
単球性樹状細胞およびT細胞にTH−1応答をプライミングするための組成物および方法
【課題】未成熟樹状細胞(DC)の成熟を誘導するため、ならびにこれらの細胞に1型免疫応答を誘導することをプライミングするための組成物および方法を提供すること。
【解決手段】本発明はまた、1型サイトカインおよび/または1型応答の産生に向かって極性化したT細胞を活性化および調製するために有用な樹状細胞集団を提供する。同様に、活性化され極性化したT細胞集団、およびこれらを作製する方法が提供される。本発明の方法はまた、未成熟樹状細胞を提供する工程;ならびに該未成熟樹状細胞を、該未成熟樹状細胞の成熟に適切な培養条件下で、有効量のBCGおよびインターフェロンγ(IFNγ)と接触させて、成熟樹状細胞集団を形成させる工程を包含する、成熟樹状細胞集団を産生するための方法である。
【解決手段】本発明はまた、1型サイトカインおよび/または1型応答の産生に向かって極性化したT細胞を活性化および調製するために有用な樹状細胞集団を提供する。同様に、活性化され極性化したT細胞集団、およびこれらを作製する方法が提供される。本発明の方法はまた、未成熟樹状細胞を提供する工程;ならびに該未成熟樹状細胞を、該未成熟樹状細胞の成熟に適切な培養条件下で、有効量のBCGおよびインターフェロンγ(IFNγ)と接触させて、成熟樹状細胞集団を形成させる工程を包含する、成熟樹状細胞集団を産生するための方法である。
【発明の詳細な説明】
【背景技術】
【0001】
(発明の背景)
抗原提示細胞(APC)は、有効な免疫応答を惹起する際に重要である。これは、抗原特異的T細胞レセプターを有するT細胞に対して抗原を提示するだけでなく、T細胞活性化に必要なシグナルも提供する。これらのシグナルは、完全に規定されていないままであるが、種々の細胞表面分子ならびにサイトカインまたは増殖因子を含む。ナイーブT細胞または非極性化T細胞の活性化に必要な因子は、記憶T細胞の再活性化に必要とされる因子とは異なり得る。APCが抗原を提示する能力およびT細胞活性化シグナルを送達する能力は両方とも、通常、補助細胞機能といわれる。単球およびB細胞はコンピテントAPCであることが示されているが、それらのインビトロでの抗原提示能力は、以前に感作されたT細胞の再活性化に制限されているようである。それゆえ、これらは、機能的にナイーブなT細胞集団も、プライミングされていないT細胞集団も直接的に活性化することができない。
【0002】
樹状細胞(DC)は、ナイーブT細胞および記憶T細胞の両方を活性化し得ると考えられる、免疫系のプロフェッショナル抗原提示細胞である。樹状細胞は、免疫治療(特に、癌の免疫治療)における使用のために、ますますエキソビボで調製されている。最適な免疫刺激特性を有する樹状細胞の調製は、エキソビボでの培養のために、これらの細胞の生物学の理解および活用を必要とする。各プロトコルに起因する種々の利点を有する、これらの細胞の培養のための種々のプロトコルが記載されている。近年のプロトコルとしては、無血清培地の使用、ならびに所望の免疫刺激特性を培養細胞に付与する成熟条件の使用が挙げられる。
【0003】
樹状細胞の成熟は、未成熟DC(これは、皮膚ランゲルハンス細胞に表現型的に類似する)を、リンパ節へと移動し得る成熟した抗原提示細胞へと転換するプロセスである。このプロセスは、この未成熟樹状細胞を特徴付ける強力な抗原取り込み能力の喪失、ならびに補助刺激細胞表面分子および種々のサイトカインの発現のアップレギュレーションをもたらす。公知の成熟プロトコルは、DCが抗原への曝露の間またはその後に遭遇すると考えられるインビボでの環境に基づく。このアプローチの最良の例は、細胞培養培地としての、単球馴化培地(MCM)の使用である。MCMは、単球を培養することによってインビトロで生成され、そして成熟因子の供給源として用いられる。成熟の原因となる、MCM中の主な成分は、炎症(誘発)性サイトカインインターロイキン1β(IL−1β)、インターロイキン6(IL−6)および腫瘍壊死因子α(TNFα)であると報告されている。他の成熟因子としては、プロスタグランジンE2(PGE2)、ポリ−dIdC、血管作用性(vasointestinal)ペプチド(VIP)、細菌リポ多糖(LPS)、ならびにマイコバクテリアまたはマイコバクテリア成分(例えば、特定の細胞壁成分)が挙げられる。
【0004】
充分に成熟した樹状細胞は、未成熟DCとは定性的および定量的に異なる。充分に成熟したDCは、より高レベルのMHCクラスI抗原およびMHCクラスII抗原、ならびにより高レベルのT細胞補助刺激分子(すなわち、CD80およびCD86)を発現する。これらの変化は、樹状細胞がT細胞を活性化する能力を増大させる。なぜなら、これらは、細胞表面の抗原密度、ならびにT細胞上の補助刺激分子の対応物(例えば、CD28など)を通るT細胞活性化シグナルの大きさを増大させるからである。さらに、成熟DCは、多量のサイトカインを産生し、このサイトカインは、T細胞応答を刺激して導く。これらのサイトカインのうちの2つは、インターロイキン10(IL−10)およびインターロイキン(IL−12)である。これらのサイトカインは、誘導されるT細胞応答の方向に対して反対の効果を有する。IL−10産生は、Th−2型応答の誘導をもたらし、一方、IL−12産生は、Th−1型応答をもたらす。後者の応答は、細胞免疫応答が所望される場合(例えば、癌免疫治療において)、特に望ましい。Th−1型応答は、細胞傷害性Tリンパ球(CTL)(これは、細胞性免疫系のエフェクターアームである)の誘導および分化をもたらす。このエフェクターアームは、腫瘍増殖と戦う際に最も有効である。IL−12はまた、ナチュラルキラー(NK)細胞の増殖を誘導し、そして抗脈管形成活性を有する。これらは両方とも、有効な抗腫瘍防御手段である。従って、IL−12を産生する樹状細胞の使用は、理論的に、免疫刺激における使用に最も適している。
【0005】
特定の樹状細胞成熟因子(例えば、細菌性リポ多糖、細菌性CpG DNA、二本鎖RNAおよびCD40リガンド)は、未成熟DCを誘導して、IL−12を産生させ、未成熟DCにTh−1応答をプライミングすると報告されている。対照的に、抗炎症分子(例えば、IL−10、TGF−β、PGE−2およびコルチコステロイド(corticosteriod))はIL−12産生を阻害し、そして細胞にTh−2応答をプライミングし得る。
【0006】
近年、樹状細胞によるIL−12産生の増強は、インターフェロンγと特定の樹状細胞成熟因子(例えば、細菌性リポ多糖(LPS)およびCD40)とを合わせることによって報告されている。しかし、LPSおよびCD40は両方とも、成熟の間に少量のIL−12を誘導する既知の能力を有する。従って、IFNγの添加は、その産生を増強するにすぎないことが可能である。インターフェロンγシグナル伝達は、Jak2−Stat1経路を使用し、これは、核へのその移動の前のStat1の701位のチロシン残基のチロシンリン酸化、およびその後のインターフェロンγ応答遺伝子の転写増強を含む。しかし、ヒト単球由来樹状細胞におけるシグナル伝達経路についてはごくわずかしか公知でない。これらの細胞におけるインターフェロンγ作用についての機構は、確立されていない。
【0007】
Mycobacterium tuberculosisの弱毒化ウシ株(Mycobacterium bovis)(現在、カルメット−ゲラン杆菌(BCG)として公知)は、癌免疫治療において用いられている。1例では、生BCGの膀胱内投与は、膀胱癌の処置に有効であることが証明されているが、この処置についての機構は、公知ではない。BCG投与の効果は、例えば、癌細胞を攻撃する免疫応答の誘導によって媒介されると仮定されている。この応答におけるBCGの特定の役割は、免疫反応性の一般化された誘導因子のものであり、免疫系に対する腫瘍抗原の提示においてアジュバント機能を有すると考えられる。
【0008】
BCGはまた、成熟マーカーCD83をアップレギュレートする能力を有する、樹状細胞に対する強力な成熟因子であることが見出されている。BCGはまた、エンドサイトーシス能力の低減を伴って、MHC分子ならびに補助刺激分子CD80およびCD86をアップレギュレートし得る。さらに、IL−12の産生が特異的に阻害されることが見出された、他のDC成熟因子について見出された結果とは対照的に、BCGまたはBCG由来リポアラビドマンナンは、サイトカイン産生を増大させることが報告されている。この前者の特性(IL−12産生の阻害)は、強力な細胞媒介性細胞傷害性応答(Th−1応答)が所望される免疫治療用の樹状細胞の成熟のためにBCGを用いることの魅力を低減する。
【発明の概要】
【発明が解決しようとする課題】
【0009】
従って、能動免疫治療におけるBCGの使用は、樹状細胞成熟を誘導する可能性を有する。しかし、樹状細胞のこのような成熟を誘導し、広範な免疫刺激を同時に提供し、そしてこれらの樹状細胞に、強固な細胞傷害性T細胞応答を有する1型(Th−1)免疫応答をプライミングさせる、組成物およびこのような組成物を用いる方法についての必要性が存在する。
【課題を解決するための手段】
【0010】
(発明の簡単な要旨)
本発明は、広範な免疫刺激を同時に提供する薬剤(すなわち、BCG)を用いて未成熟樹状細胞(DC)の成熟を誘導するための方法および組成物、ならびにこれらの細胞に抗原特異的な細胞傷害性T細胞応答をプライミングさせるための方法および組成物を提供する。1つの局面では、成熟樹状細胞集団を産生するための方法が提供され、この方法は、未成熟樹状細胞を提供する工程;ならびにこの未成熟樹状細胞を、この未成熟樹状細胞の成熟に適切な培養条件下で、有効濃度のBCGおよびインターフェロンγ(IFNγ)と接触させて、成熟樹状細胞集団を形成させる工程を包含する。この成熟樹状細胞集団は、成熟の間にBCGおよびIFNγ単独に接触させなかった未成熟樹状細胞集団よりも上昇したインターロイキン12対インターロイキン10比を生じる。この未成熟樹状細胞は、BCGおよびIFNγとの接触の前または接触の間に所定の抗原と接触され得る。この所定の抗原は、例えば、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、抗原を発現する組換え細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原(例えば、合成ペプチド抗原)、または単離抗原であり得る。
【0011】
特定の実施形態では、この方法は、単球性樹状細胞前駆体を単離する工程;およびこの前駆体を分化因子の存在下で培養して未成熟樹状細胞集団を形成する工程を必要に応じてさらに包含し得る。適切な分化因子としては、例えば、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13が挙げられる。この単球性樹状細胞前駆体は、ヒト被験体から単離され得る。特定の実施形態では、この成熟樹状細胞は、少なくとも1:1のIL−12対IL−10の比を生じる。
【0012】
別の局面では、成熟樹状細胞集団を産生するための方法が提供される。この方法は一般に、未成熟樹状細胞を提供する工程;ならびにこの未成熟樹状細胞を、この未成熟樹状細胞の成熟に適切な培養条件下で、有効量のBCGおよびインターフェロンγ(IFNγ)と接触させて、成熟樹状細胞集団を形成させる工程を包含する。得られる成熟樹状細胞集団は、1型免疫応答を生じる。この未成熟樹状細胞を、BCGおよびIFNγとの接触の前または接触の間に、所定の抗原と接触させ得る。この所定の抗原は、例えば、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、抗原を発現する組換え細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原(すなわち、合成ペプチド)、または単離抗原であり得る。
【0013】
特定の実施形態では、この方法は、単球性樹状細胞前駆体を単離する工程;およびこの前駆体を分化因子の存在下で培養して、この未成熟樹状細胞を形成させる工程を必要に応じてさらに包含し得る。適切な分化因子としては、例えば、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13が挙げられる。この単球性樹状細胞前駆体は、ヒト被験体から単離される。特定の実施形態では、この成熟樹状細胞は、少なくとも約1:1のIL−12対IL−10の比を生じる。
【0014】
なお別の局面では、T細胞を活性化するための組成物が提供される。この組成物は、成熟について適切な条件下で有効濃度のBCGおよびIFNγを用いて成熟された樹状細胞集団;および所定の抗原を含み得る。この樹状細胞集団は、成熟の間にIFNγを伴わずにBCGと接触させた成熟樹状細胞集団よりも上昇したインターロイキン12(IL−12)対インターロイキン10(IL−10)比を生じ得る。特定の実施形態では、この樹状細胞集団は、少なくとも約10:1の比のIL−12対IL−10を生じ得る。他の実施形態では、この樹状細胞集団は、成熟の間にIFNγを伴わずにBCGの存在下で培養した同様の未成熟樹状細胞集団よりも少なくとも約1:100の比のIL−12対IL−10を生じ得る。
【0015】
別の局面では、単離された未成熟樹状細胞集団が提供される。この細胞集団は、未成熟単球性樹状細胞、ならびにこの未成熟樹状細胞の成熟を誘導する有効濃度のBCGおよびIFNγを含む。得られる成熟樹状細胞は、成熟の間にIFNγを伴わずにBCGの存在下で培養した同様の未成熟樹状細胞集団よりも高いインターロイキン12(IL−12)対インターロイキン10(IL−10)を生じる。この細胞集団は、所定の抗原および/または単離されたT細胞(例えば、ナイーブT細胞)を必要に応じて含み得る。このT細胞は、単離されたリンパ球の調製物中に必要に応じて存在し得る。
【0016】
活性化T細胞を産生するための方法もまた提供される。この方法は一般に、未成熟樹状細胞を提供する工程;この未成熟樹状細胞を所定の抗原と接触させる工程;ならびにこの未成熟樹状細胞を、この未成熟樹状細胞の成熟に適切な培養条件下で、有効濃度のBCGおよびIFNγと接触させて、成熟樹状細胞を形成させる工程を包含する。この成熟樹状細胞を、ナイーブT細胞と接触させて、IFNγを産生するか、および/または1型(Th−1)応答に極性化した活性化T細胞を形成させ得る。適切な抗原としては、例えば、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、抗原を発現する組換え細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原(例えば、合成ペプチド抗原)、または単離抗原が挙げられる。
【0017】
この未成熟樹状細胞を所定の抗原、BCGおよびIFNγと同時に接触させ得るか、またはこの細胞を、BCGおよびIFNγと接触させる前にこの所定の抗原と接触させ得る。特定の実施形態では、この方法は、単球性樹状細胞前駆体を単離する工程;および分化因子の存在下でこの前駆体を培養して、この未成熟樹状細胞の形成を誘導する工程をさらに包含し得る。適切な分化因子としては、例えば、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13が挙げられる。この単球性樹状細胞前駆体は、ヒト被験体から必要に応じて単離され得る。特定の実施形態では、この未成熟樹状細胞およびT細胞は、互いに自己である。
【0018】
インターロイキン10(IL−10)よりも多くのインターロイキン12(IL−12)を産生する単離された成熟樹状細胞もまた提供される。この成熟樹状細胞は、この樹状細胞の成熟に適切な条件下で有効濃度のBCGおよびIFNγを含む組成物を用いた未成熟樹状細胞の成熟によって提供され得る。所定の抗原が、この単離された成熟樹状細胞に必要に応じて含まれ得る。所定の抗原がローディングされた、単離された成熟樹状細胞もまた提供される。この樹状細胞は、成熟の間にIFNγを伴わずにBCGの存在下で培養した同様の未成熟樹状細胞集団よりも多くの、インターロイキン10(IL−10)よりも多くのインターロイキン12(IL−12)(例えば、IL−10よりも少なくとも10倍多くのIL−12)を産生し得る。
【0019】
動物において1型(Th−1)免疫応答を産生するための方法もまた提供される。この方法は一般に、未成熟樹状細胞を提供する工程;この未成熟樹状細胞を、この未成熟樹状細胞の成熟に適切な培養条件下で、有効量のBCGおよびインターフェロンγ(IFNγ)、ならびに所定の抗原とを接触させて、成熟樹状細胞を形成させる工程を包含する。この成熟樹状細胞を動物に投与するかまたはナイーブT細胞と接触させるかのいずれかによって、インターフェロンγ(IFNγ)および/または腫瘍壊死因子α(TNFα)の産生によって特徴付けられる活性化T細胞を形成させ得る。この活性化T細胞は、特定の抗原に対する細胞傷害性T細胞応答の刺激の必要のある動物に投与され得る。適切な抗原としては、例えば、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、抗原を発現する組換え細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原(例えば、合成ペプチド抗原)、または単離抗原が挙げられる。この未成熟樹状細胞を、この所定の抗原、BCGおよびIFNγと必要に応じて同時に接触させ得るか、またはBCGおよびIFNγと接触させる前に、この未成熟樹状細胞をこの所定の抗原と接触させ得る。
【0020】
特定の実施形態では、この方法は、単球性樹状細胞前駆体をこの動物から単離する工程;およびこの前駆体を分化因子の存在下で培養して、この未成熟樹状細胞を形成させる工程をさらに包含し得る。この分化因子は、例えば、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13であり得る。
【0021】
この未成熟樹状細胞およびT細胞は、この動物に対して自己であってもよく、またはこの動物に対して同種異系であってもよい。あるいは、この未成熟樹状細胞およびT細胞は、この動物と同じMHCハプロタイプを有してもよく、またはMHCマーカーを共有してもよい。特定の実施形態では、この動物は、ヒトであっても非ヒト動物であってもよい。
例えば、本発明は以下の項目を提供する。
(項目1)
成熟樹状細胞集団を産生するための方法であって、該方法は、以下:
未成熟樹状細胞を提供する工程;ならびに
該未成熟樹状細胞を、該未成熟樹状細胞の成熟に適切な培養条件下で、有効量のBCGおよびインターフェロンγ(IFNγ)と接触させて、成熟樹状細胞集団を形成させる工程;
を包含し、ここで、該成熟樹状細胞集団は、成熟の間にBCGおよびIFNγと接触させなかった未成熟樹状細胞集団よりも上昇したインターロイキン12対インターロイキン10比を生じる、方法。
(項目2)
BCGおよびIFNγと接触させる前に、前記未成熟樹状細胞を所定の抗原と接触させる工程をさらに包含する、項目1に記載の方法。
(項目3)
前記未成熟樹状細胞を所定の抗原、BCGおよびIFNγと同時に接触させる工程をさらに包含する、項目1に記載の方法。
(項目4)
前記所定の抗原が、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原、または単離抗原である、項目2または3に記載の方法。
(項目5)
単球性樹状細胞前駆体を単離する工程;および該前駆体を分化因子の存在下で培養して前記未成熟樹状細胞を形成させる工程をさらに包含する、項目1に記載の方法。
(項目6)
前記分化因子が、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13である、項目5に記載の方法。
(項目7)
前記単球性樹状細胞前駆体が、ヒト被験体から単離される、項目5に記載の方法。
(項目8)
前記成熟樹状細胞が、少なくとも約1:1、少なくとも約10:1、少なくとも約100:1のIL−12対IL−10の比を生じる、項目1に記載の方法。
(項目9)
成熟樹状細胞集団を産生するための方法であって、該方法は、以下:
未成熟樹状細胞を提供する工程;ならびに
該未成熟樹状細胞を、該未成熟樹状細胞の成熟に適切な培養条件下で、有効量のBCGおよびインターフェロンγ(IFNγ)と接触させて、成熟樹状細胞集団を形成させる工程;
を包含し、ここで、該成熟樹状細胞集団が、1型免疫応答を生じる、方法。
(項目10)
BCGおよびIFNγと接触させる前に、前記未成熟樹状細胞を所定の抗原と接触させる工程をさらに包含する、項目9に記載の方法。
(項目11)
前記未成熟樹状細胞を所定の抗原、BCGおよびIFNγと同時に接触させる工程をさらに包含する、項目9に記載の方法。
(項目12)
前記所定の抗原が、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、抗原を発現する組換え細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原、または単離抗原である、項目10または11に記載の方法。
(項目13)
単球性樹状細胞前駆体を単離する工程;および該前駆体を分化因子の存在下で培養して前記未成熟樹状細胞を形成させる工程をさらに包含する、項目9に記載の方法。
(項目14)
前記分化因子が、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13である、項目13に記載の方法。
(項目15)
前記単球性樹状細胞前駆体が、ヒト被験体から単離される、項目13に記載の方法。
(項目16)
前記成熟樹状細胞が、少なくとも約1:1、少なくとも約10:1、少なくとも約100:1のIL−12対IL−10の比を生じる、項目9に記載の方法。
(項目17)
T細胞を活性化するための組成物であって、該組成物は、以下:
成熟に適切な条件下で有効濃度のBCGおよびIFNγを用いて成熟させた樹状細胞集団;ならびに
所定の抗原;
を含み、ここで、前記樹状細胞集団が、成熟の間にIFNγを伴わずにBCGと接触させた成熟樹状細胞集団よりも上昇したインターロイキン12(IL−12)対インターロイキン10(IL−10)比を生じる、組成物。
(項目18)
前記樹状細胞集団が、少なくとも約10:1の比のIL−12対IL−10を生じる、項目17に記載の組成物。
(項目19)
前記樹状細胞集団が、少なくとも約100:1の比のIL−12対IL−10を生じる、項目17に記載の組成物。
(項目20)
単離された未成熟樹状細胞集団であって、該細胞集団は、以下:
単離された未成熟単球性樹状細胞、ならびに該未成熟樹状細胞の成熟を誘導する有効濃度のBCGおよびIFNγを含み;ここで、得られる成熟樹状細胞が、インターロイキン10(IL−10)よりも多くのインターロイキン12(IL−12)を産生する、細胞集団。
(項目21)
所定の抗原をさらに含む、項目20に記載の細胞集団。
(項目22)
単離されたT細胞をさらに含む、項目20に記載の細胞集団。
(項目23)
前記T細胞が、ナイーブT細胞である、項目22に記載の細胞集団。
(項目24)
単離されたリンパ球をさらに含む、項目20に記載の細胞集団。
(項目25)
T細胞を産生するための方法であって、該方法は、以下:
未成熟樹状細胞を提供する工程;
該未成熟樹状細胞を所定の抗原と接触させる工程;
該未成熟樹状細胞を、該未成熟樹状細胞の成熟に適切な培養条件下で、有効濃度のBCGおよびIFNγと接触させて、成熟樹状細胞を形成させる工程;ならびに
該成熟樹状細胞をナイーブT細胞と接触させて、IFNγを産生する活性化T細胞を形成させる工程
を包含する、方法。
(項目26)
前記所定の抗原が、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、抗原を発現する組換え細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原、または単離抗原である、項目25に記載の方法。
(項目27)
前記未成熟樹状細胞が所定の抗原、BCGおよびIFNγと同時に接触される、項目25に記載の方法。
(項目28)
単球性樹状細胞前駆体を単離する工程;および該前駆体を分化因子の存在下で培養して前記未成熟樹状細胞を形成させる工程をさらに包含する、項目25に記載の方法。
(項目29)
前記分化因子が、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13である、項目28に記載の方法。
(項目30)
前記単球性樹状細胞前駆体が、ヒト被験体から単離される、項目28に記載の方法。
(項目31)
前記未成熟樹状細胞およびT細胞が、互いに対して自己である、項目25に記載の方法。
(項目32)
インターロイキン10(IL−10)より多くのインターロイキン12(IL−12)を産生する、単離された成熟樹状細胞であって、該樹状細胞の成熟に適切な条件下で、有効濃度のBCGおよびIFNγを含む組成物を用いた未成熟樹状細胞の成熟によって調製された、単離された成熟樹状細胞。
(項目33)
所定の抗原をさらに包含する、項目32に記載の単離された成熟樹状細胞。
(項目34)
所定の抗原がローディングされた単離された成熟樹状細胞であって、該樹状細胞は、インターロイキン10(IL−10)よりも多くのインターロイキン12(IL−12)を産生する、単離された成熟樹状細胞。
(項目35)
前記細胞が、IL−10よりも少なくとも10倍多くのIL−12を産生する、項目34に記載の単離された成熟樹状細胞。
(項目36)
動物内で1型免疫応答を生じるための方法であって、該方法は、以下:
未成熟樹状細胞を提供する工程;
該未成熟樹状細胞を、該未成熟樹状細胞の成熟に適切な培養条件下で、有効量のBCGおよびインターフェロンγ(IFNγ)および所定の抗原と接触させて、成熟樹状細胞を形成させる工程;
該成熟樹状細胞をナイーブT細胞と接触させて、インターロイキン10(IL−10)よりも多くのインターロイキン12(IL−12)を産生する活性化T細胞を産生させる工程;ならびに
該活性化T細胞を該動物に投与する工程
を包含する、方法。
(項目37)
前記所定の抗原が、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、抗原を発現する組換え細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原、または単離抗原である、項目36に記載の方法。
(項目38)
前記未成熟樹状細胞を所定の抗原、BCGおよびIFNγと同時に接触させる、項目36に記載の方法。
(項目39)
BCGおよびIFNγと接触させる前に、前記未成熟樹状細胞を前記所定の抗原と接触させる、項目36に記載の方法。
(項目40)
前記動物から単球性樹状細胞前駆体を単離する工程;および該前駆体を分化因子の存在下で培養して前記未成熟樹状細胞を形成させる工程をさらに包含する、項目36に記載の方法。
(項目41)
前記分化因子が、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13である、項目40に記載の方法。
(項目42)
前記未成熟樹状細胞およびT細胞が、前記動物に対して自己である、項目36に記載の方法。
(項目43)
前記未成熟樹状細胞およびT細胞が、前記動物に対して同種異系である、項目36に記載の方法。
(項目44)
前記未成熟樹状細胞およびT細胞が、前記動物と同じMHCハプロタイプを有する、項目36に記載の方法。
(項目45)
前記動物がヒトである、項目36に記載の方法。
【図面の簡単な説明】
【0022】
【図1】記載なし。
【発明を実施するための形態】
【0023】
(発明の詳細な説明)
本発明は、未成熟樹状細胞(DC)の成熟を誘導するための方法およびこれらの細胞に抗原特異的細胞傷害性T細胞応答(Th−1応答)をプライミングさせるための方法を提供する。本発明はまた、1型サイトカイン(例えば、IFNγ、TNFα、および/またはIL−2)の産生に極性化したT細胞集団を活性化および調製するために有用な樹状細胞集団を提供する。このような樹状細胞集団としては、適切な成熟条件下でBCG、IFNγおよび所定の抗原と接触させた未成熟単球性樹状細胞が挙げられる。この未成熟樹状細胞を、成熟の間または成熟前にこの抗原と接触させ得る。あるいは、抗原に(例えば、インビボで)既に曝露した未成熟単球性樹状細胞を、適切な成熟条件下でBCGおよびIFNγと接触させ得る。得られる成熟樹状細胞は、T細胞を活性化して1型応答に対してT細胞を極性化することがプライミングされる。1型応答としては、1型サイトカイン(例えば、IFNγ、および/またはIL−2)の産生、IL−10よりも多くのIL−12の産生、細胞傷害性T細胞応答、Th−1細胞の産生、および特定の型の抗体の産生が挙げられる。腫瘍壊死因子α(TNFα)もまたアップレギュレートされ得る。対照的に、2型応答は、IL−4、IL−5およびIL−10の産生、IL−12よりも多いIL−10の産生、Th2細胞の産生、ならびにCTL応答の誘導欠如によって特徴付けられる。
【0024】
関連の局面では、未成熟樹状細胞(例えば、単球性樹状細胞前駆体)についての成熟因子(これは、これらの樹状細胞に1型応答をもプライミングさせ得る)を含む組成物が提供される。成熟し、プライミングされたこのような単球性樹状細胞は、所定の抗原(すなわち、所定の外因性抗原)の主要組織適合遺伝子複合体(MHC)クラス−I提示を増加させ得る。抗原のMHCクラスI提示は、細胞傷害性Tリンパ球(CTL)の分化および標的細胞の抗原特異的CTL媒介溶解の刺激を誘導するために所望される。このような組成物は、未成熟樹状細胞を含む細胞集団と混合され得、未成熟樹状細胞を成熟させるため、ならびにこの未成熟樹状細胞とBCGとの接触によって誘導されるIL−12の阻害を転換または克服するための、BCGおよびIFNγを含む。このような組成物を接触させた未成熟樹状細胞は、成熟を経て、そして代表的には、BCG単独と接触させた未成熟樹状細胞集団と比較して、IL−10よりも多量のIL−12を産生する。
【0025】
別の局面では、被験体またはドナーから得た単球性樹状細胞前駆体を、サイトカイン(例えば、GM−CSFおよびIL−4)と接触させて、未成熟樹状細胞を獲得し得る。次いで、この未成熟樹状細胞を、BCGおよびIFNγのみと組み合わせたかまたはサイトカインと組み合わせた所定の抗原と接触させて、この樹状細胞を成熟させ得、そしてこの細胞がT細胞における1型免疫応答を誘導することをプライミングさせ得る。特定の実施形態では、MHCクラス−I抗原プロセシングが刺激され、このことは、所定の抗原を提示する細胞に対してCTL応答を惹起するために有用である。
【0026】
樹状細胞は、種々のリンパ組織および非リンパ組織に見出される抗原提示細胞の多様な集団である(Liu,Cell 106:259−62(2001);Steinman,Ann.Rev.Immunol.9:271−96(1991)を参照のこと)。樹状細胞としては、脾臓のリンパ樹状細胞、表皮のランゲルハンス細胞、および血液循環中のベール細胞が挙げられる。集合的に、樹状細胞は、それらの形態、高レベルの表面MHC−クラスII発現、ならびにT細胞、B細胞、単球、およびナチュラルキラー細胞上に発現される特定の他の表面マーカーが存在しないことに基づいて、1群として分類される。特に、単球由来樹状細胞(単球性樹状細胞ともいう)は通常、CD11c、CD80、CD86を発現し、そしてHLA−DR+であるが、CD14−である。
【0027】
対照的に、単球性樹状細胞前駆体(代表的に、単球)は通常、CD14+である。単球性樹状細胞前駆体は、これらが存在する任意の組織(特に、リンパ組織(例えば、脾臓、骨髄、リンパ節および胸腺))から入手され得る。単球性樹状細胞前駆体はまた、循環系から単離され得る。末梢血は、単球性樹状細胞前駆体の容易に利用可能な供給源である。臍帯血は、単球性樹状細胞前駆体の別の供給源である。単球性樹状細胞前駆体は、免疫応答が惹起され得る種々の生物から単離され得る。このような生物としては、動物(例えば、ヒトおよび非ヒト動物(例えば、霊長類、哺乳動物(イヌ、ネコ、マウスおよびラットが挙げられる)、鳥類(ニワトリが挙げられる)、ならびにそれらのトランスジェニック種が挙げられる)が挙げられる。
【0028】
特定の実施形態では、この単球性樹状細胞前駆体および/または未成熟樹状細胞は、健常な被験体、または免疫刺激の必要のある被験体(例えば、前立腺癌患者)、または細胞性免疫刺激が有利であり得るかもしくは所望され得る他の被験体(すなわち、細菌感染もしくはウイルス感染を有する被験体など)から単離され得る。樹状細胞前駆体および/または未成熟樹状細胞はまた、免疫刺激の必要がある、HLAが一致した被験体への投与のために、HLAが一致した健常個体から入手され得る。
【0029】
(樹状細胞前駆体および未成熟樹状細胞)
樹状細胞前駆体および未成熟樹状細胞が富化された細胞集団を、種々の供給源(血液および骨髄を含む)から単離するための方法が当該分野で公知である。例えば、樹状細胞前駆体および未成熟樹状細胞は、ヘパリン処理した血液を収集することによって、アフェレーシスもしくは白血球搬出によって、バフィコートの調製、ロゼット形成、遠心分離、密度勾配遠心分離(例えば、Ficoll(例えば、FICOLL−PAQUE(登録商標))、PERCOLL(登録商標)(透析可能ではないポリビニルピロリドン(PVP)でコーティングしたコロイド状シリカ粒子(直径15〜30mm))、スクロースなどを用いる)、細胞の示差的溶解、濾過などによって単離され得る。特定の実施形態では、白血球集団は、例えば、被験体からの血液の収集、血小板を除去するためのフィブリン除去(defribrinating)および赤血球の溶解によって調製され得る。樹状細胞前駆体および未成熟樹状細胞は、例えば、PERCOLL(登録商標)勾配を通しての遠心分離によって、単球性樹状細胞前駆体について必要に応じて富化され得る。
【0030】
樹状細胞前駆体および未成熟樹状細胞は、閉じた無菌系において必要に応じて調製され得る。本明細書中で用いられる場合、用語「閉じた無菌系」または「閉じた系」とは、非滅菌空気、周囲の空気、もしくは循環空気、または他の非無菌条件への曝露が最小であるか排除されている系をいう。樹状細胞前駆体および未成熟樹状細胞を単離するための閉じた系は、一般に、頂部が開いたチューブにおける密度勾配遠心分離、細胞の開放空気下での移植、組織培養プレートまたはシールしていないフラスコにおける細胞培養などを除外する。代表的な実施形態では、この閉じた系は、無菌でない空気への曝露を伴わずに、最初の収集容器からシール可能な組織培養容器への、樹状細胞前駆体および未成熟樹状細胞の無菌移植を可能にする。
【0031】
特定の実施形態では、単球性樹状細胞前駆体は、米国特許出願第60/307,978号(2001年7月25日出願(代理人文書番号020093−002600US)(この開示は、本明細書中に参考として援用される))に開示されるように、単球結合基材への接着によって単離される。例えば、白血球集団(例えば、白血球搬出によって単離される)を、単球性樹状細胞前駆体接着基材と接触させ得る。この白血球集団をこの基材と接触させる場合、この白血球集団中の単球性樹状細胞前駆体は優先的にこの基材に接着する。他の白血球(他の潜在的樹状細胞前駆体を含む)は、基質に対する低減した結合親和性を示し、それゆえ、この単球性樹状細胞前駆体がこの基材表面に優先的に富化されるのを可能にする。
【0032】
適切な基材としては、例えば、大きな表面積対容積比を有する基材が挙げられる。このような基材は、例えば、粒子状基材または繊維状基材であり得る。適切な粒子状基材としては、例えば、ガラス粒子、プラスチック粒子、ガラスでコーティングしたプラスチック粒子、ガラスでコーティングしたポリスチレン粒子、およびタンパク質吸収に適切な他のビーズが挙げられる。適切な繊維状基材としては、マイクロキャピラリーチューブおよび微絨毛膜が挙げられる。この粒子状基材または繊維状基材は通常、付着した単球性樹状細胞前駆体が、接着した細胞の生存率を実質的に低下させることなく、溶出されるのを可能にする。粒子状基材または繊維状基材は、この基材からの単球性樹状細胞前駆体または樹状細胞の溶出を容易にするために実質的に非多孔質であり得る。「実質的に非多孔質」の基材は、その基材中に存在する少なくとも大部分の孔が、この基材中に捕捉される細胞を最小にするために、この細胞よりも小さい基材である。
【0033】
この基材に対するこの単球性樹状細胞前駆体の付着は、結合培地の添加によって、必要に応じて増強され得る。適切な結合培地としては、例えば、サイトカイン(例えば、顆粒球/マクロファージコロニー刺激因子(GM−CSF)、インターロイキン4(IL−4)、またはインターロイキン13(IL−13))、血漿、血清(例えば、ヒト血清(例えば、自己血清もしくは同種異系血清))、精製タンパク質(例えば、血清アルブミン、二価カチオン(例えば、カルシウムイオンおよび/またはマグネシウムイオン))ならびにこの基材に対する単球性樹状細胞前駆体の特異的付着を補助するかまたはこの基材への非単球性樹状細胞前駆体の付着を妨害する、他の分子が個々にまたは任意の組み合わせで補充された、単球性樹状細胞前駆体培養培地(例えば、AIM−V(登録商標)、RPMI 1640、DMEM、X−VIVO 15(登録商標)など)が挙げられる。特定の実施形態では、血漿または血清は、熱不活化され得る。熱不活化血漿は、白血球に対して自己または異種であり得る。
【0034】
この基材への単球性樹状細胞前駆体の付着後、付着していない白血球は、単球性樹状細胞前駆体/基材複合体から分離される。任意の適切な手段を用いて、付着していない細胞を複合体から分離し得る。例えば、付着していない白血球と複合体との混合物を沈澱させ得、そして付着していない白血球および培地を傾瀉または排出し得る。あるいは、この混合物を遠心分離し得、そして付着していない白血球を含む上清を、ペレット化複合体から傾瀉または排出し得る。
【0035】
単離された樹状細胞前駆体は、分化、成熟および/または拡大のためにエキソビボで培養され得る。(本明細書中で用いられる場合、単離された未成熟樹状細胞、樹状細胞前駆体、T細胞、および他の細胞とは、ヒトの手によって、それらのネイティブな環境とは離れて存在し、それゆえ、天然の産物ではない細胞をいう。単離された細胞は、純粋な形態、半ば精製された形態、またはネイティブではない環境に存在し得る。)手短に述べると、エキソビボでの分化は代表的に、樹状細胞前駆体、または樹状細胞前駆体を有する細胞集団を、1以上の分化薬剤の存在下で培養する工程を包含する。適切な分化因子は、例えば、細胞増殖因子(例えば、サイトカイン(例えば、(GM−CSF)、インターロイキン4(IL−4)、インターロイキン13(IL−13)、および/またはそれらの組み合わせ))であり得る。特定の実施形態では、この単球性樹状細胞前駆体を分化させて、単球由来未成熟樹状細胞を形成させる。
【0036】
この樹状細胞前駆体は、適切な培養条件で培養および分化され得る。適切な組織培養培地としては、AIM−V(登録商標)、RPMI 1640、DMEM、X−VIVO 15(登録商標)などが挙げられる。この組織培養培地には、細胞の分化を促進するために、血清、アミノ酸、ビタミン、サイトカイン(例えば、GM−CSFおよび/またはIL−4)、二価カチオンなどが添加され得る。特定の実施形態では、この樹状細胞前駆体は、無血清培地中で培養され得る。このような培養条件は、必要に応じて任意の動物由来産物を排除し得る。代表的樹状細胞培養培地における代表的なサイトカインの組み合わせは、各々約500単位/mlのGM−CSFおよびIL−4である。樹状細胞前駆体は、分化して未成熟樹状細胞を形成した場合、皮膚ランゲルハンス細胞に表現型的に類似する。未成熟樹状細胞は代表的に、CD14−およびCD11c+であり、低レベルのCD86およびCD83を発現し、そして極性化されたエンドサイトーシスを介して可溶性抗原を捕捉し得る。
【0037】
未成熟樹状細胞は成熟して成熟樹状細胞を形成する。成熟DCは、抗原を取り込む能力を失い、発現がアップレギュレートされた補助刺激細胞表面分子および種々のサイトカインを提示する。特に、成熟DCは、未成熟樹状細胞よりも高いレベルのMHCクラスI抗原およびMHCクラスII抗原を発現し、そして成熟樹状細胞は、一般に、CD80+、CD83+、CD86+、およびCD14−であると同定される。より多くのMHC発現は、DC表面での抗原密度の増加をもたらし、一方、補助刺激分子CD80およびCD86のアップレギュレーションは、T細胞上の補助刺激分子(例えば、CD28)の対応物を通して、T細胞活性化シグナルを強化する。
【0038】
本発明の成熟樹状細胞は、未成熟樹状細胞を有効量または有効濃度のBCGおよびIFNγと接触させることによって、調製(すなわち、成熟)され得る。有効量のBCGは、代表的に、組織培養培地1ミリリットルあたり約105〜約107cfuの範囲に及ぶ。IFNγの有効量は代表的に、組織培養培地1ミリリットルあたり約100〜約1000Uの範囲に及ぶ。カルメットゲラン杆菌(Bacillus Calmette−Guerin;BCG)は、M.bovisの無発病性株である。本明細書中で用いられる場合、BCGは、BCG全体ならびに細胞壁構成成分、BCG由来リポアラビドマンナン、および2型免疫応答の誘導に関連した他のBCG成分をいう。BCGは、必要に応じて不活化される(例えば、熱不活化BCG、ホルマリン処理BCGなど)。
【0039】
BCGは、IL−12産生の阻害およびエンドサイトーシスによる抗原の排除に付随して、樹状細胞上の表面成熟マーカーCD83およびCD86の発現を増大させる。何らかの特定の理論に束縛されることを意図しないが、BCGによる樹状細胞成熟はまた、腫瘍壊死因子α(TNFα)のホモタイプな凝集および放出を含むと特徴付けられている(例えば、本明細書中に参考として援用される、Thurnherら,Int.J.Cancer 70:128−34(1997)を参照のこと)。IFNγおよびBCGを用いて未成熟樹状細胞を成熟させることは、IL−12のDC産生を促進し、そしてIL−10の産生を低減または阻害し、それによって、成熟樹状細胞に1型(Th−1)応答をプライミングさせる。
【0040】
この未成熟DCを、代表的に、有効量のBCGおよびIFNγと、約1時間〜約24時間にわたって接触させる。この未成熟樹状細胞は、適切な成熟培養条件において培養されて成熟し得る。適切な組織培養培地としては、AIM−V(登録商標)、RPMI 1640、DMEM、X−VIVO 15(登録商標)などが挙げられる。これらの組織培養培地には、細胞の成熟を促進するために、アミノ酸、ビタミン、サイトカイン(例えば、GM−CSFおよび/またはIL−4)、二価カチオンなどが補充され得る。代表的なサイトカインの組み合わせは、各々約500単位/mlのGM−CSFおよびIL−4である。
【0041】
樹状細胞の成熟は、当該分野で公知の方法によってモニタリングされ得る。細胞表面マーカーは、当該分野で通常のアッセイ(例えば、フローサイトメトリー、免疫組織化学など)において検出され得る。これらの細胞はまた、(例えば、ELISA、FACS、または他の免疫アッセイによって)サイトカイン産生についてモニタリングされ得る。本発明に従って成熟したDC集団では、IL−12レベルは、IL−10レベルよりも高く、1型(Th−1)応答を促進する。例えば、このDCは、1:1より大きく、約10:1または約100:1までのIL−12/IL−10比を生じ得る。成熟DCはまた、飲細胞運動によって抗原を取り込む能力を失う。飲細胞運動は、当業者には通常の取り込みアッセイによって分析され得る。樹状細胞前駆体、未成熟樹状細胞、および成熟樹状細胞は、抗原を用いてプライミングされていようとプライミングされていなかろうと、後日の使用のために凍結保存され得る。凍結保存のための方法は、当該分野で周知である。例えば、その全体が本明細書中に参考として援用される、米国特許第5,788,963号を参照のこと。
【0042】
(抗原)
本発明による、成熟しプライミングされた樹状細胞は、抗原をT細胞に対して提示し得る。成熟しプライミングされた樹状細胞は、成熟の前または成熟の間のいずれかに、未成熟樹状細胞を所定の抗原と接触させることによって形成され得る。あるいは、(例えば、単離前にインビボで)抗原と既に接触した未成熟樹状細胞を、BCGおよびIFNγを含む組成物と接触させて、1型(Th−1)応答がプライミングされた成熟樹状細胞を形成し得る。
【0043】
適切な所定の抗原は、T細胞活性化が所望される任意の抗原を含み得る。このような抗原は、例えば、細菌抗原、腫瘍特異性抗原または腫瘍関連抗原(例えば、細胞全体、腫瘍細胞溶解産物、腫瘍由来の単離抗原、融合タンパク質、リポソームなど)、ウイルス抗原、および任意の他の抗原または抗原(例えば、ペプチド抗原またはポリペプチド抗原)のフラグメントを包含し得る。特定の実施形態では、この抗原は、例えば、前立腺特異性膜抗原(PSMA)、前立腺酸性ホスファターゼ(PAP)、または前立腺特異性抗原(PSA)であり得るがこれらに限定されない(例えば、Pepsideroら,Cancer Res.40:2428−32(1980);McCormackら,Urology 45:729−44(1995)を参照のこと)。この抗原はまた、細菌細胞、細菌溶解産物、細胞溶解産物由来の膜フラグメント、または当該分野で公知の任意の他の供給源であり得る。この抗原は、組換えによって発現もしくは産生されてもよく、または化学的に合成されてすらよい。組換え抗原はまた、宿主細胞(例えば、細菌細胞、酵母細胞、昆虫細胞、脊椎動物細胞または哺乳動物細胞)の表面に発現されてもよく、溶解産物中に存在してもよく、またはこの溶解産物から精製されてもよい。
【0044】
抗原はまた、被験体由来のサンプル中に存在し得る。例えば、被験体における過剰増殖状態または他の状態に由来する組織サンプルは、抗原供給源として用いられ得る。このようなサンプルは、例えば、生検または手術切開によって入手され得る。このような抗原は、溶解産物として、または単離された調製物として用いられ得る。あるいは、被験体(例えば、癌患者)の細胞または確立された細胞株の膜調製物はまた、抗原または抗原供給源として用いられ得る。
【0045】
例示的な実施形態では、手術標本から収集された前立腺腫瘍細胞の溶解産物は、抗原供給源として用いられ得る。例えば、生検または手術切開によって得られた、癌患者自体の腫瘍のサンプルは、抗原を樹状細胞に提示するため、または抗原提示用細胞溶解産物を提供するために、直接的に用いられ得る。あるいは、癌患者の腫瘍細胞の膜調製物が用いられ得る。この腫瘍細胞は、前立腺の腫瘍細胞、肺の腫瘍細胞、卵巣の腫瘍細胞、結腸の腫瘍細胞、脳の腫瘍細胞、黒色腫、または任意の他の種類の腫瘍細胞であり得る。溶解産物および膜調製物は、当該分野で公知の方法によって、単離された腫瘍細胞から調製され得る。
【0046】
別の例示的実施形態では、精製または半精製された前立腺特異的膜抗原(PSMA、PSM抗原としても公知)は、モノクローナル抗体7E11−C.5と特異的に反応し、抗原として用いられ得る(一般的に、Horoszewiczら,Prog.Clin.Biol.Res.37:115−32(1983)、米国特許第5,162,504号;米国特許第5,788,963号;Fengら,Proc.Am.Assoc.Cancer Res.32:(Abs.1418)238(1991)を参照のこと;これらの開示は、本明細書中に参考として援用される)。なお別の例示的実施形態では、アミノ酸残基配列Leu Leu His Glu Thr Asp Ser Ala Val(配列番号1)を有する抗原性ペプチド(PSM−P1と称される)は、PSMAのアミノ酸残基4〜12に対応し、抗原として用いられ得る。あるいは、アミノ酸残基配列Ala
Leu Phe Asp Ile Glu Ser Lys Val(配列番号2)を有する抗原性ペプチド(PSM−P2と称される)は、PSMAのアミノ酸残基711〜719に対応し、抗原として用いられ得る。
【0047】
特定の実施形態では、アミノ酸残基配列Xaa Leu(またはMet)Xaa Xaa Xaa Xaa Xaa Xaa Val(またはLeu)(PSM−PXと称される)を有する抗原性ペプチド(ここで、Xaaは任意のアミノ酸残基を表す)は、抗原として用いられ得る。このペプチドは、HLA−A0201結合モチーフ、すなわち、HLA−A2患者において見出される「アンカー残基」ロイシンおよびバリンを有する9〜10アミノ酸残基の結合モチーフに似る(例えば、Greyら,Cancer Surveys 22:37−49(1995)を参照のこと)。このペプチドは、HLA−A2+患者についての抗原として用いられ得る(Central Data Analysis
Committee「Allele Frequencies」,第6.3節,Tsuji,K.ら(編),Tokyo University Press,1066−1077頁を参照のこと)。同様に、他のHLA結合モチーフに似るペプチドが用いられ得る。
【0048】
代表的に、未成熟樹状細胞は、上記のように、適切な成熟条件下で、BCG、IFNγおよび所定の抗原の存在下で培養される。必要に応じて、この未成熟樹状細胞は、GM−CSFおよびIL−4または成熟因子を含まない代表的樹状細胞培養培地中で、所定の抗原と混合され得る。この抗原との少なくとも約10分間〜2日間の培養後、この抗原は除去され得、そしてBCGおよびIFNγが補充された培養培地が添加され得る。サイトカイン(例えば、GM−CSFおよびIL−4)はまた、成熟培地に添加され得る。樹状細胞を接触させるための方法は、当該分野で一般に公知である(一般的に、SteelおよびNutman,J.Immunol.160:351−60(1998);Taoら,J.Immunol.158:4237−44(1997);DozmorovおよびMiller,Cell Immunol 178:187−96(1997);Inabaら,J Exp Med.166:182−94(1987);Macatoniaら,J Exp Med.169:1255−64(1989);De Bruijnら,Eur.J.Immunol.22:3013−20(1992)を参照のこと;これらの開示は、本明細書中に参考として援用される)。
【0049】
次いで、得られる成熟しプライミングされた樹状細胞は、T細胞(例えば、ナイーブT細胞)と共存インキュベートされる。T細胞またはT細胞サブセットは、レスポンダー細胞として使用するために種々のリンパ組織から入手され得る。このような組織としては、脾臓、リンパ節、および/または末梢血が挙げられるがこれらに限定されない。これらの細胞は、成熟しプライミングされた樹状細胞とともに、混合T細胞集団として、または精製されたT細胞サブセットとして共存培養され得る。T細胞精製は、CD2に対する抗体、CD3に対する抗体、CD4に対する抗体、CD8に対する抗体などの使用を含むがこれらに限定されない、ポジティブ選択またはネガティブ選択によって達成され得る。
【0050】
T細胞を、成熟しプライミングされた樹状細胞と接触させることによって、抗原反応性か活性化された、極性化T細胞または極性化Tリンパ球が提供される。本明細書中で用いられる場合、用語「極性化(polarized)」とは、高レベルのIFNγを産生するか、さもなければ1型(Th−1)応答を誘導するようにプライミングされているT細胞をいう。このような方法は、代表的に、未成熟樹状細胞をBCGおよびIFNγと接触させて、成熟しプライミングされた樹状細胞を調製する工程を包含する。この未成熟樹状細胞を、成熟の間または成熟前に所定の抗原と接触させ得る。この未成熟樹状細胞は、成熟の間、T細胞(例えば、ナイーブT細胞)とともに共存培養されてもよく、または、成熟および1型応答を誘導するための樹状細胞のプライミングの後にT細胞(例えば、ナイーブT細胞)と共存培養されてもよい。この未成熟樹状細胞または成熟樹状細胞は、成熟前に富化され得る。さらに、T細胞は、樹状細胞との接触前にリンパ球集団から富化され得る。特定の実施形態では、富化または精製されたCD4+T細胞集団を、この樹状細胞と接触させる。成熟し、プライミングされた樹状細胞とT細胞との共存培養は、特異的T細胞の刺激をもたらし、この特異的T細胞は、成熟して、抗原反応性CD4+T細胞または抗原反応性CD8+T細胞になる。
【0051】
別の局面では、T細胞を、1型(Th−1)T細胞応答を誘導することに向けてプライミングされた成熟樹状細胞の存在下で培養することによる、インビトロでのT細胞の再刺激のための方法が提供される。このようなT細胞は、フィーダー細胞上で必要に応じて培養され得る。成熟しプライミングされた樹状細胞は、T細胞との接触前に必要に応じて照射され得る。適切な培養条件は、1以上のサイトカイン(例えば、精製されたIL−2、コンカナバリンA刺激脾臓細胞上清、またはインターロイキン15(IL−15))を含み得る。未成熟樹状細胞、BCG、IFNγおよび所定の抗原の添加による、インビトロでのT細胞再刺激は、T細胞集団の増大を促進するために用いられ得る。
【0052】
安定な抗原特異的、極性化T細胞培養物またはT細胞株は、周期的再刺激によってインビトロで長期間にわたり維持され得る。このようにして作製されたT細胞培養物またはT細胞株は、保存され得、そして(例えば、凍結保存剤を用いた処方および凍結によって)貯蔵される場合、活性化された極性化T細胞を長期にわたって所望の間隔で再供給するために用いられ得る。
【0053】
特定の実施形態では、活性化されたCD8+T細胞またはCD4+T細胞は、本発明の方法に従って作製され得る。代表的に、抗原反応性の極性化T細胞を作製するために用いられる、成熟しプライミングされた樹状細胞は、投与されるべき被験体に対して同質遺伝子的である(例えば、この被験体から入手される)。あるいは、意図されるレシピエント被験体と同じHLAハプロタイプを有する樹状細胞は、HLAが一致したドナー由来の非癌性細胞(例えば、正常細胞)を用いてインビトロで調製され得る。特定の実施形態では、抗原反応性T細胞(CTL細胞およびTh−1細胞を含む)は、免疫刺激のための細胞供給源としてインビトロで増大される。
【0054】
(細胞集団のインビボでの投与)
本発明の別の局面では、成熟しプライミングされた樹状細胞もしくは活性化され極性化したT細胞、またはこのような細胞を含む細胞集団を、免疫刺激の必要がある被験体に投与するための方法が提供される。このような細胞集団は、成熟しプライミングされた樹状細胞集団および/または活性化され極性化したT細胞集団の両方を包含し得る。特定の実施形態では、このような方法は、樹状細胞前駆体または未成熟樹状細胞を入手し、これらの細胞を、BCG、IFNγおよび所定の抗原の存在下で分化および成熟させて、Th−1応答に対してプライミングされた成熟樹状細胞集団を形成させることによって実施される。この未成熟樹状細胞を、成熟前または成熟の間に抗原と接触させ得る。このような成熟しプライミングされた樹状細胞は、免疫刺激の必要がある被験体に直接的に投与され得る。
【0055】
関連した実施形態では、この成熟しプライミングされた樹状細胞を、被験体由来のリンパ球と接触させて、このリンパ球集団内のT細胞を刺激し得る。この活性化され極性化されたリンパ球は、必要に応じて続いて抗原反応性のCD4+T細胞および/またはCD8+T細胞の細胞培養物中でクローン増大が行われ、免疫刺激の必要がある被験体に投与され得る。特定の実施形態では、活性化され極性化されたT細胞は、この被験体に対して自己である。
【0056】
別の実施形態では、この樹状細胞、T細胞、およびレシピエント被験体は、同じMHC(HLA)ハプロタイプを有する。被験体のHLAハプロタイプを決定する方法は、当該分野で公知である。関連の実施形態では、この樹状細胞および/またはT細胞は、このレシピエント被験体に対して同種異系である。例えば、この樹状細胞は、このT細胞およびこのレシピエントに対して同種異系であり得、これは、同じMHC(HLA)ハプロタイプを有する。この同種異系細胞は代表的に、少なくとも1つのMHC対立遺伝子が一致している(例えば、少なくとも1つだが全てではないMHC対立遺伝子を共有する)。それほど代表的でない実施形態では、この樹状細胞、T細胞およびレシピエント被験体は全て、互いに対して同種異系であるが、全て、少なくとも1つの共通のMHC対立遺伝子を共通に有する。
【0057】
1つの実施形態によれば、このT細胞は、この未成熟樹状細胞を得たのと同じ被験体から得られる。インビトロでの成熟および極性化の後、この自己T細胞は、既存の免疫応答を誘発および/または増強するためにこの被験体に投与される。例えば、T細胞は、(例えば、体表面積1m2あたり約108〜109細胞の用量での)静脈内注入によって投与され得る(例えば、本明細書中に参考として援用される、Ridellら,Science 257:238−41(1992)を参照のこと)。注入は、所望の間隔で(例えば、毎月)繰り返され得る。レシピエントは、有害な影響の何らかの証拠について、T細胞注入の間およびその後、モニタリングされ得る。
【0058】
別の実施形態によれば、本発明に従ってBCGおよびIFNγを用いて成熟させた樹状細胞は、腫瘍または標的抗原を含む他の組織中に直接的に注射され得る。このような成熟細胞は、抗原を取り込んでこの抗原をT細胞に対してインビボで提示し得る。
【実施例】
【0059】
以下の実施例を、本発明の種々の局面の単に例示として提供し、そして本発明を如何様にも限定すると解釈すべきでない。
【0060】
(実施例1:異なる成熟条件下でのIL−10およびIL−12の産生:)
本実施例では、サイトカイン産生を、成熟因子BCGおよび/またはIFNγと接触させた未成熟樹状細胞集団から決定した。未成熟DCを、1%ヒト血漿を補充したOptiMEM(登録商標)培地(Gibco−BRL)の存在下で末梢血単球をプラスチックと接触させることによって調製した。未結合の単球を、洗浄によって除去した。結合した単球を、1ミリリットルあたり500U GM−CSFおよび500U IL−4の存在下で6日間にわたってX−VIVO 15(登録商標)培地中で培養した。
【0061】
第1研究では、未成熟樹状細胞を、不活化BCGの添加によって成熟させた。得られる成熟樹状細胞のサイトカイン産生を決定した。不活化BCGを、種々の濃度で、X−VIVO 15(登録商標)培地中の未成熟樹状細胞に添加し、続いて37℃にて24時間培養した。4.1×108cfu/mlストックから開始した、1ミリリットルあたりで添加したBCGの希釈を、表に指定する。サイトカイン産生を、検出されるべきサイトカインに対する抗体を用いたELISAアッセイによって決定した。手短に述べると、サイトカイン(例えば、IL−12またはIL−10)に対して特異的な抗体を用いて、固体表面のサイトカインを捕捉する。次いで、この固体表面を、このサイトカインに対する第2の標識抗体で処理して、捕捉されたサイトカインの存在を検出する。第2の抗体は、代表的に、酵素で標識されて、比色アッセイによる検出を容易にする。代表的な実験の結果を、以下の表1に下記に示す。サイトカインの量、すなわちサイトカイン産生を、pg/mlとして示す。
【0062】
【表1】

表1を言及して、この結果は、BCGの添加が、サイトカインIL−12、IL−10、またはそれらのサブユニットの産生を増大させ得るが、サイトカイン産生の相対レベルおよび絶対レベルの両方がドナー依存性であることを実証する。最大レベルの増加が、IL−12 p40およびIL−10について見られ、このことは、IL−12 p40のレベルと比較して低レベルの、観察されたIL−12 p70(p35サブユニットおよびp40サブユニットから構成される)が、IL−12 p35産生の欠除に起因することを示唆した。BCGが存在する1つの条件を除く全ての条件において、IL−12 p70レベル:IL−10レベルの比は、1以下である。このことは、BCG単独の存在下での未成熟樹状細胞の成熟が、ナイーブT細胞をTh−2 応答に向けて極性化させるようであることを示す。
【0063】
同様の条件下でIFNγを導入することの効果もまた決定した。これを、以下の表2に提示する。サイトカイン産生を上記の通りに測定した。表1と表2とを比較して、成熟因子(例えば、BCG)の存在下でのIFNγの添加が、IL−12 p70の産生を増大させたことが明らかである。特に、成熟の間でのBCGを伴うIFNγの添加は、IL−12 p35産生を増大させ、そしてIL−10産生を減少させた。その結果、IL−12 p70:IL−10の比は、BCGとともにIFNγを添加すると、いつも1より大きかった。何人かのドナーでは、および特定の条件下では、IL−12 p70産生:IL−10産生の比は、BCGを伴うIFNγの添加によって100:1より大きく増大し得る。従って、これらの結果は、驚くことに、成熟因子BCGを伴うIFNγの添加が、IL−12 p70産生を劇的に増大させ得ることを実証する。
【0064】
【表2】
(実施例2:IFNγによるIL−10のダウンレギュレーションは、用量依存性である:)
本実施例では、BCGと組み合わせたIFNγが、樹状細胞集団におけるIL−10産生をダウンレギュレートする能力を実証する。未成熟樹状細胞を、上記の通りに調製した。これらの未成熟樹状細胞を単独でインキュベートし、2つのうちの1つの濃度のBCG(1:1000希釈または1:250希釈の4.1×108cfu/mlストック)の存在下で、または1ミリリットルあたり0U〜1000Uの範囲の濃度のIFNγ単独に曝露して、成熟させた。得られる樹状細胞によるIL−10産生を、市販の抗体(例えば、R & D Systems,Minneapolis,MN製)を用いたELISA(前出)によって測定し、そしてpg/mlで報告した。コントロールでは、単独で培養した未成熟DC(BCGもIFNγも添加しない)は、検出可能なIL−10を産生しなかった。対照的に、IFNγ単独の存在下で培養したDCは、少量のIL−10(約20〜30pg/ml)を産生した。産生されたIL−10の量は、1ミリリットルあたり10U〜1000UのIFNγの範囲にわたって用量依存性ではなかった。
【0065】
対照的に、BCG単独の存在下での成熟によって産生されたDCは、顕著な量のIL−10を産生した:1:1000または1:250倍希釈のBCGストックの存在下、それぞれ、約150pg/mlまたは250pg/mlを超えるIL−10。DC成熟の間のBCGへのIFNγの添加は、IL−10産生のダウンレギュレーションを用量依存性様式でもたらした。1:1000希釈のBCGの存在下で培養したDCについては、IL−10産生は、約150pg/mlのIL−10(IFNγなし)から約20〜30pg/mlのIL−10(1000U IFNγ)へと減少した。1:250希釈のBCGの存在下で培養したDCについては、IL−10産生は、約270pg/mlのIL−10(IFNγなし)から約50pg/mlのIL−10(1000U IFNγ)へと減少した。従って、BCGおよびIFNγの存在下での未成熟DCの成熟は、IL−10産生をダウンレギュレーションし得、そしてBCG単独によって誘導されるIL−10産生の見かけの刺激を克服し得た。
【0066】
(実施例3:IFNγによるIL−12のアップレギュレーションは、用量依存性である:)
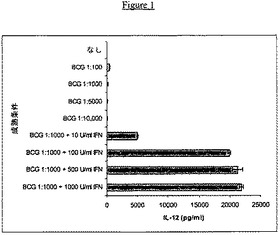
本実施例では、IFNγがIL−12産生をアップレギュレートする能力が実証された。未成熟樹状細胞を、上記のように、GM−CSFおよびIL−4の存在下で増殖させた単球の6日間培養物から誘導した。これらの未成熟DCを、種々の希釈のBCG単独を用いて、または種々の濃度のIFNγと組み合わせたBCGを用いてさらに2日間処理した。培養上清を、IL−12 p70の存在について、上記の通りに、ELISAアッセイによって試験した。
【0067】
代表的実験による結果を図1に示す。各培養物について、この結果を3連で決定した。増加する量のBCG単独に応答して、比較的低い(<1000pg/mlの)平均濃度のIL−12 p70がこれらの成熟DCによって産生された。IL−12産生の量は、BCGの量が増加するにつれて用量依存様式で減少した。対照的に、BCG(ストックの1:1000希釈物)を有するIFNγ(10U/ml)の添加によって、IL−12の量は、約5000pg/mlまで劇的に増加した。BCG(ストックの1:1000希釈物)を有するIFNγ(100U/ml)の添加によって、IFNγの量は、ほぼ20,000pg/mlまで増加した。BCGを有する500U/mlまたは1000U/mlのIFNγ(ストックの1:1000希釈物)の添加は、それぞれ、約21,000pg/mlおよび約22,000pg/mlのIL−12レベルをもたらす。
【0068】
まとめると、BCGは単独では、IL−12産生を拮抗するようであるが、BCGおよびIFNγの存在下での未成熟DCの成熟は、IL−12産生を劇的に増加させた。
【0069】
(実施例4:抗原特異的T細胞の刺激:)
本実施例では、BCGおよびIFNγの存在下で成熟させた未成熟DCは、抗原特異的T細胞によるIFNγ産生を刺激することが示された。インフルエンザAに特異的なT細胞株を、2×106細胞/mlの末梢血単核細胞(PBMC)を5%ヒト血清を補充したAIM−V(登録商標)培地中で5μg/mlのインフルエンザM1ペプチド(GILGFVFTL;配列番号3)とともにインキュベートすることによって作製した。これらの培養条件は、インフルエンザM1ペプチドに特異的なT細胞の選択的増大をもたらした。2日間の培養後、20U/ml IL−2および5ng/ml IL−15をこれらの培養物に添加した。約7〜14日間の培養後、これらのT細胞株を、サイトカインを含まない培地中に一晩配置した。
【0070】
次いで、抗原特異的T細胞を、未成熟DCとともに、BCG単独を用いて成熟させたDCとともに、またはBCGおよびIFNγを用いて成熟させたDCとともに、共存培養した。抗原特異的T細胞:DCの比は、1:1であった。T細胞およびDCを、37℃にて24時間インキュベートした。これらのDCは、インフルエンザM1ペプチドが直接的にローディングされるかまたは浸透圧的にローディングされるかのいずれかであった。手短に述べると、未成熟DCを培養フラスコから収集し、そして遠心分離によって濃縮した。浸透圧ローディングについては、これらの細胞を、小容量の高浸透圧培地中に再懸濁し、続いてPBS中の等容量のインフルエンザM1ペプチドを添加した。氷上での10分間のインキュベーションの後、これらの細胞を徹底的に洗浄した。直接的ローディングについては、これらの細胞を、等容量のX−VIVO 15(登録商標)培地およびPBS中のインフルエンザM1ペプチド中に再懸濁し、37℃で1時間にわたってインキュベートした。これらの細胞を、37℃にて2時間にわたってインキュベートして、抗原プロセシングを可能にした。
【0071】
T細胞およびDCの共存培養後、T細胞応答を、培養上清の100μlサンプルからのIFNγのELISA定量によって測定した。この結果は、CDローディングの方法にかかわらず、BCGおよびIFNγで刺激したDCが、抗原特異的T細胞の優れた刺激因子であることを示す。未成熟DCとともに共存培養したT細胞は、極めてわずかなIFNγ(<2,000pg/ml)を産生し、一方、BCG単独を用いて成熟させたDCとともに共存培養したT細胞は、IFNγの中程度の生産者であった(>5,000pg/ml)。BCGおよびIFNγを用いて成熟させたDCとともに共存培養したT細胞は、高レベルのIFNγ(浸透圧によってローディングしたDCについて>20,000pg/ml、または直接的にローディングしたDCについて>25,000pg/ml)を産生した。
【0072】
従って、BCGおよびIFNγを用いて成熟させたDCは、DCに抗原をローディングする方法とは独立して、抗原特異的T細胞のより良好な刺激因子であった。
【0073】
(実施例5:インビトロでの抗原特異的T細胞応答のデノボ作製:)
キーホールリンペットヘモシアニン(KLH)に特異的なT細胞株を、BCG、またはBCGおよびIFNγのいずれかを用いて成熟させたDCを用いて、PBMCを刺激することによって作製し、KLHまたはコントロールタンパク質を、10:1のT細胞:DC比でローディングした。T細胞および成熟させたDCに、新鮮な培地(5%ヒトAB血清、20U/ml IL−2、および5ng/ml IL−15を補充したAIM−V(登録商標)培地)を3〜4日毎に提供した。これらの細胞を、必要に応じて、より大きなフラスコへと増大させた。特定の抗原に対する前駆体頻度全体が低く、ナイーブ細胞は応答するために強力な刺激を必要とするので、刺激を、10〜21日間の間隔で3〜4回繰り返した。細胞をサイトカインを含まない培地中で一晩回復させた後に再刺激した。
【0074】
標準的な3日間のチミジン取り込みアッセイを用いて、刺激T細胞株をKLH特異的細胞増殖について試験した。刺激されたT細胞を、KLHパルス未成熟樹状細胞を用いて、種々のDC:T細胞比でインキュベートした。T細胞増殖を、1分間あたりのカウント(CPM)として測定した。刺激に応答した、細胞増殖またはサイトカインの産生を、抗原特異的応答の証拠と解釈した。抗原特異性について制御するために、ネガティブコントロールの抗原(インフルエンザAウイルス)もまたこのアッセイに含めた。
【0075】
BCG単独によって成熟させたDCを用いてT細胞を刺激する場合、T細胞増殖は一貫して低かった(<5,000CPM)。この低レベルの増殖は、DCをKLHと接触させようがインフルエンザAウイルスと接触させようが、低かった。低レベルの増殖もまた、未成熟DCを用いたインキュベーションに応答して観察された。50:1、25:1、または12.5:1の応答者:刺激者(T細胞:DC)の比で用いた3つの群のDCの間に顕著な差は観察されなかった。対照的に、BCGおよびIFNγを用いて成熟させた樹状細胞を用いてT細胞を刺激し、KLHパルスDCは、インフルエンザAでパルスした未成熟DCまたは成熟DCが誘導するよりも一貫してさらに多いT細胞増殖を誘導した(約10,000〜33,000CPM)。KLH抗原でパルスした成熟DCについては、T細胞増殖は、応答者:刺激者の比の増大に比例して増加した。
【0076】
T細胞エフェクター機能もまた、サイトカイン分泌によってモニタリングされた。KLH特異的T細胞株(上記の通りに作製された)を、BCG単独またはBCGおよびIFNγのいずれかを用いて成熟させたDCを用いて刺激した。刺激されたT細胞株を抗CD3抗体(50ng/ml)およびPMA(5ng/ml)を用いて非特異的に刺激した後、これらの細胞を、細胞内サイトカイン染色によってサイトカイン産生について試験した。サイトカイン産生を、特定のサイトカインを産生する細胞の百分率として測定した。極めて低い百分率から検出不可能な百分率(<<5%)のサンプリング細胞は、細胞内IL−2、IL−4、IL−5、またはIL−10を産生した。IL−5およびIL−10は、BCGおよびIFNγを用いて成熟させたDCによって刺激されたT細胞において検出されなかった。BCG単独によって成熟させたDCによって刺激されたT細胞は、低レベルのIFNγ(<10%)およびTNF−α(<15%)を産生した。対照的に、IFNγおよびBCGを用いて成熟させたDCを用いて刺激された有意な比率のT細胞は、IFNγ(約35%)およびTNFα(>45%)を産生した。IFNγは、IL−12産生の公知の刺激因子である。従って、BCGおよびIFNγを用いて成熟させたDCを用いてT細胞を刺激することによって、これらのT細胞は、1型(Th−1)応答に向けて極性化される。
【0077】
(実施例6:Th−1サイトカイン腫瘍壊死因子α(TNFα)の誘導:)
本実施例では、BCGおよびIFNγの組み合わせが1型サイトカイン腫瘍壊死因子α(TNFα)をアップレギュレートする能力が実証された。手短に述べると、未成熟樹状細胞を上記の通りに誘導し、そしてGM−CSFおよびIL−4の存在下で増殖させた(prown)。これらの未成熟DCを、約24時間にわたってBCG単独またはIFNγとの組み合わせのいずれかとともに培養した。続いて、タンパク質輸送インヒビター(GolgiPlugTM,PharMingen)を添加して、ゴルジ複合体から産生されたサイトカインの輸送をブロックし、そしてこれらの細胞を一晩インキュベートした。次いで、これらの細胞を、当該分野で周知の方法を用いて、収集し、透過性にし、そしてTNFαについて特異的な蛍光標識抗体またはアイソタイプコントロール抗体を用いて内部染色した。TNFαについてDCポジティブの頻度およびこれらの細胞の蛍光強度を、FACS分析によって決定した(表3)。IFNγの存在下でBCGを用いてDCの成熟は、DCがTh−1サイトカインTNFαを産生する能力を増強することが見出された。
【0078】
【表3】
(実施例7:細胞関連抗原に対する応答の誘導:)
本実施例では、BCGおよびIFNγの存在下で成熟させたDCは、BCG単独を用いて成熟させたDCと比較して、有意に高い腫瘍特異性T細胞INFγ放出および類似のレベルの抗原特異的細胞傷害性を惹起することが実証された。未成熟樹状細胞を上記の通りに単離し、そしてGM−CSFおよびIL−4の存在下で培養した。次いで、これらのDCに、緑色蛍光タンパク質(GFP)またはインフルエンザAウイルスのM1タンパク質のいずれかを発現する組換えアデノウイルスで予め感染させた腫瘍細胞(A549)全体のいずれかをローディングした。これらのDCを、BCGまたはIFNγと組み合わせたBCGのいずれかを用いて24時間後に成熟させた。腫瘍をローディングしたDCもしくはGFPまたはM1発現腫瘍細胞を用いて、自己M1特異的T細胞株を刺激した。24時間後、細胞培養上清を収集し、そして標準的IFNγ ELISAを行った。M1発現腫瘍細胞をローディングしたDCのみがIFNγ放出を刺激し得、そしてBCG+IFNγにおいて成熟させたDCは、この応答を誘導する際に、未成熟DCまたはBCG成熟DCのいずれかよりも有意により強力であった。
【0079】
【表4】
上記の実施例は、本願発明の範囲を例示するために提供されるが、本願発明の範囲を限定しない。本発明の他の改変体は、当業者に容易に明らかであり、添付の特許請求の範囲によって包含される。本明細書中に引用される全ての刊行物、特許、特許出願および他の参考文献は、それらの全体が、本明細書中に参考として援用される。
(配列表)
【背景技術】
【0001】
(発明の背景)
抗原提示細胞(APC)は、有効な免疫応答を惹起する際に重要である。これは、抗原特異的T細胞レセプターを有するT細胞に対して抗原を提示するだけでなく、T細胞活性化に必要なシグナルも提供する。これらのシグナルは、完全に規定されていないままであるが、種々の細胞表面分子ならびにサイトカインまたは増殖因子を含む。ナイーブT細胞または非極性化T細胞の活性化に必要な因子は、記憶T細胞の再活性化に必要とされる因子とは異なり得る。APCが抗原を提示する能力およびT細胞活性化シグナルを送達する能力は両方とも、通常、補助細胞機能といわれる。単球およびB細胞はコンピテントAPCであることが示されているが、それらのインビトロでの抗原提示能力は、以前に感作されたT細胞の再活性化に制限されているようである。それゆえ、これらは、機能的にナイーブなT細胞集団も、プライミングされていないT細胞集団も直接的に活性化することができない。
【0002】
樹状細胞(DC)は、ナイーブT細胞および記憶T細胞の両方を活性化し得ると考えられる、免疫系のプロフェッショナル抗原提示細胞である。樹状細胞は、免疫治療(特に、癌の免疫治療)における使用のために、ますますエキソビボで調製されている。最適な免疫刺激特性を有する樹状細胞の調製は、エキソビボでの培養のために、これらの細胞の生物学の理解および活用を必要とする。各プロトコルに起因する種々の利点を有する、これらの細胞の培養のための種々のプロトコルが記載されている。近年のプロトコルとしては、無血清培地の使用、ならびに所望の免疫刺激特性を培養細胞に付与する成熟条件の使用が挙げられる。
【0003】
樹状細胞の成熟は、未成熟DC(これは、皮膚ランゲルハンス細胞に表現型的に類似する)を、リンパ節へと移動し得る成熟した抗原提示細胞へと転換するプロセスである。このプロセスは、この未成熟樹状細胞を特徴付ける強力な抗原取り込み能力の喪失、ならびに補助刺激細胞表面分子および種々のサイトカインの発現のアップレギュレーションをもたらす。公知の成熟プロトコルは、DCが抗原への曝露の間またはその後に遭遇すると考えられるインビボでの環境に基づく。このアプローチの最良の例は、細胞培養培地としての、単球馴化培地(MCM)の使用である。MCMは、単球を培養することによってインビトロで生成され、そして成熟因子の供給源として用いられる。成熟の原因となる、MCM中の主な成分は、炎症(誘発)性サイトカインインターロイキン1β(IL−1β)、インターロイキン6(IL−6)および腫瘍壊死因子α(TNFα)であると報告されている。他の成熟因子としては、プロスタグランジンE2(PGE2)、ポリ−dIdC、血管作用性(vasointestinal)ペプチド(VIP)、細菌リポ多糖(LPS)、ならびにマイコバクテリアまたはマイコバクテリア成分(例えば、特定の細胞壁成分)が挙げられる。
【0004】
充分に成熟した樹状細胞は、未成熟DCとは定性的および定量的に異なる。充分に成熟したDCは、より高レベルのMHCクラスI抗原およびMHCクラスII抗原、ならびにより高レベルのT細胞補助刺激分子(すなわち、CD80およびCD86)を発現する。これらの変化は、樹状細胞がT細胞を活性化する能力を増大させる。なぜなら、これらは、細胞表面の抗原密度、ならびにT細胞上の補助刺激分子の対応物(例えば、CD28など)を通るT細胞活性化シグナルの大きさを増大させるからである。さらに、成熟DCは、多量のサイトカインを産生し、このサイトカインは、T細胞応答を刺激して導く。これらのサイトカインのうちの2つは、インターロイキン10(IL−10)およびインターロイキン(IL−12)である。これらのサイトカインは、誘導されるT細胞応答の方向に対して反対の効果を有する。IL−10産生は、Th−2型応答の誘導をもたらし、一方、IL−12産生は、Th−1型応答をもたらす。後者の応答は、細胞免疫応答が所望される場合(例えば、癌免疫治療において)、特に望ましい。Th−1型応答は、細胞傷害性Tリンパ球(CTL)(これは、細胞性免疫系のエフェクターアームである)の誘導および分化をもたらす。このエフェクターアームは、腫瘍増殖と戦う際に最も有効である。IL−12はまた、ナチュラルキラー(NK)細胞の増殖を誘導し、そして抗脈管形成活性を有する。これらは両方とも、有効な抗腫瘍防御手段である。従って、IL−12を産生する樹状細胞の使用は、理論的に、免疫刺激における使用に最も適している。
【0005】
特定の樹状細胞成熟因子(例えば、細菌性リポ多糖、細菌性CpG DNA、二本鎖RNAおよびCD40リガンド)は、未成熟DCを誘導して、IL−12を産生させ、未成熟DCにTh−1応答をプライミングすると報告されている。対照的に、抗炎症分子(例えば、IL−10、TGF−β、PGE−2およびコルチコステロイド(corticosteriod))はIL−12産生を阻害し、そして細胞にTh−2応答をプライミングし得る。
【0006】
近年、樹状細胞によるIL−12産生の増強は、インターフェロンγと特定の樹状細胞成熟因子(例えば、細菌性リポ多糖(LPS)およびCD40)とを合わせることによって報告されている。しかし、LPSおよびCD40は両方とも、成熟の間に少量のIL−12を誘導する既知の能力を有する。従って、IFNγの添加は、その産生を増強するにすぎないことが可能である。インターフェロンγシグナル伝達は、Jak2−Stat1経路を使用し、これは、核へのその移動の前のStat1の701位のチロシン残基のチロシンリン酸化、およびその後のインターフェロンγ応答遺伝子の転写増強を含む。しかし、ヒト単球由来樹状細胞におけるシグナル伝達経路についてはごくわずかしか公知でない。これらの細胞におけるインターフェロンγ作用についての機構は、確立されていない。
【0007】
Mycobacterium tuberculosisの弱毒化ウシ株(Mycobacterium bovis)(現在、カルメット−ゲラン杆菌(BCG)として公知)は、癌免疫治療において用いられている。1例では、生BCGの膀胱内投与は、膀胱癌の処置に有効であることが証明されているが、この処置についての機構は、公知ではない。BCG投与の効果は、例えば、癌細胞を攻撃する免疫応答の誘導によって媒介されると仮定されている。この応答におけるBCGの特定の役割は、免疫反応性の一般化された誘導因子のものであり、免疫系に対する腫瘍抗原の提示においてアジュバント機能を有すると考えられる。
【0008】
BCGはまた、成熟マーカーCD83をアップレギュレートする能力を有する、樹状細胞に対する強力な成熟因子であることが見出されている。BCGはまた、エンドサイトーシス能力の低減を伴って、MHC分子ならびに補助刺激分子CD80およびCD86をアップレギュレートし得る。さらに、IL−12の産生が特異的に阻害されることが見出された、他のDC成熟因子について見出された結果とは対照的に、BCGまたはBCG由来リポアラビドマンナンは、サイトカイン産生を増大させることが報告されている。この前者の特性(IL−12産生の阻害)は、強力な細胞媒介性細胞傷害性応答(Th−1応答)が所望される免疫治療用の樹状細胞の成熟のためにBCGを用いることの魅力を低減する。
【発明の概要】
【発明が解決しようとする課題】
【0009】
従って、能動免疫治療におけるBCGの使用は、樹状細胞成熟を誘導する可能性を有する。しかし、樹状細胞のこのような成熟を誘導し、広範な免疫刺激を同時に提供し、そしてこれらの樹状細胞に、強固な細胞傷害性T細胞応答を有する1型(Th−1)免疫応答をプライミングさせる、組成物およびこのような組成物を用いる方法についての必要性が存在する。
【課題を解決するための手段】
【0010】
(発明の簡単な要旨)
本発明は、広範な免疫刺激を同時に提供する薬剤(すなわち、BCG)を用いて未成熟樹状細胞(DC)の成熟を誘導するための方法および組成物、ならびにこれらの細胞に抗原特異的な細胞傷害性T細胞応答をプライミングさせるための方法および組成物を提供する。1つの局面では、成熟樹状細胞集団を産生するための方法が提供され、この方法は、未成熟樹状細胞を提供する工程;ならびにこの未成熟樹状細胞を、この未成熟樹状細胞の成熟に適切な培養条件下で、有効濃度のBCGおよびインターフェロンγ(IFNγ)と接触させて、成熟樹状細胞集団を形成させる工程を包含する。この成熟樹状細胞集団は、成熟の間にBCGおよびIFNγ単独に接触させなかった未成熟樹状細胞集団よりも上昇したインターロイキン12対インターロイキン10比を生じる。この未成熟樹状細胞は、BCGおよびIFNγとの接触の前または接触の間に所定の抗原と接触され得る。この所定の抗原は、例えば、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、抗原を発現する組換え細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原(例えば、合成ペプチド抗原)、または単離抗原であり得る。
【0011】
特定の実施形態では、この方法は、単球性樹状細胞前駆体を単離する工程;およびこの前駆体を分化因子の存在下で培養して未成熟樹状細胞集団を形成する工程を必要に応じてさらに包含し得る。適切な分化因子としては、例えば、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13が挙げられる。この単球性樹状細胞前駆体は、ヒト被験体から単離され得る。特定の実施形態では、この成熟樹状細胞は、少なくとも1:1のIL−12対IL−10の比を生じる。
【0012】
別の局面では、成熟樹状細胞集団を産生するための方法が提供される。この方法は一般に、未成熟樹状細胞を提供する工程;ならびにこの未成熟樹状細胞を、この未成熟樹状細胞の成熟に適切な培養条件下で、有効量のBCGおよびインターフェロンγ(IFNγ)と接触させて、成熟樹状細胞集団を形成させる工程を包含する。得られる成熟樹状細胞集団は、1型免疫応答を生じる。この未成熟樹状細胞を、BCGおよびIFNγとの接触の前または接触の間に、所定の抗原と接触させ得る。この所定の抗原は、例えば、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、抗原を発現する組換え細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原(すなわち、合成ペプチド)、または単離抗原であり得る。
【0013】
特定の実施形態では、この方法は、単球性樹状細胞前駆体を単離する工程;およびこの前駆体を分化因子の存在下で培養して、この未成熟樹状細胞を形成させる工程を必要に応じてさらに包含し得る。適切な分化因子としては、例えば、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13が挙げられる。この単球性樹状細胞前駆体は、ヒト被験体から単離される。特定の実施形態では、この成熟樹状細胞は、少なくとも約1:1のIL−12対IL−10の比を生じる。
【0014】
なお別の局面では、T細胞を活性化するための組成物が提供される。この組成物は、成熟について適切な条件下で有効濃度のBCGおよびIFNγを用いて成熟された樹状細胞集団;および所定の抗原を含み得る。この樹状細胞集団は、成熟の間にIFNγを伴わずにBCGと接触させた成熟樹状細胞集団よりも上昇したインターロイキン12(IL−12)対インターロイキン10(IL−10)比を生じ得る。特定の実施形態では、この樹状細胞集団は、少なくとも約10:1の比のIL−12対IL−10を生じ得る。他の実施形態では、この樹状細胞集団は、成熟の間にIFNγを伴わずにBCGの存在下で培養した同様の未成熟樹状細胞集団よりも少なくとも約1:100の比のIL−12対IL−10を生じ得る。
【0015】
別の局面では、単離された未成熟樹状細胞集団が提供される。この細胞集団は、未成熟単球性樹状細胞、ならびにこの未成熟樹状細胞の成熟を誘導する有効濃度のBCGおよびIFNγを含む。得られる成熟樹状細胞は、成熟の間にIFNγを伴わずにBCGの存在下で培養した同様の未成熟樹状細胞集団よりも高いインターロイキン12(IL−12)対インターロイキン10(IL−10)を生じる。この細胞集団は、所定の抗原および/または単離されたT細胞(例えば、ナイーブT細胞)を必要に応じて含み得る。このT細胞は、単離されたリンパ球の調製物中に必要に応じて存在し得る。
【0016】
活性化T細胞を産生するための方法もまた提供される。この方法は一般に、未成熟樹状細胞を提供する工程;この未成熟樹状細胞を所定の抗原と接触させる工程;ならびにこの未成熟樹状細胞を、この未成熟樹状細胞の成熟に適切な培養条件下で、有効濃度のBCGおよびIFNγと接触させて、成熟樹状細胞を形成させる工程を包含する。この成熟樹状細胞を、ナイーブT細胞と接触させて、IFNγを産生するか、および/または1型(Th−1)応答に極性化した活性化T細胞を形成させ得る。適切な抗原としては、例えば、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、抗原を発現する組換え細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原(例えば、合成ペプチド抗原)、または単離抗原が挙げられる。
【0017】
この未成熟樹状細胞を所定の抗原、BCGおよびIFNγと同時に接触させ得るか、またはこの細胞を、BCGおよびIFNγと接触させる前にこの所定の抗原と接触させ得る。特定の実施形態では、この方法は、単球性樹状細胞前駆体を単離する工程;および分化因子の存在下でこの前駆体を培養して、この未成熟樹状細胞の形成を誘導する工程をさらに包含し得る。適切な分化因子としては、例えば、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13が挙げられる。この単球性樹状細胞前駆体は、ヒト被験体から必要に応じて単離され得る。特定の実施形態では、この未成熟樹状細胞およびT細胞は、互いに自己である。
【0018】
インターロイキン10(IL−10)よりも多くのインターロイキン12(IL−12)を産生する単離された成熟樹状細胞もまた提供される。この成熟樹状細胞は、この樹状細胞の成熟に適切な条件下で有効濃度のBCGおよびIFNγを含む組成物を用いた未成熟樹状細胞の成熟によって提供され得る。所定の抗原が、この単離された成熟樹状細胞に必要に応じて含まれ得る。所定の抗原がローディングされた、単離された成熟樹状細胞もまた提供される。この樹状細胞は、成熟の間にIFNγを伴わずにBCGの存在下で培養した同様の未成熟樹状細胞集団よりも多くの、インターロイキン10(IL−10)よりも多くのインターロイキン12(IL−12)(例えば、IL−10よりも少なくとも10倍多くのIL−12)を産生し得る。
【0019】
動物において1型(Th−1)免疫応答を産生するための方法もまた提供される。この方法は一般に、未成熟樹状細胞を提供する工程;この未成熟樹状細胞を、この未成熟樹状細胞の成熟に適切な培養条件下で、有効量のBCGおよびインターフェロンγ(IFNγ)、ならびに所定の抗原とを接触させて、成熟樹状細胞を形成させる工程を包含する。この成熟樹状細胞を動物に投与するかまたはナイーブT細胞と接触させるかのいずれかによって、インターフェロンγ(IFNγ)および/または腫瘍壊死因子α(TNFα)の産生によって特徴付けられる活性化T細胞を形成させ得る。この活性化T細胞は、特定の抗原に対する細胞傷害性T細胞応答の刺激の必要のある動物に投与され得る。適切な抗原としては、例えば、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、抗原を発現する組換え細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原(例えば、合成ペプチド抗原)、または単離抗原が挙げられる。この未成熟樹状細胞を、この所定の抗原、BCGおよびIFNγと必要に応じて同時に接触させ得るか、またはBCGおよびIFNγと接触させる前に、この未成熟樹状細胞をこの所定の抗原と接触させ得る。
【0020】
特定の実施形態では、この方法は、単球性樹状細胞前駆体をこの動物から単離する工程;およびこの前駆体を分化因子の存在下で培養して、この未成熟樹状細胞を形成させる工程をさらに包含し得る。この分化因子は、例えば、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13であり得る。
【0021】
この未成熟樹状細胞およびT細胞は、この動物に対して自己であってもよく、またはこの動物に対して同種異系であってもよい。あるいは、この未成熟樹状細胞およびT細胞は、この動物と同じMHCハプロタイプを有してもよく、またはMHCマーカーを共有してもよい。特定の実施形態では、この動物は、ヒトであっても非ヒト動物であってもよい。
例えば、本発明は以下の項目を提供する。
(項目1)
成熟樹状細胞集団を産生するための方法であって、該方法は、以下:
未成熟樹状細胞を提供する工程;ならびに
該未成熟樹状細胞を、該未成熟樹状細胞の成熟に適切な培養条件下で、有効量のBCGおよびインターフェロンγ(IFNγ)と接触させて、成熟樹状細胞集団を形成させる工程;
を包含し、ここで、該成熟樹状細胞集団は、成熟の間にBCGおよびIFNγと接触させなかった未成熟樹状細胞集団よりも上昇したインターロイキン12対インターロイキン10比を生じる、方法。
(項目2)
BCGおよびIFNγと接触させる前に、前記未成熟樹状細胞を所定の抗原と接触させる工程をさらに包含する、項目1に記載の方法。
(項目3)
前記未成熟樹状細胞を所定の抗原、BCGおよびIFNγと同時に接触させる工程をさらに包含する、項目1に記載の方法。
(項目4)
前記所定の抗原が、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原、または単離抗原である、項目2または3に記載の方法。
(項目5)
単球性樹状細胞前駆体を単離する工程;および該前駆体を分化因子の存在下で培養して前記未成熟樹状細胞を形成させる工程をさらに包含する、項目1に記載の方法。
(項目6)
前記分化因子が、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13である、項目5に記載の方法。
(項目7)
前記単球性樹状細胞前駆体が、ヒト被験体から単離される、項目5に記載の方法。
(項目8)
前記成熟樹状細胞が、少なくとも約1:1、少なくとも約10:1、少なくとも約100:1のIL−12対IL−10の比を生じる、項目1に記載の方法。
(項目9)
成熟樹状細胞集団を産生するための方法であって、該方法は、以下:
未成熟樹状細胞を提供する工程;ならびに
該未成熟樹状細胞を、該未成熟樹状細胞の成熟に適切な培養条件下で、有効量のBCGおよびインターフェロンγ(IFNγ)と接触させて、成熟樹状細胞集団を形成させる工程;
を包含し、ここで、該成熟樹状細胞集団が、1型免疫応答を生じる、方法。
(項目10)
BCGおよびIFNγと接触させる前に、前記未成熟樹状細胞を所定の抗原と接触させる工程をさらに包含する、項目9に記載の方法。
(項目11)
前記未成熟樹状細胞を所定の抗原、BCGおよびIFNγと同時に接触させる工程をさらに包含する、項目9に記載の方法。
(項目12)
前記所定の抗原が、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、抗原を発現する組換え細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原、または単離抗原である、項目10または11に記載の方法。
(項目13)
単球性樹状細胞前駆体を単離する工程;および該前駆体を分化因子の存在下で培養して前記未成熟樹状細胞を形成させる工程をさらに包含する、項目9に記載の方法。
(項目14)
前記分化因子が、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13である、項目13に記載の方法。
(項目15)
前記単球性樹状細胞前駆体が、ヒト被験体から単離される、項目13に記載の方法。
(項目16)
前記成熟樹状細胞が、少なくとも約1:1、少なくとも約10:1、少なくとも約100:1のIL−12対IL−10の比を生じる、項目9に記載の方法。
(項目17)
T細胞を活性化するための組成物であって、該組成物は、以下:
成熟に適切な条件下で有効濃度のBCGおよびIFNγを用いて成熟させた樹状細胞集団;ならびに
所定の抗原;
を含み、ここで、前記樹状細胞集団が、成熟の間にIFNγを伴わずにBCGと接触させた成熟樹状細胞集団よりも上昇したインターロイキン12(IL−12)対インターロイキン10(IL−10)比を生じる、組成物。
(項目18)
前記樹状細胞集団が、少なくとも約10:1の比のIL−12対IL−10を生じる、項目17に記載の組成物。
(項目19)
前記樹状細胞集団が、少なくとも約100:1の比のIL−12対IL−10を生じる、項目17に記載の組成物。
(項目20)
単離された未成熟樹状細胞集団であって、該細胞集団は、以下:
単離された未成熟単球性樹状細胞、ならびに該未成熟樹状細胞の成熟を誘導する有効濃度のBCGおよびIFNγを含み;ここで、得られる成熟樹状細胞が、インターロイキン10(IL−10)よりも多くのインターロイキン12(IL−12)を産生する、細胞集団。
(項目21)
所定の抗原をさらに含む、項目20に記載の細胞集団。
(項目22)
単離されたT細胞をさらに含む、項目20に記載の細胞集団。
(項目23)
前記T細胞が、ナイーブT細胞である、項目22に記載の細胞集団。
(項目24)
単離されたリンパ球をさらに含む、項目20に記載の細胞集団。
(項目25)
T細胞を産生するための方法であって、該方法は、以下:
未成熟樹状細胞を提供する工程;
該未成熟樹状細胞を所定の抗原と接触させる工程;
該未成熟樹状細胞を、該未成熟樹状細胞の成熟に適切な培養条件下で、有効濃度のBCGおよびIFNγと接触させて、成熟樹状細胞を形成させる工程;ならびに
該成熟樹状細胞をナイーブT細胞と接触させて、IFNγを産生する活性化T細胞を形成させる工程
を包含する、方法。
(項目26)
前記所定の抗原が、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、抗原を発現する組換え細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原、または単離抗原である、項目25に記載の方法。
(項目27)
前記未成熟樹状細胞が所定の抗原、BCGおよびIFNγと同時に接触される、項目25に記載の方法。
(項目28)
単球性樹状細胞前駆体を単離する工程;および該前駆体を分化因子の存在下で培養して前記未成熟樹状細胞を形成させる工程をさらに包含する、項目25に記載の方法。
(項目29)
前記分化因子が、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13である、項目28に記載の方法。
(項目30)
前記単球性樹状細胞前駆体が、ヒト被験体から単離される、項目28に記載の方法。
(項目31)
前記未成熟樹状細胞およびT細胞が、互いに対して自己である、項目25に記載の方法。
(項目32)
インターロイキン10(IL−10)より多くのインターロイキン12(IL−12)を産生する、単離された成熟樹状細胞であって、該樹状細胞の成熟に適切な条件下で、有効濃度のBCGおよびIFNγを含む組成物を用いた未成熟樹状細胞の成熟によって調製された、単離された成熟樹状細胞。
(項目33)
所定の抗原をさらに包含する、項目32に記載の単離された成熟樹状細胞。
(項目34)
所定の抗原がローディングされた単離された成熟樹状細胞であって、該樹状細胞は、インターロイキン10(IL−10)よりも多くのインターロイキン12(IL−12)を産生する、単離された成熟樹状細胞。
(項目35)
前記細胞が、IL−10よりも少なくとも10倍多くのIL−12を産生する、項目34に記載の単離された成熟樹状細胞。
(項目36)
動物内で1型免疫応答を生じるための方法であって、該方法は、以下:
未成熟樹状細胞を提供する工程;
該未成熟樹状細胞を、該未成熟樹状細胞の成熟に適切な培養条件下で、有効量のBCGおよびインターフェロンγ(IFNγ)および所定の抗原と接触させて、成熟樹状細胞を形成させる工程;
該成熟樹状細胞をナイーブT細胞と接触させて、インターロイキン10(IL−10)よりも多くのインターロイキン12(IL−12)を産生する活性化T細胞を産生させる工程;ならびに
該活性化T細胞を該動物に投与する工程
を包含する、方法。
(項目37)
前記所定の抗原が、腫瘍特異性抗原、腫瘍関連抗原、ウイルス抗原、細菌抗原、腫瘍細胞、細菌細胞、抗原を発現する組換え細胞、細胞溶解産物、膜調製物、組換え産生抗原、ペプチド抗原、または単離抗原である、項目36に記載の方法。
(項目38)
前記未成熟樹状細胞を所定の抗原、BCGおよびIFNγと同時に接触させる、項目36に記載の方法。
(項目39)
BCGおよびIFNγと接触させる前に、前記未成熟樹状細胞を前記所定の抗原と接触させる、項目36に記載の方法。
(項目40)
前記動物から単球性樹状細胞前駆体を単離する工程;および該前駆体を分化因子の存在下で培養して前記未成熟樹状細胞を形成させる工程をさらに包含する、項目36に記載の方法。
(項目41)
前記分化因子が、GM−CSF、インターロイキン4、GM−CSFとインターロイキン4との組み合わせ、またはインターロイキン13である、項目40に記載の方法。
(項目42)
前記未成熟樹状細胞およびT細胞が、前記動物に対して自己である、項目36に記載の方法。
(項目43)
前記未成熟樹状細胞およびT細胞が、前記動物に対して同種異系である、項目36に記載の方法。
(項目44)
前記未成熟樹状細胞およびT細胞が、前記動物と同じMHCハプロタイプを有する、項目36に記載の方法。
(項目45)
前記動物がヒトである、項目36に記載の方法。
【図面の簡単な説明】
【0022】
【図1】記載なし。
【発明を実施するための形態】
【0023】
(発明の詳細な説明)
本発明は、未成熟樹状細胞(DC)の成熟を誘導するための方法およびこれらの細胞に抗原特異的細胞傷害性T細胞応答(Th−1応答)をプライミングさせるための方法を提供する。本発明はまた、1型サイトカイン(例えば、IFNγ、TNFα、および/またはIL−2)の産生に極性化したT細胞集団を活性化および調製するために有用な樹状細胞集団を提供する。このような樹状細胞集団としては、適切な成熟条件下でBCG、IFNγおよび所定の抗原と接触させた未成熟単球性樹状細胞が挙げられる。この未成熟樹状細胞を、成熟の間または成熟前にこの抗原と接触させ得る。あるいは、抗原に(例えば、インビボで)既に曝露した未成熟単球性樹状細胞を、適切な成熟条件下でBCGおよびIFNγと接触させ得る。得られる成熟樹状細胞は、T細胞を活性化して1型応答に対してT細胞を極性化することがプライミングされる。1型応答としては、1型サイトカイン(例えば、IFNγ、および/またはIL−2)の産生、IL−10よりも多くのIL−12の産生、細胞傷害性T細胞応答、Th−1細胞の産生、および特定の型の抗体の産生が挙げられる。腫瘍壊死因子α(TNFα)もまたアップレギュレートされ得る。対照的に、2型応答は、IL−4、IL−5およびIL−10の産生、IL−12よりも多いIL−10の産生、Th2細胞の産生、ならびにCTL応答の誘導欠如によって特徴付けられる。
【0024】
関連の局面では、未成熟樹状細胞(例えば、単球性樹状細胞前駆体)についての成熟因子(これは、これらの樹状細胞に1型応答をもプライミングさせ得る)を含む組成物が提供される。成熟し、プライミングされたこのような単球性樹状細胞は、所定の抗原(すなわち、所定の外因性抗原)の主要組織適合遺伝子複合体(MHC)クラス−I提示を増加させ得る。抗原のMHCクラスI提示は、細胞傷害性Tリンパ球(CTL)の分化および標的細胞の抗原特異的CTL媒介溶解の刺激を誘導するために所望される。このような組成物は、未成熟樹状細胞を含む細胞集団と混合され得、未成熟樹状細胞を成熟させるため、ならびにこの未成熟樹状細胞とBCGとの接触によって誘導されるIL−12の阻害を転換または克服するための、BCGおよびIFNγを含む。このような組成物を接触させた未成熟樹状細胞は、成熟を経て、そして代表的には、BCG単独と接触させた未成熟樹状細胞集団と比較して、IL−10よりも多量のIL−12を産生する。
【0025】
別の局面では、被験体またはドナーから得た単球性樹状細胞前駆体を、サイトカイン(例えば、GM−CSFおよびIL−4)と接触させて、未成熟樹状細胞を獲得し得る。次いで、この未成熟樹状細胞を、BCGおよびIFNγのみと組み合わせたかまたはサイトカインと組み合わせた所定の抗原と接触させて、この樹状細胞を成熟させ得、そしてこの細胞がT細胞における1型免疫応答を誘導することをプライミングさせ得る。特定の実施形態では、MHCクラス−I抗原プロセシングが刺激され、このことは、所定の抗原を提示する細胞に対してCTL応答を惹起するために有用である。
【0026】
樹状細胞は、種々のリンパ組織および非リンパ組織に見出される抗原提示細胞の多様な集団である(Liu,Cell 106:259−62(2001);Steinman,Ann.Rev.Immunol.9:271−96(1991)を参照のこと)。樹状細胞としては、脾臓のリンパ樹状細胞、表皮のランゲルハンス細胞、および血液循環中のベール細胞が挙げられる。集合的に、樹状細胞は、それらの形態、高レベルの表面MHC−クラスII発現、ならびにT細胞、B細胞、単球、およびナチュラルキラー細胞上に発現される特定の他の表面マーカーが存在しないことに基づいて、1群として分類される。特に、単球由来樹状細胞(単球性樹状細胞ともいう)は通常、CD11c、CD80、CD86を発現し、そしてHLA−DR+であるが、CD14−である。
【0027】
対照的に、単球性樹状細胞前駆体(代表的に、単球)は通常、CD14+である。単球性樹状細胞前駆体は、これらが存在する任意の組織(特に、リンパ組織(例えば、脾臓、骨髄、リンパ節および胸腺))から入手され得る。単球性樹状細胞前駆体はまた、循環系から単離され得る。末梢血は、単球性樹状細胞前駆体の容易に利用可能な供給源である。臍帯血は、単球性樹状細胞前駆体の別の供給源である。単球性樹状細胞前駆体は、免疫応答が惹起され得る種々の生物から単離され得る。このような生物としては、動物(例えば、ヒトおよび非ヒト動物(例えば、霊長類、哺乳動物(イヌ、ネコ、マウスおよびラットが挙げられる)、鳥類(ニワトリが挙げられる)、ならびにそれらのトランスジェニック種が挙げられる)が挙げられる。
【0028】
特定の実施形態では、この単球性樹状細胞前駆体および/または未成熟樹状細胞は、健常な被験体、または免疫刺激の必要のある被験体(例えば、前立腺癌患者)、または細胞性免疫刺激が有利であり得るかもしくは所望され得る他の被験体(すなわち、細菌感染もしくはウイルス感染を有する被験体など)から単離され得る。樹状細胞前駆体および/または未成熟樹状細胞はまた、免疫刺激の必要がある、HLAが一致した被験体への投与のために、HLAが一致した健常個体から入手され得る。
【0029】
(樹状細胞前駆体および未成熟樹状細胞)
樹状細胞前駆体および未成熟樹状細胞が富化された細胞集団を、種々の供給源(血液および骨髄を含む)から単離するための方法が当該分野で公知である。例えば、樹状細胞前駆体および未成熟樹状細胞は、ヘパリン処理した血液を収集することによって、アフェレーシスもしくは白血球搬出によって、バフィコートの調製、ロゼット形成、遠心分離、密度勾配遠心分離(例えば、Ficoll(例えば、FICOLL−PAQUE(登録商標))、PERCOLL(登録商標)(透析可能ではないポリビニルピロリドン(PVP)でコーティングしたコロイド状シリカ粒子(直径15〜30mm))、スクロースなどを用いる)、細胞の示差的溶解、濾過などによって単離され得る。特定の実施形態では、白血球集団は、例えば、被験体からの血液の収集、血小板を除去するためのフィブリン除去(defribrinating)および赤血球の溶解によって調製され得る。樹状細胞前駆体および未成熟樹状細胞は、例えば、PERCOLL(登録商標)勾配を通しての遠心分離によって、単球性樹状細胞前駆体について必要に応じて富化され得る。
【0030】
樹状細胞前駆体および未成熟樹状細胞は、閉じた無菌系において必要に応じて調製され得る。本明細書中で用いられる場合、用語「閉じた無菌系」または「閉じた系」とは、非滅菌空気、周囲の空気、もしくは循環空気、または他の非無菌条件への曝露が最小であるか排除されている系をいう。樹状細胞前駆体および未成熟樹状細胞を単離するための閉じた系は、一般に、頂部が開いたチューブにおける密度勾配遠心分離、細胞の開放空気下での移植、組織培養プレートまたはシールしていないフラスコにおける細胞培養などを除外する。代表的な実施形態では、この閉じた系は、無菌でない空気への曝露を伴わずに、最初の収集容器からシール可能な組織培養容器への、樹状細胞前駆体および未成熟樹状細胞の無菌移植を可能にする。
【0031】
特定の実施形態では、単球性樹状細胞前駆体は、米国特許出願第60/307,978号(2001年7月25日出願(代理人文書番号020093−002600US)(この開示は、本明細書中に参考として援用される))に開示されるように、単球結合基材への接着によって単離される。例えば、白血球集団(例えば、白血球搬出によって単離される)を、単球性樹状細胞前駆体接着基材と接触させ得る。この白血球集団をこの基材と接触させる場合、この白血球集団中の単球性樹状細胞前駆体は優先的にこの基材に接着する。他の白血球(他の潜在的樹状細胞前駆体を含む)は、基質に対する低減した結合親和性を示し、それゆえ、この単球性樹状細胞前駆体がこの基材表面に優先的に富化されるのを可能にする。
【0032】
適切な基材としては、例えば、大きな表面積対容積比を有する基材が挙げられる。このような基材は、例えば、粒子状基材または繊維状基材であり得る。適切な粒子状基材としては、例えば、ガラス粒子、プラスチック粒子、ガラスでコーティングしたプラスチック粒子、ガラスでコーティングしたポリスチレン粒子、およびタンパク質吸収に適切な他のビーズが挙げられる。適切な繊維状基材としては、マイクロキャピラリーチューブおよび微絨毛膜が挙げられる。この粒子状基材または繊維状基材は通常、付着した単球性樹状細胞前駆体が、接着した細胞の生存率を実質的に低下させることなく、溶出されるのを可能にする。粒子状基材または繊維状基材は、この基材からの単球性樹状細胞前駆体または樹状細胞の溶出を容易にするために実質的に非多孔質であり得る。「実質的に非多孔質」の基材は、その基材中に存在する少なくとも大部分の孔が、この基材中に捕捉される細胞を最小にするために、この細胞よりも小さい基材である。
【0033】
この基材に対するこの単球性樹状細胞前駆体の付着は、結合培地の添加によって、必要に応じて増強され得る。適切な結合培地としては、例えば、サイトカイン(例えば、顆粒球/マクロファージコロニー刺激因子(GM−CSF)、インターロイキン4(IL−4)、またはインターロイキン13(IL−13))、血漿、血清(例えば、ヒト血清(例えば、自己血清もしくは同種異系血清))、精製タンパク質(例えば、血清アルブミン、二価カチオン(例えば、カルシウムイオンおよび/またはマグネシウムイオン))ならびにこの基材に対する単球性樹状細胞前駆体の特異的付着を補助するかまたはこの基材への非単球性樹状細胞前駆体の付着を妨害する、他の分子が個々にまたは任意の組み合わせで補充された、単球性樹状細胞前駆体培養培地(例えば、AIM−V(登録商標)、RPMI 1640、DMEM、X−VIVO 15(登録商標)など)が挙げられる。特定の実施形態では、血漿または血清は、熱不活化され得る。熱不活化血漿は、白血球に対して自己または異種であり得る。
【0034】
この基材への単球性樹状細胞前駆体の付着後、付着していない白血球は、単球性樹状細胞前駆体/基材複合体から分離される。任意の適切な手段を用いて、付着していない細胞を複合体から分離し得る。例えば、付着していない白血球と複合体との混合物を沈澱させ得、そして付着していない白血球および培地を傾瀉または排出し得る。あるいは、この混合物を遠心分離し得、そして付着していない白血球を含む上清を、ペレット化複合体から傾瀉または排出し得る。
【0035】
単離された樹状細胞前駆体は、分化、成熟および/または拡大のためにエキソビボで培養され得る。(本明細書中で用いられる場合、単離された未成熟樹状細胞、樹状細胞前駆体、T細胞、および他の細胞とは、ヒトの手によって、それらのネイティブな環境とは離れて存在し、それゆえ、天然の産物ではない細胞をいう。単離された細胞は、純粋な形態、半ば精製された形態、またはネイティブではない環境に存在し得る。)手短に述べると、エキソビボでの分化は代表的に、樹状細胞前駆体、または樹状細胞前駆体を有する細胞集団を、1以上の分化薬剤の存在下で培養する工程を包含する。適切な分化因子は、例えば、細胞増殖因子(例えば、サイトカイン(例えば、(GM−CSF)、インターロイキン4(IL−4)、インターロイキン13(IL−13)、および/またはそれらの組み合わせ))であり得る。特定の実施形態では、この単球性樹状細胞前駆体を分化させて、単球由来未成熟樹状細胞を形成させる。
【0036】
この樹状細胞前駆体は、適切な培養条件で培養および分化され得る。適切な組織培養培地としては、AIM−V(登録商標)、RPMI 1640、DMEM、X−VIVO 15(登録商標)などが挙げられる。この組織培養培地には、細胞の分化を促進するために、血清、アミノ酸、ビタミン、サイトカイン(例えば、GM−CSFおよび/またはIL−4)、二価カチオンなどが添加され得る。特定の実施形態では、この樹状細胞前駆体は、無血清培地中で培養され得る。このような培養条件は、必要に応じて任意の動物由来産物を排除し得る。代表的樹状細胞培養培地における代表的なサイトカインの組み合わせは、各々約500単位/mlのGM−CSFおよびIL−4である。樹状細胞前駆体は、分化して未成熟樹状細胞を形成した場合、皮膚ランゲルハンス細胞に表現型的に類似する。未成熟樹状細胞は代表的に、CD14−およびCD11c+であり、低レベルのCD86およびCD83を発現し、そして極性化されたエンドサイトーシスを介して可溶性抗原を捕捉し得る。
【0037】
未成熟樹状細胞は成熟して成熟樹状細胞を形成する。成熟DCは、抗原を取り込む能力を失い、発現がアップレギュレートされた補助刺激細胞表面分子および種々のサイトカインを提示する。特に、成熟DCは、未成熟樹状細胞よりも高いレベルのMHCクラスI抗原およびMHCクラスII抗原を発現し、そして成熟樹状細胞は、一般に、CD80+、CD83+、CD86+、およびCD14−であると同定される。より多くのMHC発現は、DC表面での抗原密度の増加をもたらし、一方、補助刺激分子CD80およびCD86のアップレギュレーションは、T細胞上の補助刺激分子(例えば、CD28)の対応物を通して、T細胞活性化シグナルを強化する。
【0038】
本発明の成熟樹状細胞は、未成熟樹状細胞を有効量または有効濃度のBCGおよびIFNγと接触させることによって、調製(すなわち、成熟)され得る。有効量のBCGは、代表的に、組織培養培地1ミリリットルあたり約105〜約107cfuの範囲に及ぶ。IFNγの有効量は代表的に、組織培養培地1ミリリットルあたり約100〜約1000Uの範囲に及ぶ。カルメットゲラン杆菌(Bacillus Calmette−Guerin;BCG)は、M.bovisの無発病性株である。本明細書中で用いられる場合、BCGは、BCG全体ならびに細胞壁構成成分、BCG由来リポアラビドマンナン、および2型免疫応答の誘導に関連した他のBCG成分をいう。BCGは、必要に応じて不活化される(例えば、熱不活化BCG、ホルマリン処理BCGなど)。
【0039】
BCGは、IL−12産生の阻害およびエンドサイトーシスによる抗原の排除に付随して、樹状細胞上の表面成熟マーカーCD83およびCD86の発現を増大させる。何らかの特定の理論に束縛されることを意図しないが、BCGによる樹状細胞成熟はまた、腫瘍壊死因子α(TNFα)のホモタイプな凝集および放出を含むと特徴付けられている(例えば、本明細書中に参考として援用される、Thurnherら,Int.J.Cancer 70:128−34(1997)を参照のこと)。IFNγおよびBCGを用いて未成熟樹状細胞を成熟させることは、IL−12のDC産生を促進し、そしてIL−10の産生を低減または阻害し、それによって、成熟樹状細胞に1型(Th−1)応答をプライミングさせる。
【0040】
この未成熟DCを、代表的に、有効量のBCGおよびIFNγと、約1時間〜約24時間にわたって接触させる。この未成熟樹状細胞は、適切な成熟培養条件において培養されて成熟し得る。適切な組織培養培地としては、AIM−V(登録商標)、RPMI 1640、DMEM、X−VIVO 15(登録商標)などが挙げられる。これらの組織培養培地には、細胞の成熟を促進するために、アミノ酸、ビタミン、サイトカイン(例えば、GM−CSFおよび/またはIL−4)、二価カチオンなどが補充され得る。代表的なサイトカインの組み合わせは、各々約500単位/mlのGM−CSFおよびIL−4である。
【0041】
樹状細胞の成熟は、当該分野で公知の方法によってモニタリングされ得る。細胞表面マーカーは、当該分野で通常のアッセイ(例えば、フローサイトメトリー、免疫組織化学など)において検出され得る。これらの細胞はまた、(例えば、ELISA、FACS、または他の免疫アッセイによって)サイトカイン産生についてモニタリングされ得る。本発明に従って成熟したDC集団では、IL−12レベルは、IL−10レベルよりも高く、1型(Th−1)応答を促進する。例えば、このDCは、1:1より大きく、約10:1または約100:1までのIL−12/IL−10比を生じ得る。成熟DCはまた、飲細胞運動によって抗原を取り込む能力を失う。飲細胞運動は、当業者には通常の取り込みアッセイによって分析され得る。樹状細胞前駆体、未成熟樹状細胞、および成熟樹状細胞は、抗原を用いてプライミングされていようとプライミングされていなかろうと、後日の使用のために凍結保存され得る。凍結保存のための方法は、当該分野で周知である。例えば、その全体が本明細書中に参考として援用される、米国特許第5,788,963号を参照のこと。
【0042】
(抗原)
本発明による、成熟しプライミングされた樹状細胞は、抗原をT細胞に対して提示し得る。成熟しプライミングされた樹状細胞は、成熟の前または成熟の間のいずれかに、未成熟樹状細胞を所定の抗原と接触させることによって形成され得る。あるいは、(例えば、単離前にインビボで)抗原と既に接触した未成熟樹状細胞を、BCGおよびIFNγを含む組成物と接触させて、1型(Th−1)応答がプライミングされた成熟樹状細胞を形成し得る。
【0043】
適切な所定の抗原は、T細胞活性化が所望される任意の抗原を含み得る。このような抗原は、例えば、細菌抗原、腫瘍特異性抗原または腫瘍関連抗原(例えば、細胞全体、腫瘍細胞溶解産物、腫瘍由来の単離抗原、融合タンパク質、リポソームなど)、ウイルス抗原、および任意の他の抗原または抗原(例えば、ペプチド抗原またはポリペプチド抗原)のフラグメントを包含し得る。特定の実施形態では、この抗原は、例えば、前立腺特異性膜抗原(PSMA)、前立腺酸性ホスファターゼ(PAP)、または前立腺特異性抗原(PSA)であり得るがこれらに限定されない(例えば、Pepsideroら,Cancer Res.40:2428−32(1980);McCormackら,Urology 45:729−44(1995)を参照のこと)。この抗原はまた、細菌細胞、細菌溶解産物、細胞溶解産物由来の膜フラグメント、または当該分野で公知の任意の他の供給源であり得る。この抗原は、組換えによって発現もしくは産生されてもよく、または化学的に合成されてすらよい。組換え抗原はまた、宿主細胞(例えば、細菌細胞、酵母細胞、昆虫細胞、脊椎動物細胞または哺乳動物細胞)の表面に発現されてもよく、溶解産物中に存在してもよく、またはこの溶解産物から精製されてもよい。
【0044】
抗原はまた、被験体由来のサンプル中に存在し得る。例えば、被験体における過剰増殖状態または他の状態に由来する組織サンプルは、抗原供給源として用いられ得る。このようなサンプルは、例えば、生検または手術切開によって入手され得る。このような抗原は、溶解産物として、または単離された調製物として用いられ得る。あるいは、被験体(例えば、癌患者)の細胞または確立された細胞株の膜調製物はまた、抗原または抗原供給源として用いられ得る。
【0045】
例示的な実施形態では、手術標本から収集された前立腺腫瘍細胞の溶解産物は、抗原供給源として用いられ得る。例えば、生検または手術切開によって得られた、癌患者自体の腫瘍のサンプルは、抗原を樹状細胞に提示するため、または抗原提示用細胞溶解産物を提供するために、直接的に用いられ得る。あるいは、癌患者の腫瘍細胞の膜調製物が用いられ得る。この腫瘍細胞は、前立腺の腫瘍細胞、肺の腫瘍細胞、卵巣の腫瘍細胞、結腸の腫瘍細胞、脳の腫瘍細胞、黒色腫、または任意の他の種類の腫瘍細胞であり得る。溶解産物および膜調製物は、当該分野で公知の方法によって、単離された腫瘍細胞から調製され得る。
【0046】
別の例示的実施形態では、精製または半精製された前立腺特異的膜抗原(PSMA、PSM抗原としても公知)は、モノクローナル抗体7E11−C.5と特異的に反応し、抗原として用いられ得る(一般的に、Horoszewiczら,Prog.Clin.Biol.Res.37:115−32(1983)、米国特許第5,162,504号;米国特許第5,788,963号;Fengら,Proc.Am.Assoc.Cancer Res.32:(Abs.1418)238(1991)を参照のこと;これらの開示は、本明細書中に参考として援用される)。なお別の例示的実施形態では、アミノ酸残基配列Leu Leu His Glu Thr Asp Ser Ala Val(配列番号1)を有する抗原性ペプチド(PSM−P1と称される)は、PSMAのアミノ酸残基4〜12に対応し、抗原として用いられ得る。あるいは、アミノ酸残基配列Ala
Leu Phe Asp Ile Glu Ser Lys Val(配列番号2)を有する抗原性ペプチド(PSM−P2と称される)は、PSMAのアミノ酸残基711〜719に対応し、抗原として用いられ得る。
【0047】
特定の実施形態では、アミノ酸残基配列Xaa Leu(またはMet)Xaa Xaa Xaa Xaa Xaa Xaa Val(またはLeu)(PSM−PXと称される)を有する抗原性ペプチド(ここで、Xaaは任意のアミノ酸残基を表す)は、抗原として用いられ得る。このペプチドは、HLA−A0201結合モチーフ、すなわち、HLA−A2患者において見出される「アンカー残基」ロイシンおよびバリンを有する9〜10アミノ酸残基の結合モチーフに似る(例えば、Greyら,Cancer Surveys 22:37−49(1995)を参照のこと)。このペプチドは、HLA−A2+患者についての抗原として用いられ得る(Central Data Analysis
Committee「Allele Frequencies」,第6.3節,Tsuji,K.ら(編),Tokyo University Press,1066−1077頁を参照のこと)。同様に、他のHLA結合モチーフに似るペプチドが用いられ得る。
【0048】
代表的に、未成熟樹状細胞は、上記のように、適切な成熟条件下で、BCG、IFNγおよび所定の抗原の存在下で培養される。必要に応じて、この未成熟樹状細胞は、GM−CSFおよびIL−4または成熟因子を含まない代表的樹状細胞培養培地中で、所定の抗原と混合され得る。この抗原との少なくとも約10分間〜2日間の培養後、この抗原は除去され得、そしてBCGおよびIFNγが補充された培養培地が添加され得る。サイトカイン(例えば、GM−CSFおよびIL−4)はまた、成熟培地に添加され得る。樹状細胞を接触させるための方法は、当該分野で一般に公知である(一般的に、SteelおよびNutman,J.Immunol.160:351−60(1998);Taoら,J.Immunol.158:4237−44(1997);DozmorovおよびMiller,Cell Immunol 178:187−96(1997);Inabaら,J Exp Med.166:182−94(1987);Macatoniaら,J Exp Med.169:1255−64(1989);De Bruijnら,Eur.J.Immunol.22:3013−20(1992)を参照のこと;これらの開示は、本明細書中に参考として援用される)。
【0049】
次いで、得られる成熟しプライミングされた樹状細胞は、T細胞(例えば、ナイーブT細胞)と共存インキュベートされる。T細胞またはT細胞サブセットは、レスポンダー細胞として使用するために種々のリンパ組織から入手され得る。このような組織としては、脾臓、リンパ節、および/または末梢血が挙げられるがこれらに限定されない。これらの細胞は、成熟しプライミングされた樹状細胞とともに、混合T細胞集団として、または精製されたT細胞サブセットとして共存培養され得る。T細胞精製は、CD2に対する抗体、CD3に対する抗体、CD4に対する抗体、CD8に対する抗体などの使用を含むがこれらに限定されない、ポジティブ選択またはネガティブ選択によって達成され得る。
【0050】
T細胞を、成熟しプライミングされた樹状細胞と接触させることによって、抗原反応性か活性化された、極性化T細胞または極性化Tリンパ球が提供される。本明細書中で用いられる場合、用語「極性化(polarized)」とは、高レベルのIFNγを産生するか、さもなければ1型(Th−1)応答を誘導するようにプライミングされているT細胞をいう。このような方法は、代表的に、未成熟樹状細胞をBCGおよびIFNγと接触させて、成熟しプライミングされた樹状細胞を調製する工程を包含する。この未成熟樹状細胞を、成熟の間または成熟前に所定の抗原と接触させ得る。この未成熟樹状細胞は、成熟の間、T細胞(例えば、ナイーブT細胞)とともに共存培養されてもよく、または、成熟および1型応答を誘導するための樹状細胞のプライミングの後にT細胞(例えば、ナイーブT細胞)と共存培養されてもよい。この未成熟樹状細胞または成熟樹状細胞は、成熟前に富化され得る。さらに、T細胞は、樹状細胞との接触前にリンパ球集団から富化され得る。特定の実施形態では、富化または精製されたCD4+T細胞集団を、この樹状細胞と接触させる。成熟し、プライミングされた樹状細胞とT細胞との共存培養は、特異的T細胞の刺激をもたらし、この特異的T細胞は、成熟して、抗原反応性CD4+T細胞または抗原反応性CD8+T細胞になる。
【0051】
別の局面では、T細胞を、1型(Th−1)T細胞応答を誘導することに向けてプライミングされた成熟樹状細胞の存在下で培養することによる、インビトロでのT細胞の再刺激のための方法が提供される。このようなT細胞は、フィーダー細胞上で必要に応じて培養され得る。成熟しプライミングされた樹状細胞は、T細胞との接触前に必要に応じて照射され得る。適切な培養条件は、1以上のサイトカイン(例えば、精製されたIL−2、コンカナバリンA刺激脾臓細胞上清、またはインターロイキン15(IL−15))を含み得る。未成熟樹状細胞、BCG、IFNγおよび所定の抗原の添加による、インビトロでのT細胞再刺激は、T細胞集団の増大を促進するために用いられ得る。
【0052】
安定な抗原特異的、極性化T細胞培養物またはT細胞株は、周期的再刺激によってインビトロで長期間にわたり維持され得る。このようにして作製されたT細胞培養物またはT細胞株は、保存され得、そして(例えば、凍結保存剤を用いた処方および凍結によって)貯蔵される場合、活性化された極性化T細胞を長期にわたって所望の間隔で再供給するために用いられ得る。
【0053】
特定の実施形態では、活性化されたCD8+T細胞またはCD4+T細胞は、本発明の方法に従って作製され得る。代表的に、抗原反応性の極性化T細胞を作製するために用いられる、成熟しプライミングされた樹状細胞は、投与されるべき被験体に対して同質遺伝子的である(例えば、この被験体から入手される)。あるいは、意図されるレシピエント被験体と同じHLAハプロタイプを有する樹状細胞は、HLAが一致したドナー由来の非癌性細胞(例えば、正常細胞)を用いてインビトロで調製され得る。特定の実施形態では、抗原反応性T細胞(CTL細胞およびTh−1細胞を含む)は、免疫刺激のための細胞供給源としてインビトロで増大される。
【0054】
(細胞集団のインビボでの投与)
本発明の別の局面では、成熟しプライミングされた樹状細胞もしくは活性化され極性化したT細胞、またはこのような細胞を含む細胞集団を、免疫刺激の必要がある被験体に投与するための方法が提供される。このような細胞集団は、成熟しプライミングされた樹状細胞集団および/または活性化され極性化したT細胞集団の両方を包含し得る。特定の実施形態では、このような方法は、樹状細胞前駆体または未成熟樹状細胞を入手し、これらの細胞を、BCG、IFNγおよび所定の抗原の存在下で分化および成熟させて、Th−1応答に対してプライミングされた成熟樹状細胞集団を形成させることによって実施される。この未成熟樹状細胞を、成熟前または成熟の間に抗原と接触させ得る。このような成熟しプライミングされた樹状細胞は、免疫刺激の必要がある被験体に直接的に投与され得る。
【0055】
関連した実施形態では、この成熟しプライミングされた樹状細胞を、被験体由来のリンパ球と接触させて、このリンパ球集団内のT細胞を刺激し得る。この活性化され極性化されたリンパ球は、必要に応じて続いて抗原反応性のCD4+T細胞および/またはCD8+T細胞の細胞培養物中でクローン増大が行われ、免疫刺激の必要がある被験体に投与され得る。特定の実施形態では、活性化され極性化されたT細胞は、この被験体に対して自己である。
【0056】
別の実施形態では、この樹状細胞、T細胞、およびレシピエント被験体は、同じMHC(HLA)ハプロタイプを有する。被験体のHLAハプロタイプを決定する方法は、当該分野で公知である。関連の実施形態では、この樹状細胞および/またはT細胞は、このレシピエント被験体に対して同種異系である。例えば、この樹状細胞は、このT細胞およびこのレシピエントに対して同種異系であり得、これは、同じMHC(HLA)ハプロタイプを有する。この同種異系細胞は代表的に、少なくとも1つのMHC対立遺伝子が一致している(例えば、少なくとも1つだが全てではないMHC対立遺伝子を共有する)。それほど代表的でない実施形態では、この樹状細胞、T細胞およびレシピエント被験体は全て、互いに対して同種異系であるが、全て、少なくとも1つの共通のMHC対立遺伝子を共通に有する。
【0057】
1つの実施形態によれば、このT細胞は、この未成熟樹状細胞を得たのと同じ被験体から得られる。インビトロでの成熟および極性化の後、この自己T細胞は、既存の免疫応答を誘発および/または増強するためにこの被験体に投与される。例えば、T細胞は、(例えば、体表面積1m2あたり約108〜109細胞の用量での)静脈内注入によって投与され得る(例えば、本明細書中に参考として援用される、Ridellら,Science 257:238−41(1992)を参照のこと)。注入は、所望の間隔で(例えば、毎月)繰り返され得る。レシピエントは、有害な影響の何らかの証拠について、T細胞注入の間およびその後、モニタリングされ得る。
【0058】
別の実施形態によれば、本発明に従ってBCGおよびIFNγを用いて成熟させた樹状細胞は、腫瘍または標的抗原を含む他の組織中に直接的に注射され得る。このような成熟細胞は、抗原を取り込んでこの抗原をT細胞に対してインビボで提示し得る。
【実施例】
【0059】
以下の実施例を、本発明の種々の局面の単に例示として提供し、そして本発明を如何様にも限定すると解釈すべきでない。
【0060】
(実施例1:異なる成熟条件下でのIL−10およびIL−12の産生:)
本実施例では、サイトカイン産生を、成熟因子BCGおよび/またはIFNγと接触させた未成熟樹状細胞集団から決定した。未成熟DCを、1%ヒト血漿を補充したOptiMEM(登録商標)培地(Gibco−BRL)の存在下で末梢血単球をプラスチックと接触させることによって調製した。未結合の単球を、洗浄によって除去した。結合した単球を、1ミリリットルあたり500U GM−CSFおよび500U IL−4の存在下で6日間にわたってX−VIVO 15(登録商標)培地中で培養した。
【0061】
第1研究では、未成熟樹状細胞を、不活化BCGの添加によって成熟させた。得られる成熟樹状細胞のサイトカイン産生を決定した。不活化BCGを、種々の濃度で、X−VIVO 15(登録商標)培地中の未成熟樹状細胞に添加し、続いて37℃にて24時間培養した。4.1×108cfu/mlストックから開始した、1ミリリットルあたりで添加したBCGの希釈を、表に指定する。サイトカイン産生を、検出されるべきサイトカインに対する抗体を用いたELISAアッセイによって決定した。手短に述べると、サイトカイン(例えば、IL−12またはIL−10)に対して特異的な抗体を用いて、固体表面のサイトカインを捕捉する。次いで、この固体表面を、このサイトカインに対する第2の標識抗体で処理して、捕捉されたサイトカインの存在を検出する。第2の抗体は、代表的に、酵素で標識されて、比色アッセイによる検出を容易にする。代表的な実験の結果を、以下の表1に下記に示す。サイトカインの量、すなわちサイトカイン産生を、pg/mlとして示す。
【0062】
【表1】
表1を言及して、この結果は、BCGの添加が、サイトカインIL−12、IL−10、またはそれらのサブユニットの産生を増大させ得るが、サイトカイン産生の相対レベルおよび絶対レベルの両方がドナー依存性であることを実証する。最大レベルの増加が、IL−12 p40およびIL−10について見られ、このことは、IL−12 p40のレベルと比較して低レベルの、観察されたIL−12 p70(p35サブユニットおよびp40サブユニットから構成される)が、IL−12 p35産生の欠除に起因することを示唆した。BCGが存在する1つの条件を除く全ての条件において、IL−12 p70レベル:IL−10レベルの比は、1以下である。このことは、BCG単独の存在下での未成熟樹状細胞の成熟が、ナイーブT細胞をTh−2 応答に向けて極性化させるようであることを示す。
【0063】
同様の条件下でIFNγを導入することの効果もまた決定した。これを、以下の表2に提示する。サイトカイン産生を上記の通りに測定した。表1と表2とを比較して、成熟因子(例えば、BCG)の存在下でのIFNγの添加が、IL−12 p70の産生を増大させたことが明らかである。特に、成熟の間でのBCGを伴うIFNγの添加は、IL−12 p35産生を増大させ、そしてIL−10産生を減少させた。その結果、IL−12 p70:IL−10の比は、BCGとともにIFNγを添加すると、いつも1より大きかった。何人かのドナーでは、および特定の条件下では、IL−12 p70産生:IL−10産生の比は、BCGを伴うIFNγの添加によって100:1より大きく増大し得る。従って、これらの結果は、驚くことに、成熟因子BCGを伴うIFNγの添加が、IL−12 p70産生を劇的に増大させ得ることを実証する。
【0064】
【表2】
(実施例2:IFNγによるIL−10のダウンレギュレーションは、用量依存性である:)
本実施例では、BCGと組み合わせたIFNγが、樹状細胞集団におけるIL−10産生をダウンレギュレートする能力を実証する。未成熟樹状細胞を、上記の通りに調製した。これらの未成熟樹状細胞を単独でインキュベートし、2つのうちの1つの濃度のBCG(1:1000希釈または1:250希釈の4.1×108cfu/mlストック)の存在下で、または1ミリリットルあたり0U〜1000Uの範囲の濃度のIFNγ単独に曝露して、成熟させた。得られる樹状細胞によるIL−10産生を、市販の抗体(例えば、R & D Systems,Minneapolis,MN製)を用いたELISA(前出)によって測定し、そしてpg/mlで報告した。コントロールでは、単独で培養した未成熟DC(BCGもIFNγも添加しない)は、検出可能なIL−10を産生しなかった。対照的に、IFNγ単独の存在下で培養したDCは、少量のIL−10(約20〜30pg/ml)を産生した。産生されたIL−10の量は、1ミリリットルあたり10U〜1000UのIFNγの範囲にわたって用量依存性ではなかった。
【0065】
対照的に、BCG単独の存在下での成熟によって産生されたDCは、顕著な量のIL−10を産生した:1:1000または1:250倍希釈のBCGストックの存在下、それぞれ、約150pg/mlまたは250pg/mlを超えるIL−10。DC成熟の間のBCGへのIFNγの添加は、IL−10産生のダウンレギュレーションを用量依存性様式でもたらした。1:1000希釈のBCGの存在下で培養したDCについては、IL−10産生は、約150pg/mlのIL−10(IFNγなし)から約20〜30pg/mlのIL−10(1000U IFNγ)へと減少した。1:250希釈のBCGの存在下で培養したDCについては、IL−10産生は、約270pg/mlのIL−10(IFNγなし)から約50pg/mlのIL−10(1000U IFNγ)へと減少した。従って、BCGおよびIFNγの存在下での未成熟DCの成熟は、IL−10産生をダウンレギュレーションし得、そしてBCG単独によって誘導されるIL−10産生の見かけの刺激を克服し得た。
【0066】
(実施例3:IFNγによるIL−12のアップレギュレーションは、用量依存性である:)
本実施例では、IFNγがIL−12産生をアップレギュレートする能力が実証された。未成熟樹状細胞を、上記のように、GM−CSFおよびIL−4の存在下で増殖させた単球の6日間培養物から誘導した。これらの未成熟DCを、種々の希釈のBCG単独を用いて、または種々の濃度のIFNγと組み合わせたBCGを用いてさらに2日間処理した。培養上清を、IL−12 p70の存在について、上記の通りに、ELISAアッセイによって試験した。
【0067】
代表的実験による結果を図1に示す。各培養物について、この結果を3連で決定した。増加する量のBCG単独に応答して、比較的低い(<1000pg/mlの)平均濃度のIL−12 p70がこれらの成熟DCによって産生された。IL−12産生の量は、BCGの量が増加するにつれて用量依存様式で減少した。対照的に、BCG(ストックの1:1000希釈物)を有するIFNγ(10U/ml)の添加によって、IL−12の量は、約5000pg/mlまで劇的に増加した。BCG(ストックの1:1000希釈物)を有するIFNγ(100U/ml)の添加によって、IFNγの量は、ほぼ20,000pg/mlまで増加した。BCGを有する500U/mlまたは1000U/mlのIFNγ(ストックの1:1000希釈物)の添加は、それぞれ、約21,000pg/mlおよび約22,000pg/mlのIL−12レベルをもたらす。
【0068】
まとめると、BCGは単独では、IL−12産生を拮抗するようであるが、BCGおよびIFNγの存在下での未成熟DCの成熟は、IL−12産生を劇的に増加させた。
【0069】
(実施例4:抗原特異的T細胞の刺激:)
本実施例では、BCGおよびIFNγの存在下で成熟させた未成熟DCは、抗原特異的T細胞によるIFNγ産生を刺激することが示された。インフルエンザAに特異的なT細胞株を、2×106細胞/mlの末梢血単核細胞(PBMC)を5%ヒト血清を補充したAIM−V(登録商標)培地中で5μg/mlのインフルエンザM1ペプチド(GILGFVFTL;配列番号3)とともにインキュベートすることによって作製した。これらの培養条件は、インフルエンザM1ペプチドに特異的なT細胞の選択的増大をもたらした。2日間の培養後、20U/ml IL−2および5ng/ml IL−15をこれらの培養物に添加した。約7〜14日間の培養後、これらのT細胞株を、サイトカインを含まない培地中に一晩配置した。
【0070】
次いで、抗原特異的T細胞を、未成熟DCとともに、BCG単独を用いて成熟させたDCとともに、またはBCGおよびIFNγを用いて成熟させたDCとともに、共存培養した。抗原特異的T細胞:DCの比は、1:1であった。T細胞およびDCを、37℃にて24時間インキュベートした。これらのDCは、インフルエンザM1ペプチドが直接的にローディングされるかまたは浸透圧的にローディングされるかのいずれかであった。手短に述べると、未成熟DCを培養フラスコから収集し、そして遠心分離によって濃縮した。浸透圧ローディングについては、これらの細胞を、小容量の高浸透圧培地中に再懸濁し、続いてPBS中の等容量のインフルエンザM1ペプチドを添加した。氷上での10分間のインキュベーションの後、これらの細胞を徹底的に洗浄した。直接的ローディングについては、これらの細胞を、等容量のX−VIVO 15(登録商標)培地およびPBS中のインフルエンザM1ペプチド中に再懸濁し、37℃で1時間にわたってインキュベートした。これらの細胞を、37℃にて2時間にわたってインキュベートして、抗原プロセシングを可能にした。
【0071】
T細胞およびDCの共存培養後、T細胞応答を、培養上清の100μlサンプルからのIFNγのELISA定量によって測定した。この結果は、CDローディングの方法にかかわらず、BCGおよびIFNγで刺激したDCが、抗原特異的T細胞の優れた刺激因子であることを示す。未成熟DCとともに共存培養したT細胞は、極めてわずかなIFNγ(<2,000pg/ml)を産生し、一方、BCG単独を用いて成熟させたDCとともに共存培養したT細胞は、IFNγの中程度の生産者であった(>5,000pg/ml)。BCGおよびIFNγを用いて成熟させたDCとともに共存培養したT細胞は、高レベルのIFNγ(浸透圧によってローディングしたDCについて>20,000pg/ml、または直接的にローディングしたDCについて>25,000pg/ml)を産生した。
【0072】
従って、BCGおよびIFNγを用いて成熟させたDCは、DCに抗原をローディングする方法とは独立して、抗原特異的T細胞のより良好な刺激因子であった。
【0073】
(実施例5:インビトロでの抗原特異的T細胞応答のデノボ作製:)
キーホールリンペットヘモシアニン(KLH)に特異的なT細胞株を、BCG、またはBCGおよびIFNγのいずれかを用いて成熟させたDCを用いて、PBMCを刺激することによって作製し、KLHまたはコントロールタンパク質を、10:1のT細胞:DC比でローディングした。T細胞および成熟させたDCに、新鮮な培地(5%ヒトAB血清、20U/ml IL−2、および5ng/ml IL−15を補充したAIM−V(登録商標)培地)を3〜4日毎に提供した。これらの細胞を、必要に応じて、より大きなフラスコへと増大させた。特定の抗原に対する前駆体頻度全体が低く、ナイーブ細胞は応答するために強力な刺激を必要とするので、刺激を、10〜21日間の間隔で3〜4回繰り返した。細胞をサイトカインを含まない培地中で一晩回復させた後に再刺激した。
【0074】
標準的な3日間のチミジン取り込みアッセイを用いて、刺激T細胞株をKLH特異的細胞増殖について試験した。刺激されたT細胞を、KLHパルス未成熟樹状細胞を用いて、種々のDC:T細胞比でインキュベートした。T細胞増殖を、1分間あたりのカウント(CPM)として測定した。刺激に応答した、細胞増殖またはサイトカインの産生を、抗原特異的応答の証拠と解釈した。抗原特異性について制御するために、ネガティブコントロールの抗原(インフルエンザAウイルス)もまたこのアッセイに含めた。
【0075】
BCG単独によって成熟させたDCを用いてT細胞を刺激する場合、T細胞増殖は一貫して低かった(<5,000CPM)。この低レベルの増殖は、DCをKLHと接触させようがインフルエンザAウイルスと接触させようが、低かった。低レベルの増殖もまた、未成熟DCを用いたインキュベーションに応答して観察された。50:1、25:1、または12.5:1の応答者:刺激者(T細胞:DC)の比で用いた3つの群のDCの間に顕著な差は観察されなかった。対照的に、BCGおよびIFNγを用いて成熟させた樹状細胞を用いてT細胞を刺激し、KLHパルスDCは、インフルエンザAでパルスした未成熟DCまたは成熟DCが誘導するよりも一貫してさらに多いT細胞増殖を誘導した(約10,000〜33,000CPM)。KLH抗原でパルスした成熟DCについては、T細胞増殖は、応答者:刺激者の比の増大に比例して増加した。
【0076】
T細胞エフェクター機能もまた、サイトカイン分泌によってモニタリングされた。KLH特異的T細胞株(上記の通りに作製された)を、BCG単独またはBCGおよびIFNγのいずれかを用いて成熟させたDCを用いて刺激した。刺激されたT細胞株を抗CD3抗体(50ng/ml)およびPMA(5ng/ml)を用いて非特異的に刺激した後、これらの細胞を、細胞内サイトカイン染色によってサイトカイン産生について試験した。サイトカイン産生を、特定のサイトカインを産生する細胞の百分率として測定した。極めて低い百分率から検出不可能な百分率(<<5%)のサンプリング細胞は、細胞内IL−2、IL−4、IL−5、またはIL−10を産生した。IL−5およびIL−10は、BCGおよびIFNγを用いて成熟させたDCによって刺激されたT細胞において検出されなかった。BCG単独によって成熟させたDCによって刺激されたT細胞は、低レベルのIFNγ(<10%)およびTNF−α(<15%)を産生した。対照的に、IFNγおよびBCGを用いて成熟させたDCを用いて刺激された有意な比率のT細胞は、IFNγ(約35%)およびTNFα(>45%)を産生した。IFNγは、IL−12産生の公知の刺激因子である。従って、BCGおよびIFNγを用いて成熟させたDCを用いてT細胞を刺激することによって、これらのT細胞は、1型(Th−1)応答に向けて極性化される。
【0077】
(実施例6:Th−1サイトカイン腫瘍壊死因子α(TNFα)の誘導:)
本実施例では、BCGおよびIFNγの組み合わせが1型サイトカイン腫瘍壊死因子α(TNFα)をアップレギュレートする能力が実証された。手短に述べると、未成熟樹状細胞を上記の通りに誘導し、そしてGM−CSFおよびIL−4の存在下で増殖させた(prown)。これらの未成熟DCを、約24時間にわたってBCG単独またはIFNγとの組み合わせのいずれかとともに培養した。続いて、タンパク質輸送インヒビター(GolgiPlugTM,PharMingen)を添加して、ゴルジ複合体から産生されたサイトカインの輸送をブロックし、そしてこれらの細胞を一晩インキュベートした。次いで、これらの細胞を、当該分野で周知の方法を用いて、収集し、透過性にし、そしてTNFαについて特異的な蛍光標識抗体またはアイソタイプコントロール抗体を用いて内部染色した。TNFαについてDCポジティブの頻度およびこれらの細胞の蛍光強度を、FACS分析によって決定した(表3)。IFNγの存在下でBCGを用いてDCの成熟は、DCがTh−1サイトカインTNFαを産生する能力を増強することが見出された。
【0078】
【表3】
(実施例7:細胞関連抗原に対する応答の誘導:)
本実施例では、BCGおよびIFNγの存在下で成熟させたDCは、BCG単独を用いて成熟させたDCと比較して、有意に高い腫瘍特異性T細胞INFγ放出および類似のレベルの抗原特異的細胞傷害性を惹起することが実証された。未成熟樹状細胞を上記の通りに単離し、そしてGM−CSFおよびIL−4の存在下で培養した。次いで、これらのDCに、緑色蛍光タンパク質(GFP)またはインフルエンザAウイルスのM1タンパク質のいずれかを発現する組換えアデノウイルスで予め感染させた腫瘍細胞(A549)全体のいずれかをローディングした。これらのDCを、BCGまたはIFNγと組み合わせたBCGのいずれかを用いて24時間後に成熟させた。腫瘍をローディングしたDCもしくはGFPまたはM1発現腫瘍細胞を用いて、自己M1特異的T細胞株を刺激した。24時間後、細胞培養上清を収集し、そして標準的IFNγ ELISAを行った。M1発現腫瘍細胞をローディングしたDCのみがIFNγ放出を刺激し得、そしてBCG+IFNγにおいて成熟させたDCは、この応答を誘導する際に、未成熟DCまたはBCG成熟DCのいずれかよりも有意により強力であった。
【0079】
【表4】
上記の実施例は、本願発明の範囲を例示するために提供されるが、本願発明の範囲を限定しない。本発明の他の改変体は、当業者に容易に明らかであり、添付の特許請求の範囲によって包含される。本明細書中に引用される全ての刊行物、特許、特許出願および他の参考文献は、それらの全体が、本明細書中に参考として援用される。
(配列表)
【特許請求の範囲】
【請求項1】
明細書に記載の発明。
【請求項1】
明細書に記載の発明。
【図1】

【公開番号】特開2011−188872(P2011−188872A)
【公開日】平成23年9月29日(2011.9.29)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−148711(P2011−148711)
【出願日】平成23年7月4日(2011.7.4)
【分割の表示】特願2003−526345(P2003−526345)の分割
【原出願日】平成14年9月6日(2002.9.6)
【出願人】(500513619)ノースウエスト バイオセラピューティクス,インコーポレイティド (9)
【Fターム(参考)】
【公開日】平成23年9月29日(2011.9.29)
【国際特許分類】
【出願番号】特願2011−148711(P2011−148711)
【出願日】平成23年7月4日(2011.7.4)
【分割の表示】特願2003−526345(P2003−526345)の分割
【原出願日】平成14年9月6日(2002.9.6)
【出願人】(500513619)ノースウエスト バイオセラピューティクス,インコーポレイティド (9)
【Fターム(参考)】
[ Back to top ]