルフィド結合を形成することを指し示している。電流軌跡は反応の間の電流振幅の変化を示している。電流遮断85%の過渡状態に印を付している(*)。d)ジチオトレイトール(DTT)による(M113F-RL2)6(M113C-D8RL2)1-βCDの切断。矢印は、DTTによるジスルフィド結合の切断後にシクロデキストリン誘導体が113位から離脱することを指し示している。電流軌跡は切断の間の電流振幅の変化を示している。電流遮断98%の過渡状態に印を付している(*)。e)(M113F-RL2)7・βCD(τ)およびαHL(M113F-RL2)6(M113C-D8RL2)1-βCD(■)に関する単一チャネルI-V曲線。条件:25mM Tris・HCl、pH 8.0、1M KCl。
【図4】図3cの拡大図を示している。αHL(M113F-RL2)6(M113C-D8RL2)1のアミノ酸108〜120のカートゥーン表示。Phe-113残基はスティック形態で表示され、Cys-113は褐色に彩色されて同じくスティックで表示されている。
【図5】非共有結合複合体αHL(M113N-RL2)7-βCDおよび共有結合物αHL(M113N-RL2)6(T117C-D8RL3)1-βCDの特性の比較を示している。各実験は少なくとも6回繰り返した。a)突然変異型αHL細孔(M113N-RL2)7のアミノ酸108〜120のカートゥーン表示。Asn-113残基はスティック形態で表示されている。25mM Tris・HCl、pH 8.0、1M KCl中、-100mVでの、(M113N-RL2)7細孔からの単一チャネル電流の軌跡。b)(M113N-RL2)7・βCDのX線構造によって判明した、βCDが結合したαHL(M113N-RL2)7のカートゥーン表示。矢印は、βCDが細孔を自由に出入りして動くことを指し示している。この電流軌跡は、最初は占有されていなかった細孔へのβCDの結合、およびその後の解離を指し示している(平均τoff=10.7±1.5s)。c)αHL(M113N-RL2)6(T117C-D8RL3)1のアミノ酸108〜120のカートゥーン表示。Asn-113残基はスティック形態で表示され、Cys-117は褐色に彩色されて同じくスティックで表示されている。図5cの拡大図を図6に見ることができる。矢印は、βCD-PDPが細孔に入ると第二級ヒドロキシルがAsn-113の側鎖と相互作用し、Cys-117がβCD-PDPのピリジルジスルフィドと反応してジスルフィド結合を形成することを指し示している。この電流軌跡は、Cys-117とβCD-PDPのピリジルジスルフィドとの間で反応が起こった時の電流振幅の変化を示している。電流遮断80%の過渡状態に印を付している(*)。d)DTTによる(M113N-RL2)6(T117C-D8RL3)1-βCDの切断。矢印は、ジスルフィド結合の切断後にシクロデキストリン誘導体が117位から離脱することを指し示している。電流軌跡は切断の間の電流振幅の変化を示している。電流遮断90%の過渡状態に印を付している(*)。e)(M113N-RL2)7・βCD(τ)およびαHL(M113N-RL2)6(T117C-D8RL3)1-βCD(■)に関する単一チャネルI-V曲線。条件:25mM Tris・HCl、pH 8.0、1M KCl。
【図6】図5cの拡大図を示している。αHL(M113N-RL2)6(T117C-D8RL3)1のアミノ酸108〜120のカートゥーン表示。Asn-113残基はスティック形態で表示され、Cys-117は褐色に彩色されて同じくスティックで表示されている。
【図7】共有結合したβCDを含むαHL細孔による2-アダマンタンアミンの検出を示している。a)25mM Tris・HCl、pH 8.0、1M KCl中、+100mVでの、単一の(M113F-RL2)6(M113C-D8RL2)1-βCD細孔からの電流軌跡。b)シスチャンバーにおける30μMの2-アダマンタンアミンの存在下での電流軌跡。c)トランスチャンバーにおける30μMの2-アダマンタンアミンの下での電流軌跡。d)25mM Tris・HCl、pH 8.0、1M KCl中、+100mVでの、単一の(M113N-RL2)6(T117C-D8RL3)1-βCD細孔からの電流軌跡。e)シスチャンバーにおける30μMの2-アダマンタンアミンの下での電流軌跡。f)トランスチャンバーにおける30μM 2-アダマンタンアミンの下での電流軌跡。
【図8】シス-リトコール酸と(M113F-RL2)6(M113C-D8RL2)1-βCDとの長寿命結合イベントを示している。a)分子アダプター(M113F-RL2)6(M113C-D8RL2)1-βCDが結合したαHL、およびシス-リトコール酸の構造。b)25mM Tris・HCl、pH 8.0、1M KCl中、-100mVでの、シス-リトコール酸(50μM)と(M113F-RL2)6(M113C-D8RL2)1-βCDとの特徴的な結合イベント。平均τoff=8.6±7.8s(イベントの数=60)。レベル1:(M113F-RL2)6(M113C-D8RL2)1-βCD;レベル2:(M113F-RL2)6(M113C-D8RL2)1-βCD・リトコール酸、遮断92%;レベル3:(M113F-RL2)6(M113C-D8RL2)1-βCD・リトコール酸、遮断98%。
【図9】(M113F-RL2)6(M113C-D8RL2)1-βCDおよび(M113F-RL2)7がいずれも50μMのβCDの存在下でシス-リトコール酸の非存在下で閉鎖を示し、それはシス-リトコール酸結合と混同される可能性があるもののイベントはより短いことを示している。a)図6と同じ条件下でのシス-リトコール酸の非存在下における(M113F-RL2)6(M113C-D8RL2)1-βCD。レベル1:(M113F-RL2)6(M113C-D8RL2)1-βCD;レベル2:細孔の自然発生的ゲート開閉。イベントの持続時間は30〜300msの間でさまざまであった。b)25mM Tris・HCl、pH 8.0、1M KCl中、-100mVでの、50μMのβCDの存在下における(M113F-RL2)7。τoff=0.68±0.12s。レベル1:(M113F-RL2)7;レベル2:(M113F-RL2)7・βCD;レベル3:細孔複合体(M113F-RL2)7・βCDの自然発生的ゲート開閉。
【図10】25mM Tris・HCl、pH 8.0、1M KCl中、+100mVでの、(M113F-RL2)6(M113C-D8RL2)1-βCDの単一チャネル電流の変動を温度とともに示している。プロット中に示されている値は、3つの異なる実験の平均値である。単一チャネル電流は温度に直線的に依存する。I(pA)=14.307+0.5402T(℃)。
【図11a】(M113F-RL2)6(M113C-D8RL2)1-βCDおよび(M113N)6(T117C-D8RL3)1-βCDのイオン選択性を示している。シス:200mM KCl;トランス:1000mM KCl(n=3)を伴う25mM Tris・HCl、pH 8.0中で行った記録に基づく、(M113F-RL2)6(M113C-D8RL2)1( )および(M113F-RL2)6(M113C-D8RL2)1-βCD(■)に関するI-V曲線。逆転電位(Vr)には矢印を付している。
【図11b】(M113F-RL2)6(M113C-D8RL2)1-βCDおよび(M113N)6(T117C-D8RL3)1-βCDのイオン選択性を示している。シス:1000mM KCl;トランス:200mM KCl(n=3)を伴う25mM Tris・HCl、pH 8.0中で行った記録に基づく、(M113F-RL2)6(M113C-D8RL2)1( )および(M113F-RL2)6(M113C-D8RL2)1-βCD(■)に関するI-V曲線。逆転電位(Vr)には矢印を付している。
【図12】αHL-D8RL3(オクタ-アスパラギン酸尾部(octa-aspartate tail)を含む)をコードする配列を示している。この配列はSEQ ID NO:18に示されている。αHL-D8RL3をコードする配列(SEQ ID NO:18)は、実施例で用いているT117C-D8RL3をコードするSEQ ID NO:15のヌクレオチド13〜918と、SEQ ID NO:15が残基361〜363にTGC(ACGではなく)を有する点を除いて同一である。
【0010】
配列表フリーテキスト
配列の説明
SEQ ID NO:1は、野生型α-ヘモリシンの1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:2は、野生型α-ヘモリシンの1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:3は、α-ヘモリシンM113H-RL2の1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:4は、α-ヘモリシンM113H-RL2の1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:5は、α-ヘモリシンM113K-RL2の1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:6は、α-ヘモリシンM113K-RL2の1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:7は、α-ヘモリシンM113R-RL2の1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:8は、α-ヘモリシンM113R-RL2の1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:9は、実施例で用いているα-ヘモリシンM113F-RL2の1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:10は、実施例で用いているα-ヘモリシンM113F-RL2の1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:11は、実施例で用いているα-ヘモリシンM113N-RL2の1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:12は、実施例で用いているα-ヘモリシンM113N-RL2の1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:13は、実施例で用いているα-ヘモリシンM113C-D8RL2の1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:14は、実施例で用いているα-ヘモリシンM113C-D8RL2の1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:15は、実施例で用いているα-ヘモリシンT117C-D8RL3の1つのサブユニットのアミノ酸配列を示している。
SEQ ID NO:16は、実施例で用いているα-ヘモリシンT117C-D8RL3の1つのサブユニットをコードするポリヌクレオチド配列を示している。
SEQ ID NO:17は、λエキソヌクレアーゼのアミノ酸配列を示している。この配列は、集合して三量体となる3つの同一なサブユニットの1つである。
SEQ ID NO:18は、αHL-D8RL3(図12中の配列)をコードする配列を示している。
【発明を実施するための形態】
【0011】
発明の詳細な説明
開示される生成物および方法の種々の適用物は、当技術分野における具体的な要求に合わせて適応させうることが理解されるべきである。また、本明細書で用いる専門用語は特定の態様を説明することのみを目的としており、限定的であることは意図していないことも理解されるべきである。
【0012】
さらに、本明細書および添付の特許請求の範囲で用いる場合、単数形の「1つの(a)」、「1つの(an)」および「その(the)」は、その文脈で明らかに別の指示がなされない限り、複数のものに関する言及も含む。したがって、例えば、「1つの分析物(an analyte)」に対する言及は「複数の分析物(analytes)」を含み、「1つの膜貫通タンパク質細孔」に対する言及は2つまたはそれ以上のそのような細孔を含み、「1つの分子アダプター」に対する言及は2つまたはそれ以上のそのようなアダプターを含むなどである。
【0013】
本明細書で引用したすべての刊行物、特許および特許出願は、上記または下記のいずれであるかを問わず、その全体が参照により本明細書に組み入れられる。
【0014】
膜貫通タンパク質細孔
本発明は、分析物の検出に用いるための膜貫通タンパク質細孔に関する。膜貫通タンパク質細孔は、分析物との相互作用を助長する分子アダプターを含む。アダプターは、細孔と特定の向きで共有結合している。アダプターは、分析物と細孔との間の相互作用の間に、分析物が、細孔を通して流れる電流に対して、その分析物に特異的な様式で影響を及ぼすことを可能にするような向きにある。アダプターはしたがって、確率的センシングを介して分析物を検出するために細孔を用いることを可能にする向きで、細孔と共有結合している。
【0015】
本発明の膜貫通タンパク質細孔は、確率的センシングのための有用なツールである。分子アダプターの固定的な性質は、細孔から得られる信号が細孔のバレル内またはチャネル内の分析物の存在に完全に依存し、細孔からのアダプターの解離によっては影響されないことを意味する。換言すれば、アダプターの固定的な性質は、分析物が細孔と相互作用する時には常に、特徴的な電流が細孔を通して流れることを意味する。これはその結果として:(1)分析物の濃度が低い場合などのような、稀な相互作用イベントの検出;(2)分析物濃度が高い場合などのような、頻度の高い相互作用イベントの検出;(3)アダプターに対する分析物の親和性が高い場合などのような、長寿命(long-lived)相互作用イベントの検出;および(4)アダプターに対する分析物の親和性を測定しうるようにする分析物の滞留時間の測定、を可能にする、より高感度な系をもたらす。最も重要なこととして、信号に中断が全く存在しないことは、細孔の中に入ったすべての分析物が検出されることを意味する。そのような効率的な検出系は、核酸のシークエンシングを容易にする。これについて以下にさらに考察する。
【0016】
本発明の膜貫通タンパク質細孔にはほかにも利点がある。実施例の項に記載しているように、本発明の膜貫通タンパク質細孔は、最高55℃まで、または最高100℃までというような高温で機能することができる。これは、細孔が分析物を極限条件下で検出するために適していることを意味する。
【0017】
本発明の分子アダプターの固定的な性質は、それが細孔と無関係に分析物と自由に相互作用することはないことを意味する。これは干渉を減少させるともに、分析物の濃度のより正確な測定を得ることができることを意味する。
【0018】
実施例の項に記載しているように、細孔に対する分子アダプターの結合はそのイオン選択性を永続的に変化させる。このことは特定のイオン選択性を有する膜貫通タンパク質細孔の作製を可能にする。
【0019】
さらになお、本発明の分子アダプターの固定的な性質は細孔およびアダプターを一緒に保管することを可能にし、その結果、直ちに使用できる(ready-to-use)センサーの製造を可能にする。
【0020】
膜貫通タンパク質細孔
膜貫通タンパク質細孔とは、イオンが印加電圧に沿って膜の一方の側からもう一方に流れることを許容するポリペプチドのことである。細孔は好ましくは、分析物が印加電圧に沿って膜の一方の側からもう一方に流れることを許容する。
【0021】
細孔は、典型的にはオリゴマーである。細孔は好ましくは、6個、7個または8個のサブユニットといった数個の繰り返しサブユニットで構成される。細孔はより好ましくは七量体である。細孔は典型的には、イオンが流れることができるバレルまたはチャネルを含む。細孔のサブユニットは典型的には中心軸を取り囲み、ストランドを膜貫通性のβバレルもしくはチャネル、または膜貫通性のα-ヘリックス束もしくはチャネルへと導くのに寄与する。
【0022】
細孔のバレルまたはチャネルは典型的には、分析物との相互作用を助長するアミノ酸を含む。これらのアミノ酸は好ましくは、バレルまたはチャネルの狭窄部(constriction)の付近に位置する。ヌクレオチドまたは核酸の検出に用いるための細孔は、典型的には、アルギニン、リジンまたはヒスチジンといった、1つまたは複数の正に荷電したアミノ酸を含む。これらのアミノ酸は典型的には、ヌクレオチド中のリン酸基と相互作用することによって、またはヌクレオチド中の塩基とのπ-カチオン相互作用によって、細孔とヌクレオチドとの間の相互作用を助長する。ヌクレオチドまたは核酸の検出に用いるための細孔は、好ましくは、バレルまたはチャネルの狭窄部の付近に位置する、アルギニン、リジンまたはヒスチジンといった正に荷電したアミノ酸の環を有する。正に荷電した各アミノ酸は、典型的には、細孔サブユニットのそれぞれによって提供される。
【0023】
本発明に従って用いるための細孔は、β-バレル細孔またはα-ヘリックス束細孔でありうる。β-バレル細孔は、β-シートから形成されたバレルまたはチャネルを含む。適したβ-バレル細孔には、α-ヘモリシンおよびロイコシジンといったβ-毒素、ならびに恥垢菌(Mycobacterium smegmatis)ポーリンA(MspA)、外膜ポーリンF(OmpF)、外膜ポーリンG(OmpG)、外膜ホスホリパーゼA、およびナイセリア菌(Neisseria)自己輸送体リポタンパク質(NalP)といった細菌の外膜タンパク質/ポーリンが非限定的に含まれる。α-ヘリックス束細孔は、α-ヘリックスから形成されたバレルまたはチャネルを含む。適したα-ヘリックス束細孔には、内膜タンパク質およびα外膜タンパク質、例えばWZAなどが非限定的に含まれる。
【0024】
本発明に用いるために最も好ましい細孔は、α-ヘモリシンまたはその変異体である。α-ヘモリシン細孔は7個の同一なサブユニットで形成される(すなわち、それは七量体である)。α-ヘモリシンの1つのサブユニットの配列はSEQ ID NO:2に示されている。変異体は、7個のサブユニットのうち1つまたは複数がSEQ ID NO:2のものとは違いのあるアミノ酸配列を有し、かつ細孔活性を保っている七量体細孔である。変異体は、以下に考察するような、アダプターの共有結合またはアダプターの配向を助長する修飾(modification)を含んでもよい。
【0025】
変異体α-ヘモリシン中のサブユニットの1個、2個、3個、4個、5個、6個または7個が、SEQ ID NO:2のものとは違いのあるアミノ酸配列を有してもよい。変異体細孔の中の7個のサブユニットすべては同一であってもよいが典型的には異なり、これは、サブユニットの1つまたは複数が、以下に考察するようにアダプターの共有結合またはアダプターの配向を助長するように修飾されている場合には特にそうである。
【0026】
ヌクレオチドまたは核酸の検出に用いるためのα-ヘモリシンの好ましい変異体は、バレルまたはチャネルの狭窄部の付近に位置する、アルギニン、リジンまたはヒスチジンといった1つまたは複数の正に荷電したアミノ酸を有する。細孔は好ましくは、バレルまたはチャネルの狭窄部の付近に位置する、アルギニン、リジンまたはヒスチジンなどの4個、5個、6個または好ましくは7個の正に荷電したアミノ酸の環を有する。環内の各アミノ酸は、典型的には、変異体サブユニットのそれぞれによって提供される。適した変異体は、正に荷電したアミノ酸を各サブユニットの113位に含む。ヌクレオチドまたは核酸の検出に用いるための細孔は、好ましくは、SEQ ID NO:4に示されているような7個のサブユニットを含むα-ヘモリシン(M113K-RL2)7、または好ましくは、SEQ ID NO:6に示されているような7個のサブユニットを含むα-ヘモリシン(M113H-RL2)7、または最も好ましくは、SEQ ID NO:8に示されているような7個のサブユニットを含むα-ヘモリシン(M113R-RL2)7である。
【0027】
α-ヘモリシンの他の好ましい変異体は、バレルまたはチャネルの狭窄部の付近に位置する、アスパラギンなどの1つもしくは複数の非荷電アミノ酸、またはフェニルアラニンなどの1つもしくは複数の芳香族アミノ酸を有する。これには、β-CD-PDPと結合してそれを共有結合反応のために配向させること、および共有結合後のβ-CD-PDPの向きを維持することという目的がある。細孔は好ましくは、バレルまたはチャネルの狭窄部の付近に位置する、4個、5個、6個または好ましくは7個の非荷電アミノ酸または芳香族アミノ酸の環を有する。環内の各アミノ酸は、典型的には、変異体サブユニットのそれぞれによって提供される。適した変異体は、非荷電アミノ酸または芳香族アミノ酸を各サブユニットの113位に含む。本発明に従って用いるための細孔は、好ましくはα-ヘモリシン(M113F-RL2)6(M113C-D8RL2)1である。
【0028】
変異体は、生物体によって、例えばブドウ球菌(Staphylococcus bacterium)によって発現される、天然に存在する変異体であってよい。また、変異体には、組換え技術によって作製された非天然の変異体も含まれる。SEQ ID NO:2のアミノ酸配列の全長にわたって、変異体のサブユニットは好ましくは、その配列に対して、アミノ酸同一性に基づいて少なくとも50%相同であると考えられる。より好ましくは、サブユニットポリペプチドは、SEQ ID NO:2のアミノ酸配列に対して、配列全体にわたって、アミノ酸同一性に基づいて少なくとも55%、少なくとも60%、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、少なくとも85%、少なくとも90%、より好ましくは少なくとも95%、97%または99%相同であってよい。200個またはそれ以上、例えば230個、250個、270個または280個またはそれ以上の連続したアミノ酸にわたって、少なくとも80%の、例えば少なくとも85%、90%または95%のアミノ酸同一性が存在しうる(「厳密な相同性(hard homology)」)。
【0029】
アミノ酸置換を、例えば最大で1個、2個、3個、4個、5個、10個、20個または30個までの置換を、SEQ ID NO:2のアミノ酸配列に対して施すことができる。保存的置換は、例えば、以下の表1に従って施すことができる。
【0030】
(表1)保存的置換
第2列の中の同じブロック内の、好ましくは第3列の中の同じ行内のアミノ酸は、互いに置換することができる。
【0031】
SEQ ID NO:2のアミノ酸配列の1つまたは複数のアミノ酸残基を、代替的または追加的に、削除することができる。最大で1個、2個、3個、4個、5個、10個、20個もしくは30個またはそれ以上の残基を削除することができる。
【0032】
変異体は、SEQ ID NO:2の断片でできたサブユニットを含んでもよい。そのような断片は細孔形成活性を保っている。断片は、少なくとも50、100、200または250アミノ酸長でありうる。そのような断片は、キメラ性細孔を作製するために用いることができる。断片は好ましくは、SEQ ID NO:2の細孔形成ドメインを含む。
【0033】
変異体には、SEQ ID NO:2の断片または部分を含むキメラ性タンパク質細孔が含まれる。キメラ性タンパク質細孔は、それぞれがSEQ ID NO:2の断片または部分を含むサブユニットから形成される。キメラ性タンパク質細孔の細孔部分またはチャネル部分は、典型的には、SEQ ID NO:2の断片または部分によって形成される。
【0034】
1つまたは複数のアミノ酸を、代替的または追加的に、上記のポリペプチドに付加することができる。伸張は、SEQ ID NO:2のアミノ酸配列、またはそのポリペプチド変異体もしくは断片のN末端またはC末端に与えることができる。伸張は、極めて短くてもよく、例えば1〜10アミノ酸長であってもよい。または、伸張がより長く、例えば、最大で50または100アミノ酸であってもよい。担体タンパク質を、本発明によるアミノ酸配列と融合させてもよい。
【0035】
相同性を決定するためには、当技術分野における標準的な方法を用いることができる。例えば、UWGCG Packageは、相同性を計算するために、例えばそのデフォルト設定で用いることのできる、BESTFITプログラムを備えている(Devereux et al (1984) Nucleic Acids Research 12, p387-395)。PILEUPアルゴリズムおよびBLASTアルゴリズムは、例えば、Altschul S. F. (1993) J Mol Evol 36:290-300;Altschul, S.F et al (1990) J Mol Biol 215:403-10に記載されているように、相同性を計算するため、または配列を整列させるため(等価な残基または対応する配列を(典型的にはそれらのデフォルト設定で)同定することなど)に用いることができる。
【0036】
BLAST分析を行うためのソフトウエアは、National Center for Biotechnology Information(http://www.ncbi.nlm.nih.gov/)から公に入手可能である。このアルゴリズムでは、データベース配列中の同じ長さのワードとアラインメントを行った場合に何らかの正値の閾値スコアTと一致するか、またはそれを満たす、長さWの短いワードをクエリー配列中に同定することにより、高スコア配列ペア(high scoring sequence pair)(HSP)をまず同定する。Tは近隣ワードスコア閾値(neighbourhood word score threshold)と呼ばれる(Altschulら、前記)。これらの初期の近隣ワードでのヒットは、それらを含むHSPを見いだすための検索を開始するシードとなる。ワードヒットは、累積アラインメントスコアが増加する限り、各配列の両方向に対して延長される。各方向へのワードヒットの延長は以下の場合に停止する:累積アラインメントスコアが最大達成値に比べて量X低下した場合:1つもしくは複数の負スコアの残基アラインメントの蓄積のために累積スコアがゼロもしくはそれ未満になった場合;または配列のいずれかの末端に達した場合。BLASTアルゴリズムのパラメーターであるW、TおよびXは、アラインメントの感度および速度を決定する。BLASTプログラムはデフォルトとしてワード長(W)11、BLOSUM62スコアリングマトリックス(Henikoff and Henikoff (1992), Proc. Natl. Acad. Sci. USA 89: 10915-10919を参照)アラインメント(B)50、期待値(E)10、M=5、N=4および両ストランドの比較を用いる。
【0037】
BLASTアルゴリズムは、2つの配列間の類似性に関する統計分析も行う;例えば、Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90: 5873-5787を参照。BLASTアルゴリズムによって得られる類似性の指標の1つは最小合計確率(smallest sum probability)(P(N))であり、これは2つのアミノ酸配列間の一致が偶然に起こる確率の指標を与える。例えば、ある配列は、第1の配列と第2の配列との比較による最小合計確率が約1未満、好ましくは約0.1未満、より好ましくは約0.01未満、最も好ましくは約0.001未満の場合に、もう1つの配列と類似しているとみなされる。
【0038】
膜貫通タンパク質細孔は、例えば、それらの同定もしくは精製を助けるためのヒスチジン残基もしくはアスパラギン酸残基の付加によって、またはそのポリペプチドがそのような配列を天然には含まない場合に細胞からのそれらの分泌を促進するためのシグナル配列を付加することによって、修飾することができる。
【0039】
細孔は、顕在化標識(revealing label)で標識することができる。顕在化標識は、細孔を検出することを可能にする任意の適した標識であってよい。適した標識には、蛍光分子、放射性同位体、例えば125I、35Sなど、酵素、抗体、抗原、ポリヌクレオチド、およびビオチンなどのリガンドが非限定的に含まれる。
【0040】
細孔は、黄色ブドウ球菌(Staphylococcus aureus)などの細孔産生性の生物体から単離すること、または合成的に、もしくは組換え手段によって作製することができる。例えば、細孔をインビトロの翻訳および転写によって合成することができる。細孔のアミノ酸配列は、非天然のアミノ酸を含むように、または化合物の安定性を高めるように修飾することができる。細孔を合成的手段によって作製する場合には、そのようなアミノ酸を作製中に導入することができる。また、細孔を、合成的または組換え的な作製の後に改変することもできる。
【0041】
また、細孔をD-アミノ酸を用いて作製することもできる。これは、そのようなタンパク質またはペプチドを作製することに関して当技術分野で慣習的である。
【0042】
上記および下記において詳細に考察しているように、細孔は、アダプターの結合および配向を助長するための、1つまたは複数の特異的な修飾を含むことができる。細孔はまた、他の非特異的な修飾も、それらがアダプターの結合および配向と干渉しない限りは含んでよい。さまざまな非特異的な側鎖修飾が当技術分野で公知であり、細孔の側鎖に対して施すことができる。そのような修飾には、例えば、アルデヒドとの反応に続いてNaBH4による還元を行うことによるアミノ酸の還元的アルキル化、アセトイミド酸メチルによるアミド化、または無水酢酸によるアシル化が含まれる。
【0043】
組換え型の膜貫通細孔は、当技術分野で公知の標準的な方法を用いて作製することができる。細孔または細孔サブユニットをコードする核酸配列は、当技術分野における標準的な方法を用いて、単離して複製させることができる。細孔または細孔サブユニットをコードする核酸配列を、当技術分野において標準的な手法を用いて、細菌宿主細胞において発現させることもできる。細孔または細孔サブユニットは、組換え発現ベクターからのポリペプチドのインサイチュー発現によって細胞内で産生させることができる。発現ベクターは任意で、ポリペプチドの発現を制御するための誘導性プロモーターを保有する。
【0044】
細孔または細孔サブユニットをコードする核酸配列は、当技術分野における標準的な方法を用いて、単離して複製させることができる。染色体DNAを、細孔産生性の生物体、例えば黄色ブドウ球菌などから抽出する。細孔または細孔サブユニットをコードする遺伝子を、特異的プライマーを伴うPCRを用いて増幅する。増幅された配列を、続いて、クローニングベクターなどの複製可能な組換えベクター中に組み入れる。このベクターは、適合性のある宿主細胞内で核酸を複製させるために用いることができる。このように、細孔または細孔サブユニットをコードする核酸配列は、細孔または細孔サブユニットをコードするポリヌクレオチドを複製可能なベクター中に導入し、ベクターを適合性のある宿主細胞内に導入して、ベクターの複製をもたらす条件下で宿主細胞を増殖させることによって作ることができる。ベクターは宿主細胞から回収することができる。細孔または細孔サブユニットをコードするポリヌクレオチドのクローニングのために適した宿主細胞は当技術分野で公知であり、以下にさらに詳細に説明する。
【0045】
細孔または細孔サブユニットをコードする核酸配列は、適した発現ベクター中にクローニングすることができる。発現ベクター中で、細孔または細孔サブユニットをコードする核酸配列は、典型的には、宿主細胞によるコード配列の発現をもたらすことのできる制御配列と機能的に連結される。そのような発現ベクターは、細孔または細孔サブユニットを発現させるために用いることができる。
【0046】
「機能的に連結された」という用語は、記載された成分が、それらの意図した様式でそれらが機能することを可能にする関係で並列されていることを指す。コード配列と「機能的に連結された」制御配列は、制御配列と適合性のある条件下でコード配列の発現が達成されるような様式で連結されている。同じまたは異なる、細孔または細孔サブユニットの遺伝子の複数のコピーを、ベクター中に導入することができる。
【0047】
続いて、発現ベクターを適した宿主細胞内に導入することができる。このため、1つの種類のサブユニットのみを含む単量体性の細孔またはオリゴマー性の細孔の場合には、本発明の方法は、細孔または細孔サブユニットをコードする核酸配列を発現ベクター中に導入して、ベクターを適合性のある細菌宿主細胞内に導入して、細孔または細孔サブユニットをコードする核酸配列の発現をもたらす条件下で宿主細胞を増殖させることによって作製された細胞に対して実施することができる。組換え的に発現された細孔サブユニットは、宿主細胞膜の中で細孔へと自己集合すると考えられる。または、このようにして作製された組換え細孔を宿主細胞から単離して、別の膜に挿入することもできる。少なくとも2種類の異なるサブユニットを含むオリゴマー性の細孔の場合には、異なるサブユニットを上記のように異なる宿主細胞内で別々に発現させ、宿主細胞から取り出して、別の膜、例えばウサギ細胞膜などの中で細孔へと集合させることができる。
【0048】
ベクターは例えば、複製起点、任意で前記核酸配列の発現のためのプロモーター、および任意でプロモーターの調節要素(regulator)を備えた、プラスミドベクター、ウイルスベクターまたはファージベクターであってよい。ベクターは、1つまたは複数の選択マーカー遺伝子、例えばテトラサイクリン耐性遺伝子を含むことができる。プロモーターおよび他の発現調節シグナルは、それに対して発現ベクターを設計しようとする宿主細胞と適合性があるように選択することができる。T7、trc、lac、araまたはλLプロモーターが典型的には用いられる。
【0049】
宿主細胞は典型的には、細孔または細孔サブユニットを高レベルで発現する。細孔または細孔サブユニットをコードする核酸配列によって形質転換される宿主細胞は、その細胞を形質転換するために用いる発現ベクターと適合性があるように選択される。宿主細胞は典型的には細菌であり、好ましくは大腸菌(Escherichia coli)である。λ DE3溶原を有する任意の細胞、例えばC41(DE3)、BL21(DE3)、JM109(DE3)、B834(DE3)、TUNER、OrigamiおよびOrigami Bは、T7プロモーターを含むベクターを発現することができる。
【0050】
細孔または細孔サブユニットは、細孔産生性の生物体から、または上記の組換え発現後の、任意のタンパク質液体クロマトグラフィーによる精製を経て、大規模に生産することができる。典型的なタンパク質液体クロマトグラフィーシステムには、FPLC、AKTAシステム、Bio-Cadシステム、Bio-Rad BioLogicシステムおよびGilson HPLCシステムが含まれる。続いて、天然のまたは組換え的に作製された細孔または細孔サブユニットを、本発明による使用のために天然のまたは人工的な膜の中に挿入することができる。
【0051】
細孔または細孔サブユニットは、当技術分野で公知の任意の方法を用いて膜の中に挿入することができる。例えば、国際出願第PCT/GB2006/001057号(WO 2006/100484として公開)に記載された「ピック・アンド・プレース(pick and place)」法を用いて、細孔または細孔サブユニットを膜の中に導入することができる。
【0052】
分子アダプター
膜貫通細孔は、細孔と分析物との間の相互作用を助長する分子アダプターを含む。アダプターの存在は、細孔および分析物のホスト-ゲスト化学作用(host-guest chemistry)を改善する。ホスト-ゲスト化学作用の原理は当技術分野において周知である。アダプターは細孔の物理的または化学的な特性に影響を及ぼして、分析物とのその相互作用を改善する。アダプターは典型的には、細孔のバレルもしくはチャネルの電荷を改変して、または分析物と特異的に相互作用もしくは結合して、それによって細孔とのその相互作用を助長する。
【0053】
好ましくは、アダプターの少なくとも一部は、細孔のバレルまたはチャネルの内側にある。より好ましくは、アダプターのすべてが細孔のバレルまたはチャネルの内側にある。
【0054】
アダプターは、分析物と細孔との間の相互作用を媒介する。分析物は好ましくは、アダプターを介して、またはアダプターとともに細孔と可逆的に結合する。分析物は、最も好ましくは、それが膜を横切って細孔を通過する際に、アダプターを介して、またはアダプターとともに細孔と可逆的に結合する。分析物はまた、それが細孔膜を横切って細孔を通過する際に、アダプターを介して、またはアダプターとともに細孔のバレルまたはチャネルと可逆的に結合することもできる。アダプターは好ましくは、バレルまたはチャネルを、それが分析物と相互作用しうるように狭窄させる。アダプターはより好ましくは、バレルまたはチャネルを狭窄させ、細孔を通してのイオンの流れを制限する。細孔を通してのイオンの流れは、アダプターの存在が、細孔を通しての、および膜を横切ってのイオンの動きと干渉するならば制限される。細孔を通してのイオンの流れは、分析物の検出のために必要である。上記で考察したように、分析物と細孔との間の相互作用は、その分析物に特異的な様式で、細孔を通して流れるイオン電流に影響を及ぼす。アダプターは典型的には環状である。アダプターは好ましくは、細孔と同じ対称性を有する。細孔が七量体である(例えば、中心軸の周りに、14本のストランドを1つの膜貫通βバレルに提供する7個のサブユニットを有する)場合には、7回対称性(seven-fold symmetry)を有するアダプターが典型的には用いられる。同様に、細孔が六量体(例えば、中心軸の周りに、12本のストランドを1つの膜貫通βバレルに提供する6個のサブユニットを有するか、または12本のストランドのβバレルである)場合には、6回対称性を有するアダプターが典型的には用いられる。
【0055】
アダプターは典型的には、分析物と相互作用する、例えば結合する。アダプターと分析物との間の相互作用、例えば結合などは、分析物が細孔と相互作用する尤度を高める。アダプターは典型的には、ホスト-ゲスト化学作用を介して分析物と相互作用する。アダプターの一部分が、典型的には分析物と相互作用することができる。その部分は、分析物と相互作用することのできる1つまたは複数の化学基を含む。その1つまたは複数の化学基は、好ましくは、疎水性相互作用、水素結合、ファンデルワールス力、π-カチオン相互作用および/または静電力といった非共有結合相互作用によって分析物と相互作用する。分析物と相互作用することのできる1つまたは複数の化学基は、好ましくは、アミノ基および/またはヒドロキシル基を含む。アミノ基またはヒドロキシル基は、第一級、第二級または第三級炭素原子と結合していてよい。
【0056】
分析物と相互作用することのできるアダプターの部分は、好ましくは、π-カチオン相互作用を介して分析物と相互作用することのできる芳香環を含む。分析物と相互作用することのできるアダプターの部分は、より好ましくは、アミノ基またはヒドロキシル基の環、例えば、6個、7個または8個のアミノ基またはヒドロキシル基の環を含む。その部分は最も好ましくは、7個のアミノ基またはヒドロキシル基の環を含む。プロトン化アミノ基の環は、分析物中の負に荷電した基、例えばヌクレオチド中のリン酸基などと相互作用することができる。アミノ基の環は、細孔のバレルまたはチャネルの狭窄部において、正に荷電したアミノ酸の環との組み合わせで、分析物と相互作用することができる。ヒドロキシル基の環は、水素結合を介して分析物と相互作用することができる。ヒドロキシル基の環は、細孔のバレルまたはチャネルの狭窄部において、非荷電アミノ酸の環との組み合わせで、分析物と相互作用することができる。
【0057】
アダプターは、最も好ましくは、(1)バレルまたはチャネルを狭窄させ、それによって細孔を通してのイオンの流れを制限する、かつ(2)分析物と相互作用するか、または結合する。
【0058】
以下により詳細に考察するように、細孔のバレルまたはチャネルの中でのアダプターの正しい配向は、アダプターと細孔との間のホスト-ゲスト化学作用によって助長される。アダプターは好ましくは、細孔内の1つまたは複数のアミノ酸と相互作用することのできる、1つまたは複数の化学基を含む。アダプターはより好ましくは、非共有結合性の相互作用、例えば疎水性相互作用、水素結合、ファンデルワールス力、π-カチオン相互作用および/または静電力をなどを介して、細孔内の1つまたは複数のアミノ酸と相互作用することのできる、1つまたは複数の化学基を含む。細孔内の1つまたは複数のアミノ酸と相互作用することのできる化学基は、典型的にはヒドロキシルである。ヒドロキシル基は、第一級、第二級または第三級炭素原子と結合していてよい。ヒドロキシル基は、細孔内の、好ましくは細孔のバレル内またはチャネル内の非荷電アミノ酸と水素結合を形成することができる。以下により詳細に考察するように、アダプター内の1つまたは複数の化学基と、細孔内の1つまたは複数のアミノ酸との相互作用は、アダプターを正しい向きに保たせるために用いることができる。細孔内の1つまたは複数のアミノ酸と相互作用してアダプターを正しい向きに保たせる、アダプター内の1つまたは複数の化学基は、分析物と相互作用することのできる1つもしくは複数の化学基と同じでもよく、または分析物と相互作用することのできる1つもしくは複数の化学基と異なってもよい。
【0059】
細孔と分析物との間の相互作用を助長する、任意のアダプターを用いることができる。適したアダプターには、シクロデキストリン、環状ペプチドおよびカボチャ型分子が非限定的に含まれる。アダプターは好ましくは、ヘプタキス-6-アミノ-β-シクロデキストリン(am7-β-CD)または6-モノデオキシ-6-モノアミノ-β-シクロデキストリン(am1β-CD)である。以下の表2は、細孔およびアダプターの好ましい組み合わせを示している。
【0060】
(表2)細孔およびアダプターの適した組み合わせ
【0061】
好ましくは、アダプターは、ペプチド、ポリペプチド、タンパク質、核酸またはポリヌクレオチドなどのような、反復性サブユニットでできた線状ポリマーではない。より好ましくは、アダプターはプロテインキナーゼインヒビターペプチドではない。
【0062】
共有結合
アダプターは細孔と共有結合している。アダプターは、当技術分野で公知の任意の方法を用いて細孔と共有結合させることができる。アダプターは、細孔と直接結合させることができる。アダプターは好ましくは、二官能性架橋剤(bifunctional crosslinker)を用いて細孔と結合される。適した架橋剤は当技術分野において周知である。好ましい架橋剤はサクシニミジル3-(2-ピリジルジチオ)プロピオネート(SPDP)である。典型的には、アダプターは、アダプター/架橋剤複合体を細孔と共有結合させる前に二官能性架橋剤と共有結合させるが、二官能性架橋剤/細孔複合体をアダプターと結合させる前に二官能性架橋剤を細孔と共有結合させることも可能である。
【0063】
共有結合の部位は、アダプターが分析物と細孔との相互作用を助長し、それによって分析物の検出を可能にするように選択される。共有結合の部位は、分析物が、細孔を通して流れる電流に対して、その分析物に特異的な様式で影響を及ぼすように選択される。
【0064】
好ましくは、アダプターは、細孔のバレルまたはチャネルと共有結合される。アダプターは、アダプターが分析物と細孔との相互作用を助長し、それによって分析物の検出を可能にする限り、バレルまたはチャネル内の任意の部位に共有結合させることができる。好ましくは、アダプターは、アダプターとバレルまたはチャネルとの相互作用が起こる部位の近くにある、バレルまたはチャネル内のアミノ酸と共有結合される。アダプターを、アダプターとバレルまたはチャネルとの間の相互作用が起こる部位とは隔たっている、バレルまたはチャネル内のアミノ酸と共有結合させる場合には、アダプターが、アダプターとバレルまたはチャネルとの間の相互作用が起こるバレルまたはチャネル内の部位に到達しうるように、適した長さの二官能性架橋剤を用いるとよい。アダプターは、細孔と接触する際にそれが可逆的に相互作用するバレルまたはチャネル内の部位と結合していなければならないわけではない。
【0065】
細孔は好ましくは、分子アダプターと二官能性架橋剤との共有結合を助長するように修飾される。細孔のバレルまたはチャネルは、より好ましくは、分子アダプターと二官能性架橋剤との共有結合を助長するように修飾される。細孔は、当技術分野で公知の任意の方法を用いて修飾することができる。上記で考察した変異体の任意のものを、分子アダプターまたは二官能性架橋剤の共有結合を助長するために用いることができる。
【0066】
修飾は典型的には、アダプターと二官能性架橋剤との共有結合を助長するための、細孔内への、好ましくは細孔のバレルまたはチャネル内への、1つまたは複数の、例えば1個、2個、3個、4個、5個、6個または7個などのアミノ酸の導入を伴う。1つまたは複数のアミノ酸を、細孔の同じまたは異なるサブユニット中に導入することができる。共有結合を形成することのできる任意のアミノ酸、例えばシステインなどを導入することができる。アミノ酸は天然または非天然のいずれでもよい。1つまたは複数のアミノ酸は、好ましくは置換によって導入される。以下の表3は、特定の化学基を介したアダプターまたは二官能性架橋剤の共有結合を助長するために細孔内に導入することのできるアミノ酸の種類の例を示している。
【0067】
(表3)アダプターの共有結合を助長するための細孔の修飾
【0068】
好ましい態様においては、SEQ ID NO:14および16に示されているサブユニットの1つを用いて、七量体性細孔内にシステイン残基を導入する。
【0069】
アダプターの向き
アダプターは、分析物を細孔を用いて検出することを可能にする向きで細孔と共有結合される。アダプターは、分析物が、細孔を通して流れる電流に対して、その分析物に特異的な様式で影響を及ぼすような向きに置かれる。アダプターは、それが細孔および分析物のホスト-ゲスト化学作用を改善するような向きに置かれる。アダプターは、それが細孔の物理的または化学的な特性に影響を及ぼして、分析物とのその相互作用を改善するような向きに置かれる。アダプターが分析物と特異的に相互作用すること、または結合することができる場合には、アダプターは、それが分析物と特異的に相互作用すること、または結合することができるような向きに置かれる。後者の態様において、分析物と相互作用するアダプターの部分は、好ましくは、分析物がそこを通って入る細孔の末端に向けて置かれる。より好ましくは、分析物と相互作用することのできる、アダプター内の1つまたは複数の化学基は、分析物がそこを通って入る細孔の末端に向けて置かれる。基は好ましくはアミノ基またはヒドロキシル基である。分析物がそこを通って入る細孔の末端は、シス末端またはトランス末端であってよい。末端は好ましくはトランス末端である。
【0070】
アダプターの向きは、アダプターと細孔との間の共有結合および/またはホスト-ゲスト化学作用によって決定される。共有結合は、アダプターが正しい向きに置かれるように設計することができる。例えば、アダプターが細孔と共有結合する部位(例えば、アミノ酸)を、アダプターが正しい向きに置かれるように設計することもでき、および/または、二官能性架橋剤を、アダプターが正しい向きに置かれるように用いることもできる。
【0071】
好ましくは、細孔は、アダプターの配向を助長するように修飾される。細孔の修飾は、アダプターと細孔との間のホスト-ゲスト化学作用を改善し、それによってアダプターを正しい向きに保たせることができる。より好ましくは、細孔のバレルまたはチャネルは、アダプターの配向を助長するように修飾される。細孔のバレルまたはチャネルは、アダプターの配向を助長する任意の部位で修飾することができる。好ましくは、細孔のバレルまたはチャネルは、共有結合されたアダプターと細孔との間でホスト-ゲスト化学作用が起こる部位で修飾される。最も好ましくは、共有結合されたアダプターと非共有結合性の相互作用を介して相互作用することができる細孔のバレルまたはチャネル内の1つまたは複数のアミノ酸は、アダプターの配向を助長するように修飾される。修飾されるアミノ酸は、細孔と分析物との相互作用を助長する、上記で考察したアミノ酸の1つまたは複数であってよい。細孔は、当技術分野で公知の任意の方法を用いて修飾することができる。上記で考察した変異体の任意のものを、分子アダプターの配向を助長するために用いることができる。
【0072】
修飾は典型的には、アダプターの配向を助長するための、細孔内への、または好ましくはバレルまたはチャネル内への、1つまたは複数の、例えば1個、2個、3個、4個、5個、6個または7個などのアミノ酸の導入を伴う。1つまたは複数のアミノ酸を、同じまたは異なるサブユニット中に導入することができる。1つまたは複数のアミノ酸は、好ましくは、アダプターを正しい向きに保たせるアミノ酸である。1つまたは複数のアミノ酸は、より好ましくは、疎水性相互作用、水素結合、ファンデルワールス力、π-カチオン相互作用および/または静電力といった非共有結合性の相互作用を介してアダプターを正しい向きに保たせるアミノ酸である。1つまたは複数のアミノ酸は、天然または非天然のいずれでもよい。1つまたは複数のアミノ酸は、好ましくは非荷電アミノ酸である。非荷電アミノ酸はアスパラギンのように極性であってもよく、またはフェニルアラニンのように非極性であってもよい。細孔内の非荷電アミノ酸は、アダプター内のヒドロキシル基と水素結合を介して相互作用することによって、アダプターを方向づけることができる。1つまたは複数のアミノ酸は、好ましくは置換によって導入される。以下の表4は、種々の化学基を含むアダプターを正しく方向づけるために細孔内に導入することのできるアミノ酸の種類を示している。
【0073】
(表4)アダプターを配向させるための細孔の修飾
【0074】
好ましい態様において、七量体性細孔は、第一級炭素原子と結合したヒドロキシル基を含むアダプター、例えば6-モノデオキシ-6-モノアミノ-β-シクロデキストリン(am1βCD)などを細孔のシス側の方に配向させることのできる、6個のフェニルアラニン残基を含む。この七量体性細孔は、SEQ ID NO:10に示されたサブユニット6個を含むことができる。
【0075】
別の好ましい態様において、七量体性細孔は、第二級ヒドロキシル基を含むアダプター、例えば6-モノデオキシ-6-モノアミノ-β-シクロデキストリン(am1βCD)などを細孔のトランス側の方に配向させることのできる、6個のアスパラギン残基を含む。この七量体性細孔は、SEQ ID NO:12に示されたサブユニット6個を含むことができる。
【0076】
最も好ましくは、細孔は、アダプターの共有結合を助長するように、および配向を助長するように修飾される。そのような態様において、共有結合の部位と、アダプターの配向を助長するように修飾される細孔の部位との間の空間的関係は、アダプターが正しい向きに確実に保たれるように設計される。修飾は、アダプターが正しい向きに保たれる限り、細孔内の、好ましくはバレルまたはチャネル内の任意の場所に施すことができる。好ましくは、アダプターの共有結合を助長する修飾、および配向を助長する修飾は、細孔内で空間的に近接している。より好ましくは、アダプターの共有結合を助長する修飾、および配向を助長する修飾は、細孔のバレルまたはチャネル内の同一のアミノ酸の環内に作製される。環内の1つのアミノ酸をアダプターの共有結合を助長するように修飾し、一方、環内の他のアミノ酸のうち1つまたは複数、例えば1個、2個、3個、4個、5個または6個などをアダプターの配向を助長するように修飾することができる。共有結合を助長する修飾がアダプターの配向を助長する修飾から隔たっている場合には、アダプターが、アダプターの配向を助長する修飾の近くに配置され、それによって正しく配向されるように、適切な長さの二官能性架橋剤を用いるとよい。上記で考察した修飾の任意のものを用いることができる。
【0077】
最も好ましい態様においては、SEQ ID NO:10に示されているサブユニット6個およびSEQ ID NO:14に示されているサブユニット1個を用いて、またはSEQ ID NO:12に示されているサブユニット6個およびSEQ ID NO:16に示されているサブユニット1個を用いて、七量体性細孔を作製する。
【0078】
本発明の膜貫通タンパク質細孔を作製する方法
本発明はまた、本発明の膜貫通タンパク質細孔を作製する方法にも関する。本方法は、膜貫通タンパク質細孔と、細孔と分析物との間の相互作用を助長する分子アダプターとを共有結合させる段階を含む。アダプターは、当技術分野で公知の任意の方法を用いて、細孔と共有結合させることができる。
【0079】
上記で考察した細孔、アダプターおよび二官能性架橋剤の任意のものを、本方法に用いることができる。共有結合の部位は、上記で考察したように選択される。
【0080】
細孔は好ましくは、アダプターの共有結合を助長し、および/またはアダプターの配向を助長するように修飾される。これについては以上により詳細に考察している。
【0081】
本方法はまた、アダプターが、分析物を細孔を用いて検出することを可能にする向きで細孔と結合しているか否かを判定する段階も含む。これは、膜貫通タンパク質細孔を、分析物の有無を判定するために用いうるか否かを判定することを伴う。これは、以下により詳細に説明するようにして行うことができる。分析物の有無を判定することができるならば、アダプターは正しい向きにあり、本発明の膜貫通タンパク質細孔が作製されたことになる。分析物の有無を判定することができないならば、アダプターは正しくない向きにある可能性が高く、本発明の膜貫通タンパク質細孔は作製されなかったことになる。
【0082】
分析物を検出する方法
本発明はまた、分析物の有無を判定する方法にも関する。本方法は、分析物を、本発明の膜貫通タンパク質細孔と、分析物が細孔と相互作用するように接触させる段階、および相互作用の間に細孔を通過する電流を測定し、それによって分析物の有無を判定する段階を含む。本発明の膜貫通タンパク質細孔の任意のものを用いることができる。分析物を検出するために本発明の膜貫通タンパク質細孔を用いることに伴う利点については上記で考察している。
【0083】
電流がその分析物に特異的な様式で細孔を通って流れるならば(すなわち、その分析物に伴ってみられる特徴的な電流が細孔を通って流れるのが検出されたならば)、その分析物は存在する。電流がその分析物に特異的な様式で細孔を通って流れないならば、その分析物は存在しない。
【0084】
本発明はこのため、分析物の確率的センシングを伴う。本発明は、構造の類似した分析物を、それらが膜貫通タンパク質細孔を通過する電流に対して及ぼす影響の違いに基づいて識別するために用いることができる。本発明はまた、試料中の特定の分析物の濃度を測定するために用いることもできる。
【0085】
また、本発明を、バルクセンシング用途に多数のまたは数千個もの本発明の細孔を用いるセンサーに用いることもできる。
【0086】
本方法は、本発明の膜貫通タンパク質細孔を膜の中に挿入する任意の適した膜/細孔系を用いて実施することができる。本方法は典型的には、(i)本発明の膜貫通タンパク質細孔を含む人工膜、(ii)本発明の膜貫通タンパク質細孔を含む、単離された天然に存在する膜、または(iii)本発明に従って修飾された膜貫通タンパク質細孔を発現する細胞、を用いて実施される。本方法は好ましくは人工膜を用いて実施される。膜は、本発明の膜貫通タンパク質細孔に加えて、他の膜貫通タンパク質および/または膜内タンパク質、さらには他の分子も含むことができる。
【0087】
本発明の方法は、典型的にはインビトロで実施される。
【0088】
膜
膜はイオンおよび分析物の流れに対する障壁を形成する。膜は好ましくは脂質二重層である。本発明による使用のために適した脂質二重層は、当技術分野で公知の方法を用いて作製することができる。例えば、脂質二重層膜は、Montal and Mueller (1972)の方法を用いて形成させることができる。本発明の方法は、リン脂質、糖脂質、コレステロールおよびそれらの混合物を非限定的に含む任意の膜脂質から形成された脂質二重層を用いて実施することができる。脂質二重層は好ましくは、1,2-ジフィタノイル-sn-グリセロ-3-ホスホコリンから形成される。
【0089】
脂質二重層などの膜の中に細孔を挿入するための方法は、当技術分野で公知である。例えば、細孔を、脂質二重層を含む溶液中に精製形態で懸濁させ、それが脂質二重層へ拡散して、脂質二重層との結合および機能的状態への集合によって挿入されるようにすることができる。または、細孔を、M. A. Holden, H. Bayley. J. Am. Chem. Soc. 2005, 127, 6502-6503に記載された方法を用いて膜の中に直接挿入することもできる。
【0090】
細孔と分析物との間の相互作用
分析物は、膜のいずれかの側で細孔と接触させることができる。分析物は、膜のいずれかの側で細孔に導入することができる。分析物は、分析物が細孔を通って膜の他方の側へと通過することを可能にする膜の側と接触させられる。例えば、分析物は、その本来の環境においてイオンまたは低分子、例えば分析物などが細孔のバレルまたはチャネル内に入ることを可能にする細孔の末端と、イオンまたは低分子が細孔を通過しうるように、接触させられる。そのような場合、分析物は、それが膜を横切って細孔のバレルまたはチャネルを通過する際に、細孔および/またはアダプターと相互作用する。または、分析物を、分析物がアダプターを介して、またはアダプターとともに細孔と相互作用し、細孔から解離して、膜の同じ側に残ることを可能にする膜の側と接触させることもできる。本発明は、アダプターの向きが固定されている細孔を提供する。その結果として、分析物は好ましくは、アダプターが配向している方の細孔の末端と接触する。最も好ましくは、分析物は、分析物と相互作用するアダプターの部分が方向づけられている細孔の末端と接触させられる。
【0091】
分析物は、任意の様式かつ任意の部位で、細孔と相互作用することができる。上記で考察したように、分析物は好ましくは、アダプターを介して、またはアダプターとともに細孔と可逆的に結合する。分析物は最も好ましくは、それが膜を横切って細孔を通過する際に、アダプターを介して、またはアダプターとともに細孔と可逆的に結合する。分析物はまた、それが膜を横切って細孔を通過する際に、アダプターを介して、またはアダプターとともに細孔のバレルまたはチャネルと可逆的に結合することもできる。
【0092】
分析物と細孔との間の相互作用の間に、分析物は、細孔を通して流れる電流に対して、その分析物に特異的な様式で影響を及ぼす。例えば、特定の分析物は、特定の平均期間にわたって特定の程度で、細孔を通して流れる電流を減少させると考えられる。換言すれば、細孔を通して流れる電流は、特定の分析物に関して特徴的である。対照実験を、細孔を通して流れる電流に対して特定の分析物が及ぼす効果を決定するために実施することができる。続いて、試料中の特定の分析物を同定するため、または試料中に特定の分析物が存在するか否かを判定するために、被験試料に対して本発明の方法を実施したことによる結果を、そのような対照実験から得られたものと比較することができる。細孔を通して流れる電流が特定の分析物を指し示す様式で影響を受ける頻度を、試料中のその分析物の濃度を決定するために用いることができる。
【0093】
装置
本方法は、膜貫通タンパク質細孔が膜の中に挿入されている膜/細孔系を調べるために適した任意の装置を用いて実施することができる。本方法は、確率学的センシングのために適した任意の装置を用いて実施することができる。例えば、装置は、水溶液を含むチャンバーおよびチャンバーを2つの区域に分ける障壁を含む。障壁は、細孔を含む膜が形成されている開口部を有する。分析物は、分析物をチャンバー内に導入することによって細孔と接触させることができる。分析物は、チャンバーの2つの区域のいずれかの中に導入することができる。
【0094】
本発明の方法は、分析物との相互作用の間に、細孔を通過する電流を測定することを伴う。したがって、装置はまた、電位を加えて膜および細孔を横切る電気信号を測定することのできる電気回路も含む。本方法は、パッチクランプ(patch clmp)または電圧固定(voltage clamp)を用いて実施されうる。本方法は、好ましくは、電圧固定の使用を伴う。実施例は、電圧固定法を実施するための1つのやり方を開示している。
【0095】
分析物
分析物は、試料中の任意の物質でありうる。適した分析物には、金属イオン、無機塩、重合体、例えば重合酸または塩基など、色素、脱色剤、医薬品、診断用薬、娯楽薬(recreational drug)、爆発物および環境汚染物質が非限定的に含まれる。
【0096】
分析物は、細胞から分泌される分析物であってもよい。または、分析物は、本発明が実施可能となる前に分析物を細胞から抽出しなければならないような、細胞の内側に存在する分析物であってもよい。
【0097】
分析物は好ましくは、アミノ酸、ペプチド、ポリペプチドまたはタンパク質である。アミノ酸、ペプチド、ポリペプチドまたはタンパク質は天然または非天然のいずれでもありうる。ポリペプチドまたはタンパク質は、その内部に合成アミノ酸または修飾アミノ酸を含みうる。アミノ酸に対するさまざまな異なる種類の修飾が当技術分野で公知である。本発明の目的には、分析物を、当技術分野において入手可能な任意の方法によって修飾しうることが理解されるべきである。
【0098】
タンパク質は、酵素、抗体、ホルモン、増殖因子または増殖調節タンパク質、例えばサイトカインなどでありうる。サイトカインは、インターロイキン、好ましくはIFN-1、IL-2、IL-4、IL-5、IL-6、IL-10、IL-12もしくはIL-13、インターフェロン、好ましくはIL-γ、または他のサイトカイン、例えばTNF-αなどから選択することができる。タンパク質は、細菌タンパク質、真菌タンパク質、ウイルスタンパク質または寄生生物由来タンパク質であってよい。それを細孔と接触させる前に、タンパク質をアンフォールディングさせてポリペプチド鎖を形成させ、それによってそれが細孔のバレルまたはチャネルの中に入り、アダプターを介して、またはアダプターとともに細孔と相互作用することを可能にすることもできる。
【0099】
ヌクレオチドおよび核酸の検出については、以下にさらに詳細に考察している。
【0100】
分析物は任意の適した試料中に存在する。本発明は典型的には、分析物を含むことが分かっているか、または分析物を含むことが疑われる試料に対して実施される。本発明を、その実体が不明である1つまたは複数の分析物を含む試料に対して実施することもできる。または、本発明を、試料中に存在することが分かっているかまたは予想される、1つまたは複数の分析物の実体を確認するために、試料に対して実施することもできる。
【0101】
試料は生物試料であってよい。本発明は、任意の生物体または微生物から得られた、または抽出された試料に対して、インビトロで実施することができる。生物体または微生物は、典型的には原核生物または真核生物であり、典型的には、植物界、動物界、菌界、モネラ界および原生生物界という5つの界の1つに属する。本発明は、任意のウイルスから得られた、または抽出された試料に対してインビトロで実施することができる。試料は好ましくは流体試料である。試料は典型的には患者の体液を含む。試料は、尿、リンパ、唾液、粘液または羊水でありうるが、好ましくは血液、血漿または血清である。典型的には、試料はヒトが起源であるが、またはそれは、ウマ、ウシ、ヒツジもしくはブタなどの商業的な飼育動物など別の哺乳動物由来であってもよく、またはネコもしくはイヌなどのペットであってもよい。
【0102】
試料は非生物試料であってもよい。非生物試料は好ましくは流体試料である。非生物試料の例には、手術液、水、例えば飲用水、海水または河川水など、および臨床検査のための試薬が含まれる。
【0103】
試料は典型的には、アッセイされる前に、例えば遠心分離によって、または望ましくない分子もしくは細胞、例えば赤血球などを濾別する膜を通過させることによって処理される。試料は、採取次第直ちに測定してもよい。また、試料は典型的には、アッセイの前に、好ましくは-70℃未満で保存してもよい。
【0104】
条件
本発明の方法は、分析物との相互作用の間に細孔を通過する電流を測定することを伴う。膜貫通タンパク質細孔を通るイオン電流を測定するために適した条件は当技術分野で公知であり、実施例に開示されている。本方法は、膜および細孔をまたいで印加される電圧を用いて実施される。用いられる電圧は、典型的には-250mV〜+250mVである。用いられる電圧は、好ましくは、-200mV、-150mV、-100mV、-50mV、-20mVおよび0mVから選択される下限、ならびに+10mV、+20mV、+50mV、+100mV、+150mVおよび+200mVから独立に選択される上限を有する範囲にある。
【0105】
本方法は、任意のアルカリ金属塩化物塩の存在下で実施される。上記で考察した例示的な装置において、塩はチャンバー内の水溶液中に存在する。塩化カリウム(KCl)、塩化ナトリウム(NaCl)または塩化セシウム(CsCl)が、典型的には用いられる。KClが好ましい。塩濃度は、典型的には0.1〜2.5M、0.3〜1.9M、0.5〜1.8M、0.7〜1.7M、0.9〜1.6M、または1M〜1.4Mである。塩濃度は好ましくは約1Mである。
【0106】
本方法は、典型的には、緩衝液の存在下で実施される。上記で考察した例示的な装置において、緩衝液はチャンバー内の水溶液中に存在する。任意の緩衝液を本発明の方法に用いることができる。1つの適した緩衝液は、Tris-HCl緩衝液である。本方法は、典型的には4.0〜10.0、4.5〜9.5、5.0〜9.0、5.5〜8.8、6.0〜8.7、または7.0〜8.8、または7.5〜8.5のpHで実施される。用いられるpHは好ましくは約8.0である。
【0107】
本方法は典型的には、0℃〜100℃、15℃〜95℃、16℃〜90℃、17℃〜85℃、18℃〜80℃、19℃〜70℃、または20℃〜60℃で実施される。本方法は好ましくは室温で実施される。
【0108】
個々のヌクレオチドを同定する方法
本発明はまた、個々のヌクレオチドを同定する方法にも関する。本方法は、ヌクレオチドを、本発明の膜貫通タンパク質細孔と、ヌクレオチドが細孔と相互作用するように接触させる段階;および相互作用の間に細孔を通過する電流を測定し、それによってヌクレオチドの実体を決定する段階を含む。本発明はこのため、個々のヌクレオチドの確率的センシングを伴う。本発明は、構造の類似したヌクレオチドを、それらが膜貫通タンパク質細孔を通過する電流に対して及ぼす影響の違いに基づいて識別するために用いることができる。個々のヌクレオチドを、それらが細孔と相互作用する時のそれらの電流振幅から単分子レベルで同定することができる。本発明はまた、特定のヌクレオチドが試料中に存在するか否かを判定するために用いることもできる。本発明はまた、試料中の特定のヌクレオチドの濃度を測定するために用いることもできる。
【0109】
個々のヌクレオチド
本発明による個々のヌクレオチドとは、単一のヌクレオチドのことである。個々のヌクレオチドとは、別のポリヌクレオチドとヌクレオチド結合によって結合していないもののことである。ヌクレオチド結合は、ヌクレオチドのリン酸基の1つが、別のヌクレオチドの糖基(sugar group)と結合したものを含む。個々のヌクレオチドは、典型的には、少なくとも5、少なくとも10、少なくとも20、少なくとも50、少なくとも100、少なくとも200、少なくとも500、少なくとも1000、または少なくとも5000ヌクレオチドの別のポリヌクレオチド配列とヌクレオチド結合によって結合していないものである。例えば、個々のヌクレオチドは、DNAストランドまたはRNAストランドなどの標的ポリヌクレオチド配列から消化される。
【0110】
本発明の方法は、任意のヌクレオチドを同定するために用いうる。ヌクレオチドは天然または人工的のいずれであってもよい。ヌクレオチドは、典型的には、核酸塩基、糖、および少なくとも1つのリン酸基を含む。核酸塩基は、典型的には複素環式である。適した核酸塩基には、プリンおよびピリミジン、より具体的にはアデニン、グアニン、チミン、ウラシルおよびシトシンが含まれる。糖は、典型的には五炭糖である。適した糖には、リボースおよびデオキシリボースが非限定的に含まれる。ヌクレオチドは、典型的には、リボヌクレオチドまたはデオキシリボヌクレオチドである。ヌクレオチドは、典型的には、モノホスフェート、ジホスフェートまたはトリホスフェートを含む。
【0111】
適したヌクレオチドには、以下のものが非限定的に含まれる:アデノシン一リン酸(AMP)、アデノシン二リン酸(ADP)、アデノシン三リン酸(ATP)、グアノシン一リン酸(GMP)、グアノシン二リン酸(GDP)、グアノシン三リン酸(GTP)、チミジン一リン酸(TMP)、チミジン二リン酸(TDP)、チミジン三リン酸(TTP)、ウリジン一リン酸(UMP)、ウリジン二リン酸(UDP)、ウリジン三リン酸(UTP)、シチジン一リン酸(CMP)、シチジン二リン酸(CDP)、シチジン三リン酸(CTP)、サイクリックアデノシン一リン酸(cAMP)、サイクリックグアノシン一リン酸(cGMP)、デオキシアデノシン一リン酸(dAMP)、デオキシアデノシン二リン酸(dADP)、デオキシアデノシン三リン酸(dATP)、デオキシグアノシン一リン酸(dGMP)、デオキシグアノシン二リン酸(dGDP)、デオキシグアノシン三リン酸(dGTP)、デオキシチミジン一リン酸(dTMP)、デオキシチミジン二リン酸(dTDP)、デオキシチミジン三リン酸(dTTP)、デオキシウリジン一リン酸(dUMP)、デオキシウリジン二リン酸(dUDP)、デオキシウリジン三リン酸(dUTP)、デオキシシチジン一リン酸(dCMP)、デオキシシチジン二リン酸(dCDP)およびデオキシシチジン三リン酸(dCTP)。ヌクレオチドは、好ましくはAMP、TMP、GMP、UMP、dAMP、dTMP、dGMPまたはdCMPである。
【0112】
ヌクレオチドを、リボ核酸(RNA)またはデオキシリボ核酸などの核酸配列の消化によって導くこともできる。単一の核酸配列由来の個々のヌクレオチドを、その核酸の全体または一部のシークエンシングのために、連続的な様式で細孔と接触させることができる。本発明の第2の態様に従った核酸のシークエンシングについて、以下により詳細に考察する。
【0113】
ヌクレオチドは、核酸配列の消化によって導かれる場合のように、典型的には非修飾である。または、ヌクレオチドを修飾すること、または損傷させることもできる。ヌクレオチドは典型的にはメチル化される。ヌクレオチドを顕在化標識で標識することもできる。顕在化標識は、ヌクレオチドが検出されることを可能にする任意の適した標識であってよい。適した標識には、蛍光分子、放射性同位体、例えば125I、35Sなど、およびビオチンなどのリンカーが含まれる。
【0114】
ヌクレオチドは、典型的には、任意の適した生物試料中に存在する。適した生物試料については上記で考察している。
【0115】
本方法の実施
分析物の検出にかかわる、特に用いうる細孔、膜、装置および条件にかかわる上記の考察のすべては、この方法に対して等しく適用される。
【0116】
条件に関しては、用いられる電圧は、好ましくは、デオキシリボヌクレオチド5'一リン酸、例えばdAMP、dTMP、dGMPおよびdCMPなどについては約+130mVであり、リボヌクレオチド5'一リン酸、例えばAMP、TMP、GMPおよびUMPなどについては+110mVである。本方法は好ましくは、デオキシリボヌクレオチド5'一リン酸、例えばdAMP、dTMP、dGMPおよびdCMPなどについてはpH 8.0、1M KClにて+130mVで実施され、リボヌクレオチド5'一リン酸、例えばAMP、TMP、GMPおよびUMPなどについてはpH 8.0、1M KClにて+110mVで実施される。
【0117】
核酸のシークエンシングの方法
本発明はまた、標的核酸配列のシークエンシングの方法にも関する。本方法は、(a)個々のヌクレオチドを、標的配列の一方の末端から、連続移動性エキソヌクレアーゼを用いて消化する段階;(b)ヌクレオチドを、本発明の膜貫通タンパク質細孔と、ヌクレオチドが細孔と相互作用するように接触させる段階;(c)相互作用の間に細孔を通過する電流を測定し、それによってヌクレオチドの実体を決定する段階;および(d)段階(a)から(c)までを核酸配列の同じ末端で繰り返し、それによって核酸の配列を決定する段階、を含む。それ故に、本方法は、核酸のシークエンシングのための、連続的な様式での、核酸配列の各々の単一ヌクレオチドの確率的センシングを伴う。
【0118】
本発明の膜貫通タンパク質細孔は、この方法のために特に適している。核酸の有効なシークエンシングのためには、核酸中のすべてのヌクレオチドが連続的な様式で確実に同定されるようにすることが重要である。本発明の膜貫通タンパク質細孔におけるアダプターの固定的な性質は、各々の連続的なヌクレオチドが細孔と相互作用する時には常に、特徴的な電流が細孔を通して流れることを意味する。
【0119】
核酸の全体または一部のみを、この方法を用いてシークエンシングすることができる。核酸は任意の長さでありうる。例えば、核酸は、少なくとも10、少なくとも50、少なくとも100、少なくとも150、少なくとも200、少なくとも250、少なくとも300、少なくとも400、または少なくとも500ヌクレオチド長でありうる。核酸は天然または人工的のいずれであってもよい。例えば、本方法は、製造されたオリゴヌクレオチドの配列を検証するために用いることができる。本方法は典型的にはインビトロで実施される。
【0120】
この方法の段階(b)および(c)は、上記で考察したヌクレオチドを同定する方法において実施される段階と概ね同一である。分析物の検出にかかわる、特に用いうる細孔、膜、装置および条件にかかわる上記の考察のすべては、この方法に対して等しく適用される。核酸は典型的には、上記で考察したような任意の生物試料中に存在する。
【0121】
連続移動性エキソヌクレアーゼ
標的核酸のシークエンシングの方法は、核酸を連続移動性エキソヌクレアーゼと接触させて、個々のヌクレオチドを核酸の一方の末端から遊離させることを伴う。連続移動性エキソヌクレアーゼとは、典型的には、核酸の一方の末端に取り付き、配列を消化してその末端から1回に1個ずつのヌクレオチドを分離させる酵素のことである。連続移動性エキソヌクレアーゼは、核酸を5'から3'の方向に、または3'から5'の方向に消化することができる。連続移動性エキソヌクレアーゼが結合する核酸の末端は、典型的には、用いる酵素の選択を通じて、および/または当技術分野で公知の方法を用いて決定される。核酸配列の特定の末端に対する連続移動性エキソヌクレアーゼの結合を防止または助長するためには、典型的には、核酸配列のいずれかの末端でヒドロキシル基またはキャップ構造を用いるとよい。
【0122】
任意の連続移動性エキソヌクレアーゼ酵素を、本方法に用いることができる。本方法における使用のために好ましい酵素は、λエキソヌクレアーゼである。λエキソヌクレアーゼの1つのサブユニットの配列は、SEQ ID NO:17に示されている。3つの同一のサブユニットが相互作用して、三量体性エキソヌクレアーゼを形成する。λエキソヌクレアーゼの変異体は、SEQ ID NO:17のそれとは違いのあるアミノ酸配列を有し、かつ連続移動性エキソヌクレアーゼ活性を保っているポリペプチドサブユニットから形成された酵素である。これらの変異体は、上記のSEQ ID NO:2の変異体に関して上記で考察したのと同じ様式で、かつ同じ程度で、SEQ ID NO:17とは違っていてよい。変異体は、好ましくは、核酸と結合して核酸を消化する原因となるドメイン(触媒ドメイン)を含む。変異体は、好ましくは、野生型酵素と比較して、必要に応じて上昇または低下した酵素活性速度、および/またはより高い塩耐性を有する。連続移動性エキソヌクレアーゼは、膜貫通タンパク質細孔の作製に関して上記で考察した方法の任意のものを用いて作製することができる。
【0123】
標的核酸配列のシークエンシングの方法は、核酸配列を、連続移動性エキソヌクレアーゼと、上記で考察したようにヌクレオチドが各々の個々のヌクレオチドの同定を可能にする速度で核酸の末端から消化されるように、接触させることを含む。これを行うための方法は当技術分野において周知である。例えば、ポリペプチドの末端から単一のアミノ酸を連続的に消化して、それらを高速液体クロマトグラフィー(HPLC)を用いて同定しうるようにするために、エドマン分解が用いられる。類似の方法を本発明に用いることもできる。
【0124】
連続移動性エキソヌクレアーゼは、好ましくは、膜貫通タンパク質細孔と共有結合される。連続移動性エキソヌクレアーゼを細孔と共有結合させるための方法は、当技術分野において周知である。
【0125】
連続移動性エキソヌクレアーゼが機能しなければならない速度は、典型的には、野生型連続移動性エキソヌクレアーゼの至適速度よりも遅い。第2の態様の方法における連続移動性エキソヌクレアーゼの適した活性速度には、1秒当たり0.5〜1000ヌクレオチド、1秒当たり0.6〜500ヌクレオチド、1秒当たり0.7〜200ヌクレオチド、1秒当たり0.8〜100ヌクレオチド、1秒当たり0.9〜50ヌクレオチド、または1秒当たり1〜20もしくは10ヌクレオチドの消化が含まれる。速度は、好ましくは1秒当たり1、10、100、500または1000ヌクレオチドである。連続移動性エキソヌクレアーゼの適した活性速度は、さまざまなやり方で達成することができる。例えば、至適活性速度が低下している変異型連続移動性エキソヌクレアーゼを本発明に従って用いることができる。
【0126】
連続移動性エキソヌクレアーゼの活性は、典型的にはpH依存性であり、pHが低下するにつれてそれらの活性が低下する。それ故に、第2の態様の方法は、典型的には、7.5〜8.0、または7.7〜8.0というpHで実施される。用いられるpHは、好ましくは約8.0である。
【0127】
連続移動性エキソヌクレアーゼの活性速度は、典型的には、塩濃度が上昇するにつれて低下する。しかし、極めて高い塩濃度は、典型的には、酵素の活性に対して有害な影響を及ぼす。酵素の速度を制限するもう1つのやり方は、活性に有害な影響を及ぼすことなしに酵素の活性速度を低下させる塩濃度で、本方法を実施することである。例えば、本方法を0.5〜1Mの塩濃度で実施することができる。塩濃度は好ましくは約1Mである。
【0128】
キット
本発明はまた、標的核酸配列のシークエンシングの方法を実施するために用いうるキットにも関する。したがって、本キットは、核酸配列のシークエンシングのために適している。キットは、本発明の膜貫通細孔および連続移動性エキソヌクレアーゼを含む。
【0129】
キットは、上述した方法の態様の任意のものを実施することを可能にする1つまたは複数の他の試薬または器具をさらに含みうる。そのような試薬または器具には、以下の1つまたは複数が含まれる:適した緩衝液(水溶液)、対象から試料を得るための手段(例えば、注射針を含む容器または器具)、核酸配列を増幅するための手段、上記で定義した膜、または電圧固定もしくはパッチクランプの装置。試薬は、キット中に、流体試料が試薬を再懸濁させるような乾燥状態で存在しうる。キットはまた、任意で、キットを本発明の方法に用いることを可能にする説明書、または本方法を使用しうる患者に関する詳細を含みうる。キットは任意で、ヌクレオチドを含みうる。
【0130】
以下の実施例は、本発明を例示的に説明している。
【0131】
実施例
この実施例において、本発明者らは、β-シクロデキストリン(β-CD)をαHL細孔に対して、アダプターを共有結合のために配向させ、かつ反応後の細孔内腔の中でそれらを安定化するための113位の突然変異を用いて共有結合させた。このアプローチには2つの重要な成果があった。第1に、アダプターはナノポア(nanopore)から解離することができない。第2に、細孔のトランス入口(trans entrance)に面した第一級ヒドロキシルまたは第二級ヒドロキシルのいずれかを用いて、シクロデキストリンの向きを制御することができる。
【0132】
1.材料および方法
1.1 αHL細孔
α-ヘモリシン突然変異型細孔(M113F-RL2)7(WTバックグラウンド;SEQ ID NO:10)、(M113N-RL2)7(WTバックグラウンド;SEQ ID NO:12)、(M113F-RL2)6(M113C-D8RL2)1(SEQ ID NO:10および14)、(M113N)6(T117C-D8RL3)1(SEQ ID NO:12および16)を、以前に記載した通りに発現させ、集合させて精製した。
【0133】
RL2は、膜貫通βバレルをコードする配列のカセット突然変異誘発を可能とするように考案された半合成遺伝子の産物である(Cheley, S., Braha, O., Lu, X., Conlan, S., and Bayley, H. (1999) Protein Sci. 8, 1257-1267)。これは、6個のサイレント制限部位、およびコードされるポリペプチド配列中に5個の改変アミノ酸を含む(K8A、V124L、G130S、N139QおよびI142L)。D8RL2は、オクタ-アスパラギン酸尾部(octa-aspartate tail)を有するRL2である。
【0134】
RL3は、アミノ酸レベルではWT αHLポリペプチドと同一であるが、この遺伝子は、ループをコードする領域内に、カセット突然変異誘発の一助となる6個のサイレント突然変異を含む(S. Cheley, unpublished)。D8RL3は、オクタ-アスパラギン酸尾部を有するRL3である。図12は、pT7-RL3-D8の完全ポリヌクレオチド配列を示している。
【0135】
1.2 化学物質
試薬は以下の通りに入手した:1,2-ジフィタノイル-sn-グリセロ-3-ホスホコリン(Avanti Polar Lipids)、ペンタン(JT Baker)、ヘキサデカン(99+%、Sigma-Aldrich)、塩酸6-モノデオキシ-6-モノアミノ-β-シクロデキストリン(99%、Cyclolabs Budapest, Hungary)、Trizma塩基(99.9%、Sigma-Aldrich)、濃HCl(分析用試薬等級、Fisher Scientific)、塩化カリウム(99%、Sigma-Aldrich)、2-アダマンタンアミン・HCl(99%、Aldrich)、サクシニミジル3-(2-ピリジルジチオ)プロピオネート(SPDP、95%、Fluka)、トリエチルアミン(99.5%、Fluka)。
【0136】
1.3 6-モノデオキシ-6-[3-(2-ピリジルジチオ)プロピオニル]モノアミノ-β-シクロデキストリン(βCD-PDP)の合成
塩酸6-モノデオキシ-6-モノアミノ-β-シクロデキストリン(am1βCD)(11.7mg、0.01mmol)およびサクシニミジル3-(2-ピリジルジチオ)プロピオネート(SPDP、6.24mg、0.02mmol)を、5mLのMeOH/H2O中に溶解させた(1:1)。トリエチルアミン(100μL)を5分間かけて添加し、この混合物を一晩撹拌した。所望の産物は白色沈殿物として出現し、これを濾別して、冷水およびアセトンで連続して洗浄した。分析用試料は、DMSOおよび水からの再結晶化によって得られた。
【0137】
1.4 単一チャネル電流の記録
1,2-ジフィタノイル-sn-グリセロ-3-ホスホコリン(Avanti Polar Lipids)の二重層を、平面状二重層チャンバーをシスおよびトランスという2つの区画に区分するポリカーボネートフィルム(20μm厚、Goodfellow, Malvern, PAによる)中の直径100〜150μmの開口部全体にわたって形成させた。いずれの区画も緩衝液1mLを含んでいた。工学的に作製したタンパク質細孔をシス区画に添加し、それを接地させた。βCD-PDPをトランス区画に添加し、それを増幅器のヘッドステージ(head-stage)に接続した。
【0138】
別に指示する場合を除き、実験はすべて、25mM Tris・HCl、1M KCl、pH 8.0中、22.5±2℃にて±100mVで実施した。タンパク質およびβCD-PDPの新たな凍結アリコートを毎日用いた。電流はパッチクランプ増幅器(Axopatch 200B;Axon instruments, Foster City, CA)で記録し、内蔵の四極Besselフィルターにより10kHzでローパスフィルター処理を行った上で、Digiデータ 1200 A/Dコンバーター(Axon instruments)を装備したコンピュータによって20kHzでサンプリングを行った。
【0139】
1.5 データ分析
電流軌跡は、pClamp 9.0ソフトウエア(Axon Instruments)を用いて分析した。イベントはEvent Detection機能を用いて検出し、それを用いて振幅および滞留時間のヒストグラムを作成した。Origin(Microcal, Northampton, MA)を、曲線フィッティングおよびグラフ提示のために用いた。
【0140】
1.6 分子モデル
Michelle MontoyaおよびEric Gouauxによって提供された七量体性α-ヘモリシン細孔(M113F-RL2)7および(M113N-RL2)7のpdbファイルを、PyMOL(DeLano Scientific, San Carlos, CA)に表示した。βCD-PDP構造も、構築ツールを用いることによってPyMOLに表示した。
【0141】
2.結果
2.1 リンカーの設計および工学的に作製したαHL細孔のβCD-PDPによる誘導体化
(M113F-RL2)7・βCDおよび(M113N)7・βCDのX線構造(図1、Montoya and Gouaux, unpublished)から、αHL細孔のβバレル内の非共有結合的に結合したβCDの位置が判明している。(M113F-RL2)7・βCDでは、7個のPhe-113のフェニル環がβCDの7個の第一級6-ヒドロキシル基との疎水性相互作用に関与し、その結果、第一級ヒドロキシルがシス口部(cis mouth)の方を向く、細孔内でのβCDの特異的な配向をもたらす(図1b)。対照的に、(M113N-RL2)7・βCDでは、7個のAsn-113のアミド基およびLys-147のε-アミノ基がそれぞれ、βCDの7個の2-第二級ヒドロキシル基および7個の3-第二級ヒドロキシル基の水素結合距離の内部にある。この向きでは、βCDの第一級ヒドロキシル基は細孔のトランス入口の方を向く(図1c)。
【0142】
分子モデリングにより、本発明者らは、αHLバレルの内腔内の、βCDを結合させるのに適した位置を同定した。(M113F-RL2)7・βCDでは、Phe-113のフェニル環の中心とβCDの第一級ヒドロキシル基の最近接O原子との間の距離は6.5±0.5Åである(7つの位置の平均)(図1b)。(M113N-RL2)7・βCDでは、βCDの第一級ヒドロキシル基のO原子とThr-117の最近接O原子との間の距離は平均6.0±0.5Åである(図1c)。本発明者らは、6-モノデオキシ-6-モノアミノ-β-シクロデキストリン(am1βCD、これはβCDの単一の第一級ヒドロキシル基がアミノ基によって置換されている)を、位置-113または位置-117にあるシステイン残基と連結させるために、二官能性架橋剤サクシニミジル3-(2-ピリジルジチオ)プロピオネート(SPDP)を選んだ。本発明者らはまず、am1βCDをSPDPとカップリングさせてβCD-PDPを形成させた(図2)。βCD-PDPでは、リンカーの末端にあるピリジルジスルフィドを、αHL細孔内のシステイン残基の遊離チオールによって攻撃してジスルフィド結合を形成させることができる。リンカーの長さは、βCD-PDPをPyMolの中で構築することによって測定した;伸長コンフォメーションでは、am1βCDのアミド結合内のN原子と、ピリジルS原子との間の距離はおよそ7.0Åである。したがって、このリンカーは、am1βCDを、(M113F-RL2)7に見られる向きにあるβCDを用いて位置-113のシステインと連結させるため、または(M113N-RL2)7に見られる向きにあるβCDを用いて位置-117のシステインと連結させるために必要とされるよりも幾分長い。
【0143】
これらのモデリング研究に基づき、位置-113に6個のフェニルアラニン残基を有し、第7の位置に単一のシステインを有する、ヘテロ七量体性αHL細孔(M113F-RL2)6(M113C-D8RL2)1を工学的に作製した。M113Cサブユニット中のシステイン残基は、シクロデキストリンの第一級ヒドロキシルがM113Fサブユニットの6個のPhe残基の近隣に保たれるような様式で、二重層のトランス側から提示されるβCD-PDPのピリジルジスルフィドと反応しうるように設計した。反応後に、βCDは永続的な分子アダプターとしてαHL細孔の内側に係留されると考えられる。もう1つの構築物において、本発明者らは、工学的に作製した細孔(M113N-RL2)6(T117C-D8RL3)1内の117位にシステイン残基を配置した。この細孔は、M113Nサブユニット内の残基Asn-113およびLys-147と水素結合を形成した第二級ヒドロキシルの相互作用の安定化を維持しながらβCD-PDPと反応するように設計した。
【0144】
本発明者らは、PyMOLの中で、所望の共有結合複合体(M113F-RL2)6(M113C-D8RL2)1-βCDおよび(M113N-RL2)6(T117C-D8RL3)1-βCDを描き出した。(M113F-RL2)6(M113C-D8RL2)1-βCDにおいて、am1βCDのアミドN原子と、113位のシステインのS原子との間の距離は5.6Å(図3cおよび4)である。(M113N-RL2)6(T117C-D8RL3)1-βCDにおいて、am1βCDのアミドN原子と、117位のシステインのSとの間の距離は6.2Åである(図5cおよび6)。これらの値は非共有結合複合体のX線構造から求められた対応する測定値に近く(上記参照)、このことはリンカーの長さの選択が正しいことを裏づけている。
【0145】
2.2 (M113F-RL2)6(M113C-D8RL2)1-βCDの電気特性決定
単一の(M113F-RL2)6(M113C-D8RL2)1細孔をシスチャンバーから脂質二重層内に導入した場合には、本発明者らが測定したイオン電流は98±5pAであった(+100mV、25mM Tris・HCl、pH 8.0、1M KCl、n=6)。細孔との反応を観察するために、βCD-PDPをトランスチャンバーに添加して、電位を+100mVに保った。全電流の70%および95%に相当する2種類の過渡的遮断イベントが観察された。対照的に、βCDが(M113F-RL2)7と相互作用する時には、70%の電流遮断しか観察されない。このため、その第一級ヒドロキシルがPhe-113側鎖と相互作用し、リンカーがシス側に向けて突出している細孔内にβCD-PDPが結合する時には、70%の遮断が生じる可能性が非常に高い。95%の遮断は、ピリジル環がシクロデキストリンの中空部の内側に停留している細孔にβCD-PDPが入り、それによって電流の流れを低下させる時に起こる可能性がある。最終的に、5分間から1時間までの範囲にわたる期間の後には、電流レベルは70%遮断の状態に永続的に固定される。このイベントの前には、48±7msにわたって続く85%遮断がみられた(n=5)(図3cおよび4)。70%遮断状態から非修飾細孔のそれへの電流の回復は、24時間という全記録期間中に全く観察されなかった(16回の実験を通じて)。これに比して、(M113F-RL2)7と非共有的に結合したβCDの平均τoffは、同じ条件下で316±62msである。この遮断状態は、位置113でのジスルフィド結合を通じて共有結合したβCDを有するαHL細孔を表していると推定される(図3cおよび4)。この考え方に沿う形で、ジスルフィド結合はシスチャンバーへの2mM DTTの添加によって切断された。切断の間に、寿命が150±12ms(n=5)で電流遮断98%の過渡的な中間状態が観察された(図3d)。(M113F-RL2)6(M113C-D8RL2)1-βCDにおける70%遮断は、±250mVの範囲内で印加電位を上昇させたり低下させたりした後も維持された。αHL細孔内でのβCDなどの中性分子の結合は電圧依存性であり、それは電気浸透の影響によることを指摘しておきたい(Gu, L.-Q., Cheley, S., and Bayley, H. (2003) Proc.Natl. Acad. Sci. USA 100, 15498-15503)。したがって、共有結合は、非共有結合複合体が短寿命である電位でβCD解離を防止する。
【0146】
電流-電圧(I-V)曲線を、(M113F-RL2)6(M113C-D8RL2)1-βCDおよび非共有結合複合体(M113F)7・βCDに関して計測した。I-V曲線は+150〜-60mVの間では有意な違いがなかった。しかし、印加電位が-60mV未満の場合には、(M113F-RL2)6(M113C-D8RL2)1-βCDに関するI-V曲線の方が(M113F-RL2)7・βCDのそれよりも整流的であった(図3e)。
【0147】
2.3 (M113N-RL2)6(T117C-D8RL3)1の電気特性決定
単一の(M113N-RL2)6(T117C-D8RL3)1タンパク質細孔をシスチャンバーから脂質二重層内に導入した場合には、本発明者らが測定したイオン電流は-80±5pAであった(-100mV、25mM Tris・HCl、pH 8.0、1M KCl、n=6)。βCD-PDPをトランス区画に添加した後には、(M113N-RL2)7・βCDで観察された65%遮断に近い、70%の電流遮断という1種類の過渡的遮断イベントのみが観察された。1分間から20分間までの範囲にわたる期間の後には、寿命が5±2ms(n=5)で電流遮断80%という中間状態が観察され、その後には永続的な70%電流遮断が続いた(図4および5c)。(M113F-RL2)6(M113C-D8RL2)1-βCDの場合と同じように、この70%遮断は合計24時間の間には回復しなかったが(16回の実験を通じて)、一方、(M113N-RL2)7・βCDに関する平均τoffは同じ条件下で10.7±1.5sに過ぎなかった。この場合も、印加電位を±250mVの範囲内で上げ下げしても細孔の遮断解消は生じなかった。したがって、この状態は、ジスルフィド結合を通じて117位に共有結合したβCDを有するαHL細孔を表すと推定される(図4および5c)。この場合も、ジスルフィド結合はシスチャンバーへの2mM DTTの添加後に切断された(図5d)。切断の間に、電流遮断80%で寿命が300±20ms(n=5)という過渡的中間状態が観察された(図5d)。
【0148】
電流-電圧(I-V)曲線を、付加生成物(M113N-RL2)6(T117C-D8RL3)1-βCDに関して計測した。この複合体のI-V曲線は±100mVの範囲内では(M113N-RL2)7・βCDのそれに類似しており、高度の正電位および負電位では幾分逸脱する(図5e)。
【0149】
2.4 βCDを逆向きに連結させる試み
共有結合したβCDに関して提唱した向きについてさらに調べ、確認するために、本発明者らは、X線構造で立証されているように、βCD-PDPとタンパク質との間の共有結合形成が、113突然変異が通常支えると考えられるシクロデキストリンの向きとは反対の向きをいずれの場合にも生じさせる、ヘテロ七量体(M113N-RL2)6(M113C-D8RL3)1および(M113F-RL2)6(T117C-D8RL2)1を調製した(図1)。
【0150】
(M113N-RL2)6(M113C-D8RL2)1の場合には、βCD-PDPとCys-113との共有結合は、各30分ずつの10回の試行後には起こらなかった。対照的に、ヘテロ七量体(M113F-RL2)6(T117C-D8RL3)1は、永続的な80%の電流遮断によって示されたように、βCD-PDPと反応した。前述した2つの場合と同じように、結合したβCDは±250mVの電位の上げ下げに耐え、シスチャンバーへの2mM DTTの添加によって離脱させることができた(提示せず)。推定される共有結合物である(M113F-RL2)6(T117C-D8RL3)1-βCDについて、25mM Tris・HCl、pH 8.0、1M KCl中、±100mVでの電気記録によって検討した(n=3)。単一チャネル電流(24.0pA)は(M113N-RL2)6(T117C-D8RL3)1-βCDのそれ(40.0pA)よりも小さく、正電位および陰電位の両方で頻回な完全遮断を呈した(データ非提示)。加えて、(M113F-RL2)6(T117C-D8RL3)1-βCDの脇を通過する電流は、(M113N-RL2)6(T117C-D8RL3)1-βCDで観察されたものよりもはるかにノイズが多く、このために、それを有機分析物の確率的センシングのために用いることはできない(下記参照)。
【0151】
2.5 共有結合したβCDアダプターを有する工学的に作製したαHL細孔を用いた分析物検出
新たに作り出した構築物をバイオセンサー検出素子として評価するために、本発明者らはモデル分析物である2-アダマンタンアミン(2-AdNH2)の確率的センシングを行った(12,15)。上記の通りに、インサイチューで、βCDアダプターを(M113F-RL2)6(M113C-D8RL2)1または(M113N-RL2)6(T117C-D8RL3)1と結合させた。続いて、2-アダマンタンアミン・HClをシスチャンバーまたはトランスチャンバーのいずれかに添加し、過渡的な電流の準安定状態(substate)を観察した(図7)。これらのイベントは、工学的に作製したαHL細孔内での個々の2-AdNH2・βCD複合体の形成および解離を表す。平均滞留時間(τoff)は、2-AdNH2を非共有結合複合体(M113F-RL2)7・βCDおよび(M113N-RL2)7・βCDに添加した場合に観察されたものと同程度であった(以下の表5)。τoffのわずかな違いはおそらくリンカーによる干渉によって生じたものと考えられる。
【0152】
(表5)βCDおよびαHLの共有結合性および非共有結合性の付加物による、2-アダマンタンアミン(2-AdNH2)の確率的センシング
条件:25mM Tris・HCl、pH 8.0、1M KCl、+100mVで、30μMの2-AdNH2・HCl(シスまたはトランス)を伴う。個々の実験の数は括弧内に示されている。
【0153】
2.6 共有結合したアダプターを有するαHL細孔を用いた長寿命結合イベントの検討
共有結合させたアダプターを用いることで、非共有結合的に結合したアダプターを用いて検討することがより困難な、長寿命イベントおよび稀なイベントという2種類のイベントの動態を研究することが可能である。シス-リトコール酸はβCDに関して知られている最も強固な結合分子である(Yang, Z., and Breslow, R. (1997) Tetrahedron Lett. 38, 6171-6172)。このため、本発明者らはこの分子を選び、長時間にわたる結合イベントを研究するために新たに構築した細孔の試験を行った。(M113F-RL2)6(M113C-D8RL2)1-βCDを選択した理由は、分析物の非存在下でのバックグラウンドシグナルのノイズが少ないためである(例えば、図7参照)。シス-リトコール酸(50μM、図8a)を25mM Tris・HCl、pH 8.0、1M KCl中、+100mVのシスチャンバーに導入したところ(n=3)、驚いたことに、全くシグナルが観察されなかった。これはおそらく、βCDをバレル壁に結合させるリンカーによる(M113F-RL2)6(M113C-D8RL2)1-βCDのシス側に対する結合の立体障害に起因すると考えられる。シス-リトコール酸(50μM)を-100mVのトランスチャンバーに導入したところ(n=3)、本発明者らは多くの極めて長寿命のイベントを観察した。これらのイベントは98%の電流遮断を示し、各イベントの最初の200±100ms、およびイベントの最後の150±100msの間は92%の遮断であった(図8b、図9)。イベントの平均τoffは8.6±7.8sであり(イベントの数=60;範囲0.5s〜80s)、これは同じ条件下の(M113F-RL2)7・βCD複合体におけるβCDに関するτoff(0.68±0.12s)よりもかなり長寿命である、すなわち、βCDは、リトコール酸結合イベント中には非共有結合性の(M113F-RL2)7・βCDから最も高い頻度で解離すると考えられる。これは、長寿命の結合イベントを検討することに関しての、非共有結合複合体を上回る、共有結合させたアダプターの利点を示している。
【0154】
2.7 高温での、共有結合したアダプターを有するαHL細孔
本発明者らは、CDの滞留時間が減少する高温では、共有結合したCDを有するαHL細孔が、誘導体化されていない細孔よりも優れた働きをする可能性があると推測した(Kang, X., Gu, L.-Q., Cheley, S., and Bayley, H. (2005) Angew. Chem. Int. Ed. Engl., 1495-1499)。最高55℃までの(M113F)6(M113C-D8RL2)1-βCDの単一チャネル電流軌跡を得た。単一チャネル電流は温度に直線的に依存した(図10)。25mM Tris・HCl、pH 8.0、1M KCl中、+100mVでI(pA)=14.3+0.54T(℃)であり、これは非共有結合的に結合したβCDを有するαHL細孔で以前に観察されたものと同様の温度依存性である(Kang, X., Gu, L.-Q., Cheley, S., and Bayley, H. (2005) Angew. Chem. Int. Ed. Engl., 1495-1499)。これらの実験中に、(M113F-RL2)6(M113C-D8RL2)1-βCD細孔からのβCDの解離は観察されなかった。例えば、(M113F-RL2)6(M113C-D8RL2)1-βCDの脇を通過する電流は、55℃での5分間の測定中に一定に保たれたが、一方、(M113F-RL2)6(M113C-D8RL2)1と結合したβCDに関するτoffは同じ条件下で56±3.8msに過ぎなかった。これらのデータは、共有結合したCDを有するαHL細孔が、極限条件下の水溶液中での確率的センシングを可能とすることを示している。
【0155】
2.8 共有結合したアダプターを有するαHL細孔におけるイオン選択性の永続的な改変
αHL細孔内に停留した非共有結合性の分子アダプターは、イオン輸送に関する荷電選択性を劇的に改変することができる(Gu, L.-Q., Dalla Serra, M., Vincent, J. B., Vigh, G., Cheley, S., Braha, O., and Bayley, H. (2000) Proc.Natl.Acad.Sci. USA 97, 3959-3964)。例えば、pH 7.5では、WT-αHLの荷電選択性(PK+/PCl-、KCl:シス200mM、トランス1000mM)は、βCDが内腔に停留するようになった後には0.55±0.02から0.25±0.01に変化し、すなわち細孔はよりアニオン選択的になる(Gu, L.-Q., Dalla Serra, M., Vincent, J. B., Vigh, G., Cheley, S., Braha, 0., and Bayley, H. (2000) Proc.Natl.Acad.Sct. USA 97, 3959-3964)。このため、本発明者らは、βバレルの内側に共有結合したβCDは、αHL細孔のイオン選択性を永続的に改変すると予想した。本発明者らは、(M113F-RL2)6(M113C-D8RL2)1および(M113F-RL2)6(M113C-D8RL2)1-βCDを被験事例として採用し、シス/トランスおよびトランス/シスKCl勾配の両方において記録した単一チャネル電流に関してI-V曲線を作成した(図11)。非誘導体化細孔および誘導体化細孔の両方の、測定されたコンダクタンス値(g)および逆転電位(Vr)、ならびにGoldman-Hodgkin-Katz式から計算された荷電選択性(PK+/PCl-)(Hille, B. (2001) Ion channels of excitable membranes, 3rd edition,, Sinauer, Sunderland, MA, USA)を、表6(下記)に示している。
【0156】
(表6)(M113F-RL2)6(M113C-D8RL2)1および(M113F-RL2)6(M113C-D8RL2)1-βCDの逆転電位(Vr)、計算された荷電選択性(PK+/PCl-)およびコンダクタンス値(g)
各項目に関して、3回の別々の実験を行った。逆転電位(Vr)は、記述した条件下での平均値(n=3)である。荷電選択性(PK+/PCl-、n=3)およびコンダクタンス値(n=3)は、平均±SDとして引用している。PK+/PCl-は、活性および実験上のVr値(20)を用いることにより、Goldman-Hodgkin-Katz式を用いて計算した。
* 25mM Tris・HCl、pH 8.0。塩濃度(KCl)はmM単位で提示されている。
† いずれのチャンバーにおいても、+100mV、1M KCl、25mM Tris・HCl、pH 8.0
【0157】
共有結合させた分子アダプターはαHL細孔の荷電選択性を実際に改変し、この効果は永続的であった。この場合も、細孔はよりアニオン選択性となり、PK+/PCl-について観察された変化は、非共有結合的に結合したβCDで認められたものと同程度である(Gu, L.-Q., Dalla Serra, M., Vincent, J. B., Vigh, G., Cheley, S., Braha, 0., and Bayley, H. (2000) Proc.Natl.Acad.Sct. USA 97, 3959-3964)。この特徴は、イオン浸透の基礎研究およびバイオテクノロジーにおける応用のいずれについても、膜チャネルのデノボ設計において有用なはずであり、修飾された特性を有する細孔のさまざまな組み立てには大きな有用性があると考えられる(Hwang, W. L., Holden, M. A., White, S., and Bayley, H. (2007) submitted for publication;およびHolden, M. A., Needham, D., and Bayley, H. (2007) J Am Chem Soc, in press)。
【0158】
3.結論
確率的検出のための有機分析物の結合を可能にするためのαHL細孔の工学的操作はごく稀にしか成功していないが(Guan, X., Gu, L.-Q., Cheley, S., Braha, O., and Bayley, H. (2005) ChemBioChem 6, 1875-1881)、非共有結合性の分子アダプターは、絶え間なく起こる細孔との会合および解離によって課せられるある種の制約にもかかわらず、この点に関して有用であることが立証されている。後者の問題を改善するために、本発明者らは今回、βCDアダプターをαHL細孔の内側に共有結合させた。さらに、βCDの向きに関して制御して、すなわち、βCDの第一級ヒドロキシルを細孔のトランス入口またはシス入口のいずれかに面するようにさせて、結合を行わせた。共有結合は検出に欠落がないことを意味し、βCDの向きに対する制御は、ある種の分析物が2つの入口のうち一方のみを通ってβCD中空部と結合する、またはそれらが入る側に応じて異なる様式で結合するという可能性が高いことから重要である(Kang, X. F., Cheley, S., Guan, X., and Bayley, H. (2006) J Am Chem Soc 128(33),10684-10685)。共有結合させたβCDを有するαHL細孔の確率的センシングにおける有用性を例示するために、本発明者らは本明細書で、細孔を用いて、βCD内部での滞留時間が長寿命である分析物を検出しうることを示している。この場合には、βCDは通常、分析物がβCDから解離する前に細孔から解離して、信号の情報量を劣化させると考えられる(Braha, O., Webb, J., Gu, L.-Q., Kim, K., and Bayley, H. (2005) ChemPhysChem 6, 889-892)。絶え間なく起こるCDの会合および解離の影響は高温で悪化するが(Kang, X., Gu, L.-Q., Cheley, S., and Bayley, H. (2005) Angew.Chem.Int.Ed.Engl., 1495-1499)、本明細書で実証しているように共有結合はこの問題も改善する。本発明者らはまた、共有結合させたβCDを有するαHL細孔がイオン選択性を永続的に改変し、それが例えば、ナノバッテリー(nanobattery)を構築するために有用な可能性があることも示している(Hwang, W. L., Holden, M. A., White, S., and Bayley, H. (2007) submitted for publication;およびHolden, M. A., Needham, D., and Bayley, H. (2007) J Am Chem Soc, in press)。
【0159】
また、単分子DNAシークエンシング(Bayley, H. (2006) Curr Opin Chem Biol 10, 628-637)も、共有結合したβCDを含むタンパク質ナノポアの使用によって恩典を受けると考えられる。1つのアプローチでは、デオキシリボヌクレオシド5'-一リン酸を、連続移動性エキソヌクレアーゼによって個々のDNAストランドから遊離させる(Jett, J. H., Keller, R. A., Martin, J. C., Marrone, B. L., Moyzis, R. K., Ratliff, R. L., Seitzinger, N. K., Shera, E. B., and Stewart, C. C. (1989) J.Biomol.Struct.Dynam. 7, 301-309)。元々明示されたものでは、完全に置換されて転写されたDNAストランドから蛍光性塩基類似体が遊離されるが、それは遂行するのが極めて困難なことが判明している。しかし、本発明者らは最近、非共有結合性のαHL(M113R-RL2)7・am7βCD複合体(式中、am7βCDはヘプタキス-(6-デオキシ-6-アミノ)-β-シクロデキストリンである)を用いて、4種類のデオキシリボヌクレオシド5'-一リン酸を識別しうることを示した(Astier, Y., Braha, O., and Bayley, H. (2006) J Am Chem Soc 128(5), 1705-1710)。これは一歩前進ではあるが、シークエンシングを成功させるためには、エキソヌクレアーゼによって遊離させられるすべての塩基を検出素子によって捕捉して同定しなければならない。このため、分子アダプターを有するαHL細孔を用いるのであれば、共有結合させたアダプターが、シークエンシングにおける欠落を回避する助けになると考えられる。
【0160】
以下の表7〜16は、配列情報を示している。
【0161】
(表7)SEQ ID NO:1および2‐黄色ブドウ球菌由来の野生型α-ヘモリシン
SEQ ID NO:2はSEQ ID NO:1の残基1〜879によってコードされる
【0162】
(表8)SEQ ID NO:3および4‐α-ヘモリシンM113H-RL2
SEQ ID NO:4はSEQ ID NO:3の残基39〜920によってコードされる
n=a、c、gまたはt
場所5にあるXはLeuまたはPheを表す
場所7にあるXはTyr、Trp、Cys、Ser、LeuまたはPheを表す
【0163】
(表9)SEQ ID NO:5および6‐α-ヘモリシンM113K-RL2
SEQ ID NO:6はSEQ ID NO:5の残基39〜920によってコードされる
n=a、c、gまたはt
場所5にあるXはLeuまたはPheを表す
場所7にあるXはTyr、Trp、Cys、Ser、LeuまたはPheを表す
【0164】
(表10)SEQ ID NO:7および8‐α-ヘモリシンM113R-RL2
SEQ ID NO:8はSEQ ID NO:7の残基39〜920によってコードされる
n=a、c、gまたはt
場所5にあるXはLeuまたはPheを表す
場所7にあるXはTyr、Trp、Cys、Ser、LeuまたはPheを表す
【0165】
(表11)SEQ ID NO:9および10‐α-ヘモリシンM113F-RL2
SEQ ID NO:10はSEQ ID NO:9の残基39〜920によってコードされる
n=a、c、gまたはt
場所5にあるXはLeuまたはPheを表す
場所7にあるXはTyr、Trp、Cys、Ser、LeuまたはPheを表す
【0166】
(表12)SEQ ID NO:11および12‐α-ヘモリシンM113N-RL2
SEQ ID NO:12はSEQ ID NO:11の残基39〜920によってコードされる
n=a、c、gまたはt
場所5にあるXはLeuまたはPheを表す
場所7にあるXはTyr、Trp、Cys、Ser、LeuまたはPheを表す
【0167】
(表13)SEQ ID NO:13および14‐α-ヘモリシンM113C-D8RL2
SEQ ID NO:14はSEQ ID NO:13の残基39〜944によってコードされる
n=a、c、gまたはt
場所5にあるXはLeuまたはPheを表す
場所7にあるXはTyr、Trp、Cys、Ser、LeuまたはPheを表す
【0168】
(表14)SEQ ID NO:15および16‐α-ヘモリシンT117C-D8RL3
SEQ ID NO:16はSEQ ID NO:15の残基13〜918によってコードされる
【0169】
(表15)SEQ ID NO:17‐λエキソヌクレアーゼ
【0170】
(表16)SEQ ID NO:18‐α-ヘモリシン-D8RL3
【特許請求の範囲】
【請求項1】
試料中の分析物の検出に用いるための膜貫通タンパク質細孔であって、細孔と分析物との間の相互作用を助長する分子アダプターを含んでおり、該アダプターが、分析物を細孔を用いて検出することを可能にする向きで細孔と共有結合している、膜貫通タンパク質細孔。
【請求項2】
分析物と相互作用することのできるアダプターの部分が、分析物がそこを通って入る細孔の末端を向いている、請求項1記載の膜貫通タンパク質細孔。
【請求項3】
前記部分が、分析物と相互作用することのできる1つまたは複数の化学基を含む、請求項2記載の膜貫通タンパク質細孔。
【請求項4】
化学基がアミノ基またはヒドロキシル基である、請求項3記載の膜貫通タンパク質細孔。
【請求項5】
その末端がシス末端またはトランス末端である、請求項2〜4のいずれか一項記載の膜貫通タンパク質細孔。
【請求項6】
アダプターの共有結合を助長するように修飾(modify)されている、前記請求項のいずれか一項記載の膜貫通タンパク質細孔。
【請求項7】
アダプターの配向を助長するように修飾されている、前記請求項のいずれか一項記載の膜貫通タンパク質細孔。
【請求項8】
SEQ ID NO:2またはその変異体を含む、前記請求項のいずれか一項記載の膜貫通タンパク質細孔。
【請求項9】
分子アダプターがシクロデキストリンである、前記請求項のいずれか一項記載の膜貫通タンパク質細孔。
【請求項10】
シクロデキストリンが、ヘプタキス-6-アミノ-β-シクロデキストリン(am7-β-CD)または6-モノデオキシ-6-モノアミノ-β-シクロデキストリン(am1βCD)である、請求項9記載の膜貫通タンパク質細孔。
【請求項11】
分析物が個々のヌクレオチドである、前記請求項のいずれか一項記載の膜貫通細孔。
【請求項12】
アダプターが二官能性架橋剤を介して細孔と共有結合している、前記請求項のいずれか一項記載の膜貫通タンパク質細孔。
【請求項13】
以下の段階を含む、請求項1〜12のいずれか一項記載の膜貫通タンパク質細孔を作製する方法:
(a)膜貫通タンパク質細孔に、細孔と分析物との間の相互作用を助長する分子アダプターを共有結合させる段階;および
(b)該アダプターが、分析物を細孔を用いて検出することを可能にする向きで細孔と結合しているか否かを判定する段階。
【請求項14】
アダプターの共有結合を助長するため、かつ/または配向を助長するために、細孔を修飾する段階をさらに含む、請求項13記載の方法。
【請求項15】
以下の段階を含む、分析物の有無を判定する方法:
(a)分析物を、請求項1〜12のいずれか一項記載の膜貫通タンパク質細孔と、分析物が細孔と相互作用するように接触させる段階;および
(b)相互作用の間に細孔を通過する電流を測定し、それによって分析物の有無を判定する段階。
【請求項16】
以下の段階を含む、個々のヌクレオチドを同定する方法:
(a)ヌクレオチドを、請求項1〜12のいずれか一項記載の膜貫通タンパク質細孔と、ヌクレオチドが細孔と相互作用するように接触させる段階;および
(b)相互作用の間に細孔を通過する電流を測定し、それによってヌクレオチドの実体を決定する段階。
【請求項17】
以下の段階を含む、標的核酸配列のシークエンシングの方法:
(a)個々のヌクレオチドを、標的配列の一方の末端から、連続移動性(processive)エキソヌクレアーゼを用いて消化する段階;
(b)ヌクレオチドを、請求項1〜12のいずれか一項記載の膜貫通タンパク質細孔と、ヌクレオチドがアダプターと相互作用するように接触させる段階;
(c)相互作用の間に細孔を通過する電流を測定し、それによってヌクレオチドの実体を決定する段階;および
(d)段階(a)から(c)までを核酸配列の同じ末端で繰り返し、それによって核酸の配列を決定する段階。
【請求項18】
請求項1〜12のいずれか一項記載の膜貫通タンパク質細孔および連続移動性エキソヌクレアーゼを含む、核酸のシークエンシングのためのキット。
【請求項1】
試料中の分析物の検出に用いるための膜貫通タンパク質細孔であって、細孔と分析物との間の相互作用を助長する分子アダプターを含んでおり、該アダプターが、分析物を細孔を用いて検出することを可能にする向きで細孔と共有結合している、膜貫通タンパク質細孔。
【請求項2】
分析物と相互作用することのできるアダプターの部分が、分析物がそこを通って入る細孔の末端を向いている、請求項1記載の膜貫通タンパク質細孔。
【請求項3】
前記部分が、分析物と相互作用することのできる1つまたは複数の化学基を含む、請求項2記載の膜貫通タンパク質細孔。
【請求項4】
化学基がアミノ基またはヒドロキシル基である、請求項3記載の膜貫通タンパク質細孔。
【請求項5】
その末端がシス末端またはトランス末端である、請求項2〜4のいずれか一項記載の膜貫通タンパク質細孔。
【請求項6】
アダプターの共有結合を助長するように修飾(modify)されている、前記請求項のいずれか一項記載の膜貫通タンパク質細孔。
【請求項7】
アダプターの配向を助長するように修飾されている、前記請求項のいずれか一項記載の膜貫通タンパク質細孔。
【請求項8】
SEQ ID NO:2またはその変異体を含む、前記請求項のいずれか一項記載の膜貫通タンパク質細孔。
【請求項9】
分子アダプターがシクロデキストリンである、前記請求項のいずれか一項記載の膜貫通タンパク質細孔。
【請求項10】
シクロデキストリンが、ヘプタキス-6-アミノ-β-シクロデキストリン(am7-β-CD)または6-モノデオキシ-6-モノアミノ-β-シクロデキストリン(am1βCD)である、請求項9記載の膜貫通タンパク質細孔。
【請求項11】
分析物が個々のヌクレオチドである、前記請求項のいずれか一項記載の膜貫通細孔。
【請求項12】
アダプターが二官能性架橋剤を介して細孔と共有結合している、前記請求項のいずれか一項記載の膜貫通タンパク質細孔。
【請求項13】
以下の段階を含む、請求項1〜12のいずれか一項記載の膜貫通タンパク質細孔を作製する方法:
(a)膜貫通タンパク質細孔に、細孔と分析物との間の相互作用を助長する分子アダプターを共有結合させる段階;および
(b)該アダプターが、分析物を細孔を用いて検出することを可能にする向きで細孔と結合しているか否かを判定する段階。
【請求項14】
アダプターの共有結合を助長するため、かつ/または配向を助長するために、細孔を修飾する段階をさらに含む、請求項13記載の方法。
【請求項15】
以下の段階を含む、分析物の有無を判定する方法:
(a)分析物を、請求項1〜12のいずれか一項記載の膜貫通タンパク質細孔と、分析物が細孔と相互作用するように接触させる段階;および
(b)相互作用の間に細孔を通過する電流を測定し、それによって分析物の有無を判定する段階。
【請求項16】
以下の段階を含む、個々のヌクレオチドを同定する方法:
(a)ヌクレオチドを、請求項1〜12のいずれか一項記載の膜貫通タンパク質細孔と、ヌクレオチドが細孔と相互作用するように接触させる段階;および
(b)相互作用の間に細孔を通過する電流を測定し、それによってヌクレオチドの実体を決定する段階。
【請求項17】
以下の段階を含む、標的核酸配列のシークエンシングの方法:
(a)個々のヌクレオチドを、標的配列の一方の末端から、連続移動性(processive)エキソヌクレアーゼを用いて消化する段階;
(b)ヌクレオチドを、請求項1〜12のいずれか一項記載の膜貫通タンパク質細孔と、ヌクレオチドがアダプターと相互作用するように接触させる段階;
(c)相互作用の間に細孔を通過する電流を測定し、それによってヌクレオチドの実体を決定する段階;および
(d)段階(a)から(c)までを核酸配列の同じ末端で繰り返し、それによって核酸の配列を決定する段階。
【請求項18】
請求項1〜12のいずれか一項記載の膜貫通タンパク質細孔および連続移動性エキソヌクレアーゼを含む、核酸のシークエンシングのためのキット。
【図1】

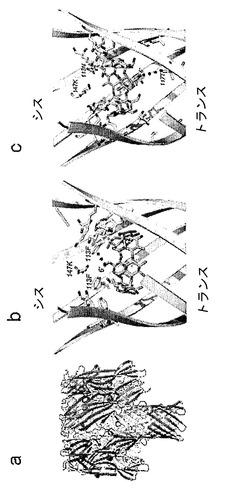
【図2】
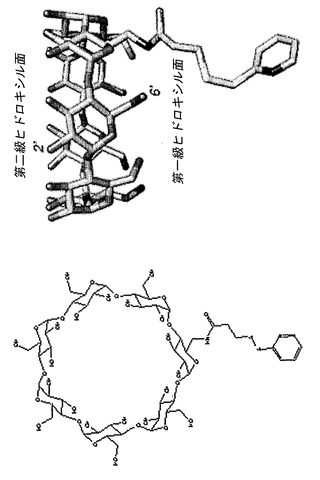
【図3】

【図4】

【図5】
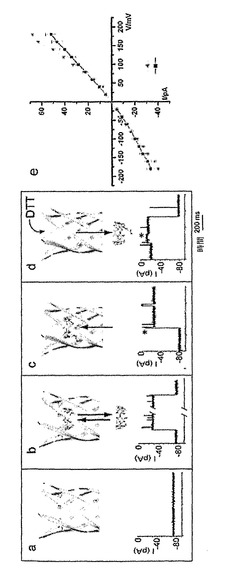
【図6】
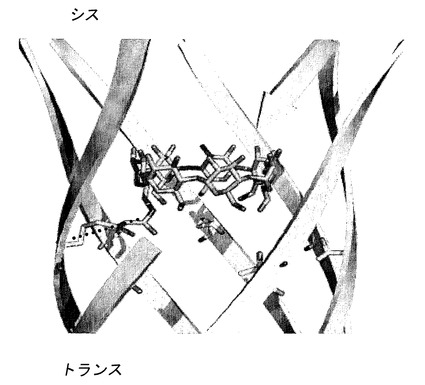
【図7】
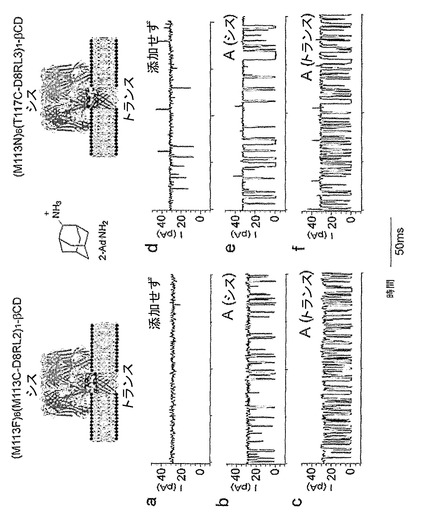
【図8】

【図9】
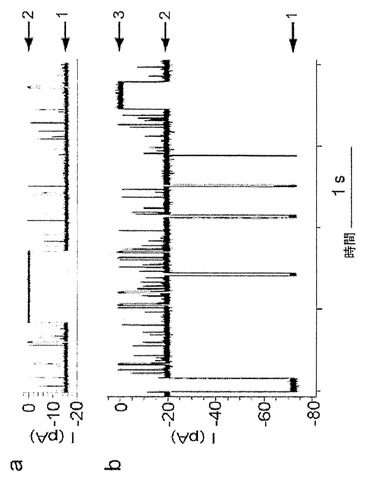
【図10】
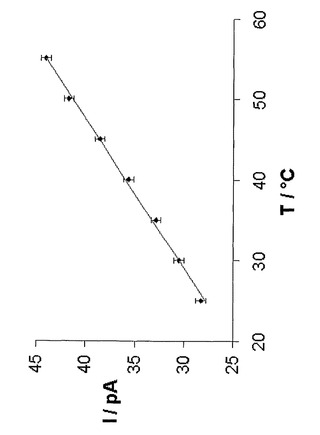
【図11a】
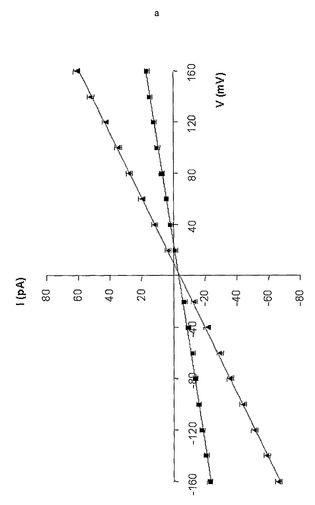
【図11b】
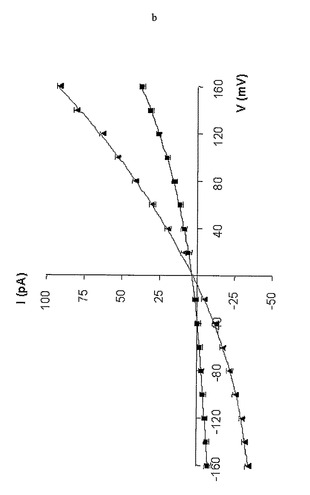
【図12】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11a】
【図11b】
【図12】
【公表番号】特表2010−539966(P2010−539966A)
【公表日】平成22年12月24日(2010.12.24)
【国際特許分類】
【出願番号】特願2010−527535(P2010−527535)
【出願日】平成20年10月6日(2008.10.6)
【国際出願番号】PCT/GB2008/003372
【国際公開番号】WO2009/044170
【国際公開日】平成21年4月9日(2009.4.9)
【出願人】(508144107)イシス イノベーション リミテッド (4)
【Fターム(参考)】
【公表日】平成22年12月24日(2010.12.24)
【国際特許分類】
【出願日】平成20年10月6日(2008.10.6)
【国際出願番号】PCT/GB2008/003372
【国際公開番号】WO2009/044170
【国際公開日】平成21年4月9日(2009.4.9)
【出願人】(508144107)イシス イノベーション リミテッド (4)
【Fターム(参考)】
[ Back to top ]