
分類不能型インフルエンザ菌の中耳炎単離物の遺伝子
【課題】分類不能型株のインフルエンザ菌(NTHi)のポリヌクレオチド配列および該ポリヌクレオチドにコードされるポリペプチドならびにその使用を提供すること。
【解決手段】本発明はまた、中耳および/または鼻咽頭へのNTHi感染中または該感染に応答して上方調節されるNTHi遺伝子に関する。本発明はまた、本発明のNTHiポリペプチドに特異的な抗体を提供する。ヒトまたは試料、例えば、血清、痰、耳液、血液、尿、リンパ液および脳脊髄液などにおいてNTHi菌を検出する方法が想定される。このような方法は、特異的ポリヌクレオチドプローブを用いてNTHiポリヌクレオチドを検出すること、または特異的抗体を用いてNTHiポリペプチドを検出することを含む。本発明ではまた、このようなNTHi菌の検出方法を用いる診断用キットが想定される。
【解決手段】本発明はまた、中耳および/または鼻咽頭へのNTHi感染中または該感染に応答して上方調節されるNTHi遺伝子に関する。本発明はまた、本発明のNTHiポリペプチドに特異的な抗体を提供する。ヒトまたは試料、例えば、血清、痰、耳液、血液、尿、リンパ液および脳脊髄液などにおいてNTHi菌を検出する方法が想定される。このような方法は、特異的ポリヌクレオチドプローブを用いてNTHiポリヌクレオチドを検出すること、または特異的抗体を用いてNTHiポリペプチドを検出することを含む。本発明ではまた、このようなNTHi菌の検出方法を用いる診断用キットが想定される。
【発明の詳細な説明】
【技術分野】
【0001】
本出願は、2005年6月16日に出願された米国仮出願第60/691,214号、および2004年3月5日に出願された米国出願第10/795,159号(これは、2003年3月6日に出願された米国仮出願第60/453,134号への優先権を主張する)への優先権を主張する。これらの全ては、それらの全体において本明細書で参考として援用される。
【0002】
(発明の分野)
本発明は、分類不能型インフルエンザ菌(NTHi)ゲノムのポリヌクレオチド配列、該ゲノム内に含まれるNTHi遺伝子および該ポリヌクレオチドにコードされるポリペプチドに関する。本発明はまた、このようなNTHiポリヌクレオチドおよびNTHiポリペプチドの使用、例えば、ワクチンおよびNTHi関連障害を処置および予防する方法に関する。本発明はまた、中耳または鼻咽頭のNTHi感染中または該感染に応答して上方調節されるNTHi遺伝子に関する。
【背景技術】
【0003】
(背景)
中耳炎(OM)は、世界中で非常によく見られる小児疾患であり、緊急治療室での小児の往診の主な原因である(非特許文献1)。最近の統計学的データにより、1990年では2,450万人の小児科医の診療所での往診がOMのためになされたことが示されており、1980年代に報告されたものの200%を超える増加を表す。長引いても死亡率と関連することは稀であるが、OMに関連する罹病率は相当である。聴力低下は、この疾患に伴う一般的な問題であり、しばしば、子供の行動、教育および言語能力の発達に影響する(非特許文献2;非特許文献3;非特許文献4)。OMの社会経済的影響もまた大きく、OMの診断およびマネージメントの直接および間接費用は、米国だけで年間50億ドルを超える(非特許文献5)。
【0004】
抗生物質療法は一般的であり、中耳腔換気用チューブの外科的留置は、滲出液の排出、感染の浄化および中耳内の液の蓄積に伴う痛みの緩和の点では成功裏であるが、多数の抗生物質耐性菌の出現およびチューブ留置に伴う侵襲性の性質により、OMのマネージメント、好ましくは予防に対するより有効で許容されるアプローチの必要性が促されている。慢性OMの外科的マネージメントは、子供に対して全身麻酔下での鼓膜を介する中耳腔換気用チューブの挿入を伴う。この手順は一般的であり(普及率は約13%である;Brightら、Am.J.Public Health,83(7):1026−8,1993)、中耳の蓄積液の排出による有痛症状の緩和の点で非常に有効であるが、処置の侵襲性の性質およびその施術(incumbent)の危険性のため、非難されている(Bermanら、Pediatrics,93(3):353−63,1994;Brightら、前掲)。;Cimons,ASM News,60:527−528;Paap,Ann.Pharmacother.,30(11):1291−7,1996)。
【0005】
ワクチン開発における進歩は、7価莢膜−コンジュゲートワクチンPREVNAR(登録商標)(EskolaおよびKilpi,Pedriatr.Infect.Dis.J.16:S72−78,2000)の最近の認可および発売によって示されるように、急性OM(AOM)の主な原因因子である肺炎連鎖球菌に関して最も進んでいる。PREVNAR(登録商標)は、浸潤性の肺炎球菌疾患に非常に有効であるが、OMに関する範囲は期待はずれ(6〜8%)であり、該ワクチンに含まれていない血清型のため、OM症例数が増加しているという報告がある(Blackら,Pedriatr.Infect.Dis J.,19:187−195;Eskolaら,Pedriatr.Infect.Dis J,19:S72−78,2000;Eskolaら,N.Engl.J.Med.344:403−409,2001;Snowら,Otol.Neurotol.,.23:1−2,2002)。滲出液を伴う慢性OMにおいてよく見られるグラム陰性病原体である分類不能型インフルエンザ菌(NTHi)に関してはほとんど進歩していない(非特許文献6;非特許文献7)。NTHiに対する有効なワクチンの開発の障害は、現時点におけるNTHi誘導型中耳疾患の病因の理解が不充分なことである。この遅滞の一因は、該疾患が、良性の鼻咽頭共生生物の一宿主免疫学的寛容から、通常無菌の中耳空間への日和見的侵入菌に対する積極防御反応のものに進行することによる、病原菌が発現するビルレンス因子と宿主の免疫応答との動態的相互作用の理解の欠如である。
【0006】
Rd菌株は、現在の小ゲノム配列決定原理のための代表例の生物体であり、インフルエンザ菌生物学の研究のための重要なモデル生物体であるが、Rd菌株は、ヘモフィルス属の構成員によって引き起こされる病原性の研究には不充分なモデルである。インフルエンザ菌のb血清型菌株は、浸潤性疾患(例えば、髄膜炎)を引き起こし、分類不能型インフルエンザ菌(NTHi)菌株は、主に、限局性呼吸器疾患、特に中耳炎(OM)、急性副鼻腔炎、市中肺炎において役割を有し、慢性閉塞性肺疾患または嚢胞性線維症の患者に重要な結果をもたらす(Kilpiら,Pediatr Infect Dis J 20:654−62,2001;Murphy,Curr Opin Infect Dis 16:129−34,2003;Romanら,J Clin Microbiol 42:1450−9,2004;Sethi,Clin Microbiol Rev 14:336−63,2001;St Geme,Vaccine 19 Suppl l:S41−50,2000)。しかしながら、Rd菌株はd血清型菌株の誘導体である。d血清型菌株が疾患と関連するのは稀である(Dainesら,J Med Microbiol 52:277−82 2003;Heathら,Pediatr Infect Dis J 20:300−5,2001;Rodriguezら,Infect Immun 71:1635−42,2003,Skoczynskaら,J Clin Microbiol 43:938−41,2005)。生物体の生物学の研究における最も有用なデータセットの一例はそのゲノム配列であるため、いくつかの調査により、b型インフルエンザ菌菌株、インフルエンザ菌Biogroup Aegyptius菌株またはRd菌株には存在しない分類不能型菌株に見られる遺伝子が同定および特性評価された(Bergmanら,Infect Immun 71:1098−108,2003;Changら,Infect Immun 68:2630−7,2000;Erdosら,Int J Pediatr Otorhinolaryngol 67:749−55.2003;Liら,Mol.Microbiol 47:1101−11,2003;McMichael & Green,Curr Opin Investig Drugs 4:953−8,2003;Pomposiello & Demple,2001;Smootら,Infect Immun 70:2694−9,2002)。
現在、どのようにしてNTHiが小児においてOMを引き起こすのかの理解は不充分である。OMの誘導に必要な推定ビルレンス因子の同定は、宿主−病原体相互作用の理解、そして最終的には潜在的なワクチン候補および化学療法の標的の同定に大きく寄与する。中耳炎のマネージメント、好ましくは予防に対するより有効で許容されるアプローチを開発する多大な必要性が存在する。ワクチン開発は、非常に有望であり、本目的を達成するための費用効果のある方法である(非特許文献8:非特許文献9)。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】Infante−RivandおよびFernandez,Epidemiol.Rev.(1993)15:444−465
【非特許文献2】Baldwin,Am.J.Otol(1993)14:601−604
【非特許文献3】Hunterら,Ann.Otol.Rhinol.Laryngol.Suppl(1994)163:59−61
【非特許文献4】Teeleら、J.Infect.Dis.(1990)162:685−694
【非特許文献5】Kaplanら,Pediatr.Infect.Dis.J.(1997)16:S9−11
【非特許文献6】Klein,Pedriatr.Infect.Dis J(1997)16:S5−8
【非特許文献7】Spinolaら,J.Infect.Dis.(1986)154:100−109
【非特許文献8】Giebank,Pedriatr.Infect.Dis J(1994)13(11):1064−8
【非特許文献9】Karmaら,Int.J.Pedritr.OtorhinolaryngoL(1995)32(Suppl.):S127−34
【発明の概要】
【課題を解決するための手段】
【0008】
(発明の概要)
本発明は、NTHi インフルエンザ86−028NP菌株のゲノム配列の同定およびキャラクタライゼーションならびにそれにコードされるポリペプチド配列を提供する。NTHiゲノム配列の3倍解析を、配列番号:1〜576として示す一連のコンティグ配列に示し(set out)、その後のゲノム配列の8倍解析を、配列番号:675〜685として示す一連の11個のコンティグ配列に示す。これらのコンティグは生データであり、当業者によって、常套的な方法を用い、重複配列を比較することにより、これらのコンティグをアセンブルし、NTHi 86−028NP菌株の完全なゲノムが構築され得る。
【0009】
NTHi 86−028NP菌株の注釈付きの完全なゲノムを、配列番号:772として示す。オープンリーディングフレームを表6に、配列番号:773〜2593として示す。得られる遺伝子産物のアミノ酸配列を、配列番号:2594〜4414として示す。該ゲノムは、1.91kbの大きさであり、Rd菌株のゲノムよりもわずかに大きい。Rd菌株のゲノムに対する総ゲノム再編成のいくつかの領域ならびに86−028NP菌株に特有のいくつかの遺伝子が同定されている。
【0010】
本発明はまた、本発明のNTHiポリペプチドに特異的な抗体を提供する。ヒトまたは試料、例えば、血清、痰、耳液、血液、尿、リンパ液および脳脊髄液などにおいてNTHi菌を検出する方法が想定される。このような方法は、特異的ポリヌクレオチドプローブを用いてNTHiポリヌクレオチドを検出すること、または特異的抗体を用いてNTHiポリペプチドを検出することを含む。本発明ではまた、このようなNTHi菌の検出方法を用いる診断用キットが想定される。
【0011】
本発明ではまた、本発明のNTHiポリペプチドまたはそのNTHiペプチドを投与することによる、免疫応答の惹起方法が想定される。このような方法は、NTHi感染によって引き起こされる疾患、例えばOMなどの処置および/または予防のために、NTHiポリペプチドまたはNTHiペプチドをワクチンとして投与することを含む。以下のNTHi遺伝子は、中耳および/または鼻咽頭感染中または該感染に応答して上方調節される。これらの遺伝子:HisB、lppB、sapA、lolA、rbsC、purE、ribB、arcB、uxuA、dsbB、ureH、licC、HI1647、ispZ、radC、mukF、glpR、ihfB、argR、cspD、HI0094、HI1163、HI1063、HI0665、HI1292、HI1064にコードされるポリペプチドおよびそのペプチドが可能なOMワクチン候補および/または化学療法の標的として想定される。NTHi HisB遺伝子は、ヌクレオチド配列配列番号:615として示され、配列番号:616で示されるアミノ酸配列をコードする。NTHi sapA遺伝子は、ヌクレオチド配列配列番号:617で示され、配列番号:618で示されるアミノ酸配列をコードする。NTHi rbsC遺伝子は、ヌクレオチド配列配列番号:619で示され、配列番号:620で示されるアミノ酸配列をコードする。NTHi purE遺伝子は、ヌクレオチド配列配列番号:621で示され、配列番号:622で示されるアミノ酸配列をコードする。NTHi ribB遺伝子は、ヌクレオチド配列配列番号:623で示され、配列番号:624で示されるアミノ酸配列をコードする。NTHi
arcB遺伝子は、ヌクレオチド配列配列番号:625で示され、配列番号:626で示されるアミノ酸配列をコードする。NTHi uxuA遺伝子は、ヌクレオチド配列配列番号:627で示され、配列番号:628で示されるアミノ酸配列をコードする。NTHi dsbB遺伝子は、ヌクレオチド配列配列番号:629で示され、配列番号:630で示されるアミノ酸配列をコードする。NTHi ureH遺伝子は、ヌクレオチド配列配列番号:631で示され、配列番号:632で示されるアミノ酸配列をコードする。NTHi licC遺伝子は、ヌクレオチド配列配列番号:633で示され、配列番号:634で示されるアミノ酸配列をコードする。NTHi HI1647遺伝子は、ヌクレオチド配列配列番号:635で示され、配列番号:636で示されるアミノ酸配列をコードする。NTHi ispZ遺伝子は、ヌクレオチド配列配列番号:637で示され、配列番号:638で示されるアミノ酸配列をコードする。NTHi radC遺伝子は、ヌクレオチド配列配列番号:639で示され、配列番号:640で示されるアミノ酸配列をコードする。NTHi mukF遺伝子は、ヌクレオチド配列配列番号:641で示され、配列番号:642で示されるアミノ酸配列をコードする。NTHi glpR遺伝子は、ヌクレオチド配列配列番号:643で示され、配列番号:644で示されるアミノ酸配列をコードする。NTHi ihfB遺伝子は、ヌクレオチド配列配列番号:645で示され、配列番号:646で示されるアミノ酸配列をコードする。NTHi argR遺伝子は、ヌクレオチド配列配列番号:647で示され、配列番号:648で示されるアミノ酸配列をコードする。NTHi cspD遺伝子は、ヌクレオチド配列配列番号:649で示され、配列番号:650で示されるアミノ酸配列をコードする。NTHi HI1163遺伝子は、ヌクレオチド配列配列番号:651で示され、配列番号:652で示されるアミノ酸配列をコードする。NTHi HI1063遺伝子は、ヌクレオチド配列配列番号:653で示され、配列番号:654で示されるアミノ酸配列をコードする。NTHi HI0665遺伝子は、ヌクレオチド配列配列番号:655で示され、配列番号:656で示されるアミノ酸配列をコードする。NTHi HI1292遺伝子は、ヌクレオチド配列配列番号:657で示され、配列番号:658で示されるアミノ酸配列をコードする。
【0012】
配列番号:1〜576、配列番号:675〜685として示されるポリヌクレオチド配列および表4と4Bに示すヌクレオチド配列に含まれる該新規なNTHi遺伝子はまた、中耳および/または鼻咽頭感染中に上方調節され、したがって、OMワクチン候補および/または化学療法の標的をコードしていると想定される。また、以下のNTHi遺伝子は、ビルレンス関連遺伝子であると想定され、したがって、有力なOMワクチンおよび/または化学療法の標的:HI1386、HI1462、HI1369、lav、HI1598をコードしていると想定される。NTHi HI1386遺伝子配列は、配列番号:659として示され、配列番号:660で示されるアミノ酸配列をコードする。NTHi HI1462遺伝子配列は、配列番号:661として示され、配列番号:662で示されるアミノ酸配列をコードする。NTHi HI1369遺伝子配列は、配列番号:665として示され、配列番号:666で示されるアミノ酸配列をコードする。NTHi lav遺伝子配列は、配列番号:663として示され、配列番号:664で示されるアミノ酸配列をコードする。NTHi HI1598遺伝子配列は、配列番号:669および配列番号:671として示され、配列番号:670および配列番号:672で示されるアミノ酸配列をコードする。ビルレンスと関連するさらなるNTHi遺伝子としては、配列番号:667および配列番号:673として示されるポリヌクレオチド配列が挙げられる。
【0013】
NTHi感染の処置または予防方法として、本発明では、NTHiポリペプチドの発現または活性を阻害する分子であって、感染中、上方調節されるか、または活性である分子を投与することが想定される。特に、本発明では、NTHi感染中に上方調節されるNTHi遺伝子に特異的に結合するアンチセンスオリゴヌクレオチドを投与することにより、NTHiタンパク質発現をモジュレートすることを含む、NTHi感染の処置または予防方法が想定され、かかる遺伝子としては、hisB、lppB、sapA、lolA、rbsC、purE、ribB、arcB、uxuA、dsbB、ureH、licC、HI1647、ispZ、radC、mukF、glpR、ihfB、argR、cspD、HI0094、HI1163、HI1063、HI0665、HI1292、HI1064が挙げられる。本発明ではまた、これらの(theses)遺伝子にコードされるタンパク質の活性をモジュレートする抗体または小分子を投与することを含む、NTHi感染の処置または予防方法が想定される。配列 配列番号:1〜576、配列番号:675〜685として示されるポリヌクレオチドおよび表4と4Bに示すヌクレオチド配列に含まれる該新規なNTHi遺伝子はまた、中耳および/または鼻咽頭感染中に上方調節され、したがって、このようなポリヌクレオチド配列に特異的に結合するアンチセンスオリゴヌクレオチドもまた想定される。
【0014】
本発明のポリヌクレオチドおよびポリペプチド
本発明は、NTHi 86−028NP菌株ゲノムの配列を提供する。該ゲノム配列の3倍解析を、本明細書において「コンティグ1〜576」で表す一連のコンティグ配列として示す。各コンティグを、その「コンティグ番号」と相関する配列同定番号 で表示する。したがって、配列番号:1〜576で示される本発明のコンティグ。これらのコンティグポリヌクレオチド配列は、常套的な方法を用いてNTHi 86−028NP菌株の完全なゲノム配列にアセンブルされ得る。NTHi菌株82−028NPゲノムの8倍配列解析が終了したら、該ゲノム配列を、本明細書において配列番号:675〜685で表す11個のコンティグに編集した。最後に、該完全なゲノムを、本明細書において配列番号:772で表す1つの核酸配列として示す。
【0015】
本発明は、配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593のコンティグ内に含まれるNTHiポリヌクレオチド配列およびオープンリーディングフレーム、ならびに表3B、表4Bおよび表5に示すヌクレオチド配列を提供する。本発明はまた、本発明のNTHiポリヌクレオチドにコードされるポリペプチド配列、例えば、配列番号:2594〜4414、表3B、表4Bおよび表5に示すアミノ酸配列などを提供する。本発明は、ストリンジェント条件下で、(a)配列番号:1〜576;配列番号:675〜685、配列番号:773〜2593のヌクレオチド配列および本明細書の表3B、表4Bおよび表5に示すヌクレオチド配列の相補配列、(b)上記の任意のポリヌクレオチドの対立遺伝子バリアントであるポリヌクレオチド;(c)上記の任意のタンパク質の種ホモログをコードするポリヌクレオチド;または(d)本発明のNTHiポリペプチドの特異的ドメインもしくは切断型を含むポリペプチドをコードするポリヌクレオチドにハイブリダイズするポリヌクレオチドを提供する。
【0016】
本発明のNTHiポリヌクレオチドはまた、上記のポリヌクレオチドと実質的に等価なヌクレオチド配列を含む。本発明によるポリヌクレオチドは、上記のNTHiポリヌクレオチドに対して、例えば、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、81%、82%、83%、84%、85%、86%、87%、88%または89%、より典型的には少なくとも90%、91%、92%、93%または94%、さらにより典型的には少なくとも95%、96%、97%、98%または99%の配列同一性を有するものであり得る。
【0017】
本発明の核酸配列の範囲内には、ストリンジェント条件下で、配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593のNTHi ヌクレオチド配列、ならびに本明細書の表3B、表4Bおよび表5に示すヌクレオチド配列、またはその相補配列にハイブリダイズする核酸配列断片が含まれ、該断片は、約5個より多くのヌクレオチド、好ましくは7個のヌクレオチド、より好ましくは9個より多くのヌクレオチド、最も好ましくは17個より多くのヌクレオチドである。例えば、本発明のポリヌクレオチドの任意の1つに選択的な(すなわち、特異的にハイブリダイズする)15、17または20個以上のヌクレオチドの断片が想定される。ポリヌクレオチドに特異的にハイブリダイズすることができるプローブにより、本発明のNTHiポリヌクレオチド配列が、同じ遺伝子ファミリー内の他のポリヌクレオチド配列と区別され得るか、またはNTHi遺伝子が他の細菌の遺伝子と区別され得、好ましくは特異的ヌクレオチド配列に基づくものである。
【0018】
用語「ストリンジェント」は、当該技術分野においてストリンジェントと一般的に理解される条件をいうために使用される。ハイブリダイゼーションのストリンジェンシーは、主に、温度、イオン強度および変性剤(例えば、ホルムアミドなど)の濃度によって決定される。ハイブリダイゼーションおよび洗浄のストリンジェント条件の例は、0.015M塩化ナトリウム、0.0015Mクエン酸ナトリウム、65〜68℃または0.015M塩化ナトリウム、0.0015Mクエン酸ナトリウムおよび50%ホルムアミド、42℃である。Sambrookら,Molecular Cloning:A Laboratory Manual,第2版,Cold Spring Harbor Laboratory,(Cold Spring Harbor,N.Y.1989)を参照のこと。よりストリンジェント条件(例えば、より高温、より低イオン強度、より高ホルムアミド、または他の変性剤など)もまた使用され得るが、ハイブリダイゼーションの速度が影響を受ける。デオキシオリゴヌクレオチドのハイブリダイゼーションが懸念される場合、さらなる例示的なストリンジェントハイブリダイゼーション条件としては、37℃(14塩基オリゴの場合)、48℃(17塩基オリゴの場合)、55℃(20塩基オリゴの場合)および60℃(23塩基オリゴの場合)での6×SSC 0.05%ピロリン酸ナトリウム中での洗浄が挙げられる。
【0019】
他の薬剤を、非特異的および/またはバックグラウンドハイブリダイゼーションを低減させる目的でハイブリダイゼーションおよび洗浄バッファー中に含めてもよい。例は、0.1%ウシ血清アルブミン、0.1%ポリビニルピロリドン、0.1%ピロリン酸ナトリウム、0.1%ドデシル硫酸ナトリウム、NaDodSO4、(SDS)、フィコール、デンハルト溶液、超音波処理サケ精子DNA(または他の非相補DNA)、および硫酸デキストランであるが、他の適当な薬剤もまた使用され得る。このような添加剤の濃度および型は、ハイブリダイゼーション条件のストリンジェンシーに実質的に影響を与えることなく、変更され得る。ハイブリダイゼーション実験は、通常、pH6.8〜7.4で行なわれるが、典型的なイオン強度条件では、ハイブリダイゼーションの速度は、ほぼpHに非依存性である。Andersonら,Nucleic Acid Hybridisation:A Practical Approach,Ch.4,IRL Press
Limited(Oxford,England)を参照のこと。ハイブリダイゼーション条件は、これらの可変量を適合させるため、および関連のある異なる配列のDNAがハイブリッドを形成するのを可能にするために、当業者によって調整され得る。
【0020】
本発明の範囲内に含まれる配列は、このような特定の配列に限定されないだけでなく、その対立遺伝子および種の異型も包含する。対立遺伝子および種の異型は、配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593に示した配列、および本明細書の表3B、表4Bおよび表5に示したヌクレオチド配列、好ましくは、そのオープンリーディングフレーム、代表的なその断片または配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593内のオープンリーディングフレーム、表3B、表4Bおよび表5に示すヌクレオチド配列と少なくとも90%同一である、好ましくは95%同一であるヌクレオチド配列を、同じ種の別の単離ヌクレオチド(isolete)由来の配列と比較することにより、常套的に決定され得る。2つの配列間の同一性および類似性を決定するための好ましいコkンピュータプログラム法としては、限定されないが、GCG プログラムパッケージ、例えば、GAP(Devereuxら,Niicl.Acid.Res.,12:387,1984;Genetics Computer Group,University of
Wisconsin,Madison,WI)、BLASTP、BLASTNおよびFASTA(Altschulら,J.Mol Biol,215:403−410,1990)が挙げられる。BLASTX プログラムは、National Center for Biotechnology Information(NCBI)および他の供給源(BLAST Manual,Altschulら NCB/NLM/NIH Bethesda,MD 20894;Altschulら、前掲)から公に入手可能である。よく知られたSmith Watermanアルゴリズムもまた、同一性を決定するために使用され得る。
【0021】
さらにまた、コドンの多様性に適合させるため、本発明は、本明細書に開示する特定のオープンリーディングフレーム(ORF)にコードされるものと同じアミノ酸配列をコードする核酸分子を包含する。すなわち、ORFのコード領域において、1つのコドンを、同じアミノ酸をコードする別のコドンと置換することが明白に想定され得る。
【0022】
本発明の単離されたポリペプチドとしては、限定されないが、配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593として示されるポリヌクレオチド配列内に含まれるヌクレオチド配列、ならびに表3B、表4Bおよび表5に示すヌクレオチド配列にコードされるアミノ酸配列を含むポリペプチド、または対応する完全長もしくは成熟タンパク質が挙げられる。本発明のポリペプチドとしては、本明細書の表3B、表4Bおよび表5に示す配列番号:616、配列番号:618、配列番号:620、配列番号:622、配列番号:624、配列番号:626、配列番号:628、配列番号:628、配列番号:630、配列番号:632、配列番号:634、配列番号:636、配列番号:638、配列番号:640、配列番号:642、配列番号:644、配列番号:646、配列番号:648、配列番号:650、配列番号:652、配列番号:654、配列番号:656、配列番号:658、配列番号:660、配列番号:662、配列番号:664、配列番号:666、配列番号:668、配列番号:670、配列番号:672、配列番号:674、配列番号:687、配列番号:689、配列番号:691、配列番号:693、配列番号:695、配列番号:697、配列番号:699、配列番号:701、配列番号:703、配列番号:705、配列番号:707、配列番号:709、配列番号:711、配列番号:713、配列番号:715、配列番号:717、配列番号:719、配列番号:721、配列番号:723、配列番号:725、配列番号:727、配列番号:729、配列番号:731、配列番号:733、配列番号:735、配列番号:737、配列番号:739、配列番号:741、配列番号:743、配列番号:745、配列番号:747、配列番号:749、配列番号:751、配列番号:753、配列番号:755、配列番号:757、配列番号:759、配列番号:761、763、配列番号:765、配列番号:767、配列番号:769または配列番号:771、配列番号:2594〜4414のアミノ酸配列が挙げられる。
【0023】
また、本発明のポリペプチドとしては、好ましくは生物学的または免疫学的活性を有し、(a)配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593として示されるヌクレオチド配列、ならびに表3B、表4Bおよび表5に示すヌクレオチド配列内に含まれるオープンリーディングフレームまたは(b)(a)のポリヌクレオチドの相補配列にストリンジェントハイブリダイゼーション条件下でハイブリダイズするポリヌクレオチドにコードされるポリペプチドが挙げられる。
【0024】
本発明はまた、本発明のアミノ酸配列の生物学的に活性または免疫学的に活性なバリアント、および生物学的および/または免疫学的活性を保持している、その「実質的な等価物」(例えば、少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、86%、87%、88%、89%、少なくとも約90%、91%、92%、93%、94%、典型的には少なくとも約95%、96%、97%、より典型的には少なくとも約98%または最も典型的には少なくとも約99%のアミノ酸同一性を有する)を提供する。対立遺伝子バリアントにコードされるポリペプチドは、配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593に示されるヌクレオチド配列、ならびに配列番号:2594〜4414として、および本明細書の表3B、表4Bと表5に示されるアミノ酸配列をコードするヌクレオチド配列内に含まれるポリヌクレオチドにコードされるポリペプチド、ならびに配列番号:2594〜4414、本明細書の表3B、表4Bおよび表5に示されるアミノ酸配列を有するポリペプチドと比べて、同様の、増大された、または低下した活性を有するものであり得る。
【0025】
NTHiペプチドとは、配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593に示されるヌクレオチド配列、または本明細書の表3B、表4Bおよび表5に示すヌクレオチド配列
にコードされるNTHiポリペプチドの断片、および配列番号:2594〜4414 本明細書の表3B、表4Bおよび表5に占められるアミノ酸配列を有するポリペプチドをいう。好ましいNTHiペプチドは、生物学的および/または免疫学的に活性である。
【0026】
さらに、本発明は、本発明のNTHi核酸断片または本発明の核酸断片の縮重バリアントにコードされる、単離されたNTHiポリペプチドまたはNTHiペプチドを提供する。用語「縮重バリアント」は、本発明の核酸断片(例えば、ORF)をヌクレオチド配列は異なるが、遺伝暗号の縮重のため、同一のNTHiポリペプチド配列をコードするヌクレオチド断片をいう。本発明の好ましい核酸断片は、タンパク質をコードするORFである。
【0027】
本発明はまた、該ポリペプチドの生物学的および/または免疫学的活性に影響を及ぼさない1つ以上の同類アミノ酸置換を有するNTHiポリペプチドを提供する。あるはまた、本発明のNTHiポリペプチドは、生物学的活性を改変するものまたは改変しないものであり得る同類アミノ酸置換を有することが想定される。用語「同類アミノ酸置換」は、天然のアミノ酸残基と、基準残基、例えば、天然に存在するアミノ酸および天然に存在しないアミノ酸との、その位置でのアミノ酸残基の極性または電荷に対して影響がほとんどまたは全くないような置換をいう。例えば、同類置換は、ポリペプチド内の無極性残基と任意の他の無極性残基との置き換えによりもたらされる。
さらに、該ポリペプチド内の任意の天然の残基はまた、「アラニンスキャニング変異誘発」法に従ってアラニンで置換され得る。天然に存在するアミノ酸は、その側差に基づき、以下のとおりに特徴づけされる:塩基性:アルギニン、リシン、ヒスチジン;酸性:グルタミン酸、アスパラギン酸;非電荷極性:グルタミン、アスパラギン、セリン、トレオニン、チロシン;および無極性:フェニルアラニン、トリプトファン、システイン、グリシン、アラニン、バリン、プロリン、メチオニン、ロイシン、ノルロイシン、イソロイシン。アミノ酸置換に関する一般規則を以下の表1に示す。
【0028】
【表1】

NTHiポリペプチドをコードするポリヌクレオチドに相補的なアンチセンスポリヌクレオチドもまた提供される。
【0029】
本発明では、本発明のポリヌクレオチドが増幅または発現のためにベクター内に挿入され得ることが想定される。発現のためには、該ポリヌクレオチドを、適切な発現制御配列、例えば、プロモーターおよびポリアデニル化シグナル配列などに作動可能に連結させる。さらに、本発明のポリヌクレオチドを含む細胞が提供される。例示的な原核生物宿主としては、大腸菌、バチルス属、ストレプトミセス属、シュードモナス属、サルモネラ属およびセラチア属などの細菌が挙げられる。
【0030】
用語「単離される」は、物質が、これが天然に存在する環境のその他の成分から取り出されており、該成分を本質的に含まないことをいう。例えば、ポリペプチドは、他の細胞性タンパク質から分離されたものであり、またはDNAは、これが天然状態で存在するゲノム内でこれとフランキングしている他のDNA分離されたものである。
【0031】
免疫応答を惹起するための抗体および方法
本発明は、NTHiポリペプチドに特有(すなわち、特異的)な抗原性エピトープに結合する抗体を提供する。また、多数のインフルエンザ菌亜型間に共通する抗原性エピトープを含むが、任意の他の抗原性エピトープに関して他と異なる抗体を提供する。該抗体は、ポリクローナル抗体、モノクローナル抗体、その特異的エピトープに結合する能力を保持している抗体断片(例えば、Fv、FabおよびF(ab)2断片)、単鎖抗体ならびにヒト抗体またはヒト化抗体であり得る。抗体は、当該技術分野において標準的な手法によって作製され得る。
【0032】
インフルエンザ菌の莢膜多糖類を模倣する抗原により、インビトロアッセイで細菌を死滅させる能力を示す抗体が生成されることが、当該技術分野において知られている。このような抗体はまた、動物モデル系において、インフルエンザ菌による抗原刺激を防御することが知られている。この研究は、莢膜多糖類(polysaccharride)に対する抗体が、おそらく、ヒトにおいて防御的免疫応答を惹起することを示す。本発明は、インフルエンザ菌を死滅させるとともに、NTHi感染からヒトを保護する能力を示す、NTHi本発明のポリペプチドおよびその断片に特異的な抗体を提供する。本発明はまた、インフルエンザ菌のビルレンスを低減させる、付着を阻害する、細胞分裂を阻害する、および/または上皮内への侵入を阻害する、あるいは、インフルエンザ菌の食作用を増強する、本発明のNTHiポリペプチドに特異的な抗体を提供する。
【0033】
インビトロ補体媒介性殺菌アッセイ系(Musherら,Infect.Immun.39:297−304,1983;Andersonら,J.Clin.Invest.51:31−38,1972)が、抗NTHi抗体の殺菌活性を測定するために使用され得る。the ability of NTHiポリペプチドがNTHiペプチドが防御的抗体応答を惹起する能力に関するさらなるデータは、感染の動物モデル、例えば、本明細書に記載のチンチラのモデル系を用いることにより得られ得る。
【0034】
また、NTHiのエピトープに対する予備形成抗体、例えば、NTHi OMP、LOSまたは非被膜タンパク質に対する抗体などの投与による受動免疫療法により、宿主に対して短期的防御を付与することが可能である。したがって、想定されるワクチン製剤は、受動免疫療法における使用のための抗体を生成させるために使用され得る。異種免疫グロブリンは、その外来免疫学的成分に対して免疫応答が誘発され得るため、ヒト免疫グロブリンがヒト用の薬に好ましい。かかる受動免疫処置は、緊急的に、特別な危険に曝された非免疫処置個体の即座の防御のために使用され得る。あるはまた、このような抗体は、抗イディオタイプ抗体の作製において使用され得、これが、こんどはNTHi エピトープに対する免疫応答を刺激するための抗原として使用され得る。
【0035】
本発明では、個体においてNTHiに対する免疫応答を惹起する方法が想定される。
この方法により、NTHi菌の死滅、細胞へのNTHi付着の阻止および/またはNTHi複製の遅滞の1種類以上を含む免疫応答が惹起される。本発明の組成物の「免疫学的用量」は、投与前に検出可能な免疫応答と比べて、または投与前の標準的な免疫応答と比べて、投与後に検出可能な体液性および/または細胞性免疫応答を生じさせるものである。本発明では、該方法によりもたらされる免疫応答が防御的および/または治療的であり得ることが想定される。一実施形態において、該方法は、本発明のNTHiタンパク質またはNTHiペプチドを含む組成物の免疫学的用量を投与する工程を含む。NTHiタンパク質またはその抗原性ペプチドは、それ自体では抗体を産生し得ないが、第1のタンパク質を安定化させ、かつ免疫学的および防御的特性を有する融合タンパク質を産生することができる補助タンパク質と融合させてもよい。したがって、有効組換えタンパク質は、好ましくは、抗原性補助タンパク質、例えば、グルタチオン−S−トランスフェラーゼ(GST)またはβ−ガラクトシダーゼ、該タンパク質を可溶化し、その産生および精製を助長する比較的大きな補助タンパク質をさらに含む。またさらに、補助タンパク質は、免疫系の全身性刺激を提供するという意味で、アジュバントとしての機能を果たすものであり得る。補助タンパク質は、第1のタンパク質のアミノ末端またはカルボキシ末端のいずれかに結合され得る。本発明により、本発明のポリヌクレオチドにコードされるNTHiポリペプチドまたはその抗原性ペプチドを含む組成物、特にワクチン組成物、および方法が提供される。
【0036】
本発明ではまた、個体において多数のインフルエンザ菌亜型に対する免疫応答を惹起する方法が想定される。この方法により、インフルエンザ菌の死滅、細胞へのインフルエンザ菌付着の阻止および/またはインフルエンザ菌複製の遅滞の1種類以上を含む免疫応答が惹起される。この方法は、多数のインフルエンザ菌亜型間に共通する抗原性エピトープを含むが、任意の他の抗原性エピトープに関して他と異なる本発明のNTHiタンパク質またはNTHiペプチドを含む組成物の免疫学的用量を投与する工程を含む。
【0037】
別の実施形態において、該方法は、本発明のNTHiタンパク質またはNTHiペプチドを発現する細胞を含む組成物の免疫学的用量を投与することを含む。また別の実施形態において、該方法は、本発明のNTHiタンパク質またはNTHiペプチドをコードするポリヌクレオチドを含む組成物の免疫学的用量を投与することを含む。該ポリヌクレオチドは、任意の他の核酸と会合されていない裸のポリヌクレオチドであってもよく、ベクター、例えば、プラスミドまたはウイルスベクター(例えば、あでの随伴ウイルスベクターまたはアデノウイルスベクター)内に存在するものであったもよい。本発明の組成物は、1種類以上のNTHiタンパク質またはNTHiペプチドを単独、またはNTHi菌に対する免疫応答を惹起する他のエピトープとの組合せで含むものであり得る。本発明の組成物はまた、1種類以上のNTHiタンパク質またはペプチドを、1種類以上の インフルエンザ菌亜型に対する免疫応答を惹起するエピトープとの組合せで含むものであり得る。該組成物の投与は、当該技術分野において標準的な経路、例えば、非経口、静脈内、経口、口腔内、経鼻、肺経由、経直腸または経膣によるものであり得る。該方法は、一人の個体に組合せで使用され得る。該方法は、個体のNTHi感染の前包埋後に使用され得る。
【0038】
中耳および/または鼻咽頭のNTHi感染の際に上方調節される遺伝子、ならびにNTHiビルレンスと関連する遺伝子を、本明細書において記載する。このようなNTHi遺伝子にコードされるポリペプチドおよびそのペプチドは、NTHi感染と関連する障害、例えばOMなどを処置または予防するための免疫応答の惹起に有用であると想定される。このような遺伝子にコードされるポリペプチドの一例としては、ヒスチジン生合成タンパク質、リポタンパク質B、ペプチドABC輸送体、ペリプラズムSapA前駆体、外膜リポタンパク質担体タンパク質前駆体、リボース輸送系パーミアーゼタンパク質、ホスホリボシルアミノイミダゾールカルボキシラーゼ触媒性サブユニット、PurE、ホスホリボシルアミノイミダゾールカルボキシラーゼ触媒性サブユニット、オルニチンカルバモイルトランスフェラーゼ、マンノン酸脱水酵素、ジスルフィドオキシドレダクターゼ、ウレアーゼアクセサリータンパク質、ホスホコリンシチジリル(cytidylyl)トランスフェラーゼ、推定ピリドキシン生合成タンパク質、一重項酸素抵抗性タンパク質、細胞内隔壁化タンパク質、DNA修復タンパク質、MukFタンパク質、グリセロール−3−ホスフェートレギュロンレセプター、組込み宿主因子βサブユニット、アルギニン抑制因子、コールドショック様タンパク質、ストレス応答タンパク質、LicA、MukF、RadAおよびHI0094、HI1163、HI0665、HI1292、HI1064 HI186、HI0352遺伝子にコードされる仮想タンパク質が挙げられる。NTHi
OMP、LOSおよび非被膜タンパク質もまた、NTHi感染と関連する障害の予防および処置のための免疫応答を惹起することが想定される。
【0039】
本発明は、個体において宿主細胞へのNTHi菌の結合を阻止する方法を含む。該方法は、NTHiの細胞付着の結合を阻止する本発明の抗体またはポリペプチドを投与することを含む。あるはまた、NTHiの細胞付着の結合を阻止する1種類以上の小分子の投与が想定される。本発明の抗体、ポリペプチドまたは小分子がNTHiの細胞付着を阻止する能力を示すために、インビトロアッセイが使用され得る。
【0040】
NTHiの細胞付着を阻止する本発明の抗体、本発明のポリペプチドおよび/または本
発明の小分子を含む医薬組成物が提供される。該医薬組成物は、前述の活性成分の1種類単独からなるものであってもよく、前述の活性成分の組合せを含むものであってもよく、細菌感染を処置するために使用されるさらなる活性成分を含むものであってもよい。医薬組成物は、医薬として有効な担体などの1種類以上の付加的成分を含んでいてもよい。医薬組成物の投与の投薬量および頻度は、標準的な手法によって決定され、例えば、個体の体重および年齢、投与経路ならびに症状の重篤度に依存する。医薬組成物の投与は、当該技術分野において標準的な経路、例えば、非経口、静脈内、経口、口腔内、経鼻、肺経由、経直腸または経膣によるものであり得る。
【0041】
また、本発明により、個体においてNTHi感染を検出するための方法が提供される。一実施形態において、該方法は、試料中の本発明のNTHiポリヌクレオチドを、該ポリヌクレオチドに特異的に結合するプライマーまたはプローブ用いて検出することを含む。該ポリヌクレオチドの検出は、例えば、ハイブリダイゼーションおよびPCRを伴う当該技術分野において常套的な数多くの手法によってなされ得る。
【0042】
本発明の抗体はまた、インフルエンザ菌感染が疑われる個体の種々の体液におけるNTHi抗原(NTHiポリペプチドおよびそのペプチド)の検出のための診断用アッセイにおける使用のための試薬を提供するために使用され得る。別の実施形態において、本発明のNTHiタンパク質およびペプチドは、種々の患者組織および体液、例えば、限定されないが:血液、血清、耳液、髄液、痰、尿、リンパ液および脳脊髄液におけるNTHiの検出のためのイムノアッセイにおける抗原として使用され得る。本発明の抗原は、当該技術分野において知られた任意のイムノアッセイ系、例えば、限定されないが:ラジオイムノアッセイ、ELISAアッセイ、サンドイッチアッセイ、沈降素反応、ゲル拡散沈降素反応、免疫拡散アッセイ、凝集アッセイ、蛍光イムノアッセイ、プロテインAイムノアッセイおよび免疫電気泳動アッセイにおいて使用され得る。
【0043】
ワクチンおよび化学療法標的
本発明の一態様は、個体にNTHi抗原タンパク質またはその抗原性ペプチドを接種することを含む、個体、特に哺乳動物における免疫学的応答を低減させるための方法に関する。
【0044】
本発明はまた、本発明の免疫学的組換えNTHiタンパク質またはNTHiペプチドを、適当な担体とともに含むワクチン製剤を提供する。ワクチン候補および/または化学療法の標的として想定されるNTHiポリペプチドおよびそのペプチドとしては、限定されないが、ヒスチジン生合成タンパク質、リポタンパク質B、ペプチドABC輸送体、ペリプラズムSapA前駆体、外膜リポタンパク質担体タンパク質前駆体、リボース輸送系パーミアーゼタンパク質、ホスホリボシルアミノイミダゾールカルボキシラーゼ触媒性サブユニット、PurE、3,4−ジヒドロキシ(dihydroxt)−2−ブトン4−ホスフェートシンターゼ、オルニチンカルバモイルトランスフェラーゼ、マンノン酸脱水酵素、ジスルフィドオキシドレダクターゼ、ウレアーゼアクセサリータンパク質、ホスホコリンシチジリルトランスフェラーゼ、推定ピリドキシン生合成タンパク質、一重項酸素抵抗性タンパク質、細胞内隔壁化タンパク質、DNA修復タンパク質、MUKFタンパク質、グリセロール−3−ホスフェートレギュロンレセプター、組込み宿主因子βサブユニット、アルギニン抑制因子、コールドショック様タンパク質、ストレス応答タンパク質、LicA、RadAならびにHI0094、HI1163、HI0665、HI1292、HI1064 HI1386、HI0352遺伝子にコードされる仮想タンパク質、配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593として示すヌクレオチド配列および本明細書の表3B、表4Bおよび表5に示すヌクレオチド配列内に存在する新規なNTHiポリヌクレオチド配列にコードされるNTHi OMP、NTHi LOSおよびNTHi非被膜タンパク質ならびにポリペプチド、ならびに配列番号:2594〜4414、本明細書の表3B、表4Bおよび表5に示すアミノ酸配列を有するポリペプチドが挙げられる。
【0045】
該タンパク質は、胃の中で分解され得るため、好ましくは、非経口、例えば、皮下、筋肉内、静脈内または皮内である投与で投与する。非経口投与に適した製剤としては、水性および非水性の滅菌注射用液剤(これは、抗酸化剤、緩衝剤、静菌剤および製剤を個体の体液(好ましくは血液)と等張性にする溶質を含んでいてもよい);ならびに水性および非水性滅菌懸濁剤(これは、懸濁剤または増粘剤を含んでいてもよい)が挙げられる。該製剤は、単位投与容器または反復投与容器、例えば、密閉アンプルおよびバイアル内に提示され得、使用直前に滅菌液状担体の添加のみが必要とされる凍結乾燥条件で保存され得る。該ワクチン製剤はまた、製剤の免疫原性を増強するためのアジュバント系(例えば、水中油型および当該技術分野において知られた他の系など)を含んでいてもよい。投薬量は、ワクチンの比活性に依存し、常套的な実験手法容易に決定され得る。
【0046】
A.ペプチドワクチン
ペプチドワクチンなどのペプチド治療用薬剤は、当該技術分野においてよく知られており、製剤分野での使用が増大している。かかるペプチド化合物の非経口投与に一貫する欠点は、急速な分解または変性であった。輸液ポンプならびにワックスまたはオイル埋入物は、治療用薬剤の長期投与のために、ペプチド様治療用薬剤の存在を延長させるためと、かかる薬剤の完全性を保持するための両方に対する取り組みにおいて使用されている。さらにまた、該ペプチド様薬剤は、理想的には、長期間(特に、該ペプチド様薬剤の各エピトープに関して)天然状態の立体構造を維持するものであるのがよく、さらに、抗原刺激された動物または免疫処置したヒトにおいて免疫学的応答が誘導されるのに適した様式で提示されるものであるのがよい。
【0047】
本発明のNTHi抗原性ペプチドは、いくつかの慣用的な様式で調製され得る。短鎖ペプチド配列は、標準的な手段を用い、化学合成によって調製され得る。特に簡便であるのは固相手法である(例えば、Eriksonら,The Proteins(1976)v.2,Academic Press,New York,p.255を参照)。自動固相合成装置が市販されている。また、配列の修飾は、適切な残基の置換、付加または欠失によって容易になされる。例えば、担体タンパク質への簡便な連結のためのスルフヒドリル基をもたらすため、システイン残基がカルボキシ末端に付加され得、またはスペーサーエレメント、例えば、さらなるグリシン残基などが、該ペプチドのC末端連結アミノ酸とその残部間の配列内に組み込まれ得る。短鎖NTHiペプチドはまた、組換え手法によって作製され得る。この長さのペプチドのコード配列は、化学的手法、例えば、Matteucciら,JAm Chem Soc,103:3185(1981)に記載のホスホトリエステル法によって容易に合成され得る。
【0048】
本明細書において想定される一部のNTHiペプチド配列は、免疫原性であるには小さすぎると考えられ得、この性質を付与するために担体物質に結合させ得る。かかる連結を作出する当該技術分野において知られた任意の方法が使用され得る。連結は、ジスルフィド結合を一方の官能基の末端に生成させ、ペプチド結合を他方に生成させるヘテロ二官能性薬剤、例えば、ジスルフィドアミド形成剤(例えば、N−スクシジミジル(succidimidyl)−3−(2−ピリジルジチオ)プロピオネート(SPDP))(例えば、Jansenら,Immun.Rev.62:185,1982を参照)およびジスルフィド結合ではなくチオエーテルを形成する二官能性カップリング剤、例えば、6−マレイミドカプロン酸、2−ブロモ酢酸、2−ヨード酢酸、4−(N−マレイミドメチル)シクロへキサン−1−カルボン酸などの反応性エステル、ならびにスクシンイミドまたは1−ヒドロキシ−2−ニトロ−4−スルホン酸と合わせ、スクシンイミジル4−(N−mareイミド−メチル)シクロへキサン−1−カルボキシレート(carobxylate)(SMCC)などのナトリウム塩にすることによりカルボキシル基を活性化させるカップリング剤を用いて形成され得る。
【0049】
B.ワクチン組成物および投与
免疫原を初回抗原刺激投与し、続いて該免疫原を1回以上の追加免疫刺激に曝露することは、有効なワクチンとするために必要であり得る(Krampら,Infect.Immun.,25:771−773,1979;Davisら,Immunology Letters,14:341−8 1986 1987)。共投与されると免疫応答が有益に増強され得るタンパク質またはポリペプチドの例としては、サイトカイン(例えば、IL−2、IL−12、GM−CSF)、サイトカイン誘導分子(例えば、Leaf)または共起刺激分子が挙げられる。ヘルパー(HTL)エピトープを細胞内標的化シグナルに連結し、CTLエピトープとは別に発現させてもよい。これにより、CTL エピトープと異なる細胞区画へのHTLエピトープの指向が可能となり得る。必要な場合は、これにより、MHCクラスII経路内へのHTLエピトープのより効率的な侵入を助長し、それによりCTL誘導を改善し得る。CTL誘導とは対照的に、ある種の疾患では、免疫抑制分子(例えば、TGF−β)の共発現によって免疫応答を特異的に低下させることが有益であり得る。
【0050】
理想的には、免疫原は2つの特性;対応する抗体の形成を刺激する能力、およびこのような抗体と特異的に反応する傾向を示すものである。免疫原は、抗体のコーミング(combing)部位によって認識され得る免疫原の最小の部分であるエピトープを1つ以上有する。特別な場合では、免疫原、免疫原の画分または免疫原が提示される条件は、所望の免疫学的応答を促すのに不充分であり、不充分な免疫性がもたらされる。これは、しばしば、ペプチドまたは他の小分子が免疫原として使用される場合である。また、免疫調節物質(例えば、サイトカイン、例えばインターロイキン)などの他の物質をワクチンと合わせてもよい。
【0051】
ワクチン分野では、免疫原ともに使用して免疫応答を増強するアジュバントと呼ばれるある種の物質の使用が認識されている。アジュバントは、アジュバントの使用なしで惹起され得るものより速くて大きな免疫応答を惹起するためにさらに使用される。また、アジュバントは、アジュバントを含めない場合に必要とされ得るより少ない免疫原を用いて免疫学的応答をもたらすため、免疫学的防御をもたらすある種の抗体サブクラスの産生を増大させるため、または免疫応答の成分(例えば、体液性、細胞性)を増強するために使用され得る。既知のアジュバントとしては、フロイントアジュバントなどの乳剤および他の油状乳剤、百日咳菌、MF59、Quillaja saponaria(QS21)由来の精製サポニン、水酸化物、リン酸塩およびミョウバンなどのアルミニウム塩、リン酸カルシウム(および他の金属塩)、水酸化アルミニウム塩などのゲル、マイコバクテリウムの産性物、例えばムラミルジペプチド、固形物質、リポソームおよびビロソームなどの粒子が挙げられる。アジュバントとして使用されることが知られている天然産物および細菌の産生物の例としては、モノホスホリルリピドA(MPL)、RC−529(合成MPL様アセチル化単糖)、OM−174(これは、大腸菌由来のリピドA誘導体である)、ホロ毒素、例えばコレラ毒素(CT)またはその誘導体の1つ、百日咳毒素(PT)および大腸菌の非耐熱性毒素(LT)またはその誘導体の1つ、ならびにCpGオリゴヌクレオチドが挙げられる。アジュバント活性は、いくつかの要素、例えば、担体効果、デポ形成、リンパ球再循環の改変、T−リンパ球の刺激、B−リンパ球の直接刺激およびマクロファージの刺激などによって影響され得る。
【0052】
ワクチンは、典型的には注射用剤として、液状の液剤もしくは懸濁剤のいずれかとして調製され、注射前に液体中で液剤もしくは懸濁剤にするのに適した固形形態もまた調製され得る。また、該調製物は乳化されたものであってもよい。免疫学的活性成分は、多くの場合、薬学的に許容され得、活性成分と適合性である賦形剤と混合される。好適な賦形剤は、例えば、水、生理食塩水、デキストロース、グリセロール、エタノールなど、およびその組合せである。また、所望の場合、ワクチンに、ワクチンの有効性を増強する少量の補助物質、例えば、湿潤剤もしくは乳化剤、pH緩衝剤またはアジュバントなどを含めてもよい。ワクチンは、注射によって非経口で、例えば、皮下または筋肉内のいずれかで慣用的に投与される。他の投与様式に適したさらなる製剤としては、坐薬および、場合によっては経口製剤が挙げられる。坐薬では、従来の結合剤および担体として、例えば、ポリアルキレン(alkalene)グリコールまたはトリグリセリドが挙げられ得る。かかる坐薬は、0.5%〜10%、好ましくは1〜2%の範囲の活性成分を含有する混合物から形成され得る。経口製剤は、例えば、医薬等級のマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなどの通常使用される賦形剤を含む。このような組成物は、液剤、懸濁剤、錠剤、丸剤、カプセル剤、徐放製剤または粉末剤の形態をとり、10%〜95%、好ましくは25〜70%の活性成分を含有する。
【0053】
また、ワクチンは、ジェット式注射器、極微針、エレクトロポレーション、ソノポレーション、マイクロカプセル封入、ポリマーまたはリポソームを用いる経皮的経路、ネブライザー、エーロゾルおよび経鼻スプレー剤用いる経粘膜経路および鼻腔内経路により投与され得る。天然または合成のポリマー、例えば、デンプン、アルギネートおよびキトサン、D−ポリL−ラクテート(PLA)、D−ポリDL−乳酸−コグリコール酸ミクロスフィア、ポリカプロラクトン、ポリオルトエステル、ポリ無水物ならびにポリホスファゼン
ポリホスファタザン(phosphatazane)などを用いるマイクロカプセル封入は、経皮的経路および経粘膜投与の両方に有用である。合成ポリオルニテート、ポリリシンおよびポリアルギニンまたは両親媒性ペプチドを含むポリマー複合体は、経皮的送達系に有用である。また、その両親媒性の性質により、リポソームは、経皮的、経粘膜および鼻腔内ワクチン送達系に想定される。ワクチン送達に使用される一般的な脂質としては、N−(1)2,3−(ジオレイル−ジヒドロキシプロピル)−N,N,N−トリメチルアンモニウム−メチル硫酸塩(DOTAP)、塩化ジオレイルオキシプロピル−トリメチルアンモニウムDOTMA、ジミスチルオキシプロピル−3−ジメチル−ヒドロキシエチルアンモニウム(DMRIE)、臭化ジメチルジオクタデシルアンモニウム(DDAB)および9N(N’,N−ジメチルアミノエタン)カルバモイル)コレステロール(DC−Chol)が挙げられる。ヘルパー脂質とリポソームの組合せにより、皮膚を通したリポソームの取込みが増強される。このようなヘルパー脂質としては、ジオレオイルホスファチジルエタノールアミン(DOPE)、ジラウトイルホスファチジルエタノールアミン(DLPE)、ジミリストイルホスファチジルエタノールアミン(DMPE)、ジパルミトイルホスファチジルエタノールアミン(DPPE)が挙げられる。また、チリのユッカ(soap tree)樹皮から誘導されるトリテルペノイドグリコシドまたはサポニン(Quillaja saponaria)およびキトサン(脱アセチル化キタン(chitan))は、鼻腔内および経粘膜ワクチン送達に有用なアジュバントとして想定される。
【0054】
該タンパク質は、中性形態または塩形態としてワクチンに製剤化され得る。薬学的に許容され得る塩としては、酸付加塩(ペプチドの遊離アミノ基と形成される)および無機酸、例えば、塩酸もしくはリン酸など)と形成されるもの、または酢酸、シュウ酸、酒石酸、マンデル酸などの有機酸が挙げられる。また、遊離カルボキシル基と形成される塩は、無機塩基、例えば、ナトリウム、カリウム、アンモニウム、カルシウムまたは第2鉄の水酸化物など、ならびにイソプロピルアミン、トリメチルアミン、2−エチルアミノエタノール、ヒスチジンおよびプロカインなどの有機塩基から誘導され得る。
【0055】
ワクチンは、投薬製剤と適合性の様式で、治療上有効で免疫原性となるような量で投与される。投与される量は、処置される被験体、被験体の免疫系が抗体を合成する能力、および所望される防御の程度に依存する。投与に必要とされる活性成分の正確な量は、担当医師の判断に依存し、各個体に対して独自である。しかしながら、適当な投薬量範囲は、個体1人あたり活性成分ほぼ数百マイクログラム程度である。また、初期投与および追加免疫刺激注射に好適なレジメン(regime)は種々であるが、初期投与後、1ないし3ヶ月間隔で後続の注射または他の投与を行なうのが典型的である。
【0056】
本明細書に記載のワクチン組成物での免疫処置の際、宿主の免疫系は、所望の抗原に特異的なCTLを大量に産生することによりワクチンに対して応答し、宿主は、後の感染に対して少なくとも部分的に免疫性となるか、または慢性感染の発症に対して抵抗性となる。本発明のNTHiポリペプチドまたはNTHiペプチドを含有するワクチン組成物は、細菌感染に易感染性あるいはそのリスクがある患者に対して、その抗原に対する免疫応答を惹起し、したがって、患者自身の免疫応答能を増強するために投与される。この使用において、この場合も、正確な量は、患者の健康状態および体重、投与様式、製剤の性質などに依存するが、一般的に、約1.0μg〜約5000/70キログラム患者、より一般的には約10〜約500mg/70kg体重の範囲である。治療目的または免疫処置目的のため、本発明のNTHiポリペプチドまたはNTHiペプチドはまた、弱毒化ウイルス宿主、例えば、ワクシニアまたは鶏痘によって発現させ得る。このアプローチは、本発明のペプチドをコードするヌクレオチド配列を発現させるためのベクターとしてワクシニアウイルスの使用を伴う。急性または慢性感染宿主または非感染宿主内に導入されると、組換えワクシニアウイルスは免疫学的ペプチドを発現し、それにより宿主CTL応答が惹起される。
【0057】
体液性免疫応答は、多くのよく知られた方法、例えば、単純放射免疫拡散アッセイ(Single Radial Immunodiffussion Assay)(SRID)、酵素イムノアッセイ(EIA)および血球凝集阻害アッセイ(HAI)などによって測定され得る。特に、SRIDでは、試験対象の免疫原を含有するゲル(例えば、アガロースなど)の層が用いられる。ゲルを切り出して凹部を作り、試験対象の血清を凹部内に入れる。ゲル内への抗体拡散により沈殿環の形成がもたらされ、その面積は試験対象の血清の抗体の濃度に比例する。EIAは、ELISA(酵素結合イムノアッセイ)としても知られ、試料中の全抗体を測定するために使用される。免疫原をマイクロタイタープレートの表面に吸着させる。試験血清を該プレートに曝露した後、酵素結合免疫グロブリン(例えば、IgGなど)に曝露する。プレートに付着した酵素の活性を、分光測光法などの任意の簡便な手段によって定量され、試験試料中に存在する免疫原に対して指向される抗体の濃度に比例する。HAIでは、免疫原、例えばウイルス系タンパク質がトリ赤血球(など)を凝集させる能力が利用される。該アッセイでは、中和抗体、すなわち、血液凝集を阻害し得る抗体が検出される。試験血清の希釈物を標準濃度の免疫原とともにインキュベートした後、赤血球を添加する。中和抗体の存在により、免疫原による赤血球の凝集が阻害される。細胞性免疫応答を測定するための試験は、遅延型過敏症の測定または標的免疫原に対するリンパ球の増殖性応答の測定を含む。
【0058】
分類不能型インフルエンザ菌(NTHi)
インフルエンザ菌は、小型の非運動性グラム陰性菌である。他のインフルエンザ菌株とは異なり、分類不能型インフルエンザ菌(NTHi)菌株は、多糖莢膜を欠き、時として「非莢膜性」と示される。NTHi菌株は、被包性菌株と遺伝学的に異なり、b型インフルエンザ単離菌よりも不均質性(heterogenous)である。NTHiは、ヒト宿主に対して複雑な一連の抗原を提示する。防御をもたらし得る可能な抗原としては、OMP、リポ多糖類、リポタンパク質、接着性タンパク質および非被膜タンパク質が挙げられる。
【0059】
ヒトは、インフルエンザ菌(influenze)の唯一の宿主である。NTHi菌株は、通常、鼻咽頭および後部口腔咽頭部を含む上気道、下気道ならびに女性の生殖管に宿する。NTHiは、ヒトにおいて広範囲の疾患、例えば、限定されないが、中耳炎、肺炎、副鼻腔炎、敗血症、心内膜炎、喉頭蓋炎、化膿性関節炎、髄膜炎、分娩後および新生児感染、分娩後および新生児セプシス、急性および慢性の卵管炎/卵管炎(salpingitis)、喉頭蓋炎(epiglottis)、心膜炎、蜂巣炎、骨髄炎、心内膜炎、胆嚢炎、腹腔内感染、尿路感染症、乳様突起炎、大動脈移植片感染症、結膜炎(conjunctitivitis)、ブラジル紫斑熱、不顕性菌血症ならびに潜在する肺疾患、例えば、慢性気管支炎、気管支拡張症(bronchietasis)および嚢胞性線維症などの増悪を引き起こす。
【0060】
NTHiの疫学的試験により、該菌株は、外膜タンパク質プロフィール(Barenlcampら,Infect.Immun.,36:535−40,1982)、酵素アロタイプ(Musserら,Infect.Immun.,52:183−191,1986)および他の一般的に使用される疫学的ツールに関して不均一であることが示された。亜型NTHiに対していくつかの試みがなされているが、方法論はいずれも完全に満足のいくものではなかった。NTHiの外膜タンパク質組成物は、ほぼ20のタンパク質からなる。すべてのNTHi菌株は、30,000および16,600ダルトンの分子量を有する2つの共通するOMPを含む。NTHi菌株は、32,000〜42,000ダルトンの範囲内の2つのOMPに基づいて亜型に分けられ得る。NTHiのリポ多糖プロフィールは、グラム陰性腸内細菌と基本的に異なり、20,000ダルトン未満の範囲における1〜4個の異なるバンドに分かれる。
【0061】
基本型のNTHi単離菌は、低継代単離菌86−028NPであり、これは、慢性中耳炎の小児から採取された。この菌株は、インビトロ(Bakaletzら,Infect.Immun.,53:331−5,1988;Holmesら,Microb.Pathog.,23:157−66,1997)ならびにチンチラOMモデル(本明細書に記載)(Bakaletzら,Vaccine,15:955−61,1997;Suzukiら,Infect.Immun.,62:1710−8,1994;DeMariaら,Infect.Immun.,64:5187−92,1996)において充分特性評価されている。86−028NP菌株は、本明細書に記載のように、中耳炎のチンチラモデルでの発現において上方調節される遺伝子、およびチンチラの中耳内でのNTHiの生存に必要な遺伝子を同定するために使用した。
【0062】
DFIストラテジー
異差蛍光誘導(DFI)ストラテジーを、本明細書において、チンチラ動物モデルのOM時に誘導されるNTHi遺伝子を同定するために使用した。感染中、菌体のビルレンスに寄与する細菌遺伝子を同定するため、いくつかの方法を開発した。かかる方法としては、インビボ発現手法(IVET)(この場合、細菌のプロモーターにより、宿主内での生存に必要とされる必須栄養素の合成に必要な遺伝子(1つまたは複数)の発現が調節される);シグニチャータグ化(signature−tagged)突然変異誘発(STM)(変異されると、微生物のビルレンス特性を改変する遺伝子のタグ特異的同定を可能にする;DNAマイクロアレイ手法(転写活性遺伝子を全体的にスクリーニングするため)およびDFI(FACS解析を用いて転写活性プロモーターを選択する(Chiangら,Annu.Rev.Microbiol.,53:129−154,1999)が挙げられる。DFIは、発現の基礎レベルとは無関係に差次的に調節される遺伝子の同定を可能にするハイスループット法であるが、インビトロでの培養に不可欠なものを排除しない。
【0063】
DFIは、多くの微生物において成功裏に利用されている。例えば、GFPレポーター系およびフローサイトメトリーを用いて、マクロファージとの相互作用の際のマイコバクテリウムの遺伝子発現が研究された(Dhandayuthapaniら,Mol.Microbiol,17:901−912,1995)。プロモータートラップ系を用いて、サルモネラ属がインビボ増殖を刺激する環境に供された場合、および培養マクロファージ様細胞によってインターナリゼーションされた場合に、その転写が増大する遺伝子が同定された(ValdiviaおよびFalkow,Mol.Microbiol,22:367−378,1996;ValdiviaおよびFalkow,Science,277:2007−2011,1997;ValdiviaおよびFalkow,Curr.Opin.Microbiol,1:359−363,1998)。また、DFIを用いて、感染を刺激する種々のインビトロ条件で培養すると、S.pneumoniaeおよびS.aureusにおいて発現されるプロモーターが同定されている(Marraら,Infect.Immun.,148:1483−1491,2002;Schneiderら,Proc.Natl Acad.Sci.U.S.A.,97:1671−1676,2000)。また、DFIを用いて、環境性刺激に応答したBacillus cereusにおける遺伝子調節(DunnおよびHandelsman,Gene,226:297−305,1999)、能力刺激ペプチドに応答したS.pneumoniaeにおける遺伝子調節(Bartilsonら,Mol.Microbiol,39:126−135,2001)、ならびに宿主細胞との相互作用時および該細胞内への侵入時のBartonella henselae(LeeおよびFalkow,Infect.Immun.,66:3964−3967,1998)、Listeria monocytogenes Wilsonら,Infect.Immun.,69:5016−5024,2001)、Brucella abortus(Eskraら,Infect.Immun.,69:7736−7742,2001)および大腸菌(Badgerら,Mol.Microbiol,36:174−182,2000)における遺伝子調節が研究された。
【0064】
DFIは、インビボ環境を刺激するために設計された感染の細胞培養モデルまたはインビトロ条件において活性なプロモーターを同定するために成功裏に使用されたが、DFIは、完全体の動物内の特定の生物学的ニッチにおいて調節されるプロモーターを同定するためには、ほとんど適用されていない。これは、おそらく、インビボ環境からの分取に伴う数多くの課題ためである。宿主の炎症性応答、播種および/または感染部位からの菌体のクリアランス、ならびに上皮細胞への細菌の付着(おそらく、バイオフィルム形成による)により、細菌は、生体動物からの回復に利用不可能となり得る。これらの要素は、とりわけ、該細菌がこのような変化を感知し、これに応答すると、微小環境の複雑さ、および遺伝子発現の不均一性に寄与する。最近、DFIを用い、該細菌を気道感染のマウスモデルおよびOMのアレチネズミ感染モデルにおいてスクリーニングすると、S.pneumoniaeにおいて発現されるプロモーターが同定された(Marraら,Infect.Immun.70:1422−33,2002;Marraら,Microbiol,148:1483−91,2002)。
【0065】
動物モデル
チンチラモデルは、広く認められたOMの実験モデルである。特に、NTHi誘導型OMのチンチラモデルは充分特性評価されており(Bakaletzら,J.Infect.Dis.,168:865−872,1993;BakaletzおよびHolmes,Clin.Diagn.Lab.Immunol.A:223−225,1997;SuzukiおよびBakaletz,Infect.Immun.,62:1710−1718,1994)、OMに対する痘苗原としてのいくつかのNTHi外膜タンパク質、外膜タンパク質の組合せ、キメラ合成ペプチドワクチン成分およびアジュバント製剤の防御的有効性を調べるために使用されている(Bakaletzら,Vaccine,15:955−961,1997;Bakaletzら,Infect.Immun.,61:2746−2762,1999;Kennedyら,Infect.Immun.,68:2756−2765,2000)。
【0066】
特に、アデノウイルスがチンチラにインフルエンザ菌誘導型中耳炎をもたらす独自のインビボモデルが存在し、これにより、NTHiの生物学的評価のための関連する細胞、組織および器官の培養系の確立が可能となった(Bakaletzら,J.Infect.Dis.,168:865−72,1993;Suzukiら,Infect.Immunity 62:1710−8,1994)。アデノウイルス感染を単独で用い、誘導された血清抗体の鼓室内への漏出が評価され(Bakaletzら,CHn.Diagnostic Lab Immunol,4(2):223−5,1997)、NTHiとともに共病原体として用い、中耳炎に対する痘苗原としての種々のNTHi外膜タンパク質、OMPの組合せ、キメラ合成ペプチドワクチン成分およびアジュバント製剤を標的化するいくつかの能動および受動免疫処置レジメンの防御的有効性が調べられた(Bakaletzら,Infect Immunity,67(6):2746−62,1999;Kennedyら,Infect Immun.,68(5):2756−65,2000;Novotnyら,Infect Immunity 68(4):2119−28,2000;Poolmanら,Vaccine 19(Suppl.1):S108−15,2000)。
【0067】
中耳のNTHi感染に応答してインビボで上方調節される遺伝子
中耳のNTHi感染に応答して差次的に調節されるプロモーターを同定するため、プロモータートラップライブラリーを構築し、分取パラメータを規定した。プロモータートラップライブラリーの一部分をチンチラの中耳内に直接接種し、OM発症を、ビデオ耳鏡検査およびティンパノメトリによって24時間および48時間の時点でモニターした。また、中耳液を感染24時間および48時間後に採取した。2色FACS解析を用い、GFPを発現している細菌を、他の細胞および滲出液に付随する残屑から単離した(isolated)。単離後、gfpmut3遺伝子の5’のインフルエンザ菌挿入配列のDNA配列を決定し、解析した。このようにした、本発明者らは、AOM時にNTHiがチンチラの中耳環境を感知し、これに応答すると上方調節される遺伝子を同定した。以下の遺伝子が同定され、これらは、NTHi感染時に上方調節されるため、NTHi感染およびビルレンスにおいて役割を果たしている可能性がある。
【0068】
実施例7において後述するように、gfp発現クローンの上記のDFI手順および続くFACS解析の後、潜在的なインビボ調節プロモーターを含む52個の候補クローンが単離された。これらのクローンが制御する遺伝子を、概要および細胞内での機能に基づいて分類し、一般的な代謝過程、環境情報的プロセッシングおよび膜輸送、膜タンパク質および仮想タンパク質を含む。これらの52個のクローンのうち8個は、NTHi86−028NP菌株に特異的配列を含む。重要なことに、3個のクローンを、1匹より多くの動物における独立したスクリーニングで単離し、それにより単離方法を確認した。
【0069】
該FACSデータを独立して確認するため、本発明者らは、候補遺伝子の相対発現を定量的RT−PCRによって調べた。親86−028NP菌株を本試験に使用した。したがって、遺伝子調節に対してプラスミドコピー数の影響がない野生型遺伝子発現を解析し、FACSによる偽陽性クローン同定の識別を可能にした。インフルエンザRd菌株で同定されたものと同様の配列を含む44個の候補クローンのうち、26個の遺伝子(60%)について、インビトロおよびインビボでの遺伝子発現の定量的比較により、NTHiがチンチラの中耳内に存在する環境的なきっかけ(cue)に応答すると、上方調節された遺伝子発現が確認された。この解析により、膜輸送、環境情報的プロセッシング、細胞性代謝、遺伝子調節ならびに未知機能を有する仮想タンパク質に関与する遺伝子の発現を駆動するインビボ調節型プロモーターが同定された。(実施例6の表4参照)。
【0070】
定量的RT−PCRにより、lolA発現の2倍増加が示された。内膜から外膜へのリポタンパク質輸送が可能となった。細菌は、中耳環境内で急速に増殖し、48時間以内に、中耳液1mlあたり5.0×108CFU NTHiに達する。細菌は、該環境を感知し、これに応答して、増殖および生存のための必須栄養素を獲得または合成する。リボース糖輸送における膜成分をコードする遺伝子rbsC(配列番号:619)は、インビトロでの細胞培養と比べ、インビボでは5倍の発現増加を示した。また、代謝過程に関与する多くの遺伝子は、インビトロでの細胞培養と比べ、インビボでは遺伝子発現の劇的な増加を示す。これらとしては、リボフラビン合成遺伝子HbB(配列番号:623)、プリンヌクレオチド生合成遺伝子purE(配列番号:621)、尿素回路によるアルギニン分解に関与するオルニチンカルバモイルトランスフェラーゼarcB(配列番号:625)、ならびにD−グルクロネートの取込みおよびグリセルアルデヒド3−リン酸への形質転換に必要とされるマンノネート(mannonate)ヒドロラーゼをコードするuxuA(配列番号:627)が挙げられる。また、程度は低いが、ヒスチジン生合成のための遺伝子(hisB;配列番号:615)、DNA修復のための遺伝子(radC;配列番号:639)および推定細胞内隔壁化膜貫通タンパク質の遺伝子(ispZ;配列番号:637)も上方調節された。
【0071】
ジスルフィド結合形成は、細菌内での多くの分泌タンパク質のフォールディングおよび合成に重要である。原核生物では、DsbAおよびDsbBがジスルフィドの形成を担う酸化的経路を構成する。DsbBはDsbAを再酸化し、これは、ジスルフィドが変性ポリペプチドに直接結合することを表す。DsbBは、酸化キノンから新たにジスルフィドを生成させることが示されている(ColletおよびBardwell,Mol.Microbiol,44:1−8,2002)。インフルエンザRd菌株では、DsbAは、形質転換能に必要とされる(Tomb,Proc.Natl.Acad.Sd.U.S.A.,89:10252−10256,1992)。ここでは、ほぼ3倍のdsbB遺伝子(配列番号:629)の転写増加が示され、中耳環境内で増殖するTHiに対するジスルフィド相互交換の重要な役割が明らかになった。
【0072】
中耳での細菌のコロニー形成は、正常な滅菌環境であり、宿主炎症性応答および続いて好中球浸潤をもたらす。細菌は、この宿主の応答に対処するため、数多くの戦略を進化させてきた。NTHiは、ヘリコバクター属において活性ウレアーゼの発現に必要とされ、酸耐性に関与することが示されている(Youngら,J.Bacterol.,178:6487−6495,1996)遺伝子のホモログであるureH(配列番号:631)遺伝子発現を増大させる(4倍)。最近、ウレアーゼ活性は、感染時に起こるpHの減少を打ち消すことにより、慢性のActinobacillus pleuropneumoniae感染において役割を果たしている可能性があることが報告された(Baltesら,Infect.Immun.,69:472−478,2000;Baltesら,Infect.Immun.,69:472−478,2001;BosseおよびMachines,Can.J.Vet.Res.,64:145−150)。中耳滲出液由来のNTHi単離菌に関するバイオタイプ解析により、87%はウレアーゼ陽性であることが示された(DeMariaら,J.Clin.Microbiol,20:1102−1104,1984)。しかしながら、NTHiビルレンスにおけるウレアーゼの役割は未知である。同様に、その産物がS.pneumoniaeのピリドキシン生合成タンパク質に対して88%の配列同一性および抗酸化剤としての機能を果たし得る推定一重項酸素抵抗性タンパク質に対して60%相同性を示す遺伝子発現の増大。ホスホリルコリン(ChoP)は、NTHiの病因に関与している(Weiserら,Infect.Immun.,65:943−950,1997)。NTHiは、phase variationによってChoP発現モジュレートし、細胞表面上のLOSを修飾する。ChoPは、抗菌性ペプチドに対する感受性を低下させることによって気道内のNTHi残留に寄与し得(Lyseckoら,Infect.Immun.,68:1664−1671,2000)、C反応性タンパク質(CRP)によって媒介される血清殺傷性に対する感受性を改変し得る(Weiserら,J.Exp.Med.,187:631−640,1998)。チンチラ鼻咽頭では、ChoP+菌株がより多くのコロニー形成を示すため、鼻咽頭および中耳腔の微小環境ではChoP+表現型が選択され得る(Tongら,Infect.Immun.,68:4593−4597,2000)。また、licC遺伝子(配列番号:633)の発現も増大された。licC遺伝子は、ホスホリルコリン誘導性LOSの生合成において役割を果たすホスホリルコリンシチジリルトランスフェラーゼをコードする(Rockら,J.Bacterol,183:4927−4931,2001)。
【0073】
また、インビボ誘導型遺伝子のうち、その産物が、続いて遺伝子発現またはDNA複製を調節する一連のものも含まれる。このような遺伝子としては、glpリプレッサーglpR(配列番号:643)によるグリセロール代謝の転写調節、アルギニン抑制因子遺伝子argR(配列番号:647)、および組込み宿主因子(IHF)βサブユニットihfB(配列番号:645)が挙げられる。IHFはヒストン様タンパク質であり、これは、特定配列のDNAに結合し、複製、部位特異的組換えおよび転写に関与するアクセサリー因子であり、多数のオペロンの活性を改変させる(Goosenおよびvan de Putte,Mol.Microbiol.16:1−7,1995)。また、CspDは、大腸菌において定常期誘導性ストレス応答の際、DNA複製を阻害し(Yamanakaら,Mol.Microbiol,39:1572−1584,2001)、mukF(配列番号:641)遺伝子タンパク質ホモログは、細胞分離前に、よりコンパクトな形態へのヌクレオチド(nucleiod)構造のリモデリングに寄与する(SawitzkeおよびAustin,Proc.Natl.Acad.ScL U.S.A.,62:1710−1718,2000)。また、本明細書に記載のDFIストラテジーにより、未知機能の遺伝子に関してインビボで誘導されるプロモーターが同定された。仮想タンパク質HI0094では、OM初期に遺伝子発現の8倍増加が示されたが、その役割は依然として未知である。HI1163(配列番号:651)は、大腸菌の推定オキシダーゼである仮想YdiJタンパク質と、58%アミノ酸同一性を示した。
【0074】
高密度トランスポゾン変異誘発ストラテジーを用いて、富化培地での増殖に不可欠なインフルエンザ菌遺伝子が同定された(Akerleyら,Proc.Natl.Acad.Sd.U.S.A.,99:966−971,2002)。6つの遺伝子が、本明細書に記載のスクリーニングにおいて同定され、これらは、Akerleyら、前掲に記載の必須遺伝子の組に含まれる(hisB、lppB、lolA、ispZ、mukFおよび未知HI0665)。最近、2つのヒト気道由来上皮細胞株と相互作用すると発現される分類不能型インフルエンザ菌の遺伝子が同定された。このような遺伝子には、代謝過程、ストレス応答、遺伝子発現、細胞外被の生合成、DNA関連プロセス、細胞分裂および未知機能のORFのコードタンパク質に関与するものが含まれた。(Ulsenら,Mol.Microbiol.,45:485−500,2002)。同様に、ストレス応答遺伝子であるcspD(配列番号:649)、プリンおよびリボフラビンの生合成に関与する遺伝子、および未知機能のvapAが、本明細書に記載のスクリーニングにおいて同定された。vapAの発現はインビトロで検出されたが、vapA遺伝子発現は、インビボでは2倍増加した。これらの特有のアプローチでは、NTHi誘導型OMにおいて上方調節され、したがって、おそらくNTHi感染およびビルレンスにおいて役割を果たしており、NTHi関連障害のワクチンおよびアンチセンス療法ならびに他の治療的処置方法の潜在的候補であり得る既知遺伝子が同定された。
また、DFIストラテジーにより、未知機能の遺伝子に関してインビボで誘導されるプロモーターの同定がもたらされた。仮想タンパク質HI0094では、OM初期に遺伝子発現において8倍増加が示されたが、その役割は依然として未知である。HI1163(配列番号:651)は、大腸菌の推定オキシダーゼである仮想YdiJタンパク質と、58%アミノ酸同一性を示した。したがって、このような仮想遺伝子は、おそらく、NTHi感染によって誘導されるOMにおいて役割を果たしている。
例えば、本発明は以下の項目を提供する。
(項目1)
配列番号:577〜579、配列番号:589〜614、配列番号:675〜685、配列番号:615、配列番号:617、配列番号:619、配列番号:621、配列番号:623、配列番号:625、配列番号:627、配列番号:629、配列番号:631、配列番号:633、配列番号:635、配列番号:637、配列番号:639、配列番号:641、配列番号:643、配列番号:645、配列番号:647、配列番号:649、配列番号:651、配列番号:653、配列番号:655、配列番号:657、配列番号:659、配列番号:661、配列番号:663、配列番号:665、配列番号:667、配列番号:669、671、配列番号:673、配列番号:686、配列番号:688、配列番号:692、配列番号:694、配列番号:696、配列番号:698、配列番号:700、配列番号:702、配列番号:704、配列番号:706、配列番号:708、配列番号:710、配列番号:712、配列番号:714、配列番号:716、配列番号:718、配列番号:720、配列番号:722、配列番号:724、配列番号:726、配列番号:728、配列番号:730、配列番号:732、配列番号:734、配列番号:736、配列番号:738、配列番号:740、配列番号:742、配列番号:744、配列番号:746、配列番号:748、配列番号:750、配列番号:752、配列番号:754、配列番号:756、配列番号:758、配列番号:760、配列番号:762、配列番号:764、配列番号:766、配列番号:768 配列番号:770または配列番号:773〜2593のいずれか1つのヌクレオチド配列を含む、単離されたポリヌクレオチド。
(項目2)
項目1に記載のヌクレオチド配列の組またはその断片にコードされるアミノ酸配列を含む、単離されたポリペプチド。
(項目3)
配列番号:616、配列番号:618、配列番号:620、配列番号:622、配列番号:624、配列番号:626、配列番号:628、配列番号:628、配列番号:630、配列番号:632、配列番号:634、配列番号:636、配列番号:638、配列番号:640、配列番号:642、配列番号:644、配列番号:646、配列番号:648、配列番号:650、配列番号:652、配列番号:654、配列番号:656、配列番号:658、配列番号:660、配列番号:662、配列番号:664、配列番号:666、配列番号:668、配列番号:670、配列番号:672、配列番号:674、配列番号:687、配列番号:689、配列番号:691、配列番号:693、配列番号:695、配列番号:697、配列番号:699、配列番号:701、配列番号:703、配列番号:705、配列番号:707、配列番号:709、配列番号:711、配列番号:713、配列番号:715、配列番号:717、配列番号:719、配列番号:721、配列番号:723、配列番号:725、配列番号:727、配列番号:729、配列番号:731、配列番号:733、配列番号:735、配列番号:737、配列番号:739、配列番号:741、配列番号:743、配列番号:745、配列番号:747、配列番号:749、配列番号:751、配列番号:753、配列番号:755、配列番号:757、配列番号:759、配列番号:761、763、配列番号:765、配列番号:767、配列番号:769、配列番号:771または配列番号:2581〜4414のいずれか1つのアミノ酸配列を含む、単離されたポリペプチド。
(項目4)
項目2または3に記載のポリペプチドおよび薬学的に許容され得る担体を含む組成物。
(項目5)
項目2もしくは3に記載のポリペプチドまたはその断片に特異的に結合する抗体。
(項目6)
項目5に記載の抗体および薬学的に許容され得る担体を含む組成物。
(項目7)
生物学的試料内のNTHi菌を検出するための方法であって、
(a)項目1に記載のポリヌクレオチドまたはその断片を生物学的試料と接触させる工程、および
(b)該試料内での該ポリヌクレオチドのハイブリダイゼーションを検出する工程、
を包含する、方法。
(項目8)
生物学的試料内のNTHi菌を検出するための方法であって、
(a)項目5に記載の抗体と生物学的試料を接触させる工程、および
(b)該試料内での該抗体の結合を検出する工程
を包含する、方法。
(項目9)
前記生物学的試料が、血清、痰、耳液、血液、尿、リンパ液および脳脊髄液からなる群より選択される、項目7または8に記載の方法。
(項目10)
NTHi菌に対する免疫応答を惹起させるための方法であって、免疫原性的に有効な用量の項目2または3に記載のポリペプチドまたはその断片を、NTHi菌感染のリスクのある患者に投与する工程を包含する、方法。
(項目11)
項目2または3に記載のポリペプチドまたはその断片および医薬用に適した担体を含むワクチン。
(項目12)
前記免疫応答が、NTHi菌の死滅、NTHi菌の複製の遅滞またはNTHi菌の結合の阻止の1種類以上を含む項目10に記載の方法。
(項目13)
前記NTHi感染が中耳内である、項目10に記載の方法。
(項目14)
NTHi菌感染を処置または予防する方法であって、項目2または3に記載のポリペプチドの発現または活性を阻害する分子を、処置または予防を必要とする患者に投与する工程を包含する、方法。
(項目15)
前記NTHiポリペプチドが、hisB、lppB、sapA、rbsC、pure、rib、arcB、uxuA、licC、ispZ、mukF、glpR、ihfB、cspD、lav、HI1647、HI0094、HI1163、HI0665、HI1292、HI1064、HI1386、HI1462、HI1369およびHI1598からなる群より選択されるNTHi遺伝子にコードされる、項目14に記載の方法。
(項目16)
前記NTHiポリペプチドが、配列番号:616、配列番号:618、配列番号:620、配列番号:622、配列番号:624、配列番号:626、配列番号:628、配列番号:634、配列番号:638、配列番号:642、配列番号:644、配列番号:646、配列番号:650、配列番号:652、配列番号:656、配列番号:658、配列番号:660、配列番号:662、配列番号:664、配列番号:666、配列番号:668、配列番号:670、配列番号:672および配列番号:674からなる群より選択されるアミノ酸配列を含む、項目14に記載の方法。
(項目17)
前記必要とする患者に投与される前記分子がアンチセンスオリゴヌクレオチドである、項目14に記載の方法。
(項目18)
前記必要とする患者に投与される前記分子が抗体である、項目14に記載の方法。
(項目19)
前記必要とする患者に投与される前記分子が小分子である、項目14に記載の方法。
(項目20)
前記NTHi感染が中耳内である、項目14に記載の方法。
【図面の簡単な説明】
【0075】
【図1】図1は、一群のインフルエンザ単離菌におけるLKP遺伝子領域を示す。86−028NP菌株の配列は、この領域において、NTHi菌株R3001の配列と同一である。これらのNTHiはともに、赤血球凝集性線毛(hemagglutinating pilus)をコードするhif遺伝子クラスターを欠く。
【図2】図2は、一群のインフルエンザ単離菌におけるrfaD領域を示す。86−028NP菌株ゲノムのrfaD領域内の遺伝子構成は、Rd菌株のゲノムのものと類似するが、調べたほとんどのNTHiのゲノムに見られるこれらの遺伝子の構成と異なる。
【図3A】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3B】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3C】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3D】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3E】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3F】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3G】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3H】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3I】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3J】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3K】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3L】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3M】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【発明を実施するための形態】
【0076】
(詳細な説明)
以下の実施例は、本発明を説明する。ここで、実施例1はNTHiゲノムの配列を示し、実施例2は同定されたコンティグおよび最初の遺伝子発見を示し、実施例3はNTHi
プロモータートラップライブラリーの構築を示し、実施例4はGFPを発現する86−028NP誘導体の解析を示し、実施例5は中耳液由来の細菌の直接標識を示し、実施例6は、急性中耳炎においてインビボで誘導されるプロモーターの同定を示し、実施例7はビルレンス関連遺伝子の同定を示し、実施例8は、NTHi特異的遺伝子配列の同定を示し、実施例9は、完全NTHi86−028NPゲノムの解析を示し、実施例10では、NTHi86−028NP菌株およびインフルエンザ菌d血清型菌株kw20のゲノムDNA配列を比較している。
【0077】
実施例1
分類不能型インフルエンザ菌のゲノムの配列
NTHi86−028NP菌株は、コロンブス小児病院において、慢性OMのため鼓膜切開およびチューブ挿入を受けた小児患者から採取された、最小限に継代された臨床用単離菌である(Bakaletzら Infection and Immunity,56(2):331−335,1988)。86−028NP菌株は、American Type Tissue Collection(Manassas,VA 20108
USA)に2002年10月16日に寄託し、受託番号PTA−4764が付与された。
【0078】
NTHiのビルレンス決定基の同定に、より広くアプローチするための取り組みにおいて、NTHi86−028NP菌株のゲノムを3倍の範囲まで配列決定した。Puregeneプロトコルを用いて86−028NP菌株から染色体DNAを調製し、Hydroshear装置(Gene Machines)により2〜4kbの大きさに切断した。切断されたDNAをエタノール沈殿させ、クレノウ酵素とT4 DNAポリメラーゼの混合物を用いて末端修復し、アガロースゲル電気泳動によってサイズ選択し、Chissoeら(Methods:a Companion to Methods ofEnzymology 3:55−65,1991)およびSambrookら(Molecular Cloning:a Laboratory Manual,第2版 Cold Springs Harbor,NY,1989)に記載のような2〜4kb断片を得た。これらの断片をベクターpUC18内に、Smal制限部位(ホスファターゼ処理済)を用いてクローニングし、大腸菌XL−I Blue内に形質転換し、アンピシリン耐性に関して選択した。挿入物を含むコロニーを、X−galを入れたLB−Ampプレート上にて青/白スクリーニングによって同定し、1.5mlのTB−Amp(TB=Terrific Broth)培養液を入れた96ディープェルプレートに移した。このディープェルプレート培養物を、一晩(18〜22時間)37℃で培養した。鋳型の調製、配列決定およびコンティグ編集を行なった。
【0079】
自動鋳型調製を、Chissoeら、前掲に記載のBeckman Biomek 2000自動ロボットワークステーションにおいて行なった。簡単には、上記で調製されたクローンの入った各96ディープェルプレートを遠心分離して細胞をペレット化し、上清みを廃棄し、細胞を−20℃で凍結させた(必要な場合)。4つの96ディープェルブロックをBiomekの台上に配置し、液体処理ロボットを用い、Chissoeら、前掲に記載の自動型の典型的なSDS−NaOH溶解プロトコルを使用して鋳型を調製した。最後にエタノール沈殿させた鋳型を、各々、50μlのddH2Oに溶解し、DNA配列決定に使用した。
【0080】
配列決定反応は、Robbins Hydra 96ロボットを用いて、鋳型(96ウェルプレートから)を384ウェルプレートに再度整列させることにより行なった。PE
Big−Dye(登録商標)ターミネーターおよび普遍的プライマー(M13フォワードおよびリバース)用いてサイクル型配列決定反応を行ない、Sephadex G50カラム上で浄化し、PE Biosystems 3700キャピラリー電気泳動DNA配列決定装置において製造業者の使用説明書に従って解析した。配列決定の読み値(8219)を576個のコンティグ(本明細書における配列番号:1〜576)に編集した。3倍配列決定の統計学的データ表2Aに示す。アセンブリ17の全特異的配列は1.74Mbである
【0081】
【表2】
続いて、NTHiゲノムの8倍配列決定解析を行なった。8倍配列決定により、NTHiゲノムを11個のコンティグに編集した。コンティグ5、8、9、10、12〜18を、本明細書において配列番号:675〜685として示す。8倍配列決定の統計学的データを表2Bに示す。
【0082】
【表3】
実施例2
コンティグの説明および所期遺伝子の発見
3倍配列解析で同定された≧5000bp長さを有する88個のコンティグのうち75個は、BLASTNにより、インフルエンザRd菌株の遺伝子と有意な類似性を示す。インフルエンザ86−028NP菌株における遺伝子順序とインフルエンザRd菌株との潜在的関連性を視覚化するため、86−028NP 3倍コンティグの組とRd遺伝子の組を、BLASTNを用いて双方向で比較した。コンティグをRd遺伝子ヒット遺伝子の座標に基づいて分類し、各コンティグを、Rayら,(Bioinformatics 17:1105−12,2001)に記載のとおりに見られる最小座標に固定(anchor)することにより、結果を遺伝子順序対コンティグの間隙(space)においてプロットした。このようにして比較し、完全に既知のゲノムと同一の遺伝子順序を有するゲノムの不完全なアセンブリにより、単調に増大する段階工程の形態が示され得る。
【0083】
BLASTXを用い、Rd菌株のゲノム内の遺伝子ならびにインフルエンザRd菌株に見られない遺伝子に対して相同性を有する配列のヒットを同定した。Rd菌株配列に対するヒットはデsータセットから除外し、その他のヒットを表3Aにまとめる。データは以下:コンティグ#(配列番号:#)、第1欄;各ヒットのEスコア、第2欄;記載のコンティグのアミノ酸翻訳部分に対して相同性を有したタンパク質の名称、第3欄;該ホモログを産性する生物体、第4欄;第3欄に示したタンパク質の各々のGenbank確認用タンパク質、第5欄;コンティグ内の対応するヌクレオチド(配列番号:で示す)のように表示する。ほとんどの場合でいくつかのホモログが同定されたが、明確にするため、相同性の大きいタンパク質を表3Aに示す。
【0084】
表3Aに示した一部の遺伝子の配列を、NTHiゲノムの8倍配列決定において同定した。表3Bに、11個のコンティグ内のこれらの遺伝子の位置、完全長オープンリーディングフレーム配列(配列番号:で特定)、オープンリーディングフレームにコードされる誘導アミノ酸配列、およびBLASTXによって同定された高い相同性を有する遺伝子(表3Aに記載)を示す。
【0085】
NTHiおよびRd菌株における比較的短い範囲の遺伝子構成を調べるため、充分報告された2つの遺伝子クラスター内の遺伝子順序を比較した。まず、赤血球凝集性線毛(LKP)遺伝子領域内に存在する遺伝子を調べた。(Mhlanga−Mutangaduraら,J Bacteriol.180(17):4693−703,1998)。線毛遺伝子クラスターは、purE遺伝子とpepN遺伝子の間に位置し、その断片のみを図1に示す。b血清型の菌株であるEaganは、hifABCDE遺伝子クラスターを含み、赤血球凝集性線毛を産生する。Rd菌株は、hicAB遺伝子ならびにhifABCDE遺伝子クラスターを欠く。一般に、これまでに調査された分類不能型菌株は、hicAB遺伝子を含んでいたが、赤血球凝集性線毛をコードするhif遺伝子は含んでいなかった。86−028NP菌株の配列(本明細書に記載)は、この領域において、NTHi菌株R3001の配列と同一である(図1)。rfaD遺伝子は、内毒素の生合成に関与する酵素をコードする。また、NTHi 2019菌株由来のrfaD遺伝子は、Nicholsらによって特性評価された(Infect Immunity 65(4):1377−86,1997)。2019菌株において、rfaD遺伝子は、内毒素生合成に関与する別の酵素をコードするrfaF遺伝子のすぐ上流に存在する。Rd菌株の遺伝子構成は異なる;rfaD遺伝子とrfaF遺伝子は、ほぼ11kbの配列によって分断されている。調査されたほとんどの分類不能型菌株は、2019菌株に見られる遺伝子構成を含んでいた。対照的に、86−028NP菌株は、Rd菌株に見られるものと同一の遺伝子構成を有する(図2)。
【0086】
現在の(current)アセンブリの大域解析により、遺伝子の含有量および順序は、Rd菌株のものと類似していることが示される。より詳細な解析によって、PasteurellaceaeおよびNTHi遺伝子の含有量と順序がRd菌株に見られるものと異なる一部の領域では、これまでにわかっていない相当な数のNTHi遺伝子が存在することが明らかになった。したがって、現在のデータにより、86−028NP菌株ゲノムは、Rdおよび非Rd様の特徴の複雑な寄せ集めを含むことが示される。
【0087】
また、DFIストラテジーにより、遺伝子発現が増大された新規なNTHi配列が同定された。他の生物体(主にグラム陰性細菌)ORFに対して相同性を有する遺伝子または遺伝子断片を含有するこのような新規なコンティグ配列の一覧を表3Aに示す。例えば、コンティグ442(配列番号:442)のヌクレオチド配列であるヌクレオチド1498〜1845は、インフルエンザRd菌株のリポタンパク質B(LppB)のアミノ酸1〜116をコードする配列に対して高度に相同である。該遺伝子は、定常期生存遺伝子urEと、最近同定され細菌リポタンパク質LppB/NlpD(これは、ビルレンスと関連している)に対して高度に相同な43kDの抗原性外膜リポタンパク質をコードする遺伝子との間に位置する(Padmalayamら,Infect.Immun.,68:4972−4979,2000)。最近、Zhangおよび協働者らによって、nlpDおよびsurE遺伝子発現は、Thermotoga maritimaにおいて細菌増殖の定常期に誘導されることが示された(Zhangら,Structure(Camb),9:1095−1106,2001)。したがって、中耳内のストレス誘導条件下では、このNTHiリポタンパク質が発現され得る。
【0088】
【表4】
【0089】
【表5】
【0090】
【表6】
【0091】
【表7】
【0092】
【表8】
【0093】
【表9】
【0094】
【表10】
【0095】
【表11】
【0096】
【表12】
【0097】
【表13】
【0098】
【表14】
【0099】
【表15】
【0100】
【表16】
【0101】
【表17】
【0102】
【表18】
【0103】
【表19】
【0104】
【表20】
【0105】
【表21】
【0106】
【表22】
【0107】
【表23】
実施例3
NTHiプロモータートラップライブラリーの構築
NTHiの潜在的ビルレンス決定基を同定するため、NTHi誘導型中耳炎(OM)のチンチラモデルの特定の1つの解剖学的ニッチにおいて、疾患進行初期に、細菌の遺伝子発現をディファレンシャル蛍光誘導(DFI)によってモニターした。NTHi86−028NP菌株のゲノムDNA断片を、プロモータートラップライブラリーを用いてプロモーターなしのgfpmut3遺伝子の上流にクローニングした。プラスミドpGZRS39Aは、Actinobacillus pleuropneumoniaeから単離されたpGZRS−1の誘導体であり、A.pleuropneumoniae−大腸菌シャトルベクターである。このプラスミドは、A.pleuropneumoniae由来の複製起点、pUC19由来のlacZa遺伝子およびTn903由来のカナマイシン耐性遺伝子を含む(Westら,Genes,160:81−86,1995)。
【0108】
プロモータートラップベクターは、GTP変異型gfpmufi遺伝子をBamHIからEcoRI方向の断片としてpGZRS−39A内にクローニングし、pRSM2167を形成することにより構築した。この変異型GTP遺伝子は、2つのアミノ酸変化S65GおよびS72Aを含み、これらにより、488nmで励起すると蛍光発光が増強される。また、この変異型は、高可溶性および発色団形成の高反応速度論を有する(Cormackら,Gene,173:33−38,1996)。このプラスミドで、エレクトロポレーションによってNTHi86−028NP菌株を形質転換し、親プラスミド86−028NP菌株/pRSM2169を作製した。
【0109】
ランダムゲノムDNA断片(実施例1に記載)を調製し、プロモータープローブベクターン内にライゲーションした。86−028NP菌株からゲノムDNAを、Puregene DNA単離キット(Gentra Systems,Minneapolis,MN)を製造業者のプロトコルに従って用いて単離した。制限障壁のため、プラスミドDNAを単離し、これをライブラリーの作製に用いることが必要であった。単離したDNAは、Sau3AI(NEB,Beverly,MA;0.25単位/μg DNA)で1時間37℃にて部分消化し、ゲル電気泳動によって分離し、Qiagenゲル抽出キットを用いて0.5〜1.5kbの大きさのDNA断片を回収した。ベクター作製のため、pRSM2167を一晩培養物から、Wizard Plus Maxiprep DNA精製系(Promega,Madison WI)を製造業者のプロトコルに従ってを用いて単離した。
【0110】
プラスミドDNAをBamHI消化によって線状化し、5’リン酸基を、ウシ腸アルカリホスファターゼ(CIAP;GibcoBRL Life Technologies)での処理によって除去した。ゲノムDNA断片を、この線状化ホスファターゼ処理ベクターを用いてライゲートし、改良プロトコル(Mitchellら,Nucleic Acids Res.,19:3625−3628,1991)に従ってエレクトロポレーション用に作製したコンピテントNP菌株内にエレクトロポレーションした。プラスミドDNAをエレクトロポレーションしてNTHi86−028NP菌株内に戻すと、形質転換効率は1000倍改善された。簡単には、細胞をOD60O=0.3まで、sBHI(脳心臓浸出物)培養液中で37℃、220rpmにて培養した。細胞を氷上で30分間冷却し、続いて、等容量の0.5×SG(1×SG:15%グリセロール、272mMスクロース)で4℃にて洗浄した。洗浄を合計3回繰返した。続いて、細胞をI×SG中で100×濃縮容量まで希釈した。BioRad Gene Pulser IIセットを200オーム、2.5kVおよび25μFで用いて細胞をエレクトロポレーションし、次いで、1mlの予備加温sBHI中で希釈し、37℃、5%CO2で2時間インキュベートし、形質転換体の一晩培養のためのチョコレート寒天上にプレーティングした。
【0111】
形質転換体を選択し、1000クローンのプール毎に、20%グリセロール(vol/vol)を含有するスキムミルク中で凍結させた。68,000構成員のgfpプロモータープローブライブラリーを作製した。ClarkeおよびCarbon(Cell,9:91−99,1976)の確率計算を用い、86−028NP菌株DNA(1.8×106bp/ゲノム)の300bp断片のライブラリー内に所与のDNA配列が提示される確率99%を得るためには、27,629個のクローンのライブラリーが必要であった。したがって、本発明のライブラリーは、86−028NPゲノムの2.5倍範囲を提示する。
【0112】
このライブラリーの質を評価するため、50個のクローンを無作為に選択して一晩チョコレート寒天上で培養し、プラスミドを単離して挿入DNAを配列した。選択されたクローンの大部分(64%)は、200〜500bpの範囲の挿入物サイズを有したが、32%は500bpを超えていた。この大部分の挿入物は、インフルエンザRd菌株の特異的オープンリーディングフレーム(ORF)に対して相同性を示し、15個のクローンは、86−028NP菌株のDNAに特有の配列を有した。Rd菌株に対して相同性を有するクローンのうち、60%は正しい向きであり、その36%は、ORFの上流に配列を含んでいた。大部分のクローンは、500bp未満の挿入物サイズを有していたが、挿入物サイズが小さいこととGFP発現の増大との間に相関性は認められなかった。実際、4個のクローンは、インビトロで軽微から中程度の蛍光を示し、そのうち3個は、200〜500塩基対の挿入物サイズを有し、1個は700塩基対より大きい挿入物を有した。
【0113】
ライブラリーの一部(ほぼ1000個のクローン)をチョコレート寒天上で培養し、PBS中に採取し、フローサイトメトリーによってGFP蛍光について解析した。挿入DNAのないプロモータートラップベクターを含む86−028NP菌株/pRSM2169と比べ、該ライブラリークローンのプールは、蛍光強度の増大を示す。したがって、該ライブラリーは、種々の活性レベルのプロモーターを有するクローンを含む。
【0114】
実施例4
GFPを発現する86−028NP誘導体の解析
gfp発現菌を同定および分取するために必要なFACSパラメータ確立するため、種々のレベルのgfp発現を示す一群の単離菌を用いた。86−028NP菌株/pRSM2169(陰性対照)を用いてバックグラウンド蛍光を評価し、したがって、観察される任意の蛍光は、gfp発現を誘導するlacZプロモーターによるものであり得る。しかしながら、この菌株は検出可能なレベルのGFPを産生せず、実際、親86−028NP菌株と比較した場合、蛍光の増大は示されない。高レベルgfp発現単離菌を、外膜タンパク質P2発現のための強力プロモーターを含有する500bp断片を、SalI−BamHI消化pRSM2167内にクローニングすることにより作製した。このプラスミドで、エレクトロポレーションにより86−028NPを形質転換し、高レベルgfp発現86−028NP菌株/pRSM2211(高度に蛍光性の対照)を作製した。この菌株は、86−028NP菌株/pRSM2169と比べて、ほぼ100倍のGFP蛍光増加を示した。中間蛍光誘導体クローン86−028NP/pKMM4B5(中間蛍光性の対照)は、FACS解析によって単離し、予備実験において、および細胞分取ための対照としての両方に使用した。gfp発現をインビトロで誘導するプロモーターを含有するDNA断片は、他の生物体のDNAに対して既知の相同性を持たない86−028NP菌株に特有である。このクローンは、86−028NP菌株/pRSM2169と比べて、ほぼ10倍の蛍光増加を示す。
【0115】
対照菌株を、チョコレート寒天上での培養から再懸濁させ、交差反応性Phycoprobe R−PE抗ヒトIgG(H+L)抗体(100μlのPBS中10μg/ml;Biomeda Corp)で30分間4℃にて標識した。3回連続洗浄して未結合抗体を除去した後、FACS解析のため、細菌を300μlのDPBS中に再懸濁した。これらの対照調製物を用い、488nmのアルゴンレーザー発光器を取り付けたCoulter Epics Eliteフローサイトメーター(Coulter Corp.)を使用して、適切なサイズおよび蛍光ゲートを設定した。細菌は、log前方角および側方散乱検出に基づいた大きさに関して、ならびに細菌のFITC/PE標識による分取に関してゲーティングさせた。分取細胞を定温sBHI中に回収し、チョコレート寒天上にプレーティングした。一晩培養後、細胞は、二次ラウンドの感染のために回収するか、または個々に選択し、一晩培養し、インビトロ培養したときの蛍光に関して個々のクローンによってスクリーニングし、20%(vol/vol)グリセロールを含有するスキムミルク中で凍結させた後、プラスミドの単離および挿入DNAの配列決定を行なった。対照菌株の分取効率を、Coulter EPICSフローサイトメーター(Coulter Corp.)を用いて確認した。
【0116】
多くのプラスミドは、抗生物質選別の非存在下では、インビトロで速やかに分離した。したがって、ここで使用したプロモータートラップベクターにこの事象の傾向があるか否かを評価するため、86−028NP菌株/pRSM2211(高度に蛍光性の対照)の単一のコロニーをチョコレート寒天上に単離し、抗生物質選別の非存在下で20回継代した。抗生物質の存在下で培養した細菌と比較した場合、蛍光強度の有意な減少は観察されなかった。また、該プラスミドは、抗生物質選別の非存在下、インビボで維持される。チンチラから採取した細菌含有中耳液を、カナマイシン含有または無含有チョコレート寒天上にプレーティングした場合、同様の細菌の計数が観察された。これらのデータは、プロモータートラップベクターが、抗生物質選別の非存在下で安定に維持されたことを示す。
【0117】
プラスミドの安定性の問題に加え、宿主−病原体相互作用を試験するためのレポーターとしてのGFPの使用に関する初期の研究により、GFPは、低毒性を有する細胞質タンパク質として継続的に合成され得、細菌の細胞表面動力学に対する影響は最小限であることが示された(Chalfieら,Science,263:802−805,1994)。高レベルgfp発現性誘導体の構築により、NTHiに対するGFP毒性の評価が可能となった。野生型菌株(86−028NP)と高GFP産生86−028NP菌株/pRSM2211両方の増殖曲線を、同様の条件下で培養した場合で比較した。増殖速度は同様であり、GFP発現は該細胞に対して毒性でなかったことを示す。
【0118】
86−028NP gfp発現性誘導体を用い、効率的な細胞分取のためのパラメータを規定した。86−028NP菌株/pRSM2169を、中間gfp発現性誘導体である86−028NP菌株/pKMM4B5と、100:1の比で混合し、全細菌集団と比べて少量割合のgfp発現クローンを含むことが予測されるインビボ環境をシミュレーションした。この混合物をFACS解析に供し、1.8%の最大蛍光集団および52%の最小蛍光集団が回収された。分取された集団のフローサイトメトリー解析により、細菌集団の65%に86−028NP菌株/pKMM4B5の富化が示され、これは、陰性集団において分取した場合では観察されなかった現象である。後続の分取ラウンドでは、この中間蛍光集団がさらに富化されることが予測され得る。陰性分取において蛍光細菌の量が減少し得ないことは、陰性分取のために設定されたゲートの大きさのためであった。GFP陰性細胞は、10%の最小蛍光集団においてゲーティングすることにより富化された。
【0119】
実施例5
中耳液からの細菌の直接標識
同様のストラテジー(実施例5に記載)を、AOM時にチンチラの中耳から採取した滲出液からの蛍光クローンの分取に適用した。本発明者らがディファレンシャル蛍光誘導(DFI)をインビボで使用可能であることは、本発明者らがgfp発現菌を分取を、非蛍光細菌、蛍光および非蛍光細胞性残屑ならびに真核生物細胞からできる能力に依存した。
【0120】
耳鏡検査またはティンパノメトリのいずれかで中耳感染の徴候のない健常な成体チンチラ(Chinchilla lanigera)を用い、ライブラリーを、インビボでのプロモーター活性についてスクリーニングした。NTHi/pRSM2169 ライブラリーの2つのプール(各々1000個のクローン)を一晩、カナマイシンを含有するチョコレート寒天上で培養した。ライブラリーを合わせ、低温の10mM滅菌PBS中で3.3×106CFU/mlまで希釈し、300μl(1.0×106CFU;500CFU/クローン)を用いて、左および右のチンチラの経水疱腔(transbullar cavity)に接種した(2000クローン/耳)。OM発症を、ビデオ耳鏡検査およびティンパノメトリによって24時間および48時間の時点でモニターした。細菌は中耳腔内で増殖し、予測どおり、48時間までに、イノキュラム用量の500倍の濃度に達した(Bakaletzら,Infect.Immunity 67:2746−62,1999)。この細菌の宿主環境への適応により炎症性応答がもたらされ、これは、紅斑、鼓膜の血管拡張および腫脹、多核白血球(PMN)の浸潤、ならびに採取した滲出液の耳鏡検査および顕微鏡検査によって観察される中耳腔内への液の蓄積によって示される。24時間および48時間後、鼓室のタッピングによって中耳液を採取し、FACS用に調製した。
【0121】
この解析を、中耳液内で採取され得る細菌に限定したことに注目することは重要である。場合によっては、FACS解析用の細菌を採取するために、中耳腔を洗浄することが必要であった。したがって、この解析には、NTHiが粘膜にゆるく結合すると上方調節される遺伝子が含まれる。NTHiは、OMのチンチラモデルの中耳腔内でバイオフィルムを形成することが観察されている(Erhlichら,JAMA,287:1710−5,2002)。本明細書に記載のプロトコルでは、プランクトン様集団から採取されるクローンを選択するため、該細菌が粘膜のバイオフィルムと結合すると遺伝子が上方調節されるクローンが採取されることは期待されない。しかしながら、中耳粘膜のホモジネーション、続いて細菌細胞の単離を行ない、本発明者らによって、このようなクローンの採取が可能となり得る。また、一部のGFP発現クローンは、滲出液において採取されるが、滲出液中に剥離細胞として、または凝集して存在する真核生物細胞に接着していることが考えられる。このような細菌は、分取効率を損なうことなく滲出液から採取するのは困難である。したがって、中耳液を粘液溶解剤で処理し、次いで遠心分離して大きな集塊および真核生物細胞を除去した後、標識した。
【0122】
チンチラの中耳液は、必要な場合には、滅菌生理食塩水で250μlまで希釈した。等容量のN−アセチル−L−システイン(0.5%;w/v)含有DPBS(pH7.4)を、粘液溶解剤として室温で5分間添加した(MiyamotoおよびBakaletz,Microb.Pathog.,21:343−356 1996)。液体を遠心分離し(300×g、5分間)、細胞性残屑、赤血球および炎症性細胞を除去した。細菌を含む上清みを新たなチューブに移した。細菌を、NTHi 86−028NP菌株由来の完全OMP調製物に対して指向されたチンチラ抗血清(1:50希釈度)とともに、45分間4℃にてインキュベートし、遠心分離(2000×g、5分間)によってペレット化し、0.05%ウシ血清アルブミンを含有する冷DPBSで2回洗浄した。続いて、細菌を交差反応性フィコプローブ(phycoprobe)R−PE抗ヒトIgG(H+L)抗体(100μlのPBS中10μg/ml;Biomeda Corp)で、30分間4℃にて標識した。3回連続洗浄して未結合抗体を除去した後、FACS解析のため、細胞を300μlのDPBS中に再懸濁した。
【0123】
実施例6
急性中耳炎においてインビボで誘導されるプロモーターの同定
プロモータートラップライブラリーで形質転換したインフルエンザ菌86−028NPを一晩、チョコレート寒天上で培養した。インビトロでgfpを発現させたプロモーターを含むクローンを選択するため、ライブラリーを1回のFACS解析(実施例6に記載のもの)に供し、低レベル量のGFPを発現するクローンのみを採取した。このクローンをプールし、チンチラの中耳に経水疱的に接種するために使用した。感染の24時間および48時間後、細菌を含む滲出液を鼓室のタッピングによって取り出した。細菌をR−PE標識抗体で間接的に標識し、蛍光タグ化細菌をゲーティングさせるが、同時に発現しているものは分取することによるFACS解析に供した。このクローンを用い、さらなる富化のために動物を再感染させた。最終回の分取後、単一のコロニー単離菌を、蛍光の消失についてインビトロでスクリーニングした。
【0124】
FACS解析によって単離されたクローン(インビボでGFP蛍光に対して陽性)(これは、インビトロでは蛍光を発しなかった)を、プラスミドの単離および挿入DNA配列の同定のために調製した。このクローンを一晩、カナマイシンを含有するチョコレート寒天プレート上で培養し、Qiaprep Miniprep Kit(Qiagen)を製造業者のプロトコルに従って用い、プラスミドの単離のために調製した。プラスミド挿入DNAは、gfpmut3遺伝子に相補的であり該挿入DNAの下流にあるプライマー5’−TGCCCATTAACATCACCATCTA−3’(配列番号:588)を用いて配列決定した。配列決定反応は、ABI prism BigDye(登録商標)ターミネーターサイクル型配列決定既製反応キット(Applied Biosystems)を製造業者のプロトコルに従って使用し、Gene Amp PCR System
9700(Applied Biosystems)用いて行なった。次いで、該配列を、96ウェルマルチスクリーンHVプレート(Millipore)内のセファデックスG−50に通すことによって精製し、続いて、ABI Prism 3100 DNA解析装置(Applied Biosystems)にて解析した。
【0125】
挿入配列を、インフルエンザRd菌株の完全にアノテートされた配列と比較した。Rd菌株に対してヌクレオチド相同性をもたない挿入物を、続いて、BLASTNおよびBLASTXアルゴリズムを用いて解析した。DNASTAR(Madison,Wise)を用い、さらなる配列解析を行なった。正しい向きであり推定ORFの5’側に配列を含む挿入配列は、NTHi菌がチンチラの中耳内に存在すると優先的に活性となる推定プロモーターを含んでいた。
【0126】
インビボで調節される推定プロモーターを有する52個のクローンを単離した。インフルエンザRd菌株で同定されたものと同様の配列を含む44個の候補クローンのうち、26個の遺伝子(60%)について、インビトロおよびインビボでの遺伝子発現の定量的比較により、NTHiがチンチラの中耳内に存在する環境的なきっかけに応答すると、上方調節された遺伝子発現が確認され、このような遺伝子を以下の表4Aにまとめる。遺伝子の発現を駆動するインビボ調節プロモーターは、膜輸送、環境情報的プロセッシング、細胞性代謝、遺伝子調節ならびに未知機能を有する仮想タンパク質に関与すると推定される。
【0127】
推定プロモーター候補のインビボでの誘導を確認するため、NTHi 86−028NP菌株を、対数増殖期中期までインビトロで、またはインビボで48時間培養した場合のメッセンジャーRNAの発現の相対量を比較した。TRIzol LS試薬(Gibco
Life Technologies)を製造業者のプロトコルに従って用い、RNAを単離した。DNAをRNA調製物から、DNA遊離キット(Ambion)を製造業者のプロトコルに従ってを用いて取り出した。DNアーゼI処理RNA試料をQiagen
RNeasyカラムに通すことによって精製した。RNAの純度および完全性を、それぞれ、260/280nmでの分光測光装置の読み値によって、およびAgilent 2100 Bioanalyzer(Agilent Technologies)において評価した。
【0128】
該FACSデータを独立して確認するため、本発明者らは、候補遺伝子の相対発現を定量的RT−PCRによって調べた。親86−028NP菌株を本試験に使用した。1工程QuantiTect SYBR Green RT−PCRキット(Qiagen)を用いるリアルタイム定量的RT−PCRにより、製造業者の使用説明書に従って転写レベルを評価した。簡単には、FACS解析で同定された推定インビボ誘導プロモーターの下流のオープンリーディングフレームに対して作製されたプライマーを用い、遺伝子特異的mRNAを逆転写し、ABI Prism 7700配列検出系(Applied Biosystems)にてRT−PCRによって増幅した。ジャイレース(gyr)の遺伝子断片を増幅することにより既知量の細菌ゲノムDNA(102〜107ゲノムコピーDNA)に対して作成した標準曲線を用い、産物の量を算出した。RNA調製物中のDNAの非存在(−RT対照)、ならびに鋳型RNAを含まない対照試料中のプライマー二量体の非存在を確認するための対照を並行して解析した。また、RT−PCR産物をゲル電気泳動によって解析し、すべての場合で、適切な塩基対サイズの単一の産物が観察された。試料間の細菌RNAの量を、gyr発現(本発明者らがインビトロで試験した種々の培養条件下で構成的に発現されることが示された)に対して標準化した。既知量の細菌ゲノムDNA(102〜107ゲノムコピーDNA)を用い、ジャイレース(gyr)の遺伝子断片を増幅させることによりRT−PCR定量の標準曲線を作成した。ジャイレースは、種々の培養条件下にてインビトロで構成的に発現され、したがって、試料間の全細菌RNAレベルを標準化するために使用した。インビボでの相対遺伝子発現を、インビトロでの遺伝子発現のものと比較し、増加倍数で示したデータを表4Aにまとめる。
【0129】
NTHiゲノムの8倍配列決定により、表4Aに示す遺伝子のほとんどの完全長オープンリーディングフレームが同定された。表4Bには、NTHiゲノム内の完全長ヌクレオチド配列および対応するアミノ酸配列を示す。便宜上、表4Bには、チンチラの中耳に存在する環境的なきっかけによる遺伝子の誘導倍率ならびに該遺伝子の産物および機能を繰り返し記載する。
【0130】
【表24】
【0131】
【表25】
【0132】
【表26】
実施例7
ビルレンス関連遺伝子の同定
多くの細菌種において、ビルレンス関連遺伝子のサブセットは、短い反復配列の複製エラーによって調節される。このような反復配列は、遺伝子の5’側またはコード配列内に存在し得、その存在は該遺伝子の発現制御の表示であり、ビルレンスとの関連性を示す。反復配列の付加または欠失により、特定のビルレンス決定基の発現または発現の欠損がもたらされる。
【0133】
NTHiインフルエンザ86−028NP菌株のコンティグの組は、短いオリゴヌクレオチド反復配列についてクエリーを行なった。該反復配列の周囲領域を解析し、該反復配列と関連する遺伝子(1つまたは複数)を同定した。表5に、同定された反復配列および各反復配列と関連するORF(BLASTによって同定)を示す。
【0134】
さらなる配列解析によって、ビルレンス関連遺伝子の完全長ヌクレオチド配列およびそのORFにコードされる対応アミノ酸配列が同定された。誘導されたアミノ酸配列は、記載のGenbank配列に対して高度に相同性である。
【0135】
【表27】
【0136】
【表28】
実施例8
NTHi特異的遺伝子配列の同定
NTHiビルレンスと関連する遺伝子もまた、NTHi菌を組織に感染させた場合の遺伝子の発現レベルと、NTHiを人工研究用培地上で培養した場合の同じ遺伝子の発現レベルを比較することにより同定された。これらの新規な遺伝子は、実施例4〜6で上記のプロモータートラップ手法を用いて同定され、続いて、既知のRdゲノムと比較すると、これらの遺伝子はNTHi 86−028NP菌株に特有であることが示された。
【0137】
このスクリーニング手順を用いて同定されたDNA配列を配列番号:577〜580として示す。これらの配列は、インフルエンザ菌Rdゲノム配列におけるホモログを有する遺伝子または遺伝子断片を含んでいなかった。これらは、チンチラの中耳内でのNTHi感染時の発現レベルから、完全に新規な配列ではないが、おそらく、これらの遺伝子の発現は、NTHiビルレンスに関与している。
【0138】
実施例9
NTHi 86−028NP菌株の完全な配列解析
ライブラリーの構築:86−028NP菌株から、Puregene試薬(Gentra Systems,Minneapolis,MN)を用いて染色体DNAを調製した。このゲノムの最初のショットガン配列決定のため、ゲノムDNAの1〜2kbおよび2〜4kbのライブラリーをpUC18において、既報のとおりにして構築した(Munsonら,Infect Immun 72:3002−10,2004)。骨格形成用(scaffolding)ライブラリーでは、ゲノムDNAを平均断片サイズ40kbに、Hamiltonシリンジを用いて手作業で切断した。末端修復後、0.7%低融点アガロースゲルを用いて、断片を分画した。30kbより大きい断片を切り出し、pEpiFOS−5へのゲル内ライゲーションを行なった。ゲルから回収したライゲーション混合物をλファージ内にインビトロでパッケージングし、EPI100細胞(Epicentre,Madison,WI)のトランスフェクションのために使用した。
。
【0139】
配列決定:配列決定のショットガン部分では、前述のPE Big−Dye(登録商標)ターミネーターおよび普遍的プライマー(M13フォワードおよびリバース)を用い、サイクル型配列決定反応を実施した(Munsonら,Infect Immun 72:3002−10,2004)。骨格形成用ライブラリーの末端を配列決定するため、プラスミドを、まず、R.E.A.L.Prep 96 Plasmid Kit(QIAGEN Inc.,Valencia,CA)を用いて精製し、次いで、TempliPhi(登録商標)DNA Amplification Kit(Amersham Biosciences Corp.,Piscataway,NJ)を用いて増幅した後、PE Big−Dye(登録商標)ターミネーターならびにpEpiFOS−5フォワードおよびリバース配列決定用プライマー(Epicentre,Madison,WI)を用いて反応を実施した。該プロジェクトの浄化(clean−up)部分のための反応は、PE Big−Dye(登録商標)ターミネーターおよびカスタムプライマー(Integrated DNA Technologies,Coralville,IA)を用いて実施した。過剰の色素ターミネーターをSephadex G50カラムで96ウェル形式にて除去し、ABI 3700またはABI 3100キャピラリー電気泳動DNA配列決定装置(Applied Biosystems,Foster City,CA)のいずれかにて配列を決定した。
【0140】
ゲノム閉鎖:骨格形成用ライブラリーおよびPCR由来の対の末端配列を用いて、コンティグを順に並べ、配列が少ない範囲の領域に配列を付加した。対のカスタムプライマー(Integrated DNA Technologies,Coralville,IA)は、各コンティグならびに配列が少ない範囲の領域に隣接する領域の両端に結合するように設計した。介在領域は、Sambrookら,Molecular Cloning:A Laboratory Manual(第3版)Cold Spring Harbor Laboratory Press.,2001に記載の標準的なPCRプロトコルにより、Taqポリメラーゼ(Roche Diagnostics,Indianapolis,IN)を用いて増幅し、両方の鎖を配列決定した。骨格形成用ライブラリー由来のクローンを鋳型として用い、リボソームのRNAオペロンおよびHMW遺伝子クラスターを完全に配列決定した。
【0141】
編集:Phred/Phrapをデータ編集に使用し、Munsonら,Infect
Immun 72:3002−10,2004に記載のデフォルト編集パラメータを用いた(Ewingら,Genome Res 8:186−94,1998;Ewingら,Genome Res 8:175−85,1988;Gordonら,Genome Res 8:195−202,1998)。507個のクローン由来の対の末端配列データを用い、DNASTAR 総合ソフトウェアSeqman IIプログラムを使用して編集データを確認した。
【0142】
データ解析:Glimmer2長orfプログラム(Delcherら,Nucleic Acids Res 27:4636−41,1999)によって同定された1178個の最長ORFの組に対して調整されたGlimmer2(v2.13)を用いてコード領域を同定した。Glimmer ORFの組を、Rd菌株プロテオーム、SwissProtデータベース、NCBI COGデータベースおよびKlEGGデータベースに対して検索することにより、類似性による自動アノテーションを行なった。Rd菌株データベースを86−028NP菌株ORFの組と、トリクロス(tricross)を用いて双方向で比較し、類似性の高信頼度領域を決定し、ゲノム組織のドットプロット比較を得た(Rayら,Bioinformatics 17:1105−12,2001)。
【0143】
自動的に推定されたアノテーション情報を、Artemis(Rodriguezら,Infect Immun 71:1635−42,2003)を用いてマニュアルでさらに操作(curate)して目的のゲノム領域を視覚化および境界画定し、カスタムFileMaker Proデータベースを作成し、次いで、これをマニュアル改定の適用および機能の割当てに関連するデータの取得に使用した。主な自動比較にはFASTA解析を使用した。80−028NP菌株のゲノムとRd菌株のゲノム間の強いシンテニーにより、大部分の遺伝子機能の自動的な割当てが可能であり、マッチングでは、アミノ酸レベルで90%以上の類似性を有していた。Rd菌株のゲノムに対してほぼ1対1の86−028NP菌株ゲノムのマッピングが、Rd菌株ORFの86−028NP菌株ゲノム配列上へのアセンブリおよび86−028NP菌株ORFのRdゲノム上への逆アセンブリ(SeqManプログラムをヌクレオチドレベルで80%同一性のアセンブリ基準で使用)によって確認された。
【0144】
マニュアルBLAST解析を用い、既知遺伝子に対して強い類似性を示さなかったORFの潜在的機能を調査した。自動割当てのマニュアル操作は、アノテーションが現在の文献に従うように行ない、自動化アルゴリズムが誤りをもたらし易いいくつかの位置(特に、HMW遺伝子クラスター、ヘモグロビン結合タンパク質およびhsd遺伝子クラスター、その高いファミリー類似性により自動割当てが混同される)を修復した。
【0145】
tRNA遺伝子をtRNAscan−SE vl.11(Loweら,Nucleic
Acids Res 25:955−64,1997)によって同定した。rRNAオペロンをRd菌株との16、23および5S rRNA類似性、ならびにこれらの遺伝子を含む隣接部のCLUSTALWアラインメントに基づいて同定し、半保存領域の境界を決定した。
【0146】
インフルエンザ菌(influennzae)86−028NP菌株の完全なゲノムDNA配列を、配列番号:772として示す。該ゲノムDNA内のオープンリーディングフレームを配列番号:773〜2593として示し、表6に示す。得られる遺伝子産物を配列番号:2581〜4414として示し、表7に示す。オープンリーディングフレームのヌクレオチドの前に示す「c」は、オープンリーディングフレームが配列番号:722として示す配列の5’−3’配列に相補的であることを示す。表6において、NTHi特異的遺伝子に印を付けている。このゲノム配列は、GenBankから受託番号CP000057(これは、引用によりその全体が本明細書に組み込まれる)として入手可能である。
【0147】
下記のオープンリーディングフレーム:配列番号:822(kdgK)、配列番号:928、配列番号:994(bolA)、配列番号:276(adhC)配列番号:1102(dusC)、配列番号:1121(merP)、配列番号:1135、配列番号:1236、配列番号:1254、配列番号:1376(lic2C)、配列番号:1431(hgpD)、配列番号:1502、配列番号:1505(ppx)、配列番号:1523(hgpC)、配列番号:1585(lex2A)、配列番号:1637配列番号:1713、配列番号:1856(mod)、配列番号:1899、配列番号:2006、配列番号:2080、配列番号:2155、配列番号:2202、配列番号:2257、配列番号:2331、配列番号:2345、配列番号:2365、配列番号:2555(metE)および配列番号:2563(pmi)は、これらのヌクレオチド配列がその配列内にフレームシフトまたは終止コドンを含むため、表6において偽遺伝子と規定している。hgpD、hgpC、lex2AおよびNTHI1769遺伝子は、リーディングフレーム内外への配列シフトを引き起こす偶発性(contingency)反復配列を含み、生じるアミノ酸配列(配列番号:3242、3332および4142)は部分翻訳物である。このような偶発性反復配列により、インフルエンザ菌において遺伝子発現が調節され、したがって、このようなタンパク質をコードする遺伝子は、1つ以上の反復配列の挿入または欠失後、正確に翻訳される。
【0148】
【表29】
【0149】
【表30】
【0150】
【表31】
【0151】
【表32】
【0152】
【表33】
【0153】
【表34】
【0154】
【表35】
【0155】
【表36】
【0156】
【表37】
【0157】
【表38】
【0158】
【表39】
【0159】
【表40】
【0160】
【表41】
【0161】
【表42】
【0162】
【表43】
【0163】
【表44】
【0164】
【表45】
【0165】
【表46】
【0166】
【表47】
【0167】
【表48】
【0168】
【表49】
【0169】
【表50】
【0170】
【表51】
【0171】
【表52】
【0172】
【表53】
【0173】
【表54】
【0174】
【表55】
【0175】
【表56】
【0176】
【表57】
【0177】
【表58】
【0178】
【表59】
【0179】
【表60】
【0180】
【表61】
【0181】
【表62】
【0182】
【表63】
【0183】
【表64】
【0184】
【表65】
【0185】
【表66】
【0186】
【表67】
【0187】
【表68】
【0188】
【表69】
【0189】
【表70】
【0190】
【表71】
【0191】
【表72】
【0192】
【表73】
【0193】
【表74】
【0194】
【表75】
【0195】
【表76】
【0196】
【表77】
【0197】
【表78】
【0198】
【表79】
【0199】
【表80】
【0200】
【表81】
【0201】
【表82】
【0202】
【表83】
【0203】
【表84】
【0204】
【表85】
【0205】
【表86】
【0206】
【表87】
【0207】
【表88】
【0208】
【表89】
【0209】
【表90】
【0210】
【表91】
【0211】
【表92】
【0212】
【表93】
【0213】
【表94】
【0214】
【表95】
【0215】
【表96】
【0216】
【表97】
【0217】
【表98】
【0218】
【表99】
【0219】
【表100】
【0220】
【表101】
【0221】
【表102】
【0222】
【表103】
【0223】
【表104】
【0224】
【表105】
【0225】
【表106】
【0226】
【表107】
【0227】
【表108】
【0228】
【表109】
【0229】
【表110】
実施例10
NTHi、86−028NP菌株およびインフルエンザRd菌株のゲノムの比較
86−028NP菌株のゲノム配列は、1,913,428bpを含む。これは、Rd菌株のゲノム(1,830,137bp)(Fleischmannら,Science
269:496−512,1995)よりほぼ4パーセント大きい。また、多数の遺伝子が86−028NP菌株に存在する(Rd菌株で1743に対して1942)。該遺伝子の相補性を、DNASTAR総合ソフトウェアのSeqmanプログラムを用いてRd菌株のものと比較した。排除値としてヌクレオチドレベルで80%同一性では、Rd菌株のゲノムには存在しない285個のORFが86−028NPゲノムにおいて同定され、86−028NP菌株ゲノムには存在しない167個のORFがRd菌株のゲノムにおいて同定された。
【0230】
86−028NP菌株は、Rd菌株と同様、6個のリボソームオペロンを有する。tRNAscan−SE vl.11を用い、20個の共通アミノ酸を表す58個のtRNA遺伝子が86−028NP菌株ゲノムにおいて同定された。tRNA−Glu、tRNA−AlaおよびtRNA−Ile遺伝子は、16SリボソームRNA遺伝子と23SリボソームRNA遺伝子の間のスペーサー領域内に位置した。UCAアンチコドンを含むtRNA遺伝子もまた同定された。このアンチコドンはオパール終止コドンに相当し、典型的には、セレノシステインを組み込むオパール抑制tRNAと関連している。tRNAは、Sec tRNA特異的伸長因子であるselB(NTHI0836)と、セレンの組込みによってセレノシステインを形成する予備としてセリンをデヒドロアラニンに変換させる酵素であるselA(NTHI0835)をコードする2つの遺伝子と隣接している(Forchhammerら,Nature 342:453−6,1989)。セレノホスフェートシンテターゼをコードするselD遺伝子(NTHI0297)もまた同定された。このセレノシステイン系の重要性は、おそらくセレノシステインコドンとして読まれるインフレームTGA終止コドンを含むギ酸デヒドロゲナーゼのαサブユニットのコード配列(NTHI0007)によって証明される。インフレームTGA終止コドンは、Rd菌株のギ酸デヒドロゲナーゼ遺伝子(GenPept受託番号P46448)の現在のアノテーションにおいて既に記載されて注釈されている。
【0231】
86−028NP菌株とRd菌株の遺伝子順序の解析に関与するゲノム間の全体比較により、大きな逆位の形態の単一の大きな再編成が示されている。この471kbの逆位は、86−028NP菌株ゲノムのほぼ25%を表し、NTHI1391およびNTHI1394(それぞれHI1218およびHI1645のホモログ)またはNTHI1949およびNTHI1950(それぞれHI1219およびHI1647のホモログ)で境界されている。HI1219およびHI1646は、Rd菌株において一部重複した遺伝子であり、cmkAおよびcmkBとアノテートされている(シチジレートキナーゼ)。1つのcmk遺伝子(NTHI1949)が、86−028NP菌株内にcmk様小断片とともに、NTHI1391とNTHI1394の間に存在する。骨格形成用ライブラリー由来のいくつかのクローンは、86−028NPゲノム内で該逆位の各末端が重複しており、本発明者らのアセンブリが認証される。この大きな逆位内には、いくつかの挿入部があり、その最大のものはほぼ13kb、27kbおよび51kbの大きさである。これらの領域は、主に仮想遺伝子および仮想保存遺伝子ならびにファージ遺伝子のいくつかのホモログを含む。例えば、27kb挿入部は、HP1−およびHP2様ファージ遺伝子の残部を含む。最大挿入部は、インテグラーゼ遺伝子のホモログと境界されている。Rd菌株では、mu様ファージがこの領域に局在している(Morganら,J Mol Biol 317:337−59,2002)。このファージは、86−028NP菌株ゲノムには存在しない。また、この大きな逆位領域内には、Rdゲノムとのシンテニーを回復させる21kb逆位が存在する。
【0232】
この大きな逆位に加え、86−028NP菌株は、Rd菌株のゲノムとの共線形成(co−linearity)と相違する他の領域を有する。これらには、Rd菌株に存在するDNAとの明白な相同性もたない配列を含む5kbより大きい9つの領域が含まれる。これらの領域のうち2つは、後述するHMW付着因子を含む。仮想遺伝子は、該特異的領域のうち6つで優位を占める。第9番目の領域はほぼ56kbの大きさである。これは、NTHI0100とNTHI0165の間に存在する。BLASTn解析により、ICEHin86−028NPと表示するこの領域内の遺伝子は、b型インフルエンザ菌プラスミド内の遺伝子ICEHin1056(Mohd−Zainら,J Bacterial
186:8114−22,2004)に対して高い相同性を有することが示された。ICEHin1056は、一連の共通コア遺伝子によって規定されるゲノムアイランドの拡張ファミリーの一構成員である(Mohd−Zainら,J Bacteriol 186:8114−22,2004)。ICEHin86−028NPは、45個のICEHm 1056 ORFのホモログを有する。これらには、主に、プラスミドの複製およびコンジュゲーションにおいて推定される役割を有するタンパク質をコードするICEHin86−028NPの5’末端付近のORF(規定のコア遺伝子を含む)、ならびに主に、膜会合性またはまたは膜輸送性のいずれかであり得ることが示唆されるモチーフを有する保存仮想タンパク質をコードする3’末端付近のORFが含まれる。注目すべきことは、ICEHin86−028NPが、ICEHin1056には見られるテトラサイクリン、クロラムフェニコールおよびβ−ラクタム耐性に関与するタンパク質をコードする遺伝子を欠くことである。ICEHin86−028NP内には、トランスポゼース、レソルバーゼおよび推定インテグラーゼ調節因子が散在しており、これは、ICEHin86−028NPが、いくつかの可動性遺伝因子に由来する複合エレメントであることを示す。
【0233】
ICEHin1506は、attP部位と表示される配列を第1の遺伝子の5’側に有する。86−028NP菌株には、このattP部位の完璧なコピーがNTHI0101の5’側に存在し、単一ヌクレオチド変化を有するこのattP部位のコピーがNTHI0164の3’側に存在する。attP部位は、ゲノムアイランドを形成する細菌染色体内への可動性遺伝因子の組込みに関与しており、おそらく、この大きな遺伝物質セクションが86−028NP菌株ゲノム内に組み込まれた状態となる機構を示唆する(Dimopoulouら,Antimicrob Agents Chemother 46:1602−3,2002)。ICEHin86−028NPは、その他の任意の関連ゲノムアイランドより少なく、86−028NP菌株のゲノム全体のG+C含量38%に近い39%のG+C含量を有する。これは、このエレメントについて長期ゲノム関連を暗示する。このエレメントとともに、膜会合タンパク質および分泌タンパク質をコードしていると考えられるICEHin1506のものに相同な遺伝子(Dimopoulouら,Antimicrob Agents Chemother 46:1602−3,2002)のその相補配列の存在は、86−028NP菌株のビルレンスの重要な暗示を有し得る
パスツレラ科(Pasteurellaceae)のいくつかの構成員、例えば、ヘモフィラス デュクレイ(Haemophilus ducreyi)、パスツレラ マルトシダ(Pasteurella multocida)およびアクチノバチラス アクチノマイセテムコミタンス(Actinobacillus actinomycetemcomitans)は、充分特性評価されたタンパク質毒素を産性する。対照的に、インフルエンザ菌は、タンパク質毒素を産性しないようであり、推定タンパク質毒素をコードする遺伝子は、86−028NP菌株ゲノムにおいて同定されなかった。インフルエンザ菌では、内毒素生合成を担うグリコシルトランスフェラーゼをコードする遺伝子、および感染過程において細菌に増強された「適応度」を付与するタンパク質をコードする遺伝子が、一般的にビルレンス決定基とみなされている。これらの遺伝子としては、付着因子、ヘムおよびヘモグロビン結合タンパク質をコードするもの、ならびに酸化的ストレスを防御するタンパク質をコードする遺伝子が挙げられる。
【0234】
偶発性遺伝子
インフルエンザ菌は、限定数の2成分調節系および他の総合調節因子を有する。Moxonおよび協働者らにより、「単純偶発性(simple contingency)遺伝子座」と称する遺伝子座が遺伝子発現調節の代替機構を提供し、したがって、該生物体が変化する環境条件に速やかに応答する能力に寄与することにより、生物体の適応度が増大すると論じられた。これらの遺伝子座は、短いタンデム反復配列を、コード領域内またはコード領域の5’側のいずれかに含む。DNA複製中、リーディングフレーム内での反復配列の付加または欠損により、リーディングフレームの改変がもたらされる。コード領域の5’側に局在する場合、反復配列の付加または欠損により、プロモーター活性の変化がもたらされる(Baylissら,Clin Invest 107:657−62,2001)。単純反復配列を含む遺伝子座は、例えば、インフルエンザ菌において広範に研究されている(Hoodら,Proc Natl Acad Sci USA 93:11121−5,1996)。以下のセクションに相変異性(phase variable)として記載する遺伝子座のいくつかは、単純反復配列を含む。
【0235】
付着因子
86−028NP菌株は、主に宿主細胞への付着において機能する産物をコードするいくつかの遺伝子を保有する(表8)。これらのうちの1つである外膜タンパク質P5は、以前に同定され、その機能が注意深く分析された(Jiangら,Infect Immun 67:187−92,1999;Kennedyら,Infect Immun 68:2756−65,2000;Novotnyら,J Immunol 171:1978−83,2003;Novotnyら,Infect Immun 68:2119−28,2000;Novotnyら,Vaccine 20:3590−7,2002;Sirakovaら,Infect Immun 62:2002−20,1994)。86−028NP菌株は、Rd菌株Actinobacillus pleuropneumoniaeおよびP.multocida由来のpilABCDのホモログである4つの遺伝子を含む遺伝子クラスターを保有する(Bakaletzら,Infect
Immun 73:1635−4,2005;Doughtyら,Vet Microbiol 72:79−90,2000;Ruffoloら,Infect Immun
65:339−43,1997 Stevensonら,Vet Microbiol
92:121−34,2003)。これらの遺伝子は、comE遺伝子および未同定遺伝子とともに、鼻咽頭組織への86−028NP菌株の付着において役割を有するIV型線毛をコードする(Kennedyら,Infect.Immun.,68:2756−2765,2000)。
【0236】
86−028NP菌株は、Rd菌株には存在しない2つの高分子量(HMW)付着因子遺伝子クラスターを保有する。該高分子量付着因子は、最初にNTHi 12菌株において特性評価され、これは、各々、3種類のタンパク質(HMWA、HMWBおよびHMWC)をコードする2つのHMW遺伝子クラスターを有する。HMWAは、該付着因子の構造的成分であり、HMWBは、膜貫通転位において役割を有する一方、HMWCは、HMWAのグリコシル化に必要とされる(Barenkampら,Infect Immun
60:1302−13,1992;Barenkampら,Infect Immun
62:3320−8;1994;Grassら,Mol.Microbiol 48:737−51,2003;St Gemeら,Mol.Microbiol 27:617−30,1998)。同様に、86−028NP菌株の2つのHMW遺伝子クラスターは、12菌株と同じ遺伝子状況でHmwA、BおよびC遺伝子のホモログを含む(Buscherら,J Bacteriol 186:4209−17,2004)。86−028NP菌株由来のHMW1AおよびHMW2Aタンパク質は、主要相違領域、例えば、HMW2A 内の41アミノ酸挿入部とC末端に向かって72%同一である。86−028NP菌株由来のHMWBとHMWCの対のタンパク質は、それぞれ99%同一である。配列ATCTTTCは、hmw1Aの上流で17回およびhmw2Aの上流で23回反復される。12菌株では、この配列の16反復配列が、各hmw遺伝子クラスターの5’側に見られる(Barenkampら,Infect Immun 60:1302−13,1992)。
【0237】
Hapは、IgAlプロテアーゼの触媒性ドメインに相同なドメインを有する自己輸送型タンパク質である。NTHI0354遺伝子は、NTHi菌株N187由来のHapと83%同一性を有するタンパク質をコードする(St Gemeら,Mol.Microbiol 14:217−3,1994)。86−028NP菌株は、HMW1およびHMW2を保有する他のNTHi菌株と同様、別のヘモフィルス属付着因子であるHiaをコードする遺伝子を欠く(Barenkampら,Mol.Microbiol 19:1215−23,1996)。また、86−028NP菌株は、本発明者らが以前に報告したように、赤血球凝集性線毛をコードするhif遺伝子クラスターを欠く(Munsonら,Infect Immun 72:3002−10,2004)。
【0238】
【表111】
表8、9および10において、「NTHI番号」は、Microbial−Pathogenesis H.influenzae 86028 NPウェブサイトおよびGenbank 受託番号CP000057で表示されるNTHi 86−028NP菌株ゲノム 内の遺伝子座タグ番号を示す。「HI番号」は、TIGR(The Institute for genomic redearc)内の対応する遺伝子座タグ番号を示す。
【0239】
リポオリゴ糖合成[[当初の出願の第13頁参照]
インフルエンザ菌リポオリゴ糖(LOS)の構造、生合成およびビルレンスにおける役割は、広範に研究されている。表9に、リポオリゴ糖生合成に関与する遺伝子の一覧を網羅する。86−028NP菌株は、LOSの七炭糖−Kdo−リピドA部分合成するのに必要とされる遺伝子の完全な相補配列を有する。lgtFおよびlpsA遺伝子は、グルコース、およびグルコースまたはガラクトースを七炭糖の第1および第3残基にそれぞれ付加するグリコシルトランスフェラーゼをコードする。これらの遺伝子はともに、86−028NP菌株ゲノム内に存在し、したがって、おそらく、炭水化物鎖は、86−028NP菌株LOSの第1七炭糖残基および第3七炭糖残基から伸長され得る(Hoodら,Microbiology 150:2089−97,2004)。b血清型菌株RM153において、lic2C遺伝子は、グルコースを第2七炭糖に付加するグリコシルトランスフェラーゼをコードする(Hoodら,Microbiology 150:2089−97,2004)。86−028NP菌株ゲノムでは、この遺伝子はフレームシフトを含む。相変異性lic2AおよびlicA遺伝子は、それぞれ、ガラクトシルトランスフェラーゼおよびコリンキナーゼをコードし、86−028NP菌株ゲノム内に存在する(Highら,Mol.Microbiol 9:1275−82,1993;Hoodら,Glycobiology 11:957−67,2001;Weiserら,Infect Immun 65:943−50,1997)。lex2B遺伝子は、b血清型DL42菌株ならびにいくつかの他の血清型分類可能菌株においてグリコシルトランスフェラーゼをコードし、86−028NP菌株ゲノム内に存在する(Griffinら,Microbiology 149:3165−75,2003;Jarosikら,Infect Immun 62:4861−7,1994)。DL42菌株のlex2B遺伝子に対して5’側は、短い相変異性lex2A遺伝子である。86−028NP菌株では、この遺伝子は、1つの4ヌクレオチド反復配列の欠損および5bp欠失のため、DL42配列(Genbank受託番号U05670)と比べるとフレーム外である。最近、Hoodおよび協働者らにより、Rd菌株において、HI0866〜HI0874を含むHmgと表示される遺伝子座が報告された(Hoodら,J Bacteriol 186:7429−39,2004)。rmlBのホモログは例外として、これらの遺伝子は、86−028NP菌株ゲノムには存在しない。これには、NTHI 2019菌株においてバイオフィルム形成に重要であることが最近示されたシアリルトランスフェラーゼをコードするsiaA遺伝子が含まれる(Greinerら,Infect Immun
72:4249−60,2004;Jonesら,J Biol Chem 277:14598−611,2002)。別のシアリルトランスフェラーゼをコードするlic3A遺伝子のホモログの2つのコピー(Hoodら,Mol.Microbiol 39:341−50,2001;Jonesら,JBiol Chem 277:14598−611,2002)、ならびに別のシアリルトランスフェラーゼをコードするlsgB遺伝子の1つのコピー(Jonesら,J Biol Chem 277:14598−611,2002)が、86−028NP菌株ゲノムにおいて同定された。
【0240】
【表112】
【0241】
【表113】
【0242】
【表114】
鉄分獲得
インフルエンザ菌株は、ヘムの中間体前駆体であるプロトポルフィリンIX(PPIX)とともにヘムまたは鉄分のいずれかの絶対要件を有する(Evansら,J Med Microbiol 7:359−65,1974;Whiteら,J Bacteriol 85:842−50,1963)。表10に、鉄分獲得に関与する遺伝子の一覧を網羅する。3種類のヘモグロビンおよびヘモグロビン−ハプトグロビン結合タンパク質HgpA、HgpBおよびHgpCがb型インフルエンザ菌HI689菌株において同定された(Jinら,Microbiology 145(Pt 4):905−14,1999;Mortonら,Infect Immun 67:2729−39,1999;Renら,Infect Immun 66:4733−41,1998)。HI689菌株では、これらの遺伝子は、CCAA4ヌクレオチド反復配列を有し、スリップストランド(slip−strand)誤対合によって調節されることが知られている。これらの遺伝子のうち2つは、86−028NP菌株内に存在する。これらはともに、CCAA反復配列を含み、hgpB遺伝子はインフレームであるが、hgpC遺伝子はフレーム外である。CCAA反復配列を含む第3の遺伝子の誘導アミノ酸配列は、hgpAと45%同一である。本発明者らは、この遺伝子をhgpDと命名した。この遺伝子はフレーム外である。ヘムおよびヘム−ヘモペキシン錯体をコードするb型インフルエンザ菌のhxuABC遺伝子のホモログ(Copeら,Infect Immun 69:2353−63,2001;Copeら,Infect Immun 66:4511−6,1998;Copeら,J Bacteriol 177:2644−53,1995)ならびにhemR受容体のホモログが同定された。86−028NP菌株はまた、ヘム結合リポタンパク質HbpAをコードする遺伝子を有する(Heathら,Pediatr Infect Dis J 20:300−5,2001)。hbpAの下流は、その産物が、推定ヘム利用タンパク質を含むクラスターであるCOG0748の構成員である仮想遺伝子であるNTHI1022である。最近b型インフルエンザ菌において同定され、一般的なヘム利用タンパク質をコードするhup遺伝子のホモログもまた同定された(Mortonら,Microbiology 150:3923−33,2004)。
【0243】
ヘム輸送系に加え、鉄輸送系wereもまた同定された。hitABC遺伝子はそれぞれ、シデロフォア欠損大腸菌株にb型インフルエンザ菌の菌株からクローニングしたHtABC遺伝子を補完することにより完結に特性評価された高度に特異的な鉄イオンABC輸送系の構成員であるFbpABCタンパク質をコードする(Andersonら,J Bacteriol 186:6220−9,2004)。tbpAB(Gray−Owenら,Infect Immun 63:1201−10,1995;Gray−Owenら,Infect Immun 63:3809−15,1995)ならびにhfeABCDと表示され、鉄取込みに関与するABC輸送系のホモログであり、かつ最初にペスト菌において特性評価された(Beardenら,J Bacteriol 180:1135−47,1998)遺伝子にコードされるトランスフェリン結合タンパク質1および2が同定された。この後者の遺伝子クラスターは、Rd菌株にも存在する。NTHI2035 は、Mn2+およびFe2+輸送体のNRAMPファミリーの推定ホモログをコードする(Richerら,J Mol Evol 57:363−7,2003)。
【0244】
上記のように、インフルエンザ菌は、インビトロでの増殖のためのヘムの供給源として、PPIXとともに鉄分を利用し得る。PPIXへの鉄の組込みを触媒する(Schlorら,Infect Immun 68:3007−9,2000)フェロキレターゼをコードするhemH遺伝子が同定された。調節因子をコードする遺伝子Furもまた同定された(Andrewsら,FEMS Microbiol Rev 27:215−37,2003;Smootら,J Med Microbiol 48:629−3,1999)。
【0245】
【表115】
【0246】
【表116】
酸化的ストレス
増殖に必要であるが、鉄分の能動的獲得は、菌体に対して有害な効果を有し得る。フェントン反応により、鉄分は過酸化水素と反応し、高度に反応性のヒドロキシル残基を生成させ得る。この生成物は、著しい効果、例えば、脂質過酸化および鉄分含有酵素とDNAの両方に対する損傷を有する(Imlay,Annu Rev Microbiol 57:395−418,2003)。ヒドロキシル残基に対する最もよく知られた防御系は、スーパーオキシドディスムターゼAおよびBからなり、これらは、高度に反応性のスーパーオキシドを過酸化水素に変換し、次いで、これは、カタラーゼによって水と酸素に変換される(Demple,Annu Rev Genet 25:315−37,1991)。86−028NP菌株およびRd菌株は、sodA遺伝子(NTHI1251)を含むが、sodB遺伝子を欠く。また、両菌株とも、カタラーゼ遺伝子hktE(NTHI1099)(Bishaiら,J Bacteriol 176:2914−21,1994)、酸化的ストレスに対する保護に関与する遺伝子の一次調節因子(Maciver & Hansen,Infect Immun 64:4618−29,1996;Pomposielloら,Trends Biotechnol 19:109−14,2001)をコードするoxyR遺伝子(NTHI0704)、およびPrx/Grxと称され、小アルキルヒドロペルオキシドに対する保護においてグルタチオン依存的役割を有するキメラペルオキシダーゼをコードする遺伝子(Pauwelsら,J Biol
Chem 278:16658−66,2003;Vergauwenら,J Bacterial 185:5555−62,2003;Vergauwenら,J Bacteriol 185:1572−81,2003)を保有する。本発明者らは、以前に、Rd菌株には存在しないP.multocidaペルオキシレドキシンTsaAのホモログをコードする遺伝子であるNTHI0212を同定した(Munsonら,Infect Immun 72:3002−10,2004)。しかしながら、86−028NP菌株は、サルモネラ属においてTsaAの還元に関与していることがわかっている(Pooleら,Biochemistry 39:6602−15,2000)専用(dedicated)アルキルヒドロペルオキシドレダクターゼであるAhpFが欠損している。酸化的ストレスに対するさらなる保護は、ftnAおよびftnB(それぞれ、NTHI1773およびNTHI1772)遺伝子にコードされるフェリチン様タンパク質によってもたらされ得る。これらのタンパク質の過剰発現により、鉄過剰負荷大腸菌fur変異型が酸化的損傷から保護されることが示された(Touatiら,J Bαcteriol 177:2305−14,1995)。仮想保存遺伝子NTHI1817は、DNA結合フェリチン様タンパク質と相同性を有するタンパク質をコードする。これは、S.entericaでは鉄の存在下および食作用中の両方において酸化的ストレスに対する保護の役割を有し、サルモネラ属感染マウスモデルにおいてビルレンスに重要である(Halseyら,Infect Immun 72:1155−8,2004)非特異的DNA結合タンパク質のDpsファミリーの構成員である。大腸菌では、Dpsは、過酸化水素によって酸化された鉄分に優先的に結合し、したがって、フェントン反応によって生成されるヒドロキシル残基の発生の抑止に重要な役割を有することが示された(Zhaoら,J Biol Chem 277:27689−96,2002)。
【0247】
分泌
Sec系に加え、86−028NP菌株は、2つのアルギニンをそのシグナルペプチド内に有するタンパク質のSec非依存的輸送に関与するTatA、BおよびC タンパク質、細胞質膜会合タンパク質をコードする遺伝子(NTHI0279、NTHI0280およびNTHI0282)を有する(Bolhuisら,J Biol Chem 276:20213−9,2001;Yenら Arch Microbiol 177:441−50,2002)。既報のように、86−028NP菌株は、自己輸送型タンパク質Lavをコードする遺伝子NTHI0585を保有する(Munsonら,Infect Immun 72:3002−10,2004)。このタンパク質はRd菌株には存在しないが、ナイセリア菌には存在し、ヘモフィルス属では、病原性菌株に限定されるようである(Davisら,J Bacteriol 183:4626−35,2001)。86−028NP菌株はまた、IgAプロテアーゼをコードする遺伝子(NTHI1164)(PoulsenらJ Bacteriol 174:2913−21,1992)、および上記のようなHap付着因子をコードする遺伝子を有する。ともに、自己輸送体の類型のタンパク質である、上記のように、HMW付着因子は、2パートナー分泌経路群のタンパク質の構成員である。
【0248】
外膜タンパク質
いくつかの外膜タンパク質(OMP)をコードする遺伝子が、他のインフルエンザ単離菌のものとの相同性によって同定された。これらとしては、主要なOMP(すべて、最初はb型インフルエンザ菌において同定);表面発現P1(NTHI0522)、ポーリンP2(NTHI0225)、ホスホモノエステラーゼおよびヘム輸送体P4(NTHI0816)、付着因子P5(NTHI1332)ならびにリポタンパク質P6(NTHI0501)が挙げられる。また、86−028NP菌株は他のヘモフィルス属菌株と、いくつかの微量OMPを共有する。これらとしては、b型インフルエンザ菌由来のD15およびトランスフェリン結合タンパク質、ならびにNTHi菌株289において同定されたOMP26のホモログ(Munsonら,Infect Immun 56:2235−42,1988;Munsonら,Infect Immun 49:544−9,1985;Munsonら,J Clin Invest 72:677−84,1983;Reidlら,J Exp Med 183:621−9,Reillyら,J Bacteriol 181:6797−805,1999;Reillyら,FEBS Lett 494:19−23,2001)が挙げられる。すべては、その後、NTHi菌株において特性評価され、潜在的なワクチン候補として解析された(Poolmanら,Vaccine 19 Suppl 1:S108−15,2000;Murphyら Curr Opin Infect Dis 16:129−34,2003;McMichaelら,Curr Opin Investig Drugs 4:953−8,2003 Crippsら, Immunol Cell Biol 81:46−51,2003;Bakaletzら Ann Otol Rhinol Laryngol Suppl 188:82−94,2002)。
【0249】
制限酵素系:
86−028NP菌株は、HindIIおよびHindIII II型制限系を欠く(Fleischmannら,Science 269:496−512.80,1995;Nwankwoら,Gene 150:75−80.104,1994,Smith,& Marley.Methods Enzymol 65:104−8,1980)。対照的に、H.aegyptiusにおいて最初に同定された(Slatkoら,Gene 74:45−50,1988)HaeII系をコードする遺伝子は、86−028NP菌株ゲノム内に存在するが、Rd菌株には存在しない。86−028NP菌株およびRd菌株はともに、メチルトランスフェラーゼ(HsdM)、配列認識タンパク質(HsdS)および制限酵素(HsdR)をコードするHsd型I制限系を有する(Robertsら,Nucleic Acids Res 31:1805−12,2003)。これらの伝子は、Rd菌株のゲノムでは隣接している(HI1285〜HI1287)。86−028NPゲノムは、各々4つの遺伝子を含有する3つのhsd様遺伝子座を含む。hsd系の1つはNTHI1838〜NTHI1843にコードされている。この遺伝子クラスターでは、NTHI1841は仮想タンパク質をコードする。第2のhsd様遺伝子座はNTHI0314〜NTHI0318にコードされている。この遺伝子クラスターでは、NTHI0316は推定アンチコドンヌクレアーゼをコードする。このhsd様系は、大腸菌のprr系に類似したものであり得る(Tyndallら,J Mol Biol 237:266−74,1994)。第3のhsd遺伝子座はNTHI0188〜NTHI0193にコードされている。この遺伝子クラスターでは、NTHI0190 は、ヘリックス−ターン−ヘリックスドメインを有する推定転写調節因子をコードする。
【技術分野】
【0001】
本出願は、2005年6月16日に出願された米国仮出願第60/691,214号、および2004年3月5日に出願された米国出願第10/795,159号(これは、2003年3月6日に出願された米国仮出願第60/453,134号への優先権を主張する)への優先権を主張する。これらの全ては、それらの全体において本明細書で参考として援用される。
【0002】
(発明の分野)
本発明は、分類不能型インフルエンザ菌(NTHi)ゲノムのポリヌクレオチド配列、該ゲノム内に含まれるNTHi遺伝子および該ポリヌクレオチドにコードされるポリペプチドに関する。本発明はまた、このようなNTHiポリヌクレオチドおよびNTHiポリペプチドの使用、例えば、ワクチンおよびNTHi関連障害を処置および予防する方法に関する。本発明はまた、中耳または鼻咽頭のNTHi感染中または該感染に応答して上方調節されるNTHi遺伝子に関する。
【背景技術】
【0003】
(背景)
中耳炎(OM)は、世界中で非常によく見られる小児疾患であり、緊急治療室での小児の往診の主な原因である(非特許文献1)。最近の統計学的データにより、1990年では2,450万人の小児科医の診療所での往診がOMのためになされたことが示されており、1980年代に報告されたものの200%を超える増加を表す。長引いても死亡率と関連することは稀であるが、OMに関連する罹病率は相当である。聴力低下は、この疾患に伴う一般的な問題であり、しばしば、子供の行動、教育および言語能力の発達に影響する(非特許文献2;非特許文献3;非特許文献4)。OMの社会経済的影響もまた大きく、OMの診断およびマネージメントの直接および間接費用は、米国だけで年間50億ドルを超える(非特許文献5)。
【0004】
抗生物質療法は一般的であり、中耳腔換気用チューブの外科的留置は、滲出液の排出、感染の浄化および中耳内の液の蓄積に伴う痛みの緩和の点では成功裏であるが、多数の抗生物質耐性菌の出現およびチューブ留置に伴う侵襲性の性質により、OMのマネージメント、好ましくは予防に対するより有効で許容されるアプローチの必要性が促されている。慢性OMの外科的マネージメントは、子供に対して全身麻酔下での鼓膜を介する中耳腔換気用チューブの挿入を伴う。この手順は一般的であり(普及率は約13%である;Brightら、Am.J.Public Health,83(7):1026−8,1993)、中耳の蓄積液の排出による有痛症状の緩和の点で非常に有効であるが、処置の侵襲性の性質およびその施術(incumbent)の危険性のため、非難されている(Bermanら、Pediatrics,93(3):353−63,1994;Brightら、前掲)。;Cimons,ASM News,60:527−528;Paap,Ann.Pharmacother.,30(11):1291−7,1996)。
【0005】
ワクチン開発における進歩は、7価莢膜−コンジュゲートワクチンPREVNAR(登録商標)(EskolaおよびKilpi,Pedriatr.Infect.Dis.J.16:S72−78,2000)の最近の認可および発売によって示されるように、急性OM(AOM)の主な原因因子である肺炎連鎖球菌に関して最も進んでいる。PREVNAR(登録商標)は、浸潤性の肺炎球菌疾患に非常に有効であるが、OMに関する範囲は期待はずれ(6〜8%)であり、該ワクチンに含まれていない血清型のため、OM症例数が増加しているという報告がある(Blackら,Pedriatr.Infect.Dis J.,19:187−195;Eskolaら,Pedriatr.Infect.Dis J,19:S72−78,2000;Eskolaら,N.Engl.J.Med.344:403−409,2001;Snowら,Otol.Neurotol.,.23:1−2,2002)。滲出液を伴う慢性OMにおいてよく見られるグラム陰性病原体である分類不能型インフルエンザ菌(NTHi)に関してはほとんど進歩していない(非特許文献6;非特許文献7)。NTHiに対する有効なワクチンの開発の障害は、現時点におけるNTHi誘導型中耳疾患の病因の理解が不充分なことである。この遅滞の一因は、該疾患が、良性の鼻咽頭共生生物の一宿主免疫学的寛容から、通常無菌の中耳空間への日和見的侵入菌に対する積極防御反応のものに進行することによる、病原菌が発現するビルレンス因子と宿主の免疫応答との動態的相互作用の理解の欠如である。
【0006】
Rd菌株は、現在の小ゲノム配列決定原理のための代表例の生物体であり、インフルエンザ菌生物学の研究のための重要なモデル生物体であるが、Rd菌株は、ヘモフィルス属の構成員によって引き起こされる病原性の研究には不充分なモデルである。インフルエンザ菌のb血清型菌株は、浸潤性疾患(例えば、髄膜炎)を引き起こし、分類不能型インフルエンザ菌(NTHi)菌株は、主に、限局性呼吸器疾患、特に中耳炎(OM)、急性副鼻腔炎、市中肺炎において役割を有し、慢性閉塞性肺疾患または嚢胞性線維症の患者に重要な結果をもたらす(Kilpiら,Pediatr Infect Dis J 20:654−62,2001;Murphy,Curr Opin Infect Dis 16:129−34,2003;Romanら,J Clin Microbiol 42:1450−9,2004;Sethi,Clin Microbiol Rev 14:336−63,2001;St Geme,Vaccine 19 Suppl l:S41−50,2000)。しかしながら、Rd菌株はd血清型菌株の誘導体である。d血清型菌株が疾患と関連するのは稀である(Dainesら,J Med Microbiol 52:277−82 2003;Heathら,Pediatr Infect Dis J 20:300−5,2001;Rodriguezら,Infect Immun 71:1635−42,2003,Skoczynskaら,J Clin Microbiol 43:938−41,2005)。生物体の生物学の研究における最も有用なデータセットの一例はそのゲノム配列であるため、いくつかの調査により、b型インフルエンザ菌菌株、インフルエンザ菌Biogroup Aegyptius菌株またはRd菌株には存在しない分類不能型菌株に見られる遺伝子が同定および特性評価された(Bergmanら,Infect Immun 71:1098−108,2003;Changら,Infect Immun 68:2630−7,2000;Erdosら,Int J Pediatr Otorhinolaryngol 67:749−55.2003;Liら,Mol.Microbiol 47:1101−11,2003;McMichael & Green,Curr Opin Investig Drugs 4:953−8,2003;Pomposiello & Demple,2001;Smootら,Infect Immun 70:2694−9,2002)。
現在、どのようにしてNTHiが小児においてOMを引き起こすのかの理解は不充分である。OMの誘導に必要な推定ビルレンス因子の同定は、宿主−病原体相互作用の理解、そして最終的には潜在的なワクチン候補および化学療法の標的の同定に大きく寄与する。中耳炎のマネージメント、好ましくは予防に対するより有効で許容されるアプローチを開発する多大な必要性が存在する。ワクチン開発は、非常に有望であり、本目的を達成するための費用効果のある方法である(非特許文献8:非特許文献9)。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】Infante−RivandおよびFernandez,Epidemiol.Rev.(1993)15:444−465
【非特許文献2】Baldwin,Am.J.Otol(1993)14:601−604
【非特許文献3】Hunterら,Ann.Otol.Rhinol.Laryngol.Suppl(1994)163:59−61
【非特許文献4】Teeleら、J.Infect.Dis.(1990)162:685−694
【非特許文献5】Kaplanら,Pediatr.Infect.Dis.J.(1997)16:S9−11
【非特許文献6】Klein,Pedriatr.Infect.Dis J(1997)16:S5−8
【非特許文献7】Spinolaら,J.Infect.Dis.(1986)154:100−109
【非特許文献8】Giebank,Pedriatr.Infect.Dis J(1994)13(11):1064−8
【非特許文献9】Karmaら,Int.J.Pedritr.OtorhinolaryngoL(1995)32(Suppl.):S127−34
【発明の概要】
【課題を解決するための手段】
【0008】
(発明の概要)
本発明は、NTHi インフルエンザ86−028NP菌株のゲノム配列の同定およびキャラクタライゼーションならびにそれにコードされるポリペプチド配列を提供する。NTHiゲノム配列の3倍解析を、配列番号:1〜576として示す一連のコンティグ配列に示し(set out)、その後のゲノム配列の8倍解析を、配列番号:675〜685として示す一連の11個のコンティグ配列に示す。これらのコンティグは生データであり、当業者によって、常套的な方法を用い、重複配列を比較することにより、これらのコンティグをアセンブルし、NTHi 86−028NP菌株の完全なゲノムが構築され得る。
【0009】
NTHi 86−028NP菌株の注釈付きの完全なゲノムを、配列番号:772として示す。オープンリーディングフレームを表6に、配列番号:773〜2593として示す。得られる遺伝子産物のアミノ酸配列を、配列番号:2594〜4414として示す。該ゲノムは、1.91kbの大きさであり、Rd菌株のゲノムよりもわずかに大きい。Rd菌株のゲノムに対する総ゲノム再編成のいくつかの領域ならびに86−028NP菌株に特有のいくつかの遺伝子が同定されている。
【0010】
本発明はまた、本発明のNTHiポリペプチドに特異的な抗体を提供する。ヒトまたは試料、例えば、血清、痰、耳液、血液、尿、リンパ液および脳脊髄液などにおいてNTHi菌を検出する方法が想定される。このような方法は、特異的ポリヌクレオチドプローブを用いてNTHiポリヌクレオチドを検出すること、または特異的抗体を用いてNTHiポリペプチドを検出することを含む。本発明ではまた、このようなNTHi菌の検出方法を用いる診断用キットが想定される。
【0011】
本発明ではまた、本発明のNTHiポリペプチドまたはそのNTHiペプチドを投与することによる、免疫応答の惹起方法が想定される。このような方法は、NTHi感染によって引き起こされる疾患、例えばOMなどの処置および/または予防のために、NTHiポリペプチドまたはNTHiペプチドをワクチンとして投与することを含む。以下のNTHi遺伝子は、中耳および/または鼻咽頭感染中または該感染に応答して上方調節される。これらの遺伝子:HisB、lppB、sapA、lolA、rbsC、purE、ribB、arcB、uxuA、dsbB、ureH、licC、HI1647、ispZ、radC、mukF、glpR、ihfB、argR、cspD、HI0094、HI1163、HI1063、HI0665、HI1292、HI1064にコードされるポリペプチドおよびそのペプチドが可能なOMワクチン候補および/または化学療法の標的として想定される。NTHi HisB遺伝子は、ヌクレオチド配列配列番号:615として示され、配列番号:616で示されるアミノ酸配列をコードする。NTHi sapA遺伝子は、ヌクレオチド配列配列番号:617で示され、配列番号:618で示されるアミノ酸配列をコードする。NTHi rbsC遺伝子は、ヌクレオチド配列配列番号:619で示され、配列番号:620で示されるアミノ酸配列をコードする。NTHi purE遺伝子は、ヌクレオチド配列配列番号:621で示され、配列番号:622で示されるアミノ酸配列をコードする。NTHi ribB遺伝子は、ヌクレオチド配列配列番号:623で示され、配列番号:624で示されるアミノ酸配列をコードする。NTHi
arcB遺伝子は、ヌクレオチド配列配列番号:625で示され、配列番号:626で示されるアミノ酸配列をコードする。NTHi uxuA遺伝子は、ヌクレオチド配列配列番号:627で示され、配列番号:628で示されるアミノ酸配列をコードする。NTHi dsbB遺伝子は、ヌクレオチド配列配列番号:629で示され、配列番号:630で示されるアミノ酸配列をコードする。NTHi ureH遺伝子は、ヌクレオチド配列配列番号:631で示され、配列番号:632で示されるアミノ酸配列をコードする。NTHi licC遺伝子は、ヌクレオチド配列配列番号:633で示され、配列番号:634で示されるアミノ酸配列をコードする。NTHi HI1647遺伝子は、ヌクレオチド配列配列番号:635で示され、配列番号:636で示されるアミノ酸配列をコードする。NTHi ispZ遺伝子は、ヌクレオチド配列配列番号:637で示され、配列番号:638で示されるアミノ酸配列をコードする。NTHi radC遺伝子は、ヌクレオチド配列配列番号:639で示され、配列番号:640で示されるアミノ酸配列をコードする。NTHi mukF遺伝子は、ヌクレオチド配列配列番号:641で示され、配列番号:642で示されるアミノ酸配列をコードする。NTHi glpR遺伝子は、ヌクレオチド配列配列番号:643で示され、配列番号:644で示されるアミノ酸配列をコードする。NTHi ihfB遺伝子は、ヌクレオチド配列配列番号:645で示され、配列番号:646で示されるアミノ酸配列をコードする。NTHi argR遺伝子は、ヌクレオチド配列配列番号:647で示され、配列番号:648で示されるアミノ酸配列をコードする。NTHi cspD遺伝子は、ヌクレオチド配列配列番号:649で示され、配列番号:650で示されるアミノ酸配列をコードする。NTHi HI1163遺伝子は、ヌクレオチド配列配列番号:651で示され、配列番号:652で示されるアミノ酸配列をコードする。NTHi HI1063遺伝子は、ヌクレオチド配列配列番号:653で示され、配列番号:654で示されるアミノ酸配列をコードする。NTHi HI0665遺伝子は、ヌクレオチド配列配列番号:655で示され、配列番号:656で示されるアミノ酸配列をコードする。NTHi HI1292遺伝子は、ヌクレオチド配列配列番号:657で示され、配列番号:658で示されるアミノ酸配列をコードする。
【0012】
配列番号:1〜576、配列番号:675〜685として示されるポリヌクレオチド配列および表4と4Bに示すヌクレオチド配列に含まれる該新規なNTHi遺伝子はまた、中耳および/または鼻咽頭感染中に上方調節され、したがって、OMワクチン候補および/または化学療法の標的をコードしていると想定される。また、以下のNTHi遺伝子は、ビルレンス関連遺伝子であると想定され、したがって、有力なOMワクチンおよび/または化学療法の標的:HI1386、HI1462、HI1369、lav、HI1598をコードしていると想定される。NTHi HI1386遺伝子配列は、配列番号:659として示され、配列番号:660で示されるアミノ酸配列をコードする。NTHi HI1462遺伝子配列は、配列番号:661として示され、配列番号:662で示されるアミノ酸配列をコードする。NTHi HI1369遺伝子配列は、配列番号:665として示され、配列番号:666で示されるアミノ酸配列をコードする。NTHi lav遺伝子配列は、配列番号:663として示され、配列番号:664で示されるアミノ酸配列をコードする。NTHi HI1598遺伝子配列は、配列番号:669および配列番号:671として示され、配列番号:670および配列番号:672で示されるアミノ酸配列をコードする。ビルレンスと関連するさらなるNTHi遺伝子としては、配列番号:667および配列番号:673として示されるポリヌクレオチド配列が挙げられる。
【0013】
NTHi感染の処置または予防方法として、本発明では、NTHiポリペプチドの発現または活性を阻害する分子であって、感染中、上方調節されるか、または活性である分子を投与することが想定される。特に、本発明では、NTHi感染中に上方調節されるNTHi遺伝子に特異的に結合するアンチセンスオリゴヌクレオチドを投与することにより、NTHiタンパク質発現をモジュレートすることを含む、NTHi感染の処置または予防方法が想定され、かかる遺伝子としては、hisB、lppB、sapA、lolA、rbsC、purE、ribB、arcB、uxuA、dsbB、ureH、licC、HI1647、ispZ、radC、mukF、glpR、ihfB、argR、cspD、HI0094、HI1163、HI1063、HI0665、HI1292、HI1064が挙げられる。本発明ではまた、これらの(theses)遺伝子にコードされるタンパク質の活性をモジュレートする抗体または小分子を投与することを含む、NTHi感染の処置または予防方法が想定される。配列 配列番号:1〜576、配列番号:675〜685として示されるポリヌクレオチドおよび表4と4Bに示すヌクレオチド配列に含まれる該新規なNTHi遺伝子はまた、中耳および/または鼻咽頭感染中に上方調節され、したがって、このようなポリヌクレオチド配列に特異的に結合するアンチセンスオリゴヌクレオチドもまた想定される。
【0014】
本発明のポリヌクレオチドおよびポリペプチド
本発明は、NTHi 86−028NP菌株ゲノムの配列を提供する。該ゲノム配列の3倍解析を、本明細書において「コンティグ1〜576」で表す一連のコンティグ配列として示す。各コンティグを、その「コンティグ番号」と相関する配列同定番号 で表示する。したがって、配列番号:1〜576で示される本発明のコンティグ。これらのコンティグポリヌクレオチド配列は、常套的な方法を用いてNTHi 86−028NP菌株の完全なゲノム配列にアセンブルされ得る。NTHi菌株82−028NPゲノムの8倍配列解析が終了したら、該ゲノム配列を、本明細書において配列番号:675〜685で表す11個のコンティグに編集した。最後に、該完全なゲノムを、本明細書において配列番号:772で表す1つの核酸配列として示す。
【0015】
本発明は、配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593のコンティグ内に含まれるNTHiポリヌクレオチド配列およびオープンリーディングフレーム、ならびに表3B、表4Bおよび表5に示すヌクレオチド配列を提供する。本発明はまた、本発明のNTHiポリヌクレオチドにコードされるポリペプチド配列、例えば、配列番号:2594〜4414、表3B、表4Bおよび表5に示すアミノ酸配列などを提供する。本発明は、ストリンジェント条件下で、(a)配列番号:1〜576;配列番号:675〜685、配列番号:773〜2593のヌクレオチド配列および本明細書の表3B、表4Bおよび表5に示すヌクレオチド配列の相補配列、(b)上記の任意のポリヌクレオチドの対立遺伝子バリアントであるポリヌクレオチド;(c)上記の任意のタンパク質の種ホモログをコードするポリヌクレオチド;または(d)本発明のNTHiポリペプチドの特異的ドメインもしくは切断型を含むポリペプチドをコードするポリヌクレオチドにハイブリダイズするポリヌクレオチドを提供する。
【0016】
本発明のNTHiポリヌクレオチドはまた、上記のポリヌクレオチドと実質的に等価なヌクレオチド配列を含む。本発明によるポリヌクレオチドは、上記のNTHiポリヌクレオチドに対して、例えば、少なくとも65%、少なくとも70%、少なくとも75%、少なくとも80%、81%、82%、83%、84%、85%、86%、87%、88%または89%、より典型的には少なくとも90%、91%、92%、93%または94%、さらにより典型的には少なくとも95%、96%、97%、98%または99%の配列同一性を有するものであり得る。
【0017】
本発明の核酸配列の範囲内には、ストリンジェント条件下で、配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593のNTHi ヌクレオチド配列、ならびに本明細書の表3B、表4Bおよび表5に示すヌクレオチド配列、またはその相補配列にハイブリダイズする核酸配列断片が含まれ、該断片は、約5個より多くのヌクレオチド、好ましくは7個のヌクレオチド、より好ましくは9個より多くのヌクレオチド、最も好ましくは17個より多くのヌクレオチドである。例えば、本発明のポリヌクレオチドの任意の1つに選択的な(すなわち、特異的にハイブリダイズする)15、17または20個以上のヌクレオチドの断片が想定される。ポリヌクレオチドに特異的にハイブリダイズすることができるプローブにより、本発明のNTHiポリヌクレオチド配列が、同じ遺伝子ファミリー内の他のポリヌクレオチド配列と区別され得るか、またはNTHi遺伝子が他の細菌の遺伝子と区別され得、好ましくは特異的ヌクレオチド配列に基づくものである。
【0018】
用語「ストリンジェント」は、当該技術分野においてストリンジェントと一般的に理解される条件をいうために使用される。ハイブリダイゼーションのストリンジェンシーは、主に、温度、イオン強度および変性剤(例えば、ホルムアミドなど)の濃度によって決定される。ハイブリダイゼーションおよび洗浄のストリンジェント条件の例は、0.015M塩化ナトリウム、0.0015Mクエン酸ナトリウム、65〜68℃または0.015M塩化ナトリウム、0.0015Mクエン酸ナトリウムおよび50%ホルムアミド、42℃である。Sambrookら,Molecular Cloning:A Laboratory Manual,第2版,Cold Spring Harbor Laboratory,(Cold Spring Harbor,N.Y.1989)を参照のこと。よりストリンジェント条件(例えば、より高温、より低イオン強度、より高ホルムアミド、または他の変性剤など)もまた使用され得るが、ハイブリダイゼーションの速度が影響を受ける。デオキシオリゴヌクレオチドのハイブリダイゼーションが懸念される場合、さらなる例示的なストリンジェントハイブリダイゼーション条件としては、37℃(14塩基オリゴの場合)、48℃(17塩基オリゴの場合)、55℃(20塩基オリゴの場合)および60℃(23塩基オリゴの場合)での6×SSC 0.05%ピロリン酸ナトリウム中での洗浄が挙げられる。
【0019】
他の薬剤を、非特異的および/またはバックグラウンドハイブリダイゼーションを低減させる目的でハイブリダイゼーションおよび洗浄バッファー中に含めてもよい。例は、0.1%ウシ血清アルブミン、0.1%ポリビニルピロリドン、0.1%ピロリン酸ナトリウム、0.1%ドデシル硫酸ナトリウム、NaDodSO4、(SDS)、フィコール、デンハルト溶液、超音波処理サケ精子DNA(または他の非相補DNA)、および硫酸デキストランであるが、他の適当な薬剤もまた使用され得る。このような添加剤の濃度および型は、ハイブリダイゼーション条件のストリンジェンシーに実質的に影響を与えることなく、変更され得る。ハイブリダイゼーション実験は、通常、pH6.8〜7.4で行なわれるが、典型的なイオン強度条件では、ハイブリダイゼーションの速度は、ほぼpHに非依存性である。Andersonら,Nucleic Acid Hybridisation:A Practical Approach,Ch.4,IRL Press
Limited(Oxford,England)を参照のこと。ハイブリダイゼーション条件は、これらの可変量を適合させるため、および関連のある異なる配列のDNAがハイブリッドを形成するのを可能にするために、当業者によって調整され得る。
【0020】
本発明の範囲内に含まれる配列は、このような特定の配列に限定されないだけでなく、その対立遺伝子および種の異型も包含する。対立遺伝子および種の異型は、配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593に示した配列、および本明細書の表3B、表4Bおよび表5に示したヌクレオチド配列、好ましくは、そのオープンリーディングフレーム、代表的なその断片または配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593内のオープンリーディングフレーム、表3B、表4Bおよび表5に示すヌクレオチド配列と少なくとも90%同一である、好ましくは95%同一であるヌクレオチド配列を、同じ種の別の単離ヌクレオチド(isolete)由来の配列と比較することにより、常套的に決定され得る。2つの配列間の同一性および類似性を決定するための好ましいコkンピュータプログラム法としては、限定されないが、GCG プログラムパッケージ、例えば、GAP(Devereuxら,Niicl.Acid.Res.,12:387,1984;Genetics Computer Group,University of
Wisconsin,Madison,WI)、BLASTP、BLASTNおよびFASTA(Altschulら,J.Mol Biol,215:403−410,1990)が挙げられる。BLASTX プログラムは、National Center for Biotechnology Information(NCBI)および他の供給源(BLAST Manual,Altschulら NCB/NLM/NIH Bethesda,MD 20894;Altschulら、前掲)から公に入手可能である。よく知られたSmith Watermanアルゴリズムもまた、同一性を決定するために使用され得る。
【0021】
さらにまた、コドンの多様性に適合させるため、本発明は、本明細書に開示する特定のオープンリーディングフレーム(ORF)にコードされるものと同じアミノ酸配列をコードする核酸分子を包含する。すなわち、ORFのコード領域において、1つのコドンを、同じアミノ酸をコードする別のコドンと置換することが明白に想定され得る。
【0022】
本発明の単離されたポリペプチドとしては、限定されないが、配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593として示されるポリヌクレオチド配列内に含まれるヌクレオチド配列、ならびに表3B、表4Bおよび表5に示すヌクレオチド配列にコードされるアミノ酸配列を含むポリペプチド、または対応する完全長もしくは成熟タンパク質が挙げられる。本発明のポリペプチドとしては、本明細書の表3B、表4Bおよび表5に示す配列番号:616、配列番号:618、配列番号:620、配列番号:622、配列番号:624、配列番号:626、配列番号:628、配列番号:628、配列番号:630、配列番号:632、配列番号:634、配列番号:636、配列番号:638、配列番号:640、配列番号:642、配列番号:644、配列番号:646、配列番号:648、配列番号:650、配列番号:652、配列番号:654、配列番号:656、配列番号:658、配列番号:660、配列番号:662、配列番号:664、配列番号:666、配列番号:668、配列番号:670、配列番号:672、配列番号:674、配列番号:687、配列番号:689、配列番号:691、配列番号:693、配列番号:695、配列番号:697、配列番号:699、配列番号:701、配列番号:703、配列番号:705、配列番号:707、配列番号:709、配列番号:711、配列番号:713、配列番号:715、配列番号:717、配列番号:719、配列番号:721、配列番号:723、配列番号:725、配列番号:727、配列番号:729、配列番号:731、配列番号:733、配列番号:735、配列番号:737、配列番号:739、配列番号:741、配列番号:743、配列番号:745、配列番号:747、配列番号:749、配列番号:751、配列番号:753、配列番号:755、配列番号:757、配列番号:759、配列番号:761、763、配列番号:765、配列番号:767、配列番号:769または配列番号:771、配列番号:2594〜4414のアミノ酸配列が挙げられる。
【0023】
また、本発明のポリペプチドとしては、好ましくは生物学的または免疫学的活性を有し、(a)配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593として示されるヌクレオチド配列、ならびに表3B、表4Bおよび表5に示すヌクレオチド配列内に含まれるオープンリーディングフレームまたは(b)(a)のポリヌクレオチドの相補配列にストリンジェントハイブリダイゼーション条件下でハイブリダイズするポリヌクレオチドにコードされるポリペプチドが挙げられる。
【0024】
本発明はまた、本発明のアミノ酸配列の生物学的に活性または免疫学的に活性なバリアント、および生物学的および/または免疫学的活性を保持している、その「実質的な等価物」(例えば、少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、86%、87%、88%、89%、少なくとも約90%、91%、92%、93%、94%、典型的には少なくとも約95%、96%、97%、より典型的には少なくとも約98%または最も典型的には少なくとも約99%のアミノ酸同一性を有する)を提供する。対立遺伝子バリアントにコードされるポリペプチドは、配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593に示されるヌクレオチド配列、ならびに配列番号:2594〜4414として、および本明細書の表3B、表4Bと表5に示されるアミノ酸配列をコードするヌクレオチド配列内に含まれるポリヌクレオチドにコードされるポリペプチド、ならびに配列番号:2594〜4414、本明細書の表3B、表4Bおよび表5に示されるアミノ酸配列を有するポリペプチドと比べて、同様の、増大された、または低下した活性を有するものであり得る。
【0025】
NTHiペプチドとは、配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593に示されるヌクレオチド配列、または本明細書の表3B、表4Bおよび表5に示すヌクレオチド配列
にコードされるNTHiポリペプチドの断片、および配列番号:2594〜4414 本明細書の表3B、表4Bおよび表5に占められるアミノ酸配列を有するポリペプチドをいう。好ましいNTHiペプチドは、生物学的および/または免疫学的に活性である。
【0026】
さらに、本発明は、本発明のNTHi核酸断片または本発明の核酸断片の縮重バリアントにコードされる、単離されたNTHiポリペプチドまたはNTHiペプチドを提供する。用語「縮重バリアント」は、本発明の核酸断片(例えば、ORF)をヌクレオチド配列は異なるが、遺伝暗号の縮重のため、同一のNTHiポリペプチド配列をコードするヌクレオチド断片をいう。本発明の好ましい核酸断片は、タンパク質をコードするORFである。
【0027】
本発明はまた、該ポリペプチドの生物学的および/または免疫学的活性に影響を及ぼさない1つ以上の同類アミノ酸置換を有するNTHiポリペプチドを提供する。あるはまた、本発明のNTHiポリペプチドは、生物学的活性を改変するものまたは改変しないものであり得る同類アミノ酸置換を有することが想定される。用語「同類アミノ酸置換」は、天然のアミノ酸残基と、基準残基、例えば、天然に存在するアミノ酸および天然に存在しないアミノ酸との、その位置でのアミノ酸残基の極性または電荷に対して影響がほとんどまたは全くないような置換をいう。例えば、同類置換は、ポリペプチド内の無極性残基と任意の他の無極性残基との置き換えによりもたらされる。
さらに、該ポリペプチド内の任意の天然の残基はまた、「アラニンスキャニング変異誘発」法に従ってアラニンで置換され得る。天然に存在するアミノ酸は、その側差に基づき、以下のとおりに特徴づけされる:塩基性:アルギニン、リシン、ヒスチジン;酸性:グルタミン酸、アスパラギン酸;非電荷極性:グルタミン、アスパラギン、セリン、トレオニン、チロシン;および無極性:フェニルアラニン、トリプトファン、システイン、グリシン、アラニン、バリン、プロリン、メチオニン、ロイシン、ノルロイシン、イソロイシン。アミノ酸置換に関する一般規則を以下の表1に示す。
【0028】
【表1】
NTHiポリペプチドをコードするポリヌクレオチドに相補的なアンチセンスポリヌクレオチドもまた提供される。
【0029】
本発明では、本発明のポリヌクレオチドが増幅または発現のためにベクター内に挿入され得ることが想定される。発現のためには、該ポリヌクレオチドを、適切な発現制御配列、例えば、プロモーターおよびポリアデニル化シグナル配列などに作動可能に連結させる。さらに、本発明のポリヌクレオチドを含む細胞が提供される。例示的な原核生物宿主としては、大腸菌、バチルス属、ストレプトミセス属、シュードモナス属、サルモネラ属およびセラチア属などの細菌が挙げられる。
【0030】
用語「単離される」は、物質が、これが天然に存在する環境のその他の成分から取り出されており、該成分を本質的に含まないことをいう。例えば、ポリペプチドは、他の細胞性タンパク質から分離されたものであり、またはDNAは、これが天然状態で存在するゲノム内でこれとフランキングしている他のDNA分離されたものである。
【0031】
免疫応答を惹起するための抗体および方法
本発明は、NTHiポリペプチドに特有(すなわち、特異的)な抗原性エピトープに結合する抗体を提供する。また、多数のインフルエンザ菌亜型間に共通する抗原性エピトープを含むが、任意の他の抗原性エピトープに関して他と異なる抗体を提供する。該抗体は、ポリクローナル抗体、モノクローナル抗体、その特異的エピトープに結合する能力を保持している抗体断片(例えば、Fv、FabおよびF(ab)2断片)、単鎖抗体ならびにヒト抗体またはヒト化抗体であり得る。抗体は、当該技術分野において標準的な手法によって作製され得る。
【0032】
インフルエンザ菌の莢膜多糖類を模倣する抗原により、インビトロアッセイで細菌を死滅させる能力を示す抗体が生成されることが、当該技術分野において知られている。このような抗体はまた、動物モデル系において、インフルエンザ菌による抗原刺激を防御することが知られている。この研究は、莢膜多糖類(polysaccharride)に対する抗体が、おそらく、ヒトにおいて防御的免疫応答を惹起することを示す。本発明は、インフルエンザ菌を死滅させるとともに、NTHi感染からヒトを保護する能力を示す、NTHi本発明のポリペプチドおよびその断片に特異的な抗体を提供する。本発明はまた、インフルエンザ菌のビルレンスを低減させる、付着を阻害する、細胞分裂を阻害する、および/または上皮内への侵入を阻害する、あるいは、インフルエンザ菌の食作用を増強する、本発明のNTHiポリペプチドに特異的な抗体を提供する。
【0033】
インビトロ補体媒介性殺菌アッセイ系(Musherら,Infect.Immun.39:297−304,1983;Andersonら,J.Clin.Invest.51:31−38,1972)が、抗NTHi抗体の殺菌活性を測定するために使用され得る。the ability of NTHiポリペプチドがNTHiペプチドが防御的抗体応答を惹起する能力に関するさらなるデータは、感染の動物モデル、例えば、本明細書に記載のチンチラのモデル系を用いることにより得られ得る。
【0034】
また、NTHiのエピトープに対する予備形成抗体、例えば、NTHi OMP、LOSまたは非被膜タンパク質に対する抗体などの投与による受動免疫療法により、宿主に対して短期的防御を付与することが可能である。したがって、想定されるワクチン製剤は、受動免疫療法における使用のための抗体を生成させるために使用され得る。異種免疫グロブリンは、その外来免疫学的成分に対して免疫応答が誘発され得るため、ヒト免疫グロブリンがヒト用の薬に好ましい。かかる受動免疫処置は、緊急的に、特別な危険に曝された非免疫処置個体の即座の防御のために使用され得る。あるはまた、このような抗体は、抗イディオタイプ抗体の作製において使用され得、これが、こんどはNTHi エピトープに対する免疫応答を刺激するための抗原として使用され得る。
【0035】
本発明では、個体においてNTHiに対する免疫応答を惹起する方法が想定される。
この方法により、NTHi菌の死滅、細胞へのNTHi付着の阻止および/またはNTHi複製の遅滞の1種類以上を含む免疫応答が惹起される。本発明の組成物の「免疫学的用量」は、投与前に検出可能な免疫応答と比べて、または投与前の標準的な免疫応答と比べて、投与後に検出可能な体液性および/または細胞性免疫応答を生じさせるものである。本発明では、該方法によりもたらされる免疫応答が防御的および/または治療的であり得ることが想定される。一実施形態において、該方法は、本発明のNTHiタンパク質またはNTHiペプチドを含む組成物の免疫学的用量を投与する工程を含む。NTHiタンパク質またはその抗原性ペプチドは、それ自体では抗体を産生し得ないが、第1のタンパク質を安定化させ、かつ免疫学的および防御的特性を有する融合タンパク質を産生することができる補助タンパク質と融合させてもよい。したがって、有効組換えタンパク質は、好ましくは、抗原性補助タンパク質、例えば、グルタチオン−S−トランスフェラーゼ(GST)またはβ−ガラクトシダーゼ、該タンパク質を可溶化し、その産生および精製を助長する比較的大きな補助タンパク質をさらに含む。またさらに、補助タンパク質は、免疫系の全身性刺激を提供するという意味で、アジュバントとしての機能を果たすものであり得る。補助タンパク質は、第1のタンパク質のアミノ末端またはカルボキシ末端のいずれかに結合され得る。本発明により、本発明のポリヌクレオチドにコードされるNTHiポリペプチドまたはその抗原性ペプチドを含む組成物、特にワクチン組成物、および方法が提供される。
【0036】
本発明ではまた、個体において多数のインフルエンザ菌亜型に対する免疫応答を惹起する方法が想定される。この方法により、インフルエンザ菌の死滅、細胞へのインフルエンザ菌付着の阻止および/またはインフルエンザ菌複製の遅滞の1種類以上を含む免疫応答が惹起される。この方法は、多数のインフルエンザ菌亜型間に共通する抗原性エピトープを含むが、任意の他の抗原性エピトープに関して他と異なる本発明のNTHiタンパク質またはNTHiペプチドを含む組成物の免疫学的用量を投与する工程を含む。
【0037】
別の実施形態において、該方法は、本発明のNTHiタンパク質またはNTHiペプチドを発現する細胞を含む組成物の免疫学的用量を投与することを含む。また別の実施形態において、該方法は、本発明のNTHiタンパク質またはNTHiペプチドをコードするポリヌクレオチドを含む組成物の免疫学的用量を投与することを含む。該ポリヌクレオチドは、任意の他の核酸と会合されていない裸のポリヌクレオチドであってもよく、ベクター、例えば、プラスミドまたはウイルスベクター(例えば、あでの随伴ウイルスベクターまたはアデノウイルスベクター)内に存在するものであったもよい。本発明の組成物は、1種類以上のNTHiタンパク質またはNTHiペプチドを単独、またはNTHi菌に対する免疫応答を惹起する他のエピトープとの組合せで含むものであり得る。本発明の組成物はまた、1種類以上のNTHiタンパク質またはペプチドを、1種類以上の インフルエンザ菌亜型に対する免疫応答を惹起するエピトープとの組合せで含むものであり得る。該組成物の投与は、当該技術分野において標準的な経路、例えば、非経口、静脈内、経口、口腔内、経鼻、肺経由、経直腸または経膣によるものであり得る。該方法は、一人の個体に組合せで使用され得る。該方法は、個体のNTHi感染の前包埋後に使用され得る。
【0038】
中耳および/または鼻咽頭のNTHi感染の際に上方調節される遺伝子、ならびにNTHiビルレンスと関連する遺伝子を、本明細書において記載する。このようなNTHi遺伝子にコードされるポリペプチドおよびそのペプチドは、NTHi感染と関連する障害、例えばOMなどを処置または予防するための免疫応答の惹起に有用であると想定される。このような遺伝子にコードされるポリペプチドの一例としては、ヒスチジン生合成タンパク質、リポタンパク質B、ペプチドABC輸送体、ペリプラズムSapA前駆体、外膜リポタンパク質担体タンパク質前駆体、リボース輸送系パーミアーゼタンパク質、ホスホリボシルアミノイミダゾールカルボキシラーゼ触媒性サブユニット、PurE、ホスホリボシルアミノイミダゾールカルボキシラーゼ触媒性サブユニット、オルニチンカルバモイルトランスフェラーゼ、マンノン酸脱水酵素、ジスルフィドオキシドレダクターゼ、ウレアーゼアクセサリータンパク質、ホスホコリンシチジリル(cytidylyl)トランスフェラーゼ、推定ピリドキシン生合成タンパク質、一重項酸素抵抗性タンパク質、細胞内隔壁化タンパク質、DNA修復タンパク質、MukFタンパク質、グリセロール−3−ホスフェートレギュロンレセプター、組込み宿主因子βサブユニット、アルギニン抑制因子、コールドショック様タンパク質、ストレス応答タンパク質、LicA、MukF、RadAおよびHI0094、HI1163、HI0665、HI1292、HI1064 HI186、HI0352遺伝子にコードされる仮想タンパク質が挙げられる。NTHi
OMP、LOSおよび非被膜タンパク質もまた、NTHi感染と関連する障害の予防および処置のための免疫応答を惹起することが想定される。
【0039】
本発明は、個体において宿主細胞へのNTHi菌の結合を阻止する方法を含む。該方法は、NTHiの細胞付着の結合を阻止する本発明の抗体またはポリペプチドを投与することを含む。あるはまた、NTHiの細胞付着の結合を阻止する1種類以上の小分子の投与が想定される。本発明の抗体、ポリペプチドまたは小分子がNTHiの細胞付着を阻止する能力を示すために、インビトロアッセイが使用され得る。
【0040】
NTHiの細胞付着を阻止する本発明の抗体、本発明のポリペプチドおよび/または本
発明の小分子を含む医薬組成物が提供される。該医薬組成物は、前述の活性成分の1種類単独からなるものであってもよく、前述の活性成分の組合せを含むものであってもよく、細菌感染を処置するために使用されるさらなる活性成分を含むものであってもよい。医薬組成物は、医薬として有効な担体などの1種類以上の付加的成分を含んでいてもよい。医薬組成物の投与の投薬量および頻度は、標準的な手法によって決定され、例えば、個体の体重および年齢、投与経路ならびに症状の重篤度に依存する。医薬組成物の投与は、当該技術分野において標準的な経路、例えば、非経口、静脈内、経口、口腔内、経鼻、肺経由、経直腸または経膣によるものであり得る。
【0041】
また、本発明により、個体においてNTHi感染を検出するための方法が提供される。一実施形態において、該方法は、試料中の本発明のNTHiポリヌクレオチドを、該ポリヌクレオチドに特異的に結合するプライマーまたはプローブ用いて検出することを含む。該ポリヌクレオチドの検出は、例えば、ハイブリダイゼーションおよびPCRを伴う当該技術分野において常套的な数多くの手法によってなされ得る。
【0042】
本発明の抗体はまた、インフルエンザ菌感染が疑われる個体の種々の体液におけるNTHi抗原(NTHiポリペプチドおよびそのペプチド)の検出のための診断用アッセイにおける使用のための試薬を提供するために使用され得る。別の実施形態において、本発明のNTHiタンパク質およびペプチドは、種々の患者組織および体液、例えば、限定されないが:血液、血清、耳液、髄液、痰、尿、リンパ液および脳脊髄液におけるNTHiの検出のためのイムノアッセイにおける抗原として使用され得る。本発明の抗原は、当該技術分野において知られた任意のイムノアッセイ系、例えば、限定されないが:ラジオイムノアッセイ、ELISAアッセイ、サンドイッチアッセイ、沈降素反応、ゲル拡散沈降素反応、免疫拡散アッセイ、凝集アッセイ、蛍光イムノアッセイ、プロテインAイムノアッセイおよび免疫電気泳動アッセイにおいて使用され得る。
【0043】
ワクチンおよび化学療法標的
本発明の一態様は、個体にNTHi抗原タンパク質またはその抗原性ペプチドを接種することを含む、個体、特に哺乳動物における免疫学的応答を低減させるための方法に関する。
【0044】
本発明はまた、本発明の免疫学的組換えNTHiタンパク質またはNTHiペプチドを、適当な担体とともに含むワクチン製剤を提供する。ワクチン候補および/または化学療法の標的として想定されるNTHiポリペプチドおよびそのペプチドとしては、限定されないが、ヒスチジン生合成タンパク質、リポタンパク質B、ペプチドABC輸送体、ペリプラズムSapA前駆体、外膜リポタンパク質担体タンパク質前駆体、リボース輸送系パーミアーゼタンパク質、ホスホリボシルアミノイミダゾールカルボキシラーゼ触媒性サブユニット、PurE、3,4−ジヒドロキシ(dihydroxt)−2−ブトン4−ホスフェートシンターゼ、オルニチンカルバモイルトランスフェラーゼ、マンノン酸脱水酵素、ジスルフィドオキシドレダクターゼ、ウレアーゼアクセサリータンパク質、ホスホコリンシチジリルトランスフェラーゼ、推定ピリドキシン生合成タンパク質、一重項酸素抵抗性タンパク質、細胞内隔壁化タンパク質、DNA修復タンパク質、MUKFタンパク質、グリセロール−3−ホスフェートレギュロンレセプター、組込み宿主因子βサブユニット、アルギニン抑制因子、コールドショック様タンパク質、ストレス応答タンパク質、LicA、RadAならびにHI0094、HI1163、HI0665、HI1292、HI1064 HI1386、HI0352遺伝子にコードされる仮想タンパク質、配列番号:1〜576、配列番号:675〜685、配列番号:773〜2593として示すヌクレオチド配列および本明細書の表3B、表4Bおよび表5に示すヌクレオチド配列内に存在する新規なNTHiポリヌクレオチド配列にコードされるNTHi OMP、NTHi LOSおよびNTHi非被膜タンパク質ならびにポリペプチド、ならびに配列番号:2594〜4414、本明細書の表3B、表4Bおよび表5に示すアミノ酸配列を有するポリペプチドが挙げられる。
【0045】
該タンパク質は、胃の中で分解され得るため、好ましくは、非経口、例えば、皮下、筋肉内、静脈内または皮内である投与で投与する。非経口投与に適した製剤としては、水性および非水性の滅菌注射用液剤(これは、抗酸化剤、緩衝剤、静菌剤および製剤を個体の体液(好ましくは血液)と等張性にする溶質を含んでいてもよい);ならびに水性および非水性滅菌懸濁剤(これは、懸濁剤または増粘剤を含んでいてもよい)が挙げられる。該製剤は、単位投与容器または反復投与容器、例えば、密閉アンプルおよびバイアル内に提示され得、使用直前に滅菌液状担体の添加のみが必要とされる凍結乾燥条件で保存され得る。該ワクチン製剤はまた、製剤の免疫原性を増強するためのアジュバント系(例えば、水中油型および当該技術分野において知られた他の系など)を含んでいてもよい。投薬量は、ワクチンの比活性に依存し、常套的な実験手法容易に決定され得る。
【0046】
A.ペプチドワクチン
ペプチドワクチンなどのペプチド治療用薬剤は、当該技術分野においてよく知られており、製剤分野での使用が増大している。かかるペプチド化合物の非経口投与に一貫する欠点は、急速な分解または変性であった。輸液ポンプならびにワックスまたはオイル埋入物は、治療用薬剤の長期投与のために、ペプチド様治療用薬剤の存在を延長させるためと、かかる薬剤の完全性を保持するための両方に対する取り組みにおいて使用されている。さらにまた、該ペプチド様薬剤は、理想的には、長期間(特に、該ペプチド様薬剤の各エピトープに関して)天然状態の立体構造を維持するものであるのがよく、さらに、抗原刺激された動物または免疫処置したヒトにおいて免疫学的応答が誘導されるのに適した様式で提示されるものであるのがよい。
【0047】
本発明のNTHi抗原性ペプチドは、いくつかの慣用的な様式で調製され得る。短鎖ペプチド配列は、標準的な手段を用い、化学合成によって調製され得る。特に簡便であるのは固相手法である(例えば、Eriksonら,The Proteins(1976)v.2,Academic Press,New York,p.255を参照)。自動固相合成装置が市販されている。また、配列の修飾は、適切な残基の置換、付加または欠失によって容易になされる。例えば、担体タンパク質への簡便な連結のためのスルフヒドリル基をもたらすため、システイン残基がカルボキシ末端に付加され得、またはスペーサーエレメント、例えば、さらなるグリシン残基などが、該ペプチドのC末端連結アミノ酸とその残部間の配列内に組み込まれ得る。短鎖NTHiペプチドはまた、組換え手法によって作製され得る。この長さのペプチドのコード配列は、化学的手法、例えば、Matteucciら,JAm Chem Soc,103:3185(1981)に記載のホスホトリエステル法によって容易に合成され得る。
【0048】
本明細書において想定される一部のNTHiペプチド配列は、免疫原性であるには小さすぎると考えられ得、この性質を付与するために担体物質に結合させ得る。かかる連結を作出する当該技術分野において知られた任意の方法が使用され得る。連結は、ジスルフィド結合を一方の官能基の末端に生成させ、ペプチド結合を他方に生成させるヘテロ二官能性薬剤、例えば、ジスルフィドアミド形成剤(例えば、N−スクシジミジル(succidimidyl)−3−(2−ピリジルジチオ)プロピオネート(SPDP))(例えば、Jansenら,Immun.Rev.62:185,1982を参照)およびジスルフィド結合ではなくチオエーテルを形成する二官能性カップリング剤、例えば、6−マレイミドカプロン酸、2−ブロモ酢酸、2−ヨード酢酸、4−(N−マレイミドメチル)シクロへキサン−1−カルボン酸などの反応性エステル、ならびにスクシンイミドまたは1−ヒドロキシ−2−ニトロ−4−スルホン酸と合わせ、スクシンイミジル4−(N−mareイミド−メチル)シクロへキサン−1−カルボキシレート(carobxylate)(SMCC)などのナトリウム塩にすることによりカルボキシル基を活性化させるカップリング剤を用いて形成され得る。
【0049】
B.ワクチン組成物および投与
免疫原を初回抗原刺激投与し、続いて該免疫原を1回以上の追加免疫刺激に曝露することは、有効なワクチンとするために必要であり得る(Krampら,Infect.Immun.,25:771−773,1979;Davisら,Immunology Letters,14:341−8 1986 1987)。共投与されると免疫応答が有益に増強され得るタンパク質またはポリペプチドの例としては、サイトカイン(例えば、IL−2、IL−12、GM−CSF)、サイトカイン誘導分子(例えば、Leaf)または共起刺激分子が挙げられる。ヘルパー(HTL)エピトープを細胞内標的化シグナルに連結し、CTLエピトープとは別に発現させてもよい。これにより、CTL エピトープと異なる細胞区画へのHTLエピトープの指向が可能となり得る。必要な場合は、これにより、MHCクラスII経路内へのHTLエピトープのより効率的な侵入を助長し、それによりCTL誘導を改善し得る。CTL誘導とは対照的に、ある種の疾患では、免疫抑制分子(例えば、TGF−β)の共発現によって免疫応答を特異的に低下させることが有益であり得る。
【0050】
理想的には、免疫原は2つの特性;対応する抗体の形成を刺激する能力、およびこのような抗体と特異的に反応する傾向を示すものである。免疫原は、抗体のコーミング(combing)部位によって認識され得る免疫原の最小の部分であるエピトープを1つ以上有する。特別な場合では、免疫原、免疫原の画分または免疫原が提示される条件は、所望の免疫学的応答を促すのに不充分であり、不充分な免疫性がもたらされる。これは、しばしば、ペプチドまたは他の小分子が免疫原として使用される場合である。また、免疫調節物質(例えば、サイトカイン、例えばインターロイキン)などの他の物質をワクチンと合わせてもよい。
【0051】
ワクチン分野では、免疫原ともに使用して免疫応答を増強するアジュバントと呼ばれるある種の物質の使用が認識されている。アジュバントは、アジュバントの使用なしで惹起され得るものより速くて大きな免疫応答を惹起するためにさらに使用される。また、アジュバントは、アジュバントを含めない場合に必要とされ得るより少ない免疫原を用いて免疫学的応答をもたらすため、免疫学的防御をもたらすある種の抗体サブクラスの産生を増大させるため、または免疫応答の成分(例えば、体液性、細胞性)を増強するために使用され得る。既知のアジュバントとしては、フロイントアジュバントなどの乳剤および他の油状乳剤、百日咳菌、MF59、Quillaja saponaria(QS21)由来の精製サポニン、水酸化物、リン酸塩およびミョウバンなどのアルミニウム塩、リン酸カルシウム(および他の金属塩)、水酸化アルミニウム塩などのゲル、マイコバクテリウムの産性物、例えばムラミルジペプチド、固形物質、リポソームおよびビロソームなどの粒子が挙げられる。アジュバントとして使用されることが知られている天然産物および細菌の産生物の例としては、モノホスホリルリピドA(MPL)、RC−529(合成MPL様アセチル化単糖)、OM−174(これは、大腸菌由来のリピドA誘導体である)、ホロ毒素、例えばコレラ毒素(CT)またはその誘導体の1つ、百日咳毒素(PT)および大腸菌の非耐熱性毒素(LT)またはその誘導体の1つ、ならびにCpGオリゴヌクレオチドが挙げられる。アジュバント活性は、いくつかの要素、例えば、担体効果、デポ形成、リンパ球再循環の改変、T−リンパ球の刺激、B−リンパ球の直接刺激およびマクロファージの刺激などによって影響され得る。
【0052】
ワクチンは、典型的には注射用剤として、液状の液剤もしくは懸濁剤のいずれかとして調製され、注射前に液体中で液剤もしくは懸濁剤にするのに適した固形形態もまた調製され得る。また、該調製物は乳化されたものであってもよい。免疫学的活性成分は、多くの場合、薬学的に許容され得、活性成分と適合性である賦形剤と混合される。好適な賦形剤は、例えば、水、生理食塩水、デキストロース、グリセロール、エタノールなど、およびその組合せである。また、所望の場合、ワクチンに、ワクチンの有効性を増強する少量の補助物質、例えば、湿潤剤もしくは乳化剤、pH緩衝剤またはアジュバントなどを含めてもよい。ワクチンは、注射によって非経口で、例えば、皮下または筋肉内のいずれかで慣用的に投与される。他の投与様式に適したさらなる製剤としては、坐薬および、場合によっては経口製剤が挙げられる。坐薬では、従来の結合剤および担体として、例えば、ポリアルキレン(alkalene)グリコールまたはトリグリセリドが挙げられ得る。かかる坐薬は、0.5%〜10%、好ましくは1〜2%の範囲の活性成分を含有する混合物から形成され得る。経口製剤は、例えば、医薬等級のマンニトール、ラクトース、デンプン、ステアリン酸マグネシウム、サッカリンナトリウム、セルロース、炭酸マグネシウムなどの通常使用される賦形剤を含む。このような組成物は、液剤、懸濁剤、錠剤、丸剤、カプセル剤、徐放製剤または粉末剤の形態をとり、10%〜95%、好ましくは25〜70%の活性成分を含有する。
【0053】
また、ワクチンは、ジェット式注射器、極微針、エレクトロポレーション、ソノポレーション、マイクロカプセル封入、ポリマーまたはリポソームを用いる経皮的経路、ネブライザー、エーロゾルおよび経鼻スプレー剤用いる経粘膜経路および鼻腔内経路により投与され得る。天然または合成のポリマー、例えば、デンプン、アルギネートおよびキトサン、D−ポリL−ラクテート(PLA)、D−ポリDL−乳酸−コグリコール酸ミクロスフィア、ポリカプロラクトン、ポリオルトエステル、ポリ無水物ならびにポリホスファゼン
ポリホスファタザン(phosphatazane)などを用いるマイクロカプセル封入は、経皮的経路および経粘膜投与の両方に有用である。合成ポリオルニテート、ポリリシンおよびポリアルギニンまたは両親媒性ペプチドを含むポリマー複合体は、経皮的送達系に有用である。また、その両親媒性の性質により、リポソームは、経皮的、経粘膜および鼻腔内ワクチン送達系に想定される。ワクチン送達に使用される一般的な脂質としては、N−(1)2,3−(ジオレイル−ジヒドロキシプロピル)−N,N,N−トリメチルアンモニウム−メチル硫酸塩(DOTAP)、塩化ジオレイルオキシプロピル−トリメチルアンモニウムDOTMA、ジミスチルオキシプロピル−3−ジメチル−ヒドロキシエチルアンモニウム(DMRIE)、臭化ジメチルジオクタデシルアンモニウム(DDAB)および9N(N’,N−ジメチルアミノエタン)カルバモイル)コレステロール(DC−Chol)が挙げられる。ヘルパー脂質とリポソームの組合せにより、皮膚を通したリポソームの取込みが増強される。このようなヘルパー脂質としては、ジオレオイルホスファチジルエタノールアミン(DOPE)、ジラウトイルホスファチジルエタノールアミン(DLPE)、ジミリストイルホスファチジルエタノールアミン(DMPE)、ジパルミトイルホスファチジルエタノールアミン(DPPE)が挙げられる。また、チリのユッカ(soap tree)樹皮から誘導されるトリテルペノイドグリコシドまたはサポニン(Quillaja saponaria)およびキトサン(脱アセチル化キタン(chitan))は、鼻腔内および経粘膜ワクチン送達に有用なアジュバントとして想定される。
【0054】
該タンパク質は、中性形態または塩形態としてワクチンに製剤化され得る。薬学的に許容され得る塩としては、酸付加塩(ペプチドの遊離アミノ基と形成される)および無機酸、例えば、塩酸もしくはリン酸など)と形成されるもの、または酢酸、シュウ酸、酒石酸、マンデル酸などの有機酸が挙げられる。また、遊離カルボキシル基と形成される塩は、無機塩基、例えば、ナトリウム、カリウム、アンモニウム、カルシウムまたは第2鉄の水酸化物など、ならびにイソプロピルアミン、トリメチルアミン、2−エチルアミノエタノール、ヒスチジンおよびプロカインなどの有機塩基から誘導され得る。
【0055】
ワクチンは、投薬製剤と適合性の様式で、治療上有効で免疫原性となるような量で投与される。投与される量は、処置される被験体、被験体の免疫系が抗体を合成する能力、および所望される防御の程度に依存する。投与に必要とされる活性成分の正確な量は、担当医師の判断に依存し、各個体に対して独自である。しかしながら、適当な投薬量範囲は、個体1人あたり活性成分ほぼ数百マイクログラム程度である。また、初期投与および追加免疫刺激注射に好適なレジメン(regime)は種々であるが、初期投与後、1ないし3ヶ月間隔で後続の注射または他の投与を行なうのが典型的である。
【0056】
本明細書に記載のワクチン組成物での免疫処置の際、宿主の免疫系は、所望の抗原に特異的なCTLを大量に産生することによりワクチンに対して応答し、宿主は、後の感染に対して少なくとも部分的に免疫性となるか、または慢性感染の発症に対して抵抗性となる。本発明のNTHiポリペプチドまたはNTHiペプチドを含有するワクチン組成物は、細菌感染に易感染性あるいはそのリスクがある患者に対して、その抗原に対する免疫応答を惹起し、したがって、患者自身の免疫応答能を増強するために投与される。この使用において、この場合も、正確な量は、患者の健康状態および体重、投与様式、製剤の性質などに依存するが、一般的に、約1.0μg〜約5000/70キログラム患者、より一般的には約10〜約500mg/70kg体重の範囲である。治療目的または免疫処置目的のため、本発明のNTHiポリペプチドまたはNTHiペプチドはまた、弱毒化ウイルス宿主、例えば、ワクシニアまたは鶏痘によって発現させ得る。このアプローチは、本発明のペプチドをコードするヌクレオチド配列を発現させるためのベクターとしてワクシニアウイルスの使用を伴う。急性または慢性感染宿主または非感染宿主内に導入されると、組換えワクシニアウイルスは免疫学的ペプチドを発現し、それにより宿主CTL応答が惹起される。
【0057】
体液性免疫応答は、多くのよく知られた方法、例えば、単純放射免疫拡散アッセイ(Single Radial Immunodiffussion Assay)(SRID)、酵素イムノアッセイ(EIA)および血球凝集阻害アッセイ(HAI)などによって測定され得る。特に、SRIDでは、試験対象の免疫原を含有するゲル(例えば、アガロースなど)の層が用いられる。ゲルを切り出して凹部を作り、試験対象の血清を凹部内に入れる。ゲル内への抗体拡散により沈殿環の形成がもたらされ、その面積は試験対象の血清の抗体の濃度に比例する。EIAは、ELISA(酵素結合イムノアッセイ)としても知られ、試料中の全抗体を測定するために使用される。免疫原をマイクロタイタープレートの表面に吸着させる。試験血清を該プレートに曝露した後、酵素結合免疫グロブリン(例えば、IgGなど)に曝露する。プレートに付着した酵素の活性を、分光測光法などの任意の簡便な手段によって定量され、試験試料中に存在する免疫原に対して指向される抗体の濃度に比例する。HAIでは、免疫原、例えばウイルス系タンパク質がトリ赤血球(など)を凝集させる能力が利用される。該アッセイでは、中和抗体、すなわち、血液凝集を阻害し得る抗体が検出される。試験血清の希釈物を標準濃度の免疫原とともにインキュベートした後、赤血球を添加する。中和抗体の存在により、免疫原による赤血球の凝集が阻害される。細胞性免疫応答を測定するための試験は、遅延型過敏症の測定または標的免疫原に対するリンパ球の増殖性応答の測定を含む。
【0058】
分類不能型インフルエンザ菌(NTHi)
インフルエンザ菌は、小型の非運動性グラム陰性菌である。他のインフルエンザ菌株とは異なり、分類不能型インフルエンザ菌(NTHi)菌株は、多糖莢膜を欠き、時として「非莢膜性」と示される。NTHi菌株は、被包性菌株と遺伝学的に異なり、b型インフルエンザ単離菌よりも不均質性(heterogenous)である。NTHiは、ヒト宿主に対して複雑な一連の抗原を提示する。防御をもたらし得る可能な抗原としては、OMP、リポ多糖類、リポタンパク質、接着性タンパク質および非被膜タンパク質が挙げられる。
【0059】
ヒトは、インフルエンザ菌(influenze)の唯一の宿主である。NTHi菌株は、通常、鼻咽頭および後部口腔咽頭部を含む上気道、下気道ならびに女性の生殖管に宿する。NTHiは、ヒトにおいて広範囲の疾患、例えば、限定されないが、中耳炎、肺炎、副鼻腔炎、敗血症、心内膜炎、喉頭蓋炎、化膿性関節炎、髄膜炎、分娩後および新生児感染、分娩後および新生児セプシス、急性および慢性の卵管炎/卵管炎(salpingitis)、喉頭蓋炎(epiglottis)、心膜炎、蜂巣炎、骨髄炎、心内膜炎、胆嚢炎、腹腔内感染、尿路感染症、乳様突起炎、大動脈移植片感染症、結膜炎(conjunctitivitis)、ブラジル紫斑熱、不顕性菌血症ならびに潜在する肺疾患、例えば、慢性気管支炎、気管支拡張症(bronchietasis)および嚢胞性線維症などの増悪を引き起こす。
【0060】
NTHiの疫学的試験により、該菌株は、外膜タンパク質プロフィール(Barenlcampら,Infect.Immun.,36:535−40,1982)、酵素アロタイプ(Musserら,Infect.Immun.,52:183−191,1986)および他の一般的に使用される疫学的ツールに関して不均一であることが示された。亜型NTHiに対していくつかの試みがなされているが、方法論はいずれも完全に満足のいくものではなかった。NTHiの外膜タンパク質組成物は、ほぼ20のタンパク質からなる。すべてのNTHi菌株は、30,000および16,600ダルトンの分子量を有する2つの共通するOMPを含む。NTHi菌株は、32,000〜42,000ダルトンの範囲内の2つのOMPに基づいて亜型に分けられ得る。NTHiのリポ多糖プロフィールは、グラム陰性腸内細菌と基本的に異なり、20,000ダルトン未満の範囲における1〜4個の異なるバンドに分かれる。
【0061】
基本型のNTHi単離菌は、低継代単離菌86−028NPであり、これは、慢性中耳炎の小児から採取された。この菌株は、インビトロ(Bakaletzら,Infect.Immun.,53:331−5,1988;Holmesら,Microb.Pathog.,23:157−66,1997)ならびにチンチラOMモデル(本明細書に記載)(Bakaletzら,Vaccine,15:955−61,1997;Suzukiら,Infect.Immun.,62:1710−8,1994;DeMariaら,Infect.Immun.,64:5187−92,1996)において充分特性評価されている。86−028NP菌株は、本明細書に記載のように、中耳炎のチンチラモデルでの発現において上方調節される遺伝子、およびチンチラの中耳内でのNTHiの生存に必要な遺伝子を同定するために使用した。
【0062】
DFIストラテジー
異差蛍光誘導(DFI)ストラテジーを、本明細書において、チンチラ動物モデルのOM時に誘導されるNTHi遺伝子を同定するために使用した。感染中、菌体のビルレンスに寄与する細菌遺伝子を同定するため、いくつかの方法を開発した。かかる方法としては、インビボ発現手法(IVET)(この場合、細菌のプロモーターにより、宿主内での生存に必要とされる必須栄養素の合成に必要な遺伝子(1つまたは複数)の発現が調節される);シグニチャータグ化(signature−tagged)突然変異誘発(STM)(変異されると、微生物のビルレンス特性を改変する遺伝子のタグ特異的同定を可能にする;DNAマイクロアレイ手法(転写活性遺伝子を全体的にスクリーニングするため)およびDFI(FACS解析を用いて転写活性プロモーターを選択する(Chiangら,Annu.Rev.Microbiol.,53:129−154,1999)が挙げられる。DFIは、発現の基礎レベルとは無関係に差次的に調節される遺伝子の同定を可能にするハイスループット法であるが、インビトロでの培養に不可欠なものを排除しない。
【0063】
DFIは、多くの微生物において成功裏に利用されている。例えば、GFPレポーター系およびフローサイトメトリーを用いて、マクロファージとの相互作用の際のマイコバクテリウムの遺伝子発現が研究された(Dhandayuthapaniら,Mol.Microbiol,17:901−912,1995)。プロモータートラップ系を用いて、サルモネラ属がインビボ増殖を刺激する環境に供された場合、および培養マクロファージ様細胞によってインターナリゼーションされた場合に、その転写が増大する遺伝子が同定された(ValdiviaおよびFalkow,Mol.Microbiol,22:367−378,1996;ValdiviaおよびFalkow,Science,277:2007−2011,1997;ValdiviaおよびFalkow,Curr.Opin.Microbiol,1:359−363,1998)。また、DFIを用いて、感染を刺激する種々のインビトロ条件で培養すると、S.pneumoniaeおよびS.aureusにおいて発現されるプロモーターが同定されている(Marraら,Infect.Immun.,148:1483−1491,2002;Schneiderら,Proc.Natl Acad.Sci.U.S.A.,97:1671−1676,2000)。また、DFIを用いて、環境性刺激に応答したBacillus cereusにおける遺伝子調節(DunnおよびHandelsman,Gene,226:297−305,1999)、能力刺激ペプチドに応答したS.pneumoniaeにおける遺伝子調節(Bartilsonら,Mol.Microbiol,39:126−135,2001)、ならびに宿主細胞との相互作用時および該細胞内への侵入時のBartonella henselae(LeeおよびFalkow,Infect.Immun.,66:3964−3967,1998)、Listeria monocytogenes Wilsonら,Infect.Immun.,69:5016−5024,2001)、Brucella abortus(Eskraら,Infect.Immun.,69:7736−7742,2001)および大腸菌(Badgerら,Mol.Microbiol,36:174−182,2000)における遺伝子調節が研究された。
【0064】
DFIは、インビボ環境を刺激するために設計された感染の細胞培養モデルまたはインビトロ条件において活性なプロモーターを同定するために成功裏に使用されたが、DFIは、完全体の動物内の特定の生物学的ニッチにおいて調節されるプロモーターを同定するためには、ほとんど適用されていない。これは、おそらく、インビボ環境からの分取に伴う数多くの課題ためである。宿主の炎症性応答、播種および/または感染部位からの菌体のクリアランス、ならびに上皮細胞への細菌の付着(おそらく、バイオフィルム形成による)により、細菌は、生体動物からの回復に利用不可能となり得る。これらの要素は、とりわけ、該細菌がこのような変化を感知し、これに応答すると、微小環境の複雑さ、および遺伝子発現の不均一性に寄与する。最近、DFIを用い、該細菌を気道感染のマウスモデルおよびOMのアレチネズミ感染モデルにおいてスクリーニングすると、S.pneumoniaeにおいて発現されるプロモーターが同定された(Marraら,Infect.Immun.70:1422−33,2002;Marraら,Microbiol,148:1483−91,2002)。
【0065】
動物モデル
チンチラモデルは、広く認められたOMの実験モデルである。特に、NTHi誘導型OMのチンチラモデルは充分特性評価されており(Bakaletzら,J.Infect.Dis.,168:865−872,1993;BakaletzおよびHolmes,Clin.Diagn.Lab.Immunol.A:223−225,1997;SuzukiおよびBakaletz,Infect.Immun.,62:1710−1718,1994)、OMに対する痘苗原としてのいくつかのNTHi外膜タンパク質、外膜タンパク質の組合せ、キメラ合成ペプチドワクチン成分およびアジュバント製剤の防御的有効性を調べるために使用されている(Bakaletzら,Vaccine,15:955−961,1997;Bakaletzら,Infect.Immun.,61:2746−2762,1999;Kennedyら,Infect.Immun.,68:2756−2765,2000)。
【0066】
特に、アデノウイルスがチンチラにインフルエンザ菌誘導型中耳炎をもたらす独自のインビボモデルが存在し、これにより、NTHiの生物学的評価のための関連する細胞、組織および器官の培養系の確立が可能となった(Bakaletzら,J.Infect.Dis.,168:865−72,1993;Suzukiら,Infect.Immunity 62:1710−8,1994)。アデノウイルス感染を単独で用い、誘導された血清抗体の鼓室内への漏出が評価され(Bakaletzら,CHn.Diagnostic Lab Immunol,4(2):223−5,1997)、NTHiとともに共病原体として用い、中耳炎に対する痘苗原としての種々のNTHi外膜タンパク質、OMPの組合せ、キメラ合成ペプチドワクチン成分およびアジュバント製剤を標的化するいくつかの能動および受動免疫処置レジメンの防御的有効性が調べられた(Bakaletzら,Infect Immunity,67(6):2746−62,1999;Kennedyら,Infect Immun.,68(5):2756−65,2000;Novotnyら,Infect Immunity 68(4):2119−28,2000;Poolmanら,Vaccine 19(Suppl.1):S108−15,2000)。
【0067】
中耳のNTHi感染に応答してインビボで上方調節される遺伝子
中耳のNTHi感染に応答して差次的に調節されるプロモーターを同定するため、プロモータートラップライブラリーを構築し、分取パラメータを規定した。プロモータートラップライブラリーの一部分をチンチラの中耳内に直接接種し、OM発症を、ビデオ耳鏡検査およびティンパノメトリによって24時間および48時間の時点でモニターした。また、中耳液を感染24時間および48時間後に採取した。2色FACS解析を用い、GFPを発現している細菌を、他の細胞および滲出液に付随する残屑から単離した(isolated)。単離後、gfpmut3遺伝子の5’のインフルエンザ菌挿入配列のDNA配列を決定し、解析した。このようにした、本発明者らは、AOM時にNTHiがチンチラの中耳環境を感知し、これに応答すると上方調節される遺伝子を同定した。以下の遺伝子が同定され、これらは、NTHi感染時に上方調節されるため、NTHi感染およびビルレンスにおいて役割を果たしている可能性がある。
【0068】
実施例7において後述するように、gfp発現クローンの上記のDFI手順および続くFACS解析の後、潜在的なインビボ調節プロモーターを含む52個の候補クローンが単離された。これらのクローンが制御する遺伝子を、概要および細胞内での機能に基づいて分類し、一般的な代謝過程、環境情報的プロセッシングおよび膜輸送、膜タンパク質および仮想タンパク質を含む。これらの52個のクローンのうち8個は、NTHi86−028NP菌株に特異的配列を含む。重要なことに、3個のクローンを、1匹より多くの動物における独立したスクリーニングで単離し、それにより単離方法を確認した。
【0069】
該FACSデータを独立して確認するため、本発明者らは、候補遺伝子の相対発現を定量的RT−PCRによって調べた。親86−028NP菌株を本試験に使用した。したがって、遺伝子調節に対してプラスミドコピー数の影響がない野生型遺伝子発現を解析し、FACSによる偽陽性クローン同定の識別を可能にした。インフルエンザRd菌株で同定されたものと同様の配列を含む44個の候補クローンのうち、26個の遺伝子(60%)について、インビトロおよびインビボでの遺伝子発現の定量的比較により、NTHiがチンチラの中耳内に存在する環境的なきっかけ(cue)に応答すると、上方調節された遺伝子発現が確認された。この解析により、膜輸送、環境情報的プロセッシング、細胞性代謝、遺伝子調節ならびに未知機能を有する仮想タンパク質に関与する遺伝子の発現を駆動するインビボ調節型プロモーターが同定された。(実施例6の表4参照)。
【0070】
定量的RT−PCRにより、lolA発現の2倍増加が示された。内膜から外膜へのリポタンパク質輸送が可能となった。細菌は、中耳環境内で急速に増殖し、48時間以内に、中耳液1mlあたり5.0×108CFU NTHiに達する。細菌は、該環境を感知し、これに応答して、増殖および生存のための必須栄養素を獲得または合成する。リボース糖輸送における膜成分をコードする遺伝子rbsC(配列番号:619)は、インビトロでの細胞培養と比べ、インビボでは5倍の発現増加を示した。また、代謝過程に関与する多くの遺伝子は、インビトロでの細胞培養と比べ、インビボでは遺伝子発現の劇的な増加を示す。これらとしては、リボフラビン合成遺伝子HbB(配列番号:623)、プリンヌクレオチド生合成遺伝子purE(配列番号:621)、尿素回路によるアルギニン分解に関与するオルニチンカルバモイルトランスフェラーゼarcB(配列番号:625)、ならびにD−グルクロネートの取込みおよびグリセルアルデヒド3−リン酸への形質転換に必要とされるマンノネート(mannonate)ヒドロラーゼをコードするuxuA(配列番号:627)が挙げられる。また、程度は低いが、ヒスチジン生合成のための遺伝子(hisB;配列番号:615)、DNA修復のための遺伝子(radC;配列番号:639)および推定細胞内隔壁化膜貫通タンパク質の遺伝子(ispZ;配列番号:637)も上方調節された。
【0071】
ジスルフィド結合形成は、細菌内での多くの分泌タンパク質のフォールディングおよび合成に重要である。原核生物では、DsbAおよびDsbBがジスルフィドの形成を担う酸化的経路を構成する。DsbBはDsbAを再酸化し、これは、ジスルフィドが変性ポリペプチドに直接結合することを表す。DsbBは、酸化キノンから新たにジスルフィドを生成させることが示されている(ColletおよびBardwell,Mol.Microbiol,44:1−8,2002)。インフルエンザRd菌株では、DsbAは、形質転換能に必要とされる(Tomb,Proc.Natl.Acad.Sd.U.S.A.,89:10252−10256,1992)。ここでは、ほぼ3倍のdsbB遺伝子(配列番号:629)の転写増加が示され、中耳環境内で増殖するTHiに対するジスルフィド相互交換の重要な役割が明らかになった。
【0072】
中耳での細菌のコロニー形成は、正常な滅菌環境であり、宿主炎症性応答および続いて好中球浸潤をもたらす。細菌は、この宿主の応答に対処するため、数多くの戦略を進化させてきた。NTHiは、ヘリコバクター属において活性ウレアーゼの発現に必要とされ、酸耐性に関与することが示されている(Youngら,J.Bacterol.,178:6487−6495,1996)遺伝子のホモログであるureH(配列番号:631)遺伝子発現を増大させる(4倍)。最近、ウレアーゼ活性は、感染時に起こるpHの減少を打ち消すことにより、慢性のActinobacillus pleuropneumoniae感染において役割を果たしている可能性があることが報告された(Baltesら,Infect.Immun.,69:472−478,2000;Baltesら,Infect.Immun.,69:472−478,2001;BosseおよびMachines,Can.J.Vet.Res.,64:145−150)。中耳滲出液由来のNTHi単離菌に関するバイオタイプ解析により、87%はウレアーゼ陽性であることが示された(DeMariaら,J.Clin.Microbiol,20:1102−1104,1984)。しかしながら、NTHiビルレンスにおけるウレアーゼの役割は未知である。同様に、その産物がS.pneumoniaeのピリドキシン生合成タンパク質に対して88%の配列同一性および抗酸化剤としての機能を果たし得る推定一重項酸素抵抗性タンパク質に対して60%相同性を示す遺伝子発現の増大。ホスホリルコリン(ChoP)は、NTHiの病因に関与している(Weiserら,Infect.Immun.,65:943−950,1997)。NTHiは、phase variationによってChoP発現モジュレートし、細胞表面上のLOSを修飾する。ChoPは、抗菌性ペプチドに対する感受性を低下させることによって気道内のNTHi残留に寄与し得(Lyseckoら,Infect.Immun.,68:1664−1671,2000)、C反応性タンパク質(CRP)によって媒介される血清殺傷性に対する感受性を改変し得る(Weiserら,J.Exp.Med.,187:631−640,1998)。チンチラ鼻咽頭では、ChoP+菌株がより多くのコロニー形成を示すため、鼻咽頭および中耳腔の微小環境ではChoP+表現型が選択され得る(Tongら,Infect.Immun.,68:4593−4597,2000)。また、licC遺伝子(配列番号:633)の発現も増大された。licC遺伝子は、ホスホリルコリン誘導性LOSの生合成において役割を果たすホスホリルコリンシチジリルトランスフェラーゼをコードする(Rockら,J.Bacterol,183:4927−4931,2001)。
【0073】
また、インビボ誘導型遺伝子のうち、その産物が、続いて遺伝子発現またはDNA複製を調節する一連のものも含まれる。このような遺伝子としては、glpリプレッサーglpR(配列番号:643)によるグリセロール代謝の転写調節、アルギニン抑制因子遺伝子argR(配列番号:647)、および組込み宿主因子(IHF)βサブユニットihfB(配列番号:645)が挙げられる。IHFはヒストン様タンパク質であり、これは、特定配列のDNAに結合し、複製、部位特異的組換えおよび転写に関与するアクセサリー因子であり、多数のオペロンの活性を改変させる(Goosenおよびvan de Putte,Mol.Microbiol.16:1−7,1995)。また、CspDは、大腸菌において定常期誘導性ストレス応答の際、DNA複製を阻害し(Yamanakaら,Mol.Microbiol,39:1572−1584,2001)、mukF(配列番号:641)遺伝子タンパク質ホモログは、細胞分離前に、よりコンパクトな形態へのヌクレオチド(nucleiod)構造のリモデリングに寄与する(SawitzkeおよびAustin,Proc.Natl.Acad.ScL U.S.A.,62:1710−1718,2000)。また、本明細書に記載のDFIストラテジーにより、未知機能の遺伝子に関してインビボで誘導されるプロモーターが同定された。仮想タンパク質HI0094では、OM初期に遺伝子発現の8倍増加が示されたが、その役割は依然として未知である。HI1163(配列番号:651)は、大腸菌の推定オキシダーゼである仮想YdiJタンパク質と、58%アミノ酸同一性を示した。
【0074】
高密度トランスポゾン変異誘発ストラテジーを用いて、富化培地での増殖に不可欠なインフルエンザ菌遺伝子が同定された(Akerleyら,Proc.Natl.Acad.Sd.U.S.A.,99:966−971,2002)。6つの遺伝子が、本明細書に記載のスクリーニングにおいて同定され、これらは、Akerleyら、前掲に記載の必須遺伝子の組に含まれる(hisB、lppB、lolA、ispZ、mukFおよび未知HI0665)。最近、2つのヒト気道由来上皮細胞株と相互作用すると発現される分類不能型インフルエンザ菌の遺伝子が同定された。このような遺伝子には、代謝過程、ストレス応答、遺伝子発現、細胞外被の生合成、DNA関連プロセス、細胞分裂および未知機能のORFのコードタンパク質に関与するものが含まれた。(Ulsenら,Mol.Microbiol.,45:485−500,2002)。同様に、ストレス応答遺伝子であるcspD(配列番号:649)、プリンおよびリボフラビンの生合成に関与する遺伝子、および未知機能のvapAが、本明細書に記載のスクリーニングにおいて同定された。vapAの発現はインビトロで検出されたが、vapA遺伝子発現は、インビボでは2倍増加した。これらの特有のアプローチでは、NTHi誘導型OMにおいて上方調節され、したがって、おそらくNTHi感染およびビルレンスにおいて役割を果たしており、NTHi関連障害のワクチンおよびアンチセンス療法ならびに他の治療的処置方法の潜在的候補であり得る既知遺伝子が同定された。
また、DFIストラテジーにより、未知機能の遺伝子に関してインビボで誘導されるプロモーターの同定がもたらされた。仮想タンパク質HI0094では、OM初期に遺伝子発現において8倍増加が示されたが、その役割は依然として未知である。HI1163(配列番号:651)は、大腸菌の推定オキシダーゼである仮想YdiJタンパク質と、58%アミノ酸同一性を示した。したがって、このような仮想遺伝子は、おそらく、NTHi感染によって誘導されるOMにおいて役割を果たしている。
例えば、本発明は以下の項目を提供する。
(項目1)
配列番号:577〜579、配列番号:589〜614、配列番号:675〜685、配列番号:615、配列番号:617、配列番号:619、配列番号:621、配列番号:623、配列番号:625、配列番号:627、配列番号:629、配列番号:631、配列番号:633、配列番号:635、配列番号:637、配列番号:639、配列番号:641、配列番号:643、配列番号:645、配列番号:647、配列番号:649、配列番号:651、配列番号:653、配列番号:655、配列番号:657、配列番号:659、配列番号:661、配列番号:663、配列番号:665、配列番号:667、配列番号:669、671、配列番号:673、配列番号:686、配列番号:688、配列番号:692、配列番号:694、配列番号:696、配列番号:698、配列番号:700、配列番号:702、配列番号:704、配列番号:706、配列番号:708、配列番号:710、配列番号:712、配列番号:714、配列番号:716、配列番号:718、配列番号:720、配列番号:722、配列番号:724、配列番号:726、配列番号:728、配列番号:730、配列番号:732、配列番号:734、配列番号:736、配列番号:738、配列番号:740、配列番号:742、配列番号:744、配列番号:746、配列番号:748、配列番号:750、配列番号:752、配列番号:754、配列番号:756、配列番号:758、配列番号:760、配列番号:762、配列番号:764、配列番号:766、配列番号:768 配列番号:770または配列番号:773〜2593のいずれか1つのヌクレオチド配列を含む、単離されたポリヌクレオチド。
(項目2)
項目1に記載のヌクレオチド配列の組またはその断片にコードされるアミノ酸配列を含む、単離されたポリペプチド。
(項目3)
配列番号:616、配列番号:618、配列番号:620、配列番号:622、配列番号:624、配列番号:626、配列番号:628、配列番号:628、配列番号:630、配列番号:632、配列番号:634、配列番号:636、配列番号:638、配列番号:640、配列番号:642、配列番号:644、配列番号:646、配列番号:648、配列番号:650、配列番号:652、配列番号:654、配列番号:656、配列番号:658、配列番号:660、配列番号:662、配列番号:664、配列番号:666、配列番号:668、配列番号:670、配列番号:672、配列番号:674、配列番号:687、配列番号:689、配列番号:691、配列番号:693、配列番号:695、配列番号:697、配列番号:699、配列番号:701、配列番号:703、配列番号:705、配列番号:707、配列番号:709、配列番号:711、配列番号:713、配列番号:715、配列番号:717、配列番号:719、配列番号:721、配列番号:723、配列番号:725、配列番号:727、配列番号:729、配列番号:731、配列番号:733、配列番号:735、配列番号:737、配列番号:739、配列番号:741、配列番号:743、配列番号:745、配列番号:747、配列番号:749、配列番号:751、配列番号:753、配列番号:755、配列番号:757、配列番号:759、配列番号:761、763、配列番号:765、配列番号:767、配列番号:769、配列番号:771または配列番号:2581〜4414のいずれか1つのアミノ酸配列を含む、単離されたポリペプチド。
(項目4)
項目2または3に記載のポリペプチドおよび薬学的に許容され得る担体を含む組成物。
(項目5)
項目2もしくは3に記載のポリペプチドまたはその断片に特異的に結合する抗体。
(項目6)
項目5に記載の抗体および薬学的に許容され得る担体を含む組成物。
(項目7)
生物学的試料内のNTHi菌を検出するための方法であって、
(a)項目1に記載のポリヌクレオチドまたはその断片を生物学的試料と接触させる工程、および
(b)該試料内での該ポリヌクレオチドのハイブリダイゼーションを検出する工程、
を包含する、方法。
(項目8)
生物学的試料内のNTHi菌を検出するための方法であって、
(a)項目5に記載の抗体と生物学的試料を接触させる工程、および
(b)該試料内での該抗体の結合を検出する工程
を包含する、方法。
(項目9)
前記生物学的試料が、血清、痰、耳液、血液、尿、リンパ液および脳脊髄液からなる群より選択される、項目7または8に記載の方法。
(項目10)
NTHi菌に対する免疫応答を惹起させるための方法であって、免疫原性的に有効な用量の項目2または3に記載のポリペプチドまたはその断片を、NTHi菌感染のリスクのある患者に投与する工程を包含する、方法。
(項目11)
項目2または3に記載のポリペプチドまたはその断片および医薬用に適した担体を含むワクチン。
(項目12)
前記免疫応答が、NTHi菌の死滅、NTHi菌の複製の遅滞またはNTHi菌の結合の阻止の1種類以上を含む項目10に記載の方法。
(項目13)
前記NTHi感染が中耳内である、項目10に記載の方法。
(項目14)
NTHi菌感染を処置または予防する方法であって、項目2または3に記載のポリペプチドの発現または活性を阻害する分子を、処置または予防を必要とする患者に投与する工程を包含する、方法。
(項目15)
前記NTHiポリペプチドが、hisB、lppB、sapA、rbsC、pure、rib、arcB、uxuA、licC、ispZ、mukF、glpR、ihfB、cspD、lav、HI1647、HI0094、HI1163、HI0665、HI1292、HI1064、HI1386、HI1462、HI1369およびHI1598からなる群より選択されるNTHi遺伝子にコードされる、項目14に記載の方法。
(項目16)
前記NTHiポリペプチドが、配列番号:616、配列番号:618、配列番号:620、配列番号:622、配列番号:624、配列番号:626、配列番号:628、配列番号:634、配列番号:638、配列番号:642、配列番号:644、配列番号:646、配列番号:650、配列番号:652、配列番号:656、配列番号:658、配列番号:660、配列番号:662、配列番号:664、配列番号:666、配列番号:668、配列番号:670、配列番号:672および配列番号:674からなる群より選択されるアミノ酸配列を含む、項目14に記載の方法。
(項目17)
前記必要とする患者に投与される前記分子がアンチセンスオリゴヌクレオチドである、項目14に記載の方法。
(項目18)
前記必要とする患者に投与される前記分子が抗体である、項目14に記載の方法。
(項目19)
前記必要とする患者に投与される前記分子が小分子である、項目14に記載の方法。
(項目20)
前記NTHi感染が中耳内である、項目14に記載の方法。
【図面の簡単な説明】
【0075】
【図1】図1は、一群のインフルエンザ単離菌におけるLKP遺伝子領域を示す。86−028NP菌株の配列は、この領域において、NTHi菌株R3001の配列と同一である。これらのNTHiはともに、赤血球凝集性線毛(hemagglutinating pilus)をコードするhif遺伝子クラスターを欠く。
【図2】図2は、一群のインフルエンザ単離菌におけるrfaD領域を示す。86−028NP菌株ゲノムのrfaD領域内の遺伝子構成は、Rd菌株のゲノムのものと類似するが、調べたほとんどのNTHiのゲノムに見られるこれらの遺伝子の構成と異なる。
【図3A】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3B】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3C】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3D】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3E】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3F】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3G】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3H】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3I】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3J】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3K】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3L】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【図3M】図3A〜3Mは、表4に示すヌクレオチド配列(配列番号:589〜614)を示し、これらは、OM感染時、上方調節されると同定された(実施例6参照)。また、既知遺伝子に対応するヌクレオチド(nt.)および配列番号:1〜576として示すコンティグ配列に対応するnt.を示す。
【発明を実施するための形態】
【0076】
(詳細な説明)
以下の実施例は、本発明を説明する。ここで、実施例1はNTHiゲノムの配列を示し、実施例2は同定されたコンティグおよび最初の遺伝子発見を示し、実施例3はNTHi
プロモータートラップライブラリーの構築を示し、実施例4はGFPを発現する86−028NP誘導体の解析を示し、実施例5は中耳液由来の細菌の直接標識を示し、実施例6は、急性中耳炎においてインビボで誘導されるプロモーターの同定を示し、実施例7はビルレンス関連遺伝子の同定を示し、実施例8は、NTHi特異的遺伝子配列の同定を示し、実施例9は、完全NTHi86−028NPゲノムの解析を示し、実施例10では、NTHi86−028NP菌株およびインフルエンザ菌d血清型菌株kw20のゲノムDNA配列を比較している。
【0077】
実施例1
分類不能型インフルエンザ菌のゲノムの配列
NTHi86−028NP菌株は、コロンブス小児病院において、慢性OMのため鼓膜切開およびチューブ挿入を受けた小児患者から採取された、最小限に継代された臨床用単離菌である(Bakaletzら Infection and Immunity,56(2):331−335,1988)。86−028NP菌株は、American Type Tissue Collection(Manassas,VA 20108
USA)に2002年10月16日に寄託し、受託番号PTA−4764が付与された。
【0078】
NTHiのビルレンス決定基の同定に、より広くアプローチするための取り組みにおいて、NTHi86−028NP菌株のゲノムを3倍の範囲まで配列決定した。Puregeneプロトコルを用いて86−028NP菌株から染色体DNAを調製し、Hydroshear装置(Gene Machines)により2〜4kbの大きさに切断した。切断されたDNAをエタノール沈殿させ、クレノウ酵素とT4 DNAポリメラーゼの混合物を用いて末端修復し、アガロースゲル電気泳動によってサイズ選択し、Chissoeら(Methods:a Companion to Methods ofEnzymology 3:55−65,1991)およびSambrookら(Molecular Cloning:a Laboratory Manual,第2版 Cold Springs Harbor,NY,1989)に記載のような2〜4kb断片を得た。これらの断片をベクターpUC18内に、Smal制限部位(ホスファターゼ処理済)を用いてクローニングし、大腸菌XL−I Blue内に形質転換し、アンピシリン耐性に関して選択した。挿入物を含むコロニーを、X−galを入れたLB−Ampプレート上にて青/白スクリーニングによって同定し、1.5mlのTB−Amp(TB=Terrific Broth)培養液を入れた96ディープェルプレートに移した。このディープェルプレート培養物を、一晩(18〜22時間)37℃で培養した。鋳型の調製、配列決定およびコンティグ編集を行なった。
【0079】
自動鋳型調製を、Chissoeら、前掲に記載のBeckman Biomek 2000自動ロボットワークステーションにおいて行なった。簡単には、上記で調製されたクローンの入った各96ディープェルプレートを遠心分離して細胞をペレット化し、上清みを廃棄し、細胞を−20℃で凍結させた(必要な場合)。4つの96ディープェルブロックをBiomekの台上に配置し、液体処理ロボットを用い、Chissoeら、前掲に記載の自動型の典型的なSDS−NaOH溶解プロトコルを使用して鋳型を調製した。最後にエタノール沈殿させた鋳型を、各々、50μlのddH2Oに溶解し、DNA配列決定に使用した。
【0080】
配列決定反応は、Robbins Hydra 96ロボットを用いて、鋳型(96ウェルプレートから)を384ウェルプレートに再度整列させることにより行なった。PE
Big−Dye(登録商標)ターミネーターおよび普遍的プライマー(M13フォワードおよびリバース)用いてサイクル型配列決定反応を行ない、Sephadex G50カラム上で浄化し、PE Biosystems 3700キャピラリー電気泳動DNA配列決定装置において製造業者の使用説明書に従って解析した。配列決定の読み値(8219)を576個のコンティグ(本明細書における配列番号:1〜576)に編集した。3倍配列決定の統計学的データ表2Aに示す。アセンブリ17の全特異的配列は1.74Mbである
【0081】
【表2】
続いて、NTHiゲノムの8倍配列決定解析を行なった。8倍配列決定により、NTHiゲノムを11個のコンティグに編集した。コンティグ5、8、9、10、12〜18を、本明細書において配列番号:675〜685として示す。8倍配列決定の統計学的データを表2Bに示す。
【0082】
【表3】
実施例2
コンティグの説明および所期遺伝子の発見
3倍配列解析で同定された≧5000bp長さを有する88個のコンティグのうち75個は、BLASTNにより、インフルエンザRd菌株の遺伝子と有意な類似性を示す。インフルエンザ86−028NP菌株における遺伝子順序とインフルエンザRd菌株との潜在的関連性を視覚化するため、86−028NP 3倍コンティグの組とRd遺伝子の組を、BLASTNを用いて双方向で比較した。コンティグをRd遺伝子ヒット遺伝子の座標に基づいて分類し、各コンティグを、Rayら,(Bioinformatics 17:1105−12,2001)に記載のとおりに見られる最小座標に固定(anchor)することにより、結果を遺伝子順序対コンティグの間隙(space)においてプロットした。このようにして比較し、完全に既知のゲノムと同一の遺伝子順序を有するゲノムの不完全なアセンブリにより、単調に増大する段階工程の形態が示され得る。
【0083】
BLASTXを用い、Rd菌株のゲノム内の遺伝子ならびにインフルエンザRd菌株に見られない遺伝子に対して相同性を有する配列のヒットを同定した。Rd菌株配列に対するヒットはデsータセットから除外し、その他のヒットを表3Aにまとめる。データは以下:コンティグ#(配列番号:#)、第1欄;各ヒットのEスコア、第2欄;記載のコンティグのアミノ酸翻訳部分に対して相同性を有したタンパク質の名称、第3欄;該ホモログを産性する生物体、第4欄;第3欄に示したタンパク質の各々のGenbank確認用タンパク質、第5欄;コンティグ内の対応するヌクレオチド(配列番号:で示す)のように表示する。ほとんどの場合でいくつかのホモログが同定されたが、明確にするため、相同性の大きいタンパク質を表3Aに示す。
【0084】
表3Aに示した一部の遺伝子の配列を、NTHiゲノムの8倍配列決定において同定した。表3Bに、11個のコンティグ内のこれらの遺伝子の位置、完全長オープンリーディングフレーム配列(配列番号:で特定)、オープンリーディングフレームにコードされる誘導アミノ酸配列、およびBLASTXによって同定された高い相同性を有する遺伝子(表3Aに記載)を示す。
【0085】
NTHiおよびRd菌株における比較的短い範囲の遺伝子構成を調べるため、充分報告された2つの遺伝子クラスター内の遺伝子順序を比較した。まず、赤血球凝集性線毛(LKP)遺伝子領域内に存在する遺伝子を調べた。(Mhlanga−Mutangaduraら,J Bacteriol.180(17):4693−703,1998)。線毛遺伝子クラスターは、purE遺伝子とpepN遺伝子の間に位置し、その断片のみを図1に示す。b血清型の菌株であるEaganは、hifABCDE遺伝子クラスターを含み、赤血球凝集性線毛を産生する。Rd菌株は、hicAB遺伝子ならびにhifABCDE遺伝子クラスターを欠く。一般に、これまでに調査された分類不能型菌株は、hicAB遺伝子を含んでいたが、赤血球凝集性線毛をコードするhif遺伝子は含んでいなかった。86−028NP菌株の配列(本明細書に記載)は、この領域において、NTHi菌株R3001の配列と同一である(図1)。rfaD遺伝子は、内毒素の生合成に関与する酵素をコードする。また、NTHi 2019菌株由来のrfaD遺伝子は、Nicholsらによって特性評価された(Infect Immunity 65(4):1377−86,1997)。2019菌株において、rfaD遺伝子は、内毒素生合成に関与する別の酵素をコードするrfaF遺伝子のすぐ上流に存在する。Rd菌株の遺伝子構成は異なる;rfaD遺伝子とrfaF遺伝子は、ほぼ11kbの配列によって分断されている。調査されたほとんどの分類不能型菌株は、2019菌株に見られる遺伝子構成を含んでいた。対照的に、86−028NP菌株は、Rd菌株に見られるものと同一の遺伝子構成を有する(図2)。
【0086】
現在の(current)アセンブリの大域解析により、遺伝子の含有量および順序は、Rd菌株のものと類似していることが示される。より詳細な解析によって、PasteurellaceaeおよびNTHi遺伝子の含有量と順序がRd菌株に見られるものと異なる一部の領域では、これまでにわかっていない相当な数のNTHi遺伝子が存在することが明らかになった。したがって、現在のデータにより、86−028NP菌株ゲノムは、Rdおよび非Rd様の特徴の複雑な寄せ集めを含むことが示される。
【0087】
また、DFIストラテジーにより、遺伝子発現が増大された新規なNTHi配列が同定された。他の生物体(主にグラム陰性細菌)ORFに対して相同性を有する遺伝子または遺伝子断片を含有するこのような新規なコンティグ配列の一覧を表3Aに示す。例えば、コンティグ442(配列番号:442)のヌクレオチド配列であるヌクレオチド1498〜1845は、インフルエンザRd菌株のリポタンパク質B(LppB)のアミノ酸1〜116をコードする配列に対して高度に相同である。該遺伝子は、定常期生存遺伝子urEと、最近同定され細菌リポタンパク質LppB/NlpD(これは、ビルレンスと関連している)に対して高度に相同な43kDの抗原性外膜リポタンパク質をコードする遺伝子との間に位置する(Padmalayamら,Infect.Immun.,68:4972−4979,2000)。最近、Zhangおよび協働者らによって、nlpDおよびsurE遺伝子発現は、Thermotoga maritimaにおいて細菌増殖の定常期に誘導されることが示された(Zhangら,Structure(Camb),9:1095−1106,2001)。したがって、中耳内のストレス誘導条件下では、このNTHiリポタンパク質が発現され得る。
【0088】
【表4】
【0089】
【表5】
【0090】
【表6】
【0091】
【表7】
【0092】
【表8】
【0093】
【表9】
【0094】
【表10】
【0095】
【表11】
【0096】
【表12】
【0097】
【表13】
【0098】
【表14】
【0099】
【表15】
【0100】
【表16】
【0101】
【表17】
【0102】
【表18】
【0103】
【表19】
【0104】
【表20】
【0105】
【表21】
【0106】
【表22】
【0107】
【表23】
実施例3
NTHiプロモータートラップライブラリーの構築
NTHiの潜在的ビルレンス決定基を同定するため、NTHi誘導型中耳炎(OM)のチンチラモデルの特定の1つの解剖学的ニッチにおいて、疾患進行初期に、細菌の遺伝子発現をディファレンシャル蛍光誘導(DFI)によってモニターした。NTHi86−028NP菌株のゲノムDNA断片を、プロモータートラップライブラリーを用いてプロモーターなしのgfpmut3遺伝子の上流にクローニングした。プラスミドpGZRS39Aは、Actinobacillus pleuropneumoniaeから単離されたpGZRS−1の誘導体であり、A.pleuropneumoniae−大腸菌シャトルベクターである。このプラスミドは、A.pleuropneumoniae由来の複製起点、pUC19由来のlacZa遺伝子およびTn903由来のカナマイシン耐性遺伝子を含む(Westら,Genes,160:81−86,1995)。
【0108】
プロモータートラップベクターは、GTP変異型gfpmufi遺伝子をBamHIからEcoRI方向の断片としてpGZRS−39A内にクローニングし、pRSM2167を形成することにより構築した。この変異型GTP遺伝子は、2つのアミノ酸変化S65GおよびS72Aを含み、これらにより、488nmで励起すると蛍光発光が増強される。また、この変異型は、高可溶性および発色団形成の高反応速度論を有する(Cormackら,Gene,173:33−38,1996)。このプラスミドで、エレクトロポレーションによってNTHi86−028NP菌株を形質転換し、親プラスミド86−028NP菌株/pRSM2169を作製した。
【0109】
ランダムゲノムDNA断片(実施例1に記載)を調製し、プロモータープローブベクターン内にライゲーションした。86−028NP菌株からゲノムDNAを、Puregene DNA単離キット(Gentra Systems,Minneapolis,MN)を製造業者のプロトコルに従って用いて単離した。制限障壁のため、プラスミドDNAを単離し、これをライブラリーの作製に用いることが必要であった。単離したDNAは、Sau3AI(NEB,Beverly,MA;0.25単位/μg DNA)で1時間37℃にて部分消化し、ゲル電気泳動によって分離し、Qiagenゲル抽出キットを用いて0.5〜1.5kbの大きさのDNA断片を回収した。ベクター作製のため、pRSM2167を一晩培養物から、Wizard Plus Maxiprep DNA精製系(Promega,Madison WI)を製造業者のプロトコルに従ってを用いて単離した。
【0110】
プラスミドDNAをBamHI消化によって線状化し、5’リン酸基を、ウシ腸アルカリホスファターゼ(CIAP;GibcoBRL Life Technologies)での処理によって除去した。ゲノムDNA断片を、この線状化ホスファターゼ処理ベクターを用いてライゲートし、改良プロトコル(Mitchellら,Nucleic Acids Res.,19:3625−3628,1991)に従ってエレクトロポレーション用に作製したコンピテントNP菌株内にエレクトロポレーションした。プラスミドDNAをエレクトロポレーションしてNTHi86−028NP菌株内に戻すと、形質転換効率は1000倍改善された。簡単には、細胞をOD60O=0.3まで、sBHI(脳心臓浸出物)培養液中で37℃、220rpmにて培養した。細胞を氷上で30分間冷却し、続いて、等容量の0.5×SG(1×SG:15%グリセロール、272mMスクロース)で4℃にて洗浄した。洗浄を合計3回繰返した。続いて、細胞をI×SG中で100×濃縮容量まで希釈した。BioRad Gene Pulser IIセットを200オーム、2.5kVおよび25μFで用いて細胞をエレクトロポレーションし、次いで、1mlの予備加温sBHI中で希釈し、37℃、5%CO2で2時間インキュベートし、形質転換体の一晩培養のためのチョコレート寒天上にプレーティングした。
【0111】
形質転換体を選択し、1000クローンのプール毎に、20%グリセロール(vol/vol)を含有するスキムミルク中で凍結させた。68,000構成員のgfpプロモータープローブライブラリーを作製した。ClarkeおよびCarbon(Cell,9:91−99,1976)の確率計算を用い、86−028NP菌株DNA(1.8×106bp/ゲノム)の300bp断片のライブラリー内に所与のDNA配列が提示される確率99%を得るためには、27,629個のクローンのライブラリーが必要であった。したがって、本発明のライブラリーは、86−028NPゲノムの2.5倍範囲を提示する。
【0112】
このライブラリーの質を評価するため、50個のクローンを無作為に選択して一晩チョコレート寒天上で培養し、プラスミドを単離して挿入DNAを配列した。選択されたクローンの大部分(64%)は、200〜500bpの範囲の挿入物サイズを有したが、32%は500bpを超えていた。この大部分の挿入物は、インフルエンザRd菌株の特異的オープンリーディングフレーム(ORF)に対して相同性を示し、15個のクローンは、86−028NP菌株のDNAに特有の配列を有した。Rd菌株に対して相同性を有するクローンのうち、60%は正しい向きであり、その36%は、ORFの上流に配列を含んでいた。大部分のクローンは、500bp未満の挿入物サイズを有していたが、挿入物サイズが小さいこととGFP発現の増大との間に相関性は認められなかった。実際、4個のクローンは、インビトロで軽微から中程度の蛍光を示し、そのうち3個は、200〜500塩基対の挿入物サイズを有し、1個は700塩基対より大きい挿入物を有した。
【0113】
ライブラリーの一部(ほぼ1000個のクローン)をチョコレート寒天上で培養し、PBS中に採取し、フローサイトメトリーによってGFP蛍光について解析した。挿入DNAのないプロモータートラップベクターを含む86−028NP菌株/pRSM2169と比べ、該ライブラリークローンのプールは、蛍光強度の増大を示す。したがって、該ライブラリーは、種々の活性レベルのプロモーターを有するクローンを含む。
【0114】
実施例4
GFPを発現する86−028NP誘導体の解析
gfp発現菌を同定および分取するために必要なFACSパラメータ確立するため、種々のレベルのgfp発現を示す一群の単離菌を用いた。86−028NP菌株/pRSM2169(陰性対照)を用いてバックグラウンド蛍光を評価し、したがって、観察される任意の蛍光は、gfp発現を誘導するlacZプロモーターによるものであり得る。しかしながら、この菌株は検出可能なレベルのGFPを産生せず、実際、親86−028NP菌株と比較した場合、蛍光の増大は示されない。高レベルgfp発現単離菌を、外膜タンパク質P2発現のための強力プロモーターを含有する500bp断片を、SalI−BamHI消化pRSM2167内にクローニングすることにより作製した。このプラスミドで、エレクトロポレーションにより86−028NPを形質転換し、高レベルgfp発現86−028NP菌株/pRSM2211(高度に蛍光性の対照)を作製した。この菌株は、86−028NP菌株/pRSM2169と比べて、ほぼ100倍のGFP蛍光増加を示した。中間蛍光誘導体クローン86−028NP/pKMM4B5(中間蛍光性の対照)は、FACS解析によって単離し、予備実験において、および細胞分取ための対照としての両方に使用した。gfp発現をインビトロで誘導するプロモーターを含有するDNA断片は、他の生物体のDNAに対して既知の相同性を持たない86−028NP菌株に特有である。このクローンは、86−028NP菌株/pRSM2169と比べて、ほぼ10倍の蛍光増加を示す。
【0115】
対照菌株を、チョコレート寒天上での培養から再懸濁させ、交差反応性Phycoprobe R−PE抗ヒトIgG(H+L)抗体(100μlのPBS中10μg/ml;Biomeda Corp)で30分間4℃にて標識した。3回連続洗浄して未結合抗体を除去した後、FACS解析のため、細菌を300μlのDPBS中に再懸濁した。これらの対照調製物を用い、488nmのアルゴンレーザー発光器を取り付けたCoulter Epics Eliteフローサイトメーター(Coulter Corp.)を使用して、適切なサイズおよび蛍光ゲートを設定した。細菌は、log前方角および側方散乱検出に基づいた大きさに関して、ならびに細菌のFITC/PE標識による分取に関してゲーティングさせた。分取細胞を定温sBHI中に回収し、チョコレート寒天上にプレーティングした。一晩培養後、細胞は、二次ラウンドの感染のために回収するか、または個々に選択し、一晩培養し、インビトロ培養したときの蛍光に関して個々のクローンによってスクリーニングし、20%(vol/vol)グリセロールを含有するスキムミルク中で凍結させた後、プラスミドの単離および挿入DNAの配列決定を行なった。対照菌株の分取効率を、Coulter EPICSフローサイトメーター(Coulter Corp.)を用いて確認した。
【0116】
多くのプラスミドは、抗生物質選別の非存在下では、インビトロで速やかに分離した。したがって、ここで使用したプロモータートラップベクターにこの事象の傾向があるか否かを評価するため、86−028NP菌株/pRSM2211(高度に蛍光性の対照)の単一のコロニーをチョコレート寒天上に単離し、抗生物質選別の非存在下で20回継代した。抗生物質の存在下で培養した細菌と比較した場合、蛍光強度の有意な減少は観察されなかった。また、該プラスミドは、抗生物質選別の非存在下、インビボで維持される。チンチラから採取した細菌含有中耳液を、カナマイシン含有または無含有チョコレート寒天上にプレーティングした場合、同様の細菌の計数が観察された。これらのデータは、プロモータートラップベクターが、抗生物質選別の非存在下で安定に維持されたことを示す。
【0117】
プラスミドの安定性の問題に加え、宿主−病原体相互作用を試験するためのレポーターとしてのGFPの使用に関する初期の研究により、GFPは、低毒性を有する細胞質タンパク質として継続的に合成され得、細菌の細胞表面動力学に対する影響は最小限であることが示された(Chalfieら,Science,263:802−805,1994)。高レベルgfp発現性誘導体の構築により、NTHiに対するGFP毒性の評価が可能となった。野生型菌株(86−028NP)と高GFP産生86−028NP菌株/pRSM2211両方の増殖曲線を、同様の条件下で培養した場合で比較した。増殖速度は同様であり、GFP発現は該細胞に対して毒性でなかったことを示す。
【0118】
86−028NP gfp発現性誘導体を用い、効率的な細胞分取のためのパラメータを規定した。86−028NP菌株/pRSM2169を、中間gfp発現性誘導体である86−028NP菌株/pKMM4B5と、100:1の比で混合し、全細菌集団と比べて少量割合のgfp発現クローンを含むことが予測されるインビボ環境をシミュレーションした。この混合物をFACS解析に供し、1.8%の最大蛍光集団および52%の最小蛍光集団が回収された。分取された集団のフローサイトメトリー解析により、細菌集団の65%に86−028NP菌株/pKMM4B5の富化が示され、これは、陰性集団において分取した場合では観察されなかった現象である。後続の分取ラウンドでは、この中間蛍光集団がさらに富化されることが予測され得る。陰性分取において蛍光細菌の量が減少し得ないことは、陰性分取のために設定されたゲートの大きさのためであった。GFP陰性細胞は、10%の最小蛍光集団においてゲーティングすることにより富化された。
【0119】
実施例5
中耳液からの細菌の直接標識
同様のストラテジー(実施例5に記載)を、AOM時にチンチラの中耳から採取した滲出液からの蛍光クローンの分取に適用した。本発明者らがディファレンシャル蛍光誘導(DFI)をインビボで使用可能であることは、本発明者らがgfp発現菌を分取を、非蛍光細菌、蛍光および非蛍光細胞性残屑ならびに真核生物細胞からできる能力に依存した。
【0120】
耳鏡検査またはティンパノメトリのいずれかで中耳感染の徴候のない健常な成体チンチラ(Chinchilla lanigera)を用い、ライブラリーを、インビボでのプロモーター活性についてスクリーニングした。NTHi/pRSM2169 ライブラリーの2つのプール(各々1000個のクローン)を一晩、カナマイシンを含有するチョコレート寒天上で培養した。ライブラリーを合わせ、低温の10mM滅菌PBS中で3.3×106CFU/mlまで希釈し、300μl(1.0×106CFU;500CFU/クローン)を用いて、左および右のチンチラの経水疱腔(transbullar cavity)に接種した(2000クローン/耳)。OM発症を、ビデオ耳鏡検査およびティンパノメトリによって24時間および48時間の時点でモニターした。細菌は中耳腔内で増殖し、予測どおり、48時間までに、イノキュラム用量の500倍の濃度に達した(Bakaletzら,Infect.Immunity 67:2746−62,1999)。この細菌の宿主環境への適応により炎症性応答がもたらされ、これは、紅斑、鼓膜の血管拡張および腫脹、多核白血球(PMN)の浸潤、ならびに採取した滲出液の耳鏡検査および顕微鏡検査によって観察される中耳腔内への液の蓄積によって示される。24時間および48時間後、鼓室のタッピングによって中耳液を採取し、FACS用に調製した。
【0121】
この解析を、中耳液内で採取され得る細菌に限定したことに注目することは重要である。場合によっては、FACS解析用の細菌を採取するために、中耳腔を洗浄することが必要であった。したがって、この解析には、NTHiが粘膜にゆるく結合すると上方調節される遺伝子が含まれる。NTHiは、OMのチンチラモデルの中耳腔内でバイオフィルムを形成することが観察されている(Erhlichら,JAMA,287:1710−5,2002)。本明細書に記載のプロトコルでは、プランクトン様集団から採取されるクローンを選択するため、該細菌が粘膜のバイオフィルムと結合すると遺伝子が上方調節されるクローンが採取されることは期待されない。しかしながら、中耳粘膜のホモジネーション、続いて細菌細胞の単離を行ない、本発明者らによって、このようなクローンの採取が可能となり得る。また、一部のGFP発現クローンは、滲出液において採取されるが、滲出液中に剥離細胞として、または凝集して存在する真核生物細胞に接着していることが考えられる。このような細菌は、分取効率を損なうことなく滲出液から採取するのは困難である。したがって、中耳液を粘液溶解剤で処理し、次いで遠心分離して大きな集塊および真核生物細胞を除去した後、標識した。
【0122】
チンチラの中耳液は、必要な場合には、滅菌生理食塩水で250μlまで希釈した。等容量のN−アセチル−L−システイン(0.5%;w/v)含有DPBS(pH7.4)を、粘液溶解剤として室温で5分間添加した(MiyamotoおよびBakaletz,Microb.Pathog.,21:343−356 1996)。液体を遠心分離し(300×g、5分間)、細胞性残屑、赤血球および炎症性細胞を除去した。細菌を含む上清みを新たなチューブに移した。細菌を、NTHi 86−028NP菌株由来の完全OMP調製物に対して指向されたチンチラ抗血清(1:50希釈度)とともに、45分間4℃にてインキュベートし、遠心分離(2000×g、5分間)によってペレット化し、0.05%ウシ血清アルブミンを含有する冷DPBSで2回洗浄した。続いて、細菌を交差反応性フィコプローブ(phycoprobe)R−PE抗ヒトIgG(H+L)抗体(100μlのPBS中10μg/ml;Biomeda Corp)で、30分間4℃にて標識した。3回連続洗浄して未結合抗体を除去した後、FACS解析のため、細胞を300μlのDPBS中に再懸濁した。
【0123】
実施例6
急性中耳炎においてインビボで誘導されるプロモーターの同定
プロモータートラップライブラリーで形質転換したインフルエンザ菌86−028NPを一晩、チョコレート寒天上で培養した。インビトロでgfpを発現させたプロモーターを含むクローンを選択するため、ライブラリーを1回のFACS解析(実施例6に記載のもの)に供し、低レベル量のGFPを発現するクローンのみを採取した。このクローンをプールし、チンチラの中耳に経水疱的に接種するために使用した。感染の24時間および48時間後、細菌を含む滲出液を鼓室のタッピングによって取り出した。細菌をR−PE標識抗体で間接的に標識し、蛍光タグ化細菌をゲーティングさせるが、同時に発現しているものは分取することによるFACS解析に供した。このクローンを用い、さらなる富化のために動物を再感染させた。最終回の分取後、単一のコロニー単離菌を、蛍光の消失についてインビトロでスクリーニングした。
【0124】
FACS解析によって単離されたクローン(インビボでGFP蛍光に対して陽性)(これは、インビトロでは蛍光を発しなかった)を、プラスミドの単離および挿入DNA配列の同定のために調製した。このクローンを一晩、カナマイシンを含有するチョコレート寒天プレート上で培養し、Qiaprep Miniprep Kit(Qiagen)を製造業者のプロトコルに従って用い、プラスミドの単離のために調製した。プラスミド挿入DNAは、gfpmut3遺伝子に相補的であり該挿入DNAの下流にあるプライマー5’−TGCCCATTAACATCACCATCTA−3’(配列番号:588)を用いて配列決定した。配列決定反応は、ABI prism BigDye(登録商標)ターミネーターサイクル型配列決定既製反応キット(Applied Biosystems)を製造業者のプロトコルに従って使用し、Gene Amp PCR System
9700(Applied Biosystems)用いて行なった。次いで、該配列を、96ウェルマルチスクリーンHVプレート(Millipore)内のセファデックスG−50に通すことによって精製し、続いて、ABI Prism 3100 DNA解析装置(Applied Biosystems)にて解析した。
【0125】
挿入配列を、インフルエンザRd菌株の完全にアノテートされた配列と比較した。Rd菌株に対してヌクレオチド相同性をもたない挿入物を、続いて、BLASTNおよびBLASTXアルゴリズムを用いて解析した。DNASTAR(Madison,Wise)を用い、さらなる配列解析を行なった。正しい向きであり推定ORFの5’側に配列を含む挿入配列は、NTHi菌がチンチラの中耳内に存在すると優先的に活性となる推定プロモーターを含んでいた。
【0126】
インビボで調節される推定プロモーターを有する52個のクローンを単離した。インフルエンザRd菌株で同定されたものと同様の配列を含む44個の候補クローンのうち、26個の遺伝子(60%)について、インビトロおよびインビボでの遺伝子発現の定量的比較により、NTHiがチンチラの中耳内に存在する環境的なきっかけに応答すると、上方調節された遺伝子発現が確認され、このような遺伝子を以下の表4Aにまとめる。遺伝子の発現を駆動するインビボ調節プロモーターは、膜輸送、環境情報的プロセッシング、細胞性代謝、遺伝子調節ならびに未知機能を有する仮想タンパク質に関与すると推定される。
【0127】
推定プロモーター候補のインビボでの誘導を確認するため、NTHi 86−028NP菌株を、対数増殖期中期までインビトロで、またはインビボで48時間培養した場合のメッセンジャーRNAの発現の相対量を比較した。TRIzol LS試薬(Gibco
Life Technologies)を製造業者のプロトコルに従って用い、RNAを単離した。DNAをRNA調製物から、DNA遊離キット(Ambion)を製造業者のプロトコルに従ってを用いて取り出した。DNアーゼI処理RNA試料をQiagen
RNeasyカラムに通すことによって精製した。RNAの純度および完全性を、それぞれ、260/280nmでの分光測光装置の読み値によって、およびAgilent 2100 Bioanalyzer(Agilent Technologies)において評価した。
【0128】
該FACSデータを独立して確認するため、本発明者らは、候補遺伝子の相対発現を定量的RT−PCRによって調べた。親86−028NP菌株を本試験に使用した。1工程QuantiTect SYBR Green RT−PCRキット(Qiagen)を用いるリアルタイム定量的RT−PCRにより、製造業者の使用説明書に従って転写レベルを評価した。簡単には、FACS解析で同定された推定インビボ誘導プロモーターの下流のオープンリーディングフレームに対して作製されたプライマーを用い、遺伝子特異的mRNAを逆転写し、ABI Prism 7700配列検出系(Applied Biosystems)にてRT−PCRによって増幅した。ジャイレース(gyr)の遺伝子断片を増幅することにより既知量の細菌ゲノムDNA(102〜107ゲノムコピーDNA)に対して作成した標準曲線を用い、産物の量を算出した。RNA調製物中のDNAの非存在(−RT対照)、ならびに鋳型RNAを含まない対照試料中のプライマー二量体の非存在を確認するための対照を並行して解析した。また、RT−PCR産物をゲル電気泳動によって解析し、すべての場合で、適切な塩基対サイズの単一の産物が観察された。試料間の細菌RNAの量を、gyr発現(本発明者らがインビトロで試験した種々の培養条件下で構成的に発現されることが示された)に対して標準化した。既知量の細菌ゲノムDNA(102〜107ゲノムコピーDNA)を用い、ジャイレース(gyr)の遺伝子断片を増幅させることによりRT−PCR定量の標準曲線を作成した。ジャイレースは、種々の培養条件下にてインビトロで構成的に発現され、したがって、試料間の全細菌RNAレベルを標準化するために使用した。インビボでの相対遺伝子発現を、インビトロでの遺伝子発現のものと比較し、増加倍数で示したデータを表4Aにまとめる。
【0129】
NTHiゲノムの8倍配列決定により、表4Aに示す遺伝子のほとんどの完全長オープンリーディングフレームが同定された。表4Bには、NTHiゲノム内の完全長ヌクレオチド配列および対応するアミノ酸配列を示す。便宜上、表4Bには、チンチラの中耳に存在する環境的なきっかけによる遺伝子の誘導倍率ならびに該遺伝子の産物および機能を繰り返し記載する。
【0130】
【表24】
【0131】
【表25】
【0132】
【表26】
実施例7
ビルレンス関連遺伝子の同定
多くの細菌種において、ビルレンス関連遺伝子のサブセットは、短い反復配列の複製エラーによって調節される。このような反復配列は、遺伝子の5’側またはコード配列内に存在し得、その存在は該遺伝子の発現制御の表示であり、ビルレンスとの関連性を示す。反復配列の付加または欠失により、特定のビルレンス決定基の発現または発現の欠損がもたらされる。
【0133】
NTHiインフルエンザ86−028NP菌株のコンティグの組は、短いオリゴヌクレオチド反復配列についてクエリーを行なった。該反復配列の周囲領域を解析し、該反復配列と関連する遺伝子(1つまたは複数)を同定した。表5に、同定された反復配列および各反復配列と関連するORF(BLASTによって同定)を示す。
【0134】
さらなる配列解析によって、ビルレンス関連遺伝子の完全長ヌクレオチド配列およびそのORFにコードされる対応アミノ酸配列が同定された。誘導されたアミノ酸配列は、記載のGenbank配列に対して高度に相同性である。
【0135】
【表27】
【0136】
【表28】
実施例8
NTHi特異的遺伝子配列の同定
NTHiビルレンスと関連する遺伝子もまた、NTHi菌を組織に感染させた場合の遺伝子の発現レベルと、NTHiを人工研究用培地上で培養した場合の同じ遺伝子の発現レベルを比較することにより同定された。これらの新規な遺伝子は、実施例4〜6で上記のプロモータートラップ手法を用いて同定され、続いて、既知のRdゲノムと比較すると、これらの遺伝子はNTHi 86−028NP菌株に特有であることが示された。
【0137】
このスクリーニング手順を用いて同定されたDNA配列を配列番号:577〜580として示す。これらの配列は、インフルエンザ菌Rdゲノム配列におけるホモログを有する遺伝子または遺伝子断片を含んでいなかった。これらは、チンチラの中耳内でのNTHi感染時の発現レベルから、完全に新規な配列ではないが、おそらく、これらの遺伝子の発現は、NTHiビルレンスに関与している。
【0138】
実施例9
NTHi 86−028NP菌株の完全な配列解析
ライブラリーの構築:86−028NP菌株から、Puregene試薬(Gentra Systems,Minneapolis,MN)を用いて染色体DNAを調製した。このゲノムの最初のショットガン配列決定のため、ゲノムDNAの1〜2kbおよび2〜4kbのライブラリーをpUC18において、既報のとおりにして構築した(Munsonら,Infect Immun 72:3002−10,2004)。骨格形成用(scaffolding)ライブラリーでは、ゲノムDNAを平均断片サイズ40kbに、Hamiltonシリンジを用いて手作業で切断した。末端修復後、0.7%低融点アガロースゲルを用いて、断片を分画した。30kbより大きい断片を切り出し、pEpiFOS−5へのゲル内ライゲーションを行なった。ゲルから回収したライゲーション混合物をλファージ内にインビトロでパッケージングし、EPI100細胞(Epicentre,Madison,WI)のトランスフェクションのために使用した。
。
【0139】
配列決定:配列決定のショットガン部分では、前述のPE Big−Dye(登録商標)ターミネーターおよび普遍的プライマー(M13フォワードおよびリバース)を用い、サイクル型配列決定反応を実施した(Munsonら,Infect Immun 72:3002−10,2004)。骨格形成用ライブラリーの末端を配列決定するため、プラスミドを、まず、R.E.A.L.Prep 96 Plasmid Kit(QIAGEN Inc.,Valencia,CA)を用いて精製し、次いで、TempliPhi(登録商標)DNA Amplification Kit(Amersham Biosciences Corp.,Piscataway,NJ)を用いて増幅した後、PE Big−Dye(登録商標)ターミネーターならびにpEpiFOS−5フォワードおよびリバース配列決定用プライマー(Epicentre,Madison,WI)を用いて反応を実施した。該プロジェクトの浄化(clean−up)部分のための反応は、PE Big−Dye(登録商標)ターミネーターおよびカスタムプライマー(Integrated DNA Technologies,Coralville,IA)を用いて実施した。過剰の色素ターミネーターをSephadex G50カラムで96ウェル形式にて除去し、ABI 3700またはABI 3100キャピラリー電気泳動DNA配列決定装置(Applied Biosystems,Foster City,CA)のいずれかにて配列を決定した。
【0140】
ゲノム閉鎖:骨格形成用ライブラリーおよびPCR由来の対の末端配列を用いて、コンティグを順に並べ、配列が少ない範囲の領域に配列を付加した。対のカスタムプライマー(Integrated DNA Technologies,Coralville,IA)は、各コンティグならびに配列が少ない範囲の領域に隣接する領域の両端に結合するように設計した。介在領域は、Sambrookら,Molecular Cloning:A Laboratory Manual(第3版)Cold Spring Harbor Laboratory Press.,2001に記載の標準的なPCRプロトコルにより、Taqポリメラーゼ(Roche Diagnostics,Indianapolis,IN)を用いて増幅し、両方の鎖を配列決定した。骨格形成用ライブラリー由来のクローンを鋳型として用い、リボソームのRNAオペロンおよびHMW遺伝子クラスターを完全に配列決定した。
【0141】
編集:Phred/Phrapをデータ編集に使用し、Munsonら,Infect
Immun 72:3002−10,2004に記載のデフォルト編集パラメータを用いた(Ewingら,Genome Res 8:186−94,1998;Ewingら,Genome Res 8:175−85,1988;Gordonら,Genome Res 8:195−202,1998)。507個のクローン由来の対の末端配列データを用い、DNASTAR 総合ソフトウェアSeqman IIプログラムを使用して編集データを確認した。
【0142】
データ解析:Glimmer2長orfプログラム(Delcherら,Nucleic Acids Res 27:4636−41,1999)によって同定された1178個の最長ORFの組に対して調整されたGlimmer2(v2.13)を用いてコード領域を同定した。Glimmer ORFの組を、Rd菌株プロテオーム、SwissProtデータベース、NCBI COGデータベースおよびKlEGGデータベースに対して検索することにより、類似性による自動アノテーションを行なった。Rd菌株データベースを86−028NP菌株ORFの組と、トリクロス(tricross)を用いて双方向で比較し、類似性の高信頼度領域を決定し、ゲノム組織のドットプロット比較を得た(Rayら,Bioinformatics 17:1105−12,2001)。
【0143】
自動的に推定されたアノテーション情報を、Artemis(Rodriguezら,Infect Immun 71:1635−42,2003)を用いてマニュアルでさらに操作(curate)して目的のゲノム領域を視覚化および境界画定し、カスタムFileMaker Proデータベースを作成し、次いで、これをマニュアル改定の適用および機能の割当てに関連するデータの取得に使用した。主な自動比較にはFASTA解析を使用した。80−028NP菌株のゲノムとRd菌株のゲノム間の強いシンテニーにより、大部分の遺伝子機能の自動的な割当てが可能であり、マッチングでは、アミノ酸レベルで90%以上の類似性を有していた。Rd菌株のゲノムに対してほぼ1対1の86−028NP菌株ゲノムのマッピングが、Rd菌株ORFの86−028NP菌株ゲノム配列上へのアセンブリおよび86−028NP菌株ORFのRdゲノム上への逆アセンブリ(SeqManプログラムをヌクレオチドレベルで80%同一性のアセンブリ基準で使用)によって確認された。
【0144】
マニュアルBLAST解析を用い、既知遺伝子に対して強い類似性を示さなかったORFの潜在的機能を調査した。自動割当てのマニュアル操作は、アノテーションが現在の文献に従うように行ない、自動化アルゴリズムが誤りをもたらし易いいくつかの位置(特に、HMW遺伝子クラスター、ヘモグロビン結合タンパク質およびhsd遺伝子クラスター、その高いファミリー類似性により自動割当てが混同される)を修復した。
【0145】
tRNA遺伝子をtRNAscan−SE vl.11(Loweら,Nucleic
Acids Res 25:955−64,1997)によって同定した。rRNAオペロンをRd菌株との16、23および5S rRNA類似性、ならびにこれらの遺伝子を含む隣接部のCLUSTALWアラインメントに基づいて同定し、半保存領域の境界を決定した。
【0146】
インフルエンザ菌(influennzae)86−028NP菌株の完全なゲノムDNA配列を、配列番号:772として示す。該ゲノムDNA内のオープンリーディングフレームを配列番号:773〜2593として示し、表6に示す。得られる遺伝子産物を配列番号:2581〜4414として示し、表7に示す。オープンリーディングフレームのヌクレオチドの前に示す「c」は、オープンリーディングフレームが配列番号:722として示す配列の5’−3’配列に相補的であることを示す。表6において、NTHi特異的遺伝子に印を付けている。このゲノム配列は、GenBankから受託番号CP000057(これは、引用によりその全体が本明細書に組み込まれる)として入手可能である。
【0147】
下記のオープンリーディングフレーム:配列番号:822(kdgK)、配列番号:928、配列番号:994(bolA)、配列番号:276(adhC)配列番号:1102(dusC)、配列番号:1121(merP)、配列番号:1135、配列番号:1236、配列番号:1254、配列番号:1376(lic2C)、配列番号:1431(hgpD)、配列番号:1502、配列番号:1505(ppx)、配列番号:1523(hgpC)、配列番号:1585(lex2A)、配列番号:1637配列番号:1713、配列番号:1856(mod)、配列番号:1899、配列番号:2006、配列番号:2080、配列番号:2155、配列番号:2202、配列番号:2257、配列番号:2331、配列番号:2345、配列番号:2365、配列番号:2555(metE)および配列番号:2563(pmi)は、これらのヌクレオチド配列がその配列内にフレームシフトまたは終止コドンを含むため、表6において偽遺伝子と規定している。hgpD、hgpC、lex2AおよびNTHI1769遺伝子は、リーディングフレーム内外への配列シフトを引き起こす偶発性(contingency)反復配列を含み、生じるアミノ酸配列(配列番号:3242、3332および4142)は部分翻訳物である。このような偶発性反復配列により、インフルエンザ菌において遺伝子発現が調節され、したがって、このようなタンパク質をコードする遺伝子は、1つ以上の反復配列の挿入または欠失後、正確に翻訳される。
【0148】
【表29】
【0149】
【表30】
【0150】
【表31】
【0151】
【表32】
【0152】
【表33】
【0153】
【表34】
【0154】
【表35】
【0155】
【表36】
【0156】
【表37】
【0157】
【表38】
【0158】
【表39】
【0159】
【表40】
【0160】
【表41】
【0161】
【表42】
【0162】
【表43】
【0163】
【表44】
【0164】
【表45】
【0165】
【表46】
【0166】
【表47】
【0167】
【表48】
【0168】
【表49】
【0169】
【表50】
【0170】
【表51】
【0171】
【表52】
【0172】
【表53】
【0173】
【表54】
【0174】
【表55】
【0175】
【表56】
【0176】
【表57】
【0177】
【表58】
【0178】
【表59】
【0179】
【表60】
【0180】
【表61】
【0181】
【表62】
【0182】
【表63】
【0183】
【表64】
【0184】
【表65】
【0185】
【表66】
【0186】
【表67】
【0187】
【表68】
【0188】
【表69】
【0189】
【表70】
【0190】
【表71】
【0191】
【表72】
【0192】
【表73】
【0193】
【表74】
【0194】
【表75】
【0195】
【表76】
【0196】
【表77】
【0197】
【表78】
【0198】
【表79】
【0199】
【表80】
【0200】
【表81】
【0201】
【表82】
【0202】
【表83】
【0203】
【表84】
【0204】
【表85】
【0205】
【表86】
【0206】
【表87】
【0207】
【表88】
【0208】
【表89】
【0209】
【表90】
【0210】
【表91】
【0211】
【表92】
【0212】
【表93】
【0213】
【表94】
【0214】
【表95】
【0215】
【表96】
【0216】
【表97】
【0217】
【表98】
【0218】
【表99】
【0219】
【表100】
【0220】
【表101】
【0221】
【表102】
【0222】
【表103】
【0223】
【表104】
【0224】
【表105】
【0225】
【表106】
【0226】
【表107】
【0227】
【表108】
【0228】
【表109】
【0229】
【表110】
実施例10
NTHi、86−028NP菌株およびインフルエンザRd菌株のゲノムの比較
86−028NP菌株のゲノム配列は、1,913,428bpを含む。これは、Rd菌株のゲノム(1,830,137bp)(Fleischmannら,Science
269:496−512,1995)よりほぼ4パーセント大きい。また、多数の遺伝子が86−028NP菌株に存在する(Rd菌株で1743に対して1942)。該遺伝子の相補性を、DNASTAR総合ソフトウェアのSeqmanプログラムを用いてRd菌株のものと比較した。排除値としてヌクレオチドレベルで80%同一性では、Rd菌株のゲノムには存在しない285個のORFが86−028NPゲノムにおいて同定され、86−028NP菌株ゲノムには存在しない167個のORFがRd菌株のゲノムにおいて同定された。
【0230】
86−028NP菌株は、Rd菌株と同様、6個のリボソームオペロンを有する。tRNAscan−SE vl.11を用い、20個の共通アミノ酸を表す58個のtRNA遺伝子が86−028NP菌株ゲノムにおいて同定された。tRNA−Glu、tRNA−AlaおよびtRNA−Ile遺伝子は、16SリボソームRNA遺伝子と23SリボソームRNA遺伝子の間のスペーサー領域内に位置した。UCAアンチコドンを含むtRNA遺伝子もまた同定された。このアンチコドンはオパール終止コドンに相当し、典型的には、セレノシステインを組み込むオパール抑制tRNAと関連している。tRNAは、Sec tRNA特異的伸長因子であるselB(NTHI0836)と、セレンの組込みによってセレノシステインを形成する予備としてセリンをデヒドロアラニンに変換させる酵素であるselA(NTHI0835)をコードする2つの遺伝子と隣接している(Forchhammerら,Nature 342:453−6,1989)。セレノホスフェートシンテターゼをコードするselD遺伝子(NTHI0297)もまた同定された。このセレノシステイン系の重要性は、おそらくセレノシステインコドンとして読まれるインフレームTGA終止コドンを含むギ酸デヒドロゲナーゼのαサブユニットのコード配列(NTHI0007)によって証明される。インフレームTGA終止コドンは、Rd菌株のギ酸デヒドロゲナーゼ遺伝子(GenPept受託番号P46448)の現在のアノテーションにおいて既に記載されて注釈されている。
【0231】
86−028NP菌株とRd菌株の遺伝子順序の解析に関与するゲノム間の全体比較により、大きな逆位の形態の単一の大きな再編成が示されている。この471kbの逆位は、86−028NP菌株ゲノムのほぼ25%を表し、NTHI1391およびNTHI1394(それぞれHI1218およびHI1645のホモログ)またはNTHI1949およびNTHI1950(それぞれHI1219およびHI1647のホモログ)で境界されている。HI1219およびHI1646は、Rd菌株において一部重複した遺伝子であり、cmkAおよびcmkBとアノテートされている(シチジレートキナーゼ)。1つのcmk遺伝子(NTHI1949)が、86−028NP菌株内にcmk様小断片とともに、NTHI1391とNTHI1394の間に存在する。骨格形成用ライブラリー由来のいくつかのクローンは、86−028NPゲノム内で該逆位の各末端が重複しており、本発明者らのアセンブリが認証される。この大きな逆位内には、いくつかの挿入部があり、その最大のものはほぼ13kb、27kbおよび51kbの大きさである。これらの領域は、主に仮想遺伝子および仮想保存遺伝子ならびにファージ遺伝子のいくつかのホモログを含む。例えば、27kb挿入部は、HP1−およびHP2様ファージ遺伝子の残部を含む。最大挿入部は、インテグラーゼ遺伝子のホモログと境界されている。Rd菌株では、mu様ファージがこの領域に局在している(Morganら,J Mol Biol 317:337−59,2002)。このファージは、86−028NP菌株ゲノムには存在しない。また、この大きな逆位領域内には、Rdゲノムとのシンテニーを回復させる21kb逆位が存在する。
【0232】
この大きな逆位に加え、86−028NP菌株は、Rd菌株のゲノムとの共線形成(co−linearity)と相違する他の領域を有する。これらには、Rd菌株に存在するDNAとの明白な相同性もたない配列を含む5kbより大きい9つの領域が含まれる。これらの領域のうち2つは、後述するHMW付着因子を含む。仮想遺伝子は、該特異的領域のうち6つで優位を占める。第9番目の領域はほぼ56kbの大きさである。これは、NTHI0100とNTHI0165の間に存在する。BLASTn解析により、ICEHin86−028NPと表示するこの領域内の遺伝子は、b型インフルエンザ菌プラスミド内の遺伝子ICEHin1056(Mohd−Zainら,J Bacterial
186:8114−22,2004)に対して高い相同性を有することが示された。ICEHin1056は、一連の共通コア遺伝子によって規定されるゲノムアイランドの拡張ファミリーの一構成員である(Mohd−Zainら,J Bacteriol 186:8114−22,2004)。ICEHin86−028NPは、45個のICEHm 1056 ORFのホモログを有する。これらには、主に、プラスミドの複製およびコンジュゲーションにおいて推定される役割を有するタンパク質をコードするICEHin86−028NPの5’末端付近のORF(規定のコア遺伝子を含む)、ならびに主に、膜会合性またはまたは膜輸送性のいずれかであり得ることが示唆されるモチーフを有する保存仮想タンパク質をコードする3’末端付近のORFが含まれる。注目すべきことは、ICEHin86−028NPが、ICEHin1056には見られるテトラサイクリン、クロラムフェニコールおよびβ−ラクタム耐性に関与するタンパク質をコードする遺伝子を欠くことである。ICEHin86−028NP内には、トランスポゼース、レソルバーゼおよび推定インテグラーゼ調節因子が散在しており、これは、ICEHin86−028NPが、いくつかの可動性遺伝因子に由来する複合エレメントであることを示す。
【0233】
ICEHin1506は、attP部位と表示される配列を第1の遺伝子の5’側に有する。86−028NP菌株には、このattP部位の完璧なコピーがNTHI0101の5’側に存在し、単一ヌクレオチド変化を有するこのattP部位のコピーがNTHI0164の3’側に存在する。attP部位は、ゲノムアイランドを形成する細菌染色体内への可動性遺伝因子の組込みに関与しており、おそらく、この大きな遺伝物質セクションが86−028NP菌株ゲノム内に組み込まれた状態となる機構を示唆する(Dimopoulouら,Antimicrob Agents Chemother 46:1602−3,2002)。ICEHin86−028NPは、その他の任意の関連ゲノムアイランドより少なく、86−028NP菌株のゲノム全体のG+C含量38%に近い39%のG+C含量を有する。これは、このエレメントについて長期ゲノム関連を暗示する。このエレメントとともに、膜会合タンパク質および分泌タンパク質をコードしていると考えられるICEHin1506のものに相同な遺伝子(Dimopoulouら,Antimicrob Agents Chemother 46:1602−3,2002)のその相補配列の存在は、86−028NP菌株のビルレンスの重要な暗示を有し得る
パスツレラ科(Pasteurellaceae)のいくつかの構成員、例えば、ヘモフィラス デュクレイ(Haemophilus ducreyi)、パスツレラ マルトシダ(Pasteurella multocida)およびアクチノバチラス アクチノマイセテムコミタンス(Actinobacillus actinomycetemcomitans)は、充分特性評価されたタンパク質毒素を産性する。対照的に、インフルエンザ菌は、タンパク質毒素を産性しないようであり、推定タンパク質毒素をコードする遺伝子は、86−028NP菌株ゲノムにおいて同定されなかった。インフルエンザ菌では、内毒素生合成を担うグリコシルトランスフェラーゼをコードする遺伝子、および感染過程において細菌に増強された「適応度」を付与するタンパク質をコードする遺伝子が、一般的にビルレンス決定基とみなされている。これらの遺伝子としては、付着因子、ヘムおよびヘモグロビン結合タンパク質をコードするもの、ならびに酸化的ストレスを防御するタンパク質をコードする遺伝子が挙げられる。
【0234】
偶発性遺伝子
インフルエンザ菌は、限定数の2成分調節系および他の総合調節因子を有する。Moxonおよび協働者らにより、「単純偶発性(simple contingency)遺伝子座」と称する遺伝子座が遺伝子発現調節の代替機構を提供し、したがって、該生物体が変化する環境条件に速やかに応答する能力に寄与することにより、生物体の適応度が増大すると論じられた。これらの遺伝子座は、短いタンデム反復配列を、コード領域内またはコード領域の5’側のいずれかに含む。DNA複製中、リーディングフレーム内での反復配列の付加または欠損により、リーディングフレームの改変がもたらされる。コード領域の5’側に局在する場合、反復配列の付加または欠損により、プロモーター活性の変化がもたらされる(Baylissら,Clin Invest 107:657−62,2001)。単純反復配列を含む遺伝子座は、例えば、インフルエンザ菌において広範に研究されている(Hoodら,Proc Natl Acad Sci USA 93:11121−5,1996)。以下のセクションに相変異性(phase variable)として記載する遺伝子座のいくつかは、単純反復配列を含む。
【0235】
付着因子
86−028NP菌株は、主に宿主細胞への付着において機能する産物をコードするいくつかの遺伝子を保有する(表8)。これらのうちの1つである外膜タンパク質P5は、以前に同定され、その機能が注意深く分析された(Jiangら,Infect Immun 67:187−92,1999;Kennedyら,Infect Immun 68:2756−65,2000;Novotnyら,J Immunol 171:1978−83,2003;Novotnyら,Infect Immun 68:2119−28,2000;Novotnyら,Vaccine 20:3590−7,2002;Sirakovaら,Infect Immun 62:2002−20,1994)。86−028NP菌株は、Rd菌株Actinobacillus pleuropneumoniaeおよびP.multocida由来のpilABCDのホモログである4つの遺伝子を含む遺伝子クラスターを保有する(Bakaletzら,Infect
Immun 73:1635−4,2005;Doughtyら,Vet Microbiol 72:79−90,2000;Ruffoloら,Infect Immun
65:339−43,1997 Stevensonら,Vet Microbiol
92:121−34,2003)。これらの遺伝子は、comE遺伝子および未同定遺伝子とともに、鼻咽頭組織への86−028NP菌株の付着において役割を有するIV型線毛をコードする(Kennedyら,Infect.Immun.,68:2756−2765,2000)。
【0236】
86−028NP菌株は、Rd菌株には存在しない2つの高分子量(HMW)付着因子遺伝子クラスターを保有する。該高分子量付着因子は、最初にNTHi 12菌株において特性評価され、これは、各々、3種類のタンパク質(HMWA、HMWBおよびHMWC)をコードする2つのHMW遺伝子クラスターを有する。HMWAは、該付着因子の構造的成分であり、HMWBは、膜貫通転位において役割を有する一方、HMWCは、HMWAのグリコシル化に必要とされる(Barenkampら,Infect Immun
60:1302−13,1992;Barenkampら,Infect Immun
62:3320−8;1994;Grassら,Mol.Microbiol 48:737−51,2003;St Gemeら,Mol.Microbiol 27:617−30,1998)。同様に、86−028NP菌株の2つのHMW遺伝子クラスターは、12菌株と同じ遺伝子状況でHmwA、BおよびC遺伝子のホモログを含む(Buscherら,J Bacteriol 186:4209−17,2004)。86−028NP菌株由来のHMW1AおよびHMW2Aタンパク質は、主要相違領域、例えば、HMW2A 内の41アミノ酸挿入部とC末端に向かって72%同一である。86−028NP菌株由来のHMWBとHMWCの対のタンパク質は、それぞれ99%同一である。配列ATCTTTCは、hmw1Aの上流で17回およびhmw2Aの上流で23回反復される。12菌株では、この配列の16反復配列が、各hmw遺伝子クラスターの5’側に見られる(Barenkampら,Infect Immun 60:1302−13,1992)。
【0237】
Hapは、IgAlプロテアーゼの触媒性ドメインに相同なドメインを有する自己輸送型タンパク質である。NTHI0354遺伝子は、NTHi菌株N187由来のHapと83%同一性を有するタンパク質をコードする(St Gemeら,Mol.Microbiol 14:217−3,1994)。86−028NP菌株は、HMW1およびHMW2を保有する他のNTHi菌株と同様、別のヘモフィルス属付着因子であるHiaをコードする遺伝子を欠く(Barenkampら,Mol.Microbiol 19:1215−23,1996)。また、86−028NP菌株は、本発明者らが以前に報告したように、赤血球凝集性線毛をコードするhif遺伝子クラスターを欠く(Munsonら,Infect Immun 72:3002−10,2004)。
【0238】
【表111】
表8、9および10において、「NTHI番号」は、Microbial−Pathogenesis H.influenzae 86028 NPウェブサイトおよびGenbank 受託番号CP000057で表示されるNTHi 86−028NP菌株ゲノム 内の遺伝子座タグ番号を示す。「HI番号」は、TIGR(The Institute for genomic redearc)内の対応する遺伝子座タグ番号を示す。
【0239】
リポオリゴ糖合成[[当初の出願の第13頁参照]
インフルエンザ菌リポオリゴ糖(LOS)の構造、生合成およびビルレンスにおける役割は、広範に研究されている。表9に、リポオリゴ糖生合成に関与する遺伝子の一覧を網羅する。86−028NP菌株は、LOSの七炭糖−Kdo−リピドA部分合成するのに必要とされる遺伝子の完全な相補配列を有する。lgtFおよびlpsA遺伝子は、グルコース、およびグルコースまたはガラクトースを七炭糖の第1および第3残基にそれぞれ付加するグリコシルトランスフェラーゼをコードする。これらの遺伝子はともに、86−028NP菌株ゲノム内に存在し、したがって、おそらく、炭水化物鎖は、86−028NP菌株LOSの第1七炭糖残基および第3七炭糖残基から伸長され得る(Hoodら,Microbiology 150:2089−97,2004)。b血清型菌株RM153において、lic2C遺伝子は、グルコースを第2七炭糖に付加するグリコシルトランスフェラーゼをコードする(Hoodら,Microbiology 150:2089−97,2004)。86−028NP菌株ゲノムでは、この遺伝子はフレームシフトを含む。相変異性lic2AおよびlicA遺伝子は、それぞれ、ガラクトシルトランスフェラーゼおよびコリンキナーゼをコードし、86−028NP菌株ゲノム内に存在する(Highら,Mol.Microbiol 9:1275−82,1993;Hoodら,Glycobiology 11:957−67,2001;Weiserら,Infect Immun 65:943−50,1997)。lex2B遺伝子は、b血清型DL42菌株ならびにいくつかの他の血清型分類可能菌株においてグリコシルトランスフェラーゼをコードし、86−028NP菌株ゲノム内に存在する(Griffinら,Microbiology 149:3165−75,2003;Jarosikら,Infect Immun 62:4861−7,1994)。DL42菌株のlex2B遺伝子に対して5’側は、短い相変異性lex2A遺伝子である。86−028NP菌株では、この遺伝子は、1つの4ヌクレオチド反復配列の欠損および5bp欠失のため、DL42配列(Genbank受託番号U05670)と比べるとフレーム外である。最近、Hoodおよび協働者らにより、Rd菌株において、HI0866〜HI0874を含むHmgと表示される遺伝子座が報告された(Hoodら,J Bacteriol 186:7429−39,2004)。rmlBのホモログは例外として、これらの遺伝子は、86−028NP菌株ゲノムには存在しない。これには、NTHI 2019菌株においてバイオフィルム形成に重要であることが最近示されたシアリルトランスフェラーゼをコードするsiaA遺伝子が含まれる(Greinerら,Infect Immun
72:4249−60,2004;Jonesら,J Biol Chem 277:14598−611,2002)。別のシアリルトランスフェラーゼをコードするlic3A遺伝子のホモログの2つのコピー(Hoodら,Mol.Microbiol 39:341−50,2001;Jonesら,JBiol Chem 277:14598−611,2002)、ならびに別のシアリルトランスフェラーゼをコードするlsgB遺伝子の1つのコピー(Jonesら,J Biol Chem 277:14598−611,2002)が、86−028NP菌株ゲノムにおいて同定された。
【0240】
【表112】
【0241】
【表113】
【0242】
【表114】
鉄分獲得
インフルエンザ菌株は、ヘムの中間体前駆体であるプロトポルフィリンIX(PPIX)とともにヘムまたは鉄分のいずれかの絶対要件を有する(Evansら,J Med Microbiol 7:359−65,1974;Whiteら,J Bacteriol 85:842−50,1963)。表10に、鉄分獲得に関与する遺伝子の一覧を網羅する。3種類のヘモグロビンおよびヘモグロビン−ハプトグロビン結合タンパク質HgpA、HgpBおよびHgpCがb型インフルエンザ菌HI689菌株において同定された(Jinら,Microbiology 145(Pt 4):905−14,1999;Mortonら,Infect Immun 67:2729−39,1999;Renら,Infect Immun 66:4733−41,1998)。HI689菌株では、これらの遺伝子は、CCAA4ヌクレオチド反復配列を有し、スリップストランド(slip−strand)誤対合によって調節されることが知られている。これらの遺伝子のうち2つは、86−028NP菌株内に存在する。これらはともに、CCAA反復配列を含み、hgpB遺伝子はインフレームであるが、hgpC遺伝子はフレーム外である。CCAA反復配列を含む第3の遺伝子の誘導アミノ酸配列は、hgpAと45%同一である。本発明者らは、この遺伝子をhgpDと命名した。この遺伝子はフレーム外である。ヘムおよびヘム−ヘモペキシン錯体をコードするb型インフルエンザ菌のhxuABC遺伝子のホモログ(Copeら,Infect Immun 69:2353−63,2001;Copeら,Infect Immun 66:4511−6,1998;Copeら,J Bacteriol 177:2644−53,1995)ならびにhemR受容体のホモログが同定された。86−028NP菌株はまた、ヘム結合リポタンパク質HbpAをコードする遺伝子を有する(Heathら,Pediatr Infect Dis J 20:300−5,2001)。hbpAの下流は、その産物が、推定ヘム利用タンパク質を含むクラスターであるCOG0748の構成員である仮想遺伝子であるNTHI1022である。最近b型インフルエンザ菌において同定され、一般的なヘム利用タンパク質をコードするhup遺伝子のホモログもまた同定された(Mortonら,Microbiology 150:3923−33,2004)。
【0243】
ヘム輸送系に加え、鉄輸送系wereもまた同定された。hitABC遺伝子はそれぞれ、シデロフォア欠損大腸菌株にb型インフルエンザ菌の菌株からクローニングしたHtABC遺伝子を補完することにより完結に特性評価された高度に特異的な鉄イオンABC輸送系の構成員であるFbpABCタンパク質をコードする(Andersonら,J Bacteriol 186:6220−9,2004)。tbpAB(Gray−Owenら,Infect Immun 63:1201−10,1995;Gray−Owenら,Infect Immun 63:3809−15,1995)ならびにhfeABCDと表示され、鉄取込みに関与するABC輸送系のホモログであり、かつ最初にペスト菌において特性評価された(Beardenら,J Bacteriol 180:1135−47,1998)遺伝子にコードされるトランスフェリン結合タンパク質1および2が同定された。この後者の遺伝子クラスターは、Rd菌株にも存在する。NTHI2035 は、Mn2+およびFe2+輸送体のNRAMPファミリーの推定ホモログをコードする(Richerら,J Mol Evol 57:363−7,2003)。
【0244】
上記のように、インフルエンザ菌は、インビトロでの増殖のためのヘムの供給源として、PPIXとともに鉄分を利用し得る。PPIXへの鉄の組込みを触媒する(Schlorら,Infect Immun 68:3007−9,2000)フェロキレターゼをコードするhemH遺伝子が同定された。調節因子をコードする遺伝子Furもまた同定された(Andrewsら,FEMS Microbiol Rev 27:215−37,2003;Smootら,J Med Microbiol 48:629−3,1999)。
【0245】
【表115】
【0246】
【表116】
酸化的ストレス
増殖に必要であるが、鉄分の能動的獲得は、菌体に対して有害な効果を有し得る。フェントン反応により、鉄分は過酸化水素と反応し、高度に反応性のヒドロキシル残基を生成させ得る。この生成物は、著しい効果、例えば、脂質過酸化および鉄分含有酵素とDNAの両方に対する損傷を有する(Imlay,Annu Rev Microbiol 57:395−418,2003)。ヒドロキシル残基に対する最もよく知られた防御系は、スーパーオキシドディスムターゼAおよびBからなり、これらは、高度に反応性のスーパーオキシドを過酸化水素に変換し、次いで、これは、カタラーゼによって水と酸素に変換される(Demple,Annu Rev Genet 25:315−37,1991)。86−028NP菌株およびRd菌株は、sodA遺伝子(NTHI1251)を含むが、sodB遺伝子を欠く。また、両菌株とも、カタラーゼ遺伝子hktE(NTHI1099)(Bishaiら,J Bacteriol 176:2914−21,1994)、酸化的ストレスに対する保護に関与する遺伝子の一次調節因子(Maciver & Hansen,Infect Immun 64:4618−29,1996;Pomposielloら,Trends Biotechnol 19:109−14,2001)をコードするoxyR遺伝子(NTHI0704)、およびPrx/Grxと称され、小アルキルヒドロペルオキシドに対する保護においてグルタチオン依存的役割を有するキメラペルオキシダーゼをコードする遺伝子(Pauwelsら,J Biol
Chem 278:16658−66,2003;Vergauwenら,J Bacterial 185:5555−62,2003;Vergauwenら,J Bacteriol 185:1572−81,2003)を保有する。本発明者らは、以前に、Rd菌株には存在しないP.multocidaペルオキシレドキシンTsaAのホモログをコードする遺伝子であるNTHI0212を同定した(Munsonら,Infect Immun 72:3002−10,2004)。しかしながら、86−028NP菌株は、サルモネラ属においてTsaAの還元に関与していることがわかっている(Pooleら,Biochemistry 39:6602−15,2000)専用(dedicated)アルキルヒドロペルオキシドレダクターゼであるAhpFが欠損している。酸化的ストレスに対するさらなる保護は、ftnAおよびftnB(それぞれ、NTHI1773およびNTHI1772)遺伝子にコードされるフェリチン様タンパク質によってもたらされ得る。これらのタンパク質の過剰発現により、鉄過剰負荷大腸菌fur変異型が酸化的損傷から保護されることが示された(Touatiら,J Bαcteriol 177:2305−14,1995)。仮想保存遺伝子NTHI1817は、DNA結合フェリチン様タンパク質と相同性を有するタンパク質をコードする。これは、S.entericaでは鉄の存在下および食作用中の両方において酸化的ストレスに対する保護の役割を有し、サルモネラ属感染マウスモデルにおいてビルレンスに重要である(Halseyら,Infect Immun 72:1155−8,2004)非特異的DNA結合タンパク質のDpsファミリーの構成員である。大腸菌では、Dpsは、過酸化水素によって酸化された鉄分に優先的に結合し、したがって、フェントン反応によって生成されるヒドロキシル残基の発生の抑止に重要な役割を有することが示された(Zhaoら,J Biol Chem 277:27689−96,2002)。
【0247】
分泌
Sec系に加え、86−028NP菌株は、2つのアルギニンをそのシグナルペプチド内に有するタンパク質のSec非依存的輸送に関与するTatA、BおよびC タンパク質、細胞質膜会合タンパク質をコードする遺伝子(NTHI0279、NTHI0280およびNTHI0282)を有する(Bolhuisら,J Biol Chem 276:20213−9,2001;Yenら Arch Microbiol 177:441−50,2002)。既報のように、86−028NP菌株は、自己輸送型タンパク質Lavをコードする遺伝子NTHI0585を保有する(Munsonら,Infect Immun 72:3002−10,2004)。このタンパク質はRd菌株には存在しないが、ナイセリア菌には存在し、ヘモフィルス属では、病原性菌株に限定されるようである(Davisら,J Bacteriol 183:4626−35,2001)。86−028NP菌株はまた、IgAプロテアーゼをコードする遺伝子(NTHI1164)(PoulsenらJ Bacteriol 174:2913−21,1992)、および上記のようなHap付着因子をコードする遺伝子を有する。ともに、自己輸送体の類型のタンパク質である、上記のように、HMW付着因子は、2パートナー分泌経路群のタンパク質の構成員である。
【0248】
外膜タンパク質
いくつかの外膜タンパク質(OMP)をコードする遺伝子が、他のインフルエンザ単離菌のものとの相同性によって同定された。これらとしては、主要なOMP(すべて、最初はb型インフルエンザ菌において同定);表面発現P1(NTHI0522)、ポーリンP2(NTHI0225)、ホスホモノエステラーゼおよびヘム輸送体P4(NTHI0816)、付着因子P5(NTHI1332)ならびにリポタンパク質P6(NTHI0501)が挙げられる。また、86−028NP菌株は他のヘモフィルス属菌株と、いくつかの微量OMPを共有する。これらとしては、b型インフルエンザ菌由来のD15およびトランスフェリン結合タンパク質、ならびにNTHi菌株289において同定されたOMP26のホモログ(Munsonら,Infect Immun 56:2235−42,1988;Munsonら,Infect Immun 49:544−9,1985;Munsonら,J Clin Invest 72:677−84,1983;Reidlら,J Exp Med 183:621−9,Reillyら,J Bacteriol 181:6797−805,1999;Reillyら,FEBS Lett 494:19−23,2001)が挙げられる。すべては、その後、NTHi菌株において特性評価され、潜在的なワクチン候補として解析された(Poolmanら,Vaccine 19 Suppl 1:S108−15,2000;Murphyら Curr Opin Infect Dis 16:129−34,2003;McMichaelら,Curr Opin Investig Drugs 4:953−8,2003 Crippsら, Immunol Cell Biol 81:46−51,2003;Bakaletzら Ann Otol Rhinol Laryngol Suppl 188:82−94,2002)。
【0249】
制限酵素系:
86−028NP菌株は、HindIIおよびHindIII II型制限系を欠く(Fleischmannら,Science 269:496−512.80,1995;Nwankwoら,Gene 150:75−80.104,1994,Smith,& Marley.Methods Enzymol 65:104−8,1980)。対照的に、H.aegyptiusにおいて最初に同定された(Slatkoら,Gene 74:45−50,1988)HaeII系をコードする遺伝子は、86−028NP菌株ゲノム内に存在するが、Rd菌株には存在しない。86−028NP菌株およびRd菌株はともに、メチルトランスフェラーゼ(HsdM)、配列認識タンパク質(HsdS)および制限酵素(HsdR)をコードするHsd型I制限系を有する(Robertsら,Nucleic Acids Res 31:1805−12,2003)。これらの伝子は、Rd菌株のゲノムでは隣接している(HI1285〜HI1287)。86−028NPゲノムは、各々4つの遺伝子を含有する3つのhsd様遺伝子座を含む。hsd系の1つはNTHI1838〜NTHI1843にコードされている。この遺伝子クラスターでは、NTHI1841は仮想タンパク質をコードする。第2のhsd様遺伝子座はNTHI0314〜NTHI0318にコードされている。この遺伝子クラスターでは、NTHI0316は推定アンチコドンヌクレアーゼをコードする。このhsd様系は、大腸菌のprr系に類似したものであり得る(Tyndallら,J Mol Biol 237:266−74,1994)。第3のhsd遺伝子座はNTHI0188〜NTHI0193にコードされている。この遺伝子クラスターでは、NTHI0190 は、ヘリックス−ターン−ヘリックスドメインを有する推定転写調節因子をコードする。
【特許請求の範囲】
【請求項1】
明細書に記載の発明。
【請求項1】
明細書に記載の発明。
【図1】

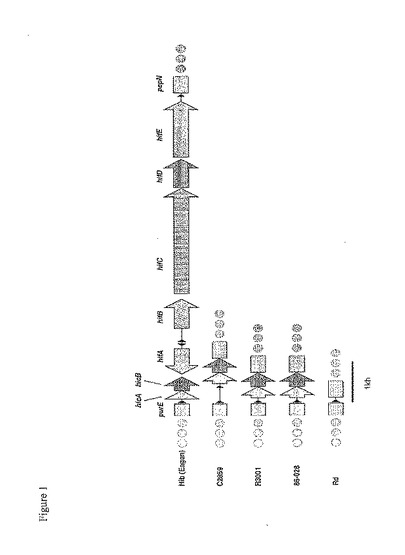
【図2】
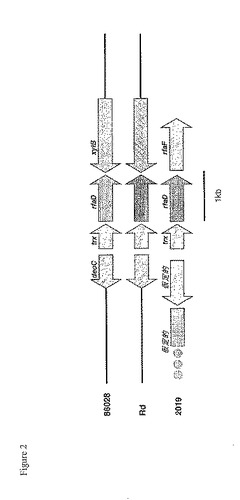
【図3A】

【図3B】

【図3C】

【図3D】

【図3E】

【図3F】

【図3G】

【図3H】

【図3I】

【図3J】

【図3K】

【図3L】

【図3M】

【図2】
【図3A】
【図3B】
【図3C】
【図3D】
【図3E】
【図3F】
【図3G】
【図3H】
【図3I】
【図3J】
【図3K】
【図3L】
【図3M】
【公開番号】特開2012−120543(P2012−120543A)
【公開日】平成24年6月28日(2012.6.28)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−62469(P2012−62469)
【出願日】平成24年3月19日(2012.3.19)
【分割の表示】特願2008−517126(P2008−517126)の分割
【原出願日】平成18年6月15日(2006.6.15)
【出願人】(502200830)ネイションワイド チルドレンズ ホスピタル, インコーポレイテッド (11)
【出願人】(507410021)ザ ボード オブ リージェンツ オブ ユニバーシティ オブ オクラホマ (2)
【Fターム(参考)】
【公開日】平成24年6月28日(2012.6.28)
【国際特許分類】
【出願番号】特願2012−62469(P2012−62469)
【出願日】平成24年3月19日(2012.3.19)
【分割の表示】特願2008−517126(P2008−517126)の分割
【原出願日】平成18年6月15日(2006.6.15)
【出願人】(502200830)ネイションワイド チルドレンズ ホスピタル, インコーポレイテッド (11)
【出願人】(507410021)ザ ボード オブ リージェンツ オブ ユニバーシティ オブ オクラホマ (2)
【Fターム(参考)】
[ Back to top ]