
反芻動物のルーメン発酵に影響を及ぼし、かつエネルギーおよびタンパク質の保持を改善する植物、植物抽出物、および植物からの天然同一成分の使用
反芻動物のルーメン発酵に影響を及ぼし、かつエネルギーおよび窒素源のアベイラビリティを増大させる、スイカズラ、ゲンチアナ・アスクレピデア、キバナリンドウ、クローブバッド、ヒナギク、オリーブ、ヒレハリソウ、ヒメヒレアザミ、シャクヤク、ヤマナラシ、サクランボ、バッコヤナギ、セイタカダイオウ、ハンニチバナ、クマコケモモ、インディアンルバーブ、エゾアカバナ、クナウチア・アルベンシス、レタス(Latuca sativa)およびネトルおよびそれらの抽出物、およびβ−ミルセンから選択される植物材料、植物抽出物、または天然同一成分の使用。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ルーメン発酵のエネルギーおよび窒素保持を改善するための、または乳酸アシドーシスを減少させるための、植物材料、植物抽出物、または天然同一精油化合物からの1つもしくはそれ以上の成分を含有する添加物の使用に関する。この化合物は、動物の飼料または飲料水によって反芻動物に適切に経口投与される。
【背景技術】
【0002】
ルーメンは、ウシ、ヒツジ、およびヤギなど反芻動物の第一胃である。その構造上および栄養上の意義は、ハンゲート(Hungate)(1966年)によって記載されている。ルーメンは、これらの動物が大部分の非反芻動物に不適切な高繊維植物材料を消化することを可能にする。ルーメンは、乳牛における150kgを超える消化物を含有する巨大な臓器であり、ここで消化のすべては微生物によって行われる。ルーメンは、動物が微生物の繊維消化活動から恩恵を受けるようにして発達した。最終生成物である、揮発性脂肪酸は、ルーメン壁にわたって吸収され、エネルギーおよびタンパク質合成に使用される。後腸発酵と比べルーメン前腸発酵の主な利点は、発酵中に形成される微生物細胞が第4胃に移動し、そこでそれらは消化され、アミノ酸が吸収されることである。したがって、後腸発酵とは違って、微生物アミノ酸は宿主動物に利用可能となる。したがって、ルーメンは、乏しい牧草地で自給する草食動物の進化との関連できわめて効率的な臓器である。
【0003】
ルーメン発酵はいくつかの欠点ももたらす。メタンは嫌気性発酵の自然の結果として生成される(ハンゲート(Hungate)、1966年)。メタンの放出は動物からのエネルギーの損失も表す。さらに、メタンは強力な温室効果ガスであり、したがってこの意味で、反芻動物は環境に損害を与える(レング(Leng)、1992年)。反芻動物はまた、食事タンパク質の利用において他の種よりも効率的でない。反芻動物からの窒素損失は、特に草食動物において例外的に高い(ノラン(Nolan)、1975年)。これは、窒素豊富な糞尿の環境に対する影響のため環境問題であると同時に経済問題である。さらに、障害のあるルーメン微生物発酵の直接の結果として生じる反芻動物に特有の消化系障害がある。
【0004】
化学的および抗生物質の飼料添加物は過去においてこれらの問題の一部を緩和するために使用されている。食物連鎖におけるかかる材料の存在は、規制当局および消費者にはますます受入れられなくなっている。したがって、反芻動物の家畜生産の問題解決は自然かつ持続可能でなければならない。
【0005】
ルーメンに入る食事タンパク質は明らかに非制御的に分解され、結果としてアンモニア形成およびその後の尿中窒素の損失がもたらされる(ノラン(Nolan)、1975年)。主な経済的損失を表す窒素保持の低い効率は、動物における代謝性侵襲を引き起こし、かつ窒素豊富な廃棄物によって環境への負担ともなる。分解過程が減少されれば、これらの問題が減少されるであろう。
【0006】
さらに、ルーメンにおけるタンパク質保持を改善するために存在する第2の可能性は、ルーメン繊毛原虫の集団の抑制である。原虫はルーメンにおける大量の細菌を消費し、それらの分解は結果として50%までのルーメン発酵からの微生物タンパク質の正味収量の減少をもたらしうる(ウォリス(Wallace)ら、1997年)。原虫が抑制されうる場合は、アンモニア形成が少なくなり、かつ食事タンパク質の補給の必要が少なくなる。
【0007】
乳酸は通常、ルーメン発酵の微量生成物にすぎない。しかし、迅速分解飼料があまりに急速に導入される場合、または濃縮物が高い割合の食事を形成する場合、揮発性脂肪酸産生がルーメンの緩衝能力を超える。これはルーメンにおけるきわめて低いpH値をもたらすため、乳酸産生細菌のみが増殖しうる(ラッセル(Russell)とヒノ(Hino)、1985年)。
【0008】
乳酸が揮発性脂肪酸よりも強い酸であると、pHはさらに低下し、正常な発酵の回復は不可能になり、そして動物は死亡する。しかし、より一般的には、高濃度の食事の状態にある反芻動物は無症状のアシドーシスにかかるが、これは苦難を伴い、生命にかかわることはないものの生産効率を減少させる。化学バッファーはこの状態を食い止めることができるが、乳酸産生細菌の増殖が抑制されればより良いであろう。
【0009】
公報国際公開第03/094628号パンフレットは、リモネン、オイゲノール、サリチル酸塩、キノリン、バニラ、チモール、およびクレゾールからなる群から選択される1つもしくはそれ以上の精油化合物の添加によって反芻動物の消化活動から発するメタン産生を削減する試みを記載している。同時にルーメンにおける微生物の活動および揮発性脂肪酸、例えばプロピオン酸の産生が許容可能レベルで維持されうる。
【発明の詳細な説明】
【0010】
本発明の目的は、全体として、かつ一括してルーメンにおける消化系障害に取組み、かつ反芻動物の成長に利用可能なエネルギーおよび窒素源の利用を促進することである。この目的は、ルーメン壁にわたって吸収され、エネルギーおよびタンパク質合成に使用される揮発性脂肪酸の形成を助けることによって、かつ食事タンパク質および微生物タンパク質の分解を減少させることによって達成されうる。他の方法は、メタン生成を制限し、乳酸産生細菌を制御または抑制することである。
【0011】
本発明によれば、前記目的は、スイカズラ、ゲンチアナ・アスクレピデア、キバナリンドウ、クローブバッド、ヒナギク、オリーブ、ヒレハリソウ、ヒメヒレアザミ、シャクヤク、ヤマナラシ、サクランボ、バッコヤナギ、セイタカダイオウ、ハンニチバナ、クマコケモモ、インディアンルバーブ、エゾアカバナ、クナウチア・アルベンシス、レタス(Latuca sativa)、およびネトルおよびそれらの抽出物、およびβ−ミルセンからなる群から選択される植物材料の1つもしくはそれ以上の成分を含有する添加物を動物飼料または飲料水によって適切に経口投与することによって達成されうる。動物に投与される成分の総量は、1日およびkg体重当り0.02mg〜20gであることが適切である。
【0012】
本発明の成分はすべて、考慮すべき欠点なしにルーメンにおける発酵の効率に実質的に寄与することが証明されている。成分の一部は原虫の活性を抑制し、その他はタンパク質分解活性、メタン生成活性、または乳酸の産生を減少させる。
【0013】
本発明は、1つもしくはそれ以上の規定された成分を含有する添加物および飼料組成物にも関する。
【0014】
成分は発酵に対して多様に影響を有するため、本発明の目的に記載の発酵を最適化する特定の化合物を組み合わせることが有利である。
【0015】
大規模な試験では、ハンニチバナ、クマコケモモ、エゾアカバナ、クナウチア・アルベンシス、およびインディアンルバーブ、およびそれらの抽出物からの植物材料からなる亜群の成分はルーメンのタンパク質分解を大幅に削減することが示された。また、これらの成分のすべては原虫の活性を阻害し、その大部分は、メタン産生、揮発性脂肪酸の量、消化率、微生物バイオマス、および/または発酵効率など、他の必須の発酵パラメータに対する有利な効果も有する。最も適切な成分は、クマコケモモ、クナウチア・アルベンシス、およびハンニチバナ、およびそれらの抽出物からの植物材料である。最も好ましい成分は、クナウチア・アルベンシスおよびその抽出物、特にメタノール抽出物からの植物材料である。
【0016】
スイカズラ、ゲンチアナ・アスクレピデア、キバナリンドウ、クローブバッド、ヒナギク、オリーブおよびヒレハリソウおよびそれらの抽出物およびβ−ミルセンからの植物材料は、その成分がルーメン液中の繊毛原虫の細菌溶解活性を抑制する別の亜群を形成する。クローブバッドも微生物バイオマスを増大させ、メタン産生を削減する。ヒナギクはメタン産生の小さな削減を示し、発酵効率および微生物バイオマス産生を改善もするが、ヒレハリソウは発酵効率を増大させ、アシドーシス効果を削減する。β−ミルセンは原虫活性に対する中程度の阻害効果を有するが、メタン産生を減少させるためにも使用される。スイカズラの花から調製される抽出物が繊毛原虫の細菌分解活性を無効にした。スイカズラの種子調製物も原虫活性を抑制したが、これは約60%にすぎない。この亜群からの好ましい成分は、ヒナギク、クローブバッド、オリーブおよびヒレハリソウおよびそれらの抽出物、特にメタノール、およびヒナギクおよびゲンチアナ・アスクレピデアの水抽出物からの植物材料である。
【0017】
ヒメヒレアザミ、シャクヤク、ヤマナラシ、サクランボ、バッコヤナギおよびセイタカダイオウおよびそれらの抽出物からの植物材料は、その成分がルーメン消化物中のメタン生成活性を減少させるために適切に使用される第3の亜群を形成する。これらの成分には重要な有害な効果が他の発酵パラメータでは確認されていない。好ましい成分は、ヒメヒレアザミ、ヤマナラシ、およびセイタカダイオウおよびそれらの抽出物であり、特に好ましいのは、ヒメヒレアザミおよびセイタカダイオウからの植物材料である。
【0018】
最後に、第4の亜群が、レタス(Latuca sativa)およびネトルおよびそれらの抽出物からの植物材料によって形成される。これらの成分は乳酸の活性および/または形成を低下させる。レタス(Latuca sativa)は、ワイルドレタス(Latuca virosa)と呼ばれることもあり、これもメタン産生および原虫活性を減少させ、揮発性脂肪酸の形成を増大させる。ネトルは追加の効果を有し、タンパク質分解および原虫活性を阻害し、かつ揮発性脂肪酸の形成を助ける。
【0019】
本発明の適切な実施形態において、添加物は、上記規定された亜群の1つ以上成分を含有し、添加物は4つの亜群すべてからの成分を含有することが有利でありうる。有利には、添加物は、
i)クナウチア・アルベンシス、クマコケモモ、およびハンニチバナおよびそれらの抽出物からの植物材料と、
ii)ヒナギク、クローブバッド、オリーブ、スイカズラおよびヒレハリソウおよびそれらの抽出物からの植物材料と、
iii)ヒメヒレアザミ、ヤマナラシ、およびセイタカダイオウおよびそれらの抽出物からの植物材料と、
iv)レタス(Latuca sativa)およびネトルおよびそれらの抽出物からの植物材料と
からなる群の各々から選択される少なくとも1つの成分を含有する。
i)〜iv)の群に属する成分の総量は適切には20〜100重量%であり、好ましくは本発明により存在する成分の総量の40〜100重量%である。
【0020】
本発明による植物材料の抽出物としては、凝結物、精油、樹脂状物質、チンキ剤、遊離物および純油が挙げられるがこれらに限定されない。植物または植物部位の抽出物の生成に際しては、水、有機溶媒、およびそれらの混合物が従来の方法に従って使用されうるが、乾燥蒸留も適用されうる。適切な有機溶媒の例は、メタノール、エタノール、プロパノール、ブタノール、酢酸エチル、メチルエチルエーテル、ジエチルエーテル、塩化メチレン、クロロホルム、四塩化炭素、ベンゼン、トルエン、石油エーテル、およびアセトンである。
【0021】
動物に投与される成分の総量は、その成分が精油化合物、植物材料の抽出物、または植物材料であるかどうかによって決まる。成分がすべてβ−ミルセン、および/または抽出物である場合には、投与量は通常、1日およびkg体重当り0.1〜50mgであるが、成分が植物材料のみからなる場合、投与される量は通常、1日およびkg体重当り0.02〜10gである。
【0022】
本発明の添加物は、本発明の成分以外の成分を含有しうる。適切には、添加物は、0.1〜100重量%、好ましくは、0.2〜90重量%の成分、および成長改善剤、香味料、吸収担体および/または他の飼料成分を含有する。好ましくは、成長改善剤、香味料、吸収担体および/または他の飼料成分の総量は、10〜75重量%の添加物である。成分が植物材料である場合、それらは適切には乾燥され、もしあれば、添加物中で他の成分と混合される前に粒子または粉末に粉砕される。
【0023】
適切な成長改善添加物および香味料の例は、クレゾール、アネトール、デカ−、ウンデカ−および/またはドデカ−ラクトン、イオノン、イロン、ジンジェロール、ピペリジン、プロピリデンファタライド(propylidene phatalide)、ブチリデンファタライド(butylidene phatalide)、カプサイシンおよび/またはタンニンである。担体は、例えば、40〜50重量%の木質繊維、8〜10重量%のステアリン、4〜5重量%のクルクマ粉末、4〜5重量%のローズマリー粉末、22〜28重量%の石灰石、1〜3重量%のアラビアゴムなどゴム、5〜50重量%の砂糖および/またはでんぷん、および5〜15重量%の水を含有しうる。他の飼料成分は、適切にはビタミン、酵素、鉱塩、粉砕穀類、タンパク質含有成分、炭水化物含有成分、ミドリング粉および/またはふすまからなる群から選択される。添加物は適切に、飼料が通常、0.4ppm〜80重量%の成分を含有するような量で本発明による飼料組成物に添加される。
【0024】
本発明による飼料組成物は通常、飼料の乾燥重量で計算され、以下の成分を含有する。
a)0〜80乾燥重量%の穀類、
b)0〜30乾燥重量%の油脂、
c)0〜85乾燥重量%の穀類以外の種類のタンパク質含有栄養物、および
d)10ppm〜40乾燥重量%の本発明の成分。
a)〜d)の総量は、好ましくは、少なくとも50乾燥重量%である。
【0025】
飼料組成物を調製する場合、添加物は、粉砕または破砕されたコムギ、オートムギ、オオムギ、トウモロコシ、およびコメなどの穀類、例えば、ナタネ、ダイズ、およびヒマワリの種に基づく植物性タンパク質源、動物性タンパク質源、糖蜜、およびさまざまな粉ミルクやホエー粉末など乳製品からなる乾燥成分と混合されうる。乾燥成分すべてを混合した後、液体成分、および加熱後に液体になる成分が添加されうる。液体成分は、場合により加熱によって液体化される油脂、例えば植物性油脂などの脂質、および/または脂肪酸などのカルボン酸からなりうる。完全な混合後、成分の粉砕度によって、粉状または粒子状の稠度が得られる。貯蔵中の分離を阻止するには、動物飼料に水を添加することが好ましく、次いで従来のペレット化、膨張、押出し工程にかけられる。余分な水は乾燥によって除去されうる。必要に応じて、結果として生じる粒状の動物飼料も破砕され、より小さな粒径にすることができる。記載された飼料組成物は通常、乾燥生草および/またはサイレージといっしょに投与される。
【0026】
飲料水サプリメントは少なくとも1乾燥重量%、適切には1〜99乾燥重量%、好ましくは10〜50乾燥重量%の成分を含有うる。1つもしくはそれ以上の成分のほかに、サプリメントは1〜99乾燥重量%の数多くの他の成分をも含有しうる。他の成分の適切な例は、鉱塩、ビタミン、健康および成長強化添加物、香味料、水溶性または水分散性担体、例えば砂糖、粉ミルク、乳副産物およびセルロース誘導体、分散剤および安定剤、例えば水溶性または水分散性ポリマー、およびそれらの混合物である。飲料水を調製する場合、サプリメントは通常、成分の濃度が1ppm〜10重量%になるような量で水に添加される。
【0027】
本発明の範囲内では、飼料組成物の懸濁液を生成することも可能である。これは、飼料が即時消費用に調製される場合に特に便利である。
【0028】
本発明をこれから以下の実施例によってさらに例示する。
【実施例】
【0029】
本実施例において多くの試験が植物材料、植物材料の抽出物、および精油化合物の試料で実行された。植物材料の試料は凍結乾燥され、粉砕され、1mmの篩に通された。試料は以下の通りであった。
【0030】
【表1】
【0031】
本発明による試料1〜22のルーメン消化物に対する効果を、ルーメン繊毛原虫の細菌分解活性、ルーメン消化物のタンパク質分解活性、ルーメン消化によって引き起こされるメタン形成、乳酸アシドーシス、および揮発性脂肪酸の形成のほか、発酵に対する他の効果に関して試験した。試験においては以下の方法を適用した。
【0032】
ルーメン繊毛原虫の細菌溶解活性
[14C]ロイシン標識セレノモナス・ルミナンチューム(Selenomonas ruminantium)(S.ルミナンチューム)の分解速度を、以前に記載されているように(ウォリス(Wallace)とマックフェルソン(McPherson)、1987年)、5mM非標識L−ロイシンの存在下に標識S.ルミナンチュームで染色ルーメン液をインビトロでインキュベートすることによって測定した。これらの条件下、大多数の細菌タンパク質分解は繊毛原虫による細菌の摂取および消化によって引き起こされる。試料は、他に記述がない限り、5mg/mlの濃度で添加された。ヒツジには混合干草、すなわち濃縮食を与えた(フルムホルツ(Frumholtz)ら、1989年)。
【0033】
抗原虫活性の継続
試料をコールマンバッファー、ルーメン液、または水とともに24時間、プレインキュベーションにかけ、次いでその後に上述した細菌分解アッセイで新鮮ルーメン液でインキュベートした。
【0034】
ルーメン消化物のタンパク質分解活性
方法1.同じヒツジからの染色ルーメン液を使用し、ウォリス(Wallace(1983年)の14C−カゼイン法を使用してタンパク質分解活性を測定した。試料を2.5mg/mlの最終濃度まで添加し、1時間のインキュベーション時間を使用した。
【0035】
方法2.ホフマン(Hoffman)ら(2003a、b)に基づく。典型的な乳牛食が与えられた瘻孔形成泌乳期ホルスタイン乳牛5頭をドナー動物として使用した。ルーメン液を朝の給餌前に給餌マットから手動で回収した。ルーメン液を100μmナイロンネットにろ過し、10%(v/v)のインキュベーションバッファーと混合した。75ml体積中緩衝化ルーメン液を、トウモロコシサイレージ450mg、オオムギ粒225mg(両方とも1mm篩に通るように粉砕)、および脱脂した未加工のダイズミール150mgとウシ血清アルブミン10mgからなるタンパク質サプリメントの基質混合物を含有する(RPT)インキュベーション瓶に分注した。試験植物が添加されていないこの基質を陰性対照としてインキュベートした。試験植物を添加した場合、それらはサイレージに置き換えた。陽性対照として使用したモネンシンをエタノール(14mg/ml)中に溶解し、この保存溶液11.25μlを3μMモネンシンの最終濃度まで充填直後に各フラスコへ注入した。すべてのフラスコを39℃下、12時間までインキュベートした。各処置の2つの複製を一定の間隔でサンプリングすると同時に、第3の補助複製をガスリーディングのために用意した。各サンプリング時間で、2×900μlを強く攪拌しながらインキュベーション瓶から回収し、SCFA、アンモニア、およびタンパク質濃度の測定のために調製した。SDS−PAGEをレムリ(Laemmli)法によって行い、総タンパク質定量をホフマン(Hoffman)ら(2002年)によるドットブロットによって行った。
【0036】
ルーメン発酵に対する一般的効果
発酵に対する一般的効果のホーエンハイム法
実験計画法:
インキュベーション系:リーディング・プレッシャー・テクニック(RPT)、75ml、総実行時間24時間
ドナー動物:非分泌乳期ホルスタイン乳牛3頭
基礎基質:牧草サイレージ0.750g(対照)
試験植物の含有レベル:10%(置換サイレージ)
ガスリーディング:2、4、6、8、10、12、16、24時間
サンプリング:SCFAおよびTDには24時間(ナイロンバッグ中NDS処置)
SCFA(900μl)およびRNA(300μl、分析最大値)には6、8、10時間
平行:2通りで3つの独立した実行(3種類のドナー動物)
【0037】
【表2】
【0038】
方法の詳細な説明:
インビトロインキュベーション
8:00時および16:00時に2つの同じ分量で給餌される維持において牧草サイレージ干草混合物を与えられた挿管乳牛からルーメン液を採取した。ルーメン液を魔法瓶に供給する前に採取し、100μmナイロンネットにろ過し、還元バッファー鉱液に添加した。すべてのステップを39℃でCO2下に行い、嫌気性状態を維持した。
【0039】
基質を10mg/ml培地の割合でインキュベートし、すなわち、基質750mgをろ過ルーメン液7.5ml(10%v/vルーメン液)を含有する緩衝化ルーメン液75mlでインキュベートした。
【0040】
ガス量または圧力をインキュベーションの2、4、6、8、10、12、16、および24時間の時点で頻繁に記録し、試料を6、8、10、および24時間後に採取した。
【0041】
分析
ガス圧はRPT系の血清瓶で記録され、実験的に測定された較正曲線を使用して体積へ変換される。各々の測定後、ガスは血清瓶から放出される。インビトロの真の消化率は、インキュベーション内容物を予め秤量したポリエステルバッグ(アンコム(Ankom)51μm孔径)に移すことによって測定された。バッグは熱シールされ、中性清浄液中で1時間煮沸され、105℃で一夜乾燥され、これを消化率の測定のために秤量した。SCFAは、GP10%SP1000 1%H3PO4、クロモソブ(Chromosob)WAW(サペルコ社(Suppelco Inc.)、Bellafonte、ペンシルベニア(PA))でパックされたステンレス鋼カラムを使用するガスクロマトグラフィーによって測定された。インキュベーション上清0.9mlに内部基準(1%メチルブトリル(Methylbutryric)酸)を含有するギ酸0.1mlを添加した。タンパク質を4℃で一夜沈殿させた。試料を遠心分離し(30,000g、10分、4℃)、上清を適切なGCバイアルへ分析のために採取した。
【0042】
RNAを改良型フェノールクロロホルムプロトコールに従って抽出した。pH5.1フェノール600μl、pH5.1バッファー270μl、SDS 30μl(20%w/v)、およびジルコニウムビーズ1.0gを試料(300μl)に添加した。試料を2×2分間、60℃で10分のインキュベーションで中断されるビーズビーター(50Hz)中で攪拌することによって細胞を溶解した。試料を氷上で10分間冷却し、クロロホルム300μlを添加した。強く、さらに10分間、室温(RT)下に振盪した後、試料を遠心分離し(10,000g、5分、4℃)、水相および有機相を分離した。水相を量的に除去し、NH4酢酸300μl(7.5M)およびイソプロパノール900μlを含有するバイアルに移した。試料を一夜、−20℃でインキュベートし、遠心分離(16,000g、10分、4℃)によって核酸を沈殿させた。上清を廃棄し、試料を1回、80%エタノール中で洗浄した。核酸をアガロースゲルに装填し、臭化エチジウムで染色した後、較正曲線に対して濃度測定的に定量化した(25〜300ng/μl)。
【0043】
発酵に対する一般的効果のリーディング法
一連の13連続発酵を毎週1回、リーディング・プレッシャー・テクニック[RPT]を使用して行い、発酵に対するこれら試験材料を含めることからの潜在的な効果、および基礎試料[トウモロコシサイレージ]の分解特性を確認した。β−ミルセン[含有レベル10および40mg g−1]を除く全試料を3通に、2つの含有レベル[100および400mg g−1DM基質]で検査した。トウモロコシサイレージを予備乾燥し、2mm篩に通るように粉砕した。全部で1.0gの混合基質[サイレージ+基質]を各々の発酵フラスコに配置した。次いで、緩衝化インキュベーション培地[90ml]を添加し、フラスコを密閉し、一夜室温下に保存した。調製ルーメン液での接種前に、フラスコ内容物を39℃に上昇させた。トウモロコシ/牧草サイレージを濃縮物[60:40]の割当量で自由に与えた泌乳期乳牛2頭から給餌前[07:00時]にルーメン液を手動で[用手圧搾内容物]得た。液体を2層のモスリンにろ過し、CO2下に39℃で使用するまで保持した。調製液10mlを各々のフラスコに添加し、これらを試験期間中39℃でインキュベートした。6つの陰性対照[緩衝化培地+ルーメン液のみ]を各々の実行に含め、直接のルーメン液効果に対するガス値および分解残留物を補正した。また、非補充トウモロコシサイレージ[1.0gフラスコ−1]をすべての実行に含めた[実行当りフラスコ6個]。試験内の結果を陽性対照値に対して表した。
【0044】
ヘッドスペースガス圧リーディングを接種後2、4、6、8、10、12、14、16、18、21、および24時間に得た。最後の測定後、発酵培地約3.0mlを各々のフラスコからサンプリングし、試験材料/含有レベルによってバルク化し、バリアン(Varian)3600GCを使用するVFA組成物のその後の分析まで[−20℃]で凍結保存した。発酵残留物をフラスコ内容物を60mlグーチろ過器[空隙率1、100〜160μm]を通じて軽真空下にろ過することによって回収した。次いで、残留物を乾燥し[24時間100℃]、秤量し、灰化し[一夜500℃]、再秤量した。測定された乾物[DM]および有機物[OM]の量を使用してiDMDおよびiOMD[それぞれ、インビトロDMおよびOM分解]を評価した。発酵ガス放出の程度[インキュベートされたmlガス g−1OM]および速度[ml h−1]を以前に導出された二次関数を使用して生成し、圧力を体積に変換した。飼料発酵効率[FE]を接種後24時間でのiDMD[g kg−1]/累積ガス[ml]放出として推定した。処置手段と陽性対照[トウモロコシサイレージのみ]との間の統計的差異をスチューデントT検定を使用して評価し、P>0.05を有意とした。
【0045】
発酵に対する一般的効果のレオン法
75%アルファルファ干草および25%オオムギからなる飼料で給餌されるルーメン挿管ヒツジ(ビタミン−ミネラルサプリメントおよび水を自由に与える)をルーメン液のドナーとして使用した。ルーメン液を朝食直前に採取した。
【0046】
50%アルファルファ干草、40%牧草の干草、および10%オオムギ粒からなるバッチ培養に使用された基質を1mm篩のハンマーミルで粉砕した。使用された基質の量は500mgであり、試験植物の含有率は10%(約50mg)であった。両方を秤量して血清瓶へ入れ、ここでろ過ルーメン液10mlおよびゲーリング(Goering)とファン・ゼースト(Van Soest)、1970年)によって記載された培地40mlを含有する緩衝化ルーメン液50mlを好気的に分散させた。これらの瓶を密閉し、39℃下にインキュベートした。インキュベーションの24時間後、確実な測定値を記録した。すなわち、
−総ガス生産(ml)、圧力トランスデューサを使用し、ガスを目盛付き注射器に収集。
−インキュベーション培地中のpH。
−インキュベーション倍地中の揮発性脂肪酸濃度、GCによる(VFAも接種時に測定し、24時間後のVFA産生を計算する)。
−DMインキュベーション残留物、焼結つぼ中で瓶内容物をろ過し、乾燥後の残留物を秤量することによって。その後、残留物の中性デタージェント繊維(NDF)内容物を測定し、非分解DNFの量を計算した。これからDMおよびNDFの消失を推定した。
【0047】
植物ごとに3つの複製をインキュベートし、ブランク(基質なし植物なし)および対照(植物なし基質500mg、および植物なし基質550mg)を使用した。
【0048】
ルーメン消化物によるメタン形成
レオンの一般法によって行われたインキュベーションをメタン形成について分析した。生産されたガスの代表的な試料をその後の分析のために採取した。サンプリングは、試料の固定および貯蔵を可能にする弁を装着した特殊なガス気密注射器を使用して行われた。試料は、ストッパーの中隔を通じて針の挿入後にヘッドスペースから直接採取され、試料を注射器に取り、弁を閉じた。この試料を総ガス生産の測定後に採取し、ガス組成物が放出されたガス中およびヘッドスペース中の残留物中で同じであると想定した。
【0049】
生産されたガス中のメタン濃度をガスクロマトグラフィーによって測定した。ガス試料(300〜500μl)をガス気密注射器から直接、対応するポートへ注入した。分析に使用されたGCパラメータ設定値は以下の通りであった。
器具:炎イオン化検出器(FID)を備えたシマズ(Shimadzu)GC−14B
カラム:60/80メッシュカルボキセン(Carboxen)1000固定相でパックした長さ2.3メートル×内径2.1mmステンレス鋼カラム
搬送ガス:ヘリウム、流量(100kPa)、一定流モード
温度:
検出器:FID。温度200℃、合成空気流量(50kPa)、H2流量(50kPa)
インジェクタ:温度200℃
カラムオーブン:170℃(定温)
【0050】
較正に使用される標準は純粋メタン(99.9%)であった。標準曲線は、メタンの注射された標準量に対してピーク面積の線形回帰をプロットすることによって確立され、反応因子を得た。
【0051】
アシドーシスアッセイ
48時間の発酵期間にわたって対照に対して、インキュベーション培地pHおよび産生される乳酸の濃度の減少が、潜在的なアシドーシス効果を消失する試験基質の能力の指標を示すモデルを使用した。本試験では粉砕コムギ[2mm]を基礎基質として使用し、試験材料を100mg g−1DM[または10mg g−1DM β−ミルセン]で含めた。前述同様に、総基質1.0gを各々のフラスコに添加した。乳酸産生を強化するために、調製したバッファー溶液を50%希釈した。これはインキュベーション培地の緩衝能力が上回り、結果として生じるpHの減少が乳酸菌および他の乳酸産生細菌の成長を促進することを確実にするためであった。ルーメン液は、使用された2頭の乳牛が高レベルの粗い粉砕コムギで早期泌乳割当量が与えられたこと除き既述されているように得られ、調製された。液体のpH測定は、接種後1時間[開始pH]、次いでさらに23および47時間後にpH電極を直接各々のフラスコへ挿入することによって行った。最後の測定の直後、液体約3.0mlを各々のフラスコから採取し、試料によってバルク化し、L[+]乳酸について酵素的に分析されるまで−20℃下に凍結保存した。
【0052】
結果は対照に対する絶対差として示されている。アシドーシスの好ましい効果が、24時間までのpH低下の低下率、高い終点pH、および低い乳酸濃度として確認された。
【0053】
結果
発酵活性の測定から得られた結果は、以下の表2a、2b、および2cに示されている。
【0054】
原虫アッセイ、方法1によるタンパク質分解アッセイ、およびメタン形成において得られた結果が同じ表に示されている。
【0055】
異なる含有率での選択された植物/植物抽出物の原虫の細菌溶解活性に対するインビトロでの影響は図1〜図7に示されている。原虫の細菌溶解活性の継続率は、特にG.アスクレピアダ(G.asclepiadea)でバッファーまたは水においてよりもルーメン液において高かった(表1)。K.アルベンシス(K.arvensis)の影響は図8に示されている。
【0056】
【表3】
【0057】
【表4】
【0058】
【表5】
【0059】
【表6】
【0060】
引用文献
ホフマン(Hoffman)EM、モイツェル(Meutzel)S、ベッカー(Becker)K(2002年)、A modified dot−blot method of protein determination applied in the tannin−protein−precipitation assay to facilitate the evaluation of tannin activity in animal feeds、Br.J.Nutr.87:421−426頁
ホフマン(Hoffman)EM、モイツェル(Meutzel)S、ベッカー(Becker)(2003年)、Effects of Moringa oleifera seed extract on rumen fermentation in vitro、Arch.Anim.Nutr.57:65−81頁
ホフマン(Hoffman)EM、モイツェル(Meutzel)S、ベッカー(Becker)(2003年)、The fermentation of soybean meal by rumen microbes in vitro reveals different kinetic features for the inactivation and the degradation of trypsin inhibitor protein、Anim.Feed Sci.Technol.106:189−197頁
ハンゲート(Hungate)RE(1966年)、The rumen and its microbes,Academic Press,New York and London Leng RA(1992)、Ruminant production and greenhouse gas emissions、Proc.NZ Soc.Anim.Prod.52(補遺):15−23頁
ナガラヤ(Nagaraja)TG、ニューボールド(Newbold)CJ、ファン・ネーベル(Van Nevel)CJ、デマイヤー(Demeyer)DI(1997年)、Manipulation of ruminal fermentation,in The rumen microbial ecosystem、ホブソン(Hobson)PNおよびスチュワート(Stewart)CS編、チャップマン・アンド・ホール(Chapman & Hall)、London、523−632頁
ノラン(Nolan)JV (1975年)、Quantitative models of nitrogen metabolism in sheep,in Digestion and Metabolism in the Ruminant、McDonald IW and Warner ACI編、University of New England Publishing Unit、Armidale、Australia、416−431頁
ラッセル(Russell)JB、ヒノ(Hino)T(1985年)、Regulation of lactate production in Streptococcus bovis:a spiraling effect that contributes to rumen acidosis、J.Dairy Sci.68:1712−1721頁
ウォリス(Wallace)RJ、マックフェルソン(McPherson)CA (1987年)、Factors affecting the rate of breakdown of bacterial protein in rumen fluid、Br.J.Nutr.58:313−323頁
ウォリス(Wallace)RJ(1983年)、Hydrolysis of 14C−labelled proteins by rumen micro−organisms and by proteolytic enzymes prepared from rumen bacteria、Br.J.Nutr.50:345−355頁
ウォリス(Wallace)RJ、オノデラ(Onodera)R、コッタ(Cotta)MA (1997年)、Metabolism of nitrogen−containing compounds, in The rumen microbial ecosystem、ホブソン(Hobson)PNおよびスチュワート(Stewart) CS編、チャップマン・アンド・ホール(Chapman & Hall)、London、283−328頁
【図面の簡単な説明】
【0061】
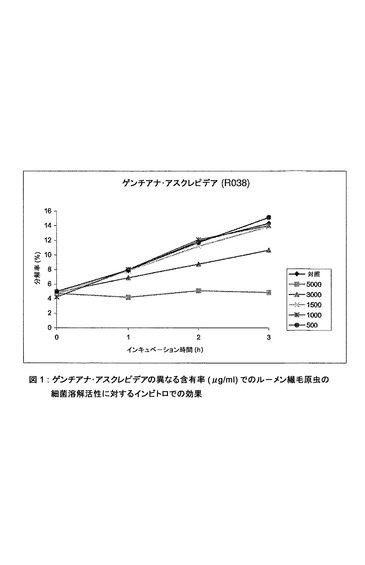
【図1】ゲンチアナ・アスクレピデアの異なる含有率(μg/ml)でのルーメン繊毛原虫の細菌溶解活性に対するインビトロでの効果。
【図2】スイカズラの異なる含有率(μg/ml)でのルーメン繊毛原虫の細菌溶解活性に対するインビトロでの効果。
【図3】クローブバッドの異なる含有率(μg/ml)でのルーメン繊毛原虫の細菌溶解活性に対するインビトロでの効果。
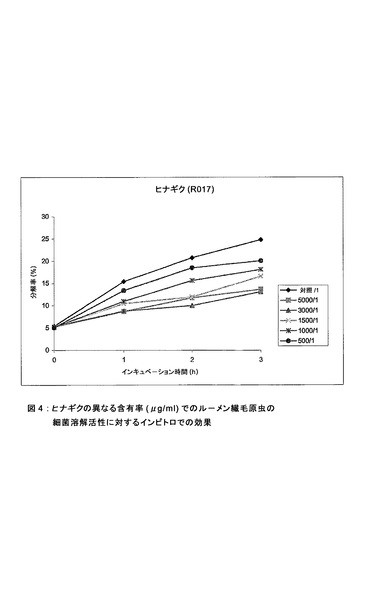
【図4】ヒナギクの異なる含有率(μg/ml)でのルーメン繊毛原虫の細菌溶解活性に対するインビトロでの効果。
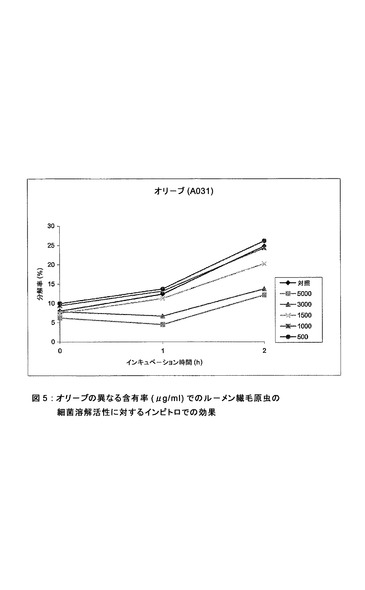
【図5】オリーブの異なる含有率(μg/ml)でのルーメン繊毛原虫の細菌溶解活性に対するインビトロでの効果。
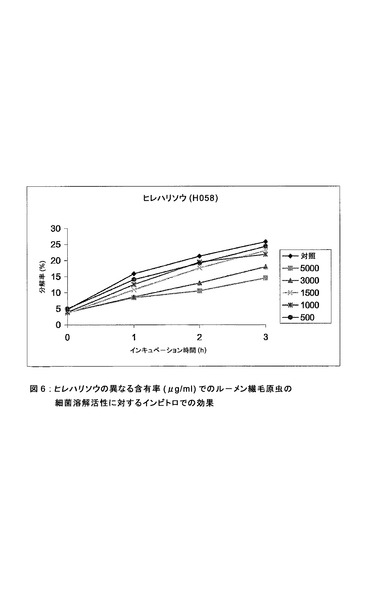
【図6】ヒレハリソウの異なる含有率(μg/ml)でのルーメン繊毛原虫の細菌溶解活性に対するインビトロでの効果。
【図7】β−ミルセンの異なる含有率(μg/ml)でのルーメン繊毛原虫の細菌溶解活性に対するインビトロでの効果。
【図8】ドットブロットアッセイによるタンパク質分解の定量により、K.アルベンシスの2種類の受入によるタンパク質分解の阻害が確認される。
【技術分野】
【0001】
本発明は、ルーメン発酵のエネルギーおよび窒素保持を改善するための、または乳酸アシドーシスを減少させるための、植物材料、植物抽出物、または天然同一精油化合物からの1つもしくはそれ以上の成分を含有する添加物の使用に関する。この化合物は、動物の飼料または飲料水によって反芻動物に適切に経口投与される。
【背景技術】
【0002】
ルーメンは、ウシ、ヒツジ、およびヤギなど反芻動物の第一胃である。その構造上および栄養上の意義は、ハンゲート(Hungate)(1966年)によって記載されている。ルーメンは、これらの動物が大部分の非反芻動物に不適切な高繊維植物材料を消化することを可能にする。ルーメンは、乳牛における150kgを超える消化物を含有する巨大な臓器であり、ここで消化のすべては微生物によって行われる。ルーメンは、動物が微生物の繊維消化活動から恩恵を受けるようにして発達した。最終生成物である、揮発性脂肪酸は、ルーメン壁にわたって吸収され、エネルギーおよびタンパク質合成に使用される。後腸発酵と比べルーメン前腸発酵の主な利点は、発酵中に形成される微生物細胞が第4胃に移動し、そこでそれらは消化され、アミノ酸が吸収されることである。したがって、後腸発酵とは違って、微生物アミノ酸は宿主動物に利用可能となる。したがって、ルーメンは、乏しい牧草地で自給する草食動物の進化との関連できわめて効率的な臓器である。
【0003】
ルーメン発酵はいくつかの欠点ももたらす。メタンは嫌気性発酵の自然の結果として生成される(ハンゲート(Hungate)、1966年)。メタンの放出は動物からのエネルギーの損失も表す。さらに、メタンは強力な温室効果ガスであり、したがってこの意味で、反芻動物は環境に損害を与える(レング(Leng)、1992年)。反芻動物はまた、食事タンパク質の利用において他の種よりも効率的でない。反芻動物からの窒素損失は、特に草食動物において例外的に高い(ノラン(Nolan)、1975年)。これは、窒素豊富な糞尿の環境に対する影響のため環境問題であると同時に経済問題である。さらに、障害のあるルーメン微生物発酵の直接の結果として生じる反芻動物に特有の消化系障害がある。
【0004】
化学的および抗生物質の飼料添加物は過去においてこれらの問題の一部を緩和するために使用されている。食物連鎖におけるかかる材料の存在は、規制当局および消費者にはますます受入れられなくなっている。したがって、反芻動物の家畜生産の問題解決は自然かつ持続可能でなければならない。
【0005】
ルーメンに入る食事タンパク質は明らかに非制御的に分解され、結果としてアンモニア形成およびその後の尿中窒素の損失がもたらされる(ノラン(Nolan)、1975年)。主な経済的損失を表す窒素保持の低い効率は、動物における代謝性侵襲を引き起こし、かつ窒素豊富な廃棄物によって環境への負担ともなる。分解過程が減少されれば、これらの問題が減少されるであろう。
【0006】
さらに、ルーメンにおけるタンパク質保持を改善するために存在する第2の可能性は、ルーメン繊毛原虫の集団の抑制である。原虫はルーメンにおける大量の細菌を消費し、それらの分解は結果として50%までのルーメン発酵からの微生物タンパク質の正味収量の減少をもたらしうる(ウォリス(Wallace)ら、1997年)。原虫が抑制されうる場合は、アンモニア形成が少なくなり、かつ食事タンパク質の補給の必要が少なくなる。
【0007】
乳酸は通常、ルーメン発酵の微量生成物にすぎない。しかし、迅速分解飼料があまりに急速に導入される場合、または濃縮物が高い割合の食事を形成する場合、揮発性脂肪酸産生がルーメンの緩衝能力を超える。これはルーメンにおけるきわめて低いpH値をもたらすため、乳酸産生細菌のみが増殖しうる(ラッセル(Russell)とヒノ(Hino)、1985年)。
【0008】
乳酸が揮発性脂肪酸よりも強い酸であると、pHはさらに低下し、正常な発酵の回復は不可能になり、そして動物は死亡する。しかし、より一般的には、高濃度の食事の状態にある反芻動物は無症状のアシドーシスにかかるが、これは苦難を伴い、生命にかかわることはないものの生産効率を減少させる。化学バッファーはこの状態を食い止めることができるが、乳酸産生細菌の増殖が抑制されればより良いであろう。
【0009】
公報国際公開第03/094628号パンフレットは、リモネン、オイゲノール、サリチル酸塩、キノリン、バニラ、チモール、およびクレゾールからなる群から選択される1つもしくはそれ以上の精油化合物の添加によって反芻動物の消化活動から発するメタン産生を削減する試みを記載している。同時にルーメンにおける微生物の活動および揮発性脂肪酸、例えばプロピオン酸の産生が許容可能レベルで維持されうる。
【発明の詳細な説明】
【0010】
本発明の目的は、全体として、かつ一括してルーメンにおける消化系障害に取組み、かつ反芻動物の成長に利用可能なエネルギーおよび窒素源の利用を促進することである。この目的は、ルーメン壁にわたって吸収され、エネルギーおよびタンパク質合成に使用される揮発性脂肪酸の形成を助けることによって、かつ食事タンパク質および微生物タンパク質の分解を減少させることによって達成されうる。他の方法は、メタン生成を制限し、乳酸産生細菌を制御または抑制することである。
【0011】
本発明によれば、前記目的は、スイカズラ、ゲンチアナ・アスクレピデア、キバナリンドウ、クローブバッド、ヒナギク、オリーブ、ヒレハリソウ、ヒメヒレアザミ、シャクヤク、ヤマナラシ、サクランボ、バッコヤナギ、セイタカダイオウ、ハンニチバナ、クマコケモモ、インディアンルバーブ、エゾアカバナ、クナウチア・アルベンシス、レタス(Latuca sativa)、およびネトルおよびそれらの抽出物、およびβ−ミルセンからなる群から選択される植物材料の1つもしくはそれ以上の成分を含有する添加物を動物飼料または飲料水によって適切に経口投与することによって達成されうる。動物に投与される成分の総量は、1日およびkg体重当り0.02mg〜20gであることが適切である。
【0012】
本発明の成分はすべて、考慮すべき欠点なしにルーメンにおける発酵の効率に実質的に寄与することが証明されている。成分の一部は原虫の活性を抑制し、その他はタンパク質分解活性、メタン生成活性、または乳酸の産生を減少させる。
【0013】
本発明は、1つもしくはそれ以上の規定された成分を含有する添加物および飼料組成物にも関する。
【0014】
成分は発酵に対して多様に影響を有するため、本発明の目的に記載の発酵を最適化する特定の化合物を組み合わせることが有利である。
【0015】
大規模な試験では、ハンニチバナ、クマコケモモ、エゾアカバナ、クナウチア・アルベンシス、およびインディアンルバーブ、およびそれらの抽出物からの植物材料からなる亜群の成分はルーメンのタンパク質分解を大幅に削減することが示された。また、これらの成分のすべては原虫の活性を阻害し、その大部分は、メタン産生、揮発性脂肪酸の量、消化率、微生物バイオマス、および/または発酵効率など、他の必須の発酵パラメータに対する有利な効果も有する。最も適切な成分は、クマコケモモ、クナウチア・アルベンシス、およびハンニチバナ、およびそれらの抽出物からの植物材料である。最も好ましい成分は、クナウチア・アルベンシスおよびその抽出物、特にメタノール抽出物からの植物材料である。
【0016】
スイカズラ、ゲンチアナ・アスクレピデア、キバナリンドウ、クローブバッド、ヒナギク、オリーブおよびヒレハリソウおよびそれらの抽出物およびβ−ミルセンからの植物材料は、その成分がルーメン液中の繊毛原虫の細菌溶解活性を抑制する別の亜群を形成する。クローブバッドも微生物バイオマスを増大させ、メタン産生を削減する。ヒナギクはメタン産生の小さな削減を示し、発酵効率および微生物バイオマス産生を改善もするが、ヒレハリソウは発酵効率を増大させ、アシドーシス効果を削減する。β−ミルセンは原虫活性に対する中程度の阻害効果を有するが、メタン産生を減少させるためにも使用される。スイカズラの花から調製される抽出物が繊毛原虫の細菌分解活性を無効にした。スイカズラの種子調製物も原虫活性を抑制したが、これは約60%にすぎない。この亜群からの好ましい成分は、ヒナギク、クローブバッド、オリーブおよびヒレハリソウおよびそれらの抽出物、特にメタノール、およびヒナギクおよびゲンチアナ・アスクレピデアの水抽出物からの植物材料である。
【0017】
ヒメヒレアザミ、シャクヤク、ヤマナラシ、サクランボ、バッコヤナギおよびセイタカダイオウおよびそれらの抽出物からの植物材料は、その成分がルーメン消化物中のメタン生成活性を減少させるために適切に使用される第3の亜群を形成する。これらの成分には重要な有害な効果が他の発酵パラメータでは確認されていない。好ましい成分は、ヒメヒレアザミ、ヤマナラシ、およびセイタカダイオウおよびそれらの抽出物であり、特に好ましいのは、ヒメヒレアザミおよびセイタカダイオウからの植物材料である。
【0018】
最後に、第4の亜群が、レタス(Latuca sativa)およびネトルおよびそれらの抽出物からの植物材料によって形成される。これらの成分は乳酸の活性および/または形成を低下させる。レタス(Latuca sativa)は、ワイルドレタス(Latuca virosa)と呼ばれることもあり、これもメタン産生および原虫活性を減少させ、揮発性脂肪酸の形成を増大させる。ネトルは追加の効果を有し、タンパク質分解および原虫活性を阻害し、かつ揮発性脂肪酸の形成を助ける。
【0019】
本発明の適切な実施形態において、添加物は、上記規定された亜群の1つ以上成分を含有し、添加物は4つの亜群すべてからの成分を含有することが有利でありうる。有利には、添加物は、
i)クナウチア・アルベンシス、クマコケモモ、およびハンニチバナおよびそれらの抽出物からの植物材料と、
ii)ヒナギク、クローブバッド、オリーブ、スイカズラおよびヒレハリソウおよびそれらの抽出物からの植物材料と、
iii)ヒメヒレアザミ、ヤマナラシ、およびセイタカダイオウおよびそれらの抽出物からの植物材料と、
iv)レタス(Latuca sativa)およびネトルおよびそれらの抽出物からの植物材料と
からなる群の各々から選択される少なくとも1つの成分を含有する。
i)〜iv)の群に属する成分の総量は適切には20〜100重量%であり、好ましくは本発明により存在する成分の総量の40〜100重量%である。
【0020】
本発明による植物材料の抽出物としては、凝結物、精油、樹脂状物質、チンキ剤、遊離物および純油が挙げられるがこれらに限定されない。植物または植物部位の抽出物の生成に際しては、水、有機溶媒、およびそれらの混合物が従来の方法に従って使用されうるが、乾燥蒸留も適用されうる。適切な有機溶媒の例は、メタノール、エタノール、プロパノール、ブタノール、酢酸エチル、メチルエチルエーテル、ジエチルエーテル、塩化メチレン、クロロホルム、四塩化炭素、ベンゼン、トルエン、石油エーテル、およびアセトンである。
【0021】
動物に投与される成分の総量は、その成分が精油化合物、植物材料の抽出物、または植物材料であるかどうかによって決まる。成分がすべてβ−ミルセン、および/または抽出物である場合には、投与量は通常、1日およびkg体重当り0.1〜50mgであるが、成分が植物材料のみからなる場合、投与される量は通常、1日およびkg体重当り0.02〜10gである。
【0022】
本発明の添加物は、本発明の成分以外の成分を含有しうる。適切には、添加物は、0.1〜100重量%、好ましくは、0.2〜90重量%の成分、および成長改善剤、香味料、吸収担体および/または他の飼料成分を含有する。好ましくは、成長改善剤、香味料、吸収担体および/または他の飼料成分の総量は、10〜75重量%の添加物である。成分が植物材料である場合、それらは適切には乾燥され、もしあれば、添加物中で他の成分と混合される前に粒子または粉末に粉砕される。
【0023】
適切な成長改善添加物および香味料の例は、クレゾール、アネトール、デカ−、ウンデカ−および/またはドデカ−ラクトン、イオノン、イロン、ジンジェロール、ピペリジン、プロピリデンファタライド(propylidene phatalide)、ブチリデンファタライド(butylidene phatalide)、カプサイシンおよび/またはタンニンである。担体は、例えば、40〜50重量%の木質繊維、8〜10重量%のステアリン、4〜5重量%のクルクマ粉末、4〜5重量%のローズマリー粉末、22〜28重量%の石灰石、1〜3重量%のアラビアゴムなどゴム、5〜50重量%の砂糖および/またはでんぷん、および5〜15重量%の水を含有しうる。他の飼料成分は、適切にはビタミン、酵素、鉱塩、粉砕穀類、タンパク質含有成分、炭水化物含有成分、ミドリング粉および/またはふすまからなる群から選択される。添加物は適切に、飼料が通常、0.4ppm〜80重量%の成分を含有するような量で本発明による飼料組成物に添加される。
【0024】
本発明による飼料組成物は通常、飼料の乾燥重量で計算され、以下の成分を含有する。
a)0〜80乾燥重量%の穀類、
b)0〜30乾燥重量%の油脂、
c)0〜85乾燥重量%の穀類以外の種類のタンパク質含有栄養物、および
d)10ppm〜40乾燥重量%の本発明の成分。
a)〜d)の総量は、好ましくは、少なくとも50乾燥重量%である。
【0025】
飼料組成物を調製する場合、添加物は、粉砕または破砕されたコムギ、オートムギ、オオムギ、トウモロコシ、およびコメなどの穀類、例えば、ナタネ、ダイズ、およびヒマワリの種に基づく植物性タンパク質源、動物性タンパク質源、糖蜜、およびさまざまな粉ミルクやホエー粉末など乳製品からなる乾燥成分と混合されうる。乾燥成分すべてを混合した後、液体成分、および加熱後に液体になる成分が添加されうる。液体成分は、場合により加熱によって液体化される油脂、例えば植物性油脂などの脂質、および/または脂肪酸などのカルボン酸からなりうる。完全な混合後、成分の粉砕度によって、粉状または粒子状の稠度が得られる。貯蔵中の分離を阻止するには、動物飼料に水を添加することが好ましく、次いで従来のペレット化、膨張、押出し工程にかけられる。余分な水は乾燥によって除去されうる。必要に応じて、結果として生じる粒状の動物飼料も破砕され、より小さな粒径にすることができる。記載された飼料組成物は通常、乾燥生草および/またはサイレージといっしょに投与される。
【0026】
飲料水サプリメントは少なくとも1乾燥重量%、適切には1〜99乾燥重量%、好ましくは10〜50乾燥重量%の成分を含有うる。1つもしくはそれ以上の成分のほかに、サプリメントは1〜99乾燥重量%の数多くの他の成分をも含有しうる。他の成分の適切な例は、鉱塩、ビタミン、健康および成長強化添加物、香味料、水溶性または水分散性担体、例えば砂糖、粉ミルク、乳副産物およびセルロース誘導体、分散剤および安定剤、例えば水溶性または水分散性ポリマー、およびそれらの混合物である。飲料水を調製する場合、サプリメントは通常、成分の濃度が1ppm〜10重量%になるような量で水に添加される。
【0027】
本発明の範囲内では、飼料組成物の懸濁液を生成することも可能である。これは、飼料が即時消費用に調製される場合に特に便利である。
【0028】
本発明をこれから以下の実施例によってさらに例示する。
【実施例】
【0029】
本実施例において多くの試験が植物材料、植物材料の抽出物、および精油化合物の試料で実行された。植物材料の試料は凍結乾燥され、粉砕され、1mmの篩に通された。試料は以下の通りであった。
【0030】
【表1】
【0031】
本発明による試料1〜22のルーメン消化物に対する効果を、ルーメン繊毛原虫の細菌分解活性、ルーメン消化物のタンパク質分解活性、ルーメン消化によって引き起こされるメタン形成、乳酸アシドーシス、および揮発性脂肪酸の形成のほか、発酵に対する他の効果に関して試験した。試験においては以下の方法を適用した。
【0032】
ルーメン繊毛原虫の細菌溶解活性
[14C]ロイシン標識セレノモナス・ルミナンチューム(Selenomonas ruminantium)(S.ルミナンチューム)の分解速度を、以前に記載されているように(ウォリス(Wallace)とマックフェルソン(McPherson)、1987年)、5mM非標識L−ロイシンの存在下に標識S.ルミナンチュームで染色ルーメン液をインビトロでインキュベートすることによって測定した。これらの条件下、大多数の細菌タンパク質分解は繊毛原虫による細菌の摂取および消化によって引き起こされる。試料は、他に記述がない限り、5mg/mlの濃度で添加された。ヒツジには混合干草、すなわち濃縮食を与えた(フルムホルツ(Frumholtz)ら、1989年)。
【0033】
抗原虫活性の継続
試料をコールマンバッファー、ルーメン液、または水とともに24時間、プレインキュベーションにかけ、次いでその後に上述した細菌分解アッセイで新鮮ルーメン液でインキュベートした。
【0034】
ルーメン消化物のタンパク質分解活性
方法1.同じヒツジからの染色ルーメン液を使用し、ウォリス(Wallace(1983年)の14C−カゼイン法を使用してタンパク質分解活性を測定した。試料を2.5mg/mlの最終濃度まで添加し、1時間のインキュベーション時間を使用した。
【0035】
方法2.ホフマン(Hoffman)ら(2003a、b)に基づく。典型的な乳牛食が与えられた瘻孔形成泌乳期ホルスタイン乳牛5頭をドナー動物として使用した。ルーメン液を朝の給餌前に給餌マットから手動で回収した。ルーメン液を100μmナイロンネットにろ過し、10%(v/v)のインキュベーションバッファーと混合した。75ml体積中緩衝化ルーメン液を、トウモロコシサイレージ450mg、オオムギ粒225mg(両方とも1mm篩に通るように粉砕)、および脱脂した未加工のダイズミール150mgとウシ血清アルブミン10mgからなるタンパク質サプリメントの基質混合物を含有する(RPT)インキュベーション瓶に分注した。試験植物が添加されていないこの基質を陰性対照としてインキュベートした。試験植物を添加した場合、それらはサイレージに置き換えた。陽性対照として使用したモネンシンをエタノール(14mg/ml)中に溶解し、この保存溶液11.25μlを3μMモネンシンの最終濃度まで充填直後に各フラスコへ注入した。すべてのフラスコを39℃下、12時間までインキュベートした。各処置の2つの複製を一定の間隔でサンプリングすると同時に、第3の補助複製をガスリーディングのために用意した。各サンプリング時間で、2×900μlを強く攪拌しながらインキュベーション瓶から回収し、SCFA、アンモニア、およびタンパク質濃度の測定のために調製した。SDS−PAGEをレムリ(Laemmli)法によって行い、総タンパク質定量をホフマン(Hoffman)ら(2002年)によるドットブロットによって行った。
【0036】
ルーメン発酵に対する一般的効果
発酵に対する一般的効果のホーエンハイム法
実験計画法:
インキュベーション系:リーディング・プレッシャー・テクニック(RPT)、75ml、総実行時間24時間
ドナー動物:非分泌乳期ホルスタイン乳牛3頭
基礎基質:牧草サイレージ0.750g(対照)
試験植物の含有レベル:10%(置換サイレージ)
ガスリーディング:2、4、6、8、10、12、16、24時間
サンプリング:SCFAおよびTDには24時間(ナイロンバッグ中NDS処置)
SCFA(900μl)およびRNA(300μl、分析最大値)には6、8、10時間
平行:2通りで3つの独立した実行(3種類のドナー動物)
【0037】
【表2】
【0038】
方法の詳細な説明:
インビトロインキュベーション
8:00時および16:00時に2つの同じ分量で給餌される維持において牧草サイレージ干草混合物を与えられた挿管乳牛からルーメン液を採取した。ルーメン液を魔法瓶に供給する前に採取し、100μmナイロンネットにろ過し、還元バッファー鉱液に添加した。すべてのステップを39℃でCO2下に行い、嫌気性状態を維持した。
【0039】
基質を10mg/ml培地の割合でインキュベートし、すなわち、基質750mgをろ過ルーメン液7.5ml(10%v/vルーメン液)を含有する緩衝化ルーメン液75mlでインキュベートした。
【0040】
ガス量または圧力をインキュベーションの2、4、6、8、10、12、16、および24時間の時点で頻繁に記録し、試料を6、8、10、および24時間後に採取した。
【0041】
分析
ガス圧はRPT系の血清瓶で記録され、実験的に測定された較正曲線を使用して体積へ変換される。各々の測定後、ガスは血清瓶から放出される。インビトロの真の消化率は、インキュベーション内容物を予め秤量したポリエステルバッグ(アンコム(Ankom)51μm孔径)に移すことによって測定された。バッグは熱シールされ、中性清浄液中で1時間煮沸され、105℃で一夜乾燥され、これを消化率の測定のために秤量した。SCFAは、GP10%SP1000 1%H3PO4、クロモソブ(Chromosob)WAW(サペルコ社(Suppelco Inc.)、Bellafonte、ペンシルベニア(PA))でパックされたステンレス鋼カラムを使用するガスクロマトグラフィーによって測定された。インキュベーション上清0.9mlに内部基準(1%メチルブトリル(Methylbutryric)酸)を含有するギ酸0.1mlを添加した。タンパク質を4℃で一夜沈殿させた。試料を遠心分離し(30,000g、10分、4℃)、上清を適切なGCバイアルへ分析のために採取した。
【0042】
RNAを改良型フェノールクロロホルムプロトコールに従って抽出した。pH5.1フェノール600μl、pH5.1バッファー270μl、SDS 30μl(20%w/v)、およびジルコニウムビーズ1.0gを試料(300μl)に添加した。試料を2×2分間、60℃で10分のインキュベーションで中断されるビーズビーター(50Hz)中で攪拌することによって細胞を溶解した。試料を氷上で10分間冷却し、クロロホルム300μlを添加した。強く、さらに10分間、室温(RT)下に振盪した後、試料を遠心分離し(10,000g、5分、4℃)、水相および有機相を分離した。水相を量的に除去し、NH4酢酸300μl(7.5M)およびイソプロパノール900μlを含有するバイアルに移した。試料を一夜、−20℃でインキュベートし、遠心分離(16,000g、10分、4℃)によって核酸を沈殿させた。上清を廃棄し、試料を1回、80%エタノール中で洗浄した。核酸をアガロースゲルに装填し、臭化エチジウムで染色した後、較正曲線に対して濃度測定的に定量化した(25〜300ng/μl)。
【0043】
発酵に対する一般的効果のリーディング法
一連の13連続発酵を毎週1回、リーディング・プレッシャー・テクニック[RPT]を使用して行い、発酵に対するこれら試験材料を含めることからの潜在的な効果、および基礎試料[トウモロコシサイレージ]の分解特性を確認した。β−ミルセン[含有レベル10および40mg g−1]を除く全試料を3通に、2つの含有レベル[100および400mg g−1DM基質]で検査した。トウモロコシサイレージを予備乾燥し、2mm篩に通るように粉砕した。全部で1.0gの混合基質[サイレージ+基質]を各々の発酵フラスコに配置した。次いで、緩衝化インキュベーション培地[90ml]を添加し、フラスコを密閉し、一夜室温下に保存した。調製ルーメン液での接種前に、フラスコ内容物を39℃に上昇させた。トウモロコシ/牧草サイレージを濃縮物[60:40]の割当量で自由に与えた泌乳期乳牛2頭から給餌前[07:00時]にルーメン液を手動で[用手圧搾内容物]得た。液体を2層のモスリンにろ過し、CO2下に39℃で使用するまで保持した。調製液10mlを各々のフラスコに添加し、これらを試験期間中39℃でインキュベートした。6つの陰性対照[緩衝化培地+ルーメン液のみ]を各々の実行に含め、直接のルーメン液効果に対するガス値および分解残留物を補正した。また、非補充トウモロコシサイレージ[1.0gフラスコ−1]をすべての実行に含めた[実行当りフラスコ6個]。試験内の結果を陽性対照値に対して表した。
【0044】
ヘッドスペースガス圧リーディングを接種後2、4、6、8、10、12、14、16、18、21、および24時間に得た。最後の測定後、発酵培地約3.0mlを各々のフラスコからサンプリングし、試験材料/含有レベルによってバルク化し、バリアン(Varian)3600GCを使用するVFA組成物のその後の分析まで[−20℃]で凍結保存した。発酵残留物をフラスコ内容物を60mlグーチろ過器[空隙率1、100〜160μm]を通じて軽真空下にろ過することによって回収した。次いで、残留物を乾燥し[24時間100℃]、秤量し、灰化し[一夜500℃]、再秤量した。測定された乾物[DM]および有機物[OM]の量を使用してiDMDおよびiOMD[それぞれ、インビトロDMおよびOM分解]を評価した。発酵ガス放出の程度[インキュベートされたmlガス g−1OM]および速度[ml h−1]を以前に導出された二次関数を使用して生成し、圧力を体積に変換した。飼料発酵効率[FE]を接種後24時間でのiDMD[g kg−1]/累積ガス[ml]放出として推定した。処置手段と陽性対照[トウモロコシサイレージのみ]との間の統計的差異をスチューデントT検定を使用して評価し、P>0.05を有意とした。
【0045】
発酵に対する一般的効果のレオン法
75%アルファルファ干草および25%オオムギからなる飼料で給餌されるルーメン挿管ヒツジ(ビタミン−ミネラルサプリメントおよび水を自由に与える)をルーメン液のドナーとして使用した。ルーメン液を朝食直前に採取した。
【0046】
50%アルファルファ干草、40%牧草の干草、および10%オオムギ粒からなるバッチ培養に使用された基質を1mm篩のハンマーミルで粉砕した。使用された基質の量は500mgであり、試験植物の含有率は10%(約50mg)であった。両方を秤量して血清瓶へ入れ、ここでろ過ルーメン液10mlおよびゲーリング(Goering)とファン・ゼースト(Van Soest)、1970年)によって記載された培地40mlを含有する緩衝化ルーメン液50mlを好気的に分散させた。これらの瓶を密閉し、39℃下にインキュベートした。インキュベーションの24時間後、確実な測定値を記録した。すなわち、
−総ガス生産(ml)、圧力トランスデューサを使用し、ガスを目盛付き注射器に収集。
−インキュベーション培地中のpH。
−インキュベーション倍地中の揮発性脂肪酸濃度、GCによる(VFAも接種時に測定し、24時間後のVFA産生を計算する)。
−DMインキュベーション残留物、焼結つぼ中で瓶内容物をろ過し、乾燥後の残留物を秤量することによって。その後、残留物の中性デタージェント繊維(NDF)内容物を測定し、非分解DNFの量を計算した。これからDMおよびNDFの消失を推定した。
【0047】
植物ごとに3つの複製をインキュベートし、ブランク(基質なし植物なし)および対照(植物なし基質500mg、および植物なし基質550mg)を使用した。
【0048】
ルーメン消化物によるメタン形成
レオンの一般法によって行われたインキュベーションをメタン形成について分析した。生産されたガスの代表的な試料をその後の分析のために採取した。サンプリングは、試料の固定および貯蔵を可能にする弁を装着した特殊なガス気密注射器を使用して行われた。試料は、ストッパーの中隔を通じて針の挿入後にヘッドスペースから直接採取され、試料を注射器に取り、弁を閉じた。この試料を総ガス生産の測定後に採取し、ガス組成物が放出されたガス中およびヘッドスペース中の残留物中で同じであると想定した。
【0049】
生産されたガス中のメタン濃度をガスクロマトグラフィーによって測定した。ガス試料(300〜500μl)をガス気密注射器から直接、対応するポートへ注入した。分析に使用されたGCパラメータ設定値は以下の通りであった。
器具:炎イオン化検出器(FID)を備えたシマズ(Shimadzu)GC−14B
カラム:60/80メッシュカルボキセン(Carboxen)1000固定相でパックした長さ2.3メートル×内径2.1mmステンレス鋼カラム
搬送ガス:ヘリウム、流量(100kPa)、一定流モード
温度:
検出器:FID。温度200℃、合成空気流量(50kPa)、H2流量(50kPa)
インジェクタ:温度200℃
カラムオーブン:170℃(定温)
【0050】
較正に使用される標準は純粋メタン(99.9%)であった。標準曲線は、メタンの注射された標準量に対してピーク面積の線形回帰をプロットすることによって確立され、反応因子を得た。
【0051】
アシドーシスアッセイ
48時間の発酵期間にわたって対照に対して、インキュベーション培地pHおよび産生される乳酸の濃度の減少が、潜在的なアシドーシス効果を消失する試験基質の能力の指標を示すモデルを使用した。本試験では粉砕コムギ[2mm]を基礎基質として使用し、試験材料を100mg g−1DM[または10mg g−1DM β−ミルセン]で含めた。前述同様に、総基質1.0gを各々のフラスコに添加した。乳酸産生を強化するために、調製したバッファー溶液を50%希釈した。これはインキュベーション培地の緩衝能力が上回り、結果として生じるpHの減少が乳酸菌および他の乳酸産生細菌の成長を促進することを確実にするためであった。ルーメン液は、使用された2頭の乳牛が高レベルの粗い粉砕コムギで早期泌乳割当量が与えられたこと除き既述されているように得られ、調製された。液体のpH測定は、接種後1時間[開始pH]、次いでさらに23および47時間後にpH電極を直接各々のフラスコへ挿入することによって行った。最後の測定の直後、液体約3.0mlを各々のフラスコから採取し、試料によってバルク化し、L[+]乳酸について酵素的に分析されるまで−20℃下に凍結保存した。
【0052】
結果は対照に対する絶対差として示されている。アシドーシスの好ましい効果が、24時間までのpH低下の低下率、高い終点pH、および低い乳酸濃度として確認された。
【0053】
結果
発酵活性の測定から得られた結果は、以下の表2a、2b、および2cに示されている。
【0054】
原虫アッセイ、方法1によるタンパク質分解アッセイ、およびメタン形成において得られた結果が同じ表に示されている。
【0055】
異なる含有率での選択された植物/植物抽出物の原虫の細菌溶解活性に対するインビトロでの影響は図1〜図7に示されている。原虫の細菌溶解活性の継続率は、特にG.アスクレピアダ(G.asclepiadea)でバッファーまたは水においてよりもルーメン液において高かった(表1)。K.アルベンシス(K.arvensis)の影響は図8に示されている。
【0056】
【表3】
【0057】
【表4】
【0058】
【表5】
【0059】
【表6】
【0060】
引用文献
ホフマン(Hoffman)EM、モイツェル(Meutzel)S、ベッカー(Becker)K(2002年)、A modified dot−blot method of protein determination applied in the tannin−protein−precipitation assay to facilitate the evaluation of tannin activity in animal feeds、Br.J.Nutr.87:421−426頁
ホフマン(Hoffman)EM、モイツェル(Meutzel)S、ベッカー(Becker)(2003年)、Effects of Moringa oleifera seed extract on rumen fermentation in vitro、Arch.Anim.Nutr.57:65−81頁
ホフマン(Hoffman)EM、モイツェル(Meutzel)S、ベッカー(Becker)(2003年)、The fermentation of soybean meal by rumen microbes in vitro reveals different kinetic features for the inactivation and the degradation of trypsin inhibitor protein、Anim.Feed Sci.Technol.106:189−197頁
ハンゲート(Hungate)RE(1966年)、The rumen and its microbes,Academic Press,New York and London Leng RA(1992)、Ruminant production and greenhouse gas emissions、Proc.NZ Soc.Anim.Prod.52(補遺):15−23頁
ナガラヤ(Nagaraja)TG、ニューボールド(Newbold)CJ、ファン・ネーベル(Van Nevel)CJ、デマイヤー(Demeyer)DI(1997年)、Manipulation of ruminal fermentation,in The rumen microbial ecosystem、ホブソン(Hobson)PNおよびスチュワート(Stewart)CS編、チャップマン・アンド・ホール(Chapman & Hall)、London、523−632頁
ノラン(Nolan)JV (1975年)、Quantitative models of nitrogen metabolism in sheep,in Digestion and Metabolism in the Ruminant、McDonald IW and Warner ACI編、University of New England Publishing Unit、Armidale、Australia、416−431頁
ラッセル(Russell)JB、ヒノ(Hino)T(1985年)、Regulation of lactate production in Streptococcus bovis:a spiraling effect that contributes to rumen acidosis、J.Dairy Sci.68:1712−1721頁
ウォリス(Wallace)RJ、マックフェルソン(McPherson)CA (1987年)、Factors affecting the rate of breakdown of bacterial protein in rumen fluid、Br.J.Nutr.58:313−323頁
ウォリス(Wallace)RJ(1983年)、Hydrolysis of 14C−labelled proteins by rumen micro−organisms and by proteolytic enzymes prepared from rumen bacteria、Br.J.Nutr.50:345−355頁
ウォリス(Wallace)RJ、オノデラ(Onodera)R、コッタ(Cotta)MA (1997年)、Metabolism of nitrogen−containing compounds, in The rumen microbial ecosystem、ホブソン(Hobson)PNおよびスチュワート(Stewart) CS編、チャップマン・アンド・ホール(Chapman & Hall)、London、283−328頁
【図面の簡単な説明】
【0061】
【図1】ゲンチアナ・アスクレピデアの異なる含有率(μg/ml)でのルーメン繊毛原虫の細菌溶解活性に対するインビトロでの効果。
【図2】スイカズラの異なる含有率(μg/ml)でのルーメン繊毛原虫の細菌溶解活性に対するインビトロでの効果。
【図3】クローブバッドの異なる含有率(μg/ml)でのルーメン繊毛原虫の細菌溶解活性に対するインビトロでの効果。
【図4】ヒナギクの異なる含有率(μg/ml)でのルーメン繊毛原虫の細菌溶解活性に対するインビトロでの効果。
【図5】オリーブの異なる含有率(μg/ml)でのルーメン繊毛原虫の細菌溶解活性に対するインビトロでの効果。
【図6】ヒレハリソウの異なる含有率(μg/ml)でのルーメン繊毛原虫の細菌溶解活性に対するインビトロでの効果。
【図7】β−ミルセンの異なる含有率(μg/ml)でのルーメン繊毛原虫の細菌溶解活性に対するインビトロでの効果。
【図8】ドットブロットアッセイによるタンパク質分解の定量により、K.アルベンシスの2種類の受入によるタンパク質分解の阻害が確認される。
【特許請求の範囲】
【請求項1】
反芻動物のルーメン発酵に影響を及ぼし、かつエネルギーおよび窒素源のアベイラビリティを増大させる1つもしくはそれ以上の植物材料、植物抽出物、または天然同一成分を含有する添加物の使用であって、
前記成分がスイカズラ(Lonicera japonica)、ゲンチアナ・アスクレピデア(Gentiana asclepidea)、キバナリンドウ(Gentiana lutea)、クローブバッド(Eugenia caryophyllata)、ヒナギク(Bellis perennis)、オリーブ(Olea europaea)、ヒレハリソウ(Symphytum officinale)、ヒメヒレアザミ(Carduus pycnocephalus)、シャクヤク(Paeoniae alba radix)、ヤマナラシ(Populus tremula)、サクランボ(Prunus avium)、バッコヤナギ(Salix caprea)、セイタカダイオウ(Rheum nobile)、ハンニチバナ(Helianthemum canum)、クマコケモモ(Arctostaphylos uva−ursi)、インディアンルバーブ(Peltiphyllum peltatum)、エゾアカバナ(Epilobium montanum)、クナウチア・アルベンシス(Knautia arvensis)、レタス(Latuca sativa)およびネトル(Urtica dioica)およびそれらの抽出物、およびβ−ミルセンからなる群から選択される植物材料であることを特徴とする使用。
【請求項2】
前記選択される成分が、ルーメン消化のタンパク質分解活性を減少させる、ハンニチバナ、クマコケモモ、エゾアカバナ、クナウチア・アルベンシス、およびインディアンルバーブおよびそれらの抽出物からの植物材料からなる亜群からの1つもしくはそれ以上の成分を含有することを特徴とする請求項1に記載の使用。
【請求項3】
前記選択される成分が、原虫の活性を抑制する、スイカズラ、ゲンチアナ・アスクレピデア、クローブバッド、ヒナギク、オリーブ、ヒレハリソウおよびそれらの抽出物、およびβ−ミルセンからの植物材料からなる群からの1つもしくはそれ以上の成分を含有することを特徴とする請求項1または2に記載の使用。
【請求項4】
前記選択される成分が、前記ルーメン消化のメタン生成活性を減少させる、ヒメヒレアザミ、シャクヤク、ヤマナラシ、サクランボ、バッコヤナギ、およびセイタカダイオウおよびそれらの抽出物からなる群からの1つもしくはそれ以上の成分を含有することを特徴とする請求項1〜3のいずれか一項に記載の使用。
【請求項5】
前記選択される成分が、乳酸の産生を減少させる、レタス(Latuca sativa)およびネトルおよびそれらの抽出物からの植物材料からなる群からの1つもしくは2つの成分を含有することを特徴とする請求項1〜4のいずれか一項に記載の使用。
【請求項6】
前記添加物が請求項2〜5に記載の亜群の少なくとも2つからの成分を含有することを特徴とする請求項2〜5に記載の使用。
【請求項7】
前記添加物が請求項2〜5に記載の全亜群からの成分を含有することを特徴とする請求項6に記載の使用。
【請求項8】
前記成分の少なくとも1つが、
i)クナウチア・アルベンシス、クマコケモモ、およびハンニチバナおよびそれらの抽出物からの植物材料からなり、
ii)ヒナギク、クローブバッド、オリーブ、スイカズラおよびヒレハリソウおよびそれらの抽出物からの植物材料からなり、
iii)ヒメヒレアザミ、ヤマナラシ、およびセイタカダイオウ、およびそれらの抽出物からの植物材料からなり、かつ
iv)レタス(Latuca sativa)およびネトルおよびそれらの抽出物からの植物材料からなる
群の各々から選択されることを特徴とする請求項7に記載の使用。
【請求項9】
前記群i)〜iv)に属する前記成分の総量が、本発明により存在する前記成分の総量の20〜100重量%であることを特徴とする請求項8に記載の使用。
【請求項10】
投与される前記成分の総量が、1日当り、かつ前記反芻動物のkg体重当り0.02mg〜20gであることを特徴とする請求項1〜7に記載の使用。
【請求項11】
前記添加物が、0.2〜90重量%の請求項1〜9のいずれか一項に記載の前記成分、および10〜75重量%の成長改善剤、香味料、吸収担体、および/または他の飼料成分を含有することを特徴とする添加物。
【請求項12】
前記添加物が、
a)0〜80乾燥重量%の穀類と、
b)0〜30乾燥重量%の油脂と、
c)穀類以外の種類の栄養物を含有する0〜85乾燥重量%のタンパク質と、
d)請求項1〜9のいずれか一項に記載の10ppm〜40乾燥重量%の成分とを含有し、a)〜d)の総量が少なくとも50乾燥重量%であることを特徴とする動物飼料。
【請求項13】
請求項1〜9のいずれか一項に記載の少なくとも1〜99乾燥重量%の前記成分と、1〜99乾燥重量%の他の成分とを含有する動物の飲料水サプリメント。
【請求項1】
反芻動物のルーメン発酵に影響を及ぼし、かつエネルギーおよび窒素源のアベイラビリティを増大させる1つもしくはそれ以上の植物材料、植物抽出物、または天然同一成分を含有する添加物の使用であって、
前記成分がスイカズラ(Lonicera japonica)、ゲンチアナ・アスクレピデア(Gentiana asclepidea)、キバナリンドウ(Gentiana lutea)、クローブバッド(Eugenia caryophyllata)、ヒナギク(Bellis perennis)、オリーブ(Olea europaea)、ヒレハリソウ(Symphytum officinale)、ヒメヒレアザミ(Carduus pycnocephalus)、シャクヤク(Paeoniae alba radix)、ヤマナラシ(Populus tremula)、サクランボ(Prunus avium)、バッコヤナギ(Salix caprea)、セイタカダイオウ(Rheum nobile)、ハンニチバナ(Helianthemum canum)、クマコケモモ(Arctostaphylos uva−ursi)、インディアンルバーブ(Peltiphyllum peltatum)、エゾアカバナ(Epilobium montanum)、クナウチア・アルベンシス(Knautia arvensis)、レタス(Latuca sativa)およびネトル(Urtica dioica)およびそれらの抽出物、およびβ−ミルセンからなる群から選択される植物材料であることを特徴とする使用。
【請求項2】
前記選択される成分が、ルーメン消化のタンパク質分解活性を減少させる、ハンニチバナ、クマコケモモ、エゾアカバナ、クナウチア・アルベンシス、およびインディアンルバーブおよびそれらの抽出物からの植物材料からなる亜群からの1つもしくはそれ以上の成分を含有することを特徴とする請求項1に記載の使用。
【請求項3】
前記選択される成分が、原虫の活性を抑制する、スイカズラ、ゲンチアナ・アスクレピデア、クローブバッド、ヒナギク、オリーブ、ヒレハリソウおよびそれらの抽出物、およびβ−ミルセンからの植物材料からなる群からの1つもしくはそれ以上の成分を含有することを特徴とする請求項1または2に記載の使用。
【請求項4】
前記選択される成分が、前記ルーメン消化のメタン生成活性を減少させる、ヒメヒレアザミ、シャクヤク、ヤマナラシ、サクランボ、バッコヤナギ、およびセイタカダイオウおよびそれらの抽出物からなる群からの1つもしくはそれ以上の成分を含有することを特徴とする請求項1〜3のいずれか一項に記載の使用。
【請求項5】
前記選択される成分が、乳酸の産生を減少させる、レタス(Latuca sativa)およびネトルおよびそれらの抽出物からの植物材料からなる群からの1つもしくは2つの成分を含有することを特徴とする請求項1〜4のいずれか一項に記載の使用。
【請求項6】
前記添加物が請求項2〜5に記載の亜群の少なくとも2つからの成分を含有することを特徴とする請求項2〜5に記載の使用。
【請求項7】
前記添加物が請求項2〜5に記載の全亜群からの成分を含有することを特徴とする請求項6に記載の使用。
【請求項8】
前記成分の少なくとも1つが、
i)クナウチア・アルベンシス、クマコケモモ、およびハンニチバナおよびそれらの抽出物からの植物材料からなり、
ii)ヒナギク、クローブバッド、オリーブ、スイカズラおよびヒレハリソウおよびそれらの抽出物からの植物材料からなり、
iii)ヒメヒレアザミ、ヤマナラシ、およびセイタカダイオウ、およびそれらの抽出物からの植物材料からなり、かつ
iv)レタス(Latuca sativa)およびネトルおよびそれらの抽出物からの植物材料からなる
群の各々から選択されることを特徴とする請求項7に記載の使用。
【請求項9】
前記群i)〜iv)に属する前記成分の総量が、本発明により存在する前記成分の総量の20〜100重量%であることを特徴とする請求項8に記載の使用。
【請求項10】
投与される前記成分の総量が、1日当り、かつ前記反芻動物のkg体重当り0.02mg〜20gであることを特徴とする請求項1〜7に記載の使用。
【請求項11】
前記添加物が、0.2〜90重量%の請求項1〜9のいずれか一項に記載の前記成分、および10〜75重量%の成長改善剤、香味料、吸収担体、および/または他の飼料成分を含有することを特徴とする添加物。
【請求項12】
前記添加物が、
a)0〜80乾燥重量%の穀類と、
b)0〜30乾燥重量%の油脂と、
c)穀類以外の種類の栄養物を含有する0〜85乾燥重量%のタンパク質と、
d)請求項1〜9のいずれか一項に記載の10ppm〜40乾燥重量%の成分とを含有し、a)〜d)の総量が少なくとも50乾燥重量%であることを特徴とする動物飼料。
【請求項13】
請求項1〜9のいずれか一項に記載の少なくとも1〜99乾燥重量%の前記成分と、1〜99乾燥重量%の他の成分とを含有する動物の飲料水サプリメント。
【図1】

【図2】

【図3】

【図4】
【図5】
【図6】
【図7】

【図8】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公表番号】特表2007−532609(P2007−532609A)
【公表日】平成19年11月15日(2007.11.15)
【国際特許分類】
【出願番号】特願2007−507805(P2007−507805)
【出願日】平成17年4月15日(2005.4.15)
【国際出願番号】PCT/EP2005/051673
【国際公開番号】WO2005/099729
【国際公開日】平成17年10月27日(2005.10.27)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【出願人】(506347296)オール−テクノロジー アイルランド リミテッド (1)
【Fターム(参考)】
【公表日】平成19年11月15日(2007.11.15)
【国際特許分類】
【出願日】平成17年4月15日(2005.4.15)
【国際出願番号】PCT/EP2005/051673
【国際公開番号】WO2005/099729
【国際公開日】平成17年10月27日(2005.10.27)
【出願人】(503220392)ディーエスエム アイピー アセッツ ビー.ブイ. (873)
【出願人】(506347296)オール−テクノロジー アイルランド リミテッド (1)
【Fターム(参考)】
[ Back to top ]