
原料植物種判定方法
【課題】加工食品に含まれる原料植物の植物種を高精度かつ定量的に判定できる方法を提供する。
【解決手段】植物ゲノムに原料植物の植物種固有のレトロトランスポゾン挿入部位が存在するか否かを検出し、この検出結果に基づいて、原料植物の植物種を判定するので、高精度かつ定量的な原料植物の植物種の判定を行うことが可能となる。
【解決手段】植物ゲノムに原料植物の植物種固有のレトロトランスポゾン挿入部位が存在するか否かを検出し、この検出結果に基づいて、原料植物の植物種を判定するので、高精度かつ定量的な原料植物の植物種の判定を行うことが可能となる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、加工食品に含まれる原料植物の植物種を判定する原料植物種判定方法に関する。
【背景技術】
【0002】
植物種の同定技術としては、例えば非特許文献1に記載のように、葉緑体DNAの遺伝子間領域の塩基配列の違いを利用して、植物種を同定する方法(PS−ID法)が知られている。また、加工食品に含まれるアレルゲン物質を定性分析・定量分析する方法としては、分析対象のアレルゲン物質に対する抗体を用いる方法が開発されている。また、コムギ、そば、落花生等といったアレルゲン物質となる植物種の遺伝子をPCRにより検出する方法が開発されている。
【0003】
また、非特許文献2には、ゲノムにおけるレトロトランスポゾンの個々の挿入位置を、制限酵素処理末端へのアダプター付加法(S-SAP:Sequence-Specific Amplification Polymorphism法)により、挿入位置近傍のゲノム配列(制限酵素認識部位)とLTR配列間の核酸増幅を区別する方法が記載されている。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Nakamura, I., N. Kameya, Y. Kato, S. Yamanaka, H. Jomori, and Y. Sato. A proposal for identifying the short ID sequence which addresses the plastid subtype of higher plants. Breeding Science 47: 385-388 (1997)
【非特許文献2】Waugh, R., K. McLean, A. J. Flavell, S. R. Pearce, A. Kumar, B. B. T. Thomas and W. Powell. Genetic distribution of Bare-1-like retrotransposable elements in the barley genome revealed by sequence-specific amplification polymorphisms (S-SAP). Mol Gen Genet 253: 687-694 (1997)
【発明の概要】
【発明が解決しようとする課題】
【0005】
しかしながら、上述のような従来技術には、以下の問題がある。
【0006】
まず、PS−ID法を用いて植物種を同定する技術では、使用する葉緑体DNAの遺伝子領域において、同一の植物種間であっても相違する塩基配列が存在するため、厳密な植物種レベルの識別が困難であるという問題がある。また、葉緑体は、1つの植物細胞内に多数存在する。さらには、この葉緑体の遺伝子領域の塩基配列のなかには、核ゲノム中の塩基配列と同一の塩基配列が存在する。それゆえ、葉緑体の遺伝子領域の塩基配列の複製数は、植物の品種間でも違いが見られることが考えられる。このため、葉緑体の遺伝子領域の塩基配列を用いるPS−ID法では、定量分析が、植物種内の品種間でもばらつきが生じることが考えられ、植物種間レベルで正確に定量比較を行うことが困難である。
【0007】
また、アレルゲン物質に対する抗体を用いる技術では、例えばコムギに対する抗体は、ライムギやエンバクにも反応するため、植物種間レベルでのアレルゲン物質の識別が困難である。また、アレルゲン物質となる植物種の遺伝子をPCRにより検出する方法では、分析対象となる植物種の遺伝子の複製数が多く、定量的な検査が困難である。
【0008】
本発明は、上記の問題点に鑑みてなされたものであり、その目的は、加工食品に含まれる原料植物の植物種を高精度かつ定量的に判定できる方法を提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、上記の課題を解決するために鋭意検討した結果、植物のゲノム中に存在する転位因子の1つであるレトロトランスポゾンの植物ゲノムへの挿入位置が原料植物の植物種固有のものであるか否かを検出する方法は、加工食品に含まれる原料植物の植物種(species)の判定においても適用できることを見出し、本発明を完成させるに至った。そして、かかる「原料植物の植物種固有のトランスポゾン挿入部位」の有無を判定し、それを定量することにより、加工食品に原料植物種が含まれているかの判定を高精度かつ定量的に行うことができることを確認して、本発明を完成させるに至った。
【0010】
すなわち、本発明の原料植物種判定方法は、上記の課題を解決するために、加工食品に含まれる原料植物の植物種を判定する原料植物種判定方法であって、植物ゲノムに原料植物の植物種固有のレトロトランスポゾン挿入部位が存在するか否かを検出し、この検出結果に基づいて、原料植物の植物種を判定することを特徴としている。
【0011】
さらに、植物ゲノムに存在する原料植物の植物種固有のレトロトランスポゾン挿入部位は、通常、その植物種に存在する全ての種類の染色体の1組には1カ所にしか存在しないため、定量的な遺伝子増幅法などを利用して定量することができる。即ち、加工食品の原料に複数の植物種が使われている場合、原料となったそれぞれの植物種について固有のレトロトランスポゾン挿入部位の量を相対的に測定し比較することで、加工食品において使用された原料植物種の相対的な割合を推定することが可能となる。したがって、原料植物の植物種固有のレトロトランスポゾン挿入部位を利用して、高精度かつ定量的な原料植物の植物種の判定を行うことが可能となる。
【0012】
本発明の原料植物種判定方法では、植物ゲノムに原料植物の植物種固有のレトロトランスポゾン挿入部位が存在するか否かの検出には、判定対象とする複数の品種のゲノムに共通して存在しているレトロトランスポゾン挿入部位における、当該レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマーと、レトロトランスポゾンの塩基配列に基づいて設計されるプライマーとを組み合わせたプライマーセットを用いて、加工食品から調製したDNAを鋳型として核酸増幅反応を行い、増幅産物の有無を確認する方法を用いることが好ましい。上記プライマーセットにより増幅される断片の長さを任意に設計できるため、短い断片が増幅されるようにプライマーセットを設計すれば、加工食品中のDNAが高度に断片化している場合でもこれを鋳型として増幅断片の有無を確認することが可能となる。また、増幅産物の有無を確認するものであるため、簡便な電気泳動により確認でき、結果の判定に熟練を要しない。また、加工食品に複数の原料植物が含まれている場合に、使用された原料植物種の割合は定量的な遺伝子増幅を用いてそれぞれの植物種に固有の挿入部位の量を相対的に測定することで推定することが可能である。
【0013】
また、本発明の原料植物種判定方法では、判定対象とする複数の品種について、ゲノム中のレトロトランスポゾン挿入部位を比較して、複数の品種のゲノムに共通して存在しているレトロトランスポゾン挿入部位を選択し、選択されたレトロトランスポゾン挿入部位について、原料植物の近縁種のゲノムに存在するか否かの確認を行い、上記確認の結果、原料植物の近縁種のゲノムに存在しないレトロトランスポゾン挿入部位を、上記原料植物の植物種固有のレトロトランスポゾン挿入部位として決定することが好ましい。例えば、S-SAP法などにより、判定対象とする原料植物の複数の品種について、ゲノム中のレトロトランスポゾン挿入部位を比較し、複数の品種のゲノムに共通して存在している挿入部位を見いだせば、この挿入部位が原料植物の近縁種のゲノムに存在するか否かを確認することにより、原料植物の植物種固有のレトロトランスポゾン挿入部位を決定することができる。
【0014】
また、本発明の原料植物種判定方法では、上記増幅産物の断片長は40bp〜600bpであることが好ましい。増幅産物の長さが短くなるようにプライマーを設計することにより、加工食品から抽出されるDNAが高度に断片化したものであっても、それを鋳型として核酸増幅反応を行うことが可能となる。
【0015】
また、本発明の原料植物種判定方法では、上記レトロトランスポゾンに隣接するゲノム領域の塩基配列は、当該領域を含むDNA断片をシーケンスすることにより得られる塩基配列であることが好ましい。
【0016】
また、本発明の原料植物種判定方法では、上記レトロトランスポゾンに隣接するゲノム領域の塩基配列は、当該領域を含むDNA断片をクローニングし、当該断片をシーケンスすることにより得られる塩基配列であることが好ましい。
【0017】
また、本発明の原料植物種判定方法の対象とする加工食品はアズキを原料に含むことが好ましく、アズキ加工食品を対象とする場合には、利用するレトロトランスポゾンはPHARE-1であることが好ましい。
【0018】
上記PHARE-1を利用してアズキ植物種の判定を行う場合、PHARE-1に隣接するゲノム領域を含むDNA断片をシーケンスすることにより得られる塩基配列として、配列番号1または2に示される塩基配列を挙げることができる。すなわち、配列番号1または2に示される塩基配列に含まれるPHARE-1に隣接するゲノム領域の塩基配列に基づいて、プライマーを設計することができる。
【0019】
上記配列番号1または2に示される塩基配列に含まれるPHARE-1に隣接するゲノム領域の塩基配列に基づいて設計されるプライマーとしては、配列番号4または5に示される塩基配列からなるプライマーを挙げることができる。また、PHARE-1の塩基配列に基づいて設計されるプライマーは配列番号3に示される塩基配列からなるプライマーを挙げることができる。なお、配列番号3〜5に示されるプライマーも本発明に含まれる。
【0020】
また、本発明の原料植物種判定キットは、上述の原料植物種判定方法を実施するためのキットであって、レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマーと、レトロトランスポゾンの塩基配列に基づいて設計されるプライマーとを含むことを特徴とプライマーを含むキットとすることにより、本発明の原料植物種判定方法をより高精度かつ定量的に実施することが可能となる。
【0021】
上記キットに含まれるプライマーとしては、配列番号3に示される塩基配列からなるプライマーと、配列番号4または5に示される塩基配列からなるプライマーのうち少なくとも1つ以上とを挙げることができる。これにより、アズキ種判定キットを構成することができる。
【発明の効果】
【0022】
本発明の原料植物種判定方法は、以上のように、植物ゲノムに原料植物の植物種固有のレトロトランスポゾン挿入部位が存在するか否かを検出し、この検出結果に基づいて、原料植物の植物種を判定するので、高精度かつ定量的な原料植物の植物種の判定を行うことが可能となる。
【0023】
さらに、本発明の原料植物種判定方法は、増幅される断片の長さを比較するものではなく、増幅産物の有無を確認するものであるため、簡便な電気泳動により正確な判定ができる。したがって、高価な分析機器を必要とせず、低コストで実施できるという効果を奏する。さらに、判定に熟練を要しないという効果を奏する。
【0024】
また、本発明の原料植物種判定キットは、本発明の原料植物種判定方法を実施するためのキットであるため、本方法をより迅速かつ簡便に実施できるという効果を奏する。
【0025】
以上のように、本発明によれば、加工食品の原料植物種を迅速、簡便かつ正確に判定することができるので、本方法を用いて加工食品の原料植物種を判定すれば、加工食品の原料植物種の不正表示を防止することができるという効果を奏する。
【図面の簡単な説明】
【0026】
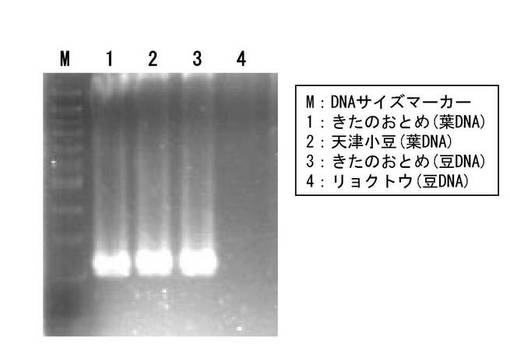
【図1】アズキ(Vigna angularis (Willd.) Ohwi & H. Ohashi)、及び該アズキと同じササゲ属(Vigna)に属する最も近縁なリョクトウ(Vigna radiate (L.) R. Wilczek)のゲノムDNAを鋳型として、Az-Copia1-LYT-D/ P1-DE-T1-5A1プライマーセットを用いたPCR増幅産物の電気泳動結果を示す画像である。
【図2】アズキ(Vigna angularis (Willd.) Ohwi & H. Ohashi)、及び該アズキと同じササゲ属(Vigna)に属する最も近縁なリョクトウ(Vigna radiate (L.) R. Wilczek)のゲノムDNAを鋳型として、Az-Copia1-LYT-D/ P1-DE-T2-4A1プライマーセットを用いたPCR増幅産物の電気泳動結果を示す画像である。
【発明を実施するための形態】
【0027】
本発明の実施の一形態について説明すれば、以下のとおりである。なお、本発明はこれに限定されるものではない。
【0028】
本発明の原料植物種判定方法は、加工食品に含まれる原料植物の植物種を判定する方法であって、植物ゲノムに原料植物の植物種固有のレトロトランスポゾン挿入部位が存在するか否かを検出することによって、原料植物の植物種を判定する方法である。
【0029】
対象とする加工食品は、原料に植物が含まれるものであって、当該加工食品から原料植物のDNAを抽出可能なものであれば特に限定されるものではない。また、原料植物体の全部または一部、あるいは原料植物から得られる収穫物(果実、種子等)に対して加熱、乾燥等の加工処理を施したものが加工食品に該当する。したがって、加工食品には最終製品のみではなく、中間加工品も含まれる。
【0030】
レトロトランスポゾンは転移因子の一種であり、進化の過程で自身の複製を作り、ゲノムに挿入するという形式で転移していた遺伝因子である。多くの植物において、多種類のレトロトランスポゾンが複製された状態で存在する。現在の植物ゲノムにおいては、レトロトランスポゾンのほとんどの配列は転移活性を失っており、ゲノムに挿入された状態で安定的に遺伝している。レトロトランスポゾンとそれに隣接するゲノム配列は、現存する植物種の共通祖先でレトロトランスポゾンの転移が生じ、その転移が生じた部位がその共通祖先において遺伝的に固定すれば、その祖先種から分岐してきた植物種の全てに存在することとなる。一方、特定の植物種の祖先がその植物種に最も近縁の植物種と種分化した後、その祖先となった集団に含まれる個体においてレトロトランスポゾンの転移が生じ、その転移が生じた部位がその祖先となった集団において遺伝的に固定すれば、その挿入部位は、染色体の変異などによりその部位が失われることがない限り、その植物種に固有でかつその植物種に属する全ての個体に存在することになる。また、種分化以前に存在していたレトロトランスポゾンの挿入部位であっても、特定の植物種においてのみ遺伝的に固定した部位は、その植物種に固有のものとなる。なお、種分化後、その種内でレトロトランスポゾンの転移が生じた場合には、ある品種ではレトロトランスポゾンの挿入が見られるゲノムの場所に、別の品種では挿入が見られないという、品種間の多型となる。
【0031】
本発明の原料植物種判定方法に利用可能なレトロトランスポゾンは、植物に存在するレトロトランスポゾンであれば特に限定されるものではない。すなわち、公知のレトロトランスポゾンのみならず、今後新たに見出されるレトロトランスポゾンも利用することができる。
【0032】
LTR型のレトロトランスポゾンは、今まで調査された範囲では、全ての高等植物に存在しており、主要な作物については、個別の配列が多数単離され、命名されている (Johns et al (1985) A low copy number, copia-like transposon in maize. EMBO J 8: 1093-1102.、Manninen and Schulman (1993) BARE-1, a copia-like retroelement in barley (Hordeum vulgare L.). Plant Mol Biol 22: 829-846.、Laten et al. (1998) SIRE-1, a copia/Ty1-like retroelement from soybean, encodes a retroviral envelope-like protein. Proc Natl Acad Sci USA 95: 6897-6902.、Kumekawa et al. (1999) Identification and characterization of novel retrotransposons of the gypsy type in rice. Mol Gen Genet 260: 593-602.)。
【0033】
LTR型のレトロトランスポゾンのうち、Ty1-Copia型に分類されるものについては、逆転写酵素領域に存在するアミノ酸保存配列をターゲットとするPCRにより、同酵素領域の配列が多数の植物から単離されている(Voytas et al. (1992) copia-like retrotransposons are ubiquitous among plants. Proc Natl Acad Sci USA 89: 7124-7128.、Hirochika et al. (1992) Retrotransposon families in rice. Mol Gen Genet 233: 209-216.)。したがって、レトロトランスポゾンの全長配列が調べられていない作物であっても、DNAデータベースにおいて既知の、あるいは、新たに逆転写酵素領域の配列を同定すれば、その延長配列を単離するTAIL(Thermal Asymmetric InterLaced)PCR(Liu et al. (1995) Efficient isolation and mapping of Arabidopsis thaliana T-DNA insert junctions by thermal asymmetric interlaced PCR. Plant Journal 8:457-463.)、Suppression PCR(Siebert et al. (1995) An improved PCR method for walking in uncloned genomic DNA. Nucleic Acids Res. 23:1087-1088.)やその他の方法(Pearse et al. (1999) Rapid isolation of plant Ty1-copia group retrotransposon LTR sequences for molecular marker studies. Plant Journal 19: 711-717.)により、末端のLTR配列までを同定することができる。
【0034】
植物ゲノムに原料植物の植物種(以下、原料植物種と記す)固有のレトロトランスポゾン挿入部位が存在するかの検出とは、植物ゲノムに、原料植物種に存在するが原料植物種以外の植物種には存在しないレトロトランスポゾンの挿入部位を検出することを意味する。上述のように、レトロトランスポゾンの挿入部位には品種間で共通したゲノムの場所に挿入が見られる。それゆえ、この共通したレトロトランスポゾンの挿入部位が、原料植物種以外の植物種(特に原料植物の近縁種)のゲノムにも存在するか否かを検討することにより、原料植物種固有のレトロトランスポゾン挿入部位を同定することができる。本発明の植物種判定方法は、原料植物種固有のレトロトランスポゾン挿入部位の有無を明らかとすることにより、加工食品における原料植物種の判定を行うものである。
【0035】
レトロトランスポゾンの挿入は、転写されたレトロトランスポゾンの塩基配列が植物ゲノムに挿入されることにより生じる。その際、レトロトランスポゾンの挿入が生じる部位は、特に規則性がない。このことから、植物ゲノムのそれぞれの部位でのレトロトランスポゾンの挿入が偶然別の細胞や個体で生じる確率は、極めて小さい。このため、ゲノムの基本構造が原料植物種と類似している近縁種であっても、レトロトランスポゾンの挿入が近縁種との共通祖先のゲノムにおいて生じたものでない限り、原料植物種のレトロトランスポゾン挿入部位が、近縁種のゲノムで見出されることは、確率的に見てゼロに等しい。
【0036】
それゆえ、レトロトランスポゾンの挿入部位のうち、その植物種において遺伝的に固定している挿入部位(全ての品種間において共通して見られる挿入部位)が、その植物種の近縁種において見出されない場合は、植物種の特異性が極めて高いものとなる。そして、このような特異性を利用して、原料植物種において遺伝的に固定しているレトロトランスポゾン挿入部位を、原料植物種を識別するマーカーとして用いることができる。
【0037】
上記植物ゲノムに原料植物種固有のレトロトランスポゾン挿入部位が存在するか否かの検出は、複数の挿入部位について行うことが好ましい。上述のように植物ゲノムには多数のレトロトランスポゾン配列が存在するため、S-SAP法などにより、特定の原料植物種のみに存在する挿入部位(原料植物種固有のレトロトランスポゾン挿入部位)を見いだせば、その部位における挿入の有無により原料植物種の有無を判定できる。また、このような植物種に固有の挿入部位を多数の植物種について見いだすことで、加工食品中に原料植物として複数の植物種が含まれる場合でも、本発明の植物種判定方法を適用できる。
【0038】
上記「原料植物種特有のレトロトランスポゾン挿入部位」の選択は、判定対象とする原料植物種における複数の品種について、ゲノム中のレトロトランスポゾン挿入部位を比較して、品種間で共通するレトロトランスポゾン挿入部位を選択すればよい。なお、ゲノム中の原料植物種特有のレトロトランスポゾン挿入部位を検出する方法については後述する。
【0039】
植物ゲノムに原料植物種特有のレトロトランスポゾン挿入部位が存在するか否かを検出する方法としては特に限定されるものではなく、公知の方法を適宜選択して用いればよいが、核酸増幅反応を用いることが好ましい。核酸増幅反応を用いる方法としては、例えば、品種間で共通して存在しているレトロトランスポゾン挿入部位について、当該レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマーと、レトロトランスポゾンの塩基配列に基づいて設計されるプライマーとを組み合わせたプライマーセットを用いて、加工食品から調製したDNAを鋳型として核酸増幅反応を行い、増幅産物の有無を確認する方法を挙げることができる。
【0040】
レトロトランスポゾンに隣接するゲノム領域はレトロトランスポゾン配列の5’側に隣接する領域でも3’側に隣接する領域でもよい。レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマー(以下「隣接プライマー」と略記する。)とレトロトランスポゾンの塩基配列に基づいて設計されるプライマー(以下「RTPプライマー」と略記する。)とは、これらを組み合わせたプライマーセットにより核酸増幅が可能な向きに設計されていればよい。
【0041】
隣接プライマーは、レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計すればよいが、当該領域の塩基配列の中で特異性の高い部分の塩基配列に基づいて設計されることが好ましい。配列特異性の高いプライマーが設計できれば、非特異的増幅産物の出現を抑制することができ、結果の判定が正確かつ容易となるからである。
【0042】
また、RTPプライマーは、レトロトランスポゾンの塩基配列のうち高度に保存されている配列部分に基づいて設計されることが好ましい。高度に保存されている配列部分に基づいてRTPプライマーを設計すれば、個々の隣接プライマーに対応するRTPプライマーを別々に設計する必要がなく、あらかじめ設計したRTPプライマーと個々の隣接プライマーとの組み合わせによりプライマーセットを構成することが可能となる。
【0043】
核酸増幅反応の鋳型DNAは加工食品から調製したDNAである。加工食品からDNAを調製する方法は、特に限定されるものではなく、公知の方法を用いればよい。例えば、「遺伝子組換え食品の検査と分析のマニュアル」(JAS 分析試験ハンドブック 2002)に記載の方法を挙げることができる。
【0044】
核酸増幅反応は、公知の核酸増幅手段を適宜選択して用いればよい。具体的には、例えば、PCR法、ICAN法、UCAN法、LAMP法、プライマーエクステンション法等を挙げることができるが、これらに限定されるものではない。中でもPCR法が、本原料品種判定方法に用いる増幅方法として好適である。
【0045】
増幅産物の有無を確認する方法としては、例えば電気泳動を挙げることができるが、これに限定されるものではない。本発明の原料品種判定方法は、増幅産物の長さの違いを検出するものではなく、目的とするおおよその大きさの増幅産物の有無を検出するものであることが大きな特徴の1つである。そのため、シーケンサー等の機器を用いて数塩基の違いを検出するような厳密な電気泳動を行う必要がなく、アガロースゲル電気泳動等の簡便な電気泳動でも正確な判定が可能である。したがって、低コストで実施でき、また、判定や機器の操作に熟練を要しないという利点がある。
【0046】
原料植物種の判定は、上述のように、品種間で共通するレトロトランスポゾンの挿入がある部位における増幅産物の有無により行う。また、加工食品に複数の原料植物が含まれている場合には、それぞれの植物種に固有の挿入部位の量を定量的な遺伝子増幅法を用いて相対的な量を測定することで使用された原料植物種の割合を推定することが可能である。
【0047】
本方法が加工食品の原料植物種判定に特に適している特徴点として、増幅産物の長さを変更することが可能であるという点を挙げることができる。すなわち、上記隣接プライマーをRTPプライマーから遠い位置に設計すれば増幅断片長は長くなり、隣接プライマーをRTPプライマーから近い位置に設計すれば増幅断片長は短くなる。例えば、RTPプライマーをレトロトランスポゾン配列の5’末端または3’末端に設計し、隣接プライマーをRTPプライマーに接するように設計すれば、理論的には、両プライマーを合わせた長さ程度の増幅断片とすることも可能である。通常加工食品から得られるDNAは断片化が進んでいるため、増幅断片長を可能な限り短く設計しておけば、高度に断片化が進んだ加工食品由来のDNAを鋳型とした場合でも原料植物種判定が可能となる。したがって、本発明の原料植物種判定方法は、DNAが劣化・断片化した加工食品の原料植物種の判定に非常に適した方法であるといえる。ただし、常に増幅断片長が最も短くなるようにプライマーを設計する必要はなく、対象とする加工食品に含まれるDNAがどの程度断片化されているかを確認し、その程度に断片化されたDNAを鋳型としても増幅可能となるようにプライマーを設計すればよい。
【0048】
具体的には、隣接プライマーは、レトロトランスポゾン配列のいずれか一方の末端から500bp以内の隣接するゲノム領域の塩基配列に基づいて設計されることが好ましい。より好ましくは、400bp以内、さらに好ましくは300bp以内である。また、隣接プライマーとRTPプライマーとの組み合わせによる増幅産物の断片長は40bp〜600bpであることが好ましい。より好ましくは40bp〜400bp、さらに好ましくは40bp〜300bpである。このような範囲内に隣接プライマーを設計し、増幅産物の長さをコントロールすれば、より多くの加工食品に本発明の原料植物種判定方法を適用することが可能となる。
【0049】
次に、ゲノム中の原料植物種特有のレトロトランスポゾン挿入部位を検出する方法について説明する。
【0050】
ゲノム中の原料植物種特有のレトロトランスポゾン挿入部位を検出するためには、判定対象の原料植物種の品種間で、レトロトランスポゾン挿入部位を正確に比較することが重要である。レトロトランスポゾン挿入部位を比較する方法は、特に限定されるものではないが、例えば、制限酵素処理末端へのアダプター付加法(Sequence-Specific Amplification Polymorphism法、非特許文献2参照、以下「S-SAP法」と略記する。)を挙げることができる。S-SAP法を用いて原料植物種特有のレトロトランスポゾン挿入部位を検出する手順の概略を以下に説明する。
【0051】
まず、判定対象の原料植物の品種からそれぞれ調製したゲノムDNAを制限酵素で完全に消化後、これにより生じた断片の末端にアダプターDNAを付加する。植物からのゲノムDNAの調製法は特に限定されるものではなく、公知の方法を用いればよい。例えば、CTAB(cetyltrimethylammonium bromide)法(早川孝彦(1997)タバコのDNA・RNA単離法.“新版植物のPCR実験プロトコール”島本功、佐々木卓治監修.秀潤社、東京,49-56.)を挙げることができる。用いる制限酵素は、次の核酸増幅によって増幅される断片に含まれるレトロトランスポゾン配列中に認識部位を持たない制限酵素であればよい。アダプターDNAは、上記制限酵素処理によりDNA断片の末端に生じた粘着末端と同じ粘着末端を片側に持つ二重鎖DNA(アダプターDNA)であればよい。結合の処理としては、一般にDNAのライゲーションに用いられる方法を用いることができる。
【0052】
次に上記アダプターDNAを付加したゲノムDNA断片を鋳型としてレトロトランスポゾンの塩基配列に基づくプライマーとアダプターの塩基配列に基づくプライマーとを用いて品種ごとに核酸増幅を行う。この際、いずれか一方のプライマーを標識しておけば、増幅断片の比較が容易となる。標識としては、蛍光標識、放射線標識等を挙げることができるがこれらに限定されるものではない。例えば、蛍光標識されたプライマーを用いた場合は、増幅産物を当該蛍光物質の検出が可能なDNAシーケンサー等によって電気泳動し、各品種から得られた増幅産物のバンドパターンを比較すればよい。全ての品種について同じ位置にバンドが検出される場合は、いずれの品種においてもゲノム中の同一部位にレトロトランスポゾンが挿入されており、多型がない、すなわち、品種間で共通していると判断できる。同じ位置にバンドが検出される品種とバンドが検出されない品種がある場合にはレトロトランスポゾンの挿入位置に品種間多型があると判断できる。このようにして、品種間で共通したと認められるレトロトランスポゾン挿入部位を選択する。
【0053】
選択したレトロトランスポゾン挿入部位について、レトロトランスポゾンに隣接するゲノム領域の塩基配列は、実際にその領域をシーケンスすることにより決定することができる。塩基配列の決定には公知の遺伝子工学的方法を用いればよい。例えば、上記S-SAP法と同様にアダプターを付加した制限酵素消化断片を鋳型としてRTPプライマーとアダプタープライマーとを用いてPCRを行い、増幅断片を適当なプラスミドベクターにクローニングする。当該ベクターを大腸菌等の適当な宿主に導入して増殖させた後、プラスミドを抽出・精製し、公知のシーケンス反応を行ってシーケンサーに供すればよい。または、上記増幅断片をクローニングせずに、ダイレクトシーケンスすることも可能である。
【0054】
以上により、決定されたレトロトランスポゾンと隣接するゲノム領域の塩基配列に基づいて、各隣接プライマーを設計することができる。
【0055】
次に、上記選択したレトロトランスポゾン挿入部位が、原料植物種と近縁種の植物のゲノムに存在するか否かを検討する。具体的には、近縁種の複数の品種や系統からそれぞれ調製したゲノムDNAを鋳型として、レトロトランスポゾンと隣接するゲノム領域の塩基配列に基づいて設計したその挿入部位のプライマーを用いてPCRなどにより核酸増幅を行う。この核酸増幅の結果、近縁種の全品種もしくは一部の品種において、増幅断片が検出された場合、上記選択したレトロトランスポゾン挿入部位が近縁種のゲノムDNAに存在すると判断できる。一方、近縁種の全品種について、増幅断片が検出されない場合、上記選択したレトロトランスポゾン挿入部位が、近縁種のゲノムDNAに存在せず、原料植物種特有のレトロトランスポゾン挿入部位であると判断できる。
【0056】
本発明者らは、本発明の原料植物種判定方法を用いて、アズキ(Vigna angularis)と、アズキと最も近縁な栽培種であるリョクトウ(Vigna radiata (L.) R. Wilczek)とについて、植物種判定の検討を行っている。そこで以下に、アズキを原料とした加工食品の原料品種判定方法について説明する。
【0057】
対象とするアズキ加工食品は、原料にアズキが含まれるものであって、当該アズキ加工食品からアズキ原料のDNAを抽出可能なものであれば特に限定されるものではない。また、アズキ原料植物体の全部または一部、あるいは原料植物から得られる収穫物(果実、種子等)に対して加熱、乾燥等の加工処理を施したものがアズキ加工食品に該当する。したがって、アズキ加工食品には最終製品のみではなく、中間加工品も含まれる。また、上記アズキ加工食品は、単一の品種の原料植物を含む加工食品に限定されるものではなく、複数の品種の原料植物を含む加工食品であってもよい。
【0058】
利用可能なレトロトランスポゾンは、アズキに存在するレトロトランスポゾンであれば特に限定されるものではない。本発明者は、前述のHirochikaらの方法(Hirochika et al. (1992) Retrotransposon families in rice. Mol Gen Genet 233: 209-216.)に従い、Ty1-Copia型レトロトランスポゾンの逆転写酵素領域に存在するアミノ酸保存配列をターゲットとするPCRにより、同酵素領域の配列をアズキ品種「しゅまり」のゲノムから単離した。このようにして単離した逆転写酵素領域の配列の延長配列をSuppression PCR(Siebert et al. (1995) An improved PCR method for walking in uncloned genomic DNA. Nucleic Acids Res. 23:1087-1088.)法により単離し、末端のLTR配列までを同定し、PHARE-1と命名した。そして、このPHARE-1を用いて、植物種判定の検討を行っている。このPHARE-1は、アズキに存在するレトロトランスポゾンの一例であり、他のレトロトランスポゾンも利用可能であることはいうまでもない。
【0059】
本発明者らは、まず、アズキ品種や系統26種類で共通すると認められるPHARE-1挿入部位を見出すために、S-SAP法を用いて、北海道立中央農業試験場遺伝資源部が保有しているアズキ品種や系統26種類について、PHARE-1の挿入部位を比較した。
【0060】
その結果、26種類のズキ品種や系統間で共通するPHARE-1の挿入部位(PHARE-1に隣接するゲノム領域を含む)が2個見出された。そして、これら2個の挿入部位をクローニングして塩基配列を決定した。これら2個のクローンの塩基配列が、配列番号1及び2に示される塩基配列である。配列番号1及び2について、レトロトランスポゾン(PHARE-1)に隣接するゲノム領域の塩基配列部分を以下に示す。
配列番号1:第1位〜第57位
配列番号2:第1位〜第57位
本発明者らは、上記配列番号1及び2に示される塩基配列のレトロトランスポゾン(PHARE-1)に隣接するゲノム領域の塩基配列に基づいて、各隣接プライマーを設計し、PHARE-1の塩基配列に基づいて、RTPプライマーを設計している。なお、発明者らが実際に使用したRTPプライマーは、配列番号3に示される塩基配列からなるプライマーであり、隣接プライマーは、配列番号4及び5に示される塩基配列からなるプライマーである。
【0061】
このようにして設計したプライマーを用いてPCRを行い、アズキ(Vigna angularis)と、アズキと最も近縁な栽培種であるリョクトウ(Vigna radiata (L.) R. Wilczek)とについて、Phare-1の挿入部位を調べた。その結果、Phareの挿入部位が、アズキ種に固有であり、近縁種であるリョクトウに存在しないことを見出した(図1、図2参照)。
【0062】
このように、配列番号3に示される塩基配列からなるRTPプライマーと配列番号4及び5に示される塩基配列からなる隣接プライマーとを用いて、本発明の原料植物種判定方法を実施すれば、加工食品中にアズキが含まれているか否かを識別することが可能である。
【0063】
本発明の原料植物種識別キットは、本発明の原料植物種判定方法を実施するためのキットであって、レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマーと、レトロトランスポゾンの塩基配列に基づいて設計されるプライマーとを含むものであればよい。キットには、プライマー以外に対象とする加工食品からDNAを抽出・精製するために用いる試薬・器具、核酸増幅反応に用いる試薬・器具、電気泳動に用いる試薬・器具等を含むことが好ましい。
【0064】
また、本発明のアズキ種識別キットは、本発明の原料植物種判定方法をアズキを含む加工食品を対象として実施するためのキットであって、上記配列番号3に示される塩基配列からなるプライマーを含み、かつ、配列番号4または5に示される塩基配列からなるプライマーのうち少なくとも1つ以上を含むものであればよい。また、キットには、プライマー以外に対象とする加工食品からDNAを抽出・精製するために用いる試薬・器具、核酸増幅反応に用いる試薬・器具、電気泳動に用いる試薬・器具等を含むことが好ましい。
【0065】
なお本発明は上述した各実施形態に限定されるものではなく、請求項に示した範囲で種々の変更が可能であり、異なる実施形態にそれぞれ開示された技術的手段を適宜組み合わせて得られる実施形態についても本発明の技術的範囲に含まれる。
【実施例1】
【0066】
アズキのレトロトランスポゾンPHARE-1のアズキゲノムへの挿入部位(P1)をPCRにより調査したところ、供試した全てのアズキ品種でその挿入部位が存在することが判明した。また、同じPCRにより、アズキ(Vigna angularis (Willd.) Ohwi & H. Ohashi)と同じササゲ属(Vigna)に属し、アズキと最も近縁な栽培種であるリョクトウ(Vigna radiata (L.) R. Wilczek)の個体において調査したところ、PHARE-1のこの挿入部位の存在を示す結果は得られなかった。
【0067】
(1)材料
本発明者が、調べた北海道立中央農業試験場遺伝資源部が保有しているアズキ品種や系統26種類を表1に示す。
【0068】
【表1】

【0069】
(2)ゲノムDNAの抽出
アズキ植物体のゲノムDNAは、CTAB(cetyltrimethylammonium bromide)法(早川孝彦(1997)タバコのDNA・RNA単離法.“新版植物のPCR実験プロトコール”島本功、佐々木卓治監修.秀潤社、東京,49-56.)を基本に抽出した。
【0070】
(3)アズキのゲノムにおけるPHARE-1挿入部位の特定
北海道立中央農業試験場遺伝資源部が保有しているアズキ品種や系統26種類について、PHARE-1のLTR配列を利用してS-SAP法による分析を行った。PHARE-1のLTR配列は、アズキ品種「しゅまり」のゲノムから、前述のHirochikaらの方法に従い、Ty1-Copia型レトロトランスポゾンの逆転写酵素領域に存在するアミノ酸保存配列をターゲットとするPCRにより同酵素領域の配列を単離し、このようにして単離した逆転写酵素領域の配列の延長配列をSuppression PCR法により単離することで、PHARE-1の末端部に存在する配列として同定したものである。このS-SAP法による電気泳動の結果から、これら26種類に全てに共通して存在すると見られるPHARE-1レトロトランスポゾン挿入部位を特定した。
【0071】
(4)アズキの品種間に共通する挿入部のPCR増幅による確認
特定した挿入部位の内の3カ所をクローニングしてその塩基配列(配列番号1及び2)を決定し、その配列における、PHARE-1レトロトランスポゾンの塩基配列とそれに隣接するそれぞれのゲノム領域の塩基配列に基づいてプライマーを設計した。設計したPHARE-1レトロトランスポゾンの塩基配列に基づくRTPプライマー(Az-Copia1-LYT-D)、及び配列番号1及び2のゲノム領域の塩基配列に基づく隣接プライマーの塩基配列は、以下の通りである。
Az-Copia1-LYT-Dプライマー(RTPプライマー):5'-GCTAAACACATAAGGGAGATTTCTCC-3'(配列番号3)
P1-DE-T1-5A1プライマー(配列番号1の塩基配列に基づく隣接プライマー):5'-CATCTAACACACCCTAAGAGATAGTC-3'(配列番号4)
P1-DE-T2-4A1プライマー(配列番号2の塩基配列に基づく隣接プライマー):5'-CCAGTAAGGATCCTTGGGTTTTCTTC-3'(配列番号5)
Az-Copia1-LYT-DプライマーとP1-DE-T1-5A1プライマー及びAz-Copia1-LYT-DプライマーとP1-DE-T2-4A1プライマーの2つのプライマー組合せについて、表1に示されたアズキ植物体から抽出したゲノムDNA溶液(20ng/μl)1μlを鋳型としてPCRを行った。PCRは、容量10μlの反応液として、E x Taq(TaKaRa)を用いて、94℃4分間の変性、94℃30秒間・59℃30秒間・72℃20秒間を40サイクル、72℃5分間の最終伸長反応のサイクリング条件で行った。増幅産物をアガロースゲル電気泳動に供し、エチジウムブロマイドで染色して観察した。その結果、これら2つのプライマー組合せのPCRにおいては、プライマーの設計位置から期待された長さのDNA断片(Az-Copia1-LYT-DプライマーとP1-DE-T1-5A1プライマーでは、219bp、Az-Copia1-LYT-DプライマーとP1-DE-T2-4A1プライマーでは、312bp)がアズキの26種類の全てで増幅した。
【0072】
(5)アズキ品種に共通する挿入部がアズキ近縁種において存在しないこと調査
配列番号1と2のようなアズキの品種には共通して存在するPHARE-1レトロトランスポゾンの挿入部位が、アズキと同じ属に属する最も近縁なマメ科作物などには存在しないことを確認するため、以下の植物について、葉または乾燥子実から上記(2)の方法によりゲノムDNAを抽出し、(4)の方法によるPCR増幅により、ゲノム中におけるその挿入部位の存在を判定した。
植物とDNAを抽出した組織:アズキ品種「きたのおとめ」の葉、アズキ系統「天津小豆」の葉、アズキ品種「きたのおとめ」の乾燥子実、リョクトウ(市販品)の乾燥子実
その結果、図1及び図2に示されるように、アズキ品種のゲノムDNAを鋳型としたPCRでは、それぞれの挿入部位の塩基配列に基づき設計したプライマーの位置から期待された長さとなるDNA断片(Az-Copia1-LYT-DプライマーとP1-DE-T1-5A1プライマーでは、219bp、Az-Copia1-LYT-DプライマーとP1-DE-T2-4A1プライマーでは、312bp)が増幅した。一方、リョクトウのゲノムDNAを鋳型としたPCRでは、いずれのPCRにおいても増幅断片が見られなかった。なお、リョクトウから抽出した同じDNAを鋳型として別のプライマーを用いてPCRを行ったところ、プライマーの設計から予想されたDNA断片の増幅がみられた。それゆえ、図1及び図2のPCRにおいてDNA断片が増幅しなかったのは、リョクトウから抽出したDNAの品質がPCR増幅に適していなかったためではなく、Az-Copia1-LYT-DプライマーとP1-DE-T1-5A1プライマーのセット、及びAz-Copia1-LYT-DプライマーとP1-DE-T2-4A1プライマーのセットにより増幅するPHARE-1の挿入部位がリョクトウには存在しないためである。この結果、これら2つのPHARE-1の挿入部位(配列番号1及び2)は、アズキに固有のものであることが分かった。
【0073】
本発明は上述した各実施形態に限定されるものではなく、請求項に示した範囲で種々の変更が可能であり、異なる実施形態にそれぞれ開示された技術的手段を適宜組み合わせて得られる実施形態についても本発明の技術的範囲に含まれる。
【産業上の利用可能性】
【0074】
本発明は、加工食品の原料植物種の判定を高精度かつ定量的にできるものであるため、食品産業に広く利用可能である。さらに、食品の不正表示や、海外で不正に栽培された品種を原料とした輸入加工食品等の検査に利用すれば、違法食品の取り締まりに大きく貢献できるものと期待される。
【技術分野】
【0001】
本発明は、加工食品に含まれる原料植物の植物種を判定する原料植物種判定方法に関する。
【背景技術】
【0002】
植物種の同定技術としては、例えば非特許文献1に記載のように、葉緑体DNAの遺伝子間領域の塩基配列の違いを利用して、植物種を同定する方法(PS−ID法)が知られている。また、加工食品に含まれるアレルゲン物質を定性分析・定量分析する方法としては、分析対象のアレルゲン物質に対する抗体を用いる方法が開発されている。また、コムギ、そば、落花生等といったアレルゲン物質となる植物種の遺伝子をPCRにより検出する方法が開発されている。
【0003】
また、非特許文献2には、ゲノムにおけるレトロトランスポゾンの個々の挿入位置を、制限酵素処理末端へのアダプター付加法(S-SAP:Sequence-Specific Amplification Polymorphism法)により、挿入位置近傍のゲノム配列(制限酵素認識部位)とLTR配列間の核酸増幅を区別する方法が記載されている。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】Nakamura, I., N. Kameya, Y. Kato, S. Yamanaka, H. Jomori, and Y. Sato. A proposal for identifying the short ID sequence which addresses the plastid subtype of higher plants. Breeding Science 47: 385-388 (1997)
【非特許文献2】Waugh, R., K. McLean, A. J. Flavell, S. R. Pearce, A. Kumar, B. B. T. Thomas and W. Powell. Genetic distribution of Bare-1-like retrotransposable elements in the barley genome revealed by sequence-specific amplification polymorphisms (S-SAP). Mol Gen Genet 253: 687-694 (1997)
【発明の概要】
【発明が解決しようとする課題】
【0005】
しかしながら、上述のような従来技術には、以下の問題がある。
【0006】
まず、PS−ID法を用いて植物種を同定する技術では、使用する葉緑体DNAの遺伝子領域において、同一の植物種間であっても相違する塩基配列が存在するため、厳密な植物種レベルの識別が困難であるという問題がある。また、葉緑体は、1つの植物細胞内に多数存在する。さらには、この葉緑体の遺伝子領域の塩基配列のなかには、核ゲノム中の塩基配列と同一の塩基配列が存在する。それゆえ、葉緑体の遺伝子領域の塩基配列の複製数は、植物の品種間でも違いが見られることが考えられる。このため、葉緑体の遺伝子領域の塩基配列を用いるPS−ID法では、定量分析が、植物種内の品種間でもばらつきが生じることが考えられ、植物種間レベルで正確に定量比較を行うことが困難である。
【0007】
また、アレルゲン物質に対する抗体を用いる技術では、例えばコムギに対する抗体は、ライムギやエンバクにも反応するため、植物種間レベルでのアレルゲン物質の識別が困難である。また、アレルゲン物質となる植物種の遺伝子をPCRにより検出する方法では、分析対象となる植物種の遺伝子の複製数が多く、定量的な検査が困難である。
【0008】
本発明は、上記の問題点に鑑みてなされたものであり、その目的は、加工食品に含まれる原料植物の植物種を高精度かつ定量的に判定できる方法を提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、上記の課題を解決するために鋭意検討した結果、植物のゲノム中に存在する転位因子の1つであるレトロトランスポゾンの植物ゲノムへの挿入位置が原料植物の植物種固有のものであるか否かを検出する方法は、加工食品に含まれる原料植物の植物種(species)の判定においても適用できることを見出し、本発明を完成させるに至った。そして、かかる「原料植物の植物種固有のトランスポゾン挿入部位」の有無を判定し、それを定量することにより、加工食品に原料植物種が含まれているかの判定を高精度かつ定量的に行うことができることを確認して、本発明を完成させるに至った。
【0010】
すなわち、本発明の原料植物種判定方法は、上記の課題を解決するために、加工食品に含まれる原料植物の植物種を判定する原料植物種判定方法であって、植物ゲノムに原料植物の植物種固有のレトロトランスポゾン挿入部位が存在するか否かを検出し、この検出結果に基づいて、原料植物の植物種を判定することを特徴としている。
【0011】
さらに、植物ゲノムに存在する原料植物の植物種固有のレトロトランスポゾン挿入部位は、通常、その植物種に存在する全ての種類の染色体の1組には1カ所にしか存在しないため、定量的な遺伝子増幅法などを利用して定量することができる。即ち、加工食品の原料に複数の植物種が使われている場合、原料となったそれぞれの植物種について固有のレトロトランスポゾン挿入部位の量を相対的に測定し比較することで、加工食品において使用された原料植物種の相対的な割合を推定することが可能となる。したがって、原料植物の植物種固有のレトロトランスポゾン挿入部位を利用して、高精度かつ定量的な原料植物の植物種の判定を行うことが可能となる。
【0012】
本発明の原料植物種判定方法では、植物ゲノムに原料植物の植物種固有のレトロトランスポゾン挿入部位が存在するか否かの検出には、判定対象とする複数の品種のゲノムに共通して存在しているレトロトランスポゾン挿入部位における、当該レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマーと、レトロトランスポゾンの塩基配列に基づいて設計されるプライマーとを組み合わせたプライマーセットを用いて、加工食品から調製したDNAを鋳型として核酸増幅反応を行い、増幅産物の有無を確認する方法を用いることが好ましい。上記プライマーセットにより増幅される断片の長さを任意に設計できるため、短い断片が増幅されるようにプライマーセットを設計すれば、加工食品中のDNAが高度に断片化している場合でもこれを鋳型として増幅断片の有無を確認することが可能となる。また、増幅産物の有無を確認するものであるため、簡便な電気泳動により確認でき、結果の判定に熟練を要しない。また、加工食品に複数の原料植物が含まれている場合に、使用された原料植物種の割合は定量的な遺伝子増幅を用いてそれぞれの植物種に固有の挿入部位の量を相対的に測定することで推定することが可能である。
【0013】
また、本発明の原料植物種判定方法では、判定対象とする複数の品種について、ゲノム中のレトロトランスポゾン挿入部位を比較して、複数の品種のゲノムに共通して存在しているレトロトランスポゾン挿入部位を選択し、選択されたレトロトランスポゾン挿入部位について、原料植物の近縁種のゲノムに存在するか否かの確認を行い、上記確認の結果、原料植物の近縁種のゲノムに存在しないレトロトランスポゾン挿入部位を、上記原料植物の植物種固有のレトロトランスポゾン挿入部位として決定することが好ましい。例えば、S-SAP法などにより、判定対象とする原料植物の複数の品種について、ゲノム中のレトロトランスポゾン挿入部位を比較し、複数の品種のゲノムに共通して存在している挿入部位を見いだせば、この挿入部位が原料植物の近縁種のゲノムに存在するか否かを確認することにより、原料植物の植物種固有のレトロトランスポゾン挿入部位を決定することができる。
【0014】
また、本発明の原料植物種判定方法では、上記増幅産物の断片長は40bp〜600bpであることが好ましい。増幅産物の長さが短くなるようにプライマーを設計することにより、加工食品から抽出されるDNAが高度に断片化したものであっても、それを鋳型として核酸増幅反応を行うことが可能となる。
【0015】
また、本発明の原料植物種判定方法では、上記レトロトランスポゾンに隣接するゲノム領域の塩基配列は、当該領域を含むDNA断片をシーケンスすることにより得られる塩基配列であることが好ましい。
【0016】
また、本発明の原料植物種判定方法では、上記レトロトランスポゾンに隣接するゲノム領域の塩基配列は、当該領域を含むDNA断片をクローニングし、当該断片をシーケンスすることにより得られる塩基配列であることが好ましい。
【0017】
また、本発明の原料植物種判定方法の対象とする加工食品はアズキを原料に含むことが好ましく、アズキ加工食品を対象とする場合には、利用するレトロトランスポゾンはPHARE-1であることが好ましい。
【0018】
上記PHARE-1を利用してアズキ植物種の判定を行う場合、PHARE-1に隣接するゲノム領域を含むDNA断片をシーケンスすることにより得られる塩基配列として、配列番号1または2に示される塩基配列を挙げることができる。すなわち、配列番号1または2に示される塩基配列に含まれるPHARE-1に隣接するゲノム領域の塩基配列に基づいて、プライマーを設計することができる。
【0019】
上記配列番号1または2に示される塩基配列に含まれるPHARE-1に隣接するゲノム領域の塩基配列に基づいて設計されるプライマーとしては、配列番号4または5に示される塩基配列からなるプライマーを挙げることができる。また、PHARE-1の塩基配列に基づいて設計されるプライマーは配列番号3に示される塩基配列からなるプライマーを挙げることができる。なお、配列番号3〜5に示されるプライマーも本発明に含まれる。
【0020】
また、本発明の原料植物種判定キットは、上述の原料植物種判定方法を実施するためのキットであって、レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマーと、レトロトランスポゾンの塩基配列に基づいて設計されるプライマーとを含むことを特徴とプライマーを含むキットとすることにより、本発明の原料植物種判定方法をより高精度かつ定量的に実施することが可能となる。
【0021】
上記キットに含まれるプライマーとしては、配列番号3に示される塩基配列からなるプライマーと、配列番号4または5に示される塩基配列からなるプライマーのうち少なくとも1つ以上とを挙げることができる。これにより、アズキ種判定キットを構成することができる。
【発明の効果】
【0022】
本発明の原料植物種判定方法は、以上のように、植物ゲノムに原料植物の植物種固有のレトロトランスポゾン挿入部位が存在するか否かを検出し、この検出結果に基づいて、原料植物の植物種を判定するので、高精度かつ定量的な原料植物の植物種の判定を行うことが可能となる。
【0023】
さらに、本発明の原料植物種判定方法は、増幅される断片の長さを比較するものではなく、増幅産物の有無を確認するものであるため、簡便な電気泳動により正確な判定ができる。したがって、高価な分析機器を必要とせず、低コストで実施できるという効果を奏する。さらに、判定に熟練を要しないという効果を奏する。
【0024】
また、本発明の原料植物種判定キットは、本発明の原料植物種判定方法を実施するためのキットであるため、本方法をより迅速かつ簡便に実施できるという効果を奏する。
【0025】
以上のように、本発明によれば、加工食品の原料植物種を迅速、簡便かつ正確に判定することができるので、本方法を用いて加工食品の原料植物種を判定すれば、加工食品の原料植物種の不正表示を防止することができるという効果を奏する。
【図面の簡単な説明】
【0026】
【図1】アズキ(Vigna angularis (Willd.) Ohwi & H. Ohashi)、及び該アズキと同じササゲ属(Vigna)に属する最も近縁なリョクトウ(Vigna radiate (L.) R. Wilczek)のゲノムDNAを鋳型として、Az-Copia1-LYT-D/ P1-DE-T1-5A1プライマーセットを用いたPCR増幅産物の電気泳動結果を示す画像である。
【図2】アズキ(Vigna angularis (Willd.) Ohwi & H. Ohashi)、及び該アズキと同じササゲ属(Vigna)に属する最も近縁なリョクトウ(Vigna radiate (L.) R. Wilczek)のゲノムDNAを鋳型として、Az-Copia1-LYT-D/ P1-DE-T2-4A1プライマーセットを用いたPCR増幅産物の電気泳動結果を示す画像である。
【発明を実施するための形態】
【0027】
本発明の実施の一形態について説明すれば、以下のとおりである。なお、本発明はこれに限定されるものではない。
【0028】
本発明の原料植物種判定方法は、加工食品に含まれる原料植物の植物種を判定する方法であって、植物ゲノムに原料植物の植物種固有のレトロトランスポゾン挿入部位が存在するか否かを検出することによって、原料植物の植物種を判定する方法である。
【0029】
対象とする加工食品は、原料に植物が含まれるものであって、当該加工食品から原料植物のDNAを抽出可能なものであれば特に限定されるものではない。また、原料植物体の全部または一部、あるいは原料植物から得られる収穫物(果実、種子等)に対して加熱、乾燥等の加工処理を施したものが加工食品に該当する。したがって、加工食品には最終製品のみではなく、中間加工品も含まれる。
【0030】
レトロトランスポゾンは転移因子の一種であり、進化の過程で自身の複製を作り、ゲノムに挿入するという形式で転移していた遺伝因子である。多くの植物において、多種類のレトロトランスポゾンが複製された状態で存在する。現在の植物ゲノムにおいては、レトロトランスポゾンのほとんどの配列は転移活性を失っており、ゲノムに挿入された状態で安定的に遺伝している。レトロトランスポゾンとそれに隣接するゲノム配列は、現存する植物種の共通祖先でレトロトランスポゾンの転移が生じ、その転移が生じた部位がその共通祖先において遺伝的に固定すれば、その祖先種から分岐してきた植物種の全てに存在することとなる。一方、特定の植物種の祖先がその植物種に最も近縁の植物種と種分化した後、その祖先となった集団に含まれる個体においてレトロトランスポゾンの転移が生じ、その転移が生じた部位がその祖先となった集団において遺伝的に固定すれば、その挿入部位は、染色体の変異などによりその部位が失われることがない限り、その植物種に固有でかつその植物種に属する全ての個体に存在することになる。また、種分化以前に存在していたレトロトランスポゾンの挿入部位であっても、特定の植物種においてのみ遺伝的に固定した部位は、その植物種に固有のものとなる。なお、種分化後、その種内でレトロトランスポゾンの転移が生じた場合には、ある品種ではレトロトランスポゾンの挿入が見られるゲノムの場所に、別の品種では挿入が見られないという、品種間の多型となる。
【0031】
本発明の原料植物種判定方法に利用可能なレトロトランスポゾンは、植物に存在するレトロトランスポゾンであれば特に限定されるものではない。すなわち、公知のレトロトランスポゾンのみならず、今後新たに見出されるレトロトランスポゾンも利用することができる。
【0032】
LTR型のレトロトランスポゾンは、今まで調査された範囲では、全ての高等植物に存在しており、主要な作物については、個別の配列が多数単離され、命名されている (Johns et al (1985) A low copy number, copia-like transposon in maize. EMBO J 8: 1093-1102.、Manninen and Schulman (1993) BARE-1, a copia-like retroelement in barley (Hordeum vulgare L.). Plant Mol Biol 22: 829-846.、Laten et al. (1998) SIRE-1, a copia/Ty1-like retroelement from soybean, encodes a retroviral envelope-like protein. Proc Natl Acad Sci USA 95: 6897-6902.、Kumekawa et al. (1999) Identification and characterization of novel retrotransposons of the gypsy type in rice. Mol Gen Genet 260: 593-602.)。
【0033】
LTR型のレトロトランスポゾンのうち、Ty1-Copia型に分類されるものについては、逆転写酵素領域に存在するアミノ酸保存配列をターゲットとするPCRにより、同酵素領域の配列が多数の植物から単離されている(Voytas et al. (1992) copia-like retrotransposons are ubiquitous among plants. Proc Natl Acad Sci USA 89: 7124-7128.、Hirochika et al. (1992) Retrotransposon families in rice. Mol Gen Genet 233: 209-216.)。したがって、レトロトランスポゾンの全長配列が調べられていない作物であっても、DNAデータベースにおいて既知の、あるいは、新たに逆転写酵素領域の配列を同定すれば、その延長配列を単離するTAIL(Thermal Asymmetric InterLaced)PCR(Liu et al. (1995) Efficient isolation and mapping of Arabidopsis thaliana T-DNA insert junctions by thermal asymmetric interlaced PCR. Plant Journal 8:457-463.)、Suppression PCR(Siebert et al. (1995) An improved PCR method for walking in uncloned genomic DNA. Nucleic Acids Res. 23:1087-1088.)やその他の方法(Pearse et al. (1999) Rapid isolation of plant Ty1-copia group retrotransposon LTR sequences for molecular marker studies. Plant Journal 19: 711-717.)により、末端のLTR配列までを同定することができる。
【0034】
植物ゲノムに原料植物の植物種(以下、原料植物種と記す)固有のレトロトランスポゾン挿入部位が存在するかの検出とは、植物ゲノムに、原料植物種に存在するが原料植物種以外の植物種には存在しないレトロトランスポゾンの挿入部位を検出することを意味する。上述のように、レトロトランスポゾンの挿入部位には品種間で共通したゲノムの場所に挿入が見られる。それゆえ、この共通したレトロトランスポゾンの挿入部位が、原料植物種以外の植物種(特に原料植物の近縁種)のゲノムにも存在するか否かを検討することにより、原料植物種固有のレトロトランスポゾン挿入部位を同定することができる。本発明の植物種判定方法は、原料植物種固有のレトロトランスポゾン挿入部位の有無を明らかとすることにより、加工食品における原料植物種の判定を行うものである。
【0035】
レトロトランスポゾンの挿入は、転写されたレトロトランスポゾンの塩基配列が植物ゲノムに挿入されることにより生じる。その際、レトロトランスポゾンの挿入が生じる部位は、特に規則性がない。このことから、植物ゲノムのそれぞれの部位でのレトロトランスポゾンの挿入が偶然別の細胞や個体で生じる確率は、極めて小さい。このため、ゲノムの基本構造が原料植物種と類似している近縁種であっても、レトロトランスポゾンの挿入が近縁種との共通祖先のゲノムにおいて生じたものでない限り、原料植物種のレトロトランスポゾン挿入部位が、近縁種のゲノムで見出されることは、確率的に見てゼロに等しい。
【0036】
それゆえ、レトロトランスポゾンの挿入部位のうち、その植物種において遺伝的に固定している挿入部位(全ての品種間において共通して見られる挿入部位)が、その植物種の近縁種において見出されない場合は、植物種の特異性が極めて高いものとなる。そして、このような特異性を利用して、原料植物種において遺伝的に固定しているレトロトランスポゾン挿入部位を、原料植物種を識別するマーカーとして用いることができる。
【0037】
上記植物ゲノムに原料植物種固有のレトロトランスポゾン挿入部位が存在するか否かの検出は、複数の挿入部位について行うことが好ましい。上述のように植物ゲノムには多数のレトロトランスポゾン配列が存在するため、S-SAP法などにより、特定の原料植物種のみに存在する挿入部位(原料植物種固有のレトロトランスポゾン挿入部位)を見いだせば、その部位における挿入の有無により原料植物種の有無を判定できる。また、このような植物種に固有の挿入部位を多数の植物種について見いだすことで、加工食品中に原料植物として複数の植物種が含まれる場合でも、本発明の植物種判定方法を適用できる。
【0038】
上記「原料植物種特有のレトロトランスポゾン挿入部位」の選択は、判定対象とする原料植物種における複数の品種について、ゲノム中のレトロトランスポゾン挿入部位を比較して、品種間で共通するレトロトランスポゾン挿入部位を選択すればよい。なお、ゲノム中の原料植物種特有のレトロトランスポゾン挿入部位を検出する方法については後述する。
【0039】
植物ゲノムに原料植物種特有のレトロトランスポゾン挿入部位が存在するか否かを検出する方法としては特に限定されるものではなく、公知の方法を適宜選択して用いればよいが、核酸増幅反応を用いることが好ましい。核酸増幅反応を用いる方法としては、例えば、品種間で共通して存在しているレトロトランスポゾン挿入部位について、当該レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマーと、レトロトランスポゾンの塩基配列に基づいて設計されるプライマーとを組み合わせたプライマーセットを用いて、加工食品から調製したDNAを鋳型として核酸増幅反応を行い、増幅産物の有無を確認する方法を挙げることができる。
【0040】
レトロトランスポゾンに隣接するゲノム領域はレトロトランスポゾン配列の5’側に隣接する領域でも3’側に隣接する領域でもよい。レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマー(以下「隣接プライマー」と略記する。)とレトロトランスポゾンの塩基配列に基づいて設計されるプライマー(以下「RTPプライマー」と略記する。)とは、これらを組み合わせたプライマーセットにより核酸増幅が可能な向きに設計されていればよい。
【0041】
隣接プライマーは、レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計すればよいが、当該領域の塩基配列の中で特異性の高い部分の塩基配列に基づいて設計されることが好ましい。配列特異性の高いプライマーが設計できれば、非特異的増幅産物の出現を抑制することができ、結果の判定が正確かつ容易となるからである。
【0042】
また、RTPプライマーは、レトロトランスポゾンの塩基配列のうち高度に保存されている配列部分に基づいて設計されることが好ましい。高度に保存されている配列部分に基づいてRTPプライマーを設計すれば、個々の隣接プライマーに対応するRTPプライマーを別々に設計する必要がなく、あらかじめ設計したRTPプライマーと個々の隣接プライマーとの組み合わせによりプライマーセットを構成することが可能となる。
【0043】
核酸増幅反応の鋳型DNAは加工食品から調製したDNAである。加工食品からDNAを調製する方法は、特に限定されるものではなく、公知の方法を用いればよい。例えば、「遺伝子組換え食品の検査と分析のマニュアル」(JAS 分析試験ハンドブック 2002)に記載の方法を挙げることができる。
【0044】
核酸増幅反応は、公知の核酸増幅手段を適宜選択して用いればよい。具体的には、例えば、PCR法、ICAN法、UCAN法、LAMP法、プライマーエクステンション法等を挙げることができるが、これらに限定されるものではない。中でもPCR法が、本原料品種判定方法に用いる増幅方法として好適である。
【0045】
増幅産物の有無を確認する方法としては、例えば電気泳動を挙げることができるが、これに限定されるものではない。本発明の原料品種判定方法は、増幅産物の長さの違いを検出するものではなく、目的とするおおよその大きさの増幅産物の有無を検出するものであることが大きな特徴の1つである。そのため、シーケンサー等の機器を用いて数塩基の違いを検出するような厳密な電気泳動を行う必要がなく、アガロースゲル電気泳動等の簡便な電気泳動でも正確な判定が可能である。したがって、低コストで実施でき、また、判定や機器の操作に熟練を要しないという利点がある。
【0046】
原料植物種の判定は、上述のように、品種間で共通するレトロトランスポゾンの挿入がある部位における増幅産物の有無により行う。また、加工食品に複数の原料植物が含まれている場合には、それぞれの植物種に固有の挿入部位の量を定量的な遺伝子増幅法を用いて相対的な量を測定することで使用された原料植物種の割合を推定することが可能である。
【0047】
本方法が加工食品の原料植物種判定に特に適している特徴点として、増幅産物の長さを変更することが可能であるという点を挙げることができる。すなわち、上記隣接プライマーをRTPプライマーから遠い位置に設計すれば増幅断片長は長くなり、隣接プライマーをRTPプライマーから近い位置に設計すれば増幅断片長は短くなる。例えば、RTPプライマーをレトロトランスポゾン配列の5’末端または3’末端に設計し、隣接プライマーをRTPプライマーに接するように設計すれば、理論的には、両プライマーを合わせた長さ程度の増幅断片とすることも可能である。通常加工食品から得られるDNAは断片化が進んでいるため、増幅断片長を可能な限り短く設計しておけば、高度に断片化が進んだ加工食品由来のDNAを鋳型とした場合でも原料植物種判定が可能となる。したがって、本発明の原料植物種判定方法は、DNAが劣化・断片化した加工食品の原料植物種の判定に非常に適した方法であるといえる。ただし、常に増幅断片長が最も短くなるようにプライマーを設計する必要はなく、対象とする加工食品に含まれるDNAがどの程度断片化されているかを確認し、その程度に断片化されたDNAを鋳型としても増幅可能となるようにプライマーを設計すればよい。
【0048】
具体的には、隣接プライマーは、レトロトランスポゾン配列のいずれか一方の末端から500bp以内の隣接するゲノム領域の塩基配列に基づいて設計されることが好ましい。より好ましくは、400bp以内、さらに好ましくは300bp以内である。また、隣接プライマーとRTPプライマーとの組み合わせによる増幅産物の断片長は40bp〜600bpであることが好ましい。より好ましくは40bp〜400bp、さらに好ましくは40bp〜300bpである。このような範囲内に隣接プライマーを設計し、増幅産物の長さをコントロールすれば、より多くの加工食品に本発明の原料植物種判定方法を適用することが可能となる。
【0049】
次に、ゲノム中の原料植物種特有のレトロトランスポゾン挿入部位を検出する方法について説明する。
【0050】
ゲノム中の原料植物種特有のレトロトランスポゾン挿入部位を検出するためには、判定対象の原料植物種の品種間で、レトロトランスポゾン挿入部位を正確に比較することが重要である。レトロトランスポゾン挿入部位を比較する方法は、特に限定されるものではないが、例えば、制限酵素処理末端へのアダプター付加法(Sequence-Specific Amplification Polymorphism法、非特許文献2参照、以下「S-SAP法」と略記する。)を挙げることができる。S-SAP法を用いて原料植物種特有のレトロトランスポゾン挿入部位を検出する手順の概略を以下に説明する。
【0051】
まず、判定対象の原料植物の品種からそれぞれ調製したゲノムDNAを制限酵素で完全に消化後、これにより生じた断片の末端にアダプターDNAを付加する。植物からのゲノムDNAの調製法は特に限定されるものではなく、公知の方法を用いればよい。例えば、CTAB(cetyltrimethylammonium bromide)法(早川孝彦(1997)タバコのDNA・RNA単離法.“新版植物のPCR実験プロトコール”島本功、佐々木卓治監修.秀潤社、東京,49-56.)を挙げることができる。用いる制限酵素は、次の核酸増幅によって増幅される断片に含まれるレトロトランスポゾン配列中に認識部位を持たない制限酵素であればよい。アダプターDNAは、上記制限酵素処理によりDNA断片の末端に生じた粘着末端と同じ粘着末端を片側に持つ二重鎖DNA(アダプターDNA)であればよい。結合の処理としては、一般にDNAのライゲーションに用いられる方法を用いることができる。
【0052】
次に上記アダプターDNAを付加したゲノムDNA断片を鋳型としてレトロトランスポゾンの塩基配列に基づくプライマーとアダプターの塩基配列に基づくプライマーとを用いて品種ごとに核酸増幅を行う。この際、いずれか一方のプライマーを標識しておけば、増幅断片の比較が容易となる。標識としては、蛍光標識、放射線標識等を挙げることができるがこれらに限定されるものではない。例えば、蛍光標識されたプライマーを用いた場合は、増幅産物を当該蛍光物質の検出が可能なDNAシーケンサー等によって電気泳動し、各品種から得られた増幅産物のバンドパターンを比較すればよい。全ての品種について同じ位置にバンドが検出される場合は、いずれの品種においてもゲノム中の同一部位にレトロトランスポゾンが挿入されており、多型がない、すなわち、品種間で共通していると判断できる。同じ位置にバンドが検出される品種とバンドが検出されない品種がある場合にはレトロトランスポゾンの挿入位置に品種間多型があると判断できる。このようにして、品種間で共通したと認められるレトロトランスポゾン挿入部位を選択する。
【0053】
選択したレトロトランスポゾン挿入部位について、レトロトランスポゾンに隣接するゲノム領域の塩基配列は、実際にその領域をシーケンスすることにより決定することができる。塩基配列の決定には公知の遺伝子工学的方法を用いればよい。例えば、上記S-SAP法と同様にアダプターを付加した制限酵素消化断片を鋳型としてRTPプライマーとアダプタープライマーとを用いてPCRを行い、増幅断片を適当なプラスミドベクターにクローニングする。当該ベクターを大腸菌等の適当な宿主に導入して増殖させた後、プラスミドを抽出・精製し、公知のシーケンス反応を行ってシーケンサーに供すればよい。または、上記増幅断片をクローニングせずに、ダイレクトシーケンスすることも可能である。
【0054】
以上により、決定されたレトロトランスポゾンと隣接するゲノム領域の塩基配列に基づいて、各隣接プライマーを設計することができる。
【0055】
次に、上記選択したレトロトランスポゾン挿入部位が、原料植物種と近縁種の植物のゲノムに存在するか否かを検討する。具体的には、近縁種の複数の品種や系統からそれぞれ調製したゲノムDNAを鋳型として、レトロトランスポゾンと隣接するゲノム領域の塩基配列に基づいて設計したその挿入部位のプライマーを用いてPCRなどにより核酸増幅を行う。この核酸増幅の結果、近縁種の全品種もしくは一部の品種において、増幅断片が検出された場合、上記選択したレトロトランスポゾン挿入部位が近縁種のゲノムDNAに存在すると判断できる。一方、近縁種の全品種について、増幅断片が検出されない場合、上記選択したレトロトランスポゾン挿入部位が、近縁種のゲノムDNAに存在せず、原料植物種特有のレトロトランスポゾン挿入部位であると判断できる。
【0056】
本発明者らは、本発明の原料植物種判定方法を用いて、アズキ(Vigna angularis)と、アズキと最も近縁な栽培種であるリョクトウ(Vigna radiata (L.) R. Wilczek)とについて、植物種判定の検討を行っている。そこで以下に、アズキを原料とした加工食品の原料品種判定方法について説明する。
【0057】
対象とするアズキ加工食品は、原料にアズキが含まれるものであって、当該アズキ加工食品からアズキ原料のDNAを抽出可能なものであれば特に限定されるものではない。また、アズキ原料植物体の全部または一部、あるいは原料植物から得られる収穫物(果実、種子等)に対して加熱、乾燥等の加工処理を施したものがアズキ加工食品に該当する。したがって、アズキ加工食品には最終製品のみではなく、中間加工品も含まれる。また、上記アズキ加工食品は、単一の品種の原料植物を含む加工食品に限定されるものではなく、複数の品種の原料植物を含む加工食品であってもよい。
【0058】
利用可能なレトロトランスポゾンは、アズキに存在するレトロトランスポゾンであれば特に限定されるものではない。本発明者は、前述のHirochikaらの方法(Hirochika et al. (1992) Retrotransposon families in rice. Mol Gen Genet 233: 209-216.)に従い、Ty1-Copia型レトロトランスポゾンの逆転写酵素領域に存在するアミノ酸保存配列をターゲットとするPCRにより、同酵素領域の配列をアズキ品種「しゅまり」のゲノムから単離した。このようにして単離した逆転写酵素領域の配列の延長配列をSuppression PCR(Siebert et al. (1995) An improved PCR method for walking in uncloned genomic DNA. Nucleic Acids Res. 23:1087-1088.)法により単離し、末端のLTR配列までを同定し、PHARE-1と命名した。そして、このPHARE-1を用いて、植物種判定の検討を行っている。このPHARE-1は、アズキに存在するレトロトランスポゾンの一例であり、他のレトロトランスポゾンも利用可能であることはいうまでもない。
【0059】
本発明者らは、まず、アズキ品種や系統26種類で共通すると認められるPHARE-1挿入部位を見出すために、S-SAP法を用いて、北海道立中央農業試験場遺伝資源部が保有しているアズキ品種や系統26種類について、PHARE-1の挿入部位を比較した。
【0060】
その結果、26種類のズキ品種や系統間で共通するPHARE-1の挿入部位(PHARE-1に隣接するゲノム領域を含む)が2個見出された。そして、これら2個の挿入部位をクローニングして塩基配列を決定した。これら2個のクローンの塩基配列が、配列番号1及び2に示される塩基配列である。配列番号1及び2について、レトロトランスポゾン(PHARE-1)に隣接するゲノム領域の塩基配列部分を以下に示す。
配列番号1:第1位〜第57位
配列番号2:第1位〜第57位
本発明者らは、上記配列番号1及び2に示される塩基配列のレトロトランスポゾン(PHARE-1)に隣接するゲノム領域の塩基配列に基づいて、各隣接プライマーを設計し、PHARE-1の塩基配列に基づいて、RTPプライマーを設計している。なお、発明者らが実際に使用したRTPプライマーは、配列番号3に示される塩基配列からなるプライマーであり、隣接プライマーは、配列番号4及び5に示される塩基配列からなるプライマーである。
【0061】
このようにして設計したプライマーを用いてPCRを行い、アズキ(Vigna angularis)と、アズキと最も近縁な栽培種であるリョクトウ(Vigna radiata (L.) R. Wilczek)とについて、Phare-1の挿入部位を調べた。その結果、Phareの挿入部位が、アズキ種に固有であり、近縁種であるリョクトウに存在しないことを見出した(図1、図2参照)。
【0062】
このように、配列番号3に示される塩基配列からなるRTPプライマーと配列番号4及び5に示される塩基配列からなる隣接プライマーとを用いて、本発明の原料植物種判定方法を実施すれば、加工食品中にアズキが含まれているか否かを識別することが可能である。
【0063】
本発明の原料植物種識別キットは、本発明の原料植物種判定方法を実施するためのキットであって、レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマーと、レトロトランスポゾンの塩基配列に基づいて設計されるプライマーとを含むものであればよい。キットには、プライマー以外に対象とする加工食品からDNAを抽出・精製するために用いる試薬・器具、核酸増幅反応に用いる試薬・器具、電気泳動に用いる試薬・器具等を含むことが好ましい。
【0064】
また、本発明のアズキ種識別キットは、本発明の原料植物種判定方法をアズキを含む加工食品を対象として実施するためのキットであって、上記配列番号3に示される塩基配列からなるプライマーを含み、かつ、配列番号4または5に示される塩基配列からなるプライマーのうち少なくとも1つ以上を含むものであればよい。また、キットには、プライマー以外に対象とする加工食品からDNAを抽出・精製するために用いる試薬・器具、核酸増幅反応に用いる試薬・器具、電気泳動に用いる試薬・器具等を含むことが好ましい。
【0065】
なお本発明は上述した各実施形態に限定されるものではなく、請求項に示した範囲で種々の変更が可能であり、異なる実施形態にそれぞれ開示された技術的手段を適宜組み合わせて得られる実施形態についても本発明の技術的範囲に含まれる。
【実施例1】
【0066】
アズキのレトロトランスポゾンPHARE-1のアズキゲノムへの挿入部位(P1)をPCRにより調査したところ、供試した全てのアズキ品種でその挿入部位が存在することが判明した。また、同じPCRにより、アズキ(Vigna angularis (Willd.) Ohwi & H. Ohashi)と同じササゲ属(Vigna)に属し、アズキと最も近縁な栽培種であるリョクトウ(Vigna radiata (L.) R. Wilczek)の個体において調査したところ、PHARE-1のこの挿入部位の存在を示す結果は得られなかった。
【0067】
(1)材料
本発明者が、調べた北海道立中央農業試験場遺伝資源部が保有しているアズキ品種や系統26種類を表1に示す。
【0068】
【表1】
【0069】
(2)ゲノムDNAの抽出
アズキ植物体のゲノムDNAは、CTAB(cetyltrimethylammonium bromide)法(早川孝彦(1997)タバコのDNA・RNA単離法.“新版植物のPCR実験プロトコール”島本功、佐々木卓治監修.秀潤社、東京,49-56.)を基本に抽出した。
【0070】
(3)アズキのゲノムにおけるPHARE-1挿入部位の特定
北海道立中央農業試験場遺伝資源部が保有しているアズキ品種や系統26種類について、PHARE-1のLTR配列を利用してS-SAP法による分析を行った。PHARE-1のLTR配列は、アズキ品種「しゅまり」のゲノムから、前述のHirochikaらの方法に従い、Ty1-Copia型レトロトランスポゾンの逆転写酵素領域に存在するアミノ酸保存配列をターゲットとするPCRにより同酵素領域の配列を単離し、このようにして単離した逆転写酵素領域の配列の延長配列をSuppression PCR法により単離することで、PHARE-1の末端部に存在する配列として同定したものである。このS-SAP法による電気泳動の結果から、これら26種類に全てに共通して存在すると見られるPHARE-1レトロトランスポゾン挿入部位を特定した。
【0071】
(4)アズキの品種間に共通する挿入部のPCR増幅による確認
特定した挿入部位の内の3カ所をクローニングしてその塩基配列(配列番号1及び2)を決定し、その配列における、PHARE-1レトロトランスポゾンの塩基配列とそれに隣接するそれぞれのゲノム領域の塩基配列に基づいてプライマーを設計した。設計したPHARE-1レトロトランスポゾンの塩基配列に基づくRTPプライマー(Az-Copia1-LYT-D)、及び配列番号1及び2のゲノム領域の塩基配列に基づく隣接プライマーの塩基配列は、以下の通りである。
Az-Copia1-LYT-Dプライマー(RTPプライマー):5'-GCTAAACACATAAGGGAGATTTCTCC-3'(配列番号3)
P1-DE-T1-5A1プライマー(配列番号1の塩基配列に基づく隣接プライマー):5'-CATCTAACACACCCTAAGAGATAGTC-3'(配列番号4)
P1-DE-T2-4A1プライマー(配列番号2の塩基配列に基づく隣接プライマー):5'-CCAGTAAGGATCCTTGGGTTTTCTTC-3'(配列番号5)
Az-Copia1-LYT-DプライマーとP1-DE-T1-5A1プライマー及びAz-Copia1-LYT-DプライマーとP1-DE-T2-4A1プライマーの2つのプライマー組合せについて、表1に示されたアズキ植物体から抽出したゲノムDNA溶液(20ng/μl)1μlを鋳型としてPCRを行った。PCRは、容量10μlの反応液として、E x Taq(TaKaRa)を用いて、94℃4分間の変性、94℃30秒間・59℃30秒間・72℃20秒間を40サイクル、72℃5分間の最終伸長反応のサイクリング条件で行った。増幅産物をアガロースゲル電気泳動に供し、エチジウムブロマイドで染色して観察した。その結果、これら2つのプライマー組合せのPCRにおいては、プライマーの設計位置から期待された長さのDNA断片(Az-Copia1-LYT-DプライマーとP1-DE-T1-5A1プライマーでは、219bp、Az-Copia1-LYT-DプライマーとP1-DE-T2-4A1プライマーでは、312bp)がアズキの26種類の全てで増幅した。
【0072】
(5)アズキ品種に共通する挿入部がアズキ近縁種において存在しないこと調査
配列番号1と2のようなアズキの品種には共通して存在するPHARE-1レトロトランスポゾンの挿入部位が、アズキと同じ属に属する最も近縁なマメ科作物などには存在しないことを確認するため、以下の植物について、葉または乾燥子実から上記(2)の方法によりゲノムDNAを抽出し、(4)の方法によるPCR増幅により、ゲノム中におけるその挿入部位の存在を判定した。
植物とDNAを抽出した組織:アズキ品種「きたのおとめ」の葉、アズキ系統「天津小豆」の葉、アズキ品種「きたのおとめ」の乾燥子実、リョクトウ(市販品)の乾燥子実
その結果、図1及び図2に示されるように、アズキ品種のゲノムDNAを鋳型としたPCRでは、それぞれの挿入部位の塩基配列に基づき設計したプライマーの位置から期待された長さとなるDNA断片(Az-Copia1-LYT-DプライマーとP1-DE-T1-5A1プライマーでは、219bp、Az-Copia1-LYT-DプライマーとP1-DE-T2-4A1プライマーでは、312bp)が増幅した。一方、リョクトウのゲノムDNAを鋳型としたPCRでは、いずれのPCRにおいても増幅断片が見られなかった。なお、リョクトウから抽出した同じDNAを鋳型として別のプライマーを用いてPCRを行ったところ、プライマーの設計から予想されたDNA断片の増幅がみられた。それゆえ、図1及び図2のPCRにおいてDNA断片が増幅しなかったのは、リョクトウから抽出したDNAの品質がPCR増幅に適していなかったためではなく、Az-Copia1-LYT-DプライマーとP1-DE-T1-5A1プライマーのセット、及びAz-Copia1-LYT-DプライマーとP1-DE-T2-4A1プライマーのセットにより増幅するPHARE-1の挿入部位がリョクトウには存在しないためである。この結果、これら2つのPHARE-1の挿入部位(配列番号1及び2)は、アズキに固有のものであることが分かった。
【0073】
本発明は上述した各実施形態に限定されるものではなく、請求項に示した範囲で種々の変更が可能であり、異なる実施形態にそれぞれ開示された技術的手段を適宜組み合わせて得られる実施形態についても本発明の技術的範囲に含まれる。
【産業上の利用可能性】
【0074】
本発明は、加工食品の原料植物種の判定を高精度かつ定量的にできるものであるため、食品産業に広く利用可能である。さらに、食品の不正表示や、海外で不正に栽培された品種を原料とした輸入加工食品等の検査に利用すれば、違法食品の取り締まりに大きく貢献できるものと期待される。
【特許請求の範囲】
【請求項1】
加工食品に含まれる原料植物の植物種を判定する原料植物種判定方法であって、
植物ゲノムに原料植物の植物種固有のレトロトランスポゾン挿入部位が存在するか否かを検出し、この検出結果に基づいて、原料植物の植物種を判定することを特徴とする原料植物種判定方法。
【請求項2】
植物ゲノムに原料植物の植物種固有のレトロトランスポゾン挿入部位が存在するか否かの検出には、
判定対象とする植物種の品種のゲノムに共通して存在しているレトロトランスポゾン挿入部位における、当該レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマーと、レトロトランスポゾンの塩基配列に基づいて設計されるプライマーとを組み合わせたプライマーセットを用いて、加工食品から調製したDNAを鋳型として核酸増幅反応を行い、増幅産物の有無を確認する方法を用いることを特徴とする請求項1に記載の原料植物種判定方法。
【請求項3】
判定対象とする植物種の品種について、ゲノム中のレトロトランスポゾン挿入部位を比較して、当該植物種の品種のゲノムに共通して存在しているレトロトランスポゾン挿入部位を選択し、選択されたレトロトランスポゾン挿入部位について、原料植物の近縁種のゲノムに存在するか否かの確認を行い、
上記確認の結果、原料植物の近縁種のゲノムに存在しないレトロトランスポゾン挿入部位を、上記原料植物の植物種固有のレトロトランスポゾン挿入部位として決定することを特徴とする請求項1または2に記載の原料植物種判定方法。
【請求項4】
上記増幅産物の断片長は40bp〜600bpであることを特徴とする請求項2または3に記載の原料植物種判定方法。
【請求項5】
上記レトロトランスポゾンに隣接するゲノム領域の塩基配列は、当該領域を含むDNA断片をシーケンスすることにより得られる塩基配列であることを特徴とする請求項2〜4の何れか1項に記載の原料植物種判定方法。
【請求項6】
上記レトロトランスポゾンに隣接するゲノム領域の塩基配列は、当該領域を含むDNA断片をクローニングし、当該断片をシーケンスすることにより得られる塩基配列であることを特徴とする請求項2〜4の何れか1項に記載の原料植物種判定方法。
【請求項7】
上記加工食品はアズキを原料に含むことを特徴とする請求項1〜6のいずれか1項に記載の原料植物種判定方法。
【請求項8】
上記レトロトランスポゾンはPHARE-1であることを特徴とする請求項7に記載の原料植物種判定方法。
【請求項9】
上記レトロトランスポゾンに隣接するゲノム領域を含むDNA断片をシーケンスすることにより得られる塩基配列は、配列番号1または2に示される塩基配列であることを特徴とする請求項8に記載の原料植物種判定方法。
【請求項10】
上記レトロトランスポゾンの塩基配列に基づいて設計されるプライマーは、配列番号3に示される塩基配列からなるプライマーであり、
上記レトロトランスポゾン挿入部位のレトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマーは、配列番号4または5に示される塩基配列からなるプライマーから選択されることを特徴とする請求項9に記載の原料植物種判定方法。
【請求項11】
請求項10に記載の原料植物種判定方法に用いるプライマーであって、配列番号3〜5に示される塩基配列からなることを特徴とするアズキ種判定用プライマー。
【請求項12】
請求項1〜6のいずれか1項に記載の原料植物種判定方法を実施するためのキットであって、レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマーと、レトロトランスポゾンの塩基配列に基づいて設計されるプライマーとを含むことを特徴とする原料植物種判定キット。
【請求項13】
請求項10に記載の原料植物種判定方法を実施するためのキットであって、配列番号3に示される塩基配列からなるプライマーを含み、かつ、配列番号4または5に示される塩基配列からなるプライマーのうち少なくとも1つ以上を含むことを特徴とするアズキ種判定キット。
【請求項1】
加工食品に含まれる原料植物の植物種を判定する原料植物種判定方法であって、
植物ゲノムに原料植物の植物種固有のレトロトランスポゾン挿入部位が存在するか否かを検出し、この検出結果に基づいて、原料植物の植物種を判定することを特徴とする原料植物種判定方法。
【請求項2】
植物ゲノムに原料植物の植物種固有のレトロトランスポゾン挿入部位が存在するか否かの検出には、
判定対象とする植物種の品種のゲノムに共通して存在しているレトロトランスポゾン挿入部位における、当該レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマーと、レトロトランスポゾンの塩基配列に基づいて設計されるプライマーとを組み合わせたプライマーセットを用いて、加工食品から調製したDNAを鋳型として核酸増幅反応を行い、増幅産物の有無を確認する方法を用いることを特徴とする請求項1に記載の原料植物種判定方法。
【請求項3】
判定対象とする植物種の品種について、ゲノム中のレトロトランスポゾン挿入部位を比較して、当該植物種の品種のゲノムに共通して存在しているレトロトランスポゾン挿入部位を選択し、選択されたレトロトランスポゾン挿入部位について、原料植物の近縁種のゲノムに存在するか否かの確認を行い、
上記確認の結果、原料植物の近縁種のゲノムに存在しないレトロトランスポゾン挿入部位を、上記原料植物の植物種固有のレトロトランスポゾン挿入部位として決定することを特徴とする請求項1または2に記載の原料植物種判定方法。
【請求項4】
上記増幅産物の断片長は40bp〜600bpであることを特徴とする請求項2または3に記載の原料植物種判定方法。
【請求項5】
上記レトロトランスポゾンに隣接するゲノム領域の塩基配列は、当該領域を含むDNA断片をシーケンスすることにより得られる塩基配列であることを特徴とする請求項2〜4の何れか1項に記載の原料植物種判定方法。
【請求項6】
上記レトロトランスポゾンに隣接するゲノム領域の塩基配列は、当該領域を含むDNA断片をクローニングし、当該断片をシーケンスすることにより得られる塩基配列であることを特徴とする請求項2〜4の何れか1項に記載の原料植物種判定方法。
【請求項7】
上記加工食品はアズキを原料に含むことを特徴とする請求項1〜6のいずれか1項に記載の原料植物種判定方法。
【請求項8】
上記レトロトランスポゾンはPHARE-1であることを特徴とする請求項7に記載の原料植物種判定方法。
【請求項9】
上記レトロトランスポゾンに隣接するゲノム領域を含むDNA断片をシーケンスすることにより得られる塩基配列は、配列番号1または2に示される塩基配列であることを特徴とする請求項8に記載の原料植物種判定方法。
【請求項10】
上記レトロトランスポゾンの塩基配列に基づいて設計されるプライマーは、配列番号3に示される塩基配列からなるプライマーであり、
上記レトロトランスポゾン挿入部位のレトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマーは、配列番号4または5に示される塩基配列からなるプライマーから選択されることを特徴とする請求項9に記載の原料植物種判定方法。
【請求項11】
請求項10に記載の原料植物種判定方法に用いるプライマーであって、配列番号3〜5に示される塩基配列からなることを特徴とするアズキ種判定用プライマー。
【請求項12】
請求項1〜6のいずれか1項に記載の原料植物種判定方法を実施するためのキットであって、レトロトランスポゾンに隣接するゲノム領域の塩基配列に基づいて設計されるプライマーと、レトロトランスポゾンの塩基配列に基づいて設計されるプライマーとを含むことを特徴とする原料植物種判定キット。
【請求項13】
請求項10に記載の原料植物種判定方法を実施するためのキットであって、配列番号3に示される塩基配列からなるプライマーを含み、かつ、配列番号4または5に示される塩基配列からなるプライマーのうち少なくとも1つ以上を含むことを特徴とするアズキ種判定キット。
【図1】

【図2】

【図2】
【公開番号】特開2010−172322(P2010−172322A)
【公開日】平成22年8月12日(2010.8.12)
【国際特許分類】
【出願番号】特願2009−21695(P2009−21695)
【出願日】平成21年2月2日(2009.2.2)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成20年度、独立行政法人農業・食品産業技術総合研究機構 プロジェクト研究「食品・農産物の表示の信頼性確保と機能性解析のための基盤技術の開発」に係る委託研究、産業技術力強化法第19条の適用を受けるもの
【出願人】(504147243)国立大学法人 岡山大学 (444)
【Fターム(参考)】
【公開日】平成22年8月12日(2010.8.12)
【国際特許分類】
【出願日】平成21年2月2日(2009.2.2)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成20年度、独立行政法人農業・食品産業技術総合研究機構 プロジェクト研究「食品・農産物の表示の信頼性確保と機能性解析のための基盤技術の開発」に係る委託研究、産業技術力強化法第19条の適用を受けるもの
【出願人】(504147243)国立大学法人 岡山大学 (444)
【Fターム(参考)】
[ Back to top ]