
分子抗原アレイ
【課題】バクテリオファージAP205ウイルス様粒子(VLP)および抗原を含む組成物、また該VLPに結合した抗原または抗原決定基を産生する方法を提供する。
【解決手段】バクテリオファージAP205のコートタンパク質の会合したVLP、および該タンパク質、さらにVLPコア粒子に有機分子の結合した組成物。該組成物は、感染症、アレルギー、癌等の疾患、障害、または病態の予防または治療に有用な免疫反応の誘導、および効率的な自己特異的免疫反応、特に抗体反応を誘導するのに有用である。
【解決手段】バクテリオファージAP205のコートタンパク質の会合したVLP、および該タンパク質、さらにVLPコア粒子に有機分子の結合した組成物。該組成物は、感染症、アレルギー、癌等の疾患、障害、または病態の予防または治療に有用な免疫反応の誘導、および効率的な自己特異的免疫反応、特に抗体反応を誘導するのに有用である。
Notice: Undefined index: DEJ in /mnt/www/gzt_disp.php on line 298
【特許請求の範囲】
【請求項1】
(a) 配列番号:1に記載のアミノ酸配列を有するタンパク質;
(b) 配列番号:3に記載のアミノ酸配列を有するタンパク質;および
(c) (a)または(b)のタンパク質の突然変異タンパク質
からなる群より選択される少なくとも1つのタンパク質を含むウイルス様粒子。
【請求項2】
タンパク質が組換え体である、請求項1記載のウイルス様粒子。
【請求項3】
突然変異タンパク質が配列番号:1に記載または配列番号:3に記載のアミノ酸配列を有し、少なくとも1つのアミノ酸残基、好ましくは3つのアミノ酸残基、より好ましくは2つのアミノ酸残基、さらにより好ましくは1つのアミノ酸残基が付加、除去、または置換され、好ましくは少なくとも1つの置換が保存的置換である、請求項1または2のいずれか一項記載のウイルス様粒子。
【請求項4】
突然変異タンパク質が配列番号:1に記載または配列番号:3に記載のアミノ酸配列を有し、少なくとも1つのシステイン残基、好ましくは2つのシステイン残基が除去または置換され、好ましくは少なくとも1つの置換が保存的置換である、請求項1〜3のいずれか一項記載のウイルス様粒子。
【請求項5】
突然変異タンパク質が配列番号:1に記載または配列番号:3に記載のアミノ酸配列を有し、少なくとも1つのリジン残基、好ましくは3つのリジン残基、より好ましくは2つのリジン残基、さらにより好ましくは1つのリジン残基が付加、除去、または置換され、好ましくは少なくとも1つの置換が保存的置換である、請求項1〜4のいずれか一項記載のウイルス様粒子。
【請求項6】
配列番号:3に記載のアミノ酸配列を有する突然変異タンパク質。
【請求項7】
配列番号:1または配列番号:3の組換えタンパク質の突然変異タンパク質。
【請求項8】
少なくとも1つのアミノ酸残基、好ましくは3つのアミノ酸残基、より好ましくは2つのアミノ酸残基、さらにより好ましくは1つのアミノ酸残基が付加、除去、または置換され、好ましくは少なくとも1つの置換が保存的置換である、請求項7記載の突然変異タンパク質。
【請求項9】
少なくとも1つのシステイン残基、好ましくは2つのシステイン残基が除去または置換され、好ましくは少なくとも1つの置換が保存的置換である、請求項7または8のいずれかに記載の突然変異タンパク質。
【請求項10】
少なくとも1つのリジン残基、好ましくは3つのリジン残基、より好ましくは2つのリジン残基、さらにより好ましくは1つのリジン残基が付加、除去、または置換され、好ましくは少なくとも1つの置換が保存的置換である、請求項7〜9のいずれか一項記載の突然変異タンパク質。
【請求項11】
少なくとも80%、好ましくは少なくとも90%、より好ましくは少なくとも95%、さらにより好ましくは99%が配列番号:2または配列番号:4のヌクレオチド配列と同一であるヌクレオチド配列を含む、AP205ウイルス様粒子を産生するためのベクター。
【請求項12】
タンパク質に融合したポリペプチドをコードするヌクレオチド配列を含む、組換えタンパク質を産生するためのベクターであって、該タンパク質が、
(a) 配列番号:1に記載のアミノ酸配列を有するタンパク質;
(b) 配列番号:3に記載のアミノ酸配列を有するタンパク質;および
(c) (a)または(b)のポリペプチドの突然変異タンパク質
からなる群より選択される組換えタンパク質を産生するための、ベクター。
【請求項13】
(a) (i) AP205ウイルス粒子;および
(ii) AP205ウイルス様粒子
からなる群より選択されるコア粒子;および
(b) 有機分子
を含み、有機分子がコア粒子に結合している組成物。
【請求項14】
有機分子およびコア粒子が、コア粒子の表面上における有機分子の規則正しくかつ反復性のアレイを形成する、請求項13記載の組成物。
【請求項15】
有機分子が、コア粒子を有機分子に連結させる第3の分子を介してコア粒子に結合している、請求項13または14のいずれかに記載の組成物。
【請求項16】
有機分子が少なくとも1つの共有結合によりコア粒子に結合し、好ましくは共有結合がペプチド結合を含む、請求項13〜15のいずれか一項記載の組成物。
【請求項17】
有機分子が少なくとも1つの共有結合によりコア粒子に結合し、好ましくは共有結合が非ペプチド結合を含む、請求項13〜16のいずれか一項記載の組成物。
【請求項18】
有機分子がハプテン、抗原、または抗原決定基を含み、または好ましくはハプテン、抗原、または抗原決定基であり、より好ましくは有機分子が抗原または抗原決定基である、請求項13〜17のいずれか一項記載の組成物。
【請求項19】
ウイルス様粒子が少なくとも1つの第1付着部位を含み、有機分子が少なくとも1つの第2付着部位を含み、その結果、好ましくは少なくとも1つの非ペプチド結合により、第2付着部位が第1付着部位と結合して規則正しくかつ反復性の抗原アレイを形成し得る、請求項13〜18のいずれか一項記載の組成物。
【請求項20】
第1付着部位がアミノ基を含み、好ましくはアミノ基であり、好ましくは第1付着部位がリジン残基を含み、好ましくはリジン残基であり、第2付着部位がスルフヒドリル基を含み、好ましくはスルフヒドリル基であり、好ましくは第2付着部位がシステイン残基を含み、好ましくはシステイン残基である、請求項13〜19のいずれか一項記載の組成物。
【請求項21】
第2付着部位が有機分子内に天然には存在しない、請求項13〜20のいずれか一項記載の組成物。
【請求項22】
組成物がアミノ酸リンカーを含み、好ましくはアミノ酸リンカーが少なくとも1つの共有結合、好ましくは少なくとも1つのペプチド結合により抗原または抗原決定基に結合している、請求項13〜21のいずれか一項記載の組成物。
【請求項23】
アミノ酸リンカーが第2付着部位を含む、請求項13〜22のいずれか一項記載の組成物。
【請求項24】
アミノ酸リンカーが
(a) CGG;
(b) N末端γ1-リンカー;
(c) N末端γ3-リンカー;
(d) Igヒンジ領域;
(e) N末端グリシンリンカー;
(f) n=0〜12およびk=0〜5である(G)kC(G)n(配列番号:93);
(g) N末端グリシン-セリンリンカー;
(h) n=0〜3、k=0〜5、m=0〜10、l=0〜2である(G)kC(G)m(S)l(GGGGS)n(配列番号:94);
(i) GGC;
(k) GGC-NH2;
(l) C末端γ1-リンカー;
(m) C末端γ3-リンカー;
(n) C末端グリシンリンカー;
(o) n=0〜12およびk=0〜5である(G)nC(G)k(配列番号:95);
(p) C末端グリシン-セリンリンカー;および
(q) n=0〜3、k=0〜5、m=0〜10、l=0〜2およびo=0〜8である(G)m(S)l(GGGGS)n(G)oC(G)k(配列番号:96)
からなる群より選択される、請求項13〜23のいずれか一項記載の組成物。
【請求項25】
有機分子が
(a) 癌細胞に対する免疫反応を誘導するのに適した有機分子;
(b) 感染症に対する免疫反応を誘導するのに適した有機分子;
(c) アレルゲンに対する免疫反応を誘導するのに適した有機分子;
(d) 自己抗原に対する改善された反応を誘導するのに適した有機分子;
(e) 家畜またはペットにおいて免疫反応を誘導するのに適した有機分子;
(f) 薬剤、ホルモン、または毒性化合物に対する反応を誘導するのに適した有機分子;および
(g) (a)〜(f)に記載の分子の断片、突然変異タンパク質、またはドメイン
からなる群より選択される、請求項13〜24のいずれか一項記載の組成物。
【請求項26】
有機分子が、
(a) 癌細胞に対する免疫反応を誘導するのに適した抗原または抗原決定基;
(b) 感染症に対する免疫反応を誘導するのに適した抗原または抗原決定基;
(c) アレルゲンに対する免疫反応を誘導するのに適した抗原または抗原決定基;
(d) 自己抗原に対する改善された反応を誘導するのに適した抗原または抗原決定基;
(e) 家畜またはペットにおいて免疫反応を誘導するのに適した抗原または抗原決定基;
(f) 薬剤、ホルモン、または毒性化合物に対する反応を誘導するのに適した抗原または抗原決定基;および
(g) (a)〜(f)に記載の分子の断片またはドメイン
からなる群より選択される抗原もしくは抗原決定基、またはそれらの断片もしくは突然変異タンパク質である、請求項13〜25のいずれか一項記載の組成物。
【請求項27】
有機分子が、
(a) HIVのポリペプチド、
(b) インフルエンザウイルスのポリペプチド、
(c) C型肝炎ウイルスのポリペプチド、
(d) トキソプラズマのポリペプチド、
(e) 熱帯熱マラリア原虫のポリペプチド、
(f) 三日熱マラリア原虫のポリペプチド、
(g) 卵形マラリア原虫のポリペプチド、
(h) 四日熱マラリア原虫のポリペプチド、
(i) 乳癌細胞のポリペプチド、
(j) 腎臓癌細胞のポリペプチド、
(k) 前立腺癌細胞のポリペプチド、
(l) 皮膚癌細胞のポリペプチド、
(m) 脳腫瘍細胞のポリペプチド、
(n) 白血病細胞のポリペプチド、
(o) 組換えプロファイリング(a recombinant profiling)、
(p) ハチ刺されアレルギーのポリペプチド、
(q) ナッツアレルギーのポリペプチド、
(r) 食物アレルギーのポリペプチド、
(s) 喘息のポリペプチド、または
(t) クラミジアのポリペプチド、
(u) Her2、
(v) GD2、
(w) EGF-R、
(x) CEA、
(y) CD52、
(z) ヒトメラノーマgp100、
(aa) ヒトメラノーマメランA/MART-1、
(bb) チロシナーゼ、
(cc) NA17-A nt、
(dd) MAGE3、
(ee) p53、および
(ff) HPV16E7;ならびに
(gg) (a)〜(z)および(aa)〜(ff)の抗原の任意の断片または突然変異タンパク質
からなる群より選択される抗原である、請求項13〜26のいずれか一項記載の組成物。
【請求項28】
抗原または抗原決定基が、
a) ホスホリパーゼA2タンパク質;
b) ヒトIgE;
c) リンホトキシン;
d) インフルエンザM2タンパク質;および
e) Der p Iペプチド
からなる群より選択されるペプチド、タンパク質、またはタンパク質もしくはペプチドの断片もしくは突然変異タンパク質である、請求項13〜27のいずれか一項記載の組成物。
【請求項29】
有機分子が抗原または抗原決定基であり、さらに抗原または抗原決定基が自己抗原もしくは抗イディオタイプ抗体、またはそのいずれかの断片である、請求項13〜28のいずれか一項記載の組成物。
【請求項30】
自己抗原が、
a) リンホトキシン;
b) リンホトキシン受容体;
c) RANKL;
d) VEGF;
e) VEGFR;
f) インターロイキン-5;
g) インターロイキン-8
h) インターロイキン-17;
h) インターロイキン-13;
i) アンジオテンシン;
k) CCL21;
l) CXCL12;
m) SDF-1;
n) MCP-1;
o) エンドグリン;
p) レジスチン;
q) GHRH;
r) LHRH;
s) TRH;
t) MIF;
u) エオタキシン;
v) ブラジキニン;
w) BLC;
x) M-CSF;
x) 腫瘍壊死因子α(TNFα);
y) アミロイドβペプチド(Aβ1-42);および
z) ヒトIgE
からなる群より選択されるタンパク質、ペプチド、またはそれらの任意の断片もしくは突然変異タンパク質である、請求項13〜29のいずれか一項記載の組成物。
【請求項31】
自己抗原が
a) リンホトキシンα(LTα);
b) リンホトキシンβ(LTβ);および
c) LTαとLTβの混合物または組み合わせ
からなる群より選択されるリンホトキシンまたはその断片である、請求項13〜30のいずれか一項記載の組成物。
【請求項32】
有機分子が、薬剤、ホルモン、または毒素に対する免疫反応を誘導するのに適した有機分子である、請求項13〜31のいずれか一項記載の組成物。
【請求項33】
有機分子が薬剤に対する免疫反応を誘導するのに適した有機分子である、請求項13〜32のいずれか一項記載の組成物。
【請求項34】
薬剤が
(a) コデイン;
(b) フェンタニル;
(c) ヘロイン;
(d) モルヒネ;
(e) アンフェタミン;
(f) コカイン;
(g) メチレンジオキシメタンフェタミン;
(h) メタンフェタミン;
(i) メチルフェニデート;
(j) ニコチン;
(k) LSD;
(l) メスカリン;
(m) シロシビン;および
(n) テトラヒドロカンナビノール
からなる群より選択される、請求項13〜33のいずれか一項記載の組成物。
【請求項35】
薬剤がニコチンである、請求項13〜34のいずれか一項記載の組成物。
【請求項36】
有機分子がホルモンに対する免疫反応を誘導するのに適している、請求項13〜35のいずれか一項記載の組成物。
【請求項37】
有機分子が、
(a) プロゲステロン;
(b) エストロゲン;
(c) テストステロン;
(d) 卵胞刺激ホルモン;
(e) メラニン刺激ホルモン;
(f) アドレナリン;および
(g) ノルアドレナリン
を含む群、好ましくはこれらからなる群より選択されるホルモンである、請求項13〜36のいずれか一項記載の組成物。
【請求項38】
有機分子が毒素に対する免疫反応を誘導するのに適している、請求項13〜37のいずれか一項記載の組成物。
【請求項39】
有機分子が、
(a) アフラトキシン;
(b) シガテラ毒;
(c) テトロドトキシン;
(d) 抗生物質;および
(e) 抗癌剤
からなる群より選択される毒素である、請求項13〜38のいずれか一項記載の組成物。
【請求項40】
a) 請求項13〜39のいずれか一項記載の組成物および
b) 許容される薬学的担体
を含む、薬学的組成物。
【請求項41】
請求項13〜39のいずれか一項記載の組成物の免疫学的に有効な量を含むワクチン組成物。
【請求項42】
請求項41記載の組成物のワクチン組成物を投与する段階を含む免疫法。
【請求項43】
アジュバントをさらに含む、請求項41記載のワクチン組成物。
【請求項44】
(a) (i) AP205ウイルス;および
(ii) AP205ウイルス様粒子;
の群より選択されるコア粒子を含む分子骨格を提供する段階;ならびに
(b) 免疫反応を誘発するのに適した有機分子を提供する段階;
(c) 任意で(a)および/または(b)内に含まれる、または別の分子として、(a)と(b)を結合する手段を提供する段階;ならびに
(d) (a)〜(c)の要素を組み合わせ、その結果、有機分子が骨格と結合して規則正しくかつ反復性の抗原アレイを形成する段階
を含む、非天然的に存在する規則正しくかつ反復性の抗原アレイを産生する方法。
【請求項45】
請求項13〜39のいずれか一項記載の組成物を個体に投与する段階を含む、個体の疾患、障害、または生理的状態を治療または予防する方法。
【請求項46】
請求項41記載のワクチン組成物を個体に投与する段階を含む、個体の疾患、障害、または生理的状態を治療または予防する方法。
【請求項47】
配列番号:125に記載のヌクレオチド配列を含む核酸分子。
【請求項48】
請求項47記載の核酸または請求項11記載のベクターを含む宿主細胞。
【請求項49】
大腸菌である、請求項48記載の宿主細胞。
【請求項50】
(a) 請求項47記載の核酸または請求項11記載のベクターを提供する段階
(b) 核酸またはベクターを宿主細胞に導入する段階;
(c) 宿主細胞において核酸またはベクターの配列を発現させて、請求項1記載のウイルス様粒子を形成し得るタンパク質または突然変異タンパク質を得る段階を含む、請求項1〜5のいずれか一項記載のウイルス様粒子を産生する方法。
【請求項51】
宿主細胞が大腸菌である、請求項50記載の方法。
【請求項1】
(a) 配列番号:1に記載のアミノ酸配列を有するタンパク質;
(b) 配列番号:3に記載のアミノ酸配列を有するタンパク質;および
(c) (a)または(b)のタンパク質の突然変異タンパク質
からなる群より選択される少なくとも1つのタンパク質を含むウイルス様粒子。
【請求項2】
タンパク質が組換え体である、請求項1記載のウイルス様粒子。
【請求項3】
突然変異タンパク質が配列番号:1に記載または配列番号:3に記載のアミノ酸配列を有し、少なくとも1つのアミノ酸残基、好ましくは3つのアミノ酸残基、より好ましくは2つのアミノ酸残基、さらにより好ましくは1つのアミノ酸残基が付加、除去、または置換され、好ましくは少なくとも1つの置換が保存的置換である、請求項1または2のいずれか一項記載のウイルス様粒子。
【請求項4】
突然変異タンパク質が配列番号:1に記載または配列番号:3に記載のアミノ酸配列を有し、少なくとも1つのシステイン残基、好ましくは2つのシステイン残基が除去または置換され、好ましくは少なくとも1つの置換が保存的置換である、請求項1〜3のいずれか一項記載のウイルス様粒子。
【請求項5】
突然変異タンパク質が配列番号:1に記載または配列番号:3に記載のアミノ酸配列を有し、少なくとも1つのリジン残基、好ましくは3つのリジン残基、より好ましくは2つのリジン残基、さらにより好ましくは1つのリジン残基が付加、除去、または置換され、好ましくは少なくとも1つの置換が保存的置換である、請求項1〜4のいずれか一項記載のウイルス様粒子。
【請求項6】
配列番号:3に記載のアミノ酸配列を有する突然変異タンパク質。
【請求項7】
配列番号:1または配列番号:3の組換えタンパク質の突然変異タンパク質。
【請求項8】
少なくとも1つのアミノ酸残基、好ましくは3つのアミノ酸残基、より好ましくは2つのアミノ酸残基、さらにより好ましくは1つのアミノ酸残基が付加、除去、または置換され、好ましくは少なくとも1つの置換が保存的置換である、請求項7記載の突然変異タンパク質。
【請求項9】
少なくとも1つのシステイン残基、好ましくは2つのシステイン残基が除去または置換され、好ましくは少なくとも1つの置換が保存的置換である、請求項7または8のいずれかに記載の突然変異タンパク質。
【請求項10】
少なくとも1つのリジン残基、好ましくは3つのリジン残基、より好ましくは2つのリジン残基、さらにより好ましくは1つのリジン残基が付加、除去、または置換され、好ましくは少なくとも1つの置換が保存的置換である、請求項7〜9のいずれか一項記載の突然変異タンパク質。
【請求項11】
少なくとも80%、好ましくは少なくとも90%、より好ましくは少なくとも95%、さらにより好ましくは99%が配列番号:2または配列番号:4のヌクレオチド配列と同一であるヌクレオチド配列を含む、AP205ウイルス様粒子を産生するためのベクター。
【請求項12】
タンパク質に融合したポリペプチドをコードするヌクレオチド配列を含む、組換えタンパク質を産生するためのベクターであって、該タンパク質が、
(a) 配列番号:1に記載のアミノ酸配列を有するタンパク質;
(b) 配列番号:3に記載のアミノ酸配列を有するタンパク質;および
(c) (a)または(b)のポリペプチドの突然変異タンパク質
からなる群より選択される組換えタンパク質を産生するための、ベクター。
【請求項13】
(a) (i) AP205ウイルス粒子;および
(ii) AP205ウイルス様粒子
からなる群より選択されるコア粒子;および
(b) 有機分子
を含み、有機分子がコア粒子に結合している組成物。
【請求項14】
有機分子およびコア粒子が、コア粒子の表面上における有機分子の規則正しくかつ反復性のアレイを形成する、請求項13記載の組成物。
【請求項15】
有機分子が、コア粒子を有機分子に連結させる第3の分子を介してコア粒子に結合している、請求項13または14のいずれかに記載の組成物。
【請求項16】
有機分子が少なくとも1つの共有結合によりコア粒子に結合し、好ましくは共有結合がペプチド結合を含む、請求項13〜15のいずれか一項記載の組成物。
【請求項17】
有機分子が少なくとも1つの共有結合によりコア粒子に結合し、好ましくは共有結合が非ペプチド結合を含む、請求項13〜16のいずれか一項記載の組成物。
【請求項18】
有機分子がハプテン、抗原、または抗原決定基を含み、または好ましくはハプテン、抗原、または抗原決定基であり、より好ましくは有機分子が抗原または抗原決定基である、請求項13〜17のいずれか一項記載の組成物。
【請求項19】
ウイルス様粒子が少なくとも1つの第1付着部位を含み、有機分子が少なくとも1つの第2付着部位を含み、その結果、好ましくは少なくとも1つの非ペプチド結合により、第2付着部位が第1付着部位と結合して規則正しくかつ反復性の抗原アレイを形成し得る、請求項13〜18のいずれか一項記載の組成物。
【請求項20】
第1付着部位がアミノ基を含み、好ましくはアミノ基であり、好ましくは第1付着部位がリジン残基を含み、好ましくはリジン残基であり、第2付着部位がスルフヒドリル基を含み、好ましくはスルフヒドリル基であり、好ましくは第2付着部位がシステイン残基を含み、好ましくはシステイン残基である、請求項13〜19のいずれか一項記載の組成物。
【請求項21】
第2付着部位が有機分子内に天然には存在しない、請求項13〜20のいずれか一項記載の組成物。
【請求項22】
組成物がアミノ酸リンカーを含み、好ましくはアミノ酸リンカーが少なくとも1つの共有結合、好ましくは少なくとも1つのペプチド結合により抗原または抗原決定基に結合している、請求項13〜21のいずれか一項記載の組成物。
【請求項23】
アミノ酸リンカーが第2付着部位を含む、請求項13〜22のいずれか一項記載の組成物。
【請求項24】
アミノ酸リンカーが
(a) CGG;
(b) N末端γ1-リンカー;
(c) N末端γ3-リンカー;
(d) Igヒンジ領域;
(e) N末端グリシンリンカー;
(f) n=0〜12およびk=0〜5である(G)kC(G)n(配列番号:93);
(g) N末端グリシン-セリンリンカー;
(h) n=0〜3、k=0〜5、m=0〜10、l=0〜2である(G)kC(G)m(S)l(GGGGS)n(配列番号:94);
(i) GGC;
(k) GGC-NH2;
(l) C末端γ1-リンカー;
(m) C末端γ3-リンカー;
(n) C末端グリシンリンカー;
(o) n=0〜12およびk=0〜5である(G)nC(G)k(配列番号:95);
(p) C末端グリシン-セリンリンカー;および
(q) n=0〜3、k=0〜5、m=0〜10、l=0〜2およびo=0〜8である(G)m(S)l(GGGGS)n(G)oC(G)k(配列番号:96)
からなる群より選択される、請求項13〜23のいずれか一項記載の組成物。
【請求項25】
有機分子が
(a) 癌細胞に対する免疫反応を誘導するのに適した有機分子;
(b) 感染症に対する免疫反応を誘導するのに適した有機分子;
(c) アレルゲンに対する免疫反応を誘導するのに適した有機分子;
(d) 自己抗原に対する改善された反応を誘導するのに適した有機分子;
(e) 家畜またはペットにおいて免疫反応を誘導するのに適した有機分子;
(f) 薬剤、ホルモン、または毒性化合物に対する反応を誘導するのに適した有機分子;および
(g) (a)〜(f)に記載の分子の断片、突然変異タンパク質、またはドメイン
からなる群より選択される、請求項13〜24のいずれか一項記載の組成物。
【請求項26】
有機分子が、
(a) 癌細胞に対する免疫反応を誘導するのに適した抗原または抗原決定基;
(b) 感染症に対する免疫反応を誘導するのに適した抗原または抗原決定基;
(c) アレルゲンに対する免疫反応を誘導するのに適した抗原または抗原決定基;
(d) 自己抗原に対する改善された反応を誘導するのに適した抗原または抗原決定基;
(e) 家畜またはペットにおいて免疫反応を誘導するのに適した抗原または抗原決定基;
(f) 薬剤、ホルモン、または毒性化合物に対する反応を誘導するのに適した抗原または抗原決定基;および
(g) (a)〜(f)に記載の分子の断片またはドメイン
からなる群より選択される抗原もしくは抗原決定基、またはそれらの断片もしくは突然変異タンパク質である、請求項13〜25のいずれか一項記載の組成物。
【請求項27】
有機分子が、
(a) HIVのポリペプチド、
(b) インフルエンザウイルスのポリペプチド、
(c) C型肝炎ウイルスのポリペプチド、
(d) トキソプラズマのポリペプチド、
(e) 熱帯熱マラリア原虫のポリペプチド、
(f) 三日熱マラリア原虫のポリペプチド、
(g) 卵形マラリア原虫のポリペプチド、
(h) 四日熱マラリア原虫のポリペプチド、
(i) 乳癌細胞のポリペプチド、
(j) 腎臓癌細胞のポリペプチド、
(k) 前立腺癌細胞のポリペプチド、
(l) 皮膚癌細胞のポリペプチド、
(m) 脳腫瘍細胞のポリペプチド、
(n) 白血病細胞のポリペプチド、
(o) 組換えプロファイリング(a recombinant profiling)、
(p) ハチ刺されアレルギーのポリペプチド、
(q) ナッツアレルギーのポリペプチド、
(r) 食物アレルギーのポリペプチド、
(s) 喘息のポリペプチド、または
(t) クラミジアのポリペプチド、
(u) Her2、
(v) GD2、
(w) EGF-R、
(x) CEA、
(y) CD52、
(z) ヒトメラノーマgp100、
(aa) ヒトメラノーマメランA/MART-1、
(bb) チロシナーゼ、
(cc) NA17-A nt、
(dd) MAGE3、
(ee) p53、および
(ff) HPV16E7;ならびに
(gg) (a)〜(z)および(aa)〜(ff)の抗原の任意の断片または突然変異タンパク質
からなる群より選択される抗原である、請求項13〜26のいずれか一項記載の組成物。
【請求項28】
抗原または抗原決定基が、
a) ホスホリパーゼA2タンパク質;
b) ヒトIgE;
c) リンホトキシン;
d) インフルエンザM2タンパク質;および
e) Der p Iペプチド
からなる群より選択されるペプチド、タンパク質、またはタンパク質もしくはペプチドの断片もしくは突然変異タンパク質である、請求項13〜27のいずれか一項記載の組成物。
【請求項29】
有機分子が抗原または抗原決定基であり、さらに抗原または抗原決定基が自己抗原もしくは抗イディオタイプ抗体、またはそのいずれかの断片である、請求項13〜28のいずれか一項記載の組成物。
【請求項30】
自己抗原が、
a) リンホトキシン;
b) リンホトキシン受容体;
c) RANKL;
d) VEGF;
e) VEGFR;
f) インターロイキン-5;
g) インターロイキン-8
h) インターロイキン-17;
h) インターロイキン-13;
i) アンジオテンシン;
k) CCL21;
l) CXCL12;
m) SDF-1;
n) MCP-1;
o) エンドグリン;
p) レジスチン;
q) GHRH;
r) LHRH;
s) TRH;
t) MIF;
u) エオタキシン;
v) ブラジキニン;
w) BLC;
x) M-CSF;
x) 腫瘍壊死因子α(TNFα);
y) アミロイドβペプチド(Aβ1-42);および
z) ヒトIgE
からなる群より選択されるタンパク質、ペプチド、またはそれらの任意の断片もしくは突然変異タンパク質である、請求項13〜29のいずれか一項記載の組成物。
【請求項31】
自己抗原が
a) リンホトキシンα(LTα);
b) リンホトキシンβ(LTβ);および
c) LTαとLTβの混合物または組み合わせ
からなる群より選択されるリンホトキシンまたはその断片である、請求項13〜30のいずれか一項記載の組成物。
【請求項32】
有機分子が、薬剤、ホルモン、または毒素に対する免疫反応を誘導するのに適した有機分子である、請求項13〜31のいずれか一項記載の組成物。
【請求項33】
有機分子が薬剤に対する免疫反応を誘導するのに適した有機分子である、請求項13〜32のいずれか一項記載の組成物。
【請求項34】
薬剤が
(a) コデイン;
(b) フェンタニル;
(c) ヘロイン;
(d) モルヒネ;
(e) アンフェタミン;
(f) コカイン;
(g) メチレンジオキシメタンフェタミン;
(h) メタンフェタミン;
(i) メチルフェニデート;
(j) ニコチン;
(k) LSD;
(l) メスカリン;
(m) シロシビン;および
(n) テトラヒドロカンナビノール
からなる群より選択される、請求項13〜33のいずれか一項記載の組成物。
【請求項35】
薬剤がニコチンである、請求項13〜34のいずれか一項記載の組成物。
【請求項36】
有機分子がホルモンに対する免疫反応を誘導するのに適している、請求項13〜35のいずれか一項記載の組成物。
【請求項37】
有機分子が、
(a) プロゲステロン;
(b) エストロゲン;
(c) テストステロン;
(d) 卵胞刺激ホルモン;
(e) メラニン刺激ホルモン;
(f) アドレナリン;および
(g) ノルアドレナリン
を含む群、好ましくはこれらからなる群より選択されるホルモンである、請求項13〜36のいずれか一項記載の組成物。
【請求項38】
有機分子が毒素に対する免疫反応を誘導するのに適している、請求項13〜37のいずれか一項記載の組成物。
【請求項39】
有機分子が、
(a) アフラトキシン;
(b) シガテラ毒;
(c) テトロドトキシン;
(d) 抗生物質;および
(e) 抗癌剤
からなる群より選択される毒素である、請求項13〜38のいずれか一項記載の組成物。
【請求項40】
a) 請求項13〜39のいずれか一項記載の組成物および
b) 許容される薬学的担体
を含む、薬学的組成物。
【請求項41】
請求項13〜39のいずれか一項記載の組成物の免疫学的に有効な量を含むワクチン組成物。
【請求項42】
請求項41記載の組成物のワクチン組成物を投与する段階を含む免疫法。
【請求項43】
アジュバントをさらに含む、請求項41記載のワクチン組成物。
【請求項44】
(a) (i) AP205ウイルス;および
(ii) AP205ウイルス様粒子;
の群より選択されるコア粒子を含む分子骨格を提供する段階;ならびに
(b) 免疫反応を誘発するのに適した有機分子を提供する段階;
(c) 任意で(a)および/または(b)内に含まれる、または別の分子として、(a)と(b)を結合する手段を提供する段階;ならびに
(d) (a)〜(c)の要素を組み合わせ、その結果、有機分子が骨格と結合して規則正しくかつ反復性の抗原アレイを形成する段階
を含む、非天然的に存在する規則正しくかつ反復性の抗原アレイを産生する方法。
【請求項45】
請求項13〜39のいずれか一項記載の組成物を個体に投与する段階を含む、個体の疾患、障害、または生理的状態を治療または予防する方法。
【請求項46】
請求項41記載のワクチン組成物を個体に投与する段階を含む、個体の疾患、障害、または生理的状態を治療または予防する方法。
【請求項47】
配列番号:125に記載のヌクレオチド配列を含む核酸分子。
【請求項48】
請求項47記載の核酸または請求項11記載のベクターを含む宿主細胞。
【請求項49】
大腸菌である、請求項48記載の宿主細胞。
【請求項50】
(a) 請求項47記載の核酸または請求項11記載のベクターを提供する段階
(b) 核酸またはベクターを宿主細胞に導入する段階;
(c) 宿主細胞において核酸またはベクターの配列を発現させて、請求項1記載のウイルス様粒子を形成し得るタンパク質または突然変異タンパク質を得る段階を含む、請求項1〜5のいずれか一項記載のウイルス様粒子を産生する方法。
【請求項51】
宿主細胞が大腸菌である、請求項50記載の方法。
【図2】


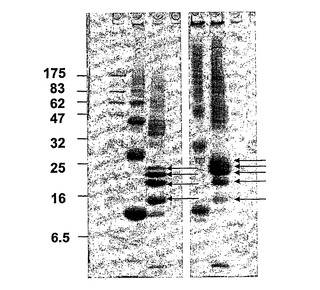
【図3】
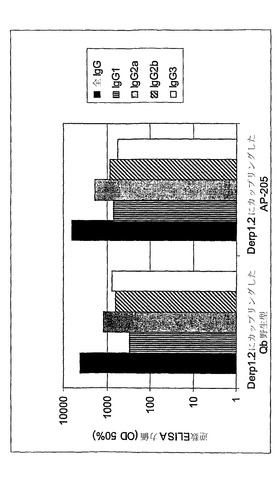
【図4A】
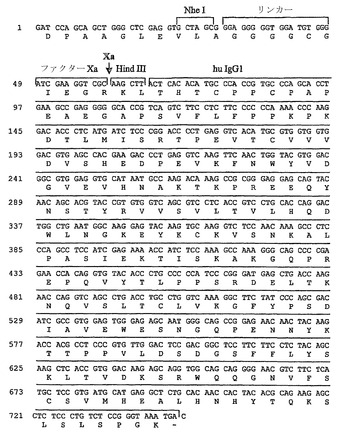
【図4B】
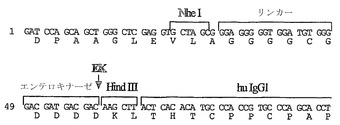
【図4C】
【図5】
【図6】
【図7】
【図8】

【図3】
【図4A】
【図4B】
【図4C】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2011−87580(P2011−87580A)
【公開日】平成23年5月6日(2011.5.6)
【国際特許分類】
【外国語出願】
【出願番号】特願2010−248015(P2010−248015)
【出願日】平成22年11月5日(2010.11.5)
【分割の表示】特願2004−520607(P2004−520607)の分割
【原出願日】平成15年7月14日(2003.7.14)
【出願人】(304042375)サイトス バイオテクノロジー アーゲー (26)
【Fターム(参考)】
【公開日】平成23年5月6日(2011.5.6)
【国際特許分類】
【出願番号】特願2010−248015(P2010−248015)
【出願日】平成22年11月5日(2010.11.5)
【分割の表示】特願2004−520607(P2004−520607)の分割
【原出願日】平成15年7月14日(2003.7.14)
【出願人】(304042375)サイトス バイオテクノロジー アーゲー (26)
【Fターム(参考)】
[ Back to top ]