
分子タグ化システム
【課題】化合物の探知、回収、および同定のための分子タグ化システムを提供すること。
【解決手段】本発明は、オリゴヌクレオチドタグの使用により分子の種類または亜集団を探知、同定、および/または分類するための方法および物質を提供する。二本鎖DNAを固相支持体に結合させる方法;集団中のポリヌクレオチドにオリゴヌクレオチドタグを結合させる方法;固相支持体およびオリゴヌクレオチドタグ相補物の均一な集団を含む、組成物;微粒子の混合物を含む組成物;微粒子に結合している複数のcDNAを含むcDNAライブラリー;微粒子に結合している複数のゲノムDNAフラグメントのライブラリーを含むゲノムDNAフラグメントのライブラリー;オリゴヌクレオチドタグのレパートリー;が、開示される。
【解決手段】本発明は、オリゴヌクレオチドタグの使用により分子の種類または亜集団を探知、同定、および/または分類するための方法および物質を提供する。二本鎖DNAを固相支持体に結合させる方法;集団中のポリヌクレオチドにオリゴヌクレオチドタグを結合させる方法;固相支持体およびオリゴヌクレオチドタグ相補物の均一な集団を含む、組成物;微粒子の混合物を含む組成物;微粒子に結合している複数のcDNAを含むcDNAライブラリー;微粒子に結合している複数のゲノムDNAフラグメントのライブラリーを含むゲノムDNAフラグメントのライブラリー;オリゴヌクレオチドタグのレパートリー;が、開示される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、一般にはオリゴヌクレオチドラベルで分子(特にポリヌクレオチド)を同定、分類、および/または探知するための方法に関し、より詳細にはオリゴヌクレオチドタグへの特異的ハイブリダイゼーションによりポリヌクレオチドを分類する方法に関する。
【背景技術】
【0002】
オリゴヌクレオチドおよびそのアナログの特異的ハイブリダイゼーションは、広く多様な研究的適用、医学的適用および産業的適用で用いられている基礎的なプロセス(診断アッセイにおける疾患関連のポリヌクレオチドの同定、新規の標的ポリヌクレオチドのクローンのスクリーニング、ポリヌクレオチド混合物のブロットにおける特異的ポリヌクレオチドの同定、特異的な標的ポリヌクレオチドの増幅、不適切に発現された遺伝子の治療的ブロッキング、DNA配列決定などを含む)である(例えば、非特許文献1〜5)。
【0003】
特異的ハイブリダイゼーションはまた、オリゴヌクレオチドタグで標識された化合物を探知、回収、および同定する方法として提案されている。例えば、複数のDNA配列決定において、オリゴヌクレオチドタグは、同じ配列決定反応において生じるDNAフラグメントからなるゲル上の電気泳動的に分離されたバンドを同定するために用いられる。このように、多くの配列決定反応由来のDNAフラグメントが、ゲルの同じレーンで分離され、次いで異なる固相物質でブロットされる。そこでは、異なる配列決定反応由来のフラグメントバンドが、相補的なタグに特異的にハイブリダイズするオリゴヌクレオチドプローブで可視化される(非特許文献6)。オリゴヌクレオチドタグの同様の用途はまた、爆発物、潜在的な汚染物質(例えば、原油)、および模造品の防止または検出のために通貨を同定するために提供されてきた(例えば、非特許文献7)にレビューされた)。さらに最近、オリゴヌクレオチドタグを用いるシステムはまた、複雑なコンビナトリアルケミカルライブラリー中の個々の分子を操作および同定する方法として(例えば、このような薬物候補物のライブラリーのスクリーニングを助けるために)提案されている(非特許文献8〜10)。
【0004】
このようなタグ化スキームをうまく実施することは、タグとその相補的プローブとの間での特異的なハイブリダイゼーションの達成における成功に大部分依存している。すなわち、オリゴヌクレオチドタグが基質をうまく同定するためには、擬陽性および擬陰性のシグナルの数を最小限にしなければならない。不幸にも、このような偽のシグナルは希ではない。なぜなら塩基対形成および塩基の積み重ね(base stacking)の自由エネルギーは二重鎖または三重鎖構造におけるヌクレオチドのあいだで広く変化するから
である。例えば、その相補物に結合したデオキシアデニン(A)およびチミジン(T)の繰り返し配列からなる二重鎖は、ミスマッチを含む部分的に相補的な標的に結合したデオキシグアニジン(G)およびデオキシシチジン(C)の繰り返し配列からなる同じ長さの二重鎖より安定性が低い可能性がある。従って、大きなコンビナトリアルケミカルライブラリー由来の所望の化合物が前者のオリゴヌクレオチドでタグ化された場合、完全にマッチしたATに富む二重鎖を検出するように設計されたハイブリダイゼーション条件下では、GCに富むオリゴヌクレオチドで標識された所望されない化合物(ミスマッチした二重鎖においてさえ)が、ATに富むタグからなる完全にマッチした二重鎖とともに検出されるという重大な可能性が存在する。非特許文献8(上記)により提案された分子タグ化システムでは、親密に関連するタグの関連するミスハイブリダイゼーションの問題は、いわゆる「無限(comma−less)」コードを用いることにより処理され、これはその相補的タグについて登録をはずれた(またはフレームシフトした)プローブが、その5またはそれ以上の3塩基ワードまたは「コドン」のおのおのについて1つまたはそれ以上のミスマッチを有する二重鎖を生じることを保証する。
【0005】
たとえ塩化テトラメチルアンモニウムのような試薬がオリゴヌクレオチド二重鎖の塩基特異的安定性の相違を無効にするために利用可能であるとしても、このような試薬の効果はしばしば限定され、そしてその存在は選択された化合物のさらなる操作(例えば、ポリメーラーゼ連鎖反応(PCR)などによる増幅)を不可能にし得るか、またはより困難にし得る。
【0006】
このような問題は、例えば多数回PCR、リバースドットブロッティングなどを介する多数のまたは複合した遺伝子座の分析における多数のハイブリダイゼーションプローブの同時使用を非常に困難にしている。その結果、例えばHLA遺伝子のような特定の座の直接配列決定は、遺伝子型の同定のための特異的ハイブリダイゼーションを用いる間接的な方法に対する信頼し得る代替法として奨励される(例えば、非特許文献11))。
【0007】
異なる固相支持体上で、クローン化されかつ同様にタグ化されたDNAを分類する能力は、特に、並行して多くのサンプルに同時に適用し得る非ゲルベースの配列決定方法と連結される場合に、このような配列決定を促進する。
【非特許文献1】Sambrookら、「Molecular Cloning:A Laboratory Manual」 第2版 Cold Spring Harbor Laboratory,New York,1989年
【非特許文献2】KellerおよびManak、「DNA Probes」 第2版 Stockton Press.New York,1993年
【非特許文献3】Milliganら、J.Med.Chem. 第36巻 p.1923−1937 1993年
【非特許文献4】Drmanacら、Science,第260巻 p.1649−1652 1993年
【非特許文献5】Bains J. DNA Sequencing and Mapping,第4巻 p.143−150 1993年
【非特許文献6】Churchら、Science,第240巻 p.185−188 1988年
【非特許文献7】Dollinger、Birkhauser,Boston,Mullisら編、「The Polymerase Chain Reaction」 p.265−274、1994年
【非特許文献8】BrennerおよびLerner. Proc.Natl.Acad.Sci.、第89巻 p.5381−5383 1992年
【非特許文献9】Alper. Science、第264巻 p.1399−1401 1994年
【非特許文献10】Needelsら、Proc.Natl.Acad.Sci.、第90巻 p.10700−10704 1993年
【非特許文献11】Gyllenstenら、Proc.Natl.Acad.Sci.,第85巻 p.7652−7656 1988年
【発明の開示】
【発明が解決しようとする課題】
【0008】
上記を考慮すると、多くのタグのレパートリーを提供するが、また天然の塩基対形成および塩基の積み重ねの自由エネルギーの差違を変化させるための特別な試薬を用いる必要はなく、擬陽性および擬陰性のシグナルの発生を最小限にするオリゴヌクレオチドベースのタグ化システムが利用可能である場合、これは有用である。このようなタグ化システムにより、コンビナトリアルケミカルライブラリーの構築および使用、DNAの大スケールのマッピングおよび配列決定、遺伝子同定、医学診断薬などを含む多くの領域での適用が見い出される。
【0009】
本発明の目的は、化合物の探知、回収、および同定のための分子タグ化システムを提供することである
【課題を解決するための手段】
【0010】
(発明の要旨)
1つの局面において、本発明は、分子集団から分子または分子亜集団を分類する方法を提供し、この方法は、以下の工程:
(a)タグのレパートリー由来のオリゴヌクレオチドタグを、分子集団中の各分子に結
合する工程であって、この工程は、(i)この集団中の実質的に同じ分子または実質的に同じ分子亜集団が、結合した同じオリゴヌクレオチドタグを有し、そしてこの集団中の実質的にすべての異なる分子または異なる分子亜集団が、結合した異なるオリゴヌクレオチドタグを結合しているように、そして(ii)このレパートリー由来の各オリゴヌクレオチドタグが複数のサブユニットを包み、そしてこの複数のサブユニットのそれぞれが3から6ヌクレオチドまたは3から6塩基対の長さを有するオリゴヌクレオチドからなり、このサブユニットが最少に交差ハイブリダイズしているセットから選択されるように行われる工程;および
(b)このオリゴヌクレオチドタグとそれぞれの相補物とを特異的にハイブリダイズすることにより、この集団からこの分子または分子亜集団を分類する工程;
を包含する。
【0011】
1つの実施形態において、上記分子または分子亜集団が、ポリヌクレオチドまたはポリヌクレオチド亜集団である。好ましい実施形態において、上記オリゴヌクレオチドの相補物が、固相支持体に結合される。より好ましい実施形態において、上記固相支持体が、上記相補物の均一な集団が結合している微粒子である。さらにより好ましい実施形態において、上記オリゴヌクレオチドタグおよび上記相補物が、一本鎖オリゴヌクレオチドである。
なおさらにより好ましい実施形態において、上記ポリヌクレオチドまたはポリヌクレオチド亜集団が、50から5000ヌクレオチドの範囲の長さを有する。なおさらにより好ましい別の実施形態において、上記微粒子が、ガラス微粒子、磁気ビーズ、グリシダルメタクリレート微粒子およびポリスチレン微粒子からなる群から選択される。なおさらにより好ましい実施形態において、上記微粒子の直径が1μmと100μmとの間である。さらにより好ましい別の実施形態において、上記固相支持体が、上記相補物の均一な集団を結合している複数の空間的に離れた表面領域を有する平面基板である。なおさらにより好ましい実施形態において、異なる上記の複数の空間的に離れた表面領域が、異なる上記相補物の均一な集団を有する。なおさらにより好ましい実施形態において、上記平面基板が、ガラス、シリコンおよびプラスチックからなる群から選択される。
【0012】
別の実施形態において、上記分子または分子亜集団が、ポリペプチドまたはポリペプチド亜集団である。好ましい実施形態において、上記オリゴヌクレオチドタグの相補物が、固相支持体に結合している。より好ましい実施形態において、上記オリゴヌクレオチドタグおよび上記相補物が、一本鎖オリゴヌクレオチドである。さらにより好ましい実施形態において、上記固相支持体が、上記相補物の均一な集団が結合している微粒子である。なおさらにより好ましい実施形態において、上記微粒子が、ガラス微粒子、磁気ビーズ、およびポリスチレン微粒子からなる群から選択される。なおさらにより好ましい実施形態において、上記固相支持体が、上記相補物の均一な集団を結合している複数の空間的に離れた表面領域を有する平面基板である。なおさらにより好ましい実施形態において、異なる上記の複数の空間的に離れた表面領域が、異なる上記相補物の均一な集団を有する。なおさらにより好ましい実施形態において、上記平面基板が、ガラス、シリコンおよびプラスチックからなる群から選択される。
【0013】
別の実施形態において、上記分子または分子亜集団が、以下の形態の線形ポリマー分子:
−(M−L)n−
を含み、ここで、リン(V)結合基であり;
Mは、1〜20個の炭素原子と、酸素、窒素、およびイオウからなる群から選択される0〜10個のヘテロ原子とを有する、直鎖分子構造、環式分子構造、または分枝有機分子構造であり;そして
nは、3〜100の範囲にある。
【0014】
好ましい実施形態において、Mは、1〜16個の炭素原子を有するアルキル、アルコキシ、アルケニル、またはアリールであるか;3〜8個の炭素原子と、酸素、窒素、およびイオウからなる群から選択される1〜3個のヘテロ原子とを有するヘテロ環である。より好ましい実施形態において、上記オリゴヌクレオチドタグの相補物は、固相支持体に結合されている。さらにより好ましい実施形態において、上記微粒子が、ガラス微粒子、磁気ビーズ、およびポリスチレン微粒子からなる群から選択される。なおさらにより好ましい別の実施形態において、上記固相支持体が、上記オリゴヌクレオチドタグの相補物の均一な集団を内部に結合している複数の離れた非重複表面領域を有する、平面基板である。なおさらにより好ましい実施形態において、異なる上記の複数の空間的に離れた表面領域が、異なる上記相補物の均一な集団を有する。
【0015】
別の局面において、本発明は、標的ポリヌクレオチドのヌクレオチド配列を決定する方法を提供し、この方法は、以下の工程:
この標的ポリヌクレオチドから、この標的ポリヌクレオチドをカバーする複数のフラグメントを生成する工程;
タグのレパートリー由来のオリゴヌクレオチドタグを、この複数のフラグメントのそれぞれに結合する工程であって、この工程は、(i)実質的にすべての同じフラグメントが、結合した同じオリゴヌクレオチドタグを有し、実質的にすべての異なるフラグメントが、異なるオリゴヌクレオチドタグを結合しているように、そして、(ii)このレパートリー由来の各オリゴヌクレオチドタグが複数のサブユニットを包含し、そしてこの複数のサブユニットのそれぞれが、3から6ヌクレオチドまたは3から6塩基対の長さを有するオリゴヌクレオチドからなり、このサブユニットが、最少に交差ハイブリダイズしているセットから選択されるように行われる、工程;
このオリゴヌクレオチドタグとそれぞれの相補物とを特異的にハイブリダイズすることによりこのフラグメントを分類する工程;
この複数のフラグメントのそれぞれの部分のヌクレオチド配列を決定する工程;および
このフラグメントの配列を対照することにより、この標的ポリヌクレオチドのヌクレオチド配列を決定する工程、
を包含する。
【0016】
1つの実施形態において、上記オリゴヌクレオチドタグおよび上記相補物が、固相支持体に結合されている。好ましい実施形態において、上記オリゴヌクレオチドタグおよび上記相補物が、一本鎖オリゴヌクレオチドである。より好ましい実施形態において、上記生成工程が、上記標的ポリヌクレオチドのランダムに重複するフラグメントを生成する。さらにより好ましい実施形態において、上記フラグメントのヌクレオチド配列を決定する工程が、単一塩基配列決定方法により、上記の複数のフラグメントについて同時に行われる。なおさらにより好ましい実施形態において、上記フラグメントのそれぞれの部分が、12から50ヌクレオチドを含む。なおさらにより好ましい実施形態において、上記フラグメントのそれぞれの部分が12から25ヌクレオチドを含む。なおさらにより好ましい実施形態において、上記標的ポリヌクレオチドが、1キロベース長と50キロベース長との間の長さである。好ましい別の実施形態において、上記固相支持体が、上記相補物の均一な集団をそれぞれ結合している複数の微粒子である。より好ましい実施形態において、上記分類工程後、上記の複数の微粒子が平面基板に固定される。さらにより好ましい実施形態において、上記の複数の微粒子が、1平方センチメートル当たり約1000微粒子から約100,000微粒子の間の密度で、上記平面基板の表面にランダムに配置される。
【0017】
別の局面において、本発明は、組成物を提供し、この組成物は:
1つまたはそれ以上の空間的に離れた領域を有する固相支持体;および
この1つまたはそれ以上の空間的に離れた領域の少なくとも1つにおいてこの固相支持
体に共有結合されたオリゴヌクレオチドタグ相補物の均一な集団であって、このオリゴヌクレオチドタグ相補物は複数のサブユニットを含み、各サブユニットは3から6ヌクレオチドの長さを有するオリゴヌクレオチドからなり、そして各サブユニットは、最少の交差ハイブリダイズしているセットから選択される、オリゴヌクレオチドタグ相補物の均一な集団、
を含む。
【0018】
1つの実施形態において、上記の複数のサブユニットが4個から10個の範囲にある。好ましい実施形態において、上記固相支持体が、単一の空間的に離れた領域を有する微粒子である。より好ましい実施形態において、上記微粒子が、ガラス微粒子、磁気ビーズおよびポリスチレン微粒子からなる群から選択される。
【0019】
別の局面において、本発明は、ポリヌクレオチド集団を分類する方法を提供し、この方法は、以下の工程:
オリゴヌクレオチドタグを、この集団の各ポリヌクレオチドに結合させる工程であって、この工程は、(i)実質的にすべての同じポリヌクレオチドが、結合した同じオリゴヌクレオチドタグを有し、実質的にすべての異なるポリヌクレオチドが、結合した異なるオリゴヌクレオチドタグを有しているように、そして(ii)各オリゴヌクレオチドタグが複数のサブユニットを包含し、そしてこの複数のサブユニットのそれぞれが、3から6ヌクレオチドの長さを有するオリゴヌクレオチドからなり、このサブユニットが、最少に交差ハイブリダイズしているセットから選択されるように行われる工程;
このオリゴヌクレオチドタグとそれぞれの相補物とを特異的にハイブリダイズすることにより、このポリヌクレオチドを分類する工程;
この分類されたポリヌクレオチドのそれぞれの部分のヌクレオチド配列を決定する工程;および
このポリヌクレオチドの配列の部分の頻度分布により、このポリヌクレオチド集団を分類する工程、
を包含する。
【0020】
1つの実施形態において、上記オリゴヌクレオチドタグの相補物が、固相支持体に共有結合される。好ましい実施形態において、上記固相支持体が微粒子であり、そして上記相補物の均一な集団が、この微粒子各々に結合される。より好ましい実施形態において、上記ポリヌクレオチドの集団が、cDNAライブラリーである。さらにより好ましい実施形態において、上記ポリヌクレオチドの部分が、12から50ヌクレオチドの範囲にある。なおさらにより好ましい実施形態において、上記ポリヌクレオチドの部分が、12から25ヌクレオチドの範囲にある。
【0021】
別の局面において、本発明は、オリゴヌクレオチドタグのレパートリーを提供し、このレパートリーは、以下の形態のオリゴヌクレオチド:
S1S2S3...Sn
からなる群から選択され、ここで、S1からSnのそれぞれは、3から6ヌクレオチドの長さを有するオリゴヌクレオチドからなり、そして最少に交差ハイブリダイズしているセットから選択されるサブユニットであり;そして
nは4から10の範囲にある。
【0022】
1つの実施形態において、上記サブユニットが、4から5ヌクレオチドの長さを有するオリゴヌクレオチドからなり、そして上記nが6から9の範囲にある。
【0023】
別の局面において、本発明は、オリゴヌクレオチドタグをポリヌクレオチドへ結合させるためのクローニングベクターのレパートリーを提供し、このレパートリーは、以下の形
態の二本鎖要素:
S1S2S3...Sn
を有し、ここで、S1からSnのそれぞれは、3から6ヌクレオチドの長さを有するオリゴヌクレオチドからなり、そして最少に交差ハイブリダイズしているセットから選択されるサブユニットであり;そして
nは4から10の範囲にある。
【0024】
1つの実施形態において、上記二本鎖要素が、上記ポリヌクレオチドを上記クローニングベクターに挿入するための1つまたはそれ以上の制限エンドヌクレアーゼ切断部位を含むポリリンカー領域に、隣接する。
【0025】
別の局面において、本発明は、ポリヌクレオチド混合物を分類する方法であって、この方法は、以下の工程:
ポリヌクレオチド混合物を含有する溶液を提供する工程であって、この混合物の各ポリヌクレオチドは、タグのレパートリー由来のオリゴヌクレオチドタグを結合しており、このレパートリー由来の各オリゴヌクレオチドタグは、複数のサブユニットを含み、この複数のサブユニットのそれぞれは、3から6ヌクレオチドの長さを有するオリゴヌクレオチドからなり、このサブユニットは、最少に交差ハイブリダイズしているセットから選択される工程;
このポリヌクレオチド混合物をサンプリングしてポリヌクレオチド亜集団を形成させる工程であって、ここで同じ配列の実質的にすべてのポリヌクレオチドは、結合した同じオリゴヌクレオチドタグを有し、異なる配列の実質的にすべてのポリヌクレオチドは異なるオリゴヌクレオチドタグを結合している、工程;および
このオリゴヌクレオチドタグとそれぞれの相補物との間に完全にマッチした二重鎖の形成を促進する条件下で、この亜集団と、このオリゴヌクレオチドタグの相補物を結合している1つまたはそれ以上の固相支持体とを接触させる工程、
を包含する。
【0026】
1つの実施形態において、上記固相支持体が、上記相補物の均一な集団を結合している微粒子である。好ましい実施形態において、上記オリゴヌクレオチドタグおよび上記相補物が、一本鎖オリゴヌクレオチドである。より好ましい実施形態において、上記ポリヌクレオチドが50から5000ヌクレオチドの範囲の長さを有する。さらにより好ましい実施形態において、上記微粒子が、ガラス微粒子、磁気ビーズ、グリシダルメタクリレート微粒子およびポリスチレン微粒子からなる群から選択される。なおさらにより好ましい実施形態において、上記固相支持体が、上記相補物の均一な集団を結合している複数の空間的に離れた表面領域を有する平面基板である。なおさらにより好ましい実施形態において、上記の異なる複数の空間的に離れた表面領域が、異なる上記相補物の均一な集団を有する。
【0027】
二本鎖DNAの末端に、予め決定された長さの一本鎖を生成させる方法であって、該方法は、以下の工程:
この二本鎖DNAの末端に5’鎖を供給する工程であって、この工程は、このような5’鎖は、3種以下のヌクレオチドからなる第1の群から選択されるヌクレオチドから構成されるように、そしてこの5’鎖のあらかじめ決定された長さが、この第1の群には存在しない種類の第2のヌクレオチドによって規定されるように、行われる工程;および
この第2のヌクレオチドのヌクレオシド三リン酸の存在下で、この二本鎖DNAの末端を、T4 DNAポリメラーゼに曝露する工程;
を包含する。
【0028】
cDNAライブラリーにおいて新規なcDNA分子を検出する方法であって、この方法
は、以下の工程:
mRNA分子集団からcDNAライブラリーを形成する工程であって、このcDNAライブラリー中の各cDNA分子は、結合したオリゴヌクレオチドを有し、この工程は、(i)実質的にすべての同じcDNA分子が、結合した同じオリゴヌクレオチドタグを有し、実質的にすべての異なるcDNA分子が、結合した異なるオリゴヌクレオチドタグを有しているように、そして(ii)各オリゴヌクレオチドタグが、複数のサブユニットを含み、そしてこの複数のサブユニットのそれぞれが、3から6ヌクレオチドの長さを有するオリゴヌクレオチドからなり、このサブユニットが、最少に交差ハイブリダイズしているセットから選択されるように行われる工程;
このオリゴヌクレオチドタグとそれぞれの相補物とを特異的にハイブリダイズすることにより、このcDNA分子を分類する工程;
この分類されたcDNA分子のそれぞれの部分のヌクレオチド配列を決定する工程;
この分類されたcDNA分子の配列の部分のヌクレオチド配列を、既知のcDNA分子の配列と比較することにより、新規なcDNA分子を同定する工程、
を包含する。
【0029】
1つの実施形態において、上記オリゴヌクレオチドタグの相補物が、固相支持体に共有結合される。好ましい実施形態において、上記固相支持体が微粒子であり、そして上記相補物の均一な集団が、この微粒子各々に結合される。より好ましい実施形態において、上記cDNA分子の部分が、12から50ヌクレオチドの範囲にある。さらにより好ましい実施形態において、上記cDNA分子の部分が、12から25ヌクレオチドの範囲にある。
【0030】
本発明の目的は、化合物の探知、回収、および同定のための分子タグ化システムを提供することである。
【0031】
本発明の別の目的は、オリゴヌクレオチドタグおよびその相補物の特異的ハイブリダイゼーションにより固相物質の表面上の同一の分子、または分子のサブクラス(特にポリヌクレオチド)を分類するための方法を提供することである。
【0032】
本発明のさらなる目的は、コンビナトリアルケミカルライブラリーを提供することであり、それらのメンバー化合物は、オリゴヌクレオチドタグおよびその相補物の特異的ハイブリダイゼーションにより同定される。
【0033】
本発明のなおさらなる目的は、同時分析および/または配列決定用の標的ポリヌクレオチドの数千ものフラグメント(特にランダムに重複するフラグメント)をタグ化および分類するためのシステムを提供することである。
【0034】
本発明の別の目的は、数百塩基対から数万塩基対の範囲の長さを有する標的ポリヌクレオチドを配列決定するための迅速かつ信頼できる方法を提供することである。
【0035】
本発明は、オリゴヌクレオチドタグの使用により分子の種類または亜集団を探知、同定、および/または分類するための方法および物質を提供することにより、これらおよび他の目的を達成する。本発明のオリゴヌクレオチドタグは多くのサブユニットからなり、各サブユニットは3から6ヌクレオチド長のオリゴヌクレオチドからなる。オリゴヌクレオチドタグのサブユニットは、最少に交差ハイブリダイズしているセット(minimally cross−hybridizing set)から選択される。このようなセットでは、このセットのサブユニットおよびこのセットの任意の他のサブユニットの相補物からなる二重鎖または三重鎖は少なくとも2つのミスマッチを含む。言い換えると、最少に交差ハイブリダイズしているセットのサブユニットは最良でも、同じセットの任意の他
のサブユニットの相補物と2つのミスマッチを有する二重鎖または三重鎖を形成する。特定の実施態様に用いられ得るオリゴヌクレオチドタグの数は、タグあたりのサブユニット数およびサブユニットの長さに依存する。その数は、一般的にタグの長さの全ての可能性のある配列の数(1つのタグがnヌクレオチド長であるタグの数は4nである)よりずっと少ない。より好ましくは、サブユニットは4から5ヌクレオチド長のオリゴヌクレオチドである。
【0036】
本発明の1つの局面において、固相支持体に結合したオリゴヌクレオチドタグの相補物は、ポリヌクレオチドをそれぞれタグを含むポリヌクレオチドの混合物から分類するために用いられる。この実施態様では、オリゴヌクレオチドタグの相補物は、同一配列の集団が特定の領域で産生されるように、微細ビーズのような固相支持体の表面上、または単一の支持体上の合成位置のアレイ(array)上の特定の位置で合成される。すなわち、各支持体(ビーズの場合)または各領域(アレイの場合)の表面は、特定の配列を有する唯一のタイプの相補物によって誘導体化される。このようなビーズ集団または領域集団は別個の配列を有する相補物のレパートリーを含み、そのレパートリーのサイズはオリゴヌクレオチドタグあたりのサブユニットの数および用いられたサブユニットの長さに依存する。同様に、分類すべきポリヌクレオチドは、同一のポリヌクレオチドが同じタグを有し、そして異なるポリヌクレオチドが異なるタグを有するようにレパートリー中のオリゴヌクレオチドタグをおのおの含む。下記でより十分に説明するように、この条件は、タグ化されたポリヌクレオチドの完全なアンサンブルからタグ化されたポリヌクレオチドの十分に小さいサンプルを取ることにより達成される。従って、支持体およびポリヌクレオチドの集団がそれぞれの相補物とオリゴヌクレオチドタグの特異的ハイブリダイゼーションを可能にする条件下で混合される場合、同一ポリヌクレオチドの亜集団は特定のビーズまたは領域上に分類される。次いでポリヌクレオチドの亜集団はミクロ生化学的技法により固相支持体上で操作され得る。
【0037】
一般的に、本発明の方法は以下の工程を包含する:(a)分子集団中の各分子にタグのレパートリー由来のオリゴヌクレオチドタグを結合して、(i)集団中の実質的に全ての同じ分子または分子の同じ亜集団は結合した同じオリゴヌクレオチドタグを有し、そして集団中の実質的に全ての異なる分子または分子の異なる亜集団は結合した異なるオリゴヌクレオチドタグを有し、ならびに(ii)レパートリー由来の各オリゴヌクレオチドタグは多数のサブユニットを含有し、そして多数の各サブユニットは3から6ヌクレオチドまたは3から6塩基対の長さを有するオリゴヌクレオチドからなり、そのサブユニットが最少に交差ハイブリダイズしているセットから選択される、工程:および(b)それらのそれぞれの相補物とオリゴヌクレオチドタグを特異的にハイブリダイズすることにより、集団の分子または分子の亜集団を分類する、工程。
【0038】
本発明の重要な局面は、並行する配列決定のためにポリヌクレオチドを分類するためのオリゴヌクレオチドタグの使用である。好ましくは、このような配列決定は以下の工程により行われる:(a)標的ポリヌクレオチドをカバーする多数のフラグメントを標的ポリヌクレオチドから生成する、工程;(b)タグのレパートリー由来のオリゴヌクレオチドタグをその多数の各フラグメントに結合して、(i)実質的に全ての同じフラグメントは、結合した同じオリゴヌクレオチドタグを有し、そして実質的に全ての異なるフラグメントは、結合した異なるオリゴヌクレオチドタグを有し、ならびに(ii)そのレパートリー由来の各オリゴヌクレオチドタグは多数のサブユニットを含有し、そして多数の各サブユニットは3から6ヌクレオチドまたは3から6塩基対の長さを有するオリゴヌクレオチドからなり、そのサブユニットが最少に交差ハイブリダイズしているセットから選択される、工程;
それらのそれぞれの相補物とオリゴヌクレオチドタグを特異的にハイブリダイズすることによりフラグメントを分類する、工程;(c)下記のように、好ましくは単一塩基配列
決定方法により、多数の各フラグメントの一部のヌクレオチド配列を決定する工程;ならびに(d)標的ポリヌクレオチドのヌクレオチド配列を、フラグメントの配列を対照することにより決定する、工程。
【0039】
本発明は、オリゴヌクレオチドで分子をタグ化または標識する現在の方法の鍵となる欠点を克服する。本発明に従ってタグの配列をコードすることにより、タグと別のタグに対する相補物との間でミスマッチした任意の二重鎖または三重鎖の安定性は、タグとそれ自身の相補物との間で完全にマッチした任意の二重鎖の安定性よりはるかに低い。従って、不正確な分類の問題は、GCに富むタグのミスマッチした二重鎖が完全にマッチしたATに富むタグよりも安定であるため、除去される。
【0040】
微細ビーズのような固相支持体と組み合わせて用いた場合、本発明は、大スケールのDNA配列決定のような大スケールの並行操作において特に有用なポリヌクレオチドを操作および分類するための容易な自動化システムを提供する。ここでは、多くの標的ポリヌクレオチドまたは単一標的ポリヌクレオチドの多くの部分が同時に配列決定および/または分析される。
【0041】
(定義)
オリゴヌクレオチドタグに関して本明細書中に用いられる「相補物」または「タグ相補物」は、オリゴヌクレオチドタグが特異的にハイブリダイズして完全にマッチした二重鎖または三重鎖を形成するオリゴヌクレオチドをいう。特異的ハイブリダイゼーションが三重鎖を生じる実施態様では、オリゴヌクレオチドタグは2本鎖または1本鎖のいずれかであることが選択され得る。従って、三重鎖が形成される場合、用語「相補物」は、1本鎖オリゴヌクレオチドタグの2本鎖相補物または2本鎖オリゴヌクレオチドタグの一本鎖相補物のいずれかを包含することを意味する。
【0042】
本明細書で用いられる用語「オリゴヌクレオチド」は、モノマー−モノマー相互作用の通常のパターン(例えば、Watson−Crick型の塩基対形成、塩基の積み重ね、Hoogsteenまたは逆Hoogsteen型の塩基対形成など)を介して標的ポリヌクレオチドに特異的に結合し得る、天然のまたは改変されたモノマーまたは連結物(デオキシリボヌクレオシド、リボヌクレオシド、それらのαアノマー型、ペプチド核酸(PNA)などを含む)の直鎖オリゴマーを含む。通常、モノマーはホスホジエステル結合またはそのアナログにより連結され、少しのモノマーユニット(例えば3〜4)から数十のモノマーユニットの大きさの範囲にわたってオリゴヌクレオチドを形成する。オリゴヌクレオチドが一連の文字(例えば、「ATGCCTG」)により示される場合はいつも、ヌクレオチドは左から右に5’→3’の順であり、他に記載されない限り、「A」はデオキシアデノシンを示し、「C」はデオキシシチジンを示し、「G」はデオキシグアノシンを示し、および「T」はチミジンを示すことが理解される。ホスホジエステル結合のアナログは、ホスホロチオエート、ホスホロジチオエート、ホスホルアニリデート、ホスホルアミデートなどを含む。通常、本発明のオリゴヌクレオチドは4つの天然のヌクレオチドを包含する;しかし、それらは非天然のヌクレオチドアナログも含有し得る。天然または非天然のヌクレオチドを有するオリゴヌクレオチドが用いられ得る場合(例えば、酵素によりプロセスされる必要がある場合)、通常天然のヌクレオチドからなるオリゴヌクレオチドが必要であることは当業者には明らかである。
【0043】
二重鎖に関して「完全にマッチした」は、各鎖中の全てのヌクレオチドが他鎖中のヌクレオチドとWatson−Crick塩基対形成を受けるように、二重鎖を作るポリヌクレオチド鎖またはオリゴヌクレオチド鎖が、互いに2本鎖構造を形成することを意味する。この用語はまた、用いられ得るヌクレオシドアナログ(例えば、デオキシイノシン、2−アミノプリン塩基を有するヌクレオシドなど)の対形成を含む。三重鎖に関しては、こ
の用語は三重鎖が完全にマッチした二重鎖および第3鎖からなり、ここで、全てのヌクレオチドが完全にマッチした二重鎖の塩基対とHoogsteenまたは逆Hoogsteen会合を受けることを意味する。逆に、二重鎖におけるタグとオリゴヌクレオチドとの間の「ミスマッチ」は、二重鎖または三重鎖におけるヌクレオチドの対またはトリプレット(triplet)がWatson−Crickおよび/またはHoogsteenおよび/または逆Hoogsteen結合を受け損なうことを意味する。
【0044】
本明細書中に用いられる「ヌクレオシド」は、2’−デオキシ型および2’−ヒドロキシル型(例えば、KornbergおよびBaker、DNA Replication、第2版(Freeman,San Francisco,1992)に記載されたように)を含む天然ヌクレオシドを含む。ヌクレオシドに関して「アナログ」は、改変された塩基部分および/または改変された糖部分を有する合成ヌクレオシド(例えば、Scheit,Nucleotide Analogs(John Wiley,New York,1980);UhlmanおよびPeyman,Chemical Reviews,90:543−584(1990)などに記載された)を、それらが特異的ハイブリダイゼーションをし得るという条件つきで含む。このようなアナログは、結合特性を増強し、退化を低減し、特異性を増すなどのように設計された合成ヌクレオシドを包含する。
【0045】
(発明の詳細な説明)
本発明は、分子(特にポリヌクレオチド)をオリゴヌクレオチドタグを用いることにより標識および分類する方法を提供する。本発明のオリゴヌクレオチドタグは、サブユニットの最少に交差ハイブリダイズしているセットから選択される多数の「ワード」またはサブユニットを包含する。このようなセットのサブユニットは、2つより少なくミスマッチしたヌクレオチドを有する同じセットの別のサブユニットの相補物と二重鎖または三重鎖を形成し得ない。従って、二重鎖を形成するレパートリーの任意の2つのオリゴヌクレオチドタグの配列は、2ヌクレオチドの異なりより「厳密(close)」でない。特定の実施態様では、レパートリーの任意の2つのオリゴヌクレオチドタグの配列は、例えばサブユニットが3つより少なくミスマッチしたヌクレオチドを有する同じセットの別のサブユニットの相補物と二重鎖を形成し得ないように最少に交差ハイブリダイズしているセットを設計することなどにより、「さらに」離れることさえあり得る。このような実施態様では、より大きい特異性が達成されるが、タグの総レパートリーはより小さい。従って、一定の長さおよびワードサイズのタグについて、所望の特異性の程度と所望のレパートリーのサイズとの間で取引がされるに違いない。本発明は、配列決定、フィンガープリンティング、または他のタイプの分析のような並行操作のためにポリヌクレオチドを標識および分類するのに特に有用である。
【0046】
(サブユニットの最少に交差ハイブリダイズしているセットからのオリゴヌクレオチドタグの構築)
任意の最少に交差ハイブリダイズしているセット用のサブユニットのヌクレオチド配列は、図3に図示した一般アルゴリズムに従い、そしてソースコードが付表Iに挙げられるプログラムminhxにより例示されたような簡単なコンピュータプログラムにより便利に計算される。minhxは、3種のヌクレオチドからなるサブユニットを有し、そして4の長さを有する全ての最少に交差ハイブリダイズしているセットを算定する。
【0047】
図3のアルゴリズムは最少に交差ハイブリダイズしているセットのサブユニットの特性、すなわちメンバー間の塩基の長さ、数の差異、および組成(例えば、2、3、または4種の塩基からなる)を最初に定義することにより実施される。一定の長さおよび組成の全ての可能性のある配列からなるテーブルMn・n=1が生じる(100)。最初のサブユニットS1を選択し、そしてテーブルの最後までi=n+1である連続するサブユニットSiと比較する(120)。連続するサブユニットが最少に交差ハイブリダイズしている
セットのメンバーであるために必要なミスマッチの数を有する場合はいつも、新たなテーブルMn+1(125)にたくわえられ、これは前述の工程120までの経路で以前に選択されたサブユニットをも含む。例えば、比較の第一セットでは、M2はS1を含む:比較の第二セットでは、M3はS1およびS2を含む:比較の第三セットでは、M4はS1、S2、およびS3を含む:など。同様に、テーブルMjにおける比較は、Sjと全ての連続サブユニットMjとの間にある。各連続するテーブルMn+1は、サブユニットが工程130までの連続する経路において除去されるので、その前の物より小さいことに気を付ける。テーブルMnの全てのサブユニットが比較された(140)後に、古いテーブルは新たなテーブルMn+1に置き換わり、そして次のラウンドの比較が開始される。選択されたサブユニットSiに比較するための連続するサブユニットを含まないテーブルMn(すなわちMn=Mn+1)が達する場合、プロセスは停止する(160)。
【0048】
好ましくは、最少に交差ハイブリダイズしているセットはセットの全ての他のサブユニットのように二重鎖安定性にほぼ等しい寄与をするサブユニットを含有する。このように、全てのサブユニットとその相補物との間で完全にマッチした二重鎖の安定性はほぼ等しい。このようなセットを選択するためのガイダンスは至適なPCRプライマーの選択および二重鎖安定性の計算についての報告された技術により提供される、例えばRychlikら、Nucleic Acids Research,17:8543−8551(1989)および18:6409−6412(1990):Breslauerら、Proc.Natl.Acad.Sci.,83:3746−3750(1986):Wetmur,Crit. Rev. Biochem. Mol. Biol.,26:227−259(1991)など。より短いタグ(例えば、約30ヌクレオチドまたはそれより少ない)について、RychlikおよびWetmurにより記載されたアルゴリズムが好ましく、より長いタグ(例えば、約30〜35ヌクレオチドまたはそれより大きい)については、Suggsら、Brown編のICN−UCLA Symp. Dev. Biol.,第23巻(Academic Press,New York,1981)の683−693頁に開示された演算法が便利に用いられ得る。明らかに、本発明の範囲内で最少に交差ハイブリダイズしているサブユニットのセットを設計するために当業者に利用可能な多くのアプローチがある。例えば、サブユニットを組み立てる場合、末端ヌクレオチドの異なる塩基の積み重ねエネルギーの影響を最小にするために、サブユニットは同じ末端ヌクレオチドを有するように提供され得る。このように、サブユニットを連結する場合、全ての隣接する末端ヌクレオチドの塩基の積み重ねエネルギーの総量は同じであり、これによりタグ融解温度の多様性が低減または除去される。
【0049】
最少に交差ハイブリダイズしているセットの好ましい実施態様は、そのサブユニットが4つの天然ヌクレオチドのうちの3つからなるセットである。下記でより詳細に述べるように、オリゴヌクレオチドタグ中のヌクレオチドの1タイプのヌクレオチドの欠損は、DNAポリメラーゼの5’→3’エクソヌクレアーゼ活性の使用により標的ポリヌクレオチドが固相支持体上にロードされることを可能にする。以下は、A、G、およびTからなる群から選択される4つのヌクレオチドをおのおの含有するサブユニットの最少に交差ハイブリダイズしているセットの例である:
【0050】
【表1】

このセットでは、各メンバーは全ての他のメンバーの相補物と3つミスマッチした塩基を有する二重鎖を形成する。
【0051】
さらなる模範的な最少に交差ハイブリダイズしているセットが下記の表Iに挙げられる。明らかに、さらなるセットは異なるグループのヌクレオチドを置換することにより、または公知の最少に交差ハイブリダイズしているセットのサブユニットを用いることにより生じ得る。
【0052】
【表2】
本発明のオリゴヌクレオチドタグおよびそれらの相補物は、自動化DNA合成機(例えば、Applied Biosystems,Inc.(Foster City,California)モデル392または394型DNA/RNA合成機)で、ホスホルアミダイトケミストリーのような標準ケミストリー(例えば、以下の参考文献で開示される)を用いて便利に合成される:BeaucageおよびIyer. Tetrahedron. 48:2223−2311(1992);Molkoら、米国特許第4,980,460号;Kosterら、米国特許第4,725,677号;Caruthersら、米国特許第4,415,732号;第4,458,066号;および第4,973,679号;など。得られたオリゴヌクレオチドが特異的ハイブリダイゼーションをし得るという条件で、別のケミストリー(例えば、ホスホロチオエート、ホスホルアミデートなど
のような非天然骨格基(backbone group)で生じる)も用いられ得る。いくつかの実施態様では、タグは酵素によりプロセスまたは操作され得る天然に存在するヌクレオチドを含有し得、対応するタグ相補物は、分類の間、より安定な二重鎖の形成を促進するペプチド核酸または同様の化合物のような非天然ヌクレオチドアナログを含有し得る。
【0053】
微粒子が支持体として用いられる場合、オリゴヌクレオチドタグおよびタグ相補物のレパートリーは、好ましくは、例えば、Shortleら、国際特許出願PCT/US93/03418に開示されるような「分裂および混合(split and mix)」技術を介するサブユニット様式(subunit−wise)合成により生じる。簡単に述べると、合成の基本ユニットは、オリゴヌクレオチドタグのサブユニットである。好ましくは、ホスホルアミダイトケミストリーを用い、3’ホスホルアミダイトオリゴヌクレオチドが最少に交差ハイブリダイズしているセット(例えば、上記に最初に挙げたセットについて、8つの4マー3’−ホスホルアミダイトがある)における各サブユニット用に調製される。合成は、Shortleらにより開示されたように、またはヌクレオシドモノマーを用いる種々のオリゴヌクレオチドライブラリーを生じるために用いられた技術(例えば、Teleniusら、Genomics,13:718−725(1992);Welshら、Nucleic Acids Research,19:5275−5279(1991);Grothuesら、Nucleic Acids Research,21:1321−1322(1993);Hartley、ヨーロッパ特許出願第90304496.4号;Lamら、Nature,354;82−84(1991);Zuckermanら、Int. J. Pept. Protein Research,40:498−507(1992);などに開示された)に直接類似してプロセスされる。一般的に、これらの技術はカップリング工程の間、成長するオリゴヌクレオチドへの活性化されたモノマーの混合物の適用を単に必要とする。
【0054】
2本鎖型のタグは、相補物鎖を別個に合成し、続いて二重鎖形成を可能にする条件下で混合することにより作製され得る。あるいは、2本鎖タグは、プライマー結合部位として働く公知のオリゴヌクレオチド配列に連結した一本鎖レパートリーを最初に合成することにより形成され得る。次いで、第2の鎖はプライマーと一本鎖レパートリーとを結合し、ポリメラーゼで伸長することにより合成される。この後者のアプローチは、Oliphantら、Gene,44:177−183(1986)に記載される。次いで、このような二重鎖タグは、本発明に従って標的ポリヌクレオチドの分類および操作用の標的ポリヌクレオチドと一緒にクローニングベクターに挿入され得る。
【0055】
特異的なハイブリダイゼーションが三重鎖形成を経て起きる実施態様では、タグ配列のコード化は、二重鎖形成タグについての原理と同じ原理に従う。しかし、サブユニット配列の選択において、さらなる制約が存在する。一般に、結合のHoogsteen型を経た第3鎖の会合は、2本鎖標的においてホモピリミジン−ホモプリントラックに沿って最も安定である。通常、塩基トリプレットは、T−A*TまたはC−G*Cモチーフ(ここで、「−」はWatson−Crick対形成を示し、そして「*」は結合のHoogsteen型を示す)を形成する。しかし、他のモチーフもまた可能である。例えば、Hoogsteen塩基対形成は、鎖の状態および組成に依存して、第3鎖(Hoogsteen鎖)と第3鎖に結合する二重鎖のプリンに富む鎖との間に平行および逆平行(antiparallel)の配向を可能にする。特定の実施態様で所望されるように、三重鎖の安定性を最大にするか、またはそうでなければ調節するための、適切な配列、配向、条件、ヌクレオシド型(例えば、リボースヌクレオシドまたはデオキシリボースヌクレオシドのどちらが用いられるか)、塩基修飾 (例えば、メチル化シトシンなど)の選択についての広範囲にわたるガイダンスが文献中に見られる。例えば、Robertsら、Proc.Natl.Acad.Sci.、88:9397−9401(1991);Rob
ertsら、Science、258:1463−1466(1992);Distefanoら、Proc.Natl.Acad.Sci.、90:1179−1183(1993);Mergnyら、Biochemistry、30:9791−9798(1991);Chengら、J.Am.Chem.Soc.、114:4465−4474(1992);BealおよびDervan、Nucleic Acids Research、20:2773−2776(1992);BealおよびDervan、J.Am.Chem.Soc.、114:4976−4982(1992);Giovannangeliら、Proc.Natl.Acad.Sci.、89:8631−8635(1992);MoserおよびDervan、Science、238:645−650(1987);McShanら、J.Biol.Chem.、267:5712−5721(1992);Yoonら、Proc.Natl.Acad.Sci.、89:3840−3844(1992);Blumeら、Nucleic Acids Research、20:1777−1784(1992);ThuongおよびHelene、Angew.Chem.Int.Ed.Engl.、32:666−690(1993)など。1本鎖タグまたは二重鎖タグをそれらの1本鎖相補物または二重鎖相補物にアニールするための条件は、周知である(例えば、Jiら、Anal.Chem.、65:1323−1328(1993))。
【0056】
本発明のオリゴヌクレオチドタグは、12から60までのヌクレオチドまたは塩基対を長さの範囲とし得る。好ましくは、オリゴヌクレオチドタグは、18から40までのヌクレオチドまたは塩基対を長さの範囲とする。さらに好ましくは、オリゴヌクレオチドタグは、25から40までのヌクレオチドまたは塩基対を長さの範囲とする。サブユニットの好ましい数およびさらに好ましい数に関して、それらの範囲は、以下のように表され得る:
【0057】
【表3】
最も好ましくは、オリゴヌクレオチドタグが1本鎖であり、そして特異的なハイブリダイゼーションが、タグ相補物とのWatson−Crick対形成により起こる。
【0058】
(分子へのタグの結合)
オリゴヌクレオチドタグは、当該分野で周知の種々の反応性官能基により、多くの異なる種類の分子に結合し得る(例えば、Haugland,Handbook of Fluorescent Probes and Research Chemicals(Molecular Probes,Inc.,Eugene,1992);Khannaら、米国特許第4,318,846号など)。表IVは、オリゴヌクレオチドタグまた
は目的の分子上に存在し得る、例となる官能基およびカウンターパート反応基を提供する。官能基およびカウンターパート反応体がともに反応する場合、いくつかの場合では活性化の後に結合基が形成される。さらに、以下により十分に記載されるように、タグは、選択された分子と同時に合成され、コンビナトリアルケミカルライブラリーを形成し得る。
【0059】
【表4】
コンビナトリアルケミカルライブラリーの生成において特に都合の良い分子の種類は、以下の形態の線形ポリマー分子を含む:
−(M−L)n−
ここで、Lはリンカー(linker)部分であり、そしてMは、広い範囲の化学構造から選択され得、不活性な非立体的に障害となるスペーサー部分として働くことから、他の成分に結合する分岐点、標識に結合するための部位:オリゴヌクレオチドに結合するための部位、あるいは治療標的にハイブリダイズまたは結合するための他の結合ポリマー:または溶解性、二重鎖および/または三重鎖形成の促進に影響を与える他の基を結合するための部位として働き得る反応性官能基を提供することまでの範囲の機能を提供するモノマー(例えば、インターカレーター、アルキル化剤など)である。このような線形ポリマー分子の配列およびその結果の組成は、BrennerおよびLerner(上記)により教示されたように、タグに結合するポリヌクレオチド内にコードされ得る。しかし、選択事象後、選択分子のタグを増幅し、次に配列決定する代わりに、タグそれ自体または付加
コードセグメントは、目的の分子の放出後、例えば、タグ中の操作された部位の制限消化により、直接的に(いわゆる以下に記載の「単一塩基」アプローチを用いて)配列決定され得る。明らかに、タグ部位の同時合成と両立できる一連のケミストリー工程により生産される任意の分子は、コンビナトリアルライブラリーの生成に用いられ得る。
【0060】
都合良く、コンビナトリアルライブラリーの生成に用いられ得る広い多様性のホスフェート結合モノマーが存在する。以下の参考文献は、本発明での使用に適する、いくつかのホスホルアミダイトおよび/または水素ホスホネートモノマーを開示し、そしてそれらの合成およびオリゴヌクレオチドへの包含に対するガイダンスを提供する:Newtonら、Nucleic Acids Research、21:1155−1162(1993);Griffinら、J.Am.Chem.Soc.、114:7976−7982(1992);Jaschkeら、Tetrahedron Letters、34:301−304(1992);Maら、国際出願PCT/CA92/00423;Zonら、国際出願PCT/US90/06630;Durandら、Nucleic Acids Research、18:6353−6359(1990);Salunkheら、J.Am.Chem.Soc.、114:8768−8772(1992);Urdeaら、米国特許第5,093,232号;Ruth、米国特許第4,948,882号;Cruickshank、米国特許第5,091,519号;Haralambidisら、Nucleic Acids Research、15:4857−4876(1987)など。さらに特に、Mは、1〜20個の炭素原子、ならびに酸素、窒素、およびイオウからなる群から選択される0〜10個のヘテロ原子を有する直鎖、環式、または分枝有機分子構造であり得る。好ましくは、Mは、1〜16個の炭素原子を有するアルキル、アルコキシ、アルケニル、またはアリール ;3〜8個の炭素原子および酸素、窒素、およびイオウからなる群から選択される1〜3個のヘテロ原子を有するヘテロ環;グリコシル;またはヌクレオシジルである。さらに好ましくは、Mは、1〜8個の炭素原子を有する、アルキル、アルコキシ、アルケニル、またはアリール;グリコシル;またはヌクレオシジルである。
【0061】
好ましくは、Lはリン(V)結合基であり、このリン結合基は、ホシホジエステル、ホスホトリエステル、メチルホスホネートまたはエチルホスホネート、ホスホロチオエート、ホスホロジチオエート、ホスホルアミデートなどであり得る。一般に、ホスホルアミダイトまたは水素ホスホネート前駆体由来の結合が好ましく、そのため本発明の線形ポリマー単位は、市販の自動化DNA合成機(例えば、Applied Biosystems,Inc.(Foster City,CA)モデル394など)で都合良く合成され得る。
【0062】
nは、MおよびLの性質に依存して顕著に変化し得る。通常、nは、約3から約100まで変化する。Mが、ヌクレオシドまたはそのアナログまたはヌクレオシドサイズのモノマーであり、そしてLがリン(V)結合の場合、その時nは約12から約100まで変化する。好ましくは、 Mが、ヌクレオシドまたはそのアナログまたはヌクレオシドサイズのモノマーであり、そしてLがリン(V)結合の場合、その時nは約12から約40まで変化する。
【0063】
ペプチドは、本発明のタグに結合する別の好ましい種類の分子である。本発明で用いられ得るペプチド−オリゴヌクレオチド結合体の合成は、Nielsenら、J.Am.Chem.Soc.、115:9812−9813(1993);Haralambidisら(上記)および国際特許出願PCT/AU88/004417;Truffertら、Tetrahedron Letters、35:2353−2356(1994);de la Torreら、 Tetrahedron Letters、35:2733−2736(1994)などの参考文献で教示されている。好ましくは、ペプチド−オ
リゴヌクレオチド結合体は、下記のように合成される。本発明に従って合成されたペプチドは、天然アミノ酸モノマーまたは非天然モノマー(天然アミノ酸のD異性体などを含む)からなり得る。
【0064】
(コンビナトリアルケミカルライブラリー)
本発明のタグを用いるコンビナトリアルケミカルライブラリーは、好ましくは、Nielsenら(上記)で開示され、そして詳細な実施態様のために図4で図解された方法により調製される。簡単にいえば、固相支持体(例えば、CPG)は、タグを合成するために用いられるケミストリーおよびいくつかの選択プロセスを経る分子を合成するために用いられるケミストリーの両方に適合する切断可能なリンカーで誘導体化される。好ましくは、タグは、上記のようなホスホルアミダイトケミストリーを用いて、およびNeilsenら(上記)により推奨される改変(すなわち、メチル保護ホスファイトおよびホスフェート部分を有するDMT−5’−O−保護3’−ホスホルアミダイト−誘導体化サブユニットが、各合成サイクルに加えられる)により合成される。ライブラリー化合物は、好ましくは、Fmocまたは同等物(連続モノマーをカップリングする官能基をマスクする保護基)を有するモノマーである。DMTおよびFmoc保護基(本明細書中でサルコシンリンカーという)の両方を用いるケミストリーに対する適切なリンカーは、Brownら、J.Chem.Soc.Chem.Commun.、1989:891−893により開示され、この開示は、参考として援用される。
【0065】
図4は、オリゴヌクレオチドタグと結合するペプチドのコンビナトリアルケミカルライブラリーを生成するためのスキームを例示する。固相支持体200は、Nielsenら(上記)により教示されるようにサルコシンリンカー205(以下の式で例示される)により誘導体化され、このリンカーは、試薬の接近を促進する伸長結合部分を有する。
【0066】
【化1】
ここで、「CPG」は、徐放性の多孔性ガラス支持体を表す。「DMT」はジメトキシトリチルを表し、そして、「Fmoc」は、9−フルオレニルメトキシカルボニルを表す。好ましい実施態様では、オリゴヌクレオチドセグメント214は、最初に合成され、そのため2本鎖の形態で、微粒子または基質の様なものの上で分類した後に、制限エンドヌクレアーゼ部位が、ライブラリー化合物を切断するために提供される。連続したサブユニットS1、S2、S3などの代替的な付加により、合成は進行してタグ212を形成し、そしてそれらの対応するライブラリー化合物モノマーA1、A2、A3などにより、合成は進行してライブラリー化合物216を形成する。「分裂および混合」技術を用ると、多様性が生まれる。
【0067】
最少に交差ハイブリダイズしているセット中のサブユニットは、ライブラリー化合物中に加えられたモノマーをコードする。従って、9ワードのセットは、9つのモノマーから構築されるライブラリー化合物を曖昧性なしにコードし得る。いくつかの曖昧さが受容可能であれば、単一のサブユニットは、1つより多いモノマーをコードし得る。
【0068】
合成が完了した後、産物は、切断および脱保護され(220)、タグライブラリー化合物225を形成し、次いで、この化合物は選択230(例えば、タンパク質のような予定された標的235への結合)を受ける。次いで、選択プロセス230から回収されたライブラリー化合物のサブセットは、固相支持体245上で、そのタグ部分(そこでは、相補
的サブユニットおよびヌクレオチドがイタリック体で示される)を介して分類される(240)。オリゴヌクレオチドスプリント242をタグ相補物250に結合し、制限部位255を形成した後、結合体は、対応する制限エンドヌクレアーゼで消化され、ライブラリー化合物(図4の実施例中のぺプチド)をオリゴヌクレオチド部分から切断する。次いで、タグの配列、それゆえライブラリー化合物の同定は、以下に記載の本発明の好ましい単一塩基配列決定技術により決定される。
【0069】
(固相支持体)
本発明で用いられる固相支持体は、広い種類の形態を有し得、例えば、微粒子、ビーズ、および膜、スライド、プレート、ミクロマシーンドチップ(micromachined chip)などが挙げられる。同様に、本発明の固相支持体は、広い種類の組成物を含み得、例えば、ガラス、プラスチック、シリコン、アルカンチオレート誘導体化金(alkanethiolate−derivatized gold)、セルロース、低架橋ポリスチレンおよび高架橋ポリスチレン、シリカゲル、ポリアミドなどが挙げられる。好ましくは、分離した粒子の集団のいずれかは、各々が同じタグ(および他はなし)への相補的配列の、均一のコーティング(または集団)を有するように用いられ、あるいは、単一支持体または数個の支持体は、各々が同じタグ(および他はなし)への相補的配列の、均一のコーティング(または集団)を含む、空間的に分離した領域で用いられる。後者の実施態様では、領域の面積は、特別な適用に従って変化し得、通常、数(例えば、3〜5)μm2から数百(例えば、100〜500)μm2の面積の領域範囲である。好ましくは、このような領域は、空間的に分離され、そのため隣接領域での事象(例えば、蛍光発光)により発生されるシグナルが、用いられる検出システムにより解明され得る。いくつかの適用では、1つより多いタグ相補物(例えば、同時配列分析、または別々にタグ化された分子を極めて近接に持ってくるための)の均一コーティングの領域を有することが、望ましくあり得る。
【0070】
タグ相補物は、その上でタグ相補物が合成される固相支持体と共に用いられ得、あるいは、別々に合成され得、そして使用のために固相支持体に結合され得る。例えば、このことは以下に開示される:Lundら、Nucleic Acids Research、16:10861−10880(1988);Albretsenら、Anal.Biochem.、189:40−50(1990);Wolfら、Nucleic Acids Research、15:2911−2926(1987);またはGhoshら、Nucleic Acids Research、15:5353−5372(1987)。好ましくは、タグ相補物は、同じ固相支持体上で合成され、そして同じ固相支持体と共に用いられ得、それは、種々の形態を含み、そして種々の結合部分を含み得る。このような支持体は、タグ相補物の均一集団が合成される領域の微粒子、またはアレイ、またはマトリックスを含み得る。広い種類の微粒子支持体が、本発明で用いられ得、例えば、徐放性の多孔性ガラス(CPG)、高架橋ポリスチレン、アクリルコポリマー、セルロース、ナイロン、デキストラン、ラテックス、ポリアクロレインなどにより作製される微粒子が挙げられ、以下の模範的な参考文献中に開示されている:Meth.Enzymol.、Section A、11−147頁、第44巻(Academic Press、New York、1976);米国特許第4,678,814号;第4,413,070号;および第4,046,720号;ならびにPon、19章、Agrawal(編)、Methods in Molecular Biology、第20巻(Humana
Press、Totowa、NJ、1993)。微粒子支持体としては、市販のヌクレオシド−誘導体化CPGおよびポリスチレンビーズ(例えば、Applied Biosystems、Foster City、CA);誘導体化磁気ビーズ;ポリエチレングリコールでグラフトされたポリスチレン(例えば、TentaGelTM、Rapp Polymere、Tubingen Germany)などがさらに挙げられる。支持体の特性(例えば、物質、多孔度、サイズ、形態など)の選択、および用いられる結合部分
のタイプは、タグを用いる際の条件に依存する。例えば、酵素による連続的なプロセスを含む適用において、酵素の立体障害を最小にし、そして基質への接近を容易にする支持体およびリンカーが好ましい。最も適切な微粒子支持体を選択する際に、考慮されるべき他の重要なファクターとしては、サイズの均一性、合成支持体としての効率、既知の表面積の度合い、および光学特性(例えば、以下により十分に説明するように、表面で多くのビーズを扱う場合、透明で平滑なビーズは、計測的な(instrumentational)利点を提供する)が挙げられる。
【0071】
微粒子表面上でタグを結合させるおよび/または合成するための模範的な結合部分は、以下に開示されている:Ponら、Biotechniques、6:768−775(1988);Webb、米国特許第4,659,774号:Baranyら、国際特許出願 PCT/US91/06103;Brownら、J.Chem.Soc.Commun.、1989:891−893;Damhaら、Nucleic Acids Research、18:3813−3821(1990);Beattieら、Clinical Chemistry、39:719−722(1993);MaskosおよびSouthern、Nucleic Acids Research、20:1679−1684(1992)など。
【0072】
上記のように、タグ相補物はまた、単一の(または数個の)固相支持体上で合成され得、タグ相補物で均一にコートされた領域のアレイを形成する。すなわち、このようなアレイ中の各領域内で、同じタグ相補物が合成される。このようなアレイを合成するための技術は、以下に開示される:McGallら、国際出願PCT/US93/03767;Peaseら、Proc.Natl.Acad.Sci.、91:5022−5026(1994);SouthernおよびMaskos、国際出願PCT/GB89/01114;MaskosおよびSouthern(上記);Southernら、Genomics、13:1008−1017(1992);およびMaskosおよびSouthern、Nucleic Acids Research、21:4663−4669(1993)。
【0073】
好ましくは、本発明は、同じタグ配列の相補物で均一にコートされた微粒子またはビーズを用いて実行される。微粒子支持体、およびそれらの表面とオリゴヌクレオチドとを共有結合または非共有結合する方法は、周知であり、以下の参考文献により例示される:BeaucageおよびIyer(上記);Gait(編)Oligonucleotide Synthesis;A Practical Approach(IRL Press、Oxford、1984);および上記の参考文献。一般に、微粒子のサイズおよび形態は重要ではない;しかし、最少の試薬およびサンプルの使用でオリゴヌクレオチドタグの多くのレパートリーの構築および操作を容易にするので、直径数μm(例えば、1〜2μm)から数百μm(例えば、200〜1000μm)の範囲のサイズの微粒子が好ましい。
【0074】
いくつかの好ましい適用では、市販の徐放性の多孔性ガラス(CPG)またはポリスチレン支持体が、本発明における固相支持体として用いられる。このような支持体は、塩基不安定性のリンカーおよび結合した最初のヌクレオチド(例えば、Applied Biosystems(Foster City、CA))を用いて有効になる。好ましくは、500オングストローム〜1000オングストロームの間のポアサイズを有する微粒子が用いられる。
【0075】
他の好ましい適用では、小孔のない微粒子が、その光学特性のために用いられ、この微粒子は、平面支持体(例えば、顕微鏡スライド)の上で多くの微粒子を探知する場合、有利に用いられ得る。特に、好ましい小孔のない微粒子は、Bangs Laborato
ries(Carmel.IN)から入手できるグリシダルメタクリレート(glycidal methacrylate)(GMA)である。このような微粒子は、種々のサイズで有用であり、そしてタグまたはタグ相補物を合成するための種々の結合基で誘導体化される。好ましくは、タグ化された微粒子の多量(massively)の平行操作のために、直径5μmのGMAビーズが用いられる。
【0076】
(標的ポリヌクレオチドの微粒子への結合)
本発明の重要な局面は、同一のポリヌクレオチド(例えば、cDNAライブラリー由来)の集団の分類、ならびに各微粒子または領域が単一種のポリヌクレオチドのみを有するような微粒子あるいは固相支持体の分離領域へのそれらの結合である。この後者の条件は、タグのレパートリーをポリヌクレオチドの集団へ連結することにより、本質的に満たされ得る。次いで、連結産物は、クローン化され、増幅され、そしてサンプリングされる。サンプルが十分小さければ、以下により十分に説明するように、得られたライブラリーの実質的に全てのタグポリヌクレオチド結合体は、唯一である。すなわち、各ポリヌクレオチドは、唯一のタグを有し、逆も同様である。次いで、ポリヌクレオチドは、タグをその相補物にハイブリダイズすることにより分類される。
【0077】
オリゴヌクレオチドタグのレパートリーは、多くの方法(例えば、直接酵素的連結による方法、増幅(例としてPCRによるもの)、タグ配列を含むプライマーの使用など)でポリヌクレオチドの集団に連結され得る。最初の連結工程は、タグ−ポリヌクレオチド結合体の非常に大きな集団を産生し、そのため単一のタグが、一般に多くの異なるポリヌクレオチドに結合する。しかし、結合体の十分小さいサンプルを採取することにより、「ダブル」を得る可能性(すなわち、2つの異なるポリヌクレオチド上の同じタグ)は、無視できる(サンプル中の同じポリヌクレオチドで異なるタグを得る可能性もあることに注目。この場合、単にプロセスされた(例えば、2回配列された)ポリヌクレオチドを生じ、そのため、通常問題はない)。以下により十分に説明したように、サンプル中にダブルを得る可能性は、ポアソン分布により評価され得る。なぜなら、サンプル中の結合体の数は大きく(例えば、千以上の桁数)、そして特別なタグを選択する可能性は小さいからである。なぜなら、タグのレパートリーは大きい(例えば、1万以上の桁)からである。一般に、サンプルが大きければ大きいほど、ダブルを得る可能性は大きくなる。従って、タグ−ポリヌクレオチド結合体の大きなサンプルを選択すること(例えば、それは、ショットガン配列決定操作における標的ポリヌクレオチド結合体の適切な適用範囲を確かめる)と最少数のダブルが存在することを確かめる小さなサンプルを選択することとの間に、設計の取引が存在する。大部分の実施態様では、ダブルの存在は、単にノイズのさらなる供給源を加えるか、または配列決定の場合、スキャニングおよびシグナルプロセシングにおける小さな複雑化要因を加えるのみである。なぜなら、多数の蛍光シグナルを与える微粒子が簡単に無視され得るからである。本明細書中で用いられる、タグを分子(特に、ヌクレオチド)に結合することに関する用語「実質的に全て」は、本質的にダブルのないタグ−分子結合体の集団を得るのに用いられるサンプリング手法の統計的な性質を反映するつもりである。タグ−分子結合体の実際の割合に関して、「実質的に全て」の意味は、タグがどのように用いられているかに依存する。好ましくは、核酸配列決定において、「実質的に全て」は、少なくともタグの80%が、結合した唯一のポリヌクレオチドを有することを意味する。より好ましくは、少なくともタグの90%が、結合した唯一のポリヌクレオチドを有することを意味する。さらにより好ましくは、少なくともタグの95%が、結合した唯一のポリヌクレオチドを有することを意味する。そして、最も好ましくは、少なくともタグの99%が、結合した唯一のポリヌクレオチドを有することを意味する。
【0078】
好ましくは、ポリヌクレオチドの集団が、メッセンジャーRNA(mRNA)からなる場合、オリゴヌクレオチドタグは、mRNAをタグ配列のプライマー含有相補物のセットで逆転写することにより、結合する。このようなプライマーの模範的なセットは、以下の
配列を有し得る:
【0079】
【化2】
ここで、「[W,W,W,C]9」は、4つのヌクレオチドの9つのサブユニットのオリゴヌクレオチドタグの配列を表し、そして「[W,W,W,C]」は、上記で挙げたサブユニット配列(すなわち、「W」はTまたはAを表す)を表す。下線を引いた部分の配列は、任意の制限エンドヌクレアーゼ部位と同一である。その部位は、用いられる場合、ビオチンを介した固相支持体への結合からポリヌクレオチドを放出させるのに用いられ得る。上記のプライマーに関して、微粒子に結合した相補物は、以下の形態を有し得る:
5’−[G,W,W,W]9TGG−リンカー−微粒子。
【0080】
逆転写後、mRNAは、例えば、RNase H 消化により除去され、そして、cDNAの第2の鎖は、例えば、以下の形態のプライマーを用いて合成される:
5’−NRRGATCYNNN−3’
ここで、NはA、T、G、またはCのいずれか一つであり、Rはプリン含有ヌクレオチドであり、そしてYはピリミジン含有ヌクレオチドである。この特定のプライマーは、得られた2本鎖DNA中にBst Yl制限部位を作り、それは、Sal I部位とともに、例えばBam HIおよびXho I部位を用いたベクター中へのクローニングを容易にする。Bst YlおよびSal I消化の後、模範的な結合体は以下の形態を有する:
【0081】
【化3】
好ましくは、リガーゼベースの配列決定の方法を用いた場合、Bst YlおよびSal
I消化フラグメントが以下の単一コピー制限部位を有するBam HI−/Xho I−消化ベクター中にクローン化される:
【0082】
【化4】
これは、以下でより十分に議論される配列決定プロセスの開始を可能とするFok I部位を添加する。
【0083】
増幅後の一本鎖のタグを露出させるための一般的な方法は、標的ポリヌクレオチド含有結合体をT4 DNAポリメラーゼまたは同様の酵素の5’→3’エキソヌクレアーゼ活性で消化することを含む。単一ヌクレオシドトリホスフェートの存在下で用いられる場合、このようなポリメラーゼは、単一ヌクレオシドトリホスフェートの相補物がテンプレート鎖に届くまで、2本鎖フラグメントの非テンプレート鎖に存在する3’陥凹(recessed)末端からヌクレオチドを切断する。このようなヌクレオチドが届いた場合、5’→3’消化は、効果的に停止する。というのは、除去活性がヌクレオチドを除去するより早い速度で、ポリメラーゼの伸長活性がヌクレオチドを付加するからである。結果として、3つのヌクレオチドで構成されるタグは、固相支持体上にロードするために容易に調製される。
【0084】
この技術はまた、標的ポリヌクレオチドの内部のFok I部位を選択的にメチル化し
、一方ポリヌクレオチドの末端で一本鎖のFok I部位をメチル化されないまま残すために用いられる。最初に、末端Fok I部位は、デオキシシチジントリホスフェートとともにポリメラーゼを用いて1本鎖とされる。次いで、フラグメントの2本鎖部分は、メチル化され、その後、1本鎖末端は、4つ全てのヌクレオシドトリホスフェートの存在下で、DNAポリメラーゼを用いて満たされ、それによりFok I部位を再生する。明らかに、この手順は、Fok I 以外のエンドヌクレアーゼに一般化され得る。
【0085】
オリゴヌクレオチドタグが、特定のハイブリダイゼーション(例えば、上記のようにそれらを1本鎖にすることにより)のために調製された後、ポリヌクレオチドは、タグとその相補物との間の完全にマッチする二重鎖の形成に有利な条件下で、タグの相補的配列を含む微粒子と混合される。これらの条件を作るための広範囲のガイダンスが文献中に存在する。このようなガイダンスを提供する例示的な参考文献としては、Wetmur、Biochemistry and Molecular Biology中のCritical Reviews、26:227−259(1991);Sambrookら、Molecular Cloning;A Laboratory Manual、第2版(Cold Spring Harbor Laboratory、New York、1989)などが挙げられる。好ましくは、ハイブリダイゼーション条件は、完全にマッチした配列のみが安定した二重鎖を形成するのに十分にストリンジェントである。このような条件下で、それらのタグにより特にハイブリダイズされたポリヌクレオチドは、微粒子に結合した相補的配列に連結される。最終的に、微粒子は洗浄され、連結されないポリヌクレオチドが除去される。
【0086】
合成支持体として通常使用されるCPG微粒子が用いられる場合、微粒子表面上でタグ相補物の密度は、代表的にいくつかの配列決定操作に必要な密度以上である。すなわち、ポリヌクレオチドを種々の酵素に結合させる連続的な処理を必要とする配列決定アプローチにおいては、空間を密にとったポリヌクレオチドは、比較的大きな酵素のポリヌクレオチドへの接近を阻害する傾向があり得る。
【0087】
このような場合、ポリヌクレオチドは、好ましくは、タグ相補物が、かなり過剰に(例えば、ポリヌクレオチドに対して10:1から100:1まで、あるいはそれ以上)存在するように微粒子と混合される。このことは、微粒子表面上におけるポリヌクレオチドの密度が、酵素の接近を阻害するほど高くないことを保証する。好ましくは、微粒子表面上のポリヌクレオチド間の平均空間は、30〜100nmの位である。標準CPG支持体およびBallotiniビーズ(固体ガラス支持体の1種)についての割合の選択におけるガイダンスは、MaskosおよびSouthern、Nucleic Acids Research、20:1679−1684(1992)中に見られる。好ましくは、配列決定の適用において、そして直径20〜50μmの範囲の標準CPGビーズが、約105のポリヌクレオチドでロードされ、そして直径5〜10μmの範囲のGMAビーズが、数万(例えば、4×104〜6×104)のポリヌクレオチドでロードされる。
【0088】
上記の方法は、以下に記載される平行配列決定方法とカップリングする場合、mRNA集団をフィンガープリントするために用いられ得る。部分配列情報は、上記の方法のような分離された微粒子に結合したcDNAの大量の(例えば、1万〜10万)サンプルから同時に得られる。部分配列の頻度分布は、種々の細胞または組織型、ならびに疾患組織(例えば、ガン)からのmRNA集団を同定し得る。このようなmRNAフィンガープリントは、病状をモニタリングおよび診断するのに有用である(例えば、国際出願 PCT/US95/21944。これは、同じ目的のための発現配列タグ(EST)の使用を記載する)。
【0089】
(単一塩基DNA配列決定)
本発明は、DNA配列決定の従来の方法、例えば、Hultmanら、Nucleic
Acids Research、17:4937−4946(1989)によって開示された方法とともに用いられ得る。しかし、複数のポリヌクレオチドの並行または同時の配列決定に関しては、以下の参考文献が挙げられる:Cheeseman、米国特許第5,302,509号; Tsienら、国際出願第WO 91/06678号; Rosenthalら、国際出願第WO 93/21340号; Canardら、Gene、148:1−6(1994); およびMetzkerら、Nucleic Acids
Research、22:4259−4267(1994)。
【0090】
本発明で用いられるのに適切であり、DNAフラグメントの電気泳動での分離を必要としないDNA配列決定の「単一塩基」方法が、国際出願第PCT/US95/03678号に記載されている。この方法は、以下の工程を包含する:(a)突出鎖を有するポリヌクレオチドの末端にプローブを連結して連結複合体を形成する工程であって、このプローブは、ポリヌクレオチドの鎖に対して相補的な突出鎖を有し、かつ、このプローブは、ヌクレアーゼ認識部位を有する工程;(b)この連結複合体から非連結プローブを除去する工程;(c)連結プローブの同定によりポリヌクレオチドの突出鎖における1つまたはそれ以上のヌクレオチドを同定する工程;(d)ヌクレアーゼで連結複合体を切断する工程;および(e)ポリヌクレオチドのヌクレオチド配列が決定されるまで(a)から(d)までの工程を繰り返す工程。以下にさらに記載するように、1つまたはそれ以上のヌクレオチドの同定は、標的ポリヌクレオチドからの連結複合体の切断前または切断後のいずれかに行われ得る。好適には、天然のタンパク質エンドヌクレアーゼが用いられる時は常に、この方法は、配列決定操作の開始時に、標的ポリヌクレオチドをメチル化する工程をさらに包含する。
【0091】
この方法の重要な特徴は、標的ポリヌクレオチドに連結されるプローブである。プローブの好適な形態は、図1aに示されている。一般に、プローブは、一方の末端10において突出鎖を有する二本鎖DNAである。プローブは、認識部位と突出末端10との間に少なくとも1つのヌクレアーゼ認識部位12およびスペーサー領域14を含む。好適には、プローブはまた、標識16を含み、これは、この特定の実施態様において、突出鎖の反対側の末端に示されている。プローブは、種々の手段により、そして種々の位置において標識され得、唯一の制限は、選択された標識手段が、連結工程またはヌクレアーゼによるプローブの認識を邪魔しないということである。
【0092】
プローブの突出鎖10が、5’末端または3’末端であるかどうかは重要ではない。しかし、標的ポリヌクレオチドおよびプローブの突出鎖が、完全に一致する二重鎖を形成し得、特異的な連結を可能にすることが重要である。もし、標的ポリヌクレオチドおよびプローブの突出鎖が異なる長さであれば、生じるギャップは、例えば、Backmanら、欧州特許出願第91100959.5号に開示されている「ギャップLCR」におけるように、連結以前にポリメラーゼによってフィルインされ得る。好適には、プローブおよび標的ポリヌクレオチドの両方の鎖が、フィルイン工程なしに連結され得るように、それぞれの突出鎖におけるヌクレオチドの数が同じである。好適には、プローブの突出鎖は2〜6のヌクレオチドの長さである。以下に示すように、突出鎖の長さが長くなるにつれて、各連結および切断サイクルの間に、標的ポリヌクレオチドへ適用されるプローブ混合物の複雑性が増す。
【0093】
プローブの相補鎖は、好都合にも、自動DNA合成機(例えば、Applied Biosystems、Inc.(Foster City、California)モデル
392または394 DNA/RNA Synthesizer)上で、標準的なケミストリーを用いて合成される。合成後、相補鎖は、組み合わされて二本鎖プローブを形成する。一般に、プローブの突出鎖は、混合物として合成されるので、すべての可能な配列
は、突出部に表される。例えば、もし、突出部が4つのヌクレオチドからなっているなら、ある実施態様において、4つの混合物が以下のように調製される:
【0094】
【化5】
ここで、「NNN」は、すべての可能な3マーを表し、「X」は、鎖の二重鎖形成部分を表す。従って、上に挙げられた4つプローブのそれぞれが、43すなわち64の異なる配列を含む;すなわち、言い換えれば、4つのプローブのそれぞれは、64の縮重を有する。例えば、X1X2...XiNNNAは、以下の配列を含む:
【0095】
【化6】
このような混合物は、例えば、Teleniusら(上記)に開示されているように、周知の技術を用いて簡単に合成される。一般に、これらの技術は、単に、縮重を導入したい位置でのカップリング工程の間、成長しているオリゴヌクレオチドへの活性化モノマーの混合物の適用を必要とするだけである。ある実施態様において、プローブの縮重を減少させることが所望され得る。これは、例えばKong Thoo Linら、Nucleic Acids Research、20:5149−5152または米国特許第5,002,867号によって教示されるように、デオキシイノシン、2−アミノプリンなどのような縮重減少アナログを用いて達成され得る。
【0096】
好適には、ホスホジエステル結合を有するオリゴヌクレオチドについては、プローブの二重鎖形成領域は、長さが約12〜約30塩基対であり、より好適には、その長さは、約15〜約25塩基対である。
【0097】
従来のリガーゼが本発明に用いられる場合、以下にさらに記載されるように、プローブの5’末端が、ある実施態様においてリン酸化され得る。5’モノホスフェートが、化学的にまたはキナーゼを用いて酵素的に(例えば、Sambrookら(上記))のいずれ
かで第2のオリゴヌクレオチドに結合され得る。化学的リン酸化は、HornおよびUrdea、Tetrahedron Lett.、27:4705(1986)に記載され、開示されたプロトコルを実施するための試薬は、市販されている(例えば、Clontech Laboratories(Palo Alto、California)からの5’Phosphate−ONTM))。従って、ある実施態様において、プローブは、以下の形態を有し得る:
【0098】
【化7】
ここで、Y’は、X’の相補ヌクレオチドであり、「p」は、モノホスフェート基である。
【0099】
上記のプローブは、種々の方法で標識され得る(放射性部分、蛍光部分、比色部分、化学ルミネセンスマーカーなどの直接または間接結合を含む)。DNAの標識およびDNAプローブの構築のための方法の多くの包括的な概説が、本発明のプローブの構築に適用可能な指針を提供している。このような概説は、Kricka編、Nonisotopic
DNA Probe Techniques (Academic Press、San Diego、1992); Haugland、Handbook of Fluorescent Probes and Research Chemicals (Molecular Probes、Inc.、Eugene、1992); KellerおよびManak、DNA Probes、第2版 (Stockton Press、New York、1993); およびEckstein編、Oligonucleotides and Analogues; A Practical Approach (IRL Press、Oxford、1991); Kessler編、Nonradioactive Labeling and Detection of Biomolecules (Springer−Verlag、Berlin、1992); Wetmur(上記); などを含む。
【0100】
好適には、プローブは、1つまたはそれ以上の蛍光色素で標識される(例えば、Menchenら、米国特許第5,188,934号; Begotら、国際出願第PCT/US90/05565号に開示されている)。
【0101】
この方法によれば、プローブは、標的ポリヌクレオチドの末端へ連結され、各連結および切断のサイクルにおいて連結複合体を形成する。連結複合体は、標的ポリヌクレオチドおよびプローブの突出鎖がアニールし、そして、少なくとも一対のプローブおよび標的の同じ向きの鎖が連結された、すなわち、互いに共有結合された後に、形成された二本鎖構造である。連結は、酵素的または化学的のいずれかで達成され得る。化学的連結方法は、当該分野において周知である(例えば、Ferrisら、Nucleosides & Nucleotides、8:407−414(1989); Shabarovaら、Nucleic Acids Research、19;4247−4251 (1991)など)。しかし、好適には、連結は、標準的なプロトコルにおいてリガーゼを用いて、酵素的に行われる。多くのリガーゼが公知であり、本発明の使用に適している(例えば、Lehman、Science、186: 790−797 (1974); Englerら、DNAリガーゼ、3−30頁、Boyer編、The Enzymes、Vol.15B (Academic Press、New York、1982)など)。好適なリガーゼは、T4 DNAリガーゼ、T7 DNA リガーゼ、E. coli DNAリガーゼ、Taqリガーゼ、Pfuリガーゼ、およびTthリガーゼを含む。これらの使用の際のプロトコルは、周知である(例えば、Sambrookら(上記); B
arany,PCR Methods and Applications、1:5−16 (1991); Marshら、Strategies、5:73−76 (1992)など)。一般に、リガーゼは、5’ホスフェート基が、接する鎖の3’水酸基への連結のために存在することを必要とする。これは、好都合に、5’ホスフェートを残すヌクレアーゼ(例えば、Fok I)を選択することによって、標的ポリヌクレオチドの少なくとも1つの鎖に対して提供される。
【0102】
非リン酸化プローブを用いる配列決定方法の実施態様において、連結工程は、(i) リガーゼで標的ポリヌクレオチドへプローブを連結させ、その結果連結複合体が、一方の鎖上にニックを有して形成される工程;(ii) 従来のプロトコルを用いて、キナーゼでニックにおいて5’水酸基をリン酸化する工程(例えば、Sambrookら(上記));および(iii) 再度連結を行い、ニックにおいて鎖を共有的に連結させる、すなわち、ニックを除去する工程を包含する。
【0103】
(微粒子表面での酵素的プロセスおよび/または結合事象の観察装置)
本発明の目的は、タグおよびその相補物の特異的ハイブリダイゼーションによって、微粒子の表面上の同一分子、特に、ポリヌクレオチドを分類することである。一旦、このような分類が行われると、分子の存在またはそれらに対して行われる操作は、タグ化された分子の性質、微粒子が、分離して、または「バッチ」で検出されるかどうか、反復された測定が所望であるかどうかなどに応じて、多くの方法で検出され得る。代表的に、分類された分子は、例えば、薬物開発において、結合のためにリガンドに曝されるか、または、例えば、ポリヌクレオチド配列決定において化学的または酵素的プロセスに供される。これらの使用の両方において、多くの微粒子上でのこのような事象またはプロセスに対応する信号を同時に観察することがしばしば望まれる。分類された分子を担持する微粒子(本明細書において「ロードされた」微粒子と称する)が、このような大規模な並行操作に役立つ(例えば、Lamら(上記)に示されている)。
【0104】
好適には、発光信号(例えば、化学ルミネセンス、蛍光など)が、事象またはプロセスを検出するために用いられるときは常に、ロードされた微粒子は、平面基板(例えば、ガラススライド)上に、(例えば、国際特許出願第PCT/US91/09217号、同第PCT/NL90/00081号および同第PCT/US95/01886号に記載されているような)走査システムでの検査のために広がっている。走査システムは、再現性よく基板を走査し、かつ、座標系によって、所定の領域に各微粒子の位置を規定し得るべきである。ポリヌクレオチド配列決定応用において、微粒子の位置同定は、連続的な走査工程において反復可能であることが重要である。
【0105】
このような走査システムは、市販の構成要素(例えば、1つまたはそれ以上の光電子増倍管、あるいは、CCDアレイおよび、例えば、蛍光信号を励起、収集および分類するための適切な光学機器を備える検出システムとともに用いられるデジタルコンピューターによって制御されるx−y移動テーブル)から構築され得る。ある実施態様において、共焦光学システムが所望であり得る。4色配列決定での使用に適切な走査システムの一例が、図5に模式的に示されている。基板300(例えば、固定微粒子を有する顕微鏡スライド)が、x−y移動テーブル302上に置かれ、これが、種々の市販のパーソナルコンピューター(例えば、Apple Computer(Cupertino、CA)から入手可能な486−ベースの機械またはPowerPC モデル7100または8100)のうちのいずれかであり得る適切にプログラムされたデジタルコンピューター304に接続され、そしてそれにより制御される。テーブル移動およびデータ収集機能のためのコンピューターソフトウェアは、National Instrumentsから入手可能なLab Windows(登録商標)のような市販の実験ソフトウェアによって提供され得る。
【0106】
基板300およびテーブル302は、光を集め、基板300に固定した微粒子へ伝達し得る1つまたはそれ以上の対物レンズ308を有する顕微鏡306と動作が連動する。好適にはレーザーである光源312からの励起光線310が、光線スプリッター314(例えば、二色性ミラー)へ指向し、これが、光線を顕微鏡306および対物レンズ308を通して再指向し、今度は、対物レンズが光線の焦点を基板300に合わせる。レンズ308は、微粒子から発光された蛍光316を集め、それを光線スプリッター314を通して信号分配光学機器318へ指向し、それが、今度は、蛍光をいくつかの蛍光特性(例えば、強度、寿命など)を電気信号に変換するための1つまたはそれ以上の適切な光学電気デバイスへ指向する。信号分配光学機器318は、帯域通過フィルター、ファイバー光学機器、回転ミラー、固定位置ミラーおよびレンズ、回折格子などのような当該分野において標準的な種々の構成要素を備える得る。図5に図示するように、信号分配光学機器318は蛍光316を4つの分離した光電子増倍管330、332、334および336へ向け、次いで、これらの出力は、プリアンプおよびフォトンカウンター350、352、354および356に向けられる。フォトンカウンターの出力は、コンピューター304によって収集され、そこで、蓄積され、分析され、ビデオ360において観察され得る。あるいは、信号分配光学機器318は、蛍光信号318をCCDアレイ上に向ける回折格子であり得る。
【0107】
走査の際の位置付けの安定性および再現性が、間隔が密接な微粒子を分離するための解像度をかなりの程度まで決定する。好適には、走査システムは、間隔が密接な(例えば、1粒子直径またはそれ以下分だけ分離されている)微粒子を解像し得るべきである。従って、ほとんどの応用に対して(例えば、CPG微粒子を用いて)、走査システムは、少なくとも10〜100μm程度に対象物を解像する能力を有するべきである。ある実施態様において、さらに高い解像度が所望であり得るが、解像度が増すにつれて、基板を完全に走査するのに必要な時間が増す。従って、ある実施態様において、速度と解像度との間で妥協が必要であり得る。走査時間の増加は、微粒子が配置していることが知られている(例えば、最初のフル走査から)位置のみを走査するシステムによって達成され得る。好適には、微粒子サイズおよび走査システム解像度が、1cm2当たり約1万〜10万の間の微粒子の密度で平面上にランダムに配置される蛍光標識された微粒子の解像度を可能にするように選択される。
【0108】
配列決定応用において、ロードされた微粒子は、種々の方法で基板の表面に固定され得る。固定は、微粒子が、顕著な損失なく、試薬露出および洗浄の連続的なサイクルを行うのに十分に強力であるべきである。基板がガラスであるとき、その表面は、市販の試薬(例えば、Pierce Chemical)を用いて、アルキルアミノリンカーで誘導体化され得、これは次に再び従来のケミストリーを用いて、アビジンに架橋され、アビジン化された表面を形成し得る。ビオチン部分は、多くの方法でロードされた微粒子に導入され得る。例えば、ポリヌクレオチドにタグを結合するために用いられるクローニングベクターの画分(例えば、10〜15パーセント)が、タグの反対側のポリヌクレオチドの末端でポリヌクレオチド挿入物に隣接して(消化において粘着末端を提供する)唯一の制限部位を含有するように操作される。この部位は、微粒子にロードするためのポリヌクレオチドおよびタグで切り出される。ロード後、ロードされたポリヌクレオチドの約10〜15パーセントが、微粒子表面から遠位の唯一の制限部位を有する。関連の制限エンドヌクレアーゼで消化後、ビオチン部分を含有している適切な二本鎖アダプターが、粘着末端に連結される。それから、得られた微粒子が、アビジン化されたガラス表面に広げられ、そこで、ビオチン−アビジン結合を介して固定される。
【0109】
あるいは、そして好適には、連結による配列決定が用いられると、最初の連結工程において、プローブの混合物が、ロードされた微粒子にアプライされる:プローブの一画分は
、配列決定方法に必要であるようなIIs型制限認識部位を含み、そしてプローブの一画分は、このような認識部位を有さないが、その代わりに、その非連結末端においてビオチン部分を含む。好適には、混合物は、約10〜15パーセントのビオチン化プローブを含む。
【0110】
さらに別の実施態様において、DNAがロードされた微粒子が、ガラス基板に塗布されると、DNAは、数時間(例えば、24時間)のインキュベーションで、ガラス表面に非特異的に吸着され、微粒子の顕著な損失なく、反復される試薬への露出および洗浄を可能とするために十分に強力な結合を形成し得る。好適には、このようなガラス基板は、フローセル (flow cell) であり、これは、ガラススライドにエッチングされた流路を含み得る。好適には、このような流路は、流体がそこを通って吸い上げられ得るように閉じられ、かつ、微粒子の単層が規定された観察領域内に閉じ込められるように、微粒子の直径と十分に近い深さを有する。
【0111】
(並行配列決定)
本発明のタグ化システムは、数キロベースまでの長さのポリヌクレオチドを配列決定するための単一塩基配列決定方法とともに用いられ得る。タグ化システムは、標的ポリヌクレオチドの何千ものフラグメントが、1つまたはそれ以上の固相支持体上に分類され、同時に配列決定され得ることを可能にする。この方法の好適な実施によると、各分類されたフラグメントの一部分が、上記のような走査システムまたは画像分析システムに関連する、顕微鏡スライドのような共通の基板に固定された何千ものロードされた微粒子のそれぞれにおいて、段階的に配列決定される。配列決定されたフラグメントの部分のサイズは、いくつかの要因、例えば、生成および分類されたフラグメントの数、標的ポリヌクレオチドの長さ、用いられた単一塩基方法の速度および正確さ、微粒子および/または同時にモニターされ得る別個の領域の数などに依存する。好適には、12〜50塩基が、各微粒子または領域で同定され、さらに好適には、18〜30塩基が、各微粒子または領域で同定される。この情報により、標的ポリヌクレオチドの配列が、重複する領域を介して12〜50塩基のフラグメントを対照することによって決定される(例えば、米国特許第5,002,867号に記載されている)。以下の引例は、所与の長さの標的ポリヌクレオチドを首尾良く再構築するために配列決定されなければならないフラグメントの部分を決定する際のさらなる指針を提供する:LanderおよびWaterman、 Genomics、2:231−239 (1988): Drmanacら、 Genomics、4:114−128 (1989): Bains、DNA Sequencing and Mapping、4: 143−150 (1993): Bains、Genomics、11:294−301 (1991): Drmanacら、J.Biomolecular Structure and Dynamics、8: 1085−1102 (1991): およびPevzner、J.Biomolecular Structure and Dynamics、7: 63−73 (1989)。好適には、標的ポリヌクレオチドの長さは、1キロベース〜50キロベースである。より好適には、長さは、10キロベース〜40キロベースである。LanderおよびWaterman(上記)は、配列決定されるフラグメントの数(すなわち、サンプルサイズ)、各フラグメントから得られた配列情報の量、および標的ポリヌクレオチドが、ギャップがない部分的な配列、すなわち、「アイランド」から再構築され得る可能性の間の相互関係に関する指針を提供する。本発明において、所与のサンプルサイズおよびフラグメント配列のサイズで得られ得る最大のポリヌクレオチドサイズを以下に示す。
【0112】
【化8】
フラグメントは、最小の重複で標的ポリヌクレオチドをカバーするフラグメントのセットを生成しようとする、いわゆる「直接的」なアプローチ、およびランダムに重複するフラグメントが生成される、いわゆる「ショットガン」アプローチを含む、種々の方法で標的ポリヌクレオチドから生成され得る。好適には、フラグメント生成についての「ショットガン」アプローチが、その簡便性および固有の縮重のために用いられる。例えば、標的ポリヌクレオチドをカバーする、ランダムに重複するフラグメントは、以下の従来の「ショットガン」配列決定プロトコルで生成される(例えば、Sambrookら(上記)に開示されている)。本明細書に用いられるような、この文脈での「カバーする(cover)」は、標的ポリヌクレオチド配列のすべての部分が、生成されたフラグメントの各サイズ範囲(例えば、全てのフラグメントは長さが、100塩基対〜200塩基対の範囲である)で表されることを意味する。簡潔には、適切なクローニングベクター(例えば、λファージ)における挿入物としての標的ポリヌクレオチドから始まって、ベクターは拡大され、精製され、そして適切な制限酵素で消化され、約10〜15μgの精製された挿入物を得る。代表的に、プロトコルは、1マイクログラムの開始DNA当たり約500〜1000のサブクローンをもたらす。挿入物は、調製ゲル電気泳動によってベクターフラグメントから分離され、従来の方法によってゲルから取り出され、そしてTE(Tris−EDTA)のような標準的な緩衝液に再懸濁される。ベクターから挿入物を切り出すために選択される制限酵素は、好適には、挿入物に適合性の粘着末端を残し、これにより、挿入物が、ランダムに重複するフラグメントを生成するために、調製において自己連結され得る。Sambrookら(上記)に説明されるように、環状化DNAは、以下に用いられる断片化方法における直鎖状DNAに比べて、フラグメントのよりよいランダム分配を与える。挿入物を例えば、T4リガーゼで従来のプロトコルを用いて、自己連結した後、生成された連結挿入物が、標準的なプロトコル(例えば、Mn++の存在下における超音波処理またはDNase I 消化)によって断片化される。断片化後、フラグメントの末端が、例えば、Sambrookら(上記)に記載されるように修復され、そして修復されたフラグメントは、ゲル電気泳動を用いて、サイズによって分離される。300〜500塩基対の範囲のフラグメントが選択され、従来の手段によってゲルから溶出され、上記のように、タグを有するベクター中に連結され、タグ−フラグメント結合体のライブラリーを形成する。
【0113】
上記のように、数千のタグ−フラグメント結合体を含有するサンプルが、ライブラリーから採取され、そして拡大され、その後、タグ−フラグメント挿入物がベクターから切り出され、そして上記のように、微粒子上のタグ相補物への特異的なハイブリダイゼーションのために調製される。標的ポリヌクレオチドのサイズに応じて、多重サンプルが、標識−フラグメントライブラリーから採取され、別々に拡大され、微粒子上にロードされ、そして配列決定される。選択された二重の数は、サンプル中に表されたタグレパートリーの画分に依存する(トリプル−−同じタグを有する3つの異なるポリヌクレオチド−−または上記を得る可能性は、安全に無視し得る)。上記のように、サンプル中のダブルの可能
性は、Poisson分布 p(ダブル)= m2e−m/2(ここで、mは、サンプル中のタグレパートリー画分である)から評価され得る。下の表Vは、所与のタグサイズ、サンプルサイズおよびレパートリーの多様性についてサンプル中でダブルを得る可能性を挙げている。
【0114】
【表5】
いずれの場合においても、次いで、ロードされた微粒子が、好適には、アビジン−ビオチン結合を介して、ガラス顕微鏡スライド上に分散、および固定される。好適には、それぞれのランダムフラグメントの少なくとも15〜20のヌクレオチドが、単一塩基方法で同時に配列決定される。次いで、標的ポリヌクレオチドの配列が、それらの重複する部分によって、アセンブリングコンティグに用いられるのと類似の、または上記の引例に開示されたようなハイブリダイゼーションにより配列決定するために開発されたような、アルゴリズムを用いて、ランダムフラグメントの部分的な配列を照合することによって再構築される。
【0115】
(本発明の方法を実施するためのキット)
本発明は、本発明の種々の実施態様を実施するためのキットを含む。好適には、本発明のキットは、固相支持体に結合されたタグ相補物のレパートリーを含む。さらに、本発明のキットは、対応するタグのレパートリー(例えば、ポリヌクレオチドを増幅して分類するためのプライマーとして、またはポリヌクレオチドを増幅して分類するために用いられ得るクローニングベクターの要素として)を含み得る。好適には、タグ相補物のレパートリーは、微粒子に結合される。キットはまた、酵素的プロセシング、検出ケミストリー(例えば、蛍光タグまたは化学ルミネセンスタグ)などのための適切な緩衝液、使用説明書、プロセシング酵素(例えば、リガーゼ、ポリメラーゼ、トランスフェラーゼなど)を含み得る。配列決定のための重要な実施態様において、キットはまた、プロセシングのためにロードされた微粒子を固定するための、アビジン化された顕微鏡スライドのような基板を含み得る。
【0116】
(cDNAライブラリーにおける新規なポリヌクレオチドの同定)
cDNAライブラリーにおける新規なポリヌクレオチドは、上記のように微粒子に結合されたcDNA分子のライブラリーを構築することによって、同定され得る。次いで、ライブラリーの大きな画分、またはライブラリー全体でさえ、部分的に並行に配列決定され得る。mRNAの単離、およびおそらく、Soaresら、Proc. Natl. A
cad. Sci.、91:9228−9232 (1994)、または同様の引例によって教示されるように、その集団の正規化の後、以下のプライマーが、従来のプロトコルを用いて、逆転写酵素で第1鎖の合成のためのポリAテールにハイブリダイズされ得る:
【0117】
【化9】
ここで、[W,W,W,C]9は、上記のようなタグを表し、「ACCAGCTGATC」は、二本鎖形態で制限部位を形成する任意の配列であり、そして「プライマー部位」は、PCRによって目的のポリヌクレオチドを増幅するためのプライマー結合部位として後に用いられる、ライブラリーのすべてのメンバーに共通の配列である。
【0118】
従来の技術による逆転写および第2鎖合成の後、二本鎖フラグメントが、上記のようにクローニングベクターに挿入され、そして増幅される。次いで、増幅されたライブラリーがサンプリングされ、そしてサンプルは増幅する。増幅されたサンプルからのクローニングベクターが単離され、そしてタグ化されたcDNAフラグメントが切り出され、そして精製される。上記のようなポリメラーゼでタグを一本鎖にした後、フラグメントはメチル化され、そして本発明によって微粒子上に分類される。好適には、上記のように、クローニングベクターは、タグ化されたcDNAが、分類および微粒子への連結後に好適な単一塩基方法による即座の配列決定を可能にする、Fok I のようなエンドヌクレアーゼで切り出され得るように構築される。
【0119】
次いで、段階的な配列決定が、本発明に従って、ライブラリー全体またはライブラリーの1つまたはそれ以上の大きな画分で同時に行われ、ついには、十分な数のヌクレオチドが、ライブラリーが由来する生物のゲノムに特有の表示のために各cDNAについて同定される。例えば、もし、ライブラリーが哺乳動物のmRNAに由来するなら、14〜15ヌクレオチドの長さのランダムに選択された配列が、典型的な哺乳動物のゲノムの2000〜3000メガベースの間で特有の表示を有すると予想される。もちろん、細菌または他の下等生物に由来するライブラリーに特有の表示には、ずっと少ないヌクレオチドの同定で十分である。好適には、少なくとも20〜30ヌクレオチドが、特有の表示を確実にし、かつ、以下に記載するように、適切なプライマーの構築を可能にするために同定される。次いで、表にされた配列は、ユニークなcDNAを同定するために、既知の配列と比較され得る。
【0120】
次いで、特有のcDNAは、従来の技術(例えば、プライム部位およびその配列が決定されたcDNAの部分に関するプライマーで製造されたPCRアンプリコンからプローブを構築する技術)によって単離される。次いで、プローブは、従来のスクリーニングプロトコルを用いてライブラリー中のcDNAを同定するために用いられ得る。
【実施例】
【0121】
(実施例I)
(pUC19由来の多重標的ポリヌクレオチドの分類)
3つの標的ポリヌクレオチドタグ結合体の混合物を、以下のように得る:まず、以下の6つのオリゴヌクレオチドを合成し、対として組み合わせ、タグ1、タグ2およびタグ3を形成する。
【0122】
【化10】
ここで、「p」はモノホスフェートを示し、wiは表Iで定義されるサブユニットを表し、用語「(**)」はそれぞれの相補物を表す。pUC19を、Sal IおよびHind IIIで消化し、大きなフラグメントを精製し、そしてタグ1、タグ2およびタグ3で別々に連結して、それぞれ、pUC19−1、 pUC19−2、 pUC19−3を形成する。この3つの組換え体を、別々に増幅し単離し、その後、pUC19−1を、Hind IIIおよびAat Iで消化し、 pUC19−2をHind IIIおよびSsp Iで消化し、pUC19−3を、Hind IIIおよびXmn Iで消化する。小さなフラグメントを従来のプロトコルを用いて単離して、それぞれ、約250、375および575塩基対の長さの3つの二本鎖フラグメントを得る。それぞれは、タグに隣接する陥没した3’鎖および反対側の末端には平滑または3’突出鎖を有する。およそ12nmolの各フラグメントを、製造者が推薦する、33μM デオキシシトシントリホスフェートを含有する反応緩衝液中で、5ユニットのT4 DNAポリメラーゼと混合する。反応混合物を37℃で30分間インキュベートし、その後、氷の上に置くことによって反応を止める。次いで、フラグメントを、従来の手段で精製する。
【0123】
CPG微粒子(粒子サイズ37〜74mm、孔サイズ500オングストローム、Pierce Chemical)を、MaskosおよびSouthen、Nucleic Acids Research、20:1679−1684 (1992)によって開示されたリンカーで誘導体化する。3つのアリコートに分離後、タグ1、タグ2およびタグ3の相補物を、従来の自動化されたDNA合成機(例えば、model 392 DNA合成機(Applied Biosystems、Foster City、CA))を用いて、微粒子上で合成する。異なって誘導体化された微粒子のそれぞれ約1mgを、別個の容器に入れる。
【0124】
pUC19−1、pUC19−2およびpUC19−3から切り出されたT4 DNAポリメラーゼ処理のフラグメントを、製造者推薦のTaq DNAリガーゼ(New England Biolabs)のための緩衝液50μLに再懸濁する。次いで、混合物を、誘導体化CPG微粒子をそれぞれ1mg含有している3つの容器に等分する。5ユニットのTaq DNAリガーゼを各容器に添加し、その後、それらを55℃で15分間インキュベートする。氷の上に置くことによって反応を止め、そして微粒子を、繰り返し遠心分離およびTEに再懸濁することによって数回洗浄する。最後に、微粒子をNde I反応緩衝液(New England Biolabs)で再懸濁し、結合したポリヌク
レオチドを消化する。微粒子からの分離後、Nde I消化によって放出されたポリヌクレオチドフラグメントを、Sequenase DNAポリメラーゼおよびフルオレセイン標識チミジントリホスフェート(Applied Biosystems、Foster City、CA)でインキュベートすることにより、蛍光標識する。次いで、そのフラグメントを、Applied Biosystems model 373 DNA配列決定機を用いて、非変性ポリアクリルアミドゲル上で別々に分析する。
【0125】
(実施例 II)
(SV40フラグメントの並行配列決定)
表Iから選択される9つの4−ヌクレオチドサブユニットからなる36マーのタグのレパートリーを、上記のような、分裂または混合手段によって、タグおよびタグ相補物を別々に合成することによって調製する。Sma I/Hind III消化M13mp19に連結可能なように、レパートリーを合成する。従って、実施例Iと同様に、1組のオリゴヌクレオチドはAの添加で開始し、9回の分裂および混合合成が続く。このオリゴヌクレオチドを、表Iのサブユニットに対応して3’−ホスホルアミダイト誘導体化4マーによりサブユニットで伸張する。次いで、Sma I認識部位(GGG) の半分、2つのCおよび5’−モノホスフェートを、例えば、Clontech Laboratories (Palo Alto、CA)から入手可能であるPhosphate−ON試薬を用いて、ヌクレオチド毎に添加することで合成を完了する。他方の組のオリゴヌクレオチドは、3つのC(Sma I認識部位の一部)および2つのGの添加で開始し、9回の分裂および混合合成が続く。このオリゴヌクレオチドを、表Iのサブユニットの相補物に対応して、3’−ホスホルアミダイト誘導体化4マーにより伸張する。Hind III認識部位および5’−モノホスフェートのヌクレオチド毎の添加により合成を完了する。合成支持体からの分離後、オリゴヌクレオチドを以下の二重鎖の形成を可能にする条件下で混合する:
【0126】
【化11】
次いで、二重鎖の混合物をSma I/Hind III消化M13mp19に連結する。タグ相補物のレパートリーを上記のようにCPG微粒子上で合成する。
【0127】
次に、Fok I部位ならびにEco RIおよびSma I部位の一部を含有する、以下のアダプターを調製する:
【0128】
【化12】
このアダプターを、上記のように、Eco RI/Sma I消化M13に連結する。
【0129】
別に、Sambrookら(上記)に記載のプロトコルに従って超音波処理により、SV40 DNAを断片化する。得られたフラグメントを標準的なプロトコルを用いて修復し、そしてサイズによって分離する。300〜500塩基対の範囲のフラグメントを選択し、上記のSma I消化M13に連結してフラグメント−タグ結合体のライブラリーを形成し、次いで、増幅する。数千の異なるフラグメント−結合体を含有するサンプルをラ
イブラリーから取り出し、さらに増幅し、そしてフラグメント−タグ挿入物を、Eco R IおよびHind IIIで消化することにより切り出す。切り出されたフラグメント−タグ結合体を、実施例Iに記載のように、デオキシシチジントリホスフェートの存在下でT4 DNAポリメラーゼで処理して、特異的なハイブリダイゼーションのためのオリゴヌクレオチドタグをCPG微粒子に曝露する。
【0130】
実施例Iに記載のように、ハイブリダイゼーションおよび連結後、ロードされた微粒子をFok Iで処理して、予め決定した配列の4−ヌクレオチド突出鎖を生成する。以下のプローブの10:1の混合物(プローブ1:プローブ2)を微粒子上のポリヌクレオチドに連結する。
【0131】
【化13】
FAMは、Appiled Biosystems から入手可能なアミノホスフェートリンカー(Aminolinker)を介してプローブ1の上段の鎖の5’−水酸基に結合されるフルオレセインを表す。ビオチンはまた、Aminolinker部分を介して結合され得、そして必要に応じて、ポリエチレンオキシドリンカー(例えば、Jaschkeら(上記))を介してさらに伸張され得る。
【0132】
次いで、ロードされた微粒子をアビジン化ガラススライドの表面上に堆積する。試薬および洗浄溶液は表面上へ送達および表面上から除去され得る。結合した微粒子を有するアビジン化スライドにより、走査型蛍光顕微鏡(例えば、Newport Model PM500−Cモーションコントローラー 、488nmの励起光を発生するSpectra−Physics Model 2020アルゴンイオンレーザー、および520nm波長域発光フィルターなどの装置を備えたZeiss Axioskop)で検査する。励起光および蛍光発光を、同じ対物レンズを通して、それぞれ、送達および収集する。二色性ミラーによって励起光および収集蛍光を分離する。二色性ミラーは、収集された蛍光を一連の帯域フィルターを介して、モニターされている蛍光物質に対応するフォトン計数デバイス(例えば、Hamamatsu model 9403−02光電子増倍管、Stanford Research Systems model SR445増幅器およびmodel SR430多重チャンネルスケーラーおよびデジタルコンピューター(例えば、486−ベースのコンピューター)を備える)へ指向する。コンピューターは、微粒子の位置を記録するスライドの二次元マップを生成する。
【0133】
最初のプローブを除去するためのFok Iでの切断後、結合微粒子上のポリヌクレオチドを、以下に記載の好適な単一塩基配列決定方法に従い、プローブ連結、洗浄、検出、切断および洗浄工程を20サイクル行う。各検出工程において、走査システムが、各微粒子で同定された塩基に対応する蛍光発光を記録する。一般に、以下の反応および洗浄を、特に記載されない限り、用いる酵素のための、製造者 (New England Biolabs) の推薦する緩衝液で実施する。標準的な緩衝液もまた、Sambrookら(上記)に記載されている。
【0134】
以下の4組の混合プローブを、標的ポリヌクレオチドへの添加用に提供する:
【0135】
【化14】
ここで、TAMRA、FAM、ROXおよびJOEは、Aminolinker IIによって結合された分光解像可能な蛍光標識である(すべて、Applied Biosystems、Inc.、Forster City、Californiaから入手可能である);太字のヌクレオチドは、Fok Iエンドヌクレアーゼのための認識部位であり、「N」は、4つのヌクレオチド、A、C、G、T.のいずれか1つを表す。TAMRA(テトラメチルローダミン)、FAM(フルオレセイン)、ROX(ローダミンX)、およびJOE(2’,7’−ジメトキシ−4’,5’−ジクロロフルオレセイン)およびそれらのオリゴヌクレオチドへの結合はまた、Fungら米国特許第4,855,225号に記載されている。
【0136】
上記のプローブを、約5モル過剰量の標的ポリヌクレオチド末端中で、以下のようにインキュベートする:プローブを、200ユニットのT4 DNAリガーゼおよびアンカー化標的ポリヌクレオチドとともに60分間16℃でT4 DNAリガーゼ緩衝液中でインキュベートする:次いで、洗浄後、標的ポリヌクレオチドを、100ユニットのT4ポリヌクレオチドキナーゼとともに30分間37℃で製造者の推薦する緩衝液中でインキュベートし、洗浄し、そして再度、200ユニットのT4 DNAリガーゼおよびアンカー化標的ポリヌクレオチドとともに30分間16℃でT4 DNAリガーゼ緩衝液中でインキュベートする。洗浄は、スライド上に洗浄緩衝液(例えば、TE)を大量に連続的に流すことによって達成される(Sambrookら(上記)に開示されている)。連結−リン酸化−連結のサイクルおよび最後の洗浄後、結合微粒子が、蛍光標識の存在について走査され、その位置および特徴が走査システムによって記録される。次いで、標識された標的ポリヌクレオチド、即ち、連結複合体を、製造者の推薦する緩衝液中で30分間37℃で、10ユニットのFor Iとともにインキュベートし、その後、TE中で洗浄する。その結果、標的ポリヌクレオチドは各鎖上で1つのヌクレオチド分短くされ、次の連結および切断サイクルのために準備される。20のヌクレオチドが同定されるまでこのプロセスを続ける。
【0137】
【表6】
【0138】
【表7】
【0139】
【表8】
【図面の簡単な説明】
【0140】
【図1a】図1a−1cは、本発明で用いられ得る「単一塩基」配列決定の好ましい方法において用いられる標識されたプローブの構造を例示する。
【図1b】図1a−1cは、本発明で用いられ得る「単一塩基」配列決定の好ましい方法において用いられる標識されたプローブの構造を例示する。
【図1c】図1a−1cは、本発明で用いられ得る「単一塩基」配列決定の好ましい方法において用いられる標識されたプローブの構造を例示する。
【図2】図2は、好ましい「単一塩基」配列決定方法において用いられた標的ポリヌクレオチドとプローブとの間で形成された連結された複合体中のヌクレアーゼ認識部位、連結部位および切断部位の相対的な位置を例示する。
【図3】図3は、最少に交差ハイブリダイズしているセットを生じるための一般的なアルゴリズムを例示するフローチャートである。
【図4】図4は、コンビナトリアルケミカルライブラリーを合成および使用するためのスキームを例示し、ここでメンバー化合物は本発明に従ってオリゴヌクレオチドタグで標識される。
【図5】図5は、本発明に従ってポリヌクレオチド配列決定のような並行操作を行うための装置を図で例示する。
【0141】
(配列表)
【0142】
【表9】
【0143】
【表10】
【0144】
【表11】
【技術分野】
【0001】
本発明は、一般にはオリゴヌクレオチドラベルで分子(特にポリヌクレオチド)を同定、分類、および/または探知するための方法に関し、より詳細にはオリゴヌクレオチドタグへの特異的ハイブリダイゼーションによりポリヌクレオチドを分類する方法に関する。
【背景技術】
【0002】
オリゴヌクレオチドおよびそのアナログの特異的ハイブリダイゼーションは、広く多様な研究的適用、医学的適用および産業的適用で用いられている基礎的なプロセス(診断アッセイにおける疾患関連のポリヌクレオチドの同定、新規の標的ポリヌクレオチドのクローンのスクリーニング、ポリヌクレオチド混合物のブロットにおける特異的ポリヌクレオチドの同定、特異的な標的ポリヌクレオチドの増幅、不適切に発現された遺伝子の治療的ブロッキング、DNA配列決定などを含む)である(例えば、非特許文献1〜5)。
【0003】
特異的ハイブリダイゼーションはまた、オリゴヌクレオチドタグで標識された化合物を探知、回収、および同定する方法として提案されている。例えば、複数のDNA配列決定において、オリゴヌクレオチドタグは、同じ配列決定反応において生じるDNAフラグメントからなるゲル上の電気泳動的に分離されたバンドを同定するために用いられる。このように、多くの配列決定反応由来のDNAフラグメントが、ゲルの同じレーンで分離され、次いで異なる固相物質でブロットされる。そこでは、異なる配列決定反応由来のフラグメントバンドが、相補的なタグに特異的にハイブリダイズするオリゴヌクレオチドプローブで可視化される(非特許文献6)。オリゴヌクレオチドタグの同様の用途はまた、爆発物、潜在的な汚染物質(例えば、原油)、および模造品の防止または検出のために通貨を同定するために提供されてきた(例えば、非特許文献7)にレビューされた)。さらに最近、オリゴヌクレオチドタグを用いるシステムはまた、複雑なコンビナトリアルケミカルライブラリー中の個々の分子を操作および同定する方法として(例えば、このような薬物候補物のライブラリーのスクリーニングを助けるために)提案されている(非特許文献8〜10)。
【0004】
このようなタグ化スキームをうまく実施することは、タグとその相補的プローブとの間での特異的なハイブリダイゼーションの達成における成功に大部分依存している。すなわち、オリゴヌクレオチドタグが基質をうまく同定するためには、擬陽性および擬陰性のシグナルの数を最小限にしなければならない。不幸にも、このような偽のシグナルは希ではない。なぜなら塩基対形成および塩基の積み重ね(base stacking)の自由エネルギーは二重鎖または三重鎖構造におけるヌクレオチドのあいだで広く変化するから
である。例えば、その相補物に結合したデオキシアデニン(A)およびチミジン(T)の繰り返し配列からなる二重鎖は、ミスマッチを含む部分的に相補的な標的に結合したデオキシグアニジン(G)およびデオキシシチジン(C)の繰り返し配列からなる同じ長さの二重鎖より安定性が低い可能性がある。従って、大きなコンビナトリアルケミカルライブラリー由来の所望の化合物が前者のオリゴヌクレオチドでタグ化された場合、完全にマッチしたATに富む二重鎖を検出するように設計されたハイブリダイゼーション条件下では、GCに富むオリゴヌクレオチドで標識された所望されない化合物(ミスマッチした二重鎖においてさえ)が、ATに富むタグからなる完全にマッチした二重鎖とともに検出されるという重大な可能性が存在する。非特許文献8(上記)により提案された分子タグ化システムでは、親密に関連するタグの関連するミスハイブリダイゼーションの問題は、いわゆる「無限(comma−less)」コードを用いることにより処理され、これはその相補的タグについて登録をはずれた(またはフレームシフトした)プローブが、その5またはそれ以上の3塩基ワードまたは「コドン」のおのおのについて1つまたはそれ以上のミスマッチを有する二重鎖を生じることを保証する。
【0005】
たとえ塩化テトラメチルアンモニウムのような試薬がオリゴヌクレオチド二重鎖の塩基特異的安定性の相違を無効にするために利用可能であるとしても、このような試薬の効果はしばしば限定され、そしてその存在は選択された化合物のさらなる操作(例えば、ポリメーラーゼ連鎖反応(PCR)などによる増幅)を不可能にし得るか、またはより困難にし得る。
【0006】
このような問題は、例えば多数回PCR、リバースドットブロッティングなどを介する多数のまたは複合した遺伝子座の分析における多数のハイブリダイゼーションプローブの同時使用を非常に困難にしている。その結果、例えばHLA遺伝子のような特定の座の直接配列決定は、遺伝子型の同定のための特異的ハイブリダイゼーションを用いる間接的な方法に対する信頼し得る代替法として奨励される(例えば、非特許文献11))。
【0007】
異なる固相支持体上で、クローン化されかつ同様にタグ化されたDNAを分類する能力は、特に、並行して多くのサンプルに同時に適用し得る非ゲルベースの配列決定方法と連結される場合に、このような配列決定を促進する。
【非特許文献1】Sambrookら、「Molecular Cloning:A Laboratory Manual」 第2版 Cold Spring Harbor Laboratory,New York,1989年
【非特許文献2】KellerおよびManak、「DNA Probes」 第2版 Stockton Press.New York,1993年
【非特許文献3】Milliganら、J.Med.Chem. 第36巻 p.1923−1937 1993年
【非特許文献4】Drmanacら、Science,第260巻 p.1649−1652 1993年
【非特許文献5】Bains J. DNA Sequencing and Mapping,第4巻 p.143−150 1993年
【非特許文献6】Churchら、Science,第240巻 p.185−188 1988年
【非特許文献7】Dollinger、Birkhauser,Boston,Mullisら編、「The Polymerase Chain Reaction」 p.265−274、1994年
【非特許文献8】BrennerおよびLerner. Proc.Natl.Acad.Sci.、第89巻 p.5381−5383 1992年
【非特許文献9】Alper. Science、第264巻 p.1399−1401 1994年
【非特許文献10】Needelsら、Proc.Natl.Acad.Sci.、第90巻 p.10700−10704 1993年
【非特許文献11】Gyllenstenら、Proc.Natl.Acad.Sci.,第85巻 p.7652−7656 1988年
【発明の開示】
【発明が解決しようとする課題】
【0008】
上記を考慮すると、多くのタグのレパートリーを提供するが、また天然の塩基対形成および塩基の積み重ねの自由エネルギーの差違を変化させるための特別な試薬を用いる必要はなく、擬陽性および擬陰性のシグナルの発生を最小限にするオリゴヌクレオチドベースのタグ化システムが利用可能である場合、これは有用である。このようなタグ化システムにより、コンビナトリアルケミカルライブラリーの構築および使用、DNAの大スケールのマッピングおよび配列決定、遺伝子同定、医学診断薬などを含む多くの領域での適用が見い出される。
【0009】
本発明の目的は、化合物の探知、回収、および同定のための分子タグ化システムを提供することである
【課題を解決するための手段】
【0010】
(発明の要旨)
1つの局面において、本発明は、分子集団から分子または分子亜集団を分類する方法を提供し、この方法は、以下の工程:
(a)タグのレパートリー由来のオリゴヌクレオチドタグを、分子集団中の各分子に結
合する工程であって、この工程は、(i)この集団中の実質的に同じ分子または実質的に同じ分子亜集団が、結合した同じオリゴヌクレオチドタグを有し、そしてこの集団中の実質的にすべての異なる分子または異なる分子亜集団が、結合した異なるオリゴヌクレオチドタグを結合しているように、そして(ii)このレパートリー由来の各オリゴヌクレオチドタグが複数のサブユニットを包み、そしてこの複数のサブユニットのそれぞれが3から6ヌクレオチドまたは3から6塩基対の長さを有するオリゴヌクレオチドからなり、このサブユニットが最少に交差ハイブリダイズしているセットから選択されるように行われる工程;および
(b)このオリゴヌクレオチドタグとそれぞれの相補物とを特異的にハイブリダイズすることにより、この集団からこの分子または分子亜集団を分類する工程;
を包含する。
【0011】
1つの実施形態において、上記分子または分子亜集団が、ポリヌクレオチドまたはポリヌクレオチド亜集団である。好ましい実施形態において、上記オリゴヌクレオチドの相補物が、固相支持体に結合される。より好ましい実施形態において、上記固相支持体が、上記相補物の均一な集団が結合している微粒子である。さらにより好ましい実施形態において、上記オリゴヌクレオチドタグおよび上記相補物が、一本鎖オリゴヌクレオチドである。
なおさらにより好ましい実施形態において、上記ポリヌクレオチドまたはポリヌクレオチド亜集団が、50から5000ヌクレオチドの範囲の長さを有する。なおさらにより好ましい別の実施形態において、上記微粒子が、ガラス微粒子、磁気ビーズ、グリシダルメタクリレート微粒子およびポリスチレン微粒子からなる群から選択される。なおさらにより好ましい実施形態において、上記微粒子の直径が1μmと100μmとの間である。さらにより好ましい別の実施形態において、上記固相支持体が、上記相補物の均一な集団を結合している複数の空間的に離れた表面領域を有する平面基板である。なおさらにより好ましい実施形態において、異なる上記の複数の空間的に離れた表面領域が、異なる上記相補物の均一な集団を有する。なおさらにより好ましい実施形態において、上記平面基板が、ガラス、シリコンおよびプラスチックからなる群から選択される。
【0012】
別の実施形態において、上記分子または分子亜集団が、ポリペプチドまたはポリペプチド亜集団である。好ましい実施形態において、上記オリゴヌクレオチドタグの相補物が、固相支持体に結合している。より好ましい実施形態において、上記オリゴヌクレオチドタグおよび上記相補物が、一本鎖オリゴヌクレオチドである。さらにより好ましい実施形態において、上記固相支持体が、上記相補物の均一な集団が結合している微粒子である。なおさらにより好ましい実施形態において、上記微粒子が、ガラス微粒子、磁気ビーズ、およびポリスチレン微粒子からなる群から選択される。なおさらにより好ましい実施形態において、上記固相支持体が、上記相補物の均一な集団を結合している複数の空間的に離れた表面領域を有する平面基板である。なおさらにより好ましい実施形態において、異なる上記の複数の空間的に離れた表面領域が、異なる上記相補物の均一な集団を有する。なおさらにより好ましい実施形態において、上記平面基板が、ガラス、シリコンおよびプラスチックからなる群から選択される。
【0013】
別の実施形態において、上記分子または分子亜集団が、以下の形態の線形ポリマー分子:
−(M−L)n−
を含み、ここで、リン(V)結合基であり;
Mは、1〜20個の炭素原子と、酸素、窒素、およびイオウからなる群から選択される0〜10個のヘテロ原子とを有する、直鎖分子構造、環式分子構造、または分枝有機分子構造であり;そして
nは、3〜100の範囲にある。
【0014】
好ましい実施形態において、Mは、1〜16個の炭素原子を有するアルキル、アルコキシ、アルケニル、またはアリールであるか;3〜8個の炭素原子と、酸素、窒素、およびイオウからなる群から選択される1〜3個のヘテロ原子とを有するヘテロ環である。より好ましい実施形態において、上記オリゴヌクレオチドタグの相補物は、固相支持体に結合されている。さらにより好ましい実施形態において、上記微粒子が、ガラス微粒子、磁気ビーズ、およびポリスチレン微粒子からなる群から選択される。なおさらにより好ましい別の実施形態において、上記固相支持体が、上記オリゴヌクレオチドタグの相補物の均一な集団を内部に結合している複数の離れた非重複表面領域を有する、平面基板である。なおさらにより好ましい実施形態において、異なる上記の複数の空間的に離れた表面領域が、異なる上記相補物の均一な集団を有する。
【0015】
別の局面において、本発明は、標的ポリヌクレオチドのヌクレオチド配列を決定する方法を提供し、この方法は、以下の工程:
この標的ポリヌクレオチドから、この標的ポリヌクレオチドをカバーする複数のフラグメントを生成する工程;
タグのレパートリー由来のオリゴヌクレオチドタグを、この複数のフラグメントのそれぞれに結合する工程であって、この工程は、(i)実質的にすべての同じフラグメントが、結合した同じオリゴヌクレオチドタグを有し、実質的にすべての異なるフラグメントが、異なるオリゴヌクレオチドタグを結合しているように、そして、(ii)このレパートリー由来の各オリゴヌクレオチドタグが複数のサブユニットを包含し、そしてこの複数のサブユニットのそれぞれが、3から6ヌクレオチドまたは3から6塩基対の長さを有するオリゴヌクレオチドからなり、このサブユニットが、最少に交差ハイブリダイズしているセットから選択されるように行われる、工程;
このオリゴヌクレオチドタグとそれぞれの相補物とを特異的にハイブリダイズすることによりこのフラグメントを分類する工程;
この複数のフラグメントのそれぞれの部分のヌクレオチド配列を決定する工程;および
このフラグメントの配列を対照することにより、この標的ポリヌクレオチドのヌクレオチド配列を決定する工程、
を包含する。
【0016】
1つの実施形態において、上記オリゴヌクレオチドタグおよび上記相補物が、固相支持体に結合されている。好ましい実施形態において、上記オリゴヌクレオチドタグおよび上記相補物が、一本鎖オリゴヌクレオチドである。より好ましい実施形態において、上記生成工程が、上記標的ポリヌクレオチドのランダムに重複するフラグメントを生成する。さらにより好ましい実施形態において、上記フラグメントのヌクレオチド配列を決定する工程が、単一塩基配列決定方法により、上記の複数のフラグメントについて同時に行われる。なおさらにより好ましい実施形態において、上記フラグメントのそれぞれの部分が、12から50ヌクレオチドを含む。なおさらにより好ましい実施形態において、上記フラグメントのそれぞれの部分が12から25ヌクレオチドを含む。なおさらにより好ましい実施形態において、上記標的ポリヌクレオチドが、1キロベース長と50キロベース長との間の長さである。好ましい別の実施形態において、上記固相支持体が、上記相補物の均一な集団をそれぞれ結合している複数の微粒子である。より好ましい実施形態において、上記分類工程後、上記の複数の微粒子が平面基板に固定される。さらにより好ましい実施形態において、上記の複数の微粒子が、1平方センチメートル当たり約1000微粒子から約100,000微粒子の間の密度で、上記平面基板の表面にランダムに配置される。
【0017】
別の局面において、本発明は、組成物を提供し、この組成物は:
1つまたはそれ以上の空間的に離れた領域を有する固相支持体;および
この1つまたはそれ以上の空間的に離れた領域の少なくとも1つにおいてこの固相支持
体に共有結合されたオリゴヌクレオチドタグ相補物の均一な集団であって、このオリゴヌクレオチドタグ相補物は複数のサブユニットを含み、各サブユニットは3から6ヌクレオチドの長さを有するオリゴヌクレオチドからなり、そして各サブユニットは、最少の交差ハイブリダイズしているセットから選択される、オリゴヌクレオチドタグ相補物の均一な集団、
を含む。
【0018】
1つの実施形態において、上記の複数のサブユニットが4個から10個の範囲にある。好ましい実施形態において、上記固相支持体が、単一の空間的に離れた領域を有する微粒子である。より好ましい実施形態において、上記微粒子が、ガラス微粒子、磁気ビーズおよびポリスチレン微粒子からなる群から選択される。
【0019】
別の局面において、本発明は、ポリヌクレオチド集団を分類する方法を提供し、この方法は、以下の工程:
オリゴヌクレオチドタグを、この集団の各ポリヌクレオチドに結合させる工程であって、この工程は、(i)実質的にすべての同じポリヌクレオチドが、結合した同じオリゴヌクレオチドタグを有し、実質的にすべての異なるポリヌクレオチドが、結合した異なるオリゴヌクレオチドタグを有しているように、そして(ii)各オリゴヌクレオチドタグが複数のサブユニットを包含し、そしてこの複数のサブユニットのそれぞれが、3から6ヌクレオチドの長さを有するオリゴヌクレオチドからなり、このサブユニットが、最少に交差ハイブリダイズしているセットから選択されるように行われる工程;
このオリゴヌクレオチドタグとそれぞれの相補物とを特異的にハイブリダイズすることにより、このポリヌクレオチドを分類する工程;
この分類されたポリヌクレオチドのそれぞれの部分のヌクレオチド配列を決定する工程;および
このポリヌクレオチドの配列の部分の頻度分布により、このポリヌクレオチド集団を分類する工程、
を包含する。
【0020】
1つの実施形態において、上記オリゴヌクレオチドタグの相補物が、固相支持体に共有結合される。好ましい実施形態において、上記固相支持体が微粒子であり、そして上記相補物の均一な集団が、この微粒子各々に結合される。より好ましい実施形態において、上記ポリヌクレオチドの集団が、cDNAライブラリーである。さらにより好ましい実施形態において、上記ポリヌクレオチドの部分が、12から50ヌクレオチドの範囲にある。なおさらにより好ましい実施形態において、上記ポリヌクレオチドの部分が、12から25ヌクレオチドの範囲にある。
【0021】
別の局面において、本発明は、オリゴヌクレオチドタグのレパートリーを提供し、このレパートリーは、以下の形態のオリゴヌクレオチド:
S1S2S3...Sn
からなる群から選択され、ここで、S1からSnのそれぞれは、3から6ヌクレオチドの長さを有するオリゴヌクレオチドからなり、そして最少に交差ハイブリダイズしているセットから選択されるサブユニットであり;そして
nは4から10の範囲にある。
【0022】
1つの実施形態において、上記サブユニットが、4から5ヌクレオチドの長さを有するオリゴヌクレオチドからなり、そして上記nが6から9の範囲にある。
【0023】
別の局面において、本発明は、オリゴヌクレオチドタグをポリヌクレオチドへ結合させるためのクローニングベクターのレパートリーを提供し、このレパートリーは、以下の形
態の二本鎖要素:
S1S2S3...Sn
を有し、ここで、S1からSnのそれぞれは、3から6ヌクレオチドの長さを有するオリゴヌクレオチドからなり、そして最少に交差ハイブリダイズしているセットから選択されるサブユニットであり;そして
nは4から10の範囲にある。
【0024】
1つの実施形態において、上記二本鎖要素が、上記ポリヌクレオチドを上記クローニングベクターに挿入するための1つまたはそれ以上の制限エンドヌクレアーゼ切断部位を含むポリリンカー領域に、隣接する。
【0025】
別の局面において、本発明は、ポリヌクレオチド混合物を分類する方法であって、この方法は、以下の工程:
ポリヌクレオチド混合物を含有する溶液を提供する工程であって、この混合物の各ポリヌクレオチドは、タグのレパートリー由来のオリゴヌクレオチドタグを結合しており、このレパートリー由来の各オリゴヌクレオチドタグは、複数のサブユニットを含み、この複数のサブユニットのそれぞれは、3から6ヌクレオチドの長さを有するオリゴヌクレオチドからなり、このサブユニットは、最少に交差ハイブリダイズしているセットから選択される工程;
このポリヌクレオチド混合物をサンプリングしてポリヌクレオチド亜集団を形成させる工程であって、ここで同じ配列の実質的にすべてのポリヌクレオチドは、結合した同じオリゴヌクレオチドタグを有し、異なる配列の実質的にすべてのポリヌクレオチドは異なるオリゴヌクレオチドタグを結合している、工程;および
このオリゴヌクレオチドタグとそれぞれの相補物との間に完全にマッチした二重鎖の形成を促進する条件下で、この亜集団と、このオリゴヌクレオチドタグの相補物を結合している1つまたはそれ以上の固相支持体とを接触させる工程、
を包含する。
【0026】
1つの実施形態において、上記固相支持体が、上記相補物の均一な集団を結合している微粒子である。好ましい実施形態において、上記オリゴヌクレオチドタグおよび上記相補物が、一本鎖オリゴヌクレオチドである。より好ましい実施形態において、上記ポリヌクレオチドが50から5000ヌクレオチドの範囲の長さを有する。さらにより好ましい実施形態において、上記微粒子が、ガラス微粒子、磁気ビーズ、グリシダルメタクリレート微粒子およびポリスチレン微粒子からなる群から選択される。なおさらにより好ましい実施形態において、上記固相支持体が、上記相補物の均一な集団を結合している複数の空間的に離れた表面領域を有する平面基板である。なおさらにより好ましい実施形態において、上記の異なる複数の空間的に離れた表面領域が、異なる上記相補物の均一な集団を有する。
【0027】
二本鎖DNAの末端に、予め決定された長さの一本鎖を生成させる方法であって、該方法は、以下の工程:
この二本鎖DNAの末端に5’鎖を供給する工程であって、この工程は、このような5’鎖は、3種以下のヌクレオチドからなる第1の群から選択されるヌクレオチドから構成されるように、そしてこの5’鎖のあらかじめ決定された長さが、この第1の群には存在しない種類の第2のヌクレオチドによって規定されるように、行われる工程;および
この第2のヌクレオチドのヌクレオシド三リン酸の存在下で、この二本鎖DNAの末端を、T4 DNAポリメラーゼに曝露する工程;
を包含する。
【0028】
cDNAライブラリーにおいて新規なcDNA分子を検出する方法であって、この方法
は、以下の工程:
mRNA分子集団からcDNAライブラリーを形成する工程であって、このcDNAライブラリー中の各cDNA分子は、結合したオリゴヌクレオチドを有し、この工程は、(i)実質的にすべての同じcDNA分子が、結合した同じオリゴヌクレオチドタグを有し、実質的にすべての異なるcDNA分子が、結合した異なるオリゴヌクレオチドタグを有しているように、そして(ii)各オリゴヌクレオチドタグが、複数のサブユニットを含み、そしてこの複数のサブユニットのそれぞれが、3から6ヌクレオチドの長さを有するオリゴヌクレオチドからなり、このサブユニットが、最少に交差ハイブリダイズしているセットから選択されるように行われる工程;
このオリゴヌクレオチドタグとそれぞれの相補物とを特異的にハイブリダイズすることにより、このcDNA分子を分類する工程;
この分類されたcDNA分子のそれぞれの部分のヌクレオチド配列を決定する工程;
この分類されたcDNA分子の配列の部分のヌクレオチド配列を、既知のcDNA分子の配列と比較することにより、新規なcDNA分子を同定する工程、
を包含する。
【0029】
1つの実施形態において、上記オリゴヌクレオチドタグの相補物が、固相支持体に共有結合される。好ましい実施形態において、上記固相支持体が微粒子であり、そして上記相補物の均一な集団が、この微粒子各々に結合される。より好ましい実施形態において、上記cDNA分子の部分が、12から50ヌクレオチドの範囲にある。さらにより好ましい実施形態において、上記cDNA分子の部分が、12から25ヌクレオチドの範囲にある。
【0030】
本発明の目的は、化合物の探知、回収、および同定のための分子タグ化システムを提供することである。
【0031】
本発明の別の目的は、オリゴヌクレオチドタグおよびその相補物の特異的ハイブリダイゼーションにより固相物質の表面上の同一の分子、または分子のサブクラス(特にポリヌクレオチド)を分類するための方法を提供することである。
【0032】
本発明のさらなる目的は、コンビナトリアルケミカルライブラリーを提供することであり、それらのメンバー化合物は、オリゴヌクレオチドタグおよびその相補物の特異的ハイブリダイゼーションにより同定される。
【0033】
本発明のなおさらなる目的は、同時分析および/または配列決定用の標的ポリヌクレオチドの数千ものフラグメント(特にランダムに重複するフラグメント)をタグ化および分類するためのシステムを提供することである。
【0034】
本発明の別の目的は、数百塩基対から数万塩基対の範囲の長さを有する標的ポリヌクレオチドを配列決定するための迅速かつ信頼できる方法を提供することである。
【0035】
本発明は、オリゴヌクレオチドタグの使用により分子の種類または亜集団を探知、同定、および/または分類するための方法および物質を提供することにより、これらおよび他の目的を達成する。本発明のオリゴヌクレオチドタグは多くのサブユニットからなり、各サブユニットは3から6ヌクレオチド長のオリゴヌクレオチドからなる。オリゴヌクレオチドタグのサブユニットは、最少に交差ハイブリダイズしているセット(minimally cross−hybridizing set)から選択される。このようなセットでは、このセットのサブユニットおよびこのセットの任意の他のサブユニットの相補物からなる二重鎖または三重鎖は少なくとも2つのミスマッチを含む。言い換えると、最少に交差ハイブリダイズしているセットのサブユニットは最良でも、同じセットの任意の他
のサブユニットの相補物と2つのミスマッチを有する二重鎖または三重鎖を形成する。特定の実施態様に用いられ得るオリゴヌクレオチドタグの数は、タグあたりのサブユニット数およびサブユニットの長さに依存する。その数は、一般的にタグの長さの全ての可能性のある配列の数(1つのタグがnヌクレオチド長であるタグの数は4nである)よりずっと少ない。より好ましくは、サブユニットは4から5ヌクレオチド長のオリゴヌクレオチドである。
【0036】
本発明の1つの局面において、固相支持体に結合したオリゴヌクレオチドタグの相補物は、ポリヌクレオチドをそれぞれタグを含むポリヌクレオチドの混合物から分類するために用いられる。この実施態様では、オリゴヌクレオチドタグの相補物は、同一配列の集団が特定の領域で産生されるように、微細ビーズのような固相支持体の表面上、または単一の支持体上の合成位置のアレイ(array)上の特定の位置で合成される。すなわち、各支持体(ビーズの場合)または各領域(アレイの場合)の表面は、特定の配列を有する唯一のタイプの相補物によって誘導体化される。このようなビーズ集団または領域集団は別個の配列を有する相補物のレパートリーを含み、そのレパートリーのサイズはオリゴヌクレオチドタグあたりのサブユニットの数および用いられたサブユニットの長さに依存する。同様に、分類すべきポリヌクレオチドは、同一のポリヌクレオチドが同じタグを有し、そして異なるポリヌクレオチドが異なるタグを有するようにレパートリー中のオリゴヌクレオチドタグをおのおの含む。下記でより十分に説明するように、この条件は、タグ化されたポリヌクレオチドの完全なアンサンブルからタグ化されたポリヌクレオチドの十分に小さいサンプルを取ることにより達成される。従って、支持体およびポリヌクレオチドの集団がそれぞれの相補物とオリゴヌクレオチドタグの特異的ハイブリダイゼーションを可能にする条件下で混合される場合、同一ポリヌクレオチドの亜集団は特定のビーズまたは領域上に分類される。次いでポリヌクレオチドの亜集団はミクロ生化学的技法により固相支持体上で操作され得る。
【0037】
一般的に、本発明の方法は以下の工程を包含する:(a)分子集団中の各分子にタグのレパートリー由来のオリゴヌクレオチドタグを結合して、(i)集団中の実質的に全ての同じ分子または分子の同じ亜集団は結合した同じオリゴヌクレオチドタグを有し、そして集団中の実質的に全ての異なる分子または分子の異なる亜集団は結合した異なるオリゴヌクレオチドタグを有し、ならびに(ii)レパートリー由来の各オリゴヌクレオチドタグは多数のサブユニットを含有し、そして多数の各サブユニットは3から6ヌクレオチドまたは3から6塩基対の長さを有するオリゴヌクレオチドからなり、そのサブユニットが最少に交差ハイブリダイズしているセットから選択される、工程:および(b)それらのそれぞれの相補物とオリゴヌクレオチドタグを特異的にハイブリダイズすることにより、集団の分子または分子の亜集団を分類する、工程。
【0038】
本発明の重要な局面は、並行する配列決定のためにポリヌクレオチドを分類するためのオリゴヌクレオチドタグの使用である。好ましくは、このような配列決定は以下の工程により行われる:(a)標的ポリヌクレオチドをカバーする多数のフラグメントを標的ポリヌクレオチドから生成する、工程;(b)タグのレパートリー由来のオリゴヌクレオチドタグをその多数の各フラグメントに結合して、(i)実質的に全ての同じフラグメントは、結合した同じオリゴヌクレオチドタグを有し、そして実質的に全ての異なるフラグメントは、結合した異なるオリゴヌクレオチドタグを有し、ならびに(ii)そのレパートリー由来の各オリゴヌクレオチドタグは多数のサブユニットを含有し、そして多数の各サブユニットは3から6ヌクレオチドまたは3から6塩基対の長さを有するオリゴヌクレオチドからなり、そのサブユニットが最少に交差ハイブリダイズしているセットから選択される、工程;
それらのそれぞれの相補物とオリゴヌクレオチドタグを特異的にハイブリダイズすることによりフラグメントを分類する、工程;(c)下記のように、好ましくは単一塩基配列
決定方法により、多数の各フラグメントの一部のヌクレオチド配列を決定する工程;ならびに(d)標的ポリヌクレオチドのヌクレオチド配列を、フラグメントの配列を対照することにより決定する、工程。
【0039】
本発明は、オリゴヌクレオチドで分子をタグ化または標識する現在の方法の鍵となる欠点を克服する。本発明に従ってタグの配列をコードすることにより、タグと別のタグに対する相補物との間でミスマッチした任意の二重鎖または三重鎖の安定性は、タグとそれ自身の相補物との間で完全にマッチした任意の二重鎖の安定性よりはるかに低い。従って、不正確な分類の問題は、GCに富むタグのミスマッチした二重鎖が完全にマッチしたATに富むタグよりも安定であるため、除去される。
【0040】
微細ビーズのような固相支持体と組み合わせて用いた場合、本発明は、大スケールのDNA配列決定のような大スケールの並行操作において特に有用なポリヌクレオチドを操作および分類するための容易な自動化システムを提供する。ここでは、多くの標的ポリヌクレオチドまたは単一標的ポリヌクレオチドの多くの部分が同時に配列決定および/または分析される。
【0041】
(定義)
オリゴヌクレオチドタグに関して本明細書中に用いられる「相補物」または「タグ相補物」は、オリゴヌクレオチドタグが特異的にハイブリダイズして完全にマッチした二重鎖または三重鎖を形成するオリゴヌクレオチドをいう。特異的ハイブリダイゼーションが三重鎖を生じる実施態様では、オリゴヌクレオチドタグは2本鎖または1本鎖のいずれかであることが選択され得る。従って、三重鎖が形成される場合、用語「相補物」は、1本鎖オリゴヌクレオチドタグの2本鎖相補物または2本鎖オリゴヌクレオチドタグの一本鎖相補物のいずれかを包含することを意味する。
【0042】
本明細書で用いられる用語「オリゴヌクレオチド」は、モノマー−モノマー相互作用の通常のパターン(例えば、Watson−Crick型の塩基対形成、塩基の積み重ね、Hoogsteenまたは逆Hoogsteen型の塩基対形成など)を介して標的ポリヌクレオチドに特異的に結合し得る、天然のまたは改変されたモノマーまたは連結物(デオキシリボヌクレオシド、リボヌクレオシド、それらのαアノマー型、ペプチド核酸(PNA)などを含む)の直鎖オリゴマーを含む。通常、モノマーはホスホジエステル結合またはそのアナログにより連結され、少しのモノマーユニット(例えば3〜4)から数十のモノマーユニットの大きさの範囲にわたってオリゴヌクレオチドを形成する。オリゴヌクレオチドが一連の文字(例えば、「ATGCCTG」)により示される場合はいつも、ヌクレオチドは左から右に5’→3’の順であり、他に記載されない限り、「A」はデオキシアデノシンを示し、「C」はデオキシシチジンを示し、「G」はデオキシグアノシンを示し、および「T」はチミジンを示すことが理解される。ホスホジエステル結合のアナログは、ホスホロチオエート、ホスホロジチオエート、ホスホルアニリデート、ホスホルアミデートなどを含む。通常、本発明のオリゴヌクレオチドは4つの天然のヌクレオチドを包含する;しかし、それらは非天然のヌクレオチドアナログも含有し得る。天然または非天然のヌクレオチドを有するオリゴヌクレオチドが用いられ得る場合(例えば、酵素によりプロセスされる必要がある場合)、通常天然のヌクレオチドからなるオリゴヌクレオチドが必要であることは当業者には明らかである。
【0043】
二重鎖に関して「完全にマッチした」は、各鎖中の全てのヌクレオチドが他鎖中のヌクレオチドとWatson−Crick塩基対形成を受けるように、二重鎖を作るポリヌクレオチド鎖またはオリゴヌクレオチド鎖が、互いに2本鎖構造を形成することを意味する。この用語はまた、用いられ得るヌクレオシドアナログ(例えば、デオキシイノシン、2−アミノプリン塩基を有するヌクレオシドなど)の対形成を含む。三重鎖に関しては、こ
の用語は三重鎖が完全にマッチした二重鎖および第3鎖からなり、ここで、全てのヌクレオチドが完全にマッチした二重鎖の塩基対とHoogsteenまたは逆Hoogsteen会合を受けることを意味する。逆に、二重鎖におけるタグとオリゴヌクレオチドとの間の「ミスマッチ」は、二重鎖または三重鎖におけるヌクレオチドの対またはトリプレット(triplet)がWatson−Crickおよび/またはHoogsteenおよび/または逆Hoogsteen結合を受け損なうことを意味する。
【0044】
本明細書中に用いられる「ヌクレオシド」は、2’−デオキシ型および2’−ヒドロキシル型(例えば、KornbergおよびBaker、DNA Replication、第2版(Freeman,San Francisco,1992)に記載されたように)を含む天然ヌクレオシドを含む。ヌクレオシドに関して「アナログ」は、改変された塩基部分および/または改変された糖部分を有する合成ヌクレオシド(例えば、Scheit,Nucleotide Analogs(John Wiley,New York,1980);UhlmanおよびPeyman,Chemical Reviews,90:543−584(1990)などに記載された)を、それらが特異的ハイブリダイゼーションをし得るという条件つきで含む。このようなアナログは、結合特性を増強し、退化を低減し、特異性を増すなどのように設計された合成ヌクレオシドを包含する。
【0045】
(発明の詳細な説明)
本発明は、分子(特にポリヌクレオチド)をオリゴヌクレオチドタグを用いることにより標識および分類する方法を提供する。本発明のオリゴヌクレオチドタグは、サブユニットの最少に交差ハイブリダイズしているセットから選択される多数の「ワード」またはサブユニットを包含する。このようなセットのサブユニットは、2つより少なくミスマッチしたヌクレオチドを有する同じセットの別のサブユニットの相補物と二重鎖または三重鎖を形成し得ない。従って、二重鎖を形成するレパートリーの任意の2つのオリゴヌクレオチドタグの配列は、2ヌクレオチドの異なりより「厳密(close)」でない。特定の実施態様では、レパートリーの任意の2つのオリゴヌクレオチドタグの配列は、例えばサブユニットが3つより少なくミスマッチしたヌクレオチドを有する同じセットの別のサブユニットの相補物と二重鎖を形成し得ないように最少に交差ハイブリダイズしているセットを設計することなどにより、「さらに」離れることさえあり得る。このような実施態様では、より大きい特異性が達成されるが、タグの総レパートリーはより小さい。従って、一定の長さおよびワードサイズのタグについて、所望の特異性の程度と所望のレパートリーのサイズとの間で取引がされるに違いない。本発明は、配列決定、フィンガープリンティング、または他のタイプの分析のような並行操作のためにポリヌクレオチドを標識および分類するのに特に有用である。
【0046】
(サブユニットの最少に交差ハイブリダイズしているセットからのオリゴヌクレオチドタグの構築)
任意の最少に交差ハイブリダイズしているセット用のサブユニットのヌクレオチド配列は、図3に図示した一般アルゴリズムに従い、そしてソースコードが付表Iに挙げられるプログラムminhxにより例示されたような簡単なコンピュータプログラムにより便利に計算される。minhxは、3種のヌクレオチドからなるサブユニットを有し、そして4の長さを有する全ての最少に交差ハイブリダイズしているセットを算定する。
【0047】
図3のアルゴリズムは最少に交差ハイブリダイズしているセットのサブユニットの特性、すなわちメンバー間の塩基の長さ、数の差異、および組成(例えば、2、3、または4種の塩基からなる)を最初に定義することにより実施される。一定の長さおよび組成の全ての可能性のある配列からなるテーブルMn・n=1が生じる(100)。最初のサブユニットS1を選択し、そしてテーブルの最後までi=n+1である連続するサブユニットSiと比較する(120)。連続するサブユニットが最少に交差ハイブリダイズしている
セットのメンバーであるために必要なミスマッチの数を有する場合はいつも、新たなテーブルMn+1(125)にたくわえられ、これは前述の工程120までの経路で以前に選択されたサブユニットをも含む。例えば、比較の第一セットでは、M2はS1を含む:比較の第二セットでは、M3はS1およびS2を含む:比較の第三セットでは、M4はS1、S2、およびS3を含む:など。同様に、テーブルMjにおける比較は、Sjと全ての連続サブユニットMjとの間にある。各連続するテーブルMn+1は、サブユニットが工程130までの連続する経路において除去されるので、その前の物より小さいことに気を付ける。テーブルMnの全てのサブユニットが比較された(140)後に、古いテーブルは新たなテーブルMn+1に置き換わり、そして次のラウンドの比較が開始される。選択されたサブユニットSiに比較するための連続するサブユニットを含まないテーブルMn(すなわちMn=Mn+1)が達する場合、プロセスは停止する(160)。
【0048】
好ましくは、最少に交差ハイブリダイズしているセットはセットの全ての他のサブユニットのように二重鎖安定性にほぼ等しい寄与をするサブユニットを含有する。このように、全てのサブユニットとその相補物との間で完全にマッチした二重鎖の安定性はほぼ等しい。このようなセットを選択するためのガイダンスは至適なPCRプライマーの選択および二重鎖安定性の計算についての報告された技術により提供される、例えばRychlikら、Nucleic Acids Research,17:8543−8551(1989)および18:6409−6412(1990):Breslauerら、Proc.Natl.Acad.Sci.,83:3746−3750(1986):Wetmur,Crit. Rev. Biochem. Mol. Biol.,26:227−259(1991)など。より短いタグ(例えば、約30ヌクレオチドまたはそれより少ない)について、RychlikおよびWetmurにより記載されたアルゴリズムが好ましく、より長いタグ(例えば、約30〜35ヌクレオチドまたはそれより大きい)については、Suggsら、Brown編のICN−UCLA Symp. Dev. Biol.,第23巻(Academic Press,New York,1981)の683−693頁に開示された演算法が便利に用いられ得る。明らかに、本発明の範囲内で最少に交差ハイブリダイズしているサブユニットのセットを設計するために当業者に利用可能な多くのアプローチがある。例えば、サブユニットを組み立てる場合、末端ヌクレオチドの異なる塩基の積み重ねエネルギーの影響を最小にするために、サブユニットは同じ末端ヌクレオチドを有するように提供され得る。このように、サブユニットを連結する場合、全ての隣接する末端ヌクレオチドの塩基の積み重ねエネルギーの総量は同じであり、これによりタグ融解温度の多様性が低減または除去される。
【0049】
最少に交差ハイブリダイズしているセットの好ましい実施態様は、そのサブユニットが4つの天然ヌクレオチドのうちの3つからなるセットである。下記でより詳細に述べるように、オリゴヌクレオチドタグ中のヌクレオチドの1タイプのヌクレオチドの欠損は、DNAポリメラーゼの5’→3’エクソヌクレアーゼ活性の使用により標的ポリヌクレオチドが固相支持体上にロードされることを可能にする。以下は、A、G、およびTからなる群から選択される4つのヌクレオチドをおのおの含有するサブユニットの最少に交差ハイブリダイズしているセットの例である:
【0050】
【表1】
このセットでは、各メンバーは全ての他のメンバーの相補物と3つミスマッチした塩基を有する二重鎖を形成する。
【0051】
さらなる模範的な最少に交差ハイブリダイズしているセットが下記の表Iに挙げられる。明らかに、さらなるセットは異なるグループのヌクレオチドを置換することにより、または公知の最少に交差ハイブリダイズしているセットのサブユニットを用いることにより生じ得る。
【0052】
【表2】
本発明のオリゴヌクレオチドタグおよびそれらの相補物は、自動化DNA合成機(例えば、Applied Biosystems,Inc.(Foster City,California)モデル392または394型DNA/RNA合成機)で、ホスホルアミダイトケミストリーのような標準ケミストリー(例えば、以下の参考文献で開示される)を用いて便利に合成される:BeaucageおよびIyer. Tetrahedron. 48:2223−2311(1992);Molkoら、米国特許第4,980,460号;Kosterら、米国特許第4,725,677号;Caruthersら、米国特許第4,415,732号;第4,458,066号;および第4,973,679号;など。得られたオリゴヌクレオチドが特異的ハイブリダイゼーションをし得るという条件で、別のケミストリー(例えば、ホスホロチオエート、ホスホルアミデートなど
のような非天然骨格基(backbone group)で生じる)も用いられ得る。いくつかの実施態様では、タグは酵素によりプロセスまたは操作され得る天然に存在するヌクレオチドを含有し得、対応するタグ相補物は、分類の間、より安定な二重鎖の形成を促進するペプチド核酸または同様の化合物のような非天然ヌクレオチドアナログを含有し得る。
【0053】
微粒子が支持体として用いられる場合、オリゴヌクレオチドタグおよびタグ相補物のレパートリーは、好ましくは、例えば、Shortleら、国際特許出願PCT/US93/03418に開示されるような「分裂および混合(split and mix)」技術を介するサブユニット様式(subunit−wise)合成により生じる。簡単に述べると、合成の基本ユニットは、オリゴヌクレオチドタグのサブユニットである。好ましくは、ホスホルアミダイトケミストリーを用い、3’ホスホルアミダイトオリゴヌクレオチドが最少に交差ハイブリダイズしているセット(例えば、上記に最初に挙げたセットについて、8つの4マー3’−ホスホルアミダイトがある)における各サブユニット用に調製される。合成は、Shortleらにより開示されたように、またはヌクレオシドモノマーを用いる種々のオリゴヌクレオチドライブラリーを生じるために用いられた技術(例えば、Teleniusら、Genomics,13:718−725(1992);Welshら、Nucleic Acids Research,19:5275−5279(1991);Grothuesら、Nucleic Acids Research,21:1321−1322(1993);Hartley、ヨーロッパ特許出願第90304496.4号;Lamら、Nature,354;82−84(1991);Zuckermanら、Int. J. Pept. Protein Research,40:498−507(1992);などに開示された)に直接類似してプロセスされる。一般的に、これらの技術はカップリング工程の間、成長するオリゴヌクレオチドへの活性化されたモノマーの混合物の適用を単に必要とする。
【0054】
2本鎖型のタグは、相補物鎖を別個に合成し、続いて二重鎖形成を可能にする条件下で混合することにより作製され得る。あるいは、2本鎖タグは、プライマー結合部位として働く公知のオリゴヌクレオチド配列に連結した一本鎖レパートリーを最初に合成することにより形成され得る。次いで、第2の鎖はプライマーと一本鎖レパートリーとを結合し、ポリメラーゼで伸長することにより合成される。この後者のアプローチは、Oliphantら、Gene,44:177−183(1986)に記載される。次いで、このような二重鎖タグは、本発明に従って標的ポリヌクレオチドの分類および操作用の標的ポリヌクレオチドと一緒にクローニングベクターに挿入され得る。
【0055】
特異的なハイブリダイゼーションが三重鎖形成を経て起きる実施態様では、タグ配列のコード化は、二重鎖形成タグについての原理と同じ原理に従う。しかし、サブユニット配列の選択において、さらなる制約が存在する。一般に、結合のHoogsteen型を経た第3鎖の会合は、2本鎖標的においてホモピリミジン−ホモプリントラックに沿って最も安定である。通常、塩基トリプレットは、T−A*TまたはC−G*Cモチーフ(ここで、「−」はWatson−Crick対形成を示し、そして「*」は結合のHoogsteen型を示す)を形成する。しかし、他のモチーフもまた可能である。例えば、Hoogsteen塩基対形成は、鎖の状態および組成に依存して、第3鎖(Hoogsteen鎖)と第3鎖に結合する二重鎖のプリンに富む鎖との間に平行および逆平行(antiparallel)の配向を可能にする。特定の実施態様で所望されるように、三重鎖の安定性を最大にするか、またはそうでなければ調節するための、適切な配列、配向、条件、ヌクレオシド型(例えば、リボースヌクレオシドまたはデオキシリボースヌクレオシドのどちらが用いられるか)、塩基修飾 (例えば、メチル化シトシンなど)の選択についての広範囲にわたるガイダンスが文献中に見られる。例えば、Robertsら、Proc.Natl.Acad.Sci.、88:9397−9401(1991);Rob
ertsら、Science、258:1463−1466(1992);Distefanoら、Proc.Natl.Acad.Sci.、90:1179−1183(1993);Mergnyら、Biochemistry、30:9791−9798(1991);Chengら、J.Am.Chem.Soc.、114:4465−4474(1992);BealおよびDervan、Nucleic Acids Research、20:2773−2776(1992);BealおよびDervan、J.Am.Chem.Soc.、114:4976−4982(1992);Giovannangeliら、Proc.Natl.Acad.Sci.、89:8631−8635(1992);MoserおよびDervan、Science、238:645−650(1987);McShanら、J.Biol.Chem.、267:5712−5721(1992);Yoonら、Proc.Natl.Acad.Sci.、89:3840−3844(1992);Blumeら、Nucleic Acids Research、20:1777−1784(1992);ThuongおよびHelene、Angew.Chem.Int.Ed.Engl.、32:666−690(1993)など。1本鎖タグまたは二重鎖タグをそれらの1本鎖相補物または二重鎖相補物にアニールするための条件は、周知である(例えば、Jiら、Anal.Chem.、65:1323−1328(1993))。
【0056】
本発明のオリゴヌクレオチドタグは、12から60までのヌクレオチドまたは塩基対を長さの範囲とし得る。好ましくは、オリゴヌクレオチドタグは、18から40までのヌクレオチドまたは塩基対を長さの範囲とする。さらに好ましくは、オリゴヌクレオチドタグは、25から40までのヌクレオチドまたは塩基対を長さの範囲とする。サブユニットの好ましい数およびさらに好ましい数に関して、それらの範囲は、以下のように表され得る:
【0057】
【表3】
最も好ましくは、オリゴヌクレオチドタグが1本鎖であり、そして特異的なハイブリダイゼーションが、タグ相補物とのWatson−Crick対形成により起こる。
【0058】
(分子へのタグの結合)
オリゴヌクレオチドタグは、当該分野で周知の種々の反応性官能基により、多くの異なる種類の分子に結合し得る(例えば、Haugland,Handbook of Fluorescent Probes and Research Chemicals(Molecular Probes,Inc.,Eugene,1992);Khannaら、米国特許第4,318,846号など)。表IVは、オリゴヌクレオチドタグまた
は目的の分子上に存在し得る、例となる官能基およびカウンターパート反応基を提供する。官能基およびカウンターパート反応体がともに反応する場合、いくつかの場合では活性化の後に結合基が形成される。さらに、以下により十分に記載されるように、タグは、選択された分子と同時に合成され、コンビナトリアルケミカルライブラリーを形成し得る。
【0059】
【表4】
コンビナトリアルケミカルライブラリーの生成において特に都合の良い分子の種類は、以下の形態の線形ポリマー分子を含む:
−(M−L)n−
ここで、Lはリンカー(linker)部分であり、そしてMは、広い範囲の化学構造から選択され得、不活性な非立体的に障害となるスペーサー部分として働くことから、他の成分に結合する分岐点、標識に結合するための部位:オリゴヌクレオチドに結合するための部位、あるいは治療標的にハイブリダイズまたは結合するための他の結合ポリマー:または溶解性、二重鎖および/または三重鎖形成の促進に影響を与える他の基を結合するための部位として働き得る反応性官能基を提供することまでの範囲の機能を提供するモノマー(例えば、インターカレーター、アルキル化剤など)である。このような線形ポリマー分子の配列およびその結果の組成は、BrennerおよびLerner(上記)により教示されたように、タグに結合するポリヌクレオチド内にコードされ得る。しかし、選択事象後、選択分子のタグを増幅し、次に配列決定する代わりに、タグそれ自体または付加
コードセグメントは、目的の分子の放出後、例えば、タグ中の操作された部位の制限消化により、直接的に(いわゆる以下に記載の「単一塩基」アプローチを用いて)配列決定され得る。明らかに、タグ部位の同時合成と両立できる一連のケミストリー工程により生産される任意の分子は、コンビナトリアルライブラリーの生成に用いられ得る。
【0060】
都合良く、コンビナトリアルライブラリーの生成に用いられ得る広い多様性のホスフェート結合モノマーが存在する。以下の参考文献は、本発明での使用に適する、いくつかのホスホルアミダイトおよび/または水素ホスホネートモノマーを開示し、そしてそれらの合成およびオリゴヌクレオチドへの包含に対するガイダンスを提供する:Newtonら、Nucleic Acids Research、21:1155−1162(1993);Griffinら、J.Am.Chem.Soc.、114:7976−7982(1992);Jaschkeら、Tetrahedron Letters、34:301−304(1992);Maら、国際出願PCT/CA92/00423;Zonら、国際出願PCT/US90/06630;Durandら、Nucleic Acids Research、18:6353−6359(1990);Salunkheら、J.Am.Chem.Soc.、114:8768−8772(1992);Urdeaら、米国特許第5,093,232号;Ruth、米国特許第4,948,882号;Cruickshank、米国特許第5,091,519号;Haralambidisら、Nucleic Acids Research、15:4857−4876(1987)など。さらに特に、Mは、1〜20個の炭素原子、ならびに酸素、窒素、およびイオウからなる群から選択される0〜10個のヘテロ原子を有する直鎖、環式、または分枝有機分子構造であり得る。好ましくは、Mは、1〜16個の炭素原子を有するアルキル、アルコキシ、アルケニル、またはアリール ;3〜8個の炭素原子および酸素、窒素、およびイオウからなる群から選択される1〜3個のヘテロ原子を有するヘテロ環;グリコシル;またはヌクレオシジルである。さらに好ましくは、Mは、1〜8個の炭素原子を有する、アルキル、アルコキシ、アルケニル、またはアリール;グリコシル;またはヌクレオシジルである。
【0061】
好ましくは、Lはリン(V)結合基であり、このリン結合基は、ホシホジエステル、ホスホトリエステル、メチルホスホネートまたはエチルホスホネート、ホスホロチオエート、ホスホロジチオエート、ホスホルアミデートなどであり得る。一般に、ホスホルアミダイトまたは水素ホスホネート前駆体由来の結合が好ましく、そのため本発明の線形ポリマー単位は、市販の自動化DNA合成機(例えば、Applied Biosystems,Inc.(Foster City,CA)モデル394など)で都合良く合成され得る。
【0062】
nは、MおよびLの性質に依存して顕著に変化し得る。通常、nは、約3から約100まで変化する。Mが、ヌクレオシドまたはそのアナログまたはヌクレオシドサイズのモノマーであり、そしてLがリン(V)結合の場合、その時nは約12から約100まで変化する。好ましくは、 Mが、ヌクレオシドまたはそのアナログまたはヌクレオシドサイズのモノマーであり、そしてLがリン(V)結合の場合、その時nは約12から約40まで変化する。
【0063】
ペプチドは、本発明のタグに結合する別の好ましい種類の分子である。本発明で用いられ得るペプチド−オリゴヌクレオチド結合体の合成は、Nielsenら、J.Am.Chem.Soc.、115:9812−9813(1993);Haralambidisら(上記)および国際特許出願PCT/AU88/004417;Truffertら、Tetrahedron Letters、35:2353−2356(1994);de la Torreら、 Tetrahedron Letters、35:2733−2736(1994)などの参考文献で教示されている。好ましくは、ペプチド−オ
リゴヌクレオチド結合体は、下記のように合成される。本発明に従って合成されたペプチドは、天然アミノ酸モノマーまたは非天然モノマー(天然アミノ酸のD異性体などを含む)からなり得る。
【0064】
(コンビナトリアルケミカルライブラリー)
本発明のタグを用いるコンビナトリアルケミカルライブラリーは、好ましくは、Nielsenら(上記)で開示され、そして詳細な実施態様のために図4で図解された方法により調製される。簡単にいえば、固相支持体(例えば、CPG)は、タグを合成するために用いられるケミストリーおよびいくつかの選択プロセスを経る分子を合成するために用いられるケミストリーの両方に適合する切断可能なリンカーで誘導体化される。好ましくは、タグは、上記のようなホスホルアミダイトケミストリーを用いて、およびNeilsenら(上記)により推奨される改変(すなわち、メチル保護ホスファイトおよびホスフェート部分を有するDMT−5’−O−保護3’−ホスホルアミダイト−誘導体化サブユニットが、各合成サイクルに加えられる)により合成される。ライブラリー化合物は、好ましくは、Fmocまたは同等物(連続モノマーをカップリングする官能基をマスクする保護基)を有するモノマーである。DMTおよびFmoc保護基(本明細書中でサルコシンリンカーという)の両方を用いるケミストリーに対する適切なリンカーは、Brownら、J.Chem.Soc.Chem.Commun.、1989:891−893により開示され、この開示は、参考として援用される。
【0065】
図4は、オリゴヌクレオチドタグと結合するペプチドのコンビナトリアルケミカルライブラリーを生成するためのスキームを例示する。固相支持体200は、Nielsenら(上記)により教示されるようにサルコシンリンカー205(以下の式で例示される)により誘導体化され、このリンカーは、試薬の接近を促進する伸長結合部分を有する。
【0066】
【化1】
ここで、「CPG」は、徐放性の多孔性ガラス支持体を表す。「DMT」はジメトキシトリチルを表し、そして、「Fmoc」は、9−フルオレニルメトキシカルボニルを表す。好ましい実施態様では、オリゴヌクレオチドセグメント214は、最初に合成され、そのため2本鎖の形態で、微粒子または基質の様なものの上で分類した後に、制限エンドヌクレアーゼ部位が、ライブラリー化合物を切断するために提供される。連続したサブユニットS1、S2、S3などの代替的な付加により、合成は進行してタグ212を形成し、そしてそれらの対応するライブラリー化合物モノマーA1、A2、A3などにより、合成は進行してライブラリー化合物216を形成する。「分裂および混合」技術を用ると、多様性が生まれる。
【0067】
最少に交差ハイブリダイズしているセット中のサブユニットは、ライブラリー化合物中に加えられたモノマーをコードする。従って、9ワードのセットは、9つのモノマーから構築されるライブラリー化合物を曖昧性なしにコードし得る。いくつかの曖昧さが受容可能であれば、単一のサブユニットは、1つより多いモノマーをコードし得る。
【0068】
合成が完了した後、産物は、切断および脱保護され(220)、タグライブラリー化合物225を形成し、次いで、この化合物は選択230(例えば、タンパク質のような予定された標的235への結合)を受ける。次いで、選択プロセス230から回収されたライブラリー化合物のサブセットは、固相支持体245上で、そのタグ部分(そこでは、相補
的サブユニットおよびヌクレオチドがイタリック体で示される)を介して分類される(240)。オリゴヌクレオチドスプリント242をタグ相補物250に結合し、制限部位255を形成した後、結合体は、対応する制限エンドヌクレアーゼで消化され、ライブラリー化合物(図4の実施例中のぺプチド)をオリゴヌクレオチド部分から切断する。次いで、タグの配列、それゆえライブラリー化合物の同定は、以下に記載の本発明の好ましい単一塩基配列決定技術により決定される。
【0069】
(固相支持体)
本発明で用いられる固相支持体は、広い種類の形態を有し得、例えば、微粒子、ビーズ、および膜、スライド、プレート、ミクロマシーンドチップ(micromachined chip)などが挙げられる。同様に、本発明の固相支持体は、広い種類の組成物を含み得、例えば、ガラス、プラスチック、シリコン、アルカンチオレート誘導体化金(alkanethiolate−derivatized gold)、セルロース、低架橋ポリスチレンおよび高架橋ポリスチレン、シリカゲル、ポリアミドなどが挙げられる。好ましくは、分離した粒子の集団のいずれかは、各々が同じタグ(および他はなし)への相補的配列の、均一のコーティング(または集団)を有するように用いられ、あるいは、単一支持体または数個の支持体は、各々が同じタグ(および他はなし)への相補的配列の、均一のコーティング(または集団)を含む、空間的に分離した領域で用いられる。後者の実施態様では、領域の面積は、特別な適用に従って変化し得、通常、数(例えば、3〜5)μm2から数百(例えば、100〜500)μm2の面積の領域範囲である。好ましくは、このような領域は、空間的に分離され、そのため隣接領域での事象(例えば、蛍光発光)により発生されるシグナルが、用いられる検出システムにより解明され得る。いくつかの適用では、1つより多いタグ相補物(例えば、同時配列分析、または別々にタグ化された分子を極めて近接に持ってくるための)の均一コーティングの領域を有することが、望ましくあり得る。
【0070】
タグ相補物は、その上でタグ相補物が合成される固相支持体と共に用いられ得、あるいは、別々に合成され得、そして使用のために固相支持体に結合され得る。例えば、このことは以下に開示される:Lundら、Nucleic Acids Research、16:10861−10880(1988);Albretsenら、Anal.Biochem.、189:40−50(1990);Wolfら、Nucleic Acids Research、15:2911−2926(1987);またはGhoshら、Nucleic Acids Research、15:5353−5372(1987)。好ましくは、タグ相補物は、同じ固相支持体上で合成され、そして同じ固相支持体と共に用いられ得、それは、種々の形態を含み、そして種々の結合部分を含み得る。このような支持体は、タグ相補物の均一集団が合成される領域の微粒子、またはアレイ、またはマトリックスを含み得る。広い種類の微粒子支持体が、本発明で用いられ得、例えば、徐放性の多孔性ガラス(CPG)、高架橋ポリスチレン、アクリルコポリマー、セルロース、ナイロン、デキストラン、ラテックス、ポリアクロレインなどにより作製される微粒子が挙げられ、以下の模範的な参考文献中に開示されている:Meth.Enzymol.、Section A、11−147頁、第44巻(Academic Press、New York、1976);米国特許第4,678,814号;第4,413,070号;および第4,046,720号;ならびにPon、19章、Agrawal(編)、Methods in Molecular Biology、第20巻(Humana
Press、Totowa、NJ、1993)。微粒子支持体としては、市販のヌクレオシド−誘導体化CPGおよびポリスチレンビーズ(例えば、Applied Biosystems、Foster City、CA);誘導体化磁気ビーズ;ポリエチレングリコールでグラフトされたポリスチレン(例えば、TentaGelTM、Rapp Polymere、Tubingen Germany)などがさらに挙げられる。支持体の特性(例えば、物質、多孔度、サイズ、形態など)の選択、および用いられる結合部分
のタイプは、タグを用いる際の条件に依存する。例えば、酵素による連続的なプロセスを含む適用において、酵素の立体障害を最小にし、そして基質への接近を容易にする支持体およびリンカーが好ましい。最も適切な微粒子支持体を選択する際に、考慮されるべき他の重要なファクターとしては、サイズの均一性、合成支持体としての効率、既知の表面積の度合い、および光学特性(例えば、以下により十分に説明するように、表面で多くのビーズを扱う場合、透明で平滑なビーズは、計測的な(instrumentational)利点を提供する)が挙げられる。
【0071】
微粒子表面上でタグを結合させるおよび/または合成するための模範的な結合部分は、以下に開示されている:Ponら、Biotechniques、6:768−775(1988);Webb、米国特許第4,659,774号:Baranyら、国際特許出願 PCT/US91/06103;Brownら、J.Chem.Soc.Commun.、1989:891−893;Damhaら、Nucleic Acids Research、18:3813−3821(1990);Beattieら、Clinical Chemistry、39:719−722(1993);MaskosおよびSouthern、Nucleic Acids Research、20:1679−1684(1992)など。
【0072】
上記のように、タグ相補物はまた、単一の(または数個の)固相支持体上で合成され得、タグ相補物で均一にコートされた領域のアレイを形成する。すなわち、このようなアレイ中の各領域内で、同じタグ相補物が合成される。このようなアレイを合成するための技術は、以下に開示される:McGallら、国際出願PCT/US93/03767;Peaseら、Proc.Natl.Acad.Sci.、91:5022−5026(1994);SouthernおよびMaskos、国際出願PCT/GB89/01114;MaskosおよびSouthern(上記);Southernら、Genomics、13:1008−1017(1992);およびMaskosおよびSouthern、Nucleic Acids Research、21:4663−4669(1993)。
【0073】
好ましくは、本発明は、同じタグ配列の相補物で均一にコートされた微粒子またはビーズを用いて実行される。微粒子支持体、およびそれらの表面とオリゴヌクレオチドとを共有結合または非共有結合する方法は、周知であり、以下の参考文献により例示される:BeaucageおよびIyer(上記);Gait(編)Oligonucleotide Synthesis;A Practical Approach(IRL Press、Oxford、1984);および上記の参考文献。一般に、微粒子のサイズおよび形態は重要ではない;しかし、最少の試薬およびサンプルの使用でオリゴヌクレオチドタグの多くのレパートリーの構築および操作を容易にするので、直径数μm(例えば、1〜2μm)から数百μm(例えば、200〜1000μm)の範囲のサイズの微粒子が好ましい。
【0074】
いくつかの好ましい適用では、市販の徐放性の多孔性ガラス(CPG)またはポリスチレン支持体が、本発明における固相支持体として用いられる。このような支持体は、塩基不安定性のリンカーおよび結合した最初のヌクレオチド(例えば、Applied Biosystems(Foster City、CA))を用いて有効になる。好ましくは、500オングストローム〜1000オングストロームの間のポアサイズを有する微粒子が用いられる。
【0075】
他の好ましい適用では、小孔のない微粒子が、その光学特性のために用いられ、この微粒子は、平面支持体(例えば、顕微鏡スライド)の上で多くの微粒子を探知する場合、有利に用いられ得る。特に、好ましい小孔のない微粒子は、Bangs Laborato
ries(Carmel.IN)から入手できるグリシダルメタクリレート(glycidal methacrylate)(GMA)である。このような微粒子は、種々のサイズで有用であり、そしてタグまたはタグ相補物を合成するための種々の結合基で誘導体化される。好ましくは、タグ化された微粒子の多量(massively)の平行操作のために、直径5μmのGMAビーズが用いられる。
【0076】
(標的ポリヌクレオチドの微粒子への結合)
本発明の重要な局面は、同一のポリヌクレオチド(例えば、cDNAライブラリー由来)の集団の分類、ならびに各微粒子または領域が単一種のポリヌクレオチドのみを有するような微粒子あるいは固相支持体の分離領域へのそれらの結合である。この後者の条件は、タグのレパートリーをポリヌクレオチドの集団へ連結することにより、本質的に満たされ得る。次いで、連結産物は、クローン化され、増幅され、そしてサンプリングされる。サンプルが十分小さければ、以下により十分に説明するように、得られたライブラリーの実質的に全てのタグポリヌクレオチド結合体は、唯一である。すなわち、各ポリヌクレオチドは、唯一のタグを有し、逆も同様である。次いで、ポリヌクレオチドは、タグをその相補物にハイブリダイズすることにより分類される。
【0077】
オリゴヌクレオチドタグのレパートリーは、多くの方法(例えば、直接酵素的連結による方法、増幅(例としてPCRによるもの)、タグ配列を含むプライマーの使用など)でポリヌクレオチドの集団に連結され得る。最初の連結工程は、タグ−ポリヌクレオチド結合体の非常に大きな集団を産生し、そのため単一のタグが、一般に多くの異なるポリヌクレオチドに結合する。しかし、結合体の十分小さいサンプルを採取することにより、「ダブル」を得る可能性(すなわち、2つの異なるポリヌクレオチド上の同じタグ)は、無視できる(サンプル中の同じポリヌクレオチドで異なるタグを得る可能性もあることに注目。この場合、単にプロセスされた(例えば、2回配列された)ポリヌクレオチドを生じ、そのため、通常問題はない)。以下により十分に説明したように、サンプル中にダブルを得る可能性は、ポアソン分布により評価され得る。なぜなら、サンプル中の結合体の数は大きく(例えば、千以上の桁数)、そして特別なタグを選択する可能性は小さいからである。なぜなら、タグのレパートリーは大きい(例えば、1万以上の桁)からである。一般に、サンプルが大きければ大きいほど、ダブルを得る可能性は大きくなる。従って、タグ−ポリヌクレオチド結合体の大きなサンプルを選択すること(例えば、それは、ショットガン配列決定操作における標的ポリヌクレオチド結合体の適切な適用範囲を確かめる)と最少数のダブルが存在することを確かめる小さなサンプルを選択することとの間に、設計の取引が存在する。大部分の実施態様では、ダブルの存在は、単にノイズのさらなる供給源を加えるか、または配列決定の場合、スキャニングおよびシグナルプロセシングにおける小さな複雑化要因を加えるのみである。なぜなら、多数の蛍光シグナルを与える微粒子が簡単に無視され得るからである。本明細書中で用いられる、タグを分子(特に、ヌクレオチド)に結合することに関する用語「実質的に全て」は、本質的にダブルのないタグ−分子結合体の集団を得るのに用いられるサンプリング手法の統計的な性質を反映するつもりである。タグ−分子結合体の実際の割合に関して、「実質的に全て」の意味は、タグがどのように用いられているかに依存する。好ましくは、核酸配列決定において、「実質的に全て」は、少なくともタグの80%が、結合した唯一のポリヌクレオチドを有することを意味する。より好ましくは、少なくともタグの90%が、結合した唯一のポリヌクレオチドを有することを意味する。さらにより好ましくは、少なくともタグの95%が、結合した唯一のポリヌクレオチドを有することを意味する。そして、最も好ましくは、少なくともタグの99%が、結合した唯一のポリヌクレオチドを有することを意味する。
【0078】
好ましくは、ポリヌクレオチドの集団が、メッセンジャーRNA(mRNA)からなる場合、オリゴヌクレオチドタグは、mRNAをタグ配列のプライマー含有相補物のセットで逆転写することにより、結合する。このようなプライマーの模範的なセットは、以下の
配列を有し得る:
【0079】
【化2】
ここで、「[W,W,W,C]9」は、4つのヌクレオチドの9つのサブユニットのオリゴヌクレオチドタグの配列を表し、そして「[W,W,W,C]」は、上記で挙げたサブユニット配列(すなわち、「W」はTまたはAを表す)を表す。下線を引いた部分の配列は、任意の制限エンドヌクレアーゼ部位と同一である。その部位は、用いられる場合、ビオチンを介した固相支持体への結合からポリヌクレオチドを放出させるのに用いられ得る。上記のプライマーに関して、微粒子に結合した相補物は、以下の形態を有し得る:
5’−[G,W,W,W]9TGG−リンカー−微粒子。
【0080】
逆転写後、mRNAは、例えば、RNase H 消化により除去され、そして、cDNAの第2の鎖は、例えば、以下の形態のプライマーを用いて合成される:
5’−NRRGATCYNNN−3’
ここで、NはA、T、G、またはCのいずれか一つであり、Rはプリン含有ヌクレオチドであり、そしてYはピリミジン含有ヌクレオチドである。この特定のプライマーは、得られた2本鎖DNA中にBst Yl制限部位を作り、それは、Sal I部位とともに、例えばBam HIおよびXho I部位を用いたベクター中へのクローニングを容易にする。Bst YlおよびSal I消化の後、模範的な結合体は以下の形態を有する:
【0081】
【化3】
好ましくは、リガーゼベースの配列決定の方法を用いた場合、Bst YlおよびSal
I消化フラグメントが以下の単一コピー制限部位を有するBam HI−/Xho I−消化ベクター中にクローン化される:
【0082】
【化4】
これは、以下でより十分に議論される配列決定プロセスの開始を可能とするFok I部位を添加する。
【0083】
増幅後の一本鎖のタグを露出させるための一般的な方法は、標的ポリヌクレオチド含有結合体をT4 DNAポリメラーゼまたは同様の酵素の5’→3’エキソヌクレアーゼ活性で消化することを含む。単一ヌクレオシドトリホスフェートの存在下で用いられる場合、このようなポリメラーゼは、単一ヌクレオシドトリホスフェートの相補物がテンプレート鎖に届くまで、2本鎖フラグメントの非テンプレート鎖に存在する3’陥凹(recessed)末端からヌクレオチドを切断する。このようなヌクレオチドが届いた場合、5’→3’消化は、効果的に停止する。というのは、除去活性がヌクレオチドを除去するより早い速度で、ポリメラーゼの伸長活性がヌクレオチドを付加するからである。結果として、3つのヌクレオチドで構成されるタグは、固相支持体上にロードするために容易に調製される。
【0084】
この技術はまた、標的ポリヌクレオチドの内部のFok I部位を選択的にメチル化し
、一方ポリヌクレオチドの末端で一本鎖のFok I部位をメチル化されないまま残すために用いられる。最初に、末端Fok I部位は、デオキシシチジントリホスフェートとともにポリメラーゼを用いて1本鎖とされる。次いで、フラグメントの2本鎖部分は、メチル化され、その後、1本鎖末端は、4つ全てのヌクレオシドトリホスフェートの存在下で、DNAポリメラーゼを用いて満たされ、それによりFok I部位を再生する。明らかに、この手順は、Fok I 以外のエンドヌクレアーゼに一般化され得る。
【0085】
オリゴヌクレオチドタグが、特定のハイブリダイゼーション(例えば、上記のようにそれらを1本鎖にすることにより)のために調製された後、ポリヌクレオチドは、タグとその相補物との間の完全にマッチする二重鎖の形成に有利な条件下で、タグの相補的配列を含む微粒子と混合される。これらの条件を作るための広範囲のガイダンスが文献中に存在する。このようなガイダンスを提供する例示的な参考文献としては、Wetmur、Biochemistry and Molecular Biology中のCritical Reviews、26:227−259(1991);Sambrookら、Molecular Cloning;A Laboratory Manual、第2版(Cold Spring Harbor Laboratory、New York、1989)などが挙げられる。好ましくは、ハイブリダイゼーション条件は、完全にマッチした配列のみが安定した二重鎖を形成するのに十分にストリンジェントである。このような条件下で、それらのタグにより特にハイブリダイズされたポリヌクレオチドは、微粒子に結合した相補的配列に連結される。最終的に、微粒子は洗浄され、連結されないポリヌクレオチドが除去される。
【0086】
合成支持体として通常使用されるCPG微粒子が用いられる場合、微粒子表面上でタグ相補物の密度は、代表的にいくつかの配列決定操作に必要な密度以上である。すなわち、ポリヌクレオチドを種々の酵素に結合させる連続的な処理を必要とする配列決定アプローチにおいては、空間を密にとったポリヌクレオチドは、比較的大きな酵素のポリヌクレオチドへの接近を阻害する傾向があり得る。
【0087】
このような場合、ポリヌクレオチドは、好ましくは、タグ相補物が、かなり過剰に(例えば、ポリヌクレオチドに対して10:1から100:1まで、あるいはそれ以上)存在するように微粒子と混合される。このことは、微粒子表面上におけるポリヌクレオチドの密度が、酵素の接近を阻害するほど高くないことを保証する。好ましくは、微粒子表面上のポリヌクレオチド間の平均空間は、30〜100nmの位である。標準CPG支持体およびBallotiniビーズ(固体ガラス支持体の1種)についての割合の選択におけるガイダンスは、MaskosおよびSouthern、Nucleic Acids Research、20:1679−1684(1992)中に見られる。好ましくは、配列決定の適用において、そして直径20〜50μmの範囲の標準CPGビーズが、約105のポリヌクレオチドでロードされ、そして直径5〜10μmの範囲のGMAビーズが、数万(例えば、4×104〜6×104)のポリヌクレオチドでロードされる。
【0088】
上記の方法は、以下に記載される平行配列決定方法とカップリングする場合、mRNA集団をフィンガープリントするために用いられ得る。部分配列情報は、上記の方法のような分離された微粒子に結合したcDNAの大量の(例えば、1万〜10万)サンプルから同時に得られる。部分配列の頻度分布は、種々の細胞または組織型、ならびに疾患組織(例えば、ガン)からのmRNA集団を同定し得る。このようなmRNAフィンガープリントは、病状をモニタリングおよび診断するのに有用である(例えば、国際出願 PCT/US95/21944。これは、同じ目的のための発現配列タグ(EST)の使用を記載する)。
【0089】
(単一塩基DNA配列決定)
本発明は、DNA配列決定の従来の方法、例えば、Hultmanら、Nucleic
Acids Research、17:4937−4946(1989)によって開示された方法とともに用いられ得る。しかし、複数のポリヌクレオチドの並行または同時の配列決定に関しては、以下の参考文献が挙げられる:Cheeseman、米国特許第5,302,509号; Tsienら、国際出願第WO 91/06678号; Rosenthalら、国際出願第WO 93/21340号; Canardら、Gene、148:1−6(1994); およびMetzkerら、Nucleic Acids
Research、22:4259−4267(1994)。
【0090】
本発明で用いられるのに適切であり、DNAフラグメントの電気泳動での分離を必要としないDNA配列決定の「単一塩基」方法が、国際出願第PCT/US95/03678号に記載されている。この方法は、以下の工程を包含する:(a)突出鎖を有するポリヌクレオチドの末端にプローブを連結して連結複合体を形成する工程であって、このプローブは、ポリヌクレオチドの鎖に対して相補的な突出鎖を有し、かつ、このプローブは、ヌクレアーゼ認識部位を有する工程;(b)この連結複合体から非連結プローブを除去する工程;(c)連結プローブの同定によりポリヌクレオチドの突出鎖における1つまたはそれ以上のヌクレオチドを同定する工程;(d)ヌクレアーゼで連結複合体を切断する工程;および(e)ポリヌクレオチドのヌクレオチド配列が決定されるまで(a)から(d)までの工程を繰り返す工程。以下にさらに記載するように、1つまたはそれ以上のヌクレオチドの同定は、標的ポリヌクレオチドからの連結複合体の切断前または切断後のいずれかに行われ得る。好適には、天然のタンパク質エンドヌクレアーゼが用いられる時は常に、この方法は、配列決定操作の開始時に、標的ポリヌクレオチドをメチル化する工程をさらに包含する。
【0091】
この方法の重要な特徴は、標的ポリヌクレオチドに連結されるプローブである。プローブの好適な形態は、図1aに示されている。一般に、プローブは、一方の末端10において突出鎖を有する二本鎖DNAである。プローブは、認識部位と突出末端10との間に少なくとも1つのヌクレアーゼ認識部位12およびスペーサー領域14を含む。好適には、プローブはまた、標識16を含み、これは、この特定の実施態様において、突出鎖の反対側の末端に示されている。プローブは、種々の手段により、そして種々の位置において標識され得、唯一の制限は、選択された標識手段が、連結工程またはヌクレアーゼによるプローブの認識を邪魔しないということである。
【0092】
プローブの突出鎖10が、5’末端または3’末端であるかどうかは重要ではない。しかし、標的ポリヌクレオチドおよびプローブの突出鎖が、完全に一致する二重鎖を形成し得、特異的な連結を可能にすることが重要である。もし、標的ポリヌクレオチドおよびプローブの突出鎖が異なる長さであれば、生じるギャップは、例えば、Backmanら、欧州特許出願第91100959.5号に開示されている「ギャップLCR」におけるように、連結以前にポリメラーゼによってフィルインされ得る。好適には、プローブおよび標的ポリヌクレオチドの両方の鎖が、フィルイン工程なしに連結され得るように、それぞれの突出鎖におけるヌクレオチドの数が同じである。好適には、プローブの突出鎖は2〜6のヌクレオチドの長さである。以下に示すように、突出鎖の長さが長くなるにつれて、各連結および切断サイクルの間に、標的ポリヌクレオチドへ適用されるプローブ混合物の複雑性が増す。
【0093】
プローブの相補鎖は、好都合にも、自動DNA合成機(例えば、Applied Biosystems、Inc.(Foster City、California)モデル
392または394 DNA/RNA Synthesizer)上で、標準的なケミストリーを用いて合成される。合成後、相補鎖は、組み合わされて二本鎖プローブを形成する。一般に、プローブの突出鎖は、混合物として合成されるので、すべての可能な配列
は、突出部に表される。例えば、もし、突出部が4つのヌクレオチドからなっているなら、ある実施態様において、4つの混合物が以下のように調製される:
【0094】
【化5】
ここで、「NNN」は、すべての可能な3マーを表し、「X」は、鎖の二重鎖形成部分を表す。従って、上に挙げられた4つプローブのそれぞれが、43すなわち64の異なる配列を含む;すなわち、言い換えれば、4つのプローブのそれぞれは、64の縮重を有する。例えば、X1X2...XiNNNAは、以下の配列を含む:
【0095】
【化6】
このような混合物は、例えば、Teleniusら(上記)に開示されているように、周知の技術を用いて簡単に合成される。一般に、これらの技術は、単に、縮重を導入したい位置でのカップリング工程の間、成長しているオリゴヌクレオチドへの活性化モノマーの混合物の適用を必要とするだけである。ある実施態様において、プローブの縮重を減少させることが所望され得る。これは、例えばKong Thoo Linら、Nucleic Acids Research、20:5149−5152または米国特許第5,002,867号によって教示されるように、デオキシイノシン、2−アミノプリンなどのような縮重減少アナログを用いて達成され得る。
【0096】
好適には、ホスホジエステル結合を有するオリゴヌクレオチドについては、プローブの二重鎖形成領域は、長さが約12〜約30塩基対であり、より好適には、その長さは、約15〜約25塩基対である。
【0097】
従来のリガーゼが本発明に用いられる場合、以下にさらに記載されるように、プローブの5’末端が、ある実施態様においてリン酸化され得る。5’モノホスフェートが、化学的にまたはキナーゼを用いて酵素的に(例えば、Sambrookら(上記))のいずれ
かで第2のオリゴヌクレオチドに結合され得る。化学的リン酸化は、HornおよびUrdea、Tetrahedron Lett.、27:4705(1986)に記載され、開示されたプロトコルを実施するための試薬は、市販されている(例えば、Clontech Laboratories(Palo Alto、California)からの5’Phosphate−ONTM))。従って、ある実施態様において、プローブは、以下の形態を有し得る:
【0098】
【化7】
ここで、Y’は、X’の相補ヌクレオチドであり、「p」は、モノホスフェート基である。
【0099】
上記のプローブは、種々の方法で標識され得る(放射性部分、蛍光部分、比色部分、化学ルミネセンスマーカーなどの直接または間接結合を含む)。DNAの標識およびDNAプローブの構築のための方法の多くの包括的な概説が、本発明のプローブの構築に適用可能な指針を提供している。このような概説は、Kricka編、Nonisotopic
DNA Probe Techniques (Academic Press、San Diego、1992); Haugland、Handbook of Fluorescent Probes and Research Chemicals (Molecular Probes、Inc.、Eugene、1992); KellerおよびManak、DNA Probes、第2版 (Stockton Press、New York、1993); およびEckstein編、Oligonucleotides and Analogues; A Practical Approach (IRL Press、Oxford、1991); Kessler編、Nonradioactive Labeling and Detection of Biomolecules (Springer−Verlag、Berlin、1992); Wetmur(上記); などを含む。
【0100】
好適には、プローブは、1つまたはそれ以上の蛍光色素で標識される(例えば、Menchenら、米国特許第5,188,934号; Begotら、国際出願第PCT/US90/05565号に開示されている)。
【0101】
この方法によれば、プローブは、標的ポリヌクレオチドの末端へ連結され、各連結および切断のサイクルにおいて連結複合体を形成する。連結複合体は、標的ポリヌクレオチドおよびプローブの突出鎖がアニールし、そして、少なくとも一対のプローブおよび標的の同じ向きの鎖が連結された、すなわち、互いに共有結合された後に、形成された二本鎖構造である。連結は、酵素的または化学的のいずれかで達成され得る。化学的連結方法は、当該分野において周知である(例えば、Ferrisら、Nucleosides & Nucleotides、8:407−414(1989); Shabarovaら、Nucleic Acids Research、19;4247−4251 (1991)など)。しかし、好適には、連結は、標準的なプロトコルにおいてリガーゼを用いて、酵素的に行われる。多くのリガーゼが公知であり、本発明の使用に適している(例えば、Lehman、Science、186: 790−797 (1974); Englerら、DNAリガーゼ、3−30頁、Boyer編、The Enzymes、Vol.15B (Academic Press、New York、1982)など)。好適なリガーゼは、T4 DNAリガーゼ、T7 DNA リガーゼ、E. coli DNAリガーゼ、Taqリガーゼ、Pfuリガーゼ、およびTthリガーゼを含む。これらの使用の際のプロトコルは、周知である(例えば、Sambrookら(上記); B
arany,PCR Methods and Applications、1:5−16 (1991); Marshら、Strategies、5:73−76 (1992)など)。一般に、リガーゼは、5’ホスフェート基が、接する鎖の3’水酸基への連結のために存在することを必要とする。これは、好都合に、5’ホスフェートを残すヌクレアーゼ(例えば、Fok I)を選択することによって、標的ポリヌクレオチドの少なくとも1つの鎖に対して提供される。
【0102】
非リン酸化プローブを用いる配列決定方法の実施態様において、連結工程は、(i) リガーゼで標的ポリヌクレオチドへプローブを連結させ、その結果連結複合体が、一方の鎖上にニックを有して形成される工程;(ii) 従来のプロトコルを用いて、キナーゼでニックにおいて5’水酸基をリン酸化する工程(例えば、Sambrookら(上記));および(iii) 再度連結を行い、ニックにおいて鎖を共有的に連結させる、すなわち、ニックを除去する工程を包含する。
【0103】
(微粒子表面での酵素的プロセスおよび/または結合事象の観察装置)
本発明の目的は、タグおよびその相補物の特異的ハイブリダイゼーションによって、微粒子の表面上の同一分子、特に、ポリヌクレオチドを分類することである。一旦、このような分類が行われると、分子の存在またはそれらに対して行われる操作は、タグ化された分子の性質、微粒子が、分離して、または「バッチ」で検出されるかどうか、反復された測定が所望であるかどうかなどに応じて、多くの方法で検出され得る。代表的に、分類された分子は、例えば、薬物開発において、結合のためにリガンドに曝されるか、または、例えば、ポリヌクレオチド配列決定において化学的または酵素的プロセスに供される。これらの使用の両方において、多くの微粒子上でのこのような事象またはプロセスに対応する信号を同時に観察することがしばしば望まれる。分類された分子を担持する微粒子(本明細書において「ロードされた」微粒子と称する)が、このような大規模な並行操作に役立つ(例えば、Lamら(上記)に示されている)。
【0104】
好適には、発光信号(例えば、化学ルミネセンス、蛍光など)が、事象またはプロセスを検出するために用いられるときは常に、ロードされた微粒子は、平面基板(例えば、ガラススライド)上に、(例えば、国際特許出願第PCT/US91/09217号、同第PCT/NL90/00081号および同第PCT/US95/01886号に記載されているような)走査システムでの検査のために広がっている。走査システムは、再現性よく基板を走査し、かつ、座標系によって、所定の領域に各微粒子の位置を規定し得るべきである。ポリヌクレオチド配列決定応用において、微粒子の位置同定は、連続的な走査工程において反復可能であることが重要である。
【0105】
このような走査システムは、市販の構成要素(例えば、1つまたはそれ以上の光電子増倍管、あるいは、CCDアレイおよび、例えば、蛍光信号を励起、収集および分類するための適切な光学機器を備える検出システムとともに用いられるデジタルコンピューターによって制御されるx−y移動テーブル)から構築され得る。ある実施態様において、共焦光学システムが所望であり得る。4色配列決定での使用に適切な走査システムの一例が、図5に模式的に示されている。基板300(例えば、固定微粒子を有する顕微鏡スライド)が、x−y移動テーブル302上に置かれ、これが、種々の市販のパーソナルコンピューター(例えば、Apple Computer(Cupertino、CA)から入手可能な486−ベースの機械またはPowerPC モデル7100または8100)のうちのいずれかであり得る適切にプログラムされたデジタルコンピューター304に接続され、そしてそれにより制御される。テーブル移動およびデータ収集機能のためのコンピューターソフトウェアは、National Instrumentsから入手可能なLab Windows(登録商標)のような市販の実験ソフトウェアによって提供され得る。
【0106】
基板300およびテーブル302は、光を集め、基板300に固定した微粒子へ伝達し得る1つまたはそれ以上の対物レンズ308を有する顕微鏡306と動作が連動する。好適にはレーザーである光源312からの励起光線310が、光線スプリッター314(例えば、二色性ミラー)へ指向し、これが、光線を顕微鏡306および対物レンズ308を通して再指向し、今度は、対物レンズが光線の焦点を基板300に合わせる。レンズ308は、微粒子から発光された蛍光316を集め、それを光線スプリッター314を通して信号分配光学機器318へ指向し、それが、今度は、蛍光をいくつかの蛍光特性(例えば、強度、寿命など)を電気信号に変換するための1つまたはそれ以上の適切な光学電気デバイスへ指向する。信号分配光学機器318は、帯域通過フィルター、ファイバー光学機器、回転ミラー、固定位置ミラーおよびレンズ、回折格子などのような当該分野において標準的な種々の構成要素を備える得る。図5に図示するように、信号分配光学機器318は蛍光316を4つの分離した光電子増倍管330、332、334および336へ向け、次いで、これらの出力は、プリアンプおよびフォトンカウンター350、352、354および356に向けられる。フォトンカウンターの出力は、コンピューター304によって収集され、そこで、蓄積され、分析され、ビデオ360において観察され得る。あるいは、信号分配光学機器318は、蛍光信号318をCCDアレイ上に向ける回折格子であり得る。
【0107】
走査の際の位置付けの安定性および再現性が、間隔が密接な微粒子を分離するための解像度をかなりの程度まで決定する。好適には、走査システムは、間隔が密接な(例えば、1粒子直径またはそれ以下分だけ分離されている)微粒子を解像し得るべきである。従って、ほとんどの応用に対して(例えば、CPG微粒子を用いて)、走査システムは、少なくとも10〜100μm程度に対象物を解像する能力を有するべきである。ある実施態様において、さらに高い解像度が所望であり得るが、解像度が増すにつれて、基板を完全に走査するのに必要な時間が増す。従って、ある実施態様において、速度と解像度との間で妥協が必要であり得る。走査時間の増加は、微粒子が配置していることが知られている(例えば、最初のフル走査から)位置のみを走査するシステムによって達成され得る。好適には、微粒子サイズおよび走査システム解像度が、1cm2当たり約1万〜10万の間の微粒子の密度で平面上にランダムに配置される蛍光標識された微粒子の解像度を可能にするように選択される。
【0108】
配列決定応用において、ロードされた微粒子は、種々の方法で基板の表面に固定され得る。固定は、微粒子が、顕著な損失なく、試薬露出および洗浄の連続的なサイクルを行うのに十分に強力であるべきである。基板がガラスであるとき、その表面は、市販の試薬(例えば、Pierce Chemical)を用いて、アルキルアミノリンカーで誘導体化され得、これは次に再び従来のケミストリーを用いて、アビジンに架橋され、アビジン化された表面を形成し得る。ビオチン部分は、多くの方法でロードされた微粒子に導入され得る。例えば、ポリヌクレオチドにタグを結合するために用いられるクローニングベクターの画分(例えば、10〜15パーセント)が、タグの反対側のポリヌクレオチドの末端でポリヌクレオチド挿入物に隣接して(消化において粘着末端を提供する)唯一の制限部位を含有するように操作される。この部位は、微粒子にロードするためのポリヌクレオチドおよびタグで切り出される。ロード後、ロードされたポリヌクレオチドの約10〜15パーセントが、微粒子表面から遠位の唯一の制限部位を有する。関連の制限エンドヌクレアーゼで消化後、ビオチン部分を含有している適切な二本鎖アダプターが、粘着末端に連結される。それから、得られた微粒子が、アビジン化されたガラス表面に広げられ、そこで、ビオチン−アビジン結合を介して固定される。
【0109】
あるいは、そして好適には、連結による配列決定が用いられると、最初の連結工程において、プローブの混合物が、ロードされた微粒子にアプライされる:プローブの一画分は
、配列決定方法に必要であるようなIIs型制限認識部位を含み、そしてプローブの一画分は、このような認識部位を有さないが、その代わりに、その非連結末端においてビオチン部分を含む。好適には、混合物は、約10〜15パーセントのビオチン化プローブを含む。
【0110】
さらに別の実施態様において、DNAがロードされた微粒子が、ガラス基板に塗布されると、DNAは、数時間(例えば、24時間)のインキュベーションで、ガラス表面に非特異的に吸着され、微粒子の顕著な損失なく、反復される試薬への露出および洗浄を可能とするために十分に強力な結合を形成し得る。好適には、このようなガラス基板は、フローセル (flow cell) であり、これは、ガラススライドにエッチングされた流路を含み得る。好適には、このような流路は、流体がそこを通って吸い上げられ得るように閉じられ、かつ、微粒子の単層が規定された観察領域内に閉じ込められるように、微粒子の直径と十分に近い深さを有する。
【0111】
(並行配列決定)
本発明のタグ化システムは、数キロベースまでの長さのポリヌクレオチドを配列決定するための単一塩基配列決定方法とともに用いられ得る。タグ化システムは、標的ポリヌクレオチドの何千ものフラグメントが、1つまたはそれ以上の固相支持体上に分類され、同時に配列決定され得ることを可能にする。この方法の好適な実施によると、各分類されたフラグメントの一部分が、上記のような走査システムまたは画像分析システムに関連する、顕微鏡スライドのような共通の基板に固定された何千ものロードされた微粒子のそれぞれにおいて、段階的に配列決定される。配列決定されたフラグメントの部分のサイズは、いくつかの要因、例えば、生成および分類されたフラグメントの数、標的ポリヌクレオチドの長さ、用いられた単一塩基方法の速度および正確さ、微粒子および/または同時にモニターされ得る別個の領域の数などに依存する。好適には、12〜50塩基が、各微粒子または領域で同定され、さらに好適には、18〜30塩基が、各微粒子または領域で同定される。この情報により、標的ポリヌクレオチドの配列が、重複する領域を介して12〜50塩基のフラグメントを対照することによって決定される(例えば、米国特許第5,002,867号に記載されている)。以下の引例は、所与の長さの標的ポリヌクレオチドを首尾良く再構築するために配列決定されなければならないフラグメントの部分を決定する際のさらなる指針を提供する:LanderおよびWaterman、 Genomics、2:231−239 (1988): Drmanacら、 Genomics、4:114−128 (1989): Bains、DNA Sequencing and Mapping、4: 143−150 (1993): Bains、Genomics、11:294−301 (1991): Drmanacら、J.Biomolecular Structure and Dynamics、8: 1085−1102 (1991): およびPevzner、J.Biomolecular Structure and Dynamics、7: 63−73 (1989)。好適には、標的ポリヌクレオチドの長さは、1キロベース〜50キロベースである。より好適には、長さは、10キロベース〜40キロベースである。LanderおよびWaterman(上記)は、配列決定されるフラグメントの数(すなわち、サンプルサイズ)、各フラグメントから得られた配列情報の量、および標的ポリヌクレオチドが、ギャップがない部分的な配列、すなわち、「アイランド」から再構築され得る可能性の間の相互関係に関する指針を提供する。本発明において、所与のサンプルサイズおよびフラグメント配列のサイズで得られ得る最大のポリヌクレオチドサイズを以下に示す。
【0112】
【化8】
フラグメントは、最小の重複で標的ポリヌクレオチドをカバーするフラグメントのセットを生成しようとする、いわゆる「直接的」なアプローチ、およびランダムに重複するフラグメントが生成される、いわゆる「ショットガン」アプローチを含む、種々の方法で標的ポリヌクレオチドから生成され得る。好適には、フラグメント生成についての「ショットガン」アプローチが、その簡便性および固有の縮重のために用いられる。例えば、標的ポリヌクレオチドをカバーする、ランダムに重複するフラグメントは、以下の従来の「ショットガン」配列決定プロトコルで生成される(例えば、Sambrookら(上記)に開示されている)。本明細書に用いられるような、この文脈での「カバーする(cover)」は、標的ポリヌクレオチド配列のすべての部分が、生成されたフラグメントの各サイズ範囲(例えば、全てのフラグメントは長さが、100塩基対〜200塩基対の範囲である)で表されることを意味する。簡潔には、適切なクローニングベクター(例えば、λファージ)における挿入物としての標的ポリヌクレオチドから始まって、ベクターは拡大され、精製され、そして適切な制限酵素で消化され、約10〜15μgの精製された挿入物を得る。代表的に、プロトコルは、1マイクログラムの開始DNA当たり約500〜1000のサブクローンをもたらす。挿入物は、調製ゲル電気泳動によってベクターフラグメントから分離され、従来の方法によってゲルから取り出され、そしてTE(Tris−EDTA)のような標準的な緩衝液に再懸濁される。ベクターから挿入物を切り出すために選択される制限酵素は、好適には、挿入物に適合性の粘着末端を残し、これにより、挿入物が、ランダムに重複するフラグメントを生成するために、調製において自己連結され得る。Sambrookら(上記)に説明されるように、環状化DNAは、以下に用いられる断片化方法における直鎖状DNAに比べて、フラグメントのよりよいランダム分配を与える。挿入物を例えば、T4リガーゼで従来のプロトコルを用いて、自己連結した後、生成された連結挿入物が、標準的なプロトコル(例えば、Mn++の存在下における超音波処理またはDNase I 消化)によって断片化される。断片化後、フラグメントの末端が、例えば、Sambrookら(上記)に記載されるように修復され、そして修復されたフラグメントは、ゲル電気泳動を用いて、サイズによって分離される。300〜500塩基対の範囲のフラグメントが選択され、従来の手段によってゲルから溶出され、上記のように、タグを有するベクター中に連結され、タグ−フラグメント結合体のライブラリーを形成する。
【0113】
上記のように、数千のタグ−フラグメント結合体を含有するサンプルが、ライブラリーから採取され、そして拡大され、その後、タグ−フラグメント挿入物がベクターから切り出され、そして上記のように、微粒子上のタグ相補物への特異的なハイブリダイゼーションのために調製される。標的ポリヌクレオチドのサイズに応じて、多重サンプルが、標識−フラグメントライブラリーから採取され、別々に拡大され、微粒子上にロードされ、そして配列決定される。選択された二重の数は、サンプル中に表されたタグレパートリーの画分に依存する(トリプル−−同じタグを有する3つの異なるポリヌクレオチド−−または上記を得る可能性は、安全に無視し得る)。上記のように、サンプル中のダブルの可能
性は、Poisson分布 p(ダブル)= m2e−m/2(ここで、mは、サンプル中のタグレパートリー画分である)から評価され得る。下の表Vは、所与のタグサイズ、サンプルサイズおよびレパートリーの多様性についてサンプル中でダブルを得る可能性を挙げている。
【0114】
【表5】
いずれの場合においても、次いで、ロードされた微粒子が、好適には、アビジン−ビオチン結合を介して、ガラス顕微鏡スライド上に分散、および固定される。好適には、それぞれのランダムフラグメントの少なくとも15〜20のヌクレオチドが、単一塩基方法で同時に配列決定される。次いで、標的ポリヌクレオチドの配列が、それらの重複する部分によって、アセンブリングコンティグに用いられるのと類似の、または上記の引例に開示されたようなハイブリダイゼーションにより配列決定するために開発されたような、アルゴリズムを用いて、ランダムフラグメントの部分的な配列を照合することによって再構築される。
【0115】
(本発明の方法を実施するためのキット)
本発明は、本発明の種々の実施態様を実施するためのキットを含む。好適には、本発明のキットは、固相支持体に結合されたタグ相補物のレパートリーを含む。さらに、本発明のキットは、対応するタグのレパートリー(例えば、ポリヌクレオチドを増幅して分類するためのプライマーとして、またはポリヌクレオチドを増幅して分類するために用いられ得るクローニングベクターの要素として)を含み得る。好適には、タグ相補物のレパートリーは、微粒子に結合される。キットはまた、酵素的プロセシング、検出ケミストリー(例えば、蛍光タグまたは化学ルミネセンスタグ)などのための適切な緩衝液、使用説明書、プロセシング酵素(例えば、リガーゼ、ポリメラーゼ、トランスフェラーゼなど)を含み得る。配列決定のための重要な実施態様において、キットはまた、プロセシングのためにロードされた微粒子を固定するための、アビジン化された顕微鏡スライドのような基板を含み得る。
【0116】
(cDNAライブラリーにおける新規なポリヌクレオチドの同定)
cDNAライブラリーにおける新規なポリヌクレオチドは、上記のように微粒子に結合されたcDNA分子のライブラリーを構築することによって、同定され得る。次いで、ライブラリーの大きな画分、またはライブラリー全体でさえ、部分的に並行に配列決定され得る。mRNAの単離、およびおそらく、Soaresら、Proc. Natl. A
cad. Sci.、91:9228−9232 (1994)、または同様の引例によって教示されるように、その集団の正規化の後、以下のプライマーが、従来のプロトコルを用いて、逆転写酵素で第1鎖の合成のためのポリAテールにハイブリダイズされ得る:
【0117】
【化9】
ここで、[W,W,W,C]9は、上記のようなタグを表し、「ACCAGCTGATC」は、二本鎖形態で制限部位を形成する任意の配列であり、そして「プライマー部位」は、PCRによって目的のポリヌクレオチドを増幅するためのプライマー結合部位として後に用いられる、ライブラリーのすべてのメンバーに共通の配列である。
【0118】
従来の技術による逆転写および第2鎖合成の後、二本鎖フラグメントが、上記のようにクローニングベクターに挿入され、そして増幅される。次いで、増幅されたライブラリーがサンプリングされ、そしてサンプルは増幅する。増幅されたサンプルからのクローニングベクターが単離され、そしてタグ化されたcDNAフラグメントが切り出され、そして精製される。上記のようなポリメラーゼでタグを一本鎖にした後、フラグメントはメチル化され、そして本発明によって微粒子上に分類される。好適には、上記のように、クローニングベクターは、タグ化されたcDNAが、分類および微粒子への連結後に好適な単一塩基方法による即座の配列決定を可能にする、Fok I のようなエンドヌクレアーゼで切り出され得るように構築される。
【0119】
次いで、段階的な配列決定が、本発明に従って、ライブラリー全体またはライブラリーの1つまたはそれ以上の大きな画分で同時に行われ、ついには、十分な数のヌクレオチドが、ライブラリーが由来する生物のゲノムに特有の表示のために各cDNAについて同定される。例えば、もし、ライブラリーが哺乳動物のmRNAに由来するなら、14〜15ヌクレオチドの長さのランダムに選択された配列が、典型的な哺乳動物のゲノムの2000〜3000メガベースの間で特有の表示を有すると予想される。もちろん、細菌または他の下等生物に由来するライブラリーに特有の表示には、ずっと少ないヌクレオチドの同定で十分である。好適には、少なくとも20〜30ヌクレオチドが、特有の表示を確実にし、かつ、以下に記載するように、適切なプライマーの構築を可能にするために同定される。次いで、表にされた配列は、ユニークなcDNAを同定するために、既知の配列と比較され得る。
【0120】
次いで、特有のcDNAは、従来の技術(例えば、プライム部位およびその配列が決定されたcDNAの部分に関するプライマーで製造されたPCRアンプリコンからプローブを構築する技術)によって単離される。次いで、プローブは、従来のスクリーニングプロトコルを用いてライブラリー中のcDNAを同定するために用いられ得る。
【実施例】
【0121】
(実施例I)
(pUC19由来の多重標的ポリヌクレオチドの分類)
3つの標的ポリヌクレオチドタグ結合体の混合物を、以下のように得る:まず、以下の6つのオリゴヌクレオチドを合成し、対として組み合わせ、タグ1、タグ2およびタグ3を形成する。
【0122】
【化10】
ここで、「p」はモノホスフェートを示し、wiは表Iで定義されるサブユニットを表し、用語「(**)」はそれぞれの相補物を表す。pUC19を、Sal IおよびHind IIIで消化し、大きなフラグメントを精製し、そしてタグ1、タグ2およびタグ3で別々に連結して、それぞれ、pUC19−1、 pUC19−2、 pUC19−3を形成する。この3つの組換え体を、別々に増幅し単離し、その後、pUC19−1を、Hind IIIおよびAat Iで消化し、 pUC19−2をHind IIIおよびSsp Iで消化し、pUC19−3を、Hind IIIおよびXmn Iで消化する。小さなフラグメントを従来のプロトコルを用いて単離して、それぞれ、約250、375および575塩基対の長さの3つの二本鎖フラグメントを得る。それぞれは、タグに隣接する陥没した3’鎖および反対側の末端には平滑または3’突出鎖を有する。およそ12nmolの各フラグメントを、製造者が推薦する、33μM デオキシシトシントリホスフェートを含有する反応緩衝液中で、5ユニットのT4 DNAポリメラーゼと混合する。反応混合物を37℃で30分間インキュベートし、その後、氷の上に置くことによって反応を止める。次いで、フラグメントを、従来の手段で精製する。
【0123】
CPG微粒子(粒子サイズ37〜74mm、孔サイズ500オングストローム、Pierce Chemical)を、MaskosおよびSouthen、Nucleic Acids Research、20:1679−1684 (1992)によって開示されたリンカーで誘導体化する。3つのアリコートに分離後、タグ1、タグ2およびタグ3の相補物を、従来の自動化されたDNA合成機(例えば、model 392 DNA合成機(Applied Biosystems、Foster City、CA))を用いて、微粒子上で合成する。異なって誘導体化された微粒子のそれぞれ約1mgを、別個の容器に入れる。
【0124】
pUC19−1、pUC19−2およびpUC19−3から切り出されたT4 DNAポリメラーゼ処理のフラグメントを、製造者推薦のTaq DNAリガーゼ(New England Biolabs)のための緩衝液50μLに再懸濁する。次いで、混合物を、誘導体化CPG微粒子をそれぞれ1mg含有している3つの容器に等分する。5ユニットのTaq DNAリガーゼを各容器に添加し、その後、それらを55℃で15分間インキュベートする。氷の上に置くことによって反応を止め、そして微粒子を、繰り返し遠心分離およびTEに再懸濁することによって数回洗浄する。最後に、微粒子をNde I反応緩衝液(New England Biolabs)で再懸濁し、結合したポリヌク
レオチドを消化する。微粒子からの分離後、Nde I消化によって放出されたポリヌクレオチドフラグメントを、Sequenase DNAポリメラーゼおよびフルオレセイン標識チミジントリホスフェート(Applied Biosystems、Foster City、CA)でインキュベートすることにより、蛍光標識する。次いで、そのフラグメントを、Applied Biosystems model 373 DNA配列決定機を用いて、非変性ポリアクリルアミドゲル上で別々に分析する。
【0125】
(実施例 II)
(SV40フラグメントの並行配列決定)
表Iから選択される9つの4−ヌクレオチドサブユニットからなる36マーのタグのレパートリーを、上記のような、分裂または混合手段によって、タグおよびタグ相補物を別々に合成することによって調製する。Sma I/Hind III消化M13mp19に連結可能なように、レパートリーを合成する。従って、実施例Iと同様に、1組のオリゴヌクレオチドはAの添加で開始し、9回の分裂および混合合成が続く。このオリゴヌクレオチドを、表Iのサブユニットに対応して3’−ホスホルアミダイト誘導体化4マーによりサブユニットで伸張する。次いで、Sma I認識部位(GGG) の半分、2つのCおよび5’−モノホスフェートを、例えば、Clontech Laboratories (Palo Alto、CA)から入手可能であるPhosphate−ON試薬を用いて、ヌクレオチド毎に添加することで合成を完了する。他方の組のオリゴヌクレオチドは、3つのC(Sma I認識部位の一部)および2つのGの添加で開始し、9回の分裂および混合合成が続く。このオリゴヌクレオチドを、表Iのサブユニットの相補物に対応して、3’−ホスホルアミダイト誘導体化4マーにより伸張する。Hind III認識部位および5’−モノホスフェートのヌクレオチド毎の添加により合成を完了する。合成支持体からの分離後、オリゴヌクレオチドを以下の二重鎖の形成を可能にする条件下で混合する:
【0126】
【化11】
次いで、二重鎖の混合物をSma I/Hind III消化M13mp19に連結する。タグ相補物のレパートリーを上記のようにCPG微粒子上で合成する。
【0127】
次に、Fok I部位ならびにEco RIおよびSma I部位の一部を含有する、以下のアダプターを調製する:
【0128】
【化12】
このアダプターを、上記のように、Eco RI/Sma I消化M13に連結する。
【0129】
別に、Sambrookら(上記)に記載のプロトコルに従って超音波処理により、SV40 DNAを断片化する。得られたフラグメントを標準的なプロトコルを用いて修復し、そしてサイズによって分離する。300〜500塩基対の範囲のフラグメントを選択し、上記のSma I消化M13に連結してフラグメント−タグ結合体のライブラリーを形成し、次いで、増幅する。数千の異なるフラグメント−結合体を含有するサンプルをラ
イブラリーから取り出し、さらに増幅し、そしてフラグメント−タグ挿入物を、Eco R IおよびHind IIIで消化することにより切り出す。切り出されたフラグメント−タグ結合体を、実施例Iに記載のように、デオキシシチジントリホスフェートの存在下でT4 DNAポリメラーゼで処理して、特異的なハイブリダイゼーションのためのオリゴヌクレオチドタグをCPG微粒子に曝露する。
【0130】
実施例Iに記載のように、ハイブリダイゼーションおよび連結後、ロードされた微粒子をFok Iで処理して、予め決定した配列の4−ヌクレオチド突出鎖を生成する。以下のプローブの10:1の混合物(プローブ1:プローブ2)を微粒子上のポリヌクレオチドに連結する。
【0131】
【化13】
FAMは、Appiled Biosystems から入手可能なアミノホスフェートリンカー(Aminolinker)を介してプローブ1の上段の鎖の5’−水酸基に結合されるフルオレセインを表す。ビオチンはまた、Aminolinker部分を介して結合され得、そして必要に応じて、ポリエチレンオキシドリンカー(例えば、Jaschkeら(上記))を介してさらに伸張され得る。
【0132】
次いで、ロードされた微粒子をアビジン化ガラススライドの表面上に堆積する。試薬および洗浄溶液は表面上へ送達および表面上から除去され得る。結合した微粒子を有するアビジン化スライドにより、走査型蛍光顕微鏡(例えば、Newport Model PM500−Cモーションコントローラー 、488nmの励起光を発生するSpectra−Physics Model 2020アルゴンイオンレーザー、および520nm波長域発光フィルターなどの装置を備えたZeiss Axioskop)で検査する。励起光および蛍光発光を、同じ対物レンズを通して、それぞれ、送達および収集する。二色性ミラーによって励起光および収集蛍光を分離する。二色性ミラーは、収集された蛍光を一連の帯域フィルターを介して、モニターされている蛍光物質に対応するフォトン計数デバイス(例えば、Hamamatsu model 9403−02光電子増倍管、Stanford Research Systems model SR445増幅器およびmodel SR430多重チャンネルスケーラーおよびデジタルコンピューター(例えば、486−ベースのコンピューター)を備える)へ指向する。コンピューターは、微粒子の位置を記録するスライドの二次元マップを生成する。
【0133】
最初のプローブを除去するためのFok Iでの切断後、結合微粒子上のポリヌクレオチドを、以下に記載の好適な単一塩基配列決定方法に従い、プローブ連結、洗浄、検出、切断および洗浄工程を20サイクル行う。各検出工程において、走査システムが、各微粒子で同定された塩基に対応する蛍光発光を記録する。一般に、以下の反応および洗浄を、特に記載されない限り、用いる酵素のための、製造者 (New England Biolabs) の推薦する緩衝液で実施する。標準的な緩衝液もまた、Sambrookら(上記)に記載されている。
【0134】
以下の4組の混合プローブを、標的ポリヌクレオチドへの添加用に提供する:
【0135】
【化14】
ここで、TAMRA、FAM、ROXおよびJOEは、Aminolinker IIによって結合された分光解像可能な蛍光標識である(すべて、Applied Biosystems、Inc.、Forster City、Californiaから入手可能である);太字のヌクレオチドは、Fok Iエンドヌクレアーゼのための認識部位であり、「N」は、4つのヌクレオチド、A、C、G、T.のいずれか1つを表す。TAMRA(テトラメチルローダミン)、FAM(フルオレセイン)、ROX(ローダミンX)、およびJOE(2’,7’−ジメトキシ−4’,5’−ジクロロフルオレセイン)およびそれらのオリゴヌクレオチドへの結合はまた、Fungら米国特許第4,855,225号に記載されている。
【0136】
上記のプローブを、約5モル過剰量の標的ポリヌクレオチド末端中で、以下のようにインキュベートする:プローブを、200ユニットのT4 DNAリガーゼおよびアンカー化標的ポリヌクレオチドとともに60分間16℃でT4 DNAリガーゼ緩衝液中でインキュベートする:次いで、洗浄後、標的ポリヌクレオチドを、100ユニットのT4ポリヌクレオチドキナーゼとともに30分間37℃で製造者の推薦する緩衝液中でインキュベートし、洗浄し、そして再度、200ユニットのT4 DNAリガーゼおよびアンカー化標的ポリヌクレオチドとともに30分間16℃でT4 DNAリガーゼ緩衝液中でインキュベートする。洗浄は、スライド上に洗浄緩衝液(例えば、TE)を大量に連続的に流すことによって達成される(Sambrookら(上記)に開示されている)。連結−リン酸化−連結のサイクルおよび最後の洗浄後、結合微粒子が、蛍光標識の存在について走査され、その位置および特徴が走査システムによって記録される。次いで、標識された標的ポリヌクレオチド、即ち、連結複合体を、製造者の推薦する緩衝液中で30分間37℃で、10ユニットのFor Iとともにインキュベートし、その後、TE中で洗浄する。その結果、標的ポリヌクレオチドは各鎖上で1つのヌクレオチド分短くされ、次の連結および切断サイクルのために準備される。20のヌクレオチドが同定されるまでこのプロセスを続ける。
【0137】
【表6】
【0138】
【表7】
【0139】
【表8】
【図面の簡単な説明】
【0140】
【図1a】図1a−1cは、本発明で用いられ得る「単一塩基」配列決定の好ましい方法において用いられる標識されたプローブの構造を例示する。
【図1b】図1a−1cは、本発明で用いられ得る「単一塩基」配列決定の好ましい方法において用いられる標識されたプローブの構造を例示する。
【図1c】図1a−1cは、本発明で用いられ得る「単一塩基」配列決定の好ましい方法において用いられる標識されたプローブの構造を例示する。
【図2】図2は、好ましい「単一塩基」配列決定方法において用いられた標的ポリヌクレオチドとプローブとの間で形成された連結された複合体中のヌクレアーゼ認識部位、連結部位および切断部位の相対的な位置を例示する。
【図3】図3は、最少に交差ハイブリダイズしているセットを生じるための一般的なアルゴリズムを例示するフローチャートである。
【図4】図4は、コンビナトリアルケミカルライブラリーを合成および使用するためのスキームを例示し、ここでメンバー化合物は本発明に従ってオリゴヌクレオチドタグで標識される。
【図5】図5は、本発明に従ってポリヌクレオチド配列決定のような並行操作を行うための装置を図で例示する。
【0141】
(配列表)
【0142】
【表9】
【0143】
【表10】
【0144】
【表11】
【特許請求の範囲】
【請求項1】
1つまたはそれ以上の固相支持体上にポリヌクレオチドの集団からのポリヌクレオチドを分類する方法であって、該方法は、以下の工程:
(a)最少に交差ハイブリダイズしているセットから選択されるタグのレパートリー由来のオリゴヌクレオチドタグを、ポリヌクレオチドの集団中の各ポリヌクレオチドに結合させる工程であって、該集団中の実質的にすべての異なるポリヌクレオチドが、結合された異なるオリゴヌクレオチドタグを有してタグ化ポリヌクレオチドの集団を形成する、工程;および
(b)該タグ化ポリヌクレオチドの該オリゴヌクレオチドタグとそれぞれの相補物とを特異的にハイブリダイズすることにより該タグ化ポリヌクレオチドを分類する工程であって、該相補物のそれぞれが、1つまたはそれ以上の固相支持体の空間的に離れている領域において均一な集団として結合されている、工程、
を包含する、方法。
【請求項1】
1つまたはそれ以上の固相支持体上にポリヌクレオチドの集団からのポリヌクレオチドを分類する方法であって、該方法は、以下の工程:
(a)最少に交差ハイブリダイズしているセットから選択されるタグのレパートリー由来のオリゴヌクレオチドタグを、ポリヌクレオチドの集団中の各ポリヌクレオチドに結合させる工程であって、該集団中の実質的にすべての異なるポリヌクレオチドが、結合された異なるオリゴヌクレオチドタグを有してタグ化ポリヌクレオチドの集団を形成する、工程;および
(b)該タグ化ポリヌクレオチドの該オリゴヌクレオチドタグとそれぞれの相補物とを特異的にハイブリダイズすることにより該タグ化ポリヌクレオチドを分類する工程であって、該相補物のそれぞれが、1つまたはそれ以上の固相支持体の空間的に離れている領域において均一な集団として結合されている、工程、
を包含する、方法。
【図1a】

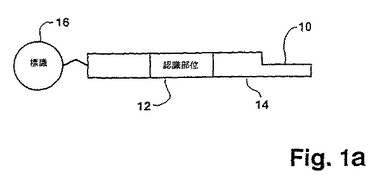
【図1b】
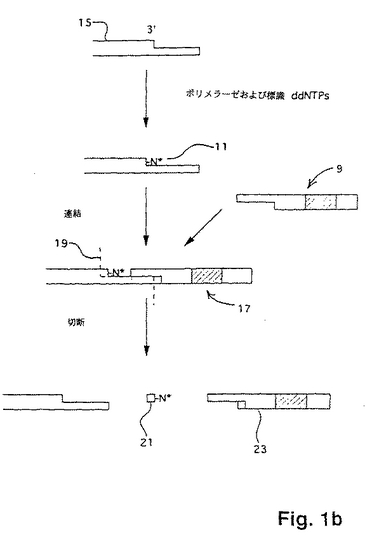
【図1c】
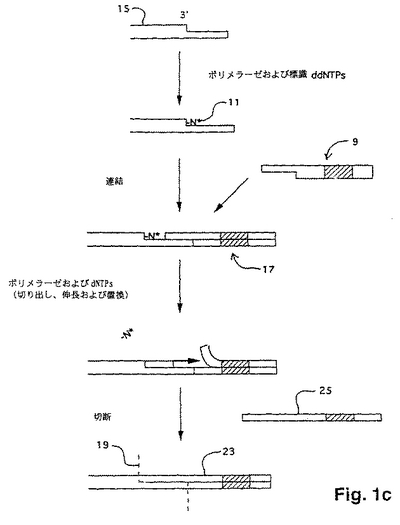
【図2】
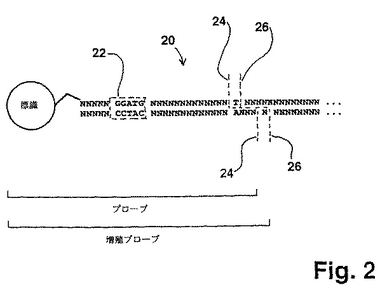
【図3】
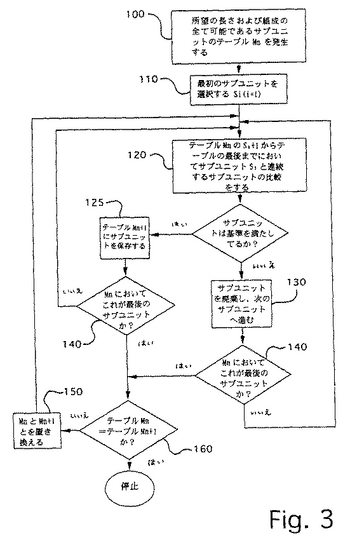
【図4】
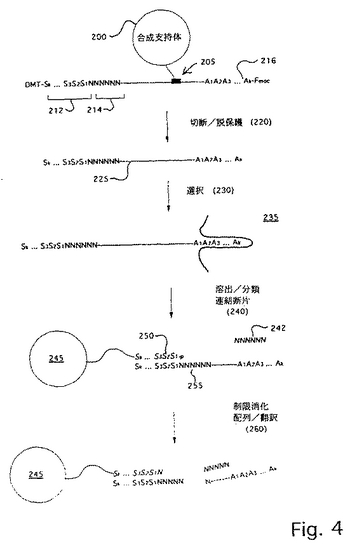
【図5】
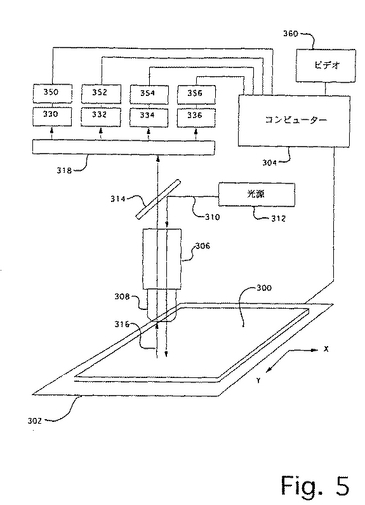
【図1b】
【図1c】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2006−320321(P2006−320321A)
【公開日】平成18年11月30日(2006.11.30)
【国際特許分類】
【出願番号】特願2006−128002(P2006−128002)
【出願日】平成18年5月1日(2006.5.1)
【分割の表示】特願2004−261709(P2004−261709)の分割
【原出願日】平成7年10月12日(1995.10.12)
【出願人】(598114930)ソレクサ・インコーポレイテッド (10)
【氏名又は名称原語表記】Solexa,Inc.
【住所又は居所原語表記】3832 Bay Center Place, Hayward, California 94545 U.S.A.
【Fターム(参考)】
【公開日】平成18年11月30日(2006.11.30)
【国際特許分類】
【出願日】平成18年5月1日(2006.5.1)
【分割の表示】特願2004−261709(P2004−261709)の分割
【原出願日】平成7年10月12日(1995.10.12)
【出願人】(598114930)ソレクサ・インコーポレイテッド (10)
【氏名又は名称原語表記】Solexa,Inc.
【住所又は居所原語表記】3832 Bay Center Place, Hayward, California 94545 U.S.A.
【Fターム(参考)】
[ Back to top ]