
半数体植物胚および倍加半数体植物胚を作出する方法
【課題】半数体植物胚を作出するための方法を提供すること。
【解決手段】本発明は、半数体植物胚を作出するための方法に関し、この方法は、細胞分裂誘発分子を含む小胞子または花粉を提供すること;小胞子または花粉を、半数体胚を作製すべき植物体の胚嚢細胞、特に卵細胞に受粉させること;および半数体植物胚を得るために、小胞子または花粉が胚嚢細胞、特に卵細胞中にまたはその近くに細胞分裂誘発分子を放出することを可能にして、その分裂をトリガーすることを含む。倍加半数体植物胚を作出すべき場合、染色体数の倍加は、受粉後のある段階の、特に細胞分裂中にまたは胚を得た後に起こる。本発明はさらに、このようにして得られた胚、これから再生された植物体、およびその子孫に関する。
【解決手段】本発明は、半数体植物胚を作出するための方法に関し、この方法は、細胞分裂誘発分子を含む小胞子または花粉を提供すること;小胞子または花粉を、半数体胚を作製すべき植物体の胚嚢細胞、特に卵細胞に受粉させること;および半数体植物胚を得るために、小胞子または花粉が胚嚢細胞、特に卵細胞中にまたはその近くに細胞分裂誘発分子を放出することを可能にして、その分裂をトリガーすることを含む。倍加半数体植物胚を作出すべき場合、染色体数の倍加は、受粉後のある段階の、特に細胞分裂中にまたは胚を得た後に起こる。本発明はさらに、このようにして得られた胚、これから再生された植物体、およびその子孫に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、半数体植物胚および倍加半数体植物胚を作出する新たな方法に関する。本発明はさらに、このようにして得られた植物胚、これから再生された植物体、ならびにこうした植物体の子孫、細胞、組織および種子に関する。
【背景技術】
【0002】
1964年にGuhaとMaheshwari(Nature 204:497)が半数体胞子から植物体を再生できることを発見して以来、他の種で類似の知見を得るために多くの研究が行われてきた(例えば、「In vitro Haploid production in Higher plants」Vol.1、2、3、4、5、Eds:S.Jain、S.SoporyおよびR.Veilleux(1996)Kluwer Academic Publishersを参照されたい)。
【0003】
現代の最新植物育種では、倍加半数体(DH)の使用は、遺伝的純系の作製を迅速化するための、さらには同義遺伝子/複対立遺伝子によってコードされる形質など困難な形質を評価しモニタリングするための、極めて貴重なツールとなっている。
【0004】
作物育種における倍加半数体の作出および使用は多く種でよく知られている(例えば、Thomas W.ら(2003)、In:Doubled haploid production in crop plants.A Manual.Eds.M.Maluszynski、K.Kasha、B.ForsterおよびI.Szarejko.Kluwer Academic Publishers、pp337−349を参照されたい)。一般に、倍加半数体は、雄性器官由来の胞子から得ることができる。この場合、胞子は「小胞子」と称され、in vitro培養は「小胞子培養」と呼ばれる。倍加半数体は、雌性器官または「大胞子」から得ることもできる。対応するin vitro培養は一般に「雌性発生」と称される。
【0005】
典型的な小胞子培養は、古くからアブラナ属で十分に確立されている(例えば、Kellerら(1984)In:K.Giles、S.Sen(eds.)、Plant Cell Culture in Crop Improvement pp169−183.Plenum Pub.Corp.、New Yorkを参照されたい)。典型的な雌性発生培養はサトウダイコンで知られている(例えば、Hosemans D.およびBossoutrot,Z.Pflanzenzuchtg.91:74−77(1983)を参照されたい)。キュウリにおいても雌性発生は極めて十分に確立された技法である(EP0374755を参照されたい)。
【0006】
雌性発生に取り組む方法は、放射線照射された花粉を利用した単為生殖による卵細胞の胚形成の誘発に基づく。よく知られている一例は、メロンで公開されており、現在いくつかの育種会社で常法に従い応用されている(Sauton AおよびR.Dumas de Vaulx、Agronomie 7:141−148(1987)を参照されたい)。単為生殖では、未受精卵から新たな植物体が発生する。この技法の成功率は低い。
【0007】
30年を超える研究を経て利用可能な技術および経験が豊富に得られたにもかかわらず、多く技法の成功は従順な遺伝子型に限定される。これは、DHの使用による膨大な利益を存分に利用できていないことを意味する。一部の遺伝子型の反応性の変動は別として、トマト、ワタなどいくつかの作物種では、いまだにDHの誘導が困難である。
【発明の概要】
【発明が解決しようとする課題】
【0008】
したがって、本発明の目的は、半数体植物胚または倍加半数体植物胚を作出するための新たな方法を提供することである。
【課題を解決するための手段】
【0009】
本発明をもたらした研究において、驚くべきことに、細胞分裂誘発分子を含む花粉または小胞子を卵細胞に受粉することにより、卵細胞を、胚形成を遂げるように誘導できることが見出された。次いで、この卵細胞を、受精なしで胚を形成するようにトリガーする。花粉により卵細胞は実際に受精していないので、二倍体接合体は形成されず、生じた胚は半数体のままである。受粉後のある段階の細胞分裂中に、自然な染色体倍加を起こして、部分的にまたは完全にDHである胚を得ることができる。染色体倍加は、例えば、コルヒチンなどの知られている化学的手段によって誘発することもできる。
【0010】
したがって、本発明は、半数体植物胚を作出するための方法に関し、この方法は、
a)細胞分裂誘発分子を含む小胞子または花粉を提供する工程と、
b)小胞子または花粉を、半数体胚を作製すべき植物体の胚嚢細胞、特に卵細胞に受粉させる工程と、
c)半数体植物胚を得るために、小胞子または花粉が胚嚢細胞、特に卵細胞中にまたはその近くに細胞分裂誘発分子を放出することを可能にして、その分裂をトリガーする工程と
を含む。
【0011】
代替の実施形態では、本発明は、倍加半数体植物胚を作出するための方法に関し、この方法は、
a)細胞分裂誘発分子を含む小胞子または花粉を提供する工程と、
b)小胞子または花粉を、倍加半数体胚を作製すべき植物体の胚嚢細胞、特に卵細胞に受粉させる工程と、
c)植物胚を得るために、小胞子または花粉が胚嚢細胞、特に卵細胞中にまたはその近くに細胞分裂誘発分子を放出することを可能にして、その分裂をトリガーする工程と
を含み、染色体数の倍加が受粉後のある段階の、特に細胞分裂中にまたは胚を得た後に起こる。
【0012】
したがって、本発明は、胚嚢細胞または卵細胞の細胞分裂をトリガーするためのビヒクルとしての花粉または小胞子の使用に関する。
【図面の簡単な説明】
【0013】
【図1】アラビドプシスp35S:AP2mutの花序を示す図である。
【図2】パーティクルガン法により得られたGUS陽性の花粉を示す図である。
【図3】パーティクルガン法による花粉の形質転換の代表的な結果を示す図である。
【図4】トマトの15の様々な開花期を示す図である。
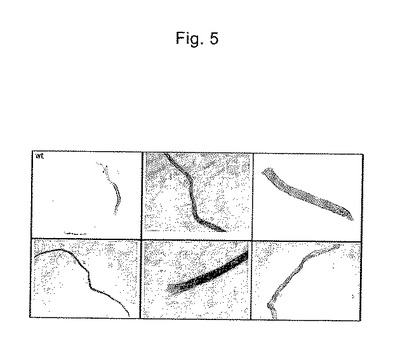
【図5】GUS陽性の花粉管の代表例を示す図である。
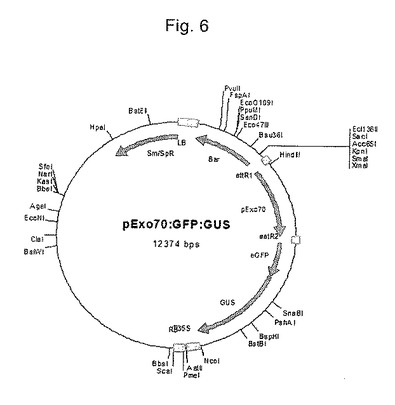
【図6】DNAプラスミドpExo70::GFP:GUSの構造を示す図である。
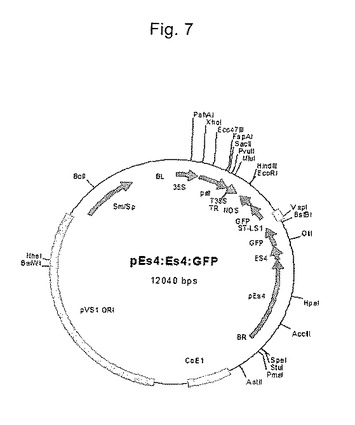
【図7】pES4::ES4:GFP構築物の構造を示す図である。
【発明を実施するための形態】
【0014】
第1の実施形態では、細胞分裂誘発分子は、花粉中で、例えば、プラスミド上に存在する核酸から一過性に発現される。細胞分裂誘発分子は、核酸またはタンパク質のいずれかとすることができ、プラスミドからの構成的発現により花粉または小胞子中で産生される。このようにして産生された細胞分裂誘発分子は、受粉後に卵細胞中に放出される。
【0015】
第2の実施形態では、細胞分裂誘発分子は、花粉ゲノム中に安定に組み込まれている核酸から発現される。細胞分裂誘発分子は、核酸またはタンパク質のいずれかとすることができ、構成的発現により花粉または小胞子中で産生される。このようにして産生された細胞分裂誘発分子は、受粉後の胚嚢または卵細胞中に放出される。
【0016】
第3の実施形態では、細胞分裂誘発分子は、胚嚢細胞特異的または卵細胞特異的プロモーターの制御下にある核酸からの発現により胚嚢細胞または卵細胞中で産生される。この核酸は、核酸を含む花粉または小胞子を受粉させることにより胚嚢細胞または卵細胞中に入る。胚嚢細胞または卵細胞中で組織特異的プロモーターが作動し、したがって細胞分裂誘発分子の産生がもたらされる。
【0017】
第4の実施形態では、花粉または小胞子は、直接形質転換されるのではなく、細胞分裂誘発分子をコードする核酸を運ぶトランスジェニック植物体で形成される。このようなトランスジェニック花粉または小胞子は、構成的プロモーター、または花粉特異的もしくは小胞子特異的プロモーター、または胚嚢細胞特異的もしくは卵細胞特異的プロモーターの制御下にあってよい。第1の場合、転写物は、植物体にとって有害でないことが好ましい。第2の場合、トランスジーンは花粉または小胞子のみで発現され、細胞分裂誘発分子は花粉または小胞子のみで産生される。第3の場合、トランスジーンは、受粉後に核酸が卵細胞または胚嚢細胞に入ると発現される。
【0018】
こうしたすべての実施形態では、花粉による卵細胞の実際の受精は回避しなればならない。これは、放射線照射を用いて、または花粉および卵細胞について異なる種を使用することによって達成することができる。タンパク質は、放射線照射によって損傷を受けない、または実質的に損傷を受けないので、放射線照射は、タンパク質が花粉中で産生される実施形態に特に適している。核酸を花粉または小胞子から胚嚢または卵細胞に移すべき場合、卵細胞供与植物体とは別の種を使用することが好ましい。
【0019】
本発明の特定の実施形態によれば、細胞分裂誘発分子を含む小胞子または花粉は、核酸による形質転換によって得られる。形質転換は、任意の適当な方法で、例えば、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)を用いて、または微粒子銃(バイオリスティック法)を用いて実施することができる。
【0020】
こうした形質転換技法はよく知られている。アグロバクテリウム・ツメファシエンスを用いた植物細胞の形質転換は十分に確立されており、例えば、De la Rivaら、EJB Vol.1(3)(1998)およびBent、Plant Physiol.124:1540−1547(2000)にその総説がある。
【0021】
近年、植物の遺伝的形質転換は、アグロバクテリウム属だけに限定されず、他の細菌も植物を形質転換する潜在能力を有することが発見された(参照により本明細書に組み込まれているBroothaertsら、Nature 433、629−633(2005))。植物に関連するこうした共生細菌は、不活性化(disarmed)Tiプラスミドおよび適当なバイナリーベクターの両方の獲得により、遺伝子導入に対してコンピテントにされた。このような形質転換系も本発明における使用に適している。
【0022】
バイオリスティック形質転換も当分野の技術者によく知られており、このような用途のためのツールは数年前から市販されている(Ralph Book、In:QiagenNews、Issue No.5、1997)。
【0023】
本発明において使用するのに適した技法はまた、例えば、アンティリヌム・マユス(Antirrhinum majus)における小胞子のレベルでのDNAのデリバリーおよびその一過性発現が示されている、Barinovaら(J Exp Bot.53(371):1119−29(2002))に、またはアルファルファ(メディカゴ・サティバL.(Medicago sativa L.))に関するRamaiahら(Current Science 73:674−682(1997))に記載されている。
【0024】
タバコにおけるバイオリスティック微粒子銃を用いた小胞子または花粉の形質転換のための方法は、Baubak Bajoghli(Matrikel number:9802743、University of Vienna、Experimentelle Genetic III.Plant Biotechnology by Alisher Touraev、July 2001)で見出すことができる。Van der Leede−Plegtら、Transgenic Research 4(2):77−86(1995)は、微粒子銃を用いたタバコ(ニコチアナ・グルティノサ(Nicotiana glutinosa))の花粉中へのDNAの直接的なデリバリーについて記載している。こうした技法および他の技法は、本発明において使用するための花粉または小胞子の形質転換に使用することができる。
【0025】
したがって、特定の実施形態では、花粉および小胞子は、核酸の存在に基づいて細胞分裂誘発分子を含む。導入される核酸は、細胞分裂誘発分子それ自体とすることができ、または細胞分裂誘発分子をコードすることができる。後者の場合、この誘発分子はタンパク質またはペプチドである。前者の場合、この誘発分子は核酸である。この核酸は、それ自体誘導性とすることができ、または他の核酸が発現するのを遮断することができる。例えば、この核酸は、Kip関連タンパク質ファミリーのメンバーまたは網膜芽細胞腫に対するRNAiとすることができ、またはそれをコードすることができる(例えば、Park Jら、Plant Journal 42:153−163(2005)を参照されたい)。網膜芽細胞腫タンパク質は、植物における細胞増殖、分化および核内倍加を調節する。
【0026】
あるいは、この核酸は、細胞分裂誘発分子の前駆体、または細胞分裂誘発分子を産生する酵素をコードすることができる。核酸がこの酵素をコードする場合、これは、誘発分子であるかまたはこれを直接産生する酵素であってよく、あるいは最終的に誘発分子が得られる経路の部分である酵素であってよい。本明細書では、「細胞分裂誘発分子」は、細胞分裂を直接的または間接的にトリガーするあらゆる分子を包含するものとする。
【0027】
本発明は、細胞分裂誘発分子が、形質転換された花粉または小胞子を用いて胚嚢または卵細胞に送達される原理に基づく。細胞分裂を作動させることができる遺伝子構築物または分子は、それ自体知られており、本発明の新たな方法で使用することができる。
【0028】
本発明に従って使用することができる遺伝子の例は、栄養成長を継続するようにプログラム化された体細胞(栄養細胞)が、転写因子をコードする(1種または複数の)遺伝子でこの細胞を形質転換することにより胚発生に容易に転換することを開示するStoneら(PNAS 98、11806−11811(2001))に記載されている。別の例は、Baby Boom(Boutilierら、The Plant Cell 14、1737−1749(2002))またはleafy cotyledon(Stone Sら、PNAS 25:11806−11811(2001))である。こうした遺伝子および他の遺伝子は、本発明の細胞分裂誘発分子をコードするために使用することができる。
【0029】
Zuoら(The Plant Journal 30:1−12(2002))では、体細胞胚形成が、いわゆるWuschel遺伝子を過剰発現させることによりアラビドプシスのあらゆる組織で植物ホルモンなしで誘発されることが見出されたことについて記載されている。この技術はUS2003/0082813にも記載されている。本発明との相違は、US2003/0082813では植物細胞がWuschel DNA配列により一過性ではなく、安定に形質転換されることである。次いで、この安定に導入されたWuschel遺伝子は、安定に組み込まれている組織で過剰発現され、この組織中で細胞分裂および胚形成が誘発される。
【0030】
本発明によれば、Wuschel遺伝子の発現産物であってよい細胞分裂誘発分子をコードする核酸は、細胞分裂を遂げるべき細胞のゲノム中に安定に組み込まれていない。細胞分裂誘発分子をコードする核酸は、卵細胞特異的プロモーターの制御下にあるので、分裂細胞中で一過性に発現されるか、または花粉中で発現され、その後、コードされる細胞分裂誘発分子が、細胞分裂を開始するようにトリガーすべき細胞(卵細胞または胚嚢細胞)中に放出されるかのいずれかである。したがって、花粉または小胞子は、分子それ自体(コード配列ではなく、Wuschel遺伝子発現産物)を、細胞分裂を開始するようにトリガーすべき細胞中に導入するためのビヒクルである。
【0031】
卵細胞を誘発する遺伝子の適用は、上述の遺伝子などの遺伝子の異所性および一過性発現に限定されないが、iaaM、iaaHなどのホルモンを産生することができる酵素をコードする遺伝子(Thomashowら、(1986)Science 231、616−618を参照されたい)および細胞周期タンパク質をコードする遺伝子を使用することによっても類似の結果を得ることができる。
【0032】
加えて、卵細胞の分裂および胚形成の誘発をさらに最適化するために遺伝子の組合せを使用することができる。細胞周期遺伝子の一過性発現も卵細胞の分裂を引き起こすことができる(細胞周期遺伝子の総説については、Murray A(Cell 116:221−234(2004))を参照されたい)。特に、サイクリンEおよびDの単独でのまたは組み合わせての一過性発現を使用して卵細胞分裂をトリガーすることができる。
【0033】
本発明によれば、核酸は、小胞子または花粉中で一過性にもしくはゲノム中への安定な組込みの後のいずれかで発現されるか、または卵細胞中で卵細胞特異的プロモーターから一過性に発現されるかのいずれかである。このように、得られた胚における構成的発現は回避される。
【0034】
一過性発現は、組織特異的または細胞特異的に起こり得る。代替の実施形態では、一過性発現は、核酸を組織特異的または誘導性プロモーターの制御下に置くことによって得られる。組織特異的プロモーターは、適切には、花粉特異的または小胞子特異的プロモーターである。花粉特異的プロモーターはよく知られており、一過性発現は単子葉植物種および双子葉植物種の両方で示されている。こうした型のプロモーターの例は、例えば、Twell,Dら、Development 109(3):705−713(1990);Hamilton,Dら、Plant Mol.Biol.18:211−218(1992)に記載されている。
【0035】
したがって、小胞子(例えば、アグロバクテリウム形質転換またはバイオリスティック形質転換により)または花粉粒を形質転換する従来の方法を使用することにより、Baby Boom、Wuschel、leafy cotyledon、サイクリン、サイクリン依存性キナーゼ(CDK)、E2F(高等真核生物における転写因子ファミリーのメンバー;Zheng,Nら、Genes Dev.13:666(1999))、DP(Magyar Zら、FEBS Lett.486(1):79−87(2000))などのような遺伝子の一過性発現をもたらす本発明によれば、形質転換された小胞子または花粉粒を雌受容体に受粉すると、卵細胞の分裂を誘発することが可能である。
【0036】
好ましくは、花粉または小胞子の雄原核は不活性化または破壊される。このように、卵細胞の受精は確かに回避される。誘発分子を不必要に損傷しないためには、雄原核の不活性化または破壊は、花粉または小胞子の形質転換の前に実施することが好ましい。不活性化または破壊は、適切には放射線照射を用いて行われる。
【0037】
花粉核の放射線照射は、雄原核を分解するためのよく知られている方法であるが、放射線照射量に依存して、これは、花粉管が形成され、卵細胞中に放出することを妨害しない。Grantら(New Zealand Journal of Botany 18、339−341(1980))ではこの技法について記載されている。
【0038】
放射線照射され、続いて形質転換された花粉/小胞子細胞は、その後、同じ種または前記花粉/小胞子細胞の花粉放出が起こり得る種由来の植物体の雌ずいに移される。異種の受粉の一例は、花粉供与植物体としてのナス科および受容植物体としてのトマトに属する種の使用である。他の例は、de Martinis,Dら、Planta 214(5):806−812(2002)およびDore Cら、Plant Cell Reports 15:758−761(1996)に記載されている。一般に、異種の受粉に適している種は同じ科に属するものである。
【0039】
種に依存して、分裂に由来した種子は、卵細胞または卵巣から収穫することができる。あるいは、胚珠培養では、発生中の胚を生存させることが必要であり得る。
【0040】
したがって、基本的に、不活性化された花粉粒は、卵細胞分裂および胚形成を誘発することができるシグナル分子を一過性に運ぶ。この分子は一過性であるので、卵細胞DNAは安定に形質転換されない。
【0041】
したがって、特定の実施形態では、本発明は、雄原核を不活性化するために放射線照射によって不活性化された小胞子または花粉粒中で一過性に存在し発現される細胞分裂分子(タンパク質、DNA、RNA)の利用に関し、それにより前記細胞分裂分子は、花粉管により卵細胞中にまたはその近くに放出されるとその作用を発揮する。
【0042】
別の実施形態では、花粉または小胞子の供与植物体は、細胞分裂誘発分子をコードする遺伝子で安定に形質転換されるが、卵細胞誘発分子の発現は一過性である。この詳細な実施形態では、花粉または小胞子を提供する植物体は、そのゲノムDNAが、好ましくは胚嚢細胞特異的または卵細胞特異的プロモーターの制御下にある細胞分裂分子をコードする遺伝子または遺伝子構築物を運ぶように安定に形質転換される。次いで、この遺伝子または遺伝子構築物は、胚嚢細胞中に、特に卵細胞中にまたはその近くに放出されると発現される。生じた細胞分裂誘発分子は細胞分裂をトリガーする。
【0043】
受粉の前に、花粉または小胞子を放射線照射して雄原核を不活性化する。あるいは、花粉または小胞子は、前記花粉/小胞子細胞の花粉放出が起こり得る別の種の雌ずいに移される。この方法の利点は、細胞分裂誘発分子をコードする核酸が雄原核に運ばれ、最終的に精細胞中に行き着くことである。好ましくは、細胞分裂誘発分子は、供与植物体に、結果として精細胞に複数コピー存在する。花粉または小胞子の発生への遺伝子構築物の存在の干渉を回避するために、花粉または小胞子が胚嚢細胞、特に卵細胞中に移された場合のみ遺伝子または遺伝子構築物の発現を可能にする誘導性または特異的プロモーター、好ましくは胚嚢細胞特異的または卵細胞特異的プロモーターが使用される。
【0044】
本発明はさらに、本発明の方法を用いて得られる半数体胚および倍加半数体胚、ならびにこのような半数体胚または倍加半数体胚から再生された植物体、このような植物体の子孫、ならびにこのような植物体またはその子孫由来の種子、細胞、組織、小胞子および卵細胞に関する。
【0045】
本願明細書では、卵細胞という用語は、読みやすくする目的で単独で使用されることがあるが、この場合「胚嚢細胞、特に卵細胞」と解釈するものとする。
【0046】
本発明はさらに、例示目的にすぎず、本発明を限定するものと決して解釈されない続く実施例で明らかにされよう。
【実施例】
【0047】
実施例1
微粒子銃を用いたアラビドプシス花粉の形質転換
DNAプラスミドpCAMBIA 1301およびpExo70::GFP:GUSを使用して1μmの金粒子をコーティングした。pCAMBIA 1301は、800ヌクレオチドのCaMV35Sプロモーターによって調節されるGUSを含有するバイナリーベクターである(Robertsら、pCAMBIA Vector release manual version 3.05(1998))。pExo70::GFP:GUSは、β−グルクロニダーゼ(GUS)および緑色蛍光タンパク質(GFP)を含有する(図6)。
【0048】
アラビドプシスp35S:AP2mutの3つの花序をペトリ皿の中央に置いた(図1)。ペトリ皿をパーティクルガンに設置し、コーティングされた金粒子を3回撃ち込んだ。銃の撃ち込みから2日後に発現を調べた。図2は、このようにして得られたGUS陽性の花粉を示す。
【0049】
同じ実験をペトリ皿でパーティクルガンにおけるレベル3で繰り返した。2200psiの圧力で2回撃ち込んだ。図3A〜Dは、この実験の代表的な結果を示す。
【0050】
この実験から、微粒子銃を使用して花粉を形質転換することができるという結果になる。
【0051】
実施例2
微粒子銃の撃込み後のin vitroでの花粉発芽
成熟した花粉は静止状態である。花粉粒が雌株の柱頭に付着した後、柱頭からの水移動による再水和から花粉発芽の過程が始まる。この実施例では、微粒子銃の撃込み後、in vitroで花粉を発芽させる。
【0052】
図4は、トマトの15の様々な開花期を示す。段階1、段階5および段階14の花粉を本実験に使用した。段階1の花粉は完全に成熟しており、段階5の花粉も成熟している。段階14は、単核性の後期/二核性の初期である。
【0053】
段階1、段階5および段階14の花の花粉を200μlのNLN13培地(13%ショ糖を補充したNLN培地(Lichter R.、Z.Pflanzenzuecht 105:427−437(1982)))中に分離した。この200μlをgenescreenメンブレン上にスポットし、5分間乾燥させる。次いで、このメンブレンを1/2 MS寒天平板上に置き、Exo70::GFP:GUSでコーティングされた1μmの金粒子を2200psiで撃ち込む。撃込み後、メンブレンは6ウェルタイタープレートに置く。段階1および段階5の花粉は、1.5mlの発芽培地A(Clarke)(20mM MES、0.07%Ca(NO3)2・H2O、0.02%MgSO4・7H2O、0.01%KNO3、0.01%H3BO3、2%ショ糖および15%PEG4000)中でインキュベートする。段階14の小胞子は、NLN13培地中でインキュベートする。3時間インキュベートした後、1.5mlの2×GUS染色バッファーを添加し、試料を37℃で一晩置いた。図5は、GUS陽性の花粉管の代表例を示す。形質転換後でも花粉は管を形成できることが明らかである。
【0054】
実施例3
細胞分裂刺激因子を運ぶ花粉を受粉した後の卵細胞の細胞分裂を誘発するためのこうした花粉の調製
実施例1および2から、GUSを用いたモデル系で花粉を形質転換できることが実証される。ここで、花粉を放射線照射した後の、細胞分裂誘発分子BabyBoom(BBM)(Boutilierら、2002、前掲)によるトマト花粉の形質転換法を記載する。
【0055】
CaMV35Sプロモーター::GFP構築物を運ぶ安定に形質転換された植物体由来の花粉を使用した。この構築物を可視の非破壊マーカーとして使用して、有性事象に由来する胚および内乳と、本発明の方法に由来する胚とを判別する。CaMV35Sプロモーターは胚および内乳で活性であるが、胚珠で不活性であり、したがって受精に由来する胚のみを特徴づける。花粉供与体として使用した植物体は、このCaMV35Sプロモーター::GFP構築物に関してホモ接合である。
【0056】
この花粉を放射線照射し、少数の花粉でも卵細胞を受精させ、受粉により正常な接合胚形成を誘発できるように放射線照射量を選択した。これは、子房から果実への成長も刺激したが、この果実は、種子を正常な数の10%未満含有しており、有性生殖過程は、完全には無効にされなかったが、著しく影響を受けたことが示された。
【0057】
アラビドプシスExo70遺伝子(Atg28640)のプロモーター配列によって駆動されるBBM遺伝子による形質転換は、実施例2に記載の通り微粒子銃で実施する。構築物は、BBMと融合したEXO70プロモーター配列(pExo70)を含有する。
【0058】
実施例4
形質転換された花粉の受粉および胚形成
トマトの花を除雄し、実施例3で得られる形質転換された花粉を受粉させた。受粉後、子房が拡大し、果実様体を形成した。若い果実様構造は、植物体で2〜4週間維持された。植物体を気候条件下(昼22℃、夜18℃)で生育した。果実を収穫し、GFP発現(CaMV−35S::GFP)についてイメージングして受精に由来する胚を除去した。
【0059】
胚珠の一部が本発明による単為発生を開始し、GFP蛍光を示さずに胚様構造を生じた。生存した胚をNeal,CAおよびTopoleski,LD(J.Amer.Soc.Hort.Sci.108(3):434−438(1983)に記載の通り培地上でさらにインキュベートした。未成熟の胚の25%〜50%を生存可能な小植物に再生することができた。この小植物はいずれもトランスジェニックではなく、したがって母系起源の胚を示す。
【0060】
加えて、雄性特異的分子マーカーを使用して胚の起源をさらに確認した(Vos P.ら、Nucleic Acids Research、23:4407−4414(1995))。
【0061】
半数体胚と二倍体胚を判別するために、DNA倍数性レベルの測定を、De Laat,Aら、Plant Breeding 99:303−307(1987)に記載の方法に従いフローサイトメトリーによって実施した。得られた小植物の大多数は倍加半数体であると思われた。
【0062】
実施例5
卵細胞特異的プロモーターにより駆動された発現
アラビドプシス由来の花粉を、トウモロコシ由来のpES4プロモーターの制御下にあるレポーター遺伝子GFPを含むpES4::ES4:GFP構築物(図7参照)で実施例1に記載の通り形質転換した(Cordts,Sら、The Plant J.25:103−114(2001))。形質転換された花粉で蛍光は観察されなかった。
【0063】
形質転換された花粉をアラビドプシス植物に受粉した後、蛍光が卵細胞中で特異的に検出可能であった。この実験から、pES4プロモーターは、卵細胞中で活性であり、細胞分裂誘発分子の発現を作動させるために使用できることが実証される。
【技術分野】
【0001】
本発明は、半数体植物胚および倍加半数体植物胚を作出する新たな方法に関する。本発明はさらに、このようにして得られた植物胚、これから再生された植物体、ならびにこうした植物体の子孫、細胞、組織および種子に関する。
【背景技術】
【0002】
1964年にGuhaとMaheshwari(Nature 204:497)が半数体胞子から植物体を再生できることを発見して以来、他の種で類似の知見を得るために多くの研究が行われてきた(例えば、「In vitro Haploid production in Higher plants」Vol.1、2、3、4、5、Eds:S.Jain、S.SoporyおよびR.Veilleux(1996)Kluwer Academic Publishersを参照されたい)。
【0003】
現代の最新植物育種では、倍加半数体(DH)の使用は、遺伝的純系の作製を迅速化するための、さらには同義遺伝子/複対立遺伝子によってコードされる形質など困難な形質を評価しモニタリングするための、極めて貴重なツールとなっている。
【0004】
作物育種における倍加半数体の作出および使用は多く種でよく知られている(例えば、Thomas W.ら(2003)、In:Doubled haploid production in crop plants.A Manual.Eds.M.Maluszynski、K.Kasha、B.ForsterおよびI.Szarejko.Kluwer Academic Publishers、pp337−349を参照されたい)。一般に、倍加半数体は、雄性器官由来の胞子から得ることができる。この場合、胞子は「小胞子」と称され、in vitro培養は「小胞子培養」と呼ばれる。倍加半数体は、雌性器官または「大胞子」から得ることもできる。対応するin vitro培養は一般に「雌性発生」と称される。
【0005】
典型的な小胞子培養は、古くからアブラナ属で十分に確立されている(例えば、Kellerら(1984)In:K.Giles、S.Sen(eds.)、Plant Cell Culture in Crop Improvement pp169−183.Plenum Pub.Corp.、New Yorkを参照されたい)。典型的な雌性発生培養はサトウダイコンで知られている(例えば、Hosemans D.およびBossoutrot,Z.Pflanzenzuchtg.91:74−77(1983)を参照されたい)。キュウリにおいても雌性発生は極めて十分に確立された技法である(EP0374755を参照されたい)。
【0006】
雌性発生に取り組む方法は、放射線照射された花粉を利用した単為生殖による卵細胞の胚形成の誘発に基づく。よく知られている一例は、メロンで公開されており、現在いくつかの育種会社で常法に従い応用されている(Sauton AおよびR.Dumas de Vaulx、Agronomie 7:141−148(1987)を参照されたい)。単為生殖では、未受精卵から新たな植物体が発生する。この技法の成功率は低い。
【0007】
30年を超える研究を経て利用可能な技術および経験が豊富に得られたにもかかわらず、多く技法の成功は従順な遺伝子型に限定される。これは、DHの使用による膨大な利益を存分に利用できていないことを意味する。一部の遺伝子型の反応性の変動は別として、トマト、ワタなどいくつかの作物種では、いまだにDHの誘導が困難である。
【発明の概要】
【発明が解決しようとする課題】
【0008】
したがって、本発明の目的は、半数体植物胚または倍加半数体植物胚を作出するための新たな方法を提供することである。
【課題を解決するための手段】
【0009】
本発明をもたらした研究において、驚くべきことに、細胞分裂誘発分子を含む花粉または小胞子を卵細胞に受粉することにより、卵細胞を、胚形成を遂げるように誘導できることが見出された。次いで、この卵細胞を、受精なしで胚を形成するようにトリガーする。花粉により卵細胞は実際に受精していないので、二倍体接合体は形成されず、生じた胚は半数体のままである。受粉後のある段階の細胞分裂中に、自然な染色体倍加を起こして、部分的にまたは完全にDHである胚を得ることができる。染色体倍加は、例えば、コルヒチンなどの知られている化学的手段によって誘発することもできる。
【0010】
したがって、本発明は、半数体植物胚を作出するための方法に関し、この方法は、
a)細胞分裂誘発分子を含む小胞子または花粉を提供する工程と、
b)小胞子または花粉を、半数体胚を作製すべき植物体の胚嚢細胞、特に卵細胞に受粉させる工程と、
c)半数体植物胚を得るために、小胞子または花粉が胚嚢細胞、特に卵細胞中にまたはその近くに細胞分裂誘発分子を放出することを可能にして、その分裂をトリガーする工程と
を含む。
【0011】
代替の実施形態では、本発明は、倍加半数体植物胚を作出するための方法に関し、この方法は、
a)細胞分裂誘発分子を含む小胞子または花粉を提供する工程と、
b)小胞子または花粉を、倍加半数体胚を作製すべき植物体の胚嚢細胞、特に卵細胞に受粉させる工程と、
c)植物胚を得るために、小胞子または花粉が胚嚢細胞、特に卵細胞中にまたはその近くに細胞分裂誘発分子を放出することを可能にして、その分裂をトリガーする工程と
を含み、染色体数の倍加が受粉後のある段階の、特に細胞分裂中にまたは胚を得た後に起こる。
【0012】
したがって、本発明は、胚嚢細胞または卵細胞の細胞分裂をトリガーするためのビヒクルとしての花粉または小胞子の使用に関する。
【図面の簡単な説明】
【0013】
【図1】アラビドプシスp35S:AP2mutの花序を示す図である。
【図2】パーティクルガン法により得られたGUS陽性の花粉を示す図である。
【図3】パーティクルガン法による花粉の形質転換の代表的な結果を示す図である。
【図4】トマトの15の様々な開花期を示す図である。
【図5】GUS陽性の花粉管の代表例を示す図である。
【図6】DNAプラスミドpExo70::GFP:GUSの構造を示す図である。
【図7】pES4::ES4:GFP構築物の構造を示す図である。
【発明を実施するための形態】
【0014】
第1の実施形態では、細胞分裂誘発分子は、花粉中で、例えば、プラスミド上に存在する核酸から一過性に発現される。細胞分裂誘発分子は、核酸またはタンパク質のいずれかとすることができ、プラスミドからの構成的発現により花粉または小胞子中で産生される。このようにして産生された細胞分裂誘発分子は、受粉後に卵細胞中に放出される。
【0015】
第2の実施形態では、細胞分裂誘発分子は、花粉ゲノム中に安定に組み込まれている核酸から発現される。細胞分裂誘発分子は、核酸またはタンパク質のいずれかとすることができ、構成的発現により花粉または小胞子中で産生される。このようにして産生された細胞分裂誘発分子は、受粉後の胚嚢または卵細胞中に放出される。
【0016】
第3の実施形態では、細胞分裂誘発分子は、胚嚢細胞特異的または卵細胞特異的プロモーターの制御下にある核酸からの発現により胚嚢細胞または卵細胞中で産生される。この核酸は、核酸を含む花粉または小胞子を受粉させることにより胚嚢細胞または卵細胞中に入る。胚嚢細胞または卵細胞中で組織特異的プロモーターが作動し、したがって細胞分裂誘発分子の産生がもたらされる。
【0017】
第4の実施形態では、花粉または小胞子は、直接形質転換されるのではなく、細胞分裂誘発分子をコードする核酸を運ぶトランスジェニック植物体で形成される。このようなトランスジェニック花粉または小胞子は、構成的プロモーター、または花粉特異的もしくは小胞子特異的プロモーター、または胚嚢細胞特異的もしくは卵細胞特異的プロモーターの制御下にあってよい。第1の場合、転写物は、植物体にとって有害でないことが好ましい。第2の場合、トランスジーンは花粉または小胞子のみで発現され、細胞分裂誘発分子は花粉または小胞子のみで産生される。第3の場合、トランスジーンは、受粉後に核酸が卵細胞または胚嚢細胞に入ると発現される。
【0018】
こうしたすべての実施形態では、花粉による卵細胞の実際の受精は回避しなればならない。これは、放射線照射を用いて、または花粉および卵細胞について異なる種を使用することによって達成することができる。タンパク質は、放射線照射によって損傷を受けない、または実質的に損傷を受けないので、放射線照射は、タンパク質が花粉中で産生される実施形態に特に適している。核酸を花粉または小胞子から胚嚢または卵細胞に移すべき場合、卵細胞供与植物体とは別の種を使用することが好ましい。
【0019】
本発明の特定の実施形態によれば、細胞分裂誘発分子を含む小胞子または花粉は、核酸による形質転換によって得られる。形質転換は、任意の適当な方法で、例えば、アグロバクテリウム・ツメファシエンス(Agrobacterium tumefaciens)を用いて、または微粒子銃(バイオリスティック法)を用いて実施することができる。
【0020】
こうした形質転換技法はよく知られている。アグロバクテリウム・ツメファシエンスを用いた植物細胞の形質転換は十分に確立されており、例えば、De la Rivaら、EJB Vol.1(3)(1998)およびBent、Plant Physiol.124:1540−1547(2000)にその総説がある。
【0021】
近年、植物の遺伝的形質転換は、アグロバクテリウム属だけに限定されず、他の細菌も植物を形質転換する潜在能力を有することが発見された(参照により本明細書に組み込まれているBroothaertsら、Nature 433、629−633(2005))。植物に関連するこうした共生細菌は、不活性化(disarmed)Tiプラスミドおよび適当なバイナリーベクターの両方の獲得により、遺伝子導入に対してコンピテントにされた。このような形質転換系も本発明における使用に適している。
【0022】
バイオリスティック形質転換も当分野の技術者によく知られており、このような用途のためのツールは数年前から市販されている(Ralph Book、In:QiagenNews、Issue No.5、1997)。
【0023】
本発明において使用するのに適した技法はまた、例えば、アンティリヌム・マユス(Antirrhinum majus)における小胞子のレベルでのDNAのデリバリーおよびその一過性発現が示されている、Barinovaら(J Exp Bot.53(371):1119−29(2002))に、またはアルファルファ(メディカゴ・サティバL.(Medicago sativa L.))に関するRamaiahら(Current Science 73:674−682(1997))に記載されている。
【0024】
タバコにおけるバイオリスティック微粒子銃を用いた小胞子または花粉の形質転換のための方法は、Baubak Bajoghli(Matrikel number:9802743、University of Vienna、Experimentelle Genetic III.Plant Biotechnology by Alisher Touraev、July 2001)で見出すことができる。Van der Leede−Plegtら、Transgenic Research 4(2):77−86(1995)は、微粒子銃を用いたタバコ(ニコチアナ・グルティノサ(Nicotiana glutinosa))の花粉中へのDNAの直接的なデリバリーについて記載している。こうした技法および他の技法は、本発明において使用するための花粉または小胞子の形質転換に使用することができる。
【0025】
したがって、特定の実施形態では、花粉および小胞子は、核酸の存在に基づいて細胞分裂誘発分子を含む。導入される核酸は、細胞分裂誘発分子それ自体とすることができ、または細胞分裂誘発分子をコードすることができる。後者の場合、この誘発分子はタンパク質またはペプチドである。前者の場合、この誘発分子は核酸である。この核酸は、それ自体誘導性とすることができ、または他の核酸が発現するのを遮断することができる。例えば、この核酸は、Kip関連タンパク質ファミリーのメンバーまたは網膜芽細胞腫に対するRNAiとすることができ、またはそれをコードすることができる(例えば、Park Jら、Plant Journal 42:153−163(2005)を参照されたい)。網膜芽細胞腫タンパク質は、植物における細胞増殖、分化および核内倍加を調節する。
【0026】
あるいは、この核酸は、細胞分裂誘発分子の前駆体、または細胞分裂誘発分子を産生する酵素をコードすることができる。核酸がこの酵素をコードする場合、これは、誘発分子であるかまたはこれを直接産生する酵素であってよく、あるいは最終的に誘発分子が得られる経路の部分である酵素であってよい。本明細書では、「細胞分裂誘発分子」は、細胞分裂を直接的または間接的にトリガーするあらゆる分子を包含するものとする。
【0027】
本発明は、細胞分裂誘発分子が、形質転換された花粉または小胞子を用いて胚嚢または卵細胞に送達される原理に基づく。細胞分裂を作動させることができる遺伝子構築物または分子は、それ自体知られており、本発明の新たな方法で使用することができる。
【0028】
本発明に従って使用することができる遺伝子の例は、栄養成長を継続するようにプログラム化された体細胞(栄養細胞)が、転写因子をコードする(1種または複数の)遺伝子でこの細胞を形質転換することにより胚発生に容易に転換することを開示するStoneら(PNAS 98、11806−11811(2001))に記載されている。別の例は、Baby Boom(Boutilierら、The Plant Cell 14、1737−1749(2002))またはleafy cotyledon(Stone Sら、PNAS 25:11806−11811(2001))である。こうした遺伝子および他の遺伝子は、本発明の細胞分裂誘発分子をコードするために使用することができる。
【0029】
Zuoら(The Plant Journal 30:1−12(2002))では、体細胞胚形成が、いわゆるWuschel遺伝子を過剰発現させることによりアラビドプシスのあらゆる組織で植物ホルモンなしで誘発されることが見出されたことについて記載されている。この技術はUS2003/0082813にも記載されている。本発明との相違は、US2003/0082813では植物細胞がWuschel DNA配列により一過性ではなく、安定に形質転換されることである。次いで、この安定に導入されたWuschel遺伝子は、安定に組み込まれている組織で過剰発現され、この組織中で細胞分裂および胚形成が誘発される。
【0030】
本発明によれば、Wuschel遺伝子の発現産物であってよい細胞分裂誘発分子をコードする核酸は、細胞分裂を遂げるべき細胞のゲノム中に安定に組み込まれていない。細胞分裂誘発分子をコードする核酸は、卵細胞特異的プロモーターの制御下にあるので、分裂細胞中で一過性に発現されるか、または花粉中で発現され、その後、コードされる細胞分裂誘発分子が、細胞分裂を開始するようにトリガーすべき細胞(卵細胞または胚嚢細胞)中に放出されるかのいずれかである。したがって、花粉または小胞子は、分子それ自体(コード配列ではなく、Wuschel遺伝子発現産物)を、細胞分裂を開始するようにトリガーすべき細胞中に導入するためのビヒクルである。
【0031】
卵細胞を誘発する遺伝子の適用は、上述の遺伝子などの遺伝子の異所性および一過性発現に限定されないが、iaaM、iaaHなどのホルモンを産生することができる酵素をコードする遺伝子(Thomashowら、(1986)Science 231、616−618を参照されたい)および細胞周期タンパク質をコードする遺伝子を使用することによっても類似の結果を得ることができる。
【0032】
加えて、卵細胞の分裂および胚形成の誘発をさらに最適化するために遺伝子の組合せを使用することができる。細胞周期遺伝子の一過性発現も卵細胞の分裂を引き起こすことができる(細胞周期遺伝子の総説については、Murray A(Cell 116:221−234(2004))を参照されたい)。特に、サイクリンEおよびDの単独でのまたは組み合わせての一過性発現を使用して卵細胞分裂をトリガーすることができる。
【0033】
本発明によれば、核酸は、小胞子または花粉中で一過性にもしくはゲノム中への安定な組込みの後のいずれかで発現されるか、または卵細胞中で卵細胞特異的プロモーターから一過性に発現されるかのいずれかである。このように、得られた胚における構成的発現は回避される。
【0034】
一過性発現は、組織特異的または細胞特異的に起こり得る。代替の実施形態では、一過性発現は、核酸を組織特異的または誘導性プロモーターの制御下に置くことによって得られる。組織特異的プロモーターは、適切には、花粉特異的または小胞子特異的プロモーターである。花粉特異的プロモーターはよく知られており、一過性発現は単子葉植物種および双子葉植物種の両方で示されている。こうした型のプロモーターの例は、例えば、Twell,Dら、Development 109(3):705−713(1990);Hamilton,Dら、Plant Mol.Biol.18:211−218(1992)に記載されている。
【0035】
したがって、小胞子(例えば、アグロバクテリウム形質転換またはバイオリスティック形質転換により)または花粉粒を形質転換する従来の方法を使用することにより、Baby Boom、Wuschel、leafy cotyledon、サイクリン、サイクリン依存性キナーゼ(CDK)、E2F(高等真核生物における転写因子ファミリーのメンバー;Zheng,Nら、Genes Dev.13:666(1999))、DP(Magyar Zら、FEBS Lett.486(1):79−87(2000))などのような遺伝子の一過性発現をもたらす本発明によれば、形質転換された小胞子または花粉粒を雌受容体に受粉すると、卵細胞の分裂を誘発することが可能である。
【0036】
好ましくは、花粉または小胞子の雄原核は不活性化または破壊される。このように、卵細胞の受精は確かに回避される。誘発分子を不必要に損傷しないためには、雄原核の不活性化または破壊は、花粉または小胞子の形質転換の前に実施することが好ましい。不活性化または破壊は、適切には放射線照射を用いて行われる。
【0037】
花粉核の放射線照射は、雄原核を分解するためのよく知られている方法であるが、放射線照射量に依存して、これは、花粉管が形成され、卵細胞中に放出することを妨害しない。Grantら(New Zealand Journal of Botany 18、339−341(1980))ではこの技法について記載されている。
【0038】
放射線照射され、続いて形質転換された花粉/小胞子細胞は、その後、同じ種または前記花粉/小胞子細胞の花粉放出が起こり得る種由来の植物体の雌ずいに移される。異種の受粉の一例は、花粉供与植物体としてのナス科および受容植物体としてのトマトに属する種の使用である。他の例は、de Martinis,Dら、Planta 214(5):806−812(2002)およびDore Cら、Plant Cell Reports 15:758−761(1996)に記載されている。一般に、異種の受粉に適している種は同じ科に属するものである。
【0039】
種に依存して、分裂に由来した種子は、卵細胞または卵巣から収穫することができる。あるいは、胚珠培養では、発生中の胚を生存させることが必要であり得る。
【0040】
したがって、基本的に、不活性化された花粉粒は、卵細胞分裂および胚形成を誘発することができるシグナル分子を一過性に運ぶ。この分子は一過性であるので、卵細胞DNAは安定に形質転換されない。
【0041】
したがって、特定の実施形態では、本発明は、雄原核を不活性化するために放射線照射によって不活性化された小胞子または花粉粒中で一過性に存在し発現される細胞分裂分子(タンパク質、DNA、RNA)の利用に関し、それにより前記細胞分裂分子は、花粉管により卵細胞中にまたはその近くに放出されるとその作用を発揮する。
【0042】
別の実施形態では、花粉または小胞子の供与植物体は、細胞分裂誘発分子をコードする遺伝子で安定に形質転換されるが、卵細胞誘発分子の発現は一過性である。この詳細な実施形態では、花粉または小胞子を提供する植物体は、そのゲノムDNAが、好ましくは胚嚢細胞特異的または卵細胞特異的プロモーターの制御下にある細胞分裂分子をコードする遺伝子または遺伝子構築物を運ぶように安定に形質転換される。次いで、この遺伝子または遺伝子構築物は、胚嚢細胞中に、特に卵細胞中にまたはその近くに放出されると発現される。生じた細胞分裂誘発分子は細胞分裂をトリガーする。
【0043】
受粉の前に、花粉または小胞子を放射線照射して雄原核を不活性化する。あるいは、花粉または小胞子は、前記花粉/小胞子細胞の花粉放出が起こり得る別の種の雌ずいに移される。この方法の利点は、細胞分裂誘発分子をコードする核酸が雄原核に運ばれ、最終的に精細胞中に行き着くことである。好ましくは、細胞分裂誘発分子は、供与植物体に、結果として精細胞に複数コピー存在する。花粉または小胞子の発生への遺伝子構築物の存在の干渉を回避するために、花粉または小胞子が胚嚢細胞、特に卵細胞中に移された場合のみ遺伝子または遺伝子構築物の発現を可能にする誘導性または特異的プロモーター、好ましくは胚嚢細胞特異的または卵細胞特異的プロモーターが使用される。
【0044】
本発明はさらに、本発明の方法を用いて得られる半数体胚および倍加半数体胚、ならびにこのような半数体胚または倍加半数体胚から再生された植物体、このような植物体の子孫、ならびにこのような植物体またはその子孫由来の種子、細胞、組織、小胞子および卵細胞に関する。
【0045】
本願明細書では、卵細胞という用語は、読みやすくする目的で単独で使用されることがあるが、この場合「胚嚢細胞、特に卵細胞」と解釈するものとする。
【0046】
本発明はさらに、例示目的にすぎず、本発明を限定するものと決して解釈されない続く実施例で明らかにされよう。
【実施例】
【0047】
実施例1
微粒子銃を用いたアラビドプシス花粉の形質転換
DNAプラスミドpCAMBIA 1301およびpExo70::GFP:GUSを使用して1μmの金粒子をコーティングした。pCAMBIA 1301は、800ヌクレオチドのCaMV35Sプロモーターによって調節されるGUSを含有するバイナリーベクターである(Robertsら、pCAMBIA Vector release manual version 3.05(1998))。pExo70::GFP:GUSは、β−グルクロニダーゼ(GUS)および緑色蛍光タンパク質(GFP)を含有する(図6)。
【0048】
アラビドプシスp35S:AP2mutの3つの花序をペトリ皿の中央に置いた(図1)。ペトリ皿をパーティクルガンに設置し、コーティングされた金粒子を3回撃ち込んだ。銃の撃ち込みから2日後に発現を調べた。図2は、このようにして得られたGUS陽性の花粉を示す。
【0049】
同じ実験をペトリ皿でパーティクルガンにおけるレベル3で繰り返した。2200psiの圧力で2回撃ち込んだ。図3A〜Dは、この実験の代表的な結果を示す。
【0050】
この実験から、微粒子銃を使用して花粉を形質転換することができるという結果になる。
【0051】
実施例2
微粒子銃の撃込み後のin vitroでの花粉発芽
成熟した花粉は静止状態である。花粉粒が雌株の柱頭に付着した後、柱頭からの水移動による再水和から花粉発芽の過程が始まる。この実施例では、微粒子銃の撃込み後、in vitroで花粉を発芽させる。
【0052】
図4は、トマトの15の様々な開花期を示す。段階1、段階5および段階14の花粉を本実験に使用した。段階1の花粉は完全に成熟しており、段階5の花粉も成熟している。段階14は、単核性の後期/二核性の初期である。
【0053】
段階1、段階5および段階14の花の花粉を200μlのNLN13培地(13%ショ糖を補充したNLN培地(Lichter R.、Z.Pflanzenzuecht 105:427−437(1982)))中に分離した。この200μlをgenescreenメンブレン上にスポットし、5分間乾燥させる。次いで、このメンブレンを1/2 MS寒天平板上に置き、Exo70::GFP:GUSでコーティングされた1μmの金粒子を2200psiで撃ち込む。撃込み後、メンブレンは6ウェルタイタープレートに置く。段階1および段階5の花粉は、1.5mlの発芽培地A(Clarke)(20mM MES、0.07%Ca(NO3)2・H2O、0.02%MgSO4・7H2O、0.01%KNO3、0.01%H3BO3、2%ショ糖および15%PEG4000)中でインキュベートする。段階14の小胞子は、NLN13培地中でインキュベートする。3時間インキュベートした後、1.5mlの2×GUS染色バッファーを添加し、試料を37℃で一晩置いた。図5は、GUS陽性の花粉管の代表例を示す。形質転換後でも花粉は管を形成できることが明らかである。
【0054】
実施例3
細胞分裂刺激因子を運ぶ花粉を受粉した後の卵細胞の細胞分裂を誘発するためのこうした花粉の調製
実施例1および2から、GUSを用いたモデル系で花粉を形質転換できることが実証される。ここで、花粉を放射線照射した後の、細胞分裂誘発分子BabyBoom(BBM)(Boutilierら、2002、前掲)によるトマト花粉の形質転換法を記載する。
【0055】
CaMV35Sプロモーター::GFP構築物を運ぶ安定に形質転換された植物体由来の花粉を使用した。この構築物を可視の非破壊マーカーとして使用して、有性事象に由来する胚および内乳と、本発明の方法に由来する胚とを判別する。CaMV35Sプロモーターは胚および内乳で活性であるが、胚珠で不活性であり、したがって受精に由来する胚のみを特徴づける。花粉供与体として使用した植物体は、このCaMV35Sプロモーター::GFP構築物に関してホモ接合である。
【0056】
この花粉を放射線照射し、少数の花粉でも卵細胞を受精させ、受粉により正常な接合胚形成を誘発できるように放射線照射量を選択した。これは、子房から果実への成長も刺激したが、この果実は、種子を正常な数の10%未満含有しており、有性生殖過程は、完全には無効にされなかったが、著しく影響を受けたことが示された。
【0057】
アラビドプシスExo70遺伝子(Atg28640)のプロモーター配列によって駆動されるBBM遺伝子による形質転換は、実施例2に記載の通り微粒子銃で実施する。構築物は、BBMと融合したEXO70プロモーター配列(pExo70)を含有する。
【0058】
実施例4
形質転換された花粉の受粉および胚形成
トマトの花を除雄し、実施例3で得られる形質転換された花粉を受粉させた。受粉後、子房が拡大し、果実様体を形成した。若い果実様構造は、植物体で2〜4週間維持された。植物体を気候条件下(昼22℃、夜18℃)で生育した。果実を収穫し、GFP発現(CaMV−35S::GFP)についてイメージングして受精に由来する胚を除去した。
【0059】
胚珠の一部が本発明による単為発生を開始し、GFP蛍光を示さずに胚様構造を生じた。生存した胚をNeal,CAおよびTopoleski,LD(J.Amer.Soc.Hort.Sci.108(3):434−438(1983)に記載の通り培地上でさらにインキュベートした。未成熟の胚の25%〜50%を生存可能な小植物に再生することができた。この小植物はいずれもトランスジェニックではなく、したがって母系起源の胚を示す。
【0060】
加えて、雄性特異的分子マーカーを使用して胚の起源をさらに確認した(Vos P.ら、Nucleic Acids Research、23:4407−4414(1995))。
【0061】
半数体胚と二倍体胚を判別するために、DNA倍数性レベルの測定を、De Laat,Aら、Plant Breeding 99:303−307(1987)に記載の方法に従いフローサイトメトリーによって実施した。得られた小植物の大多数は倍加半数体であると思われた。
【0062】
実施例5
卵細胞特異的プロモーターにより駆動された発現
アラビドプシス由来の花粉を、トウモロコシ由来のpES4プロモーターの制御下にあるレポーター遺伝子GFPを含むpES4::ES4:GFP構築物(図7参照)で実施例1に記載の通り形質転換した(Cordts,Sら、The Plant J.25:103−114(2001))。形質転換された花粉で蛍光は観察されなかった。
【0063】
形質転換された花粉をアラビドプシス植物に受粉した後、蛍光が卵細胞中で特異的に検出可能であった。この実験から、pES4プロモーターは、卵細胞中で活性であり、細胞分裂誘発分子の発現を作動させるために使用できることが実証される。
【特許請求の範囲】
【請求項1】
半数体植物胚を作出するための方法であって、
a)細胞分裂誘発分子を含む小胞子または花粉を提供する工程と、
b)小胞子または花粉を、半数体胚を作製すべき植物体の胚嚢細胞、特に卵細胞に受粉させる工程と、
c)小胞子または花粉が胚嚢細胞、特に卵細胞中にまたはその近くに細胞分裂誘発分子を放出することを可能にして、その分裂をトリガーして、半数体植物胚を得る工程とを含む方法。
【請求項2】
倍加半数体植物胚を作出するための方法であって、
a)細胞分裂誘発分子を含む小胞子または花粉を提供する工程と、
b)小胞子または花粉を、倍加半数体胚を作製すべき植物体の胚嚢細胞、特に卵細胞に受粉させる工程と、
c)小胞子または花粉が胚嚢細胞、特に卵細胞中にまたはその近くに細胞分裂誘発分子を放出することを可能にして、その分裂をトリガーして、植物胚を得る工程とを含み、染色体数の倍加が受粉後のある段階の、特に細胞分裂中にまたは胚を得た後に起こる方法。
【請求項3】
染色体数の倍加が自然に起こる、請求項2に記載の方法。
【請求項4】
染色体数の倍加が薬品処理を用いて、特にコルヒチンを用いて行われる、請求項2に記載の方法。
【請求項5】
細胞分裂誘発分子を含む小胞子または花粉が、核酸による小胞子または花粉の形質転換によって得ることができる、請求項1〜4いずれか一項記載の方法。
【請求項6】
細胞分裂誘発分子を含む小胞子または花粉が、細胞分裂誘発分子をコードする核酸で形質転換された植物体から得ることができる、請求項1〜4いずれか一項記載の方法。
【請求項7】
形質転換がアグロバクテリウム・ツメファシエンスまたはバイオリスティック法を用いて行われる、請求項5または6に記載の方法。
【請求項8】
核酸が、細胞分裂誘発分子であるか、または細胞分裂誘発分子、細胞分裂誘発分子の前駆体、もしくは細胞分裂誘発分子を産生する酵素をコードする、請求項5、6または7に記載の方法。
【請求項9】
細胞分裂誘発分子である核酸が、細胞分裂を阻害または停止する遺伝子の発現を遮断するRNAiである、請求項5、6または7に記載の方法。
【請求項10】
細胞分裂を阻害または停止する遺伝子が、網膜芽細胞腫(Rb)またはKip関連タンパク質(KRP)ファミリーのメンバーである、請求項9に記載の方法。
【請求項11】
細胞分裂誘発分子が、Baby Boom、Leafy cotyledon、WUSCHEL、サイクリン、サイクリン依存性キナーゼ(CDK)、E2FおよびDPからなる群から選択される、請求項5、6または7に記載の方法。
【請求項12】
核酸が小胞子または花粉中で一過性に発現される、請求項5、6または7に記載の方法。
【請求項13】
一過性発現が、核酸を組織特異的または誘導性プロモーターの制御下に置くことによって得られる、請求項12に記載の方法。
【請求項14】
組織特異的プロモーターが花粉特異的または小胞子特異的プロモーターである、請求項13に記載の方法。
【請求項15】
核酸が、小胞子または花粉中では安定に組み込まれているが、胚嚢細胞または卵細胞中では一過性に発現される、請求項5、6または7に記載の方法。
【請求項16】
一過性発現が、核酸を花粉または小胞子中では活性のない組織特異的または誘導性プロモーターの制御下に置くことによって得られる、請求項15に記載の方法。
【請求項17】
組織特異的プロモーターが胚嚢細胞特異的または卵細胞特異的プロモーターである、請求項13または16に記載の方法。
【請求項18】
細胞分裂誘発分子をコードする核酸が安定に組み込まれているか、または一過性に発現される小胞子もしくは花粉が、胚嚢細胞または卵細胞を提供する受容植物体とは別の種に属する供与植物体由来である、請求項1〜17いずれか一項記載の方法。
【請求項19】
花粉または小胞子の雄原核が不活性化または破壊されている、請求項1〜17いずれか一項記載の方法。
【請求項20】
雄原核の不活性化または破壊が花粉または小胞子の形質転換の前に行われる、請求項19に記載の方法。
【請求項21】
雄原核の不活性化または破壊が放射線照射を用いて行われる、請求項19または20に記載の方法。
【請求項22】
請求項1または請求項5〜21いずれか一項記載の方法を用いて得ることができる半数体植物胚。
【請求項23】
請求項2〜20いずれか一項記載の方法を用いて得られる倍加半数体植物胚。
【請求項24】
請求項22または23に記載の植物胚から再生された植物体。
【請求項25】
請求項24に記載の植物体の子孫。
【請求項26】
請求項24または25に記載の植物体の種子。
【請求項27】
請求項24または25に記載の植物体由来の細胞。
【請求項28】
花粉、小胞子、胚嚢細胞および卵細胞から選択される、請求項27に記載の細胞。
【請求項29】
請求項24または25に記載の植物体由来の組織。
【請求項30】
胚嚢組織である、請求項29に記載の組織。
【請求項1】
半数体植物胚を作出するための方法であって、
a)細胞分裂誘発分子を含む小胞子または花粉を提供する工程と、
b)小胞子または花粉を、半数体胚を作製すべき植物体の胚嚢細胞、特に卵細胞に受粉させる工程と、
c)小胞子または花粉が胚嚢細胞、特に卵細胞中にまたはその近くに細胞分裂誘発分子を放出することを可能にして、その分裂をトリガーして、半数体植物胚を得る工程とを含む方法。
【請求項2】
倍加半数体植物胚を作出するための方法であって、
a)細胞分裂誘発分子を含む小胞子または花粉を提供する工程と、
b)小胞子または花粉を、倍加半数体胚を作製すべき植物体の胚嚢細胞、特に卵細胞に受粉させる工程と、
c)小胞子または花粉が胚嚢細胞、特に卵細胞中にまたはその近くに細胞分裂誘発分子を放出することを可能にして、その分裂をトリガーして、植物胚を得る工程とを含み、染色体数の倍加が受粉後のある段階の、特に細胞分裂中にまたは胚を得た後に起こる方法。
【請求項3】
染色体数の倍加が自然に起こる、請求項2に記載の方法。
【請求項4】
染色体数の倍加が薬品処理を用いて、特にコルヒチンを用いて行われる、請求項2に記載の方法。
【請求項5】
細胞分裂誘発分子を含む小胞子または花粉が、核酸による小胞子または花粉の形質転換によって得ることができる、請求項1〜4いずれか一項記載の方法。
【請求項6】
細胞分裂誘発分子を含む小胞子または花粉が、細胞分裂誘発分子をコードする核酸で形質転換された植物体から得ることができる、請求項1〜4いずれか一項記載の方法。
【請求項7】
形質転換がアグロバクテリウム・ツメファシエンスまたはバイオリスティック法を用いて行われる、請求項5または6に記載の方法。
【請求項8】
核酸が、細胞分裂誘発分子であるか、または細胞分裂誘発分子、細胞分裂誘発分子の前駆体、もしくは細胞分裂誘発分子を産生する酵素をコードする、請求項5、6または7に記載の方法。
【請求項9】
細胞分裂誘発分子である核酸が、細胞分裂を阻害または停止する遺伝子の発現を遮断するRNAiである、請求項5、6または7に記載の方法。
【請求項10】
細胞分裂を阻害または停止する遺伝子が、網膜芽細胞腫(Rb)またはKip関連タンパク質(KRP)ファミリーのメンバーである、請求項9に記載の方法。
【請求項11】
細胞分裂誘発分子が、Baby Boom、Leafy cotyledon、WUSCHEL、サイクリン、サイクリン依存性キナーゼ(CDK)、E2FおよびDPからなる群から選択される、請求項5、6または7に記載の方法。
【請求項12】
核酸が小胞子または花粉中で一過性に発現される、請求項5、6または7に記載の方法。
【請求項13】
一過性発現が、核酸を組織特異的または誘導性プロモーターの制御下に置くことによって得られる、請求項12に記載の方法。
【請求項14】
組織特異的プロモーターが花粉特異的または小胞子特異的プロモーターである、請求項13に記載の方法。
【請求項15】
核酸が、小胞子または花粉中では安定に組み込まれているが、胚嚢細胞または卵細胞中では一過性に発現される、請求項5、6または7に記載の方法。
【請求項16】
一過性発現が、核酸を花粉または小胞子中では活性のない組織特異的または誘導性プロモーターの制御下に置くことによって得られる、請求項15に記載の方法。
【請求項17】
組織特異的プロモーターが胚嚢細胞特異的または卵細胞特異的プロモーターである、請求項13または16に記載の方法。
【請求項18】
細胞分裂誘発分子をコードする核酸が安定に組み込まれているか、または一過性に発現される小胞子もしくは花粉が、胚嚢細胞または卵細胞を提供する受容植物体とは別の種に属する供与植物体由来である、請求項1〜17いずれか一項記載の方法。
【請求項19】
花粉または小胞子の雄原核が不活性化または破壊されている、請求項1〜17いずれか一項記載の方法。
【請求項20】
雄原核の不活性化または破壊が花粉または小胞子の形質転換の前に行われる、請求項19に記載の方法。
【請求項21】
雄原核の不活性化または破壊が放射線照射を用いて行われる、請求項19または20に記載の方法。
【請求項22】
請求項1または請求項5〜21いずれか一項記載の方法を用いて得ることができる半数体植物胚。
【請求項23】
請求項2〜20いずれか一項記載の方法を用いて得られる倍加半数体植物胚。
【請求項24】
請求項22または23に記載の植物胚から再生された植物体。
【請求項25】
請求項24に記載の植物体の子孫。
【請求項26】
請求項24または25に記載の植物体の種子。
【請求項27】
請求項24または25に記載の植物体由来の細胞。
【請求項28】
花粉、小胞子、胚嚢細胞および卵細胞から選択される、請求項27に記載の細胞。
【請求項29】
請求項24または25に記載の植物体由来の組織。
【請求項30】
胚嚢組織である、請求項29に記載の組織。
【図1】


【図2】

【図3】

【図4】

【図5】
【図6】
【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2013−74892(P2013−74892A)
【公開日】平成25年4月25日(2013.4.25)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−257445(P2012−257445)
【出願日】平成24年11月26日(2012.11.26)
【分割の表示】特願2008−514018(P2008−514018)の分割
【原出願日】平成18年5月31日(2006.5.31)
【出願人】(500502222)ライク・ズワーン・ザードテールト・アン・ザードハンデル・ベスローテン・フェンノートシャップ (19)
【Fターム(参考)】
【公開日】平成25年4月25日(2013.4.25)
【国際特許分類】
【出願番号】特願2012−257445(P2012−257445)
【出願日】平成24年11月26日(2012.11.26)
【分割の表示】特願2008−514018(P2008−514018)の分割
【原出願日】平成18年5月31日(2006.5.31)
【出願人】(500502222)ライク・ズワーン・ザードテールト・アン・ザードハンデル・ベスローテン・フェンノートシャップ (19)
【Fターム(参考)】
[ Back to top ]