
化膿連鎖球菌システインプロテアーゼの組換え発現およびその免疫原性組成物
本発明は一般に分子生物学、臨床細菌学、およびタンパク質折り畳みの分野に関する。より詳しくは、本発明は、宿主細胞における可溶性成熟化膿連鎖球菌外毒素B(SpeB)ポリペプチドを組換え発現させるための方法に関する。

【発明の詳細な説明】
【発明の詳細な説明】
【0001】
(技術分野)
本発明は、分子生物学、臨床細菌学、およびタンパク質折り畳みの分野に関する。より詳しくは、本発明は、宿主細胞における成熟化膿連鎖球菌外毒素B(SpeB)ポリペプチドを組換え発現させるための方法に関する。
【0002】
(従来技術)
A群連鎖球菌(group A streptococci)(GAS)とも呼ばれる化膿連鎖球菌は、ヒトの一般的なグラム陽性細菌病原体である。化膿連鎖球菌は、咽頭炎、膿痂疹、および敗血症を含むヒトにおけるさまざまな病状を引き起こす。感染後には、リウマチ熱および急性糸球体腎炎など自己免疫合併症もヒトにおいて発生する。化膿連鎖球菌は、猩紅熱、壊疽性筋膜炎、および毒性ショックなど重篤な急性疾患も引き起こす。
【0003】
A群連鎖球菌によって引き起こされる咽頭炎は、一般的に「連鎖球菌性咽頭炎」と呼ばれ、季節によって一般医療における全呼び出しの少なくとも16%を占める(非特許文献1)。A群連鎖球菌は、壊疽性筋膜炎に関与する毒性ショックの北米および他の四大陸における最近の再流行の原因でもある(非特許文献2)。
【0004】
連鎖球菌感染は現在、抗生物質療法によって治療されている。しかし、治療された者の25−30%は再発性疾患を有し、かつ/または粘膜分泌物中の微生物を削減する。毒性ショックおよび重篤な感染症の抗生物質による治療は多くの場合、無効であり、かつ死亡率が50%を越えうる(非特許文献3)。ペニシリンが重篤な侵襲性の連鎖球菌感染をうまく治療しえないことは、大きな接種原が迅速に固定相に達し、かつペニシリンがゆっくり増殖する細菌に対してあまり有効ではない現象によって生じる(非特許文献4)。したがって、依然として、連鎖球菌感染を予防し、または改善する有効な手段の継続的必要性がある。さらに具体的には、連鎖球菌感染を予防する免疫原性組成物に有用である抗原(または免疫原)を同定または開発する必要性が存在する。現在、免疫原として考えられているかかるポリペプチド抗原が、連鎖球菌システインプロテアーゼ、連鎖球菌プロテイナーゼ、またはストレプトペイン(streptopain)としても知られる化膿連鎖球菌外毒素B(SpeB)である。
【0005】
化膿連鎖球菌外毒素B(SpeB)は、40kDa不活性プレ−プロ−酵素(すなわち、チモーゲン)として発現され(非特許文献5、非特許文献6)、27アミノ酸NH2−末端シグナル配列を有し、そして118アミノ酸プロ−ペプチド配列(アミノ酸28−145)、および253アミノ酸成熟配列(アミノ酸146−398)が続く。分泌すると、40kDa SpeBチモーゲンは自己触媒活性化を受け、結果として12kDa NH2−末端プロ−ペプチドの除去および成熟28kDa活性SpeB酵素の形成が生じる。この活性酵素への変換機序は、望ましくないタンパク質分解を阻止し、かつタンパク質分解活性の空間的および時間的調節を可能にする(非特許文献7)。
【0006】
酵素のシステインエンドペプチダーゼ基のメンバーとして、SpeBは活性部位でCys−His対を含有する(非特許文献8、非特許文献9、非特許文献10)。192位での単一のシステイン残基のセリンへ(以下、「C192S」)の置換は結果としてSpeB酵素の検出可能なタンパク質分解活性の喪失が生じ、40kDa SpeBチモーゲンの28kDa成熟SpeB形態へのプロセッシングを阻止する(非特許文献11、非特許文献12、非特許文献13)。
【0007】
以前の試験の結果は、C192S SpeB変異株の成熟形態が野生型SpeB酵素への最大の阻害活性による抗体の生成に必要であることを示した(非特許文献12)。これは、免疫化のためにC192S SpeB変異株のNH2−末端切断、28kDa成熟形態を生成する必要を示す。しかし、成熟C192S SpeBの組換え発現(すなわち、そのNH2−末端プロ配列の欠損)の結果として大腸菌における不溶性タンパク質の蓄積が生じる。
【0008】
可溶性成熟C192S SpeB変異株を生成するための1つの方法は、40kDa SpeBチモーゲンの制限タンパク質分解によっている。例えば、成熟C192S SpeB変異株を生成する40kDa C192S SpeB変異株チモーゲンの制限タンパク質分解は、エラスターゼ、ペプシン、サーモリシン(非特許文献12)、およびパパインを含む一部のプロテアーゼを使用して達成されている。これらのデータのほか、トリプシンおよびスブチリシンを利用する非特許文献6によって公開されたデータは、所望の28kDa成熟C192S SpeB変異株が、さまざまなプロテイナーゼによる40kDa C192S SpeB変異株チモーゲンの処理によって有効に生成されることを示す。
【0009】
しかし、この方法には大規模生成に対するいくつかの制限を有する。第一に、成熟プロテアーゼの最終生成物収量は2つの連続精製ステップに対する要件、完全長チモーゲンに対する第1と、加工成熟プロテアーゼに対する第2の要件のため低い。第二に、特に大規模に、制限タンパク質分解反応のコンシステンシーおよび再現性と関係がある難点がある。最後に、切断に使用される酵素的に活性の外来プロテアーゼによる最終生成物の固有の汚染リスクがある。かかる汚染は、反応が樹脂固定化プロテアーゼで行われる場合でも回避することはきわめて困難である。
【0010】
したがって、依然として、哺乳類宿主における連鎖球菌感染に対して有効な免疫原性組成物に対する技術上の必要性がある。したがって、成熟SpeBが哺乳類宿主に投与された場合でも免疫原性である、成熟SpeBポリペプチドを生成し、または発現させる方法を特定することがきわめて望ましい。また、成熟SpeBポリペプチドの免疫原性形態を生成し、または発現させるかかる方法が、SpeB収量の減少、タンパク質分解コンシステンシー/再現性の制限、および外来酵素汚染のリスクなど前記大規模制限を回避することも望ましい。
【非特許文献1】ホープ・シンプソン(Hope−Simpson)、1981年
【非特許文献2】スティーブンス(Stevens)、1992年
【非特許文献3】デービス(Davies)ら、1996年
【非特許文献4】スティーブンス(Stevens)ら、1993年
【非特許文献5】ショセ(Chaussee)ら、1993年
【非特許文献6】リュー(Liu)とエリオット(Elliot)、1965年
【非特許文献7】カーン(Khan)とジェイムズ(James)、1998年
【非特許文献8】リュー(Liu)ら、1965年
【非特許文献9】リュー(Liu)、1965年
【非特許文献10】タイ(Tai)ら、1976年
【非特許文献11】グッバ(Gubba)ら、1998年
【非特許文献12】マツカ(Matsuka)ら、1999年
【非特許文献13】ムサー(Musser)、1996年
【0011】
(発明の開示)
本発明は、概して、宿主細胞における成熟化膿連鎖球菌外毒素B(SpeB)ポリペプチドを組換え発現させる方法、およびその免疫原性組成物に関する。より詳しくは、本発明は、宿主細胞における12kDa SpeBプロ−ペプチドおよび28kDa成熟SpeBポリペプチドを共発現させ、ここで成熟SpeBポリペプチドが宿主細胞において可溶性である新たな方法に関する。
【0012】
したがって、一部の実施形態においては、本発明は、宿主細胞における成熟化膿連鎖球菌外毒素B(SpeB)ポリペプチドを組換え発現させるための方法であって、この方法は、(i)(a)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列と(b)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列とを含んで成る、ポリシストロニックプラスミドで宿主細胞を形質転換し、形質導入し、トランスフェクトし、または感染させるステップと、(ii)宿主細胞によって成熟SpeBポリペプチドおよびSpeBプロ−ポリペプチドドメインの発現を可能にする条件下で宿主細胞を培養するステップとを含んで成り、かつ成熟SpeBポリペプチドが宿主細胞において可溶性である方法に関する。したがって、本発明のポリシストロニックプラスミド系においては、単一のプロモーター(例えば、T7プロモーター)がポリシストロニックmRNA転写物の発現を推進するが、ここでポリシストロニックmRNAはその正確なリーディングフレームにおける2つもしくはそれ以上のポリペプチド(例えば、SpeBプロ−ポリペプチドドメインおよび成熟SpeBポリペプチド)をコード化する。一部の実施形態においては、SpeBプロ−ポリペプチドドメインは配列番号2のアミノ酸残基28〜145を含んで成るポリペプチドとしてさらに規定され、かつ成熟SpeBポリペプチドは配列番号2のアミノ酸残基146〜398を含んで成るポリペプチドとしてさらに規定される。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192でのシステインはセリンによって置換される。別の実施形態においては、成熟SpeBポリペプチドは哺乳類宿主において免疫原性である。さらに別の実施形態においては、成熟SpeBポリペプチドに特異的な抗体は野生型SpeBポリペプチドと交差反応し、かつSpeBポリペプチド活性を無力化する。一部の好ましい実施形態においては、プラスミドはT7プロモーター含有プラスミドである。特定の実施形態においては、T7プロモーター含有プラスミドはpET、pRSET、pCRT7−CTTOPO、およびpIVeXより成る群から選択される。別の好ましい実施形態においては、宿主細胞は細菌細胞である。一部の実施形態においては、細菌宿主細胞は大腸菌である。さらに他の実施形態においては、大腸菌は、BLR(DE3)、BLR(DE3)pLysS、AD494(DE3)、AD494(DE3)pLysS、BL21(DE3)、BL21(DE3)pLysS、BL21(DE3)pLysE、BL21(DE3)pLacI、BL21trxB(DE3)、BL21trxB(DE3)pLysS、HMS174(DE3)、HMS174(DE3)pLysS、HMS174(DE3)pLysE、Origami(DE3)、Origami(DE3)pLysS、Origami(DE3)pLysE、Origami(DE3)pLacI、OrigamiB(DE3)、OrigamiB(DE3)pLysS、OrigamiB(DE3)pLysE、OrigamiB(DE3)pLacI、Rosetta(DE3)、Rosetta(DE3)pLysS、Rosetta(DE3)pLysE、Rosetta(DE3)pLacI、Tuner(DE3)、Tuner(DE3)pLysS、およびTuner(DE3)pLacIより成る群から選択される菌株である。
【0013】
他の実施形態においては、本発明は、(a)(i)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列を含んで成るプラスミドおよび(ii)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るプラスミドで宿主細胞を形質転換し、形質導入し、トランスフェクトし、または感染させるステップと、(b)SpeBプロ−ポリペプチドドメインおよび成熟SpeBポリペプチドを共発現させるのに適切な条件下で宿主細胞を培養するステップとを含み、成熟SpeBポリペプチドが宿主細胞において可溶性である、宿主細胞において成熟SpeBポリペプチドを組換え発現させるための方法に関する。一部の実施形態においては、SpeBプロ−ポリペプチドドメインは配列番号2のアミノ酸残基28〜145を含んで成るポリペプチドとしてさらに規定され、かつ成熟SpeBポリペプチドは配列番号2のアミノ酸残基146〜398を含んで成るポリペプチドとしてさらに規定される。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192でのシステインはセリンによって置換される。他の実施形態においては、成熟SpeBポリペプチドは哺乳類宿主において免疫原性である。さらに他の実施形態においては、成熟SpeBポリペプチドに特異的な抗体は野生型SpeBポリペプチドと交差反応し、かつSpeBポリペプチド活性を無力化する。別の好ましい実施形態においては、プラスミドはT7プロモーター含有プラスミドである。特定の実施形態においては、T7プロモーター含有プラスミドはpET、pRSET、pCRT7−CTTOPO、およびpIVeXより成る群から選択される。さらに他の実施形態においては、宿主細胞は細菌細胞である。1つの好ましい実施形態においては、宿主細胞は大腸菌であり、ここで大腸菌は、BLR(DE3)、BLR(DE3)pLysS、AD494(DE3)、AD494(DE3)pLysS、BL21(DE3)、BL21(DE3)pLysS、BL21(DE3)pLysE、BL21(DE3)pLacI、BL21trxB(DE3)、BL21trxB(DE3)pLysS、HMS174(DE3)、HMS174(DE3)pLysS、HMS174(DE3)pLysE、Origami(DE3)、Origami(DE3)pLysS、Origami(DE3)pLysE、Origami(DE3)pLacI、OrigamiB(DE3)、OrigamiB(DE3)pLysS、OrigamiB(DE3)pLysE、OrigamiB(DE3)pLacI、Rosetta(DE3)、Rosetta(DE3)pLysS、Rosetta(DE3)pLysE、Rosetta(DE3)pLacI、Tuner(DE3)、Tuner(DE3)pLysS、およびTuner(DE3)pLacIより成る群から選択される菌株である。
【0014】
別の実施形態においては、本発明は、(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るプラスミドを宿主細胞において組換え発現させ、ここで前記SpeBポリペプチドが宿主細胞において不溶性ポリペプチド集合体を形成するステップと、(b)ポリペプチド集合体を可溶化し、可溶化ポリペプチドが非ネイティブ成熟SpeBポリペプチドとして規定されるステップと、(c)シャペロンタンパク質の存在下に非ネイティブ成熟SpeBポリペプチドを再折り畳みし、非ネイティブ成熟SpeBポリペプチドがネイティブ成熟SpeBポリペプチドへ折り畳まれるステップと、(d)ネイティブSpeBポリペプチドを回収するステップとを含んで成る成熟SpeBポリペプチドを生成するための方法に関する。好ましい実施形態においては、シャペロンタンパク質は、GroEL、GroEL/GroES、PPI、PDI、およびSpeBプロ−ポリペプチドドメインより成る群から選択される。特定の実施形態においては、シャペロンタンパク質は、配列番号2のアミノ酸残基28〜145を含んで成るSpeBプロ−ポリペプチドドメインである。好ましい実施形態においては、成熟SpeBは、配列番号2のアミノ酸残基146〜398を含んで成るポリペプチドである。別の好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192におけるシステインがセリンによって置換される。さらに別の実施形態においては、不溶性ポリペプチド集合体は封入体としてさらに規定される。他の実施形態においては、ポリペプチドの可溶化が尿素、塩化グアニジニウム(guanidinium chloride)、ドデシル硫酸ナトリウム(SDS)、および熱などより成る群から選択される変性剤である。
【0015】
さらに他の実施形態においては、本発明は、宿主細胞において(i)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(ii)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成るポリシストロニックプラスミドを発現させるステップを含んで成り、成熟SpeBポリペプチドが宿主細胞において可溶性である、宿主細胞において成熟SpeBポリペプチドを組換え発現させるための方法に関する。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192におけるシステインがセリンによって置換される。特定の実施形態においては、プラスミドは、GroESポリペプチドをコード化するポリヌクレオチドをさらに含んで成る。
【0016】
さらに他の実施形態においては、本発明は、(a)(i)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(ii)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成るポリシストロニックプラスミドで宿主細胞を形質転換し、形質導入し、トランスフェクトし、または感染させるステップと、(b)成熟SpeBポリペプチドおよび前記GroELポリペプチドを発現させるのに適切な条件下で宿主細胞を培養し、ここで成熟SpeBポリペプチドが宿主細胞に可溶性であるステップと、(c)ネイティブ成熟SpeBポリペプチドを回収するステップとを含んで成る成熟SpeBポリペプチドを生成するために方法に関する。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192におけるシステインがセリンによって置換される。
【0017】
一部の他の実施形態においては、本発明は、本発明に記載された方法の1つもしくはそれ以上に従って生成される成熟SpeBポリペプチドに関する。他の実施形態においては、本発明は、本発明の方法の1つに従って生成されるSpeBポリペプチドを含んで成る免疫原性組成物に関する。さらに他の実施形態においては、本発明は、化膿連鎖球菌に対して哺乳類の対象を免疫化する方法に関し、この方法は、対象に免疫原性量の免疫原性組成物を投与するステップを含んで成り、ここで免疫原性組成物は本発明の方法に従って生成される成熟SpeBポリペプチドを含んで成る。
【0018】
他の実施形態においては、本発明は、(a)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列と、(b)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列とを含んで成り、成熟SpeBポリペプチドが宿主細胞において発現されると可溶性である、ポリシストロニックプラスミドに関する。一部の実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192でのシステインはセリンによって置換される。一部の他の実施形態においては、プラスミドはT7プロモーター含有プラスミドである。
【0019】
一部の他の実施形態においては、本発明は、(a)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列を含んで成るプラスミド、および(b)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るプラスミドに関し、成熟SpeBポリペプチドは宿主細胞において発現されると可溶性である。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192でのシステインはセリンによって置換される。プラスミドはT7プロモーター含有プラスミドである。
【0020】
別の実施形態においては、本発明は、(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(b)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成り、成熟SpeBポリペプチドが宿主細胞で発現されると可溶性であるポリシストロニックプラスミドに関する。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192におけるシステインはセリンによって置換される。一部の他の実施形態においては、プラスミドはT7プロモーター含有プラスミドである。
【0021】
さらに別の実施形態においては、本発明は、(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(b)GroELポリペプチドをコード化するポリヌクレオチド配列と、(c)GroESポリペプチドをコード化するポリヌクレオチド配列とを含んで成り、成熟SpeBポリペプチドが宿主細胞で発現されると可溶性であるポリシストロニックプラスミドに関する。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192におけるシステインはセリンによって置換される。他の実施形態においては、プラスミドはT7プロモーター含有プラスミドである。
【0022】
さらに他の実施形態においては、(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(b)GroEL、GroES、PDI、およびPPIより成る群から選択される1つもしくはそれ以上のポリペプチドをコード化するポリヌクレオチド配列とを含んで成り、成熟SpeBポリペプチドが宿主細胞で発現されると可溶性であるポリシストロニックプラスミドに関する。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192におけるシステインがセリンによって置換される。他の実施形態においては、プラスミドはT7プロモーター含有プラスミドである。
【0023】
別の実施形態においては、本発明は、本発明のプラスミドにより形質転換され、形質導入され、トランスフェクトされ、または感染された宿主細胞を提供する。
【0024】
本発明の他の特徴および利点は、以下の詳細な説明、その好ましい実施形態、および請求の範囲から明らかであろう。
【0025】
(図面の簡単な記載)
図1Aおよび1Bは組換え成熟SpeBとプロ配列ドメインの相互作用を示す。図1Aはペプシン生成成熟C192S SpeB(黒丸)、および成熟野生型SpeB(黒四角)の濃度増大を示し、これらはプロ配列ドメイン(黒記号)、またはリゾチーム(白記号)含有マイクロタイタープレートでインキュベートされ、かつ実施例1によって示されているようにELISAで分析された。データは、各々二重に行われた3つの実験を代表する。図1Bは実施例1に示されているようにバイオコア(Biocore)3000の使用によって行われた、プロ配列ドメインと成熟SpeBポリペプチドとの相互作用の実時間分析を示す。結果は表面プラスモン共鳴(相対反応)として表されている。
【0026】
図2Aおよび2Bは成熟SpeBのプロ配列ドメイン介在阻害を示す。示されているのは、レゾルフィン標識カゼイン(図2A)またはシステインプロテアーゼパパイン(図2B)を使用した成熟野生型SpeBの阻害であり、SpeBプロ配列ドメイン(黒四角)、またはリゾチーム(白三角)を使用した基質が実施例1に示されているように分析された。
【0027】
図3は変性成熟SpeBの再折り畳みに対する組換えプロ配列ドメインの効果を示す。変性成熟SpeBを、PBS、0.5Mアルギニン(黒丸)、PBS、0.5Mアルギニン、20μMプロテアーゼ阻害剤E−64(白丸)、PBS(黒四角)、およびPBS、20μM E−64(白四角)を含有する増大濃度のプロ配列ドメイン中に迅速に希釈した。反応を実施例1に示されているように行い、レゾルフィン標識カゼイン切断アッセイを使用してプロテアーゼ活性の存在について評価した。
【0028】
図4は2−プラスミドベース発現構成物を示す略図である。2−プラスミドベース発現ベクターの特徴としては、図のように、KanR−カナマイシン耐性遺伝子(aph(3’)−la)、AmpR−アンピシリン耐性遺伝子(b−ラクタマーゼ)、複製のOri−origin、laclq−lacレプレッサ、T7ベースプロモーター(T7またはT7lac)、およびT7ターミネータが挙げられる。
【0029】
図5Aおよび5Bは、各々、ポリシストロニック発現系および合成リンカー領域を示す図表である。ポリシストロニック発現ベクター(図5A)の特徴としては、図のように、KanR−カナマイシン耐性遺伝子(aph(3’)−la)、複製のOri−origin、laclq−lacレプレッサ、T7lacプロモーター、およびT7ターミネータが挙げられる。分析されたリンカー領域の塩基組成が示されている(図5B)。各リンカー領域のヌクレオチド番号(5nt、10nt、20nt、40nt)は、プロ配列ドメインの遺伝子工学的TAA翻訳停止コドン(PSD停止)(太字テキスト)と、最適化シャイン・ダルガノリボソーム結合部位(SD)との間の塩基の数を示す。第2のシストロンの翻訳ATG開始コドンは太字イタリックで示されている。
【0030】
図6Aおよび6Bはポリシストロニック発現系における可溶性成熟SpeBおよびプロ配列ドメインレベルの評価および相対定量を示す。可溶性フラクションにおけるプロ配列ドメイン、および成熟SpeB発現に対する5nt、10nt、20nt、40ntリンカー含有C192S SpeBポリシストロニック系のSDS−PAGE評価が行われた(図6A)。発現タンパク質の定量は、分子動力学パーソナル濃度計を使用して行われ、スキャン面の測定は発現レベルとして示された(図6B)。
【0031】
図7はポリシストロニックcDNAの定量的PCR分析を示す。cDNAおよび−RT制御は実施例1に記載されているように調製され分析された。すべてのCt値はKanR mRNA発現に標準化された。
【0032】
図8は成熟C192S SpeBの熱変性を示す。発現C192S SpeBチモーゲンのペプシン切断によって、または2−プラスミドおよびポリシストロニック共発現系によって生成された精製成熟C192S SpeBの変性曲線は、実施例1に示されているように分析された。
【0033】
図9は成熟野生型SpeBの熱誘発変性を示す。自己触媒的プロセシング(すなわち、チモーゲン生成)によって、または2−プラスミドおよびポリシストロニック共発現系によって生成された精製成熟野生型SpeBの変性曲線は、実施例1に示されているように分析された。
【0034】
図10は組換え成熟SpeBの操作モル濃度の評価を示す。自動触媒(黒四角)のほか、2−プラスミド(黒丸)およびポリシストロニック(黒三角)共発現によって生成された当量(0.12μM)の精製成熟野生型SpeBが、実施例1に記載されているようにレゾルフィン標識カゼイン切断アッセイを使用して評価された。25℃下に1時間のインキュベーション時間を切断反応用に使用した。ポリシストロニック発現によって生成された精製成熟C192S SpeB(白ダイヤ)が対照として評価された。
【0035】
図11は野生型SpeBタンパク質分解活性の抗体介在阻害を示す。2−プラスミド(黒ダイヤ)、またはポリシストロニック系(白四角)によって生成された成熟C192S SpeBに対して生成される抗血清の増大量が、レゾルフィン標識カゼイン切断アッセイを使用して成熟野生型SpeBのタンパク質分解活性を特異的に阻害する能力について評価された。37℃下に2時間のインキュベーション時間を切断反応用に使用した。免疫前血清が分析用の陰性対照(トリ−スター)が使用された。
【0036】
(発明の詳細な記載)
以下に記載される本発明は、免疫原性組成物で使用する化膿連鎖球菌外毒素B(以下、「SpeB」)を生成する方法に対する技術上の必要性に対処する。一部の好ましい実施形態においては、本発明は、免疫原性組成物で使用する成熟SpeBを生成する方法に関する。より詳しくは、以下に記載される本発明は、宿主細胞における成熟SpeBポリペプチドを組換え発現させる方法に対する技術上の必要性に対処し、ここで発現成熟SpeBポリペプチドは宿主細胞において可溶性である。好ましい実施形態においては、本発明の方法に従って生成される成熟SpeBポリペプチドは、哺乳類宿主に投与されると免疫原性である。
【0037】
以下に規定される、化膿連鎖球菌で発現される「SpeBチモーゲン」は、配列番号2のアミノ酸1−398を含んで成る40kDaプレ−プロ−ポリペプチドである。SpeBチモーゲンは、27アミノ酸NH2−末端シグナル(「プレ」)配列を有する酵素的に不活性な(すなわち、チモーゲン)、40kDa、プレ−プロ−ポリペプチドとして発現され、そして118アミノ酸プロ−ポリペプチド(「プロ」)配列、および253アミノ酸成熟ポリペプチド配列が続く。したがって、以下に規定されているように、SpeBチモーゲンの「プレ」配列は、配列番号2のアミノ酸1−27を含んで成る。同様に、以下に規定されているように、SpeBチモーゲンの「プロ配列」、「プロ配列ドメイン」、「プロ−ポリペプチド配列」、および「プロ−ポリペプチドドメイン」は同じ意味で使用され、ここでプロ配列は配列番号2のアミノ酸28−145を含んで成る。
【0038】
ネイティブ細菌(すなわち、化膿連鎖球菌)によって分泌されると、40kDa SpeBチモーゲンは自己触媒活性化を受け、結果として12kDa NH2−末端プロ−ポリペプチド配列(すなわち、配列番号2のアミノ酸8−145)の除去および成熟28kDaタンパク質分解活性SpeBポリペプチド(または酵素)の形成が生じる。以下に規定されているように、「成熟SpeB」ポリペプチドは、配列番号2のアミノ酸146−398を含んで成る28kDaポリペプチドであり、ここで成熟SpeBポリペプチドはシステインプロテアーゼ活性を有する。以下に規定されているように、「成熟SpeB」ポリペプチドおよび「成熟野生型SpeB」ポリペプチドは同じ意味で使用され、その両方は配列番号2のアミノ酸146−398を含んで成る野生型28kDaポリペプチドを指し、ここでポリペプチドはシステインプロテアーゼ活性を有する。
【0039】
SpeB活性部位は、配列番号2のアミノ酸残基192および340でCys−His対を含有する。192位での単一のシステイン残基のセリン残基(以下、「C192S」または「C192S変異株」)へのアミノ酸置換(すなわち、突然変異)は、成熟C192S SpeBポリペプチドの検出可能なタンパク質分解活性の喪失をもたらす。したがって、以下に規定されているように、「C192S SpeBチモーゲン」または「C192S SpeB変異株」は、配列番号2のアミノ酸28−398を含んで成り、ここで192位でのシステインアミノ酸残基はセリンアミノ酸残基に変異されている。以下に規定されているように、「成熟C192S SpeB」ポリペプチドは、配列番号2のアミノ酸146−398を含んで成る28kDaポリペプチドであり、ここで192位でのシステインアミノ酸残基はセリンアミノ酸残基に変異されており、ここで成熟C192S SpeBポリペプチドは成熟野生型SpeBポリペプチドに対するシステインプロテアーゼ活性を有さない。
【0040】
したがって、一部の実施形態においては、本発明は、成熟SpeBポリペプチド、かつより好ましくは、成熟C192S SpeBポリペプチドを含んで成る免疫原性組成物に関する。例えば、C192S SpeB変異株による以前の免疫学的試験では、成熟C192S SpeBポリペプチド(すなわち、NH2−末端プロ−ポリペプチド配列の欠損)が、成熟野生型SpeBポリペプチドへの最大の阻害活性(すなわち、交差再活性)による抗体の生成に必要であることが明らかにされている(マツカ(Matsuka)ら、1999年)。したがって、本明細書では、化膿連鎖球菌感染に対して哺乳類を免疫化するために有効な免疫原性組成物が、少なくとも成熟C192S SpeBまたは成熟野生型SpeBポリペプチド抗原を含んで成ることが考えられている。しかし、大腸菌宿主細胞における成熟SpeBポリペプチドの組換え発現が、もっぱら大腸菌における不溶性SpeBポリペプチド集合体の生成をもたらすことが当技術分野で周知である(マツカ(Matsuka)ら、1999年)。
【0041】
したがって、特定の実施形態においては、本発明は、「成熟」および「可溶性」である宿主細胞におけるSpeBポリペプチドの発現の困難さを克服する方法に関する。例えば、本発明の1つの実施形態においては、成熟C192S SpeBポリペプチド(または成熟野生型SpeB)をコード化するプラスミドと、宿主細胞におけるプロ−ポリペプチドドメイン(すなわち、配列番号2のアミノ酸28−145)をコード化するプラスミドとの共発現が、宿主細胞における可溶性、成熟C192S SpeBポリペプチド(または成熟野生型SpeB)の有効な発現をもたらすことが明らかにされている(例えば、実施例3を参照)。同様に、ポリシストロニックプラスミドが、成熟C192S SpeBポリペプチド(または成熟野生型SpeB)とプロ−ポリペプチド配列(配列番号2のアミノ酸28−145)の両方をコード化する、宿主細胞におけるポリシストロニックプラスミドの組換え共発現も、」宿主細胞における可溶性、成熟C192S SpeBポリペプチド(または成熟野生型SpeB)の発現をもたらす(例えば、実施例4を参照)。
【0042】
したがって、以下に記載される本発明は、宿主細胞において可溶性成熟C192S SpeBポリペプチド(または成熟野生型SpeB)を発現させ、可溶性成熟C192S SpeBポリペプチドが化膿連鎖球菌感染に対して哺乳類を免疫化するための免疫原性組成物において特に有用である、新たな方法、およびその新たな組成物を提供する。したがって、一部の好ましい実施形態においては、本発明は、成熟C192S SpeBポリペプチド(または成熟野生型SpeB)をコード化するプラスミドと、宿主細胞におけるプロ−ポリペプチドドメイン(すなわち、配列番号2のアミノ酸28−145)をコード化するプラスミドを共発現させ、ここで成熟SpeBポリペプチドが宿主細胞において可溶性である方法に関する。一部の他の好ましい実施形態においては、本発明は、宿主細胞におけるポリシストロニックプラスミドを発現させる方法に関し、ここでポリシストロニックプラスミドは、成熟C192S SpeBポリペプチド(または成熟野生型SpeB)とプロ−ポリペプチド配列(配列番号2のアミノ酸28−145)の両方をコード化し、ここで成熟SpeBポリペプチドは宿主細胞において可溶性である。
【0043】
本発明は、可溶性成熟SpeBの生成を調節するプロ配列ドメインの要件を明らかにし、プロ配列ドメインが成熟SpeBポリペプチドの適切な折り畳みに対する分子内シャペロンとして機能することを示す。成熟野生型SpeBポリペプチドまたは成熟C192S SpeBのいずれかとのプロ配列ドメインの関連性は、それぞれ、約11nmおよび34nMの解離定数(Kd)を有する(実施例2)。これらの結合値は、プロ配列ドメインと成熟SpeBポリペプチドドメインとの間の高い親和性を示す。また、プロ配列ドメインの分子シャペロン活性は、尿素変性成熟SpeBを使用してインビトロで明らかにされている(実施例2)。
【0044】
したがって、特定の実施形態においては、本発明は、不溶性成熟SpeB集合体のタンパク質補助折り畳みの方法に関する。例えば、本発明の1つの実施形態は、(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るプラスミドを宿主細胞において組換え発現させ、ここでSpeBポリペプチドが宿主細胞において不溶性ポリペプチド集合体を形成するステップと、(b)ポリペプチド集合体を可溶化し、可溶化ポリペプチドが非ネイティブ成熟SpeBポリペプチドとして規定されるステップと、(c)1つもしくはそれ以上のシャペロンタンパク質の存在下に非ネイティブ成熟SpeBポリペプチドを再折り畳みし、非ネイティブ成熟SpeBポリペプチドがネイティブ成熟SpeBポリペプチドへ折り畳まれるステップと、(d)ネイティブ成熟SpeBポリペプチドを回収するステップとを含んで成る成熟SpeBポリペプチドを生成するための方法に関する。
【0045】
同様に、他の実施形態においては、本発明は、成熟SpeBポリペプチドが1つもしくはそれ以上の分子シャペロンタンパク質の存在下に発現される、タンパク質補助折り畳みの方法に関する。例えば、本発明は、(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(b)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成るポリシストロニックプラスミドを宿主細胞において組換え発現させ、ここで成熟SpeBポリペプチドが宿主細胞において可溶性であり、かつ(c)ネイティブ成熟SpeBポリペプチドを回収するステップを含んで成る、宿主細胞において成熟SpeBポリペプチドを発現するための方法を提供する。
【0046】
A.SpeB抗原をコード化するポリヌクレオチド
本発明の単離および精製化膿連鎖球菌ポリヌクレオチドは、成熟SpeBポリペプチド抗原の生成において使用される。さらに具体的には、一部の実施形態においては、本発明のポリヌクレオチドは成熟SpeBポリペプチドおよびSpeBプロ−ポリペプチドドメインをコード化し、ここで成熟SpeBポリペプチドは宿主細胞において発現されると可溶性である。したがって、本発明の一部の実施形態においては、ポリヌクレオチドは配列番号2のアミノ酸146〜398を含んで成る成熟SpeBポリペプチドをコード化し、第2のポリヌクレオチドは配列番号2のアミノ酸28〜145を含んで成るプロ−ポリペプチドドメインをコード化する。好ましい実施形態においては、本発明のポリヌクレオチドは成熟C192SpeBポリペプチドをコード化し、ここで成熟C192SSpeBポリペプチドは配列番号2のアミノ酸146〜398を含んで成り、ここで配列番号2のアミノ酸残基192でのシステインはセリン残基に変異されている。一部の好ましい実施形態においては、成熟SpeBポリペプチドをコード化するポリヌクレオチドは配列番号1のヌクレオチド436〜1197を含んで成り、成熟C192S SpeBポリペプチドをコード化するポリヌクレオチドは配列番号1のヌクレオチド436〜1197を含んで成り、ここで配列番号2のアミノ酸残基192はセリンアミノ酸であり、かつプロ−ポリペプチドドメインをコード化するポリヌクレオチドは配列番号1のヌクレオチド82〜435を含んで成る。
【0047】
したがって、特定の実施形態においては、本発明のポリヌクレオチドはDNA分子であり、ここでDNAはゲノムDNA、プラスミドDNA、またはcDNAでありうる。好ましい実施形態においては、本発明のポリヌクレオチドは組換えcDNAポリヌクレオチドである。別の好ましい実施形態においては、成熟SpeBポリペプチドをコード化するポリヌクレオチドは第1のプラスミドベクターに含まれており、かつSpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチドは第2のプラスミドベクターに含まれており、ここで両方のベクターは宿主細胞において共発現される。別の好ましい実施形態においては、成熟SpeBポリペプチドをコード化するポリヌクレオチドおよびSpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチドはポリシストロニック発現構成物に含まれている。セクションE(実施例4および図9)に記載されているように、本発明のポリシストロニック構成物は、5’〜3’の方向でSpeBプロ−ポリペプチドドメインを第1のシストロンとして含んで成り、そして翻訳エンハンサーおよび最適化シャイン・ダルガノリボソーム結合部位および成熟SpeBを第2のシストトロンとして含んで成る合成リンカーが続く。別の実施形態においては、本発明のポリシストロニック構成物は、5’〜3’の方向で成熟SpeBを第1のシストロンとして含んで成り、そして翻訳エンハンサーおよび最適化シャイン・ダルガノリボソーム結合部位およびSpeBプロ−ポリペプチドドメインを第2のシストロンとして含んで成る合成リンカーが続く。
【0048】
以下で使用される「ポリヌクレオチド」という語は、ホスホジエステル結合によって結合されたヌクレオチドの配列を意味する。ポリヌクレオチドは以下、5’〜3’の方向で示されている。本発明のポリヌクレオチドは、約10〜約数十万の塩基対を含んで成る。好ましくは、ポリヌクレオチドは約10〜約3,000の塩基対を含んで成る。特定のポリヌクレオチドの好ましい長さは以下に記載されている。
【0049】
本発明のポリヌクレオチドは、デオキシリボ核酸(DNA)分子、リボ核酸(RNA)分子、またはヌクレオチド類似体を使用して生成されるDNAもしくはRNAの類似体である。核酸分子は一本鎖また二本鎖であるが、好ましくは、二本鎖DNAである。ポリヌクレオチドはDNA分子である場合、その分子は遺伝子、cDNA分子、またはゲノムDNA分子である。ヌクレオチド塩基は、単一の文字表記によって以下に示されている。すなわち、アデニン(A)、グアニン(G)、チミン(T)、シトシン(C)、イノシン(I)およびウラシル(U)。
【0050】
「単離」は、天然状態から「人が作ったものによって」変化されていることを意味する。「単離」組成物または物質は、その最初の環境から変更もしくは除去、または変更および除去されているものである。例えば、生きた動物に天然に存在するポリヌクレオチドまたはポリペプチドは「単離」されず、その天然状態の共存する物質から分離された同じポリヌクレオチドまたはポリペプチドは、その語が以下で使用されるように「単離」である。
【0051】
好ましくは、「単離」ポリヌクレオチドは、核酸が由来する生物のゲノムDNAにおける核酸を天然にフランキングする配列(すなわち、核酸の5’および3’末端に位置した配列)を含まない。例えば、さまざまな実施形態においては、単離SpeB核酸分子は、核酸が由来する細胞のゲノムDNAにおける核酸分子を天然にフランキングする約5kb、4kb、3kb、2kb、1kb、0.5kbまたは0.1kb未満のヌクレオチド配列を含有する。しかし、SpeB核酸分子はタンパク質コードまたは調節配列に融合し、それでも単離とみなされる。
【0052】
本発明のSpeBポリヌクレオチドは、標準クローニングおよびスクリーニング法を使用し、mRNAに由来するcDNAライブラリーから得られる。本発明のポリヌクレオチドは、ゲノムDNAライブラリー(例えば、化膿連鎖球菌ライブラリー)などの天然源からも得られ、または公知および市販の方法を使用して合成される。
【0053】
成熟SpeBおよび/またはSpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチドの相同分子種および対立遺伝子変異体は、当技術分野で周知の方法を使用して容易に同定される。ポリヌクレオチドの対立遺伝子変異体および相同分子種は、配列番号1に示されているヌクレオチド配列、またはこのヌクレオチド配列の断片の一般的に少なくとも約70−75%、より一般的に少なくとも約80−85%、かつ最も一般的に少なくとも約90−95%もしくはそれ以上の相同のヌクレオチド配列を含んで成る。かかる核酸分子は、好ましくは、ストリンジェントな条件下で、配列番号1に示されているヌクレオチド配列、またはこのヌクレオチド配列の断片にハイブリッド可能であることが容易に同定される。
【0054】
成熟SpeBポリペプチドおよびSpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチドは、宿主細胞における可溶性成熟SpeBポリペプチドまたはその断片の組換え生成のために使用される。ポリヌクレオチドは、成熟SpeBポリペプチドのコード配列および/またはSpeBプロ−ポリペプチドドメインのコード配列を含みうる。ポリヌクレオチドは、転写、非翻訳配列、スプライシングシグナル、プロモーター/エンハンサー配列、リボソーム結合部位、およびポリアデニル化シグナルなど非コード5’および3’配列も含有しうる。
【0055】
したがって、一部の実施形態においては、本発明によって提供されるポリペプチド配列情報(すなわち、配列番号1)は、以下に開示される選択ポリヌクレオチドの遺伝子配列に特異的にハイブリッドする能力を有する比較的短いDNA(またはRNA)オリゴヌクレオチド配列の調製を可能にする。以下で使用される「オリゴヌクレオチド」という語は、2個もしくはそれ以上のデオキシリボヌクレオチドまたはリボヌクレオチド、通常、3個(3)超、および通常10個(10)超、さらに100個(100)もしくはそれ以上(ただし好ましくは、20〜30個)までから成る分子として規定される。正確なサイズは、多くの因子によって決まり、言い換えると、オリゴヌクレオチドの最終の機能または使用によって決まる。したがって、本発明の特定の実施形態においては、適切な長さの核酸プローブが選択ヌクレオチド配列の検討に基づき調製される。かかる核酸プローブが成熟SpeBポリペプチドまたはSpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチドに特異的にハイブリッドする能力は、さまざまな実施形態においてそれらに特定の有用性を与える。最も重要なことだが、これらのプローブは所定のサンプルにおける相補配列の存在を検出するためのさまざまなアッセイにおいて使用される。
【0056】
一部の実施形態においては、オリゴヌクレオチドプライマーを使用することが有利である。これらのプライマーは、化学合成、DNA複製、逆転写、またはその組合せを含む任意のやり方で生成されうる。かかるプライマーの配列は、ポリメラーゼ連鎖反応(PCR)技術を使用して原核細胞からSpeBポリペプチドをコード化するSpeBポリヌクレオチドの規定部分を検出、増幅、または変異で使用するための本発明のポリヌクレオチドを使用して指定される。
【0057】
一部の実施形態においては、本発明のポリヌクレオチドをハイブリッド形成を検出するための適切な標識と組合せて使用することが有利である。多種多様の適切な標識は、検出可能シグナルを与える能力があるアビジン/ビオチンなど、放射性、酵素、または他のリガンドを含めて当技術分野で周知である。
【0058】
本発明による利点の一部を提供するために、ハイブリダイゼーション試験またはアッセイに使用される好ましい核酸配列としては、配列番号2のポリペプチドをコード化するポリヌクレオチドの少なくとも10個〜約18個のヌクレオチド長ストレッチに相補的であるプローブ分子が挙げられる。長さが少なくとも10個のヌクレオチドのサイズは、断片が安定かつ選択的である二重分子を形成するのに十分な長さであることを確実にするために役立つ。しかし、ハイブリッドの安定性および選択性を増大させ、それによって得られる特異的ハイブリッド分子の質および程度を改善するために、長さが10個超の塩基のストレッチにわたって相補配列を有する分子が一般に好ましい。かかる断片は、例えば、化学的手段によって断片を直接合成することによって、(参照することにより本明細書で援用される米国特許第4,683,202号明細書)のPCR技術など核酸再生成技術の適用によって、または適切な挿入および適切な制限酵素部位を含有する組換えプラスミドから選択DNA断片を切除することによって容易に調製される。
【0059】
したがって、本発明のポリヌクレオチドプローブ分子は、遺伝子の相補ストレッチで二重分子を選択的に形成するその能力のために使用される。想定される適用によって、ターゲット配列のためにプローブの各種程度の選択性を達成するのに各種条件のハイブリダイゼーションのストリンジェンシーを使用することが望まれる(以下の表1を参照)。高度の選択性を必要とする用途のために、通常、比較的ストリンジェントな条件を使用してハイブリッドを形成することが望まれる。一部の用途、例えば、基礎を成すテンプレートにハイブリッドされる変異株プライマー鎖を使用する変異株を調製することが望まれ、または相同ポリペプチドコード配列を他の細胞、機能的同等物などから単離することが求められる場合には、ヘテロ2本鎖形成を可能にするあまり厳しくないハイブリダイゼーション条件が通常、必要とされる(表1を参照)。それによって、クロスハイブリダイゼーション種は、対照ハイブリダイゼーションに対する陽性ハイブリダイゼーションシグナルとして容易に同定される。ともかく、一般に、条件は、温度の上昇と同じやり方でハイブリッド二本鎖の不安定化に使用される増加量のホルムアミドの添加によって、より厳しく与えられることが予想される。したがって、ハイブリダイゼーション条件は容易に操作され、したがって、一般に所望の結果によって選択される方法となる。
【0060】
本発明は、低ストリンジェントな条件、より好ましくは、ストリンジェントな条件、かつ最も好ましくは、高ストリンジェントなな条件下で、以下に記載されたポリヌクレオチドにハイブリッド可能なポリヌクレオチドも含む。ストリンジェントな条件の例は以下の表1に示されている。すなわち、高ストリンジェントな条件は、少なくとも、例えば、条件A−Fのストリンジェンシーであり、ストリンジェントな条件は、少なくとも、例えば、条件G−Lのストリンジェンシーであり、低ストリンジェントな条件は、少なくとも、例えば、条件M−Rのストリンジェンシーである。
【0061】
【表1】
【0062】
(bp)I:ハイブリッド長さは、ハイブリダイズするポリヌクレオチドののハイブリッド領域について予想される。ポリヌクレオチドを未知の配列のターゲットポリヌクレオチドにハイブリダイズする場合、ハイブリッド長さはポリヌクレオチドのハイブリダイゼーションのものと想定される。既知の配列のポリヌクレオチドがハイブリダイズされる場合、ハイブリッド長さはポリヌクレオチドの配列を調整し、最適な配列相補性の領域または各領域を同定することによって決定される。
バッファーH:SSPE(1×SSPEは、0.15M NaCl、10mM NaH2PO4、および1.25mM EDTA、pH7.4である)は、ハイブリダイゼーションおよび洗浄バッファー中SSC(1×SSCは0.15M NaCl、および15mM クエン酸ナトリウム)に代替され、洗浄はハイブリダイゼーションの完了後15分間行われる。
TB〜TR:長さが50未満の塩基対であることが予想されるハイブリッドのハイブリダイゼーション温度は、ハイブリッドの融解温度(Tm)よりも5−10℃低くあるべきであり、ここでTmは以下の等式に従って決定される。長さが18未満の塩基対のハイブリッドは、Tm(℃)=2(A+T塩基の#)+4(G+C塩基の#)。長さが18〜49の塩基対のハイブリッドでは、Tm(℃)=81.5+16.6(log10[Na+])+0.41(%G+C)−(600/N)であり、ここでNはハイブリッド中の塩基の数であり、[Na+]はハイブリダイゼーションバッファー中のナトリウムイオンの濃度である(1×SSCの[Na+]=0.165M)。
【0063】
ポリヌクレオチドハイブリダイゼーションのストリンジェントな条件の追加の例は、参照することにより以下に援用される、サムブルック(Sambrook)ら、1989年、Molecular Cloning:A Laboratory Manual、コールド・スプリング・ハーバー・ラボラトリー・プレス(Cold Spring Harbor Laboratory Press)、コールド・スプリング・ハーバー(Cold Spring Harbor)、ニューヨーク州、第9章および第11章、およびオーズベル(Ausubel)ら、1995年、Current Protocols in Molecular Biology、ジョン・ワイリー・アンド・サンズ(John Wiley & Sons,Inc.)編、第2.10節および第6.3−6.4節に示されている。
【0064】
B.成熟SPEBポリペプチド抗原
特定の実施形態においては、本発明は、成熟SpeBポリペプチドおよび/または成熟C192S SpeBポリペプチドを含んで成る免疫原を提供する。一部の実施形態においては、これらの免疫原は化膿連鎖球菌感染に対して哺乳類宿主を免疫化するための免疫原性組成物において使用される。好ましい実施形態においては、免疫原は、高率の異種化膿連鎖球菌株に対する保護(すなわち、交差保護)を与える成熟C192S SpeBポリペプチドである。
【0065】
抗原または免疫原が通常、免疫原性に基づき規定される。免疫原性は、体液性および/または細胞介在免疫応答を誘発する能力として規定される。したがって、「抗原」または「免疫原」という語は、以下に規定されているように、体液性および/または細胞介在免疫応答を誘発する能力を有する分子である。
【0066】
したがって、1つの実施形態においては、本発明は、少なくとも化膿連鎖球菌成熟SpeBポリペプチドおよび/または成熟C192S SpeBポリペプチドを含んで成る免疫原性組成物に関する。好ましい実施形態においては、成熟SpeBポリペプチド抗原は、配列番号2のアミノ酸146〜398を含んで成る。別の好ましい実施形態においては、成熟C192S SpeBポリペプチド抗原は配列番号2のアミノ酸146〜398を含んで成り、ここで配列番号2の192位でのアミノ酸はシステインからセリン残基に変異されている。本発明の一部の他の実施形態においては、成熟SpeBポリペプチドおよび/または成熟C192S SpeBポリペプチドを含んで成る免疫原性組成物は、1つまたは追加のポリペプチド抗原(すなわち、SpeB以外のポリペプチド)をさらに含んで成り、ここで1つもしくはそれ以上の追加のポリペプチド抗原は化膿連鎖球菌抗原または他の感染性細菌および/またはウイルスからの抗原である。
【0067】
本発明によるSpeBポリペプチドの生物学的同等物または変異体は、SpeBチモーゲン、C192S SpeBチモーゲン、成熟SpeBポリペプチド、成熟C192S SpeBポリペプチド、およびSpeBプロ−ポリペプチドドメインより成る群から選択される化膿連鎖球菌ポリペプチドと実質的な相同を含有するポリペプチドを包含する。例えば、成熟SpeBポリペプチドおよび成熟C192S SpeBポリペプチドの生物学的同等物または変異体も、成熟SpeBポリペプチドが免疫原性反応を引き出す能力を維持する限り、配列番号2の成熟SpeBポリペプチドが修飾されているポリペプチドを含む。一般に、成熟SpeBポリペプチドの機能的な生物学的同等物または変異体は天然アミノ酸配列変異体であり、ここで成熟SpeBポリペプチドは免疫原性反応を引き出す能力を維持する。
【0068】
修飾および変更が本発明の成熟SpeBポリペプチドの構造においてなされ、それでもSpeB免疫原性特性を有する分子が得られる。ポリペプチドの生物学的機能活性を規定するのはポリペプチドの相互作用的能力および性質であるため、一部のアミノ酸配列置換がポリペプチド配列(または、もちろん、その基礎を成すDNAコード配列)においてなされ、それでも同様の特性を有するポリペプチドが得られる。
【0069】
アミノ酸置換は一般に、アミノ酸側鎖置換、例えば、その疎水性、親水性、電荷、サイズなどの相対類似性に基づく(例えば、以下、参照することによりその全体が援用される、カイト(Kyte)とドリトル(Doolittle)、1982年、および米国特許第4,554,101号明細書を参照)。さまざまな上述の特性を考慮に入れる例示の置換は当業者には公知であり、アルギニンおよびリジン、グルタミン酸塩およびアスパラギン酸塩、セリンおよびトレオニン、グルタミンおよびアスパラギン、およびバリン、ロイシンおよびイソロイシンを含む(以下の表2を参照)。したがって、本発明では、上記されているように成熟SpeBポリペプチドの機能的または生物学的同等物が意図されている。
【0070】
【表2】
【0071】
本発明の特定の実施形態においては、多価または組合せ免疫原性組成物が提供される。組合せ免疫原性組成物が、1つもしくはそれ以上の追加の抗原とともに、1つもしくはそれ以上の本発明のポリペプチド(例えば、成熟C192S SpeB)、またはその断片(例えば、SpeBエピトープ断片)を含むことによって提供される。特に、組合せ免疫原性組成物は1つもしくはそれ以上の成熟SpeBポリペプチド、またはその断片を、1つもしくはそれ以上のポリペプチド、ポリペプチド断片、糖質、オリゴ糖類、脂質、リポオリゴ糖類、多糖類、オリゴ糖類−タンパク質コンジュゲート、多糖類−タンパク質コンジュゲート、ペプチド−タンパク質コンジュゲート、オリゴ糖類−ペプチドコンジュゲート、多糖類−ペプチドコンジュゲート、タンパク質−タンパク質コンジュゲート、リポオリゴ糖類−タンパク質コンジュゲート、または多糖類−タンパク質コンジュゲートと組合わせることによって提供される。
【0072】
したがって、一部の実施形態においては、上記の1つもしくはそれ以上の抗原は、化学的付着(すなわち、接合)によって抗原担体タンパク質にコンジュゲートされる。ポリペプチドを担体タンパク質に接合させる手段は当技術分野で周知であり、グルタルアルデヒド、m−マレイミドベンコイル−N−ヒドロキシスクシニミドエステル、カルボジイミド、およびビス−ビアゾチ化(biazotized)ベンジジンを含む。
【0073】
例示の従来のタンパク質担体は、大腸菌DnaKタンパク質、ガラクトキナーゼ(galK)、ユビキチン、α−接合因子、β−ガラクトシダーゼ、およびインフルエンザNS−1タンパク質を含むがこれらに限定されない。トキソイド(すなわち、その毒性活性を削減するように十分な修飾による天然毒素をコード化する配列)、例えば、ジフテリアトキソイドおよび破傷風トキソイド、その各毒素、およびこれらタンパク質の突然変異型も担体として使用されうる。例示の担体タンパク質はジフテリア毒素CRM197である(ジフテリア毒素の非毒性形態、参照することによりその全体が本明細書で援用される、米国特許第5,614,382号明細書を参照)。他の担体としては、緑膿菌(Pseudomonas)の外毒素A、大腸菌の易熱性毒素、コレラ菌(Vibrio cholera)、およびロタウイルス粒子(ロタウイルスおよびVP6粒子を含む)が挙げられる。あるいは、担体タンパク質または他の免疫原性タンパク質の断片またはエピトープが使用されうる。例えば、ハプテンが細菌毒素のT細胞エピトープに結合されうる(米国特許第5,785,973号明細書を参照。同様に、さまざなな細菌熱ショックタンパク質、例えば、マイコバクテリアhsp−70が使用されうる。グルタチオン−S−トランスフェラーゼ(GST)は別の有用な担体である。当業者は、これに関して使用される適切な担体を容易に選択することができる。
【0074】
一部の実施形態においては、本発明は、不溶性成熟SpeB集合体のタンパク質補助折り畳みの方法に関する。同様に、他の実施形態においては、本発明はタンパク質補助折り畳みの方法に関し、ここで成熟SpeBポリペプチドは、1つもしくはそれ以上の分子シャペロンタンパク質の存在下に発現される。
【0075】
例えば、1つの実施形態においては、本発明は、(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るプラスミドを宿主細胞において組換え発現させ、ここでSpeBポリペプチドが宿主細胞において不溶性ポリペプチド集合体を形成するステップと、(b)ポリペプチド集合体を可溶化し、可溶化ポリペプチドが非ネイティブ成熟SpeBポリペプチドとして規定されるステップと、(c)シャペロンタンパク質の存在下に非ネイティブ成熟SpeBポリペプチドを再折り畳みし、非ネイティブ成熟SpeBポリペプチドがネイティブ成熟SpeBポリペプチドへ折り畳まれるステップと、(d)前記ネイティブ成熟SpeBポリペプチドを回収するステップとを含んで成る成熟SpeBポリペプチドを生成するための方法に関する。一部の他の実施形態においては、本発明は、(i)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(ii)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成るポリシストロニックプラスミドを含んで成る宿主細胞における成熟SpeBポリペプチドを組換え発現させ、ここで成熟SpeBポリペプチドが宿主細胞において可溶性である方法を提供する。
【0076】
したがって、特定の実施形態においては、本発明は、成熟SpeBポリペプチドの折り畳みを支援する分子シャペロンタンパク質を提供する。分子シャペロンは当技術分野で周知であり、トリガー因子(TF)などのリボソーム結合タンパク質、Hsp70、DnaK、Hsp40、DnaJ、GrpEなどのシャペロンのHsp70ファミリー、およびGroEL、GroES、Hsp60、Hsp10などのシャペロンのシャペロニンファミリーを含むがこれらに限定されない(クレイトン(Creighton)、1993年、ハルトル(Hartl)とヘイヤー・ハルトル(Hayer−Hartl)、2002年)。本発明の使用に考えられている分子シャペロンタンパク質のポリヌクレオチドおよびポリペプチド配列は当技術分野で周知であり(例えば、各々参照することによりその全体が本明細書で援用される、米国特許第6,159,708号明細書、米国特許第6,010,879号明細書、米国特許第5,776,724号明細書、およびロリマー(Lorimer)とボールドウィン(Baldwin)、Methods in Enzymology、1998年を参照)、折り畳み/再折り畳み要件、補因子などが、所定の分子シャペロンタンパク質に対して以下に記載されている。
【0077】
したがって、非限定的実施例によって、大腸菌からの分子シャペロニンGroELはシャペロンの熱ショックタンパク質60(Hsp60)クラスのメンバーであり、大腸菌GroEオペロンからのGroESとともに発現される。GroELは、集合の傾向があるポリペプチド中間体の濃度およびオフ経路集合率を減少させる折り畳まれていないタンパク質(例えば、成熟SpeB)を結合することによってタンパク質折り畳み反応を支援し、それによってネイティブな立体構造(例えば、適切に折り畳まれた可溶性成熟SpeB)に有利に分配する。補シャペロニンGroES、およびATP、K+、およびMg2+などの補因子は、GroEL介在ポリペプチド折り畳み反応の収量をさらに増大させることが知られている。したがって、特定の実施形態においては、当業者は、シャペロン介在(または補助)タンパク質折り畳みを改善または強化するために当技術分野で周知の成分、補因子、追加のシャペロンタンパク質などを含めるであろう。
【0078】
機序は所定のシャペロンファミリーまたはクラスで変動しうるが、すべてのシャペロンタンパク質(以下に記載されるPDIおよびPPIを除き)によって共有される基礎を成す特徴は、その非ネイティブ立体構造においてタンパク質を結合する能力である。以下に規定されているように、本発明の「分子シャペロン」または「シャペロン」タンパク質は、インビボおよび/またはインビトロでのポリペプチドの折り畳みを補助するタンパク質である。
【0079】
本明細書では分子シャペロン「型」タンパク質としても考えられているのが、タンパク質ジスフィルドイソメラーゼ(PDI)およびペプチジル−プロリルcis/transイソメラーゼ(PPI)であり、これらはタンパク質ジスフィド結合形成およびプロリンcis/trans異性化をそれぞれ触媒する(ロリマー(Lorimer)とボールドウィン(Baldwin)、Methods in Enzymology、1998年を参照)。例えば、化膿連鎖球菌におけるSpeBの発現がRopAタンパク質を必要とすることが、化膿連鎖球菌突然変異によって明らかにされた(リヨン(Lyon)ら、1998年)。RopAタンパク質は、リボソーム上で新生ポリペプチドに結合し、GroELに結合し、かつペプチジル−プロリルイソメラーゼ活性を有することが知られている分子シャペロンである。したがって、本発明の一部の他の実施形態においては、成熟SpeBポリペプチドを生成するための方法は、非ネイティブ成熟SpeBポリペプチドを再折り畳みし、またはPPI、特にRopA PPIの存在下に成熟SpeBを発現させるステップを含んで成る。
【0080】
C.組換え発現ベクターおよび宿主細胞
別の実施形態においては、本発明は、SpeBプロ−ポリペプチドドメインおよび成熟SpeBポリペプチドをコード化するポリヌクレオチドを含んで成る発現ベクターに関する。一部の実施形態においては、本発明の発現ベクターはポリシストロニック核酸配列を含んで成り、ここで1つのシストロンがSpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチドを含んで成り、第2のシストロンが成熟SpeBポリペプチドをコード化するポリヌクレオチドを含んで成る。一部の他の実施形態においては、本発明の発現ベクターは、SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチドを含んで成り、第2の発現ベクターは成熟SpeBポリペプチドをコード化するポリヌクレオチドを含んで成る。1つの好ましい実施形態においては、本発明の発現ベクターはプラスミド構成物である。別の好ましい実施形態においては、本発明の発現ベクターは表3、実施例1に記載されているプラスミドである。
【0081】
本明細書で使用される「ベクター」という語は、それが結合されている別の核酸を輸送する能力がある核酸分子を指す。ベクターの1つの型は「プラスミド」であり、追加のDNA部分がライゲーションされているこれは環状二本鎖DNAループを指す。別の型のベクターはウイルスベクターであり、ここでは追加のDNA部分がウイルスゲノムへライゲーションされている。一部のベクターは、それらが導入される宿主細胞(例えば、複製の細菌源を有する細菌ベクターおよびエピソーム哺乳類ベクター)において自己複製が可能である。さらに、一部のベクターは、それらが操作的に結合される遺伝子の発現を方向づけることが可能である。かかるベクターは、本明細書では「発現ベクター」と呼ばれる。一般に、組換えDNA技術において有用性の発現ベクターはしばしばプラスミドの形態である。本明細書においては、「プラスミド」および「ベクター」は、プラスミドがベクターの最も一般的に使用される形態であるため同じ意味で使用される。しかし、本発明では、同等の機能を果たすウイルスベクター(例えば、複製欠損レトロウイルス、アデノウイルス、およびアデノ関連性ウイルス)など他の形態のかかる発現ベクターを含むことは意図されている。
【0082】
原核生物におけるタンパク質の発現は、融合または非融合タンパク質のいずれかの発現を方向づける構成的または誘導的プロモーターを含有するベクターにより大腸菌において最も多く行われる。融合ベクターは、その中でコード化されたタンパク質、組換えタンパク質のアミノまたはカルボキシ末端に多くのアミノ酸を添加する。
【0083】
一部の実施形態においては、宿主細胞が、(i)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列、および(ii)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るポリシストロニックプラスミドで形質転換され、形質導入され、トランスフェクトされ、または感染される。本明細書で規定されているように、「ポリシストロニックmRNA」は、2つもしくはそれ以上のポリペプチドをコードする。したがって、以下に規定されているように、本発明の「ポリシストロニックポリヌクレオチド」、「ポリシストロニックcDNA」、または「ポリシストロニックプラスミド」は、ポリシストロニックmRNAをコードし、次いで2つもしくはそれ以上のポリペプチドをコード化する。
【0084】
適切な誘導的、非融合、大腸菌発現ベクターの例としては、pTrc(アマン(Amann)ら、1988年)、pET IId(スタディール(Studier)ら、1990年)、pET、pRSET、pCRT7−CTTOPO、およびpIVeXが挙げられる。pTrcベクターからのターゲット遺伝子発現は、ハイブリッドtrp−lac融合プロモーターからの宿主RNAポリメラーゼ転写に依拠する。pET IIdベクターからのターゲット遺伝子発現は、共発現ウイルスRNAポリメラーゼT7gnlによって介在されるT7gn1β−lac融合プロモーターからの転写に依拠する。このウイルスポリメラーゼは、lacUV5プロモーターの転写制御下にT7gnl遺伝子を有する常在プロファージからの宿主菌株BL21(DE3)またはHMSI74(DE3)によって供給される。一部の実施形態においては、pRK5、pCMVBlue、pCMV−LIC、pAPL400−023、pAPL400−087、およびpAPL400−088などヒトCMVまたはシミアンCMVプロモーターを含んで成るプラスミドベクターも考えられている。
【0085】
大腸菌における組換えタンパク質発現を最大限にする1つの方法は、組換えタンパク質をタンパク質分解的に切断する能力障害による宿主細菌におけるタンパク質の発現である。別の方法は、発現ベクターへ挿入される核酸の核酸配列を変化させ、各アミノ酸に対する個々のコドンが大腸菌において優先的に利用されるものとなることである。本発明の核酸配列のかかる変化は、標準のDNA突然変異または合成法によって行われる。
【0086】
プロモーターは、通常、転写の開始点(すなわち、転写開始部位)の前(上流)で約100個のヌクレオチド対内のDNA分子の領域である。その領域は通常、異なる遺伝子における同様のそれぞれの位置に配置されている数種類のDNA配列要素を含有する。本明細書で使用される「プロモーター」という語は、技術上、上流プロモーター領域またはプロモーター領域と呼ばれるものを含む。
【0087】
本発明の別の態様は、本発明の組換え発現ベクターが導入されている宿主細胞に関する。「宿主細胞」、「遺伝子組換え宿主細胞」、および「組換え宿主細胞」は、本明細書では同じ意味で使用される。かかる語は特定の対象の細胞だけではなく、かかる細胞の子孫または潜在的子孫を指すことが理解される。特定の修飾が突然変異または環境的影響により後世において起こりうるため、かかる子孫は、実際には、親細胞と同一ではありえないが、それでも本明細書で使用される語の範囲内に含まれる。宿主細胞は原核または真核生物細胞であるが、好ましくは、原核細胞である。例えば、SpeBポリペプチドは大腸菌、および化膿連鎖球菌など細菌細胞で発現される。他の実施形態においては、SpeBポリペプチドは昆虫細胞(例えば、Sf9、高5およびSf21細胞)、酵母(例えば、P.パストリス(P.pastoris)、P.メタノリカ(P.methanolica)、S.ポンベ(S.pombe)、およびS.セレヴィシエ(S.cerevisiae))または哺乳類細胞(例えば、チャイニーズハムスター卵巣細胞(CHO)、Cos−1、CV−1、HeLa、NIH3T3、PER−C6、およびNSOにおいて発現される。他の適切な宿主細胞は当業者に周知である。
【0088】
培養における原核祝細胞など本発明の宿主細胞が、SpeBポリペプチドを生成する(すなわち、発現させる)ために使用される。1つの実施形態においては、方法は、成熟SpeBポリペプチドおよびSpeBプロ−ポリペプチドドメインが生成されるまで適切な培地で(成熟SpeBポリペプチドおよびSpeBプロ−ポリペプチドドメインをコード化する組換え発現ベクターが導入されている)本発明の宿主細胞を培養するステップを含んで成る。別の実施形態においては、方法は、成熟SpeBポリペプチドを培地または宿主細胞から単離するステップをさらに含んで成る。
【0089】
ベクターDNAは、従来のトランスフォーメーション、形質導入、感染、またはトランスフェクション法によって原核または真核細胞へ導入される。本明細書で使用される「トランスフォーメーション」、「感染」、および「トランスフェクション」は、リン酸カルシウムまたは塩化カルシウム共沈殿、DEAE−デキストラン介在トランスフェクション、リポフェクション、感染、またはエレクトロポレーションを含む宿主細胞へ外来核酸(例えば、DNA)を導入する技術上認められたさまざまな方法を指すことが意図されている。宿主細胞を形質転換し、感染し、トランスフェクトする適切な方法は、サムブルック(Sambrook)ら(「Molecular Cloning:A Laboratory Manual」、第2版、コールド・スプリング・ハーバー・ラボラトリー(Cold Spring Harbor Laboratory)、コールド・スプリング・ハーバー・ラボラトリー・プレス(Cold Spring Harbor Laboratory Press)、コールド・スプリング・ハーバー(Cold Spring Harbor)、ニューヨーク州、1989年)、および他の実験マニュアルに見出される。
【0090】
最も広く使用されている方法は、リン酸カルシウムまたはDEAE−デキストランのいずれかによって介在されるトランスフェクションである。機序は依然として不明であるが、トランスフェクトされたDNAはエンドサイトーシスによって細胞の細胞質に入り、核に輸送されると考えられている。細胞型によって、培養細胞の90%までの集団がつねにトランスフェクトされる。その高い効率により、リン酸カルシウムまたはDEAE−デキストランによって介在されるトランスフェクションは、多数の細胞における外来DNAの一時的発現を必要とする実験に選択される方法である。リン酸カルシウム介在トランスフェクションは、通常、宿主細胞ゲノムへの頭−尾タンデムで配置される外来DNAのコピーを組込む細胞系を確立するためにも使用される。
【0091】
発現ベクターのコード配列は、操作的に転写終了領域へ結合される。RNAポリメラーゼは、ポリアデニル化が起こる部位を通じてコード化DNA配列を転写する。通常、ポリアデニル化部位の数百塩基対下流に位置したDNA配列は転写を終了させる役目をする。それらのDNA配列は本明細書では転写終了領域と呼ばれる。それらの領域は転写メッセンジャーRNA(mRNA)の有効なポリアデニル化に必要とされる。転写終了領域は当技術分野で周知である。例示の転写終了領域は、SV40またはプロタミン遺伝子のポリアデニル化シグナルを含んで成る。
【0092】
本発明のDNA分子、遺伝子、またはポリヌクレオチドは、当技術分野で周知である多くの方法によってベクターへ組入れられる。例えば、ベクターpUC18は有用性が明らかにされている。同様に、関連ベクターM13mp18およびM13mp19は、特に、ジデオキシ配列決定の実施における本発明の一部の実施形態において使用される。
【0093】
好ましい実施形態においては、本発明の組換え宿主細胞は原核宿主細胞である。好ましくは、本発明の組換え宿主細胞は大腸菌のDH5α菌株の細菌細胞である。一般に、原核細胞が、DNA配列の初期クローニングおよび本発明において有用であるベクターの構成のために好ましい。例えば、大腸菌K12菌株はとりわけ有益である。使用される他の微生物菌株としては、大腸菌Bおよび大腸菌x(E.colix)1976(ATCC番号31537)が挙げられる。これらの例は、もちろん、限定ではなく例示が目的とされている。
【0094】
原核生物も発現のために使用される。上記の菌株のほか、W3110(ATCC番号273325)、BLR(DE3)、BLR(DE3)pLysS、AD494(DE3)、AD494(DE3)pLysS、BL21(DE3)、BL21(DE3)pLysS、BL21(DE3)pLysE、BL21(DE3)pLacI、BL21trxB(DE3)、BL21trxB(DE3)pLysS、 HMS174(DE3)、HMS174(DE3)pLysS、HMS174(DE3)pLysE、Origami(DE3)、Origami(DE3)pLysS、Origami(DE3)pLysE、Origami(DE3)pLacI、OrigamiB(DE3)、OrigamiB(DE3)pLysS、OrigamiB(DE3)pLysE、OrigamiB(DE3)pLacI、Rosetta(DE3)、Rosetta(DE3)pLysS、Rosetta(DE3)pLysE、Rosetta(DE3)pLacI、Tuner(DE3)、Tuner(DE3)pLysS、およびTuner(DE3)pLacIなどの大腸菌株、枯草菌(Bacillus subtilis)などの桿菌、またはネズミチフス菌(Salmonella typhimurium)もしくは霊菌(Serratia marcesans)、およびさまざまなシュードモナス(Pseudomonas)種などの腸内細菌科が使用される。
【0095】
一般に、宿主細胞と共存できる種由来であるレプリコンおよび制御配列を含有するプラスミドベクターが、これらの宿主といっしょに使用される。ベクターは通常、複製部位、および形質転換された細胞における表現型選択を提供する能力がある標識配列を有する。例えば、大腸菌は、大腸菌種由来のプラスミド、pBR322を使用して形質転換される(ボリバー(Bolivar)ら、1977年)。pBR322は、アンピシリンおよびテトラサイクリン耐性の遺伝子を含有し、したがって形質転換された細胞を同定する容易な手段を提供する。pBRプラスミド、もしくは他の微生物プラスミドまたはファージは、それぞれのポリペプチドの発現のための微生物によって使用されるプロモーターも含有しなければならず、または含有するように修飾される必要がある。
【0096】
組換えDNA構成において最も一般的に使用されるプロモーターとしては、β−ラクタマーゼ(ペニシリナーゼ)およびラクトースプロモーター系(チャン(Chang)ら、1978年、イタクラ(Itakura)ら、1977年、ゲデル(Goeddel)ら、1979年、ゲデル(Goeddel)ら、1980年)、トリプトファン(TRP)プロモーター系(欧州出願第EP0036776号明細書、シーブウェンリスト(Siebwenlist)ら、1980年)、およびT7またはT7lacプロモーター系が挙げられる。これらは最も一般的に使用されているが、他の微生物プロモーターが発見され、かつ利用されており、それらのヌクレオチド配列に関する詳細は公開されており、当業者が機能的プロモーターをプラスミドベクターへ導入することが可能である(シーブウェンリスト(Siebwenlist)ら、1980年)。
【0097】
トランスフェクション後、細胞は培養条件下でSpeBポリペプチドの発現に十分な時間にわたって維持される。培養条件は当技術分野で周知であり、イオン組成および濃度、温度、pHなどを含む。通常、トランスフェクトされた細胞は培養培地で培養条件下で維持される。さまざまな細胞型のための適切な培地は当技術分野で周知である。好ましい実施形態においては、温度は約20℃〜約50℃、より好ましくは、約30℃〜約40℃、かつさらに好ましくは、約37℃である。
【0098】
pHは、約6.0の値〜約8.0の値、より好ましくは、約6.8の値〜約7.8の値、かつ最も好ましくは、約7.4である。オスモル濃度は、好ましくは、約200ミリオスモル/リットル(mosm/L)〜約400mosm/L、より好ましくは、約290mosm/L〜約310mosm/Lである。コード化ポリペプチドのトランスフェクションおよび発現のための他の生物学的条件は当技術分野で周知である。
【0099】
トランスフェクトされた細胞は、SpeBポリペプチドの発現に十分な時間にわたって維持される。適切な時間は、とりわけ、使用される細胞型によって決まり、当業者によって容易に決定可能である。通常、維持時間は、約2〜約14日である。
【0100】
組換えSpeBポリペプチドは、トランスフェクトされた細胞またはその細胞が培養されている培地のいずれかより回収され、または収集される。回収は、SpeBポリペプチドの単離および精製を含んで成る。ポリペプチドのための単離および精製法は当技術分野で周知であり、沈殿、ろ過、クロマトグラフィー、電気泳動などの方法を含む。
【0101】
さらに別の実施形態においては、本発明のSpeBポリペプチドは、細胞を含まない翻訳系などインビトロタンパク質翻訳系によって生成される(例えば、ベトン(Betton)、2003年、ブラウン(Braun)ら、2002年、ジェルムツス(Jermutus)ら、1998年、キグワ(Kigawa)ら、1999年、キム(Kim)ら、1996年、およびスピリン(Spirin)ら、1988年を参照)。例えば、各々参照することによりその全体が本明細書で援用される、米国特許第6,399,323号明細書、および米国特許第5,478,730号明細書は、細胞を含まない(インビトロ)翻訳系におけるポリペプチダーゼの調製または生成のための方法、条件などを記載している。
【0102】
D.免疫原性組成物
一部の好ましい実施形態においては、本発明は、成熟SpeBポリペプチド免疫原(すなわち、成熟野生型または成熟C192S)および生理的に許容される担体を含んで成る成熟SpeB免疫原性組成物を提供する。より好ましくは、免疫原性組成物は、配列番号2のアミノ酸146−398を含んで成る成熟野生型SpeBポリペプチド、または配列番号2のアミノ酸146−398を含んで成る成熟C192S SpeBポリペプチドを含んで成り、ここで配列番号2のシステインアミノ酸192はセリンに変異されている。本発明のさらに他の実施形態においては、多価または組合せ免疫原性組成物が提供される。組合せ免疫原性組成物が、化膿連鎖球菌および/または他の細菌種からの1つもしくはそれ以上の追加の抗原とともに、1つもしくはそれ以上の本発明のポリペプチド(例えば、成熟C192S SpeB)を含むことによって提供される。特に、組合せ免疫原性組成物が、本発明の1つもしくはそれ以上の成熟SpeBポリペプチドを、1つもしくはそれ以上のポリペプチド、ポリペプチド断片、糖質、オリゴ糖類、脂質、リポオリゴ糖類、多糖類、オリゴ糖類−タンパク質コンジュゲート、多糖類−タンパク質コンジュゲート、ペプチド−タンパク質コンジュゲート、オリゴ糖類−ペプチドコンジュゲート、多糖類−ペプチドコンジュゲート、タンパク質−タンパク質コンジュゲート、リポオリゴ糖類−タンパク質コンジュゲート、または多糖類−タンパク質コンジュゲートと組合わせることによって提供される。
【0103】
本発明の成熟SpeBポリペプチドは、哺乳類対象、例えば、ヒトへの投与に適切な免疫原性組成物へ組入れられる。かかる組成物は通常、「免疫原性」組成物および薬剤として許容される担体を含んで成る。以下で使用される「薬剤として許容される担体」という言葉は、薬剤投与と適合するすべての溶剤、分散剤、コーティング剤、抗菌剤および抗真菌剤、等張剤および吸収遅延剤などを含むことが意図されている。薬剤活性物質のためのかかる溶剤および薬剤の使用は当技術分野で周知である。従来の溶剤または薬剤が活性化合物と不適合である場合を除き、かかる溶剤は本発明の組成物において使用される。補助活性化合物も組成物へ組入れられる。
【0104】
本発明の免疫原性組成物は、その投与の意図された経路と適合性であるように製剤化される。投与経路の例としては、非経口(例えば、静脈内、皮内、皮下、筋内、腹腔内)、粘膜(例えば、経口、直腸、鼻内、口腔、膣、呼吸器)、および経皮(局所)が挙げられる。非経口、皮内、または皮下適用に使用される溶液または懸濁液としては、以下の要素、すなわち、注射用の水、食塩水、固定油、ポリエチレングリコール、グリセリン、プロピレングリコール、または他の合成溶剤など滅菌希釈剤、ベンジルアルコールまたはメチルパラベンなど抗菌剤、アスコルビン酸または重亜硫酸ナトリウムなど抗酸化剤、エチレンジアミン四酢酸などキレート化剤、酢酸塩、クエン酸塩、またはリン酸塩などバッファー、および塩化ナトリウムまたはデキストロースなど張性の調整剤が挙げられる。pHは、塩酸または水酸化ナトリウムなど酸または塩酸で調整される。非経口製剤は、アンプル、使い捨て注射器、もしくはガラスまたはプラスチック製のマルチドーズ用バイアルに入れられる。
【0105】
滅菌注射溶液は、活性化合物(例えば、成熟SpeB)を、必要に応じて上記列挙した成分の1つまたは組合せとともに適切な溶剤中で必要な量において組入れた後、ろ過滅菌によって調製される。一般に、分散剤は、基礎分散溶剤および上記列挙したものからの必要な他の成分を含有する滅菌ビヒクルへ活性化合物を組入れることによって調製される。滅菌注射溶液用の滅菌粉末の場合、調製の好ましい方法は真空乾燥および凍結乾燥であり、これによりその以前に滅菌ろ過した溶液から活性成分+追加の所望の成分の粉末が得られる。
【0106】
経口組成物は一般に不活性希釈剤または食用担体を含む。それらはゼラチンカプセルに入れられ、または錠剤の中へ圧縮される。治療用の経口投与のために、活性化合物は賦形剤とともに組入れられ、錠剤、トローチ、またはカプセルの形で使用される。経口組成物は、口内洗浄剤として使用される液体担体を使用しても調製され、ここで液体担体中の化合物は経口適用され、局所使用され、吐出され、または燕下される。製剤適合性結合剤、および/または佐剤物質が組成物の一部として含まれる。錠剤、ピル、カプセル、トローチなどは、以下の成分のいずれか、または類似した性質の化合物のいずれかを含有する。すなわち、微結晶性セルロース、トラガカントゴム、またはゼラチンなど結合剤、でんぷんまたはラクトースなど賦形剤、アルギン酸、プリモゲル(Primogel)、またはコーンスターチなど崩壊剤、ステアリン酸マグネシウムまたはステロート(Sterotes)など潤滑剤、コロイド状二酸化ケイ素など流動促進剤、スクロースまたはサッカリンなど甘味剤、またはペパーミント、サルチル酸メチル、またはオレンジ香味料など香料添加剤。
【0107】
吸入による投与のために、化合物は、適切な推進剤、例えば、炭酸ガスなどのガスを含有する加圧容器またはディスペンサー、またはネブライザーからのエアロゾルスプレーの形で送達される。全身投与も粘膜または経皮手段による。粘膜または経皮投与のために、浸透されるバリアに適切な浸透剤が製剤で使用される。かかる浸透剤は一般に当技術分野で周知であり、例えば、粘膜投与のための洗浄剤、胆汁塩、およびフシジン酸誘導体を含む。粘膜投与は、鼻腔用スプレーまたは坐剤を使用して行われる。経皮投与のために、活性化合物は軟膏、膏薬、ゲル、または一般に当技術分野で周知であるクリームへ処方される。
【0108】
化合物は坐剤(例えば、カカオバターおよび他のグリセリドなど従来の坐剤の基剤による)または直腸送達用の停留浣腸の形でも調製される。
【0109】
1つの実施形態においては、活性化合物は、インプラントおよびマイクロカプセル化送達系を含む、制御放出製剤など体内からの急速な排泄に対して化合物を保護する担体とともに調製される。
【0110】
生体分解性、生体適合性ポリマー、例えば、エチレン酢酸ビニル、ポリ無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル、およびポリ乳酸などが使用される。かかる製剤の調製方法は当業者には明らかであろう。材料は、アルザ・コーポレーション(Alza Corporation)およびノバ・ファルマセウティカルズ(Nova Pharmaceuticals,Inc.)からも市販されている。リポソーム懸濁液(ウイルス抗原に対するモノクローナル抗体による感染細胞にターゲティングされたリポソームを含む)も薬剤として許容される担体として使用される。これらは、例えば、以下で参照することにより援用される、米国特許第4,522,811号明細書に記載されているように、当業者に周知の方法に従って調製される。
【0111】
投与を容易にし、投与量を均一にするために、用量単位剤形で経口または非経口組成物を処方することが特に有利である。以下で使用される用量単位剤形は、治療される対象のための単位用量として適した物理的に分離した単位を指し、各単位は、必要な薬剤担体とともに所望の治療効果を生じるように計算された所定量の活性化合物を含有する。本発明の用量単位剤形の仕様は、活性化合物の独自の特徴および達成される特定の治療効果、ならびに個人の治療用にかかる活性化合物を配合する技術上固有の制限によって決定され、かつ直接それらに依存している。
【0112】
薬剤として許容されるビヒクルは、副作用を引き起こすことがなく、例えば、活性化合物の投与を促進し、体内におけるその寿命および/またはその有効性を増大し、溶液中の溶解度を増大し、あるいは、その保存を強化することを可能にする薬剤または免疫原性組成物へ入る化合物または化合物の組合せを指定すると理解されている。これら薬剤として許容されるビヒクルは公知であり、選択された活性化合物の性質および投与方法に従って当業者によって適合されるであろう。
【0113】
本発明の免疫原性原性は、1つもしくはそれ以上の佐剤をさらに含んで成りうる。「佐剤」は、抗原の免疫原性を強化するのに役立つ物質である。したがって、佐剤はしばしば免疫応答を高め、当業者には公知である。本発明で意図されている佐剤の例は、リン酸アルミニウムおよび水酸化アルミニウムなどアルミニウム塩(alum)、ヒト結核菌(Mycobacterium tuberculosis)、百日咳菌(Bordetella pertussis)、細菌性リポ多糖、アミノアルキルグルコサミンリン酸化合物(AGP)、もしくはその誘導体または類似体を含むがこれらに限定されず、これらはコリザ(Corixa)(ハミルトン(Hamilton)、モンタナ州(MT))から市販されており、かつ米国特許第6,113,918号明細書に記載されており、かかるAGPの1つは、2−[(R)−3−テトラデカノイルオキシテトラデカノイルアミノ]エチル2−デオキシ−4−O−ホスホノ−3−O−[(R)−3−テトラデカノイルオキシテトラデカノイル(tetradecanoyoxytetradecanoyl)]−2−[(R)−3−テトラデカノイルオキシテトラデカノイルアミノ(tetradecanoyoxytetradecanoylamino)]−b−D−グルコピラノシドであり、これは529としても知られる(以前はRC529として知られる)が、これは安定乳剤または水性剤形として処方されており、米国特許第4,912,094号明細書に記載されているMPL(商標)(3−O−脱アシル化モノホスホリル脂質A)(コリザ(Corixa))、CpGモチーフ(米国特許第6,207,646号明細書)を含有するオリゴヌクレオチドなど合成ポリヌクレオチド、ポリペプチド、米国特許第5,057,540号明細書に記載されている、Quil AまたはSTIMULON(商標)QS−21(アンチジェニックス(Antigenics)、フラミンガム(Framingham)、マサシューセッツ州)などサポニン、百日咳毒素(PT)、または大腸菌易熱性毒素(LT)、特にLT−K63、LT−R72、CT−S109、PT−K9/G129、例えば、国際特許公開国際公開第93/13302号パンフレットおよび同第92/19265号パンフレットを参照、コレラ毒素(野生型または突然変異体のいずれか、例えば、ここでアミノ酸29位でのグルタミン酸が別のアミノ酸、好ましくは、公開された国際特許出願番号国際公開第00/18434号パンフレットに従って、ヒスチジンで置換されている)。さまざまなサイトカインおよびリンフォカインが佐剤としての使用に適している。かかる佐剤の1つが、米国特許第5,078,996号明細書に記載されているヌクレオチド配列を有する、顆粒球マクロファージコロニー刺激因子(GM−CSF)である。GM−CSF cDNAを含有するプラスミドは、大腸菌へ形質転換されており、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection)(ATCC)、1081University Boulevard,Manassas、バージニア州(VA)20110−2209により、登録番号39900で寄託されている。サイトカインインターロイキン−12(IL−12)は、米国特許第5,723,127号明細書に記載されている別の佐剤である。他のサイトカインまたはリンフォカインは、インターロイキン1−α、1−β、2、4、5、6、7、8、10、13、14、15、16、17、および 18、インターフェロン−α、β、およびγ、顆粒球コロニー刺激因子、および腫瘍壊死因子αおよびβを含むがこれらに限定されない免疫修飾活性を有することが示されており、佐剤としての使用に適している。
【0114】
本発明の組成物は通常、標準、公知、非毒性の生理的に許容される担体、佐剤、および所望のビヒクルを含有する用量単位製剤で非経口投与される。以下で使用される非経口という語は、静脈内、皮下、皮内、筋内、動脈内注射、または注入技術を含む。
【0115】
注射製剤、例えば、滅菌注射水性または油性懸濁液は、適切な分散または湿潤剤および懸濁剤を使用して既知の技術に従って処方される。滅菌注射製剤は、例えば、1,3−ブタンジオール中の溶液として、非毒性の非経口的に許容される希釈剤または溶剤中の注射溶液または懸濁液でもある。
【0116】
使用されうる許容されるビヒクルおよび溶剤としては、水、リンガー溶液、および生理食塩液が挙げられる。また、滅菌、固定油が従来、溶剤または懸濁化剤として使用されている。このために、合成モノ−またはジ−グリセリドを含む無菌性固定油が使用されている。また、オレイン酸など脂肪酸が注射剤の調製に使用されている。
【0117】
好ましい担体としては、リン酸塩、乳酸塩、トリス(Tris)などによる中性食塩水バッファーが挙げられる。ウイルスベクターの投与に際しては、ベクターは十分に精製され、欠陥干渉アデノウイルス粒子または内毒素および他の発熱源など、望ましくない汚染物を実質的に含まないようにし、ベクター構成物が投与される個人において有害反応を引き起こさないようにする。ベクターを精製する好ましい手段には、塩化セシウム勾配遠心分離など浮遊密度勾配の使用が含まれる。
【0118】
担体はリポソームでもある。送達ビヒクルとしてリポソームを使用する手段は当技術分野で周知である。
【0119】
特定の実施形態においては、本発明の免疫原性組成物は、遺伝子発現を制御する調節配列と操作的に関連する本発明のポリヌクレオチド配列を含んで成る。目的とするポリヌクレオチド配列は、DNAの発現を促進する調節要素、すなわち、プロモーターおよび/またはエンハンサー要素の制御下にプラスミドなど発現ベクターへ工学的に入れられる。好ましい実施形態においては、ヒトサイトメガロウイルス介在早期プロモーター/エンハンサーが使用される(米国特許第5,168,062号明細書)。プロモーターは細胞特異的であり、かつ所定の細胞でのみポリヌクレオチドの実質的な転写を可能にしうる。
【0120】
ポリヌクレオチドは、「裸の」DNA(米国特許第5,580,859号明細書)として宿主へ直接導入されるか、またはブピビカインおよび他の局所麻酔薬(米国特許第5,593,972号明細書)およびカチオンポリアミン(米国特許第6,127,170号明細書)など免疫化を促進する薬剤で組成物中に処方される。
【0121】
このポリヌクレオチド免疫化方法においては、本発明のポリペプチドはインビボでの一過性に発現され、宿主の染色体に挿入または組込まれる遺伝物質はない。この方法は、目標が目的とする遺伝物資を染色体へ挿入または組込む遺伝子療法とは区別されるべきである。免疫化によって投与されるポリヌクレオチドが宿主における形質転換表現型を生じさせないことを確認するアッセイが使用される(米国特許第6,168,918号明細書)。
【0122】
すべての特許および刊行物は、参照することにより本明細書で援用される。
E.実施例
以下の実施例は、他の方法で詳細に記載されている場合を除き、当業者に公知であり、かつルーチンである標準の方法を使用して行われる。以下の実施例は、例示目的に示されており、決して本発明の範囲を限定するものとして考えるべきではない。
【0123】
実施例1
材料と方法
培地と試薬。大腸菌BLR(DE3)(ノバーゲン(Novagen)、カリフォルニア州(CA))を発現試験用に使用した。適切な抗生物質選択条件下で細菌をルリア寒天培地(LB)で生育させた。アンピシリンを濃度100μg/mLで使用し、カナミシンを50μg/mLで使用した。ZeroBluntTOPクローニングベクターpCR−Blunt(インビトロジェン(InVitrogen)、カールズバッド(Carlsbad)、カリフォルニア州(CA))をPCR生成断片のクローニング用に使用した。Century−Plus RNA Markers(商標)、Millennium RNA Markers(商標)、RNAlater(商標)、RNAqueous−Midi(商標)、DNA−free(商標)、ULTRAhyb(商標)、NorthernMax(商標)、およびRETROscript(商標)をアンビオン(Ambion)(オースチン(Austin)、テキサス州(TX))から得た。GeneScreen(商標)ハイブリダイゼーション膜、蛍光−N6−dATP、Renaissance(登録商標)Antifluorescein−APコンジュゲートポリクローナル抗体、およびCDP−Star(登録商標)をパーキンエルマー・ライフ・サイエンシス(PerkinElmer Life Sciences)、ボストン(Boston)、マサチューセッツ州(MA)から得た。すべての制限酵素をニュー・イングランド・バイオラブス(New England Biolabs)(べバリー(Beverly)、マサチューセッツ州(MA))から入手した。
【0124】
ポリメラーゼ連鎖反応。特に断りのない限り、PCR増幅を以下の反応条件を使用して最終容量50μLで行った。すなわち、0.2mM dNTPs、1.0mM DTT、0.8μM各プライマー、10U熱安定ポリメラーゼ、および1×熱安定ポリメラーゼバッファー。クローニングおよび突然変異反応のために、Pwoポリメラーゼ(ベーリンガー・マンハイム(Boehringer Mannheim)、インディアポリス(Indianapolis)、インディアナ州(IN))を使用したが、他のすべて増幅ではTaqポリメラーゼ(アプライド・バイオシステムズ(Applied Biosystems)、フォスターシティー(Foster City)、カリフォルニア州(CA))を使用した。増幅は、94℃で30秒間、55℃で30秒間、および72℃で30秒間の25サイクルで構成された。
【0125】
プラスミド構成物。成熟SpeBコード領域に対応する770bp断片を、それぞれNco IおよびBamH I部位を含有するフォワード (5’CCATGGAACCAGTTGTTAAATCTCTCC3’)(配列番号3)プライマーおよびリバース(5’GGATCCTAAGGTTTGATGCCTACAACAGC3’)(配列番号4)プライマー(下線)を使用してPCR増幅した。フォワードプライマーは、NcoIクローニング部位内にネストしたATG翻訳開始コドンを含有するように設計され、結果として発現タンパク質のN末端へ追加のメチオニン残基が生じ、第1の残基をグルタミン(CAA)からグルタミン酸塩(GAA)へ変更した。同様に、プロ配列ドメイン(アミノ酸28−146)を含む367bp断片を、クローニングのための工学的NcoI(フォワード)部位およびBamHI(リバース)部位とともに、フォワード(5’CCATGGATCAAAACTTTGCTCGTAACG3’)(配列番号5)プライマー、およびリバース(5’GGATCCTTATTTAATCTCAGCGGTACCAGC3’)(配列番号6)プライマーを使用して増幅した。リバースプライマーにおけるBamHI部位のすぐ3’方向に終止コドンを挿入し、発現組換えタンパク質の翻訳終了を指令した。PCR反応では、以前に記載されているように(マツカ(Matsuka)ら、1999年)、単一システインを192位でセリン置換(C192S)にコード化するTGT〜AGT突然変異によるSpeBチモーゲンを含有するプラスミドベースのテンプレートを使用した。PCR産物をpCR−Bluntヘサブクローン化し、その後にNcoIおよびBamH Iを使用して制限消化によって切除した。770bp成熟C192S SpeBおよび367bpプロ配列ドメインコード断片をアガロースゲル電気泳動によって精製し、それぞれ、NcoIおよびBamHI制限部位を使用してpET28aおよびpET3dへライゲートした。結果として生じる発現プラスミド、pLP681およびpLP682を標準方法を使用して大腸菌BLR(DE3)へ同時形質転換した。生成細菌発現菌株を2−プラスミドベースの共発現分析に利用した。
【0126】
成熟野生型SpeB発現構成物を、単一のA〜T塩基変更(太字)とともに、重複フォワード
(配列番号7)プライマー、およびリバース
(配列番号8)プライマーを使用する突然変異PCRを使用して生成し、PCRのためのテンプレートとして使用したpLP681におけるC192S突然変異を復帰した。突然変異生成は、ワイナー(Weiner)ら(1994年)によって記載された方法の変形で構成され、以下のサイクル条件を使用した。すなわち、94℃で1分、94℃で15秒および68℃で10分の16サイクルの後、94℃で15秒および68℃で10分の12サイクルとともに、68℃で各サイクルで15秒増分で増加させる延長時間により、結果として68℃で13分の最終延長時間が生じた。PCR反応を大腸菌へのトランスフォーメーション前にDpnIで切断した。結果として生じるクローンpLP680を使用前に配列決定し、所望の突然変異を検証した。プラスミドをpLP682で大腸菌BLR(DE3)へ同時形質転換し、2−プラスミドベース共発現試験で使用した。
【0127】
ポリシストロニック発現ベクターを構成するために、pLP682からのSpeBプロ配列ドメインコード領域をNcoIおよびBamHIを使用する制限消化によって切断した。367bp断片をアガロースゲル電気泳動によって精製し、やはりNco IおよびBamH Iで制限したpET28aへライゲートし、結果としてpLP688が生じた。
【0128】
長さを増大させる4種類のリンカー領域に引き続いて、以下のフォワードプライマーを使用するPCRによって成熟SpeBコード領域を生成した。すなわち、
すべてのフォワードプライマーは、5’末端(下線)上にBglII制限部位、最適化シャイン・ダルガノ部位(太字イタリック)、および翻訳開始コドン(太字)を含んだ。翻訳終止コドンへの5’直後にXhoI部位(下線)を含有するリバースプライマー(5’CTCGAGCTAAGGTTTGATGCCTA−CAACAGC3’)(配列番号13)をすべてのリンカーベースの増幅反応のために使用した。
【0129】
上記のプライマーを使用し、5nt、10nt、または20ntリンカー+成熟C192S SpeBコード領域を増幅し、pLP681をPCRのためのテンプレートとして利用した。同様に、pLP680をテンプレートとして使用し、20ntリンカー+成熟野生型SpeB PCR産物を生成した。PCR断片をpCR−Bluntへサブクローン化し、BglIIおよびXhoIによる制限消化によって切除し、アガロースゲル電気泳動によって精製した。単離後、BamHIおよびXhoI制限部位を使用して断片をpLP688へライゲートし、pLP683、pLP684、pLP685、およびpLP687ポリシストロニック発現構成物を生成した。同一のやり方で、上記のプライマーによりPCRのためのテンプレートとしてpLP685を使用してpLP686を合成した。インビボ分析のために生成され、使用されたすべての発現プラスミドの明細が表3に示されている。表3に示されているすべての発現は、特に断りのない限り、pET28aバックグラウンドにおけるものである。
【0130】
【表3】
【0131】
組換えタンパク質の発現。ポリシストロニックおよび2−プラスミドベースの共発現試験のために、適切な抗生物質を含有するLB200mLを所望の細菌発現ショックで接種し、37℃下に一夜、生育させた。一夜培養物を培地を含有する新鮮抗生物質2Lへ1:10に希釈し、25℃下に約0.6のOD600に生育させ、25℃下に16時間、1mM IPTGで誘発した。遠心分離で細胞を収集し、結果として生じる細胞ペレットを使用するまで−20℃下に保存した。誘発前後のサンプルをタンパク質発現およびRNA単離の分析用に採取した。RNA分析用のサンプルを使用するまで−70℃下にRNAlater(商標)で保存した。
【0132】
タンパク質発現を超音波処理による細胞融解後のSDS−PAGE分析によって評価した。不溶性材料を遠心分離によってペレット化し、可溶性上清フラクションを回収した。細胞残屑をPBS 1mL中に再懸濁する前にPBSで2回洗浄した。すべての可溶性および不溶性フラクションを細胞融解前のOD600の細菌培養に基づき標準化した。クマシーブルー染色によって、かつ標準方法によるPVDF膜への移動後のウェスタンブロット分析によってタンパク質発現を視覚化した。SpeBのチモーゲン形態に対して生成されたポリクローナル抗体を使用してブロットをプローブし、発現プロ配列ドメインおよび成熟28kDa SpeBの同時の検出を可能にした。
【0133】
RNAの単離。総RNAをRNAqueous−Midi(商標)を使用して細菌培養物から単離した。RNAlater(商標)で保存した誘発後サンプルを室温下に解凍し、RNAqueous(商標)溶解/結合バッファー1mL中に再懸濁する前にペレット化した。次いで、溶解物を説明マニュアルに規定されているように処理した。単離RNAをLiClを使用して沈殿させ、DNA−free(商標)で2回、メーカーの仕様とおり処理し、汚染ゲノムまたはプラスミドDNAを除去した。260nmでの吸光度の分析によってRNAを定量し、吸光度比A260nm/A280nmの測定によって純度を評価した。RNAサンプル(1μg)を、以前に記載されているように適切なプライマー使用してPCRによって分析し、汚染プラスミドDNAが存在しないことを検証した。また、すべてのサンプルをPCR反応に添加したpLP685プラスミド1ngのスパイク回収分析によって残留DNAseについて評価した。DNAおよびDNase汚染がないことが証明されたサンプルのみその後の分析に使用した。
【0134】
ノーザンブロットハイブリダイゼーション。総RNA(5μg)を、メーカーによって規定されているようにNorthernMax(商標)を使用して変性条件下で1%アガロースゲル上で断片化した。ミレニアムおよびセンチュリーRNAマーカー(各2μg)をEtBrで前染色し、これを使用してRNAサイズを評価した。サンプルをナイロン膜(GeneScreen(商標))に移し、UV架橋し、80℃下に2時間乾燥させ、RNA断片化に使用されるホルムアルデヒド反応を逆転させた。RNA標準をUVによって視覚し、断片位置をハイブリダイゼーション前にナイロン膜上に示した。膜を2×SSC(0.3M Nacl、0.03Mクエン酸ナトリウム)で前湿潤させ、42℃下に2時間、ULTRAhyp(商標)で前ハイブリダイゼーションした。ノーザン分析用のプローブをテンプレートとしてpLP685、および上記で詳述されているように、プロ配列ドメイン(367bp)および成熟SpeB配列(770bp)に特異的な一連のプライマーを使用してPCR合成した。蛍光−N6−dATP(10μM)をPCR反応で使用し、無作為に標識成熟SpeBおよびプロ配列ドメインDNAプローブを生成した。結果として生じるプローブをアガロースゲル電気泳動によって精製し、各々の32pgを変性させ、42℃下に一夜、ULTRAhyp(商標)での膜ハイブリダイゼーションに使用した。ハイブリダイゼーション後、膜を2回過剰な2×SSCで10分間、室温下、2×SSC、1%SDSで2回20分間、42℃下、および0.2×SSC、0.1%SDSで2回20分間、42℃下に洗浄した。膜をBLOTTOでブロックし、アルカリホスファターゼ−コンジュゲート抗フルオレセインポリクローナル抗体、およびCDP−Star(登録商標)を、メーカーの仕様のとおりに使用して展開した。ブロットをBioMax MR−2オートラジオグラフフィルム(イーストマン・コダック(Eastman Kodak)、ロチェスター(Rochester)、ニューヨーク州(NY))に5分間、室温下に露出し、信号検出させた。
【0135】
cDNAの合成。RT−PCR用のRETROscript(商標)ファースト・ストランド(First Strand)合成キット、および無作為十量体をメーカーによって規定された条件下で使用する逆転写によって総RNAの2μgからcDNAを調製した。陰性対照サンプルを同一の条件を使用し、ただし逆転写酵素(−RT)の非存在下に生成した。反応産物をPCRおよび定量的PCR分析の使用前にヌクレアーゼを含まない水中に1:200で希釈した。
【0136】
cDNAの定量的PCR(qPCR)分析。プロ配列ドメイン、成熟SpeB、およびpET28aコード化カナマイシン耐性遺伝子(KanR)に特異的なプライマーおよびプローブを、プライマー発現ソフトウェア(アプライド・バイオシステムズ(Applied Biosystems)、フォスターシティー(Foster City)、カリフォルニア州(CA))を使用して設計した。定量的PCR反応を以下の条件下で行った。すなわち、300nMの各プライマー(フォワードおよびリバース)、200nM FAM/TAMRAプローブ、2×TaqManユニバーサルPCRマスターミックス(アプライド・バイオシステムズ(Applied Biosystems)、および1μL希釈cDNAまたは陰性対照サンプル。サイクル条件として、50℃で2分間、95℃で10分間、95℃で15秒間および60℃で1分間の40サイクルをABI7000配列検出システムで使用して最終容量25μLで反応を行った。KanR cDNAレベルを内部制御として使用し、全反応をKanR閾値サイクル(Ct)に基づき標準化した。結果は標準化Ct値として表されている。
【0137】
cDNAのPCR分析。希釈cDNAおよび陰性対照サンプルをPCRによって分析し、発現プラスミドから生じたmRNA転写をさらに評価した。プロ配列ドメイン、成熟SpeB、およびプロ配列ドメインフォワード(5’CCATGGATCAAAACTTTGCTCGTAACG3’)(配列番号14)プライマー、および成熟プロテアーゼリバース(5’CTCGAGCTAAGGTTTGATGCCTACAACAGC3’)(配列番号15)プライマーに特異的な上記のPCRプライマーからの第3のセットを使用し、それぞれ、プロ配列ドメイン、成熟SpeB、および完全長SpeBチモーゲン(1119bp)、およびポリシストロニックcDNA(1145−1180bp)を増幅した。1μLの各希釈サンプルを、増幅サイクルを30に増大したことを除き、上記の条件下で使用し、PCR産物をアガロースゲル電気泳動によって分析した。
【0138】
SpeBプロ配列ドメインの精製。溶解バッファー(20mMトリス、pH7.2、10mM MgCl2、10μg/μL DNase)中、15mLバッファー/g細胞の割合で細胞を再懸濁し、M110−Yマイクロフルイダイザー(Microfluidizer)(マイクロフルイディクス(Microfluidics)、ニュートン(Newton)、マサチューセッツ州(MA))を使用して溶解した。細胞残屑を遠心分離でペレット化し、可溶性フラクションを回収し、50mMグリシン−HCl(pH3.2)に対して4℃下に一夜、透析した。透析は、大腸菌タンパク質の顕著な沈殿によって達成された。沈殿物質を遠心分離で除去し、回収上清を100mM酢酸ナトリウム(pH4.5)に対する透析前にpH4.5に変更した。サンプルをSP−セファロース陽イオン交換カラムへ供給し、組換えプロ配列ドメインタンパク質を100mM酢酸ナトリウム(pH4.5)、1M塩化ナトリウムの勾配で溶出した。プロ配列ドメイン含有フラクションをプールし、SDS−PAGE電気泳動によって純度をチェックし、タンパク質濃度をBCAアッセイによって測定した(スミス(Smith)ら、1985年)。
【0139】
組換え発現成熟野生型SpeBの精製。溶解バッファー(20mMトリス、pH7.2、10mM MgCl2、10μg/μL DNase)中、15mLバッファー/g細胞の割合で細胞を再懸濁し、顕微溶液化によって溶解した。細胞残屑を遠心分離で除去し、不溶性成熟野生型SpeB含有フラクションを回収した。ペレットを処理し(50mMトリス、pH8.0、150mM NaCl、1mM EDTA、2%トリトンX−100)4℃下に一夜、室温下に1.5時間、室温下に3.5時間、すべて静かに攪拌して3連続洗浄を行った。ペレットを室温下に一夜、攪拌して20mMトリス、pH8.0、0.8M尿素中に溶解し、100mM酢酸ナトリウム、pH4.5、8M尿素で平衡したSP−セファロースカラム上にサンプルを塗布する前に酢酸ナトリウムでpHを4.5に変更した。組換え成熟野生型SpeBタンパク質を0〜750mM勾配の塩化ナトリウムで溶出し、SDS−PAGE電気泳動によって純度を分析し、タンパク質濃度をBCAによって測定した。精製、変性タンパク質をインビトロ再折り畳み実験に利用した。
【0140】
共発現成熟SpeBの精製。ポリシストロニックまたは2−プラスミド系のいずれかによって共発現された組換え成熟野生型SpeBおよび成熟C192S SpeBの精製方法は同一であった。溶解バッファー(20mMトリス、pH7.2、10mM MgCl2、10μg/μL DNase)中、15mLバッファー/g細胞の割合で誘発細菌細胞ペレットを再懸濁し、顕微溶液化によって溶解した。細胞残屑を遠心分離でペレット化し、可溶性フラクションを回収し、50mMグリシン−HCl(pH3.2)に対して4℃下に一夜、透析した。透析は、大腸菌タンパク質の顕著な沈殿によって達成され、pHを4.5に迅速に変更する前に遠心分離で浄化した。サンプルを10M尿素で1:1に希釈し、100mM酢酸ナトリウム(pH4.5)、5M尿素で平衡したSP−セファロースカラム上に供給した。カラムからの非結合物質をA280nmがベースラインに達するまで平衡バッファーで洗浄した。カラムを100mM酢酸ナトリウム(pH4.5)で洗浄し、尿素を除去し、組換え成熟プロテアーゼを0〜1.0M塩化ナトリウムの勾配で溶出した。回収タンパク質をPBS(pH7.4)に対して透析し、SDS−PAGEゲル電気泳動によって純度を評価し、タンパク質濃度をBCAによって測定した。
【0141】
ポリクローナル抗血清の生成。精製成熟C192S SpeB(ポリシストロニックまたは2−プラスミドベースの共発現によって生成)を抗血清の生成のために使用した。スイス・ウェブスター(Swiss Webster)マウスを0、4、6週にMPL 50mgおよびAIPO4 100μgを佐剤として使用して精製タンパク質5μgで免疫化した。
【0142】
成熟SpeBのプロ配列ドメインとの相互作用。SpeBプロ配列ドメイン/成熟SpeB相互作用の解離定数(Kd)を酵素免疫吸着測定法(ELISA)またはバイオコア(Biocore)3000装置を使用して測定した。ELISA分析のために、20μM E−64で阻害した増大濃度のペプシン生成組換え成熟C192S(マツカ(Matsuka)ら、1999年)および組換え成熟野生型SpeBを、精製プロ配列ドメイン、またはリゾチーム(陰性対照)のいずれかでコーティングしたマイクロタイターウェルでインキュベートした。プレートをTBS、pH7.4、0.05%Tween20で洗浄し、検出されたSpeBを405nmでの吸光度を測定することによって成熟SpeBに対して生成された親和性精製ポリクローナル抗体と結合した。結果として生じる吸光度(A)の濃度依存増大を等式ΔA=Amax+[L]/Kd+[L]に適合したが、ここでKdは解離定数であり、[L]は遊離リガンドの濃度である。
【0143】
プロ配列ドメインのペプシン生成組換え成熟C192S SpeBの実時間相互作用をバイオコア(Biocore)3000(バイオコア(Biocore)、Piscataway、ニュージャージー州(NJ))を使用する表面プラスモン共鳴(SPR)によって明らかにした。精製プロ配列ドメインをメーカーの仕様に従って活性化カルボキシメチルデキストランでコーティングされたバイオセンサチップに共有結合した。結合実験をTBS(pH7.4)、0.05%Tween20中、25℃下に行った。増大濃度のペプシン生成組換え成熟C192S SpeBを固定化プロ配列ドメインに添加し、その結合を実時間でモニタリングした。結合過程のセンソグラムを器具とともに供給されたソフトウェアを使用して分析した。
【0144】
SpeBプロ配列ドメインの阻害活性の評価。成熟SpeB(0.1μM)を、増大濃度のプロ配列ドメイン、またはリゾチーム(陰性対照)を利用し、レゾルフィン標識カゼイン基質(0.4%)の存在下、PBS(pH7.4)、10mM DTT中25℃下に1時間インキュベートした。インキュベーション後、非消化基質を2%トリクロロ酢酸を使用する沈殿によって除去し、浄化上清フラクションにおける放出レゾルフィン標識ペプチドの吸光度を574nmで分光光度法により測定した。また、密接に関係したシステインプロテアーゼ(パパイン)を陰性対照と同一条件下で分析し、プロ配列ドメイン/成熟SpeB相互作用の特異性を明らかにした。
【0145】
変性成熟SpeBのインビトロ再折り畳み。100mM酢酸ナトリウム(pH4.5)、8M尿素中の組換え変性成熟SpeBを、0.5Mアルギニンを含有するPBS(pH7.4)、またはPBS(pH7.4)のいずれかにより迅速に希釈した(1:20v/v)。希釈は、示されているように増大濃度の精製プロ配列ドメインの存在下に20μM E−64阻害剤の有無により行った。希釈後、10mM DTTを各反応物に添加し、サンプルを4℃下に24時間インキュベートした。反応物におけるSpeBの最終濃度は5μMであった。インキュベーション後、100μL反応物アリコートをレゾルフィン標識カゼイン切断アッセイを使用して評価し、再折り畳みSpeBの活性を評価した。
【0146】
成熟SpeBのカゼイン分解活性。酵素のタンパク質分解活性を評価するために、指示量のSpeBを、規定されているように、0.4%レゾルフィン標識カゼイン、PBS(pH7.4)、10mM DTT(2%)の存在下にインキュベートした。非消化基質をトリクロロ酢酸沈殿(2%)によって除去し、サンプルを遠心分離によって浄化し、上清フラクションにおける放出レゾルフィン標識ペプチドの吸光度を574nmで分光光度法により測定した。
【0147】
成熟SpeBの熱誘発変性。組換え発現成熟SpeBの融解をTBS(pH7.4)中のタンパク質サンプルを加熱することによって評価すると同時に、SLM AB2蛍光分光光度計を使用し、280nmでの励起により、350nm/320nmでの固有の蛍光強度の比をモニタリングした。
【0148】
実施例2
SpeBプロ配列ドメインの阻害およびシャペロン活性
そのNH2末端プロ配列ドメインを欠く成熟SpeBの組換え発現は、もっぱら大腸菌における不溶性タンパク質の生成をもたらす(マツカ(Matsuka)ら、1999年)。可溶性成熟SpeBを支配するプロ配列ドメインの要件は、ドメインが分子内シャペロンとして機能し、タンパク質の適切な折り畳みを方向づけることを示す。かかる活性を検査するために、SpeBプロ配列ドメイン、40kDa SpeBチモーゲンのほか、成熟野生型SpeBおよび成熟C192S SpeBを特徴づけのための大腸菌から発現させ、精製した(データは示さず)。プロ配列ドメインと成熟SpeBタンパク質の結合を、ELISA(図1A)、およびバイオコア3000(図1B)を使用する表面プラスモン共鳴(SPR)を使用して調査した。実施例1で規定された計算パラメータを使用することにより、プロ配列ドメインの野生型およびC192S成熟SpeBとの相互作用が、それぞれ、11nmおよび34nMのKdを有することが推定された。SPRによって測定されるプロ配列ドメインおよび成熟C192S SpeBの相互作用の実時間分析は、11nMのKdを有することが推定された。両方の方法によって測定される結合値は、プロ配列ドメインと成熟SpeBドメインとの間の高い親和性を示す。
【0149】
プロ配列ドメインの成熟SpeBとの結合は、プロテアーゼ活性の阻害をもたらす。これは基質としてレゾルフィン標識カゼインを使用するカゼイン分解切断アッセイを使用して明らかにされた(図2A)。プロ配列ドメイン、またはリゾチーム制御を、0.1μMの成熟SpeBを使用する0〜100μMの阻害剤濃度範囲にわたって分析した。結果は、プロ配列ドメインが0.3μMの濃度で最大のプロテアーゼ活性の半分を阻害し(IC50=0.3μM)、阻害剤としてのリゾチームの添加は同じ濃度範囲にわたって成熟SpeBの活性に対する影響はないことを示した。同一条件下での密接に関係したシステインプロテアーゼ(パパイン)の分析は、パパインプロテアーゼ活性に対する影響を示さなかった(図2B)。かかる結果は、SpeBとプロ配列ドメインとの間の相互作用の特異性をさらに明らかにし、かつプロ配列ドメインの分子内阻害活性がインサイツでのプロテアーゼ活性の調節に役立ちうることを示す。
【0150】
プロ配列ドメインの分子内シャペロン活性は、尿素変性成熟システインプロテアーゼを使用してインビトロで明らかにされた。増大濃度のプロ配列ドメインの存在下に再折り畳みされた変性成熟プロテアーゼは、レゾルフィン標識カゼイン基質の切断によってモニタリングされるように回収プロテアーゼ活性の顕著な増大を明らかにした(図3)。反応における不可逆性システインプロテアーゼ阻害剤(E−64)の添加は基質切断を阻止し、確認されたカゼイン分解切断は特に再折り畳み成熟SpeBの活性に関与することを示した。これらのデータは、SpeBプロ配列ドメインが分子内シャペロンとして作用し、成熟SpeBの折り畳みを方向づけることを示す。さらに、結果は、2つの領域が適切な折り畳みを方向づけるために共有結合される必要がないことをはっきり示す。
【0151】
実施例3
SPEBプロ配列ドメインおよび成熟SPEBの2−プラスミドベースの共発現
インビトロでの成熟SpeBの正確な再折り畳みを方向づけるプロ配列ドメインの能力は、インビボでの2つのタンパク質の独立した共発現が潜在的に正確に折り畳まれる成熟SpeBの生成がもたらされることを示す。したがって、本実施例では、2−プラスミド共発現系が開発され、この場合、一方のプラスミドはプロ配列ドメイン(pLP682)をコード化し、かつ他方は成熟SpeBポリペプチド(pLP680またはpLP681)をコード化する(図4)。pLP680もしくはpLP681のいずれかのみで形質転換され、またはpLP682で同時形質転換された大腸菌を使用し、SDS−PAGEおよびウェスタンブロットを使用してタンパク質発現を調査した。成熟野生型または成熟C192S SpeB構成物のいずれかによる唯一の発現(データは示さず)は、優先的に不溶性28kDa成熟SpeBの生成をもたらした。これは、プロ配列ドメイン非存在下の成熟SpeBの発現が、不正確に折り畳まれるタンパク質の生成をもたらすことを示す。対照的に、プロ配列ドメインとともに成熟SpeBポリペプチドのインビボ共発現は、細胞の可溶性フラクションの両方のタンパク質の実質的レベルの生成をもたらし、両方のタンパク質の独立した共発現がSpeBの適切な折り畳みを促進することを示す。SpeBチモーゲンに対するポリクローナル抗体を利用するウェスタンブロット分析では、プロ配列ドメインおよび成熟SpeBポリペプチドの同時の検出が可能であった(データは示さず)。ブロット分析では、発現タンパク質の同一性が確認され、さらにSDS−PAGEによって証明される結果が検証された。これらの結果によりインビトロデータが再確認され、プロ配列ドメインおよび成熟SpeBポリペプチドが成熟SpeBの適切な折り畳みを方向づけるために共有結合される必要がないことを示した。
【0152】
実施例4
SPEBプロ配列ドメインおよび成熟SPEBのポリシストロニックベースの共発現
実施例3に記載されているように、プロ配列ドメインおよび成熟SpeBの2−プラスミドベースの共発現は、可溶性に正確に折り畳まれたSpeBを生成するかかる方法の有用性を明らかにした。成熟SpeBの大規模生成のために、プロ配列ドメインと成熟SpeBポリペプチドの独立した共発現のためのポリシストロニック発現系が開発された。したがって、この発現系は、第1のシストロンとしてのプロ配列ドメインに引き続き、翻訳エンハンサーおよび最適化シャイン・ダルガノリボソーム結合部位を含有する合成リンカー(バリック(Barrick)ら、1994年、カリー(Curry)とトミッチ(Tomich)、1998年、リングクイスト(Ringquist)ら、1992年)5’の第2のシストロンが設計された(図5)。第1のシストロンの翻訳終止コドンと第2のシストロンのシャイン・ダルガノとの間の合成リンカーの増大する長さ(5nt、10nt、20nt、40nt)を2つのタンパク質の発現レベルの違いについて調査した。リンカー領域は、転写RNAにおける2つのシストロン間の第2の構造を最小限にするように設計され、非制限リボソームの流れおよび第2のシストロンでの翻訳の再開始を可能にした。それぞれ、5nt、10nt、20nt、および40ntリンカーを含有するポリシストロニック成熟C192S SpeB発現構成物pLP683、pLP684、pLP685、およびpLP686のほか、成熟野生型SpeBの20ntポリシストロニック発現構成物(pLP687)を利用する実験を行った。
【0153】
pLP685およびpLP687ポリシストロニック構成物を使用するタンパク質発現の分析では、プロ配列ドメイン(12kDa)および成熟SpeB(28kDa)の両方の生成が確認された(データは示さず)。2−プラスミド系で示されているように、大腸菌における両方のタンパク質の同時かつ独立の共発現は、優先的に細胞の可溶性フラクションにおける成熟SpeBの生成をもたらした。4つのC192Sポリシストロニック構成物のそれぞれの誘発培養からの全細胞溶解物の可溶性フラクションもSDS−PAGEによって分析された(図6A)。発現のレベルを定量するために、濃度計を使用してゲルを分析し、28kDa成熟SpeB、または12kDaプロ配列ドメインに対応する各バンドの面積を測定した(図6B)。各構成物によってプロ配列ドメインのレベルにはほとんど違いは確認されなかったが、40ntリンカー含有ポリシストロンからの不溶性成熟SpeB発現の顕著な減少が確認された。興味深いことに、119ntリンカー領域を含有する構成物からの可溶性成熟SpeB発現は、5nt、10nt、および20ntリンカーポリシストロンのものと同等のレベルをはっきり示した(データは示さず)。
【0154】
実施例5
ポリシストロニックmRNA転写の分析および転写レベルの評価
総RNAをすべてのポリシストロニック構成物、野生型SpeBチモーゲン、成熟野生型SpeB、成熟C192S SpeB、およびプロ配列ドメインの誘発培養物から単離した。単離RNAのノーザンブロット分析を行い、20ntリンカー含有構成物、および規定されているように適切な陽性対照から生じた転写のサイズを評価した(データは示さず)。得られた結果により、対照からの成熟野生型SpeB(約892塩基)、成熟C192S SpeB(約892塩基)およびプロ配列ドメイン(約487塩基)mRNA転写はすべて予想されたサイズで移動することが明らかにされた。野生型およびC192S 20ntリンカー含有ポリシストロニック発現系の両方からのmRNA転写の検出は、野生型SpeBチモーゲン対照のもの(約1246塩基)よりわずかに高い同等のサイズ(約1287塩基)で移動する転写シグナルを示した。かかる結果は、予測されるように、各々の完全長、ポリシストロニックmRNAの生成と一致する。成熟SpeBおよびプロ配列ドメイン対照で確認されるものと同等の、これらサンプルにおけるより小さい転写の検出の欠如は、SpeBの発現がポリシストロニック転写の生成に直接帰するものであり、多様なmRNA種の存在の結果によるものではないことを示す。
【0155】
すべてのポリシストロニック種から生じる転写の完全長の性質もcDNAのPCR分析によって検証された。ポリシストロニックサンプルおよび単一のシストロン対照からの総RNAを使用し、対応するcDNAを生成し、かつ希釈cDNAサンプルをPCRによって分析した。逆転写酵素(−RT)を除くすべての反応成分を含む各サンプルの陰性対照を生成し、単離RNA内の潜在的なプラスミドDNA汚染を評価した。プロ配列ドメイン(367bp)、成熟SpeB(770bp)、および完全長プロ配列/成熟SpeB(1119−1180bp)コード領域に特異的な一連のプライマーを使用し、cDNAサンプル、−RT陰性対照、およびpLP685陽性対照を検査した(データは示さず)。プロ配列ドメインのPCR増幅により、その発現構成物におけるプロ配列ドメインヌクレオチド配列を含有するすべてのcDNAサンプルにおける予想されるサイズの産物が生じた。この配列を欠くサンプル、成熟野生型SpeBおよび成熟C192S SpeB発現構成物は、増幅産物を生成しなかった。成熟プロテアーゼコード領域の増幅について、プロ配列ドメイン発現系のものを除くすべてのcDNAサンプルは陽性バンドを生成した。完全長プロ配列/成熟SpeB領域の増幅により、ポリシストロニックおよびSpeBチモーゲンサンプルのみについてPCR産物が生じたが、成熟野生型SpeB、成熟C192S SpeB、およびプロ配列ドメイン発現系からのサンプルは、予測されるように産物生成の欠如を証明した。すべてのサンプルの−RT陰性対照は増幅産物を生成せず、mRNAの逆転写に由来するcDNAサンプルで確認された陽性シグナルの生成を示し、汚染DNAの増幅を示さなかった。
【0156】
異なる発現系のmRNA転写レベルの検査は、cDNAサンプルおよび−RT対照の定量的PCR分析によって測定された。プロ配列および成熟SpeB領域の各々に特異的な一連のプライマー/プローブを使用し、培養物間の転写レベルを比較した。カナマイシン耐性をコード化する遺伝子がすべての発現構成物上に存在することを考えて、KanRmRNAを分析中の内標準として使用し、発現サンプル間のプラスミドコピー数の潜在的な違いに対して制御した。結果は、各構成物の誘発中に生成されたmRNA転写のレベルが同等であることをはっきり示す(図7)。より重要なことには、データは、プロ配列のmRNAレベルが成熟SpeBのものと同等であることを示す。これらの結果は、ポリシストロニック構成物間の可溶性成熟SpeBの確認された量の明らかな違いが、転写の早過ぎる終了によるものではないことを示す。
【0157】
実施例6
共発現成熟SpeBの特徴づけ
2−プラスミドベースおよびポリシストロニック系の両方によって生成された精製成熟SpeBをSDS−PAGEによって分析し(データは示さず)、すべてのタンパク質を熱誘発展開実験にかけた(図8および図9)。融解(すなわち、変性)曲線を上昇温度範囲(0−90℃)にわたってタンパク質サンプルを加熱すると同時に、固有の蛍光の比の変化をモニタリングすることによって得た。2−プラスミドおよびポリシストロニック共発現C192S(図8)の両方、または野生型(図9)成熟SpeBによる変性曲線は、明確な移行部をはっきり示し、SpeBチモーゲンとはそれぞれ対応する発現のものと同様の中間点を有し、インビトロ切断(パパイン生成)、または自己触媒(野生型SpeBチモーゲン)によって加工された。かかる結果は、いずれかの共発現系によって生成される組換えタンパク質が、対応する組換え発現および加工チモーゲンと同様に展開されることを示す。
【0158】
すべての組換え成熟野生型SpeBの固有のプロテアーゼ活性をその操作的モル濃度の測定によって評価した(図10)。各々について、同等濃度のSpeB(0.12μM)を、レゾルフィン標識カゼイン基質の添加前に、不可逆性システインプロテアーゼ阻害剤、E−64の増大量の存在下にプレインキュベートした。標識ペプチドの放出を分光光度法で測定し、結果を阻害剤濃度の関数としてプロットした。2−プラスミド(0.121μM)、ポリシストロニック(0.124μM)、および自己触媒加工(0.124μM)成熟SpeBポリペプチドについて得られた値は、それらが酵素的に区別不可能であり、タンパク質濃度に基づく0.12μMの予想値と同等であることを示した。
【0159】
以前のデータは、パパインによるインビトロ切断によって生成された成熟C192S SpeBに対して生成された抗体が、野生型SpeBのタンパク質分解活性を阻害する能力があることを示した(マツカ(Matsuka)ら、1999年)。いずれにかの共発現系によって生成される組換えタンパク質が同様の活性を発揮するを判定するために、2−プラスミドまたは20ntポリシストロニック系のいずれかによって生成された成熟C192S SpeBで免疫化したマウスからの増大濃度の抗血清をカゼイン分解切断アッセイを使用して分析した(図11)。結果は、基質の加水分解が、プレ免疫対照に対して、いずれかの共発現系によって生成されるタンパク質で免疫化した動物からの血清の増大量の存在下に阻害されることを示す。さらに、各々について証明された阻害のレベルは、SpeBチモーゲンとして発現され、かつペプシンを使用してインビトロで加工された成熟C192S SpeBに対して生成された血清について以前に報告されたものと同等である(マツカ(Matsuka)ら、1999年)。
【0160】
【0161】
【0162】
【0163】
【0164】
【図面の簡単な説明】
【0165】
【図1】組換え成熟SpeBとプロ配列ドメインの相互作用を示す。
【図2】成熟SpeBのプロ配列ドメイン介在阻害を示す。
【図3】変性成熟SpeBの再折り畳みに対する組換えプロ配列ドメインの効果を示す。
【図4】2−プラスミドベース発現構成物を示す略図である。
【図5】ポリシストロニック発現系および合成リンカー領域を示す図表である。
【図6】ポリシストロニック発現系における可溶性成熟SpeBおよびプロ配列ドメインレベルの評価および相対定量を示す。
【図7】ポリシストロニックcDNAの定量的PCR分析を示す。
【図8】成熟C192S SpeBの熱変性を示す。
【図9】成熟野生型SpeBの熱誘発変性を示す。
【図10】組換え成熟SpeBの操作モル濃度の評価を示す。
【図11】野生型SpeBタンパク質分解活性の抗体介在阻害を示す。
【発明の詳細な説明】
【0001】
(技術分野)
本発明は、分子生物学、臨床細菌学、およびタンパク質折り畳みの分野に関する。より詳しくは、本発明は、宿主細胞における成熟化膿連鎖球菌外毒素B(SpeB)ポリペプチドを組換え発現させるための方法に関する。
【0002】
(従来技術)
A群連鎖球菌(group A streptococci)(GAS)とも呼ばれる化膿連鎖球菌は、ヒトの一般的なグラム陽性細菌病原体である。化膿連鎖球菌は、咽頭炎、膿痂疹、および敗血症を含むヒトにおけるさまざまな病状を引き起こす。感染後には、リウマチ熱および急性糸球体腎炎など自己免疫合併症もヒトにおいて発生する。化膿連鎖球菌は、猩紅熱、壊疽性筋膜炎、および毒性ショックなど重篤な急性疾患も引き起こす。
【0003】
A群連鎖球菌によって引き起こされる咽頭炎は、一般的に「連鎖球菌性咽頭炎」と呼ばれ、季節によって一般医療における全呼び出しの少なくとも16%を占める(非特許文献1)。A群連鎖球菌は、壊疽性筋膜炎に関与する毒性ショックの北米および他の四大陸における最近の再流行の原因でもある(非特許文献2)。
【0004】
連鎖球菌感染は現在、抗生物質療法によって治療されている。しかし、治療された者の25−30%は再発性疾患を有し、かつ/または粘膜分泌物中の微生物を削減する。毒性ショックおよび重篤な感染症の抗生物質による治療は多くの場合、無効であり、かつ死亡率が50%を越えうる(非特許文献3)。ペニシリンが重篤な侵襲性の連鎖球菌感染をうまく治療しえないことは、大きな接種原が迅速に固定相に達し、かつペニシリンがゆっくり増殖する細菌に対してあまり有効ではない現象によって生じる(非特許文献4)。したがって、依然として、連鎖球菌感染を予防し、または改善する有効な手段の継続的必要性がある。さらに具体的には、連鎖球菌感染を予防する免疫原性組成物に有用である抗原(または免疫原)を同定または開発する必要性が存在する。現在、免疫原として考えられているかかるポリペプチド抗原が、連鎖球菌システインプロテアーゼ、連鎖球菌プロテイナーゼ、またはストレプトペイン(streptopain)としても知られる化膿連鎖球菌外毒素B(SpeB)である。
【0005】
化膿連鎖球菌外毒素B(SpeB)は、40kDa不活性プレ−プロ−酵素(すなわち、チモーゲン)として発現され(非特許文献5、非特許文献6)、27アミノ酸NH2−末端シグナル配列を有し、そして118アミノ酸プロ−ペプチド配列(アミノ酸28−145)、および253アミノ酸成熟配列(アミノ酸146−398)が続く。分泌すると、40kDa SpeBチモーゲンは自己触媒活性化を受け、結果として12kDa NH2−末端プロ−ペプチドの除去および成熟28kDa活性SpeB酵素の形成が生じる。この活性酵素への変換機序は、望ましくないタンパク質分解を阻止し、かつタンパク質分解活性の空間的および時間的調節を可能にする(非特許文献7)。
【0006】
酵素のシステインエンドペプチダーゼ基のメンバーとして、SpeBは活性部位でCys−His対を含有する(非特許文献8、非特許文献9、非特許文献10)。192位での単一のシステイン残基のセリンへ(以下、「C192S」)の置換は結果としてSpeB酵素の検出可能なタンパク質分解活性の喪失が生じ、40kDa SpeBチモーゲンの28kDa成熟SpeB形態へのプロセッシングを阻止する(非特許文献11、非特許文献12、非特許文献13)。
【0007】
以前の試験の結果は、C192S SpeB変異株の成熟形態が野生型SpeB酵素への最大の阻害活性による抗体の生成に必要であることを示した(非特許文献12)。これは、免疫化のためにC192S SpeB変異株のNH2−末端切断、28kDa成熟形態を生成する必要を示す。しかし、成熟C192S SpeBの組換え発現(すなわち、そのNH2−末端プロ配列の欠損)の結果として大腸菌における不溶性タンパク質の蓄積が生じる。
【0008】
可溶性成熟C192S SpeB変異株を生成するための1つの方法は、40kDa SpeBチモーゲンの制限タンパク質分解によっている。例えば、成熟C192S SpeB変異株を生成する40kDa C192S SpeB変異株チモーゲンの制限タンパク質分解は、エラスターゼ、ペプシン、サーモリシン(非特許文献12)、およびパパインを含む一部のプロテアーゼを使用して達成されている。これらのデータのほか、トリプシンおよびスブチリシンを利用する非特許文献6によって公開されたデータは、所望の28kDa成熟C192S SpeB変異株が、さまざまなプロテイナーゼによる40kDa C192S SpeB変異株チモーゲンの処理によって有効に生成されることを示す。
【0009】
しかし、この方法には大規模生成に対するいくつかの制限を有する。第一に、成熟プロテアーゼの最終生成物収量は2つの連続精製ステップに対する要件、完全長チモーゲンに対する第1と、加工成熟プロテアーゼに対する第2の要件のため低い。第二に、特に大規模に、制限タンパク質分解反応のコンシステンシーおよび再現性と関係がある難点がある。最後に、切断に使用される酵素的に活性の外来プロテアーゼによる最終生成物の固有の汚染リスクがある。かかる汚染は、反応が樹脂固定化プロテアーゼで行われる場合でも回避することはきわめて困難である。
【0010】
したがって、依然として、哺乳類宿主における連鎖球菌感染に対して有効な免疫原性組成物に対する技術上の必要性がある。したがって、成熟SpeBが哺乳類宿主に投与された場合でも免疫原性である、成熟SpeBポリペプチドを生成し、または発現させる方法を特定することがきわめて望ましい。また、成熟SpeBポリペプチドの免疫原性形態を生成し、または発現させるかかる方法が、SpeB収量の減少、タンパク質分解コンシステンシー/再現性の制限、および外来酵素汚染のリスクなど前記大規模制限を回避することも望ましい。
【非特許文献1】ホープ・シンプソン(Hope−Simpson)、1981年
【非特許文献2】スティーブンス(Stevens)、1992年
【非特許文献3】デービス(Davies)ら、1996年
【非特許文献4】スティーブンス(Stevens)ら、1993年
【非特許文献5】ショセ(Chaussee)ら、1993年
【非特許文献6】リュー(Liu)とエリオット(Elliot)、1965年
【非特許文献7】カーン(Khan)とジェイムズ(James)、1998年
【非特許文献8】リュー(Liu)ら、1965年
【非特許文献9】リュー(Liu)、1965年
【非特許文献10】タイ(Tai)ら、1976年
【非特許文献11】グッバ(Gubba)ら、1998年
【非特許文献12】マツカ(Matsuka)ら、1999年
【非特許文献13】ムサー(Musser)、1996年
【0011】
(発明の開示)
本発明は、概して、宿主細胞における成熟化膿連鎖球菌外毒素B(SpeB)ポリペプチドを組換え発現させる方法、およびその免疫原性組成物に関する。より詳しくは、本発明は、宿主細胞における12kDa SpeBプロ−ペプチドおよび28kDa成熟SpeBポリペプチドを共発現させ、ここで成熟SpeBポリペプチドが宿主細胞において可溶性である新たな方法に関する。
【0012】
したがって、一部の実施形態においては、本発明は、宿主細胞における成熟化膿連鎖球菌外毒素B(SpeB)ポリペプチドを組換え発現させるための方法であって、この方法は、(i)(a)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列と(b)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列とを含んで成る、ポリシストロニックプラスミドで宿主細胞を形質転換し、形質導入し、トランスフェクトし、または感染させるステップと、(ii)宿主細胞によって成熟SpeBポリペプチドおよびSpeBプロ−ポリペプチドドメインの発現を可能にする条件下で宿主細胞を培養するステップとを含んで成り、かつ成熟SpeBポリペプチドが宿主細胞において可溶性である方法に関する。したがって、本発明のポリシストロニックプラスミド系においては、単一のプロモーター(例えば、T7プロモーター)がポリシストロニックmRNA転写物の発現を推進するが、ここでポリシストロニックmRNAはその正確なリーディングフレームにおける2つもしくはそれ以上のポリペプチド(例えば、SpeBプロ−ポリペプチドドメインおよび成熟SpeBポリペプチド)をコード化する。一部の実施形態においては、SpeBプロ−ポリペプチドドメインは配列番号2のアミノ酸残基28〜145を含んで成るポリペプチドとしてさらに規定され、かつ成熟SpeBポリペプチドは配列番号2のアミノ酸残基146〜398を含んで成るポリペプチドとしてさらに規定される。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192でのシステインはセリンによって置換される。別の実施形態においては、成熟SpeBポリペプチドは哺乳類宿主において免疫原性である。さらに別の実施形態においては、成熟SpeBポリペプチドに特異的な抗体は野生型SpeBポリペプチドと交差反応し、かつSpeBポリペプチド活性を無力化する。一部の好ましい実施形態においては、プラスミドはT7プロモーター含有プラスミドである。特定の実施形態においては、T7プロモーター含有プラスミドはpET、pRSET、pCRT7−CTTOPO、およびpIVeXより成る群から選択される。別の好ましい実施形態においては、宿主細胞は細菌細胞である。一部の実施形態においては、細菌宿主細胞は大腸菌である。さらに他の実施形態においては、大腸菌は、BLR(DE3)、BLR(DE3)pLysS、AD494(DE3)、AD494(DE3)pLysS、BL21(DE3)、BL21(DE3)pLysS、BL21(DE3)pLysE、BL21(DE3)pLacI、BL21trxB(DE3)、BL21trxB(DE3)pLysS、HMS174(DE3)、HMS174(DE3)pLysS、HMS174(DE3)pLysE、Origami(DE3)、Origami(DE3)pLysS、Origami(DE3)pLysE、Origami(DE3)pLacI、OrigamiB(DE3)、OrigamiB(DE3)pLysS、OrigamiB(DE3)pLysE、OrigamiB(DE3)pLacI、Rosetta(DE3)、Rosetta(DE3)pLysS、Rosetta(DE3)pLysE、Rosetta(DE3)pLacI、Tuner(DE3)、Tuner(DE3)pLysS、およびTuner(DE3)pLacIより成る群から選択される菌株である。
【0013】
他の実施形態においては、本発明は、(a)(i)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列を含んで成るプラスミドおよび(ii)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るプラスミドで宿主細胞を形質転換し、形質導入し、トランスフェクトし、または感染させるステップと、(b)SpeBプロ−ポリペプチドドメインおよび成熟SpeBポリペプチドを共発現させるのに適切な条件下で宿主細胞を培養するステップとを含み、成熟SpeBポリペプチドが宿主細胞において可溶性である、宿主細胞において成熟SpeBポリペプチドを組換え発現させるための方法に関する。一部の実施形態においては、SpeBプロ−ポリペプチドドメインは配列番号2のアミノ酸残基28〜145を含んで成るポリペプチドとしてさらに規定され、かつ成熟SpeBポリペプチドは配列番号2のアミノ酸残基146〜398を含んで成るポリペプチドとしてさらに規定される。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192でのシステインはセリンによって置換される。他の実施形態においては、成熟SpeBポリペプチドは哺乳類宿主において免疫原性である。さらに他の実施形態においては、成熟SpeBポリペプチドに特異的な抗体は野生型SpeBポリペプチドと交差反応し、かつSpeBポリペプチド活性を無力化する。別の好ましい実施形態においては、プラスミドはT7プロモーター含有プラスミドである。特定の実施形態においては、T7プロモーター含有プラスミドはpET、pRSET、pCRT7−CTTOPO、およびpIVeXより成る群から選択される。さらに他の実施形態においては、宿主細胞は細菌細胞である。1つの好ましい実施形態においては、宿主細胞は大腸菌であり、ここで大腸菌は、BLR(DE3)、BLR(DE3)pLysS、AD494(DE3)、AD494(DE3)pLysS、BL21(DE3)、BL21(DE3)pLysS、BL21(DE3)pLysE、BL21(DE3)pLacI、BL21trxB(DE3)、BL21trxB(DE3)pLysS、HMS174(DE3)、HMS174(DE3)pLysS、HMS174(DE3)pLysE、Origami(DE3)、Origami(DE3)pLysS、Origami(DE3)pLysE、Origami(DE3)pLacI、OrigamiB(DE3)、OrigamiB(DE3)pLysS、OrigamiB(DE3)pLysE、OrigamiB(DE3)pLacI、Rosetta(DE3)、Rosetta(DE3)pLysS、Rosetta(DE3)pLysE、Rosetta(DE3)pLacI、Tuner(DE3)、Tuner(DE3)pLysS、およびTuner(DE3)pLacIより成る群から選択される菌株である。
【0014】
別の実施形態においては、本発明は、(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るプラスミドを宿主細胞において組換え発現させ、ここで前記SpeBポリペプチドが宿主細胞において不溶性ポリペプチド集合体を形成するステップと、(b)ポリペプチド集合体を可溶化し、可溶化ポリペプチドが非ネイティブ成熟SpeBポリペプチドとして規定されるステップと、(c)シャペロンタンパク質の存在下に非ネイティブ成熟SpeBポリペプチドを再折り畳みし、非ネイティブ成熟SpeBポリペプチドがネイティブ成熟SpeBポリペプチドへ折り畳まれるステップと、(d)ネイティブSpeBポリペプチドを回収するステップとを含んで成る成熟SpeBポリペプチドを生成するための方法に関する。好ましい実施形態においては、シャペロンタンパク質は、GroEL、GroEL/GroES、PPI、PDI、およびSpeBプロ−ポリペプチドドメインより成る群から選択される。特定の実施形態においては、シャペロンタンパク質は、配列番号2のアミノ酸残基28〜145を含んで成るSpeBプロ−ポリペプチドドメインである。好ましい実施形態においては、成熟SpeBは、配列番号2のアミノ酸残基146〜398を含んで成るポリペプチドである。別の好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192におけるシステインがセリンによって置換される。さらに別の実施形態においては、不溶性ポリペプチド集合体は封入体としてさらに規定される。他の実施形態においては、ポリペプチドの可溶化が尿素、塩化グアニジニウム(guanidinium chloride)、ドデシル硫酸ナトリウム(SDS)、および熱などより成る群から選択される変性剤である。
【0015】
さらに他の実施形態においては、本発明は、宿主細胞において(i)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(ii)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成るポリシストロニックプラスミドを発現させるステップを含んで成り、成熟SpeBポリペプチドが宿主細胞において可溶性である、宿主細胞において成熟SpeBポリペプチドを組換え発現させるための方法に関する。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192におけるシステインがセリンによって置換される。特定の実施形態においては、プラスミドは、GroESポリペプチドをコード化するポリヌクレオチドをさらに含んで成る。
【0016】
さらに他の実施形態においては、本発明は、(a)(i)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(ii)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成るポリシストロニックプラスミドで宿主細胞を形質転換し、形質導入し、トランスフェクトし、または感染させるステップと、(b)成熟SpeBポリペプチドおよび前記GroELポリペプチドを発現させるのに適切な条件下で宿主細胞を培養し、ここで成熟SpeBポリペプチドが宿主細胞に可溶性であるステップと、(c)ネイティブ成熟SpeBポリペプチドを回収するステップとを含んで成る成熟SpeBポリペプチドを生成するために方法に関する。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192におけるシステインがセリンによって置換される。
【0017】
一部の他の実施形態においては、本発明は、本発明に記載された方法の1つもしくはそれ以上に従って生成される成熟SpeBポリペプチドに関する。他の実施形態においては、本発明は、本発明の方法の1つに従って生成されるSpeBポリペプチドを含んで成る免疫原性組成物に関する。さらに他の実施形態においては、本発明は、化膿連鎖球菌に対して哺乳類の対象を免疫化する方法に関し、この方法は、対象に免疫原性量の免疫原性組成物を投与するステップを含んで成り、ここで免疫原性組成物は本発明の方法に従って生成される成熟SpeBポリペプチドを含んで成る。
【0018】
他の実施形態においては、本発明は、(a)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列と、(b)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列とを含んで成り、成熟SpeBポリペプチドが宿主細胞において発現されると可溶性である、ポリシストロニックプラスミドに関する。一部の実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192でのシステインはセリンによって置換される。一部の他の実施形態においては、プラスミドはT7プロモーター含有プラスミドである。
【0019】
一部の他の実施形態においては、本発明は、(a)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列を含んで成るプラスミド、および(b)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るプラスミドに関し、成熟SpeBポリペプチドは宿主細胞において発現されると可溶性である。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192でのシステインはセリンによって置換される。プラスミドはT7プロモーター含有プラスミドである。
【0020】
別の実施形態においては、本発明は、(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(b)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成り、成熟SpeBポリペプチドが宿主細胞で発現されると可溶性であるポリシストロニックプラスミドに関する。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192におけるシステインはセリンによって置換される。一部の他の実施形態においては、プラスミドはT7プロモーター含有プラスミドである。
【0021】
さらに別の実施形態においては、本発明は、(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(b)GroELポリペプチドをコード化するポリヌクレオチド配列と、(c)GroESポリペプチドをコード化するポリヌクレオチド配列とを含んで成り、成熟SpeBポリペプチドが宿主細胞で発現されると可溶性であるポリシストロニックプラスミドに関する。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192におけるシステインはセリンによって置換される。他の実施形態においては、プラスミドはT7プロモーター含有プラスミドである。
【0022】
さらに他の実施形態においては、(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(b)GroEL、GroES、PDI、およびPPIより成る群から選択される1つもしくはそれ以上のポリペプチドをコード化するポリヌクレオチド配列とを含んで成り、成熟SpeBポリペプチドが宿主細胞で発現されると可溶性であるポリシストロニックプラスミドに関する。好ましい実施形態においては、成熟SpeBポリペプチドのアミノ酸残基192におけるシステインがセリンによって置換される。他の実施形態においては、プラスミドはT7プロモーター含有プラスミドである。
【0023】
別の実施形態においては、本発明は、本発明のプラスミドにより形質転換され、形質導入され、トランスフェクトされ、または感染された宿主細胞を提供する。
【0024】
本発明の他の特徴および利点は、以下の詳細な説明、その好ましい実施形態、および請求の範囲から明らかであろう。
【0025】
(図面の簡単な記載)
図1Aおよび1Bは組換え成熟SpeBとプロ配列ドメインの相互作用を示す。図1Aはペプシン生成成熟C192S SpeB(黒丸)、および成熟野生型SpeB(黒四角)の濃度増大を示し、これらはプロ配列ドメイン(黒記号)、またはリゾチーム(白記号)含有マイクロタイタープレートでインキュベートされ、かつ実施例1によって示されているようにELISAで分析された。データは、各々二重に行われた3つの実験を代表する。図1Bは実施例1に示されているようにバイオコア(Biocore)3000の使用によって行われた、プロ配列ドメインと成熟SpeBポリペプチドとの相互作用の実時間分析を示す。結果は表面プラスモン共鳴(相対反応)として表されている。
【0026】
図2Aおよび2Bは成熟SpeBのプロ配列ドメイン介在阻害を示す。示されているのは、レゾルフィン標識カゼイン(図2A)またはシステインプロテアーゼパパイン(図2B)を使用した成熟野生型SpeBの阻害であり、SpeBプロ配列ドメイン(黒四角)、またはリゾチーム(白三角)を使用した基質が実施例1に示されているように分析された。
【0027】
図3は変性成熟SpeBの再折り畳みに対する組換えプロ配列ドメインの効果を示す。変性成熟SpeBを、PBS、0.5Mアルギニン(黒丸)、PBS、0.5Mアルギニン、20μMプロテアーゼ阻害剤E−64(白丸)、PBS(黒四角)、およびPBS、20μM E−64(白四角)を含有する増大濃度のプロ配列ドメイン中に迅速に希釈した。反応を実施例1に示されているように行い、レゾルフィン標識カゼイン切断アッセイを使用してプロテアーゼ活性の存在について評価した。
【0028】
図4は2−プラスミドベース発現構成物を示す略図である。2−プラスミドベース発現ベクターの特徴としては、図のように、KanR−カナマイシン耐性遺伝子(aph(3’)−la)、AmpR−アンピシリン耐性遺伝子(b−ラクタマーゼ)、複製のOri−origin、laclq−lacレプレッサ、T7ベースプロモーター(T7またはT7lac)、およびT7ターミネータが挙げられる。
【0029】
図5Aおよび5Bは、各々、ポリシストロニック発現系および合成リンカー領域を示す図表である。ポリシストロニック発現ベクター(図5A)の特徴としては、図のように、KanR−カナマイシン耐性遺伝子(aph(3’)−la)、複製のOri−origin、laclq−lacレプレッサ、T7lacプロモーター、およびT7ターミネータが挙げられる。分析されたリンカー領域の塩基組成が示されている(図5B)。各リンカー領域のヌクレオチド番号(5nt、10nt、20nt、40nt)は、プロ配列ドメインの遺伝子工学的TAA翻訳停止コドン(PSD停止)(太字テキスト)と、最適化シャイン・ダルガノリボソーム結合部位(SD)との間の塩基の数を示す。第2のシストロンの翻訳ATG開始コドンは太字イタリックで示されている。
【0030】
図6Aおよび6Bはポリシストロニック発現系における可溶性成熟SpeBおよびプロ配列ドメインレベルの評価および相対定量を示す。可溶性フラクションにおけるプロ配列ドメイン、および成熟SpeB発現に対する5nt、10nt、20nt、40ntリンカー含有C192S SpeBポリシストロニック系のSDS−PAGE評価が行われた(図6A)。発現タンパク質の定量は、分子動力学パーソナル濃度計を使用して行われ、スキャン面の測定は発現レベルとして示された(図6B)。
【0031】
図7はポリシストロニックcDNAの定量的PCR分析を示す。cDNAおよび−RT制御は実施例1に記載されているように調製され分析された。すべてのCt値はKanR mRNA発現に標準化された。
【0032】
図8は成熟C192S SpeBの熱変性を示す。発現C192S SpeBチモーゲンのペプシン切断によって、または2−プラスミドおよびポリシストロニック共発現系によって生成された精製成熟C192S SpeBの変性曲線は、実施例1に示されているように分析された。
【0033】
図9は成熟野生型SpeBの熱誘発変性を示す。自己触媒的プロセシング(すなわち、チモーゲン生成)によって、または2−プラスミドおよびポリシストロニック共発現系によって生成された精製成熟野生型SpeBの変性曲線は、実施例1に示されているように分析された。
【0034】
図10は組換え成熟SpeBの操作モル濃度の評価を示す。自動触媒(黒四角)のほか、2−プラスミド(黒丸)およびポリシストロニック(黒三角)共発現によって生成された当量(0.12μM)の精製成熟野生型SpeBが、実施例1に記載されているようにレゾルフィン標識カゼイン切断アッセイを使用して評価された。25℃下に1時間のインキュベーション時間を切断反応用に使用した。ポリシストロニック発現によって生成された精製成熟C192S SpeB(白ダイヤ)が対照として評価された。
【0035】
図11は野生型SpeBタンパク質分解活性の抗体介在阻害を示す。2−プラスミド(黒ダイヤ)、またはポリシストロニック系(白四角)によって生成された成熟C192S SpeBに対して生成される抗血清の増大量が、レゾルフィン標識カゼイン切断アッセイを使用して成熟野生型SpeBのタンパク質分解活性を特異的に阻害する能力について評価された。37℃下に2時間のインキュベーション時間を切断反応用に使用した。免疫前血清が分析用の陰性対照(トリ−スター)が使用された。
【0036】
(発明の詳細な記載)
以下に記載される本発明は、免疫原性組成物で使用する化膿連鎖球菌外毒素B(以下、「SpeB」)を生成する方法に対する技術上の必要性に対処する。一部の好ましい実施形態においては、本発明は、免疫原性組成物で使用する成熟SpeBを生成する方法に関する。より詳しくは、以下に記載される本発明は、宿主細胞における成熟SpeBポリペプチドを組換え発現させる方法に対する技術上の必要性に対処し、ここで発現成熟SpeBポリペプチドは宿主細胞において可溶性である。好ましい実施形態においては、本発明の方法に従って生成される成熟SpeBポリペプチドは、哺乳類宿主に投与されると免疫原性である。
【0037】
以下に規定される、化膿連鎖球菌で発現される「SpeBチモーゲン」は、配列番号2のアミノ酸1−398を含んで成る40kDaプレ−プロ−ポリペプチドである。SpeBチモーゲンは、27アミノ酸NH2−末端シグナル(「プレ」)配列を有する酵素的に不活性な(すなわち、チモーゲン)、40kDa、プレ−プロ−ポリペプチドとして発現され、そして118アミノ酸プロ−ポリペプチド(「プロ」)配列、および253アミノ酸成熟ポリペプチド配列が続く。したがって、以下に規定されているように、SpeBチモーゲンの「プレ」配列は、配列番号2のアミノ酸1−27を含んで成る。同様に、以下に規定されているように、SpeBチモーゲンの「プロ配列」、「プロ配列ドメイン」、「プロ−ポリペプチド配列」、および「プロ−ポリペプチドドメイン」は同じ意味で使用され、ここでプロ配列は配列番号2のアミノ酸28−145を含んで成る。
【0038】
ネイティブ細菌(すなわち、化膿連鎖球菌)によって分泌されると、40kDa SpeBチモーゲンは自己触媒活性化を受け、結果として12kDa NH2−末端プロ−ポリペプチド配列(すなわち、配列番号2のアミノ酸8−145)の除去および成熟28kDaタンパク質分解活性SpeBポリペプチド(または酵素)の形成が生じる。以下に規定されているように、「成熟SpeB」ポリペプチドは、配列番号2のアミノ酸146−398を含んで成る28kDaポリペプチドであり、ここで成熟SpeBポリペプチドはシステインプロテアーゼ活性を有する。以下に規定されているように、「成熟SpeB」ポリペプチドおよび「成熟野生型SpeB」ポリペプチドは同じ意味で使用され、その両方は配列番号2のアミノ酸146−398を含んで成る野生型28kDaポリペプチドを指し、ここでポリペプチドはシステインプロテアーゼ活性を有する。
【0039】
SpeB活性部位は、配列番号2のアミノ酸残基192および340でCys−His対を含有する。192位での単一のシステイン残基のセリン残基(以下、「C192S」または「C192S変異株」)へのアミノ酸置換(すなわち、突然変異)は、成熟C192S SpeBポリペプチドの検出可能なタンパク質分解活性の喪失をもたらす。したがって、以下に規定されているように、「C192S SpeBチモーゲン」または「C192S SpeB変異株」は、配列番号2のアミノ酸28−398を含んで成り、ここで192位でのシステインアミノ酸残基はセリンアミノ酸残基に変異されている。以下に規定されているように、「成熟C192S SpeB」ポリペプチドは、配列番号2のアミノ酸146−398を含んで成る28kDaポリペプチドであり、ここで192位でのシステインアミノ酸残基はセリンアミノ酸残基に変異されており、ここで成熟C192S SpeBポリペプチドは成熟野生型SpeBポリペプチドに対するシステインプロテアーゼ活性を有さない。
【0040】
したがって、一部の実施形態においては、本発明は、成熟SpeBポリペプチド、かつより好ましくは、成熟C192S SpeBポリペプチドを含んで成る免疫原性組成物に関する。例えば、C192S SpeB変異株による以前の免疫学的試験では、成熟C192S SpeBポリペプチド(すなわち、NH2−末端プロ−ポリペプチド配列の欠損)が、成熟野生型SpeBポリペプチドへの最大の阻害活性(すなわち、交差再活性)による抗体の生成に必要であることが明らかにされている(マツカ(Matsuka)ら、1999年)。したがって、本明細書では、化膿連鎖球菌感染に対して哺乳類を免疫化するために有効な免疫原性組成物が、少なくとも成熟C192S SpeBまたは成熟野生型SpeBポリペプチド抗原を含んで成ることが考えられている。しかし、大腸菌宿主細胞における成熟SpeBポリペプチドの組換え発現が、もっぱら大腸菌における不溶性SpeBポリペプチド集合体の生成をもたらすことが当技術分野で周知である(マツカ(Matsuka)ら、1999年)。
【0041】
したがって、特定の実施形態においては、本発明は、「成熟」および「可溶性」である宿主細胞におけるSpeBポリペプチドの発現の困難さを克服する方法に関する。例えば、本発明の1つの実施形態においては、成熟C192S SpeBポリペプチド(または成熟野生型SpeB)をコード化するプラスミドと、宿主細胞におけるプロ−ポリペプチドドメイン(すなわち、配列番号2のアミノ酸28−145)をコード化するプラスミドとの共発現が、宿主細胞における可溶性、成熟C192S SpeBポリペプチド(または成熟野生型SpeB)の有効な発現をもたらすことが明らかにされている(例えば、実施例3を参照)。同様に、ポリシストロニックプラスミドが、成熟C192S SpeBポリペプチド(または成熟野生型SpeB)とプロ−ポリペプチド配列(配列番号2のアミノ酸28−145)の両方をコード化する、宿主細胞におけるポリシストロニックプラスミドの組換え共発現も、」宿主細胞における可溶性、成熟C192S SpeBポリペプチド(または成熟野生型SpeB)の発現をもたらす(例えば、実施例4を参照)。
【0042】
したがって、以下に記載される本発明は、宿主細胞において可溶性成熟C192S SpeBポリペプチド(または成熟野生型SpeB)を発現させ、可溶性成熟C192S SpeBポリペプチドが化膿連鎖球菌感染に対して哺乳類を免疫化するための免疫原性組成物において特に有用である、新たな方法、およびその新たな組成物を提供する。したがって、一部の好ましい実施形態においては、本発明は、成熟C192S SpeBポリペプチド(または成熟野生型SpeB)をコード化するプラスミドと、宿主細胞におけるプロ−ポリペプチドドメイン(すなわち、配列番号2のアミノ酸28−145)をコード化するプラスミドを共発現させ、ここで成熟SpeBポリペプチドが宿主細胞において可溶性である方法に関する。一部の他の好ましい実施形態においては、本発明は、宿主細胞におけるポリシストロニックプラスミドを発現させる方法に関し、ここでポリシストロニックプラスミドは、成熟C192S SpeBポリペプチド(または成熟野生型SpeB)とプロ−ポリペプチド配列(配列番号2のアミノ酸28−145)の両方をコード化し、ここで成熟SpeBポリペプチドは宿主細胞において可溶性である。
【0043】
本発明は、可溶性成熟SpeBの生成を調節するプロ配列ドメインの要件を明らかにし、プロ配列ドメインが成熟SpeBポリペプチドの適切な折り畳みに対する分子内シャペロンとして機能することを示す。成熟野生型SpeBポリペプチドまたは成熟C192S SpeBのいずれかとのプロ配列ドメインの関連性は、それぞれ、約11nmおよび34nMの解離定数(Kd)を有する(実施例2)。これらの結合値は、プロ配列ドメインと成熟SpeBポリペプチドドメインとの間の高い親和性を示す。また、プロ配列ドメインの分子シャペロン活性は、尿素変性成熟SpeBを使用してインビトロで明らかにされている(実施例2)。
【0044】
したがって、特定の実施形態においては、本発明は、不溶性成熟SpeB集合体のタンパク質補助折り畳みの方法に関する。例えば、本発明の1つの実施形態は、(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るプラスミドを宿主細胞において組換え発現させ、ここでSpeBポリペプチドが宿主細胞において不溶性ポリペプチド集合体を形成するステップと、(b)ポリペプチド集合体を可溶化し、可溶化ポリペプチドが非ネイティブ成熟SpeBポリペプチドとして規定されるステップと、(c)1つもしくはそれ以上のシャペロンタンパク質の存在下に非ネイティブ成熟SpeBポリペプチドを再折り畳みし、非ネイティブ成熟SpeBポリペプチドがネイティブ成熟SpeBポリペプチドへ折り畳まれるステップと、(d)ネイティブ成熟SpeBポリペプチドを回収するステップとを含んで成る成熟SpeBポリペプチドを生成するための方法に関する。
【0045】
同様に、他の実施形態においては、本発明は、成熟SpeBポリペプチドが1つもしくはそれ以上の分子シャペロンタンパク質の存在下に発現される、タンパク質補助折り畳みの方法に関する。例えば、本発明は、(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(b)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成るポリシストロニックプラスミドを宿主細胞において組換え発現させ、ここで成熟SpeBポリペプチドが宿主細胞において可溶性であり、かつ(c)ネイティブ成熟SpeBポリペプチドを回収するステップを含んで成る、宿主細胞において成熟SpeBポリペプチドを発現するための方法を提供する。
【0046】
A.SpeB抗原をコード化するポリヌクレオチド
本発明の単離および精製化膿連鎖球菌ポリヌクレオチドは、成熟SpeBポリペプチド抗原の生成において使用される。さらに具体的には、一部の実施形態においては、本発明のポリヌクレオチドは成熟SpeBポリペプチドおよびSpeBプロ−ポリペプチドドメインをコード化し、ここで成熟SpeBポリペプチドは宿主細胞において発現されると可溶性である。したがって、本発明の一部の実施形態においては、ポリヌクレオチドは配列番号2のアミノ酸146〜398を含んで成る成熟SpeBポリペプチドをコード化し、第2のポリヌクレオチドは配列番号2のアミノ酸28〜145を含んで成るプロ−ポリペプチドドメインをコード化する。好ましい実施形態においては、本発明のポリヌクレオチドは成熟C192SpeBポリペプチドをコード化し、ここで成熟C192SSpeBポリペプチドは配列番号2のアミノ酸146〜398を含んで成り、ここで配列番号2のアミノ酸残基192でのシステインはセリン残基に変異されている。一部の好ましい実施形態においては、成熟SpeBポリペプチドをコード化するポリヌクレオチドは配列番号1のヌクレオチド436〜1197を含んで成り、成熟C192S SpeBポリペプチドをコード化するポリヌクレオチドは配列番号1のヌクレオチド436〜1197を含んで成り、ここで配列番号2のアミノ酸残基192はセリンアミノ酸であり、かつプロ−ポリペプチドドメインをコード化するポリヌクレオチドは配列番号1のヌクレオチド82〜435を含んで成る。
【0047】
したがって、特定の実施形態においては、本発明のポリヌクレオチドはDNA分子であり、ここでDNAはゲノムDNA、プラスミドDNA、またはcDNAでありうる。好ましい実施形態においては、本発明のポリヌクレオチドは組換えcDNAポリヌクレオチドである。別の好ましい実施形態においては、成熟SpeBポリペプチドをコード化するポリヌクレオチドは第1のプラスミドベクターに含まれており、かつSpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチドは第2のプラスミドベクターに含まれており、ここで両方のベクターは宿主細胞において共発現される。別の好ましい実施形態においては、成熟SpeBポリペプチドをコード化するポリヌクレオチドおよびSpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチドはポリシストロニック発現構成物に含まれている。セクションE(実施例4および図9)に記載されているように、本発明のポリシストロニック構成物は、5’〜3’の方向でSpeBプロ−ポリペプチドドメインを第1のシストロンとして含んで成り、そして翻訳エンハンサーおよび最適化シャイン・ダルガノリボソーム結合部位および成熟SpeBを第2のシストトロンとして含んで成る合成リンカーが続く。別の実施形態においては、本発明のポリシストロニック構成物は、5’〜3’の方向で成熟SpeBを第1のシストロンとして含んで成り、そして翻訳エンハンサーおよび最適化シャイン・ダルガノリボソーム結合部位およびSpeBプロ−ポリペプチドドメインを第2のシストロンとして含んで成る合成リンカーが続く。
【0048】
以下で使用される「ポリヌクレオチド」という語は、ホスホジエステル結合によって結合されたヌクレオチドの配列を意味する。ポリヌクレオチドは以下、5’〜3’の方向で示されている。本発明のポリヌクレオチドは、約10〜約数十万の塩基対を含んで成る。好ましくは、ポリヌクレオチドは約10〜約3,000の塩基対を含んで成る。特定のポリヌクレオチドの好ましい長さは以下に記載されている。
【0049】
本発明のポリヌクレオチドは、デオキシリボ核酸(DNA)分子、リボ核酸(RNA)分子、またはヌクレオチド類似体を使用して生成されるDNAもしくはRNAの類似体である。核酸分子は一本鎖また二本鎖であるが、好ましくは、二本鎖DNAである。ポリヌクレオチドはDNA分子である場合、その分子は遺伝子、cDNA分子、またはゲノムDNA分子である。ヌクレオチド塩基は、単一の文字表記によって以下に示されている。すなわち、アデニン(A)、グアニン(G)、チミン(T)、シトシン(C)、イノシン(I)およびウラシル(U)。
【0050】
「単離」は、天然状態から「人が作ったものによって」変化されていることを意味する。「単離」組成物または物質は、その最初の環境から変更もしくは除去、または変更および除去されているものである。例えば、生きた動物に天然に存在するポリヌクレオチドまたはポリペプチドは「単離」されず、その天然状態の共存する物質から分離された同じポリヌクレオチドまたはポリペプチドは、その語が以下で使用されるように「単離」である。
【0051】
好ましくは、「単離」ポリヌクレオチドは、核酸が由来する生物のゲノムDNAにおける核酸を天然にフランキングする配列(すなわち、核酸の5’および3’末端に位置した配列)を含まない。例えば、さまざまな実施形態においては、単離SpeB核酸分子は、核酸が由来する細胞のゲノムDNAにおける核酸分子を天然にフランキングする約5kb、4kb、3kb、2kb、1kb、0.5kbまたは0.1kb未満のヌクレオチド配列を含有する。しかし、SpeB核酸分子はタンパク質コードまたは調節配列に融合し、それでも単離とみなされる。
【0052】
本発明のSpeBポリヌクレオチドは、標準クローニングおよびスクリーニング法を使用し、mRNAに由来するcDNAライブラリーから得られる。本発明のポリヌクレオチドは、ゲノムDNAライブラリー(例えば、化膿連鎖球菌ライブラリー)などの天然源からも得られ、または公知および市販の方法を使用して合成される。
【0053】
成熟SpeBおよび/またはSpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチドの相同分子種および対立遺伝子変異体は、当技術分野で周知の方法を使用して容易に同定される。ポリヌクレオチドの対立遺伝子変異体および相同分子種は、配列番号1に示されているヌクレオチド配列、またはこのヌクレオチド配列の断片の一般的に少なくとも約70−75%、より一般的に少なくとも約80−85%、かつ最も一般的に少なくとも約90−95%もしくはそれ以上の相同のヌクレオチド配列を含んで成る。かかる核酸分子は、好ましくは、ストリンジェントな条件下で、配列番号1に示されているヌクレオチド配列、またはこのヌクレオチド配列の断片にハイブリッド可能であることが容易に同定される。
【0054】
成熟SpeBポリペプチドおよびSpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチドは、宿主細胞における可溶性成熟SpeBポリペプチドまたはその断片の組換え生成のために使用される。ポリヌクレオチドは、成熟SpeBポリペプチドのコード配列および/またはSpeBプロ−ポリペプチドドメインのコード配列を含みうる。ポリヌクレオチドは、転写、非翻訳配列、スプライシングシグナル、プロモーター/エンハンサー配列、リボソーム結合部位、およびポリアデニル化シグナルなど非コード5’および3’配列も含有しうる。
【0055】
したがって、一部の実施形態においては、本発明によって提供されるポリペプチド配列情報(すなわち、配列番号1)は、以下に開示される選択ポリヌクレオチドの遺伝子配列に特異的にハイブリッドする能力を有する比較的短いDNA(またはRNA)オリゴヌクレオチド配列の調製を可能にする。以下で使用される「オリゴヌクレオチド」という語は、2個もしくはそれ以上のデオキシリボヌクレオチドまたはリボヌクレオチド、通常、3個(3)超、および通常10個(10)超、さらに100個(100)もしくはそれ以上(ただし好ましくは、20〜30個)までから成る分子として規定される。正確なサイズは、多くの因子によって決まり、言い換えると、オリゴヌクレオチドの最終の機能または使用によって決まる。したがって、本発明の特定の実施形態においては、適切な長さの核酸プローブが選択ヌクレオチド配列の検討に基づき調製される。かかる核酸プローブが成熟SpeBポリペプチドまたはSpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチドに特異的にハイブリッドする能力は、さまざまな実施形態においてそれらに特定の有用性を与える。最も重要なことだが、これらのプローブは所定のサンプルにおける相補配列の存在を検出するためのさまざまなアッセイにおいて使用される。
【0056】
一部の実施形態においては、オリゴヌクレオチドプライマーを使用することが有利である。これらのプライマーは、化学合成、DNA複製、逆転写、またはその組合せを含む任意のやり方で生成されうる。かかるプライマーの配列は、ポリメラーゼ連鎖反応(PCR)技術を使用して原核細胞からSpeBポリペプチドをコード化するSpeBポリヌクレオチドの規定部分を検出、増幅、または変異で使用するための本発明のポリヌクレオチドを使用して指定される。
【0057】
一部の実施形態においては、本発明のポリヌクレオチドをハイブリッド形成を検出するための適切な標識と組合せて使用することが有利である。多種多様の適切な標識は、検出可能シグナルを与える能力があるアビジン/ビオチンなど、放射性、酵素、または他のリガンドを含めて当技術分野で周知である。
【0058】
本発明による利点の一部を提供するために、ハイブリダイゼーション試験またはアッセイに使用される好ましい核酸配列としては、配列番号2のポリペプチドをコード化するポリヌクレオチドの少なくとも10個〜約18個のヌクレオチド長ストレッチに相補的であるプローブ分子が挙げられる。長さが少なくとも10個のヌクレオチドのサイズは、断片が安定かつ選択的である二重分子を形成するのに十分な長さであることを確実にするために役立つ。しかし、ハイブリッドの安定性および選択性を増大させ、それによって得られる特異的ハイブリッド分子の質および程度を改善するために、長さが10個超の塩基のストレッチにわたって相補配列を有する分子が一般に好ましい。かかる断片は、例えば、化学的手段によって断片を直接合成することによって、(参照することにより本明細書で援用される米国特許第4,683,202号明細書)のPCR技術など核酸再生成技術の適用によって、または適切な挿入および適切な制限酵素部位を含有する組換えプラスミドから選択DNA断片を切除することによって容易に調製される。
【0059】
したがって、本発明のポリヌクレオチドプローブ分子は、遺伝子の相補ストレッチで二重分子を選択的に形成するその能力のために使用される。想定される適用によって、ターゲット配列のためにプローブの各種程度の選択性を達成するのに各種条件のハイブリダイゼーションのストリンジェンシーを使用することが望まれる(以下の表1を参照)。高度の選択性を必要とする用途のために、通常、比較的ストリンジェントな条件を使用してハイブリッドを形成することが望まれる。一部の用途、例えば、基礎を成すテンプレートにハイブリッドされる変異株プライマー鎖を使用する変異株を調製することが望まれ、または相同ポリペプチドコード配列を他の細胞、機能的同等物などから単離することが求められる場合には、ヘテロ2本鎖形成を可能にするあまり厳しくないハイブリダイゼーション条件が通常、必要とされる(表1を参照)。それによって、クロスハイブリダイゼーション種は、対照ハイブリダイゼーションに対する陽性ハイブリダイゼーションシグナルとして容易に同定される。ともかく、一般に、条件は、温度の上昇と同じやり方でハイブリッド二本鎖の不安定化に使用される増加量のホルムアミドの添加によって、より厳しく与えられることが予想される。したがって、ハイブリダイゼーション条件は容易に操作され、したがって、一般に所望の結果によって選択される方法となる。
【0060】
本発明は、低ストリンジェントな条件、より好ましくは、ストリンジェントな条件、かつ最も好ましくは、高ストリンジェントなな条件下で、以下に記載されたポリヌクレオチドにハイブリッド可能なポリヌクレオチドも含む。ストリンジェントな条件の例は以下の表1に示されている。すなわち、高ストリンジェントな条件は、少なくとも、例えば、条件A−Fのストリンジェンシーであり、ストリンジェントな条件は、少なくとも、例えば、条件G−Lのストリンジェンシーであり、低ストリンジェントな条件は、少なくとも、例えば、条件M−Rのストリンジェンシーである。
【0061】
【表1】
【0062】
(bp)I:ハイブリッド長さは、ハイブリダイズするポリヌクレオチドののハイブリッド領域について予想される。ポリヌクレオチドを未知の配列のターゲットポリヌクレオチドにハイブリダイズする場合、ハイブリッド長さはポリヌクレオチドのハイブリダイゼーションのものと想定される。既知の配列のポリヌクレオチドがハイブリダイズされる場合、ハイブリッド長さはポリヌクレオチドの配列を調整し、最適な配列相補性の領域または各領域を同定することによって決定される。
バッファーH:SSPE(1×SSPEは、0.15M NaCl、10mM NaH2PO4、および1.25mM EDTA、pH7.4である)は、ハイブリダイゼーションおよび洗浄バッファー中SSC(1×SSCは0.15M NaCl、および15mM クエン酸ナトリウム)に代替され、洗浄はハイブリダイゼーションの完了後15分間行われる。
TB〜TR:長さが50未満の塩基対であることが予想されるハイブリッドのハイブリダイゼーション温度は、ハイブリッドの融解温度(Tm)よりも5−10℃低くあるべきであり、ここでTmは以下の等式に従って決定される。長さが18未満の塩基対のハイブリッドは、Tm(℃)=2(A+T塩基の#)+4(G+C塩基の#)。長さが18〜49の塩基対のハイブリッドでは、Tm(℃)=81.5+16.6(log10[Na+])+0.41(%G+C)−(600/N)であり、ここでNはハイブリッド中の塩基の数であり、[Na+]はハイブリダイゼーションバッファー中のナトリウムイオンの濃度である(1×SSCの[Na+]=0.165M)。
【0063】
ポリヌクレオチドハイブリダイゼーションのストリンジェントな条件の追加の例は、参照することにより以下に援用される、サムブルック(Sambrook)ら、1989年、Molecular Cloning:A Laboratory Manual、コールド・スプリング・ハーバー・ラボラトリー・プレス(Cold Spring Harbor Laboratory Press)、コールド・スプリング・ハーバー(Cold Spring Harbor)、ニューヨーク州、第9章および第11章、およびオーズベル(Ausubel)ら、1995年、Current Protocols in Molecular Biology、ジョン・ワイリー・アンド・サンズ(John Wiley & Sons,Inc.)編、第2.10節および第6.3−6.4節に示されている。
【0064】
B.成熟SPEBポリペプチド抗原
特定の実施形態においては、本発明は、成熟SpeBポリペプチドおよび/または成熟C192S SpeBポリペプチドを含んで成る免疫原を提供する。一部の実施形態においては、これらの免疫原は化膿連鎖球菌感染に対して哺乳類宿主を免疫化するための免疫原性組成物において使用される。好ましい実施形態においては、免疫原は、高率の異種化膿連鎖球菌株に対する保護(すなわち、交差保護)を与える成熟C192S SpeBポリペプチドである。
【0065】
抗原または免疫原が通常、免疫原性に基づき規定される。免疫原性は、体液性および/または細胞介在免疫応答を誘発する能力として規定される。したがって、「抗原」または「免疫原」という語は、以下に規定されているように、体液性および/または細胞介在免疫応答を誘発する能力を有する分子である。
【0066】
したがって、1つの実施形態においては、本発明は、少なくとも化膿連鎖球菌成熟SpeBポリペプチドおよび/または成熟C192S SpeBポリペプチドを含んで成る免疫原性組成物に関する。好ましい実施形態においては、成熟SpeBポリペプチド抗原は、配列番号2のアミノ酸146〜398を含んで成る。別の好ましい実施形態においては、成熟C192S SpeBポリペプチド抗原は配列番号2のアミノ酸146〜398を含んで成り、ここで配列番号2の192位でのアミノ酸はシステインからセリン残基に変異されている。本発明の一部の他の実施形態においては、成熟SpeBポリペプチドおよび/または成熟C192S SpeBポリペプチドを含んで成る免疫原性組成物は、1つまたは追加のポリペプチド抗原(すなわち、SpeB以外のポリペプチド)をさらに含んで成り、ここで1つもしくはそれ以上の追加のポリペプチド抗原は化膿連鎖球菌抗原または他の感染性細菌および/またはウイルスからの抗原である。
【0067】
本発明によるSpeBポリペプチドの生物学的同等物または変異体は、SpeBチモーゲン、C192S SpeBチモーゲン、成熟SpeBポリペプチド、成熟C192S SpeBポリペプチド、およびSpeBプロ−ポリペプチドドメインより成る群から選択される化膿連鎖球菌ポリペプチドと実質的な相同を含有するポリペプチドを包含する。例えば、成熟SpeBポリペプチドおよび成熟C192S SpeBポリペプチドの生物学的同等物または変異体も、成熟SpeBポリペプチドが免疫原性反応を引き出す能力を維持する限り、配列番号2の成熟SpeBポリペプチドが修飾されているポリペプチドを含む。一般に、成熟SpeBポリペプチドの機能的な生物学的同等物または変異体は天然アミノ酸配列変異体であり、ここで成熟SpeBポリペプチドは免疫原性反応を引き出す能力を維持する。
【0068】
修飾および変更が本発明の成熟SpeBポリペプチドの構造においてなされ、それでもSpeB免疫原性特性を有する分子が得られる。ポリペプチドの生物学的機能活性を規定するのはポリペプチドの相互作用的能力および性質であるため、一部のアミノ酸配列置換がポリペプチド配列(または、もちろん、その基礎を成すDNAコード配列)においてなされ、それでも同様の特性を有するポリペプチドが得られる。
【0069】
アミノ酸置換は一般に、アミノ酸側鎖置換、例えば、その疎水性、親水性、電荷、サイズなどの相対類似性に基づく(例えば、以下、参照することによりその全体が援用される、カイト(Kyte)とドリトル(Doolittle)、1982年、および米国特許第4,554,101号明細書を参照)。さまざまな上述の特性を考慮に入れる例示の置換は当業者には公知であり、アルギニンおよびリジン、グルタミン酸塩およびアスパラギン酸塩、セリンおよびトレオニン、グルタミンおよびアスパラギン、およびバリン、ロイシンおよびイソロイシンを含む(以下の表2を参照)。したがって、本発明では、上記されているように成熟SpeBポリペプチドの機能的または生物学的同等物が意図されている。
【0070】
【表2】
【0071】
本発明の特定の実施形態においては、多価または組合せ免疫原性組成物が提供される。組合せ免疫原性組成物が、1つもしくはそれ以上の追加の抗原とともに、1つもしくはそれ以上の本発明のポリペプチド(例えば、成熟C192S SpeB)、またはその断片(例えば、SpeBエピトープ断片)を含むことによって提供される。特に、組合せ免疫原性組成物は1つもしくはそれ以上の成熟SpeBポリペプチド、またはその断片を、1つもしくはそれ以上のポリペプチド、ポリペプチド断片、糖質、オリゴ糖類、脂質、リポオリゴ糖類、多糖類、オリゴ糖類−タンパク質コンジュゲート、多糖類−タンパク質コンジュゲート、ペプチド−タンパク質コンジュゲート、オリゴ糖類−ペプチドコンジュゲート、多糖類−ペプチドコンジュゲート、タンパク質−タンパク質コンジュゲート、リポオリゴ糖類−タンパク質コンジュゲート、または多糖類−タンパク質コンジュゲートと組合わせることによって提供される。
【0072】
したがって、一部の実施形態においては、上記の1つもしくはそれ以上の抗原は、化学的付着(すなわち、接合)によって抗原担体タンパク質にコンジュゲートされる。ポリペプチドを担体タンパク質に接合させる手段は当技術分野で周知であり、グルタルアルデヒド、m−マレイミドベンコイル−N−ヒドロキシスクシニミドエステル、カルボジイミド、およびビス−ビアゾチ化(biazotized)ベンジジンを含む。
【0073】
例示の従来のタンパク質担体は、大腸菌DnaKタンパク質、ガラクトキナーゼ(galK)、ユビキチン、α−接合因子、β−ガラクトシダーゼ、およびインフルエンザNS−1タンパク質を含むがこれらに限定されない。トキソイド(すなわち、その毒性活性を削減するように十分な修飾による天然毒素をコード化する配列)、例えば、ジフテリアトキソイドおよび破傷風トキソイド、その各毒素、およびこれらタンパク質の突然変異型も担体として使用されうる。例示の担体タンパク質はジフテリア毒素CRM197である(ジフテリア毒素の非毒性形態、参照することによりその全体が本明細書で援用される、米国特許第5,614,382号明細書を参照)。他の担体としては、緑膿菌(Pseudomonas)の外毒素A、大腸菌の易熱性毒素、コレラ菌(Vibrio cholera)、およびロタウイルス粒子(ロタウイルスおよびVP6粒子を含む)が挙げられる。あるいは、担体タンパク質または他の免疫原性タンパク質の断片またはエピトープが使用されうる。例えば、ハプテンが細菌毒素のT細胞エピトープに結合されうる(米国特許第5,785,973号明細書を参照。同様に、さまざなな細菌熱ショックタンパク質、例えば、マイコバクテリアhsp−70が使用されうる。グルタチオン−S−トランスフェラーゼ(GST)は別の有用な担体である。当業者は、これに関して使用される適切な担体を容易に選択することができる。
【0074】
一部の実施形態においては、本発明は、不溶性成熟SpeB集合体のタンパク質補助折り畳みの方法に関する。同様に、他の実施形態においては、本発明はタンパク質補助折り畳みの方法に関し、ここで成熟SpeBポリペプチドは、1つもしくはそれ以上の分子シャペロンタンパク質の存在下に発現される。
【0075】
例えば、1つの実施形態においては、本発明は、(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るプラスミドを宿主細胞において組換え発現させ、ここでSpeBポリペプチドが宿主細胞において不溶性ポリペプチド集合体を形成するステップと、(b)ポリペプチド集合体を可溶化し、可溶化ポリペプチドが非ネイティブ成熟SpeBポリペプチドとして規定されるステップと、(c)シャペロンタンパク質の存在下に非ネイティブ成熟SpeBポリペプチドを再折り畳みし、非ネイティブ成熟SpeBポリペプチドがネイティブ成熟SpeBポリペプチドへ折り畳まれるステップと、(d)前記ネイティブ成熟SpeBポリペプチドを回収するステップとを含んで成る成熟SpeBポリペプチドを生成するための方法に関する。一部の他の実施形態においては、本発明は、(i)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(ii)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成るポリシストロニックプラスミドを含んで成る宿主細胞における成熟SpeBポリペプチドを組換え発現させ、ここで成熟SpeBポリペプチドが宿主細胞において可溶性である方法を提供する。
【0076】
したがって、特定の実施形態においては、本発明は、成熟SpeBポリペプチドの折り畳みを支援する分子シャペロンタンパク質を提供する。分子シャペロンは当技術分野で周知であり、トリガー因子(TF)などのリボソーム結合タンパク質、Hsp70、DnaK、Hsp40、DnaJ、GrpEなどのシャペロンのHsp70ファミリー、およびGroEL、GroES、Hsp60、Hsp10などのシャペロンのシャペロニンファミリーを含むがこれらに限定されない(クレイトン(Creighton)、1993年、ハルトル(Hartl)とヘイヤー・ハルトル(Hayer−Hartl)、2002年)。本発明の使用に考えられている分子シャペロンタンパク質のポリヌクレオチドおよびポリペプチド配列は当技術分野で周知であり(例えば、各々参照することによりその全体が本明細書で援用される、米国特許第6,159,708号明細書、米国特許第6,010,879号明細書、米国特許第5,776,724号明細書、およびロリマー(Lorimer)とボールドウィン(Baldwin)、Methods in Enzymology、1998年を参照)、折り畳み/再折り畳み要件、補因子などが、所定の分子シャペロンタンパク質に対して以下に記載されている。
【0077】
したがって、非限定的実施例によって、大腸菌からの分子シャペロニンGroELはシャペロンの熱ショックタンパク質60(Hsp60)クラスのメンバーであり、大腸菌GroEオペロンからのGroESとともに発現される。GroELは、集合の傾向があるポリペプチド中間体の濃度およびオフ経路集合率を減少させる折り畳まれていないタンパク質(例えば、成熟SpeB)を結合することによってタンパク質折り畳み反応を支援し、それによってネイティブな立体構造(例えば、適切に折り畳まれた可溶性成熟SpeB)に有利に分配する。補シャペロニンGroES、およびATP、K+、およびMg2+などの補因子は、GroEL介在ポリペプチド折り畳み反応の収量をさらに増大させることが知られている。したがって、特定の実施形態においては、当業者は、シャペロン介在(または補助)タンパク質折り畳みを改善または強化するために当技術分野で周知の成分、補因子、追加のシャペロンタンパク質などを含めるであろう。
【0078】
機序は所定のシャペロンファミリーまたはクラスで変動しうるが、すべてのシャペロンタンパク質(以下に記載されるPDIおよびPPIを除き)によって共有される基礎を成す特徴は、その非ネイティブ立体構造においてタンパク質を結合する能力である。以下に規定されているように、本発明の「分子シャペロン」または「シャペロン」タンパク質は、インビボおよび/またはインビトロでのポリペプチドの折り畳みを補助するタンパク質である。
【0079】
本明細書では分子シャペロン「型」タンパク質としても考えられているのが、タンパク質ジスフィルドイソメラーゼ(PDI)およびペプチジル−プロリルcis/transイソメラーゼ(PPI)であり、これらはタンパク質ジスフィド結合形成およびプロリンcis/trans異性化をそれぞれ触媒する(ロリマー(Lorimer)とボールドウィン(Baldwin)、Methods in Enzymology、1998年を参照)。例えば、化膿連鎖球菌におけるSpeBの発現がRopAタンパク質を必要とすることが、化膿連鎖球菌突然変異によって明らかにされた(リヨン(Lyon)ら、1998年)。RopAタンパク質は、リボソーム上で新生ポリペプチドに結合し、GroELに結合し、かつペプチジル−プロリルイソメラーゼ活性を有することが知られている分子シャペロンである。したがって、本発明の一部の他の実施形態においては、成熟SpeBポリペプチドを生成するための方法は、非ネイティブ成熟SpeBポリペプチドを再折り畳みし、またはPPI、特にRopA PPIの存在下に成熟SpeBを発現させるステップを含んで成る。
【0080】
C.組換え発現ベクターおよび宿主細胞
別の実施形態においては、本発明は、SpeBプロ−ポリペプチドドメインおよび成熟SpeBポリペプチドをコード化するポリヌクレオチドを含んで成る発現ベクターに関する。一部の実施形態においては、本発明の発現ベクターはポリシストロニック核酸配列を含んで成り、ここで1つのシストロンがSpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチドを含んで成り、第2のシストロンが成熟SpeBポリペプチドをコード化するポリヌクレオチドを含んで成る。一部の他の実施形態においては、本発明の発現ベクターは、SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチドを含んで成り、第2の発現ベクターは成熟SpeBポリペプチドをコード化するポリヌクレオチドを含んで成る。1つの好ましい実施形態においては、本発明の発現ベクターはプラスミド構成物である。別の好ましい実施形態においては、本発明の発現ベクターは表3、実施例1に記載されているプラスミドである。
【0081】
本明細書で使用される「ベクター」という語は、それが結合されている別の核酸を輸送する能力がある核酸分子を指す。ベクターの1つの型は「プラスミド」であり、追加のDNA部分がライゲーションされているこれは環状二本鎖DNAループを指す。別の型のベクターはウイルスベクターであり、ここでは追加のDNA部分がウイルスゲノムへライゲーションされている。一部のベクターは、それらが導入される宿主細胞(例えば、複製の細菌源を有する細菌ベクターおよびエピソーム哺乳類ベクター)において自己複製が可能である。さらに、一部のベクターは、それらが操作的に結合される遺伝子の発現を方向づけることが可能である。かかるベクターは、本明細書では「発現ベクター」と呼ばれる。一般に、組換えDNA技術において有用性の発現ベクターはしばしばプラスミドの形態である。本明細書においては、「プラスミド」および「ベクター」は、プラスミドがベクターの最も一般的に使用される形態であるため同じ意味で使用される。しかし、本発明では、同等の機能を果たすウイルスベクター(例えば、複製欠損レトロウイルス、アデノウイルス、およびアデノ関連性ウイルス)など他の形態のかかる発現ベクターを含むことは意図されている。
【0082】
原核生物におけるタンパク質の発現は、融合または非融合タンパク質のいずれかの発現を方向づける構成的または誘導的プロモーターを含有するベクターにより大腸菌において最も多く行われる。融合ベクターは、その中でコード化されたタンパク質、組換えタンパク質のアミノまたはカルボキシ末端に多くのアミノ酸を添加する。
【0083】
一部の実施形態においては、宿主細胞が、(i)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列、および(ii)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るポリシストロニックプラスミドで形質転換され、形質導入され、トランスフェクトされ、または感染される。本明細書で規定されているように、「ポリシストロニックmRNA」は、2つもしくはそれ以上のポリペプチドをコードする。したがって、以下に規定されているように、本発明の「ポリシストロニックポリヌクレオチド」、「ポリシストロニックcDNA」、または「ポリシストロニックプラスミド」は、ポリシストロニックmRNAをコードし、次いで2つもしくはそれ以上のポリペプチドをコード化する。
【0084】
適切な誘導的、非融合、大腸菌発現ベクターの例としては、pTrc(アマン(Amann)ら、1988年)、pET IId(スタディール(Studier)ら、1990年)、pET、pRSET、pCRT7−CTTOPO、およびpIVeXが挙げられる。pTrcベクターからのターゲット遺伝子発現は、ハイブリッドtrp−lac融合プロモーターからの宿主RNAポリメラーゼ転写に依拠する。pET IIdベクターからのターゲット遺伝子発現は、共発現ウイルスRNAポリメラーゼT7gnlによって介在されるT7gn1β−lac融合プロモーターからの転写に依拠する。このウイルスポリメラーゼは、lacUV5プロモーターの転写制御下にT7gnl遺伝子を有する常在プロファージからの宿主菌株BL21(DE3)またはHMSI74(DE3)によって供給される。一部の実施形態においては、pRK5、pCMVBlue、pCMV−LIC、pAPL400−023、pAPL400−087、およびpAPL400−088などヒトCMVまたはシミアンCMVプロモーターを含んで成るプラスミドベクターも考えられている。
【0085】
大腸菌における組換えタンパク質発現を最大限にする1つの方法は、組換えタンパク質をタンパク質分解的に切断する能力障害による宿主細菌におけるタンパク質の発現である。別の方法は、発現ベクターへ挿入される核酸の核酸配列を変化させ、各アミノ酸に対する個々のコドンが大腸菌において優先的に利用されるものとなることである。本発明の核酸配列のかかる変化は、標準のDNA突然変異または合成法によって行われる。
【0086】
プロモーターは、通常、転写の開始点(すなわち、転写開始部位)の前(上流)で約100個のヌクレオチド対内のDNA分子の領域である。その領域は通常、異なる遺伝子における同様のそれぞれの位置に配置されている数種類のDNA配列要素を含有する。本明細書で使用される「プロモーター」という語は、技術上、上流プロモーター領域またはプロモーター領域と呼ばれるものを含む。
【0087】
本発明の別の態様は、本発明の組換え発現ベクターが導入されている宿主細胞に関する。「宿主細胞」、「遺伝子組換え宿主細胞」、および「組換え宿主細胞」は、本明細書では同じ意味で使用される。かかる語は特定の対象の細胞だけではなく、かかる細胞の子孫または潜在的子孫を指すことが理解される。特定の修飾が突然変異または環境的影響により後世において起こりうるため、かかる子孫は、実際には、親細胞と同一ではありえないが、それでも本明細書で使用される語の範囲内に含まれる。宿主細胞は原核または真核生物細胞であるが、好ましくは、原核細胞である。例えば、SpeBポリペプチドは大腸菌、および化膿連鎖球菌など細菌細胞で発現される。他の実施形態においては、SpeBポリペプチドは昆虫細胞(例えば、Sf9、高5およびSf21細胞)、酵母(例えば、P.パストリス(P.pastoris)、P.メタノリカ(P.methanolica)、S.ポンベ(S.pombe)、およびS.セレヴィシエ(S.cerevisiae))または哺乳類細胞(例えば、チャイニーズハムスター卵巣細胞(CHO)、Cos−1、CV−1、HeLa、NIH3T3、PER−C6、およびNSOにおいて発現される。他の適切な宿主細胞は当業者に周知である。
【0088】
培養における原核祝細胞など本発明の宿主細胞が、SpeBポリペプチドを生成する(すなわち、発現させる)ために使用される。1つの実施形態においては、方法は、成熟SpeBポリペプチドおよびSpeBプロ−ポリペプチドドメインが生成されるまで適切な培地で(成熟SpeBポリペプチドおよびSpeBプロ−ポリペプチドドメインをコード化する組換え発現ベクターが導入されている)本発明の宿主細胞を培養するステップを含んで成る。別の実施形態においては、方法は、成熟SpeBポリペプチドを培地または宿主細胞から単離するステップをさらに含んで成る。
【0089】
ベクターDNAは、従来のトランスフォーメーション、形質導入、感染、またはトランスフェクション法によって原核または真核細胞へ導入される。本明細書で使用される「トランスフォーメーション」、「感染」、および「トランスフェクション」は、リン酸カルシウムまたは塩化カルシウム共沈殿、DEAE−デキストラン介在トランスフェクション、リポフェクション、感染、またはエレクトロポレーションを含む宿主細胞へ外来核酸(例えば、DNA)を導入する技術上認められたさまざまな方法を指すことが意図されている。宿主細胞を形質転換し、感染し、トランスフェクトする適切な方法は、サムブルック(Sambrook)ら(「Molecular Cloning:A Laboratory Manual」、第2版、コールド・スプリング・ハーバー・ラボラトリー(Cold Spring Harbor Laboratory)、コールド・スプリング・ハーバー・ラボラトリー・プレス(Cold Spring Harbor Laboratory Press)、コールド・スプリング・ハーバー(Cold Spring Harbor)、ニューヨーク州、1989年)、および他の実験マニュアルに見出される。
【0090】
最も広く使用されている方法は、リン酸カルシウムまたはDEAE−デキストランのいずれかによって介在されるトランスフェクションである。機序は依然として不明であるが、トランスフェクトされたDNAはエンドサイトーシスによって細胞の細胞質に入り、核に輸送されると考えられている。細胞型によって、培養細胞の90%までの集団がつねにトランスフェクトされる。その高い効率により、リン酸カルシウムまたはDEAE−デキストランによって介在されるトランスフェクションは、多数の細胞における外来DNAの一時的発現を必要とする実験に選択される方法である。リン酸カルシウム介在トランスフェクションは、通常、宿主細胞ゲノムへの頭−尾タンデムで配置される外来DNAのコピーを組込む細胞系を確立するためにも使用される。
【0091】
発現ベクターのコード配列は、操作的に転写終了領域へ結合される。RNAポリメラーゼは、ポリアデニル化が起こる部位を通じてコード化DNA配列を転写する。通常、ポリアデニル化部位の数百塩基対下流に位置したDNA配列は転写を終了させる役目をする。それらのDNA配列は本明細書では転写終了領域と呼ばれる。それらの領域は転写メッセンジャーRNA(mRNA)の有効なポリアデニル化に必要とされる。転写終了領域は当技術分野で周知である。例示の転写終了領域は、SV40またはプロタミン遺伝子のポリアデニル化シグナルを含んで成る。
【0092】
本発明のDNA分子、遺伝子、またはポリヌクレオチドは、当技術分野で周知である多くの方法によってベクターへ組入れられる。例えば、ベクターpUC18は有用性が明らかにされている。同様に、関連ベクターM13mp18およびM13mp19は、特に、ジデオキシ配列決定の実施における本発明の一部の実施形態において使用される。
【0093】
好ましい実施形態においては、本発明の組換え宿主細胞は原核宿主細胞である。好ましくは、本発明の組換え宿主細胞は大腸菌のDH5α菌株の細菌細胞である。一般に、原核細胞が、DNA配列の初期クローニングおよび本発明において有用であるベクターの構成のために好ましい。例えば、大腸菌K12菌株はとりわけ有益である。使用される他の微生物菌株としては、大腸菌Bおよび大腸菌x(E.colix)1976(ATCC番号31537)が挙げられる。これらの例は、もちろん、限定ではなく例示が目的とされている。
【0094】
原核生物も発現のために使用される。上記の菌株のほか、W3110(ATCC番号273325)、BLR(DE3)、BLR(DE3)pLysS、AD494(DE3)、AD494(DE3)pLysS、BL21(DE3)、BL21(DE3)pLysS、BL21(DE3)pLysE、BL21(DE3)pLacI、BL21trxB(DE3)、BL21trxB(DE3)pLysS、 HMS174(DE3)、HMS174(DE3)pLysS、HMS174(DE3)pLysE、Origami(DE3)、Origami(DE3)pLysS、Origami(DE3)pLysE、Origami(DE3)pLacI、OrigamiB(DE3)、OrigamiB(DE3)pLysS、OrigamiB(DE3)pLysE、OrigamiB(DE3)pLacI、Rosetta(DE3)、Rosetta(DE3)pLysS、Rosetta(DE3)pLysE、Rosetta(DE3)pLacI、Tuner(DE3)、Tuner(DE3)pLysS、およびTuner(DE3)pLacIなどの大腸菌株、枯草菌(Bacillus subtilis)などの桿菌、またはネズミチフス菌(Salmonella typhimurium)もしくは霊菌(Serratia marcesans)、およびさまざまなシュードモナス(Pseudomonas)種などの腸内細菌科が使用される。
【0095】
一般に、宿主細胞と共存できる種由来であるレプリコンおよび制御配列を含有するプラスミドベクターが、これらの宿主といっしょに使用される。ベクターは通常、複製部位、および形質転換された細胞における表現型選択を提供する能力がある標識配列を有する。例えば、大腸菌は、大腸菌種由来のプラスミド、pBR322を使用して形質転換される(ボリバー(Bolivar)ら、1977年)。pBR322は、アンピシリンおよびテトラサイクリン耐性の遺伝子を含有し、したがって形質転換された細胞を同定する容易な手段を提供する。pBRプラスミド、もしくは他の微生物プラスミドまたはファージは、それぞれのポリペプチドの発現のための微生物によって使用されるプロモーターも含有しなければならず、または含有するように修飾される必要がある。
【0096】
組換えDNA構成において最も一般的に使用されるプロモーターとしては、β−ラクタマーゼ(ペニシリナーゼ)およびラクトースプロモーター系(チャン(Chang)ら、1978年、イタクラ(Itakura)ら、1977年、ゲデル(Goeddel)ら、1979年、ゲデル(Goeddel)ら、1980年)、トリプトファン(TRP)プロモーター系(欧州出願第EP0036776号明細書、シーブウェンリスト(Siebwenlist)ら、1980年)、およびT7またはT7lacプロモーター系が挙げられる。これらは最も一般的に使用されているが、他の微生物プロモーターが発見され、かつ利用されており、それらのヌクレオチド配列に関する詳細は公開されており、当業者が機能的プロモーターをプラスミドベクターへ導入することが可能である(シーブウェンリスト(Siebwenlist)ら、1980年)。
【0097】
トランスフェクション後、細胞は培養条件下でSpeBポリペプチドの発現に十分な時間にわたって維持される。培養条件は当技術分野で周知であり、イオン組成および濃度、温度、pHなどを含む。通常、トランスフェクトされた細胞は培養培地で培養条件下で維持される。さまざまな細胞型のための適切な培地は当技術分野で周知である。好ましい実施形態においては、温度は約20℃〜約50℃、より好ましくは、約30℃〜約40℃、かつさらに好ましくは、約37℃である。
【0098】
pHは、約6.0の値〜約8.0の値、より好ましくは、約6.8の値〜約7.8の値、かつ最も好ましくは、約7.4である。オスモル濃度は、好ましくは、約200ミリオスモル/リットル(mosm/L)〜約400mosm/L、より好ましくは、約290mosm/L〜約310mosm/Lである。コード化ポリペプチドのトランスフェクションおよび発現のための他の生物学的条件は当技術分野で周知である。
【0099】
トランスフェクトされた細胞は、SpeBポリペプチドの発現に十分な時間にわたって維持される。適切な時間は、とりわけ、使用される細胞型によって決まり、当業者によって容易に決定可能である。通常、維持時間は、約2〜約14日である。
【0100】
組換えSpeBポリペプチドは、トランスフェクトされた細胞またはその細胞が培養されている培地のいずれかより回収され、または収集される。回収は、SpeBポリペプチドの単離および精製を含んで成る。ポリペプチドのための単離および精製法は当技術分野で周知であり、沈殿、ろ過、クロマトグラフィー、電気泳動などの方法を含む。
【0101】
さらに別の実施形態においては、本発明のSpeBポリペプチドは、細胞を含まない翻訳系などインビトロタンパク質翻訳系によって生成される(例えば、ベトン(Betton)、2003年、ブラウン(Braun)ら、2002年、ジェルムツス(Jermutus)ら、1998年、キグワ(Kigawa)ら、1999年、キム(Kim)ら、1996年、およびスピリン(Spirin)ら、1988年を参照)。例えば、各々参照することによりその全体が本明細書で援用される、米国特許第6,399,323号明細書、および米国特許第5,478,730号明細書は、細胞を含まない(インビトロ)翻訳系におけるポリペプチダーゼの調製または生成のための方法、条件などを記載している。
【0102】
D.免疫原性組成物
一部の好ましい実施形態においては、本発明は、成熟SpeBポリペプチド免疫原(すなわち、成熟野生型または成熟C192S)および生理的に許容される担体を含んで成る成熟SpeB免疫原性組成物を提供する。より好ましくは、免疫原性組成物は、配列番号2のアミノ酸146−398を含んで成る成熟野生型SpeBポリペプチド、または配列番号2のアミノ酸146−398を含んで成る成熟C192S SpeBポリペプチドを含んで成り、ここで配列番号2のシステインアミノ酸192はセリンに変異されている。本発明のさらに他の実施形態においては、多価または組合せ免疫原性組成物が提供される。組合せ免疫原性組成物が、化膿連鎖球菌および/または他の細菌種からの1つもしくはそれ以上の追加の抗原とともに、1つもしくはそれ以上の本発明のポリペプチド(例えば、成熟C192S SpeB)を含むことによって提供される。特に、組合せ免疫原性組成物が、本発明の1つもしくはそれ以上の成熟SpeBポリペプチドを、1つもしくはそれ以上のポリペプチド、ポリペプチド断片、糖質、オリゴ糖類、脂質、リポオリゴ糖類、多糖類、オリゴ糖類−タンパク質コンジュゲート、多糖類−タンパク質コンジュゲート、ペプチド−タンパク質コンジュゲート、オリゴ糖類−ペプチドコンジュゲート、多糖類−ペプチドコンジュゲート、タンパク質−タンパク質コンジュゲート、リポオリゴ糖類−タンパク質コンジュゲート、または多糖類−タンパク質コンジュゲートと組合わせることによって提供される。
【0103】
本発明の成熟SpeBポリペプチドは、哺乳類対象、例えば、ヒトへの投与に適切な免疫原性組成物へ組入れられる。かかる組成物は通常、「免疫原性」組成物および薬剤として許容される担体を含んで成る。以下で使用される「薬剤として許容される担体」という言葉は、薬剤投与と適合するすべての溶剤、分散剤、コーティング剤、抗菌剤および抗真菌剤、等張剤および吸収遅延剤などを含むことが意図されている。薬剤活性物質のためのかかる溶剤および薬剤の使用は当技術分野で周知である。従来の溶剤または薬剤が活性化合物と不適合である場合を除き、かかる溶剤は本発明の組成物において使用される。補助活性化合物も組成物へ組入れられる。
【0104】
本発明の免疫原性組成物は、その投与の意図された経路と適合性であるように製剤化される。投与経路の例としては、非経口(例えば、静脈内、皮内、皮下、筋内、腹腔内)、粘膜(例えば、経口、直腸、鼻内、口腔、膣、呼吸器)、および経皮(局所)が挙げられる。非経口、皮内、または皮下適用に使用される溶液または懸濁液としては、以下の要素、すなわち、注射用の水、食塩水、固定油、ポリエチレングリコール、グリセリン、プロピレングリコール、または他の合成溶剤など滅菌希釈剤、ベンジルアルコールまたはメチルパラベンなど抗菌剤、アスコルビン酸または重亜硫酸ナトリウムなど抗酸化剤、エチレンジアミン四酢酸などキレート化剤、酢酸塩、クエン酸塩、またはリン酸塩などバッファー、および塩化ナトリウムまたはデキストロースなど張性の調整剤が挙げられる。pHは、塩酸または水酸化ナトリウムなど酸または塩酸で調整される。非経口製剤は、アンプル、使い捨て注射器、もしくはガラスまたはプラスチック製のマルチドーズ用バイアルに入れられる。
【0105】
滅菌注射溶液は、活性化合物(例えば、成熟SpeB)を、必要に応じて上記列挙した成分の1つまたは組合せとともに適切な溶剤中で必要な量において組入れた後、ろ過滅菌によって調製される。一般に、分散剤は、基礎分散溶剤および上記列挙したものからの必要な他の成分を含有する滅菌ビヒクルへ活性化合物を組入れることによって調製される。滅菌注射溶液用の滅菌粉末の場合、調製の好ましい方法は真空乾燥および凍結乾燥であり、これによりその以前に滅菌ろ過した溶液から活性成分+追加の所望の成分の粉末が得られる。
【0106】
経口組成物は一般に不活性希釈剤または食用担体を含む。それらはゼラチンカプセルに入れられ、または錠剤の中へ圧縮される。治療用の経口投与のために、活性化合物は賦形剤とともに組入れられ、錠剤、トローチ、またはカプセルの形で使用される。経口組成物は、口内洗浄剤として使用される液体担体を使用しても調製され、ここで液体担体中の化合物は経口適用され、局所使用され、吐出され、または燕下される。製剤適合性結合剤、および/または佐剤物質が組成物の一部として含まれる。錠剤、ピル、カプセル、トローチなどは、以下の成分のいずれか、または類似した性質の化合物のいずれかを含有する。すなわち、微結晶性セルロース、トラガカントゴム、またはゼラチンなど結合剤、でんぷんまたはラクトースなど賦形剤、アルギン酸、プリモゲル(Primogel)、またはコーンスターチなど崩壊剤、ステアリン酸マグネシウムまたはステロート(Sterotes)など潤滑剤、コロイド状二酸化ケイ素など流動促進剤、スクロースまたはサッカリンなど甘味剤、またはペパーミント、サルチル酸メチル、またはオレンジ香味料など香料添加剤。
【0107】
吸入による投与のために、化合物は、適切な推進剤、例えば、炭酸ガスなどのガスを含有する加圧容器またはディスペンサー、またはネブライザーからのエアロゾルスプレーの形で送達される。全身投与も粘膜または経皮手段による。粘膜または経皮投与のために、浸透されるバリアに適切な浸透剤が製剤で使用される。かかる浸透剤は一般に当技術分野で周知であり、例えば、粘膜投与のための洗浄剤、胆汁塩、およびフシジン酸誘導体を含む。粘膜投与は、鼻腔用スプレーまたは坐剤を使用して行われる。経皮投与のために、活性化合物は軟膏、膏薬、ゲル、または一般に当技術分野で周知であるクリームへ処方される。
【0108】
化合物は坐剤(例えば、カカオバターおよび他のグリセリドなど従来の坐剤の基剤による)または直腸送達用の停留浣腸の形でも調製される。
【0109】
1つの実施形態においては、活性化合物は、インプラントおよびマイクロカプセル化送達系を含む、制御放出製剤など体内からの急速な排泄に対して化合物を保護する担体とともに調製される。
【0110】
生体分解性、生体適合性ポリマー、例えば、エチレン酢酸ビニル、ポリ無水物、ポリグリコール酸、コラーゲン、ポリオルトエステル、およびポリ乳酸などが使用される。かかる製剤の調製方法は当業者には明らかであろう。材料は、アルザ・コーポレーション(Alza Corporation)およびノバ・ファルマセウティカルズ(Nova Pharmaceuticals,Inc.)からも市販されている。リポソーム懸濁液(ウイルス抗原に対するモノクローナル抗体による感染細胞にターゲティングされたリポソームを含む)も薬剤として許容される担体として使用される。これらは、例えば、以下で参照することにより援用される、米国特許第4,522,811号明細書に記載されているように、当業者に周知の方法に従って調製される。
【0111】
投与を容易にし、投与量を均一にするために、用量単位剤形で経口または非経口組成物を処方することが特に有利である。以下で使用される用量単位剤形は、治療される対象のための単位用量として適した物理的に分離した単位を指し、各単位は、必要な薬剤担体とともに所望の治療効果を生じるように計算された所定量の活性化合物を含有する。本発明の用量単位剤形の仕様は、活性化合物の独自の特徴および達成される特定の治療効果、ならびに個人の治療用にかかる活性化合物を配合する技術上固有の制限によって決定され、かつ直接それらに依存している。
【0112】
薬剤として許容されるビヒクルは、副作用を引き起こすことがなく、例えば、活性化合物の投与を促進し、体内におけるその寿命および/またはその有効性を増大し、溶液中の溶解度を増大し、あるいは、その保存を強化することを可能にする薬剤または免疫原性組成物へ入る化合物または化合物の組合せを指定すると理解されている。これら薬剤として許容されるビヒクルは公知であり、選択された活性化合物の性質および投与方法に従って当業者によって適合されるであろう。
【0113】
本発明の免疫原性原性は、1つもしくはそれ以上の佐剤をさらに含んで成りうる。「佐剤」は、抗原の免疫原性を強化するのに役立つ物質である。したがって、佐剤はしばしば免疫応答を高め、当業者には公知である。本発明で意図されている佐剤の例は、リン酸アルミニウムおよび水酸化アルミニウムなどアルミニウム塩(alum)、ヒト結核菌(Mycobacterium tuberculosis)、百日咳菌(Bordetella pertussis)、細菌性リポ多糖、アミノアルキルグルコサミンリン酸化合物(AGP)、もしくはその誘導体または類似体を含むがこれらに限定されず、これらはコリザ(Corixa)(ハミルトン(Hamilton)、モンタナ州(MT))から市販されており、かつ米国特許第6,113,918号明細書に記載されており、かかるAGPの1つは、2−[(R)−3−テトラデカノイルオキシテトラデカノイルアミノ]エチル2−デオキシ−4−O−ホスホノ−3−O−[(R)−3−テトラデカノイルオキシテトラデカノイル(tetradecanoyoxytetradecanoyl)]−2−[(R)−3−テトラデカノイルオキシテトラデカノイルアミノ(tetradecanoyoxytetradecanoylamino)]−b−D−グルコピラノシドであり、これは529としても知られる(以前はRC529として知られる)が、これは安定乳剤または水性剤形として処方されており、米国特許第4,912,094号明細書に記載されているMPL(商標)(3−O−脱アシル化モノホスホリル脂質A)(コリザ(Corixa))、CpGモチーフ(米国特許第6,207,646号明細書)を含有するオリゴヌクレオチドなど合成ポリヌクレオチド、ポリペプチド、米国特許第5,057,540号明細書に記載されている、Quil AまたはSTIMULON(商標)QS−21(アンチジェニックス(Antigenics)、フラミンガム(Framingham)、マサシューセッツ州)などサポニン、百日咳毒素(PT)、または大腸菌易熱性毒素(LT)、特にLT−K63、LT−R72、CT−S109、PT−K9/G129、例えば、国際特許公開国際公開第93/13302号パンフレットおよび同第92/19265号パンフレットを参照、コレラ毒素(野生型または突然変異体のいずれか、例えば、ここでアミノ酸29位でのグルタミン酸が別のアミノ酸、好ましくは、公開された国際特許出願番号国際公開第00/18434号パンフレットに従って、ヒスチジンで置換されている)。さまざまなサイトカインおよびリンフォカインが佐剤としての使用に適している。かかる佐剤の1つが、米国特許第5,078,996号明細書に記載されているヌクレオチド配列を有する、顆粒球マクロファージコロニー刺激因子(GM−CSF)である。GM−CSF cDNAを含有するプラスミドは、大腸菌へ形質転換されており、アメリカン・タイプ・カルチャー・コレクション(American Type Culture Collection)(ATCC)、1081University Boulevard,Manassas、バージニア州(VA)20110−2209により、登録番号39900で寄託されている。サイトカインインターロイキン−12(IL−12)は、米国特許第5,723,127号明細書に記載されている別の佐剤である。他のサイトカインまたはリンフォカインは、インターロイキン1−α、1−β、2、4、5、6、7、8、10、13、14、15、16、17、および 18、インターフェロン−α、β、およびγ、顆粒球コロニー刺激因子、および腫瘍壊死因子αおよびβを含むがこれらに限定されない免疫修飾活性を有することが示されており、佐剤としての使用に適している。
【0114】
本発明の組成物は通常、標準、公知、非毒性の生理的に許容される担体、佐剤、および所望のビヒクルを含有する用量単位製剤で非経口投与される。以下で使用される非経口という語は、静脈内、皮下、皮内、筋内、動脈内注射、または注入技術を含む。
【0115】
注射製剤、例えば、滅菌注射水性または油性懸濁液は、適切な分散または湿潤剤および懸濁剤を使用して既知の技術に従って処方される。滅菌注射製剤は、例えば、1,3−ブタンジオール中の溶液として、非毒性の非経口的に許容される希釈剤または溶剤中の注射溶液または懸濁液でもある。
【0116】
使用されうる許容されるビヒクルおよび溶剤としては、水、リンガー溶液、および生理食塩液が挙げられる。また、滅菌、固定油が従来、溶剤または懸濁化剤として使用されている。このために、合成モノ−またはジ−グリセリドを含む無菌性固定油が使用されている。また、オレイン酸など脂肪酸が注射剤の調製に使用されている。
【0117】
好ましい担体としては、リン酸塩、乳酸塩、トリス(Tris)などによる中性食塩水バッファーが挙げられる。ウイルスベクターの投与に際しては、ベクターは十分に精製され、欠陥干渉アデノウイルス粒子または内毒素および他の発熱源など、望ましくない汚染物を実質的に含まないようにし、ベクター構成物が投与される個人において有害反応を引き起こさないようにする。ベクターを精製する好ましい手段には、塩化セシウム勾配遠心分離など浮遊密度勾配の使用が含まれる。
【0118】
担体はリポソームでもある。送達ビヒクルとしてリポソームを使用する手段は当技術分野で周知である。
【0119】
特定の実施形態においては、本発明の免疫原性組成物は、遺伝子発現を制御する調節配列と操作的に関連する本発明のポリヌクレオチド配列を含んで成る。目的とするポリヌクレオチド配列は、DNAの発現を促進する調節要素、すなわち、プロモーターおよび/またはエンハンサー要素の制御下にプラスミドなど発現ベクターへ工学的に入れられる。好ましい実施形態においては、ヒトサイトメガロウイルス介在早期プロモーター/エンハンサーが使用される(米国特許第5,168,062号明細書)。プロモーターは細胞特異的であり、かつ所定の細胞でのみポリヌクレオチドの実質的な転写を可能にしうる。
【0120】
ポリヌクレオチドは、「裸の」DNA(米国特許第5,580,859号明細書)として宿主へ直接導入されるか、またはブピビカインおよび他の局所麻酔薬(米国特許第5,593,972号明細書)およびカチオンポリアミン(米国特許第6,127,170号明細書)など免疫化を促進する薬剤で組成物中に処方される。
【0121】
このポリヌクレオチド免疫化方法においては、本発明のポリペプチドはインビボでの一過性に発現され、宿主の染色体に挿入または組込まれる遺伝物質はない。この方法は、目標が目的とする遺伝物資を染色体へ挿入または組込む遺伝子療法とは区別されるべきである。免疫化によって投与されるポリヌクレオチドが宿主における形質転換表現型を生じさせないことを確認するアッセイが使用される(米国特許第6,168,918号明細書)。
【0122】
すべての特許および刊行物は、参照することにより本明細書で援用される。
E.実施例
以下の実施例は、他の方法で詳細に記載されている場合を除き、当業者に公知であり、かつルーチンである標準の方法を使用して行われる。以下の実施例は、例示目的に示されており、決して本発明の範囲を限定するものとして考えるべきではない。
【0123】
実施例1
材料と方法
培地と試薬。大腸菌BLR(DE3)(ノバーゲン(Novagen)、カリフォルニア州(CA))を発現試験用に使用した。適切な抗生物質選択条件下で細菌をルリア寒天培地(LB)で生育させた。アンピシリンを濃度100μg/mLで使用し、カナミシンを50μg/mLで使用した。ZeroBluntTOPクローニングベクターpCR−Blunt(インビトロジェン(InVitrogen)、カールズバッド(Carlsbad)、カリフォルニア州(CA))をPCR生成断片のクローニング用に使用した。Century−Plus RNA Markers(商標)、Millennium RNA Markers(商標)、RNAlater(商標)、RNAqueous−Midi(商標)、DNA−free(商標)、ULTRAhyb(商標)、NorthernMax(商標)、およびRETROscript(商標)をアンビオン(Ambion)(オースチン(Austin)、テキサス州(TX))から得た。GeneScreen(商標)ハイブリダイゼーション膜、蛍光−N6−dATP、Renaissance(登録商標)Antifluorescein−APコンジュゲートポリクローナル抗体、およびCDP−Star(登録商標)をパーキンエルマー・ライフ・サイエンシス(PerkinElmer Life Sciences)、ボストン(Boston)、マサチューセッツ州(MA)から得た。すべての制限酵素をニュー・イングランド・バイオラブス(New England Biolabs)(べバリー(Beverly)、マサチューセッツ州(MA))から入手した。
【0124】
ポリメラーゼ連鎖反応。特に断りのない限り、PCR増幅を以下の反応条件を使用して最終容量50μLで行った。すなわち、0.2mM dNTPs、1.0mM DTT、0.8μM各プライマー、10U熱安定ポリメラーゼ、および1×熱安定ポリメラーゼバッファー。クローニングおよび突然変異反応のために、Pwoポリメラーゼ(ベーリンガー・マンハイム(Boehringer Mannheim)、インディアポリス(Indianapolis)、インディアナ州(IN))を使用したが、他のすべて増幅ではTaqポリメラーゼ(アプライド・バイオシステムズ(Applied Biosystems)、フォスターシティー(Foster City)、カリフォルニア州(CA))を使用した。増幅は、94℃で30秒間、55℃で30秒間、および72℃で30秒間の25サイクルで構成された。
【0125】
プラスミド構成物。成熟SpeBコード領域に対応する770bp断片を、それぞれNco IおよびBamH I部位を含有するフォワード (5’CCATGGAACCAGTTGTTAAATCTCTCC3’)(配列番号3)プライマーおよびリバース(5’GGATCCTAAGGTTTGATGCCTACAACAGC3’)(配列番号4)プライマー(下線)を使用してPCR増幅した。フォワードプライマーは、NcoIクローニング部位内にネストしたATG翻訳開始コドンを含有するように設計され、結果として発現タンパク質のN末端へ追加のメチオニン残基が生じ、第1の残基をグルタミン(CAA)からグルタミン酸塩(GAA)へ変更した。同様に、プロ配列ドメイン(アミノ酸28−146)を含む367bp断片を、クローニングのための工学的NcoI(フォワード)部位およびBamHI(リバース)部位とともに、フォワード(5’CCATGGATCAAAACTTTGCTCGTAACG3’)(配列番号5)プライマー、およびリバース(5’GGATCCTTATTTAATCTCAGCGGTACCAGC3’)(配列番号6)プライマーを使用して増幅した。リバースプライマーにおけるBamHI部位のすぐ3’方向に終止コドンを挿入し、発現組換えタンパク質の翻訳終了を指令した。PCR反応では、以前に記載されているように(マツカ(Matsuka)ら、1999年)、単一システインを192位でセリン置換(C192S)にコード化するTGT〜AGT突然変異によるSpeBチモーゲンを含有するプラスミドベースのテンプレートを使用した。PCR産物をpCR−Bluntヘサブクローン化し、その後にNcoIおよびBamH Iを使用して制限消化によって切除した。770bp成熟C192S SpeBおよび367bpプロ配列ドメインコード断片をアガロースゲル電気泳動によって精製し、それぞれ、NcoIおよびBamHI制限部位を使用してpET28aおよびpET3dへライゲートした。結果として生じる発現プラスミド、pLP681およびpLP682を標準方法を使用して大腸菌BLR(DE3)へ同時形質転換した。生成細菌発現菌株を2−プラスミドベースの共発現分析に利用した。
【0126】
成熟野生型SpeB発現構成物を、単一のA〜T塩基変更(太字)とともに、重複フォワード
(配列番号7)プライマー、およびリバース
(配列番号8)プライマーを使用する突然変異PCRを使用して生成し、PCRのためのテンプレートとして使用したpLP681におけるC192S突然変異を復帰した。突然変異生成は、ワイナー(Weiner)ら(1994年)によって記載された方法の変形で構成され、以下のサイクル条件を使用した。すなわち、94℃で1分、94℃で15秒および68℃で10分の16サイクルの後、94℃で15秒および68℃で10分の12サイクルとともに、68℃で各サイクルで15秒増分で増加させる延長時間により、結果として68℃で13分の最終延長時間が生じた。PCR反応を大腸菌へのトランスフォーメーション前にDpnIで切断した。結果として生じるクローンpLP680を使用前に配列決定し、所望の突然変異を検証した。プラスミドをpLP682で大腸菌BLR(DE3)へ同時形質転換し、2−プラスミドベース共発現試験で使用した。
【0127】
ポリシストロニック発現ベクターを構成するために、pLP682からのSpeBプロ配列ドメインコード領域をNcoIおよびBamHIを使用する制限消化によって切断した。367bp断片をアガロースゲル電気泳動によって精製し、やはりNco IおよびBamH Iで制限したpET28aへライゲートし、結果としてpLP688が生じた。
【0128】
長さを増大させる4種類のリンカー領域に引き続いて、以下のフォワードプライマーを使用するPCRによって成熟SpeBコード領域を生成した。すなわち、
すべてのフォワードプライマーは、5’末端(下線)上にBglII制限部位、最適化シャイン・ダルガノ部位(太字イタリック)、および翻訳開始コドン(太字)を含んだ。翻訳終止コドンへの5’直後にXhoI部位(下線)を含有するリバースプライマー(5’CTCGAGCTAAGGTTTGATGCCTA−CAACAGC3’)(配列番号13)をすべてのリンカーベースの増幅反応のために使用した。
【0129】
上記のプライマーを使用し、5nt、10nt、または20ntリンカー+成熟C192S SpeBコード領域を増幅し、pLP681をPCRのためのテンプレートとして利用した。同様に、pLP680をテンプレートとして使用し、20ntリンカー+成熟野生型SpeB PCR産物を生成した。PCR断片をpCR−Bluntへサブクローン化し、BglIIおよびXhoIによる制限消化によって切除し、アガロースゲル電気泳動によって精製した。単離後、BamHIおよびXhoI制限部位を使用して断片をpLP688へライゲートし、pLP683、pLP684、pLP685、およびpLP687ポリシストロニック発現構成物を生成した。同一のやり方で、上記のプライマーによりPCRのためのテンプレートとしてpLP685を使用してpLP686を合成した。インビボ分析のために生成され、使用されたすべての発現プラスミドの明細が表3に示されている。表3に示されているすべての発現は、特に断りのない限り、pET28aバックグラウンドにおけるものである。
【0130】
【表3】
【0131】
組換えタンパク質の発現。ポリシストロニックおよび2−プラスミドベースの共発現試験のために、適切な抗生物質を含有するLB200mLを所望の細菌発現ショックで接種し、37℃下に一夜、生育させた。一夜培養物を培地を含有する新鮮抗生物質2Lへ1:10に希釈し、25℃下に約0.6のOD600に生育させ、25℃下に16時間、1mM IPTGで誘発した。遠心分離で細胞を収集し、結果として生じる細胞ペレットを使用するまで−20℃下に保存した。誘発前後のサンプルをタンパク質発現およびRNA単離の分析用に採取した。RNA分析用のサンプルを使用するまで−70℃下にRNAlater(商標)で保存した。
【0132】
タンパク質発現を超音波処理による細胞融解後のSDS−PAGE分析によって評価した。不溶性材料を遠心分離によってペレット化し、可溶性上清フラクションを回収した。細胞残屑をPBS 1mL中に再懸濁する前にPBSで2回洗浄した。すべての可溶性および不溶性フラクションを細胞融解前のOD600の細菌培養に基づき標準化した。クマシーブルー染色によって、かつ標準方法によるPVDF膜への移動後のウェスタンブロット分析によってタンパク質発現を視覚化した。SpeBのチモーゲン形態に対して生成されたポリクローナル抗体を使用してブロットをプローブし、発現プロ配列ドメインおよび成熟28kDa SpeBの同時の検出を可能にした。
【0133】
RNAの単離。総RNAをRNAqueous−Midi(商標)を使用して細菌培養物から単離した。RNAlater(商標)で保存した誘発後サンプルを室温下に解凍し、RNAqueous(商標)溶解/結合バッファー1mL中に再懸濁する前にペレット化した。次いで、溶解物を説明マニュアルに規定されているように処理した。単離RNAをLiClを使用して沈殿させ、DNA−free(商標)で2回、メーカーの仕様とおり処理し、汚染ゲノムまたはプラスミドDNAを除去した。260nmでの吸光度の分析によってRNAを定量し、吸光度比A260nm/A280nmの測定によって純度を評価した。RNAサンプル(1μg)を、以前に記載されているように適切なプライマー使用してPCRによって分析し、汚染プラスミドDNAが存在しないことを検証した。また、すべてのサンプルをPCR反応に添加したpLP685プラスミド1ngのスパイク回収分析によって残留DNAseについて評価した。DNAおよびDNase汚染がないことが証明されたサンプルのみその後の分析に使用した。
【0134】
ノーザンブロットハイブリダイゼーション。総RNA(5μg)を、メーカーによって規定されているようにNorthernMax(商標)を使用して変性条件下で1%アガロースゲル上で断片化した。ミレニアムおよびセンチュリーRNAマーカー(各2μg)をEtBrで前染色し、これを使用してRNAサイズを評価した。サンプルをナイロン膜(GeneScreen(商標))に移し、UV架橋し、80℃下に2時間乾燥させ、RNA断片化に使用されるホルムアルデヒド反応を逆転させた。RNA標準をUVによって視覚し、断片位置をハイブリダイゼーション前にナイロン膜上に示した。膜を2×SSC(0.3M Nacl、0.03Mクエン酸ナトリウム)で前湿潤させ、42℃下に2時間、ULTRAhyp(商標)で前ハイブリダイゼーションした。ノーザン分析用のプローブをテンプレートとしてpLP685、および上記で詳述されているように、プロ配列ドメイン(367bp)および成熟SpeB配列(770bp)に特異的な一連のプライマーを使用してPCR合成した。蛍光−N6−dATP(10μM)をPCR反応で使用し、無作為に標識成熟SpeBおよびプロ配列ドメインDNAプローブを生成した。結果として生じるプローブをアガロースゲル電気泳動によって精製し、各々の32pgを変性させ、42℃下に一夜、ULTRAhyp(商標)での膜ハイブリダイゼーションに使用した。ハイブリダイゼーション後、膜を2回過剰な2×SSCで10分間、室温下、2×SSC、1%SDSで2回20分間、42℃下、および0.2×SSC、0.1%SDSで2回20分間、42℃下に洗浄した。膜をBLOTTOでブロックし、アルカリホスファターゼ−コンジュゲート抗フルオレセインポリクローナル抗体、およびCDP−Star(登録商標)を、メーカーの仕様のとおりに使用して展開した。ブロットをBioMax MR−2オートラジオグラフフィルム(イーストマン・コダック(Eastman Kodak)、ロチェスター(Rochester)、ニューヨーク州(NY))に5分間、室温下に露出し、信号検出させた。
【0135】
cDNAの合成。RT−PCR用のRETROscript(商標)ファースト・ストランド(First Strand)合成キット、および無作為十量体をメーカーによって規定された条件下で使用する逆転写によって総RNAの2μgからcDNAを調製した。陰性対照サンプルを同一の条件を使用し、ただし逆転写酵素(−RT)の非存在下に生成した。反応産物をPCRおよび定量的PCR分析の使用前にヌクレアーゼを含まない水中に1:200で希釈した。
【0136】
cDNAの定量的PCR(qPCR)分析。プロ配列ドメイン、成熟SpeB、およびpET28aコード化カナマイシン耐性遺伝子(KanR)に特異的なプライマーおよびプローブを、プライマー発現ソフトウェア(アプライド・バイオシステムズ(Applied Biosystems)、フォスターシティー(Foster City)、カリフォルニア州(CA))を使用して設計した。定量的PCR反応を以下の条件下で行った。すなわち、300nMの各プライマー(フォワードおよびリバース)、200nM FAM/TAMRAプローブ、2×TaqManユニバーサルPCRマスターミックス(アプライド・バイオシステムズ(Applied Biosystems)、および1μL希釈cDNAまたは陰性対照サンプル。サイクル条件として、50℃で2分間、95℃で10分間、95℃で15秒間および60℃で1分間の40サイクルをABI7000配列検出システムで使用して最終容量25μLで反応を行った。KanR cDNAレベルを内部制御として使用し、全反応をKanR閾値サイクル(Ct)に基づき標準化した。結果は標準化Ct値として表されている。
【0137】
cDNAのPCR分析。希釈cDNAおよび陰性対照サンプルをPCRによって分析し、発現プラスミドから生じたmRNA転写をさらに評価した。プロ配列ドメイン、成熟SpeB、およびプロ配列ドメインフォワード(5’CCATGGATCAAAACTTTGCTCGTAACG3’)(配列番号14)プライマー、および成熟プロテアーゼリバース(5’CTCGAGCTAAGGTTTGATGCCTACAACAGC3’)(配列番号15)プライマーに特異的な上記のPCRプライマーからの第3のセットを使用し、それぞれ、プロ配列ドメイン、成熟SpeB、および完全長SpeBチモーゲン(1119bp)、およびポリシストロニックcDNA(1145−1180bp)を増幅した。1μLの各希釈サンプルを、増幅サイクルを30に増大したことを除き、上記の条件下で使用し、PCR産物をアガロースゲル電気泳動によって分析した。
【0138】
SpeBプロ配列ドメインの精製。溶解バッファー(20mMトリス、pH7.2、10mM MgCl2、10μg/μL DNase)中、15mLバッファー/g細胞の割合で細胞を再懸濁し、M110−Yマイクロフルイダイザー(Microfluidizer)(マイクロフルイディクス(Microfluidics)、ニュートン(Newton)、マサチューセッツ州(MA))を使用して溶解した。細胞残屑を遠心分離でペレット化し、可溶性フラクションを回収し、50mMグリシン−HCl(pH3.2)に対して4℃下に一夜、透析した。透析は、大腸菌タンパク質の顕著な沈殿によって達成された。沈殿物質を遠心分離で除去し、回収上清を100mM酢酸ナトリウム(pH4.5)に対する透析前にpH4.5に変更した。サンプルをSP−セファロース陽イオン交換カラムへ供給し、組換えプロ配列ドメインタンパク質を100mM酢酸ナトリウム(pH4.5)、1M塩化ナトリウムの勾配で溶出した。プロ配列ドメイン含有フラクションをプールし、SDS−PAGE電気泳動によって純度をチェックし、タンパク質濃度をBCAアッセイによって測定した(スミス(Smith)ら、1985年)。
【0139】
組換え発現成熟野生型SpeBの精製。溶解バッファー(20mMトリス、pH7.2、10mM MgCl2、10μg/μL DNase)中、15mLバッファー/g細胞の割合で細胞を再懸濁し、顕微溶液化によって溶解した。細胞残屑を遠心分離で除去し、不溶性成熟野生型SpeB含有フラクションを回収した。ペレットを処理し(50mMトリス、pH8.0、150mM NaCl、1mM EDTA、2%トリトンX−100)4℃下に一夜、室温下に1.5時間、室温下に3.5時間、すべて静かに攪拌して3連続洗浄を行った。ペレットを室温下に一夜、攪拌して20mMトリス、pH8.0、0.8M尿素中に溶解し、100mM酢酸ナトリウム、pH4.5、8M尿素で平衡したSP−セファロースカラム上にサンプルを塗布する前に酢酸ナトリウムでpHを4.5に変更した。組換え成熟野生型SpeBタンパク質を0〜750mM勾配の塩化ナトリウムで溶出し、SDS−PAGE電気泳動によって純度を分析し、タンパク質濃度をBCAによって測定した。精製、変性タンパク質をインビトロ再折り畳み実験に利用した。
【0140】
共発現成熟SpeBの精製。ポリシストロニックまたは2−プラスミド系のいずれかによって共発現された組換え成熟野生型SpeBおよび成熟C192S SpeBの精製方法は同一であった。溶解バッファー(20mMトリス、pH7.2、10mM MgCl2、10μg/μL DNase)中、15mLバッファー/g細胞の割合で誘発細菌細胞ペレットを再懸濁し、顕微溶液化によって溶解した。細胞残屑を遠心分離でペレット化し、可溶性フラクションを回収し、50mMグリシン−HCl(pH3.2)に対して4℃下に一夜、透析した。透析は、大腸菌タンパク質の顕著な沈殿によって達成され、pHを4.5に迅速に変更する前に遠心分離で浄化した。サンプルを10M尿素で1:1に希釈し、100mM酢酸ナトリウム(pH4.5)、5M尿素で平衡したSP−セファロースカラム上に供給した。カラムからの非結合物質をA280nmがベースラインに達するまで平衡バッファーで洗浄した。カラムを100mM酢酸ナトリウム(pH4.5)で洗浄し、尿素を除去し、組換え成熟プロテアーゼを0〜1.0M塩化ナトリウムの勾配で溶出した。回収タンパク質をPBS(pH7.4)に対して透析し、SDS−PAGEゲル電気泳動によって純度を評価し、タンパク質濃度をBCAによって測定した。
【0141】
ポリクローナル抗血清の生成。精製成熟C192S SpeB(ポリシストロニックまたは2−プラスミドベースの共発現によって生成)を抗血清の生成のために使用した。スイス・ウェブスター(Swiss Webster)マウスを0、4、6週にMPL 50mgおよびAIPO4 100μgを佐剤として使用して精製タンパク質5μgで免疫化した。
【0142】
成熟SpeBのプロ配列ドメインとの相互作用。SpeBプロ配列ドメイン/成熟SpeB相互作用の解離定数(Kd)を酵素免疫吸着測定法(ELISA)またはバイオコア(Biocore)3000装置を使用して測定した。ELISA分析のために、20μM E−64で阻害した増大濃度のペプシン生成組換え成熟C192S(マツカ(Matsuka)ら、1999年)および組換え成熟野生型SpeBを、精製プロ配列ドメイン、またはリゾチーム(陰性対照)のいずれかでコーティングしたマイクロタイターウェルでインキュベートした。プレートをTBS、pH7.4、0.05%Tween20で洗浄し、検出されたSpeBを405nmでの吸光度を測定することによって成熟SpeBに対して生成された親和性精製ポリクローナル抗体と結合した。結果として生じる吸光度(A)の濃度依存増大を等式ΔA=Amax+[L]/Kd+[L]に適合したが、ここでKdは解離定数であり、[L]は遊離リガンドの濃度である。
【0143】
プロ配列ドメインのペプシン生成組換え成熟C192S SpeBの実時間相互作用をバイオコア(Biocore)3000(バイオコア(Biocore)、Piscataway、ニュージャージー州(NJ))を使用する表面プラスモン共鳴(SPR)によって明らかにした。精製プロ配列ドメインをメーカーの仕様に従って活性化カルボキシメチルデキストランでコーティングされたバイオセンサチップに共有結合した。結合実験をTBS(pH7.4)、0.05%Tween20中、25℃下に行った。増大濃度のペプシン生成組換え成熟C192S SpeBを固定化プロ配列ドメインに添加し、その結合を実時間でモニタリングした。結合過程のセンソグラムを器具とともに供給されたソフトウェアを使用して分析した。
【0144】
SpeBプロ配列ドメインの阻害活性の評価。成熟SpeB(0.1μM)を、増大濃度のプロ配列ドメイン、またはリゾチーム(陰性対照)を利用し、レゾルフィン標識カゼイン基質(0.4%)の存在下、PBS(pH7.4)、10mM DTT中25℃下に1時間インキュベートした。インキュベーション後、非消化基質を2%トリクロロ酢酸を使用する沈殿によって除去し、浄化上清フラクションにおける放出レゾルフィン標識ペプチドの吸光度を574nmで分光光度法により測定した。また、密接に関係したシステインプロテアーゼ(パパイン)を陰性対照と同一条件下で分析し、プロ配列ドメイン/成熟SpeB相互作用の特異性を明らかにした。
【0145】
変性成熟SpeBのインビトロ再折り畳み。100mM酢酸ナトリウム(pH4.5)、8M尿素中の組換え変性成熟SpeBを、0.5Mアルギニンを含有するPBS(pH7.4)、またはPBS(pH7.4)のいずれかにより迅速に希釈した(1:20v/v)。希釈は、示されているように増大濃度の精製プロ配列ドメインの存在下に20μM E−64阻害剤の有無により行った。希釈後、10mM DTTを各反応物に添加し、サンプルを4℃下に24時間インキュベートした。反応物におけるSpeBの最終濃度は5μMであった。インキュベーション後、100μL反応物アリコートをレゾルフィン標識カゼイン切断アッセイを使用して評価し、再折り畳みSpeBの活性を評価した。
【0146】
成熟SpeBのカゼイン分解活性。酵素のタンパク質分解活性を評価するために、指示量のSpeBを、規定されているように、0.4%レゾルフィン標識カゼイン、PBS(pH7.4)、10mM DTT(2%)の存在下にインキュベートした。非消化基質をトリクロロ酢酸沈殿(2%)によって除去し、サンプルを遠心分離によって浄化し、上清フラクションにおける放出レゾルフィン標識ペプチドの吸光度を574nmで分光光度法により測定した。
【0147】
成熟SpeBの熱誘発変性。組換え発現成熟SpeBの融解をTBS(pH7.4)中のタンパク質サンプルを加熱することによって評価すると同時に、SLM AB2蛍光分光光度計を使用し、280nmでの励起により、350nm/320nmでの固有の蛍光強度の比をモニタリングした。
【0148】
実施例2
SpeBプロ配列ドメインの阻害およびシャペロン活性
そのNH2末端プロ配列ドメインを欠く成熟SpeBの組換え発現は、もっぱら大腸菌における不溶性タンパク質の生成をもたらす(マツカ(Matsuka)ら、1999年)。可溶性成熟SpeBを支配するプロ配列ドメインの要件は、ドメインが分子内シャペロンとして機能し、タンパク質の適切な折り畳みを方向づけることを示す。かかる活性を検査するために、SpeBプロ配列ドメイン、40kDa SpeBチモーゲンのほか、成熟野生型SpeBおよび成熟C192S SpeBを特徴づけのための大腸菌から発現させ、精製した(データは示さず)。プロ配列ドメインと成熟SpeBタンパク質の結合を、ELISA(図1A)、およびバイオコア3000(図1B)を使用する表面プラスモン共鳴(SPR)を使用して調査した。実施例1で規定された計算パラメータを使用することにより、プロ配列ドメインの野生型およびC192S成熟SpeBとの相互作用が、それぞれ、11nmおよび34nMのKdを有することが推定された。SPRによって測定されるプロ配列ドメインおよび成熟C192S SpeBの相互作用の実時間分析は、11nMのKdを有することが推定された。両方の方法によって測定される結合値は、プロ配列ドメインと成熟SpeBドメインとの間の高い親和性を示す。
【0149】
プロ配列ドメインの成熟SpeBとの結合は、プロテアーゼ活性の阻害をもたらす。これは基質としてレゾルフィン標識カゼインを使用するカゼイン分解切断アッセイを使用して明らかにされた(図2A)。プロ配列ドメイン、またはリゾチーム制御を、0.1μMの成熟SpeBを使用する0〜100μMの阻害剤濃度範囲にわたって分析した。結果は、プロ配列ドメインが0.3μMの濃度で最大のプロテアーゼ活性の半分を阻害し(IC50=0.3μM)、阻害剤としてのリゾチームの添加は同じ濃度範囲にわたって成熟SpeBの活性に対する影響はないことを示した。同一条件下での密接に関係したシステインプロテアーゼ(パパイン)の分析は、パパインプロテアーゼ活性に対する影響を示さなかった(図2B)。かかる結果は、SpeBとプロ配列ドメインとの間の相互作用の特異性をさらに明らかにし、かつプロ配列ドメインの分子内阻害活性がインサイツでのプロテアーゼ活性の調節に役立ちうることを示す。
【0150】
プロ配列ドメインの分子内シャペロン活性は、尿素変性成熟システインプロテアーゼを使用してインビトロで明らかにされた。増大濃度のプロ配列ドメインの存在下に再折り畳みされた変性成熟プロテアーゼは、レゾルフィン標識カゼイン基質の切断によってモニタリングされるように回収プロテアーゼ活性の顕著な増大を明らかにした(図3)。反応における不可逆性システインプロテアーゼ阻害剤(E−64)の添加は基質切断を阻止し、確認されたカゼイン分解切断は特に再折り畳み成熟SpeBの活性に関与することを示した。これらのデータは、SpeBプロ配列ドメインが分子内シャペロンとして作用し、成熟SpeBの折り畳みを方向づけることを示す。さらに、結果は、2つの領域が適切な折り畳みを方向づけるために共有結合される必要がないことをはっきり示す。
【0151】
実施例3
SPEBプロ配列ドメインおよび成熟SPEBの2−プラスミドベースの共発現
インビトロでの成熟SpeBの正確な再折り畳みを方向づけるプロ配列ドメインの能力は、インビボでの2つのタンパク質の独立した共発現が潜在的に正確に折り畳まれる成熟SpeBの生成がもたらされることを示す。したがって、本実施例では、2−プラスミド共発現系が開発され、この場合、一方のプラスミドはプロ配列ドメイン(pLP682)をコード化し、かつ他方は成熟SpeBポリペプチド(pLP680またはpLP681)をコード化する(図4)。pLP680もしくはpLP681のいずれかのみで形質転換され、またはpLP682で同時形質転換された大腸菌を使用し、SDS−PAGEおよびウェスタンブロットを使用してタンパク質発現を調査した。成熟野生型または成熟C192S SpeB構成物のいずれかによる唯一の発現(データは示さず)は、優先的に不溶性28kDa成熟SpeBの生成をもたらした。これは、プロ配列ドメイン非存在下の成熟SpeBの発現が、不正確に折り畳まれるタンパク質の生成をもたらすことを示す。対照的に、プロ配列ドメインとともに成熟SpeBポリペプチドのインビボ共発現は、細胞の可溶性フラクションの両方のタンパク質の実質的レベルの生成をもたらし、両方のタンパク質の独立した共発現がSpeBの適切な折り畳みを促進することを示す。SpeBチモーゲンに対するポリクローナル抗体を利用するウェスタンブロット分析では、プロ配列ドメインおよび成熟SpeBポリペプチドの同時の検出が可能であった(データは示さず)。ブロット分析では、発現タンパク質の同一性が確認され、さらにSDS−PAGEによって証明される結果が検証された。これらの結果によりインビトロデータが再確認され、プロ配列ドメインおよび成熟SpeBポリペプチドが成熟SpeBの適切な折り畳みを方向づけるために共有結合される必要がないことを示した。
【0152】
実施例4
SPEBプロ配列ドメインおよび成熟SPEBのポリシストロニックベースの共発現
実施例3に記載されているように、プロ配列ドメインおよび成熟SpeBの2−プラスミドベースの共発現は、可溶性に正確に折り畳まれたSpeBを生成するかかる方法の有用性を明らかにした。成熟SpeBの大規模生成のために、プロ配列ドメインと成熟SpeBポリペプチドの独立した共発現のためのポリシストロニック発現系が開発された。したがって、この発現系は、第1のシストロンとしてのプロ配列ドメインに引き続き、翻訳エンハンサーおよび最適化シャイン・ダルガノリボソーム結合部位を含有する合成リンカー(バリック(Barrick)ら、1994年、カリー(Curry)とトミッチ(Tomich)、1998年、リングクイスト(Ringquist)ら、1992年)5’の第2のシストロンが設計された(図5)。第1のシストロンの翻訳終止コドンと第2のシストロンのシャイン・ダルガノとの間の合成リンカーの増大する長さ(5nt、10nt、20nt、40nt)を2つのタンパク質の発現レベルの違いについて調査した。リンカー領域は、転写RNAにおける2つのシストロン間の第2の構造を最小限にするように設計され、非制限リボソームの流れおよび第2のシストロンでの翻訳の再開始を可能にした。それぞれ、5nt、10nt、20nt、および40ntリンカーを含有するポリシストロニック成熟C192S SpeB発現構成物pLP683、pLP684、pLP685、およびpLP686のほか、成熟野生型SpeBの20ntポリシストロニック発現構成物(pLP687)を利用する実験を行った。
【0153】
pLP685およびpLP687ポリシストロニック構成物を使用するタンパク質発現の分析では、プロ配列ドメイン(12kDa)および成熟SpeB(28kDa)の両方の生成が確認された(データは示さず)。2−プラスミド系で示されているように、大腸菌における両方のタンパク質の同時かつ独立の共発現は、優先的に細胞の可溶性フラクションにおける成熟SpeBの生成をもたらした。4つのC192Sポリシストロニック構成物のそれぞれの誘発培養からの全細胞溶解物の可溶性フラクションもSDS−PAGEによって分析された(図6A)。発現のレベルを定量するために、濃度計を使用してゲルを分析し、28kDa成熟SpeB、または12kDaプロ配列ドメインに対応する各バンドの面積を測定した(図6B)。各構成物によってプロ配列ドメインのレベルにはほとんど違いは確認されなかったが、40ntリンカー含有ポリシストロンからの不溶性成熟SpeB発現の顕著な減少が確認された。興味深いことに、119ntリンカー領域を含有する構成物からの可溶性成熟SpeB発現は、5nt、10nt、および20ntリンカーポリシストロンのものと同等のレベルをはっきり示した(データは示さず)。
【0154】
実施例5
ポリシストロニックmRNA転写の分析および転写レベルの評価
総RNAをすべてのポリシストロニック構成物、野生型SpeBチモーゲン、成熟野生型SpeB、成熟C192S SpeB、およびプロ配列ドメインの誘発培養物から単離した。単離RNAのノーザンブロット分析を行い、20ntリンカー含有構成物、および規定されているように適切な陽性対照から生じた転写のサイズを評価した(データは示さず)。得られた結果により、対照からの成熟野生型SpeB(約892塩基)、成熟C192S SpeB(約892塩基)およびプロ配列ドメイン(約487塩基)mRNA転写はすべて予想されたサイズで移動することが明らかにされた。野生型およびC192S 20ntリンカー含有ポリシストロニック発現系の両方からのmRNA転写の検出は、野生型SpeBチモーゲン対照のもの(約1246塩基)よりわずかに高い同等のサイズ(約1287塩基)で移動する転写シグナルを示した。かかる結果は、予測されるように、各々の完全長、ポリシストロニックmRNAの生成と一致する。成熟SpeBおよびプロ配列ドメイン対照で確認されるものと同等の、これらサンプルにおけるより小さい転写の検出の欠如は、SpeBの発現がポリシストロニック転写の生成に直接帰するものであり、多様なmRNA種の存在の結果によるものではないことを示す。
【0155】
すべてのポリシストロニック種から生じる転写の完全長の性質もcDNAのPCR分析によって検証された。ポリシストロニックサンプルおよび単一のシストロン対照からの総RNAを使用し、対応するcDNAを生成し、かつ希釈cDNAサンプルをPCRによって分析した。逆転写酵素(−RT)を除くすべての反応成分を含む各サンプルの陰性対照を生成し、単離RNA内の潜在的なプラスミドDNA汚染を評価した。プロ配列ドメイン(367bp)、成熟SpeB(770bp)、および完全長プロ配列/成熟SpeB(1119−1180bp)コード領域に特異的な一連のプライマーを使用し、cDNAサンプル、−RT陰性対照、およびpLP685陽性対照を検査した(データは示さず)。プロ配列ドメインのPCR増幅により、その発現構成物におけるプロ配列ドメインヌクレオチド配列を含有するすべてのcDNAサンプルにおける予想されるサイズの産物が生じた。この配列を欠くサンプル、成熟野生型SpeBおよび成熟C192S SpeB発現構成物は、増幅産物を生成しなかった。成熟プロテアーゼコード領域の増幅について、プロ配列ドメイン発現系のものを除くすべてのcDNAサンプルは陽性バンドを生成した。完全長プロ配列/成熟SpeB領域の増幅により、ポリシストロニックおよびSpeBチモーゲンサンプルのみについてPCR産物が生じたが、成熟野生型SpeB、成熟C192S SpeB、およびプロ配列ドメイン発現系からのサンプルは、予測されるように産物生成の欠如を証明した。すべてのサンプルの−RT陰性対照は増幅産物を生成せず、mRNAの逆転写に由来するcDNAサンプルで確認された陽性シグナルの生成を示し、汚染DNAの増幅を示さなかった。
【0156】
異なる発現系のmRNA転写レベルの検査は、cDNAサンプルおよび−RT対照の定量的PCR分析によって測定された。プロ配列および成熟SpeB領域の各々に特異的な一連のプライマー/プローブを使用し、培養物間の転写レベルを比較した。カナマイシン耐性をコード化する遺伝子がすべての発現構成物上に存在することを考えて、KanRmRNAを分析中の内標準として使用し、発現サンプル間のプラスミドコピー数の潜在的な違いに対して制御した。結果は、各構成物の誘発中に生成されたmRNA転写のレベルが同等であることをはっきり示す(図7)。より重要なことには、データは、プロ配列のmRNAレベルが成熟SpeBのものと同等であることを示す。これらの結果は、ポリシストロニック構成物間の可溶性成熟SpeBの確認された量の明らかな違いが、転写の早過ぎる終了によるものではないことを示す。
【0157】
実施例6
共発現成熟SpeBの特徴づけ
2−プラスミドベースおよびポリシストロニック系の両方によって生成された精製成熟SpeBをSDS−PAGEによって分析し(データは示さず)、すべてのタンパク質を熱誘発展開実験にかけた(図8および図9)。融解(すなわち、変性)曲線を上昇温度範囲(0−90℃)にわたってタンパク質サンプルを加熱すると同時に、固有の蛍光の比の変化をモニタリングすることによって得た。2−プラスミドおよびポリシストロニック共発現C192S(図8)の両方、または野生型(図9)成熟SpeBによる変性曲線は、明確な移行部をはっきり示し、SpeBチモーゲンとはそれぞれ対応する発現のものと同様の中間点を有し、インビトロ切断(パパイン生成)、または自己触媒(野生型SpeBチモーゲン)によって加工された。かかる結果は、いずれかの共発現系によって生成される組換えタンパク質が、対応する組換え発現および加工チモーゲンと同様に展開されることを示す。
【0158】
すべての組換え成熟野生型SpeBの固有のプロテアーゼ活性をその操作的モル濃度の測定によって評価した(図10)。各々について、同等濃度のSpeB(0.12μM)を、レゾルフィン標識カゼイン基質の添加前に、不可逆性システインプロテアーゼ阻害剤、E−64の増大量の存在下にプレインキュベートした。標識ペプチドの放出を分光光度法で測定し、結果を阻害剤濃度の関数としてプロットした。2−プラスミド(0.121μM)、ポリシストロニック(0.124μM)、および自己触媒加工(0.124μM)成熟SpeBポリペプチドについて得られた値は、それらが酵素的に区別不可能であり、タンパク質濃度に基づく0.12μMの予想値と同等であることを示した。
【0159】
以前のデータは、パパインによるインビトロ切断によって生成された成熟C192S SpeBに対して生成された抗体が、野生型SpeBのタンパク質分解活性を阻害する能力があることを示した(マツカ(Matsuka)ら、1999年)。いずれにかの共発現系によって生成される組換えタンパク質が同様の活性を発揮するを判定するために、2−プラスミドまたは20ntポリシストロニック系のいずれかによって生成された成熟C192S SpeBで免疫化したマウスからの増大濃度の抗血清をカゼイン分解切断アッセイを使用して分析した(図11)。結果は、基質の加水分解が、プレ免疫対照に対して、いずれかの共発現系によって生成されるタンパク質で免疫化した動物からの血清の増大量の存在下に阻害されることを示す。さらに、各々について証明された阻害のレベルは、SpeBチモーゲンとして発現され、かつペプシンを使用してインビトロで加工された成熟C192S SpeBに対して生成された血清について以前に報告されたものと同等である(マツカ(Matsuka)ら、1999年)。
【0160】
【0161】
【0162】
【0163】
【0164】
【図面の簡単な説明】
【0165】
【図1】組換え成熟SpeBとプロ配列ドメインの相互作用を示す。
【図2】成熟SpeBのプロ配列ドメイン介在阻害を示す。
【図3】変性成熟SpeBの再折り畳みに対する組換えプロ配列ドメインの効果を示す。
【図4】2−プラスミドベース発現構成物を示す略図である。
【図5】ポリシストロニック発現系および合成リンカー領域を示す図表である。
【図6】ポリシストロニック発現系における可溶性成熟SpeBおよびプロ配列ドメインレベルの評価および相対定量を示す。
【図7】ポリシストロニックcDNAの定量的PCR分析を示す。
【図8】成熟C192S SpeBの熱変性を示す。
【図9】成熟野生型SpeBの熱誘発変性を示す。
【図10】組換え成熟SpeBの操作モル濃度の評価を示す。
【図11】野生型SpeBタンパク質分解活性の抗体介在阻害を示す。
【特許請求の範囲】
【請求項1】
宿主細胞における成熟化膿連鎖球菌外毒素B(Streptococcus pyogenes exotoxin B)(SpeB)ポリペプチドを組換え発現させるための方法であって、(a)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列と、(b)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列とを含むポリシストロニックプラスミドで宿主細胞を形質転換し、形質導入し、トランスフェクトし、または感染させ、該宿主細胞によって該成熟SpeBポリペプチドおよび該SpeBプロ−ポリペプチドドメインの発現を可能にする条件下で該宿主細胞を培養することを含む、その成熟SpeBポリペプチドが該宿主細胞において可溶性であるところの、方法。
【請求項2】
前記SpeBプロ−ポリペプチドドメインが、配列番号2のアミノ酸残基28〜145を含んで成るポリペプチドとしてさらに規定される、請求項1に記載の方法。
【請求項3】
前記成熟SpeBポリペプチドが、配列番号2のアミノ酸残基146〜398を含んで成るポリペプチドとしてさらに規定される、請求項1に記載の方法。
【請求項4】
前記成熟SpeBポリペプチドのアミノ酸残基192での前記システインがセリンによって置換される、請求項1に記載の方法。
【請求項5】
前記成熟SpeBポリペプチドが哺乳類宿主において免疫原性である、請求項1に記載の方法。
【請求項6】
前記成熟SpeBポリペプチドに特異的な抗体が野生型SpeBポリペプチドと交差反応し、かつSpeBポリペプチド活性を無力化する、請求項1に記載の方法。
【請求項7】
前記プラスミドがT7プロモーター含有プラスミドである、請求項1に記載の方法。
【請求項8】
前記プラスミドが、pET、pRSET、pCRT7−CTTOPO、およびpIVeXより成る群から選択される、請求項7に記載の方法。
【請求項9】
前記宿主細胞が細菌細胞である、請求項1に記載の方法。
【請求項10】
前記宿主細胞が大腸菌(E.coli)である、請求項9に記載の方法。
【請求項11】
前記大腸菌が、BLR(DE3)、BLR(DE3)pLysS、AD494(DE3)、AD494(DE3)pLysS、BL21(DE3)、BL21(DE3)pLysS、BL21(DE3)pLysE、BL21(DE3)pLacI、BL21trxB(DE3)、BL21trxB(DE3)pLysS、HMS174(DE3)、HMS174(DE3)pLysS、HMS174(DE3)pLysE、Origami(DE3)、Origami(DE3)pLysS、Origami(DE3)pLysE、Origami(DE3)pLacI、OrigamiB(DE3)、OrigamiB(DE3)pLysS、OrigamiB(DE3)pLysE、OrigamiB(DE3)pLacI、Rosetta(DE3)、Rosetta(DE3)pLysS、Rosetta(DE3)pLysE、Rosetta(DE3)pLacI、Tuner(DE3)、Tuner(DE3)pLysS、およびTuner(DE3)pLacIより成る群から選択される菌株である、請求項10に記載の方法。
【請求項12】
宿主細胞における成熟SpeBポリペプチドを組換え発現させるための方法であって、(a)(i)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列を含んで成るプラスミドおよび(ii)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るプラスミドで宿主細胞を形質転換し、形質導入し、トランスフェクトし、または感染させ、
(b)前記SpeBプロ−ポリペプチドドメインおよび前記成熟SpeBポリペプチドを共発現させるのに適切な条件下で宿主細胞を培養すること
を含む、前記成熟SpeBポリペプチドが宿主細胞において可溶性であるところの、方法。
【請求項13】
前記SpeBプロ−ポリペプチドドメインが、配列番号2のアミノ酸残基28〜145を含んで成るポリペプチドとしてさらに規定される、請求項12に記載の方法。
【請求項14】
前記成熟SpeBポリペプチドが、配列番号2のアミノ酸残基146〜398を含んで成るポリペプチドとしてさらに規定される、請求項12に記載の方法。
【請求項15】
前記成熟SpeBポリペプチドのアミノ酸残基192での前記システインがセリンによって置換される、請求項12に記載の方法。
【請求項16】
前記成熟SpeBポリペプチドが哺乳類宿主において免疫原性である、請求項12に記載の方法。
【請求項17】
前記成熟SpeBポリペプチドに特異的な抗体が野生型SpeBポリペプチドと交差反応し、かつSpeBポリペプチド活性を無力化する、請求項12に記載の方法。
【請求項18】
前記プラスミドがT7プロモーター含有プラスミドである、請求項12に記載の方法。
【請求項19】
前記プラスミドが、pET、pRSET、pCRT7−CTTOPO、およびpIVeXより成る群から選択される、請求項18に記載の方法。
【請求項20】
前記宿主細胞が細菌細胞である、請求項12に記載の方法。
【請求項21】
前記宿主細胞が大腸菌(E.coli)である、請求項20に記載の方法。
【請求項22】
前記大腸菌が、BLR(DE3)、BLR(DE3)pLysS、AD494(DE3)、AD494(DE3)pLysS、BL21(DE3)、BL21(DE3)pLysS、BL21(DE3)pLysE、BL21(DE3)pLacI、BL21trxB(DE3)、BL21trxB(DE3)pLysS、HMS174(DE3)、HMS174(DE3)pLysS、HMS174(DE3)pLysE、Origami(DE3)、Origami(DE3)pLysS、Origami(DE3)pLysE、Origami(DE3)pLacI、OrigamiB(DE3)、OrigamiB(DE3)pLysS、OrigamiB(DE3)pLysE、OrigamiB(DE3)pLacI、Rosetta(DE3)、Rosetta(DE3)pLysS、Rosetta(DE3)pLysE、Rosetta(DE3)pLacI、Tuner(DE3)、Tuner(DE3)pLysS、およびTuner(DE3)pLacIより成る群から選択される菌株である、請求項21に記載の方法。
【請求項23】
成熟SpeBポリペプチドを生成するための方法であって、
(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含むプラスミドを宿主細胞において組換え発現させ、ここで前記SpeBポリペプチドが前記宿主細胞において不溶性ポリペプチド集合体を形成し、
(b)前記ポリペプチド集合体を可溶化し、ここで前記可溶化ポリペプチドが非ネイティブ成熟SpeBポリペプチドとして規定され、
(c)1つもしくはそれ以上のシャペロンタンパク質の存在下に前記非ネイティブ成熟SpeBポリペプチドを再折り畳みし、ここで前記非ネイティブ成熟SpeBポリペプチドがネイティブ成熟SpeBポリペプチドへ折り畳まれ、
(d)前記ネイティブ成熟SpeBポリペプチドを回収すること
を含んで成る方法。
【請求項24】
前記1つもしくはそれ以上のシャペロンタンパク質が、GroEL、GroEL/GroES、ペプチジル−プロリルイソメラーゼ(PPI)、ペプチドジスルフィドイソメラーゼ(PDI)およびSpeBプロ−ポリペプチドドメインより成る群から選択される、請求項23に記載の方法。
【請求項25】
前記シャペロンタンパク質が、配列番号2のアミノ酸残基28〜145を含んで成るSpeBプロ−ポリペプチドドメインである、請求項23に記載の方法。
【請求項26】
前記成熟SpeBが、配列番号2のアミノ酸残基146〜398を含んで成るポリペプチドである、請求項23に記載の方法。
【請求項27】
前記成熟SpeBポリペプチドのアミノ酸残基192における前記システインがセリンによって置換される、請求項26に記載の方法。
【請求項28】
前記不溶性ポリペプチド集合体が封入体としてさらに規定される、請求項23に記載の方法。
【請求項29】
前記ポリペプチドの可溶化が尿素、塩化グアニジニウム、および熱より成る群から選択される変性剤である、請求項23に記載の方法。
【請求項30】
宿主細胞において(i)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(ii)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成るポリシストロニックプラスミドを発現させることを含む、前記成熟SpeBポリペプチドが前記宿主細胞において可溶性である、宿主細胞において成熟SpeBポリペプチドを組換え発現させるための方法。
【請求項31】
前記成熟SpeBポリペプチドのアミノ酸残基192における前記システインがセリンによって置換される、請求項30に記載の方法。
【請求項32】
前記プラスミドが、GroESポリペプチドをコード化するポリヌクレオチドをさらに含んで成る、請求項30に記載の方法。
【請求項33】
成熟SpeBポリペプチドを生成するための方法であって、
(a)(i)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(ii)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成るポリシストロニックプラスミドで宿主細胞を形質転換し、形質導入し、トランスフェクトし、または感染させ、
(b)前記成熟SpeBポリペプチドおよび前記GroELポリペプチドを発現させるのに適切な条件下で前記宿主細胞を培養し、ここで前記成熟SpeBポリペプチドが前記宿主細胞において可溶性であり、
(c)前記ネイティブ成熟SpeBポリペプチドを回収すること
を含んで成る方法。
【請求項34】
前記成熟SpeBポリペプチドのアミノ酸残基192における前記システインがセリンによって置換される、請求項33に記載の方法。
【請求項35】
請求項1に記載の方法に従って生成される成熟SpeBポリペプチド。
【請求項36】
請求項12に記載の方法に従って生成される成熟SpeBポリペプチド。
【請求項37】
請求項23に記載の方法に従って生成される成熟SpeBポリペプチド。
【請求項38】
請求項30に記載の方法に従って生成される成熟SpeBポリペプチド。
【請求項39】
請求項33に記載の方法に従って生成される成熟SpeBポリペプチド。
【請求項40】
請求項35に記載のSpeBポリペプチドを含んで成る免疫原性組成物。
【請求項41】
請求項36に記載のSpeBポリペプチドを含んで成る免疫原性組成物。
【請求項42】
請求項37に記載のSpeBポリペプチドを含んで成る免疫原性組成物。
【請求項43】
請求項38に記載のSpeBポリペプチドを含んで成る免疫原性組成物。
【請求項44】
請求項39に記載のSpeBポリペプチドを含んで成る免疫原性組成物。
【請求項45】
請求項40に記載の免疫原性量の組成物を哺乳類対象に投与することを含む化膿連鎖球菌に対して前記対象を免疫化する方法。
【請求項46】
請求項41に記載の免疫原性量の組成物を哺乳類対象に投与することを含む化膿連鎖球菌に対して前記対象を免疫化する方法。
【請求項47】
請求項42に記載の免疫原性量の組成物を哺乳類対象に投与することを含む化膿連鎖球菌に対して前記対象を免疫化する方法。
【請求項48】
請求項43に記載の免疫原性量の組成物を哺乳類対象に投与することを含む化膿連鎖球菌に対して前記対象を免疫化する方法。
【請求項49】
請求項44に記載の免疫原性量の組成物を哺乳類対象に投与することを含む化膿連鎖球菌に対して前記対象を免疫化する方法。
【請求項50】
(a)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列と、(b)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列とを含む、ここで前記成熟SpeBポリペプチドが宿主細胞において発現されると可溶性である、ポリシストロニックプラスミド。
【請求項51】
前記成熟SpeBポリペプチドのアミノ酸残基192での前記システインがセリンによって置換される、請求項50に記載のプラスミド。
【請求項52】
前記プラスミドがT7プロモーター含有プラスミドである、請求項50に記載のプラスミド。
【請求項53】
SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列を含んで成るプラスミド、および成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るプラスミドであって、ここで前記成熟SpeBポリペプチドが宿主細胞において発現されると可溶性であるところの、プラスミド。
【請求項54】
前記成熟SpeBポリペプチドのアミノ酸残基192での前記システインがセリンによって置換される、請求項53に記載のプラスミド。
【請求項55】
前記プラスミドがT7プロモーター含有プラスミドである、請求項53に記載のプラスミド。
【請求項56】
(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(b)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成り、ここで前記成熟SpeBポリペプチドが宿主細胞で発現されると可溶性であるところの、ポリシストロニックプラスミド。
【請求項57】
前記成熟SpeBポリペプチドのアミノ酸残基192における前記システインがセリンによって置換される、請求項56に記載のプラスミド。
【請求項58】
前記プラスミドがT7プロモーター含有プラスミドである、請求項56に記載のプラスミド。
【請求項59】
(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(b)GroELポリペプチドをコード化するポリヌクレオチド配列と、(c)GroESポリペプチドをコード化するポリヌクレオチド配列とを含んで成り、ここで前記成熟SpeBポリペプチドが宿主細胞で発現されると可溶性であるところの、ポリシストロニックプラスミド。
【請求項60】
前記成熟SpeBポリペプチドのアミノ酸残基192における前記システインがセリンによって置換される、請求項59に記載のプラスミド。
【請求項61】
前記プラスミドがT7プロモーター含有プラスミドである、請求項59に記載のプラスミド。
【請求項62】
(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(b)GroEL、GroES、SpeBプロ−ポリペプチドドメイン、PDI、およびPPIより成る群から選択される1つもしくはそれ以上のポリペプチドをコード化するポリヌクレオチド配列とを含んで成り、ここで前記成熟SpeBポリペプチドが宿主細胞で発現されると可溶性であるところの、ポリシストロニックプラスミド。
【請求項63】
前記成熟SpeBポリペプチドのアミノ酸残基192における前記システインがセリンによって置換される、請求項62に記載のプラスミド。
【請求項64】
前記プラスミドがT7プロモーター含有プラスミドである、請求項62に記載のプラスミド。
【請求項65】
請求項50に記載のプラスミドで形質転換され、形質導入され、トランスフェクトされ、または感染される宿主細胞。
【請求項66】
請求項53に記載のプラスミドで形質転換され、形質導入され、トランスフェクトされ、または感染される宿主細胞。
【請求項67】
請求項56に記載のプラスミドで形質転換され、形質導入され、トランスフェクトされ、または感染される宿主細胞。
【請求項68】
請求項59に記載のプラスミドで形質転換され、形質導入され、トランスフェクトされ、または感染される宿主細胞。
【請求項69】
請求項62に記載のプラスミドで形質転換され、形質導入され、トランスフェクトされ、または感染される宿主細胞。
【請求項70】
前記PPIが化膿連鎖球菌RopA PPIである、請求項24に記載の方法。
【請求項71】
前記PPIが化膿連鎖球菌RopA PPIである、請求項62に記載のプラスミド。
【請求項1】
宿主細胞における成熟化膿連鎖球菌外毒素B(Streptococcus pyogenes exotoxin B)(SpeB)ポリペプチドを組換え発現させるための方法であって、(a)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列と、(b)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列とを含むポリシストロニックプラスミドで宿主細胞を形質転換し、形質導入し、トランスフェクトし、または感染させ、該宿主細胞によって該成熟SpeBポリペプチドおよび該SpeBプロ−ポリペプチドドメインの発現を可能にする条件下で該宿主細胞を培養することを含む、その成熟SpeBポリペプチドが該宿主細胞において可溶性であるところの、方法。
【請求項2】
前記SpeBプロ−ポリペプチドドメインが、配列番号2のアミノ酸残基28〜145を含んで成るポリペプチドとしてさらに規定される、請求項1に記載の方法。
【請求項3】
前記成熟SpeBポリペプチドが、配列番号2のアミノ酸残基146〜398を含んで成るポリペプチドとしてさらに規定される、請求項1に記載の方法。
【請求項4】
前記成熟SpeBポリペプチドのアミノ酸残基192での前記システインがセリンによって置換される、請求項1に記載の方法。
【請求項5】
前記成熟SpeBポリペプチドが哺乳類宿主において免疫原性である、請求項1に記載の方法。
【請求項6】
前記成熟SpeBポリペプチドに特異的な抗体が野生型SpeBポリペプチドと交差反応し、かつSpeBポリペプチド活性を無力化する、請求項1に記載の方法。
【請求項7】
前記プラスミドがT7プロモーター含有プラスミドである、請求項1に記載の方法。
【請求項8】
前記プラスミドが、pET、pRSET、pCRT7−CTTOPO、およびpIVeXより成る群から選択される、請求項7に記載の方法。
【請求項9】
前記宿主細胞が細菌細胞である、請求項1に記載の方法。
【請求項10】
前記宿主細胞が大腸菌(E.coli)である、請求項9に記載の方法。
【請求項11】
前記大腸菌が、BLR(DE3)、BLR(DE3)pLysS、AD494(DE3)、AD494(DE3)pLysS、BL21(DE3)、BL21(DE3)pLysS、BL21(DE3)pLysE、BL21(DE3)pLacI、BL21trxB(DE3)、BL21trxB(DE3)pLysS、HMS174(DE3)、HMS174(DE3)pLysS、HMS174(DE3)pLysE、Origami(DE3)、Origami(DE3)pLysS、Origami(DE3)pLysE、Origami(DE3)pLacI、OrigamiB(DE3)、OrigamiB(DE3)pLysS、OrigamiB(DE3)pLysE、OrigamiB(DE3)pLacI、Rosetta(DE3)、Rosetta(DE3)pLysS、Rosetta(DE3)pLysE、Rosetta(DE3)pLacI、Tuner(DE3)、Tuner(DE3)pLysS、およびTuner(DE3)pLacIより成る群から選択される菌株である、請求項10に記載の方法。
【請求項12】
宿主細胞における成熟SpeBポリペプチドを組換え発現させるための方法であって、(a)(i)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列を含んで成るプラスミドおよび(ii)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るプラスミドで宿主細胞を形質転換し、形質導入し、トランスフェクトし、または感染させ、
(b)前記SpeBプロ−ポリペプチドドメインおよび前記成熟SpeBポリペプチドを共発現させるのに適切な条件下で宿主細胞を培養すること
を含む、前記成熟SpeBポリペプチドが宿主細胞において可溶性であるところの、方法。
【請求項13】
前記SpeBプロ−ポリペプチドドメインが、配列番号2のアミノ酸残基28〜145を含んで成るポリペプチドとしてさらに規定される、請求項12に記載の方法。
【請求項14】
前記成熟SpeBポリペプチドが、配列番号2のアミノ酸残基146〜398を含んで成るポリペプチドとしてさらに規定される、請求項12に記載の方法。
【請求項15】
前記成熟SpeBポリペプチドのアミノ酸残基192での前記システインがセリンによって置換される、請求項12に記載の方法。
【請求項16】
前記成熟SpeBポリペプチドが哺乳類宿主において免疫原性である、請求項12に記載の方法。
【請求項17】
前記成熟SpeBポリペプチドに特異的な抗体が野生型SpeBポリペプチドと交差反応し、かつSpeBポリペプチド活性を無力化する、請求項12に記載の方法。
【請求項18】
前記プラスミドがT7プロモーター含有プラスミドである、請求項12に記載の方法。
【請求項19】
前記プラスミドが、pET、pRSET、pCRT7−CTTOPO、およびpIVeXより成る群から選択される、請求項18に記載の方法。
【請求項20】
前記宿主細胞が細菌細胞である、請求項12に記載の方法。
【請求項21】
前記宿主細胞が大腸菌(E.coli)である、請求項20に記載の方法。
【請求項22】
前記大腸菌が、BLR(DE3)、BLR(DE3)pLysS、AD494(DE3)、AD494(DE3)pLysS、BL21(DE3)、BL21(DE3)pLysS、BL21(DE3)pLysE、BL21(DE3)pLacI、BL21trxB(DE3)、BL21trxB(DE3)pLysS、HMS174(DE3)、HMS174(DE3)pLysS、HMS174(DE3)pLysE、Origami(DE3)、Origami(DE3)pLysS、Origami(DE3)pLysE、Origami(DE3)pLacI、OrigamiB(DE3)、OrigamiB(DE3)pLysS、OrigamiB(DE3)pLysE、OrigamiB(DE3)pLacI、Rosetta(DE3)、Rosetta(DE3)pLysS、Rosetta(DE3)pLysE、Rosetta(DE3)pLacI、Tuner(DE3)、Tuner(DE3)pLysS、およびTuner(DE3)pLacIより成る群から選択される菌株である、請求項21に記載の方法。
【請求項23】
成熟SpeBポリペプチドを生成するための方法であって、
(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含むプラスミドを宿主細胞において組換え発現させ、ここで前記SpeBポリペプチドが前記宿主細胞において不溶性ポリペプチド集合体を形成し、
(b)前記ポリペプチド集合体を可溶化し、ここで前記可溶化ポリペプチドが非ネイティブ成熟SpeBポリペプチドとして規定され、
(c)1つもしくはそれ以上のシャペロンタンパク質の存在下に前記非ネイティブ成熟SpeBポリペプチドを再折り畳みし、ここで前記非ネイティブ成熟SpeBポリペプチドがネイティブ成熟SpeBポリペプチドへ折り畳まれ、
(d)前記ネイティブ成熟SpeBポリペプチドを回収すること
を含んで成る方法。
【請求項24】
前記1つもしくはそれ以上のシャペロンタンパク質が、GroEL、GroEL/GroES、ペプチジル−プロリルイソメラーゼ(PPI)、ペプチドジスルフィドイソメラーゼ(PDI)およびSpeBプロ−ポリペプチドドメインより成る群から選択される、請求項23に記載の方法。
【請求項25】
前記シャペロンタンパク質が、配列番号2のアミノ酸残基28〜145を含んで成るSpeBプロ−ポリペプチドドメインである、請求項23に記載の方法。
【請求項26】
前記成熟SpeBが、配列番号2のアミノ酸残基146〜398を含んで成るポリペプチドである、請求項23に記載の方法。
【請求項27】
前記成熟SpeBポリペプチドのアミノ酸残基192における前記システインがセリンによって置換される、請求項26に記載の方法。
【請求項28】
前記不溶性ポリペプチド集合体が封入体としてさらに規定される、請求項23に記載の方法。
【請求項29】
前記ポリペプチドの可溶化が尿素、塩化グアニジニウム、および熱より成る群から選択される変性剤である、請求項23に記載の方法。
【請求項30】
宿主細胞において(i)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(ii)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成るポリシストロニックプラスミドを発現させることを含む、前記成熟SpeBポリペプチドが前記宿主細胞において可溶性である、宿主細胞において成熟SpeBポリペプチドを組換え発現させるための方法。
【請求項31】
前記成熟SpeBポリペプチドのアミノ酸残基192における前記システインがセリンによって置換される、請求項30に記載の方法。
【請求項32】
前記プラスミドが、GroESポリペプチドをコード化するポリヌクレオチドをさらに含んで成る、請求項30に記載の方法。
【請求項33】
成熟SpeBポリペプチドを生成するための方法であって、
(a)(i)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(ii)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成るポリシストロニックプラスミドで宿主細胞を形質転換し、形質導入し、トランスフェクトし、または感染させ、
(b)前記成熟SpeBポリペプチドおよび前記GroELポリペプチドを発現させるのに適切な条件下で前記宿主細胞を培養し、ここで前記成熟SpeBポリペプチドが前記宿主細胞において可溶性であり、
(c)前記ネイティブ成熟SpeBポリペプチドを回収すること
を含んで成る方法。
【請求項34】
前記成熟SpeBポリペプチドのアミノ酸残基192における前記システインがセリンによって置換される、請求項33に記載の方法。
【請求項35】
請求項1に記載の方法に従って生成される成熟SpeBポリペプチド。
【請求項36】
請求項12に記載の方法に従って生成される成熟SpeBポリペプチド。
【請求項37】
請求項23に記載の方法に従って生成される成熟SpeBポリペプチド。
【請求項38】
請求項30に記載の方法に従って生成される成熟SpeBポリペプチド。
【請求項39】
請求項33に記載の方法に従って生成される成熟SpeBポリペプチド。
【請求項40】
請求項35に記載のSpeBポリペプチドを含んで成る免疫原性組成物。
【請求項41】
請求項36に記載のSpeBポリペプチドを含んで成る免疫原性組成物。
【請求項42】
請求項37に記載のSpeBポリペプチドを含んで成る免疫原性組成物。
【請求項43】
請求項38に記載のSpeBポリペプチドを含んで成る免疫原性組成物。
【請求項44】
請求項39に記載のSpeBポリペプチドを含んで成る免疫原性組成物。
【請求項45】
請求項40に記載の免疫原性量の組成物を哺乳類対象に投与することを含む化膿連鎖球菌に対して前記対象を免疫化する方法。
【請求項46】
請求項41に記載の免疫原性量の組成物を哺乳類対象に投与することを含む化膿連鎖球菌に対して前記対象を免疫化する方法。
【請求項47】
請求項42に記載の免疫原性量の組成物を哺乳類対象に投与することを含む化膿連鎖球菌に対して前記対象を免疫化する方法。
【請求項48】
請求項43に記載の免疫原性量の組成物を哺乳類対象に投与することを含む化膿連鎖球菌に対して前記対象を免疫化する方法。
【請求項49】
請求項44に記載の免疫原性量の組成物を哺乳類対象に投与することを含む化膿連鎖球菌に対して前記対象を免疫化する方法。
【請求項50】
(a)SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列と、(b)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列とを含む、ここで前記成熟SpeBポリペプチドが宿主細胞において発現されると可溶性である、ポリシストロニックプラスミド。
【請求項51】
前記成熟SpeBポリペプチドのアミノ酸残基192での前記システインがセリンによって置換される、請求項50に記載のプラスミド。
【請求項52】
前記プラスミドがT7プロモーター含有プラスミドである、請求項50に記載のプラスミド。
【請求項53】
SpeBプロ−ポリペプチドドメインをコード化するポリヌクレオチド配列を含んで成るプラスミド、および成熟SpeBポリペプチドをコード化するポリヌクレオチド配列を含んで成るプラスミドであって、ここで前記成熟SpeBポリペプチドが宿主細胞において発現されると可溶性であるところの、プラスミド。
【請求項54】
前記成熟SpeBポリペプチドのアミノ酸残基192での前記システインがセリンによって置換される、請求項53に記載のプラスミド。
【請求項55】
前記プラスミドがT7プロモーター含有プラスミドである、請求項53に記載のプラスミド。
【請求項56】
(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(b)GroELポリペプチドをコード化するポリヌクレオチド配列とを含んで成り、ここで前記成熟SpeBポリペプチドが宿主細胞で発現されると可溶性であるところの、ポリシストロニックプラスミド。
【請求項57】
前記成熟SpeBポリペプチドのアミノ酸残基192における前記システインがセリンによって置換される、請求項56に記載のプラスミド。
【請求項58】
前記プラスミドがT7プロモーター含有プラスミドである、請求項56に記載のプラスミド。
【請求項59】
(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(b)GroELポリペプチドをコード化するポリヌクレオチド配列と、(c)GroESポリペプチドをコード化するポリヌクレオチド配列とを含んで成り、ここで前記成熟SpeBポリペプチドが宿主細胞で発現されると可溶性であるところの、ポリシストロニックプラスミド。
【請求項60】
前記成熟SpeBポリペプチドのアミノ酸残基192における前記システインがセリンによって置換される、請求項59に記載のプラスミド。
【請求項61】
前記プラスミドがT7プロモーター含有プラスミドである、請求項59に記載のプラスミド。
【請求項62】
(a)成熟SpeBポリペプチドをコード化するポリヌクレオチド配列と、(b)GroEL、GroES、SpeBプロ−ポリペプチドドメイン、PDI、およびPPIより成る群から選択される1つもしくはそれ以上のポリペプチドをコード化するポリヌクレオチド配列とを含んで成り、ここで前記成熟SpeBポリペプチドが宿主細胞で発現されると可溶性であるところの、ポリシストロニックプラスミド。
【請求項63】
前記成熟SpeBポリペプチドのアミノ酸残基192における前記システインがセリンによって置換される、請求項62に記載のプラスミド。
【請求項64】
前記プラスミドがT7プロモーター含有プラスミドである、請求項62に記載のプラスミド。
【請求項65】
請求項50に記載のプラスミドで形質転換され、形質導入され、トランスフェクトされ、または感染される宿主細胞。
【請求項66】
請求項53に記載のプラスミドで形質転換され、形質導入され、トランスフェクトされ、または感染される宿主細胞。
【請求項67】
請求項56に記載のプラスミドで形質転換され、形質導入され、トランスフェクトされ、または感染される宿主細胞。
【請求項68】
請求項59に記載のプラスミドで形質転換され、形質導入され、トランスフェクトされ、または感染される宿主細胞。
【請求項69】
請求項62に記載のプラスミドで形質転換され、形質導入され、トランスフェクトされ、または感染される宿主細胞。
【請求項70】
前記PPIが化膿連鎖球菌RopA PPIである、請求項24に記載の方法。
【請求項71】
前記PPIが化膿連鎖球菌RopA PPIである、請求項62に記載のプラスミド。
【図1】

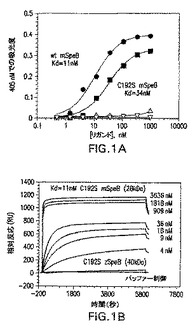
【図2】
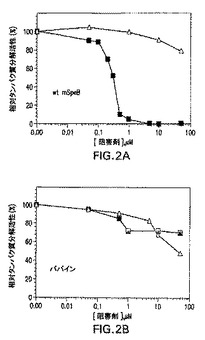
【図3】
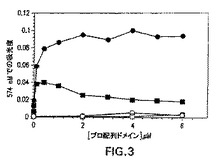
【図4】
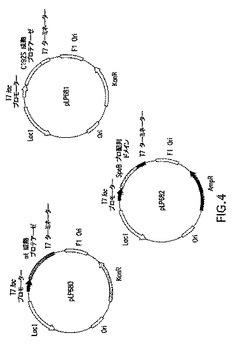
【図5】
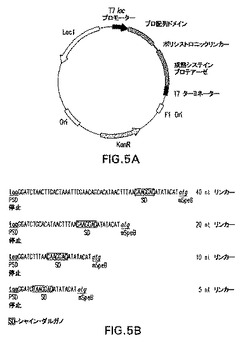
【図6】
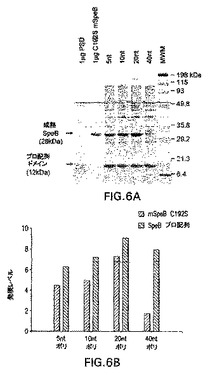
【図7】
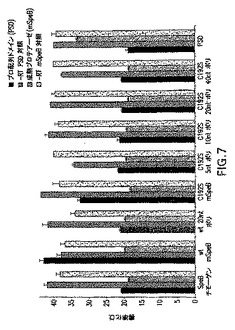
【図8】
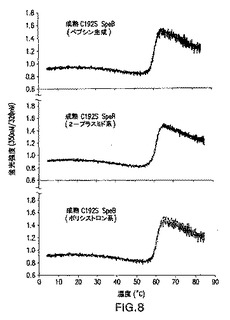
【図9】
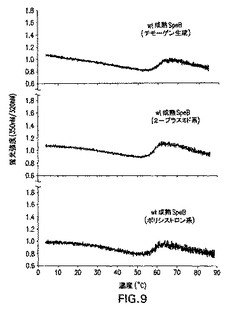
【図10】
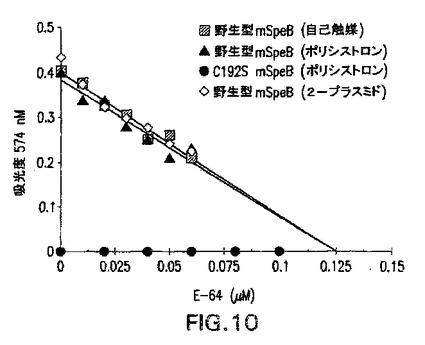
【図11】
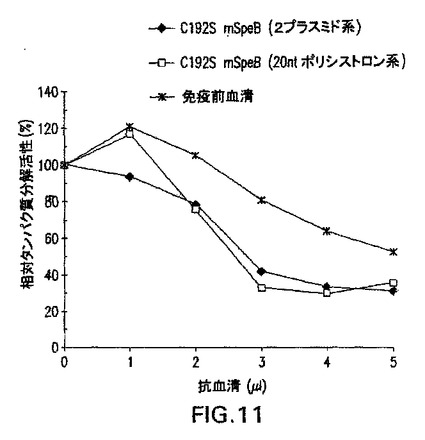
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公表番号】特表2007−536900(P2007−536900A)
【公表日】平成19年12月20日(2007.12.20)
【国際特許分類】
【出願番号】特願2006−518866(P2006−518866)
【出願日】平成16年7月7日(2004.7.7)
【国際出願番号】PCT/US2004/021714
【国際公開番号】WO2005/007690
【国際公開日】平成17年1月27日(2005.1.27)
【出願人】(591000791)ワイス・ホールディングズ・コーポレイション (43)
【氏名又は名称原語表記】Wyeth Holdings Corporation
【Fターム(参考)】
【公表日】平成19年12月20日(2007.12.20)
【国際特許分類】
【出願日】平成16年7月7日(2004.7.7)
【国際出願番号】PCT/US2004/021714
【国際公開番号】WO2005/007690
【国際公開日】平成17年1月27日(2005.1.27)
【出願人】(591000791)ワイス・ホールディングズ・コーポレイション (43)
【氏名又は名称原語表記】Wyeth Holdings Corporation
【Fターム(参考)】
[ Back to top ]