
動物及びヒト病原体に対して使用するためのバシラスサブティリス由来の抗菌化合物
動物及びヒトの病原菌に対して使用するためのBacillus subtilis由来の抗微生物化合物。Bacillus subtilisの新規な菌株は家禽の胃腸管から単離され、Clostridium perfringens、Clostridium difficile、Campylobacter jejuni、 Campylobacter coli、及びStreptococcus pneumoniaeに優れた阻止作用を有する因子を生じることが見いだされた。この因子又はこれらの因子は、加熱処理後十分な生存性及び抗微生物活性を保持する。本発明は、C. perfringensを含む病原微生物の処置方法を提供する。
【発明の詳細な説明】
【発明の詳細な説明】
【0001】
発明の背景
本発明は一般的に抗菌化合物に関し、さらに具体的には、動物及びヒト病原体に対して使用するためのBacillus subtilis PB6由来の抗菌化合物に関する。
【0002】
Clostridium perfringensによりもたらされる壊疽性腸炎、腸性毒血症は、家禽の死滅をもたらす腸壁の壊疽性損傷の進展に導く(Paulus and Ruckebusch, 1996; Tsai and Tung, 1981)。それは複雑で部分的に知られていない疫学及び病因の多因性疾患でもある(Kaldhusdal, 2000)。細菌であるC. perfringensは家禽の胃腸管中で見いだされる(Tshirdewahn等, 1991)が、しかし壊疽性腸炎の発生は偶発的である(Cowen等, 1987)。それにもかかわらず、C. perfringensで汚染された食品は、鶏の壊疽性腸炎の発生に影響を与えてきた(Kaldhusdal, 2000)。研究は、健常な鶏は胃腸管中には比較的に少ない数のC. perfringensしか有しないが、一方、細菌の濃度の増加が壊疽性腸炎症状をもたらすことがある(Craven等, 1999)。
【0003】
壊疽性腸炎を罹患している家禽を処置するのにバシトラシン、リノコマイシン及びその他の成長促進抗生物質が一般に使用される(Craven 等, 1999)。しかし、鶏又は七面鳥由来のC. perfringensの抗生物質−抵抗性株の単離が原因で(Devriese等, 1993; Kondo, 1988; Watkins等, 1997)、家禽健康当局及び生産者は、抗生物質を置き換えるプロバイオティック製品の開発と応用に興味が増加している。プロバイオティクスは、宿主の胃腸の細菌のバランスを改良することにより宿主に好影響を与える生存細菌供給サプルメントとして定義される。何人かの研究者は、腸内細菌のこの標準化は次の利点を与えると信じている:(a)競合的除外(定着部位の拮抗とも称される)による病原体に対する保護;(b)一定の栄養素及び酵素的/脱毒素化反応の提供;(c)組織形態形成及び蠕動活性の関連性;並びに(d)宿主の免疫系及び内分泌系の相互作用である。さらに、家禽の壊疽性腸炎の上昇及び多くの国による種々の飼料用抗生物質の漸進的禁止に照らして(Council of the European Communities, 1998)、動物の能力を増強させるために代替成長促進剤(alternative growth promoters: AGP)へ向けてのシフトがある。イーストや乳酸菌のような有用な微生物は、過去20年間動物生産に使用されてきた。プロバイオティク特性を持つ細菌は、腸管中の病原菌と置き換わる(replacing or displacing)ことにより健康な腸ミクロフロラを維持する主要な機能を有する。プロバイオティックスは、動物腸内の病原菌に対して競合的除外(Tournot, 1989)及び拮抗作用(Fuller, 1989)により「正常な」腸内ミクロフロラを維持できることが知られている。概して、プロバイオティク微生物は腸内で大量に増殖でき、したがって、病原菌の繁殖を抑制する。ヒト又は動物に使用できるプロバイオティク微生物には、Bacillus spp., Lactobacillus spp., Enterococcus spp.及びStreptococcus spp等がある(Lee 等,1999)。胃腸管の異なる部位でコロニーを形成できる種々の微生物が報告されている(Jin 等, 1997)。概して、微生物の濃度の最も高い盲腸に比較して、胃腸管の十二指腸部が最も低い細菌ミクロフロラの集団を示す(Mead 及びAdams, 1975; Salanitro等, 1974)。Lactobacillus spp., Streptococcus spp.及びEscherichia coliのような微生物が、十二指腸部、小腸中の空腸及び回腸部にから単離された(Shapiro及びSarles, 1949)。盲腸の一般的な微生物集団は、Streptococcus spp., Staphylococcus spp., Lactobacillus spp., Fusobacterium spp.及びBacteroidesのような絶対嫌気性生物を含む(Barnes等, 1972; Mead, 1997)。
【0004】
鶏腸管の種々のセグメント由来の有用細菌が単離され、C. perfringensに対して可能性のあるプロバイオティックスとしてスクリーニングされた。以前の研究によれば、胃腸管の上皮細胞を接着し、コロニー化するプロバイオティックス微生物の能力は特定の動物源からの特定部位の単離物に大きく依存することを示した(Barrow 等, 1980; Reid, 1999; Fuller, 1973; Wesney 及びTannock, 1979)。動物の飼料に微生物プロバイオティックスの使用に関連する多くの有利な作用がある。これらの有利な作用には、病原菌E. coli (Watkins et al., 1982), Campylobacter jejuni (Morishita等, 1997) 及びSalmonella enteritidis (Pascual等, 1999)の競合的除外等があり、有用な腸ミクロフロラの成長と生存力を増強し(Hosoi等, 2000)、鶏に栄養素の消化と吸収の改善をもたらす(Ratcliff, 2000; Scheinbach, 1998; Sissons, 1989; Thomke及びElwinger, 1998)。
【0005】
プロバイオティク細菌を単離し特徴付けるために使用されるその他の基準には、胆汁及び酸安定性(Hoa 等, 2000; Huis In't Veld及びShortt, 1996)、抗生物質の産生(Salminen等, 1996)、並びに安全性に合致し又は一般に安全と認められる(generally recognized as safe :GRAS)状態(Donohue 及びSalminen, 1996; SCAN, 2000)等がある。多数の乳酸菌(単一若しくは組合せ)が、病原微生物に向けて種々の程度の抗微生物活性を呈することが示された(Harris等, 1989; Motlagh等, 1991)。加えて、乳酸菌の成長可能な培養物又は発酵抽出物が、多くの腸の疾病の特性である、置換された内因性腸ミクロフロラの処置に有用である(Charteris等, 1997; Drake 等, 1996)。このような細菌は、上皮組織に接着することにより、酸や胆汁条件で生き残ることができ、腸管に少なくとも一時的にコロニーを形成する。それらは、豚、鶏及び子牛の成長速度を改良し、飼料に有用であることが報告された(Hale及びNewton, 1979; Tortuero, 1973; Schwab等, 1980)。さらに、これらの有用な細菌を含有する飼料を与えた豚及び子牛において下痢の発生に顕著な減少が観察された(Lee, 1999)。乳酸菌培養物も、豚のE. coliからの腸毒素の作用を中和するものと思われる(Mitchell及びKenworthy, 1976)。その他の乳酸菌の有用な作用にはC. perfringensを含む有害細菌の置換、細菌性尿素分解酵素活性の産生、ビタミンの合成、免疫系における刺激作用、及び消化に対する貢献等がある(Hofacre等, 1998)。以前の研究により、Lactobacillus rhamnosus (Alander等, 1999; Asensio 等, 1976; Silva 等, 1987), L. plantarum (Andersson, 1986; West及びWarner, 1988), Lactococcus lactis ssp. lactis (Motlagh等, 1991; Spelhaug及びHarlander, 1989), 並びにPediococcus pentosaceus (Fleming 等, 1975; Graham及びMcKay, 1985)がClostridium sppに対して殺細菌性であったことを示す。
【0006】
顕著な抗菌活性を持つ微生物ペプチドは、一般的に、動物、植物、微生物(Sahl, 1985)及び非−無菌食物(Muriana, 1993)から単離される。それらは、小さく、カチオン性であり、3000〜6000ダルトンの分子質量を有する(Roller, 1991)。前駆体ペプチドの翻訳後修飾(post-translational modification)は、分子内チオエーテル橋を導入し、Pep 5、ナイシン、及びサブチリンのようなカチオン性ペプチドにすることが示されている(Gross及びMorell, 1971; Kordel 及びStahl 1986; Kordel等, 1989)。これらのペプチドは、食物に配合するとき化学的に合成された防腐剤よりも重要な潜在的安全性利点を提供するが、多くのペプチドは生産者菌株の病原特性のため適切でない。コリシン(Konisky, 1982)、エピデルミン及びPep 5 (Ersfeld-Dressen 等, 1984; Horner等, 1989)のようなペプチドは、クリームや軟膏で局所適用に有用であるが、生産者菌株の特質のために食物に使用することが承認されそうにない(Roller, 1991)。
【0007】
本発明は、C. perfringensに対して抗菌活性を示すBacillus subtilis PB6の細胞又は発酵抽出物に関する。我々の実験室における研究は、Bacillus subtilis PB6の発酵抽出物は高熱、酸性条件、高濃度の胆汁塩、及び溶媒中での抽出可能性の条件下で安定なタンパク質特質の抗菌因子を含むことを示した。Bacillus sppによるバクテリオシンの産生が報告され、最も特性化されたバクテリオシンは、B. subtilisのサブチリン(Jansen及びHirschmann, 1944)、B megateriumのメガシン(Von Tersch及びCarlton, 1983)、B. licheniformisのリケニン(Pattnaik等, 2001), B. thuringiensisトチシン(Paik等, 1997),並びにB. cereus の数バクテリオシン(Naclerio等, 1993; Paik等, 2000)である。病原微生物の広範囲のスペクトルに対するこれらのバクテリオシンの積極的なスクリーニングにかかわらず、C. perfringensにおけるBacillus spp.の細胞又は代謝の影響を決定する研究はなされていない。我々の研究は、Bacillus subtilis PB6の発酵抽出物がC. difficile、Campylobacter jejuni、Campylobacter coli及びStreptococcus pneunoniaeに対する抑制的であることも確認した。
【0008】
発明の概要
本発明は、家禽の胃腸管から単離された新規な細菌株から構成され、Clostridium
sppを抑制するためにそれを使用する。具体的には、Bacillus subtilis PB6が、Clostridium perfringensにおける優れた抑制作用を示し、家禽の壊疽性腸炎をもたらすクロスト−リジウムエンテロトキシンの産生を制限する。本発明は、家禽に供給される汚染動物用飼料中のC. perfringensの絶滅を(これに限定されない)含む治療法を提供する。本発明は、Bacillus subtilis PB6の耐熱性菌株並びに動物用飼料のペレット化プロセス中の高温に耐えることのできる熱安定性抗クロストリジウム因子を提供することにも関する。Bacillus subtilis PB6及びそれらの抗クロストリジウム因子の細胞は、熱処理後、生存能力と抗微生物活性を十分に保持する。加えて、本発明は、家禽の胃腸管の異なるpHに対して安定であるBacillus subtilis PB6の菌株並びに抗クロストリジウム因子を提供する。本発明は、感染した家禽の下部腸管中へのBacillus subtilis PB6並びに抗クロストリジウム因子の通路を確実にし、それにより、C. perfringensをそれぞれ置換及び/又は分離並びに抑制できる。
【0009】
本発明は、Clostridium difficile, Campylobacter jejuni, Campylobacter coli, 及びStreptococcus pneumoniaeの絶滅(これらに限定されないが)を含むヒト病原の絶滅におけるBacillus subtilis PB6及びその抗クロストリジウム因子の適用を拡張することにも関する。
【0010】
図面の簡単な説明
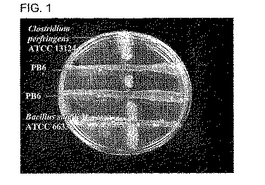
図1は、アンタゴニスト分析を示すペトリ皿の写真である、ここで、垂直縞はインジケーター生物、Clostridium perfringens ATCC 13124であり、水平縞はBacillus subtilis PB6及びBacillus subtilis ATCC 6633であり、前者は健常鶏の腸管から単離された。
【0011】
図2は、Bacillus subtilis PB6の消化された核酸の態様を示すRiboPrintの写真である。このリボタイプ化技術は、DNAフラグメントを産生する細菌から抽出されたDNAを消化するEcoR I, Pst I 及びPvu IIのような制限酵素を使用する。RRNA遺伝子のオペロンを使用する特異性プローブは、次いで、これらのDNAフラグメントを検出するために使用され、したがって、オリジナルBacillus subtilis PB6である菌株を確認する。
【0012】
好適な実施態様の詳細な記述
材料及び方法
鶏の腸管から単離した想定Bacillus sppの確認
健常な鶏の腸管をシンガポールのAgri-food & Veterinary Authority(AVA)により認証された地方マーケットから入手した。得られた新鮮な腸管を4片、すなわち、十二指腸、空腸、回腸及び盲腸に切り分けた。各片からの内容物を試験管中に集めた。これらの試験管は、0.6%イースト抽出物(Oxoid Ltd, UK) (TSBYE) (1 リットルTBSYE は30gのTryptic soyブロス粉末及び6 gのイースト抽出物を含有し、121℃のオートクレーブで20分間殺菌)を含む無菌Tryptic Soy Broth (Becton, Dickinson & Co, MD)を含有し、前記内容物を入れた試験管を80℃で20分間加熱した。加熱処理後、各試験管からの部分をTSBYE (TSAYEと称される)を含有する寒天上にストリークし(1リットルのTSAYEは30 g Tryptic soyブロス粉末、6 g イースト抽出物、及び10 g寒天を含み、121℃のオートクレーブで20分間殺菌)、37℃で18〜22時間インキュベートした。4つの四半部からのランダムコロニーを10mlの無菌TSBYE中でインキュベートし、37℃で18〜22時間インキュベートした。
【0013】
胞子形成体について選択するために、全ての培養物を100℃で30分間加熱処理に付し、成長細胞を除去した。次いで、培養物をTSAYE上にストリークし、37℃で18〜22時間インキュベートした。
【0014】
TSAYE寒天プレートからの代表的なコロニーをグラム染色し、顕微鏡的に調べ、鶏の腸管から単離した微生物の形態を決定した。さらに、TSAYE寒天プレートからの代表的なコロニーを5% Malachite Green溶液を使用して染色し、胞子形成体について顕微鏡的に調べた。
【0015】
腸管から単離されたすべての細菌を想定上同定するために生化学試験を行った。生化学試験キットであるAPI 50 CH (bio Merieux)を使用して、49種の炭水化物の発酵形態に基づく種々の細菌を同定した。鶏腸管から単離されたすべての細菌の発酵形態を確立した後、API 50 CHB/Lを使用して細菌の残りからBacillus subtilis PB6を同定した。炭水化物発酵形態を、デーベースに対してAPILAB Plusソフトウエアを使用してコンピューター計算をし、ここで、等しいか99%以上が同定率がよく一致すると認定される。同定率が80未満の場合一致するとされない。
【0016】
アンタゴニスト分析
TSBYEブロス中、37℃でインキュベートしてBacillus subtilis PB6の想定菌株を増殖させた。Clostridium perfringens ATCC 13124をインジケーター微生物として使用し、鶏の腸管から単離した微生物に対してスクリーニングした。C. perfringensの単離したコロニーをAnaerogen Pak (Oxoid)を使用する嫌気性条件で37℃でチオグリコレートブロス中に接種した。無菌布交換を使用してTSAYE寒天表面上にC. perfringensの一夜培養物をストリーク(垂直)した。次いで、Bacillus subtilis PB6の想定菌株を含む一夜培養物をC. perfringensのストリーク線を2分する同一の寒天プレートを交差させてストリークした。すべての接種したプレートを5%二酸化炭素条件下37℃でインキュベートした。24時間インキュベート後、インジケーター細菌に対する試験菌のアンタゴニスト作用を、他の菌に対する一微生物の抑制作用を示すストリーク線の交差点を囲む綺麗な領域の出現により観察できる。
【0017】
Bacillus subtilis PB6の細胞、胞子及び発酵抽出物の熱処理
Bacillus subtilis PB6の想定細胞又は胞子を、100rpmに設定した振とう式インキュベーター中、37℃で18時間一夜増殖をさせた。1ml容量の一夜培養物を試験管に入れた9mlの緩衝化ペプトン水(buffered peptone water: BPW)中に加え、90℃及び100℃に2、5及び10分各々加熱処理を行った。同様に、Bacillus subtilis PB6の発酵抽出物又は濾過物を70℃、80℃、90℃、100℃及び121℃で15分間加熱した。完全でしかも均一な加熱を確実にするために、水浴中の水位を、加熱用溶媒の位置よりも上に保った。加熱後、90℃又は100℃の異なる加熱時間の各試験管を直ちに氷水浴中に入れ、細胞のそれ以上の破壊を防止した。
【0018】
Bacillus subtilis PB6の胞子におけるpH2の影響
pH2の酸性溶液を200mlの脱イオン水中に0.2mlの10MHClを加えることにより調製した。胞子懸濁液を、無菌脱イオン水(pH6)中に1000倍に稀釈し、80℃で20分間予備加熱した。次いで、胞子懸濁液を酸性溶液(pH2)中でインキュベートし、40℃90分間インキュベートした。生存細胞数を0、30、60、90−分間隔で決定した。
【0019】
Bacillus subtilis PB6の胞子におけるpH6及び0.75%胆汁の影響
0.75%胆汁濃度を酸性溶液(pH2)中に加え、NaOH(12M)を使用して最終pHを6.0に調節した。砂嚢及び小腸のペレット化条件及びpH条件をシミュレートするために、予備加熱処理した(80℃、20分間)胞子懸濁物をHCl溶液(pH 2)中に加え、40℃で90分間インキュベートした。インキュベート90分後、pH2溶液の内容物を0.75%胆汁溶液(pH6)を含有する別のフラスコに移し、次いで、40℃でさらに90分間インキュベートした。最終的に、0.75%胆汁溶液(pH6)からのアリコットを取り、BPW中に0、30、60及び90分の時稀釈し、生存可能な細胞についてTSAYE上に塗抹標本作製(spread-plated)をした。
【0020】
生存細胞数
Bacillus subtilis PB6の生長細胞及び胞子の双方を緩衝化ペプトン水(BPW)中で稀釈し、1リットル当たり6gのイースト抽出物で補足したTryptic Soy Agar(TSAYE)上で平板培養し、多くの生存細胞を確認した。細胞懸濁物を80℃で20分間加熱処理をし、無菌的に緩衝化ペプトン水(BPW)中に稀釈してから、生存細胞の計数のためにTSAYE上に塗抹標本を作製した。同様に、非加熱の胞子懸濁物も稀釈し、生存細胞の計数のためにTSAYE上に塗抹標本を作製した。すべての培地平板を30℃で18時間インキュベートした。熱不活性化検討に関して、生存細胞総数を加熱時間に関してプロットし、D−値(分)をこれらのグラフの勾配-1から誘導した。D−値は、特定の温度(℃)で1-logまで細菌集団を減少させるのに必要な分で計った時間として定義される。
【0021】
Well Diffusion Assay
修正寒天−well diffusion法(Tagg 及びMcGiven 1971)を使用して、発酵抽出物からの濾過物又はBacillus subtilis PB6の濾過物の抗クロストリジウム活性を評価した。C. perfringens ATCC 13124 又はC. difficileの一夜培養物を日常的抗クロストリジウム活性分析のための指標菌株として使用した。チオグリコレート寒天を45℃まで調節してから、それに指標菌株の一夜培養物を接種した。次いで、この混合物の20ml容量を各無菌ペトリ皿上に注ぎ、室温で1時間かけて凝固させた。平板計数用寒天中にウエル(直径0.75cmx深さ1.0cm)を穴空け装置により無菌的に作製した。次いで、発酵抽出物又はBacillus subtilis PB6の濾過物を含有する容量100μlの試料を各ウエル中に入れた。同様にして、Streptococcus pneumoniae、Campylobacter jejuni、Campylobacter coli、及びHelicobacter pyloriの一夜培養物を適当な寒天培地中に混合し、Bacillus subtilis PB6の細胞又は発酵ブロスの作用を試験するために指標菌として使用した。すべての培養用平板を37℃で18時間インキュベートした。
【0022】
抗微生物化合物の特性
抗微生物化合物の産生のため、TSBYE中でBacillus subtilis PB6を好気的に100rpmで振とうしながら37℃で18時間かけて増殖させた。0.22μm(Sartorius)濾過ディスクを使用して培養物から細菌細胞を取り除いた。濾過物の幾らかを集め、4℃で一夜貯蔵し、濾過物上の冷蔵温度の作用を観察した。集めた濾過物を、プロナーゼ、37℃で1時間について(最終濃度が1mg/mlの)ペプシン、37℃で12時間について(最終濃度が1mg/mlの)トリプシン、及び(最終濃度が0.5mg/mlの)カタラーゼで処理した。
【0023】
動物試行設計(Animal trial design)
401日齢の健常ブロイラー(Wuxi Broiler Breeding Group Co. Ltd.)を、対照を含めてランダムに2処置に分けた。各処置は、開始期間(0〜21日)の間、1反復当たり雌雄混合した40羽を用いて5反復から構成された。21日目に12雄鳥及び12雌鳥を各反復からランダムに選択し、残りの試行期間の間分離し、最終期間(22〜42日)の間、1処置当たり10反復(1反復当たり12羽)だった。
【0024】
鳥の管理
食餌は、開始食餌と最終食餌それぞれについて20%及び18%の粗タンパク質含量を調合した。処置では、抗微生物化合物を含むPB6濾過物の3 kg/T 食事を食餌中に含ませた。両食餌の算出代謝可能エネルギーは、おおよそ2860kcal/kgだった。昼間は自然光にし、夜間は薄暗い光を与え、継続的な食事を確保した。水は任意に与えた。
【0025】
統計分析
SPSS及びDuncan's Multiple Rangeを使用して、処置間の変動及び統計的差の分析を行った。
【0026】
結 果
推定Bacillus subtilis PB6の同定
API生化学試験を使用して、PB6をBacillus spp. (Bacillus megaterium)として92% IDで同定した(表1)。PB6の発酵態様をAPIのLABデータベースと比較した。ここで、菌株の同一性を同一性の百分率(%ID)として表現した。これは、データベース中のすべての他の分類群と比較した分類に相当する態様の接近度の計算を基にする(表1)。さらに、ribotyping techniqueを使用して PB6をBacillus subtilisとして確認した(図2)。
【0027】
【表1】

【0028】
次いで、Bacillus subtilis PB6を、ribotyping techniqueを使用して確認した(図2)。
Clostridium perfringensに対するアンタゴニストスクリーニング
Bacillus subtilis PB6の細胞及び発酵抽出物はC. perfringens、C. difficile、 Campylobacter jejuni、Campylobacter coli、及びStreptococcus pneumoniaeに対して抗微生物活性を示した(表2)。
【0029】
表3はWell Diffusion Assayからのデータを示す。ここでは、Bacillus subtilis PB6の培養物を24時間記載した温度で増殖させ、それから、濾過物を集め、指標微生物としてClostridium perfringens ATCC 13124を播種した寒天のウエル中に入れた。
【0030】
【表2】
【0031】
【表3】
【0032】
Bacillus subtilis PB6の細胞、胞子及び発酵抽出物の熱処理
Bacillus subtilis PB6の生長細胞を90℃で2〜10分間加熱処理したとき、生存細胞数のおおよそ5〜6log減少が観察された(データは示さず)。試験した他のBacillus菌株と比較すると、Bacillus subtilis PB6の生長細胞のD−値は0.44分であり、最も高い熱抵抗性を示した(表4)。Bacillus subtilis PB6生長細胞の100℃におけるD−値は0.41分だった(表4)。Bacillus subtilis PB6の胞子についての90℃及び100℃におけるD−値はそれぞれ24及び1.07分だった(表4)。
【0033】
【表4】
【0034】
Bacillus subtilis PB6の抗微生物化合物の安定性
Bacillus subtilis PB6の発酵抽出物内の抗クロストリジウム因子が、70℃、80℃、90℃、100℃及び121℃で15分間並びに4℃で一夜貯蔵後も依然として活性である(表5)。
【0035】
【表5】
【0036】
Bacillus subtilis PB6の発酵抽出物内の抗クロストリジウム因子は、トリプシンで処理後も依然として活性がある(表6)。未処理濾過物を含むウエルは4℃又は25℃のいずれかで貯蔵した。後者は対照として使用した。
【0037】
【表6】
【0038】
抗微生物化合物は、カタラーゼ及びプロナーゼのような一定範囲のプロテアーゼ、並びにペプシンを用いる処理に対して安定だった(表7)。TSBYE (pH 7.0)、pH6.3のTSBYEを含有するウエルと、未処理濾過物及び過酸化水素は対照として使用した。
【0039】
【表7】
【0040】
Bacillus subtilis PB6の胞子におけるpH及び胆汁の影響
Bacillus subtilis PB6の予備加熱した胞子(80℃、20分)を酸性化した溶液(pH2)中に入れ、40℃で90分間インキュベートすると、無視できるほどの減少または主要な減少でない生存細胞が観察された(表8)。
【0041】
【表8】
【0042】
酸性溶液(pH2)中で90分インキュベート後、Bacillus胞子発芽の百分率がなお98%だった(表8)。Bacillus subtilis PB6の胞子は、pH2で処理し、次いで0.75%の胆汁を含有するpH6の溶液に加え、90分間インキュベートすると、生存ししかも発芽する(28%)ことが見いだされた(表9)。
【0043】
【表9】
【0044】
ブロイラーのFCRにおけるPB6濾過物の影響
ブロイラーの飼料換算率(feed conversion ration:FCR)における抗微生物化合物を含有する3 kg/T PB6濾過物の添加の影響を検討した。結果(表10)は、抗微生物化合物が、PB6抗微生物化合物を添加していない対照と比較して、開始期間及び最終期間の双方で鳥のFCRを改良したことを示す。
【0045】
【表10】
【0046】
前述は本発明の例証的実施態様を含む。本明細書中に記載した前述の実施態様と方法は、当業者の能力、経験、及び好適に基づき変動できる。一定の順序で方法の工程を単に列挙することは、必然的に方法の工程の順序にいかなる制限も構成しない。前述及び図面は本発明を単に説明し且つ例証するものであり、特許請求の範囲が制限することを除いて、制限しない。開示を受けた当業者は、発明の範囲から逸脱しないで修正及び変動をなすことができる。
【図面の簡単な説明】
【0047】
【図1】図1は、アンタゴニスト分析を示すペトリ皿の写真である、ここで、垂直縞はインジケーター生物、Clostridium perfringens ATCC 13124であり、水平縞はBacillus subtilis PB6及びBacillus subtilis ATCC 6633であり、前者は健常鶏の腸管から単離された。
【図2】図2は、Bacillus subtilis PB6の消化された核酸の態様を示すRiboPrintの写真である。このリボタイプ化技術は、DNAフラグメントを産生する細菌から抽出されたDNAを消化するEcoR I, Pst I 及びPvu IIのような制限酵素を使用する。RRNA遺伝子のオペロンを使用する特異性プローブは、次いで、これらのDNAフラグメントを検出するために使用され、したがって、オリジナルBacillus subtilis PB6である菌株を確認する。
【発明の詳細な説明】
【0001】
発明の背景
本発明は一般的に抗菌化合物に関し、さらに具体的には、動物及びヒト病原体に対して使用するためのBacillus subtilis PB6由来の抗菌化合物に関する。
【0002】
Clostridium perfringensによりもたらされる壊疽性腸炎、腸性毒血症は、家禽の死滅をもたらす腸壁の壊疽性損傷の進展に導く(Paulus and Ruckebusch, 1996; Tsai and Tung, 1981)。それは複雑で部分的に知られていない疫学及び病因の多因性疾患でもある(Kaldhusdal, 2000)。細菌であるC. perfringensは家禽の胃腸管中で見いだされる(Tshirdewahn等, 1991)が、しかし壊疽性腸炎の発生は偶発的である(Cowen等, 1987)。それにもかかわらず、C. perfringensで汚染された食品は、鶏の壊疽性腸炎の発生に影響を与えてきた(Kaldhusdal, 2000)。研究は、健常な鶏は胃腸管中には比較的に少ない数のC. perfringensしか有しないが、一方、細菌の濃度の増加が壊疽性腸炎症状をもたらすことがある(Craven等, 1999)。
【0003】
壊疽性腸炎を罹患している家禽を処置するのにバシトラシン、リノコマイシン及びその他の成長促進抗生物質が一般に使用される(Craven 等, 1999)。しかし、鶏又は七面鳥由来のC. perfringensの抗生物質−抵抗性株の単離が原因で(Devriese等, 1993; Kondo, 1988; Watkins等, 1997)、家禽健康当局及び生産者は、抗生物質を置き換えるプロバイオティック製品の開発と応用に興味が増加している。プロバイオティクスは、宿主の胃腸の細菌のバランスを改良することにより宿主に好影響を与える生存細菌供給サプルメントとして定義される。何人かの研究者は、腸内細菌のこの標準化は次の利点を与えると信じている:(a)競合的除外(定着部位の拮抗とも称される)による病原体に対する保護;(b)一定の栄養素及び酵素的/脱毒素化反応の提供;(c)組織形態形成及び蠕動活性の関連性;並びに(d)宿主の免疫系及び内分泌系の相互作用である。さらに、家禽の壊疽性腸炎の上昇及び多くの国による種々の飼料用抗生物質の漸進的禁止に照らして(Council of the European Communities, 1998)、動物の能力を増強させるために代替成長促進剤(alternative growth promoters: AGP)へ向けてのシフトがある。イーストや乳酸菌のような有用な微生物は、過去20年間動物生産に使用されてきた。プロバイオティク特性を持つ細菌は、腸管中の病原菌と置き換わる(replacing or displacing)ことにより健康な腸ミクロフロラを維持する主要な機能を有する。プロバイオティックスは、動物腸内の病原菌に対して競合的除外(Tournot, 1989)及び拮抗作用(Fuller, 1989)により「正常な」腸内ミクロフロラを維持できることが知られている。概して、プロバイオティク微生物は腸内で大量に増殖でき、したがって、病原菌の繁殖を抑制する。ヒト又は動物に使用できるプロバイオティク微生物には、Bacillus spp., Lactobacillus spp., Enterococcus spp.及びStreptococcus spp等がある(Lee 等,1999)。胃腸管の異なる部位でコロニーを形成できる種々の微生物が報告されている(Jin 等, 1997)。概して、微生物の濃度の最も高い盲腸に比較して、胃腸管の十二指腸部が最も低い細菌ミクロフロラの集団を示す(Mead 及びAdams, 1975; Salanitro等, 1974)。Lactobacillus spp., Streptococcus spp.及びEscherichia coliのような微生物が、十二指腸部、小腸中の空腸及び回腸部にから単離された(Shapiro及びSarles, 1949)。盲腸の一般的な微生物集団は、Streptococcus spp., Staphylococcus spp., Lactobacillus spp., Fusobacterium spp.及びBacteroidesのような絶対嫌気性生物を含む(Barnes等, 1972; Mead, 1997)。
【0004】
鶏腸管の種々のセグメント由来の有用細菌が単離され、C. perfringensに対して可能性のあるプロバイオティックスとしてスクリーニングされた。以前の研究によれば、胃腸管の上皮細胞を接着し、コロニー化するプロバイオティックス微生物の能力は特定の動物源からの特定部位の単離物に大きく依存することを示した(Barrow 等, 1980; Reid, 1999; Fuller, 1973; Wesney 及びTannock, 1979)。動物の飼料に微生物プロバイオティックスの使用に関連する多くの有利な作用がある。これらの有利な作用には、病原菌E. coli (Watkins et al., 1982), Campylobacter jejuni (Morishita等, 1997) 及びSalmonella enteritidis (Pascual等, 1999)の競合的除外等があり、有用な腸ミクロフロラの成長と生存力を増強し(Hosoi等, 2000)、鶏に栄養素の消化と吸収の改善をもたらす(Ratcliff, 2000; Scheinbach, 1998; Sissons, 1989; Thomke及びElwinger, 1998)。
【0005】
プロバイオティク細菌を単離し特徴付けるために使用されるその他の基準には、胆汁及び酸安定性(Hoa 等, 2000; Huis In't Veld及びShortt, 1996)、抗生物質の産生(Salminen等, 1996)、並びに安全性に合致し又は一般に安全と認められる(generally recognized as safe :GRAS)状態(Donohue 及びSalminen, 1996; SCAN, 2000)等がある。多数の乳酸菌(単一若しくは組合せ)が、病原微生物に向けて種々の程度の抗微生物活性を呈することが示された(Harris等, 1989; Motlagh等, 1991)。加えて、乳酸菌の成長可能な培養物又は発酵抽出物が、多くの腸の疾病の特性である、置換された内因性腸ミクロフロラの処置に有用である(Charteris等, 1997; Drake 等, 1996)。このような細菌は、上皮組織に接着することにより、酸や胆汁条件で生き残ることができ、腸管に少なくとも一時的にコロニーを形成する。それらは、豚、鶏及び子牛の成長速度を改良し、飼料に有用であることが報告された(Hale及びNewton, 1979; Tortuero, 1973; Schwab等, 1980)。さらに、これらの有用な細菌を含有する飼料を与えた豚及び子牛において下痢の発生に顕著な減少が観察された(Lee, 1999)。乳酸菌培養物も、豚のE. coliからの腸毒素の作用を中和するものと思われる(Mitchell及びKenworthy, 1976)。その他の乳酸菌の有用な作用にはC. perfringensを含む有害細菌の置換、細菌性尿素分解酵素活性の産生、ビタミンの合成、免疫系における刺激作用、及び消化に対する貢献等がある(Hofacre等, 1998)。以前の研究により、Lactobacillus rhamnosus (Alander等, 1999; Asensio 等, 1976; Silva 等, 1987), L. plantarum (Andersson, 1986; West及びWarner, 1988), Lactococcus lactis ssp. lactis (Motlagh等, 1991; Spelhaug及びHarlander, 1989), 並びにPediococcus pentosaceus (Fleming 等, 1975; Graham及びMcKay, 1985)がClostridium sppに対して殺細菌性であったことを示す。
【0006】
顕著な抗菌活性を持つ微生物ペプチドは、一般的に、動物、植物、微生物(Sahl, 1985)及び非−無菌食物(Muriana, 1993)から単離される。それらは、小さく、カチオン性であり、3000〜6000ダルトンの分子質量を有する(Roller, 1991)。前駆体ペプチドの翻訳後修飾(post-translational modification)は、分子内チオエーテル橋を導入し、Pep 5、ナイシン、及びサブチリンのようなカチオン性ペプチドにすることが示されている(Gross及びMorell, 1971; Kordel 及びStahl 1986; Kordel等, 1989)。これらのペプチドは、食物に配合するとき化学的に合成された防腐剤よりも重要な潜在的安全性利点を提供するが、多くのペプチドは生産者菌株の病原特性のため適切でない。コリシン(Konisky, 1982)、エピデルミン及びPep 5 (Ersfeld-Dressen 等, 1984; Horner等, 1989)のようなペプチドは、クリームや軟膏で局所適用に有用であるが、生産者菌株の特質のために食物に使用することが承認されそうにない(Roller, 1991)。
【0007】
本発明は、C. perfringensに対して抗菌活性を示すBacillus subtilis PB6の細胞又は発酵抽出物に関する。我々の実験室における研究は、Bacillus subtilis PB6の発酵抽出物は高熱、酸性条件、高濃度の胆汁塩、及び溶媒中での抽出可能性の条件下で安定なタンパク質特質の抗菌因子を含むことを示した。Bacillus sppによるバクテリオシンの産生が報告され、最も特性化されたバクテリオシンは、B. subtilisのサブチリン(Jansen及びHirschmann, 1944)、B megateriumのメガシン(Von Tersch及びCarlton, 1983)、B. licheniformisのリケニン(Pattnaik等, 2001), B. thuringiensisトチシン(Paik等, 1997),並びにB. cereus の数バクテリオシン(Naclerio等, 1993; Paik等, 2000)である。病原微生物の広範囲のスペクトルに対するこれらのバクテリオシンの積極的なスクリーニングにかかわらず、C. perfringensにおけるBacillus spp.の細胞又は代謝の影響を決定する研究はなされていない。我々の研究は、Bacillus subtilis PB6の発酵抽出物がC. difficile、Campylobacter jejuni、Campylobacter coli及びStreptococcus pneunoniaeに対する抑制的であることも確認した。
【0008】
発明の概要
本発明は、家禽の胃腸管から単離された新規な細菌株から構成され、Clostridium
sppを抑制するためにそれを使用する。具体的には、Bacillus subtilis PB6が、Clostridium perfringensにおける優れた抑制作用を示し、家禽の壊疽性腸炎をもたらすクロスト−リジウムエンテロトキシンの産生を制限する。本発明は、家禽に供給される汚染動物用飼料中のC. perfringensの絶滅を(これに限定されない)含む治療法を提供する。本発明は、Bacillus subtilis PB6の耐熱性菌株並びに動物用飼料のペレット化プロセス中の高温に耐えることのできる熱安定性抗クロストリジウム因子を提供することにも関する。Bacillus subtilis PB6及びそれらの抗クロストリジウム因子の細胞は、熱処理後、生存能力と抗微生物活性を十分に保持する。加えて、本発明は、家禽の胃腸管の異なるpHに対して安定であるBacillus subtilis PB6の菌株並びに抗クロストリジウム因子を提供する。本発明は、感染した家禽の下部腸管中へのBacillus subtilis PB6並びに抗クロストリジウム因子の通路を確実にし、それにより、C. perfringensをそれぞれ置換及び/又は分離並びに抑制できる。
【0009】
本発明は、Clostridium difficile, Campylobacter jejuni, Campylobacter coli, 及びStreptococcus pneumoniaeの絶滅(これらに限定されないが)を含むヒト病原の絶滅におけるBacillus subtilis PB6及びその抗クロストリジウム因子の適用を拡張することにも関する。
【0010】
図面の簡単な説明
図1は、アンタゴニスト分析を示すペトリ皿の写真である、ここで、垂直縞はインジケーター生物、Clostridium perfringens ATCC 13124であり、水平縞はBacillus subtilis PB6及びBacillus subtilis ATCC 6633であり、前者は健常鶏の腸管から単離された。
【0011】
図2は、Bacillus subtilis PB6の消化された核酸の態様を示すRiboPrintの写真である。このリボタイプ化技術は、DNAフラグメントを産生する細菌から抽出されたDNAを消化するEcoR I, Pst I 及びPvu IIのような制限酵素を使用する。RRNA遺伝子のオペロンを使用する特異性プローブは、次いで、これらのDNAフラグメントを検出するために使用され、したがって、オリジナルBacillus subtilis PB6である菌株を確認する。
【0012】
好適な実施態様の詳細な記述
材料及び方法
鶏の腸管から単離した想定Bacillus sppの確認
健常な鶏の腸管をシンガポールのAgri-food & Veterinary Authority(AVA)により認証された地方マーケットから入手した。得られた新鮮な腸管を4片、すなわち、十二指腸、空腸、回腸及び盲腸に切り分けた。各片からの内容物を試験管中に集めた。これらの試験管は、0.6%イースト抽出物(Oxoid Ltd, UK) (TSBYE) (1 リットルTBSYE は30gのTryptic soyブロス粉末及び6 gのイースト抽出物を含有し、121℃のオートクレーブで20分間殺菌)を含む無菌Tryptic Soy Broth (Becton, Dickinson & Co, MD)を含有し、前記内容物を入れた試験管を80℃で20分間加熱した。加熱処理後、各試験管からの部分をTSBYE (TSAYEと称される)を含有する寒天上にストリークし(1リットルのTSAYEは30 g Tryptic soyブロス粉末、6 g イースト抽出物、及び10 g寒天を含み、121℃のオートクレーブで20分間殺菌)、37℃で18〜22時間インキュベートした。4つの四半部からのランダムコロニーを10mlの無菌TSBYE中でインキュベートし、37℃で18〜22時間インキュベートした。
【0013】
胞子形成体について選択するために、全ての培養物を100℃で30分間加熱処理に付し、成長細胞を除去した。次いで、培養物をTSAYE上にストリークし、37℃で18〜22時間インキュベートした。
【0014】
TSAYE寒天プレートからの代表的なコロニーをグラム染色し、顕微鏡的に調べ、鶏の腸管から単離した微生物の形態を決定した。さらに、TSAYE寒天プレートからの代表的なコロニーを5% Malachite Green溶液を使用して染色し、胞子形成体について顕微鏡的に調べた。
【0015】
腸管から単離されたすべての細菌を想定上同定するために生化学試験を行った。生化学試験キットであるAPI 50 CH (bio Merieux)を使用して、49種の炭水化物の発酵形態に基づく種々の細菌を同定した。鶏腸管から単離されたすべての細菌の発酵形態を確立した後、API 50 CHB/Lを使用して細菌の残りからBacillus subtilis PB6を同定した。炭水化物発酵形態を、デーベースに対してAPILAB Plusソフトウエアを使用してコンピューター計算をし、ここで、等しいか99%以上が同定率がよく一致すると認定される。同定率が80未満の場合一致するとされない。
【0016】
アンタゴニスト分析
TSBYEブロス中、37℃でインキュベートしてBacillus subtilis PB6の想定菌株を増殖させた。Clostridium perfringens ATCC 13124をインジケーター微生物として使用し、鶏の腸管から単離した微生物に対してスクリーニングした。C. perfringensの単離したコロニーをAnaerogen Pak (Oxoid)を使用する嫌気性条件で37℃でチオグリコレートブロス中に接種した。無菌布交換を使用してTSAYE寒天表面上にC. perfringensの一夜培養物をストリーク(垂直)した。次いで、Bacillus subtilis PB6の想定菌株を含む一夜培養物をC. perfringensのストリーク線を2分する同一の寒天プレートを交差させてストリークした。すべての接種したプレートを5%二酸化炭素条件下37℃でインキュベートした。24時間インキュベート後、インジケーター細菌に対する試験菌のアンタゴニスト作用を、他の菌に対する一微生物の抑制作用を示すストリーク線の交差点を囲む綺麗な領域の出現により観察できる。
【0017】
Bacillus subtilis PB6の細胞、胞子及び発酵抽出物の熱処理
Bacillus subtilis PB6の想定細胞又は胞子を、100rpmに設定した振とう式インキュベーター中、37℃で18時間一夜増殖をさせた。1ml容量の一夜培養物を試験管に入れた9mlの緩衝化ペプトン水(buffered peptone water: BPW)中に加え、90℃及び100℃に2、5及び10分各々加熱処理を行った。同様に、Bacillus subtilis PB6の発酵抽出物又は濾過物を70℃、80℃、90℃、100℃及び121℃で15分間加熱した。完全でしかも均一な加熱を確実にするために、水浴中の水位を、加熱用溶媒の位置よりも上に保った。加熱後、90℃又は100℃の異なる加熱時間の各試験管を直ちに氷水浴中に入れ、細胞のそれ以上の破壊を防止した。
【0018】
Bacillus subtilis PB6の胞子におけるpH2の影響
pH2の酸性溶液を200mlの脱イオン水中に0.2mlの10MHClを加えることにより調製した。胞子懸濁液を、無菌脱イオン水(pH6)中に1000倍に稀釈し、80℃で20分間予備加熱した。次いで、胞子懸濁液を酸性溶液(pH2)中でインキュベートし、40℃90分間インキュベートした。生存細胞数を0、30、60、90−分間隔で決定した。
【0019】
Bacillus subtilis PB6の胞子におけるpH6及び0.75%胆汁の影響
0.75%胆汁濃度を酸性溶液(pH2)中に加え、NaOH(12M)を使用して最終pHを6.0に調節した。砂嚢及び小腸のペレット化条件及びpH条件をシミュレートするために、予備加熱処理した(80℃、20分間)胞子懸濁物をHCl溶液(pH 2)中に加え、40℃で90分間インキュベートした。インキュベート90分後、pH2溶液の内容物を0.75%胆汁溶液(pH6)を含有する別のフラスコに移し、次いで、40℃でさらに90分間インキュベートした。最終的に、0.75%胆汁溶液(pH6)からのアリコットを取り、BPW中に0、30、60及び90分の時稀釈し、生存可能な細胞についてTSAYE上に塗抹標本作製(spread-plated)をした。
【0020】
生存細胞数
Bacillus subtilis PB6の生長細胞及び胞子の双方を緩衝化ペプトン水(BPW)中で稀釈し、1リットル当たり6gのイースト抽出物で補足したTryptic Soy Agar(TSAYE)上で平板培養し、多くの生存細胞を確認した。細胞懸濁物を80℃で20分間加熱処理をし、無菌的に緩衝化ペプトン水(BPW)中に稀釈してから、生存細胞の計数のためにTSAYE上に塗抹標本を作製した。同様に、非加熱の胞子懸濁物も稀釈し、生存細胞の計数のためにTSAYE上に塗抹標本を作製した。すべての培地平板を30℃で18時間インキュベートした。熱不活性化検討に関して、生存細胞総数を加熱時間に関してプロットし、D−値(分)をこれらのグラフの勾配-1から誘導した。D−値は、特定の温度(℃)で1-logまで細菌集団を減少させるのに必要な分で計った時間として定義される。
【0021】
Well Diffusion Assay
修正寒天−well diffusion法(Tagg 及びMcGiven 1971)を使用して、発酵抽出物からの濾過物又はBacillus subtilis PB6の濾過物の抗クロストリジウム活性を評価した。C. perfringens ATCC 13124 又はC. difficileの一夜培養物を日常的抗クロストリジウム活性分析のための指標菌株として使用した。チオグリコレート寒天を45℃まで調節してから、それに指標菌株の一夜培養物を接種した。次いで、この混合物の20ml容量を各無菌ペトリ皿上に注ぎ、室温で1時間かけて凝固させた。平板計数用寒天中にウエル(直径0.75cmx深さ1.0cm)を穴空け装置により無菌的に作製した。次いで、発酵抽出物又はBacillus subtilis PB6の濾過物を含有する容量100μlの試料を各ウエル中に入れた。同様にして、Streptococcus pneumoniae、Campylobacter jejuni、Campylobacter coli、及びHelicobacter pyloriの一夜培養物を適当な寒天培地中に混合し、Bacillus subtilis PB6の細胞又は発酵ブロスの作用を試験するために指標菌として使用した。すべての培養用平板を37℃で18時間インキュベートした。
【0022】
抗微生物化合物の特性
抗微生物化合物の産生のため、TSBYE中でBacillus subtilis PB6を好気的に100rpmで振とうしながら37℃で18時間かけて増殖させた。0.22μm(Sartorius)濾過ディスクを使用して培養物から細菌細胞を取り除いた。濾過物の幾らかを集め、4℃で一夜貯蔵し、濾過物上の冷蔵温度の作用を観察した。集めた濾過物を、プロナーゼ、37℃で1時間について(最終濃度が1mg/mlの)ペプシン、37℃で12時間について(最終濃度が1mg/mlの)トリプシン、及び(最終濃度が0.5mg/mlの)カタラーゼで処理した。
【0023】
動物試行設計(Animal trial design)
401日齢の健常ブロイラー(Wuxi Broiler Breeding Group Co. Ltd.)を、対照を含めてランダムに2処置に分けた。各処置は、開始期間(0〜21日)の間、1反復当たり雌雄混合した40羽を用いて5反復から構成された。21日目に12雄鳥及び12雌鳥を各反復からランダムに選択し、残りの試行期間の間分離し、最終期間(22〜42日)の間、1処置当たり10反復(1反復当たり12羽)だった。
【0024】
鳥の管理
食餌は、開始食餌と最終食餌それぞれについて20%及び18%の粗タンパク質含量を調合した。処置では、抗微生物化合物を含むPB6濾過物の3 kg/T 食事を食餌中に含ませた。両食餌の算出代謝可能エネルギーは、おおよそ2860kcal/kgだった。昼間は自然光にし、夜間は薄暗い光を与え、継続的な食事を確保した。水は任意に与えた。
【0025】
統計分析
SPSS及びDuncan's Multiple Rangeを使用して、処置間の変動及び統計的差の分析を行った。
【0026】
結 果
推定Bacillus subtilis PB6の同定
API生化学試験を使用して、PB6をBacillus spp. (Bacillus megaterium)として92% IDで同定した(表1)。PB6の発酵態様をAPIのLABデータベースと比較した。ここで、菌株の同一性を同一性の百分率(%ID)として表現した。これは、データベース中のすべての他の分類群と比較した分類に相当する態様の接近度の計算を基にする(表1)。さらに、ribotyping techniqueを使用して PB6をBacillus subtilisとして確認した(図2)。
【0027】
【表1】
【0028】
次いで、Bacillus subtilis PB6を、ribotyping techniqueを使用して確認した(図2)。
Clostridium perfringensに対するアンタゴニストスクリーニング
Bacillus subtilis PB6の細胞及び発酵抽出物はC. perfringens、C. difficile、 Campylobacter jejuni、Campylobacter coli、及びStreptococcus pneumoniaeに対して抗微生物活性を示した(表2)。
【0029】
表3はWell Diffusion Assayからのデータを示す。ここでは、Bacillus subtilis PB6の培養物を24時間記載した温度で増殖させ、それから、濾過物を集め、指標微生物としてClostridium perfringens ATCC 13124を播種した寒天のウエル中に入れた。
【0030】
【表2】
【0031】
【表3】
【0032】
Bacillus subtilis PB6の細胞、胞子及び発酵抽出物の熱処理
Bacillus subtilis PB6の生長細胞を90℃で2〜10分間加熱処理したとき、生存細胞数のおおよそ5〜6log減少が観察された(データは示さず)。試験した他のBacillus菌株と比較すると、Bacillus subtilis PB6の生長細胞のD−値は0.44分であり、最も高い熱抵抗性を示した(表4)。Bacillus subtilis PB6生長細胞の100℃におけるD−値は0.41分だった(表4)。Bacillus subtilis PB6の胞子についての90℃及び100℃におけるD−値はそれぞれ24及び1.07分だった(表4)。
【0033】
【表4】
【0034】
Bacillus subtilis PB6の抗微生物化合物の安定性
Bacillus subtilis PB6の発酵抽出物内の抗クロストリジウム因子が、70℃、80℃、90℃、100℃及び121℃で15分間並びに4℃で一夜貯蔵後も依然として活性である(表5)。
【0035】
【表5】
【0036】
Bacillus subtilis PB6の発酵抽出物内の抗クロストリジウム因子は、トリプシンで処理後も依然として活性がある(表6)。未処理濾過物を含むウエルは4℃又は25℃のいずれかで貯蔵した。後者は対照として使用した。
【0037】
【表6】
【0038】
抗微生物化合物は、カタラーゼ及びプロナーゼのような一定範囲のプロテアーゼ、並びにペプシンを用いる処理に対して安定だった(表7)。TSBYE (pH 7.0)、pH6.3のTSBYEを含有するウエルと、未処理濾過物及び過酸化水素は対照として使用した。
【0039】
【表7】
【0040】
Bacillus subtilis PB6の胞子におけるpH及び胆汁の影響
Bacillus subtilis PB6の予備加熱した胞子(80℃、20分)を酸性化した溶液(pH2)中に入れ、40℃で90分間インキュベートすると、無視できるほどの減少または主要な減少でない生存細胞が観察された(表8)。
【0041】
【表8】
【0042】
酸性溶液(pH2)中で90分インキュベート後、Bacillus胞子発芽の百分率がなお98%だった(表8)。Bacillus subtilis PB6の胞子は、pH2で処理し、次いで0.75%の胆汁を含有するpH6の溶液に加え、90分間インキュベートすると、生存ししかも発芽する(28%)ことが見いだされた(表9)。
【0043】
【表9】
【0044】
ブロイラーのFCRにおけるPB6濾過物の影響
ブロイラーの飼料換算率(feed conversion ration:FCR)における抗微生物化合物を含有する3 kg/T PB6濾過物の添加の影響を検討した。結果(表10)は、抗微生物化合物が、PB6抗微生物化合物を添加していない対照と比較して、開始期間及び最終期間の双方で鳥のFCRを改良したことを示す。
【0045】
【表10】
【0046】
前述は本発明の例証的実施態様を含む。本明細書中に記載した前述の実施態様と方法は、当業者の能力、経験、及び好適に基づき変動できる。一定の順序で方法の工程を単に列挙することは、必然的に方法の工程の順序にいかなる制限も構成しない。前述及び図面は本発明を単に説明し且つ例証するものであり、特許請求の範囲が制限することを除いて、制限しない。開示を受けた当業者は、発明の範囲から逸脱しないで修正及び変動をなすことができる。
【図面の簡単な説明】
【0047】
【図1】図1は、アンタゴニスト分析を示すペトリ皿の写真である、ここで、垂直縞はインジケーター生物、Clostridium perfringens ATCC 13124であり、水平縞はBacillus subtilis PB6及びBacillus subtilis ATCC 6633であり、前者は健常鶏の腸管から単離された。
【図2】図2は、Bacillus subtilis PB6の消化された核酸の態様を示すRiboPrintの写真である。このリボタイプ化技術は、DNAフラグメントを産生する細菌から抽出されたDNAを消化するEcoR I, Pst I 及びPvu IIのような制限酵素を使用する。RRNA遺伝子のオペロンを使用する特異性プローブは、次いで、これらのDNAフラグメントを検出するために使用され、したがって、オリジナルBacillus subtilis PB6である菌株を確認する。
【特許請求の範囲】
【請求項1】
細菌の増殖を阻止する鶏の胃腸管から単離されたある細菌から産生された化合物。
【請求項2】
前記ある細菌がプロバイオティク微生物の構成要素である、請求項1に記載の化合物。
【請求項3】
前記化合物がBacillus subtilis PB6により産生される、請求項1に記載の化合物。
【請求項4】
前記化合物が動物及びヒトの病原菌の増殖を阻止する、請求項1に記載の化合物。
【請求項5】
前記動物病原菌が、Clostridium、Campylobacter、Streptococcus、及びHelicobacterを含む属からの種から選択されるがこれらに限定されない、請求項1に記載の化合物。
【請求項6】
前記化合物を使用して家禽中の壊疽性腸炎の症状をなくする、請求項1に記載の化合物。
【請求項7】
前記化合物を使用して家禽のFCRすなわち飼料換算率を上昇させる、請求項1に記載の化合物。
【請求項8】
前記化合物が、熱、プロテアーゼ、pH、溶媒、胆汁塩及び温度に対して安定である、請求項1に記載の化合物。
【請求項9】
細菌類Bacillus subtilis PB6が熱、温度及び胆汁塩に対して安定である、請求項1に記載の化合物。
【請求項10】
細菌類Bacillus subtilis PB6を動物及びヒトの双方に対してプロバイオティクとして使用できる、請求項1に記載の化合物。
【請求項11】
前記化合物を動物及びヒトの腸炎を治療するために使用する、請求項1に記載の化合物。
【請求項1】
細菌の増殖を阻止する鶏の胃腸管から単離されたある細菌から産生された化合物。
【請求項2】
前記ある細菌がプロバイオティク微生物の構成要素である、請求項1に記載の化合物。
【請求項3】
前記化合物がBacillus subtilis PB6により産生される、請求項1に記載の化合物。
【請求項4】
前記化合物が動物及びヒトの病原菌の増殖を阻止する、請求項1に記載の化合物。
【請求項5】
前記動物病原菌が、Clostridium、Campylobacter、Streptococcus、及びHelicobacterを含む属からの種から選択されるがこれらに限定されない、請求項1に記載の化合物。
【請求項6】
前記化合物を使用して家禽中の壊疽性腸炎の症状をなくする、請求項1に記載の化合物。
【請求項7】
前記化合物を使用して家禽のFCRすなわち飼料換算率を上昇させる、請求項1に記載の化合物。
【請求項8】
前記化合物が、熱、プロテアーゼ、pH、溶媒、胆汁塩及び温度に対して安定である、請求項1に記載の化合物。
【請求項9】
細菌類Bacillus subtilis PB6が熱、温度及び胆汁塩に対して安定である、請求項1に記載の化合物。
【請求項10】
細菌類Bacillus subtilis PB6を動物及びヒトの双方に対してプロバイオティクとして使用できる、請求項1に記載の化合物。
【請求項11】
前記化合物を動物及びヒトの腸炎を治療するために使用する、請求項1に記載の化合物。
【図1】

【図2】

【図2】
【公表番号】特表2006−514019(P2006−514019A)
【公表日】平成18年4月27日(2006.4.27)
【国際特許分類】
【出願番号】特願2004−557192(P2004−557192)
【出願日】平成15年11月12日(2003.11.12)
【国際出願番号】PCT/US2003/036094
【国際公開番号】WO2004/050832
【国際公開日】平成16年6月17日(2004.6.17)
【出願人】(500462328)ケミン、インダストリーズ、インコーポレーテッド (7)
【氏名又は名称原語表記】KEMIN INDUSTRIES, INC.
【Fターム(参考)】
【公表日】平成18年4月27日(2006.4.27)
【国際特許分類】
【出願日】平成15年11月12日(2003.11.12)
【国際出願番号】PCT/US2003/036094
【国際公開番号】WO2004/050832
【国際公開日】平成16年6月17日(2004.6.17)
【出願人】(500462328)ケミン、インダストリーズ、インコーポレーテッド (7)
【氏名又は名称原語表記】KEMIN INDUSTRIES, INC.
【Fターム(参考)】
[ Back to top ]