
受容体相互作用検出方法およびサイクリックAMPセンサータンパク質
【課題】受容体の空間的な配置を明らかにして受容体の相互作用に関する各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞において特定の受容体の相互作用を再現性良く観察することができる受容体相互作用検出方法等を提供することを課題とする。
【解決手段】本実施例において、ルシフェラーゼおよびサイクリックAMP結合タンパク質を含む細胞を作製し、作製した細胞に当該細胞外から、ルシフェリンを添加し、ルシフェリンが添加された後の細胞にセロトニンやドーパミン、フォルスコリンを添加しながら細胞の発光量を測定し、測定した発光量に基づいて細胞内におけるサイクリックAMPの濃度を解析し、解析したサイクリックAMPの濃度に基づいて、D1受容体と5HT2A受容体との相互作用を検出する。
【解決手段】本実施例において、ルシフェラーゼおよびサイクリックAMP結合タンパク質を含む細胞を作製し、作製した細胞に当該細胞外から、ルシフェリンを添加し、ルシフェリンが添加された後の細胞にセロトニンやドーパミン、フォルスコリンを添加しながら細胞の発光量を測定し、測定した発光量に基づいて細胞内におけるサイクリックAMPの濃度を解析し、解析したサイクリックAMPの濃度に基づいて、D1受容体と5HT2A受容体との相互作用を検出する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、細胞内の互いに異なる受容体同士の相互作用(例えばD1受容体と5HT2A受容体の相互作用)を、一方の受容体(例えばD1受容体)の活性(例えばサイクリックAMPの濃度)を一細胞ごとに非侵襲的に解析して検出する受容体相互作用検出方法、および互いに異なる受容体同士の相互作用を検出するためのサイクリックAMPセンサータンパク質に関するものである。
【背景技術】
【0002】
多くのホルモンや神経伝達物質は、Gタンパク質共役型受容体に働き、細胞内セカンドメッセンジャーを介して様々な細胞活動を引き起こしている。Gタンパク質共役型受容体に働く神経伝達物質は、ヒトの情動や気分、依存、認知、学習、記憶などの広範囲に亘る脳機能の調節に関わっており、なかでも、モノアミンおよびその受容体は、躁うつ病や統合失調症、気分障害、自閉症、注意欠陥/多動性障害などの精神疾患に対する治療の標的とされている。このような脳の高次機能は、脳内の伝達物質およびその受容体の時間的および空間的な濃度分布によって制御されている。したがって、特定の情動活動に関する情報伝達機構を解析するにあたって、脳内のGタンパク質共役型受容体の時間的および空間的な変化および相互作用をリアルタイムに捉えることが必要である。
【0003】
これまでに、脳内の受容体の時間的および空間的な変化および相互作用に関する研究において、情動活動に対する効果を示す薬物の存在が明らかにされ、その作用点がモノアミンのGタンパク質共役型受容体であることが解明されてきた経緯がある。モノアミンのGタンパク質共役型受容体が関与する従来のシグナル伝達の研究では、主にモデル動物を用いた行動薬理学的な手法で解析されてきた。
【0004】
例えば、Bodyらは、5HT2A受容体やD1受容体、D2受容体のサブタイプに対する選択的阻害剤を用いて、ラットの代表的なオペラント条件付けであるタイミング行動を制御する方法を開示している(非特許文献1参照)。タイミング行動に対するd−アンフェタミンの効果が、5HT2A受容体およびD1受容体に対する選択的阻害剤で阻害されることから、タイミング行動に対するd−アンフェタミンの作用には5HT2A受容体およびD1受容体が関わっていることを明らかにしている。
【0005】
また、例えば、Auclairらは、5HT2A受容体に対する選択的阻害剤を用いて、ラットにおけるd−アンフェタミン誘導性の自発運動活性を制御する方法を開示している(非特許文献2参照)。d−アンフェタミンは、脳内においてドーパミン遊離を起こさせ、ドーパミン誘導性の自発運動活性の上昇を引き起こす。5HT2A受容体に対する選択的阻害剤が、ドーパミン遊離を抑制せずにドーパミン誘導性の自発運動活性の上昇を抑制することを示しており、ドーパミン受容体と5HT2A受容体との相互作用を明らかにしている。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Body S., Cheung T.H.C., Bezzina G., Asgari K., Fone K.C.F., Glennon J.C., Bradshaw C.M., Szababi E., “Effects of d−amphatamine and DOI (2,5−dimethoxy−4−iodoamphetamine) on timing behavior: intaraction between D1 and 5−HT2A receptors”, Psycopharmacology, Vol.189, pp331−343, 2006
【非特許文献2】Auclair A., Blanc G., Glowinski J., Tassin J.−P., “Role of serotonin2A receptors in d−amphetamine−induced release of dopamine: comparison with previous data on alpha1b−adrenergic receptors”, Journal of Neurochemistry, vol.91, pp318−326, 2004
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、モデル動物を用いた行動薬理学的な手法を用いて異なるGタンパク質共役型受容体の時間的および空間的な相互作用を解析する従来の方法では、受容体の空間的な配置が不明のまま相互作用の解析を行っているため、受容体同士の相互作用を正確には示していない、という問題点があった。
【0008】
本発明は、上記問題点に鑑みてなされたもので、受容体の空間的な配置を明らかにして受容体の相互作用に関する各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞において特定の受容体の相互作用を再現性良く観察することができる受容体相互作用検出方法および受容体の相互作用を検出するためのサイクリックAMPセンサータンパク質を提供することを目的とする。
【課題を解決するための手段】
【0009】
上述した課題を解決し、目的を達成するために、本発明にかかる受容体相互作用検出方法は、細胞において第1の受容体と第2の受容体との相互作用を検出する受容体相互作用検出方法であって、前記第2の受容体の活性を示す指標となるレポータを含む前記細胞を作製する作製工程と、前記作製工程で作製した前記細胞に当該細胞外から、発光させるための所定の処置を行うと共に前記第1の受容体に結合する所定の受容体結合物質を添加する処置および添加工程と、前記処置および添加工程が行われた後の前記細胞の発光量を測定する測定工程と、前記測定工程で測定した前記発光量に基づいて前記細胞内における前記第2の受容体の活性を解析する解析工程と、前記解析工程で解析した前記第2の受容体の活性に基づいて、前記第1の受容体と前記第2の受容体との前記相互作用を検出する検出工程と、を含むことを特徴とする。ここで、レポータとは、遺伝子の発現に応じて光を発生し得る蛍光タンパク質や発光タンパク質を包含するものであり、具体的にはサイクリックAMP(adenosine monophosphate)センサータンパク質などである。また、発光タンパク質とは、具体的には生物発光タンパク質などである。また、発光させるための所定の処置とは、例えば発光基質を添加する(または予め細胞等の試料中に供給しておく)ことや励起光を照射することなどである。生物発光共鳴エネルギー転移(いわゆるBRET)による蛍光(例えばGFP、YFP等)を測定する場合には、生物発光用の基質を添加等することによって励起光の照射は不要である。
【0010】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記作製工程において、前記レポータは、サイクリックAMPセンサータンパク質であり、前記解析工程において、前記第2の受容体の活性はサイクリックAMPの濃度で解析すること、を特徴とする。
【0011】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記サイクリックAMPセンサータンパク質は、前記細胞内のサイクリックAMPと相互作用して発光するものであること、を特徴とする。
【0012】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記サイクリックAMPセンサータンパク質は、ホタルのルシフェラーゼおよびPKA(protein kinase A)のサイクリックAMP結合領域とからなる融合タンパク質であること、を特徴とする。
【0013】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記サイクリックAMP結合領域が、マウスのPKA−RIα由来であることを特徴とする。
【0014】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記サイクリックAMP結合領域が、220から250のアミノ酸残基の長さを有することを特徴とする。
【0015】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記サイクリックAMP結合領域が、少なくとも2つのサイクリックAMP結合部位を含むことを特徴とする。
【0016】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記サイクリックAMPセンサータンパク質は、前記ルシフェラーゼのN末端側断片、前記ルシフェラーゼのC末端側断片および前記サイクリックAMP結合領域を含み、前記N末端側断片と前記C末端側断片とが前記サイクリックAMPセンサータンパク質分子内で結合すると、前記ルシフェラーゼの発光酵素活性が回復することを特徴とする。
【0017】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記サイクリックAMP結合領域が、配列番号37または39に記載のアミノ酸配列を含むことを特徴とする。
【0018】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記作製工程において、前記細胞は前記第1の受容体と前記第2の受容体とを含有するものであること、を特徴とする。
【0019】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記作製工程において、前記細胞は前記第1の受容体の遺伝子と前記第2の受容体の遺伝子とが導入されたものであること、を特徴とする。
【0020】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記測定工程は、前記第2の受容体の活性に依存する発光標識の光子量を測定する光子量測定工程をさらに含むこと、を特徴とする。
【0021】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記測定工程は、前記第2の受容体の活性に依存する発光標識を撮像する撮像工程をさらに含むこと、を特徴とする。
【0022】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記撮像工程は、複数の異なる前記細胞に対して同時に実行され、前記解析工程は、前記撮像工程で撮像した複数の発光画像を前記細胞ごとに照合する照合工程をさらに含むこと、を特徴とする。
【0023】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記測定工程は繰り返し実行されること、を特徴とする。
【0024】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記第1の受容体は5HT2A受容体であり、前記第2の受容体はD1受容体であること、を特徴とする。
【0025】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記処置および添加工程において、前記受容体結合物質はセロトニンであること、を特徴とする。
【0026】
また、本発明はサイクリックAMPセンサータンパク質に関するものであり、本発明にかかるサイクリックAMPセンサータンパク質は、第1の受容体と第2の受容体との相互作用を検出するためのものであること、を特徴とする。
【0027】
また、本発明にかかるサイクリックAMPセンサータンパク質は、前記に記載のサイクリックAMPセンサータンパク質において、サイクリックAMP(adenosine monophosphate)と相互作用して発光するものであること、を特徴とする。
【0028】
また、本発明にかかるサイクリックAMPセンサータンパク質は、前記に記載のサイクリックAMPセンサータンパク質において、前記第1の受容体は5HT2A受容体であり、前記第2の受容体はD1受容体であること、を特徴とする。
【発明の効果】
【0029】
この発明によれば、第2の受容体の活性を示す指標となるレポータを含む細胞を作製し、作製した細胞に当該細胞外から、発光させるための所定の処置を行うと共に第1の受容体に結合する所定の受容体結合物質を添加し、当該処置および添加が行われた後の細胞の発光量を測定し、測定した発光量に基づいて細胞内における第2の受容体の活性を解析し、解析した第2の受容体の活性に基づいて、第1の受容体と第2の受容体との相互作用を検出するので、受容体の空間的な配置を明らかにして受容体の相互作用に関する各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞において特定の受容体の相互作用を再現性良く観察することができるという効果を奏する。
【0030】
この発明によれば、レポータはサイクリックAMPセンサータンパク質であり、第2の受容体の活性はサイクリックAMPの濃度で解析するので、生物発光によるサイクリックAMP検出原理を利用して受容体の空間的な配置を明らかにして受容体の相互作用に関する各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞において特定の受容体の相互作用を再現性良く観察することができるという効果を奏する。
【0031】
この発明によれば、サイクリックAMPセンサータンパク質は、細胞内のサイクリックAMPと相互作用して発光するものであるので、サイクリックAMPの存在下で相互作用が強化された発光によりサイクリックAMPの濃度を検出することができるという効果を奏する。
【0032】
この発明によれば、サイクリックAMPセンサータンパク質は、ホタルのルシフェラーゼおよびPKA(protein kinase A)のサイクリックAMP結合領域とからなる融合タンパク質であるので、発光量を測ればサイクリックAMP濃度を算出することができるという効果を奏する。
【0033】
この発明によれば、サイクリックAMP結合領域が、マウスのPKA−RIα由来であるので、哺乳類由来の細胞において使用することができるという効果を奏する。
【0034】
この発明によれば、サイクリックAMP結合領域が、220から250のアミノ酸残基の長さを有するので、ルシフェラーゼの活性をなるべく維持したまま融合タンパク質を作製できるという効果を奏する。
【0035】
この発明によれば、サイクリックAMP結合領域が、少なくとも2つのサイクリックAMP結合部位を含むので、サイクリックAMP濃度の変化によって、ルシフェラーゼ活性を変化させることができるという効果を奏する。
【0036】
この発明によれば、サイクリックAMPセンサータンパク質は、ルシフェラーゼのN末端側断片、ルシフェラーゼのC末端側断片およびサイクリックAMP結合領域を含み、N末端側断片とC末端側断片とが前記サイクリックAMPセンサータンパク質分子内で結合すると、ルシフェラーゼの発光酵素活性が回復するので、サイクリックAMP結合領域の構造変化により発光強度を変化させることができるという効果を奏する。
【0037】
この発明によれば、サイクリックAMP結合領域が、配列番号37または39に記載のアミノ酸配列を含むので、条件に応じてサイクリックAMPセンサータンパク質として最適なものを選択できるという効果を奏する。
【0038】
この発明によれば、細胞は第1の受容体と第2の受容体とを含有するものであるので、解析対象となる受容体を人工的に含めた扱い易い細胞を用いて実験をやり易くすることができるという効果を奏する。
【0039】
この発明によれば、細胞は第1の受容体の遺伝子と第2の受容体の遺伝子とが導入されたものであるので、解析対象となる受容体を人工的に含めた扱い易い細胞を用いて実験をやり易くすることができる他、自然に近い応答を得ることができるという効果を奏する。
【0040】
この発明によれば、第2の受容体の活性に依存する発光標識の光子量を測定するので、フォトンカウンティングの技術を利用して細胞全体または細胞集団全体からの発光量を得ることができるという効果を奏する。
【0041】
この発明によれば、第2の受容体の活性に依存する発光標識を撮像するので、発光顕微鏡を利用して個々の細胞からの発光量を得ることができるという効果を奏する。
【0042】
この発明によれば、撮像を複数の異なる細胞に対して同時に実行し、撮像した複数の発光画像を細胞ごとに照合するので、細胞一つ一つからの発光量を個別に且つ同時に得ることができるという効果を奏する。
【0043】
この発明によれば、発光量の測定を繰り返し実行するので、発光量の経時変化を観察することができるという効果を奏する。
【0044】
この発明によれば、第1の受容体は5HT2A受容体であり、第2の受容体はD1受容体であるので、D1受容体および5HT2A受容体の空間的な配置を明らかにしてD1受容体と5HT2A受容体の相互作用に関する各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞においてD1受容体と5HT2A受容体の相互作用を再現性良く観察することができるという効果を奏する。
【0045】
この発明によれば、受容体結合物質はセロトニンであるので、5HT2A受容体の活性を観察することができるという効果を奏する。
【0046】
この発明によれば、サイクリックAMPセンサータンパク質は、第1の受容体と第2の受容体との相互作用を検出するためのものであるので、当該サイクリックAMPセンサータンパク質を用いれば、受容体の空間的な配置を明らかにして受容体の相互作用に関する各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞において特定の受容体の相互作用を再現性良く観察することができるという効果を奏する。
【0047】
この発明によれば、サイクリックAMPセンサータンパク質はサイクリックAMP(adenosine monophosphate)と相互作用して発光するものであるので、当該サイクリックAMPセンサータンパク質を用いれば、サイクリックAMPの存在下で相互作用が強化された発光によりサイクリックAMP濃度を検出することができるという効果を奏する。
【0048】
この発明によれば、サイクリックAMPセンサータンパク質はD1受容体と5HT2A受容体との相互作用を検出するためのものであるので、当該サイクリックAMPセンサータンパク質を用いれば、D1受容体および5HT2A受容体の空間的な配置を明らかにしてD1受容体と5HT2A受容体の相互作用に関する各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞においてD1受容体と5HT2A受容体の相互作用を再現性良く観察することができるという効果を奏する。
【図面の簡単な説明】
【0049】
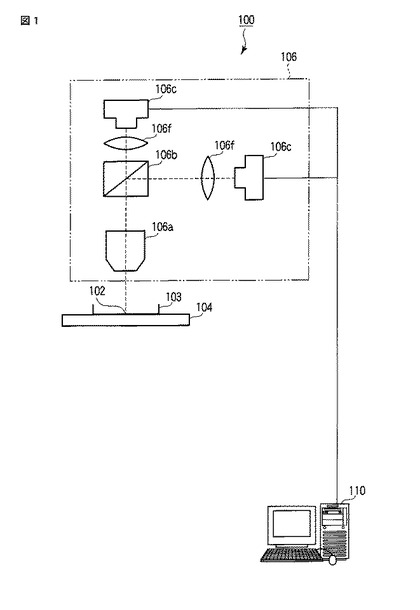
【図1】図1は、発光観察システム100の全体構成の一例を示す図である。
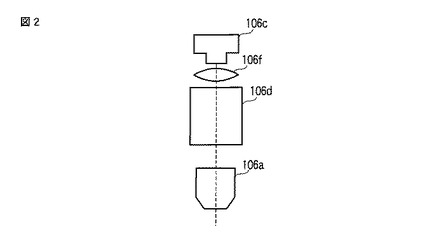
【図2】図2は、発光観察システム100の発光画像撮像ユニット106の構成の一例を示す図である。
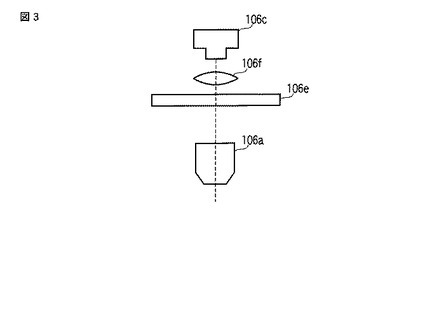
【図3】図3は、発光観察システム100の発光画像撮像ユニット106の構成の一例を示す図である。
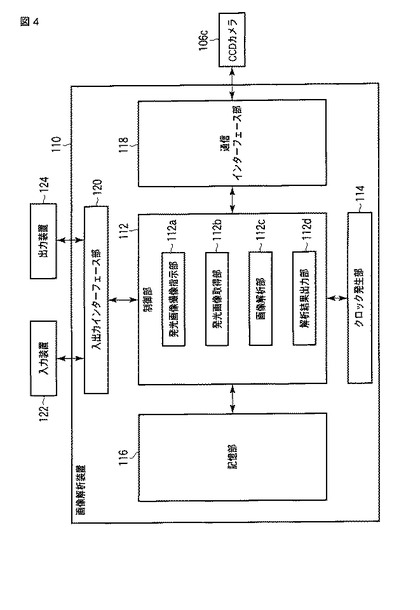
【図4】図4は、発光観察システム100の画像解析装置110の構成の一例を示すブロック図である。
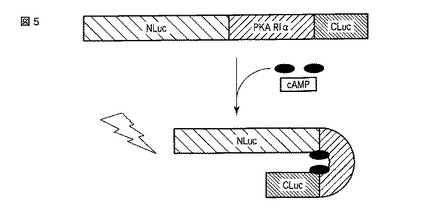
【図5】図5は、スプリットルシフェラーゼを用いたサイクリックAMPの検出原理を示す図である。
【図6】図6は、サイクリックAMPセンサータンパク質、5HT2A受容体およびD1受容体が導入されたHEK293細胞を含むセロトニン刺激前の発光画像を示す図である。
【図7】図7は、サイクリックAMPセンサータンパク質、5HT2A受容体およびD1受容体が導入されたHEK293細胞を含むセロトニン刺激5分後の発光画像を示す図である。
【図8】図8は、サイクリックAMPセンサータンパク質、5HT2A受容体およびD1受容体が導入されたHEK293細胞を含むドーパミン刺激5分後の発光画像を示す図である。
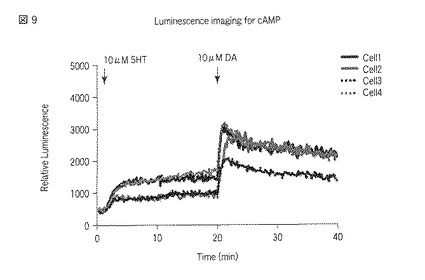
【図9】図9は、5HT2A受容体およびD1受容体が導入された、選択されたHEK293細胞におけるセロトニン刺激およびドーパミン刺激によるサイクリックAMPセンサータンパク質の発光強度の経時変化を示した図である。
【図10】図10は、サイクリックAMPセンサータンパク質、5HT2A受容体およびD2受容体が導入されたHEK293細胞を含むセロトニン刺激前の発光画像を示す図である。
【図11】図11は、サイクリックAMPセンサータンパク質、5HT2A受容体およびD2受容体が導入されたHEK293細胞を含むセロトニン刺激5分後の発光画像を示す図である。
【図12】図12は、サイクリックAMPセンサータンパク質、5HT2A受容体およびD2受容体が導入されたHEK293細胞を含むドーパミン刺激5分後の発光画像を示す図である。
【図13】図13は、サイクリックAMPセンサータンパク質、5HT2A受容体およびD2受容体が導入されたHEK293細胞を含むフォルスコリン刺激5分後の発光画像を示す図である。
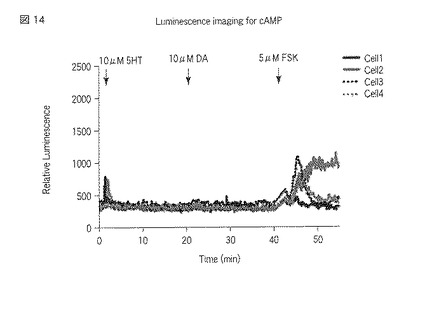
【図14】図14は、5HT2A受容体およびD2受容体が導入された、選択されたHEK293細胞におけるセロトニン刺激、ドーパミン刺激およびフォルスコリン刺激によるサイクリックAMPセンサータンパク質の発光強度の経時変化を示した図である。
【図15】図15は、サイクリックAMPセンサータンパク質および5HT2A受容体が導入されたHEK293細胞を含むセロトニン刺激前の発光画像を示す図である。
【図16】図16は、サイクリックAMPセンサータンパク質および5HT2A受容体が導入されたHEK293細胞を含むセロトニン刺激5分後の発光画像を示す図である。
【図17】図17は、サイクリックAMPセンサータンパク質および5HT2A受容体が導入されたHEK293細胞を含むフォルスコリン刺激5分後の発光画像を示す図である。
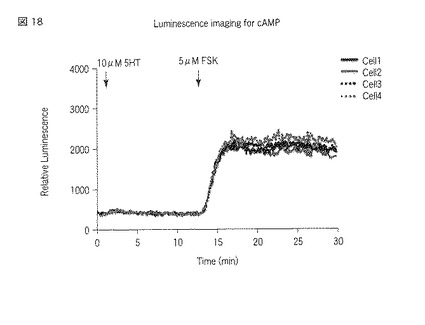
【図18】図18は、5HT2A受容体が導入された、選択されたHEK293細胞におけるセロトニン刺激およびドーパミン刺激によるサイクリックAMPセンサータンパク質の発光強度の経時変化を示した図である。
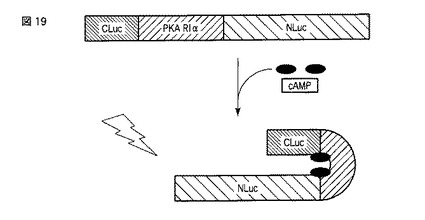
【図19】図19は、変異型のスプリットルシフェラーゼを用いたサイクリックAMPの検出原理を示す図である。
【図20】図20は、cpGL4_C_RIα_αAB_Nおよび、5HT2A受容体、D1受容体を導入したHEK293細胞において、セロトニン刺激前の発光画像を示す図である。
【図21】図21は、cpGL4_C_RIα_αAB_Nおよび、5HT2A受容体、D1受容体を導入したHEK293細胞において、セロトニン刺激5分後の発光画像を示す図である。
【図22】図22は、cpGL4_C_RIα_αAB_Nおよび、5HT2A受容体、D1受容体を導入したHEK293細胞において、ドーパミン刺激5分後の発光画像を示す図である。
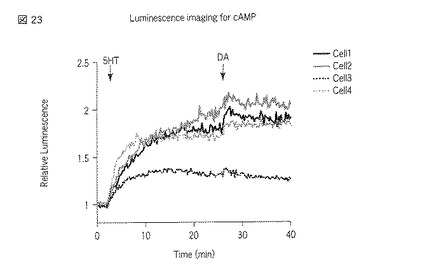
【図23】図23は、5HT2A受容体、D1受容体を導入したHEK293細胞において、選択した細胞でのセロトニンおよびドーパミン刺激によるcpGL4_C_RIα_αAB_Nの発光強度の経時変化を示した図である。
【図24】図24は、cpGL4_C_RIα_αAB_Nおよび、5HT2A受容体を導入したHEK293細胞において、セロトニン刺激前の発光画像を示す図である。
【図25】図25は、cpGL4_C_RIα_αAB_Nおよび、5HT2A受容体を導入したHEK293細胞において、セロトニン刺激5分後の発光画像を示す図である。
【図26】図26は、cpGL4_C_RIα_αAB_Nおよび、5HT2A受容体を導入したHEK293細胞において、ドーパミン刺激5分後の発光画像を示す図である。
【図27】図27は、5HT2A受容体を導入したHEK293細胞において、選択した細胞でのセロトニンおよびドーパミン刺激によるcpGL4_C_RIα_αAB_Nの発光強度の経時変化を示した図である。
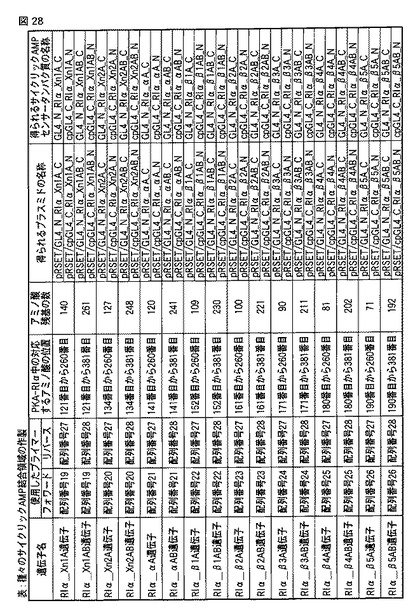
【図28】図28は、作製した種々のサイクリックAMPセンサータンパク質の詳細を示す表である。
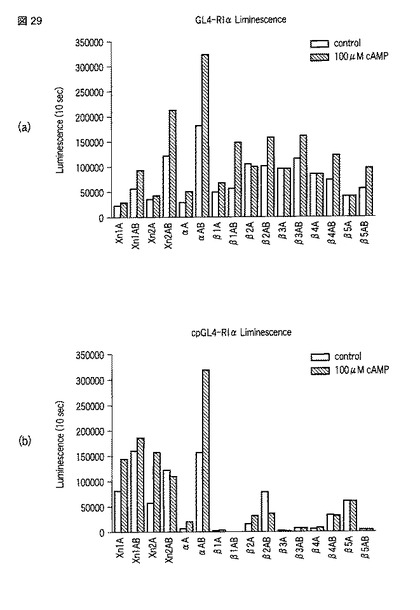
【図29】図29は、種々のサイクリックAMPセンサータンパク質の活性を比較した図である。
【発明を実施するための形態】
【0050】
以下に、本発明にかかる受容体相互作用検出方法および受容体の相互作用を検出するためのサイクリックAMPセンサータンパク質の実施の形態を図面に基づいて詳細に説明する。なお、本実施の形態により本発明が限定されるものではない。ここで、本実施の形態では、発光タンパク質(具体的には生物発光タンパク質)を用いてD1受容体と5HT2A受容体との相互作用を発光画像で検出する場合を一例として説明するが、蛍光タンパク質や他の受容体同士やフォトンカウンティングに対しても同様に実施することができる。
【0051】
[構成]
まず、本発明にかかる受容体相互作用検出方法で用いる発光観察システム100の構成について、図1、図2および図3を参照して説明する。図1は、発光観察システム100の全体構成の一例を示す図である。
【0052】
図1に示すように、発光観察システム100は、細胞102を収納した容器103(具体的にはシャーレ、スライドガラス、マイクロプレート、ゲル支持体、微粒子担体など)と、容器103を配置するステージ104と、微弱な発光を測定するための発光画像撮像ユニット106と、画像解析装置110と、で構成されている。発光画像撮像ユニット106をステージ104の下側に配置してもよい。これにより、カバー(図示せず)の開閉によるサンプル上方からの外乱光を完全に遮断でき、その結果、発光画像のS/N比を増すことができる。発光画像撮像ユニット106は、レーザー走査式の光学系であってもよい。
【0053】
細胞102は、例えば、サイクリックAMP(adenosine monophosphate)濃度に依存して発光するよう発光標識された生きた細胞である。ここで、サイクリックAMP濃度に依存して発光するタンパク質(発光タンパク質)として、N末側発光酵素とC末側発光酵素、およびサイクリックAMP結合タンパク質との融合タンパク質の他に、例えば国際公開第2007/120522号または特表2007−508014号公報に開示されている融合タンパク質を用いてもよい。また、ここで、細胞102に、当該融合タンパク質が発現されるよう構成した、当該融合タンパク質をコードするポリヌクレオチドを含む発現ベクターを細胞102に導入することにより、当該融合タンパク質が細胞102内で発現されるよう構成してもよい。なお、細胞102には、当該細胞102外から所定の発光基質(例えばルシフェリンなど)および所定の刺激(例えば薬物刺激など)が与えられる。
【0054】
発光画像撮像ユニット106は、具体的には正立型の発光顕微鏡であり、細胞102の発光画像を撮像する。発光画像撮像ユニット106は、図示の如く、対物レンズ106aと、ダイクロイックミラー106bと、CCD(charge−coupled device)カメラ106cと、結像レンズ106fと、で構成されている。対物レンズ106aは、具体的には(開口数/倍率)2の値が0.01以上のものが好ましい。ダイクロイックミラー106bは、細胞102から発せられた発光を色別に分離し、2色の発光を用いて発光量や発光強度を色別に測定する場合に用いる。CCDカメラ106cは、対物レンズ106a、ダイクロイックミラー106bおよび結像レンズ106fを介して当該CCDカメラ106cのチップ面に投影された細胞102の発光画像および明視野画像を撮る。また、CCDカメラ106cは、画像解析装置110と有線または無線で通信可能に接続される。ここで、細胞102が撮像範囲中に複数存在する場合、CCDカメラ106cは、当該撮像範囲中に含まれる複数の細胞102の発光画像および明視野画像を撮像してもよい。結像レンズ106fは、対物レンズ106aおよびダイクロイックミラー106bを介して当該結像レンズ106fに入射した像(具体的には細胞102を含む像)を結像する。なお、図1では、ダイクロイックミラー106bで分離した2つの発光に対応する発光画像を2台のCCDカメラ106cで別々に撮像する場合の一例を示しており、1つの発光を用いる場合には、発光画像撮像ユニット106は、対物レンズ106a、1台のCCDカメラ106cおよび結像レンズ106fで構成されてもよい。
【0055】
ここで、2色の発光を用いて発光量や発光強度を色別に測定する場合、発光画像撮像ユニット106は、図2に示すように、対物レンズ106aと、CCDカメラ106cと、スプリットイメージユニット106dと、結像レンズ106fと、で構成されてもよい。そして、CCDカメラ106cは、スプリットイメージユニット106dおよび結像レンズ106fを介して当該CCDカメラ106cのチップ面に投影された細胞102の発光画像(スプリットイメージ)および明視野像を撮像してもよい。スプリットイメージユニット106dは、細胞102から発せられた発光を色別に分離し、ダイクロイックミラー106bと同様、2色の発光を用いて発光量や発光強度を色別に測定する場合に用いる。
【0056】
また、複数色の発光を用いて発光量や発光強度を色別に測定する場合(つまり、多色の発光を用いる場合)、発光画像撮像ユニット106は、図3に示すように、対物レンズ106aと、CCDカメラ106cと、フィルターホイール106eと、結像レンズ106fと、で構成されてもよい。そして、CCDカメラ106cは、フィルターホイール106eおよび結像レンズ106fを介して当該CCDカメラ106cのチップ面に投影された細胞102の発光画像および明視野画像を撮像してもよい。フィルターホイール106eは、細胞102から発せられた発光をフィルタ交換によって色別に分離し、複数色の発光を用いて発光量や発光強度を色別に測定する場合に用いる。
【0057】
図1に戻り、画像解析装置110は、具体的にはパーソナルコンピュータである。そして、画像解析装置110は、図4に示すように、大別して、制御部112と、システムの時刻を計時するクロック発生部114と、記憶部116と、通信インターフェース部118と、入出力インターフェース部120と、入力装置122と、出力装置124と、で構成されており、これら各部はバスを介して接続されている。
【0058】
記憶部116は、ストレージ手段であり、具体的には、RAM(random access memory)やROM(read−only memory)等のメモリ装置、ハードディスクのような固定ディスク装置、フレキシブルディスク、光ディスク等を用いることができる。そして、記憶部116は制御部112の各部の処理により得られたデータなどを記憶する。通信インターフェース部118は、画像解析装置110と、CCDカメラ106cと、の間における通信を媒介する。すなわち、通信インターフェース部118は他の端末と有線または無線の通信回線を介してデータを通信する機能を有する。入出力インターフェース部120は、入力装置122や出力装置124に接続する。ここで、出力装置124には、モニタ(家庭用テレビを含む)の他、スピーカやプリンタを用いることができる(なお、以下で、出力装置124をモニターとして記載する場合がある。)。また、入力装置122には、キーボードやマウスやマイクの他、マウスと協働してポインティングデバイス機能を実現するモニターを用いることができる。
【0059】
制御部112は、OS(Operating System)等の制御プログラムや各種の処理手順等を規定したプログラムや所要データを格納するための内部メモリを有し、これらのプログラムに基づいて種々の処理を実行する。そして、制御部112は、大別して、発光画像撮像指示部112aと、発光画像取得部112bと、画像解析部112cと、解析結果出力部112dと、で構成されている。
【0060】
発光画像撮像指示部112aは、通信インターフェース部118を介して、CCDカメラ106cへ発光画像および明視野画像の撮像を指示する。発光画像取得部112bは、CCDカメラ106cで撮像した発光画像および明視野画像を、通信インターフェース部118を介して取得する。制御部112は、発光画像撮像指示部112aを制御して、CCDカメラ106cに細胞102の発光画像および明視野画像を繰り返し撮像させ、発光画像取得部112bを制御して、撮像した発光画像および明視野画像を、撮像される度に又は全撮像が終了後纏めて取得する。
【0061】
画像解析部112cは、発光画像取得部112bで取得した複数の発光画像に基づいて、細胞102から発せられる発光の発光強度を経時的に測定する。解析結果出力部112dは、画像解析部112cでの解析結果を出力装置124に出力する。具体的には、解析結果出力部112dは、画像解析部112cで得られた、細胞102から発せられる発光の発光強度に関する時系列データを、グラフ化して出力装置124に表示する。
【0062】
[受容体相互作用検出方法]
本発明は、細胞において第1の受容体と第2の受容体との相互作用を検出する受容体相互作用検出方法であって、前記第2の受容体の活性を示す指標となるレポータを含む前記細胞を作製する作製工程と、前記作製工程で作製した前記細胞に当該細胞外から、発光させるための所定の処置を行うと共に前記第1の受容体に結合する所定の受容体結合物質を添加する処置および添加工程と、前記処置および添加工程が行われた後の前記細胞の発光量を測定する測定工程と、前記測定工程で測定した前記発光量に基づいて前記細胞内における前記第2の受容体の活性を解析する解析工程と、前記解析工程で解析した前記第2の受容体の活性に基づいて、前記第1の受容体と前記第2の受容体との前記相互作用を検出する検出工程とを含むことを特徴とする受容体相互作用検出方法に関する。
【0063】
作製工程では、レポータを含む細胞が作製される。このレポータとは、後述するようなサイクリックAMPセンサータンパク質であってよい。対象となる細胞の種類に限定はなく、細菌細胞、酵母細胞、植物細胞および動物細胞等を使用できる。動物細胞が使用される場合、特に哺乳細胞が使用され、例えばマウスの細胞、サルの細胞およびヒトの細胞が使用される。細胞に対してレポータを導入する方法に特別な限定はなく、既知の導入方法が使用できる。細胞は、生体組織の一部として培養された細胞集合体であってもよいし、生体組織が属している臓器等や生命個体の形態でもよい。レポータがタンパク質である場合、1つの方法は、当該タンパク質をコードする塩基配列を含む又は当該配列から成る核酸を細胞に導入し、その後細胞内で当該タンパク質を発現させる方法である。例えば、当該核酸を含む発現ベクターを、リン酸カルシウム法、リポフェクション法またはエレクトロポレーション法等によって細胞内に導入し、発現ベクターから当該タンパク質を発現させることができる。ベクターから発現させるのではなく、当該核酸がゲノムに組み込まれた細胞を作製した後、ゲノムから当該タンパク質を発現させることもできる。別の方法は、細胞外で精製した当該タンパク質を細胞内に直接導入する方法である。例えば、マイクロインジェクション法によって当該タンパク質を細胞内に直接注入することができる。または、当該タンパク質を含む培養液にて細胞をインキュベートさせて、エンドサイトーシスによって当該タンパク質を細胞に取り込ませることができる。
【0064】
処置および添加工程では、発光させるための所定の処置および第1の受容体に結合する所定の受容体結合物質の添加が行われる。発光させるための所定の処置とは、例えば発光基質を添加することや励起光を照射することなどである。発光基質を添加する方法に特別な限定はなく、適宜選択できる。好ましくは、発光基質は測定の直前に添加される。発光基質の添加量に特別な限定はないが、好ましくは、発光酵素が発光反応を生じるために十分な量で添加される。第1の受容体に結合する所定の受容体結合物質の添加によって、当該受容体は活性化される。第1の受容体は例えば5HT2A受容体であってよく、受容体結合物質はセロトニン、例えば5HTとすることができる。
【0065】
測定工程では、処置および添加工程によって細胞から生じる発光が測定される。測定はイメージングまたはフォトカウンティングによって行うことができる。イメージングを行う場合、発光反応が生じている細胞の画像が取得されるので、細胞ごとの発光を測定するのに都合が良い。撮像は任意のタイミングで行うことができるが、好ましくは、基質添加から一定時間後に行われる。特に複数の測定を繰り返す場合、測定結果のばらつきを抑えるために、複数の測定を通じて同一のタイミングで撮像することが好ましい。撮像時間は条件に応じて最良の時間が適宜選択される。発光強度が小さい場合、撮像時間(顕微鏡を使用する場合、露光時間)は長くされ、逆に発光強度が大きすぎる場合、撮像時間は短くされる。撮像は、発光反応に係る波長の光のみを取得して行うこともできるが、それと同時に明視野画像または別の波長の光による画像を取得してもよい。例えば、細胞にサイクリックAMPセンサータンパク質と同時にプローブ(例えば蛍光または発光タンパク質)を融合した特定のタンパク質を導入し、サイクリックAMPセンサータンパク質に由来する光と同時に、当該特定のタンパク質からの光を検出してもよい。
【0066】
解析工程では、第2の受容体の活性が解析される。測定工程において得られた測定結果に基づき、発光が生じたかどうかが判定される。発光が生じた場合、第2の受容体が活性化したと判定される。好ましくは、第2の受容体として、D1受容体またはD2受容体を選択することができる。D1受容体は、リガンドが結合するとアデニル酸シクラーゼを活性化し、細胞内のサイクリックAMP濃度を上昇させる作用を有する。一方、D2受容体は、リガンドが結合するとホスホジエステラーゼを活性化し、細胞内のサイクリックAMP濃度を低下させる作用を有する。第2の受容体は、アデニル酸シクラーゼを活性化する作用を有するか否かが判明している受容体であることが好ましい。
【0067】
検出工程では、第2の受容体の活性の解析結果に基づいて、第1の受容体と第2の受容体との前記相互作用が検出される。処置および添加工程において第1の受容体に受容体結合物質を与えた結果、解析工程において第2の受容体の活性が認められた場合、第1の受容体と第2の受容体との相互作用に対して肯定的な結論が得られる。一方、解析工程において第2の受容体の活性が認められなかった場合、第1の受容体と第2の受容体との相互作用に対して否定的な結論が得られる。
【0068】
[サイクリックAMPセンサータンパク質]
次に、サイクリックAMPセンサータンパク質について補足的に説明する。
【0069】
本発明において、第2の受容体の活性を示す指標となるレポータとして、サイクリックAMPセンサータンパク質を使用することができる。当該サイクリックAMPセンサータンパク質として、ホタルのルシフェラーゼおよびPKAのサイクリックAMP結合領域とからなる融合タンパク質を使用することができる。
【0070】
サイクリックAMP結合領域としては、マウスのPKA−RIα由来のものを使用することができる。ここで、PKA−RIαとは、PKAの制御サブユニットαのことを意味する。PKA−RIαは、サイクリックAMP結合ドメインを有しており、サイクリックAMPに結合する能力を有する。
【0071】
サイクリックAMP結合領域の長さに関して、そのアミノ酸残基の数を120から250としてもよい。また、アミノ酸残基の数を200から250、220から250、230から250、230から245、または230から240としてもよい。例えば、アミノ酸残基数を230または241としてもよい。
【0072】
サイクリックAMP結合領域としては、少なくとも2つのサイクリックAMP結合部位を含むものを使用することができる。「サイクリックAMP結合部位」とは、サイクリックAMP1分子が結合することができるサイクリックAMP結合領域中の部位として定義される。サイクリックAMP結合部位には、前述のPKA−RIαのサイクリックAMP結合ドメインが含まれる。また、上記定義を満たせば、サイクリックAMP結合ドメインとして特定または認識されていない部位であってもサイクリックAMP結合部位に含まれる。
【0073】
本発明に係る方法において、ルシフェラーゼのN末端側断片、ルシフェラーゼのC末端側断片およびサイクリックAMP結合領域を含むサイクリックAMPセンサータンパク質であって、N末端側断片とC末端側断片とがサイクリックAMPセンサータンパク質分子内で結合すると、ルシフェラーゼの発光酵素活性が回復するサイクリックAMPセンサータンパク質を使用することができる。「ルシフェラーゼのN末端側断片」とは、C末端から複数のアミノ酸が欠失したルシフェラーゼ断片であって、発光酵素活性を失ったルシフェラーゼ断片を意味する。「ルシフェラーゼのC末端側断片」とは、N末端から複数のアミノ酸が欠失したルシフェラーゼ断片であって、発光酵素活性を失ったルシフェラーゼ断片を意味する。サイクリックAMPセンサータンパク質がそれらの断片およびサイクリックAMP結合領域を「含む」とは、サイクリックAMPセンサータンパク質が、ルシフェラーゼのN末端側断片、ルシフェラーゼのC末端側断片、サイクリックAMP結合領域および任意にその他のポリペプチドから成る融合タンパク質であることを意味する。したがって、これらの構成要素が側鎖または修飾基として含まれるわけではない。このようなサイクリックAMPセンサータンパク質の例は、図5に示されるような、サイクリックAMPセンサータンパク質のN末端側にルシフェラーゼのN末端側断片が存在し、サイクリックAMPセンサータンパク質のC末端側にルシフェラーゼのC末端側断片が存在し、これらの断片の間にPKA−RIα由来のサイクリックAMP結合領域が存在するサイクリックAMPセンサータンパク質である。また、別の例は、図19に示されるような、サイクリックAMPセンサータンパク質のN末端側にルシフェラーゼのC末端側断片が存在し、サイクリックAMPセンサータンパク質のC末端側にルシフェラーゼのN末端側断片が存在し、これらの断片の間にPKA−RIα由来のサイクリックAMP結合領域が存在するサイクリックAMPセンサータンパク質である。
【0074】
N末端側断片は、例えばルシフェラーゼの1番目から400番目のアミノ酸を含む断片であってよく、C末端側断片は、例えばルシフェラーゼの401番目から550番目のアミノ酸を含むものであってよい。特に、N末端側断片がルシフェラーゼの1番目から416番目のアミノ酸を含み、C末端側断片がルシフェラーゼの399番目から550番目のアミノ酸を含むことが好ましい。このように、N末端側断片およびC末端側断片は、それらの配列の一部が重複していてもよい。また、N末端側断片にもC末端側断片にも含まれないルシフェラーゼ配列部分が存在してもよい。
【0075】
サイクリックAMPセンサータンパク質内において、ルシフェラーゼがN末端側断片とC末端側断片とに分かれたことにより、通常の環境下(例えばサイクリックAMPが存在していない環境下)では、サイクリックAMPセンサータンパク質はルシフェラーゼの活性を有さない。しかし、特定の環境下(例えばサイクリックAMPが存在する環境下)では、サイクリックAMPセンサータンパク質はルシフェラーゼの活性を回復する。この回復は、サイクリックAMP結合領域の機能に起因する。例えば、マウスのPKA−RIα由来のサイクリックAMP結合領域を選択した場合、図5および19に示されるように、サイクリックAMP結合領域にサイクリックAMPが結合すると、サイクリックAMPセンサータンパク質全体の立体構造が変化し、結果としてN末端側断片とC末端側断片とが接近する。これらの2つの断片が協同してルシフェラーゼ活性を奏する。
【0076】
「N末端側断片とC末端側断片とが前記サイクリックAMPセンサータンパク質分子内で結合すると発光酵素活性が回復する」という表現において、N末端側断片とC末端側断片とが「結合」する態様に限定はなく、共有結合または非共有結合等の化学的結合であってもよいが、N末端側断片とC末端側断片とがルシフェラーゼの本来の機能を果たしうる程度に「接近」または「接触」している状態であることが好ましい。あるいは、N末端側断片とC末端側断片とが「会合」する状態であってもよい。すなわち、当該表現において使用される「結合」という用語は、「接近」、「接触」または「会合」という意味を含む。
【0077】
また、上記表現において、N末端側断片とC末端側断片との結合は、サイクリックAMPに応答して生じる結合であってよい。より具体的には、サイクリックAMP結合領域にサイクリックAMP分子が結合することで、サイクリックAMP結合領域の構造の変化が生じ、その結果生じるN末端側断片とC末端側断片との結合または接近であってよい。
【0078】
なお、サイクリックAMP結合領域としては、好ましくは配列番号37または39に示されるアミノ酸配列を含むポリペプチドである。
【実施例】
【0079】
[実施例1]
本実施の形態における実施例について、以下に図5〜図9を用いて説明する。ここで、図5はスプリットルシフェラーゼアッセイ法を用いたサイクリックAMPの検出原理を示す図である。
【0080】
スプリットルシフェラーゼアッセイとは、ホタルルシフェラーゼやウミシイタケルシフェラーゼ等のルシフェラーゼを特定の位置で分割して発光酵素活性(生物発光能)を失活させた後、そのフラグメント(断片)であるN末側発光酵素とC末側発光酵素を再構成して発光酵素活性を回復させる手法であり、タンパク質間相互作用の検出やタンパク質の細胞内小器官移行検出等に用いられる。
【0081】
図5には、N末側発光酵素(NLuc)とC末側発光酵素(CLuc)との非共有結合的相互作用を起こさせるタンパク質として、サイクリックAMP結合タンパク質を用いた例を示している。すなわち、細胞内で発現させた、N末側発光酵素とC末側発光酵素およびサイクリックAMP結合タンパク質との融合タンパク質における当該サイクリックAMP結合タンパク質にサイクリックAMPが結合すると、当該サイクリックAMP結合タンパク質の立体構造が変化してN末側発光酵素とC末側発光酵素とが結合し、発光酵素活性が回復して検出可能なシグナルが発せられる。本実施例では、当該スプリットルシフェラーゼアッセイ系において、サイクリックAMP結合タンパク質にPKA(protein kinase A)の制御サブユニットIα(PKA−RIα)のサイクリックAMP結合ドメインを用いた例について説明する。ここで、本実施例における実験の流れ(手順)は、以下の通りである。
【0082】
[前準備その1:N末側発光酵素遺伝子とC末側発光酵素遺伝子およびPKA−RIαのサイクリックAMP結合ドメイン遺伝子のクローニングと、融合タンパク質発現プラスミドの作製]
[手順1]
N末側発光酵素(NLuc)遺伝子とC末側発光酵素(CLuc)遺伝子の作製のために、PCR(polymerase chain reaction)に用いる合成オリゴDNA(deoxyribonucleic acid)を以下に示す配列で調製した。
[NLuc遺伝子作製用合成オリゴDNA配列]
NLuc_Fw(配列番号1):5’−TGTGGATCCAGCCACCATGGAAGATGCCAA−3’
NLuc_Rv(配列番号2):5’−CAGCTCGAGGTCCTTGTCGATGAGAGCGTT−3’
[CLuc遺伝子作製用合成オリゴDNA配列]
CLuc_Fw(配列番号3):5’−ATCAGATCTGGCTACGTTAACAACCCCGAG−3’
CLuc_Rv(配列番号4):5’−CTAGAATTCTTACACGGCGATCTTGCCGCC−3’
【0083】
また、PKA−RIαのcAMP結合ドメイン遺伝子のクローニングのために、PCRに用いる合成オリゴDNAを以下に示す配列で調製した。
PKA_RIA_β1_Fw(配列番号5):5’−ATTCTCGAGGCTATGTTTCCAGTCTCCTTT−3’
PKA_RIA_B_Rv(配列番号6):5’−GGAAGATCTGACGGACAGGGACACGAAGCT−3’
【0084】
[手順2]
pGL4.10(プロメガ(株)製)を鋳型として、N末側ルシフェラーゼ遺伝子(NLuc:GL4.10遺伝子の1番目から416番目のアミノ酸を含む。)およびC末側ルシフェラーゼ遺伝子(CLuc:GL4.10遺伝子の399番目から550番目のアミノ酸を含む。)を、上記合成オリゴDNA(配列番号1のDNA配列と配列番号2のDNA配列のペア、および配列番号3のDNA配列と配列番号4のDNA配列のペア)をプライマーとしてPCRにより増幅した。
【0085】
[手順3]
PKAの制御サブユニットIα(PKA−RIα)に2ヶ所あるサイクリックAMP結合ドメインのAとBの両方を含む領域を、マウスのPKA制御サブユニットIα(PKA−RIα)のcDNAを鋳型とし、上記合成オリゴDNAをプライマーとしてPCRにより増幅した。配列番号5のDNA配列と配列番号6のDNA配列のプライマーを用いて、RIα_β1AB遺伝子(マウスのPKA−RIα遺伝子の152番目から381番目のアミノ酸配列に対応する領域を含む。配列番号39)をPCRにより増幅した。
【0086】
[手順4:融合タンパク質遺伝子発現プラスミドの作製]
PCRで増幅させたNLuc遺伝子、CLuc遺伝子およびRIα_β1AB遺伝子を、動物細胞発現用プラスミドであるpcDNA3.1(インビトロジェン社製)のBamHI部位とEcoRI部位の間に挿入し、動物細胞発現用プラスミドpcDNA/GL4_N_RIα_β1AB_Cを作製した。
【0087】
[前準備その2:ヒトD1受容体遺伝子とD2受容体遺伝子および5HT2A受容体遺伝子のクローニング]
[手順1]
ヒトD1受容体遺伝子とD2受容体遺伝子および5HT2A受容体遺伝子の作製のために、PCRに用いる合成オリゴDNAを以下に示す配列で調製した。
[ヒトD1受容体遺伝子作製用合成オリゴDNA配列]
HU_D1R_Fw(配列番号7):5’−GCCACCATGAGGACTCTGAACACCTCTGCC−3’
HU_D1R_Rv(配列番号8):5’−TCAGGTTGGGTGCTGACCGTTTTGTGTGAT−3’
[ヒトD2受容体遺伝子作製用合成オリゴDNA配列]
HU_D2R_Fw(配列番号9):5’−GCCACCATGGATCCACTGAATCTGTCCTGG−3’
HU_D2R_Rv(配列番号10):5’−TCAGCAGTGGAGGATCTTCAGGAAGGCCTT−3’
[ヒト5HT2A受容体遺伝子作製用合成オリゴDNA配列]
HU_5HT2AR_Fw(配列番号11):5’−GCCACCATGGATATTCTTTGTGAAGAAAAT−3’
HU_5HT2AR_Rv(配列番号12):5’−TCACACACAGCTCACCTTTTCATTCACTCC−3’
【0088】
[手順2]
ヒト脳cDNAライブラリ(タカラバイオ(株)製)を鋳型として、D1受容体遺伝子(ヒトのD1受容体遺伝子の全長に対応する領域を含む。)D2受容体遺伝子(ヒトのD2受容体遺伝子の全長に対応する領域を含む。)および5HT2A受容体遺伝子(ヒトの5HT2A受容体遺伝子の全長に対応する領域を含む。)を、上記合成オリゴDNAをプライマーとしてPCRにより増幅した。
【0089】
[手順3]
PCRで増幅させたD1受容体遺伝子、D2受容体遺伝子および5HT2A受容体遺伝子をpBluescriptIIベクターにサブクローニングした後、作製したDNAの配列をDNAシーケンシングにより確認した。
【0090】
[手順4]
それぞれの遺伝子を哺乳類細胞の発現ベクターpcDNA3.1(+)(インビトロジェン(株)製)へ組み込み、ヒトD1受容体発現用プラスミドpcDNA/D1RおよびヒトD2受容体発現用プラスミドpcDNA/D2Rおよびヒト5HT2A受容体発現用プラスミドpcDNA/5HT2ARを作製した。
【0091】
[実験:ヒトD1受容体、D2受容体および5HT2A受容体を含有するHEK293細胞でのcAMP濃度変動の発光イメージング]
[手順1:HEK293細胞の培養]
ATCC(American Type Culture Collection)社より入手したHEK293細胞を、5% CO2インキュベーター内で、10% Fetal Bovine Serumおよび1× Nonessential amino acidsを添加したEarle's MEM/培地(GIBCO社製)で培養した。
【0092】
[手順2:タンパク質発現プラスミドの導入]
HEK293細胞を、直径35mmガラスボトムディッシュに2×105/dishの細胞密度で播種し、5% CO2インキュベーター内で一晩培養し、サイクリックAMPセンサータンパク質発現用プラスミドpcDNA/GL4_N_RIα_β1AB_C、ヒトD1受容体発現用プラスミドpcDNA/D1R(またはヒトD2受容体発現用プラスミドpcDNA/D2R)およびヒト5HT2A受容体発現用プラスミドpcDNA/5HT2ARを混合し、FuGENE HD(ロシュ社製)を用いてトランスフェクションを行い、5% CO2インキュベーター内で一晩培養した。なお、ヒトD1受容体のアミノ酸配列は具体的には配列番号29のアミノ酸配列であり、ヒト5HT2A受容体のアミノ酸配列は具体的には配列番号30のアミノ酸配列である。
【0093】
[手順3:発光画像の撮像]
培地中にルシフェリン2mM(プロメガ社製)を加えて1時間静置してから、培養ディッシュを発光顕微鏡“LV(LUMINOVIEW)−200”(オリンパス社製)にセットし、20秒間隔で発光画像のタイムラプス撮影を行った。発光観察条件として、対物レンズの倍率は40倍、露出時間は10秒、ビニングは2×2とした。CCDカメラ106cとしてEM−CCDカメラiXon(アンドール社製)を用い、画像解析装置110として構成したパーソナルコンピュータに発光画像を取り込んだ。
【0094】
[手順4:ドーパミン刺激による発光画像の撮像]
タイムラプス撮影開始から1分後以降に、セロトニン(5HT:最終濃度10μM)、ドーパミン(DA:最終濃度10μM)およびフォルスコリン(FSK:最終濃度5μM)で順次刺激を行い、引き続き発光画像のタイムラプス撮影を行った。
【0095】
[手順5:発光量の経時変化のグラフ表示]
刺激前に撮影した各々の発光画像(図6、図10および図15参照)に対して図示の如く複数のROI(Region of Interest:関心領域)を指定し、また刺激後に撮影した各々の発光画像(図7、図8、図11〜図13および図16〜図17参照)に対して図示の如く複数のROIを指定した。指定した各ROIの発光強度を各々の発光画像に基づいて測定し、当該測定した発光強度の経時変化をグラフ(図9、図14および図18参照)で表示した。発光画像の解析は、画像解析部112cとして機能するMetamorphソフトウェア(ユニバーサルイメージング社製)を用いて行った。なお、図9、図14および図18において、Cell1〜Cell4のそれぞれに対応する各グラフは、図6〜図8、図10〜図13および図15〜図17における番号1〜番号4のそれぞれに対応する各ROIの発光強度の経時変化を示している。
【0096】
[実験の結果]
以上の実験の結果、図6〜図9に示すように、5HT2A受容体またはD1受容体、D2受容体への刺激に対する細胞内のサイクリックAMP応答の変化をシングルセルレベル(一細胞レベル)で検出することができた。また、図9に示されている個々の細胞における発光強度の変化を見ると、5HT2A受容体とD1受容体との組合せにおいて、5HT2A受容体への刺激に対する細胞内サイクリックAMP濃度の上昇が観察された。しかし、図11〜図14および図16〜図18に示すように、5HT2A受容体とD2受容体との組合せにおいておよび5HT2A受容体のみにおいては、5HT2A受容体への刺激に対する細胞内サイクリックAMP濃度の上昇が観察されなかった。セロトニン受容体とドーパミン受容体との相互作用のこれまでの研究から、個体において、D1受容体を介すると考えられている行動が5HT2A受容体を介して引き起こされる、ということが知られている。本実験により得られた結果は、個体において観察した反応を細胞レベルで説明することが出来ることを示している。
【0097】
[本実施例のまとめ]
以上、本実施例によれば、細胞内の情報伝達に影響を及ぼすことなく、各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞においての5HT2A受容体とD1受容体との相互作用を再現性良く経時的に観察することができる。
【0098】
[他の実施の形態]
上述した実施の形態においては、主に、互いに結合することにより発光酵素活性が回復するよう分割させたN末側発光酵素とC末側発光酵素およびPKA−RIαのサイクリックAMP結合ドメインからなる融合タンパク質を、サイクリックAMPセンサータンパク質として説明を行ったが、本発明はこれに限られず、N末側発光酵素とC末側発光酵素およびPKA−RIαのサイクリックAMP結合ドメインからなる融合タンパク質に替えて、国際公開第2007/120522号および特表2007−508014号公報に開示されているサイクリックAMPセンサータンパク質を用いてもよい。
【0099】
[実施例2]
実施例1とは異なる実施形態に係るサイクリックAMPセンサータンパク質を作製し、その有用性を検討した。具体的には、図19に示されるように、サイクリックAMPセンサータンパク質のN末端側にC末端側断片が配置され、C末端側にN末端側断片が配置され、それらの間にサイクリックAMP結合領域が配置されたサイクリックAMPセンサータンパク質を作製した。すなわち、端的に表現すると、実施例1に係るサイクリックAMPセンサータンパク質に対して、C末端側断片とN末端側断片とを置き換えたサイクリックAMPセンサータンパク質を作製した。なお、本実施例に係るサイクリックAMPセンサータンパク質並びにその作製のために使用される遺伝子およびプライマー等について、それらの名称に「変異型」または「cp(circular permutaion)」等を付して区別した。
【0100】
[前準備:変異型ルシフェラーゼ作成用のN末側発光酵素遺伝子とC末側発光酵素遺伝子、およびPKA−RIαのサイクリックAMP結合ドメイン遺伝子のクローニングと、変異型ルシフェラーゼ融合タンパク質発現プラスミドの作製]
[手順1]
変異型ルシフェラーゼ作成用のN末側発光酵素(cpNLuc)遺伝子とC末側発光酵素(cpCLuc)遺伝子の作製のために、PCR(polymerase chain reaction)に用いる合成オリゴDNA(deoxyribonucleic acid)を以下に示す配列で調製した。
[cpNLuc遺伝子作製用合成オリゴDNA配列]
cpNLuc_Fw(配列番号13):5’−ATCAGATCTGAAGATGCCAAAAACATTAAG−3’
cpNLuc_Rv(配列番号14):5’−CTAGAATTCTTAGTCCTTGTCGATGAGAGC−3’
[cpCLuc遺伝子作製用合成オリゴDNA配列]
cpCLuc_Fw(配列番号15):5’−TGTGGATCCAGCCACCATGAGCGGCTACGTTAACAACCCC−3’
cpCLuc_Rv(配列番号16):5’−CAGCTCGAGCACGGCGATCTTGCCGCCCTT−3’
【0101】
また、PKA−RIαのcAMP結合ドメイン遺伝子のクローニングのために、PCRに用いる合成オリゴDNAを以下に示す配列で調製した。
PKA_RIA_α_Fw(配列番号17):5’−TTTCTCGAGCTTGATGATAACGAGAGAAGT−3’
PKA_RIA_B_Rv(配列番号18):5’−GGAAGATCTGACGGACAGGGACACGAAGCT−3’
【0102】
[手順2]
市販のルシフェラーゼ遺伝子であるpGL4.10(プロメガ(株)製)を鋳型として、変異型のN末側ルシフェラーゼ遺伝子(cpNLuc:GL4.10遺伝子の1番目から416番目のアミノ酸を含む。)および変異型のC末側ルシフェラーゼ遺伝子(cpCLuc:GL4.10遺伝子の399番目から550番目のアミノ酸を含む。)を、上記合成オリゴDNA(配列番号13のDNA配列と配列番号14のDNA配列のペア、および配列番号15のDNA配列と配列番号16のDNA配列のペア)をプライマーとしてPCRにより増幅した。
【0103】
[手順3]
PKAの制御サブユニットIα(PKA−RIα)に2ヶ所あるサイクリックAMP結合ドメインのAとBの両方を含む領域を、マウスのPKA制御サブユニットIα(PKA−RIα)のcDNAを鋳型とし、上記合成オリゴDNAをプライマーとしてPCRにより増幅した。配列番号17のDNA配列と配列番号18のDNA配列のプライマーを用いて、RIα_αAB遺伝子(マウスのPKA−RIα遺伝子の141番目から381番目のアミノ酸配列に対応する領域を含む。)(塩基配列は配列番号37に示される)をPCRにより増幅した。
【0104】
[手順4:融合タンパク質遺伝子発現プラスミドの作製]
PCRで増幅させた、cpNLuc遺伝子、cpCLuc遺伝子およびRIα_αAB遺伝子をそれぞれ含む核酸を、制限酵素にて適宜処理した後、動物細胞発現用プラスミドであるpcDNA3.1(インビトロジェン社製)のBamHI部位とEcoRI部位の間に挿入し、動物細胞発現用プラスミドpcDNA/cpGL4_C_RIα_αAB_Nを作製した。
【0105】
[実験:ヒトD1受容体、D2受容体および5HT2A受容体を含有するHEK293細胞でのcAMP濃度変動の発光イメージング]
[手順1:HEK293細胞の培養]
ATCC(American Type Culture Collection)社より入手したHEK293細胞を、5% CO2インキュベーター内で、10% Fetal Bovine Serumおよび1× Nonessential amino acidsを添加したEarle's MEM/培地(GIBCO社製)で培養した。
【0106】
[手順2:タンパク質発現プラスミドの導入]
HEK293細胞を、直径35mmガラスボトムディッシュに2×105/dishの細胞密度で播種し、5% CO2インキュベーター内で一晩培養し、サイクリックAMPセンサータンパク質発現用プラスミドpcDNA/cpGL4_C_RIα_αAB_N、ヒトD1受容体発現用プラスミドpcDNA/D1R(またはヒトD2受容体発現用プラスミドpcDNA/D2R)およびヒト5HT2A受容体発現用プラスミドpcDNA/5HT2ARを混合し、FuGENE HD(ロシュ社製)を用いてトランスフェクションを行い、5% CO2インキュベーター内で一晩培養した。なお、pcDNA/D1R、pcDNA/D2RおよびpcDNA/5HT2ARは、実施例1と同様に作製したものを使用した。
【0107】
[手順3:発光画像の撮像]
培地中にルシフェリン2mM(プロメガ社製)を加えて1時間静置してから、培養ディッシュを発光顕微鏡“LV(LUMINOVIEW)−200”(オリンパス社製)にセットし、20秒間隔で発光画像のタイムラプス撮影を行った。発光観察条件として、対物レンズの倍率は40倍、露出時間は10秒、ビニングは2×2とした。CCDカメラ106cとしてEM−CCDカメラiXon(アンドール社製)を用い、画像解析装置110として構成したパーソナルコンピュータに発光画像を取り込んだ。
【0108】
[手順4:セロトニン、ドーパミン刺激による発光画像の撮像]
タイムラプス撮影開始から1分後以降に、セロトニン(5HT:最終濃度10μM)、ドーパミン(DA:最終濃度10μM)で順次刺激を行い、引き続き発光画像のタイムラプス撮影を行った。
【0109】
[手順5:発光量の経時変化のグラフ表示]
刺激前に撮影した各々の発光画像(図20および図24参照)に対して図示の如く複数のROI(Region of Interest:関心領域)を指定し、また刺激後に撮影した各々の発光画像(図21、図22、図25および図26参照)に対して図示の如く複数のROIを指定した。指定した各ROIの発光強度を各々の発光画像に基づいて測定し、当該測定した発光強度の経時変化をグラフ(図23、および図27参照)で表示した。発光画像の解析は、画像解析部112cとして機能するMetamorphソフトウェア(ユニバーサルイメージング社製)を用いて行った。なお、図23および図27において、Cell1〜Cell4のそれぞれに対応する各グラフは、図20〜図22および図24〜図26における番号1〜番号4のそれぞれに対応する各ROIの発光強度の経時変化を示している。
【0110】
[実験の結果]
以上の実験の結果、図6〜図9および図20〜図23に示すように、5HT2A受容体またはD1受容体、D2受容体への刺激に対する細胞内のサイクリックAMP応答の変化をシングルセルレベル(一細胞レベル)で検出することができた。また、図9および図23に示されている個々の細胞における発光強度の変化を見ると、5HT2A受容体とD1受容体との組合せにおいて、5HT2A受容体への刺激に対する細胞内サイクリックAMP濃度の上昇が観察された。しかし、図11〜図14および図16〜図18、図24〜図27に示すように、5HT2A受容体とD2受容体との組合せにおいておよび5HT2A受容体のみにおいては、5HT2A受容体への刺激に対する細胞内サイクリックAMP濃度の上昇が観察されなかった。セロトニン受容体とドーパミン受容体との相互作用のこれまでの研究から、個体において、D1受容体を介すると考えられている行動が5HT2A受容体を介して引き起こされる、ということが知られている。本実験により得られた結果は、個体において観察した反応を細胞レベルで説明することが出来ることを示している。
【0111】
[本実施例のまとめ]
以上、本実施例によれば、細胞内の情報伝達に影響を及ぼすことなく、各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞においての5HT2A受容体とD1受容体との相互作用を再現性良く経時的に観察することができる。この実施例において使用するルシフェラーゼ遺伝子やルシフェリン等は、他の市販試薬であってもよいし、生物発光等を発生する生物から抽出等されたものであってもよい。
【0112】
[他の実施の形態]
上述した実施の形態においては、主に、互いに結合することにより発光酵素活性が回復するよう分割させたN末側発光酵素とC末側発光酵素、および、PKA−RIαのサイクリックAMP結合ドメインからなる融合タンパク質をサイクリックAMPセンサータンパク質として説明を行ったが、本発明はこれに限られず、N末側発光酵素とC末側発光酵素、および、PKA−RIαのサイクリックAMP結合ドメインからなる融合タンパク質に替えて特許文献1および特許文献2に開示されているサイクリックAMPセンサータンパク質を用いてもよい。
【0113】
[実施例3]
種々のサイクリックAMPセンサータンパク質を作製し、それらの有用性を検討した。具体的には、長さおよび範囲をかえた16通りのサイクリックAMP結合領域を作製し、それぞれについて、N末端側断片、サイクリックAMP結合領域およびC末端側断片の順に配置したもの、ならびに、C末端側断片、サイクリックAMP結合領域およびN末端側断片の順に配置したものの計32種のサイクリックAMPセンサータンパク質を作製した。それら全てについて、サイクリックAMPの存在下および非存在下における酵素活性を測定した。
【0114】
[手順1]
スプリットルシフェラーゼ作成用のN末側発光酵素(NLuc)遺伝子とC末側発光酵素(CLuc)遺伝子、および、変異型スプリットルシフェラーゼ作成用のN末側発光酵素(cpNLuc)遺伝子とC末側発光酵素(cpCLuc)遺伝子の作製のために、PCR(polymerase chain reaction)に用いる合成オリゴDNA(deoxyribonucleic acid)を以下に示す配列で調製した。
[NLuc遺伝子作製用合成オリゴDNA配列]
NLuc_Fw (配列番号1): 5’ − TGTGGATCCAGCCACCATGGAAGATGCCAA − 3’
NLuc_Rv (配列番号2): 5’ − CAGCTCGAGGTCCTTGTCGATGAGAGCGTT − 3’
[CLuc遺伝子作製用合成オリゴDNA配列]
CLuc_Fw (配列番号3): 5’ − ATCAGATCTGGCTACGTTAACAACCCCGAG − 3’
CLuc_Rv (配列番号4): 5’ − CTAGAATTCTTACACGGCGATCTTGCCGCC − 3’
[cpNLuc遺伝子作製用合成オリゴDNA配列]
cpNLuc_Fw(配列番号13):5’−ATCAGATCTGAAGATGCCAAAAACATTAAG−3’
cpNLuc_Rv(配列番号14):5’−CTAGAATTCTTAGTCCTTGTCGATGAGAGC−3’
[cpCLuc遺伝子作製用合成オリゴDNA配列]
cpCLuc_Fw(配列番号15):5’−TGTGGATCCAGCCACCATGAGCGGCTACGTTAACAACCCC−3’
cpCLuc_Rv(配列番号16):5’−CAGCTCGAGCACGGCGATCTTGCCGCCCTT−3’
【0115】
また、PKA−RIαのcAMP結合ドメイン遺伝子のクローニングのために、PCRに用いる合成オリゴDNAを以下に示す配列で調製した。
PKA_RIA_Xn1_Fw (配列番号19): 5’− ATTCTCGAGGATTATAAGACAATGGCTGCT −3’
PKA_RIA_Xn2_Fw (配列番号20): 5’− GCCCTCGAGAAGAATGTGCTGTTTTCACAC −3’
PKA_RIA_α_Fw (配列番号21): 5’− TTTCTCGAGCTTGATGATAACGAGAGAAGT −3’
PKA_RIA_β1_Fw (配列番号22): 5’− ATTCTCGAGGCTATGTTTCCAGTCTCCTTT −3’
PKA_RIA_β2_Fw (配列番号23): 5’− TTTCTCGAGGGAGAGACGGTTATTCAGCAA −3’
PKA_RIA_β3_Fw (配列番号24): 5’− GGTCTCGAGGGGGATAACTTCTATGTGATT −3’
PKA_RIA_β4_Fw (配列番号25): 5’− ATTCTCGAGGGAGAAATGGATGTCTATGTC −3’
PKA_RIA_β5_Fw (配列番号26): 5’− AATCTCGAGTGGGCAACCAGTGTTGGGGAA −3’
PKA_RIA_A_Rv (配列番号27): 5’− CTCAGATCTGTCCAGAGACTCTAAAATAGA −3’
PKA_RIA_B_Rv (配列番号28): 5’− GGAAGATCTGACGGACAGGGACACGAAGCT −3’
【0116】
[手順2]
そして、pGL4.10(プロメガ(株)製)を鋳型として、以下の表1のプライマーセットを使用し、ルシフェラーゼ遺伝子の断片を含む核酸をPCRによって増幅した。
【0117】
【表1】

[手順3]
PKAの制御サブユニットIα(PKA−RIα)にはcAMP結合ドメインがAとBの2ヶ所あり、Aのみを含む領域、または、AおよびBの両方を含む領域を、マウスのPKA制御サブユニットIα(PKA−RIα)のcDNAを鋳型とし、上記合成オリゴDNAをプライマーとしてPCRにより増幅した。使用したプライマーと増幅された遺伝子との関係を図28に示す(図中、「遺伝子名」「使用したプライマー」「PKA−RIα中の対するアミノ酸の位置」「アミノ酸残基の数」が参照される)。
【0118】
なお、配列番号31には、PKAの制御サブユニットIα(PKA−RIα)全体のアミノ酸配列が示される。配列番号32には、RIα−Xn1A遺伝子に対応するアミノ酸配列が記載される。配列番号33には、RIα−Xn1AB遺伝子に対応するアミノ酸配列が記載される。配列番号34には、RIα−Xn2A遺伝子に対応するアミノ酸配列が記載される。配列番号35には、RIα−Xn2AB遺伝子に対応するアミノ酸配列が記載される。配列番号36には、RIα−αA遺伝子に対応するアミノ酸配列が記載される。配列番号37には、RIα−αAB遺伝子に対応するアミノ酸配列が記載される。配列番号38には、RIα−β1A遺伝子に対応するアミノ酸配列が記載される。配列番号39には、RIα−β1AB遺伝子に対応するアミノ酸配列が記載される。配列番号40には、RIα−β2A遺伝子に対応するアミノ酸配列が記載される。配列番号41には、RIα−β2AB遺伝子に対応するアミノ酸配列が記載される。配列番号42には、RIα−β3A遺伝子に対応するアミノ酸配列が記載される。配列番号43には、RIα−β3AB遺伝子に対応するアミノ酸配列が記載される。配列番号44には、RIα−β4A遺伝子に対応するアミノ酸配列が記載される。配列番号45には、RIα−β4AB遺伝子に対応するアミノ酸配列が記載される。配列番号46には、RIα−β5A遺伝子に対応するアミノ酸配列が記載される。配列番号47には、RIα−β5AB遺伝子に対応するアミノ酸配列が記載される。
【0119】
[手順4](原核生物発現用プラスミドpRSET/GL4_N_C、および、pRSET/cpGL4_C_Nの作製)
PCRによって増幅したNLuc遺伝子を含む断片を、原核生物発現用プラスミドであるpRSET2B(インビトロジェン社製)内のBamHI部位とXhoI部位との間に挿入した後、CLuc遺伝子を含む断片を、BglII部位とEcoRI部位との間に挿入することで、原核生物発現用プラスミド(pRSET/GL4_N_C)を作製した。
【0120】
また、PCRによって増幅したcpCLuc遺伝子を含む断片を、原核生物発現用プラスミドであるpRSET2B(インビトロジェン社製)内のBamHI部位とXhoI部位との間に挿入した後、cpNLuc遺伝子を含む断片を、BglII部位とEcoRI部位との間に挿入し、原核生物発現用プラスミド(pRSET/cpGL4_C_N)を作製した。
【0121】
[手順5](融合タンパク質遺伝子発現プラスミドの作製)
手順3によって作製したPKA−RIα由来の種々の遺伝子を含む核酸を、手順4で作製したpRSET/GL4_N_CベクターまたはpRSET/cpGL4_C_Nベクター内のXhoI部位とBglII部位の間に挿入した。このようにして得られたプラスミドを図28に示す(図中、「得られるプラスミドの名称」が参照される)。
【0122】
[手順6](融合タンパク質遺伝子の大腸菌での発現および精製)
以上のように作製した融合タンパク質発現プラスミドを、大腸菌JM109(DE3)株にトランスフォームし、The QIAexpressionist(キアゲン社製)を用いて、キットのマニュアルに従って融合タンパク質を精製した。精製手順を簡略に説明すると、上記原核生物発現用プラスミドを大腸菌JM109(DE3)株にトランスフォームして、5mLのLB培地中に植菌した。一晩培養した後、遠心分離によって集菌し、集菌した大腸菌を溶解バッファー(50mM NaH2PO4, 300mM NaCl, 10mM イミダゾール, pH8.0)500μLで再懸濁してリゾチームを添加し(1mg/mL)、4℃で60分間インキュベートした。細胞を静かにボルテックスして溶解し、15,000xgで10分間遠心分離して上清を新しいチューブに移した。50% Ni−NTAレジン懸濁液 50μLを添加し、4℃で60分間静かに混合した。15,000xgで10秒間遠心してレジンを沈殿させ、レジンを洗浄バッファー(50mM NaH2PO4, 300mM NaCl, 20mM イミダゾール, pH8.0) 500μLで2回洗浄した。溶出バッファー(50mM NaH2PO4, 300mM NaCl, 250mM イミダゾール, pH8.0) 50μLで融合タンパク質を2回溶出し、融合タンパク質を得た。得られた融合タンパク質を図28に示す(図中、「得られるサイクリックAMPセンサータンパク質の名称」が参照される)。
【0123】
[手順7](融合タンパク質遺伝子の特性測定)
上記融合タンパク質の発光活性をcAMPの存在下および非存在下でそれぞれ測定した。測定方法を簡単に説明すると、上述の融合タンパク質溶液20μLに、リン酸緩衝液(PBS)20μLを加えた。さらに、4mMcAMP溶液を1μL加え(終濃度 100μM)、ネガティブコントロールにはPBSを同量加えた。Bright−Glo(プロメガ社製)を40μL加えて、Luminescencer JNR−II(アトー社製)で発光量(10秒間の積算)を測定した。
【0124】
[結果]
上記の通り、精製した融合タンパク質をPBSに希釈し、cAMP(終濃度100μM)の有無での発光量を測定した(図29)。その結果、cAMPが存在することで発光活性が上昇する融合タンパク質が存在することがわかった。cAMPの有無による活性の差が特に大きかったのは、GL4_N_RIα_β1AB_C(図29a中「β1AB」)およびcpGL4_C_RIα_αAB_Nであった((図29b中「αAB」)。
【産業上の利用可能性】
【0125】
以上のように、本発明にかかる受容体相互作用検出方法およびサイクリックAMPセンサータンパク質は、バイオ、製薬、医療など様々な分野で好適に用いることができる。
【0126】
また、以上のように、本発明にかかる5HT2A受容体とD1受容体の相互作用を検出するための発光タンパク質および解析方法は、バイオ、製薬、医療など様々な分野で好適に用いることができる。
【符号の説明】
【0127】
100 発光観察システム
103 容器(シャーレ)
104 ステージ
106 発光画像撮像ユニット
106a 対物レンズ(発光観察用)
106b ダイクロイックミラー
106c CCDカメラ
106d スプリットイメージユニット
106e フィルターホイール
106f 結像レンズ
110 画像解析装置
112 制御部
112a 発光画像撮像指示部
112b 発光画像取得部
112c 画像解析部
112d 解析結果出力部
114 クロック発生部
116 記憶部
118 通信インターフェース部
120 入出力インターフェース部
122 入力装置
124 出力装置
【技術分野】
【0001】
本発明は、細胞内の互いに異なる受容体同士の相互作用(例えばD1受容体と5HT2A受容体の相互作用)を、一方の受容体(例えばD1受容体)の活性(例えばサイクリックAMPの濃度)を一細胞ごとに非侵襲的に解析して検出する受容体相互作用検出方法、および互いに異なる受容体同士の相互作用を検出するためのサイクリックAMPセンサータンパク質に関するものである。
【背景技術】
【0002】
多くのホルモンや神経伝達物質は、Gタンパク質共役型受容体に働き、細胞内セカンドメッセンジャーを介して様々な細胞活動を引き起こしている。Gタンパク質共役型受容体に働く神経伝達物質は、ヒトの情動や気分、依存、認知、学習、記憶などの広範囲に亘る脳機能の調節に関わっており、なかでも、モノアミンおよびその受容体は、躁うつ病や統合失調症、気分障害、自閉症、注意欠陥/多動性障害などの精神疾患に対する治療の標的とされている。このような脳の高次機能は、脳内の伝達物質およびその受容体の時間的および空間的な濃度分布によって制御されている。したがって、特定の情動活動に関する情報伝達機構を解析するにあたって、脳内のGタンパク質共役型受容体の時間的および空間的な変化および相互作用をリアルタイムに捉えることが必要である。
【0003】
これまでに、脳内の受容体の時間的および空間的な変化および相互作用に関する研究において、情動活動に対する効果を示す薬物の存在が明らかにされ、その作用点がモノアミンのGタンパク質共役型受容体であることが解明されてきた経緯がある。モノアミンのGタンパク質共役型受容体が関与する従来のシグナル伝達の研究では、主にモデル動物を用いた行動薬理学的な手法で解析されてきた。
【0004】
例えば、Bodyらは、5HT2A受容体やD1受容体、D2受容体のサブタイプに対する選択的阻害剤を用いて、ラットの代表的なオペラント条件付けであるタイミング行動を制御する方法を開示している(非特許文献1参照)。タイミング行動に対するd−アンフェタミンの効果が、5HT2A受容体およびD1受容体に対する選択的阻害剤で阻害されることから、タイミング行動に対するd−アンフェタミンの作用には5HT2A受容体およびD1受容体が関わっていることを明らかにしている。
【0005】
また、例えば、Auclairらは、5HT2A受容体に対する選択的阻害剤を用いて、ラットにおけるd−アンフェタミン誘導性の自発運動活性を制御する方法を開示している(非特許文献2参照)。d−アンフェタミンは、脳内においてドーパミン遊離を起こさせ、ドーパミン誘導性の自発運動活性の上昇を引き起こす。5HT2A受容体に対する選択的阻害剤が、ドーパミン遊離を抑制せずにドーパミン誘導性の自発運動活性の上昇を抑制することを示しており、ドーパミン受容体と5HT2A受容体との相互作用を明らかにしている。
【先行技術文献】
【非特許文献】
【0006】
【非特許文献1】Body S., Cheung T.H.C., Bezzina G., Asgari K., Fone K.C.F., Glennon J.C., Bradshaw C.M., Szababi E., “Effects of d−amphatamine and DOI (2,5−dimethoxy−4−iodoamphetamine) on timing behavior: intaraction between D1 and 5−HT2A receptors”, Psycopharmacology, Vol.189, pp331−343, 2006
【非特許文献2】Auclair A., Blanc G., Glowinski J., Tassin J.−P., “Role of serotonin2A receptors in d−amphetamine−induced release of dopamine: comparison with previous data on alpha1b−adrenergic receptors”, Journal of Neurochemistry, vol.91, pp318−326, 2004
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、モデル動物を用いた行動薬理学的な手法を用いて異なるGタンパク質共役型受容体の時間的および空間的な相互作用を解析する従来の方法では、受容体の空間的な配置が不明のまま相互作用の解析を行っているため、受容体同士の相互作用を正確には示していない、という問題点があった。
【0008】
本発明は、上記問題点に鑑みてなされたもので、受容体の空間的な配置を明らかにして受容体の相互作用に関する各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞において特定の受容体の相互作用を再現性良く観察することができる受容体相互作用検出方法および受容体の相互作用を検出するためのサイクリックAMPセンサータンパク質を提供することを目的とする。
【課題を解決するための手段】
【0009】
上述した課題を解決し、目的を達成するために、本発明にかかる受容体相互作用検出方法は、細胞において第1の受容体と第2の受容体との相互作用を検出する受容体相互作用検出方法であって、前記第2の受容体の活性を示す指標となるレポータを含む前記細胞を作製する作製工程と、前記作製工程で作製した前記細胞に当該細胞外から、発光させるための所定の処置を行うと共に前記第1の受容体に結合する所定の受容体結合物質を添加する処置および添加工程と、前記処置および添加工程が行われた後の前記細胞の発光量を測定する測定工程と、前記測定工程で測定した前記発光量に基づいて前記細胞内における前記第2の受容体の活性を解析する解析工程と、前記解析工程で解析した前記第2の受容体の活性に基づいて、前記第1の受容体と前記第2の受容体との前記相互作用を検出する検出工程と、を含むことを特徴とする。ここで、レポータとは、遺伝子の発現に応じて光を発生し得る蛍光タンパク質や発光タンパク質を包含するものであり、具体的にはサイクリックAMP(adenosine monophosphate)センサータンパク質などである。また、発光タンパク質とは、具体的には生物発光タンパク質などである。また、発光させるための所定の処置とは、例えば発光基質を添加する(または予め細胞等の試料中に供給しておく)ことや励起光を照射することなどである。生物発光共鳴エネルギー転移(いわゆるBRET)による蛍光(例えばGFP、YFP等)を測定する場合には、生物発光用の基質を添加等することによって励起光の照射は不要である。
【0010】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記作製工程において、前記レポータは、サイクリックAMPセンサータンパク質であり、前記解析工程において、前記第2の受容体の活性はサイクリックAMPの濃度で解析すること、を特徴とする。
【0011】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記サイクリックAMPセンサータンパク質は、前記細胞内のサイクリックAMPと相互作用して発光するものであること、を特徴とする。
【0012】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記サイクリックAMPセンサータンパク質は、ホタルのルシフェラーゼおよびPKA(protein kinase A)のサイクリックAMP結合領域とからなる融合タンパク質であること、を特徴とする。
【0013】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記サイクリックAMP結合領域が、マウスのPKA−RIα由来であることを特徴とする。
【0014】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記サイクリックAMP結合領域が、220から250のアミノ酸残基の長さを有することを特徴とする。
【0015】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記サイクリックAMP結合領域が、少なくとも2つのサイクリックAMP結合部位を含むことを特徴とする。
【0016】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記サイクリックAMPセンサータンパク質は、前記ルシフェラーゼのN末端側断片、前記ルシフェラーゼのC末端側断片および前記サイクリックAMP結合領域を含み、前記N末端側断片と前記C末端側断片とが前記サイクリックAMPセンサータンパク質分子内で結合すると、前記ルシフェラーゼの発光酵素活性が回復することを特徴とする。
【0017】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記サイクリックAMP結合領域が、配列番号37または39に記載のアミノ酸配列を含むことを特徴とする。
【0018】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記作製工程において、前記細胞は前記第1の受容体と前記第2の受容体とを含有するものであること、を特徴とする。
【0019】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記作製工程において、前記細胞は前記第1の受容体の遺伝子と前記第2の受容体の遺伝子とが導入されたものであること、を特徴とする。
【0020】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記測定工程は、前記第2の受容体の活性に依存する発光標識の光子量を測定する光子量測定工程をさらに含むこと、を特徴とする。
【0021】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記測定工程は、前記第2の受容体の活性に依存する発光標識を撮像する撮像工程をさらに含むこと、を特徴とする。
【0022】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記撮像工程は、複数の異なる前記細胞に対して同時に実行され、前記解析工程は、前記撮像工程で撮像した複数の発光画像を前記細胞ごとに照合する照合工程をさらに含むこと、を特徴とする。
【0023】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記測定工程は繰り返し実行されること、を特徴とする。
【0024】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記第1の受容体は5HT2A受容体であり、前記第2の受容体はD1受容体であること、を特徴とする。
【0025】
また、本発明にかかる受容体相互作用検出方法は、前記に記載の受容体相互作用検出方法において、前記処置および添加工程において、前記受容体結合物質はセロトニンであること、を特徴とする。
【0026】
また、本発明はサイクリックAMPセンサータンパク質に関するものであり、本発明にかかるサイクリックAMPセンサータンパク質は、第1の受容体と第2の受容体との相互作用を検出するためのものであること、を特徴とする。
【0027】
また、本発明にかかるサイクリックAMPセンサータンパク質は、前記に記載のサイクリックAMPセンサータンパク質において、サイクリックAMP(adenosine monophosphate)と相互作用して発光するものであること、を特徴とする。
【0028】
また、本発明にかかるサイクリックAMPセンサータンパク質は、前記に記載のサイクリックAMPセンサータンパク質において、前記第1の受容体は5HT2A受容体であり、前記第2の受容体はD1受容体であること、を特徴とする。
【発明の効果】
【0029】
この発明によれば、第2の受容体の活性を示す指標となるレポータを含む細胞を作製し、作製した細胞に当該細胞外から、発光させるための所定の処置を行うと共に第1の受容体に結合する所定の受容体結合物質を添加し、当該処置および添加が行われた後の細胞の発光量を測定し、測定した発光量に基づいて細胞内における第2の受容体の活性を解析し、解析した第2の受容体の活性に基づいて、第1の受容体と第2の受容体との相互作用を検出するので、受容体の空間的な配置を明らかにして受容体の相互作用に関する各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞において特定の受容体の相互作用を再現性良く観察することができるという効果を奏する。
【0030】
この発明によれば、レポータはサイクリックAMPセンサータンパク質であり、第2の受容体の活性はサイクリックAMPの濃度で解析するので、生物発光によるサイクリックAMP検出原理を利用して受容体の空間的な配置を明らかにして受容体の相互作用に関する各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞において特定の受容体の相互作用を再現性良く観察することができるという効果を奏する。
【0031】
この発明によれば、サイクリックAMPセンサータンパク質は、細胞内のサイクリックAMPと相互作用して発光するものであるので、サイクリックAMPの存在下で相互作用が強化された発光によりサイクリックAMPの濃度を検出することができるという効果を奏する。
【0032】
この発明によれば、サイクリックAMPセンサータンパク質は、ホタルのルシフェラーゼおよびPKA(protein kinase A)のサイクリックAMP結合領域とからなる融合タンパク質であるので、発光量を測ればサイクリックAMP濃度を算出することができるという効果を奏する。
【0033】
この発明によれば、サイクリックAMP結合領域が、マウスのPKA−RIα由来であるので、哺乳類由来の細胞において使用することができるという効果を奏する。
【0034】
この発明によれば、サイクリックAMP結合領域が、220から250のアミノ酸残基の長さを有するので、ルシフェラーゼの活性をなるべく維持したまま融合タンパク質を作製できるという効果を奏する。
【0035】
この発明によれば、サイクリックAMP結合領域が、少なくとも2つのサイクリックAMP結合部位を含むので、サイクリックAMP濃度の変化によって、ルシフェラーゼ活性を変化させることができるという効果を奏する。
【0036】
この発明によれば、サイクリックAMPセンサータンパク質は、ルシフェラーゼのN末端側断片、ルシフェラーゼのC末端側断片およびサイクリックAMP結合領域を含み、N末端側断片とC末端側断片とが前記サイクリックAMPセンサータンパク質分子内で結合すると、ルシフェラーゼの発光酵素活性が回復するので、サイクリックAMP結合領域の構造変化により発光強度を変化させることができるという効果を奏する。
【0037】
この発明によれば、サイクリックAMP結合領域が、配列番号37または39に記載のアミノ酸配列を含むので、条件に応じてサイクリックAMPセンサータンパク質として最適なものを選択できるという効果を奏する。
【0038】
この発明によれば、細胞は第1の受容体と第2の受容体とを含有するものであるので、解析対象となる受容体を人工的に含めた扱い易い細胞を用いて実験をやり易くすることができるという効果を奏する。
【0039】
この発明によれば、細胞は第1の受容体の遺伝子と第2の受容体の遺伝子とが導入されたものであるので、解析対象となる受容体を人工的に含めた扱い易い細胞を用いて実験をやり易くすることができる他、自然に近い応答を得ることができるという効果を奏する。
【0040】
この発明によれば、第2の受容体の活性に依存する発光標識の光子量を測定するので、フォトンカウンティングの技術を利用して細胞全体または細胞集団全体からの発光量を得ることができるという効果を奏する。
【0041】
この発明によれば、第2の受容体の活性に依存する発光標識を撮像するので、発光顕微鏡を利用して個々の細胞からの発光量を得ることができるという効果を奏する。
【0042】
この発明によれば、撮像を複数の異なる細胞に対して同時に実行し、撮像した複数の発光画像を細胞ごとに照合するので、細胞一つ一つからの発光量を個別に且つ同時に得ることができるという効果を奏する。
【0043】
この発明によれば、発光量の測定を繰り返し実行するので、発光量の経時変化を観察することができるという効果を奏する。
【0044】
この発明によれば、第1の受容体は5HT2A受容体であり、第2の受容体はD1受容体であるので、D1受容体および5HT2A受容体の空間的な配置を明らかにしてD1受容体と5HT2A受容体の相互作用に関する各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞においてD1受容体と5HT2A受容体の相互作用を再現性良く観察することができるという効果を奏する。
【0045】
この発明によれば、受容体結合物質はセロトニンであるので、5HT2A受容体の活性を観察することができるという効果を奏する。
【0046】
この発明によれば、サイクリックAMPセンサータンパク質は、第1の受容体と第2の受容体との相互作用を検出するためのものであるので、当該サイクリックAMPセンサータンパク質を用いれば、受容体の空間的な配置を明らかにして受容体の相互作用に関する各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞において特定の受容体の相互作用を再現性良く観察することができるという効果を奏する。
【0047】
この発明によれば、サイクリックAMPセンサータンパク質はサイクリックAMP(adenosine monophosphate)と相互作用して発光するものであるので、当該サイクリックAMPセンサータンパク質を用いれば、サイクリックAMPの存在下で相互作用が強化された発光によりサイクリックAMP濃度を検出することができるという効果を奏する。
【0048】
この発明によれば、サイクリックAMPセンサータンパク質はD1受容体と5HT2A受容体との相互作用を検出するためのものであるので、当該サイクリックAMPセンサータンパク質を用いれば、D1受容体および5HT2A受容体の空間的な配置を明らかにしてD1受容体と5HT2A受容体の相互作用に関する各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞においてD1受容体と5HT2A受容体の相互作用を再現性良く観察することができるという効果を奏する。
【図面の簡単な説明】
【0049】
【図1】図1は、発光観察システム100の全体構成の一例を示す図である。
【図2】図2は、発光観察システム100の発光画像撮像ユニット106の構成の一例を示す図である。
【図3】図3は、発光観察システム100の発光画像撮像ユニット106の構成の一例を示す図である。
【図4】図4は、発光観察システム100の画像解析装置110の構成の一例を示すブロック図である。
【図5】図5は、スプリットルシフェラーゼを用いたサイクリックAMPの検出原理を示す図である。
【図6】図6は、サイクリックAMPセンサータンパク質、5HT2A受容体およびD1受容体が導入されたHEK293細胞を含むセロトニン刺激前の発光画像を示す図である。
【図7】図7は、サイクリックAMPセンサータンパク質、5HT2A受容体およびD1受容体が導入されたHEK293細胞を含むセロトニン刺激5分後の発光画像を示す図である。
【図8】図8は、サイクリックAMPセンサータンパク質、5HT2A受容体およびD1受容体が導入されたHEK293細胞を含むドーパミン刺激5分後の発光画像を示す図である。
【図9】図9は、5HT2A受容体およびD1受容体が導入された、選択されたHEK293細胞におけるセロトニン刺激およびドーパミン刺激によるサイクリックAMPセンサータンパク質の発光強度の経時変化を示した図である。
【図10】図10は、サイクリックAMPセンサータンパク質、5HT2A受容体およびD2受容体が導入されたHEK293細胞を含むセロトニン刺激前の発光画像を示す図である。
【図11】図11は、サイクリックAMPセンサータンパク質、5HT2A受容体およびD2受容体が導入されたHEK293細胞を含むセロトニン刺激5分後の発光画像を示す図である。
【図12】図12は、サイクリックAMPセンサータンパク質、5HT2A受容体およびD2受容体が導入されたHEK293細胞を含むドーパミン刺激5分後の発光画像を示す図である。
【図13】図13は、サイクリックAMPセンサータンパク質、5HT2A受容体およびD2受容体が導入されたHEK293細胞を含むフォルスコリン刺激5分後の発光画像を示す図である。
【図14】図14は、5HT2A受容体およびD2受容体が導入された、選択されたHEK293細胞におけるセロトニン刺激、ドーパミン刺激およびフォルスコリン刺激によるサイクリックAMPセンサータンパク質の発光強度の経時変化を示した図である。
【図15】図15は、サイクリックAMPセンサータンパク質および5HT2A受容体が導入されたHEK293細胞を含むセロトニン刺激前の発光画像を示す図である。
【図16】図16は、サイクリックAMPセンサータンパク質および5HT2A受容体が導入されたHEK293細胞を含むセロトニン刺激5分後の発光画像を示す図である。
【図17】図17は、サイクリックAMPセンサータンパク質および5HT2A受容体が導入されたHEK293細胞を含むフォルスコリン刺激5分後の発光画像を示す図である。
【図18】図18は、5HT2A受容体が導入された、選択されたHEK293細胞におけるセロトニン刺激およびドーパミン刺激によるサイクリックAMPセンサータンパク質の発光強度の経時変化を示した図である。
【図19】図19は、変異型のスプリットルシフェラーゼを用いたサイクリックAMPの検出原理を示す図である。
【図20】図20は、cpGL4_C_RIα_αAB_Nおよび、5HT2A受容体、D1受容体を導入したHEK293細胞において、セロトニン刺激前の発光画像を示す図である。
【図21】図21は、cpGL4_C_RIα_αAB_Nおよび、5HT2A受容体、D1受容体を導入したHEK293細胞において、セロトニン刺激5分後の発光画像を示す図である。
【図22】図22は、cpGL4_C_RIα_αAB_Nおよび、5HT2A受容体、D1受容体を導入したHEK293細胞において、ドーパミン刺激5分後の発光画像を示す図である。
【図23】図23は、5HT2A受容体、D1受容体を導入したHEK293細胞において、選択した細胞でのセロトニンおよびドーパミン刺激によるcpGL4_C_RIα_αAB_Nの発光強度の経時変化を示した図である。
【図24】図24は、cpGL4_C_RIα_αAB_Nおよび、5HT2A受容体を導入したHEK293細胞において、セロトニン刺激前の発光画像を示す図である。
【図25】図25は、cpGL4_C_RIα_αAB_Nおよび、5HT2A受容体を導入したHEK293細胞において、セロトニン刺激5分後の発光画像を示す図である。
【図26】図26は、cpGL4_C_RIα_αAB_Nおよび、5HT2A受容体を導入したHEK293細胞において、ドーパミン刺激5分後の発光画像を示す図である。
【図27】図27は、5HT2A受容体を導入したHEK293細胞において、選択した細胞でのセロトニンおよびドーパミン刺激によるcpGL4_C_RIα_αAB_Nの発光強度の経時変化を示した図である。
【図28】図28は、作製した種々のサイクリックAMPセンサータンパク質の詳細を示す表である。
【図29】図29は、種々のサイクリックAMPセンサータンパク質の活性を比較した図である。
【発明を実施するための形態】
【0050】
以下に、本発明にかかる受容体相互作用検出方法および受容体の相互作用を検出するためのサイクリックAMPセンサータンパク質の実施の形態を図面に基づいて詳細に説明する。なお、本実施の形態により本発明が限定されるものではない。ここで、本実施の形態では、発光タンパク質(具体的には生物発光タンパク質)を用いてD1受容体と5HT2A受容体との相互作用を発光画像で検出する場合を一例として説明するが、蛍光タンパク質や他の受容体同士やフォトンカウンティングに対しても同様に実施することができる。
【0051】
[構成]
まず、本発明にかかる受容体相互作用検出方法で用いる発光観察システム100の構成について、図1、図2および図3を参照して説明する。図1は、発光観察システム100の全体構成の一例を示す図である。
【0052】
図1に示すように、発光観察システム100は、細胞102を収納した容器103(具体的にはシャーレ、スライドガラス、マイクロプレート、ゲル支持体、微粒子担体など)と、容器103を配置するステージ104と、微弱な発光を測定するための発光画像撮像ユニット106と、画像解析装置110と、で構成されている。発光画像撮像ユニット106をステージ104の下側に配置してもよい。これにより、カバー(図示せず)の開閉によるサンプル上方からの外乱光を完全に遮断でき、その結果、発光画像のS/N比を増すことができる。発光画像撮像ユニット106は、レーザー走査式の光学系であってもよい。
【0053】
細胞102は、例えば、サイクリックAMP(adenosine monophosphate)濃度に依存して発光するよう発光標識された生きた細胞である。ここで、サイクリックAMP濃度に依存して発光するタンパク質(発光タンパク質)として、N末側発光酵素とC末側発光酵素、およびサイクリックAMP結合タンパク質との融合タンパク質の他に、例えば国際公開第2007/120522号または特表2007−508014号公報に開示されている融合タンパク質を用いてもよい。また、ここで、細胞102に、当該融合タンパク質が発現されるよう構成した、当該融合タンパク質をコードするポリヌクレオチドを含む発現ベクターを細胞102に導入することにより、当該融合タンパク質が細胞102内で発現されるよう構成してもよい。なお、細胞102には、当該細胞102外から所定の発光基質(例えばルシフェリンなど)および所定の刺激(例えば薬物刺激など)が与えられる。
【0054】
発光画像撮像ユニット106は、具体的には正立型の発光顕微鏡であり、細胞102の発光画像を撮像する。発光画像撮像ユニット106は、図示の如く、対物レンズ106aと、ダイクロイックミラー106bと、CCD(charge−coupled device)カメラ106cと、結像レンズ106fと、で構成されている。対物レンズ106aは、具体的には(開口数/倍率)2の値が0.01以上のものが好ましい。ダイクロイックミラー106bは、細胞102から発せられた発光を色別に分離し、2色の発光を用いて発光量や発光強度を色別に測定する場合に用いる。CCDカメラ106cは、対物レンズ106a、ダイクロイックミラー106bおよび結像レンズ106fを介して当該CCDカメラ106cのチップ面に投影された細胞102の発光画像および明視野画像を撮る。また、CCDカメラ106cは、画像解析装置110と有線または無線で通信可能に接続される。ここで、細胞102が撮像範囲中に複数存在する場合、CCDカメラ106cは、当該撮像範囲中に含まれる複数の細胞102の発光画像および明視野画像を撮像してもよい。結像レンズ106fは、対物レンズ106aおよびダイクロイックミラー106bを介して当該結像レンズ106fに入射した像(具体的には細胞102を含む像)を結像する。なお、図1では、ダイクロイックミラー106bで分離した2つの発光に対応する発光画像を2台のCCDカメラ106cで別々に撮像する場合の一例を示しており、1つの発光を用いる場合には、発光画像撮像ユニット106は、対物レンズ106a、1台のCCDカメラ106cおよび結像レンズ106fで構成されてもよい。
【0055】
ここで、2色の発光を用いて発光量や発光強度を色別に測定する場合、発光画像撮像ユニット106は、図2に示すように、対物レンズ106aと、CCDカメラ106cと、スプリットイメージユニット106dと、結像レンズ106fと、で構成されてもよい。そして、CCDカメラ106cは、スプリットイメージユニット106dおよび結像レンズ106fを介して当該CCDカメラ106cのチップ面に投影された細胞102の発光画像(スプリットイメージ)および明視野像を撮像してもよい。スプリットイメージユニット106dは、細胞102から発せられた発光を色別に分離し、ダイクロイックミラー106bと同様、2色の発光を用いて発光量や発光強度を色別に測定する場合に用いる。
【0056】
また、複数色の発光を用いて発光量や発光強度を色別に測定する場合(つまり、多色の発光を用いる場合)、発光画像撮像ユニット106は、図3に示すように、対物レンズ106aと、CCDカメラ106cと、フィルターホイール106eと、結像レンズ106fと、で構成されてもよい。そして、CCDカメラ106cは、フィルターホイール106eおよび結像レンズ106fを介して当該CCDカメラ106cのチップ面に投影された細胞102の発光画像および明視野画像を撮像してもよい。フィルターホイール106eは、細胞102から発せられた発光をフィルタ交換によって色別に分離し、複数色の発光を用いて発光量や発光強度を色別に測定する場合に用いる。
【0057】
図1に戻り、画像解析装置110は、具体的にはパーソナルコンピュータである。そして、画像解析装置110は、図4に示すように、大別して、制御部112と、システムの時刻を計時するクロック発生部114と、記憶部116と、通信インターフェース部118と、入出力インターフェース部120と、入力装置122と、出力装置124と、で構成されており、これら各部はバスを介して接続されている。
【0058】
記憶部116は、ストレージ手段であり、具体的には、RAM(random access memory)やROM(read−only memory)等のメモリ装置、ハードディスクのような固定ディスク装置、フレキシブルディスク、光ディスク等を用いることができる。そして、記憶部116は制御部112の各部の処理により得られたデータなどを記憶する。通信インターフェース部118は、画像解析装置110と、CCDカメラ106cと、の間における通信を媒介する。すなわち、通信インターフェース部118は他の端末と有線または無線の通信回線を介してデータを通信する機能を有する。入出力インターフェース部120は、入力装置122や出力装置124に接続する。ここで、出力装置124には、モニタ(家庭用テレビを含む)の他、スピーカやプリンタを用いることができる(なお、以下で、出力装置124をモニターとして記載する場合がある。)。また、入力装置122には、キーボードやマウスやマイクの他、マウスと協働してポインティングデバイス機能を実現するモニターを用いることができる。
【0059】
制御部112は、OS(Operating System)等の制御プログラムや各種の処理手順等を規定したプログラムや所要データを格納するための内部メモリを有し、これらのプログラムに基づいて種々の処理を実行する。そして、制御部112は、大別して、発光画像撮像指示部112aと、発光画像取得部112bと、画像解析部112cと、解析結果出力部112dと、で構成されている。
【0060】
発光画像撮像指示部112aは、通信インターフェース部118を介して、CCDカメラ106cへ発光画像および明視野画像の撮像を指示する。発光画像取得部112bは、CCDカメラ106cで撮像した発光画像および明視野画像を、通信インターフェース部118を介して取得する。制御部112は、発光画像撮像指示部112aを制御して、CCDカメラ106cに細胞102の発光画像および明視野画像を繰り返し撮像させ、発光画像取得部112bを制御して、撮像した発光画像および明視野画像を、撮像される度に又は全撮像が終了後纏めて取得する。
【0061】
画像解析部112cは、発光画像取得部112bで取得した複数の発光画像に基づいて、細胞102から発せられる発光の発光強度を経時的に測定する。解析結果出力部112dは、画像解析部112cでの解析結果を出力装置124に出力する。具体的には、解析結果出力部112dは、画像解析部112cで得られた、細胞102から発せられる発光の発光強度に関する時系列データを、グラフ化して出力装置124に表示する。
【0062】
[受容体相互作用検出方法]
本発明は、細胞において第1の受容体と第2の受容体との相互作用を検出する受容体相互作用検出方法であって、前記第2の受容体の活性を示す指標となるレポータを含む前記細胞を作製する作製工程と、前記作製工程で作製した前記細胞に当該細胞外から、発光させるための所定の処置を行うと共に前記第1の受容体に結合する所定の受容体結合物質を添加する処置および添加工程と、前記処置および添加工程が行われた後の前記細胞の発光量を測定する測定工程と、前記測定工程で測定した前記発光量に基づいて前記細胞内における前記第2の受容体の活性を解析する解析工程と、前記解析工程で解析した前記第2の受容体の活性に基づいて、前記第1の受容体と前記第2の受容体との前記相互作用を検出する検出工程とを含むことを特徴とする受容体相互作用検出方法に関する。
【0063】
作製工程では、レポータを含む細胞が作製される。このレポータとは、後述するようなサイクリックAMPセンサータンパク質であってよい。対象となる細胞の種類に限定はなく、細菌細胞、酵母細胞、植物細胞および動物細胞等を使用できる。動物細胞が使用される場合、特に哺乳細胞が使用され、例えばマウスの細胞、サルの細胞およびヒトの細胞が使用される。細胞に対してレポータを導入する方法に特別な限定はなく、既知の導入方法が使用できる。細胞は、生体組織の一部として培養された細胞集合体であってもよいし、生体組織が属している臓器等や生命個体の形態でもよい。レポータがタンパク質である場合、1つの方法は、当該タンパク質をコードする塩基配列を含む又は当該配列から成る核酸を細胞に導入し、その後細胞内で当該タンパク質を発現させる方法である。例えば、当該核酸を含む発現ベクターを、リン酸カルシウム法、リポフェクション法またはエレクトロポレーション法等によって細胞内に導入し、発現ベクターから当該タンパク質を発現させることができる。ベクターから発現させるのではなく、当該核酸がゲノムに組み込まれた細胞を作製した後、ゲノムから当該タンパク質を発現させることもできる。別の方法は、細胞外で精製した当該タンパク質を細胞内に直接導入する方法である。例えば、マイクロインジェクション法によって当該タンパク質を細胞内に直接注入することができる。または、当該タンパク質を含む培養液にて細胞をインキュベートさせて、エンドサイトーシスによって当該タンパク質を細胞に取り込ませることができる。
【0064】
処置および添加工程では、発光させるための所定の処置および第1の受容体に結合する所定の受容体結合物質の添加が行われる。発光させるための所定の処置とは、例えば発光基質を添加することや励起光を照射することなどである。発光基質を添加する方法に特別な限定はなく、適宜選択できる。好ましくは、発光基質は測定の直前に添加される。発光基質の添加量に特別な限定はないが、好ましくは、発光酵素が発光反応を生じるために十分な量で添加される。第1の受容体に結合する所定の受容体結合物質の添加によって、当該受容体は活性化される。第1の受容体は例えば5HT2A受容体であってよく、受容体結合物質はセロトニン、例えば5HTとすることができる。
【0065】
測定工程では、処置および添加工程によって細胞から生じる発光が測定される。測定はイメージングまたはフォトカウンティングによって行うことができる。イメージングを行う場合、発光反応が生じている細胞の画像が取得されるので、細胞ごとの発光を測定するのに都合が良い。撮像は任意のタイミングで行うことができるが、好ましくは、基質添加から一定時間後に行われる。特に複数の測定を繰り返す場合、測定結果のばらつきを抑えるために、複数の測定を通じて同一のタイミングで撮像することが好ましい。撮像時間は条件に応じて最良の時間が適宜選択される。発光強度が小さい場合、撮像時間(顕微鏡を使用する場合、露光時間)は長くされ、逆に発光強度が大きすぎる場合、撮像時間は短くされる。撮像は、発光反応に係る波長の光のみを取得して行うこともできるが、それと同時に明視野画像または別の波長の光による画像を取得してもよい。例えば、細胞にサイクリックAMPセンサータンパク質と同時にプローブ(例えば蛍光または発光タンパク質)を融合した特定のタンパク質を導入し、サイクリックAMPセンサータンパク質に由来する光と同時に、当該特定のタンパク質からの光を検出してもよい。
【0066】
解析工程では、第2の受容体の活性が解析される。測定工程において得られた測定結果に基づき、発光が生じたかどうかが判定される。発光が生じた場合、第2の受容体が活性化したと判定される。好ましくは、第2の受容体として、D1受容体またはD2受容体を選択することができる。D1受容体は、リガンドが結合するとアデニル酸シクラーゼを活性化し、細胞内のサイクリックAMP濃度を上昇させる作用を有する。一方、D2受容体は、リガンドが結合するとホスホジエステラーゼを活性化し、細胞内のサイクリックAMP濃度を低下させる作用を有する。第2の受容体は、アデニル酸シクラーゼを活性化する作用を有するか否かが判明している受容体であることが好ましい。
【0067】
検出工程では、第2の受容体の活性の解析結果に基づいて、第1の受容体と第2の受容体との前記相互作用が検出される。処置および添加工程において第1の受容体に受容体結合物質を与えた結果、解析工程において第2の受容体の活性が認められた場合、第1の受容体と第2の受容体との相互作用に対して肯定的な結論が得られる。一方、解析工程において第2の受容体の活性が認められなかった場合、第1の受容体と第2の受容体との相互作用に対して否定的な結論が得られる。
【0068】
[サイクリックAMPセンサータンパク質]
次に、サイクリックAMPセンサータンパク質について補足的に説明する。
【0069】
本発明において、第2の受容体の活性を示す指標となるレポータとして、サイクリックAMPセンサータンパク質を使用することができる。当該サイクリックAMPセンサータンパク質として、ホタルのルシフェラーゼおよびPKAのサイクリックAMP結合領域とからなる融合タンパク質を使用することができる。
【0070】
サイクリックAMP結合領域としては、マウスのPKA−RIα由来のものを使用することができる。ここで、PKA−RIαとは、PKAの制御サブユニットαのことを意味する。PKA−RIαは、サイクリックAMP結合ドメインを有しており、サイクリックAMPに結合する能力を有する。
【0071】
サイクリックAMP結合領域の長さに関して、そのアミノ酸残基の数を120から250としてもよい。また、アミノ酸残基の数を200から250、220から250、230から250、230から245、または230から240としてもよい。例えば、アミノ酸残基数を230または241としてもよい。
【0072】
サイクリックAMP結合領域としては、少なくとも2つのサイクリックAMP結合部位を含むものを使用することができる。「サイクリックAMP結合部位」とは、サイクリックAMP1分子が結合することができるサイクリックAMP結合領域中の部位として定義される。サイクリックAMP結合部位には、前述のPKA−RIαのサイクリックAMP結合ドメインが含まれる。また、上記定義を満たせば、サイクリックAMP結合ドメインとして特定または認識されていない部位であってもサイクリックAMP結合部位に含まれる。
【0073】
本発明に係る方法において、ルシフェラーゼのN末端側断片、ルシフェラーゼのC末端側断片およびサイクリックAMP結合領域を含むサイクリックAMPセンサータンパク質であって、N末端側断片とC末端側断片とがサイクリックAMPセンサータンパク質分子内で結合すると、ルシフェラーゼの発光酵素活性が回復するサイクリックAMPセンサータンパク質を使用することができる。「ルシフェラーゼのN末端側断片」とは、C末端から複数のアミノ酸が欠失したルシフェラーゼ断片であって、発光酵素活性を失ったルシフェラーゼ断片を意味する。「ルシフェラーゼのC末端側断片」とは、N末端から複数のアミノ酸が欠失したルシフェラーゼ断片であって、発光酵素活性を失ったルシフェラーゼ断片を意味する。サイクリックAMPセンサータンパク質がそれらの断片およびサイクリックAMP結合領域を「含む」とは、サイクリックAMPセンサータンパク質が、ルシフェラーゼのN末端側断片、ルシフェラーゼのC末端側断片、サイクリックAMP結合領域および任意にその他のポリペプチドから成る融合タンパク質であることを意味する。したがって、これらの構成要素が側鎖または修飾基として含まれるわけではない。このようなサイクリックAMPセンサータンパク質の例は、図5に示されるような、サイクリックAMPセンサータンパク質のN末端側にルシフェラーゼのN末端側断片が存在し、サイクリックAMPセンサータンパク質のC末端側にルシフェラーゼのC末端側断片が存在し、これらの断片の間にPKA−RIα由来のサイクリックAMP結合領域が存在するサイクリックAMPセンサータンパク質である。また、別の例は、図19に示されるような、サイクリックAMPセンサータンパク質のN末端側にルシフェラーゼのC末端側断片が存在し、サイクリックAMPセンサータンパク質のC末端側にルシフェラーゼのN末端側断片が存在し、これらの断片の間にPKA−RIα由来のサイクリックAMP結合領域が存在するサイクリックAMPセンサータンパク質である。
【0074】
N末端側断片は、例えばルシフェラーゼの1番目から400番目のアミノ酸を含む断片であってよく、C末端側断片は、例えばルシフェラーゼの401番目から550番目のアミノ酸を含むものであってよい。特に、N末端側断片がルシフェラーゼの1番目から416番目のアミノ酸を含み、C末端側断片がルシフェラーゼの399番目から550番目のアミノ酸を含むことが好ましい。このように、N末端側断片およびC末端側断片は、それらの配列の一部が重複していてもよい。また、N末端側断片にもC末端側断片にも含まれないルシフェラーゼ配列部分が存在してもよい。
【0075】
サイクリックAMPセンサータンパク質内において、ルシフェラーゼがN末端側断片とC末端側断片とに分かれたことにより、通常の環境下(例えばサイクリックAMPが存在していない環境下)では、サイクリックAMPセンサータンパク質はルシフェラーゼの活性を有さない。しかし、特定の環境下(例えばサイクリックAMPが存在する環境下)では、サイクリックAMPセンサータンパク質はルシフェラーゼの活性を回復する。この回復は、サイクリックAMP結合領域の機能に起因する。例えば、マウスのPKA−RIα由来のサイクリックAMP結合領域を選択した場合、図5および19に示されるように、サイクリックAMP結合領域にサイクリックAMPが結合すると、サイクリックAMPセンサータンパク質全体の立体構造が変化し、結果としてN末端側断片とC末端側断片とが接近する。これらの2つの断片が協同してルシフェラーゼ活性を奏する。
【0076】
「N末端側断片とC末端側断片とが前記サイクリックAMPセンサータンパク質分子内で結合すると発光酵素活性が回復する」という表現において、N末端側断片とC末端側断片とが「結合」する態様に限定はなく、共有結合または非共有結合等の化学的結合であってもよいが、N末端側断片とC末端側断片とがルシフェラーゼの本来の機能を果たしうる程度に「接近」または「接触」している状態であることが好ましい。あるいは、N末端側断片とC末端側断片とが「会合」する状態であってもよい。すなわち、当該表現において使用される「結合」という用語は、「接近」、「接触」または「会合」という意味を含む。
【0077】
また、上記表現において、N末端側断片とC末端側断片との結合は、サイクリックAMPに応答して生じる結合であってよい。より具体的には、サイクリックAMP結合領域にサイクリックAMP分子が結合することで、サイクリックAMP結合領域の構造の変化が生じ、その結果生じるN末端側断片とC末端側断片との結合または接近であってよい。
【0078】
なお、サイクリックAMP結合領域としては、好ましくは配列番号37または39に示されるアミノ酸配列を含むポリペプチドである。
【実施例】
【0079】
[実施例1]
本実施の形態における実施例について、以下に図5〜図9を用いて説明する。ここで、図5はスプリットルシフェラーゼアッセイ法を用いたサイクリックAMPの検出原理を示す図である。
【0080】
スプリットルシフェラーゼアッセイとは、ホタルルシフェラーゼやウミシイタケルシフェラーゼ等のルシフェラーゼを特定の位置で分割して発光酵素活性(生物発光能)を失活させた後、そのフラグメント(断片)であるN末側発光酵素とC末側発光酵素を再構成して発光酵素活性を回復させる手法であり、タンパク質間相互作用の検出やタンパク質の細胞内小器官移行検出等に用いられる。
【0081】
図5には、N末側発光酵素(NLuc)とC末側発光酵素(CLuc)との非共有結合的相互作用を起こさせるタンパク質として、サイクリックAMP結合タンパク質を用いた例を示している。すなわち、細胞内で発現させた、N末側発光酵素とC末側発光酵素およびサイクリックAMP結合タンパク質との融合タンパク質における当該サイクリックAMP結合タンパク質にサイクリックAMPが結合すると、当該サイクリックAMP結合タンパク質の立体構造が変化してN末側発光酵素とC末側発光酵素とが結合し、発光酵素活性が回復して検出可能なシグナルが発せられる。本実施例では、当該スプリットルシフェラーゼアッセイ系において、サイクリックAMP結合タンパク質にPKA(protein kinase A)の制御サブユニットIα(PKA−RIα)のサイクリックAMP結合ドメインを用いた例について説明する。ここで、本実施例における実験の流れ(手順)は、以下の通りである。
【0082】
[前準備その1:N末側発光酵素遺伝子とC末側発光酵素遺伝子およびPKA−RIαのサイクリックAMP結合ドメイン遺伝子のクローニングと、融合タンパク質発現プラスミドの作製]
[手順1]
N末側発光酵素(NLuc)遺伝子とC末側発光酵素(CLuc)遺伝子の作製のために、PCR(polymerase chain reaction)に用いる合成オリゴDNA(deoxyribonucleic acid)を以下に示す配列で調製した。
[NLuc遺伝子作製用合成オリゴDNA配列]
NLuc_Fw(配列番号1):5’−TGTGGATCCAGCCACCATGGAAGATGCCAA−3’
NLuc_Rv(配列番号2):5’−CAGCTCGAGGTCCTTGTCGATGAGAGCGTT−3’
[CLuc遺伝子作製用合成オリゴDNA配列]
CLuc_Fw(配列番号3):5’−ATCAGATCTGGCTACGTTAACAACCCCGAG−3’
CLuc_Rv(配列番号4):5’−CTAGAATTCTTACACGGCGATCTTGCCGCC−3’
【0083】
また、PKA−RIαのcAMP結合ドメイン遺伝子のクローニングのために、PCRに用いる合成オリゴDNAを以下に示す配列で調製した。
PKA_RIA_β1_Fw(配列番号5):5’−ATTCTCGAGGCTATGTTTCCAGTCTCCTTT−3’
PKA_RIA_B_Rv(配列番号6):5’−GGAAGATCTGACGGACAGGGACACGAAGCT−3’
【0084】
[手順2]
pGL4.10(プロメガ(株)製)を鋳型として、N末側ルシフェラーゼ遺伝子(NLuc:GL4.10遺伝子の1番目から416番目のアミノ酸を含む。)およびC末側ルシフェラーゼ遺伝子(CLuc:GL4.10遺伝子の399番目から550番目のアミノ酸を含む。)を、上記合成オリゴDNA(配列番号1のDNA配列と配列番号2のDNA配列のペア、および配列番号3のDNA配列と配列番号4のDNA配列のペア)をプライマーとしてPCRにより増幅した。
【0085】
[手順3]
PKAの制御サブユニットIα(PKA−RIα)に2ヶ所あるサイクリックAMP結合ドメインのAとBの両方を含む領域を、マウスのPKA制御サブユニットIα(PKA−RIα)のcDNAを鋳型とし、上記合成オリゴDNAをプライマーとしてPCRにより増幅した。配列番号5のDNA配列と配列番号6のDNA配列のプライマーを用いて、RIα_β1AB遺伝子(マウスのPKA−RIα遺伝子の152番目から381番目のアミノ酸配列に対応する領域を含む。配列番号39)をPCRにより増幅した。
【0086】
[手順4:融合タンパク質遺伝子発現プラスミドの作製]
PCRで増幅させたNLuc遺伝子、CLuc遺伝子およびRIα_β1AB遺伝子を、動物細胞発現用プラスミドであるpcDNA3.1(インビトロジェン社製)のBamHI部位とEcoRI部位の間に挿入し、動物細胞発現用プラスミドpcDNA/GL4_N_RIα_β1AB_Cを作製した。
【0087】
[前準備その2:ヒトD1受容体遺伝子とD2受容体遺伝子および5HT2A受容体遺伝子のクローニング]
[手順1]
ヒトD1受容体遺伝子とD2受容体遺伝子および5HT2A受容体遺伝子の作製のために、PCRに用いる合成オリゴDNAを以下に示す配列で調製した。
[ヒトD1受容体遺伝子作製用合成オリゴDNA配列]
HU_D1R_Fw(配列番号7):5’−GCCACCATGAGGACTCTGAACACCTCTGCC−3’
HU_D1R_Rv(配列番号8):5’−TCAGGTTGGGTGCTGACCGTTTTGTGTGAT−3’
[ヒトD2受容体遺伝子作製用合成オリゴDNA配列]
HU_D2R_Fw(配列番号9):5’−GCCACCATGGATCCACTGAATCTGTCCTGG−3’
HU_D2R_Rv(配列番号10):5’−TCAGCAGTGGAGGATCTTCAGGAAGGCCTT−3’
[ヒト5HT2A受容体遺伝子作製用合成オリゴDNA配列]
HU_5HT2AR_Fw(配列番号11):5’−GCCACCATGGATATTCTTTGTGAAGAAAAT−3’
HU_5HT2AR_Rv(配列番号12):5’−TCACACACAGCTCACCTTTTCATTCACTCC−3’
【0088】
[手順2]
ヒト脳cDNAライブラリ(タカラバイオ(株)製)を鋳型として、D1受容体遺伝子(ヒトのD1受容体遺伝子の全長に対応する領域を含む。)D2受容体遺伝子(ヒトのD2受容体遺伝子の全長に対応する領域を含む。)および5HT2A受容体遺伝子(ヒトの5HT2A受容体遺伝子の全長に対応する領域を含む。)を、上記合成オリゴDNAをプライマーとしてPCRにより増幅した。
【0089】
[手順3]
PCRで増幅させたD1受容体遺伝子、D2受容体遺伝子および5HT2A受容体遺伝子をpBluescriptIIベクターにサブクローニングした後、作製したDNAの配列をDNAシーケンシングにより確認した。
【0090】
[手順4]
それぞれの遺伝子を哺乳類細胞の発現ベクターpcDNA3.1(+)(インビトロジェン(株)製)へ組み込み、ヒトD1受容体発現用プラスミドpcDNA/D1RおよびヒトD2受容体発現用プラスミドpcDNA/D2Rおよびヒト5HT2A受容体発現用プラスミドpcDNA/5HT2ARを作製した。
【0091】
[実験:ヒトD1受容体、D2受容体および5HT2A受容体を含有するHEK293細胞でのcAMP濃度変動の発光イメージング]
[手順1:HEK293細胞の培養]
ATCC(American Type Culture Collection)社より入手したHEK293細胞を、5% CO2インキュベーター内で、10% Fetal Bovine Serumおよび1× Nonessential amino acidsを添加したEarle's MEM/培地(GIBCO社製)で培養した。
【0092】
[手順2:タンパク質発現プラスミドの導入]
HEK293細胞を、直径35mmガラスボトムディッシュに2×105/dishの細胞密度で播種し、5% CO2インキュベーター内で一晩培養し、サイクリックAMPセンサータンパク質発現用プラスミドpcDNA/GL4_N_RIα_β1AB_C、ヒトD1受容体発現用プラスミドpcDNA/D1R(またはヒトD2受容体発現用プラスミドpcDNA/D2R)およびヒト5HT2A受容体発現用プラスミドpcDNA/5HT2ARを混合し、FuGENE HD(ロシュ社製)を用いてトランスフェクションを行い、5% CO2インキュベーター内で一晩培養した。なお、ヒトD1受容体のアミノ酸配列は具体的には配列番号29のアミノ酸配列であり、ヒト5HT2A受容体のアミノ酸配列は具体的には配列番号30のアミノ酸配列である。
【0093】
[手順3:発光画像の撮像]
培地中にルシフェリン2mM(プロメガ社製)を加えて1時間静置してから、培養ディッシュを発光顕微鏡“LV(LUMINOVIEW)−200”(オリンパス社製)にセットし、20秒間隔で発光画像のタイムラプス撮影を行った。発光観察条件として、対物レンズの倍率は40倍、露出時間は10秒、ビニングは2×2とした。CCDカメラ106cとしてEM−CCDカメラiXon(アンドール社製)を用い、画像解析装置110として構成したパーソナルコンピュータに発光画像を取り込んだ。
【0094】
[手順4:ドーパミン刺激による発光画像の撮像]
タイムラプス撮影開始から1分後以降に、セロトニン(5HT:最終濃度10μM)、ドーパミン(DA:最終濃度10μM)およびフォルスコリン(FSK:最終濃度5μM)で順次刺激を行い、引き続き発光画像のタイムラプス撮影を行った。
【0095】
[手順5:発光量の経時変化のグラフ表示]
刺激前に撮影した各々の発光画像(図6、図10および図15参照)に対して図示の如く複数のROI(Region of Interest:関心領域)を指定し、また刺激後に撮影した各々の発光画像(図7、図8、図11〜図13および図16〜図17参照)に対して図示の如く複数のROIを指定した。指定した各ROIの発光強度を各々の発光画像に基づいて測定し、当該測定した発光強度の経時変化をグラフ(図9、図14および図18参照)で表示した。発光画像の解析は、画像解析部112cとして機能するMetamorphソフトウェア(ユニバーサルイメージング社製)を用いて行った。なお、図9、図14および図18において、Cell1〜Cell4のそれぞれに対応する各グラフは、図6〜図8、図10〜図13および図15〜図17における番号1〜番号4のそれぞれに対応する各ROIの発光強度の経時変化を示している。
【0096】
[実験の結果]
以上の実験の結果、図6〜図9に示すように、5HT2A受容体またはD1受容体、D2受容体への刺激に対する細胞内のサイクリックAMP応答の変化をシングルセルレベル(一細胞レベル)で検出することができた。また、図9に示されている個々の細胞における発光強度の変化を見ると、5HT2A受容体とD1受容体との組合せにおいて、5HT2A受容体への刺激に対する細胞内サイクリックAMP濃度の上昇が観察された。しかし、図11〜図14および図16〜図18に示すように、5HT2A受容体とD2受容体との組合せにおいておよび5HT2A受容体のみにおいては、5HT2A受容体への刺激に対する細胞内サイクリックAMP濃度の上昇が観察されなかった。セロトニン受容体とドーパミン受容体との相互作用のこれまでの研究から、個体において、D1受容体を介すると考えられている行動が5HT2A受容体を介して引き起こされる、ということが知られている。本実験により得られた結果は、個体において観察した反応を細胞レベルで説明することが出来ることを示している。
【0097】
[本実施例のまとめ]
以上、本実施例によれば、細胞内の情報伝達に影響を及ぼすことなく、各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞においての5HT2A受容体とD1受容体との相互作用を再現性良く経時的に観察することができる。
【0098】
[他の実施の形態]
上述した実施の形態においては、主に、互いに結合することにより発光酵素活性が回復するよう分割させたN末側発光酵素とC末側発光酵素およびPKA−RIαのサイクリックAMP結合ドメインからなる融合タンパク質を、サイクリックAMPセンサータンパク質として説明を行ったが、本発明はこれに限られず、N末側発光酵素とC末側発光酵素およびPKA−RIαのサイクリックAMP結合ドメインからなる融合タンパク質に替えて、国際公開第2007/120522号および特表2007−508014号公報に開示されているサイクリックAMPセンサータンパク質を用いてもよい。
【0099】
[実施例2]
実施例1とは異なる実施形態に係るサイクリックAMPセンサータンパク質を作製し、その有用性を検討した。具体的には、図19に示されるように、サイクリックAMPセンサータンパク質のN末端側にC末端側断片が配置され、C末端側にN末端側断片が配置され、それらの間にサイクリックAMP結合領域が配置されたサイクリックAMPセンサータンパク質を作製した。すなわち、端的に表現すると、実施例1に係るサイクリックAMPセンサータンパク質に対して、C末端側断片とN末端側断片とを置き換えたサイクリックAMPセンサータンパク質を作製した。なお、本実施例に係るサイクリックAMPセンサータンパク質並びにその作製のために使用される遺伝子およびプライマー等について、それらの名称に「変異型」または「cp(circular permutaion)」等を付して区別した。
【0100】
[前準備:変異型ルシフェラーゼ作成用のN末側発光酵素遺伝子とC末側発光酵素遺伝子、およびPKA−RIαのサイクリックAMP結合ドメイン遺伝子のクローニングと、変異型ルシフェラーゼ融合タンパク質発現プラスミドの作製]
[手順1]
変異型ルシフェラーゼ作成用のN末側発光酵素(cpNLuc)遺伝子とC末側発光酵素(cpCLuc)遺伝子の作製のために、PCR(polymerase chain reaction)に用いる合成オリゴDNA(deoxyribonucleic acid)を以下に示す配列で調製した。
[cpNLuc遺伝子作製用合成オリゴDNA配列]
cpNLuc_Fw(配列番号13):5’−ATCAGATCTGAAGATGCCAAAAACATTAAG−3’
cpNLuc_Rv(配列番号14):5’−CTAGAATTCTTAGTCCTTGTCGATGAGAGC−3’
[cpCLuc遺伝子作製用合成オリゴDNA配列]
cpCLuc_Fw(配列番号15):5’−TGTGGATCCAGCCACCATGAGCGGCTACGTTAACAACCCC−3’
cpCLuc_Rv(配列番号16):5’−CAGCTCGAGCACGGCGATCTTGCCGCCCTT−3’
【0101】
また、PKA−RIαのcAMP結合ドメイン遺伝子のクローニングのために、PCRに用いる合成オリゴDNAを以下に示す配列で調製した。
PKA_RIA_α_Fw(配列番号17):5’−TTTCTCGAGCTTGATGATAACGAGAGAAGT−3’
PKA_RIA_B_Rv(配列番号18):5’−GGAAGATCTGACGGACAGGGACACGAAGCT−3’
【0102】
[手順2]
市販のルシフェラーゼ遺伝子であるpGL4.10(プロメガ(株)製)を鋳型として、変異型のN末側ルシフェラーゼ遺伝子(cpNLuc:GL4.10遺伝子の1番目から416番目のアミノ酸を含む。)および変異型のC末側ルシフェラーゼ遺伝子(cpCLuc:GL4.10遺伝子の399番目から550番目のアミノ酸を含む。)を、上記合成オリゴDNA(配列番号13のDNA配列と配列番号14のDNA配列のペア、および配列番号15のDNA配列と配列番号16のDNA配列のペア)をプライマーとしてPCRにより増幅した。
【0103】
[手順3]
PKAの制御サブユニットIα(PKA−RIα)に2ヶ所あるサイクリックAMP結合ドメインのAとBの両方を含む領域を、マウスのPKA制御サブユニットIα(PKA−RIα)のcDNAを鋳型とし、上記合成オリゴDNAをプライマーとしてPCRにより増幅した。配列番号17のDNA配列と配列番号18のDNA配列のプライマーを用いて、RIα_αAB遺伝子(マウスのPKA−RIα遺伝子の141番目から381番目のアミノ酸配列に対応する領域を含む。)(塩基配列は配列番号37に示される)をPCRにより増幅した。
【0104】
[手順4:融合タンパク質遺伝子発現プラスミドの作製]
PCRで増幅させた、cpNLuc遺伝子、cpCLuc遺伝子およびRIα_αAB遺伝子をそれぞれ含む核酸を、制限酵素にて適宜処理した後、動物細胞発現用プラスミドであるpcDNA3.1(インビトロジェン社製)のBamHI部位とEcoRI部位の間に挿入し、動物細胞発現用プラスミドpcDNA/cpGL4_C_RIα_αAB_Nを作製した。
【0105】
[実験:ヒトD1受容体、D2受容体および5HT2A受容体を含有するHEK293細胞でのcAMP濃度変動の発光イメージング]
[手順1:HEK293細胞の培養]
ATCC(American Type Culture Collection)社より入手したHEK293細胞を、5% CO2インキュベーター内で、10% Fetal Bovine Serumおよび1× Nonessential amino acidsを添加したEarle's MEM/培地(GIBCO社製)で培養した。
【0106】
[手順2:タンパク質発現プラスミドの導入]
HEK293細胞を、直径35mmガラスボトムディッシュに2×105/dishの細胞密度で播種し、5% CO2インキュベーター内で一晩培養し、サイクリックAMPセンサータンパク質発現用プラスミドpcDNA/cpGL4_C_RIα_αAB_N、ヒトD1受容体発現用プラスミドpcDNA/D1R(またはヒトD2受容体発現用プラスミドpcDNA/D2R)およびヒト5HT2A受容体発現用プラスミドpcDNA/5HT2ARを混合し、FuGENE HD(ロシュ社製)を用いてトランスフェクションを行い、5% CO2インキュベーター内で一晩培養した。なお、pcDNA/D1R、pcDNA/D2RおよびpcDNA/5HT2ARは、実施例1と同様に作製したものを使用した。
【0107】
[手順3:発光画像の撮像]
培地中にルシフェリン2mM(プロメガ社製)を加えて1時間静置してから、培養ディッシュを発光顕微鏡“LV(LUMINOVIEW)−200”(オリンパス社製)にセットし、20秒間隔で発光画像のタイムラプス撮影を行った。発光観察条件として、対物レンズの倍率は40倍、露出時間は10秒、ビニングは2×2とした。CCDカメラ106cとしてEM−CCDカメラiXon(アンドール社製)を用い、画像解析装置110として構成したパーソナルコンピュータに発光画像を取り込んだ。
【0108】
[手順4:セロトニン、ドーパミン刺激による発光画像の撮像]
タイムラプス撮影開始から1分後以降に、セロトニン(5HT:最終濃度10μM)、ドーパミン(DA:最終濃度10μM)で順次刺激を行い、引き続き発光画像のタイムラプス撮影を行った。
【0109】
[手順5:発光量の経時変化のグラフ表示]
刺激前に撮影した各々の発光画像(図20および図24参照)に対して図示の如く複数のROI(Region of Interest:関心領域)を指定し、また刺激後に撮影した各々の発光画像(図21、図22、図25および図26参照)に対して図示の如く複数のROIを指定した。指定した各ROIの発光強度を各々の発光画像に基づいて測定し、当該測定した発光強度の経時変化をグラフ(図23、および図27参照)で表示した。発光画像の解析は、画像解析部112cとして機能するMetamorphソフトウェア(ユニバーサルイメージング社製)を用いて行った。なお、図23および図27において、Cell1〜Cell4のそれぞれに対応する各グラフは、図20〜図22および図24〜図26における番号1〜番号4のそれぞれに対応する各ROIの発光強度の経時変化を示している。
【0110】
[実験の結果]
以上の実験の結果、図6〜図9および図20〜図23に示すように、5HT2A受容体またはD1受容体、D2受容体への刺激に対する細胞内のサイクリックAMP応答の変化をシングルセルレベル(一細胞レベル)で検出することができた。また、図9および図23に示されている個々の細胞における発光強度の変化を見ると、5HT2A受容体とD1受容体との組合せにおいて、5HT2A受容体への刺激に対する細胞内サイクリックAMP濃度の上昇が観察された。しかし、図11〜図14および図16〜図18、図24〜図27に示すように、5HT2A受容体とD2受容体との組合せにおいておよび5HT2A受容体のみにおいては、5HT2A受容体への刺激に対する細胞内サイクリックAMP濃度の上昇が観察されなかった。セロトニン受容体とドーパミン受容体との相互作用のこれまでの研究から、個体において、D1受容体を介すると考えられている行動が5HT2A受容体を介して引き起こされる、ということが知られている。本実験により得られた結果は、個体において観察した反応を細胞レベルで説明することが出来ることを示している。
【0111】
[本実施例のまとめ]
以上、本実施例によれば、細胞内の情報伝達に影響を及ぼすことなく、各細胞からの正確な定量的結果を得ることができ、その結果、個々の細胞においての5HT2A受容体とD1受容体との相互作用を再現性良く経時的に観察することができる。この実施例において使用するルシフェラーゼ遺伝子やルシフェリン等は、他の市販試薬であってもよいし、生物発光等を発生する生物から抽出等されたものであってもよい。
【0112】
[他の実施の形態]
上述した実施の形態においては、主に、互いに結合することにより発光酵素活性が回復するよう分割させたN末側発光酵素とC末側発光酵素、および、PKA−RIαのサイクリックAMP結合ドメインからなる融合タンパク質をサイクリックAMPセンサータンパク質として説明を行ったが、本発明はこれに限られず、N末側発光酵素とC末側発光酵素、および、PKA−RIαのサイクリックAMP結合ドメインからなる融合タンパク質に替えて特許文献1および特許文献2に開示されているサイクリックAMPセンサータンパク質を用いてもよい。
【0113】
[実施例3]
種々のサイクリックAMPセンサータンパク質を作製し、それらの有用性を検討した。具体的には、長さおよび範囲をかえた16通りのサイクリックAMP結合領域を作製し、それぞれについて、N末端側断片、サイクリックAMP結合領域およびC末端側断片の順に配置したもの、ならびに、C末端側断片、サイクリックAMP結合領域およびN末端側断片の順に配置したものの計32種のサイクリックAMPセンサータンパク質を作製した。それら全てについて、サイクリックAMPの存在下および非存在下における酵素活性を測定した。
【0114】
[手順1]
スプリットルシフェラーゼ作成用のN末側発光酵素(NLuc)遺伝子とC末側発光酵素(CLuc)遺伝子、および、変異型スプリットルシフェラーゼ作成用のN末側発光酵素(cpNLuc)遺伝子とC末側発光酵素(cpCLuc)遺伝子の作製のために、PCR(polymerase chain reaction)に用いる合成オリゴDNA(deoxyribonucleic acid)を以下に示す配列で調製した。
[NLuc遺伝子作製用合成オリゴDNA配列]
NLuc_Fw (配列番号1): 5’ − TGTGGATCCAGCCACCATGGAAGATGCCAA − 3’
NLuc_Rv (配列番号2): 5’ − CAGCTCGAGGTCCTTGTCGATGAGAGCGTT − 3’
[CLuc遺伝子作製用合成オリゴDNA配列]
CLuc_Fw (配列番号3): 5’ − ATCAGATCTGGCTACGTTAACAACCCCGAG − 3’
CLuc_Rv (配列番号4): 5’ − CTAGAATTCTTACACGGCGATCTTGCCGCC − 3’
[cpNLuc遺伝子作製用合成オリゴDNA配列]
cpNLuc_Fw(配列番号13):5’−ATCAGATCTGAAGATGCCAAAAACATTAAG−3’
cpNLuc_Rv(配列番号14):5’−CTAGAATTCTTAGTCCTTGTCGATGAGAGC−3’
[cpCLuc遺伝子作製用合成オリゴDNA配列]
cpCLuc_Fw(配列番号15):5’−TGTGGATCCAGCCACCATGAGCGGCTACGTTAACAACCCC−3’
cpCLuc_Rv(配列番号16):5’−CAGCTCGAGCACGGCGATCTTGCCGCCCTT−3’
【0115】
また、PKA−RIαのcAMP結合ドメイン遺伝子のクローニングのために、PCRに用いる合成オリゴDNAを以下に示す配列で調製した。
PKA_RIA_Xn1_Fw (配列番号19): 5’− ATTCTCGAGGATTATAAGACAATGGCTGCT −3’
PKA_RIA_Xn2_Fw (配列番号20): 5’− GCCCTCGAGAAGAATGTGCTGTTTTCACAC −3’
PKA_RIA_α_Fw (配列番号21): 5’− TTTCTCGAGCTTGATGATAACGAGAGAAGT −3’
PKA_RIA_β1_Fw (配列番号22): 5’− ATTCTCGAGGCTATGTTTCCAGTCTCCTTT −3’
PKA_RIA_β2_Fw (配列番号23): 5’− TTTCTCGAGGGAGAGACGGTTATTCAGCAA −3’
PKA_RIA_β3_Fw (配列番号24): 5’− GGTCTCGAGGGGGATAACTTCTATGTGATT −3’
PKA_RIA_β4_Fw (配列番号25): 5’− ATTCTCGAGGGAGAAATGGATGTCTATGTC −3’
PKA_RIA_β5_Fw (配列番号26): 5’− AATCTCGAGTGGGCAACCAGTGTTGGGGAA −3’
PKA_RIA_A_Rv (配列番号27): 5’− CTCAGATCTGTCCAGAGACTCTAAAATAGA −3’
PKA_RIA_B_Rv (配列番号28): 5’− GGAAGATCTGACGGACAGGGACACGAAGCT −3’
【0116】
[手順2]
そして、pGL4.10(プロメガ(株)製)を鋳型として、以下の表1のプライマーセットを使用し、ルシフェラーゼ遺伝子の断片を含む核酸をPCRによって増幅した。
【0117】
【表1】
[手順3]
PKAの制御サブユニットIα(PKA−RIα)にはcAMP結合ドメインがAとBの2ヶ所あり、Aのみを含む領域、または、AおよびBの両方を含む領域を、マウスのPKA制御サブユニットIα(PKA−RIα)のcDNAを鋳型とし、上記合成オリゴDNAをプライマーとしてPCRにより増幅した。使用したプライマーと増幅された遺伝子との関係を図28に示す(図中、「遺伝子名」「使用したプライマー」「PKA−RIα中の対するアミノ酸の位置」「アミノ酸残基の数」が参照される)。
【0118】
なお、配列番号31には、PKAの制御サブユニットIα(PKA−RIα)全体のアミノ酸配列が示される。配列番号32には、RIα−Xn1A遺伝子に対応するアミノ酸配列が記載される。配列番号33には、RIα−Xn1AB遺伝子に対応するアミノ酸配列が記載される。配列番号34には、RIα−Xn2A遺伝子に対応するアミノ酸配列が記載される。配列番号35には、RIα−Xn2AB遺伝子に対応するアミノ酸配列が記載される。配列番号36には、RIα−αA遺伝子に対応するアミノ酸配列が記載される。配列番号37には、RIα−αAB遺伝子に対応するアミノ酸配列が記載される。配列番号38には、RIα−β1A遺伝子に対応するアミノ酸配列が記載される。配列番号39には、RIα−β1AB遺伝子に対応するアミノ酸配列が記載される。配列番号40には、RIα−β2A遺伝子に対応するアミノ酸配列が記載される。配列番号41には、RIα−β2AB遺伝子に対応するアミノ酸配列が記載される。配列番号42には、RIα−β3A遺伝子に対応するアミノ酸配列が記載される。配列番号43には、RIα−β3AB遺伝子に対応するアミノ酸配列が記載される。配列番号44には、RIα−β4A遺伝子に対応するアミノ酸配列が記載される。配列番号45には、RIα−β4AB遺伝子に対応するアミノ酸配列が記載される。配列番号46には、RIα−β5A遺伝子に対応するアミノ酸配列が記載される。配列番号47には、RIα−β5AB遺伝子に対応するアミノ酸配列が記載される。
【0119】
[手順4](原核生物発現用プラスミドpRSET/GL4_N_C、および、pRSET/cpGL4_C_Nの作製)
PCRによって増幅したNLuc遺伝子を含む断片を、原核生物発現用プラスミドであるpRSET2B(インビトロジェン社製)内のBamHI部位とXhoI部位との間に挿入した後、CLuc遺伝子を含む断片を、BglII部位とEcoRI部位との間に挿入することで、原核生物発現用プラスミド(pRSET/GL4_N_C)を作製した。
【0120】
また、PCRによって増幅したcpCLuc遺伝子を含む断片を、原核生物発現用プラスミドであるpRSET2B(インビトロジェン社製)内のBamHI部位とXhoI部位との間に挿入した後、cpNLuc遺伝子を含む断片を、BglII部位とEcoRI部位との間に挿入し、原核生物発現用プラスミド(pRSET/cpGL4_C_N)を作製した。
【0121】
[手順5](融合タンパク質遺伝子発現プラスミドの作製)
手順3によって作製したPKA−RIα由来の種々の遺伝子を含む核酸を、手順4で作製したpRSET/GL4_N_CベクターまたはpRSET/cpGL4_C_Nベクター内のXhoI部位とBglII部位の間に挿入した。このようにして得られたプラスミドを図28に示す(図中、「得られるプラスミドの名称」が参照される)。
【0122】
[手順6](融合タンパク質遺伝子の大腸菌での発現および精製)
以上のように作製した融合タンパク質発現プラスミドを、大腸菌JM109(DE3)株にトランスフォームし、The QIAexpressionist(キアゲン社製)を用いて、キットのマニュアルに従って融合タンパク質を精製した。精製手順を簡略に説明すると、上記原核生物発現用プラスミドを大腸菌JM109(DE3)株にトランスフォームして、5mLのLB培地中に植菌した。一晩培養した後、遠心分離によって集菌し、集菌した大腸菌を溶解バッファー(50mM NaH2PO4, 300mM NaCl, 10mM イミダゾール, pH8.0)500μLで再懸濁してリゾチームを添加し(1mg/mL)、4℃で60分間インキュベートした。細胞を静かにボルテックスして溶解し、15,000xgで10分間遠心分離して上清を新しいチューブに移した。50% Ni−NTAレジン懸濁液 50μLを添加し、4℃で60分間静かに混合した。15,000xgで10秒間遠心してレジンを沈殿させ、レジンを洗浄バッファー(50mM NaH2PO4, 300mM NaCl, 20mM イミダゾール, pH8.0) 500μLで2回洗浄した。溶出バッファー(50mM NaH2PO4, 300mM NaCl, 250mM イミダゾール, pH8.0) 50μLで融合タンパク質を2回溶出し、融合タンパク質を得た。得られた融合タンパク質を図28に示す(図中、「得られるサイクリックAMPセンサータンパク質の名称」が参照される)。
【0123】
[手順7](融合タンパク質遺伝子の特性測定)
上記融合タンパク質の発光活性をcAMPの存在下および非存在下でそれぞれ測定した。測定方法を簡単に説明すると、上述の融合タンパク質溶液20μLに、リン酸緩衝液(PBS)20μLを加えた。さらに、4mMcAMP溶液を1μL加え(終濃度 100μM)、ネガティブコントロールにはPBSを同量加えた。Bright−Glo(プロメガ社製)を40μL加えて、Luminescencer JNR−II(アトー社製)で発光量(10秒間の積算)を測定した。
【0124】
[結果]
上記の通り、精製した融合タンパク質をPBSに希釈し、cAMP(終濃度100μM)の有無での発光量を測定した(図29)。その結果、cAMPが存在することで発光活性が上昇する融合タンパク質が存在することがわかった。cAMPの有無による活性の差が特に大きかったのは、GL4_N_RIα_β1AB_C(図29a中「β1AB」)およびcpGL4_C_RIα_αAB_Nであった((図29b中「αAB」)。
【産業上の利用可能性】
【0125】
以上のように、本発明にかかる受容体相互作用検出方法およびサイクリックAMPセンサータンパク質は、バイオ、製薬、医療など様々な分野で好適に用いることができる。
【0126】
また、以上のように、本発明にかかる5HT2A受容体とD1受容体の相互作用を検出するための発光タンパク質および解析方法は、バイオ、製薬、医療など様々な分野で好適に用いることができる。
【符号の説明】
【0127】
100 発光観察システム
103 容器(シャーレ)
104 ステージ
106 発光画像撮像ユニット
106a 対物レンズ(発光観察用)
106b ダイクロイックミラー
106c CCDカメラ
106d スプリットイメージユニット
106e フィルターホイール
106f 結像レンズ
110 画像解析装置
112 制御部
112a 発光画像撮像指示部
112b 発光画像取得部
112c 画像解析部
112d 解析結果出力部
114 クロック発生部
116 記憶部
118 通信インターフェース部
120 入出力インターフェース部
122 入力装置
124 出力装置
【特許請求の範囲】
【請求項1】
細胞において第1の受容体と第2の受容体との相互作用を検出する受容体相互作用検出方法であって、
前記第2の受容体の活性を示す指標となるレポータを含む前記細胞を作製する作製工程と、
前記作製工程で作製した前記細胞に当該細胞外から、発光させるための所定の処置を行うと共に前記第1の受容体に結合する所定の受容体結合物質を添加する処置および添加工程と、
前記処置および添加工程が行われた後の前記細胞の発光量を測定する測定工程と、
前記測定工程で測定した前記発光量に基づいて前記細胞内における前記第2の受容体の活性を解析する解析工程と、
前記解析工程で解析した前記第2の受容体の活性に基づいて、前記第1の受容体と前記第2の受容体との前記相互作用を検出する検出工程と、
を含むことを特徴とする受容体相互作用検出方法。
【請求項2】
前記作製工程において、前記レポータは、サイクリックAMPセンサータンパク質であり、
前記解析工程において、前記第2の受容体の活性はサイクリックAMPの濃度であること、
を特徴とする請求項1に記載の受容体相互作用検出方法。
【請求項3】
前記サイクリックAMPセンサータンパク質は、前記細胞内のサイクリックAMPと相互作用して発光するものであること、
を特徴とする請求項2に記載の受容体相互作用検出方法。
【請求項4】
前記サイクリックAMPセンサータンパク質は、ホタルのルシフェラーゼおよびPKA(protein kinase A)のサイクリックAMP結合領域とからなる融合タンパク質であること、
を特徴とする請求項2または3に記載の受容体相互作用検出方法。
【請求項5】
前記サイクリックAMP結合領域が、マウスのPKA−RIα由来であることを特徴とする請求項4に記載の受容体相互作用検出方法。
【請求項6】
前記サイクリックAMP結合領域が、220から250のアミノ酸残基の長さを有することを特徴とする請求項4または5に記載の受容体相互作用検出方法。
【請求項7】
前記サイクリックAMP結合領域が、少なくとも2つのサイクリックAMP結合部位を含むことを特徴とする請求項4から6の何れか1項に記載の受容体相互作用検出方法。
【請求項8】
前記サイクリックAMPセンサータンパク質は、前記ルシフェラーゼのN末端側断片、前記ルシフェラーゼのC末端側断片および前記サイクリックAMP結合領域を含み、前記N末端側断片と前記C末端側断片とが前記サイクリックAMPセンサータンパク質分子内で結合すると、前記ルシフェラーゼの発光酵素活性が回復することを特徴とする請求項4から7の何れか1項に記載の受容体相互作用検出方法。
【請求項9】
前記サイクリックAMP結合領域が、配列番号37または39に記載のアミノ酸配列を含むことを特徴とする請求項4から8の何れか1項に記載の受容体相互作用検出方法。
【請求項10】
前記作製工程において、前記細胞は前記第1の受容体と前記第2の受容体とを含有するものであること、
を特徴とする請求項1から9のいずれか1つに記載の受容体相互作用検出方法。
【請求項11】
前記作製工程において、前記細胞は前記第1の受容体の遺伝子と前記第2の受容体の遺伝子とが導入されたものであること、
を特徴とする請求項1から9のいずれか1つに記載の受容体相互作用検出方法。
【請求項12】
前記測定工程は、前記第2の受容体の活性に依存する発光標識の光子量を測定する光子量測定工程をさらに含むこと、
を特徴とする請求項1から11のいずれか1つに記載の受容体相互作用検出方法。
【請求項13】
前記測定工程は、前記第2の受容体の活性に依存する発光標識を撮像する撮像工程をさらに含むこと、
を特徴とする請求項1から11のいずれか1つに記載の受容体相互作用検出方法。
【請求項14】
前記撮像工程は、複数の異なる前記細胞に対して同時に実行され、
前記解析工程は、前記撮像工程で撮像した複数の発光画像を前記細胞ごとに照合する照合工程をさらに含むこと、
を特徴とする請求項13に記載の受容体相互作用検出方法。
【請求項15】
前記測定工程は繰り返し実行されること、
を特徴とする請求項1から14のいずれか1つに記載の受容体相互作用検出方法。
【請求項16】
前記第1の受容体は5HT2A受容体であり、前記第2の受容体はD1受容体であること、
を特徴とする請求項1から15のいずれか1つに記載の受容体相互作用検出方法。
【請求項17】
前記処置および添加工程において、前記受容体結合物質はセロトニンであること、
を特徴とする請求項16に記載の受容体相互作用検出方法。
【請求項18】
第1の受容体と第2の受容体との相互作用を検出するためのものであること、
を特徴とするサイクリックAMPセンサータンパク質。
【請求項19】
サイクリックAMP(adenosine monophosphate)と相互作用して発光するものであること、
を特徴とする請求項18に記載のサイクリックAMPセンサータンパク質。
【請求項20】
前記第1の受容体は5HT2A受容体であり、前記第2の受容体はD1受容体であること、
を特徴とする請求項18または19に記載のサイクリックAMPセンサータンパク質。
【請求項1】
細胞において第1の受容体と第2の受容体との相互作用を検出する受容体相互作用検出方法であって、
前記第2の受容体の活性を示す指標となるレポータを含む前記細胞を作製する作製工程と、
前記作製工程で作製した前記細胞に当該細胞外から、発光させるための所定の処置を行うと共に前記第1の受容体に結合する所定の受容体結合物質を添加する処置および添加工程と、
前記処置および添加工程が行われた後の前記細胞の発光量を測定する測定工程と、
前記測定工程で測定した前記発光量に基づいて前記細胞内における前記第2の受容体の活性を解析する解析工程と、
前記解析工程で解析した前記第2の受容体の活性に基づいて、前記第1の受容体と前記第2の受容体との前記相互作用を検出する検出工程と、
を含むことを特徴とする受容体相互作用検出方法。
【請求項2】
前記作製工程において、前記レポータは、サイクリックAMPセンサータンパク質であり、
前記解析工程において、前記第2の受容体の活性はサイクリックAMPの濃度であること、
を特徴とする請求項1に記載の受容体相互作用検出方法。
【請求項3】
前記サイクリックAMPセンサータンパク質は、前記細胞内のサイクリックAMPと相互作用して発光するものであること、
を特徴とする請求項2に記載の受容体相互作用検出方法。
【請求項4】
前記サイクリックAMPセンサータンパク質は、ホタルのルシフェラーゼおよびPKA(protein kinase A)のサイクリックAMP結合領域とからなる融合タンパク質であること、
を特徴とする請求項2または3に記載の受容体相互作用検出方法。
【請求項5】
前記サイクリックAMP結合領域が、マウスのPKA−RIα由来であることを特徴とする請求項4に記載の受容体相互作用検出方法。
【請求項6】
前記サイクリックAMP結合領域が、220から250のアミノ酸残基の長さを有することを特徴とする請求項4または5に記載の受容体相互作用検出方法。
【請求項7】
前記サイクリックAMP結合領域が、少なくとも2つのサイクリックAMP結合部位を含むことを特徴とする請求項4から6の何れか1項に記載の受容体相互作用検出方法。
【請求項8】
前記サイクリックAMPセンサータンパク質は、前記ルシフェラーゼのN末端側断片、前記ルシフェラーゼのC末端側断片および前記サイクリックAMP結合領域を含み、前記N末端側断片と前記C末端側断片とが前記サイクリックAMPセンサータンパク質分子内で結合すると、前記ルシフェラーゼの発光酵素活性が回復することを特徴とする請求項4から7の何れか1項に記載の受容体相互作用検出方法。
【請求項9】
前記サイクリックAMP結合領域が、配列番号37または39に記載のアミノ酸配列を含むことを特徴とする請求項4から8の何れか1項に記載の受容体相互作用検出方法。
【請求項10】
前記作製工程において、前記細胞は前記第1の受容体と前記第2の受容体とを含有するものであること、
を特徴とする請求項1から9のいずれか1つに記載の受容体相互作用検出方法。
【請求項11】
前記作製工程において、前記細胞は前記第1の受容体の遺伝子と前記第2の受容体の遺伝子とが導入されたものであること、
を特徴とする請求項1から9のいずれか1つに記載の受容体相互作用検出方法。
【請求項12】
前記測定工程は、前記第2の受容体の活性に依存する発光標識の光子量を測定する光子量測定工程をさらに含むこと、
を特徴とする請求項1から11のいずれか1つに記載の受容体相互作用検出方法。
【請求項13】
前記測定工程は、前記第2の受容体の活性に依存する発光標識を撮像する撮像工程をさらに含むこと、
を特徴とする請求項1から11のいずれか1つに記載の受容体相互作用検出方法。
【請求項14】
前記撮像工程は、複数の異なる前記細胞に対して同時に実行され、
前記解析工程は、前記撮像工程で撮像した複数の発光画像を前記細胞ごとに照合する照合工程をさらに含むこと、
を特徴とする請求項13に記載の受容体相互作用検出方法。
【請求項15】
前記測定工程は繰り返し実行されること、
を特徴とする請求項1から14のいずれか1つに記載の受容体相互作用検出方法。
【請求項16】
前記第1の受容体は5HT2A受容体であり、前記第2の受容体はD1受容体であること、
を特徴とする請求項1から15のいずれか1つに記載の受容体相互作用検出方法。
【請求項17】
前記処置および添加工程において、前記受容体結合物質はセロトニンであること、
を特徴とする請求項16に記載の受容体相互作用検出方法。
【請求項18】
第1の受容体と第2の受容体との相互作用を検出するためのものであること、
を特徴とするサイクリックAMPセンサータンパク質。
【請求項19】
サイクリックAMP(adenosine monophosphate)と相互作用して発光するものであること、
を特徴とする請求項18に記載のサイクリックAMPセンサータンパク質。
【請求項20】
前記第1の受容体は5HT2A受容体であり、前記第2の受容体はD1受容体であること、
を特徴とする請求項18または19に記載のサイクリックAMPセンサータンパク質。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】

【図7】

【図8】

【図9】
【図10】

【図11】

【図12】

【図13】

【図14】
【図15】

【図16】

【図17】

【図18】
【図19】
【図20】

【図21】

【図22】

【図23】
【図24】

【図25】

【図26】

【図27】
【図28】
【図29】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【公開番号】特開2010−243485(P2010−243485A)
【公開日】平成22年10月28日(2010.10.28)
【国際特許分類】
【出願番号】特願2010−63122(P2010−63122)
【出願日】平成22年3月18日(2010.3.18)
【出願人】(000000376)オリンパス株式会社 (11,466)
【Fターム(参考)】
【公開日】平成22年10月28日(2010.10.28)
【国際特許分類】
【出願日】平成22年3月18日(2010.3.18)
【出願人】(000000376)オリンパス株式会社 (11,466)
【Fターム(参考)】
[ Back to top ]