
単球由来多能性細胞MOMC
【課題】
必要十分量を低侵襲的かつ簡便に安定供給でき、細胞移植に際して拒絶反応の問題がなく、骨、軟骨、骨格筋、脂肪などの間葉系細胞、血管内皮細胞、心筋細胞、神経細胞など多種類の細胞へと分化する能力を有する多能性細胞や、かかる多能性細胞から分化誘導された間葉系細胞、心筋細胞、血管内皮細胞、神経細胞や、これらを有効成分とする治療剤・治療方法を提供すること。
【解決手段】
末梢血単核細胞(PBMC)をフィブロネクチンでコートしたプラスチックプレート上で7〜10日間培養し、出現する線維芽細胞様形態を呈する細胞群は循環CD14+単球由来で、CD14+CD45+CD34+I型コラーゲン+という独特のフェノタイプを有し、特定の培養条件により骨、軟骨、骨格筋、脂肪などの間葉系細胞、血管内皮細胞、心筋細胞、神経細胞へと分化する能力を有する。
必要十分量を低侵襲的かつ簡便に安定供給でき、細胞移植に際して拒絶反応の問題がなく、骨、軟骨、骨格筋、脂肪などの間葉系細胞、血管内皮細胞、心筋細胞、神経細胞など多種類の細胞へと分化する能力を有する多能性細胞や、かかる多能性細胞から分化誘導された間葉系細胞、心筋細胞、血管内皮細胞、神経細胞や、これらを有効成分とする治療剤・治療方法を提供すること。
【解決手段】
末梢血単核細胞(PBMC)をフィブロネクチンでコートしたプラスチックプレート上で7〜10日間培養し、出現する線維芽細胞様形態を呈する細胞群は循環CD14+単球由来で、CD14+CD45+CD34+I型コラーゲン+という独特のフェノタイプを有し、特定の培養条件により骨、軟骨、骨格筋、脂肪などの間葉系細胞、血管内皮細胞、心筋細胞、神経細胞へと分化する能力を有する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、間葉系細胞、心筋細胞、血管内皮細胞等の中胚葉系細胞や神経細胞に分化しうるCD14+及びCD34+の単球由来多能性細胞や、該単球由来多能性細胞から分化誘導された間葉系細胞、心筋細胞、血管内皮細胞等の中胚葉系細胞・組織と神経細胞・組織や、これらを有効成分とする治療剤や、これらを投与する治療方法に関する。
【背景技術】
【0002】
末梢血単球は骨髄造血幹細胞に由来し、マクロファージ、樹状細胞、破骨細胞、中枢神経系の小グリア細胞、肝臓のクッパー細胞等の数種の食細胞に分化することが知られている(例えば、非特許文献1〜4参照。)。単球の各種食細胞への分化は、種々の成長因子からのシグナリングによって調節されている。すなわち、マクロファージへの分化はM−CSF、樹状細胞への分化はGM−CSF及びIL−4、破骨細胞への分化はNFリガンドの受容体活性因子(RANKL)やM−CSF(例えば、非特許文献2、3、5参照。)からのシグナリングによってそれぞれ調節されている。つい最近まで単球の分化能は、組織食細胞に限定的であると考えられてきた。しかし近年の研究成果から、種々の血管由来因子を組合わせた条件下でのインビトロ培養を行うことにより、ヒト単球が血管内皮様細胞へと分化する能力を示すことが明らかになった(例えば、非特許文献6、7参照。)。さらに、肉芽腫のインビトロモデルにおける単球分化プロセスにおいて骨特異的アルカリフォスファターゼが発現されることが報告さている(例えば、非特許文献8参照。)。しかし、単球の分化能は、まだ完全に解明されておらず、単球が食細胞以外の細胞型への分化能を有しているかどうかはよく知られていなかった。
【0003】
他方、多くの成人組織には、自己複製能、及び不可逆的最終分化を遂げる娘細胞産生能を有する幹細胞群が存在していることが明らかになっている(例えば、非特許文献9参照)。最も特徴づけが進んでいるのが造血幹細胞とその子孫細胞であるが、幹細胞は、間葉系、神経、及び造血幹細胞等、殆どの臓器において同定されている(例えば、非特許文献10〜12参照。)。間葉系幹細胞(MSC)は、骨髄における接着線維芽様細胞として同定されており、骨、軟骨、脂肪、筋肉、及び骨髄間質等の間葉組織系統への分化能を有している(例えば、非特許文献10参照。)。MSCと似た形態及びフェノタイプ上の特徴並びに分化能を有する間葉系前駆細胞が、臍帯血(例えば、非特許文献13参照。)、及び胎児(例えば、非特許文献14参照。)や成人(例えば、非特許文献15参照。)の末梢血において、著しく低頻度ではあるが存在していることが近年報告されている。しかし、MSC及び循環MSC様細胞においては、各種造血幹マーカー及び幹細胞/血管内皮マーカーCD34が発現されることはない(例えば、非特許文献10、13〜15参照。)。
【0004】
上記のように、組織再生における将来的な治療的介入に対する供給源候補として、種々の出生後の組織特異的幹細胞及び胚幹(ES)細胞の分析が現在進められている(例えば、非特許文献9参照。)。骨形成不全症のヒト小児の場合(例えば、非特許文献16参照。)と同様に動物モデルへ骨髄由来MSCを移植すると、かかるMSCが多くの臓器に移入し、組織特異的系統に従って分化することが報告されている(例えば、非特許文献17、18参照。)。しかしMSCは、ヒト成人骨髄では、稀少細胞群であり(0.01%〜0.001%)、再生治療に十分数量のMSCを増殖させることは、技術的に困難であり、高価であり、また長時間を要するものであった(例えば、非特許文献19参照。)。ES細胞は胚芽細胞由来の多分化能細胞であり、インビトロで未分化のまま無限増殖することができ、インビボで殆どの細胞型へと分化誘導される(例えば、非特許文献20参照。)。ES細胞はヒトから単離されたものだが、研究や治療に用いる際には倫理上の問題という足枷がある(例えば、非特許文献21参照。)。
【0005】
出生後のヒト末梢血における血管内皮細胞(例えば、非特許文献22参照。)、平滑筋細胞(例えば、非特許文献23参照。)、及び間葉系細胞(例えば、非特許文献15参照。)等の血管内皮あるいは間葉系細胞型への分化能をもつ前駆細胞が数種類、報告されている。内皮前駆細胞及び平滑筋前駆細胞をインビトロで増殖させるためには、成長因子をいくつか組み合わせて用いることが必要である(例えば、非特許文献22、23参照。)。間葉系前駆細胞は、外因性成長因子不在下における20%ウシ胎児血清(FBS)添加培地での増殖が可能であるが、PBMC培養における間葉系前駆細胞の発生は、CD14+細胞の除去によって影響されないことが報告されている(例えば、非特許文献15参照。)。しかし、これら血管内皮あるいは間葉系前駆細胞は、CD14、CD45、CD34及びI型コラーゲンに対して陽性というフェノタイプ上の特徴を有していない。
【0006】
【非特許文献1】Bioessays, 17, 977-986, 1995
【非特許文献2】Blood, 98, 2544-5254, 2001
【非特許文献3】BMC Immunol., 3, 15, 2002
【非特許文献4】Microsc Res Tech., 39, 350-364, 1997
【非特許文献5】J Exp Med., 190, 1741-1754, 1999
【非特許文献6】Differentiation, 65, 287-300, 2000
【非特許文献7】Cardiovascular Res., 49, 671-680, 2001
【非特許文献8】Immunobiology, 202, 68-81, 2000
【非特許文献9】Science, 287, 1442-1446, 2000
【非特許文献10】Science, 284, 143-147, 1999
【非特許文献11】Science, 287, 1433-1438, 2000
【非特許文献12】J. Hepatol., 29, 676-682, 1998
【非特許文献13】Br. J. Haematol., 109, 235-242, 2000
【非特許文献14】Blood, 98, 2396-2402, 2001
【非特許文献15】Arthritis Res., 2, 477-488, 2000
【非特許文献16】Nat. Med., 5, 309-313, 1999
【非特許文献17】Nat. Med., 6, 1282-1286, 2000
【非特許文献18】Science, 279, 1528-1530, 1998
【非特許文献19】Stem Cells, 19, 180-192, 2001
【非特許文献20】Trends Biotechnol., 18, 53-57, 2000
【非特許文献21】Science, 287, 1397, 2000
【非特許文献22】Science, 275, 964-967, 1997
【非特許文献23】Circulation, 106, 1199-1204, 2002
【発明の開示】
【発明が解決しようとする課題】
【0007】
現在の医学に残された大きな課題は疾患、外傷などによる臓器の欠失や機能障害の克服といわれている。このような状態に対する治療として現時点で施行しうる唯一の方法は臓器移植であるが、脳死判定やドナー供給の問題のため現実的な治療法として普及するには未だ多くの障害がある。一方、最近の幹細胞および発生生物学の進歩に伴って臓器再生を目指した再生医学が注目され、21世紀に進むべき医療の方向性として期待されている。動物実験レベルでは胚性幹細胞(ES細胞)の移植により臓器の機能回復が報告されているが、ヒトへの応用を考えた場合には拒絶反応やES細胞使用による倫理的な問題で行き詰まっているのが現状である。また、成人の各種組織幹細胞(間葉系、血管、肝など)も生体内にきわめて少数しか存在せず、その単離は技術的に難しく、移植に十分な細胞数を得ることは現時点で困難とされている。したがって、ES細胞や組織幹細胞を用いた再生医療が実地医療に応用されるためには解決しなければならない多くの問題が山積しており、特に、細胞移植による再生医療が現実のものとなるためには、分化能を有する細胞の安定供給が必要不可欠であると考えられる。
【0008】
本発明の課題は、必要十分量を低侵襲的かつ簡便に安定供給でき、細胞移植に際してドナー確保や拒絶反応の問題がなく、倫理的な問題も少ない、骨、軟骨、骨格筋、脂肪などの間葉系細胞、血管内皮細胞、心筋細胞、神経細胞など多種類の細胞へと分化する能力を有する多能性細胞や、かかる多能性細胞から分化誘導された間葉系細胞、心筋細胞、血管内皮細胞等の中胚葉系前駆細胞・中胚葉系細胞・中胚葉系組織と神経細胞・組織や、これらを有効成分とする治療剤や、これらを投与する治療方法を提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、末梢血単核細胞(PBMC)をフィブロネクチンでコートしたプラスチックプレート上で7〜10日間培養すると線維芽細胞様の細胞が出現することを観察した。この線維芽細胞様形態を呈するヒト細胞群の由来および生理機能に興味を持って解析したところ、これらの細胞は循環CD14+単球由来で、CD14+CD45+CD34+I型コラーゲン+という独特のフェノタイプを有し、特定の培養条件により骨、軟骨、骨格筋、脂肪などの間葉系細胞、血管内皮細胞、心筋細胞、神経細胞へと分化する能力を有することを見い出し、単球由来多能性細胞(monocyte-derived multipotent cell;MOMC)と命名した。このように、循環単球が食細胞だけでなく種々の間葉系細胞等への分化能を有する多能性前駆細胞であることを初めて明らかにした本発明者らによる知見に基づいて、本発明を完成するに至った。
【0010】
すなわち本発明は、単球に由来し、CD14とCD34とを発現する単球由来多能性細胞(請求項1)や、単球に由来し、CD14とCD34とCD45とI型コラーゲンとを発現する単球由来多能性細胞(請求項2)や、間葉系組織へと分化誘導する条件下での培養により、間葉系細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞(請求項3)や、間葉系細胞が、骨芽細胞、骨格筋芽細胞、軟骨細胞又は脂肪細胞であることを特徴とする請求項3記載の単球由来多能性細胞(請求項4)や、培養心筋細胞との共培養等の心筋へと分化誘導する条件下での培養により、心筋細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞(請求項5)や、培養神経細胞との共培養等の神経へと分化誘導する条件下での培養により、神経細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞(請求項6)や、血管内皮細胞を維持する条件下での培養等の血管内皮へと分化誘導する条件下での培養により、血管内皮細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞(請求項7)や、中胚葉系細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞(請求項8)に関する。
【0011】
また本発明は、請求項1〜8のいずれか記載の単球由来多能性細胞を間葉系組織へと分化誘導する条件下で培養することにより誘導された間葉系前駆細胞、間葉系細胞又は間葉系組織(請求項9)や、間葉系細胞が、骨芽細胞、骨格筋芽細胞、軟骨細胞又は脂肪細胞であることを特徴とする請求項9記載の間葉系前駆細胞、間葉系細胞又は間葉系組織(請求項10)や、請求項1〜8のいずれか記載の単球由来多能性細胞を培養心筋細胞との共培養等の心筋へと分化誘導する条件下での培養により誘導された心筋前駆細胞、心筋細胞又は心筋組織(請求項11)や、請求項1〜8のいずれか記載の単球由来多能性細胞を培養神経細胞との共培養等の神経へと分化誘導する条件下での培養により誘導された神経前駆細胞、神経細胞又は神経組織(請求項12)や、請求項1〜8のいずれか記載の単球由来多能性細胞を血管内皮細胞を維持する条件下での培養等の血管内皮へと分化誘導する条件下での培養により誘導された血管内皮前駆細胞、血管内皮細胞又は血管内皮組織(請求項13)や、請求項1〜8のいずれか記載の単球由来多能性細胞から分化・誘導された中胚葉系前駆細胞、中胚葉系細胞又は中胚葉系組織(請求項14)に関する。
【0012】
さらに本発明は、請求項1〜8のいずれか記載の単球由来多能性細胞、並びに/又は、前記単球由来多能性細胞から分化・誘導された中胚葉系前駆細胞、中胚葉系細胞及び/若しくは中胚葉系組織を有効成分とする治療剤(請求項15)や請求項1〜8のいずれか記載の単球由来多能性細胞、並びに/又は、前記単球由来多能性細胞から分化・誘導された神経前駆細胞、神経細胞及び/若しくは神経組織を有効成分とする治療剤(請求項16)に関する。
【発明の効果】
【0013】
本発明によると、細胞治療や再生医療に極めて有用な、骨、軟骨、骨格筋、脂肪などの間葉系細胞、血管内皮細胞、心筋細胞、神経細胞など多種類の細胞へと分化する能力を有する単球由来の多能性細胞MOMCを得ることができる。そして、単球は末梢血から簡単に大きな侵襲なく採取することができる上、単球は末梢血単核球の約20%を占め、必要十分量の細胞を安定供給できる。単球からMOMCへの分化誘導は簡単、迅速、安価に行え、特殊な装置を必要としない。また、細胞移植に自己の細胞を用いることができることから、ドナー確保や拒絶反応の問題がなく、倫理的な問題も少ない。本発明は、単球/食細胞系に関する伝統的な生物学的知見を覆すものであり、単球の分化能並びに単球の生体恒常性維持及び病態誘導に果たす役割を理解する上で、大きく貢献するものである。
【発明を実施するための最良の形態】
【0014】
本発明の単球由来多能性細胞(MOMC)としては、単球に由来し、CD14とCD34とを発現する細胞又は細胞群や単球に由来し、CD14とCD34とCD45とI型コラーゲンとを発現する細胞又は細胞群であれば特に制限されるものではなく、上記単球の起源としてはマウス、ラット、イヌ、ブタ、サル、ヒトなど特に限定されないが、ヒトを好適に例示することができ、ヒト単球の場合、ドナー由来でもよいが自己由来が特に好ましい。また、単球の供給源としては末梢血あるいは骨髄由来の単球、又は造血幹細胞からex vivoで分化させた単球を例示することができ、ここで単球とはCD14又はCD11b陽性と定義される。
【0015】
上記CD14とCD45とは単球及び単球由来細胞のマーカーとして、CD34は幹細胞のマーカーとして、I型コラーゲンは間葉系細胞のマーカーとして知られている。本発明の単球由来多能性細胞としては、幹細胞マーカーとしてのCD105、Sca−1や、間葉系細胞のマーカーとしてのIII型コラーゲン、フィブロネクチンや、血管内皮マーカーとしてのVEカドヘリン、Flt−1を発現するものが好ましい。かかるMOMCは、上記タンパク質の発現パターンから、単球、マクロファージ、樹状細胞とは異なる細胞であり、間葉系細胞、血管内皮細胞、幹細胞の特徴をあわせ持つ新規の細胞群といえる。
【0016】
本発明のMOMCは循環CD14+単球に由来するが、このことは以下にいくつか列挙する。第一に、MOMCは、単球系統マーカーであるCD14、CD13、CD11b、CD11c及びCD64に対して陽性である。第二に、フィブロネクチン上で培養した末梢血接着細胞のフェノタイプ分析を連続的に行った結果、CD14+接着細胞上におけるCD34の発現が増加していた。第三に、CD14+細胞を除去することにより、PBMCからのMOMC発生は殆ど完全に阻害されたが、CD34又はCD105陽性細胞を除去しても阻害されなかった。最後に、標識CD14+細胞を用いた研究から、CD14+単球においてはCD34発現がアップレギュレートされ、またCD14-細胞群の中にPBMC培養中に急速に増殖する細胞がないことが明らかになった。
【0017】
本発明のMOMCとしては、中胚葉系細胞にMSCを分化させることが知られている誘導条件下で中胚葉系細胞に分化しうる多能性を備えた細胞、具体的には、間葉系組織へと分化誘導する条件下での培養により骨芽細胞、骨格筋芽細胞、軟骨細胞、脂肪細胞、骨髄間質細胞、平滑筋細胞等の間葉系細胞に分化しうる多能性や、培養心筋細胞との共培養等の心筋へと分化誘導する条件下での培養により心筋細胞に分化しうる多能性や、血管内皮細胞を維持する条件下での培養等の血管内皮へと分化誘導する条件下での培養により血管内皮細胞に分化しうる多能性の他、培養神経細胞との共培養等の神経へと分化誘導する条件下での培養により、外胚葉系細胞である神経細胞に分化しうる多能性などを備えた細胞が好ましい。
【0018】
また、本発明は、本発明のMOMCを、中胚葉系細胞にMSCを分化させることが知られている誘導条件下で分化・誘導された中胚葉系前駆細胞、中胚葉系細胞又は中胚葉系組織、具体的には、MOMCを間葉系組織へと分化誘導する条件下で培養することにより誘導された骨芽細胞、骨格筋芽細胞、軟骨細胞、脂肪細胞等の間葉系前駆細胞、間葉系細胞又は間葉系組織や、MOMCを培養心筋細胞と共培養等の心筋へと分化誘導する条件下での培養により誘導された心筋前駆細胞、心筋細胞又は心筋組織や、MOMCを血管内皮細胞を維持する条件下での培養等の血管内皮へと分化誘導する条件下での培養により誘導された血管内皮前駆細胞、血管内皮細胞又は血管内皮組織の他、MOMCを培養神経細胞との共培養等の神経へと分化誘導する条件下での培養により誘導された外胚葉系の神経前駆細胞、神経細胞又は神経組織に関する。
【0019】
骨芽細胞への分化誘導条件下で培養することにより誘導された骨芽細胞としては、類円形の形態となり、アリザリン赤陽性のカルシウム沈着、アルカリフォスファターゼ染色陽性、細胞内Ca濃度の増加、骨芽細胞に特異的な骨シアロタンパク質II、オステオカルシン遺伝子を発現する細胞を好適に例示することができる。軟骨細胞への分化誘導条件下で培養することにより誘導された軟骨細胞としては、細胞質に富むやや大型の類円形の形態となり、軟骨細胞に特異的なII型及びX型コラーゲンを発現する細胞を好適に例示することができる。骨格筋細胞への分化誘導条件下で培養することにより誘導された骨格筋細胞としては、紡錘形の細長い形態をとり、骨格筋特異的アクチン、ミオシンを発現する細胞を好適に例示することができる。脂肪細胞への分化誘導条件下で培養することにより誘導された脂肪細胞としては、オイル赤染色陽性の脂肪滴が出現し、PPARγ、aP2遺伝子の発現が増加する細胞を好適に例示することができる。
【0020】
心筋へと分化誘導する条件下での培養により誘導された心筋前駆細胞や心筋細胞としては、ネスチン及び心筋特異的転写因子Nkx2.5、eHANDを発現し、心筋細胞構造蛋白であるミオシン軽鎖の遺伝子を発現する細胞を好適に例示することができる。神経へと分化誘導する条件下での培養により誘導された神経前駆細胞や神経細胞としては、ネスチン及び神経細胞特異的転写因子Neuro D、Neurogenin 2を発現し、その後神経マーカーのHu、Neu Nを発現する細胞を好適に例示することができる。血管内皮へと分化誘導する条件下での培養により誘導された血管内皮前駆細胞や血管内皮細胞としては、小突起を有する多型性の形態となり、血管内皮に特異的なマーカータンパク質KDR、vWF、eNOSを発現する細胞を好適に例示することができる。
【0021】
上記のように、MOMCは、主としてMSCを中胚葉系細胞に分化させることが知られている誘導条件下において間葉系細胞等の中胚葉系細胞への分化能を有しているの他、MOMCを神経へと分化誘導する条件下での培養により誘導された外胚葉系の神経細胞への分化能を有している。さらにMOMCは、系統特異的転写因子の発現タイミングという点において、MSC分化と同じ段階を追って各種間葉系細胞等の中胚葉系細胞などへと分化する。例えば、Cbfa1/Runx2の発現に続いて、シアロタンパク質II及びオステオカルシンが発現され(Cell, 108, 17-29, 2002)、SkM−アクチン及びミオシンが発現される前に、MyoDが発現される(Front Biosci., 5, D750-767, 2000)。かかる知見は、各種間葉系統への分化プロセスが、MOMCとMSCで共通していることを示すものである。
【0022】
本発明のMOMCの作製方法としては、末梢血単核細胞(PBMC)をフィブロネクチン上でインビトロ培養、例えば、フィブロネクチンでコートしたプラスチックプレート上で培養、好ましくは5〜14日間、特に好ましくは7〜10日間培養し、CD14とCD34とを発現する線維芽細胞様細胞を採取する方法であれば特に制限されるものではないが、PBMCを外因性成長因子不在下で培養することが好ましい。上記フィブロネクチンのかわりにコラーゲンやラミニンを用いてもよいが、単球のMOMCへの分化にはこれらフィブロネクチン等の他に、CD14-細胞由来の液性因子が必要とされる。上記PBMCの起源としてはマウス、ラット、イヌ、ブタ、サル等の実験動物又はヒトなど特に限定されないが、ヒトPBMCを好適に例示することができる。また、ヒトPBMCは、ヒト静脈血から常法により単離することができる。上記培養方法としては特に制限されないが、37℃、5%CO2、湿気下にて密度104〜107/ml、例えば2×106/mlで培養し、2〜4日毎に、特に好ましくは3日毎に非接着細胞を除去するとともに、新鮮培地を補充する培養方法を挙げることができる。このようにして得られる本発明のMOMCは、その元来のフェノタイプを5世代継代するまで維持しつつ、培養によって増殖させることが可能である。
【0023】
本発明は、本発明のMOMC、並びに/又は、前記MOMCから分化・誘導された中胚葉系前駆細胞、中胚葉系細胞及び/若しくは中胚葉系組織、具体的には、MOMCを間葉系組織へと分化誘導する条件下で培養することにより誘導された骨芽前駆細胞、骨格筋芽前駆細胞、軟骨前駆細胞、脂肪前駆細胞等の間葉系前駆細胞、骨芽細胞、骨格筋芽細胞、軟骨細胞、脂肪細胞等の間葉系細胞、骨、軟骨、筋肉、脂肪等の間葉系組織や、MOMCを培養心筋細胞と共培養等の心筋へと分化誘導する条件下での培養により誘導された心筋前駆細胞、心筋細胞、心筋組織や、MOMCを血管内皮細胞を維持する条件下での培養等の血管内皮へと分化誘導する条件下での培養により誘導された血管内皮前駆細胞、血管内皮細胞、血管内皮組織等を有効成分とする治療剤や、本発明のMOMC、並びに/又は、前記MOMCから分化・誘導された神経前駆細胞、神経細胞及び/若しくは神経組織等を有効成分とする治療剤に関する。また、本発明のMOMC、並びに/又は、前記MOMCから分化・誘導された、前記中胚葉系前駆細胞、中胚葉系細胞及び/若しくは中胚葉系組織や、前記神経前駆細胞、神経細胞及び/若しくは神経組織等を投与、例えば障害、欠損部位およびその近傍へ直接注入、又は末梢血へ投与することを特徴とする治療方法に関する。MOMC又は分化誘導処理したMOMCのいずれが治療剤として適しているかは、細胞の種類や疾病の種類や投与方法を考慮して適宜決定することが好ましい。また、MOMCは遺伝子導入が比較的簡単な細胞あることから、ヒトへの細胞移植の前に特定の遺伝子を導入した後に組織再生治療等に用いることもできる。例えば、ある種の先天性疾患で骨の形成に障害がある場合に、その遺伝子を是正した上で移植することや、あるいは、特定の蛋白(サイトカイン、成長因子、ホルモンなど)を産生するように調製した上で移植することもできる。
【0024】
前記のとおり、種々の間葉系組織等の中胚葉系組織や神経組織への分化能をもつことからMOMCには、先天性疾患、変性疾患及び外傷に対する組織再建治療における細胞供給源として有用である。具体的に本発明の治療剤や治療方法の対象となる疾患又は病態としては、骨形成不全、骨折、関節リウマチなど変性疾患による骨破壊や、軟骨が破壊される関節リウマチ、変形性関節症や、ジストロフィーなどの先天性疾患や筋炎などの後天性疾患による筋萎縮や、心筋梗塞や心筋症による心筋障害や、脳梗塞、パーキンソン病などの脳障害、脊髄損傷などの外傷や、動脈硬化や膠原病による血管障害を挙げることができ、便宜上、豊胸をはじめとした美容形成等も本発明の治療剤や治療方法に含まれる。このようなMOMC又は分化誘導処理したMOMCを使用する細胞治療には、現在提案されている組織特異的幹細胞及びES細胞を用いる再生治療等にはない大きな利点がある。すなわち、血液採取という低侵襲的に患者本人から大量の単球を採取できることから、循環単球は比較的容易に入手可能な自己細胞供給源ということができ、さらに、単球からMOMCを作製することは技術的に簡便で迅速に実施でき、また、ES細胞を使用する際の倫理上のジレンマも回避することができる。
【実施例】
【0025】
以下、実施例により本発明をより具体的に説明するが、本発明の技術的範囲はこれらの例示に限定されるものではない。
[方法と材料]
実施例1(MOMCの培養)
成人健常ドナーから得たヘパリン添加静脈血を、Lymphoprep(Nycomed Pharma AS社製)を用いた密度勾配遠心分離にかけてPBMCを単離した。血液検体は全て、倫理委員会が定める書面によるインフォームドコンセントを被験者が提出した後に採血したものである。単離した細胞をリン酸緩衝液(PBS)で2回洗浄し、10%熱不活化FBS(JRH Biosciences社製)添加低グルコースDMEMに懸濁した。PBMCを、外因性成長因子不在下、37℃、5%CO2、湿気下の諸条件のもと、フィブロネクチンでコートしたプラスチックプレート上にて密度2×106/mlで培養し、培養3日目に非接着細胞を除去した。3日毎に新鮮培地に交換しながら、4週間まで細胞を培養した。培養7〜10日目に接着細胞をMOMCとして回収し、以下のアッセイに用いたり、或いはフィブロネクチンでコートしたプレートに新たに移し替えて同一培養条件下で10世代まで維持した。
【0026】
MOMCの由来を考察するため、CD14+、CD34+、又はCD105/エンドグリン/SH2+細胞を除去したPBMCをフィブロネクチンでコートしたプレート上で7日間培養した。CD14+又はCD34+細胞は、磁気ビーズを結合させた抗CD14モノクローナル抗体又は抗CD34モノクローナル抗体(DynaBeads社製)を用い、製造者のマニュアルに従って磁気分離を行うことにより除去した。CD105+細胞を除去したPBMCは、先ず抗CD105モノクローナル抗体(Immunotech社製)共存下でインキュベートし、次に磁気ビーズを結合させたヤギ抗マウスIgG抗体(Dynal社製)共存下でインキュベートすることにより調製した。アイソタイプが一致するマウスモノクローナル抗体共存下、さらにビーズ結合抗マウスIgG抗体共存下でインキュベートしたモック処理PBMCを調製し、コントロールとした。フローサイトメトリーにより解析したところ、CD14+細胞を除去したPBMC画分中のCD14+細胞の割合は、一貫して0.5%未満であり、CD34+細胞除去画分中におけるCD34+細胞並びにCD105+細胞除去画分中におけるCD105+細胞の割合は、0.01%未満だった。1cm3あたりの付着細胞数を算定し、その結果を無処理のPBMC培養における付着細胞数に対する割合で表した。
【0027】
抗CD14モノクローナル抗体結合磁気ビーズ(CD14 MicroBeads;Miltenyi Biotech社製)を用い、さらに製造者マニュアルに従ってMACS(磁気細胞分離装置)によるカラム分離を行うことにより、PBMCから循環CD14+単球及びCD14-細胞を分離する実験も実施した。フローサイトメトリー分析の結果、単球画分には98%を越えるCD14+細胞が、またCD14-細胞画分には0.5%未満のCD14+細胞がそれぞれ存在していた。PKH67グリーン(Sigma社製)で単球を標識し、フィブロネクチンでコートしたプラスチックプレート又はコーティングを施していないプラスチックプレート上で非標識CD14-細胞と7日間の共培養を行った(比率1:4)。PKH67標識単球もフィブロネクチンでコートしたプレート上で単独培養した。
【0028】
実施例2(マクロファージ及び樹状細胞の調製)
プラスチックプレート上、20%FBS及び4ng/mlのM−CSF(R&D Systems社製)を添加したM199培地において接着PBMCを7日間培養し、マクロファージを調製した(Differentiation, 65, 287-300, 2000)。プラスチックに接着したPBMCを連続培養条件下において成熟させることにより、成熟単球由来樹状細胞を得た(J Immunol Methods, 196, 121-135, 1996)。その概要は、50ng/mlのGM−CSF及び50ng/mlのIL−4を含む10%FBSを添加したRPMI1640培地上で接着細胞を7日間培養した(材料は全てPeproTech社製)。次に幼若樹状細胞を、50ng/mlのTNF−α(PeproTech社製)共存下で3日間インキュベートした。フローサイトメトリー分析の結果、マクロファージ画分には、CD14+CD80+細胞が98%以上含まれており、樹状細胞画分には、CD83+HLA−DR+細胞が95%以上、CD14+細胞が1%未満の割合で含まれていた。
【0029】
実施例3(細胞株)
健常ドナーに皮膚生検を行ってヒト皮膚線維芽細胞を得て、その初代培養を樹立し、10%FBS添加低グルコースDMEM中で維持した。初代ヒト筋芽細胞は、臨床的に筋炎が疑われるが組織学上は正常な患者の筋肉生検を培養することにより作製した(J Cell Biol., 144, 631-643, 1999)。ヒト骨肉腫細胞株MG−63、横紋筋肉腫細胞株RD、及び軟骨肉腫細胞株OUMS−27は、日本ヒューマンサイエンス研究資源バンク(日本、大阪府)から入手し、10%FBS添加低グルコースDMEMで維持した。
【0030】
実施例4(MOMCの間葉系細胞へのインビトロ分化)
PBMCから新たに作製したMOMC、数代にわたり継代培養されたMOMC、又は低温保存されていたMOMCのいずれかを、新たにフィブロネクチンをコートしたプラスチックプレート、又はチャンバースライドに移して10%FBS添加高グルコースDMEM(Hyclone Laboratories社製)中でセミコンフルエントになるまで生育させた。次に、下記の種々の間葉系細胞型にMSCを分化させることが知られている誘導条件下(Science,284, 143-147, 1999, J Cell Biol., 144, 631-643, 1999, J Cell Biochem., 64, 295-312, 1997, Muscle Nerve,18, 1417-1426, 1995, Tissue Eng., 4, 415-428, 1998, Arthritis Rheum., 44, 85-95, 2001, J Biol Chem., 275, 9645-9652, 2000)で、上記細胞を培養した。コントロールとして、PBMCから新たに単離した単球、マクロファージ及び皮膚線維芽細胞も同一条件下で処理した。
【0031】
骨形成に関しては、100nMのデキサメサゾン、10mMのβ−グリセロホスフェート及び50μMのアスコルビン酸を含む骨形成誘導培地(Clonetics社製)上で接着細胞を培養した。1週間に2回培地を交換することを3週間継続した。
【0032】
筋形成に関しては、接着細胞を10μMの5−アザシチジン(Sigma社製)で24時間処理した。PBSで細胞を洗浄し、5%ウマ血清(Life Technologies社製)、50mMのヒドロコルチゾン(Sigma社製)、及び4ng/mlの塩基性線維芽細胞増殖因子(Sigma社製)を含む培地で培養した。1週間に2回培地を交換することを3週間継続した。
【0033】
軟骨形成に関しては、接着単層細胞をTGF−β(R&D Systems社製)存在下、無血清培地上で3週間培養した。TGF−βは1日おきに培養培地に加え、その最終濃度を10ng/mlとした。
【0034】
脂肪形成に関しては、1μMのデキサメサゾン、0.5mMのメチル−イソブチルキサンチン、10μg/mlのインスリン、及び100mMのインドメタシン(全てSigma社製)を添加した脂肪誘導培地にて細胞をインキュベートした。72時間後に上記培地を、10μg/mlのインスリンを単独添加した維持培地と交換して24時間おいた。脂肪誘導培地で細胞を3回処理し、さらに1週間維持培地において維持した。
【0035】
実施例5(MOMCの心筋細胞へのインビトロ分化)
MOMCを8日間ウィスターラット胎児由来の培養心筋細胞とともに共培養を行った。
培養細胞を4%パラホルムアルデヒドで固定した後、ヒト特異的抗ネスチン(nestin)抗体(Chemicon社)により免疫染色(DAB発色)を行った。
【0036】
MOMCを蛍光色素PKH67(Sigma社)により細胞膜を標識した後に、ウィスターラット培養心筋細胞と7〜10日間、共培養を行った。培養細胞を4%パラホルムアルデヒドで固定した後、抗Nkx2.5抗体、抗eHAND抗体(Santa Cruz社)、抗CD45抗体(DAKO社)により、二重蛍光免疫染色を行った。
【0037】
MOMCをウィスターラット培養心筋細胞と3日、6日、9日、12日間共培養を行った後、mRNAを抽出した。心筋細胞構造蛋白であるミオシン軽鎖(MLC2v)に対するヒト特異的PCRプライマー(TGACAAGAACGATCTGAGAG、CAGGTTCTTGTAGTCCAAGT)を作成し、RT−PCRを行った。
【0038】
実施例6(MOMCの神経細胞へのインビトロ分化)
MOMCを8日間ウィスターラット培養神経細胞とともに共培養を行った。培養細胞を4%パラホルムアルデヒドで固定した後、ヒト特異的抗ネスチン抗体により免疫染色(DAB発色)を行った。
【0039】
MOMCを蛍光色素PKH67により細胞膜を標識した後に、ウィスターラット培養神経細胞と4日間、共培養を行った。培養細胞を4%パラホルムアルデヒドで固定した後、抗NeuroD抗体(Santa Cruz社)、抗CD45抗体(DAKO社)により、二重蛍光免疫染色を行った。
【0040】
MOMCをウィスターラット培養神経細胞と3日間、共培養を行った。培養細胞を4%パラホルムアルデヒドで固定した後、ヒト特異的抗ネスチン抗体、抗Neurogenin2抗体(Chemicon社)により、二重蛍光免疫染色を行った。
【0041】
MOMCを蛍光色素PKH67により細胞膜を標識した後に、ウィスターラット培養神経細胞と9〜10日間、共培養を行った。培養細胞を4%パラホルムアルデヒドで固定した後、抗Hu抗体、抗NeuN抗体(Chemicon社)により二重蛍光免疫免疫染色を行った。
【0042】
実施例7(MOMCの血管内皮細胞へのインビトロ分化)
MOMCを付着細胞として血管内皮細胞維持培地EBM−2(Clonetics社)中で7〜10日間培養した。培養細胞を10%中性緩衝ホルマリンで固定した後、抗CD34抗体(Calbiochem-Novabiochem社)、抗vWF抗体(Dako社)、抗eNOS抗体(Bechton-Dickinson社)、抗VEGFR2/KDR/Flk−1抗体(Sigma社)により免疫染色(DAB発色)を行った。
【0043】
EBM−2培地中で7日間培養したMOMCからRNAを抽出しcDNAへ逆転写した。血管内皮細胞に発現されるFlt−1、VEGFR2/KDR/Flk−1、CD31、CD144、vWFおよびCD34、CD45、CD14、GAPDHに対する特異的プライマーを用いてPCRを行った。コントロールとして分化誘導をかける前のMOMC、培養さい帯静脈血管内皮細胞(HUVEC)由来のRNAを用いた。
【0044】
実施例8(フローサイトメトリー分析)
蛍光細胞染色は以下の手順で行った。2mMのEDTA共存下における氷上インキュベーションにより、接着細胞をプラスチックプレートから脱離させ、正常マウス又はラットの血清を用い、4℃下で10分間ブロックした。抗HLA−DR抗体、抗CD11c抗体(BD Pharmingen社製)、抗CD11b/Mac−1抗体、抗CD14抗体、抗CD29抗体、抗CD34抗体、抗CD44抗体、抗CD83抗体、抗CD105/エンドグリン/SH2抗体、抗CD117/c−kit抗体(Immunotech社製)、抗CD34抗体、抗CD133抗体(MiltenyiBiotech社製)、抗HLAクラスI抗体、抗HLA−DR抗体、抗CD31/PECAM−1抗体、抗Flt−1/VEGFR1抗体、抗Flk−1/VEGFR2抗体(Sigma社製)、抗CD40抗体、抗CD54抗体、抗CD80抗体、抗CD86抗体(Ancell社製)、抗CD144/VE−カドヘリン抗体、抗I型コラーゲン抗体(Chemicon International社製)を含むマウスモノクローナル抗体、又はラット抗Sca−1モノクローナル抗体(Cedarlane Laboratories社製)に、FITC、フィコエリトリン(PE)若しくはPC5を結合させて、又は結合させないで、上記細胞を染色した。非結合モノクローナル抗体を用いた場合には、FITC若しくはPEを結合したヤギ抗マウス抗体若しくはラットIgG F(ab’)2抗体(Immunotech社製)を2次抗体として使用した。IntraPrepTM浸透処理試薬(Immunotech社製)により細胞の浸透処理及び固定化を行い、細胞内染色を行った。細胞の分析は、CellQuestソフトウエアを使用したFACSRCaliburフローサイトメーター(Becton Dickinson社製)により行った。フォワード及びサイドスキャッターでのゲーティングにより可視化細胞を同定し、データは対数ヒストグラム又はドットプロットとして示した。
【0045】
実施例9(免疫組織化学分析)
サイトスピン法により単球及びマクロファージ、樹状細胞をスライド上に塗抹し、これら細胞以外の細胞型は、フィブロネクチンでコートしたチャンバースライド上で培養したが、フィブロネクチン染色用には、コラーゲンI型をコートしたチャンバースライドを用いた。これらの細胞を10%ホルマリンで固定し、3%過酸化物を用い、外因性ペルオキシダーゼ活性を5分間抑制した。抗CD45抗体、抗ビメンチン抗体、抗骨格筋特異的アクチン抗体(SkM−アクチン)(Dako社製)、抗CD34抗体(Calbiochem-Novabiochem社製)、抗I型コラーゲン抗体(Chemicon社製)、抗III型コラーゲン抗体、抗フィブロネクチン抗体(Sigma社製)、抗II型コラーゲン抗体(ICN Biomedicals社製)、抗骨格筋特異的ミオシン重鎖抗体(SkM−MHC)(Zymed Laboratories社製)を含むマウスモノクローナル抗体又はラット抗Sca−1モノクローナル抗体共存下にて30分間スライドをインキュベートし、さらにビオチン標識抗マウス抗体若しくは抗ラットIgG抗体共存下でのインキュベーションを行った。3,3’−ジアミノベンジジンを基質として用い、ストレプトアビジン−ホースラディッシュペルオキシダーゼ複合物(ニチレイ社製)を室温で10分間反応させることにより、抗体−ビオチン結合体を検出した。核をヘマトキシリンで対比染色した。上記1次抗体の代りに正常マウス若しくはラットのIgG抗体(Dako社製)共存下でインキュベートした細胞を負のコントロールとした。
【0046】
蛍光二重染色は以下の方法で行った。4%パラホルムアルデヒドで細胞を固定し、ヤギ抗PEBP2αA抗体若しくは抗Sox9ポリクローナル抗体(Santa Cruz社製)共存下でインキュベートし、次にAlexa FluorR568ヤギ特異的IgG抗体(Molecular Probes社製)共存下でインキュベートし、さらにFITC結合マウス抗CD45モノクローナル抗体(Dako社製)共存化でインキュベートした。同様にマウス抗MyoD抗体(Dako社製)又は抗ペルオキシゾーム増殖活性化受容体γ(PPARγ遺伝子)モノクローナル抗体(Santa Cruz社製)で染色し、その後テトラメチルローダミン・イソチオシアネート異性体Rで標識したマウス特異的IgG抗体(Dako社製)共存化でインキュベートし、さらにFITC結合抗CD45モノクローナル抗体共存化でインキュベートした。共焦点レーザー蛍光顕微鏡(LSM5 PASCAL;Carl-Zeiss社製)を用いて、これらの細胞を観察した。所定のマーカーに対して染色陽性を示した細胞の割合を数値化するために、一培養あたり少なくとも300個の細胞を低倍率で鏡検した。
【0047】
実施例10(アセチル化LDL(Ac−LDL)の摂取)
2.5μg/mlのDil−Ac−LDL(Molecular Probes社製)共存化で接着細胞を1時間培養し、Ac−LDL摂取量をフローサイトメトリーで調べた。
【0048】
実施例11(アルカリフォスファターゼ染色)
10%ホルマリンに細胞を固定し、0.2mg/mlのナフトールAS−TRホスフェート及び0.5mg/mlのFast Red RC(全てSigma社製)を含む溶液中で10分間インキュベートした。
【0049】
実施例12(細胞内カルシウムの検出)
細胞内におけるカルシウム沈着を調べるため、細胞を10%ホルマリンに固定し、2%アリザリンレッドS(Sigma社製)で3分間染色し、蒸留水で入念に洗浄した。市販のキット(Sigma社製)を使い、細胞内カルシウム濃度を測定した(J Biol Chem., 275, 9645-9652, 2000)。ウシ血清アルブミンを標準とするブラッドフォード・プロテイン・アッセイキット(Bio-Rad Laboratories社製)を使用し、細胞抽出物におけるタンパク質含量も測定した。カルシウム濃度は、タンパク質含量1μgあたりのμg数として表示した。
【0050】
実施例13(オイル-レッドO染色)
0.2%グルタルアルデヒドに細胞を5分間固定し、60%イソプロパノールで洗浄し、0.1%オイル-レッドO(Sigma社製)で10分間覆った。60%イソプロパノール、次いで蒸留水による洗浄後、細胞のヘマトキシリン対比染色を行った。
【0051】
実施例14(透過電子顕微鏡)
培養後のMOMCを直ちに2.5%グルタルアルデヒドに固定し、2%四酸化オスミウムで2次固定し、エタノール及び酸化プロピレンの一連使用により脱水してエポキシ樹脂に包埋した。LKBウルトラトーム上で、細胞をダイアモンドナイフで薄切片化した。灰色から銀色の範囲の切片を150−メッシュグリッド上に回収し、酢酸ウラニル及び酢酸鉛で二重染色を施し、JEOL-1200EXII電子顕微鏡下で観察した(Jeol社製)。
【0052】
実施例15(細胞増殖研究)
MOMCの増殖は、文献(Blood, 71, 1201-1210, 1988)記載の方法に従い、BrdUで標識することにより検出した。以下簡潔に説明する。10μMのBrdU(Sigma社製)存在下で、染色するまでの2時間MOMCを培養した。Carnoyの固定剤(メタノール/酢酸)で−20℃で30分間固定した後、細胞を風乾し、2N−HClで1時間処理してDNAを変性させ、さらに10分間0.1Mのホウ酸塩(pH8.5)で中和した。これらの細胞をマウス抗BrdUモノクローナル抗体(Chemicon International社製)共存下でインキュベートし、ビオチン−ストレプトアビジン−ペルオキシダーゼ複合体で染色した。核はヘマトキシリンで対比染色した。上記の1次抗体の代りに、アイソタイプが一致するマウスのコントロールモノクローナル抗体共存下でインキュベートした細胞を負のコントロールとした。非固定細胞を、ヨウ化プロピジウム(Sigma社製)共存下で30分間インキュベートして、蛍光顕微鏡観察を行い、アポトーシスを起こした細胞を検出した。
【0053】
文献記載(J Exp Med., 183, 2313-2328, 1996)の手法に従い、5−カルボキシフルオレセイン・ジアセテート・スクシンイミジル・エステル(CFSE)で細胞を標識し、細胞分裂の様子を考察した。フィブロネクチンでコートしたプレート上で1、3、5日間、CFSE標識単球と非標識CD14-細胞を共培養し、接着細胞を回収してPC5標識抗CD14モノクローナル抗体で染色した。CFSE標識MOMCについても、1、3、及び5日間の培養を行った。CFSE標識による発色強度をフローサイトメトリーにより解析した。
【0054】
実施例16(RT−PCR)
RNeasyキット(Qiagen社製)を用いて分化誘導処理を行った場合と行わなかった場合の双方において、MOMCから全RNAを抽出した。末梢血CD14+単球、並びにマクロファージ、樹状細胞、皮膚線維芽細胞、筋芽細胞、骨肉腫、横紋筋肉腫及び軟骨肉腫を含む種々の培養細胞からも全RNAを抽出した。ヒト筋肉及び脂肪組織由来の全RNAサンプルは、Clonetech Laboratories社から購入した。Molonyマウス白血病ウイルス・リバーストランスクリプターゼ(Takara社製)を用い、オリゴ−dTをプライマーとして、全RNAから一本鎖cDNAを合成した。次にこのcDNA(50ngの全RNAに相当)を、配列番号3〜34に示される表1記載の各種特異的プライマーを用いたPCR増幅法にかけた。このPCR産物を2%アガロースゲル電気泳動分析にかけ、臭化エチジウム染色により可視化した。
【0055】
【表1】

【0056】
実施例17(統計分析)
全ての比較における統計有意は、Mann−WhitneyのU検定により試験した。
【0057】
[結果]
実施例18(MOMCの作製)
フィブロネクチンでコートしたプレート上にて、10%FBSを単独添加したDMEM中でPBMCを培養すると、一部の細胞が直ちにプレートに付着した。円形細胞の小クラスターが24時間以内に発生し、そこから細胞突起が伸張した。培養5日目に線維芽細胞様形態を呈する接着細胞が出現した。さらに3日間培養すると、この線維芽細胞様細胞は、培養中の優勢細胞となった(図1a)。かかる線維芽細胞はクラスター周辺部に高頻度で観察された(図1b)。線維芽細胞様細胞は、培養14日目頃までは徐々にその数を増やした。それ以降、増殖能は次第に消失していったが、細胞は4週間まで生存していた。ドナー50名から得たPBMC108個を培養したところ、7日目に0.3〜1×107個の接着細胞が得られた。フローサイトメトリー分析の結果、培養7日目に回収した細胞は単一のフェノタイプを示し(95%以上がホモジニアス)、CD14、CD45、CD34、及びI型コラーゲンに対して陽性だった(図2)。このフェノタイプは独特であり、単球/マクロファージ(CD14+、CD45+、CD34-及びI型コラーゲン-)、血管内皮前駆細胞(CD14-、CD45-、CD34+及びI型コラーゲン-)(Science, 275, 964-967, 1997)、及び間葉系前駆細胞(CD14-、CD45-、CD34-、及びI型コラーゲン+)(Arthritis Res., 2, 477-488, 2000)を含む既知の末梢血由来接着細胞のフェノタイプとは異なっている。少なくとも50名のドナーから得た細胞は、同じフェノタイプを有していた。7日目に、新たにフィブロネクチンでコートしたプレートに細胞を移し替え、同一条件下で培養したところ、殆ど全ての細胞が、細長い線維芽細胞様の形態を現した(図1c)。これらの細胞は、5世代まで増殖することができ、細胞増殖は継代直後に最も盛んになるようだった。しかし5世代継代以降は、細胞増殖速度が徐々に遅くなり、8世代継代を超えると、その増殖能は消失した。特別な処理を施さない限りは、培養期間中に細胞が成熟間葉系細胞へ自然に分化することはなかったが、高いコンフルエンシーとなるように細胞を播種すると、複数の核をもつ細胞の出現が僅かに認められた。インビトロ培養でPBMCから得られたこれらの線維芽細胞様細胞を、以下の知見に基づき、MOMCと名付けた。
【0058】
電子顕微鏡観察の結果、MOMCは、多数の細胞小器官をもつ紡錐形の形態をしていることがわかった(図1d−g)。多数の一次リソソーム、細胞表面にみられる偽足状の突起、また迷路状の細胞小胞は、マクロファージ類及びその他の食細胞類に固有に見られる所見である。MOMCにはまた、突出して束をなす中間フィラメント、よく発達した二次リソソーム、伸張して分岐したミトコンドリアといった間葉系由来細胞によく見られる特徴が認められる。殆ど全てのMOMCに僅かな脂質滴が観察された。また、血管内皮細胞特有の棒状微小管に似た構造も数多く認められた。これらの超微形態所見は、食細胞、間葉系細胞及び血管内皮細胞の特徴を備えたものである。
【0059】
実施例19(循環系におけるMOMCの由来)
フィブロネクチンでコートしたプレート上でPBMCを培養して得られた接着細胞を、フローサイトメトリーにかけ、その表面におけるCD14及びCD34の発現を連続的に調べた(図3a)。1時間後にプレートに付着した細胞の大多数はCD14+及びCD34-だったが、接着CD14+細胞上におけるCD34の発現は次第にアップレギュレートされていった。培養7日後には、殆ど全ての接着細胞がCD14及びCD34に対して陽性を示した。末梢血には、CD34+血管内皮細胞前駆細胞(Science, 275, 964-967, 1997)及びCD105/エンドグリン/SH2+間葉系前駆細胞(Science, 284, 143-147, 1999, Arthritis Res., 2, 477-488, 2000, Biochem Biophys Res Commun., 265, 134-139, 1999)が含まれているため、本発明者らは、PBMCからCD14、CD34又はCD105に対する陽性細胞を除去することによりもたらされるMOMCのインビトロ誘導における効果を考察した。図3bに示したとおり、CD14+単球の除去によりMOMCの出現は、ほぼ完全に阻害されたが、CD34+又はCD105+細胞を除去しても何の効果も生じなかった。MOMCが循環単球由来であることをさらに確認すべく、PBMCからCD14+単球を分離し、PKH67で標識し、非標識CD14-細胞共存下においてフィブロネクチンでコートしたプレート上で培養した。図3cに示すように、培養一週間後にPKH67標識単球はCD34を発現した。以上の知見を総合すると、MOMCが循環CD14+単球由来であることが示唆される。しかし、培養3日目に回収された非接着細胞にもCD14+細胞が有意比率で含まれており、CD14+単球の一部のサブセットがフィブロネクチンへの付着能を有し、MOMCに分化することが示唆される。
【0060】
PKH67標識単球単独でフィブロネクチン上における培養を行うと、線維芽細胞様に変化したのは僅かな細胞のみで、CD34の発現も培養7日目の時点でアップレギュレートされていなかった(図3c)。この場合、単球におけるCD34発現を誘導するには、CD14-細胞の培養上清を加えるだけで十分であった。さらに、フィブロネクチンコーティング処理を施していないプレート上で、CD14-細胞とPKH67標識単球を共培養すると、培養7日目にプレートに付着していた細胞数は低く、また、それら付着した細胞はCD34を発現していなかった(図3c)。このことから、循環単球がMOMCに分化するには末梢血CD14-細胞由来の液性因子(1若しくは2種以上)及びフィブロネクチンへの結合が必須であると考えられる。
【0061】
MOMCの分化プロセスにおいて、CD14+単球が増殖するか否かを考察するため、CD14+単球のCFSE標識を行い、非標識CD14-細胞とフィブロネクチン上で共培養した。連続的に接着細胞を回収してCFSE標識の発色強度やCD14の発現をフローサイトメトリーにより調べた(図3d)。殆ど全てのフィブロネクチン付着単球が、主に培養24時間以内に増殖していたが、それ以降の培養期間においてもゆっくりとではあるが増殖していた。接着細胞は殆ど例外なくCFSE標識CD14+単球であり、CD14-細胞画分において接着細胞が増殖する様子は観察されず、混入したCD14-細胞群中の特異的前駆細胞が生育する結果として、MOMCが発生するのではないことが示唆される。
【0062】
実施例20(MOMCのフェノタイプ)
MOMCにおける種々の細胞表面分子及び細胞内分子の発現を、フローサイトメトリー及び免疫組織化学法により調べ(図2及び図4)、そのタンパク質発現プロフィールを、単球、マクロファージ及び樹状細胞のものと比較した(表2)。MOMCは、造血及び単球系統マーカー遺伝子(CD45、CD14、CD13、CD11b、CD11c及びCD64)を発現したが、樹状細胞マーカー遺伝子(CD1a及びCD83)は発現しなかった。MOMC上においてHLAクラスI、HLA−DR、及び共刺激分子(CD40及びCD86)が発現されることから、MOMCは抗原提示細胞として、抗原特異的T細胞活性の誘導能を有していることが強く示唆される。MOMCは、造血幹/血管内皮細胞マーカー遺伝子CD34、及び間葉系幹/血管内皮細胞マーカー遺伝子CD105/エンドグリン/SH2を発現した(Biochem Biophys Res Commun., 265, 134-139, 1999)。またMOMCは、幹細胞マーカー遺伝子Sca−1を発現したが、別の幹細胞マーカー遺伝子であるCD117/c−kit及びCD133は発現しなかった。またMOMCは、血管内皮細胞マーカー遺伝子であるCD144/VE−カドヘリン及びFlt−1/VEGFR1陽性だったが、Flk−1/VEGFR2及びvWFの発現は認められなかった。MOMCは、一般に間葉系由来細胞によって産生される細胞外マトリックスタンパク質であるI型及びIII型コラーゲン、フィブロネクチン及びビメンチン陽性だった。これらのタンパク質発現プロフィールは、5世代継代まで変化しなかった。MOMCは単球及び単球由来食細胞とは異なるフェノタイプを示した。特に、幹細胞マーカー遺伝子(CD34、Sca−1及びCD105)、血管内皮マーカー遺伝子(CD144/VE−カドヘリン及びFlt−1/VEGFR1)、及び間葉系マーカー遺伝子(I及びIII型コラーゲン及びフィブロネクチン)の発現は、MOMC独特の特徴だった。従って、MOMCは、食細胞、血管内皮細胞、間葉系細胞及び幹細胞のフェノタイプを併せもつ細胞であるとの認識が成立する。
【0063】
【表2】
【0064】
実施例21(MOMCの増殖能)
培養中にMOMCが増加する様子が認められた。この観察結果が細胞分裂に基づくものかを考察すべく、MOMCにおける分裂細胞の比率をBrdU染色によって連続的に調べた(図5a)。半数近くの接着細胞が、継代1日後にBrdUによって染色されたが、BrdU+細胞は培養5日目に著減した。1、3、5、7及び10日目に、全接着細胞におけるBrdU+細胞の割合を計算したところ、BrdU+細胞の比率は、培養1日目に最大を示し、以後経時的に減少した。ヨウ化プロピジウム染色陽性細胞は、どの時点においても1%未満だった。CFSE標識を行って調べた結果、MOMCが継代後に活発かつ同調的に分裂している様子が認められ、優勢的に増殖している細胞部分はなかった(図5c)。以上の知見は、MOMCが培養中における増殖能を有し、また、主に継代直後に増殖することを示唆するものである。
【0065】
実施例22(MOMCの間葉系細胞系統へのインビトロ分化)
MOMCは、間葉系細胞にみられるいくつかの形態及びフェノタイプ上の特徴を備えていたことから、MOMCは、いくつかの間葉系統へと分化誘導していく能力を有しているとの仮説をたてた。この仮説を確かめるため、MSCを骨、骨格筋、軟骨及び脂肪へと分化誘導することが知られている各種条件下でMOMCを培養した。
【0066】
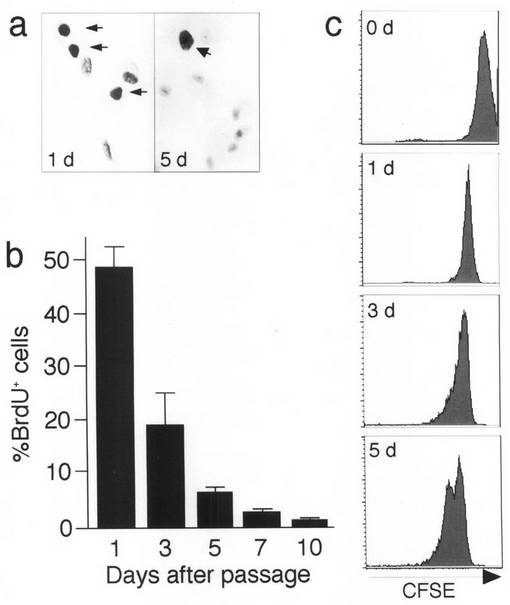
骨への分化誘導処理を施したMOMCの形態は、紡錐形から類円形へと変化した。殆ど全ての接着細胞でカルシウム沈着が生じているのが、アリザリンレッド染色から観察され(図6a)、それら接着細胞はアルカリフォスファターゼを発現していた(図6b)。この過程で、細胞内カルシウム含量が顕著な増加を示した(図6c)。骨形成誘導後、MOMCは、成熟骨細胞が産生する骨シアロタンパク質II、オステオカルシン(Calcif Tissue Int., 62, 74-82, 1998)や、骨特異的転写因子オステリックス(Cell, 108, 17-29, 2002)のmRNAを発現した。他方、かかる誘導処理後、CD34、CD45及びCD14の発現は消失した(図6d)。
【0067】
MOMCを5−アザシチジンで処理し、筋への分化誘導条件下で3週間培養したところ、細胞は細長くなったものの筋細胞のような複数核をもつ細胞は出現しなかった。この時点で、検体によってその割合は異なるが、45〜60%の接着細胞においてSkM−アクチン及びSkM−MHCの発現が誘導されていた(図6e及びf)。RT−PCRを行った結果、筋特異的転写因子マイオジェニンのmRNA及びSkM−MHCのmRNAが誘導後に検出された(図6g)。CD34、CD14及びCD45の発現は減少したが消失はしなかった。免疫組織化学分析の結果、CD34は殆ど全ての接着細胞で発現したが、CD14及びCD45は、SkM−MHCを発現しなかった細胞でその発現をみた(データは示さず)。培養した筋芽細胞、筋組織、さらには横紋筋肉腫細胞株においてさえ検出可能レベルのCD34が発現され、これは原始筋細胞のサブセットにおけるCD34発現と一致するものだった(J Cell Biol., 150, 1085-1100, 2000)。
【0068】
軟骨形成を誘導するため、MOMCをTGF−β1の存在下、マイクロマス懸濁液中で培養した。これは、MSCにおいて軟骨細胞の分化を誘導するための標準的な手法である(Science, 284, 143-147, 1999, Tissue Eng., 4, 415-428, 1998, Arthritis Rheum., 44, 85-95, 2001)。しかし、プレート上での液滴マイクロマス、及び円錐管中でのペレットマイクロマスによるいくつかの別のプロトコールに従った場合においても、MOMCは1週間以内に死滅した。そこで、TGF−β1存在下で単層MOMCを3週間培養した。図6hに示すように、関節軟骨特有のII型コラーゲンは、無処理のMOMCで弱く発現していたが、その発現は誘導処理後に顕著にアップレギュレートされた。RT−PCRの結果からさらに、誘導処理後に軟骨細胞特異的II型及びX型コラーゲンの発現がアップレギュレートされていることが明らかになった(図6i)。CD45及びCD14の発現は消失したが、CD34の発現は誘導処理後も保持された。他方、CD34は軟骨肉腫細胞株においても発現がみられた。
【0069】
電子顕微鏡観察の結果、MOMCには脂質滴が僅かに認められたが(図1d、f及びg)、誘導処理後に脂質液胞が出現し、その大きさ、数量ともに経時的に増加した。これら脂質液胞はオイル-レッドOで染色された(図6j)。この誘導処理の結果、検体によって割合は異なるが、50〜80%の接着細胞が、この系統にコミットしていた。PPARγ遺伝子のmRNA及び脂肪酸結合タンパク質aP2のmRNAは、MOMCによって弱く発現されたが、これらの遺伝子発現は上記誘導処理後、顕著にアップレギュレートされた(図6k)。CD45及びCD14のmRNA発現は消失したが、CD34発現は、誘導処理後も保持された。他方、CD34は脂肪組織においても発現がみられた。
【0070】
PBMCから新たに産生されたMOMC、5世代まで継代培養したMOMC、又は低温保存されたMOMCにおいても同様に間葉系細胞系統への分化が観察された。さらに、ドナー5名から得たMOMCも同様の多分化能を有していた。成熟間葉系細胞であるヒト皮膚線維芽細胞株2種と、新たに単離したCD14+単球及びマクロファージを同一誘導条件下でそれぞれ培養した。3週間後、皮膚線維芽細胞は、上記条件下において外見は健常だったものの分化傾向は示さなかった。1週間後、上記培養条件下にあった循環単球及びマクロファージは、何ら外見上の分化傾向をみせないままに脱離していた。系統特異的分化は、培養3週間後に付着細胞の半分以上で観察された。しかし付着細胞数は、培養当初の細胞数より20〜50%減少し、MOMCの多くが誘導処理の過程において脱離していったと考えられた。
【0071】
ある系統への分化が各種誘導処理特異的であるかどうかを調べるために、各処理における培養物をアリザリンレッド、オイル-レッドOで交叉染色し、また、SkM−MHC若しくはII型コラーゲン免疫染色も行った。これらの細胞は目的とする系統に特異的な染色に対してのみ陽性で、それ以外の染色に対してはすべて陰性だった(データは示さず)。さらに、インビトロアッセイにおいて、目的とする系統以外の系統に一部のサブセットが分化する可能性を探るため、3週間分化誘導処理した細胞を高感度RT−PCRにかけて、その発現が骨形成(オステオカルシン)、筋形成(SkM−MHC)、軟骨形成(II型コラーゲン)、又は脂肪形成(aP2)系統に限定的な複数遺伝子の転写物を増幅した。系統特異的遺伝子の発現は、目的とする系統に特異的であり(データは示さず)、分化が、施した処理に特異的に起きることが示唆された。
【0072】
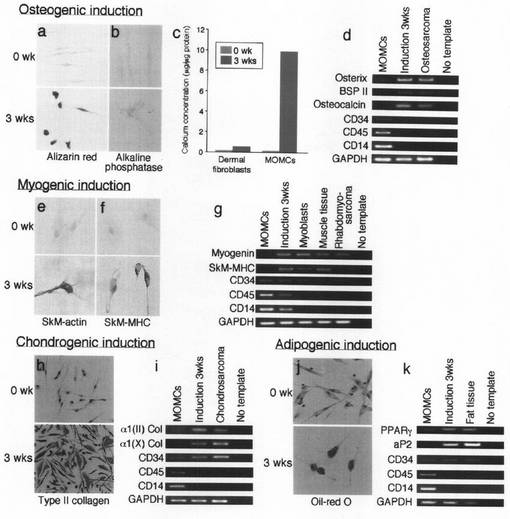
分化した間葉系細胞が、MOMC画分に混入したMSCとしての分化能をもつ細胞に由来する可能性を除外するために、誘導処理を行う前にMACS分離システムにより、MOMCからCD14+細胞をポジティブに精製した。その結果は予想どおり、かかる選択処理をしないMOMCにおけると同様に、骨形成、筋形成、軟骨形成、及び脂肪形成への分化が認められた。また、CD34又はCD105/エンドグリン/SH2を発現する細胞を、PBMC培養開始時に除去しても、間葉系細胞への分化には影響がなかった。さらに、系統特異的転写因子であるCbfa1/Runx2(Cell, 89, 755-764, 1997)、MyoD(Front Biosci., 5, D750-767, 2000)、Sox−9(Osteoarthritis Cartilage, 8, 309-334, 2000)、及びPPARγ(J Biol Chem., 276, 37731-37734, 2001)の、誘導処理1週間後におけるMOMC上での発現を調べた。この時点でMOMCはCD45を発現しているので、骨形成、筋形成、軟骨形成、及び脂肪形成誘導処理を行ったMOMCに二重染色を施し、CD45及び個々の転写因子の発現を調べた。図7に示すように、1週間にわたる骨形成分化を経たMOMCは、細胞膜及び細胞質にCD45を発現し、核にCbfa1/Runx2を発現した。同様に、CD45/MyoDとCD45/Sox−9の同時発現が、それぞれ1週間の筋形成及び軟骨形成分化誘導を受けたMOMCで認められた。PPARγは、無処理MOMCの核において弱く発現されていたが、1週間の脂肪形成誘導後、PPARγの発現は増加し、CD45の発現が減少した。これらの知見は、特異的分化誘導条件下でCD45+造血細胞の系統特異的分化が生じることを示唆するものである。免疫組織化学法及びRT−PCR法による測定の結果、PPARγ以外の上記系統特異的転写因子の発現は、誘導処理3週間後に消滅していることがわかった(データは示さず)。これらの知見より、MOMCが、誘導処理過程の初期分化段階において、上記系統特異的転写因子を一過性に発現することが示唆される。
【0073】
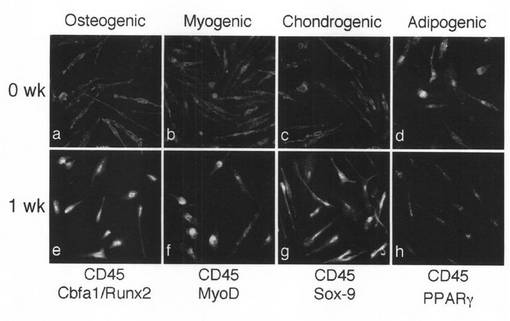
実施例23(MOMCの心筋細胞へのインビトロ分化)
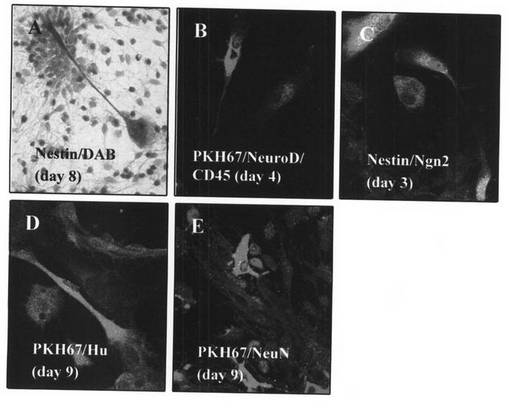
神経、心筋前駆細胞に発現するマーカーであるネスチン(Brown)が、共培養8日目のMOMCに発現し、周囲ラット培養心筋細胞との結合が観察された(図8A)。
【0074】
PKH67(Green)で標識されたMOMCが心筋細胞特異的転写因子であるNkx2.5、eHAND(Red/Alexa568:Molecular Probe社)を発現し、同時に血球系マーカーであるCD45(Blue/Alexa660:Molecular Probe社)を発現していた(図8B、図8C)。これはヒト末梢血血球細胞由来のMOMCの心筋細胞への分化過程を示すものである。
【0075】
ヒト特異的PCRプライマーを用いたRT−PCRでは、陽性コントロールのヒト心筋では心筋細胞構造蛋白であるミオシン軽鎖(MLC2v)の発現が認められ、陰性コントロールのラット心筋、共培養前のMOMCでは発現が認められない。共培養後12日目のMOMCにおいてヒトMLC2vの発現が認められた(図8D)。
【0076】
これらのことから、MOMCをラット心筋細胞と共培養することにより、MOMCの心筋細胞への分化が誘導され、共培養8−10日目には心筋前駆細胞のマーカーが発現され、12−14日目には心筋構造蛋白が発現されることが証明された。
【0077】
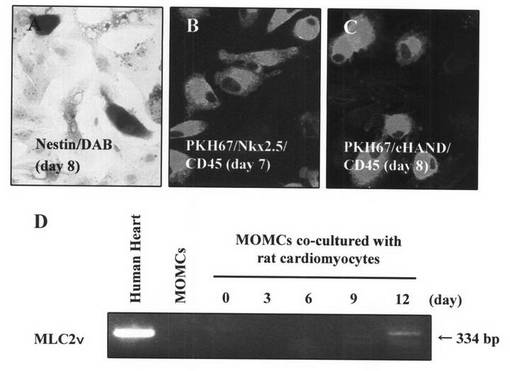
実施例24(MOMCの神経細胞へのインビトロ分化)
神経、心筋前駆細胞に発現するマーカーであるネスチン(Brown)が、共培養8日目のMOMCに発現し、周囲ラット培養神経細胞へ向かう神経突起の伸長が観察された(図9A)。
【0078】
共培養4日目にはPKH67(Green)で標識されたMOMCが神経細胞特異的転写因子であるNeuroD(Red/Alexa568:Molecular Probe社)を発現し、同時に血球系マーカーであるCD45(Blue/Alexa660:Molecular Probe社)を発現していた(図9B)。これはヒト末梢血血球細胞由来のMOMCの神経細胞への分化過程を示すものである。
【0079】
共培養3日目にはMOMCがネスチン(Green/Alexa488:Molecular Probe社)を発現すると同時に神経細胞特異的転写因子であるNeurogenin2 (Red/Alexa568:Molecular Probe社)を発現していた(図9C)。このことから、MOMCが神経細胞への分化が決定された神経細胞の前駆細胞であることが示された。
【0080】
ウィスターラット培養神経細胞と9日間の共培養では、KH67(Green)で標識されたMOMCにおいて成熟した神経のマーカーであるHu、NeuN(Red/Alexa568:Molecular Probe社)の発現が認められた(図9D、図9E)。このことからMOMCが成熟した神経細胞まで分化することが証明された。
【0081】
実施例25(MOMCの血管内皮細胞へのインビトロ分化)
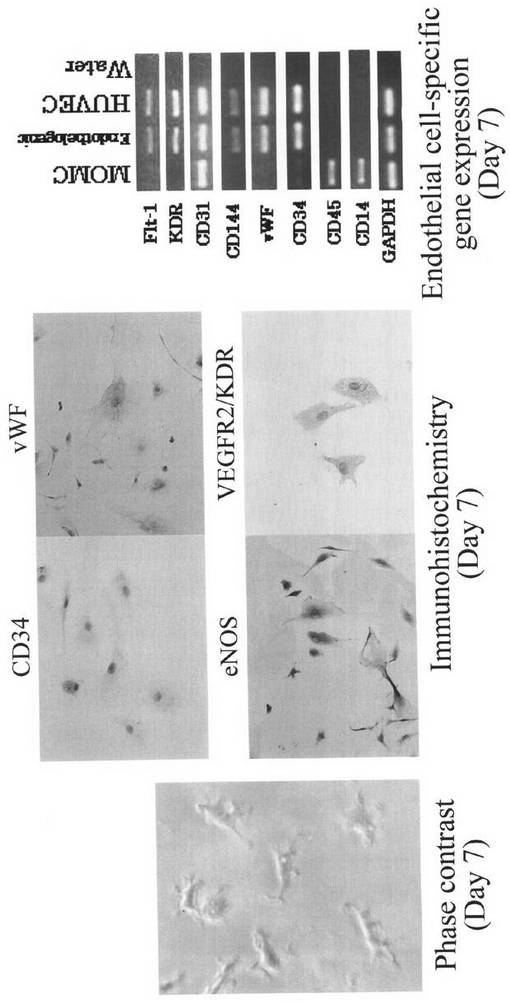
EBM−2培地で7日間分化誘導したMOMCは紡錘形から多数の突起を有する形態に変化し、もともと発現していなかった血管内皮細胞に特異的なvWF、eNOS、VEGFR2/KDR/Flk−1を発現した。
【0082】
RT−PCRによる遺伝子発現解析では、EBM−2培地で7日間分化を誘導したMOMCで血管内皮に特徴的なFlt−1、VEGFR2/KDR/Flk−1、CD31、CD144、vWFの遺伝子発現を認めた。Flt−1とCD31はMOMCで発現がみられたが、他の遺伝子は誘導後に発現された。分化誘導後にCD34の発現は増強し、CD45、CD14の発現は消失した。
【0083】
これらの成績から、MOMCを血管内皮細胞維持培地で培養することにより、血管内皮細胞への分化が誘導されることが証明された。
【図面の簡単な説明】
【0084】
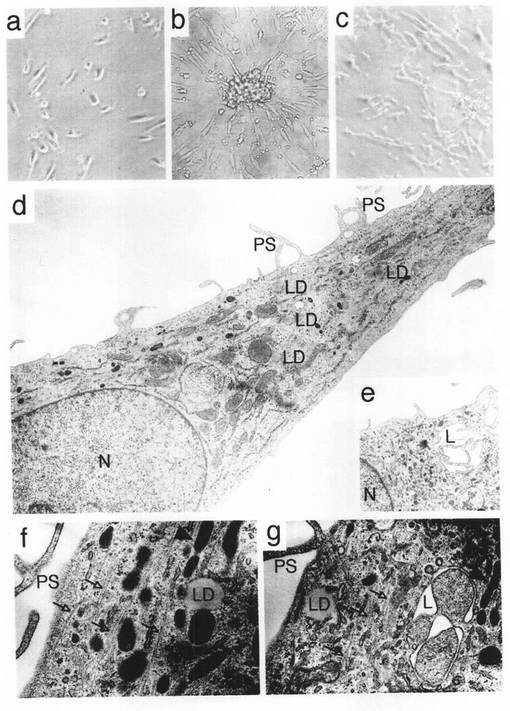
【図1】本発明のMOMCの形態を示す図である。(a,b)PBMCをフィブロネクチン上、10%FBS添加低グルコースDMEMにおいて7日間培養し、位相差顕微鏡下での観察を行った。原倍率は、aが×80、bが×40である。フィブロネクチンでコートした新たなプレートにMOMCを移し替え、24時間培養した。(c)位相差顕微鏡観察結果を示す画像である(×40)。(d、e)電子顕微鏡観察結果を示す画像である(d及びe、×5000;f及びg、×30000)。中間フィラメントの束を矢印で示し、棒状微小管に似た構造物を矢頭で示した。Lは迷路状の細胞小胞、LDは脂質滴、Nは核、PSは偽足をそれぞれ示している。
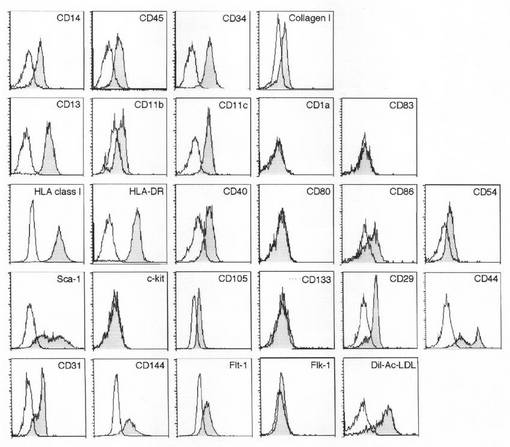
【図2】本発明のMOMCのフローサイトメトリー分析結果を示す図である。PBMCをフィブロネクチンでコートしたプレート上で培養し、7日目に接着細胞を回収し、図中に示した一連のモノクローナル抗体で染色してフローサイトメトリー分析を行った。斜線を施したヒストグラムは、目的分子の発現を示している。白抜きのヒストグラムは、アイソタイプが一致するコントロールモノクローナル抗体で染色したコントロールを示している。この結果は、少なくとも3回実施した実験から得たものである。
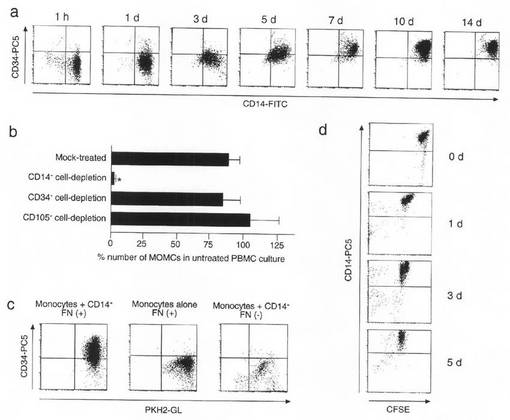
【図3】本発明のMOMCが循環CD14+単球由来であることを説明する図である。(a)PBMCをフィブロネクチン上で、1時間、1日、3日、5日、7日、10日及び14日間、それぞれ培養した。接着細胞を回収し、FITC結合抗CD14抗体及びPC5結合抗CD34モノクローナル抗体で染色し、フローサイトメトリーにかけた。この結果は、個別に3回行った実験から得たものである。(b)CD14+細胞、CD34+細胞又はCD105+細胞を消失させたPBMC、並びにモック処理を施した細胞を、フィブロネクチン上で7日間培養した。1cm3あたりに付着した細胞数を数え、その結果を無処理PBMC培養における付着細胞数に対する割合として表した。この結果は、3名のドナーから得た数値の平均及びSDである。図中のアステリスクは、モック処理PBMC培養と比較したときの有意差を示す。(c)MACS(磁気細胞分離装置)でソーティングしたCD14+単球を、PKH67染色し、非標識CD14-細胞の存在/非存在下(割合1:4)にて、フィブロネクチンでコートした、若しくはコーティングを施していないプラスチックプレート上で7日間培養した。接着細胞を回収し、PC5結合抗CD34モノクローナル抗体で染色し、フローサイトメトリー分析を行った。この結果は、3回の実験から得たものである。(d)MACSでソーティングしたCD14+単球のCFSE染色を行い、フィブロネクチン上で、0、1、3、5日間それぞれ培養した。接着細胞を回収し、PC5結合抗CD14モノクローナル抗体で染色した。これらの細胞のフローサイトメトリー分析を行った。この結果は、実験を3回実施して得たものである。
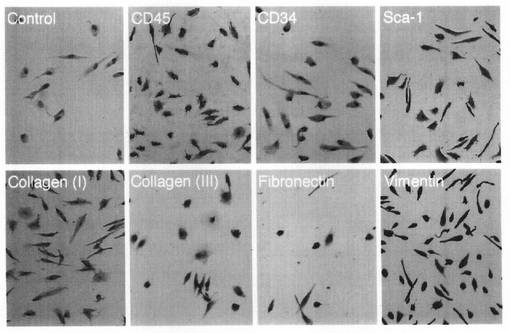
【図4】本発明のMOMCの組織免疫化学分析結果を示す図である。PBMCをフィブロネクチンでコートしたプレート上で7日間培養して得られたMOMCを、フィブロネクチンでコートしたチャンバースライド上に移した(I型コラーゲンをコートしたスライドは、フィブロネクチン染色のためにだけ用いた)。培養24時間後に、上記スライドを10%ホルマリンに固定し、図中に示したモノクローナル抗体でそれぞれ染色した。ヘマトキシリンで核を対比染色した。原倍率は、×100である。この結果は、少なくとも3回実施した実験から得たものである。
【図5】本発明のMOMCが増殖する様子を示した図である。フィブロネクチンでコートしたプレート上、PBMCを7日間培養して得られたMOMCを、フィブロネクチンでコートしたチャンバースライド上に移した。MOMCをさらに1、3、5、7、10日間培養し、BrdU染色を行った。核は、ヘマトキシリンで対比染色した。(a)1日目及び5日目における代表的な図である。矢印は、BrdU染色陽性の核を示す。(b)BrdU染色の結果をみるために少なくとも200個の細胞を数え、1、3、5、7、10日間培養した各スライド上のBrdU+細胞数を算定した。結果は、5回行った実験から得た結果の平均及びSDである。(c)MOMCをCFSEで標識し、フィブロネクチンでコートした新たなプレート上で、0、1、3、5日間培養した。接着細胞を回収し、フローサイトメトリー分析にかけた。この結果は3回実施した実験から得たものである。
【図6】本発明のMOMCが、骨、筋、軟骨、及び脂肪細胞へと分化する様子を示す図である。3週間の骨形成誘導を行う前後のMOMCをアリザリン赤で染色し(a、倍率×100)、又はアルカリフォスファターゼ染色を行った(b、×100)。骨形成誘導処理前後のMOMC及び線維芽細胞における細胞内カルシウム沈着量を測定し、タンパク質含量1μgあたりのμg数として表した(c)。3週間の骨形成誘導前後のMOMCと、骨肉腫細胞株におけるオステリックス、骨シアロタンパク質II(BSP II)、オステオカルシン、CD34、CD45、CD14及びGAPDHのmRNA発現を調べた(d)。3週間の筋形成誘導前後のMOMCを染色し、SkM−アクチン(e、×200)又はSkM−MHC(f、×200)染色を調べた。3週間の筋形成誘導前後のMOMCと、筋芽細胞、筋組織及び横紋筋肉腫細胞株におけるマイオジェニン、SkM−MHC、CD34、CD45、CD14及びGAPDHのmRNA発現を調べた(g)。3週間の軟骨形成誘導前後のMOMCを染色し、II型コラーゲン染色を調べた(h、×40)。3週間の軟骨形成誘導前後のMOMCと、軟骨肉腫細胞株におけるα1(II型)及びα1(X型)コラーゲン、CD34、CD45、CD14並びにGAPDHのmRNA発現を調べた(i)。3週間の脂肪形成誘導前後のMOMCをオイル-レッドOで染色した(j、×200)。3週間の脂肪形成誘導前後のMOMCと、脂肪組織におけるPPARγ、aP2、CD34、CD45、CD14及びGAPDHのmRNA発現を調べた(k)。この結果は、少なくとも5回行った実験から得たものである。
【図7】本発明のMOMCを、それぞれ一週間、間葉系へ分化させた後の、CD45(緑)と組織特異的転写因子(赤)の共発現の様子を示す図である。誘導処理前のMOMC(a−d)と、骨形成(e)、筋形成(f)、軟骨形成(g)若しくは脂肪形成(h)の各誘導処理を1週間行ったMOMCにおいてみられるCD45と、Cbfa1/Runx2(a及びe)、MyoD(b及びf)、Sox−9(c及びg)若しくはPPARγ(d及びh)のそれぞれとの共局在性を免疫組織化学法で調べた。共焦点レーザー蛍光顕微鏡下(原倍率×200)で細胞観察を行った。この結果は、3回行った実験から得られたものである。
【図8】本発明のMOMCが、心筋細胞へと分化する様子を示す図である。8日間ウィスターラット培養心筋細胞とともに共培養を行ったMOMCに発現したネスチン(茶)を免疫染色した(A;×200)。MOMCを蛍光色素PKH67(緑)により細胞膜を標識した後に、ウィスターラット培養心筋細胞と7日間共培養を行い、心筋細胞特異的転写因子であるNkx2.5(赤)と、血球系マーカーであるCD45(青)の二重蛍光免疫染色を行った(B;×200)。MOMCを蛍光色素PKH67(緑)により細胞膜を標識した後に、ウィスターラット培養心筋細胞と8日間共培養を行い、心筋細胞特異的転写因子であるeHAND(赤)と、血球系マーカーであるCD45(青)の二重蛍光免疫染色を行った(C;×200)。MOMCをウィスターラット培養心筋細胞と3日、6日、9日、12日間、共培養を行い、心筋細胞構造蛋白であるミオシン軽鎖(MLC2v)のmRNA発現を調べた(D)。この結果は、3回行った実験から得られたものである。
【図9】本発明のMOMCが、神経細胞へと分化する様子を示す図である。8日間ウィスターラット培養神経細胞とともに共培養を行ったMOMCに発現した、神経・心筋前駆細胞に発現するマーカーであるネスチン(茶)を免疫染色した(A;×200)。MOMCを蛍光色素PKH67(緑)により細胞膜を標識した後に、ウィスターラット培養神経細胞と4日間共培養を行い、神経細胞特異的転写因子であるNeuroD(赤)と、血球系マーカーであるCD45(青)の二重蛍光免疫染色を行った(B;×200)。MOMCをウィスターラット培養神経細胞と3日間共培養を行い、ネスチン(茶)と神経細胞特異的転写因子であるNeurogenin2(赤)の二重蛍光免疫染色を行った(C;×200)。MOMCをウィスターラット培養神経細胞と3日間共培養を行い、PKH67(緑)と成熟した神経のマーカーであるHu(赤)の二重蛍光免疫染色を行った(D;×200)。MOMCをウィスターラット培養神経細胞と9日間共培養を行い、PKH67(緑)と成熟した神経のマーカーであるNeuN(赤)の二重蛍光免疫染色を行った(E;×200)。この結果は、3回行った実験から得られたものである。
【図10】本発明のMOMCが、血管内皮細胞へと分化する様子を示す図である。血管内皮細胞維持培地EBM−2培地で7日間分化誘導したMOMCは紡錘形から多数の突起を有する形態に変化した(左;×200)。EBM−2培地で7日間分化誘導したMOMCに発現した、CD34と血管内皮細胞に特異的なvWF、eNOS、VEGFR2/KDR/Flk−1を免疫染色(茶)した(中;×200)。MOMCをEBM−2培地中で7日間培養し、血管内皮細胞に発現されるFlt−1、VEGFR2/KDR/Flk−1、CD31、CD144、vWFおよびCD34、CD45、CD14、GAPDHのmRNA発現を調べた(右)。この結果は、3回行った実験から得られたものである。
【技術分野】
【0001】
本発明は、間葉系細胞、心筋細胞、血管内皮細胞等の中胚葉系細胞や神経細胞に分化しうるCD14+及びCD34+の単球由来多能性細胞や、該単球由来多能性細胞から分化誘導された間葉系細胞、心筋細胞、血管内皮細胞等の中胚葉系細胞・組織と神経細胞・組織や、これらを有効成分とする治療剤や、これらを投与する治療方法に関する。
【背景技術】
【0002】
末梢血単球は骨髄造血幹細胞に由来し、マクロファージ、樹状細胞、破骨細胞、中枢神経系の小グリア細胞、肝臓のクッパー細胞等の数種の食細胞に分化することが知られている(例えば、非特許文献1〜4参照。)。単球の各種食細胞への分化は、種々の成長因子からのシグナリングによって調節されている。すなわち、マクロファージへの分化はM−CSF、樹状細胞への分化はGM−CSF及びIL−4、破骨細胞への分化はNFリガンドの受容体活性因子(RANKL)やM−CSF(例えば、非特許文献2、3、5参照。)からのシグナリングによってそれぞれ調節されている。つい最近まで単球の分化能は、組織食細胞に限定的であると考えられてきた。しかし近年の研究成果から、種々の血管由来因子を組合わせた条件下でのインビトロ培養を行うことにより、ヒト単球が血管内皮様細胞へと分化する能力を示すことが明らかになった(例えば、非特許文献6、7参照。)。さらに、肉芽腫のインビトロモデルにおける単球分化プロセスにおいて骨特異的アルカリフォスファターゼが発現されることが報告さている(例えば、非特許文献8参照。)。しかし、単球の分化能は、まだ完全に解明されておらず、単球が食細胞以外の細胞型への分化能を有しているかどうかはよく知られていなかった。
【0003】
他方、多くの成人組織には、自己複製能、及び不可逆的最終分化を遂げる娘細胞産生能を有する幹細胞群が存在していることが明らかになっている(例えば、非特許文献9参照)。最も特徴づけが進んでいるのが造血幹細胞とその子孫細胞であるが、幹細胞は、間葉系、神経、及び造血幹細胞等、殆どの臓器において同定されている(例えば、非特許文献10〜12参照。)。間葉系幹細胞(MSC)は、骨髄における接着線維芽様細胞として同定されており、骨、軟骨、脂肪、筋肉、及び骨髄間質等の間葉組織系統への分化能を有している(例えば、非特許文献10参照。)。MSCと似た形態及びフェノタイプ上の特徴並びに分化能を有する間葉系前駆細胞が、臍帯血(例えば、非特許文献13参照。)、及び胎児(例えば、非特許文献14参照。)や成人(例えば、非特許文献15参照。)の末梢血において、著しく低頻度ではあるが存在していることが近年報告されている。しかし、MSC及び循環MSC様細胞においては、各種造血幹マーカー及び幹細胞/血管内皮マーカーCD34が発現されることはない(例えば、非特許文献10、13〜15参照。)。
【0004】
上記のように、組織再生における将来的な治療的介入に対する供給源候補として、種々の出生後の組織特異的幹細胞及び胚幹(ES)細胞の分析が現在進められている(例えば、非特許文献9参照。)。骨形成不全症のヒト小児の場合(例えば、非特許文献16参照。)と同様に動物モデルへ骨髄由来MSCを移植すると、かかるMSCが多くの臓器に移入し、組織特異的系統に従って分化することが報告されている(例えば、非特許文献17、18参照。)。しかしMSCは、ヒト成人骨髄では、稀少細胞群であり(0.01%〜0.001%)、再生治療に十分数量のMSCを増殖させることは、技術的に困難であり、高価であり、また長時間を要するものであった(例えば、非特許文献19参照。)。ES細胞は胚芽細胞由来の多分化能細胞であり、インビトロで未分化のまま無限増殖することができ、インビボで殆どの細胞型へと分化誘導される(例えば、非特許文献20参照。)。ES細胞はヒトから単離されたものだが、研究や治療に用いる際には倫理上の問題という足枷がある(例えば、非特許文献21参照。)。
【0005】
出生後のヒト末梢血における血管内皮細胞(例えば、非特許文献22参照。)、平滑筋細胞(例えば、非特許文献23参照。)、及び間葉系細胞(例えば、非特許文献15参照。)等の血管内皮あるいは間葉系細胞型への分化能をもつ前駆細胞が数種類、報告されている。内皮前駆細胞及び平滑筋前駆細胞をインビトロで増殖させるためには、成長因子をいくつか組み合わせて用いることが必要である(例えば、非特許文献22、23参照。)。間葉系前駆細胞は、外因性成長因子不在下における20%ウシ胎児血清(FBS)添加培地での増殖が可能であるが、PBMC培養における間葉系前駆細胞の発生は、CD14+細胞の除去によって影響されないことが報告されている(例えば、非特許文献15参照。)。しかし、これら血管内皮あるいは間葉系前駆細胞は、CD14、CD45、CD34及びI型コラーゲンに対して陽性というフェノタイプ上の特徴を有していない。
【0006】
【非特許文献1】Bioessays, 17, 977-986, 1995
【非特許文献2】Blood, 98, 2544-5254, 2001
【非特許文献3】BMC Immunol., 3, 15, 2002
【非特許文献4】Microsc Res Tech., 39, 350-364, 1997
【非特許文献5】J Exp Med., 190, 1741-1754, 1999
【非特許文献6】Differentiation, 65, 287-300, 2000
【非特許文献7】Cardiovascular Res., 49, 671-680, 2001
【非特許文献8】Immunobiology, 202, 68-81, 2000
【非特許文献9】Science, 287, 1442-1446, 2000
【非特許文献10】Science, 284, 143-147, 1999
【非特許文献11】Science, 287, 1433-1438, 2000
【非特許文献12】J. Hepatol., 29, 676-682, 1998
【非特許文献13】Br. J. Haematol., 109, 235-242, 2000
【非特許文献14】Blood, 98, 2396-2402, 2001
【非特許文献15】Arthritis Res., 2, 477-488, 2000
【非特許文献16】Nat. Med., 5, 309-313, 1999
【非特許文献17】Nat. Med., 6, 1282-1286, 2000
【非特許文献18】Science, 279, 1528-1530, 1998
【非特許文献19】Stem Cells, 19, 180-192, 2001
【非特許文献20】Trends Biotechnol., 18, 53-57, 2000
【非特許文献21】Science, 287, 1397, 2000
【非特許文献22】Science, 275, 964-967, 1997
【非特許文献23】Circulation, 106, 1199-1204, 2002
【発明の開示】
【発明が解決しようとする課題】
【0007】
現在の医学に残された大きな課題は疾患、外傷などによる臓器の欠失や機能障害の克服といわれている。このような状態に対する治療として現時点で施行しうる唯一の方法は臓器移植であるが、脳死判定やドナー供給の問題のため現実的な治療法として普及するには未だ多くの障害がある。一方、最近の幹細胞および発生生物学の進歩に伴って臓器再生を目指した再生医学が注目され、21世紀に進むべき医療の方向性として期待されている。動物実験レベルでは胚性幹細胞(ES細胞)の移植により臓器の機能回復が報告されているが、ヒトへの応用を考えた場合には拒絶反応やES細胞使用による倫理的な問題で行き詰まっているのが現状である。また、成人の各種組織幹細胞(間葉系、血管、肝など)も生体内にきわめて少数しか存在せず、その単離は技術的に難しく、移植に十分な細胞数を得ることは現時点で困難とされている。したがって、ES細胞や組織幹細胞を用いた再生医療が実地医療に応用されるためには解決しなければならない多くの問題が山積しており、特に、細胞移植による再生医療が現実のものとなるためには、分化能を有する細胞の安定供給が必要不可欠であると考えられる。
【0008】
本発明の課題は、必要十分量を低侵襲的かつ簡便に安定供給でき、細胞移植に際してドナー確保や拒絶反応の問題がなく、倫理的な問題も少ない、骨、軟骨、骨格筋、脂肪などの間葉系細胞、血管内皮細胞、心筋細胞、神経細胞など多種類の細胞へと分化する能力を有する多能性細胞や、かかる多能性細胞から分化誘導された間葉系細胞、心筋細胞、血管内皮細胞等の中胚葉系前駆細胞・中胚葉系細胞・中胚葉系組織と神経細胞・組織や、これらを有効成分とする治療剤や、これらを投与する治療方法を提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、末梢血単核細胞(PBMC)をフィブロネクチンでコートしたプラスチックプレート上で7〜10日間培養すると線維芽細胞様の細胞が出現することを観察した。この線維芽細胞様形態を呈するヒト細胞群の由来および生理機能に興味を持って解析したところ、これらの細胞は循環CD14+単球由来で、CD14+CD45+CD34+I型コラーゲン+という独特のフェノタイプを有し、特定の培養条件により骨、軟骨、骨格筋、脂肪などの間葉系細胞、血管内皮細胞、心筋細胞、神経細胞へと分化する能力を有することを見い出し、単球由来多能性細胞(monocyte-derived multipotent cell;MOMC)と命名した。このように、循環単球が食細胞だけでなく種々の間葉系細胞等への分化能を有する多能性前駆細胞であることを初めて明らかにした本発明者らによる知見に基づいて、本発明を完成するに至った。
【0010】
すなわち本発明は、単球に由来し、CD14とCD34とを発現する単球由来多能性細胞(請求項1)や、単球に由来し、CD14とCD34とCD45とI型コラーゲンとを発現する単球由来多能性細胞(請求項2)や、間葉系組織へと分化誘導する条件下での培養により、間葉系細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞(請求項3)や、間葉系細胞が、骨芽細胞、骨格筋芽細胞、軟骨細胞又は脂肪細胞であることを特徴とする請求項3記載の単球由来多能性細胞(請求項4)や、培養心筋細胞との共培養等の心筋へと分化誘導する条件下での培養により、心筋細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞(請求項5)や、培養神経細胞との共培養等の神経へと分化誘導する条件下での培養により、神経細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞(請求項6)や、血管内皮細胞を維持する条件下での培養等の血管内皮へと分化誘導する条件下での培養により、血管内皮細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞(請求項7)や、中胚葉系細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞(請求項8)に関する。
【0011】
また本発明は、請求項1〜8のいずれか記載の単球由来多能性細胞を間葉系組織へと分化誘導する条件下で培養することにより誘導された間葉系前駆細胞、間葉系細胞又は間葉系組織(請求項9)や、間葉系細胞が、骨芽細胞、骨格筋芽細胞、軟骨細胞又は脂肪細胞であることを特徴とする請求項9記載の間葉系前駆細胞、間葉系細胞又は間葉系組織(請求項10)や、請求項1〜8のいずれか記載の単球由来多能性細胞を培養心筋細胞との共培養等の心筋へと分化誘導する条件下での培養により誘導された心筋前駆細胞、心筋細胞又は心筋組織(請求項11)や、請求項1〜8のいずれか記載の単球由来多能性細胞を培養神経細胞との共培養等の神経へと分化誘導する条件下での培養により誘導された神経前駆細胞、神経細胞又は神経組織(請求項12)や、請求項1〜8のいずれか記載の単球由来多能性細胞を血管内皮細胞を維持する条件下での培養等の血管内皮へと分化誘導する条件下での培養により誘導された血管内皮前駆細胞、血管内皮細胞又は血管内皮組織(請求項13)や、請求項1〜8のいずれか記載の単球由来多能性細胞から分化・誘導された中胚葉系前駆細胞、中胚葉系細胞又は中胚葉系組織(請求項14)に関する。
【0012】
さらに本発明は、請求項1〜8のいずれか記載の単球由来多能性細胞、並びに/又は、前記単球由来多能性細胞から分化・誘導された中胚葉系前駆細胞、中胚葉系細胞及び/若しくは中胚葉系組織を有効成分とする治療剤(請求項15)や請求項1〜8のいずれか記載の単球由来多能性細胞、並びに/又は、前記単球由来多能性細胞から分化・誘導された神経前駆細胞、神経細胞及び/若しくは神経組織を有効成分とする治療剤(請求項16)に関する。
【発明の効果】
【0013】
本発明によると、細胞治療や再生医療に極めて有用な、骨、軟骨、骨格筋、脂肪などの間葉系細胞、血管内皮細胞、心筋細胞、神経細胞など多種類の細胞へと分化する能力を有する単球由来の多能性細胞MOMCを得ることができる。そして、単球は末梢血から簡単に大きな侵襲なく採取することができる上、単球は末梢血単核球の約20%を占め、必要十分量の細胞を安定供給できる。単球からMOMCへの分化誘導は簡単、迅速、安価に行え、特殊な装置を必要としない。また、細胞移植に自己の細胞を用いることができることから、ドナー確保や拒絶反応の問題がなく、倫理的な問題も少ない。本発明は、単球/食細胞系に関する伝統的な生物学的知見を覆すものであり、単球の分化能並びに単球の生体恒常性維持及び病態誘導に果たす役割を理解する上で、大きく貢献するものである。
【発明を実施するための最良の形態】
【0014】
本発明の単球由来多能性細胞(MOMC)としては、単球に由来し、CD14とCD34とを発現する細胞又は細胞群や単球に由来し、CD14とCD34とCD45とI型コラーゲンとを発現する細胞又は細胞群であれば特に制限されるものではなく、上記単球の起源としてはマウス、ラット、イヌ、ブタ、サル、ヒトなど特に限定されないが、ヒトを好適に例示することができ、ヒト単球の場合、ドナー由来でもよいが自己由来が特に好ましい。また、単球の供給源としては末梢血あるいは骨髄由来の単球、又は造血幹細胞からex vivoで分化させた単球を例示することができ、ここで単球とはCD14又はCD11b陽性と定義される。
【0015】
上記CD14とCD45とは単球及び単球由来細胞のマーカーとして、CD34は幹細胞のマーカーとして、I型コラーゲンは間葉系細胞のマーカーとして知られている。本発明の単球由来多能性細胞としては、幹細胞マーカーとしてのCD105、Sca−1や、間葉系細胞のマーカーとしてのIII型コラーゲン、フィブロネクチンや、血管内皮マーカーとしてのVEカドヘリン、Flt−1を発現するものが好ましい。かかるMOMCは、上記タンパク質の発現パターンから、単球、マクロファージ、樹状細胞とは異なる細胞であり、間葉系細胞、血管内皮細胞、幹細胞の特徴をあわせ持つ新規の細胞群といえる。
【0016】
本発明のMOMCは循環CD14+単球に由来するが、このことは以下にいくつか列挙する。第一に、MOMCは、単球系統マーカーであるCD14、CD13、CD11b、CD11c及びCD64に対して陽性である。第二に、フィブロネクチン上で培養した末梢血接着細胞のフェノタイプ分析を連続的に行った結果、CD14+接着細胞上におけるCD34の発現が増加していた。第三に、CD14+細胞を除去することにより、PBMCからのMOMC発生は殆ど完全に阻害されたが、CD34又はCD105陽性細胞を除去しても阻害されなかった。最後に、標識CD14+細胞を用いた研究から、CD14+単球においてはCD34発現がアップレギュレートされ、またCD14-細胞群の中にPBMC培養中に急速に増殖する細胞がないことが明らかになった。
【0017】
本発明のMOMCとしては、中胚葉系細胞にMSCを分化させることが知られている誘導条件下で中胚葉系細胞に分化しうる多能性を備えた細胞、具体的には、間葉系組織へと分化誘導する条件下での培養により骨芽細胞、骨格筋芽細胞、軟骨細胞、脂肪細胞、骨髄間質細胞、平滑筋細胞等の間葉系細胞に分化しうる多能性や、培養心筋細胞との共培養等の心筋へと分化誘導する条件下での培養により心筋細胞に分化しうる多能性や、血管内皮細胞を維持する条件下での培養等の血管内皮へと分化誘導する条件下での培養により血管内皮細胞に分化しうる多能性の他、培養神経細胞との共培養等の神経へと分化誘導する条件下での培養により、外胚葉系細胞である神経細胞に分化しうる多能性などを備えた細胞が好ましい。
【0018】
また、本発明は、本発明のMOMCを、中胚葉系細胞にMSCを分化させることが知られている誘導条件下で分化・誘導された中胚葉系前駆細胞、中胚葉系細胞又は中胚葉系組織、具体的には、MOMCを間葉系組織へと分化誘導する条件下で培養することにより誘導された骨芽細胞、骨格筋芽細胞、軟骨細胞、脂肪細胞等の間葉系前駆細胞、間葉系細胞又は間葉系組織や、MOMCを培養心筋細胞と共培養等の心筋へと分化誘導する条件下での培養により誘導された心筋前駆細胞、心筋細胞又は心筋組織や、MOMCを血管内皮細胞を維持する条件下での培養等の血管内皮へと分化誘導する条件下での培養により誘導された血管内皮前駆細胞、血管内皮細胞又は血管内皮組織の他、MOMCを培養神経細胞との共培養等の神経へと分化誘導する条件下での培養により誘導された外胚葉系の神経前駆細胞、神経細胞又は神経組織に関する。
【0019】
骨芽細胞への分化誘導条件下で培養することにより誘導された骨芽細胞としては、類円形の形態となり、アリザリン赤陽性のカルシウム沈着、アルカリフォスファターゼ染色陽性、細胞内Ca濃度の増加、骨芽細胞に特異的な骨シアロタンパク質II、オステオカルシン遺伝子を発現する細胞を好適に例示することができる。軟骨細胞への分化誘導条件下で培養することにより誘導された軟骨細胞としては、細胞質に富むやや大型の類円形の形態となり、軟骨細胞に特異的なII型及びX型コラーゲンを発現する細胞を好適に例示することができる。骨格筋細胞への分化誘導条件下で培養することにより誘導された骨格筋細胞としては、紡錘形の細長い形態をとり、骨格筋特異的アクチン、ミオシンを発現する細胞を好適に例示することができる。脂肪細胞への分化誘導条件下で培養することにより誘導された脂肪細胞としては、オイル赤染色陽性の脂肪滴が出現し、PPARγ、aP2遺伝子の発現が増加する細胞を好適に例示することができる。
【0020】
心筋へと分化誘導する条件下での培養により誘導された心筋前駆細胞や心筋細胞としては、ネスチン及び心筋特異的転写因子Nkx2.5、eHANDを発現し、心筋細胞構造蛋白であるミオシン軽鎖の遺伝子を発現する細胞を好適に例示することができる。神経へと分化誘導する条件下での培養により誘導された神経前駆細胞や神経細胞としては、ネスチン及び神経細胞特異的転写因子Neuro D、Neurogenin 2を発現し、その後神経マーカーのHu、Neu Nを発現する細胞を好適に例示することができる。血管内皮へと分化誘導する条件下での培養により誘導された血管内皮前駆細胞や血管内皮細胞としては、小突起を有する多型性の形態となり、血管内皮に特異的なマーカータンパク質KDR、vWF、eNOSを発現する細胞を好適に例示することができる。
【0021】
上記のように、MOMCは、主としてMSCを中胚葉系細胞に分化させることが知られている誘導条件下において間葉系細胞等の中胚葉系細胞への分化能を有しているの他、MOMCを神経へと分化誘導する条件下での培養により誘導された外胚葉系の神経細胞への分化能を有している。さらにMOMCは、系統特異的転写因子の発現タイミングという点において、MSC分化と同じ段階を追って各種間葉系細胞等の中胚葉系細胞などへと分化する。例えば、Cbfa1/Runx2の発現に続いて、シアロタンパク質II及びオステオカルシンが発現され(Cell, 108, 17-29, 2002)、SkM−アクチン及びミオシンが発現される前に、MyoDが発現される(Front Biosci., 5, D750-767, 2000)。かかる知見は、各種間葉系統への分化プロセスが、MOMCとMSCで共通していることを示すものである。
【0022】
本発明のMOMCの作製方法としては、末梢血単核細胞(PBMC)をフィブロネクチン上でインビトロ培養、例えば、フィブロネクチンでコートしたプラスチックプレート上で培養、好ましくは5〜14日間、特に好ましくは7〜10日間培養し、CD14とCD34とを発現する線維芽細胞様細胞を採取する方法であれば特に制限されるものではないが、PBMCを外因性成長因子不在下で培養することが好ましい。上記フィブロネクチンのかわりにコラーゲンやラミニンを用いてもよいが、単球のMOMCへの分化にはこれらフィブロネクチン等の他に、CD14-細胞由来の液性因子が必要とされる。上記PBMCの起源としてはマウス、ラット、イヌ、ブタ、サル等の実験動物又はヒトなど特に限定されないが、ヒトPBMCを好適に例示することができる。また、ヒトPBMCは、ヒト静脈血から常法により単離することができる。上記培養方法としては特に制限されないが、37℃、5%CO2、湿気下にて密度104〜107/ml、例えば2×106/mlで培養し、2〜4日毎に、特に好ましくは3日毎に非接着細胞を除去するとともに、新鮮培地を補充する培養方法を挙げることができる。このようにして得られる本発明のMOMCは、その元来のフェノタイプを5世代継代するまで維持しつつ、培養によって増殖させることが可能である。
【0023】
本発明は、本発明のMOMC、並びに/又は、前記MOMCから分化・誘導された中胚葉系前駆細胞、中胚葉系細胞及び/若しくは中胚葉系組織、具体的には、MOMCを間葉系組織へと分化誘導する条件下で培養することにより誘導された骨芽前駆細胞、骨格筋芽前駆細胞、軟骨前駆細胞、脂肪前駆細胞等の間葉系前駆細胞、骨芽細胞、骨格筋芽細胞、軟骨細胞、脂肪細胞等の間葉系細胞、骨、軟骨、筋肉、脂肪等の間葉系組織や、MOMCを培養心筋細胞と共培養等の心筋へと分化誘導する条件下での培養により誘導された心筋前駆細胞、心筋細胞、心筋組織や、MOMCを血管内皮細胞を維持する条件下での培養等の血管内皮へと分化誘導する条件下での培養により誘導された血管内皮前駆細胞、血管内皮細胞、血管内皮組織等を有効成分とする治療剤や、本発明のMOMC、並びに/又は、前記MOMCから分化・誘導された神経前駆細胞、神経細胞及び/若しくは神経組織等を有効成分とする治療剤に関する。また、本発明のMOMC、並びに/又は、前記MOMCから分化・誘導された、前記中胚葉系前駆細胞、中胚葉系細胞及び/若しくは中胚葉系組織や、前記神経前駆細胞、神経細胞及び/若しくは神経組織等を投与、例えば障害、欠損部位およびその近傍へ直接注入、又は末梢血へ投与することを特徴とする治療方法に関する。MOMC又は分化誘導処理したMOMCのいずれが治療剤として適しているかは、細胞の種類や疾病の種類や投与方法を考慮して適宜決定することが好ましい。また、MOMCは遺伝子導入が比較的簡単な細胞あることから、ヒトへの細胞移植の前に特定の遺伝子を導入した後に組織再生治療等に用いることもできる。例えば、ある種の先天性疾患で骨の形成に障害がある場合に、その遺伝子を是正した上で移植することや、あるいは、特定の蛋白(サイトカイン、成長因子、ホルモンなど)を産生するように調製した上で移植することもできる。
【0024】
前記のとおり、種々の間葉系組織等の中胚葉系組織や神経組織への分化能をもつことからMOMCには、先天性疾患、変性疾患及び外傷に対する組織再建治療における細胞供給源として有用である。具体的に本発明の治療剤や治療方法の対象となる疾患又は病態としては、骨形成不全、骨折、関節リウマチなど変性疾患による骨破壊や、軟骨が破壊される関節リウマチ、変形性関節症や、ジストロフィーなどの先天性疾患や筋炎などの後天性疾患による筋萎縮や、心筋梗塞や心筋症による心筋障害や、脳梗塞、パーキンソン病などの脳障害、脊髄損傷などの外傷や、動脈硬化や膠原病による血管障害を挙げることができ、便宜上、豊胸をはじめとした美容形成等も本発明の治療剤や治療方法に含まれる。このようなMOMC又は分化誘導処理したMOMCを使用する細胞治療には、現在提案されている組織特異的幹細胞及びES細胞を用いる再生治療等にはない大きな利点がある。すなわち、血液採取という低侵襲的に患者本人から大量の単球を採取できることから、循環単球は比較的容易に入手可能な自己細胞供給源ということができ、さらに、単球からMOMCを作製することは技術的に簡便で迅速に実施でき、また、ES細胞を使用する際の倫理上のジレンマも回避することができる。
【実施例】
【0025】
以下、実施例により本発明をより具体的に説明するが、本発明の技術的範囲はこれらの例示に限定されるものではない。
[方法と材料]
実施例1(MOMCの培養)
成人健常ドナーから得たヘパリン添加静脈血を、Lymphoprep(Nycomed Pharma AS社製)を用いた密度勾配遠心分離にかけてPBMCを単離した。血液検体は全て、倫理委員会が定める書面によるインフォームドコンセントを被験者が提出した後に採血したものである。単離した細胞をリン酸緩衝液(PBS)で2回洗浄し、10%熱不活化FBS(JRH Biosciences社製)添加低グルコースDMEMに懸濁した。PBMCを、外因性成長因子不在下、37℃、5%CO2、湿気下の諸条件のもと、フィブロネクチンでコートしたプラスチックプレート上にて密度2×106/mlで培養し、培養3日目に非接着細胞を除去した。3日毎に新鮮培地に交換しながら、4週間まで細胞を培養した。培養7〜10日目に接着細胞をMOMCとして回収し、以下のアッセイに用いたり、或いはフィブロネクチンでコートしたプレートに新たに移し替えて同一培養条件下で10世代まで維持した。
【0026】
MOMCの由来を考察するため、CD14+、CD34+、又はCD105/エンドグリン/SH2+細胞を除去したPBMCをフィブロネクチンでコートしたプレート上で7日間培養した。CD14+又はCD34+細胞は、磁気ビーズを結合させた抗CD14モノクローナル抗体又は抗CD34モノクローナル抗体(DynaBeads社製)を用い、製造者のマニュアルに従って磁気分離を行うことにより除去した。CD105+細胞を除去したPBMCは、先ず抗CD105モノクローナル抗体(Immunotech社製)共存下でインキュベートし、次に磁気ビーズを結合させたヤギ抗マウスIgG抗体(Dynal社製)共存下でインキュベートすることにより調製した。アイソタイプが一致するマウスモノクローナル抗体共存下、さらにビーズ結合抗マウスIgG抗体共存下でインキュベートしたモック処理PBMCを調製し、コントロールとした。フローサイトメトリーにより解析したところ、CD14+細胞を除去したPBMC画分中のCD14+細胞の割合は、一貫して0.5%未満であり、CD34+細胞除去画分中におけるCD34+細胞並びにCD105+細胞除去画分中におけるCD105+細胞の割合は、0.01%未満だった。1cm3あたりの付着細胞数を算定し、その結果を無処理のPBMC培養における付着細胞数に対する割合で表した。
【0027】
抗CD14モノクローナル抗体結合磁気ビーズ(CD14 MicroBeads;Miltenyi Biotech社製)を用い、さらに製造者マニュアルに従ってMACS(磁気細胞分離装置)によるカラム分離を行うことにより、PBMCから循環CD14+単球及びCD14-細胞を分離する実験も実施した。フローサイトメトリー分析の結果、単球画分には98%を越えるCD14+細胞が、またCD14-細胞画分には0.5%未満のCD14+細胞がそれぞれ存在していた。PKH67グリーン(Sigma社製)で単球を標識し、フィブロネクチンでコートしたプラスチックプレート又はコーティングを施していないプラスチックプレート上で非標識CD14-細胞と7日間の共培養を行った(比率1:4)。PKH67標識単球もフィブロネクチンでコートしたプレート上で単独培養した。
【0028】
実施例2(マクロファージ及び樹状細胞の調製)
プラスチックプレート上、20%FBS及び4ng/mlのM−CSF(R&D Systems社製)を添加したM199培地において接着PBMCを7日間培養し、マクロファージを調製した(Differentiation, 65, 287-300, 2000)。プラスチックに接着したPBMCを連続培養条件下において成熟させることにより、成熟単球由来樹状細胞を得た(J Immunol Methods, 196, 121-135, 1996)。その概要は、50ng/mlのGM−CSF及び50ng/mlのIL−4を含む10%FBSを添加したRPMI1640培地上で接着細胞を7日間培養した(材料は全てPeproTech社製)。次に幼若樹状細胞を、50ng/mlのTNF−α(PeproTech社製)共存下で3日間インキュベートした。フローサイトメトリー分析の結果、マクロファージ画分には、CD14+CD80+細胞が98%以上含まれており、樹状細胞画分には、CD83+HLA−DR+細胞が95%以上、CD14+細胞が1%未満の割合で含まれていた。
【0029】
実施例3(細胞株)
健常ドナーに皮膚生検を行ってヒト皮膚線維芽細胞を得て、その初代培養を樹立し、10%FBS添加低グルコースDMEM中で維持した。初代ヒト筋芽細胞は、臨床的に筋炎が疑われるが組織学上は正常な患者の筋肉生検を培養することにより作製した(J Cell Biol., 144, 631-643, 1999)。ヒト骨肉腫細胞株MG−63、横紋筋肉腫細胞株RD、及び軟骨肉腫細胞株OUMS−27は、日本ヒューマンサイエンス研究資源バンク(日本、大阪府)から入手し、10%FBS添加低グルコースDMEMで維持した。
【0030】
実施例4(MOMCの間葉系細胞へのインビトロ分化)
PBMCから新たに作製したMOMC、数代にわたり継代培養されたMOMC、又は低温保存されていたMOMCのいずれかを、新たにフィブロネクチンをコートしたプラスチックプレート、又はチャンバースライドに移して10%FBS添加高グルコースDMEM(Hyclone Laboratories社製)中でセミコンフルエントになるまで生育させた。次に、下記の種々の間葉系細胞型にMSCを分化させることが知られている誘導条件下(Science,284, 143-147, 1999, J Cell Biol., 144, 631-643, 1999, J Cell Biochem., 64, 295-312, 1997, Muscle Nerve,18, 1417-1426, 1995, Tissue Eng., 4, 415-428, 1998, Arthritis Rheum., 44, 85-95, 2001, J Biol Chem., 275, 9645-9652, 2000)で、上記細胞を培養した。コントロールとして、PBMCから新たに単離した単球、マクロファージ及び皮膚線維芽細胞も同一条件下で処理した。
【0031】
骨形成に関しては、100nMのデキサメサゾン、10mMのβ−グリセロホスフェート及び50μMのアスコルビン酸を含む骨形成誘導培地(Clonetics社製)上で接着細胞を培養した。1週間に2回培地を交換することを3週間継続した。
【0032】
筋形成に関しては、接着細胞を10μMの5−アザシチジン(Sigma社製)で24時間処理した。PBSで細胞を洗浄し、5%ウマ血清(Life Technologies社製)、50mMのヒドロコルチゾン(Sigma社製)、及び4ng/mlの塩基性線維芽細胞増殖因子(Sigma社製)を含む培地で培養した。1週間に2回培地を交換することを3週間継続した。
【0033】
軟骨形成に関しては、接着単層細胞をTGF−β(R&D Systems社製)存在下、無血清培地上で3週間培養した。TGF−βは1日おきに培養培地に加え、その最終濃度を10ng/mlとした。
【0034】
脂肪形成に関しては、1μMのデキサメサゾン、0.5mMのメチル−イソブチルキサンチン、10μg/mlのインスリン、及び100mMのインドメタシン(全てSigma社製)を添加した脂肪誘導培地にて細胞をインキュベートした。72時間後に上記培地を、10μg/mlのインスリンを単独添加した維持培地と交換して24時間おいた。脂肪誘導培地で細胞を3回処理し、さらに1週間維持培地において維持した。
【0035】
実施例5(MOMCの心筋細胞へのインビトロ分化)
MOMCを8日間ウィスターラット胎児由来の培養心筋細胞とともに共培養を行った。
培養細胞を4%パラホルムアルデヒドで固定した後、ヒト特異的抗ネスチン(nestin)抗体(Chemicon社)により免疫染色(DAB発色)を行った。
【0036】
MOMCを蛍光色素PKH67(Sigma社)により細胞膜を標識した後に、ウィスターラット培養心筋細胞と7〜10日間、共培養を行った。培養細胞を4%パラホルムアルデヒドで固定した後、抗Nkx2.5抗体、抗eHAND抗体(Santa Cruz社)、抗CD45抗体(DAKO社)により、二重蛍光免疫染色を行った。
【0037】
MOMCをウィスターラット培養心筋細胞と3日、6日、9日、12日間共培養を行った後、mRNAを抽出した。心筋細胞構造蛋白であるミオシン軽鎖(MLC2v)に対するヒト特異的PCRプライマー(TGACAAGAACGATCTGAGAG、CAGGTTCTTGTAGTCCAAGT)を作成し、RT−PCRを行った。
【0038】
実施例6(MOMCの神経細胞へのインビトロ分化)
MOMCを8日間ウィスターラット培養神経細胞とともに共培養を行った。培養細胞を4%パラホルムアルデヒドで固定した後、ヒト特異的抗ネスチン抗体により免疫染色(DAB発色)を行った。
【0039】
MOMCを蛍光色素PKH67により細胞膜を標識した後に、ウィスターラット培養神経細胞と4日間、共培養を行った。培養細胞を4%パラホルムアルデヒドで固定した後、抗NeuroD抗体(Santa Cruz社)、抗CD45抗体(DAKO社)により、二重蛍光免疫染色を行った。
【0040】
MOMCをウィスターラット培養神経細胞と3日間、共培養を行った。培養細胞を4%パラホルムアルデヒドで固定した後、ヒト特異的抗ネスチン抗体、抗Neurogenin2抗体(Chemicon社)により、二重蛍光免疫染色を行った。
【0041】
MOMCを蛍光色素PKH67により細胞膜を標識した後に、ウィスターラット培養神経細胞と9〜10日間、共培養を行った。培養細胞を4%パラホルムアルデヒドで固定した後、抗Hu抗体、抗NeuN抗体(Chemicon社)により二重蛍光免疫免疫染色を行った。
【0042】
実施例7(MOMCの血管内皮細胞へのインビトロ分化)
MOMCを付着細胞として血管内皮細胞維持培地EBM−2(Clonetics社)中で7〜10日間培養した。培養細胞を10%中性緩衝ホルマリンで固定した後、抗CD34抗体(Calbiochem-Novabiochem社)、抗vWF抗体(Dako社)、抗eNOS抗体(Bechton-Dickinson社)、抗VEGFR2/KDR/Flk−1抗体(Sigma社)により免疫染色(DAB発色)を行った。
【0043】
EBM−2培地中で7日間培養したMOMCからRNAを抽出しcDNAへ逆転写した。血管内皮細胞に発現されるFlt−1、VEGFR2/KDR/Flk−1、CD31、CD144、vWFおよびCD34、CD45、CD14、GAPDHに対する特異的プライマーを用いてPCRを行った。コントロールとして分化誘導をかける前のMOMC、培養さい帯静脈血管内皮細胞(HUVEC)由来のRNAを用いた。
【0044】
実施例8(フローサイトメトリー分析)
蛍光細胞染色は以下の手順で行った。2mMのEDTA共存下における氷上インキュベーションにより、接着細胞をプラスチックプレートから脱離させ、正常マウス又はラットの血清を用い、4℃下で10分間ブロックした。抗HLA−DR抗体、抗CD11c抗体(BD Pharmingen社製)、抗CD11b/Mac−1抗体、抗CD14抗体、抗CD29抗体、抗CD34抗体、抗CD44抗体、抗CD83抗体、抗CD105/エンドグリン/SH2抗体、抗CD117/c−kit抗体(Immunotech社製)、抗CD34抗体、抗CD133抗体(MiltenyiBiotech社製)、抗HLAクラスI抗体、抗HLA−DR抗体、抗CD31/PECAM−1抗体、抗Flt−1/VEGFR1抗体、抗Flk−1/VEGFR2抗体(Sigma社製)、抗CD40抗体、抗CD54抗体、抗CD80抗体、抗CD86抗体(Ancell社製)、抗CD144/VE−カドヘリン抗体、抗I型コラーゲン抗体(Chemicon International社製)を含むマウスモノクローナル抗体、又はラット抗Sca−1モノクローナル抗体(Cedarlane Laboratories社製)に、FITC、フィコエリトリン(PE)若しくはPC5を結合させて、又は結合させないで、上記細胞を染色した。非結合モノクローナル抗体を用いた場合には、FITC若しくはPEを結合したヤギ抗マウス抗体若しくはラットIgG F(ab’)2抗体(Immunotech社製)を2次抗体として使用した。IntraPrepTM浸透処理試薬(Immunotech社製)により細胞の浸透処理及び固定化を行い、細胞内染色を行った。細胞の分析は、CellQuestソフトウエアを使用したFACSRCaliburフローサイトメーター(Becton Dickinson社製)により行った。フォワード及びサイドスキャッターでのゲーティングにより可視化細胞を同定し、データは対数ヒストグラム又はドットプロットとして示した。
【0045】
実施例9(免疫組織化学分析)
サイトスピン法により単球及びマクロファージ、樹状細胞をスライド上に塗抹し、これら細胞以外の細胞型は、フィブロネクチンでコートしたチャンバースライド上で培養したが、フィブロネクチン染色用には、コラーゲンI型をコートしたチャンバースライドを用いた。これらの細胞を10%ホルマリンで固定し、3%過酸化物を用い、外因性ペルオキシダーゼ活性を5分間抑制した。抗CD45抗体、抗ビメンチン抗体、抗骨格筋特異的アクチン抗体(SkM−アクチン)(Dako社製)、抗CD34抗体(Calbiochem-Novabiochem社製)、抗I型コラーゲン抗体(Chemicon社製)、抗III型コラーゲン抗体、抗フィブロネクチン抗体(Sigma社製)、抗II型コラーゲン抗体(ICN Biomedicals社製)、抗骨格筋特異的ミオシン重鎖抗体(SkM−MHC)(Zymed Laboratories社製)を含むマウスモノクローナル抗体又はラット抗Sca−1モノクローナル抗体共存下にて30分間スライドをインキュベートし、さらにビオチン標識抗マウス抗体若しくは抗ラットIgG抗体共存下でのインキュベーションを行った。3,3’−ジアミノベンジジンを基質として用い、ストレプトアビジン−ホースラディッシュペルオキシダーゼ複合物(ニチレイ社製)を室温で10分間反応させることにより、抗体−ビオチン結合体を検出した。核をヘマトキシリンで対比染色した。上記1次抗体の代りに正常マウス若しくはラットのIgG抗体(Dako社製)共存下でインキュベートした細胞を負のコントロールとした。
【0046】
蛍光二重染色は以下の方法で行った。4%パラホルムアルデヒドで細胞を固定し、ヤギ抗PEBP2αA抗体若しくは抗Sox9ポリクローナル抗体(Santa Cruz社製)共存下でインキュベートし、次にAlexa FluorR568ヤギ特異的IgG抗体(Molecular Probes社製)共存下でインキュベートし、さらにFITC結合マウス抗CD45モノクローナル抗体(Dako社製)共存化でインキュベートした。同様にマウス抗MyoD抗体(Dako社製)又は抗ペルオキシゾーム増殖活性化受容体γ(PPARγ遺伝子)モノクローナル抗体(Santa Cruz社製)で染色し、その後テトラメチルローダミン・イソチオシアネート異性体Rで標識したマウス特異的IgG抗体(Dako社製)共存化でインキュベートし、さらにFITC結合抗CD45モノクローナル抗体共存化でインキュベートした。共焦点レーザー蛍光顕微鏡(LSM5 PASCAL;Carl-Zeiss社製)を用いて、これらの細胞を観察した。所定のマーカーに対して染色陽性を示した細胞の割合を数値化するために、一培養あたり少なくとも300個の細胞を低倍率で鏡検した。
【0047】
実施例10(アセチル化LDL(Ac−LDL)の摂取)
2.5μg/mlのDil−Ac−LDL(Molecular Probes社製)共存化で接着細胞を1時間培養し、Ac−LDL摂取量をフローサイトメトリーで調べた。
【0048】
実施例11(アルカリフォスファターゼ染色)
10%ホルマリンに細胞を固定し、0.2mg/mlのナフトールAS−TRホスフェート及び0.5mg/mlのFast Red RC(全てSigma社製)を含む溶液中で10分間インキュベートした。
【0049】
実施例12(細胞内カルシウムの検出)
細胞内におけるカルシウム沈着を調べるため、細胞を10%ホルマリンに固定し、2%アリザリンレッドS(Sigma社製)で3分間染色し、蒸留水で入念に洗浄した。市販のキット(Sigma社製)を使い、細胞内カルシウム濃度を測定した(J Biol Chem., 275, 9645-9652, 2000)。ウシ血清アルブミンを標準とするブラッドフォード・プロテイン・アッセイキット(Bio-Rad Laboratories社製)を使用し、細胞抽出物におけるタンパク質含量も測定した。カルシウム濃度は、タンパク質含量1μgあたりのμg数として表示した。
【0050】
実施例13(オイル-レッドO染色)
0.2%グルタルアルデヒドに細胞を5分間固定し、60%イソプロパノールで洗浄し、0.1%オイル-レッドO(Sigma社製)で10分間覆った。60%イソプロパノール、次いで蒸留水による洗浄後、細胞のヘマトキシリン対比染色を行った。
【0051】
実施例14(透過電子顕微鏡)
培養後のMOMCを直ちに2.5%グルタルアルデヒドに固定し、2%四酸化オスミウムで2次固定し、エタノール及び酸化プロピレンの一連使用により脱水してエポキシ樹脂に包埋した。LKBウルトラトーム上で、細胞をダイアモンドナイフで薄切片化した。灰色から銀色の範囲の切片を150−メッシュグリッド上に回収し、酢酸ウラニル及び酢酸鉛で二重染色を施し、JEOL-1200EXII電子顕微鏡下で観察した(Jeol社製)。
【0052】
実施例15(細胞増殖研究)
MOMCの増殖は、文献(Blood, 71, 1201-1210, 1988)記載の方法に従い、BrdUで標識することにより検出した。以下簡潔に説明する。10μMのBrdU(Sigma社製)存在下で、染色するまでの2時間MOMCを培養した。Carnoyの固定剤(メタノール/酢酸)で−20℃で30分間固定した後、細胞を風乾し、2N−HClで1時間処理してDNAを変性させ、さらに10分間0.1Mのホウ酸塩(pH8.5)で中和した。これらの細胞をマウス抗BrdUモノクローナル抗体(Chemicon International社製)共存下でインキュベートし、ビオチン−ストレプトアビジン−ペルオキシダーゼ複合体で染色した。核はヘマトキシリンで対比染色した。上記の1次抗体の代りに、アイソタイプが一致するマウスのコントロールモノクローナル抗体共存下でインキュベートした細胞を負のコントロールとした。非固定細胞を、ヨウ化プロピジウム(Sigma社製)共存下で30分間インキュベートして、蛍光顕微鏡観察を行い、アポトーシスを起こした細胞を検出した。
【0053】
文献記載(J Exp Med., 183, 2313-2328, 1996)の手法に従い、5−カルボキシフルオレセイン・ジアセテート・スクシンイミジル・エステル(CFSE)で細胞を標識し、細胞分裂の様子を考察した。フィブロネクチンでコートしたプレート上で1、3、5日間、CFSE標識単球と非標識CD14-細胞を共培養し、接着細胞を回収してPC5標識抗CD14モノクローナル抗体で染色した。CFSE標識MOMCについても、1、3、及び5日間の培養を行った。CFSE標識による発色強度をフローサイトメトリーにより解析した。
【0054】
実施例16(RT−PCR)
RNeasyキット(Qiagen社製)を用いて分化誘導処理を行った場合と行わなかった場合の双方において、MOMCから全RNAを抽出した。末梢血CD14+単球、並びにマクロファージ、樹状細胞、皮膚線維芽細胞、筋芽細胞、骨肉腫、横紋筋肉腫及び軟骨肉腫を含む種々の培養細胞からも全RNAを抽出した。ヒト筋肉及び脂肪組織由来の全RNAサンプルは、Clonetech Laboratories社から購入した。Molonyマウス白血病ウイルス・リバーストランスクリプターゼ(Takara社製)を用い、オリゴ−dTをプライマーとして、全RNAから一本鎖cDNAを合成した。次にこのcDNA(50ngの全RNAに相当)を、配列番号3〜34に示される表1記載の各種特異的プライマーを用いたPCR増幅法にかけた。このPCR産物を2%アガロースゲル電気泳動分析にかけ、臭化エチジウム染色により可視化した。
【0055】
【表1】
【0056】
実施例17(統計分析)
全ての比較における統計有意は、Mann−WhitneyのU検定により試験した。
【0057】
[結果]
実施例18(MOMCの作製)
フィブロネクチンでコートしたプレート上にて、10%FBSを単独添加したDMEM中でPBMCを培養すると、一部の細胞が直ちにプレートに付着した。円形細胞の小クラスターが24時間以内に発生し、そこから細胞突起が伸張した。培養5日目に線維芽細胞様形態を呈する接着細胞が出現した。さらに3日間培養すると、この線維芽細胞様細胞は、培養中の優勢細胞となった(図1a)。かかる線維芽細胞はクラスター周辺部に高頻度で観察された(図1b)。線維芽細胞様細胞は、培養14日目頃までは徐々にその数を増やした。それ以降、増殖能は次第に消失していったが、細胞は4週間まで生存していた。ドナー50名から得たPBMC108個を培養したところ、7日目に0.3〜1×107個の接着細胞が得られた。フローサイトメトリー分析の結果、培養7日目に回収した細胞は単一のフェノタイプを示し(95%以上がホモジニアス)、CD14、CD45、CD34、及びI型コラーゲンに対して陽性だった(図2)。このフェノタイプは独特であり、単球/マクロファージ(CD14+、CD45+、CD34-及びI型コラーゲン-)、血管内皮前駆細胞(CD14-、CD45-、CD34+及びI型コラーゲン-)(Science, 275, 964-967, 1997)、及び間葉系前駆細胞(CD14-、CD45-、CD34-、及びI型コラーゲン+)(Arthritis Res., 2, 477-488, 2000)を含む既知の末梢血由来接着細胞のフェノタイプとは異なっている。少なくとも50名のドナーから得た細胞は、同じフェノタイプを有していた。7日目に、新たにフィブロネクチンでコートしたプレートに細胞を移し替え、同一条件下で培養したところ、殆ど全ての細胞が、細長い線維芽細胞様の形態を現した(図1c)。これらの細胞は、5世代まで増殖することができ、細胞増殖は継代直後に最も盛んになるようだった。しかし5世代継代以降は、細胞増殖速度が徐々に遅くなり、8世代継代を超えると、その増殖能は消失した。特別な処理を施さない限りは、培養期間中に細胞が成熟間葉系細胞へ自然に分化することはなかったが、高いコンフルエンシーとなるように細胞を播種すると、複数の核をもつ細胞の出現が僅かに認められた。インビトロ培養でPBMCから得られたこれらの線維芽細胞様細胞を、以下の知見に基づき、MOMCと名付けた。
【0058】
電子顕微鏡観察の結果、MOMCは、多数の細胞小器官をもつ紡錐形の形態をしていることがわかった(図1d−g)。多数の一次リソソーム、細胞表面にみられる偽足状の突起、また迷路状の細胞小胞は、マクロファージ類及びその他の食細胞類に固有に見られる所見である。MOMCにはまた、突出して束をなす中間フィラメント、よく発達した二次リソソーム、伸張して分岐したミトコンドリアといった間葉系由来細胞によく見られる特徴が認められる。殆ど全てのMOMCに僅かな脂質滴が観察された。また、血管内皮細胞特有の棒状微小管に似た構造も数多く認められた。これらの超微形態所見は、食細胞、間葉系細胞及び血管内皮細胞の特徴を備えたものである。
【0059】
実施例19(循環系におけるMOMCの由来)
フィブロネクチンでコートしたプレート上でPBMCを培養して得られた接着細胞を、フローサイトメトリーにかけ、その表面におけるCD14及びCD34の発現を連続的に調べた(図3a)。1時間後にプレートに付着した細胞の大多数はCD14+及びCD34-だったが、接着CD14+細胞上におけるCD34の発現は次第にアップレギュレートされていった。培養7日後には、殆ど全ての接着細胞がCD14及びCD34に対して陽性を示した。末梢血には、CD34+血管内皮細胞前駆細胞(Science, 275, 964-967, 1997)及びCD105/エンドグリン/SH2+間葉系前駆細胞(Science, 284, 143-147, 1999, Arthritis Res., 2, 477-488, 2000, Biochem Biophys Res Commun., 265, 134-139, 1999)が含まれているため、本発明者らは、PBMCからCD14、CD34又はCD105に対する陽性細胞を除去することによりもたらされるMOMCのインビトロ誘導における効果を考察した。図3bに示したとおり、CD14+単球の除去によりMOMCの出現は、ほぼ完全に阻害されたが、CD34+又はCD105+細胞を除去しても何の効果も生じなかった。MOMCが循環単球由来であることをさらに確認すべく、PBMCからCD14+単球を分離し、PKH67で標識し、非標識CD14-細胞共存下においてフィブロネクチンでコートしたプレート上で培養した。図3cに示すように、培養一週間後にPKH67標識単球はCD34を発現した。以上の知見を総合すると、MOMCが循環CD14+単球由来であることが示唆される。しかし、培養3日目に回収された非接着細胞にもCD14+細胞が有意比率で含まれており、CD14+単球の一部のサブセットがフィブロネクチンへの付着能を有し、MOMCに分化することが示唆される。
【0060】
PKH67標識単球単独でフィブロネクチン上における培養を行うと、線維芽細胞様に変化したのは僅かな細胞のみで、CD34の発現も培養7日目の時点でアップレギュレートされていなかった(図3c)。この場合、単球におけるCD34発現を誘導するには、CD14-細胞の培養上清を加えるだけで十分であった。さらに、フィブロネクチンコーティング処理を施していないプレート上で、CD14-細胞とPKH67標識単球を共培養すると、培養7日目にプレートに付着していた細胞数は低く、また、それら付着した細胞はCD34を発現していなかった(図3c)。このことから、循環単球がMOMCに分化するには末梢血CD14-細胞由来の液性因子(1若しくは2種以上)及びフィブロネクチンへの結合が必須であると考えられる。
【0061】
MOMCの分化プロセスにおいて、CD14+単球が増殖するか否かを考察するため、CD14+単球のCFSE標識を行い、非標識CD14-細胞とフィブロネクチン上で共培養した。連続的に接着細胞を回収してCFSE標識の発色強度やCD14の発現をフローサイトメトリーにより調べた(図3d)。殆ど全てのフィブロネクチン付着単球が、主に培養24時間以内に増殖していたが、それ以降の培養期間においてもゆっくりとではあるが増殖していた。接着細胞は殆ど例外なくCFSE標識CD14+単球であり、CD14-細胞画分において接着細胞が増殖する様子は観察されず、混入したCD14-細胞群中の特異的前駆細胞が生育する結果として、MOMCが発生するのではないことが示唆される。
【0062】
実施例20(MOMCのフェノタイプ)
MOMCにおける種々の細胞表面分子及び細胞内分子の発現を、フローサイトメトリー及び免疫組織化学法により調べ(図2及び図4)、そのタンパク質発現プロフィールを、単球、マクロファージ及び樹状細胞のものと比較した(表2)。MOMCは、造血及び単球系統マーカー遺伝子(CD45、CD14、CD13、CD11b、CD11c及びCD64)を発現したが、樹状細胞マーカー遺伝子(CD1a及びCD83)は発現しなかった。MOMC上においてHLAクラスI、HLA−DR、及び共刺激分子(CD40及びCD86)が発現されることから、MOMCは抗原提示細胞として、抗原特異的T細胞活性の誘導能を有していることが強く示唆される。MOMCは、造血幹/血管内皮細胞マーカー遺伝子CD34、及び間葉系幹/血管内皮細胞マーカー遺伝子CD105/エンドグリン/SH2を発現した(Biochem Biophys Res Commun., 265, 134-139, 1999)。またMOMCは、幹細胞マーカー遺伝子Sca−1を発現したが、別の幹細胞マーカー遺伝子であるCD117/c−kit及びCD133は発現しなかった。またMOMCは、血管内皮細胞マーカー遺伝子であるCD144/VE−カドヘリン及びFlt−1/VEGFR1陽性だったが、Flk−1/VEGFR2及びvWFの発現は認められなかった。MOMCは、一般に間葉系由来細胞によって産生される細胞外マトリックスタンパク質であるI型及びIII型コラーゲン、フィブロネクチン及びビメンチン陽性だった。これらのタンパク質発現プロフィールは、5世代継代まで変化しなかった。MOMCは単球及び単球由来食細胞とは異なるフェノタイプを示した。特に、幹細胞マーカー遺伝子(CD34、Sca−1及びCD105)、血管内皮マーカー遺伝子(CD144/VE−カドヘリン及びFlt−1/VEGFR1)、及び間葉系マーカー遺伝子(I及びIII型コラーゲン及びフィブロネクチン)の発現は、MOMC独特の特徴だった。従って、MOMCは、食細胞、血管内皮細胞、間葉系細胞及び幹細胞のフェノタイプを併せもつ細胞であるとの認識が成立する。
【0063】
【表2】
【0064】
実施例21(MOMCの増殖能)
培養中にMOMCが増加する様子が認められた。この観察結果が細胞分裂に基づくものかを考察すべく、MOMCにおける分裂細胞の比率をBrdU染色によって連続的に調べた(図5a)。半数近くの接着細胞が、継代1日後にBrdUによって染色されたが、BrdU+細胞は培養5日目に著減した。1、3、5、7及び10日目に、全接着細胞におけるBrdU+細胞の割合を計算したところ、BrdU+細胞の比率は、培養1日目に最大を示し、以後経時的に減少した。ヨウ化プロピジウム染色陽性細胞は、どの時点においても1%未満だった。CFSE標識を行って調べた結果、MOMCが継代後に活発かつ同調的に分裂している様子が認められ、優勢的に増殖している細胞部分はなかった(図5c)。以上の知見は、MOMCが培養中における増殖能を有し、また、主に継代直後に増殖することを示唆するものである。
【0065】
実施例22(MOMCの間葉系細胞系統へのインビトロ分化)
MOMCは、間葉系細胞にみられるいくつかの形態及びフェノタイプ上の特徴を備えていたことから、MOMCは、いくつかの間葉系統へと分化誘導していく能力を有しているとの仮説をたてた。この仮説を確かめるため、MSCを骨、骨格筋、軟骨及び脂肪へと分化誘導することが知られている各種条件下でMOMCを培養した。
【0066】
骨への分化誘導処理を施したMOMCの形態は、紡錐形から類円形へと変化した。殆ど全ての接着細胞でカルシウム沈着が生じているのが、アリザリンレッド染色から観察され(図6a)、それら接着細胞はアルカリフォスファターゼを発現していた(図6b)。この過程で、細胞内カルシウム含量が顕著な増加を示した(図6c)。骨形成誘導後、MOMCは、成熟骨細胞が産生する骨シアロタンパク質II、オステオカルシン(Calcif Tissue Int., 62, 74-82, 1998)や、骨特異的転写因子オステリックス(Cell, 108, 17-29, 2002)のmRNAを発現した。他方、かかる誘導処理後、CD34、CD45及びCD14の発現は消失した(図6d)。
【0067】
MOMCを5−アザシチジンで処理し、筋への分化誘導条件下で3週間培養したところ、細胞は細長くなったものの筋細胞のような複数核をもつ細胞は出現しなかった。この時点で、検体によってその割合は異なるが、45〜60%の接着細胞においてSkM−アクチン及びSkM−MHCの発現が誘導されていた(図6e及びf)。RT−PCRを行った結果、筋特異的転写因子マイオジェニンのmRNA及びSkM−MHCのmRNAが誘導後に検出された(図6g)。CD34、CD14及びCD45の発現は減少したが消失はしなかった。免疫組織化学分析の結果、CD34は殆ど全ての接着細胞で発現したが、CD14及びCD45は、SkM−MHCを発現しなかった細胞でその発現をみた(データは示さず)。培養した筋芽細胞、筋組織、さらには横紋筋肉腫細胞株においてさえ検出可能レベルのCD34が発現され、これは原始筋細胞のサブセットにおけるCD34発現と一致するものだった(J Cell Biol., 150, 1085-1100, 2000)。
【0068】
軟骨形成を誘導するため、MOMCをTGF−β1の存在下、マイクロマス懸濁液中で培養した。これは、MSCにおいて軟骨細胞の分化を誘導するための標準的な手法である(Science, 284, 143-147, 1999, Tissue Eng., 4, 415-428, 1998, Arthritis Rheum., 44, 85-95, 2001)。しかし、プレート上での液滴マイクロマス、及び円錐管中でのペレットマイクロマスによるいくつかの別のプロトコールに従った場合においても、MOMCは1週間以内に死滅した。そこで、TGF−β1存在下で単層MOMCを3週間培養した。図6hに示すように、関節軟骨特有のII型コラーゲンは、無処理のMOMCで弱く発現していたが、その発現は誘導処理後に顕著にアップレギュレートされた。RT−PCRの結果からさらに、誘導処理後に軟骨細胞特異的II型及びX型コラーゲンの発現がアップレギュレートされていることが明らかになった(図6i)。CD45及びCD14の発現は消失したが、CD34の発現は誘導処理後も保持された。他方、CD34は軟骨肉腫細胞株においても発現がみられた。
【0069】
電子顕微鏡観察の結果、MOMCには脂質滴が僅かに認められたが(図1d、f及びg)、誘導処理後に脂質液胞が出現し、その大きさ、数量ともに経時的に増加した。これら脂質液胞はオイル-レッドOで染色された(図6j)。この誘導処理の結果、検体によって割合は異なるが、50〜80%の接着細胞が、この系統にコミットしていた。PPARγ遺伝子のmRNA及び脂肪酸結合タンパク質aP2のmRNAは、MOMCによって弱く発現されたが、これらの遺伝子発現は上記誘導処理後、顕著にアップレギュレートされた(図6k)。CD45及びCD14のmRNA発現は消失したが、CD34発現は、誘導処理後も保持された。他方、CD34は脂肪組織においても発現がみられた。
【0070】
PBMCから新たに産生されたMOMC、5世代まで継代培養したMOMC、又は低温保存されたMOMCにおいても同様に間葉系細胞系統への分化が観察された。さらに、ドナー5名から得たMOMCも同様の多分化能を有していた。成熟間葉系細胞であるヒト皮膚線維芽細胞株2種と、新たに単離したCD14+単球及びマクロファージを同一誘導条件下でそれぞれ培養した。3週間後、皮膚線維芽細胞は、上記条件下において外見は健常だったものの分化傾向は示さなかった。1週間後、上記培養条件下にあった循環単球及びマクロファージは、何ら外見上の分化傾向をみせないままに脱離していた。系統特異的分化は、培養3週間後に付着細胞の半分以上で観察された。しかし付着細胞数は、培養当初の細胞数より20〜50%減少し、MOMCの多くが誘導処理の過程において脱離していったと考えられた。
【0071】
ある系統への分化が各種誘導処理特異的であるかどうかを調べるために、各処理における培養物をアリザリンレッド、オイル-レッドOで交叉染色し、また、SkM−MHC若しくはII型コラーゲン免疫染色も行った。これらの細胞は目的とする系統に特異的な染色に対してのみ陽性で、それ以外の染色に対してはすべて陰性だった(データは示さず)。さらに、インビトロアッセイにおいて、目的とする系統以外の系統に一部のサブセットが分化する可能性を探るため、3週間分化誘導処理した細胞を高感度RT−PCRにかけて、その発現が骨形成(オステオカルシン)、筋形成(SkM−MHC)、軟骨形成(II型コラーゲン)、又は脂肪形成(aP2)系統に限定的な複数遺伝子の転写物を増幅した。系統特異的遺伝子の発現は、目的とする系統に特異的であり(データは示さず)、分化が、施した処理に特異的に起きることが示唆された。
【0072】
分化した間葉系細胞が、MOMC画分に混入したMSCとしての分化能をもつ細胞に由来する可能性を除外するために、誘導処理を行う前にMACS分離システムにより、MOMCからCD14+細胞をポジティブに精製した。その結果は予想どおり、かかる選択処理をしないMOMCにおけると同様に、骨形成、筋形成、軟骨形成、及び脂肪形成への分化が認められた。また、CD34又はCD105/エンドグリン/SH2を発現する細胞を、PBMC培養開始時に除去しても、間葉系細胞への分化には影響がなかった。さらに、系統特異的転写因子であるCbfa1/Runx2(Cell, 89, 755-764, 1997)、MyoD(Front Biosci., 5, D750-767, 2000)、Sox−9(Osteoarthritis Cartilage, 8, 309-334, 2000)、及びPPARγ(J Biol Chem., 276, 37731-37734, 2001)の、誘導処理1週間後におけるMOMC上での発現を調べた。この時点でMOMCはCD45を発現しているので、骨形成、筋形成、軟骨形成、及び脂肪形成誘導処理を行ったMOMCに二重染色を施し、CD45及び個々の転写因子の発現を調べた。図7に示すように、1週間にわたる骨形成分化を経たMOMCは、細胞膜及び細胞質にCD45を発現し、核にCbfa1/Runx2を発現した。同様に、CD45/MyoDとCD45/Sox−9の同時発現が、それぞれ1週間の筋形成及び軟骨形成分化誘導を受けたMOMCで認められた。PPARγは、無処理MOMCの核において弱く発現されていたが、1週間の脂肪形成誘導後、PPARγの発現は増加し、CD45の発現が減少した。これらの知見は、特異的分化誘導条件下でCD45+造血細胞の系統特異的分化が生じることを示唆するものである。免疫組織化学法及びRT−PCR法による測定の結果、PPARγ以外の上記系統特異的転写因子の発現は、誘導処理3週間後に消滅していることがわかった(データは示さず)。これらの知見より、MOMCが、誘導処理過程の初期分化段階において、上記系統特異的転写因子を一過性に発現することが示唆される。
【0073】
実施例23(MOMCの心筋細胞へのインビトロ分化)
神経、心筋前駆細胞に発現するマーカーであるネスチン(Brown)が、共培養8日目のMOMCに発現し、周囲ラット培養心筋細胞との結合が観察された(図8A)。
【0074】
PKH67(Green)で標識されたMOMCが心筋細胞特異的転写因子であるNkx2.5、eHAND(Red/Alexa568:Molecular Probe社)を発現し、同時に血球系マーカーであるCD45(Blue/Alexa660:Molecular Probe社)を発現していた(図8B、図8C)。これはヒト末梢血血球細胞由来のMOMCの心筋細胞への分化過程を示すものである。
【0075】
ヒト特異的PCRプライマーを用いたRT−PCRでは、陽性コントロールのヒト心筋では心筋細胞構造蛋白であるミオシン軽鎖(MLC2v)の発現が認められ、陰性コントロールのラット心筋、共培養前のMOMCでは発現が認められない。共培養後12日目のMOMCにおいてヒトMLC2vの発現が認められた(図8D)。
【0076】
これらのことから、MOMCをラット心筋細胞と共培養することにより、MOMCの心筋細胞への分化が誘導され、共培養8−10日目には心筋前駆細胞のマーカーが発現され、12−14日目には心筋構造蛋白が発現されることが証明された。
【0077】
実施例24(MOMCの神経細胞へのインビトロ分化)
神経、心筋前駆細胞に発現するマーカーであるネスチン(Brown)が、共培養8日目のMOMCに発現し、周囲ラット培養神経細胞へ向かう神経突起の伸長が観察された(図9A)。
【0078】
共培養4日目にはPKH67(Green)で標識されたMOMCが神経細胞特異的転写因子であるNeuroD(Red/Alexa568:Molecular Probe社)を発現し、同時に血球系マーカーであるCD45(Blue/Alexa660:Molecular Probe社)を発現していた(図9B)。これはヒト末梢血血球細胞由来のMOMCの神経細胞への分化過程を示すものである。
【0079】
共培養3日目にはMOMCがネスチン(Green/Alexa488:Molecular Probe社)を発現すると同時に神経細胞特異的転写因子であるNeurogenin2 (Red/Alexa568:Molecular Probe社)を発現していた(図9C)。このことから、MOMCが神経細胞への分化が決定された神経細胞の前駆細胞であることが示された。
【0080】
ウィスターラット培養神経細胞と9日間の共培養では、KH67(Green)で標識されたMOMCにおいて成熟した神経のマーカーであるHu、NeuN(Red/Alexa568:Molecular Probe社)の発現が認められた(図9D、図9E)。このことからMOMCが成熟した神経細胞まで分化することが証明された。
【0081】
実施例25(MOMCの血管内皮細胞へのインビトロ分化)
EBM−2培地で7日間分化誘導したMOMCは紡錘形から多数の突起を有する形態に変化し、もともと発現していなかった血管内皮細胞に特異的なvWF、eNOS、VEGFR2/KDR/Flk−1を発現した。
【0082】
RT−PCRによる遺伝子発現解析では、EBM−2培地で7日間分化を誘導したMOMCで血管内皮に特徴的なFlt−1、VEGFR2/KDR/Flk−1、CD31、CD144、vWFの遺伝子発現を認めた。Flt−1とCD31はMOMCで発現がみられたが、他の遺伝子は誘導後に発現された。分化誘導後にCD34の発現は増強し、CD45、CD14の発現は消失した。
【0083】
これらの成績から、MOMCを血管内皮細胞維持培地で培養することにより、血管内皮細胞への分化が誘導されることが証明された。
【図面の簡単な説明】
【0084】
【図1】本発明のMOMCの形態を示す図である。(a,b)PBMCをフィブロネクチン上、10%FBS添加低グルコースDMEMにおいて7日間培養し、位相差顕微鏡下での観察を行った。原倍率は、aが×80、bが×40である。フィブロネクチンでコートした新たなプレートにMOMCを移し替え、24時間培養した。(c)位相差顕微鏡観察結果を示す画像である(×40)。(d、e)電子顕微鏡観察結果を示す画像である(d及びe、×5000;f及びg、×30000)。中間フィラメントの束を矢印で示し、棒状微小管に似た構造物を矢頭で示した。Lは迷路状の細胞小胞、LDは脂質滴、Nは核、PSは偽足をそれぞれ示している。
【図2】本発明のMOMCのフローサイトメトリー分析結果を示す図である。PBMCをフィブロネクチンでコートしたプレート上で培養し、7日目に接着細胞を回収し、図中に示した一連のモノクローナル抗体で染色してフローサイトメトリー分析を行った。斜線を施したヒストグラムは、目的分子の発現を示している。白抜きのヒストグラムは、アイソタイプが一致するコントロールモノクローナル抗体で染色したコントロールを示している。この結果は、少なくとも3回実施した実験から得たものである。
【図3】本発明のMOMCが循環CD14+単球由来であることを説明する図である。(a)PBMCをフィブロネクチン上で、1時間、1日、3日、5日、7日、10日及び14日間、それぞれ培養した。接着細胞を回収し、FITC結合抗CD14抗体及びPC5結合抗CD34モノクローナル抗体で染色し、フローサイトメトリーにかけた。この結果は、個別に3回行った実験から得たものである。(b)CD14+細胞、CD34+細胞又はCD105+細胞を消失させたPBMC、並びにモック処理を施した細胞を、フィブロネクチン上で7日間培養した。1cm3あたりに付着した細胞数を数え、その結果を無処理PBMC培養における付着細胞数に対する割合として表した。この結果は、3名のドナーから得た数値の平均及びSDである。図中のアステリスクは、モック処理PBMC培養と比較したときの有意差を示す。(c)MACS(磁気細胞分離装置)でソーティングしたCD14+単球を、PKH67染色し、非標識CD14-細胞の存在/非存在下(割合1:4)にて、フィブロネクチンでコートした、若しくはコーティングを施していないプラスチックプレート上で7日間培養した。接着細胞を回収し、PC5結合抗CD34モノクローナル抗体で染色し、フローサイトメトリー分析を行った。この結果は、3回の実験から得たものである。(d)MACSでソーティングしたCD14+単球のCFSE染色を行い、フィブロネクチン上で、0、1、3、5日間それぞれ培養した。接着細胞を回収し、PC5結合抗CD14モノクローナル抗体で染色した。これらの細胞のフローサイトメトリー分析を行った。この結果は、実験を3回実施して得たものである。
【図4】本発明のMOMCの組織免疫化学分析結果を示す図である。PBMCをフィブロネクチンでコートしたプレート上で7日間培養して得られたMOMCを、フィブロネクチンでコートしたチャンバースライド上に移した(I型コラーゲンをコートしたスライドは、フィブロネクチン染色のためにだけ用いた)。培養24時間後に、上記スライドを10%ホルマリンに固定し、図中に示したモノクローナル抗体でそれぞれ染色した。ヘマトキシリンで核を対比染色した。原倍率は、×100である。この結果は、少なくとも3回実施した実験から得たものである。
【図5】本発明のMOMCが増殖する様子を示した図である。フィブロネクチンでコートしたプレート上、PBMCを7日間培養して得られたMOMCを、フィブロネクチンでコートしたチャンバースライド上に移した。MOMCをさらに1、3、5、7、10日間培養し、BrdU染色を行った。核は、ヘマトキシリンで対比染色した。(a)1日目及び5日目における代表的な図である。矢印は、BrdU染色陽性の核を示す。(b)BrdU染色の結果をみるために少なくとも200個の細胞を数え、1、3、5、7、10日間培養した各スライド上のBrdU+細胞数を算定した。結果は、5回行った実験から得た結果の平均及びSDである。(c)MOMCをCFSEで標識し、フィブロネクチンでコートした新たなプレート上で、0、1、3、5日間培養した。接着細胞を回収し、フローサイトメトリー分析にかけた。この結果は3回実施した実験から得たものである。
【図6】本発明のMOMCが、骨、筋、軟骨、及び脂肪細胞へと分化する様子を示す図である。3週間の骨形成誘導を行う前後のMOMCをアリザリン赤で染色し(a、倍率×100)、又はアルカリフォスファターゼ染色を行った(b、×100)。骨形成誘導処理前後のMOMC及び線維芽細胞における細胞内カルシウム沈着量を測定し、タンパク質含量1μgあたりのμg数として表した(c)。3週間の骨形成誘導前後のMOMCと、骨肉腫細胞株におけるオステリックス、骨シアロタンパク質II(BSP II)、オステオカルシン、CD34、CD45、CD14及びGAPDHのmRNA発現を調べた(d)。3週間の筋形成誘導前後のMOMCを染色し、SkM−アクチン(e、×200)又はSkM−MHC(f、×200)染色を調べた。3週間の筋形成誘導前後のMOMCと、筋芽細胞、筋組織及び横紋筋肉腫細胞株におけるマイオジェニン、SkM−MHC、CD34、CD45、CD14及びGAPDHのmRNA発現を調べた(g)。3週間の軟骨形成誘導前後のMOMCを染色し、II型コラーゲン染色を調べた(h、×40)。3週間の軟骨形成誘導前後のMOMCと、軟骨肉腫細胞株におけるα1(II型)及びα1(X型)コラーゲン、CD34、CD45、CD14並びにGAPDHのmRNA発現を調べた(i)。3週間の脂肪形成誘導前後のMOMCをオイル-レッドOで染色した(j、×200)。3週間の脂肪形成誘導前後のMOMCと、脂肪組織におけるPPARγ、aP2、CD34、CD45、CD14及びGAPDHのmRNA発現を調べた(k)。この結果は、少なくとも5回行った実験から得たものである。
【図7】本発明のMOMCを、それぞれ一週間、間葉系へ分化させた後の、CD45(緑)と組織特異的転写因子(赤)の共発現の様子を示す図である。誘導処理前のMOMC(a−d)と、骨形成(e)、筋形成(f)、軟骨形成(g)若しくは脂肪形成(h)の各誘導処理を1週間行ったMOMCにおいてみられるCD45と、Cbfa1/Runx2(a及びe)、MyoD(b及びf)、Sox−9(c及びg)若しくはPPARγ(d及びh)のそれぞれとの共局在性を免疫組織化学法で調べた。共焦点レーザー蛍光顕微鏡下(原倍率×200)で細胞観察を行った。この結果は、3回行った実験から得られたものである。
【図8】本発明のMOMCが、心筋細胞へと分化する様子を示す図である。8日間ウィスターラット培養心筋細胞とともに共培養を行ったMOMCに発現したネスチン(茶)を免疫染色した(A;×200)。MOMCを蛍光色素PKH67(緑)により細胞膜を標識した後に、ウィスターラット培養心筋細胞と7日間共培養を行い、心筋細胞特異的転写因子であるNkx2.5(赤)と、血球系マーカーであるCD45(青)の二重蛍光免疫染色を行った(B;×200)。MOMCを蛍光色素PKH67(緑)により細胞膜を標識した後に、ウィスターラット培養心筋細胞と8日間共培養を行い、心筋細胞特異的転写因子であるeHAND(赤)と、血球系マーカーであるCD45(青)の二重蛍光免疫染色を行った(C;×200)。MOMCをウィスターラット培養心筋細胞と3日、6日、9日、12日間、共培養を行い、心筋細胞構造蛋白であるミオシン軽鎖(MLC2v)のmRNA発現を調べた(D)。この結果は、3回行った実験から得られたものである。
【図9】本発明のMOMCが、神経細胞へと分化する様子を示す図である。8日間ウィスターラット培養神経細胞とともに共培養を行ったMOMCに発現した、神経・心筋前駆細胞に発現するマーカーであるネスチン(茶)を免疫染色した(A;×200)。MOMCを蛍光色素PKH67(緑)により細胞膜を標識した後に、ウィスターラット培養神経細胞と4日間共培養を行い、神経細胞特異的転写因子であるNeuroD(赤)と、血球系マーカーであるCD45(青)の二重蛍光免疫染色を行った(B;×200)。MOMCをウィスターラット培養神経細胞と3日間共培養を行い、ネスチン(茶)と神経細胞特異的転写因子であるNeurogenin2(赤)の二重蛍光免疫染色を行った(C;×200)。MOMCをウィスターラット培養神経細胞と3日間共培養を行い、PKH67(緑)と成熟した神経のマーカーであるHu(赤)の二重蛍光免疫染色を行った(D;×200)。MOMCをウィスターラット培養神経細胞と9日間共培養を行い、PKH67(緑)と成熟した神経のマーカーであるNeuN(赤)の二重蛍光免疫染色を行った(E;×200)。この結果は、3回行った実験から得られたものである。
【図10】本発明のMOMCが、血管内皮細胞へと分化する様子を示す図である。血管内皮細胞維持培地EBM−2培地で7日間分化誘導したMOMCは紡錘形から多数の突起を有する形態に変化した(左;×200)。EBM−2培地で7日間分化誘導したMOMCに発現した、CD34と血管内皮細胞に特異的なvWF、eNOS、VEGFR2/KDR/Flk−1を免疫染色(茶)した(中;×200)。MOMCをEBM−2培地中で7日間培養し、血管内皮細胞に発現されるFlt−1、VEGFR2/KDR/Flk−1、CD31、CD144、vWFおよびCD34、CD45、CD14、GAPDHのmRNA発現を調べた(右)。この結果は、3回行った実験から得られたものである。
【特許請求の範囲】
【請求項1】
単球に由来し、CD14とCD34とを発現する単球由来多能性細胞。
【請求項2】
単球に由来し、CD14とCD34とCD45とI型コラーゲンとを発現する単球由来多能性細胞。
【請求項3】
間葉系組織へと分化誘導する条件下での培養により、間葉系細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞。
【請求項4】
間葉系細胞が、骨芽細胞、骨格筋芽細胞、軟骨細胞又は脂肪細胞であることを特徴とする請求項3記載の単球由来多能性細胞。
【請求項5】
培養心筋細胞との共培養等の心筋へと分化誘導する条件下での培養により、心筋細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞。
【請求項6】
培養神経細胞との共培養等の神経へと分化誘導する条件下での培養により、神経細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞。
【請求項7】
血管内皮細胞を維持する条件下での培養等の血管内皮へと分化誘導する条件下での培養により、血管内皮細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞。
【請求項8】
中胚葉系細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞。
【請求項9】
請求項1〜8のいずれか記載の単球由来多能性細胞を間葉系組織へと分化誘導する条件下で培養することにより誘導された間葉系前駆細胞、間葉系細胞又は間葉系組織。
【請求項10】
間葉系細胞が、骨芽細胞、骨格筋芽細胞、軟骨細胞又は脂肪細胞であることを特徴とする請求項9記載の間葉系前駆細胞、間葉系細胞又は間葉系組織。
【請求項11】
請求項1〜8のいずれか記載の単球由来多能性細胞を培養心筋細胞との共培養等の心筋へと分化誘導する条件下での培養により誘導された心筋前駆細胞、心筋細胞又は心筋組織。
【請求項12】
請求項1〜8のいずれか記載の単球由来多能性細胞を培養神経細胞との共培養等の神経へと分化誘導する条件下での培養により誘導された神経前駆細胞、神経細胞又は神経組織。
【請求項13】
請求項1〜8のいずれか記載の単球由来多能性細胞を血管内皮細胞を維持する条件下での培養等の血管内皮へと分化誘導する条件下での培養により誘導された血管内皮前駆細胞、血管内皮細胞又は血管内皮組織。
【請求項14】
請求項1〜8のいずれか記載の単球由来多能性細胞から分化・誘導された中胚葉系前駆細胞、中胚葉系細胞又は中胚葉系組織。
【請求項15】
請求項1〜8のいずれか記載の単球由来多能性細胞、並びに/又は、前記単球由来多能性細胞から分化・誘導された中胚葉系前駆細胞、中胚葉系細胞及び/若しくは中胚葉系組織を有効成分とする治療剤。
【請求項16】
請求項1〜8のいずれか記載の単球由来多能性細胞、並びに/又は、前記単球由来多能性細胞から分化・誘導された神経前駆細胞、神経細胞及び/若しくは神経組織を有効成分とする治療剤。
【請求項1】
単球に由来し、CD14とCD34とを発現する単球由来多能性細胞。
【請求項2】
単球に由来し、CD14とCD34とCD45とI型コラーゲンとを発現する単球由来多能性細胞。
【請求項3】
間葉系組織へと分化誘導する条件下での培養により、間葉系細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞。
【請求項4】
間葉系細胞が、骨芽細胞、骨格筋芽細胞、軟骨細胞又は脂肪細胞であることを特徴とする請求項3記載の単球由来多能性細胞。
【請求項5】
培養心筋細胞との共培養等の心筋へと分化誘導する条件下での培養により、心筋細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞。
【請求項6】
培養神経細胞との共培養等の神経へと分化誘導する条件下での培養により、神経細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞。
【請求項7】
血管内皮細胞を維持する条件下での培養等の血管内皮へと分化誘導する条件下での培養により、血管内皮細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞。
【請求項8】
中胚葉系細胞に分化しうることを特徴とする請求項1又は2記載の単球由来多能性細胞。
【請求項9】
請求項1〜8のいずれか記載の単球由来多能性細胞を間葉系組織へと分化誘導する条件下で培養することにより誘導された間葉系前駆細胞、間葉系細胞又は間葉系組織。
【請求項10】
間葉系細胞が、骨芽細胞、骨格筋芽細胞、軟骨細胞又は脂肪細胞であることを特徴とする請求項9記載の間葉系前駆細胞、間葉系細胞又は間葉系組織。
【請求項11】
請求項1〜8のいずれか記載の単球由来多能性細胞を培養心筋細胞との共培養等の心筋へと分化誘導する条件下での培養により誘導された心筋前駆細胞、心筋細胞又は心筋組織。
【請求項12】
請求項1〜8のいずれか記載の単球由来多能性細胞を培養神経細胞との共培養等の神経へと分化誘導する条件下での培養により誘導された神経前駆細胞、神経細胞又は神経組織。
【請求項13】
請求項1〜8のいずれか記載の単球由来多能性細胞を血管内皮細胞を維持する条件下での培養等の血管内皮へと分化誘導する条件下での培養により誘導された血管内皮前駆細胞、血管内皮細胞又は血管内皮組織。
【請求項14】
請求項1〜8のいずれか記載の単球由来多能性細胞から分化・誘導された中胚葉系前駆細胞、中胚葉系細胞又は中胚葉系組織。
【請求項15】
請求項1〜8のいずれか記載の単球由来多能性細胞、並びに/又は、前記単球由来多能性細胞から分化・誘導された中胚葉系前駆細胞、中胚葉系細胞及び/若しくは中胚葉系組織を有効成分とする治療剤。
【請求項16】
請求項1〜8のいずれか記載の単球由来多能性細胞、並びに/又は、前記単球由来多能性細胞から分化・誘導された神経前駆細胞、神経細胞及び/若しくは神経組織を有効成分とする治療剤。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2006−14741(P2006−14741A)
【公開日】平成18年1月19日(2006.1.19)
【国際特許分類】
【出願番号】特願2005−228860(P2005−228860)
【出願日】平成17年8月5日(2005.8.5)
【分割の表示】特願2003−74573(P2003−74573)の分割
【原出願日】平成15年3月18日(2003.3.18)
【出願人】(899000079)学校法人慶應義塾 (742)
【Fターム(参考)】
【公開日】平成18年1月19日(2006.1.19)
【国際特許分類】
【出願日】平成17年8月5日(2005.8.5)
【分割の表示】特願2003−74573(P2003−74573)の分割
【原出願日】平成15年3月18日(2003.3.18)
【出願人】(899000079)学校法人慶應義塾 (742)
【Fターム(参考)】
[ Back to top ]