
医薬の製造のためのプロテアーゼまたはプロテアーゼ阻害剤の使用
本発明は、SDF−1活性および/または濃度が発症および/または経過に関与する疾患を治療するための医薬の製造における、カテプシンK阻害剤(CTKI)またはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子の使用に関する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、発症および/または経過へのSDF−1の関与によって特徴付けられる疾患を治療するための医薬の製造におけるカテプシンK(CTK)またはカテプシンK阻害剤(CTKI)の使用に関する。
【背景技術】
【0002】
血液中を循環する形態学的に認識可能でありかつ機能的に有能である細胞としては、赤血球、好中球性、好酸性、および好塩基性顆粒、B−、T−、非B−、非T−リンパ球、および血小板が挙げられる。これらの成熟造血細胞は、赤血球系についての赤芽球、顆粒球系についての骨髄芽球、前骨髄球および骨髄球、ならびに血小板についての巨核球のような、それぞれの系統についての形態学的に認識可能な分裂する前駆細胞によって、必要に応じて、置換されるかまたはこれらに由来する。前記前駆細胞は、2つの主要なサブグループへ簡略化して分類され得るより始原的な細胞から生じる:幹細胞および前駆細胞[レビューについては、(1)を参照のこと]。
【0003】
CXCL12またはストロマ細胞由来因子1(SDF−1)は、リンパ球、単球、およびCD34−陽性造血前駆細胞を休止させるための強力な化学誘引物質である(2)。SDF−1は、既知のリガンドとしてこのケモカインのみを有する、1つの受容体、CXCR4へのみ結合する。
【0004】
SDF−1は、低濃度で、白血球インテグリンの活性化、キャピラリー内皮細胞上への白血球の捕捉(arrest)、およびこれらの細胞の溢出へと導く(3)。SDF−1は、B細胞リンパ球産生および骨髄細胞骨髄造血を制御し(4)、そして骨髄移植のために重要である(13)。それは、CD4+T細胞生存を促進し、そしてサイトカインおよびT細胞受容体媒介刺激のためにこれらの細胞をプライムする(5)。SDF−1/CXCR4複合体の機能的活性は、白血球エラスターゼによって媒介されるタンパク質分解によりインビトロで阻害され得ることが示され、そしてこれは、造血細胞前駆体のホーミングが調節される機構であり得る(6)。実際に、インビボでの顆粒球コロニー刺激因子(G−CSF)によって誘発される末梢血への骨髄および他の器官からのCD34+造血細胞前駆体の放出(動員)は、好中球エステラーゼによるSDF−1のタンパク質分解による不活性化に依存する(41)。
【0005】
LevesqueおよびPetitによって実証されたように、カテプシンGおよびエラスターゼはG−CSF誘発動員(G-CSF induced mobilization)においてSDF−1を切断することが公知である。
【0006】
2つの増殖因子(サイトカイン)、顆粒球コロニー刺激因子(G−CSF)および顆粒球マクロファージコロニー刺激因子(GM−CSF)は、(たとえば、骨髄移植および集中的癌化学療法後)重篤な好中球減少症を有する患者における熱および感染を予防するために広く使用される。しかし、サイトカイン療法は高価であり;発熱性好中球減少症の危険性が>=30%である場合に、G−CSFの費用は正当化される。一般的に、G−CSFが化学療法の完了の約24時間後に投与される場合に、最大の臨床的利益が生じる。5μg/kg/日(皮下)の投与量がしばしば有効である。G−CSFおよびGM−CSFは、自家骨髄移植または集中的化学療法を受ける患者において、>500/μlへの好中球カウントの戻りを促進する。
【0007】
先天性、周期性、および特発性好中球減少症における好中球産生は、G−CSF 3〜10μg/kg/日(皮下)の投与によって改善され得る。この治療は、口内炎および他のタイプの口腔咽頭の炎症、熱、および蜂巣炎ならびに他の文書化された細菌感染を有さない患者において示される。利益は滞留され、そして患者は、その効果を失うことなく、数ヶ月または数年間、毎日または隔日G−CSFに維持され得る。長期間のG−CSFはまた、脊髄形成異常症、HIVおよびAIDSならびに自己免疫疾患を含む、他の状況において、好中球産生および好中球動員を誘発して好中球減少症を予防するために使用されている。一般的に、特に重篤な好中球減少症を有さない患者について、この療法の臨床的利益はあまり明確でないが、好中球カウントは増加する。特異体質の薬物反応によって引き起こされる好中球減少症を有する患者もまた、特に遅延した回復が予想される場合、G−CSFから利益を得るかもしれない。しかし、これまでのところ、制御されない試行のみが、後者の状況において報告されている。
【0008】
SDF−1およびその受容体CXCR4の相互作用は、マウス胚発生のあいだのBMの幹細胞播種において重要な役割を果たす(10、11)。以前、本発明者らは、レシピエントとして免疫不全NOD/SCIDマウスを使用して、マウス骨髄の高レベル多系統再生(multilineage repopulation)および短期間インビボ移動(short term in vivo migration)(ホーミング)の両方がSDF−1/CXCR4相互作用に依存することを示すことが出来た(12〜15)。これらのデータのサポートにおいて、ヒトCD34+細胞における高レベルのCXCR4発現またはインビトロでの高SDF−1誘発指向性運動性(directional motility)のいずれかが、CD34+細胞の正の選択を伴う、同種異系および自家臨床移植の両方においてより早い回復と関連することが示されている(16、17)。
【0009】
他の初期に作用するサイトカイン(early acting cytokines)との相乗効果で低濃度のSDF−1は、ヒトCD34+細胞とマウス幹細胞および前駆細胞との両方の増殖を増強し、これは前駆細胞生存におけるこのケモカインの役割を示唆し(25〜29)、一方、高濃度のSDF−1は、移植されたNOD/SCIDマウスの連続再生(serial repopulation)を可能にする原始ヒト胎児肝臓CD34+細胞およびヒト長期培養始原細胞(LTCIC)の増殖との停止を誘発する(30、31)ことが十分に実証されている。
【0010】
CXCR4発現は、サイトカイン、ケモカイン、間質細胞、接着分子、およびタンパク質分解酵素のような環境因子によって調節される、動的プロセスである(18)。ヒト起源の造血幹細胞および前駆細胞において、CXCR4は、短期間(約40時間)インビトロサイトカイン培養(13、19)またはタンパク質分解酵素(たとえば、MMP−2およびMMP−920)での臍帯血(CB)CD34+の刺激よって、細胞内プールからアップレギュレートされ得る。これは、引き続いて、SDF−1勾配へのそれらのインビトロ移動ならびに移植されたNOD/SCIDおよび連続的に移植されたb2mnull NOD/SCIDにおけるそれらのインビボホーミングおよび再生能を増強し、幹細胞自己再生および発生を運動性とリンクする。最近の報告は、サイトカインカクテルとのより長い培養期間が、ヒトCB CD34+に富む細胞における細胞表面CXCCR4発現の減少(22)を生じさせることを実証し、そして減少された再生が、より長い期間インビトロで培養されたヒト前駆体で実証された。最近、本発明者らは、CB CD34+/CXCR4−ソート細胞は低レベルの細胞内CXCR4を有することを示し、これは、短期間のインビトロサイトカイン刺激に続いて、迅速に細胞表面上に機能的に発現され、移植されたNOD/SCIDマウスのSDF−1依存性ホーミングおよび再生を媒介し得る(15)。
【0011】
SDF−1の不活性化と関連するタンパク質分解切断は、MMP−2およびMMP−9、カテプシンGおよびエラスターゼを含むいくつかの分解酵素によって実証される(Petitら Natl Immunol. 3:687 2002, Valenzuela-Femandezら JBC 277:15677, 2002, Levesqueら JCI 111 :187, 2003 および McQuibbanら JBC 276: 43503, 2001)。
【0012】
ヒトおよびマウス幹細胞24の指向性移動を媒介することにおけるそれらの中心的役割に加えて、SDF−1/CXCR4相互作用はまた、他の幹細胞機能に関与する。重要なことに、SDF−1/CXCR4相互作用はまた、BMにおける幹細胞および前駆細胞の滞留に関与する(10、32、33)。この仮説はまた、マウスBM腔へ直接注入されたヒト造血幹細胞(HSC)の固定におけるSDF−1/CXCR4相互作用の関与を実証する他の研究によって確認されている(34、35)。これらの相互作用の干渉は、BMから循環へのヒトおよびマウスの両方の前駆体の放出/動員を誘発する(36〜41)。
【0013】
骨は、骨を生成する骨芽細胞および骨を再吸収する破骨細胞の活性により平衡化される恒常的な再構築を受けている。骨粗しょう症、パジェット病、特定の形態の関節炎、および骨転移のような、種々の骨疾患は、過剰な破骨細胞媒介骨再吸収によって特徴付けられる。RANKL(NKκBリガンドの受容体アクチベータ)およびその対応のRANK受容体は、破骨細胞分化および活性化において重要な役割を果たし、そしてオステオプロテゲリン(OPG)は、RANKLの生理学的阻害剤である。
【0014】
マトリクス分解は、主に、システインプロテアーゼ、CTKの活性に起因する(7、8)。CTKは、破骨細胞において高度に発現されそして骨再吸収(破骨細胞が骨を分解するプロセス)に関連する、リソソームシステインプロテアーゼである。I型コラーゲンは、有機骨量(organic bone mass)の90〜95%を構成し(Krane, S. M., and Simon, L.(1994) in Scientific American Medicine (Rubenstein, E. および Federman, D. D., eds), Vol. 3 , pp. 1-26, Scientific American, Inc., New York)、そしてCTKについての主要な生物学的基質を示す(9)。全ての哺乳動物コラゲナーゼの中で、CTKは、それらの三重らせん構造内の複数の部位で間質コラーゲンを切断し得る唯一のプロテアーゼである(Garnero, P., Borel, O., Byrjalsen, I., Ferreras, M., Drake, F. H., McQueney, M. S., Foged, N. T., Delmas, P. D., および Delaisse, J. M. (1998) J. Biol. Chem. 273, 32347-32352, 6Kafienah, W., Bromme, D., Buttle, D. J., Croucher, L. J., および Hollander, A. P. (1998) Biochem. J. 331, 727-732)。CTK活性の欠乏は、常染色体劣性骨格形成異常濃化異骨症(autosomal recessive skelら dysplasia pycnodysostosis)を有する患者において観察されるように(GeIb, B. D., Shi, G. P., Chapman, H. A., and Desnick, R. J. (1996) Science 273, 1236-1238)、破骨細胞内のリソソーム中の未消化の膠原原線維の蓄積へ導く(Everts, V., Aronson, D. C, および Beertsen, W. (1985) Calcif. Tissue Int. 37, 25-31)。CTK突然変異体の分析によって、コラーゲン分解異常は、CTKのタンパク質分解活性の損失と必ずしも連結されないことが判明した。前記プロテアーゼの活性部位から離れている、1つの疾患を引き起こす変異、Y212Cは、CTKの全体的なタンパク質分解活性に少しだけ影響を与えたが、そのコラゲナーゼ活性を完全に排除した(Hou, W.-S., Bromme, D., Zhao, Y., Mehler, E., Dushey, C, Weinstein, H., Miranda, C. S., Fraga, C, Greig, F., Carey, J., Rimoin, D. L., Desnick, R. J., および GeIb, B. D. (1999) J. Clin. Invest. 103, 731-738)。この観察は、CTKの触媒活性に加えて、他の特徴が、CTKによるコラーゲンの加水分解のために必要とされることを示した。骨−および軟骨耐性グリコサミノグルカンは、特に、カテプシンLまたはマトリックスメタロプロテアーゼIでは観察されない効果である、CTKによるI型およびII型の間質コラーゲンの分解を増強する(Li, Z., Hou, W. S., and Bromine, D. (2000) Biochemistry 39, 529-536)。この知見は、CTKのコラゲナーゼ活性が、CTKタンパク質およびグリコサミノグルカンの特異的な相互作用を必要とすることを示唆した。しかし、CTKがコラーゲンを分解する機構は、理解しにくいままである。Li Zら(J Biol Chem. 2002 Aug 9;277(32):28669-76. Epub 2002 May 30)は、CTKの膠原溶解性活性はCTKタンパク質とコンドロイチン硫酸との新規のオリゴマー複合体の形成に依存することを実証した。したがって、現在、CTK阻害剤は、骨再吸収を阻害するために使用されている。
【発明の開示】
【0015】
本発明は、SDF−1活性および/または濃度が発症および/または経過に関与する疾患を治療するための医薬の製造における、カテプシンK阻害剤(CTKI)またはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体(circularly permutated derivative)、塩または誘導因子の使用に関する。
【0016】
本発明の1局面において、CTKは、癌、炎症および感染症のような、SDF−1活性によって引き起こされる/悪化される疾患において使用される。
【0017】
本発明の一実施形態において、前記疾患はアレルギー性気道疾患である。
【0018】
本発明のさらなる実施形態において、前記疾患は慢性関節リウマチである。
【0019】
本発明のさらなる実施形態において、前記疾患は動脈硬化症である。
【0020】
本発明のさらなる実施形態において、前記疾患は、癌、たとえば、前立腺癌、腎臓癌、神経芽腫、神経膠腫、膵臓癌、結腸癌、乳癌、白血病、たとえば、急性リンパ芽球性白血病(ALL)および急性骨髄性白血病(AML)である。
【0021】
本発明のさらなる実施形態において、CTKは、好ましくはCXCR4を発現する癌細胞においいて、癌転移を予防するために使用される。
【0022】
本発明の別の実施形態において、CTKIは、HIVのような、SDF−1活性によって予防/緩和される疾患において使用される。
【0023】
さらに、本発明は、必要とする対象において幹細胞の動員を誘発するための医薬の製造における、CTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子の使用を提供する。
【0024】
一実施形態において、必要とする前記対象は、たとえば骨髄移植または癌化学療法に続いて生じる、重篤な好中球減少症に苦しむ。
【0025】
また、本発明は、たとえば必要とする対象において骨髄、肝臓および腎臓のような器官の再生を増強するための、必要とする対象において骨髄中の幹細胞の滞留を増大させるための医薬の製造におけるCTKIの使用を提供する。
【0026】
本発明はまた、有効量のCTKIまたはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を投与することを含む、SDF−1活性および/または濃度によってその発症および経過が影響される疾患を治療する方法を提供する。
【0027】
別の局面において、本発明は、有効量のCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を哺乳動物へ投与することを含む、哺乳動物における癌を治療する方法を提供する。
【0028】
本発明の好ましい実施形態において、前記癌細胞は、CXCケモカイン受容体4(CXCR4)を発現する。
【0029】
別の好ましい実施形態において、本発明の方法は、転移を予防するために使用される。
【0030】
さらに、本発明は、必要とする対象へ有効量のCTKIまたはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を投与することを含む、組織への多能性幹細胞の標的化を調節する方法を提供する。
【0031】
本発明の一実施形態において、CTKIが、標的組織への前記細胞(たとえば、正常な造血細胞)の標的化を増大させるために、該標的組織へ投与される。
【0032】
本発明の好ましい実施形態において、前記細胞がインビボで患者にあり、かつ治療的に有効量のCXCR4アゴニスト、たとえば、SDF−1、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子が、癌患者のような、必要とする該患者へ投与される。
【0033】
別の好ましい実施形態において、前記患者は自家または同種異系骨髄あるいは末梢血幹細胞移植を必要とし、および/または前記患者は細胞毒性薬で治療される。
【0034】
別の実施形態において、本発明は、CTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子が、標的組織への前記細胞(たとえば、新生細胞)の標的化を減少させるために、該標的組織へ投与される方法を提供する。
【0035】
別の局面において、本発明は、造血細胞へ有効量のCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を投与することを含む、造血細胞増殖の速度を減少させる方法を提供する。
【0036】
本発明の一実施形態において、前記造血細胞は、造血幹細胞および造血前駆細胞からなる群から選択される。
【0037】
本発明のさらなる実施形態において、前記細胞はインビボで患者にあり、かつ治療的に有効量のCXCR4アゴニスト、たとえば、SDF−1、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子が必要とする該患者へ投与される。
【0038】
本発明のさらなる実施形態において、前記患者は、自家または同種異系骨髄あるいは末梢血幹細胞移植を必要とする。
【0039】
本発明のさらなる実施形態において、前記患者は、癌を有し、そして細胞毒性薬で治療される。
【0040】
別の局面において、本発明は、CTKをSDF−1と接触させること、SDF−1の活性を測定すること、およびCTKによるSDF−1活性の阻害を防止または逆転し得る化合物を単離することを含む、CTKアンタゴニストを同定する方法を提供する。
【0041】
本発明はまた、CTKをSDF−1と接触させること、SDF−1の完全性を測定すること、およびCTKによるSDF−1活性の分解を防止し得る化合物を単離することを含む、CTKアンタゴニストを同定する方法を提供する。
【0042】
本発明は、本発明の方法によって得られるアンタゴニストに関する。
【0043】
本発明は、例示のためのみに、添付の図面を参照して本明細書に記載される。ここで詳細な図面を特に参照しているが、示される詳細は、例示のためにそして本発明の好ましい実施形態の例示的議論のためにのみ示され、そして本発明の原理および概念的局面の最も有用でありかつ容易に理解される説明であると思われるものを提供するために提示されることが強調される。この点において、本発明の基本的理解のために必要とするよりもより詳細に本発明の構造的詳細を示す試みは成されず、図面を考慮しての説明により、本発明のいくつかの形態が実際にどのように具体化され得るかが当業者に明らかとなる。
【発明を実施するための最良の形態】
【0044】
本発明は、ケモカインストロマ細胞由来因子(SDF−1)が発症および/または経過に関与する疾患を治療するための医薬の製造における、CTK阻害剤(CTKI)またはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子の使用に関する。
【0045】
本発明は、CTKがSDF−1活性を特異的に阻害し得るという知見に基づく。本発明者らは、ストレスシグナルが以下の事象を誘発することを本明細書中において示す:BM SDF−1の一時的増加、破骨細胞活性化、CTK活性化の増加、CTK分解によって媒介されるBM SDF−1濃度の減少、および細胞動員。
【0046】
本発明の原理および作用は、図面および付随の説明を参照してよりよく理解され得る。
【0047】
本発明の少なくとも1つの実施形態を詳細に説明する前に、以下の説明に記載されるかまたは実施例によって例示される詳細へ本発明がその適用で限定されないことが理解される。本発明は、他の実施形態あるいは種々の方法で実施または実行され得る。また、本明細書中で使用される表現および用語は、説明のためであり、そして限定としてみなされるべきでないことが理解される。
【0048】
下記に示される知見は、炎症および損傷のようなストレスシグナルが、破骨細胞活性化および骨髄から末梢血(PB)への前駆体動員を誘発することを実証する。本発明者らは、ストレスシグナルが以下の事象を誘発することを示す:BM SDF−1の一時的増加、破骨細胞活性化、CTK活性化の増加、CTK分解によって媒介されるBM SDF−1濃度の減少、および細胞動員。
【0049】
また、得られた結果は、外から投与されたSDF−1、あるいはG−CSFおよび/またはHGFの機能により産生されたSDF−1、あるいはストレスシグナル(たとえば、炎症または損傷)が、破骨細胞活性化、BM CTKの増加、SDF−1の減少、および前駆体動員を直接誘発することを示す。
【0050】
本発明者らは、インビボでのRANKL(破骨細胞活性化の誘導因子)投与が機能性CXCR4(SDF−1)を必要とする動員を誘発することを実証した。
【0051】
本願において得られた結果は驚くべきことであり、何故ならば、Takamatsuら(Blood. 1998 Nov l;92(9):3465-73. 1998)の結果とは対照的に、結果は、破骨細胞活性化が骨髄から循環への幹/前駆体細胞動員を媒介すること示すためである。CTK、すなわち主要な破骨細胞プロテイナーゼは、そのアミノ末端の特異的かつ迅速な切断によりSDF−1活性化を阻害することが実証された。
【0052】
したがって、本知見は、SDF−1活性を調節するために、カテプシンK阻害剤(CTKI)またはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を使用するという可能性を提供する。
【0053】
多くの疾患の発症または経過は、SDF−1濃度および/または活性のアップまたはダウンレギュレーションに伴って生じる。したがって、CTKIまたはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子は、発症および/または経過へのSDF−1の関与によって特徴付けられる疾患を治療するための医薬の製造において使用され得る。
【0054】
SDF−1およびその受容体CXCR4は、ヒト免疫不全ウイルス1型病態生理学、白血球輸送、炎症、造血、胚形成、新脈管形成、および癌転移において重要な役割を果たす。本発明者らの知見に基づいて、本発明者らは、CTK媒介タンパク質分解が、SDF−l/CXCR4依存細胞シグナル伝達を不活性化しかつ停止させることを示唆する。
【0055】
本発明の一局面において、カテプシンK阻害剤(CTKI)またはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子が、腫瘍、感染症および炎症性疾患のような、SDF−1活性によって引き起こされる/悪化される疾患において使用される。
【0056】
増加する証拠は、アレルギー性気道疾患(Gonzalo ら, [J immunol 165:499-508, 2000], Lukacs ら, [Am. J Pathol. 160:1353-1360, 2002])、慢性関節リウマチ Buckley ら, [J immunol 165:3423-3429, 2000]、および動脈硬化症(Abi-Younes ら, [Circ Res 86:131-138, 2000]のようないくつかの疾患における、腫瘍、感染性および炎症性プロセスの病因に、SDF−1/CXCR4シグナル伝達系を関係させ、ここでSDF−1はアップレギュレートされる。
【0057】
CXCRまたはSDF−1の異常発現は、前立腺癌(Taichman ら. Cancer Res 2002, 62:1832-7)、腎臓癌(Br J Cancer 2002, 86:1250-6)、神経芽腫(J Immunol. 2001, 167:4747-57)、神経膠腫(Zhou ら. J Biol. Chem. 2002, 277:49481, Salmaggi ら J Neurooncol. 2004 May;67(3):305-17.)、膵臓癌(Clin. Cancer Res 2000,6:3530)、結腸癌(Zeelenberg ら. Cancer Res 2003, 63:3833-9)および乳癌(Muller ら. Nature 2001, 410:50-6)のような固形癌において観察されている。
【0058】
大抵の白血病細胞は、接着、生存および増殖の増加を伴ってSDF−1に応答する(Juarez ら Histol Histopathol. 2004, 19 (1): 299-309)。
【0059】
SDF−1はまた、プレBALL細胞(pre-BALL cells)の増殖および生存に関与する(Nishii ら. Br J Haematol. 1999 Jun;105(3):701-10)。Tavorら(Cancer Research 64, 2817-2824, 2004)は、急性骨髄性白血病(AML)細胞がSDF−1を発現すること、および内因性SDF−1を遮断することによりAML細胞生存が減少することを示した。
【0060】
悪性神経膠腫におけるSDF−1は、新脈管形成および内皮細胞と腫瘍細胞とのあいだのクロストークにおける役割を有しているようである(Salmaggi ら J Neurooncol. 2004 May;67(3):305-17)。したがって、CTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を用いてSDF−1活性を阻害することは、臨床的に有益である。
【0061】
SDF−1およびCXCR4は、腫瘍細胞の転移先を決定する。CXCR4はヒト乳癌細胞によって発現され、そしてSDF−1は、マウスモデルにおける転移の最初の目的地である器官において高く、このケモカインシグナル伝達が浸潤反応を誘発することを示している(Muller ら., [Nature 410:50-56, 2001])。したがって、CTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を用いてSDF−1活性を遮断することは、CXCケモカイン受容体4を発現する腫瘍細胞における転移阻害のために有益である。
【0062】
得られた結果は、必要に応じて、破骨細胞活性化および/またはCTK活性を誘発することにより、動員を誘発することができることを実証する。あるいは、動員は、破骨細胞活性化および/またはCTK活性を阻害することによって防止され得る。
【0063】
本発明の一実施形態において、CTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子が、重篤な好中球減少症において前駆体の動員を誘発するために使用される。たとえば、骨髄移植後および/または集中的な癌化学療法後に生じる重篤な好中球減少症において。
【0064】
本発明の別の実施形態において、破骨細胞活性化が、重篤な好中球減少症において前駆体の動員を誘発するために使用される。
【0065】
本発明の別の局面において、CTKIは、SDF−1活性によって予防/緩和される疾患において使用される。
【0066】
SDF−1は、リンパ器官、肝臓、肺、および内皮細胞を囲む間葉細胞を含む種々の組織において産生されることが公知である(Tachibana ら. 1998. Nature 393:591.)。
【0067】
CTKは、比較的に組織特異的であり、そして破骨細胞に加えて、マクロファージ、大動脈平滑筋細胞、甲状腺上皮細胞および肺上皮細胞に存在する。これらの細胞で占められる器官において、CTKIは、SDF−1アゴニストとして機能すると予想される。たとえば、骨髄におけるCTKの活性の阻害は、この酵素によるSDF−1分解および引き続いての動員を防止する。
【0068】
したがって、別の実施形態において、CTKIは、再生(repopulation)を増強するためおよび欠点のある造血(defective hematopoiesis)を回復させるために、前駆体の動員を阻害するためおよびしたがって骨髄における前駆体の滞留を増大させるために使用される。
【0069】
本発明のこの局面にしたがう幹細胞/前駆体は、好ましくは、治療される対象から得られる。しかし、幹細胞/前駆体はまた、同系(syngeneic)、同種異系(allogeneic)から、そしてより好ましくはないが、異種(xenogeneic)ドナーから得られ得る。
【0070】
同種異系または異種幹細胞が使用される場合、レシピエント対象および/または細胞は、好ましくは、移植片対宿主および宿主対移植片拒絶反応を防止するように処理されることが理解される。免疫抑制プロトコルは、当該分野において周知であり、そして米国特許第6,447,765号に開示されている。
【0071】
CXCR4は、T細胞指向性(tropic)ヒト免疫不全ウイルス1についてのコレセプター(coreceptor)として役立ち得る(Oberlin ら, [Nature 382:833-835, 1996])。SDF−1は、この受容体のダウンレギュレーションの支配および促進を両方することによって、CXCR4へ結合することについてヒト免疫不全ウイルス(HIV)と拮抗することが示されており;したがって、それは、このウイルスに対する宿主防衛において役割を果たし得る(Amara ら, [J Exp Med 186:1390-146, 1997])。
【0072】
したがって、本発明の一実施形態において、CTKIは、HIVのようなSDF−1活性によって予防/緩和される疾患において使用される。
【0073】
一局面において、本発明は、有効量のCTKIまたはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、循環的に置換された誘導体、塩または誘導因子を含む、SDF−1濃度および/または活性のアップまたはダウンレギュレーションによって影響される疾患を治療する方法を提供する。
【0074】
一実施形態において、本発明は、哺乳動物における腫瘍細胞の転移を阻害する方法を提供し、ここで該腫瘍細胞はCXCケモカイン受容体4(CXCR4)を発現し、該方法は、該腫瘍の転移を阻害するに十分な量のCTKを該哺乳動物へ投与することを含む。
【0075】
一局面において、本発明は、対象内の組織への多能性幹細胞(たとえば、正常な前駆体)の標的化を調節するために医薬の製造のための、CTKIまたはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、循環的に置換された誘導体、塩または誘導因子の使用を提供する。
【0076】
1実施形態において、本発明は、標的組織におけるCTK活性を阻害することを含む、骨髄のような標的細胞への正常な前駆細胞の標的化を増大させることに関する。
【0077】
別の実施形態において、本発明は、標的組織におけるCTK活性を増大させることを含む、標的組織(たとえば、骨髄)への新生細胞の標的化の減少に関する。
【0078】
CTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、循環的に置換された誘導体、塩または誘導因子は、SDF−1阻害剤として機能し、したがって、それは、白血球輸送、炎症、造血、造血細胞の増殖、胚形成、新脈管形成、および癌転移においてSDF−1活性を阻害する。
【0079】
CTK阻害剤はSDF−1アゴニストとして機能し、したがって、それは、BM SDF−1を増加させる(たとえば、照射)またはSDF−1が骨髄において増加される条件を有する治療または薬剤が投与された患者において、骨髄における造血細胞の増殖を増強する。
【0080】
したがって、一実施形態において、本発明は、造血細胞へ有効量のCTK阻害剤を投与することを含む、正常な造血細胞増殖の速度を増加させる方法に関する。
【0081】
本発明の好ましい実施形態において、前記造血細胞は、造血幹細胞および造血前駆細胞からなる群から選択される。
【0082】
別の好ましい実施形態において、前記細胞は患者中に存在し、そしてSDF−1は、このような治療の必要がある患者において投与されるかまたは誘導される。
【0083】
さらに好ましい実施形態において、このような必要がある前記患者は癌を有する。
【0084】
さらなる好ましい実施形態において、前記患者は、自家または同種異系骨髄または末梢血幹細胞移植の必要性を有する。
【0085】
別の好ましい実施形態において、前記患者は細胞毒性薬で治療され、ここで、CTK阻害剤の前記有効量は、該細胞毒性薬に対する正常細胞の感受性を減少させるに十分である。
【0086】
本発明の一局面において、CTK阻害剤はSDF−1アゴニストとして機能し、したがって、それは、BM SDF−1を増加させる(たとえば、照射)またはSDF−1が骨髄において増加される条件を有する治療または薬剤が投与された患者において、骨髄への造血細胞の移植を増強する。
【0087】
本発明の一局面において、CTKはSDF−1アンタゴニストとして機能し、したがって、それは、骨髄から末梢血への前駆体動員へ導くSDF−1分解を媒介する。
【0088】
本発明の一局面によれば、CTKをSDF−1と接触させること、SDF−1の活性または完全性を測定すること、およびSDF−1活性の阻害または分解を防止または逆転し得る化合物を単離することを含む、CTKアンタゴニストを同定する方法が提供される。
【0089】
一実施形態において、測定されるSDF−1の活性は移動である。本発明は、本方法によって得られ得るCTKアンタゴニストをも意図する。
【0090】
別の局面において、本発明は、CXCR4が該癌細胞において発現されるおよび/またはSDF−1が該癌細胞によって分泌される癌患者の治療方法に関し、該方法は、該患者への治療的に有効な量のCTKの投与を含む。
【0091】
本発明の一実施形態において、癌のタイプは、SDF−1を発現する細胞を含む器官へ典型的に転移するものである。
【0092】
さらなる実施形態において、前記癌は、典型的に、皮膚、肝臓、脳、および肺に転移する。
【0093】
さらなる実施形態において、前記腫瘍細胞は、リンパ腫細胞、神経芽腫細胞、肺癌細胞、血管肉腫細胞、白血病細胞、神経膠腫細胞、または黒色腫細胞、乳癌細胞または前立腺癌細胞から選択される。
【0094】
本明細書中で使用される場合、句「幹細胞」は、特定の特別な機能を有する他の細胞タイプ(即ち、「完全に分化した」細胞)へ分化し得る細胞をいう。
【0095】
本明細書中で使用される場合、「トランスジェニック細胞」は、導入された遺伝子または断片を有する細胞である。
【0096】
用語「動員」は、たとえばサイトカインG−CSFによって誘発される、定常状態ホメオタシスの不均衡に起因して、細胞が骨髄から循環へ放出されるプロセスをいう。
【0097】
用語「前駆細胞」は、コロニー形成し得る、未熟な未分化の造血細胞の不均一な集団をいう。前駆体は幹細胞について富化されるが、それらはまた、真の幹細胞を特徴付ける長期間の骨髄再生をすることが出来ないより成熟した細胞も含む。用語「TRAP+MNC細胞」は、酵素TRAP(酒石酸抵抗性酸性フォスファターゼ)を発現する多核細胞である、活性化された破骨細胞をいう。
【0098】
用語「ストレスシグナル」は、サイトカイニン、ケモカイン、タンパク質分解酵素および接着分子のアップまたはダウンレギュレーションによって達成される、定常状態条件の重篤な不均衡によって誘発される危険状況をいう。例としては、炎症、照射、毒性薬物、サイトカイン刺激または欠乏が挙げられる。
【0099】
用語「CTKの阻害剤」は、本発明の文脈内において、CTK産生および/または機能が弱められ、減少され、あるいは部分的に、実質的にまたは完全に防止または遮断される様式で、CTK産生、発現および/または機能をダウンレギュレートするいずれの分子をもいう。用語「CTK阻害剤」は、CTK産生の阻害剤およびCTK機能の阻害剤を含むように意図される。
【0100】
産生の阻害剤は、合成、またはCTKのプロセッシングに悪影響を与えるいかなる分子でもあり得る。本発明にしたがって考慮される阻害剤は、たとえば、CTKの遺伝子発現の抑制因子、CTK mRNAの転写を減少もしくは防止するかまたは該mRNAの分解へ導くアンチセンスmRNA、正しい折り畳みを妨げるかまたは部分的にもしくは実質的にCTKの分泌を防止するタンパク質、あるいはCTKを分解するプロテアーゼなどであり得る。
【0101】
CTK機能の阻害剤は、たとえば、CTKアゴニストであり得る。アゴニストは、その基質へのCTK結合を担うCTKまたはCTK結合部位を部分的にまたは実質的に中和するに十分な親和性および特異性で、CTK分子自体へ結合し得るかまたはこれを隔離し得る。
【0102】
本明細書中で使用される場合、句「発現または活性を阻害する」は、CTKの発現(転写および/または翻訳)あるいは活性(たとえば、酵素的またはリガンド結合)を部分的にまたは完全に阻害することをいう。いくつかの異なるアプローチが、CTKの活性をダウンレギュレートするために使用され得る。
【0103】
たとえば、CTK活性を阻害することは、CTKに特異的に結合し得る抗体または抗体フラグメントのような薬剤によって達成され得る。好ましくは、該抗体は、CTKの少なくとも1つのエピトープへ特異的に結合する。本明細書中で使用される場合、用語「エピトープ」は、抗体のパラトープが結合する抗原上のいかなる抗原決定基をもいう。
【0104】
エピトープ決定基は、通常、アミノ酸あるいは炭水化物側鎖のような分子の化学的に活性な表面グルーピング(groupings)からなり、そして通常、特定の3次元構造的特徴および特定の電荷特性を有する。CTKの好ましいエピトープは、SDF−1とのCTKの会合の触媒部位または領域を含むものである。
【0105】
用語「抗体」は、本明細書中で使用される場合、インタクトな分子、ならびにFab、F(ab’)2、およびFvのような機能性フラグメントを含む。これらの機能性抗体フラグメントは、以下のように規定される:(1)Fab、抗体分子の一価の抗原結合フラグメントを含み、酵素パパインで完全抗体(whole antibody)を消化してインタクトな軽鎖および1つの重鎖の部分を得ることによって作製され得る、フラグメント;(2)Fab’、ペプシンで完全抗体を処理し、続いて還元して、インタクトな軽鎖および重鎖の部分を得ることによって得られ得る抗体分子のフラグメント;2つのFab’フラグメントが、1抗体分子当たり得られる;(3)F(ab’)2、引き続いての還元無しに酵素ペプシンで完全抗体を処理することによって得られ得る抗体のフラグメント;F(ab’)2は、2つのジスルフィド結合によって一緒に滞留された2つのFab’フラグメントの2量体である;(4)Fv、2つの鎖として発現される軽鎖の可変領域および重鎖の可変領域を含む遺伝子操作されたフラグメントとして規定される;ならびに(5)一本鎖抗体(「SCA」)、遺伝子的に融合された一本鎖分子のような、好適なポリペプチドリンカーによって結合された、軽鎖の可変領域および重鎖の可変領域を含む遺伝子操作された分子。
【0106】
ポリクローナルおよびモノクローナル抗体ならびにそれらのフラグメントを作製する方法は、当分野において周知である(たとえば、Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, New York, 1988を参照のこと。これは、参照により本明細書に組み込まれる)。CTKへ結合する市販のポリクローナルおよびモノクローナル抗体は、本発明における使用に好適である。
【0107】
本発明にしたがう抗体フラグメントは、抗体のタンパク質分解性加水分解あるいは該フラグメントをコードするDNAの大腸菌または哺乳動物細胞(たとえば、チャイニーズハムスター卵巣細胞培養または他のタンパク質発現系)における発現によって作製され得る。抗体フラグメントは、従来法による完全抗体のペプシンまたはパパイン消化によって得られ得る。たとえば、抗体フラグメントは、ペプシンでの抗体の酵素的切断によって作製され得、F(ab’)2という5Sフラグメントを提供する。このフラグメントは、さらに、チオール還元剤および必要に応じてジスルフィド結合の切断から生じるスルフヒドリル基についてのブロック基を使用して切断され得、3.5S Fab’一価フラグメントが作製される。あるいは、ペプシンを使用する酵素的切断によって、直接、2つの一価Fab’フラグメントおよびFcフラグメントが作製される。これらの方法は、たとえば、Goldenberg、米国特許第4,036,945号および第4,331,647号、ならびにそれらに含まれる参照文献によって記載されており、これらはそれらの全体が参照により本明細書中に組み込まれる。また、Porter, R. R. [Biochem. J. 73: 119-126 (1959)]を参照のこと。抗体を切断する他の方法、たとえば、一価軽−重鎖フラグメントを形成するための重鎖の分離、フラグメントのさらなる切断、または他の酵素的、化学的、もしくは遺伝学的技術もまた、インタクトな抗体によって認識される抗原へフラグメントが結合する限りにおいて、使用され得る。
【0108】
Fvフラグメントは、VHおよびVL鎖の会合を含む。この会合は、Inbarら[Proc. Nat'l Acad. Sci. USA 69:2659- 62 (1972)]において記載されるように、非共有結合的であり得る。あるいは、可変鎖は、分子間ジスルフィド結合によって連結されるか、またはグルタルアルデヒドのような化学物質によって架橋され得る。好ましくは、Fvフラグメントは、ペプチドリンカーによって連結されたVHおよびVL鎖を含む。これらの一本鎖抗原結合タンパク質(scFv)は、オリゴヌクレオチドによって連結されたVHおよびVLドメインをコードするDNA配列を含む構造遺伝子を構築することによって作製される。該構造遺伝子は、発現ベクターへ挿入され、引き続いてこれは大腸菌のような宿主へ導入される。
【0109】
抗体フラグメントの形態は、単一の相補性決定領域(CDR)をコードするペプチドである。CDRペプチド(「最小認識単位」)は、問題の抗体のCDRをコードする遺伝子を構築することによって得られ得る。このような遺伝子は、たとえば、ポリメラーゼ連鎖反応を使用して抗体産生細胞のRNAから可変領域を合成することによって、作製される。たとえば、LarrickおよびFry [Methods, 2: 106-10 (1991)]を参照のこと。
【0110】
非ヒト(たとえば、マウス)抗体のヒト化形態は、非ヒト免疫グロブリンから誘導される最小配列を含む、免疫グロブリン、その免疫グロブリン鎖またはフラグメント(たとえば、Fv、Fab、Fab’、F(ab’).sub.2または抗体の他の抗原結合サブ配列)のキメラ分子である。ヒト化抗体としては、ヒト免疫グロブリン(レシピエント抗体)が挙げられ、ここで、該レシピエントの相補性決定領域(CDR)由来の残基は、所望の特異性、親和性および能力(capacity)を有するマウス、ラットまたはウサギのような非ヒト種のCDR由来の残基(ドナー抗体)によって置換されている。場合によっては、ヒト免疫グロブリンのFvフレームワーク残基は、対応の非ヒト残基によって置換される。ヒト化抗体はまた、レシピエント抗体においてもまたは取り込まれた(imported)CDRもしくはフレームワーク配列においても見られない残基を含み得る。一般的に、ヒト化抗体は、少なくとも1つ、そして典型的には2つの、可変ドメインの実質的に全てを含み、ここで、全てまたは実質的に全てのCDR領域は、非ヒト免疫グロブリンのそれらに対応し、そして全てまたは実質的に全てのFR領域は、ヒト免疫グロブリン共通配列のそれらである。ヒト化抗体はまた、最適には、免疫グロブリン定常領域(Fc)(典型的には、ヒト免疫グロブリンのそれ)の少なくとも部分を含む[Jones ら, Nature, 321 :522-525 (1986); Riechmann ら, Nature, 332:323-329 (1988);およびPresta, Curr. Op. Struct. Biol., 2:593-596 (1992)]。
【0111】
非ヒト抗体をヒト化するための方法は、当該分野において周知である。一般的に、ヒト化抗体は、非ヒトである供給源からそれへ導入された1またはそれ以上のアミノ酸残基を有する。それらの非ヒトアミノ酸残基は、しばしば、インポート残基(import residues)と呼ばれ、これらは典型的にインポート可変ドメインから取られる。ヒト化は、齧歯類CDRまたはCDR配列をヒト抗体の対応の配列の代わりに使用することによって、Winterおよび共同研究者[Jones ら, Nature, 321 :522-525 (1986);Riechmann ら, Nature 332:323-327 (1988);Verhoeyen ら, Science, 239:1534-1536 (1988)]の方法にしたがって本質的に行われ得る。したがって、このようなヒト化抗体は、キメラ抗体であり(米国特許第4,816,567号)、ここで、実質的に1つ未満のインタクトなヒト可変ドメインは、非ヒト種由来の対応の配列によって置換されている。実際には、ヒト化抗体は、典型的に、いくつかのCDR残基および恐らくいくつかのFR残基が齧歯類抗体における類似部位由来の残基によって置換されているヒト抗体である。
【0112】
ヒト抗体はまた、ファージディスプレイライブラリ[Hoogenboom および Winter, J. MoI. Biol., 227:381 (1991);Marks ら, J. MoI. Biol., 222:581 (1991)]を含む、当該分野において公知の種々の技術を使用して作製され得る。ColeらおよびBoernerらの技術もまた、ヒトモノクローナル抗体の作製のために利用可能である(Cole ら, Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p.77 (1985)およびBoerner ら, J. Immunol., 147(l):86-95 (1991))。同様に、ヒト抗体は、内因性免疫グロブリン遺伝子が部分的にまたは完全に不活性化されているトランスジェニック動物(たとえば、マウス)へのヒト免疫グロブリン遺伝子座の導入によって作製され得る。挑戦に際して、ヒト抗体作製が観察され、これは、遺伝子再配列、集合(assembly)、および抗体レパートリーを含む、全ての点でヒトにおいて見られるものに非常に似ている。このアプローチは、たとえば、米国特許第5,545,807号;第5,545,806号;第5,569,825号;第5,625,126号;第5,633,425号;第5,661,016号、および以下の科学刊行物において記載されている:Marks ら, Bio/Technology 10: 779-783 (1992);Lonberg ら, Nature 368: 856-859 (1994);Morrison, Nature 368 812-13 (1994);Fishwild ら, Nature Biotechnology 14, 845- 51 (1996);Neuberger, Nature Biotechnology 14: 826 (1996);およびLonberg and Huszar, Intern. Rev. Immunol. 13, 65-93 (1995)。
【0113】
CTK活性の阻害はまた、CTKの公知のペプチド阻害剤を使用することによって、またはCTKの触媒部位と相互作用し得るポリペプチド配列から誘導される任意のペプチド阻害剤(たとえば、基質アナログ)によって、行われ得る。さらなる阻害剤を同定するために使用され得る好適な生化学/分子アプローチの記載は、本明細書以下に提供される。
【0114】
CTKのさらなる阻害剤は、Blanchardら(Structure 7:1125-1133, 1999)およびWattら(Structure 7:1135-1143, 1999)によって記載されるCTKの3次元分子構造についてならびにChouら(FEBS 419:49-54, 1997)によって作製されたその基質結合モデルについて、分子設計アプローチを使用して同定され得る。
【0115】
CTKを細胞内小器官/位置へ再配置し、そしてそれをその生物学的効果を示すことが出来ないようにするタンパク質によって、CTK活性はまた阻害され得る。CTK細胞の発現のダウンレギュレーションは、いくつかの分子アプローチのいずれか1つを使用して行われ得る。
【0116】
たとえば、CTK転写は、低分子干渉RNA(siRNA)分子を使用することによって、RNA干渉を介して阻害され得る。RNA干渉は、2工程プロセスであり;開始工程と呼ばれる第1工程は、恐らくDicer(dsRNA特異的リボヌクレアーゼのRNアーゼIIIファミリーのメンバー)[これは、ATP依存性様式で(直接またはトランスジーンまたはウイルスを介して導入される)dsRNAを処理する(切断する)]の作用によって、インプットされたdsRNAが21〜23ヌクレオチド(nt)の低分子干渉RNA(siRNA)へ消化される。連続する切断事象は、各々が2−ヌクレオチド3’オーバーハングを有する、19〜21bp二重鎖(siRNA)へ前記RNAを分解する[Hutvagner and Zamore Curr. Opin. Genetics and Development 12:225-232 (2002);およびBernstein Nature 409:363- 366 (2001)]。
【0117】
前記エフェクター工程において、前記siRNAデュプレックスは、ヌクレアーゼ複合体へ結合し、RNA誘導型サイレンシング複合体(RISC)を形成する。siRNAデュプレックスのATP依存性の巻戻しが、RISCの活性化のために必要とされる。活性RISCは、次いで、塩基対相互作用によって相同転写物(homologous transcript)を標的化し、そして前記mRNAをsiRNAの3’末端から12のヌクレオチドフラグメントへ切断する[Hutvagner and Zamore Curr. Opin. Genetics and Development 12:225-232 (2002);Hammond ら. (2001) Nat. Rev. Gen. 2:110-119 (2001);およびSharp Genes. Dev. 15:485-90 (2001)]。切断機構は依然として解明されていないが、研究は、各RISCが単一siRNAおよびRNアーゼを含むことを示す(Hutvagner and Zamore, Curr. Opin. Genetics and Development 12:225-232, 2002)。
【0118】
RNAiの顕著な効力のために、RNAi経路内の増幅工程が示唆されている。増幅は、より多くのsiRNAを生じるインプットdsRNAのコピーによって、あるいは形成されるsiRNAの複製によって生じ得る。あるいはまたはさらに、増幅は、RISCの複数の代謝回転事象によって行われ得る[Hammond ら. Nat. Rev. Gen. 2:110-119 (2001), Sharp Genes. Dev. 15:485-90 (2001);Hutvagner and Zamore Curr. Opin. Genetics and Development 12:225-232 (2002)]。RNAiについてのより多くの情報については、以下のレビューを参照のこと:Tuschl ChemBiochem. 2:239-245 (2001); Cullen Nat. Immunol. 3:597-599 (2002);およびBrantl Biochem. Biophys. Act. 1575:15-25 (2002)。
【0119】
本発明での使用に好適なRNAi分子の合成は、以下のように行われ得る。先ず、CTK mRNA配列を、AAジヌクレオチド配列についてAUG開始コドンの下流でスキャンする。各AAおよび3’隣接19ヌクレオチドの出現を、可能性のあるsiRNA標的部位として記録する。好ましくは、siRNA標的部位は、非翻訳領域(LITR)は調節タンパク質結合部位により富んでいるので、オープンリーディングフレームから選択される。UTR結合タンパク質および/または翻訳開始複合体は、siRNAエンドヌクレアーゼ複合体の結合に干渉し得る[Tuschl ChemBiochem. 2:239-245]。しかし、非翻訳領域に指向されるsiRNAはまた有効であり得ることが理解され、何故ならば、GAPDHについて実証されているからであり、ここで、5’UTRで向けられるsiRNAは、細胞性GAPDH mRNAの約90%減少を媒介し、そして完全にタンパク質レベルを廃した(abolished)(www.ambion.com/techlib/tn/91/912.html)。
【0120】
次に、NCBIサーバ(www.ncbi.nlm. nih.gov/BLAST/)から利用可能なBLASTソフトウエアのような任意の配列アラインメントソフトウエアを使用して、可能性のある標的部位を好適なゲノムデータベース(たとえば、ヒト、マウス、ラットなど)と比較する。他のコード配列に対して有意な相同性を示す推定標的部位を除去する。
【0121】
資格(qualifying)標的配列は、siRNA合成のためのテンプレートとして選択される。好ましい配列は低G/C含有量を含むものであり、何故ならば、これらは、55%を越えるG/C含有量を有するものと比較した場合に、遺伝子サイレンシングを媒介することにおいてより有効であると判明したためである。いくつかの標的部位は、好ましくは、評価のために該標的遺伝子の長さに沿って選択される。選択されるsiRNAのよりよい評価のために、ネガティブコントロールが、結合(conjunction)において好ましくは使用される。siRNAのネガティブコントロールは、好ましくは、該siRNAと同一のヌクレオチド組成を含むが、該ゲノムに対して有意な相同性を欠いている。したがって、前記siRNAのスクランブルされたヌクレオチド配列が、それが他の遺伝子に対していずれの有意な相同性を示さない限りにおいて、好ましくは使用される。
【0122】
CTKをダウンレギュレートし得る別の薬剤は、CTKのDNA配列またはmRNA転写物を特異的に切断し得るDNA酵素(DNAzyme)分子である。DNA酵素は、一本鎖標的配列および二本鎖標的配列の両方を切断し得る一本鎖ポリヌクレオチドである(Breaker, R.R. および Joyce, G. Chemistry and Biology 1995;2:655; Santoro, S.W. & Joyce, G.F. Proc. Natl, Acad. Sci. USA 1997;943:4262)。DNA酵素の一般的なモデル(「10−23」モデル)が提案されている。「10−23」DNA酵素は、各々7〜9のデオキシリボヌクレオチドの2つの基質認識ドメインが両側に並ぶ、15のデオキシリボヌクレオチドの触媒ドメインを有する。このタイプのDNA酵素は、プリン:ピリミジン接合部でその基質RNAを効率的に切断し得る(Santoro, S.W. & Joyce, G.F. Proc. Natl, Acad. Sci. USA 199;DNA酵素のレビューについては、Khachigian, LM [Curr Opin MoI Ther 4: 119-21 (2002)]を参照のこと)。
【0123】
一本鎖および二本鎖標的切断部位を認識する合成の人工DNA酵素の構築および増幅の例は、Joyceらの米国特許第6,326,174号に開示されている。ヒトウロキナーゼ受容体に指向された類似の設計のDNA酵素は、ウロキナーゼ受容体発現を阻害しそしてインビボで首尾良く結腸癌細胞転移を阻害することが最近観察された(Itohら, 20002, Abstract 409, Ann Meeting Am Soc Gen Ther www.asgt.org)。別の適用において、bcr−abl癌遺伝子に相補的なDNA酵素は、白血病細胞における該癌遺伝子発現を阻害し、そしてCMLおよびALLのケースにおける自家骨髄移植における再発率を減少させることに成功した。
【0124】
CTK発現の阻害はまた、CTKをコードするmRNA転写物と特異的にハイブリダイズしそれによってCTK転写物の翻訳を特異的に阻害し得るアンチセンスポリヌクレオチドを使用することによって行われ得る。
【0125】
CTK発現を効率的に阻害するために使用され得るアンチセンス分子の設計は、アンチセンスアプローチにとって重要な2つの局面を考慮しながら、行われなければならない。第1局面は、好適な細胞の細胞質への前記オリゴヌクレオチドの送達であり、一方、第2局面は、その翻訳を阻害する様式で細胞内の該設計されたmRNAへ特異的に結合するオリゴヌクレオチドの設計である。
【0126】
先行技術は、広範囲の細胞タイプにオリゴヌクレオチドを効率的に送達するために使用され得る、多数の送達戦略を教示する。[たとえば、Luft, J MoI Med 76: 75-6, 1998;Kronenwett ら, Blood 91 : 852-62, 1998;Rajur ら, Bioconjug Chem 8: 935-40, 1997;Lavigne ら, Biochem Biophys Res Commun 237: 566-71, 1997;およびAoki ら, Biochem Biophys Res Commun 231 : 540-5), 1997を参照のこと]。
【0127】
さらに、前記標的mRNAおよび前記オリゴヌクレオチドの両方における構造変化のエネルギー論の原因となる熱力学サイクルに基づく、最も高い予想される結合親和性を有する配列を同定するためのアルゴリズムもまた、利用可能である[たとえば、Walton ら. Biotechnol Bioeng 65: 1-9, 1999を参照のこと]。
【0128】
このようなアルゴリズムは、細胞におけるアンチセンスアプローチを実行するために首尾良く使用されてきた。たとえば、Waltonらによって開発されたアルゴリズムは、科学者が、ウサギベータ−グロビン(RBG)およびマウス腫瘍壊死因子−α(TNFアルファ)転写物について首尾良くアンチセンスオリゴヌクレオチドを設計することを可能にした。同一の研究グループは、より最近、動的PCR技術によって評価した場合、細胞培養物において3つのモデル標的mRNA(ヒト乳酸デヒドロゲナーゼAおよびBならびにラットgp130)に対して合理的に選択されたオリゴヌクレオチドのアンチセンス活性は、ほとんど全てのケース(ホスホジエステルおよびホスホロチオエートオリゴヌクレオチド化学で2つの細胞タイプにおける3つの異なる標的に対する試験を含む)において有効であることが判明したことを報告した。
【0129】
さらに、インビトロ系を使用して特異的オリゴヌクレオチドの有効性を設計および予想するためのいくつかのアプローチもまた公表された(Matveeva ら, Nature Biotechnology 16, 1374-1375, 1998)。
【0130】
いくつかの臨床試験は、アンチセンスオリゴヌクレオチドの安全性、実行可能性および活性を実証した。たとえば、癌の治療に好適なアンチセンスオリゴヌクレオチドが首尾良く使用され(Holmund ら, Curr Opin MoI Ther 1 :372-85, 1999)、一方、c−myb遺伝子、p53およびBcl−2を標的化するアンチセンスオリゴヌクレオチドによる血液悪性腫瘍の治療は、臨床試験に入っており、そして患者によって許容されることが示された(Gerwitz Curr Opin MoI Ther 1 :297-306, 1999)。
【0131】
より最近、ヒトヘパラナーゼ遺伝子発現のアンチセンス媒介抑制が、マウスモデルにおけるヒト癌細胞の胸膜転移を抑制すると報告された(Uno ら., Cancer Res 61 :7855-60, 2001)。
【0132】
したがって、現在の総意は、上述されるように、非常に正確なアンチセンス設計アルゴリズムおよび多種多様のオリゴヌクレオチド送達系の作製へ導いたアンチセンス技術の分野における最近の進展は、当業者が、過度な試行錯誤実験に頼る必要無しに、公知の配列の発現をダウンレギュレートするに好適なアンチセンスアプローチを設計および実行することを可能にしたということである。
【0133】
アンチセンス配列は、CTKをコードする転写物を切断し得るリボザイム配列を含み得、それによって、機能性CTKへのそれらの転写物の翻訳を防止する。このようなリボザイムは、固相オリゴヌクレオチド合成を使用して容易に合成可能である。
【0134】
リボザイムは、問題のタンパク質をコードするmRNAの切断による遺伝子発現の配列特異的阻害のためにますます使用されている[Welch ら., "Expression of ribozymes in gene transfer systems to modulate target RNA levels." Curr Opin Biotechnol. 1998 Oct;9(5):486-96]。何らかの特定の標的RNAを切断するようにリボザイムを設計する可能性のために、基礎研究および治療適用の両方においてそれらを価値あるツールにした。治療学分野において、リボザイムは、感染症におけるウイルスRNA、癌における優性癌遺伝子、および遺伝性疾患における特定の体細胞突然変異を標的化するために開発された[Welch ら, "Ribozyme gene therapy for hepatitis C virus infection." Clin Diagn Virol. 1998 JuI 15;10(2- 3): 163-71.]。最も顕著には、HIV患者についてのいくつかのリボザイム遺伝子療法プロトコルが、既に臨床第1相試験にある。より最近では、リボザイムは、トランスジェニック動物、遺伝子標的確認、および経路解明のために使用されている。いくつかのリボザイムが臨床試験の種々の段階にある。ANGIOZYMEは、ヒト臨床試験において研究される最初の化学合成されたリボザイムであった。ANGIOZYMEは、新脈管形成経路における重要な成分である、VEGF−r(血管内皮成長因子受容体)の形成を特異的に阻害する。Ribozyme Pharmaceuticals, Inc.、ならびに他の企業は、動物も出るにおける抗新脈管形成療法の重要性を実証した。HEPTAZYME、C型肝炎ウイルス(HCV)RNAを選択的に破壊するように設計されたリボザイムは、細胞培養アッセイにおいてC型肝炎ウイルスRNAを減少させることにおいて有効であると判った(Ribozyme Pharmaceuticals, Incorporated−ウェブホームページ)。
【0135】
上記の発現可能な阻害剤(たとえば、抗体フラグメント、アンチセンスなど)は組換え技術を使用して合成され得そしてたとえば注入によって直接提供され得るが、このような分子はまた、哺乳動物細胞において構成的組織特異的または誘発性転写を指向するために好適なプロモータ配列の転写制御下に配置された、該阻害剤をコードするポリヌクレオチド配列を含む発現ベクターを使用することによって細胞において直接発現され得る。
【0136】
本発明での使用に好適な構成的プロモータは、大抵の環境条件および大抵のタイプの細胞(たとえば、サイトメガロウイルス(CMV)およびラウス肉腫ウイルス(RSV))下で機能性である(即ち、転写を指向し得る)配列を含む。本発明の使用に好適な組織特異的プロモータは、造血細胞において機能性である配列を含み、例としては、たとえば、ClarkおよびGordon(Leukoc Biol. 63:153-68, 1998);Steinら(Cancer 15::2899-902, 2000);ならびにHormasら(Curr Top. Microbiol. Immunol. 211 :159-64, 1996)によって記載されるプロモータが挙げられる。本発明での使用に好適な誘導性プロモータとしては、たとえば、テトラサイクリン誘導性プロモータ(Srour ら, hromb. Haemost. 90: 398-405, 2003)が挙げられる。
【0137】
本発明の発現ベクターは、原核生物、真核生物、または好ましくは両方(たとえば、シャトルベクターにおける複製および組込みにこのベクターを好適にするさらなる配列を含む。典型的なクローニングベクターは、転写および翻訳開始配列(たとえば、プロモータ、エンハンサー)ならびに転写および翻訳ターミネータ(たとえば、ポリアデニル化シグナル)を含む。
【0138】
真核生物プロモータは、典型的に、2タイプの認識配列、TATAボックスおよび上流プロモータエレメントを含む。転写開始部位の25〜30塩基対上流に配置される、TATAボックスは、RNA合成を開始するようにRNAポリメラーゼに指令することに関与すると考えられる。他の上流プロモータエレメントは、転写開始速度を決定する。
【0139】
エンハンサーエレメントは、同種または異種プロモータから1,000倍まで転写を刺激し得る。エンハンサーは、転写開始部位から下流または上流に配置される場合、活性である。ウイルスから誘導される多くのエンハンサーエレメントは、広い宿主範囲を有し、そして種々の組織において活性である。たとえば、SV40初期遺伝子エンハンサーは、多くの細胞タイプについて好適である。本発明に好適である他のエンハンサー/プロモータ組み合わせとしては、以下から誘導されるものが挙げられる:ポリオーマウイルス、ヒトまたはマウスサイトメガロウイルス(CMV)、マウス白血病ウイルス、マウスまたはラウス肉腫ウイルスおよびHIVのようなレトロウイルス由来の長い末端反復配列(long term repeat)が挙げられる。たとえば、Enhancers and Eukaryotic Expression, Cold Spring Harbor Press, Cold Spring Harbor, N.Y. 1983を参照のこと。これは、参照により本明細書中に組み込まれる。
【0140】
ポリアデニル化配列はまた、Scfvのようなポリペプチド阻害剤の翻訳効率を増加させるために発現ベクターへ添加され得る。2つの異なる配列エレメントが、正確かつ効率的なポリアデニル化のために必要とされる:ポリアデニル化部位から下流に配置されるGUまたはUに富む配列、および11〜30ヌクレオチド上流に配置される、6ヌクレオチドの高度に保存された配列、AAUAAA。本発明に好適である終止およびポリアデニル化シグナルは、SV40から誘導されるものが挙げられる。
【0141】
既に述べたエレメントに加えて、本発明の発現ベクターは、典型的に、クローン化核酸の発現レベルを増大させるかまたは組換えDNAを有する細胞の同定を促進するように意図された他の特殊エレメントを含む。たとえば、多数の動物ウイルスは、許容性細胞タイプにおけるウイルスゲノムの余分な(extra)染色体複製を促進するDNA配列を含む。これらのウイルスレプリコンを有するプラスミドは、プラスミド上に保有される遺伝子によってまたは宿主細胞のゲノムで好適な因子が提供される限り、エピソーム複製される。
【0142】
前記ベクターは、真核細胞のレプリコンを含んでも含まなくてもよい。真核細胞レプリコンが存在する場合、前記ベクターは、好適な選択可能なマーカーを使用して、真核細胞中において増幅可能である。前記ベクターは真核細胞レプリコンを含まない場合、エピソーム性の増幅は可能でない。その代わりに、組換えDNAは、遺伝子操作された細胞のゲノムへ組み込まれ、ここで、前記プロモータは所望の核酸の発現を指令する。
【0143】
本発明の発現ベクターは、さらに、以下を可能にする追加のポリヌクレオチドを含み得る:たとえば、内部リボソーム侵入部位(IRES)およびプロモータキメラポリペプチドのゲノム組込みのための配列のような、単一のmRNAからのいくつかのタンパク質の翻訳。
【0144】
哺乳動物発現ベクターの例としては、以下が挙げられるがこれらに限定されない:pcDNA3、pcDNA3.1(+/−)、pGL3、pZeoSV2(+/−)、ρSecTag2、pDisplay、pEF/myc/cyto、pCMV/myc/cyto、pCR3.1、pSinRep5、DH26S、DHBB、pNMTl、pNMT41、pNMTSl(これらは、Invitrogenから入手可能)、pCI(これは、Promegaから入手可能)、pMbac、pPbac、pBK−RSVおよびpBK−CMV(これらは、Strategeneから入手可能)、pTRES(これは、Clontechから入手可能)、ならびにそれらの誘導体。
【0145】
レトロウイルスのような真核生物ウイルス由来の調節エレメントを含む発現ベクターもまた、本発明によって使用され得る。SV40ベクターとしては、pSVT7およびpMT2が挙げられる。ウシ乳頭腫ウイルスから誘導されるベクターとしてはpBV−IMTHAが挙げられ、そしてエプスタイン・バー・ウイルスから誘導されるベクターとしてはpHEBOおよびp2O5が挙げられる。他の例示的なベクターとしては、pMSG、pAV009/A+、pMTO10/A+、pMAMneo−5、バキュロウイルスpDSVE、ならびにSV−40初期プロモータ、SV−40後期プロモータ、メタロチオネインプロモータ、マウス乳腺癌ウイルスプロモータ、ラウス肉腫ウイルスプロモータ、ポリヘドリンプロモータ、または真核生物細胞における発現に有効と示される他のプロモータの指令下でタンパク質の発現を可能にする任意の他のベクターが挙げられる。
【0146】
ウイルスは、多くの場合において宿主防衛機構を回避するように進化した、非常に特殊な感染体である。典型的に、ウイルスは、特定の細胞タイプにおいて感染および繁殖する。ウイルスベクターの標的特異性は、その天然の特異性を使用し、所定の細胞タイプを特異的に標的化しそして感染された細胞へ組換え遺伝子を導入する。したがって、本発明によって使用されるベクターのタイプは、形質転換される細胞タイプに依存する。形質転換される細胞タイプにしたがって好適なベクターを選択する能力は、十分に当業者の能力内にあり、したがって選択考慮の一般的な説明は本明細書中において提供されない。たとえば、骨髄細胞は、ヒトT細胞白血病ウイルス1型(HTLV−I)を使用して標的化され得る。
【0147】
組換えウイルスベクターは、CTK阻害剤のインビボ発現のために有用であり、何故ならば、それらは、側方感染(lateral infection)および標的特異性のような利点を提供するからである。側方感染は、たとえばレトロウイルスのライフサイクルにおいて固有であり、そして単一の感染された細胞が多くの子孫ビリオン(これらは、出芽しそして隣接する細胞に感染する)を産生するプロセスである。結果は、その大部分が元々のウイルス粒子によって最初感染されなかった、広い領域が迅速に感染されるということである。これは、感染因子が娘子孫(daughter progeny)によってのみ広がる垂直感染とは対照的である。側方に広がることができないウイルスベクターもまた、産生され得る。この特徴は、所望の目的が限定数の標的化細胞へのみ特定の遺伝子を導入することである場合に、有用であり得る。
【0148】
造血細胞特異的発現に好適である構築物のさらなる記載は、Malikら(Blood 15: 86:2993-3005, 1995)に提供される。
【0149】
十分でない量の阻害剤を発現するか、または阻害剤の発現について通常サイレントな細胞における、CTKの内因性阻害剤の内因性産生を誘発および/または増強するためのベクターの使用もまた、本発明にしたがって考慮される。前記ベクターは、前記阻害剤を発現することが望まれる細胞中で機能性である調節配列を含むかもしれない。このような調節配列は、プロモータまたはエンハンサーを含む。次いで、調節配列は、相同組換えによってゲノムの正しい遺伝子座へ導入され、このようにして、該調節配列と該遺伝子(この発現が、誘発または増強されるように要求される)とを作動可能に連結する。前記技術は、通常、「内因性遺伝子活性化」(EGA)と呼ばれ、そしてそれは、たとえばWO91/09955に記載されている。
【0150】
同一の技術を使用して、即ち、CTKの遺伝子座へ、CTK発現のダウンレギュレーションもしくは防止を生じさせる負の調節エレメント(たとえば、サイレンシングエレメント)を導入することによって、CTK発現を停止させることが可能であることが、当業者よって理解される。CTK発現のこのようなダウンレギュレーションまたはサイレンシングは、疾患を予防および/または治療するためのCTKIの使用と同一の効果を有することを、当業者は理解する。種々の方法が、本発明の発現ベクターを造血細胞へ導入するために使用され得る。このような方法は、一般的に、以下において記載されており:Sambrookら, Molecular Cloning: A Laboratory Manual, Cold Springs Harbor Laboratory, New York (1989, 1992);Ausubelら, Current Protocols in Molecular Biology, John Wiley and Sons, Baltimore, Md. (1989);Changら, Somatic Gene Therapy, CRC Press, Ann Arbor, Mich. (1995);Vega ら, Gene Targeting, CRC Press, Ann Arbor Mich. (1995);Vectors: A Survey of Molecular Cloning Vectors and Their Uses, Butterworths, Boston Mass. (1988);およびGilboaら[Biotechniques 4 (6): 504-512, 1986]、そしてたとえば、組換えウイルスベクターを用いての安定的または一時的トランスフェクション、リポフェクション、エレクトロポレーションおよび感染を含む。さらに、陽性−陰性の選択方法については、米国特許第5,464,764号および第5,487,992号を参照のこと。
【0151】
ウイルス感染による核酸の導入は、リポフェクションおよびエレクトロポレーションのような他の方法に対していくつかの利点を提供し、何故ならば、より高いトランスフェクション効率がウイルスの感染性に起因して得られ得るためである。
【0152】
前記阻害剤を発現するために使用される発現構築物は、好ましくは、これらの細胞中での独占的かつ制御可能な発現を可能にする様式で、構築されそして造血細胞へ導入されることが理解される。たとえば、もっぱら造血細胞を形質転換し得るウイルス発現ベクターを使用することによってあるいはこのような細胞をエクスビボで形質転換することによって、そして該発現構築物(上記の例を参照のこと)において誘発性プロモータ配列を使用することによって、これらの細胞中での独占的かつ制御可能な発現が達成され得る。
【0153】
このような発現戦略は、白血病治療の文脈において使用される場合に特に有利であり、何故ならば、それは、造血(hematopoesis)に対する正確な制御およびしたがって造血細胞数の調節を可能にするためである。
【0154】
本明細書に記載されるように、CTK発現または活性化のダウンレギュレーションは、ダウンレギュレーション剤へ培養造血細胞を暴露することによってインビトロで、あるいは対象へこのような薬剤を投与することによってインビボで行われてもよい。これらのインビボおよびインビトロアプローチは、CTK阻害剤の以下の非限定的な例を処理するために使用され得る:ペプチド阻害剤、たとえば、Mu−Leu−hph−us−ph(Palmer J Med. Chem 1995, 38:3193-3196)、非ペプチド阻害剤、たとえば、SB−462795、SB−357114、462795(Glaxo Swithkline's)、E64、AAR494、アンチセンス(Inui らJ Biol Chem. 1997 Mar 28;272(13):8109-12.)、NH連結されたアリール/ヘテロアリールCTKI(Robichaud ら Bioorg Med Chem Lett. 2004 Aug 16;14(16):4291-5)、AAE581(Novartis)。
CTK阻害剤はまた、以下に開示されている:WO00/55126およびWO01/49288、Taveresら(J Med Chem 47:5049, 2004, Jmed Chem 29:47(3):588-99, 2004)、Catalaiioら(Bioorg Med Chem Lett.2004 9;14(3):719-22, and 14(1) 275-8)、Robichaudら(J Med Chem 14;46(17):3709-27, 2003)、Settiら(Bioorg Med Chem Lett 13(12):2051-3, 2003)、Altmannら(Bioorg Med Chem Lett 13(12): 1997-2001, 2003)、Chenら(J Org Chem, 68(7):2633-8, 2003)。
【0155】
本明細書中の用語「ポリペプチドおよびタンパク質」は、互換可能である。本発明はまた、本発明のCTKタンパク質のムテインに関し、このムテインは、CTKの天然に生じる配列のみを本質的に有するCTKタンパク質と同一の生物学的活性を本質的に滞留する。このような「ムテイン」は、この種の修飾が該タンパク質自体に対して該タンパク質ムテインの生物学的活性を実質的に変化させないように、約20および10アミノ酸残基までが、それぞれ、CTKタンパク質において、欠失されているか、付加されているか、または他のものによって置換されていてもよいものであり得る。
【0156】
これらのムテインは、公知の合成および/または部位特異的突然変異誘発技術によって、あるいはその好適な任意の他の公知の技術によって作製される。
【0157】
このようなムテインはいずれも、好ましくは、基本であるCTKのそれの実質的に複製的であるアミノ酸配列を有し、その結果、それと実質的に類似の活性を有する。したがって、下記実施例に記載の生物学的活性試験へこのようなムテインを供することを含む、慣用実験によって、どの所定のムテインが本発明の基本タンパク質と実質的に同一の活性を有するかが決定され得る。
【0158】
本発明にしたがって使用され得るCTKタンパク質のムテイン、またはその核酸コードは、置換ペプチドまたはポリヌクレオチドのような実質的にCTK対応配列の有限セットを含み、これらは、本明細書中で示される技術およびガイダンスに基づいて、過度な実験無しに、当業者によって慣用的に得られ得る。タンパク質化学および構造の詳細な説明については、Schulz, G.E. ら, Principles of Protein Structure, Springer- Verlag, New York, 1978;およびCreighton, T.E., Proteins: Structure and Molecular Properties, W.H. Freeman & Co., San Francisco, 1983を参照のこと。これらは参照により本明細書中に組み込まれる。コドン選択のような、ヌクレオチド配列置換の表示については、以下を参照のこと:Ausubelら, Current Protocols in Molecular Biology, Greene Publications and Wiley Interscience, New York, NY, 1987-1995;Sambrookら, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1989。
【0159】
本発明にしたがうムテインについての好ましい変化は、「保存的」置換と知られているものである。天然に生じるCTK配列を本質的に有するタンパク質におけるそれらの保存的アミノ酸置換は、あるグループ内の同義のアミノ酸を含み得、このグループは、該グループのメンバー間の置換が分子の生物学的機能を保存するという十分に類似の物理化学的特性を有する。Grantham, Science, Vol. 185, pp. 862-864 (1974)を参照のこと。特に、前記挿入または欠失が数個のアミノ酸(たとえば、50以下、そして好ましくは20CTK以下)のみを含みかつ機能性構造(functional conformation)に重要であるアミノ酸(たとえば、システイン残基)を除去または置換しない場合、アミノ酸の挿入および欠失もまた、その機能を変化させることなく、上記で定義される配列において成され得ることが明らかである[Anfinsen, "Principles That Govern The Folding of Protein Chains", Science, Vol. 181, pp. 223- 230 (1973)]。このような欠失および/または挿入によって作製されるムテインは、本発明の範囲内に入る。好ましくは、前記同義のアミノ酸基は、表Aに規定されるものである。より好ましくは、前記同義のアミノ酸は、表Bに規定されるものであり;そして最も好ましくは、前記同義のアミノ酸は、表Cに規定されるものである。
【0160】
【表1】
【0161】
【表2】
【0162】
【表3】
【0163】
本発明における使用のための前記タンパク質のムテインを得るために使用され得る、タンパク質におけるアミノ酸置換体の作製例としては、以下に記載のような任意の公知の方法工程が挙げられる:米国特許第RE 33,653号、第4,959,314号、第4,588,585号および第4,737,462号(Markら);第5,116,943号(Kothsら);第4,965,195号(Namenら);第4,879,111号(Chongら);および第5,017,691号(Leeら);ならびに米国特許第4,904,584号(Strawら)に記載のリシン置換タンパク質。
【0164】
本発明の別の好ましい実施形態において、本発明における使用のためのCTKタンパク質の任意のムテインは、本発明の上述のCTKタンパク質のそれに本質的に対応するアミノ酸配列を有する。用語「に本質的に対応する」は、特にCTKに対するその能力が関連する場合、その基本的特徴に影響を与えない基本タンパク質の配列に対して僅かな変化を有するムテインを含むように意図される。用語「に本質的に対応する」に入ると一般的に考えられるタイプの変化は、本発明のCTKタンパク質をコードするDNAの従来の突然変異誘発技術から生じるものであり、ほんの僅かな修飾を生じ、たとえば化学誘引物質に対する幹細胞の感度を増大する所望の活性についてスクリーニングする。
【0165】
本発明はまた、CTK変異体を含む。好ましいCTK変異体は、少なくとも80%アミノ酸同一性を有するものであり、より好ましいCTK変異体は、少なくとも90%アミノ酸同一性を有するものであり、そして最も好ましい変異体は、CTKアミノ酸配列に対して少なくとも95%同一性を有するものである。
【0166】
用語「配列同一性」は、本明細書中で使用される場合、Clustal−Xプログラム[これは、ClustalWマルチプル配列アラインメントプログラム(Thompsonら,1994)についてのウインドウズインターフェースである]を使用する低相同性領域のリファイン(refinement)を伴う、HanksおよびQuinn(1991)にしたがうアラインメントによって該アミノ酸配列が比較されることを意味する。Clustal−Xプログラムは、ftp://ftp-igbmc.u- strasbg.fr/pub/clustalx/でインターネットから入手可能である。当然ながら、このリンクが休止となった場合、当業者であれば、過度な実験無しに、標準的なインターネット検索技術を使用して他のリンクにあるこのプログラムのバージョンを見つけ出し得ることが理解される。特に述べない限り、本願の有効出願日現在で、本明細書中において言及されるプログラムの最新バージョンが、本発明を実施するために使用されるものである。
【0167】
「配列同一性」を測定するための別の方法は、以下である。−12のギャップオープンペナルティー(ギャップの最初のゼロ)および−4のギャップエクステンションペナルティー(該ギャップにおける追加の連続するゼロの各々当たり)を有するデフォルト(BLOSUM62)マトリクス(値−4〜+11)を使用し、Genetic Computing Group’s GDAP(グローバルなアラインメントプログラム)のバージョン9を使用して、配列を整列する。整列後、パーセンテージ同一性が、請求される配列におけるアミノ酸の数のパーセンテージとして一致の数を表現することによって計算される。
【0168】
本発明にしたがうムテインとしては、ストリンジェント条件下でDNAまたはRNAへハイブリダイズしそして本発明にしたがうCTKタンパク質をコードする核酸(たとえば、DNAまたはRNA)[CTKをコードする天然に生じる配列の本質的に全てと、遺伝暗号の縮重の理由で該天然に誘導されるヌクレオチド配列とはそのヌクレオチド配列が異なり得る配列(即ち、多少異なる核酸配列は、その縮重に起因して、同一のアミノ酸配列を依然としてコードし得る)とを含む]によってコードされるものが挙げられる。
用語「ハイブリダイゼーション」は、本明細書中で使用される場合、核酸の鎖が、塩基対形成により相補鎖と結合するいずれのプロセスをも含む(Coombs J, 1994, Dictionary of Biotechnology, stokton Press, New York NY)。「増幅」は、核酸配列のさらなるコピーの産生と規定され、そして一般的に、当該分野において周知のポリメラーゼ連鎖反応技術を使用して行われる(Dieffenbach および Dveksler, 1995, PCR Primer, a Laboratory Manual, Cold Spring Harbor Press, Plainview NY)。
【0169】
「ストリンジェンシー」は、典型的に、約Tm−5℃(プローブの融点より5℃下)からTmより約20℃〜25℃下までの範囲で生じる。
【0170】
用語「ストリンジェント条件」は、当業者が慣習的に「ストリンジェント」と呼ぶ、ハイブリダイゼーションおよび引き続いての洗浄条件をいう。Ausubelら, Current Protocols in Molecular Biology, Greene Publications and Wiley Interscience, New York, NY, 1987-1995;Sambrookら, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1989を参照のこと。
【0171】
本明細書中で使用される場合、ストリンジェンシー条件は、ハイブリダイゼーション実験において使用される温度、一価カチオンのモル濃度およびハイブリダイゼーション溶液中のホルムアミドのパーセンテージの関数である。所定のセットの条件と関連するストリンジェンシーの程度を決定するために、先ず、DNA−DNAハイブリッドの融点Tmとして表される100%同一性のハイブリッドの安定性を決定するためにMeinkothら(1984)の等式が使用される:
Tm=81.5C+16.6(LogM)+0.41(%GC)−0.61(%ホルム(form))−500/L
式中、Mは一価カチオンのモル濃度であり、%GCはDNA中のGおよびCヌクレオチドのパーセンテージであり、%ホルムはハイブリダイゼーション溶液中のホルムアミドのパーセンテージであり、そしてLは塩基対のハイブリッドの長さである。100%同一性ハイブリッドについて計算されるものからTmが低下される各1Cについて、許容されるミスマッチの量は、約1%増加される。したがって、特定の塩およびホルムアミド濃度で所定のハイブリダイゼーション実験のために使用されるTmが、Meinkothの等式にしたがう100%ハイブリッドについて計算されるTmより10C下である場合、ハイブリダイゼーションは、約10%ミスマッチまで存在しても生じる。
【0172】
本明細書中で使用される場合、「高度にストリンジェントな条件」は、上記式によって計算されるかまたは実際に測定される、標的配列との完全な二本鎖について存在するTmよりも多くて10C下であるTmを提供するものである。「中程度にストリンジェントな条件」は、上記式によって計算されるかまたは実際に測定される、標的配列との完全な二本鎖について存在するTmより多くて20C下であるTmを提供するものである。限定されること無しに、高度にストリンジェントな(前記ハイブリッドの計算または測定Tmより5〜10C下)および中程度にストリンジェントな(前記ハイブリッドの計算または測定Tmより15〜20C下)条件の例は、前記ハイブリッドの計算Tm未満の好適な温度で、2×SSC(標準クエン酸食塩水)および0.5%SDS(ドデシル硫酸ナトリウム)の洗浄溶液を使用する。特に、使用されるハイブリダイゼーション条件が、安定なハイブリッドと共にあまり安定でないハイブリッドが形成することを可能にするものである場合、前記条件の最終的なストリンジェンシーは、主として洗浄条件に起因する。より高いストリンジェンシーでの洗浄条件は、次いで、前記あまり安定でないハイブリッドを除去する。上述の高度にストリンジェントな〜中程度にストリンジェントな洗浄条件で使用され得る、典型的なハイブリダイゼーション条件は、前記Tmの約20〜25C下の温度での、6×SSC(または6×SSPE(標準食塩水−リン酸塩−EDTA)、5×Denhardt試薬、0.5% SDS、100&マイクロ(100 µ);g/ml変性断片化サケ精子DNAの溶液中におけるハイブリダイゼーションである。混合プローブが使用される場合、SSCの代わりに塩化テトラメチルアンモニウム(TMAC)を使用することが好ましい(Ausubel, 1987, 1999)。成人幹細胞は、骨髄吸引のような外科手術を使用して得られ得るか、またはNexell Therapeutics Inc. Irvine, CA, USAから入手可能なもののような市販のシステムを使用して収穫され得る。本発明によって使用される幹細胞は、好ましくは、対象の循環へのHSCの放出に対する化学療法またはサイトカイン刺激を使用する、幹細胞動員手順を使用して、回収される(即ち、採取される)。動員は骨髄手術よりもより多くのHSCおよび前駆細胞を得ることが知られているので、幹細胞は、好ましくはこの手順を使用して回収される。
【0173】
CTKの「アイソフォーム」は、SDF−1または選択的スプライシングによって産生され得るそのフラグメントを分解し得るタンパク質である。用語「円順列誘導体」は、本明細書中で使用される場合、線形分子をいい、ここで、末端が直接的にまたはリンカーを介して一緒に結合されて環状分子が作製され、次いで該環状分子が別の位置で開裂されて、元の分子の末端とは異なる末端を有する新たな線形分子が作製されている。円順列物(circular permutations)としては、その構造が、環状化され次いで開裂されている分子と等しい分子が挙げられる。したがって、円順列分子は、線形分子として新たに合成されてもよく、そして環状化および開裂工程を経由しなくてもよい。円順列誘導体の製造は、WO95/27732に記載されている。
【0174】
なおさらなる実施形態において、本発明にしたがう物質は、免疫グロブリン融合を含み、即ち、本発明にしたがう分子は、免疫グロブリンの全部または一部へ融合される。免疫グロブリン融合タンパク質を作製するための方法は、当該分野において周知である(たとえば、WO01/03737に記載されるもの)。当業者であれば、本発明の得られる融合タンパク質はCTKの生物学的活性を滞留することを理解する。得られる融合タンパク質は、体液中の延長された滞留時間(半減期)、増加された特異的活性、増加された発現レベル、または該融合タンパク質の促進された精製のような、改善された特性を理想的には有する。
【0175】
好ましくは、本発明にしたがう前記物質は、Ig分子の定常領域へ融合される。それは、たとえば、ヒトIgG1のCH2およびCH3ドメインのような、重鎖領域へ融合され得る。Ig分子の他のアイソフォームもまた、本発明にしたがう融合タンパク質の作製に好適である:たとえば、アイソフォームIgG2またはIgG4、あるいはIgMまたはIgAのような他のIgクラス。融合タンパク質は、単量体または多量体、ヘテロ−もしくはホモ多量体であってもよい。
【0176】
本発明にしたがう物質の機能性誘導体は、前記タンパク質の特性(たとえば、安定性、半減期、バイオアベイラビリティ、人体による許容性、または免疫原性)を改善するためにポリマーへ複合体化され得る。したがって、本発明の好ましい実施形態は、アミノ酸残基上の1またはそれ以上の部分として生じる1以上の官能基へ結合された少なくとも1つの部分を含む本発明にしたがう物質の機能性誘導体に関する。
【0177】
非常に好ましい実施形態は、ポリエチレングリコール(PEG)へ結合された本発明の物質に関する。PEG化は、たとえばWO92/13095に記載されるもののような、公知の方法によって行われ得る。
【0178】
本発明にしたがう「活性画分」は、たとえば、CTKのフラグメントであり得る。用語フラグメントは、前記分子の任意のサブセット、即ち、所望の生物学的活性を滞留するより短いペプチドをいう。フラグメントは、CTK分子のいずれかの末端からアミノ酸を除去し、そして得られたフラグメントを、SDF−1を分解するその特性について試験することによって、容易に得られ得る。ポリペプチドのN末端またはC末端のいずれかから1つのアミノ酸を除去するためのプロテアーゼは公知であり、そしてしたがって、所望の生物学的活性を滞留するフラグメントを決定することは、慣用的実験のみを含む。
【0179】
CTK、そのムテインおよび融合タンパク質の活性画分として、本発明は、前記タンパク質分子のポリペプチド鎖の任意のフラグメントもしくは前駆体のみ、またはそこへ結合された関連分子または残基(たとえば、糖またはリン酸残基)、あるいはそれら自体による前記タンパク質分子または前記糖残基の凝集体をさらに含み、但し、前記画分は、CTKと実質的に類似の活性を有し、たとえばSDF−1を分解する。
【0180】
用語「塩」は、本明細書中において、CTK分子またはそのアナログのアミノ酸基の酸付加塩およびカルボキシル基の塩の両方をいう。カルボキシル基の塩は、当該分野において公知の手段によって形成されてもよく、そして無機塩、たとえば、ナトリウム、カルシウム、アンモニウム、鉄(ferric)または亜鉛塩など、および有機塩基との塩、たとえば、トリエタノールアミン、アルギニンもしくはリシン、ピペリジン、プロカインのようなアミンと形成されるものを含む。酸付加塩としては、たとえば、鉱酸(たとえば、塩酸または硫酸)との塩、および有機酸(たとえば、酢酸またはシュウ酸)との塩が挙げられる。当然ながら、いずれのこのような塩も、CTKの生物学的活性、たとえばSDF−1を分解する能力を滞留しなければならない。
【0181】
幹細胞動員は、CTKまたは破骨細胞活性化のみであるいは多数の分子と組み合わせて誘発され得る。例としては、以下が挙げられるがこれらに限定されない:サイトカイン、たとえば、顆粒球コロニー刺激因子(G−CSF)、顆粒球マクロファージコロニー刺激因子(GM−CSF)、インターロイキン(IL)−7、IL−3、IL−12、幹細胞因子(SCF)、およびflt−3リガンド;ケモカイン、たとえば、IL−8、Mip−1α、Groβ,またはSDF−1;ならびに化学療法薬 シクロホスファミド(Cy)およびパクリタキセル。これらの分子は速度論および効果で相違することが理解されるが、現在公知の実施形態によれば、G−CSFが、幹細胞を動員するために、単独でまたはたとえばシクロホスファミドと組み合わせて好ましくは使用される。典型的に、G−CSFは、5〜10日間、5〜10μg/kgの用量で毎日投与される。幹細胞を動員する方法は、米国特許第6,447,766号および第6,162,427号に開示されている。ヒト胚幹細胞は、ヒト胚盤胞から単離され得る。ヒト胚盤胞は、典型的に、ヒト体内着床前胚または体外受精(IVF)胚から得られる。あるいは、単一細胞ヒト胚は、胚盤胞段階へ拡張され得る。ヒトES細胞の単離のために、透明帯が胚盤胞から除去され、そして内細胞塊(ICM)が免疫手術によって単離され、ここで、栄養外胚葉細胞が溶解され、そして穏やかなピペッティングによりインタクトなICMから除去される。次いで、ICMを、その成長(outgrowth)を可能にする好適な培地を含む組織培養フラスコ中にプレートする。9〜15日後、ICM誘導化成長を、機械的分離または酵素的分解のいずれかによって集団(clumps)に分離し、次いで該細胞を、新鮮な組織培養培地上に再度プレートする。未分化形態を実証するコロニーを、マイクロピペットによって個々に選択し、集団へ機械的に分離し、そして再度プレートする。次いで、得られるES細胞を1〜2週間毎に定期的に分ける。ヒトES細胞の作製方法についてのさらなる詳細については、Thomsonら[米国特許第5,843,780号;Science 282: 1145, 1998;Curr. Top. Dev. Biol. 38: 133, 1998;Proc. Natl. Acad. Sci. USA 92: 7844, 1995];Bongsoら[Hum Reprod 4: 706, 1989];Gardnerら[Fertil. Steril. 69: 84, 1998]を参照のこと。
【0182】
市販の幹細胞もまた、本発明のこの局面にしたがって使用され得ることが理解される。ヒトES細胞は、NIHヒト胚幹細胞登録(NIH human embryonic stem cells registry)(<http://escr.nih.gov>)から購入され得る。市販の胚幹細胞株の非限定的な例は、BGOl、BG02、BG03、BG04、CY12、CY30、CY92、CYlO、TE03、およびTE32である。
【0183】
ヒトEG細胞は、当業者に公知の実験室技術を使用して、妊娠の約8〜11週のヒト胎児から得られる始原生殖細胞から回収され得る。生殖堤を分離し、そして小さな塊に切断し、その後これを機械的分離により細胞へ分離する。次いで、ES細胞を、好適な培地を含む組織培養フラスコ中で成長させる。ES細胞と一致する細胞形態が観察されるまで、典型的に7〜30日または1〜4継代後まで、培地を毎日交換して、前記細胞を培養する。ES細胞を作製する方法についてのさらなる詳細については、Shamblottら[Proc. Natl. Acad. Sci. USA 95: 13726, 1998]および米国特許第6,090,622号を参照のこと。
【0184】
多分化能を示す幹細胞集団の富化が好ましくは行われ得ることが理解される。したがって、たとえば、上記に概述されるように、CD34+幹細胞は、以下にさらに説明されるようなFACSまたはアフィニティーカラムを使用して制御され得る。
【0185】
増殖条件下での幹細胞の培養がまた、幹細胞の数が処置における使用には少なすぎる場合、行われてもよい。幹細胞の培養は、米国特許第6,511,958号、第6,436,704号、第6,280,718号、第6,258,597号、第6,184,035号、第6,132708号および第5,837,5739号に記載されている。
【0186】
たとえば腫瘍細胞中のCXCR4受容体レベルの分析は、フローサイトメトリーによって検出され得る。このアプローチは、液体媒体中の励起源を過ぎて流れる個々の細胞をスキャンする機器を使用する。前記技術は、可視光および蛍光発光の測定に基づく、単一生存(または死)細胞についての迅速で定量的なマルチパラメータ分析を提供し得る。この基本プロトコルは以下に焦点を合わせる:蛍光標識抗体および特異的細胞結合分子(specific cell-associated molecules)に結合するリガンドによって作製される蛍光強度を測定する。蛍光活性化細胞選別装置を使用して細胞集団を単離するために、本発明の幹細胞を、R&D, 614 McKinley Place NE Minneapolis, MNから市販される抗CXCR4と接触させる。
【0187】
走化性受容体発現のレベルを定量的に評価するための他の細胞学的または生化学的方法としては、以下が挙げられるがこれらに限定されない:標識された(たとえば、放射標識された)ケモカインを使用する結合分析、ウェスタンブロット分析、細胞表面ビオチン化、および免疫蛍光染色。前記受容体発現レベルもまた前記mRNAレベルで測定され得ることが理解される。たとえば、CXCR4 mRNAは、特異的プローブへのハイブリダイゼーションによって細胞中において検出されてもよい。このようなプローブは、クローン化DNAまたはそのフラグメント、RNA(典型的に、インビトロ転写により作製される)、またはオリゴヌクレオチドプローブ(通常、固相合成によって作製される)であり得る。特異的ハイブリダイゼーションに好適なプローブを使用および作製する方法は、周知であり、そして当該分野において使用されている。問題の走化性受容体のmRNAへ特異的にハイブリダイズするプライマーを使用する、増幅反応[たとえば、PCR、"PCR Protocols: A Guide To Methods And Applications", Academic Press, San Diego, CA (1990)]を使用して、mRNAレベルの定量もまた行われ得る。
【0188】
mRNA検出アッセイにおける精度を改善するために、種々のコントロールが、通常使用され得る。たとえば、偽ハイブリダイゼーションを評価するために、サンプルは、無関係のプローブへハイブリダイズされそしてハイブリダイゼーション前にRNアーゼAで処理され得る。
【0189】
走化性受容体発現を測定するために使用され得る。たとえば、走化性剤(たとえば、SDF−1)の勾配を使用しそして膜を介しての該走化性剤への幹細胞移動が続く、機能アッセイもまた、増加した走化性を示す幹細胞を同定および単離するために使用され得る。前記細胞が十分なレベルの前記走化性受容体(たとえば、CXCR4)を発現しない場合、該細胞の大部分は膜上に存在するままである。しかし、本発明の化学誘引物質受容体の増加した発現時に、細胞は、膜を通って移動し、そして走化性プレートのウエルの底部に定着する(実施例セクションの実施例3を参照のこと)。機能ホーミングアッセイ(functional homing assay)もまた、本発明の方法によって使用され得ることが理解される。このようなアッセイは、Kollet (2001) Blood 97:3283-3291(参照文献12)に記載されている。
【0190】
本発明にしたがう好ましい個々の対象は、イヌ、ネコ、ヒツジ、ブタ、ウマ、ウシおよび好ましくはヒトのような哺乳動物である。
【0191】
本発明のさらなる目的、利点、および新規の特徴は、限定される意図は無い以下の実施例の検討の際に、当業者に明らかとなる。さらに、上記で示されそして下記特許請求の範囲セクションにおいて特許請求されるような本発明の種々の実施形態および局面の各々は、下記の実施例に実験的サポートが見られる。
【実施例】
【0192】
上記説明と共に本発明を非限定的に示す以下の実施例がここで参照される。
【0193】
一般的に、本明細書中で使用される命名および本発明において使用される実験室手順は、分子、生化学、微生物学および組換えDNA技術を含む。このような技術は、文献において十分に説明されている。たとえば、以下を参照のこと:"Molecular Cloning: A Laboratory Manual" Sambrook ら, (1989);"Current Protocols in Molecular Biology" Volumes I-III Ausubel, R. M., ed. (1994);Ausubel ら., "Current Protocols in Molecular Biology", John Wiley および Sons, Baltimore, Maryland (1989);Perbal, "A Practical Guide to Molecular Cloning", John Wiley & Sons, New York (1988);Watson ら, "Recombinant DNA", Scientific American Books, New York;Birren ら. (eds) "Genome Analysis: A Laboratory Manual Series", VoIs. 1-4, Cold Spring Harbor Laboratory Press, New York (1998);米国特許第4,666,828号、第4,683,202号、第4,801,531号、第5,192,659号および第5,272,057号に記載される方法;"Cell Biology: A Laboratory Handbook", Volumes I-III Cellis, J. E., ed. (1994);"Current Protocols in Immunology" Volumes I-III Coligaii J. E., ed. (1994);Stites ら. (eds), "Basic and Clinical Immunology" (8th Edition), Appleton & Lange, Norwalk, CT (1994);Mishell and Shiigi (eds), "Selected Methods in Cellular Immunology", W. H. Freeman and Co., New York (1980);利用可能な免疫学的検定法は、特許文献および科学文献に広く記載されており、たとえば、米国特許第3,791,932号、第3,839,153号、第3,850,752号、第3,850,578号、第3,853,987号、第3,867,517号、第3,879,262号、第3,901,654号、第3,935,074号、第3,984,533号、第3,996,345号、第4,034,074号、第4,098,876号、第4,879,219号、第5,011,771号および第5,281,521号を参照のこと;"Oligonucleotide Synthesis" Gait, M. J., ed. (1984); "Nucleic Acid Hybridization" Hames, B. D., and Higgins S. J., eds. (1985);"Transcription and Translation" Hames, B. D., および Higgins S. J., Eds. (1984);"Animal Cell Culture" Freshney, R. I., ed. (1986);"Immobilized Cells and Enzymes" IRL Press, (1986);"A Practical Guide to Molecular Cloning" Perbal, B., (1984)および"Methods in Enzymology" Vol. 1-317, Academic Press;"PCR Protocols: A Guide To Methods And Applications", Academic Press, San Diego, CA (1990);Marshak ら, "Strategies for Protein Purification and Characterization - A Laboratory Course Manual" CSHL Press (1996);これらの全ては、本明細書中に完全に記載されているかのように、参照により組み込まれる。他の一般的な参照文献は、本明細書にわたって提供される。その中の手順は、当該分野において周知であると考えられ、そして読み手の便宜のために提供される。その中に含まれる全ての情報は、参照により本明細書中に組み込まれる。
【0194】
実施例1:
LPS誘発炎症は破骨細胞活性化をもたらす。
骨破壊は、慢性関節リウマチおよび歯周炎を含むいくつかの慢性炎症性疾患の病理学的特徴である。この種の炎症誘発される骨量減少は、破骨細胞活性化および増加した数の骨再吸収性破骨細胞の誘発から生じる。
【0195】
本発明者らは、骨髄(BM)中の破骨細胞活性化に対する、内毒素 リポ多糖類(LPS)投与によって誘発される炎症の効果を研究した。
【0196】
LPS(Sigma)を、250μg/マウスの単回皮下注射でBalb/cマウスへ投与し、そしてコントロール群においてLPSの代わりに生理食塩水を注射した。LPS投与の5日後、マウスを犠牲にし、骨を固定し、脱灰し、パラフィン包埋し、そして切片化し、BMを収穫し、そしてTRAP+破骨細胞(活性化破骨細胞)のレベルをモニターした。
【0197】
本発明者らは、TRAP+破骨細胞(Sigma製のTRAP染色用のキットを使用して、赤色染色により検出される)の急激な増加を見た。このアッセイにおいて、LPS処理したマウスのBMにおける組織TRAPの固有活性を測定するために、基質は外から提供される(図1AおよびB)。
【0198】
次に、本発明者らは、SDF−1、肝細胞成長因子(HGF)および破骨細胞特異的プロテアーゼCTKのような骨髄媒介物(mediators)の転写に対するLPS媒介炎症の効果を研究した。
【0199】
各マウスの1つの大腿を、RNA抽出のためにTrireagentでフラッシュした(flush)。1μg RNAをcDNAの作製のために使用した。半定量RT−PCRを使用して、マウスSDF−1、HGFおよびCTKの転写を測定した。以下のプライマーを使用した:SDF−1(Ponomaryov, 2000, J. Clin. Invest. 106:1331):左プライマー(配列番号1)5’−GGACGCCAAGGTCGTCGCCGTG−3’、右プライマー(配列番号2)5’−TTGCATCTCCCACGGATGTCAG−3’。HGF(Weinstein, 2001, Moll Cell boil, 21(15):5122):左プライマー(配列番号3)5’−TGCCAGAAAGATATCCCGAC−3’、右プライマー(配列番号4)5’−AACTCGGATGTTTGGGTCAG−3’。CTK:左プライマー(配列番号5)5’−GGCCTCTCTTGGTGTCCATA−3’、右プライマー(配列番号6)5’−TCTGCTGCACGTATTGGAAG−3’。
【0200】
本発明者らは、LPS処置マウスのBM中におけるSDF−1転写の減少ならびにHGFおよびCTK転写の急激な増加を観察した(図1C)。
【0201】
したがって、得られた結果は、LPS誘発炎症(5日処置による)は、骨髄中の以下の変化を媒介することを実証する:破骨細胞活性化の増加、CTKおよびHGF転写の増加、ならびにSDF−1転写の減少。
【0202】
実施例2:
LPS誘発炎症はBM SDF−1濃度の減少および前駆体動員をもたらす。
前の実施例において、本発明者らは、炎症が骨髄中のSDF−1の減少を媒介することを示した。本発明者らは、さらに以下の実験を行い、LPS媒介炎症に続いてBM中のSDF−1濃度の減少が存在するかどうかを研究した。したがって、本発明者らは、LPS処理マウス対未処理マウスにおいて、BM中のSDF−1濃度をモニターした(図2A)。
【0203】
実施例1に記載されるようにマウスに5日間LPSを投与し、そして骨髄中のSDF−1濃度をモニターした(Petitら2002によって記載されるようなELISAによる)。
【0204】
図2Aに概述される結果は、LPS媒介炎症(5日間処理による)が、未処理コントロール群における1.2からLPS処理群における0.4ng/mgタンパク質まで、BM SDF−1濃度の減少(約66%)を誘発することを示す。
【0205】
周知の動員剤である顆粒球コロニー刺激因子(G−CSF)による、BMから循環への前駆体の動員は、BM SDF−1濃度についての減少によって先行されるので(Petit ら Nature immunology (3) NO 7 687, 2002)、本発明者らは、LPS媒介炎症およびBM中のSDF−1減少に動員が続くのかどうかを研究した(図2B)。マウスに5日間LPSを投与し(実施例1に記載される通り)、そして末梢血(PB)中の前駆体増加を測定した(実施例9に記載される通り)。
【0206】
本発明者らは、LPS処理マウス(5日間)のPBにおいて、コントロール未処理マウスのPBにおける前駆体のレベルと比較して前駆体の増加したレベル(約95%より高い)を見た(図2B)。たとえば、5×105播種細胞当たり約100前駆体コロニーがLPSおよびコントロール群において見られ、対して、未処理群において5×105播種細胞当たり5コロニー。
【0207】
LPS処理マウスにおけるPB中で検出された前駆体増加は、骨髄からの細胞の放出または動員から生じることを検証するために、LPS処理後まもなく(LPS処理の16時間後)血球計を使用して、フィコール化(ficolled)WBC(白血球)の数をカウントすることによって、単核細胞(MNC)をBMおよびPB中において、ならびに未処理マウスのBMおよびPB中においてモニターした(図3)。
【0208】
本発明者らは、LPS媒介炎症は、それぞれの未処理マウスと比較した場合、BM SDF−1濃度の減少(約55%)、BM中のMNC細胞の減少(約53%)、ならびにPB中のMNC細胞の増殖(約78%)を誘発することが判った(図3)。
【0209】
したがって、LPS投与の16時間後、BM SDF−1濃度の減少およびBMからPBへの細胞の放出が観察される。
【0210】
実施例3:
機能性CXCR4はLPS媒介動員のために必要とされる。
G−CSFは、BM SDF−1を減少させそしてCXCR4をアップレギュレートすることによって、幹細胞動員を誘発することが知られている(Petitら2002)。本発明者らは、実験を行い、前駆体のLPS媒介動員がCXCR4/SDF−1相互作用を必要とするかどうかを測定した。
【0211】
本発明者らは、LPS投与の16時間後、LPS処理マウスのBMおよびPBにおける、ならびにコントロール未処理マウスにおける、CXCR4発現をモニターした。本発明者らは、LPS処理マウスのBMおよびPBの両方において、フローサイトメトリーによって検出されるような(Petit 2002)CXCR4発現のレベルに対する有意な増加を見(図4A)、これは、前駆体動員の有意な増加と関連した(図4B)。
【0212】
結果は、LPSによる前駆細胞の動員は、SDF−1/CXCR4相互作用を含むことを示す。
【0213】
機能性CXCR4がLPS媒介動員のために必要とされることを確認するために、マウスをLPSのみで処理し(16時間)、LPSおよび抗CXCR4抗体[抗ラットCXCR4、これはまた、マウスCXCR4(Torrey Pines Biolabs, CA)に対して有効であり、500mcl PBS中10mcg]で共処理し、あるいは未処理のままにし、そしてPB中またはBM中の白血球(WBC)を、処理の16時間後に血球計を使用してモニターした(図5)。
【0214】
本発明者らは、LPS媒介動員は、機能性CXCR4の非存在下で減少されることが判った(図5)。したがって、結果は、機能性CXCR4およびCXCR4/SDF−1相互作用は、LPS媒介動員のために必要とすることを確認する。
【0215】
したがって、G−CSF(Petitら2002)によって誘発される動員におけるのと同様に、LPS媒介動員は、骨髄中のSDF−1の減少およびCXCR4の増加を含む。
【0216】
LPS媒介動員が続く骨髄中のSDF−1の減少は、恐らく、SDF−1分解に起因する。
【0217】
実施例1〜3に示される結果によれば、炎症は、BM破骨細胞活性化、CTKおよびHGF発現、SDF−1減少ならびに前駆体動員を誘発する。G−CSF媒介動員と同様に、炎症媒介動員は、BM SDF−1減少を含み、そして機能性CXCR4を必要とすることもまた示された。
【0218】
実施例4:
損傷は動員を引き起こす。
本発明者らは、実験を行い、制御された出血(体重の1%の単回の出血)によって誘発された損傷のモデルにおける細胞動員を研究した。
【0219】
以下のパラメータを、出血の誘発後にモニターした:A− 前駆体の動員、B− 骨髄中のSDF−1レベル、およびC− 血中のCXCR4発現(図6)。
【0220】
本発明者らは、出血開始の7〜14日後、BM SDF−1濃度の有意かつ安定な減少を観察した。CXCR4発現の一時的増加が、出血の3〜7日後に見られた(図6)。PB中の前駆体の一時的増加が、出血の3〜7日後に検出された(図6)。
【0221】
したがって、制御された出血によって引き起こされる損傷は、BM SDF−1濃度の安定な減少、ならびにPB CXCR4および前駆体動員の一時的増加を誘発する。
【0222】
実施例5:
損傷は破骨細胞活性化に影響を与える。
本発明者らは、BM中の破骨細胞活性化に対する、制御された出血によって誘発される損傷の効果を調べた。
【0223】
実験設定は、制御された出血の誘発(実施例4における通り)、ならびに処理マウス対未処理マウスにおけるBM中の破骨細胞の活性化マーカー、TRA+をモニターすること(実施例1における通り)を含んだ。
【0224】
本発明者らは、出血誘発の7日後、BM中の劇的な破骨細胞活性化を検出することができた(図7)。
【0225】
実施例1〜5における上記の知見は、炎症および損傷のようなストレスシグナルが、骨髄からPBへの前駆体動員を誘発することを実証する。ストレスシグナル動員の誘発に含まれるのは、G−CSF媒介動員と同様に、BM SDF−1濃度の減少、およびBM中のCXCR4発現の増加である。予想外なことに、本発明者らは、ストレスシグナル媒介動員は破骨細胞活性化を伴うことを見出した。
【0226】
実施例6:
破骨細胞活性化を媒介する因子。
本発明者らは、実験を行い、細胞動員および運動性(motility)の媒介物(mediators)、たとえば、G−CSF、SDF−1およびHGF(Lapidot 2002 Exp Hematol, SDF-1 , Aiuti 1997, J Exp Med、およびKollet ら JCI 112:160-169 (2003))が破骨細胞活性化を誘発し得るかどうかをインビトロで研究した。
【0227】
破骨細胞前駆体は、骨芽細胞との細胞−細胞接触によって刺激され、活性化多核化TRAP+骨吸収細胞となる。インビトロで活性化破骨細胞を得るために、破骨細胞活性化のために必要とされる、それらのM−CSF産生およびRANKL発現を増強するビタミンD3およびPG2Eと共に、頭蓋冠(clvarian)骨芽細胞をインキュベートする。次いで、破骨細胞前駆体を含むBM細胞を、5日間、前記骨芽細胞単層上に播種し、活性化多核化破骨細胞を得る。
【0228】
原発性マウス骨芽細胞およびBM(上記に詳述されるようなocl前駆体を含有する)の共培養物を、G−CSF(50ng/ml)、HGF(50ng/ml)、またはSDF−1(10および100ng/ml)の存在下で播種した。5日後、前記培養物中のTRAP+多核化破骨細胞を染色し(実施例1に記載される通り)、そして5日後カウントした。
【0229】
本発明者らは、媒介物G−CSF、HGFおよびSDF−1のいずれかでの処理が、活性化TRAP+破骨細胞のレベルを有意に増加させたことを観察した(図8Aそれぞれ、および8B要旨)。
【0230】
本発明者らは、さらに実験を行い、上記の媒介物のどれが破骨細胞を直接活性化するかを研究した。
【0231】
骨芽細胞を、G−CSF、HGFと共にインキュベートしたかまたは未処理のままとし、そしてSDF−1転写および発現を、このような細胞において測定した(図9)。
【0232】
原発性頭蓋冠骨芽細胞を、SDF−1(PeproTech,100ng/nl)、HGF(PeproTech,50ng/ml)およびG−CSF(Filgrastin,Roche,50ng/ml)の存在下で3日間培養した。条件化培地を回収し、実施例2に記載されるように、ELISAによりSDF−1濃度を測定した。SDF−1転写および発現の測定は、実施例2における通り。
【0233】
本発明者らは、G−CSFおよびHGFの両方が、骨芽細胞においてSDF−1転写およびSDF−1産生を誘発することを示し(それぞれ、図9Aおよび9B)、したがって、SDF−1は、G−CSFおよび/またはHGFによって骨芽細胞において誘発される因子であるようであり、そしてこれは、破骨細胞活性化を直接誘発する。
【0234】
本発明者らは、原発性マウス破骨細胞の培養物を染色し、そして破骨細胞がCXCR4を発現することを見出した(示さず)。
【0235】
得られた結果は、G−CSFおよびHGFが骨芽細胞においてSDF−1発現を誘発すること、ならびにSDF−1が破骨細胞表面上のCXCR4に結合しそして破骨細胞活性化へ導くことを示す。本発明者らは、SDF−1媒介破骨細胞活性化が前駆体動員を誘発すると仮説を立てる。
【0236】
本発明者らは、以下の実験を行い、SDF−1が破骨細胞活性化および造血前駆細胞(HPC)動員を誘発するという本発明者らの仮説をインビボで検証した。Balb/cマウスに、10mcg SDF−1または1.5mcg HGFあるいは両方を、5連日間、皮下注射し、そして破骨細胞活性化(実施例1におけるようにTRAP+染色によって測定した)および末梢血への動員(実施例1におけるように半固体培養物においてアッセイした)を測定した。
【0237】
本発明者らは、SDF−1およびHGF投与が、インビボで、破骨細胞活性化(図10A)および前駆体の動員(図10B)の両方を誘発することが判った。
【0238】
全てにおいて、得られた結果は、外から投与されたSDF−1、あるいはG−CSFおよび/またはHGFの作用によって産生されたSDF−1、あるいはストレスシグナル(たとえば、炎症または損傷)が、破骨細胞活性化および前駆体動員を直接誘発することを示す。
【0239】
実施例7:
NFカッパBリガンドの受容体アクチベータ(RANKL)による直接破骨細胞活性化は、インビボ造血幹細胞動員を誘発する
RANKLは、破骨細胞分化因子である(SUDAらEndocr Rev 20:345, 1999)。本発明者らは、RANKLによる破骨細胞の活性化が以下を誘発するかどうかを試験した:
A− 骨髄中におけるCTKおよびMMP−9のような活性化破骨細胞により典型的に分泌されるプロテアーゼの発現(Delaisse, Clin Chim Acta Feb 15;291(2):223-34 2002);ならびに
B− 前駆体動員。
【0240】
マウスに、以下の通りにRANKLを投与した:3日間、5mcgの2回毎日皮下注射、そしてRANKLの非存在下でのもう2日後。以下のパラメータを、処理マウスおよびコントロール未処理マウスの両方においてモニターした:TRAP+活性化破骨細胞(図11A)、骨髄中における破骨細胞プロテアーゼMMP−9(図11B)およびCTK(図11C)の誘発、ならびに動員(図12)。
【0241】
本発明者は、RANKL投与が、骨内膜(幹細胞および前駆細胞を含むことが公知の、骨を結合するBM領域)に沿ったTRAP+活性破骨細胞の形成、骨髄中におけるMMP−9およびCTK発現の誘発(それぞれ、図11A、BおよびC)ならびに前駆体の動員(図12A)を誘発したことを観察した。MMP阻害剤またはCXCR4中和抗体と共のRANKLでの以下の処理に続いて、損なわれた動員が見られた(図12B)。したがって、RANKLによって誘発される、破骨細胞活性化による動員は、CXCR4/SDF−1相互作用およびMMP2/9機能(function)を必要とする(図12B)。
【0242】
上記は、RANKL投与は、インビボで、機能性CXCR4/SDF-1相互作用を必要とする破骨細胞活性化および動員を誘発したことを実証する。
【0243】
実施例8:
破骨細胞活性化による動員の機構。
本発明者らは、前の実施例において、ストレスシグナルによる動員は、BM骨芽細胞による一時的SDF−1産生を含み、このような産生されたSDF−1は、表面破骨細胞CXCR4へ結合し、それらの活性化へ導くことを実証した。次に、破骨細胞活性化は、SDF−1分解およびBM SDF−1濃度減少を誘発する。
【0244】
実施例1において得られた結果を考慮して、本発明者らは、特異的SDF−1分解が、主要な破骨細胞骨吸収酵素、CTKによって誘発されるとい仮説を立てた。したがって、本発明者らは、SDF−1の活性に対するCTKの効果を研究した。
【0245】
本発明者らは、SDF−1についてのインビトロ機能アッセイを使用し(実施例10)、SDF−1活性に対するCTKの効果を調べた。SDF−1への白血病性Pre B ALL G2(比較的低濃度のSDF−1へ非常によく移動する細胞、Spiegel, Blood, 2004)の移動について成った。
【0246】
手短に述べると、SDF−1(125ng)を、37℃で約16時間、PBSで調節された100ul反応体積でCTK(ug/ml)と共にインキュベートした。いくつかのサンプルにおいて、プロテアーゼ阻害剤(システイン阻害剤E−64を含有する、PI、1ul、Sigma)を、37℃で1.5時間CTKと共にプレインキュベートし、その後、SDF−1を添加した。前記白血病性Pre B ALL G2細胞をトランスウエル(transwells)上に加え、そしてCTK処理および未処理SDF−1への移動を測定した。
【0247】
本発明者らは、SDF−1の走化性活性はCTKによって廃止されるが、プロテアーゼ阻害剤(PI)の存在下では廃止されないと判った(図13)。
【0248】
得られた結果は、CTKがSDF−1活性を阻害することを実証する。
【0249】
本発明者らは、さらに、分のタイムスケールでのCTKによるSDF−1切断の速度論を研究した(図14)。SDF−1のサンプル(20ng)を、37℃で0、15、30、60、120および240分間、CTK(1.76 ug/ml)と共にインキュベートした。回収後、サンプルを、SDS−PAGEへ供し、そしてブロットし、そして抗SDF−1ポリクローナルAb(R&D)またはモノクローナルAb Kl 5C(これは、該ケモカインのアミノ酸末端部分へ特異的に結合する)で検出した。
【0250】
得られた結果は、CTKによる迅速かつ部位特異的なSDF−1切断を示す。
【0251】
したがって、CTK、主要な破骨細胞プロテイナーゼは、そのアミノ酸末端の特異的かつ迅速な切断によってSDF−1活性を阻害することが実証された。
【0252】
実施例9:
CFUアッセイ
血液循環におけるマウス前駆体(mouse progenitors)のレベルを検出するために、半固体培養を、以前に記載されるように行った(Petit, Nat Immunol, 2002)。手短に述べると、マウスBM細胞(3x105細胞/ml)を、0.9%メチルセルロース(Sigma)、30%FCS、5x10-5% 2ME、50ng/ml SCF、5ng/ml IL−3、5ng/ml GM−CSF(R&D)、および2u/mlエリスロポイエチン(Orto Bio Tech, Don Mills, Canada)においてプレートした。前記培養物を、5%CO2を含有する湿気のある環境において37℃でインキュベートし、そして7日後、形態学的基準によって、倒立顕微鏡法によりスコア化した。
【0253】
実施例10:
移動アッセイ(Migration assay)
125ng/ml SDF−1αを、37℃で一晩CTK(1mcg/ml)とインキュベートするかまたは未処理に保った。CTKの酵素活性を阻害するために、1および2mclのプロテアーゼ阻害剤(Sigma)を、1.5時間CTKと共にインキュベートし、その後SDF−1へ添加した。10%FCSを補充したRPMI(600μl)を、試験されたSDF−1と共に、Costar24ウエルトランスウエル(Costar 24-wells transwell)(Corning(孔サイズ5μm),NY)の下部チャンバへ添加した。100μl培地中の1×105pre B ALL G2細胞を、上部チャンバへ加え、そして37℃で4時間移動させた。移動細胞を下部チャンバから回収し、そしてFACSCaliburを使用して60秒間カウントした。コントロール自然移動を、下部チャンバにおいてSDF−1α無しに行った。
【0254】
実施例11:
動物。
8〜10週齢のBalb/cマウスをHarlan Rehovotから購入した。全ての実験は、ワイツマン・インスティテュートの動物ケア委員会(the animal care committee of the Weizmann Institute)によって承認された。
【図面の簡単な説明】
【0255】
【図1】LPS誘発炎症が破骨細胞活性化をもたらすことを示す。Balb/cマウスに、LPSの単回注射(250mcg/マウス)をIPで注射した。5日後、骨を採取し、そして活性化された破骨細胞を、赤色での酵素TRAP染色により検出した(A)。BMを採取し、そして20ng/mlのマウスM−SCFおよび可溶性マウスRANKL(これらは、破骨細胞分化および活性化を誘発する)の存在下で播種し、アッセイを破骨細胞BM前駆体のレベルを定量するために使用した(B)。SDF−1、HGFおよびカテプシンK(CTK)の発現を、RT−PCRで評価した(C)。
【図2】LPS誘発炎症がSDF−1分解および前駆体動員をもたらすことを示す。BM液および末梢血細胞を、図1に記載のマウスから収集した。(A)ELISAによって測定したBM液中のSDF−1濃度。(B)半固体培養物(semi solid culture)において測定されたPB前駆体によって増加されたコロニーの数。
【図3】LPS誘発炎症が、16時間以内にSDF−1の減少およびBMからの細胞放出(egress)をもたらすことを示す。LPSの単回注射(250mcg)で処理したマウスを16時間後に殺した。BM SDF−1濃度をELISAによって測定し(A)、そしてBMおよび血液循環中の単核細胞の数を血球計によって測定した(B)。
【図4】SDF−1/CXCR4相互作用が、前駆体のLPS誘発動員に関与することを示す。LPSの単回注射(250mcg)16時間後、Balb/cマウスを犠牲死させ、そしてBMおよびPBを採取した。CXCR4発現を、フローサイトメトリーによって測定し(A)、そして循環中の前駆体のレベルを半固体培養物において測定した(B)。
【図5】機能性CXCR4およびMMP 2/9がLPS媒介動員(LPS-mediated mobilization)のために必要とされることを示す。Balb/cマウスにLPSの単回注射(250mcg)を注射した。MMP 2/9阻害剤(100mcg)および抗CXCR4(10ug)を2回注射した:LPS注射の2時間前および2時間後。LPS注射の16時間後、BMおよびPBを採取し、そして血中(A)およびBM中(B)の白血球の数を、血球計を使用して評価した。
【図6】制御された出血によって誘発されたストレスシグナルに応答しての、SDF−1レベルの減少およびCXCR4発現の増加を示す。Balb/cマウスを、体重1%の単回出血(single bleeding)によって処理した。3、7、10および14日後、BMおよびPBを採取した。BM中のSDF−1レベルをELISAによって測定した(A)。PB細胞によるCXCR4発現をフローサイトメトリーにより定量し(B)、そして動員を示す循環前駆体のレベルを半固体培養物において測定した(C)。
【図7】制御された出血が破骨細胞活性化を誘発することを示す。図6に記載のマウスの大腿を固定し、脱灰し、パラフィン包埋し、そして切片化した。TRAP染色を使用して、赤色の活性化破骨細胞を検出した。
【図8】G−CSF、HGFおよびSDF−1でのインビトロ刺激がOcl活性化を誘発することを示す。マウス原発性頭蓋冠骨芽細胞を、ビタミンD3およびPG2Eの存在下で成長させ、M−CSFの分泌およびRANKLの発現を増強した。BM細胞を、G−CSF(50ng/ml)、HGF(50ng/ml)、SDF−1(10および100ng/ml)有りまたは無しで、骨芽細胞単層上に播種した。成熟活性化破骨細胞を、TRAP染色により、5日後に検出した。(A)代表的TRAP染色。(B)Aの要旨。
【図9】G−CSFおよびHGFに応答しての、原発性マウス骨芽細胞によるSDF−1分泌の増加を示す。マウス原発性頭蓋冠骨芽細胞を、50ng/ml G−CSFおよびHGFの存在下で3日間培養した。(A)RT−PCRにより測定した、SDF−1 mRNAの発現。(B)ELISAにより測定された、処理した骨芽細胞によって分泌されたSDF−1のレベル。
【図10】サイトカインSDF−1およびHGFが、インビボで破骨細胞活性化およびHPC動員を誘発することを示す。Balb/cマウスに、SDF−1(10mcg)およびHGF(1.5mcg)の5連日注射を注射した。処理マウスの骨を、赤色で成熟活性破骨細胞を示すTRAPについて染色した(A)。末梢血液細胞を半固体培養物によって、動員を示す循環前駆体のレベルについて測定した(B)。
【図11】Ocl分化因子RANKLが、骨内膜に沿うTRAP+活性化破骨細胞のインビボ形成を誘発することを示す。Balb/cマウスに、可溶性マウスRANKL(1日当たり2回注射、皮下(subcoutaneous)注射当たり5mcg)を注射した。処理マウスの骨を、成熟活性化破骨細胞を示すTRAP(赤色で)について染色した(A)。BM液中のMMP−9のレベルをザイモグラフィーアッセイ(zymography assay)により測定した(B)。CTK mRNAの発現をRT−PCRにより検出した(C)。処理マウスの骨を、成熟活性化破骨細胞を示すTRAP(赤色で)について染色した。BM液中のMP−9のレベルをザイモグラフィーアッセイにより測定した(B)。CTK mRNAの発現をRT−PCRにより検出した(C)。
【図12】RANKL処理が、CXCR4およびMMP2/9によって媒介される、前駆体動員を誘発することを示す。(A)5日間注射したSDF−1(10mcg)、HGF(1.5mcg)および最初の3日間注射したsRANKL(2×5mcg)によって誘発された、半固体培養物において測定した前駆体動員。(B)RANKL誘発動員は、最後の2日間注射した、抗CXCR4抗体(10mcg)およびMMP2/9(100mcg)の投与によって阻害される。(C)処理マウスのBM中の破骨細胞前駆体を、M−CSFおよびsRANKLと共に4日間BM細胞を培養することによって測定し、続いてTRAP染色し、成熟活性破骨細胞を検出した。
【図13】SDF−1の走化性活性が、CTK(主要な破骨細胞再吸収酵素)によって廃止されることを示す。SDF−1(125ng)を、37℃で16時間、1mcg/ml CTK(反応における最終濃度)と共にインキュベートした。プロテアーゼ阻害剤を、SDF−1を添加する前に1.5時間、CTKと共にイキュベートした。処理したSDF−1へのPre B ALL G2細胞の移動は、その走化性活性を示す。
【図14】SDF−1がCTKによりN末端で切断されることを示す。SDF−1(20ng)を、37℃で、0、15、30および60分間、CTK(1.76ug/ml)と共にインキュベートした。サンプルをSDS−PAGEへ供し、ブロットし、そして抗SDF−1ポリクローナルAbまたはモノクローナルAb K15C(これは、前記ケモカインのアミノ末端部へ特異的に結合する)で検出した。
【技術分野】
【0001】
本発明は、発症および/または経過へのSDF−1の関与によって特徴付けられる疾患を治療するための医薬の製造におけるカテプシンK(CTK)またはカテプシンK阻害剤(CTKI)の使用に関する。
【背景技術】
【0002】
血液中を循環する形態学的に認識可能でありかつ機能的に有能である細胞としては、赤血球、好中球性、好酸性、および好塩基性顆粒、B−、T−、非B−、非T−リンパ球、および血小板が挙げられる。これらの成熟造血細胞は、赤血球系についての赤芽球、顆粒球系についての骨髄芽球、前骨髄球および骨髄球、ならびに血小板についての巨核球のような、それぞれの系統についての形態学的に認識可能な分裂する前駆細胞によって、必要に応じて、置換されるかまたはこれらに由来する。前記前駆細胞は、2つの主要なサブグループへ簡略化して分類され得るより始原的な細胞から生じる:幹細胞および前駆細胞[レビューについては、(1)を参照のこと]。
【0003】
CXCL12またはストロマ細胞由来因子1(SDF−1)は、リンパ球、単球、およびCD34−陽性造血前駆細胞を休止させるための強力な化学誘引物質である(2)。SDF−1は、既知のリガンドとしてこのケモカインのみを有する、1つの受容体、CXCR4へのみ結合する。
【0004】
SDF−1は、低濃度で、白血球インテグリンの活性化、キャピラリー内皮細胞上への白血球の捕捉(arrest)、およびこれらの細胞の溢出へと導く(3)。SDF−1は、B細胞リンパ球産生および骨髄細胞骨髄造血を制御し(4)、そして骨髄移植のために重要である(13)。それは、CD4+T細胞生存を促進し、そしてサイトカインおよびT細胞受容体媒介刺激のためにこれらの細胞をプライムする(5)。SDF−1/CXCR4複合体の機能的活性は、白血球エラスターゼによって媒介されるタンパク質分解によりインビトロで阻害され得ることが示され、そしてこれは、造血細胞前駆体のホーミングが調節される機構であり得る(6)。実際に、インビボでの顆粒球コロニー刺激因子(G−CSF)によって誘発される末梢血への骨髄および他の器官からのCD34+造血細胞前駆体の放出(動員)は、好中球エステラーゼによるSDF−1のタンパク質分解による不活性化に依存する(41)。
【0005】
LevesqueおよびPetitによって実証されたように、カテプシンGおよびエラスターゼはG−CSF誘発動員(G-CSF induced mobilization)においてSDF−1を切断することが公知である。
【0006】
2つの増殖因子(サイトカイン)、顆粒球コロニー刺激因子(G−CSF)および顆粒球マクロファージコロニー刺激因子(GM−CSF)は、(たとえば、骨髄移植および集中的癌化学療法後)重篤な好中球減少症を有する患者における熱および感染を予防するために広く使用される。しかし、サイトカイン療法は高価であり;発熱性好中球減少症の危険性が>=30%である場合に、G−CSFの費用は正当化される。一般的に、G−CSFが化学療法の完了の約24時間後に投与される場合に、最大の臨床的利益が生じる。5μg/kg/日(皮下)の投与量がしばしば有効である。G−CSFおよびGM−CSFは、自家骨髄移植または集中的化学療法を受ける患者において、>500/μlへの好中球カウントの戻りを促進する。
【0007】
先天性、周期性、および特発性好中球減少症における好中球産生は、G−CSF 3〜10μg/kg/日(皮下)の投与によって改善され得る。この治療は、口内炎および他のタイプの口腔咽頭の炎症、熱、および蜂巣炎ならびに他の文書化された細菌感染を有さない患者において示される。利益は滞留され、そして患者は、その効果を失うことなく、数ヶ月または数年間、毎日または隔日G−CSFに維持され得る。長期間のG−CSFはまた、脊髄形成異常症、HIVおよびAIDSならびに自己免疫疾患を含む、他の状況において、好中球産生および好中球動員を誘発して好中球減少症を予防するために使用されている。一般的に、特に重篤な好中球減少症を有さない患者について、この療法の臨床的利益はあまり明確でないが、好中球カウントは増加する。特異体質の薬物反応によって引き起こされる好中球減少症を有する患者もまた、特に遅延した回復が予想される場合、G−CSFから利益を得るかもしれない。しかし、これまでのところ、制御されない試行のみが、後者の状況において報告されている。
【0008】
SDF−1およびその受容体CXCR4の相互作用は、マウス胚発生のあいだのBMの幹細胞播種において重要な役割を果たす(10、11)。以前、本発明者らは、レシピエントとして免疫不全NOD/SCIDマウスを使用して、マウス骨髄の高レベル多系統再生(multilineage repopulation)および短期間インビボ移動(short term in vivo migration)(ホーミング)の両方がSDF−1/CXCR4相互作用に依存することを示すことが出来た(12〜15)。これらのデータのサポートにおいて、ヒトCD34+細胞における高レベルのCXCR4発現またはインビトロでの高SDF−1誘発指向性運動性(directional motility)のいずれかが、CD34+細胞の正の選択を伴う、同種異系および自家臨床移植の両方においてより早い回復と関連することが示されている(16、17)。
【0009】
他の初期に作用するサイトカイン(early acting cytokines)との相乗効果で低濃度のSDF−1は、ヒトCD34+細胞とマウス幹細胞および前駆細胞との両方の増殖を増強し、これは前駆細胞生存におけるこのケモカインの役割を示唆し(25〜29)、一方、高濃度のSDF−1は、移植されたNOD/SCIDマウスの連続再生(serial repopulation)を可能にする原始ヒト胎児肝臓CD34+細胞およびヒト長期培養始原細胞(LTCIC)の増殖との停止を誘発する(30、31)ことが十分に実証されている。
【0010】
CXCR4発現は、サイトカイン、ケモカイン、間質細胞、接着分子、およびタンパク質分解酵素のような環境因子によって調節される、動的プロセスである(18)。ヒト起源の造血幹細胞および前駆細胞において、CXCR4は、短期間(約40時間)インビトロサイトカイン培養(13、19)またはタンパク質分解酵素(たとえば、MMP−2およびMMP−920)での臍帯血(CB)CD34+の刺激よって、細胞内プールからアップレギュレートされ得る。これは、引き続いて、SDF−1勾配へのそれらのインビトロ移動ならびに移植されたNOD/SCIDおよび連続的に移植されたb2mnull NOD/SCIDにおけるそれらのインビボホーミングおよび再生能を増強し、幹細胞自己再生および発生を運動性とリンクする。最近の報告は、サイトカインカクテルとのより長い培養期間が、ヒトCB CD34+に富む細胞における細胞表面CXCCR4発現の減少(22)を生じさせることを実証し、そして減少された再生が、より長い期間インビトロで培養されたヒト前駆体で実証された。最近、本発明者らは、CB CD34+/CXCR4−ソート細胞は低レベルの細胞内CXCR4を有することを示し、これは、短期間のインビトロサイトカイン刺激に続いて、迅速に細胞表面上に機能的に発現され、移植されたNOD/SCIDマウスのSDF−1依存性ホーミングおよび再生を媒介し得る(15)。
【0011】
SDF−1の不活性化と関連するタンパク質分解切断は、MMP−2およびMMP−9、カテプシンGおよびエラスターゼを含むいくつかの分解酵素によって実証される(Petitら Natl Immunol. 3:687 2002, Valenzuela-Femandezら JBC 277:15677, 2002, Levesqueら JCI 111 :187, 2003 および McQuibbanら JBC 276: 43503, 2001)。
【0012】
ヒトおよびマウス幹細胞24の指向性移動を媒介することにおけるそれらの中心的役割に加えて、SDF−1/CXCR4相互作用はまた、他の幹細胞機能に関与する。重要なことに、SDF−1/CXCR4相互作用はまた、BMにおける幹細胞および前駆細胞の滞留に関与する(10、32、33)。この仮説はまた、マウスBM腔へ直接注入されたヒト造血幹細胞(HSC)の固定におけるSDF−1/CXCR4相互作用の関与を実証する他の研究によって確認されている(34、35)。これらの相互作用の干渉は、BMから循環へのヒトおよびマウスの両方の前駆体の放出/動員を誘発する(36〜41)。
【0013】
骨は、骨を生成する骨芽細胞および骨を再吸収する破骨細胞の活性により平衡化される恒常的な再構築を受けている。骨粗しょう症、パジェット病、特定の形態の関節炎、および骨転移のような、種々の骨疾患は、過剰な破骨細胞媒介骨再吸収によって特徴付けられる。RANKL(NKκBリガンドの受容体アクチベータ)およびその対応のRANK受容体は、破骨細胞分化および活性化において重要な役割を果たし、そしてオステオプロテゲリン(OPG)は、RANKLの生理学的阻害剤である。
【0014】
マトリクス分解は、主に、システインプロテアーゼ、CTKの活性に起因する(7、8)。CTKは、破骨細胞において高度に発現されそして骨再吸収(破骨細胞が骨を分解するプロセス)に関連する、リソソームシステインプロテアーゼである。I型コラーゲンは、有機骨量(organic bone mass)の90〜95%を構成し(Krane, S. M., and Simon, L.(1994) in Scientific American Medicine (Rubenstein, E. および Federman, D. D., eds), Vol. 3 , pp. 1-26, Scientific American, Inc., New York)、そしてCTKについての主要な生物学的基質を示す(9)。全ての哺乳動物コラゲナーゼの中で、CTKは、それらの三重らせん構造内の複数の部位で間質コラーゲンを切断し得る唯一のプロテアーゼである(Garnero, P., Borel, O., Byrjalsen, I., Ferreras, M., Drake, F. H., McQueney, M. S., Foged, N. T., Delmas, P. D., および Delaisse, J. M. (1998) J. Biol. Chem. 273, 32347-32352, 6Kafienah, W., Bromme, D., Buttle, D. J., Croucher, L. J., および Hollander, A. P. (1998) Biochem. J. 331, 727-732)。CTK活性の欠乏は、常染色体劣性骨格形成異常濃化異骨症(autosomal recessive skelら dysplasia pycnodysostosis)を有する患者において観察されるように(GeIb, B. D., Shi, G. P., Chapman, H. A., and Desnick, R. J. (1996) Science 273, 1236-1238)、破骨細胞内のリソソーム中の未消化の膠原原線維の蓄積へ導く(Everts, V., Aronson, D. C, および Beertsen, W. (1985) Calcif. Tissue Int. 37, 25-31)。CTK突然変異体の分析によって、コラーゲン分解異常は、CTKのタンパク質分解活性の損失と必ずしも連結されないことが判明した。前記プロテアーゼの活性部位から離れている、1つの疾患を引き起こす変異、Y212Cは、CTKの全体的なタンパク質分解活性に少しだけ影響を与えたが、そのコラゲナーゼ活性を完全に排除した(Hou, W.-S., Bromme, D., Zhao, Y., Mehler, E., Dushey, C, Weinstein, H., Miranda, C. S., Fraga, C, Greig, F., Carey, J., Rimoin, D. L., Desnick, R. J., および GeIb, B. D. (1999) J. Clin. Invest. 103, 731-738)。この観察は、CTKの触媒活性に加えて、他の特徴が、CTKによるコラーゲンの加水分解のために必要とされることを示した。骨−および軟骨耐性グリコサミノグルカンは、特に、カテプシンLまたはマトリックスメタロプロテアーゼIでは観察されない効果である、CTKによるI型およびII型の間質コラーゲンの分解を増強する(Li, Z., Hou, W. S., and Bromine, D. (2000) Biochemistry 39, 529-536)。この知見は、CTKのコラゲナーゼ活性が、CTKタンパク質およびグリコサミノグルカンの特異的な相互作用を必要とすることを示唆した。しかし、CTKがコラーゲンを分解する機構は、理解しにくいままである。Li Zら(J Biol Chem. 2002 Aug 9;277(32):28669-76. Epub 2002 May 30)は、CTKの膠原溶解性活性はCTKタンパク質とコンドロイチン硫酸との新規のオリゴマー複合体の形成に依存することを実証した。したがって、現在、CTK阻害剤は、骨再吸収を阻害するために使用されている。
【発明の開示】
【0015】
本発明は、SDF−1活性および/または濃度が発症および/または経過に関与する疾患を治療するための医薬の製造における、カテプシンK阻害剤(CTKI)またはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体(circularly permutated derivative)、塩または誘導因子の使用に関する。
【0016】
本発明の1局面において、CTKは、癌、炎症および感染症のような、SDF−1活性によって引き起こされる/悪化される疾患において使用される。
【0017】
本発明の一実施形態において、前記疾患はアレルギー性気道疾患である。
【0018】
本発明のさらなる実施形態において、前記疾患は慢性関節リウマチである。
【0019】
本発明のさらなる実施形態において、前記疾患は動脈硬化症である。
【0020】
本発明のさらなる実施形態において、前記疾患は、癌、たとえば、前立腺癌、腎臓癌、神経芽腫、神経膠腫、膵臓癌、結腸癌、乳癌、白血病、たとえば、急性リンパ芽球性白血病(ALL)および急性骨髄性白血病(AML)である。
【0021】
本発明のさらなる実施形態において、CTKは、好ましくはCXCR4を発現する癌細胞においいて、癌転移を予防するために使用される。
【0022】
本発明の別の実施形態において、CTKIは、HIVのような、SDF−1活性によって予防/緩和される疾患において使用される。
【0023】
さらに、本発明は、必要とする対象において幹細胞の動員を誘発するための医薬の製造における、CTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子の使用を提供する。
【0024】
一実施形態において、必要とする前記対象は、たとえば骨髄移植または癌化学療法に続いて生じる、重篤な好中球減少症に苦しむ。
【0025】
また、本発明は、たとえば必要とする対象において骨髄、肝臓および腎臓のような器官の再生を増強するための、必要とする対象において骨髄中の幹細胞の滞留を増大させるための医薬の製造におけるCTKIの使用を提供する。
【0026】
本発明はまた、有効量のCTKIまたはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を投与することを含む、SDF−1活性および/または濃度によってその発症および経過が影響される疾患を治療する方法を提供する。
【0027】
別の局面において、本発明は、有効量のCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を哺乳動物へ投与することを含む、哺乳動物における癌を治療する方法を提供する。
【0028】
本発明の好ましい実施形態において、前記癌細胞は、CXCケモカイン受容体4(CXCR4)を発現する。
【0029】
別の好ましい実施形態において、本発明の方法は、転移を予防するために使用される。
【0030】
さらに、本発明は、必要とする対象へ有効量のCTKIまたはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を投与することを含む、組織への多能性幹細胞の標的化を調節する方法を提供する。
【0031】
本発明の一実施形態において、CTKIが、標的組織への前記細胞(たとえば、正常な造血細胞)の標的化を増大させるために、該標的組織へ投与される。
【0032】
本発明の好ましい実施形態において、前記細胞がインビボで患者にあり、かつ治療的に有効量のCXCR4アゴニスト、たとえば、SDF−1、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子が、癌患者のような、必要とする該患者へ投与される。
【0033】
別の好ましい実施形態において、前記患者は自家または同種異系骨髄あるいは末梢血幹細胞移植を必要とし、および/または前記患者は細胞毒性薬で治療される。
【0034】
別の実施形態において、本発明は、CTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子が、標的組織への前記細胞(たとえば、新生細胞)の標的化を減少させるために、該標的組織へ投与される方法を提供する。
【0035】
別の局面において、本発明は、造血細胞へ有効量のCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を投与することを含む、造血細胞増殖の速度を減少させる方法を提供する。
【0036】
本発明の一実施形態において、前記造血細胞は、造血幹細胞および造血前駆細胞からなる群から選択される。
【0037】
本発明のさらなる実施形態において、前記細胞はインビボで患者にあり、かつ治療的に有効量のCXCR4アゴニスト、たとえば、SDF−1、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子が必要とする該患者へ投与される。
【0038】
本発明のさらなる実施形態において、前記患者は、自家または同種異系骨髄あるいは末梢血幹細胞移植を必要とする。
【0039】
本発明のさらなる実施形態において、前記患者は、癌を有し、そして細胞毒性薬で治療される。
【0040】
別の局面において、本発明は、CTKをSDF−1と接触させること、SDF−1の活性を測定すること、およびCTKによるSDF−1活性の阻害を防止または逆転し得る化合物を単離することを含む、CTKアンタゴニストを同定する方法を提供する。
【0041】
本発明はまた、CTKをSDF−1と接触させること、SDF−1の完全性を測定すること、およびCTKによるSDF−1活性の分解を防止し得る化合物を単離することを含む、CTKアンタゴニストを同定する方法を提供する。
【0042】
本発明は、本発明の方法によって得られるアンタゴニストに関する。
【0043】
本発明は、例示のためのみに、添付の図面を参照して本明細書に記載される。ここで詳細な図面を特に参照しているが、示される詳細は、例示のためにそして本発明の好ましい実施形態の例示的議論のためにのみ示され、そして本発明の原理および概念的局面の最も有用でありかつ容易に理解される説明であると思われるものを提供するために提示されることが強調される。この点において、本発明の基本的理解のために必要とするよりもより詳細に本発明の構造的詳細を示す試みは成されず、図面を考慮しての説明により、本発明のいくつかの形態が実際にどのように具体化され得るかが当業者に明らかとなる。
【発明を実施するための最良の形態】
【0044】
本発明は、ケモカインストロマ細胞由来因子(SDF−1)が発症および/または経過に関与する疾患を治療するための医薬の製造における、CTK阻害剤(CTKI)またはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子の使用に関する。
【0045】
本発明は、CTKがSDF−1活性を特異的に阻害し得るという知見に基づく。本発明者らは、ストレスシグナルが以下の事象を誘発することを本明細書中において示す:BM SDF−1の一時的増加、破骨細胞活性化、CTK活性化の増加、CTK分解によって媒介されるBM SDF−1濃度の減少、および細胞動員。
【0046】
本発明の原理および作用は、図面および付随の説明を参照してよりよく理解され得る。
【0047】
本発明の少なくとも1つの実施形態を詳細に説明する前に、以下の説明に記載されるかまたは実施例によって例示される詳細へ本発明がその適用で限定されないことが理解される。本発明は、他の実施形態あるいは種々の方法で実施または実行され得る。また、本明細書中で使用される表現および用語は、説明のためであり、そして限定としてみなされるべきでないことが理解される。
【0048】
下記に示される知見は、炎症および損傷のようなストレスシグナルが、破骨細胞活性化および骨髄から末梢血(PB)への前駆体動員を誘発することを実証する。本発明者らは、ストレスシグナルが以下の事象を誘発することを示す:BM SDF−1の一時的増加、破骨細胞活性化、CTK活性化の増加、CTK分解によって媒介されるBM SDF−1濃度の減少、および細胞動員。
【0049】
また、得られた結果は、外から投与されたSDF−1、あるいはG−CSFおよび/またはHGFの機能により産生されたSDF−1、あるいはストレスシグナル(たとえば、炎症または損傷)が、破骨細胞活性化、BM CTKの増加、SDF−1の減少、および前駆体動員を直接誘発することを示す。
【0050】
本発明者らは、インビボでのRANKL(破骨細胞活性化の誘導因子)投与が機能性CXCR4(SDF−1)を必要とする動員を誘発することを実証した。
【0051】
本願において得られた結果は驚くべきことであり、何故ならば、Takamatsuら(Blood. 1998 Nov l;92(9):3465-73. 1998)の結果とは対照的に、結果は、破骨細胞活性化が骨髄から循環への幹/前駆体細胞動員を媒介すること示すためである。CTK、すなわち主要な破骨細胞プロテイナーゼは、そのアミノ末端の特異的かつ迅速な切断によりSDF−1活性化を阻害することが実証された。
【0052】
したがって、本知見は、SDF−1活性を調節するために、カテプシンK阻害剤(CTKI)またはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を使用するという可能性を提供する。
【0053】
多くの疾患の発症または経過は、SDF−1濃度および/または活性のアップまたはダウンレギュレーションに伴って生じる。したがって、CTKIまたはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子は、発症および/または経過へのSDF−1の関与によって特徴付けられる疾患を治療するための医薬の製造において使用され得る。
【0054】
SDF−1およびその受容体CXCR4は、ヒト免疫不全ウイルス1型病態生理学、白血球輸送、炎症、造血、胚形成、新脈管形成、および癌転移において重要な役割を果たす。本発明者らの知見に基づいて、本発明者らは、CTK媒介タンパク質分解が、SDF−l/CXCR4依存細胞シグナル伝達を不活性化しかつ停止させることを示唆する。
【0055】
本発明の一局面において、カテプシンK阻害剤(CTKI)またはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子が、腫瘍、感染症および炎症性疾患のような、SDF−1活性によって引き起こされる/悪化される疾患において使用される。
【0056】
増加する証拠は、アレルギー性気道疾患(Gonzalo ら, [J immunol 165:499-508, 2000], Lukacs ら, [Am. J Pathol. 160:1353-1360, 2002])、慢性関節リウマチ Buckley ら, [J immunol 165:3423-3429, 2000]、および動脈硬化症(Abi-Younes ら, [Circ Res 86:131-138, 2000]のようないくつかの疾患における、腫瘍、感染性および炎症性プロセスの病因に、SDF−1/CXCR4シグナル伝達系を関係させ、ここでSDF−1はアップレギュレートされる。
【0057】
CXCRまたはSDF−1の異常発現は、前立腺癌(Taichman ら. Cancer Res 2002, 62:1832-7)、腎臓癌(Br J Cancer 2002, 86:1250-6)、神経芽腫(J Immunol. 2001, 167:4747-57)、神経膠腫(Zhou ら. J Biol. Chem. 2002, 277:49481, Salmaggi ら J Neurooncol. 2004 May;67(3):305-17.)、膵臓癌(Clin. Cancer Res 2000,6:3530)、結腸癌(Zeelenberg ら. Cancer Res 2003, 63:3833-9)および乳癌(Muller ら. Nature 2001, 410:50-6)のような固形癌において観察されている。
【0058】
大抵の白血病細胞は、接着、生存および増殖の増加を伴ってSDF−1に応答する(Juarez ら Histol Histopathol. 2004, 19 (1): 299-309)。
【0059】
SDF−1はまた、プレBALL細胞(pre-BALL cells)の増殖および生存に関与する(Nishii ら. Br J Haematol. 1999 Jun;105(3):701-10)。Tavorら(Cancer Research 64, 2817-2824, 2004)は、急性骨髄性白血病(AML)細胞がSDF−1を発現すること、および内因性SDF−1を遮断することによりAML細胞生存が減少することを示した。
【0060】
悪性神経膠腫におけるSDF−1は、新脈管形成および内皮細胞と腫瘍細胞とのあいだのクロストークにおける役割を有しているようである(Salmaggi ら J Neurooncol. 2004 May;67(3):305-17)。したがって、CTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を用いてSDF−1活性を阻害することは、臨床的に有益である。
【0061】
SDF−1およびCXCR4は、腫瘍細胞の転移先を決定する。CXCR4はヒト乳癌細胞によって発現され、そしてSDF−1は、マウスモデルにおける転移の最初の目的地である器官において高く、このケモカインシグナル伝達が浸潤反応を誘発することを示している(Muller ら., [Nature 410:50-56, 2001])。したがって、CTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を用いてSDF−1活性を遮断することは、CXCケモカイン受容体4を発現する腫瘍細胞における転移阻害のために有益である。
【0062】
得られた結果は、必要に応じて、破骨細胞活性化および/またはCTK活性を誘発することにより、動員を誘発することができることを実証する。あるいは、動員は、破骨細胞活性化および/またはCTK活性を阻害することによって防止され得る。
【0063】
本発明の一実施形態において、CTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子が、重篤な好中球減少症において前駆体の動員を誘発するために使用される。たとえば、骨髄移植後および/または集中的な癌化学療法後に生じる重篤な好中球減少症において。
【0064】
本発明の別の実施形態において、破骨細胞活性化が、重篤な好中球減少症において前駆体の動員を誘発するために使用される。
【0065】
本発明の別の局面において、CTKIは、SDF−1活性によって予防/緩和される疾患において使用される。
【0066】
SDF−1は、リンパ器官、肝臓、肺、および内皮細胞を囲む間葉細胞を含む種々の組織において産生されることが公知である(Tachibana ら. 1998. Nature 393:591.)。
【0067】
CTKは、比較的に組織特異的であり、そして破骨細胞に加えて、マクロファージ、大動脈平滑筋細胞、甲状腺上皮細胞および肺上皮細胞に存在する。これらの細胞で占められる器官において、CTKIは、SDF−1アゴニストとして機能すると予想される。たとえば、骨髄におけるCTKの活性の阻害は、この酵素によるSDF−1分解および引き続いての動員を防止する。
【0068】
したがって、別の実施形態において、CTKIは、再生(repopulation)を増強するためおよび欠点のある造血(defective hematopoiesis)を回復させるために、前駆体の動員を阻害するためおよびしたがって骨髄における前駆体の滞留を増大させるために使用される。
【0069】
本発明のこの局面にしたがう幹細胞/前駆体は、好ましくは、治療される対象から得られる。しかし、幹細胞/前駆体はまた、同系(syngeneic)、同種異系(allogeneic)から、そしてより好ましくはないが、異種(xenogeneic)ドナーから得られ得る。
【0070】
同種異系または異種幹細胞が使用される場合、レシピエント対象および/または細胞は、好ましくは、移植片対宿主および宿主対移植片拒絶反応を防止するように処理されることが理解される。免疫抑制プロトコルは、当該分野において周知であり、そして米国特許第6,447,765号に開示されている。
【0071】
CXCR4は、T細胞指向性(tropic)ヒト免疫不全ウイルス1についてのコレセプター(coreceptor)として役立ち得る(Oberlin ら, [Nature 382:833-835, 1996])。SDF−1は、この受容体のダウンレギュレーションの支配および促進を両方することによって、CXCR4へ結合することについてヒト免疫不全ウイルス(HIV)と拮抗することが示されており;したがって、それは、このウイルスに対する宿主防衛において役割を果たし得る(Amara ら, [J Exp Med 186:1390-146, 1997])。
【0072】
したがって、本発明の一実施形態において、CTKIは、HIVのようなSDF−1活性によって予防/緩和される疾患において使用される。
【0073】
一局面において、本発明は、有効量のCTKIまたはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、循環的に置換された誘導体、塩または誘導因子を含む、SDF−1濃度および/または活性のアップまたはダウンレギュレーションによって影響される疾患を治療する方法を提供する。
【0074】
一実施形態において、本発明は、哺乳動物における腫瘍細胞の転移を阻害する方法を提供し、ここで該腫瘍細胞はCXCケモカイン受容体4(CXCR4)を発現し、該方法は、該腫瘍の転移を阻害するに十分な量のCTKを該哺乳動物へ投与することを含む。
【0075】
一局面において、本発明は、対象内の組織への多能性幹細胞(たとえば、正常な前駆体)の標的化を調節するために医薬の製造のための、CTKIまたはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、循環的に置換された誘導体、塩または誘導因子の使用を提供する。
【0076】
1実施形態において、本発明は、標的組織におけるCTK活性を阻害することを含む、骨髄のような標的細胞への正常な前駆細胞の標的化を増大させることに関する。
【0077】
別の実施形態において、本発明は、標的組織におけるCTK活性を増大させることを含む、標的組織(たとえば、骨髄)への新生細胞の標的化の減少に関する。
【0078】
CTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、循環的に置換された誘導体、塩または誘導因子は、SDF−1阻害剤として機能し、したがって、それは、白血球輸送、炎症、造血、造血細胞の増殖、胚形成、新脈管形成、および癌転移においてSDF−1活性を阻害する。
【0079】
CTK阻害剤はSDF−1アゴニストとして機能し、したがって、それは、BM SDF−1を増加させる(たとえば、照射)またはSDF−1が骨髄において増加される条件を有する治療または薬剤が投与された患者において、骨髄における造血細胞の増殖を増強する。
【0080】
したがって、一実施形態において、本発明は、造血細胞へ有効量のCTK阻害剤を投与することを含む、正常な造血細胞増殖の速度を増加させる方法に関する。
【0081】
本発明の好ましい実施形態において、前記造血細胞は、造血幹細胞および造血前駆細胞からなる群から選択される。
【0082】
別の好ましい実施形態において、前記細胞は患者中に存在し、そしてSDF−1は、このような治療の必要がある患者において投与されるかまたは誘導される。
【0083】
さらに好ましい実施形態において、このような必要がある前記患者は癌を有する。
【0084】
さらなる好ましい実施形態において、前記患者は、自家または同種異系骨髄または末梢血幹細胞移植の必要性を有する。
【0085】
別の好ましい実施形態において、前記患者は細胞毒性薬で治療され、ここで、CTK阻害剤の前記有効量は、該細胞毒性薬に対する正常細胞の感受性を減少させるに十分である。
【0086】
本発明の一局面において、CTK阻害剤はSDF−1アゴニストとして機能し、したがって、それは、BM SDF−1を増加させる(たとえば、照射)またはSDF−1が骨髄において増加される条件を有する治療または薬剤が投与された患者において、骨髄への造血細胞の移植を増強する。
【0087】
本発明の一局面において、CTKはSDF−1アンタゴニストとして機能し、したがって、それは、骨髄から末梢血への前駆体動員へ導くSDF−1分解を媒介する。
【0088】
本発明の一局面によれば、CTKをSDF−1と接触させること、SDF−1の活性または完全性を測定すること、およびSDF−1活性の阻害または分解を防止または逆転し得る化合物を単離することを含む、CTKアンタゴニストを同定する方法が提供される。
【0089】
一実施形態において、測定されるSDF−1の活性は移動である。本発明は、本方法によって得られ得るCTKアンタゴニストをも意図する。
【0090】
別の局面において、本発明は、CXCR4が該癌細胞において発現されるおよび/またはSDF−1が該癌細胞によって分泌される癌患者の治療方法に関し、該方法は、該患者への治療的に有効な量のCTKの投与を含む。
【0091】
本発明の一実施形態において、癌のタイプは、SDF−1を発現する細胞を含む器官へ典型的に転移するものである。
【0092】
さらなる実施形態において、前記癌は、典型的に、皮膚、肝臓、脳、および肺に転移する。
【0093】
さらなる実施形態において、前記腫瘍細胞は、リンパ腫細胞、神経芽腫細胞、肺癌細胞、血管肉腫細胞、白血病細胞、神経膠腫細胞、または黒色腫細胞、乳癌細胞または前立腺癌細胞から選択される。
【0094】
本明細書中で使用される場合、句「幹細胞」は、特定の特別な機能を有する他の細胞タイプ(即ち、「完全に分化した」細胞)へ分化し得る細胞をいう。
【0095】
本明細書中で使用される場合、「トランスジェニック細胞」は、導入された遺伝子または断片を有する細胞である。
【0096】
用語「動員」は、たとえばサイトカインG−CSFによって誘発される、定常状態ホメオタシスの不均衡に起因して、細胞が骨髄から循環へ放出されるプロセスをいう。
【0097】
用語「前駆細胞」は、コロニー形成し得る、未熟な未分化の造血細胞の不均一な集団をいう。前駆体は幹細胞について富化されるが、それらはまた、真の幹細胞を特徴付ける長期間の骨髄再生をすることが出来ないより成熟した細胞も含む。用語「TRAP+MNC細胞」は、酵素TRAP(酒石酸抵抗性酸性フォスファターゼ)を発現する多核細胞である、活性化された破骨細胞をいう。
【0098】
用語「ストレスシグナル」は、サイトカイニン、ケモカイン、タンパク質分解酵素および接着分子のアップまたはダウンレギュレーションによって達成される、定常状態条件の重篤な不均衡によって誘発される危険状況をいう。例としては、炎症、照射、毒性薬物、サイトカイン刺激または欠乏が挙げられる。
【0099】
用語「CTKの阻害剤」は、本発明の文脈内において、CTK産生および/または機能が弱められ、減少され、あるいは部分的に、実質的にまたは完全に防止または遮断される様式で、CTK産生、発現および/または機能をダウンレギュレートするいずれの分子をもいう。用語「CTK阻害剤」は、CTK産生の阻害剤およびCTK機能の阻害剤を含むように意図される。
【0100】
産生の阻害剤は、合成、またはCTKのプロセッシングに悪影響を与えるいかなる分子でもあり得る。本発明にしたがって考慮される阻害剤は、たとえば、CTKの遺伝子発現の抑制因子、CTK mRNAの転写を減少もしくは防止するかまたは該mRNAの分解へ導くアンチセンスmRNA、正しい折り畳みを妨げるかまたは部分的にもしくは実質的にCTKの分泌を防止するタンパク質、あるいはCTKを分解するプロテアーゼなどであり得る。
【0101】
CTK機能の阻害剤は、たとえば、CTKアゴニストであり得る。アゴニストは、その基質へのCTK結合を担うCTKまたはCTK結合部位を部分的にまたは実質的に中和するに十分な親和性および特異性で、CTK分子自体へ結合し得るかまたはこれを隔離し得る。
【0102】
本明細書中で使用される場合、句「発現または活性を阻害する」は、CTKの発現(転写および/または翻訳)あるいは活性(たとえば、酵素的またはリガンド結合)を部分的にまたは完全に阻害することをいう。いくつかの異なるアプローチが、CTKの活性をダウンレギュレートするために使用され得る。
【0103】
たとえば、CTK活性を阻害することは、CTKに特異的に結合し得る抗体または抗体フラグメントのような薬剤によって達成され得る。好ましくは、該抗体は、CTKの少なくとも1つのエピトープへ特異的に結合する。本明細書中で使用される場合、用語「エピトープ」は、抗体のパラトープが結合する抗原上のいかなる抗原決定基をもいう。
【0104】
エピトープ決定基は、通常、アミノ酸あるいは炭水化物側鎖のような分子の化学的に活性な表面グルーピング(groupings)からなり、そして通常、特定の3次元構造的特徴および特定の電荷特性を有する。CTKの好ましいエピトープは、SDF−1とのCTKの会合の触媒部位または領域を含むものである。
【0105】
用語「抗体」は、本明細書中で使用される場合、インタクトな分子、ならびにFab、F(ab’)2、およびFvのような機能性フラグメントを含む。これらの機能性抗体フラグメントは、以下のように規定される:(1)Fab、抗体分子の一価の抗原結合フラグメントを含み、酵素パパインで完全抗体(whole antibody)を消化してインタクトな軽鎖および1つの重鎖の部分を得ることによって作製され得る、フラグメント;(2)Fab’、ペプシンで完全抗体を処理し、続いて還元して、インタクトな軽鎖および重鎖の部分を得ることによって得られ得る抗体分子のフラグメント;2つのFab’フラグメントが、1抗体分子当たり得られる;(3)F(ab’)2、引き続いての還元無しに酵素ペプシンで完全抗体を処理することによって得られ得る抗体のフラグメント;F(ab’)2は、2つのジスルフィド結合によって一緒に滞留された2つのFab’フラグメントの2量体である;(4)Fv、2つの鎖として発現される軽鎖の可変領域および重鎖の可変領域を含む遺伝子操作されたフラグメントとして規定される;ならびに(5)一本鎖抗体(「SCA」)、遺伝子的に融合された一本鎖分子のような、好適なポリペプチドリンカーによって結合された、軽鎖の可変領域および重鎖の可変領域を含む遺伝子操作された分子。
【0106】
ポリクローナルおよびモノクローナル抗体ならびにそれらのフラグメントを作製する方法は、当分野において周知である(たとえば、Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, New York, 1988を参照のこと。これは、参照により本明細書に組み込まれる)。CTKへ結合する市販のポリクローナルおよびモノクローナル抗体は、本発明における使用に好適である。
【0107】
本発明にしたがう抗体フラグメントは、抗体のタンパク質分解性加水分解あるいは該フラグメントをコードするDNAの大腸菌または哺乳動物細胞(たとえば、チャイニーズハムスター卵巣細胞培養または他のタンパク質発現系)における発現によって作製され得る。抗体フラグメントは、従来法による完全抗体のペプシンまたはパパイン消化によって得られ得る。たとえば、抗体フラグメントは、ペプシンでの抗体の酵素的切断によって作製され得、F(ab’)2という5Sフラグメントを提供する。このフラグメントは、さらに、チオール還元剤および必要に応じてジスルフィド結合の切断から生じるスルフヒドリル基についてのブロック基を使用して切断され得、3.5S Fab’一価フラグメントが作製される。あるいは、ペプシンを使用する酵素的切断によって、直接、2つの一価Fab’フラグメントおよびFcフラグメントが作製される。これらの方法は、たとえば、Goldenberg、米国特許第4,036,945号および第4,331,647号、ならびにそれらに含まれる参照文献によって記載されており、これらはそれらの全体が参照により本明細書中に組み込まれる。また、Porter, R. R. [Biochem. J. 73: 119-126 (1959)]を参照のこと。抗体を切断する他の方法、たとえば、一価軽−重鎖フラグメントを形成するための重鎖の分離、フラグメントのさらなる切断、または他の酵素的、化学的、もしくは遺伝学的技術もまた、インタクトな抗体によって認識される抗原へフラグメントが結合する限りにおいて、使用され得る。
【0108】
Fvフラグメントは、VHおよびVL鎖の会合を含む。この会合は、Inbarら[Proc. Nat'l Acad. Sci. USA 69:2659- 62 (1972)]において記載されるように、非共有結合的であり得る。あるいは、可変鎖は、分子間ジスルフィド結合によって連結されるか、またはグルタルアルデヒドのような化学物質によって架橋され得る。好ましくは、Fvフラグメントは、ペプチドリンカーによって連結されたVHおよびVL鎖を含む。これらの一本鎖抗原結合タンパク質(scFv)は、オリゴヌクレオチドによって連結されたVHおよびVLドメインをコードするDNA配列を含む構造遺伝子を構築することによって作製される。該構造遺伝子は、発現ベクターへ挿入され、引き続いてこれは大腸菌のような宿主へ導入される。
【0109】
抗体フラグメントの形態は、単一の相補性決定領域(CDR)をコードするペプチドである。CDRペプチド(「最小認識単位」)は、問題の抗体のCDRをコードする遺伝子を構築することによって得られ得る。このような遺伝子は、たとえば、ポリメラーゼ連鎖反応を使用して抗体産生細胞のRNAから可変領域を合成することによって、作製される。たとえば、LarrickおよびFry [Methods, 2: 106-10 (1991)]を参照のこと。
【0110】
非ヒト(たとえば、マウス)抗体のヒト化形態は、非ヒト免疫グロブリンから誘導される最小配列を含む、免疫グロブリン、その免疫グロブリン鎖またはフラグメント(たとえば、Fv、Fab、Fab’、F(ab’).sub.2または抗体の他の抗原結合サブ配列)のキメラ分子である。ヒト化抗体としては、ヒト免疫グロブリン(レシピエント抗体)が挙げられ、ここで、該レシピエントの相補性決定領域(CDR)由来の残基は、所望の特異性、親和性および能力(capacity)を有するマウス、ラットまたはウサギのような非ヒト種のCDR由来の残基(ドナー抗体)によって置換されている。場合によっては、ヒト免疫グロブリンのFvフレームワーク残基は、対応の非ヒト残基によって置換される。ヒト化抗体はまた、レシピエント抗体においてもまたは取り込まれた(imported)CDRもしくはフレームワーク配列においても見られない残基を含み得る。一般的に、ヒト化抗体は、少なくとも1つ、そして典型的には2つの、可変ドメインの実質的に全てを含み、ここで、全てまたは実質的に全てのCDR領域は、非ヒト免疫グロブリンのそれらに対応し、そして全てまたは実質的に全てのFR領域は、ヒト免疫グロブリン共通配列のそれらである。ヒト化抗体はまた、最適には、免疫グロブリン定常領域(Fc)(典型的には、ヒト免疫グロブリンのそれ)の少なくとも部分を含む[Jones ら, Nature, 321 :522-525 (1986); Riechmann ら, Nature, 332:323-329 (1988);およびPresta, Curr. Op. Struct. Biol., 2:593-596 (1992)]。
【0111】
非ヒト抗体をヒト化するための方法は、当該分野において周知である。一般的に、ヒト化抗体は、非ヒトである供給源からそれへ導入された1またはそれ以上のアミノ酸残基を有する。それらの非ヒトアミノ酸残基は、しばしば、インポート残基(import residues)と呼ばれ、これらは典型的にインポート可変ドメインから取られる。ヒト化は、齧歯類CDRまたはCDR配列をヒト抗体の対応の配列の代わりに使用することによって、Winterおよび共同研究者[Jones ら, Nature, 321 :522-525 (1986);Riechmann ら, Nature 332:323-327 (1988);Verhoeyen ら, Science, 239:1534-1536 (1988)]の方法にしたがって本質的に行われ得る。したがって、このようなヒト化抗体は、キメラ抗体であり(米国特許第4,816,567号)、ここで、実質的に1つ未満のインタクトなヒト可変ドメインは、非ヒト種由来の対応の配列によって置換されている。実際には、ヒト化抗体は、典型的に、いくつかのCDR残基および恐らくいくつかのFR残基が齧歯類抗体における類似部位由来の残基によって置換されているヒト抗体である。
【0112】
ヒト抗体はまた、ファージディスプレイライブラリ[Hoogenboom および Winter, J. MoI. Biol., 227:381 (1991);Marks ら, J. MoI. Biol., 222:581 (1991)]を含む、当該分野において公知の種々の技術を使用して作製され得る。ColeらおよびBoernerらの技術もまた、ヒトモノクローナル抗体の作製のために利用可能である(Cole ら, Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, p.77 (1985)およびBoerner ら, J. Immunol., 147(l):86-95 (1991))。同様に、ヒト抗体は、内因性免疫グロブリン遺伝子が部分的にまたは完全に不活性化されているトランスジェニック動物(たとえば、マウス)へのヒト免疫グロブリン遺伝子座の導入によって作製され得る。挑戦に際して、ヒト抗体作製が観察され、これは、遺伝子再配列、集合(assembly)、および抗体レパートリーを含む、全ての点でヒトにおいて見られるものに非常に似ている。このアプローチは、たとえば、米国特許第5,545,807号;第5,545,806号;第5,569,825号;第5,625,126号;第5,633,425号;第5,661,016号、および以下の科学刊行物において記載されている:Marks ら, Bio/Technology 10: 779-783 (1992);Lonberg ら, Nature 368: 856-859 (1994);Morrison, Nature 368 812-13 (1994);Fishwild ら, Nature Biotechnology 14, 845- 51 (1996);Neuberger, Nature Biotechnology 14: 826 (1996);およびLonberg and Huszar, Intern. Rev. Immunol. 13, 65-93 (1995)。
【0113】
CTK活性の阻害はまた、CTKの公知のペプチド阻害剤を使用することによって、またはCTKの触媒部位と相互作用し得るポリペプチド配列から誘導される任意のペプチド阻害剤(たとえば、基質アナログ)によって、行われ得る。さらなる阻害剤を同定するために使用され得る好適な生化学/分子アプローチの記載は、本明細書以下に提供される。
【0114】
CTKのさらなる阻害剤は、Blanchardら(Structure 7:1125-1133, 1999)およびWattら(Structure 7:1135-1143, 1999)によって記載されるCTKの3次元分子構造についてならびにChouら(FEBS 419:49-54, 1997)によって作製されたその基質結合モデルについて、分子設計アプローチを使用して同定され得る。
【0115】
CTKを細胞内小器官/位置へ再配置し、そしてそれをその生物学的効果を示すことが出来ないようにするタンパク質によって、CTK活性はまた阻害され得る。CTK細胞の発現のダウンレギュレーションは、いくつかの分子アプローチのいずれか1つを使用して行われ得る。
【0116】
たとえば、CTK転写は、低分子干渉RNA(siRNA)分子を使用することによって、RNA干渉を介して阻害され得る。RNA干渉は、2工程プロセスであり;開始工程と呼ばれる第1工程は、恐らくDicer(dsRNA特異的リボヌクレアーゼのRNアーゼIIIファミリーのメンバー)[これは、ATP依存性様式で(直接またはトランスジーンまたはウイルスを介して導入される)dsRNAを処理する(切断する)]の作用によって、インプットされたdsRNAが21〜23ヌクレオチド(nt)の低分子干渉RNA(siRNA)へ消化される。連続する切断事象は、各々が2−ヌクレオチド3’オーバーハングを有する、19〜21bp二重鎖(siRNA)へ前記RNAを分解する[Hutvagner and Zamore Curr. Opin. Genetics and Development 12:225-232 (2002);およびBernstein Nature 409:363- 366 (2001)]。
【0117】
前記エフェクター工程において、前記siRNAデュプレックスは、ヌクレアーゼ複合体へ結合し、RNA誘導型サイレンシング複合体(RISC)を形成する。siRNAデュプレックスのATP依存性の巻戻しが、RISCの活性化のために必要とされる。活性RISCは、次いで、塩基対相互作用によって相同転写物(homologous transcript)を標的化し、そして前記mRNAをsiRNAの3’末端から12のヌクレオチドフラグメントへ切断する[Hutvagner and Zamore Curr. Opin. Genetics and Development 12:225-232 (2002);Hammond ら. (2001) Nat. Rev. Gen. 2:110-119 (2001);およびSharp Genes. Dev. 15:485-90 (2001)]。切断機構は依然として解明されていないが、研究は、各RISCが単一siRNAおよびRNアーゼを含むことを示す(Hutvagner and Zamore, Curr. Opin. Genetics and Development 12:225-232, 2002)。
【0118】
RNAiの顕著な効力のために、RNAi経路内の増幅工程が示唆されている。増幅は、より多くのsiRNAを生じるインプットdsRNAのコピーによって、あるいは形成されるsiRNAの複製によって生じ得る。あるいはまたはさらに、増幅は、RISCの複数の代謝回転事象によって行われ得る[Hammond ら. Nat. Rev. Gen. 2:110-119 (2001), Sharp Genes. Dev. 15:485-90 (2001);Hutvagner and Zamore Curr. Opin. Genetics and Development 12:225-232 (2002)]。RNAiについてのより多くの情報については、以下のレビューを参照のこと:Tuschl ChemBiochem. 2:239-245 (2001); Cullen Nat. Immunol. 3:597-599 (2002);およびBrantl Biochem. Biophys. Act. 1575:15-25 (2002)。
【0119】
本発明での使用に好適なRNAi分子の合成は、以下のように行われ得る。先ず、CTK mRNA配列を、AAジヌクレオチド配列についてAUG開始コドンの下流でスキャンする。各AAおよび3’隣接19ヌクレオチドの出現を、可能性のあるsiRNA標的部位として記録する。好ましくは、siRNA標的部位は、非翻訳領域(LITR)は調節タンパク質結合部位により富んでいるので、オープンリーディングフレームから選択される。UTR結合タンパク質および/または翻訳開始複合体は、siRNAエンドヌクレアーゼ複合体の結合に干渉し得る[Tuschl ChemBiochem. 2:239-245]。しかし、非翻訳領域に指向されるsiRNAはまた有効であり得ることが理解され、何故ならば、GAPDHについて実証されているからであり、ここで、5’UTRで向けられるsiRNAは、細胞性GAPDH mRNAの約90%減少を媒介し、そして完全にタンパク質レベルを廃した(abolished)(www.ambion.com/techlib/tn/91/912.html)。
【0120】
次に、NCBIサーバ(www.ncbi.nlm. nih.gov/BLAST/)から利用可能なBLASTソフトウエアのような任意の配列アラインメントソフトウエアを使用して、可能性のある標的部位を好適なゲノムデータベース(たとえば、ヒト、マウス、ラットなど)と比較する。他のコード配列に対して有意な相同性を示す推定標的部位を除去する。
【0121】
資格(qualifying)標的配列は、siRNA合成のためのテンプレートとして選択される。好ましい配列は低G/C含有量を含むものであり、何故ならば、これらは、55%を越えるG/C含有量を有するものと比較した場合に、遺伝子サイレンシングを媒介することにおいてより有効であると判明したためである。いくつかの標的部位は、好ましくは、評価のために該標的遺伝子の長さに沿って選択される。選択されるsiRNAのよりよい評価のために、ネガティブコントロールが、結合(conjunction)において好ましくは使用される。siRNAのネガティブコントロールは、好ましくは、該siRNAと同一のヌクレオチド組成を含むが、該ゲノムに対して有意な相同性を欠いている。したがって、前記siRNAのスクランブルされたヌクレオチド配列が、それが他の遺伝子に対していずれの有意な相同性を示さない限りにおいて、好ましくは使用される。
【0122】
CTKをダウンレギュレートし得る別の薬剤は、CTKのDNA配列またはmRNA転写物を特異的に切断し得るDNA酵素(DNAzyme)分子である。DNA酵素は、一本鎖標的配列および二本鎖標的配列の両方を切断し得る一本鎖ポリヌクレオチドである(Breaker, R.R. および Joyce, G. Chemistry and Biology 1995;2:655; Santoro, S.W. & Joyce, G.F. Proc. Natl, Acad. Sci. USA 1997;943:4262)。DNA酵素の一般的なモデル(「10−23」モデル)が提案されている。「10−23」DNA酵素は、各々7〜9のデオキシリボヌクレオチドの2つの基質認識ドメインが両側に並ぶ、15のデオキシリボヌクレオチドの触媒ドメインを有する。このタイプのDNA酵素は、プリン:ピリミジン接合部でその基質RNAを効率的に切断し得る(Santoro, S.W. & Joyce, G.F. Proc. Natl, Acad. Sci. USA 199;DNA酵素のレビューについては、Khachigian, LM [Curr Opin MoI Ther 4: 119-21 (2002)]を参照のこと)。
【0123】
一本鎖および二本鎖標的切断部位を認識する合成の人工DNA酵素の構築および増幅の例は、Joyceらの米国特許第6,326,174号に開示されている。ヒトウロキナーゼ受容体に指向された類似の設計のDNA酵素は、ウロキナーゼ受容体発現を阻害しそしてインビボで首尾良く結腸癌細胞転移を阻害することが最近観察された(Itohら, 20002, Abstract 409, Ann Meeting Am Soc Gen Ther www.asgt.org)。別の適用において、bcr−abl癌遺伝子に相補的なDNA酵素は、白血病細胞における該癌遺伝子発現を阻害し、そしてCMLおよびALLのケースにおける自家骨髄移植における再発率を減少させることに成功した。
【0124】
CTK発現の阻害はまた、CTKをコードするmRNA転写物と特異的にハイブリダイズしそれによってCTK転写物の翻訳を特異的に阻害し得るアンチセンスポリヌクレオチドを使用することによって行われ得る。
【0125】
CTK発現を効率的に阻害するために使用され得るアンチセンス分子の設計は、アンチセンスアプローチにとって重要な2つの局面を考慮しながら、行われなければならない。第1局面は、好適な細胞の細胞質への前記オリゴヌクレオチドの送達であり、一方、第2局面は、その翻訳を阻害する様式で細胞内の該設計されたmRNAへ特異的に結合するオリゴヌクレオチドの設計である。
【0126】
先行技術は、広範囲の細胞タイプにオリゴヌクレオチドを効率的に送達するために使用され得る、多数の送達戦略を教示する。[たとえば、Luft, J MoI Med 76: 75-6, 1998;Kronenwett ら, Blood 91 : 852-62, 1998;Rajur ら, Bioconjug Chem 8: 935-40, 1997;Lavigne ら, Biochem Biophys Res Commun 237: 566-71, 1997;およびAoki ら, Biochem Biophys Res Commun 231 : 540-5), 1997を参照のこと]。
【0127】
さらに、前記標的mRNAおよび前記オリゴヌクレオチドの両方における構造変化のエネルギー論の原因となる熱力学サイクルに基づく、最も高い予想される結合親和性を有する配列を同定するためのアルゴリズムもまた、利用可能である[たとえば、Walton ら. Biotechnol Bioeng 65: 1-9, 1999を参照のこと]。
【0128】
このようなアルゴリズムは、細胞におけるアンチセンスアプローチを実行するために首尾良く使用されてきた。たとえば、Waltonらによって開発されたアルゴリズムは、科学者が、ウサギベータ−グロビン(RBG)およびマウス腫瘍壊死因子−α(TNFアルファ)転写物について首尾良くアンチセンスオリゴヌクレオチドを設計することを可能にした。同一の研究グループは、より最近、動的PCR技術によって評価した場合、細胞培養物において3つのモデル標的mRNA(ヒト乳酸デヒドロゲナーゼAおよびBならびにラットgp130)に対して合理的に選択されたオリゴヌクレオチドのアンチセンス活性は、ほとんど全てのケース(ホスホジエステルおよびホスホロチオエートオリゴヌクレオチド化学で2つの細胞タイプにおける3つの異なる標的に対する試験を含む)において有効であることが判明したことを報告した。
【0129】
さらに、インビトロ系を使用して特異的オリゴヌクレオチドの有効性を設計および予想するためのいくつかのアプローチもまた公表された(Matveeva ら, Nature Biotechnology 16, 1374-1375, 1998)。
【0130】
いくつかの臨床試験は、アンチセンスオリゴヌクレオチドの安全性、実行可能性および活性を実証した。たとえば、癌の治療に好適なアンチセンスオリゴヌクレオチドが首尾良く使用され(Holmund ら, Curr Opin MoI Ther 1 :372-85, 1999)、一方、c−myb遺伝子、p53およびBcl−2を標的化するアンチセンスオリゴヌクレオチドによる血液悪性腫瘍の治療は、臨床試験に入っており、そして患者によって許容されることが示された(Gerwitz Curr Opin MoI Ther 1 :297-306, 1999)。
【0131】
より最近、ヒトヘパラナーゼ遺伝子発現のアンチセンス媒介抑制が、マウスモデルにおけるヒト癌細胞の胸膜転移を抑制すると報告された(Uno ら., Cancer Res 61 :7855-60, 2001)。
【0132】
したがって、現在の総意は、上述されるように、非常に正確なアンチセンス設計アルゴリズムおよび多種多様のオリゴヌクレオチド送達系の作製へ導いたアンチセンス技術の分野における最近の進展は、当業者が、過度な試行錯誤実験に頼る必要無しに、公知の配列の発現をダウンレギュレートするに好適なアンチセンスアプローチを設計および実行することを可能にしたということである。
【0133】
アンチセンス配列は、CTKをコードする転写物を切断し得るリボザイム配列を含み得、それによって、機能性CTKへのそれらの転写物の翻訳を防止する。このようなリボザイムは、固相オリゴヌクレオチド合成を使用して容易に合成可能である。
【0134】
リボザイムは、問題のタンパク質をコードするmRNAの切断による遺伝子発現の配列特異的阻害のためにますます使用されている[Welch ら., "Expression of ribozymes in gene transfer systems to modulate target RNA levels." Curr Opin Biotechnol. 1998 Oct;9(5):486-96]。何らかの特定の標的RNAを切断するようにリボザイムを設計する可能性のために、基礎研究および治療適用の両方においてそれらを価値あるツールにした。治療学分野において、リボザイムは、感染症におけるウイルスRNA、癌における優性癌遺伝子、および遺伝性疾患における特定の体細胞突然変異を標的化するために開発された[Welch ら, "Ribozyme gene therapy for hepatitis C virus infection." Clin Diagn Virol. 1998 JuI 15;10(2- 3): 163-71.]。最も顕著には、HIV患者についてのいくつかのリボザイム遺伝子療法プロトコルが、既に臨床第1相試験にある。より最近では、リボザイムは、トランスジェニック動物、遺伝子標的確認、および経路解明のために使用されている。いくつかのリボザイムが臨床試験の種々の段階にある。ANGIOZYMEは、ヒト臨床試験において研究される最初の化学合成されたリボザイムであった。ANGIOZYMEは、新脈管形成経路における重要な成分である、VEGF−r(血管内皮成長因子受容体)の形成を特異的に阻害する。Ribozyme Pharmaceuticals, Inc.、ならびに他の企業は、動物も出るにおける抗新脈管形成療法の重要性を実証した。HEPTAZYME、C型肝炎ウイルス(HCV)RNAを選択的に破壊するように設計されたリボザイムは、細胞培養アッセイにおいてC型肝炎ウイルスRNAを減少させることにおいて有効であると判った(Ribozyme Pharmaceuticals, Incorporated−ウェブホームページ)。
【0135】
上記の発現可能な阻害剤(たとえば、抗体フラグメント、アンチセンスなど)は組換え技術を使用して合成され得そしてたとえば注入によって直接提供され得るが、このような分子はまた、哺乳動物細胞において構成的組織特異的または誘発性転写を指向するために好適なプロモータ配列の転写制御下に配置された、該阻害剤をコードするポリヌクレオチド配列を含む発現ベクターを使用することによって細胞において直接発現され得る。
【0136】
本発明での使用に好適な構成的プロモータは、大抵の環境条件および大抵のタイプの細胞(たとえば、サイトメガロウイルス(CMV)およびラウス肉腫ウイルス(RSV))下で機能性である(即ち、転写を指向し得る)配列を含む。本発明の使用に好適な組織特異的プロモータは、造血細胞において機能性である配列を含み、例としては、たとえば、ClarkおよびGordon(Leukoc Biol. 63:153-68, 1998);Steinら(Cancer 15::2899-902, 2000);ならびにHormasら(Curr Top. Microbiol. Immunol. 211 :159-64, 1996)によって記載されるプロモータが挙げられる。本発明での使用に好適な誘導性プロモータとしては、たとえば、テトラサイクリン誘導性プロモータ(Srour ら, hromb. Haemost. 90: 398-405, 2003)が挙げられる。
【0137】
本発明の発現ベクターは、原核生物、真核生物、または好ましくは両方(たとえば、シャトルベクターにおける複製および組込みにこのベクターを好適にするさらなる配列を含む。典型的なクローニングベクターは、転写および翻訳開始配列(たとえば、プロモータ、エンハンサー)ならびに転写および翻訳ターミネータ(たとえば、ポリアデニル化シグナル)を含む。
【0138】
真核生物プロモータは、典型的に、2タイプの認識配列、TATAボックスおよび上流プロモータエレメントを含む。転写開始部位の25〜30塩基対上流に配置される、TATAボックスは、RNA合成を開始するようにRNAポリメラーゼに指令することに関与すると考えられる。他の上流プロモータエレメントは、転写開始速度を決定する。
【0139】
エンハンサーエレメントは、同種または異種プロモータから1,000倍まで転写を刺激し得る。エンハンサーは、転写開始部位から下流または上流に配置される場合、活性である。ウイルスから誘導される多くのエンハンサーエレメントは、広い宿主範囲を有し、そして種々の組織において活性である。たとえば、SV40初期遺伝子エンハンサーは、多くの細胞タイプについて好適である。本発明に好適である他のエンハンサー/プロモータ組み合わせとしては、以下から誘導されるものが挙げられる:ポリオーマウイルス、ヒトまたはマウスサイトメガロウイルス(CMV)、マウス白血病ウイルス、マウスまたはラウス肉腫ウイルスおよびHIVのようなレトロウイルス由来の長い末端反復配列(long term repeat)が挙げられる。たとえば、Enhancers and Eukaryotic Expression, Cold Spring Harbor Press, Cold Spring Harbor, N.Y. 1983を参照のこと。これは、参照により本明細書中に組み込まれる。
【0140】
ポリアデニル化配列はまた、Scfvのようなポリペプチド阻害剤の翻訳効率を増加させるために発現ベクターへ添加され得る。2つの異なる配列エレメントが、正確かつ効率的なポリアデニル化のために必要とされる:ポリアデニル化部位から下流に配置されるGUまたはUに富む配列、および11〜30ヌクレオチド上流に配置される、6ヌクレオチドの高度に保存された配列、AAUAAA。本発明に好適である終止およびポリアデニル化シグナルは、SV40から誘導されるものが挙げられる。
【0141】
既に述べたエレメントに加えて、本発明の発現ベクターは、典型的に、クローン化核酸の発現レベルを増大させるかまたは組換えDNAを有する細胞の同定を促進するように意図された他の特殊エレメントを含む。たとえば、多数の動物ウイルスは、許容性細胞タイプにおけるウイルスゲノムの余分な(extra)染色体複製を促進するDNA配列を含む。これらのウイルスレプリコンを有するプラスミドは、プラスミド上に保有される遺伝子によってまたは宿主細胞のゲノムで好適な因子が提供される限り、エピソーム複製される。
【0142】
前記ベクターは、真核細胞のレプリコンを含んでも含まなくてもよい。真核細胞レプリコンが存在する場合、前記ベクターは、好適な選択可能なマーカーを使用して、真核細胞中において増幅可能である。前記ベクターは真核細胞レプリコンを含まない場合、エピソーム性の増幅は可能でない。その代わりに、組換えDNAは、遺伝子操作された細胞のゲノムへ組み込まれ、ここで、前記プロモータは所望の核酸の発現を指令する。
【0143】
本発明の発現ベクターは、さらに、以下を可能にする追加のポリヌクレオチドを含み得る:たとえば、内部リボソーム侵入部位(IRES)およびプロモータキメラポリペプチドのゲノム組込みのための配列のような、単一のmRNAからのいくつかのタンパク質の翻訳。
【0144】
哺乳動物発現ベクターの例としては、以下が挙げられるがこれらに限定されない:pcDNA3、pcDNA3.1(+/−)、pGL3、pZeoSV2(+/−)、ρSecTag2、pDisplay、pEF/myc/cyto、pCMV/myc/cyto、pCR3.1、pSinRep5、DH26S、DHBB、pNMTl、pNMT41、pNMTSl(これらは、Invitrogenから入手可能)、pCI(これは、Promegaから入手可能)、pMbac、pPbac、pBK−RSVおよびpBK−CMV(これらは、Strategeneから入手可能)、pTRES(これは、Clontechから入手可能)、ならびにそれらの誘導体。
【0145】
レトロウイルスのような真核生物ウイルス由来の調節エレメントを含む発現ベクターもまた、本発明によって使用され得る。SV40ベクターとしては、pSVT7およびpMT2が挙げられる。ウシ乳頭腫ウイルスから誘導されるベクターとしてはpBV−IMTHAが挙げられ、そしてエプスタイン・バー・ウイルスから誘導されるベクターとしてはpHEBOおよびp2O5が挙げられる。他の例示的なベクターとしては、pMSG、pAV009/A+、pMTO10/A+、pMAMneo−5、バキュロウイルスpDSVE、ならびにSV−40初期プロモータ、SV−40後期プロモータ、メタロチオネインプロモータ、マウス乳腺癌ウイルスプロモータ、ラウス肉腫ウイルスプロモータ、ポリヘドリンプロモータ、または真核生物細胞における発現に有効と示される他のプロモータの指令下でタンパク質の発現を可能にする任意の他のベクターが挙げられる。
【0146】
ウイルスは、多くの場合において宿主防衛機構を回避するように進化した、非常に特殊な感染体である。典型的に、ウイルスは、特定の細胞タイプにおいて感染および繁殖する。ウイルスベクターの標的特異性は、その天然の特異性を使用し、所定の細胞タイプを特異的に標的化しそして感染された細胞へ組換え遺伝子を導入する。したがって、本発明によって使用されるベクターのタイプは、形質転換される細胞タイプに依存する。形質転換される細胞タイプにしたがって好適なベクターを選択する能力は、十分に当業者の能力内にあり、したがって選択考慮の一般的な説明は本明細書中において提供されない。たとえば、骨髄細胞は、ヒトT細胞白血病ウイルス1型(HTLV−I)を使用して標的化され得る。
【0147】
組換えウイルスベクターは、CTK阻害剤のインビボ発現のために有用であり、何故ならば、それらは、側方感染(lateral infection)および標的特異性のような利点を提供するからである。側方感染は、たとえばレトロウイルスのライフサイクルにおいて固有であり、そして単一の感染された細胞が多くの子孫ビリオン(これらは、出芽しそして隣接する細胞に感染する)を産生するプロセスである。結果は、その大部分が元々のウイルス粒子によって最初感染されなかった、広い領域が迅速に感染されるということである。これは、感染因子が娘子孫(daughter progeny)によってのみ広がる垂直感染とは対照的である。側方に広がることができないウイルスベクターもまた、産生され得る。この特徴は、所望の目的が限定数の標的化細胞へのみ特定の遺伝子を導入することである場合に、有用であり得る。
【0148】
造血細胞特異的発現に好適である構築物のさらなる記載は、Malikら(Blood 15: 86:2993-3005, 1995)に提供される。
【0149】
十分でない量の阻害剤を発現するか、または阻害剤の発現について通常サイレントな細胞における、CTKの内因性阻害剤の内因性産生を誘発および/または増強するためのベクターの使用もまた、本発明にしたがって考慮される。前記ベクターは、前記阻害剤を発現することが望まれる細胞中で機能性である調節配列を含むかもしれない。このような調節配列は、プロモータまたはエンハンサーを含む。次いで、調節配列は、相同組換えによってゲノムの正しい遺伝子座へ導入され、このようにして、該調節配列と該遺伝子(この発現が、誘発または増強されるように要求される)とを作動可能に連結する。前記技術は、通常、「内因性遺伝子活性化」(EGA)と呼ばれ、そしてそれは、たとえばWO91/09955に記載されている。
【0150】
同一の技術を使用して、即ち、CTKの遺伝子座へ、CTK発現のダウンレギュレーションもしくは防止を生じさせる負の調節エレメント(たとえば、サイレンシングエレメント)を導入することによって、CTK発現を停止させることが可能であることが、当業者よって理解される。CTK発現のこのようなダウンレギュレーションまたはサイレンシングは、疾患を予防および/または治療するためのCTKIの使用と同一の効果を有することを、当業者は理解する。種々の方法が、本発明の発現ベクターを造血細胞へ導入するために使用され得る。このような方法は、一般的に、以下において記載されており:Sambrookら, Molecular Cloning: A Laboratory Manual, Cold Springs Harbor Laboratory, New York (1989, 1992);Ausubelら, Current Protocols in Molecular Biology, John Wiley and Sons, Baltimore, Md. (1989);Changら, Somatic Gene Therapy, CRC Press, Ann Arbor, Mich. (1995);Vega ら, Gene Targeting, CRC Press, Ann Arbor Mich. (1995);Vectors: A Survey of Molecular Cloning Vectors and Their Uses, Butterworths, Boston Mass. (1988);およびGilboaら[Biotechniques 4 (6): 504-512, 1986]、そしてたとえば、組換えウイルスベクターを用いての安定的または一時的トランスフェクション、リポフェクション、エレクトロポレーションおよび感染を含む。さらに、陽性−陰性の選択方法については、米国特許第5,464,764号および第5,487,992号を参照のこと。
【0151】
ウイルス感染による核酸の導入は、リポフェクションおよびエレクトロポレーションのような他の方法に対していくつかの利点を提供し、何故ならば、より高いトランスフェクション効率がウイルスの感染性に起因して得られ得るためである。
【0152】
前記阻害剤を発現するために使用される発現構築物は、好ましくは、これらの細胞中での独占的かつ制御可能な発現を可能にする様式で、構築されそして造血細胞へ導入されることが理解される。たとえば、もっぱら造血細胞を形質転換し得るウイルス発現ベクターを使用することによってあるいはこのような細胞をエクスビボで形質転換することによって、そして該発現構築物(上記の例を参照のこと)において誘発性プロモータ配列を使用することによって、これらの細胞中での独占的かつ制御可能な発現が達成され得る。
【0153】
このような発現戦略は、白血病治療の文脈において使用される場合に特に有利であり、何故ならば、それは、造血(hematopoesis)に対する正確な制御およびしたがって造血細胞数の調節を可能にするためである。
【0154】
本明細書に記載されるように、CTK発現または活性化のダウンレギュレーションは、ダウンレギュレーション剤へ培養造血細胞を暴露することによってインビトロで、あるいは対象へこのような薬剤を投与することによってインビボで行われてもよい。これらのインビボおよびインビトロアプローチは、CTK阻害剤の以下の非限定的な例を処理するために使用され得る:ペプチド阻害剤、たとえば、Mu−Leu−hph−us−ph(Palmer J Med. Chem 1995, 38:3193-3196)、非ペプチド阻害剤、たとえば、SB−462795、SB−357114、462795(Glaxo Swithkline's)、E64、AAR494、アンチセンス(Inui らJ Biol Chem. 1997 Mar 28;272(13):8109-12.)、NH連結されたアリール/ヘテロアリールCTKI(Robichaud ら Bioorg Med Chem Lett. 2004 Aug 16;14(16):4291-5)、AAE581(Novartis)。
CTK阻害剤はまた、以下に開示されている:WO00/55126およびWO01/49288、Taveresら(J Med Chem 47:5049, 2004, Jmed Chem 29:47(3):588-99, 2004)、Catalaiioら(Bioorg Med Chem Lett.2004 9;14(3):719-22, and 14(1) 275-8)、Robichaudら(J Med Chem 14;46(17):3709-27, 2003)、Settiら(Bioorg Med Chem Lett 13(12):2051-3, 2003)、Altmannら(Bioorg Med Chem Lett 13(12): 1997-2001, 2003)、Chenら(J Org Chem, 68(7):2633-8, 2003)。
【0155】
本明細書中の用語「ポリペプチドおよびタンパク質」は、互換可能である。本発明はまた、本発明のCTKタンパク質のムテインに関し、このムテインは、CTKの天然に生じる配列のみを本質的に有するCTKタンパク質と同一の生物学的活性を本質的に滞留する。このような「ムテイン」は、この種の修飾が該タンパク質自体に対して該タンパク質ムテインの生物学的活性を実質的に変化させないように、約20および10アミノ酸残基までが、それぞれ、CTKタンパク質において、欠失されているか、付加されているか、または他のものによって置換されていてもよいものであり得る。
【0156】
これらのムテインは、公知の合成および/または部位特異的突然変異誘発技術によって、あるいはその好適な任意の他の公知の技術によって作製される。
【0157】
このようなムテインはいずれも、好ましくは、基本であるCTKのそれの実質的に複製的であるアミノ酸配列を有し、その結果、それと実質的に類似の活性を有する。したがって、下記実施例に記載の生物学的活性試験へこのようなムテインを供することを含む、慣用実験によって、どの所定のムテインが本発明の基本タンパク質と実質的に同一の活性を有するかが決定され得る。
【0158】
本発明にしたがって使用され得るCTKタンパク質のムテイン、またはその核酸コードは、置換ペプチドまたはポリヌクレオチドのような実質的にCTK対応配列の有限セットを含み、これらは、本明細書中で示される技術およびガイダンスに基づいて、過度な実験無しに、当業者によって慣用的に得られ得る。タンパク質化学および構造の詳細な説明については、Schulz, G.E. ら, Principles of Protein Structure, Springer- Verlag, New York, 1978;およびCreighton, T.E., Proteins: Structure and Molecular Properties, W.H. Freeman & Co., San Francisco, 1983を参照のこと。これらは参照により本明細書中に組み込まれる。コドン選択のような、ヌクレオチド配列置換の表示については、以下を参照のこと:Ausubelら, Current Protocols in Molecular Biology, Greene Publications and Wiley Interscience, New York, NY, 1987-1995;Sambrookら, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1989。
【0159】
本発明にしたがうムテインについての好ましい変化は、「保存的」置換と知られているものである。天然に生じるCTK配列を本質的に有するタンパク質におけるそれらの保存的アミノ酸置換は、あるグループ内の同義のアミノ酸を含み得、このグループは、該グループのメンバー間の置換が分子の生物学的機能を保存するという十分に類似の物理化学的特性を有する。Grantham, Science, Vol. 185, pp. 862-864 (1974)を参照のこと。特に、前記挿入または欠失が数個のアミノ酸(たとえば、50以下、そして好ましくは20CTK以下)のみを含みかつ機能性構造(functional conformation)に重要であるアミノ酸(たとえば、システイン残基)を除去または置換しない場合、アミノ酸の挿入および欠失もまた、その機能を変化させることなく、上記で定義される配列において成され得ることが明らかである[Anfinsen, "Principles That Govern The Folding of Protein Chains", Science, Vol. 181, pp. 223- 230 (1973)]。このような欠失および/または挿入によって作製されるムテインは、本発明の範囲内に入る。好ましくは、前記同義のアミノ酸基は、表Aに規定されるものである。より好ましくは、前記同義のアミノ酸は、表Bに規定されるものであり;そして最も好ましくは、前記同義のアミノ酸は、表Cに規定されるものである。
【0160】
【表1】
【0161】
【表2】
【0162】
【表3】
【0163】
本発明における使用のための前記タンパク質のムテインを得るために使用され得る、タンパク質におけるアミノ酸置換体の作製例としては、以下に記載のような任意の公知の方法工程が挙げられる:米国特許第RE 33,653号、第4,959,314号、第4,588,585号および第4,737,462号(Markら);第5,116,943号(Kothsら);第4,965,195号(Namenら);第4,879,111号(Chongら);および第5,017,691号(Leeら);ならびに米国特許第4,904,584号(Strawら)に記載のリシン置換タンパク質。
【0164】
本発明の別の好ましい実施形態において、本発明における使用のためのCTKタンパク質の任意のムテインは、本発明の上述のCTKタンパク質のそれに本質的に対応するアミノ酸配列を有する。用語「に本質的に対応する」は、特にCTKに対するその能力が関連する場合、その基本的特徴に影響を与えない基本タンパク質の配列に対して僅かな変化を有するムテインを含むように意図される。用語「に本質的に対応する」に入ると一般的に考えられるタイプの変化は、本発明のCTKタンパク質をコードするDNAの従来の突然変異誘発技術から生じるものであり、ほんの僅かな修飾を生じ、たとえば化学誘引物質に対する幹細胞の感度を増大する所望の活性についてスクリーニングする。
【0165】
本発明はまた、CTK変異体を含む。好ましいCTK変異体は、少なくとも80%アミノ酸同一性を有するものであり、より好ましいCTK変異体は、少なくとも90%アミノ酸同一性を有するものであり、そして最も好ましい変異体は、CTKアミノ酸配列に対して少なくとも95%同一性を有するものである。
【0166】
用語「配列同一性」は、本明細書中で使用される場合、Clustal−Xプログラム[これは、ClustalWマルチプル配列アラインメントプログラム(Thompsonら,1994)についてのウインドウズインターフェースである]を使用する低相同性領域のリファイン(refinement)を伴う、HanksおよびQuinn(1991)にしたがうアラインメントによって該アミノ酸配列が比較されることを意味する。Clustal−Xプログラムは、ftp://ftp-igbmc.u- strasbg.fr/pub/clustalx/でインターネットから入手可能である。当然ながら、このリンクが休止となった場合、当業者であれば、過度な実験無しに、標準的なインターネット検索技術を使用して他のリンクにあるこのプログラムのバージョンを見つけ出し得ることが理解される。特に述べない限り、本願の有効出願日現在で、本明細書中において言及されるプログラムの最新バージョンが、本発明を実施するために使用されるものである。
【0167】
「配列同一性」を測定するための別の方法は、以下である。−12のギャップオープンペナルティー(ギャップの最初のゼロ)および−4のギャップエクステンションペナルティー(該ギャップにおける追加の連続するゼロの各々当たり)を有するデフォルト(BLOSUM62)マトリクス(値−4〜+11)を使用し、Genetic Computing Group’s GDAP(グローバルなアラインメントプログラム)のバージョン9を使用して、配列を整列する。整列後、パーセンテージ同一性が、請求される配列におけるアミノ酸の数のパーセンテージとして一致の数を表現することによって計算される。
【0168】
本発明にしたがうムテインとしては、ストリンジェント条件下でDNAまたはRNAへハイブリダイズしそして本発明にしたがうCTKタンパク質をコードする核酸(たとえば、DNAまたはRNA)[CTKをコードする天然に生じる配列の本質的に全てと、遺伝暗号の縮重の理由で該天然に誘導されるヌクレオチド配列とはそのヌクレオチド配列が異なり得る配列(即ち、多少異なる核酸配列は、その縮重に起因して、同一のアミノ酸配列を依然としてコードし得る)とを含む]によってコードされるものが挙げられる。
用語「ハイブリダイゼーション」は、本明細書中で使用される場合、核酸の鎖が、塩基対形成により相補鎖と結合するいずれのプロセスをも含む(Coombs J, 1994, Dictionary of Biotechnology, stokton Press, New York NY)。「増幅」は、核酸配列のさらなるコピーの産生と規定され、そして一般的に、当該分野において周知のポリメラーゼ連鎖反応技術を使用して行われる(Dieffenbach および Dveksler, 1995, PCR Primer, a Laboratory Manual, Cold Spring Harbor Press, Plainview NY)。
【0169】
「ストリンジェンシー」は、典型的に、約Tm−5℃(プローブの融点より5℃下)からTmより約20℃〜25℃下までの範囲で生じる。
【0170】
用語「ストリンジェント条件」は、当業者が慣習的に「ストリンジェント」と呼ぶ、ハイブリダイゼーションおよび引き続いての洗浄条件をいう。Ausubelら, Current Protocols in Molecular Biology, Greene Publications and Wiley Interscience, New York, NY, 1987-1995;Sambrookら, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY, 1989を参照のこと。
【0171】
本明細書中で使用される場合、ストリンジェンシー条件は、ハイブリダイゼーション実験において使用される温度、一価カチオンのモル濃度およびハイブリダイゼーション溶液中のホルムアミドのパーセンテージの関数である。所定のセットの条件と関連するストリンジェンシーの程度を決定するために、先ず、DNA−DNAハイブリッドの融点Tmとして表される100%同一性のハイブリッドの安定性を決定するためにMeinkothら(1984)の等式が使用される:
Tm=81.5C+16.6(LogM)+0.41(%GC)−0.61(%ホルム(form))−500/L
式中、Mは一価カチオンのモル濃度であり、%GCはDNA中のGおよびCヌクレオチドのパーセンテージであり、%ホルムはハイブリダイゼーション溶液中のホルムアミドのパーセンテージであり、そしてLは塩基対のハイブリッドの長さである。100%同一性ハイブリッドについて計算されるものからTmが低下される各1Cについて、許容されるミスマッチの量は、約1%増加される。したがって、特定の塩およびホルムアミド濃度で所定のハイブリダイゼーション実験のために使用されるTmが、Meinkothの等式にしたがう100%ハイブリッドについて計算されるTmより10C下である場合、ハイブリダイゼーションは、約10%ミスマッチまで存在しても生じる。
【0172】
本明細書中で使用される場合、「高度にストリンジェントな条件」は、上記式によって計算されるかまたは実際に測定される、標的配列との完全な二本鎖について存在するTmよりも多くて10C下であるTmを提供するものである。「中程度にストリンジェントな条件」は、上記式によって計算されるかまたは実際に測定される、標的配列との完全な二本鎖について存在するTmより多くて20C下であるTmを提供するものである。限定されること無しに、高度にストリンジェントな(前記ハイブリッドの計算または測定Tmより5〜10C下)および中程度にストリンジェントな(前記ハイブリッドの計算または測定Tmより15〜20C下)条件の例は、前記ハイブリッドの計算Tm未満の好適な温度で、2×SSC(標準クエン酸食塩水)および0.5%SDS(ドデシル硫酸ナトリウム)の洗浄溶液を使用する。特に、使用されるハイブリダイゼーション条件が、安定なハイブリッドと共にあまり安定でないハイブリッドが形成することを可能にするものである場合、前記条件の最終的なストリンジェンシーは、主として洗浄条件に起因する。より高いストリンジェンシーでの洗浄条件は、次いで、前記あまり安定でないハイブリッドを除去する。上述の高度にストリンジェントな〜中程度にストリンジェントな洗浄条件で使用され得る、典型的なハイブリダイゼーション条件は、前記Tmの約20〜25C下の温度での、6×SSC(または6×SSPE(標準食塩水−リン酸塩−EDTA)、5×Denhardt試薬、0.5% SDS、100&マイクロ(100 µ);g/ml変性断片化サケ精子DNAの溶液中におけるハイブリダイゼーションである。混合プローブが使用される場合、SSCの代わりに塩化テトラメチルアンモニウム(TMAC)を使用することが好ましい(Ausubel, 1987, 1999)。成人幹細胞は、骨髄吸引のような外科手術を使用して得られ得るか、またはNexell Therapeutics Inc. Irvine, CA, USAから入手可能なもののような市販のシステムを使用して収穫され得る。本発明によって使用される幹細胞は、好ましくは、対象の循環へのHSCの放出に対する化学療法またはサイトカイン刺激を使用する、幹細胞動員手順を使用して、回収される(即ち、採取される)。動員は骨髄手術よりもより多くのHSCおよび前駆細胞を得ることが知られているので、幹細胞は、好ましくはこの手順を使用して回収される。
【0173】
CTKの「アイソフォーム」は、SDF−1または選択的スプライシングによって産生され得るそのフラグメントを分解し得るタンパク質である。用語「円順列誘導体」は、本明細書中で使用される場合、線形分子をいい、ここで、末端が直接的にまたはリンカーを介して一緒に結合されて環状分子が作製され、次いで該環状分子が別の位置で開裂されて、元の分子の末端とは異なる末端を有する新たな線形分子が作製されている。円順列物(circular permutations)としては、その構造が、環状化され次いで開裂されている分子と等しい分子が挙げられる。したがって、円順列分子は、線形分子として新たに合成されてもよく、そして環状化および開裂工程を経由しなくてもよい。円順列誘導体の製造は、WO95/27732に記載されている。
【0174】
なおさらなる実施形態において、本発明にしたがう物質は、免疫グロブリン融合を含み、即ち、本発明にしたがう分子は、免疫グロブリンの全部または一部へ融合される。免疫グロブリン融合タンパク質を作製するための方法は、当該分野において周知である(たとえば、WO01/03737に記載されるもの)。当業者であれば、本発明の得られる融合タンパク質はCTKの生物学的活性を滞留することを理解する。得られる融合タンパク質は、体液中の延長された滞留時間(半減期)、増加された特異的活性、増加された発現レベル、または該融合タンパク質の促進された精製のような、改善された特性を理想的には有する。
【0175】
好ましくは、本発明にしたがう前記物質は、Ig分子の定常領域へ融合される。それは、たとえば、ヒトIgG1のCH2およびCH3ドメインのような、重鎖領域へ融合され得る。Ig分子の他のアイソフォームもまた、本発明にしたがう融合タンパク質の作製に好適である:たとえば、アイソフォームIgG2またはIgG4、あるいはIgMまたはIgAのような他のIgクラス。融合タンパク質は、単量体または多量体、ヘテロ−もしくはホモ多量体であってもよい。
【0176】
本発明にしたがう物質の機能性誘導体は、前記タンパク質の特性(たとえば、安定性、半減期、バイオアベイラビリティ、人体による許容性、または免疫原性)を改善するためにポリマーへ複合体化され得る。したがって、本発明の好ましい実施形態は、アミノ酸残基上の1またはそれ以上の部分として生じる1以上の官能基へ結合された少なくとも1つの部分を含む本発明にしたがう物質の機能性誘導体に関する。
【0177】
非常に好ましい実施形態は、ポリエチレングリコール(PEG)へ結合された本発明の物質に関する。PEG化は、たとえばWO92/13095に記載されるもののような、公知の方法によって行われ得る。
【0178】
本発明にしたがう「活性画分」は、たとえば、CTKのフラグメントであり得る。用語フラグメントは、前記分子の任意のサブセット、即ち、所望の生物学的活性を滞留するより短いペプチドをいう。フラグメントは、CTK分子のいずれかの末端からアミノ酸を除去し、そして得られたフラグメントを、SDF−1を分解するその特性について試験することによって、容易に得られ得る。ポリペプチドのN末端またはC末端のいずれかから1つのアミノ酸を除去するためのプロテアーゼは公知であり、そしてしたがって、所望の生物学的活性を滞留するフラグメントを決定することは、慣用的実験のみを含む。
【0179】
CTK、そのムテインおよび融合タンパク質の活性画分として、本発明は、前記タンパク質分子のポリペプチド鎖の任意のフラグメントもしくは前駆体のみ、またはそこへ結合された関連分子または残基(たとえば、糖またはリン酸残基)、あるいはそれら自体による前記タンパク質分子または前記糖残基の凝集体をさらに含み、但し、前記画分は、CTKと実質的に類似の活性を有し、たとえばSDF−1を分解する。
【0180】
用語「塩」は、本明細書中において、CTK分子またはそのアナログのアミノ酸基の酸付加塩およびカルボキシル基の塩の両方をいう。カルボキシル基の塩は、当該分野において公知の手段によって形成されてもよく、そして無機塩、たとえば、ナトリウム、カルシウム、アンモニウム、鉄(ferric)または亜鉛塩など、および有機塩基との塩、たとえば、トリエタノールアミン、アルギニンもしくはリシン、ピペリジン、プロカインのようなアミンと形成されるものを含む。酸付加塩としては、たとえば、鉱酸(たとえば、塩酸または硫酸)との塩、および有機酸(たとえば、酢酸またはシュウ酸)との塩が挙げられる。当然ながら、いずれのこのような塩も、CTKの生物学的活性、たとえばSDF−1を分解する能力を滞留しなければならない。
【0181】
幹細胞動員は、CTKまたは破骨細胞活性化のみであるいは多数の分子と組み合わせて誘発され得る。例としては、以下が挙げられるがこれらに限定されない:サイトカイン、たとえば、顆粒球コロニー刺激因子(G−CSF)、顆粒球マクロファージコロニー刺激因子(GM−CSF)、インターロイキン(IL)−7、IL−3、IL−12、幹細胞因子(SCF)、およびflt−3リガンド;ケモカイン、たとえば、IL−8、Mip−1α、Groβ,またはSDF−1;ならびに化学療法薬 シクロホスファミド(Cy)およびパクリタキセル。これらの分子は速度論および効果で相違することが理解されるが、現在公知の実施形態によれば、G−CSFが、幹細胞を動員するために、単独でまたはたとえばシクロホスファミドと組み合わせて好ましくは使用される。典型的に、G−CSFは、5〜10日間、5〜10μg/kgの用量で毎日投与される。幹細胞を動員する方法は、米国特許第6,447,766号および第6,162,427号に開示されている。ヒト胚幹細胞は、ヒト胚盤胞から単離され得る。ヒト胚盤胞は、典型的に、ヒト体内着床前胚または体外受精(IVF)胚から得られる。あるいは、単一細胞ヒト胚は、胚盤胞段階へ拡張され得る。ヒトES細胞の単離のために、透明帯が胚盤胞から除去され、そして内細胞塊(ICM)が免疫手術によって単離され、ここで、栄養外胚葉細胞が溶解され、そして穏やかなピペッティングによりインタクトなICMから除去される。次いで、ICMを、その成長(outgrowth)を可能にする好適な培地を含む組織培養フラスコ中にプレートする。9〜15日後、ICM誘導化成長を、機械的分離または酵素的分解のいずれかによって集団(clumps)に分離し、次いで該細胞を、新鮮な組織培養培地上に再度プレートする。未分化形態を実証するコロニーを、マイクロピペットによって個々に選択し、集団へ機械的に分離し、そして再度プレートする。次いで、得られるES細胞を1〜2週間毎に定期的に分ける。ヒトES細胞の作製方法についてのさらなる詳細については、Thomsonら[米国特許第5,843,780号;Science 282: 1145, 1998;Curr. Top. Dev. Biol. 38: 133, 1998;Proc. Natl. Acad. Sci. USA 92: 7844, 1995];Bongsoら[Hum Reprod 4: 706, 1989];Gardnerら[Fertil. Steril. 69: 84, 1998]を参照のこと。
【0182】
市販の幹細胞もまた、本発明のこの局面にしたがって使用され得ることが理解される。ヒトES細胞は、NIHヒト胚幹細胞登録(NIH human embryonic stem cells registry)(<http://escr.nih.gov>)から購入され得る。市販の胚幹細胞株の非限定的な例は、BGOl、BG02、BG03、BG04、CY12、CY30、CY92、CYlO、TE03、およびTE32である。
【0183】
ヒトEG細胞は、当業者に公知の実験室技術を使用して、妊娠の約8〜11週のヒト胎児から得られる始原生殖細胞から回収され得る。生殖堤を分離し、そして小さな塊に切断し、その後これを機械的分離により細胞へ分離する。次いで、ES細胞を、好適な培地を含む組織培養フラスコ中で成長させる。ES細胞と一致する細胞形態が観察されるまで、典型的に7〜30日または1〜4継代後まで、培地を毎日交換して、前記細胞を培養する。ES細胞を作製する方法についてのさらなる詳細については、Shamblottら[Proc. Natl. Acad. Sci. USA 95: 13726, 1998]および米国特許第6,090,622号を参照のこと。
【0184】
多分化能を示す幹細胞集団の富化が好ましくは行われ得ることが理解される。したがって、たとえば、上記に概述されるように、CD34+幹細胞は、以下にさらに説明されるようなFACSまたはアフィニティーカラムを使用して制御され得る。
【0185】
増殖条件下での幹細胞の培養がまた、幹細胞の数が処置における使用には少なすぎる場合、行われてもよい。幹細胞の培養は、米国特許第6,511,958号、第6,436,704号、第6,280,718号、第6,258,597号、第6,184,035号、第6,132708号および第5,837,5739号に記載されている。
【0186】
たとえば腫瘍細胞中のCXCR4受容体レベルの分析は、フローサイトメトリーによって検出され得る。このアプローチは、液体媒体中の励起源を過ぎて流れる個々の細胞をスキャンする機器を使用する。前記技術は、可視光および蛍光発光の測定に基づく、単一生存(または死)細胞についての迅速で定量的なマルチパラメータ分析を提供し得る。この基本プロトコルは以下に焦点を合わせる:蛍光標識抗体および特異的細胞結合分子(specific cell-associated molecules)に結合するリガンドによって作製される蛍光強度を測定する。蛍光活性化細胞選別装置を使用して細胞集団を単離するために、本発明の幹細胞を、R&D, 614 McKinley Place NE Minneapolis, MNから市販される抗CXCR4と接触させる。
【0187】
走化性受容体発現のレベルを定量的に評価するための他の細胞学的または生化学的方法としては、以下が挙げられるがこれらに限定されない:標識された(たとえば、放射標識された)ケモカインを使用する結合分析、ウェスタンブロット分析、細胞表面ビオチン化、および免疫蛍光染色。前記受容体発現レベルもまた前記mRNAレベルで測定され得ることが理解される。たとえば、CXCR4 mRNAは、特異的プローブへのハイブリダイゼーションによって細胞中において検出されてもよい。このようなプローブは、クローン化DNAまたはそのフラグメント、RNA(典型的に、インビトロ転写により作製される)、またはオリゴヌクレオチドプローブ(通常、固相合成によって作製される)であり得る。特異的ハイブリダイゼーションに好適なプローブを使用および作製する方法は、周知であり、そして当該分野において使用されている。問題の走化性受容体のmRNAへ特異的にハイブリダイズするプライマーを使用する、増幅反応[たとえば、PCR、"PCR Protocols: A Guide To Methods And Applications", Academic Press, San Diego, CA (1990)]を使用して、mRNAレベルの定量もまた行われ得る。
【0188】
mRNA検出アッセイにおける精度を改善するために、種々のコントロールが、通常使用され得る。たとえば、偽ハイブリダイゼーションを評価するために、サンプルは、無関係のプローブへハイブリダイズされそしてハイブリダイゼーション前にRNアーゼAで処理され得る。
【0189】
走化性受容体発現を測定するために使用され得る。たとえば、走化性剤(たとえば、SDF−1)の勾配を使用しそして膜を介しての該走化性剤への幹細胞移動が続く、機能アッセイもまた、増加した走化性を示す幹細胞を同定および単離するために使用され得る。前記細胞が十分なレベルの前記走化性受容体(たとえば、CXCR4)を発現しない場合、該細胞の大部分は膜上に存在するままである。しかし、本発明の化学誘引物質受容体の増加した発現時に、細胞は、膜を通って移動し、そして走化性プレートのウエルの底部に定着する(実施例セクションの実施例3を参照のこと)。機能ホーミングアッセイ(functional homing assay)もまた、本発明の方法によって使用され得ることが理解される。このようなアッセイは、Kollet (2001) Blood 97:3283-3291(参照文献12)に記載されている。
【0190】
本発明にしたがう好ましい個々の対象は、イヌ、ネコ、ヒツジ、ブタ、ウマ、ウシおよび好ましくはヒトのような哺乳動物である。
【0191】
本発明のさらなる目的、利点、および新規の特徴は、限定される意図は無い以下の実施例の検討の際に、当業者に明らかとなる。さらに、上記で示されそして下記特許請求の範囲セクションにおいて特許請求されるような本発明の種々の実施形態および局面の各々は、下記の実施例に実験的サポートが見られる。
【実施例】
【0192】
上記説明と共に本発明を非限定的に示す以下の実施例がここで参照される。
【0193】
一般的に、本明細書中で使用される命名および本発明において使用される実験室手順は、分子、生化学、微生物学および組換えDNA技術を含む。このような技術は、文献において十分に説明されている。たとえば、以下を参照のこと:"Molecular Cloning: A Laboratory Manual" Sambrook ら, (1989);"Current Protocols in Molecular Biology" Volumes I-III Ausubel, R. M., ed. (1994);Ausubel ら., "Current Protocols in Molecular Biology", John Wiley および Sons, Baltimore, Maryland (1989);Perbal, "A Practical Guide to Molecular Cloning", John Wiley & Sons, New York (1988);Watson ら, "Recombinant DNA", Scientific American Books, New York;Birren ら. (eds) "Genome Analysis: A Laboratory Manual Series", VoIs. 1-4, Cold Spring Harbor Laboratory Press, New York (1998);米国特許第4,666,828号、第4,683,202号、第4,801,531号、第5,192,659号および第5,272,057号に記載される方法;"Cell Biology: A Laboratory Handbook", Volumes I-III Cellis, J. E., ed. (1994);"Current Protocols in Immunology" Volumes I-III Coligaii J. E., ed. (1994);Stites ら. (eds), "Basic and Clinical Immunology" (8th Edition), Appleton & Lange, Norwalk, CT (1994);Mishell and Shiigi (eds), "Selected Methods in Cellular Immunology", W. H. Freeman and Co., New York (1980);利用可能な免疫学的検定法は、特許文献および科学文献に広く記載されており、たとえば、米国特許第3,791,932号、第3,839,153号、第3,850,752号、第3,850,578号、第3,853,987号、第3,867,517号、第3,879,262号、第3,901,654号、第3,935,074号、第3,984,533号、第3,996,345号、第4,034,074号、第4,098,876号、第4,879,219号、第5,011,771号および第5,281,521号を参照のこと;"Oligonucleotide Synthesis" Gait, M. J., ed. (1984); "Nucleic Acid Hybridization" Hames, B. D., and Higgins S. J., eds. (1985);"Transcription and Translation" Hames, B. D., および Higgins S. J., Eds. (1984);"Animal Cell Culture" Freshney, R. I., ed. (1986);"Immobilized Cells and Enzymes" IRL Press, (1986);"A Practical Guide to Molecular Cloning" Perbal, B., (1984)および"Methods in Enzymology" Vol. 1-317, Academic Press;"PCR Protocols: A Guide To Methods And Applications", Academic Press, San Diego, CA (1990);Marshak ら, "Strategies for Protein Purification and Characterization - A Laboratory Course Manual" CSHL Press (1996);これらの全ては、本明細書中に完全に記載されているかのように、参照により組み込まれる。他の一般的な参照文献は、本明細書にわたって提供される。その中の手順は、当該分野において周知であると考えられ、そして読み手の便宜のために提供される。その中に含まれる全ての情報は、参照により本明細書中に組み込まれる。
【0194】
実施例1:
LPS誘発炎症は破骨細胞活性化をもたらす。
骨破壊は、慢性関節リウマチおよび歯周炎を含むいくつかの慢性炎症性疾患の病理学的特徴である。この種の炎症誘発される骨量減少は、破骨細胞活性化および増加した数の骨再吸収性破骨細胞の誘発から生じる。
【0195】
本発明者らは、骨髄(BM)中の破骨細胞活性化に対する、内毒素 リポ多糖類(LPS)投与によって誘発される炎症の効果を研究した。
【0196】
LPS(Sigma)を、250μg/マウスの単回皮下注射でBalb/cマウスへ投与し、そしてコントロール群においてLPSの代わりに生理食塩水を注射した。LPS投与の5日後、マウスを犠牲にし、骨を固定し、脱灰し、パラフィン包埋し、そして切片化し、BMを収穫し、そしてTRAP+破骨細胞(活性化破骨細胞)のレベルをモニターした。
【0197】
本発明者らは、TRAP+破骨細胞(Sigma製のTRAP染色用のキットを使用して、赤色染色により検出される)の急激な増加を見た。このアッセイにおいて、LPS処理したマウスのBMにおける組織TRAPの固有活性を測定するために、基質は外から提供される(図1AおよびB)。
【0198】
次に、本発明者らは、SDF−1、肝細胞成長因子(HGF)および破骨細胞特異的プロテアーゼCTKのような骨髄媒介物(mediators)の転写に対するLPS媒介炎症の効果を研究した。
【0199】
各マウスの1つの大腿を、RNA抽出のためにTrireagentでフラッシュした(flush)。1μg RNAをcDNAの作製のために使用した。半定量RT−PCRを使用して、マウスSDF−1、HGFおよびCTKの転写を測定した。以下のプライマーを使用した:SDF−1(Ponomaryov, 2000, J. Clin. Invest. 106:1331):左プライマー(配列番号1)5’−GGACGCCAAGGTCGTCGCCGTG−3’、右プライマー(配列番号2)5’−TTGCATCTCCCACGGATGTCAG−3’。HGF(Weinstein, 2001, Moll Cell boil, 21(15):5122):左プライマー(配列番号3)5’−TGCCAGAAAGATATCCCGAC−3’、右プライマー(配列番号4)5’−AACTCGGATGTTTGGGTCAG−3’。CTK:左プライマー(配列番号5)5’−GGCCTCTCTTGGTGTCCATA−3’、右プライマー(配列番号6)5’−TCTGCTGCACGTATTGGAAG−3’。
【0200】
本発明者らは、LPS処置マウスのBM中におけるSDF−1転写の減少ならびにHGFおよびCTK転写の急激な増加を観察した(図1C)。
【0201】
したがって、得られた結果は、LPS誘発炎症(5日処置による)は、骨髄中の以下の変化を媒介することを実証する:破骨細胞活性化の増加、CTKおよびHGF転写の増加、ならびにSDF−1転写の減少。
【0202】
実施例2:
LPS誘発炎症はBM SDF−1濃度の減少および前駆体動員をもたらす。
前の実施例において、本発明者らは、炎症が骨髄中のSDF−1の減少を媒介することを示した。本発明者らは、さらに以下の実験を行い、LPS媒介炎症に続いてBM中のSDF−1濃度の減少が存在するかどうかを研究した。したがって、本発明者らは、LPS処理マウス対未処理マウスにおいて、BM中のSDF−1濃度をモニターした(図2A)。
【0203】
実施例1に記載されるようにマウスに5日間LPSを投与し、そして骨髄中のSDF−1濃度をモニターした(Petitら2002によって記載されるようなELISAによる)。
【0204】
図2Aに概述される結果は、LPS媒介炎症(5日間処理による)が、未処理コントロール群における1.2からLPS処理群における0.4ng/mgタンパク質まで、BM SDF−1濃度の減少(約66%)を誘発することを示す。
【0205】
周知の動員剤である顆粒球コロニー刺激因子(G−CSF)による、BMから循環への前駆体の動員は、BM SDF−1濃度についての減少によって先行されるので(Petit ら Nature immunology (3) NO 7 687, 2002)、本発明者らは、LPS媒介炎症およびBM中のSDF−1減少に動員が続くのかどうかを研究した(図2B)。マウスに5日間LPSを投与し(実施例1に記載される通り)、そして末梢血(PB)中の前駆体増加を測定した(実施例9に記載される通り)。
【0206】
本発明者らは、LPS処理マウス(5日間)のPBにおいて、コントロール未処理マウスのPBにおける前駆体のレベルと比較して前駆体の増加したレベル(約95%より高い)を見た(図2B)。たとえば、5×105播種細胞当たり約100前駆体コロニーがLPSおよびコントロール群において見られ、対して、未処理群において5×105播種細胞当たり5コロニー。
【0207】
LPS処理マウスにおけるPB中で検出された前駆体増加は、骨髄からの細胞の放出または動員から生じることを検証するために、LPS処理後まもなく(LPS処理の16時間後)血球計を使用して、フィコール化(ficolled)WBC(白血球)の数をカウントすることによって、単核細胞(MNC)をBMおよびPB中において、ならびに未処理マウスのBMおよびPB中においてモニターした(図3)。
【0208】
本発明者らは、LPS媒介炎症は、それぞれの未処理マウスと比較した場合、BM SDF−1濃度の減少(約55%)、BM中のMNC細胞の減少(約53%)、ならびにPB中のMNC細胞の増殖(約78%)を誘発することが判った(図3)。
【0209】
したがって、LPS投与の16時間後、BM SDF−1濃度の減少およびBMからPBへの細胞の放出が観察される。
【0210】
実施例3:
機能性CXCR4はLPS媒介動員のために必要とされる。
G−CSFは、BM SDF−1を減少させそしてCXCR4をアップレギュレートすることによって、幹細胞動員を誘発することが知られている(Petitら2002)。本発明者らは、実験を行い、前駆体のLPS媒介動員がCXCR4/SDF−1相互作用を必要とするかどうかを測定した。
【0211】
本発明者らは、LPS投与の16時間後、LPS処理マウスのBMおよびPBにおける、ならびにコントロール未処理マウスにおける、CXCR4発現をモニターした。本発明者らは、LPS処理マウスのBMおよびPBの両方において、フローサイトメトリーによって検出されるような(Petit 2002)CXCR4発現のレベルに対する有意な増加を見(図4A)、これは、前駆体動員の有意な増加と関連した(図4B)。
【0212】
結果は、LPSによる前駆細胞の動員は、SDF−1/CXCR4相互作用を含むことを示す。
【0213】
機能性CXCR4がLPS媒介動員のために必要とされることを確認するために、マウスをLPSのみで処理し(16時間)、LPSおよび抗CXCR4抗体[抗ラットCXCR4、これはまた、マウスCXCR4(Torrey Pines Biolabs, CA)に対して有効であり、500mcl PBS中10mcg]で共処理し、あるいは未処理のままにし、そしてPB中またはBM中の白血球(WBC)を、処理の16時間後に血球計を使用してモニターした(図5)。
【0214】
本発明者らは、LPS媒介動員は、機能性CXCR4の非存在下で減少されることが判った(図5)。したがって、結果は、機能性CXCR4およびCXCR4/SDF−1相互作用は、LPS媒介動員のために必要とすることを確認する。
【0215】
したがって、G−CSF(Petitら2002)によって誘発される動員におけるのと同様に、LPS媒介動員は、骨髄中のSDF−1の減少およびCXCR4の増加を含む。
【0216】
LPS媒介動員が続く骨髄中のSDF−1の減少は、恐らく、SDF−1分解に起因する。
【0217】
実施例1〜3に示される結果によれば、炎症は、BM破骨細胞活性化、CTKおよびHGF発現、SDF−1減少ならびに前駆体動員を誘発する。G−CSF媒介動員と同様に、炎症媒介動員は、BM SDF−1減少を含み、そして機能性CXCR4を必要とすることもまた示された。
【0218】
実施例4:
損傷は動員を引き起こす。
本発明者らは、実験を行い、制御された出血(体重の1%の単回の出血)によって誘発された損傷のモデルにおける細胞動員を研究した。
【0219】
以下のパラメータを、出血の誘発後にモニターした:A− 前駆体の動員、B− 骨髄中のSDF−1レベル、およびC− 血中のCXCR4発現(図6)。
【0220】
本発明者らは、出血開始の7〜14日後、BM SDF−1濃度の有意かつ安定な減少を観察した。CXCR4発現の一時的増加が、出血の3〜7日後に見られた(図6)。PB中の前駆体の一時的増加が、出血の3〜7日後に検出された(図6)。
【0221】
したがって、制御された出血によって引き起こされる損傷は、BM SDF−1濃度の安定な減少、ならびにPB CXCR4および前駆体動員の一時的増加を誘発する。
【0222】
実施例5:
損傷は破骨細胞活性化に影響を与える。
本発明者らは、BM中の破骨細胞活性化に対する、制御された出血によって誘発される損傷の効果を調べた。
【0223】
実験設定は、制御された出血の誘発(実施例4における通り)、ならびに処理マウス対未処理マウスにおけるBM中の破骨細胞の活性化マーカー、TRA+をモニターすること(実施例1における通り)を含んだ。
【0224】
本発明者らは、出血誘発の7日後、BM中の劇的な破骨細胞活性化を検出することができた(図7)。
【0225】
実施例1〜5における上記の知見は、炎症および損傷のようなストレスシグナルが、骨髄からPBへの前駆体動員を誘発することを実証する。ストレスシグナル動員の誘発に含まれるのは、G−CSF媒介動員と同様に、BM SDF−1濃度の減少、およびBM中のCXCR4発現の増加である。予想外なことに、本発明者らは、ストレスシグナル媒介動員は破骨細胞活性化を伴うことを見出した。
【0226】
実施例6:
破骨細胞活性化を媒介する因子。
本発明者らは、実験を行い、細胞動員および運動性(motility)の媒介物(mediators)、たとえば、G−CSF、SDF−1およびHGF(Lapidot 2002 Exp Hematol, SDF-1 , Aiuti 1997, J Exp Med、およびKollet ら JCI 112:160-169 (2003))が破骨細胞活性化を誘発し得るかどうかをインビトロで研究した。
【0227】
破骨細胞前駆体は、骨芽細胞との細胞−細胞接触によって刺激され、活性化多核化TRAP+骨吸収細胞となる。インビトロで活性化破骨細胞を得るために、破骨細胞活性化のために必要とされる、それらのM−CSF産生およびRANKL発現を増強するビタミンD3およびPG2Eと共に、頭蓋冠(clvarian)骨芽細胞をインキュベートする。次いで、破骨細胞前駆体を含むBM細胞を、5日間、前記骨芽細胞単層上に播種し、活性化多核化破骨細胞を得る。
【0228】
原発性マウス骨芽細胞およびBM(上記に詳述されるようなocl前駆体を含有する)の共培養物を、G−CSF(50ng/ml)、HGF(50ng/ml)、またはSDF−1(10および100ng/ml)の存在下で播種した。5日後、前記培養物中のTRAP+多核化破骨細胞を染色し(実施例1に記載される通り)、そして5日後カウントした。
【0229】
本発明者らは、媒介物G−CSF、HGFおよびSDF−1のいずれかでの処理が、活性化TRAP+破骨細胞のレベルを有意に増加させたことを観察した(図8Aそれぞれ、および8B要旨)。
【0230】
本発明者らは、さらに実験を行い、上記の媒介物のどれが破骨細胞を直接活性化するかを研究した。
【0231】
骨芽細胞を、G−CSF、HGFと共にインキュベートしたかまたは未処理のままとし、そしてSDF−1転写および発現を、このような細胞において測定した(図9)。
【0232】
原発性頭蓋冠骨芽細胞を、SDF−1(PeproTech,100ng/nl)、HGF(PeproTech,50ng/ml)およびG−CSF(Filgrastin,Roche,50ng/ml)の存在下で3日間培養した。条件化培地を回収し、実施例2に記載されるように、ELISAによりSDF−1濃度を測定した。SDF−1転写および発現の測定は、実施例2における通り。
【0233】
本発明者らは、G−CSFおよびHGFの両方が、骨芽細胞においてSDF−1転写およびSDF−1産生を誘発することを示し(それぞれ、図9Aおよび9B)、したがって、SDF−1は、G−CSFおよび/またはHGFによって骨芽細胞において誘発される因子であるようであり、そしてこれは、破骨細胞活性化を直接誘発する。
【0234】
本発明者らは、原発性マウス破骨細胞の培養物を染色し、そして破骨細胞がCXCR4を発現することを見出した(示さず)。
【0235】
得られた結果は、G−CSFおよびHGFが骨芽細胞においてSDF−1発現を誘発すること、ならびにSDF−1が破骨細胞表面上のCXCR4に結合しそして破骨細胞活性化へ導くことを示す。本発明者らは、SDF−1媒介破骨細胞活性化が前駆体動員を誘発すると仮説を立てる。
【0236】
本発明者らは、以下の実験を行い、SDF−1が破骨細胞活性化および造血前駆細胞(HPC)動員を誘発するという本発明者らの仮説をインビボで検証した。Balb/cマウスに、10mcg SDF−1または1.5mcg HGFあるいは両方を、5連日間、皮下注射し、そして破骨細胞活性化(実施例1におけるようにTRAP+染色によって測定した)および末梢血への動員(実施例1におけるように半固体培養物においてアッセイした)を測定した。
【0237】
本発明者らは、SDF−1およびHGF投与が、インビボで、破骨細胞活性化(図10A)および前駆体の動員(図10B)の両方を誘発することが判った。
【0238】
全てにおいて、得られた結果は、外から投与されたSDF−1、あるいはG−CSFおよび/またはHGFの作用によって産生されたSDF−1、あるいはストレスシグナル(たとえば、炎症または損傷)が、破骨細胞活性化および前駆体動員を直接誘発することを示す。
【0239】
実施例7:
NFカッパBリガンドの受容体アクチベータ(RANKL)による直接破骨細胞活性化は、インビボ造血幹細胞動員を誘発する
RANKLは、破骨細胞分化因子である(SUDAらEndocr Rev 20:345, 1999)。本発明者らは、RANKLによる破骨細胞の活性化が以下を誘発するかどうかを試験した:
A− 骨髄中におけるCTKおよびMMP−9のような活性化破骨細胞により典型的に分泌されるプロテアーゼの発現(Delaisse, Clin Chim Acta Feb 15;291(2):223-34 2002);ならびに
B− 前駆体動員。
【0240】
マウスに、以下の通りにRANKLを投与した:3日間、5mcgの2回毎日皮下注射、そしてRANKLの非存在下でのもう2日後。以下のパラメータを、処理マウスおよびコントロール未処理マウスの両方においてモニターした:TRAP+活性化破骨細胞(図11A)、骨髄中における破骨細胞プロテアーゼMMP−9(図11B)およびCTK(図11C)の誘発、ならびに動員(図12)。
【0241】
本発明者は、RANKL投与が、骨内膜(幹細胞および前駆細胞を含むことが公知の、骨を結合するBM領域)に沿ったTRAP+活性破骨細胞の形成、骨髄中におけるMMP−9およびCTK発現の誘発(それぞれ、図11A、BおよびC)ならびに前駆体の動員(図12A)を誘発したことを観察した。MMP阻害剤またはCXCR4中和抗体と共のRANKLでの以下の処理に続いて、損なわれた動員が見られた(図12B)。したがって、RANKLによって誘発される、破骨細胞活性化による動員は、CXCR4/SDF−1相互作用およびMMP2/9機能(function)を必要とする(図12B)。
【0242】
上記は、RANKL投与は、インビボで、機能性CXCR4/SDF-1相互作用を必要とする破骨細胞活性化および動員を誘発したことを実証する。
【0243】
実施例8:
破骨細胞活性化による動員の機構。
本発明者らは、前の実施例において、ストレスシグナルによる動員は、BM骨芽細胞による一時的SDF−1産生を含み、このような産生されたSDF−1は、表面破骨細胞CXCR4へ結合し、それらの活性化へ導くことを実証した。次に、破骨細胞活性化は、SDF−1分解およびBM SDF−1濃度減少を誘発する。
【0244】
実施例1において得られた結果を考慮して、本発明者らは、特異的SDF−1分解が、主要な破骨細胞骨吸収酵素、CTKによって誘発されるとい仮説を立てた。したがって、本発明者らは、SDF−1の活性に対するCTKの効果を研究した。
【0245】
本発明者らは、SDF−1についてのインビトロ機能アッセイを使用し(実施例10)、SDF−1活性に対するCTKの効果を調べた。SDF−1への白血病性Pre B ALL G2(比較的低濃度のSDF−1へ非常によく移動する細胞、Spiegel, Blood, 2004)の移動について成った。
【0246】
手短に述べると、SDF−1(125ng)を、37℃で約16時間、PBSで調節された100ul反応体積でCTK(ug/ml)と共にインキュベートした。いくつかのサンプルにおいて、プロテアーゼ阻害剤(システイン阻害剤E−64を含有する、PI、1ul、Sigma)を、37℃で1.5時間CTKと共にプレインキュベートし、その後、SDF−1を添加した。前記白血病性Pre B ALL G2細胞をトランスウエル(transwells)上に加え、そしてCTK処理および未処理SDF−1への移動を測定した。
【0247】
本発明者らは、SDF−1の走化性活性はCTKによって廃止されるが、プロテアーゼ阻害剤(PI)の存在下では廃止されないと判った(図13)。
【0248】
得られた結果は、CTKがSDF−1活性を阻害することを実証する。
【0249】
本発明者らは、さらに、分のタイムスケールでのCTKによるSDF−1切断の速度論を研究した(図14)。SDF−1のサンプル(20ng)を、37℃で0、15、30、60、120および240分間、CTK(1.76 ug/ml)と共にインキュベートした。回収後、サンプルを、SDS−PAGEへ供し、そしてブロットし、そして抗SDF−1ポリクローナルAb(R&D)またはモノクローナルAb Kl 5C(これは、該ケモカインのアミノ酸末端部分へ特異的に結合する)で検出した。
【0250】
得られた結果は、CTKによる迅速かつ部位特異的なSDF−1切断を示す。
【0251】
したがって、CTK、主要な破骨細胞プロテイナーゼは、そのアミノ酸末端の特異的かつ迅速な切断によってSDF−1活性を阻害することが実証された。
【0252】
実施例9:
CFUアッセイ
血液循環におけるマウス前駆体(mouse progenitors)のレベルを検出するために、半固体培養を、以前に記載されるように行った(Petit, Nat Immunol, 2002)。手短に述べると、マウスBM細胞(3x105細胞/ml)を、0.9%メチルセルロース(Sigma)、30%FCS、5x10-5% 2ME、50ng/ml SCF、5ng/ml IL−3、5ng/ml GM−CSF(R&D)、および2u/mlエリスロポイエチン(Orto Bio Tech, Don Mills, Canada)においてプレートした。前記培養物を、5%CO2を含有する湿気のある環境において37℃でインキュベートし、そして7日後、形態学的基準によって、倒立顕微鏡法によりスコア化した。
【0253】
実施例10:
移動アッセイ(Migration assay)
125ng/ml SDF−1αを、37℃で一晩CTK(1mcg/ml)とインキュベートするかまたは未処理に保った。CTKの酵素活性を阻害するために、1および2mclのプロテアーゼ阻害剤(Sigma)を、1.5時間CTKと共にインキュベートし、その後SDF−1へ添加した。10%FCSを補充したRPMI(600μl)を、試験されたSDF−1と共に、Costar24ウエルトランスウエル(Costar 24-wells transwell)(Corning(孔サイズ5μm),NY)の下部チャンバへ添加した。100μl培地中の1×105pre B ALL G2細胞を、上部チャンバへ加え、そして37℃で4時間移動させた。移動細胞を下部チャンバから回収し、そしてFACSCaliburを使用して60秒間カウントした。コントロール自然移動を、下部チャンバにおいてSDF−1α無しに行った。
【0254】
実施例11:
動物。
8〜10週齢のBalb/cマウスをHarlan Rehovotから購入した。全ての実験は、ワイツマン・インスティテュートの動物ケア委員会(the animal care committee of the Weizmann Institute)によって承認された。
【図面の簡単な説明】
【0255】
【図1】LPS誘発炎症が破骨細胞活性化をもたらすことを示す。Balb/cマウスに、LPSの単回注射(250mcg/マウス)をIPで注射した。5日後、骨を採取し、そして活性化された破骨細胞を、赤色での酵素TRAP染色により検出した(A)。BMを採取し、そして20ng/mlのマウスM−SCFおよび可溶性マウスRANKL(これらは、破骨細胞分化および活性化を誘発する)の存在下で播種し、アッセイを破骨細胞BM前駆体のレベルを定量するために使用した(B)。SDF−1、HGFおよびカテプシンK(CTK)の発現を、RT−PCRで評価した(C)。
【図2】LPS誘発炎症がSDF−1分解および前駆体動員をもたらすことを示す。BM液および末梢血細胞を、図1に記載のマウスから収集した。(A)ELISAによって測定したBM液中のSDF−1濃度。(B)半固体培養物(semi solid culture)において測定されたPB前駆体によって増加されたコロニーの数。
【図3】LPS誘発炎症が、16時間以内にSDF−1の減少およびBMからの細胞放出(egress)をもたらすことを示す。LPSの単回注射(250mcg)で処理したマウスを16時間後に殺した。BM SDF−1濃度をELISAによって測定し(A)、そしてBMおよび血液循環中の単核細胞の数を血球計によって測定した(B)。
【図4】SDF−1/CXCR4相互作用が、前駆体のLPS誘発動員に関与することを示す。LPSの単回注射(250mcg)16時間後、Balb/cマウスを犠牲死させ、そしてBMおよびPBを採取した。CXCR4発現を、フローサイトメトリーによって測定し(A)、そして循環中の前駆体のレベルを半固体培養物において測定した(B)。
【図5】機能性CXCR4およびMMP 2/9がLPS媒介動員(LPS-mediated mobilization)のために必要とされることを示す。Balb/cマウスにLPSの単回注射(250mcg)を注射した。MMP 2/9阻害剤(100mcg)および抗CXCR4(10ug)を2回注射した:LPS注射の2時間前および2時間後。LPS注射の16時間後、BMおよびPBを採取し、そして血中(A)およびBM中(B)の白血球の数を、血球計を使用して評価した。
【図6】制御された出血によって誘発されたストレスシグナルに応答しての、SDF−1レベルの減少およびCXCR4発現の増加を示す。Balb/cマウスを、体重1%の単回出血(single bleeding)によって処理した。3、7、10および14日後、BMおよびPBを採取した。BM中のSDF−1レベルをELISAによって測定した(A)。PB細胞によるCXCR4発現をフローサイトメトリーにより定量し(B)、そして動員を示す循環前駆体のレベルを半固体培養物において測定した(C)。
【図7】制御された出血が破骨細胞活性化を誘発することを示す。図6に記載のマウスの大腿を固定し、脱灰し、パラフィン包埋し、そして切片化した。TRAP染色を使用して、赤色の活性化破骨細胞を検出した。
【図8】G−CSF、HGFおよびSDF−1でのインビトロ刺激がOcl活性化を誘発することを示す。マウス原発性頭蓋冠骨芽細胞を、ビタミンD3およびPG2Eの存在下で成長させ、M−CSFの分泌およびRANKLの発現を増強した。BM細胞を、G−CSF(50ng/ml)、HGF(50ng/ml)、SDF−1(10および100ng/ml)有りまたは無しで、骨芽細胞単層上に播種した。成熟活性化破骨細胞を、TRAP染色により、5日後に検出した。(A)代表的TRAP染色。(B)Aの要旨。
【図9】G−CSFおよびHGFに応答しての、原発性マウス骨芽細胞によるSDF−1分泌の増加を示す。マウス原発性頭蓋冠骨芽細胞を、50ng/ml G−CSFおよびHGFの存在下で3日間培養した。(A)RT−PCRにより測定した、SDF−1 mRNAの発現。(B)ELISAにより測定された、処理した骨芽細胞によって分泌されたSDF−1のレベル。
【図10】サイトカインSDF−1およびHGFが、インビボで破骨細胞活性化およびHPC動員を誘発することを示す。Balb/cマウスに、SDF−1(10mcg)およびHGF(1.5mcg)の5連日注射を注射した。処理マウスの骨を、赤色で成熟活性破骨細胞を示すTRAPについて染色した(A)。末梢血液細胞を半固体培養物によって、動員を示す循環前駆体のレベルについて測定した(B)。
【図11】Ocl分化因子RANKLが、骨内膜に沿うTRAP+活性化破骨細胞のインビボ形成を誘発することを示す。Balb/cマウスに、可溶性マウスRANKL(1日当たり2回注射、皮下(subcoutaneous)注射当たり5mcg)を注射した。処理マウスの骨を、成熟活性化破骨細胞を示すTRAP(赤色で)について染色した(A)。BM液中のMMP−9のレベルをザイモグラフィーアッセイ(zymography assay)により測定した(B)。CTK mRNAの発現をRT−PCRにより検出した(C)。処理マウスの骨を、成熟活性化破骨細胞を示すTRAP(赤色で)について染色した。BM液中のMP−9のレベルをザイモグラフィーアッセイにより測定した(B)。CTK mRNAの発現をRT−PCRにより検出した(C)。
【図12】RANKL処理が、CXCR4およびMMP2/9によって媒介される、前駆体動員を誘発することを示す。(A)5日間注射したSDF−1(10mcg)、HGF(1.5mcg)および最初の3日間注射したsRANKL(2×5mcg)によって誘発された、半固体培養物において測定した前駆体動員。(B)RANKL誘発動員は、最後の2日間注射した、抗CXCR4抗体(10mcg)およびMMP2/9(100mcg)の投与によって阻害される。(C)処理マウスのBM中の破骨細胞前駆体を、M−CSFおよびsRANKLと共に4日間BM細胞を培養することによって測定し、続いてTRAP染色し、成熟活性破骨細胞を検出した。
【図13】SDF−1の走化性活性が、CTK(主要な破骨細胞再吸収酵素)によって廃止されることを示す。SDF−1(125ng)を、37℃で16時間、1mcg/ml CTK(反応における最終濃度)と共にインキュベートした。プロテアーゼ阻害剤を、SDF−1を添加する前に1.5時間、CTKと共にイキュベートした。処理したSDF−1へのPre B ALL G2細胞の移動は、その走化性活性を示す。
【図14】SDF−1がCTKによりN末端で切断されることを示す。SDF−1(20ng)を、37℃で、0、15、30および60分間、CTK(1.76ug/ml)と共にインキュベートした。サンプルをSDS−PAGEへ供し、ブロットし、そして抗SDF−1ポリクローナルAbまたはモノクローナルAb K15C(これは、前記ケモカインのアミノ末端部へ特異的に結合する)で検出した。
【特許請求の範囲】
【請求項1】
SDF−1活性および/または濃度が発症および/または経過に関与する疾患を治療するための医薬の製造における、カテプシンK阻害剤(CTKI)またはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子の使用。
【請求項2】
前記疾患がSDF−1活性によって引き起こされる/悪化される請求項1記載のCTKの使用。
【請求項3】
前記疾患が癌、炎症および感染症から選択される請求項2記載の使用。
【請求項4】
前記疾患がアレルギー性気道疾患である請求項3記載の使用。
【請求項5】
前記疾患が慢性関節リウマチである請求項3記載の使用。
【請求項6】
前記疾患が動脈硬化症である請求項3記載の使用。
【請求項7】
前記疾患が癌である請求項3記載の使用。
【請求項8】
前記疾患が前立腺癌である請求項7記載の使用。
【請求項9】
前記疾患が腎臓癌である請求項7記載の使用。
【請求項10】
前記疾患が神経芽腫である請求項7記載の使用。
【請求項11】
前記疾患が神経膠腫である請求項7記載の使用。
【請求項12】
前記疾患が膵臓癌である請求項7記載の使用。
【請求項13】
前記疾患が結腸癌である請求項7記載の使用。
【請求項14】
前記疾患が乳癌である請求項7記載の使用。
【請求項15】
転移を予防するための請求項7記載の使用。
【請求項16】
前記癌細胞がCXCR4を発現する請求項15記載の使用。
【請求項17】
前記疾患が白血病である請求項7記載の使用。
【請求項18】
前記疾患が急性リンパ芽球性白血病(ALL)である請求項17記載の使用。
【請求項19】
前記疾患が急性骨髄性白血病(AML)である請求項17記載の使用。
【請求項20】
前記疾患がSDF−1活性によって予防/緩和される請求項1記載のCTKIの使用。
【請求項21】
前記疾患がHIVである請求項20記載の使用。
【請求項22】
必要とする対象において幹細胞の動員を誘導するための医薬の製造における、CTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子の使用。
【請求項23】
前記必要とする対象が重篤な好中球減少症に苦しむ請求項22記載の使用。
【請求項24】
前記好中球減少症が骨髄移植の際に生じる請求項23記載の使用。
【請求項25】
前記好中球減少症が癌化学療法の際に生じる請求項23記載の使用。
【請求項26】
必要とする対象において骨髄中の幹細胞の滞留を増大させるための医薬の製造におけるCTKIの使用。
【請求項27】
必要とする対象において器官の再生を増強するための請求項26記載の使用。
【請求項28】
前記器官が骨髄である請求項27記載の使用。
【請求項29】
有効量のCTKIまたはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を投与することを含む、SDF−1活性および/または濃度によってその発症および経過が影響される疾患を治療する方法。
【請求項30】
有効量のCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を投与することを含む、哺乳動物における癌を治療する方法。
【請求項31】
前記癌細胞がCXCケモカイン受容体4(CXCR4)を発現する請求項29記載の方法。
【請求項32】
転移を予防するための請求項30記載の方法。
【請求項33】
必要とする対象へ有効量のCTKIまたはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を投与することを含む、組織への多能性幹細胞の標的化を調節する方法。
【請求項34】
前記標的組織への前記細胞の標的化を増大させるために、CTKIを標的組織へ投与する請求項33記載の方法。
【請求項35】
前記細胞が正常な造血細胞である請求項34記載の方法。
【請求項36】
前記造血細胞が造血幹細胞および造血前駆細胞からなる群から選択される請求項35記載の方法。
【請求項37】
前記細胞がインビボで患者にあり、かつ治療的に有効量のCXCR4アゴニストをこのような治療を必要とする該患者へ投与する請求項36記載の方法。
【請求項38】
前記CXCR4アゴニストがSDF−1、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子である請求項36記載の方法。
【請求項39】
前記患者が癌を有する請求項38記載の方法。
【請求項40】
前記患者が自家もしくは同種異系骨髄または末梢血幹細胞移植を必要とする請求項39記載の方法。
【請求項41】
前記患者を細胞毒性薬で治療することをさらに含む請求項40記載の方法。
【請求項42】
有効量のCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子が、標的組織への前記細胞の標的化を減少させるために、該標的組織へ投与される請求項33記載の方法。
【請求項43】
前記細胞が新生細胞である請求項42記載の方法。
【請求項44】
造血細胞へ有効量のCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を投与することを含む、造血細胞増殖の速度を減少させる方法。
【請求項45】
前記造血細胞が造血幹細胞および造血前駆細胞からなる群から選択される請求項44記載の方法。
【請求項46】
前記細胞がインビボで患者にあり、かつ治療的に有効量のCXCR4アゴニストをこのような治療を必要とする該患者へ投与する請求項45記載の方法。
【請求項47】
前記患者が自家または同種異系骨髄あるいは末梢血幹細胞移植を必要とする請求項46記載の方法。
【請求項48】
前記患者が癌を有する請求項47記載の方法。
【請求項49】
前記患者を細胞毒性薬で治療することをさらに含む請求項48記載の方法。
【請求項50】
CTKをSDF−1と接触させること、SDF−1の活性を測定すること、およびCTKによるSDF−1活性の阻害を防止または逆転し得る化合物を単離することを含む、CTKアンタゴニストを同定する方法。
【請求項51】
CTKをSDF−1と接触させること、SDF−1の完全性を測定すること、およびCTKによるSDF−1活性の分解を防止し得る化合物を単離することを含む、CTKアンタゴニストを同定する方法。
【請求項52】
請求項43または44記載の方法によって得られるアンタゴニスト。
【請求項1】
SDF−1活性および/または濃度が発症および/または経過に関与する疾患を治療するための医薬の製造における、カテプシンK阻害剤(CTKI)またはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子の使用。
【請求項2】
前記疾患がSDF−1活性によって引き起こされる/悪化される請求項1記載のCTKの使用。
【請求項3】
前記疾患が癌、炎症および感染症から選択される請求項2記載の使用。
【請求項4】
前記疾患がアレルギー性気道疾患である請求項3記載の使用。
【請求項5】
前記疾患が慢性関節リウマチである請求項3記載の使用。
【請求項6】
前記疾患が動脈硬化症である請求項3記載の使用。
【請求項7】
前記疾患が癌である請求項3記載の使用。
【請求項8】
前記疾患が前立腺癌である請求項7記載の使用。
【請求項9】
前記疾患が腎臓癌である請求項7記載の使用。
【請求項10】
前記疾患が神経芽腫である請求項7記載の使用。
【請求項11】
前記疾患が神経膠腫である請求項7記載の使用。
【請求項12】
前記疾患が膵臓癌である請求項7記載の使用。
【請求項13】
前記疾患が結腸癌である請求項7記載の使用。
【請求項14】
前記疾患が乳癌である請求項7記載の使用。
【請求項15】
転移を予防するための請求項7記載の使用。
【請求項16】
前記癌細胞がCXCR4を発現する請求項15記載の使用。
【請求項17】
前記疾患が白血病である請求項7記載の使用。
【請求項18】
前記疾患が急性リンパ芽球性白血病(ALL)である請求項17記載の使用。
【請求項19】
前記疾患が急性骨髄性白血病(AML)である請求項17記載の使用。
【請求項20】
前記疾患がSDF−1活性によって予防/緩和される請求項1記載のCTKIの使用。
【請求項21】
前記疾患がHIVである請求項20記載の使用。
【請求項22】
必要とする対象において幹細胞の動員を誘導するための医薬の製造における、CTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子の使用。
【請求項23】
前記必要とする対象が重篤な好中球減少症に苦しむ請求項22記載の使用。
【請求項24】
前記好中球減少症が骨髄移植の際に生じる請求項23記載の使用。
【請求項25】
前記好中球減少症が癌化学療法の際に生じる請求項23記載の使用。
【請求項26】
必要とする対象において骨髄中の幹細胞の滞留を増大させるための医薬の製造におけるCTKIの使用。
【請求項27】
必要とする対象において器官の再生を増強するための請求項26記載の使用。
【請求項28】
前記器官が骨髄である請求項27記載の使用。
【請求項29】
有効量のCTKIまたはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を投与することを含む、SDF−1活性および/または濃度によってその発症および経過が影響される疾患を治療する方法。
【請求項30】
有効量のCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を投与することを含む、哺乳動物における癌を治療する方法。
【請求項31】
前記癌細胞がCXCケモカイン受容体4(CXCR4)を発現する請求項29記載の方法。
【請求項32】
転移を予防するための請求項30記載の方法。
【請求項33】
必要とする対象へ有効量のCTKIまたはCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を投与することを含む、組織への多能性幹細胞の標的化を調節する方法。
【請求項34】
前記標的組織への前記細胞の標的化を増大させるために、CTKIを標的組織へ投与する請求項33記載の方法。
【請求項35】
前記細胞が正常な造血細胞である請求項34記載の方法。
【請求項36】
前記造血細胞が造血幹細胞および造血前駆細胞からなる群から選択される請求項35記載の方法。
【請求項37】
前記細胞がインビボで患者にあり、かつ治療的に有効量のCXCR4アゴニストをこのような治療を必要とする該患者へ投与する請求項36記載の方法。
【請求項38】
前記CXCR4アゴニストがSDF−1、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子である請求項36記載の方法。
【請求項39】
前記患者が癌を有する請求項38記載の方法。
【請求項40】
前記患者が自家もしくは同種異系骨髄または末梢血幹細胞移植を必要とする請求項39記載の方法。
【請求項41】
前記患者を細胞毒性薬で治療することをさらに含む請求項40記載の方法。
【請求項42】
有効量のCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子が、標的組織への前記細胞の標的化を減少させるために、該標的組織へ投与される請求項33記載の方法。
【請求項43】
前記細胞が新生細胞である請求項42記載の方法。
【請求項44】
造血細胞へ有効量のCTK、そのムテイン、アイソフォーム、融合タンパク質、機能性誘導体、活性画分、円順列誘導体、塩または誘導因子を投与することを含む、造血細胞増殖の速度を減少させる方法。
【請求項45】
前記造血細胞が造血幹細胞および造血前駆細胞からなる群から選択される請求項44記載の方法。
【請求項46】
前記細胞がインビボで患者にあり、かつ治療的に有効量のCXCR4アゴニストをこのような治療を必要とする該患者へ投与する請求項45記載の方法。
【請求項47】
前記患者が自家または同種異系骨髄あるいは末梢血幹細胞移植を必要とする請求項46記載の方法。
【請求項48】
前記患者が癌を有する請求項47記載の方法。
【請求項49】
前記患者を細胞毒性薬で治療することをさらに含む請求項48記載の方法。
【請求項50】
CTKをSDF−1と接触させること、SDF−1の活性を測定すること、およびCTKによるSDF−1活性の阻害を防止または逆転し得る化合物を単離することを含む、CTKアンタゴニストを同定する方法。
【請求項51】
CTKをSDF−1と接触させること、SDF−1の完全性を測定すること、およびCTKによるSDF−1活性の分解を防止し得る化合物を単離することを含む、CTKアンタゴニストを同定する方法。
【請求項52】
請求項43または44記載の方法によって得られるアンタゴニスト。
【図1】

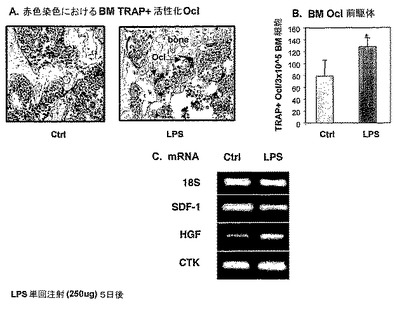
【図2】
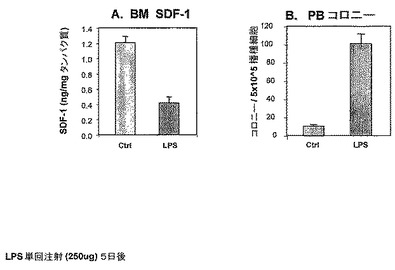
【図3】
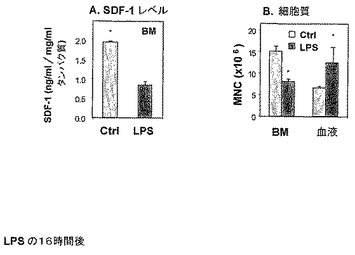
【図4】
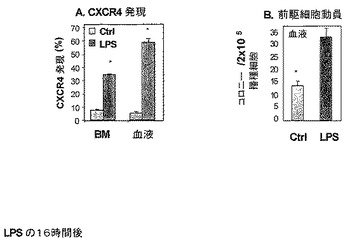
【図5】
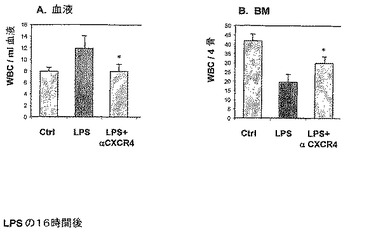
【図6】
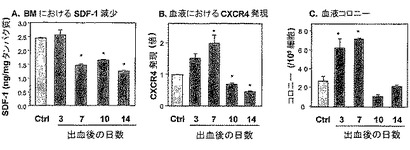
【図7】
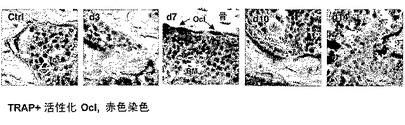
【図8】

【図9】
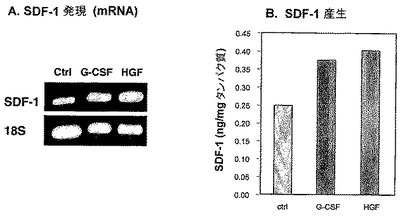
【図10】
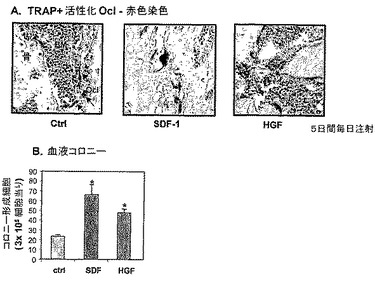
【図11】

【図12】
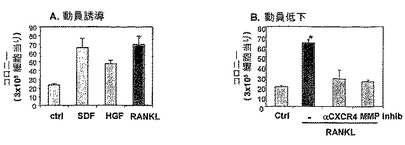
【図13】

【図14】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【公表番号】特表2008−520554(P2008−520554A)
【公表日】平成20年6月19日(2008.6.19)
【国際特許分類】
【出願番号】特願2007−538612(P2007−538612)
【出願日】平成17年10月27日(2005.10.27)
【国際出願番号】PCT/IL2005/001121
【国際公開番号】WO2006/048862
【国際公開日】平成18年5月11日(2006.5.11)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.ウィンドウズ
【出願人】(500018608)イエダ リサーチ アンド ディベロップメント カンパニー リミテッド (35)
【Fターム(参考)】
【公表日】平成20年6月19日(2008.6.19)
【国際特許分類】
【出願日】平成17年10月27日(2005.10.27)
【国際出願番号】PCT/IL2005/001121
【国際公開番号】WO2006/048862
【国際公開日】平成18年5月11日(2006.5.11)
【公序良俗違反の表示】
(特許庁注:以下のものは登録商標)
1.ウィンドウズ
【出願人】(500018608)イエダ リサーチ アンド ディベロップメント カンパニー リミテッド (35)
【Fターム(参考)】
[ Back to top ]