
単鎖組換えT細胞レセプター
TCRα鎖定常領域細胞外配列のN末端に融合したTCRα鎖可変領域配列により構成されるαセグメント、TCRβ鎖定常領域細胞外配列のN末端に融合したTCRβ鎖可変領域により構成されるβセグメント、及びαセグメントのC末端をβセグメントのN末端に又はその逆に連結するリンカー配列を含む単鎖T細胞レセプター(scTCR)。α及びβセグメントの定常領域細胞外配列は、ジスルフィド結合により連結し、リンカー配列の長さ及びジスルフィド結合の位置は、α及びβセグメントの可変領域配列が、天然のαβT細胞レセプター中と実質的に同様に相互に配向するような長さ及び位置である。2以上のこのようなscTCRの複合体及び治療及び種々のスクリーニング適用におけるscTCRの使用もまた開示される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、単鎖T細胞レセプター(TCR)に関する。
【背景技術】
【0002】
天然のTCR
例えばWO99/60120に記載されているように、TCRは、T細胞による特異的な主要組織適合性複合体(MHC)−ペプチド複合体の認識を媒介し、そしてそれ自体、免疫系の細胞兵器の働きに必須である。
【0003】
抗体及びTCRは、特異的な様式で抗原を認識する唯2つの分子型であり、したがってTCRは、MHCにおいて提示される特定ペプチド抗原(この外来ペプチドは、細胞内の異常の唯一の徴候であることが多い)の唯一のレセプターである。T細胞認識は、T細胞及び抗原提示細胞(APC)が直接物理的接触をしているときに起こり、抗原特異的TCRとpMHC複合体との結びつきにより開始される。
【0004】
天然(native)のTCRは、シグナル伝達媒介に関与するCD3複合体の不変タンパク質と会合する、免疫グロブリンスーパーファミリーのヘテロ二量体細胞表面タンパク質である。TCRはαβ及びγδ形態で存在し、これらの形態は、構造的に類似するが、全く異なる解剖学的所在及びおそらくは機能を有する。MHCクラスI及びクラスIIリガンドもまた、免疫グロブリンスーパーファミリータンパク質であるが、APC細胞表面で多様な編成(array)の短いペプチドフラグメントを提示することを可能にする高度に多形性のペプチド結合部位を有して抗原提示に特化している。
【0005】
2つのさらなるクラスのタンパク質が、TCRリガンドとして機能し得ることが知られている。(1)CD1抗原は、その遺伝子が古典的なMHCクラスI及びクラスII抗原とは異なる染色体に位置するMHCクラスI関連分子である。CD1分子は、従来のクラスI及びクラスII-MHC−pep複合体と類似の様式で、ペプチド部分及び非ペプチド部分(例えば、脂質、糖脂質)をT細胞に提示することができる(例えば、Barclayら(1997) The Leucocyte Antigen Factsbook 第2版,Academic Press、及びBauer(1997) Eur J Immunol 27(6) 1366-1373を参照)。(2)細菌性スーパー抗原は、クラスII MHC分子とTCRのサブセットとの両方に結合できる可溶性毒素である(Fraser(1989) Nature 339 221-233)。多くのスーパー抗原が1又は2のVβセグメントに対して特異性を示す一方、その他のものはより無差別な結合を示す。ともかく、スーパー抗原は、多クローン性様式でT細胞のサブセットを刺激する能力により、増強した免疫応答を誘発することができる。
【0006】
天然のヘテロ二量体αβTCRの細胞外部分は2つのポリペプチドからなり、その各個が、膜近位定常ドメイン及び膜遠位可変ドメインを有する(図1を参照)。定常ドメイン及び可変ドメインの各個が、鎖内ジスルフィド結合を含む。可変ドメインは、抗体の相補性決定領域(CDR)に類似する高度多形性ループを含む。TCRのCDR3はMHCにより提示されたペプチドと相互作用し、CDR1及びCDR2はペプチド及びMHCと相互作用する。TCR配列の多様性は、連結される可変(V)、多様性(V)、連結(J)及び定常遺伝子の体細胞再編成(somatic rearrangement)を介して生じる。機能的なα鎖ポリペプチドは、再編成されたV−J−C領域により形成される一方、β鎖はV−D−J−C領域からなる。細胞外定常ドメインは、膜近位領域及び免疫グロブリン領域を有する。単一のα鎖定常ドメイン(TRACとして知られる)及び2つの異なるβ定常ドメイン(TRBC1及びTRBC2として知られる)が存在する(IMGT命名法)。これらβ定常ドメイン間には4つのアミノ酸変化が存在し、そのうちの3つは、本発明の単鎖TCRを作成するために使用するドメイン内にある。これらの変化は全て、TRBC1及びTRBC2のエキソン1内にあり(N4K5→K4N5及びF37→Y(IMGT番号付法、差異TRBC1→TRBC2))、2つのTRCβ鎖定常領域間の最後のアミノ酸変化は、TRBC1及びTRBC2のエキソン3にある(V1→E)。TCR細胞外ドメインの各個の範囲はいくらか変化し得る。しかし、当業者は、The T Cell Receptor Facts Book,Lefranc & Lefranc,Academic Press発行,2001のような参考文献を使用して、ドメイン境界の位置を容易に決定できる。
【0007】
単鎖TCR
単鎖TCR(scTCR)は、アミノ酸単鎖からなる人工構築物であり、これは天然のヘテロ二量体TRCと同様に、MHC−ペプチド複合体に結合する。不運にも、α及びβ鎖をその両方が1つのオープンリーディングフレームで発現するように単に連結することによって機能的なα/βアナログscTCRを作成しようとする試みは、成功しなかった。これは、おそらく、α−β可溶ドメイン対合の当然の不安定性による。
【0008】
したがって、α及びβ鎖のいずれか又は両方の種々の短縮化を使用する特別な技法が、scTCRの作成に必要とされてきた。これらの方式は、非常に限られた範囲のscTCR配列に対してのみ適用可能であるようである。Hooら(1992)PNAS.89(10):4759-63は、25アミノ酸リンカーと連結した短縮型β及びα鎖と細菌細胞周辺質発現(bacterial periplasmic expression)とを使用する2C T細胞クローンからの単鎖形式でのマウスTCRの発現を報告する(Schodinら(1996)Mol.Immunol.33(9):819-29もまた参照)。この設計はまた、Hollerら(2000)PNAS.97(10):5387-92により報告されたm6単鎖TCRの基礎をなし、m6単鎖TCRは2C scTCRから誘導され、同じH2-Ld制限アロエピトープ(alloepitope)に結合する。Shustaら(2000)Nature Biotechnology 18:754-759は、酵母展示実験(yeast display experiment)における単鎖2C TCR構築物の使用を報告し、これは増強した熱安定性及び溶解性を有する変異TCRを生じた。この報告はまた、これら展示2C TCRがコグネイトpMHCを発現する細胞に選択的に結合することができることを証明した。Khandekarら(1997)J.Biol.Chem.272(51):32190-7は、マウスD10 TCRについて、このscTCRをMBPに融合して細菌細胞質で発現したが、同様な設計を報告している(Hare1ら(1999)Nat.Struct.Biol.6(6):574-81もまた参照)。Hilyardら(1994)PNAS.91(19):9057-61は、Vα−リンカー−Vβ設計を使用し細菌細胞周辺質で発現させた、インフルエンザマトリクスタンパク質−HLA-A2に特異的なヒトscTCRを報告している。
【0009】
Chungら(1994)PNAS.91(26)12654-8は、Vα−リンカー−Vβ−Cβ設計及び哺乳動物細胞株の表面での発現を使用するヒトscTCRの作成を報告している。この報告は、scTCRのペプチド−HLA特異的結合に対しては何らの言及も含んでいない。Plaksinら(1997)J.Immunol.158(5):2218-27は、HIV gp12-H-2Ddエピトープに特異的なマウスscTCRを作成するための同様なVα−リンカー−Vβ−Cβ設計を報告している。
【0010】
治療用途
疾患プロセスにより影響される細胞に局在できるターゲッティング部分(targeting moiety)の必要性が存在する。このようなターゲッティング部分は、自己免疫疾患の原因である免疫系の「指向性を誤った」作用を直接的に遮断するためか又はガン性細胞に細胞毒性物質(cytotoxic agent)を送達する手段としてのいずれかに利用し得る。
【0011】
理想的には、これらの適用に適切な分子は、関係する疾患プロセスに直接関与する細胞マーカーに対して特異的親和性を必要とする。この目的のためには抗体が使用されてきた。
【0012】
スクリーニング用途
多くの重要な細胞相互作用及び細胞応答(TCR媒介免疫系を含む)は、細胞表面レセプターと、他の細胞の表面に提示されるリガンドとの間で行なわれる接触により制御される。これらのタイプの特異的分子接触は、ヒト体内での正確な生化学的調節に決定的に重要であり、したがって精力的に研究されている。多くの場合、このような研究の目標は、疾患を予防し又は疾患と戦うために、細胞応答を調整する手段を考案することである。
【0013】
したがって、ある程度の特異性でヒトレセプター又はリガンド分子と結合する化合物を同定するために用いる方法は、新たな疾患治療学の発見及び開発のための先導として重要である。特に、特定のレセプター−リガンド相互作用に干渉する化合物は、治療剤又は治療キャリアとしての直接の可能性を有する。
【0014】
非常に大きな化合物ライブラリの比較的容易で対費用効率の高い作成を可能にするコンビナトリアル・ケミストリーにおける進歩は、化合物試験の範囲を莫大に増大させた。今や、スクリーニングプログラムの限界は、ほとんどの場合、用いることができるアッセイの性質、適切なレセプター及びリガンド分子の作成及びこれらアッセイが高スループットスクリーニング法にどのくらい十分に適合させることができるかにある。
【発明の開示】
【課題を解決するための手段】
【0015】
発明の簡単な説明
本発明は、アミノ酸単鎖の残基間のジスルフィド結合の存在により特徴付けられる新たなクラスのα/βアナログscTCRを利用可能にする。この結合は、その分子のα領域とβ領域との間の対合の安定性に寄与する。このようなTCRは、スクリーニング目的又は治療目的に有用である。
【0016】
発明の詳細な説明
本発明は、TCRα鎖定常領域細胞外配列のN末端に融合したTCRα鎖可変領域配列により構成されるαセグメントと、TCRβ鎖定常領域細胞外配列のN末端に融合したTCRβ鎖可変領域配列により構成されるβセグメントと、αセグメントのC末端をβセグメントのN末端を連結するか又はその逆であるリンカー配列とを含み、α及びβセグメントの定常領域細胞外配列は、ジスルフィド結合により連結し、リンカー配列の長さ及びジスルフィド結合の位置は、α及びβセグメントの可変領域配列が天然のαβT細胞レセプター中と実質的に同様に相互に配向するような長さ及び位置である単鎖T細胞レセプター(scTCR)を提供する。
【0017】
本発明のscTCRにおいて、α及びβセグメントの可変領域配列が天然のαβT細胞レセプター中と実質的に同様に相互に配向するという要件は、その分子が関係するTCRリガンド(pMHC複合体、CD1−抗原複合体、スーパー抗原又はスーパー抗原/pMHC複合体)に結合することを確認することによって検証する−結合すればこの要件を満たす。pMHC複合体との相互作用は、BIAcore 3000(商標)又はBIAcore 2000(商標)装置を使用して測定することができる。本明細書中の実施例3又はWO 99/6120はそれぞれ、MHC−ペプチド複合体とのTCR結合を分析するために必要となる方法の詳細な説明を提供する。これらの方法は、TCR/CD1及びTCR/スーパー抗原相互作用の研究に等しく適用可能である。これらの方法をTCR/CD1相互作用の研究に適用するためには、可溶形態のCD1が必要であり、その作成は、Bauer(1997)Eur J Immunol 27(6) 1366-1373)に記載されている。
【0018】
α及びβセグメント
α及びβセグメント中に存在する定常領域細胞外配列は、好ましくは、ヒトTCRのものに対応し、α及びβセグメント中に存在する可変領域配列も同様である。しかし、このような配列間の対応は、アミノ酸レベルで1:1である必要はない。全体として結果的に、天然のαβT細胞レセプター中と同様にα及びβセグメントの可変領域配列が相互に配向し、ペプチド−MHC結合機能性が維持されれば、対応するヒトTCR配列に対する、N若しくはC短縮並びに/又はアミノ酸の欠失及び/若しくは置換は許容される。特に、α及びβセグメント中に存在する定常領域細胞外配列は、scTCRが結合するペプチド−MHC複合体との接触に直接関与しないので、天然のTCRの細胞外定常ドメイン配列より短くてもよいし、天然のTCRの細胞外定常ドメイン配列に対する置換又は欠失を含んでもよい。
【0019】
αセグメント中に存在する定常領域細胞外配列は、TCRα鎖の細胞外定常Igドメインに対応する配列を含んでもよく、及び/又はβセグメント中に存在する定常領域細胞外配列は、TCRβ鎖の細胞外定常Igドメインに対応する配列を含んでもよい。
【0020】
本発明の1つの実施形態では、αセグメントは、TCRα鎖の定常領域の実質的に全ての細胞外ドメインのN末端に融合した、TCRα鎖の実質的に全ての可変領域に対応し;且つ/又はβセグメントは、TCRβ鎖の定常領域の実質的に全ての細胞外ドメインのN末端に融合した、TCRβ鎖の実質的に全ての可変領域に対応する。
【0021】
別の実施形態では、α及びβセグメント中に存在する定常領域細胞外配列は、TCRの天然の鎖間ジスルフィド結合を形成するシステイン残基を排除するように、C末端で短縮化された天然のTCRのα及びβ鎖の定常領域に対応する。或いは、これらシステイン残基は、天然のジスルフィド結合が欠失するように、別のアミノ酸残基(例えばセリン又はアラニン)で置換されてもよい。加えて、天然のTCRβ鎖は、対合していないシステイン残基を含むが、その残基は、本発明のscTCRのβ配列から欠失されていてもよいし、本発明のscTCRのβ配列中で非システイン残基で置換されていてもよい。
【0022】
本発明の1つの特定の実施形態では、α及びβセグメント中に存在するTCRα及びβ鎖可変領域配列は、併せて、第1のTCRの機能的な可変ドメインに対応してもよく、α及びβセグメント中に存在するTCRα及びβ鎖定常領域細胞外配列は、第2のTCRのそれに対応してもよく、第1及び第2のTCRは同じ種に由来する。したがって、α及びβセグメント中に存在するα及びβ鎖可変領域配列は、第1のヒトTCRのものに対応してもよく、α及びβ鎖定常領域細胞外配列は、第2のヒトTCRのものに対応してもよい。例えば、A6 Tax sTCR定常領域細胞外配列は、異種の可変ドメインが融合できるフレームワークとして使用することができる。
【0023】
本発明の別の実施形態では、α及びβセグメント中に存在するTCRα及びβ鎖可変領域配列は、併せて、第1のTCRの機能的な可変ドメインに対応し、α及びβセグメント中に存在するTCRα及びβ鎖定常領域細胞外配列は、第2のTCRのそれに対応し、第1及び第2のTCRは異なる種に由来する。この実施形態では、α及びβセグメント中に存在するTCRα及びβ鎖可変領域配列は、併せて、ヒトTCRの機能的可変ドメインに対応し、α及びβセグメント中に存在するTCRα及びβ鎖定常領域細胞外配列は、マウスTCRのものに対応する。本発明のこのような実施形態は、scTCRが免疫原性である可能性が高い非ヒト定常領域配列を含み、したがって標的細胞上に局在するとdTCRに対する免疫応答全体を増強する可能性が高いという利点を有する。異常細胞(例えばガン細胞)に対する免疫応答は、このように増強され得る。
【0024】
リンカー
本発明では、リンカー配列は、ポリペプチド単鎖を形成するために、α及びβセグメントを連結する。リンカー配列は、例えば、式−P−AA−P−(式中、Pはプロリンであり、AAはアミノ酸がグリシン及びセリンであるアミノ酸配列を表す)を有してもよい。
【0025】
scTCRがMHC−ペプチド複合体に結合するために、α及びβセグメントは、その可変領域配列がそのような結合に関して配向するように、対合していなければならない。したがって、リンカーは、αセグメントのC末端とβセグメントのN末端との間又はその逆の間の距離を橋渡しするに十分な長さを有するべきである。他方で、万一にもN末端可変領域配列でリンカーの末端部が、scTCRと標的ペプチド−MHC複合体との結合を遮断し又は減少させないように、好ましくは、過度なリンカー長は回避すべきである。
【0026】
例えば、α及びβセグメント中に存在する定常領域細胞外配列が、TCRの天然の鎖間ジスルフィド結合を形成するシステイン残基を排除するように、C末端で短縮化された天然のTCRのα及びβ鎖の定常領域に対応し、リンカー配列がαセグメントのC末端とβセグメントのN末端を連結する場合、リンカーは、26〜41アミノ酸、例えば29、30、31又は32アミノ酸からなってもよく、特定のリンカーは、式−PGGG−(SGGGG)5−P−(式中、Pはプロリンであり、Gはグリシンであり、Sはセリンである)を有する。
【0027】
ジスルフィド結合
本発明のscTCRの基本的に特徴付ける性状は、α及びβセグメントの定常領域細胞外配列間のジスルフィド結合である。この結合は、天然の二量体αβTCR中に存在する天然の鎖間ジスルフィド結合に対応してもよいし、天然のTCR中に対応物(counterpart)を有さない、α及びβセグメントの定常領域細胞外配列中に特異的に組み込まれたシステイン間のものであってもよい。いくつかの場合では、天然のもの及び天然のものでない両方のジスルフィド結合が、本発明のscTCRにおいて望ましくあり得る。
【0028】
ジスルフィド結合の位置は、α及びβセグメントの可変領域配列が天然のαβT細胞レセプター中と実質的に同様に相互に配向するという要件に従属する。
ジスルフィド結合は、α及びβセグメント上の非システイン残基をシステインに変異させ、その変異した残基間にジスルフィド結合を形成させることにより形成してもよい。天然の残基の代わりに導入されたシステイン残基間にジスルフィド結合を形成することができるように、天然のTCR中でそれぞれのβ炭素が約6Å(0.6nm)以下、好ましくは3.5Å(0.35nm)〜5.9Å(0.59nm)の範囲で離れている残基が好ましい。ジスルフィド結合が定常免疫グロブリン領域中の残基間であれば、その結合は膜近位領域の残基間であり得るが好ましい。システインを導入してジスルフィド結合を形成することができる好ましい部位は、TCRα鎖についてはTRAC*01の、TCRβ鎖についてはTRBC1*01又はTRBC2*01のエキソン1中の以下の残基である:
【表1】

【0029】
今や、システイン残基に変異させて本発明に従うscTCR中に新たな鎖間ジスルフィド結合を形成させることができるヒトTCR中の残基が同定されたので、当業者は他の種のTCRを同じ方法で変異させてその種のscTCRを作成することができる。ヒトにおいては、当業者は、単に、それぞれのTCR鎖中の以下のモチーフを探索して変異させるべき残基(影付きの残基がシステインへの変異のための残基である)を同定することのみが必要である。
α鎖Thr48:DSDVYITDKTVLDMRSMDFK(TRAC*01遺伝子のエキソン1のアミノ酸39〜58)
α鎖Thr45:QSKDSDVYITDKTVLDMRSM(TRAC*01遺伝子のエキソン1のアミノ酸36〜55)
α鎖Tyr10:DIQNPDPAVYQLRDSKSSDK(TRAC*01遺伝子のエキソン1のアミノ酸1〜20)
α鎖Ser15:DPAVYQLRDSKSSDKSVCLF(TRAC*01遺伝子のエキソン1のアミノ酸6〜25)
β鎖Ser57:NGKEVHSGVSTDPQPLKEQP(TRBC1*01及びTRBC2*01遺伝子のエキソン1のアミノ酸48〜67)
β鎖Ser77:ALNDSRYALSSRLRVSATFW(TRBC1*01及びTRBC2*01遺伝子のエキソン1のアミノ酸68〜87)
β鎖Ser17:PPEVAVFEPSEAEISHTQKA(TRBC1*01及びTRBC2*01遺伝子のエキソン1のアミノ酸8〜27)
β鎖Asp59:KEVHSGVSTDPQPLKEQPAL(TRBC1*01及びTRBC2*01遺伝子のエキソン1のアミノ酸50〜69)
β鎖Glu15:VFPPEVAVFEPSEAEISHTQ(TRBC1*01及びTRBC2*01遺伝子のエキソン1のアミノ酸6〜25)
【0030】
他の種においては、TCR鎖は、上記モチーフに対して100%同一性を有する領域を有さなくてもよい。しかし、当業者は、上記モチーフを使用してTCRα又はβ鎖の等価部分、したがってシステインに変異させるべき残基を同定することができる。整列技法(alignment technique)をこの点に関して使用してもよい。例えば、変異のためのTCR配列の関係部分を位置決めするために、European Bioinformatics Instituteのウェブサイト(http://www.ebi.ac.uk/index.html)で入手可能であるClustalWを使用して上記モチーフと特定のTCR鎖配列を比較することができる。
【0031】
本発明は、その範囲に、αβアナログscTCR及び他の哺乳動物(マウス、ラット、ブタ、ヤギ及びヒツジを含むがこれらに限定されない)のそれを含む。上記のようにヒト/非ヒトキメラscTCRもまた含まれる。上記のように、当業者は、システイン残基を導入して鎖間ジスルフィド結合を形成させることが可能である上記のヒト部位と等価な部位を決定することができる。例えば、以下は、マウスCα及びCβ可溶性ドメインのアミノ酸配列を、システインに変異させてTCR鎖間ジスルフィド結合を形成させることができる上記のヒト残基に等価なマウス残基を示すモチーフと共に示す(ここで、関係する残基には影が付されている)。
マウスCα可溶性ドメイン:
PYIQNPEPAVYQLKDPRSQDSTLCLFTDFDSQINVPKTMESGTFITDKTVLDMKAMDSKSNGAIAWSNQTSFTCQDIFKETNATYPSSDVP
マウスCβ可溶性ドメイン:
EDLRNVTPPKVSLFEPSKAEIANKQKATLVCLARGFFPDHVELSWWVNGREVHSGVSTDPQAYKESNYSYCLSSRLRVSATFWHNPRNHFRCQVQFHGLSEEDKWPEGSPKPVTQNISAEAWGRAD
ヒトα鎖Thr48のマウス等価物:ESGTFITDKTVLDMKAMDSK
ヒトα鎖Thr45のマウス等価物:KTMESGTFITDKTVLDMKAM
ヒトα鎖Tyr10のマウス等価物:YIQNPEPAVYQLKDPRSQDS
ヒトα鎖Ser15のマウス等価物:AVYQLKDPRSQDSTLCLFTD
ヒトβ鎖Ser57のマウス等価物:NGREVHSGVSTDPQAYKESN
ヒトβ鎖Ser77のマウス等価物:KESNYSYCLSSRLRVSATFW
ヒトβ鎖Ser17のマウス等価物:PPKVSLFEPSKAEIANKQKA
ヒトβ鎖Asp59のマウス等価物:REVHSGVSTDPQAYKESNYS
ヒトβ鎖Glu15のマウス等価物:VTPPKVSLFEPSKAEIANKQ
【0032】
上記のように、A6 Tax sTCR細胞外定常領域は、それに異種可変ドメインを融合させることが可能であるフレームワークとして使用することができる。異種可変領域配列は、ジスルフィド結合と定常領域配列のN末端との間の任意の点で定常領域配列に連結することが好ましい。A6 Tax TCRα及びβ定常領域配列の場合、ジスルフィド結合は、アミノ酸残基158及び172でそれぞれ導入されたシステイン残基間で形成され得る。したがって、異種α及びβ鎖可変領域配列の付着点が、それぞれ残基159又は173とα又はβ定常領域配列のN末端との間であれば好ましい。
【0033】
さらなる観点
本発明のscTCR(好ましくはヒトである)は、実質的に純粋な形態で又は精製若しくは単離された調製物として提供され得る。例えば、これは、他のタンパク質を実質的に含まない形態で提供され得る。
【0034】
本発明の複数のscTCRは多価複合体で提供され得る。したがって、本発明は、1つの観点では、多価T細胞レセプター(TCR)複合体を提供し、これは複数の本明細書中に記載のような可溶性T細胞レセプターを含む。複数の可溶性TCRの各個は、好ましくは、同一である。
【0035】
本発明の多価複合体において、scTCRは、多量体の形態であってもよいし、及び/又は脂質二重層(例えばリポソーム)上に存在するか若しくはこれと会合していてもよい。
【0036】
最も単純な形態では、本発明に従う多価scTCR複合体は、好ましくはリンカー分子を介して、互いに会合(例えば、共有結合又は別の結合)した2又は3又は4又はそれ以上のT細胞レセプター分子の多量体を含む。適切なリンカー分子には、アビジン、ストレプトアビジン、ニュートラアビジン(neutravidin)及びエキストラアビジン(extravidin)(これらの各個は、ビオチンに関して4つの結合部位を有する)のような多価付着分子が含まれが、これらに限定されない。したがって、ビオチン化TCR分子は、複数のTCR結合部位を有するT細胞レセプター多量体に形成することができる。多量体中のTCR分子の数は、その多量体を作成するために使用したリンカー分子の量に対するTCRの量に依存し、また他のビオチン化分子の存否にも依存する。好ましい多量体は、二量体、三量体又は四量体のTCR複合体である。
【0037】
TCR四量体より相当大きい構造は、特異的MHC−ペプチド複合体を発現する細胞を追跡又は標的することに使用してもよい。好ましくは、構造は、直径が10nm〜10μmの範囲である。各構造は、その構造上の2以上のTCR分子が細胞上の2以上のMHC−ペプチド複合体に同時に結合することを可能にし、したがってその細胞に関する多量体結合部分のアビディティを増大させるように十分に離れた距離で複数のscTCR分子を展示し得る。
【0038】
1又は複数のscTCRを有する複合体を形成するための本発明での使用に適切な構造は、膜構造(例えばリポソーム)及び固体構造(好ましくはビーズのような粒子、例えばラテックスビーズである)を含む。T細胞レセプター分子で外部を被覆され得る他の構造もまた適切である。好ましくは、構造は、個々のT細胞レセプター分子でよりむしろ、T細胞レセプター多量体で被覆される。
【0039】
リポソームの場合、T細胞レセプター分子又はその多量体は、膜に付着していてもよいし、膜とその他の会合をしていてもよい。このための技法は当業者に周知である。
【0040】
標識又は別の部分(例えば、毒性部分又は治療的部分)が、本発明の多価scTCR複合体に含まれてもよい。例えば、標識又は他の部分は、混合分子多量体に含まれてもよい。このような多量体分子の例は、3つのscTCR分子と1つのペルオキシダーゼ分子とを含む四量体である。これは、TCR及び当該酵素を3:1のモル比で混合して四量体複合体を生成し、正確な比の分子を含まない複合体から所望の複合体を分離することにより達成され得る。これらの混合分子は、立体障害がその分子の所望の機能を損なわないか又は有意には損なわないという条件で、分子の任意の組合せを含み得る。ストレプトアビジン分子上の結合部位の位置は、立体障害を起こす可能性がないので、混合四量体に適切である。
【0041】
さらなる観点では、本発明は、MHC−ペプチド複合体を検出するための方法を提供し、この方法は、
a.本発明のcTCRを提供すること
b.scTCRをMHC−ペプチド複合体と接触させること;及び
scTCRのMHC−ペプチド複合体との結合を検出すること
を含む。
【0042】
治療用途
本発明のscTCR(又はその多価複合体)は、択一的又は追加的に、例えば細胞殺傷に使用するための毒性部分又は免疫刺激物質(immunostimulating agent)(例えばインターロイキン又はサイトカイン)であり得る治療物質(therapeutic agent)と会合(例えば、共有結合又は別の結合)していてもよい。本発明の多価scTCR複合体は、非多量体のT細胞レセプターへテロ二量体と比べて、TCRリガンド(例えばpMHC複合体又はCD1分子)に関して増強した結合能力を有し得る。したがって、本発明に従う多価scTCR複合体は、特定の抗原を提示する細胞をインビトロ又はインビボで追跡又は標的するために特に有用であり、またそのような用途を有するさらに多価のTCR複合体の製造のための中間体としても有用である。したがって、scTCR又は多価scTCR複合体は、インビボでの使用のために薬学的に受容可能な製剤で提供され得る。
【0043】
本発明はまた、標的細胞に治療物質を送達するための方法を提供し、この方法は、潜在的な標的細胞を本発明に従うscTCR又は多価scTCR複合体と、標的細胞へのscTCR又は多価scTCR複合体の付着を可能にする条件下で接触させることを含み、当該scTCR又は多価scTCR複合体は、MHC−ペプチド複合体に特異的であり、これに治療物質を会合させる。
【0044】
特に、可溶性scTCR又は多価scTCR複合体を使用して、特定の抗原を提示する細胞の位置に治療物質を送達することができる。このことは、多くの状況で、特に腫瘍に対して有用である。治療物質は、その効果を局所的にではあるが、治療物質が結合する細胞上に限らずに発揮するように送達され得る。したがって、1つの特定の戦略は、腫瘍抗原に特異的なT細胞レセプター又は多価scTCR複合体に連結した抗腫瘍分子を考案する。
【0045】
多くの治療物質、例えば放射活性化合物、酵素(例えばパーフォリン)又は化学療法剤(例えばシスプラチン)がこの用途に用いられ得る。確実に所望の位置で毒性効果が発揮されるために、毒素は、それがゆっくりと放出されるように、ストレプトアビジンに連結したリポソーム内にあり得る。このことが、体内での輸送の間の損傷効果を防止し、関係する抗原提示細胞とのscTCRの結合後に毒素が最大効果を有することを確実にする。
【0046】
他の適切な治療物質には、
・小分子細胞毒性物質、すなわち、哺乳動物細胞を殺傷する能力を有する、分子量700ダルトン未満の化合物。このような化合物はまた、細胞毒性効果を有することができる毒性金属を含み得る。さらに、これら小分子細胞毒性物質はまた、プロドラッグ、すなわち、生理学的条件下で崩壊又は変換して細胞毒性物質を放出する化合物を含む。このような物質の例には、シスプラチン、メイタンシン(maytansine)誘導体、ラケルマイシン(rachelmycin)、カリケアマイシン(calicheamicin)、ドセタキセル、エトポシド、ゲムシタビン、イホスファミド、イリノテカン、メルファラン、ミトキサントロン、ソルフィマーソディウムホトフィリンII(sorfimer sodiumphotofrin II)、テモゾロマイド(temozolmide)、トポテカン、トリメトレキサート(trimetreate)、グルクロナート、オーリスタチンE(auristatin E)、ビンクリスチン及びドキソルビシンが含まれる;
・ペプチド細胞毒素、すなわち、哺乳動物細胞を殺傷する能力を有するタンパク質又はそのフラグメント。例には、リシン、ジフテリア毒素、シュードモナス細菌外毒素A、DNAアーゼ及びRNAアーゼが含まれる;
・放射性核種、すなわち、1以上のα若しくはβ粒子又はγ線の同時放射を伴って崩壊する元素の不安定同位体。例には、ヨウ素131、レニウム186、インジウム111、イットリウム90、ビスマス210及び213、アクチニウム225及びアスタチン213が含まれる;
・プロドラッグ、例えば抗体を指向する酵素プロドラッグ;
・免疫増強剤(immuno-stimulant)、すなわち、免疫応答を刺激する部分。例には、サイトカイン(例えばIL-2)、ケモカイン(例えばIL-8)、血小板第4因子、メラノーマ増殖刺激タンパク質など、抗体又はそのフラグメント、補体活性化剤、異種タンパク質ドメイン、同種タンパク質ドメイン、ウイルス性/細菌性タンパク質ドメイン及びウイルス性/細菌性ペプチドが含まれる。
【0047】
本発明の可溶性scTCR又は多価scTCR複合体は、プロドラッグを薬物に変換し得る酵素に連結されてもよい。これは、プロドラッグが薬物を必要とする(すなわち、scTCRにより標的にされる)部位でのみ薬物に変換することを可能にする。
【0048】
本発明に従うscTCRに適切なMHC−ペプチド標的の例には、ウイルス性エピトープ、例えば、HTLV-1エピトープ(例えばHLA-A2により制限されたTaxペプチド;HTLV-1は白血病に関連する)、HIVエピトープ、EBVエピトープ、CMVエピトープ;メラノーマエピトープ(例えばMAGE-1 HLA-A1制限エピトープ)及び他のガン特異的エピトープ(例えばHLA-A2により制限された腎臓細胞ガン腫関連抗原G250);並びに自己免疫疾患(例えば慢性関節リウマチ)に関連するエピトープが含まれるが、これらの限定されない。さらに、本発明での使用に適切な疾患関連pMHC標的は、HLA Factbook(Barclay編,Academic Press)に掲載されており、他にも多くのものが同定されている。
【0049】
scTCRの特異性による限局化薬物送達は、多くの疾患処置を増強することができる可能性が高い。
【0050】
そのための薬物が存在するウイルス性疾患、例えばHIV、SIV、EBV、CMVは、その薬物が感染細胞の近位で放出又は活性化されればその恩恵を受ける。ガンについては、腫瘍又は転移の近位に限局することにより、毒素又は免疫増強剤の効果が増強される。自己免疫疾患では、より長い期間にわたってより局所的な効果を有する一方で、被験者の免疫能力全体には最小限にしか影響しない免疫抑制薬物が徐々に放出され得る。移植拒絶の予防では、免疫抑制薬物の効果は、同じ方法で最適化され得る。ワクチン送達のためには、ワクチン抗原は抗原提示細胞の近くに限局され、したがって抗原の効力を増強し得る。この方法はまた、画像化目的にも適用することができる。
【0051】
本発明のscTCRは、特異的リガンド(例えばpMHC)への結合によりT細胞活性化を調整し、それによりT細胞活性化を阻害するために使用してもよい。T細胞媒介炎症及び/又は組織損傷を含む自己免疫疾患(例えばI型糖尿病)は、このアプローチに適する。関係するpMHCにより提示される特異的ペプチドエピトープの知識が、この用途に必要である。
【0052】
本発明に従う医薬は、通常、一般には薬学的に受容可能なキャリアを含む、滅菌の薬学組成物の部分として供給される。この薬学組成物は、(これを患者に投与する望ましい方法に依存して)任意の適切な形態であり得る。これは、単位投薬剤形で提供されてもよく、一般には密封容器で提供され、キットの部分として提供されてもよい。このようなキットは、(必ずしもではないが)通常、使用のための指示書を含む。これは、複数の単位投薬剤形を含み得る。
【0053】
薬学組成物は、任意の適切な経路、例えば、経口(口腔粘膜又は舌下を含む)、直腸、鼻、局所(口腔粘膜、舌下又は経皮を含む)、膣又は非経口(皮下、筋肉内、静脈内又は皮内)経路による投与に適合され得る。このような組成物は、薬学の分野において公知の任意の方法により、例えば滅菌条件下で活性成分をキャリア又は賦形剤と混合することにより製造し得る。
【0054】
スクリーニング用途
本発明のscTCRは、TCR媒介細胞免疫系の調節物質(modulator)(阻害剤を含む)を同定するために設計されたスクリーニング法で利用可能である。
当業者に公知であるように、この型のタンパク質−タンパク質相互作用スクリーニングのための適切な基礎を提供する多くのアッセイ形式が存在する。
【0055】
増幅ルミネッセンス近接ホモジニアスアッセイ(Amplified Luminescent Proximity Homogeneous Assay)システム(例えばAlphaScreen(商標))は、レセプター及びリガンドタンパク質が付着することができるヒドロゲルの層で被覆された「ドナー」及び「アクセプター」ビーズの使用に基づく。これらレセプター及びリガンド分子の間の相互作用は、ビーズを近接状態にする。これらビーズがレーザ光に曝されると、「ドナー」ビーズ中の光増感物質は、周囲の酸素をより励起された一重項状態に変換する。一重項状態の酸素分子は、拡散して、「アクセプター」ビーズ中の化学発光物質と反応し、この化学発光物質は、さらに、同じビーズに含まれる蛍光体を活性化する。続いて、蛍光体は520〜620nmの光を発光し、これは、レセプター−リガンド相互作用が生じたことをシグナル伝達する。レセプター−リガンド相互作用の阻害剤の存在により、このシグナルは低減される。
【0056】
表面プラズモン共鳴(SPR)は、一方の結合パートナー(通常レセプター)が「チップ」(センサ表面)に固定され、他方の結合パートナー(通常リガンド)(これは可溶であり、チップ上を流される)の結合が検出される界面光学アッセイである。リガンドの結合は、結果的に、チップ表面近くのタンパク質濃度の増大を生じ、このことはその領域の屈折率の変化を引き起こす。チップの表面は、屈折率の変化が、表面プラズモン共鳴により検出され得るように構成される(ここで、表面プラズモン共鳴とは光学現象であり、これにより、ある入射角の光は、薄金属フィルムで、振動する表面電荷密度の波(表面プラズモン)の共鳴励起に起因して、減弱強度の屈折ビームを生じる)。この共鳴は、金属フィルムの裏側での屈折率の変化に非常に敏感であり、固体されたタンパク質と可溶タンパク質との間の結合を検出するために使用されるのが、このシグナルである。分子相互作用のSPR検出及びデータ分析の簡便な使用を可能にするシステムは市販されている。例は、Iasys(商標)装置(Fisons)及びBiacore(商標)装置である。
【0057】
他の界面光学アッセイには、全内部反射蛍光(TIRF)、共振ミラー(RM)及び回折格子カプラセンサ(optical grating coupler sensor)(GCS)が含まれ、これは、Woodbury及びVenton(J.Chromatog.B.725 113-137(1999))でより詳細に議論されている。シンチレーション近接アッセイ(scintillation proximity assay)(SPA)は、CD28とB7との間の低親和性相互作用の阻害剤について化合物ライブラリをスクリーニングするために使用されてきた(おそらく約4μMのKd(Van der Merweら、J.Exp.Med.185:393-403 (1997),Jenhら、Anal Biochem 165(2) 287-93 (1998)))。SPAは、指示表面上に固定された閃光物質(scintillant)にエネルギーを転移させる特定の放射活性同位体からのβ粒子放出を利用する放射活性アッセイである。溶液中の短期(short range)のβ粒子により、確実に、β粒子が閃光物質に近接して発射されたときにのみ閃光が生じる。タンパク質−タンパク質相互作用の検出に適用される場合、一方の相互作用パートナーを放射性同位体で標識し、他方は閃光物質を含むビーズに結合するか、又は閃光物質と共に表面に被覆するかのいずれかである。アッセイを最適に設定することが可能な場合、放射性同位体は、閃光物質に対して、2つのタンパク質間の結合が起きたときにのみ光子発射が活性化されるに十分に近くに運ばれる。
【0058】
本発明のさらなる観点は、scTCRと、MHC−ペプチド複合体、CD1−抗原複合体、スーパー抗原及びMHC−ペプチド/スーパー抗原複合体から選択されるTCRリガンドとの間の相互作用の阻害剤を同定する方法であり、これは、scTCRをscTCRリガンド結合パートナーと、試験化合物の存在下及び非存在下で接触させること、及び試験化合物の存在により、リガンドへのscTCRの結合が減少するかどうかを測定することを含み、この減少が阻害剤を同定するとする。
【0059】
本発明の最後の観点は、scTCRと、MHC−ペプチド複合体、CD1−抗原複合体、スーパー抗原及びMHC−ペプチド/スーパー抗原複合体から選択されるTCRリガンドとの間の相互作用の潜在的な阻害剤を同定する方法であり、これは、scTCR又はscTCRリガンド結合パートナーを試験化合物と接触させること、及び試験化合物が、scTCR及び/又はリガンドに結合するかどうかを測定することを含み、この結合が潜在的阻害剤を同定するとする。本発明のこの観点は、界面光学アッセイ(例えば、BIAcore(商標)システムを使用して実施される界面光学アッセイ)における特別な有用性を見出し得る。
【0060】
本発明の各観点の好ましい性状は、必要な変更を加えて、その他の観点の各個についての性状と同様である。明細書中に言及した先行技術文献は、法が許す最大範囲で明細書中に援用する。
【実施例】
【0061】
本発明は以下の実施例においてさらに記述するが、実施例は、いかなる意味においても、本発明の範囲を限定するものではない。
【0062】
以下に添付図面について言及する。
図1a及び1bは、それぞれ、システインコドンを導入するように変異された、可溶性A6 TCRのα及びβ鎖の核酸配列を示す。影は導入したシステインコドンを示す。
図2aは、新規なジスルフィド鎖間結合を形成させるために使用したT48→C変異(下線)を含む、A6 TCRα鎖細胞外アミノ酸配列を示す。図2bは、新規なジスルフィド鎖間結合を形成させるために使用したS57→C変異(下線)を含む、A6 TCRβ鎖細胞外アミノ酸配列を示す。
図3は、Gly/Serリンカー(30マー)のDNA及びアミノ酸配列を示す。
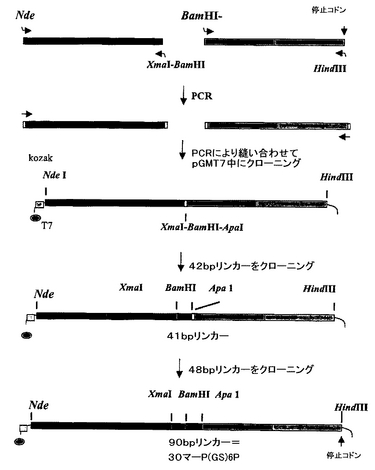
図4は、scDiS A6 TCRを作成するために使用したクローニング戦略をまとめる。
図5aは、scDiS A6 TCRのDNA配列を示す。
図5bは、scDiS A6 TCRのアミノ酸配列を示す。
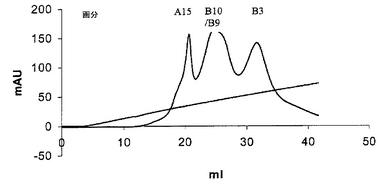
図6は、直線で示されるような0〜500mM NaCl勾配を使用するPOROS 50HQイオン交換カラムからのscDiS A6 TCRタンパク質の溶出を説明する。
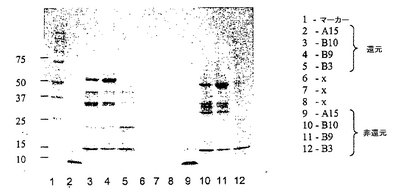
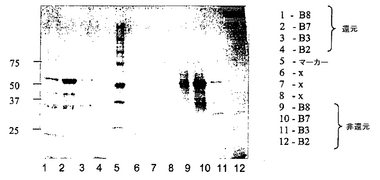
図7は、図6で説明したカラムラン(column run)からの画分A15、B10、B9及びB3の還元SDS-PAGEゲル(クーマシー染色)及び非還元SDS-PAGE(クーマシー染色)ゲルの両方の結果を示す。画分B9及びB10は、明らかに、予想したサイズのscDiS A6 TCRに対応するタンパク質を含む。
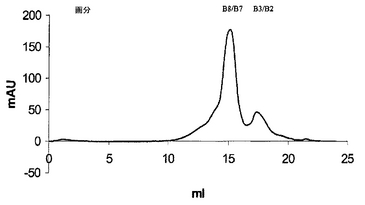
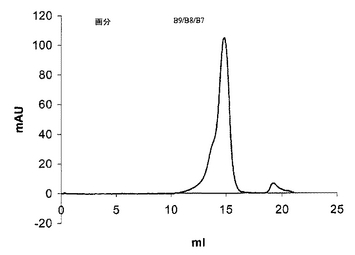
図8は、図6に示したイオン交換カラムランからの画分B10〜B7のSuperdex 200ゲル濾過カラムからのscDiS A6 TCR溶出の溶出を説明する。
図9は、図8で説明したゲル濾過カラムランからの画分B8、B7、B3及びB2の還元SDS-PAGEゲル(クーマシー染色)及び非還元SDS-PAGE(クーマシー染色)ゲルの両方の結果を示す。画分B7は、明らかに、予想したサイズのscDiS A6 TCRに対応するタンパク質を含む。
図10は、図8に示したゲル濾過ランの濃縮画分B9〜B6のBIAcore緩衝液中への最後のゲル濾過ランである。scDiS A6 TCRは単一の主要ピークとして溶出する。
図11は、HLA-A2 TAXへのscDiS A6 TCRの結合についてのBIAcoreデータである。
【0063】
実施例1−新規な鎖間ジスルフィド結合の形成に必要なシステイン残基を導入するためのプライマーの設計及びA6 Tax TCRα及びβ鎖の変異誘発
TRAC*01中のエキソン1のA6 Taxスレオニン48をシステインに変異させるために、以下のプライマーを設計した(変異を小文字で示す):
5'-C ACA GAC AAA tgT GTG CTA GAC AT
5'-AT GTC TAG CAC Aca TTT GTC TGT G
TRBC1*01及びTRBC2*01の両方でエキソン1のA6 Taxセリン57をシステインに変異させるために、以下のプライマーを設計した(変異を小文字で示す):
5'-C AGT GGG GTC tGC ACA GAC CC
5'-GG GTC TGT GCa GAC CCC ACT G
【0064】
PCR変異誘発:
A6 Tax TCRα又はβ鎖の遺伝子を含む発現プラスミドを、それぞれα鎖プライマー又はβ鎖プライマーを使用して、以下のように変異させた。100ngのプラスミドを5μlの10mM dNTP、25μlの10×Pfu緩衝液(Stratagene)、10単位のPfuポリメラーゼ(Stratagene)と混合し、最終容量をH2Oで240μlに調整した。48μlのこの混合物に、50μlの最終反応溶液中で0.2μMの最終濃度が得られるように希釈したプライマーを補充した。95℃にて30秒間の最初の変性工程の後、反応混合物を、Hybaid PCR express PCR装置において、15回の変性(95℃、30秒)、アニーリング(55℃、60秒)及び伸長(73℃、8分)に付した。次いで、産物を10単位のDpnI制限酵素(New England Biolabs)で37℃にて5時間消化した。10μlの消化反応物を、コンピテントXL1-Blue細菌に形質転換し、37℃にて18時間増殖させた。一つのコロニーを採取し、5mlのTYP+アンピシリン(16g/l細菌トリプトン、16g/l酵母抽出物、5g/l NaCl、2.5g/l K2HPO4、100mg/lアンピシリン)中で一晩増殖させた。プラスミドDNAを、製造業者の指示に従ってQiagenミニプレップカラムで精製し、配列を、オックスフォード大学生化学部の配列決定設備で自動配列決定により検証した。それぞれの変異した核酸及びアミノ酸配列を、α鎖については図1a及び2aに、β鎖については図1b及び2bに示す。
【0065】
実施例2−新規ジスルフィド鎖間結合を組み込んだ単鎖A6 TCRの設計、発現及び試験
α鎖配列の末端部から停止コドン(TAA)を除去した以外は、実施例1で製造し、図1a及び1bに示したような新規ジスルフィドの形成に必要な追加のシステイン残基を組み込んだ変異A6 TCRα及びβ鎖のDNA配列を含む発現ベクターを、以下のように、単鎖A6 TCRの作成の基礎として使用した。
【0066】
scDis A6 TCRは、TCRα鎖のC末端とβ鎖のN末端との間に、30アミノ酸リンカー配列を含む。図3は、このリンカーのDNA及びアミノ酸配列を示す。scDis A6 TCRを作成するために用いたクローニング戦略を図4にまとめる。
【0067】
簡潔には、A6 dsTCRのα及びβ鎖を、図4に示すような制限部位を含むプライマー、すなわち:
α5'プライマー:ccaaggccatatgcagaaggaagtggagcagaactct
α3'プライマー:ttgggcccgccggatccgcccccgggggaactttctgggctgggg
β5'プライマー:tcccccgggggcggatccggcgggcccaacgctggtgtcactcag
β3'プライマー:gggaagcttagtctgctctaccccaggcctcg
を使用するPCRにより増幅させた。
【0068】
このように作成した2つのフラグメントを、5'α及び3'βプライマーを使用してPCRにより縫い合わせて(PCR stitched)、部位XmaI−BamHI−ApaIを含む短いリンカーを有する単鎖TCRを得た。このフラグメントをpGMT7中にクローニングした。次いで、完全長リンカーを二段階で挿入した。最初に、42bpフラグメントを、XmaI及びBamHI部位を使用して挿入した:
5'-CC GGG GGT GGC TCT GGC GGT GGC GGT TCA GGC GGT GGC G -3'
3'- C CCA CCG AGA CCG CCA CCG CCA AGT CCG CCA CCG CCT AG-5'。
【0069】
第2に、48bpフラグメントを、BamHI及びApaI部位を使用して挿入して、α鎖の3'末端とβ鎖の5'末端との間に90bpリンカーを作成した。48bpフラグメントは、以下のオリゴの混合物のPCR伸長により作成した:
5'- GC GGA TCC GGC GGT GGC GGT TCG GGT GGC GGT GGC TC-3'
3'- CCA AGC CCA CCG CCA CCG AGT CCG CCA CCG CCC GGG TG -5'。
【0070】
この伸長の産物を、BamHI及びApaIで消化し、42bpリンカーフラグメントを含む消化プラスミド中に連結した。
scDis A6 TCRの完全なDNA及びアミノ酸配列を、それぞれ、図5a及び5bに示す。
【0071】
単鎖ジスルフィド連結A6 TCRの発現及び精製:
単鎖ジスルフィド連結A6 TCRを含む発現プラスミドを、E.coliのBL21pLysS株に形質転換し、1つのアンピシリン耐性コロニーを37℃にてTYP(アンピシリン100μg/ml)培地中でOD600が0.4になるまで増殖させた後、0.5mM IPTGでタンパク質発現を誘導した。誘導の3時間後に、Beckman J-6B中での4000rpmにて30分間の遠心分離によって細胞を採集した。細胞ペレットを、50mM Tris-HCI、25%(w/v)スクロース、1mM NaEDTA、0.1%(w/v)アジ化Na、10mM DTT(pH8.0)を含む緩衝液に再懸濁した。一晩の凍結−解凍工程の後、再懸濁した細胞を、Milsonix XL2020超音波処理器中で標準の12mm径プローブを使用して1分間のバーストで合計約10分間超音波処理した。封入体ペレットを、Beckman J2-21遠心分離器で13000rpmにて30分間の遠心分離により回収した。次いで、3回の界面活性剤洗浄を行なって細胞残渣及び膜成分を除去した。各回で、封入体ペレットをTriton緩衝液(50mM Tris-HCI、0.5%Triton-X100、200mM NaCI、10mM NaEDTA、0.1%(w/v)アジ化Na、2mM DTT(pH8.0))中でホモジナイズした後、Beckman J2-21中で13000rpmにて15分間の遠心分離によりペレット化した。次いで、界面活性剤及び塩を、以下の緩衝液中での同様な洗浄により除去した:50mM Tris-HCl、1mM NaEDTA、0.1%(w/v)アジ化Na、2mM DTT(pH8.0)。最後に、封入体を30mgのアリコートに分け、−70℃で凍結させた。封入体タンパク質の収率を、6Mグアニジン−HClでの可溶化及びBradford顔料結合アッセイ(PerBio)での測定により定量化した。
【0072】
約15mgの可溶化封入体鎖を凍結ストックから解凍した。封入体を、6Mグアニジン溶液中5mg/mlの最終濃度まで希釈し、DTT(2Mストック)を10mMの最終濃度まで加えた。混合物を37℃にて30分間インキュベートした。1リットルのリフォールディング緩衝液(100mM Tris(pH8.5)、400mM L-アルギニン、2mM EDTA、5mM還元グルタチオン、0.5mM酸化グルタチオン、5M尿素、0.2mM PMSF)を調製し、5℃±3℃にて激しく撹拌した。レドックスカップル(redox couple)(2-メルカプトエチルアミン及びシスタミン(それぞれ6.6mM及び3.7mMの最終濃度まで)を添加して約5分後に、変性TCR鎖を添加した。次いで、タンパク質を、5℃±3℃にて撹拌しながら、約5時間±15分間リフォールディングさせた。次いで、リフォールディング体(refold)を、2回、最初は10リットルの100mM尿素に対して、第2回目は10リットルの100mM尿素、10mM Tris(pH8.0)に対して透析した。リフォールディング工程及び透析工程は共に6〜8℃で行なった。
【0073】
図6のようにAkta精製器(Pharmacia)を使用して、透析したリフォールディング体をPOROS 50HQアニオン交換カラムに充填し、50カラム容量にわたる0〜500mM NaCIの勾配で結合タンパク質を溶出させることにより、 scTCRを分解産物及び不純物から分離した。ピーク画分を4℃で保存し、クーマシー染色SDS-PAGEにより分析した(図7)後、プールして濃縮した。次いで、sTCRを精製し、HBS-EP緩衝液(10mM HEPES(pH7.4)、150mM NaCl、3.5mM EDTA、0.05% nonidet p40)中で予め平衡化したSuperdex 200HRゲル濾過カラムを使用して特徴付けた(図8)。ピーク画分を4℃で保存し、クーマシー染色SDS-PAGEにより分析した(図9)後、プールして濃縮した。最後に、BIAcore緩衝液中に精製タンパク質を生成するために、濃縮画分B9〜B6を別のゲル濾過工程を通して行なった(図10)。約50kDaの相対分子量で溶出するピークをプールした。これを濃縮した後、BIAcore表面プラズモン共鳴分析により特徴付けた。
【0074】
実施例3−HLA-A2 TaxとのscTCRの結合のBIAcore表面プラズモン共鳴による特徴付け
表面プラズモン共鳴バイオセンサ(BIAcore 3000(商標))を使用して、A6 scTCRとそのペプチド−MHCリガンド(HLA-A2 Tax)との結合を分析した。これは、半配向様式でストレプトアビジン被覆結合表面に固定した単一pMHC複合体(下記に説明)を作成し、同時に4つまでの異なるpMHC(別々のフローセルに固定された)に対する可溶性T細胞レセプターの結合の効率的な試験を可能にすることによって促進させた。HLA複合体の手動での注入により、固定したクラスI分子を正確なレベルで容易に操作することが可能になった。
【0075】
このような固定化複合体は、T細胞レセプター及びコレセプターCD8ααの両方と結合でき、この両方とも可溶相に注入され得る。
【0076】
ビオチン化クラスI HLA-A2−Tax複合体を、インビトロで、構成成分のサブユニットタンパク質及び合成ペプチドを含む細菌発現封入体からリフォールディングさせ、続いて精製し、インビトロで酵素的にビオチン化した(O'Callaghanら(1999)Anal.Biochem.266:9-15)。適切な構築物中でHLA重鎖の膜貫通ドメイン及び細胞質ドメインを置換するC末端ビオチン化タグを有するHLA重鎖を発現させた。HLA軽鎖又はβ2-ミクログロブリンもまた、〜500mg/リットル細菌培養物のレベルで、適切な構築物からE.coli中で封入体として発現させた。
【0077】
E. coli細胞を溶解し、封入体を約80%の純度まで精製する。封入体からのタンパク質を6Mグアニジン−HCl、50mM Tris(pH8.1)、100mM NaCl、10mM DTT、10mM EDTA中で変性させ、5℃より低いリフォールド緩衝液中に変性タンパク質の単一パルス(single pulse of denatured protein)を添加することにより、30mg/リットル重鎖、30mg/リットルβ2mの濃度で、0.4M L-アルギニン−HCl、100mM Tris(pH8.1)、3.7mMシスタミン、mMシステアミン、4mg/mlペプチド(例えばtax 11-19)中にリフォールディングさせた。リフォールディングは少なくとも1時間4℃で完了させた。
【0078】
緩衝液を、10容量の10mM Tris(pH8.1)での透析により交換した。溶液のイオン強度を十分に減少させるために、2回の緩衝液交換が必要であった。次いで、タンパク質溶液を、1.5μm酢酸セルロースフィルターを通して濾過し、POROS 50HQアニオン交換カラム(8ml床容量)に充填した。タンパク質を、直線の0〜500mM NaCl勾配で溶出させた。HLA-A2−ペプチド複合体は約250mM NaClで溶出した。ピーク画分を収集し、プロテアーゼインヒビターのカクテル(Calbiochem)を加え、画分を氷上で冷却した。
【0079】
ビオチン化タグを付したHLA複合体を、10mM Tris(pH8.1)、5mM NaCl中に、同じ緩衝液中で平衡化したPharmacia迅速脱塩カラムを使用して緩衝液交換をした。溶出の際、即座に、タンパク質含有画分を氷上で冷却し、プロテアーゼインヒビターカクテル(Calbiochem)を加えた。次いで、ビオチン化試薬を加えた:1mMビオチン、5mM ATP(pH8に緩衝化)、7.5mM MgCl2及び5μg/ml BirA酵素(O’Callaghanら(1999)Anal.Biochem.266:9-15に従って精製)。次いで、混合物を室温にて一晩インキュベートした。
【0080】
ゲル濾過クロマトグラフィーを使用してビオチン化HLA複合体を精製した。Pharmacia Superdex 75 HR 10/30カラムを濾過PBSで予め平衡化し、1mlのビオチン化反応混合物を充填し、カラムをPBSで0.5ml/分にて展開した。ビオチン化HLA複合体は、約15mlで単一ピークとして溶出した。タンパク質を含む画分をプールし、氷上で冷却し、プロテアーゼインヒビターカクテルを加えた。クーマシー結合アッセイ(PerBio)を使用してタンパク質濃度を測定し、ビオチン化HLA複合体のアリコートを−20℃で凍結保存した。標準的なアミンカップリング法によりストレプトアビジンを固定化した。
【0081】
新規な鎖間結合を含むA6 Tax scTCRとそのリガンド/MHC複合体又は無関係のHLA−ペプチド組合せ(これらの製造は上記で説明)との間の相互作用を、BIAcore 3000(商標)表面プラズモン共鳴(SPR)バイオセンサで分析した。 SPRは、レセプターリガンド相互作用を検出しその親和性及び動力学的パラメータを分析するために使用することができる原理である、小さなフローセル内のセンサ表面近くで、応答単位(RU)で表される屈折率の変化を測定する。β2mに架橋したビオチンとフローセルの活性化表面に化学的に架橋したストレプトアビジンとの間の結合を介して、個々のHLA−ペプチド複合体を別々のフローセルに固定化することにより、プローブフローセルを準備した。次いで、異なるフローセルの表面上にscTCRを一定流速で通過させ、そうしている間のSPR応答を測定することにより、アッセイを実施した。ペプチド−HLA複合体上に一定流速で異なる濃度で可溶性sTCRを注入してバックグランドの共鳴を規定した。これらコントロールの測定値を、特異ペプチド−HLA複合体を用いて得られた値から減算し、これを使用して解離定数Kd(Price及びDwek、Principles and Problems in Physical Chemistry for Biochemists(第2版)1979,Clarendon Press、Oxford)として表される結合親和性を算出した。
【0082】
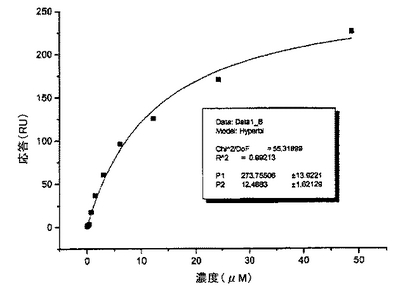
scDiS A6 TCRのBIAcore分析により、この分子がそのコグネイトリガンド(HLA-A2 TAX)に、12.4±1.62μMのkdで特異的に結合することが示された。
【図面の簡単な説明】
【0083】
【図1】a及びbは、それぞれ、システインコドンを導入するように変異された、可溶性A6 TCRのα及びβ鎖の核酸配列を示す。
【図2】aは、新規なジスルフィド鎖間結合を形成させるために使用したT48→C変異(下線)を含む、A6 TCRα鎖細胞外アミノ酸配列を示す。bは、新規なジスルフィド鎖間結合を形成させるために使用したS57→C変異(下線)を含む、A6 TCRβ鎖細胞外アミノ酸配列を示す。
【図3】Gly/Serリンカー(30マー)のDNA及びアミノ酸配列を示す。
【図4】scDiS A6 TCRを作成するために使用したクローニング戦略をまとめる。
【図5】aはscDiS A6 TCRのDNA配列を示す。bはscDiS A6 TCRのアミノ酸配列を示す。
【図6】直線で示されるような0〜500mM NaCl勾配を使用するPOROS 50HQイオン交換カラムからのscDiS A6 TCRタンパク質の溶出を説明する。
【図7】図6で説明したカラムラン(column run)からの画分A15、B10、B9及びB3の還元SDS-PAGEゲル(クーマシー染色)及び非還元SDS-PAGE(クーマシー染色)ゲルの両方の結果を示す。画分B9及びB10は、明らかに、予想したサイズのscDiS A6 TCRに対応するタンパク質を含む。
【図8】図6に示したイオン交換カラムランからの画分B10〜B7のSuperdex 200ゲル濾過カラムからのscDiS A6 TCR溶出の溶出を説明する。
【図9】図8で説明したゲル濾過カラムランからの画分B8、B7、B3及びB2の還元SDS-PAGEゲル(クーマシー染色)及び非還元SDS-PAGE(クーマシー染色)ゲルの両方の結果を示す。画分B7は、明らかに、予想したサイズのscDiS A6 TCRに対応するタンパク質を含む。
【図10】図8に示したゲル濾過ランの濃縮画分B9〜B6のBIAcore緩衝液中への最後のゲル濾過ランである。scDiS A6 TCRは単一の主要ピークとして溶出する。
【図11】図11は、HLA-A2 TAXへのscDiS A6 TCRの結合についてのBIAcoreデータである。
【技術分野】
【0001】
本発明は、単鎖T細胞レセプター(TCR)に関する。
【背景技術】
【0002】
天然のTCR
例えばWO99/60120に記載されているように、TCRは、T細胞による特異的な主要組織適合性複合体(MHC)−ペプチド複合体の認識を媒介し、そしてそれ自体、免疫系の細胞兵器の働きに必須である。
【0003】
抗体及びTCRは、特異的な様式で抗原を認識する唯2つの分子型であり、したがってTCRは、MHCにおいて提示される特定ペプチド抗原(この外来ペプチドは、細胞内の異常の唯一の徴候であることが多い)の唯一のレセプターである。T細胞認識は、T細胞及び抗原提示細胞(APC)が直接物理的接触をしているときに起こり、抗原特異的TCRとpMHC複合体との結びつきにより開始される。
【0004】
天然(native)のTCRは、シグナル伝達媒介に関与するCD3複合体の不変タンパク質と会合する、免疫グロブリンスーパーファミリーのヘテロ二量体細胞表面タンパク質である。TCRはαβ及びγδ形態で存在し、これらの形態は、構造的に類似するが、全く異なる解剖学的所在及びおそらくは機能を有する。MHCクラスI及びクラスIIリガンドもまた、免疫グロブリンスーパーファミリータンパク質であるが、APC細胞表面で多様な編成(array)の短いペプチドフラグメントを提示することを可能にする高度に多形性のペプチド結合部位を有して抗原提示に特化している。
【0005】
2つのさらなるクラスのタンパク質が、TCRリガンドとして機能し得ることが知られている。(1)CD1抗原は、その遺伝子が古典的なMHCクラスI及びクラスII抗原とは異なる染色体に位置するMHCクラスI関連分子である。CD1分子は、従来のクラスI及びクラスII-MHC−pep複合体と類似の様式で、ペプチド部分及び非ペプチド部分(例えば、脂質、糖脂質)をT細胞に提示することができる(例えば、Barclayら(1997) The Leucocyte Antigen Factsbook 第2版,Academic Press、及びBauer(1997) Eur J Immunol 27(6) 1366-1373を参照)。(2)細菌性スーパー抗原は、クラスII MHC分子とTCRのサブセットとの両方に結合できる可溶性毒素である(Fraser(1989) Nature 339 221-233)。多くのスーパー抗原が1又は2のVβセグメントに対して特異性を示す一方、その他のものはより無差別な結合を示す。ともかく、スーパー抗原は、多クローン性様式でT細胞のサブセットを刺激する能力により、増強した免疫応答を誘発することができる。
【0006】
天然のヘテロ二量体αβTCRの細胞外部分は2つのポリペプチドからなり、その各個が、膜近位定常ドメイン及び膜遠位可変ドメインを有する(図1を参照)。定常ドメイン及び可変ドメインの各個が、鎖内ジスルフィド結合を含む。可変ドメインは、抗体の相補性決定領域(CDR)に類似する高度多形性ループを含む。TCRのCDR3はMHCにより提示されたペプチドと相互作用し、CDR1及びCDR2はペプチド及びMHCと相互作用する。TCR配列の多様性は、連結される可変(V)、多様性(V)、連結(J)及び定常遺伝子の体細胞再編成(somatic rearrangement)を介して生じる。機能的なα鎖ポリペプチドは、再編成されたV−J−C領域により形成される一方、β鎖はV−D−J−C領域からなる。細胞外定常ドメインは、膜近位領域及び免疫グロブリン領域を有する。単一のα鎖定常ドメイン(TRACとして知られる)及び2つの異なるβ定常ドメイン(TRBC1及びTRBC2として知られる)が存在する(IMGT命名法)。これらβ定常ドメイン間には4つのアミノ酸変化が存在し、そのうちの3つは、本発明の単鎖TCRを作成するために使用するドメイン内にある。これらの変化は全て、TRBC1及びTRBC2のエキソン1内にあり(N4K5→K4N5及びF37→Y(IMGT番号付法、差異TRBC1→TRBC2))、2つのTRCβ鎖定常領域間の最後のアミノ酸変化は、TRBC1及びTRBC2のエキソン3にある(V1→E)。TCR細胞外ドメインの各個の範囲はいくらか変化し得る。しかし、当業者は、The T Cell Receptor Facts Book,Lefranc & Lefranc,Academic Press発行,2001のような参考文献を使用して、ドメイン境界の位置を容易に決定できる。
【0007】
単鎖TCR
単鎖TCR(scTCR)は、アミノ酸単鎖からなる人工構築物であり、これは天然のヘテロ二量体TRCと同様に、MHC−ペプチド複合体に結合する。不運にも、α及びβ鎖をその両方が1つのオープンリーディングフレームで発現するように単に連結することによって機能的なα/βアナログscTCRを作成しようとする試みは、成功しなかった。これは、おそらく、α−β可溶ドメイン対合の当然の不安定性による。
【0008】
したがって、α及びβ鎖のいずれか又は両方の種々の短縮化を使用する特別な技法が、scTCRの作成に必要とされてきた。これらの方式は、非常に限られた範囲のscTCR配列に対してのみ適用可能であるようである。Hooら(1992)PNAS.89(10):4759-63は、25アミノ酸リンカーと連結した短縮型β及びα鎖と細菌細胞周辺質発現(bacterial periplasmic expression)とを使用する2C T細胞クローンからの単鎖形式でのマウスTCRの発現を報告する(Schodinら(1996)Mol.Immunol.33(9):819-29もまた参照)。この設計はまた、Hollerら(2000)PNAS.97(10):5387-92により報告されたm6単鎖TCRの基礎をなし、m6単鎖TCRは2C scTCRから誘導され、同じH2-Ld制限アロエピトープ(alloepitope)に結合する。Shustaら(2000)Nature Biotechnology 18:754-759は、酵母展示実験(yeast display experiment)における単鎖2C TCR構築物の使用を報告し、これは増強した熱安定性及び溶解性を有する変異TCRを生じた。この報告はまた、これら展示2C TCRがコグネイトpMHCを発現する細胞に選択的に結合することができることを証明した。Khandekarら(1997)J.Biol.Chem.272(51):32190-7は、マウスD10 TCRについて、このscTCRをMBPに融合して細菌細胞質で発現したが、同様な設計を報告している(Hare1ら(1999)Nat.Struct.Biol.6(6):574-81もまた参照)。Hilyardら(1994)PNAS.91(19):9057-61は、Vα−リンカー−Vβ設計を使用し細菌細胞周辺質で発現させた、インフルエンザマトリクスタンパク質−HLA-A2に特異的なヒトscTCRを報告している。
【0009】
Chungら(1994)PNAS.91(26)12654-8は、Vα−リンカー−Vβ−Cβ設計及び哺乳動物細胞株の表面での発現を使用するヒトscTCRの作成を報告している。この報告は、scTCRのペプチド−HLA特異的結合に対しては何らの言及も含んでいない。Plaksinら(1997)J.Immunol.158(5):2218-27は、HIV gp12-H-2Ddエピトープに特異的なマウスscTCRを作成するための同様なVα−リンカー−Vβ−Cβ設計を報告している。
【0010】
治療用途
疾患プロセスにより影響される細胞に局在できるターゲッティング部分(targeting moiety)の必要性が存在する。このようなターゲッティング部分は、自己免疫疾患の原因である免疫系の「指向性を誤った」作用を直接的に遮断するためか又はガン性細胞に細胞毒性物質(cytotoxic agent)を送達する手段としてのいずれかに利用し得る。
【0011】
理想的には、これらの適用に適切な分子は、関係する疾患プロセスに直接関与する細胞マーカーに対して特異的親和性を必要とする。この目的のためには抗体が使用されてきた。
【0012】
スクリーニング用途
多くの重要な細胞相互作用及び細胞応答(TCR媒介免疫系を含む)は、細胞表面レセプターと、他の細胞の表面に提示されるリガンドとの間で行なわれる接触により制御される。これらのタイプの特異的分子接触は、ヒト体内での正確な生化学的調節に決定的に重要であり、したがって精力的に研究されている。多くの場合、このような研究の目標は、疾患を予防し又は疾患と戦うために、細胞応答を調整する手段を考案することである。
【0013】
したがって、ある程度の特異性でヒトレセプター又はリガンド分子と結合する化合物を同定するために用いる方法は、新たな疾患治療学の発見及び開発のための先導として重要である。特に、特定のレセプター−リガンド相互作用に干渉する化合物は、治療剤又は治療キャリアとしての直接の可能性を有する。
【0014】
非常に大きな化合物ライブラリの比較的容易で対費用効率の高い作成を可能にするコンビナトリアル・ケミストリーにおける進歩は、化合物試験の範囲を莫大に増大させた。今や、スクリーニングプログラムの限界は、ほとんどの場合、用いることができるアッセイの性質、適切なレセプター及びリガンド分子の作成及びこれらアッセイが高スループットスクリーニング法にどのくらい十分に適合させることができるかにある。
【発明の開示】
【課題を解決するための手段】
【0015】
発明の簡単な説明
本発明は、アミノ酸単鎖の残基間のジスルフィド結合の存在により特徴付けられる新たなクラスのα/βアナログscTCRを利用可能にする。この結合は、その分子のα領域とβ領域との間の対合の安定性に寄与する。このようなTCRは、スクリーニング目的又は治療目的に有用である。
【0016】
発明の詳細な説明
本発明は、TCRα鎖定常領域細胞外配列のN末端に融合したTCRα鎖可変領域配列により構成されるαセグメントと、TCRβ鎖定常領域細胞外配列のN末端に融合したTCRβ鎖可変領域配列により構成されるβセグメントと、αセグメントのC末端をβセグメントのN末端を連結するか又はその逆であるリンカー配列とを含み、α及びβセグメントの定常領域細胞外配列は、ジスルフィド結合により連結し、リンカー配列の長さ及びジスルフィド結合の位置は、α及びβセグメントの可変領域配列が天然のαβT細胞レセプター中と実質的に同様に相互に配向するような長さ及び位置である単鎖T細胞レセプター(scTCR)を提供する。
【0017】
本発明のscTCRにおいて、α及びβセグメントの可変領域配列が天然のαβT細胞レセプター中と実質的に同様に相互に配向するという要件は、その分子が関係するTCRリガンド(pMHC複合体、CD1−抗原複合体、スーパー抗原又はスーパー抗原/pMHC複合体)に結合することを確認することによって検証する−結合すればこの要件を満たす。pMHC複合体との相互作用は、BIAcore 3000(商標)又はBIAcore 2000(商標)装置を使用して測定することができる。本明細書中の実施例3又はWO 99/6120はそれぞれ、MHC−ペプチド複合体とのTCR結合を分析するために必要となる方法の詳細な説明を提供する。これらの方法は、TCR/CD1及びTCR/スーパー抗原相互作用の研究に等しく適用可能である。これらの方法をTCR/CD1相互作用の研究に適用するためには、可溶形態のCD1が必要であり、その作成は、Bauer(1997)Eur J Immunol 27(6) 1366-1373)に記載されている。
【0018】
α及びβセグメント
α及びβセグメント中に存在する定常領域細胞外配列は、好ましくは、ヒトTCRのものに対応し、α及びβセグメント中に存在する可変領域配列も同様である。しかし、このような配列間の対応は、アミノ酸レベルで1:1である必要はない。全体として結果的に、天然のαβT細胞レセプター中と同様にα及びβセグメントの可変領域配列が相互に配向し、ペプチド−MHC結合機能性が維持されれば、対応するヒトTCR配列に対する、N若しくはC短縮並びに/又はアミノ酸の欠失及び/若しくは置換は許容される。特に、α及びβセグメント中に存在する定常領域細胞外配列は、scTCRが結合するペプチド−MHC複合体との接触に直接関与しないので、天然のTCRの細胞外定常ドメイン配列より短くてもよいし、天然のTCRの細胞外定常ドメイン配列に対する置換又は欠失を含んでもよい。
【0019】
αセグメント中に存在する定常領域細胞外配列は、TCRα鎖の細胞外定常Igドメインに対応する配列を含んでもよく、及び/又はβセグメント中に存在する定常領域細胞外配列は、TCRβ鎖の細胞外定常Igドメインに対応する配列を含んでもよい。
【0020】
本発明の1つの実施形態では、αセグメントは、TCRα鎖の定常領域の実質的に全ての細胞外ドメインのN末端に融合した、TCRα鎖の実質的に全ての可変領域に対応し;且つ/又はβセグメントは、TCRβ鎖の定常領域の実質的に全ての細胞外ドメインのN末端に融合した、TCRβ鎖の実質的に全ての可変領域に対応する。
【0021】
別の実施形態では、α及びβセグメント中に存在する定常領域細胞外配列は、TCRの天然の鎖間ジスルフィド結合を形成するシステイン残基を排除するように、C末端で短縮化された天然のTCRのα及びβ鎖の定常領域に対応する。或いは、これらシステイン残基は、天然のジスルフィド結合が欠失するように、別のアミノ酸残基(例えばセリン又はアラニン)で置換されてもよい。加えて、天然のTCRβ鎖は、対合していないシステイン残基を含むが、その残基は、本発明のscTCRのβ配列から欠失されていてもよいし、本発明のscTCRのβ配列中で非システイン残基で置換されていてもよい。
【0022】
本発明の1つの特定の実施形態では、α及びβセグメント中に存在するTCRα及びβ鎖可変領域配列は、併せて、第1のTCRの機能的な可変ドメインに対応してもよく、α及びβセグメント中に存在するTCRα及びβ鎖定常領域細胞外配列は、第2のTCRのそれに対応してもよく、第1及び第2のTCRは同じ種に由来する。したがって、α及びβセグメント中に存在するα及びβ鎖可変領域配列は、第1のヒトTCRのものに対応してもよく、α及びβ鎖定常領域細胞外配列は、第2のヒトTCRのものに対応してもよい。例えば、A6 Tax sTCR定常領域細胞外配列は、異種の可変ドメインが融合できるフレームワークとして使用することができる。
【0023】
本発明の別の実施形態では、α及びβセグメント中に存在するTCRα及びβ鎖可変領域配列は、併せて、第1のTCRの機能的な可変ドメインに対応し、α及びβセグメント中に存在するTCRα及びβ鎖定常領域細胞外配列は、第2のTCRのそれに対応し、第1及び第2のTCRは異なる種に由来する。この実施形態では、α及びβセグメント中に存在するTCRα及びβ鎖可変領域配列は、併せて、ヒトTCRの機能的可変ドメインに対応し、α及びβセグメント中に存在するTCRα及びβ鎖定常領域細胞外配列は、マウスTCRのものに対応する。本発明のこのような実施形態は、scTCRが免疫原性である可能性が高い非ヒト定常領域配列を含み、したがって標的細胞上に局在するとdTCRに対する免疫応答全体を増強する可能性が高いという利点を有する。異常細胞(例えばガン細胞)に対する免疫応答は、このように増強され得る。
【0024】
リンカー
本発明では、リンカー配列は、ポリペプチド単鎖を形成するために、α及びβセグメントを連結する。リンカー配列は、例えば、式−P−AA−P−(式中、Pはプロリンであり、AAはアミノ酸がグリシン及びセリンであるアミノ酸配列を表す)を有してもよい。
【0025】
scTCRがMHC−ペプチド複合体に結合するために、α及びβセグメントは、その可変領域配列がそのような結合に関して配向するように、対合していなければならない。したがって、リンカーは、αセグメントのC末端とβセグメントのN末端との間又はその逆の間の距離を橋渡しするに十分な長さを有するべきである。他方で、万一にもN末端可変領域配列でリンカーの末端部が、scTCRと標的ペプチド−MHC複合体との結合を遮断し又は減少させないように、好ましくは、過度なリンカー長は回避すべきである。
【0026】
例えば、α及びβセグメント中に存在する定常領域細胞外配列が、TCRの天然の鎖間ジスルフィド結合を形成するシステイン残基を排除するように、C末端で短縮化された天然のTCRのα及びβ鎖の定常領域に対応し、リンカー配列がαセグメントのC末端とβセグメントのN末端を連結する場合、リンカーは、26〜41アミノ酸、例えば29、30、31又は32アミノ酸からなってもよく、特定のリンカーは、式−PGGG−(SGGGG)5−P−(式中、Pはプロリンであり、Gはグリシンであり、Sはセリンである)を有する。
【0027】
ジスルフィド結合
本発明のscTCRの基本的に特徴付ける性状は、α及びβセグメントの定常領域細胞外配列間のジスルフィド結合である。この結合は、天然の二量体αβTCR中に存在する天然の鎖間ジスルフィド結合に対応してもよいし、天然のTCR中に対応物(counterpart)を有さない、α及びβセグメントの定常領域細胞外配列中に特異的に組み込まれたシステイン間のものであってもよい。いくつかの場合では、天然のもの及び天然のものでない両方のジスルフィド結合が、本発明のscTCRにおいて望ましくあり得る。
【0028】
ジスルフィド結合の位置は、α及びβセグメントの可変領域配列が天然のαβT細胞レセプター中と実質的に同様に相互に配向するという要件に従属する。
ジスルフィド結合は、α及びβセグメント上の非システイン残基をシステインに変異させ、その変異した残基間にジスルフィド結合を形成させることにより形成してもよい。天然の残基の代わりに導入されたシステイン残基間にジスルフィド結合を形成することができるように、天然のTCR中でそれぞれのβ炭素が約6Å(0.6nm)以下、好ましくは3.5Å(0.35nm)〜5.9Å(0.59nm)の範囲で離れている残基が好ましい。ジスルフィド結合が定常免疫グロブリン領域中の残基間であれば、その結合は膜近位領域の残基間であり得るが好ましい。システインを導入してジスルフィド結合を形成することができる好ましい部位は、TCRα鎖についてはTRAC*01の、TCRβ鎖についてはTRBC1*01又はTRBC2*01のエキソン1中の以下の残基である:
【表1】
【0029】
今や、システイン残基に変異させて本発明に従うscTCR中に新たな鎖間ジスルフィド結合を形成させることができるヒトTCR中の残基が同定されたので、当業者は他の種のTCRを同じ方法で変異させてその種のscTCRを作成することができる。ヒトにおいては、当業者は、単に、それぞれのTCR鎖中の以下のモチーフを探索して変異させるべき残基(影付きの残基がシステインへの変異のための残基である)を同定することのみが必要である。
α鎖Thr48:DSDVYITDKTVLDMRSMDFK(TRAC*01遺伝子のエキソン1のアミノ酸39〜58)
α鎖Thr45:QSKDSDVYITDKTVLDMRSM(TRAC*01遺伝子のエキソン1のアミノ酸36〜55)
α鎖Tyr10:DIQNPDPAVYQLRDSKSSDK(TRAC*01遺伝子のエキソン1のアミノ酸1〜20)
α鎖Ser15:DPAVYQLRDSKSSDKSVCLF(TRAC*01遺伝子のエキソン1のアミノ酸6〜25)
β鎖Ser57:NGKEVHSGVSTDPQPLKEQP(TRBC1*01及びTRBC2*01遺伝子のエキソン1のアミノ酸48〜67)
β鎖Ser77:ALNDSRYALSSRLRVSATFW(TRBC1*01及びTRBC2*01遺伝子のエキソン1のアミノ酸68〜87)
β鎖Ser17:PPEVAVFEPSEAEISHTQKA(TRBC1*01及びTRBC2*01遺伝子のエキソン1のアミノ酸8〜27)
β鎖Asp59:KEVHSGVSTDPQPLKEQPAL(TRBC1*01及びTRBC2*01遺伝子のエキソン1のアミノ酸50〜69)
β鎖Glu15:VFPPEVAVFEPSEAEISHTQ(TRBC1*01及びTRBC2*01遺伝子のエキソン1のアミノ酸6〜25)
【0030】
他の種においては、TCR鎖は、上記モチーフに対して100%同一性を有する領域を有さなくてもよい。しかし、当業者は、上記モチーフを使用してTCRα又はβ鎖の等価部分、したがってシステインに変異させるべき残基を同定することができる。整列技法(alignment technique)をこの点に関して使用してもよい。例えば、変異のためのTCR配列の関係部分を位置決めするために、European Bioinformatics Instituteのウェブサイト(http://www.ebi.ac.uk/index.html)で入手可能であるClustalWを使用して上記モチーフと特定のTCR鎖配列を比較することができる。
【0031】
本発明は、その範囲に、αβアナログscTCR及び他の哺乳動物(マウス、ラット、ブタ、ヤギ及びヒツジを含むがこれらに限定されない)のそれを含む。上記のようにヒト/非ヒトキメラscTCRもまた含まれる。上記のように、当業者は、システイン残基を導入して鎖間ジスルフィド結合を形成させることが可能である上記のヒト部位と等価な部位を決定することができる。例えば、以下は、マウスCα及びCβ可溶性ドメインのアミノ酸配列を、システインに変異させてTCR鎖間ジスルフィド結合を形成させることができる上記のヒト残基に等価なマウス残基を示すモチーフと共に示す(ここで、関係する残基には影が付されている)。
マウスCα可溶性ドメイン:
PYIQNPEPAVYQLKDPRSQDSTLCLFTDFDSQINVPKTMESGTFITDKTVLDMKAMDSKSNGAIAWSNQTSFTCQDIFKETNATYPSSDVP
マウスCβ可溶性ドメイン:
EDLRNVTPPKVSLFEPSKAEIANKQKATLVCLARGFFPDHVELSWWVNGREVHSGVSTDPQAYKESNYSYCLSSRLRVSATFWHNPRNHFRCQVQFHGLSEEDKWPEGSPKPVTQNISAEAWGRAD
ヒトα鎖Thr48のマウス等価物:ESGTFITDKTVLDMKAMDSK
ヒトα鎖Thr45のマウス等価物:KTMESGTFITDKTVLDMKAM
ヒトα鎖Tyr10のマウス等価物:YIQNPEPAVYQLKDPRSQDS
ヒトα鎖Ser15のマウス等価物:AVYQLKDPRSQDSTLCLFTD
ヒトβ鎖Ser57のマウス等価物:NGREVHSGVSTDPQAYKESN
ヒトβ鎖Ser77のマウス等価物:KESNYSYCLSSRLRVSATFW
ヒトβ鎖Ser17のマウス等価物:PPKVSLFEPSKAEIANKQKA
ヒトβ鎖Asp59のマウス等価物:REVHSGVSTDPQAYKESNYS
ヒトβ鎖Glu15のマウス等価物:VTPPKVSLFEPSKAEIANKQ
【0032】
上記のように、A6 Tax sTCR細胞外定常領域は、それに異種可変ドメインを融合させることが可能であるフレームワークとして使用することができる。異種可変領域配列は、ジスルフィド結合と定常領域配列のN末端との間の任意の点で定常領域配列に連結することが好ましい。A6 Tax TCRα及びβ定常領域配列の場合、ジスルフィド結合は、アミノ酸残基158及び172でそれぞれ導入されたシステイン残基間で形成され得る。したがって、異種α及びβ鎖可変領域配列の付着点が、それぞれ残基159又は173とα又はβ定常領域配列のN末端との間であれば好ましい。
【0033】
さらなる観点
本発明のscTCR(好ましくはヒトである)は、実質的に純粋な形態で又は精製若しくは単離された調製物として提供され得る。例えば、これは、他のタンパク質を実質的に含まない形態で提供され得る。
【0034】
本発明の複数のscTCRは多価複合体で提供され得る。したがって、本発明は、1つの観点では、多価T細胞レセプター(TCR)複合体を提供し、これは複数の本明細書中に記載のような可溶性T細胞レセプターを含む。複数の可溶性TCRの各個は、好ましくは、同一である。
【0035】
本発明の多価複合体において、scTCRは、多量体の形態であってもよいし、及び/又は脂質二重層(例えばリポソーム)上に存在するか若しくはこれと会合していてもよい。
【0036】
最も単純な形態では、本発明に従う多価scTCR複合体は、好ましくはリンカー分子を介して、互いに会合(例えば、共有結合又は別の結合)した2又は3又は4又はそれ以上のT細胞レセプター分子の多量体を含む。適切なリンカー分子には、アビジン、ストレプトアビジン、ニュートラアビジン(neutravidin)及びエキストラアビジン(extravidin)(これらの各個は、ビオチンに関して4つの結合部位を有する)のような多価付着分子が含まれが、これらに限定されない。したがって、ビオチン化TCR分子は、複数のTCR結合部位を有するT細胞レセプター多量体に形成することができる。多量体中のTCR分子の数は、その多量体を作成するために使用したリンカー分子の量に対するTCRの量に依存し、また他のビオチン化分子の存否にも依存する。好ましい多量体は、二量体、三量体又は四量体のTCR複合体である。
【0037】
TCR四量体より相当大きい構造は、特異的MHC−ペプチド複合体を発現する細胞を追跡又は標的することに使用してもよい。好ましくは、構造は、直径が10nm〜10μmの範囲である。各構造は、その構造上の2以上のTCR分子が細胞上の2以上のMHC−ペプチド複合体に同時に結合することを可能にし、したがってその細胞に関する多量体結合部分のアビディティを増大させるように十分に離れた距離で複数のscTCR分子を展示し得る。
【0038】
1又は複数のscTCRを有する複合体を形成するための本発明での使用に適切な構造は、膜構造(例えばリポソーム)及び固体構造(好ましくはビーズのような粒子、例えばラテックスビーズである)を含む。T細胞レセプター分子で外部を被覆され得る他の構造もまた適切である。好ましくは、構造は、個々のT細胞レセプター分子でよりむしろ、T細胞レセプター多量体で被覆される。
【0039】
リポソームの場合、T細胞レセプター分子又はその多量体は、膜に付着していてもよいし、膜とその他の会合をしていてもよい。このための技法は当業者に周知である。
【0040】
標識又は別の部分(例えば、毒性部分又は治療的部分)が、本発明の多価scTCR複合体に含まれてもよい。例えば、標識又は他の部分は、混合分子多量体に含まれてもよい。このような多量体分子の例は、3つのscTCR分子と1つのペルオキシダーゼ分子とを含む四量体である。これは、TCR及び当該酵素を3:1のモル比で混合して四量体複合体を生成し、正確な比の分子を含まない複合体から所望の複合体を分離することにより達成され得る。これらの混合分子は、立体障害がその分子の所望の機能を損なわないか又は有意には損なわないという条件で、分子の任意の組合せを含み得る。ストレプトアビジン分子上の結合部位の位置は、立体障害を起こす可能性がないので、混合四量体に適切である。
【0041】
さらなる観点では、本発明は、MHC−ペプチド複合体を検出するための方法を提供し、この方法は、
a.本発明のcTCRを提供すること
b.scTCRをMHC−ペプチド複合体と接触させること;及び
scTCRのMHC−ペプチド複合体との結合を検出すること
を含む。
【0042】
治療用途
本発明のscTCR(又はその多価複合体)は、択一的又は追加的に、例えば細胞殺傷に使用するための毒性部分又は免疫刺激物質(immunostimulating agent)(例えばインターロイキン又はサイトカイン)であり得る治療物質(therapeutic agent)と会合(例えば、共有結合又は別の結合)していてもよい。本発明の多価scTCR複合体は、非多量体のT細胞レセプターへテロ二量体と比べて、TCRリガンド(例えばpMHC複合体又はCD1分子)に関して増強した結合能力を有し得る。したがって、本発明に従う多価scTCR複合体は、特定の抗原を提示する細胞をインビトロ又はインビボで追跡又は標的するために特に有用であり、またそのような用途を有するさらに多価のTCR複合体の製造のための中間体としても有用である。したがって、scTCR又は多価scTCR複合体は、インビボでの使用のために薬学的に受容可能な製剤で提供され得る。
【0043】
本発明はまた、標的細胞に治療物質を送達するための方法を提供し、この方法は、潜在的な標的細胞を本発明に従うscTCR又は多価scTCR複合体と、標的細胞へのscTCR又は多価scTCR複合体の付着を可能にする条件下で接触させることを含み、当該scTCR又は多価scTCR複合体は、MHC−ペプチド複合体に特異的であり、これに治療物質を会合させる。
【0044】
特に、可溶性scTCR又は多価scTCR複合体を使用して、特定の抗原を提示する細胞の位置に治療物質を送達することができる。このことは、多くの状況で、特に腫瘍に対して有用である。治療物質は、その効果を局所的にではあるが、治療物質が結合する細胞上に限らずに発揮するように送達され得る。したがって、1つの特定の戦略は、腫瘍抗原に特異的なT細胞レセプター又は多価scTCR複合体に連結した抗腫瘍分子を考案する。
【0045】
多くの治療物質、例えば放射活性化合物、酵素(例えばパーフォリン)又は化学療法剤(例えばシスプラチン)がこの用途に用いられ得る。確実に所望の位置で毒性効果が発揮されるために、毒素は、それがゆっくりと放出されるように、ストレプトアビジンに連結したリポソーム内にあり得る。このことが、体内での輸送の間の損傷効果を防止し、関係する抗原提示細胞とのscTCRの結合後に毒素が最大効果を有することを確実にする。
【0046】
他の適切な治療物質には、
・小分子細胞毒性物質、すなわち、哺乳動物細胞を殺傷する能力を有する、分子量700ダルトン未満の化合物。このような化合物はまた、細胞毒性効果を有することができる毒性金属を含み得る。さらに、これら小分子細胞毒性物質はまた、プロドラッグ、すなわち、生理学的条件下で崩壊又は変換して細胞毒性物質を放出する化合物を含む。このような物質の例には、シスプラチン、メイタンシン(maytansine)誘導体、ラケルマイシン(rachelmycin)、カリケアマイシン(calicheamicin)、ドセタキセル、エトポシド、ゲムシタビン、イホスファミド、イリノテカン、メルファラン、ミトキサントロン、ソルフィマーソディウムホトフィリンII(sorfimer sodiumphotofrin II)、テモゾロマイド(temozolmide)、トポテカン、トリメトレキサート(trimetreate)、グルクロナート、オーリスタチンE(auristatin E)、ビンクリスチン及びドキソルビシンが含まれる;
・ペプチド細胞毒素、すなわち、哺乳動物細胞を殺傷する能力を有するタンパク質又はそのフラグメント。例には、リシン、ジフテリア毒素、シュードモナス細菌外毒素A、DNAアーゼ及びRNAアーゼが含まれる;
・放射性核種、すなわち、1以上のα若しくはβ粒子又はγ線の同時放射を伴って崩壊する元素の不安定同位体。例には、ヨウ素131、レニウム186、インジウム111、イットリウム90、ビスマス210及び213、アクチニウム225及びアスタチン213が含まれる;
・プロドラッグ、例えば抗体を指向する酵素プロドラッグ;
・免疫増強剤(immuno-stimulant)、すなわち、免疫応答を刺激する部分。例には、サイトカイン(例えばIL-2)、ケモカイン(例えばIL-8)、血小板第4因子、メラノーマ増殖刺激タンパク質など、抗体又はそのフラグメント、補体活性化剤、異種タンパク質ドメイン、同種タンパク質ドメイン、ウイルス性/細菌性タンパク質ドメイン及びウイルス性/細菌性ペプチドが含まれる。
【0047】
本発明の可溶性scTCR又は多価scTCR複合体は、プロドラッグを薬物に変換し得る酵素に連結されてもよい。これは、プロドラッグが薬物を必要とする(すなわち、scTCRにより標的にされる)部位でのみ薬物に変換することを可能にする。
【0048】
本発明に従うscTCRに適切なMHC−ペプチド標的の例には、ウイルス性エピトープ、例えば、HTLV-1エピトープ(例えばHLA-A2により制限されたTaxペプチド;HTLV-1は白血病に関連する)、HIVエピトープ、EBVエピトープ、CMVエピトープ;メラノーマエピトープ(例えばMAGE-1 HLA-A1制限エピトープ)及び他のガン特異的エピトープ(例えばHLA-A2により制限された腎臓細胞ガン腫関連抗原G250);並びに自己免疫疾患(例えば慢性関節リウマチ)に関連するエピトープが含まれるが、これらの限定されない。さらに、本発明での使用に適切な疾患関連pMHC標的は、HLA Factbook(Barclay編,Academic Press)に掲載されており、他にも多くのものが同定されている。
【0049】
scTCRの特異性による限局化薬物送達は、多くの疾患処置を増強することができる可能性が高い。
【0050】
そのための薬物が存在するウイルス性疾患、例えばHIV、SIV、EBV、CMVは、その薬物が感染細胞の近位で放出又は活性化されればその恩恵を受ける。ガンについては、腫瘍又は転移の近位に限局することにより、毒素又は免疫増強剤の効果が増強される。自己免疫疾患では、より長い期間にわたってより局所的な効果を有する一方で、被験者の免疫能力全体には最小限にしか影響しない免疫抑制薬物が徐々に放出され得る。移植拒絶の予防では、免疫抑制薬物の効果は、同じ方法で最適化され得る。ワクチン送達のためには、ワクチン抗原は抗原提示細胞の近くに限局され、したがって抗原の効力を増強し得る。この方法はまた、画像化目的にも適用することができる。
【0051】
本発明のscTCRは、特異的リガンド(例えばpMHC)への結合によりT細胞活性化を調整し、それによりT細胞活性化を阻害するために使用してもよい。T細胞媒介炎症及び/又は組織損傷を含む自己免疫疾患(例えばI型糖尿病)は、このアプローチに適する。関係するpMHCにより提示される特異的ペプチドエピトープの知識が、この用途に必要である。
【0052】
本発明に従う医薬は、通常、一般には薬学的に受容可能なキャリアを含む、滅菌の薬学組成物の部分として供給される。この薬学組成物は、(これを患者に投与する望ましい方法に依存して)任意の適切な形態であり得る。これは、単位投薬剤形で提供されてもよく、一般には密封容器で提供され、キットの部分として提供されてもよい。このようなキットは、(必ずしもではないが)通常、使用のための指示書を含む。これは、複数の単位投薬剤形を含み得る。
【0053】
薬学組成物は、任意の適切な経路、例えば、経口(口腔粘膜又は舌下を含む)、直腸、鼻、局所(口腔粘膜、舌下又は経皮を含む)、膣又は非経口(皮下、筋肉内、静脈内又は皮内)経路による投与に適合され得る。このような組成物は、薬学の分野において公知の任意の方法により、例えば滅菌条件下で活性成分をキャリア又は賦形剤と混合することにより製造し得る。
【0054】
スクリーニング用途
本発明のscTCRは、TCR媒介細胞免疫系の調節物質(modulator)(阻害剤を含む)を同定するために設計されたスクリーニング法で利用可能である。
当業者に公知であるように、この型のタンパク質−タンパク質相互作用スクリーニングのための適切な基礎を提供する多くのアッセイ形式が存在する。
【0055】
増幅ルミネッセンス近接ホモジニアスアッセイ(Amplified Luminescent Proximity Homogeneous Assay)システム(例えばAlphaScreen(商標))は、レセプター及びリガンドタンパク質が付着することができるヒドロゲルの層で被覆された「ドナー」及び「アクセプター」ビーズの使用に基づく。これらレセプター及びリガンド分子の間の相互作用は、ビーズを近接状態にする。これらビーズがレーザ光に曝されると、「ドナー」ビーズ中の光増感物質は、周囲の酸素をより励起された一重項状態に変換する。一重項状態の酸素分子は、拡散して、「アクセプター」ビーズ中の化学発光物質と反応し、この化学発光物質は、さらに、同じビーズに含まれる蛍光体を活性化する。続いて、蛍光体は520〜620nmの光を発光し、これは、レセプター−リガンド相互作用が生じたことをシグナル伝達する。レセプター−リガンド相互作用の阻害剤の存在により、このシグナルは低減される。
【0056】
表面プラズモン共鳴(SPR)は、一方の結合パートナー(通常レセプター)が「チップ」(センサ表面)に固定され、他方の結合パートナー(通常リガンド)(これは可溶であり、チップ上を流される)の結合が検出される界面光学アッセイである。リガンドの結合は、結果的に、チップ表面近くのタンパク質濃度の増大を生じ、このことはその領域の屈折率の変化を引き起こす。チップの表面は、屈折率の変化が、表面プラズモン共鳴により検出され得るように構成される(ここで、表面プラズモン共鳴とは光学現象であり、これにより、ある入射角の光は、薄金属フィルムで、振動する表面電荷密度の波(表面プラズモン)の共鳴励起に起因して、減弱強度の屈折ビームを生じる)。この共鳴は、金属フィルムの裏側での屈折率の変化に非常に敏感であり、固体されたタンパク質と可溶タンパク質との間の結合を検出するために使用されるのが、このシグナルである。分子相互作用のSPR検出及びデータ分析の簡便な使用を可能にするシステムは市販されている。例は、Iasys(商標)装置(Fisons)及びBiacore(商標)装置である。
【0057】
他の界面光学アッセイには、全内部反射蛍光(TIRF)、共振ミラー(RM)及び回折格子カプラセンサ(optical grating coupler sensor)(GCS)が含まれ、これは、Woodbury及びVenton(J.Chromatog.B.725 113-137(1999))でより詳細に議論されている。シンチレーション近接アッセイ(scintillation proximity assay)(SPA)は、CD28とB7との間の低親和性相互作用の阻害剤について化合物ライブラリをスクリーニングするために使用されてきた(おそらく約4μMのKd(Van der Merweら、J.Exp.Med.185:393-403 (1997),Jenhら、Anal Biochem 165(2) 287-93 (1998)))。SPAは、指示表面上に固定された閃光物質(scintillant)にエネルギーを転移させる特定の放射活性同位体からのβ粒子放出を利用する放射活性アッセイである。溶液中の短期(short range)のβ粒子により、確実に、β粒子が閃光物質に近接して発射されたときにのみ閃光が生じる。タンパク質−タンパク質相互作用の検出に適用される場合、一方の相互作用パートナーを放射性同位体で標識し、他方は閃光物質を含むビーズに結合するか、又は閃光物質と共に表面に被覆するかのいずれかである。アッセイを最適に設定することが可能な場合、放射性同位体は、閃光物質に対して、2つのタンパク質間の結合が起きたときにのみ光子発射が活性化されるに十分に近くに運ばれる。
【0058】
本発明のさらなる観点は、scTCRと、MHC−ペプチド複合体、CD1−抗原複合体、スーパー抗原及びMHC−ペプチド/スーパー抗原複合体から選択されるTCRリガンドとの間の相互作用の阻害剤を同定する方法であり、これは、scTCRをscTCRリガンド結合パートナーと、試験化合物の存在下及び非存在下で接触させること、及び試験化合物の存在により、リガンドへのscTCRの結合が減少するかどうかを測定することを含み、この減少が阻害剤を同定するとする。
【0059】
本発明の最後の観点は、scTCRと、MHC−ペプチド複合体、CD1−抗原複合体、スーパー抗原及びMHC−ペプチド/スーパー抗原複合体から選択されるTCRリガンドとの間の相互作用の潜在的な阻害剤を同定する方法であり、これは、scTCR又はscTCRリガンド結合パートナーを試験化合物と接触させること、及び試験化合物が、scTCR及び/又はリガンドに結合するかどうかを測定することを含み、この結合が潜在的阻害剤を同定するとする。本発明のこの観点は、界面光学アッセイ(例えば、BIAcore(商標)システムを使用して実施される界面光学アッセイ)における特別な有用性を見出し得る。
【0060】
本発明の各観点の好ましい性状は、必要な変更を加えて、その他の観点の各個についての性状と同様である。明細書中に言及した先行技術文献は、法が許す最大範囲で明細書中に援用する。
【実施例】
【0061】
本発明は以下の実施例においてさらに記述するが、実施例は、いかなる意味においても、本発明の範囲を限定するものではない。
【0062】
以下に添付図面について言及する。
図1a及び1bは、それぞれ、システインコドンを導入するように変異された、可溶性A6 TCRのα及びβ鎖の核酸配列を示す。影は導入したシステインコドンを示す。
図2aは、新規なジスルフィド鎖間結合を形成させるために使用したT48→C変異(下線)を含む、A6 TCRα鎖細胞外アミノ酸配列を示す。図2bは、新規なジスルフィド鎖間結合を形成させるために使用したS57→C変異(下線)を含む、A6 TCRβ鎖細胞外アミノ酸配列を示す。
図3は、Gly/Serリンカー(30マー)のDNA及びアミノ酸配列を示す。
図4は、scDiS A6 TCRを作成するために使用したクローニング戦略をまとめる。
図5aは、scDiS A6 TCRのDNA配列を示す。
図5bは、scDiS A6 TCRのアミノ酸配列を示す。
図6は、直線で示されるような0〜500mM NaCl勾配を使用するPOROS 50HQイオン交換カラムからのscDiS A6 TCRタンパク質の溶出を説明する。
図7は、図6で説明したカラムラン(column run)からの画分A15、B10、B9及びB3の還元SDS-PAGEゲル(クーマシー染色)及び非還元SDS-PAGE(クーマシー染色)ゲルの両方の結果を示す。画分B9及びB10は、明らかに、予想したサイズのscDiS A6 TCRに対応するタンパク質を含む。
図8は、図6に示したイオン交換カラムランからの画分B10〜B7のSuperdex 200ゲル濾過カラムからのscDiS A6 TCR溶出の溶出を説明する。
図9は、図8で説明したゲル濾過カラムランからの画分B8、B7、B3及びB2の還元SDS-PAGEゲル(クーマシー染色)及び非還元SDS-PAGE(クーマシー染色)ゲルの両方の結果を示す。画分B7は、明らかに、予想したサイズのscDiS A6 TCRに対応するタンパク質を含む。
図10は、図8に示したゲル濾過ランの濃縮画分B9〜B6のBIAcore緩衝液中への最後のゲル濾過ランである。scDiS A6 TCRは単一の主要ピークとして溶出する。
図11は、HLA-A2 TAXへのscDiS A6 TCRの結合についてのBIAcoreデータである。
【0063】
実施例1−新規な鎖間ジスルフィド結合の形成に必要なシステイン残基を導入するためのプライマーの設計及びA6 Tax TCRα及びβ鎖の変異誘発
TRAC*01中のエキソン1のA6 Taxスレオニン48をシステインに変異させるために、以下のプライマーを設計した(変異を小文字で示す):
5'-C ACA GAC AAA tgT GTG CTA GAC AT
5'-AT GTC TAG CAC Aca TTT GTC TGT G
TRBC1*01及びTRBC2*01の両方でエキソン1のA6 Taxセリン57をシステインに変異させるために、以下のプライマーを設計した(変異を小文字で示す):
5'-C AGT GGG GTC tGC ACA GAC CC
5'-GG GTC TGT GCa GAC CCC ACT G
【0064】
PCR変異誘発:
A6 Tax TCRα又はβ鎖の遺伝子を含む発現プラスミドを、それぞれα鎖プライマー又はβ鎖プライマーを使用して、以下のように変異させた。100ngのプラスミドを5μlの10mM dNTP、25μlの10×Pfu緩衝液(Stratagene)、10単位のPfuポリメラーゼ(Stratagene)と混合し、最終容量をH2Oで240μlに調整した。48μlのこの混合物に、50μlの最終反応溶液中で0.2μMの最終濃度が得られるように希釈したプライマーを補充した。95℃にて30秒間の最初の変性工程の後、反応混合物を、Hybaid PCR express PCR装置において、15回の変性(95℃、30秒)、アニーリング(55℃、60秒)及び伸長(73℃、8分)に付した。次いで、産物を10単位のDpnI制限酵素(New England Biolabs)で37℃にて5時間消化した。10μlの消化反応物を、コンピテントXL1-Blue細菌に形質転換し、37℃にて18時間増殖させた。一つのコロニーを採取し、5mlのTYP+アンピシリン(16g/l細菌トリプトン、16g/l酵母抽出物、5g/l NaCl、2.5g/l K2HPO4、100mg/lアンピシリン)中で一晩増殖させた。プラスミドDNAを、製造業者の指示に従ってQiagenミニプレップカラムで精製し、配列を、オックスフォード大学生化学部の配列決定設備で自動配列決定により検証した。それぞれの変異した核酸及びアミノ酸配列を、α鎖については図1a及び2aに、β鎖については図1b及び2bに示す。
【0065】
実施例2−新規ジスルフィド鎖間結合を組み込んだ単鎖A6 TCRの設計、発現及び試験
α鎖配列の末端部から停止コドン(TAA)を除去した以外は、実施例1で製造し、図1a及び1bに示したような新規ジスルフィドの形成に必要な追加のシステイン残基を組み込んだ変異A6 TCRα及びβ鎖のDNA配列を含む発現ベクターを、以下のように、単鎖A6 TCRの作成の基礎として使用した。
【0066】
scDis A6 TCRは、TCRα鎖のC末端とβ鎖のN末端との間に、30アミノ酸リンカー配列を含む。図3は、このリンカーのDNA及びアミノ酸配列を示す。scDis A6 TCRを作成するために用いたクローニング戦略を図4にまとめる。
【0067】
簡潔には、A6 dsTCRのα及びβ鎖を、図4に示すような制限部位を含むプライマー、すなわち:
α5'プライマー:ccaaggccatatgcagaaggaagtggagcagaactct
α3'プライマー:ttgggcccgccggatccgcccccgggggaactttctgggctgggg
β5'プライマー:tcccccgggggcggatccggcgggcccaacgctggtgtcactcag
β3'プライマー:gggaagcttagtctgctctaccccaggcctcg
を使用するPCRにより増幅させた。
【0068】
このように作成した2つのフラグメントを、5'α及び3'βプライマーを使用してPCRにより縫い合わせて(PCR stitched)、部位XmaI−BamHI−ApaIを含む短いリンカーを有する単鎖TCRを得た。このフラグメントをpGMT7中にクローニングした。次いで、完全長リンカーを二段階で挿入した。最初に、42bpフラグメントを、XmaI及びBamHI部位を使用して挿入した:
5'-CC GGG GGT GGC TCT GGC GGT GGC GGT TCA GGC GGT GGC G -3'
3'- C CCA CCG AGA CCG CCA CCG CCA AGT CCG CCA CCG CCT AG-5'。
【0069】
第2に、48bpフラグメントを、BamHI及びApaI部位を使用して挿入して、α鎖の3'末端とβ鎖の5'末端との間に90bpリンカーを作成した。48bpフラグメントは、以下のオリゴの混合物のPCR伸長により作成した:
5'- GC GGA TCC GGC GGT GGC GGT TCG GGT GGC GGT GGC TC-3'
3'- CCA AGC CCA CCG CCA CCG AGT CCG CCA CCG CCC GGG TG -5'。
【0070】
この伸長の産物を、BamHI及びApaIで消化し、42bpリンカーフラグメントを含む消化プラスミド中に連結した。
scDis A6 TCRの完全なDNA及びアミノ酸配列を、それぞれ、図5a及び5bに示す。
【0071】
単鎖ジスルフィド連結A6 TCRの発現及び精製:
単鎖ジスルフィド連結A6 TCRを含む発現プラスミドを、E.coliのBL21pLysS株に形質転換し、1つのアンピシリン耐性コロニーを37℃にてTYP(アンピシリン100μg/ml)培地中でOD600が0.4になるまで増殖させた後、0.5mM IPTGでタンパク質発現を誘導した。誘導の3時間後に、Beckman J-6B中での4000rpmにて30分間の遠心分離によって細胞を採集した。細胞ペレットを、50mM Tris-HCI、25%(w/v)スクロース、1mM NaEDTA、0.1%(w/v)アジ化Na、10mM DTT(pH8.0)を含む緩衝液に再懸濁した。一晩の凍結−解凍工程の後、再懸濁した細胞を、Milsonix XL2020超音波処理器中で標準の12mm径プローブを使用して1分間のバーストで合計約10分間超音波処理した。封入体ペレットを、Beckman J2-21遠心分離器で13000rpmにて30分間の遠心分離により回収した。次いで、3回の界面活性剤洗浄を行なって細胞残渣及び膜成分を除去した。各回で、封入体ペレットをTriton緩衝液(50mM Tris-HCI、0.5%Triton-X100、200mM NaCI、10mM NaEDTA、0.1%(w/v)アジ化Na、2mM DTT(pH8.0))中でホモジナイズした後、Beckman J2-21中で13000rpmにて15分間の遠心分離によりペレット化した。次いで、界面活性剤及び塩を、以下の緩衝液中での同様な洗浄により除去した:50mM Tris-HCl、1mM NaEDTA、0.1%(w/v)アジ化Na、2mM DTT(pH8.0)。最後に、封入体を30mgのアリコートに分け、−70℃で凍結させた。封入体タンパク質の収率を、6Mグアニジン−HClでの可溶化及びBradford顔料結合アッセイ(PerBio)での測定により定量化した。
【0072】
約15mgの可溶化封入体鎖を凍結ストックから解凍した。封入体を、6Mグアニジン溶液中5mg/mlの最終濃度まで希釈し、DTT(2Mストック)を10mMの最終濃度まで加えた。混合物を37℃にて30分間インキュベートした。1リットルのリフォールディング緩衝液(100mM Tris(pH8.5)、400mM L-アルギニン、2mM EDTA、5mM還元グルタチオン、0.5mM酸化グルタチオン、5M尿素、0.2mM PMSF)を調製し、5℃±3℃にて激しく撹拌した。レドックスカップル(redox couple)(2-メルカプトエチルアミン及びシスタミン(それぞれ6.6mM及び3.7mMの最終濃度まで)を添加して約5分後に、変性TCR鎖を添加した。次いで、タンパク質を、5℃±3℃にて撹拌しながら、約5時間±15分間リフォールディングさせた。次いで、リフォールディング体(refold)を、2回、最初は10リットルの100mM尿素に対して、第2回目は10リットルの100mM尿素、10mM Tris(pH8.0)に対して透析した。リフォールディング工程及び透析工程は共に6〜8℃で行なった。
【0073】
図6のようにAkta精製器(Pharmacia)を使用して、透析したリフォールディング体をPOROS 50HQアニオン交換カラムに充填し、50カラム容量にわたる0〜500mM NaCIの勾配で結合タンパク質を溶出させることにより、 scTCRを分解産物及び不純物から分離した。ピーク画分を4℃で保存し、クーマシー染色SDS-PAGEにより分析した(図7)後、プールして濃縮した。次いで、sTCRを精製し、HBS-EP緩衝液(10mM HEPES(pH7.4)、150mM NaCl、3.5mM EDTA、0.05% nonidet p40)中で予め平衡化したSuperdex 200HRゲル濾過カラムを使用して特徴付けた(図8)。ピーク画分を4℃で保存し、クーマシー染色SDS-PAGEにより分析した(図9)後、プールして濃縮した。最後に、BIAcore緩衝液中に精製タンパク質を生成するために、濃縮画分B9〜B6を別のゲル濾過工程を通して行なった(図10)。約50kDaの相対分子量で溶出するピークをプールした。これを濃縮した後、BIAcore表面プラズモン共鳴分析により特徴付けた。
【0074】
実施例3−HLA-A2 TaxとのscTCRの結合のBIAcore表面プラズモン共鳴による特徴付け
表面プラズモン共鳴バイオセンサ(BIAcore 3000(商標))を使用して、A6 scTCRとそのペプチド−MHCリガンド(HLA-A2 Tax)との結合を分析した。これは、半配向様式でストレプトアビジン被覆結合表面に固定した単一pMHC複合体(下記に説明)を作成し、同時に4つまでの異なるpMHC(別々のフローセルに固定された)に対する可溶性T細胞レセプターの結合の効率的な試験を可能にすることによって促進させた。HLA複合体の手動での注入により、固定したクラスI分子を正確なレベルで容易に操作することが可能になった。
【0075】
このような固定化複合体は、T細胞レセプター及びコレセプターCD8ααの両方と結合でき、この両方とも可溶相に注入され得る。
【0076】
ビオチン化クラスI HLA-A2−Tax複合体を、インビトロで、構成成分のサブユニットタンパク質及び合成ペプチドを含む細菌発現封入体からリフォールディングさせ、続いて精製し、インビトロで酵素的にビオチン化した(O'Callaghanら(1999)Anal.Biochem.266:9-15)。適切な構築物中でHLA重鎖の膜貫通ドメイン及び細胞質ドメインを置換するC末端ビオチン化タグを有するHLA重鎖を発現させた。HLA軽鎖又はβ2-ミクログロブリンもまた、〜500mg/リットル細菌培養物のレベルで、適切な構築物からE.coli中で封入体として発現させた。
【0077】
E. coli細胞を溶解し、封入体を約80%の純度まで精製する。封入体からのタンパク質を6Mグアニジン−HCl、50mM Tris(pH8.1)、100mM NaCl、10mM DTT、10mM EDTA中で変性させ、5℃より低いリフォールド緩衝液中に変性タンパク質の単一パルス(single pulse of denatured protein)を添加することにより、30mg/リットル重鎖、30mg/リットルβ2mの濃度で、0.4M L-アルギニン−HCl、100mM Tris(pH8.1)、3.7mMシスタミン、mMシステアミン、4mg/mlペプチド(例えばtax 11-19)中にリフォールディングさせた。リフォールディングは少なくとも1時間4℃で完了させた。
【0078】
緩衝液を、10容量の10mM Tris(pH8.1)での透析により交換した。溶液のイオン強度を十分に減少させるために、2回の緩衝液交換が必要であった。次いで、タンパク質溶液を、1.5μm酢酸セルロースフィルターを通して濾過し、POROS 50HQアニオン交換カラム(8ml床容量)に充填した。タンパク質を、直線の0〜500mM NaCl勾配で溶出させた。HLA-A2−ペプチド複合体は約250mM NaClで溶出した。ピーク画分を収集し、プロテアーゼインヒビターのカクテル(Calbiochem)を加え、画分を氷上で冷却した。
【0079】
ビオチン化タグを付したHLA複合体を、10mM Tris(pH8.1)、5mM NaCl中に、同じ緩衝液中で平衡化したPharmacia迅速脱塩カラムを使用して緩衝液交換をした。溶出の際、即座に、タンパク質含有画分を氷上で冷却し、プロテアーゼインヒビターカクテル(Calbiochem)を加えた。次いで、ビオチン化試薬を加えた:1mMビオチン、5mM ATP(pH8に緩衝化)、7.5mM MgCl2及び5μg/ml BirA酵素(O’Callaghanら(1999)Anal.Biochem.266:9-15に従って精製)。次いで、混合物を室温にて一晩インキュベートした。
【0080】
ゲル濾過クロマトグラフィーを使用してビオチン化HLA複合体を精製した。Pharmacia Superdex 75 HR 10/30カラムを濾過PBSで予め平衡化し、1mlのビオチン化反応混合物を充填し、カラムをPBSで0.5ml/分にて展開した。ビオチン化HLA複合体は、約15mlで単一ピークとして溶出した。タンパク質を含む画分をプールし、氷上で冷却し、プロテアーゼインヒビターカクテルを加えた。クーマシー結合アッセイ(PerBio)を使用してタンパク質濃度を測定し、ビオチン化HLA複合体のアリコートを−20℃で凍結保存した。標準的なアミンカップリング法によりストレプトアビジンを固定化した。
【0081】
新規な鎖間結合を含むA6 Tax scTCRとそのリガンド/MHC複合体又は無関係のHLA−ペプチド組合せ(これらの製造は上記で説明)との間の相互作用を、BIAcore 3000(商標)表面プラズモン共鳴(SPR)バイオセンサで分析した。 SPRは、レセプターリガンド相互作用を検出しその親和性及び動力学的パラメータを分析するために使用することができる原理である、小さなフローセル内のセンサ表面近くで、応答単位(RU)で表される屈折率の変化を測定する。β2mに架橋したビオチンとフローセルの活性化表面に化学的に架橋したストレプトアビジンとの間の結合を介して、個々のHLA−ペプチド複合体を別々のフローセルに固定化することにより、プローブフローセルを準備した。次いで、異なるフローセルの表面上にscTCRを一定流速で通過させ、そうしている間のSPR応答を測定することにより、アッセイを実施した。ペプチド−HLA複合体上に一定流速で異なる濃度で可溶性sTCRを注入してバックグランドの共鳴を規定した。これらコントロールの測定値を、特異ペプチド−HLA複合体を用いて得られた値から減算し、これを使用して解離定数Kd(Price及びDwek、Principles and Problems in Physical Chemistry for Biochemists(第2版)1979,Clarendon Press、Oxford)として表される結合親和性を算出した。
【0082】
scDiS A6 TCRのBIAcore分析により、この分子がそのコグネイトリガンド(HLA-A2 TAX)に、12.4±1.62μMのkdで特異的に結合することが示された。
【図面の簡単な説明】
【0083】
【図1】a及びbは、それぞれ、システインコドンを導入するように変異された、可溶性A6 TCRのα及びβ鎖の核酸配列を示す。
【図2】aは、新規なジスルフィド鎖間結合を形成させるために使用したT48→C変異(下線)を含む、A6 TCRα鎖細胞外アミノ酸配列を示す。bは、新規なジスルフィド鎖間結合を形成させるために使用したS57→C変異(下線)を含む、A6 TCRβ鎖細胞外アミノ酸配列を示す。
【図3】Gly/Serリンカー(30マー)のDNA及びアミノ酸配列を示す。
【図4】scDiS A6 TCRを作成するために使用したクローニング戦略をまとめる。
【図5】aはscDiS A6 TCRのDNA配列を示す。bはscDiS A6 TCRのアミノ酸配列を示す。
【図6】直線で示されるような0〜500mM NaCl勾配を使用するPOROS 50HQイオン交換カラムからのscDiS A6 TCRタンパク質の溶出を説明する。
【図7】図6で説明したカラムラン(column run)からの画分A15、B10、B9及びB3の還元SDS-PAGEゲル(クーマシー染色)及び非還元SDS-PAGE(クーマシー染色)ゲルの両方の結果を示す。画分B9及びB10は、明らかに、予想したサイズのscDiS A6 TCRに対応するタンパク質を含む。
【図8】図6に示したイオン交換カラムランからの画分B10〜B7のSuperdex 200ゲル濾過カラムからのscDiS A6 TCR溶出の溶出を説明する。
【図9】図8で説明したゲル濾過カラムランからの画分B8、B7、B3及びB2の還元SDS-PAGEゲル(クーマシー染色)及び非還元SDS-PAGE(クーマシー染色)ゲルの両方の結果を示す。画分B7は、明らかに、予想したサイズのscDiS A6 TCRに対応するタンパク質を含む。
【図10】図8に示したゲル濾過ランの濃縮画分B9〜B6のBIAcore緩衝液中への最後のゲル濾過ランである。scDiS A6 TCRは単一の主要ピークとして溶出する。
【図11】図11は、HLA-A2 TAXへのscDiS A6 TCRの結合についてのBIAcoreデータである。
【特許請求の範囲】
【請求項1】
TCRα鎖定常領域細胞外配列のN末端に融合したTCRα鎖可変領域配列により構成されるαセグメント、
TCRβ鎖定常領域細胞外配列のN末端に融合したTCRβ鎖可変領域により構成されるβセグメント、及び
αセグメントのC末端をβセグメントのN末端に又はその逆に連結するリンカー配列
を含み、
α及びβセグメントの定常領域細胞外配列はジスルフィド結合により連結し、
リンカー配列の長さ及びジスルフィド結合の位置は、α及びβセグメントの可変領域が天然のαβT細胞レセプター中と実質的に同様に相互に配向するような長さ及び位置である単鎖T細胞レセプター(scTCR)。
【請求項2】
α及びβセグメントの定常領域細胞外配列を連結するジスルフィド結合が、天然のαβT細胞レセプター中に等価物がないものである請求項1に記載のscTCR。
【請求項3】
ジスルフィド結合が、TCRα及びβ鎖の細胞外定常Igドメイン中に存在する配列に対応するα及びβセグメントのサブ配列中のアミノ酸残基を連結する請求項1又は2に記載のscTCR。
【請求項4】
ジスルフィド結合が、そのβ炭素原子がTCRα及びβ鎖の細胞外定常Igドメインの対応する配列中で0.6nm未満離れているアミノ酸残基から置換されたシステイン残基を連結する請求項3に記載のscTCR。
【請求項5】
αセグメント中に存在する定常領域細胞外配列が、TCRα鎖の細胞外定常Igドメインに対応する配列を含み、及び/又はβセグメント中に存在する定常領域細胞外配列が、TCRβ鎖の細胞外定常Igドメインに対応する配列を含む請求項3又は4に記載のscTCR。
【請求項6】
(a)αセグメントが、TCRα鎖のα鎖定常領域の細胞外ドメインのN末端に融合したTCRの可変領域であり、及び/又は(b)βセグメントが、TCRβ鎖の定常領域の細胞外ドメインのN末端に融合したTCRβ鎖の可変領域である請求項1〜5のいずれか1項に記載のscTCR。
【請求項7】
α及びβセグメント中に存在する定常領域細胞外配列が、TCRの天然の鎖間ジスルフィド結合を形成するシステイン残基が除去されるように、C末端で短縮化された天然のTCRのα及びβ鎖の定常領域に対応する請求項1〜5のいずれか1項に記載のscTCR。
【請求項8】
α及びβセグメント中に存在する定常領域細胞外配列が、天然の鎖間ジスルフィド結合を形成するシステイン残基が別のアミノ酸残基で置換された天然のTCRのα及びβ鎖の定常領域に対応する請求項1〜5のいずれか1項に記載のscTCR。
【請求項9】
システイン残基がセリン又はアラニンで置換されている請求項8に記載のscTCR。
【請求項10】
リンカー配列が、式-P-AA-P-(式中、Pはプロリンであり、AAはアミノ酸がグリシン及びセリンであるアミノ酸配列を表す)を有する請求項1〜9のいずれか1項に記載のscTCR。
【請求項11】
リンカー配列がαドメインのC末端をβドメインのN末端に連結している請求項1〜10のいずれか1項に記載のscTCR。
【請求項12】
リンカー配列が26〜41アミノ酸からなる請求項11に記載のscTCR。
【請求項13】
リンカー配列が29、30、31又は32アミノ酸からなる請求項11に記載のscTCR。
【請求項14】
リンカー配列が33、34、35又は36アミノ酸からなる請求項11に記載のscTCR。
【請求項15】
リンカー配列が-PGGG-(SGGGG)5-P-(ここで、Pはプロリンであり、Gはグリシンであり、Sはセリンである)である請求項11に記載のscTCR。
【請求項16】
リンカー配列が-PGGG-(SGGGG)6-P-(ここで、Pはプロリンであり、Gはグリシンであり、Sはセリンである)である請求項11に記載のscTCR。
【請求項17】
天然のTCRβ鎖中に存在する対合していないシステイン残基が存在しない請求項1〜16のいずれか1項に記載のscTCR。
【請求項18】
αセグメントの定常領域細胞外配列がTRAC*01に対応する配列を含み、βセグメントがTRBC1*01又はTRBC2*01に対応する配列を含み、天然のものではないジスルフィド結合がTRAC*01のエキソン1のThr48及びTRBC1*01又はTRBC2*01のエキソン1のSer57から置換されたシステイン残基間にある請求項1〜17のいずれか1項に記載のscTCR。
【請求項19】
ジスルフィド結合がTRAC*01のエキソン1のThr45及びTRBC1*01又はTRBC2*01のエキソン1のSer77から置換されたシステイン残基を連結している請求項1〜18のいずれか1項に記載のscTCR。
【請求項20】
ジスルフィド結合がTRAC*01のエキソン1のThr10及びTRBC1*01又はTRBC2*01のエキソン1のSer17から置換されたシステイン残基を連結している請求項1〜18のいずれか1項に記載のscTCR。
【請求項21】
ジスルフィド結合がTRAC*01のエキソン1のThr45及びTRBC1*01又はTRBC2*01のエキソン1のAsp59から置換されたシステイン残基を連結している請求項1〜18のいずれか1項に記載のscTCR。
【請求項22】
ジスルフィド結合がTRAC*01のエキソン1のSer15及びTRBC1*01又はTRBC2*01のエキソン1のGlu15から置換されたシステイン残基を連結している請求項1〜18のいずれか1項に記載のscTCR。
【請求項23】
α及びβセグメント中に存在するTCRα及びβ鎖可変領域配列が、併せて、第1のTCRの機能的可変ドメインに対応し、α及びβセグメント中に存在するTCRα及びβ鎖定常領域細胞外配列が、第2のTCRのα及びβ鎖定常領域細胞外配列に対応し、第1及び第2のTCRは同じ種に由来する請求項1〜22のいずれか1項に記載のscTCR。
【請求項24】
α及びβセグメント中に存在するTCRα及びβ鎖可変領域配列が、併せて、第1のTCRの機能的可変ドメインに対応し、α及びβセグメント中に存在するTCRα及びβ鎖定常領域細胞外配列が、第2のTCRのα及びβ鎖定常領域細胞外配列に対応し、第1及び第2のTCRは異なる種に由来する請求項1〜22のいずれか1項に記載のscTCR。
【請求項25】
α及びβセグメント中に存在するTCRα及びβ鎖可変領域配列が、併せて、ヒトTCRの機能的可変ドメインに対応し、α及びβセグメント中に存在するTCRα及びβ鎖定常領域細胞外配列が、マウスTCRのα及びβ鎖定常領域細胞外配列に対応する請求項24に記載のscTCR。
【請求項26】
TCRがペプチドMHC複合体に結合するものである請求項1〜24のいずれか1項に記載のscTCR。
【請求項27】
TCRがCD1−抗原複合体に結合するものである請求項25に記載のscTCR。
【請求項28】
TCRがスーパー抗原又はペプチド−MHC/スーパー抗原複合体に結合するものである請求項1〜24のいずれか1項に記載のscTCR。
【請求項29】
複数の請求項1〜28のいずれか1項に記載のsTCRを含む多価T細胞レセプター(TCR)複合体。
【請求項30】
治療物質に共有結合している請求項1〜28のいずれか1項に記載のscTCR又は請求項30に記載の複合体。
【請求項31】
粒子又はビーズに付着した1又は複数の請求項1〜28のいずれか1項又は請求項30に記載のscTCR。
【請求項32】
請求項1〜31のいずれか1項に記載のscTCRおよび薬学的に受容可能なキャリアを含む組成物。
【請求項33】
1又は複数の請求項1〜29のいずれか1項に記載のscTCRを用意し;scTCRをTCRリガンドと接触させ;及びscTCRとリガンドとの結合を検出することを含む、MHC−ペプチド複合体、CD1−抗原複合体、スーパー抗原及びMHC−ペプチド/スーパー抗原複合体から選択されるTCRリガンドを検出する方法。
【請求項34】
scTCRをscTCRリガンド結合パートナーと試験化合物の存在下及び非存在下で接触させ、及び試験化合物の存在によりscTCRとTCRリガンドとの結合が減少するかどうかを測定することを含み、その減少が阻害物質を同定するとされる、1または複数の請求項1〜28のいずれか1項に記載のscTCRと、MHC−ペプチド複合体、CD1−抗原複合体、スーパー抗原及びMHC−ペプチド/スーパー抗原複合体から選択されるTCRリガンドとの間の相互作用の阻害剤を同定する方法。
【請求項35】
scTCR又はscTCRリガンド結合パートナーと試験化合物を接触させ、及び試験化合物がscTCR及び/又はTCRリガンドに結合するかどうかを測定することを含み、その結合が潜在的阻害剤を同定するとされる、1または複数の請求項1〜28のいずれか1項に記載のscTCRと、MHC−ペプチド複合体、CD1−抗原複合体、スーパー抗原及びMHC−ペプチド/スーパー抗原複合体から選択されるTCRリガンドとの間の相互作用の潜在的阻害剤を同定する方法。
【請求項36】
請求項1〜28のいずれか1項に記載のscTCRをコードする配列又はこれと相補的な配列を含む核酸分子。
【請求項37】
請求項36に記載の核酸分子を含むベクター。
【請求項1】
TCRα鎖定常領域細胞外配列のN末端に融合したTCRα鎖可変領域配列により構成されるαセグメント、
TCRβ鎖定常領域細胞外配列のN末端に融合したTCRβ鎖可変領域により構成されるβセグメント、及び
αセグメントのC末端をβセグメントのN末端に又はその逆に連結するリンカー配列
を含み、
α及びβセグメントの定常領域細胞外配列はジスルフィド結合により連結し、
リンカー配列の長さ及びジスルフィド結合の位置は、α及びβセグメントの可変領域が天然のαβT細胞レセプター中と実質的に同様に相互に配向するような長さ及び位置である単鎖T細胞レセプター(scTCR)。
【請求項2】
α及びβセグメントの定常領域細胞外配列を連結するジスルフィド結合が、天然のαβT細胞レセプター中に等価物がないものである請求項1に記載のscTCR。
【請求項3】
ジスルフィド結合が、TCRα及びβ鎖の細胞外定常Igドメイン中に存在する配列に対応するα及びβセグメントのサブ配列中のアミノ酸残基を連結する請求項1又は2に記載のscTCR。
【請求項4】
ジスルフィド結合が、そのβ炭素原子がTCRα及びβ鎖の細胞外定常Igドメインの対応する配列中で0.6nm未満離れているアミノ酸残基から置換されたシステイン残基を連結する請求項3に記載のscTCR。
【請求項5】
αセグメント中に存在する定常領域細胞外配列が、TCRα鎖の細胞外定常Igドメインに対応する配列を含み、及び/又はβセグメント中に存在する定常領域細胞外配列が、TCRβ鎖の細胞外定常Igドメインに対応する配列を含む請求項3又は4に記載のscTCR。
【請求項6】
(a)αセグメントが、TCRα鎖のα鎖定常領域の細胞外ドメインのN末端に融合したTCRの可変領域であり、及び/又は(b)βセグメントが、TCRβ鎖の定常領域の細胞外ドメインのN末端に融合したTCRβ鎖の可変領域である請求項1〜5のいずれか1項に記載のscTCR。
【請求項7】
α及びβセグメント中に存在する定常領域細胞外配列が、TCRの天然の鎖間ジスルフィド結合を形成するシステイン残基が除去されるように、C末端で短縮化された天然のTCRのα及びβ鎖の定常領域に対応する請求項1〜5のいずれか1項に記載のscTCR。
【請求項8】
α及びβセグメント中に存在する定常領域細胞外配列が、天然の鎖間ジスルフィド結合を形成するシステイン残基が別のアミノ酸残基で置換された天然のTCRのα及びβ鎖の定常領域に対応する請求項1〜5のいずれか1項に記載のscTCR。
【請求項9】
システイン残基がセリン又はアラニンで置換されている請求項8に記載のscTCR。
【請求項10】
リンカー配列が、式-P-AA-P-(式中、Pはプロリンであり、AAはアミノ酸がグリシン及びセリンであるアミノ酸配列を表す)を有する請求項1〜9のいずれか1項に記載のscTCR。
【請求項11】
リンカー配列がαドメインのC末端をβドメインのN末端に連結している請求項1〜10のいずれか1項に記載のscTCR。
【請求項12】
リンカー配列が26〜41アミノ酸からなる請求項11に記載のscTCR。
【請求項13】
リンカー配列が29、30、31又は32アミノ酸からなる請求項11に記載のscTCR。
【請求項14】
リンカー配列が33、34、35又は36アミノ酸からなる請求項11に記載のscTCR。
【請求項15】
リンカー配列が-PGGG-(SGGGG)5-P-(ここで、Pはプロリンであり、Gはグリシンであり、Sはセリンである)である請求項11に記載のscTCR。
【請求項16】
リンカー配列が-PGGG-(SGGGG)6-P-(ここで、Pはプロリンであり、Gはグリシンであり、Sはセリンである)である請求項11に記載のscTCR。
【請求項17】
天然のTCRβ鎖中に存在する対合していないシステイン残基が存在しない請求項1〜16のいずれか1項に記載のscTCR。
【請求項18】
αセグメントの定常領域細胞外配列がTRAC*01に対応する配列を含み、βセグメントがTRBC1*01又はTRBC2*01に対応する配列を含み、天然のものではないジスルフィド結合がTRAC*01のエキソン1のThr48及びTRBC1*01又はTRBC2*01のエキソン1のSer57から置換されたシステイン残基間にある請求項1〜17のいずれか1項に記載のscTCR。
【請求項19】
ジスルフィド結合がTRAC*01のエキソン1のThr45及びTRBC1*01又はTRBC2*01のエキソン1のSer77から置換されたシステイン残基を連結している請求項1〜18のいずれか1項に記載のscTCR。
【請求項20】
ジスルフィド結合がTRAC*01のエキソン1のThr10及びTRBC1*01又はTRBC2*01のエキソン1のSer17から置換されたシステイン残基を連結している請求項1〜18のいずれか1項に記載のscTCR。
【請求項21】
ジスルフィド結合がTRAC*01のエキソン1のThr45及びTRBC1*01又はTRBC2*01のエキソン1のAsp59から置換されたシステイン残基を連結している請求項1〜18のいずれか1項に記載のscTCR。
【請求項22】
ジスルフィド結合がTRAC*01のエキソン1のSer15及びTRBC1*01又はTRBC2*01のエキソン1のGlu15から置換されたシステイン残基を連結している請求項1〜18のいずれか1項に記載のscTCR。
【請求項23】
α及びβセグメント中に存在するTCRα及びβ鎖可変領域配列が、併せて、第1のTCRの機能的可変ドメインに対応し、α及びβセグメント中に存在するTCRα及びβ鎖定常領域細胞外配列が、第2のTCRのα及びβ鎖定常領域細胞外配列に対応し、第1及び第2のTCRは同じ種に由来する請求項1〜22のいずれか1項に記載のscTCR。
【請求項24】
α及びβセグメント中に存在するTCRα及びβ鎖可変領域配列が、併せて、第1のTCRの機能的可変ドメインに対応し、α及びβセグメント中に存在するTCRα及びβ鎖定常領域細胞外配列が、第2のTCRのα及びβ鎖定常領域細胞外配列に対応し、第1及び第2のTCRは異なる種に由来する請求項1〜22のいずれか1項に記載のscTCR。
【請求項25】
α及びβセグメント中に存在するTCRα及びβ鎖可変領域配列が、併せて、ヒトTCRの機能的可変ドメインに対応し、α及びβセグメント中に存在するTCRα及びβ鎖定常領域細胞外配列が、マウスTCRのα及びβ鎖定常領域細胞外配列に対応する請求項24に記載のscTCR。
【請求項26】
TCRがペプチドMHC複合体に結合するものである請求項1〜24のいずれか1項に記載のscTCR。
【請求項27】
TCRがCD1−抗原複合体に結合するものである請求項25に記載のscTCR。
【請求項28】
TCRがスーパー抗原又はペプチド−MHC/スーパー抗原複合体に結合するものである請求項1〜24のいずれか1項に記載のscTCR。
【請求項29】
複数の請求項1〜28のいずれか1項に記載のsTCRを含む多価T細胞レセプター(TCR)複合体。
【請求項30】
治療物質に共有結合している請求項1〜28のいずれか1項に記載のscTCR又は請求項30に記載の複合体。
【請求項31】
粒子又はビーズに付着した1又は複数の請求項1〜28のいずれか1項又は請求項30に記載のscTCR。
【請求項32】
請求項1〜31のいずれか1項に記載のscTCRおよび薬学的に受容可能なキャリアを含む組成物。
【請求項33】
1又は複数の請求項1〜29のいずれか1項に記載のscTCRを用意し;scTCRをTCRリガンドと接触させ;及びscTCRとリガンドとの結合を検出することを含む、MHC−ペプチド複合体、CD1−抗原複合体、スーパー抗原及びMHC−ペプチド/スーパー抗原複合体から選択されるTCRリガンドを検出する方法。
【請求項34】
scTCRをscTCRリガンド結合パートナーと試験化合物の存在下及び非存在下で接触させ、及び試験化合物の存在によりscTCRとTCRリガンドとの結合が減少するかどうかを測定することを含み、その減少が阻害物質を同定するとされる、1または複数の請求項1〜28のいずれか1項に記載のscTCRと、MHC−ペプチド複合体、CD1−抗原複合体、スーパー抗原及びMHC−ペプチド/スーパー抗原複合体から選択されるTCRリガンドとの間の相互作用の阻害剤を同定する方法。
【請求項35】
scTCR又はscTCRリガンド結合パートナーと試験化合物を接触させ、及び試験化合物がscTCR及び/又はTCRリガンドに結合するかどうかを測定することを含み、その結合が潜在的阻害剤を同定するとされる、1または複数の請求項1〜28のいずれか1項に記載のscTCRと、MHC−ペプチド複合体、CD1−抗原複合体、スーパー抗原及びMHC−ペプチド/スーパー抗原複合体から選択されるTCRリガンドとの間の相互作用の潜在的阻害剤を同定する方法。
【請求項36】
請求項1〜28のいずれか1項に記載のscTCRをコードする配列又はこれと相補的な配列を含む核酸分子。
【請求項37】
請求項36に記載の核酸分子を含むベクター。
【図3】


【図4】
【図5】

【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公表番号】特表2006−502741(P2006−502741A)
【公表日】平成18年1月26日(2006.1.26)
【国際特許分類】
【出願番号】特願2005−500992(P2005−500992)
【出願日】平成15年10月3日(2003.10.3)
【国際出願番号】PCT/GB2003/004310
【国際公開番号】WO2004/033685
【国際公開日】平成16年4月22日(2004.4.22)
【出願人】(505121578)アヴィデックス リミテッド (17)
【氏名又は名称原語表記】AVIDEX LTD
【住所又は居所原語表記】57C Milton Park,Abingdon OX14 4RX,United Kingdom
【Fターム(参考)】
【公表日】平成18年1月26日(2006.1.26)
【国際特許分類】
【出願日】平成15年10月3日(2003.10.3)
【国際出願番号】PCT/GB2003/004310
【国際公開番号】WO2004/033685
【国際公開日】平成16年4月22日(2004.4.22)
【出願人】(505121578)アヴィデックス リミテッド (17)
【氏名又は名称原語表記】AVIDEX LTD
【住所又は居所原語表記】57C Milton Park,Abingdon OX14 4RX,United Kingdom
【Fターム(参考)】
[ Back to top ]