
分枝酵素の活性を改変した小麦、ならびにそれから得たデンプンおよびデンプン含有品
相対的に多量のアミロースを含有することができ、SBEIIa活性のレベルを低下させた小麦。ゲノムA中にSBEIIa変異遺伝子を有する小麦。さらに、この小麦のSBEIIb活性のレベルを低下させてもよい。本発明の小麦粒は、アミロペクチン合成経路に障害があるにもかかわらず、非しわ型のフェノタイプとなることができ、相対的に多量のアミロースを含有することもできる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、相対的にアミロース含有量が高い穀粒デンプンを有する小麦植物に関する。本発明はまた、胚乳中のデンプン分枝酵素IIa(SBEIIa)の活性を低下させた小麦、およびこのような植物を得る方法にも関する。本発明はまた、穀粒およびデンプン、ならびにそれから得た食料品および非食料品にも関する。
【背景技術】
【0002】
穀類では、成熟した穀粒の約45〜65%重量をデンプンが占めている。デンプンは、2つのタイプの分子、すなわちアミロースおよびアミロペクチンからなっている。アミロースは、α−1,4結合したグルコシド鎖からなるほぼ直鎖状の分子であり、一方、アミロペクチンは直鎖同士を結合するα−1,6グルコシド結合で高度に枝分かれしている。
【0003】
高等植物の胚乳におけるデンプンの合成は、4つの重要なステップを触媒する一揃いの酵素群によって行われる。第1に、ADP−グルコースピロホスホリラーゼが、G−1−PおよびATPからADP−グルコースを合成することによって、デンプンのモノマー前駆物質を活性化する。第2に、活性化されたグルコシル基供与体、すなわちADP−グルコースが、デンプン合成酵素によって、既存のα−1,4結合の非還元末端に移される。第3に、デンプン分枝酵素が、α−1,4結合したグルカン部分を切断することにより分岐点を導入し、続いて切断した鎖を受け入れ先の鎖に移して新しいα−1,6結合を形成させる。デンプン分枝酵素は、α−ポリグルカンにα−1,6結合を導入できる唯一の酵素であり、したがって、アミロペクチンの形成において極めて重要な役割を果たしている。最後に、作用機序は解明されていないが、デンプン脱分枝酵素が分枝結合の一部を除去する(Myers他、2000年)。
【0004】
少なくともこれら4種の酵素活性が高等植物における通常のデンプン粒合成に必要であることは明らかであり、4種の酵素活性のそれぞれについて複数のアイソフォームが高等植物の胚乳中で発見されており、変異解析に基づいて(Wang他、1998年、Buleon他、1998年)、または遺伝子導入手法を用いて遺伝子発現レベルを改変することによって(Abel他、1996年、Jobling他、1999年、Scwall他、2000年)、個々のアイソフォームの具体的な役割が提示されてきた。しかし、デンプン生合成に対する各酵素活性の各アイソフォームの正確な寄与は依然として分かっておらず、これらの寄与が種間で著しく異なるかどうかも分かっていない。穀類の胚乳には、ADP−グルコースピロホスホリラーゼの2種のアイソフォームが存在する。すなわちアミロプラスト内部に1つの型、細胞質に1つの型が存在している(Denyer他、1996年、Thorbjornsen他、1996年)。各型は、2種のサブユニットからなっている。トウモロコシのしわ型(shrunken)(sh2)変異体および脆性(brittle)(bt2)変異体は、それぞれ大サブユニットおよび小サブユニット中の損傷を示している(GirouxおよびHannah、1994年)。穀類の胚乳には次の4クラスのデンプン合成酵素が存在している。デンプン粒の内部にもっぱら局在しているアイソフォーム、すなわち顆粒結合型デンプン合成酵素(GBSS)、顆粒と可溶性画分の間に分配されている2種の型(SSI(Li他、1999年a);SSII(Li他、1999年b))、ならびに可溶性画分中に全体がある4番目の型、すなわちSSIII(Cao他、2000年、Li他、1999年b、Li他、2000年)である。GBSSはアミロース合成に不可欠であることが示されており(Shure他、1983年)、SSIIおよびSSIIIの変異によりアミロペクチン構造が変えられることが示されている(Gao他、1998年、Craig他、1998年)。SSI活性の役割を定義する変異は記載されていない。
【0005】
分枝酵素の3種の型、すなわち、分枝酵素I(SBEI)、分枝酵素IIa(SBEIIa)、分枝酵素IIb(SBEIIb)は、穀類の胚乳で発現される(HedmanおよびBoyer、1982年、BoyerおよびPreiss、1978年、Mizuno他、1992年、Sun他、1997年)。コメ(NakamuraおよびYamanouchi、1992年)、トウモロコシ(Baba他、1991年;Fisher他、1993年;Gao他、1997年)、および小麦(Repellin他、1997年;Nair他、1997年;Rahman他、1997年)のゲノム配列およびcDNA配列は特徴を明らかにされている。配列アラインメントにより、ヌクレオチドレベルでもアミノ酸レベルでも配列の類似性の程度が高いことが判明しており、SBEIクラス、SBEIIaクラス、およびSBEIIbクラスへの分類が可能である。SBEIIaおよびSBEIIbは一般に、特に遺伝子の中心領域において、互いに約80%の配列同一性を示す。SBEIIaおよびSBEIIbは、その発現パターンによって区別することもできる。SBEIIbは一般に胚乳で特異的に発現されるが、SBEIIaは植物のすべての組織中に存在する。
【0006】
小麦胚乳では、SBEI(Morell他、1997年)はもっぱら可溶性画分に存在し、一方、SBEIIaおよびSBEIIbは、可溶性画分にもデンプン粒結合画分にも存在している(Rahman他、1995年)。トウモロコシおよびコメでは、高アミロースのフェノタイプは、アミロース増量(ae)遺伝子としても知られているSBEIIb遺伝子の損傷に起因することが示されている(BoyerおよびPreiss、1981年、Mizuno他、1993年;Nishi他、2001年)。これらSBEIIb変異体では、胚乳のデンプン粒が異常な形態を示し、アミロース含有量はかなり高くなり、残存するアミロペクチンの分岐頻度は少なくなり、短鎖(<重合度17、特に重合度8〜12)の比率が低くなっていた。さらに、デンプンの糊化温度は高くなっていた。加えて、アミロースとアミロペクチンの「中間体」と定義される物質がかなり溜まっていた(Boyer他、1980年、Takeda、他、1993年b)。一方、突然変異誘発(Mu)挿入因子によってSBEIIa遺伝子で突然変異が起こり、その結果としてSBEIIaタンパク質発現を欠いているトウモロコシ植物は、葉のデンプンでは分岐が変わっていたが、胚乳デンプンの分岐については野生型植物との区別が不可能であった(Blauth他、2001年)。同様に、SBEIIa活性が欠損したコメ植物でも、胚乳(Nakamura 2002年)のアミロペクチン鎖の性質に有意な変化は起きなかった(Nakamura 2002年)。トウモロコシおよびコメのいずれにおいても、SBEIIa遺伝子およびSBEIIb遺伝子はゲノム中で互いに結合していない。
【0007】
トウモロコシでは、dull1変異が起こると、変化の程度は遺伝的背景に応じて変わるが、デンプン含有量が少なくなり、胚乳のアミロースレベルが高くなり、残存アミロペクチンの分岐の程度が高くなる(ShannonおよびGarwood、1984年)。この変異に対応する遺伝子は、トランスポゾン突然変異誘発遺伝子(Mu)を用いるトランスポゾン標識法によって同定および単離され、デンプン合成酵素II(SSII)と呼ばれる酵素をコードしていることが示されている(Gao他、1998年)。現在この酵素は、穀類のSSIIIファミリーのメンバーとして認識されている(Li他、2003年)。変異胚乳では、dull1変異に伴ってSBEIIa活性のレベルが低下していた。他の穀類では、これに対応する変異は報告されていない。これらの研究結果が他の穀類、例えば小麦に関連性があるかどうかは分かっていない。
【0008】
WO94/09144では、センス遺伝子およびアンチセンス遺伝子を使用してトウモロコシのデンプン合成酵素(SS)とSBEとの天然の比率を変えることを示唆している。しかし、提案されている分子戦略を実証するデータは何も提示されておらず、SBEIIa活性を特異的に低下させることは示唆されていない。
【0009】
ジャガイモでは、その後の研究によりいくらかの性質の変化が確認されたものの(Safford他、1998年)、SBEIのみをダウンレギュレーションしてもデンプン構造に最小限の影響しか与えない(Filpse他、1996年)。しかし、ジャガイモでSBEIIおよびSBEIを一緒にダウンレギュレーションすると、SBEIIのみをダウンレギュレーションした場合よりもはるかに相対的アミロース含有量が増加した(Schwall他、2000年)。
【0010】
高等植物には2種類の型の脱分枝酵素が存在しており、その基質特異性に基づいて、イソアミラーゼ型脱分枝酵素およびプルラナーゼ型脱分枝酵素と定義されている(Myers他、2000年)。トウモロコシおよびコメにおけるシュガリー(sugary)−1変異は、双方の脱分枝酵素の欠損に関連付けられている(James他、1995年、Kubo他、1999年)が、原因となる変異は、イソアミラーゼ型脱分枝酵素遺伝子と同じ場所に位置している。穀類からクローン化されている代表的なデンプン生合成遺伝子を表1に挙げる。
表1.穀類で明らかにされているデンプン分枝酵素遺伝子
【0011】
【表1】

【0012】
デンプンは、食品工業、製紙工業、および化学工業で広く使用されている。デンプンの物理的構造は、食料品または非食料品もしくは工業製品用のデンプンの栄養特性および取扱特性に大きな影響を与え得る。アミロペクチン鎖長の分布、結晶性の程度およびタイプ、ならびに糊化温度、粘度、膨潤体積などの特性を含めて、いくつかの特質をデンプン構造の指標とみなすことができる。アミロペクチン鎖長の変化は、結晶性の変化、糊化、またはアミロペクチンの老化の指標とすることができる。
【0013】
デンプン組成物、特にアミロース含有量が高いことに関連付けることができるレジスタントスターチと呼ばれる型は、腸の健康、特に大腸の健康に重要な意味をもっている。したがって、腸の健康を促進する手段として食品に使用するために、トウモロコシなどいくつかの穀物において高アミロースデンプンの開発が行われてきた。レジスタントスターチの有益な効果は、大腸に栄養物が供給され、そこで大腸内の腸内微生物叢にエネルギー源が与えられ、このエネルギー源が発酵されて、特に短鎖脂肪酸が形成されることにより得られる。これらの短鎖脂肪酸は、結腸細胞に栄養物を提供し、一部の栄養物の大腸からの摂取を増進し、結腸の生理活性を促進する。一般に、レジスタントスターチまたは他の食物繊維が提供されない場合は、結腸は代謝的に比較的不活発である。
【0014】
化学的方法あるいは別法で改変されたデンプンは、食品に利用することができ、改変されていない供給源によっては通常得られない機能性を与えるが、そのような改変は、改変に伴うプロセスが原因となって、価値のある他の成分を改変しまたは望ましくない感覚をもたらす傾向がある。したがって、未改変の形態で食品に使用できる構成成分からなる供給源を提供することが好ましい。
【0015】
全世界で毎年生産される小麦の量は、穀物作物のうちで最も多い。小麦のデンプン構造で分かっている変異は、トウモロコシまたはコメで得られる変異と比べて限定されており、小麦の形質転換効率が他の穀類での効率に遅れをとっていること、ならびにパン小麦が6倍体の性質を有することがその理由の一部である。Triticum aestivum(普通小麦)には3種のゲノムが存在するため、2倍体種では変異をより容易に確認できるのに対して、個々のゲノムの変異を隠すことによる緩衝効果がある。トウモロコシまたはコメのアミロース増量フェノタイプに対応するSBEIIbの変異体は、小麦では明らかにされていない。SBEIIa変異またはSBEIIb変異によって与えられる小麦のフェノタイプは分かっていない。モチ性遺伝子(GBSS、ZhaoおよびSharp、1998年)の変異体およびSGP−1タンパク質を完全に欠いた変異体(Yamamori他、2000年)が知られている。後者の変異体は、タンパク質電気泳動での分析によりSGP−1(SSII)タンパク質のゲノムA、ゲノムB、およびゲノムDの個々の型を欠く系統を交配することによって作製された。SSII ヌル型の種子の調査により、変異によってアミロペクチン構造が変化し、デンプン粒が変形し、相対的アミロース含有量がデンプンの約30〜37%に増え、野生型のレベルより約8%多くなることが示された(Yamamori他、2000年)。比色測定、電流滴定(いずれもヨウ素結合について行う)、およびコンカナバリンA法によってアミロースを測定した。SSII ヌル変異体から得たデンプンは、等価な非変異型の植物から得たデンプンと比べて、低い糊化温度を示した。SSII−ヌル型穀粒のデンプン含有量は、野生型での60%から50%未満にまで減少していた。
【0016】
WO99/14314では、小麦関連の2倍体植物であるAegilops tauschii(タルホ小麦)からのSBEIIa遺伝子の単離について記載しているが、改変デンプンを含む小麦は作製しなかった。
【0017】
WO00/15810では、小麦のSBEIIb遺伝子のcDNAクローニングについて記載している。彼らは、アミロースレベルを変えた小麦植物を得ることはせず、アミロースを少なくとも50%含むデンプンを有する小麦を教示しなかった。
【0018】
WO01/62934も、小麦のSBEIIb遺伝子について記載し、小麦植物に分枝酵素活性の阻害物質を導入することを示唆しているが、アミロースを少なくとも50%含むデンプンを有する小麦を教示していない。
【0019】
WO01/32886は、小麦胚乳に存在するSBEIの1つの型をコードしているcDNAを明らかにした。コードされているポリペプチドは、A型のデンプン粒に優先的に関連していることが発見された。彼らは、SBEI活性を抑制することも、改変されたデンプン粒の形態を示すことも、小麦のアミロース量を増やすこともしなかった。
【0020】
したがって、アミロース比率が約50%より高いデンプンを有する小麦は知られていない。高アミロースのトウモロコシ変種およびオオムギ変種が知られているが、これらの穀類から作られた製品は、小麦が好ましい穀類である製品に関しては、例えばパン、パスタ、またはヌードルではアミロース含有量が非常に多い小麦と比べて不利である。
【0021】
これらのタイプの高アミロースデンプンは有用であるが、アミロース含有量のより高い小麦デンプンが、特にデンプン合成の改善および他の特質、例えば収穫後改変の必要が減ることと関連している場合は好ましい。このようなデンプン製品はまた、消化に対して比較的抵抗性があり、健康面でより大きな利益をもたらす。
【0022】
当業者なら、本明細書で記述する本発明が、具体的に記述するもの以外の変更や修正を受けることが分かるであろう。本明細書で記述する本発明が、このような変形法および改良法のすべてを含むことを理解すべきである。本発明はまた、本明細書で参照または指示するこのようなステップ、特徴、組成物、および化合物のすべても個別にまたは合わせて含み、また、前記ステップまたは特徴のうちのいずれか2つ以上からなる任意の組合せおよびすべての組合せも含む。
【0023】
本明細書を通じて、文脈上別の意味で解釈する必要がない限り、「含む(comprise)」という単語および「comprises」や「comprising」などの変形は、記載した整数(integer)もしくはステップ、または整数(intergers)もしくはステップの群を包含するが、他の整数(integer)およびステップ、ならびに整数(integers)およびステップの群のどれも除外しないことを意味しているものと理解されたい。本発明は、本明細書で記述する個々の実施形態によって範囲を限定されるべきではなく、これらの実施形態は例証するためのものであるにすぎない。本明細書で記述するように、機能的に等価な製品、組成物、および方法が本発明の範囲に含まれることは明らかである。
【0024】
本明細書中で本発明者らが参照した刊行物の書誌詳細は、明細書の最後にまとめている。本明細書で記述する参考文献は、その全体を参照により本明細書に組み込む。従来技術に関する本明細書の参考文献は、任意の1種または複数の従来技術の文献を含めて、前記の従来技術がオーストラリアで周知の一般的知識であり、あるいはオーストラリアで周知の一般的知識の一部を形成していることを認めるものまたは示唆するものとしてみなすべきではない。
【0025】
本明細書では、「に由来する、から得た(derived from)」という用語は、特定の完全体(integer)または完全体群(integers)が、指定した種に由来するものであるが、必ずしも指定の供給源から直接得られているわけではないことを示すと理解すべきである。
【0026】
本明細書で参照するヌクレオチド残基の記号は、IUPAC−IUB生化学命名法委員会により推奨される記号であり、Aはアデニンを示し、Cはシトシンを示し、Gはグアニンを示し、Tはチミジンを示す。
【発明の開示】
【0027】
<発明の概要>
【0028】
第1の態様で、本発明は、穀粒のデンプン中のアミロース比率が少なくとも50%である小麦植物から得た穀粒を提供する。この小麦植物は、SBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低くてよく、SBEIIaとSBEIIbの両方の遺伝子発現、酵素活性、またはその双方のレベルが低いのが好ましい。この穀粒は、遺伝的変異、すなわちSBEIIa遺伝子発現、酵素活性、またはその双方を阻害するSBEIIa遺伝子の変異、あるいはSBEIIa遺伝子発現、酵素活性、またはその双方を阻害する導入核酸のいずれかを含んでよい。さらに、この穀粒はSBEIに同様の遺伝的変異を含んでもよい。この穀粒は、ADPグルコースピロホスホリラーゼ、GBSS、SSI、SSII、SSIII、イソアミラーゼ型脱分枝酵素、及びプルラナーゼ型脱分枝酵素からなる群から選択されるタンパク質および/または酵素活性を、改変されたレベルでさらに含んでもよい。この穀粒は、導入遺伝子を含んでよく、また、この導入遺伝子は、アンチセンスRNA分子、コサプレッションRNA分子、リボザイムRNA分子、または二重鎖RNA分子をコードしてよい。この導入遺伝子によって、SBEIIaをコードしているmRNAの発現レベルが低められることが好ましい。この穀粒は、SBEIIa遺伝子に変異を含んでよく、1つの形態では少なくとも1つのゲノムにSBEIIa遺伝子のヌル変異があり、ゲノムのうちの2つもしくは3つにヌル変異があってもよい。穀粒のデンプン中のアミロース比率は、少なくとも40%、50%、55%、60%、70%、または80%でよい。別の形態では、穀粒内部のデンプン粒の少なくとも50%は、偏光下で観察すると非複屈折性に見える。穀粒は、非しわ型でよく、平均重量は少なくとも36mgまたは40mgである。代わりの形態では、殻のない状態での穀粒のデンプン含有量は、少なくとも25%(w/w)または少なくとも35%(w/w)であり、野生型穀粒のデンプン含有量の少なくとも90%でよい。穀粒は、全粒、玄穀、製粉した穀粒、挽き割りにした穀粒、圧扁した穀粒、精白した穀粒、粉砕した穀粒、またはパーボイルド穀粒でよい。
【0029】
第1の態様の別の形態で、本発明は、デンプンと、野生型の穀粒に比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルを低下させる遺伝的変異とを含む、小麦植物から得た穀粒を提供する。この遺伝的変異は、SBEIIa遺伝子の変異またはSBEIIa遺伝子の発現の阻害物質をコードする導入核酸を含み、穀粒のデンプン中のアミロース比率は少なくとも30%である。
【0030】
第2の態様では、本発明は、それだけには限らないが、小麦粉、全粒小麦粉、セモリナ粉を含めて第1の態様の穀粒に由来する製粉製品または本発明の穀粒から得たデンプン、あるいはこのような小麦粉、全粒小麦粉、セモリナ粉、デンプンを組み込んだ食料品、あるいは穀粒を圧扁した製品、薄く削った製品、または押し出した製品を提供する。この製品には、別の供給源から得た小麦粉、全粒小麦粉、セモリナ粉、またはデンプンと混合した、本発明の第1の態様の穀粒から得た小麦粉、全粒小麦粉、セモリナ粉、またはデンプンを含めてよい。
【0031】
第3の態様では、本発明は、第1の態様の小麦植物の穀粒から得たデンプン粒またはデンプンを提供する。第3の態様の特定の形態では、さらに、小麦植物の胚乳のSBEIIa酵素活性レベルが低められている。
【0032】
第4の態様では、本発明は、本発明の第3の態様に記載のデンプンと食品成分または水を含む組成物にあると言える。この態様は、食品、非食品組成物及びデンプンに他のデンプンまたはデンプン含有品とを混合したものを含む。
【0033】
第5の態様では、本発明は、上記の本発明の第4の態様に記載のデンプン粒および別の食品成分または水を含む組成物を提供する。
【0034】
第6の態様では、本発明は、先の態様に記載の穀粒またはデンプン粒もしくはデンプンを製造するのに使用できる小麦植物を提供する。小麦植物は、トランスジェニックでも非トランスジェニックでもよく、それから得られる穀粒も同様である。
【0035】
第7の態様では、本発明は、i)親となる小麦植物または小麦種子に遺伝的変異を導入するステップと、ii)野生型の植物または種子と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップとを含む、穀粒のデンプン中のアミロース比率が少なくとも50%であるデンプンを含有する穀粒を結実することができる小麦植物を作製する方法を提供する。
【0036】
第7の態様の第2の形態では、本発明は、i)SBEIIa遺伝子の変異またはSBEIIa遺伝子の発現の阻害物質をコードする導入核酸を含む遺伝的変異を、親となる小麦植物または小麦種子に導入するステップと、ii)野生型の穀粒と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップとを含む、穀粒のデンプン中のアミロース比率が少なくとも30%であるデンプンを含有する穀粒を結実することができる小麦植物を製造する方法を提供する。遺伝的変異を導入するステップは、SBEIIa遺伝子発現の阻害物質を発現する外来の核酸の導入を含んでもよく、親となる小麦植物の突然変異誘発を含んでもよい。
【0037】
第7の態様の第3の形態では、本発明は、i)SBEIIa遺伝子の変異を含む遺伝的変異を、親となる小麦植物または小麦種子に導入するステップと、ii)野生型の穀粒と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップと、iii)SBEIIb遺伝子の変異を含む遺伝的変異を、親となる小麦植物または小麦種子に導入するステップと、iv)野生型の穀粒と比べて胚乳のSBEIIbの遺伝子発現、SBEIIaの酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップと、v)胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低い植物を、胚乳のSBEIIb遺伝子発現、SBEIIb酵素活性、またはその双方のレベルが低い植物と交配させるステップと、SBEIIaとSBEIIbの双方の遺伝子発現、酵素活性、またはその双方が低い小麦植物を同定するステップとを含む、穀粒を結実することができる小麦植物を作製する方法を提供する。
【0038】
第7の態様の第4の形態では、本発明は、a)小麦のAゲノム、Bゲノム、またはDゲノムから発現されるSBEIIa活性が低下した小麦植物または穀粒を同定し、b)前記小麦植物またはステップa)の穀粒から得た小麦植物を、SBEIIa活性の低い第2の小麦植物と交配させるステップ、あるいは、c)SBEIIa酵素活性の低い植物をSBEIIb酵素活性の低い植物と交配させ、SBEIIaとSBEIIbの双方の活性が低い小麦植物を同定するステップを含む、その穀粒のデンプン中の相対的アミロース含有量が少なくとも50%であり、好ましくは胚乳のSBEIIa酵素活性が低活性である小麦植物を作製する方法を提供する。第7の態様の植物は、Triticum aestivum ssp.aestivumであることが好ましい。
【0039】
第8の態様では、上記に示した方法によって植物を改変し特性を改変したデンプンを抽出してなる、改変デンプンの作製方法を提供する。
【0040】
第9の態様では、本発明は、小麦のSBEIIb遺伝子、またはSBEIIa遺伝子にそれぞれ結合する分子マーカーを用いて小麦植物または小麦種子の集団をスクリーニングするステップと、結合した分子マーカーの有無に基づいて植物または種子を同定するステップとを含む、SBEIIa遺伝子またはSBEIIb遺伝子の変異に関して小麦植物または小麦種子を同定する方法を提供する。
【0041】
第9の態様の第2の形態では、本発明は、小麦のSBEIIbタンパク質、またはSBEIIaタンパク質にそれぞれ特異的な抗体を用いて小麦植物または小麦種子の集団をスクリーニングするステップと、抗体結合の有無に基づいて植物または種子を同定するステップとを含む、SBEIIa遺伝子またはSBEIIb遺伝子の変異に関して小麦植物または小麦種子を同定する方法を提供する。
【0042】
第10の態様では、本発明は、SBEIIa遺伝子が染色体2Aの長腕から欠けている変異、あるいは染色体2Aの長腕上のSBEIIa遺伝子が、野生型穀粒と比べて穀粒の胚乳のSBEIIaタンパク質、SBEIIa酵素活性、またはその双方を低下させる変異を有する変異を含む、小麦植物から得た穀粒を提供する。この変異はSBEIIa遺伝子のヌル変異でも、SBEIIa遺伝子の少なくとも一部の欠失でもよい。穀粒は、SBEIIb遺伝子が染色体2Aの長腕から欠けている変異、あるいは染色体2Aの長腕上のSBEIIb遺伝子が、野生型穀粒と比べて穀粒の胚乳のSBEIIbタンパク質、SBEIIb酵素活性、またはその双方を低下させる変異を有する変異をさらに含んでもよい。遺伝子欠失によって、染色体2Aの長腕上のSBEIIa遺伝子およびSBEIIb遺伝子の双方の発現を中断させてもよい。
【0043】
植物は、野生型穀粒と比べて染色体2Bの長腕上のSBEIIa遺伝子にコードされたデンプン分枝酵素活性を低下させる遺伝的変異をさらに含むことができるデュラム小麦植物でよい。別の遺伝的変異は、染色体2Bの長腕からのSBEIIa遺伝子の欠失、または野生型穀粒と比べて穀粒の胚乳のSBEIIa酵素活性を低下させる染色体2Bの長腕上のSBEIIa遺伝子の変異を含んでよい。
【0044】
植物は、野生型穀粒と比べて染色体2B、染色体2D、またはその双方の長腕上のSBEIIa遺伝子にコードされたデンプン分枝酵素活性を低下させる遺伝的変異をおそらくはさらに含むであろうTriticum aestivum ssp.aestivum(普通小麦の亜種)であってもよい。さらなる遺伝的変異は、前記染色体のうちの少なくとも1つからのSBEIIa遺伝子の欠失、または野生型穀粒と比べて穀粒の胚乳のSBEIIa酵素活性を低下させる前記染色体のうちの少なくとも1つのSBEIIa遺伝子の変異を含む。
【0045】
植物には、SBEIIa遺伝子発現、活性またはその双方の阻害物質をコードする核酸を導入してよい。SBEIIa酵素活性のレベルは、野生型穀粒と比べて少なくとも40%低下させることができる。穀粒のデンプン中のアミロース比率は、少なくとも30%または少なくとも50%でよい。穀粒は、非しわ型でよく、平均重量は少なくとも36mgでよい。穀粒から得たデンプン粒の少なくとも50%が、偏光下で観察すると非複屈折性に見えるものでよい。本発明の一形態では、殻のない状態での穀粒のデンプン含有量は、少なくとも25%(w/w)であり、あるいは、デンプン含有量は野生型穀粒のデンプン含有量の少なくとも90%である。
【0046】
本発明の形態のうちのいずれかに記載の穀粒のアミロペクチンでは、アミロペクチンをイソアミラーゼで脱分枝した後に測定すると、野生型穀粒のアミロペクチンと比べて重合度が4〜12の鎖長部分の比率が低下していてもよい。
【0047】
穀粒は、SBEIタンパク質、SBEI酵素活性、またはその双方を低レベルで含んでもよく、また、ADPグルコースピロホスホリラーゼ、GBSS、SSI、SSII、SSIII、イソアミラーゼ型脱分枝酵素、プルラナーゼ型脱分枝酵素、およびこれらの任意の組合せからなる群から選択される、野生型穀粒と比べてレベルを変えられた酵素をさらに含んでもよい。
【0048】
本発明のこの第10の態様の形態は、穀粒、穀粒から抽出されたデンプン粒、および、例えば小麦粉、全粒小麦粉、またはセモリナ粉など穀粒から製造された製品またそのデンプンを包含する。
【0049】
<小麦のSBEIIaの改変>
本発明は、小麦胚乳のSBEIIa活性を低下させると、修飾されたデンプンが産生され、特に小麦穀粒の相対的なアミロースレベルが高くなるという研究結果に基づいている。この予想外の結果は、SBEIIaの変異によってアミロペクチン/アミロースの特性が改変されなかったトウモロコシおよびコメでの研究結果と対照的である(Blauth他、2001年、Nakamura、2002年)。別の実施形態では、SBEIIb活性ならびにSBEIIa活性を低下させるなど、1種または複数の追加のデンプン生合成酵素活性を改変する。これら2種の活性をコードする遺伝子の変異には、SBEIIaおよびSBEIIbが、トウモロコシおよびコメでは関連していないのとは対照的に、小麦では密接に関連しているという驚くべき研究結果が役立つ。本発明者らは、予想外に、SBEIIa活性およびSBEIIb活性のレベルが低い小麦植物の穀粒が非しわ型であるということも発見した。
【0050】
<小麦植物を作製する方法>
一態様では、本発明は、穀粒中のデンプンを改変した小麦植物を作製する方法、特にデンプン中のアミロース比率を少なくとも30%に増加させる方法を提供する。通常、6倍体の小麦およびデュラム小麦では、デンプン中のアミロース比率は、約18〜約30%の範囲であり、一部の変異体(SGP−1欠損)では最大約35%である。一実施形態では、本発明の方法は、アミロースを少なくとも30%含むデンプンを有する穀粒を結実する小麦植物を提供するために、親となる小麦植物または小麦種子に遺伝的変異を導入するステップを含む。本明細書で定義するデンプン中のアミロース比率は、重量/重量(w/w)、すなわち、穀粒から得たデンプンの重量に対するパーセンテージとしてアミロース重量を示した値に基づく。別の実施形態では、デンプン中のアミロース比率は、少なくとも40%、少なくとも50%、少なくとも55%、少なくとも60%、少なくとも65%、少なくとも70%、または少なくとも75%である(それぞれw/w)。本発明の別の実施形態では、この方法により、少なくとも80%または少なくとも90%(w/w)のアミロース比率が得られる。
【0051】
別の実施形態では、この方法は、小麦胚乳のデンプン分枝酵素IIa(SBEIIa)タンパク質、酵素活性またはその双方のレベルを変えること、好ましくは低下させることを含む。すなわち、小麦植物に遺伝的変異が導入されると、直接的にまたは間接的にSBEIIaのレベルが変更され、その結果として本明細書に記述するようにデンプンが修飾される。先の実施形態と相互に排他的ではない別の実施形態では、この方法は、小麦胚乳のSBEIIa遺伝子の発現レベルを変えること、好ましくは低下させることを含み、あるいは、この方法は、胚乳のSBEIIa活性を低下させるような小麦SBEIIa遺伝子の変異を含む。SBEIIa遺伝子発現のレベルまたは他の遺伝子のレベルの低下は、核酸、例えば阻害性分子をコードする導入遺伝子の導入によって実現することができる。阻害性分子の例としては、アンチセンスRNA分子、コサプレッションRNA分子、リボザイムRNA分子、または二重鎖RNA分子が挙げられる。
【0052】
本明細書では、「改変すること」、「増加させること」、「増加された」、「低下させること」、「低下した」、「阻害された」などの用語は、比較的な用語、すなわち野生型または未改変の状態と比較しての用語とみなす。「タンパク質のレベル」とは、特定のタンパク質、例えばSBEIIaの量を意味し、これは、例えば、ウェスタンブロット分析や他の免疫学的方法など当技術分野で既知の任意の方法によって測定することができる。「酵素活性のレベル」とは、酵素アッセイで測定した特定の酵素の量を意味する。変異体の酵素活性のレベルは改変されているがタンパク質自体の発現レベル(量)は改変されていないことがあると理解されよう。逆に言えば、タンパク質の量を変えることはできるが、程度の差はあれ活性なタンパク質が産生される場合は、その活性は同じままである。例えば、その酵素をコードする遺伝子が不活性化されるようなときには、量および活性の双方の低下も起こり得る。一部の実施形態では、タンパク質または活性のレベルは、未修飾の小麦の胚乳におけるタンパク質または活性のレベルと比べて、少なくとも40%または少なくとも60%、あるいは少なくとも75%、少なくとも90%、または少なくとも95%低下している。タンパク質活性、酵素活性、または遺伝子発現のレベルの低下は、穀粒のどの発育段階でも、特に発育中の胚乳でデンプンが合成される時期である穀粒の登熟期の間に起こってもよく、成熟に至るまでの穀粒発育のすべての段階で起こってもよい。
【0053】
本明細書では、本質的にα−グルコピラノース単位で構成された多糖類を「デンプン」と定義する。デンプンは、小麦中の主要な貯蔵炭水化物であり、アミロプラスト中で合成され、顆粒中で形成および貯蔵される。デンプンは、ほぼ直鎖状のα−1,4−D−グルコピラノースポリマーであるアミロースと、主にα−1,4結合したα−D−グルコピラノース単位にα−1,6結合分枝が付いた短鎖を有するアミロペクチンとを含む。野生型植物から得た小麦デンプンは、アミロースを最大約20%〜30%、アミロペクチンを最大約70%〜80%含む。これら2種の分子の別の有意な差は、分子量である。アミロースはらせん状の構造で分子量が104〜106ダルトンであり、一方、アミロペクチンの分子量は107〜108ダルトンである。近年の研究により、アミロース中に最大約0.1%のα−1,6−グリコシドの枝分かれ部位が存在することがあることが示され、したがって、「ほぼ直鎖状」と表現されている。本明細書では、α−1,4結合のグルコシド(グルコピラノース)単位およびアミロース様の長鎖アミロペクチン(「中間体」または「アミロース様アミロペクチン」と呼ばれることがある。Takeda他、1993年b;Fergason、1994年)から構成されるほぼ直鎖状の分子を含めて、「アミロース」と定義する。例えば90%(w/v)DMSOを用いてのサイズ排除高速液体クロマトグラフィー(HPLC)、コンカナバリンA法(Megazyme社、アイルランド)を含めて当技術分野で既知の方法のいずれかを用いるか、好ましくは例えば実施例1で記述するヨード滴定法によってアミロース含有量を決定することができる。HPLC法は、デンプンの脱分枝を伴うものでも(BateyおよびCurtin、1996年)、脱分枝を伴わないものでもよい。穀粒重量およびアミロース含有量から、穀粒当たりのアミロース蓄積量を計算し、トランスジェニック系統とコントロールの系統とを比較することができる。
【0054】
別の実施形態では、この方法は、当技術分野で既知の任意の方法を用いて小麦胚乳中のSBEIIaの量または活性を決定するステップを含む。ある実施形態では、例えば、ウェスタンブロット法やELISA検定法などの免疫検出法によってタンパク質のレベルを測定し、または、それに対応するmRNAのレベルを、ノーザンブロットハイブリダイゼーション分析や逆転写ポリメラーゼ連鎖反応法(RT−PCR)など当技術分野で公知の方法によって測定する。別の実施形態では、この方法は、胚乳中のSBEIIaタンパク質または酵素活性のレベルが変えられている小麦植物または穀粒を選択またはスクリーニングするステップを含む。選択ステップは、SBEIIa活性またはタンパク質のレベルの低下に基づいてもよく、あるいは、アミロース比率の上昇やアミロペクチン比率の低下など小麦植物の穀粒のフェノタイプ、見た目のフェノタイプ、例えばしわ型穀粒、またはデンプン粒の特性の変化に基づいてもよい。
【0055】
本発明は、本明細書で記述する方法のうちのいずれかを用いて、直接的にデンプンの特性を測定して、または間接的に、例えば、植物またはその穀粒中の遺伝的変異の存在を検出することによって、穀粒のデンプンの特性が変えられた小麦植物を同定する方法を含むと理解されるであろう。植物は、小麦植物の集団の植物、例えば、小麦育種の植物などでよい。
【0056】
SBE活性は、酵素アッセイ、例えばホスホリラーゼ刺激法(BoyerおよびPreiss、1978年)によって直接測定することができる。この検定法では、ホスホリラーゼaによるメタノール不溶性ポリマー(α−D−グルカン)へのグルコース1−リン酸の取込に対するSBEの刺激を測定する。SBE活性は、グルカンポリマーの枝分かれの結果として生じるグルカン−ポリヨウ素錯体の吸光度の減少を測定するヨウ素染色法によって測定することができる。SBE活性は、イソアミラーゼによる分解の後、基質として還元されたアミロースから生成される還元末端を測定する分枝結合検定法によって検定することもできる(Takeda他、1993年a)。SBEI活性またはSBEIIb活性がない状態で活性を測定することが好ましい。SBEのアイソフォームは、様々な基質特異性を示す。例えば、SBEIはアミロースを枝分かれさせる活性が高く、一方、SBEIIaおよびSBEIIbは、アミロペクチン基質を枝分かれさせる割合が高い。アイソフォームは、移動されるグルカン鎖の長さに基づいて区別することもできる。本明細書で記述するものなど特異的な抗体を用いて、SBEタンパク質を測定してもよい。SBEII活性は、穀粒の発育中に、発育中の胚乳で測定してもよく、あるいは、等価であるが改変されていない穀粒中にタンパク質がなお存在し、免疫学的方法で検定することができる成熟穀粒で測定してもよい。
【0057】
別の態様では、本発明は、穀粒のデンプン中のアミロース比率が少なくとも50%になるように、小麦胚乳中の複数のデンプン生合成酵素の活性を改変し、好ましくは低下させる方法であって、前記酵素のうちの1種がSBEIIaである方法を提供する。いくつかの実施形態では、SBEIIaおよびSBEIIbの双方のタンパク質または酵素活性のレベルが低められ、あるいは、SBEIIa、SBEIIb、およびSBEIの3種すべてのレベルが低められる。SBEIIaとともに改変することができる他のデンプン生合成酵素は、SSI、SSII、SSIIIである。デンプン脱分枝酵素は、例えばイソアミラーゼまたはプルラナーゼの活性を改変することもできる。SBEIIaが改変される限りにおいて、上記の酵素の任意の組合せでも提供される。別の実施形態では、植物の胚乳以外の組織に存在する1種または複数のデンプン生合成酵素の活性が改変される。例えば、主に胚乳での発現を目的としたSBEIIa阻害分子をコードする導入遺伝子によって活性の損失が起こると、それを補うために、葉のSBEIまたはSBEIIの活性が増大されることがある。改変は、例えば、量の増加または減少でも、発現時期の変化でもよい。あるいは、SBEIIaの低下とともに、1種または複数のデンプン生合成酵素を過剰発現させることによって、デンプン合成をさらに改良することができる。このような酵素をコードする遺伝子は、様々な供給源のうちのいずれかから、例えば、細菌または小麦以外の他の供給源から得てもよく、また、触媒特性を変えるように、例えば、酵素の温度依存性を変えるように修飾することができる(例えば、WO94/09144を参照のこと)。
【0058】
高アミロースのフェノタイプは、SBEIIa遺伝子の発現、またはSBEIIa遺伝子およびSBEIIb遺伝子を部分的にまたは全体的に阻害することによって実現することができる。1つまたは複数の遺伝子が阻害される程度によって、小麦穀粒で作られるデンプンの特質がある程度決められることになる。修飾された小麦胚乳から抽出されたタンパク質に対して一連のゲル電気泳動法のいずれかを実施すると、その特質とSBEIIa活性および/またはSBEIIb活性に対する修飾の程度が明らかになる。修飾は、SBEIIa活性および/またはSBEIIb活性の低下、酵素活性の完全な消失、または胚乳内のSBEIIbまたは他の酵素の分布の変化として起こることがある。これらの検定を実施するために、例えば、Rahman他により1995年に概説されているように、小麦胚乳からデンプンを抽出し、その中のタンパク質を分析することができる。SDSポリアクリルアミドゲル電気泳動(PAGE)やイムノブロッティングなど当技術分野で公知の技術を、可溶性画分およびデンプン粒画分に対して実施し、結果を用いてSBEIIa酵素および/またはSBEIIb酵素に修飾が起こっている植物または穀粒を同定する。
【0059】
<小麦植物>
別の態様では、本発明は、デンプン中のアミロース比率が少なくとも30%である穀粒を結実することができる小麦植物を提供する。別の実施形態では、アミロース比率は、少なくとも40%、少なくとも50%、少なくとも55%、少なくとも60%、少なくとも65%、少なくとも70%、または少なくとも80%である。別の実施形態では、その穀粒がデンプン中にこれらのレベルのうちのいずれかのアミロースを含む小麦植物は、野生型穀粒と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルを低下させる遺伝的変異を含む。好ましい実施形態では、この遺伝的変異は、SBEIIa遺伝子の変異またはSBEIIa遺伝子発現の阻害物質をコードする導入核酸を含む。この阻害物質は、アンチセンスRNA分子、コサプレッションRNA分子、リボザイムRNA分子、二重鎖RNA分子、あるいはSBEIIaの発現および/または活性を阻害する類似の分子を含んでよい。
【0060】
本明細書では、例えば、Triticum aestivum L.ssp.aestivum(一般的な小麦またはパン小麦)、Triticum aestivumの他の亜種、Triticum turgidumL.ssp.durum(マカロニ小麦または硬質小麦としても知られているデュラム小麦)、Triticum monococcum L.ssp.monococcum(栽培型のヒトツブ小麦または小型のスペルト小麦)、Triticum timopheevi ssp.timopheevi、Triticum turgigum L.ssp.dicoccon(栽培型のエンマー小麦)、およびTriticum turgidumの他の亜種を含めて(Feldman)、商業的に栽培されている普通小麦(Triticum)属の種に属する任意の植物を小麦植物と定義する。小麦は、AABBDD型のゲノムを有する6倍体小麦でも、AABB型のゲノムを有する4倍体小麦でもよい。本発明による小麦の遺伝的変異は、ライムギおよびオオムギを含めた一部の関連種にハイブリダイゼーションによって伝達することができるため、本発明は、パン小麦とライムギの交配種であるライ小麦を含めて、このようにして形成された交配種も含む。特定の実施形態では、小麦植物は、Triticum aestivum種のものであり、好ましくは、亜種aestivumのものである。あるいは、Triticum aestivumからデュラム小麦に変異または導入遺伝子を容易に移すことができるため、小麦は、Triticum turgidum L.ssp.durumであることが好ましい。
【0061】
本発明は、野生型植物から抽出したデンプンと比べてアミロースを高比率で含むデンプンを有する穀粒を結実することができる、胚乳中のSBEIIaタンパク質、酵素活性、またはその双方のレベルを低下させた小麦植物も提供する。SBEIIaのレベルの低下は、穀粒の発育過程の少なくとも一段階で起こってもよく、成熟に至るまでの全過程で起こってもよい。別の実施形態では、胚乳中のSBEIIaのレベルは、野生型と比べて少なくとも50%、少なくとも75%、少なくとも90%、または少なくとも95%低められる。「野生型」という用語は、遺伝学の分野におけるの通常の意味を指し、本明細書で教示するような修飾をしていない小麦の栽培品種または遺伝子型を含む。
【0062】
本発明は、遺伝子型および/またはフェノタイプにおいて、親の小麦植物の望ましい形質を有する子孫植物および子孫穀粒も提供する。本発明の範囲は、培養組織や培養細胞などのような望ましい形質を有する植物を作製するのに使用できる、小麦植物の増殖性の材料のどれにでも及ぶ。
【0063】
本発明は、SBEIIa活性の低下に加えて、SBEIIbまたは他のデンプン生合成酵素を改変し、好ましくは低下させた小麦植物も含む。SBEIIa活性およびSBEIIb活性の低下した植物は、SBEIIaの低下した植物とSBEIIbの低下した植物とを交配することによって、あるいは、SBEIIa遺伝子およびSBEIIb遺伝子の双方の発現を阻害する分子をコードする導入遺伝子を導入することによって作製することができる。本明細書で示すように、小麦のSBEIIa遺伝子およびSBEIIb遺伝子は密に結びついているため、小麦のゲノムのうちの1つによりコードされているSBEIIaアイソフォームおよびSBEIIbアイソフォームを欠いている変種を同定し、そのような変種を交配して少なくとも2つのゲノムによってコードされているアイソフォームを低下させた植物を作製することによって、両方の活性を低下させた植物を作製することもできる。
【0064】
本発明は、他の遺伝的背景または前述の小麦植物と交配できる他種における遺伝的変異または変異も含む。改変された(変異した)植物を、より望ましい遺伝的背景を有する植物と交配することができる。最初の交配後、適切な回数だけ戻し交配を実施して、望ましくない遺伝的背景を取り除くことができる。望ましい遺伝的背景として、商業用の収穫高および農業生産力や非生物ストレスへの耐性など他の特質を与える遺伝子の適切な組合せを挙げることができる。遺伝的背景には、改変デンプンを生合成または修飾する他の遺伝子、例えば、原因となる遺伝子が分かっていないしわ型胚乳を有する他の小麦系統に由来する遺伝子も含まれ得る。
【0065】
植物は、トランスジェニックでも非トランスジェニックでもよい。
【0066】
本発明は、SBEIIa遺伝子が染色体2Aの長腕(2AL)から欠けている変異、あるいは染色体2Aの長腕上のSBEIIa遺伝子が、野生型穀粒と比べて穀粒の胚乳のSBEIIa酵素活性を低下させる変異を有する変異を含む小麦植物も提供する。2400種の小麦の登録品種を徹底してスクリーニングしたが、本発明者らは、天然に存在するこのような植物を発見できなかった。このことは、2AL上の機能的なSBEIIa遺伝子を保持するための選択が自然に起こっている可能性があることを示唆している。しかし、突然変異誘発後には、このような植物を作製し、同定することができた。これらの植物は、一部の市場で望ましいとされる非トランスジェニックである。これらの植物は、パン小麦でも、デュラム小麦でも、他の小麦でもよい。好ましい実施形態では、この小麦植物は、2AL染色体上のSBEIIa遺伝子の少なくとも一部の欠失を含み、この欠失はSBEIIb遺伝子の少なくとも一部に及ぶことがある。当技術分野で理解されているように、パン小麦などの6倍体小麦は、一般にゲノムA、ゲノムB、ゲノムDと呼ばれる3種のゲノムを含み、一方、デュラム小麦などの4倍体小麦は、一般にゲノムAおよびゲノムBと呼ばれる2種のゲノムを含む。各ゲノムは7対の染色体を含み、これらは減数分裂の進行中に細胞学的方法によって観察することができる。これらの染色体は、通常、大きさに従い最大のものから最小のものの順序で示され、したがって、染色体2は各ゲノムで2番目に大きい染色体である。各染色体には、染色体2上に非対称的に配置されたセントロメアがあり、それゆえに染色体2の2つの腕は、「短腕」および「長腕」と呼ばれている。本明細書では、この用語の標準的な意味に従って、長腕に沿ったセントロメアと先端の間の染色体部分を「染色体2Aの長腕」と定義する。「染色体2Bの長腕」および「染色体2Dの長腕」という用語は、それぞれ小麦のゲノムBまたはゲノムDの染色体2に関するという点を除いて、同様に定義される。
【0067】
本発明者らは、小麦の染色体2上でSBEIIa遺伝子およびSBEIIb遺伝子が密に結びついしていることを発見した。特定の実施形態では、この小麦植物は、少なくともSBEIIa遺伝子の変異を含む染色体腕2ALを過半数(>50%)含む。すなわち、本質的には、少なくともAゲノムのSBEIIa遺伝子に変異を含む染色体2ALが存在している。2ALの存在は、例えば、in situハイブリダイゼーション法(実施例9を参照のこと)などの細胞学的技術によって、または2ALに特異的な分子マーカーを用いることによって、確認することができる。好ましい実施形態では、この小麦植物は、前記変異に関してホモ接合性である。この変異は、ヌル変異でよい。この変異は、遺伝子欠失でよい。
【0068】
特定の実施形態では、欠失対立遺伝子は、MLT2B8植物またはMLT2D1植物に由来するものである。これらの植物の変異したSBEIIa対立遺伝子は2AL染色体上に存在するため、交配によって、これらの対立遺伝子をパン小麦またはデュラム小麦の変種に導入することができる。したがって、本発明は、このような植物、およびそれらから得られる穀粒およびデンプン製品を含む。これらの対立遺伝子を、他の有用なデンプン生合成遺伝子もしくは対立遺伝子、または他の有用な遺伝形質と組み合わせてよい。
【0069】
本発明の範囲が、このような小麦植物、またはこのような植物から得られる穀粒を作製または同定する方法に及ぶことは明らかである。
【0070】
<穀粒>
本発明は、野生型小麦穀粒から抽出したデンプンと比べて改変されたデンプンを含む小麦穀粒も提供する。本明細書では、ほぼ成熟した穀粒を穀粒と定義する。これには、商業的環境で収穫された穀粒が含まれる。一実施形態では、改変デンプンは、少なくとも一部には小麦穀粒の胚乳の発育中にSBEIIa活性を低下させた結果物である。先の実施形態と相互に排他的ではない別の実施形態では、この穀粒のアミロース比率(全デンプンに対するパーセンテージ)は上昇している。これは、野生型植物から得た穀粒と比べた、デンプン中のアミロペクチンの減少率として測定することができる。野生型小麦デンプンは、アミロースを約20〜30%、アミロペクチンを約70〜80%有する。本発明の穀粒は、好ましくはアミロースを少なくとも50%(w/w)含むデンプンを含有する。別の実施形態では、SBEIIa活性およびSBEIIb活性の双方が、胚乳の発育中に低められる。別の実施形態では、SBEIの活性も低められる。別の実施形態では、当技術分野でよく理解されている方法によって測定したアミロース比率がは、穀粒のデンプンの少なくとも55%、少なくとも60%、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、または少なくとも90%(各w/w)である。アミロースレベルの上昇は、デンプン粒の形態が異常になること、または光学顕微鏡もしくは当技術分野で既知の他の方法で観察すると顆粒の複屈折性が失われていることによって、証明することができる。特定の実施形態では、ヨード滴定法によって、例えば、MorrisonおよびLaignelet(1983年)の方法のような分光光度的方法でよい、または高速液体クロマトグラフィー(HPLC、例えば、BateyおよびCurtin、1996年)によって、アミロース比率を測定する。
【0071】
別の実施形態では、この小麦穀粒は、例えば、糊化温度の上昇もしくは低下、糊化の進行中もしくは糊化後の膨潤特性の変化、粘度の変化、アミロペクチンの鎖長分布の変化、またはこれらの任意の組合せなどの物理的特質が改変されたデンプンを含む。糊化温度の上昇または低下は、糊化の第1ピーク、第2ピーク、またはその双方に対するものでよい。例えば糊化エンタルピーなどデンプンの1種または複数の特性は未変化でよい。示差走査熱量計で測定した糊化の第1ピーク(最高点)の温度は、野生型穀粒から得た対応するデンプンでの第1ピークの温度と比べて、少なくとも3℃または5℃、好ましくは少なくとも7℃または8℃、より好ましくは少なくとも10℃上昇していてよい。特定の実施形態では、温度上昇は、3℃〜12℃の範囲である。
【0072】
穀粒は、しわ型でも非しわ型でもよく、好ましくは非しわ型のフェノタイプを有する。本明細書では、穀粒の大多数、好ましくは個々の穀粒の少なくとも90%が、丸いフェノタイプまたは十分に充実したフェノタイプを示す場合、「非しわ型」と定義する。これは、通常、正常レベルまたはほぼ正常なレベルのデンプン蓄積に関連付けられている。一方、本明細書では、「しわ型」フェノタイプという用語は、穀粒の大多数、具体的には少なくとも穀粒の90%のデンプン蓄積量が減少している場合を意味する。それぞれ野生型穀粒と比べて、少ししわ型の穀粒とは、平均デンプン含有量が少なくとも30%に低下しているものであり、中程度のしわ型穀粒とは、平均デンプン含有量が少なくとも50%に低下しているものであり、高度のしわ型穀粒とは、平均デンプン含有量が少なくとも70%に低下しているものである。しわの程度は、成熟穀粒の重量に対するパーセンテージとして、相対的デンプン含有量によって測定することもできる。改変されていない屋外栽培の小麦穀粒のデンプン含有量は約65%であるのに対し、しわ型穀粒のデンプン含有量は50%未満に減少している。
【0073】
別の実施形態では、穀粒の平均重量は、少なくとも36mgまたは40mgである。穀粒の平均重量は、数が分かっている穀粒、すなわち代表試料となるひとかたまりの穀粒の重量を測定し、穀粒の数で全重量を割ることによって、決定する。デンプン含有量、平均重量、および非しわ型フェノタイプなど穀粒の特質が野生型レベルに近いものが、穀粒の商業的製造に望ましいことが理解されよう。
【0074】
本発明は、小麦粉、粗挽き粉、練り粉、または穀粒からもしくは穀粒を用いて製造した他の製品も提供する。これらは、例えば、分別または漂白による加工をしなくても加工をしてもよい。本発明は、本発明の小麦植物から得た、食品製造に有用な小麦穀粒をさらに提供する。さらに、本発明は、他の方法で加工された穀粒も含む。したがって、穀粒は、製粉しても、粉砕しても、圧扁しても、精白しても、粗挽きもしくは挽き割りにしても、パーボイルドしても(ポレンタ)よく、例えばクスクス粉でもよい。
【0075】
<デンプン>
別の態様では、本発明は、本明細書で記述する小麦植物の穀粒から得た、アミロース比率が高くアミロペクチン比率が低いデンプンを提供する。好ましい実施形態では、デンプンは、野生型小麦に比べて胚乳のSBEIIaタンパク質、SBEIIa酵素活性、またはその双方のレベルが低下した小麦植物の穀粒から得られる。別の実施形態では、野生型小麦に比べて、SBEIIa活性およびSBEIIb活性の双方が低く、または、SBEIIa、SBEIIb、およびSBEIの3種すべてのレベルが低い。
【0076】
別の態様では、本発明は、本明細書で記述する小麦植物の穀粒から得た、アミロースを少なくとも50%、少なくとも55%、少なくとも60%、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、または少なくとも90%含むデンプンを提供する。このデンプンは、少なくとも部分的に精製されている。すなわち、このデンプンは、穀粒の少なくとも1種の他の成分から分離されている。精製デンプンは、デンプンをタンパク質、油、および繊維から分離することを伴う精白プロセス、例えば湿式粉砕プロセスによって穀粒から得ることができる。精白プロセスの最初の製品は、デンプン粒の混合物または組成物であり、したがって、本発明は、本明細書で記述する加工デンプンを含むこのような顆粒を包含する。
【0077】
デンプンの糊化温度は上昇していても、低下していてもよいが、上昇していることが好ましい。特定の実施形態では、DSCによる測定によれば、第1ピークの立ち上がりの温度または第1ピークの最高点の温度のうちの少なくとも1つが、野生型小麦穀粒から抽出したデンプンと比べて、少なくとも3℃、少なくとも5℃、少なくとも7℃、または少なくとも10℃上昇している。特定の実施形態では、温度上昇の範囲は3℃〜12℃である。特に留意すべきことには、糊化温度は、第1ピークの立ち上がりの温度が低下するとともに、ピーク最高点の温度が上昇していることがある。先の実施形態と相互に排他的ではない別の実施形態では、DSCにより測定すると、デンプンの糊化温度の第1ピークは変わっているが、アミロースと脂質の高熱分解に対応する第2ピークの温度はほぼ変わっていない。別の実施形態では、デンプンの糊化エンタルピーは減少しており、例えば、対応する野生型小麦デンプンのエンタルピーと比べて、少なくとも25%または少なくとも40%減少している。
【0078】
別の実施形態では、このデンプンは、特異的な物理的特質によって示される改変された構造を有するレジスタントスターチを高レベルで含む。このような特質には、消化酵素に物理的に接近しにくい特質が含まれることがあり、これは、デンプン粒の形態が変わっていること、相当量のデンプン結合脂質が存在すること、結晶化度の変化、アミロペクチン鎖長分布の変化、またはこれらの任意の組合せが原因となっている可能性がある。高アミロース比率も、レジスタントスターチのレベルに寄与している。
【0079】
本発明は、好ましくは高レベルのレジスタントスターチとともに、増量した食物繊維を含む、例示的な小麦植物の穀粒から得たデンプンも提供する。
【0080】
本発明の範囲が、本明細書で記述する小麦デンプンを製造する方法に及ぶことは明らかである。一実施形態では、本方法は、本明細書で記述する小麦穀粒を得るステップと、その穀粒からデンプンを抽出するステップを含む。この小麦穀粒は、本明細書で記述する小麦植物を栽培し、穀粒を収穫することによって得ても、穀粒の生産者または穀粒の輸入業者から入手してもよい。
【0081】
<遺伝子活性の低減方法>
SBEIIa、SBEIIbその他のデンプン生合成または修飾遺伝子の発現および/または活性は、1つまたは複数の遺伝的変異を小麦植物に導入することにより変わることがある。本明細書において用いる場合、「遺伝的変異」とは、この文脈において、対象とする遺伝子の発現または活性に影響を及ぼす、小麦植物ゲノムにおける遺伝性の任意の変化を意味する。遺伝的変異としては、点変異、挿入、置換、逆位、重複、転座および好ましくは欠失などの突然変異ならびに1つまたは複数の導入遺伝子のゲノムへの導入を含む。
【0082】
本明細書において用いる場合、「核酸分子」および「核酸配列」という語句は、ヌクレオチドのポリマーを指し、これは一本鎖でもよい二本鎖でもよい。それは、例えば、ゲノムDNAまたはcDNAなどのDNA、RNA、mRNAまたはこれらの任意の組合せを含み得る。小麦細胞への導入のために、送達性または安定性を改善するべく核酸分子を化学的に修飾してもよいし、ウイルスベクターなどのベクターの一部として保護してもよい。核酸分子はクローン技術によって得てもよいし、周知の技術である技術によって合成してもよい。核酸分子は、コード鎖もしくは非コード鎖(アンチセンス)または、例えば、逆位反復コンストラクトにおけるような、これらの組合せを含み得る。遺伝子に「対応する」核酸配列と言う場合、「対応する」という用語は、ヌクレオチド配列が、参照遺伝子またはその指示された部分と同じヌクレオチド配列を有するか、通常のワトソン−クリック型塩基対において正確に相補的であるヌクレオチド配列を有するか、そのような配列のRNA(例えばmRNA)等価物であるか、遺伝子のmRNAに由来するcDNAであるような、ヌクレオチド配列関係を指す。
【0083】
ヌクレオチド配列は、5'から3'方向へ、標準的な1文字ヌクレオチド省略形を使用して一本鎖配列によって表す。「相補性」は、塩基対合によってアニールされる2本の一本鎖核酸分子または配列の関係を記述するものである。例えば、5'−GACT−3'は、その相補鎖5'−AGTC−3に対合する。「相同性」または「相同的」は、文脈によって2つ以上のヌクレオチド配列または2つ以上のポリペプチド配列の間の配列の類似性または同一性を指す。「%同一性」という用語は、例えば、CLUSTAL VアルゴリズムまたはBlastnもしくはBLAST2配列プログラム(これらは米国国立バイオテクノロジー情報センター(National Center for Biotechnology Information)から入手可能で、インターネット上ではhttp://www.ncbi.nlm.nih.gov/BLAST/で利用可能であり、好ましくはデフォルトのパラメータ設定とする)などの標準的なものとなっているアルゴリズムを使用してアラインメントを行った2つのヌクレオチド配列間のヌクレオチドの合致の%を指す。同様にして、「%同一性」はポリペプチド配列を指すこともある。
【0084】
本明細書において、SBEIIa、SBEIIbその他のデンプン生合成支配遺伝子またはアンチセンス、コサプレッション、リボザイム、二重鎖RNA分子その他をコードする遺伝子などを含め「遺伝子」という場合、その最も幅広い文脈で解するものとし、プロモーターと転写ターミネーター−ポリアデニル化配列などの制御領域を伴う転写領域を有する古典的なゲノム遺伝子を含む。転写領域は転写されたが、翻訳されなかった配列(非翻訳配列、UTR)を含み、状況に応じて、タンパク質コード領域もしくはイントロン(これは成熟したRNAを形成するために除かれる)またはこれらの任意の組合せを含んでもよい。「遺伝子」は、エキソンに対応するcDNAから得られる形態およびRNAゲノムで見られるようなRNA遺伝子を含む。「遺伝子」という用語は、機能的な産物の全部または一部をコードする合成または融合分子を記述するためにも用いられる。
【0085】
細胞、好ましくは、小麦細胞中に存在する場合、「遺伝子」は、「生物活性」分子または「遺伝子産物」(これはRNAでもポリペプチドでもよい。)の「発現」を指示する。この過程は、最も一般的にはRNAを生産するための転写とタンパク質を生産するための翻訳による。これらの産物は、その後細胞内で修飾されてもよい。RNAは、例えば、ポリアデニル化、スプライシング、キャッピング、21〜23のヌクレオチド断片へのダイシング、核からの輸送またはタンパク質に対する共有結合もしくは非共有結合的な相互作用によって修飾されてもよい。例えば、タンパク質はリン酸化、グリコシル化または脂質化によって修飾されてもよい。本明細書において用いる場合、これらの過程はすべて「遺伝子の発現」その他の用語に包含される。
【0086】
本明細書において用いる場合、「小麦SBEIIa遺伝子」および「小麦SBEIIb遺伝子」という用語ならびに関連語は、それぞれ、SBEIIaまたはSBEIIb酵素をコードする小麦から同定された遺伝子、および他の小麦の種類に存在する相同遺伝子を指す。これらは表1に列挙する遺伝子配列を含むが、これらに限定されるものではない。SBEIIaとSBEIIb遺伝子の配列には、異なる小麦の種類からの自然変異があると思われる。相同遺伝子は、当業者であれば容易に認識できる。SBEIIb遺伝子またはタンパク質についてと同様に、相同的なSBEIIa遺伝子またはタンパク質の間の配列同一性の程度は、少なくとも90%であると考えられる。
【0087】
本発明で用いる遺伝子は、天然に存在するSBEIIa、SBEIIbその他に由来してもよいし、標準的な組み換え技術によってその他のデンプン生合成支配遺伝子に由来するものでもよい。本明細書において用いる場合、「組み換え核酸分子」その他の用語は、天然に存在しない配列であるか、本来別個の2以上の配列セグメントの人工的組合せによって作られる配列を有する配列を指す。この人工的組合せは、化学合成によって、または、より一般的に、例えば、周知の技術である遺伝子工学技術によって、単離された核酸セグメントを人工的に操作することによって作られるものでもよい。「組み換え」という用語は、単に核酸の一部の追加、置換または欠失のみによって変更された核酸を含む。多くの場合、組み換え核酸は、プロモーター配列に機能的に連結された核酸配列を含んでもよい。そのような組み換え核酸は、例えば、細胞を形質転換するのに用いられるベクターの一部でもよい。
【0088】
一般に、遺伝子は1つまたは複数のヌクレオチド置換、欠失および/または添加(例えばコドン修飾)を生じる突然変異生成に付してもよい。そのような遺伝子のヌクレオチド挿入型の誘導体は、5'および3'端融合ならびに1つまたは複数のヌクレオチドの内部配列挿入を含む。挿入型のヌクレオチド配列変異形では、1つまたは複数のヌクレオチドがヌクレオチド配列で予め定められたサイトに導入されるが、ランダムな挿入を行い結果として生じる産物を適宜スクリーニングすることも可能である。欠失型変異形は、配列から1つまたは複数のヌクレオチドの除去によって特徴付けられる。置換型のヌクレオチド変異形は、配列の少なくとも1つのヌクレオチドが取り除かれ、異なるヌクレオチドがその位置に挿入されたものである。置換がコドンによって定義されるアミノ酸を変えないという点で、そのような置換は「サイレント」でもよい。あるいは、保存的な置換基は、1つのアミノ酸を別の同様に働くアミノ酸に変えるように設計される。典型的な置換は、以下に従ってなされるものである:
【0089】
保存的なアミノ酸置換に適する残基
元の残基 置換例
Ala Ser
Arg Lys
Asn Gln;His
Asp Glu
Cys Ser
Gln Asn
Glu Asp
Gly Ala
His Asn;Gln
Ile Leu;Val
Leu Ile;Val
Lys Arg;Gln;Glu
Met Leu;Ile
Phe Met;Leu;Tyr
Ser Thr
Thr Ser
Trp Tyr
Tyr Trp;Phe
Val Ile;Leu
【0090】
<導入遺伝子>
SBElla、SBEIIbまたは他デンプン生合成または修飾遺伝子の発現および/または活性は、1つまたは複数の導入遺伝子を小麦植物にもたらすことによって変更され得る。本明細書で言う「導入遺伝子」は、バイオテクノロジーの技術分野における通常の意味を有し、組換えDNAまたはRNA技術によって製造されたか変更され、対象とする有機体または細胞(好ましくは小麦細胞)に導入された遺伝子配列を含む。導入遺伝子は、有機体または細胞に由来する遺伝子配列(例えば、アンチセンス配列)を含んでもよい。導入遺伝子は、典型的には、前記有機体または細胞に由来しない外来性の核酸を含む。「トランスジェニック」とは、導入遺伝子を含む有機体または細胞を指す。「非トランスジェニック」とは、ゲノム中にいかなる導入遺伝子も存在しないことを指す。安定した継承のために、好ましくは導入遺伝子を有機体または細胞のゲノムに組み込む。
【0091】
当業者は、遺伝子またはそれに対する相補配列の発現のためには、前記遺伝子が、細胞内でプロモーター配列との機能的な関係に置かれる必要があると承知している。本明細書の目的のためのプロモーターの選択は、必要な発現レベルおよび/または発現させるべき組織、器官および種によって、特に胚乳特異的プロモーターによって変わり得る。
【0092】
プロモーター配列の制御調節の下に核酸分子を置くことは、発現がプロモーター配列によって制御されるように前記分子を位置付けることを意味する。プロモーターは、通常(但し、必須ではない)、それが調節する核酸分子の上流に、または5'末端に位置する。さらにまた、プロモーターを含む調節要素は、通常、遺伝子の転写開始部位の2kb以内に位置する。異種プロモーター/構造遺伝子組合せの構築では、一般に、その自然の条件下でプロモーターとそれが制御する遺伝子(すなわち、それからプロモーターが誘導される遺伝子)との間の距離とほぼ同じ距離に、遺伝子転写開始部位からプロモーターを隔てて配置するのが好ましい。当技術分野において知られているように、この距離が若干変化してもプロモーター機能を損なうことなく対応できる。同様に、その制御下に置かれる異種遺伝子に関しての調節配列要素の好ましい位置は、その自然の条件下での要素(すなわち、それから誘導される遺伝子)の位置によって定義される。また、当技術分野で知られているように、この距離においても若干の変化は生じ得る。
【0093】
本発明の遺伝子コンストラクトにおける使用に適したプロモーターの例としては、ウイルス、イースト、カビ、バクテリア、昆虫、鳥、哺乳類および植物の遺伝子に由来するプロモーターが挙げられ、好ましくは植物細胞で機能し得るもの、より好ましくは小麦の胚乳で発現し得るものが挙げられる。プロモーターは構成的に、または発現が起こる組織によって異なるように発現を調節するものでよい。あるいは、発現は発現が起こる発育時期に関して、または、外刺激(例えば生理的ストレスまたは温度)に応じて異なってもよい。
【0094】
SBEIIaその他のデンプン生合成支配遺伝子活性を低減する方法は、導入遺伝子を小麦の再生可能な細胞に導入して、形質転換細胞からトランスジェニック小麦植物を再生させるステップを含み得る。アミロペクチンの合成に関係する分枝酵素はSBEI、SBEIIaとSBEIIbを含み、本発明はSBEIIaのみの、または、SBEIIbまたはSBEI発現の変更と組み合わせた発現低減を包含する。したがって、導入遺伝子(複数可)はこれらの遺伝子のうちの2つ以上を不活性化してもよい。さらに、導入遺伝子(例えば、二重鎖RNA、アンチセンスまたはリボザイムRNAをコードするもの。下記参照)が直接SBEllbまたはSBEI遺伝子形質発現を標的とするという点で、SBEIIbやSBEIの失活は直接的でもよいし、あるいは、それはSBEIIbまたはSBEIの発現に間接的に変更をもたらすものでもよい。例えば、導入遺伝子RNAは配列同一性または塩基の対合に関してSBEIIa遺伝子/RNAのみを標的としてもよいが、胚乳におけるタンパク質の安定性または分布を変えることによって、SBEIIbまたはSBEI活性の低減をもたらすものでもよい。さらに、本発明の形態は、SBEIIaの変更された活性と1つまたは複数の他のアミロペクチン合成酵素(これらの酵素はSSI、SSII、SSIIIおよびイソアミラーゼまたはプルラナーゼなどの脱分枝酵素を含んでもよい。)の変更の組合せにある。これらのいずれかまたはすべての発現は、導入遺伝子の導入によって変更され得る。
【0095】
小麦のアミロペクチン合成遺伝子についてはいくつかのDNAの塩基配列が知られており、それらのいずれでも小麦における遺伝子失活のための導入遺伝子を設計するためのベースとすることができる。これらのものとしてはSBEIIa(GenBankアクセッション番号Y11282、AF338431およびAF338432)およびSBEIIb(WO00/15810、WO01/62934)が挙げられる。小麦のSBEI遺伝子は、Rahman他(1997年)およびRahman他(1999年)に記載されている。SBEIのためTriticum tauschii配列(これは小麦DゲノムSBEI遺伝子との相同性が高い。)は、WO99/14314に見ることができる。小麦のSBEIのためのcDNA配列は、GenBankデータベースにおいてアクセッション番号AF076679でアクセスすることができる。大麦その他の密接に関連する種から得られる他のアミロペクチン合成遺伝子のホモログは、小麦において遺伝子形質発現レベルを修飾するのに用いることもできる。そのような遺伝子またはその断片は、PCR増幅または標識化プローブへのハイブリダイゼーションを含む当技術分野では周知である方法によって得ることができる。
【0096】
本明細書において用いる場合、「ストリンジェントなハイブリダイゼーション条件」は、少なくとも90%、好ましくは少なくとも95%の配列同一性がプローブと標的配列の間にあるならば、ハイブリダイゼーションが通常、起こることを意味する。ストリンジェントなハイブリダイゼーション条件の例は、50%ホルムアミド、5×SSC(1×SSC=150mM NaCl(15mMクエン酸トリナトリウム))、50mMリン酸ナトリウム(pH7.6)、5×デンハルト溶液、10%硫酸デキストランおよびサケ精子DNAなどの変性剪断キャリアDNA20μg/mlからなる溶液中で一夜インキュベーションした後、ハイブリダイゼーションの支持体をおよそ65℃で0.1×SSCで洗浄するというものである。他のハイブリダイゼーションおよび洗浄条件はよく知られており、例えば、Sambrook他、Molecular Cloning:A Laboratory Manual、第2版、Cold Spring Harbor、NY(1989年)、特に第11章に例示されている。
【0097】
導入遺伝子コンストラクトの調製に用いるホモログの領域は、対応する小麦遺伝子と少なくとも85%、好ましくは少なくとも90%、さらに好ましくは95〜100%の同一性を適当な領域内に有すべきである。また、導入遺伝子は、小麦胚乳内で発現されるアミロペクチン合成遺伝子を特異的に標的とし、植物中、他の場所でのアミロペクチン合成にはより小さなまたは最小限の影響しか与えないことが好ましい。これは、導入遺伝子中の胚乳特異的プロモーターなどの適当な調節配列の使用によって達成してもよい。
【0098】
<アンチセンス>
小麦などの植物において遺伝子活性に変更を加える、特に低減する遺伝子工学アプローチは、当技術分野において周知である。これらの方法は、標的遺伝子のRNAと相補的でハイブリダイズすることができる適当なアンチセンス分子の発現のための、遺伝子コンストラクトの導入を含む。アンチセンス分子は標的遺伝子のmRNAの翻訳または処理または安定性に干渉し、それによって遺伝子の発現を不活性化すると考えられる。アンチセンス配列を考案する方法は周知の技術であり、これらの例は米国特許第5190131号、欧州特許明細書第0467349−A1号、欧州特許明細書第0223399−A1号および欧州特許明細書第0240208号に見ることができ、これらは参照によって本明細書に組み込まれる。植物におけるアンチセンス法の使用は、Bourque(1995年)とSenior(1998年)によって概説されている。Bourqueは、植物システムにおいてアンチセンス配列を使った遺伝子失活の多数の例を列挙している。彼女も、部分的な阻害がシステムにおいてかなりの変化を確かにもたらすであろうから、酵素活性の100%の抑制を達成することは必要ではないと述べている。Senior(1998年)は、アンチセンス法は、現在、植物で遺伝子形質発現を操作することの非常に安定した技術であると述べている。
【0099】
小麦SBElla、SBEIIb、SBEIその他のデンプン生合成または修飾遺伝子についてのアンチセンス分子は、小麦mRNA配列に基づくことができるか、他の種(例えば、大麦)から得られる相同なDNAまたはmRNA配列に由来し得る。アンチセンス配列は、これらの遺伝子のいずれかの転写物の全部または一部に、または、それらの発現(例えば、それらのスプライシング)の制御を及ぼす配列について対応すればよい。アンチセンス配列は、小麦SBEIIaその他の遺伝子の標的とされたコード領域または5'−非翻訳領域(UTR)もしくは3'−UTRまたはこれらの組合せに対応してもよい。それは、ある程度、イントロン配列(それは転写の間または後に除かれ得る)と相補的でもよいが好ましくは標的遺伝子のエキソン配列のみに相補的である。UTRは一般に違いがより大きいことからすれば、これらの領域を標的とすることは、遺伝子抑制により大きな特異性をもたらす。特定の実施形態において、アンチセンス配列の長さは、遺伝子RNA配列の相補鎖と対応する少なくとも19の隣接するヌクレオチド、少なくとも50、少なくとも100、少なくとも200、少なくとも500または少なくとも1000のヌクレオチドである。すべての遺伝子転写物と相補的な全長配列を用いてもよい。特定の実施形態において、アンチセンス配列の長さは、100〜2000ヌクレオチドである。さらなる実施形態において、標的とされた転写物に相補的なアンチセンス配列の配列同一性の程度は、少なくとも85%、少なくとも90%または95〜100%である。アンチセンスRNA分子は、もちろん、分子を安定させるために機能してもよい無関係な配列を含んでもよい。
【0100】
<コサプレッション>
使用し得る別の分子生物学的製剤アプローチは、コサプレッションである。コサプレッションのメカニズムは、十分理解されていないが、転写後遺伝子サイレンシング(PTGS)を含むと考えられており、この点ではアンチセンス抑制の多くの例と、非常に類似している。それは、その発現のためには、プロモーターに関してセンス方向に植物内に遺伝子の余分なコピーまたはその断片を導入することが必要である。センス断片のサイズ、標的遺伝子領域との対応および標的遺伝子への配列同一性の程度は、上述のアンチセンス配列に関してと同様である。若干の例では、遺伝子配列のさらなるコピーは、標的植物遺伝子の発現の妨害をする。コサプレッションアプローチを実行する方法については、WO97/20936および欧州特許明細書第0465572号を参照されたい。
【0101】
<二重鎖RNA媒介遺伝子サイレンシング>
遺伝的変異を小麦植物に導入するために使用され得るさらなる方法は、二重鎖あるいは二本鎖RNAによって媒介される遺伝子サイレンシングである。この方法も、PTGSを含む。この方法では、失活させるべき標的遺伝子に相同で少なくとも部分的に二本鎖であるRNA産物(複数可)の合成を指示するDNAを導入する。したがって、DNAは、RNAに転写されるときハイブリダイズして二本鎖RNA領域を形成することができるセンスとアンチセンス配列からなる。好ましい実施形態において、センス配列とアンチセンス配列は、RNAに転写されるとき、スプライシングで除かれるイントロンからなるスペーサー領域によって分離されている。この構成は、遺伝子サイレンシングをより高効率にすることが示された。二本鎖領域は、1または2のDNA領域から転写される1または2のRNA分子を含んでもよい。二本鎖分子の存在は、二本鎖RNAおよびさらには標的植物遺伝子からの相同なRNA転写物も破壊する内在性植物システムからの応答を誘発し、標的遺伝子の活性を能率的に低減するか排除する。この技術を実施する方法については、オーストラリア特許明細書第99/29514−A号およびWO99/53050を参照されたい。特定の実施形態において、ハイブリダイズするセンス配列およびアンチセンス配列の長さは、少なくとも19の隣接するヌクレオチド、少なくとも30、少なくとも50、少なくとも100、少なくとも200、少なくとも500または少なくとも1000のヌクレオチドである。すべての遺伝子転写物と対応する全長配列を用いてもよい。特定の実施形態においては、長さは範囲100〜2000ヌクレオチドである。さらなる実施形態において、センス配列およびアンチセンス配列の標的とされた転写物への配列同一性の程度は、少なくとも85%、少なくとも90%または95〜100%である。RNA分子は、もちろん、分子を安定させるために機能してもよい無関係な配列を含んでもよい。RNA分子は、RNAポリメラーゼIIまたはRNAポリメラーゼIIIプロモーターの制御下に発現されるものでもよい。後者の例は、tRNAまたはsnRNAプロモーターを含む。二本鎖RNA分子は、互いに連結された複数の遺伝子からの配列を含んでもよく、それによって複数の遺伝子を標的としてもよい。
【0102】
<リボザイム>
小麦の遺伝子形質発現の望ましい失活をもたらす遺伝的変異は、1つまたは複数のリボザイムをコードする核酸分子を含む。リボザイムは、1またはしばしば2つのハイブリダイズしている配列によって定義される特定のサイトで他のRNA分子を切断することができる、酵素または触媒機能を有するRNA分子である。RNAの切断により、標的遺伝子の発現を不活性化する。リボザイムはアンチセンス分子の働きをしてもよく、それが遺伝子失活に寄与してもよい。リボザイムは、ハイブリダイズしている配列の間で1つまたは複数の触媒領域(好ましくはハンマーヘッドまたはヘアピンタイプ)を含む。RNAseP、Group IまたはIIイントロンおよびデルタ型肝炎ウイルスタイプを含め、他のリボザイムのモチーフを用いてもよい。欧州特許明細書第0321201号および米国特許第6,221,661号が参照される。例えばWegener他(1994年)によって、遺伝子導入植物の遺伝子を不活性化するリボザイムの使用が示されている。
【0103】
<遺伝子コンストラクト/ベクター>
本発明は、遺伝子阻害分子をコードするRNAまたはDNA、好ましくはDNAを含む単離された核酸分子も提供する。ある実施形態において、核酸分子は、小麦SBEIIa遺伝子配列を標的とし、小麦穀粒の胚乳のその発現を失活させるアンチセンス、センス(コサプレッション)、二本鎖RNAまたはリボザイム分子をコードする。本発明はまた、プロモーター、エンハンサーおよび転写末端またはポリアデニル化配列などの1つまたは複数の調整要素を含む単離された核酸分子を含むかコードする遺伝子のコンストラクトを提供する。そのような要素は当技術分野において周知である。植物、特に小麦などの単子葉植物において、遺伝子のコンストラクトは、導入遺伝子の発現を助けるイントロン配列を含んでもよい。「イントロン」という用語は、その通常の意味では、転写されるが、タンパク質をコードせず、翻訳前にRNAからスプライシングにより除かれる遺伝子の部分を意味するように使われる。導入遺伝子が翻訳産物をコードするか、そうでなく転写領域のどこかをコードするならば、イントロンは5'−UTRまたはコード領域に組み入れられてもよい。
【0104】
本発明はさらに遺伝子のコンストラクトを含むベクター(例えば、プラスミドベクター)を提供する。「ベクター」という用語は、in vitroまたはin vivo発現が可能な発現ベクターおよび1つの細胞または有機体から別のもう1つまで転移され得る形質転換ベクターを含む。例えば、大腸菌またはアグロバクテリウムなどの原核細胞において、ベクターは細胞内での複製をもたらす配列を含む。特定の実施形態において、ベクターは少なくとも1つのT−DNA境界配列によって定まるT−DNA配列(これは小麦細胞に導入することができる。)を含むバイナリーベクターである。本発明は、さらに、ベクターを含む細胞、例えば、未熟な胚の胚盤の細胞などの再生可能な細胞でもよいアグロバクテリウムまたは小麦細胞を提供する。あるいは、細胞は導入遺伝子を含む形質転換された小麦細胞でもよい。
【0105】
<プロモーター/ターミネーター>
別の実施形態において、本発明の導入遺伝子または他の遺伝子コンストラクトは、小麦の胚乳内で調節されたか構成的な発現をもたらし得る転写開始領域(プロモーター)を含む。プロモーターは組織特異的で、選択的に、または、もっぱら胚乳の発現を与えるものでもよい。プロモーターは、(高分子グルテニンプロモーター、小麦SSIプロモーター、小麦SBEIIプロモーター、小麦GBSSプロモーターなどの)胚乳特異的または(ユビキチンプロモーターまたはCaMV35Sまたは強化された35Sプロモーターなどの)胚乳に非特異的なプロモーターのいずれから選択してもよい。プロモーターは、温度、光またはストレスなどの要因によって調整されてもよい。通常、プロモーターは、発現されるべき遺伝子配列の5'をもたらされる。コンストラクトは、nos3'またはocs3'ポリアデニル化領域または転写ターミネーターなどの転写を強化する他の要素も含んでもよい。例示したDNA領域は、適当な選択可能なマーカー遺伝子配列と他の要素を含むベクターに、または、ベクターがこれらの配列を含んで同時形質転換され得るベクターに取り込まれる。
【0106】
<小麦の形質転換法>
小麦などの単子葉植物の形質転換法、すなわち、外来性の核酸の導入によって遺伝的変異を植物に導入し、プロトプラストまたは未熟な植物胚から植物を再生する方法は、当技術分野において知られており、例えば、Becker他、1994年、Cheng他、1997年、He他、1994年、Hess他、1990年、Nehra他、1994年、Vasil他、1992年、Vasil他、1993年、Weeks他、1993年、Weir他、2001年、オーストラリア特許出願番号第75460/94号、欧州特許出願番号第709462号、WO93/04178、WO89/12012、WO94/13822およびWO99/14314を参照されたい。望ましいヌクレオチド配列または遺伝子のコンストラクトおよび選択可能なマーカーを運ぶベクターは、組織栽培された植物または外植体の再生可能な小麦細胞またはプロトプラストなどの適当な植物システムに導入すればよい。選択可能なマーカー遺伝子は、小麦細胞に抗生物質または除草剤耐性を提供してもよいし、マンノースなどの基質の利用性を可能にするものでもよい。選択可能なマーカーは、好ましくはアスラム、ジェネテシン(geneticin)またはハイグロマイシン耐性を小麦細胞に与える。再生可能な小麦細胞は、好ましくは、未熟な胚の胚盤、成熟した胚、これらに由来するカルスまたは分裂組織に由来する。
【0107】
形質転換植物は選択可能なマーカー遺伝子を含んでもよいし、あるいは、そのような遺伝子は再生の間か後に、例えば、ゲノムからの選択可能なマーカー遺伝子の切除により、または、SBEIIa阻害導入遺伝子から離れた選択可能なマーカー遺伝子の分離によって除去してもよい。
【0108】
導入遺伝子または突然変異が染色体に融和した植物は、例えば、導入遺伝子またはフェノタイプ観察に特有の適当な核酸プローブを使うことによりスクリーニングすることができる。いくつかの方法のいずれも、形質転換植物の存在を決定するために使用され得る。例えば、ポリメラーゼチェーンリアクション法(PCR)は形質転換植物に特有な配列を増幅するのに用い得る(ゲル電気泳動その他の方法による増幅産物の検出を行う)。DNAは従来法を使用して植物から抽出してもよく、PCR反応は、形質転換植物と非形質転換植物を識別するプライマーを使って実行してもよい。例えば、コンストラクトに読み込まれる形質転換ベクターからDNA領域を増幅するプライマーを設計してもよいし、逆方向プライマーは対象とすべき遺伝子から設計してもよい。植物がうまく形質転換されたならば、これらのプライマーは断片を増幅するのみである。陽性の形質転換体を確かめる別の方法は、当技術分野において周知であるサザンブロットハイブリダイゼーションによる。形質転換植物または変異体は、非形質転換植物または野生型植物からそのフェノタイプ(例えば、選択可能なマーカー遺伝子の存在によって、または免疫学的方法による特定のタンパク質の存在によって、または、タンパク質の欠如(例えば、ELISA分析アッセイまたはウェスタンブロット分析によって検出されるものとしての胚乳のSBEIIaタンパク質の欠如)によって与えられる)によって識別すなわち区別される。そのような植物をスクリーニングする際に使われる徴候は、穀粒のフェノタイプの特徴の観察(例えば、縮んだ穀物の視覚検査または測定によって)、増加したアミロース含有量の試験または顕微鏡で複屈折の存在について調べることによることもできる。
【0109】
<変異>
小麦胚乳においてSBEIIa酵素その他のデンプン生合成酵素の活性の低減をもたらす遺伝的変異の導入は、それぞれの遺伝子内における適当な突然変異にもよって達成されてもよいし、遺伝子の調節配列内における適当な突然変異にもよって達成されてもよい。本明細書の文脈では、「誘発突然変異」は化学産物、放射線または生物ベースの突然変異生成(例えば、トランスポゾンまたはT−DNA挿入)の結果でもよい人工的に誘発された遺伝的変異である。遺伝子が阻害される程度は、ある程度、生産されるデンプンの特徴を決定する。突然変異はトランケーションまたはヌル変異でもよく、これらがデンプンの性質に重要な影響を及ぼすことは知られている。しかし、変更されたデンプン構造は、デンプンまたは小麦穀粒において対象とする特徴をもたらすのに十分にアミロペクチン合成酵素活性を低減する漏出突然変異からも生じるであろう。他の染色体再編成も有効であり得、これらは挿入、欠失、逆位、重複または点変異を含んでもよい。本明細書で用いられる「ヌル変異」は、対象とすべき遺伝子の活性の完全かまたは完全に近い損失をもたらす突然変異を指し、例えば、遺伝子活性がもはや見出されないものを指す。
【0110】
SBEIIa遺伝子は、染色体2の長腕に位置する。この遺伝子その他の遺伝子への突然変異(特に欠失突然変異)は、対象とすべき遺伝子(例えば、SBEIIa遺伝子または二重突然変異体の場合には、おそらく結合されたSBEIIb遺伝子にまで拡大される。)に集中させることが好ましい。この文脈における遺伝子は、転写領域とともにプロモーター領域と転写終了/ポリアデニル化信号を含む。転写領域は、タンパク質コード領域(複数可)ならびにmRNAの5'非翻訳および3'非翻訳領域、さらに存在し得る任意のイントロン領域を含む。遺伝子への突然変異は、遺伝子の任意の領域におけるまたは領域の組合せでもよく、1つのヌクレオチドのみの変更(例えば、コード領域のフレームシフト突然変異)からすべての遺伝子の欠失に及び得る。遺伝的変異のためにホモ接合性である植物が好ましい。
【0111】
欠失は、100または2、300程度、おそらくは500キロベース単位のサイズに制限してもよい。特定の実施形態において、欠失は少なくとも2、3000キロベースまたは5000キロベース未満まで拡がる。本発明は、それぞれのゲノムの染色体2の長腕の多くを含むより大きな欠失を含んでもよいが、染色体2の長腕は多数の他の遺伝子をその上に集中させて有するので、小麦植物の生命力に対し影響を与えるため、これは好ましくない。したがって、大きな欠損が起こる場合は、これらは植物生命力に対し悪影響を与え、それゆえに、その商業的な生存能力に対し悪影響を与えるため、少なくとも染色体2の長腕の大多数が存在することが望ましい。好ましい実施形態において、染色体2Aの長腕の大部分は存在する。
【0112】
突然変異生成は、化学または放射線手段、例えば、種子のEMSまたはアジ化ナトリウム(ZwarおよびChandler、1995年)処理またはガンマ照射によって達成できる。変異体の単離は、突然変異を起こす植物または種をスクリーニングすることによって達成してもよい。例えば、小麦の突然変異集団は、穀粒のハイアミロース含有量および/または通常より長いアミロペクチン鎖分布またはELISAによるSBEIIaタンパク質の損失、または、変更された穀粒形状(Green他、1997年)についてスクリーニングされ得る。スクリーニングは、好ましくは、既にSBE活性のうちの1つが欠如している小麦遺伝子型、例えば、SBEIIb陰性のバックグラウンドでなされる。そのような突然変異は、次いで、望ましい遺伝的バックグラウンドを有する植物との間で交配し、当初の望ましくない親のバックグラウンドを消し去るために適当な数の戻し交配を実行することによって、望ましい遺伝的バックグラウンドに導入し得る。
【0113】
別の実施形態において、突然変異は小麦においてSBEIIaとSBEIIb遺伝子の発現または活性に影響を及ぼす。そのような突然変異の確認は、トウモロコシまたはコメと対照的に2つの遺伝子が小麦では密接に結びついているという予想外の知見によって容易になる。1つの遺伝子の欠失は他の遺伝子に容易に及ぶことがあり、無効な対立遺伝子(ヌル変異)を両方の遺伝子にもたらす。この知見は、小麦の少なくとも1つのゲノムの両方の遺伝子の変異体である自然な変異体のスクリーニングを容易にし、2つまたは3つのゲノムにおいて両方の遺伝子に複合的な突然変異のある小麦を生産するためのスクリーニングを可能にする。そのような小麦は、小麦穀粒の高アミロースな非トランスジェニック源と産物を提供する。
【0114】
SBEIIaまたはアミロペクチン合成に関与している他の酵素をコードする遺伝子の突然変異は、通常、アミロース含有量の比率の増大を引き起こす。個々の穀粒当たりのアミロースの量はアミロペクチンからアミロースまで炭素の流れが変更される結果として増大し得るが、あるいは、穀粒当たりのデンプン生産高の顕著な減少があるならば、それは減少することもある。いずれにせよ、デンプン中パーセンテージとしてのアミロースの相対的レベルは増加する。
【0115】
変形したデンプン粒を有する種がハイアミロース大麦(Morell他、2003年)および、デンプン中におよそ90%のアミロースを有する低アミロペクチン(LAPS)トウモロコシ(Sidebottom他、1998年)で報告されている。
【0116】
複屈折は、2つの方向に光を屈折させる物質の能力である;これは、偏光顕微鏡で見たとき、「マルタ十字」と呼ばれる暗い十字架を各々のデンプン粒上に生み出す。複屈折は、顆粒内のポリマーの構造組織の秩序程度の指標である(ThomasおよびAtwell、1999年)。デンプン粒の複屈折の損失は、通常、アミロース含有量の増加とよく相関している。
【0117】
<食糧生産への適性>
別の面で、本発明は食糧生産に役立つ小麦(穀粒が相対的に高いアミロース含有量を有し低減されたアミロペクチン含有量を有するデンプンを含有する。)を提供する。好ましくは、穀粒が得られる小麦植物は、成長の間、胚乳で低減されたSBEIIa活性レベルを有する。本発明の小麦植物は、食糧生産のために、特に、商業的食糧生産のために役立つ。そのような食糧生産は、商業的食糧生産の成分となし得る小麦粉、生地その他の産物の製造を含んでもよい。
【0118】
小麦の望ましい遺伝的バックグラウンドは、農業生産高および他の特徴の考慮を含む。そうした特徴には冬小麦種または春小麦種、農業的な成績、耐病性と非生物的なストレス耐性を有するのが望ましいか否かを含んでもよい。オーストラリアでは、バクスター、ケネディ、ジャンズ(Janz)、フレーム、ロゼッルラ(Rosella)、カドゥー(Cadoux)、ダイアモンドバード(Diamondbird)その他一般に栽培されている種類などの小麦栽培品種に変更されたデンプンの特徴を交配してみたいであろう。提供される例はオーストラリアの生産領域に特有であるが、他の変種は他の栽培地域に適している。本発明の小麦変種は、少なくともいくつかの栽培地域における、対応する野生型変種の80%以上の生産量を与えることが好ましく、90%以上がより好ましく、95%以上がさらにより好ましい。生産量は、制御された野外試験ですぐに計測できる。
【0119】
さらなる実施形態において、穀粒のデンプン含有量は、少なくともおよそ25%、35%、45%または55%〜65%(w/w)である。商業的に栽培される野生型小麦は、通常55〜65%の範囲のデンプン含有量を有し、ある程度は成長させる栽培品種に依存する。あるいは、本発明の穀粒は、等価であるが改変されていない小麦由来の穀粒の少なくとも90%のデンプン含有量を有する。野生型より低いデンプン含有量は、おそらくはアミロペクチンレベルが低減された結果である。デンプン含有量がたとえ低くても、ハイアミロース産物に比較的高い価値があるため、穀粒は商業食糧生産に依然有用となり得る。他の望ましい特徴は、穀粒を挽く能力、特に穀粒硬度を含む。より高価値の小麦植物を作る別の面は、穀粒からのデンプン抽出の程度であり、抽出率が高いほどより有用である。穀粒の形状は植物の商業的な有用性に影響を与え得るもう1つの特徴でもあり、このように、穀粒の形状は穀粒を挽く上での便利さまたはその反対の影響を及ぼし得る。例えば、細長い穀粒形態では、それを挽いて、処理するのが難しくなる可能性がある。
【0120】
より実の詰まった穀粒はより大きな生産量を達成する観点から望ましく、高レベルのアミロースを有するデンプンの生産または、変更された鎖長分布を有する改変されたデンプン中でのデンプン生産などのような本発明の特定の利点は成し遂げられ得る。このように、穀粒は好ましくは非縮小フェノタイプを有する。しかし、本発明の他の面は、より充填度の低い穀粒によって、より良好に達成されることもあり得る。このように、アリューロン層または胚またはタンパク質のデンプンへの割合はより充填度の低い穀粒でより高いこともあり、それによって、アリューロン層またはタンパク質といった有益な構成要素がより高い小麦粉その他の産物がもたらされる。高いアリューロン層産物はこのように葉酸塩などの特定のビタミン分がより高くてもよいし、あるいは、それはカルシウムなどの特定のミネラル分がより高くてもよく、より高い抵抗性デンプンレベルと組み合わさって、大腸におけるミネラル分の強化摂取に相乗的効果をもたらし得る。
【0121】
デンプンは、標準分析法、例えば、Schulman他(1991年)の方法を用いて小麦穀粒から容易に分離される。工業的スケールでは、湿式または乾式ミリングを使うことができる。デンプン粒サイズは、より大きなA顆粒をより小さなB顆粒から分離するデンプン加工産業で重要である。本発明の小麦植物の粒から得られるデンプンは、相対的に高いアミロース含有量を有する。
【0122】
<変化したデンプンの物理的特徴>
糊化は、過剰量の水中における分子秩序の熱による崩壊(破壊)であり、粒膨潤、結晶化温度、複屈折の喪失、粘性の増大とデンプン可溶化などの特性の同時的かつ非可逆的変化を伴う。トウモロコシのae(アミロース増量体)変異体からのハイアミロースデンプンは、通常のトウモロコシより高い温度を示した(Fuwa他、1999年、Krueger他、1987年)。他方、デンプン合成酵素IIa活性が欠如している大麦sex6変異体からのデンプンは低い糊化温度を有し、対照植物と比較したとき、糊化ピークに対するエンタルピーは減少した(Morell他、2003年)。
【0123】
本発明の別の面では、示差走査熱量測定で測定した際にデンプンは変更された糊化温度を有する。これは野生型植物由来のデンプンと比較して増加することも減少することもある。変更された糊化温度は、相対的に高いアミロース含有量に加わる場合もある。示差走査熱量測定で測定した場合、野生型小麦デンプンの糊化温度は、典型的には、最初のピーク(開始温度と定義される)の温度が約61℃(Rahman他、2000年)である。
【0124】
デンプンは、野生型デンプンと比較した過剰の熱水によるその膨張率によって特徴付けることができる。体積膨張は、典型的にはデンプンまたは小麦粉と過剰の水を混合し、加熱(典型的には90℃以上)することで測定される。次いで、試料を遠心によって集め、沈殿した物質の質量を試料の乾燥重量によって割ることで膨張体積を表す。食品調製物、特に水和する食品調製物のデンプン含有量を増やすよう求められる場合は、低い膨張特性が有用である。
【0125】
本発明の選択された形の小麦のデンプン構造は、結晶化度が小麦から分離される通常のデンプンと比較して低減するという点でも異なり得る。デンプンの結晶化度の低減は、官能特性の強化と結びつくとも考えられ、よりスムーズな舌触りに寄与する。このように、デンプンはさらに、1つまたは複数のアミロペクチン合成酵素の活性レベルの低減から生じる低減された結晶化度を示してもよい。結晶化度は、典型的にX線結晶学によって調べられる。
【0126】
変更されたアミロペクチン構造の1つの基準は、デンプンの鎖長分布または重合度である。鎖長分布は、イソアミラーゼ脱分枝の後、蛍光ラベル糖鎖電気泳動(Fluorophore−assisted carbohydrate electrophoresis:FACE)を用いて決定され得る。本発明のデンプンのアミロペクチンは、脱分枝した時点で、野生型植物からのデンプンの分布より大きい5〜60の範囲の鎖長分布を有し得る。より長い鎖長のデンプンは、枝分れの頻度が相応して減少する。したがって、デンプンは依然存在するアミロペクチン中により長いアミロペクチン鎖長の分布を有してもよい。
【0127】
<食物特性>
デンプンは人間の食餌における主要な炭水化物源であり、本発明とそれに由来する産物の穀粒は食物調製に用いることができる。例えば家畜生産において、または、ペットフードにおいて、食物は人または動物によって消費され得る。変更された小麦植物に由来する穀粒は食品加工の方法においてすぐに使うことができ、したがって、本発明は、前述の小麦植物を処理した穀粒または全粒から得られる、挽いたか、粉砕したか、粗挽きしたか、球状にしたか、ロール状にした穀粒または製品(小麦粉を含む。)を含む。これらの製品は、次いで、様々な食品、例えば、パン、ケーキ、ビスケットなどのデンプン質製品、または増粘剤または結着剤などの食品添加物中で、または、飲物、麺、パスタまたはインスタントスープに用いてもよい。本発明の穀粒に由来する穀粒または産物は、特に朝食用シリアルにおいて、または、押出加工された製品として望ましい。本発明のハイアミロースデンプンは、菓子類産業において有用なハイストレングスゲル類を形成するのに用いることもでき、または、より低温で短時間の硬化時間を可能にする。それらは、例えば、深揚げポテトその他の食品において油吸収を減らすためのコーティングとしても用い得る。
【0128】
<食物繊維>
本明細書において、食物繊維は、健康な人間の小腸では吸収されずに大腸に入る炭水化物および炭水化物消化産物である。これは、抵抗性デンプン、β−グルカンならびに他の可溶性および不溶性炭水化物ポリマーを含む。それは、少なくとも部分的には、常駐微生物相によって大腸内で発酵可能な炭水化物部分を含むことを目的とする。
【0129】
本発明のデンプンは、好ましくは比較的高いレベルの食物繊維、特にアミロースを含む。本発明の穀粒の食物繊維含有量は、単に胚乳のアミロース含有量の相対的な増加によって生じてもよいし、そうでなくてもよい。
【0130】
本発明の実施形態は、高レベルの食物繊維との組合せにおいて、アリューロン層と胚の組合せに起因してもよい。具体的には、アリューロンまたは胚の相対的レベルがより高い状態で存在する場合に、これが起こるのでもよい。小麦穀粒がわずかに縮小する場合、胚乳は量が低減されて存在し、アリューロン層と胚は比較的に増大した量で存在する。このように、小麦は高いレジスタントスターチと組み合わせて比較的高いレベルのある種の有益な要素またはビタミンを有し、そのような要素としては、二価カチオン、生物が利用可能なCa++と葉酸塩などのビタミン、あるいは、トコフェロールやトコトリエノールなどの抗酸化剤を含む。粉砕製品の1つの具体的形状は、アリューロン層が粉砕製品に含まれるものでもよい。具体的な粉砕プロセスは、粉砕製品中のアリューロン層の量を強化するように行ってもよい。このように、アリューロン層および胚を含むように粉砕または他の加工をした穀粒に由来する製品はいずれも、別の源からこれらの要素を加える必要がなく、さらなる栄養的な利益を有する。
【0131】
<レジスタントスターチ>
レジスタントスターチは、健康な人間の小腸に吸収されずに大腸に入るデンプンおよびデンプン消化産物の合計と定義される。したがって、レジスタントスターチは、消化されて小腸で吸収される産物を除外する。レジスタントスターチは物理的に接触しがたいデンプン(RS1形)、抵抗性顆粒(RS2)、老化デンプン(RS3)、化学修飾デンプン(RS4)を含む。食物で消費されるとき、本発明のデンプンの変更されたデンプン構造と特にハイアミロースレベルはレジスタントスターチの増加を引き起こす。デンプンは消化するにはいくぶん接触しがたいRS1形でもよい。V−複合体結晶度で測定されるデンプン−脂質会合体もレジスタントスターチのレベルに寄与しそうである。
【0132】
本発明の1つの利点は、それが特定の栄養的な利益である産物を提供し、さらに、デンプンまたは小麦穀粒の他の構成要素を修飾する必要なしにそのような産物を提供するという点であることが理解されるであろう。しかし、デンプンまたは穀粒の他の成分への修飾は望ましい場合もあり、本発明はそのような修飾された構成成分を包含する。修飾の方法はよく知られており、抵抗性形を増やすために、従来法によるデンプンその他の成分の抽出とデンプンの修飾を含む。デンプンは、熱および/または水分での処理によって、物理的に(例えば、ボールミリング)、酵素処理で(例えば、α−またはβ−アミラーゼ、プルラナーゼその他)、化学加水分解(液体またはガス状の試薬を使用して湿式または乾式で)、酸化、二官能基試薬(例えばナトリウムトリメタリン酸、オキシ塩化リン)を用いた架橋またはカルボキシメチル化によって修飾され得る。
【0133】
<血糖指数>
血糖指数(GI)は、デンプンを含む食品の消化率に関するものであって、運動時、血中グルコース濃度に関する白パンまたはブドウ糖の効果を試験食物の効果と比較するものである。血糖指数は、食事後の血清グルコース濃度と血中グルコースホメオスタシスのためにインシュリンの必要量に関する食物の影響可能性の目安である。本発明の食品によって提供される1つの重要な特徴は、血糖インデックスが減少することである。さらにまた、食品は最終的な消化レベルが低く、したがって比較的低カロリーとなし得る。低カロリー産物は、粉砕小麦穀粒から作り出される小麦粉の含有に基づくものでもよい。そのような食品は、お腹を満たし、腸の健康を強化し、食後の血清グルコースと脂質濃度を減らすとともに低カロリー食品を提供するという効果を有し得る。
【0134】
<食品以外の応用>
本発明は、アミロースレベルを上昇させ、アミロペクチンを低減させた修飾または改善されたデンプンを提供し、これらの特性は種々の産業の必要条件のいずれをも満たす。デンプンは、フィルム、紙、織物、段ボールおよび接着性産業(Young、1984年)で、例えば、サイズ剤として、広く非食品産業において使われている。小麦デンプンは、ブドウ糖シロップの生産のための、または、エタノール産生のための基質として用いてもよい。改変されていないデンプンの物性は若干のアプリケーションでその有用性を制限し、高価であるか他の不利益があることがあり得る化学修飾がしばしば必要となる。特に他の物性と組み合わせてアミロペクチン含有量を低減したために、本発明は収穫後改変の必要がより少ないデンプンを提供する。例えば、糊化温度、デンプンの剪断応力抵抗性、フィルム強さおよび/または水抵抗ならびに本発明の穀粒から製造される産物は、変更してもよい。デンプンは、ポリスチレンや他のパッキング材料の代わりとして生分解性のある緩い充填パッキング材料を調製するのに用いてもよい。
【0135】
本発明の各面に関して様々な説明を行ってきたが、本発明が本発明の2つ以上の面の組合せによるでもよいことが理解されるであろう。
【実施例】
【0136】
<実施例1.材料と方法>
炭水化物の決定と分析
デンプンは、Schulman他(1991年)の方法を使用して小麦穀粒から分離した。デンプン含有量は、Megazyme(Bray、Co Wicklow、アイルランド共和国)によって供給される全デンプン分析キットを使用して測定した。次いで、デンプン含有量を対照植物と比較する。総穀粒重量からのデンプン重量を引いて穀粒の全非デンプン含有量を求め、総重量の低減がデンプン含有量の低減によるかどうか確定する。
【0137】
デンプン試料のアミロース含有量は、以下の通り、MorrisonおよびLaignelet(1983年)の比色(ヨウ素還元)方法をわずかに変更して測定した。およそ2mgのデンプンを、ふたのゴムワッシャーと嵌まる2mlのネジキャップ付チューブに、正確に秤量した(0.1mgまで正確)。脂質を取り除くために、85%(v/v)メタノール1mlをデンプンと混合し、時折ボルテックスにかけながら65℃水浴中で1時間チューブを加熱した。13,000g、5分間の遠心の後、上清を慎重に取り除き、抽出工程を繰り返した。次いで、デンプンを65℃で1時間乾燥して、2mgデンプン(上記のように秤量)につき1mlの尿素−ジメチルスルホキシド溶液(UDMSO;1容積の6M尿素量に対し9容積のジメチルスルホキシド)を用いて溶解させた。混合物は直ちにボルテックスにかけ激しく混合し、デンプンを完全に溶解させるために断続的にボルテックスにかけつつ、95℃水浴中で1時間インキュベートした。デンプン−UDMSO溶液(50μl)のアリコートを、水ml当たり2mgのヨウ素と20mgのヨウ化カリウムを含む20μlのI2−KI試薬で処理した。混合物を水で全量1mlとした。200μlをマイクロプレートに移し、Emax Precision Microplate Reader(Molecular Devices、米国)を用いて吸光度を読むことにより650nmでの混合物の吸光度を測定した。0〜100%のアミロースと100%〜0%のアミロペクチンを含む標準試料をポテトアミロースとコーン(またはポテト)アミロペクチン(Sigma)から作り、試験試料と同様に処理した。アミロース含有量(パーセンテージアミロース)は標準試料の吸光度に由来する回帰式を使用して吸光度値から決定した。脱分枝したデンプンのアミラーゼ/アミロペクチン比の分析は、Case他(1998年)によって行ってもよいし、またはBateyおよびCurtin(1996年)によって記載されるように脱分枝デンプンを分離するHPLC法によって行ってもよい。
【0138】
デンプン試料の脱分枝後に、デンプン鎖長の分布は、Morell他(1998年)によるキャピラリー電気泳動ユニットを用いて蛍光ラベル糖鎖電気泳動(Fluorophore−assisted carbohydrate electrophoresis:FACE)によって分析し得る。デンプン試料の糊化温度プロフィールは、Pyris 1示差走査熱量計(Perkin Elmer、Norwalk、コネチカット州、米国)で測定し得る。デンプン溶液の粘度は、例えば、Batey他(1997年)が報告したような条件を用いてRapid−Visco−Analyser(RVA、Newport Scientific Pty Ltd、Warriewood、シドニー)上で測定し得る。測定し得るパラメータとしては、ピーク粘度(最大熱ペースト粘度)、維持強さ、最終的な粘性および糊化温度が挙げられる。小麦粉またはデンプンの膨潤量は、Konik−Rose他(2001年)の方法によって測定され得る。水の取り込みは、小麦粉またはデンプン試料を水と混ぜる前後、所定の温度で試料を量り、糊化した材料を回収することで測定される。
【0139】
β−Glucanレベルは、Megazyme(Bray、Co Wicklow、アイルランド共和国)によって供給されるキットを使用して測定され得る。
【0140】
胚乳のタンパク質発現の分析
胚乳の特定のタンパク質発現は、ウェスタンブロット手順によって分析した。胚乳はすべての母体組織から切り離し、およそ0.2mgの試料を、5mM EDTA、20%グリセリン、5mM DTTおよび1mMペファブロック(Pefabloc)を含む50mM KPi緩衝液(42mM K2HPO4と8mM KH2PO4)、pH7.5の600μl中でホモジナイズした。粉砕試料を13、000gで10分間遠心処理し、上清アリコートを使用まで−80℃で凍結した。総タンパク評価のために、0.25mg/mlBSA標準液0、20、40、60、80および100μlアリコートを用いてBSA標準曲線を準備した。試料(3μl)は蒸留水で全量100μlとし、1mlのCoomassie Plus Protein試薬を各々に加えた。吸光度はブランクとして標準曲線から0 BSA試料を用い、595nmで5分後に計測し、試料のタンパク質レベルを測定した。各々の胚乳から20μg総タンパクを含む試料を、0.34MTris−HCl(pH8.8)、アクリルアミド(8.0%)、過硫酸アンモニウム(0.06%)およびTEMED(0.1%)を含む8%非変性ポリアクリルアミドゲル上で泳動させた。電気泳動後、タンパク質はMorell他(1997年)によるニトロセルロース膜に移し、SBEIIaまたはSBEIIb特異抗体と免疫反応させた。
【0141】
<実施例2.小麦SBEIIAとSBEIIB発現変更のための遺伝子コンストラクト>
小麦のSBEIIaまたはSBEIIb遺伝子の発現を低減すべく二重鎖RNA(dsRNA)コンストラクトを作成した。そのようなコンストラクトにおいて、SBEIIaまたはSBEIIb遺伝子の一部に対応する望ましい核酸配列は、発現したRNAが塩基対合可能で二重鎖または二本鎖RNAを形成し得る相補領域を含むように、プロモーターに対しセンスとアンチセンスの両方に存在した。センスとアンチセンス配列の間のスペーサー領域は、形質転換植物においてRNAの一部として転写されるとき、堅い「ヘアピン」二重構造を作るためにスプライシングにより除かれるイントロン配列からなるものとした。イントロンを含むことで、二重鎖RNAコンストラクトによって与えられる遺伝子サイレンシングの効率が増すことが分かっている(Smith他、2000年)。望ましい核酸を、高分子量グルテニン(HMWG)プロモーター配列(Dx5サブユニット遺伝子、アクセッション番号X12928、Anderson他、1989年のプロモーター)とアグロバクテリウム由来ノパリンシンターゼ遺伝子(nos3')からのターミネーター配列に連結した。これは、dsRNA配列の胚乳特異的発現をもたらした。
【0142】
SBEIIa二重鎖RNAコンストラクトは、小麦SBEIIa遺伝子(GenBankアクセッション番号AF338431、図1参照)からPCRによって増幅される1536bpのヌクレオチド配列を含んでいた。これは以下を含んでいた;エキソン1と2の全体とエキソン3の一部を含み(図1のヌクレオチド位置1058〜1336、1664〜1761および2038〜2219)EcoRIとKpnIの制限部位を各側に有する468bp配列(断片1)、SBEIIaのエキソン3と4の一部およびイントロン3の全部を含み(図1のヌクレオチド位置2220〜2731)KpnIとSacIの部位を各側に有する512bp配列(断片2)ならびにSBEIIaの完全なエキソン1、2および3を含み(図1のヌクレオチド位置1058〜1336、1664〜1761および2038〜2279)BamHIとSacIの部位を各側に有する528bp配列(断片3)。次いで、断片3の配列が断片1に対してアンチセンス方向において断片2にライゲートされるように、断片1、2と3をライゲートした。最初に、HMWGプロモーター配列とnos3'ターミネーターを含むベクターpDVO3000内で二重鎖RNAコンストラクトを生成した。ベクターpDVO3000の遺伝子コンストラクトはpDVO3−IIaと名付け、二重鎖RNA遺伝子はds−SBEIIaとした。
【0143】
SBEIIb二重鎖RNAコンストラクトについての戦略も同様とした。SBEIIbコンストラクトは、小麦SBEIIb遺伝子(配列は、図2で概説)からPCRによって増幅される1607bpの断片を含んでいた。これは、エキソン1と2の全体とエキソン3の一部を含み(図2のヌクレオチド位置489〜640、789〜934および1598〜1769)EcoRIとKpnIの制限部位を各側に有する471bp配列(断片1)、SBEIIbのエキソン3と4の一部およびイントロン3の全部を含み(図2のヌクレオチド位置1770〜2364)KpnIとSacIの制限部位を各側に有する589bp配列(断片2)ならびにSBEIIbの完全なエキソン1、2および3を含み(図2のヌクレオチド位置489〜640、789〜934および1598〜1872)BamHIとSacIの制限部位を各側に有する528bp配列(断片3)を含んでいた。次いで、断片3の配列が断片1に対してアンチセンス方向において断片2にライゲートされるように、断片1、2と3をライゲートした。ベクターpDVO3000のSBEIIb二重鎖RNA遺伝子コンストラクトはpDVO3−IIbと名付け、二重鎖RNA遺伝子はds−SBEIIbとした。コンストラクトを図3に模式的に示す。
【0144】
次いで、ds−RNA発現カセットの各々を制限酵素−Xholで切り離し、バイナリー形質転換ベクターpGB53およびpBIOS340に挿入した。pGB53は、コメアクチンプロモーターによってドライブされるアスラム耐性(sul)をコードする遺伝子の導入によってpSBll(Komari他、1996年)から作られ、対象とすべき遺伝子の導入のためにT−DNA右側境界に隣接して特有のXhoIサイトを残した。同様に、pBIOS340はカナマイシンとジェネテシン耐性をコードするnptII遺伝子の導入によってpSBl(Komari他、1996年)から作られ、コメアクチンプロモーターによってドライブされ、ここでも、右側境界に隣接して特有のXhoIサイトを残した。pGB53とpBIOS340のSBEIIaコンストラクトはそれぞれpCL51とpCL59と名付け、pGB53とpBIOS340中のSBEIIbコンストラクトはそれぞれpCL54とpCL60とした。
【0145】
<実施例3:小麦の形質転換>
virプラスミドpAL4404およびpSBlを運ぶ無害化したAgrobacteilum tumefaciens株LBA4404に小麦の形質転換のための遺伝子コンストラクトをエレクトロポーレーションによって導入し、続いてスペクチノマイシンを含む培地上で選択した。形質転換したアグロバクテリウム株を、2日間固化させたYEP培地上27℃でインキュベートした。次いでバクテリアを回収して、小麦接種のため、650nmで光学濃度2.4まで400mMアセトシリンゴンを含むTSIM1(100mg/lmyo−イノシトール、10g/lブドウ糖、50mg/l MES緩衝液pH5.5によるMS培地)に再懸濁させた。
【0146】
小麦植物(品種NB1、Nickerson Seeds Ltd、Rothwell、Lincs.から得られる春小麦種)を、1日16時間の日照とするために明かりを補って温室内で22/15℃日/夜の温度で成長させた。50cmのひこばえ茎を含むように開花期後およそ14日で(胚:長さおよそ1mm)でひこばえを収穫した。次いで、止め葉以外、すべての葉をひこばえから取り除き、止め葉は清浄化してカビ胞子による汚染を除いた。次いで、各々の小穂の包穎と最初の2つの小花から外穎を慎重に取り除いて未熟種子を露出させた。一般に、各々の小穂にはこれらの2粒の種のみが見つかった。この手順を、花序の全長に沿って実行した。次いで、穂には簡単な表面殺菌を行うために70%IMSを吹き付けた。
【0147】
アグロバクテリウム懸濁液(1μl)を、10μlハミルトン注射器を使って、未熟種子内においてほぼ胚盤:胚乳の界面の位置で、露出した種子すべてに接種した。次いで、ひこばえを水中に置き、種の乾燥を防ぐために半透明のビニール袋で覆い、明かりをつけたインキュベーター中に23℃で3日間、日照1日当たり16時間、45_Em−2s−1PARで置いた。同時培養3日の後、接種した未熟種子を取り除き、70%エチルアルコール(30秒)、次いで20%漂白剤(Domestos、20分)で表面を殺菌し、無菌蒸留水で完全に洗浄した。未熟な胚を無菌的に単離し、W3培地(MSに20g/lショ糖と2mg/l 2,4−Dを補い、6g/l Type Iアガロース(Sigma)で固めたもの)に置き、150mg/lタイメンチン(Timentin)(W3T)を加え、胚盤(プレート当たり20胚)を一番上とした。培養物を、25℃で光(1当たり16時間の日照、80_Em−2s−1PAR)中に置いた。胚上の胚軸の成長を単離後およそ5日後に評価し、カルス生産を改善するために必要な場合、軸は取り除いた。胚は4週間W3T上で維持し、単離2週間後、新しい培地に移し、胚形成能力を評価した。
【0148】
4週成長させた後、接種した胚に由来するカルスは、W3T媒地上に植菌した非接種胚から得られる対照カルスと非常に類似していた。バクテリアの存在は、接種した胚に由来するカルスの胚形成能力を実質的に低減させたようには見えなかった。胚形成カルスを、2mg/lアスラム(pGB53誘導体を使用した)または25mg/l(pBIOS340誘導体)および150mg/lタイメンチン(W32AT)を含むW3培地に移した。カルスをさらに2週間、この媒地上で維持し、次いで、各カルスを2mm−サイズの小片に分け、W32AT上に再植菌した。バイナリーベクターコンストラクトなしでLBA4404の接種に由来する対照胚は、選択培地上で形質転換カルスを生じなかった。
【0149】
さらに2週間培養後、胚形成性カルスの成長についてすべての組織を評価した:4週後にも選択培地上で継続的な成長の徴候を示しているすべてのカルスが、再生培地(40g/lマルトースと150mg/lタイメンチン(pH5.8)を加えたRMT−MS、6g/lアガロース(Sigma 1型)で固化)に移された。新芽をこの培地上で4週間再生し、次いで、芽の伸長と発根のために150mg/lタイメンチンを加えたMS30に移した。次いで、幼若植物を土壌混合物に移し、2週間噴霧ベンチに保持し、最後に温室に移した。
【0150】
pCL54またはpCL60(ds−SBEIIb)を使っている合計3217個の胚およびpCL51またはpCL59(ds−SBEIIa)を使っている2010個の胚をこの方法によって処理し、61個の植物をIIb形質転換のためにカルスから再生し、31個の植物をIIa形質転換のためにカルスから再生した。選択培地上の生存状況は、それらが遺伝子コンストラクトによって首尾よく形質転換されていることを示唆した。選択可能なマーカー遺伝子で形質転換した植物の大多数(すべてではない)は、SBEIIaまたはSBEIIb阻害遺伝子を一体化するものと期待される;これらは以下の例に記載するように容易に識別することができた。
【0151】
実験から得られた良好な再生可能性を伴う複数の安定した組み込みイベントの回復は、本明細書で用いられる種接種形質転換法が小麦について報告されている他の方法と同じように効率的なことを示した。株AGL1などの代わりのアグロバクテリウム株またはハイグロマイシン抵抗性をコードする遺伝子などの選択可能なマーカーも本明細書方法に用い得る。
【0152】
<実施例4.小麦形質転換体の分析>
形質転換は、以下の方法の1つまたは複数で測定した:
導入遺伝子の1つまたは複数についてのPCR分析。PCR分析は、StaceyおよびIsaac(1994年)によって記載されるミニプレップ法を使用し、新鮮な葉資料1〜2cm2から抽出されるゲノムDNAについて行った。PCR反応は、SBEIIa遺伝子からの断片(462bp)を増幅すべく設計された、例えば、プライマーSBEIIa−For:5'−CCCGCTGCTTTCGCTCATTTTG−3'[配列番号9]およびSBEIIa−Rev:5'−GACTACCGGAGCTCCCACCTTC−3'[配列番号10]を用いて、またはSBEIIb(505bp)についてSBEIIb−DupFor5'−AGATGTGAATGGCTGCTTGCTG−3'[配列番号11]およびSBEIIb−DupRev5'−CAGGTCGACCATATGGGAGAGC−3'[配列番号12]を用いて行った。反応条件は以下の通りとした:「ホットスタート」(94C、3分)後、30サイクルの変性(95℃、30秒)、アニーリング(55℃、30秒)、伸長(73℃、2分)後、73℃(5分)で1サイクル。
【0153】
サザンブロットハイブリダイゼーション分析は、凍結乾燥した粉砕組織(StaceyおよびIsaac、1994年)から、より大規模(9ml)に抽出したDNAに対して実行する。DNA試料は0.2mg/mlに調整し、HindIII、EcoRIおよびKpnIなどの制限酵素で消化した。StaceyおよびIsaac(1994年)に記載されるように、制限酵素消化、ゲル電気泳動と真空ブロッティングを実行する。ds−SBEIIコンストラクトのイントロン3領域を含むジゴキシゲニンで標識したプローブは、McCreeryおよびHelentjaris(1994年)の方法によってPCRにより製造される。サザンブロットへのプローブのハイブリダイゼーションおよび化学発光による検出は、McCreeryおよびHelentjaris(1994年)の方法によって実行する。
【0154】
PCR分析の結果を表2にまとめる。PCRによって示される導入遺伝子が陽性だった植物は、ds−SBEIIaについて27の独立した形質転換イベントとds−SBEIIbについての61の独立したイベントを含んでいた。
表2.SBEIIaとSBEIIbRNA二重鎖コンストラクトによる小麦の形質転換
【0155】
【表2】
【0156】
<実施例5.二重鎖RNAコンストラクトで形質転換した植物から得た穀粒の分析>
デンプン顆粒形態
T0形質転換小麦植物から得た成熟したT1種子からのデンプン顆粒の形態を、光学顕微鏡法によって観察した。ds−SBEIIaで独立に形質転換した25本のT0植物とds−SBEIIbで独立に形質転換した12本の植物の各々からのそれぞれ10個の穀粒を分析した。各胚乳はデンプン顆粒を放出させるために穏やかに押しつぶし、それを水中に分散して、光学顕微鏡の下で視覚化した。分析した25のds−SBEIIa系統のうち、12個は変形顆粒(例えば、図4参照)を含む穀粒を有していたが、視覚的観察では異なる種子では変形レベルは様々であった。これに対し、12のds−SBEIIb系統のいずれも、光学顕微鏡法下で観察したとき胚乳中のデンプン顆粒の顕著な変形は示さなかった。結果を、表3と4にまとめて示す。
表3.ds−SBEIIaトランスジェニック小麦系統のT1種子のデンプン顆粒形態
【0157】
【表3】
【0158】
*各系統からの10粒の種子のデンプン顆粒形態を観察した。
10粒すべての種が通常の顆粒形態を有する場合は、形態を+として示し、ひどく変形した種子がある場合は−で、また、若干の異常がある(すなわち、少なくともいくつかの種子にある程度の変形があるが、ひどい変形のものはない)場合は+/−とした。
【0159】
表4.ds−SBEIIbトランスジェニック小麦系統のT1種のデンプン顆粒形態。
【0160】
【表4】
【0161】
+は、各系統からの10粒の種子すべてが正常なデンプン顆粒形状を有していたことを示す。
偏光下のデンプン顆粒を観察したところ、ds−SBEIIa穀粒については、変形顆粒(図5)のため、複屈折が著しく低減していたことが分かった。複屈折の損失は、系統50.lbからの種子では顆粒の94%で観察され、一方、同じ系統の別の種子からの正常な顆粒が完全な複屈折(表5)を示しており、変形フェノタイプと相関している。正常な顆粒による種子は、導入遺伝子が不足している分離体であると思われ、したがって、発現型的には正常である。
表5.ds−SBEIIaトランスジェニック小麦系統50.1bのT1種子からのデンプン顆粒の複屈折
【0162】
【表5】
【0163】
+通常のデンプン顆粒、−ひどく変形した顆粒、+/−若干の変形のある顆粒
【0164】
光学顕微鏡観察の結果は、デンプン顆粒の走査型電子顕微鏡(SEM)によって確認する。このために、精製したデンプンに金でスパッタリングして、室温、15kVで走査する。
【0165】
穀粒重量
ds−SBEIIa形質転換植物(温室で等しい条件下で生育)からの個々の穀粒を秤量した(表6)。植物50.1b、58.2a、61.2aおよび109からのひどく変形した顆粒を有する穀粒は、同じ条件下で生育した野生型植物の粒と比較して、平均重量で顕著な低減はなかった。したがって、デンプン生産は、大きく変形したデンプン顆粒を有する種子においてであっても実質的にされる低減するようには見えなかった。このデータは、胚乳中のSBEIIa活性の低減した野外生育小麦の収穫高がだいたい普通であることも示唆している。
表6.ds−SBEIIaトランスジェニック小麦系統からのT1種子の穀粒重量
【0166】
【表6】
【0167】
T2トランスジェニック小麦胚乳のSBEIIaとSBEIIbタンパク質の分析
5つの独立した形質転換系統を代表して13ds−SBEIIa形質転換T1植物からと、3つの独立した形質転換系統を代表して9ds−SBEIIaからの種子(T2)を、胚乳内のSBEIIaとSBEIIbタンパク質発現について、非変性PAGEとウェスタンブロッティングによって分析した。先に述べたように、ds−SBEIIb系統はすべて正常の顆粒形態を有するが、ds−SBEIIa植物はすべて異常なデンプン顆粒形態を有する系統からのものであった。SBEIIaの検出のために用いた抗体は、ウサギ由来の3KLHであり、これはアミノ酸配列AASPGKVLVPDESDDLGC[配列番号13]を有し、成熟SBEIIaのN末端からの配列に対応する合成ペプチドに対して誘導したものであり、使用に際して1:5000に希釈した。SBEIIbの検出のために用いた抗体はR6であり、これはアミノ酸配列AGGPSGEVMIGC[配列番号14]を有し、成熟SBEIIbのN末端からの推定配列に対応する合成ペプチドに対して誘導したものであり、使用前に1:6000に希釈した。使用した二次抗体は、GAR−ホースラディッシュペルオキダーゼ複合体とした(1:3000希釈)。免疫反応バンドは、アマシャムECL−検出システムを使用して明らかにした。
【0168】
植物の中には導入遺伝子についてヘテロ接合のものもあると予想されたため、22本のT1植物の各々から7粒の成長中の穀粒(開花後15日)の各々から胚乳を分析した。13のds−SBEIIa植物のうちの12は、胚乳中にSBEIIaタンパク質レベルの低減を示すT2子孫を生じた。1系統からの全7粒の種子(50.3x.9)はSBEIIaを完全に欠いているようであったが、他の4本の植物からの全7粒の種はSBEIIaの明らかに低減した発現を示した(表7)。これらは、導入遺伝子についてホモ接合性である系統を表しているであろう。7系統はSBEIIaが存在しない点またはSBEIIaレベルが低減している点、または場合によってはタンパク質が明らかには低減してない点で分かれており、これらの系統はおそらく導入遺伝子についてのヘテロ接合体を表している。13系統(50.3x.6)は野生型発現についてホモ接合性だった(表7)。
表7.T2 ds−SBEIIaトランスジェニック小麦系統からの胚乳タンパク質のウェスタンブロット分析
【0169】
【表7】
【0170】
試験した9のds−SBEIIbトランスジェニック系統のうち、3つ(110.16b.2、110.16b.5および110.16b.19)は一様に、7粒の子孫種子のいずれにおいてもSBEIIb発現は示さなかったが、2は野生型発現について均一、残る4は発現なし、発現低減または野生型に分離した(表7)。種子からの胚は、T2植物とT3種子を生じさせるべく成長させてもよく(胚レスキュー)、これらは導入遺伝子に関してT2種子の遺伝子の状態を確かめるためにPCRとタンパク質発現分析でスクリーニングする。
【0171】
これらのデータは、二重鎖RNAコンストラクトが小麦胚乳のSBEIIa遺伝子およびSBEIIb遺伝子の発現低減に関して有効なことを示す。データはまた、SBEIIb発現のみの低減はデンプン顆粒形態を実質的に変化させなかったことをも示す。
【0172】
ds−SBEIIa導入遺伝子を含み、SBEIIaタンパク質が欠如しているトランスジェニック種子におけるSBEIIb遺伝子の発現とds−SBEIIbを含む種子のSBEIIa遺伝子の発現の発現も、ウェスタンブロット法によって分析した。予想外にも、ds−SBEIIaを含むトランスジェニック種子では、SBEIIbが大きく減少した。しかし、ds−SBEIIbについてトランスジェニックな種子では、逆の影響は観察されなかった。SBEIIbがds−SBEIIbによって完全にサイレンスにされた種子でSBEIIa発現は変わらなかった。SBEIIbの発現は、二重鎖コンストラクトのために使われる領域の遺伝子間での配列相同性のためにds−SBEIIaコンストラクトによって抑制された可能性があり、SBEIIbの活性は、いくつかの他のメカニズムによってds−SBEIIa導入遺伝子によって低減された可能性がある。
【0173】
SBEIIa遺伝子およびSBEIIb遺伝子の発現レベルは、mRNAレベルにおいても、ノーザンハイブリダイゼーションまたはRT−PCR法などの標準的な技術によって(例えば、非保存領域由来のプローブを用い、または遺伝子のうちの1つにおいて特異的なサイトにハイブリダイズする他のサイト、例えば、3'非翻訳領域ではハイブリダイズしないプライマー対を用いて)具体的に決定し得る。そのような領域またはサイトは、2つの遺伝子配列の比較によって、容易に確認できる。
【0174】
<実施例6.形質転換小麦のデンプン分析>
トランスジェニック小麦穀粒中でのアミロースおよびアミロペクチンレベル
実施例1に記載したように、6つのプールしたT1種子試料からのデンプンのアミロース含有量を測定した。プールした種子試料は、以下のようにしてトランスジェニック小麦系統から得た:
【0175】
プール1−ds−SBEIIaトランスジェニック系統85.2cからデンプン変形顆粒を有していた種子
プール2−ds−SBEIIaトランスジェニック系統85.1aから正常顆粒を有していた種子
プール3−ds−SBEIIbトランスジェニック系統110.18aから正常顆粒を有していた種子
プール4−ds−SBEIIaトランスジェニック系統58.1a、58.2aおよび61.2a(一緒にプール)から変形顆粒を有していた種子
プール5−ds−SBEIIaトランスジェニック系統83.1bから正常顆粒を有していた種子
プール6−ds−SBEIIbトランスジェニック系統75.3xから正常顆粒を有していた種子
【0176】
各々の分析は、デンプン試料の4つの複製を用いて行った。これらの分析のために、吸光度をアミロース含有量に変換するのに用いられる回帰式は、Y=57.548x−8.793であった(ここで、Yはアミロース含有量(%)であり、xは吸光度)。
【0177】
結果を表8に示す。変形デンプン顆粒の存在は、明らかにアミロース含有量の増加を伴っていた。ds−SBEIIaトランスジェニック系統(プール1および4)からの変形顆粒を有する穀粒由来のデンプンは50%を超えるアミロース含有量を有していたのに対し、他のデンプンプール(正常なデンプン顆粒を有する穀粒に由来する)は21〜26%の範囲でアミロース含有量を有していた。これはSBEIIb(表8)の発現が低減された系統IIb 110.18aからのデンプンを含んでおり、このことは、小麦中においてSBEIIbのみ失活させても、穀粒デンプン中でのアミロースレベルは実質的に増加しないことを示唆する。
表8.トランスジェニック小麦系統のヨウ素還元法によって推定されるアミロース含有量
【0178】
【表8】
【0179】
分析の第2セットは、プール 4とSSII(Yamamori他、2000年)が欠如している小麦からのデンプンおよびSSIIaの変異体である大麦系統M292からの試料を用いてヨウ素還元法によって行った。プール4小麦種子(ds−SBEIIaトランスジェニック系統)について測定されたアミロース含有量は、小麦および大麦のSSII変異体からのデンプンのそれより相当に高かった。
【0180】
これはこれらの穀粒中のデンプンのアミロペクチン含有量が相当に、すなわち、野生型のおよそ75%から、50%未満またはさらには20%未満にまで低減されることを意味する。
【0181】
ds−SBEIIaとds−SBEIIb導入遺伝子を含む系統は、上述の遺伝子導入植物を交配することによって発生する。そのような子孫の穀粒デンプン中のアミロース含有量は、ds−SBEIIaのみを含む植物由来のデンプンのアミロース含有量以上に、例えば75または80%まで増加し、SBEIIaに加えてSBEIIbの抑制がさらにアミロースレベルを上昇させることを示している。
【0182】
<実施例7.A、BおよびD ゲノムからのSBEIIaの比較>
小麦cDNAのコンストラクトおよびゲノムライブラリー
小麦胚乳cDNAとゲノムライブラリーは、ファージベクター(Sambrook他、1989年)における標準分析法によって作成した。2つのcDNAライブラリー(一方は栽培品種Rosella(Rahman他、1999年)由来のRNAから、他方は栽培品種Wyuna(Rahman他、2001年)から作成した。RosellaライブラリーはベクターZAPII中にあってEcoRIとNotIプライマーを用い、他方、Wyunaライブラリーは試薬に伴って与えられるプロトコルに従ってZipLoxベクター(Life Technology)中に存在した。ライブラリーの力価は、大腸菌のY1090(ZL)株で試験して2×106pfuの力価であった。ゲノムライブラリーは、A.tauschii変異体10097由来のDNAから作成した。DNAをSau3Aで消化し、部分装填lambdaGEM12ベクター(Promega)にライゲートした。クローンフラグメントは、SacIまたはXhoI消化で放出できた。T.aestivum DNAのゲノムライブラリーは、Turner他(1999年)に記載されている。
【0183】
SBEIIa cDNA配列の隔離
小麦SBEI遺伝子配列のプローブを使って低ストリンジェンシーで(Rahman他、2001年)で、栽培品種Rosellaから調製されるライブラリーからcDNAを分離した。得られた最も長いクローン(sbe9と命名)の配列決定を行ったところ、SBEIIaタイプの配列(Genbank AF338432.1)をコードすることが分かった。その後、栽培品種Wyuna(Rahman他、2001年)から調製した胚乳cDNAライブラリーから、sbe9の位置536〜890に対応するプローブを用いて3つのクローンを分離した。ライブラリースクリーニングの条件は、25%ホルムアミド、5×SSC、0.1%SDS、10×デンハルト溶液、100μg/mlサケ精液DNAで42℃、16時間のハイブリダイゼーション後、2×SSC、0.1%SDS、65℃、3×1時間(中程度のストリンジェンシー)で洗浄とした。クローンを配列決定することにより、3つの異なる配列が得られ、これらを以下、sr995およびsr997(図6)と表す。
【0184】
これらのcDNA配列を調べることにより、様々な配列が小麦胚乳で発現していることが示されたが、これらは異なる小麦ゲノムからのSBEIIa転写物と対応しているようである。他の既知の小麦SBEIIa cDNA配列による配列とのPILEUP比較は、sr995とsr996配列が、D−ゲノム配列wSBE−D1(sr854)(図7)に由来するmRNA配列とクラスターになっていることを示し、sr995およびsr996がDゲノムSBEIIaからの転写物を表していることを示唆した。Y11282配列(Nair他、(1997年))とクラスターになっているSr997は、それらがおそらく同じゲノム(AゲノムかBゲノム)に由来するものであることを示している。前述のsbe9(AF338432.1)はおそらくY11282と同じゲノムに由来するものであろうが、選択的スプライシングイベントを表し、5'末端の近くで1つのエキソンがスプライシングで除かれたものと一致している。
【0185】
小麦T.aestivumのA、BおよびDゲノムからのSBEIIa遺伝子との区別
遺伝子またはRNA転写配列における相違は、遺伝子レベルにせよRNAレベルにせよ、突然変異スクリーニングのためのA、BおよびDゲノム特異的プライマー設計の基礎として用い得る。例えば、図6は、Genbank アクセッションY11282を含むcDNA由来のSBEIIaヌクレオチド配列とcDNA sbe9(AF338432.1)、sr997およびsr995の部分配列と比較している。ゲノム配列はT.aestivum由来のSBEIIa遺伝子に利用可能である(例えば、表1参照)。ゲノム配列は、A、BおよびDゲノムに割り当てられた。比較は染色体の多型を示し、そのいずれもが配列を分子的手段によって識別するのに用いることができる。
【0186】
エキソン5領域に基づくフォワードプライマー(5'−ATCACTTACCGAGAATGGG−3')[配列番号15]およびエキソン6領域に基づくリバースプライマー(5'−CTGCATTTGGATTCCAATTG−3')[配列番号16]をA、BおよびDゲノム由来の産物間の識別をするのに用いた。そのようなプライマーは、A、BまたはDゲノム(下記参照)由来のSBEIIa遺伝子の1つまたは複数の変異体である変種またはアクセッションをスクリーニングするためにPCR反応で用いてもよい。
【0187】
SBEIIb遺伝子を小麦のA、BおよびDゲノムから区別するためにPCRベースのマーカーも開発した。例えば、プライマー対ARA19F(5'−CACCCATTGTAATTGGGTACACTG−3')[配列番号17]とARA15R(5'−TCCATGCCTCCTTCGTGTTCATCA−3')[配列番号18]を用いたPCR反応の後、制限酵素RsaIで増幅産物の消化を行うことで、SBEIIb遺伝子と3つのゲノムを区別することができた。
【0188】
cDNA配列の違いを、推定タンパク質配列に反映する。例えば、Dゲノム(sr854)とAまたはBゲノム(Y11282)ポリペプチドに対して推定したノーカットアミノ酸配列を図8で比較する。1つまたは複数のゲノムに特異的な活性が欠如している小麦の種類をスクリーニングするために、SBEIIaタンパク質に対するゲノム特異的抗体を産生するために使うことのできた領域688〜698と735〜6とでは顕著な差異が明らかである。他の違いは、図8のアミノ酸位置1〜54と対応する輸送ペプチド配列で起こる。
【0189】
<実施例8.1つまたは複数のSBEII遺伝子における小麦の種類変異体の識別>
BおよびDゲノム中のSBEIIbヌル変異の識別
300のオーストラリアの小麦の種類を含む1500の小麦寄託種の合計、オーストラリア冬穀物コレクション(AWCC、Australian Winter Cereal Collection、Tamworth、NSW Australia)からの900の小麦寄託種と300の小麦生産地種を、プライマーARA19F(上記参照)とARA23R(5'−CTGCGCATAAATCCAAACTTCTCG−3')[配列番号19]を用いて多形イントロン3領域に対応するSBEIIbマーカーを用いたPCR増幅でスクリーニングした。PCR増幅は、上述の条件を用いた。増幅産物を制限酵素RsaIで消化して、ポリアクリルアミドゲル上で電気泳動にかけた。3つの系統(Aus12745、Aus17340およびAus10103)はDゲノムマーカーが欠けており、2つの系統(Aus12509およびAus12565)はBゲノムマーカー(図9)が欠けていた。これらの系統は、BまたはDゲノムのためのSBEIIb遺伝子において推定されるヌル変異体を表す。
【0190】
PCRの結果を確認するために、ヌル変異体系統からDNA上で、サザンブロットハイブリダイゼーション分析を行った。標準分析法によって植物から調製したDNAをHindIIIで消化して、1%アガロースゲル上で電気泳動にかけ、Hybond N+ナイロン膜(Amersham)上にブロットした。
【0191】
Megaprime DNAラベリングシステム(Amersham Pharmacia Biotech UK Ltd)を使用して、SBEIIb(位置2019〜2391、図2参照)遺伝子のイントロン3領域から放射性ラベリングしたプローブを生成し、厳しいハイブリダイゼーション条件下に使用した。Aus17340とAus10103は、Dゲノムから〜4.8kBバンドを失っており、Aus12509とAus12565は、Bゲノムから〜3.4kBを失っていた(図10)。したがって、これらの系統は、それぞれ、DまたはBゲノムSBEIIb遺伝子についてのヌル変異体であることが確認された。
【0192】
B+D2倍ヌル変異体の世代
以下の交配を実行した:
Aus17340a×Aus12509
Aus17340b×Aus12509
Aus17340a×Aus12565
Aus17340b×Aus12565
Aus12745 ×Aus12509
Aus12745 ×Aus12565
【0193】
Aus17340aとAus17340bは、同じAus17340栽培品種の2つの異なるバイオタイプであり、両方とも、DゲノムSBEIIb遺伝子マーカーについて無効であることを確認した。F1植物を自家受粉し、F2子孫はBとD両方のゲノムSBEIIb遺伝子の変異体(2倍ヌル変異体)である植物のためのPCR方法によってスクリーニングした。プライマー対AR2b19cF(5'−CTATGCCAATTGAACAACAATGC−3'[配列番号20]とAR2b23cR(5'−CGTGTTCATCAATGTCTGAACG−3'[配列番号21](これは、ARA19F/ARA23Rと同じ領域を増幅する)を用いたPCR増幅後、制限酵素Rsalを用いた消化によってSBEIIb突然変異の分離を観察した。典型的分離パターンを図11に示す。カイ二乗分析で、交配Aus17340a×Aus12509とAus17340a×Aus12565の分離パターンが9:3:3:1(表9)の期待される比率に当て嵌まることが分かった。分離は、他の交配では非常に歪んでいた。
表9.BとDサブゲノムにおけるSBEIIbヌル変異体間の交配のF2集団のカイ二乗分析
【0194】
【表9】
【0195】
表中のχ2(9:3:3:1)の値(0.05)、df3=7.81
【0196】
アルビノ植物は親の系統にかかわりなくすべて交配で検出され、変異体葉緑素関連の遺伝子も集団内で分離していたことを示した。分析した24のアルビノ植物のうち、23はB+D二重ヌル変異体であり、1は野生型であるように見えた。B+D二重ヌル変異を伴う5つの正常に見える緑色植物を、試験した718の系統から確認した。これらのうちの3はAus17340b×Aus12509(BD219、BD303、BD341)、1つはAus17340a×Aus12509(BD54)、1つはAus17340b×Aus12565(BD636)に由来するものであった。この結果から、BおよびDゲノムSBEIIb遺伝子における突然変異は、2つの突然変異した遺伝子位置が一緒になった場合、アルビノフェノタイプを与えている葉緑素関連遺伝子における突然変異と密接な関連があることが明らかになった。しかし、極めて低い頻度ではあるものの、正常のB+D二重ヌル変異体系統をもたらす、SBED遺伝子と葉緑素関連遺伝子の間の組み換え事象が確認された。これは、2つの遺伝子が密接に結合されるが、分離され得ることを示す。
【0197】
<実施例9.SBEIIaとSBEIIbは小麦中で結合している>
BACクローンの単離
A.tauschii meyeri品種(Moullet他、1999年)から作られる大きな挿入コスミドバイナリーコスミド(BAC)ライブラリーをSBEIIb遺伝子のイントロン3領域(位置2019〜2391、図2)でプローブし、SBEIIb遺伝子を含むBACを単離した。4つのポジティブなクローンを単離して、BAC−4、−5、−9および−12と称した。これらがSBEIIb遺伝子を含んでいることを確認するために、これらのクローンからDNAを抽出し、HindIIIまたはEcoRlで消化し、同じプローブ(図12)を使ってサザンブロットハイブリダイゼーションを実行した。クローンBAC−5は、EcoRlを伴う〜7.5kBのサイズの1つの強いハイブリダイズバンドと、HindIIIを伴う〜6.1、3.6、2.3および1.7のサイズの4つのバンドを示した(図12)。これは、BAC−5上でのSBEIIbの存在を示した。BAC−5上での遺伝子の3'の存在を調べるため、SBEIIbcDNA配列に基づく、エキソン17を増幅するように設計された特定のプライマー(AR2b3pr2F、5'−GGATATGTATGATTTCATGG−3'[配列番号22]およびAR2b3pr2R、5'−CCATAAAGTTAAGATAACCC−3')[配列番号23]、および20(AR2b3pr1F、5'−GACATCAGACCACCAGTACG−3')[配列番号24]およびAR2b3pr1R、5'CTTCCCAGGCTTTAAACAGC−3')[配列番号25]を使ってPCR増幅を行った。プライマーのセットは両方とも、エキソン17については128bp、エキソン20については145bpサイズの期待される産物を増幅し、BAC−5がSBEIIbの3'末端を含むことを示した。これは、エキソン20からPCR産物を配列決定することによってさらに確かめられた。
【0198】
SBEIIbに加えてSBEIIa遺伝子の存在についてもBAC−5を調べた。プライマーAR2akpnIF5'−GGTACCGCAGAAAATATACGAGATTGACCC−3'[配列番号26]を使ったヌクレオチド配列決定反応は、SBEIIa遺伝子のイントロン3領域に対応する配列を与え、wSBEII−Dlの位置2265〜2478(図1)に由来する配列と同じであった。この結果は、SBEIIaがBAC−5上にも存在することを示唆し、SBEIIaとSBEIIbがおそらく小麦において密接に結合しているであろうことを意味した。
【0199】
蛍光in situハイブリッド形成(FISH)
Turner他(1999年)によって記載されたようにして、wSBE II−D1ゲノムクローンF2(Rahman他、2001年)とwSBEII−D2クローン(Rahman他、2001年)とのin situハイブリダイゼーションを、Aegilops tauschiiと小麦からの染色体潰砕物に対して実行した。ハイブリダイズした染色体の同一性を、染色体識別(Mukai他、1990年)のために使われる反復塩基配列である二重標識withpSc119.2によって確認した。wSBEIIクローンの両方とも染色体2(図13)の先端領域にハイブリダイズし、小麦における2つのSBEII遺伝子の近くを示した。
【0200】
小麦SBEIIbヌル変異体はSBEIIaについての変異体でもある
先に述べたように確認したSBEIIbヌル変異体を、プライマーSr913F(5'−ATCACTTACCGAGAATGGG−3')[配列番号27]およびE6R(5'−CTGCATTTGGATTCCAATTG−3')[配列番号28]を用いて、SBEIIa遺伝子における突然変異についてスクリーニングした。これらのプライマーは、wSBE II−D1のイントロン5領域を増幅して、A、BおよびDゲノム上でSBEIIa遺伝子を識別するように設計した。
【0201】
SBEIIb Bゲノムヌル変異体Aus12565およびAus12509は、SBEIIa遺伝子のBゲノムヌル変異体でもあると分かった。同様に、SBEIIb、Aus17340およびAus10103のDゲノムヌル変異体は、SBEIIaのDゲノムヌル変異体でもあった。さらにまた、SBEIIb、BD341およびBD636のB+Dゲノム二重突然変異体系統は、SBEIIa遺伝子のB+Dゲノム二重ヌル変異体でもあった。データは、SBEIIaとSBEIIbが、コメとトウモロコシとは対照的に、小麦においては密接に結合することを証明し、上述のこれらの遺伝子のBおよびDゲノムコピーについての突然変異が欠失突然変異を表すことを示す。
【0202】
3重ヌルSBEIIa小麦変異体
上述の方法は、SBEIIaおよび/またはSBEIIbのA−ゲノム変異体を単離させるのに用いてもよい。例えば、SBEIIaおよび/またはSBEIIbに密接に結合するBAC−5領域は、遺伝子中でA−ゲノム突然変異をスクリーニングするためにプローブまたはPCRプライマーの設計のために使われる。A−ゲノム変異体は、B+D二重ヌル系統と交配してA+B+D3重ヌル系統を生産する。あるいは、B+Dゲノム二重ヌル変異体の突然変異生成は照射その他の手段で実行され、三重ヌル変異体は、SBEIIa活性を完全に欠いており、状況に応じてSBEIIb活性が特定される。非常に高いアミロース濃度を備えた非トランスジェニック小麦品種が提供される。
【0203】
<実施例10.小麦におけるSBEIIA遺伝子の突然変異>
SBEIIaの活性低減に至る小麦におけるSBEIIa遺伝子の突然変異は、ガンマ線照射または化学的突然変異誘発、例えばエチルメタンスルホン酸エステル(EMS)を用いて達成する。ガンマ線誘発突然変異のために、種子は60Co源(ZikiryaevaおよびKasimov(1972年)から20〜50kRの量の照射を受ける。EMS突然変異生成は、Mullins他(1999年)に従って種子をEMS(0.03%、v/v)で処理して行う。B+D二重ヌルバックグラウンドでは、変異体穀粒は増加したアミロース含有量または変更したデンプン顆粒形態に基づいて特定され、上述の方法によって確認される。SBEIIb活性を保持するSBEIIaの変異体は再変異してもよく、子孫は、SBEIIaに加えてSBEIIb活性の喪失についてスクリーニングでき、または、SBEIIa変異体はSBEIIb変異体と交配して、突然変異を結合して、胚乳において実質的にSBEII活性が欠如している種類の非トランスジェニック小麦を生産する。
【0204】
<実施例11.SGP−1小麦変異体はSBEIIa活性およびSBEIIb活性において低減している>
小麦(Triticum aestivum)のA、BおよびDゲノムからのデンプン合成酵素II(SSII)遺伝子は、糖−顆粒タンパク質(Starch Granule Proteins−1;SGP−1)としても知られている100−105kDaのポリペプチドをコードする。SSII(SGP−1)は、それぞれ染色体7B、7Aおよび7D(Denyer他、1995年;YamamoriおよびEndo1996年)短腕上遺伝子の相同的集合によってコードされるおよその分子量が100、104および105kDaである3つのポリペプチドからなる。Yamamori他(2000年)は、タンパク質電気泳動による分析で、A、BおよびDゲノムの特異的なSGP−1タンパク質の形態を欠いている系統を交配することによってSGP−1ヌル小麦を生産した。SGP−1ヌルな種子の検査は、突然変異がアミロペクチン構造の変化をもたらし、アミロース含有量を上昇させ、デンプン顆粒を変形させることを示した(Yamamori他、2000年)。そのうえ、成熟した穀粒についての電気泳動実験では、顆粒結合SBEII(SGP−2)とSSI(SGP−3)のレベルがかなり減少することが分かった。SGP−1ヌル系統をもたらす突然変異(複数可)の分子的基礎は知られていなかった。
【0205】
我々は、さらに、成熟穀粒からのデンプン顆粒において完全にSGP−1を欠いている小麦系統を特徴付けるために実験を行った。SSII遺伝子がSGP−1ヌルな小麦のA、BおよびDゲノムの各々に存在するかどうか決定するために、DNAをSGP−1ヌルな小麦と野生型(対照)cvChinese Spring小麦から抽出し、Bゲノムについてはプライマーの組合せssIIa(5'−CCAAGTACCAGTGGTGAACGC−3'[配列番号29]およびssIIb(5'−CGGTGGGATCCAACGGCCC−3'[配列番号30]を用いたPCRによって、AおよびDゲノムについてはssIIaとssIIc(5'−CATGTGAGCTAGCTTTCGCCC−3'[配列番号31]を用いたPCRによって分析した。増幅した領域は、wSSIIA(GenBankアクセッション番号AF155217)またはwSSIIBまたはwSSIIDの対応する領域である位置2472〜2821bpの間にあった。A、BおよびDゲノム産物の明確な識別が可能であったので、増幅した領域はエキソン8の部分を構成して選ばれた。増幅は、94℃で30秒、60℃で1分および702℃で2分の35サイクルを用いて行った。SGP−1ヌルな小麦のA、BおよびDゲノム産物から生産されるPCR断片は、Chinese springから作り出した対応する断片と同じサイズであった。イソアミラーゼとSSI遺伝子(それはSSIIに最も密接に位置するデンプン生合成支配遺伝子であって、SSII(Li他、2002年)の両側に位置する)の遺伝子セグメントのPCR増幅は、これらの遺伝子がSGP−1ヌルな小麦のA、BおよびDゲノムの各々について増幅され得ることを示した。したがって、SGP−1ヌルなフェノタイプは、染色体7短腕上でこれら遺伝子のいずれの欠失にも起因しなかった。
【0206】
走査型電子顕微鏡によって調べた場合、SGP−1ヌルな発育中種子からのデンプン顆粒は、開花後10日から成熟まで明らかに変形した。キャピラリー電気泳動法によって調べた場合、変異体中の脱分枝したデンプンの鎖長分布は、より短いチェーン(重合度8まで)の比率の増加を示す一方、重合度9−22の比率の減少を示す。
【0207】
SGP−I胚乳におけるデンプン合成酵素の発現と分枝酵素
デンプン顆粒中におけるデンプン合成酵素と分枝酵素の発現を、SGP−1ヌルにおいて調べ、野生型栽培品種Chinese Springのそれらと比較した。SGP−1ヌル系統では、種子発育のステージに関係なく、SSII(図14)の欠如に加え顆粒中のSBEIIとSSIの量においてほぼ90%〜96%の顕著な低減があった。特異的抗体の使用は、顆粒から得られるSBEIIバンドがChinese Springではおよそ1:3の比率のSBEIIaとSBEIIbを含むことを示した。SGP−1ヌル変異体においては、量があまりにも低かったため、相対的な比率を、抗体を用いて測定することはできなかった。また、穀粒成長の初期からGBSS Iレベルの低減があった。SGP−1変異体においては、SBEIIaとSBEIIbを含むデンプン顆粒結合ポリペプチドのレベル低減があることは、明らかである。デンプン−顆粒結合ポリペプチド(SBEIIおよびSSI)の低減は、SGP−1ヌル(Yamamori他、2000年)を産生するのに用いられる小麦系統の穀粒では観察されなかった、影響が特にSSIIの欠如に起因することを示唆している。
【0208】
分枝酵素とデンプン合成酵素は、胚乳を成長させる溶液段階でも分析した。溶解性SBEIIbの相対量はChinese SpringとSGP−1ヌル系統で同様であり、変異体の溶解段階にあるSBEIIaの量の低減があった(図15)。しかし、これはSGP−1ヌルな系統の系図に部分的に原因したものでもよい。
【0209】
これらのデータは、SBEIIa活性がSSII遺伝子中の突然変異によって多形質発現的に減少し得ることを証明する。SSIIのみの突然変異が、デンプン中の相対的アミロースレベルを50%未満にさせるが、それはSBEIIa以外の遺伝子の突然変異がSBEII突然変異と組合せ可能であり、アミロースレベルを増やし、変化したデンプンを生じることを示唆する。
【0210】
<実施例12.SBEIのノックアウト−多数のアイソフォーム>
陰イオン交換クロマトグラフィーを通しての小麦デンプン分枝酵素の精製は、活性を3つのピークに分解した(図16、Morell他、1997年)。栽培品種Chinese Spring(CS)からの胚乳抽出は、ポリクローンナル抗体である抗−WBE−1(タンパク質のピーク1(図16B)のN末端配列に対応するアミノ酸配列を有する合成ペプチドに対して誘導されたもの)を用い、非変性PAGE上で4つのSBEIポリペプチドの存在を明らかにした。CSヌルの同染色体−四染色体系統の分析では、これらのポリペプチドが染色体7上にコード化されていることが分かった。免疫ブロットの上のバンドはA(Aバンド)、B(Bバンド)とD(DiとDiiバンド)ゲノムに割り当てられ、それぞれ、活性はA−メジャー活性、B−メジャー活性およびD−メジャー活性と名付けられた。陰イオン交換クロマトグラフィーによって得られる活性なピークを表している精製画分の免疫ブロット分析では、最初のピークがSBEI AメジャーおよびDメジャー活性を含み、第2のピークはSBEI B−メジャー活性を含むことが分かった(図16C)。
【0211】
染色体7上で大きなSBEI活性をコードする遺伝子の位置は、3つのよく特徴が分かっている関連した遺伝子、wSBEI−D2、wSBEI−D3およびwSBEI−D4の決定された位置と一致している。SBEI−メジャーの推定したタンパク質配列は、それがこれらの遺伝子の最後にコードされていることを示した(wSBEI−D4(Rahman他、1997年、Suzuki他、2003年)。第4のSBEI遺伝子の存在は、サザンブロットハイブリダイゼーションデータ(Suzuki他、2003年)に基づいて提案された。
【0212】
SBEIメジャーのヌル変異体の特定
1つまたは複数のアイソフォームの発現が欠如しているヌル変異体を同定するため、非変性ゲル電気泳動の後、SBEI−メジャー免疫ブロット検出によって小麦生殖細胞質のコレクションをスクリーニングした。上述の抗wSBEI抗体を用いた。分析した182のオーストラリア6倍体小麦のアクセッションのうち、SBEI−Dメジャーを発現しない13系統、SBEI−Bメジャーを欠く16系統、SBEI−Aを欠く10系統、およびAとBアイソフォームの両方ともを欠く2つ(BindawarraとVectis)が同定された。これらの系統は、それぞれのゲノムSBEI遺伝子のヌル変異を有すると考えられる。SBEI−メジャー遺伝子のヌル変異の頻度(〜23%)はGBSS遺伝子(22%)(Boggini他、2001年)のそれと同様だった。
【0213】
SBEI−メジャー三重ヌル系統の生成
免疫ブロット分析から、栽培品種BindwarraとVectisがA−メジャーおよびB−メジャーSBEI活性を欠いていることは明らかであり、栽培品種CadouxがD−メジャーSBEI活性を欠くものとして同定された。Vectis×Cadouxの交配から得られる185系統のF2子孫集団を、イムノブロッティングでスクリーニングした。しかし、3つの活性のすべてを欠いている系統は得られず、そのような子孫は生存能力が低いか、あるいは、ゲノム間に何等かの相互作用があることが示唆された。そこで、B−メジャーおよびD−メジャー活性を欠いている子孫系統VC3.1.11を、A−メジャー活性を欠き遺伝子操作したChinese Spring(CS7AL−15)と交配した。二重ハプロイド系統をプライマーARBE1CF(5'−GGGCAAACGGAATCTGATCC−3')[配列番号32]およびARA9R(5'−CCAGATCGTATATCGGAAGGTCG−3')[配列番号33]を用いてPCRとイムノブロッティングの両方でスクリーニングを行い、160のうち2系統(A113とD13)は、非変性ゲル類の上でイムノブロッティングによって判断すると、SBEIメジャー活性を完全に欠いていた。図17は、A113(レーン6)を含む倍加半数体系統の代表的分離パターンを示す。
【0214】
系統A113の胚乳に残存SBE活性がないか調べた。野生型小麦変種(D28)は、SBEI活性の2つのピークを示した。一方、A113抽出物は、最初のピークは与えたが、活性の第2のピークは完全に欠いていた。この活性を含む精製画分から得られるアミノ酸配列は、A113中ではタンパク質のSBEIタイプの存在を示した。しかし、この画分は、非変性ジェル中では抗WBEI抗体との反応を示さなかった。A113中での枝分れ活性は、SBEIIタイプ酵素のものであり得る約80kDaのタンパク質に対応する(それはポテトSBEとトウモロコシSBEII抗体で交差反応した)。
【0215】
これらのデータは、SBEI変異体系統が小麦で発生し得ることを示す。SBEIIaと状況に応じてSBEIIb突然変異によるSBEI突然変異の組合せは、穀粒デンプン中に非常に高いアミロースレベルを有する小麦植物を生産する。
【0216】
<実施例13.SBEII遺伝子中に突然変異を有する染色体2Aを含む変異体小麦の識別>
SBEIIaまたはSBEIIb遺伝子中に突然変異を有する小麦系統を特定するべく、2400の6倍体小麦アクセッションをA、BまたはDゲノム中、SBEIIbのヌル変異についてスクリーニングした。プライマーAR2bl9cF/AR2b23cRを各系統の小麦植物のゲノムDNA試料へのPCR反応で使用し、RsaIで増幅産物の消化を行いゲル電気泳動した。このマーカーはイントロン3領域(小麦SBEIIb遺伝子(図2)のヌクレオチド位置2085〜2336)を増幅して、SBEIIbに特異的だった。上の実施例で記載したように、このスクリーニングでは3つのDゲノムSBEII−ヌル変異体と2つのBゲノムSBEII−ヌル変異体の識別がなされた。SBEIIbと対応しているAゲノムバンドを欠いた変異体系統は検出されなかった。これは、変異体SBEIIb遺伝子を有する染色体2Aを含む小麦系統が自然に存在しないことを示唆した。
【0217】
Tony PriorとRohit Mago(CSIRO)によって生成されたガンマ系統線(60Co源)誘導変異体小麦集団を用いて小麦SBEII中の誘発突然変異をスクリーニングした。小麦集団は、Gabo1BL.1RSとVeery 3とを交配させたF2子孫から生成した。この集団からの合計2694粒の変異体種を、プライマーAR2bl9cFとAR2b23cRを用い先に述べたようにPCR反応でスクリーニングした。1本の植物に由来し、SBEIIb Aゲノムアレルを欠く2個の種子(MLT2B8およびMLT2D1と命名)が同定された(図18)。集団内の種子で、BまたはDゲノム中にSBEIIbのヌル変異を含むものは特定されなかった。
【0218】
上の実施例で示すように、SBEIIaとSBEIIb遺伝子は、小麦では染色体2の長腕上で密接に結合している。したがって、我々は、プライマーSr913F/E6Rを用いたPCR反応で、AゲノムSBEIIa遺伝子の有無についてこれらの種子からのDNAを調べた。これらのプライマーはwSBEII−D1のイントロン5(ヌクレオチド位置2959〜3189、図1[配列番号1])領域を増幅する。増幅後、産物を5%のシークエンシングジェル(ABI Prism DNAシーケンサー)上で電気泳動にかけた。蛍光ラベリングした産物を、ソフトウェアGenescanを用いて分析した。走査プロフィールは変異体種子MLT2B8およびMLT2D1の両方についての増幅産物がAゲノムSBEIIa遺伝子に対応している産物を欠いていることを示し、両方の種子がSBEIIbに加えてAゲノムSBEIIaについてヌルなアレルを有することを示した。
【0219】
これらの種子において、SBEIIaに特異的なAゲノムマーカーであるARIIaAF(5'−GCAAAAGCCAGATCATAAATTTAGAGC−3')[配列番号34]およびAゲノムSBEIIa遺伝子の遺伝子産物のみを増幅するARIIaAR(5'−CTTCCAATTCATTGTTAATGGTCACAC−3')[配列番号35]を用いてヌル変異をさらに確認した(3131 wSBE II−DA1(図1)へのヌクレオチド位置3024)。この一組のプライマーは変種Chinese Springからの植物材料から110bp産物を増幅する一方、この産物は2粒の推定上の変異体種には明らかに欠けていた。これは陰性対照dt2AS(それは染色体2Aの長腕の欠けているChinese Springの染色体操作系統である)に関してと同じである。SBEIIa遺伝子とSBEIIb遺伝子が染色体2の長腕に置かれるので、この系統は両方の遺伝子のAゲノムアレルを欠いており、それゆえに、陰性対照(図19)として用いることができた。
【0220】
変異体種子MLT2B8およびMLT2D1(SBEIIaとSBEIIbに対するAゲノム変異体として識別される)からの胚を育てて植物とする。これら植物由来の種から得られるデンプンは、アミロース含有量、鎖長および他の特性がAゲノム上のSBEIIaとSBEIIb両方のヌル変異がデンプン特性に影響を及ぼすかどうか決定するために分析される。
【0221】
先に述べたように、BとDゲノムの両方にSBEIIaとSBEIIb遺伝子の突然変異を有する5系統が発生した。これらのうち、系統BD 219とBD 636は、温室で成長して、Aヌル変異体系統MLT2B8 とMLT2Dとが交配される。倍加半数体集団は、ホモ接合の三重ヌル変異体植物を提供するために、これらの交配のF1種子から生成される。そのような三重ヌル変異体植物は、8に1の頻度で、倍加半数体集団内に生じるはずである。Aゲノムヌル変異は、類似の交配によってBゲノム突然変異またはDゲノム突然変異と生み合わせることができる。さらなる交配において、ヌルなアレルのいずれかを農学的または他の特徴のために、任意の適当な遺伝的バックグラウンドにも導入することができる。
【0222】
以下の交配は、AゲノムSBEIIaとSBEIIbに突然変異を有するデュラム小麦(例えば品種Wollaroi)を産生するために実行される:
1)Wollaroi×MLT2B8またはMLT2D1(WollaroiバックグラウンドにおいてA−ゲノム SBEIIa/SBEIIbヌルデュラムを生じるため)
2)A−ゲノムヌルなデュラム(Wollaroi)×B−ゲノムヌルSBEIIa/SBEIIb小麦系統(AB2倍ヌルSBEIIa/SBEIIbデュラム(Wollaroi)を生じるため)
あるいは、
1)Wollaroi×B−ゲノムヌル小麦系統(B−ゲノムヌルなデュラムを生じるために)(Wollaroi)
2)B−ゲノムヌルなWollaroi×Aヌル小麦系統(AB2倍のヌルSBEIIa/SBEIIbデュラムを生じるために)。
これらの交配は、高アミロース6倍体小麦のそれと類似している健康に有利な特定の最終用途を有する高アミロースデュラムコムギの生成をもたらす。
【0223】
<実施例14.穀粒中の高アミロース含有量のセファロース2Bカラム分別法による確認>
SBEIIa/SBEIIb阻害遺伝子コンストラクトを含むトランスジェニック小麦植物の穀粒におけるデンプンのアミロース含有量をセファロースカラム分別法により決定する。この方法では、デンプン分子は、その分子量に基づいてカラム上で分離した。次いで、分離した画分を、デンプン分析キット(Sigma社)を利用し供給元の説明書に従って分析した。
【0224】
デンプンおよそ10mgを、37℃で30分間、インキュベーションすることにより、3.0mlの1NNaOH(脱ガス後)に溶解した。未溶解成分を落とすため15分遠心処理した。上清を、1ml/分のポンプ速度でセファロースCL2Bカラムにロードした。緩衝液として10mM NaOHを用い、各2.5ml50画分を集めた。画分9〜50のpHは、1MHC1の35μlで4.5に調整した。各試料のアリコート(250μl)を管に移し、250μlのデンプン試薬(デンプン分析評価キット、Sigma)を添加した。対照は以下のものを含んでいた。デンプン試薬(250μl)と水(250μl)のみを含むブランクのデンプン分析評価試薬、500μlの水のみを含むブランクのブドウ糖分析評価試薬、250μlデンプン試料と250μl水のみを含むブランク試料と250μlのデンプン試薬と250μlデンプン試料のみを含む試料テスト。試料と対照は60℃で60分、次いで、各200μlを新しい管に移し、その後、1mlのブドウ糖試薬(デンプン分析評価キット、Sigma)を添加して30分間、37℃でインキュベートする。デンプンの量(mg)を測定するのには、各画分においてキット添付の説明書に従って340nmの吸光度を用いた。
【0225】
デンプン試料のクロマトグラムは、セファロースカラムから溶出する2つのピークを明らかにした。各試料のアミロース含有量(第2のピーク)を、両方のピークの範囲内のデンプンの総量のパーセンテージとして計算した。
【0226】
この方法を用い、ds−SBEIIaトランスジェニック系統Acc.144087のアミロース含有量(それは導入遺伝子についてはホモ接合性であることが示された)は78%になるように計算され、ds−SBEIIbトランスジェニック系統Acc.144008(イベントIIb 110.16bからのホモ接合のトランスジェニック系統)のそれは23%であると見積った(図20)。比較において、88.47%および27.29%のこれらの系統に対してヨード還元的定法によりアミロース含有量を求めた(表10)。
【0227】
糊化温度、ペースト粘度および膨潤体積などのデンプンなどの機能的な特性を、示差走査熱量測定(DSC)、Rapid Visco Analyser(RVA)およびデンプン膨潤力試験によってそれぞれ測定した。これらのデンプンの構造は、X線結晶学と粒度分析によって分析される。
表10.ヨード還元的定法により評価した小麦トランスジェニック系統のアミロース含有量
【0228】
【表10】
【0229】
<実施例15.鎖長分布分析>
デンプン試料の鎖長分布は、デンプンのイソアミラーゼ脱分枝後、蛍光ラベル糖鎖電気泳動(Fluorophore−assisted carbohydrate electrophoresis:FACE)により測定した。トランスジェニック種子から由来のデンプン中のうち重合度6−11、重合度12−30および重合度31−60からの鎖長のパーセンテージを、非トランスジェニックの対照と比較して表11に示す。高アミローストランスジェニック系統由来のデンプンについて正規化した鎖長分布を同遺伝子型の非形質転換対照由来のデンプンについて正規化した鎖長分布から差し引いたモル差を、図21に示す。
表11.小麦トランスジェニック系統由来のイソアミラーゼ脱分枝デンプンの鎖長分布
【0230】
【表11】
【0231】
ds−SBEIIbトランスジェニック種由来のデンプンにおける重合度4−12の鎖長は、非形質転換種子またはds−SBEIIaトランスジェニック種子由来のデンプンと比較して顕著に低い比率であった。重合度>13の鎖長の比率は、他と比較してds−SBEIIaトランスジェニック種子でより高かった。これらの結果は、SBEIIaが小麦デンプンにおいて重合度4−12のより短い鎖の合成に選択的に関与しているという可能性を示唆する。しかし、SSIIa変異体由来のデンプンにおいては、より短い鎖長の比率がアミロースで増加していた。
【0232】
<実施例16.トランスジェニック小麦由来のデンプンの特性>
ds−SBEIIaおよびds−SBEIIbトランスジェニック系統からの糊化温度を含むデンプンの物性を、Perkin Elmer Diamond示差走査熱量計を使って分析した。各デンプンおよそ20mgを、1:2の比率で水に混合し、すなわち、含水率66.7%として、DSCパンに封入した。1分当たり10℃の加熱速度を用いて、0から150℃まで試験試料と参照試料を加熱した。データは、装置で利用できるソフトウェアを使用して分析した。
【0233】
各デンプンについてサーモグラムDSCトレースにおいて2つの吸熱ピークが観察された。最初のピークは、デンプンの糊化の間の結晶構造の崩壊を意味した。第2のピークは、アミロース−脂質解離の吸熱を表した。ds−SBEIIaトランスジェニック系統からのデンプンの糊化ピーク温度は非形質転換対照デンプンについてのピーク温度と比較しておよそ7〜10℃の増加を示し、ds−SBEIIbトランスジェニック系統からのデンプンと比較しておよそ3〜7℃増加した。
表12.示差走査熱量計(DSC)で測定したトランスジェニック小麦デンプンの熱的性質
【0234】
【表12】
【0235】
これらの系統では、非形質転換対照およびds−SBEIIbトランスジェニック系統と比較して、およそ16〜19℃の糊化(最初のピーク)終了温度の著しい増加が観察された。糊化の開始温度は、ds−SBEIIaトランスジェニック系統で対照またはds−SBEIIbトランスジェニック系統より早く現れた。Ng他(1997年)は、アミロース増量(ae)トウモロコシデンプンの糊化開始温度が通常のトウモロコシデンプンのそれと類似していると報告したが、aeデンプンでは正常のデンプンと比較してピークの糊化温度が顕著に増加していた。ds−SBEIIaトランスジェニック系統からのデンプンの糊化エンタルピーは、対照およびds−SBEIIb系統のそれより顕著に低かった。これは、糊化ピーク面積の顕著な低下(これはds−SBEIIaトランスジェニック系統におけるアミロペクチン量の低減を表している)を反映しているようである。アミロース−脂質分離ピークでは、トランスジェニック系統のいずれでも著しい変化は観察されなかった。したがって、我々は、この新しい一揃いの特性を有するデンプンを得た。
【0236】
<参考文献>
Abel et al., (1996). The Plant Journal 10, 981-991.
Anderson et al., (1989). Nucl Acids Res 17, 461-462.
Baba et al., (1991). Biochem Biophys Res Commun 181: 87-94.
Batey and Curtin. (1996). Starch 48, 338-344.
Batey et al., (1997). Cereal Chemistry 74, 497-501.
Becker et al., (1994). Plant J. 5: 299-307.
Blauth et al., (2001). Plant Physiology 125, 1396-1405.
Bourque. (1995). Plant Science 105, 125-149.
Boyer and Preiss, (1978). Carbohydrate Research 61, 321-334.
Boyer and Preiss, (1981). Plant Physiology 67, 1141-1145.
Boyer et al., (1980). Starch 32, 217-222.
Buleon et al., (1998). International Journal of Biological Macromolecules 23, 85-112.
Cao et al., (2000). Archives. of Biochemistry and Biophysics. 373, 135-146.
Case et al., (1998). Journal of Cereal Science 27, 301-314.
Cheng et al., (1997). Plant Physiol 115: 971-980.
Craig et al., (1998). Plant Cell 10, 413-426.
Denyer et al., (1995). Planta 196: 256-265.
Denyer et al., (1996). Plant Physiology 112, 779-785.
Feldman. (***) pp 3-56 in The World Wheat Book, A history of wheat breeding. Eds Bonjean and Angus, Lavoisier Publishing, Paris.
Fergason. 1994. pp 55-77 in "Speciality Corns" eds , CRC Press Inc.
Filpse et al.,(1996). Planta 198, 340.
Fisher et al., (1993). Plant Physiol 102:1045-1046.
Fisher et al., (1996). Plant Physiol 110: 611-619.
Fuwa et al., (1999). Starch/Starke. 51, 147-151.
Gao et al., (1997). Plant Physiol 114: 69-78.
Gao et al., (1998). Plant Cell 10, 399-412.
Giroux and Hannah. (1994). Molecular and General Genetics 243, 400-408.
Green et al., (1997). Plant Physiology 114, 203-212.
He et al., (1994). Plant Cell Reports 14: 192-196.
Hedman and Boyer, (1982). Biochemical Genetics 20, 483-492.
Hess et al., (1990). Plant Science 72: 233-244.
James et al., (1995). Plant Cell 7, 417-429.
Jobling et al., (1999). Plant Journal 18, 163-171.
Komari et al., (1996). Plant Journal 10:165-174.
Konik-Rose et al (2001) Starch 53, 14-20.
Krueger et al., (1987). Cereal Chemistry 64, 187-190.
Kubo et al., (1999). Plant physiology. 121, 399-409.
Li et al., (1999a). Plant physiology. 120, 1147-1155.
Li et al., (1999b). Theoretical and Applied Genetics 98, 1208-1216.
Li et al., (2000). Plant Physiology 123, 613-624.
Li et al., (2003). Funct Integr Genomics 3:76-85.
Maniatis et al., (1982). Molecular cloning: a laboratory manual. Cold Spring Harbour Laboratory Press New York.
McCreery and Helentjaris (1994). Methods in Molecular Biology, Vol. 28: Protocols for nucleic acid analysis by non-radioactive probes, 67-71, Humana Press Inc., Totawa, NJ.
Mizuno et al., (1993). Journal of Biological Chemistry 268, 19084-19091.
Mizuno et al., (1992). Journal of Biochemistry 112, 643-651.
Morell et al., (1997). Plant Physiology 113, 201-208.
Morell et al., (1998). Electrophoresis 19, 2603-2611.
Morell et al., (2003). Plant J. 34: 173-185.
Morrison and Laignelet (1983). Journal of Cereal Science 1:9-20.
Mullins et al., (1999). European Journal of Plant Pathology 105: 465-475.
Myers et al., (2000). Plant Physiology 122, 989-997.
Nakamura (2002). Plant Cell Physiology 43, 718-725.
Nakamura and Yamanouchi (1992). Plant Physiol 99: 1265-1266.
Nair et al., (1997). Plant Sci 122: 153-163.
Nehra et al., (1994). Plant J. 5: 285-297.
Ng et al., (1997) Cereal Chemistry 74: 288-292.
Nishi et al., (2001). Plant Physiology 127, 459-472.
Rahman et al., (1995). Australian Journal of Plant Physiology 22, 793-803.
Rahman et al., (1997). Genome 40: 465-474.
Rahman et al., (1999). Theor Appl Genet. 98: 156-163.
Rahman et al., (2000). J Cereal Sci 31: 91-110.
Rahman et al., (2001). Plant Physiol 125: 1314-1324.
Repellin et al., (1997). Plant Gene Reg 97-094
Safford et al., (1998). Carbohydrate Polymers 35, 155-168.
Schulman and Kammiovirta, (1991). Starch 43, 387-389.
Schwall et al., (2000). Nature Biotechnology 18, 551-554.
Senior (1998). Biotechnology and Genetic Engineering Reviews 15, 79-119.
Shannon and Garwood, (1984). In Starch: Chemistry and Technology, Whistler et al., eds, Academic Press, Orlando, FL, pp25-86.
Shure et al., (1983). Cell 35, 225-233.
Sidebottom et al., (1998). Journal of Cereal Science 27, 279-287.
Stacey and Isaac (1994). Methods in Molecular Biology, Vol. 28: Protocols for nucleic acid analysis by non-radioactive probes, pp 9-15, Humana Press Inc., Totawa, NJ.
Sun et al., (1997). The New Phytologist 137 , 215-215.
Takeda et al., (1993a). Carbohydrate Research 240, 253-262.
Takeda et al., (1993b). Carbohydrate Research 246, 273-281.
Thomas and Atwell 1999 Starches Eagen Press, St Paul, Minnesota, USA pp: 13-24.
Thorbjornsen et al., (1996). Plant Journal 10, 243-250.
Vasil et al., (1992). Bio/Technology 10: 667-674.
Vasil et al., (1993). Bio/Technology 11: 1553-1558.
Wang et al., (1998). Journal of Experimental Botany 49, 481-502.
Weeks et al., (1993). Plant Physiol 102: 1077-1084.
Wegener et al., 1994. Mol. Gen Genet. 245, 465-470.
Weir et al., (2001). Aust J Plant Physiol 28: 807-818.
Yamamori and Endo, (1996). Theoretical and Applied Genetics 93, 275-281.
Yamamori et al., (2000). Theor.Appl.Genet. 101, 21-29
Young. (1984). in Whistler et al. (eds), Academic Press, Orlando, FL, chap 8.
Zhao and Sharp, (1998). Plant Breeding 117: 488-490.
Zikiryaeva and Kasimov, (1972). Uzbekskii Biologicheskii Zhurnal 6, 18-20.
Zwar and Chandler, (1995). Planta 197, 39-48.
【図面の簡単な説明】
【0237】
【図1】A.tauschii(タルホ小麦)から得たデンプン分枝酵素IIa遺伝子(wSBEII−D1)の配列を示す表である[配列番号1]。6倍体の小麦(T.aestivum(普通小麦))のゲノムDのSBEIIa遺伝子に対応している。
【図2】T.aestivum(普通小麦)から得た小麦のSBEIIb遺伝子配列(wbe2bゲノム)の一部を示す表である[配列番号2]。
【図3】二重鎖RNAコンストラクトの概略図である。A:使用されている遺伝子要素の順序は、センス方向にプロモーター、SBEIIaまたはSBEIIb遺伝子配列(エキソン1、2および3)、アンチセンス方向に、イントロン(イントロン3)、SBEIIaまたはSBEIIb遺伝子配列(エキソン1、2、3および4)、および転写ターミネーター/ポリアデニル化配列であった。B:ds−SBEIIaおよびds−SBEIIb遺伝子の転写物は、センス配列とアンチセンス配列のハイブリダイゼーションにより形成された二本鎖部分を有する「ヘアピン」RNA構造を形成する。ヌクレオチドのGTおよびAGにはさまれたイントロン配列はスプライシングされる。
【図4】光学顕微鏡で観察したデンプン粒の写真である。A)は、ds−SBEIIaトランスジェニック系統83.1bに由来する野生型デンプン粒を含む小麦種子でから得たデンプン粒であり、B)は、ds−SBEIIaトランスジェニック系統50.1bに由来する変形デンプン粒を含む小麦種子から得たデンプン粒である。
【図5】図4の小麦種子から得たデンプン粒の複屈折性を偏光下で可視化した写真である。
【図6】小麦SBEIIaのcDNA配列の一部分を比較する表である。sbe9は、AF338432.1の一部に対応する。Y11282[配列番号3]、sr997[配列番号4]、sr995[配列番号5]、sbe9[配列番号6]の配列の一部を示している。
【図7】初めの63個のアミノ酸に基づき小麦のSBEIIa配列の一部分をパイルアップ(PILEUP)比較した図である。クローンに対応する遺伝子の推定されるゲノム位置を示している。
【図8】D−ゲノムポリペプチド(sr854)[配列番号7]の推測されるアミノ酸配列をAゲノムまたはBゲノム(y11282)[配列番号8]の産生物と比較した表である。トランジット配列(位置1〜54)はイタリック体で示している。
【図9】様々な小麦登録種から得たSBEIIb遺伝子のイントロン3領域をPCR増幅した結果を示す図である(レーン1〜11)。PCR増幅は、プライマーARA19FおよびARA23Rを用い、次にRsa1で消化して行った。ゲノムA、B、およびDに対応するバンドを矢印で示している。レーン3(Aus17340)およびレーン5(Aus10103)は、ゲノムDに特異的なマーカーを欠いており、一方、レーン8(Aus12509)およびレーン9(Aus12565)は、ゲノムBのマーカーを欠いている。
【図10】SBEIIbのイントロン3領域に由来するプローブを用いて、小麦登録種由来のHindIIIで消化したDNAをサザンハイブリダイゼーションした結果を示す写真である。各レーンは、1)Aus12565、2)Aus12509、3)Aus10103、4)CSDT2DL−4、5)Aus12530(デュラム小麦)、6)CSDT2BL−9、7)Aus6323、8)CSDT2DS、9)Aus17340、10)Aus12745、11)CSDT2DL−4、12)Aegilops tauschii(タルホ小麦)に対応している。
【図11】SBEIIbのイントロン3領域のPCR増幅により、Aus17340aとAus12509を交配したF2集団をスクリーニングした結果を示す図である。PCR増幅は、プライマーAR2b19cFおよびAR2b23cRを用い、次にRsaIで消化して行った。レーン8はゲノムBのマーカーとゲノムDのマーカーの双方を欠いており、したがってBD54系統は、BDの二重ヌル(double−ヌル)系統を示している。
【図12】SBEIIbのイントロン3領域に由来するプローブを用いて、HindIII(レーン1〜4)およびEcoR1(レーン5〜8)で消化したBACクローンをサザンハイブリダイゼーションした結果を示す写真である。各レーンは、1)BAC4、2)BAC5、3)BAC9、4)BAC12、5)BAC4、6)BAC5、7)BAC9、8)BAC12に対応している。
【図13】A)A.tauschii(タルホ小麦)染色体(大きな写真および挿入写真の下の方)および小麦染色体(挿入写真の上の方)に対してwSBEII−DA1プローブおよび反復DNA配列プローブ(pSc 119.2)を用いて行った蛍光in situハイブリダイゼーション(FISH)法の結果を示す写真である。 B)小麦染色体に対してSBEIIbプローブを用いて行った蛍光in situハイブリダイゼーション(FISH)法の結果を示す写真である。
【図14】指定した種子発育のいくつかの段階(開花の10、15、25日後、M=成熟)の野生型Chinese Spring(CS)系統およびSGP−1 ヌル小麦系統の顆粒結合タンパク質をSDS−ポリアクリルアミドゲル電気泳動(PAGE)により分析した結果を示す図である。銀染色ゲルの画像のタンパク質バンド強度を測定した。成熟したCS種子のGBSSのバンド強度を基準の100とし、指定した発育段階における他の酵素の量を、成熟CSのGBSSに対するパーセンテージとして表している。a)がGBSS、b)がSSI、c)がSBEIIである。黒い棒グラフは、SGP−1 ヌルを示す。CSおよびSGP−1 ヌル系統から得た顆粒結合タンパク質の電気泳動画像の例を示す。
【図15】可溶性画分中のSBEIIaおよびSBEIIbの相対量を示すグラフである。SDS−PAGEのイムノブロットをスキャンし、画像のタンパク質バンド強度を測定した。SBEIIaとSBEIIbの融合タンパク質を標準としてゲル上で使用して、タンパク質の量を概算した。
【図16】A.小麦(cv Rosella)胚乳分枝酵素活性を陰イオン交換クロマトグラフィーにより分析した結果を示すグラフである。胚乳の可溶性タンパク質を硫酸アンモニウムで分画し、セファクリル(Sephacryl)S−200カラム、続いてResource Q陰イオン交換カラムに添加してクロマトグラフィー分析をした。B)小麦胚乳SBEIの抗−WBE1抗体を用いて免疫検出分析を行い、非変性ポリアクリルアミドゲル電気泳動(PAGE)で分離させた結果を示す写真である。AおよびBとして示すSBEIタンパク質バンドは、それぞれゲノムAおよびゲノムBに由来する産生物であり、DiおよびDiiは、ゲノムDに由来する産生物である。レーン1はCS、レーン2はN7BT7A、レーン3はN7AT7B、レーン4はN7DT7Aから得た抽出物に対応するC)抗−WBE1抗体を用いた陰イオン交換クロマトグラムで活性なピークを示す、精製画分の免疫検出分析の結果を示す写真である。レーン1は、胚乳の粗製な可溶性抽出物であり、レーン2の画分はピーク1を示し、レーン3の画分はピーク2を示している。
【図17】抗WBEI抗体を用いた免疫検出によりSBEIアイソフォームを分離するために、VC3.1.11とCS7AL−15の交配種の倍加半数体の子孫をスクリーニングした結果を示す写真である。レーン1〜14は、倍加半数体の子孫系統に対応している。レーン6は、A113と呼ばれる三重(triple)ヌル型SBEI変異系統であり、レーン7は、D28と呼ばれるSBEIアイソフォームが正常な系統である。
【図18】Veery3とGabo 1BL.1RSの交配種のガンマ線で誘発した突然変異種子から得たDNA(レーン1〜6)を、プライマーAR2b19cF/AR2b23cRを用いてPCR増幅した結果を示す写真である。レーン2は、変異種子MLT2B8を示し、レーン7は、Chinese Springを示す。
【図19】小麦SBEIIa遺伝子のゲノムA特異的プライマー、すなわちARIIaAF/ARIIaARを用いて、小麦系統から得たDNAをPCR増幅した結果を示す写真である。レーン1〜5は、順に、CS、MLT2B8、MLT2D1、Dt2AS、およびBD219(ゲノムBとゲノムDの双方のSBEIIaおよびSBEIIb植物がヌル変異体である植物)である。
【図20】デンプンアッセイキット(Sigma社)を用いて、小麦系統a)Acc144008およびb)Acc144087から得たデンプンをセファロース(Sepharose)CL2Bゲルクロマトグラムにより分析した結果を示すグラフである。
【図21】小麦のトランスジェニック系統から得たデンプンの鎖長の特性を、形質転換されていないコントロールのNB1(小麦)の鎖長特性に対して比較した結果を示すグラフである。形質転換されていないコントロールから得たデンプンの個々の少糖類の全重量のパーセンテージを、トランスジェニック系統から得たデンプンの対応する値から引いている。サンプルの凡例は、085(黒の菱形)、025(黒の三角)、008(白丸)である。
【技術分野】
【0001】
本発明は、相対的にアミロース含有量が高い穀粒デンプンを有する小麦植物に関する。本発明はまた、胚乳中のデンプン分枝酵素IIa(SBEIIa)の活性を低下させた小麦、およびこのような植物を得る方法にも関する。本発明はまた、穀粒およびデンプン、ならびにそれから得た食料品および非食料品にも関する。
【背景技術】
【0002】
穀類では、成熟した穀粒の約45〜65%重量をデンプンが占めている。デンプンは、2つのタイプの分子、すなわちアミロースおよびアミロペクチンからなっている。アミロースは、α−1,4結合したグルコシド鎖からなるほぼ直鎖状の分子であり、一方、アミロペクチンは直鎖同士を結合するα−1,6グルコシド結合で高度に枝分かれしている。
【0003】
高等植物の胚乳におけるデンプンの合成は、4つの重要なステップを触媒する一揃いの酵素群によって行われる。第1に、ADP−グルコースピロホスホリラーゼが、G−1−PおよびATPからADP−グルコースを合成することによって、デンプンのモノマー前駆物質を活性化する。第2に、活性化されたグルコシル基供与体、すなわちADP−グルコースが、デンプン合成酵素によって、既存のα−1,4結合の非還元末端に移される。第3に、デンプン分枝酵素が、α−1,4結合したグルカン部分を切断することにより分岐点を導入し、続いて切断した鎖を受け入れ先の鎖に移して新しいα−1,6結合を形成させる。デンプン分枝酵素は、α−ポリグルカンにα−1,6結合を導入できる唯一の酵素であり、したがって、アミロペクチンの形成において極めて重要な役割を果たしている。最後に、作用機序は解明されていないが、デンプン脱分枝酵素が分枝結合の一部を除去する(Myers他、2000年)。
【0004】
少なくともこれら4種の酵素活性が高等植物における通常のデンプン粒合成に必要であることは明らかであり、4種の酵素活性のそれぞれについて複数のアイソフォームが高等植物の胚乳中で発見されており、変異解析に基づいて(Wang他、1998年、Buleon他、1998年)、または遺伝子導入手法を用いて遺伝子発現レベルを改変することによって(Abel他、1996年、Jobling他、1999年、Scwall他、2000年)、個々のアイソフォームの具体的な役割が提示されてきた。しかし、デンプン生合成に対する各酵素活性の各アイソフォームの正確な寄与は依然として分かっておらず、これらの寄与が種間で著しく異なるかどうかも分かっていない。穀類の胚乳には、ADP−グルコースピロホスホリラーゼの2種のアイソフォームが存在する。すなわちアミロプラスト内部に1つの型、細胞質に1つの型が存在している(Denyer他、1996年、Thorbjornsen他、1996年)。各型は、2種のサブユニットからなっている。トウモロコシのしわ型(shrunken)(sh2)変異体および脆性(brittle)(bt2)変異体は、それぞれ大サブユニットおよび小サブユニット中の損傷を示している(GirouxおよびHannah、1994年)。穀類の胚乳には次の4クラスのデンプン合成酵素が存在している。デンプン粒の内部にもっぱら局在しているアイソフォーム、すなわち顆粒結合型デンプン合成酵素(GBSS)、顆粒と可溶性画分の間に分配されている2種の型(SSI(Li他、1999年a);SSII(Li他、1999年b))、ならびに可溶性画分中に全体がある4番目の型、すなわちSSIII(Cao他、2000年、Li他、1999年b、Li他、2000年)である。GBSSはアミロース合成に不可欠であることが示されており(Shure他、1983年)、SSIIおよびSSIIIの変異によりアミロペクチン構造が変えられることが示されている(Gao他、1998年、Craig他、1998年)。SSI活性の役割を定義する変異は記載されていない。
【0005】
分枝酵素の3種の型、すなわち、分枝酵素I(SBEI)、分枝酵素IIa(SBEIIa)、分枝酵素IIb(SBEIIb)は、穀類の胚乳で発現される(HedmanおよびBoyer、1982年、BoyerおよびPreiss、1978年、Mizuno他、1992年、Sun他、1997年)。コメ(NakamuraおよびYamanouchi、1992年)、トウモロコシ(Baba他、1991年;Fisher他、1993年;Gao他、1997年)、および小麦(Repellin他、1997年;Nair他、1997年;Rahman他、1997年)のゲノム配列およびcDNA配列は特徴を明らかにされている。配列アラインメントにより、ヌクレオチドレベルでもアミノ酸レベルでも配列の類似性の程度が高いことが判明しており、SBEIクラス、SBEIIaクラス、およびSBEIIbクラスへの分類が可能である。SBEIIaおよびSBEIIbは一般に、特に遺伝子の中心領域において、互いに約80%の配列同一性を示す。SBEIIaおよびSBEIIbは、その発現パターンによって区別することもできる。SBEIIbは一般に胚乳で特異的に発現されるが、SBEIIaは植物のすべての組織中に存在する。
【0006】
小麦胚乳では、SBEI(Morell他、1997年)はもっぱら可溶性画分に存在し、一方、SBEIIaおよびSBEIIbは、可溶性画分にもデンプン粒結合画分にも存在している(Rahman他、1995年)。トウモロコシおよびコメでは、高アミロースのフェノタイプは、アミロース増量(ae)遺伝子としても知られているSBEIIb遺伝子の損傷に起因することが示されている(BoyerおよびPreiss、1981年、Mizuno他、1993年;Nishi他、2001年)。これらSBEIIb変異体では、胚乳のデンプン粒が異常な形態を示し、アミロース含有量はかなり高くなり、残存するアミロペクチンの分岐頻度は少なくなり、短鎖(<重合度17、特に重合度8〜12)の比率が低くなっていた。さらに、デンプンの糊化温度は高くなっていた。加えて、アミロースとアミロペクチンの「中間体」と定義される物質がかなり溜まっていた(Boyer他、1980年、Takeda、他、1993年b)。一方、突然変異誘発(Mu)挿入因子によってSBEIIa遺伝子で突然変異が起こり、その結果としてSBEIIaタンパク質発現を欠いているトウモロコシ植物は、葉のデンプンでは分岐が変わっていたが、胚乳デンプンの分岐については野生型植物との区別が不可能であった(Blauth他、2001年)。同様に、SBEIIa活性が欠損したコメ植物でも、胚乳(Nakamura 2002年)のアミロペクチン鎖の性質に有意な変化は起きなかった(Nakamura 2002年)。トウモロコシおよびコメのいずれにおいても、SBEIIa遺伝子およびSBEIIb遺伝子はゲノム中で互いに結合していない。
【0007】
トウモロコシでは、dull1変異が起こると、変化の程度は遺伝的背景に応じて変わるが、デンプン含有量が少なくなり、胚乳のアミロースレベルが高くなり、残存アミロペクチンの分岐の程度が高くなる(ShannonおよびGarwood、1984年)。この変異に対応する遺伝子は、トランスポゾン突然変異誘発遺伝子(Mu)を用いるトランスポゾン標識法によって同定および単離され、デンプン合成酵素II(SSII)と呼ばれる酵素をコードしていることが示されている(Gao他、1998年)。現在この酵素は、穀類のSSIIIファミリーのメンバーとして認識されている(Li他、2003年)。変異胚乳では、dull1変異に伴ってSBEIIa活性のレベルが低下していた。他の穀類では、これに対応する変異は報告されていない。これらの研究結果が他の穀類、例えば小麦に関連性があるかどうかは分かっていない。
【0008】
WO94/09144では、センス遺伝子およびアンチセンス遺伝子を使用してトウモロコシのデンプン合成酵素(SS)とSBEとの天然の比率を変えることを示唆している。しかし、提案されている分子戦略を実証するデータは何も提示されておらず、SBEIIa活性を特異的に低下させることは示唆されていない。
【0009】
ジャガイモでは、その後の研究によりいくらかの性質の変化が確認されたものの(Safford他、1998年)、SBEIのみをダウンレギュレーションしてもデンプン構造に最小限の影響しか与えない(Filpse他、1996年)。しかし、ジャガイモでSBEIIおよびSBEIを一緒にダウンレギュレーションすると、SBEIIのみをダウンレギュレーションした場合よりもはるかに相対的アミロース含有量が増加した(Schwall他、2000年)。
【0010】
高等植物には2種類の型の脱分枝酵素が存在しており、その基質特異性に基づいて、イソアミラーゼ型脱分枝酵素およびプルラナーゼ型脱分枝酵素と定義されている(Myers他、2000年)。トウモロコシおよびコメにおけるシュガリー(sugary)−1変異は、双方の脱分枝酵素の欠損に関連付けられている(James他、1995年、Kubo他、1999年)が、原因となる変異は、イソアミラーゼ型脱分枝酵素遺伝子と同じ場所に位置している。穀類からクローン化されている代表的なデンプン生合成遺伝子を表1に挙げる。
表1.穀類で明らかにされているデンプン分枝酵素遺伝子
【0011】
【表1】
【0012】
デンプンは、食品工業、製紙工業、および化学工業で広く使用されている。デンプンの物理的構造は、食料品または非食料品もしくは工業製品用のデンプンの栄養特性および取扱特性に大きな影響を与え得る。アミロペクチン鎖長の分布、結晶性の程度およびタイプ、ならびに糊化温度、粘度、膨潤体積などの特性を含めて、いくつかの特質をデンプン構造の指標とみなすことができる。アミロペクチン鎖長の変化は、結晶性の変化、糊化、またはアミロペクチンの老化の指標とすることができる。
【0013】
デンプン組成物、特にアミロース含有量が高いことに関連付けることができるレジスタントスターチと呼ばれる型は、腸の健康、特に大腸の健康に重要な意味をもっている。したがって、腸の健康を促進する手段として食品に使用するために、トウモロコシなどいくつかの穀物において高アミロースデンプンの開発が行われてきた。レジスタントスターチの有益な効果は、大腸に栄養物が供給され、そこで大腸内の腸内微生物叢にエネルギー源が与えられ、このエネルギー源が発酵されて、特に短鎖脂肪酸が形成されることにより得られる。これらの短鎖脂肪酸は、結腸細胞に栄養物を提供し、一部の栄養物の大腸からの摂取を増進し、結腸の生理活性を促進する。一般に、レジスタントスターチまたは他の食物繊維が提供されない場合は、結腸は代謝的に比較的不活発である。
【0014】
化学的方法あるいは別法で改変されたデンプンは、食品に利用することができ、改変されていない供給源によっては通常得られない機能性を与えるが、そのような改変は、改変に伴うプロセスが原因となって、価値のある他の成分を改変しまたは望ましくない感覚をもたらす傾向がある。したがって、未改変の形態で食品に使用できる構成成分からなる供給源を提供することが好ましい。
【0015】
全世界で毎年生産される小麦の量は、穀物作物のうちで最も多い。小麦のデンプン構造で分かっている変異は、トウモロコシまたはコメで得られる変異と比べて限定されており、小麦の形質転換効率が他の穀類での効率に遅れをとっていること、ならびにパン小麦が6倍体の性質を有することがその理由の一部である。Triticum aestivum(普通小麦)には3種のゲノムが存在するため、2倍体種では変異をより容易に確認できるのに対して、個々のゲノムの変異を隠すことによる緩衝効果がある。トウモロコシまたはコメのアミロース増量フェノタイプに対応するSBEIIbの変異体は、小麦では明らかにされていない。SBEIIa変異またはSBEIIb変異によって与えられる小麦のフェノタイプは分かっていない。モチ性遺伝子(GBSS、ZhaoおよびSharp、1998年)の変異体およびSGP−1タンパク質を完全に欠いた変異体(Yamamori他、2000年)が知られている。後者の変異体は、タンパク質電気泳動での分析によりSGP−1(SSII)タンパク質のゲノムA、ゲノムB、およびゲノムDの個々の型を欠く系統を交配することによって作製された。SSII ヌル型の種子の調査により、変異によってアミロペクチン構造が変化し、デンプン粒が変形し、相対的アミロース含有量がデンプンの約30〜37%に増え、野生型のレベルより約8%多くなることが示された(Yamamori他、2000年)。比色測定、電流滴定(いずれもヨウ素結合について行う)、およびコンカナバリンA法によってアミロースを測定した。SSII ヌル変異体から得たデンプンは、等価な非変異型の植物から得たデンプンと比べて、低い糊化温度を示した。SSII−ヌル型穀粒のデンプン含有量は、野生型での60%から50%未満にまで減少していた。
【0016】
WO99/14314では、小麦関連の2倍体植物であるAegilops tauschii(タルホ小麦)からのSBEIIa遺伝子の単離について記載しているが、改変デンプンを含む小麦は作製しなかった。
【0017】
WO00/15810では、小麦のSBEIIb遺伝子のcDNAクローニングについて記載している。彼らは、アミロースレベルを変えた小麦植物を得ることはせず、アミロースを少なくとも50%含むデンプンを有する小麦を教示しなかった。
【0018】
WO01/62934も、小麦のSBEIIb遺伝子について記載し、小麦植物に分枝酵素活性の阻害物質を導入することを示唆しているが、アミロースを少なくとも50%含むデンプンを有する小麦を教示していない。
【0019】
WO01/32886は、小麦胚乳に存在するSBEIの1つの型をコードしているcDNAを明らかにした。コードされているポリペプチドは、A型のデンプン粒に優先的に関連していることが発見された。彼らは、SBEI活性を抑制することも、改変されたデンプン粒の形態を示すことも、小麦のアミロース量を増やすこともしなかった。
【0020】
したがって、アミロース比率が約50%より高いデンプンを有する小麦は知られていない。高アミロースのトウモロコシ変種およびオオムギ変種が知られているが、これらの穀類から作られた製品は、小麦が好ましい穀類である製品に関しては、例えばパン、パスタ、またはヌードルではアミロース含有量が非常に多い小麦と比べて不利である。
【0021】
これらのタイプの高アミロースデンプンは有用であるが、アミロース含有量のより高い小麦デンプンが、特にデンプン合成の改善および他の特質、例えば収穫後改変の必要が減ることと関連している場合は好ましい。このようなデンプン製品はまた、消化に対して比較的抵抗性があり、健康面でより大きな利益をもたらす。
【0022】
当業者なら、本明細書で記述する本発明が、具体的に記述するもの以外の変更や修正を受けることが分かるであろう。本明細書で記述する本発明が、このような変形法および改良法のすべてを含むことを理解すべきである。本発明はまた、本明細書で参照または指示するこのようなステップ、特徴、組成物、および化合物のすべても個別にまたは合わせて含み、また、前記ステップまたは特徴のうちのいずれか2つ以上からなる任意の組合せおよびすべての組合せも含む。
【0023】
本明細書を通じて、文脈上別の意味で解釈する必要がない限り、「含む(comprise)」という単語および「comprises」や「comprising」などの変形は、記載した整数(integer)もしくはステップ、または整数(intergers)もしくはステップの群を包含するが、他の整数(integer)およびステップ、ならびに整数(integers)およびステップの群のどれも除外しないことを意味しているものと理解されたい。本発明は、本明細書で記述する個々の実施形態によって範囲を限定されるべきではなく、これらの実施形態は例証するためのものであるにすぎない。本明細書で記述するように、機能的に等価な製品、組成物、および方法が本発明の範囲に含まれることは明らかである。
【0024】
本明細書中で本発明者らが参照した刊行物の書誌詳細は、明細書の最後にまとめている。本明細書で記述する参考文献は、その全体を参照により本明細書に組み込む。従来技術に関する本明細書の参考文献は、任意の1種または複数の従来技術の文献を含めて、前記の従来技術がオーストラリアで周知の一般的知識であり、あるいはオーストラリアで周知の一般的知識の一部を形成していることを認めるものまたは示唆するものとしてみなすべきではない。
【0025】
本明細書では、「に由来する、から得た(derived from)」という用語は、特定の完全体(integer)または完全体群(integers)が、指定した種に由来するものであるが、必ずしも指定の供給源から直接得られているわけではないことを示すと理解すべきである。
【0026】
本明細書で参照するヌクレオチド残基の記号は、IUPAC−IUB生化学命名法委員会により推奨される記号であり、Aはアデニンを示し、Cはシトシンを示し、Gはグアニンを示し、Tはチミジンを示す。
【発明の開示】
【0027】
<発明の概要>
【0028】
第1の態様で、本発明は、穀粒のデンプン中のアミロース比率が少なくとも50%である小麦植物から得た穀粒を提供する。この小麦植物は、SBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低くてよく、SBEIIaとSBEIIbの両方の遺伝子発現、酵素活性、またはその双方のレベルが低いのが好ましい。この穀粒は、遺伝的変異、すなわちSBEIIa遺伝子発現、酵素活性、またはその双方を阻害するSBEIIa遺伝子の変異、あるいはSBEIIa遺伝子発現、酵素活性、またはその双方を阻害する導入核酸のいずれかを含んでよい。さらに、この穀粒はSBEIに同様の遺伝的変異を含んでもよい。この穀粒は、ADPグルコースピロホスホリラーゼ、GBSS、SSI、SSII、SSIII、イソアミラーゼ型脱分枝酵素、及びプルラナーゼ型脱分枝酵素からなる群から選択されるタンパク質および/または酵素活性を、改変されたレベルでさらに含んでもよい。この穀粒は、導入遺伝子を含んでよく、また、この導入遺伝子は、アンチセンスRNA分子、コサプレッションRNA分子、リボザイムRNA分子、または二重鎖RNA分子をコードしてよい。この導入遺伝子によって、SBEIIaをコードしているmRNAの発現レベルが低められることが好ましい。この穀粒は、SBEIIa遺伝子に変異を含んでよく、1つの形態では少なくとも1つのゲノムにSBEIIa遺伝子のヌル変異があり、ゲノムのうちの2つもしくは3つにヌル変異があってもよい。穀粒のデンプン中のアミロース比率は、少なくとも40%、50%、55%、60%、70%、または80%でよい。別の形態では、穀粒内部のデンプン粒の少なくとも50%は、偏光下で観察すると非複屈折性に見える。穀粒は、非しわ型でよく、平均重量は少なくとも36mgまたは40mgである。代わりの形態では、殻のない状態での穀粒のデンプン含有量は、少なくとも25%(w/w)または少なくとも35%(w/w)であり、野生型穀粒のデンプン含有量の少なくとも90%でよい。穀粒は、全粒、玄穀、製粉した穀粒、挽き割りにした穀粒、圧扁した穀粒、精白した穀粒、粉砕した穀粒、またはパーボイルド穀粒でよい。
【0029】
第1の態様の別の形態で、本発明は、デンプンと、野生型の穀粒に比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルを低下させる遺伝的変異とを含む、小麦植物から得た穀粒を提供する。この遺伝的変異は、SBEIIa遺伝子の変異またはSBEIIa遺伝子の発現の阻害物質をコードする導入核酸を含み、穀粒のデンプン中のアミロース比率は少なくとも30%である。
【0030】
第2の態様では、本発明は、それだけには限らないが、小麦粉、全粒小麦粉、セモリナ粉を含めて第1の態様の穀粒に由来する製粉製品または本発明の穀粒から得たデンプン、あるいはこのような小麦粉、全粒小麦粉、セモリナ粉、デンプンを組み込んだ食料品、あるいは穀粒を圧扁した製品、薄く削った製品、または押し出した製品を提供する。この製品には、別の供給源から得た小麦粉、全粒小麦粉、セモリナ粉、またはデンプンと混合した、本発明の第1の態様の穀粒から得た小麦粉、全粒小麦粉、セモリナ粉、またはデンプンを含めてよい。
【0031】
第3の態様では、本発明は、第1の態様の小麦植物の穀粒から得たデンプン粒またはデンプンを提供する。第3の態様の特定の形態では、さらに、小麦植物の胚乳のSBEIIa酵素活性レベルが低められている。
【0032】
第4の態様では、本発明は、本発明の第3の態様に記載のデンプンと食品成分または水を含む組成物にあると言える。この態様は、食品、非食品組成物及びデンプンに他のデンプンまたはデンプン含有品とを混合したものを含む。
【0033】
第5の態様では、本発明は、上記の本発明の第4の態様に記載のデンプン粒および別の食品成分または水を含む組成物を提供する。
【0034】
第6の態様では、本発明は、先の態様に記載の穀粒またはデンプン粒もしくはデンプンを製造するのに使用できる小麦植物を提供する。小麦植物は、トランスジェニックでも非トランスジェニックでもよく、それから得られる穀粒も同様である。
【0035】
第7の態様では、本発明は、i)親となる小麦植物または小麦種子に遺伝的変異を導入するステップと、ii)野生型の植物または種子と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップとを含む、穀粒のデンプン中のアミロース比率が少なくとも50%であるデンプンを含有する穀粒を結実することができる小麦植物を作製する方法を提供する。
【0036】
第7の態様の第2の形態では、本発明は、i)SBEIIa遺伝子の変異またはSBEIIa遺伝子の発現の阻害物質をコードする導入核酸を含む遺伝的変異を、親となる小麦植物または小麦種子に導入するステップと、ii)野生型の穀粒と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップとを含む、穀粒のデンプン中のアミロース比率が少なくとも30%であるデンプンを含有する穀粒を結実することができる小麦植物を製造する方法を提供する。遺伝的変異を導入するステップは、SBEIIa遺伝子発現の阻害物質を発現する外来の核酸の導入を含んでもよく、親となる小麦植物の突然変異誘発を含んでもよい。
【0037】
第7の態様の第3の形態では、本発明は、i)SBEIIa遺伝子の変異を含む遺伝的変異を、親となる小麦植物または小麦種子に導入するステップと、ii)野生型の穀粒と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップと、iii)SBEIIb遺伝子の変異を含む遺伝的変異を、親となる小麦植物または小麦種子に導入するステップと、iv)野生型の穀粒と比べて胚乳のSBEIIbの遺伝子発現、SBEIIaの酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップと、v)胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低い植物を、胚乳のSBEIIb遺伝子発現、SBEIIb酵素活性、またはその双方のレベルが低い植物と交配させるステップと、SBEIIaとSBEIIbの双方の遺伝子発現、酵素活性、またはその双方が低い小麦植物を同定するステップとを含む、穀粒を結実することができる小麦植物を作製する方法を提供する。
【0038】
第7の態様の第4の形態では、本発明は、a)小麦のAゲノム、Bゲノム、またはDゲノムから発現されるSBEIIa活性が低下した小麦植物または穀粒を同定し、b)前記小麦植物またはステップa)の穀粒から得た小麦植物を、SBEIIa活性の低い第2の小麦植物と交配させるステップ、あるいは、c)SBEIIa酵素活性の低い植物をSBEIIb酵素活性の低い植物と交配させ、SBEIIaとSBEIIbの双方の活性が低い小麦植物を同定するステップを含む、その穀粒のデンプン中の相対的アミロース含有量が少なくとも50%であり、好ましくは胚乳のSBEIIa酵素活性が低活性である小麦植物を作製する方法を提供する。第7の態様の植物は、Triticum aestivum ssp.aestivumであることが好ましい。
【0039】
第8の態様では、上記に示した方法によって植物を改変し特性を改変したデンプンを抽出してなる、改変デンプンの作製方法を提供する。
【0040】
第9の態様では、本発明は、小麦のSBEIIb遺伝子、またはSBEIIa遺伝子にそれぞれ結合する分子マーカーを用いて小麦植物または小麦種子の集団をスクリーニングするステップと、結合した分子マーカーの有無に基づいて植物または種子を同定するステップとを含む、SBEIIa遺伝子またはSBEIIb遺伝子の変異に関して小麦植物または小麦種子を同定する方法を提供する。
【0041】
第9の態様の第2の形態では、本発明は、小麦のSBEIIbタンパク質、またはSBEIIaタンパク質にそれぞれ特異的な抗体を用いて小麦植物または小麦種子の集団をスクリーニングするステップと、抗体結合の有無に基づいて植物または種子を同定するステップとを含む、SBEIIa遺伝子またはSBEIIb遺伝子の変異に関して小麦植物または小麦種子を同定する方法を提供する。
【0042】
第10の態様では、本発明は、SBEIIa遺伝子が染色体2Aの長腕から欠けている変異、あるいは染色体2Aの長腕上のSBEIIa遺伝子が、野生型穀粒と比べて穀粒の胚乳のSBEIIaタンパク質、SBEIIa酵素活性、またはその双方を低下させる変異を有する変異を含む、小麦植物から得た穀粒を提供する。この変異はSBEIIa遺伝子のヌル変異でも、SBEIIa遺伝子の少なくとも一部の欠失でもよい。穀粒は、SBEIIb遺伝子が染色体2Aの長腕から欠けている変異、あるいは染色体2Aの長腕上のSBEIIb遺伝子が、野生型穀粒と比べて穀粒の胚乳のSBEIIbタンパク質、SBEIIb酵素活性、またはその双方を低下させる変異を有する変異をさらに含んでもよい。遺伝子欠失によって、染色体2Aの長腕上のSBEIIa遺伝子およびSBEIIb遺伝子の双方の発現を中断させてもよい。
【0043】
植物は、野生型穀粒と比べて染色体2Bの長腕上のSBEIIa遺伝子にコードされたデンプン分枝酵素活性を低下させる遺伝的変異をさらに含むことができるデュラム小麦植物でよい。別の遺伝的変異は、染色体2Bの長腕からのSBEIIa遺伝子の欠失、または野生型穀粒と比べて穀粒の胚乳のSBEIIa酵素活性を低下させる染色体2Bの長腕上のSBEIIa遺伝子の変異を含んでよい。
【0044】
植物は、野生型穀粒と比べて染色体2B、染色体2D、またはその双方の長腕上のSBEIIa遺伝子にコードされたデンプン分枝酵素活性を低下させる遺伝的変異をおそらくはさらに含むであろうTriticum aestivum ssp.aestivum(普通小麦の亜種)であってもよい。さらなる遺伝的変異は、前記染色体のうちの少なくとも1つからのSBEIIa遺伝子の欠失、または野生型穀粒と比べて穀粒の胚乳のSBEIIa酵素活性を低下させる前記染色体のうちの少なくとも1つのSBEIIa遺伝子の変異を含む。
【0045】
植物には、SBEIIa遺伝子発現、活性またはその双方の阻害物質をコードする核酸を導入してよい。SBEIIa酵素活性のレベルは、野生型穀粒と比べて少なくとも40%低下させることができる。穀粒のデンプン中のアミロース比率は、少なくとも30%または少なくとも50%でよい。穀粒は、非しわ型でよく、平均重量は少なくとも36mgでよい。穀粒から得たデンプン粒の少なくとも50%が、偏光下で観察すると非複屈折性に見えるものでよい。本発明の一形態では、殻のない状態での穀粒のデンプン含有量は、少なくとも25%(w/w)であり、あるいは、デンプン含有量は野生型穀粒のデンプン含有量の少なくとも90%である。
【0046】
本発明の形態のうちのいずれかに記載の穀粒のアミロペクチンでは、アミロペクチンをイソアミラーゼで脱分枝した後に測定すると、野生型穀粒のアミロペクチンと比べて重合度が4〜12の鎖長部分の比率が低下していてもよい。
【0047】
穀粒は、SBEIタンパク質、SBEI酵素活性、またはその双方を低レベルで含んでもよく、また、ADPグルコースピロホスホリラーゼ、GBSS、SSI、SSII、SSIII、イソアミラーゼ型脱分枝酵素、プルラナーゼ型脱分枝酵素、およびこれらの任意の組合せからなる群から選択される、野生型穀粒と比べてレベルを変えられた酵素をさらに含んでもよい。
【0048】
本発明のこの第10の態様の形態は、穀粒、穀粒から抽出されたデンプン粒、および、例えば小麦粉、全粒小麦粉、またはセモリナ粉など穀粒から製造された製品またそのデンプンを包含する。
【0049】
<小麦のSBEIIaの改変>
本発明は、小麦胚乳のSBEIIa活性を低下させると、修飾されたデンプンが産生され、特に小麦穀粒の相対的なアミロースレベルが高くなるという研究結果に基づいている。この予想外の結果は、SBEIIaの変異によってアミロペクチン/アミロースの特性が改変されなかったトウモロコシおよびコメでの研究結果と対照的である(Blauth他、2001年、Nakamura、2002年)。別の実施形態では、SBEIIb活性ならびにSBEIIa活性を低下させるなど、1種または複数の追加のデンプン生合成酵素活性を改変する。これら2種の活性をコードする遺伝子の変異には、SBEIIaおよびSBEIIbが、トウモロコシおよびコメでは関連していないのとは対照的に、小麦では密接に関連しているという驚くべき研究結果が役立つ。本発明者らは、予想外に、SBEIIa活性およびSBEIIb活性のレベルが低い小麦植物の穀粒が非しわ型であるということも発見した。
【0050】
<小麦植物を作製する方法>
一態様では、本発明は、穀粒中のデンプンを改変した小麦植物を作製する方法、特にデンプン中のアミロース比率を少なくとも30%に増加させる方法を提供する。通常、6倍体の小麦およびデュラム小麦では、デンプン中のアミロース比率は、約18〜約30%の範囲であり、一部の変異体(SGP−1欠損)では最大約35%である。一実施形態では、本発明の方法は、アミロースを少なくとも30%含むデンプンを有する穀粒を結実する小麦植物を提供するために、親となる小麦植物または小麦種子に遺伝的変異を導入するステップを含む。本明細書で定義するデンプン中のアミロース比率は、重量/重量(w/w)、すなわち、穀粒から得たデンプンの重量に対するパーセンテージとしてアミロース重量を示した値に基づく。別の実施形態では、デンプン中のアミロース比率は、少なくとも40%、少なくとも50%、少なくとも55%、少なくとも60%、少なくとも65%、少なくとも70%、または少なくとも75%である(それぞれw/w)。本発明の別の実施形態では、この方法により、少なくとも80%または少なくとも90%(w/w)のアミロース比率が得られる。
【0051】
別の実施形態では、この方法は、小麦胚乳のデンプン分枝酵素IIa(SBEIIa)タンパク質、酵素活性またはその双方のレベルを変えること、好ましくは低下させることを含む。すなわち、小麦植物に遺伝的変異が導入されると、直接的にまたは間接的にSBEIIaのレベルが変更され、その結果として本明細書に記述するようにデンプンが修飾される。先の実施形態と相互に排他的ではない別の実施形態では、この方法は、小麦胚乳のSBEIIa遺伝子の発現レベルを変えること、好ましくは低下させることを含み、あるいは、この方法は、胚乳のSBEIIa活性を低下させるような小麦SBEIIa遺伝子の変異を含む。SBEIIa遺伝子発現のレベルまたは他の遺伝子のレベルの低下は、核酸、例えば阻害性分子をコードする導入遺伝子の導入によって実現することができる。阻害性分子の例としては、アンチセンスRNA分子、コサプレッションRNA分子、リボザイムRNA分子、または二重鎖RNA分子が挙げられる。
【0052】
本明細書では、「改変すること」、「増加させること」、「増加された」、「低下させること」、「低下した」、「阻害された」などの用語は、比較的な用語、すなわち野生型または未改変の状態と比較しての用語とみなす。「タンパク質のレベル」とは、特定のタンパク質、例えばSBEIIaの量を意味し、これは、例えば、ウェスタンブロット分析や他の免疫学的方法など当技術分野で既知の任意の方法によって測定することができる。「酵素活性のレベル」とは、酵素アッセイで測定した特定の酵素の量を意味する。変異体の酵素活性のレベルは改変されているがタンパク質自体の発現レベル(量)は改変されていないことがあると理解されよう。逆に言えば、タンパク質の量を変えることはできるが、程度の差はあれ活性なタンパク質が産生される場合は、その活性は同じままである。例えば、その酵素をコードする遺伝子が不活性化されるようなときには、量および活性の双方の低下も起こり得る。一部の実施形態では、タンパク質または活性のレベルは、未修飾の小麦の胚乳におけるタンパク質または活性のレベルと比べて、少なくとも40%または少なくとも60%、あるいは少なくとも75%、少なくとも90%、または少なくとも95%低下している。タンパク質活性、酵素活性、または遺伝子発現のレベルの低下は、穀粒のどの発育段階でも、特に発育中の胚乳でデンプンが合成される時期である穀粒の登熟期の間に起こってもよく、成熟に至るまでの穀粒発育のすべての段階で起こってもよい。
【0053】
本明細書では、本質的にα−グルコピラノース単位で構成された多糖類を「デンプン」と定義する。デンプンは、小麦中の主要な貯蔵炭水化物であり、アミロプラスト中で合成され、顆粒中で形成および貯蔵される。デンプンは、ほぼ直鎖状のα−1,4−D−グルコピラノースポリマーであるアミロースと、主にα−1,4結合したα−D−グルコピラノース単位にα−1,6結合分枝が付いた短鎖を有するアミロペクチンとを含む。野生型植物から得た小麦デンプンは、アミロースを最大約20%〜30%、アミロペクチンを最大約70%〜80%含む。これら2種の分子の別の有意な差は、分子量である。アミロースはらせん状の構造で分子量が104〜106ダルトンであり、一方、アミロペクチンの分子量は107〜108ダルトンである。近年の研究により、アミロース中に最大約0.1%のα−1,6−グリコシドの枝分かれ部位が存在することがあることが示され、したがって、「ほぼ直鎖状」と表現されている。本明細書では、α−1,4結合のグルコシド(グルコピラノース)単位およびアミロース様の長鎖アミロペクチン(「中間体」または「アミロース様アミロペクチン」と呼ばれることがある。Takeda他、1993年b;Fergason、1994年)から構成されるほぼ直鎖状の分子を含めて、「アミロース」と定義する。例えば90%(w/v)DMSOを用いてのサイズ排除高速液体クロマトグラフィー(HPLC)、コンカナバリンA法(Megazyme社、アイルランド)を含めて当技術分野で既知の方法のいずれかを用いるか、好ましくは例えば実施例1で記述するヨード滴定法によってアミロース含有量を決定することができる。HPLC法は、デンプンの脱分枝を伴うものでも(BateyおよびCurtin、1996年)、脱分枝を伴わないものでもよい。穀粒重量およびアミロース含有量から、穀粒当たりのアミロース蓄積量を計算し、トランスジェニック系統とコントロールの系統とを比較することができる。
【0054】
別の実施形態では、この方法は、当技術分野で既知の任意の方法を用いて小麦胚乳中のSBEIIaの量または活性を決定するステップを含む。ある実施形態では、例えば、ウェスタンブロット法やELISA検定法などの免疫検出法によってタンパク質のレベルを測定し、または、それに対応するmRNAのレベルを、ノーザンブロットハイブリダイゼーション分析や逆転写ポリメラーゼ連鎖反応法(RT−PCR)など当技術分野で公知の方法によって測定する。別の実施形態では、この方法は、胚乳中のSBEIIaタンパク質または酵素活性のレベルが変えられている小麦植物または穀粒を選択またはスクリーニングするステップを含む。選択ステップは、SBEIIa活性またはタンパク質のレベルの低下に基づいてもよく、あるいは、アミロース比率の上昇やアミロペクチン比率の低下など小麦植物の穀粒のフェノタイプ、見た目のフェノタイプ、例えばしわ型穀粒、またはデンプン粒の特性の変化に基づいてもよい。
【0055】
本発明は、本明細書で記述する方法のうちのいずれかを用いて、直接的にデンプンの特性を測定して、または間接的に、例えば、植物またはその穀粒中の遺伝的変異の存在を検出することによって、穀粒のデンプンの特性が変えられた小麦植物を同定する方法を含むと理解されるであろう。植物は、小麦植物の集団の植物、例えば、小麦育種の植物などでよい。
【0056】
SBE活性は、酵素アッセイ、例えばホスホリラーゼ刺激法(BoyerおよびPreiss、1978年)によって直接測定することができる。この検定法では、ホスホリラーゼaによるメタノール不溶性ポリマー(α−D−グルカン)へのグルコース1−リン酸の取込に対するSBEの刺激を測定する。SBE活性は、グルカンポリマーの枝分かれの結果として生じるグルカン−ポリヨウ素錯体の吸光度の減少を測定するヨウ素染色法によって測定することができる。SBE活性は、イソアミラーゼによる分解の後、基質として還元されたアミロースから生成される還元末端を測定する分枝結合検定法によって検定することもできる(Takeda他、1993年a)。SBEI活性またはSBEIIb活性がない状態で活性を測定することが好ましい。SBEのアイソフォームは、様々な基質特異性を示す。例えば、SBEIはアミロースを枝分かれさせる活性が高く、一方、SBEIIaおよびSBEIIbは、アミロペクチン基質を枝分かれさせる割合が高い。アイソフォームは、移動されるグルカン鎖の長さに基づいて区別することもできる。本明細書で記述するものなど特異的な抗体を用いて、SBEタンパク質を測定してもよい。SBEII活性は、穀粒の発育中に、発育中の胚乳で測定してもよく、あるいは、等価であるが改変されていない穀粒中にタンパク質がなお存在し、免疫学的方法で検定することができる成熟穀粒で測定してもよい。
【0057】
別の態様では、本発明は、穀粒のデンプン中のアミロース比率が少なくとも50%になるように、小麦胚乳中の複数のデンプン生合成酵素の活性を改変し、好ましくは低下させる方法であって、前記酵素のうちの1種がSBEIIaである方法を提供する。いくつかの実施形態では、SBEIIaおよびSBEIIbの双方のタンパク質または酵素活性のレベルが低められ、あるいは、SBEIIa、SBEIIb、およびSBEIの3種すべてのレベルが低められる。SBEIIaとともに改変することができる他のデンプン生合成酵素は、SSI、SSII、SSIIIである。デンプン脱分枝酵素は、例えばイソアミラーゼまたはプルラナーゼの活性を改変することもできる。SBEIIaが改変される限りにおいて、上記の酵素の任意の組合せでも提供される。別の実施形態では、植物の胚乳以外の組織に存在する1種または複数のデンプン生合成酵素の活性が改変される。例えば、主に胚乳での発現を目的としたSBEIIa阻害分子をコードする導入遺伝子によって活性の損失が起こると、それを補うために、葉のSBEIまたはSBEIIの活性が増大されることがある。改変は、例えば、量の増加または減少でも、発現時期の変化でもよい。あるいは、SBEIIaの低下とともに、1種または複数のデンプン生合成酵素を過剰発現させることによって、デンプン合成をさらに改良することができる。このような酵素をコードする遺伝子は、様々な供給源のうちのいずれかから、例えば、細菌または小麦以外の他の供給源から得てもよく、また、触媒特性を変えるように、例えば、酵素の温度依存性を変えるように修飾することができる(例えば、WO94/09144を参照のこと)。
【0058】
高アミロースのフェノタイプは、SBEIIa遺伝子の発現、またはSBEIIa遺伝子およびSBEIIb遺伝子を部分的にまたは全体的に阻害することによって実現することができる。1つまたは複数の遺伝子が阻害される程度によって、小麦穀粒で作られるデンプンの特質がある程度決められることになる。修飾された小麦胚乳から抽出されたタンパク質に対して一連のゲル電気泳動法のいずれかを実施すると、その特質とSBEIIa活性および/またはSBEIIb活性に対する修飾の程度が明らかになる。修飾は、SBEIIa活性および/またはSBEIIb活性の低下、酵素活性の完全な消失、または胚乳内のSBEIIbまたは他の酵素の分布の変化として起こることがある。これらの検定を実施するために、例えば、Rahman他により1995年に概説されているように、小麦胚乳からデンプンを抽出し、その中のタンパク質を分析することができる。SDSポリアクリルアミドゲル電気泳動(PAGE)やイムノブロッティングなど当技術分野で公知の技術を、可溶性画分およびデンプン粒画分に対して実施し、結果を用いてSBEIIa酵素および/またはSBEIIb酵素に修飾が起こっている植物または穀粒を同定する。
【0059】
<小麦植物>
別の態様では、本発明は、デンプン中のアミロース比率が少なくとも30%である穀粒を結実することができる小麦植物を提供する。別の実施形態では、アミロース比率は、少なくとも40%、少なくとも50%、少なくとも55%、少なくとも60%、少なくとも65%、少なくとも70%、または少なくとも80%である。別の実施形態では、その穀粒がデンプン中にこれらのレベルのうちのいずれかのアミロースを含む小麦植物は、野生型穀粒と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルを低下させる遺伝的変異を含む。好ましい実施形態では、この遺伝的変異は、SBEIIa遺伝子の変異またはSBEIIa遺伝子発現の阻害物質をコードする導入核酸を含む。この阻害物質は、アンチセンスRNA分子、コサプレッションRNA分子、リボザイムRNA分子、二重鎖RNA分子、あるいはSBEIIaの発現および/または活性を阻害する類似の分子を含んでよい。
【0060】
本明細書では、例えば、Triticum aestivum L.ssp.aestivum(一般的な小麦またはパン小麦)、Triticum aestivumの他の亜種、Triticum turgidumL.ssp.durum(マカロニ小麦または硬質小麦としても知られているデュラム小麦)、Triticum monococcum L.ssp.monococcum(栽培型のヒトツブ小麦または小型のスペルト小麦)、Triticum timopheevi ssp.timopheevi、Triticum turgigum L.ssp.dicoccon(栽培型のエンマー小麦)、およびTriticum turgidumの他の亜種を含めて(Feldman)、商業的に栽培されている普通小麦(Triticum)属の種に属する任意の植物を小麦植物と定義する。小麦は、AABBDD型のゲノムを有する6倍体小麦でも、AABB型のゲノムを有する4倍体小麦でもよい。本発明による小麦の遺伝的変異は、ライムギおよびオオムギを含めた一部の関連種にハイブリダイゼーションによって伝達することができるため、本発明は、パン小麦とライムギの交配種であるライ小麦を含めて、このようにして形成された交配種も含む。特定の実施形態では、小麦植物は、Triticum aestivum種のものであり、好ましくは、亜種aestivumのものである。あるいは、Triticum aestivumからデュラム小麦に変異または導入遺伝子を容易に移すことができるため、小麦は、Triticum turgidum L.ssp.durumであることが好ましい。
【0061】
本発明は、野生型植物から抽出したデンプンと比べてアミロースを高比率で含むデンプンを有する穀粒を結実することができる、胚乳中のSBEIIaタンパク質、酵素活性、またはその双方のレベルを低下させた小麦植物も提供する。SBEIIaのレベルの低下は、穀粒の発育過程の少なくとも一段階で起こってもよく、成熟に至るまでの全過程で起こってもよい。別の実施形態では、胚乳中のSBEIIaのレベルは、野生型と比べて少なくとも50%、少なくとも75%、少なくとも90%、または少なくとも95%低められる。「野生型」という用語は、遺伝学の分野におけるの通常の意味を指し、本明細書で教示するような修飾をしていない小麦の栽培品種または遺伝子型を含む。
【0062】
本発明は、遺伝子型および/またはフェノタイプにおいて、親の小麦植物の望ましい形質を有する子孫植物および子孫穀粒も提供する。本発明の範囲は、培養組織や培養細胞などのような望ましい形質を有する植物を作製するのに使用できる、小麦植物の増殖性の材料のどれにでも及ぶ。
【0063】
本発明は、SBEIIa活性の低下に加えて、SBEIIbまたは他のデンプン生合成酵素を改変し、好ましくは低下させた小麦植物も含む。SBEIIa活性およびSBEIIb活性の低下した植物は、SBEIIaの低下した植物とSBEIIbの低下した植物とを交配することによって、あるいは、SBEIIa遺伝子およびSBEIIb遺伝子の双方の発現を阻害する分子をコードする導入遺伝子を導入することによって作製することができる。本明細書で示すように、小麦のSBEIIa遺伝子およびSBEIIb遺伝子は密に結びついているため、小麦のゲノムのうちの1つによりコードされているSBEIIaアイソフォームおよびSBEIIbアイソフォームを欠いている変種を同定し、そのような変種を交配して少なくとも2つのゲノムによってコードされているアイソフォームを低下させた植物を作製することによって、両方の活性を低下させた植物を作製することもできる。
【0064】
本発明は、他の遺伝的背景または前述の小麦植物と交配できる他種における遺伝的変異または変異も含む。改変された(変異した)植物を、より望ましい遺伝的背景を有する植物と交配することができる。最初の交配後、適切な回数だけ戻し交配を実施して、望ましくない遺伝的背景を取り除くことができる。望ましい遺伝的背景として、商業用の収穫高および農業生産力や非生物ストレスへの耐性など他の特質を与える遺伝子の適切な組合せを挙げることができる。遺伝的背景には、改変デンプンを生合成または修飾する他の遺伝子、例えば、原因となる遺伝子が分かっていないしわ型胚乳を有する他の小麦系統に由来する遺伝子も含まれ得る。
【0065】
植物は、トランスジェニックでも非トランスジェニックでもよい。
【0066】
本発明は、SBEIIa遺伝子が染色体2Aの長腕(2AL)から欠けている変異、あるいは染色体2Aの長腕上のSBEIIa遺伝子が、野生型穀粒と比べて穀粒の胚乳のSBEIIa酵素活性を低下させる変異を有する変異を含む小麦植物も提供する。2400種の小麦の登録品種を徹底してスクリーニングしたが、本発明者らは、天然に存在するこのような植物を発見できなかった。このことは、2AL上の機能的なSBEIIa遺伝子を保持するための選択が自然に起こっている可能性があることを示唆している。しかし、突然変異誘発後には、このような植物を作製し、同定することができた。これらの植物は、一部の市場で望ましいとされる非トランスジェニックである。これらの植物は、パン小麦でも、デュラム小麦でも、他の小麦でもよい。好ましい実施形態では、この小麦植物は、2AL染色体上のSBEIIa遺伝子の少なくとも一部の欠失を含み、この欠失はSBEIIb遺伝子の少なくとも一部に及ぶことがある。当技術分野で理解されているように、パン小麦などの6倍体小麦は、一般にゲノムA、ゲノムB、ゲノムDと呼ばれる3種のゲノムを含み、一方、デュラム小麦などの4倍体小麦は、一般にゲノムAおよびゲノムBと呼ばれる2種のゲノムを含む。各ゲノムは7対の染色体を含み、これらは減数分裂の進行中に細胞学的方法によって観察することができる。これらの染色体は、通常、大きさに従い最大のものから最小のものの順序で示され、したがって、染色体2は各ゲノムで2番目に大きい染色体である。各染色体には、染色体2上に非対称的に配置されたセントロメアがあり、それゆえに染色体2の2つの腕は、「短腕」および「長腕」と呼ばれている。本明細書では、この用語の標準的な意味に従って、長腕に沿ったセントロメアと先端の間の染色体部分を「染色体2Aの長腕」と定義する。「染色体2Bの長腕」および「染色体2Dの長腕」という用語は、それぞれ小麦のゲノムBまたはゲノムDの染色体2に関するという点を除いて、同様に定義される。
【0067】
本発明者らは、小麦の染色体2上でSBEIIa遺伝子およびSBEIIb遺伝子が密に結びついしていることを発見した。特定の実施形態では、この小麦植物は、少なくともSBEIIa遺伝子の変異を含む染色体腕2ALを過半数(>50%)含む。すなわち、本質的には、少なくともAゲノムのSBEIIa遺伝子に変異を含む染色体2ALが存在している。2ALの存在は、例えば、in situハイブリダイゼーション法(実施例9を参照のこと)などの細胞学的技術によって、または2ALに特異的な分子マーカーを用いることによって、確認することができる。好ましい実施形態では、この小麦植物は、前記変異に関してホモ接合性である。この変異は、ヌル変異でよい。この変異は、遺伝子欠失でよい。
【0068】
特定の実施形態では、欠失対立遺伝子は、MLT2B8植物またはMLT2D1植物に由来するものである。これらの植物の変異したSBEIIa対立遺伝子は2AL染色体上に存在するため、交配によって、これらの対立遺伝子をパン小麦またはデュラム小麦の変種に導入することができる。したがって、本発明は、このような植物、およびそれらから得られる穀粒およびデンプン製品を含む。これらの対立遺伝子を、他の有用なデンプン生合成遺伝子もしくは対立遺伝子、または他の有用な遺伝形質と組み合わせてよい。
【0069】
本発明の範囲が、このような小麦植物、またはこのような植物から得られる穀粒を作製または同定する方法に及ぶことは明らかである。
【0070】
<穀粒>
本発明は、野生型小麦穀粒から抽出したデンプンと比べて改変されたデンプンを含む小麦穀粒も提供する。本明細書では、ほぼ成熟した穀粒を穀粒と定義する。これには、商業的環境で収穫された穀粒が含まれる。一実施形態では、改変デンプンは、少なくとも一部には小麦穀粒の胚乳の発育中にSBEIIa活性を低下させた結果物である。先の実施形態と相互に排他的ではない別の実施形態では、この穀粒のアミロース比率(全デンプンに対するパーセンテージ)は上昇している。これは、野生型植物から得た穀粒と比べた、デンプン中のアミロペクチンの減少率として測定することができる。野生型小麦デンプンは、アミロースを約20〜30%、アミロペクチンを約70〜80%有する。本発明の穀粒は、好ましくはアミロースを少なくとも50%(w/w)含むデンプンを含有する。別の実施形態では、SBEIIa活性およびSBEIIb活性の双方が、胚乳の発育中に低められる。別の実施形態では、SBEIの活性も低められる。別の実施形態では、当技術分野でよく理解されている方法によって測定したアミロース比率がは、穀粒のデンプンの少なくとも55%、少なくとも60%、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、または少なくとも90%(各w/w)である。アミロースレベルの上昇は、デンプン粒の形態が異常になること、または光学顕微鏡もしくは当技術分野で既知の他の方法で観察すると顆粒の複屈折性が失われていることによって、証明することができる。特定の実施形態では、ヨード滴定法によって、例えば、MorrisonおよびLaignelet(1983年)の方法のような分光光度的方法でよい、または高速液体クロマトグラフィー(HPLC、例えば、BateyおよびCurtin、1996年)によって、アミロース比率を測定する。
【0071】
別の実施形態では、この小麦穀粒は、例えば、糊化温度の上昇もしくは低下、糊化の進行中もしくは糊化後の膨潤特性の変化、粘度の変化、アミロペクチンの鎖長分布の変化、またはこれらの任意の組合せなどの物理的特質が改変されたデンプンを含む。糊化温度の上昇または低下は、糊化の第1ピーク、第2ピーク、またはその双方に対するものでよい。例えば糊化エンタルピーなどデンプンの1種または複数の特性は未変化でよい。示差走査熱量計で測定した糊化の第1ピーク(最高点)の温度は、野生型穀粒から得た対応するデンプンでの第1ピークの温度と比べて、少なくとも3℃または5℃、好ましくは少なくとも7℃または8℃、より好ましくは少なくとも10℃上昇していてよい。特定の実施形態では、温度上昇は、3℃〜12℃の範囲である。
【0072】
穀粒は、しわ型でも非しわ型でもよく、好ましくは非しわ型のフェノタイプを有する。本明細書では、穀粒の大多数、好ましくは個々の穀粒の少なくとも90%が、丸いフェノタイプまたは十分に充実したフェノタイプを示す場合、「非しわ型」と定義する。これは、通常、正常レベルまたはほぼ正常なレベルのデンプン蓄積に関連付けられている。一方、本明細書では、「しわ型」フェノタイプという用語は、穀粒の大多数、具体的には少なくとも穀粒の90%のデンプン蓄積量が減少している場合を意味する。それぞれ野生型穀粒と比べて、少ししわ型の穀粒とは、平均デンプン含有量が少なくとも30%に低下しているものであり、中程度のしわ型穀粒とは、平均デンプン含有量が少なくとも50%に低下しているものであり、高度のしわ型穀粒とは、平均デンプン含有量が少なくとも70%に低下しているものである。しわの程度は、成熟穀粒の重量に対するパーセンテージとして、相対的デンプン含有量によって測定することもできる。改変されていない屋外栽培の小麦穀粒のデンプン含有量は約65%であるのに対し、しわ型穀粒のデンプン含有量は50%未満に減少している。
【0073】
別の実施形態では、穀粒の平均重量は、少なくとも36mgまたは40mgである。穀粒の平均重量は、数が分かっている穀粒、すなわち代表試料となるひとかたまりの穀粒の重量を測定し、穀粒の数で全重量を割ることによって、決定する。デンプン含有量、平均重量、および非しわ型フェノタイプなど穀粒の特質が野生型レベルに近いものが、穀粒の商業的製造に望ましいことが理解されよう。
【0074】
本発明は、小麦粉、粗挽き粉、練り粉、または穀粒からもしくは穀粒を用いて製造した他の製品も提供する。これらは、例えば、分別または漂白による加工をしなくても加工をしてもよい。本発明は、本発明の小麦植物から得た、食品製造に有用な小麦穀粒をさらに提供する。さらに、本発明は、他の方法で加工された穀粒も含む。したがって、穀粒は、製粉しても、粉砕しても、圧扁しても、精白しても、粗挽きもしくは挽き割りにしても、パーボイルドしても(ポレンタ)よく、例えばクスクス粉でもよい。
【0075】
<デンプン>
別の態様では、本発明は、本明細書で記述する小麦植物の穀粒から得た、アミロース比率が高くアミロペクチン比率が低いデンプンを提供する。好ましい実施形態では、デンプンは、野生型小麦に比べて胚乳のSBEIIaタンパク質、SBEIIa酵素活性、またはその双方のレベルが低下した小麦植物の穀粒から得られる。別の実施形態では、野生型小麦に比べて、SBEIIa活性およびSBEIIb活性の双方が低く、または、SBEIIa、SBEIIb、およびSBEIの3種すべてのレベルが低い。
【0076】
別の態様では、本発明は、本明細書で記述する小麦植物の穀粒から得た、アミロースを少なくとも50%、少なくとも55%、少なくとも60%、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、または少なくとも90%含むデンプンを提供する。このデンプンは、少なくとも部分的に精製されている。すなわち、このデンプンは、穀粒の少なくとも1種の他の成分から分離されている。精製デンプンは、デンプンをタンパク質、油、および繊維から分離することを伴う精白プロセス、例えば湿式粉砕プロセスによって穀粒から得ることができる。精白プロセスの最初の製品は、デンプン粒の混合物または組成物であり、したがって、本発明は、本明細書で記述する加工デンプンを含むこのような顆粒を包含する。
【0077】
デンプンの糊化温度は上昇していても、低下していてもよいが、上昇していることが好ましい。特定の実施形態では、DSCによる測定によれば、第1ピークの立ち上がりの温度または第1ピークの最高点の温度のうちの少なくとも1つが、野生型小麦穀粒から抽出したデンプンと比べて、少なくとも3℃、少なくとも5℃、少なくとも7℃、または少なくとも10℃上昇している。特定の実施形態では、温度上昇の範囲は3℃〜12℃である。特に留意すべきことには、糊化温度は、第1ピークの立ち上がりの温度が低下するとともに、ピーク最高点の温度が上昇していることがある。先の実施形態と相互に排他的ではない別の実施形態では、DSCにより測定すると、デンプンの糊化温度の第1ピークは変わっているが、アミロースと脂質の高熱分解に対応する第2ピークの温度はほぼ変わっていない。別の実施形態では、デンプンの糊化エンタルピーは減少しており、例えば、対応する野生型小麦デンプンのエンタルピーと比べて、少なくとも25%または少なくとも40%減少している。
【0078】
別の実施形態では、このデンプンは、特異的な物理的特質によって示される改変された構造を有するレジスタントスターチを高レベルで含む。このような特質には、消化酵素に物理的に接近しにくい特質が含まれることがあり、これは、デンプン粒の形態が変わっていること、相当量のデンプン結合脂質が存在すること、結晶化度の変化、アミロペクチン鎖長分布の変化、またはこれらの任意の組合せが原因となっている可能性がある。高アミロース比率も、レジスタントスターチのレベルに寄与している。
【0079】
本発明は、好ましくは高レベルのレジスタントスターチとともに、増量した食物繊維を含む、例示的な小麦植物の穀粒から得たデンプンも提供する。
【0080】
本発明の範囲が、本明細書で記述する小麦デンプンを製造する方法に及ぶことは明らかである。一実施形態では、本方法は、本明細書で記述する小麦穀粒を得るステップと、その穀粒からデンプンを抽出するステップを含む。この小麦穀粒は、本明細書で記述する小麦植物を栽培し、穀粒を収穫することによって得ても、穀粒の生産者または穀粒の輸入業者から入手してもよい。
【0081】
<遺伝子活性の低減方法>
SBEIIa、SBEIIbその他のデンプン生合成または修飾遺伝子の発現および/または活性は、1つまたは複数の遺伝的変異を小麦植物に導入することにより変わることがある。本明細書において用いる場合、「遺伝的変異」とは、この文脈において、対象とする遺伝子の発現または活性に影響を及ぼす、小麦植物ゲノムにおける遺伝性の任意の変化を意味する。遺伝的変異としては、点変異、挿入、置換、逆位、重複、転座および好ましくは欠失などの突然変異ならびに1つまたは複数の導入遺伝子のゲノムへの導入を含む。
【0082】
本明細書において用いる場合、「核酸分子」および「核酸配列」という語句は、ヌクレオチドのポリマーを指し、これは一本鎖でもよい二本鎖でもよい。それは、例えば、ゲノムDNAまたはcDNAなどのDNA、RNA、mRNAまたはこれらの任意の組合せを含み得る。小麦細胞への導入のために、送達性または安定性を改善するべく核酸分子を化学的に修飾してもよいし、ウイルスベクターなどのベクターの一部として保護してもよい。核酸分子はクローン技術によって得てもよいし、周知の技術である技術によって合成してもよい。核酸分子は、コード鎖もしくは非コード鎖(アンチセンス)または、例えば、逆位反復コンストラクトにおけるような、これらの組合せを含み得る。遺伝子に「対応する」核酸配列と言う場合、「対応する」という用語は、ヌクレオチド配列が、参照遺伝子またはその指示された部分と同じヌクレオチド配列を有するか、通常のワトソン−クリック型塩基対において正確に相補的であるヌクレオチド配列を有するか、そのような配列のRNA(例えばmRNA)等価物であるか、遺伝子のmRNAに由来するcDNAであるような、ヌクレオチド配列関係を指す。
【0083】
ヌクレオチド配列は、5'から3'方向へ、標準的な1文字ヌクレオチド省略形を使用して一本鎖配列によって表す。「相補性」は、塩基対合によってアニールされる2本の一本鎖核酸分子または配列の関係を記述するものである。例えば、5'−GACT−3'は、その相補鎖5'−AGTC−3に対合する。「相同性」または「相同的」は、文脈によって2つ以上のヌクレオチド配列または2つ以上のポリペプチド配列の間の配列の類似性または同一性を指す。「%同一性」という用語は、例えば、CLUSTAL VアルゴリズムまたはBlastnもしくはBLAST2配列プログラム(これらは米国国立バイオテクノロジー情報センター(National Center for Biotechnology Information)から入手可能で、インターネット上ではhttp://www.ncbi.nlm.nih.gov/BLAST/で利用可能であり、好ましくはデフォルトのパラメータ設定とする)などの標準的なものとなっているアルゴリズムを使用してアラインメントを行った2つのヌクレオチド配列間のヌクレオチドの合致の%を指す。同様にして、「%同一性」はポリペプチド配列を指すこともある。
【0084】
本明細書において、SBEIIa、SBEIIbその他のデンプン生合成支配遺伝子またはアンチセンス、コサプレッション、リボザイム、二重鎖RNA分子その他をコードする遺伝子などを含め「遺伝子」という場合、その最も幅広い文脈で解するものとし、プロモーターと転写ターミネーター−ポリアデニル化配列などの制御領域を伴う転写領域を有する古典的なゲノム遺伝子を含む。転写領域は転写されたが、翻訳されなかった配列(非翻訳配列、UTR)を含み、状況に応じて、タンパク質コード領域もしくはイントロン(これは成熟したRNAを形成するために除かれる)またはこれらの任意の組合せを含んでもよい。「遺伝子」は、エキソンに対応するcDNAから得られる形態およびRNAゲノムで見られるようなRNA遺伝子を含む。「遺伝子」という用語は、機能的な産物の全部または一部をコードする合成または融合分子を記述するためにも用いられる。
【0085】
細胞、好ましくは、小麦細胞中に存在する場合、「遺伝子」は、「生物活性」分子または「遺伝子産物」(これはRNAでもポリペプチドでもよい。)の「発現」を指示する。この過程は、最も一般的にはRNAを生産するための転写とタンパク質を生産するための翻訳による。これらの産物は、その後細胞内で修飾されてもよい。RNAは、例えば、ポリアデニル化、スプライシング、キャッピング、21〜23のヌクレオチド断片へのダイシング、核からの輸送またはタンパク質に対する共有結合もしくは非共有結合的な相互作用によって修飾されてもよい。例えば、タンパク質はリン酸化、グリコシル化または脂質化によって修飾されてもよい。本明細書において用いる場合、これらの過程はすべて「遺伝子の発現」その他の用語に包含される。
【0086】
本明細書において用いる場合、「小麦SBEIIa遺伝子」および「小麦SBEIIb遺伝子」という用語ならびに関連語は、それぞれ、SBEIIaまたはSBEIIb酵素をコードする小麦から同定された遺伝子、および他の小麦の種類に存在する相同遺伝子を指す。これらは表1に列挙する遺伝子配列を含むが、これらに限定されるものではない。SBEIIaとSBEIIb遺伝子の配列には、異なる小麦の種類からの自然変異があると思われる。相同遺伝子は、当業者であれば容易に認識できる。SBEIIb遺伝子またはタンパク質についてと同様に、相同的なSBEIIa遺伝子またはタンパク質の間の配列同一性の程度は、少なくとも90%であると考えられる。
【0087】
本発明で用いる遺伝子は、天然に存在するSBEIIa、SBEIIbその他に由来してもよいし、標準的な組み換え技術によってその他のデンプン生合成支配遺伝子に由来するものでもよい。本明細書において用いる場合、「組み換え核酸分子」その他の用語は、天然に存在しない配列であるか、本来別個の2以上の配列セグメントの人工的組合せによって作られる配列を有する配列を指す。この人工的組合せは、化学合成によって、または、より一般的に、例えば、周知の技術である遺伝子工学技術によって、単離された核酸セグメントを人工的に操作することによって作られるものでもよい。「組み換え」という用語は、単に核酸の一部の追加、置換または欠失のみによって変更された核酸を含む。多くの場合、組み換え核酸は、プロモーター配列に機能的に連結された核酸配列を含んでもよい。そのような組み換え核酸は、例えば、細胞を形質転換するのに用いられるベクターの一部でもよい。
【0088】
一般に、遺伝子は1つまたは複数のヌクレオチド置換、欠失および/または添加(例えばコドン修飾)を生じる突然変異生成に付してもよい。そのような遺伝子のヌクレオチド挿入型の誘導体は、5'および3'端融合ならびに1つまたは複数のヌクレオチドの内部配列挿入を含む。挿入型のヌクレオチド配列変異形では、1つまたは複数のヌクレオチドがヌクレオチド配列で予め定められたサイトに導入されるが、ランダムな挿入を行い結果として生じる産物を適宜スクリーニングすることも可能である。欠失型変異形は、配列から1つまたは複数のヌクレオチドの除去によって特徴付けられる。置換型のヌクレオチド変異形は、配列の少なくとも1つのヌクレオチドが取り除かれ、異なるヌクレオチドがその位置に挿入されたものである。置換がコドンによって定義されるアミノ酸を変えないという点で、そのような置換は「サイレント」でもよい。あるいは、保存的な置換基は、1つのアミノ酸を別の同様に働くアミノ酸に変えるように設計される。典型的な置換は、以下に従ってなされるものである:
【0089】
保存的なアミノ酸置換に適する残基
元の残基 置換例
Ala Ser
Arg Lys
Asn Gln;His
Asp Glu
Cys Ser
Gln Asn
Glu Asp
Gly Ala
His Asn;Gln
Ile Leu;Val
Leu Ile;Val
Lys Arg;Gln;Glu
Met Leu;Ile
Phe Met;Leu;Tyr
Ser Thr
Thr Ser
Trp Tyr
Tyr Trp;Phe
Val Ile;Leu
【0090】
<導入遺伝子>
SBElla、SBEIIbまたは他デンプン生合成または修飾遺伝子の発現および/または活性は、1つまたは複数の導入遺伝子を小麦植物にもたらすことによって変更され得る。本明細書で言う「導入遺伝子」は、バイオテクノロジーの技術分野における通常の意味を有し、組換えDNAまたはRNA技術によって製造されたか変更され、対象とする有機体または細胞(好ましくは小麦細胞)に導入された遺伝子配列を含む。導入遺伝子は、有機体または細胞に由来する遺伝子配列(例えば、アンチセンス配列)を含んでもよい。導入遺伝子は、典型的には、前記有機体または細胞に由来しない外来性の核酸を含む。「トランスジェニック」とは、導入遺伝子を含む有機体または細胞を指す。「非トランスジェニック」とは、ゲノム中にいかなる導入遺伝子も存在しないことを指す。安定した継承のために、好ましくは導入遺伝子を有機体または細胞のゲノムに組み込む。
【0091】
当業者は、遺伝子またはそれに対する相補配列の発現のためには、前記遺伝子が、細胞内でプロモーター配列との機能的な関係に置かれる必要があると承知している。本明細書の目的のためのプロモーターの選択は、必要な発現レベルおよび/または発現させるべき組織、器官および種によって、特に胚乳特異的プロモーターによって変わり得る。
【0092】
プロモーター配列の制御調節の下に核酸分子を置くことは、発現がプロモーター配列によって制御されるように前記分子を位置付けることを意味する。プロモーターは、通常(但し、必須ではない)、それが調節する核酸分子の上流に、または5'末端に位置する。さらにまた、プロモーターを含む調節要素は、通常、遺伝子の転写開始部位の2kb以内に位置する。異種プロモーター/構造遺伝子組合せの構築では、一般に、その自然の条件下でプロモーターとそれが制御する遺伝子(すなわち、それからプロモーターが誘導される遺伝子)との間の距離とほぼ同じ距離に、遺伝子転写開始部位からプロモーターを隔てて配置するのが好ましい。当技術分野において知られているように、この距離が若干変化してもプロモーター機能を損なうことなく対応できる。同様に、その制御下に置かれる異種遺伝子に関しての調節配列要素の好ましい位置は、その自然の条件下での要素(すなわち、それから誘導される遺伝子)の位置によって定義される。また、当技術分野で知られているように、この距離においても若干の変化は生じ得る。
【0093】
本発明の遺伝子コンストラクトにおける使用に適したプロモーターの例としては、ウイルス、イースト、カビ、バクテリア、昆虫、鳥、哺乳類および植物の遺伝子に由来するプロモーターが挙げられ、好ましくは植物細胞で機能し得るもの、より好ましくは小麦の胚乳で発現し得るものが挙げられる。プロモーターは構成的に、または発現が起こる組織によって異なるように発現を調節するものでよい。あるいは、発現は発現が起こる発育時期に関して、または、外刺激(例えば生理的ストレスまたは温度)に応じて異なってもよい。
【0094】
SBEIIaその他のデンプン生合成支配遺伝子活性を低減する方法は、導入遺伝子を小麦の再生可能な細胞に導入して、形質転換細胞からトランスジェニック小麦植物を再生させるステップを含み得る。アミロペクチンの合成に関係する分枝酵素はSBEI、SBEIIaとSBEIIbを含み、本発明はSBEIIaのみの、または、SBEIIbまたはSBEI発現の変更と組み合わせた発現低減を包含する。したがって、導入遺伝子(複数可)はこれらの遺伝子のうちの2つ以上を不活性化してもよい。さらに、導入遺伝子(例えば、二重鎖RNA、アンチセンスまたはリボザイムRNAをコードするもの。下記参照)が直接SBEllbまたはSBEI遺伝子形質発現を標的とするという点で、SBEIIbやSBEIの失活は直接的でもよいし、あるいは、それはSBEIIbまたはSBEIの発現に間接的に変更をもたらすものでもよい。例えば、導入遺伝子RNAは配列同一性または塩基の対合に関してSBEIIa遺伝子/RNAのみを標的としてもよいが、胚乳におけるタンパク質の安定性または分布を変えることによって、SBEIIbまたはSBEI活性の低減をもたらすものでもよい。さらに、本発明の形態は、SBEIIaの変更された活性と1つまたは複数の他のアミロペクチン合成酵素(これらの酵素はSSI、SSII、SSIIIおよびイソアミラーゼまたはプルラナーゼなどの脱分枝酵素を含んでもよい。)の変更の組合せにある。これらのいずれかまたはすべての発現は、導入遺伝子の導入によって変更され得る。
【0095】
小麦のアミロペクチン合成遺伝子についてはいくつかのDNAの塩基配列が知られており、それらのいずれでも小麦における遺伝子失活のための導入遺伝子を設計するためのベースとすることができる。これらのものとしてはSBEIIa(GenBankアクセッション番号Y11282、AF338431およびAF338432)およびSBEIIb(WO00/15810、WO01/62934)が挙げられる。小麦のSBEI遺伝子は、Rahman他(1997年)およびRahman他(1999年)に記載されている。SBEIのためTriticum tauschii配列(これは小麦DゲノムSBEI遺伝子との相同性が高い。)は、WO99/14314に見ることができる。小麦のSBEIのためのcDNA配列は、GenBankデータベースにおいてアクセッション番号AF076679でアクセスすることができる。大麦その他の密接に関連する種から得られる他のアミロペクチン合成遺伝子のホモログは、小麦において遺伝子形質発現レベルを修飾するのに用いることもできる。そのような遺伝子またはその断片は、PCR増幅または標識化プローブへのハイブリダイゼーションを含む当技術分野では周知である方法によって得ることができる。
【0096】
本明細書において用いる場合、「ストリンジェントなハイブリダイゼーション条件」は、少なくとも90%、好ましくは少なくとも95%の配列同一性がプローブと標的配列の間にあるならば、ハイブリダイゼーションが通常、起こることを意味する。ストリンジェントなハイブリダイゼーション条件の例は、50%ホルムアミド、5×SSC(1×SSC=150mM NaCl(15mMクエン酸トリナトリウム))、50mMリン酸ナトリウム(pH7.6)、5×デンハルト溶液、10%硫酸デキストランおよびサケ精子DNAなどの変性剪断キャリアDNA20μg/mlからなる溶液中で一夜インキュベーションした後、ハイブリダイゼーションの支持体をおよそ65℃で0.1×SSCで洗浄するというものである。他のハイブリダイゼーションおよび洗浄条件はよく知られており、例えば、Sambrook他、Molecular Cloning:A Laboratory Manual、第2版、Cold Spring Harbor、NY(1989年)、特に第11章に例示されている。
【0097】
導入遺伝子コンストラクトの調製に用いるホモログの領域は、対応する小麦遺伝子と少なくとも85%、好ましくは少なくとも90%、さらに好ましくは95〜100%の同一性を適当な領域内に有すべきである。また、導入遺伝子は、小麦胚乳内で発現されるアミロペクチン合成遺伝子を特異的に標的とし、植物中、他の場所でのアミロペクチン合成にはより小さなまたは最小限の影響しか与えないことが好ましい。これは、導入遺伝子中の胚乳特異的プロモーターなどの適当な調節配列の使用によって達成してもよい。
【0098】
<アンチセンス>
小麦などの植物において遺伝子活性に変更を加える、特に低減する遺伝子工学アプローチは、当技術分野において周知である。これらの方法は、標的遺伝子のRNAと相補的でハイブリダイズすることができる適当なアンチセンス分子の発現のための、遺伝子コンストラクトの導入を含む。アンチセンス分子は標的遺伝子のmRNAの翻訳または処理または安定性に干渉し、それによって遺伝子の発現を不活性化すると考えられる。アンチセンス配列を考案する方法は周知の技術であり、これらの例は米国特許第5190131号、欧州特許明細書第0467349−A1号、欧州特許明細書第0223399−A1号および欧州特許明細書第0240208号に見ることができ、これらは参照によって本明細書に組み込まれる。植物におけるアンチセンス法の使用は、Bourque(1995年)とSenior(1998年)によって概説されている。Bourqueは、植物システムにおいてアンチセンス配列を使った遺伝子失活の多数の例を列挙している。彼女も、部分的な阻害がシステムにおいてかなりの変化を確かにもたらすであろうから、酵素活性の100%の抑制を達成することは必要ではないと述べている。Senior(1998年)は、アンチセンス法は、現在、植物で遺伝子形質発現を操作することの非常に安定した技術であると述べている。
【0099】
小麦SBElla、SBEIIb、SBEIその他のデンプン生合成または修飾遺伝子についてのアンチセンス分子は、小麦mRNA配列に基づくことができるか、他の種(例えば、大麦)から得られる相同なDNAまたはmRNA配列に由来し得る。アンチセンス配列は、これらの遺伝子のいずれかの転写物の全部または一部に、または、それらの発現(例えば、それらのスプライシング)の制御を及ぼす配列について対応すればよい。アンチセンス配列は、小麦SBEIIaその他の遺伝子の標的とされたコード領域または5'−非翻訳領域(UTR)もしくは3'−UTRまたはこれらの組合せに対応してもよい。それは、ある程度、イントロン配列(それは転写の間または後に除かれ得る)と相補的でもよいが好ましくは標的遺伝子のエキソン配列のみに相補的である。UTRは一般に違いがより大きいことからすれば、これらの領域を標的とすることは、遺伝子抑制により大きな特異性をもたらす。特定の実施形態において、アンチセンス配列の長さは、遺伝子RNA配列の相補鎖と対応する少なくとも19の隣接するヌクレオチド、少なくとも50、少なくとも100、少なくとも200、少なくとも500または少なくとも1000のヌクレオチドである。すべての遺伝子転写物と相補的な全長配列を用いてもよい。特定の実施形態において、アンチセンス配列の長さは、100〜2000ヌクレオチドである。さらなる実施形態において、標的とされた転写物に相補的なアンチセンス配列の配列同一性の程度は、少なくとも85%、少なくとも90%または95〜100%である。アンチセンスRNA分子は、もちろん、分子を安定させるために機能してもよい無関係な配列を含んでもよい。
【0100】
<コサプレッション>
使用し得る別の分子生物学的製剤アプローチは、コサプレッションである。コサプレッションのメカニズムは、十分理解されていないが、転写後遺伝子サイレンシング(PTGS)を含むと考えられており、この点ではアンチセンス抑制の多くの例と、非常に類似している。それは、その発現のためには、プロモーターに関してセンス方向に植物内に遺伝子の余分なコピーまたはその断片を導入することが必要である。センス断片のサイズ、標的遺伝子領域との対応および標的遺伝子への配列同一性の程度は、上述のアンチセンス配列に関してと同様である。若干の例では、遺伝子配列のさらなるコピーは、標的植物遺伝子の発現の妨害をする。コサプレッションアプローチを実行する方法については、WO97/20936および欧州特許明細書第0465572号を参照されたい。
【0101】
<二重鎖RNA媒介遺伝子サイレンシング>
遺伝的変異を小麦植物に導入するために使用され得るさらなる方法は、二重鎖あるいは二本鎖RNAによって媒介される遺伝子サイレンシングである。この方法も、PTGSを含む。この方法では、失活させるべき標的遺伝子に相同で少なくとも部分的に二本鎖であるRNA産物(複数可)の合成を指示するDNAを導入する。したがって、DNAは、RNAに転写されるときハイブリダイズして二本鎖RNA領域を形成することができるセンスとアンチセンス配列からなる。好ましい実施形態において、センス配列とアンチセンス配列は、RNAに転写されるとき、スプライシングで除かれるイントロンからなるスペーサー領域によって分離されている。この構成は、遺伝子サイレンシングをより高効率にすることが示された。二本鎖領域は、1または2のDNA領域から転写される1または2のRNA分子を含んでもよい。二本鎖分子の存在は、二本鎖RNAおよびさらには標的植物遺伝子からの相同なRNA転写物も破壊する内在性植物システムからの応答を誘発し、標的遺伝子の活性を能率的に低減するか排除する。この技術を実施する方法については、オーストラリア特許明細書第99/29514−A号およびWO99/53050を参照されたい。特定の実施形態において、ハイブリダイズするセンス配列およびアンチセンス配列の長さは、少なくとも19の隣接するヌクレオチド、少なくとも30、少なくとも50、少なくとも100、少なくとも200、少なくとも500または少なくとも1000のヌクレオチドである。すべての遺伝子転写物と対応する全長配列を用いてもよい。特定の実施形態においては、長さは範囲100〜2000ヌクレオチドである。さらなる実施形態において、センス配列およびアンチセンス配列の標的とされた転写物への配列同一性の程度は、少なくとも85%、少なくとも90%または95〜100%である。RNA分子は、もちろん、分子を安定させるために機能してもよい無関係な配列を含んでもよい。RNA分子は、RNAポリメラーゼIIまたはRNAポリメラーゼIIIプロモーターの制御下に発現されるものでもよい。後者の例は、tRNAまたはsnRNAプロモーターを含む。二本鎖RNA分子は、互いに連結された複数の遺伝子からの配列を含んでもよく、それによって複数の遺伝子を標的としてもよい。
【0102】
<リボザイム>
小麦の遺伝子形質発現の望ましい失活をもたらす遺伝的変異は、1つまたは複数のリボザイムをコードする核酸分子を含む。リボザイムは、1またはしばしば2つのハイブリダイズしている配列によって定義される特定のサイトで他のRNA分子を切断することができる、酵素または触媒機能を有するRNA分子である。RNAの切断により、標的遺伝子の発現を不活性化する。リボザイムはアンチセンス分子の働きをしてもよく、それが遺伝子失活に寄与してもよい。リボザイムは、ハイブリダイズしている配列の間で1つまたは複数の触媒領域(好ましくはハンマーヘッドまたはヘアピンタイプ)を含む。RNAseP、Group IまたはIIイントロンおよびデルタ型肝炎ウイルスタイプを含め、他のリボザイムのモチーフを用いてもよい。欧州特許明細書第0321201号および米国特許第6,221,661号が参照される。例えばWegener他(1994年)によって、遺伝子導入植物の遺伝子を不活性化するリボザイムの使用が示されている。
【0103】
<遺伝子コンストラクト/ベクター>
本発明は、遺伝子阻害分子をコードするRNAまたはDNA、好ましくはDNAを含む単離された核酸分子も提供する。ある実施形態において、核酸分子は、小麦SBEIIa遺伝子配列を標的とし、小麦穀粒の胚乳のその発現を失活させるアンチセンス、センス(コサプレッション)、二本鎖RNAまたはリボザイム分子をコードする。本発明はまた、プロモーター、エンハンサーおよび転写末端またはポリアデニル化配列などの1つまたは複数の調整要素を含む単離された核酸分子を含むかコードする遺伝子のコンストラクトを提供する。そのような要素は当技術分野において周知である。植物、特に小麦などの単子葉植物において、遺伝子のコンストラクトは、導入遺伝子の発現を助けるイントロン配列を含んでもよい。「イントロン」という用語は、その通常の意味では、転写されるが、タンパク質をコードせず、翻訳前にRNAからスプライシングにより除かれる遺伝子の部分を意味するように使われる。導入遺伝子が翻訳産物をコードするか、そうでなく転写領域のどこかをコードするならば、イントロンは5'−UTRまたはコード領域に組み入れられてもよい。
【0104】
本発明はさらに遺伝子のコンストラクトを含むベクター(例えば、プラスミドベクター)を提供する。「ベクター」という用語は、in vitroまたはin vivo発現が可能な発現ベクターおよび1つの細胞または有機体から別のもう1つまで転移され得る形質転換ベクターを含む。例えば、大腸菌またはアグロバクテリウムなどの原核細胞において、ベクターは細胞内での複製をもたらす配列を含む。特定の実施形態において、ベクターは少なくとも1つのT−DNA境界配列によって定まるT−DNA配列(これは小麦細胞に導入することができる。)を含むバイナリーベクターである。本発明は、さらに、ベクターを含む細胞、例えば、未熟な胚の胚盤の細胞などの再生可能な細胞でもよいアグロバクテリウムまたは小麦細胞を提供する。あるいは、細胞は導入遺伝子を含む形質転換された小麦細胞でもよい。
【0105】
<プロモーター/ターミネーター>
別の実施形態において、本発明の導入遺伝子または他の遺伝子コンストラクトは、小麦の胚乳内で調節されたか構成的な発現をもたらし得る転写開始領域(プロモーター)を含む。プロモーターは組織特異的で、選択的に、または、もっぱら胚乳の発現を与えるものでもよい。プロモーターは、(高分子グルテニンプロモーター、小麦SSIプロモーター、小麦SBEIIプロモーター、小麦GBSSプロモーターなどの)胚乳特異的または(ユビキチンプロモーターまたはCaMV35Sまたは強化された35Sプロモーターなどの)胚乳に非特異的なプロモーターのいずれから選択してもよい。プロモーターは、温度、光またはストレスなどの要因によって調整されてもよい。通常、プロモーターは、発現されるべき遺伝子配列の5'をもたらされる。コンストラクトは、nos3'またはocs3'ポリアデニル化領域または転写ターミネーターなどの転写を強化する他の要素も含んでもよい。例示したDNA領域は、適当な選択可能なマーカー遺伝子配列と他の要素を含むベクターに、または、ベクターがこれらの配列を含んで同時形質転換され得るベクターに取り込まれる。
【0106】
<小麦の形質転換法>
小麦などの単子葉植物の形質転換法、すなわち、外来性の核酸の導入によって遺伝的変異を植物に導入し、プロトプラストまたは未熟な植物胚から植物を再生する方法は、当技術分野において知られており、例えば、Becker他、1994年、Cheng他、1997年、He他、1994年、Hess他、1990年、Nehra他、1994年、Vasil他、1992年、Vasil他、1993年、Weeks他、1993年、Weir他、2001年、オーストラリア特許出願番号第75460/94号、欧州特許出願番号第709462号、WO93/04178、WO89/12012、WO94/13822およびWO99/14314を参照されたい。望ましいヌクレオチド配列または遺伝子のコンストラクトおよび選択可能なマーカーを運ぶベクターは、組織栽培された植物または外植体の再生可能な小麦細胞またはプロトプラストなどの適当な植物システムに導入すればよい。選択可能なマーカー遺伝子は、小麦細胞に抗生物質または除草剤耐性を提供してもよいし、マンノースなどの基質の利用性を可能にするものでもよい。選択可能なマーカーは、好ましくはアスラム、ジェネテシン(geneticin)またはハイグロマイシン耐性を小麦細胞に与える。再生可能な小麦細胞は、好ましくは、未熟な胚の胚盤、成熟した胚、これらに由来するカルスまたは分裂組織に由来する。
【0107】
形質転換植物は選択可能なマーカー遺伝子を含んでもよいし、あるいは、そのような遺伝子は再生の間か後に、例えば、ゲノムからの選択可能なマーカー遺伝子の切除により、または、SBEIIa阻害導入遺伝子から離れた選択可能なマーカー遺伝子の分離によって除去してもよい。
【0108】
導入遺伝子または突然変異が染色体に融和した植物は、例えば、導入遺伝子またはフェノタイプ観察に特有の適当な核酸プローブを使うことによりスクリーニングすることができる。いくつかの方法のいずれも、形質転換植物の存在を決定するために使用され得る。例えば、ポリメラーゼチェーンリアクション法(PCR)は形質転換植物に特有な配列を増幅するのに用い得る(ゲル電気泳動その他の方法による増幅産物の検出を行う)。DNAは従来法を使用して植物から抽出してもよく、PCR反応は、形質転換植物と非形質転換植物を識別するプライマーを使って実行してもよい。例えば、コンストラクトに読み込まれる形質転換ベクターからDNA領域を増幅するプライマーを設計してもよいし、逆方向プライマーは対象とすべき遺伝子から設計してもよい。植物がうまく形質転換されたならば、これらのプライマーは断片を増幅するのみである。陽性の形質転換体を確かめる別の方法は、当技術分野において周知であるサザンブロットハイブリダイゼーションによる。形質転換植物または変異体は、非形質転換植物または野生型植物からそのフェノタイプ(例えば、選択可能なマーカー遺伝子の存在によって、または免疫学的方法による特定のタンパク質の存在によって、または、タンパク質の欠如(例えば、ELISA分析アッセイまたはウェスタンブロット分析によって検出されるものとしての胚乳のSBEIIaタンパク質の欠如)によって与えられる)によって識別すなわち区別される。そのような植物をスクリーニングする際に使われる徴候は、穀粒のフェノタイプの特徴の観察(例えば、縮んだ穀物の視覚検査または測定によって)、増加したアミロース含有量の試験または顕微鏡で複屈折の存在について調べることによることもできる。
【0109】
<変異>
小麦胚乳においてSBEIIa酵素その他のデンプン生合成酵素の活性の低減をもたらす遺伝的変異の導入は、それぞれの遺伝子内における適当な突然変異にもよって達成されてもよいし、遺伝子の調節配列内における適当な突然変異にもよって達成されてもよい。本明細書の文脈では、「誘発突然変異」は化学産物、放射線または生物ベースの突然変異生成(例えば、トランスポゾンまたはT−DNA挿入)の結果でもよい人工的に誘発された遺伝的変異である。遺伝子が阻害される程度は、ある程度、生産されるデンプンの特徴を決定する。突然変異はトランケーションまたはヌル変異でもよく、これらがデンプンの性質に重要な影響を及ぼすことは知られている。しかし、変更されたデンプン構造は、デンプンまたは小麦穀粒において対象とする特徴をもたらすのに十分にアミロペクチン合成酵素活性を低減する漏出突然変異からも生じるであろう。他の染色体再編成も有効であり得、これらは挿入、欠失、逆位、重複または点変異を含んでもよい。本明細書で用いられる「ヌル変異」は、対象とすべき遺伝子の活性の完全かまたは完全に近い損失をもたらす突然変異を指し、例えば、遺伝子活性がもはや見出されないものを指す。
【0110】
SBEIIa遺伝子は、染色体2の長腕に位置する。この遺伝子その他の遺伝子への突然変異(特に欠失突然変異)は、対象とすべき遺伝子(例えば、SBEIIa遺伝子または二重突然変異体の場合には、おそらく結合されたSBEIIb遺伝子にまで拡大される。)に集中させることが好ましい。この文脈における遺伝子は、転写領域とともにプロモーター領域と転写終了/ポリアデニル化信号を含む。転写領域は、タンパク質コード領域(複数可)ならびにmRNAの5'非翻訳および3'非翻訳領域、さらに存在し得る任意のイントロン領域を含む。遺伝子への突然変異は、遺伝子の任意の領域におけるまたは領域の組合せでもよく、1つのヌクレオチドのみの変更(例えば、コード領域のフレームシフト突然変異)からすべての遺伝子の欠失に及び得る。遺伝的変異のためにホモ接合性である植物が好ましい。
【0111】
欠失は、100または2、300程度、おそらくは500キロベース単位のサイズに制限してもよい。特定の実施形態において、欠失は少なくとも2、3000キロベースまたは5000キロベース未満まで拡がる。本発明は、それぞれのゲノムの染色体2の長腕の多くを含むより大きな欠失を含んでもよいが、染色体2の長腕は多数の他の遺伝子をその上に集中させて有するので、小麦植物の生命力に対し影響を与えるため、これは好ましくない。したがって、大きな欠損が起こる場合は、これらは植物生命力に対し悪影響を与え、それゆえに、その商業的な生存能力に対し悪影響を与えるため、少なくとも染色体2の長腕の大多数が存在することが望ましい。好ましい実施形態において、染色体2Aの長腕の大部分は存在する。
【0112】
突然変異生成は、化学または放射線手段、例えば、種子のEMSまたはアジ化ナトリウム(ZwarおよびChandler、1995年)処理またはガンマ照射によって達成できる。変異体の単離は、突然変異を起こす植物または種をスクリーニングすることによって達成してもよい。例えば、小麦の突然変異集団は、穀粒のハイアミロース含有量および/または通常より長いアミロペクチン鎖分布またはELISAによるSBEIIaタンパク質の損失、または、変更された穀粒形状(Green他、1997年)についてスクリーニングされ得る。スクリーニングは、好ましくは、既にSBE活性のうちの1つが欠如している小麦遺伝子型、例えば、SBEIIb陰性のバックグラウンドでなされる。そのような突然変異は、次いで、望ましい遺伝的バックグラウンドを有する植物との間で交配し、当初の望ましくない親のバックグラウンドを消し去るために適当な数の戻し交配を実行することによって、望ましい遺伝的バックグラウンドに導入し得る。
【0113】
別の実施形態において、突然変異は小麦においてSBEIIaとSBEIIb遺伝子の発現または活性に影響を及ぼす。そのような突然変異の確認は、トウモロコシまたはコメと対照的に2つの遺伝子が小麦では密接に結びついているという予想外の知見によって容易になる。1つの遺伝子の欠失は他の遺伝子に容易に及ぶことがあり、無効な対立遺伝子(ヌル変異)を両方の遺伝子にもたらす。この知見は、小麦の少なくとも1つのゲノムの両方の遺伝子の変異体である自然な変異体のスクリーニングを容易にし、2つまたは3つのゲノムにおいて両方の遺伝子に複合的な突然変異のある小麦を生産するためのスクリーニングを可能にする。そのような小麦は、小麦穀粒の高アミロースな非トランスジェニック源と産物を提供する。
【0114】
SBEIIaまたはアミロペクチン合成に関与している他の酵素をコードする遺伝子の突然変異は、通常、アミロース含有量の比率の増大を引き起こす。個々の穀粒当たりのアミロースの量はアミロペクチンからアミロースまで炭素の流れが変更される結果として増大し得るが、あるいは、穀粒当たりのデンプン生産高の顕著な減少があるならば、それは減少することもある。いずれにせよ、デンプン中パーセンテージとしてのアミロースの相対的レベルは増加する。
【0115】
変形したデンプン粒を有する種がハイアミロース大麦(Morell他、2003年)および、デンプン中におよそ90%のアミロースを有する低アミロペクチン(LAPS)トウモロコシ(Sidebottom他、1998年)で報告されている。
【0116】
複屈折は、2つの方向に光を屈折させる物質の能力である;これは、偏光顕微鏡で見たとき、「マルタ十字」と呼ばれる暗い十字架を各々のデンプン粒上に生み出す。複屈折は、顆粒内のポリマーの構造組織の秩序程度の指標である(ThomasおよびAtwell、1999年)。デンプン粒の複屈折の損失は、通常、アミロース含有量の増加とよく相関している。
【0117】
<食糧生産への適性>
別の面で、本発明は食糧生産に役立つ小麦(穀粒が相対的に高いアミロース含有量を有し低減されたアミロペクチン含有量を有するデンプンを含有する。)を提供する。好ましくは、穀粒が得られる小麦植物は、成長の間、胚乳で低減されたSBEIIa活性レベルを有する。本発明の小麦植物は、食糧生産のために、特に、商業的食糧生産のために役立つ。そのような食糧生産は、商業的食糧生産の成分となし得る小麦粉、生地その他の産物の製造を含んでもよい。
【0118】
小麦の望ましい遺伝的バックグラウンドは、農業生産高および他の特徴の考慮を含む。そうした特徴には冬小麦種または春小麦種、農業的な成績、耐病性と非生物的なストレス耐性を有するのが望ましいか否かを含んでもよい。オーストラリアでは、バクスター、ケネディ、ジャンズ(Janz)、フレーム、ロゼッルラ(Rosella)、カドゥー(Cadoux)、ダイアモンドバード(Diamondbird)その他一般に栽培されている種類などの小麦栽培品種に変更されたデンプンの特徴を交配してみたいであろう。提供される例はオーストラリアの生産領域に特有であるが、他の変種は他の栽培地域に適している。本発明の小麦変種は、少なくともいくつかの栽培地域における、対応する野生型変種の80%以上の生産量を与えることが好ましく、90%以上がより好ましく、95%以上がさらにより好ましい。生産量は、制御された野外試験ですぐに計測できる。
【0119】
さらなる実施形態において、穀粒のデンプン含有量は、少なくともおよそ25%、35%、45%または55%〜65%(w/w)である。商業的に栽培される野生型小麦は、通常55〜65%の範囲のデンプン含有量を有し、ある程度は成長させる栽培品種に依存する。あるいは、本発明の穀粒は、等価であるが改変されていない小麦由来の穀粒の少なくとも90%のデンプン含有量を有する。野生型より低いデンプン含有量は、おそらくはアミロペクチンレベルが低減された結果である。デンプン含有量がたとえ低くても、ハイアミロース産物に比較的高い価値があるため、穀粒は商業食糧生産に依然有用となり得る。他の望ましい特徴は、穀粒を挽く能力、特に穀粒硬度を含む。より高価値の小麦植物を作る別の面は、穀粒からのデンプン抽出の程度であり、抽出率が高いほどより有用である。穀粒の形状は植物の商業的な有用性に影響を与え得るもう1つの特徴でもあり、このように、穀粒の形状は穀粒を挽く上での便利さまたはその反対の影響を及ぼし得る。例えば、細長い穀粒形態では、それを挽いて、処理するのが難しくなる可能性がある。
【0120】
より実の詰まった穀粒はより大きな生産量を達成する観点から望ましく、高レベルのアミロースを有するデンプンの生産または、変更された鎖長分布を有する改変されたデンプン中でのデンプン生産などのような本発明の特定の利点は成し遂げられ得る。このように、穀粒は好ましくは非縮小フェノタイプを有する。しかし、本発明の他の面は、より充填度の低い穀粒によって、より良好に達成されることもあり得る。このように、アリューロン層または胚またはタンパク質のデンプンへの割合はより充填度の低い穀粒でより高いこともあり、それによって、アリューロン層またはタンパク質といった有益な構成要素がより高い小麦粉その他の産物がもたらされる。高いアリューロン層産物はこのように葉酸塩などの特定のビタミン分がより高くてもよいし、あるいは、それはカルシウムなどの特定のミネラル分がより高くてもよく、より高い抵抗性デンプンレベルと組み合わさって、大腸におけるミネラル分の強化摂取に相乗的効果をもたらし得る。
【0121】
デンプンは、標準分析法、例えば、Schulman他(1991年)の方法を用いて小麦穀粒から容易に分離される。工業的スケールでは、湿式または乾式ミリングを使うことができる。デンプン粒サイズは、より大きなA顆粒をより小さなB顆粒から分離するデンプン加工産業で重要である。本発明の小麦植物の粒から得られるデンプンは、相対的に高いアミロース含有量を有する。
【0122】
<変化したデンプンの物理的特徴>
糊化は、過剰量の水中における分子秩序の熱による崩壊(破壊)であり、粒膨潤、結晶化温度、複屈折の喪失、粘性の増大とデンプン可溶化などの特性の同時的かつ非可逆的変化を伴う。トウモロコシのae(アミロース増量体)変異体からのハイアミロースデンプンは、通常のトウモロコシより高い温度を示した(Fuwa他、1999年、Krueger他、1987年)。他方、デンプン合成酵素IIa活性が欠如している大麦sex6変異体からのデンプンは低い糊化温度を有し、対照植物と比較したとき、糊化ピークに対するエンタルピーは減少した(Morell他、2003年)。
【0123】
本発明の別の面では、示差走査熱量測定で測定した際にデンプンは変更された糊化温度を有する。これは野生型植物由来のデンプンと比較して増加することも減少することもある。変更された糊化温度は、相対的に高いアミロース含有量に加わる場合もある。示差走査熱量測定で測定した場合、野生型小麦デンプンの糊化温度は、典型的には、最初のピーク(開始温度と定義される)の温度が約61℃(Rahman他、2000年)である。
【0124】
デンプンは、野生型デンプンと比較した過剰の熱水によるその膨張率によって特徴付けることができる。体積膨張は、典型的にはデンプンまたは小麦粉と過剰の水を混合し、加熱(典型的には90℃以上)することで測定される。次いで、試料を遠心によって集め、沈殿した物質の質量を試料の乾燥重量によって割ることで膨張体積を表す。食品調製物、特に水和する食品調製物のデンプン含有量を増やすよう求められる場合は、低い膨張特性が有用である。
【0125】
本発明の選択された形の小麦のデンプン構造は、結晶化度が小麦から分離される通常のデンプンと比較して低減するという点でも異なり得る。デンプンの結晶化度の低減は、官能特性の強化と結びつくとも考えられ、よりスムーズな舌触りに寄与する。このように、デンプンはさらに、1つまたは複数のアミロペクチン合成酵素の活性レベルの低減から生じる低減された結晶化度を示してもよい。結晶化度は、典型的にX線結晶学によって調べられる。
【0126】
変更されたアミロペクチン構造の1つの基準は、デンプンの鎖長分布または重合度である。鎖長分布は、イソアミラーゼ脱分枝の後、蛍光ラベル糖鎖電気泳動(Fluorophore−assisted carbohydrate electrophoresis:FACE)を用いて決定され得る。本発明のデンプンのアミロペクチンは、脱分枝した時点で、野生型植物からのデンプンの分布より大きい5〜60の範囲の鎖長分布を有し得る。より長い鎖長のデンプンは、枝分れの頻度が相応して減少する。したがって、デンプンは依然存在するアミロペクチン中により長いアミロペクチン鎖長の分布を有してもよい。
【0127】
<食物特性>
デンプンは人間の食餌における主要な炭水化物源であり、本発明とそれに由来する産物の穀粒は食物調製に用いることができる。例えば家畜生産において、または、ペットフードにおいて、食物は人または動物によって消費され得る。変更された小麦植物に由来する穀粒は食品加工の方法においてすぐに使うことができ、したがって、本発明は、前述の小麦植物を処理した穀粒または全粒から得られる、挽いたか、粉砕したか、粗挽きしたか、球状にしたか、ロール状にした穀粒または製品(小麦粉を含む。)を含む。これらの製品は、次いで、様々な食品、例えば、パン、ケーキ、ビスケットなどのデンプン質製品、または増粘剤または結着剤などの食品添加物中で、または、飲物、麺、パスタまたはインスタントスープに用いてもよい。本発明の穀粒に由来する穀粒または産物は、特に朝食用シリアルにおいて、または、押出加工された製品として望ましい。本発明のハイアミロースデンプンは、菓子類産業において有用なハイストレングスゲル類を形成するのに用いることもでき、または、より低温で短時間の硬化時間を可能にする。それらは、例えば、深揚げポテトその他の食品において油吸収を減らすためのコーティングとしても用い得る。
【0128】
<食物繊維>
本明細書において、食物繊維は、健康な人間の小腸では吸収されずに大腸に入る炭水化物および炭水化物消化産物である。これは、抵抗性デンプン、β−グルカンならびに他の可溶性および不溶性炭水化物ポリマーを含む。それは、少なくとも部分的には、常駐微生物相によって大腸内で発酵可能な炭水化物部分を含むことを目的とする。
【0129】
本発明のデンプンは、好ましくは比較的高いレベルの食物繊維、特にアミロースを含む。本発明の穀粒の食物繊維含有量は、単に胚乳のアミロース含有量の相対的な増加によって生じてもよいし、そうでなくてもよい。
【0130】
本発明の実施形態は、高レベルの食物繊維との組合せにおいて、アリューロン層と胚の組合せに起因してもよい。具体的には、アリューロンまたは胚の相対的レベルがより高い状態で存在する場合に、これが起こるのでもよい。小麦穀粒がわずかに縮小する場合、胚乳は量が低減されて存在し、アリューロン層と胚は比較的に増大した量で存在する。このように、小麦は高いレジスタントスターチと組み合わせて比較的高いレベルのある種の有益な要素またはビタミンを有し、そのような要素としては、二価カチオン、生物が利用可能なCa++と葉酸塩などのビタミン、あるいは、トコフェロールやトコトリエノールなどの抗酸化剤を含む。粉砕製品の1つの具体的形状は、アリューロン層が粉砕製品に含まれるものでもよい。具体的な粉砕プロセスは、粉砕製品中のアリューロン層の量を強化するように行ってもよい。このように、アリューロン層および胚を含むように粉砕または他の加工をした穀粒に由来する製品はいずれも、別の源からこれらの要素を加える必要がなく、さらなる栄養的な利益を有する。
【0131】
<レジスタントスターチ>
レジスタントスターチは、健康な人間の小腸に吸収されずに大腸に入るデンプンおよびデンプン消化産物の合計と定義される。したがって、レジスタントスターチは、消化されて小腸で吸収される産物を除外する。レジスタントスターチは物理的に接触しがたいデンプン(RS1形)、抵抗性顆粒(RS2)、老化デンプン(RS3)、化学修飾デンプン(RS4)を含む。食物で消費されるとき、本発明のデンプンの変更されたデンプン構造と特にハイアミロースレベルはレジスタントスターチの増加を引き起こす。デンプンは消化するにはいくぶん接触しがたいRS1形でもよい。V−複合体結晶度で測定されるデンプン−脂質会合体もレジスタントスターチのレベルに寄与しそうである。
【0132】
本発明の1つの利点は、それが特定の栄養的な利益である産物を提供し、さらに、デンプンまたは小麦穀粒の他の構成要素を修飾する必要なしにそのような産物を提供するという点であることが理解されるであろう。しかし、デンプンまたは穀粒の他の成分への修飾は望ましい場合もあり、本発明はそのような修飾された構成成分を包含する。修飾の方法はよく知られており、抵抗性形を増やすために、従来法によるデンプンその他の成分の抽出とデンプンの修飾を含む。デンプンは、熱および/または水分での処理によって、物理的に(例えば、ボールミリング)、酵素処理で(例えば、α−またはβ−アミラーゼ、プルラナーゼその他)、化学加水分解(液体またはガス状の試薬を使用して湿式または乾式で)、酸化、二官能基試薬(例えばナトリウムトリメタリン酸、オキシ塩化リン)を用いた架橋またはカルボキシメチル化によって修飾され得る。
【0133】
<血糖指数>
血糖指数(GI)は、デンプンを含む食品の消化率に関するものであって、運動時、血中グルコース濃度に関する白パンまたはブドウ糖の効果を試験食物の効果と比較するものである。血糖指数は、食事後の血清グルコース濃度と血中グルコースホメオスタシスのためにインシュリンの必要量に関する食物の影響可能性の目安である。本発明の食品によって提供される1つの重要な特徴は、血糖インデックスが減少することである。さらにまた、食品は最終的な消化レベルが低く、したがって比較的低カロリーとなし得る。低カロリー産物は、粉砕小麦穀粒から作り出される小麦粉の含有に基づくものでもよい。そのような食品は、お腹を満たし、腸の健康を強化し、食後の血清グルコースと脂質濃度を減らすとともに低カロリー食品を提供するという効果を有し得る。
【0134】
<食品以外の応用>
本発明は、アミロースレベルを上昇させ、アミロペクチンを低減させた修飾または改善されたデンプンを提供し、これらの特性は種々の産業の必要条件のいずれをも満たす。デンプンは、フィルム、紙、織物、段ボールおよび接着性産業(Young、1984年)で、例えば、サイズ剤として、広く非食品産業において使われている。小麦デンプンは、ブドウ糖シロップの生産のための、または、エタノール産生のための基質として用いてもよい。改変されていないデンプンの物性は若干のアプリケーションでその有用性を制限し、高価であるか他の不利益があることがあり得る化学修飾がしばしば必要となる。特に他の物性と組み合わせてアミロペクチン含有量を低減したために、本発明は収穫後改変の必要がより少ないデンプンを提供する。例えば、糊化温度、デンプンの剪断応力抵抗性、フィルム強さおよび/または水抵抗ならびに本発明の穀粒から製造される産物は、変更してもよい。デンプンは、ポリスチレンや他のパッキング材料の代わりとして生分解性のある緩い充填パッキング材料を調製するのに用いてもよい。
【0135】
本発明の各面に関して様々な説明を行ってきたが、本発明が本発明の2つ以上の面の組合せによるでもよいことが理解されるであろう。
【実施例】
【0136】
<実施例1.材料と方法>
炭水化物の決定と分析
デンプンは、Schulman他(1991年)の方法を使用して小麦穀粒から分離した。デンプン含有量は、Megazyme(Bray、Co Wicklow、アイルランド共和国)によって供給される全デンプン分析キットを使用して測定した。次いで、デンプン含有量を対照植物と比較する。総穀粒重量からのデンプン重量を引いて穀粒の全非デンプン含有量を求め、総重量の低減がデンプン含有量の低減によるかどうか確定する。
【0137】
デンプン試料のアミロース含有量は、以下の通り、MorrisonおよびLaignelet(1983年)の比色(ヨウ素還元)方法をわずかに変更して測定した。およそ2mgのデンプンを、ふたのゴムワッシャーと嵌まる2mlのネジキャップ付チューブに、正確に秤量した(0.1mgまで正確)。脂質を取り除くために、85%(v/v)メタノール1mlをデンプンと混合し、時折ボルテックスにかけながら65℃水浴中で1時間チューブを加熱した。13,000g、5分間の遠心の後、上清を慎重に取り除き、抽出工程を繰り返した。次いで、デンプンを65℃で1時間乾燥して、2mgデンプン(上記のように秤量)につき1mlの尿素−ジメチルスルホキシド溶液(UDMSO;1容積の6M尿素量に対し9容積のジメチルスルホキシド)を用いて溶解させた。混合物は直ちにボルテックスにかけ激しく混合し、デンプンを完全に溶解させるために断続的にボルテックスにかけつつ、95℃水浴中で1時間インキュベートした。デンプン−UDMSO溶液(50μl)のアリコートを、水ml当たり2mgのヨウ素と20mgのヨウ化カリウムを含む20μlのI2−KI試薬で処理した。混合物を水で全量1mlとした。200μlをマイクロプレートに移し、Emax Precision Microplate Reader(Molecular Devices、米国)を用いて吸光度を読むことにより650nmでの混合物の吸光度を測定した。0〜100%のアミロースと100%〜0%のアミロペクチンを含む標準試料をポテトアミロースとコーン(またはポテト)アミロペクチン(Sigma)から作り、試験試料と同様に処理した。アミロース含有量(パーセンテージアミロース)は標準試料の吸光度に由来する回帰式を使用して吸光度値から決定した。脱分枝したデンプンのアミラーゼ/アミロペクチン比の分析は、Case他(1998年)によって行ってもよいし、またはBateyおよびCurtin(1996年)によって記載されるように脱分枝デンプンを分離するHPLC法によって行ってもよい。
【0138】
デンプン試料の脱分枝後に、デンプン鎖長の分布は、Morell他(1998年)によるキャピラリー電気泳動ユニットを用いて蛍光ラベル糖鎖電気泳動(Fluorophore−assisted carbohydrate electrophoresis:FACE)によって分析し得る。デンプン試料の糊化温度プロフィールは、Pyris 1示差走査熱量計(Perkin Elmer、Norwalk、コネチカット州、米国)で測定し得る。デンプン溶液の粘度は、例えば、Batey他(1997年)が報告したような条件を用いてRapid−Visco−Analyser(RVA、Newport Scientific Pty Ltd、Warriewood、シドニー)上で測定し得る。測定し得るパラメータとしては、ピーク粘度(最大熱ペースト粘度)、維持強さ、最終的な粘性および糊化温度が挙げられる。小麦粉またはデンプンの膨潤量は、Konik−Rose他(2001年)の方法によって測定され得る。水の取り込みは、小麦粉またはデンプン試料を水と混ぜる前後、所定の温度で試料を量り、糊化した材料を回収することで測定される。
【0139】
β−Glucanレベルは、Megazyme(Bray、Co Wicklow、アイルランド共和国)によって供給されるキットを使用して測定され得る。
【0140】
胚乳のタンパク質発現の分析
胚乳の特定のタンパク質発現は、ウェスタンブロット手順によって分析した。胚乳はすべての母体組織から切り離し、およそ0.2mgの試料を、5mM EDTA、20%グリセリン、5mM DTTおよび1mMペファブロック(Pefabloc)を含む50mM KPi緩衝液(42mM K2HPO4と8mM KH2PO4)、pH7.5の600μl中でホモジナイズした。粉砕試料を13、000gで10分間遠心処理し、上清アリコートを使用まで−80℃で凍結した。総タンパク評価のために、0.25mg/mlBSA標準液0、20、40、60、80および100μlアリコートを用いてBSA標準曲線を準備した。試料(3μl)は蒸留水で全量100μlとし、1mlのCoomassie Plus Protein試薬を各々に加えた。吸光度はブランクとして標準曲線から0 BSA試料を用い、595nmで5分後に計測し、試料のタンパク質レベルを測定した。各々の胚乳から20μg総タンパクを含む試料を、0.34MTris−HCl(pH8.8)、アクリルアミド(8.0%)、過硫酸アンモニウム(0.06%)およびTEMED(0.1%)を含む8%非変性ポリアクリルアミドゲル上で泳動させた。電気泳動後、タンパク質はMorell他(1997年)によるニトロセルロース膜に移し、SBEIIaまたはSBEIIb特異抗体と免疫反応させた。
【0141】
<実施例2.小麦SBEIIAとSBEIIB発現変更のための遺伝子コンストラクト>
小麦のSBEIIaまたはSBEIIb遺伝子の発現を低減すべく二重鎖RNA(dsRNA)コンストラクトを作成した。そのようなコンストラクトにおいて、SBEIIaまたはSBEIIb遺伝子の一部に対応する望ましい核酸配列は、発現したRNAが塩基対合可能で二重鎖または二本鎖RNAを形成し得る相補領域を含むように、プロモーターに対しセンスとアンチセンスの両方に存在した。センスとアンチセンス配列の間のスペーサー領域は、形質転換植物においてRNAの一部として転写されるとき、堅い「ヘアピン」二重構造を作るためにスプライシングにより除かれるイントロン配列からなるものとした。イントロンを含むことで、二重鎖RNAコンストラクトによって与えられる遺伝子サイレンシングの効率が増すことが分かっている(Smith他、2000年)。望ましい核酸を、高分子量グルテニン(HMWG)プロモーター配列(Dx5サブユニット遺伝子、アクセッション番号X12928、Anderson他、1989年のプロモーター)とアグロバクテリウム由来ノパリンシンターゼ遺伝子(nos3')からのターミネーター配列に連結した。これは、dsRNA配列の胚乳特異的発現をもたらした。
【0142】
SBEIIa二重鎖RNAコンストラクトは、小麦SBEIIa遺伝子(GenBankアクセッション番号AF338431、図1参照)からPCRによって増幅される1536bpのヌクレオチド配列を含んでいた。これは以下を含んでいた;エキソン1と2の全体とエキソン3の一部を含み(図1のヌクレオチド位置1058〜1336、1664〜1761および2038〜2219)EcoRIとKpnIの制限部位を各側に有する468bp配列(断片1)、SBEIIaのエキソン3と4の一部およびイントロン3の全部を含み(図1のヌクレオチド位置2220〜2731)KpnIとSacIの部位を各側に有する512bp配列(断片2)ならびにSBEIIaの完全なエキソン1、2および3を含み(図1のヌクレオチド位置1058〜1336、1664〜1761および2038〜2279)BamHIとSacIの部位を各側に有する528bp配列(断片3)。次いで、断片3の配列が断片1に対してアンチセンス方向において断片2にライゲートされるように、断片1、2と3をライゲートした。最初に、HMWGプロモーター配列とnos3'ターミネーターを含むベクターpDVO3000内で二重鎖RNAコンストラクトを生成した。ベクターpDVO3000の遺伝子コンストラクトはpDVO3−IIaと名付け、二重鎖RNA遺伝子はds−SBEIIaとした。
【0143】
SBEIIb二重鎖RNAコンストラクトについての戦略も同様とした。SBEIIbコンストラクトは、小麦SBEIIb遺伝子(配列は、図2で概説)からPCRによって増幅される1607bpの断片を含んでいた。これは、エキソン1と2の全体とエキソン3の一部を含み(図2のヌクレオチド位置489〜640、789〜934および1598〜1769)EcoRIとKpnIの制限部位を各側に有する471bp配列(断片1)、SBEIIbのエキソン3と4の一部およびイントロン3の全部を含み(図2のヌクレオチド位置1770〜2364)KpnIとSacIの制限部位を各側に有する589bp配列(断片2)ならびにSBEIIbの完全なエキソン1、2および3を含み(図2のヌクレオチド位置489〜640、789〜934および1598〜1872)BamHIとSacIの制限部位を各側に有する528bp配列(断片3)を含んでいた。次いで、断片3の配列が断片1に対してアンチセンス方向において断片2にライゲートされるように、断片1、2と3をライゲートした。ベクターpDVO3000のSBEIIb二重鎖RNA遺伝子コンストラクトはpDVO3−IIbと名付け、二重鎖RNA遺伝子はds−SBEIIbとした。コンストラクトを図3に模式的に示す。
【0144】
次いで、ds−RNA発現カセットの各々を制限酵素−Xholで切り離し、バイナリー形質転換ベクターpGB53およびpBIOS340に挿入した。pGB53は、コメアクチンプロモーターによってドライブされるアスラム耐性(sul)をコードする遺伝子の導入によってpSBll(Komari他、1996年)から作られ、対象とすべき遺伝子の導入のためにT−DNA右側境界に隣接して特有のXhoIサイトを残した。同様に、pBIOS340はカナマイシンとジェネテシン耐性をコードするnptII遺伝子の導入によってpSBl(Komari他、1996年)から作られ、コメアクチンプロモーターによってドライブされ、ここでも、右側境界に隣接して特有のXhoIサイトを残した。pGB53とpBIOS340のSBEIIaコンストラクトはそれぞれpCL51とpCL59と名付け、pGB53とpBIOS340中のSBEIIbコンストラクトはそれぞれpCL54とpCL60とした。
【0145】
<実施例3:小麦の形質転換>
virプラスミドpAL4404およびpSBlを運ぶ無害化したAgrobacteilum tumefaciens株LBA4404に小麦の形質転換のための遺伝子コンストラクトをエレクトロポーレーションによって導入し、続いてスペクチノマイシンを含む培地上で選択した。形質転換したアグロバクテリウム株を、2日間固化させたYEP培地上27℃でインキュベートした。次いでバクテリアを回収して、小麦接種のため、650nmで光学濃度2.4まで400mMアセトシリンゴンを含むTSIM1(100mg/lmyo−イノシトール、10g/lブドウ糖、50mg/l MES緩衝液pH5.5によるMS培地)に再懸濁させた。
【0146】
小麦植物(品種NB1、Nickerson Seeds Ltd、Rothwell、Lincs.から得られる春小麦種)を、1日16時間の日照とするために明かりを補って温室内で22/15℃日/夜の温度で成長させた。50cmのひこばえ茎を含むように開花期後およそ14日で(胚:長さおよそ1mm)でひこばえを収穫した。次いで、止め葉以外、すべての葉をひこばえから取り除き、止め葉は清浄化してカビ胞子による汚染を除いた。次いで、各々の小穂の包穎と最初の2つの小花から外穎を慎重に取り除いて未熟種子を露出させた。一般に、各々の小穂にはこれらの2粒の種のみが見つかった。この手順を、花序の全長に沿って実行した。次いで、穂には簡単な表面殺菌を行うために70%IMSを吹き付けた。
【0147】
アグロバクテリウム懸濁液(1μl)を、10μlハミルトン注射器を使って、未熟種子内においてほぼ胚盤:胚乳の界面の位置で、露出した種子すべてに接種した。次いで、ひこばえを水中に置き、種の乾燥を防ぐために半透明のビニール袋で覆い、明かりをつけたインキュベーター中に23℃で3日間、日照1日当たり16時間、45_Em−2s−1PARで置いた。同時培養3日の後、接種した未熟種子を取り除き、70%エチルアルコール(30秒)、次いで20%漂白剤(Domestos、20分)で表面を殺菌し、無菌蒸留水で完全に洗浄した。未熟な胚を無菌的に単離し、W3培地(MSに20g/lショ糖と2mg/l 2,4−Dを補い、6g/l Type Iアガロース(Sigma)で固めたもの)に置き、150mg/lタイメンチン(Timentin)(W3T)を加え、胚盤(プレート当たり20胚)を一番上とした。培養物を、25℃で光(1当たり16時間の日照、80_Em−2s−1PAR)中に置いた。胚上の胚軸の成長を単離後およそ5日後に評価し、カルス生産を改善するために必要な場合、軸は取り除いた。胚は4週間W3T上で維持し、単離2週間後、新しい培地に移し、胚形成能力を評価した。
【0148】
4週成長させた後、接種した胚に由来するカルスは、W3T媒地上に植菌した非接種胚から得られる対照カルスと非常に類似していた。バクテリアの存在は、接種した胚に由来するカルスの胚形成能力を実質的に低減させたようには見えなかった。胚形成カルスを、2mg/lアスラム(pGB53誘導体を使用した)または25mg/l(pBIOS340誘導体)および150mg/lタイメンチン(W32AT)を含むW3培地に移した。カルスをさらに2週間、この媒地上で維持し、次いで、各カルスを2mm−サイズの小片に分け、W32AT上に再植菌した。バイナリーベクターコンストラクトなしでLBA4404の接種に由来する対照胚は、選択培地上で形質転換カルスを生じなかった。
【0149】
さらに2週間培養後、胚形成性カルスの成長についてすべての組織を評価した:4週後にも選択培地上で継続的な成長の徴候を示しているすべてのカルスが、再生培地(40g/lマルトースと150mg/lタイメンチン(pH5.8)を加えたRMT−MS、6g/lアガロース(Sigma 1型)で固化)に移された。新芽をこの培地上で4週間再生し、次いで、芽の伸長と発根のために150mg/lタイメンチンを加えたMS30に移した。次いで、幼若植物を土壌混合物に移し、2週間噴霧ベンチに保持し、最後に温室に移した。
【0150】
pCL54またはpCL60(ds−SBEIIb)を使っている合計3217個の胚およびpCL51またはpCL59(ds−SBEIIa)を使っている2010個の胚をこの方法によって処理し、61個の植物をIIb形質転換のためにカルスから再生し、31個の植物をIIa形質転換のためにカルスから再生した。選択培地上の生存状況は、それらが遺伝子コンストラクトによって首尾よく形質転換されていることを示唆した。選択可能なマーカー遺伝子で形質転換した植物の大多数(すべてではない)は、SBEIIaまたはSBEIIb阻害遺伝子を一体化するものと期待される;これらは以下の例に記載するように容易に識別することができた。
【0151】
実験から得られた良好な再生可能性を伴う複数の安定した組み込みイベントの回復は、本明細書で用いられる種接種形質転換法が小麦について報告されている他の方法と同じように効率的なことを示した。株AGL1などの代わりのアグロバクテリウム株またはハイグロマイシン抵抗性をコードする遺伝子などの選択可能なマーカーも本明細書方法に用い得る。
【0152】
<実施例4.小麦形質転換体の分析>
形質転換は、以下の方法の1つまたは複数で測定した:
導入遺伝子の1つまたは複数についてのPCR分析。PCR分析は、StaceyおよびIsaac(1994年)によって記載されるミニプレップ法を使用し、新鮮な葉資料1〜2cm2から抽出されるゲノムDNAについて行った。PCR反応は、SBEIIa遺伝子からの断片(462bp)を増幅すべく設計された、例えば、プライマーSBEIIa−For:5'−CCCGCTGCTTTCGCTCATTTTG−3'[配列番号9]およびSBEIIa−Rev:5'−GACTACCGGAGCTCCCACCTTC−3'[配列番号10]を用いて、またはSBEIIb(505bp)についてSBEIIb−DupFor5'−AGATGTGAATGGCTGCTTGCTG−3'[配列番号11]およびSBEIIb−DupRev5'−CAGGTCGACCATATGGGAGAGC−3'[配列番号12]を用いて行った。反応条件は以下の通りとした:「ホットスタート」(94C、3分)後、30サイクルの変性(95℃、30秒)、アニーリング(55℃、30秒)、伸長(73℃、2分)後、73℃(5分)で1サイクル。
【0153】
サザンブロットハイブリダイゼーション分析は、凍結乾燥した粉砕組織(StaceyおよびIsaac、1994年)から、より大規模(9ml)に抽出したDNAに対して実行する。DNA試料は0.2mg/mlに調整し、HindIII、EcoRIおよびKpnIなどの制限酵素で消化した。StaceyおよびIsaac(1994年)に記載されるように、制限酵素消化、ゲル電気泳動と真空ブロッティングを実行する。ds−SBEIIコンストラクトのイントロン3領域を含むジゴキシゲニンで標識したプローブは、McCreeryおよびHelentjaris(1994年)の方法によってPCRにより製造される。サザンブロットへのプローブのハイブリダイゼーションおよび化学発光による検出は、McCreeryおよびHelentjaris(1994年)の方法によって実行する。
【0154】
PCR分析の結果を表2にまとめる。PCRによって示される導入遺伝子が陽性だった植物は、ds−SBEIIaについて27の独立した形質転換イベントとds−SBEIIbについての61の独立したイベントを含んでいた。
表2.SBEIIaとSBEIIbRNA二重鎖コンストラクトによる小麦の形質転換
【0155】
【表2】
【0156】
<実施例5.二重鎖RNAコンストラクトで形質転換した植物から得た穀粒の分析>
デンプン顆粒形態
T0形質転換小麦植物から得た成熟したT1種子からのデンプン顆粒の形態を、光学顕微鏡法によって観察した。ds−SBEIIaで独立に形質転換した25本のT0植物とds−SBEIIbで独立に形質転換した12本の植物の各々からのそれぞれ10個の穀粒を分析した。各胚乳はデンプン顆粒を放出させるために穏やかに押しつぶし、それを水中に分散して、光学顕微鏡の下で視覚化した。分析した25のds−SBEIIa系統のうち、12個は変形顆粒(例えば、図4参照)を含む穀粒を有していたが、視覚的観察では異なる種子では変形レベルは様々であった。これに対し、12のds−SBEIIb系統のいずれも、光学顕微鏡法下で観察したとき胚乳中のデンプン顆粒の顕著な変形は示さなかった。結果を、表3と4にまとめて示す。
表3.ds−SBEIIaトランスジェニック小麦系統のT1種子のデンプン顆粒形態
【0157】
【表3】
【0158】
*各系統からの10粒の種子のデンプン顆粒形態を観察した。
10粒すべての種が通常の顆粒形態を有する場合は、形態を+として示し、ひどく変形した種子がある場合は−で、また、若干の異常がある(すなわち、少なくともいくつかの種子にある程度の変形があるが、ひどい変形のものはない)場合は+/−とした。
【0159】
表4.ds−SBEIIbトランスジェニック小麦系統のT1種のデンプン顆粒形態。
【0160】
【表4】
【0161】
+は、各系統からの10粒の種子すべてが正常なデンプン顆粒形状を有していたことを示す。
偏光下のデンプン顆粒を観察したところ、ds−SBEIIa穀粒については、変形顆粒(図5)のため、複屈折が著しく低減していたことが分かった。複屈折の損失は、系統50.lbからの種子では顆粒の94%で観察され、一方、同じ系統の別の種子からの正常な顆粒が完全な複屈折(表5)を示しており、変形フェノタイプと相関している。正常な顆粒による種子は、導入遺伝子が不足している分離体であると思われ、したがって、発現型的には正常である。
表5.ds−SBEIIaトランスジェニック小麦系統50.1bのT1種子からのデンプン顆粒の複屈折
【0162】
【表5】
【0163】
+通常のデンプン顆粒、−ひどく変形した顆粒、+/−若干の変形のある顆粒
【0164】
光学顕微鏡観察の結果は、デンプン顆粒の走査型電子顕微鏡(SEM)によって確認する。このために、精製したデンプンに金でスパッタリングして、室温、15kVで走査する。
【0165】
穀粒重量
ds−SBEIIa形質転換植物(温室で等しい条件下で生育)からの個々の穀粒を秤量した(表6)。植物50.1b、58.2a、61.2aおよび109からのひどく変形した顆粒を有する穀粒は、同じ条件下で生育した野生型植物の粒と比較して、平均重量で顕著な低減はなかった。したがって、デンプン生産は、大きく変形したデンプン顆粒を有する種子においてであっても実質的にされる低減するようには見えなかった。このデータは、胚乳中のSBEIIa活性の低減した野外生育小麦の収穫高がだいたい普通であることも示唆している。
表6.ds−SBEIIaトランスジェニック小麦系統からのT1種子の穀粒重量
【0166】
【表6】
【0167】
T2トランスジェニック小麦胚乳のSBEIIaとSBEIIbタンパク質の分析
5つの独立した形質転換系統を代表して13ds−SBEIIa形質転換T1植物からと、3つの独立した形質転換系統を代表して9ds−SBEIIaからの種子(T2)を、胚乳内のSBEIIaとSBEIIbタンパク質発現について、非変性PAGEとウェスタンブロッティングによって分析した。先に述べたように、ds−SBEIIb系統はすべて正常の顆粒形態を有するが、ds−SBEIIa植物はすべて異常なデンプン顆粒形態を有する系統からのものであった。SBEIIaの検出のために用いた抗体は、ウサギ由来の3KLHであり、これはアミノ酸配列AASPGKVLVPDESDDLGC[配列番号13]を有し、成熟SBEIIaのN末端からの配列に対応する合成ペプチドに対して誘導したものであり、使用に際して1:5000に希釈した。SBEIIbの検出のために用いた抗体はR6であり、これはアミノ酸配列AGGPSGEVMIGC[配列番号14]を有し、成熟SBEIIbのN末端からの推定配列に対応する合成ペプチドに対して誘導したものであり、使用前に1:6000に希釈した。使用した二次抗体は、GAR−ホースラディッシュペルオキダーゼ複合体とした(1:3000希釈)。免疫反応バンドは、アマシャムECL−検出システムを使用して明らかにした。
【0168】
植物の中には導入遺伝子についてヘテロ接合のものもあると予想されたため、22本のT1植物の各々から7粒の成長中の穀粒(開花後15日)の各々から胚乳を分析した。13のds−SBEIIa植物のうちの12は、胚乳中にSBEIIaタンパク質レベルの低減を示すT2子孫を生じた。1系統からの全7粒の種子(50.3x.9)はSBEIIaを完全に欠いているようであったが、他の4本の植物からの全7粒の種はSBEIIaの明らかに低減した発現を示した(表7)。これらは、導入遺伝子についてホモ接合性である系統を表しているであろう。7系統はSBEIIaが存在しない点またはSBEIIaレベルが低減している点、または場合によってはタンパク質が明らかには低減してない点で分かれており、これらの系統はおそらく導入遺伝子についてのヘテロ接合体を表している。13系統(50.3x.6)は野生型発現についてホモ接合性だった(表7)。
表7.T2 ds−SBEIIaトランスジェニック小麦系統からの胚乳タンパク質のウェスタンブロット分析
【0169】
【表7】
【0170】
試験した9のds−SBEIIbトランスジェニック系統のうち、3つ(110.16b.2、110.16b.5および110.16b.19)は一様に、7粒の子孫種子のいずれにおいてもSBEIIb発現は示さなかったが、2は野生型発現について均一、残る4は発現なし、発現低減または野生型に分離した(表7)。種子からの胚は、T2植物とT3種子を生じさせるべく成長させてもよく(胚レスキュー)、これらは導入遺伝子に関してT2種子の遺伝子の状態を確かめるためにPCRとタンパク質発現分析でスクリーニングする。
【0171】
これらのデータは、二重鎖RNAコンストラクトが小麦胚乳のSBEIIa遺伝子およびSBEIIb遺伝子の発現低減に関して有効なことを示す。データはまた、SBEIIb発現のみの低減はデンプン顆粒形態を実質的に変化させなかったことをも示す。
【0172】
ds−SBEIIa導入遺伝子を含み、SBEIIaタンパク質が欠如しているトランスジェニック種子におけるSBEIIb遺伝子の発現とds−SBEIIbを含む種子のSBEIIa遺伝子の発現の発現も、ウェスタンブロット法によって分析した。予想外にも、ds−SBEIIaを含むトランスジェニック種子では、SBEIIbが大きく減少した。しかし、ds−SBEIIbについてトランスジェニックな種子では、逆の影響は観察されなかった。SBEIIbがds−SBEIIbによって完全にサイレンスにされた種子でSBEIIa発現は変わらなかった。SBEIIbの発現は、二重鎖コンストラクトのために使われる領域の遺伝子間での配列相同性のためにds−SBEIIaコンストラクトによって抑制された可能性があり、SBEIIbの活性は、いくつかの他のメカニズムによってds−SBEIIa導入遺伝子によって低減された可能性がある。
【0173】
SBEIIa遺伝子およびSBEIIb遺伝子の発現レベルは、mRNAレベルにおいても、ノーザンハイブリダイゼーションまたはRT−PCR法などの標準的な技術によって(例えば、非保存領域由来のプローブを用い、または遺伝子のうちの1つにおいて特異的なサイトにハイブリダイズする他のサイト、例えば、3'非翻訳領域ではハイブリダイズしないプライマー対を用いて)具体的に決定し得る。そのような領域またはサイトは、2つの遺伝子配列の比較によって、容易に確認できる。
【0174】
<実施例6.形質転換小麦のデンプン分析>
トランスジェニック小麦穀粒中でのアミロースおよびアミロペクチンレベル
実施例1に記載したように、6つのプールしたT1種子試料からのデンプンのアミロース含有量を測定した。プールした種子試料は、以下のようにしてトランスジェニック小麦系統から得た:
【0175】
プール1−ds−SBEIIaトランスジェニック系統85.2cからデンプン変形顆粒を有していた種子
プール2−ds−SBEIIaトランスジェニック系統85.1aから正常顆粒を有していた種子
プール3−ds−SBEIIbトランスジェニック系統110.18aから正常顆粒を有していた種子
プール4−ds−SBEIIaトランスジェニック系統58.1a、58.2aおよび61.2a(一緒にプール)から変形顆粒を有していた種子
プール5−ds−SBEIIaトランスジェニック系統83.1bから正常顆粒を有していた種子
プール6−ds−SBEIIbトランスジェニック系統75.3xから正常顆粒を有していた種子
【0176】
各々の分析は、デンプン試料の4つの複製を用いて行った。これらの分析のために、吸光度をアミロース含有量に変換するのに用いられる回帰式は、Y=57.548x−8.793であった(ここで、Yはアミロース含有量(%)であり、xは吸光度)。
【0177】
結果を表8に示す。変形デンプン顆粒の存在は、明らかにアミロース含有量の増加を伴っていた。ds−SBEIIaトランスジェニック系統(プール1および4)からの変形顆粒を有する穀粒由来のデンプンは50%を超えるアミロース含有量を有していたのに対し、他のデンプンプール(正常なデンプン顆粒を有する穀粒に由来する)は21〜26%の範囲でアミロース含有量を有していた。これはSBEIIb(表8)の発現が低減された系統IIb 110.18aからのデンプンを含んでおり、このことは、小麦中においてSBEIIbのみ失活させても、穀粒デンプン中でのアミロースレベルは実質的に増加しないことを示唆する。
表8.トランスジェニック小麦系統のヨウ素還元法によって推定されるアミロース含有量
【0178】
【表8】
【0179】
分析の第2セットは、プール 4とSSII(Yamamori他、2000年)が欠如している小麦からのデンプンおよびSSIIaの変異体である大麦系統M292からの試料を用いてヨウ素還元法によって行った。プール4小麦種子(ds−SBEIIaトランスジェニック系統)について測定されたアミロース含有量は、小麦および大麦のSSII変異体からのデンプンのそれより相当に高かった。
【0180】
これはこれらの穀粒中のデンプンのアミロペクチン含有量が相当に、すなわち、野生型のおよそ75%から、50%未満またはさらには20%未満にまで低減されることを意味する。
【0181】
ds−SBEIIaとds−SBEIIb導入遺伝子を含む系統は、上述の遺伝子導入植物を交配することによって発生する。そのような子孫の穀粒デンプン中のアミロース含有量は、ds−SBEIIaのみを含む植物由来のデンプンのアミロース含有量以上に、例えば75または80%まで増加し、SBEIIaに加えてSBEIIbの抑制がさらにアミロースレベルを上昇させることを示している。
【0182】
<実施例7.A、BおよびD ゲノムからのSBEIIaの比較>
小麦cDNAのコンストラクトおよびゲノムライブラリー
小麦胚乳cDNAとゲノムライブラリーは、ファージベクター(Sambrook他、1989年)における標準分析法によって作成した。2つのcDNAライブラリー(一方は栽培品種Rosella(Rahman他、1999年)由来のRNAから、他方は栽培品種Wyuna(Rahman他、2001年)から作成した。RosellaライブラリーはベクターZAPII中にあってEcoRIとNotIプライマーを用い、他方、Wyunaライブラリーは試薬に伴って与えられるプロトコルに従ってZipLoxベクター(Life Technology)中に存在した。ライブラリーの力価は、大腸菌のY1090(ZL)株で試験して2×106pfuの力価であった。ゲノムライブラリーは、A.tauschii変異体10097由来のDNAから作成した。DNAをSau3Aで消化し、部分装填lambdaGEM12ベクター(Promega)にライゲートした。クローンフラグメントは、SacIまたはXhoI消化で放出できた。T.aestivum DNAのゲノムライブラリーは、Turner他(1999年)に記載されている。
【0183】
SBEIIa cDNA配列の隔離
小麦SBEI遺伝子配列のプローブを使って低ストリンジェンシーで(Rahman他、2001年)で、栽培品種Rosellaから調製されるライブラリーからcDNAを分離した。得られた最も長いクローン(sbe9と命名)の配列決定を行ったところ、SBEIIaタイプの配列(Genbank AF338432.1)をコードすることが分かった。その後、栽培品種Wyuna(Rahman他、2001年)から調製した胚乳cDNAライブラリーから、sbe9の位置536〜890に対応するプローブを用いて3つのクローンを分離した。ライブラリースクリーニングの条件は、25%ホルムアミド、5×SSC、0.1%SDS、10×デンハルト溶液、100μg/mlサケ精液DNAで42℃、16時間のハイブリダイゼーション後、2×SSC、0.1%SDS、65℃、3×1時間(中程度のストリンジェンシー)で洗浄とした。クローンを配列決定することにより、3つの異なる配列が得られ、これらを以下、sr995およびsr997(図6)と表す。
【0184】
これらのcDNA配列を調べることにより、様々な配列が小麦胚乳で発現していることが示されたが、これらは異なる小麦ゲノムからのSBEIIa転写物と対応しているようである。他の既知の小麦SBEIIa cDNA配列による配列とのPILEUP比較は、sr995とsr996配列が、D−ゲノム配列wSBE−D1(sr854)(図7)に由来するmRNA配列とクラスターになっていることを示し、sr995およびsr996がDゲノムSBEIIaからの転写物を表していることを示唆した。Y11282配列(Nair他、(1997年))とクラスターになっているSr997は、それらがおそらく同じゲノム(AゲノムかBゲノム)に由来するものであることを示している。前述のsbe9(AF338432.1)はおそらくY11282と同じゲノムに由来するものであろうが、選択的スプライシングイベントを表し、5'末端の近くで1つのエキソンがスプライシングで除かれたものと一致している。
【0185】
小麦T.aestivumのA、BおよびDゲノムからのSBEIIa遺伝子との区別
遺伝子またはRNA転写配列における相違は、遺伝子レベルにせよRNAレベルにせよ、突然変異スクリーニングのためのA、BおよびDゲノム特異的プライマー設計の基礎として用い得る。例えば、図6は、Genbank アクセッションY11282を含むcDNA由来のSBEIIaヌクレオチド配列とcDNA sbe9(AF338432.1)、sr997およびsr995の部分配列と比較している。ゲノム配列はT.aestivum由来のSBEIIa遺伝子に利用可能である(例えば、表1参照)。ゲノム配列は、A、BおよびDゲノムに割り当てられた。比較は染色体の多型を示し、そのいずれもが配列を分子的手段によって識別するのに用いることができる。
【0186】
エキソン5領域に基づくフォワードプライマー(5'−ATCACTTACCGAGAATGGG−3')[配列番号15]およびエキソン6領域に基づくリバースプライマー(5'−CTGCATTTGGATTCCAATTG−3')[配列番号16]をA、BおよびDゲノム由来の産物間の識別をするのに用いた。そのようなプライマーは、A、BまたはDゲノム(下記参照)由来のSBEIIa遺伝子の1つまたは複数の変異体である変種またはアクセッションをスクリーニングするためにPCR反応で用いてもよい。
【0187】
SBEIIb遺伝子を小麦のA、BおよびDゲノムから区別するためにPCRベースのマーカーも開発した。例えば、プライマー対ARA19F(5'−CACCCATTGTAATTGGGTACACTG−3')[配列番号17]とARA15R(5'−TCCATGCCTCCTTCGTGTTCATCA−3')[配列番号18]を用いたPCR反応の後、制限酵素RsaIで増幅産物の消化を行うことで、SBEIIb遺伝子と3つのゲノムを区別することができた。
【0188】
cDNA配列の違いを、推定タンパク質配列に反映する。例えば、Dゲノム(sr854)とAまたはBゲノム(Y11282)ポリペプチドに対して推定したノーカットアミノ酸配列を図8で比較する。1つまたは複数のゲノムに特異的な活性が欠如している小麦の種類をスクリーニングするために、SBEIIaタンパク質に対するゲノム特異的抗体を産生するために使うことのできた領域688〜698と735〜6とでは顕著な差異が明らかである。他の違いは、図8のアミノ酸位置1〜54と対応する輸送ペプチド配列で起こる。
【0189】
<実施例8.1つまたは複数のSBEII遺伝子における小麦の種類変異体の識別>
BおよびDゲノム中のSBEIIbヌル変異の識別
300のオーストラリアの小麦の種類を含む1500の小麦寄託種の合計、オーストラリア冬穀物コレクション(AWCC、Australian Winter Cereal Collection、Tamworth、NSW Australia)からの900の小麦寄託種と300の小麦生産地種を、プライマーARA19F(上記参照)とARA23R(5'−CTGCGCATAAATCCAAACTTCTCG−3')[配列番号19]を用いて多形イントロン3領域に対応するSBEIIbマーカーを用いたPCR増幅でスクリーニングした。PCR増幅は、上述の条件を用いた。増幅産物を制限酵素RsaIで消化して、ポリアクリルアミドゲル上で電気泳動にかけた。3つの系統(Aus12745、Aus17340およびAus10103)はDゲノムマーカーが欠けており、2つの系統(Aus12509およびAus12565)はBゲノムマーカー(図9)が欠けていた。これらの系統は、BまたはDゲノムのためのSBEIIb遺伝子において推定されるヌル変異体を表す。
【0190】
PCRの結果を確認するために、ヌル変異体系統からDNA上で、サザンブロットハイブリダイゼーション分析を行った。標準分析法によって植物から調製したDNAをHindIIIで消化して、1%アガロースゲル上で電気泳動にかけ、Hybond N+ナイロン膜(Amersham)上にブロットした。
【0191】
Megaprime DNAラベリングシステム(Amersham Pharmacia Biotech UK Ltd)を使用して、SBEIIb(位置2019〜2391、図2参照)遺伝子のイントロン3領域から放射性ラベリングしたプローブを生成し、厳しいハイブリダイゼーション条件下に使用した。Aus17340とAus10103は、Dゲノムから〜4.8kBバンドを失っており、Aus12509とAus12565は、Bゲノムから〜3.4kBを失っていた(図10)。したがって、これらの系統は、それぞれ、DまたはBゲノムSBEIIb遺伝子についてのヌル変異体であることが確認された。
【0192】
B+D2倍ヌル変異体の世代
以下の交配を実行した:
Aus17340a×Aus12509
Aus17340b×Aus12509
Aus17340a×Aus12565
Aus17340b×Aus12565
Aus12745 ×Aus12509
Aus12745 ×Aus12565
【0193】
Aus17340aとAus17340bは、同じAus17340栽培品種の2つの異なるバイオタイプであり、両方とも、DゲノムSBEIIb遺伝子マーカーについて無効であることを確認した。F1植物を自家受粉し、F2子孫はBとD両方のゲノムSBEIIb遺伝子の変異体(2倍ヌル変異体)である植物のためのPCR方法によってスクリーニングした。プライマー対AR2b19cF(5'−CTATGCCAATTGAACAACAATGC−3'[配列番号20]とAR2b23cR(5'−CGTGTTCATCAATGTCTGAACG−3'[配列番号21](これは、ARA19F/ARA23Rと同じ領域を増幅する)を用いたPCR増幅後、制限酵素Rsalを用いた消化によってSBEIIb突然変異の分離を観察した。典型的分離パターンを図11に示す。カイ二乗分析で、交配Aus17340a×Aus12509とAus17340a×Aus12565の分離パターンが9:3:3:1(表9)の期待される比率に当て嵌まることが分かった。分離は、他の交配では非常に歪んでいた。
表9.BとDサブゲノムにおけるSBEIIbヌル変異体間の交配のF2集団のカイ二乗分析
【0194】
【表9】
【0195】
表中のχ2(9:3:3:1)の値(0.05)、df3=7.81
【0196】
アルビノ植物は親の系統にかかわりなくすべて交配で検出され、変異体葉緑素関連の遺伝子も集団内で分離していたことを示した。分析した24のアルビノ植物のうち、23はB+D二重ヌル変異体であり、1は野生型であるように見えた。B+D二重ヌル変異を伴う5つの正常に見える緑色植物を、試験した718の系統から確認した。これらのうちの3はAus17340b×Aus12509(BD219、BD303、BD341)、1つはAus17340a×Aus12509(BD54)、1つはAus17340b×Aus12565(BD636)に由来するものであった。この結果から、BおよびDゲノムSBEIIb遺伝子における突然変異は、2つの突然変異した遺伝子位置が一緒になった場合、アルビノフェノタイプを与えている葉緑素関連遺伝子における突然変異と密接な関連があることが明らかになった。しかし、極めて低い頻度ではあるものの、正常のB+D二重ヌル変異体系統をもたらす、SBED遺伝子と葉緑素関連遺伝子の間の組み換え事象が確認された。これは、2つの遺伝子が密接に結合されるが、分離され得ることを示す。
【0197】
<実施例9.SBEIIaとSBEIIbは小麦中で結合している>
BACクローンの単離
A.tauschii meyeri品種(Moullet他、1999年)から作られる大きな挿入コスミドバイナリーコスミド(BAC)ライブラリーをSBEIIb遺伝子のイントロン3領域(位置2019〜2391、図2)でプローブし、SBEIIb遺伝子を含むBACを単離した。4つのポジティブなクローンを単離して、BAC−4、−5、−9および−12と称した。これらがSBEIIb遺伝子を含んでいることを確認するために、これらのクローンからDNAを抽出し、HindIIIまたはEcoRlで消化し、同じプローブ(図12)を使ってサザンブロットハイブリダイゼーションを実行した。クローンBAC−5は、EcoRlを伴う〜7.5kBのサイズの1つの強いハイブリダイズバンドと、HindIIIを伴う〜6.1、3.6、2.3および1.7のサイズの4つのバンドを示した(図12)。これは、BAC−5上でのSBEIIbの存在を示した。BAC−5上での遺伝子の3'の存在を調べるため、SBEIIbcDNA配列に基づく、エキソン17を増幅するように設計された特定のプライマー(AR2b3pr2F、5'−GGATATGTATGATTTCATGG−3'[配列番号22]およびAR2b3pr2R、5'−CCATAAAGTTAAGATAACCC−3')[配列番号23]、および20(AR2b3pr1F、5'−GACATCAGACCACCAGTACG−3')[配列番号24]およびAR2b3pr1R、5'CTTCCCAGGCTTTAAACAGC−3')[配列番号25]を使ってPCR増幅を行った。プライマーのセットは両方とも、エキソン17については128bp、エキソン20については145bpサイズの期待される産物を増幅し、BAC−5がSBEIIbの3'末端を含むことを示した。これは、エキソン20からPCR産物を配列決定することによってさらに確かめられた。
【0198】
SBEIIbに加えてSBEIIa遺伝子の存在についてもBAC−5を調べた。プライマーAR2akpnIF5'−GGTACCGCAGAAAATATACGAGATTGACCC−3'[配列番号26]を使ったヌクレオチド配列決定反応は、SBEIIa遺伝子のイントロン3領域に対応する配列を与え、wSBEII−Dlの位置2265〜2478(図1)に由来する配列と同じであった。この結果は、SBEIIaがBAC−5上にも存在することを示唆し、SBEIIaとSBEIIbがおそらく小麦において密接に結合しているであろうことを意味した。
【0199】
蛍光in situハイブリッド形成(FISH)
Turner他(1999年)によって記載されたようにして、wSBE II−D1ゲノムクローンF2(Rahman他、2001年)とwSBEII−D2クローン(Rahman他、2001年)とのin situハイブリダイゼーションを、Aegilops tauschiiと小麦からの染色体潰砕物に対して実行した。ハイブリダイズした染色体の同一性を、染色体識別(Mukai他、1990年)のために使われる反復塩基配列である二重標識withpSc119.2によって確認した。wSBEIIクローンの両方とも染色体2(図13)の先端領域にハイブリダイズし、小麦における2つのSBEII遺伝子の近くを示した。
【0200】
小麦SBEIIbヌル変異体はSBEIIaについての変異体でもある
先に述べたように確認したSBEIIbヌル変異体を、プライマーSr913F(5'−ATCACTTACCGAGAATGGG−3')[配列番号27]およびE6R(5'−CTGCATTTGGATTCCAATTG−3')[配列番号28]を用いて、SBEIIa遺伝子における突然変異についてスクリーニングした。これらのプライマーは、wSBE II−D1のイントロン5領域を増幅して、A、BおよびDゲノム上でSBEIIa遺伝子を識別するように設計した。
【0201】
SBEIIb Bゲノムヌル変異体Aus12565およびAus12509は、SBEIIa遺伝子のBゲノムヌル変異体でもあると分かった。同様に、SBEIIb、Aus17340およびAus10103のDゲノムヌル変異体は、SBEIIaのDゲノムヌル変異体でもあった。さらにまた、SBEIIb、BD341およびBD636のB+Dゲノム二重突然変異体系統は、SBEIIa遺伝子のB+Dゲノム二重ヌル変異体でもあった。データは、SBEIIaとSBEIIbが、コメとトウモロコシとは対照的に、小麦においては密接に結合することを証明し、上述のこれらの遺伝子のBおよびDゲノムコピーについての突然変異が欠失突然変異を表すことを示す。
【0202】
3重ヌルSBEIIa小麦変異体
上述の方法は、SBEIIaおよび/またはSBEIIbのA−ゲノム変異体を単離させるのに用いてもよい。例えば、SBEIIaおよび/またはSBEIIbに密接に結合するBAC−5領域は、遺伝子中でA−ゲノム突然変異をスクリーニングするためにプローブまたはPCRプライマーの設計のために使われる。A−ゲノム変異体は、B+D二重ヌル系統と交配してA+B+D3重ヌル系統を生産する。あるいは、B+Dゲノム二重ヌル変異体の突然変異生成は照射その他の手段で実行され、三重ヌル変異体は、SBEIIa活性を完全に欠いており、状況に応じてSBEIIb活性が特定される。非常に高いアミロース濃度を備えた非トランスジェニック小麦品種が提供される。
【0203】
<実施例10.小麦におけるSBEIIA遺伝子の突然変異>
SBEIIaの活性低減に至る小麦におけるSBEIIa遺伝子の突然変異は、ガンマ線照射または化学的突然変異誘発、例えばエチルメタンスルホン酸エステル(EMS)を用いて達成する。ガンマ線誘発突然変異のために、種子は60Co源(ZikiryaevaおよびKasimov(1972年)から20〜50kRの量の照射を受ける。EMS突然変異生成は、Mullins他(1999年)に従って種子をEMS(0.03%、v/v)で処理して行う。B+D二重ヌルバックグラウンドでは、変異体穀粒は増加したアミロース含有量または変更したデンプン顆粒形態に基づいて特定され、上述の方法によって確認される。SBEIIb活性を保持するSBEIIaの変異体は再変異してもよく、子孫は、SBEIIaに加えてSBEIIb活性の喪失についてスクリーニングでき、または、SBEIIa変異体はSBEIIb変異体と交配して、突然変異を結合して、胚乳において実質的にSBEII活性が欠如している種類の非トランスジェニック小麦を生産する。
【0204】
<実施例11.SGP−1小麦変異体はSBEIIa活性およびSBEIIb活性において低減している>
小麦(Triticum aestivum)のA、BおよびDゲノムからのデンプン合成酵素II(SSII)遺伝子は、糖−顆粒タンパク質(Starch Granule Proteins−1;SGP−1)としても知られている100−105kDaのポリペプチドをコードする。SSII(SGP−1)は、それぞれ染色体7B、7Aおよび7D(Denyer他、1995年;YamamoriおよびEndo1996年)短腕上遺伝子の相同的集合によってコードされるおよその分子量が100、104および105kDaである3つのポリペプチドからなる。Yamamori他(2000年)は、タンパク質電気泳動による分析で、A、BおよびDゲノムの特異的なSGP−1タンパク質の形態を欠いている系統を交配することによってSGP−1ヌル小麦を生産した。SGP−1ヌルな種子の検査は、突然変異がアミロペクチン構造の変化をもたらし、アミロース含有量を上昇させ、デンプン顆粒を変形させることを示した(Yamamori他、2000年)。そのうえ、成熟した穀粒についての電気泳動実験では、顆粒結合SBEII(SGP−2)とSSI(SGP−3)のレベルがかなり減少することが分かった。SGP−1ヌル系統をもたらす突然変異(複数可)の分子的基礎は知られていなかった。
【0205】
我々は、さらに、成熟穀粒からのデンプン顆粒において完全にSGP−1を欠いている小麦系統を特徴付けるために実験を行った。SSII遺伝子がSGP−1ヌルな小麦のA、BおよびDゲノムの各々に存在するかどうか決定するために、DNAをSGP−1ヌルな小麦と野生型(対照)cvChinese Spring小麦から抽出し、Bゲノムについてはプライマーの組合せssIIa(5'−CCAAGTACCAGTGGTGAACGC−3'[配列番号29]およびssIIb(5'−CGGTGGGATCCAACGGCCC−3'[配列番号30]を用いたPCRによって、AおよびDゲノムについてはssIIaとssIIc(5'−CATGTGAGCTAGCTTTCGCCC−3'[配列番号31]を用いたPCRによって分析した。増幅した領域は、wSSIIA(GenBankアクセッション番号AF155217)またはwSSIIBまたはwSSIIDの対応する領域である位置2472〜2821bpの間にあった。A、BおよびDゲノム産物の明確な識別が可能であったので、増幅した領域はエキソン8の部分を構成して選ばれた。増幅は、94℃で30秒、60℃で1分および702℃で2分の35サイクルを用いて行った。SGP−1ヌルな小麦のA、BおよびDゲノム産物から生産されるPCR断片は、Chinese springから作り出した対応する断片と同じサイズであった。イソアミラーゼとSSI遺伝子(それはSSIIに最も密接に位置するデンプン生合成支配遺伝子であって、SSII(Li他、2002年)の両側に位置する)の遺伝子セグメントのPCR増幅は、これらの遺伝子がSGP−1ヌルな小麦のA、BおよびDゲノムの各々について増幅され得ることを示した。したがって、SGP−1ヌルなフェノタイプは、染色体7短腕上でこれら遺伝子のいずれの欠失にも起因しなかった。
【0206】
走査型電子顕微鏡によって調べた場合、SGP−1ヌルな発育中種子からのデンプン顆粒は、開花後10日から成熟まで明らかに変形した。キャピラリー電気泳動法によって調べた場合、変異体中の脱分枝したデンプンの鎖長分布は、より短いチェーン(重合度8まで)の比率の増加を示す一方、重合度9−22の比率の減少を示す。
【0207】
SGP−I胚乳におけるデンプン合成酵素の発現と分枝酵素
デンプン顆粒中におけるデンプン合成酵素と分枝酵素の発現を、SGP−1ヌルにおいて調べ、野生型栽培品種Chinese Springのそれらと比較した。SGP−1ヌル系統では、種子発育のステージに関係なく、SSII(図14)の欠如に加え顆粒中のSBEIIとSSIの量においてほぼ90%〜96%の顕著な低減があった。特異的抗体の使用は、顆粒から得られるSBEIIバンドがChinese Springではおよそ1:3の比率のSBEIIaとSBEIIbを含むことを示した。SGP−1ヌル変異体においては、量があまりにも低かったため、相対的な比率を、抗体を用いて測定することはできなかった。また、穀粒成長の初期からGBSS Iレベルの低減があった。SGP−1変異体においては、SBEIIaとSBEIIbを含むデンプン顆粒結合ポリペプチドのレベル低減があることは、明らかである。デンプン−顆粒結合ポリペプチド(SBEIIおよびSSI)の低減は、SGP−1ヌル(Yamamori他、2000年)を産生するのに用いられる小麦系統の穀粒では観察されなかった、影響が特にSSIIの欠如に起因することを示唆している。
【0208】
分枝酵素とデンプン合成酵素は、胚乳を成長させる溶液段階でも分析した。溶解性SBEIIbの相対量はChinese SpringとSGP−1ヌル系統で同様であり、変異体の溶解段階にあるSBEIIaの量の低減があった(図15)。しかし、これはSGP−1ヌルな系統の系図に部分的に原因したものでもよい。
【0209】
これらのデータは、SBEIIa活性がSSII遺伝子中の突然変異によって多形質発現的に減少し得ることを証明する。SSIIのみの突然変異が、デンプン中の相対的アミロースレベルを50%未満にさせるが、それはSBEIIa以外の遺伝子の突然変異がSBEII突然変異と組合せ可能であり、アミロースレベルを増やし、変化したデンプンを生じることを示唆する。
【0210】
<実施例12.SBEIのノックアウト−多数のアイソフォーム>
陰イオン交換クロマトグラフィーを通しての小麦デンプン分枝酵素の精製は、活性を3つのピークに分解した(図16、Morell他、1997年)。栽培品種Chinese Spring(CS)からの胚乳抽出は、ポリクローンナル抗体である抗−WBE−1(タンパク質のピーク1(図16B)のN末端配列に対応するアミノ酸配列を有する合成ペプチドに対して誘導されたもの)を用い、非変性PAGE上で4つのSBEIポリペプチドの存在を明らかにした。CSヌルの同染色体−四染色体系統の分析では、これらのポリペプチドが染色体7上にコード化されていることが分かった。免疫ブロットの上のバンドはA(Aバンド)、B(Bバンド)とD(DiとDiiバンド)ゲノムに割り当てられ、それぞれ、活性はA−メジャー活性、B−メジャー活性およびD−メジャー活性と名付けられた。陰イオン交換クロマトグラフィーによって得られる活性なピークを表している精製画分の免疫ブロット分析では、最初のピークがSBEI AメジャーおよびDメジャー活性を含み、第2のピークはSBEI B−メジャー活性を含むことが分かった(図16C)。
【0211】
染色体7上で大きなSBEI活性をコードする遺伝子の位置は、3つのよく特徴が分かっている関連した遺伝子、wSBEI−D2、wSBEI−D3およびwSBEI−D4の決定された位置と一致している。SBEI−メジャーの推定したタンパク質配列は、それがこれらの遺伝子の最後にコードされていることを示した(wSBEI−D4(Rahman他、1997年、Suzuki他、2003年)。第4のSBEI遺伝子の存在は、サザンブロットハイブリダイゼーションデータ(Suzuki他、2003年)に基づいて提案された。
【0212】
SBEIメジャーのヌル変異体の特定
1つまたは複数のアイソフォームの発現が欠如しているヌル変異体を同定するため、非変性ゲル電気泳動の後、SBEI−メジャー免疫ブロット検出によって小麦生殖細胞質のコレクションをスクリーニングした。上述の抗wSBEI抗体を用いた。分析した182のオーストラリア6倍体小麦のアクセッションのうち、SBEI−Dメジャーを発現しない13系統、SBEI−Bメジャーを欠く16系統、SBEI−Aを欠く10系統、およびAとBアイソフォームの両方ともを欠く2つ(BindawarraとVectis)が同定された。これらの系統は、それぞれのゲノムSBEI遺伝子のヌル変異を有すると考えられる。SBEI−メジャー遺伝子のヌル変異の頻度(〜23%)はGBSS遺伝子(22%)(Boggini他、2001年)のそれと同様だった。
【0213】
SBEI−メジャー三重ヌル系統の生成
免疫ブロット分析から、栽培品種BindwarraとVectisがA−メジャーおよびB−メジャーSBEI活性を欠いていることは明らかであり、栽培品種CadouxがD−メジャーSBEI活性を欠くものとして同定された。Vectis×Cadouxの交配から得られる185系統のF2子孫集団を、イムノブロッティングでスクリーニングした。しかし、3つの活性のすべてを欠いている系統は得られず、そのような子孫は生存能力が低いか、あるいは、ゲノム間に何等かの相互作用があることが示唆された。そこで、B−メジャーおよびD−メジャー活性を欠いている子孫系統VC3.1.11を、A−メジャー活性を欠き遺伝子操作したChinese Spring(CS7AL−15)と交配した。二重ハプロイド系統をプライマーARBE1CF(5'−GGGCAAACGGAATCTGATCC−3')[配列番号32]およびARA9R(5'−CCAGATCGTATATCGGAAGGTCG−3')[配列番号33]を用いてPCRとイムノブロッティングの両方でスクリーニングを行い、160のうち2系統(A113とD13)は、非変性ゲル類の上でイムノブロッティングによって判断すると、SBEIメジャー活性を完全に欠いていた。図17は、A113(レーン6)を含む倍加半数体系統の代表的分離パターンを示す。
【0214】
系統A113の胚乳に残存SBE活性がないか調べた。野生型小麦変種(D28)は、SBEI活性の2つのピークを示した。一方、A113抽出物は、最初のピークは与えたが、活性の第2のピークは完全に欠いていた。この活性を含む精製画分から得られるアミノ酸配列は、A113中ではタンパク質のSBEIタイプの存在を示した。しかし、この画分は、非変性ジェル中では抗WBEI抗体との反応を示さなかった。A113中での枝分れ活性は、SBEIIタイプ酵素のものであり得る約80kDaのタンパク質に対応する(それはポテトSBEとトウモロコシSBEII抗体で交差反応した)。
【0215】
これらのデータは、SBEI変異体系統が小麦で発生し得ることを示す。SBEIIaと状況に応じてSBEIIb突然変異によるSBEI突然変異の組合せは、穀粒デンプン中に非常に高いアミロースレベルを有する小麦植物を生産する。
【0216】
<実施例13.SBEII遺伝子中に突然変異を有する染色体2Aを含む変異体小麦の識別>
SBEIIaまたはSBEIIb遺伝子中に突然変異を有する小麦系統を特定するべく、2400の6倍体小麦アクセッションをA、BまたはDゲノム中、SBEIIbのヌル変異についてスクリーニングした。プライマーAR2bl9cF/AR2b23cRを各系統の小麦植物のゲノムDNA試料へのPCR反応で使用し、RsaIで増幅産物の消化を行いゲル電気泳動した。このマーカーはイントロン3領域(小麦SBEIIb遺伝子(図2)のヌクレオチド位置2085〜2336)を増幅して、SBEIIbに特異的だった。上の実施例で記載したように、このスクリーニングでは3つのDゲノムSBEII−ヌル変異体と2つのBゲノムSBEII−ヌル変異体の識別がなされた。SBEIIbと対応しているAゲノムバンドを欠いた変異体系統は検出されなかった。これは、変異体SBEIIb遺伝子を有する染色体2Aを含む小麦系統が自然に存在しないことを示唆した。
【0217】
Tony PriorとRohit Mago(CSIRO)によって生成されたガンマ系統線(60Co源)誘導変異体小麦集団を用いて小麦SBEII中の誘発突然変異をスクリーニングした。小麦集団は、Gabo1BL.1RSとVeery 3とを交配させたF2子孫から生成した。この集団からの合計2694粒の変異体種を、プライマーAR2bl9cFとAR2b23cRを用い先に述べたようにPCR反応でスクリーニングした。1本の植物に由来し、SBEIIb Aゲノムアレルを欠く2個の種子(MLT2B8およびMLT2D1と命名)が同定された(図18)。集団内の種子で、BまたはDゲノム中にSBEIIbのヌル変異を含むものは特定されなかった。
【0218】
上の実施例で示すように、SBEIIaとSBEIIb遺伝子は、小麦では染色体2の長腕上で密接に結合している。したがって、我々は、プライマーSr913F/E6Rを用いたPCR反応で、AゲノムSBEIIa遺伝子の有無についてこれらの種子からのDNAを調べた。これらのプライマーはwSBEII−D1のイントロン5(ヌクレオチド位置2959〜3189、図1[配列番号1])領域を増幅する。増幅後、産物を5%のシークエンシングジェル(ABI Prism DNAシーケンサー)上で電気泳動にかけた。蛍光ラベリングした産物を、ソフトウェアGenescanを用いて分析した。走査プロフィールは変異体種子MLT2B8およびMLT2D1の両方についての増幅産物がAゲノムSBEIIa遺伝子に対応している産物を欠いていることを示し、両方の種子がSBEIIbに加えてAゲノムSBEIIaについてヌルなアレルを有することを示した。
【0219】
これらの種子において、SBEIIaに特異的なAゲノムマーカーであるARIIaAF(5'−GCAAAAGCCAGATCATAAATTTAGAGC−3')[配列番号34]およびAゲノムSBEIIa遺伝子の遺伝子産物のみを増幅するARIIaAR(5'−CTTCCAATTCATTGTTAATGGTCACAC−3')[配列番号35]を用いてヌル変異をさらに確認した(3131 wSBE II−DA1(図1)へのヌクレオチド位置3024)。この一組のプライマーは変種Chinese Springからの植物材料から110bp産物を増幅する一方、この産物は2粒の推定上の変異体種には明らかに欠けていた。これは陰性対照dt2AS(それは染色体2Aの長腕の欠けているChinese Springの染色体操作系統である)に関してと同じである。SBEIIa遺伝子とSBEIIb遺伝子が染色体2の長腕に置かれるので、この系統は両方の遺伝子のAゲノムアレルを欠いており、それゆえに、陰性対照(図19)として用いることができた。
【0220】
変異体種子MLT2B8およびMLT2D1(SBEIIaとSBEIIbに対するAゲノム変異体として識別される)からの胚を育てて植物とする。これら植物由来の種から得られるデンプンは、アミロース含有量、鎖長および他の特性がAゲノム上のSBEIIaとSBEIIb両方のヌル変異がデンプン特性に影響を及ぼすかどうか決定するために分析される。
【0221】
先に述べたように、BとDゲノムの両方にSBEIIaとSBEIIb遺伝子の突然変異を有する5系統が発生した。これらのうち、系統BD 219とBD 636は、温室で成長して、Aヌル変異体系統MLT2B8 とMLT2Dとが交配される。倍加半数体集団は、ホモ接合の三重ヌル変異体植物を提供するために、これらの交配のF1種子から生成される。そのような三重ヌル変異体植物は、8に1の頻度で、倍加半数体集団内に生じるはずである。Aゲノムヌル変異は、類似の交配によってBゲノム突然変異またはDゲノム突然変異と生み合わせることができる。さらなる交配において、ヌルなアレルのいずれかを農学的または他の特徴のために、任意の適当な遺伝的バックグラウンドにも導入することができる。
【0222】
以下の交配は、AゲノムSBEIIaとSBEIIbに突然変異を有するデュラム小麦(例えば品種Wollaroi)を産生するために実行される:
1)Wollaroi×MLT2B8またはMLT2D1(WollaroiバックグラウンドにおいてA−ゲノム SBEIIa/SBEIIbヌルデュラムを生じるため)
2)A−ゲノムヌルなデュラム(Wollaroi)×B−ゲノムヌルSBEIIa/SBEIIb小麦系統(AB2倍ヌルSBEIIa/SBEIIbデュラム(Wollaroi)を生じるため)
あるいは、
1)Wollaroi×B−ゲノムヌル小麦系統(B−ゲノムヌルなデュラムを生じるために)(Wollaroi)
2)B−ゲノムヌルなWollaroi×Aヌル小麦系統(AB2倍のヌルSBEIIa/SBEIIbデュラムを生じるために)。
これらの交配は、高アミロース6倍体小麦のそれと類似している健康に有利な特定の最終用途を有する高アミロースデュラムコムギの生成をもたらす。
【0223】
<実施例14.穀粒中の高アミロース含有量のセファロース2Bカラム分別法による確認>
SBEIIa/SBEIIb阻害遺伝子コンストラクトを含むトランスジェニック小麦植物の穀粒におけるデンプンのアミロース含有量をセファロースカラム分別法により決定する。この方法では、デンプン分子は、その分子量に基づいてカラム上で分離した。次いで、分離した画分を、デンプン分析キット(Sigma社)を利用し供給元の説明書に従って分析した。
【0224】
デンプンおよそ10mgを、37℃で30分間、インキュベーションすることにより、3.0mlの1NNaOH(脱ガス後)に溶解した。未溶解成分を落とすため15分遠心処理した。上清を、1ml/分のポンプ速度でセファロースCL2Bカラムにロードした。緩衝液として10mM NaOHを用い、各2.5ml50画分を集めた。画分9〜50のpHは、1MHC1の35μlで4.5に調整した。各試料のアリコート(250μl)を管に移し、250μlのデンプン試薬(デンプン分析評価キット、Sigma)を添加した。対照は以下のものを含んでいた。デンプン試薬(250μl)と水(250μl)のみを含むブランクのデンプン分析評価試薬、500μlの水のみを含むブランクのブドウ糖分析評価試薬、250μlデンプン試料と250μl水のみを含むブランク試料と250μlのデンプン試薬と250μlデンプン試料のみを含む試料テスト。試料と対照は60℃で60分、次いで、各200μlを新しい管に移し、その後、1mlのブドウ糖試薬(デンプン分析評価キット、Sigma)を添加して30分間、37℃でインキュベートする。デンプンの量(mg)を測定するのには、各画分においてキット添付の説明書に従って340nmの吸光度を用いた。
【0225】
デンプン試料のクロマトグラムは、セファロースカラムから溶出する2つのピークを明らかにした。各試料のアミロース含有量(第2のピーク)を、両方のピークの範囲内のデンプンの総量のパーセンテージとして計算した。
【0226】
この方法を用い、ds−SBEIIaトランスジェニック系統Acc.144087のアミロース含有量(それは導入遺伝子についてはホモ接合性であることが示された)は78%になるように計算され、ds−SBEIIbトランスジェニック系統Acc.144008(イベントIIb 110.16bからのホモ接合のトランスジェニック系統)のそれは23%であると見積った(図20)。比較において、88.47%および27.29%のこれらの系統に対してヨード還元的定法によりアミロース含有量を求めた(表10)。
【0227】
糊化温度、ペースト粘度および膨潤体積などのデンプンなどの機能的な特性を、示差走査熱量測定(DSC)、Rapid Visco Analyser(RVA)およびデンプン膨潤力試験によってそれぞれ測定した。これらのデンプンの構造は、X線結晶学と粒度分析によって分析される。
表10.ヨード還元的定法により評価した小麦トランスジェニック系統のアミロース含有量
【0228】
【表10】
【0229】
<実施例15.鎖長分布分析>
デンプン試料の鎖長分布は、デンプンのイソアミラーゼ脱分枝後、蛍光ラベル糖鎖電気泳動(Fluorophore−assisted carbohydrate electrophoresis:FACE)により測定した。トランスジェニック種子から由来のデンプン中のうち重合度6−11、重合度12−30および重合度31−60からの鎖長のパーセンテージを、非トランスジェニックの対照と比較して表11に示す。高アミローストランスジェニック系統由来のデンプンについて正規化した鎖長分布を同遺伝子型の非形質転換対照由来のデンプンについて正規化した鎖長分布から差し引いたモル差を、図21に示す。
表11.小麦トランスジェニック系統由来のイソアミラーゼ脱分枝デンプンの鎖長分布
【0230】
【表11】
【0231】
ds−SBEIIbトランスジェニック種由来のデンプンにおける重合度4−12の鎖長は、非形質転換種子またはds−SBEIIaトランスジェニック種子由来のデンプンと比較して顕著に低い比率であった。重合度>13の鎖長の比率は、他と比較してds−SBEIIaトランスジェニック種子でより高かった。これらの結果は、SBEIIaが小麦デンプンにおいて重合度4−12のより短い鎖の合成に選択的に関与しているという可能性を示唆する。しかし、SSIIa変異体由来のデンプンにおいては、より短い鎖長の比率がアミロースで増加していた。
【0232】
<実施例16.トランスジェニック小麦由来のデンプンの特性>
ds−SBEIIaおよびds−SBEIIbトランスジェニック系統からの糊化温度を含むデンプンの物性を、Perkin Elmer Diamond示差走査熱量計を使って分析した。各デンプンおよそ20mgを、1:2の比率で水に混合し、すなわち、含水率66.7%として、DSCパンに封入した。1分当たり10℃の加熱速度を用いて、0から150℃まで試験試料と参照試料を加熱した。データは、装置で利用できるソフトウェアを使用して分析した。
【0233】
各デンプンについてサーモグラムDSCトレースにおいて2つの吸熱ピークが観察された。最初のピークは、デンプンの糊化の間の結晶構造の崩壊を意味した。第2のピークは、アミロース−脂質解離の吸熱を表した。ds−SBEIIaトランスジェニック系統からのデンプンの糊化ピーク温度は非形質転換対照デンプンについてのピーク温度と比較しておよそ7〜10℃の増加を示し、ds−SBEIIbトランスジェニック系統からのデンプンと比較しておよそ3〜7℃増加した。
表12.示差走査熱量計(DSC)で測定したトランスジェニック小麦デンプンの熱的性質
【0234】
【表12】
【0235】
これらの系統では、非形質転換対照およびds−SBEIIbトランスジェニック系統と比較して、およそ16〜19℃の糊化(最初のピーク)終了温度の著しい増加が観察された。糊化の開始温度は、ds−SBEIIaトランスジェニック系統で対照またはds−SBEIIbトランスジェニック系統より早く現れた。Ng他(1997年)は、アミロース増量(ae)トウモロコシデンプンの糊化開始温度が通常のトウモロコシデンプンのそれと類似していると報告したが、aeデンプンでは正常のデンプンと比較してピークの糊化温度が顕著に増加していた。ds−SBEIIaトランスジェニック系統からのデンプンの糊化エンタルピーは、対照およびds−SBEIIb系統のそれより顕著に低かった。これは、糊化ピーク面積の顕著な低下(これはds−SBEIIaトランスジェニック系統におけるアミロペクチン量の低減を表している)を反映しているようである。アミロース−脂質分離ピークでは、トランスジェニック系統のいずれでも著しい変化は観察されなかった。したがって、我々は、この新しい一揃いの特性を有するデンプンを得た。
【0236】
<参考文献>
Abel et al., (1996). The Plant Journal 10, 981-991.
Anderson et al., (1989). Nucl Acids Res 17, 461-462.
Baba et al., (1991). Biochem Biophys Res Commun 181: 87-94.
Batey and Curtin. (1996). Starch 48, 338-344.
Batey et al., (1997). Cereal Chemistry 74, 497-501.
Becker et al., (1994). Plant J. 5: 299-307.
Blauth et al., (2001). Plant Physiology 125, 1396-1405.
Bourque. (1995). Plant Science 105, 125-149.
Boyer and Preiss, (1978). Carbohydrate Research 61, 321-334.
Boyer and Preiss, (1981). Plant Physiology 67, 1141-1145.
Boyer et al., (1980). Starch 32, 217-222.
Buleon et al., (1998). International Journal of Biological Macromolecules 23, 85-112.
Cao et al., (2000). Archives. of Biochemistry and Biophysics. 373, 135-146.
Case et al., (1998). Journal of Cereal Science 27, 301-314.
Cheng et al., (1997). Plant Physiol 115: 971-980.
Craig et al., (1998). Plant Cell 10, 413-426.
Denyer et al., (1995). Planta 196: 256-265.
Denyer et al., (1996). Plant Physiology 112, 779-785.
Feldman. (***) pp 3-56 in The World Wheat Book, A history of wheat breeding. Eds Bonjean and Angus, Lavoisier Publishing, Paris.
Fergason. 1994. pp 55-77 in "Speciality Corns" eds , CRC Press Inc.
Filpse et al.,(1996). Planta 198, 340.
Fisher et al., (1993). Plant Physiol 102:1045-1046.
Fisher et al., (1996). Plant Physiol 110: 611-619.
Fuwa et al., (1999). Starch/Starke. 51, 147-151.
Gao et al., (1997). Plant Physiol 114: 69-78.
Gao et al., (1998). Plant Cell 10, 399-412.
Giroux and Hannah. (1994). Molecular and General Genetics 243, 400-408.
Green et al., (1997). Plant Physiology 114, 203-212.
He et al., (1994). Plant Cell Reports 14: 192-196.
Hedman and Boyer, (1982). Biochemical Genetics 20, 483-492.
Hess et al., (1990). Plant Science 72: 233-244.
James et al., (1995). Plant Cell 7, 417-429.
Jobling et al., (1999). Plant Journal 18, 163-171.
Komari et al., (1996). Plant Journal 10:165-174.
Konik-Rose et al (2001) Starch 53, 14-20.
Krueger et al., (1987). Cereal Chemistry 64, 187-190.
Kubo et al., (1999). Plant physiology. 121, 399-409.
Li et al., (1999a). Plant physiology. 120, 1147-1155.
Li et al., (1999b). Theoretical and Applied Genetics 98, 1208-1216.
Li et al., (2000). Plant Physiology 123, 613-624.
Li et al., (2003). Funct Integr Genomics 3:76-85.
Maniatis et al., (1982). Molecular cloning: a laboratory manual. Cold Spring Harbour Laboratory Press New York.
McCreery and Helentjaris (1994). Methods in Molecular Biology, Vol. 28: Protocols for nucleic acid analysis by non-radioactive probes, 67-71, Humana Press Inc., Totawa, NJ.
Mizuno et al., (1993). Journal of Biological Chemistry 268, 19084-19091.
Mizuno et al., (1992). Journal of Biochemistry 112, 643-651.
Morell et al., (1997). Plant Physiology 113, 201-208.
Morell et al., (1998). Electrophoresis 19, 2603-2611.
Morell et al., (2003). Plant J. 34: 173-185.
Morrison and Laignelet (1983). Journal of Cereal Science 1:9-20.
Mullins et al., (1999). European Journal of Plant Pathology 105: 465-475.
Myers et al., (2000). Plant Physiology 122, 989-997.
Nakamura (2002). Plant Cell Physiology 43, 718-725.
Nakamura and Yamanouchi (1992). Plant Physiol 99: 1265-1266.
Nair et al., (1997). Plant Sci 122: 153-163.
Nehra et al., (1994). Plant J. 5: 285-297.
Ng et al., (1997) Cereal Chemistry 74: 288-292.
Nishi et al., (2001). Plant Physiology 127, 459-472.
Rahman et al., (1995). Australian Journal of Plant Physiology 22, 793-803.
Rahman et al., (1997). Genome 40: 465-474.
Rahman et al., (1999). Theor Appl Genet. 98: 156-163.
Rahman et al., (2000). J Cereal Sci 31: 91-110.
Rahman et al., (2001). Plant Physiol 125: 1314-1324.
Repellin et al., (1997). Plant Gene Reg 97-094
Safford et al., (1998). Carbohydrate Polymers 35, 155-168.
Schulman and Kammiovirta, (1991). Starch 43, 387-389.
Schwall et al., (2000). Nature Biotechnology 18, 551-554.
Senior (1998). Biotechnology and Genetic Engineering Reviews 15, 79-119.
Shannon and Garwood, (1984). In Starch: Chemistry and Technology, Whistler et al., eds, Academic Press, Orlando, FL, pp25-86.
Shure et al., (1983). Cell 35, 225-233.
Sidebottom et al., (1998). Journal of Cereal Science 27, 279-287.
Stacey and Isaac (1994). Methods in Molecular Biology, Vol. 28: Protocols for nucleic acid analysis by non-radioactive probes, pp 9-15, Humana Press Inc., Totawa, NJ.
Sun et al., (1997). The New Phytologist 137 , 215-215.
Takeda et al., (1993a). Carbohydrate Research 240, 253-262.
Takeda et al., (1993b). Carbohydrate Research 246, 273-281.
Thomas and Atwell 1999 Starches Eagen Press, St Paul, Minnesota, USA pp: 13-24.
Thorbjornsen et al., (1996). Plant Journal 10, 243-250.
Vasil et al., (1992). Bio/Technology 10: 667-674.
Vasil et al., (1993). Bio/Technology 11: 1553-1558.
Wang et al., (1998). Journal of Experimental Botany 49, 481-502.
Weeks et al., (1993). Plant Physiol 102: 1077-1084.
Wegener et al., 1994. Mol. Gen Genet. 245, 465-470.
Weir et al., (2001). Aust J Plant Physiol 28: 807-818.
Yamamori and Endo, (1996). Theoretical and Applied Genetics 93, 275-281.
Yamamori et al., (2000). Theor.Appl.Genet. 101, 21-29
Young. (1984). in Whistler et al. (eds), Academic Press, Orlando, FL, chap 8.
Zhao and Sharp, (1998). Plant Breeding 117: 488-490.
Zikiryaeva and Kasimov, (1972). Uzbekskii Biologicheskii Zhurnal 6, 18-20.
Zwar and Chandler, (1995). Planta 197, 39-48.
【図面の簡単な説明】
【0237】
【図1】A.tauschii(タルホ小麦)から得たデンプン分枝酵素IIa遺伝子(wSBEII−D1)の配列を示す表である[配列番号1]。6倍体の小麦(T.aestivum(普通小麦))のゲノムDのSBEIIa遺伝子に対応している。
【図2】T.aestivum(普通小麦)から得た小麦のSBEIIb遺伝子配列(wbe2bゲノム)の一部を示す表である[配列番号2]。
【図3】二重鎖RNAコンストラクトの概略図である。A:使用されている遺伝子要素の順序は、センス方向にプロモーター、SBEIIaまたはSBEIIb遺伝子配列(エキソン1、2および3)、アンチセンス方向に、イントロン(イントロン3)、SBEIIaまたはSBEIIb遺伝子配列(エキソン1、2、3および4)、および転写ターミネーター/ポリアデニル化配列であった。B:ds−SBEIIaおよびds−SBEIIb遺伝子の転写物は、センス配列とアンチセンス配列のハイブリダイゼーションにより形成された二本鎖部分を有する「ヘアピン」RNA構造を形成する。ヌクレオチドのGTおよびAGにはさまれたイントロン配列はスプライシングされる。
【図4】光学顕微鏡で観察したデンプン粒の写真である。A)は、ds−SBEIIaトランスジェニック系統83.1bに由来する野生型デンプン粒を含む小麦種子でから得たデンプン粒であり、B)は、ds−SBEIIaトランスジェニック系統50.1bに由来する変形デンプン粒を含む小麦種子から得たデンプン粒である。
【図5】図4の小麦種子から得たデンプン粒の複屈折性を偏光下で可視化した写真である。
【図6】小麦SBEIIaのcDNA配列の一部分を比較する表である。sbe9は、AF338432.1の一部に対応する。Y11282[配列番号3]、sr997[配列番号4]、sr995[配列番号5]、sbe9[配列番号6]の配列の一部を示している。
【図7】初めの63個のアミノ酸に基づき小麦のSBEIIa配列の一部分をパイルアップ(PILEUP)比較した図である。クローンに対応する遺伝子の推定されるゲノム位置を示している。
【図8】D−ゲノムポリペプチド(sr854)[配列番号7]の推測されるアミノ酸配列をAゲノムまたはBゲノム(y11282)[配列番号8]の産生物と比較した表である。トランジット配列(位置1〜54)はイタリック体で示している。
【図9】様々な小麦登録種から得たSBEIIb遺伝子のイントロン3領域をPCR増幅した結果を示す図である(レーン1〜11)。PCR増幅は、プライマーARA19FおよびARA23Rを用い、次にRsa1で消化して行った。ゲノムA、B、およびDに対応するバンドを矢印で示している。レーン3(Aus17340)およびレーン5(Aus10103)は、ゲノムDに特異的なマーカーを欠いており、一方、レーン8(Aus12509)およびレーン9(Aus12565)は、ゲノムBのマーカーを欠いている。
【図10】SBEIIbのイントロン3領域に由来するプローブを用いて、小麦登録種由来のHindIIIで消化したDNAをサザンハイブリダイゼーションした結果を示す写真である。各レーンは、1)Aus12565、2)Aus12509、3)Aus10103、4)CSDT2DL−4、5)Aus12530(デュラム小麦)、6)CSDT2BL−9、7)Aus6323、8)CSDT2DS、9)Aus17340、10)Aus12745、11)CSDT2DL−4、12)Aegilops tauschii(タルホ小麦)に対応している。
【図11】SBEIIbのイントロン3領域のPCR増幅により、Aus17340aとAus12509を交配したF2集団をスクリーニングした結果を示す図である。PCR増幅は、プライマーAR2b19cFおよびAR2b23cRを用い、次にRsaIで消化して行った。レーン8はゲノムBのマーカーとゲノムDのマーカーの双方を欠いており、したがってBD54系統は、BDの二重ヌル(double−ヌル)系統を示している。
【図12】SBEIIbのイントロン3領域に由来するプローブを用いて、HindIII(レーン1〜4)およびEcoR1(レーン5〜8)で消化したBACクローンをサザンハイブリダイゼーションした結果を示す写真である。各レーンは、1)BAC4、2)BAC5、3)BAC9、4)BAC12、5)BAC4、6)BAC5、7)BAC9、8)BAC12に対応している。
【図13】A)A.tauschii(タルホ小麦)染色体(大きな写真および挿入写真の下の方)および小麦染色体(挿入写真の上の方)に対してwSBEII−DA1プローブおよび反復DNA配列プローブ(pSc 119.2)を用いて行った蛍光in situハイブリダイゼーション(FISH)法の結果を示す写真である。 B)小麦染色体に対してSBEIIbプローブを用いて行った蛍光in situハイブリダイゼーション(FISH)法の結果を示す写真である。
【図14】指定した種子発育のいくつかの段階(開花の10、15、25日後、M=成熟)の野生型Chinese Spring(CS)系統およびSGP−1 ヌル小麦系統の顆粒結合タンパク質をSDS−ポリアクリルアミドゲル電気泳動(PAGE)により分析した結果を示す図である。銀染色ゲルの画像のタンパク質バンド強度を測定した。成熟したCS種子のGBSSのバンド強度を基準の100とし、指定した発育段階における他の酵素の量を、成熟CSのGBSSに対するパーセンテージとして表している。a)がGBSS、b)がSSI、c)がSBEIIである。黒い棒グラフは、SGP−1 ヌルを示す。CSおよびSGP−1 ヌル系統から得た顆粒結合タンパク質の電気泳動画像の例を示す。
【図15】可溶性画分中のSBEIIaおよびSBEIIbの相対量を示すグラフである。SDS−PAGEのイムノブロットをスキャンし、画像のタンパク質バンド強度を測定した。SBEIIaとSBEIIbの融合タンパク質を標準としてゲル上で使用して、タンパク質の量を概算した。
【図16】A.小麦(cv Rosella)胚乳分枝酵素活性を陰イオン交換クロマトグラフィーにより分析した結果を示すグラフである。胚乳の可溶性タンパク質を硫酸アンモニウムで分画し、セファクリル(Sephacryl)S−200カラム、続いてResource Q陰イオン交換カラムに添加してクロマトグラフィー分析をした。B)小麦胚乳SBEIの抗−WBE1抗体を用いて免疫検出分析を行い、非変性ポリアクリルアミドゲル電気泳動(PAGE)で分離させた結果を示す写真である。AおよびBとして示すSBEIタンパク質バンドは、それぞれゲノムAおよびゲノムBに由来する産生物であり、DiおよびDiiは、ゲノムDに由来する産生物である。レーン1はCS、レーン2はN7BT7A、レーン3はN7AT7B、レーン4はN7DT7Aから得た抽出物に対応するC)抗−WBE1抗体を用いた陰イオン交換クロマトグラムで活性なピークを示す、精製画分の免疫検出分析の結果を示す写真である。レーン1は、胚乳の粗製な可溶性抽出物であり、レーン2の画分はピーク1を示し、レーン3の画分はピーク2を示している。
【図17】抗WBEI抗体を用いた免疫検出によりSBEIアイソフォームを分離するために、VC3.1.11とCS7AL−15の交配種の倍加半数体の子孫をスクリーニングした結果を示す写真である。レーン1〜14は、倍加半数体の子孫系統に対応している。レーン6は、A113と呼ばれる三重(triple)ヌル型SBEI変異系統であり、レーン7は、D28と呼ばれるSBEIアイソフォームが正常な系統である。
【図18】Veery3とGabo 1BL.1RSの交配種のガンマ線で誘発した突然変異種子から得たDNA(レーン1〜6)を、プライマーAR2b19cF/AR2b23cRを用いてPCR増幅した結果を示す写真である。レーン2は、変異種子MLT2B8を示し、レーン7は、Chinese Springを示す。
【図19】小麦SBEIIa遺伝子のゲノムA特異的プライマー、すなわちARIIaAF/ARIIaARを用いて、小麦系統から得たDNAをPCR増幅した結果を示す写真である。レーン1〜5は、順に、CS、MLT2B8、MLT2D1、Dt2AS、およびBD219(ゲノムBとゲノムDの双方のSBEIIaおよびSBEIIb植物がヌル変異体である植物)である。
【図20】デンプンアッセイキット(Sigma社)を用いて、小麦系統a)Acc144008およびb)Acc144087から得たデンプンをセファロース(Sepharose)CL2Bゲルクロマトグラムにより分析した結果を示すグラフである。
【図21】小麦のトランスジェニック系統から得たデンプンの鎖長の特性を、形質転換されていないコントロールのNB1(小麦)の鎖長特性に対して比較した結果を示すグラフである。形質転換されていないコントロールから得たデンプンの個々の少糖類の全重量のパーセンテージを、トランスジェニック系統から得たデンプンの対応する値から引いている。サンプルの凡例は、085(黒の菱形)、025(黒の三角)、008(白丸)である。
【特許請求の範囲】
【請求項1】
デンプンを含む小麦植物から得た穀粒であって、穀粒のデンプン中のアミロース比率が少なくとも50%である穀粒。
【請求項2】
野生型穀粒と比べて低レベルの胚乳のSBEIIaタンパク質、SBEIIa酵素活性、またはその双方を含む、請求項1に記載の穀粒。
【請求項3】
野生型穀粒と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルを低下させる遺伝的変異を含み、遺伝的変異がSBEIIa遺伝子の変異またはSBEIIa遺伝子発現の阻害物質をコードする導入核酸を含む、請求項1または2のいずれか一項に記載の穀粒。
【請求項4】
デンプンと、野生型穀粒に比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルを低下させる遺伝的変異とを含み、遺伝的変異がSBEIIa遺伝子の変異またはSBEIIa遺伝子発現の阻害物質をコードする導入核酸を含む、小麦植物から得た穀粒であって、穀粒のデンプン中のアミロース比率が少なくとも30%である穀粒。
【請求項5】
前記遺伝的変異が、野生型穀粒と比べて胚乳のSBEIIaタンパク質、SBEIIa酵素活性、またはその双方のレベルを低下させるSBEIIa遺伝子の変異を含む、請求項4に記載の穀粒。
【請求項6】
少なくとも1種のSBEIIa遺伝子中にヌル変異を含む、請求項1から5までのいずれか一項に記載の穀粒。
【請求項7】
前記植物が6倍体であり、穀粒が2種または3種のSBEIIa遺伝子中にヌル変異を含む、請求項6に記載の穀粒。
【請求項8】
SBEIIa遺伝子発現の阻害物質をコードする導入遺伝子を含む、請求項1、2または4のいずれか一項に記載の穀粒。
【請求項9】
前記導入遺伝子がアンチセンスRNA分子、コサプレッションRNA分子、リボザイムRNA分子、または二重鎖RNA分子をコードする、請求項8に記載の穀粒。
【請求項10】
野生型穀粒と比べて低レベルのSBEIIbタンパク質、酵素活性、またはその双方を含む、請求項1から9のいずれか一項に記載の穀粒。
【請求項11】
野生型穀粒と比べて低レベルのSBEIタンパク質、酵素活性、またはその双方を含む、請求項1から10のいずれか一項に記載の穀粒。
【請求項12】
野生型穀粒と比べて改変されたレベルの酵素を含み、前記酵素がADPグルコースピロホスホリラーゼ、GBSS、SSI、SSII、SSIII、イソアミラーゼ型脱分枝酵素、プルラナーゼ型脱分枝酵素、およびこれらの任意の組合せからなる群から選択される、請求項1から11のいずれか一項に記載の穀粒。
【請求項13】
穀粒のデンプン中のアミロース比率が少なくとも40%である、請求項4または5のいずれかに記載の穀粒。
【請求項14】
非しわ型であり、または平均重量が少なくとも約36mgであり、あるいはその両方を満たす、請求項1から13のいずれか一項に記載の穀粒。
【請求項15】
穀粒から得たデンプン粒のうちの少なくとも50%が偏光下で観察すると非複屈折性に見える、請求項1から14のいずれか一項に記載の穀粒。
【請求項16】
殻のない状態での穀粒のデンプン含有量が少なくとも25%(w/w)であるか、またはデンプン含有量が野生型穀粒のデンプン含有量の少なくとも90%であるか、あるいはその両方を満たす、請求項1から15のいずれか一項に記載の穀粒。
【請求項17】
全粒、製粉した穀粒、粉砕した穀粒、精白した穀粒、圧扁した穀粒、粗挽きした穀粒、パーボイルドした穀粒、または挽き割りにした穀粒である、請求項1から16のいずれか一項に記載の穀粒。
【請求項18】
請求項1から17のいずれか一項に記載の穀粒を結実することができる小麦植物。
【請求項19】
請求項1から17のいずれか一項に記載の穀粒から抽出したデンプン粒。
【請求項20】
前記顆粒のデンプン中のアミロース比率が少なくとも50%であるデンプンを含む請求項19に記載のデンプン粒。
【請求項21】
請求項1から17のいずれか一項に記載の穀粒から抽出したデンプン。
【請求項22】
請求項1から17のいずれか一項に記載の穀粒、小麦粉、全粒小麦粉、セモリナ粉、請求項1から17のいずれか一項に記載の穀粒から得たデンプン、またはこれらの任意の組合せを含む製品。
【請求項23】
前記小麦粉、全粒小麦粉、セモリナ粉、またはデンプンを別の供給源から得た小麦粉、全粒小麦粉、セモリナ粉、またはデンプンと混合した請求項22に記載の製品。
【請求項24】
非食料品である請求項22に記載の製品。
【請求項25】
請求項21に記載のデンプンおよび別の食品成分または水を含む組成物。
【請求項26】
i)親となる小麦植物または小麦種子に遺伝的変異を導入するステップと、
ii)野生型の植物または種子と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップとを含む、
穀粒を結実することができる小麦植物を作製する方法であって、穀粒がデンプンを含有し、穀粒のデンプン中のアミロース比率が少なくとも50%である小麦植物の作製方法。
【請求項27】
i)SBEIIa遺伝子の変異またはSBEIIa遺伝子の発現の阻害物質をコードする導入核酸を含む遺伝的変異を、親となる小麦植物または小麦種子に導入するステップと、
ii)野生型の穀粒と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップとを含む、
穀粒を結実することができる小麦植物を作製する方法であって、穀粒がデンプンを含有し、穀粒のデンプン中のアミロース比率が少なくとも30%である小麦植物の作製方法。
【請求項28】
前記遺伝的変異を導入するステップが、SBEIIa遺伝子発現の阻害物質を発現する外来の核酸の導入を含む、請求項26または27に記載の小麦植物の作製方法。
【請求項29】
前記遺伝的変異を導入するステップが、化学薬剤または放射線による親小麦植物または小麦種子の突然変異誘発を含む、請求項26または27のいずれか一項に記載の小麦植物の作製方法。
【請求項30】
i)SBEIIa遺伝子の変異を含む遺伝的変異を、親となる小麦植物または小麦種子に導入するステップと、
ii)野生型の穀粒と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップと、
iii)SBEIIb遺伝子の変異を含む遺伝的変異を、親となる小麦植物または小麦種子に導入するステップと、
iv)野生型の穀粒と比べて胚乳のSBEIIb遺伝子発現、SBEIIb酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップと、
v)胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低い植物を、胚乳のSBEIIb遺伝子発現、SBEIIb酵素活性、またはその双方のレベルが低い植物と交配するステップとを含む、
穀粒を結実することができる小麦植物を作製する方法であって、SBEIIaとSBEIIbの双方の遺伝子発現、酵素活性、またはその双方が低い小麦植物を同定するステップとを含む、小麦植物を作製する方法。
【請求項31】
請求項1から17のいずれか一項に記載の穀粒からデンプンを抽出するステップを含む、改変した小麦デンプンを製造する方法。
【請求項32】
Triticum aestivum ssp.aestivumである、請求項18に記載の小麦植物。
【請求項33】
小麦のSBEIIb遺伝子またはSBEIIa遺伝子にそれぞれ結合する分子マーカーを用いて小麦植物または小麦種子の集団をスクリーニングするステップと、結合した分子マーカーの有無に基づいて植物または種子を同定するステップとを含む、SBEIIa遺伝子またはSBEIIb遺伝子の変異を有する小麦植物または小麦種子を同定する方法。
【請求項34】
小麦のSBEIIbタンパク質またはSBEIIaタンパク質にそれぞれ特異的な抗体を用いて小麦植物または小麦種子の集団をスクリーニングするステップと、抗体結合の有無に基づいて植物または種子を同定するステップとを含む、SBEIIa遺伝子またはSBEIIb遺伝子の変異を有する小麦植物または小麦種子を同定する方法。
【請求項35】
SBEIIa遺伝子が染色体2Aの長腕から欠けている変異、あるいは染色体2Aの長腕上のSBEIIa遺伝子が、野生型穀粒と比べて穀粒の胚乳のSBEIIaタンパク質、SBEIIa酵素活性、またはその双方を低下させる変異を有する変異を含む、小麦植物から得た穀粒。
【請求項36】
前記変異がSBEIIa遺伝子のヌル変異である、請求項35に記載の穀粒。
【請求項37】
前記変異がSBEIIa遺伝子の少なくとも一部分の欠失である、請求項35または36のいずれかに記載の穀粒。
【請求項38】
SBEIIb遺伝子が染色体2Aの長腕から欠けている変異、あるいは染色体2Aの長腕上のSBEIIb遺伝子が、野生型穀粒と比べて前記穀粒の胚乳のSBEIIbタンパク質、SBEIIb酵素活性、またはその双方を低下させる変異を有する変異をさらに含む、請求項35から37のいずれか一項に記載の穀粒。
【請求項39】
欠失が、染色体2Aの長腕上のSBEIIa遺伝子およびSBEIIb遺伝子の双方の発現を阻害する、請求項35から38のいずれか一項に記載の穀粒。
【請求項40】
前記植物がデュラム小麦植物である、請求項35から39のいずれか一項に記載の穀粒。
【請求項41】
野生型穀粒と比べて染色体2Bの長腕上のSBEIIa遺伝子によってコードされているデンプン分枝酵素活性を低下させる遺伝的変異をさらに含む、請求項40に記載の穀粒。
【請求項42】
前記遺伝的変異が、染色体2Bの長腕からのSBEIIa遺伝子の不在、または野生型穀粒と比べて前記穀粒の胚乳のSBEIIa酵素活性を低下させる染色体2Bの長腕上のSBEIIa遺伝子の変異を含む、請求項41に記載の穀粒。
【請求項43】
前記植物がTriticum aestivum ssp aestivumである、請求項35から39のいずれか一項に記載の穀粒。
【請求項44】
野生型穀粒と比べて染色体2B、染色体2D、または両方の染色体の長腕上のSBEIIa遺伝子によってコードされているデンプン分枝酵素活性を低下させる遺伝的変異を含む、請求項43に記載の穀粒。
【請求項45】
前記遺伝的変異が、前記染色体のうちの少なくとも1つからのSBEIIa遺伝子の欠失、または野生型穀粒と比べて前記穀粒の胚乳のSBEIIa酵素活性を低下させる前記染色体のうちの少なくとも1つのSBEIIa遺伝子の変異を含む、請求項44に記載の穀粒。
【請求項46】
前記遺伝的変異が、SBEIIa遺伝子発現、活性またはその双方の阻害物質をコードする導入核酸を含む、請求項41または44のいずれか一項に記載の穀粒。
【請求項47】
前記SBEIIa酵素活性のレベルが野生型穀粒と比べて少なくとも40%低下している、請求項35から46のいずれか一項に記載の穀粒。
【請求項48】
穀粒のデンプン中のアミロース比率が少なくとも30%である、請求項35から47のいずれか一項に記載の穀粒。
【請求項49】
穀粒のデンプン中のアミロース比率が少なくとも50%である、請求項48に記載の穀粒。
【請求項50】
穀粒のアミロペクチンの重合度が4から12の鎖長部分の比率は、アミロペクチンをイソアミラーゼで脱分枝した後に測定すると、野生型穀粒のアミロペクチンと比べて低下している、請求項1から17、および請求項35から49のいずれか一項に記載の穀粒。
【請求項51】
非しわ型である、請求項35から50のいずれか一項に記載の穀粒。
【請求項52】
平均重量が少なくとも約36mgである、請求項35から51のいずれか一項に記載の穀粒。
【請求項53】
穀粒から得たデンプン粒の少なくとも50%が、偏光下で観察すると非複屈折性に見える、請求項48から52のいずれか一項に記載の穀粒。
【請求項54】
殻のない状態での穀粒のデンプン含有量が少なくとも25%(w/w)であり、または、デンプン含有量が野生型穀粒のデンプン含有量の少なくとも90%である、請求項46から53のいずれか一項に記載の穀粒。
【請求項55】
全粒、製粉した穀粒、粉砕した穀粒、精白した穀粒、圧扁した穀粒、粗挽きした穀粒、パーボイルドした穀粒、または挽き割りにした穀粒である、請求項35から54のいずれか一項に記載の穀粒。
【請求項56】
低レベルのSBEIタンパク質、SBEI酵素活性、またはその双方をさらに含む、請求項35から55のいずれか一項に記載の穀粒。
【請求項57】
野生型穀粒と比べて改変したレベルの酵素をさらに含む穀粒であって、前記酵素が、ADPグルコースピロホスホリラーゼ、GBSS、SSI、SSII、SSIII、イソアミラーゼ型脱分枝酵素、プルラナーゼ型脱分枝酵素、およびこれらの任意の組合せからなる群から選択される、請求項35から56のいずれか一項に記載の穀粒。
【請求項58】
請求項35から57のいずれか一項に記載の穀粒を結実することができる小麦植物。
【請求項59】
請求項35から57のいずれか一項に記載の穀粒から抽出したデンプン粒。
【請求項60】
前記顆粒のデンプン中のアミロース比率が少なくとも50%である、デンプンを含む請求項59に記載のデンプン粒。
【請求項61】
請求項35から57のいずれか一項に記載の穀粒から抽出したデンプン。
【請求項62】
請求項35から57のいずれか一項に記載の穀粒、あるいは請求項35から57のいずれか一項に記載の穀粒から製造した小麦粉、全粒小麦粉、セモリナ粉、またはデンプンを含む製品。
【請求項63】
前記小麦粉、全粒小麦粉、セモリナ粉、またはデンプンが、別の供給源から得た小麦粉、全粒小麦粉、セモリナ粉、またはデンプンと混合される、請求項62に記載の製品。
【請求項64】
非食料品である、請求項62に記載の製品。
【請求項65】
請求項61に記載のデンプンおよび別の食品成分または水を含む組成物。
【請求項66】
(i)小麦植物を育てるステップと(ii)前記小麦植物から穀粒を収穫するステップとを含む穀粒を製造する方法であって、前記穀粒が、SBEIIa遺伝子が染色体2Aの長腕から欠けている変異、あるいは染色体2Aの長腕上のSBEIIa遺伝子が、野生型穀粒と比べて前記穀粒の胚乳のSBEIIa酵素活性を低下させる変異を有する変異を含む穀粒を製造する方法。
【請求項67】
(i)請求項35から57のいずれか一項に記載の穀粒を得るステップと、(ii)前記穀粒からデンプンを抽出するステップとを含む、デンプンを製造する方法。
【請求項68】
(i)小麦植物を栽培するステップと、(ii)前記小麦植物から穀粒を収穫するステップとを含む、請求項1から17、請求項35から57のいずれか一項に記載の穀粒を製造する方法。
【請求項1】
デンプンを含む小麦植物から得た穀粒であって、穀粒のデンプン中のアミロース比率が少なくとも50%である穀粒。
【請求項2】
野生型穀粒と比べて低レベルの胚乳のSBEIIaタンパク質、SBEIIa酵素活性、またはその双方を含む、請求項1に記載の穀粒。
【請求項3】
野生型穀粒と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルを低下させる遺伝的変異を含み、遺伝的変異がSBEIIa遺伝子の変異またはSBEIIa遺伝子発現の阻害物質をコードする導入核酸を含む、請求項1または2のいずれか一項に記載の穀粒。
【請求項4】
デンプンと、野生型穀粒に比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルを低下させる遺伝的変異とを含み、遺伝的変異がSBEIIa遺伝子の変異またはSBEIIa遺伝子発現の阻害物質をコードする導入核酸を含む、小麦植物から得た穀粒であって、穀粒のデンプン中のアミロース比率が少なくとも30%である穀粒。
【請求項5】
前記遺伝的変異が、野生型穀粒と比べて胚乳のSBEIIaタンパク質、SBEIIa酵素活性、またはその双方のレベルを低下させるSBEIIa遺伝子の変異を含む、請求項4に記載の穀粒。
【請求項6】
少なくとも1種のSBEIIa遺伝子中にヌル変異を含む、請求項1から5までのいずれか一項に記載の穀粒。
【請求項7】
前記植物が6倍体であり、穀粒が2種または3種のSBEIIa遺伝子中にヌル変異を含む、請求項6に記載の穀粒。
【請求項8】
SBEIIa遺伝子発現の阻害物質をコードする導入遺伝子を含む、請求項1、2または4のいずれか一項に記載の穀粒。
【請求項9】
前記導入遺伝子がアンチセンスRNA分子、コサプレッションRNA分子、リボザイムRNA分子、または二重鎖RNA分子をコードする、請求項8に記載の穀粒。
【請求項10】
野生型穀粒と比べて低レベルのSBEIIbタンパク質、酵素活性、またはその双方を含む、請求項1から9のいずれか一項に記載の穀粒。
【請求項11】
野生型穀粒と比べて低レベルのSBEIタンパク質、酵素活性、またはその双方を含む、請求項1から10のいずれか一項に記載の穀粒。
【請求項12】
野生型穀粒と比べて改変されたレベルの酵素を含み、前記酵素がADPグルコースピロホスホリラーゼ、GBSS、SSI、SSII、SSIII、イソアミラーゼ型脱分枝酵素、プルラナーゼ型脱分枝酵素、およびこれらの任意の組合せからなる群から選択される、請求項1から11のいずれか一項に記載の穀粒。
【請求項13】
穀粒のデンプン中のアミロース比率が少なくとも40%である、請求項4または5のいずれかに記載の穀粒。
【請求項14】
非しわ型であり、または平均重量が少なくとも約36mgであり、あるいはその両方を満たす、請求項1から13のいずれか一項に記載の穀粒。
【請求項15】
穀粒から得たデンプン粒のうちの少なくとも50%が偏光下で観察すると非複屈折性に見える、請求項1から14のいずれか一項に記載の穀粒。
【請求項16】
殻のない状態での穀粒のデンプン含有量が少なくとも25%(w/w)であるか、またはデンプン含有量が野生型穀粒のデンプン含有量の少なくとも90%であるか、あるいはその両方を満たす、請求項1から15のいずれか一項に記載の穀粒。
【請求項17】
全粒、製粉した穀粒、粉砕した穀粒、精白した穀粒、圧扁した穀粒、粗挽きした穀粒、パーボイルドした穀粒、または挽き割りにした穀粒である、請求項1から16のいずれか一項に記載の穀粒。
【請求項18】
請求項1から17のいずれか一項に記載の穀粒を結実することができる小麦植物。
【請求項19】
請求項1から17のいずれか一項に記載の穀粒から抽出したデンプン粒。
【請求項20】
前記顆粒のデンプン中のアミロース比率が少なくとも50%であるデンプンを含む請求項19に記載のデンプン粒。
【請求項21】
請求項1から17のいずれか一項に記載の穀粒から抽出したデンプン。
【請求項22】
請求項1から17のいずれか一項に記載の穀粒、小麦粉、全粒小麦粉、セモリナ粉、請求項1から17のいずれか一項に記載の穀粒から得たデンプン、またはこれらの任意の組合せを含む製品。
【請求項23】
前記小麦粉、全粒小麦粉、セモリナ粉、またはデンプンを別の供給源から得た小麦粉、全粒小麦粉、セモリナ粉、またはデンプンと混合した請求項22に記載の製品。
【請求項24】
非食料品である請求項22に記載の製品。
【請求項25】
請求項21に記載のデンプンおよび別の食品成分または水を含む組成物。
【請求項26】
i)親となる小麦植物または小麦種子に遺伝的変異を導入するステップと、
ii)野生型の植物または種子と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップとを含む、
穀粒を結実することができる小麦植物を作製する方法であって、穀粒がデンプンを含有し、穀粒のデンプン中のアミロース比率が少なくとも50%である小麦植物の作製方法。
【請求項27】
i)SBEIIa遺伝子の変異またはSBEIIa遺伝子の発現の阻害物質をコードする導入核酸を含む遺伝的変異を、親となる小麦植物または小麦種子に導入するステップと、
ii)野生型の穀粒と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップとを含む、
穀粒を結実することができる小麦植物を作製する方法であって、穀粒がデンプンを含有し、穀粒のデンプン中のアミロース比率が少なくとも30%である小麦植物の作製方法。
【請求項28】
前記遺伝的変異を導入するステップが、SBEIIa遺伝子発現の阻害物質を発現する外来の核酸の導入を含む、請求項26または27に記載の小麦植物の作製方法。
【請求項29】
前記遺伝的変異を導入するステップが、化学薬剤または放射線による親小麦植物または小麦種子の突然変異誘発を含む、請求項26または27のいずれか一項に記載の小麦植物の作製方法。
【請求項30】
i)SBEIIa遺伝子の変異を含む遺伝的変異を、親となる小麦植物または小麦種子に導入するステップと、
ii)野生型の穀粒と比べて胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップと、
iii)SBEIIb遺伝子の変異を含む遺伝的変異を、親となる小麦植物または小麦種子に導入するステップと、
iv)野生型の穀粒と比べて胚乳のSBEIIb遺伝子発現、SBEIIb酵素活性、またはその双方のレベルが低下している、親小麦植物または小麦種子の子孫の植物または種子を同定するステップと、
v)胚乳のSBEIIa遺伝子発現、SBEIIa酵素活性、またはその双方のレベルが低い植物を、胚乳のSBEIIb遺伝子発現、SBEIIb酵素活性、またはその双方のレベルが低い植物と交配するステップとを含む、
穀粒を結実することができる小麦植物を作製する方法であって、SBEIIaとSBEIIbの双方の遺伝子発現、酵素活性、またはその双方が低い小麦植物を同定するステップとを含む、小麦植物を作製する方法。
【請求項31】
請求項1から17のいずれか一項に記載の穀粒からデンプンを抽出するステップを含む、改変した小麦デンプンを製造する方法。
【請求項32】
Triticum aestivum ssp.aestivumである、請求項18に記載の小麦植物。
【請求項33】
小麦のSBEIIb遺伝子またはSBEIIa遺伝子にそれぞれ結合する分子マーカーを用いて小麦植物または小麦種子の集団をスクリーニングするステップと、結合した分子マーカーの有無に基づいて植物または種子を同定するステップとを含む、SBEIIa遺伝子またはSBEIIb遺伝子の変異を有する小麦植物または小麦種子を同定する方法。
【請求項34】
小麦のSBEIIbタンパク質またはSBEIIaタンパク質にそれぞれ特異的な抗体を用いて小麦植物または小麦種子の集団をスクリーニングするステップと、抗体結合の有無に基づいて植物または種子を同定するステップとを含む、SBEIIa遺伝子またはSBEIIb遺伝子の変異を有する小麦植物または小麦種子を同定する方法。
【請求項35】
SBEIIa遺伝子が染色体2Aの長腕から欠けている変異、あるいは染色体2Aの長腕上のSBEIIa遺伝子が、野生型穀粒と比べて穀粒の胚乳のSBEIIaタンパク質、SBEIIa酵素活性、またはその双方を低下させる変異を有する変異を含む、小麦植物から得た穀粒。
【請求項36】
前記変異がSBEIIa遺伝子のヌル変異である、請求項35に記載の穀粒。
【請求項37】
前記変異がSBEIIa遺伝子の少なくとも一部分の欠失である、請求項35または36のいずれかに記載の穀粒。
【請求項38】
SBEIIb遺伝子が染色体2Aの長腕から欠けている変異、あるいは染色体2Aの長腕上のSBEIIb遺伝子が、野生型穀粒と比べて前記穀粒の胚乳のSBEIIbタンパク質、SBEIIb酵素活性、またはその双方を低下させる変異を有する変異をさらに含む、請求項35から37のいずれか一項に記載の穀粒。
【請求項39】
欠失が、染色体2Aの長腕上のSBEIIa遺伝子およびSBEIIb遺伝子の双方の発現を阻害する、請求項35から38のいずれか一項に記載の穀粒。
【請求項40】
前記植物がデュラム小麦植物である、請求項35から39のいずれか一項に記載の穀粒。
【請求項41】
野生型穀粒と比べて染色体2Bの長腕上のSBEIIa遺伝子によってコードされているデンプン分枝酵素活性を低下させる遺伝的変異をさらに含む、請求項40に記載の穀粒。
【請求項42】
前記遺伝的変異が、染色体2Bの長腕からのSBEIIa遺伝子の不在、または野生型穀粒と比べて前記穀粒の胚乳のSBEIIa酵素活性を低下させる染色体2Bの長腕上のSBEIIa遺伝子の変異を含む、請求項41に記載の穀粒。
【請求項43】
前記植物がTriticum aestivum ssp aestivumである、請求項35から39のいずれか一項に記載の穀粒。
【請求項44】
野生型穀粒と比べて染色体2B、染色体2D、または両方の染色体の長腕上のSBEIIa遺伝子によってコードされているデンプン分枝酵素活性を低下させる遺伝的変異を含む、請求項43に記載の穀粒。
【請求項45】
前記遺伝的変異が、前記染色体のうちの少なくとも1つからのSBEIIa遺伝子の欠失、または野生型穀粒と比べて前記穀粒の胚乳のSBEIIa酵素活性を低下させる前記染色体のうちの少なくとも1つのSBEIIa遺伝子の変異を含む、請求項44に記載の穀粒。
【請求項46】
前記遺伝的変異が、SBEIIa遺伝子発現、活性またはその双方の阻害物質をコードする導入核酸を含む、請求項41または44のいずれか一項に記載の穀粒。
【請求項47】
前記SBEIIa酵素活性のレベルが野生型穀粒と比べて少なくとも40%低下している、請求項35から46のいずれか一項に記載の穀粒。
【請求項48】
穀粒のデンプン中のアミロース比率が少なくとも30%である、請求項35から47のいずれか一項に記載の穀粒。
【請求項49】
穀粒のデンプン中のアミロース比率が少なくとも50%である、請求項48に記載の穀粒。
【請求項50】
穀粒のアミロペクチンの重合度が4から12の鎖長部分の比率は、アミロペクチンをイソアミラーゼで脱分枝した後に測定すると、野生型穀粒のアミロペクチンと比べて低下している、請求項1から17、および請求項35から49のいずれか一項に記載の穀粒。
【請求項51】
非しわ型である、請求項35から50のいずれか一項に記載の穀粒。
【請求項52】
平均重量が少なくとも約36mgである、請求項35から51のいずれか一項に記載の穀粒。
【請求項53】
穀粒から得たデンプン粒の少なくとも50%が、偏光下で観察すると非複屈折性に見える、請求項48から52のいずれか一項に記載の穀粒。
【請求項54】
殻のない状態での穀粒のデンプン含有量が少なくとも25%(w/w)であり、または、デンプン含有量が野生型穀粒のデンプン含有量の少なくとも90%である、請求項46から53のいずれか一項に記載の穀粒。
【請求項55】
全粒、製粉した穀粒、粉砕した穀粒、精白した穀粒、圧扁した穀粒、粗挽きした穀粒、パーボイルドした穀粒、または挽き割りにした穀粒である、請求項35から54のいずれか一項に記載の穀粒。
【請求項56】
低レベルのSBEIタンパク質、SBEI酵素活性、またはその双方をさらに含む、請求項35から55のいずれか一項に記載の穀粒。
【請求項57】
野生型穀粒と比べて改変したレベルの酵素をさらに含む穀粒であって、前記酵素が、ADPグルコースピロホスホリラーゼ、GBSS、SSI、SSII、SSIII、イソアミラーゼ型脱分枝酵素、プルラナーゼ型脱分枝酵素、およびこれらの任意の組合せからなる群から選択される、請求項35から56のいずれか一項に記載の穀粒。
【請求項58】
請求項35から57のいずれか一項に記載の穀粒を結実することができる小麦植物。
【請求項59】
請求項35から57のいずれか一項に記載の穀粒から抽出したデンプン粒。
【請求項60】
前記顆粒のデンプン中のアミロース比率が少なくとも50%である、デンプンを含む請求項59に記載のデンプン粒。
【請求項61】
請求項35から57のいずれか一項に記載の穀粒から抽出したデンプン。
【請求項62】
請求項35から57のいずれか一項に記載の穀粒、あるいは請求項35から57のいずれか一項に記載の穀粒から製造した小麦粉、全粒小麦粉、セモリナ粉、またはデンプンを含む製品。
【請求項63】
前記小麦粉、全粒小麦粉、セモリナ粉、またはデンプンが、別の供給源から得た小麦粉、全粒小麦粉、セモリナ粉、またはデンプンと混合される、請求項62に記載の製品。
【請求項64】
非食料品である、請求項62に記載の製品。
【請求項65】
請求項61に記載のデンプンおよび別の食品成分または水を含む組成物。
【請求項66】
(i)小麦植物を育てるステップと(ii)前記小麦植物から穀粒を収穫するステップとを含む穀粒を製造する方法であって、前記穀粒が、SBEIIa遺伝子が染色体2Aの長腕から欠けている変異、あるいは染色体2Aの長腕上のSBEIIa遺伝子が、野生型穀粒と比べて前記穀粒の胚乳のSBEIIa酵素活性を低下させる変異を有する変異を含む穀粒を製造する方法。
【請求項67】
(i)請求項35から57のいずれか一項に記載の穀粒を得るステップと、(ii)前記穀粒からデンプンを抽出するステップとを含む、デンプンを製造する方法。
【請求項68】
(i)小麦植物を栽培するステップと、(ii)前記小麦植物から穀粒を収穫するステップとを含む、請求項1から17、請求項35から57のいずれか一項に記載の穀粒を製造する方法。
【図1】


【図1】

【図1】

【図1】

【図2】

【図2】

【図3】
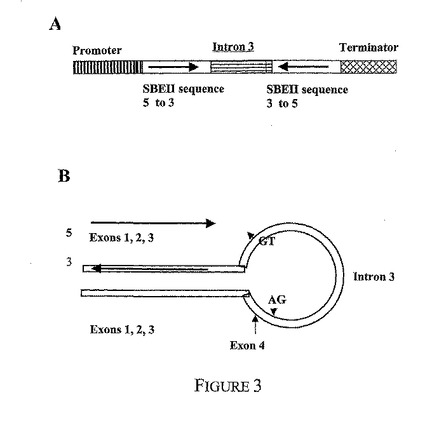
【図4】
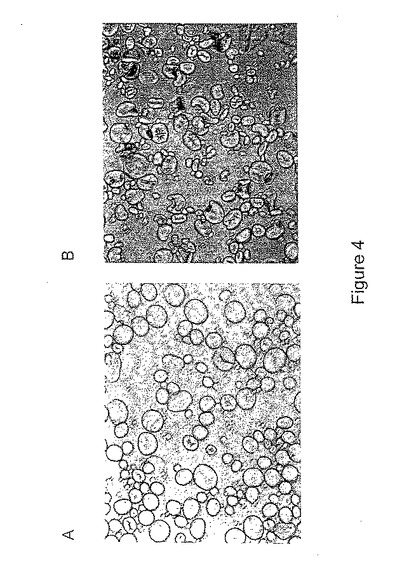
【図5】
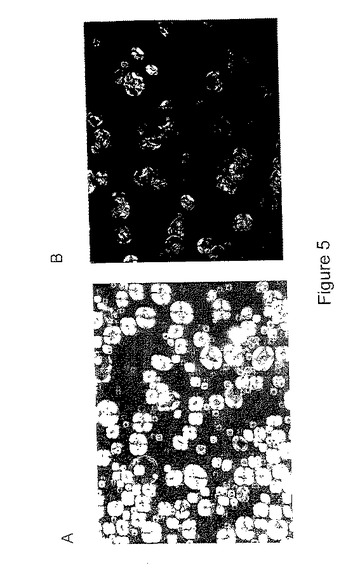
【図6】

【図7】
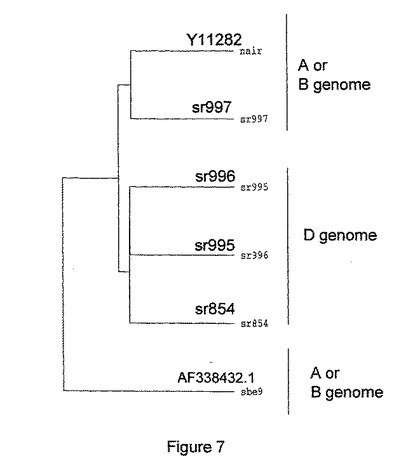
【図8】

【図8】

【図8】

【図8】

【図9】
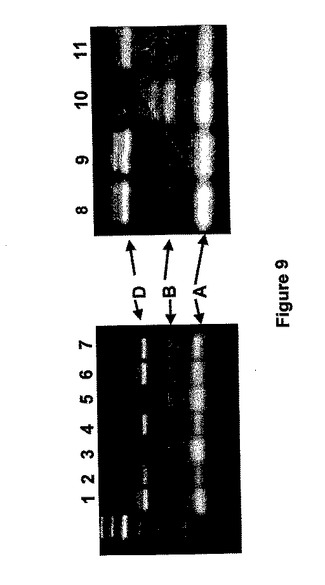
【図10】
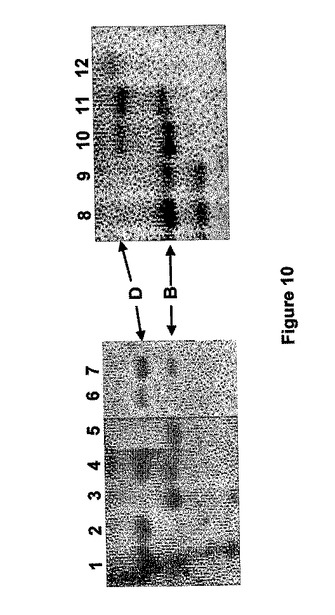
【図11】
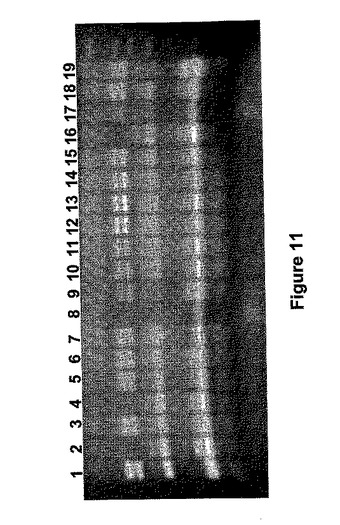
【図12】
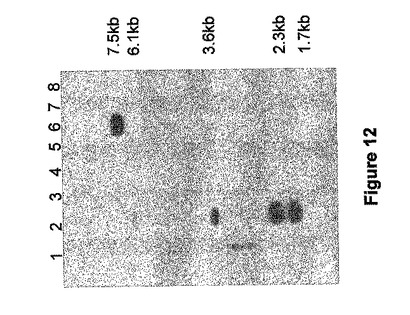
【図13】
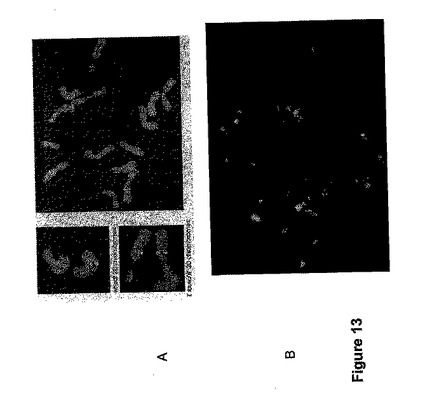
【図14】
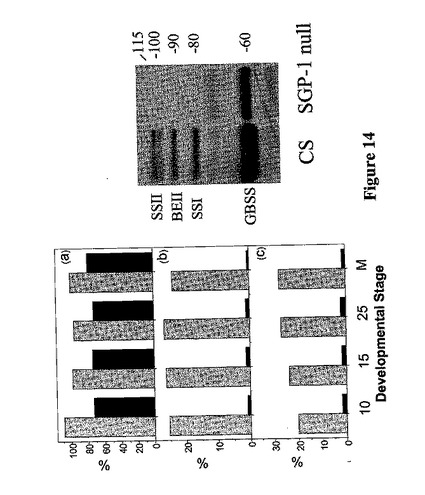
【図15】
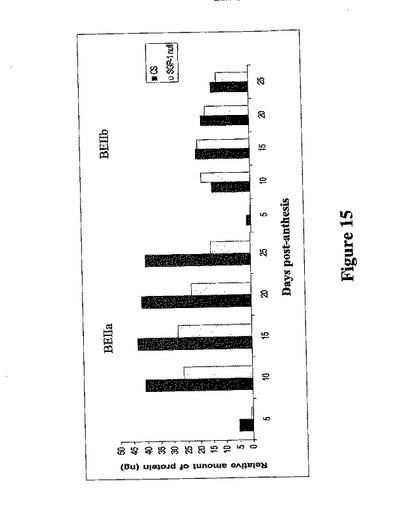
【図16】
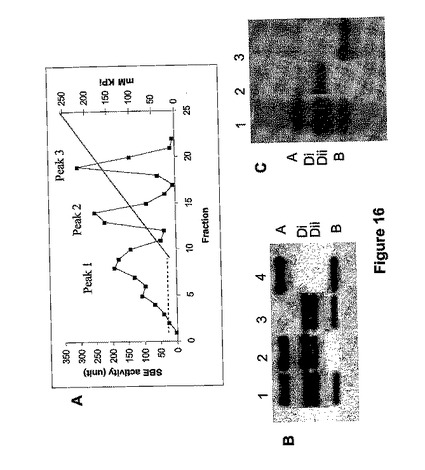
【図17】
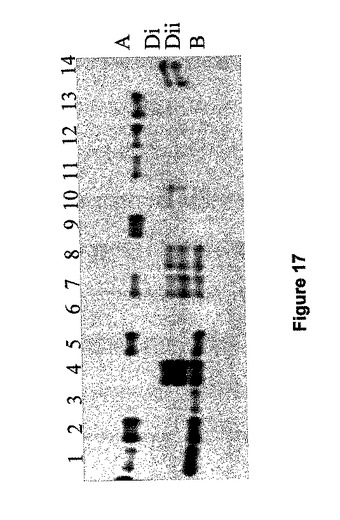
【図18】
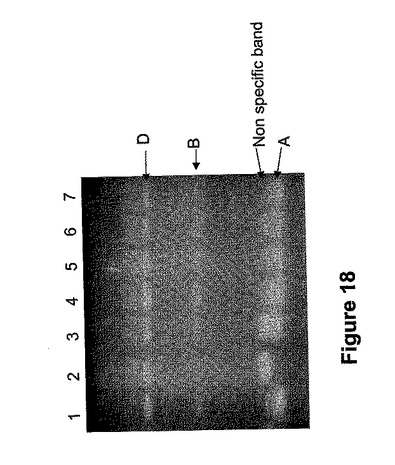
【図19】
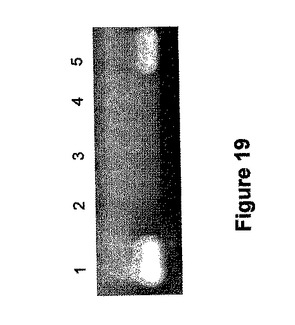
【図20】
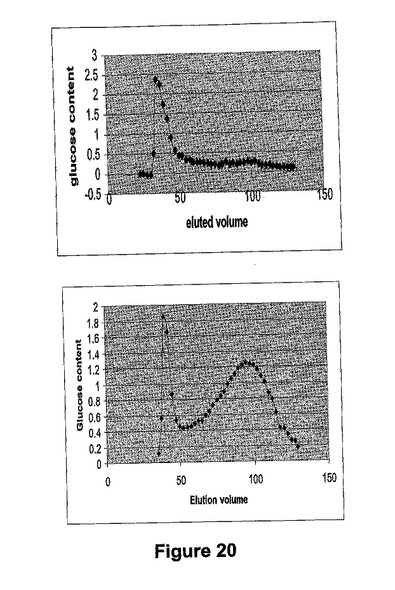
【図21】
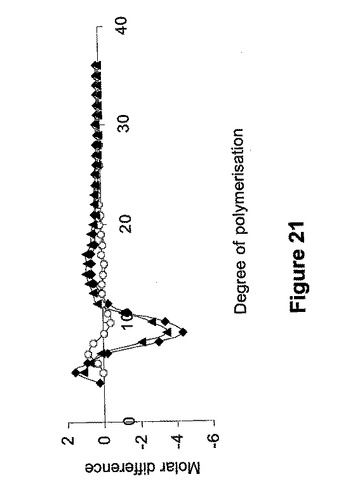
【図1】
【図1】
【図1】
【図2】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図8】
【図8】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【公表番号】特表2007−504803(P2007−504803A)
【公表日】平成19年3月8日(2007.3.8)
【国際特許分類】
【出願番号】特願2006−517898(P2006−517898)
【出願日】平成16年6月30日(2004.6.30)
【国際出願番号】PCT/AU2004/000901
【国際公開番号】WO2005/001098
【国際公開日】平成17年1月6日(2005.1.6)
【出願人】(598152079)コモンウェルス サイエンティフィック アンド インダストリアル リサーチ オーガナイゼイション (16)
【出願人】(501359630)バイオジェーマ (2)
【Fターム(参考)】
【公表日】平成19年3月8日(2007.3.8)
【国際特許分類】
【出願日】平成16年6月30日(2004.6.30)
【国際出願番号】PCT/AU2004/000901
【国際公開番号】WO2005/001098
【国際公開日】平成17年1月6日(2005.1.6)
【出願人】(598152079)コモンウェルス サイエンティフィック アンド インダストリアル リサーチ オーガナイゼイション (16)
【出願人】(501359630)バイオジェーマ (2)
【Fターム(参考)】
[ Back to top ]