
受容体チロシンキナーゼであるFLT4ならびに診断および治療におけるその使用
【課題】リンパ組織、とくにたとえばリンパ管腫、転移性リンパ節ならびに炎症性疾患、感染性疾患および免疫性疾患などの疾患状態のリンパ管および高内皮性小静脈における変化を検出するための診断道具を提供すること。
【解決手段】モノクローナル抗FLT4抗体は、リンパ管および高内皮性小静脈の内皮細胞に対する特異的マーカーとして、リンパ組織、とくにたとえばリンパ管腫、転移性リンパ節ならびに炎症性疾患、感染性疾患および免疫性疾患などの疾患状態のリンパ管および高内皮性小静脈における変化を検出するための診断道具として有用である。
【解決手段】モノクローナル抗FLT4抗体は、リンパ管および高内皮性小静脈の内皮細胞に対する特異的マーカーとして、リンパ組織、とくにたとえばリンパ管腫、転移性リンパ節ならびに炎症性疾患、感染性疾患および免疫性疾患などの疾患状態のリンパ管および高内皮性小静脈における変化を検出するための診断道具として有用である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は概して、受容体チロシンキナーゼ、核酸プローブおよび前記受容体を特異的に認識する抗体、ならびに動物およびヒト組織中のリンパ管と高内皮性小静脈(high endothelial venules)(HEV)を、そして培養物中(in culture)のリンパ内皮細胞(lymphatic endothelial cells)を同定するための前記プローブおよび抗体の使用に関する。さらに詳しくは、本発明は受容体チロシンキナーゼであるFLT4に特異的な抗体、リンパ管におけるFLT4の発現を同定する方法ならびに究極的には、たとえば炎症性疾患、感染性疾患および免疫性疾患、転移性リンパ節ならびにリンパ管腫などのリンパ組織における変化を含む動物およびヒトにおける疾患の状態の診断ならびに治療に関する。
【背景技術】
【0002】
いくつかの疾患と同様、胚の脈管形成および血管形成(embryonic vasculogenesis and angiogenesis)、血液凝固ならびに創傷の治癒および再生などの脈管系の生理機能は、血管に並ぶ脈管内皮(vascular endothelium)に関係している。脈管枝(vascular tree)の発達は血管形成を経ておこり、いくつかの理論によると、リンパ系の形成は、静脈からの発芽(sprouting)により動脈および静脈の発達(development)後すぐに開始する(1、2)。
【0003】
胎児期(fetal period)ののち、新生血管形成に関連した脈管形成の期間を除いて、内皮細胞は非常にゆっくりと増殖する。脈管形成を刺激する成長因子は、特異的な内皮細胞表面受容体であるチロシンキナーゼを介してそれらの効果を及ぼす。
【0004】
ヒト赤白血病細胞系からクローニングした受容体チロシンキナーゼであるFLT4のcDNAのタンパク質産物は、Nを介してグリコシル化(N−glycosylated)され、その細胞外ドメインには7つの免疫グロブリン様ループを含んでいる。FLT4の細胞質チロシンキナーゼドメインは、FLT1およびKDRの対応するドメインとアミノ酸レベルで約80%同一であり、血小板由来成長因子に対する受容体、コロニー刺激因子−1に対する受容体、幹細胞因子に対する受容体および受容体FLT3と約60%同一である(3)。
【0005】
FLT4の生物学的機能は現在までのところ知られていないが、その制限された発現のパターンは前記機能が脈管内皮に関係しているかもしれないということを示した。本発明者らが以前にえた結果から、ジャーナル・オブ・エクスペリメンタル・メディスン(J.Exp.Med.)、178巻、2077〜2088頁(1993年)にカイパイネン(Kaipainen)らが開示したように、いくつかの胎児の器官の発達中の管(vessels)の内皮細胞におけるFLT4のmRNAの発現が明らかになった。受容体FLT4、FLT1およびKDR/FLK−1のmRNAシグナルは、重複(overlapping)を示したが、研究した組織においては異なる発現パターンを示した(4)。これらのデータは、この遺伝子ファミリーによってコードされる受容体チロシンキナーゼは血管の成長の調節および/または分化において異なる機能を有するかもしれないことを示唆している。
【0006】
リンパ系の主要な機能は、組織から流体が戻るようにし、多くの脈管外物質(extravascular substances)を血液へ戻すことである。さらに、成熟過程のあいだ、リンパ球は血液を離れ、リンパ系器官および他の組織を通り移動してリンパ管に入り、胸管を通り血液に戻る。専門化した小静脈である高内皮性小静脈(HEV)は再びリンパ球に結合し、組織への管外遊出を引きおこす。したがって、リンパ管、とくにリンパ節は免疫学における重要な役割を果たしており、異なる腫瘍の転移が発達する部位でもある。
【0007】
20世紀以来、リンパ系の胚起源(embryonic origin)に関する3つの異なる理論が提起されている。しかしながら、リンパ管に対して利用できる特異的マーカーは存在しなかったため、本発明より前は、リンパ管を同定することは困難であった。
【0008】
リンパ管は、リンパ管造影法を用いてもっとも一般的に研究される。リンパ管造影法において、エックス線造影剤はリンパ管に直接注射される。前記造影剤はリンパ系の輸出ドレナージ管(efferent drainage vessels)に沿って分配される。前記造影剤はリンパ節に集められ、半年間はそこにとどまり、そのあいだにエックス線解析によりリンパ節の大きさと構造を追跡することができる。この診断は、リンパ節に転移を有する癌患者およびたとえばリンパ腫などのリンパの悪性疾患においてとくに重要である。
【発明の概要】
【0009】
本発明は、FLT4ペプチドおよびその他の構築物、ならびにリンパ内皮細胞に対する特異的マーカーとしてのFLT4の用途に関する。
【0010】
また本発明は、核酸プローブおよびFLT4を特異的に認識する抗体、とくにモノクローナル抗体ならびに前記抗体を含む組成物に関する。さらに本出願では、組織、とくにリンパ組織およびリンパ内皮細胞における受容体FLT4を検出し、その量を測定する診断目的への前記モノクローナル抗体の使用が開示される。
【0011】
好ましい態様において、本発明は受容体FLT4を特異的に認識するモノクローナル抗体を提供する。より明確には、本発明は9D9F9と名付けられたモノクローナル抗体を提供する。モノクローナル抗体9D9F9を産生するハイブリドーマ細胞系は、ブダペスト条約の規定によりドイチェ・サムルング・フォン・ミクロオルガニスメン・ウント・ツエルクルツレン・ゲゼルシャフト・ミット・ベシュレンクテル・ハフツング(Deutsche Sammlung von Mikroorganismen und Zellkulturen Gmb H)(DSM)に寄託されている(DSM受託番号ACC2210)。
【0012】
また検出しうるマーカーで標識したモノクローナル抗体も提供する。本明細書で使用するとき、「検出しうるマーカー」という用語は当業者に知られたあらゆる検出しうるマーカーを包含する。しかしながら、本発明の好ましい態様における検出しうるマーカーは、放射性同位体、蛍光色素、色素、酵素およびビオチンからなる群より選ばれる。本発明の目的に対して、適当な放射性同位体は125Iおよび131Iを含むがこれらに限定されるものではない。
【0013】
本発明のモノクローナル抗体は、細胞サンプル中の受容体FLT4の存在を検出する方法に使用しうる。前記方法は、細胞サンプルを本発明のモノクローナル抗体にさらす工程および前記モノクローナル抗体の受容体FLT4への結合を検出する工程からなる。
【0014】
したがって、本発明の別の面は、細胞サンプル中の受容体FLT4の存在を決定する方法であって、
(a)細胞サンプルを本発明のモノクローナル抗体にさらす工程、および
(b)前記モノクローナル抗体の受容体FLT4への結合を検出する工程
からなる方法に関する。
【0015】
細胞混合物を本発明のモノクローナル抗体にさらすことは、蛍光により活性化された細胞の選別(fluorescence-activated cell sorting)のばあいは、溶液中で行なうことができ、たとえば生検材料などの固形組織標本上でも行なうこともでき、またカラムクロマトグラフィーまたは直接免疫接着(direct immune adherence)を用いるばあいは、固体担体に固定化されたモノクローナル抗体を用いて行なうことができる。モノクローナル抗体にさらされる細胞混合物は、血液細胞または組織細胞のいかなる溶液であってもよい。好ましくは、細胞混合物はリンパ内皮細胞を含むかまたは含むことが疑われる正常または病的な組織に由来する。細胞混合物を前記モノクローナル抗体にさらしたのち、受容体FLT4を有する細胞は前記モノクローナル抗体と結合し、抗体−受容体FLT4複合体を形成するであろう。抗体−受容体FLT4複合体の存在、したがって受容体FLT4の存在は、当該技術分野において既知の方法によって検出することができる。これらの方法はたとえば蛍光抗体法、FACS分析、ELISA、IRMA(サンドイッチ型免疫化学分析法)、免疫組織化学的方法、125I標識およびオートラジオグラフィーを用いたRIAなどの当該技術分野において標準的な免疫組織化学的方法を含む。
【0016】
本発明はまた映像化しうる試薬に接合(conjugated)したモノクローナル抗体も提供する。本明細書で使用するとき、「映像化しうる試薬」という用語は放射性同位体を含むが、これに限定されるものではない。好ましい放射性同位体はテクネチウム99mである。
【0017】
特定の態様において、本発明は組織サンプルおよび生物中のリンパ管およびその内皮細胞をモニターする方法に関する。さらに本発明はリンパ組織、とくにリンパ管の状態(炎症、感染、外傷(traumas)、成長、新形成など)を描く臨床的検出方法および生物中のリンパ管、したがってリンパ系血管新生(lymphatic vascularization)を検出する方法を提供する。
【0018】
さらに具体的には、本発明は、転移性癌、炎症状態、感染状態および免疫状態に関係したFLT4の発現によって特徴付けられるリンパ系の変化(lymphatic changes)を検出し、同定する方法を提供する。前記方法は、
(a)リンパ系の変化が疑われる組織および/または体液サンプルをうる工程、
(b)前記サンプルを、FLT4に特異的なモノクローナル抗体とその抗原の複合体を形成するのに適した条件下で、FLT4に特異的なモノクローナル抗体と接触させる工程、および
(c)形成されたあらゆる複合体の存在を検出する工程
からなる。
【0019】
前記方法により検出されうる組織は、FLT4を含むリンパ系細胞(lymphatic cells)または受容体FLT4を発現している細胞を有するあらゆる正常な、前癌性のまたは癌性の固形腫瘍組織である。本発明の1つの態様において、モノクローナル抗体は本明細書に記載されたような検出しうるマーカーで標識される。本発明の方法は様々な形態の癌、とくにリンパ節における転移およびリンパ管腫に加えて、たとえばリンパ腫などのその他のリンパの悪性疾患の検出および識別に有用である。
【0020】
ヒト患者におけるリンパ管、高内皮性小静脈またはリンパ節の存在を映像化する方法も本発明によって提供される。前記方法は標識した抗体を投与し、リンパ管またはリンパ節においてFLT4を発現している細胞が存在する部位を映像化することにより検出することからなる。
【0021】
本発明はさらにリンパ血管新生ならびに炎症状態、感染状態および免疫状態におけるFTL4の機能を刺激するまたは拮抗する方法に関する。前記方法は、このような反応に関与するFLT4内皮細胞部位、とくにFLT4の機能が、たとえば転移性癌、リンパ腫、炎症性疾患(慢性または急性)、感染性疾患および免疫性疾患などの疾患に関連している部位をブロックするのに充分な量のFLT4に結合する化合物を投与することによりFLT4が介在するリンパ血管新生を阻害することからなる。
【図面の簡単な説明】
【0022】
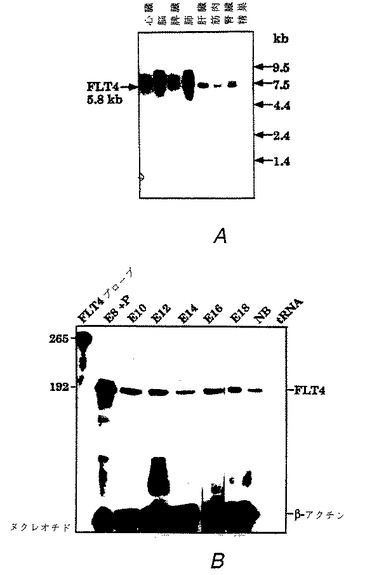
【図1】図1はマウス組織におけるFLT4のmRNAの発現を表わし、Aは成体のマウスの表示した組織から単離したポリアデニル化RNA(polyadenylated RNA)のハイブリダイゼーションを表わす。FLT4のmRNAのバンドの大きさはkbで示す。Bは様々な存胎齢(E8〜E18)のマウス胚および新生マウス(1日)から単離したRNAのRNアーゼプロテクション分析(RNA se protection analysis)を表わしている。サンプルE8+Pは胎盤も含んでいる。プローブおよび保護されたFLT4断片の大きさはヌクレオチドで示し、β−アクチンを対照として使用した。
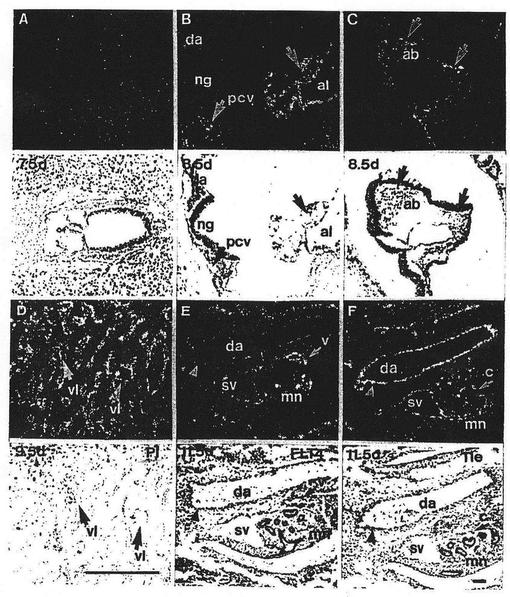
【図2】図2は交尾後7.5日、8.5日および11.5日の胚におけるFLT4のmRNAの発現を表わしている。インサイチュ・オートラジオグラムの暗視野(darkfield)および明視野(brightfield)の顕微鏡写真を示す。FLT4のmRNAの発現は、7.5日の胚(A)においては検出することができなかった。交尾後8.5日のマウス胚のFLT4の発現を(B)および(C)に示す。矢印は、後主静脈の内皮(cv)、(B)における尿膜(al)および(C)の頭部間葉の血管芽細胞(ab)におけるFLT4陽性細胞を示している。交尾後8.5日の胎盤において、FLT4転写物は、静脈裂孔(venous lacunae)(vl)の内皮細胞において見ることができた(D)。パネルEおよびFは、交尾後11.5日の胚におけるFLT4およびTieのハイブリダイゼーションシグナルの比較を示している。発達中の胸部大動脈および後腎(mn)領域を示す(20×)。胸部大動脈はFLT4に対して陰性であるがTieのmRNAに対しては陽性であるのに対して、両方のプローブはともに心内膜下静脈(subendocardial vein)(sv)の内皮とハイブリダイズすることに注意すべきである。またFLT4のプローブは、後腎静脈(v)からシグナルを出すが、Tieは後腎毛細管(metanephric capillaries)(c、矢印)と大部分がハイブリダイズする。daは胸部大動脈を、ngは神経溝を示す。スケールバーは30μmである。
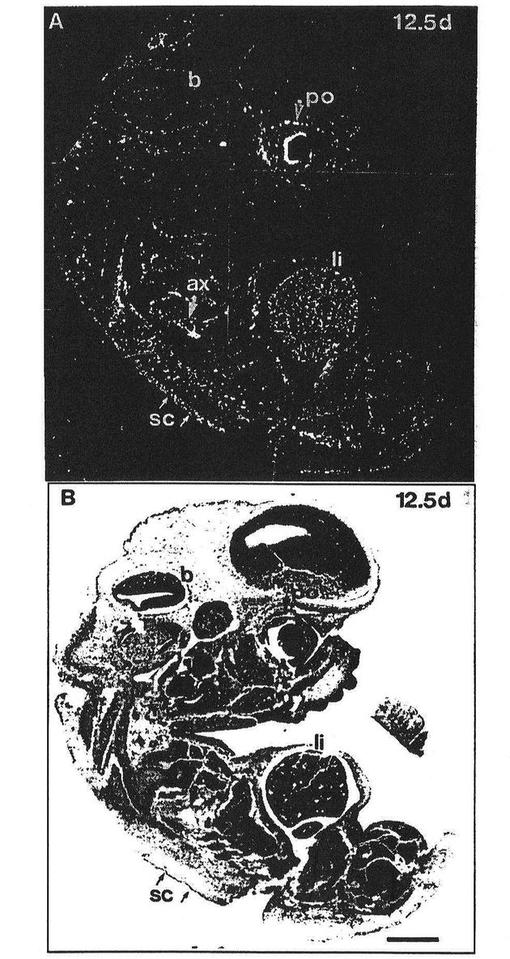
【図3】図3は交尾後12.5日の胚におけるFLT4のmRNAの発現を表わしている。腋窩面(axillar plane)を通る矢状切断図を示す。FLT4のmRNAは、腋窩(ax)の拡張された管(dilated vessels)、眼窩周辺(po)領域の叢様パターン、脊椎傍組織(矢じり)および皮下(sc)組織において顕著であった。bは脳を、liは肝臓を示す。スケールバーは5μmである。
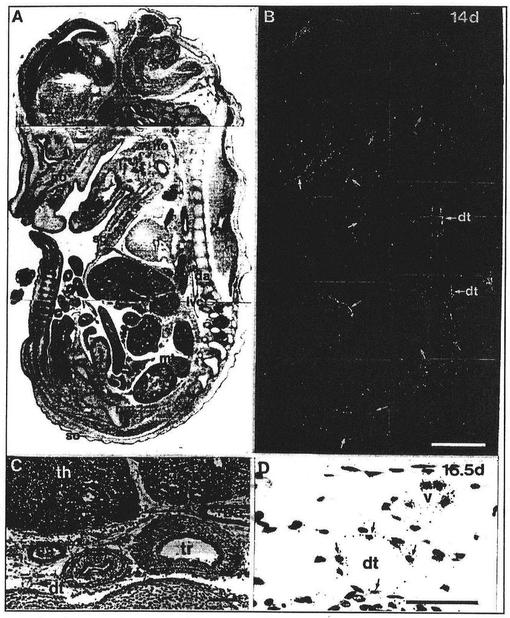
【図4】図4は交尾後14日および16.5日の胚におけるFLT4を表わしている。パネルAおよびBは、正中矢状切断図の明視野および暗視野の映像を示している。poは眼窩領域を、ljは下顎を、neは頸領域を、scは皮下組織を、mtは腸間膜を、aoは大動脈を、dtは胸管を示す。(C)はヘマトキシリン−エオシン染色した16.5日の胚の横断面を示す。thは胸腺を、trは気管を、eは食道を、caは頸動脈を、baは腕頭の動脈を、dtは胸郭を示す。(D)は胸管領域の高拡大図(40x)を示し、オートラジオグラフィーの粒子が内皮細胞全体に見ることができる。また、前記写真の上の部分における小管(v)は陽性である。スケールバーは10μm(A〜C)および1μm(D)である。
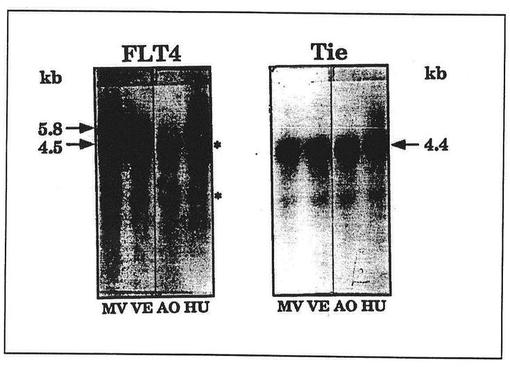
【図5】図5は培養した内皮細胞におけるFLT4およびTieのmRNA発現の比較を表わしている。ヒトの包皮の微小管(MV)、大腿の静脈(VE)、大動脈(AO)および臍帯の静脈(HU)の内皮細胞由来のポリアデニル化RNAのノーザンブロット分析を示す。比較のために、受容体チロシンキナーゼであるTieのmRNAのハイブリダイゼーションシグナルを示す。プローブのリボソームRNAへの非特異的結合の結果生じたバンドをアスタリスクで印をつけた。
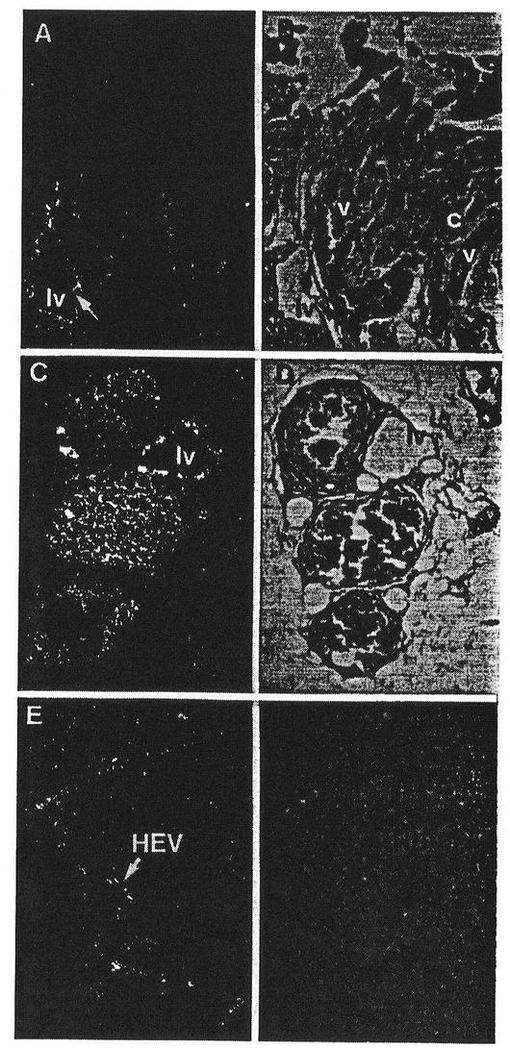
【図6】図6は成人のヒト腸間膜(A、B)、肺(C、D)および扁桃(E、F)のリンパ管におけるFLT4を表わしている。AおよびCにおけるリンパ管のみがFLT4シグナルを出し、静脈、毛細管および動脈ではFLT4のmRNAに対して陰性であることに注意すべきである。扁桃において、シグナルはいくらかのHEVの内皮において見出された。スケールバーは200μmである。
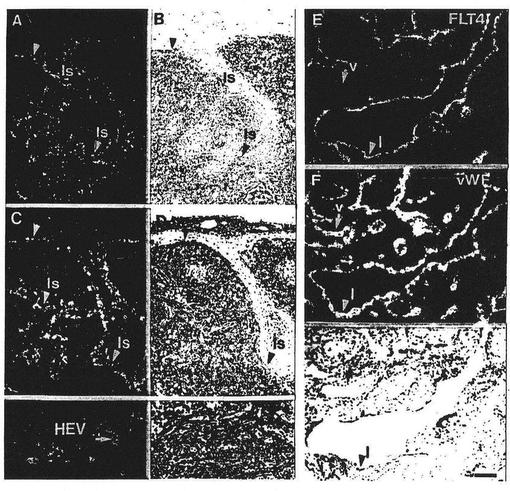
【図7】図7は正常(A、B)および転移性(C、D)リンパ節ならびにリンパ管腫(E〜G)におけるFLT4のmRNAを表わしている。矢じりは、FLT4陽性であるリンパ洞およびHEVに印をつけている。FLT4およびフォンビルブラント因子のシグナルの比較では、両方がリンパ内皮において示されるが、毛細管(c)および静脈(v)の内皮においてはフォンビルブラント因子のシグナルのみが示された。スケールバーは10μm(A〜D)および100μm(E〜G)である。
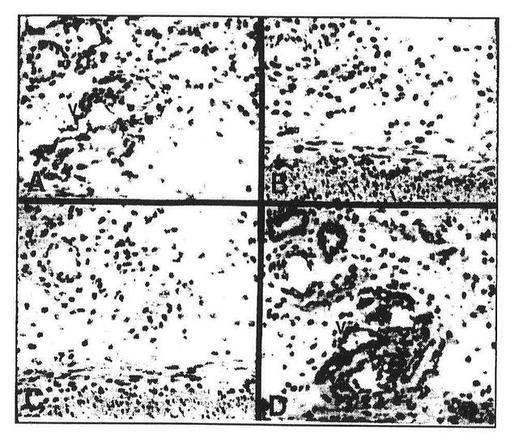
【図8】図8はイムノペルオキシダーゼ染色によって検出された胎児の腸間膜管(mesenterial vessels)におけるFLT4の発現を表している。親和性により精製した抗FLT4抗体(A)、抗原でブロックした抗血清(B)、あらかじめ免疫した血清(C)および第VIII因子関連抗原に対して特異的な抗血清(D)を用いて断片を染色した。染色はいくらかのものに制限されるが、すべての管(v)には制限されないことに注意すべきである。スケールバーは0.05mmである。
【発明を実施するための形態】
【0023】
リンパ組織、とくに転移性癌および免疫性疾患状態に関連するリンパ節における変化を同定することの重要性を認識して、本発明者らはFLT4がリンパ管内皮を検出する特異的なマーカーであり、したがってヒトの病的状態におけるリンパ系の変化に対するマーカーとして有用であることを示した。
【0024】
本発明者らは以前に、FLT1およびKDRと比較したFLT4の発現パターンは、妊娠18週目のヒト胎児の組織において大きく異なることを示した(4)。発達におけるFLT4の役割を理解するために、本発明者らはマウスFLT4の部分的なcDNAをクローニングした。これらのプローブをインサイチュ・ハイブリダイゼーションに使用して、マウスの発達におけるFLT4のmRNAの発現を分析し、FLT4はリンパ系の脈管形成中に発現することを見出した。FLT4はいくらかの高内皮性小静脈(HEV)と同様に、正常および病的な状態の成人の組織のリンパ内皮細胞において見出されたので、前記の知見の妥当性は正常および病的な成人の組織においても確認された。
【0025】
マウスFLT4のcDNA断片のクローニングにより、前記断片から推定されるアミノ酸配列は対応するヒトの配列とほとんど同一であることが示された(研究した両方の区域におけるアミノ酸の同一性は約96%)。マウスFLT4のcDNAの同一性のさらなる証拠はノーザンハイブリダイゼーションによりえられ、そこでは両方の種に由来するプローブが典型的なマウス組織から5.8kbのmRNAシグナルを出した。成体マウスの様々な組織から単離したRNAの解析は、肝臓、肺、心臓、脾臓および腎臓におけるFLT4の発現を示し、脳および精巣においてはまったくないかまたはわずかのハイブリダイゼーションを示した。このパターンは先にガランド(Galland)らにより報告されたパターンと類似している(5)。RNアーゼプロテクション(RNase protection)の結果により、FLT4遺伝子は交尾後8.5日の胚から始まって、マウスの発達中必要であることが示唆され、相対的な発現レベルはかなり安定しているようであった。
【0026】
インサイチュ・ハイブリダイゼーションのために、細胞外ドメイン配列をコードするマウスFLT4のcDNA断片を2つ選んだ。このことによりハイブリダイゼーションパターンを関連の受容体FLK−1およびFLT−1のパターンと明確に区別することができる。このことは細胞外領域におけるFLT4との配列同一性が非常に低い程度のものでしかないことを示している(6、7、8、9)。内皮の受容体チロシンキナーゼであるFLK−1、FLT−1、TieおよびTekの遺伝子と同様に、FLT4の遺伝子は、交尾後7.5日の胚では発現しなかった。交尾後8.5日の胚においては、強いFLT4シグナルが尿膜、頭部間葉(head mesenchyme)の血管芽細胞および主静脈に局在した。対照的に、胸部大動脈(dorsal aorta)、心臓の心内膜および卵黄包の血管芽細胞は、Tie、Tek、FLK−1ならびにFLT−1、TieおよびTekに対してとは異なり、FLT4に対して陰性であった(10、8)。FLT4発現の静脈系(venous system)への限定は、11.5日のマウス胚由来のサンプルにおいてよりいっそう明確になり、ここではTieのmRNAは動脈においても発現した。交尾後12.5日の胚においては、FLT4シグナルは発達中の静脈性内皮および推定リンパ内皮を装飾したが、内皮の受容体チロシンキナーゼであるTieに対してとは異なり、動脈内皮は陰性であった。発達の後半段階中、FLT4のmRNAは、発達中のリンパ管を表わす血液細胞を欠いた脈管叢(vascular plexuses)に限定されるようになった。成人の組織において、リンパ内皮およびいくらかの高内皮性小静脈のみがFLT4のmRNAを発現した。増加した発現が転移性リンパ節およびリンパ管腫におけるリンパ洞および高内皮性小静脈においておこった。
【0027】
マウス胚からえたデータの解釈が困難であり、ヒトのリンパ系は非常によく定義されているため、ヒト内皮を研究した。また、様々な内皮から確立した細胞を、FLT4の発現の特異性がインビトロ条件において持続するかどうかをみるために、細胞培養中で研究することができた。内皮細胞系はインビトロ培養において分化した特徴(differentiated features)を失うことが知られている。それゆえ、前記内皮細胞系がFLT4に対して陰性であることは予期されないことではなかった。培養した大動脈の内皮細胞もまたFLT4のmRNAを欠いていた。しかしながら、微小脈管構造ならびに大腿および臍帯の静脈から成長したヒト内皮細胞からシグナルはえられた。したがって、少なくともFLT4発現の特異性のいくらかは細胞培養中で維持されていた。
【0028】
成人の組織のインサイチュ・ハイブリダイゼーション分析は、発達中のマウス胚においてみられたFLT4のリンパ系への限定を確認した。FLT4の発現はリンパ内皮およびヒトリンパ節のリンパ洞においてみられた。興味深いことに、立方体様内皮(cuboidal endothelium)を有し、白血球のリンパ節への取り引き(trafficking)における機能を示すいくらかのHEVもまたFLT4陽性であった。さらに、パラレル・ハイブリダイゼーション(parallel hybridization)分析により、正常なリンパ節と比較して、転移性のこれらの構造においてはFLT4のmRNAレベルが上ったことが示された。またFLT4は、結合組織基質(connective tissue stroma)および成長している、内皮に並んだリンパチャネル(endothelial-lined lymphatic channels)からなる良性腫瘍であるリンパ管腫において非常に目立った。FLT4のmRNAは、前記腫瘍のリンパ内皮に限定され、前記腫瘍の動脈、静脈および毛細管には存在しなかった。ヒトの肺において、本発明者らは、この組織における唯一のFLT4に陽性な管であるリンパ構造を同定することができた。
【0029】
前記の結果は、FLT4は成人の組織におけるリンパ管およびいくらかの高内皮性小静脈に対する新規なマーカーであることを示唆している。また前記の結果は、リンパ管の静脈起源(venous origin)についての理論を支持する。成長因子の受容体としてのFLT4は前記の管の分化および機能に関係しているかもしれない。
【0030】
本発明に係るFLT4結合化合物(FLT4−binding compounds)を組合わせた結果は、とくに可視化することができる放射性の、電子的に密集した(electron-dense)またはその他のリポーター物質につないだ本発明の抗体を使用することによって、リンパ内皮を選択的に標識することができる。受容体FLT4の内在化を誘導するモノクローナル抗体を含む物質をリンパ系に注射し、それによってあらかじめ定義した(predefined)分子をリンパ内皮に運ぶことが可能であろう。また、前記の本発明に係るFLT4結合化合物を高内皮性小静脈、とくに受容体FLT4を高いレベルで発現する活性化HEVの検出に使用することが可能であろう。本発明者らが知る限り、リンパ内皮に対する特異的なマーカーは現在入手できない。
【0031】
以下の実施例は本発明を説明するためのみに与えられるものであり、いかなるばあいも本発明の範囲を限定するものではない。
【実施例】
【0032】
実施例1
マウスFLT4のcDNAプローブのクローニング
129SVマウス由来のイフィックス(登録商標)ツー ゲノムライブラリー(iFIX(登録商標) II genomic library)(ストラタジーン(Stratagene))の約106のプラークを、細胞外ドメインを含むS2.5ヒト受容体FLT4のcDNA断片を用いてスクリーニングした(3)。2.5kbのBam HI断片を陽性のプラークからサブクローニングし、両方の末端から配列決定を行なった。前記サブクローニング物から、ポリメラーゼ連鎖反応を使用して増幅し、マウスFLT4のcDNA配列のヌクレオチド1745〜2049を含むエクソン断片をpBluescript KSII+/−ベクター(ストラタジーン)へクローニングした(9)。
【0033】
ヌクレオチド1〜192を含む第2の断片も同様にクローニングした。
【0034】
実施例2
マウス組織におけるFLT4のmRNAの分析
コムチュンスキー(Chomczynski)らにしたがい、発達中の胚(交尾後8〜18日のマウスおよび1日齢のマウス)から全RNAを単離した(11)。交尾後8日の胚由来のサンプルは胎盤も含んでいた。
【0035】
RNアーゼプロテクション分析のために、RNAプローブを、アンチセンス配向(antisense orientation)のために[32P]−UTPおよびT7ポリメラーゼを使用して、実施例1にしたがってえた直線状FLT4プラスミドから生成した。使用したβ−アクチンプローブは公開されているマウスβ−アクチン配列のヌクレオチド1188〜1279に対応する(12)。6%ポリアクリルアミド/7M尿素ゲル中で精製したのち、標識した転写物を30μgの全RNAと52℃で一晩ハイブリダイズさせた。ハイブリダイズしなかったRNAを、RNアーゼA(10U/ml)およびT1(1μg/ml)を用い37℃、pH7.5で1時間かけて消化した。RNアーゼを、プロテイナーゼKを用いた37℃での15分間の消化により不活性化し、サンプルを6%ポリアクリルアミド/7M尿素ゲル中で分析した。
【0036】
本実験において分析したFLT4の発現パターンにより、肺、肝臓、心臓、腎臓、骨格筋および脾臓からは非常に弱いmRNAシグナルがえられたが、精巣および脳には明白に特異的なシグナルはないということが示された(図1A)。RNアーゼプロテクション法による、マウス発生の異なる時期(phase)に採取した一連のRNAの分析によって、FLT4のmRNAは、交尾後8日から新生マウスまで終始シグナル強度における大きな変動なしに発現することが示された(図1B)。
【0037】
実施例3
マウス胚におけるFLT4に対するインサイチュ(in situ)・ハイブリダイゼーション
FLT4転写物を細胞および組織に首尾よくもどすために、交尾後7.5日および8.5日のマウス胚の切片を、標識したFLT4のRNAとハイブリダイズさせた。マウス胚はCBAおよびNMRIマウスの交配に由来した。妊娠したマウスを頸部の脱臼(cervical dislocation)により屠殺し、胚をただちに凍結するかまたはリン酸緩衝液を経て4%パラホルムアルデヒド中へ移した。胚および単離したマウスの器官を4℃で18時間かけて固定し、脱水し、パラフィン中に包埋し、ついで6μm切片に切出した。
【0038】
192ヌクレオチドおよび349ヌクレオチドのRNAプローブ(アンチセンスおよびセンス)(実施例1参照)を、[35S]−UTPを使用して、直線状プラスミドから生成した。切片のインサイチュ・ハイブリダイゼーションを、ウィルキンソン(Wilkinson)らの文献(13、14)にしたがい、以下に示す改変、1)パラフィンワックス中への包埋前に、トルエンの代わりにキシレンを使用し、2)6μm切片を切出し、2%の3−トリエトキシシリルプロピルアミンを用いて前処理したガラススライド表面上のジエチルピロ炭酸塩で処理した水の層に置き、3)プローブのアルカリ加水分解を省略し、4)ハイ・ストリンジェンシー・ウオッシュ(high stringency wash)を、30mM DTTおよび1×SSCを含む溶液中、65℃で80分間かけて行なうという改変を加えて行なった。切片をNTB−2エマルジョン(コダック(Kodak))でおおい、4℃で貯蔵した。スライドを14日間さらし、現像し(developed)、ヘマトキシリンを用いて染色した。センス鎖およびRNアーゼAで処理した切片を用いた対照のハイブリダイゼーションはバックグラウンドをこえる特異的なシグナルを出さなかった。
【0039】
図2Aおよび2Bに示したように、FLT4のmRNAは交尾後7.5日のマウス胚においては発現しなかったが、発生8.5日の後主静脈(cv)においては明るいシグナルが検出された。対照的に、発達中の心臓(データは示さず)および胸部大動脈(da)はFLT4陰性であった。胚外の組織において、FLT4は尿膜(パネルBにおけるal)においては顕著に発現したが、発達中の卵黄包の血島は陰性であった(データは示さず)。一方、頭部の間葉の血管芽細胞(ab)は強いFLT4陽性であった(C)。発達中の胎盤において、FLT4シグナルは、末梢の洞様毛細血管静脈(peripheral sinusoidal veins)において最初に見られた(データは示さず)。交尾後9.5日の胎盤において、静脈の裂孔(venous lacunae)の内皮(Dにおけるvl)およびレイチャート膜(Reichert´s membrane)に部分的に融合した巨細胞(データは示さず)はFLT4のmRNAを発現した。
【0040】
したがって、FLT4の発現は、もっとも初期の内皮細胞前駆体である血管芽細胞において非常に顕著であるが、交尾後8.5日のマウス胚の特定の管のみに限定されるように思われる。受容体Tieは発達中のマウス胚のすべての内皮細胞において発現し、したがって前記細胞に対するマーカーを提供することが知られている。とくに、Tieプローブとは対照的に、FLT4プローブは、交尾後11.5日のマウス胚の動脈性内皮、たとえば発達中の胸部大動脈(図2のE、Fにおけるda)または頸動脈(データは示さず)の内皮細胞とハイブリダイズしたとしても非常に弱かった。その代わりに、FLT4シグナルは発達中の静脈において非常に顕著であった。たとえば、FLT4シグナルは発達中の後腎を囲む静脈(Eにおけるv、sv)において検出され、Tieプローブは後腎内の毛細管(c)を顕著に認識した(F)。
【0041】
図3から理解されうるように、FLT4のmRNAは交尾後12.5日のマウス胚のいくつかの領域に分布し、とくに腋窩領域(ax)の拡張された管において顕著である。類似したFLT4陽性の管構造が、頸静脈域における正中矢状切断図において見られた(データは示さず)。FLT4を発現している管の叢様パターンは、眼窩周辺領域(po)および発生中の椎骨の周囲(vb)において見られた。また、発達中の皮膚のちょうど下のFLT4陽性脈管網(FLT4−positive vascular network)が明らかになった(sc)。より弱い毛細管シグナルが発達中の脳(b)を含むいくつかの領域からえられた。FLT4のmRNAは、尾の領域と同様、頸の領域の小管、発達中の鼻の小管および発達中の舌の基底部(base)においても検出することができた(データは示さず)。そのうえ、肝臓(li)は斑点様のパターンでFLT4のmRNAに対して強い陽性であった。
【0042】
さらなる発達のあいだ、FLT4のRNAは胚の特定の管にさらに限定されるようになったと思われる。交尾後14.5日のマウス胚はこの限定された発現パターンを精密に示している(図4A、B)。図4の正中矢状切断図において、最も顕著なFLT4のシグナルは、前記切断図の前方部分における発達中の脊柱に沿って見られる。前記シグナルは、発達のこの時点において形成された最大のリンパ管である胸管(dt)の内皮細胞に由来していると思われた。対照的に、胸部大動脈(da)および下大静脈(vc)は陰性であった。また腸間膜領域の拡張された管もFLT4に対して強い陽性であった。さらに、交尾後12.5日のマウス胚において、頸(ne)の領域と同様、眼窩周辺(po)、下顎(lj)における解剖学的な境界線に沿った管の網(vessel networks)はFLT4陽性の内皮を含んでいた。類似した構造は心膜空間(pericardial space)および皮下(sc)組織のいたる所に存在した。とくに、FLT4陰性の管とは対照的に、すべてのFLT4陽性の管は、それらの管腔(lumen)に血液細胞を欠いていた。前記発現パターンは、発達のこの時点においてリンパ管の内皮にFLT4が限定されることを示唆している。本発明者らがFLT4発現を見たさらなる部位は、発達中の骨髄(bm)の洞様毛細血管にあった。
【0043】
FLT4プローブとハイブリダイズした交尾後16.5日の胚の上部胸郭(upper thorax)の横径切断写真を図4のパネルCおよびDに示す。Cに示される切片を、ヘマトキシリン−エオシンを用いて染色し、前記区域における異なる型の管を可視化した。前記区域は頸の動脈(ca)、腕頭の動脈(ba)、大静脈(vc)およびサイズが小さく筋肉および結合組織の周囲には存在しない胸管(矢印)を含んでいる。胸管領域の拡大図をパネルDに示し、ここではFLT4のオートラジオグラフィーの粒子を見ることができる。周辺の小管(v)と同様、胸管の内皮細胞はFLT4プローブとハイブリダイズする。
【0044】
実施例4
培養した内皮細胞におけるFLT4のmRNAの分析
実施例3に記載したインサイチュ・ハイブリダイゼーションの結果により、FLT4は静脈の内皮細胞ならびに後にはリンパ管およびいくらかの静脈の内皮細胞では発現するが、動脈の内皮では発現しないことが示された。前記のような調節がインビトロで維持されるかどうかを見るために、本発明者らはノーザンブロッティングおよびハイブリダイゼーション分析を使用して培養した内皮細胞を研究した。
【0045】
ヒトの大動脈、大腿静脈、臍帯静脈および包皮微小管由来の内皮細胞を単離し、培養して、ファン ヒンスバーグ(Van Hinsberg)によりすでに記載されたように特徴づけを行なった(15、16)。前記内皮細胞を、ポリアデニル化RNAを単離するために、5〜8回継代(分裂比(split ratio)1:3)したのち、コンフルエントな密度(confluent density)で使用した。
【0046】
内皮細胞系EA・hy926、BCEおよびLEIIはFLT4を発現しなかった(データは示さず)。しかしながら、培養したヒトの微小管、静脈および臍帯の静脈の内皮細胞は、FLT4に特異的な5.8kbおよび4.5kbのmRNAに関して陽性であり、一方大動脈の内皮細胞は陰性であった(図5)。対照的に、別の内皮の受容体チロシンキナーゼの遺伝子であるTieは、研究したすべての型の内皮細胞において4.4kbのmRNAとして発現した。
【0047】
実施例5
成人組織におけるFLT4のmRNA
実施例3においてえられた結果により、FLT4のmRNAは、発達中、リンパ管の内皮に大規模に制限されるようになることが示された。ヒトにおけるこの知見の潜在的な重要性のため、本発明者らは、ヒトFLT4プローブを使用して成人組織におけるFLT4も研究した。使用したヒトFLT4プローブは、cDNAの塩基対1〜595を含むEcoRI−SphI断片であった(3)。フォンビルブラント因子プローブは塩基対1〜2334を含むEcoRI−HindIII断片であった(17)。
【0048】
本発明者らは、組織病理学的診断に送るための、慣用の操作で固定された材料を使用した。正常な肺組織を類表皮癌におかされた左の肺の下葉の切除によりえた。腸間膜および腸間膜のリンパ節を、結腸の腺癌を有する患者からえた。異常な大きさであったので、睡液腺に隣接した正常なリンパ節を摘出した。2人の患者由来の扁桃およびその2つの垂(appendixes)は診断上の変化はなかった。2つのリンパ管筋腫瘍(lymphangiomyomas)および3つの嚢胞性リンパ管腫を研究し類似した結果をえた。
【0049】
ヒト組織について、ヒト組織は組織病理学的診断のために10%ホルマリンを用いて固定した慣用サンプルであり、正常なインサイチュでの手順は単にバックグラウンドのみを与えるが、プロテイナーゼKの代わりのマイクロ波処理は特異的ハイブリダイゼーションを可能にした(18、19)。
【0050】
腸間膜において、肺および垂のリンパ内皮(lv)はFLT4シグナルを出すが、静脈(v)、動脈(a)および毛細管(c)は陰性であった(図6A〜Dおよびデータは示さず)。FLT4がHEVで発現するかどうかを研究するために、扁桃を研究した。実際に扁桃について、FLT4に特異的なオートラジオグラフィーの粒子が、いくらかのHEVにおいては検出された(E、F)。
【0051】
実施例6
正常および転移性リンパ節ならびにリンパ管腫におけるFLT4のmRNAの分析
ヒト腸間膜リンパ節の一部(実施例5参照)をFLT4の発現について分析した。結果を図7に示す。
【0052】
FLT4はリンパ洞(ls)ならびに輸入(afferent)および輸出(efferent)のリンパ管において発現する(データは示さず)。同じパターンが腺癌の転移を含むリンパ節において見られる(C、D)。正常および転移性リンパ節の両方におけるいくつかのHEVも陽性であった。パネルEにおいて、FLT4の発現は嚢胞性リンパ管腫において見られる(Fにおけるヘマトキシリン−エオシンで染色した切片と比較した)。とくに、FLT4のリンパ内皮に対する特異性は、すべての血管におけるフォンビルブラント因子に対するインサイチュでのシグナルとの比較により明らかである(F)。
【0053】
実施例7
胎児の内皮細胞におけるFLT4の局在化
短型(short form)のカルボキシ末端側の40のアミノ酸をコードするFLT4のcDNA断片を、657bpのEcoRI断片として、読み取り枠(frame)中にグルタチオン−S−トランスフェラーゼをコードする領域を有するpGEX−1IT細菌性発現ベクター(ファルマシア(Pharmacia))にクローニングした。えられたGST−FLT4融合タンパク質を大腸菌中で産生し、グルタチオン−セファロース 4B カラムを使用したアフィニティークロマトグラフィーにより精製した。精製したタンパク質を凍結乾燥し、PBS中に溶解し、フロイントアジュバントと混合し、ウサギの免疫に使用した。抗血清は4回のブースター免疫ののちのものを使用した。
【0054】
17週齢および20週齢のヒト胎児由来の組織を、プロスタグランジンを用いて誘導した合法の流産よりえた。研究はヘルシンキ大学中央病院の倫理委員会の承認を受けた。存胎齢は胎児の脚の長さより推定した。胎児の組織をティシュー−テック(Tissue-Tek)(マイルス(Miles))中に包埋し、直ちに凍結し、−70℃で貯蔵した。
【0055】
抗FLT4抗血清をGST−セファロースカラムに交差吸収(cross-absorbed)させて抗GST抗体を除去し、ついでGST−FLT4アフィニティークロマトグラフィーにより精製した。厚さ6μmのいくつかの組織のクリオスタット切片(cryostat sections)をアセトンを用いて固定し、メタノール中の0.3%H2O2で30分間処理し内因性ペルオキシダーゼ活性をブロックした。洗浄後、切片を5%正常ブタ血清とともにインキュベートした。ついで切片をFLT4に対する抗体とともにインキュベートし、洗浄して、結合した抗体をペルオキシダーゼが結合したブタ抗ウサギIgGを用いて検出し、続いて基質として0.2%の3,3−ジアミノベンジジン(アマシャム(Amersham))を使用してペルオキシダーゼ活性に対する染色を行なった。切片をマイヤーのヘマトキシリン(Meyer´s hematoxylin)中で対比染色した。
【0056】
ヒト胎児の腸間膜の抗FLT4イムノペルオキシダーゼ染色は、いくつかの管の内皮中のFLT4タンパク質を示したが(図8A)、抗原でブロックした抗FLT4抗体(B)およびあらかじめ免疫した血清(C)は陰性であった。比較のために、図8Dは、脈管の内皮細胞に特異的な第VIII因子関連抗原に対する抗血清を使用した染色の結果を示す。
【0057】
実施例8
FLT4に対するモノクローナル抗体の産生
融合I:
4月齢のBalb/c雄マウスを、フロイント完全アジュバントを用いて乳化した濃縮培地(150μg/マウス)中の組換えにより産生したFLT4タンパク質(実施例7参照)の腹腔内注射により免疫した。3〜4週間おきに150μgのブースター注射をし、さらに3週間の間隔をおいたのち最終ブースター(PBS中の10μgのFLT4の腹腔内投与)を行なった。最終ブースター投与から4日後、マウスを屠殺し、マウス脾リンパ様細胞(lymphoid cells)をSP2/0プラスマ細胞腫細胞とそれぞれ2:1の割合で融合させた。
【0058】
融合細胞を、20%ウシ胎児血清およびHAT補足物(HAT supplement)(ヒポキサンチン−アミノプテリン−チミジン、ギブコ(GIBCO)、043−01060H、50倍希釈)を含むイーエックス−セル320(EX-Cell 320)培地(セララブ(SERALAB))を添加した96ウェル培養プレート(ヌンク(NUNC))中からえた。細胞を5%CO2雰囲気下、+37℃で培養した。10日後、HAT補足培地をHT補足細胞培養培地(ギブコ、043−01065H、50倍希釈)に交換した。HT培地はアミノプテリンを含まないことを除くとHAT培地と同一である。
【0059】
3週間後、特異的な抗体の産生を、実施例10に記載されている抗原特異的免疫蛍光分析法(antigen-specific immunofluorometric assay)であるIFMAにより決定した。マスタークローンを、スタスツェウスキー(Staszewski)ら、エール・ジャーナル・オブ・バイオロジー・アンド・メディシン(Yale Journal of Biology and Medicine)、57巻、865〜868頁(1984年)に記載された限界希釈(limited dilutions)によりクローニングした。陽性クローンを24ウェル組織培養プレート(ヌンク)上で増やし、再クローニングし、前記と同様の方法により再試験を行なった。陽性のクローンを蛍光活性化細胞選別法(FACS)により試験した。
【0060】
安定したクローンは、おそらくクラスIgAに属するIgを産生した1つのクローンを除いて、IgG1クラスに属する免疫グロブリンを分泌した。モノクローナル抗体のサブクラスを、IFMAにおいて、マウスのサブクラスに対するラットのモノクローナル抗体をビオチン接合物(biotin conjugate)(セロテック(SEROTEC))として使用して決定した。
【0061】
腹水(ascites fluid)中にモノクローナル抗体を産生するためにBalb/cマウスを使用した。プリスタン(pristane)(2,6,10,14−テトラメチルペンタデカン、98%、アルドリッチ−ケミエ(ALDRICH-CHEMIE)、デー(D)−7924、シュタインハイム(Steinheim)、カタログ番号 T2,280−2)でマウスを前処理したのち、前記ハイブリドーマを前記マウスに腹腔内注射した。ハイブリドーマ細胞注射の約2週間前に、プリスタン0.5mlを(静脈内)注射した。注射した細胞量はマウス1匹あたり約7.5〜9×106であった。ハイブリドーマ注射後10〜14日後、腹水を集めた。
【0062】
融合II:
2月齢のBalb/cマウス(雌)を、フロイント完全アジュバントを用いて乳化した組換えにより産生したFLT4タンパク質(実施例7参照)(20μg/マウス)の腹腔内注射により免疫した。3〜4週間おきに20μgのブースター注射をし、さらに3週間の間隔をおいたのち最終ブースター(PBS中に10μgのFLT4、静脈内投与)を行なった。最終ブースター投与から4日後、マウスを屠殺し、マウス脾リンパ様細胞をSP2/0プラスマ細胞腫細胞とそれぞれ2:1の割合で融合させた。
【0063】
融合細胞を、20%ウシ胎児血清およびHAT補足物(ヒポキサンチン−アミノプテリン−チミジン、ギブコ ビーアールエル(GIBCO BRL)21060−017、1:50に希釈)を含むオプティエムイーエム(OptiMEM)1(グルタマックス(Glutamax)1を有する、51985−026、ギブコ ビーアールエル)を添加した96ウェル培養プレート(ファルコン(FALCON))中からえた。細胞を5%CO2雰囲気下、+37℃で培養した。10日後、HAT補足培地をHT補足細胞培養培地(ギブコ ビーアールエル、41065−012、1:50倍に希釈)に交換した。HT培地はアミノプテリンを含まないことを除くとHAT培地と同一である。
【0064】
3週間後、特異的な抗体の産生を、実施例9に記載されている抗原特異的免疫蛍光分析法(IFMA)により決定した。マスタークローンを、スタスツェウスキーら、エール・ジャーナル・オブ・バイオロジー・アンド・メディシン、57巻、865〜868頁(1984年)に記載された限界希釈によりクローニングした。陽性クローンを24ウェル組織培養プレート(ファルコン)上で増やし、再クローニングし、前記と同様の方法により再試験を行なった。陽性のクローンを蛍光活性化細胞選別法(FACS)により試験した。
【0065】
2E11および6B2クローンは、IgG1クラスに属する免疫グロブリンを分泌し、2B12クローンはサブクラスIgMに属するIgを産生した。マウスのサブクラスのIgG1を、IFMAにおいて、マウスのサブクラスの重鎖に対するラットモノクローナル抗体をビオチン接合物(セロテック)として使用して決定し、マウスのサブクラスのIgMを、マウスモノクローナル抗体アイソタイピング・キット(Mouse Monoclonal Antibody Isotyping Kit)(ディップスティック フォーマット(Dipstick Format))(19663−012、ライフ テクノロジーズ社(Life Technologies Inc.))を用いて決定した。
【0066】
実施例9
FLT4に対するモノクローナル抗体の特異性
融合Iの抗体:
実施例7に記載したFLT4の細胞外ドメインを、アナリティカル・バイオケミストリー(Anal.Biochem.)、176巻(2)、319〜325頁(1989年)に記載されたムッカラ(Mukkala)らの方法に、以下に示す改変:250倍をこえるモル量のイソチオシアネートDTTA−Eu(N1キレート(N1 chelate)、ワラック(Wallac)、フィンランド共和国)をFLT4溶液(PBS中に0.5mg/ml)に添加し、pHを0.5mol/L炭酸ナトリウム緩衝液、pH9.8を添加することにより約9に調整したことを加えて標識化した。標識化を+4℃で一晩行なった。結合しなかった標識を、溶離液としてのTSA緩衝液(0.15mol/lのNaClを含む50mmol/L Tris−HCl、pH7.8)とともにPD−10(ファルマシア、スウェーデン王国)を用いて除去した。
【0067】
精製後、標識されたFLT4に1mg/mlウシ血清アルブミン(BSA)を添加し、標識物を+4℃で貯蔵した。既知のEuCl3標準の蛍光に対する蛍光の割合を測定することにより決定したFLT4の1分子あたりに取り込まれたユウロピウムイオン数は1.9であった(ヘミッラ(Hemmilae)ら、アナリティカル・バイオケミストリー、137巻、335〜343頁(1984年))。
【0068】
実施例8において産生した抗体を、ウサギ抗マウスIg(Z259、ダコパッツ(DAKOPATTS))で被覆したマイクロタイトレーション・ストリップ・ウェル(microtitration strip wells)(ヌンク(NUNC)、ポリソーブ(polysorb))を用いたサンドイッチ型免疫蛍光分析法を使用してスクリーニングした。あらかじめ被覆したウェルをデルフィア(DELFIA)洗浄溶液を用いたプレートウォッシュ(Platewash)1296−024(ワラック)により一度洗浄した。デルフィア分析緩衝液を、細胞培養上清および予備的スクリーニング分析(preliminary screening assays)における正の対照として使用した脾摘出したマウスの血清(1:1000〜1:100000の希釈)の希釈緩衝液として使用した。
【0069】
+4℃で一晩(または室温で2時間)のインキュベーションを、プレートシェイク振とう機(Plateshake shaker)(1296−001、ワラック)上での5分間の振とうにより開始し、続いて前記洗浄溶液を用いて4回洗浄した。
【0070】
ユウロピウムで標識したFLT4を、分析緩衝液100μl中、1:500の希釈で添加した。プレートシェイク振とう機上で5分間および室温(RT)で1時間インキュベートしたのち、前記と同様にストリップを洗浄した。
【0071】
エンハンスメント溶液(Enhancement solution)(デルフィア)を200μl/ウェルの濃度で添加した。ついで、プレートを、プレートシェイク振とう機上で5分間振とうし、アルクス(ARCUS)−1230(ワラック)により蛍光強度を10〜15分間測定した(レブグレン(Loevgren)ら、コリンス ダブリュー ピー(Collins W.P.)(編)、オルターナティブ・イムノアッセイズ(Alternative Immunoassays)、ジョン・ウィレー・アンド・ソンズ社(John Wiley & Sons Ltd.)、203〜216頁(1985年))。
【0072】
えられたFLT4に対するモノクローナル抗体および対応するFACSの結果を表1に要約する。
【0073】
【表1】

【0074】
抗FLT4の9D9F9と名付けられた1つのクローンは、IFMAにより免疫グロブリンクラスIgG1に属することが決定されたモノクローナル抗体を安定して分泌した。ハイブリドーマ9d9f9は、1995年3月23日に、ドイツ連邦共和国、3300 ブラウンシュヴァイク(Braunschweig)、マシェロデル ヴェク 1ベー(Mascheroder Weg 1b)のドイツ微生物および細胞培養コレクション(German Collection of Microorganisms and Cell Cultures)、ヒトおよび動物細胞ならびにウイルス部門(Department of Human and Animal Cell Cultures and Viruses)に寄託され、受託番号ACC2210を与えられている。
【0075】
融合IIの抗体:
実施例7に記載したFLT4の細胞外ドメインを、アナリティカル・バイオケミストリー、176巻(2)、319〜325頁(1989年)に記載されたムッカラらの方法に、以下に示す改変:250倍をこえるモル量のイソチオシアネートDTTA−Eu(N1キレート、ワラック、フィンランド共和国)をFLT4溶液(PBS中に0.5mg/ml)に添加し、pHを0.5mol/L炭酸ナトリウム緩衝液、pH9.8を添加することにより約9に調整したことを加えて標識化した。標識化を+4℃で一晩行なった。結合しなかった標識は、溶離液としてのTSA緩衝液(0.15mol/LのNaClを含む50mmol/L Tris−HCl、pH7.8)とともにPD−10(ファルマシア)を用いて除去した。
【0076】
精製後、標識されたFLT4に1mg/mlウシ血清アルブミン(BSA)を添加し、標識物を+4℃で貯蔵した。既知のEuCl3標準の蛍光に対する蛍光の割合を測定することにより決定したFLT4の1分子あたりに取り込まれたユウロピウムイオン数は1.9であった(ヘミッリ(Hemmil)ら、アナリティカル・バイオケミストリー、137巻、335〜343頁(1984年))。
【0077】
実施例8において産生した抗体を、ウサギ抗マウスIg(Z259、ダコ(DAKO))で被覆したマイクロタイトレーション・ウェル(ヌンク、ポリソーブ)を用いたFLT4特異的IFMAを使用してスクリーニングした。あらかじめ被覆したウェルをデルフィア・プレート・ウォッシュにより洗浄溶液(ワラック)を用いて一度洗浄した。
【0078】
デルフィア分析緩衝液を、細胞培養上清(予備的スクリーニングにおいて1:2の希釈)および正の対照として使用した脾摘出したマウスの血清(1:1000〜1:100000の希釈)の希釈緩衝液として使用した。標準として、精製した抗FLT4の9D9F9(マウスのサブクラスIgG1)を1.0ng/ml〜250ng/mlのあいだの濃度で使用した。サンプルをはじめに室温下、プレート・シェイク(ワラック)上で5分間振とうし、ついで+4℃で約18時間インキュベートした。はじめにフレーム(frames)を4回洗浄し、ついでユウロピウムで標識したFLT4(分析緩衝液100μl中、1:2000)を添加し、最終的に前記フレームを室温下で1時間インキュベートした。前記のように洗浄したのち、エンハンスメント溶液(200μl/ウェル、ワラック)を添加し、プレートをプレート・シェイク上で5分間振とうした。蛍光強度をアルクス−1230(ワラック)により測定した。
【0079】
えられたFLT4に対するモノクローナル抗体および対応する結果を表2に要約する。
【0080】
抗FLT4抗体の定量の標準曲線を、親和性により精製した抗FLT4である9D9F9を使用して作成した。直線の範囲は1.0ng/ml〜250ng/mlに達した。
【0081】
細胞表面上に全長のFLT4を発現するpLTRFLT4構築物を同時にトランスフェクト(cotransfected)したNIH3T3細胞の細胞溶解物を、6.5%SDS−PAGE中で電気泳動し、タンパク質をニトロセルロース硝酸塩(nitrocellulose nitrate)膜(0.45μm、シュレイチャー アンド シュエル(SCHLEICHER & SCHUELL))上に移し、モノクローナル抗体細胞培養上清(1:10に希釈、50mmol/L TRIS−4%メタノールを含む40mmol/Lグリシン緩衝液、0.04%SDS)を用いてイムノブロットした。モノクローナル抗体の特異性をエイチアールピー(HRP)接合ウサギ抗マウスIg(P161、ダコ、150mmol/L食塩水および5%ミルク粉末を含む20mmol/L TRIS緩衝液、pH7.5中、1:1000に希釈)およびイーシーエル(ECL)(エンハンスド ケミルミネッセンス(Enhanced chemiluminescence)、アマシャム)とのインキュベーションを使用して検出した。
【0082】
【表2】
【0083】
前記より明らかなように、本発明に係る抗体は、リンパ管、リンパ内皮細胞、高内皮性小静脈、リンパ管腫、転移性リンパ節およびその他のリンパ系の疾患状態の診断ならびに同定、転移の蔓延(metastatic spread)の検出およびモニタリングに、リンパ管および高内皮性小静脈の内皮細胞の刺激ならびに阻害に、内皮細胞への分子の選択的な導入に、そしてリンパ管およびリンパ管の疾患状態の映像化において有用である。本明細書において請求した主題の他の用途は当業者にとって明らかである。
【0084】
[参考文献]
1. Sabin,F.R.1909.リンパ系全体としての形態を考慮した、ヒト胚のリンパ系。Am.J.Anat.9:43.
2. van der Putte,S.C.J.1975.ヒトにおけるリンパ系の発達。Adv.Anat.Embryol.Cell Biol.51:3.
3. Pajusola,K.,O.Aprelikova,J.Korhonen,A.Kaipainen,L.Pertovaara,R.Alitalo,and K.Alitalo.1992.受容体チロシンキナーゼであるFLT4は7つの免疫グロブリン様ループを含み、多数のヒト組織および細胞系で発現する。Cancer Res.52:5738.
4. Kaipainen,A.,J.Korhonen,K.Pajusola,O.Aprelikova,M.G.Persico,B.I.Terman,and K.Alitalo.1993.受容体チロシンキナーゼである関連したFLT4、FLT1およびKDRは、ヒト胎児の内皮細胞において異なる発現パターンを示す。J.Exp.Med.178:2077.
5. Galland,F.,A.Karamysheva,M.-J.Pebusque,J.-P.Borg,R.Rottapel,P.Dubreuil,O.Rosnet,and D.Birnbaum.1993.FLT4遺伝子は、脈管の内皮成長因子受容体に関係するトランスメンブレン型チロシンキナーゼをコードする。Oncogene.8:1233.
6. Millauer,B.,S.Wizigmann-Voos,H.Schnuerch,R.Martinez,N.-P.H.Moller,W.Risau,and A.Ullrich.1993.高親和性のVEGFの結合および発達上の発現は、脈管形成および血管形成の主要なレギュレーターとしてのFlk−1を示唆する。Cell.72:835.
7. Yamaguchi,T.P.,D.Dumont,R.A.Conlon,M.L.Breitman,and J.Rossant.1993.flt関連チロシンキナーゼであるflk−1は内皮細胞前駆体に対する早期のマーカーである。Development.118:489.
8. Peters,K.G.,C.De Vries,and L.T.Williams.1993.脈管内皮の成長因子の受容体の胚形成および組織修復中の発現は、内皮の分化および血管の成長における役割を示唆する。Proc.Natl.Acad.Sci.USA 90:8915.
9. Finnerty,H.,K.Kelleher,G.Morris E.,K.Bean,D.Merberg,R.Kritz,J.Morris C.,H.Sookdeo,K.J.Turner,and C.R.Wood 1993.マウスのFLTおよびFLT4の分子クローニング。Oncogene.8:2293.
10. Korhonen,J.,A.Polvi,J.Partanen,and K.Alitalo.1993.マウスの受容体チロシンキナーゼであるtie遺伝子:胚の血管形成中の発現。Oncogene.8:395.
11. Chomczynski,P.,and N.Sacchi.1987.酸性グアニジンチオシアネート−フェノール−クロロホルム抽出によるRNA単離の単一工程方法。Anal.Biochem.162:156.
12. Tokunaga,K.,H.Taniguchi,K.Yoda,M.Shimizu,and S.Sakiyama.1986.マウスの細胞骨格βに作用するmRNAに対する全長のcDNAのヌクレオチド配列。Nucleic.Acid.Res.14:2829.
13. Wilkinson,D.G.,J.A.Bailes,J.E.Champion,andA.P.McMahon.1987.交尾後(post coitum)8〜10日のマウスの発達の分子解析により胚のグロビン発現のみに変化が検出される。Development.99:493.
14. Wilkinson,D.G.,J.A.Bailes,and A.P.McMahon.1987.プロトオンコジーンであるint−1の発現は、発達中のマウスの胚の特定の神経細胞に限定される。Cell.50:79.
15. Van Hinsberg,V.W.M.,D.Binnema,M.A.Scheffer,E.D.Sprengers,T.Kooistra,and D.C.Rijken.1987.成人のヒト血管由来の連続的に増殖した内皮細胞によるプラスミノーゲン活性化因子および阻害因子の産生。Arteriosclerosis.7:389.
16. Van Hinsberg,V.W.M.,M.A.Scheffer,and T.Kooistra.1987.ヒト包皮微小管の内皮細胞によるプラスミノーゲン活性化因子およびPA阻害因子−1産生に対するトロンビンの作用。Thromb.Haemostas.57:148.
17. Bonthron,D.T.,E.C.Orr,L.M.Mitsock,D.Ginsberg,R.I.Handin,and S.H.Orkin.1986.プレ−プロ−フォンビルブラント因子のcDNAのヌクレオチド配列。Nucleic Acids Res.141:7125.
18. Shi,E.,M.Kan,J.Xu,and W.L.McKeehan.1991.(繊維芽細胞)成長因子結合型の16キロダルトンのヘパリンのあるものは、受容体に依存した内在化ののち、安定な40キロダルトンの複合体として生じる。J.Biol.Chem.266:5774.
19. Catoretti,G.,M.H.G.Becker,G.Key,M.Duchrow,C.Schlueter,J.Galle,and J.Gerdest.1992.Ki−67抗原の組換え部分に対するモノクローナル抗体(MIB1およびMIB3)は、マイクロ波で処理したホルマリン固定化パラフィン切片中の増殖している細胞を検出した。J.of Pathol.168:357.
【0085】
[配列の表示]
(1)一般情報
(i)出願人:
(A)名称:ヘルシンキ ユニバシティ ライセンシング リミテッド オケサ ユキチュア
(A)名称:オリオン コーポレーション
(ii)発明の名称:受容体チロシンキナーゼであるFLT4ならびに診断および治療におけるその用途
(iii)配列の数:1
(iv)コンピュータ読取フォーム:
(A)ソフトウェア:PatentIn Release 1.0、バージョン1.25(ヨーロッパ特許局)
(v)優先権データ:
(A)出願番号:アメリカ合衆国、08/257754
(B)出願日:1994年6月9日
(2)配列番号:1
【技術分野】
【0001】
本発明は概して、受容体チロシンキナーゼ、核酸プローブおよび前記受容体を特異的に認識する抗体、ならびに動物およびヒト組織中のリンパ管と高内皮性小静脈(high endothelial venules)(HEV)を、そして培養物中(in culture)のリンパ内皮細胞(lymphatic endothelial cells)を同定するための前記プローブおよび抗体の使用に関する。さらに詳しくは、本発明は受容体チロシンキナーゼであるFLT4に特異的な抗体、リンパ管におけるFLT4の発現を同定する方法ならびに究極的には、たとえば炎症性疾患、感染性疾患および免疫性疾患、転移性リンパ節ならびにリンパ管腫などのリンパ組織における変化を含む動物およびヒトにおける疾患の状態の診断ならびに治療に関する。
【背景技術】
【0002】
いくつかの疾患と同様、胚の脈管形成および血管形成(embryonic vasculogenesis and angiogenesis)、血液凝固ならびに創傷の治癒および再生などの脈管系の生理機能は、血管に並ぶ脈管内皮(vascular endothelium)に関係している。脈管枝(vascular tree)の発達は血管形成を経ておこり、いくつかの理論によると、リンパ系の形成は、静脈からの発芽(sprouting)により動脈および静脈の発達(development)後すぐに開始する(1、2)。
【0003】
胎児期(fetal period)ののち、新生血管形成に関連した脈管形成の期間を除いて、内皮細胞は非常にゆっくりと増殖する。脈管形成を刺激する成長因子は、特異的な内皮細胞表面受容体であるチロシンキナーゼを介してそれらの効果を及ぼす。
【0004】
ヒト赤白血病細胞系からクローニングした受容体チロシンキナーゼであるFLT4のcDNAのタンパク質産物は、Nを介してグリコシル化(N−glycosylated)され、その細胞外ドメインには7つの免疫グロブリン様ループを含んでいる。FLT4の細胞質チロシンキナーゼドメインは、FLT1およびKDRの対応するドメインとアミノ酸レベルで約80%同一であり、血小板由来成長因子に対する受容体、コロニー刺激因子−1に対する受容体、幹細胞因子に対する受容体および受容体FLT3と約60%同一である(3)。
【0005】
FLT4の生物学的機能は現在までのところ知られていないが、その制限された発現のパターンは前記機能が脈管内皮に関係しているかもしれないということを示した。本発明者らが以前にえた結果から、ジャーナル・オブ・エクスペリメンタル・メディスン(J.Exp.Med.)、178巻、2077〜2088頁(1993年)にカイパイネン(Kaipainen)らが開示したように、いくつかの胎児の器官の発達中の管(vessels)の内皮細胞におけるFLT4のmRNAの発現が明らかになった。受容体FLT4、FLT1およびKDR/FLK−1のmRNAシグナルは、重複(overlapping)を示したが、研究した組織においては異なる発現パターンを示した(4)。これらのデータは、この遺伝子ファミリーによってコードされる受容体チロシンキナーゼは血管の成長の調節および/または分化において異なる機能を有するかもしれないことを示唆している。
【0006】
リンパ系の主要な機能は、組織から流体が戻るようにし、多くの脈管外物質(extravascular substances)を血液へ戻すことである。さらに、成熟過程のあいだ、リンパ球は血液を離れ、リンパ系器官および他の組織を通り移動してリンパ管に入り、胸管を通り血液に戻る。専門化した小静脈である高内皮性小静脈(HEV)は再びリンパ球に結合し、組織への管外遊出を引きおこす。したがって、リンパ管、とくにリンパ節は免疫学における重要な役割を果たしており、異なる腫瘍の転移が発達する部位でもある。
【0007】
20世紀以来、リンパ系の胚起源(embryonic origin)に関する3つの異なる理論が提起されている。しかしながら、リンパ管に対して利用できる特異的マーカーは存在しなかったため、本発明より前は、リンパ管を同定することは困難であった。
【0008】
リンパ管は、リンパ管造影法を用いてもっとも一般的に研究される。リンパ管造影法において、エックス線造影剤はリンパ管に直接注射される。前記造影剤はリンパ系の輸出ドレナージ管(efferent drainage vessels)に沿って分配される。前記造影剤はリンパ節に集められ、半年間はそこにとどまり、そのあいだにエックス線解析によりリンパ節の大きさと構造を追跡することができる。この診断は、リンパ節に転移を有する癌患者およびたとえばリンパ腫などのリンパの悪性疾患においてとくに重要である。
【発明の概要】
【0009】
本発明は、FLT4ペプチドおよびその他の構築物、ならびにリンパ内皮細胞に対する特異的マーカーとしてのFLT4の用途に関する。
【0010】
また本発明は、核酸プローブおよびFLT4を特異的に認識する抗体、とくにモノクローナル抗体ならびに前記抗体を含む組成物に関する。さらに本出願では、組織、とくにリンパ組織およびリンパ内皮細胞における受容体FLT4を検出し、その量を測定する診断目的への前記モノクローナル抗体の使用が開示される。
【0011】
好ましい態様において、本発明は受容体FLT4を特異的に認識するモノクローナル抗体を提供する。より明確には、本発明は9D9F9と名付けられたモノクローナル抗体を提供する。モノクローナル抗体9D9F9を産生するハイブリドーマ細胞系は、ブダペスト条約の規定によりドイチェ・サムルング・フォン・ミクロオルガニスメン・ウント・ツエルクルツレン・ゲゼルシャフト・ミット・ベシュレンクテル・ハフツング(Deutsche Sammlung von Mikroorganismen und Zellkulturen Gmb H)(DSM)に寄託されている(DSM受託番号ACC2210)。
【0012】
また検出しうるマーカーで標識したモノクローナル抗体も提供する。本明細書で使用するとき、「検出しうるマーカー」という用語は当業者に知られたあらゆる検出しうるマーカーを包含する。しかしながら、本発明の好ましい態様における検出しうるマーカーは、放射性同位体、蛍光色素、色素、酵素およびビオチンからなる群より選ばれる。本発明の目的に対して、適当な放射性同位体は125Iおよび131Iを含むがこれらに限定されるものではない。
【0013】
本発明のモノクローナル抗体は、細胞サンプル中の受容体FLT4の存在を検出する方法に使用しうる。前記方法は、細胞サンプルを本発明のモノクローナル抗体にさらす工程および前記モノクローナル抗体の受容体FLT4への結合を検出する工程からなる。
【0014】
したがって、本発明の別の面は、細胞サンプル中の受容体FLT4の存在を決定する方法であって、
(a)細胞サンプルを本発明のモノクローナル抗体にさらす工程、および
(b)前記モノクローナル抗体の受容体FLT4への結合を検出する工程
からなる方法に関する。
【0015】
細胞混合物を本発明のモノクローナル抗体にさらすことは、蛍光により活性化された細胞の選別(fluorescence-activated cell sorting)のばあいは、溶液中で行なうことができ、たとえば生検材料などの固形組織標本上でも行なうこともでき、またカラムクロマトグラフィーまたは直接免疫接着(direct immune adherence)を用いるばあいは、固体担体に固定化されたモノクローナル抗体を用いて行なうことができる。モノクローナル抗体にさらされる細胞混合物は、血液細胞または組織細胞のいかなる溶液であってもよい。好ましくは、細胞混合物はリンパ内皮細胞を含むかまたは含むことが疑われる正常または病的な組織に由来する。細胞混合物を前記モノクローナル抗体にさらしたのち、受容体FLT4を有する細胞は前記モノクローナル抗体と結合し、抗体−受容体FLT4複合体を形成するであろう。抗体−受容体FLT4複合体の存在、したがって受容体FLT4の存在は、当該技術分野において既知の方法によって検出することができる。これらの方法はたとえば蛍光抗体法、FACS分析、ELISA、IRMA(サンドイッチ型免疫化学分析法)、免疫組織化学的方法、125I標識およびオートラジオグラフィーを用いたRIAなどの当該技術分野において標準的な免疫組織化学的方法を含む。
【0016】
本発明はまた映像化しうる試薬に接合(conjugated)したモノクローナル抗体も提供する。本明細書で使用するとき、「映像化しうる試薬」という用語は放射性同位体を含むが、これに限定されるものではない。好ましい放射性同位体はテクネチウム99mである。
【0017】
特定の態様において、本発明は組織サンプルおよび生物中のリンパ管およびその内皮細胞をモニターする方法に関する。さらに本発明はリンパ組織、とくにリンパ管の状態(炎症、感染、外傷(traumas)、成長、新形成など)を描く臨床的検出方法および生物中のリンパ管、したがってリンパ系血管新生(lymphatic vascularization)を検出する方法を提供する。
【0018】
さらに具体的には、本発明は、転移性癌、炎症状態、感染状態および免疫状態に関係したFLT4の発現によって特徴付けられるリンパ系の変化(lymphatic changes)を検出し、同定する方法を提供する。前記方法は、
(a)リンパ系の変化が疑われる組織および/または体液サンプルをうる工程、
(b)前記サンプルを、FLT4に特異的なモノクローナル抗体とその抗原の複合体を形成するのに適した条件下で、FLT4に特異的なモノクローナル抗体と接触させる工程、および
(c)形成されたあらゆる複合体の存在を検出する工程
からなる。
【0019】
前記方法により検出されうる組織は、FLT4を含むリンパ系細胞(lymphatic cells)または受容体FLT4を発現している細胞を有するあらゆる正常な、前癌性のまたは癌性の固形腫瘍組織である。本発明の1つの態様において、モノクローナル抗体は本明細書に記載されたような検出しうるマーカーで標識される。本発明の方法は様々な形態の癌、とくにリンパ節における転移およびリンパ管腫に加えて、たとえばリンパ腫などのその他のリンパの悪性疾患の検出および識別に有用である。
【0020】
ヒト患者におけるリンパ管、高内皮性小静脈またはリンパ節の存在を映像化する方法も本発明によって提供される。前記方法は標識した抗体を投与し、リンパ管またはリンパ節においてFLT4を発現している細胞が存在する部位を映像化することにより検出することからなる。
【0021】
本発明はさらにリンパ血管新生ならびに炎症状態、感染状態および免疫状態におけるFTL4の機能を刺激するまたは拮抗する方法に関する。前記方法は、このような反応に関与するFLT4内皮細胞部位、とくにFLT4の機能が、たとえば転移性癌、リンパ腫、炎症性疾患(慢性または急性)、感染性疾患および免疫性疾患などの疾患に関連している部位をブロックするのに充分な量のFLT4に結合する化合物を投与することによりFLT4が介在するリンパ血管新生を阻害することからなる。
【図面の簡単な説明】
【0022】
【図1】図1はマウス組織におけるFLT4のmRNAの発現を表わし、Aは成体のマウスの表示した組織から単離したポリアデニル化RNA(polyadenylated RNA)のハイブリダイゼーションを表わす。FLT4のmRNAのバンドの大きさはkbで示す。Bは様々な存胎齢(E8〜E18)のマウス胚および新生マウス(1日)から単離したRNAのRNアーゼプロテクション分析(RNA se protection analysis)を表わしている。サンプルE8+Pは胎盤も含んでいる。プローブおよび保護されたFLT4断片の大きさはヌクレオチドで示し、β−アクチンを対照として使用した。
【図2】図2は交尾後7.5日、8.5日および11.5日の胚におけるFLT4のmRNAの発現を表わしている。インサイチュ・オートラジオグラムの暗視野(darkfield)および明視野(brightfield)の顕微鏡写真を示す。FLT4のmRNAの発現は、7.5日の胚(A)においては検出することができなかった。交尾後8.5日のマウス胚のFLT4の発現を(B)および(C)に示す。矢印は、後主静脈の内皮(cv)、(B)における尿膜(al)および(C)の頭部間葉の血管芽細胞(ab)におけるFLT4陽性細胞を示している。交尾後8.5日の胎盤において、FLT4転写物は、静脈裂孔(venous lacunae)(vl)の内皮細胞において見ることができた(D)。パネルEおよびFは、交尾後11.5日の胚におけるFLT4およびTieのハイブリダイゼーションシグナルの比較を示している。発達中の胸部大動脈および後腎(mn)領域を示す(20×)。胸部大動脈はFLT4に対して陰性であるがTieのmRNAに対しては陽性であるのに対して、両方のプローブはともに心内膜下静脈(subendocardial vein)(sv)の内皮とハイブリダイズすることに注意すべきである。またFLT4のプローブは、後腎静脈(v)からシグナルを出すが、Tieは後腎毛細管(metanephric capillaries)(c、矢印)と大部分がハイブリダイズする。daは胸部大動脈を、ngは神経溝を示す。スケールバーは30μmである。
【図3】図3は交尾後12.5日の胚におけるFLT4のmRNAの発現を表わしている。腋窩面(axillar plane)を通る矢状切断図を示す。FLT4のmRNAは、腋窩(ax)の拡張された管(dilated vessels)、眼窩周辺(po)領域の叢様パターン、脊椎傍組織(矢じり)および皮下(sc)組織において顕著であった。bは脳を、liは肝臓を示す。スケールバーは5μmである。
【図4】図4は交尾後14日および16.5日の胚におけるFLT4を表わしている。パネルAおよびBは、正中矢状切断図の明視野および暗視野の映像を示している。poは眼窩領域を、ljは下顎を、neは頸領域を、scは皮下組織を、mtは腸間膜を、aoは大動脈を、dtは胸管を示す。(C)はヘマトキシリン−エオシン染色した16.5日の胚の横断面を示す。thは胸腺を、trは気管を、eは食道を、caは頸動脈を、baは腕頭の動脈を、dtは胸郭を示す。(D)は胸管領域の高拡大図(40x)を示し、オートラジオグラフィーの粒子が内皮細胞全体に見ることができる。また、前記写真の上の部分における小管(v)は陽性である。スケールバーは10μm(A〜C)および1μm(D)である。
【図5】図5は培養した内皮細胞におけるFLT4およびTieのmRNA発現の比較を表わしている。ヒトの包皮の微小管(MV)、大腿の静脈(VE)、大動脈(AO)および臍帯の静脈(HU)の内皮細胞由来のポリアデニル化RNAのノーザンブロット分析を示す。比較のために、受容体チロシンキナーゼであるTieのmRNAのハイブリダイゼーションシグナルを示す。プローブのリボソームRNAへの非特異的結合の結果生じたバンドをアスタリスクで印をつけた。
【図6】図6は成人のヒト腸間膜(A、B)、肺(C、D)および扁桃(E、F)のリンパ管におけるFLT4を表わしている。AおよびCにおけるリンパ管のみがFLT4シグナルを出し、静脈、毛細管および動脈ではFLT4のmRNAに対して陰性であることに注意すべきである。扁桃において、シグナルはいくらかのHEVの内皮において見出された。スケールバーは200μmである。
【図7】図7は正常(A、B)および転移性(C、D)リンパ節ならびにリンパ管腫(E〜G)におけるFLT4のmRNAを表わしている。矢じりは、FLT4陽性であるリンパ洞およびHEVに印をつけている。FLT4およびフォンビルブラント因子のシグナルの比較では、両方がリンパ内皮において示されるが、毛細管(c)および静脈(v)の内皮においてはフォンビルブラント因子のシグナルのみが示された。スケールバーは10μm(A〜D)および100μm(E〜G)である。
【図8】図8はイムノペルオキシダーゼ染色によって検出された胎児の腸間膜管(mesenterial vessels)におけるFLT4の発現を表している。親和性により精製した抗FLT4抗体(A)、抗原でブロックした抗血清(B)、あらかじめ免疫した血清(C)および第VIII因子関連抗原に対して特異的な抗血清(D)を用いて断片を染色した。染色はいくらかのものに制限されるが、すべての管(v)には制限されないことに注意すべきである。スケールバーは0.05mmである。
【発明を実施するための形態】
【0023】
リンパ組織、とくに転移性癌および免疫性疾患状態に関連するリンパ節における変化を同定することの重要性を認識して、本発明者らはFLT4がリンパ管内皮を検出する特異的なマーカーであり、したがってヒトの病的状態におけるリンパ系の変化に対するマーカーとして有用であることを示した。
【0024】
本発明者らは以前に、FLT1およびKDRと比較したFLT4の発現パターンは、妊娠18週目のヒト胎児の組織において大きく異なることを示した(4)。発達におけるFLT4の役割を理解するために、本発明者らはマウスFLT4の部分的なcDNAをクローニングした。これらのプローブをインサイチュ・ハイブリダイゼーションに使用して、マウスの発達におけるFLT4のmRNAの発現を分析し、FLT4はリンパ系の脈管形成中に発現することを見出した。FLT4はいくらかの高内皮性小静脈(HEV)と同様に、正常および病的な状態の成人の組織のリンパ内皮細胞において見出されたので、前記の知見の妥当性は正常および病的な成人の組織においても確認された。
【0025】
マウスFLT4のcDNA断片のクローニングにより、前記断片から推定されるアミノ酸配列は対応するヒトの配列とほとんど同一であることが示された(研究した両方の区域におけるアミノ酸の同一性は約96%)。マウスFLT4のcDNAの同一性のさらなる証拠はノーザンハイブリダイゼーションによりえられ、そこでは両方の種に由来するプローブが典型的なマウス組織から5.8kbのmRNAシグナルを出した。成体マウスの様々な組織から単離したRNAの解析は、肝臓、肺、心臓、脾臓および腎臓におけるFLT4の発現を示し、脳および精巣においてはまったくないかまたはわずかのハイブリダイゼーションを示した。このパターンは先にガランド(Galland)らにより報告されたパターンと類似している(5)。RNアーゼプロテクション(RNase protection)の結果により、FLT4遺伝子は交尾後8.5日の胚から始まって、マウスの発達中必要であることが示唆され、相対的な発現レベルはかなり安定しているようであった。
【0026】
インサイチュ・ハイブリダイゼーションのために、細胞外ドメイン配列をコードするマウスFLT4のcDNA断片を2つ選んだ。このことによりハイブリダイゼーションパターンを関連の受容体FLK−1およびFLT−1のパターンと明確に区別することができる。このことは細胞外領域におけるFLT4との配列同一性が非常に低い程度のものでしかないことを示している(6、7、8、9)。内皮の受容体チロシンキナーゼであるFLK−1、FLT−1、TieおよびTekの遺伝子と同様に、FLT4の遺伝子は、交尾後7.5日の胚では発現しなかった。交尾後8.5日の胚においては、強いFLT4シグナルが尿膜、頭部間葉(head mesenchyme)の血管芽細胞および主静脈に局在した。対照的に、胸部大動脈(dorsal aorta)、心臓の心内膜および卵黄包の血管芽細胞は、Tie、Tek、FLK−1ならびにFLT−1、TieおよびTekに対してとは異なり、FLT4に対して陰性であった(10、8)。FLT4発現の静脈系(venous system)への限定は、11.5日のマウス胚由来のサンプルにおいてよりいっそう明確になり、ここではTieのmRNAは動脈においても発現した。交尾後12.5日の胚においては、FLT4シグナルは発達中の静脈性内皮および推定リンパ内皮を装飾したが、内皮の受容体チロシンキナーゼであるTieに対してとは異なり、動脈内皮は陰性であった。発達の後半段階中、FLT4のmRNAは、発達中のリンパ管を表わす血液細胞を欠いた脈管叢(vascular plexuses)に限定されるようになった。成人の組織において、リンパ内皮およびいくらかの高内皮性小静脈のみがFLT4のmRNAを発現した。増加した発現が転移性リンパ節およびリンパ管腫におけるリンパ洞および高内皮性小静脈においておこった。
【0027】
マウス胚からえたデータの解釈が困難であり、ヒトのリンパ系は非常によく定義されているため、ヒト内皮を研究した。また、様々な内皮から確立した細胞を、FLT4の発現の特異性がインビトロ条件において持続するかどうかをみるために、細胞培養中で研究することができた。内皮細胞系はインビトロ培養において分化した特徴(differentiated features)を失うことが知られている。それゆえ、前記内皮細胞系がFLT4に対して陰性であることは予期されないことではなかった。培養した大動脈の内皮細胞もまたFLT4のmRNAを欠いていた。しかしながら、微小脈管構造ならびに大腿および臍帯の静脈から成長したヒト内皮細胞からシグナルはえられた。したがって、少なくともFLT4発現の特異性のいくらかは細胞培養中で維持されていた。
【0028】
成人の組織のインサイチュ・ハイブリダイゼーション分析は、発達中のマウス胚においてみられたFLT4のリンパ系への限定を確認した。FLT4の発現はリンパ内皮およびヒトリンパ節のリンパ洞においてみられた。興味深いことに、立方体様内皮(cuboidal endothelium)を有し、白血球のリンパ節への取り引き(trafficking)における機能を示すいくらかのHEVもまたFLT4陽性であった。さらに、パラレル・ハイブリダイゼーション(parallel hybridization)分析により、正常なリンパ節と比較して、転移性のこれらの構造においてはFLT4のmRNAレベルが上ったことが示された。またFLT4は、結合組織基質(connective tissue stroma)および成長している、内皮に並んだリンパチャネル(endothelial-lined lymphatic channels)からなる良性腫瘍であるリンパ管腫において非常に目立った。FLT4のmRNAは、前記腫瘍のリンパ内皮に限定され、前記腫瘍の動脈、静脈および毛細管には存在しなかった。ヒトの肺において、本発明者らは、この組織における唯一のFLT4に陽性な管であるリンパ構造を同定することができた。
【0029】
前記の結果は、FLT4は成人の組織におけるリンパ管およびいくらかの高内皮性小静脈に対する新規なマーカーであることを示唆している。また前記の結果は、リンパ管の静脈起源(venous origin)についての理論を支持する。成長因子の受容体としてのFLT4は前記の管の分化および機能に関係しているかもしれない。
【0030】
本発明に係るFLT4結合化合物(FLT4−binding compounds)を組合わせた結果は、とくに可視化することができる放射性の、電子的に密集した(electron-dense)またはその他のリポーター物質につないだ本発明の抗体を使用することによって、リンパ内皮を選択的に標識することができる。受容体FLT4の内在化を誘導するモノクローナル抗体を含む物質をリンパ系に注射し、それによってあらかじめ定義した(predefined)分子をリンパ内皮に運ぶことが可能であろう。また、前記の本発明に係るFLT4結合化合物を高内皮性小静脈、とくに受容体FLT4を高いレベルで発現する活性化HEVの検出に使用することが可能であろう。本発明者らが知る限り、リンパ内皮に対する特異的なマーカーは現在入手できない。
【0031】
以下の実施例は本発明を説明するためのみに与えられるものであり、いかなるばあいも本発明の範囲を限定するものではない。
【実施例】
【0032】
実施例1
マウスFLT4のcDNAプローブのクローニング
129SVマウス由来のイフィックス(登録商標)ツー ゲノムライブラリー(iFIX(登録商標) II genomic library)(ストラタジーン(Stratagene))の約106のプラークを、細胞外ドメインを含むS2.5ヒト受容体FLT4のcDNA断片を用いてスクリーニングした(3)。2.5kbのBam HI断片を陽性のプラークからサブクローニングし、両方の末端から配列決定を行なった。前記サブクローニング物から、ポリメラーゼ連鎖反応を使用して増幅し、マウスFLT4のcDNA配列のヌクレオチド1745〜2049を含むエクソン断片をpBluescript KSII+/−ベクター(ストラタジーン)へクローニングした(9)。
【0033】
ヌクレオチド1〜192を含む第2の断片も同様にクローニングした。
【0034】
実施例2
マウス組織におけるFLT4のmRNAの分析
コムチュンスキー(Chomczynski)らにしたがい、発達中の胚(交尾後8〜18日のマウスおよび1日齢のマウス)から全RNAを単離した(11)。交尾後8日の胚由来のサンプルは胎盤も含んでいた。
【0035】
RNアーゼプロテクション分析のために、RNAプローブを、アンチセンス配向(antisense orientation)のために[32P]−UTPおよびT7ポリメラーゼを使用して、実施例1にしたがってえた直線状FLT4プラスミドから生成した。使用したβ−アクチンプローブは公開されているマウスβ−アクチン配列のヌクレオチド1188〜1279に対応する(12)。6%ポリアクリルアミド/7M尿素ゲル中で精製したのち、標識した転写物を30μgの全RNAと52℃で一晩ハイブリダイズさせた。ハイブリダイズしなかったRNAを、RNアーゼA(10U/ml)およびT1(1μg/ml)を用い37℃、pH7.5で1時間かけて消化した。RNアーゼを、プロテイナーゼKを用いた37℃での15分間の消化により不活性化し、サンプルを6%ポリアクリルアミド/7M尿素ゲル中で分析した。
【0036】
本実験において分析したFLT4の発現パターンにより、肺、肝臓、心臓、腎臓、骨格筋および脾臓からは非常に弱いmRNAシグナルがえられたが、精巣および脳には明白に特異的なシグナルはないということが示された(図1A)。RNアーゼプロテクション法による、マウス発生の異なる時期(phase)に採取した一連のRNAの分析によって、FLT4のmRNAは、交尾後8日から新生マウスまで終始シグナル強度における大きな変動なしに発現することが示された(図1B)。
【0037】
実施例3
マウス胚におけるFLT4に対するインサイチュ(in situ)・ハイブリダイゼーション
FLT4転写物を細胞および組織に首尾よくもどすために、交尾後7.5日および8.5日のマウス胚の切片を、標識したFLT4のRNAとハイブリダイズさせた。マウス胚はCBAおよびNMRIマウスの交配に由来した。妊娠したマウスを頸部の脱臼(cervical dislocation)により屠殺し、胚をただちに凍結するかまたはリン酸緩衝液を経て4%パラホルムアルデヒド中へ移した。胚および単離したマウスの器官を4℃で18時間かけて固定し、脱水し、パラフィン中に包埋し、ついで6μm切片に切出した。
【0038】
192ヌクレオチドおよび349ヌクレオチドのRNAプローブ(アンチセンスおよびセンス)(実施例1参照)を、[35S]−UTPを使用して、直線状プラスミドから生成した。切片のインサイチュ・ハイブリダイゼーションを、ウィルキンソン(Wilkinson)らの文献(13、14)にしたがい、以下に示す改変、1)パラフィンワックス中への包埋前に、トルエンの代わりにキシレンを使用し、2)6μm切片を切出し、2%の3−トリエトキシシリルプロピルアミンを用いて前処理したガラススライド表面上のジエチルピロ炭酸塩で処理した水の層に置き、3)プローブのアルカリ加水分解を省略し、4)ハイ・ストリンジェンシー・ウオッシュ(high stringency wash)を、30mM DTTおよび1×SSCを含む溶液中、65℃で80分間かけて行なうという改変を加えて行なった。切片をNTB−2エマルジョン(コダック(Kodak))でおおい、4℃で貯蔵した。スライドを14日間さらし、現像し(developed)、ヘマトキシリンを用いて染色した。センス鎖およびRNアーゼAで処理した切片を用いた対照のハイブリダイゼーションはバックグラウンドをこえる特異的なシグナルを出さなかった。
【0039】
図2Aおよび2Bに示したように、FLT4のmRNAは交尾後7.5日のマウス胚においては発現しなかったが、発生8.5日の後主静脈(cv)においては明るいシグナルが検出された。対照的に、発達中の心臓(データは示さず)および胸部大動脈(da)はFLT4陰性であった。胚外の組織において、FLT4は尿膜(パネルBにおけるal)においては顕著に発現したが、発達中の卵黄包の血島は陰性であった(データは示さず)。一方、頭部の間葉の血管芽細胞(ab)は強いFLT4陽性であった(C)。発達中の胎盤において、FLT4シグナルは、末梢の洞様毛細血管静脈(peripheral sinusoidal veins)において最初に見られた(データは示さず)。交尾後9.5日の胎盤において、静脈の裂孔(venous lacunae)の内皮(Dにおけるvl)およびレイチャート膜(Reichert´s membrane)に部分的に融合した巨細胞(データは示さず)はFLT4のmRNAを発現した。
【0040】
したがって、FLT4の発現は、もっとも初期の内皮細胞前駆体である血管芽細胞において非常に顕著であるが、交尾後8.5日のマウス胚の特定の管のみに限定されるように思われる。受容体Tieは発達中のマウス胚のすべての内皮細胞において発現し、したがって前記細胞に対するマーカーを提供することが知られている。とくに、Tieプローブとは対照的に、FLT4プローブは、交尾後11.5日のマウス胚の動脈性内皮、たとえば発達中の胸部大動脈(図2のE、Fにおけるda)または頸動脈(データは示さず)の内皮細胞とハイブリダイズしたとしても非常に弱かった。その代わりに、FLT4シグナルは発達中の静脈において非常に顕著であった。たとえば、FLT4シグナルは発達中の後腎を囲む静脈(Eにおけるv、sv)において検出され、Tieプローブは後腎内の毛細管(c)を顕著に認識した(F)。
【0041】
図3から理解されうるように、FLT4のmRNAは交尾後12.5日のマウス胚のいくつかの領域に分布し、とくに腋窩領域(ax)の拡張された管において顕著である。類似したFLT4陽性の管構造が、頸静脈域における正中矢状切断図において見られた(データは示さず)。FLT4を発現している管の叢様パターンは、眼窩周辺領域(po)および発生中の椎骨の周囲(vb)において見られた。また、発達中の皮膚のちょうど下のFLT4陽性脈管網(FLT4−positive vascular network)が明らかになった(sc)。より弱い毛細管シグナルが発達中の脳(b)を含むいくつかの領域からえられた。FLT4のmRNAは、尾の領域と同様、頸の領域の小管、発達中の鼻の小管および発達中の舌の基底部(base)においても検出することができた(データは示さず)。そのうえ、肝臓(li)は斑点様のパターンでFLT4のmRNAに対して強い陽性であった。
【0042】
さらなる発達のあいだ、FLT4のRNAは胚の特定の管にさらに限定されるようになったと思われる。交尾後14.5日のマウス胚はこの限定された発現パターンを精密に示している(図4A、B)。図4の正中矢状切断図において、最も顕著なFLT4のシグナルは、前記切断図の前方部分における発達中の脊柱に沿って見られる。前記シグナルは、発達のこの時点において形成された最大のリンパ管である胸管(dt)の内皮細胞に由来していると思われた。対照的に、胸部大動脈(da)および下大静脈(vc)は陰性であった。また腸間膜領域の拡張された管もFLT4に対して強い陽性であった。さらに、交尾後12.5日のマウス胚において、頸(ne)の領域と同様、眼窩周辺(po)、下顎(lj)における解剖学的な境界線に沿った管の網(vessel networks)はFLT4陽性の内皮を含んでいた。類似した構造は心膜空間(pericardial space)および皮下(sc)組織のいたる所に存在した。とくに、FLT4陰性の管とは対照的に、すべてのFLT4陽性の管は、それらの管腔(lumen)に血液細胞を欠いていた。前記発現パターンは、発達のこの時点においてリンパ管の内皮にFLT4が限定されることを示唆している。本発明者らがFLT4発現を見たさらなる部位は、発達中の骨髄(bm)の洞様毛細血管にあった。
【0043】
FLT4プローブとハイブリダイズした交尾後16.5日の胚の上部胸郭(upper thorax)の横径切断写真を図4のパネルCおよびDに示す。Cに示される切片を、ヘマトキシリン−エオシンを用いて染色し、前記区域における異なる型の管を可視化した。前記区域は頸の動脈(ca)、腕頭の動脈(ba)、大静脈(vc)およびサイズが小さく筋肉および結合組織の周囲には存在しない胸管(矢印)を含んでいる。胸管領域の拡大図をパネルDに示し、ここではFLT4のオートラジオグラフィーの粒子を見ることができる。周辺の小管(v)と同様、胸管の内皮細胞はFLT4プローブとハイブリダイズする。
【0044】
実施例4
培養した内皮細胞におけるFLT4のmRNAの分析
実施例3に記載したインサイチュ・ハイブリダイゼーションの結果により、FLT4は静脈の内皮細胞ならびに後にはリンパ管およびいくらかの静脈の内皮細胞では発現するが、動脈の内皮では発現しないことが示された。前記のような調節がインビトロで維持されるかどうかを見るために、本発明者らはノーザンブロッティングおよびハイブリダイゼーション分析を使用して培養した内皮細胞を研究した。
【0045】
ヒトの大動脈、大腿静脈、臍帯静脈および包皮微小管由来の内皮細胞を単離し、培養して、ファン ヒンスバーグ(Van Hinsberg)によりすでに記載されたように特徴づけを行なった(15、16)。前記内皮細胞を、ポリアデニル化RNAを単離するために、5〜8回継代(分裂比(split ratio)1:3)したのち、コンフルエントな密度(confluent density)で使用した。
【0046】
内皮細胞系EA・hy926、BCEおよびLEIIはFLT4を発現しなかった(データは示さず)。しかしながら、培養したヒトの微小管、静脈および臍帯の静脈の内皮細胞は、FLT4に特異的な5.8kbおよび4.5kbのmRNAに関して陽性であり、一方大動脈の内皮細胞は陰性であった(図5)。対照的に、別の内皮の受容体チロシンキナーゼの遺伝子であるTieは、研究したすべての型の内皮細胞において4.4kbのmRNAとして発現した。
【0047】
実施例5
成人組織におけるFLT4のmRNA
実施例3においてえられた結果により、FLT4のmRNAは、発達中、リンパ管の内皮に大規模に制限されるようになることが示された。ヒトにおけるこの知見の潜在的な重要性のため、本発明者らは、ヒトFLT4プローブを使用して成人組織におけるFLT4も研究した。使用したヒトFLT4プローブは、cDNAの塩基対1〜595を含むEcoRI−SphI断片であった(3)。フォンビルブラント因子プローブは塩基対1〜2334を含むEcoRI−HindIII断片であった(17)。
【0048】
本発明者らは、組織病理学的診断に送るための、慣用の操作で固定された材料を使用した。正常な肺組織を類表皮癌におかされた左の肺の下葉の切除によりえた。腸間膜および腸間膜のリンパ節を、結腸の腺癌を有する患者からえた。異常な大きさであったので、睡液腺に隣接した正常なリンパ節を摘出した。2人の患者由来の扁桃およびその2つの垂(appendixes)は診断上の変化はなかった。2つのリンパ管筋腫瘍(lymphangiomyomas)および3つの嚢胞性リンパ管腫を研究し類似した結果をえた。
【0049】
ヒト組織について、ヒト組織は組織病理学的診断のために10%ホルマリンを用いて固定した慣用サンプルであり、正常なインサイチュでの手順は単にバックグラウンドのみを与えるが、プロテイナーゼKの代わりのマイクロ波処理は特異的ハイブリダイゼーションを可能にした(18、19)。
【0050】
腸間膜において、肺および垂のリンパ内皮(lv)はFLT4シグナルを出すが、静脈(v)、動脈(a)および毛細管(c)は陰性であった(図6A〜Dおよびデータは示さず)。FLT4がHEVで発現するかどうかを研究するために、扁桃を研究した。実際に扁桃について、FLT4に特異的なオートラジオグラフィーの粒子が、いくらかのHEVにおいては検出された(E、F)。
【0051】
実施例6
正常および転移性リンパ節ならびにリンパ管腫におけるFLT4のmRNAの分析
ヒト腸間膜リンパ節の一部(実施例5参照)をFLT4の発現について分析した。結果を図7に示す。
【0052】
FLT4はリンパ洞(ls)ならびに輸入(afferent)および輸出(efferent)のリンパ管において発現する(データは示さず)。同じパターンが腺癌の転移を含むリンパ節において見られる(C、D)。正常および転移性リンパ節の両方におけるいくつかのHEVも陽性であった。パネルEにおいて、FLT4の発現は嚢胞性リンパ管腫において見られる(Fにおけるヘマトキシリン−エオシンで染色した切片と比較した)。とくに、FLT4のリンパ内皮に対する特異性は、すべての血管におけるフォンビルブラント因子に対するインサイチュでのシグナルとの比較により明らかである(F)。
【0053】
実施例7
胎児の内皮細胞におけるFLT4の局在化
短型(short form)のカルボキシ末端側の40のアミノ酸をコードするFLT4のcDNA断片を、657bpのEcoRI断片として、読み取り枠(frame)中にグルタチオン−S−トランスフェラーゼをコードする領域を有するpGEX−1IT細菌性発現ベクター(ファルマシア(Pharmacia))にクローニングした。えられたGST−FLT4融合タンパク質を大腸菌中で産生し、グルタチオン−セファロース 4B カラムを使用したアフィニティークロマトグラフィーにより精製した。精製したタンパク質を凍結乾燥し、PBS中に溶解し、フロイントアジュバントと混合し、ウサギの免疫に使用した。抗血清は4回のブースター免疫ののちのものを使用した。
【0054】
17週齢および20週齢のヒト胎児由来の組織を、プロスタグランジンを用いて誘導した合法の流産よりえた。研究はヘルシンキ大学中央病院の倫理委員会の承認を受けた。存胎齢は胎児の脚の長さより推定した。胎児の組織をティシュー−テック(Tissue-Tek)(マイルス(Miles))中に包埋し、直ちに凍結し、−70℃で貯蔵した。
【0055】
抗FLT4抗血清をGST−セファロースカラムに交差吸収(cross-absorbed)させて抗GST抗体を除去し、ついでGST−FLT4アフィニティークロマトグラフィーにより精製した。厚さ6μmのいくつかの組織のクリオスタット切片(cryostat sections)をアセトンを用いて固定し、メタノール中の0.3%H2O2で30分間処理し内因性ペルオキシダーゼ活性をブロックした。洗浄後、切片を5%正常ブタ血清とともにインキュベートした。ついで切片をFLT4に対する抗体とともにインキュベートし、洗浄して、結合した抗体をペルオキシダーゼが結合したブタ抗ウサギIgGを用いて検出し、続いて基質として0.2%の3,3−ジアミノベンジジン(アマシャム(Amersham))を使用してペルオキシダーゼ活性に対する染色を行なった。切片をマイヤーのヘマトキシリン(Meyer´s hematoxylin)中で対比染色した。
【0056】
ヒト胎児の腸間膜の抗FLT4イムノペルオキシダーゼ染色は、いくつかの管の内皮中のFLT4タンパク質を示したが(図8A)、抗原でブロックした抗FLT4抗体(B)およびあらかじめ免疫した血清(C)は陰性であった。比較のために、図8Dは、脈管の内皮細胞に特異的な第VIII因子関連抗原に対する抗血清を使用した染色の結果を示す。
【0057】
実施例8
FLT4に対するモノクローナル抗体の産生
融合I:
4月齢のBalb/c雄マウスを、フロイント完全アジュバントを用いて乳化した濃縮培地(150μg/マウス)中の組換えにより産生したFLT4タンパク質(実施例7参照)の腹腔内注射により免疫した。3〜4週間おきに150μgのブースター注射をし、さらに3週間の間隔をおいたのち最終ブースター(PBS中の10μgのFLT4の腹腔内投与)を行なった。最終ブースター投与から4日後、マウスを屠殺し、マウス脾リンパ様細胞(lymphoid cells)をSP2/0プラスマ細胞腫細胞とそれぞれ2:1の割合で融合させた。
【0058】
融合細胞を、20%ウシ胎児血清およびHAT補足物(HAT supplement)(ヒポキサンチン−アミノプテリン−チミジン、ギブコ(GIBCO)、043−01060H、50倍希釈)を含むイーエックス−セル320(EX-Cell 320)培地(セララブ(SERALAB))を添加した96ウェル培養プレート(ヌンク(NUNC))中からえた。細胞を5%CO2雰囲気下、+37℃で培養した。10日後、HAT補足培地をHT補足細胞培養培地(ギブコ、043−01065H、50倍希釈)に交換した。HT培地はアミノプテリンを含まないことを除くとHAT培地と同一である。
【0059】
3週間後、特異的な抗体の産生を、実施例10に記載されている抗原特異的免疫蛍光分析法(antigen-specific immunofluorometric assay)であるIFMAにより決定した。マスタークローンを、スタスツェウスキー(Staszewski)ら、エール・ジャーナル・オブ・バイオロジー・アンド・メディシン(Yale Journal of Biology and Medicine)、57巻、865〜868頁(1984年)に記載された限界希釈(limited dilutions)によりクローニングした。陽性クローンを24ウェル組織培養プレート(ヌンク)上で増やし、再クローニングし、前記と同様の方法により再試験を行なった。陽性のクローンを蛍光活性化細胞選別法(FACS)により試験した。
【0060】
安定したクローンは、おそらくクラスIgAに属するIgを産生した1つのクローンを除いて、IgG1クラスに属する免疫グロブリンを分泌した。モノクローナル抗体のサブクラスを、IFMAにおいて、マウスのサブクラスに対するラットのモノクローナル抗体をビオチン接合物(biotin conjugate)(セロテック(SEROTEC))として使用して決定した。
【0061】
腹水(ascites fluid)中にモノクローナル抗体を産生するためにBalb/cマウスを使用した。プリスタン(pristane)(2,6,10,14−テトラメチルペンタデカン、98%、アルドリッチ−ケミエ(ALDRICH-CHEMIE)、デー(D)−7924、シュタインハイム(Steinheim)、カタログ番号 T2,280−2)でマウスを前処理したのち、前記ハイブリドーマを前記マウスに腹腔内注射した。ハイブリドーマ細胞注射の約2週間前に、プリスタン0.5mlを(静脈内)注射した。注射した細胞量はマウス1匹あたり約7.5〜9×106であった。ハイブリドーマ注射後10〜14日後、腹水を集めた。
【0062】
融合II:
2月齢のBalb/cマウス(雌)を、フロイント完全アジュバントを用いて乳化した組換えにより産生したFLT4タンパク質(実施例7参照)(20μg/マウス)の腹腔内注射により免疫した。3〜4週間おきに20μgのブースター注射をし、さらに3週間の間隔をおいたのち最終ブースター(PBS中に10μgのFLT4、静脈内投与)を行なった。最終ブースター投与から4日後、マウスを屠殺し、マウス脾リンパ様細胞をSP2/0プラスマ細胞腫細胞とそれぞれ2:1の割合で融合させた。
【0063】
融合細胞を、20%ウシ胎児血清およびHAT補足物(ヒポキサンチン−アミノプテリン−チミジン、ギブコ ビーアールエル(GIBCO BRL)21060−017、1:50に希釈)を含むオプティエムイーエム(OptiMEM)1(グルタマックス(Glutamax)1を有する、51985−026、ギブコ ビーアールエル)を添加した96ウェル培養プレート(ファルコン(FALCON))中からえた。細胞を5%CO2雰囲気下、+37℃で培養した。10日後、HAT補足培地をHT補足細胞培養培地(ギブコ ビーアールエル、41065−012、1:50倍に希釈)に交換した。HT培地はアミノプテリンを含まないことを除くとHAT培地と同一である。
【0064】
3週間後、特異的な抗体の産生を、実施例9に記載されている抗原特異的免疫蛍光分析法(IFMA)により決定した。マスタークローンを、スタスツェウスキーら、エール・ジャーナル・オブ・バイオロジー・アンド・メディシン、57巻、865〜868頁(1984年)に記載された限界希釈によりクローニングした。陽性クローンを24ウェル組織培養プレート(ファルコン)上で増やし、再クローニングし、前記と同様の方法により再試験を行なった。陽性のクローンを蛍光活性化細胞選別法(FACS)により試験した。
【0065】
2E11および6B2クローンは、IgG1クラスに属する免疫グロブリンを分泌し、2B12クローンはサブクラスIgMに属するIgを産生した。マウスのサブクラスのIgG1を、IFMAにおいて、マウスのサブクラスの重鎖に対するラットモノクローナル抗体をビオチン接合物(セロテック)として使用して決定し、マウスのサブクラスのIgMを、マウスモノクローナル抗体アイソタイピング・キット(Mouse Monoclonal Antibody Isotyping Kit)(ディップスティック フォーマット(Dipstick Format))(19663−012、ライフ テクノロジーズ社(Life Technologies Inc.))を用いて決定した。
【0066】
実施例9
FLT4に対するモノクローナル抗体の特異性
融合Iの抗体:
実施例7に記載したFLT4の細胞外ドメインを、アナリティカル・バイオケミストリー(Anal.Biochem.)、176巻(2)、319〜325頁(1989年)に記載されたムッカラ(Mukkala)らの方法に、以下に示す改変:250倍をこえるモル量のイソチオシアネートDTTA−Eu(N1キレート(N1 chelate)、ワラック(Wallac)、フィンランド共和国)をFLT4溶液(PBS中に0.5mg/ml)に添加し、pHを0.5mol/L炭酸ナトリウム緩衝液、pH9.8を添加することにより約9に調整したことを加えて標識化した。標識化を+4℃で一晩行なった。結合しなかった標識を、溶離液としてのTSA緩衝液(0.15mol/lのNaClを含む50mmol/L Tris−HCl、pH7.8)とともにPD−10(ファルマシア、スウェーデン王国)を用いて除去した。
【0067】
精製後、標識されたFLT4に1mg/mlウシ血清アルブミン(BSA)を添加し、標識物を+4℃で貯蔵した。既知のEuCl3標準の蛍光に対する蛍光の割合を測定することにより決定したFLT4の1分子あたりに取り込まれたユウロピウムイオン数は1.9であった(ヘミッラ(Hemmilae)ら、アナリティカル・バイオケミストリー、137巻、335〜343頁(1984年))。
【0068】
実施例8において産生した抗体を、ウサギ抗マウスIg(Z259、ダコパッツ(DAKOPATTS))で被覆したマイクロタイトレーション・ストリップ・ウェル(microtitration strip wells)(ヌンク(NUNC)、ポリソーブ(polysorb))を用いたサンドイッチ型免疫蛍光分析法を使用してスクリーニングした。あらかじめ被覆したウェルをデルフィア(DELFIA)洗浄溶液を用いたプレートウォッシュ(Platewash)1296−024(ワラック)により一度洗浄した。デルフィア分析緩衝液を、細胞培養上清および予備的スクリーニング分析(preliminary screening assays)における正の対照として使用した脾摘出したマウスの血清(1:1000〜1:100000の希釈)の希釈緩衝液として使用した。
【0069】
+4℃で一晩(または室温で2時間)のインキュベーションを、プレートシェイク振とう機(Plateshake shaker)(1296−001、ワラック)上での5分間の振とうにより開始し、続いて前記洗浄溶液を用いて4回洗浄した。
【0070】
ユウロピウムで標識したFLT4を、分析緩衝液100μl中、1:500の希釈で添加した。プレートシェイク振とう機上で5分間および室温(RT)で1時間インキュベートしたのち、前記と同様にストリップを洗浄した。
【0071】
エンハンスメント溶液(Enhancement solution)(デルフィア)を200μl/ウェルの濃度で添加した。ついで、プレートを、プレートシェイク振とう機上で5分間振とうし、アルクス(ARCUS)−1230(ワラック)により蛍光強度を10〜15分間測定した(レブグレン(Loevgren)ら、コリンス ダブリュー ピー(Collins W.P.)(編)、オルターナティブ・イムノアッセイズ(Alternative Immunoassays)、ジョン・ウィレー・アンド・ソンズ社(John Wiley & Sons Ltd.)、203〜216頁(1985年))。
【0072】
えられたFLT4に対するモノクローナル抗体および対応するFACSの結果を表1に要約する。
【0073】
【表1】
【0074】
抗FLT4の9D9F9と名付けられた1つのクローンは、IFMAにより免疫グロブリンクラスIgG1に属することが決定されたモノクローナル抗体を安定して分泌した。ハイブリドーマ9d9f9は、1995年3月23日に、ドイツ連邦共和国、3300 ブラウンシュヴァイク(Braunschweig)、マシェロデル ヴェク 1ベー(Mascheroder Weg 1b)のドイツ微生物および細胞培養コレクション(German Collection of Microorganisms and Cell Cultures)、ヒトおよび動物細胞ならびにウイルス部門(Department of Human and Animal Cell Cultures and Viruses)に寄託され、受託番号ACC2210を与えられている。
【0075】
融合IIの抗体:
実施例7に記載したFLT4の細胞外ドメインを、アナリティカル・バイオケミストリー、176巻(2)、319〜325頁(1989年)に記載されたムッカラらの方法に、以下に示す改変:250倍をこえるモル量のイソチオシアネートDTTA−Eu(N1キレート、ワラック、フィンランド共和国)をFLT4溶液(PBS中に0.5mg/ml)に添加し、pHを0.5mol/L炭酸ナトリウム緩衝液、pH9.8を添加することにより約9に調整したことを加えて標識化した。標識化を+4℃で一晩行なった。結合しなかった標識は、溶離液としてのTSA緩衝液(0.15mol/LのNaClを含む50mmol/L Tris−HCl、pH7.8)とともにPD−10(ファルマシア)を用いて除去した。
【0076】
精製後、標識されたFLT4に1mg/mlウシ血清アルブミン(BSA)を添加し、標識物を+4℃で貯蔵した。既知のEuCl3標準の蛍光に対する蛍光の割合を測定することにより決定したFLT4の1分子あたりに取り込まれたユウロピウムイオン数は1.9であった(ヘミッリ(Hemmil)ら、アナリティカル・バイオケミストリー、137巻、335〜343頁(1984年))。
【0077】
実施例8において産生した抗体を、ウサギ抗マウスIg(Z259、ダコ(DAKO))で被覆したマイクロタイトレーション・ウェル(ヌンク、ポリソーブ)を用いたFLT4特異的IFMAを使用してスクリーニングした。あらかじめ被覆したウェルをデルフィア・プレート・ウォッシュにより洗浄溶液(ワラック)を用いて一度洗浄した。
【0078】
デルフィア分析緩衝液を、細胞培養上清(予備的スクリーニングにおいて1:2の希釈)および正の対照として使用した脾摘出したマウスの血清(1:1000〜1:100000の希釈)の希釈緩衝液として使用した。標準として、精製した抗FLT4の9D9F9(マウスのサブクラスIgG1)を1.0ng/ml〜250ng/mlのあいだの濃度で使用した。サンプルをはじめに室温下、プレート・シェイク(ワラック)上で5分間振とうし、ついで+4℃で約18時間インキュベートした。はじめにフレーム(frames)を4回洗浄し、ついでユウロピウムで標識したFLT4(分析緩衝液100μl中、1:2000)を添加し、最終的に前記フレームを室温下で1時間インキュベートした。前記のように洗浄したのち、エンハンスメント溶液(200μl/ウェル、ワラック)を添加し、プレートをプレート・シェイク上で5分間振とうした。蛍光強度をアルクス−1230(ワラック)により測定した。
【0079】
えられたFLT4に対するモノクローナル抗体および対応する結果を表2に要約する。
【0080】
抗FLT4抗体の定量の標準曲線を、親和性により精製した抗FLT4である9D9F9を使用して作成した。直線の範囲は1.0ng/ml〜250ng/mlに達した。
【0081】
細胞表面上に全長のFLT4を発現するpLTRFLT4構築物を同時にトランスフェクト(cotransfected)したNIH3T3細胞の細胞溶解物を、6.5%SDS−PAGE中で電気泳動し、タンパク質をニトロセルロース硝酸塩(nitrocellulose nitrate)膜(0.45μm、シュレイチャー アンド シュエル(SCHLEICHER & SCHUELL))上に移し、モノクローナル抗体細胞培養上清(1:10に希釈、50mmol/L TRIS−4%メタノールを含む40mmol/Lグリシン緩衝液、0.04%SDS)を用いてイムノブロットした。モノクローナル抗体の特異性をエイチアールピー(HRP)接合ウサギ抗マウスIg(P161、ダコ、150mmol/L食塩水および5%ミルク粉末を含む20mmol/L TRIS緩衝液、pH7.5中、1:1000に希釈)およびイーシーエル(ECL)(エンハンスド ケミルミネッセンス(Enhanced chemiluminescence)、アマシャム)とのインキュベーションを使用して検出した。
【0082】
【表2】
【0083】
前記より明らかなように、本発明に係る抗体は、リンパ管、リンパ内皮細胞、高内皮性小静脈、リンパ管腫、転移性リンパ節およびその他のリンパ系の疾患状態の診断ならびに同定、転移の蔓延(metastatic spread)の検出およびモニタリングに、リンパ管および高内皮性小静脈の内皮細胞の刺激ならびに阻害に、内皮細胞への分子の選択的な導入に、そしてリンパ管およびリンパ管の疾患状態の映像化において有用である。本明細書において請求した主題の他の用途は当業者にとって明らかである。
【0084】
[参考文献]
1. Sabin,F.R.1909.リンパ系全体としての形態を考慮した、ヒト胚のリンパ系。Am.J.Anat.9:43.
2. van der Putte,S.C.J.1975.ヒトにおけるリンパ系の発達。Adv.Anat.Embryol.Cell Biol.51:3.
3. Pajusola,K.,O.Aprelikova,J.Korhonen,A.Kaipainen,L.Pertovaara,R.Alitalo,and K.Alitalo.1992.受容体チロシンキナーゼであるFLT4は7つの免疫グロブリン様ループを含み、多数のヒト組織および細胞系で発現する。Cancer Res.52:5738.
4. Kaipainen,A.,J.Korhonen,K.Pajusola,O.Aprelikova,M.G.Persico,B.I.Terman,and K.Alitalo.1993.受容体チロシンキナーゼである関連したFLT4、FLT1およびKDRは、ヒト胎児の内皮細胞において異なる発現パターンを示す。J.Exp.Med.178:2077.
5. Galland,F.,A.Karamysheva,M.-J.Pebusque,J.-P.Borg,R.Rottapel,P.Dubreuil,O.Rosnet,and D.Birnbaum.1993.FLT4遺伝子は、脈管の内皮成長因子受容体に関係するトランスメンブレン型チロシンキナーゼをコードする。Oncogene.8:1233.
6. Millauer,B.,S.Wizigmann-Voos,H.Schnuerch,R.Martinez,N.-P.H.Moller,W.Risau,and A.Ullrich.1993.高親和性のVEGFの結合および発達上の発現は、脈管形成および血管形成の主要なレギュレーターとしてのFlk−1を示唆する。Cell.72:835.
7. Yamaguchi,T.P.,D.Dumont,R.A.Conlon,M.L.Breitman,and J.Rossant.1993.flt関連チロシンキナーゼであるflk−1は内皮細胞前駆体に対する早期のマーカーである。Development.118:489.
8. Peters,K.G.,C.De Vries,and L.T.Williams.1993.脈管内皮の成長因子の受容体の胚形成および組織修復中の発現は、内皮の分化および血管の成長における役割を示唆する。Proc.Natl.Acad.Sci.USA 90:8915.
9. Finnerty,H.,K.Kelleher,G.Morris E.,K.Bean,D.Merberg,R.Kritz,J.Morris C.,H.Sookdeo,K.J.Turner,and C.R.Wood 1993.マウスのFLTおよびFLT4の分子クローニング。Oncogene.8:2293.
10. Korhonen,J.,A.Polvi,J.Partanen,and K.Alitalo.1993.マウスの受容体チロシンキナーゼであるtie遺伝子:胚の血管形成中の発現。Oncogene.8:395.
11. Chomczynski,P.,and N.Sacchi.1987.酸性グアニジンチオシアネート−フェノール−クロロホルム抽出によるRNA単離の単一工程方法。Anal.Biochem.162:156.
12. Tokunaga,K.,H.Taniguchi,K.Yoda,M.Shimizu,and S.Sakiyama.1986.マウスの細胞骨格βに作用するmRNAに対する全長のcDNAのヌクレオチド配列。Nucleic.Acid.Res.14:2829.
13. Wilkinson,D.G.,J.A.Bailes,J.E.Champion,andA.P.McMahon.1987.交尾後(post coitum)8〜10日のマウスの発達の分子解析により胚のグロビン発現のみに変化が検出される。Development.99:493.
14. Wilkinson,D.G.,J.A.Bailes,and A.P.McMahon.1987.プロトオンコジーンであるint−1の発現は、発達中のマウスの胚の特定の神経細胞に限定される。Cell.50:79.
15. Van Hinsberg,V.W.M.,D.Binnema,M.A.Scheffer,E.D.Sprengers,T.Kooistra,and D.C.Rijken.1987.成人のヒト血管由来の連続的に増殖した内皮細胞によるプラスミノーゲン活性化因子および阻害因子の産生。Arteriosclerosis.7:389.
16. Van Hinsberg,V.W.M.,M.A.Scheffer,and T.Kooistra.1987.ヒト包皮微小管の内皮細胞によるプラスミノーゲン活性化因子およびPA阻害因子−1産生に対するトロンビンの作用。Thromb.Haemostas.57:148.
17. Bonthron,D.T.,E.C.Orr,L.M.Mitsock,D.Ginsberg,R.I.Handin,and S.H.Orkin.1986.プレ−プロ−フォンビルブラント因子のcDNAのヌクレオチド配列。Nucleic Acids Res.141:7125.
18. Shi,E.,M.Kan,J.Xu,and W.L.McKeehan.1991.(繊維芽細胞)成長因子結合型の16キロダルトンのヘパリンのあるものは、受容体に依存した内在化ののち、安定な40キロダルトンの複合体として生じる。J.Biol.Chem.266:5774.
19. Catoretti,G.,M.H.G.Becker,G.Key,M.Duchrow,C.Schlueter,J.Galle,and J.Gerdest.1992.Ki−67抗原の組換え部分に対するモノクローナル抗体(MIB1およびMIB3)は、マイクロ波で処理したホルマリン固定化パラフィン切片中の増殖している細胞を検出した。J.of Pathol.168:357.
【0085】
[配列の表示]
(1)一般情報
(i)出願人:
(A)名称:ヘルシンキ ユニバシティ ライセンシング リミテッド オケサ ユキチュア
(A)名称:オリオン コーポレーション
(ii)発明の名称:受容体チロシンキナーゼであるFLT4ならびに診断および治療におけるその用途
(iii)配列の数:1
(iv)コンピュータ読取フォーム:
(A)ソフトウェア:PatentIn Release 1.0、バージョン1.25(ヨーロッパ特許局)
(v)優先権データ:
(A)出願番号:アメリカ合衆国、08/257754
(B)出願日:1994年6月9日
(2)配列番号:1
【特許請求の範囲】
【請求項1】
受容体チロシンキナーゼFLT4の細胞外ドメインに対するモノクローナル抗体の、転移性癌、リンパ腫、リンパ管腫、炎症性疾患(慢性または急性)、感染性疾患および免疫性疾患からなる群より選ばれる疾患と関連するFLT−4が介在するリンパ血管新生を阻害するための医薬組成物の製造における使用。
【請求項2】
前記モノクローナル抗体がDSM ACC 2210として寄託されているハイブリドーマ細胞系により産生されるものである請求項1記載の使用。
【請求項1】
受容体チロシンキナーゼFLT4の細胞外ドメインに対するモノクローナル抗体の、転移性癌、リンパ腫、リンパ管腫、炎症性疾患(慢性または急性)、感染性疾患および免疫性疾患からなる群より選ばれる疾患と関連するFLT−4が介在するリンパ血管新生を阻害するための医薬組成物の製造における使用。
【請求項2】
前記モノクローナル抗体がDSM ACC 2210として寄託されているハイブリドーマ細胞系により産生されるものである請求項1記載の使用。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2012−193191(P2012−193191A)
【公開日】平成24年10月11日(2012.10.11)
【国際特許分類】
【出願番号】特願2012−138421(P2012−138421)
【出願日】平成24年6月20日(2012.6.20)
【分割の表示】特願2009−23515(P2009−23515)の分割
【原出願日】平成7年6月9日(1995.6.9)
【出願人】(508088775)ベジェニクス ピーティーワイ リミテッド (5)
【氏名又は名称原語表記】VEGENICS PTY LTD
【Fターム(参考)】
【公開日】平成24年10月11日(2012.10.11)
【国際特許分類】
【出願日】平成24年6月20日(2012.6.20)
【分割の表示】特願2009−23515(P2009−23515)の分割
【原出願日】平成7年6月9日(1995.6.9)
【出願人】(508088775)ベジェニクス ピーティーワイ リミテッド (5)
【氏名又は名称原語表記】VEGENICS PTY LTD
【Fターム(参考)】
[ Back to top ]