
分節化されたマイナス鎖RNAウイルスの検出のための方法および組成物
本発明は、インフルエンザウイルスのごとき分節化されたマイナス鎖RNAウイルスの存在、不存在または量の検出のための組成物および方法を提供する。好ましくは、アンチセンス配向における、分節化されたマイナス鎖RNAウイルスの5’UTRおよび3'UTRならびにレポーター遺伝子のオープンリーディングフレームを含む人工セグメントを含む遺伝子改変された脊椎動物細胞を、分節化されたマイナス鎖ウイルスを含むことが疑われる生物学的試料と接触させる。分節化されたマイナス鎖RNAウイルスでの細胞の感染の結果、そのレポーター遺伝子によりコード化されたポリペプチドを発現する。また、本発明の遺伝子改変された細胞は、人工セグメントをコードする組換えDNAを含むことができる。その組換えDNAは、人工セグメントの転写を指令するRNAポリメラーゼI用のプロモーターを含むことができる。

【発明の詳細な説明】
【発明の詳細な説明】
【0001】
発明の分野
本願は、一般的にウイルス学の分野、より詳細には、生物学的試料中のマイナス鎖の分節化されたRNAウイルスを検出および定量する方法に関する。また、本願は、かかる方法に用いるために改変された細胞に関する。
【0002】
背景
マイナス鎖RNAウイルスは、ヒトおよび動物において多数の疾患の原因である。かくして、生物学的試料におけるマイナス鎖RNAウイルスの検出および定量化は、臨床医および研究者の双方に重要である。
【0003】
マイナス鎖RNAウイルスは、分節化されたRNAまたは分節化されていないRNAを含むことができる。分節化されていないマイナス鎖RNAウイルスにおいて、全ウイルスゲノムは、単一のネガティブセンスRNA分子により含まれるが、分節化されたマイナス鎖RNAウイルスにおいて、そのウイルスゲノムは複数のネガティブセンスのRNAセグメントにより含まれる。分節化されたマイナス鎖RNAウイルスのうち、Arenaviridae、BunyaviridaeおよびOrthomyxoviridaeは、各々、2、3および6〜8個のセグメントを含む。vRNAの各セグメントは、5'から3'の順序にて、ウイルスポリペプチドをコードする少なくとも1つのアンチセンスのオープンリーディングフレーム配列の5'非翻訳部位(「UTR」)、および3'UTRを含む。UTRは、RNA依存性RNAポリメラーゼのごとき分節化されたマイナス鎖RNA複製および/または遺伝子発現と関連した少なくとも1つの蛋白質により認識され、および/またはそれに結合する。セグメントによりコード化された蛋白質の発現は、セグメントが5'および3'末端配列を正確に規定することを必要とすることは今や一般的に受け入れられている。
【0004】
Olivoらの米国特許第5,591,579号および第5,851,757号は、生物学的試料における分節化されていないRNAウイルスの存在を検出するための細胞系および方法を開示する。その細胞系は、細胞内DNA依存性RNAポリメラーゼにより認識できるプロモーターを含むDNA分子で形質転換される。構造上不完全なRNAウイルスのcDNAの転写を指令できるプロモーターは、レポーターcDNAに連結される。ウイルス酵素のある種のトランス活性化に寄与する活性関連ウイルスが供されない限りは、DNA依存性RNAポリメラーゼにより転写されたRNA分子は、レポーターcDNAの翻訳を引き起こすことができない。
【0005】
Olivoらの米国特許第6,270,958号は、生物学的試料中の分節化されていないマイナス鎖RNAウイルスの検出のための診断アッセイを開示する。その開示されたアッセイにおいて、遺伝子改変された細胞は、マイナス鎖RNAウイルスのミニゲノムまたはミニアンチゲノムを発現する。ミニゲノムまたはミニアンチゲノムは、レポーター遺伝子またはその相補体、およびウイルスRNAからのフランキング配列を含む。また、該細胞は、各々がウイルスのヌクレオカプシド蛋白質の発現を指令するプロモーターを含む組換えDNA配列を含む。これらの細胞系におけるレポーター遺伝子の発現は、ヌクレオカプシド蛋白質の存在ならびにウイルスでの細胞の感染に依存する。
【0006】
分節化されたマイナス鎖RNAウイルスのセグメントによりコード化されたポリペプチドの発現が、正確に規定された3'および5'末端配列を必要とするために、研究者は、RNAポリメラーゼI(Pol I)の生化学的特性を利用して、分節化されたマイナス鎖RNAウイルスのセグメントのcDNAの発現を調べた。Pol Iは、正確なヌクレオチド配列にて転写を開始および終了すると考えられる。この点に関してZobelら, Nucleic Acids Research 21: 3607-3614,1993およびNeumannら, Virology 202: 477-479,1994は、マウスリボソームRNA(rDNA)遺伝子に由来するPol I(「Pol Iプロモーター」)により認識されるプロモーターを含む組換えDNA構築体を生成した。これらの組換えDNA構築体におけるPol Iプロモーターは、Pol I転写開始部位、インフルエンザウイルスRNAセグメントの5’非翻訳領域(UTR)、クロラムフェニコールアセチル基転移酵素(CAT)コード配列に相補的な配列(赤血球凝集素cDNAを置き換える)、および3’UTRを含むインフルエンザウイルスセグメント、ならびにPol Iポリメラーゼにより認識される転写ターミネーター配列(Pol Iターミネーター)を含む作動可能に連結したDNA配列の転写(恐らくPol Iによる)を指令する。このDNA構築体でトランスフェクトしたインフルエンザウイルスに感染した細胞は、CAT活性を発現し、これはウイルスでコード化されたRNA依存性RNAポリメラーゼは、テンプレートとしてPol I転写体を用いることができ、その結果、翻訳可能なmRNAが活性CAT酵素をコードすることを示す。
【0007】
これらおよび同様の試験が、インフルエンザウイルスセグメントのcDNAを含む組換えDNAを用いて、インフルエンザウイルスの完全な生成に導いた。この連結において、Neumannら、 Proceedings of the National Academy of Sciences USA 96: 9345-9350,1999,およびHoffmannら, Proceedings of the National Academy of Sciences USA 97: 6108-6113,2000は、クローン化されたcDNAからの完全なインフルエンザA型ウイルスの生成についての「逆遺伝学」DNA感染系を開示する。これらの試験において、細胞は、第1のシリーズの、各々がvRNAセグメントのcDNAに作動可能に連結したPol Iプロモーターを含むDNA配列を含むプラスミド、ならびに第2のシリーズの、各々がウイルス蛋白をコードするcDNAに作動可能に連結されたPol IIプロモーターを含むプラスミドでトランスフェクトされた。この系において転写されたウイルス蛋白質は、ウイルスの核タンパク質(NP)およびポリメラーゼ複合蛋白質(蛋白質PA、PB1およびPB2を含む)を含んだ。
【0008】
発明の概要
本発明は、分節化されたマイナス鎖RNAウイルスを含むことが疑われる生物学的試料における少なくとも1つの分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出または決定するための組成物および方法を包含する。また、本発明は、分節化されたマイナス鎖RNAウイルスに対して活性な抗ウイルス化合物をスクリーニングするための組成物および方法を包含する。
【0009】
1つの具体例において、本発明は、あるポリペプチドをコードするレポーター遺伝子を含む組換えRNA分子を含む、遺伝子改変された脊椎動物細胞を提供する。好ましい具体例において、脊椎動物細胞は哺乳動物細胞で有り得る。ある具体例において、細胞は、存在、不存在または量の検出または決定が求められる分節化されたマイナス鎖RNAウイルスのウイルスRNA依存性RNAポリメラーゼを含まない。レポーター遺伝子を含む翻訳可能なmRNAの形成は、分節化されたマイナス鎖ウイルスのRNA依存性RNAポリメラーゼの細胞へのその導入を必要とする。
【0010】
結果的に、もう一つの具体例において、本発明は、分節化されたマイナス鎖RNAウイルスを含むことが疑われる生物学的試料中の分節化されたマイナス鎖ウイルスの存在、不存在または量を検出または決定する方法を提供する。該方法は、生物学的試料と前記のごとき組換えRNA分子を含む遺伝子改変された脊椎動物細胞とを接触させ、次いで、レポーター遺伝子によりコード化されたポリペプチドの不存在、存在または量を検出することを含む。
【0011】
もう一つの具体例において、本発明は、発現が分節化されたマイナス鎖ウイルスのウイルスRNA依存性RNAポリメラーゼの細胞内の存在に依存するレポーター遺伝子を含む組換えDNA分子を含む遺伝子改変された脊椎動物細胞を提供する。かくして、また、本発明は、DNA分子を含む細胞内の発現が分節化されたマイナス鎖ウイルスのウイルスRNA依存性RNAポリメラーゼの細胞内の存在に依存するレポーター遺伝子を含む単離された組換えDNA分子を提供する。
【0012】
結果的に、もう一つの具体例において、本発明は、分節化されたマイナス鎖RNAウイルスを含むことが疑われる生物学的試料における分節化されたマイナス鎖ウイルスの存在、不存在または量を検出または決定するもう一つの方法を提供する。該方法は、生物学的試料と、前記のごとき組換えDNA分子を含む遺伝子改変された脊椎動物細胞とを接触させ、次いで、レポーター遺伝子によりコード化されたポリペプチドの不存在、存在または量を検出することを含む。
【0013】
もう一つの具体例において、本発明は、生物学的試料における、第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出または決定するための遺伝子改変された脊椎動物細胞を提供する。この具体例の細胞は、発現が第1の分節化されたマイナス鎖RNAウイルスのウイルスRNAの細胞内の存在に依存する第1のレポーター遺伝子を含む第1の組換えRNA分子、ならびに発現が第2の分節化されたマイナス鎖RNAウイルスのウイルスRNAの細胞内の存在に依存する第2のレポーター遺伝子を含む第2の組換えRNA分子を含む。
【0014】
結果的に、もう一つの具体例において、本発明は、生物学的試料における、第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出または決定する方法を提供する。該方法は、生物学的試料と、前記のごとき第1の組換えRNA分子および第2の組換えRNA分子を含む遺伝子改変された脊椎動物細胞とを接触させ、次いで、第1のレポーター遺伝子によりコード化された第1のポリペプチドの不存在、存在または量、ならびに第2のレポーター遺伝子によりコード化された第2のポリペプチドの不存在、存在または量を検出することを含む。
【0015】
もう一つの具体例において、本発明は、生物学的試料における、第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出または決定するための遺伝子改変された脊椎動物細胞を提供する。この具体例の細胞は、発現が第1の分節化されたマイナス鎖RNAウイルスのウイルスRNAの細胞内の存在に依存する第1のレポーター遺伝子を含む第1の組換えDNA分子、ならびに発現が第2の分節化されたマイナス鎖RNAウイルスのウイルスRNAの細胞内の存在に依存する第2のレポーター遺伝子を含む第2の組換えDNA分子を含む。
【0016】
結果的に、もう一つの具体例において、本発明は、生物学的試料における、第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出または決定する方法を提供する。該方法は、生物学的試料と、前記のごとき第1の組換えDNA分子および第2の組換えDNA分子を含む遺伝子改変された脊椎動物細胞とを接触させ、次いで、第1のレポーター遺伝子によりコード化される第1のポリペプチドの不存在、存在または量ならびに第2のレポーター遺伝子によりコード化される第2のポリペプチドの不存在、存在または量を検出することを含む。
【0017】
さらにもう一つの具体例において、本発明は、インフルエンザウイルスを含むことが疑われる生物学的試料において、インフルエンザA型ウイルス、インフルエンザB型ウイルスまたはどちらでもないウイルスの感染の間の鑑別診断を決定する方法を提供する。該方法は、生物学的試料と、遺伝子改変された細胞とを接触させることを含み、ここに、実質的に複数の遺伝子改変された細胞は、インフルエンザA型ウイルスのセグメントの少なくとも1つのUTRを含む組換えRNA分子、およびレポーター遺伝子を含む。この具体例において、レポーター遺伝子ポリペプチド発現は、インフルエンザA型またはインフルエンザB型ウイルスのウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存する。その細胞内のレポーター遺伝子発現の定量分析は、生物学的試料のウイルス含量:インフルエンザA型ウイルスを含む生物学的試料と接触した細胞集団は、感染していない対照培養物のそれより実質的に大きい量のレポーター遺伝子産物を生成し、一方、インフルエンザB型ウイルスを含む生物学的試料と接触した細胞集団は、感染していない対照培養物のそれよりわずかに大きな量のレポーター遺伝子産物を生成し、一方、病原体ウイルスを含まないか、またはインフルエンザA型ウイルスもしくはインフルエンザB型ウイルス以外の病原体を含む生物学的試料と接触した細胞集団は、感染していない対照培養物のそれほど著しく大きくない量のレポーター遺伝子産物を生成することを明らかにする。
【0018】
もう一つの具体例において、本発明は、包装化または使用説明書、および単離組換えDNA分子を含み、単離組換えDNA分子が蛋白質発現が分節化されたマイナス鎖ウイルスのウイルスRNA依存性RNAポリメラーゼの細胞内の存在に依存するレポーター遺伝子を含むキットを提供する。
【0019】
もう一つの具体例において、本発明は、分節化されたマイナス鎖RNAウイルスに指向された抗ウイルス薬についての定量的スクリーニング方法を提供する。該方法は、候補抗ウイルス化合物と、前記のごときレポーター遺伝子を含む人工セグメントを含む遺伝子改変された細胞とを接触させ、ここに、その細胞は、ウイルスRNA依存性RNAポリメラーゼを含むウイルスタンパク質複合体も含み、次いで、そのレポーター遺伝子ポリペプチドの発現レベルを測定することを含む。ある具体例において、タンパク質複合体は、細胞により含まれた複数のプラスミドの発現により提供でき、各プラスミドは、タンパク質複合体を含む少なくとも1つのポリペプチドをコードする。もう一つの具体例において、タンパク質複合体は、分節化されたマイナス鎖RNAウイルスによる細胞の感染により提供できる。候補抗ウイルス化合物の効力は、その化合物と接触した細胞(または細胞集団)内のレポーター遺伝子ポリペプチドの発現レベルを、その化合物と接触していない対照の細胞または細胞集団のそれと比較することにより決定される。
【0020】
もう一つの具体例において、本発明は、分節化されたマイナス鎖RNAウイルスに指向される抗ウイルス薬についての定量的スクリーニング方法を提供する。該方法は、候補抗ウイルス化合物と、ウイルスに感染した細胞とを接触させ;次いで、その細胞により生成されたウイルスの存在、不存在または量を検出することを含む。その検出は、前記のごとく、レポーター遺伝子を含む人工セグメントを含む遺伝子改変された細胞(または細胞の集団)と、候補抗ウイルス化合物と接触した細胞により生成されたウイルスとを接触させることを含む。候補抗ウイルス化合物の効力は、遺伝子改変された細胞または細胞集団によるレポーター遺伝子ポリペプチドの発現レベルを、候補抗ウイルス化合物と接触していない感染細胞により生成されたウイルスと接触した対照の細胞または細胞集団のそれと比較することにより決定される。
【0021】
詳細な記載
本発明により、分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出する方法および組成物が提供される。本発明は、遺伝子改変された細胞により供された分節化されたマイナス鎖RNAウイルスの人工セグメントが、細胞が分節化されたマイナス鎖RNAウイルスのRNA依存性RNAポリメラーゼをさらに含む場合、レポーター遺伝子を含む翻訳可能なmRNAの形成にアクセス可能であるという発明者らの観察に基づいている。ある具体例において、遺伝子改変された細胞は、RNA依存性RNAポリメラーゼのポリペプチド成分をコードする1以上の核酸をさらに含む細胞に基づくRNA依存性RNAポリメラーゼを含む。いくつかの具体例において、RNA依存性RNAポリメラーゼをコードする核酸は、例えば、分節化されたマイナス鎖RNAウイルスでの細胞の感染により供されるウイルスゲノムRNAとなることができる。レポーター遺伝子を含む翻訳可能なmRNAの形成は、感染に先立ち、細胞内のウイルスヌクレオカプシド蛋白質の不存在下でさえ生じることができる。他の具体例において、RNA依存性RNAポリメラーゼをコードする核酸は、1以上の組換えDNA分子になることができる。組換えDNA分子は、例えば、プラスミドまたは複数のプラスミドのごとき、例えば、ベクターまたは複数のベクターで有り得、ここに、そのベクターは、RNA依存性RNAポリメラーゼを含むウイルスタンパク質複合体に寄与するウイルスのポリペプチドをコードするcDNAを含む。組換えDNA分子は、ウイルスのヌクレオカプシド蛋白質のcDNAを含む1以上のベクターをさらに含むことができる。加えて、本発明は、レポーター遺伝子の産物の発現が、分節化されたマイナス鎖RNAウイルスのRNA依存性RNAポリメラーゼの不存在下、バックグラウンドレベルを著しく超えて検出されないという発明者らの観察に基づいている。宿主細胞にRNA依存性RNAポリメラーゼを導入する方法が、分節化されたマイナス鎖RNAウイルスにより細胞の感染を含むために、該方法および組成物は、生物学的試料における1以上の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出または診断することを可能とする。
【0022】
本明細書に記載された方法および組成物は、当業者によく知れた実験技術を利用し、Sambrook, J. , Fritsch, E. F. and Maniatis, T. (1989) Molecular Cloning: A Laboratory Manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NYのごとき実験マニュアルに見出すことができる。
【0023】
本発明による遺伝子改変された細胞は、組換え核酸を含む脊椎動物細胞であり、検出すべき分節化されたマイナス鎖RNAウイルスによる感染に感受性である。分節化されたマイナス鎖RNAウイルスは、例えば、アレナウイルス、ブンヤウイルスまたはオルトミクソウイルスで有り得る。オルトミクソウイルスは、例えば、インフルエンザA型ウイルス、インフルエンザB型ウイルスまたはインフルエンザC型ウイルスで有り得る。本発明の遺伝子改変された細胞は、予め形成された人工セグメントRNAからの翻訳可能なレポーター遺伝子mRNAの効率的な合成に必要な蛋白質が発現されるように、感染させる分節化されたマイナス鎖RNAウイルスのゲノムが、細胞に入ることができ、その複製サイクルにおいて少なくともはるかに十分に進むことができるならば、感染に感受性である。好ましくは、分節化されたマイナス鎖RNAウイルスでの細胞の感染は、十分に増殖性であり、すなわち、子孫ウイルスが、感染細胞により生成され、次いで、細胞から細胞へ広がる。さらに、ある具体例において、遺伝子改変された細胞は、好ましくは、人工セグメントが組換えDNA鋳型から転写できるように、(例えば、ヒトまたはマウス起源の)Pol Iプロモーターが機能するものである。特別の分節化されたマイナス鎖RNAウイルスによる感染に感受性の細胞系が、公知の感受性細胞系についての文献を検索することにより、および/または単にルーチン実験を必要とするよく知られた手順を用いる候補細胞系をスクリーニングすることにより容易に同定できると当業者ならば理解するであろう。分節化されたマイナス鎖RNAウイルスに感受性の細胞の非限定の例は、以下を含む:293T腎臓、LA−4、MRC5、A549、HEp2、2fTGHおよびSTATI−欠損2fTGH(U3A)のごときヒト細胞、ならびにベイビーハムスター腎臓(BHK)、CV−1、ベロ(Vero)、LLC−MK1、サル初代腎臓およびチャイニーズハムスター卵巣(CHO)のごとき非ヒト細胞。ヒト293T細胞は、ヒトを含めた哺乳動物を感染させる分節化されたマイナス鎖RNAウイルスを検出するための宿主細胞としての使用のために好ましい。感受性細胞は当該技術分野においてよく知られ、American Type Culture Collection, 10801 University Boulevard, Manassas, VA 20110-2209のごときソースから入手可能である。本明細書に記載された組換えDNA分子を含む本発明の遺伝子改変された細胞は、安定的にトランスフェクトした、あるいは一過性にトランスフェクトした細胞で有り得る。また、ある態様において、細胞により含まれた組換えDNAは、ベクター、好ましくはプラスミドにより含むことができる。また、ベクターは、当該技術分野においてよく知られた方法により用いられる、例えば、選択マーカーまたは複製開始点のごとき、組換えDNA分子の選択、維持または増殖に有用な他の配列を含むことができる(例えば、Sambrookら、前記)。
【0024】
また、本発明は、分節化されたマイナス鎖RNAウイルスの人工セグメントまたはその相補体を含む遺伝子改変された脊椎動物細胞を提供する。本発明の人工セグメントは、分節化されたマイナス鎖RNAウイルスのセグメントの5'UTR、レポーター遺伝子のオープンリーディングフレーム、および分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRを含む一本鎖RNAである。好ましくは、少なくとも1つの、好ましくは、双方の5'UTRおよび3'UTRは、その存在、不存在または量が、生物学的試料において検出されるべきある種の分節化されたマイナス鎖RNAウイルスからのものである。その5'UTRおよび3'UTRは、同一または異なるセグメントからのもので有り得る。好ましくは、その5'UTRおよび3'UTRは、その存在、不存在または量が生物学的試料において検出されるべき分節化されたマイナス鎖RNAウイルスの同一セグメントからのものである。好ましくは、人工セグメントのレポーター遺伝子のオープンリーディングフレームは、アンチセンス配置にあって、人工セグメントの相補体のレポーター遺伝子のオープンリーディングフレームは、センス配置にある。ウイルスRNA依存性RNAポリメラーゼの不存在下、宿主細胞は、翻訳可能なウイルスmRNA分子を提供も、そのセグメントに相補的なRNAを合成することもできない。しかしながら、ウイルスRNA依存性RNAポリメラーゼの存在下、宿主細胞は、翻訳可能なウイルスmRNA分子を提供することができ、鋳型として相補鎖を用いて、各ケースにおいて、人工セグメントおよびその相補体の両方を合成も複製もできる。ウイルスRNA依存性RNAポリメラーゼを含む宿主細胞において、人工セグメントのUTRを認識し、ウイルスRNA依存性RNAポリメラーゼにより転写開始部位として機能できる。人工セグメントにおいて、レポーター遺伝子のオープンリーディングフレームは、好ましくはアンチセンス配向にある。理論により拘束されることなくして、ウイルスRNA依存性RNAポリメラーゼのごとき1以上のウイルス蛋白質は、その人工セグメントのUTRを認識し、それに結合すると考えられる。ウイルス蛋白質は、宿主細胞の感染に際してウイルスにより宿主細胞に導入できる。さらに、鋳型として人工セグメントを利用することにより、RNA依存性RNAポリメラーゼは、人工セグメント(「ポジティブセンス」鎖)に相補的なRNA鎖の合成を触媒し、ポジティブセンス鎖は、人工セグメントの合成用の鋳型として用いることができると考えられる。また、さらに、RNA依存性RNAポリメラーゼは、キャップされたポリアデニル化されたmRNAに処理できる人工mRNAの合成用鋳型としてポジティブセンス鎖を利用する(例えば、Mikulasovaら, Acta Virol. 44: 273-282,2000参照)。センス配向にてレポーター遺伝子配列を含むmRNAは、細胞による翻訳に利用可能である結果、レポーター遺伝子によりコード化されたポリペプチドを生成する。従って、分節化されたマイナス鎖RNAウイルスでの細胞の感染は、例えば、その細胞と診断テスト試料との接触により供されるウイルスが、ウイルスRNA依存性RNAポリメラーゼを含むものを含めたウイルスのポリペプチドの発現に導き、従って、レポーター遺伝子ポリペプチドの発現に導く。加えて、遺伝子改変された細胞または細胞集団により生成されたポリペプチドの量は、ウイルスのレベルを飽和させるまで細胞(または細胞集団)を感染させる、分節化されたマイナス鎖RNAウイルスの量に単調に関連し、それにより、生物学的試料中のウイルスレベルの定量化のための手段を提供する。レポーター遺伝子をコード化するmRNAが、ウイルスRNA依存性RNAポリメラーゼ不存在下で宿主細胞内で合成されないので、本発明の感染していない遺伝子改変された細胞のレポーター遺伝子ポリペプチドの量は、人工セグメントを含まない対照細胞のものほど著しく大きくない。
【0025】
その存在、不存在または量が検出されるべき分節化されたマイナス鎖RNAウイルスは、アレナウイルス、ブンヤウイルスまたはオルトミクソウイルスのごときいずれかの分節化されたマイナス鎖RNAウイルスで有り得、好ましくは、オルトミクソウイルス、より好ましくは、インフルエンザウイルスである。インフルエンザウイルスは、インフルエンザA型ウイルス、インフルエンザB型ウイルスまたはインフルエンザC型ウイルスで有り得、好ましくは、インフルエンザA型ウイルスまたはインフルエンザB型ウイルスである。好ましくは、分節化されたマイナス鎖RNAウイルスの不存在、存在または量を検出するために用いた人工セグメントは、インフルエンザウイルス、好ましくは、インフルエンザA型ウイルスのセグメントからの5'UTR、好ましくは、インフルエンザA型ウイルスのNPセグメントの5'UTRを含む。好ましくは、その人工セグメントは、インフルエンザウイルス、好ましくは、インフルエンザA型ウイルスのセグメントからの3'UTR、好ましくは、インフルエンザA型ウイルスのNPセグメントの3'UTRを含む。好ましくは、その5'UTRおよび3'UTRは、同一ウイルス、好ましくは、インフルエンザA型ウイルスからのものであり、好ましくは、双方がインフルエンザA型ウイルスのNPセグメントからのものである。
【0026】
本発明のレポーター遺伝子は、細胞内で発現した場合、検出可能な産物をコードするいずれかの核酸配列(DNAまたはRNA)である。レポーター遺伝子は、例えば、cDNA配列、ゲノムDNA配列または合成配列のごときオープンリーディングフレームを含むDNAまたはRNA配列で有り得る。また、レポーター遺伝子は、イントロンを含むことができる。レポーター遺伝子核酸配列におけるイントロンの存在は、ウイルスRNA依存性RNAポリメラーゼでの細胞の刺激に際して、宿主細胞内でコード化されたポリペプチドの発現レベルを増加できると考えられる。レポーター遺伝子産物は、好ましくは、細胞内で容易にアッセイまたは検出できるポリペプチドである。ポリペプチドのアッセイまたは検出は、例えば、レポーター遺伝子ポリペプチドに指向された抗体を用いる免疫検出;例えば、蛍光発生的、色素生産性、化学発光もしくは放射性の基質を用いる酵素活性検出;または蛋白質蛍光のごときいずれかの公知の検出方法によることができる。レポーター遺伝子産物の非限定の例は、クロラムフェニコールアセチルトランスフェラーゼ、β-ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質(GFP)および分泌アルカリフォスファターゼを含む。好ましくは、レポーター遺伝子産物は、緑色蛍光タンパク質のごとき本質的に蛍光性であるポリペプチドであるか、またはホタルルシフェラーゼのごとき化学発光の基質が利用可能である酵素である。好ましくは、ポリペプチドの不存在、存在または量の検出は、そのポリペプチドまたはその基質により発光した光量子シグナルの不存在、存在または量の検出を含む。光量子シグナルは、例えば、緑色蛍光タンパク質のごとき蛍光蛋白質からの蛍光発光、または例えば、ホタルルシフェラーゼまたはウミシイタケルシフェラーゼのごとき化学発光の基質が利用できる酵素からの化学発光で有り得る。光シグナルの検出または定量化は、当該技術分野において知られたいずれかの検出または定量化の方法、例えば、蛍光顕微鏡法、フローサイトメトリーによることができる。「緑色蛍光蛋白質」なる用語は、Aequorea victoria (Chalfieら, Science, 263:802-805, 1994)からのGFPの自然発生野生型配列、ならびに蛍光変異体およびその誘導体(色に拘わらず)、例えば、Tsienらへの米国特許第6,054,321号、Chalfieらへの米国特許第6,146、826号、Michaelsへの米国特許第6,096、865号に記載された蛍光蛋白質を含むもののごとき蛍光ポリペプチドを含む。ルシフェラーゼなる用語は、ホタルルシフェラーゼおよびウミシイタケルシフェラーゼのごとき自然発生の化学発光基質を有する酵素を含む。
【0027】
ある具体例において、遺伝子改変された細胞は、人工セグメントをコードする組換えDNA配列を含む。遺伝子改変された細胞は、当該技術分野において知られたいずれかの方法により生成できる。いくつかの具体例において、遺伝子改変された細胞は、本発明の組換えDNAでの細胞のトランスフェクションにより生成できる。トランスフェクションは、当該技術分野においてよく知られた方法を用いて達成できる。好ましくは、トランスフェクションは、TransITR-LT1トランスフェクション試薬(Mirus Corporation, Madison, WI)のごときトランスフェクション剤を用いて達成される。組換えDNAは、人工セグメントをコード化する組換えDNA配列を含む単離組換えDNA分子として提供できる。単離組換えDNA分子は、組換えプラスミドまたは組換えウイルスのごときベクターを含むことができる。組換えウイルスは、例えば、組換えバクテリオファージまたは組換え脊椎動物ウイルスで有り得る。
【0028】
ある具体例において、これらの具体例の組換えDNA分子は、5'から3'の順序にて、DNA依存性RNAポリメラーゼ用のプロモーター、DNA依存性RNAポリメラーゼ用の転写開始部位、人工セグメントまたはその相補体をコード化するDNA配列、ならびにDNA依存性RNAポリメラーゼ用の転写終結部位を含む。組換えDNA配列の転写のための本発明の遺伝子改変された細胞により用いられたDNA依存性RNAポリメラーゼは、細胞に内因的なDNA依存性RNAポリメラーゼ、または例えば、Olivoらへの特許第6,270,958号に開示されたDNA依存性RNAポリメラーゼのごとき異種のDNA依存性RNAポリメラーゼで有り得る。好ましくは、そのDNA依存性RNAポリメラーゼは、細胞に内因的なDNA依存性RNAポリメラーゼであり、より好ましくは、RNAポリメラーゼIである。従って、そのプロモーターは、脊椎動物細胞内で機能できるいずれかのRNAポリメラーゼ用のプロモーターで有り得、好ましくは、細胞に内因的なRNAポリメラーゼ用のプロモーター、好ましくは、RNAポリメラーゼI用のプロモーターである。プロモーターは、好ましくは、Pol I転写開始部位である転写開始部位に作動可能に連結される。Pol I転写開始部位は、Pol Iが、例えば、配列5'-CCAATAA-3'のごとき転写を開始するために認識できるいずれかの配列で有り得る。この配列がPol Iプロモーターに続く場合、転写は、この配列の直ぐ3'のヌクレオチドにて開始されると考えられる。Pol Iが正確に規定された配列にて転写を開始するために、Pol I転写開始部位は、好ましくは、DNAの転写体が、5'末端が人工セグメントをコードするcDNAの5'UTRの5'端と正確にまたはその5'端の約10のヌクレオチド内で一致する人工セグメントを含むにように配置される。好ましくは、5'末端は、5'UTRの5'端と正確に一致する。別法として、人工セグメントがプロモーターに関してポジティブセンスの配置にあるならば、DNAの転写体は、5'末端が人工セグメントをコードするcDNAの3'UTRの相補体の5'端と正確にまたはその5'端の約10のヌクレオチド内で一致する人工セグメントを含む。好ましくは、5'末端は、3'UTRの5'端と正確に一致する。人工セグメントをコードするcDNAは、分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRのcDNA、ポリペプチドをコードするレポーター遺伝子のオープンリーディングフレームのDNA配列、および分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRのcDNAを含む。レポーター遺伝子のオープンリーディングフレームのDNA配列は、組換えDNA分子により含まれたプロモーターに関してセンスまたはアンチセンス配置のいずれかで有り得、好ましくはプロモーターに関してアンチセンス配置にある。人工セグメントをコード化する配列は、プロモーターに関してセンスまたはアンチセンス配向に有り得る。かくして、レポーター遺伝子DNA配列は、プロモーターに関してセンスまたはアンチセンス配向のいずれかでも有り得る。さらに、いくつかの具体例において、レポーター遺伝子DNA配列もイントロンを含むことができる。
【0029】
組換えDNA分子はさらに転写終結部位を含む。「転写終結部位」は、例えば、転写終了のためのDNA依存性RNAポリメラーゼ、自己切断リボザイムのごとき触媒的なRNA配列をコード化する配列または直線DNAの末端配列のごとき規定された3’末端を有するRNA配列に転写または処理できるいずれかのDNA配列を含むように本明細書に規定される。転写終結部位は、好ましくは、RNAポリメラーゼIにより認識された転写終結部位である。好ましい具体例において、Pol I転写終結部位は、配列5'-GGGGGG-3'により描写できる。別法の具体例において、転写終結部位は、自己切断リボザイムのごとき触媒的なRNA配列をコードする配列を含むことができる。本発明の転写終結部位は、転写に際して、DNAから転写されたRNAの3'末端が人工セグメントの3'UTRの3'端に正確にあるか、またはその3’端の約10ヌクレオチド内にあるようにDNA配列内に位置する。好ましい具体例において、転写に際して、DNAから転写されたRNAの3'末端は、正確に3'UTRの3'端にある。別法として、人工セグメントがプロモーターに関してポジティブセンスの配置にあるならば、本発明の転写終結部位は、転写に際して、DNAから転写されたRNAの3'末端が、人工セグメントの相補体の5'UTRの相補体の3'端に正確にあるか、またはその3'端の約10ヌクレオチド内にあるようにDNA配列中に位置する。好ましい具体例において、そのDNAから転写されたRNAの3'末端は、5'UTRの相補体の3'端に正確にある。
【0030】
ある具体例において、組換えDNA分子は、例えば、組換えプラスミドまたは組換えウイルスのごときベクターである。好ましくは、ベクターは組換えプラスミドである。プラスミドの非限定の例は、pHH21NPUTR-lucおよびpHH21NPUTR-GFPを含み、ここに、各々、DNA依存性RNAポリメラーゼ用のプロモーターはPol Iプロモーターであり、そのUTRはインフルエンザA型ウイルスのNPセグメントからのもであって、レポーター遺伝子はホタルルシフェラーゼおよび緑色蛍光タンパク質をコードする。かかるベクターは、当業者によく知られた材料および方法を用いて構築できる(例えば、Sambrookら、前記)。非限定の例において、かかるベクターは、プラスミドプラスミドpHH21のごときPol Iプロモーターを含むプラスミドを用いて構築できる(Neumann, G.ら, Proceedings of the National Academy of Sciences USA 96: 9345-9350,1999 ; Neumann, G.ら, Virology 202: 477-479,1994)。
【0031】
理論により拘束されることなくして、その配列構造のために、本発明のいくつかの具体例における組換えDNAは、Pol IのごときRNAポリメラーゼにより転写されて、分節化されたマイナス鎖ウイルスのセグメントまたはセグメント群のものに同一またはほとんど同一である5'および3'末端配列を有するRNA転写体を形成し、それにより、人工セグメントを形成すると考えられる。従って、人工セグメントは、生物学的試料により含まれたウイルスによる感染に先立ち細胞内に蓄積する。分節化されていないウイルスの検出方法と異なり、本発明の方法による分節化されたウイルスの検出は、ウイルス感染に先立ち、またはウイルス感染に加えて、ウイルス蛋白質の細胞内の発現を必要としない。しかしながら、遺伝子改変された細胞がヌクレオカプシド蛋白質のごときウイルス蛋白質をさらに含むならば、レポーターポリペプチドの発現レベルは増強できる。例えば、かかるウイルス蛋白質は、ウイルス蛋白質用のcDNAに作動可能に連結されたDNA依存性RNAポリメラーゼ(RNAポリメラーゼIIのごとき)用のプロモーターを各々含む1以上のプラスミドでの細胞のトランスフェクションを介して細胞に供することができる。
【0032】
もう一つの具体例において、本発明は、生物学的試料における第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出するための組成物および方法を提供する。この具体例の遺伝子改変された細胞は、2つの人工セグメントまたはそれらの相補体を含む。第1の人工セグメントは、第1の分節化されたマイナス鎖RNAウイルスの5'UTR、レポーターポリペプチドをコードする第1の核酸配列、および第1の分節化されたマイナス鎖RNAウイルスの第1の3'UTRを含む。第2の人工セグメントは、第2の分節化されたマイナス鎖RNAウイルスの5'UTR、レポーターポリペプチドをコードする第2の核酸配列、および第2の分節化されたマイナス鎖RNAウイルスの第2の3'UTRを含む。第1および第2の人工セグメントの双方について、レポーターポリペプチドをコードする核酸配列は、センスまたはアンチセンス配置で有り得、好ましくはアンチセンス配置にある。これらの具体例における細胞は、前記のごとき人工セグメントの転写用の配列に作動可能に連結したDNA依存性RNAポリメラーゼ用のプロモーターを各々含む第1の組換えDNA分子および第2の組換えDNA分子のいずれもさらに含むことができる。あるいは、単一の組換えDNA分子は、DNA依存性RNAポリメラーゼ用の2つのプロモーターを含むことができ、各プロモーターは、前記のごとき2つの人工セグメントのうちの1つの転写のための配列に作動可能に連結される。これらの具体例において、ウイルスは、非限定の例において、アレナウイルス、ブンヤウイルス、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスから選択された1対のウイルスのごときいずれかの1対の分節化されたマイナス鎖ウイルスで有り得る。2つのウイルスを含む生物学的試料との接触に際して、レポーター遺伝子を含むmRNAは各ウイルスの量に比例して形成される。従って、レポーター遺伝子は比例して発現され、生物学的試料のウイルス含量を反映する。例えば、この具体例の遺伝子改変された細胞は、インフルエンザA型からのUTRを含む第1の人工セグメント、およびインフルエンザB型からのUTRを含む第2の人工セグメントを含むことができ、人工セグメントは、異なったレポーター遺伝子を含む。その細胞と、インフルエンザA型およびインフルエンザB型ウイルスの双方を含む生物学的試料との接触に際して、細胞により作成されたレポーター遺伝子ポリペプチドの量は、その標本におけるウイルス量に比例するであろう。好ましくは、これらの具体例のレポーター遺伝子によりコード化されたポリペプチドは、光発光ポリペプチドであるか、または光発光基質が利用可能である酵素、好ましくは、ホタルルシフェラーゼ、ウミシイタケルシフェラーゼおよび緑色蛍光タンパク質よりなる群から選択される酵素である。
【0033】
さらにもう一つの具体例において、本発明は、インフルエンザウイルスを含むことが疑われる生物学的試料における、インフルエンザA型ウイルス、インフルエンザB型ウイルスまたはどちらでもないウイルスの感染の間の鑑別診断を決定する方法を提供する。該方法は、その生物学的試料と、遺伝子改変された細胞の集団とを接触させることを含み、ここに、実質的に複数の遺伝子改変された細胞は、インフルエンザA型ウイルスのセグメントの少なくとも1つのUTRおよびポリペプチドをコードするレポーター遺伝子を含む人工セグメントを含む。人工セグメントを含む「実質的に複数」の遺伝子改変された細胞は、人工セグメントを含む細胞の少なくとも約50%、好ましくは少なくとも約75%、より好ましくは少なくとも約90%で有り得る。レポーター遺伝子は、好ましくはアンチセンス配置にある。これらの細胞の人工セグメントの5'UTRおよび3'UTRの片方または双方は、好ましくは、インフルエンザA型ウイルスのセグメントのUTR、より好ましくはNPセグメントである。レポーター遺伝子ポリペプチドの発現は、インフルエンザウイルスA型ウイルスまたはインフルエンザB型ウイルスのウイルスRNA依存性RNAポリメラーゼの細胞内の存在に依存する。好ましい具体例において、レポーター遺伝子によりコード化されたポリペプチドは、光発光により検出できる蛋白質、例えば、緑色蛍光タンパク質またはルシフェラーゼ蛋白質である。これらの具体例において、細胞集団中のレポーター遺伝子発現の定量的測定は、生物学的試料のウイルス含量:インフルエンザA型ウイルスを含む生物学的試料と接触した細胞集団が、感染していない対照培養物のそれより実質的に大きな量のレポーター遺伝子産物を生成し、一方、インフルエンザB型ウイルスを含む生物学的試料と接触した細胞集団は、感染していない対照培養物よりわずかに大きな量のレポーター遺伝子産物を生成することを明らかにする。いくつかの具体例において、レポーター遺伝子発現の定量分析は、細胞集団で全体として発現されたレポーター遺伝子ポリペプチドの測定を含むことができる。もう一つの具体例において、定量分析は、例えば、レポーター遺伝子およびフローサイトメトリーまたは蛍光標示式細胞分取器を用いて、細胞ごとのベースでレポーター遺伝子ポリペプチドレベルを測定することを含むことができる。かくして、いくつかの具体例において、インフルエンザA型ウイルス、インフルエンザB型またはどちらでもないウイルスを含むことが知られた試料を用いて、これらのウイルスに応じた培養によりレポーター遺伝子発現レベルを確立できる。これらの発現レベルは、インフルエンザウイルスを含むことが疑われる生物学的試料により含まれたウイルスのタイプを決定するためのガイダンスを提供できる。例えば、レポーター遺伝子ポリペプチドを発現する細胞のパーセンテージは、そのウイルスのうちの1つに感染した対照培養物を用いて測定されたパーセンテージを反映するであろう。かくして、例えば、その細胞集団と接触した生物学的試料は、大多数の細胞中のレポーターポリペプチドの発現に導く、例えば、少なくとも約80%の細胞、または少なくとも約90%の細胞は、インフルエンザA型ウイルスを含むと診断でき、一方、その細胞集団と接触した生物学的試料は、少数の細胞、例えば、約10%の細胞におけるレポーターポリペプチドの発現に導き、インフルエンザB型ウイルスを含むと診断することができる。さらに、その細胞集団と接触した生物学的試料は、約3%未満の細胞におけるレポーターポリペプチドの発現に導き、どちらのウイルスも含まないと診断できる。
【0034】
もう一つの具体例において、本発明は、包装化および単離組換えDNA分子を含むキットを提供し、単離組換え分子は、その分子でトランスフェクトされた細胞の発現が分節化されたマイナス鎖ウイルスのウイルスRNAの細胞内の存在に依存するレポーター遺伝子を含む。この具体例において、単離組換えDNA分子は、5'から3'の順序にて、DNA依存性RNAポリメラーゼ用のプロモーター、DNA依存性RNAポリメラーゼ用の転写開始部位、分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRのcDNA、ポリペプチドをコードするレポーター遺伝子のオープンリーディングフレームのDNA配列、分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRのcDNAならびにDNA依存性RNAポリメラーゼ用の転写終結部位を含む。この具体例のレポーター遺伝子のオープンリーディングフレームのDNA配列は、センスまたはアンチセンス配向で有り得、好ましくはアンチセンス配向にある。また、この具体例のキットは、前記されたもののごとき分節化されたマイナス鎖RNAウイルスによる感染に感受性の細胞または細胞集団を含むことができる。その細胞または細胞集団は、単離組換えDNA分子で感染した遺伝子改変された細胞で有り得る。あるいは、その細胞または細胞集団は、非トランスフェクトで有り得る。キットの使用者は、当業者によく知られた標準的な方法を用いて、そのDNAで細胞または細胞集団をトランスフェクトし、次いで、そのトランスフェクトした細胞を用いて、分節化されたマイナス鎖RNAウイルスの存在、不存在または量を測定できる。トランスフェクションは、Sambrookら、前記に記載されたもののごとき当業者によく知られた標準的手順により行うことができる。
【0035】
もう一つの具体例において、本発明は、分節化されたマイナス鎖RNAウイルスの増殖または遺伝子発現に指向された抗ウイルス薬についての定量的スクリーニング方法を提供する。そのウイルスは、好ましくは、インフルエンザA型ウイルスのごときインフルエンザウイルスである。定量的スクリーニング方法は、 (a)候補抗ウイルス化合物と、(i)前記のレポーター遺伝子ポリペプチドのコード化した配列を含む人工セグメントおよび(ii)分節化されたマイナス鎖RNAウイルスのRNA依存性RNAポリメラーゼを含むタンパク質複合体を含む遺伝子改変された細胞または細胞集団とを接触させ、次いで、(b)細胞または細胞集団により生成されたポリペプチドの量を検出することを含む。ある具体例において、分節化されたマイナス鎖RNAウイルスのRNA依存性RNAポリメラーゼを含むタンパク質複合体は、1以上の核酸により細胞または細胞集団において提供でき、各核酸は、分節化されたマイナス鎖RNAウイルスの複製複合体の少なくとも1つのポリペプチド成分をコードする。他の具体例において、タンパク質複合体は、そのウイルスでの細胞の感染、またはウイルスRNAでの細胞のトランスフェクションにより細胞に提供できる。候補抗ウイルス化合物の効力は、その化合物と接触した細胞または細胞集団のレポーター遺伝子ポリペプチドの発現レベルをその化合物と接触しない対照の細胞または細胞集団のそれに比較することにより決定される。レポーター遺伝子ポリペプチドの発現レベルは、細胞により発現されたレポーター遺伝子ポリペプチドの合計量の測定により決定できる。いくつかの具体例において、レポーター遺伝子ポリペプチドの発現は、レポーター遺伝子ポリペプチドを発現する細胞のパーセンテージの測定により決定できる。非限定の例において、レポーター遺伝子ポリペプチドがGFPまたはルシフェラーゼのいずれかである場合、フローサイトメトリーを用いて、細胞数−対−レポーター遺伝子ポリペプチド発現レベルの2次元プロットを提供できる。標準的な分析技術を用いて、候補化合物の効力を決定でき、例えば、集めたデータを用いて、候補化合物のレポーター遺伝子発現(ED50)の50%の低下を供する有効用量を決定できる。この定量的スクリーニング方法は、迅速な結果を提供し、実施するのは簡単である。
【0036】
もう一つの具体例において、本発明は、分節化されたマイナス鎖RNAウイルスに指向された抗ウイルス薬についてのウイルス生成アッセイ(viral yield assay)を提供する。この具体例において、第1の細胞(または細胞集団)は、候補抗ウイルス化合物と接触する。また、第1の細胞はウイルスに感染する。第1の細胞は、ウイルスによる増殖感染に感受性であるタイプである。次いで、細胞または細胞集団によるウイルスの生成は、候補抗ウイルス化合物との感染および接触後、第1の細胞または細胞集団により生成されたウイルスの試料(もしあれば)を含む生成試料を用いて測定される。その化合物の抗ウイルスの活性は、生成試料と、第2の細胞(または細胞集団)とを接触させることにより決定され、ここに、第2の細胞または細胞集団は、発現が前記のごとく、ウイルスのRNA依存性RNAポリメラーゼの細胞内における存在に依存するポリペプチドをコードするレポーター遺伝子を含む人工セグメントを含む遺伝子改変された細胞または細胞集団である。(対照と比較した)レポーター遺伝子の発現レベルの低下は、候補抗ウイルス化合物による抗ウイルス活性を示す。
【0037】
以下の実施例は、例示的な目的だけに提供され、本発明の範囲を限定するものと解釈されるべきでない。その実施例は、本発明についてのより良好な理解およびその利点のより良好な認識を可能にするであろう。
【0038】
実施例1
本実施例は、プラスミドpHH21 NP UTR HIルシフェラーゼの構造および機能化を示す。
【0039】
このプラスミド(図1)は、インフルエンザA/WSN/33のセグメント5のUTRにより隣接して挟まれたルシフェラーゼORFをコードする。このプラスミドを含む細胞において、転写は、プラスミドがPol Iにより転写される場合、ネガティブまたはゲノムのセンスRNA転写体が生成されるように、RNAポリメラーゼIプロモーターおよびターミネーターにより制御される。このプラスミドにおいて、ルシフェラーゼ・レポーターポリペプチドをコードするDNA配列は、ヒトRNAポリメラーゼI発現ベクターにより含まれたインフルエンザA型ウイルスNPセグメントのcDNAのオープンリーディングフレームを交換する(plasmid pHH21 ; Neumann, G.ら, Proceedings of the National Academy of Sciences USA 96: 9345-9350,1999 ; Neumann,G.ら, Virology 202: 477-479,1994)。そのプラスミドの機能領域に対応するヌクレオチド配列を示す。このプラスミドでトランスフェクトし、インフルエンザA型ウイルスに感染した細胞は、ルシフェラーゼを発現できる。同様に、このプラスミドでトランスフェクトし、さらにウイルス蛋白質PA、PB1、PB2およびNPをコードするプラスミドでトランスフェクトした細胞も、ルシフェラーゼを発現できる。
【0040】
実施例2
この実施例はインフルエンザA型ウイルス検出プロセスを示す。
【0041】
この実施例(図2)において、トランスフェクトした細胞は、RNAポリメラーゼI酵素を用いて、ネガティブまたはゲノムのセンスRNA転写体を生成する。インフルエンザA型ウイルスでの感染またはインフルエンザA型ウイルスのポリメラーゼ複合体の発現に際して、転写体は、全長のポジティブセンスRNAまたは5’キャップされたポリアデニル化mRNAに複製される。ポジティブセンスRNAを複製して、さらなるウイルスネガティブセンスRNAを形成し、一方、そのmRNAを宿主細胞機構によりレポーター蛋白質に翻訳する。ルシフェラーゼ・レポーターポリペプチドは、当該技術分野において知られた標準的な方法により検出および定量できる。
【0042】
実施例3
この実施例はインフルエンザA型ウイルス検出手順を示す。
【0043】
この手順において、細胞をそのpHH21ルシフェラーゼまたは同様のプラスミドでトランスフェクトする(図3)。次いで、細胞を安定的にトランスフェクトした亜集団について選択するか、または直接的に用いることができる。次いで、生物学的試料(例えば、臨床検査試料またはウイルス試料)を検出可能なシグナルの生成に十分であると期待される期間細胞と共にインキュベートする。インキュベーション時間は少なくとも約6時間で有り得る。好ましくは、そのインキュベーション時間は、少なくとも約6時間から約14日まで、少なくとも約6時間から約120時間まで、または少なくとも約6時間から約24時間までで有り得る。試料に対する曝露後、その細胞を当該技術分野において知られた標準的な方法によりレポーター遺伝子ポリペプチドの発現について分析する。
【0044】
実施例4
この実施例は、人工のインフルエンザA型セグメントからのレポーター遺伝子の発現を示す。
【0045】
この実施例において、ホタルルシフェラーゼ遺伝子を、そのNPセグメントからのUTRを含むヒトRNAポリメラーゼI発現ベクターpHH21におけるNPオープンリーディングフレームに代えて、クローン化した。得られたプラスミドのpHH21NPUTR-lucは、インフルエンザウイルスRNAポリメラーゼ複合体により転写できる機能的なインフルエンザウイルス人工セグメントRNAを発現させ、それにより、ウイルス感染の存在下、ルシフェラーゼ発現を可能とする。ヒト293T細胞をpHH21NPUTR-lucでトランスフェクトし、5の感染多重度(MOI)にてインフルエンザウイルス株WSN/33でトランスフェクトさせたか、またはインフルエンザウイルス複製およびヌクレオカプシド蛋白質(ウイルスポリペプチドPA、PB1、PB2およびNPを各々コードするpPA、pPBl、pPB2およびpNP)用の発現ベクターで共トランスフェクトさせた。トランスフェクションの42時間後(または、pHH21NPUTR-luc単独でトランスフェクトした細胞については、インフルエンザウイルスによる感染の18時間後)に、細胞を溶解し、GLOWキット(Promega, Madison WI)を用いて、ホタルルシフェラーゼ活性につきアッセイした。感染の不存在下のそのプラスミドでのトランスフェクションは、16,000を超えるのシグナル対ノイズの非常に高いシグナル(>2×106 LCPS)を生じさせた(図4、第3レーン)。しかしながら、人工セグメント(pHH21NPUTR-luc)を含むプラスミドだけでトランスフェクトした細胞のウイルス感染も、レポーター遺伝子発現を促すのに十分であった(図4、第2レーン)。
【0046】
実施例5
この実施例は、インフルエンザA型ウイルス活性の指標としてのGFPレポーター遺伝子を示す。
【0047】
蛍光ベースの読取りを用いたインフルエンザA型ウイルス活性についてのスクリーニングを促すために、インフルエンザウイルスA/WSN/33のNPセグメントのUTRにより隣接して挟まれたGFPオープンリーディングフレームを含むcDNAを構築した。このcDNAを、ヒトRNAポリメラーゼIプロモーターおよびターミネーターを介して、人工セグメント、NP UTR-GFPの転写を駆動するそのpHH21ベクターにクローン化した。293T細胞をNP UTR-GFPでトランスフェクトした。トランスフェクションの24時間後に、細胞をインフルエンザA型ウイルスに感染させた。感染の18時間後、細胞をGFP発現につき分析した。多量のGFPが、ウイルス感染細胞の100%に存在した (図5A;黒色、満たされたヒストグラム)が、偽感染させた細胞は、ほとんどないまたは全くGFPを発現しなかった(図5A;白色、破線ヒストグラム)。これらのデータは、NP UTR-GFPを発現している細胞におけるGFPの感染依存性の生成を示す。この感染依存性の方法は、ウイルス検出系を可能とする。
【0048】
このレポーター遺伝子構築体が、感染に独立したプロセスにおいて機能できるかを決定するために、293T細胞を、pNP-UTR-GFPと共に、RNA複製および転写(PA、PB1、PB2およびNP)に必要なインフルエンザORFを発現するプラスミドでトランスフェクトした。外因性のORFを含まないプラスミドは、対照としてトランスフェクションに含まれた。4つのウイルスのポリメラーゼ蛋白質のすべてをコードするcDNAでトランスフェクトした細胞の中で、GFPは、細胞の約35%で検出され(図5B、黒色、満たされたヒストグラム)、一方、4つのウイルスのポリメラーゼ発現プラスミドのうちの3つだけでトランスフェクトした細胞は、GFPのかなりの発現を示さなかった(図5B、白色、破線ヒストグラム)。これらのデータは、NP-UTR-GFPからのGFPレポーター遺伝子活性が感染に独立したアッセイにおいて検出可能であることを示す。かくして、図3におけるデータは、ポリメラーゼが感染またはトランスフェクションにより送達されるかのウイルスRNA依存性RNAポリメラーゼ活性を定量するためのツールとしてのインフルエンザA型ウイルスUTRにより隣接して挟まれたレポーター遺伝子の使用を示す。
【0049】
実施例6
この実施例は、インフルエンザC型ウイルスではなくインフルエンザA型ウイルスおよびインフルエンザB型ウイルスによるレポーター遺伝子発現の誘導を示す。
【0050】
インフルエンザA型ウイルスNP-UTR-GFP転写体が、オルソミクソウイルスの他のメンバーのポリメラーゼ複合体により認識できるかを決定するために、pHH21NPUTR-GFPトランスフェクトした細胞を、インフルエンザA型(WSN/33株)、インフルエンザB型(Yamagata/59株)またはインフルエンザC型(Jerusalem/99株)に感染させた。インフルエンザA型/WSN/33の感染細胞は、99%を超えるGFPポジティブ細胞を示し、一方、インフルエンザB/Yamagata/59はわずか9.8%のGFPポジティブ細胞を示した。さらに、GFPポジティブインフルエンザA/WSN/33-感染細胞における平均チャネル蛍光は2963であり、一方、インフルエンザB/Yamagata/59-感染細胞における対応する集団は、わずか88の平均チャネル蛍光を有した。これらのデータは、GFPレポーターがインフルエンザA型ウイルス感染細胞において高度に発現されるが、インフルエンザB型ウイルス感染細胞においてはそれが単にわずかに発現されることを示す。偽感染およびンフルエンザC型感染した培養物の双方は、2.5%のGFPのポジティブ細胞を有し、それはそのレポーターがインフルエンザC型感染細胞において発現されなかったことを示す。これは、NP UTR GFP転写体がインフルエンザC型ウイルスポリメラーゼ複合体により認識も転写もされなかったことを示唆する。このアッセイを診断的に用いて、生物学的試料、例えば、インフルエンザの症状を示すヒト患者からの血液試料がインフルエンザA型ウイルス、インフルエンザB型ウイルスまたはどちらでもないウイルスを含むかどうかを決定できる。
【0051】
本発明の範囲を逸脱することなく、種々の変更が、前記の方法および組成物に行うことができるので、前記の記載に含まれたすべての内容は例示として解釈され、限定する意味に解釈されるものではないことを意図する。
【0052】
本明細書に引用されたすべての引用文献は、ここに出典明示してその全てを本明細書の一部とみなす。本明細書の引用文献の説明は、それらの著者によりなされた主張を単に概括することを意図し、いずれの参考文献も特許性に関連する先行技術を構成するという承認はなされていない。出願人は、引用された参考文献の正確さおよび適切さを試みるための権利を留保する。
【図面の簡単な説明】
【0053】
【図1】図1は、プラスミドpHH21 NP UTR HIルシフェラーゼの地図を示す。
【図2】図2は、人工セグメントを用いるインフルエンザA型ウイルス検出を示す。
【図3】図3は、インフルエンザA型ウイルス検出手順のブロックダイヤグラムを示す。
【図4】図4は、インフルエンザA型ウイルスのセグメントからのUTRおよびルシフェラーゼORFを含む人工セグメントを含む遺伝子改変された細胞によるホタルルシフェラーゼ発現を示す。
【図5】図5は、インフルエンザA型ウイルスのセグメントからのUTRおよびGFP ORFを含む人工セグメントを含む遺伝子改変された細胞による緑色蛍光タンパク質発現を示す。
【発明の詳細な説明】
【0001】
発明の分野
本願は、一般的にウイルス学の分野、より詳細には、生物学的試料中のマイナス鎖の分節化されたRNAウイルスを検出および定量する方法に関する。また、本願は、かかる方法に用いるために改変された細胞に関する。
【0002】
背景
マイナス鎖RNAウイルスは、ヒトおよび動物において多数の疾患の原因である。かくして、生物学的試料におけるマイナス鎖RNAウイルスの検出および定量化は、臨床医および研究者の双方に重要である。
【0003】
マイナス鎖RNAウイルスは、分節化されたRNAまたは分節化されていないRNAを含むことができる。分節化されていないマイナス鎖RNAウイルスにおいて、全ウイルスゲノムは、単一のネガティブセンスRNA分子により含まれるが、分節化されたマイナス鎖RNAウイルスにおいて、そのウイルスゲノムは複数のネガティブセンスのRNAセグメントにより含まれる。分節化されたマイナス鎖RNAウイルスのうち、Arenaviridae、BunyaviridaeおよびOrthomyxoviridaeは、各々、2、3および6〜8個のセグメントを含む。vRNAの各セグメントは、5'から3'の順序にて、ウイルスポリペプチドをコードする少なくとも1つのアンチセンスのオープンリーディングフレーム配列の5'非翻訳部位(「UTR」)、および3'UTRを含む。UTRは、RNA依存性RNAポリメラーゼのごとき分節化されたマイナス鎖RNA複製および/または遺伝子発現と関連した少なくとも1つの蛋白質により認識され、および/またはそれに結合する。セグメントによりコード化された蛋白質の発現は、セグメントが5'および3'末端配列を正確に規定することを必要とすることは今や一般的に受け入れられている。
【0004】
Olivoらの米国特許第5,591,579号および第5,851,757号は、生物学的試料における分節化されていないRNAウイルスの存在を検出するための細胞系および方法を開示する。その細胞系は、細胞内DNA依存性RNAポリメラーゼにより認識できるプロモーターを含むDNA分子で形質転換される。構造上不完全なRNAウイルスのcDNAの転写を指令できるプロモーターは、レポーターcDNAに連結される。ウイルス酵素のある種のトランス活性化に寄与する活性関連ウイルスが供されない限りは、DNA依存性RNAポリメラーゼにより転写されたRNA分子は、レポーターcDNAの翻訳を引き起こすことができない。
【0005】
Olivoらの米国特許第6,270,958号は、生物学的試料中の分節化されていないマイナス鎖RNAウイルスの検出のための診断アッセイを開示する。その開示されたアッセイにおいて、遺伝子改変された細胞は、マイナス鎖RNAウイルスのミニゲノムまたはミニアンチゲノムを発現する。ミニゲノムまたはミニアンチゲノムは、レポーター遺伝子またはその相補体、およびウイルスRNAからのフランキング配列を含む。また、該細胞は、各々がウイルスのヌクレオカプシド蛋白質の発現を指令するプロモーターを含む組換えDNA配列を含む。これらの細胞系におけるレポーター遺伝子の発現は、ヌクレオカプシド蛋白質の存在ならびにウイルスでの細胞の感染に依存する。
【0006】
分節化されたマイナス鎖RNAウイルスのセグメントによりコード化されたポリペプチドの発現が、正確に規定された3'および5'末端配列を必要とするために、研究者は、RNAポリメラーゼI(Pol I)の生化学的特性を利用して、分節化されたマイナス鎖RNAウイルスのセグメントのcDNAの発現を調べた。Pol Iは、正確なヌクレオチド配列にて転写を開始および終了すると考えられる。この点に関してZobelら, Nucleic Acids Research 21: 3607-3614,1993およびNeumannら, Virology 202: 477-479,1994は、マウスリボソームRNA(rDNA)遺伝子に由来するPol I(「Pol Iプロモーター」)により認識されるプロモーターを含む組換えDNA構築体を生成した。これらの組換えDNA構築体におけるPol Iプロモーターは、Pol I転写開始部位、インフルエンザウイルスRNAセグメントの5’非翻訳領域(UTR)、クロラムフェニコールアセチル基転移酵素(CAT)コード配列に相補的な配列(赤血球凝集素cDNAを置き換える)、および3’UTRを含むインフルエンザウイルスセグメント、ならびにPol Iポリメラーゼにより認識される転写ターミネーター配列(Pol Iターミネーター)を含む作動可能に連結したDNA配列の転写(恐らくPol Iによる)を指令する。このDNA構築体でトランスフェクトしたインフルエンザウイルスに感染した細胞は、CAT活性を発現し、これはウイルスでコード化されたRNA依存性RNAポリメラーゼは、テンプレートとしてPol I転写体を用いることができ、その結果、翻訳可能なmRNAが活性CAT酵素をコードすることを示す。
【0007】
これらおよび同様の試験が、インフルエンザウイルスセグメントのcDNAを含む組換えDNAを用いて、インフルエンザウイルスの完全な生成に導いた。この連結において、Neumannら、 Proceedings of the National Academy of Sciences USA 96: 9345-9350,1999,およびHoffmannら, Proceedings of the National Academy of Sciences USA 97: 6108-6113,2000は、クローン化されたcDNAからの完全なインフルエンザA型ウイルスの生成についての「逆遺伝学」DNA感染系を開示する。これらの試験において、細胞は、第1のシリーズの、各々がvRNAセグメントのcDNAに作動可能に連結したPol Iプロモーターを含むDNA配列を含むプラスミド、ならびに第2のシリーズの、各々がウイルス蛋白をコードするcDNAに作動可能に連結されたPol IIプロモーターを含むプラスミドでトランスフェクトされた。この系において転写されたウイルス蛋白質は、ウイルスの核タンパク質(NP)およびポリメラーゼ複合蛋白質(蛋白質PA、PB1およびPB2を含む)を含んだ。
【0008】
発明の概要
本発明は、分節化されたマイナス鎖RNAウイルスを含むことが疑われる生物学的試料における少なくとも1つの分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出または決定するための組成物および方法を包含する。また、本発明は、分節化されたマイナス鎖RNAウイルスに対して活性な抗ウイルス化合物をスクリーニングするための組成物および方法を包含する。
【0009】
1つの具体例において、本発明は、あるポリペプチドをコードするレポーター遺伝子を含む組換えRNA分子を含む、遺伝子改変された脊椎動物細胞を提供する。好ましい具体例において、脊椎動物細胞は哺乳動物細胞で有り得る。ある具体例において、細胞は、存在、不存在または量の検出または決定が求められる分節化されたマイナス鎖RNAウイルスのウイルスRNA依存性RNAポリメラーゼを含まない。レポーター遺伝子を含む翻訳可能なmRNAの形成は、分節化されたマイナス鎖ウイルスのRNA依存性RNAポリメラーゼの細胞へのその導入を必要とする。
【0010】
結果的に、もう一つの具体例において、本発明は、分節化されたマイナス鎖RNAウイルスを含むことが疑われる生物学的試料中の分節化されたマイナス鎖ウイルスの存在、不存在または量を検出または決定する方法を提供する。該方法は、生物学的試料と前記のごとき組換えRNA分子を含む遺伝子改変された脊椎動物細胞とを接触させ、次いで、レポーター遺伝子によりコード化されたポリペプチドの不存在、存在または量を検出することを含む。
【0011】
もう一つの具体例において、本発明は、発現が分節化されたマイナス鎖ウイルスのウイルスRNA依存性RNAポリメラーゼの細胞内の存在に依存するレポーター遺伝子を含む組換えDNA分子を含む遺伝子改変された脊椎動物細胞を提供する。かくして、また、本発明は、DNA分子を含む細胞内の発現が分節化されたマイナス鎖ウイルスのウイルスRNA依存性RNAポリメラーゼの細胞内の存在に依存するレポーター遺伝子を含む単離された組換えDNA分子を提供する。
【0012】
結果的に、もう一つの具体例において、本発明は、分節化されたマイナス鎖RNAウイルスを含むことが疑われる生物学的試料における分節化されたマイナス鎖ウイルスの存在、不存在または量を検出または決定するもう一つの方法を提供する。該方法は、生物学的試料と、前記のごとき組換えDNA分子を含む遺伝子改変された脊椎動物細胞とを接触させ、次いで、レポーター遺伝子によりコード化されたポリペプチドの不存在、存在または量を検出することを含む。
【0013】
もう一つの具体例において、本発明は、生物学的試料における、第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出または決定するための遺伝子改変された脊椎動物細胞を提供する。この具体例の細胞は、発現が第1の分節化されたマイナス鎖RNAウイルスのウイルスRNAの細胞内の存在に依存する第1のレポーター遺伝子を含む第1の組換えRNA分子、ならびに発現が第2の分節化されたマイナス鎖RNAウイルスのウイルスRNAの細胞内の存在に依存する第2のレポーター遺伝子を含む第2の組換えRNA分子を含む。
【0014】
結果的に、もう一つの具体例において、本発明は、生物学的試料における、第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出または決定する方法を提供する。該方法は、生物学的試料と、前記のごとき第1の組換えRNA分子および第2の組換えRNA分子を含む遺伝子改変された脊椎動物細胞とを接触させ、次いで、第1のレポーター遺伝子によりコード化された第1のポリペプチドの不存在、存在または量、ならびに第2のレポーター遺伝子によりコード化された第2のポリペプチドの不存在、存在または量を検出することを含む。
【0015】
もう一つの具体例において、本発明は、生物学的試料における、第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出または決定するための遺伝子改変された脊椎動物細胞を提供する。この具体例の細胞は、発現が第1の分節化されたマイナス鎖RNAウイルスのウイルスRNAの細胞内の存在に依存する第1のレポーター遺伝子を含む第1の組換えDNA分子、ならびに発現が第2の分節化されたマイナス鎖RNAウイルスのウイルスRNAの細胞内の存在に依存する第2のレポーター遺伝子を含む第2の組換えDNA分子を含む。
【0016】
結果的に、もう一つの具体例において、本発明は、生物学的試料における、第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出または決定する方法を提供する。該方法は、生物学的試料と、前記のごとき第1の組換えDNA分子および第2の組換えDNA分子を含む遺伝子改変された脊椎動物細胞とを接触させ、次いで、第1のレポーター遺伝子によりコード化される第1のポリペプチドの不存在、存在または量ならびに第2のレポーター遺伝子によりコード化される第2のポリペプチドの不存在、存在または量を検出することを含む。
【0017】
さらにもう一つの具体例において、本発明は、インフルエンザウイルスを含むことが疑われる生物学的試料において、インフルエンザA型ウイルス、インフルエンザB型ウイルスまたはどちらでもないウイルスの感染の間の鑑別診断を決定する方法を提供する。該方法は、生物学的試料と、遺伝子改変された細胞とを接触させることを含み、ここに、実質的に複数の遺伝子改変された細胞は、インフルエンザA型ウイルスのセグメントの少なくとも1つのUTRを含む組換えRNA分子、およびレポーター遺伝子を含む。この具体例において、レポーター遺伝子ポリペプチド発現は、インフルエンザA型またはインフルエンザB型ウイルスのウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存する。その細胞内のレポーター遺伝子発現の定量分析は、生物学的試料のウイルス含量:インフルエンザA型ウイルスを含む生物学的試料と接触した細胞集団は、感染していない対照培養物のそれより実質的に大きい量のレポーター遺伝子産物を生成し、一方、インフルエンザB型ウイルスを含む生物学的試料と接触した細胞集団は、感染していない対照培養物のそれよりわずかに大きな量のレポーター遺伝子産物を生成し、一方、病原体ウイルスを含まないか、またはインフルエンザA型ウイルスもしくはインフルエンザB型ウイルス以外の病原体を含む生物学的試料と接触した細胞集団は、感染していない対照培養物のそれほど著しく大きくない量のレポーター遺伝子産物を生成することを明らかにする。
【0018】
もう一つの具体例において、本発明は、包装化または使用説明書、および単離組換えDNA分子を含み、単離組換えDNA分子が蛋白質発現が分節化されたマイナス鎖ウイルスのウイルスRNA依存性RNAポリメラーゼの細胞内の存在に依存するレポーター遺伝子を含むキットを提供する。
【0019】
もう一つの具体例において、本発明は、分節化されたマイナス鎖RNAウイルスに指向された抗ウイルス薬についての定量的スクリーニング方法を提供する。該方法は、候補抗ウイルス化合物と、前記のごときレポーター遺伝子を含む人工セグメントを含む遺伝子改変された細胞とを接触させ、ここに、その細胞は、ウイルスRNA依存性RNAポリメラーゼを含むウイルスタンパク質複合体も含み、次いで、そのレポーター遺伝子ポリペプチドの発現レベルを測定することを含む。ある具体例において、タンパク質複合体は、細胞により含まれた複数のプラスミドの発現により提供でき、各プラスミドは、タンパク質複合体を含む少なくとも1つのポリペプチドをコードする。もう一つの具体例において、タンパク質複合体は、分節化されたマイナス鎖RNAウイルスによる細胞の感染により提供できる。候補抗ウイルス化合物の効力は、その化合物と接触した細胞(または細胞集団)内のレポーター遺伝子ポリペプチドの発現レベルを、その化合物と接触していない対照の細胞または細胞集団のそれと比較することにより決定される。
【0020】
もう一つの具体例において、本発明は、分節化されたマイナス鎖RNAウイルスに指向される抗ウイルス薬についての定量的スクリーニング方法を提供する。該方法は、候補抗ウイルス化合物と、ウイルスに感染した細胞とを接触させ;次いで、その細胞により生成されたウイルスの存在、不存在または量を検出することを含む。その検出は、前記のごとく、レポーター遺伝子を含む人工セグメントを含む遺伝子改変された細胞(または細胞の集団)と、候補抗ウイルス化合物と接触した細胞により生成されたウイルスとを接触させることを含む。候補抗ウイルス化合物の効力は、遺伝子改変された細胞または細胞集団によるレポーター遺伝子ポリペプチドの発現レベルを、候補抗ウイルス化合物と接触していない感染細胞により生成されたウイルスと接触した対照の細胞または細胞集団のそれと比較することにより決定される。
【0021】
詳細な記載
本発明により、分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出する方法および組成物が提供される。本発明は、遺伝子改変された細胞により供された分節化されたマイナス鎖RNAウイルスの人工セグメントが、細胞が分節化されたマイナス鎖RNAウイルスのRNA依存性RNAポリメラーゼをさらに含む場合、レポーター遺伝子を含む翻訳可能なmRNAの形成にアクセス可能であるという発明者らの観察に基づいている。ある具体例において、遺伝子改変された細胞は、RNA依存性RNAポリメラーゼのポリペプチド成分をコードする1以上の核酸をさらに含む細胞に基づくRNA依存性RNAポリメラーゼを含む。いくつかの具体例において、RNA依存性RNAポリメラーゼをコードする核酸は、例えば、分節化されたマイナス鎖RNAウイルスでの細胞の感染により供されるウイルスゲノムRNAとなることができる。レポーター遺伝子を含む翻訳可能なmRNAの形成は、感染に先立ち、細胞内のウイルスヌクレオカプシド蛋白質の不存在下でさえ生じることができる。他の具体例において、RNA依存性RNAポリメラーゼをコードする核酸は、1以上の組換えDNA分子になることができる。組換えDNA分子は、例えば、プラスミドまたは複数のプラスミドのごとき、例えば、ベクターまたは複数のベクターで有り得、ここに、そのベクターは、RNA依存性RNAポリメラーゼを含むウイルスタンパク質複合体に寄与するウイルスのポリペプチドをコードするcDNAを含む。組換えDNA分子は、ウイルスのヌクレオカプシド蛋白質のcDNAを含む1以上のベクターをさらに含むことができる。加えて、本発明は、レポーター遺伝子の産物の発現が、分節化されたマイナス鎖RNAウイルスのRNA依存性RNAポリメラーゼの不存在下、バックグラウンドレベルを著しく超えて検出されないという発明者らの観察に基づいている。宿主細胞にRNA依存性RNAポリメラーゼを導入する方法が、分節化されたマイナス鎖RNAウイルスにより細胞の感染を含むために、該方法および組成物は、生物学的試料における1以上の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出または診断することを可能とする。
【0022】
本明細書に記載された方法および組成物は、当業者によく知れた実験技術を利用し、Sambrook, J. , Fritsch, E. F. and Maniatis, T. (1989) Molecular Cloning: A Laboratory Manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NYのごとき実験マニュアルに見出すことができる。
【0023】
本発明による遺伝子改変された細胞は、組換え核酸を含む脊椎動物細胞であり、検出すべき分節化されたマイナス鎖RNAウイルスによる感染に感受性である。分節化されたマイナス鎖RNAウイルスは、例えば、アレナウイルス、ブンヤウイルスまたはオルトミクソウイルスで有り得る。オルトミクソウイルスは、例えば、インフルエンザA型ウイルス、インフルエンザB型ウイルスまたはインフルエンザC型ウイルスで有り得る。本発明の遺伝子改変された細胞は、予め形成された人工セグメントRNAからの翻訳可能なレポーター遺伝子mRNAの効率的な合成に必要な蛋白質が発現されるように、感染させる分節化されたマイナス鎖RNAウイルスのゲノムが、細胞に入ることができ、その複製サイクルにおいて少なくともはるかに十分に進むことができるならば、感染に感受性である。好ましくは、分節化されたマイナス鎖RNAウイルスでの細胞の感染は、十分に増殖性であり、すなわち、子孫ウイルスが、感染細胞により生成され、次いで、細胞から細胞へ広がる。さらに、ある具体例において、遺伝子改変された細胞は、好ましくは、人工セグメントが組換えDNA鋳型から転写できるように、(例えば、ヒトまたはマウス起源の)Pol Iプロモーターが機能するものである。特別の分節化されたマイナス鎖RNAウイルスによる感染に感受性の細胞系が、公知の感受性細胞系についての文献を検索することにより、および/または単にルーチン実験を必要とするよく知られた手順を用いる候補細胞系をスクリーニングすることにより容易に同定できると当業者ならば理解するであろう。分節化されたマイナス鎖RNAウイルスに感受性の細胞の非限定の例は、以下を含む:293T腎臓、LA−4、MRC5、A549、HEp2、2fTGHおよびSTATI−欠損2fTGH(U3A)のごときヒト細胞、ならびにベイビーハムスター腎臓(BHK)、CV−1、ベロ(Vero)、LLC−MK1、サル初代腎臓およびチャイニーズハムスター卵巣(CHO)のごとき非ヒト細胞。ヒト293T細胞は、ヒトを含めた哺乳動物を感染させる分節化されたマイナス鎖RNAウイルスを検出するための宿主細胞としての使用のために好ましい。感受性細胞は当該技術分野においてよく知られ、American Type Culture Collection, 10801 University Boulevard, Manassas, VA 20110-2209のごときソースから入手可能である。本明細書に記載された組換えDNA分子を含む本発明の遺伝子改変された細胞は、安定的にトランスフェクトした、あるいは一過性にトランスフェクトした細胞で有り得る。また、ある態様において、細胞により含まれた組換えDNAは、ベクター、好ましくはプラスミドにより含むことができる。また、ベクターは、当該技術分野においてよく知られた方法により用いられる、例えば、選択マーカーまたは複製開始点のごとき、組換えDNA分子の選択、維持または増殖に有用な他の配列を含むことができる(例えば、Sambrookら、前記)。
【0024】
また、本発明は、分節化されたマイナス鎖RNAウイルスの人工セグメントまたはその相補体を含む遺伝子改変された脊椎動物細胞を提供する。本発明の人工セグメントは、分節化されたマイナス鎖RNAウイルスのセグメントの5'UTR、レポーター遺伝子のオープンリーディングフレーム、および分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRを含む一本鎖RNAである。好ましくは、少なくとも1つの、好ましくは、双方の5'UTRおよび3'UTRは、その存在、不存在または量が、生物学的試料において検出されるべきある種の分節化されたマイナス鎖RNAウイルスからのものである。その5'UTRおよび3'UTRは、同一または異なるセグメントからのもので有り得る。好ましくは、その5'UTRおよび3'UTRは、その存在、不存在または量が生物学的試料において検出されるべき分節化されたマイナス鎖RNAウイルスの同一セグメントからのものである。好ましくは、人工セグメントのレポーター遺伝子のオープンリーディングフレームは、アンチセンス配置にあって、人工セグメントの相補体のレポーター遺伝子のオープンリーディングフレームは、センス配置にある。ウイルスRNA依存性RNAポリメラーゼの不存在下、宿主細胞は、翻訳可能なウイルスmRNA分子を提供も、そのセグメントに相補的なRNAを合成することもできない。しかしながら、ウイルスRNA依存性RNAポリメラーゼの存在下、宿主細胞は、翻訳可能なウイルスmRNA分子を提供することができ、鋳型として相補鎖を用いて、各ケースにおいて、人工セグメントおよびその相補体の両方を合成も複製もできる。ウイルスRNA依存性RNAポリメラーゼを含む宿主細胞において、人工セグメントのUTRを認識し、ウイルスRNA依存性RNAポリメラーゼにより転写開始部位として機能できる。人工セグメントにおいて、レポーター遺伝子のオープンリーディングフレームは、好ましくはアンチセンス配向にある。理論により拘束されることなくして、ウイルスRNA依存性RNAポリメラーゼのごとき1以上のウイルス蛋白質は、その人工セグメントのUTRを認識し、それに結合すると考えられる。ウイルス蛋白質は、宿主細胞の感染に際してウイルスにより宿主細胞に導入できる。さらに、鋳型として人工セグメントを利用することにより、RNA依存性RNAポリメラーゼは、人工セグメント(「ポジティブセンス」鎖)に相補的なRNA鎖の合成を触媒し、ポジティブセンス鎖は、人工セグメントの合成用の鋳型として用いることができると考えられる。また、さらに、RNA依存性RNAポリメラーゼは、キャップされたポリアデニル化されたmRNAに処理できる人工mRNAの合成用鋳型としてポジティブセンス鎖を利用する(例えば、Mikulasovaら, Acta Virol. 44: 273-282,2000参照)。センス配向にてレポーター遺伝子配列を含むmRNAは、細胞による翻訳に利用可能である結果、レポーター遺伝子によりコード化されたポリペプチドを生成する。従って、分節化されたマイナス鎖RNAウイルスでの細胞の感染は、例えば、その細胞と診断テスト試料との接触により供されるウイルスが、ウイルスRNA依存性RNAポリメラーゼを含むものを含めたウイルスのポリペプチドの発現に導き、従って、レポーター遺伝子ポリペプチドの発現に導く。加えて、遺伝子改変された細胞または細胞集団により生成されたポリペプチドの量は、ウイルスのレベルを飽和させるまで細胞(または細胞集団)を感染させる、分節化されたマイナス鎖RNAウイルスの量に単調に関連し、それにより、生物学的試料中のウイルスレベルの定量化のための手段を提供する。レポーター遺伝子をコード化するmRNAが、ウイルスRNA依存性RNAポリメラーゼ不存在下で宿主細胞内で合成されないので、本発明の感染していない遺伝子改変された細胞のレポーター遺伝子ポリペプチドの量は、人工セグメントを含まない対照細胞のものほど著しく大きくない。
【0025】
その存在、不存在または量が検出されるべき分節化されたマイナス鎖RNAウイルスは、アレナウイルス、ブンヤウイルスまたはオルトミクソウイルスのごときいずれかの分節化されたマイナス鎖RNAウイルスで有り得、好ましくは、オルトミクソウイルス、より好ましくは、インフルエンザウイルスである。インフルエンザウイルスは、インフルエンザA型ウイルス、インフルエンザB型ウイルスまたはインフルエンザC型ウイルスで有り得、好ましくは、インフルエンザA型ウイルスまたはインフルエンザB型ウイルスである。好ましくは、分節化されたマイナス鎖RNAウイルスの不存在、存在または量を検出するために用いた人工セグメントは、インフルエンザウイルス、好ましくは、インフルエンザA型ウイルスのセグメントからの5'UTR、好ましくは、インフルエンザA型ウイルスのNPセグメントの5'UTRを含む。好ましくは、その人工セグメントは、インフルエンザウイルス、好ましくは、インフルエンザA型ウイルスのセグメントからの3'UTR、好ましくは、インフルエンザA型ウイルスのNPセグメントの3'UTRを含む。好ましくは、その5'UTRおよび3'UTRは、同一ウイルス、好ましくは、インフルエンザA型ウイルスからのものであり、好ましくは、双方がインフルエンザA型ウイルスのNPセグメントからのものである。
【0026】
本発明のレポーター遺伝子は、細胞内で発現した場合、検出可能な産物をコードするいずれかの核酸配列(DNAまたはRNA)である。レポーター遺伝子は、例えば、cDNA配列、ゲノムDNA配列または合成配列のごときオープンリーディングフレームを含むDNAまたはRNA配列で有り得る。また、レポーター遺伝子は、イントロンを含むことができる。レポーター遺伝子核酸配列におけるイントロンの存在は、ウイルスRNA依存性RNAポリメラーゼでの細胞の刺激に際して、宿主細胞内でコード化されたポリペプチドの発現レベルを増加できると考えられる。レポーター遺伝子産物は、好ましくは、細胞内で容易にアッセイまたは検出できるポリペプチドである。ポリペプチドのアッセイまたは検出は、例えば、レポーター遺伝子ポリペプチドに指向された抗体を用いる免疫検出;例えば、蛍光発生的、色素生産性、化学発光もしくは放射性の基質を用いる酵素活性検出;または蛋白質蛍光のごときいずれかの公知の検出方法によることができる。レポーター遺伝子産物の非限定の例は、クロラムフェニコールアセチルトランスフェラーゼ、β-ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質(GFP)および分泌アルカリフォスファターゼを含む。好ましくは、レポーター遺伝子産物は、緑色蛍光タンパク質のごとき本質的に蛍光性であるポリペプチドであるか、またはホタルルシフェラーゼのごとき化学発光の基質が利用可能である酵素である。好ましくは、ポリペプチドの不存在、存在または量の検出は、そのポリペプチドまたはその基質により発光した光量子シグナルの不存在、存在または量の検出を含む。光量子シグナルは、例えば、緑色蛍光タンパク質のごとき蛍光蛋白質からの蛍光発光、または例えば、ホタルルシフェラーゼまたはウミシイタケルシフェラーゼのごとき化学発光の基質が利用できる酵素からの化学発光で有り得る。光シグナルの検出または定量化は、当該技術分野において知られたいずれかの検出または定量化の方法、例えば、蛍光顕微鏡法、フローサイトメトリーによることができる。「緑色蛍光蛋白質」なる用語は、Aequorea victoria (Chalfieら, Science, 263:802-805, 1994)からのGFPの自然発生野生型配列、ならびに蛍光変異体およびその誘導体(色に拘わらず)、例えば、Tsienらへの米国特許第6,054,321号、Chalfieらへの米国特許第6,146、826号、Michaelsへの米国特許第6,096、865号に記載された蛍光蛋白質を含むもののごとき蛍光ポリペプチドを含む。ルシフェラーゼなる用語は、ホタルルシフェラーゼおよびウミシイタケルシフェラーゼのごとき自然発生の化学発光基質を有する酵素を含む。
【0027】
ある具体例において、遺伝子改変された細胞は、人工セグメントをコードする組換えDNA配列を含む。遺伝子改変された細胞は、当該技術分野において知られたいずれかの方法により生成できる。いくつかの具体例において、遺伝子改変された細胞は、本発明の組換えDNAでの細胞のトランスフェクションにより生成できる。トランスフェクションは、当該技術分野においてよく知られた方法を用いて達成できる。好ましくは、トランスフェクションは、TransITR-LT1トランスフェクション試薬(Mirus Corporation, Madison, WI)のごときトランスフェクション剤を用いて達成される。組換えDNAは、人工セグメントをコード化する組換えDNA配列を含む単離組換えDNA分子として提供できる。単離組換えDNA分子は、組換えプラスミドまたは組換えウイルスのごときベクターを含むことができる。組換えウイルスは、例えば、組換えバクテリオファージまたは組換え脊椎動物ウイルスで有り得る。
【0028】
ある具体例において、これらの具体例の組換えDNA分子は、5'から3'の順序にて、DNA依存性RNAポリメラーゼ用のプロモーター、DNA依存性RNAポリメラーゼ用の転写開始部位、人工セグメントまたはその相補体をコード化するDNA配列、ならびにDNA依存性RNAポリメラーゼ用の転写終結部位を含む。組換えDNA配列の転写のための本発明の遺伝子改変された細胞により用いられたDNA依存性RNAポリメラーゼは、細胞に内因的なDNA依存性RNAポリメラーゼ、または例えば、Olivoらへの特許第6,270,958号に開示されたDNA依存性RNAポリメラーゼのごとき異種のDNA依存性RNAポリメラーゼで有り得る。好ましくは、そのDNA依存性RNAポリメラーゼは、細胞に内因的なDNA依存性RNAポリメラーゼであり、より好ましくは、RNAポリメラーゼIである。従って、そのプロモーターは、脊椎動物細胞内で機能できるいずれかのRNAポリメラーゼ用のプロモーターで有り得、好ましくは、細胞に内因的なRNAポリメラーゼ用のプロモーター、好ましくは、RNAポリメラーゼI用のプロモーターである。プロモーターは、好ましくは、Pol I転写開始部位である転写開始部位に作動可能に連結される。Pol I転写開始部位は、Pol Iが、例えば、配列5'-CCAATAA-3'のごとき転写を開始するために認識できるいずれかの配列で有り得る。この配列がPol Iプロモーターに続く場合、転写は、この配列の直ぐ3'のヌクレオチドにて開始されると考えられる。Pol Iが正確に規定された配列にて転写を開始するために、Pol I転写開始部位は、好ましくは、DNAの転写体が、5'末端が人工セグメントをコードするcDNAの5'UTRの5'端と正確にまたはその5'端の約10のヌクレオチド内で一致する人工セグメントを含むにように配置される。好ましくは、5'末端は、5'UTRの5'端と正確に一致する。別法として、人工セグメントがプロモーターに関してポジティブセンスの配置にあるならば、DNAの転写体は、5'末端が人工セグメントをコードするcDNAの3'UTRの相補体の5'端と正確にまたはその5'端の約10のヌクレオチド内で一致する人工セグメントを含む。好ましくは、5'末端は、3'UTRの5'端と正確に一致する。人工セグメントをコードするcDNAは、分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRのcDNA、ポリペプチドをコードするレポーター遺伝子のオープンリーディングフレームのDNA配列、および分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRのcDNAを含む。レポーター遺伝子のオープンリーディングフレームのDNA配列は、組換えDNA分子により含まれたプロモーターに関してセンスまたはアンチセンス配置のいずれかで有り得、好ましくはプロモーターに関してアンチセンス配置にある。人工セグメントをコード化する配列は、プロモーターに関してセンスまたはアンチセンス配向に有り得る。かくして、レポーター遺伝子DNA配列は、プロモーターに関してセンスまたはアンチセンス配向のいずれかでも有り得る。さらに、いくつかの具体例において、レポーター遺伝子DNA配列もイントロンを含むことができる。
【0029】
組換えDNA分子はさらに転写終結部位を含む。「転写終結部位」は、例えば、転写終了のためのDNA依存性RNAポリメラーゼ、自己切断リボザイムのごとき触媒的なRNA配列をコード化する配列または直線DNAの末端配列のごとき規定された3’末端を有するRNA配列に転写または処理できるいずれかのDNA配列を含むように本明細書に規定される。転写終結部位は、好ましくは、RNAポリメラーゼIにより認識された転写終結部位である。好ましい具体例において、Pol I転写終結部位は、配列5'-GGGGGG-3'により描写できる。別法の具体例において、転写終結部位は、自己切断リボザイムのごとき触媒的なRNA配列をコードする配列を含むことができる。本発明の転写終結部位は、転写に際して、DNAから転写されたRNAの3'末端が人工セグメントの3'UTRの3'端に正確にあるか、またはその3’端の約10ヌクレオチド内にあるようにDNA配列内に位置する。好ましい具体例において、転写に際して、DNAから転写されたRNAの3'末端は、正確に3'UTRの3'端にある。別法として、人工セグメントがプロモーターに関してポジティブセンスの配置にあるならば、本発明の転写終結部位は、転写に際して、DNAから転写されたRNAの3'末端が、人工セグメントの相補体の5'UTRの相補体の3'端に正確にあるか、またはその3'端の約10ヌクレオチド内にあるようにDNA配列中に位置する。好ましい具体例において、そのDNAから転写されたRNAの3'末端は、5'UTRの相補体の3'端に正確にある。
【0030】
ある具体例において、組換えDNA分子は、例えば、組換えプラスミドまたは組換えウイルスのごときベクターである。好ましくは、ベクターは組換えプラスミドである。プラスミドの非限定の例は、pHH21NPUTR-lucおよびpHH21NPUTR-GFPを含み、ここに、各々、DNA依存性RNAポリメラーゼ用のプロモーターはPol Iプロモーターであり、そのUTRはインフルエンザA型ウイルスのNPセグメントからのもであって、レポーター遺伝子はホタルルシフェラーゼおよび緑色蛍光タンパク質をコードする。かかるベクターは、当業者によく知られた材料および方法を用いて構築できる(例えば、Sambrookら、前記)。非限定の例において、かかるベクターは、プラスミドプラスミドpHH21のごときPol Iプロモーターを含むプラスミドを用いて構築できる(Neumann, G.ら, Proceedings of the National Academy of Sciences USA 96: 9345-9350,1999 ; Neumann, G.ら, Virology 202: 477-479,1994)。
【0031】
理論により拘束されることなくして、その配列構造のために、本発明のいくつかの具体例における組換えDNAは、Pol IのごときRNAポリメラーゼにより転写されて、分節化されたマイナス鎖ウイルスのセグメントまたはセグメント群のものに同一またはほとんど同一である5'および3'末端配列を有するRNA転写体を形成し、それにより、人工セグメントを形成すると考えられる。従って、人工セグメントは、生物学的試料により含まれたウイルスによる感染に先立ち細胞内に蓄積する。分節化されていないウイルスの検出方法と異なり、本発明の方法による分節化されたウイルスの検出は、ウイルス感染に先立ち、またはウイルス感染に加えて、ウイルス蛋白質の細胞内の発現を必要としない。しかしながら、遺伝子改変された細胞がヌクレオカプシド蛋白質のごときウイルス蛋白質をさらに含むならば、レポーターポリペプチドの発現レベルは増強できる。例えば、かかるウイルス蛋白質は、ウイルス蛋白質用のcDNAに作動可能に連結されたDNA依存性RNAポリメラーゼ(RNAポリメラーゼIIのごとき)用のプロモーターを各々含む1以上のプラスミドでの細胞のトランスフェクションを介して細胞に供することができる。
【0032】
もう一つの具体例において、本発明は、生物学的試料における第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出するための組成物および方法を提供する。この具体例の遺伝子改変された細胞は、2つの人工セグメントまたはそれらの相補体を含む。第1の人工セグメントは、第1の分節化されたマイナス鎖RNAウイルスの5'UTR、レポーターポリペプチドをコードする第1の核酸配列、および第1の分節化されたマイナス鎖RNAウイルスの第1の3'UTRを含む。第2の人工セグメントは、第2の分節化されたマイナス鎖RNAウイルスの5'UTR、レポーターポリペプチドをコードする第2の核酸配列、および第2の分節化されたマイナス鎖RNAウイルスの第2の3'UTRを含む。第1および第2の人工セグメントの双方について、レポーターポリペプチドをコードする核酸配列は、センスまたはアンチセンス配置で有り得、好ましくはアンチセンス配置にある。これらの具体例における細胞は、前記のごとき人工セグメントの転写用の配列に作動可能に連結したDNA依存性RNAポリメラーゼ用のプロモーターを各々含む第1の組換えDNA分子および第2の組換えDNA分子のいずれもさらに含むことができる。あるいは、単一の組換えDNA分子は、DNA依存性RNAポリメラーゼ用の2つのプロモーターを含むことができ、各プロモーターは、前記のごとき2つの人工セグメントのうちの1つの転写のための配列に作動可能に連結される。これらの具体例において、ウイルスは、非限定の例において、アレナウイルス、ブンヤウイルス、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスから選択された1対のウイルスのごときいずれかの1対の分節化されたマイナス鎖ウイルスで有り得る。2つのウイルスを含む生物学的試料との接触に際して、レポーター遺伝子を含むmRNAは各ウイルスの量に比例して形成される。従って、レポーター遺伝子は比例して発現され、生物学的試料のウイルス含量を反映する。例えば、この具体例の遺伝子改変された細胞は、インフルエンザA型からのUTRを含む第1の人工セグメント、およびインフルエンザB型からのUTRを含む第2の人工セグメントを含むことができ、人工セグメントは、異なったレポーター遺伝子を含む。その細胞と、インフルエンザA型およびインフルエンザB型ウイルスの双方を含む生物学的試料との接触に際して、細胞により作成されたレポーター遺伝子ポリペプチドの量は、その標本におけるウイルス量に比例するであろう。好ましくは、これらの具体例のレポーター遺伝子によりコード化されたポリペプチドは、光発光ポリペプチドであるか、または光発光基質が利用可能である酵素、好ましくは、ホタルルシフェラーゼ、ウミシイタケルシフェラーゼおよび緑色蛍光タンパク質よりなる群から選択される酵素である。
【0033】
さらにもう一つの具体例において、本発明は、インフルエンザウイルスを含むことが疑われる生物学的試料における、インフルエンザA型ウイルス、インフルエンザB型ウイルスまたはどちらでもないウイルスの感染の間の鑑別診断を決定する方法を提供する。該方法は、その生物学的試料と、遺伝子改変された細胞の集団とを接触させることを含み、ここに、実質的に複数の遺伝子改変された細胞は、インフルエンザA型ウイルスのセグメントの少なくとも1つのUTRおよびポリペプチドをコードするレポーター遺伝子を含む人工セグメントを含む。人工セグメントを含む「実質的に複数」の遺伝子改変された細胞は、人工セグメントを含む細胞の少なくとも約50%、好ましくは少なくとも約75%、より好ましくは少なくとも約90%で有り得る。レポーター遺伝子は、好ましくはアンチセンス配置にある。これらの細胞の人工セグメントの5'UTRおよび3'UTRの片方または双方は、好ましくは、インフルエンザA型ウイルスのセグメントのUTR、より好ましくはNPセグメントである。レポーター遺伝子ポリペプチドの発現は、インフルエンザウイルスA型ウイルスまたはインフルエンザB型ウイルスのウイルスRNA依存性RNAポリメラーゼの細胞内の存在に依存する。好ましい具体例において、レポーター遺伝子によりコード化されたポリペプチドは、光発光により検出できる蛋白質、例えば、緑色蛍光タンパク質またはルシフェラーゼ蛋白質である。これらの具体例において、細胞集団中のレポーター遺伝子発現の定量的測定は、生物学的試料のウイルス含量:インフルエンザA型ウイルスを含む生物学的試料と接触した細胞集団が、感染していない対照培養物のそれより実質的に大きな量のレポーター遺伝子産物を生成し、一方、インフルエンザB型ウイルスを含む生物学的試料と接触した細胞集団は、感染していない対照培養物よりわずかに大きな量のレポーター遺伝子産物を生成することを明らかにする。いくつかの具体例において、レポーター遺伝子発現の定量分析は、細胞集団で全体として発現されたレポーター遺伝子ポリペプチドの測定を含むことができる。もう一つの具体例において、定量分析は、例えば、レポーター遺伝子およびフローサイトメトリーまたは蛍光標示式細胞分取器を用いて、細胞ごとのベースでレポーター遺伝子ポリペプチドレベルを測定することを含むことができる。かくして、いくつかの具体例において、インフルエンザA型ウイルス、インフルエンザB型またはどちらでもないウイルスを含むことが知られた試料を用いて、これらのウイルスに応じた培養によりレポーター遺伝子発現レベルを確立できる。これらの発現レベルは、インフルエンザウイルスを含むことが疑われる生物学的試料により含まれたウイルスのタイプを決定するためのガイダンスを提供できる。例えば、レポーター遺伝子ポリペプチドを発現する細胞のパーセンテージは、そのウイルスのうちの1つに感染した対照培養物を用いて測定されたパーセンテージを反映するであろう。かくして、例えば、その細胞集団と接触した生物学的試料は、大多数の細胞中のレポーターポリペプチドの発現に導く、例えば、少なくとも約80%の細胞、または少なくとも約90%の細胞は、インフルエンザA型ウイルスを含むと診断でき、一方、その細胞集団と接触した生物学的試料は、少数の細胞、例えば、約10%の細胞におけるレポーターポリペプチドの発現に導き、インフルエンザB型ウイルスを含むと診断することができる。さらに、その細胞集団と接触した生物学的試料は、約3%未満の細胞におけるレポーターポリペプチドの発現に導き、どちらのウイルスも含まないと診断できる。
【0034】
もう一つの具体例において、本発明は、包装化および単離組換えDNA分子を含むキットを提供し、単離組換え分子は、その分子でトランスフェクトされた細胞の発現が分節化されたマイナス鎖ウイルスのウイルスRNAの細胞内の存在に依存するレポーター遺伝子を含む。この具体例において、単離組換えDNA分子は、5'から3'の順序にて、DNA依存性RNAポリメラーゼ用のプロモーター、DNA依存性RNAポリメラーゼ用の転写開始部位、分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRのcDNA、ポリペプチドをコードするレポーター遺伝子のオープンリーディングフレームのDNA配列、分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRのcDNAならびにDNA依存性RNAポリメラーゼ用の転写終結部位を含む。この具体例のレポーター遺伝子のオープンリーディングフレームのDNA配列は、センスまたはアンチセンス配向で有り得、好ましくはアンチセンス配向にある。また、この具体例のキットは、前記されたもののごとき分節化されたマイナス鎖RNAウイルスによる感染に感受性の細胞または細胞集団を含むことができる。その細胞または細胞集団は、単離組換えDNA分子で感染した遺伝子改変された細胞で有り得る。あるいは、その細胞または細胞集団は、非トランスフェクトで有り得る。キットの使用者は、当業者によく知られた標準的な方法を用いて、そのDNAで細胞または細胞集団をトランスフェクトし、次いで、そのトランスフェクトした細胞を用いて、分節化されたマイナス鎖RNAウイルスの存在、不存在または量を測定できる。トランスフェクションは、Sambrookら、前記に記載されたもののごとき当業者によく知られた標準的手順により行うことができる。
【0035】
もう一つの具体例において、本発明は、分節化されたマイナス鎖RNAウイルスの増殖または遺伝子発現に指向された抗ウイルス薬についての定量的スクリーニング方法を提供する。そのウイルスは、好ましくは、インフルエンザA型ウイルスのごときインフルエンザウイルスである。定量的スクリーニング方法は、 (a)候補抗ウイルス化合物と、(i)前記のレポーター遺伝子ポリペプチドのコード化した配列を含む人工セグメントおよび(ii)分節化されたマイナス鎖RNAウイルスのRNA依存性RNAポリメラーゼを含むタンパク質複合体を含む遺伝子改変された細胞または細胞集団とを接触させ、次いで、(b)細胞または細胞集団により生成されたポリペプチドの量を検出することを含む。ある具体例において、分節化されたマイナス鎖RNAウイルスのRNA依存性RNAポリメラーゼを含むタンパク質複合体は、1以上の核酸により細胞または細胞集団において提供でき、各核酸は、分節化されたマイナス鎖RNAウイルスの複製複合体の少なくとも1つのポリペプチド成分をコードする。他の具体例において、タンパク質複合体は、そのウイルスでの細胞の感染、またはウイルスRNAでの細胞のトランスフェクションにより細胞に提供できる。候補抗ウイルス化合物の効力は、その化合物と接触した細胞または細胞集団のレポーター遺伝子ポリペプチドの発現レベルをその化合物と接触しない対照の細胞または細胞集団のそれに比較することにより決定される。レポーター遺伝子ポリペプチドの発現レベルは、細胞により発現されたレポーター遺伝子ポリペプチドの合計量の測定により決定できる。いくつかの具体例において、レポーター遺伝子ポリペプチドの発現は、レポーター遺伝子ポリペプチドを発現する細胞のパーセンテージの測定により決定できる。非限定の例において、レポーター遺伝子ポリペプチドがGFPまたはルシフェラーゼのいずれかである場合、フローサイトメトリーを用いて、細胞数−対−レポーター遺伝子ポリペプチド発現レベルの2次元プロットを提供できる。標準的な分析技術を用いて、候補化合物の効力を決定でき、例えば、集めたデータを用いて、候補化合物のレポーター遺伝子発現(ED50)の50%の低下を供する有効用量を決定できる。この定量的スクリーニング方法は、迅速な結果を提供し、実施するのは簡単である。
【0036】
もう一つの具体例において、本発明は、分節化されたマイナス鎖RNAウイルスに指向された抗ウイルス薬についてのウイルス生成アッセイ(viral yield assay)を提供する。この具体例において、第1の細胞(または細胞集団)は、候補抗ウイルス化合物と接触する。また、第1の細胞はウイルスに感染する。第1の細胞は、ウイルスによる増殖感染に感受性であるタイプである。次いで、細胞または細胞集団によるウイルスの生成は、候補抗ウイルス化合物との感染および接触後、第1の細胞または細胞集団により生成されたウイルスの試料(もしあれば)を含む生成試料を用いて測定される。その化合物の抗ウイルスの活性は、生成試料と、第2の細胞(または細胞集団)とを接触させることにより決定され、ここに、第2の細胞または細胞集団は、発現が前記のごとく、ウイルスのRNA依存性RNAポリメラーゼの細胞内における存在に依存するポリペプチドをコードするレポーター遺伝子を含む人工セグメントを含む遺伝子改変された細胞または細胞集団である。(対照と比較した)レポーター遺伝子の発現レベルの低下は、候補抗ウイルス化合物による抗ウイルス活性を示す。
【0037】
以下の実施例は、例示的な目的だけに提供され、本発明の範囲を限定するものと解釈されるべきでない。その実施例は、本発明についてのより良好な理解およびその利点のより良好な認識を可能にするであろう。
【0038】
実施例1
本実施例は、プラスミドpHH21 NP UTR HIルシフェラーゼの構造および機能化を示す。
【0039】
このプラスミド(図1)は、インフルエンザA/WSN/33のセグメント5のUTRにより隣接して挟まれたルシフェラーゼORFをコードする。このプラスミドを含む細胞において、転写は、プラスミドがPol Iにより転写される場合、ネガティブまたはゲノムのセンスRNA転写体が生成されるように、RNAポリメラーゼIプロモーターおよびターミネーターにより制御される。このプラスミドにおいて、ルシフェラーゼ・レポーターポリペプチドをコードするDNA配列は、ヒトRNAポリメラーゼI発現ベクターにより含まれたインフルエンザA型ウイルスNPセグメントのcDNAのオープンリーディングフレームを交換する(plasmid pHH21 ; Neumann, G.ら, Proceedings of the National Academy of Sciences USA 96: 9345-9350,1999 ; Neumann,G.ら, Virology 202: 477-479,1994)。そのプラスミドの機能領域に対応するヌクレオチド配列を示す。このプラスミドでトランスフェクトし、インフルエンザA型ウイルスに感染した細胞は、ルシフェラーゼを発現できる。同様に、このプラスミドでトランスフェクトし、さらにウイルス蛋白質PA、PB1、PB2およびNPをコードするプラスミドでトランスフェクトした細胞も、ルシフェラーゼを発現できる。
【0040】
実施例2
この実施例はインフルエンザA型ウイルス検出プロセスを示す。
【0041】
この実施例(図2)において、トランスフェクトした細胞は、RNAポリメラーゼI酵素を用いて、ネガティブまたはゲノムのセンスRNA転写体を生成する。インフルエンザA型ウイルスでの感染またはインフルエンザA型ウイルスのポリメラーゼ複合体の発現に際して、転写体は、全長のポジティブセンスRNAまたは5’キャップされたポリアデニル化mRNAに複製される。ポジティブセンスRNAを複製して、さらなるウイルスネガティブセンスRNAを形成し、一方、そのmRNAを宿主細胞機構によりレポーター蛋白質に翻訳する。ルシフェラーゼ・レポーターポリペプチドは、当該技術分野において知られた標準的な方法により検出および定量できる。
【0042】
実施例3
この実施例はインフルエンザA型ウイルス検出手順を示す。
【0043】
この手順において、細胞をそのpHH21ルシフェラーゼまたは同様のプラスミドでトランスフェクトする(図3)。次いで、細胞を安定的にトランスフェクトした亜集団について選択するか、または直接的に用いることができる。次いで、生物学的試料(例えば、臨床検査試料またはウイルス試料)を検出可能なシグナルの生成に十分であると期待される期間細胞と共にインキュベートする。インキュベーション時間は少なくとも約6時間で有り得る。好ましくは、そのインキュベーション時間は、少なくとも約6時間から約14日まで、少なくとも約6時間から約120時間まで、または少なくとも約6時間から約24時間までで有り得る。試料に対する曝露後、その細胞を当該技術分野において知られた標準的な方法によりレポーター遺伝子ポリペプチドの発現について分析する。
【0044】
実施例4
この実施例は、人工のインフルエンザA型セグメントからのレポーター遺伝子の発現を示す。
【0045】
この実施例において、ホタルルシフェラーゼ遺伝子を、そのNPセグメントからのUTRを含むヒトRNAポリメラーゼI発現ベクターpHH21におけるNPオープンリーディングフレームに代えて、クローン化した。得られたプラスミドのpHH21NPUTR-lucは、インフルエンザウイルスRNAポリメラーゼ複合体により転写できる機能的なインフルエンザウイルス人工セグメントRNAを発現させ、それにより、ウイルス感染の存在下、ルシフェラーゼ発現を可能とする。ヒト293T細胞をpHH21NPUTR-lucでトランスフェクトし、5の感染多重度(MOI)にてインフルエンザウイルス株WSN/33でトランスフェクトさせたか、またはインフルエンザウイルス複製およびヌクレオカプシド蛋白質(ウイルスポリペプチドPA、PB1、PB2およびNPを各々コードするpPA、pPBl、pPB2およびpNP)用の発現ベクターで共トランスフェクトさせた。トランスフェクションの42時間後(または、pHH21NPUTR-luc単独でトランスフェクトした細胞については、インフルエンザウイルスによる感染の18時間後)に、細胞を溶解し、GLOWキット(Promega, Madison WI)を用いて、ホタルルシフェラーゼ活性につきアッセイした。感染の不存在下のそのプラスミドでのトランスフェクションは、16,000を超えるのシグナル対ノイズの非常に高いシグナル(>2×106 LCPS)を生じさせた(図4、第3レーン)。しかしながら、人工セグメント(pHH21NPUTR-luc)を含むプラスミドだけでトランスフェクトした細胞のウイルス感染も、レポーター遺伝子発現を促すのに十分であった(図4、第2レーン)。
【0046】
実施例5
この実施例は、インフルエンザA型ウイルス活性の指標としてのGFPレポーター遺伝子を示す。
【0047】
蛍光ベースの読取りを用いたインフルエンザA型ウイルス活性についてのスクリーニングを促すために、インフルエンザウイルスA/WSN/33のNPセグメントのUTRにより隣接して挟まれたGFPオープンリーディングフレームを含むcDNAを構築した。このcDNAを、ヒトRNAポリメラーゼIプロモーターおよびターミネーターを介して、人工セグメント、NP UTR-GFPの転写を駆動するそのpHH21ベクターにクローン化した。293T細胞をNP UTR-GFPでトランスフェクトした。トランスフェクションの24時間後に、細胞をインフルエンザA型ウイルスに感染させた。感染の18時間後、細胞をGFP発現につき分析した。多量のGFPが、ウイルス感染細胞の100%に存在した (図5A;黒色、満たされたヒストグラム)が、偽感染させた細胞は、ほとんどないまたは全くGFPを発現しなかった(図5A;白色、破線ヒストグラム)。これらのデータは、NP UTR-GFPを発現している細胞におけるGFPの感染依存性の生成を示す。この感染依存性の方法は、ウイルス検出系を可能とする。
【0048】
このレポーター遺伝子構築体が、感染に独立したプロセスにおいて機能できるかを決定するために、293T細胞を、pNP-UTR-GFPと共に、RNA複製および転写(PA、PB1、PB2およびNP)に必要なインフルエンザORFを発現するプラスミドでトランスフェクトした。外因性のORFを含まないプラスミドは、対照としてトランスフェクションに含まれた。4つのウイルスのポリメラーゼ蛋白質のすべてをコードするcDNAでトランスフェクトした細胞の中で、GFPは、細胞の約35%で検出され(図5B、黒色、満たされたヒストグラム)、一方、4つのウイルスのポリメラーゼ発現プラスミドのうちの3つだけでトランスフェクトした細胞は、GFPのかなりの発現を示さなかった(図5B、白色、破線ヒストグラム)。これらのデータは、NP-UTR-GFPからのGFPレポーター遺伝子活性が感染に独立したアッセイにおいて検出可能であることを示す。かくして、図3におけるデータは、ポリメラーゼが感染またはトランスフェクションにより送達されるかのウイルスRNA依存性RNAポリメラーゼ活性を定量するためのツールとしてのインフルエンザA型ウイルスUTRにより隣接して挟まれたレポーター遺伝子の使用を示す。
【0049】
実施例6
この実施例は、インフルエンザC型ウイルスではなくインフルエンザA型ウイルスおよびインフルエンザB型ウイルスによるレポーター遺伝子発現の誘導を示す。
【0050】
インフルエンザA型ウイルスNP-UTR-GFP転写体が、オルソミクソウイルスの他のメンバーのポリメラーゼ複合体により認識できるかを決定するために、pHH21NPUTR-GFPトランスフェクトした細胞を、インフルエンザA型(WSN/33株)、インフルエンザB型(Yamagata/59株)またはインフルエンザC型(Jerusalem/99株)に感染させた。インフルエンザA型/WSN/33の感染細胞は、99%を超えるGFPポジティブ細胞を示し、一方、インフルエンザB/Yamagata/59はわずか9.8%のGFPポジティブ細胞を示した。さらに、GFPポジティブインフルエンザA/WSN/33-感染細胞における平均チャネル蛍光は2963であり、一方、インフルエンザB/Yamagata/59-感染細胞における対応する集団は、わずか88の平均チャネル蛍光を有した。これらのデータは、GFPレポーターがインフルエンザA型ウイルス感染細胞において高度に発現されるが、インフルエンザB型ウイルス感染細胞においてはそれが単にわずかに発現されることを示す。偽感染およびンフルエンザC型感染した培養物の双方は、2.5%のGFPのポジティブ細胞を有し、それはそのレポーターがインフルエンザC型感染細胞において発現されなかったことを示す。これは、NP UTR GFP転写体がインフルエンザC型ウイルスポリメラーゼ複合体により認識も転写もされなかったことを示唆する。このアッセイを診断的に用いて、生物学的試料、例えば、インフルエンザの症状を示すヒト患者からの血液試料がインフルエンザA型ウイルス、インフルエンザB型ウイルスまたはどちらでもないウイルスを含むかどうかを決定できる。
【0051】
本発明の範囲を逸脱することなく、種々の変更が、前記の方法および組成物に行うことができるので、前記の記載に含まれたすべての内容は例示として解釈され、限定する意味に解釈されるものではないことを意図する。
【0052】
本明細書に引用されたすべての引用文献は、ここに出典明示してその全てを本明細書の一部とみなす。本明細書の引用文献の説明は、それらの著者によりなされた主張を単に概括することを意図し、いずれの参考文献も特許性に関連する先行技術を構成するという承認はなされていない。出願人は、引用された参考文献の正確さおよび適切さを試みるための権利を留保する。
【図面の簡単な説明】
【0053】
【図1】図1は、プラスミドpHH21 NP UTR HIルシフェラーゼの地図を示す。
【図2】図2は、人工セグメントを用いるインフルエンザA型ウイルス検出を示す。
【図3】図3は、インフルエンザA型ウイルス検出手順のブロックダイヤグラムを示す。
【図4】図4は、インフルエンザA型ウイルスのセグメントからのUTRおよびルシフェラーゼORFを含む人工セグメントを含む遺伝子改変された細胞によるホタルルシフェラーゼ発現を示す。
【図5】図5は、インフルエンザA型ウイルスのセグメントからのUTRおよびGFP ORFを含む人工セグメントを含む遺伝子改変された細胞による緑色蛍光タンパク質発現を示す。
【特許請求の範囲】
【請求項1】
ウイルスを含むことが疑われる生物学的試料中の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出する方法であって、
a)生物学的試料と、ポリペプチドをコードするレポーター遺伝子を含む組換えRNA分子を含む遺伝子改変された脊椎動物細胞とを接触させ、ここに、そのポリペプチドの発現は、ウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存し;次いで、
b)レポーター遺伝子によりコード化されたポリペプチドの不存在、存在または量を検出することを特徴とする該方法。
【請求項2】
分節化されたマイナス鎖RNAウイルスが、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスよりなる群から選択されることを特徴とする請求項1記載の方法。
【請求項3】
RNA分子が、人工セグメントまたはその相補体であり、人工セグメントが、さらにセグメントの5'UTRおよびセグメントの3'UTRを含み、ここに、該5'UTRおよび3'UTRのうちの少なくとも1つは、ウイルスのUTRであることを特徴とする請求項1記載の方法。
【請求項4】
5'UTRを含む人工セグメントが、インフルエンザA型ウイルスのNPセグメントの5'UTRを含むことを特徴とする請求項3記載の方法。
【請求項5】
人工セグメントの3'UTRが、インフルエンザA型ウイルスのNPセグメントの3'UTRを含むことを特徴とする請求項3記載の方法。
【請求項6】
レポーター遺伝子によりコード化されたポリペプチドが、クロラムフェニコールアセチルトランスフェラーゼ、β−ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質および分泌アルカリフォスファターゼよりなる群から選択されることを特徴とする請求項1記載の方法。
【請求項7】
レポーター遺伝子によりコード化されたポリペプチドが、緑色蛍光タンパク質およびホタルルシフェラーゼよりなる群から選択されることを特徴とする請求項6記載の方法。
【請求項8】
レポーター遺伝子によりコード化されたポリペプチドの不存在、存在または量の検出が、ポリペプチドにより発光された光量子シグナルの不存在、存在または量を検出することを特徴とする請求項1記載の方法。
【請求項9】
遺伝子改変された脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択されることを特徴とする請求項1記載の方法。
【請求項10】
分節化されたマイナス鎖RNAウイルスを含むことが疑われる生物学的試料中の分節化されたマイナス鎖ウイルスの存在、不存在または量を検出する方法であって、
a)生物学的試料と、ポリペプチドをコードするレポーター遺伝子を含む組換えDNA分子を含む遺伝子改変された脊椎動物細胞とを接触させ、ここに、該ポリペプチドの発現は、ウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存し;次いで、
b)レポーター遺伝子によりコード化されたポリペプチドの不存在、存在または量を検出することを特徴とする該方法。
【請求項11】
組換えDNA分子が、5'から3'の順序にて、
DNA依存性RNAポリメラーゼ用のプロモーター;
DNA依存性RNAポリメラーゼ用の転写開始部位;
人工セグメントをコード化するDNA配列;および
転写終結部位を含むことを特徴とする請求項10記載の方法。
【請求項12】
DNA依存性RNAポリメラーゼ用のプロモーターが、RNAポリメラーゼI用のプロモーターであることを特徴とする請求項11記載の方法。
【請求項13】
転写開始部位が、RNAポリメラーゼI用の転写開始部位であることを特徴とする請求項11記載の方法。
【請求項14】
人工セグメントをコードするDNA配列が、人工セグメントの転写がアンチセンス配向にあるようにそのプロモーターに作動可能に連結されることを特徴とする請求項11記載の方法。
【請求項15】
人工セグメントをコード化するDNAが、人工セグメントの転写がセンス配向にあるようにそのプロモーターに作動可能に連結される請求項11記載の方法。
【請求項16】
人工セグメントが、マイナス鎖RNAウイルスのセグメントの5'UTRのcDNA、レポーター遺伝子、およびマイナス鎖RNAウイルスのセグメントの3'UTRのcDNAを含み、ここに、3'UTRおよび5'UTRの少なくとも1つは、生物学的試料を含むことが疑われるウイルスのUTRであることを特徴とする請求項11記載の方法。
【請求項17】
3'UTRおよび5'UTRの少なくとも1つが、インフルエンザA型ウイルスのNPセグメントのUTRであることを特徴とする請求項16記載の方法。
【請求項18】
分節化されたマイナス鎖ウイルスが、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスよりなる群から選択されるウイルスであることを特徴とする請求項10記載の方法。
【請求項19】
レポーター遺伝子が、クロラムフェニコールアセチルトランスフェラーゼ、β-ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質および分泌アルカリフォスファターゼβ-よりなる群から選択されるポリペプチドをコードすることを特徴とする請求項11記載の方法。
【請求項20】
ポリペプチドが、緑色蛍光タンパク質およびホタルルシフェラーゼよりなる群から選択されることを特徴とする請求項26記載の方法。
【請求項21】
転写終結シグナルが、RNAポリメラーゼIの転写終結シグナルであることを特徴とする請求項11記載の方法。
【請求項22】
転写終結部位が、自己切断リボザイムをコードする配列を含むことを特徴とする請求項11記載の方法。
【請求項23】
遺伝子改変された脊椎動物細胞が、安定的にトランスフェクトした遺伝子改変された脊椎動物細胞であることを特徴とする請求項10記載の方法。
【請求項24】
遺伝子改変された脊椎動物細胞が、一過性にトランスフェクトした遺伝子改変された脊椎動物細胞であることを特徴とする請求項10記載の方法。
【請求項25】
遺伝子改変された脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択される感染細胞であることを特徴とする請求項10記載の方法。
【請求項26】
生物学的試料中のインフルエンザA型、インフルエンザB型およびいずれでもないウイルス感染間の鑑別診断を決定する方法であって、
a)生物学的試料と、遺伝子改変された細胞の集団とを接触させ、ここに、実質的に複数の遺伝子改変された細胞は、ポリペプチドをコードするレポーター遺伝子を含む組換えRNA分子を含み、ここに、該ポリペプチドの発現は、インフルエンザウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存し;次いで
b)遺伝子改変された細胞の集団におけるレポーター遺伝子の発現を測定することを特徴とする該方法。
【請求項27】
組換えRNAが、インフルエンザA型ウイルスの人工セグメントであり、人工セグメントが、インフルエンザA型ウイルスのセグメントの5'UTR、ポリペプチドをコードするレポーター遺伝子のオープンリーディングフレームおよびインフルエンザA型ウイルスのセグメントの3'UTRを含むことを特徴とする請求項26記載の方法。
【請求項28】
遺伝子改変された細胞の集団におけるレポーター遺伝子の発現の測定が、レポーター遺伝子を発現する細胞のパーセンテージを決定することを特徴とする請求項26記載の方法。
【請求項29】
遺伝子改変された細胞の集団におけるレポーター遺伝子の発現の測定が、平均シグナル強度を決定することを特徴とする請求項26記載の方法。
【請求項30】
レポーター遺伝子が、クロラムフェニコールアセチルトランスフェラーゼ、β−ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質および分泌アルカリフォスファターゼよりなる群から選択されることを特徴とする請求項26記載の方法。
【請求項31】
遺伝子改変された細胞の集団におけるレポーター遺伝子の発現の測定が、ルシフェラーゼレポーター遺伝子ポリペプチドまたはGFPレポーター遺伝子ポリペプチドからの光量子シグナルを測定することを特徴とする請求項30記載の方法。
【請求項32】
ポリペプチドをコードするレポーター遺伝子が、センス配向にあることを特徴とする請求項26記載の方法。
【請求項33】
ポリペプチドをコードするレポーター遺伝子が、アンチセンス配向にあることを特徴とする請求項26記載の方法。
【請求項34】
遺伝子改変された脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択されることを特徴とする請求項26記載の方法。
【請求項35】
実質的に複数の遺伝子改変された細胞が、細胞の少なくとも約50%を占めることを特徴とする請求項26記載の方法。
【請求項36】
実質的に複数の遺伝子改変された細胞が、細胞の少なくとも約75%を占めることを特徴とする請求項26記載の方法。
【請求項37】
実質的に複数の遺伝子改変された細胞が、細胞の少なくとも約90%を占めることを特徴とする請求項26記載の方法。
【請求項38】
生物学的試料中の第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出する方法であって、
a)生物学的試料と、遺伝子改変された脊椎動物細胞とを接触させ、該細胞が、第1のレポーター遺伝子を含む第1の組換えRNA分子、ここに、その発現が、第1の分節化されたマイナス鎖ウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存し、ならびに第2のレポーター遺伝子を含む第2の組換えRNA分子、ここに、その発現が、第2の分節化されたマイナス鎖ウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存し;次いで、
b)第1のレポーター遺伝子によりコード化された第1のポリペプチドの不存在、存在もしくは量、または第2のレポーター遺伝子によりコード化された第2のポリペプチドの不存在、存在もしくは量を検出することを特徴とする該方法。
【請求項39】
第1の組換えRNA分子が、第1の分節化されたマイナス鎖RNAウイルスの5'UTR、第1のレポーター遺伝子のオープンリーディングフレームおよび第1の分節化されたマイナス鎖RNAウイルスの3'UTRを含む第1の人工セグメントであって、第2の組換えRNA分子が、第2の分節化されたマイナス鎖RNAウイルスの5'UTR、第2のレポーター遺伝子のオープンリーディングフレームおよび第2の分節化されたマイナス鎖RNAウイルスの3'UTRを含む第2の人工セグメントであることを特徴とする請求項38記載の方法。
【請求項40】
第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスが各々、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスよりなる群から選択される請求項38記載の方法。
【請求項41】
第1のレポーター遺伝子および第2のレポーター遺伝子が各々、クロラムフェニコールアセチルトランスフェラーゼ、β-ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質および分泌アルカリフォスファターゼβ-よりなる群から選択されるポリペプチドをコードすることを特徴とする請求項44記載の方法。
【請求項42】
第1のレポーター遺伝子および第2のレポーター遺伝子が各々、ホタルルシフェラーゼ、ウミシイタケルシフェラーゼおよび緑色蛍光タンパク質よりなる群から選択されるポリペプチドをコードすることを特徴とする請求項41記載の方法。
【請求項43】
第1のレポーター遺伝子および第2のレポーター遺伝子の少なくとも1つのオープンリーディングフレームが、アンチセンス配向にあることを特徴とする請求項38記載の方法。
【請求項44】
第1のレポーター遺伝子および第2のレポーター遺伝子の少なくとも1つのオープンリーディングフレームが、センス配向にあることを特徴とする請求項38記載の方法。
【請求項45】
遺伝子改変された脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞、またチャイニーズハムスター卵巣細胞よりなる群から選択されるトランスフェクト細胞である請求項38記載の方法。
【請求項46】
第1もしくは第2の分節化されたマイナス鎖RNAウイルスを含むことが疑われる生物学的試料中の第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出する方法であって、
a)生物学的試料と、その発現が第1の分節化されたマイナス鎖ウイルスのウイルスRNAの細胞内の存在に依存する第1のレポーター遺伝子を含む第1の組換えDNA分子、ならびにその発現が第2の分節化されたマイナス鎖ウイルスのウイルスRNAの細胞内の存在に依存する第2のレポーター遺伝子を含む第2の組換えDNA分子を含む遺伝子改変された脊椎動物細胞とを接触させ;次いで、
b)第1および第2のレポーター遺伝子によりコード化されたポリペプチドの不存在、存在または量を検出することを特徴とする該方法。
【請求項47】
第1の組換えRNA分子が、5'から3'の順序にて:
a)DNA依存性RNAポリメラーゼ用のプロモーター;
b)DNA依存性RNAポリメラーゼ用の転写開始部位;
c)第1の分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRのcDNA、第1のレポーター遺伝子のオープンリーディングフレームのDNA配列および第1の分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRのcDNAを含む第1の人工セグメント;
d)DNA依存性RNAポリメラーゼ用の転写終結部位を含み、
第2の組換えRNA分子が、5'から3'の順序にて:
e)DNA依存性RNAポリメラーゼ用のプロモーター;
f)DNA依存性RNAポリメラーゼ用の転写開始部位;
g)第2の分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRのcDNA、第2のレポーター遺伝子のオープンリーディングフレームのDNA配列、および第2の分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRのcDNAを含む第2の人工セグメント;および
h)DNA依存性RNAポリメラーゼ用の転写終結部位を含むことを特徴とする請求項46記載の方法。
【請求項48】
第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスが各々、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスよりなる群から選択されることを特徴とする請求項46記載の方法。
【請求項49】
DNA依存性RNAポリメラーゼ用の各プロモーターが、RNAポリメラーゼI用のプロモーターであることを特徴とする請求項47記載の方法。
【請求項50】
DNA依存性RNAポリメラーゼ用の各転写開始部位が、RNAポリメラーゼI用の転写開始部位であることを特徴とする請求項47記載の方法。
【請求項51】
各転写終結部位が、自己切断リボザイムをコードする配列を含むことを特徴とする請求項47記載の方法。
【請求項52】
各転写終結部位が、RNAポリメラーゼI終止部位であることを特徴とする請求項47記載の方法。
【請求項53】
遺伝子改変された脊椎動物細胞が、
DNA依存性RNAポリメラーゼ用の第1のプロモーター;
DNA依存性RNAポリメラーゼ用の第1の転写開始部位;
第1の分節化されたマイナス鎖RNAウイルスの第1の5'UTRおよび第1の3'UTRならびに第1のレポーター遺伝子のオープンリーディングフレームを含む第1の人工セグメントをコードする第1のDNA配列;
第1の転写終結部位;
DNA依存性RNAポリメラーゼ用の第2のプロモーター;
DNA依存性RNAポリメラーゼ用の第2の転写開始部位;
第2の分節化されたマイナス鎖RNAウイルスの第2の5'UTRおよび第2の3'UTRならびに第2のレポーター遺伝子のオープンリーディングフレームを含む第2の人工セグメントをコードする第2のDNA配列;ならびに
第2の転写終結部位を含む組換えDNA分子を含むことを特徴とする請求項46記載の方法。
【請求項54】
遺伝子改変された脊椎動物細胞が、安定的にトランスフェクトした遺伝子改変された脊椎動物細胞であることを特徴とする請求項46記載の方法。
【請求項55】
遺伝子改変された脊椎動物細胞が、一過性にトランスフェクトした遺伝子改変された脊椎動物細胞であることを特徴とする請求項46記載の方法。
【請求項56】
遺伝子改変された脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択される感染細胞であることを特徴とする請求項46記載の方法。
【請求項57】
使用説明書および単離された組換えDNA分子を含み、ここに、該単離された組換えDNA分子は、その分子でトランスフェクトした細胞における発現が、分節化されたマイナス鎖RNAウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存するレポーター遺伝子を含む、分節化されたマイナス鎖ウイルスの存在、不存在または量を検出するためのキット。
【請求項58】
単離組換えDNA分子が、5'から3'の順序にて:
a)DNA依存性RNAポリメラーゼ用のプロモーター;
b)DNA依存性RNAポリメラーゼ用の転写開始部位;
c)ポリペプチドをコードするレポーター遺伝子のオープンリーディングフレームを含むDNA配列を含む人工セグメントをコード化する配列;および
d)DNA依存性RNAポリメラーゼ用の転写終結部位
を含む請求項57記載のキット。
【請求項59】
分節化されたマイナス鎖RNAウイルスが、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスよりなる群から選択される請求項57記載のキット。
【請求項60】
DNA依存性RNAポリメラーゼ用のプロモーターが、RNAポリメラーゼI用のプロモーターである請求項57記載のキット。
【請求項61】
DNA依存性RNAポリメラーゼ用の転写開始部位が、RNAポリメラーゼI用の転写開始部位である請求項57記載のキット。
【請求項62】
人工セグメントが、分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRのcDNA、および分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRのcDNAをさらに含む請求項57記載のキット。
【請求項63】
分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRのcDNA、および分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRのcDNAの少なくとも1つが、インフルエンザA型ウイルスのNPセグメントのUTRのcDNAである請求項62記載のキット。
【請求項64】
レポーター遺伝子のオープンリーディングフレームのDNA配列が、クロラムフェニコールアセチルトランスフェラーゼ、β-ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質および分泌アルカリフォスファターゼよりなる群から選択されるポリペプチドをコードする請求項57記載のキット。
【請求項65】
ポリペプチドが、緑色蛍光タンパク質およびホタルルシフェラーゼよりなる群から選択される請求項64記載のキット。
【請求項66】
レポーター遺伝子のオープンリーディングフレームのDNA配列が、アンチセンス配向にある請求項57記載のキット。
【請求項67】
レポーター遺伝子のオープンリーディングフレームのDNA配列が、センス配向にある請求項57記載のキット。
【請求項68】
転写終結シグナルが、RNAポリメラーゼI用の転写終結シグナルである請求項57記載のキット。
【請求項69】
転写終結シグナルが、リボザイム配列を含む請求項57記載のキット。
【請求項70】
キットが、さらに脊椎動物細胞を含む請求項57記載のキット。
【請求項71】
脊椎動物細胞が、哺乳動物細胞である請求項57記載のキット。
【請求項72】
脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択される請求項71記載のキット。
【請求項73】
単離組換えDNA分子が、脊椎動物細胞により含まれる請求項72記載のキット。
【請求項74】
脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択される請求項80記載のキット。
【請求項75】
分節化されたマイナス鎖RNAウイルスに指向された抗ウイルス薬についての定量的スクリーニング方法であって、
a)候補抗ウイルス化合物と、遺伝子改変された細胞または細胞集団とを接触させ、ここに、該細胞または細胞集団は、その発現がそのウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存するポリペプチドをコードするレポーター遺伝子を含む組換えRNA分子を含み、さらに分節化されたマイナス鎖RNAウイルスのRNA依存性RNAポリメラーゼを含み;次いで、
b)細胞または細胞集団により生成されたポリペプチドの量を検出する
ことを特徴とする該方法。
【請求項76】
分節化されたマイナス鎖RNAウイルスが、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスよりなる群から選択されることを特徴とする請求項75記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項77】
組換えRNA分子が、分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRおよび分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRをさらに含むことを特徴とする請求項75記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項78】
人工セグメントの5'UTRが、インフルエンザA型ウイルスのNPセグメントの5'UTRを含むことを特徴とする請求項76記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項79】
人工セグメントの3'UTRが、インフルエンザA型ウイルスのNPセグメントの3'UTRを含むことを特徴とする請求項76記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項80】
レポーター遺伝子によりコード化されたポリペプチドが、クロラムフェニコールアセチルトランスフェラーゼ、β-ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質および分泌アルカリフォスファターゼよりなる群から選択されることを特徴とする請求項75記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項81】
レポーター遺伝子によりコード化されたポリペプチドが、緑色蛍光タンパク質およびホタルルシフェラーゼよりなる群から選択され、レポーター遺伝子によりコード化されたポリペプチドの量の検出が、ポリペプチドまたはその基質により発光された光量子シグナルの量による検出を含むことを特徴とする請求項75記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項82】
遺伝子改変された脊椎動物細胞が、安定的にトランスフェクトした遺伝子改変された脊椎動物細胞であり、該細胞が組換えRNAをコード化する組換えDNAをさらに含むことを特徴とする請求項75記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項83】
遺伝子改変された脊椎動物細胞が、一過性にトランスフェクトした遺伝子改変された脊椎動物細胞であり、該細胞が、組換えRNAをコード化する組換えDNAをさらに含む請求項75記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項84】
遺伝子改変された脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択されるトランスフェクト細胞であることを特徴とする請求項75記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項85】
候補抗ウイルス化合物と、そのウイルスによる増殖感染に感受性である第1の細胞とを接触させ;
第1の細胞を分節化されたマイナス鎖RNAウイルスで感染させ、それにより生成試料を供し;
第2の細胞とその生成試料とを接触させ、ここに、第2が、その発現が該ウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存するポリペプチドをコードするレポーター遺伝子を含む組換えRNA分子を含み;次いで、
第2の細胞により生成されたポリペプチドの量を検出することを含む、
分節化されたマイナス鎖RNAウイルスに指向された抗−ウイルス薬のための定量的スクリーニング方法。
【請求項86】
分節化されたマイナス鎖RNAウイルスが、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスよりなる群から選択されることを特徴とする請求項84記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項87】
組換えRNA分子が、分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRおよび分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRをさらに含む人工セグメントであることを特徴とする請求項84記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項88】
人工セグメントの5'UTRが、インフルエンザA型ウイルスのNPセグメントの5'UTRを含むことを特徴とする請求項86記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項89】
人工セグメントの3'UTRが、インフルエンザA型ウイルスのNPセグメントの3'UTRを含むことを特徴とする請求項86記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項90】
レポーター遺伝子によりコード化されたポリペプチドが、クロラムフェニコールアセチルトランスフェラーゼ、β-ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質および分泌アルカリフォスファターゼよりなる群から選択されることを特徴とする請求項84記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項91】
レポーター遺伝子によりコード化されたポリペプチドが、緑色蛍光タンパク質およびホタルルシフェラーゼよりなる群から選択され、レポーター遺伝子によりコード化されたポリペプチドの量の検出が、ポリペプチドまたはその基質により発光された光量子シグナルの量による検出を含むことを特徴とする請求項84記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項92】
遺伝子改変された脊椎動物細胞が、安定的にトランスフェクトした遺伝子改変された脊椎動物細胞であり、該細胞が組換えRNAをコード化する組換えDNAをさらに含むことを特徴とする請求項84記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項93】
遺伝子改変された脊椎動物細胞が、一過性にトランスフェクトした遺伝子改変された脊椎動物細胞であり、該細胞が、組換えRNAをコード化する組換えDNAをさらに含む請求項84記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項94】
遺伝子改変された脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択されるトランスフェクト細胞であることを特徴とする請求項84記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項1】
ウイルスを含むことが疑われる生物学的試料中の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出する方法であって、
a)生物学的試料と、ポリペプチドをコードするレポーター遺伝子を含む組換えRNA分子を含む遺伝子改変された脊椎動物細胞とを接触させ、ここに、そのポリペプチドの発現は、ウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存し;次いで、
b)レポーター遺伝子によりコード化されたポリペプチドの不存在、存在または量を検出することを特徴とする該方法。
【請求項2】
分節化されたマイナス鎖RNAウイルスが、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスよりなる群から選択されることを特徴とする請求項1記載の方法。
【請求項3】
RNA分子が、人工セグメントまたはその相補体であり、人工セグメントが、さらにセグメントの5'UTRおよびセグメントの3'UTRを含み、ここに、該5'UTRおよび3'UTRのうちの少なくとも1つは、ウイルスのUTRであることを特徴とする請求項1記載の方法。
【請求項4】
5'UTRを含む人工セグメントが、インフルエンザA型ウイルスのNPセグメントの5'UTRを含むことを特徴とする請求項3記載の方法。
【請求項5】
人工セグメントの3'UTRが、インフルエンザA型ウイルスのNPセグメントの3'UTRを含むことを特徴とする請求項3記載の方法。
【請求項6】
レポーター遺伝子によりコード化されたポリペプチドが、クロラムフェニコールアセチルトランスフェラーゼ、β−ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質および分泌アルカリフォスファターゼよりなる群から選択されることを特徴とする請求項1記載の方法。
【請求項7】
レポーター遺伝子によりコード化されたポリペプチドが、緑色蛍光タンパク質およびホタルルシフェラーゼよりなる群から選択されることを特徴とする請求項6記載の方法。
【請求項8】
レポーター遺伝子によりコード化されたポリペプチドの不存在、存在または量の検出が、ポリペプチドにより発光された光量子シグナルの不存在、存在または量を検出することを特徴とする請求項1記載の方法。
【請求項9】
遺伝子改変された脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択されることを特徴とする請求項1記載の方法。
【請求項10】
分節化されたマイナス鎖RNAウイルスを含むことが疑われる生物学的試料中の分節化されたマイナス鎖ウイルスの存在、不存在または量を検出する方法であって、
a)生物学的試料と、ポリペプチドをコードするレポーター遺伝子を含む組換えDNA分子を含む遺伝子改変された脊椎動物細胞とを接触させ、ここに、該ポリペプチドの発現は、ウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存し;次いで、
b)レポーター遺伝子によりコード化されたポリペプチドの不存在、存在または量を検出することを特徴とする該方法。
【請求項11】
組換えDNA分子が、5'から3'の順序にて、
DNA依存性RNAポリメラーゼ用のプロモーター;
DNA依存性RNAポリメラーゼ用の転写開始部位;
人工セグメントをコード化するDNA配列;および
転写終結部位を含むことを特徴とする請求項10記載の方法。
【請求項12】
DNA依存性RNAポリメラーゼ用のプロモーターが、RNAポリメラーゼI用のプロモーターであることを特徴とする請求項11記載の方法。
【請求項13】
転写開始部位が、RNAポリメラーゼI用の転写開始部位であることを特徴とする請求項11記載の方法。
【請求項14】
人工セグメントをコードするDNA配列が、人工セグメントの転写がアンチセンス配向にあるようにそのプロモーターに作動可能に連結されることを特徴とする請求項11記載の方法。
【請求項15】
人工セグメントをコード化するDNAが、人工セグメントの転写がセンス配向にあるようにそのプロモーターに作動可能に連結される請求項11記載の方法。
【請求項16】
人工セグメントが、マイナス鎖RNAウイルスのセグメントの5'UTRのcDNA、レポーター遺伝子、およびマイナス鎖RNAウイルスのセグメントの3'UTRのcDNAを含み、ここに、3'UTRおよび5'UTRの少なくとも1つは、生物学的試料を含むことが疑われるウイルスのUTRであることを特徴とする請求項11記載の方法。
【請求項17】
3'UTRおよび5'UTRの少なくとも1つが、インフルエンザA型ウイルスのNPセグメントのUTRであることを特徴とする請求項16記載の方法。
【請求項18】
分節化されたマイナス鎖ウイルスが、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスよりなる群から選択されるウイルスであることを特徴とする請求項10記載の方法。
【請求項19】
レポーター遺伝子が、クロラムフェニコールアセチルトランスフェラーゼ、β-ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質および分泌アルカリフォスファターゼβ-よりなる群から選択されるポリペプチドをコードすることを特徴とする請求項11記載の方法。
【請求項20】
ポリペプチドが、緑色蛍光タンパク質およびホタルルシフェラーゼよりなる群から選択されることを特徴とする請求項26記載の方法。
【請求項21】
転写終結シグナルが、RNAポリメラーゼIの転写終結シグナルであることを特徴とする請求項11記載の方法。
【請求項22】
転写終結部位が、自己切断リボザイムをコードする配列を含むことを特徴とする請求項11記載の方法。
【請求項23】
遺伝子改変された脊椎動物細胞が、安定的にトランスフェクトした遺伝子改変された脊椎動物細胞であることを特徴とする請求項10記載の方法。
【請求項24】
遺伝子改変された脊椎動物細胞が、一過性にトランスフェクトした遺伝子改変された脊椎動物細胞であることを特徴とする請求項10記載の方法。
【請求項25】
遺伝子改変された脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択される感染細胞であることを特徴とする請求項10記載の方法。
【請求項26】
生物学的試料中のインフルエンザA型、インフルエンザB型およびいずれでもないウイルス感染間の鑑別診断を決定する方法であって、
a)生物学的試料と、遺伝子改変された細胞の集団とを接触させ、ここに、実質的に複数の遺伝子改変された細胞は、ポリペプチドをコードするレポーター遺伝子を含む組換えRNA分子を含み、ここに、該ポリペプチドの発現は、インフルエンザウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存し;次いで
b)遺伝子改変された細胞の集団におけるレポーター遺伝子の発現を測定することを特徴とする該方法。
【請求項27】
組換えRNAが、インフルエンザA型ウイルスの人工セグメントであり、人工セグメントが、インフルエンザA型ウイルスのセグメントの5'UTR、ポリペプチドをコードするレポーター遺伝子のオープンリーディングフレームおよびインフルエンザA型ウイルスのセグメントの3'UTRを含むことを特徴とする請求項26記載の方法。
【請求項28】
遺伝子改変された細胞の集団におけるレポーター遺伝子の発現の測定が、レポーター遺伝子を発現する細胞のパーセンテージを決定することを特徴とする請求項26記載の方法。
【請求項29】
遺伝子改変された細胞の集団におけるレポーター遺伝子の発現の測定が、平均シグナル強度を決定することを特徴とする請求項26記載の方法。
【請求項30】
レポーター遺伝子が、クロラムフェニコールアセチルトランスフェラーゼ、β−ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質および分泌アルカリフォスファターゼよりなる群から選択されることを特徴とする請求項26記載の方法。
【請求項31】
遺伝子改変された細胞の集団におけるレポーター遺伝子の発現の測定が、ルシフェラーゼレポーター遺伝子ポリペプチドまたはGFPレポーター遺伝子ポリペプチドからの光量子シグナルを測定することを特徴とする請求項30記載の方法。
【請求項32】
ポリペプチドをコードするレポーター遺伝子が、センス配向にあることを特徴とする請求項26記載の方法。
【請求項33】
ポリペプチドをコードするレポーター遺伝子が、アンチセンス配向にあることを特徴とする請求項26記載の方法。
【請求項34】
遺伝子改変された脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択されることを特徴とする請求項26記載の方法。
【請求項35】
実質的に複数の遺伝子改変された細胞が、細胞の少なくとも約50%を占めることを特徴とする請求項26記載の方法。
【請求項36】
実質的に複数の遺伝子改変された細胞が、細胞の少なくとも約75%を占めることを特徴とする請求項26記載の方法。
【請求項37】
実質的に複数の遺伝子改変された細胞が、細胞の少なくとも約90%を占めることを特徴とする請求項26記載の方法。
【請求項38】
生物学的試料中の第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出する方法であって、
a)生物学的試料と、遺伝子改変された脊椎動物細胞とを接触させ、該細胞が、第1のレポーター遺伝子を含む第1の組換えRNA分子、ここに、その発現が、第1の分節化されたマイナス鎖ウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存し、ならびに第2のレポーター遺伝子を含む第2の組換えRNA分子、ここに、その発現が、第2の分節化されたマイナス鎖ウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存し;次いで、
b)第1のレポーター遺伝子によりコード化された第1のポリペプチドの不存在、存在もしくは量、または第2のレポーター遺伝子によりコード化された第2のポリペプチドの不存在、存在もしくは量を検出することを特徴とする該方法。
【請求項39】
第1の組換えRNA分子が、第1の分節化されたマイナス鎖RNAウイルスの5'UTR、第1のレポーター遺伝子のオープンリーディングフレームおよび第1の分節化されたマイナス鎖RNAウイルスの3'UTRを含む第1の人工セグメントであって、第2の組換えRNA分子が、第2の分節化されたマイナス鎖RNAウイルスの5'UTR、第2のレポーター遺伝子のオープンリーディングフレームおよび第2の分節化されたマイナス鎖RNAウイルスの3'UTRを含む第2の人工セグメントであることを特徴とする請求項38記載の方法。
【請求項40】
第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスが各々、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスよりなる群から選択される請求項38記載の方法。
【請求項41】
第1のレポーター遺伝子および第2のレポーター遺伝子が各々、クロラムフェニコールアセチルトランスフェラーゼ、β-ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質および分泌アルカリフォスファターゼβ-よりなる群から選択されるポリペプチドをコードすることを特徴とする請求項44記載の方法。
【請求項42】
第1のレポーター遺伝子および第2のレポーター遺伝子が各々、ホタルルシフェラーゼ、ウミシイタケルシフェラーゼおよび緑色蛍光タンパク質よりなる群から選択されるポリペプチドをコードすることを特徴とする請求項41記載の方法。
【請求項43】
第1のレポーター遺伝子および第2のレポーター遺伝子の少なくとも1つのオープンリーディングフレームが、アンチセンス配向にあることを特徴とする請求項38記載の方法。
【請求項44】
第1のレポーター遺伝子および第2のレポーター遺伝子の少なくとも1つのオープンリーディングフレームが、センス配向にあることを特徴とする請求項38記載の方法。
【請求項45】
遺伝子改変された脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞、またチャイニーズハムスター卵巣細胞よりなる群から選択されるトランスフェクト細胞である請求項38記載の方法。
【請求項46】
第1もしくは第2の分節化されたマイナス鎖RNAウイルスを含むことが疑われる生物学的試料中の第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスの存在、不存在または量を検出する方法であって、
a)生物学的試料と、その発現が第1の分節化されたマイナス鎖ウイルスのウイルスRNAの細胞内の存在に依存する第1のレポーター遺伝子を含む第1の組換えDNA分子、ならびにその発現が第2の分節化されたマイナス鎖ウイルスのウイルスRNAの細胞内の存在に依存する第2のレポーター遺伝子を含む第2の組換えDNA分子を含む遺伝子改変された脊椎動物細胞とを接触させ;次いで、
b)第1および第2のレポーター遺伝子によりコード化されたポリペプチドの不存在、存在または量を検出することを特徴とする該方法。
【請求項47】
第1の組換えRNA分子が、5'から3'の順序にて:
a)DNA依存性RNAポリメラーゼ用のプロモーター;
b)DNA依存性RNAポリメラーゼ用の転写開始部位;
c)第1の分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRのcDNA、第1のレポーター遺伝子のオープンリーディングフレームのDNA配列および第1の分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRのcDNAを含む第1の人工セグメント;
d)DNA依存性RNAポリメラーゼ用の転写終結部位を含み、
第2の組換えRNA分子が、5'から3'の順序にて:
e)DNA依存性RNAポリメラーゼ用のプロモーター;
f)DNA依存性RNAポリメラーゼ用の転写開始部位;
g)第2の分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRのcDNA、第2のレポーター遺伝子のオープンリーディングフレームのDNA配列、および第2の分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRのcDNAを含む第2の人工セグメント;および
h)DNA依存性RNAポリメラーゼ用の転写終結部位を含むことを特徴とする請求項46記載の方法。
【請求項48】
第1の分節化されたマイナス鎖RNAウイルスおよび第2の分節化されたマイナス鎖RNAウイルスが各々、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスよりなる群から選択されることを特徴とする請求項46記載の方法。
【請求項49】
DNA依存性RNAポリメラーゼ用の各プロモーターが、RNAポリメラーゼI用のプロモーターであることを特徴とする請求項47記載の方法。
【請求項50】
DNA依存性RNAポリメラーゼ用の各転写開始部位が、RNAポリメラーゼI用の転写開始部位であることを特徴とする請求項47記載の方法。
【請求項51】
各転写終結部位が、自己切断リボザイムをコードする配列を含むことを特徴とする請求項47記載の方法。
【請求項52】
各転写終結部位が、RNAポリメラーゼI終止部位であることを特徴とする請求項47記載の方法。
【請求項53】
遺伝子改変された脊椎動物細胞が、
DNA依存性RNAポリメラーゼ用の第1のプロモーター;
DNA依存性RNAポリメラーゼ用の第1の転写開始部位;
第1の分節化されたマイナス鎖RNAウイルスの第1の5'UTRおよび第1の3'UTRならびに第1のレポーター遺伝子のオープンリーディングフレームを含む第1の人工セグメントをコードする第1のDNA配列;
第1の転写終結部位;
DNA依存性RNAポリメラーゼ用の第2のプロモーター;
DNA依存性RNAポリメラーゼ用の第2の転写開始部位;
第2の分節化されたマイナス鎖RNAウイルスの第2の5'UTRおよび第2の3'UTRならびに第2のレポーター遺伝子のオープンリーディングフレームを含む第2の人工セグメントをコードする第2のDNA配列;ならびに
第2の転写終結部位を含む組換えDNA分子を含むことを特徴とする請求項46記載の方法。
【請求項54】
遺伝子改変された脊椎動物細胞が、安定的にトランスフェクトした遺伝子改変された脊椎動物細胞であることを特徴とする請求項46記載の方法。
【請求項55】
遺伝子改変された脊椎動物細胞が、一過性にトランスフェクトした遺伝子改変された脊椎動物細胞であることを特徴とする請求項46記載の方法。
【請求項56】
遺伝子改変された脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択される感染細胞であることを特徴とする請求項46記載の方法。
【請求項57】
使用説明書および単離された組換えDNA分子を含み、ここに、該単離された組換えDNA分子は、その分子でトランスフェクトした細胞における発現が、分節化されたマイナス鎖RNAウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存するレポーター遺伝子を含む、分節化されたマイナス鎖ウイルスの存在、不存在または量を検出するためのキット。
【請求項58】
単離組換えDNA分子が、5'から3'の順序にて:
a)DNA依存性RNAポリメラーゼ用のプロモーター;
b)DNA依存性RNAポリメラーゼ用の転写開始部位;
c)ポリペプチドをコードするレポーター遺伝子のオープンリーディングフレームを含むDNA配列を含む人工セグメントをコード化する配列;および
d)DNA依存性RNAポリメラーゼ用の転写終結部位
を含む請求項57記載のキット。
【請求項59】
分節化されたマイナス鎖RNAウイルスが、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスよりなる群から選択される請求項57記載のキット。
【請求項60】
DNA依存性RNAポリメラーゼ用のプロモーターが、RNAポリメラーゼI用のプロモーターである請求項57記載のキット。
【請求項61】
DNA依存性RNAポリメラーゼ用の転写開始部位が、RNAポリメラーゼI用の転写開始部位である請求項57記載のキット。
【請求項62】
人工セグメントが、分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRのcDNA、および分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRのcDNAをさらに含む請求項57記載のキット。
【請求項63】
分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRのcDNA、および分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRのcDNAの少なくとも1つが、インフルエンザA型ウイルスのNPセグメントのUTRのcDNAである請求項62記載のキット。
【請求項64】
レポーター遺伝子のオープンリーディングフレームのDNA配列が、クロラムフェニコールアセチルトランスフェラーゼ、β-ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質および分泌アルカリフォスファターゼよりなる群から選択されるポリペプチドをコードする請求項57記載のキット。
【請求項65】
ポリペプチドが、緑色蛍光タンパク質およびホタルルシフェラーゼよりなる群から選択される請求項64記載のキット。
【請求項66】
レポーター遺伝子のオープンリーディングフレームのDNA配列が、アンチセンス配向にある請求項57記載のキット。
【請求項67】
レポーター遺伝子のオープンリーディングフレームのDNA配列が、センス配向にある請求項57記載のキット。
【請求項68】
転写終結シグナルが、RNAポリメラーゼI用の転写終結シグナルである請求項57記載のキット。
【請求項69】
転写終結シグナルが、リボザイム配列を含む請求項57記載のキット。
【請求項70】
キットが、さらに脊椎動物細胞を含む請求項57記載のキット。
【請求項71】
脊椎動物細胞が、哺乳動物細胞である請求項57記載のキット。
【請求項72】
脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択される請求項71記載のキット。
【請求項73】
単離組換えDNA分子が、脊椎動物細胞により含まれる請求項72記載のキット。
【請求項74】
脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択される請求項80記載のキット。
【請求項75】
分節化されたマイナス鎖RNAウイルスに指向された抗ウイルス薬についての定量的スクリーニング方法であって、
a)候補抗ウイルス化合物と、遺伝子改変された細胞または細胞集団とを接触させ、ここに、該細胞または細胞集団は、その発現がそのウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存するポリペプチドをコードするレポーター遺伝子を含む組換えRNA分子を含み、さらに分節化されたマイナス鎖RNAウイルスのRNA依存性RNAポリメラーゼを含み;次いで、
b)細胞または細胞集団により生成されたポリペプチドの量を検出する
ことを特徴とする該方法。
【請求項76】
分節化されたマイナス鎖RNAウイルスが、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスよりなる群から選択されることを特徴とする請求項75記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項77】
組換えRNA分子が、分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRおよび分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRをさらに含むことを特徴とする請求項75記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項78】
人工セグメントの5'UTRが、インフルエンザA型ウイルスのNPセグメントの5'UTRを含むことを特徴とする請求項76記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項79】
人工セグメントの3'UTRが、インフルエンザA型ウイルスのNPセグメントの3'UTRを含むことを特徴とする請求項76記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項80】
レポーター遺伝子によりコード化されたポリペプチドが、クロラムフェニコールアセチルトランスフェラーゼ、β-ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質および分泌アルカリフォスファターゼよりなる群から選択されることを特徴とする請求項75記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項81】
レポーター遺伝子によりコード化されたポリペプチドが、緑色蛍光タンパク質およびホタルルシフェラーゼよりなる群から選択され、レポーター遺伝子によりコード化されたポリペプチドの量の検出が、ポリペプチドまたはその基質により発光された光量子シグナルの量による検出を含むことを特徴とする請求項75記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項82】
遺伝子改変された脊椎動物細胞が、安定的にトランスフェクトした遺伝子改変された脊椎動物細胞であり、該細胞が組換えRNAをコード化する組換えDNAをさらに含むことを特徴とする請求項75記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項83】
遺伝子改変された脊椎動物細胞が、一過性にトランスフェクトした遺伝子改変された脊椎動物細胞であり、該細胞が、組換えRNAをコード化する組換えDNAをさらに含む請求項75記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項84】
遺伝子改変された脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択されるトランスフェクト細胞であることを特徴とする請求項75記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項85】
候補抗ウイルス化合物と、そのウイルスによる増殖感染に感受性である第1の細胞とを接触させ;
第1の細胞を分節化されたマイナス鎖RNAウイルスで感染させ、それにより生成試料を供し;
第2の細胞とその生成試料とを接触させ、ここに、第2が、その発現が該ウイルスのRNA依存性RNAポリメラーゼの細胞内の存在に依存するポリペプチドをコードするレポーター遺伝子を含む組換えRNA分子を含み;次いで、
第2の細胞により生成されたポリペプチドの量を検出することを含む、
分節化されたマイナス鎖RNAウイルスに指向された抗−ウイルス薬のための定量的スクリーニング方法。
【請求項86】
分節化されたマイナス鎖RNAウイルスが、インフルエンザA型ウイルス、インフルエンザB型ウイルスおよびインフルエンザC型ウイルスよりなる群から選択されることを特徴とする請求項84記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項87】
組換えRNA分子が、分節化されたマイナス鎖RNAウイルスのセグメントの5'UTRおよび分節化されたマイナス鎖RNAウイルスのセグメントの3'UTRをさらに含む人工セグメントであることを特徴とする請求項84記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項88】
人工セグメントの5'UTRが、インフルエンザA型ウイルスのNPセグメントの5'UTRを含むことを特徴とする請求項86記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項89】
人工セグメントの3'UTRが、インフルエンザA型ウイルスのNPセグメントの3'UTRを含むことを特徴とする請求項86記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項90】
レポーター遺伝子によりコード化されたポリペプチドが、クロラムフェニコールアセチルトランスフェラーゼ、β-ガラクトシダーゼ、β-グルコロニダーゼ、ウミシイタケルシフェラーゼ、ホタルルシフェラーゼ、緑色蛍光タンパク質および分泌アルカリフォスファターゼよりなる群から選択されることを特徴とする請求項84記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項91】
レポーター遺伝子によりコード化されたポリペプチドが、緑色蛍光タンパク質およびホタルルシフェラーゼよりなる群から選択され、レポーター遺伝子によりコード化されたポリペプチドの量の検出が、ポリペプチドまたはその基質により発光された光量子シグナルの量による検出を含むことを特徴とする請求項84記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項92】
遺伝子改変された脊椎動物細胞が、安定的にトランスフェクトした遺伝子改変された脊椎動物細胞であり、該細胞が組換えRNAをコード化する組換えDNAをさらに含むことを特徴とする請求項84記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項93】
遺伝子改変された脊椎動物細胞が、一過性にトランスフェクトした遺伝子改変された脊椎動物細胞であり、該細胞が、組換えRNAをコード化する組換えDNAをさらに含む請求項84記載の抗ウイルス薬についての定量的スクリーニング方法。
【請求項94】
遺伝子改変された脊椎動物細胞が、293T腎臓細胞、LA−4細胞、MRC5細胞、A549細胞、CV−1細胞、ベロ細胞、LLC−MK1細胞、HEp2細胞、2fTGH細胞、U3A細胞、BHK細胞、サル初代腎臓細胞およびチャイニーズハムスター卵巣細胞よりなる群から選択されるトランスフェクト細胞であることを特徴とする請求項84記載の抗ウイルス薬についての定量的スクリーニング方法。
【図1】

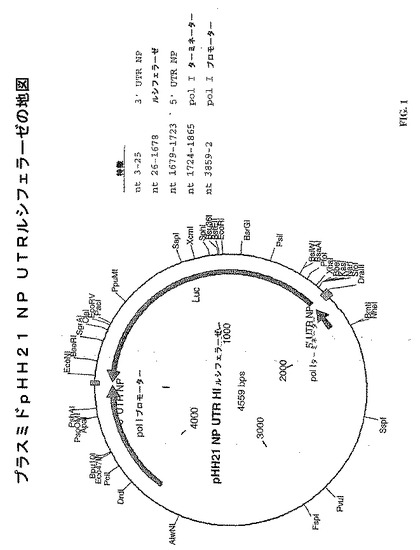
【図2】
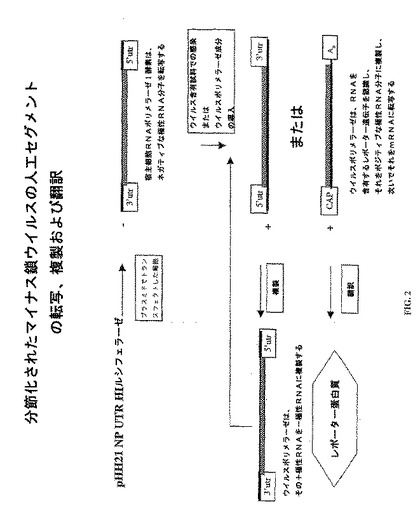
【図3】
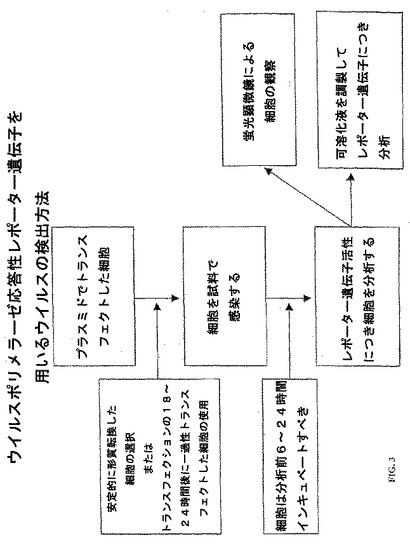
【図4】
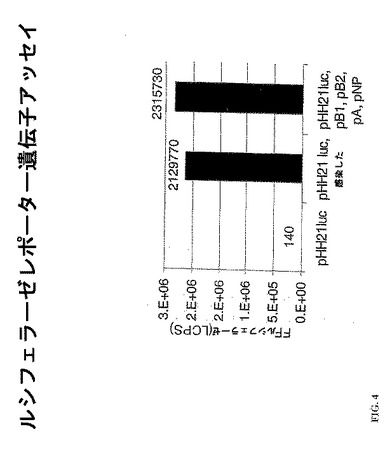
【図5】
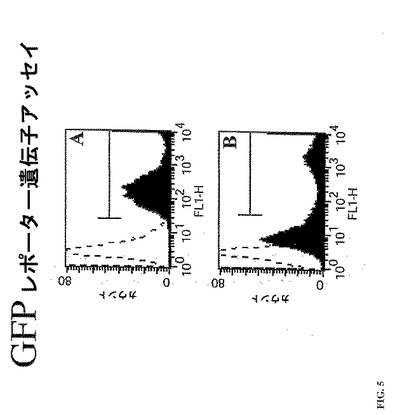
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2007−524404(P2007−524404A)
【公表日】平成19年8月30日(2007.8.30)
【国際特許分類】
【出願番号】特願2006−538136(P2006−538136)
【出願日】平成16年10月22日(2004.10.22)
【国際出願番号】PCT/US2004/035168
【国際公開番号】WO2005/044986
【国際公開日】平成17年5月19日(2005.5.19)
【出願人】(597025806)ワシントン・ユニバーシティ (26)
【氏名又は名称原語表記】Washington University School of Medicine
【出願人】(506151464)アパス・リミテッド・ライアビリティ・カンパニー (1)
【氏名又は名称原語表記】APATH LLC
【Fターム(参考)】
【公表日】平成19年8月30日(2007.8.30)
【国際特許分類】
【出願日】平成16年10月22日(2004.10.22)
【国際出願番号】PCT/US2004/035168
【国際公開番号】WO2005/044986
【国際公開日】平成17年5月19日(2005.5.19)
【出願人】(597025806)ワシントン・ユニバーシティ (26)
【氏名又は名称原語表記】Washington University School of Medicine
【出願人】(506151464)アパス・リミテッド・ライアビリティ・カンパニー (1)
【氏名又は名称原語表記】APATH LLC
【Fターム(参考)】
[ Back to top ]