
原核細胞由来の組み換えN−グリコシル化タンパク質
【課題】原核生物中で産生できるN−グリコシル化効率最適化タンパク質、ならびに該タンパク質の産生方法、および該タンパク質の抗原性・安定性・生物活性を改変するためのN−グリカンの組み換えタンパク質へのより効果的な導入方法、さらにその表面上に該組換えN−グリコシル化タンパク質を効率的にディスプレイする宿主細胞の提供。
【解決手段】1つまたは複数のN−グリコシル化効率最適化アミノ酸配列を含む組み換えN−グリコシル化タンパク質、これらのタンパク質をコードする核酸、ならびに相当するベクターおよび宿主細胞。また、医薬品調製のための、前記タンパク質、核酸、ベクター、および宿主細胞の使用。さらに、該タンパク質を産生するための方法。
【解決手段】1つまたは複数のN−グリコシル化効率最適化アミノ酸配列を含む組み換えN−グリコシル化タンパク質、これらのタンパク質をコードする核酸、ならびに相当するベクターおよび宿主細胞。また、医薬品調製のための、前記タンパク質、核酸、ベクター、および宿主細胞の使用。さらに、該タンパク質を産生するための方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、1つまたは複数の導入されたNグリコシル化された最適化アミノ酸コンセンサス配列を含む組み換えN−グリコシル化タンパク質、これらのタンパク質をコードする核酸、ならびに相当するベクターおよび宿主細胞に関する。さらに、本発明は、医薬品を調製するための、前記タンパク質、核酸、ベクター、および宿主細胞の使用に関する。さらに、本発明は、前記タンパク質を産生するための方法を提供する。
【背景技術】
【0002】
発明の背景
N−結合タンパク質のグリコシル化は、真核生物の小胞体で起こる本質的かつ保存されたプロセスである。それは、タンパク質のフォールディング、オリゴマー化、安定性、品質管理、選別、ならびに分泌および膜タンパク質の輸送にとって重要である(ヘレニウス,A(Helenius,A.)およびアービィ,M.(Aebi,M.)(2004年)、「Roles of N−linked glycans in the endoplasmic reticulum.」「Annu.Rev.Biochem.」73、1019〜1049)。
【0003】
タンパク質のグリコシル化は、タンパク質の抗原性、安定性、および半減期に対して重大な影響を有する。さらに、グリコシル化は、クロマトグラフィ、例えば、タンパク質のグリコシル化された部分と相互作用する、固相に結合されたレクチンリガンドを用いるアフィニティクロマトグラフィによるタンパク質の精製を助けることができる。したがって、生物学的および薬学的に有用なグリコシル化パターンを提供するために、真核生物細胞において、組換えにより多くのグリコシル化タンパク質を産生することは、確立された手法である。
【0004】
ごく近年、細菌、すなわち食物性の病原菌カンピロバクター ジェジュニ(Campylobacter jejuni)も、そのタンパク質をN−グリコシル化することができることが実証されている(シマンスキー(Szymanski)ら(1999年)、「Evidence for a system of general protein glycosylation in Campylobacter jejuni.」「Mol.Microbiol.」32、1022〜1030)。グリコシル化のために必要とされる機構は、いわゆるpgl遺伝子座にクラスター化される12の遺伝子によってコードされる。N−グリコシル化の破壊は、C.ジェジュニ(C.jejuni)の侵入および病原性に影響を及ぼすが、大抵の真核性の生物において見られるように致死的ではない(ブルダP.(Burda P.)およびM.アービィ(M.Aebi.)(1999年)、「The dolichol pathway of N−linked glycosylation.」「Biochim Biophys Acta」1426(2):239〜57)。大腸菌(E.coli)中でpgl遺伝子座とアクセプター糖タンパク質を組換えにより同時に発現させることによって、C.ジェジュニ(C.jejuni)タンパク質のN−グリコシル化を元に戻すことが可能である(ワッカー(Wacker)ら(2002年)「N−linked glycosylation in Campylobacter jejuni and its functional transfer into E coli.」「Science」298、1790〜1793)。
【0005】
欧州特許出願公開第03 702 276 1号明細書(欧州特許第1 481 057号明細書)、すなわち本発明の発明者の以前の発明は、(i)脂質担体上へのオリゴ糖の構築のための特異的なグリコシルトランスフェラーゼ、(ii)コンセンサス配列「N−X−S/T」(式中、Xは、プロリン以外の任意のアミノ酸であり得る)を含む組み換え標的タンパク質、ならびに(iii)前記オリゴ糖を、標的タンパク質のコンセンサス配列に共有結合的に連結させるC.ジェジュニ(C.jejuni)のオリゴサッカリルトランスフェラーゼ(OTアーゼ)、をコードする核酸が導入される原核生物を教示する。前記原核生物は、特定のグリコシルトランスフェラーゼのタイプによって定義される、特定の構造をもつN−グリカンを産生する。
【0006】
たとえ、タンパク質中の既知のN−グリコシル化コンセンサス配列の存在によって、C.ジェジュニ(C.jejuni)のオリゴサッカリルトランスフェラーゼ(OTアーゼ)を含む原核生物における組み換え標的タンパク質のN−グリコシル化が確かに可能になったとしても、いくつかの標的タンパク質のN−グリコシル化は、しばしば非効率的である。
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明の目的は、原核生物中でin vivoで産生することができる、N−グリコシル化の効率が最適化タンパク質、ならびにこうしたタンパク質を産生するための手段および方法を提供することである。本発明の別の目的は、前記タンパク質の抗原性、安定性、生物学的、予防的、および/または治療的活性を改変するための、N−グリカンの組み換えタンパク質へのより効果的な導入を意図する。さらなる目的は、その表面上に本発明の組み換えN−グリコシル化タンパク質を効率的にディスプレイする宿主細胞の提供である。
【課題を解決するための手段】
【0008】
第1の態様では、本発明は、1つまたは複数の下記N−グリコシル化された最適化アミノ酸配列を含む、組み換えN−グリコシル化タンパク質を提供する:
D/E−X−N−Z−S/T(最適化コンセンサス配列)
式中、XおよびZは、Pro以外の任意の天然のアミノ酸であり得、かつ少なくとも1つの前記N−グリコシル化された部分的なアミノ酸コンセンサス配列が導入される。
【発明の効果】
【0009】
驚くべきことに、特定の部分的なアミノ酸配列(最適化コンセンサス配列)をタンパク質に導入することで、カンピロバクター(Campylobacter)種、好ましくはC.ジェジュニ(C.jejuni)由来のオリゴサッカリルトランスフェラーゼ(OST、OTアーゼ)によって、これらが導入された位置で、非常に効率的にN−グリコシル化されたタンパク質がもたらされることが判明した。
【図面の簡単な説明】
【0010】
【図1】構築物AからCより得られるLipタンパク質のN−グリコシル化を説明する図である(実施例1を参照のこと)。C.ジェジュニ(C.jejuni)由来の機能性のpglオペロン(ワッカー(Wacker)ら、2002年、上記)、および、構築物A(レーン2)、B(レーン1)、およびC(レーン3)をコードするプラスミド、または突然変異D121Aを伴う構築物Cのミュータント(レーン4)を保有する大腸菌(E.coli)Top 10細胞。タンパク質を発現させ、周辺質抽出物から精製した。SDS−PAGE、および精製されたタンパク質の分画を染色しているクーマシーブリリアントブルーが示される。
【図2】プラスミドから前記タンパク質を発現する、CLM24細胞(フェルドマン(Feldman)ら、(2005年)、「Engineering N−linked protein glycosylation with diverse O antigen lipopolysaccharide structures in Escherichia coli.」「Proc.Natl.Acad.Sci.U S A」102、3016〜3021)、またはTop 10細胞(パネルE、レーン1〜6)、またはSCM7細胞(アライモ,C.(Alaimo、C.)、キャトレイン,I.(Catrein、I.)、モルフ,L.(Morf,L.)、マロルダ,C.L.(Marolda,C.L.)、カレワエルト,N.(Callewaert,N.)、ヴァルヴァノ,M.A.(Valvano、M.A.)、フェルドマン,M.F.(Feldman,M.F.)、アービィ,M.(Aebi,M.)(2006年)「Two distinct but interchangeable mechanisms for flipping of lipid−linked oligosaccharides.」「EMBO journal」25、967〜976)(パネルE、レーン7,8)中で、C.ジェジュニ(C.jejuni)pglオペロン(ワッカー(Wacker)ら、2002年、上記)による配列特異的なN−グリコシル化について分析される、様々なタンパク質のN−グリコシル化分析を示す図である。ニトロセルロース膜に移され、特異的な抗血清を用いて視覚化された、SDS−PAGE分離された周辺質抽出物が示される。パネルA〜Dでは、上のパネルが、抗AcrA抗血清(ワッカー(Wacker)ら、2002年、上記;ニタ−ラザー,M(Nita−Lazar,M)ワッカー,M.(Wacker,M.)、シェッグ,B.(Schegg、B.)、アンバー,S.(Amber,S.)、およびアービィ,M(Aebi,M.)(2005年)、「The N−X−S/T consensus sequence is required but not sufficient for bacterial N−linked protein glycosylation.」「Glycobiology」15、361〜367)を用いて探索される免疫ブロットを示すのに対し、下のパネルは、R12抗血清(ワッカー(Wacker)ら、2002年、上記)を用いて探索される免疫ブロットを示す。+および−は、細胞中の機能性または変異型のpglオペロンの存在を示す。パネルAは、pelBシグナル配列と、ヘキサヒスチジンタグを伴う可溶性の野生型AcrA(レーン1、2)、AcrA−N273Q(レーン3、4)、およびAcrA−D121A(レーン5)のサンプルを含有する。パネルB:AcrA(レーン1、2)、AcrA−T145D(レーン3)、AcrA−N123Q−N273Q−T145D(レーン4、5)。パネルC:AcrA−F115D−T145D(レーン1、2)、AcrA−N123Q−N273Q−N272D(レーン3、4)。パネルD:AcrA−N273Q(レーン1、2)、AcrA−N273Q−F122P(レーン3、4)。パネルE:CtxB(レーン1、2)、CtxB−W88D(レーン3、4)、CtxB−Q56/DSNIT(レーン5、6)、およびCtxB−W88D−Q56/DSNIT。
【図3】OmpH1中の複数のグリコシル化部位の操作を示す図である。ΔwaaL株SCM6を、プラスミドpACYCpgl(全pgl遺伝子座をコードする)と、野生型OmpH1(レーン1)、OmpH1N139s−myc(レーン2)、OmpH1KGN→NIT、HFGDD→DSNIT−myc(レーン3)、OmpH1RGD→NIT、HFGDD→DSNIT−myc(レーン4)、OmpH1KGN→NIT、RGD→NIT−myc(レーン5)、OmpH1KGN→NIT、RGD→NIT、HFGDD→DSNIT−myc(レーン6)、またはOmpH1RGD→NIT V83T−myc(レーン7)を発現するプラスミドを用いて同時形質転換させた。細胞を好気的に増殖させ、分析の前に3時間、0.5%アラビノースで誘導させた。全細胞溶解物を、材料および方法のセクションに述べた通りに、培養物の光学濃度を均一にした後にTCA沈殿させた。15% SDS−PAGEによってタンパク質を分離し、PVDF膜上に移した。最初のパネル;抗mycタグ抗体を用いて探索された全細胞溶解物の免疫ブロット。下のパネル;グリカン特異的な抗血清を用いて探索された全細胞溶解物の免疫ブロット。非グリコシル化およびグリコシル化されたOmpH1の位置は、右側に示される。
【図4】様々なOmpH1変異体を発現する細胞の蛍光顕微鏡検査を示す図である。野生型OmpH1およびその変異体のための発現プラスミドを含有する大腸菌(E.coli)株CLM24またはSCM6の培養物を、等しくOD600の0.25/mlにした。細胞を、リン酸緩衝食塩水(PBS)(pH 7.4)で2回洗浄し、100μl細胞懸濁液を、ゼラチンを塗ったスライドガラス上に滴下し、加湿チャンバー内で30分間、室温(RT)でインキュベートした。全細胞免疫蛍光標識化におけるその後のステップはすべて、加湿チャンバー内で室温で行った。結合していない細胞を除去し、残留物を、室温で30分間、4%パラホルムアルデヒドを含有するPBSで固定した。重要なことに、パラホルムアルデヒドは、細胞を透過させないが、膜が無損傷であることによって区画化が保たれると考えられる。固定された細胞を、PBSで2回洗浄し、5% BSA(PBS中)を含有するブロッキング緩衝液に再懸濁した。ブロック後、細胞を、抗mycモノクローナルマウスIgG(1:50、カルバイオケム(Calbiochem))および/または抗グリカン抗血清(1:4000)と共に、5% BSAを含有する100μlのPBS中で、1時間インキュベートした。細胞を、100μlのPBSで3回、各5分間洗浄し、FITCと結合させた抗ウサギ二次抗体(1:250、ジャクソンイムノリサーチラボラトリーズ(Jackson Immunoresearch Laboratories))、および/またはCy3と結合させた抗マウス抗体(1:250、ジャクソンイムノリサーチラボラトリーズ(Jackson Immunoresearch Laboratories))と共に、5% BSAを含有する100μlのPBS中で1時間インキュベートした。必要とされる場合、細菌のDNAを染色するために、4,6−ジアミノ−2−フェニルインドール(DAPI)(シグマ(Sigma))(0.5μg/ml)を、二次抗体インキュベーション時に加えた。二次抗体を、PBSで細胞から洗い落とし、ベクタシールド(vectashield)(ベクターラボラトリーズ(Vector Laboratories))マウンティングメディアを使用することによって、カバースリップをスライドに載せ,マニキュア液を用いて密閉した。Axioplan2顕微鏡(カールツァイス(Carl Zeiss))を使用することによって、蛍光顕微鏡検査を実施した。Adobe Photoshop(バージョンCS2)を使用することによって、画像を組み合わせた。SCM6細胞は、以下を発現する:OmpH1(パネルA)、OmpH1N139S(パネルB)、OmpH1C20S(パネルC)、OmpH1KGN→NIT、HFGDD→DSNIT(パネルD)、OmpH1RGD→NIT、HFGDD→DSNIT(パネルE)、OmpH1KGN→NIT、RGD→NIT(パネルF)、OmpH1V83T、KGN→NIT(パネルG)、およびOmpH1KGN→NIT、RGD→NIT、HFGDD→DSNIT(パネルH)。第1列は、黒色バックグラウンド上に灰色のトーンで表された、第2、3、および4列の画像の組合せである。第2列:DAPI染色による灰色のトーン中の青色蛍光、第3列:グリカン特異的な蛍光による緑色蛍光、第4列:抗myc染色による赤色蛍光。
【発明を実施するための形態】
【0011】
本発明において使用される用語「部分的なアミノ酸配列」はまた、「最適化コンセンサス配列」とも称される。最適化コンセンサス配列は、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来のオリゴサッカリルトランスフェラーゼ(OST、OTアーゼ)によって、従来技術で知られている通常のコンセンサス配列「N−X−S/T」よりもかなり効率的にN−グリコシル化される。
【0012】
一般に、用語「組み換えN−グリコシル化タンパク質」は、前記タンパク質をコードする核酸を天然には含まない宿主細胞中で産生される任意の異種ポリペプチドまたはオリゴペプチドを指す。本発明において、この用語は、任意の宿主細胞、例えば真核または原核生物の宿主細胞、好ましくは原核の宿主細胞、例えばエシェリキア亜種(Escherichia ssp.)、カンピロバクター亜種(Campylobacter ssp.)、サルモネラ亜種(Salmonella ssp.)、シゲラ亜種(Shigella ssp.)、ヘリコバクター亜種(Helicobacter ssp.)、シュードモナス亜種(Pseudomonas ssp.)、バチルス亜種(Bacillus ssp.)、より好ましくは、大腸菌(Escherichia coli)、カンピロバクター ジェジュニ(Campylobacter jejuni)、サルモネラ チフィムリウム(Salmonella typhimurium)などの中で、組換えによって産生されるタンパク質を指す。ここでは、前記タンパク質をコードする核酸が、前記宿主細胞に導入される。また、コードされたタンパク質は、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来のOTアーゼ、天然に存在する、あるいは組換えにより前記宿主細胞に導入される前記トランスフェラーゼ酵素によってN−グリコシル化される。
【0013】
アミノ酸について国際的に認められた1文字記号によれば、省略形D、E、N、S、およびTはそれぞれ、アスパラギン酸、グルタミン酸、アスパラギン、セリン、およびトレオニンを表す。本発明のタンパク質は、1つまたは複数の最適化コンセンサス配列D/E−X−N−Z−S/Tが導入され、かつN−グリコシル化されるという点で、天然または従来技術のタンパク質とは異なる。したがって、本発明のタンパク質は、天然に存在するC.ジェジュニ(C.jejuni)タンパク質(これも、最適化コンセンサス配列を含有するが、任意のさらなる(導入された)最適化コンセンサス配列は含まない)とは異なる。
【0014】
最適化コンセンサス配列の導入は、1つまたは複数のアミノ酸の付加、欠損、および/または置換によって達成することができる。最適化コンセンサス配列を導入するための、1つまたは複数のアミノ酸の付加、欠損、および/または置換は、固相支持型の化学ペプチド合成などの、当業者によく知られている化学合成戦略によって達成することができる。あるいは、より大きなポリペプチドにとって好ましいことだが、本発明のタンパク質は、標準の組み換え技術によって調製することができる。
【0015】
本発明のタンパク質は、高効率で、かつ、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロンを含む任意の原核の宿主中で産生され得るという利点を有する。本発明の態様および実施形態を実践するためのカンピロバクター種由来の好ましい代替のOTアーゼは、カンピロバクター コリ(Campylobacter coli)およびカンピロバクター ラリ(Campylobacter lari)である(シマンスキーC.M.(Szymanski C.M.)およびレン B.W.(Wren B.W.)(2005年)、「Protein glycosylation in bacterial mucosal pathogens.」「Nat.Rev.Microbiol.」3:225〜237を参照のこと)。前記原核宿主がカンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)である場合、機能性のpglオペロンは、天然に存在するものであり得る。しかし、当分野で以前に実証された通り、また、上述した通り、pglオペロンは、細胞に導入することができ、前記新規の細胞の環境中で機能性のままでいることが可能である。
【0016】
用語「カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロン」は、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)の機能性のオリゴサッカリルトランスフェラーゼ(OTアーゼ)、および脂質担体上にオリゴ糖を構築することが可能な1種または複数の特異的なグリコシルトランスフェラーゼをコードする核酸のクラスターを指すものとする。ここでは、前記オリゴ糖は、OTアーゼによって、脂質担体から、1つまたは複数の最適化アミノ酸配列:D/E−XN−Z−S/Tを有する標的タンパク質に転移することができる。用語「カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロン」は、本発明において、必ずしも単一の転写単位としてのオペロンを指すというわけではないことを理解するべきである。この用語は、単に、ある宿主細胞における、組み換えタンパク質のN−グリコシル化のための機能性の成分の存在を要するだけである。これらの成分は、1つまたは複数の別々のmRNAとして転写されてもよく、一緒にあるいは別々に調節されてもよい。例えば、この用語はまた、ある宿主細胞中のゲノムDNAおよびプラスミド中に位置する機能性の成分も包含する。効率のためには、機能性のpglオペロンのすべての成分が、調節されおよび同時に発現されることが好ましい。
【0017】
機能性のオリゴサッカリルトランスフェラーゼ(OTアーゼ)だけは、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)に由来するべきであること、また、脂質担体上にオリゴ糖を構築することが可能な1つまたは複数の特異的なグリコシルトランスフェラーゼは、宿主細胞に由来するものであっても組換えにより前記宿主細胞に導入されるものであってもよいが、唯一の機能的制約は、前記グリコシルトランスフェラーゼによって構築されるオリゴ糖が、OTアーゼによって、脂質担体から、1つまたは複数の最適化コンセンサス配列を有する標的タンパク質に転移可能であるということであることを認識することは重要である。したがって、特異的なグリコシルトランスフェラーゼを天然に含む宿主細胞の選択および/または前記宿主中に天然に存在する特異的なグリコシルトランスフェラーゼを無能力化することおよび異種特異的なグリコシルトランスフェラーゼの導入によって、当業者は、本発明のタンパク質中の最適化N−グリコシル化コンセンサス部位に結合されたN−グリカンを変えることが可能となる。
【0018】
上記の結果として、本発明は、本発明のタンパク質上のN−グリカン−パターンの個々の設計を提供する。したがって、タンパク質は、生物学、薬学、および精製の要件に合わせるために、そのN−グリカンパターンにおいて個別化させることができる。
【0019】
好ましい実施形態では、本発明のタンパク質は、1つ、さらに2つ以上、好ましくは少なくとも2つ、好ましくは少なくとも3つ、より好ましくは少なくとも5つの前記N−グリコシル化された最適化アミノ酸配列を含んでよい。
【0020】
本発明のタンパク質中の1つまたは複数のN−グリコシル化された最適化アミノ酸配列の存在は、その抗原性を増大させること、その安定性を増大させること、その生物活性に影響を及ぼすこと、その生物学的半減期を延長させること、および/またはその精製を単純化させることのために好都合であり得る。
【0021】
最適化コンセンサス配列は、位置XおよびZに、プロリン以外の任意のアミノ酸を含んでよい。用語「任意のアミノ酸」は、最適化コンセンサス配列を、OTアーゼによってN−グリコシル化させることが可能となる、一般的および稀な天然のアミノ酸、ならびに合成アミノ酸誘導体および類似体を包含するものとする。天然に存在する一般的および稀なアミノ酸が、XおよびZに対して好ましい。XとZは、同じであっても異なってもよい。
【0022】
XおよびZが、本発明のタンパク質中の最適化コンセンサス配列各々について異なっていてもよいことに注意されたい。
【0023】
OTアーゼによる転移のために、脂質担体上にオリゴ糖を構築する場合、最適化コンセンサス配列に結合されるN−グリカンは、特異的なグリコシルトランスフェラーゼおよびその相互作用によって決定されることとなる。当業者は、所望の宿主細胞中に存在する特異的なグリコシルトランスフェラーゼのタイプおよび量を変化させることによって、N−グリカンを設計することができる。
【0024】
N−グリカンは、本明細書では、Nグリコシド結合を介してタンパク質中のアスパラギン残基のε−アミド窒素と連結された、様々な組成の単糖、オリゴ糖、または多糖と定義される。好ましくは、OTアーゼによって転移されるN−グリカンは、グラム陰性または陽性細菌の細胞膜に存在するウンデカプレノール−ピロリン酸脂質−アンカー上に構築される。これは、O抗原、O多糖、およびペプチドグリカンの合成に関与する(ビュック,T.D.(Bugg,T.D.)およびブランディッシュ,P.E.(Brandish,P.E.)(1994年)、「From peptidoglycan to glycoproteins:common features of lipid−linked oligosaccharide biosynthesis.」「FEM Microbiol Lett」119、255〜262;ヴァルヴァノ,M.A.(Valvano,M.A.)(2003年)、「Export of O−specific lipopolysaccharide.」「Front Biosci」8、s452〜471)。
【0025】
好ましい実施形態では、本発明の組み換えタンパク質は、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来のN−グリカンの群から選択される1種または複数のN−グリカンを含み、N−グリカンは、グラム陰性細菌中のO多糖形成O抗原に転移されるオリゴ糖および多糖、あるいはグラム陽性細菌由来の莢膜多糖、好ましくは:緑膿菌(P.aeruginosa)O9、O11;大腸菌(E.coli)O7、O9、O16、O157、および志賀赤痢菌(Shigella dysenteriae)O1ならびに多糖構造に影響を及ぼすグリコシルトランスフェラーゼおよびエピメラーゼを挿入するあるいは欠損させることによって得られるそれらの操作された変異体から得られる。
【0026】
さらなる好ましい実施形態では、本発明の組み換えタンパク質は、2つ以上の異なるN−グリカンを含む。
【0027】
例えば、同じタンパク質上の異なるN−グリカンは、早いまたは遅いプロモーターを使用して、あるいは個々の特異的なグリコシルトランスフェラーゼのプロモーター活性を開始する、静まらせる、強化する、および/または低下させるための因子を導入して、特異的なグリコシルトランスフェラーゼの発現のタイミングを制御することによって調製することができる。それらの活性を制御する適切なプロモーターおよび因子は、当分野でごく普通に利用可能なものであるので、これ以上は論じない。
【0028】
本発明の組み換えタンパク質の起源に対する制約はない。好ましくは、前記タンパク質は、哺乳類、細菌、ウイルス、菌類、または植物のタンパク質から得られる。より好ましくは、タンパク質は、哺乳類、最も好ましくはヒトのタンパク質から得られる。本発明の抗原性の組み換えタンパク質を調製するために、好ましくはワクチン中で活性成分を使用するためには、組み換えタンパク質は、細菌、ウイルス、または菌類のタンパク質から得られることが好ましい。
【0029】
さらなる好ましい実施形態では、本発明は、組み換えタンパク質を提供する。ここでは、タンパク質および/またはN−グリカンは、治療および/または予防的に活性である。少なくとも1つの最適化N−グリコシル化されたコンセンサス配列を導入することによって、タンパク質における治療および/または予防的活性を改変する、さらには導入することができる。より好ましい実施形態では、これは、免疫原的に活性であるタンパク質および/またはN−グリカンである。この場合、導入されるN−グリコシル化は、タンパク質の生物活性に対する改変効果を有していてもよく、および/または新規の抗原性部位を導入し、および/または分解ステップを避けるためにタンパク質をマスキングしてもよく、および/または半減期を増大する。
【0030】
本発明の組み換えタンパク質を、宿主細胞、好ましくは細菌、より好ましくはグラム陰性細菌の外膜および/または表面に効率的にターゲティングすることができる。表面ディスプレイおよび/または外膜局在化を助けるために、本発明の組み換えタンパク質は、細菌、好ましくはグラム陰性細菌の外膜および/または細胞表面に前記組み換えタンパク質をターゲティングすることが可能な少なくとも1つのポリペプチド配列をさらに含むことが好ましい。
【0031】
好ましい実施形態では、本発明の組み換えタンパク質は、前記ターゲティングするポリペプチド配列が、タイプIIシグナルペプチド(パエトゼル,M.(Paetzel,M.)、カリア,A.(Karia、A.)、ストリナドカ,N.C.(Strynadka,N.C.)、およびダルビー,R.E.(Dalbey,R.E.)、2002年、「Signal peptidases.」「Chem Rev」102:4549〜4580)、または外膜タンパク質(ウェメラス,H.(Wemerus,H.)およびシュタール,S.(Stahl,S.)2004年、「Biotechnological applications for surface−engineered bacteria.」「Biotechnol Appl Biochem」40:209〜228に概説)からなる群から選択される、好ましくはC.ジェジュニ(C.jejuni)由来のOmpH1、C.ジェジュニ(C.jejuni)由来のJlpA、大腸菌(E.coli)由来の外膜タンパク質、好ましくはOmpS、OmpC、OmpA、OprF、PhoE、LamB、Lpp’OmpA(表面ディスプレイ技術のための融合タンパク質、フランシスコ,J.A.(Francisco,J.A.)、エアハート,C.F.(Earhart,C.F.)、およびジョルジオ,G.(Georgiou,G.、1992年、「Transport and anchoring of beta−lactamase to the external surface of Escherichia coli.」「Proc Natl Acad Sci U S A」89:2713〜2717を参照のこと)、および緑膿菌(Pseudomonas aeruginosa)由来のlnpタンパク質の全長タンパク質またはシグナルペプチドからなる群から選択されるものである。
【0032】
異なる態様では、本発明は、本発明の組み換えタンパク質をコードする核酸に関する。好ましくは、前記核酸は、mRNA、DNA、またはPNA、より好ましくは、mRNAまたはDNA、最も好ましくはDNAである。核酸は、前記タンパク質をコードする配列、また、それに加えて制御配列(例えば、プロモーター、エンハンサー、終止コドン、開始コドン)などの他の配列、および、挙げられた制御配列などを介して組み換えタンパク質の発現を調節するために必要とされる遺伝子を含む可能性がある。用語「本発明の組み換えタンパク質をコードする核酸」は、前記コード配列と、場合によっては任意のさらなる核酸配列(カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロンを含有する宿主細胞中で、この核酸が本発明の組み換えタンパク質を産生することが可能な限り、配列情報を問わない)とを含む核酸を指す。より好ましくは、本発明は、プロモーターに作動可能に連結される、好ましくは、原核生物の既知の誘導性の恒常的プロモーターからなる群から選択されるプロモーター、より好ましくは、テトラサイクリンプロモーター、アラビノースプロモーター、サリチル酸プロモーター、lac、trc、およびtacプロモーター(バネイクス,F.(Baneyx、F.)(1999年)、「Recombinant protein expression in Escherichia coli.」「Curr Opin Biotechnol」10、411〜421;ビルマン−ヤコブ,H.(Billman−Jacobe,H.)(1996年)、「Expression in bacteria other than Escherichia coli.」「Curr Opin Biotechnol」7、500〜504)に連結される、単離および精製された核酸を提供する。前記作動可能に連結された核酸は、例えばワクチン接種のために使用することができる。
【0033】
さらに、本発明の別の態様は、本発明の核酸および/またはベクターを含む宿主細胞に関する。宿主細胞のタイプは、それがC.ジェジュニ(C.jejuni)由来の機能性のpglオペロン、および本発明の組み換え標的タンパク質をコードする1種または複数の核酸に適応する限り、制限されない。好ましい宿主細胞は、原核生物の宿主細胞、より好ましくは細菌、最も好ましくは以下からなる群から選択されるものである:エシェリキア亜種(Escherichia ssp.)、カンピロバクター亜種(Campylobacter ssp.)、サルモネラ亜種(Salmonella ssp.)、シゲラ亜種(Shigella ssp.)、ヘリコバクター亜種(Helicobacter ssp.)、シュードモナス亜種(Pseudomonas ssp.)、バチルス亜種(Bacillus ssp.)、好ましくは大腸菌(E.coli)、より好ましくは、大腸菌(E.coli)株Top10、W3110、CLM24、BL21、SCM6、およびSCM7(Feldman(フェルドマン)ら(2005年)、「Engineering N−linked protein glycosylation with diverse O antigen lipopolysaccharide structures in Escherichia coli.」「Proc Natl. Acad. Sci.U S A」102、3016〜3021;アライモ,C.(Alaimo、C.)、キャトレイン,I.(Catrein、I.)、モルフ,L.(Morf,L.)、マロルダ,C.L.(Marolda,C.L.)、カレワエルト,N.(Callewaert,N.)、ヴァルヴァノ,M.A.(Valvano、M.A.)、フェルドマン,M.F.(Feldman,M.F.)、アービィ,M.(Aebi,M.)(2006年)、「Two distinct but interchangeable mechanisms for flipping of lipid−linked oligosacchardes.」「EMBO journal」25、967〜976)、およびS.エンテリカ(S.enterica)株SL3261(サルモネラエンテリカ(Salmonela Enterica)sv.ティフィリウム(Typhimurium)LT2(デルタ)aroA、ヘイセト,S.K.(Hoiseth,S.K.)およびストッカー,B.A.(Stocker,B.A.)、1981年、「Aromatic−dependent Salmonella typhimurium are non−virulent and effective as live vaccines.」「Nature」291:238〜239を参照のこと)、SL3749(サルモネラエンテリカ(Salmonella enterica)sv.ティフィリウム(Typhimurium)LT2 waaL、カニュク(Kaniuk)ら、「J.Biol.Chem.」279:36470〜36480を参照のこと)、およびSL326ΔwaaL。
【0034】
より好ましい実施形態では、本発明の宿主細胞は、本発明の組み換えタンパク質の外膜および/または表面ディスプレイへのターゲティングに有用であるもの、好ましくは、前記宿主細胞が、組み換えグラム陰性細菌であって:
i)以下をコードするヌクレオチド配列を含む遺伝子型
a)脂質担体上へのオリゴ糖の構築のための少なくとも1種の天然または組み換え型の特異的なグリコシルトランスフェラーゼ、
b)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の少なくとも1種の天然または組み換え型の原核生物オリゴサッカリルトランスフェラーゼ(OTアーゼ)、
c)本発明の少なくとも1種の組み換えタンパク質、好ましくは、ターゲティングポリペプチドをさらに含むタンパク質、および
ii)グラム陰性細菌の外膜内および/または上に位置する、本発明の組み換えN−グリコシル化タンパク質を含む表現型、
を有するグラム陰性細菌である。
【0035】
上記形態のための宿主細胞は、好ましくは、以下からなる群から選択される:エシェリキア亜種(Escherichia ssp.)、カンピロバクター亜種(Campylobacter ssp.)、シゲラ亜種(Shigella ssp.)、ヘリコバクター亜種(Helicobacter ssp.)、およびシュードモナス亜種(Pseudomonas ssp.)、サルモネラ亜種(Salmonella ssp.)、好ましくは大腸菌(E.coli)、より好ましくは、大腸菌(E.coli)株Top10、W3110、CLM24、BL21、SCM6、およびSCM7、ならびにS.エンテリカ(S.enterica)株SL3261、SL3749、およびSL3261ΔwaaL.(ヘイセト,S.K.(Hoiseth,S.K.)およびストッカー,B.A.(Stocker,B.A.)、1981年、「Aromatic−dependent Salmonella typhimurium are non−virulent and effective as live vaccines.」「Nature」291:238〜239を参照のこと)、SL3749(カニュク,N.A.(Kaniuk,N.A.)、ビノグラードフ,E.(Vinogradov,E.)、およびホイットフィールド,C.(Whitfield,C.)2004年、「Investigation of the structural requirements in the lipopolysaccharide core acceptor for ligation of O antigens in the genus Salmonella: WaaL“ligase”is not the sole determinant of acceptor specificity.」「J Biol Chem」279:36470〜36480)。
【0036】
本発明の好ましいタンパク質は、それ自身で、および/または導入されたN−グリコシル化部位に起因して、治療または予防的活性を有する可能性があるので、これらは、医薬品の調製のために使用することができる。本発明を実施するのためのタンパク質のタイプは、限定されないので、数例を挙げると、その中に最適化されたN−グリコシル化されたコンセンサス配列が導入された、EPO、IFN−アルファ、TNFアルファ、IgG、IgM、IgA、インターロイキン、サイトカイン;C.ジェジュニ(C.jejuni)タンパク質のようなワクチン接種のためのウイルスおよび細菌のタンパク質など(HisJ(Cj0734c)、AcrA(Cj0367c)、OmpH1(Cj0982c)、ジフテリア毒素(CRM197)、コレラ毒素、緑膿菌エキソプロテイン(exoprotein)など)の本発明のタンパク質が、医薬品を調製するために有用である(ウィシンスカ,A(Wyszynska,A.)、ラズコー,A.(Raczko,A.)、リス,M.(Lis,M.)、およびジャグシン−クリニカ(Jagusztyn−Krynicka),E.K.(2004年)、「Oral immunization of chickens with avirulent Salmonella vaccine strain carrying C.jejuni 72Dz/92 cjaA gene elicits specific humoral immune response associated with protection against challenge with wild−type Campylobacter.」「Vaccine 22」、1379〜1389)。
【0037】
さらに、本発明の核酸および/またはベクターも、医薬品の調製のために、好ましくは遺伝子治療で使用するために有用である。
【0038】
さらに、本発明の宿主細胞、好ましくは、細菌(好ましくはグラム陰性細菌、より好ましくは上記に列挙したグラム陰性細菌のうちの1種)の外膜内および/または上に位置する本発明のN−グリコシル化された組み換えタンパク質を含む表現型を有するものが、医薬品の調製のために特に有用である。
【0039】
より好ましくは、本発明のタンパク質は、その必要がある対象の治療および/または予防用ワクチン接種のための医薬品の調製のために使用される。
【0040】
より好ましい実施形態では、本発明は、好ましくは遺伝子治療による、その必要がある対象の治療および/または予防用ワクチン接種のための医薬品の調製のための、本発明の核酸および/またはベクターの使用に関する。
【0041】
ディスプレイされたN−グリコシル化タンパク質が、宿主細胞の表面上に多量に存在し、免疫細胞、特に、その親水性N−グリカンによって十分に利用可能であるので、また、宿主細胞が、アジュバントのそれ以上の効果を有する(生存する場合、さらにある程度複製し、そのワクチン接種効果を増幅してもよい)ので、前記N−グリコシル化された組み換えタンパク質をディスプレイする本発明の宿主細胞は、ワクチンを調製するために特に有用である。
【0042】
好ましくは、本発明の医療態様を実践するための宿主細胞は、弱毒化されたあるいは殺された宿主細胞である。
【0043】
医薬品、好ましくはワクチンを調製することに対する本発明の宿主細胞の使用の別の利点は、それが、細胞の成分に起因するIgA抗体を誘導することである。
【0044】
好ましくは、前記宿主細胞は、動物(好ましくは哺乳類、げっ歯類、ヒツジ、ウマ、イヌ、ウシ、またはヒト)中にIgA抗体を誘導するために、本発明に従って使用される。
【0045】
ワクチン接種を必要とする前記対象は、鳥類、哺乳類、または魚類、好ましくは哺乳類、より好ましくは、ウシ、ヒツジ、ウマ、イヌ、ネコ、およびヒトからなる群から選択される哺乳類、最も好ましくはヒトであることが好ましい。家禽類もまた、好ましい。
【0046】
本発明のさらなる態様は、本発明の少なくとも1種のタンパク質、少なくとも1種の核酸、少なくとも1種のベクター、および/または少なくとも1種の宿主細胞を含む薬剤組成物に関する。タンパク質または宿主細胞、好ましくは弱毒化されたあるいは殺された宿主細胞を含む医薬品の調製、および、遺伝子治療のための核酸および/またはベクターを含む医薬品の調製は、当分野でよく知られている。最終の薬剤組成物のための調製スキーム、ならびにその投与の方式および詳細は、用いられるタンパク質、宿主細胞、核酸、および/またはベクターに依存することとなる。
【0047】
好ましい実施形態では、本発明の薬剤組成物は、薬学的に許容される賦形剤、希釈剤、および/または補助剤を含む。
【0048】
本発明は、以下のうちの少なくとも1種を含む薬剤組成物を提供する:(i)本発明の組み換えタンパク質である/をコードする/を発現する組み換えタンパク質、宿主細胞、核酸、および/または組み換えベクター、および(ii)薬学的に許容される賦形剤、希釈剤、および/または補助剤。
【0049】
適切な賦形剤、希釈剤、および/または補助剤は、当分野でよく知られている。賦形剤または希釈剤は、活性成分のための媒体または媒質として働いてよい固体、半固体、または液体材料であってよい。組成物を調製する分野における当業者であれば、選択される産物の特定の特性、治療される疾患または状態、疾患または状態の段階、および他の関連する状況に応じて、投与の適切な形および様式を容易に選択することができる(「Remington’s Pharmaceutical Sciences」、マックパブリッシング(Mack Publishing Co.)(1990年))。薬学的に許容される希釈剤または賦形剤の割合および性質は、選択される薬学的に活性な化合物の溶解性および化学的性質、選択される投与経路、および標準の薬務によって決定される。製剤は、経口、非経口、または局所使用に適合されてもよく、また、錠剤、カプセル、座薬、液剤、懸濁剤などの形で患者に投与されてもよい。本発明の薬学的に活性な化合物は、それ自体で有効でありながら、安定性、結晶化の都合、溶解性の増大などの目的で、酸付加塩または塩基付加塩などの薬学的に許容されるその塩の形で調製および投与することもできる。
【0050】
本発明のさらなる態様は、N−結合グリコシル化タンパク質を産生するための方法であって、
a)以下をコードする核酸を含む組み換え生物、好ましくは原核生物を提供するステップと、
i)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロン、および
ii)1つまたは複数の下記N−グリコシル化された最適化アミノ酸コンセンサス配列を含む少なくとも1種の組み換え標的タンパク質であって:
D/E−X−N−Z−S/T
式中、XおよびZは、Pro以外の任意の天然のアミノ酸であり得、少なくとも1つの前記N−グリコシル化された最適化アミノ酸コンセンサス配列が導入される組み換え標的タンパク質、および
b)標的タンパク質の産生およびN−グリコシル化に適した方式で、組み換え生物を培養するステップと、
を含む方法に関する。
【0051】
好ましくは、標的タンパク質は、本発明の上記の組み換えタンパク質のうちの1つである。
【0052】
本発明の好ましい方法では、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロンは、
OTアーゼによって標的タンパク質に転移されることとなる、脂質担体上へのオリゴ糖の構築のための、
i)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の組み換えOTアーゼ、および
OTアーゼによって標的タンパク質に転移されることとなる、脂質担体上へのオリゴ糖の構築のための、
ii)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の組み換え型および/または天然の特異的なグリコシルトランスフェラーゼ、および/または
iii)カンピロバクター種以外の種から得られる組み換え型および/または天然の特異的なグリコシルトランスフェラーゼ、
をコードする核酸を含む。
【0053】
さらに、好ましい実施形態では、本発明は、本発明の宿主細胞を調製するための方法であって、
i)グラム陰性細菌を提供するステップと、
ii)以下をコードする少なくとも1つのヌクレオチド配列を前記細菌に導入するステップと、
a)脂質担体上へのオリゴ糖の構築のための少なくとも1種の組み換え型の特異的なグリコシルトランスフェラーゼ、および/または
b)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の少なくとも1種の組み換えオリゴサッカリルトランスフェラーゼ(OTアーゼ)、および/または
c)1つまたは複数の下記N−グリコシル化された最適化アミノ酸コンセンサス配列を含む少なくとも1種の組み換えタンパク質であって:
D/E−X−N−Z−S/T
式中、XおよびZは、Pro以外の任意の天然のアミノ酸であり得、少なくとも1つの前記N−グリコシル化された最適化アミノ酸コンセンサス配列が導入される組み換えタンパク質、
iii)c)のヌクレオチド配列によってコードされる少なくとも1種の組み換えN−グリコシル化タンパク質が、グラム陰性細菌の外膜内および/または上に位置されるまで、前記細菌を培養するステップと、
を含む方法に関する。
【0054】
上記好ましい方法を実施するために、組み換え型の原核生物または宿主細胞は、好ましくは、エシェリキア亜種(Escherichia ssp.)、カンピロバクター亜種(Campylobacter ssp.)、サルモネラ亜種(Salmonella ssp.)、シゲラ亜種(Shigella ssp.)、ヘリコバクター亜種(Helicobacter ssp.)、シュードモナス亜種(Pseudomonas ssp.)、バチルス亜種(Bacillus ssp.)、好ましくは大腸菌(Escherichia coli)、好ましくは大腸菌(E.coli)株Top10、W3110、CLM24、BL21、SCM6、およびSCM7、ならびにS.エンテリカ(S.enterica)株SL3261、SL3749、およびSL3261ΔwaaLからなる細菌の群から選択される。
【0055】
本発明の組み換えタンパク質を産生、単離、および/または精製するための別の好ましい方法は、次のステップを含む:
a)請求項15または16に記載の宿主細胞を培養するステップ、
b)前記組み換えグラム陰性細菌の外膜を除去するステップ、および
c)前記組み換えタンパク質を回収するステップ。
【0056】
細胞、好ましくは原核細胞、より好ましくはグラム陰性細菌細胞の外膜を除去するための例示的な方法は、適切な酵素処理方法、浸透圧衝撃、界面活性剤可溶化、およびフレンチプレス法である。
【0057】
最も好ましくは、本発明は、カンピロバクター種以外の種、好ましくはC.ジェジュニ(C.jejuni)から得られる組み換え型または天然の特異的なグリコシルトランスフェラーゼが、前記宿主細胞中で機能的に発現させることが可能である細菌、古細菌、および/または真核生物に由来するグリコシルトランスフェラーゼおよびエピメラーゼの群から選択される方法に関する。
【0058】
以下の実施例は、本発明をさらに説明する役割を果たすものであり、いずれにせよその範囲を限定するものではない。
【実施例】
【0059】
実施例
N−グリコシル化を最適化するためのモデルタンパク質としてのAcrAの選択
N−グリコシル化のためのアクセプタータンパク質の要件を最適化するために、C.ジェジュニ(C.jejuni)糖タンパク質AcrA(Cj0367c)に対して、詳細な研究を実施した。AcrAは、350アミノ酸残基の周辺質リポタンパク質である。脂質に固定されるのではなく周辺質に分泌されることが、グリコシル化のための必要条件であることが示されている(ニタ−ラザー(Nita−Lazar)ら、2005年、上記)。大腸菌(E.Coli)中で発現される場合、搬出のためのシグナルは、天然のAcrAシグナル配列または異種起源のPelBシグナルであり得る。5つの潜在的なN−結合グリコシル化シークオン(sequon)(N117、N123、N147、N273、N274)のうち、2つの同じものが、C.ジェジュニ(C.jejuni)および大腸菌中で使用される(N123およびN273(ニタ−ラザー(Nita−Lazar)ら、2005年、上記))。AcrAは、詳細な構造情報が利用可能であるC.ジェジュニ(C.jejuni)の唯一の周辺質N−糖タンパク質であるので、モデルとして選択された。近年では、AcrA相同体、すなわちグラム陰性細菌緑膿菌(P.aeruginosa)由来のMexAタンパク質の結晶構造が公開されている(ヒギンズ(Higgins)ら(2004年)、「Structure of the periplasmic component of a bacterial drug efflux pump.」「Proc.Natl.Acad.Sci.U S A」101、9994〜9999)。どちらのタンパク質も、いわゆる周辺質排出ポンプタンパク質のメンバーである(PEP、(ジョンソン,J.M.(Johnson,J.M.)およびチャーチ,G.M.(Church,G.M.)(1999年))「Alignment and structure prediction of divergent protein families:periplasmic and outer membrane proteins of bacterial efflux pumps.」「J.Mol.Biol.287、695〜715))。細長い分子は、3つの直線的に配置されたサブドメイン:αらせんの逆平行のコイルドコイルを含有し、これは、リポイルドメインによって塩基で結びつけられ、これに、6本鎖のβバレル構造ドメインが続く。N末端の23〜28残基およびC末端における95〜101残基は、結晶中に組み込まれていない。MexAタンパク質とAcrAタンパク質の配列は、29.3%同一であり、50%類似である。したがって、2つのタンパク質はおそらく、全体的に類似の折りたたみを示す。
【0060】
実施例1 グリコシル化を誘発する第1のペプチド配列の解明
緑膿菌のMexAと類似である、したがってまた、C.ジェジュニ(C.jejuni)のAcrA中にあるリポイルドメインは、大腸菌において個々に発現させることができるコンパクトなタンパク質を形成することが知られている(バーグ,A.(Berg,A.)、およびデコック,A.(de Kok,A.)(1997年)、「2−Oxo acid dehydrogenase multienzyme complexes.」「The central role of the lipoyl domain.Biol.Chem.」378、617〜634によって概説される)。どのアクセプターペプチド配列が、大腸菌におけるpgl機構によるN−グリコシル化に必要とされるのかを確認するために、AcrAのリポイルドメインを取り出した。これを、異なる長さのペプチドを周辺質に輸送し、それらをin vivoでpgl機構に与えるための分子骨格として使用した。
【0061】
したがって、リポイルドメイン(Lip)をコードするプラスミドを構築し、OmpA(チェ,J.H.(Choi,J.H.)およびリー,S.Y.(Lee,S.Y.)(2004年)、「Secretory and extracellular production of recombinant proteins using Escherichia coli.」「Appl Microbiol Biotechnol」64、625〜635)のシグナル配列にN末端を、ヘキサヒスチジンタグにC末端を融合させた。クローニングを行い、アラビノースプロモーターの制御下で、遺伝子発現を行った。Lipドメイン境界については、MexA中でのリポイルドメイン部分のドメイン境界と同じ位置で出現すると考えられるアミノ酸位を選択した。N−グリカンを受け入れるその能力について、異なるペプチドを試験するために、配列のストレッチを、Lipドメインの2つのハンマーヘッド様部分の間に挿入した。ストレッチは、C.ジェジュニ(C.jejuni)AcrAのN−グリコシル化部位N123を含む配列からなるものであった。得られたオープンリーディングフレームは、OmpAシグナル配列をコードする配列、AcrAのN末端基ハンマーヘッド様部分(D60〜D95、アミノ酸の番号付けは、成熟したAcrAポリペプチドの配列番号付けに関連する)、AcrAの天然のN123グリコシル化部位を含有する別のストレッチ(下記参照)、AcrA−Lip(L167−D210)のC末端のハンマーヘッド様部分、およびC末端のhis−タグからなるものであった。
【0062】
プラスミドの構築は、標準の分子生物学技術によって達成された。異なる長さのAcrAの天然のN123グリコシル化部位を含有する3つのストレッチを、Lipの中間に挿入し、3つの異なるORFをもたらした:
構築物Aは、次のタンパク質配列をもたらすA118−S130を含有する:
【化1】

構築物Bは、次のタンパク質配列をもたらすF122−E138を含有する:
【化2】
構築物Cは、次のタンパク質配列をもたらすD121−A127を含有する:
【化3】
下線が引かれた配列のストレッチは、OmpAシグナルペプチドを示し、単独で下線が引かれた残基は、クローン化のために、あるいはタンパク質を分解に対して耐性にするために導入されたものである。太字:AcrAのN123に相当するグリコシル化部位。イタリック体:ヘキサ−ヒスチジンタグ。相当する遺伝子を、プラスミドpEC415(シュルツ,H.(Schulz,H.)、ヘンネック,H.(Hennecke,H.)、およびトニー−マイヤー,L.(Thony−Meyer,L.)(1998年)、「Prototype of a heme chaperone essential for cytochrome c maturation.」「Science」281、1997〜1200)の主鎖中のアラビノースプロモーターの制御下で発現させた。
【0063】
3つのストレッチのうちのどれがLipタンパク質のグリコシル化を誘発したかを確認するために、タンパク質発現実験を実施した。pACYCpglまたはpACYCpglmut(ワッカー(Wacker)ら、2002年、上記)と、構築物A、B、またはCをコードするプラスミドを保有する大腸菌Top10細胞(インビトロジェン(Invitrogen)、米国カリフォルニア州カールズバッド(Carlsbad,CA,USA))を、アンピシリンおよびクロラムフェニコールを含有するLB媒質中で37℃で0.5ODまで増殖させた。誘導のために1/1000体積の20%アラビノース(w/v)溶液を加え、さらに2時間細胞を増殖させた。細胞を、遠心分離によってその後収集し、20mMトリス/HCl(pH 8.5)、20%スクロース(w/v)、1mM EDTA、1mM PMSF、および1g/l(w/v)リゾチーム中に再懸濁させ、4℃で1時間保温した。球状の芽球(spheroblasts)をペレット化すると、周辺質抽出物が得られ、これを1/9体積(v/v)の10×緩衝液A(3M NaCl、0.5MのTris/HCl(pH 8.0)、および0.1M イミダゾール)で希釈し、MgSO4を加えて2.5mMにした。Ni−アフィニティ精製は、緩衝液A中で、アマシャムファルマシアバイオテック(Amersham Pharmacia Biotech)(スウェーデン、ウプサラ(Uppsala,Sweden))からの1ml Ni−セファロース(Sepharose)カラム上で実施した。タンパク質は、0.25M イミダゾールを含有する緩衝液Aで溶出させた。
【0064】
図1は、クーマシーブリリアントブルー染色した、Ni−精製された周辺質抽出物からのピーク溶出分画のSDS−PAGEゲルを示す。発現分析によって、構築物Bが、突出した単一のタンパク質種を産生することが示された(図1、レーン1)。構築物AとCは両方とも、突出したタンパク質に加えて、電気泳動移動度がより遅い第2のタンパク質バンドをもたらす(図1、レーン2および3)。より重いタンパク質種が確かにグリコシル化されたことが、MALDI−TOF/TOFによって証明された(図示せず)。構築物B中では失われたが、AおよびCでは存在する唯一のアミノ酸は、D121、すなわちグリコシル化されたN123に対して2つN末端方向に位置するアスパラギン酸残基であった。このことは、D121が、OTアーゼによるグリコシル化のために重要な役割を果たすことを実証している。D121がグリコシル化に不可欠なものであるということを検証するために、それは構築物C発現分析においてアラニンへ突然変異され、1つだけのタンパク質バンドをもたらし(図1、レーン4)、その結果、D121がグリコシル化に重要であることを示した。さらに、人工のペプチドディスプレイタンパク質がグリコシル化され得るという事実は、D/E−X−N−Y−S/Tタイプの短いペプチドが、C.ジェジュニ(C.jejuni)が担うN−グリコシル化が起こるためのすべての情報を含有することを示す。
【0065】
実施例2 実施例1の検証;AcrA−D121Aは、N123でグリコシル化されない
ペプチドディスプレイ手法による発見を確認するために、アスパラギン酸からアラニンへの突然変異を、位置121(D121A、すなわちグリコシル化されたN123よりも2つ前の残基)に挿入し、AcrAタンパク質の全長の可溶性バージョンにおいて、部位N123が大腸菌中で依然としてグリコシル化され得るかどうかを試験した。試験を行うために、このAcrA−D121Aを発現させ、そのグリコシル化状態を分析した。分析のために、操作されたAcrAを使用した。これは、周辺質への分泌のためのPelBシグナル配列(チェ(Choi)およびリー(Lee)、2004年、上記)、および、精製のためのC末端のヘキサヒスチジンタグを含有するという点で、本来のC.ジェジュニ(C.jejuni)遺伝子とは異なっていた。このAcrA変異体は、脂質に固定された未変性タンパク質として分泌され、シグナルペプチド切断され、グリコシル化されることが示されている(ニタ−ラザー(Nita−Lazar)ら、2005年、上記))。以下は、可溶性のAcrAタンパク質のタンパク質配列である。
【化4】
【0066】
下線が引かれた残基は、PelBシグナルペプチドであり、イタリック体は、ヘキサ−ヒスチジンタグであり、太字は、N123およびN273の2つの天然のグリコシル化部位である。pEC415プラスミド(シュルツ(Schulz)ら、1998年)中に上記のタンパク質に対するORFを含有するプラスミドを、pAcAperを産生するために構築した。
【0067】
AcrAおよびそのミュータント(下記参照)のグリコシル化状態を試験するアッセイは、以下の通りであった:その活性または不活性の形の、プラスミドが担うpglオペロン(pACYCpglまたはpACYCpglmut、(ワッカー(Wacker)ら、2002年、上記)を参照のこと)と、AcrAをコードするプラスミド(pAcrAper)を含有する、指数的に増殖する大腸菌CLM24(フェルドマン(Feldman)ら、2005年、上記)細胞において、0.02%アラビノースを用いて、AcrAの発現を誘導した。4時間の誘導の後、周辺質抽出物を、上述した通りに調製し、抗AcrA抗血清またはR12抗血清のいずれかを用いる、SDS−PAGE、電気移動(electrotransfer)、および免疫検出によって分析した。R12抗血清は、C.ジェジュニ(C.jejuni)N−グリカンを含有するタンパク質に対して特異的である(ワッカー(Wacker)ら、2002年、上記)。
【0068】
図2Aの最初の2本のレーンは、機能性のpglオペロンが存在する場合と存在しない場合のAcrAを示す。機能性のpglオペロンが存在しない場合には、1つのバンドのみが現れ、存在する場合には3つのバンドが現れる(図2A、上のパネル)。これらは、非グリコシル化AcrA(レーン1)、ならびに、非グリコシル化、モノグリコシル化、およびジグリコシル化AcrA(レーン2)に相当する。レーン2中の2つのより重いタンパク質がグリコシル化されたことを、R12ウェスタンブロットによって確認した(レーン2、下のパネル)。ミュータントAcrA−N273Qを同じ方法で発現させた場合、機能性のグリコシル化pglオペロンが存在する場合には、モノグリコシル化されたAcrAだけが検出された(レーン3)。機能性のpgl遺伝子座が存在しない場合には、非グリコシル化AcrAが検出された(レーン4)。ミュータントAcrA−D121Aの分析は、2つのバンドのみをもたらし、これらのうちの1つは、レーン3においてAcrA−N273Qで観察されるように、グリコシル化されたものであった(レーン5)。これは、D121が、位置123〜125での効率的なグリコシル化のために必須であることを意味する。
【0069】
実施例3 人工のグリコシル化部位のAcrAへの導入
アスパラギン酸残基の導入が、グリコシル化部位をもたらすことができるかどうか試験するために、可溶性のAcrAの位置N117およびN147の使用されないグリコシル化部位の−2位置の残基がアスパラギン酸(F115D、T145D)と交換されたAcrAミュータントを産生した。改変されたグリコシル化部位を、実施例2に記載されるのと同じアッセイによってグリコシル化することができるかどうかを、その後試験した。両方の突然変異を、AcrAの可溶性のバージョンの野生型配列に、あるいは、(どちらも使用されるグリコシル化部位を欠損させた)(N123QおよびN273Q)二重のミュータント中に、個々に挿入した。4時間誘導させた培養物の周辺質抽出物を調製し、SDSページによって分離し、ウェスタンブロッティングによって分析した(図2B)。対照として、野生型のサンプルをグリコシル化し、非グリコシル化されたAcrAを、同じゲル上に流した(レーン1および2)。T145D突然変異は、天然には使用されないグリコシル化シークオンN147−S149の−2位置に影響を及ぼした。AcrA−T145Dを発現させると、抗AcrA抗血清を用いるウェスタンブロッティングは、4つのバンドをもたらし、それらの最も高いものは、レーン2における2重にグリコシル化されたタンパク質よりも電気泳動移動度が遅かった(図2Bにおけるレーン3)。R12ブロットによって、第4のバンドは、3重にグリコシル化されたAcrAであることが確認された。抗AcrAに対する低い強度にもかかわらず、最も重いバンドは、グリコシル化特異的なR12抗血清を用いると最も強いシグナルを示した。同じミュータントAcrA−T145Dが、天然のN−グリコシル化配列(AcrA−N123Q−N273Q−T145D)の非存在下で発現された場合、機能性のpglオペロンの存在下で、モノグリコシル化されたAcrAのみが検出され(図2B、レーン4)、機能性のpglオペロンの非存在下では検出されなかった(レーン5)。これは、レーン4中のより重いバンドが、グリコシル化されたことを実証する。したがって、単純にT145D突然変異を導入することによって、最適化グリコシル化部位がもたらされた(DFNNS)。
【0070】
アスパラギン酸残基を−2位置に挿入することによって、グリコシル化部位を導入することが可能であることをさらに確認するために、天然には使用されない部位N117−119およびN274−T276を、N−グリコシル化を最適化するために変化させた。この目的のために、さらなるミュータントを産生した(図2C)。上記のシステムにおけるAcrA−F115D−T145Dの発現は、抗AcrA抗血清を用いて検出される5つのタンパク質種をもたらした(レーン2)。これは、同じAcrA分子上に起こる4つのグリコシル化の指標となる。C.ジェジュニ(C.jejuni)N−グリカン特異的なR12抗血清を用いる検出を実施すると、5つのバンドのラダーが検出された。最も低いかすかなバンドは、非グリコシル化AcrAであり、これは、グリコシル化の非存在下でも存在するからであり(レーン1)、最も高いものは、4重にグリコシル化されたAcrA中の5つの抗原性の決定因子におそらく起因する、強いシグナルをもたらす。したがって、2つの導入された部位(N117およびN147で)および2つの天然に使用される部位(N123およびN273)が使用され、pgl機構によってグリコシル化される。pglオペロンを含むおよび含まないAcrA−N123Q−N273Q−N272Dの発現は、第3の人工的に導入されたグリコシル化部位、N274(DNNST)も、pglオペロンによって認識されたことを実証した(図2C、レーン3および4)。
【0071】
上記の実験によって、C.ジェジュニ(C.jejuni)のOTアーゼによって認識される細菌のN−グリコシル化部位は、部分的に真核性のものと同じコンセンサスからなるが(N−X−S/T、X≠P)、さらに、効率を増大するために、−2位置におけるアスパラギン酸が必要とされるという知見が確認される。さらに、これらの実験は、こうした最適化コンセンサス配列を組換えにより導入することによって、所望の部位でタンパク質をグリコシル化することが可能性であることを実証する。
【0072】
実施例4 最適化N−グリコシル化配列における位置−1の確認
細菌のグリコシル化部位における−1の位置が、真核生物における+1の位置と同じ制限を示すかどうか試験するために、さらなる実験を実施した(インペリアリ,B.(Imperiali,B.)およびシャノン,K.L.(Shannon,K.L.)(1991年)、「Differences between Asn−Xaa−Thr−containing peptides:a comparison of solution conformation and substrate behaviour with oligosaccharyl−transferase.」「Biochemistry」30、4374〜4380;ラッド,P.M.(Rudd,P.M.)およびドゥウェック,R.A.(Dwek,R.A.)(1997年)、「Glycosylation:heterogeneity and the 3D structure of proteins.」、「Crit.Rev.Biochem.Mol.Biol.」32、1〜100)。+1のプロリン残基は、グリコシル化が抑制されるように、ペプチドを制限すると考えられる。類似の効果が、−1の位置でも観察されることができるかどうか試験するために、第2の天然の部位がノックアウトされた点変異体(AcrA−N273Q−F122P)において、第1の天然に使用される部位のその位置に、プロリン残基を導入した。AcrA−N273Qの対照発現は、機能性のpglオペロンが存在する場合には、モノグリコシル化されたタンパク質を示した(図2D、レーン1および2)。しかし、AcrA−N273Q−F122Pは、グリコシル化されなかった(図2D、レーン3および4)。これは、プロリンは、これがアスパラギンと−2位置の負に荷電する残基との間に残基を構成する場合、細菌のN−グリコシル化を抑制することを示している。
【0073】
C.ジェジュニ(C.jejuni)pgl機構によってグリコシル化されることが知られているすべての部位の配列アラインメントは、これらがすべて、−2位置にDまたはEを含むことを示す(ニタ−ラザー(Nita−Lazar)ら、2005年、上記;ワッカー(Wacker)ら、2002年、上記;ヤング(Young)ら、(2002年)、「Structure of the N−linked glycan present on a multiple glycoproteins in the Gram−negative bacterium, Campylobacter jejuni.」「J.Biol.Chem.」277、42530〜42539)。したがって、細菌のグリコシル化コンセンサス配列は、−2位置の負に荷電するアミノ酸によって最適化され、D/E−X−N−Z−S/T(式中、XおよびZ≠P)をもたらす可能性があることが確立された。
【0074】
実施例5 非C.ジェジュニ(C.jejuni)タンパク質のN−グリコシル化
第1の配列要件(最適化コンセンサス配列)が、細菌におけるN−グリコシル化に十分であることを実証するために、非C.ジェジュニ(C.jejuni)タンパク質を、上記の戦略を適用することによってグリコシル化させることができるかどうかを試験した。コレラ毒素Bサブユニット(CtxB)を、グリコシル化標的として用いた。相当する遺伝子を、それが、まさに実施例1における構築物AからCと同様の、N末端上のOmpAシグナル配列のコード配列とC末端のヘキサヒスチジンタグを含有するような方式で、コレラ菌から増幅させた。得られたDNAを、実施例1で用いられたプラスミド中の構築物Aと置き換えるようにクローン化した。W88のDへの点突然変異、またはW88の後のD挿入は、最適化グリコシル化部位(DNNKT)をもたらした。野生型、ならびにシグナル配列およびhis−タグを含有するW88D CtxBタンパク質を、C.ジェジュニ(C.jejuni)由来の機能性のpgl遺伝子座の存在下および非存在下で、大腸菌Top 10および他の細胞型中で発現させた。Top 10細胞由来の周辺質抽出物を、SDS−PAGE、電気移動、およびCtxB抗血清を用いる連続的な免疫ブロット法によって分析すると、CtxB W88Dだけは、pgl遺伝子座バックグラウンドにおいて、より高い、したがって、グリコシル化されたバンドを産生していた(図2E、レーン3と4を比較せよ)。コンセンサス配列(DSNIT)はまた、CtxBのG54またはQ56(後者は、CtxB−Q56/DSNITと表される)、すなわち、CtxBのガングリオシドGM1結合活性に寄与することが報告されたループのうちの1つの中のものを置き換えることによって挿入された。図2Eのレーン5および6は、操作されたタンパク質(Top 10細胞中に発現されるQ56ではなくペプチド配列DSNITを含有する構築物によって例示される)が、上で述べたのと同様に分析した場合、グリコシル化を欠く細胞ではなくグリコシル化コンピテントな細胞において、より移動性が低い、したがって、グリコシル化されたバンドをもたらしたことを実証している。2つの操作、すなわち、W88の後のDの挿入ならびにDSNITのQ56の置き換えを含むCxtBは、SCM7細胞(アライモ(Alaimo)ら、「EMBO journal」25:967〜976(2006年))において2重にグリコシル化されたことも実証された(パネルE、レーン7および8)。レーン7に示される2重にグリコシル化されたタンパク質CtxBを、Ni2+アフィニティ精製し、標準のプロトコルによるゲル内トリプシン処理の後、ESI−MS/MSによって分析した。予測されるグリコペプチドが検出され、細菌のN−グリコシル化はまた、本発明の細菌のN−グリコシル化のための最適化コンセンサス配列を突然変異させる、あるいは挿入することによって、非C.ジェジュニ(C.jejuni)タンパク質も対象とすることが可能であることが確認された(図示せず)。本発明を実践するための他の適切な例示的な大腸菌(E.coli)株の例は、W3110、CLM24、BL21(ストラタジーン(Stratagene)、米国カリフォルニア州、ラホーヤ(La Jolla,CA,USA))、SCM6、およびSCM7である。
【0075】
ここで使用されるCtxBタンパク質のアミノ酸配列を、以下に示す(組み換えOmpAシグナル配列は下線部、ヘキサ−ヒスチジンタグはイタリック体、W88は太字):
【化5】
【0076】
実施例6 C.ジェジュニ(C.jejuni)外膜タンパク質(OmpH1)への人工のN−グリコシル化部位の導入
細菌におけるN−グリコシル化の潜在的な用途は、遺伝子型に表現型を連結し、それによって特定の遺伝子突然変異について選択するための、細菌の宿主細胞の表面上のグリカンのディスプレイである。N−グリカンを、外膜タンパク質上に提示することが可能であることを実証するために、OmpH1タンパク質を、それが、本発明の複数の最適化コンセンサス部位を含有するように設計した。既知の結晶構造から推測される通り、この部位をタンパク質のループ領域に設計した(ミュラー,A.(Muller,A.)、トーマス,G.H.(Thomas,G.H.)、ホーラー、R.(Horler,R.)、ブラニガン,J.A.(Brannigan,J.A.)、ブラゴヴァ,E.(Blagova,E.)、レヴディコフ,V.M.(Levdikov,V.M.)、フォッグ,M.J.(Fogg,M.J.)、ウィルソン,K.S.(Wilson,K.S.)、およびウィルキンソン,A.J(Wilkinson,A.J.)2005年、「An ATP−binding cassette−type cysteine transporter in Campylobacter jejuni inferred from the structure of an extracytoplasmic solute receptor protein.」「Mol.Microbiol.」57:143〜155)。以前の実験では、最良のグリコシル化配列は、突然変異V83T、K59N−G60I−N61T、R190N−G191I−D192T、およびH263D−F264S−G265N−D266I−D267Tによって産生されたことが示されている。表面ディスプレイについては、最大にN−グリカン特異的なサンプルを確立するために、それらの導入された部位の異なる組み合わせを評価することが所望された。こうした組み合わせを、プラスミド構築物をコードする野生型OmpH1中に作り出し、AcrAについて述べられたのと同様の方式で試験した。図3は、既存の野生型シークオンに加えて、複数のグリコシル化シークオンを有する様々なOmpH1変異体の分析を示す。OmpH1変異体は、3つ(レーン3、4、5および7)、および4つ(レーン6)のグリコシル化シークオンを伴って産生された。グリコシル化シークオンを1つだけ有する野生型OmpH1と、グリコシル化のための重要なアスパラギンを欠くミュータントもまた、この実験に含まれた。ここで試験されるすべての変異体は、高レベルのグリコシル化効率を実証するものに限られるわけではなく、あらゆるグリコシル化シークオンが利用された。結果は、カンピロバクターN−グリカン特異的な免疫血清を用いて確認した(図3、下のパネル)。
【0077】
以下は、カンピロバクタージェジュニ(Campylobacter jejuni)(81−176株)のOmpH1タンパク質のタンパク質配列である。付着されたmycタグは、イタリック体である。
【化6】
タンパク質中の天然のグリコシル化部位は太字であり、シグナル配列には下線が引かれている。
【0078】
実施例7 大腸菌細胞の外膜上のOmpH1上のC.ジェジュニ(C.jejuni)由来のN−グリカンの表面ディスプレイ
複数のグリコシル化されたOmpH1変異体を、細菌細胞の表面上にディスプレイさせることができるかどうかという問題に答えるために、様々なOmpH1変異体を発現している細菌のCLM24またはSCM6(これは、SCM7ΔwaaLである)細胞に、免疫蛍光法を実施した。野生型OmpH1と、グリコシル化のための重要なアスパラギンを欠くミュータントを、実験に含めた。さらに、周辺質内にタンパク質を保持する、したがって、この実験における対象としての役割を果たすために、C20Sミュータントを構築した。パラホルムアルデヒドで処理した細胞に対して免疫染色を行った。パラホルムアルデヒドは、細胞の構造または区画化を壊さずに細胞を固定する。FITCおよびCy3に結合させた相当する二次抗体と組み合わせて、c−Myc−およびN−グリカン−特異的な免疫血清を使用して、細菌細胞表面上のタンパク質(赤色蛍光)およびN−グリカン(緑色)をそれぞれ検出した。さらに、4,6−ジアミノ−2−フェニルインドール(DAPI、青色)を用いて、細菌のDNAを染色し、細菌細胞と細胞残屑を明確に区別した。野生型OmpH1を発現している細胞が染色された場合、タンパク質ならびにN−グリカンに特異的な免疫蛍光が検出された(図4A)。重要なアスパラギンN139Sを欠くミュータントが、抗−Myc−とN−グリカン−の両方に特異的な免疫血清で染色された場合、グリカン特異的なシグナルではなく、タンパク質のみに特異的なシグナルが得られ(パネル4B)、これは、N−グリカン特異的な免疫血清の特異性を示した。タンパク質が、C20Sミュータントにおいて見られるように周辺質内に保持された場合、タンパク質特異的な赤い免疫蛍光は検出されず、これは、抗体が細胞内に拡散することが不可能であっても、任意の表面現象を検出するのに十分コンピテントであったことを示していた(4Cパネル)。次に、グリコシル化において異なる複数のOmpH1変異体を発現している細胞が染色された:OmpH1KGN→NIT、HFGDD→DSNIT(パネル4D)、OmpH1RGD→NIT、HFGDD→DSNIT(パネル4E)、OmpH1KGN→NIT、RGD→NIT(パネル4F)、OmpH1V83T、KGN→NIT(パネル4G)、およびOmpH1KGN→NIT、RGD→NIT、HFGDD→DSNIT(パネル4H)。すべてのOmpH1変異体は、2重に染色され、これは、細菌の表面上のグリコシル化タンパク質の存在を示していた。図4は、グレースケールで表したものであり、第1行は、同じ列の他の画像を組合せた画像である。
【技術分野】
【0001】
本発明は、1つまたは複数の導入されたNグリコシル化された最適化アミノ酸コンセンサス配列を含む組み換えN−グリコシル化タンパク質、これらのタンパク質をコードする核酸、ならびに相当するベクターおよび宿主細胞に関する。さらに、本発明は、医薬品を調製するための、前記タンパク質、核酸、ベクター、および宿主細胞の使用に関する。さらに、本発明は、前記タンパク質を産生するための方法を提供する。
【背景技術】
【0002】
発明の背景
N−結合タンパク質のグリコシル化は、真核生物の小胞体で起こる本質的かつ保存されたプロセスである。それは、タンパク質のフォールディング、オリゴマー化、安定性、品質管理、選別、ならびに分泌および膜タンパク質の輸送にとって重要である(ヘレニウス,A(Helenius,A.)およびアービィ,M.(Aebi,M.)(2004年)、「Roles of N−linked glycans in the endoplasmic reticulum.」「Annu.Rev.Biochem.」73、1019〜1049)。
【0003】
タンパク質のグリコシル化は、タンパク質の抗原性、安定性、および半減期に対して重大な影響を有する。さらに、グリコシル化は、クロマトグラフィ、例えば、タンパク質のグリコシル化された部分と相互作用する、固相に結合されたレクチンリガンドを用いるアフィニティクロマトグラフィによるタンパク質の精製を助けることができる。したがって、生物学的および薬学的に有用なグリコシル化パターンを提供するために、真核生物細胞において、組換えにより多くのグリコシル化タンパク質を産生することは、確立された手法である。
【0004】
ごく近年、細菌、すなわち食物性の病原菌カンピロバクター ジェジュニ(Campylobacter jejuni)も、そのタンパク質をN−グリコシル化することができることが実証されている(シマンスキー(Szymanski)ら(1999年)、「Evidence for a system of general protein glycosylation in Campylobacter jejuni.」「Mol.Microbiol.」32、1022〜1030)。グリコシル化のために必要とされる機構は、いわゆるpgl遺伝子座にクラスター化される12の遺伝子によってコードされる。N−グリコシル化の破壊は、C.ジェジュニ(C.jejuni)の侵入および病原性に影響を及ぼすが、大抵の真核性の生物において見られるように致死的ではない(ブルダP.(Burda P.)およびM.アービィ(M.Aebi.)(1999年)、「The dolichol pathway of N−linked glycosylation.」「Biochim Biophys Acta」1426(2):239〜57)。大腸菌(E.coli)中でpgl遺伝子座とアクセプター糖タンパク質を組換えにより同時に発現させることによって、C.ジェジュニ(C.jejuni)タンパク質のN−グリコシル化を元に戻すことが可能である(ワッカー(Wacker)ら(2002年)「N−linked glycosylation in Campylobacter jejuni and its functional transfer into E coli.」「Science」298、1790〜1793)。
【0005】
欧州特許出願公開第03 702 276 1号明細書(欧州特許第1 481 057号明細書)、すなわち本発明の発明者の以前の発明は、(i)脂質担体上へのオリゴ糖の構築のための特異的なグリコシルトランスフェラーゼ、(ii)コンセンサス配列「N−X−S/T」(式中、Xは、プロリン以外の任意のアミノ酸であり得る)を含む組み換え標的タンパク質、ならびに(iii)前記オリゴ糖を、標的タンパク質のコンセンサス配列に共有結合的に連結させるC.ジェジュニ(C.jejuni)のオリゴサッカリルトランスフェラーゼ(OTアーゼ)、をコードする核酸が導入される原核生物を教示する。前記原核生物は、特定のグリコシルトランスフェラーゼのタイプによって定義される、特定の構造をもつN−グリカンを産生する。
【0006】
たとえ、タンパク質中の既知のN−グリコシル化コンセンサス配列の存在によって、C.ジェジュニ(C.jejuni)のオリゴサッカリルトランスフェラーゼ(OTアーゼ)を含む原核生物における組み換え標的タンパク質のN−グリコシル化が確かに可能になったとしても、いくつかの標的タンパク質のN−グリコシル化は、しばしば非効率的である。
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明の目的は、原核生物中でin vivoで産生することができる、N−グリコシル化の効率が最適化タンパク質、ならびにこうしたタンパク質を産生するための手段および方法を提供することである。本発明の別の目的は、前記タンパク質の抗原性、安定性、生物学的、予防的、および/または治療的活性を改変するための、N−グリカンの組み換えタンパク質へのより効果的な導入を意図する。さらなる目的は、その表面上に本発明の組み換えN−グリコシル化タンパク質を効率的にディスプレイする宿主細胞の提供である。
【課題を解決するための手段】
【0008】
第1の態様では、本発明は、1つまたは複数の下記N−グリコシル化された最適化アミノ酸配列を含む、組み換えN−グリコシル化タンパク質を提供する:
D/E−X−N−Z−S/T(最適化コンセンサス配列)
式中、XおよびZは、Pro以外の任意の天然のアミノ酸であり得、かつ少なくとも1つの前記N−グリコシル化された部分的なアミノ酸コンセンサス配列が導入される。
【発明の効果】
【0009】
驚くべきことに、特定の部分的なアミノ酸配列(最適化コンセンサス配列)をタンパク質に導入することで、カンピロバクター(Campylobacter)種、好ましくはC.ジェジュニ(C.jejuni)由来のオリゴサッカリルトランスフェラーゼ(OST、OTアーゼ)によって、これらが導入された位置で、非常に効率的にN−グリコシル化されたタンパク質がもたらされることが判明した。
【図面の簡単な説明】
【0010】
【図1】構築物AからCより得られるLipタンパク質のN−グリコシル化を説明する図である(実施例1を参照のこと)。C.ジェジュニ(C.jejuni)由来の機能性のpglオペロン(ワッカー(Wacker)ら、2002年、上記)、および、構築物A(レーン2)、B(レーン1)、およびC(レーン3)をコードするプラスミド、または突然変異D121Aを伴う構築物Cのミュータント(レーン4)を保有する大腸菌(E.coli)Top 10細胞。タンパク質を発現させ、周辺質抽出物から精製した。SDS−PAGE、および精製されたタンパク質の分画を染色しているクーマシーブリリアントブルーが示される。
【図2】プラスミドから前記タンパク質を発現する、CLM24細胞(フェルドマン(Feldman)ら、(2005年)、「Engineering N−linked protein glycosylation with diverse O antigen lipopolysaccharide structures in Escherichia coli.」「Proc.Natl.Acad.Sci.U S A」102、3016〜3021)、またはTop 10細胞(パネルE、レーン1〜6)、またはSCM7細胞(アライモ,C.(Alaimo、C.)、キャトレイン,I.(Catrein、I.)、モルフ,L.(Morf,L.)、マロルダ,C.L.(Marolda,C.L.)、カレワエルト,N.(Callewaert,N.)、ヴァルヴァノ,M.A.(Valvano、M.A.)、フェルドマン,M.F.(Feldman,M.F.)、アービィ,M.(Aebi,M.)(2006年)「Two distinct but interchangeable mechanisms for flipping of lipid−linked oligosaccharides.」「EMBO journal」25、967〜976)(パネルE、レーン7,8)中で、C.ジェジュニ(C.jejuni)pglオペロン(ワッカー(Wacker)ら、2002年、上記)による配列特異的なN−グリコシル化について分析される、様々なタンパク質のN−グリコシル化分析を示す図である。ニトロセルロース膜に移され、特異的な抗血清を用いて視覚化された、SDS−PAGE分離された周辺質抽出物が示される。パネルA〜Dでは、上のパネルが、抗AcrA抗血清(ワッカー(Wacker)ら、2002年、上記;ニタ−ラザー,M(Nita−Lazar,M)ワッカー,M.(Wacker,M.)、シェッグ,B.(Schegg、B.)、アンバー,S.(Amber,S.)、およびアービィ,M(Aebi,M.)(2005年)、「The N−X−S/T consensus sequence is required but not sufficient for bacterial N−linked protein glycosylation.」「Glycobiology」15、361〜367)を用いて探索される免疫ブロットを示すのに対し、下のパネルは、R12抗血清(ワッカー(Wacker)ら、2002年、上記)を用いて探索される免疫ブロットを示す。+および−は、細胞中の機能性または変異型のpglオペロンの存在を示す。パネルAは、pelBシグナル配列と、ヘキサヒスチジンタグを伴う可溶性の野生型AcrA(レーン1、2)、AcrA−N273Q(レーン3、4)、およびAcrA−D121A(レーン5)のサンプルを含有する。パネルB:AcrA(レーン1、2)、AcrA−T145D(レーン3)、AcrA−N123Q−N273Q−T145D(レーン4、5)。パネルC:AcrA−F115D−T145D(レーン1、2)、AcrA−N123Q−N273Q−N272D(レーン3、4)。パネルD:AcrA−N273Q(レーン1、2)、AcrA−N273Q−F122P(レーン3、4)。パネルE:CtxB(レーン1、2)、CtxB−W88D(レーン3、4)、CtxB−Q56/DSNIT(レーン5、6)、およびCtxB−W88D−Q56/DSNIT。
【図3】OmpH1中の複数のグリコシル化部位の操作を示す図である。ΔwaaL株SCM6を、プラスミドpACYCpgl(全pgl遺伝子座をコードする)と、野生型OmpH1(レーン1)、OmpH1N139s−myc(レーン2)、OmpH1KGN→NIT、HFGDD→DSNIT−myc(レーン3)、OmpH1RGD→NIT、HFGDD→DSNIT−myc(レーン4)、OmpH1KGN→NIT、RGD→NIT−myc(レーン5)、OmpH1KGN→NIT、RGD→NIT、HFGDD→DSNIT−myc(レーン6)、またはOmpH1RGD→NIT V83T−myc(レーン7)を発現するプラスミドを用いて同時形質転換させた。細胞を好気的に増殖させ、分析の前に3時間、0.5%アラビノースで誘導させた。全細胞溶解物を、材料および方法のセクションに述べた通りに、培養物の光学濃度を均一にした後にTCA沈殿させた。15% SDS−PAGEによってタンパク質を分離し、PVDF膜上に移した。最初のパネル;抗mycタグ抗体を用いて探索された全細胞溶解物の免疫ブロット。下のパネル;グリカン特異的な抗血清を用いて探索された全細胞溶解物の免疫ブロット。非グリコシル化およびグリコシル化されたOmpH1の位置は、右側に示される。
【図4】様々なOmpH1変異体を発現する細胞の蛍光顕微鏡検査を示す図である。野生型OmpH1およびその変異体のための発現プラスミドを含有する大腸菌(E.coli)株CLM24またはSCM6の培養物を、等しくOD600の0.25/mlにした。細胞を、リン酸緩衝食塩水(PBS)(pH 7.4)で2回洗浄し、100μl細胞懸濁液を、ゼラチンを塗ったスライドガラス上に滴下し、加湿チャンバー内で30分間、室温(RT)でインキュベートした。全細胞免疫蛍光標識化におけるその後のステップはすべて、加湿チャンバー内で室温で行った。結合していない細胞を除去し、残留物を、室温で30分間、4%パラホルムアルデヒドを含有するPBSで固定した。重要なことに、パラホルムアルデヒドは、細胞を透過させないが、膜が無損傷であることによって区画化が保たれると考えられる。固定された細胞を、PBSで2回洗浄し、5% BSA(PBS中)を含有するブロッキング緩衝液に再懸濁した。ブロック後、細胞を、抗mycモノクローナルマウスIgG(1:50、カルバイオケム(Calbiochem))および/または抗グリカン抗血清(1:4000)と共に、5% BSAを含有する100μlのPBS中で、1時間インキュベートした。細胞を、100μlのPBSで3回、各5分間洗浄し、FITCと結合させた抗ウサギ二次抗体(1:250、ジャクソンイムノリサーチラボラトリーズ(Jackson Immunoresearch Laboratories))、および/またはCy3と結合させた抗マウス抗体(1:250、ジャクソンイムノリサーチラボラトリーズ(Jackson Immunoresearch Laboratories))と共に、5% BSAを含有する100μlのPBS中で1時間インキュベートした。必要とされる場合、細菌のDNAを染色するために、4,6−ジアミノ−2−フェニルインドール(DAPI)(シグマ(Sigma))(0.5μg/ml)を、二次抗体インキュベーション時に加えた。二次抗体を、PBSで細胞から洗い落とし、ベクタシールド(vectashield)(ベクターラボラトリーズ(Vector Laboratories))マウンティングメディアを使用することによって、カバースリップをスライドに載せ,マニキュア液を用いて密閉した。Axioplan2顕微鏡(カールツァイス(Carl Zeiss))を使用することによって、蛍光顕微鏡検査を実施した。Adobe Photoshop(バージョンCS2)を使用することによって、画像を組み合わせた。SCM6細胞は、以下を発現する:OmpH1(パネルA)、OmpH1N139S(パネルB)、OmpH1C20S(パネルC)、OmpH1KGN→NIT、HFGDD→DSNIT(パネルD)、OmpH1RGD→NIT、HFGDD→DSNIT(パネルE)、OmpH1KGN→NIT、RGD→NIT(パネルF)、OmpH1V83T、KGN→NIT(パネルG)、およびOmpH1KGN→NIT、RGD→NIT、HFGDD→DSNIT(パネルH)。第1列は、黒色バックグラウンド上に灰色のトーンで表された、第2、3、および4列の画像の組合せである。第2列:DAPI染色による灰色のトーン中の青色蛍光、第3列:グリカン特異的な蛍光による緑色蛍光、第4列:抗myc染色による赤色蛍光。
【発明を実施するための形態】
【0011】
本発明において使用される用語「部分的なアミノ酸配列」はまた、「最適化コンセンサス配列」とも称される。最適化コンセンサス配列は、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来のオリゴサッカリルトランスフェラーゼ(OST、OTアーゼ)によって、従来技術で知られている通常のコンセンサス配列「N−X−S/T」よりもかなり効率的にN−グリコシル化される。
【0012】
一般に、用語「組み換えN−グリコシル化タンパク質」は、前記タンパク質をコードする核酸を天然には含まない宿主細胞中で産生される任意の異種ポリペプチドまたはオリゴペプチドを指す。本発明において、この用語は、任意の宿主細胞、例えば真核または原核生物の宿主細胞、好ましくは原核の宿主細胞、例えばエシェリキア亜種(Escherichia ssp.)、カンピロバクター亜種(Campylobacter ssp.)、サルモネラ亜種(Salmonella ssp.)、シゲラ亜種(Shigella ssp.)、ヘリコバクター亜種(Helicobacter ssp.)、シュードモナス亜種(Pseudomonas ssp.)、バチルス亜種(Bacillus ssp.)、より好ましくは、大腸菌(Escherichia coli)、カンピロバクター ジェジュニ(Campylobacter jejuni)、サルモネラ チフィムリウム(Salmonella typhimurium)などの中で、組換えによって産生されるタンパク質を指す。ここでは、前記タンパク質をコードする核酸が、前記宿主細胞に導入される。また、コードされたタンパク質は、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来のOTアーゼ、天然に存在する、あるいは組換えにより前記宿主細胞に導入される前記トランスフェラーゼ酵素によってN−グリコシル化される。
【0013】
アミノ酸について国際的に認められた1文字記号によれば、省略形D、E、N、S、およびTはそれぞれ、アスパラギン酸、グルタミン酸、アスパラギン、セリン、およびトレオニンを表す。本発明のタンパク質は、1つまたは複数の最適化コンセンサス配列D/E−X−N−Z−S/Tが導入され、かつN−グリコシル化されるという点で、天然または従来技術のタンパク質とは異なる。したがって、本発明のタンパク質は、天然に存在するC.ジェジュニ(C.jejuni)タンパク質(これも、最適化コンセンサス配列を含有するが、任意のさらなる(導入された)最適化コンセンサス配列は含まない)とは異なる。
【0014】
最適化コンセンサス配列の導入は、1つまたは複数のアミノ酸の付加、欠損、および/または置換によって達成することができる。最適化コンセンサス配列を導入するための、1つまたは複数のアミノ酸の付加、欠損、および/または置換は、固相支持型の化学ペプチド合成などの、当業者によく知られている化学合成戦略によって達成することができる。あるいは、より大きなポリペプチドにとって好ましいことだが、本発明のタンパク質は、標準の組み換え技術によって調製することができる。
【0015】
本発明のタンパク質は、高効率で、かつ、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロンを含む任意の原核の宿主中で産生され得るという利点を有する。本発明の態様および実施形態を実践するためのカンピロバクター種由来の好ましい代替のOTアーゼは、カンピロバクター コリ(Campylobacter coli)およびカンピロバクター ラリ(Campylobacter lari)である(シマンスキーC.M.(Szymanski C.M.)およびレン B.W.(Wren B.W.)(2005年)、「Protein glycosylation in bacterial mucosal pathogens.」「Nat.Rev.Microbiol.」3:225〜237を参照のこと)。前記原核宿主がカンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)である場合、機能性のpglオペロンは、天然に存在するものであり得る。しかし、当分野で以前に実証された通り、また、上述した通り、pglオペロンは、細胞に導入することができ、前記新規の細胞の環境中で機能性のままでいることが可能である。
【0016】
用語「カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロン」は、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)の機能性のオリゴサッカリルトランスフェラーゼ(OTアーゼ)、および脂質担体上にオリゴ糖を構築することが可能な1種または複数の特異的なグリコシルトランスフェラーゼをコードする核酸のクラスターを指すものとする。ここでは、前記オリゴ糖は、OTアーゼによって、脂質担体から、1つまたは複数の最適化アミノ酸配列:D/E−XN−Z−S/Tを有する標的タンパク質に転移することができる。用語「カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロン」は、本発明において、必ずしも単一の転写単位としてのオペロンを指すというわけではないことを理解するべきである。この用語は、単に、ある宿主細胞における、組み換えタンパク質のN−グリコシル化のための機能性の成分の存在を要するだけである。これらの成分は、1つまたは複数の別々のmRNAとして転写されてもよく、一緒にあるいは別々に調節されてもよい。例えば、この用語はまた、ある宿主細胞中のゲノムDNAおよびプラスミド中に位置する機能性の成分も包含する。効率のためには、機能性のpglオペロンのすべての成分が、調節されおよび同時に発現されることが好ましい。
【0017】
機能性のオリゴサッカリルトランスフェラーゼ(OTアーゼ)だけは、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)に由来するべきであること、また、脂質担体上にオリゴ糖を構築することが可能な1つまたは複数の特異的なグリコシルトランスフェラーゼは、宿主細胞に由来するものであっても組換えにより前記宿主細胞に導入されるものであってもよいが、唯一の機能的制約は、前記グリコシルトランスフェラーゼによって構築されるオリゴ糖が、OTアーゼによって、脂質担体から、1つまたは複数の最適化コンセンサス配列を有する標的タンパク質に転移可能であるということであることを認識することは重要である。したがって、特異的なグリコシルトランスフェラーゼを天然に含む宿主細胞の選択および/または前記宿主中に天然に存在する特異的なグリコシルトランスフェラーゼを無能力化することおよび異種特異的なグリコシルトランスフェラーゼの導入によって、当業者は、本発明のタンパク質中の最適化N−グリコシル化コンセンサス部位に結合されたN−グリカンを変えることが可能となる。
【0018】
上記の結果として、本発明は、本発明のタンパク質上のN−グリカン−パターンの個々の設計を提供する。したがって、タンパク質は、生物学、薬学、および精製の要件に合わせるために、そのN−グリカンパターンにおいて個別化させることができる。
【0019】
好ましい実施形態では、本発明のタンパク質は、1つ、さらに2つ以上、好ましくは少なくとも2つ、好ましくは少なくとも3つ、より好ましくは少なくとも5つの前記N−グリコシル化された最適化アミノ酸配列を含んでよい。
【0020】
本発明のタンパク質中の1つまたは複数のN−グリコシル化された最適化アミノ酸配列の存在は、その抗原性を増大させること、その安定性を増大させること、その生物活性に影響を及ぼすこと、その生物学的半減期を延長させること、および/またはその精製を単純化させることのために好都合であり得る。
【0021】
最適化コンセンサス配列は、位置XおよびZに、プロリン以外の任意のアミノ酸を含んでよい。用語「任意のアミノ酸」は、最適化コンセンサス配列を、OTアーゼによってN−グリコシル化させることが可能となる、一般的および稀な天然のアミノ酸、ならびに合成アミノ酸誘導体および類似体を包含するものとする。天然に存在する一般的および稀なアミノ酸が、XおよびZに対して好ましい。XとZは、同じであっても異なってもよい。
【0022】
XおよびZが、本発明のタンパク質中の最適化コンセンサス配列各々について異なっていてもよいことに注意されたい。
【0023】
OTアーゼによる転移のために、脂質担体上にオリゴ糖を構築する場合、最適化コンセンサス配列に結合されるN−グリカンは、特異的なグリコシルトランスフェラーゼおよびその相互作用によって決定されることとなる。当業者は、所望の宿主細胞中に存在する特異的なグリコシルトランスフェラーゼのタイプおよび量を変化させることによって、N−グリカンを設計することができる。
【0024】
N−グリカンは、本明細書では、Nグリコシド結合を介してタンパク質中のアスパラギン残基のε−アミド窒素と連結された、様々な組成の単糖、オリゴ糖、または多糖と定義される。好ましくは、OTアーゼによって転移されるN−グリカンは、グラム陰性または陽性細菌の細胞膜に存在するウンデカプレノール−ピロリン酸脂質−アンカー上に構築される。これは、O抗原、O多糖、およびペプチドグリカンの合成に関与する(ビュック,T.D.(Bugg,T.D.)およびブランディッシュ,P.E.(Brandish,P.E.)(1994年)、「From peptidoglycan to glycoproteins:common features of lipid−linked oligosaccharide biosynthesis.」「FEM Microbiol Lett」119、255〜262;ヴァルヴァノ,M.A.(Valvano,M.A.)(2003年)、「Export of O−specific lipopolysaccharide.」「Front Biosci」8、s452〜471)。
【0025】
好ましい実施形態では、本発明の組み換えタンパク質は、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来のN−グリカンの群から選択される1種または複数のN−グリカンを含み、N−グリカンは、グラム陰性細菌中のO多糖形成O抗原に転移されるオリゴ糖および多糖、あるいはグラム陽性細菌由来の莢膜多糖、好ましくは:緑膿菌(P.aeruginosa)O9、O11;大腸菌(E.coli)O7、O9、O16、O157、および志賀赤痢菌(Shigella dysenteriae)O1ならびに多糖構造に影響を及ぼすグリコシルトランスフェラーゼおよびエピメラーゼを挿入するあるいは欠損させることによって得られるそれらの操作された変異体から得られる。
【0026】
さらなる好ましい実施形態では、本発明の組み換えタンパク質は、2つ以上の異なるN−グリカンを含む。
【0027】
例えば、同じタンパク質上の異なるN−グリカンは、早いまたは遅いプロモーターを使用して、あるいは個々の特異的なグリコシルトランスフェラーゼのプロモーター活性を開始する、静まらせる、強化する、および/または低下させるための因子を導入して、特異的なグリコシルトランスフェラーゼの発現のタイミングを制御することによって調製することができる。それらの活性を制御する適切なプロモーターおよび因子は、当分野でごく普通に利用可能なものであるので、これ以上は論じない。
【0028】
本発明の組み換えタンパク質の起源に対する制約はない。好ましくは、前記タンパク質は、哺乳類、細菌、ウイルス、菌類、または植物のタンパク質から得られる。より好ましくは、タンパク質は、哺乳類、最も好ましくはヒトのタンパク質から得られる。本発明の抗原性の組み換えタンパク質を調製するために、好ましくはワクチン中で活性成分を使用するためには、組み換えタンパク質は、細菌、ウイルス、または菌類のタンパク質から得られることが好ましい。
【0029】
さらなる好ましい実施形態では、本発明は、組み換えタンパク質を提供する。ここでは、タンパク質および/またはN−グリカンは、治療および/または予防的に活性である。少なくとも1つの最適化N−グリコシル化されたコンセンサス配列を導入することによって、タンパク質における治療および/または予防的活性を改変する、さらには導入することができる。より好ましい実施形態では、これは、免疫原的に活性であるタンパク質および/またはN−グリカンである。この場合、導入されるN−グリコシル化は、タンパク質の生物活性に対する改変効果を有していてもよく、および/または新規の抗原性部位を導入し、および/または分解ステップを避けるためにタンパク質をマスキングしてもよく、および/または半減期を増大する。
【0030】
本発明の組み換えタンパク質を、宿主細胞、好ましくは細菌、より好ましくはグラム陰性細菌の外膜および/または表面に効率的にターゲティングすることができる。表面ディスプレイおよび/または外膜局在化を助けるために、本発明の組み換えタンパク質は、細菌、好ましくはグラム陰性細菌の外膜および/または細胞表面に前記組み換えタンパク質をターゲティングすることが可能な少なくとも1つのポリペプチド配列をさらに含むことが好ましい。
【0031】
好ましい実施形態では、本発明の組み換えタンパク質は、前記ターゲティングするポリペプチド配列が、タイプIIシグナルペプチド(パエトゼル,M.(Paetzel,M.)、カリア,A.(Karia、A.)、ストリナドカ,N.C.(Strynadka,N.C.)、およびダルビー,R.E.(Dalbey,R.E.)、2002年、「Signal peptidases.」「Chem Rev」102:4549〜4580)、または外膜タンパク質(ウェメラス,H.(Wemerus,H.)およびシュタール,S.(Stahl,S.)2004年、「Biotechnological applications for surface−engineered bacteria.」「Biotechnol Appl Biochem」40:209〜228に概説)からなる群から選択される、好ましくはC.ジェジュニ(C.jejuni)由来のOmpH1、C.ジェジュニ(C.jejuni)由来のJlpA、大腸菌(E.coli)由来の外膜タンパク質、好ましくはOmpS、OmpC、OmpA、OprF、PhoE、LamB、Lpp’OmpA(表面ディスプレイ技術のための融合タンパク質、フランシスコ,J.A.(Francisco,J.A.)、エアハート,C.F.(Earhart,C.F.)、およびジョルジオ,G.(Georgiou,G.、1992年、「Transport and anchoring of beta−lactamase to the external surface of Escherichia coli.」「Proc Natl Acad Sci U S A」89:2713〜2717を参照のこと)、および緑膿菌(Pseudomonas aeruginosa)由来のlnpタンパク質の全長タンパク質またはシグナルペプチドからなる群から選択されるものである。
【0032】
異なる態様では、本発明は、本発明の組み換えタンパク質をコードする核酸に関する。好ましくは、前記核酸は、mRNA、DNA、またはPNA、より好ましくは、mRNAまたはDNA、最も好ましくはDNAである。核酸は、前記タンパク質をコードする配列、また、それに加えて制御配列(例えば、プロモーター、エンハンサー、終止コドン、開始コドン)などの他の配列、および、挙げられた制御配列などを介して組み換えタンパク質の発現を調節するために必要とされる遺伝子を含む可能性がある。用語「本発明の組み換えタンパク質をコードする核酸」は、前記コード配列と、場合によっては任意のさらなる核酸配列(カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロンを含有する宿主細胞中で、この核酸が本発明の組み換えタンパク質を産生することが可能な限り、配列情報を問わない)とを含む核酸を指す。より好ましくは、本発明は、プロモーターに作動可能に連結される、好ましくは、原核生物の既知の誘導性の恒常的プロモーターからなる群から選択されるプロモーター、より好ましくは、テトラサイクリンプロモーター、アラビノースプロモーター、サリチル酸プロモーター、lac、trc、およびtacプロモーター(バネイクス,F.(Baneyx、F.)(1999年)、「Recombinant protein expression in Escherichia coli.」「Curr Opin Biotechnol」10、411〜421;ビルマン−ヤコブ,H.(Billman−Jacobe,H.)(1996年)、「Expression in bacteria other than Escherichia coli.」「Curr Opin Biotechnol」7、500〜504)に連結される、単離および精製された核酸を提供する。前記作動可能に連結された核酸は、例えばワクチン接種のために使用することができる。
【0033】
さらに、本発明の別の態様は、本発明の核酸および/またはベクターを含む宿主細胞に関する。宿主細胞のタイプは、それがC.ジェジュニ(C.jejuni)由来の機能性のpglオペロン、および本発明の組み換え標的タンパク質をコードする1種または複数の核酸に適応する限り、制限されない。好ましい宿主細胞は、原核生物の宿主細胞、より好ましくは細菌、最も好ましくは以下からなる群から選択されるものである:エシェリキア亜種(Escherichia ssp.)、カンピロバクター亜種(Campylobacter ssp.)、サルモネラ亜種(Salmonella ssp.)、シゲラ亜種(Shigella ssp.)、ヘリコバクター亜種(Helicobacter ssp.)、シュードモナス亜種(Pseudomonas ssp.)、バチルス亜種(Bacillus ssp.)、好ましくは大腸菌(E.coli)、より好ましくは、大腸菌(E.coli)株Top10、W3110、CLM24、BL21、SCM6、およびSCM7(Feldman(フェルドマン)ら(2005年)、「Engineering N−linked protein glycosylation with diverse O antigen lipopolysaccharide structures in Escherichia coli.」「Proc Natl. Acad. Sci.U S A」102、3016〜3021;アライモ,C.(Alaimo、C.)、キャトレイン,I.(Catrein、I.)、モルフ,L.(Morf,L.)、マロルダ,C.L.(Marolda,C.L.)、カレワエルト,N.(Callewaert,N.)、ヴァルヴァノ,M.A.(Valvano、M.A.)、フェルドマン,M.F.(Feldman,M.F.)、アービィ,M.(Aebi,M.)(2006年)、「Two distinct but interchangeable mechanisms for flipping of lipid−linked oligosacchardes.」「EMBO journal」25、967〜976)、およびS.エンテリカ(S.enterica)株SL3261(サルモネラエンテリカ(Salmonela Enterica)sv.ティフィリウム(Typhimurium)LT2(デルタ)aroA、ヘイセト,S.K.(Hoiseth,S.K.)およびストッカー,B.A.(Stocker,B.A.)、1981年、「Aromatic−dependent Salmonella typhimurium are non−virulent and effective as live vaccines.」「Nature」291:238〜239を参照のこと)、SL3749(サルモネラエンテリカ(Salmonella enterica)sv.ティフィリウム(Typhimurium)LT2 waaL、カニュク(Kaniuk)ら、「J.Biol.Chem.」279:36470〜36480を参照のこと)、およびSL326ΔwaaL。
【0034】
より好ましい実施形態では、本発明の宿主細胞は、本発明の組み換えタンパク質の外膜および/または表面ディスプレイへのターゲティングに有用であるもの、好ましくは、前記宿主細胞が、組み換えグラム陰性細菌であって:
i)以下をコードするヌクレオチド配列を含む遺伝子型
a)脂質担体上へのオリゴ糖の構築のための少なくとも1種の天然または組み換え型の特異的なグリコシルトランスフェラーゼ、
b)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の少なくとも1種の天然または組み換え型の原核生物オリゴサッカリルトランスフェラーゼ(OTアーゼ)、
c)本発明の少なくとも1種の組み換えタンパク質、好ましくは、ターゲティングポリペプチドをさらに含むタンパク質、および
ii)グラム陰性細菌の外膜内および/または上に位置する、本発明の組み換えN−グリコシル化タンパク質を含む表現型、
を有するグラム陰性細菌である。
【0035】
上記形態のための宿主細胞は、好ましくは、以下からなる群から選択される:エシェリキア亜種(Escherichia ssp.)、カンピロバクター亜種(Campylobacter ssp.)、シゲラ亜種(Shigella ssp.)、ヘリコバクター亜種(Helicobacter ssp.)、およびシュードモナス亜種(Pseudomonas ssp.)、サルモネラ亜種(Salmonella ssp.)、好ましくは大腸菌(E.coli)、より好ましくは、大腸菌(E.coli)株Top10、W3110、CLM24、BL21、SCM6、およびSCM7、ならびにS.エンテリカ(S.enterica)株SL3261、SL3749、およびSL3261ΔwaaL.(ヘイセト,S.K.(Hoiseth,S.K.)およびストッカー,B.A.(Stocker,B.A.)、1981年、「Aromatic−dependent Salmonella typhimurium are non−virulent and effective as live vaccines.」「Nature」291:238〜239を参照のこと)、SL3749(カニュク,N.A.(Kaniuk,N.A.)、ビノグラードフ,E.(Vinogradov,E.)、およびホイットフィールド,C.(Whitfield,C.)2004年、「Investigation of the structural requirements in the lipopolysaccharide core acceptor for ligation of O antigens in the genus Salmonella: WaaL“ligase”is not the sole determinant of acceptor specificity.」「J Biol Chem」279:36470〜36480)。
【0036】
本発明の好ましいタンパク質は、それ自身で、および/または導入されたN−グリコシル化部位に起因して、治療または予防的活性を有する可能性があるので、これらは、医薬品の調製のために使用することができる。本発明を実施するのためのタンパク質のタイプは、限定されないので、数例を挙げると、その中に最適化されたN−グリコシル化されたコンセンサス配列が導入された、EPO、IFN−アルファ、TNFアルファ、IgG、IgM、IgA、インターロイキン、サイトカイン;C.ジェジュニ(C.jejuni)タンパク質のようなワクチン接種のためのウイルスおよび細菌のタンパク質など(HisJ(Cj0734c)、AcrA(Cj0367c)、OmpH1(Cj0982c)、ジフテリア毒素(CRM197)、コレラ毒素、緑膿菌エキソプロテイン(exoprotein)など)の本発明のタンパク質が、医薬品を調製するために有用である(ウィシンスカ,A(Wyszynska,A.)、ラズコー,A.(Raczko,A.)、リス,M.(Lis,M.)、およびジャグシン−クリニカ(Jagusztyn−Krynicka),E.K.(2004年)、「Oral immunization of chickens with avirulent Salmonella vaccine strain carrying C.jejuni 72Dz/92 cjaA gene elicits specific humoral immune response associated with protection against challenge with wild−type Campylobacter.」「Vaccine 22」、1379〜1389)。
【0037】
さらに、本発明の核酸および/またはベクターも、医薬品の調製のために、好ましくは遺伝子治療で使用するために有用である。
【0038】
さらに、本発明の宿主細胞、好ましくは、細菌(好ましくはグラム陰性細菌、より好ましくは上記に列挙したグラム陰性細菌のうちの1種)の外膜内および/または上に位置する本発明のN−グリコシル化された組み換えタンパク質を含む表現型を有するものが、医薬品の調製のために特に有用である。
【0039】
より好ましくは、本発明のタンパク質は、その必要がある対象の治療および/または予防用ワクチン接種のための医薬品の調製のために使用される。
【0040】
より好ましい実施形態では、本発明は、好ましくは遺伝子治療による、その必要がある対象の治療および/または予防用ワクチン接種のための医薬品の調製のための、本発明の核酸および/またはベクターの使用に関する。
【0041】
ディスプレイされたN−グリコシル化タンパク質が、宿主細胞の表面上に多量に存在し、免疫細胞、特に、その親水性N−グリカンによって十分に利用可能であるので、また、宿主細胞が、アジュバントのそれ以上の効果を有する(生存する場合、さらにある程度複製し、そのワクチン接種効果を増幅してもよい)ので、前記N−グリコシル化された組み換えタンパク質をディスプレイする本発明の宿主細胞は、ワクチンを調製するために特に有用である。
【0042】
好ましくは、本発明の医療態様を実践するための宿主細胞は、弱毒化されたあるいは殺された宿主細胞である。
【0043】
医薬品、好ましくはワクチンを調製することに対する本発明の宿主細胞の使用の別の利点は、それが、細胞の成分に起因するIgA抗体を誘導することである。
【0044】
好ましくは、前記宿主細胞は、動物(好ましくは哺乳類、げっ歯類、ヒツジ、ウマ、イヌ、ウシ、またはヒト)中にIgA抗体を誘導するために、本発明に従って使用される。
【0045】
ワクチン接種を必要とする前記対象は、鳥類、哺乳類、または魚類、好ましくは哺乳類、より好ましくは、ウシ、ヒツジ、ウマ、イヌ、ネコ、およびヒトからなる群から選択される哺乳類、最も好ましくはヒトであることが好ましい。家禽類もまた、好ましい。
【0046】
本発明のさらなる態様は、本発明の少なくとも1種のタンパク質、少なくとも1種の核酸、少なくとも1種のベクター、および/または少なくとも1種の宿主細胞を含む薬剤組成物に関する。タンパク質または宿主細胞、好ましくは弱毒化されたあるいは殺された宿主細胞を含む医薬品の調製、および、遺伝子治療のための核酸および/またはベクターを含む医薬品の調製は、当分野でよく知られている。最終の薬剤組成物のための調製スキーム、ならびにその投与の方式および詳細は、用いられるタンパク質、宿主細胞、核酸、および/またはベクターに依存することとなる。
【0047】
好ましい実施形態では、本発明の薬剤組成物は、薬学的に許容される賦形剤、希釈剤、および/または補助剤を含む。
【0048】
本発明は、以下のうちの少なくとも1種を含む薬剤組成物を提供する:(i)本発明の組み換えタンパク質である/をコードする/を発現する組み換えタンパク質、宿主細胞、核酸、および/または組み換えベクター、および(ii)薬学的に許容される賦形剤、希釈剤、および/または補助剤。
【0049】
適切な賦形剤、希釈剤、および/または補助剤は、当分野でよく知られている。賦形剤または希釈剤は、活性成分のための媒体または媒質として働いてよい固体、半固体、または液体材料であってよい。組成物を調製する分野における当業者であれば、選択される産物の特定の特性、治療される疾患または状態、疾患または状態の段階、および他の関連する状況に応じて、投与の適切な形および様式を容易に選択することができる(「Remington’s Pharmaceutical Sciences」、マックパブリッシング(Mack Publishing Co.)(1990年))。薬学的に許容される希釈剤または賦形剤の割合および性質は、選択される薬学的に活性な化合物の溶解性および化学的性質、選択される投与経路、および標準の薬務によって決定される。製剤は、経口、非経口、または局所使用に適合されてもよく、また、錠剤、カプセル、座薬、液剤、懸濁剤などの形で患者に投与されてもよい。本発明の薬学的に活性な化合物は、それ自体で有効でありながら、安定性、結晶化の都合、溶解性の増大などの目的で、酸付加塩または塩基付加塩などの薬学的に許容されるその塩の形で調製および投与することもできる。
【0050】
本発明のさらなる態様は、N−結合グリコシル化タンパク質を産生するための方法であって、
a)以下をコードする核酸を含む組み換え生物、好ましくは原核生物を提供するステップと、
i)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロン、および
ii)1つまたは複数の下記N−グリコシル化された最適化アミノ酸コンセンサス配列を含む少なくとも1種の組み換え標的タンパク質であって:
D/E−X−N−Z−S/T
式中、XおよびZは、Pro以外の任意の天然のアミノ酸であり得、少なくとも1つの前記N−グリコシル化された最適化アミノ酸コンセンサス配列が導入される組み換え標的タンパク質、および
b)標的タンパク質の産生およびN−グリコシル化に適した方式で、組み換え生物を培養するステップと、
を含む方法に関する。
【0051】
好ましくは、標的タンパク質は、本発明の上記の組み換えタンパク質のうちの1つである。
【0052】
本発明の好ましい方法では、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロンは、
OTアーゼによって標的タンパク質に転移されることとなる、脂質担体上へのオリゴ糖の構築のための、
i)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の組み換えOTアーゼ、および
OTアーゼによって標的タンパク質に転移されることとなる、脂質担体上へのオリゴ糖の構築のための、
ii)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の組み換え型および/または天然の特異的なグリコシルトランスフェラーゼ、および/または
iii)カンピロバクター種以外の種から得られる組み換え型および/または天然の特異的なグリコシルトランスフェラーゼ、
をコードする核酸を含む。
【0053】
さらに、好ましい実施形態では、本発明は、本発明の宿主細胞を調製するための方法であって、
i)グラム陰性細菌を提供するステップと、
ii)以下をコードする少なくとも1つのヌクレオチド配列を前記細菌に導入するステップと、
a)脂質担体上へのオリゴ糖の構築のための少なくとも1種の組み換え型の特異的なグリコシルトランスフェラーゼ、および/または
b)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の少なくとも1種の組み換えオリゴサッカリルトランスフェラーゼ(OTアーゼ)、および/または
c)1つまたは複数の下記N−グリコシル化された最適化アミノ酸コンセンサス配列を含む少なくとも1種の組み換えタンパク質であって:
D/E−X−N−Z−S/T
式中、XおよびZは、Pro以外の任意の天然のアミノ酸であり得、少なくとも1つの前記N−グリコシル化された最適化アミノ酸コンセンサス配列が導入される組み換えタンパク質、
iii)c)のヌクレオチド配列によってコードされる少なくとも1種の組み換えN−グリコシル化タンパク質が、グラム陰性細菌の外膜内および/または上に位置されるまで、前記細菌を培養するステップと、
を含む方法に関する。
【0054】
上記好ましい方法を実施するために、組み換え型の原核生物または宿主細胞は、好ましくは、エシェリキア亜種(Escherichia ssp.)、カンピロバクター亜種(Campylobacter ssp.)、サルモネラ亜種(Salmonella ssp.)、シゲラ亜種(Shigella ssp.)、ヘリコバクター亜種(Helicobacter ssp.)、シュードモナス亜種(Pseudomonas ssp.)、バチルス亜種(Bacillus ssp.)、好ましくは大腸菌(Escherichia coli)、好ましくは大腸菌(E.coli)株Top10、W3110、CLM24、BL21、SCM6、およびSCM7、ならびにS.エンテリカ(S.enterica)株SL3261、SL3749、およびSL3261ΔwaaLからなる細菌の群から選択される。
【0055】
本発明の組み換えタンパク質を産生、単離、および/または精製するための別の好ましい方法は、次のステップを含む:
a)請求項15または16に記載の宿主細胞を培養するステップ、
b)前記組み換えグラム陰性細菌の外膜を除去するステップ、および
c)前記組み換えタンパク質を回収するステップ。
【0056】
細胞、好ましくは原核細胞、より好ましくはグラム陰性細菌細胞の外膜を除去するための例示的な方法は、適切な酵素処理方法、浸透圧衝撃、界面活性剤可溶化、およびフレンチプレス法である。
【0057】
最も好ましくは、本発明は、カンピロバクター種以外の種、好ましくはC.ジェジュニ(C.jejuni)から得られる組み換え型または天然の特異的なグリコシルトランスフェラーゼが、前記宿主細胞中で機能的に発現させることが可能である細菌、古細菌、および/または真核生物に由来するグリコシルトランスフェラーゼおよびエピメラーゼの群から選択される方法に関する。
【0058】
以下の実施例は、本発明をさらに説明する役割を果たすものであり、いずれにせよその範囲を限定するものではない。
【実施例】
【0059】
実施例
N−グリコシル化を最適化するためのモデルタンパク質としてのAcrAの選択
N−グリコシル化のためのアクセプタータンパク質の要件を最適化するために、C.ジェジュニ(C.jejuni)糖タンパク質AcrA(Cj0367c)に対して、詳細な研究を実施した。AcrAは、350アミノ酸残基の周辺質リポタンパク質である。脂質に固定されるのではなく周辺質に分泌されることが、グリコシル化のための必要条件であることが示されている(ニタ−ラザー(Nita−Lazar)ら、2005年、上記)。大腸菌(E.Coli)中で発現される場合、搬出のためのシグナルは、天然のAcrAシグナル配列または異種起源のPelBシグナルであり得る。5つの潜在的なN−結合グリコシル化シークオン(sequon)(N117、N123、N147、N273、N274)のうち、2つの同じものが、C.ジェジュニ(C.jejuni)および大腸菌中で使用される(N123およびN273(ニタ−ラザー(Nita−Lazar)ら、2005年、上記))。AcrAは、詳細な構造情報が利用可能であるC.ジェジュニ(C.jejuni)の唯一の周辺質N−糖タンパク質であるので、モデルとして選択された。近年では、AcrA相同体、すなわちグラム陰性細菌緑膿菌(P.aeruginosa)由来のMexAタンパク質の結晶構造が公開されている(ヒギンズ(Higgins)ら(2004年)、「Structure of the periplasmic component of a bacterial drug efflux pump.」「Proc.Natl.Acad.Sci.U S A」101、9994〜9999)。どちらのタンパク質も、いわゆる周辺質排出ポンプタンパク質のメンバーである(PEP、(ジョンソン,J.M.(Johnson,J.M.)およびチャーチ,G.M.(Church,G.M.)(1999年))「Alignment and structure prediction of divergent protein families:periplasmic and outer membrane proteins of bacterial efflux pumps.」「J.Mol.Biol.287、695〜715))。細長い分子は、3つの直線的に配置されたサブドメイン:αらせんの逆平行のコイルドコイルを含有し、これは、リポイルドメインによって塩基で結びつけられ、これに、6本鎖のβバレル構造ドメインが続く。N末端の23〜28残基およびC末端における95〜101残基は、結晶中に組み込まれていない。MexAタンパク質とAcrAタンパク質の配列は、29.3%同一であり、50%類似である。したがって、2つのタンパク質はおそらく、全体的に類似の折りたたみを示す。
【0060】
実施例1 グリコシル化を誘発する第1のペプチド配列の解明
緑膿菌のMexAと類似である、したがってまた、C.ジェジュニ(C.jejuni)のAcrA中にあるリポイルドメインは、大腸菌において個々に発現させることができるコンパクトなタンパク質を形成することが知られている(バーグ,A.(Berg,A.)、およびデコック,A.(de Kok,A.)(1997年)、「2−Oxo acid dehydrogenase multienzyme complexes.」「The central role of the lipoyl domain.Biol.Chem.」378、617〜634によって概説される)。どのアクセプターペプチド配列が、大腸菌におけるpgl機構によるN−グリコシル化に必要とされるのかを確認するために、AcrAのリポイルドメインを取り出した。これを、異なる長さのペプチドを周辺質に輸送し、それらをin vivoでpgl機構に与えるための分子骨格として使用した。
【0061】
したがって、リポイルドメイン(Lip)をコードするプラスミドを構築し、OmpA(チェ,J.H.(Choi,J.H.)およびリー,S.Y.(Lee,S.Y.)(2004年)、「Secretory and extracellular production of recombinant proteins using Escherichia coli.」「Appl Microbiol Biotechnol」64、625〜635)のシグナル配列にN末端を、ヘキサヒスチジンタグにC末端を融合させた。クローニングを行い、アラビノースプロモーターの制御下で、遺伝子発現を行った。Lipドメイン境界については、MexA中でのリポイルドメイン部分のドメイン境界と同じ位置で出現すると考えられるアミノ酸位を選択した。N−グリカンを受け入れるその能力について、異なるペプチドを試験するために、配列のストレッチを、Lipドメインの2つのハンマーヘッド様部分の間に挿入した。ストレッチは、C.ジェジュニ(C.jejuni)AcrAのN−グリコシル化部位N123を含む配列からなるものであった。得られたオープンリーディングフレームは、OmpAシグナル配列をコードする配列、AcrAのN末端基ハンマーヘッド様部分(D60〜D95、アミノ酸の番号付けは、成熟したAcrAポリペプチドの配列番号付けに関連する)、AcrAの天然のN123グリコシル化部位を含有する別のストレッチ(下記参照)、AcrA−Lip(L167−D210)のC末端のハンマーヘッド様部分、およびC末端のhis−タグからなるものであった。
【0062】
プラスミドの構築は、標準の分子生物学技術によって達成された。異なる長さのAcrAの天然のN123グリコシル化部位を含有する3つのストレッチを、Lipの中間に挿入し、3つの異なるORFをもたらした:
構築物Aは、次のタンパク質配列をもたらすA118−S130を含有する:
【化1】
構築物Bは、次のタンパク質配列をもたらすF122−E138を含有する:
【化2】
構築物Cは、次のタンパク質配列をもたらすD121−A127を含有する:
【化3】
下線が引かれた配列のストレッチは、OmpAシグナルペプチドを示し、単独で下線が引かれた残基は、クローン化のために、あるいはタンパク質を分解に対して耐性にするために導入されたものである。太字:AcrAのN123に相当するグリコシル化部位。イタリック体:ヘキサ−ヒスチジンタグ。相当する遺伝子を、プラスミドpEC415(シュルツ,H.(Schulz,H.)、ヘンネック,H.(Hennecke,H.)、およびトニー−マイヤー,L.(Thony−Meyer,L.)(1998年)、「Prototype of a heme chaperone essential for cytochrome c maturation.」「Science」281、1997〜1200)の主鎖中のアラビノースプロモーターの制御下で発現させた。
【0063】
3つのストレッチのうちのどれがLipタンパク質のグリコシル化を誘発したかを確認するために、タンパク質発現実験を実施した。pACYCpglまたはpACYCpglmut(ワッカー(Wacker)ら、2002年、上記)と、構築物A、B、またはCをコードするプラスミドを保有する大腸菌Top10細胞(インビトロジェン(Invitrogen)、米国カリフォルニア州カールズバッド(Carlsbad,CA,USA))を、アンピシリンおよびクロラムフェニコールを含有するLB媒質中で37℃で0.5ODまで増殖させた。誘導のために1/1000体積の20%アラビノース(w/v)溶液を加え、さらに2時間細胞を増殖させた。細胞を、遠心分離によってその後収集し、20mMトリス/HCl(pH 8.5)、20%スクロース(w/v)、1mM EDTA、1mM PMSF、および1g/l(w/v)リゾチーム中に再懸濁させ、4℃で1時間保温した。球状の芽球(spheroblasts)をペレット化すると、周辺質抽出物が得られ、これを1/9体積(v/v)の10×緩衝液A(3M NaCl、0.5MのTris/HCl(pH 8.0)、および0.1M イミダゾール)で希釈し、MgSO4を加えて2.5mMにした。Ni−アフィニティ精製は、緩衝液A中で、アマシャムファルマシアバイオテック(Amersham Pharmacia Biotech)(スウェーデン、ウプサラ(Uppsala,Sweden))からの1ml Ni−セファロース(Sepharose)カラム上で実施した。タンパク質は、0.25M イミダゾールを含有する緩衝液Aで溶出させた。
【0064】
図1は、クーマシーブリリアントブルー染色した、Ni−精製された周辺質抽出物からのピーク溶出分画のSDS−PAGEゲルを示す。発現分析によって、構築物Bが、突出した単一のタンパク質種を産生することが示された(図1、レーン1)。構築物AとCは両方とも、突出したタンパク質に加えて、電気泳動移動度がより遅い第2のタンパク質バンドをもたらす(図1、レーン2および3)。より重いタンパク質種が確かにグリコシル化されたことが、MALDI−TOF/TOFによって証明された(図示せず)。構築物B中では失われたが、AおよびCでは存在する唯一のアミノ酸は、D121、すなわちグリコシル化されたN123に対して2つN末端方向に位置するアスパラギン酸残基であった。このことは、D121が、OTアーゼによるグリコシル化のために重要な役割を果たすことを実証している。D121がグリコシル化に不可欠なものであるということを検証するために、それは構築物C発現分析においてアラニンへ突然変異され、1つだけのタンパク質バンドをもたらし(図1、レーン4)、その結果、D121がグリコシル化に重要であることを示した。さらに、人工のペプチドディスプレイタンパク質がグリコシル化され得るという事実は、D/E−X−N−Y−S/Tタイプの短いペプチドが、C.ジェジュニ(C.jejuni)が担うN−グリコシル化が起こるためのすべての情報を含有することを示す。
【0065】
実施例2 実施例1の検証;AcrA−D121Aは、N123でグリコシル化されない
ペプチドディスプレイ手法による発見を確認するために、アスパラギン酸からアラニンへの突然変異を、位置121(D121A、すなわちグリコシル化されたN123よりも2つ前の残基)に挿入し、AcrAタンパク質の全長の可溶性バージョンにおいて、部位N123が大腸菌中で依然としてグリコシル化され得るかどうかを試験した。試験を行うために、このAcrA−D121Aを発現させ、そのグリコシル化状態を分析した。分析のために、操作されたAcrAを使用した。これは、周辺質への分泌のためのPelBシグナル配列(チェ(Choi)およびリー(Lee)、2004年、上記)、および、精製のためのC末端のヘキサヒスチジンタグを含有するという点で、本来のC.ジェジュニ(C.jejuni)遺伝子とは異なっていた。このAcrA変異体は、脂質に固定された未変性タンパク質として分泌され、シグナルペプチド切断され、グリコシル化されることが示されている(ニタ−ラザー(Nita−Lazar)ら、2005年、上記))。以下は、可溶性のAcrAタンパク質のタンパク質配列である。
【化4】
【0066】
下線が引かれた残基は、PelBシグナルペプチドであり、イタリック体は、ヘキサ−ヒスチジンタグであり、太字は、N123およびN273の2つの天然のグリコシル化部位である。pEC415プラスミド(シュルツ(Schulz)ら、1998年)中に上記のタンパク質に対するORFを含有するプラスミドを、pAcAperを産生するために構築した。
【0067】
AcrAおよびそのミュータント(下記参照)のグリコシル化状態を試験するアッセイは、以下の通りであった:その活性または不活性の形の、プラスミドが担うpglオペロン(pACYCpglまたはpACYCpglmut、(ワッカー(Wacker)ら、2002年、上記)を参照のこと)と、AcrAをコードするプラスミド(pAcrAper)を含有する、指数的に増殖する大腸菌CLM24(フェルドマン(Feldman)ら、2005年、上記)細胞において、0.02%アラビノースを用いて、AcrAの発現を誘導した。4時間の誘導の後、周辺質抽出物を、上述した通りに調製し、抗AcrA抗血清またはR12抗血清のいずれかを用いる、SDS−PAGE、電気移動(electrotransfer)、および免疫検出によって分析した。R12抗血清は、C.ジェジュニ(C.jejuni)N−グリカンを含有するタンパク質に対して特異的である(ワッカー(Wacker)ら、2002年、上記)。
【0068】
図2Aの最初の2本のレーンは、機能性のpglオペロンが存在する場合と存在しない場合のAcrAを示す。機能性のpglオペロンが存在しない場合には、1つのバンドのみが現れ、存在する場合には3つのバンドが現れる(図2A、上のパネル)。これらは、非グリコシル化AcrA(レーン1)、ならびに、非グリコシル化、モノグリコシル化、およびジグリコシル化AcrA(レーン2)に相当する。レーン2中の2つのより重いタンパク質がグリコシル化されたことを、R12ウェスタンブロットによって確認した(レーン2、下のパネル)。ミュータントAcrA−N273Qを同じ方法で発現させた場合、機能性のグリコシル化pglオペロンが存在する場合には、モノグリコシル化されたAcrAだけが検出された(レーン3)。機能性のpgl遺伝子座が存在しない場合には、非グリコシル化AcrAが検出された(レーン4)。ミュータントAcrA−D121Aの分析は、2つのバンドのみをもたらし、これらのうちの1つは、レーン3においてAcrA−N273Qで観察されるように、グリコシル化されたものであった(レーン5)。これは、D121が、位置123〜125での効率的なグリコシル化のために必須であることを意味する。
【0069】
実施例3 人工のグリコシル化部位のAcrAへの導入
アスパラギン酸残基の導入が、グリコシル化部位をもたらすことができるかどうか試験するために、可溶性のAcrAの位置N117およびN147の使用されないグリコシル化部位の−2位置の残基がアスパラギン酸(F115D、T145D)と交換されたAcrAミュータントを産生した。改変されたグリコシル化部位を、実施例2に記載されるのと同じアッセイによってグリコシル化することができるかどうかを、その後試験した。両方の突然変異を、AcrAの可溶性のバージョンの野生型配列に、あるいは、(どちらも使用されるグリコシル化部位を欠損させた)(N123QおよびN273Q)二重のミュータント中に、個々に挿入した。4時間誘導させた培養物の周辺質抽出物を調製し、SDSページによって分離し、ウェスタンブロッティングによって分析した(図2B)。対照として、野生型のサンプルをグリコシル化し、非グリコシル化されたAcrAを、同じゲル上に流した(レーン1および2)。T145D突然変異は、天然には使用されないグリコシル化シークオンN147−S149の−2位置に影響を及ぼした。AcrA−T145Dを発現させると、抗AcrA抗血清を用いるウェスタンブロッティングは、4つのバンドをもたらし、それらの最も高いものは、レーン2における2重にグリコシル化されたタンパク質よりも電気泳動移動度が遅かった(図2Bにおけるレーン3)。R12ブロットによって、第4のバンドは、3重にグリコシル化されたAcrAであることが確認された。抗AcrAに対する低い強度にもかかわらず、最も重いバンドは、グリコシル化特異的なR12抗血清を用いると最も強いシグナルを示した。同じミュータントAcrA−T145Dが、天然のN−グリコシル化配列(AcrA−N123Q−N273Q−T145D)の非存在下で発現された場合、機能性のpglオペロンの存在下で、モノグリコシル化されたAcrAのみが検出され(図2B、レーン4)、機能性のpglオペロンの非存在下では検出されなかった(レーン5)。これは、レーン4中のより重いバンドが、グリコシル化されたことを実証する。したがって、単純にT145D突然変異を導入することによって、最適化グリコシル化部位がもたらされた(DFNNS)。
【0070】
アスパラギン酸残基を−2位置に挿入することによって、グリコシル化部位を導入することが可能であることをさらに確認するために、天然には使用されない部位N117−119およびN274−T276を、N−グリコシル化を最適化するために変化させた。この目的のために、さらなるミュータントを産生した(図2C)。上記のシステムにおけるAcrA−F115D−T145Dの発現は、抗AcrA抗血清を用いて検出される5つのタンパク質種をもたらした(レーン2)。これは、同じAcrA分子上に起こる4つのグリコシル化の指標となる。C.ジェジュニ(C.jejuni)N−グリカン特異的なR12抗血清を用いる検出を実施すると、5つのバンドのラダーが検出された。最も低いかすかなバンドは、非グリコシル化AcrAであり、これは、グリコシル化の非存在下でも存在するからであり(レーン1)、最も高いものは、4重にグリコシル化されたAcrA中の5つの抗原性の決定因子におそらく起因する、強いシグナルをもたらす。したがって、2つの導入された部位(N117およびN147で)および2つの天然に使用される部位(N123およびN273)が使用され、pgl機構によってグリコシル化される。pglオペロンを含むおよび含まないAcrA−N123Q−N273Q−N272Dの発現は、第3の人工的に導入されたグリコシル化部位、N274(DNNST)も、pglオペロンによって認識されたことを実証した(図2C、レーン3および4)。
【0071】
上記の実験によって、C.ジェジュニ(C.jejuni)のOTアーゼによって認識される細菌のN−グリコシル化部位は、部分的に真核性のものと同じコンセンサスからなるが(N−X−S/T、X≠P)、さらに、効率を増大するために、−2位置におけるアスパラギン酸が必要とされるという知見が確認される。さらに、これらの実験は、こうした最適化コンセンサス配列を組換えにより導入することによって、所望の部位でタンパク質をグリコシル化することが可能性であることを実証する。
【0072】
実施例4 最適化N−グリコシル化配列における位置−1の確認
細菌のグリコシル化部位における−1の位置が、真核生物における+1の位置と同じ制限を示すかどうか試験するために、さらなる実験を実施した(インペリアリ,B.(Imperiali,B.)およびシャノン,K.L.(Shannon,K.L.)(1991年)、「Differences between Asn−Xaa−Thr−containing peptides:a comparison of solution conformation and substrate behaviour with oligosaccharyl−transferase.」「Biochemistry」30、4374〜4380;ラッド,P.M.(Rudd,P.M.)およびドゥウェック,R.A.(Dwek,R.A.)(1997年)、「Glycosylation:heterogeneity and the 3D structure of proteins.」、「Crit.Rev.Biochem.Mol.Biol.」32、1〜100)。+1のプロリン残基は、グリコシル化が抑制されるように、ペプチドを制限すると考えられる。類似の効果が、−1の位置でも観察されることができるかどうか試験するために、第2の天然の部位がノックアウトされた点変異体(AcrA−N273Q−F122P)において、第1の天然に使用される部位のその位置に、プロリン残基を導入した。AcrA−N273Qの対照発現は、機能性のpglオペロンが存在する場合には、モノグリコシル化されたタンパク質を示した(図2D、レーン1および2)。しかし、AcrA−N273Q−F122Pは、グリコシル化されなかった(図2D、レーン3および4)。これは、プロリンは、これがアスパラギンと−2位置の負に荷電する残基との間に残基を構成する場合、細菌のN−グリコシル化を抑制することを示している。
【0073】
C.ジェジュニ(C.jejuni)pgl機構によってグリコシル化されることが知られているすべての部位の配列アラインメントは、これらがすべて、−2位置にDまたはEを含むことを示す(ニタ−ラザー(Nita−Lazar)ら、2005年、上記;ワッカー(Wacker)ら、2002年、上記;ヤング(Young)ら、(2002年)、「Structure of the N−linked glycan present on a multiple glycoproteins in the Gram−negative bacterium, Campylobacter jejuni.」「J.Biol.Chem.」277、42530〜42539)。したがって、細菌のグリコシル化コンセンサス配列は、−2位置の負に荷電するアミノ酸によって最適化され、D/E−X−N−Z−S/T(式中、XおよびZ≠P)をもたらす可能性があることが確立された。
【0074】
実施例5 非C.ジェジュニ(C.jejuni)タンパク質のN−グリコシル化
第1の配列要件(最適化コンセンサス配列)が、細菌におけるN−グリコシル化に十分であることを実証するために、非C.ジェジュニ(C.jejuni)タンパク質を、上記の戦略を適用することによってグリコシル化させることができるかどうかを試験した。コレラ毒素Bサブユニット(CtxB)を、グリコシル化標的として用いた。相当する遺伝子を、それが、まさに実施例1における構築物AからCと同様の、N末端上のOmpAシグナル配列のコード配列とC末端のヘキサヒスチジンタグを含有するような方式で、コレラ菌から増幅させた。得られたDNAを、実施例1で用いられたプラスミド中の構築物Aと置き換えるようにクローン化した。W88のDへの点突然変異、またはW88の後のD挿入は、最適化グリコシル化部位(DNNKT)をもたらした。野生型、ならびにシグナル配列およびhis−タグを含有するW88D CtxBタンパク質を、C.ジェジュニ(C.jejuni)由来の機能性のpgl遺伝子座の存在下および非存在下で、大腸菌Top 10および他の細胞型中で発現させた。Top 10細胞由来の周辺質抽出物を、SDS−PAGE、電気移動、およびCtxB抗血清を用いる連続的な免疫ブロット法によって分析すると、CtxB W88Dだけは、pgl遺伝子座バックグラウンドにおいて、より高い、したがって、グリコシル化されたバンドを産生していた(図2E、レーン3と4を比較せよ)。コンセンサス配列(DSNIT)はまた、CtxBのG54またはQ56(後者は、CtxB−Q56/DSNITと表される)、すなわち、CtxBのガングリオシドGM1結合活性に寄与することが報告されたループのうちの1つの中のものを置き換えることによって挿入された。図2Eのレーン5および6は、操作されたタンパク質(Top 10細胞中に発現されるQ56ではなくペプチド配列DSNITを含有する構築物によって例示される)が、上で述べたのと同様に分析した場合、グリコシル化を欠く細胞ではなくグリコシル化コンピテントな細胞において、より移動性が低い、したがって、グリコシル化されたバンドをもたらしたことを実証している。2つの操作、すなわち、W88の後のDの挿入ならびにDSNITのQ56の置き換えを含むCxtBは、SCM7細胞(アライモ(Alaimo)ら、「EMBO journal」25:967〜976(2006年))において2重にグリコシル化されたことも実証された(パネルE、レーン7および8)。レーン7に示される2重にグリコシル化されたタンパク質CtxBを、Ni2+アフィニティ精製し、標準のプロトコルによるゲル内トリプシン処理の後、ESI−MS/MSによって分析した。予測されるグリコペプチドが検出され、細菌のN−グリコシル化はまた、本発明の細菌のN−グリコシル化のための最適化コンセンサス配列を突然変異させる、あるいは挿入することによって、非C.ジェジュニ(C.jejuni)タンパク質も対象とすることが可能であることが確認された(図示せず)。本発明を実践するための他の適切な例示的な大腸菌(E.coli)株の例は、W3110、CLM24、BL21(ストラタジーン(Stratagene)、米国カリフォルニア州、ラホーヤ(La Jolla,CA,USA))、SCM6、およびSCM7である。
【0075】
ここで使用されるCtxBタンパク質のアミノ酸配列を、以下に示す(組み換えOmpAシグナル配列は下線部、ヘキサ−ヒスチジンタグはイタリック体、W88は太字):
【化5】
【0076】
実施例6 C.ジェジュニ(C.jejuni)外膜タンパク質(OmpH1)への人工のN−グリコシル化部位の導入
細菌におけるN−グリコシル化の潜在的な用途は、遺伝子型に表現型を連結し、それによって特定の遺伝子突然変異について選択するための、細菌の宿主細胞の表面上のグリカンのディスプレイである。N−グリカンを、外膜タンパク質上に提示することが可能であることを実証するために、OmpH1タンパク質を、それが、本発明の複数の最適化コンセンサス部位を含有するように設計した。既知の結晶構造から推測される通り、この部位をタンパク質のループ領域に設計した(ミュラー,A.(Muller,A.)、トーマス,G.H.(Thomas,G.H.)、ホーラー、R.(Horler,R.)、ブラニガン,J.A.(Brannigan,J.A.)、ブラゴヴァ,E.(Blagova,E.)、レヴディコフ,V.M.(Levdikov,V.M.)、フォッグ,M.J.(Fogg,M.J.)、ウィルソン,K.S.(Wilson,K.S.)、およびウィルキンソン,A.J(Wilkinson,A.J.)2005年、「An ATP−binding cassette−type cysteine transporter in Campylobacter jejuni inferred from the structure of an extracytoplasmic solute receptor protein.」「Mol.Microbiol.」57:143〜155)。以前の実験では、最良のグリコシル化配列は、突然変異V83T、K59N−G60I−N61T、R190N−G191I−D192T、およびH263D−F264S−G265N−D266I−D267Tによって産生されたことが示されている。表面ディスプレイについては、最大にN−グリカン特異的なサンプルを確立するために、それらの導入された部位の異なる組み合わせを評価することが所望された。こうした組み合わせを、プラスミド構築物をコードする野生型OmpH1中に作り出し、AcrAについて述べられたのと同様の方式で試験した。図3は、既存の野生型シークオンに加えて、複数のグリコシル化シークオンを有する様々なOmpH1変異体の分析を示す。OmpH1変異体は、3つ(レーン3、4、5および7)、および4つ(レーン6)のグリコシル化シークオンを伴って産生された。グリコシル化シークオンを1つだけ有する野生型OmpH1と、グリコシル化のための重要なアスパラギンを欠くミュータントもまた、この実験に含まれた。ここで試験されるすべての変異体は、高レベルのグリコシル化効率を実証するものに限られるわけではなく、あらゆるグリコシル化シークオンが利用された。結果は、カンピロバクターN−グリカン特異的な免疫血清を用いて確認した(図3、下のパネル)。
【0077】
以下は、カンピロバクタージェジュニ(Campylobacter jejuni)(81−176株)のOmpH1タンパク質のタンパク質配列である。付着されたmycタグは、イタリック体である。
【化6】
タンパク質中の天然のグリコシル化部位は太字であり、シグナル配列には下線が引かれている。
【0078】
実施例7 大腸菌細胞の外膜上のOmpH1上のC.ジェジュニ(C.jejuni)由来のN−グリカンの表面ディスプレイ
複数のグリコシル化されたOmpH1変異体を、細菌細胞の表面上にディスプレイさせることができるかどうかという問題に答えるために、様々なOmpH1変異体を発現している細菌のCLM24またはSCM6(これは、SCM7ΔwaaLである)細胞に、免疫蛍光法を実施した。野生型OmpH1と、グリコシル化のための重要なアスパラギンを欠くミュータントを、実験に含めた。さらに、周辺質内にタンパク質を保持する、したがって、この実験における対象としての役割を果たすために、C20Sミュータントを構築した。パラホルムアルデヒドで処理した細胞に対して免疫染色を行った。パラホルムアルデヒドは、細胞の構造または区画化を壊さずに細胞を固定する。FITCおよびCy3に結合させた相当する二次抗体と組み合わせて、c−Myc−およびN−グリカン−特異的な免疫血清を使用して、細菌細胞表面上のタンパク質(赤色蛍光)およびN−グリカン(緑色)をそれぞれ検出した。さらに、4,6−ジアミノ−2−フェニルインドール(DAPI、青色)を用いて、細菌のDNAを染色し、細菌細胞と細胞残屑を明確に区別した。野生型OmpH1を発現している細胞が染色された場合、タンパク質ならびにN−グリカンに特異的な免疫蛍光が検出された(図4A)。重要なアスパラギンN139Sを欠くミュータントが、抗−Myc−とN−グリカン−の両方に特異的な免疫血清で染色された場合、グリカン特異的なシグナルではなく、タンパク質のみに特異的なシグナルが得られ(パネル4B)、これは、N−グリカン特異的な免疫血清の特異性を示した。タンパク質が、C20Sミュータントにおいて見られるように周辺質内に保持された場合、タンパク質特異的な赤い免疫蛍光は検出されず、これは、抗体が細胞内に拡散することが不可能であっても、任意の表面現象を検出するのに十分コンピテントであったことを示していた(4Cパネル)。次に、グリコシル化において異なる複数のOmpH1変異体を発現している細胞が染色された:OmpH1KGN→NIT、HFGDD→DSNIT(パネル4D)、OmpH1RGD→NIT、HFGDD→DSNIT(パネル4E)、OmpH1KGN→NIT、RGD→NIT(パネル4F)、OmpH1V83T、KGN→NIT(パネル4G)、およびOmpH1KGN→NIT、RGD→NIT、HFGDD→DSNIT(パネル4H)。すべてのOmpH1変異体は、2重に染色され、これは、細菌の表面上のグリコシル化タンパク質の存在を示していた。図4は、グレースケールで表したものであり、第1行は、同じ列の他の画像を組合せた画像である。
【特許請求の範囲】
【請求項1】
1つまたは複数の下記N−グリコシル化された最適化アミノ酸コンセンサス配列を含む組み換えN−グリコシル化タンパク質であって:
D/E−X−N−Z−S/T
式中、XおよびZは、Pro以外の任意の天然のアミノ酸であり得、かつ少なくとも1つの前記N−グリコシル化された最適化アミノ酸コンセンサス配列が導入される、組み換えN−グリコシル化タンパク質。
【請求項2】
少なくとも2つ、好ましくは少なくとも3つ、より好ましくは少なくとも5つの前記N−グリコシル化された最適化アミノ酸コンセンサス配列を含む請求項1に記載の組み換えタンパク質。
【請求項3】
タンパク質が、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来のN−グリカンの群から選択される1つまたは複数のN−グリカンを含み、前記N−グリカンは、グラム陰性細菌中のO多糖形成O抗原に転移されるオリゴ糖および多糖、あるいは、グラム陽性細菌由来の莢膜多糖、好ましくは:緑膿菌(P.aeruginosa)O9、O11;大腸菌(E.coli)O7、O9、O16、O157、および志賀赤痢菌(Shigella dysenteriae)O1から得られる、請求項1または2に記載の組み換えタンパク質。
【請求項4】
タンパク質が、2つ以上の異なるN−グリカンを含む、請求項1から3のいずれか一項に記載の組み換えタンパク質。
【請求項5】
タンパク質が、哺乳類、細菌、ウイルス、菌類、または植物タンパク質由来である、請求項1から4のいずれか一項に記載の組み換えタンパク質。
【請求項6】
タンパク質および/またはN−グリカンが、治療的および/または予防的に活性である、請求項1から5のいずれか一項に記載の組み換えタンパク質。
【請求項7】
タンパク質および/またはN−グリカンが、免疫原的に活性である、請求項1から6のいずれか一項に記載の組み換えタンパク質。
【請求項8】
前記組み換えタンパク質を、グラム陰性細菌の外膜にターゲティングすることが可能な少なくとも1つのポリペプチド配列をさらに含む、請求項1から7のいずれか一項に記載の組み換えタンパク質。
【請求項9】
前記ターゲティングポリペプチド配列が、好ましくはC.ジェジュニ(C.jejuni)由来のOmpH1、C.ジェジュニ(C.jejuni)由来のJlpA、大腸菌(E.coli)由来の外膜タンパク質(好ましくはOmpS、OmpC、OmpA、OprF、PhoE、LamB、Lpp;OmpA)、および緑膿菌(Pseudomonas aeruginosa)由来のlnpタンパク質の全長タンパク質またはシグナルペプチドからなる群から選択される、タイプIIシグナルペプチドまたは外膜タンパク質からなる群から選択される、請求項8に記載の組み換えタンパク質。
【請求項10】
請求項1から9のいずれか一項に記載のタンパク質をコードする核酸。
【請求項11】
請求項10に記載の核酸を含むベクター。
【請求項12】
請求項10に記載の核酸および/または請求項11に記載のベクターを含む宿主細胞。
【請求項13】
宿主細胞が、原核性または真核性、好ましくは原核性である、請求項12に記載の宿主細胞。
【請求項14】
宿主細胞が、エシェリキア亜種(Escherichia ssp.)、カンピロバクター亜種(Campylobacter ssp.)、サルモネラ亜種(Salmonella ssp.)、シゲラ亜種(Shigella ssp.)、ヘリコバクター亜種(Helicobacter ssp.)、シュードモナス亜種(Pseudomonas ssp.)、バチルス亜種(Bacillus ssp.)、好ましくは、大腸菌(Escherichia coli)、より好ましくは、大腸菌(E.coli)株Top10、W3110、CLM24、BL21、SCM6、およびSCM7、ならびにS.エンテリカ(S.enterica)株SL3261、SL3749、およびSL3261ΔwaaLからなる細菌の群から選択される、請求項13に記載の宿主細胞。
【請求項15】
前記宿主細胞が、
i)以下をコードするヌクレオチド配列を含む遺伝子型、
a)脂質担体上へのオリゴ糖の構築のための少なくとも1種の天然または組み換え型の特異的なグリコシルトランスフェラーゼ、
b)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の、少なくとも1種の天然または組み換え型のオリゴサッカリルトランスフェラーゼ(OTアーゼ)、
c)請求項1から9のいずれか一項、好ましくは、請求項8または9に記載の少なくとも1種の組み換えタンパク質、および
ii)グラム陰性細菌の外膜内および/または上に位置する請求項1から9のいずれか一項に記載の組み換えN−グリコシル化タンパク質を含む表現型、
を有する組み換えグラム陰性細菌である、請求項12から14のいずれか一項に記載の宿主細胞。
【請求項16】
エシェリキア亜種(Escherichia ssp.)、カンピロバクター亜種(Campylobacter ssp.)、シゲラ亜種(Shigella ssp)、ヘリコバクター亜種(Helicobacter ssp.)、およびシュードモナス亜種(Pseudomonas ssp.)、サルモネラ亜種(Salmonella ssp.)、好ましくは、大腸菌(E.coli)、より好ましくは、大腸菌(E.coli)株Top10、W3110、CLM24、BL21、SCM6、およびSCM7、ならびにS.エンテリカ(S.enterica)株SL3261、SL3749、およびSL3261ΔwaaLからなる群から選択される、請求項15に記載の宿主細胞。
【請求項17】
医薬品の調製のための、請求項1から9のいずれか一項に記載の組み換えタンパク質の使用。
【請求項18】
医薬品の調製のための、請求項10に記載の核酸および/または請求項11に記載のベクターの使用。
【請求項19】
医薬品の調製のための、請求項15または16に記載の宿主細胞の使用。
【請求項20】
その必要性のある対象の治療および/または予防用ワクチン接種のための医薬品の調製のための、請求項17に記載のタンパク質の使用。
【請求項21】
その必要性のある対象の治療および/または予防用ワクチン接種のための医薬品の調製のための、請求項18に記載の核酸および/またはベクターの使用。
【請求項22】
その必要性のある対象の治療および/または予防用ワクチン接種のための医薬品の調製のための、請求項15または16に記載の宿主細胞の使用。
【請求項23】
動物(好ましくは哺乳類、より好ましくはげっ歯類、ヒツジ、ウマ、イヌ、ウシ、またはヒト)においてIgA抗体を誘導するための、請求項19または22に記載の宿主細胞の使用。
【請求項24】
請求項1から9のいずれか一項に記載の少なくとも1種のタンパク質を含む薬剤組成物。
【請求項25】
少なくとも1種の請求項10に記載の核酸および/または請求項11に記載のベクターを含む薬剤組成物。
【請求項26】
請求項16または17に記載の少なくとも1種の宿主細胞を含む薬剤組成物。
【請求項27】
薬学的に許容される賦形剤、希釈剤、および/または補助剤をさらに含む、請求項24から26のいずれか一項に記載の薬剤組成物。
【請求項28】
N−結合グリコシル化タンパク質を産生するための方法であって
a)以下をコードする核酸を含む組み換え生物、好ましくは原核生物を提供するステップ:
i)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロン、および
ii)1つまたは複数の下記N−グリコシル化された最適化アミノ酸コンセンサス配列を含む、少なくとも1種の組み換え標的タンパク質であって:
D/E−X−N−Z−S/T
式中、XおよびZは、Pro以外の任意の天然のアミノ酸であり得、かつ少なくとも1つの前記N−グリコシル化された最適化アミノ酸コンセンサス配列が導入される組み換え標的タンパク質、および
b)標的タンパク質の産生およびN−グリコシル化に適した方式で、組み換え生物を培養するステップ、
を含む方法。
【請求項29】
標的タンパク質が、請求項1から9のいずれか一項に記載の組み換えタンパク質である、請求項28に記載の方法。
【請求項30】
カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロンが、
OTアーゼによって標的タンパク質に転移されることとなる、脂質担体上へのオリゴ糖の構築のための、
i)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の組み換えOTアーゼ、および
ii)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni))由来の組み換え型および/または天然の特異的なグリコシルトランスフェラーゼ、および/または
iii)カンピロバクター種以外の種由来の組み換え型および/または天然の特異的なグリコシルトランスフェラーゼ、
をコードする核酸を含む、請求項28または29に記載の方法。
【請求項31】
請求項15または16に記載の宿主細胞を調製するための方法であって、
i)グラム陰性細菌を提供するステップと、
ii)以下をコードする少なくとも1つのヌクレオチド配列を前記細菌に導入するステップと、
a)脂質担体上へのオリゴ糖の構築のための少なくとも1種の組み換え型の特異的なグリコシルトランスフェラーゼ、および/または
b)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の少なくとも1種の組み換えオリゴサッカリルトランスフェラーゼ(OTアーゼ)、および/または
c)1つまたは複数の下記N−グリコシル化された最適化アミノ酸コンセンサス配列を含む少なくとも1種の組み換えタンパク質であって:
D/E−X−N−Z−S/T
式中、XおよびZは、Pro以外の任意の天然のアミノ酸であり得、かつ少なくとも1つの前記N−グリコシル化された最適化アミノ酸コンセンサス配列が導入される組み換えタンパク質、
iii)c)のヌクレオチド配列によってコードされる少なくとも1種の組み換えN−グリコシル化タンパク質が、グラム陰性細菌の外膜内および/または上に位置されるまで、前記細菌を培養するステップと、
を含む方法。
【請求項32】
組み換え型の原核生物または宿主細胞が、エシェリキア亜種(Escherichia ssp)、カンピロバクター亜種(Campylobacter ssp.)、サルモネラ亜種(Salmonella ssp.)、シゲラ亜種(Shigella ssp.)、ヘリコバクター亜種(Helicobacter ssp.)、シュードモナス亜種(Pseudomonas ssp.)、バチルス亜種(Bacillus ssp.)、好ましくは、大腸菌(Escherichia coli)、好ましくは、大腸菌(E.coli)株Top10、W3110、CLM24、BL21、SCM6、およびSCM7、ならびにS.エンテリカ(S.enterica)株SL3261、SL3749、およびSL3261ΔwaaLからなる細菌の群から選択される、請求項28から31のいずれか一項に記載の方法。
【請求項33】
請求項1から9のいずれか一項に記載の組み換えタンパク質を産生、単離、および/または精製するための方法であって、
a)請求項15または16に記載の宿主細胞を培養するステップと、
b)前記組み換えグラム陰性細菌の外膜を除去するステップと、
c)前記組み換えタンパク質を回収するステップと、
を含む方法。
【請求項1】
1つまたは複数の下記N−グリコシル化された最適化アミノ酸コンセンサス配列を含む組み換えN−グリコシル化タンパク質であって:
D/E−X−N−Z−S/T
式中、XおよびZは、Pro以外の任意の天然のアミノ酸であり得、かつ少なくとも1つの前記N−グリコシル化された最適化アミノ酸コンセンサス配列が導入される、組み換えN−グリコシル化タンパク質。
【請求項2】
少なくとも2つ、好ましくは少なくとも3つ、より好ましくは少なくとも5つの前記N−グリコシル化された最適化アミノ酸コンセンサス配列を含む請求項1に記載の組み換えタンパク質。
【請求項3】
タンパク質が、カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来のN−グリカンの群から選択される1つまたは複数のN−グリカンを含み、前記N−グリカンは、グラム陰性細菌中のO多糖形成O抗原に転移されるオリゴ糖および多糖、あるいは、グラム陽性細菌由来の莢膜多糖、好ましくは:緑膿菌(P.aeruginosa)O9、O11;大腸菌(E.coli)O7、O9、O16、O157、および志賀赤痢菌(Shigella dysenteriae)O1から得られる、請求項1または2に記載の組み換えタンパク質。
【請求項4】
タンパク質が、2つ以上の異なるN−グリカンを含む、請求項1から3のいずれか一項に記載の組み換えタンパク質。
【請求項5】
タンパク質が、哺乳類、細菌、ウイルス、菌類、または植物タンパク質由来である、請求項1から4のいずれか一項に記載の組み換えタンパク質。
【請求項6】
タンパク質および/またはN−グリカンが、治療的および/または予防的に活性である、請求項1から5のいずれか一項に記載の組み換えタンパク質。
【請求項7】
タンパク質および/またはN−グリカンが、免疫原的に活性である、請求項1から6のいずれか一項に記載の組み換えタンパク質。
【請求項8】
前記組み換えタンパク質を、グラム陰性細菌の外膜にターゲティングすることが可能な少なくとも1つのポリペプチド配列をさらに含む、請求項1から7のいずれか一項に記載の組み換えタンパク質。
【請求項9】
前記ターゲティングポリペプチド配列が、好ましくはC.ジェジュニ(C.jejuni)由来のOmpH1、C.ジェジュニ(C.jejuni)由来のJlpA、大腸菌(E.coli)由来の外膜タンパク質(好ましくはOmpS、OmpC、OmpA、OprF、PhoE、LamB、Lpp;OmpA)、および緑膿菌(Pseudomonas aeruginosa)由来のlnpタンパク質の全長タンパク質またはシグナルペプチドからなる群から選択される、タイプIIシグナルペプチドまたは外膜タンパク質からなる群から選択される、請求項8に記載の組み換えタンパク質。
【請求項10】
請求項1から9のいずれか一項に記載のタンパク質をコードする核酸。
【請求項11】
請求項10に記載の核酸を含むベクター。
【請求項12】
請求項10に記載の核酸および/または請求項11に記載のベクターを含む宿主細胞。
【請求項13】
宿主細胞が、原核性または真核性、好ましくは原核性である、請求項12に記載の宿主細胞。
【請求項14】
宿主細胞が、エシェリキア亜種(Escherichia ssp.)、カンピロバクター亜種(Campylobacter ssp.)、サルモネラ亜種(Salmonella ssp.)、シゲラ亜種(Shigella ssp.)、ヘリコバクター亜種(Helicobacter ssp.)、シュードモナス亜種(Pseudomonas ssp.)、バチルス亜種(Bacillus ssp.)、好ましくは、大腸菌(Escherichia coli)、より好ましくは、大腸菌(E.coli)株Top10、W3110、CLM24、BL21、SCM6、およびSCM7、ならびにS.エンテリカ(S.enterica)株SL3261、SL3749、およびSL3261ΔwaaLからなる細菌の群から選択される、請求項13に記載の宿主細胞。
【請求項15】
前記宿主細胞が、
i)以下をコードするヌクレオチド配列を含む遺伝子型、
a)脂質担体上へのオリゴ糖の構築のための少なくとも1種の天然または組み換え型の特異的なグリコシルトランスフェラーゼ、
b)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の、少なくとも1種の天然または組み換え型のオリゴサッカリルトランスフェラーゼ(OTアーゼ)、
c)請求項1から9のいずれか一項、好ましくは、請求項8または9に記載の少なくとも1種の組み換えタンパク質、および
ii)グラム陰性細菌の外膜内および/または上に位置する請求項1から9のいずれか一項に記載の組み換えN−グリコシル化タンパク質を含む表現型、
を有する組み換えグラム陰性細菌である、請求項12から14のいずれか一項に記載の宿主細胞。
【請求項16】
エシェリキア亜種(Escherichia ssp.)、カンピロバクター亜種(Campylobacter ssp.)、シゲラ亜種(Shigella ssp)、ヘリコバクター亜種(Helicobacter ssp.)、およびシュードモナス亜種(Pseudomonas ssp.)、サルモネラ亜種(Salmonella ssp.)、好ましくは、大腸菌(E.coli)、より好ましくは、大腸菌(E.coli)株Top10、W3110、CLM24、BL21、SCM6、およびSCM7、ならびにS.エンテリカ(S.enterica)株SL3261、SL3749、およびSL3261ΔwaaLからなる群から選択される、請求項15に記載の宿主細胞。
【請求項17】
医薬品の調製のための、請求項1から9のいずれか一項に記載の組み換えタンパク質の使用。
【請求項18】
医薬品の調製のための、請求項10に記載の核酸および/または請求項11に記載のベクターの使用。
【請求項19】
医薬品の調製のための、請求項15または16に記載の宿主細胞の使用。
【請求項20】
その必要性のある対象の治療および/または予防用ワクチン接種のための医薬品の調製のための、請求項17に記載のタンパク質の使用。
【請求項21】
その必要性のある対象の治療および/または予防用ワクチン接種のための医薬品の調製のための、請求項18に記載の核酸および/またはベクターの使用。
【請求項22】
その必要性のある対象の治療および/または予防用ワクチン接種のための医薬品の調製のための、請求項15または16に記載の宿主細胞の使用。
【請求項23】
動物(好ましくは哺乳類、より好ましくはげっ歯類、ヒツジ、ウマ、イヌ、ウシ、またはヒト)においてIgA抗体を誘導するための、請求項19または22に記載の宿主細胞の使用。
【請求項24】
請求項1から9のいずれか一項に記載の少なくとも1種のタンパク質を含む薬剤組成物。
【請求項25】
少なくとも1種の請求項10に記載の核酸および/または請求項11に記載のベクターを含む薬剤組成物。
【請求項26】
請求項16または17に記載の少なくとも1種の宿主細胞を含む薬剤組成物。
【請求項27】
薬学的に許容される賦形剤、希釈剤、および/または補助剤をさらに含む、請求項24から26のいずれか一項に記載の薬剤組成物。
【請求項28】
N−結合グリコシル化タンパク質を産生するための方法であって
a)以下をコードする核酸を含む組み換え生物、好ましくは原核生物を提供するステップ:
i)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロン、および
ii)1つまたは複数の下記N−グリコシル化された最適化アミノ酸コンセンサス配列を含む、少なくとも1種の組み換え標的タンパク質であって:
D/E−X−N−Z−S/T
式中、XおよびZは、Pro以外の任意の天然のアミノ酸であり得、かつ少なくとも1つの前記N−グリコシル化された最適化アミノ酸コンセンサス配列が導入される組み換え標的タンパク質、および
b)標的タンパク質の産生およびN−グリコシル化に適した方式で、組み換え生物を培養するステップ、
を含む方法。
【請求項29】
標的タンパク質が、請求項1から9のいずれか一項に記載の組み換えタンパク質である、請求項28に記載の方法。
【請求項30】
カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の機能性のpglオペロンが、
OTアーゼによって標的タンパク質に転移されることとなる、脂質担体上へのオリゴ糖の構築のための、
i)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の組み換えOTアーゼ、および
ii)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni))由来の組み換え型および/または天然の特異的なグリコシルトランスフェラーゼ、および/または
iii)カンピロバクター種以外の種由来の組み換え型および/または天然の特異的なグリコシルトランスフェラーゼ、
をコードする核酸を含む、請求項28または29に記載の方法。
【請求項31】
請求項15または16に記載の宿主細胞を調製するための方法であって、
i)グラム陰性細菌を提供するステップと、
ii)以下をコードする少なくとも1つのヌクレオチド配列を前記細菌に導入するステップと、
a)脂質担体上へのオリゴ糖の構築のための少なくとも1種の組み換え型の特異的なグリコシルトランスフェラーゼ、および/または
b)カンピロバクター種、好ましくはC.ジェジュニ(C.jejuni)由来の少なくとも1種の組み換えオリゴサッカリルトランスフェラーゼ(OTアーゼ)、および/または
c)1つまたは複数の下記N−グリコシル化された最適化アミノ酸コンセンサス配列を含む少なくとも1種の組み換えタンパク質であって:
D/E−X−N−Z−S/T
式中、XおよびZは、Pro以外の任意の天然のアミノ酸であり得、かつ少なくとも1つの前記N−グリコシル化された最適化アミノ酸コンセンサス配列が導入される組み換えタンパク質、
iii)c)のヌクレオチド配列によってコードされる少なくとも1種の組み換えN−グリコシル化タンパク質が、グラム陰性細菌の外膜内および/または上に位置されるまで、前記細菌を培養するステップと、
を含む方法。
【請求項32】
組み換え型の原核生物または宿主細胞が、エシェリキア亜種(Escherichia ssp)、カンピロバクター亜種(Campylobacter ssp.)、サルモネラ亜種(Salmonella ssp.)、シゲラ亜種(Shigella ssp.)、ヘリコバクター亜種(Helicobacter ssp.)、シュードモナス亜種(Pseudomonas ssp.)、バチルス亜種(Bacillus ssp.)、好ましくは、大腸菌(Escherichia coli)、好ましくは、大腸菌(E.coli)株Top10、W3110、CLM24、BL21、SCM6、およびSCM7、ならびにS.エンテリカ(S.enterica)株SL3261、SL3749、およびSL3261ΔwaaLからなる細菌の群から選択される、請求項28から31のいずれか一項に記載の方法。
【請求項33】
請求項1から9のいずれか一項に記載の組み換えタンパク質を産生、単離、および/または精製するための方法であって、
a)請求項15または16に記載の宿主細胞を培養するステップと、
b)前記組み換えグラム陰性細菌の外膜を除去するステップと、
c)前記組み換えタンパク質を回収するステップと、
を含む方法。
【図1】

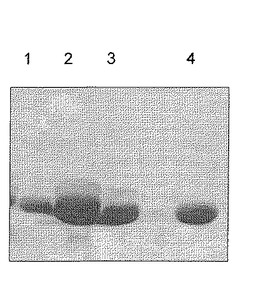
【図2】
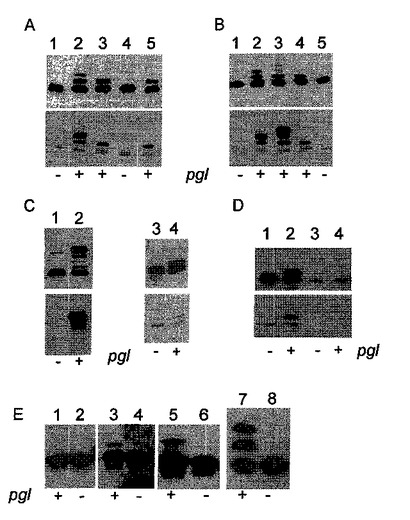
【図3】
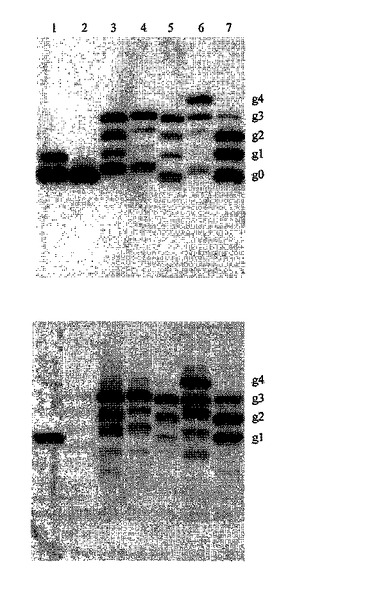
【図4】

【図2】
【図3】
【図4】
【公開番号】特開2012−100678(P2012−100678A)
【公開日】平成24年5月31日(2012.5.31)
【国際特許分類】
【外国語出願】
【出願番号】特願2012−1342(P2012−1342)
【出願日】平成24年1月6日(2012.1.6)
【分割の表示】特願2008−510495(P2008−510495)の分割
【原出願日】平成18年5月10日(2006.5.10)
【出願人】(507372121)
【Fターム(参考)】
【公開日】平成24年5月31日(2012.5.31)
【国際特許分類】
【出願番号】特願2012−1342(P2012−1342)
【出願日】平成24年1月6日(2012.1.6)
【分割の表示】特願2008−510495(P2008−510495)の分割
【原出願日】平成18年5月10日(2006.5.10)
【出願人】(507372121)
【Fターム(参考)】
[ Back to top ]