
可変性リンパ球受容体、その関連ポリペプチドおよび核酸、ならびにその使用
【課題】可変性リンパ球受容体(VLR)に関連する組成物および方法の提供。
【解決手段】可変性リンパ球受容体(VLR)に関連する組成物および方法が開示される。本発明は、N末端ロイシンリッチリピート(LRRNT)、1つまたは複数のロイシンリッチリピート(LRR)(本明細書では内部LRRと呼ぶ)、C末端ロイシンリッチリピート(LRRCT)、および連結ペプチドを含む単離されたポリペプチドを提供するが、このとき連結ペプチドはαヘリックスを含んでいる。ポリペプチドの長さは、約130という少数のアミノ酸または約225という多数のアミノ酸を含むことができる。
【解決手段】可変性リンパ球受容体(VLR)に関連する組成物および方法が開示される。本発明は、N末端ロイシンリッチリピート(LRRNT)、1つまたは複数のロイシンリッチリピート(LRR)(本明細書では内部LRRと呼ぶ)、C末端ロイシンリッチリピート(LRRCT)、および連結ペプチドを含む単離されたポリペプチドを提供するが、このとき連結ペプチドはαヘリックスを含んでいる。ポリペプチドの長さは、約130という少数のアミノ酸または約225という多数のアミノ酸を含むことができる。
【発明の詳細な説明】
【技術分野】
【0001】
本出願は、2004年5月21日に出願した米国特許仮出願第60/573,563号の利益を要求する。これは本明細書中でその全体が参考として援用される。
【0002】
本発明は、NIH/NIAIS助成金AI39816およびHG02526−01、ならびにNSF助成金MCB−0317460およびIBN−0321461の下での政府の支援を受けて成された。政府は、本発明において特定の権利を有する。
【背景技術】
【0003】
(発明の背景)
有顎類脊椎動物における適応免疫応答は、抗原が特異的リンパ球受容体によって認識されると開始される。抗原受容体多様性は、免疫グロブリン(Ig)およびT細胞受容体(TCR)の遺伝子座における可変性遺伝子セグメント、多様性遺伝子セグメントおよび結合遺伝子セグメントの組み換えによって生成される。この組み換え再配列は、処理されていない抗原に対する抗体ならびに主要組織適合遺伝子複合体(MHC)クラスIおよびII分子の先端内で提示された抗原フラグメントを認識するTCRの膨大なレバートリーを生成する。そこでクローン性が多様なリンパ球は、主要造血組織および胸腺内で幹細胞前駆体から分化する、Igを有するB細胞およびTCRを有するT細胞の形態で、脊椎動物の適応免疫の基礎を形成している。この組み換え免疫系の重要なエレメントは全有顎類脊椎動物において保存されており、多重遺伝子のTCR遺伝子座およびIg遺伝子座は、最も基本的な有顎類の代表であるサメ、ガンギエイ、およびエイにおいてさえ顕著に複雑である(非特許文献1;非特許文献2;非特許文献3)。
【0004】
早期の脊椎動物の分散(radiation)から生き残っている極めて少数の末裔の無顎類脊椎動物であるヤツメウナギおよびメクラウナギにおいても適応免疫についての豊富な証拠がある(非特許文献4)。これらの無顎類については、体液性タイプおよび細胞媒介性タイプの免疫応答が報告されている。例えば、ヤツメウナギは一次抗原刺激に応答して特異的循環性アグルチニンを産生し、追加免疫後にはより高度のアグルチニンレベルを生成し(非特許文献5;非特許文献6;非特許文献7;非特許文献8;非特許文献9;非特許文献10)、加速された速度で第2セットの表皮同種移植片を拒絶し(非特許文献11;非特許文献12;非特許文献9;非特許文献13)、そして遅延型の過敏性反応を示す(非特許文献5;非特許文献9)。無顎類の適応免疫応答は、有顎類脊椎動物のリンパ造血組織および血液中で見いだされるリンパ球に形態学的に似ている細胞に起因していた(非特許文献5;非特許文献11;非特許文献12;非特許文献14;非特許文献15;非特許文献9;非特許文献16;非特許文献17;非特許文献18;非特許文献13;非特許文献19;非特許文献20)。それらの哺乳動物対応物と同様に、ヤツメウナギリンパ球は他の血球タイプよりはるかに照射感受性であり(非特許文献9)、抗原刺激に応答して凝集かつ増殖し(非特許文献5;非特許文献14;非特許文献15)、そしてPU.1/Spi−BおよびIkarosなどの哺乳動物リンパ球分化に関係している転写因子を発現する(非特許文献21;非特許文献22;非特許文献23;非特許文献24)。しかし驚くべきことに、Ig遺伝子、TCR遺伝子、およびMHC遺伝子は、以前には無顎類脊椎動物または無脊椎動物の尾索類Ciona intestinalisのゲノム配列内では同定されていなかった(非特許文献25)。本発明は、新規なリンパ球受容体および新規なリンパ球受容体をコードする核酸に関する。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Rast,J.P.,Michele K.Anderson,M.K.,Strong,S.J.,Luer,C,Litman,R.T.,および Litman,G.W.α,β,g,and δ T Cell Antigen Receptor Genes Arose Early in Vertebrate Phylogeny.Immunity(1997)6:1−11
【非特許文献2】Flajnik MFおよびKasahara M、Comparative genomics of the MHC:glimpses into the evolution of the adaptive immune system.Immunity(2001)15:351−62
【非特許文献3】Flajnik MF、Comparative analyses of immunoglobulin genes:surprises and portents.Nat.Rev.Immunol(2002)2:688−98
【非特許文献4】Forey PLおよびJanvier P、Agnathans and the origin of jawed vertebrates.Nature(1993)361:129−134
【非特許文献5】Finstad J,Papermaster BWおよびGood RA、Evolution of the immune response.III.Morphologic studies of the thymus and organized lymphoid tissue.Lab Invest.(1964)13:490−512
【非特許文献6】Marchalonis JJおよびEdelman GM、Phylogenetic origins of antibody structure.3.Antibodies in the primary immune response of the sea lamprey,Petromyzon marinus.J.Exp.Med(1968)127:891−914
【非特許文献7】Litman GW,Frommel D,Finstad FJ,Howell J,Pollara BWおよびGood RA、The evolution of the immune response.VIII.Structural studies of the lamprey immunoglobulin.J.Immunol(1970)105:1278−85
【非特許文献8】Pollara B,Litman GW,Finstad J,Howell JおよびGood RA、The evolution of the immune response.VII.Antibody to human”O”cells and properties of the immunoglobulin in lamprey.J.Immunol(1970)105:738−45
【非特許文献9】Good,R.A.,Finstad,J.&Litman,G.W.、The biology of lampreys II:Immunology(編者 Hardisty,M.V.&Potter,LC.)(1972)405−432(Academic Press,London)
【非特許文献10】Hagen M,Filosa MFおよびYouson JH、The immune response in adult sea lamprey{Petromyzon marinus L.):the effect of temperature.Comp.Biochem.Physiol(1985)82:207−10
【非特許文献11】Finstad J,Papermaster BWおよびGood RA、Evolution of the immune response.JJ.Morphologic studies of the thymus and organized lymphoid tissue.Lab Invest.(1964)13:490−512
【非特許文献12】Perey DY,Finstad J,Pollara BおよびGood RA、Evolution of the immune response.VI.First and second set skin homograft rejections in primitive fishes.Lab.Invest.(1968)19:591−7
【非特許文献13】Fujii TおよびHayakawa I、A histological and electron−microscopic study of the cell types involved in rejection of skin allografts in ammocoetes.Cell Tissue Res.(1983)231:301−12
【非特許文献14】Cooper AJ、Ammocoete lymphoid cell populations in vitro.:4th Leukocyte Culture Conference.O.R.Mclntyre(編)New York Appleton Century−Crofts(1971)pp.137− 47
【非特許文献15】Piavis GWおよびHiatt JL、Blood cell lineage in the sea lamprey Petromyzon marinus(Pisces:Petromyzontidae).Copeia(1971)4:722−8
【非特許文献16】Kilarski WおよびPlytycz B、The presence of plasma cells in the lamprey(Agnatha).Dev.Comp.Immunol(1981)5:361−6
【非特許文献17】Zapata A,Ardavin CF,Gomariz RPおよびLeceta J、Plasma cells in the ammocoete of Petromyzon marinus.Cell Tissue Res.(1981)221:203−8
【非特許文献18】Fujii T、Electron microscopy of the leukocytes of the typhlosole in ammocoetes,with special attention to the antibody−producing cells.J.Morphol(1982)173:87−100
【非特許文献19】Ardavin CFおよびZapata A、Ultrastructure and changes during metamorphosis of the lympho−hemopoietic tissue of the larval anadromous sea lamprey Petromyzon marinus.Dev.Comp.Immunol.(1987)11:79−93
【非特許文献20】Mayer WE,Uinuk−Ool T,Tichy H,Gartland LA,Klein JおよびCooper MD、Isolation and characterization of lymphocyte−like cells from a lamprey.Proc.Natl.Acad.Sci,USA(2002a)99:14350−5
【非特許文献21】Haire RN,Miracle AL,Rast JPおよびLitman GW、Members of the Ikaros gene family are present in early representative vertebrates.J.Immunol.(2000)165:306−12
【非特許文献22】Shintani S,Terzic J,Sato A,Saraga−Babic M,O’hUigin C,Tichy HおよびKlein J、Do lampreys have lymphocytes? The Spi evidence.Proc.Natl.Acad.Sci.,USA(2000)97:7417−22
【非特許文献23】Anderson MK,Sun X,Miracle AL,Litman GWおよびRothenberg EV Evolution of hematopoiesis: Three members of the PU.1 transcription factor family in a cartilaginous fish,Raja eglanteria.Proc.Natl.Acad.Sci USA(2001)98:553−8
【非特許文献24】Mayer WE,O’Huigin C,Tichy H,Terzic JおよびSaraga−Babic M、Identification of two Ikaros−like transcription factors in lamprey.Scand.J.Immunol(2002b)55:162−70
【非特許文献25】Azumi K et al.,Genomic analysis of immunity in a Urochordate and the emergence of the vertebrate immune system:”waiting for Godot”Immunogenetics(2003)55:570−81
【発明の概要】
【課題を解決するための手段】
【0006】
(発明の概要)
本明細書に具体的かつ広範囲に記載する本発明の目的によると、本発明は、1つの態様では、新規なリンパ球受容体またはそのフラグメントを含むポリペプチドに関する。本発明は、前記リンパ球受容体またはフラグメントをコードする核酸にさらに関する。前記ポリペプチドおよび核酸を作製かつ使用する方法もまた提供される。そのような使用には、広範囲の精製、治療および診断方法が含まれる。
【0007】
本発明の追加の長所は、一部は以下の説明の中に記載され、そして一部は本説明から明白になるか、または本発明の実践によって学習することができるであろう。本発明の長所は、添付の特許請求の範囲の中で特別に指摘された要素および組み合わせによって実現かつ達成されるであろう。上記の一般的説明および下記に詳述する説明の両方は、代表的かつ説明的なものに過ぎず、特許請求されるように本発明を限定するものではないことを理解されたい。
例えば、本願発明は以下の項目を提供する。
(項目1)
1つのN末端ロイシンリッチリピート(LRRNT)、1つまたは複数のロイシンリッチリピート(LRR)、1つのC末端ロイシンリッチリピート(LRRCT)、および1つの連結ペプチドを含む単離されたポリペプチドであって、前記連結ペプチドがαヘリックスを含む、ポリペプチド。
(項目2)
前記連結ペプチドがLRRCTへ連結している、項目1に記載のポリペプチド。
(項目3)
柄領域およびグリコシル−ホスファチジル−イノシトールアンカーをさらに含む、項目1に記載のポリペプチド。
(項目4)
疎水性尾部をさらに含む、項目3に記載のポリペプチド。
(項目5)
前記柄領域がトレオニン−プロリンリッチ領域を含む、項目3に記載のポリペプチド。
(項目6)
1つのシグナルペプチドをさらに含む、項目1に記載のポリペプチド。
(項目7)
1〜9個のLRRが存在する、前記LRRNTに隣接するLRR1を備える、項目1に記載のポリペプチド。
(項目8)
LRR1が約20個未満のアミノ酸を含む、項目7に記載のポリペプチド。
(項目9)
LRR1が約18個のアミノ酸を含む、項目7に記載のポリペプチド。
(項目10)
LRR2〜9の各々が約25個未満のアミノ酸を含む、項目7に記載のポリペプチド。
(項目11)
前記LRRNTが約40個未満のアミノ酸を含む、項目1に記載のポリペプチド。
(項目12)
前記LRRNTが配列番号157のアミノ酸配列を含む、項目11に記載のポリペプチド。
(項目13)
前記LRRNTが1つまたは複数の保存的アミノ酸置換を有する配列番号157のアミノ酸配列を含む、項目12に記載のポリペプチド。
(項目14)
前記LRRCTが約60個未満のアミノ酸を含む、項目1に記載のポリペプチド。
(項目15)
前記LRRCTが配列番号158のアミノ酸配列を含む、項目14に記載のポリペプチド。
(項目16)
前記LRRCTが1つまたは複数の保存的アミノ酸置換を有する配列番号158のアミノ酸配列を含む、項目14に記載のポリペプチド。
(項目17)
前記連結ペプチドが約15個未満のアミノ酸を含む、項目1に記載のポリペプチド。
(項目18)
前記LRRのアミノ酸配列が相互に相違する、そしてLRRNTおよびLRRCTと相違する、項目1に記載のポリペプチド。
(項目19)
前記ポリペプチドが約130〜約225アミノ酸長である、項目1に記載のポリペプチド。
(項目20)
前記ポリペプチドが選択的に作用因子へ結合する、項目1に記載のポリペプチド。
(項目21)
前記作用因子が病原体である、項目20に記載のポリペプチド。
(項目22)
前記病原体が細菌である、項目21に記載のポリペプチド。
(項目23)
前記作用因子が毒素である、項目20に記載のポリペプチド。
(項目24)
項目1に記載のポリペプチドをコードする単離された核酸。
(項目25)
発現制御配列に機能的に連結した項目24に記載の核酸を含む発現ベクター。
(項目26)
項目25記載のベクターを含む培養された細胞。
(項目27)
前記ポリペプチドが固体支持体へ結合している、項目1に記載のポリペプチド。
(項目28)
前記固体支持体が可動固体支持体である、項目27に記載のポリペプチド。
(項目29)
前記固体支持体がカラムである、項目27に記載のポリペプチド。
(項目30)
前記固体支持体がチップである、項目27に記載のポリペプチド。
(項目31)
前記固体支持体がマルチウエルプレートである、項目27に記載のポリペプチド。
(項目32)
前記LRRがポリペプチド全体で高度に可変性である、項目1記載の複数のポリペプチド。
(項目33)
前記複数が固体支持体へ結合している、項目32に記載の複数のポリペプチド。
(項目34)
前記固体支持体が可動固体支持体である、項目32に記載の複数のポリペプチド。
(項目35)
前記固体支持体がカラムである、項目32に記載の複数のポリペプチド。
(項目36)
前記固体支持体がチップである、項目32に記載の複数のポリペプチド。
(項目37)
前記固体支持体がマルチウエルプレートである、項目32に記載の複数のポリペプチド。
(項目38)
サンプル中の作用因子を検出する方法であって、
a.前記サンプルを項目20記載のポリペプチドとを、前記サンプル中の前記作用因子へ前記ポリペプチドが結合するための条件下でと接触させる工程と;
b.前記サンプル中の前記作用因子へ結合したポリペプチドを検出する工程であって、結合したポリペプチドが前記サンプル中の作用因子を指示する工程と、を含む方法。
(項目39)
前記ポリペプチドが検出可能な成分で標識される、項目38に記載の方法。
(項目40)
前記サンプル中の前記作用因子の定量をさらに含む、項目38記載の方法。
(項目41)
作用因子の活性を遮断する方法であって、前記作用因子を項目20に記載の前記ポリペプチドとを、前記ポリペプチドが前記作用因子に結合するための条件下で接触させる工程を含み、前記ポリペプチドの前記作用因子への結合が前記作用因子の活性を遮断する方法。
(項目42)
接触させる工程がインビトロである、項目38に記載の方法。
(項目43)
接触させる工程がインビボである、項目38に記載の方法。
(項目44)
被験者に有効量の項目1に記載のポリペプチドを投与する工程、および前記被験者において結合したポリペプチドの所在位置を検出する工程、を含むイメージング法。
(項目45)
前記ポリペプチドが検出可能な成分で標識される、項目44に記載の方法。
(項目46)
サンプルからの作用因子を精製する方法であって、
a.前記サンプルを項目20記載のポリペプチドと、前記ポリペプチドが作用因子に結合してポリペプチド/作用因子複合体を形成する条件下で、接触させる工程と;
b.前記サンプルから前記ポリペプチド/作用因子複合体を単離する工程と;
c.前記ポリペプチド/作用因子複合体から前記作用因子を単離する工程と、を含む方法。
(項目47)
被験者における病原性作用を減少または予防する方法であって、前記被験者に有効量の項目21のポリペプチドを投与する工程を含む方法。
(項目48)
ポリペプチドを製造する方法であって、前記ポリペプチドの発現を許容する条件下で項目26に記載の細胞を培養する工程と、前記ポリペプチドを細胞または細胞の培地から精製する工程と、を含む方法。
(項目49)
被験者における1つまたは複数の可変性リンパ球受容体をスクリーニングする方法であって、被験者において、1つのN末端ロイシンリッチリピート(LRRNT)、1つまたは複数のロイシンリッチリピート(LRR)、1つのC末端ロイシンリッチリピート(LRRCT)、および1つの連結ペプチドを含む1つまたは複数のポリペプチドを同定する工程を含み、このとき前記連結ペプチドがαヘリックスを含む、方法。
(項目50)
遺伝子の機能を決定する方法であって、遺伝子を発現する細胞の細胞質内へ前記遺伝子のタンパク質産物に対して特異的なポリペプチドを導入する工程、および機能が知られていない遺伝子のタンパク質産物の消失に起因する作用を監視する工程、を含む方法。
【図面の簡単な説明】
【0008】
本明細書に組み込まれていてその一部を構成する添付の図面は、本発明のいくつかの実施形態を図示しており、下記の説明とともに、本発明の原理を説明するために役立つ。
【図1】図1は、ヤツメウナギの白血球およびVLRを示した図である。図1aは、抗原/マイトジェンカクテルを用いた免疫賦活前後の白血球についての光散乱分析を示している。図1bは、選別された免疫賦活白血球:小リンパ球(R1)、大リンパ球(R2)または骨髄性細胞(R3)を示している。Wright−Giemsa染色、100×。スケールバー=10μm。図1cは、VLRおよびGAPDH(コントロール)の視覚的なノーザンブロットを示している。免疫賦活および非免疫賦活幼生の組織から、または造血器官および血液由来の選別された細胞から増幅させたcDNAが示されている。図1dは、VLRスティックモデル:シグナルペプチド、N末端LRR、9個のLRR、連結ペプチド、C末端LRR、トレオニン−プロリンリッチ柄部、GPIアンカーおよび疎水性尾部を示している(クローン12.26、417残基、AY577974)。図1eは、細菌GPI−ホスホリパーゼCを用いて処理した(+PLC)または用いずに処理した(−PLC)マウス胸腺腫細胞内で発現したエピトープタグ付きVLRおよびFcγRIIb(コントロール)の細胞表面発現を示している。図1fは、2回転させて見たVLR多様性領域の3Dモデルを示している(クローン12.26)。
【図2】図2は、2匹のヤツメウナギの幼生におけるVLR多様性についての調査を示した図である。リンパ球からPCR増幅した20の多様性領域のアライメント。PCRプライマーは、全VLR配列内に保存された領域内に所在した:LRRNTの5’側およびLRRCTの3’側近くのシグナルペプチド。ドナー動物およびクローン数が指示されている。LRRモチーフの位置もまた示されている。黒色:100%同一性;灰色:60〜99%同一性;白色:60%同一性。配列1.3〜2.10は、本明細書では各々配列番号1〜20に対応する。
【図3】図3は、13匹の個々の幼生におけるVLRタンパク質多様性の評価を示した図である。cDNAおよびゲノムPCRクローン由来の112のVLR多様性領域の遺伝子距離デンドログラム(樹系図)。幼生数およびクローン数(例、6.20=ドナー6、クローン20)は、免疫賦活ドナー(N=27)については赤色および非免疫賦活ドナー(N=41)については緑色で指示されている。星印(*)は、1つの単離体由来の2個のVLR(9.16S、9.16L)を含む単一の細胞単離体(N=12)由来のクローンを示している;そしてコントロール10細胞プール由来のクローンは10Cと表示されている(N=4)。ゲノムDNA由来の成熟VLR配列は青色である(N=28;血液#10、12;死体#11、13)。全セットについての平均多様性は1.36±0.03であり、13の固体からの配列群内で0.28〜0.54の範囲である。
【図4】図4は、VLRのN末端またはC末端のプローブを用いてハイブリダイズした制限酵素消化DNAのVLRゲノムブロットを示した図である。図4(a)は、3匹のヤツメウナギのブロットを示している(血液DNA #10、12;死体#13)。動物13だけが多型BamHIパターンを示した。図4bは、10匹のヤツメウナギからプールされた赤血球からゲノムの広がりを示している。パルスフィールドブロット・ハイブリダイゼーションは両方のプローブに対して適合するパターンを示し、さらなる350kbのNotI N末端バンドは5’gVLR重複に対応する。
【図5】図5は、VLR遺伝子座のゲノム組織を示した図である。図5aは、20kbにわたってオーバーラップするクローンPAC16(44kb)およびPAC3(33kb)から融合された57kbのgVLRコンティーグ(AY577941)内で同定されたモチーフを示している。点線はPAC挿入物を表している;赤いバーは、N末端およびC末端プローブを指示している。図5bは、PAC4(58kb、AY577942)が11.7kbにわたってgVLRコンティーグとアライメントすることを示している(ヌクレオチド45,882〜57,609)。1〜3個のLRRのカセットは順方向または逆方向で配置されている:8個はgVLRコンティーグ内であり、17個はPAC4内である。図5cは、gVLRのLR−PCR分析を示している。血液(#10)または死体(#13)由来のDNAはプライマーgVLR.F1+gVLR.R1を用いて増幅させた(図5aおよび図5eに示した)。PAC16アンプリコンがコントロールとして機能した。約20kbのバンドは生殖細胞系VLRに対応し、約8kbのバンドは成熟VLRに対応する。図5dは、成熟VLRのリンパ球特異的再配列を示している。選別された100個のリンパ球または赤血球のプール由来のLR−PCR。約14kbのバンドは生殖細胞系VLRに対応し、約〜1kbのバンドはリンパ球DNAからのみ増幅された成熟VLRに対応する。図5eは、8kbの成熟VLRアンプリコンを例示している。
【図6】図6は、ESTクローンから予測された22個のVLRタンパク質の複合アライメントを示した図である(シングルパス5’配列、一部不完全なC末端)。黒色:完全同一性;黄色:80〜99%同一性;緑色:60〜79%同一性;白色:60%未満の同一性。LyEST3090〜LyEST5266に対するアミノ酸配列は、各々配列番号21〜42に対応する。
【図7】図7は、代表的なVLR(cDNAクローンLyEST2913、AY578059)のORFを示した図である。開始メチオニンはヌクレオチド118〜120にあり、終止コドンはヌクレオチド937〜939にある。生殖細胞系VLRのエクソン2およびエクソン4内に保存されたヌクレオチド配列は赤色で表示されている;エクソン3に対応する多様な5’LRRCTは緑色で表示されている。構造モチーフはタンパク質配列の上方に表示されている;GPI開裂部位は青色で表示されている。図示したアミノ酸配列は配列番号43に対応し、図示した核酸配列は配列番号156に対応する。
【図8−1】図8は、13匹のヤツメウナギからPCR増幅させた112のVLR多様性領域の複合アライメントを示した図である。免疫賦活ヤツメウナギおよび非免疫賦活ヤツメウナギからのゲノムクローンおよびRT−PCRクローン。非免疫賦活動物:#1〜4と指定された動物(N=41)、#8と指定された動物由来の選別された単リンパ球(N=4)および#8.10Cと指定された動物由来の10個の細胞のプール由来のクローン(N=4);免疫賦活動物:2個のVLR(9.16S、9.16L)を備える1つの単離体を含む、#5〜7と指定された動物(N=27)および#9と指定された動物由来の選別された単リンパ球(N=8);成熟VLR:#10〜13(N=28)または死体(#11、13)と指定された血液から抽出した幼生ゲノムDNA。黒色:80〜100%同一性;黄色:60〜79%;緑色:40〜59%;白色:<40%。アライメントの上部から、1.1に対するアミノ酸配列は配列番号13に対応し、アミノ酸配列7.27〜4.7は配列番号45〜52に対応し、アミノ酸配列1.5は配列番号12に対応し、アミノ酸配列4.14は配列番号54に対応し、アミノ酸配列1.7は配列番号8に対応し、アミノ酸配列3.15は配列番号56に対応し、アミノ酸配列2.1は配列番号5に対応し、アミノ酸配列2.2は配列番号10に対応し、アミノ酸配列2.7は配列番号11に対応し、アミノ酸配列4.8〜6.22は配列番号60〜65に対応し、アミノ酸配列2.4は配列番号3に対応し、アミノ酸配列1.8は配列番号2に対応し、アミノ酸配列7.3〜6.21は配列番号68〜72に対応し、アミノ酸配列1.2は配列番号5に対応し、アミノ酸配列2.14は配列番号6に対応し、アミノ酸配列3.7は配列番号75に対応し、アミノ酸配列1.6は配列番号7に対応し、アミノ酸配列5.3は配列番号77に対応し、アミノ酸配列10.1は配列番号78に対応し、アミノ酸配列2.14は配列番号4に対応し、アミノ酸配列1.3は配列番号1に対応し、アミノ酸配列6.16〜7.26は配列番号81〜119に対応し、アミノ酸配列2.15は配列番号14に対応し、アミノ酸配列2.8は配列番号17に対応し、アミノ酸配列5.6〜7.33は配列番号122〜125に対応し、アミノ酸配列1.10は配列番号19に対応し、アミノ酸配列2.10は配列番号20に対応し、アミノ酸配列1.4は配列番号15に対応し、アミノ酸配列12.19〜4.3は配列番号129〜132に対応し、アミノ酸配列1.9は配列番号16に対応し、アミノ酸配列5.5〜3.3は配列番号134〜144に対応し、アミノ酸配列2.13は配列番号18に対応し、およびアミノ酸配列3.6〜3.9は配列番号146〜155に対応する。
【図8−2】(図8−1の続き)
【図9】図9は、進化的に保存された無顎類VLRを示した図である。沿岸メクラウナギ(Eptatretus burgeri)、太平洋メクラウナギ(E.stoutii)、ウミヤツメ(Petromyzon marinus;GenBank受託番号AY577946)、アメリカカワヤツメ(Lampetra appendix)および北部カワヤツメ(Ichthyomyzon fossor)を表しているVLRアミノ酸配列。青色の影:100%同一性;黄色:60〜99%同一性;緑色:40〜59%同一性;赤色:疎水性尾部領域。
【図10】図10は、太平洋メクラウナギVLR多様性領域(LRRNTからLRRCT)間の遺伝子距離を示した図である。5匹の動物からの、PCR増幅リンパ球様cDNAクローンまたは血液ゲノムPCRアンプリコンから予測されるタンパク質。スケールバーは5%アミノ酸多様性を表している。A.VLR−A(N=139)。B.VLR−B(N=70)。緑色:非免疫賦活;赤色:免疫賦活;青色:ゲノム成熟VLR;星印−関連配列。
【図11】図11は、メクラウナギVLR遺伝子座を示した図である。図11Aは、太平洋メクラウナギVLR−Aを示している。図11Bは、沿岸メクラウナギVLR−Aを示している。図11Cは、太平洋メクラウナギVLR−Bを示している。図11Dは、沿岸メクラウナギVLR−Bを示している。4個のBACクローンからの挿入物の配列であり、占められていないギャップがマーキングされている。逆方向または順方向にあるVLR生殖細胞系遺伝子およびフランキングカセットの位置はキロベース単位で指示されている(図は一定尺度から外れている)。GenScan遺伝子予測は青色で表示されている:太平洋メクラウナギ生殖細胞系VLR−A遺伝子から上流の非関連性LRR遺伝子ならびに沿岸メクラウナギVLR−Aおよび太平洋メクラウナギVLR−B遺伝子座における2つの隣接するトランスポザーゼORF。
【図12】図12は、無顎類VLR遺伝子、転写産物および系統発生を示した図である。図12Aは、太平洋メクラウナギおよびウミヤツメの生殖細胞系および成熟VLR遺伝子の略図を示している。着色したバーは、コーディング領域を示している;ヌクレオチド単位のサイズ;メクラウナギVLRを増幅させるために使用したPCRプライマーの位置(表5)は矢印で指示され、F(順方向)、R(逆方向)と指示されている。図12Bは、リンパ球様転写産物(RT−PCR)または血液ゲノムDNAからPCR増幅させた太平洋メクラウナギVLRを示している。アガロースゲル画像;左には分子量マーカーが表示されている(キロベース);生殖細胞系および成熟VLRアンプリコンの位置は右側に表示されている。図12Cは、無顎類VLRの系統発生的分析を示している。メクラウナギおよびヤツメウナギVLRタンパク質の近隣結合系統樹(図9の配列と同一配列);ブートストラップ値が指示されている。スケールバーは10%アミノ酸多様性を表している。図12Dは、無顎類VLRの進化のモデルを示している。
【図13】図13は、沿岸メクラウナギVLR多様性領域(LRRNTからLRRCT)間の遺伝子距離を示した図である。タンパク質は、3匹の動物の白血球cDNAクローンまたはゲノムDNA由来の成熟VLRアンプリコンから予測した。スケールバーは5%アミノ酸多様性を表している。A.VLR−A(N=66)。B.VLR−B(N=18)。赤色:メクラウナギ#7;緑色:#8;青色:メクラウナギ#9由来のゲノム成熟VLR。
【発明を実施するための形態】
【0009】
(詳細な説明)
最古の脊椎動物の中で現代に生きる代表であるウミヤツメPetromyzon marinusの脊椎動物免疫系の原始エレメントを対象にリンパに集中した調査が開始された。ヤツメウナギ造血組織由来のリンパ球様細胞によって発現した転写産物の初期の分析は、免疫系分子の数種のホモログを同定したが(Mayer et al.,2002a;Uinuk−Ool et al.,2002;Uinuk−Ool et al.,2003)、有顎類脊椎動物が特異的適応免疫のために使用する基本的なIgスーパーファミリー受容体エレメントは同定されなかった。血流中に存在する活性化リンパ芽球は適応応答に含まれる遺伝子を発現する可能性が高いと判断して、本試験は、免疫賦活ヤツメウナギ幼生由来リンパ球のトランスクリプトームの調査から開始した。この研究により、本明細書に記載する新規なタイプの高可変性リンパ球受容体が明らかになった。
【0010】
本発明は、本明細書に含まれた本発明の好ましい実施形態および実施例についての下記の詳細な説明および図面ならびにそれらについての上記や下記の説明を参照することによってより容易に理解することができる。
【0011】
本発明の化合物、組成物、製品、装置、および/または方法を開示および説明する前に、当然ながらそれらは変動することがあるので、本発明は、他に特に規定しない限り、特定の合成方法、特定の組み換えバイオテクノロジー方法にも限定されず、他に規定しない限り特定の試薬にも限定されないことを理解されたい。さらにまた、本明細書で使用する用語は特定の実施形態を説明するために使用されたに過ぎず、限定的であることは意図されていないことも理解されたい。
本明細書および添付の特許請求項において使用する単数形の「1つの」および「その」には、状況が他の場合を明確に指さない限り、複数対象が含まれる。そこで、例えば「1つの医薬担体」との言及には、2つ以上のそのような担体などの混合物が含まれる。
【0012】
本明細書での範囲は、「約」1つの特定値から、および/または「約」別の特定値までを表すことができる。そのような範囲が明示される場合は、別の実施形態は、1つの特定値から他の特定値までを含んでいる。同様に、先行詞「約」の使用によって近似値として数値が明示される場合は、特定値が別の実施形態を形成することを理解されたい。さらに、範囲各々の終点は、他方の終点と関連して、および他方の終点からは独立してのどちらにおいても重要であることを理解されたい。
【0013】
「任意の」または「任意で」は、引き続いて記載される事象または状況が発生しても発生しなくてもよく、そしてその説明は前記事象または状況が発生する場合およびそれが発生しない場合を含んでいることを意味する。
【0014】
本明細書で使用する「ポリペプチド」、「タンパク質」、および「ペプチド」は、アミノ酸配列について言及するために互換的に使用される。
【0015】
本発明は、1〜12個のロイシンリッチリピートを含み、適応免疫において機能できる体細胞性再配列が可能なポリペプチドである可変性リンパ球受容体(VLR)に関する。
【0016】
本発明は、N末端ロイシンリッチリピート(LRRNT)、1つまたは複数のロイシンリッチリピート(LRR)(本明細書では内部LRRと呼ぶ)、C末端ロイシンリッチリピート(LRRCT)、および連結ペプチドを含む単離されたポリペプチドを提供するが、このとき連結ペプチドはαヘリックスを含んでいる。ポリペプチドの長さは、約130という少数のアミノ酸または約225という多数のアミノ酸を含むことができる。ポリペプチドおよびコーディング核酸の一般構造および特定配列の例は図に示されている。さらに様々な領域(シグナルペプチド、LRRNT、LRR、LRRCT、連結ペプチド、柄部および疎水性尾部を含む)の極めて多数の例は図の中に見いだすことができる。
【0017】
任意で、連結ペプチドはLRRCTのN末端側に位置しており、より具体的には内部LRRおよびLRRCTの間に位置している。連結ペプチドは、内部LRRおよびLRRCTへ連結させることができる。したがって、本明細書ではLRRNT、1つまたは複数の内部LRR、連結ペプチド、およびLRRCTをその順序で含むポリペプチドが開示される。さらに、LRRNTおよびLRRCT間の内部LRR領域が、1個、2個、3個、4個、5個、6個、7個、8個または9個のロイシンリッチリピートを含むポリペプチドが開示され、LRR1はLRRNTに隣接して位置するかまたは近位に位置する。本明細書で使用するようにLRR1、LRR2、LRR3、LRR4、LRR5、LRR6、LRR7、LRR8、またはLRR9は、LRRNTからLRRCTへ連続的に進むとみなされる。そこで本明細書には、LRRNT、LRR1、LRR1〜2、LRR1〜3、LRR1〜4、LRR1〜5、LRR1〜6、LRR1〜7、LRR1〜8、またはLRR1〜9、連結ペプチド、およびLRRCTをその順序で含むポリペプチドが開示される。
【0018】
ロイシンリッチリピート(LRR)は、タンパク質対タンパク質相互作用に典型的に関係する短い配列モチーフであり、LRRは複数のロイシン残基を含んでいる。LRRは、例えば2位、5位、7位、12位、16位、21位および24位でロイシンまたは他の脂肪族残基を含有する。しかし、ロイシンまたは他の脂肪族残基は2位、5位、7位、12位、16位、21位および24位での残基の場所に加えて、またはそれらの代わりに他の位置に存在し得ると理解されており、本明細書では企図されている。例えば、ロイシンは2位ではなく3位に存在し得る。構造的には、モチーフがβシート構造を形成することもまた理解されたい。そこで、例えばLRRNT、5個のLRR、LRRCT、および連結ペプチドを含む開示されたポリペプチドは、7つのβシート構造および連結ペプチドのαヘリックスを含むであろう。
【0019】
各LRRの長さおよび配列は、ポリペプチド内の他のLRRならびにLRRNTおよびLRRCTとは相違することがあると理解されたい。例えば、本発明の1つの実施形態は、LRRNT、1〜9個のLRR、連結ペプチド、およびLRRCTを含むポリペプチドであるが、このとき第1内部LRRはLRR1であり、LRR1は約20個未満のアミノ酸を含んでいる。さらにLRR1が約18個のアミノ酸を含むポリペプチドが開示される。任意で、このポリペプチドはさらにLRR2〜9を含むが、このときLRR2〜9は各々約25未満のアミノ酸である。さらにLRR2〜9が各々約24個のアミノ酸を含むポリペプチドが開示される。LRR1〜9は、長さおよび特異的アミノ酸配列の両方において所定のポリペプチド内で同一であっても、または相互から相違していてもよい。
【0020】
LRRNTおよびLRRCTと指される末端LRRは、典型的には各内部LRRより長い。LRRNTおよびLRRCTは、不変領域(類似の可変性リンパ球受容体と比較してポリペプチドの残りに比してほとんど変化を有していない領域)を含む。可変領域は、特異性を備える受容体を提供するが、不変領域および受容体にわたる一般構造類似性は保護免疫機能を維持するのに役立つ。ポリペプチドはLRRNTを含むことができ、このときLRRNTは約40未満のアミノ酸を含んでいる。そこで、LRRNTは、1つまたは複数の保存的アミノ酸置換の存在下または不在下で、任意でアミノ酸配列CPSQCSC(配列番号157)、CPSRCSC(配列番号307)、CPAQCSC(配列番号308)、CPSQCLC(配列番号309)、CPSQCPC(配列番号310)、NGATCKK(配列番号311)、またはNEALCKK(配列番号312)を含んでいる。さらにまた、LRRCTが約60アミノ酸長未満、および任意で40〜60アミノ酸長であるLRRCTを含むポリペプチドが開示される。詳細には、特に、LRRCTが保存的アミノ酸置換の存在下または不在下で、アミノ酸配列TNTPVRAVTEASTSPSKCP(配列番号158)、SGKPVRSIICP(配列番号313)、SSKAVLDVTEEEAAEDCV(配列番号314)、またはQSKAVLEITEKDAASDCV(配列番号315)を含むポリペプチドが開示される。
【0021】
すべてのペプチド、ポリペプチド、およびタンパク質と同様に、ペプチド、ポリペプチド、またはタンパク質の性質または機能を変化させないLRRCTおよびLRRNTのアミノ酸配列における置換が発生する可能性があると理解されたい。そのような置換は、保存的アミノ酸置換を含んでおり、下記でより詳細に考察する。
【0022】
開示された組成物は、連結ペプチドも含むことができる。典型的には、そのようなペプチドは15アミノ酸長未満の短いペプチドであり、αヘリックスを含んでいる。そこで、例えばαヘリックスを含む10アミノ酸長、11アミノ酸長、12アミノ酸長、13アミノ酸長、14アミノ酸長、および15アミノ酸長の連結ペプチドが特異的に開示される。連結ペプチドは、ポリペプチドの構造的成分を連結させるために機能すると理解されたい。さらに、ポリペプチドの連結ペプチドは、LRRCTへ連結できると理解されたい。
【0023】
本発明のポリペプチドは、可溶形または膜結合形を含むことができる。ポリペプチドを可溶形または膜結合形にさせる多数の機序が存在する。例えば、膜貫通ドメインを有していないポリペプチドは細胞によって直接的に分泌されることがある。または、ポリペプチドは、膜表面上にそのポリペプチドを維持するグリコシル−ホスファチジル−イノシトール(GPI)アンカーを含むことができる。このため、本明細書ではGPIアンカーを含むポリペプチドが開示される。表面へ結合したポリペプチドを維持するための他の機構は、当技術分野において知られている。例えば、本ポリペプチドは、膜の脂質二重層との共有相互作用を形成するシングルパスまたはマルチパス膜貫通領域を通して疎水層へ結合させることができる。または、本ポリペプチドは、表面タンパク質との非共有相互作用を通して表面へ結合させることができる。
【0024】
本発明のポリペプチドは、表面結合ポリペプチドであってよい。細胞表面へのトラフィッキングは、細胞の表面へポリペプチドを送達するための細胞内輸送機構についてのインジケータを提供するシグナルペプチドによって実施することができる。そこで、本発明のまた別の実施形態では、本発明のポリペプチドは本ポリペプチドのN末端のシグナルペプチドを含んでいる。
【0025】
本ポリペプチドが疎水性尾部を含むことができることは理解されており、本明細書で企図されている。
【0026】
本ポリペプチドは、柄領域を含むことができる。柄領域は、トレオニン−プロリンリッチ領域を含んでおり、GPIアンカーおよび疎水性尾部とともに、任意で本ポリペプチドの膜結合形で存在する。
【0027】
本発明のポリペプチドの例には、配列番号1〜43、45〜52、54、56、60〜65、68〜72、75、77〜78、81〜119、122〜125、129〜132、134〜144、および146〜155のアミノ酸配列を含むポリペプチドが含まれる。配列には、GenBank受託番号AY577941〜AY578059およびCK988414〜CK988652が含まれる。配列番号1〜20のアミノ酸配列を含む配列は、全長VLRの例を表している。配列番号43のアミノ酸配列を含む配列は、シグナルペプチドを備える全長VLRの例である。さらなる全長VLRおよびそのアミノ酸配列を含むフラグメントは図の中に見いだすことができる。本発明のポリペプチドについて本明細書で教示される構造に基づいて、これらの配列は1つの属のポリペプチドの例であることは理解されるであろう。本発明は全長VLRおよびそのフラグメントを含んでいると理解されたい。
【0028】
開示された組成物を調製するために使用される構成成分ならびに本明細書に開示した方法の中で使用される組成物自体が開示される。これらやその他の材料は本明細書に開示されており、これらの材料の組み合わせ、サブセット、相互作用、グループなどが開示される場合は、これらの化合物の各々の様々な個別および集合的組み合わせおよび順列についての具体的な参照は明示的には開示されないかもしれないが、各々は本明細書に詳細に企図かつ説明されていると理解されたい。例えば、特定のポリペプチドが開示かつ考察され、多数のポリペプチドへ加えることのできる多数の修飾が考察される場合は、ポリペプチドのありとあらゆる組み合わせおよび順列ならびにそれの反対が特別に指示されない限り可能である修飾が特別に企図されている。そこで、1クラスの分子A、B、およびCが開示され、ならびに1クラスの分子D、E、およびFと組み合わせ分子の例A〜Dが開示される場合は、各々が個別に言及されていない場合でさえ、各々が個別に、そしてA−E、A−F、B−D、B−E、B−F、C−D、C−E、およびC−Fの集合的に企図された意味の組み合わせが開示されると考えられる。同様に、これらの任意のサブセットまたは組み合わせもまた開示されている。そこで、例えば、A−E、B−F、およびC−Eのサブグループが開示されると考えられる。この概念は、本明細書に開示した組成物の製造方法および使用方法における工程を含むがそれらに限定されない本出願のすべての態様に当てはまる。そこで、実施できる様々な追加の工程が存在する場合は、これらの追加の工程の各々は本明細書に開示した方法の任意の特定実施形態または実施形態の組み合わせにより実施できると理解されたい。
【0029】
本発明のポリペプチドは、所望の機能を有する。本明細書に記載したポリペプチドは、抗体が選択的に抗原または作用因子へ結合するのとほとんど同様に、抗原または作用因子に選択的へ結合する。ポリペプチドは、任意に可変性リンパ球受容体(天然型または非天然型)またはそのフラグメントもしくは変異体である。用語「可変性リンパ球受容体」は、本明細書では広範囲の意味で使用され、同様に用語「抗体」は様々な特異性を有する様々なバージョンを含んでいる。ポリペプチドは、本明細書に記載したインビトロアッセイを使用してまたは類似方法によってそれらの所望の活性について試験され、その後にそれらの治療的、診断的、またはその他の精製活性が公知の試験方法にしたがって試験される。
【0030】
本発明のポリペプチドは、細胞外物質(例、病原体)または抗原へ結合できる。作用因子または抗原には、ペプチド、ポリペプチド、脂質、糖脂質、およびタンパク質を含むことができるが、それらに限定されない。作用因子または抗原は、病原生物を含むがそれらに限定されない様々な起源に由来することがある。作用因子または抗原への結合は、選択的であると理解されたい。「選択的に結合」または「特異的に結合」は、他の抗原または作用因子を部分的または完全に除外して1つの作用因子または抗原へ結合することを意味する。「結合」は、アッセイ方法のバックグラウンドの少なくとも約1.5倍での検出可能な結合を意味する。選択的または特異的結合については、そのような検出可能な結合は所定の抗原または作用因子については検出できるが、コントロール抗原または作用因子については検出できない。そこで、例えばウイルス、細菌、真菌、または原生動物の抗原または作用因子に選択的に結合するポリペプチドが開示される。
【0031】
ポリペプチドが詳細に開示されるが、前記ポリペプチドは作用因子へ結合し、このとき前記作用因子は病原性作用因子である。さらに、病原性作用因子に選択的に結合する本発明のポリペプチドが開示されるが、このとき前記病原体はウイルスである。多数のウイルスが存在することが知られている。ウイルスは、単純ヘルペスウイルス1型、単純ヘルペスウイルス2型、サイトメガロウイルス、エプスタイン・バーウイルス、水疱・帯状疱疹ウイルス、ヒトヘルペスウイルス6型、ヒトヘルペスウイルス7型、ヒトヘルペスウイルス8型、痘瘡ウイルス、水疱性口内炎ウイルス、A型肝炎ウイルス、B型肝炎ウイルス、C型肝炎ウイルス、D型肝炎ウイルス、E型肝炎ウイルス、ライノウイルス、コロナウイルス、インフルエンザウイルスA型、インフルエンザウイルスB型、麻疹ウイルス、ポリオーマウイルス、ヒトパピローマウイルス、RS(呼吸系合胞体)ウイルス、アデノウイルス、コクサッキーウイルス、デング熱ウイルス、流行性耳下腺炎ウイルス、ポリオウイルス、狂犬病ウイルス、ラウス肉腫ウイルス、黄熱病ウイルス、エボラウイルス、マールブルグウイルス、ラッサ熱ウイルス、東部ウマ脳炎ウイルス、日本脳炎ウイルス、セントルイス脳炎ウイルス、マレーバレー熱(Murray Valley fever)ウイルス、西ナイルウイルス、リフトバレー熱ウイルス、ロタウイルスA型、ロタウイルスB型、ロタウイルスC型、シンドビスウイルス、サル免疫不全ウイルス、ヒトT細胞白血球ウイルス1型、ハンタウイルス、風疹ウイルス、サル免疫不全ウイルス、ヒト免疫不全ウイルス1型、およびヒト免疫不全ウイルス2型からなるウイルス群から選択できる。
【0032】
さらにまた、病原体が細菌である本発明のポリペプチドが開示される。多数の細菌が存在することが知られている。本明細書では、病原体に選択的に結合するポリペプチドが特別に企図かつ開示されるが、このとき前記病原体は結核菌(M.tuberculosis)、ウシ型結核菌(M.bovis)、ウシ型結核菌株BCG、BCG亜株、トリ型結核菌(M.avium)、マイコバクテリウム・イントラセルラーレ(M.intracellulare)、マイコバクテリウム・アフリカヌム(M.africanum)、マイコバクテリウム・カンサシイ(M.kansasii)、マイコバクテリウム・マリナム(M.marinum)、マイコバクテリウム・ウルセランス(M.ulcerans)、トリ型結核菌亜種パラ結核菌(paratuberculosis)、ノカルジア・アステオロイデス(Nocardia asteroides)、その他のノカルジア種、レジオネラ・ニューモフィラ菌(Legionella pneumophila)、その他のレジオネラ種、ネズミチフス菌(Salmonella typhi)、その他のサルモネラ種、赤痢菌種(Shigella species)、ペスト菌(Yersinia pestis)、パスツレラ・ヘモリチカ(Pasteurella haemolytica)、動物パスツレラ症病原菌(Pasteurella multocida)、その他のパスツレラ種、アクチノバチルス胸膜肺炎菌(Actinobacillus pleuropneumoniae)、リステリア・モノサイトゲネス(Listeria monocytogenes)、リステリア・イヴァノビ(Listeria ivanovii)、ブルセラ・アボルツス(Brucella abortus)、その他のブルセラ種、コードリア・ルミナンチウム(Cowdria ruminantium)、クラミジア肺炎菌(Chlamydia pneumoniae)、トラコーマ・クラミジア(Chlamydia trachomatis)、オーム病病原体(Chlamydia psittaci)、コクシエラ・バーネッティイ(Coxiella burnetti)、その他のリケッチア(Rickettsial)種、エールリヒア(Ehrlichia)種、黄色ブドウ球菌(Staphylococcus
aureus)、表皮ブドウ球菌(Staphylococcus epidermidis)、化膿性連鎖球菌(Streptococcus pyogenes)、ストレプトコッカス・アガラクティエ(Streptococcus agalactiae)、炭疽菌(Bacillus anthracis)、大腸菌(Escherichia
coli)、コレラ菌(Vibrio cholerae)、カンピロバクター(Campylobacter)種、髄膜炎菌(Neiserria meningitidis)、淋菌(Neiserria gonorrhea)、緑膿菌(Pseudomonas aeruginosa)、その他のシュードモナス種、インフルエンザ菌(Haemophilus influenzae)、軟性下疳菌(Haemophilus ducreyi)、その他のヘモフィルス(Hemophilus)種、破傷風菌(Clostridium tetani)、その他のクロストリジウム(Clostridium)種、エルシニア・エンテロコリチカ(Yersinia enterocolitica)、およびその他のエルシニア種からなる細菌リストから選択される細菌である。
【0033】
さらに、病原体に選択的に結合する本発明のポリペプチドが開示されるが、前記病原体は原生動物または他の寄生虫である。多数の寄生虫性感染症が存在することが知られている。本明細書では病原体に選択的に結合するポリペプチドが特別に企図かつ開示されるが、前記病原体は、トキソプラズマ原虫(Toxoplasma gondii)、熱帯熱マラリア原虫(Plasmodium falciparum)、三日熱マラリア原虫(Plasmodium vivax)、四日熱マラリア原虫(Plasmodium malariae)、その他のプラズモジウム(Plasmodium)種、トリパノソーマ・ブルース(Trypanosoma brucei)、トリパノソーマ・クルーズ(Trypanosoma cruzi)、森林型熱帯リーシュマニア(Leishmania major)、その他のリーシュマニア(Leishmania)種、マンソン住血吸虫(Schistosoma mansoni)、その他の住血吸虫(Schistosoma)種、および赤痢アメーバ(Entamoeba histolytica)からなる群から選択される寄生虫性感染である。
【0034】
さらに、病原体に選択的に結合する本発明のポリペプチドが開示されるが、前記病原体は真菌である。多数の真菌が存在することが知られている。本明細書には特別にポリペプチドが企図かつ開示されているが、このとき病原体は、カンジダ・アルビカンス(Candida albicans)、クリプトコックス・ネオホルマンス(Cryptococcus neoformans)、ヒストプラズマ・カプスラーツ(Histoplasma capsulatum)、アスペルギルス・フミガーツス(Aspergillus fumigatus)、コクシジオイデス・イミティス(Coccidiodes
immitis)、ブラジル・パラコクシジオイデス(Paracoccidiodes brasiliensis)、ブラストミセス・デルマティディス(Blastomyces dermitidis)、カリニ肺炎(Pneomocystis carnii)、ペニシリウム症(Penicillium marneffi)、およびアルテルナリア・アルテルナータ(Alternaria alternata)からなる真菌群から選択される真菌である。
【0035】
本発明のポリペプチドは、毒素にもまた選択的に結合できる。本明細書では、「毒素」は、それ(毒素)が接触した任意の細胞を効果的に破壊する任意の化学的または生物学的物質を意味する。毒素の著明な例には、リシン、百日咳毒素、サリン、細菌内毒素、中毒性ショック症候群毒素1、コレラ毒素、およびヘビ毒毒素が挙げられる。そこで、毒素へ結合するポリペプチドが特別に開示される。
【0036】
本明細書に記載したポリペプチドは、所望の機能が維持される限り修飾および変化させることができる。任意の変異体および誘導体または本明細書に開示した遺伝子およびタンパク質から発生する可能性のある変異体および誘導体を規定する1つの方法は、特異的な既知配列との相同性によって変異体および誘導体を規定することによると理解されたい。例えば、配列番号1は、本発明の任意の数の核酸によってコードされたポリペプチドの特定アミノ酸配列を規定している。特に開示されているのは、上記の配列に対して少なくとも70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、または99パーセントの相同性を有する本明細書に開示されたこれらや他の遺伝子およびタンパク質の変異体である。当業者であれば、2つのタンパク質または核酸(例えば、遺伝子)の相同性を決定する方法を容易に理解できる。例えば、相同性は、相同性がその最高レベルにあるように2つの配列をアライメントした後に計算できる。
【0037】
一般に、任意の公知の変異体および誘導体または本明細書に開示した遺伝子およびタンパク質から発生する可能性のある変異体および誘導体を規定する1つの方法は、特異的な既知配列との相同性によって変異体および誘導体を規定することによる方法であると理解されたい。本明細書に開示した特定配列のこの同一性については、本明細書の他の場所においても考察する。一般に、本明細書に開示した遺伝子およびタンパク質の変異体は、典型的には、上記の配列または天然配列に対して少なくとも約70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、または99パーセントの相同性を有する。当業者であれば、2つのタンパク質または核酸(例えば、遺伝子)の相同性を決定する方法を容易に理解できる。例えば、相同性は、相同性がその最高レベルにあるように2つの配列をアライメントした後に計算できる。
【0038】
相同性を計算するまた別の方法は、発表されたアルゴリズムによって実施できる。比較のための配列の最適アライメントは、SmithおよびWaterman Adv.Appl.Math.2:482(1981)の局所相同性アルゴリズムによって、NeedlemanおよびWunsch,J.MoL Biol.48:443(1970)の相同性アライメントアルゴリズムによって、PearsonおよびLipman,Proc.Natl.Acad.Sci.U.S.A.85:2444(1988)の類似性検索方法によって、これらのアルゴリズムのコンピュータを使用した実行(GAP、BESTFIT、FASTA、およびTFASTA、Wisconsin Genetics Software Package,Genetics Computer Group,575 Science Dr.、ワイオミング州マディソン)によって、または目視検査によって実施できる。
【0039】
核酸についての同一タイプの相同性は、例えば少なくとも核酸アライメントに関する材料について参照して本明細書に組み込まれるZuker,M.Science 244:48−52,1989,Jaeger et al.Proc.Natl.Acad.Sci.USA 86:7706−7710,1989,Jaeger et al.,Methods Enzymol.183:281−306,1989の中で開示されたアルゴリズムによって入手できる。典型的にはこれらの方法のいずれかを使用できること、そして所定の場合にはこれらの様々な方法の結果は相違することがあるが、当業者であれば、これらの方法の少なくとも1つを用いて同一性が見いだされると、それらの配列は規定された同一性を有しており、本明細書に開示されていると理解することができることが理解されよう。
【0040】
例えば、本明細書で使用するような他の配列との特定パーセンテージの相同性を有すると言及された配列は、任意の1つまたは複数の上記に記載した計算方法によって計算された言及された相同性を有する配列を意味する。例えば、第1配列は、第1配列がZuker計算法を用いて第2配列と80%相同性を有すると計算された場合は、たとえ第1配列が他の任意の計算方法によって計算した場合に第2配列との80%相同性を有していない場合でさえ、本明細書に規定したように第2配列との80%相同性を有する。また別の例として、第1配列は、第1配列がZuker計算法およびPearsonおよびLipman計算法の両方を用いて第2配列と80%相同性を有すると計算された場合は、たとえ第1配列がSmithおよびWaterman計算法、NeedlemanおよびWunsch計算法、Jaeger計算法、または任意の他の計算方法によって計算した場合に第2配列との80%相同性を有していない場合でさえ、本明細書に規定したように第2配列との80%相同性を有する。さらにまた別の例として、第1配列は、第1配列が各計算法を用いて第2配列と80%相同性を有すると計算された場合は、本明細書に規定したように第2配列との80%相同性を有する(だが、実際に様々な計算法はしばしば相違する計算相同性パーセンテージを生じさせるであろう)。
【0041】
タンパク質の変異体および誘導体は、当業者には明確に理解されており、アミノ酸配列修飾を含むことができる。例えば、アミノ酸配列の修飾は、3つのクラス:置換、挿入または欠失変異体の1つまたは複数に分類される。挿入には、単一または複数アミノ酸残基のアミノおよび/またはカルボキシル末端融合ならびに配列内挿入が含まれる。挿入は、通常は、アミノまたはカルボキシル末端融合の挿入より少ない挿入、例えばおよそ1〜4個の残基であろう。実施例に記載された誘導体などの免疫原性融合タンパク質誘導体は、標的配列へ免疫原性を付与するために十分に大きなポリペプチドをインビトロ架橋結合によって融合させる工程によって、またはその融合をコードするDNAを用いて形質転換された組み換え細胞培養によって作製される。欠失は、タンパク質配列からの1つまたは複数のアミノ酸残基の除去を特徴とする。典型的には、約2〜6未満の残基がタンパク質分子内の任意の1つの部位で欠失している。これらの変異体は、通常はタンパク質をコードするDNA内のヌクレオチドの部位特異的突然変異によって調製され、それによってその変異体をコードするDNAが産生され、その後に組み換え細胞培養内でDNAが発現させられる。知られている配列を有するDNA内の規定部位で置換突然変異を作製するための技術、例えばM13プライマー突然変異誘発およびPCR突然変異誘発はよく知られている。アミノ酸置換は、典型的には単一残基の置換であるが、1回に多数の相違する場所で発生してもよい;挿入は、通常はおよそ約1〜10アミノ酸残基であろう;また欠失は約1〜30残基の範囲に及ぶであろう。欠失または挿入は、好ましくは隣接対、すなわち2残基の欠失または2残基の挿入で作製される。置換、欠失、挿入またはそれらの任意の組み合わせは、最終構築物に達するために組合わせることができる。突然変異は、配列をリーディングフレームの外に配置してはならず、好ましくは二次mRNA構造を生成できる相補的領域を作製しないであろう。置換変異体は、少なくとも1つの残基が取り除かれており、異なる残基がその場所に挿入されている変異体である。そのような置換は、一般の表1および2にしたがって作製され、保存的置換と言われる。
【0042】
【表1】

【0043】
【表2】
機能または免疫学的同一性の実質的な変化は、表2に記載の置換より保存的ではない置換を選択することによって、すなわち(a)例えばシートまたはヘリックス立体構造のような、置換領域におけるポリペプチド骨格の構造、(b)標的部位での分子の電荷もしくは疎水性、または(c)側鎖のかさ、を維持する作用がより大きく相違する残基を選択する工程によってなされる。一般にタンパク質特性における最大の変化を生じさせると予測される置換は、(a)例えばセリルもしくはトレオニルのような親水性残基が、例えばロイシル、イソロイシル、フェニルアラニル、バリルもしくはアラニルのような疎水性残基と(によって)置換されている;(b)システインもしくはプロリンが任意の他の残基と(によって)置換されている;(c)例えばリシル、アルギニル、もしくはヒスチジルのような陽性側鎖を有する残基が、例えばグルタミルもしくはアスパルチルのような陰性残基と(によって)置換されている;(d)例えばフェニルアラニンのようなかさの大きな側鎖を有する残基が、この場合にはグリシンのような側鎖を有していない残基と(によって)置換されている、または(e)硫酸化および/またはグリコシル化のための部位数を増加させる、ことによる置換であろう。
【0044】
例えば、1つのアミノ酸残基と生物学的および/または化学的に類似する別のアミノ酸残基との置換は、当業者には保存的置換として知られている。例えば、保存的置換は1つの疎水性残基と別の疎水性残基とを置換させ、または1つの極性残基と他の極性残基とを置換させるであろう。置換には、例えば、GIy、Ala;Val、Ile、Leu;Asp、Glu;Asn、Gln;Ser、Thr;Lys、Arg;およびPhe、Tyrなどの組み合わせが挙げられる。各々明示的に開示された配列のそのような保存的に置換された変形は、本明細書に提供したモザイクポリペプチドの範囲内に含まれる。
【0045】
置換または欠失突然変異誘発は、N−グリコシル化(Asn−X−Thr/Ser)またはO−グリコシル化(SerまたはThr)のための部位を挿入するために使用できる。システインまたはその他の易変残基の欠失もまた望ましいことがある。潜在的タンパク質分解部位、例えばArgの欠失または置換は、例えば塩基性残基の1つを欠失させる工程によって、または1つをグルタミニル残基もしくはヒスチジル残基で置換する工程によって遂行される。
【0046】
所定の翻訳後誘導体化は、発現したポリペプチド上の組み換え宿主細胞の作用の結果である。グルタミニル残基およびアスパラギニル残基は、しばしば対応するグルタミル残基およびアスパルチル残基へ翻訳後に脱アミド化される。または、これらの残基は弱酸性条件下では脱アミド化される。その他の翻訳後修飾にはプロリンおよびリシンの水酸化、セリルもしくはトレオニル残基のヒドロキシル基のリン酸化、リシン側鎖、アルギニン側鎖、およびヒスチジン側鎖のo−アミノ基のメチル化(T.E.Creighton,Proteins:Structure and Molecular Properties,W.H.Freeman & Co.,San Francisco pp79−86[1983])、N末端アミンのアセチル化、および一部の例では、C末端カルボキシルのアミド化が挙げられる。
【0047】
本明細書で使用する用語「可変性リンパ球受容体」は、被験者に投与されると減少した免疫原性を有するように修飾されているポリペプチドも意味することがある。例えば、ヒトアミノ酸配列は、抗体がヒト化されるのとほとんど同様に、ヒト被験者にとって低免疫原性のバージョンにするためにポリペプチドに挿入または追加することができる。多数の非ヒト可変性リンパ球受容体(例、ヤツメウナギ、マウス、ラット、またはウサギに由来するもの)は、ヒトにおいて天然抗原性である可能性があり、したがってヒトへ投与すると望ましくない免疫応答を発生させることがある。このため、本発明の方法における修飾されたポリペプチドの使用は、ヒトに投与されたポリペプチドが望ましくない免疫応答を惹起する可能性を低下させるために役立つことがある。
【0048】
修飾技術は、可変性リンパ球受容体分子の1つまたは複数のポリペプチド領域をコードするDNA配列を操作するための組み換えDNAテクノロジーの使用を含むことができる。したがって、可変性リンパ球受容体(またはそのフラグメント)のヒト化形は、キメラ可変性リンパ球受容体、好ましくはヒト(レシピエント)アミノ酸配列に組み込まれた、非ヒト(ドナー)可変性リンパ球受容体由来の抗原(作用因子)結合部位を含有する可変性リンパ球受容体の抗原(作用因子)結合部分である。
【0049】
これらのタンパク質配列、その変異体およびフラグメントをコードできる核酸もまた開示されることを理解されたい。これには、特異的タンパク質配列に関連するすべての縮重配列、すなわち1つの特定タンパク質配列をコードする配列を有するすべての核酸、ならびに本明細書に開示したタンパク質配列の変異体および誘導体をコードする縮重核酸を含むすべての核酸が含まれるであろう。そこで、各特定核酸配列は本明細書に書き出されない可能性があるが、ありとあらゆる配列が開示されたタンパク質配列を通して本明細書に実際に開示かつ記載されていると理解されたい。
【0050】
ヒト化可変性リンパ球受容体はさらにまた、レシピエントの可変性リンパ球受容体にも取込まれたヒト配列内にも見いだされないアミノ酸配列を含有することがある。
【0051】
本発明のポリペプチドは、融合タンパク質を作製するためにも使用できる。これらのポリペプチドは、融合タンパク質におけるターゲッティング機能に役立つことができる。そこで本発明のポリペプチドは、第2成分へ、組み換え技術によってコンジュゲートするか、さもなければ連結することができる。第2成分は、例えば細胞殺滅が所望であれば、毒素を含むことができる。そこで、例えば原生動物に選択的に結合するポリペプチドは、原生動物を標的とすることができ、融合タンパク質の毒素成分は細胞を殺滅できる。同様に、本発明のポリペプチドは送達機能を実行できる。そこで第2成分は治療薬であってよい。
【0052】
本発明のポリペプチドは、検出可能なタグに連結させることができる。「検出可能なタグ」は、インビボまたはインビトロでイメージング法または検出法を用いて視認できる任意のタグである。検出可能なタグは、放射線不透過性物質、放射標識、化学発光標識、蛍光標識、または磁気標識であってよい。検出可能なタグは、γ線エミッター、β線エミッター、α線エミッター、γ線エミッター、ポジトロンエミッター、X線エミッターおよび蛍光エミッターからなる群から選択できる。適切な蛍光化合物には、フルオレセインナトリウム、フルオレセインイソチオシアネート、フィコエリトリン、およびテキサスレッドスルホニルクロリド、アロフィコシアニン(APC)、Cy5−PE、CY7−APC、およびカスケードイエローが挙げられる。
【0053】
標識のために適合する放射性同位体には、ヨウ素131、ヨウ素123、ヨウ素125、ヨウ素126、ヨウ素133、臭素77、インジウム111、インジウム113m、ガリウム67、ガリウム68、ルテニウム95、ルテニウム97、ルテニウム103、ルテニウム105、水銀107、水銀203、レニウム99m、レニウム105、レニウム101、テルル121m、テルル122m、テルル125m、ツリウム165、ツリウム167、ツリウム168、テクネチウム99mおよびフッ素18が挙げられる。
【0054】
任意で、検出可能なタグは、組織化学的技術、ELISA様アッセイ、共焦点顕微鏡、蛍光検出法、セルソーティング法、核磁気共鳴法、ラジオイムノシンチグラフィー、X線検査、陽電子放射断層撮影法、コンピュータ体軸断層撮影法、磁気共鳴イメージング法、および超音波検査法を用いて視認できる。
【0055】
または、ポリペプチドをビオチニル化して、蛍光標識ストレプトアビジンのようなその後に検出可能な標識を使用するとポリペプチドを間接的に検出できる。ビオチンは、当技術分野において知られている数種の技術のいずれかによって検出される。例えば、ビオチンは蛍光標識アビジンと結合する工程によって検出可能であり、アビジンは各結合事象に関連するシグナルを増加させるためにフィコエリトリンまたは連鎖蛍光標識を用いて標識される。
【0056】
任意で、ポリペプチドはスライド、培養皿、マルチウエルプレート、カラム、チップ、アレイまたは安定性ビーズなどの固体支持体へ結合させられる。「アレイ」には、マイクロプレートまたはスライドなどの1つまたは複数のマルチウエルアレイ化手段が挙げられる。
【0057】
任意で、ポリペプチドは、例えばセルソーティングテクノロジーを用いてソーティングできるビーズなどの可動性固体支持体へ結合させられる。「可動性固体支持体」とは、1セットの識別可能に標識されたマイクロスフェアまたはビーズを意味する。好ましくは、マイクロスフェアは、ポリスチレン・ジビニルベンゼンビーズである。特異的蛍光色素でマークされ、特異的蛍光プロフィールを有するマイクロスフェアのセットは、例えばLuminex Corporation(テキサス州オースティン)から市販で入手できる。
【0058】
本発明は、さらに複数の本発明のポリペプチドを提供する。任意で、ポリペプチドのLRRは、ポリペプチド全体にわたり高度に可変性である。そこで、大多数は、内部LRRの変動性に基づいて、相違する結合特異性を備えるポリペプチドを含むことができる。
【0059】
さらにまた、本発明のポリペプチドを備える容器または本発明のポリペプチドを備える固定性固体支持体もしくは可動性固体支持体を含むキットもまた提供される。任意で、ポリペプチドは固体支持体またはキットへ結合させられる。任意で、キットは、ポリペプチド、固体支持体、およびポリペプチドを固体支持体へ結合させるための連結手段を含有している。
【0060】
本発明は、本発明のポリペプチドをコードする単離された核酸を提供する。そのような核酸の1つの例は、配列番号156のヌクレオチド配列(代表的VLRのORF)を含んでいる。VLRまたはそのフラグメントをコードする核酸の他の例には、配列番号44、配列番号53〜55、配列番号57〜59、配列番号66〜67、配列番号73〜74、配列番号76、配列番号79〜80、および配列番号172〜302が挙げられる。VLR遺伝子に関連する種々の配列には、Genbank受託番号AY57791〜AY578059、AY964719〜AY964931、AY965520〜AY965612、AY965658〜AY965681、およびCK988414〜CK988652を有する様々な配列がある。これらの配列は、全体が、ならびにその中に含まれる個別のサブ配列(領域またはフラグメント)が参照として本明細書に援用される。
【0061】
そのような核酸配列は、一例として核酸の属によって提供されるが、限定することは意図されていない。さらに、これらの核酸を含む発現ベクターも提供されるが、このとき核酸は発現制御配列へ機能的に連結している。さらに発現ベクターを含む培養細胞も提供される。そのような発現ベクターおよび培養細胞を使用すると、本発明のポリペプチドを作製できる。
【0062】
本明細書では、例えばVLRまたはそのフラグメントもしくは変異体をコードする核酸を含む、核酸に基づく様々な分子が開示されている。開示された核酸は、ヌクレオチド、ヌクレオチドアナログ、またはヌクレオチド置換基から作製される。
【0063】
ヌクレオチドアナログは、塩基、糖、またはリン酸塩成分のいずれかへの一部のタイプの修飾を含有するヌクレオチドである。塩基成分に対する修飾は、A、C、G、およびT/Uの天然および合成修飾、ならびにウラシル−5−イル(.psi.)、ヒポキサンチン−9−イル(I)、および2−アミノアデニン−9−イルなどの様々なプリンもしくはピリミジン塩基の天然および合成修飾を含む。修飾された塩基には、5−メチルシトシン(5−me−C)、5−ヒドロキシメチルシトシン、キサンチン、ヒポキサンチン、2−アミノアデニン、アデニンおよびグアニンの6−メチルおよびその他のアルキル誘導体、アデニンおよびグアニンの2−プロピルおよびその他のアルキル誘導体、2−チオウラシル、2−チオチミンおよび2−チオシトシン、5−ハロウラシルおよび5−ハロシトシン、5−プロピニルウラシルおよび5−プロピニルシトシン、6−アゾウラシル、6−アゾシトシンおよび6−アゾチミン、5−ウラシル(プソイドウラシル)、4−チオウラシル、8−ハロ、8−アミノ、8−チオール、8−チオアルキル、8−ヒドロキシルならびに他の8−置換のアデニンおよびグアニン、5−ハロ、特に5−ブロモ、5−トリフルオロメチルおよび他の5−置換のウラシルおよびシトシン、7−メチルグアニンおよび7−メチルアデニン、8−アザグアニンおよび8−アザアデニン、7−デアザグアニンおよび7−デアザアデニン、ならびに3−デアザグアニンおよび3−デアザアデニンが挙げられるが、それらに限定されない。追加の塩基修飾は、例えば米国特許第3,687,808号、Englisch et al.,Angewandte Chemie,International Edition,1991,30,613、およびSanghvi,Y.S.,Chapter 15,Antisense Research and Applications,pages 289−302,Crooke,S.T.およびLebleu,B.編,CRC Press,1993に見いだすことができる。5−置換ピリミジン、6−アザピリミジンなどの所定のヌクレオチドアナログならびに2−アミノプロピルアデニン、5−プロピニルウラシルおよび5−プロピニルシトシン、5−メチルシトシンを含むN−2、N−6およびO−6置換プリンは、二本鎖形成の安定性を増加させることができる。多くの場合、塩基修飾は例えば、二本鎖安定性上昇などの独特の特性を達成するために、2’−O−メトキシエチルなどの糖修飾と組み合わせることができる。広範囲の塩基修飾について詳述している第4,845,205号;第5,130,302号;第5,134,066号;第5,175,273号;第5,367,066号;第5,432,272号;第5,457,187号;第5,459,255号;第5,484,908号;第5,502,177号;第5,525,711号;第5,552,540号;第5,587,469号;第5,594,121号;第5,596,091号;第5,614,617号;および第5,681,941号などの多数の米国特許がある。これらの各特許は、参照して本明細書に組み込まれる。
【0064】
ヌクレオチドアナログは、さらに糖成分の修飾を含むことができる。糖部分の修飾は、リボースおよびデオキシリボースの天然修飾ならびに合成修飾を含むであろう。糖修飾は、2’位での以下の修飾を含むがそれらに限定されない:OH;F;O−、S−、もしくはN−アルキル;O−、S−、もしくはN−アルケニル;O−、S−もしくはN−アルキニル;またはO−アルキル−O−アルキル。このときアルキル、アルケニルおよびアルキニルは置換または未置換C1〜C10アルキルまたはC2〜C10アルケニルおよびアルキニルであってよい。2’糖修飾には−O[(CH2)nO]mCH3、−O(CH2)nOCH3、−O(CH2)nNH2、−O(CH2)nCH3、−O(CH2)n−ONH2、および−O(CH2)nON[(CH2)nCH3)]2(式中、nおよびmは1〜約10である)が含まれるがそれらに限定されない。
【0065】
2’位でのその他の糖修飾には:C1〜C10低級アルキル、置換低級アルキル、アルカリル、アラルキル、O−アルカリルもしくはO−アラルキル、SH、SCH3、OCN、Cl、Br、CN、CF3、OCF3、SOCH3、SO2CH3、ONO2、NO2、N3、NH2、ヘテロシクロアルキル、ヘテロシクロアルカリル、アミノアルキルアミノ、ポリアルキルアミノ、置換シリル、RNA開裂基、レポーター基、インターカレーター、オリゴヌクレオチドの薬物動態特性を改良するための基、またはオリゴヌクレオチドの薬力学的特性を改良するための基、および類似の特性を有する他の置換基が含まれるがそれらに限定されない。類似の修飾は、糖の上の他の位置、特に3’末端ヌクレオチド上または2’〜5’結合オリゴヌクレオチド内の糖の3’位、および5’末端ヌクレオチドの5’位で行なうことができる。修飾された糖は、例えばCH2およびSなどの架橋環状酸素での修飾を含有する糖も含むであろう。ヌクレオチド糖アナログは、ペントフラノシル糖の代わりにシクロブチル成分などの糖ミメティックもまた有する可能性がある。そのような修飾された糖構造の調製を教示する第4,981,957号;第5,118,800号;第5,319,080号;第5,359,044号;第5,393,878号;第5,446,137号;第5,466,786号;第5,514,785号;第5,519,134号;第5,567,811号;第5,576,427号;第5,591,722号;第5,597,909号;第5,610,300号;第5,627,053号;第5,639,873号;第5,646,265号;第5,658,873号;第5,670,633号;および第5,700,920号などの多数の米国特許が存在しており、それらは各々全体として参照して本明細書に組み込まれる。
【0066】
ヌクレオチドアナログは、さらにリン酸塩成分で修飾することもできる。修飾されたリン酸塩成分には、2つのヌクレオチド間の連結がホスホロチオエート、キラルホスホロチオエート、ホスホロジチオエート、ホスホトリエステル、アミノアルキルホスホトリエステル、メチルならびに3’−アルキレンホスホネートおよびキラルホスホネートを含む他のアルキルホスホネート、ホスフィネート、3’−アミノホスホルアミデートおよびアミノアルキルホスホルアミデート、チオホスホルアミデートを含むホスホルアミデート、チオノアルキルホスホネート、チオノアルキルホスホトリエステル、およびボラノホスフェートを含むように修飾できる成分が含まれるがそれらに限定されない。これらのホスフェートまたは2つのヌクレオチド間の修飾ホスフェート結合は3’−5’結合または2’−5’結合を通して行なうことができ、この結合は3’−5’から5’−3’または2’−5’から5’−2’などの逆極性を含有できることを理解されたい。様々な塩、混合塩および遊離酸形もまた含まれる。多数の米国特許が修飾リン酸エステルを含有するヌクレオチドの作製方法および使用方法を教示しており、それらにはその各々が参照して本明細書に組み込まれる第3,687,808号;第4,469,863号;第4,476,301号;第5,023,243号;第5,177,196号;第5,188,897号;第5,264,423号;第5,276,019号;第5,278,302号;第5,286,717号;第5,321,131号;第5,399,676号;第5,405,939号;第5,453,496号;第5,455,233号;第5,466,677号;第5,476,925号;第5,519,126号;第5,536,821号;第5,541,306号;第5,550,111号;第5,563,253号;第5,571,799号;第5,587,361号;および第5,625,050号が含まれるがそれらに限定されない。
【0067】
ヌクレオチドアナログは必ずしも単一修飾を含有している必要はなく、成分の1つの中または相違する成分間に複数の修飾を含有していてよいことも理解されたい。
【0068】
ヌクレオチド置換基は、ヌクレオチドと類似の機能的特性を有する分子であるが、ペプチド核酸(PNA)などはリン酸エステル成分を含有していない。ヌクレオチド置換基は、Watson−Crick様式またはHoogsteen様式で核酸を認識するであろうが、リン酸エステル成分以外の成分を通して連結される分子である。ヌクレオチド置換基は、適切な標的核酸と相互作用すると二本鎖型構造に適合することができる。
【0069】
ヌクレオチド置換基は、リン酸エステル成分および/または糖成分が置換されているヌクレオチドまたはヌクレオチドアナログである。ヌクレオチド置換基は、標準的なリン原子を含有していない。リン酸エステルに対する置換基は、例えば、短鎖アルキルもしくはシクロアルキルヌクレオシド間結合、混合ヘテロ原子およびアルキルもしくはシクロアルキルヌクレオシド間結合、または1つまたは複数の短鎖ヘテロ原子もしくは複素環ヌクレオシド間結合であってよい。これらには、モルホリノ結合(一部にはヌクレオシドの糖部分から形成される);シロキサン骨格;硫化物、スルホキシドおよびスルホン骨格;ホルムアセチルおよびチオホルムアセチル骨格;メチレンホルムアセチルおよびチオホルムアセチル骨格;アルケン含有骨格;スルファメート骨格;メチレンイミノおよびメチレンヒドラジノ骨格;スルホン酸エステル骨格およびスルホンアミド骨格;アミド骨格;ならびに混合N、O、SおよびCH2成分部分を有するその他が含まれる。多数の米国特許がリン酸エステル置換基の作製方法および使用方法を教示しており、それらにはその各々が参照して本明細書に組み込まれる第5,034,506号;第5,166,315号;第5,185,444号;第5,214,134号;第5,216,141号;第5,235,033号;第5,264,562号;第5,264,564号;第5,405,938号;第5,434,257号;第5,466,677号;第5,470,967号;第5,489,677号;第5,541,307号;第5,561,225号;第5,596,086号;第5,602,240号;第5,610,289号;第5,602,240号;第5,608,046号;第5,610,289号;第5,618,704号;第5,623,070号;第5,663,312号;第5,633,360号;第5,677,437号;および第5,677,439号が含まれるがそれらに限定されない。
【0070】
さらにヌクレオチド置換基ではヌクレオチドの糖およびリン酸部分の両方の成分が、例えばアミド型結合(アミノエチルグリシン)(PNA)によって置換されてよいことを理解されたい。各々が参照して本明細書に組み込まれる米国特許第5,539,082号;第5,714,331号;および第5,719,262号は、PNA分子の製造方法および使用方法を教示している。(さらに、Nielsen et al.,Science,1991,254,1497−1500も参照されたい)。
【0071】
例えば細胞取り込みを強化させるために、他のタイプの分子(コンジュゲート)をヌクレオチドまたはヌクレオチドアナログへ結合させることも可能である。コンジュゲートは、ヌクレオチドまたはヌクレオチドアナログへ化学的に結合させることができる。そのようなコンジュゲートには、コレステロール成分などの脂質成分(Letsinger et al.,Proc.Natl.Acad.Sci.USA,1989,86,6553−6556)、コール酸(Manoharan et al.,Bioorg.Med.Chem.Let.,1994,4,1053−1060)、チオエーテル、例えばヘキシル−S−トリチルチオール(Manoharan et al.,Ann.N.Y.Acad.Sci.,1992,660,306−309;Manoharan et al.,Bioorg.Med.Chem.Let.,1993,3,2765−2770)、チオコレステロール(Oberhauser et al.,Nucl.Acids Res.,1992,20,533−538)、脂肪族鎖、例えばドデカンジオールもしくはウンデシル残基(Saison−Behmoaras et al.,EMBO
J,1991,10,1111−1118;Kabanov et al,FEBS Lett.,1990,259,327−330;Svinarchuk et al.,Biochimie,1993,75,49−54)、リン脂質、例えばジ−ヘキサデシル−rac−グリセロールもしくはトリエチルアンモニウム1,2−ジ−O−ヘキサデシル−rac−グリセロ−3−H−ホスホン酸塩(Manoharan et al.,Tetrahedron Lett.,1995,36,3651−3654;Shea
et al.,Nucl.Acids Res.,1990,18,3777−3783)、ポリアミンもしくはポリエチレングリコール鎖(Manoharan et al.,Nucleosides & Nucleotides,1995,14,969−973)、またはアダマンタン酢酸(Manoharan et al.,Tetrahedron Lett.,1995,36,3651−3654)、パルミチル成分(Mishra et al.,Biochem.Biophys.Acta,1995,1264,229−237)、またはオクタデシルアミンもしくはヘキシルアミノ−カルボニル−オキシコレステロール成分(Crooke et al.,J.Pharmacol.Exp.Ther.,1996,277,923−937が含まれるがそれらに限定されない。多数の米国特許がそのようなコンジュゲートの調製を教示しており、それらには各々が参照して本明細書に組み込まれる米国特許第4,828,979号;第4,948,882号;第5,218,105号;第5,525,465号;第5,541,313号;第5,545,730号;第5,552,538号;第5,578,717,第5,580,731号;第5,580,731号;第5,591,584号;第5,109,124号;第5,118,802号;第5,138,045号;第5,414,077号;第5,486,603号;第5,512,439号;第5,578,718号;第5,608,046号;第4,587,044号;第4,605,735号;第4,667,025号;第4,762,779号;第4,789,737号;第4,824,941号;第4,835,263号;第4,876,335号;第4,904,582号;第4,958,013号;第5,082,830号;第5,112,963号;第5,214,136号;第5,082,830号;第5,112,963号;第5,214,136号;第5,245,022号;第5,254,469号;第5,258,506号;第5,262,536号;第5,272,250号;第5,292,873号;第5,317,098号;第5,371,241号、第5,391,723号;第5,416,203号、第5,451,463号;第5,510,475号;第5,512,667号;第5,514,785号;第5,565,552号;第5,567,810号;第5,574,142号;第5,585,481号;第5,587,371号;第5,595,726号;第5,597,696号;第5,599,923号;第5,599,928号;および第5,688,941号が含まれるがそれらに限定されない。
【0072】
VLR遺伝子、または匹敵する遺伝子と相互作用できるプライマーおよびプローブを含む組成物が開示される。所定の実施形態では、プライマーはDNA増幅反応を支援するために使用される。典型的には、プライマーは配列特異的方法で延長させることができるであろう。配列特異的方法でのプライマーの伸長には、それにプライマーがハイブリダイズされる、さもなければ関連付けられる核酸分子の配列および/または組成がプライマーの伸長によって生成する生成物の組成または配列を指示する、または影響を及ぼす任意の方法が含まれる。このため配列特異的方法でのプライマーの伸長には、PCR、DNAシーケンシング、DNA伸長、DNA重合、RNA転写、または逆転写が含まれるがそれらに限定されない。配列特異的方法でプライマーを増幅させる技術および条件が好ましい。所定の実施形態では、プライマーはPCRまたは直接的シーケンシングなどのDNA増幅反応のために使用される。所定の実施形態では、プライマーはさらにまた非酵素的技術を用いて伸長させることもできることを理解されたいが、このとき例えばプライマーを伸長させるために使用されるヌクレオチドもしくはオリゴヌクレオチドは、それらが配列特異的方法でプライマーを伸長させるように化学的に反応するように修飾させられる。
【0073】
所定の実施形態においてVLR遺伝子と相互作用するためのプライマーもしくはプローブのサイズは、DNA増幅などのプライマーの所望の酵素的操作またはプローブもしくはプライマーの単純なハイブリダイゼーションを支援する任意のサイズであってよい。典型的なVLRプライマーもしくはプローブは、少なくとも6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、125、150、175、200、225、250、275、300、325、350、375、400、425、450、475、500、550、600、650、700、750、800、850、900、950、1000、1250、1500、1750、2000、2250、2500、2750、3000、3500、または4000ヌクレオチド長であろう。
【0074】
本発明のポリペプチドおよび核酸は、様々な技術において使用できる。例えば、本ポリペプチドは選択された作用因子を検出するため、選択された作用因子の活性を遮断するため、作用因子を精製するために、イメージングツールとして、および治療薬として使用できる。
【0075】
本明細書では、サンプル中の作用因子を検出する方法であって、ポリペプチドがサンプル中の作用因子へ結合できる条件下でサンプルをポリペプチドと接触させる工程、およびサンプル中の作用因子へ結合したポリペプチドを検出する工程、を含む方法が提供される。結合したポリペプチドは、サンプル中に作用因子が含まれることを示す。検出方法は、当技術分野においてよく知られている。例えば、ポリペプチドは、上述したように検出可能なタグを用いて標識できる。検出方法を使用すると、サンプル中の作用因子の存在または不在を記録することができる。しかし、検出方法はさらに定量方法と結合することができる。インビトロアッセイ法には、標準物質に比較して量を確定するために使用できる知られている作用因子量のコントロールサンプルとの比較に基づいて作用因子の定量を可能にするELISAなどの比色アッセイ法が含まれる。この方法は、さらに、放出された放射線に基づいて定量を可能にする放射分析アッセイおよび蛍光アッセイまたは上述した可視化および定量の任意の手段を含むことができる。
【0076】
サンプルは、任意の生物学的サンプルを含む、試験される任意のサンプルであってよい。サンプルには、液体サンプル(水、血液、尿など)、組織サンプル、培養サンプル、細胞サンプルなどを含むことができる。
【0077】
本発明のポリペプチドは、さらにまた遮断抗体に匹敵して、それへ結合する作用因子の活性を遮断するために使用することもできる。そこで、作用因子の活性を遮断する方法であって、作用因子にポリペプチドが結合できる条件下でその作用因子を本発明のポリペプチドと接触させる工程を含む方法もまた開示される。作用因子へのポリペプチドの結合は、その作用因子の活性を遮断する。接触させるステップは、インビボまたはインビトロのいずれであってもよい。そこで、例えば、サンプルの汚染を減少させるために、毒素へ結合するポリペプチドをサンプルに添加して、毒素活性を遮断することができる。
【0078】
本発明のポリペプチドは、さらにまた拮抗抗体に匹敵して、それへ結合する任意の作用因子の活性を促進するために使用することもできる。そこで、作用因子の活性を促進する方法であって、作用因子にポリペプチドが結合できる条件下でその作用因子を本発明のポリペプチドと接触させる工程を含む方法もまた開示される。作用因子へのポリペプチドの結合は、その作用因子の活性を促進する。
【0079】
本明細書に開示したポリペプチドは、知られていない機能を備える遺伝子の機能を決定するために使用することができる。そこで、開示されたポリペプチドをタンパク質ノックダウンアッセイにおいて使用する方法が開示される。例えば、開示されたポリペプチドは、機能が知られていない遺伝子を含む細胞の細胞質内で発現させることができる。RNA転写産物が細胞の細胞質内で翻訳されている場合は、開示されたポリペプチドは問題の遺伝子のタンパク質産物へ結合することができる。タンパク質発現の消失が細胞に及ぼす作用を監視することによって、タンパク質機能を決定することができる。そこで、機能が知られていない遺伝子産物に対して特異的なポリペプチドが特別に開示される。さらに、遺伝子の機能を決定する方法であって、遺伝子のタンパク質産物に対して特異的なポリペプチドを、その遺伝子を発現する細胞の細胞質内へ導入する工程、および機能が知られていない遺伝子のタンパク質産物の消失に起因する作用を監視する工程、を含む方法もまた開示される。
【0080】
本発明のポリペプチドは、イメージング法においても使用できる。例えば、本発明は、被験者に有効量のポリペプチドを投与する工程、および被験者において結合したポリペプチドの所在位置を検出する工程、を含むイメージング法を提供する。イメージング法の例は、上述されている。
【0081】
本発明は、さらに精製方法を提供する。本明細書では、サンプルから作用因子を精製する方法であって、ポリペプチドがそのサンプルへ結合してポリペプチド/作用因子複合体を形成する条件下でサンプルをポリペプチドと接触させる工程;およびその作用因子をポリペプチド/作用因子複合体から単離する工程、を含む方法が開示される。例えば、ポリペプチドはカラムへ結合させ、サンプルは、サンプル内の作用因子が結合したポリペプチドへ結合できる条件下でカラムを通過させることができる。作用因子は、引き続いて所望の溶離剤中でカラムから溶出させることができる。精製方法は、研究方法として、および商業的方法として有用であろう。例えば、そのような方法は、薬理学的化合物から汚染物質を除去する際に有用であろう。
【0082】
本ポリペプチドは、治療方法においても使用できる。例えば、本明細書では、被験者における病原性作用を減少させる、または予防する方法であって、病原体へ結合する有効量のポリペプチドを被験者に投与する工程を含む方法が提供される。さらにまた、有害作用を減少させられる、または有効な作用を促進できるように作用因子の活性を遮断または促進する方法も提供される。
【0083】
本明細書では、本発明のポリペプチドまたは核酸および医薬上許容される担体を含む組成物が提供される。本発明の組成物は、インビボで投与することもできる。組成物は、経口、非経口(例、静脈内)、筋肉内注射、腹腔内注射、経皮的、体外、局所的などによって投与されてよいが、典型的には局所的鼻腔内投与または吸入剤による投与が好ましい。本明細書で使用する「局所的鼻腔内投与」は、鼻孔の一方または両方を通して鼻および鼻腔内への組成物の送達を意味しており、スプレー機構もしくは滴下機構、または核酸もしくはベクターのエーロゾル化を通しての送達を含むことができる。後者は、大量の動物を同時に治療しなければならない場合には有効なことがある。吸入剤による組成物の投与は、スプレーまたは滴下機構による送達を介して鼻または口を通して行なうことができる。送達は、挿管を通して呼吸器系の任意の領域(例、肺)へ直接的に行なうこともできる。必要とされる組成物の正確な量は、被験者の種、年齢、体重および全身状態、治療されるアレルギー性障害の重症度、使用される特定の核酸またはベクター、投与様式などに依存して、被験者毎に相違するであろう。そこで、各組成物について正確な量を規定するのは不可能である。しかし、適切な量は、当業者が本明細書の教示を前提に日常的な実験器具を使用さえすれば決定することができる。
【0084】
使用する場合、組成物の非経口投与は一般に注射を特徴とする。注射剤は、液剤もしくは懸濁剤、注射前に液体中の懸濁液にするために適合する固体形、またはエマルジョンとしてのいずれかの便宜的形態で調製できる。非経口投与のために最近改良されたアプローチは、一定用量が維持されるような徐放系または持続放出系の使用が含まれる。例えば、参照して本明細書に組み込まれる米国特許第3,610,795号を参照されたい。
【0085】
物質は、(例えば、微粒子、リポソーム、または細胞内に組み込まれて)溶液、懸濁液中にあってよい。これらは、抗体、受容体、または受容体リガンドを介して特定細胞タイプを標的とすることができる。下記の参考文献は、腫瘍組織へ特異的タンパク質をターゲッティングするためのこのテクノロジーの使用例である(Senter,et al.,Bioconjugate Chem.,2:447−451,(1991);Bagshawe,K.D.,Br.J.Cancer,60:275−281,(1989);Bagshawe,et al.,Br.J.Cancer,58:700−703,(1988);Senter,et al.,Bioconjugate Chem.,4:3−9,(1993);Battelli,et al.,Cancer Immunol.Immunother.,35:421−425,(1992);PieterszおよびMcKenzie,Immunolog.Reviews,129:57−80,(1992);ならびにRoffler,et al.,Biochem.Pharmacol,42:2062−2065,(1991))。「ステルス」などのビヒクルおよびその他の抗体結合リポソーム(大腸癌を標的とする脂質媒介性薬物を含む)、細胞特異的リガンドを通してのDNAの受容体媒介性ターゲッティング、リンパ球指向性腫瘍ターゲッティング、およびインビボのマウス神経膠腫細胞に対する高度に特異的な治療的レトロウイルスターゲッティング。下記の参考文献は、腫瘍組織へ特異的タンパク質をターゲッティングするためのこのテクノロジーの使用例である(Hughes et al.,Cancer Research,49:6214−6220,(1989);およびLitzingerおよびHuang,Biochimica et Biophysica Acta,1104:179−187,(1992))。一般に、受容体は、構成的またはリガンド誘導性のいずれかで、エンドサイトーシスの経路に含まれている。これらの受容体は、クラスリン被覆ピット内でクラスターを形成し、クラスリン被覆小胞を介して細胞内に進入し、その中で受容体が分類される酸性化エンドソームを通過し、そして次に細胞表面へ再循環する、細胞内に蓄積される、またはリソソーム内で分解される、のいずれかとなる。内在化経路は、栄養素取り込み、活性化タンパク質の除去、高分子のクリアランス、ウイルスおよび毒素の日和見進入、リガンドの解離および分解、ならびに受容体レベル調節などの様々な機能に役立つ。多数の受容体は、細胞タイプ、受容体濃度、リガンドのタイプ、リガンドの価数、およびリガンドの濃度に依存して、2つ以上の細胞内経路にしたがう。受容体媒介性エンドサイトーシスの分子および細胞機序は精査されている(BrownおよびGreene,DNA and Cell Biology 10:6,399−409 (1991))。
【0086】
「医薬上許容される」は、生物学的またはその他の点で不都合ではない物質を意味しており、すなわちその物質は、何の不都合な生物学的作用を誘発することなく、またはそれが含有される医薬組成物の他の成分のいずれかと有害な方法で相互作用することなく本発明のポリペプチドと一緒に被験者に投与することができる。担体は、当業者にはよく知られているように、当然ながら有効成分の分解を最小限に抑えるように、そして被験者における有害な副作用を最小限に抑えるように選択されるであろう。適切な担体およびそれらの調製物は、Remington:The Science and Practice
of Pharmacy(第19版)編者A.R.Gennaro,Mack Publishing Company,Easton,PA 1995に記載されている。典型的には、調製物を等張性にするために、調製物中では適切な量の医薬上許容される塩が使用される。医薬上許容される担体の例には、食塩液、リンゲル液およびデキストロース液が含まれるがそれらに限定されない。溶液のpHは、好ましくは約5〜約8、より好ましくは約7〜約7.5である。その他の担体には、可変性リンパ球受容体を含有する固体疎水性ポリマーの半透性マトリックスなどの徐放性調製物が含まれるが、そのマトリックスは成形製品、例えばフィルム、リポソームまたは微粒子の形状にある。当業者には、例えば投与経路、投与される可変性リンパ球受容体の濃度に依存して所定の担体がより好ましいことは明白であろう。
【0087】
医薬担体は、当業者には知られている。これらは、最も典型的には無菌水、食塩液、および生理的pHにある緩衝液などの溶液を含むヒトに薬物を投与するための標準的担体であろう。本組成物は、例えば筋肉内または皮下投与することができる。その他の化合物は、当業者によって使用される標準方法にしたがって投与されるであろう。
【0088】
本医薬組成物は、精選分子に加えて、担体、増粘剤、希釈剤、バッファー、保存剤、界面活性剤などを含むことができる。本医薬組成物は、さらに抗菌物質、抗炎症薬、麻酔薬などの1つまたは複数の有効成分を含むことができる。
【0089】
非経口投与のための本調製物には、無菌の水性または非水性液剤、懸濁剤、およびエマルジョンが含まれる。非水性溶媒の例は、プロピレングリコール、ポリエチレングリコール、オリーブ油などの植物油、およびオレイン酸エチルなどの注射用有機エステルである。水性担体には、水、食塩液および緩衝溶媒を含むアルコール性/水性の溶液、エマルジョンもしくは懸濁液が含まれる。非経口ビヒクルには、塩化ナトリウム溶液、リンガーデキストロース、デキストロースおよび塩化ナトリウム、乳酸加リンガー液、または固定油が含まれる。静注用ビヒクルには、液体および栄養補充剤、電解質補充剤(リンガーデキストロースをベースとする補充剤など)などが含まれる。例えば抗菌物質、抗酸化剤、キレート剤、および不活性ガスなどの保存剤およびその他の添加剤もまた存在していてよい。
【0090】
局所適用のための本調製物は、軟膏、ローション剤、クリーム剤、ゲル剤、滴剤、坐剤、スプレー剤、液剤および散剤を含むことができる。従来型医薬担体、水性、粉末状または油性基剤、増粘剤などが必要になる、または望ましいことがある。
【0091】
経口投与のための本組成物には、散剤もしくは顆粒剤、水もしくは非水性溶媒中の懸濁剤もしくは液剤、カプセル剤、サシェ剤、または錠剤が含まれる。増粘剤、矯味剤、希釈剤、乳化剤、分散助剤または結合剤が望ましいことがある。
【0092】
本組成物の一部は、もしかすると塩酸塩、臭化水素酸、過塩素酸、硝酸、チオシアン酸、硫酸、およびリン酸などの無機酸と、ギ酸、酢酸、プロピオン酸、グリコール酸、乳酸、ピルビン酸、シュウ酸、マロン酸、コハク酸、マレイン酸、およびフマル酸などの有機酸との反応によって、または水酸化ナトリウム、水酸化アンモニウム、水酸化カリウムなどの無機塩基、およびモノ−、ジ−、トリアルキルおよびアリールエミンならびに置換されたエタノールアミンなどの有機塩基との反応によって生成される医薬上許容される酸付加塩もしくは塩基付加塩として投与できる。
【0093】
組成物を投与するための用量範囲は、症状や障害に有効である所望の作用を生じさせるために十分な量である。用量は、望ましくない交差反応、アナフィラキシー反応などの有害な副作用を惹起するほど大量であってはならない。一般に、用量は、患者における年齢、状態、性別および疾患の程度に伴って変動するが、当業者であれば決定することができる。用量は、万一何らかの禁忌がある場合に個々の医師によって調整することができる。用量は変動することがあり、1日もしくは数日にわたって、1回または複数回の投与で投与することができる。
【0094】
本発明の可変性リンパ球受容体および可変性リンパ球受容体フラグメントおよび変異体は、さらにまた患者もしくは被験者に、その患者もしくは被験者自身の細胞が核酸を取り込んで、コードされた可変性リンパ球受容体もしくは可変性リンパ球受容体フラグメントを生成かつ分泌するように、可変性リンパ球受容体または可変性リンパ球受容体フラグメントもしくは変異体をコードする核酸調製物(例、DNAもしくはRNA)として投与することもできる。
【0095】
インビトロまたはインビボのいずれかで、核酸を細胞に送達するために使用できる多数の組成物および方法がある。これらの方法および組成物は、大きく分けると次の2つのクラスに分類できる:ウイルスベースの送達系および非ウイルスベースの送達系。例えば、核酸は、エレクトロポレーション、リポフェクション、リン酸カルシウム沈降法、プラスミド、ウイルスベクター、ウイルス核酸、ファージ核酸、ファージ、コスミドなどの多数の直接送達系を通して、またはカチオン性リポソームなどの細胞もしくは担体中の遺伝物質の輸送を介して送達できる。ウイルスベクター、化学的トランスフェクタントを含むトランスフェクションのための適切な手段、またはエレクトロポレーションおよびDNAの直接拡散法などの物理−機械的方法は、例えばWolff,J.A.,et al.,Science,247,1465−1468,(1990);およびWolff,J.A.Nature,352,815−818,(1991)によって記載されている。そのような方法は当技術分野においてよく知られており、本明細書に記載した組成物および方法と一緒に使用するために容易に適合させることができる。所定の場合には、これらの方法は大きなDNA分子とともに特異的に機能するように修飾されるであろう。さらに、これらの方法を使用すると、担体のターゲッティング特性を使用することによって、所定の疾患および細胞集団を標的とすることができる。
【0096】
トランスファーベクターは、核酸を細胞内へ送達するために、または遺伝子を送達するための一般的戦略の一部として、例えば組み換えレトロウイルスまたはアデノウイルスの一部として使用される任意のヌクレオチド構造(例、プラスミド)であってよい(Ram
et al.Cancer Res.53:83−88,(1993))。本明細書で使用するプラスミドまたはウイルスベクターは、VLRなどの開示された核酸を細胞内へ分解させずに輸送する作用因子であり、その中にそれが送達される細胞内での遺伝子の発現を生じさせるプロモーターが含まれる。ウイルスベクターは、例えば、アデノウイルス、アデノ関連ウイルス、ヘルペスウイルス、ワクシニアウイルス、ポリオウイルス、AIDSウイルス、ニューロン栄養ウイルス、シンドビスウイルス、およびHIV骨格を備えるウイルスを含むその他のRNAウイルスである。さらに好ましいのは、それらをベクターとして使用するために適合させるこれらのウイルスの特性を共有する任意のウイルスファミリーである。レトロウイルスには、マウスマロネー白血病ウイルス(MMLV)、およびベクターとしてのMMLVの所望の特性を発現するレトロウイルスが含まれる。レトロウイルスベクターは、他のウイルスベクターより大きな遺伝的ペイロード、すなわち導入遺伝子またはマーカー遺伝子を運ぶことができ、このために一般的に使用されるベクターである。しかし、それらは非増殖細胞中では有用ではない。アデノウイルスベクターは比較的安定性であり、作業するのが容易であり、高い力価を有し、エーロゾル調製物中で送達することができ、非分割細胞をトランスフェクトすることができる。ポックスウイルスベクターは大きく、遺伝子を挿入するための幾つかの部位を有しており、それらは熱安定性で、室温で保管することができる。好ましい実施形態は、ウイルス抗原によって惹起される、宿主生体の免疫応答を抑制できるように組み換えられているウイルスベクターである。このタイプの好ましいベクターは、インターロイキン8または10のためのコーディング領域を有するであろう。
【0097】
ウイルスベクターは、細胞内へ遺伝子を導入するために化学的または物理的方法より高度のトランスアクション能力(遺伝子を導入する能力)を有する可能性がある。典型的には、ウイルスベクターは、非構造的初期遺伝子、構造的後期遺伝子、RNAポリメラーゼIII転写産物、複製およびエンカプシデーションのために必要な逆方向末端反復、およびウイルスゲノムの転写および複製を制御するためのプロモーターを含有する。ベクターとして組み換えられると、典型的にはウイルスから1つまたは複数の初期遺伝子が除去され、遺伝子もしくは遺伝子/プロモーターカセットが除去されたウイルスDNAの代わりにウイルスゲノム内に挿入される。このタイプの構築体は、約8kbまでの外来遺伝物質を運ぶことができる。除去された初期遺伝子の必要な機能は、典型的には初期遺伝子の遺伝子産物をトランスで発現するように組み換えられている細胞系によって補充される。
【0098】
レトロウイルスは、任意のタイプ、サブファミリー、属、または向性を含むレトロウイルスのウイルスファミリーに属する動物ウイルスである。レトロウイルスベクターは、一般に、参照して本明細書組み込まれるVerma,I.M.,Retroviral vectors for gene transfer.In Microbiology−1985,American Society for Microbiology,pp.229−232,Washington,(1985)の中に記載されている。遺伝子療法のためにレトロウイルスベクターを使用する方法の例は、それらの教示が参照して本明細書に組み込まれる米国特許第4,868,116号および第4,980,286号;PCT国際特許公開WO90/02806およびWO89/07136;ならびにMulligan,(Science 260:926−932(1993))に記載されている。
【0099】
レトロウイルスは、本質的に核酸カーゴ内にパッキングされているパッケージである。核酸カーゴは、それとともにパッケージングシグナルを運ぶが、これは複製された娘分子がパッケージコート内に効率的にパッケージングされることを保証する。パッケージングシグナルに加えて、複製、および複製されたウイルスのパッケージングのためにシス形に必要とされる多数の分子が存在する。典型的には、レトロウイルスゲノムは、タンパク質コートを作製する際に関係しているgag、pol、およびenv遺伝子を含有している。標的細胞へ輸送されるべき異質DNAによって典型的に置換されるのはgag、pol、およびenv遺伝子である。レトロウイルスベクターは、典型的にはパッケージコート内に組み込むためのパッケージングシグナル、gag転写ユニットの開始をシグナルする配列、逆転写のtRNAプライマーへ結合するためのプライマー結合部位を含む逆転写のために必要なエレメント、DNA合成中のRNAストランドのスイッチを誘導する末端反復配列、DNA合成の第2ストランドの合成のための開始部位として機能する3’LTRに対して5’のプリンリッチ配列、および宿主ゲノム内に挿入するためにDNA状態のレトロウイルスの挿入を可能にするLTRの両末端近くの特異的配列を含有する。gag、pol、およびenv遺伝子の除去は、約8kbの異質配列をウイルスゲノム内に挿入し、逆転写させ、複製後には新規なレトロウイルス粒子内にパッケージングすることを可能にする。この核酸の量は、各転写産物のサイズに依存して、1つから多数までの遺伝子を送達するために十分である。挿入物内に他の遺伝子と一緒に陽性または陰性いずれかの選択可能なマーカーを含めることが好ましい。
【0100】
大多数のレトロウイルスベクター内の複製機構およびパッケージングタンパク質は除去されているので(gag、pol、およびenv)、これらのベクターは典型的にはそれらをパッケージング細胞系内へ配置することによって生成される。パッケージング細胞系は、複製およびパッケージング機構を含有するが、任意のパッケージングシグナルが欠けているレトロウイルスを用いてトランスフェクトまたは形質転換されている細胞系である。精選DNAを運ぶベクターがこれらの細胞系内にトランスフェクトされると、当該遺伝子を含有するベクターが複製され、ヘルパー細胞によってシス形で提供される機構によって新規なレトロウイルス粒子内にパッケージングされる。この機構のためのゲノムは、それらに必要なシグナルが欠如しているためにパッケージングされない。
【0101】
複製欠損アデノウイルスの構築については記載されている(Berkner et al.,J.Virology 61:1213−1220(1987);Massie et al.,Mol.Cell.Biol.6:2872−2883(1986);Haj−Ahmad et al.,J.Virology 57:267−274(1986);Davidson et al.,J.Virology 61:1226−1239(1987);Zhang “Generation and identification of recombinant adenovirus by liposome−mediated transfection and PCR analysis” BioTechniques 15:868−872(1993))。これらのウイルスをベクターとして使用する有益性は、それらが初期感染細胞内で複製できるが、新規な感染性ウイルス粒子を形成することはできないために、それらが他の細胞タイプへ拡大できる程度が限定されることにある。組み換えアデノウイルスは、気道上皮、肝細胞、血管内皮、CNS実質、および多数の他の組織部位へ直接的なインビボ送達後に高い効率の遺伝子導入を達成することが証明されている(Morsy,J.Clin.Invest.92:1580−1586(1993);Kirshenbaum,J Clin.Invest.92:381−387(1993);Roessler,J.Clin.Invest.92:1085−1092(1993);Moullier,Nature Genetics 4:154−159(1993);La Salle,Science 259:988−990(1993);Gomez−Foix,J.Biol.Chem.267:25129−25134(1992);Rich,Human Gene Therapy 4:461−476(1993);Zabner,Nature Genetics 6:75−83(1994);Guzman,Circulation Research 73:1201−1207(1993);Bout,Human Gene Therapy 5:3−10(1994);Zabner,Cell 75:207−216(1993);Caillaud,Eur.J.Neuroscience 5:1287−1291(1993);およびRagot,J.Gen.Virology 74:501−507(1993))。組み換えアデノウイルスは、特異的細胞表面受容体へ結合することによって遺伝子形質導入を達成するが、その後にウイルスは、野生型または複製欠損アデノウイルスと同一方法で受容体媒介性エンドサイトーシスによって内在化される(ChardonnetおよびDales,Virolog
y 40:462−477(1970);BrownおよびBurlingham,J Virology 12:386−396(1973);SvenssonおよびPersson,J.Virology 55:442−449(1985);Seth,et
al.,J.Virol.51:650−655(1984);Seth,et al.,Mol.Cell.Biol.4:1528−1533(1984);Varga et al.,J Virology 65:6061−6070(1991);Wickham et al.,Cell 73:309−319(1993))。
【0102】
ウイルスベクターは、E1遺伝子が除去されているアデノウイルスをベースとするウイルスベクターであってよく、これらのバイロン(viron)はヒト293細胞系などの細胞系において生成される。また別の好ましい実施形態では、E1およびE3遺伝子の両方がアデノウイルスゲノムから除去される。
【0103】
また別のタイプのウイルスベクターは、アデノ関連ウイルス(AAV)をベースとしている。この欠陥のあるパルボウイルスは、多数の細胞タイプを感染させることができ、ヒトにとって非病原性であるので好ましい。AAV型ベクターは約4〜5kbを輸送することができ、野生型AAVは19番染色体中へ安定性で挿入できることが知られている。この部位特異的組み込み特性を含有するベクターが好ましい。このタイプのベクターの特に好ましい実施形態は、Avigen(カリフォルニア州サンフランシスコ)によって製造された、単純ヘルペスウイルスチミジンキナーゼ遺伝子、HSV−tk、および/または緑色蛍光タンパク質(GFP)をコードする遺伝子などのマーカー遺伝子を含有することのできるP4.1Cベクターである。
【0104】
また別のタイプのAAVウイルスでは、AAVは、異種遺伝子に機能的に連結した細胞特異的発現を指令するプロモーターを含有する少なくとも1つのカセットを挟んでいる1対の逆方向末端反復(ITR)を含有している。この状況における異種とは、AAVまたはB19パルボウイルスにとって自然ではない任意のヌクレオチド配列または遺伝子を意味する。
【0105】
典型的には、AAVおよびB19コーディング領域は、安全な非細胞毒性ベクターを生じさせるように欠失させられている。AAV ITR、またはそれらの修飾は、感染性および部位特異的組み込みを付与するが、細胞毒性は付与せず、そしてこのプロモーターは細胞特異的発現を指令する。AAVベクターに関連する物質については、本明細書に参照して組み込まれる米国特許第6,261,834号に記載されている。
【0106】
そこで本発明のベクターは、実質的毒性を伴わずに哺乳動物染色体内へ組み込むことのできるDNA分子を提供する。
【0107】
ウイルスおよびレトロウイルス内に挿入された遺伝子は、通常は、所望の遺伝子産物の発現を制御するのに役立つプロモーター、および/またはエンハンサーを含有している。プロモーターは、一般に、転写開始部位に関して相当に固定された場所にある場合に機能するDNAの配列である。プロモーターは、RNAポリメラーゼと転写因子との基本的相互作用のために必要とされるコアエレメントを含有しており、上流エレメントおよび応答エレメントを含有していてよい。
【0108】
大きなヒトヘルペスウイルスを用いた分子的遺伝学実験は、それによって大きな異種DNAフラグメントをクローン化し、増殖させ、そしてヘルペスウイルスを用いた感染のために許容される細胞内で確立することができる手段を提供してきた(Sun et al.,Nature genetics 8:33−41,1994;CotterおよびRobertson,.Curr Opin Mol Ther 5:633−644,1999)。これらの大きなDNAウイルス(単純ヘルペスウイルス(HSV)およびエプスタイン・バーウイルス(EBV))は、特定細胞へ150kbを超えるヒト異種DNAのフラグメントを送達する能力を有している。EBV組み換え体は、エピソームDNAとしての感染したB細胞内に大きなDNAの塊を維持することができる。330kbまでのヒトゲノム挿入物を運ぶ個別クローンは、遺伝的に安定性であると思われた。これらのエピソームを維持するためには、EBVによる感染中に構成的に発現した特異的EBV核タンパク質EBNA1を必要とする。さらに、これらのベクターは、大量のタンパク質をインビトロで一時的に生成できるトランスフェクションのために使用できる。ヘルペスウイルスアンプリコン系もまた、220kbを超えるDNA破片をパッケージングし、DNAをエピソームとして安定性で維持できる細胞を感染させるために使用されている。
【0109】
その他の有用な系には、例えば、複製および宿主限定非複製ワクシニアウイルスベクターが含まれる。
【0110】
開示された組成物は、様々な方法で標的細胞へ送達することができる。例えば、本組成物はエレクトロポレーション、またはリポフェクション、またはリン酸カルシウム沈降法を通して送達できる。選択される送達機構は、一部には標的とされる細胞のタイプ、およびその送達が例えばインビボまたはインビトロのどちらで発生するのかに依存するであろう。
【0111】
そこで、本組成物は、開示されたベクターに加えて、例えばカチオン性リポソーム(例、DOTMA、DOPE、DC−コレステロール)またはアニオン性リポソームなどのリポソームのような脂質を含むことができる。リポソームは、所望であれば特定細胞をターゲッティングすることを促進するためのタンパク質をさらに含むことができる。化合物およびカチオン性リポソームを含む組成物の投与は、標的器官へ輸入性の血液に投与する、または気道の細胞へターゲッティングするために気道内へ吸入させることができる。リポソームに関しては、例えば、Brigham et al.Am.J.Resp.Cell.Mol.Biol.1:95−100 (1989);Felgner et al.Proc.Natl.Acad.Sci USA 84:7413−7417 (1987);米国特許第4,897,355号を参照されたい。さらに、化合物はマクロファージなどの特定細胞タイプへターゲッティングできる、またはマイクロカプセルからの化合物の拡散もしくは化合物の送達が特定の速度もしくは用量のために設計されているマイクロカプセルの構成成分として投与できる。
【0112】
投与および被験者の細胞内への外因性DNAの取り込み(すなわち、遺伝子形質導入またはトランスフェクション)を含む上述した方法では、細胞への組成物の送達は様々な機構を介して行なうことができる。1つの例として、送達は、市販で入手できるLIPOFECTIN、LIPOFECTAMINE(GIBCO−BRL,Inc.、メリーランド州ゲーサーズバーグ)、SUPERFECT(Qiagen,Inc.、独国ヒルデン)およびTRANSFECTAM(Promega Biotec,Inc.,、ワイオミング州マディソン)などのリポソーム調製物、ならびに当技術分野において標準的な方法にしたがって開発された他のリポソームを使用して、リポソームを介して行なうことができる。さらに、本発明の核酸またはベクターは、エレクトロポレーションによってインビボで送達することができ、そのためのテクノロジーはGenetronics,Inc.(カリフォルニア州サンディエゴ)から、ならびにSONOPORATION装置(ImaRx Pharmaceutical Corp.、アリゾナ州トゥーソン)によって入手できる。
【0113】
物質は、(例えば、微粒子、リポソーム、または細胞内に組み込まれて)溶液や懸濁液中に含まれていてよい。これらは、VLR、抗体、受容体、または受容体リガンドを介して特定細胞タイプを標的とすることができる。下記の参考文献は、腫瘍組織へ特異的タンパク質をターゲッティングするためのこのテクノロジーの使用例である(Senter,et al.,Bioconjugate Chem.,2:447−451,(1991);Bagshawe,K.D.,Br.J.Cancer,60:275−281,(1989);Bagshawe,et al.,Br.J.Cancer,58:700−703,(1988);Senter,et al.,Bioconjugate Chem.,4:3−9,(1993);Battelli,et al.,Cancer Immunol.Immunother.,35:421−425,(1992);PieterszおよびMcKenzie,Immunolog.Reviews,129:57−80,(1992);およびRoffler,et al.,Biochem.Pharmacol,42:2062−2065,(1991))。これらの技術は、様々な他の特定細胞タイプに対して使用できる。「ステルス」などのビヒクルおよびその他の抗体またはVLR結合リポソーム(大腸癌を標的とする脂質媒介性薬物を含む)、細胞特異的リガンドを通してのDNAの受容体媒介性ターゲッティング、リンパ球指向性腫瘍ターゲッティング、およびインビボのマウス神経膠腫細胞に対する高度に特異的な治療的レトロウイルスターゲッティング。下記の参考文献は、腫瘍組織へ特異的タンパク質をターゲッティングするためのこのテクノロジーの使用例である(Hughes et al.,Cancer Research,49:6214−6220,(1989);およびLitzingerおよびHuang,Biochimica et Biophysica Acta,1104:179−187,(1992))。一般に、受容体は、構成的またはリガンド誘導性のいずれかで、エンドサイトーシスの経路に含まれている。これらの受容体は、クラスリン被覆ピット内でクラスターを形成し、クラスリン被覆小胞を介して細胞内に進入し、その中で受容体が分類される酸性化エンドソームを通過し、そして次に細胞表面へ再循環する、細胞内に蓄積される、またはリソソーム内で分解される、のいずれかとなる。内在化経路は、栄養素取り込み、活性化タンパク質の除去、高分子のクリアランス、ウイルスおよび毒素の日和見進入、リガンドの解離および分解、ならびに受容体レベル調節などの様々な機能に役立つ。多数の受容体は、細胞タイプ、受容体濃度、リガンドのタイプ、リガンドの価数、およびリガンドの濃度に依存して、2つ以上の細胞内経路にしたがう。受容体媒介性エンドサイトーシスの分子および細胞機序は精査されている(BrownおよびGreene,DNA and Cell Biology
10:6,399−409 (1991))。
【0114】
宿主細胞ゲノム内に組み込まれるべき細胞へ送達される核酸は、典型的には組み込み配列を含有している。これらの配列は、特にウイルスをベースとする系が使用される場合は、ウイルス関連配列であることが多い。これらのウイルス組み込み系は、送達系内に含有される核酸を宿主ゲノム内に組み込むことができるように、リポソームなどの非核酸をベースとする送達系を用いて送達されるべき核酸内に組み込むこともできる。
【0115】
その他の宿主ゲノム内に組み込むための一般的技術には、例えば、宿主ゲノムとの同種組み換えを促進するように設計された系が含まれる。これらの系は、典型的には、送達された核酸が宿主ゲノム内に組み込まれるのを誘発するように、ベクター核酸と標的核酸との組み換えが発生する宿主細胞ゲノム内で標的配列と十分な相同性を有する、発現する核酸に隣接する配列に依存している。同種組み換えを促進するために必要なこれらの系および方法は、当業者には知られている。
【0116】
上述したように、本組成物は医薬上許容される担体中で投与することができ、当技術分野においてよく知られている様々な機構(例、裸のDNAの取り込み、リポソーム融合、遺伝子銃を介してのDNAの筋肉内注射、エンドサイトーシスなど)によって、被験者の細胞へインビボおよび/またはエックスビボで送達することができる。
【0117】
エクソビボ法を使用する場合は、細胞または組織を取り出し、当技術分野においてよく知られている標準プロトコールにしたがって体外で維持することができる。本組成物は、例えばリン酸カルシウム媒介性遺伝子送達、エレクトロポレーション、マイクロインジェクションまたはプロテオリポソームなどの任意の遺伝子輸送機構を介して細胞内へ導入することができる。形質導入された細胞は、次に(例えば医薬上許容される担体中で)注入する、または細胞もしくは組織タイプにとって標準的方法によって被験者に同位置へ逆移植することができる。被験者に様々な細胞を移植または注入するための標準方法は知られている。
【0118】
細胞へ送達される核酸は、典型的には発現制御系を含有している。例えば、ウイルスおよびレトロウイルス内に挿入された遺伝子は、通常は、所望の遺伝子産物の発現を制御するのに役立つようにプロモーター、および/またはエンハンサーを含有している。プロモーターは、一般に、転写開始部位に関して比較的固定された場所にある場合に機能するDNAの配列である。プロモーターは、RNAポリメラーゼと転写因子との基本的相互作用のために必要とされるコアエレメントを含有しており、上流エレメントおよび応答エレメントを含有していてよい。
【0119】
哺乳動物宿主細胞中のベクターからの転写を制御する好ましいプロモーターは、例えば、ポリオーマウイルス、サルウイルス40(SV40)、アデノウイルス、レトロウイルス、B型肝炎ウイルスおよび最も好ましくはサイトメガロウイルスなどのウイルスゲノムの様々な起源から、または異種哺乳動物プロモーター、例えばβアクチンプロモーターから入手できる。SV40ウイルスの初期および後期プロモーターは、便宜的には、SV40ウイルス複製起点も含有するSV40制限フラグメントとして入手される(Fiers
et al.,Nature,273:113(1978))。ヒトサイトメガロウイルスの前初期プロモーターは、便宜的にはHindIII E制限フラグメントとして入手される(Greenway,P.J.et al.,Gene 18:355−360(1982))。当然ながら、宿主細胞または関連種由来のプロモーターもまたこの場合に有用である。
【0120】
エンハンサーは、一般に転写開始部位から固定されない距離で機能し、転写ユニットに対して5’(Laimins,L.et al.,Proc.Natl.Acad.Sci.78:993(1981))または3’(Lusky,M.L.,et al.,Mol.Cell Bio.3:1108(1983))のいずれかであってよいDNAの配列を意味する。さらに、エンハンサーはイントロン内(Banerji,J.L.et
al.,Cell 33:729(1983))ならびにコーディング配列自体の内側(Osborne,T.F.,et al.,Mol.Cell Bio.4:1293(1984))にあってよい。それらは通常は10〜300塩基対長であり、それらはシス形で機能する。エンハンサーは、隣接プロモーターからの転写を増加させるように機能する。エンハンサーは、さらにまた転写の調節を媒介する応答エレメントを含有することが多い。プロモーターは、さらにまた転写の調節を媒介する応答エレメントを含有することができる。エンハンサーは、しばしば遺伝子の発現の調節を決定する。多くのエンハンサー配列は哺乳動物遺伝子から現在では知られているが(グロビン、エラスターゼ、アルブミン、−フェトプロテインおよびインスリン)、典型的には一般的発現のためには真核細胞ウイルス由来のエンハンサーが使用されるであろう。好ましい例は、複製起点の後期側(100〜270塩基対)上のSV40エンハンサー、サイトメガロウイルス初期プロモーターエンハンサー、複製起点の後期側上のポリオーマエンハンサー、およびアデノウイルスエンハンサーである。
【0121】
プロモーターおよび/またはエンハンサーは、それらの機能を始動させる光線または特異的化学事象のいずれかによって特異的に活性化することができる。系は、テトラサイクリンおよびデキサメタゾンなどの試薬によって調節できる。ガンマ線照射などの照射への曝露、またはアルキル化化学療法薬によってウイルスベクター遺伝子発現を強化する方法もある。
【0122】
所定の実施形態では、プロモーターおよび/またはエンハンサー領域は、転写されるべき転写ユニットの領域の発現を最大限にするための構成的プロモーターおよび/またはエンハンサーとして機能することができる。所定の構築物では、プロモーターおよび/またはエンハンサー領域は、特定時間に特定タイプの細胞中でしか発現しないとはいえ、全真核細胞タイプで活性な可能性がある。このタイプの好ましいプロモーターは、CMVプロモーター(650塩基)である。その他の好ましいプロモーターは、SV40プロモーター、サイトメガロウイルス(全長プロモーター)、およびレトロウイルスベクターLTFである。
【0123】
真核宿主細胞(酵母、真菌、昆虫、植物、動物、ヒトまたは有核細胞)中で使用される発現ベクターは、さらにまたmRNA発現に影響を及ぼすことのできる転写の終結のために必要な配列を含有することができる。これらの領域は、組織因子タンパク質をコードするmRNAの非翻訳部分におけるポリアデニル化セグメントとして転写される。3’非翻訳領域には、転写終結部位も含まれる。転写ユニットがさらにポリアデニル化領域も含有することが好ましい。この領域の1つの利点は、転写されたユニットがプロセッシングされてmRNAと同様に輸送される可能性をそれが増加させることにある。発現構築物中のポリアデニル化シグナルの同定および使用は、明白に確立されている。導入遺伝子構築物中で同種ポリアデニル化シグナルが使用されるのが好ましい。所定の転写ユニットでは、ポリアデニル化領域はSV40初期ポリアデニル化シグナルから引き出され、約400塩基からなる。さらにまた、転写されたユニットは他の標準配列を単独で、または上記の配列と組み合わせて含有し、その構築物からの発現、またはその構築物の安定性を改良するのが好ましい。
【0124】
ウイルスベクターは、マーカー産物をコードする核酸配列を含有することができる。このマーカー産物は、遺伝子が細胞に送達されているかどうか、送達されている場合は発現しているかどうかを決定するために使用される。好ましいマーカー遺伝子は、β−ガラクトシダーゼをコードする大腸菌lacZ遺伝子、および緑色蛍光タンパク質である。
【0125】
一部の実施形態では、マーカーは選択可能なマーカーであってよい。哺乳動物細胞のために適切な選択可能なマーカーの例は、ジヒドロ葉酸還元酵素(DHFR)、チミジンキナーゼ、ネオマイシン、ネオマイシンアナログG418、ハイドロマイシン、およびピューロマイシンである。そのような選択可能なマーカーを哺乳動物宿主細胞内へ移すのに成功すると、形質転換された哺乳動物宿主細胞は、選択的圧力下に置かれたときに生存することができる。選択的レジメンには広く使用されている2つの明確なカテゴリーが存在する。第1カテゴリーは細胞の代謝、および補給された培地とは無関係に増殖する能力が欠如する突然変異細胞系の使用に基づいている。2つの例は、CHO DHFR細胞およびマウスLTK細胞である。これらの細胞は、チミジンまたはヒポキサンチンなどの栄養素を添加せずに増殖する能力が欠如している。これらの細胞は完全なヌクレオチド合成経路のために必要な所定の遺伝子が欠如しているので、それらは欠けているヌクレオチドが補給培地中に提供されない限り生存することはできない。培地への補給の代替策は、各遺伝子が欠けている細胞中へ無傷DHFRまたはTK遺伝子を導入すること、これによりそれらの増殖要件を変化させることである。DHFRまたはTK遺伝子を用いて形質転換されなかった個々の細胞は、非補給培地中では生存することができないであろう。
【0126】
第2カテゴリーは、任意の細胞タイプで使用される選択スキームに関する、突然変異細胞系の使用を必要としない優性選択である。これらのスキームは、典型的には宿主細胞の増殖を停止させるための薬剤を使用する。新規な遺伝子を有するそれらの細胞は、薬物耐性を備えるタンパク質を発現し、その選択を生き延びるであろう。そのような優性選択の例は、薬物のネオマイシン(Southern P.and Berg,P.,J.Molec.Appl.Genet.1:327 (1982))、ミコフェノール酸(Mulligan,R.C.and Berg,P.Science 209:1422 (1980))、またはハイグロマイシン(Sugden,B.et al.,Mol.Cell.Biol.5:410−413 (1985))を使用する。これら3つの例は、適切な薬剤G418もしくはネオマイシン(ゲネシチン)、xgpt(ミコフェノール酸)またはハイグロマイシン各々に対する耐性を備える真核生物制御下の細菌遺伝子を使用する。その他には、ネオマイシンアナログG418およびpuramycinが含まれる。
【0127】
被験者の細胞内への外因性DNAの投与および取り込み(すなわち遺伝子形質導入またはトランスフェクション)を含んでいる上述した方法では、本発明の核酸は裸のDNAもしくはRNAの形状にあってよい、または、当業者には明白であるように、それによって抗体をコードするDNAフラグメントがプロモーターの転写調節下となるように、これらの核酸は核酸を細胞へ送達するためのベクター内に含まれていてよい。ベクターは、アデノウイルスベクター(Quantum Biotechnologies,Inc.(カナダ国ケベック州ラバル)などの市販で入手できる調製物であってよい。細胞への核酸またはベクターの送達は、様々な機構を介して行なわれる。1つの例として、送達は、市販で入手できるLIPOFECTIN、LIPOFECTAMINE(GIBCO−BRL,Inc.、メリーランド州ゲーサーズバーグ)、SUPERFECT(Qiagen,Inc.、独国ヒルデン)およびTRANSFECTAM(Promega Biotec,Inc.,、ワイオミング州マディソン)などのリポソーム調製物、ならびに当技術分野において標準的な方法にしたがって開発された他のリポソームを使用して、リポソームを介して行なうことができる。さらに、本発明の核酸またはベクターは、エレクトロポレーションによってインビボで送達することができ、そのためのテクノロジーはGenetronics,Inc.(San Diego,CA)から、ならびにSONOPORATION装置(ImaRx Pharmaceutical Corp.、アリゾナ州トゥーソン)によって入手できる。
【0128】
1つの例として、ベクターの送達は、組み換えレトロウイルスゲノムをパッケージできるレトロウイルスベクター系などのウイルス系によって行なうことができる(例えば、Pastan et al.,Proc.Natl.Acad.Sci.U.S.A.85:4486,1988;Miller et al.,Mol.Cell.Biol.6:2895,1986を参照されたい)。組み換えレトロウイルスは、感染させ、それによって感染した細胞へ本発明の広範な中和抗体をコードする核酸(またはその活性フラグメント)を送達するために使用できる。変化した核酸を哺乳動物細胞中へ導入する正確な方法は、当然ながらレトロウイルスベクターの使用に限定されない。この方法のためには、アデノウイルスベクター(Mitani et al.,Hum.Gene Ther.5:941−948,1994)、アデノ関連ウイルス(AAV)ベクター(Goodman et al.,Blood 84:1492−1500,1994)、レンチウイルスベクター(Naidini et al.,Science 272:263−267,1996)、シュードタイプ化レトロウイルスベクター(Agrawal et al.,Exper.Hematol.24:738−747,1996)の使用を含む他の技術を広汎に利用できる。リポソーム送達および受容体媒介性ならびにその他のエンドサイトーシス機構などの物理的形質導入技術もまた使用できる(例えば、Schwartzenberger et al.,Blood 87:472−478,1996を参照されたい)。本発明は、これらやその他の一般に使用される遺伝子導入法のいずれかと結び付けて使用できる。
【0129】
1つの例として、本発明の抗体をコードする核酸がアデノウイルスベクター内で被験者の細胞へ送達される場合は、ヒトへアデノウイルスを投与するための用量は注射1回当たり約107〜109プラーク形成単位(pfu)の範囲に及んでよいが、注射1回当たり1012pfuという高用量でもよい(Crystal,Hum.Gene Ther.8:985−1001,1997;AlvarezおよびCuriel,Hum.Gene Ther.8:597−613,1997)。被験者は単回注射を受けることができる。または追加の注射が必要な場合は、6カ月間隔(または熟練の医師によって決定される他の適切な時間間隔)で無期限の期間にわたり、および/または治療の有効性が確定されるまで繰り返すことができる。
使用する場合、本発明の核酸またはベクターの非経口投与は一般に注射を特徴とする。注射剤は、液剤もしくは懸濁剤、注射前に液体中の懸濁液にするために適合する固体形、またはエマルジョンとしてのいずれかの従来の形態で調製できる。非経口投与のために最近改訂されたアプローチは、一定用量が維持されるような徐放系または持続放出系の使用が含まれる。例えば、参照して本明細書に組み込まれる米国特許第3,610,795号を参照されたい。適切な調製物および治療用化合物の様々な投与経路についてのこれ以上の考察については、例えば、Remington:The Science and Practice of Pharmacy(第19版)編者A.R.Gennaro,Mack Publishing Company,Easton,PA 1995を参照されたい。
【0130】
本発明は、さらに本発明のポリペプチドの製造方法であって、ポリペプチドをコードする核酸を含むベクターを含む細胞を培養する工程と、およびそのポリペプチドを細胞から、または培地から精製する工程と、を含む方法を提供する。さらにまた、タンパク質合成技術を使用した本発明のポリペプチドの製造方法が提供される。
【0131】
さらに、被験者における1つまたは複数の可変性リンパ球受容体をスクリーニングする方法であって、被験者においてN末端ロイシンリッチリピート(LRRNT)、1つまたは複数のロイシンリッチリピート(LRR)、C末端ロイシンリッチリピート(LRRCT)、および連結ペプチドを含む1つまたは複数のポリペプチドを同定する工程を含む方法が開示されるが、このとき連結ペプチドはαヘリックスを含んでいる。
【実施例】
【0132】
下記の実施例は、当業者に、本明細書で特許請求した化合物、組成物、製品、装置および/または方法の製造方法および評価方法に関する完全な開示および説明を提供できるように提示するものであり、純粋に本発明の例示となることを意図しており、本発明者らが本発明であると見なす本発明の範囲を限定することは意図されていない。数字(例、量、温度など)に関して正確さを保証できるように努力はしてきたが、いくらかの誤差および偏差が含まれる可能性がある。他に特に記載しない限り、部は重量部であり、温度は℃または周囲温度であり、そして圧力は大気圧またはほぼ大気圧である。
【0133】
(実施例1:ウミヤツメにおける可変性リンパ球受容体)
(免疫賦活された血液リンパ球由来の転写産物の分析)
活性化リンパ球のトランスクリプトームを調査するために、生きている大腸菌、ヒツジ赤血球、フィトヘマグルチニンおよびアメリカヤマゴボウマイトジェンを含有する抗原/マイトジェンカクテルの腹腔内注射によって、ヤツメウナギの幼生を1週間間隔で2〜4回免疫賦活した。第2回追加免疫賦活から3日後の末梢白血球中の大リンパ球の分画は、非免疫賦活個体に比して13倍大きく、骨髄細胞の分画もまた6倍大きかった(図1a)。小リンパ球に比較して、大リンパ球はサイズがほぼ2倍であり、広範囲のアズール親和性細胞質を有し、顕著な核小体を特徴とした(図1b)。これらの細胞を選別し、ヤツメウナギ活性化骨髄細胞または赤血球由来のcDNAに対する差引きによって活性化リンパ球のメッセージが富裕なcDNAライブラリーを構築するために使用した。
【0134】
差引きされたライブラリーからの1,507クローン中で同定された最も豊富な配列群から、非免疫賦活リンパ球転写産物の調査からの1セットの52個のLRRを含有する発現配列タグ(EST)とともにクラスター化した、可変数の様々なロイシンリッチリピート(LRR)モチーフを備える319種のタンパク質が予測された。3’末端配列をパージングした後、1セットの239種の固有に多様なLRRタンパク質が同定され、それらのうちの22種が239〜304アミノ酸のオープンリーディングフレーム(ORF)の大多数または全部をコードしていた(図6)。これらのヤツメウナギタンパク質は、これらの239の配列各々が固有であり、それらの転写産物は主としてリンパ球によって発現することが見いだされたために、暫定的に可変性リンパ球受容体(VLR)と命名された(図1c)。造血細胞組織由来のリンパ球は非免疫賦活動物において最高VLRレベルを示し、そして免疫賦活は大リンパ球によるVLR転写の増強を生じさせた。これらのVLRの基本的組成は、保存されたシグナルペプチド、N末端LRR(LRRNT)、可変数の多様なLRR、および連結ペプチドを含んでおり、その後にC末端LRR(LRRCT)ならびにトレオニンリッチおよびプラリンリッチ柄部、全体的グリコシル−ホスファチジル−イノシトール(GPI)−アンカー部位および疎水性尾部から構成される保存されたC末端が続いた(図1dおよび図7)。エピトープタグ付きVLRをコードするレトロウイルス構築物が哺乳動物細胞系内にトランスフェクトされると、免疫蛍光分析により、タンパク質の細胞表面局在化が確証され、細菌性GPI特異的ホスホリパーゼCを用いた処置は、細胞表面発現のレベルを有意に減少させ(図1e)、上清中へVLRタンパク質を放出した。3次元構造モデルを生成するために、11個のLRRからなる最長VLR配列を関連LRRタンパク質の結晶構造座標上に通した(Schwede et al.,2000)。このモデルは、Toll様受容体(TLR)エクトドメインのために予測されたモデル(Bell et al.,2003)に類似して、9つのβシートがLRRNTおよびLRRCTによって両端上でキャップされる凹状ソレノイド構造(図1f)を提供する。
【0135】
(VLRレパートリーは個々のヤツメウナギにおいて高度に多様である)
個々のヤツメウナギにおいてRT−PCRによってVLR多様性について調査した。3匹の免疫賦活幼生および4匹の非免疫賦活幼生由来の白血球mRNAを、VLR多様性領域に隣接するプライマーを用いて増幅させた。動物1匹に付き約10クローンのシーケンシングにより69個の固有のVLRが生じ、1匹の個体から同一のクローンは2個しか生じなかった。2匹の動物由来の20個のVLRの可変性配列は、タンパク質多様性を示している(図2;図3および図8に含まれた全セット)。134〜214アミノ酸のサイズ変動は、主としてLRRモジュール数における差に起因した。各配列は、LRRNT、18アミノ酸LRR1、ほぼ変わりなく24アミノ酸長の1〜9個のLRR、13アミノ酸の連結ペプチドおよびC末端LRRCTを含有する;LRRNTは、30〜38アミノ酸を有し、LRRCTは48〜58アミノ酸を有する。顕著な配列多様性の領域は各LRRモチーフから明白であるが、LRRNTにおける最初から7個の残基およびLRRCTにおける最終20個の残基はほぼ不変である。
【0136】
個々のリンパ球のレベルでのVLR多様性を評価するために、全ORFに隣接するプライマーを用いたRT−PCRを使用した。単細胞単離体を免疫賦活および非免疫賦活幼生の血液から選別した。非免疫賦活動物からの6回の単細胞反応および免疫賦活幼生からの7回の反応から入手したPCR産物の分析は、リンパ球13個中12個が単一VLRを発現し(図3)、10個の非免疫賦活細胞のコントロールプールからのVLRクローン6個中5個が固有であることを証明した。1個の細胞単離体は2個のVLR(9.16S、9.16L)を生じたが、この単離体が2個のリンパ球を含有していた可能性を排除することはできない。3個のVLRは、短縮されたタンパク質を予測させるインフレーム終止コドンを有していた。興味深いことに、免疫賦活幼生由来の5個のリンパ球(9.1=9.16S;9.2=9.16L;9.7=9.9)間では、同一VLRの組み合わせが同定された。追加の3匹の免疫賦活幼生(#5〜7)由来の血液サンプルの分析では固有のVLRしか明らかにならなかった(N=27)。これらの所見は、多様なVLRの単一対立遺伝子発現を示唆しており、VLRを有するリンパ球のクローン性増殖についての予備的証拠を提供する。
【0137】
(VLR遺伝子座の複雑性)
保存されたC末端プローブを用いたゲノムブロット・ハイブリダイゼーションにより単一バンドが明らかになった(図4a)。保存された5’UTRおよびシグナルペプチドからなるN末端プローブは、そのブロットが2つの追加のBamHIハンドを示した個体を除いて、使用した制限酵素に依存して、2〜3つのバンドと反応した。さらに、ゲノムパルスフィールドCHEFブロットにより、全6個の消化物中でC末端プローブとの単一ハイブリダイゼーションバンドが明らかになったが、他方N末端プローブは1つの追加の350kbのNotIバンドとのマッチングパターンを生じた(図4b)。これらの所見は、100〜150kbのゲノム内に含有される生殖細胞系VLR遺伝子(gVLR)のN末端およびC末端を備える単一VLR遺伝子座を示している(図4b;PacI消化物)。この遺伝子座を詳細に特性付けるために、これらのプローブを使用して、成体の赤血球DNAから構築された大きな挿入物ウミヤツメP1細菌人工染色体(PAC)ライブラリーをスクリーニングした。両方のプローブを用いてハイブリダイズした5つのPACの分析では、単一の14kbのVLR遺伝子(gVLR)アンプリコンが、ORFフランキングプライマーを用いたロングレンジPCR(LR−PCR)によって同定された。PCR産物の制限酵素分析では、同一のEcoRIバンドおよび2つの対立遺伝子BamHIパターンが明らかになった。2つのgVLR対立遺伝子を表しているPACクローンは、各々33および44kb挿入物を備えるPAC3およびPAC16であるとシーケンシングされた。それらの配列はgVLRを含有する20kbの領域にわたりオーバーラップしていた;PAC16は、gVLRから25kb上流に伸長し、PAC3は18kb下流に伸長した。PAC3および16の間のオーバーラップ領域は、PAC16のgVLRにおける短い欠失を除いて、ほぼ同一であった(24、43および78塩基対)。これらの配列は、このためPAC3のわずかに長い配列を保存しているgVLRコンティーグ内へ融合させられた(図5a)。
【0138】
PAC3/16コンティーグ内のgVLRは4つのエクソンからなる。第1は5’UTRの一部を含有する;エクソン2は5’UTRの残り、シグナルペプチドおよびLRRNTの5’側半分を含有する;エクソン3はLRRCTの5’側半分を含有し、そしてエクソン4はLRRCTの3’側半分、C末端および3’UTRをコードする。標準的な真核細胞スプライス部位は5’UTRイントロンでしか同定されなかったが、他方gVLR内の他のエクソン/イントロン境界はcDNA配列とのアライメントによって決定された。注目すべきことに、gVLR配列は、3’LRRNT、LRR1または24アミノ酸LRRのいずれも含有していなかった。このgVLRから上流では、順方向または逆方向のいずれかに配置されたLRRNT、LRR1およびLRRを含む、一重または二重の可変性LRRモジュールの6つのカセットが同定された。これらのLRRカセットはコンティーグの最初の6kbに及んだが、他方2つの多様な5’LRRCTカセットはgVLRから7kb下流に位置した。
【0139】
また別のクローンのPAC4は、N末端プローブを用いた場合しかハイブリダイズしなかったが、LRRNTおよびLRR1コンセンサスプライマーを用いるPCRによって同定された複数のLRRをコードすることが見いだされた。全挿入物は58kb長であり(図5b)、この配列は小さなギャップ(PAC4内の210〜738塩基対の4つのギャップおよびPAC3/16コンティーグ内の25〜55塩基対の8つのギャップ)を備える11.7kbのgVLRコンティーグとオーバーラップした。このオーバーラップはgVLRエクソン2と3の間の介在配列内に伸長したが、PAC4の553塩基対の末端配列は固有であった。総計30の、1〜3個の多様なLRRモジュールの17のカセットは、部分gVLRから15kb上流に位置したPAC4内の31kb領域内でコードされた。PAC3/16gVLRコンティーグとPAC4配列との比較により、追加の1〜5kbの領域が90%を超える同一性を備えることが明らかになったが、これらは非関連配列によって中断された。PAC4は、5’フランクおよび約半分のgVLRを含む約12kbの重複、または高度に相違するVLR対立遺伝子のいずれかを表すことができよう。これらの可能性を区別するために、これらのPAC挿入物からのgVLR内の制限部位のマップに対するN末端プローブ(図4a)とゲノムハイブリダイゼーションのパターンを比較した。ブロットパターンおよび制限マップは、ブロット内の2kbバンドとは相違していたPAC4からの5.7kbのHindIIIフラグメントを除いて、全フラグメントについて匹敵していた(図4a)。3つのブロットされたゲノムおよびPACライブラリーからのゲノム間のそのような限定された変動性を考えると、PAC4は多型性gVLR対立遺伝子を表している可能性はないと思われる。限定されたVLR対立遺伝子変動は、マイクロサテライト遺伝子座においてさえ低対立遺伝子多様性の他の証拠と一致しており(Bryan et al.,2003)、北米五大湖内のウミヤツメ集団およびその他の淡水に生息する集団が高度に近親交配されていることを示している。そこでこの分析は、単一ヤツメウナギgVLR遺伝子座がgVLRのN末端半分の追加のコピーを有していることを示している。
【0140】
(体細胞性gVLR再配列は多様な成熟VLRを生成する)
VLR多様性領域に隣接するプライマーを用いたPCR増幅によって幼生DNAサンプルを分析すると、6個の固有のイントロンレスVLR ORFが得られた(図3、動物番号#10、12)。この興味のある所見にしたがって、ORFフランキングプライマーを用いた幼生DNAサンプルのPCR増幅は、5’UTRイントロンを含む1.5〜2kbのVLRクローンを生成し、これはクローン14個中13個の固有の配列を明らかにした(#10、11)。これらのゲノムPCRクローンは連続したVLR ORFを含有していたので、それらは「不完全」生殖細胞系VLRからそれらを区別するために暫定的に成熟VLRと名付けられた。配列分析は、これらの成熟VLRがN末端プローブとハイブリダイズする1〜1.3kbの多型性EcoRIバンドを生成することを示唆したが、これらのバンドはリンパ球DNAブロット内でしか観察されなかった(図4cが含まれる)。これらの観察は、ペレット化された赤血球または幼生全身から抽出されたヤツメウナギDNAサンプルが成熟VLRを含有しているが、これらのサンプルからのDNAブロットで検出されるほど十分に豊富なのは生殖細胞系VLRのコピーだけであることを指摘している。
【0141】
この不可解な謎を解決するために、ヤツメウナギリンパ球内での体細胞性遺伝子再配列が、生殖細胞系gVLRからの非コーディングDNAを上流および下流カセットからの多様なLRRと置換して、小さな成熟VLRを生成すると理論付けた。この仮説を試験するために、生殖細胞系gVLRにわたるPCR増幅のための、上流の約3kbおよび下流フランクの約3kbを含むプライマーを設計した(図5a)。幼生DNAサンプルからのLR−PCR増幅は、PAC16プラスミドからのgVLRアンプリコンに類似する約20kbの小さなバンドにプラスして約8kbの追加の顕著なバンドを生じた(図5c)。2匹の幼生サンプルからの8kbアンプリコンの配列分析は、クローン10個中9個が固有の成熟VLRをコードすることを明らかにし(図3)、それらのフランクはgVLRのフランクと同一であった(図5d)。全28個の固有の成熟VLRは、4つの幼生DNAサンプルからのPCR産物間で同定された。リンパ球DNAは、おそらくこれらのDNAサンプルを抽出するために使用されたペレット化赤血球または幼生全身の小さな分画としての、これらの成熟VLRにとってのテンプレートであった。明らかに、LR−PCR中にはリンパ球成熟VLRのより短いテンプレートが優先的に増幅された。gVLR ORFに隣接するプライマーを用いて増幅した場合に、1.5〜2kbの成熟VLRおよび14kbのgVLRの2つのアンプリコンを生じさせる類似のPCRバイアスが観察された。
【0142】
そこでヤツメウナギにおける適応免疫応答を惹起するであろうリンパ球受容体についての探索は、有顎類脊椎動物のIgおよびTCRとは完全に相違する可変性リンパ球受容体の系を同定する。VLRは、複数のLRRモジュールおよびGPIアンカーを介してリンパ球血漿膜へ結合させられている不変柄領域からなる。N末端およびC末端LRRのフランキングチップは不変であり、介在LRRの数および配列における変化は顕著なVLR多様性の一因となる。潜在的VLR多様性は354の固有配列中345を備えて膨大であり、免疫賦活リンパ球由来の同一VLRは3対に過ぎず、ほぼ同一のVLRは3個であった。したがってVLRは、この代表的無顎類に多様なリンパ球受容体レバートリーを与えている。
【0143】
これらの高度に多様なVLRは、病原体の認識において役割を果たす。多様なLRRモジュールを特徴とするタンパク質は、それらが並外れた広範囲のリガンドアレイと相互作用する傾向を示すために、動物および植物の主要な本質的免疫受容体である。動物TLRは、ウイルス、細菌、真菌および原生動物上で保存されたエピトープの認識に関係しており、炎症反応において最高点に達するシグナル伝達カスケードを活性化する(Beutler,2004)。可溶形でも見いだされるGPI固定LRRタンパク質であるCD14は、細菌リポ多糖およびリン脂質へ結合して、TLR4受容体とのシグナリング複合体を形成する(Landmann,2000)。さらにまた別のサイトゾルLRRタンパク質の哺乳動物ファミリーであるNBS−LRRは、細胞内病原体を認識する(Chamaillard et al.,2003)。植物疾患耐性遺伝子は、数百種ものNBS−LRRタンパク質、LRR受容体様キナーゼおよびLRR受容体様タンパク質を含む大きな多重遺伝子ファミリーのメンバーであり、それらの多くは抗病原体反応の特異的活性化に関係することが証明されている(Jones et al.,2004)。顕著な多様性を備える抗原結合VLRは、ヤツメウナギにおいて観察される適応免疫応答を媒介する。リンパ球の表面へのVLRのGPI固定は、これらの受容体のGPI特異的ホスホリパーゼ放出を許容し(Ikezawa 2002)、VLRに予想免疫系における表面受容体および体液性アグルチニンの両方としての二重機能性を付与する。
【0144】
ゲノムPACクローンのシーケンシングにより、シグナルペプチド、5’LRRNT、5’LRRCT、3’LRRCTおよびC末端だけをコードする4つのエクソンからなる生殖細胞系gVLRが同定された。gVLRは5’LRRCTを除いて多様性LRRモジュールを欠如しており、これは修飾しないと高度に多様なVLRメッセージをコードできないであろうことを示唆していた。しかし、複数の多様なLRRカセットはgVLRの上流および下流で見いだされ、これらは成熟VLR遺伝子を組み立てるためにgVLR内へ挿入するために利用できよう。成熟VLRは上流および下流LRRカセットを備える生殖細胞系gVLR内の非コーディングDNAの体細胞性置換を通して生成されるという仮説を試験するために、LR−PCRを使用して生殖細胞系および成熟VLR遺伝子両方の存在を検出した。gVLRからの約20kbの予想された産物は2匹のヤツメウナギのゲノムDNA、そしてさらに、多様なセットの成熟VLRをコードすることが見いだされた成熟VLRからの予測8kbのアンプリコンから入手された。さらに、少数の例では、候補LRRドナーはVLR配列に対する同一性に基づいてgVLR隣接カセット間で同定することができ、そしてgVLR 5’LRRNTおよび3’LRRCT内の高度に保存された配列は遺伝子変換プロセスのための固定領域として潜在的に機能できるであろう。VLRは、体細胞性DNA再配列の機構によって生成される。
【0145】
非減数分裂性DNA再配列は、他の系から知られている。例えば、表面構成成分をコードする遺伝子の再配列は、慢性感染中の免疫認識から逃れるために数種の病原体によって使用される戦略である。淋菌(Neisseria gonorrhoeae)のピリンにおける抗原性変化は、pilE遺伝子座と複数のサイレントなpilSコピーとの間の非可逆的組み換えを含んでおり(Hamrick,2001)、ライム病ボレリア属スピロヘータ類(Borrelia spirochaetes)における抗原性変化は1アレイの15のサイレントカセットとvlsE発現部位間の遺伝子変換によって生成される(Wang et al.,2003)。同様に原生動物ブルーストリパノソーマ(Trypanosoma brucei)は反復されたDNA再配列によるそれらの変異体表面コート糖タンパク質の発現を交互に繰り返し(Donelson,2003)、ならびにマラリア原虫熱帯熱マラリア原虫(Plasmodium falciparum)および腸常在菌であるランブル鞭毛虫(Giardia lamblia)は複数の表面抗原遺伝子間で頻回に切り替わる。宿主と寄生虫との間の進化上の武器競争(arm race)において、脊椎動物は生殖細胞系受容体の体細胞性再配列によって感染性疾患に対抗するために類似の戦略を採用した。多様なリンパ球抗原受容体は、有顎類では対のトランスポザーゼ様RAG1およびRAG2のカットアンドペースト活性によって(Schluter et al.,1999)および無顎類ではいまだ特性付けられていない機構によって組み立てられる。
【0146】
ヤツメウナギVLR系の特徴は、2つの顕著な相違とともに、有顎類脊椎動物リンパ球のIgおよびTCRとの類似性を有する。第1に、ヤツメウナギVLRはLRRモジュールからなるが、他方有顎類抗原受容体はIgドメインからなる。ヤツメウナギの免疫は、祖先の生殖細胞系をコードするLRR受容体の多様性を、その生殖細胞系VLR遺伝子を体細胞性で多様化したものである可変性リンパ球LRR受容体の系と置換する、段階的進化プロセスを受ける。これとは対照的に、有顎類脊椎動物の組み換えリンパ球受容体のコア成分としてのIgドメインは、Igスーパーファミリーメンバーが有顎類脊椎動物以外の動物における病原体または同種移植片の任意のタイプの免疫認識において何らかの役割を果たすことは証明されていないので、それらの祖先からの興味深い追跡不能な進化的推進力である。(Kaufman,2002)。第2に、ヤツメウナギ中のMHC分子の存在についての証拠は見いだされていない。有顎類脊椎動物においては、抗原ペプチドをT細胞へ効率的に提示させるためには多型性MHC分子が必須であるが、他方近親交配されたMHCホモ接合体は疾患耐性の損傷に苦しむと思われる(Penn et al.,2002;Grimholt et al.,2003)。ヤツメウナギは五大湖内の近親交配集団として成長するので、これはそれらのVLR系が多型性構成成分とは無関係に機能するように進化してきた可能性を示している。
【0147】
(動物)
ウミヤツメの幼生(体長8〜13cm)はミシガン湖の支流(Lamprey Services、ミシガン州ラジントン)、またはヒューロン湖の支流(Hammond Bay Biological Station、ミシガン州ミラーズバーグ)からであった。免疫賦活のための幼生は鎮静させ(100mg/L MS222;Sigma)、107大腸菌BL21(DE3)、107ヒツジ赤血球、50μgのフィトヘマグルチニンおよび25μgのヤマゴボウマイトジェン(Sigma)を含有する75μLの0.67×PBSを腹腔内注射した。免疫賦活は1週間間隔で2または4回繰返し、細胞は最終免疫賦活の3〜4日後に採取した。尾を切断した幼生から採血し、0.57×PBSおよび30mM EDTAを用いて1:1に希釈した。バフィコート白血球を50gでの5分間の遠心によって収集した。MoFlo血球計算器を用いて(Mayer et al.,2002a)に記載されたように細胞を選別した。
【0148】
(差引きした免疫賦活白血球cDNAライブラリー)
Super SMART PCR cDNA合成法(BD Biosciences)を1週間間隔で4回免疫賦活した幼生から選別した大リンパ球、骨髄細胞および赤血球からのmRNAとともに使用した。2回の反応において活性化リンパ球cDNAは、骨髄細胞または赤血球のcDNAに対して差引きした(PCR−Select,BD Biosciences)。差引きした産物をpGEM−T Easy(Promega)内でクローン化し、1,507配列について分析した。
【0149】
【表3】
(VLR RT−PCR)
非免疫賦活幼生(#1〜4)、または1週間間隔で2回免疫賦活した幼生(#5〜7)からのバフィコート白血球を300gで5分間かけてペレット化した。第1鎖cDNAは50ngのランダムヘキサマー(Superscript III;Invitrogen)を用いてプライミングした。VLR多様性領域は、LRR.F1+LRR.R1を用いるExpand High Fidelity(Roche)を使用して増幅させた(表3)。熱サイクルは次のとおりであった:94℃で1分間、次に94℃で30秒間、59℃で30秒間、72℃で1分間の35サイクル。動物1匹当たり10〜12クローンをシーケンシングした。
【0150】
(VLR単細胞RT−PCR)
非免疫賦活幼生(#8)、および1週間間隔で2回免疫賦活した幼生(#9)のバフィコート由来の単一リンパ球、または10−細胞プールを0.2mLのTRIzol(Invitrogen)内へ選別した。第1鎖cDNAはLRR_C.R2を用いてプライミングした。VLRは、最初はAdvantage II(BD Biosciences)を用いるSlit.F+LRR_C.R2および次にExpand High Fidelityを用いるLRR_N.F1+LRR_C.R1の2ラウンドのネステッドPCRによって増幅させた。サイクリングパラメータは次のとおりであった:94℃で1分間、次に94℃で30秒間、60℃で30秒間、72℃で1分間の40サイクル。ベクタープライマーを用いたコロニーPCRによって、12個の細胞各々からの6コロニー内で単一サイズ挿入物が明らかになり、それらのうちの3つをシーケンシングした。細胞9.16からのコロニーは2サイズであることが明らかになり、3つの短い挿入物および3つの長い挿入物をシーケンシングした。10個の非免疫賦活細胞プールからは、6個のクローンをシーケンシングした。
【0151】
(ゲノムDNAおよびゲノムPCR)
ゲノムDNAは、幼生全身の1/3、50gで5分間かけてペレット化した0.25mLの血液からの赤血球、または107の選別されたリンパ球から単離した。PCRは、Expand Long Template(Roche)を用いて400ngのgDNAから実施した。VLR多様性領域は、LRR.F1+LRR.R1を用いて幼生#10および12から増幅させた。成熟VLRは、Slit.F+LRR_C.R2またはLRR_N.F1+LRR_C.R1を用いて、動物#10および11から増幅させた。gVLRを超える増幅は、gVLR.F1+gVLR.R1を用いて動物#10および13からであった。8kbバンドは、pCR−XL(Invitrogen)内でクローン化し、M13.Forward、M13.Reverse、Slit.FおよびLRR_C.R2を用いてシーケンシングした。
【0152】
(実際のノーザンブロットおよびDNAブロット)
実際のノーザンは、推奨されたように調製した(Super SMARTマニュアル)。20サイクルで増幅させたcDNAは、幼生尾部、肝臓ならびに非免疫賦活動物の血液、盲樋および腎臓から選別したリンパ球から、または1週間間隔4回免疫賦活した幼生の血液から選別した小および大リンパ球、骨髄細胞および赤血球からであった。
【0153】
1レーン当たり10μgの幼生#10、12および13からのゲノムDNAは、BamHI、EcoRIまたはHindIII(Roche)を用いて消化した;5μgのリンパ球DNAはEcoRIを用いて消化した。パルスフィールドCHEFブロットのために、10匹の幼生からの赤血球をアガロース中に包埋し、1レーン当たり20μgのDNAはAscI、FseI、NotI、PacI、PmeI、またはSfiIを用いて消化した。
【0154】
次の32P標識プローブを使用した:Slit.F+Slit.Rを用いてクローンLRR−2913からPCR増幅させたVLR N末端プローブ(196塩基対)、およびLRR_C.F1+LRR_C.R1を用いて増幅させたC末端プローブ(208塩基対);GAPDH.F+GAPDH.Rを用いてクローンPmGAPDHから増幅させたGAPDHプローブ(314塩基対)。
【0155】
(PACライブラリーおよびクローン)
pCYPAC6(AF133437)中のアレイ化ウミヤツメPACライブラリーは、部分MboI消化物を用いて1匹のミシガン湖成体の赤血球DNAから構築した。6×104クローンは、1〜2倍のゲノム被度を有する65kbの平均挿入物を有していた。ライブラリーは、N末端およびC末端プローブ両方を用いてスクリーニングした。陽性クローンのプラスミドは、EcoRI消化し、ブロッティングし、そしてN末端またはC末端プローブのどちらかとハイブリダイズした。5種のPACは両方のプローブとハイブリダイズし(2、3、15、16、17)、5種のPACはN末端プローブとのみハイブリダイズした(4、9、14、35、42、43)。
【0156】
gVLRは、Slit.F+LRR_C.R2を用いてPAC2、3、15、16および17のプラスミドからのExpand Long Templateを用いて増幅させた。全PCR産物は、2セットのBamHIパターン(PAC2、3および15〜17)を含む14kbであった。PAC3、4および16はMcGill University(カナダ国ケベック州)でシーケンシングされた。
【0157】
(VLR GPIアンカー)
LRRNTから終止コドンまでのVLR挿入物は、Dis_LRR.F+Dis_LRR.R1を用いるExpand High Fidelityを使用してクローンLRR−2913から増幅させ、pDisplay(Invitrogen)内のIgκシグナルペプチドおよびヘマグルチニンエピトープへ融合させた。表面局在化およびVLR GPIアンカーは、BW1547細胞、またはmFcγRIIbを発現するコントロール中で分析した。細胞は、30℃で45分間にわたり1単位/mLの細菌性GPI特異的ホスホリパーゼC(Sigma)を用いて処理した。エピトープタグ付けタンパク質は、抗HA−タグmAb 12CA5を用いて表面染色した。
【0158】
(配列分析)
配列変動性は、MEGA2.1 UPGMA(Kumar et al.,2001)を用いて推定した。GPIアンカー部位は:http://129.194.185.165/dgpi/によって同定した。SWISS−MODEL VLR 3D構造は:http://cubic.bioc.columbia.edu/predictprotein/submit_meta.htmlによって同定した。クローン12.26からの残基22〜319は、CD42a(1m10.pdb)およびNOGO−66受容体(lp8t.pdb)の結晶座標上に進ませた。
【0159】
(実施例2:メクラウナギにおける可変性リンパ球受容体)
(円口類VLRホモログ)
VLRの2つの明確なタイプであるVLR−AおよびVLR−Bは、沿岸メクラウナギであるEptatretus burgeriの12,000個の白血球cDNAクローンから発現した配列タグ間で同定された(Suzuki et al,2004B)。適合するVLRは次に、太平洋メクラウナギE.stoutiiのリンパ球様細胞の転写産物からRT−PCRによってクローニングした。図9は、メクラウナギVLR−AおよびVLR−B、ウミヤツメVLR(Petromyzon marinus)および2つの非寄生性ヤツメウナギのVLR、アメリカカワヤツメ(Lampetra appendix)および北部カワヤツメ(Ichthyomyzon fossor)のアミノ酸配列のアライメントを描出している。これらのVLRは類似の構造的ドメインを共有している:シグナルペプチド(SP)、N末端LRR(LRRNT)、18残基LRR1、その後に可変数の24残基LRR、13残基連結ペプチド(CP)およびC末端LRR(LRRCT)が続く。C末端の最初では、ヤツメウナギVLRおよびメクラウナギVLR−Bはトレオニン/プロリンリッチ領域を有しているが、この領域はメクラウナギVLR−A内では明確には保存されていない。全VLRタンパク質は、グリコシル−ホスファチジル−イノシトール(GPI)細胞表面膜アンカーを加えるためにタンパク質を修飾するために必要とされる疎水性尾部領域で終了する。ウミヤツメVLRと同様に、メクラウナギVLR−AはGPI固定タンパク質であると予測されたが、ω開裂部位は同定されなかった(DGPI http://129.194.185.165/dgpi/);VLR−BについてのC末端疎水性プロファイルもまたGPI修飾の予測因子である。
【0160】
メクラウナギVLRの転写産物はリンパ球様細胞中では豊富であるが、それらの光散乱特性によって選別された骨髄細胞または赤血球中では豊富ではない。VLR−A転写産物レベルは、白血球サンプル中のVLR−Bレベルより約3倍高かった。太平洋メクラウナギの両方のVLRタイプは高度に異質であり(図10AおよびB)、これは可変数の24残基のLRRモジュールおよび顕著なLRR配列多様性を示していた。匹敵する多様性は沿岸メクラウナギからのVLR−A配列(N=66)およびVLR−B配列(N=18)について観察された(図13)。興味深いことに、同一であった、または1〜2個の残基しか相違していなかった2〜4個のVLR−Aクローンの5つのクラスターは、抗原とマイトジェンとのカクテルの4回にわたる週1回の注射が投与されたメクラウナギ#5(図10Aにおいて星印で表示した)からの40の転写産物間で見いだされた。このメクラウナギからのVLR−A転写産物の30%が関連配列のクラスターから構成されるという所見は、リンパ球を有するVLR−Aのクローン性増殖を示唆している。1〜2のアミノ酸置換を備えるクローンは、体細胞性超変異を通しての追加のVLR多様化を反映している。
【0161】
固有の配列である太平洋メクラウナギVLR−Aのデータセット(N=130)から、24残基LRRの転写産物による2〜6コピーが明らかになる(N=527;平均4)。VLR−Bデータセット(N=69)内には、1〜6コピーの24残基LRRが存在したが(N=195;平均2.8)、他方129のウミヤツメVLR(19;GenBank受託番号AY577943〜AY578059)のデータセット内には1〜9コピーの24残基LRRが存在した(N=325;平均2.5)。そして、これらのVLRの個別構成成分を、確実なアライメントのために種間で相違し過ぎるLRRNTおよびLRRCTを除いて(表4;328個のLRR1ドメイン、328個のCPドメイン、および24残基LRRの1,047個の単一ドメイン)、近隣結合系統樹において個別に分析した。
【0162】
【表4】
これらのクラスターは、ほぼ排他的に同一タイプおよび種起源、すなわち太平洋メクラウナギVLR−A、VLR−B、またはウミヤツメVLRクラスタリングであった。相違するVLRタイプ間に同一LRRドメインの例はなかった。しかし、メクラウナギVLR−Aクラスター内およびヤツメウナギVLRクラスター内のLRR1およびCPドメインの大部分は同一であった(表4)。これとは対照的に、メクラウナギVLR−B内のLRR1ドメインは98%固有であった;24残基LRRのセットは、さらに主として固有配列から構成された:97%はメクラウナギVLR−B中で、90%はVLR−A中で、および83%はウミヤツメVLR中で固有であった。この顕著に高度の多様性は、24残基LRRタイプ各々について引き出されたコンセンサス配列が少なくとも10フレームワーク残基を共有することを前提にすると特に注目に値する。
【0163】
(メクラウナギVLR遺伝子)
太平洋メクラウナギおよび沿岸メクラウナギのVLR遺伝子座のゲノム組織は、細菌性人工染色体(BAC)ライブラリーから単離した大きな挿入物のゲノムクローンの配列から決定したが、BACは各VLRタイプに対して1つであった(図11)。メクラウナギゲノム内では、各gVLRについて1コピーしか同定されなかった。遺伝子座の配列および組織は、両方の種でほぼ同一であり、gVLR−AおよびgVLR−B間で相当に保存されている。メクラウナギgVLRは、5’非翻訳領域(UTR)で始まり、2つのコーディング領域が続く(図12A)。ウミヤツメgVLRにおけると同様に、5’UTRは、太平洋メクラウナギgVLR−A内では6.4kb長およびgVLR−B中では220塩基対であるイントロンによって分割されている。メクラウナギgVLR内の第1コーディング領域は、gVLR−A内のシグナルペプチドおよびLRRNTドメインならびにgVLR−B内の23残基シグナルペプチド中の残基1〜13だけをコードする。次に、gVLR−AおよびgVLR−Bに対して各々171および211塩基対の短い介在配列が存在する。第2コーディング領域は、LRRCTの5’末端をコードするヤツメウナギ領域が欠けていること以外は、ウミヤツメgVLR中と同様に、LRRCTの3’末端およびC末端からなる。メクラウナギgVLRは、開始コドンから終止コドンまでがgVLR−A内では671塩基対そしてgVLR−B内では410塩基対とコンパクトである。
【0164】
メクラウナギgVLR遺伝子座は、生殖細胞系遺伝子から約20〜40kb下流に位置する多様なLRRモチーフをコードするカセットを有している(図11)。VLR−A遺伝子座では、多様なCPドメインの6〜8末端残基およびgVLR−A 3’LRRCTとの4残基の同一オーバーラップを含む5’LRRCTをコードするカセットが存在する。さらに下流には、gVLR−Aに比較して逆方向に配置された2つの多様なLRRおよび次に逆方向不完全5’LRRCTを含むカセットが存在する。gVLR−B遺伝子座には、シグナルペプチドの残基7〜23および5’LRRNT、次に多様なCPドメインおよび5’LRRCT、1つの逆方向LRRをコードするカセットがあり、さらにより下流では、LRRの12末端残基および8個の近位残基からなる別の逆方向LRRカセットが存在する。gVLRから約50kb上流および約70kb下流にわたるフランキングDNA内では他の多様なLRRモジュールは同定されなかった。しかし、リンパ球様細胞からのゲノムPCRアンプリコンのサンプル:2匹の動物からの35個の固有の成熟VLR−Aおよび38個のVLR−B配列中で同定された成熟VLR遺伝子の欠如している構成成分を提供するために多様なLRRエレメントがゲノム内のどこかに存在すると思われる(図10)。そこで、メクラウナギ成熟VLR遺伝子は、ヤツメウナギのための場合と同様に、体細胞性組み換えを通して組み立てられなければならない。
【0165】
メクラウナギリンパ球様細胞中の生殖細胞系VLR遺伝子は、遺伝子再配列の前に能動的に転写される。VLR−A生殖細胞系転写産物のPCRアンプリコンは、約0.7kb長およびVLR−Bについては約0.5kbであるが(図12B、RT−PCR;PCRプライマーの位置は図12Aに示されている)、他方より大きなアンプリコンは再配列された成熟VLR遺伝子からの転写産物に対応して、VLR−AおよびVLR−Bに対して各々約1.1および約0.8kbであった。血中ゲノムDNAからの対応するPCRアンプリコンは、生殖細胞系遺伝子に対しては約0.7kbならびに成熟VLR−A遺伝子および成熟VLR−B遺伝子に対して約1.1kbである(図12B、ゲノムPCR)。生殖細胞系および成熟VLR遺伝子からの転写産物中では、5’イントロンはスプライシングされて出され、対応するゲノムPCRアンプリコンより短いRT−PCR産物が産生する(図12BにおけるVLR−Bを参照;gVLR−Aアンプリコンは6.4kbイントロンを含んでいない)。しかし、コーディングエクソン間の介在配列は、それらにコンセンサスな真核細胞のスプライス部位が欠けているために、生殖細胞系転写産物中に保持されている。それに対して生殖細胞系スイッチ領域転写が必須である哺乳動物抗体クラススイッチ組み換えにおける場合と同様に、gVLR再配列のために生殖細胞系転写が必要とされることがある(Bottaro et al,1994;Hein et al.,1998)。
【0166】
(VLR系統発生学)
無顎類VLRタンパク質の系統発生学的分析により、ヤツメウナギVLR配列、メクラウナギVLR−A配列およびVLR−B配列によって各々構成される3つの別個のクラスターが明らかになる(図12C)。メクラウナギVLR−BおよびヤツメウナギVLRは、メクラウナギVLR−Aとの分科とは別個の分科でクラスターを形成している。VLR多様性領域であるLRRNT〜LRRCTまたはLRR1〜CPだけがアライメントされた場合は同一樹形が見られた。そこでメクラウナギVLR−Aは祖先遺伝子の重複によって発生した(図12D)か、あるいは4億9,900万年±3,800万年前のカンブリア紀においてメクラウナギとヤツメウナギ系統に分裂した後にVLR−Aオルソログを消失した(Hedges et al.,2001)。ヤツメウナギVLR−Aオルソログが存在するが、それは極めて低レベルで発現する、または非リンパ系細胞中では発現しないために、ヤツメウナギリンパ球様細胞由来の18,000を超えるcDNA配列中では検出されなかった可能性もある(Pancer et al.,2004)。
【0167】
現存する円口類の両方種類におけるVLRの存在は、脊椎動物が先行の分子認識系を発達させるための強力な進化圧力の指標である。この分析は、カンブリア紀における4,000万年未満以内に、LRRまたはIgいずれかの遺伝子フラグメントが組み換えアッセンブリーを受けてリンパ球受容体の多様なレパートリーを生成する2つの根本的に相違する系が無顎類および有顎類において進化したことを示唆している。この進化のシナリオは、多くの興味深い疑問を引き起こすが、それらの1つは2つの適応免疫戦略が収束進化を意味するかどうか、または一方が他方の先祖であるかどうかという問題に関する。VLRが先駆脊椎動物免疫受容体であったのか、または個別に進化したVLRおよびIgの再配列であったのかは、現存および絶滅無顎類および有顎類脊椎動物の群間での系統発生的関係が明確に解決された場合にしか明らかにならないであろう(Mallatt et al.,2003;Meyer et al.,2003)。しかしこの点について、現代の無顎類の両方の種類におけるVLRの存在は、円口類の単一系統起源に好都合な追加の分子的証拠を提供している。
【0168】
(動物)
太平洋メクラウナギEptatretus stoutii(体長30〜60cm)の生きている標本は、Marinus Scientific(カリフォルニア州ロングビーチ)から購入し、12℃の人工海水(Oceanic System、テキサス州ダラス)中で2カ月間維持した。アメリカカワヤツメ(Lampetra appendix)および北部カワヤツメ(Ichthyomyzon fossor)の幼生(体長:15〜20cm)は、五大湖の支流から入手した(Lamprey Services、ミシガン州ラジントン)。
【0169】
メクラウナギはpH=7に緩衝した0.5g/LのMS222(Sigma、ミズーリ州セントルイス)中へ15分間浸漬することにより鎮静し、その後に0.5mLのメクラウナギPBS中の抗原/マイトジェンカクテル(1Lに付き:28gのNaCl、0.2gのKCL、1.44gのNa2HPO4、0.24gのKH2PO4、pH=7.4、1オスモル)を用いて腹腔内注射した。このカクテルは、109の生きている大腸菌TG1細菌、109のヒツジ赤血球(Colorado Serum Company、コロラド州デンバー)ならびに各100μgのフィトヘマグルチニンおよびヤマゴボウマイトジェン(Sigma)を含有していた。免疫賦活は1週間間隔で繰返し、第4回目の免疫賦活から4日後にシリンジを用いて尾血洞から血液を採取し、30mM EDTAを含有するメクラウナギPBSを用いて1:1に希釈した。50×gでの5分間の遠心後に採取したバフィコート白血球は報告されたように(Newton et al.,1994;Raison et al.,1994)、MoFlo血球計算器(Cytomation,コロラド州フォートコリンズ)を用いて光散乱特性によって選別した。
【0170】
(メクラウナギVLR)
沿岸メクラウナギEptatretus burgeriのVLRホモログは、非免疫賦活動物#7、8の白血球RNAから発現した配列タグのデータベースに対するBLASTクエリーとしてヤツメウナギVLRを用いて同定した(Suzuki et al.,2004B)。両方のストランド上の有意な適合を有するクローンを:64個のVLR−A cDNAクローンおよび15個のVLR−B cDNAクローン上でシーケンシングした。太平洋メクラウナギに対しては、3匹の非免疫賦活個体(#1〜3、6)からの分離されていない血球およびバフィコート白血球、および2匹の免疫賦活動物(#4、5)からのバフィコート白血球を血液ゲノムDNAおよび白血球RNAを抽出するために使用した。RNAの抽出はTRIzol試薬(Invitrogen、カリフォルニア州カールズバッド)を用いて行い、ポリA RNAはDynabeads mRNA精製キット(Dynal Biotech、ニューヨーク州レークサクセス)を用いて選択した。第1ストランドcDNA合成は、Superscript III First Strand cDNA合成キット(Invitrogen)を用いて、VLR−Aについては20pmoleのHgVLRA.F1(表5)、またはVLR−BについてはHgVLRB.F1を用いてプライムし、生成物はカラム精製した(QIAquick PCR精製;QIAGEN、カリフォルニア州バレンシア)。
【0171】
【表5】
VLRは次に、10pmole/μLでフォワードプライマーおよびリバースプライマーのセット(F1またはF2およびR1またはR2)各1μL、5μLの10×バッファー、36.25μLのDDW、5μLのcDNAもしくはゲノムDNA(250ng)および0.75μLの伸長酵素を含有する50μL反応液中で、Expand High Fidelity PCR(Roche Applied Science、インディアナ州インディアナポリス)を用いて、cDNAまたはゲノムDNAからPCR増幅させた。反応液は、94℃で1分間の1サイクル、次に94℃で30秒間、58℃で30秒間および72℃で1分間の35サイクル、および最後に72℃で7分間の伸長を用いて増幅させた。生成物をカラム精製し、pCRII−TOPO(Invitrogen)中でクローン化し、挿入物をシーケンシングした。太平洋メクラウナギについては、109個のVLR−A RT−PCRクローンをシーケンシングし(4個はインフレーム終止コドンを含有していた)、そして36個のゲノム成熟VLR−Aアンプリコンをシーケンシングした(2個がインフレーム終止コドンを含有していた)。VLR−Bについては、37個のRT−PCRクローンをシーケンシングし(1個はインフレーム終止コドンを含有していた)、そして38個のゲノム成熟VLR−Bアンプリコンをシーケンシングした(4個がインフレーム終止コドンを含有していた)。沿岸メクラウナギ#9からの肝臓ゲノムDNA(Suzuki et al.,2004B)をPCRクローニングのために使用し、成熟VLRについてシーケンシングした:4個の成熟VLR−Aアンプリコン(2個はインフレーム終止コドンを含有していた)および3個の成熟VLR−Bアンプリコン。
【0172】
(非寄生性ヤツメウナギVLR)
第1鎖cDNAは、リバースプライマーVLR_3UT.R(ウミヤツメ3’UTRプライマー、表5)を用いて上述したように合成した。アメリカカワヤツメについてはフォワードプライマーおよびリバースプライマーはSlit.F(ウミヤツメ5’UTRプライマー)であり、北部カワヤツメについてはLRR_N.F1(また別のウミヤツメ5’UTRプライマー)であった。アメリカカワヤツメの総計13個および北部カワヤツメの7個の固有のVLRクローンをシーケンシングした。
【0173】
(BACライブラリーおよびクローン)
沿岸メクラウナギBACライブラリー(Suzuki et al.,2004A)は、上述したようにVLRプライマーを用いるPCRによってスクリーニングした(F1またはF2およびR1またはR2)。太平洋メクラウナギBACライブラリー(VMRC23)は、ベクターpCCBACE1(Epicentre Technologies、ワイオミング州マディソン)内の単一標本からの赤血球DNAのEcoRI部分消化物から構築した。このライブラリーは、約184,000個の組み換え体からなり、メクラウナギゲノムの約5×の有効範囲を含んでいる。全ライブラリーは5’および3’のVLR−AプローブおよびVLR−Bプローブを用いたハイブリダイゼーションによってスクリーニングし、陽性クローンはPCRによって正しいことを確証された。太平洋および沿岸メクラウナギ由来の各VLRタイプについて1つのBACを約10×有効範囲でシーケンシングし、コンティーグに組み立てた(Macrogen、韓国ソウル)。挿入物の不完全配列の場合は、コンティーグ内にギャップを占めないgVLRおよびLRRカセットを含有する部分だけを含めた:Eb_gVLR−A、43,362bp;Eb_gVLR−B、92,072bp;Es_gVLR−A、81,648bp;Es_gVLR−B,76,730bp。
【0174】
(配列分析)
隣接結合およびUPGMA樹は、MEGA3 Molecular Evolutionary Genetics Analysisからのプログラムを用いてペアワイズ欠失選択を用いて構築した(Kumar et al.,2004)。BAC挿入物中の遺伝子の予測は、ftp://ftp.ncbi.nlm.nih.gov/blast/executables/およびGenScanサーバー:genes.mit.edu/GENSCAN.htmlからダウンロードしたローカルBLASTを使用することによって遂行した。
【0175】
本出願を通して、様々な刊行物が参照されている。これらの刊行物の開示は、本発明が関係する技術の現在の状態をより十分に記載するために、これにより全体として本出願に組み込まれる。開示した参考文献は、その参考文献が依拠している文章の中で考察されている、その中に含有される物質については、本明細書に個別かつ詳細に参照して組み込まれる。
【0176】
当業者には、本発明において、本発明の範囲または精神から逸脱せずに様々な修飾および変化を加えられることは明白であろう。本発明のその他の実施形態は、当業者には本明細書に開示した本明細書の説明および実践の考察から明白であろう。本明細書および実施例は例示に過ぎないと見なされ、本発明の真の範囲および精神は添付の特許請求の範囲によって指示されることが意図されている。
【0177】
【表6】
【0178】
【表7】
【0179】
【表8】
【0180】
【表9】
【0181】
【表10】
【数1】
【数2】
【数3】
【数4】
【数5】
【数6】
【数7】
【数8】
【数9】
【数10】
【数11】
【数12】
【数13】
【数14】
【数15】
【数16】
【数17】
【数18】
【数19】
【数20】
【数21】
【数22】
【数23】
【数24】
【数25】
【数26】
【数27】
【数28】
【数29】
【数30】
【数31】
【数32】
【数33】
【数34】
【数35】
【数36】
【数37】
【数38】
【数39】
【数40】
【数41】
【数42】
【数43】
【数44】
【数45】
【数46】
【数47】
【数48】
【数49】
【数50】
【数51】
【数52】
【数53】
【数54】
【数55】
【数56】
【数57】
【数58】
【数59】
【数60】
【数61】
【数62】
【数63】
【数64】
【数65】
【数66】
【数67】
【数68】
【数69】
【数70】
【数71】
【数72】
【数73】
【数74】
【数75】
【数76】
【数77】
【数78】
【数79】
【数80】
【数81】
【数82】
【数83】
【数84】
【数85】
【数86】
【数87】
【数88】
【数89】
【数90】
【数91】
【数92】
【数93】
【数94】
【数95】
【数96】
【数97】
【数98】
【数99】
【数100】
【数101】
【数102】
【数103】
【数104】
【数105】
【数106】
【数107】
【数108】
【数109】
【数110】
【数111】
【数112】
【数113】
【数114】
【数115】
【数116】
【数117】
【数118】
【数119】
【数120】
【数121】
【数122】
【数123】
【数124】
【数125】
【数126】
【数127】
【数128】
【数129】
【数130】
【数131】
【数132】
【数133】
【数134】
【数135】
【数136】
【数137】
【数138】
【数139】
【数140】
【数141】
【数142】
【数143】
【数144】
【数145】
【数146】
【数147】
【数148】
【数149】
【数150】
【数151】
【数152】
【数153】
【数154】
【数155】
【数156】
【数157】
【数158】
【技術分野】
【0001】
本出願は、2004年5月21日に出願した米国特許仮出願第60/573,563号の利益を要求する。これは本明細書中でその全体が参考として援用される。
【0002】
本発明は、NIH/NIAIS助成金AI39816およびHG02526−01、ならびにNSF助成金MCB−0317460およびIBN−0321461の下での政府の支援を受けて成された。政府は、本発明において特定の権利を有する。
【背景技術】
【0003】
(発明の背景)
有顎類脊椎動物における適応免疫応答は、抗原が特異的リンパ球受容体によって認識されると開始される。抗原受容体多様性は、免疫グロブリン(Ig)およびT細胞受容体(TCR)の遺伝子座における可変性遺伝子セグメント、多様性遺伝子セグメントおよび結合遺伝子セグメントの組み換えによって生成される。この組み換え再配列は、処理されていない抗原に対する抗体ならびに主要組織適合遺伝子複合体(MHC)クラスIおよびII分子の先端内で提示された抗原フラグメントを認識するTCRの膨大なレバートリーを生成する。そこでクローン性が多様なリンパ球は、主要造血組織および胸腺内で幹細胞前駆体から分化する、Igを有するB細胞およびTCRを有するT細胞の形態で、脊椎動物の適応免疫の基礎を形成している。この組み換え免疫系の重要なエレメントは全有顎類脊椎動物において保存されており、多重遺伝子のTCR遺伝子座およびIg遺伝子座は、最も基本的な有顎類の代表であるサメ、ガンギエイ、およびエイにおいてさえ顕著に複雑である(非特許文献1;非特許文献2;非特許文献3)。
【0004】
早期の脊椎動物の分散(radiation)から生き残っている極めて少数の末裔の無顎類脊椎動物であるヤツメウナギおよびメクラウナギにおいても適応免疫についての豊富な証拠がある(非特許文献4)。これらの無顎類については、体液性タイプおよび細胞媒介性タイプの免疫応答が報告されている。例えば、ヤツメウナギは一次抗原刺激に応答して特異的循環性アグルチニンを産生し、追加免疫後にはより高度のアグルチニンレベルを生成し(非特許文献5;非特許文献6;非特許文献7;非特許文献8;非特許文献9;非特許文献10)、加速された速度で第2セットの表皮同種移植片を拒絶し(非特許文献11;非特許文献12;非特許文献9;非特許文献13)、そして遅延型の過敏性反応を示す(非特許文献5;非特許文献9)。無顎類の適応免疫応答は、有顎類脊椎動物のリンパ造血組織および血液中で見いだされるリンパ球に形態学的に似ている細胞に起因していた(非特許文献5;非特許文献11;非特許文献12;非特許文献14;非特許文献15;非特許文献9;非特許文献16;非特許文献17;非特許文献18;非特許文献13;非特許文献19;非特許文献20)。それらの哺乳動物対応物と同様に、ヤツメウナギリンパ球は他の血球タイプよりはるかに照射感受性であり(非特許文献9)、抗原刺激に応答して凝集かつ増殖し(非特許文献5;非特許文献14;非特許文献15)、そしてPU.1/Spi−BおよびIkarosなどの哺乳動物リンパ球分化に関係している転写因子を発現する(非特許文献21;非特許文献22;非特許文献23;非特許文献24)。しかし驚くべきことに、Ig遺伝子、TCR遺伝子、およびMHC遺伝子は、以前には無顎類脊椎動物または無脊椎動物の尾索類Ciona intestinalisのゲノム配列内では同定されていなかった(非特許文献25)。本発明は、新規なリンパ球受容体および新規なリンパ球受容体をコードする核酸に関する。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Rast,J.P.,Michele K.Anderson,M.K.,Strong,S.J.,Luer,C,Litman,R.T.,および Litman,G.W.α,β,g,and δ T Cell Antigen Receptor Genes Arose Early in Vertebrate Phylogeny.Immunity(1997)6:1−11
【非特許文献2】Flajnik MFおよびKasahara M、Comparative genomics of the MHC:glimpses into the evolution of the adaptive immune system.Immunity(2001)15:351−62
【非特許文献3】Flajnik MF、Comparative analyses of immunoglobulin genes:surprises and portents.Nat.Rev.Immunol(2002)2:688−98
【非特許文献4】Forey PLおよびJanvier P、Agnathans and the origin of jawed vertebrates.Nature(1993)361:129−134
【非特許文献5】Finstad J,Papermaster BWおよびGood RA、Evolution of the immune response.III.Morphologic studies of the thymus and organized lymphoid tissue.Lab Invest.(1964)13:490−512
【非特許文献6】Marchalonis JJおよびEdelman GM、Phylogenetic origins of antibody structure.3.Antibodies in the primary immune response of the sea lamprey,Petromyzon marinus.J.Exp.Med(1968)127:891−914
【非特許文献7】Litman GW,Frommel D,Finstad FJ,Howell J,Pollara BWおよびGood RA、The evolution of the immune response.VIII.Structural studies of the lamprey immunoglobulin.J.Immunol(1970)105:1278−85
【非特許文献8】Pollara B,Litman GW,Finstad J,Howell JおよびGood RA、The evolution of the immune response.VII.Antibody to human”O”cells and properties of the immunoglobulin in lamprey.J.Immunol(1970)105:738−45
【非特許文献9】Good,R.A.,Finstad,J.&Litman,G.W.、The biology of lampreys II:Immunology(編者 Hardisty,M.V.&Potter,LC.)(1972)405−432(Academic Press,London)
【非特許文献10】Hagen M,Filosa MFおよびYouson JH、The immune response in adult sea lamprey{Petromyzon marinus L.):the effect of temperature.Comp.Biochem.Physiol(1985)82:207−10
【非特許文献11】Finstad J,Papermaster BWおよびGood RA、Evolution of the immune response.JJ.Morphologic studies of the thymus and organized lymphoid tissue.Lab Invest.(1964)13:490−512
【非特許文献12】Perey DY,Finstad J,Pollara BおよびGood RA、Evolution of the immune response.VI.First and second set skin homograft rejections in primitive fishes.Lab.Invest.(1968)19:591−7
【非特許文献13】Fujii TおよびHayakawa I、A histological and electron−microscopic study of the cell types involved in rejection of skin allografts in ammocoetes.Cell Tissue Res.(1983)231:301−12
【非特許文献14】Cooper AJ、Ammocoete lymphoid cell populations in vitro.:4th Leukocyte Culture Conference.O.R.Mclntyre(編)New York Appleton Century−Crofts(1971)pp.137− 47
【非特許文献15】Piavis GWおよびHiatt JL、Blood cell lineage in the sea lamprey Petromyzon marinus(Pisces:Petromyzontidae).Copeia(1971)4:722−8
【非特許文献16】Kilarski WおよびPlytycz B、The presence of plasma cells in the lamprey(Agnatha).Dev.Comp.Immunol(1981)5:361−6
【非特許文献17】Zapata A,Ardavin CF,Gomariz RPおよびLeceta J、Plasma cells in the ammocoete of Petromyzon marinus.Cell Tissue Res.(1981)221:203−8
【非特許文献18】Fujii T、Electron microscopy of the leukocytes of the typhlosole in ammocoetes,with special attention to the antibody−producing cells.J.Morphol(1982)173:87−100
【非特許文献19】Ardavin CFおよびZapata A、Ultrastructure and changes during metamorphosis of the lympho−hemopoietic tissue of the larval anadromous sea lamprey Petromyzon marinus.Dev.Comp.Immunol.(1987)11:79−93
【非特許文献20】Mayer WE,Uinuk−Ool T,Tichy H,Gartland LA,Klein JおよびCooper MD、Isolation and characterization of lymphocyte−like cells from a lamprey.Proc.Natl.Acad.Sci,USA(2002a)99:14350−5
【非特許文献21】Haire RN,Miracle AL,Rast JPおよびLitman GW、Members of the Ikaros gene family are present in early representative vertebrates.J.Immunol.(2000)165:306−12
【非特許文献22】Shintani S,Terzic J,Sato A,Saraga−Babic M,O’hUigin C,Tichy HおよびKlein J、Do lampreys have lymphocytes? The Spi evidence.Proc.Natl.Acad.Sci.,USA(2000)97:7417−22
【非特許文献23】Anderson MK,Sun X,Miracle AL,Litman GWおよびRothenberg EV Evolution of hematopoiesis: Three members of the PU.1 transcription factor family in a cartilaginous fish,Raja eglanteria.Proc.Natl.Acad.Sci USA(2001)98:553−8
【非特許文献24】Mayer WE,O’Huigin C,Tichy H,Terzic JおよびSaraga−Babic M、Identification of two Ikaros−like transcription factors in lamprey.Scand.J.Immunol(2002b)55:162−70
【非特許文献25】Azumi K et al.,Genomic analysis of immunity in a Urochordate and the emergence of the vertebrate immune system:”waiting for Godot”Immunogenetics(2003)55:570−81
【発明の概要】
【課題を解決するための手段】
【0006】
(発明の概要)
本明細書に具体的かつ広範囲に記載する本発明の目的によると、本発明は、1つの態様では、新規なリンパ球受容体またはそのフラグメントを含むポリペプチドに関する。本発明は、前記リンパ球受容体またはフラグメントをコードする核酸にさらに関する。前記ポリペプチドおよび核酸を作製かつ使用する方法もまた提供される。そのような使用には、広範囲の精製、治療および診断方法が含まれる。
【0007】
本発明の追加の長所は、一部は以下の説明の中に記載され、そして一部は本説明から明白になるか、または本発明の実践によって学習することができるであろう。本発明の長所は、添付の特許請求の範囲の中で特別に指摘された要素および組み合わせによって実現かつ達成されるであろう。上記の一般的説明および下記に詳述する説明の両方は、代表的かつ説明的なものに過ぎず、特許請求されるように本発明を限定するものではないことを理解されたい。
例えば、本願発明は以下の項目を提供する。
(項目1)
1つのN末端ロイシンリッチリピート(LRRNT)、1つまたは複数のロイシンリッチリピート(LRR)、1つのC末端ロイシンリッチリピート(LRRCT)、および1つの連結ペプチドを含む単離されたポリペプチドであって、前記連結ペプチドがαヘリックスを含む、ポリペプチド。
(項目2)
前記連結ペプチドがLRRCTへ連結している、項目1に記載のポリペプチド。
(項目3)
柄領域およびグリコシル−ホスファチジル−イノシトールアンカーをさらに含む、項目1に記載のポリペプチド。
(項目4)
疎水性尾部をさらに含む、項目3に記載のポリペプチド。
(項目5)
前記柄領域がトレオニン−プロリンリッチ領域を含む、項目3に記載のポリペプチド。
(項目6)
1つのシグナルペプチドをさらに含む、項目1に記載のポリペプチド。
(項目7)
1〜9個のLRRが存在する、前記LRRNTに隣接するLRR1を備える、項目1に記載のポリペプチド。
(項目8)
LRR1が約20個未満のアミノ酸を含む、項目7に記載のポリペプチド。
(項目9)
LRR1が約18個のアミノ酸を含む、項目7に記載のポリペプチド。
(項目10)
LRR2〜9の各々が約25個未満のアミノ酸を含む、項目7に記載のポリペプチド。
(項目11)
前記LRRNTが約40個未満のアミノ酸を含む、項目1に記載のポリペプチド。
(項目12)
前記LRRNTが配列番号157のアミノ酸配列を含む、項目11に記載のポリペプチド。
(項目13)
前記LRRNTが1つまたは複数の保存的アミノ酸置換を有する配列番号157のアミノ酸配列を含む、項目12に記載のポリペプチド。
(項目14)
前記LRRCTが約60個未満のアミノ酸を含む、項目1に記載のポリペプチド。
(項目15)
前記LRRCTが配列番号158のアミノ酸配列を含む、項目14に記載のポリペプチド。
(項目16)
前記LRRCTが1つまたは複数の保存的アミノ酸置換を有する配列番号158のアミノ酸配列を含む、項目14に記載のポリペプチド。
(項目17)
前記連結ペプチドが約15個未満のアミノ酸を含む、項目1に記載のポリペプチド。
(項目18)
前記LRRのアミノ酸配列が相互に相違する、そしてLRRNTおよびLRRCTと相違する、項目1に記載のポリペプチド。
(項目19)
前記ポリペプチドが約130〜約225アミノ酸長である、項目1に記載のポリペプチド。
(項目20)
前記ポリペプチドが選択的に作用因子へ結合する、項目1に記載のポリペプチド。
(項目21)
前記作用因子が病原体である、項目20に記載のポリペプチド。
(項目22)
前記病原体が細菌である、項目21に記載のポリペプチド。
(項目23)
前記作用因子が毒素である、項目20に記載のポリペプチド。
(項目24)
項目1に記載のポリペプチドをコードする単離された核酸。
(項目25)
発現制御配列に機能的に連結した項目24に記載の核酸を含む発現ベクター。
(項目26)
項目25記載のベクターを含む培養された細胞。
(項目27)
前記ポリペプチドが固体支持体へ結合している、項目1に記載のポリペプチド。
(項目28)
前記固体支持体が可動固体支持体である、項目27に記載のポリペプチド。
(項目29)
前記固体支持体がカラムである、項目27に記載のポリペプチド。
(項目30)
前記固体支持体がチップである、項目27に記載のポリペプチド。
(項目31)
前記固体支持体がマルチウエルプレートである、項目27に記載のポリペプチド。
(項目32)
前記LRRがポリペプチド全体で高度に可変性である、項目1記載の複数のポリペプチド。
(項目33)
前記複数が固体支持体へ結合している、項目32に記載の複数のポリペプチド。
(項目34)
前記固体支持体が可動固体支持体である、項目32に記載の複数のポリペプチド。
(項目35)
前記固体支持体がカラムである、項目32に記載の複数のポリペプチド。
(項目36)
前記固体支持体がチップである、項目32に記載の複数のポリペプチド。
(項目37)
前記固体支持体がマルチウエルプレートである、項目32に記載の複数のポリペプチド。
(項目38)
サンプル中の作用因子を検出する方法であって、
a.前記サンプルを項目20記載のポリペプチドとを、前記サンプル中の前記作用因子へ前記ポリペプチドが結合するための条件下でと接触させる工程と;
b.前記サンプル中の前記作用因子へ結合したポリペプチドを検出する工程であって、結合したポリペプチドが前記サンプル中の作用因子を指示する工程と、を含む方法。
(項目39)
前記ポリペプチドが検出可能な成分で標識される、項目38に記載の方法。
(項目40)
前記サンプル中の前記作用因子の定量をさらに含む、項目38記載の方法。
(項目41)
作用因子の活性を遮断する方法であって、前記作用因子を項目20に記載の前記ポリペプチドとを、前記ポリペプチドが前記作用因子に結合するための条件下で接触させる工程を含み、前記ポリペプチドの前記作用因子への結合が前記作用因子の活性を遮断する方法。
(項目42)
接触させる工程がインビトロである、項目38に記載の方法。
(項目43)
接触させる工程がインビボである、項目38に記載の方法。
(項目44)
被験者に有効量の項目1に記載のポリペプチドを投与する工程、および前記被験者において結合したポリペプチドの所在位置を検出する工程、を含むイメージング法。
(項目45)
前記ポリペプチドが検出可能な成分で標識される、項目44に記載の方法。
(項目46)
サンプルからの作用因子を精製する方法であって、
a.前記サンプルを項目20記載のポリペプチドと、前記ポリペプチドが作用因子に結合してポリペプチド/作用因子複合体を形成する条件下で、接触させる工程と;
b.前記サンプルから前記ポリペプチド/作用因子複合体を単離する工程と;
c.前記ポリペプチド/作用因子複合体から前記作用因子を単離する工程と、を含む方法。
(項目47)
被験者における病原性作用を減少または予防する方法であって、前記被験者に有効量の項目21のポリペプチドを投与する工程を含む方法。
(項目48)
ポリペプチドを製造する方法であって、前記ポリペプチドの発現を許容する条件下で項目26に記載の細胞を培養する工程と、前記ポリペプチドを細胞または細胞の培地から精製する工程と、を含む方法。
(項目49)
被験者における1つまたは複数の可変性リンパ球受容体をスクリーニングする方法であって、被験者において、1つのN末端ロイシンリッチリピート(LRRNT)、1つまたは複数のロイシンリッチリピート(LRR)、1つのC末端ロイシンリッチリピート(LRRCT)、および1つの連結ペプチドを含む1つまたは複数のポリペプチドを同定する工程を含み、このとき前記連結ペプチドがαヘリックスを含む、方法。
(項目50)
遺伝子の機能を決定する方法であって、遺伝子を発現する細胞の細胞質内へ前記遺伝子のタンパク質産物に対して特異的なポリペプチドを導入する工程、および機能が知られていない遺伝子のタンパク質産物の消失に起因する作用を監視する工程、を含む方法。
【図面の簡単な説明】
【0008】
本明細書に組み込まれていてその一部を構成する添付の図面は、本発明のいくつかの実施形態を図示しており、下記の説明とともに、本発明の原理を説明するために役立つ。
【図1】図1は、ヤツメウナギの白血球およびVLRを示した図である。図1aは、抗原/マイトジェンカクテルを用いた免疫賦活前後の白血球についての光散乱分析を示している。図1bは、選別された免疫賦活白血球:小リンパ球(R1)、大リンパ球(R2)または骨髄性細胞(R3)を示している。Wright−Giemsa染色、100×。スケールバー=10μm。図1cは、VLRおよびGAPDH(コントロール)の視覚的なノーザンブロットを示している。免疫賦活および非免疫賦活幼生の組織から、または造血器官および血液由来の選別された細胞から増幅させたcDNAが示されている。図1dは、VLRスティックモデル:シグナルペプチド、N末端LRR、9個のLRR、連結ペプチド、C末端LRR、トレオニン−プロリンリッチ柄部、GPIアンカーおよび疎水性尾部を示している(クローン12.26、417残基、AY577974)。図1eは、細菌GPI−ホスホリパーゼCを用いて処理した(+PLC)または用いずに処理した(−PLC)マウス胸腺腫細胞内で発現したエピトープタグ付きVLRおよびFcγRIIb(コントロール)の細胞表面発現を示している。図1fは、2回転させて見たVLR多様性領域の3Dモデルを示している(クローン12.26)。
【図2】図2は、2匹のヤツメウナギの幼生におけるVLR多様性についての調査を示した図である。リンパ球からPCR増幅した20の多様性領域のアライメント。PCRプライマーは、全VLR配列内に保存された領域内に所在した:LRRNTの5’側およびLRRCTの3’側近くのシグナルペプチド。ドナー動物およびクローン数が指示されている。LRRモチーフの位置もまた示されている。黒色:100%同一性;灰色:60〜99%同一性;白色:60%同一性。配列1.3〜2.10は、本明細書では各々配列番号1〜20に対応する。
【図3】図3は、13匹の個々の幼生におけるVLRタンパク質多様性の評価を示した図である。cDNAおよびゲノムPCRクローン由来の112のVLR多様性領域の遺伝子距離デンドログラム(樹系図)。幼生数およびクローン数(例、6.20=ドナー6、クローン20)は、免疫賦活ドナー(N=27)については赤色および非免疫賦活ドナー(N=41)については緑色で指示されている。星印(*)は、1つの単離体由来の2個のVLR(9.16S、9.16L)を含む単一の細胞単離体(N=12)由来のクローンを示している;そしてコントロール10細胞プール由来のクローンは10Cと表示されている(N=4)。ゲノムDNA由来の成熟VLR配列は青色である(N=28;血液#10、12;死体#11、13)。全セットについての平均多様性は1.36±0.03であり、13の固体からの配列群内で0.28〜0.54の範囲である。
【図4】図4は、VLRのN末端またはC末端のプローブを用いてハイブリダイズした制限酵素消化DNAのVLRゲノムブロットを示した図である。図4(a)は、3匹のヤツメウナギのブロットを示している(血液DNA #10、12;死体#13)。動物13だけが多型BamHIパターンを示した。図4bは、10匹のヤツメウナギからプールされた赤血球からゲノムの広がりを示している。パルスフィールドブロット・ハイブリダイゼーションは両方のプローブに対して適合するパターンを示し、さらなる350kbのNotI N末端バンドは5’gVLR重複に対応する。
【図5】図5は、VLR遺伝子座のゲノム組織を示した図である。図5aは、20kbにわたってオーバーラップするクローンPAC16(44kb)およびPAC3(33kb)から融合された57kbのgVLRコンティーグ(AY577941)内で同定されたモチーフを示している。点線はPAC挿入物を表している;赤いバーは、N末端およびC末端プローブを指示している。図5bは、PAC4(58kb、AY577942)が11.7kbにわたってgVLRコンティーグとアライメントすることを示している(ヌクレオチド45,882〜57,609)。1〜3個のLRRのカセットは順方向または逆方向で配置されている:8個はgVLRコンティーグ内であり、17個はPAC4内である。図5cは、gVLRのLR−PCR分析を示している。血液(#10)または死体(#13)由来のDNAはプライマーgVLR.F1+gVLR.R1を用いて増幅させた(図5aおよび図5eに示した)。PAC16アンプリコンがコントロールとして機能した。約20kbのバンドは生殖細胞系VLRに対応し、約8kbのバンドは成熟VLRに対応する。図5dは、成熟VLRのリンパ球特異的再配列を示している。選別された100個のリンパ球または赤血球のプール由来のLR−PCR。約14kbのバンドは生殖細胞系VLRに対応し、約〜1kbのバンドはリンパ球DNAからのみ増幅された成熟VLRに対応する。図5eは、8kbの成熟VLRアンプリコンを例示している。
【図6】図6は、ESTクローンから予測された22個のVLRタンパク質の複合アライメントを示した図である(シングルパス5’配列、一部不完全なC末端)。黒色:完全同一性;黄色:80〜99%同一性;緑色:60〜79%同一性;白色:60%未満の同一性。LyEST3090〜LyEST5266に対するアミノ酸配列は、各々配列番号21〜42に対応する。
【図7】図7は、代表的なVLR(cDNAクローンLyEST2913、AY578059)のORFを示した図である。開始メチオニンはヌクレオチド118〜120にあり、終止コドンはヌクレオチド937〜939にある。生殖細胞系VLRのエクソン2およびエクソン4内に保存されたヌクレオチド配列は赤色で表示されている;エクソン3に対応する多様な5’LRRCTは緑色で表示されている。構造モチーフはタンパク質配列の上方に表示されている;GPI開裂部位は青色で表示されている。図示したアミノ酸配列は配列番号43に対応し、図示した核酸配列は配列番号156に対応する。
【図8−1】図8は、13匹のヤツメウナギからPCR増幅させた112のVLR多様性領域の複合アライメントを示した図である。免疫賦活ヤツメウナギおよび非免疫賦活ヤツメウナギからのゲノムクローンおよびRT−PCRクローン。非免疫賦活動物:#1〜4と指定された動物(N=41)、#8と指定された動物由来の選別された単リンパ球(N=4)および#8.10Cと指定された動物由来の10個の細胞のプール由来のクローン(N=4);免疫賦活動物:2個のVLR(9.16S、9.16L)を備える1つの単離体を含む、#5〜7と指定された動物(N=27)および#9と指定された動物由来の選別された単リンパ球(N=8);成熟VLR:#10〜13(N=28)または死体(#11、13)と指定された血液から抽出した幼生ゲノムDNA。黒色:80〜100%同一性;黄色:60〜79%;緑色:40〜59%;白色:<40%。アライメントの上部から、1.1に対するアミノ酸配列は配列番号13に対応し、アミノ酸配列7.27〜4.7は配列番号45〜52に対応し、アミノ酸配列1.5は配列番号12に対応し、アミノ酸配列4.14は配列番号54に対応し、アミノ酸配列1.7は配列番号8に対応し、アミノ酸配列3.15は配列番号56に対応し、アミノ酸配列2.1は配列番号5に対応し、アミノ酸配列2.2は配列番号10に対応し、アミノ酸配列2.7は配列番号11に対応し、アミノ酸配列4.8〜6.22は配列番号60〜65に対応し、アミノ酸配列2.4は配列番号3に対応し、アミノ酸配列1.8は配列番号2に対応し、アミノ酸配列7.3〜6.21は配列番号68〜72に対応し、アミノ酸配列1.2は配列番号5に対応し、アミノ酸配列2.14は配列番号6に対応し、アミノ酸配列3.7は配列番号75に対応し、アミノ酸配列1.6は配列番号7に対応し、アミノ酸配列5.3は配列番号77に対応し、アミノ酸配列10.1は配列番号78に対応し、アミノ酸配列2.14は配列番号4に対応し、アミノ酸配列1.3は配列番号1に対応し、アミノ酸配列6.16〜7.26は配列番号81〜119に対応し、アミノ酸配列2.15は配列番号14に対応し、アミノ酸配列2.8は配列番号17に対応し、アミノ酸配列5.6〜7.33は配列番号122〜125に対応し、アミノ酸配列1.10は配列番号19に対応し、アミノ酸配列2.10は配列番号20に対応し、アミノ酸配列1.4は配列番号15に対応し、アミノ酸配列12.19〜4.3は配列番号129〜132に対応し、アミノ酸配列1.9は配列番号16に対応し、アミノ酸配列5.5〜3.3は配列番号134〜144に対応し、アミノ酸配列2.13は配列番号18に対応し、およびアミノ酸配列3.6〜3.9は配列番号146〜155に対応する。
【図8−2】(図8−1の続き)
【図9】図9は、進化的に保存された無顎類VLRを示した図である。沿岸メクラウナギ(Eptatretus burgeri)、太平洋メクラウナギ(E.stoutii)、ウミヤツメ(Petromyzon marinus;GenBank受託番号AY577946)、アメリカカワヤツメ(Lampetra appendix)および北部カワヤツメ(Ichthyomyzon fossor)を表しているVLRアミノ酸配列。青色の影:100%同一性;黄色:60〜99%同一性;緑色:40〜59%同一性;赤色:疎水性尾部領域。
【図10】図10は、太平洋メクラウナギVLR多様性領域(LRRNTからLRRCT)間の遺伝子距離を示した図である。5匹の動物からの、PCR増幅リンパ球様cDNAクローンまたは血液ゲノムPCRアンプリコンから予測されるタンパク質。スケールバーは5%アミノ酸多様性を表している。A.VLR−A(N=139)。B.VLR−B(N=70)。緑色:非免疫賦活;赤色:免疫賦活;青色:ゲノム成熟VLR;星印−関連配列。
【図11】図11は、メクラウナギVLR遺伝子座を示した図である。図11Aは、太平洋メクラウナギVLR−Aを示している。図11Bは、沿岸メクラウナギVLR−Aを示している。図11Cは、太平洋メクラウナギVLR−Bを示している。図11Dは、沿岸メクラウナギVLR−Bを示している。4個のBACクローンからの挿入物の配列であり、占められていないギャップがマーキングされている。逆方向または順方向にあるVLR生殖細胞系遺伝子およびフランキングカセットの位置はキロベース単位で指示されている(図は一定尺度から外れている)。GenScan遺伝子予測は青色で表示されている:太平洋メクラウナギ生殖細胞系VLR−A遺伝子から上流の非関連性LRR遺伝子ならびに沿岸メクラウナギVLR−Aおよび太平洋メクラウナギVLR−B遺伝子座における2つの隣接するトランスポザーゼORF。
【図12】図12は、無顎類VLR遺伝子、転写産物および系統発生を示した図である。図12Aは、太平洋メクラウナギおよびウミヤツメの生殖細胞系および成熟VLR遺伝子の略図を示している。着色したバーは、コーディング領域を示している;ヌクレオチド単位のサイズ;メクラウナギVLRを増幅させるために使用したPCRプライマーの位置(表5)は矢印で指示され、F(順方向)、R(逆方向)と指示されている。図12Bは、リンパ球様転写産物(RT−PCR)または血液ゲノムDNAからPCR増幅させた太平洋メクラウナギVLRを示している。アガロースゲル画像;左には分子量マーカーが表示されている(キロベース);生殖細胞系および成熟VLRアンプリコンの位置は右側に表示されている。図12Cは、無顎類VLRの系統発生的分析を示している。メクラウナギおよびヤツメウナギVLRタンパク質の近隣結合系統樹(図9の配列と同一配列);ブートストラップ値が指示されている。スケールバーは10%アミノ酸多様性を表している。図12Dは、無顎類VLRの進化のモデルを示している。
【図13】図13は、沿岸メクラウナギVLR多様性領域(LRRNTからLRRCT)間の遺伝子距離を示した図である。タンパク質は、3匹の動物の白血球cDNAクローンまたはゲノムDNA由来の成熟VLRアンプリコンから予測した。スケールバーは5%アミノ酸多様性を表している。A.VLR−A(N=66)。B.VLR−B(N=18)。赤色:メクラウナギ#7;緑色:#8;青色:メクラウナギ#9由来のゲノム成熟VLR。
【発明を実施するための形態】
【0009】
(詳細な説明)
最古の脊椎動物の中で現代に生きる代表であるウミヤツメPetromyzon marinusの脊椎動物免疫系の原始エレメントを対象にリンパに集中した調査が開始された。ヤツメウナギ造血組織由来のリンパ球様細胞によって発現した転写産物の初期の分析は、免疫系分子の数種のホモログを同定したが(Mayer et al.,2002a;Uinuk−Ool et al.,2002;Uinuk−Ool et al.,2003)、有顎類脊椎動物が特異的適応免疫のために使用する基本的なIgスーパーファミリー受容体エレメントは同定されなかった。血流中に存在する活性化リンパ芽球は適応応答に含まれる遺伝子を発現する可能性が高いと判断して、本試験は、免疫賦活ヤツメウナギ幼生由来リンパ球のトランスクリプトームの調査から開始した。この研究により、本明細書に記載する新規なタイプの高可変性リンパ球受容体が明らかになった。
【0010】
本発明は、本明細書に含まれた本発明の好ましい実施形態および実施例についての下記の詳細な説明および図面ならびにそれらについての上記や下記の説明を参照することによってより容易に理解することができる。
【0011】
本発明の化合物、組成物、製品、装置、および/または方法を開示および説明する前に、当然ながらそれらは変動することがあるので、本発明は、他に特に規定しない限り、特定の合成方法、特定の組み換えバイオテクノロジー方法にも限定されず、他に規定しない限り特定の試薬にも限定されないことを理解されたい。さらにまた、本明細書で使用する用語は特定の実施形態を説明するために使用されたに過ぎず、限定的であることは意図されていないことも理解されたい。
本明細書および添付の特許請求項において使用する単数形の「1つの」および「その」には、状況が他の場合を明確に指さない限り、複数対象が含まれる。そこで、例えば「1つの医薬担体」との言及には、2つ以上のそのような担体などの混合物が含まれる。
【0012】
本明細書での範囲は、「約」1つの特定値から、および/または「約」別の特定値までを表すことができる。そのような範囲が明示される場合は、別の実施形態は、1つの特定値から他の特定値までを含んでいる。同様に、先行詞「約」の使用によって近似値として数値が明示される場合は、特定値が別の実施形態を形成することを理解されたい。さらに、範囲各々の終点は、他方の終点と関連して、および他方の終点からは独立してのどちらにおいても重要であることを理解されたい。
【0013】
「任意の」または「任意で」は、引き続いて記載される事象または状況が発生しても発生しなくてもよく、そしてその説明は前記事象または状況が発生する場合およびそれが発生しない場合を含んでいることを意味する。
【0014】
本明細書で使用する「ポリペプチド」、「タンパク質」、および「ペプチド」は、アミノ酸配列について言及するために互換的に使用される。
【0015】
本発明は、1〜12個のロイシンリッチリピートを含み、適応免疫において機能できる体細胞性再配列が可能なポリペプチドである可変性リンパ球受容体(VLR)に関する。
【0016】
本発明は、N末端ロイシンリッチリピート(LRRNT)、1つまたは複数のロイシンリッチリピート(LRR)(本明細書では内部LRRと呼ぶ)、C末端ロイシンリッチリピート(LRRCT)、および連結ペプチドを含む単離されたポリペプチドを提供するが、このとき連結ペプチドはαヘリックスを含んでいる。ポリペプチドの長さは、約130という少数のアミノ酸または約225という多数のアミノ酸を含むことができる。ポリペプチドおよびコーディング核酸の一般構造および特定配列の例は図に示されている。さらに様々な領域(シグナルペプチド、LRRNT、LRR、LRRCT、連結ペプチド、柄部および疎水性尾部を含む)の極めて多数の例は図の中に見いだすことができる。
【0017】
任意で、連結ペプチドはLRRCTのN末端側に位置しており、より具体的には内部LRRおよびLRRCTの間に位置している。連結ペプチドは、内部LRRおよびLRRCTへ連結させることができる。したがって、本明細書ではLRRNT、1つまたは複数の内部LRR、連結ペプチド、およびLRRCTをその順序で含むポリペプチドが開示される。さらに、LRRNTおよびLRRCT間の内部LRR領域が、1個、2個、3個、4個、5個、6個、7個、8個または9個のロイシンリッチリピートを含むポリペプチドが開示され、LRR1はLRRNTに隣接して位置するかまたは近位に位置する。本明細書で使用するようにLRR1、LRR2、LRR3、LRR4、LRR5、LRR6、LRR7、LRR8、またはLRR9は、LRRNTからLRRCTへ連続的に進むとみなされる。そこで本明細書には、LRRNT、LRR1、LRR1〜2、LRR1〜3、LRR1〜4、LRR1〜5、LRR1〜6、LRR1〜7、LRR1〜8、またはLRR1〜9、連結ペプチド、およびLRRCTをその順序で含むポリペプチドが開示される。
【0018】
ロイシンリッチリピート(LRR)は、タンパク質対タンパク質相互作用に典型的に関係する短い配列モチーフであり、LRRは複数のロイシン残基を含んでいる。LRRは、例えば2位、5位、7位、12位、16位、21位および24位でロイシンまたは他の脂肪族残基を含有する。しかし、ロイシンまたは他の脂肪族残基は2位、5位、7位、12位、16位、21位および24位での残基の場所に加えて、またはそれらの代わりに他の位置に存在し得ると理解されており、本明細書では企図されている。例えば、ロイシンは2位ではなく3位に存在し得る。構造的には、モチーフがβシート構造を形成することもまた理解されたい。そこで、例えばLRRNT、5個のLRR、LRRCT、および連結ペプチドを含む開示されたポリペプチドは、7つのβシート構造および連結ペプチドのαヘリックスを含むであろう。
【0019】
各LRRの長さおよび配列は、ポリペプチド内の他のLRRならびにLRRNTおよびLRRCTとは相違することがあると理解されたい。例えば、本発明の1つの実施形態は、LRRNT、1〜9個のLRR、連結ペプチド、およびLRRCTを含むポリペプチドであるが、このとき第1内部LRRはLRR1であり、LRR1は約20個未満のアミノ酸を含んでいる。さらにLRR1が約18個のアミノ酸を含むポリペプチドが開示される。任意で、このポリペプチドはさらにLRR2〜9を含むが、このときLRR2〜9は各々約25未満のアミノ酸である。さらにLRR2〜9が各々約24個のアミノ酸を含むポリペプチドが開示される。LRR1〜9は、長さおよび特異的アミノ酸配列の両方において所定のポリペプチド内で同一であっても、または相互から相違していてもよい。
【0020】
LRRNTおよびLRRCTと指される末端LRRは、典型的には各内部LRRより長い。LRRNTおよびLRRCTは、不変領域(類似の可変性リンパ球受容体と比較してポリペプチドの残りに比してほとんど変化を有していない領域)を含む。可変領域は、特異性を備える受容体を提供するが、不変領域および受容体にわたる一般構造類似性は保護免疫機能を維持するのに役立つ。ポリペプチドはLRRNTを含むことができ、このときLRRNTは約40未満のアミノ酸を含んでいる。そこで、LRRNTは、1つまたは複数の保存的アミノ酸置換の存在下または不在下で、任意でアミノ酸配列CPSQCSC(配列番号157)、CPSRCSC(配列番号307)、CPAQCSC(配列番号308)、CPSQCLC(配列番号309)、CPSQCPC(配列番号310)、NGATCKK(配列番号311)、またはNEALCKK(配列番号312)を含んでいる。さらにまた、LRRCTが約60アミノ酸長未満、および任意で40〜60アミノ酸長であるLRRCTを含むポリペプチドが開示される。詳細には、特に、LRRCTが保存的アミノ酸置換の存在下または不在下で、アミノ酸配列TNTPVRAVTEASTSPSKCP(配列番号158)、SGKPVRSIICP(配列番号313)、SSKAVLDVTEEEAAEDCV(配列番号314)、またはQSKAVLEITEKDAASDCV(配列番号315)を含むポリペプチドが開示される。
【0021】
すべてのペプチド、ポリペプチド、およびタンパク質と同様に、ペプチド、ポリペプチド、またはタンパク質の性質または機能を変化させないLRRCTおよびLRRNTのアミノ酸配列における置換が発生する可能性があると理解されたい。そのような置換は、保存的アミノ酸置換を含んでおり、下記でより詳細に考察する。
【0022】
開示された組成物は、連結ペプチドも含むことができる。典型的には、そのようなペプチドは15アミノ酸長未満の短いペプチドであり、αヘリックスを含んでいる。そこで、例えばαヘリックスを含む10アミノ酸長、11アミノ酸長、12アミノ酸長、13アミノ酸長、14アミノ酸長、および15アミノ酸長の連結ペプチドが特異的に開示される。連結ペプチドは、ポリペプチドの構造的成分を連結させるために機能すると理解されたい。さらに、ポリペプチドの連結ペプチドは、LRRCTへ連結できると理解されたい。
【0023】
本発明のポリペプチドは、可溶形または膜結合形を含むことができる。ポリペプチドを可溶形または膜結合形にさせる多数の機序が存在する。例えば、膜貫通ドメインを有していないポリペプチドは細胞によって直接的に分泌されることがある。または、ポリペプチドは、膜表面上にそのポリペプチドを維持するグリコシル−ホスファチジル−イノシトール(GPI)アンカーを含むことができる。このため、本明細書ではGPIアンカーを含むポリペプチドが開示される。表面へ結合したポリペプチドを維持するための他の機構は、当技術分野において知られている。例えば、本ポリペプチドは、膜の脂質二重層との共有相互作用を形成するシングルパスまたはマルチパス膜貫通領域を通して疎水層へ結合させることができる。または、本ポリペプチドは、表面タンパク質との非共有相互作用を通して表面へ結合させることができる。
【0024】
本発明のポリペプチドは、表面結合ポリペプチドであってよい。細胞表面へのトラフィッキングは、細胞の表面へポリペプチドを送達するための細胞内輸送機構についてのインジケータを提供するシグナルペプチドによって実施することができる。そこで、本発明のまた別の実施形態では、本発明のポリペプチドは本ポリペプチドのN末端のシグナルペプチドを含んでいる。
【0025】
本ポリペプチドが疎水性尾部を含むことができることは理解されており、本明細書で企図されている。
【0026】
本ポリペプチドは、柄領域を含むことができる。柄領域は、トレオニン−プロリンリッチ領域を含んでおり、GPIアンカーおよび疎水性尾部とともに、任意で本ポリペプチドの膜結合形で存在する。
【0027】
本発明のポリペプチドの例には、配列番号1〜43、45〜52、54、56、60〜65、68〜72、75、77〜78、81〜119、122〜125、129〜132、134〜144、および146〜155のアミノ酸配列を含むポリペプチドが含まれる。配列には、GenBank受託番号AY577941〜AY578059およびCK988414〜CK988652が含まれる。配列番号1〜20のアミノ酸配列を含む配列は、全長VLRの例を表している。配列番号43のアミノ酸配列を含む配列は、シグナルペプチドを備える全長VLRの例である。さらなる全長VLRおよびそのアミノ酸配列を含むフラグメントは図の中に見いだすことができる。本発明のポリペプチドについて本明細書で教示される構造に基づいて、これらの配列は1つの属のポリペプチドの例であることは理解されるであろう。本発明は全長VLRおよびそのフラグメントを含んでいると理解されたい。
【0028】
開示された組成物を調製するために使用される構成成分ならびに本明細書に開示した方法の中で使用される組成物自体が開示される。これらやその他の材料は本明細書に開示されており、これらの材料の組み合わせ、サブセット、相互作用、グループなどが開示される場合は、これらの化合物の各々の様々な個別および集合的組み合わせおよび順列についての具体的な参照は明示的には開示されないかもしれないが、各々は本明細書に詳細に企図かつ説明されていると理解されたい。例えば、特定のポリペプチドが開示かつ考察され、多数のポリペプチドへ加えることのできる多数の修飾が考察される場合は、ポリペプチドのありとあらゆる組み合わせおよび順列ならびにそれの反対が特別に指示されない限り可能である修飾が特別に企図されている。そこで、1クラスの分子A、B、およびCが開示され、ならびに1クラスの分子D、E、およびFと組み合わせ分子の例A〜Dが開示される場合は、各々が個別に言及されていない場合でさえ、各々が個別に、そしてA−E、A−F、B−D、B−E、B−F、C−D、C−E、およびC−Fの集合的に企図された意味の組み合わせが開示されると考えられる。同様に、これらの任意のサブセットまたは組み合わせもまた開示されている。そこで、例えば、A−E、B−F、およびC−Eのサブグループが開示されると考えられる。この概念は、本明細書に開示した組成物の製造方法および使用方法における工程を含むがそれらに限定されない本出願のすべての態様に当てはまる。そこで、実施できる様々な追加の工程が存在する場合は、これらの追加の工程の各々は本明細書に開示した方法の任意の特定実施形態または実施形態の組み合わせにより実施できると理解されたい。
【0029】
本発明のポリペプチドは、所望の機能を有する。本明細書に記載したポリペプチドは、抗体が選択的に抗原または作用因子へ結合するのとほとんど同様に、抗原または作用因子に選択的へ結合する。ポリペプチドは、任意に可変性リンパ球受容体(天然型または非天然型)またはそのフラグメントもしくは変異体である。用語「可変性リンパ球受容体」は、本明細書では広範囲の意味で使用され、同様に用語「抗体」は様々な特異性を有する様々なバージョンを含んでいる。ポリペプチドは、本明細書に記載したインビトロアッセイを使用してまたは類似方法によってそれらの所望の活性について試験され、その後にそれらの治療的、診断的、またはその他の精製活性が公知の試験方法にしたがって試験される。
【0030】
本発明のポリペプチドは、細胞外物質(例、病原体)または抗原へ結合できる。作用因子または抗原には、ペプチド、ポリペプチド、脂質、糖脂質、およびタンパク質を含むことができるが、それらに限定されない。作用因子または抗原は、病原生物を含むがそれらに限定されない様々な起源に由来することがある。作用因子または抗原への結合は、選択的であると理解されたい。「選択的に結合」または「特異的に結合」は、他の抗原または作用因子を部分的または完全に除外して1つの作用因子または抗原へ結合することを意味する。「結合」は、アッセイ方法のバックグラウンドの少なくとも約1.5倍での検出可能な結合を意味する。選択的または特異的結合については、そのような検出可能な結合は所定の抗原または作用因子については検出できるが、コントロール抗原または作用因子については検出できない。そこで、例えばウイルス、細菌、真菌、または原生動物の抗原または作用因子に選択的に結合するポリペプチドが開示される。
【0031】
ポリペプチドが詳細に開示されるが、前記ポリペプチドは作用因子へ結合し、このとき前記作用因子は病原性作用因子である。さらに、病原性作用因子に選択的に結合する本発明のポリペプチドが開示されるが、このとき前記病原体はウイルスである。多数のウイルスが存在することが知られている。ウイルスは、単純ヘルペスウイルス1型、単純ヘルペスウイルス2型、サイトメガロウイルス、エプスタイン・バーウイルス、水疱・帯状疱疹ウイルス、ヒトヘルペスウイルス6型、ヒトヘルペスウイルス7型、ヒトヘルペスウイルス8型、痘瘡ウイルス、水疱性口内炎ウイルス、A型肝炎ウイルス、B型肝炎ウイルス、C型肝炎ウイルス、D型肝炎ウイルス、E型肝炎ウイルス、ライノウイルス、コロナウイルス、インフルエンザウイルスA型、インフルエンザウイルスB型、麻疹ウイルス、ポリオーマウイルス、ヒトパピローマウイルス、RS(呼吸系合胞体)ウイルス、アデノウイルス、コクサッキーウイルス、デング熱ウイルス、流行性耳下腺炎ウイルス、ポリオウイルス、狂犬病ウイルス、ラウス肉腫ウイルス、黄熱病ウイルス、エボラウイルス、マールブルグウイルス、ラッサ熱ウイルス、東部ウマ脳炎ウイルス、日本脳炎ウイルス、セントルイス脳炎ウイルス、マレーバレー熱(Murray Valley fever)ウイルス、西ナイルウイルス、リフトバレー熱ウイルス、ロタウイルスA型、ロタウイルスB型、ロタウイルスC型、シンドビスウイルス、サル免疫不全ウイルス、ヒトT細胞白血球ウイルス1型、ハンタウイルス、風疹ウイルス、サル免疫不全ウイルス、ヒト免疫不全ウイルス1型、およびヒト免疫不全ウイルス2型からなるウイルス群から選択できる。
【0032】
さらにまた、病原体が細菌である本発明のポリペプチドが開示される。多数の細菌が存在することが知られている。本明細書では、病原体に選択的に結合するポリペプチドが特別に企図かつ開示されるが、このとき前記病原体は結核菌(M.tuberculosis)、ウシ型結核菌(M.bovis)、ウシ型結核菌株BCG、BCG亜株、トリ型結核菌(M.avium)、マイコバクテリウム・イントラセルラーレ(M.intracellulare)、マイコバクテリウム・アフリカヌム(M.africanum)、マイコバクテリウム・カンサシイ(M.kansasii)、マイコバクテリウム・マリナム(M.marinum)、マイコバクテリウム・ウルセランス(M.ulcerans)、トリ型結核菌亜種パラ結核菌(paratuberculosis)、ノカルジア・アステオロイデス(Nocardia asteroides)、その他のノカルジア種、レジオネラ・ニューモフィラ菌(Legionella pneumophila)、その他のレジオネラ種、ネズミチフス菌(Salmonella typhi)、その他のサルモネラ種、赤痢菌種(Shigella species)、ペスト菌(Yersinia pestis)、パスツレラ・ヘモリチカ(Pasteurella haemolytica)、動物パスツレラ症病原菌(Pasteurella multocida)、その他のパスツレラ種、アクチノバチルス胸膜肺炎菌(Actinobacillus pleuropneumoniae)、リステリア・モノサイトゲネス(Listeria monocytogenes)、リステリア・イヴァノビ(Listeria ivanovii)、ブルセラ・アボルツス(Brucella abortus)、その他のブルセラ種、コードリア・ルミナンチウム(Cowdria ruminantium)、クラミジア肺炎菌(Chlamydia pneumoniae)、トラコーマ・クラミジア(Chlamydia trachomatis)、オーム病病原体(Chlamydia psittaci)、コクシエラ・バーネッティイ(Coxiella burnetti)、その他のリケッチア(Rickettsial)種、エールリヒア(Ehrlichia)種、黄色ブドウ球菌(Staphylococcus
aureus)、表皮ブドウ球菌(Staphylococcus epidermidis)、化膿性連鎖球菌(Streptococcus pyogenes)、ストレプトコッカス・アガラクティエ(Streptococcus agalactiae)、炭疽菌(Bacillus anthracis)、大腸菌(Escherichia
coli)、コレラ菌(Vibrio cholerae)、カンピロバクター(Campylobacter)種、髄膜炎菌(Neiserria meningitidis)、淋菌(Neiserria gonorrhea)、緑膿菌(Pseudomonas aeruginosa)、その他のシュードモナス種、インフルエンザ菌(Haemophilus influenzae)、軟性下疳菌(Haemophilus ducreyi)、その他のヘモフィルス(Hemophilus)種、破傷風菌(Clostridium tetani)、その他のクロストリジウム(Clostridium)種、エルシニア・エンテロコリチカ(Yersinia enterocolitica)、およびその他のエルシニア種からなる細菌リストから選択される細菌である。
【0033】
さらに、病原体に選択的に結合する本発明のポリペプチドが開示されるが、前記病原体は原生動物または他の寄生虫である。多数の寄生虫性感染症が存在することが知られている。本明細書では病原体に選択的に結合するポリペプチドが特別に企図かつ開示されるが、前記病原体は、トキソプラズマ原虫(Toxoplasma gondii)、熱帯熱マラリア原虫(Plasmodium falciparum)、三日熱マラリア原虫(Plasmodium vivax)、四日熱マラリア原虫(Plasmodium malariae)、その他のプラズモジウム(Plasmodium)種、トリパノソーマ・ブルース(Trypanosoma brucei)、トリパノソーマ・クルーズ(Trypanosoma cruzi)、森林型熱帯リーシュマニア(Leishmania major)、その他のリーシュマニア(Leishmania)種、マンソン住血吸虫(Schistosoma mansoni)、その他の住血吸虫(Schistosoma)種、および赤痢アメーバ(Entamoeba histolytica)からなる群から選択される寄生虫性感染である。
【0034】
さらに、病原体に選択的に結合する本発明のポリペプチドが開示されるが、前記病原体は真菌である。多数の真菌が存在することが知られている。本明細書には特別にポリペプチドが企図かつ開示されているが、このとき病原体は、カンジダ・アルビカンス(Candida albicans)、クリプトコックス・ネオホルマンス(Cryptococcus neoformans)、ヒストプラズマ・カプスラーツ(Histoplasma capsulatum)、アスペルギルス・フミガーツス(Aspergillus fumigatus)、コクシジオイデス・イミティス(Coccidiodes
immitis)、ブラジル・パラコクシジオイデス(Paracoccidiodes brasiliensis)、ブラストミセス・デルマティディス(Blastomyces dermitidis)、カリニ肺炎(Pneomocystis carnii)、ペニシリウム症(Penicillium marneffi)、およびアルテルナリア・アルテルナータ(Alternaria alternata)からなる真菌群から選択される真菌である。
【0035】
本発明のポリペプチドは、毒素にもまた選択的に結合できる。本明細書では、「毒素」は、それ(毒素)が接触した任意の細胞を効果的に破壊する任意の化学的または生物学的物質を意味する。毒素の著明な例には、リシン、百日咳毒素、サリン、細菌内毒素、中毒性ショック症候群毒素1、コレラ毒素、およびヘビ毒毒素が挙げられる。そこで、毒素へ結合するポリペプチドが特別に開示される。
【0036】
本明細書に記載したポリペプチドは、所望の機能が維持される限り修飾および変化させることができる。任意の変異体および誘導体または本明細書に開示した遺伝子およびタンパク質から発生する可能性のある変異体および誘導体を規定する1つの方法は、特異的な既知配列との相同性によって変異体および誘導体を規定することによると理解されたい。例えば、配列番号1は、本発明の任意の数の核酸によってコードされたポリペプチドの特定アミノ酸配列を規定している。特に開示されているのは、上記の配列に対して少なくとも70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、または99パーセントの相同性を有する本明細書に開示されたこれらや他の遺伝子およびタンパク質の変異体である。当業者であれば、2つのタンパク質または核酸(例えば、遺伝子)の相同性を決定する方法を容易に理解できる。例えば、相同性は、相同性がその最高レベルにあるように2つの配列をアライメントした後に計算できる。
【0037】
一般に、任意の公知の変異体および誘導体または本明細書に開示した遺伝子およびタンパク質から発生する可能性のある変異体および誘導体を規定する1つの方法は、特異的な既知配列との相同性によって変異体および誘導体を規定することによる方法であると理解されたい。本明細書に開示した特定配列のこの同一性については、本明細書の他の場所においても考察する。一般に、本明細書に開示した遺伝子およびタンパク質の変異体は、典型的には、上記の配列または天然配列に対して少なくとも約70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、または99パーセントの相同性を有する。当業者であれば、2つのタンパク質または核酸(例えば、遺伝子)の相同性を決定する方法を容易に理解できる。例えば、相同性は、相同性がその最高レベルにあるように2つの配列をアライメントした後に計算できる。
【0038】
相同性を計算するまた別の方法は、発表されたアルゴリズムによって実施できる。比較のための配列の最適アライメントは、SmithおよびWaterman Adv.Appl.Math.2:482(1981)の局所相同性アルゴリズムによって、NeedlemanおよびWunsch,J.MoL Biol.48:443(1970)の相同性アライメントアルゴリズムによって、PearsonおよびLipman,Proc.Natl.Acad.Sci.U.S.A.85:2444(1988)の類似性検索方法によって、これらのアルゴリズムのコンピュータを使用した実行(GAP、BESTFIT、FASTA、およびTFASTA、Wisconsin Genetics Software Package,Genetics Computer Group,575 Science Dr.、ワイオミング州マディソン)によって、または目視検査によって実施できる。
【0039】
核酸についての同一タイプの相同性は、例えば少なくとも核酸アライメントに関する材料について参照して本明細書に組み込まれるZuker,M.Science 244:48−52,1989,Jaeger et al.Proc.Natl.Acad.Sci.USA 86:7706−7710,1989,Jaeger et al.,Methods Enzymol.183:281−306,1989の中で開示されたアルゴリズムによって入手できる。典型的にはこれらの方法のいずれかを使用できること、そして所定の場合にはこれらの様々な方法の結果は相違することがあるが、当業者であれば、これらの方法の少なくとも1つを用いて同一性が見いだされると、それらの配列は規定された同一性を有しており、本明細書に開示されていると理解することができることが理解されよう。
【0040】
例えば、本明細書で使用するような他の配列との特定パーセンテージの相同性を有すると言及された配列は、任意の1つまたは複数の上記に記載した計算方法によって計算された言及された相同性を有する配列を意味する。例えば、第1配列は、第1配列がZuker計算法を用いて第2配列と80%相同性を有すると計算された場合は、たとえ第1配列が他の任意の計算方法によって計算した場合に第2配列との80%相同性を有していない場合でさえ、本明細書に規定したように第2配列との80%相同性を有する。また別の例として、第1配列は、第1配列がZuker計算法およびPearsonおよびLipman計算法の両方を用いて第2配列と80%相同性を有すると計算された場合は、たとえ第1配列がSmithおよびWaterman計算法、NeedlemanおよびWunsch計算法、Jaeger計算法、または任意の他の計算方法によって計算した場合に第2配列との80%相同性を有していない場合でさえ、本明細書に規定したように第2配列との80%相同性を有する。さらにまた別の例として、第1配列は、第1配列が各計算法を用いて第2配列と80%相同性を有すると計算された場合は、本明細書に規定したように第2配列との80%相同性を有する(だが、実際に様々な計算法はしばしば相違する計算相同性パーセンテージを生じさせるであろう)。
【0041】
タンパク質の変異体および誘導体は、当業者には明確に理解されており、アミノ酸配列修飾を含むことができる。例えば、アミノ酸配列の修飾は、3つのクラス:置換、挿入または欠失変異体の1つまたは複数に分類される。挿入には、単一または複数アミノ酸残基のアミノおよび/またはカルボキシル末端融合ならびに配列内挿入が含まれる。挿入は、通常は、アミノまたはカルボキシル末端融合の挿入より少ない挿入、例えばおよそ1〜4個の残基であろう。実施例に記載された誘導体などの免疫原性融合タンパク質誘導体は、標的配列へ免疫原性を付与するために十分に大きなポリペプチドをインビトロ架橋結合によって融合させる工程によって、またはその融合をコードするDNAを用いて形質転換された組み換え細胞培養によって作製される。欠失は、タンパク質配列からの1つまたは複数のアミノ酸残基の除去を特徴とする。典型的には、約2〜6未満の残基がタンパク質分子内の任意の1つの部位で欠失している。これらの変異体は、通常はタンパク質をコードするDNA内のヌクレオチドの部位特異的突然変異によって調製され、それによってその変異体をコードするDNAが産生され、その後に組み換え細胞培養内でDNAが発現させられる。知られている配列を有するDNA内の規定部位で置換突然変異を作製するための技術、例えばM13プライマー突然変異誘発およびPCR突然変異誘発はよく知られている。アミノ酸置換は、典型的には単一残基の置換であるが、1回に多数の相違する場所で発生してもよい;挿入は、通常はおよそ約1〜10アミノ酸残基であろう;また欠失は約1〜30残基の範囲に及ぶであろう。欠失または挿入は、好ましくは隣接対、すなわち2残基の欠失または2残基の挿入で作製される。置換、欠失、挿入またはそれらの任意の組み合わせは、最終構築物に達するために組合わせることができる。突然変異は、配列をリーディングフレームの外に配置してはならず、好ましくは二次mRNA構造を生成できる相補的領域を作製しないであろう。置換変異体は、少なくとも1つの残基が取り除かれており、異なる残基がその場所に挿入されている変異体である。そのような置換は、一般の表1および2にしたがって作製され、保存的置換と言われる。
【0042】
【表1】
【0043】
【表2】
機能または免疫学的同一性の実質的な変化は、表2に記載の置換より保存的ではない置換を選択することによって、すなわち(a)例えばシートまたはヘリックス立体構造のような、置換領域におけるポリペプチド骨格の構造、(b)標的部位での分子の電荷もしくは疎水性、または(c)側鎖のかさ、を維持する作用がより大きく相違する残基を選択する工程によってなされる。一般にタンパク質特性における最大の変化を生じさせると予測される置換は、(a)例えばセリルもしくはトレオニルのような親水性残基が、例えばロイシル、イソロイシル、フェニルアラニル、バリルもしくはアラニルのような疎水性残基と(によって)置換されている;(b)システインもしくはプロリンが任意の他の残基と(によって)置換されている;(c)例えばリシル、アルギニル、もしくはヒスチジルのような陽性側鎖を有する残基が、例えばグルタミルもしくはアスパルチルのような陰性残基と(によって)置換されている;(d)例えばフェニルアラニンのようなかさの大きな側鎖を有する残基が、この場合にはグリシンのような側鎖を有していない残基と(によって)置換されている、または(e)硫酸化および/またはグリコシル化のための部位数を増加させる、ことによる置換であろう。
【0044】
例えば、1つのアミノ酸残基と生物学的および/または化学的に類似する別のアミノ酸残基との置換は、当業者には保存的置換として知られている。例えば、保存的置換は1つの疎水性残基と別の疎水性残基とを置換させ、または1つの極性残基と他の極性残基とを置換させるであろう。置換には、例えば、GIy、Ala;Val、Ile、Leu;Asp、Glu;Asn、Gln;Ser、Thr;Lys、Arg;およびPhe、Tyrなどの組み合わせが挙げられる。各々明示的に開示された配列のそのような保存的に置換された変形は、本明細書に提供したモザイクポリペプチドの範囲内に含まれる。
【0045】
置換または欠失突然変異誘発は、N−グリコシル化(Asn−X−Thr/Ser)またはO−グリコシル化(SerまたはThr)のための部位を挿入するために使用できる。システインまたはその他の易変残基の欠失もまた望ましいことがある。潜在的タンパク質分解部位、例えばArgの欠失または置換は、例えば塩基性残基の1つを欠失させる工程によって、または1つをグルタミニル残基もしくはヒスチジル残基で置換する工程によって遂行される。
【0046】
所定の翻訳後誘導体化は、発現したポリペプチド上の組み換え宿主細胞の作用の結果である。グルタミニル残基およびアスパラギニル残基は、しばしば対応するグルタミル残基およびアスパルチル残基へ翻訳後に脱アミド化される。または、これらの残基は弱酸性条件下では脱アミド化される。その他の翻訳後修飾にはプロリンおよびリシンの水酸化、セリルもしくはトレオニル残基のヒドロキシル基のリン酸化、リシン側鎖、アルギニン側鎖、およびヒスチジン側鎖のo−アミノ基のメチル化(T.E.Creighton,Proteins:Structure and Molecular Properties,W.H.Freeman & Co.,San Francisco pp79−86[1983])、N末端アミンのアセチル化、および一部の例では、C末端カルボキシルのアミド化が挙げられる。
【0047】
本明細書で使用する用語「可変性リンパ球受容体」は、被験者に投与されると減少した免疫原性を有するように修飾されているポリペプチドも意味することがある。例えば、ヒトアミノ酸配列は、抗体がヒト化されるのとほとんど同様に、ヒト被験者にとって低免疫原性のバージョンにするためにポリペプチドに挿入または追加することができる。多数の非ヒト可変性リンパ球受容体(例、ヤツメウナギ、マウス、ラット、またはウサギに由来するもの)は、ヒトにおいて天然抗原性である可能性があり、したがってヒトへ投与すると望ましくない免疫応答を発生させることがある。このため、本発明の方法における修飾されたポリペプチドの使用は、ヒトに投与されたポリペプチドが望ましくない免疫応答を惹起する可能性を低下させるために役立つことがある。
【0048】
修飾技術は、可変性リンパ球受容体分子の1つまたは複数のポリペプチド領域をコードするDNA配列を操作するための組み換えDNAテクノロジーの使用を含むことができる。したがって、可変性リンパ球受容体(またはそのフラグメント)のヒト化形は、キメラ可変性リンパ球受容体、好ましくはヒト(レシピエント)アミノ酸配列に組み込まれた、非ヒト(ドナー)可変性リンパ球受容体由来の抗原(作用因子)結合部位を含有する可変性リンパ球受容体の抗原(作用因子)結合部分である。
【0049】
これらのタンパク質配列、その変異体およびフラグメントをコードできる核酸もまた開示されることを理解されたい。これには、特異的タンパク質配列に関連するすべての縮重配列、すなわち1つの特定タンパク質配列をコードする配列を有するすべての核酸、ならびに本明細書に開示したタンパク質配列の変異体および誘導体をコードする縮重核酸を含むすべての核酸が含まれるであろう。そこで、各特定核酸配列は本明細書に書き出されない可能性があるが、ありとあらゆる配列が開示されたタンパク質配列を通して本明細書に実際に開示かつ記載されていると理解されたい。
【0050】
ヒト化可変性リンパ球受容体はさらにまた、レシピエントの可変性リンパ球受容体にも取込まれたヒト配列内にも見いだされないアミノ酸配列を含有することがある。
【0051】
本発明のポリペプチドは、融合タンパク質を作製するためにも使用できる。これらのポリペプチドは、融合タンパク質におけるターゲッティング機能に役立つことができる。そこで本発明のポリペプチドは、第2成分へ、組み換え技術によってコンジュゲートするか、さもなければ連結することができる。第2成分は、例えば細胞殺滅が所望であれば、毒素を含むことができる。そこで、例えば原生動物に選択的に結合するポリペプチドは、原生動物を標的とすることができ、融合タンパク質の毒素成分は細胞を殺滅できる。同様に、本発明のポリペプチドは送達機能を実行できる。そこで第2成分は治療薬であってよい。
【0052】
本発明のポリペプチドは、検出可能なタグに連結させることができる。「検出可能なタグ」は、インビボまたはインビトロでイメージング法または検出法を用いて視認できる任意のタグである。検出可能なタグは、放射線不透過性物質、放射標識、化学発光標識、蛍光標識、または磁気標識であってよい。検出可能なタグは、γ線エミッター、β線エミッター、α線エミッター、γ線エミッター、ポジトロンエミッター、X線エミッターおよび蛍光エミッターからなる群から選択できる。適切な蛍光化合物には、フルオレセインナトリウム、フルオレセインイソチオシアネート、フィコエリトリン、およびテキサスレッドスルホニルクロリド、アロフィコシアニン(APC)、Cy5−PE、CY7−APC、およびカスケードイエローが挙げられる。
【0053】
標識のために適合する放射性同位体には、ヨウ素131、ヨウ素123、ヨウ素125、ヨウ素126、ヨウ素133、臭素77、インジウム111、インジウム113m、ガリウム67、ガリウム68、ルテニウム95、ルテニウム97、ルテニウム103、ルテニウム105、水銀107、水銀203、レニウム99m、レニウム105、レニウム101、テルル121m、テルル122m、テルル125m、ツリウム165、ツリウム167、ツリウム168、テクネチウム99mおよびフッ素18が挙げられる。
【0054】
任意で、検出可能なタグは、組織化学的技術、ELISA様アッセイ、共焦点顕微鏡、蛍光検出法、セルソーティング法、核磁気共鳴法、ラジオイムノシンチグラフィー、X線検査、陽電子放射断層撮影法、コンピュータ体軸断層撮影法、磁気共鳴イメージング法、および超音波検査法を用いて視認できる。
【0055】
または、ポリペプチドをビオチニル化して、蛍光標識ストレプトアビジンのようなその後に検出可能な標識を使用するとポリペプチドを間接的に検出できる。ビオチンは、当技術分野において知られている数種の技術のいずれかによって検出される。例えば、ビオチンは蛍光標識アビジンと結合する工程によって検出可能であり、アビジンは各結合事象に関連するシグナルを増加させるためにフィコエリトリンまたは連鎖蛍光標識を用いて標識される。
【0056】
任意で、ポリペプチドはスライド、培養皿、マルチウエルプレート、カラム、チップ、アレイまたは安定性ビーズなどの固体支持体へ結合させられる。「アレイ」には、マイクロプレートまたはスライドなどの1つまたは複数のマルチウエルアレイ化手段が挙げられる。
【0057】
任意で、ポリペプチドは、例えばセルソーティングテクノロジーを用いてソーティングできるビーズなどの可動性固体支持体へ結合させられる。「可動性固体支持体」とは、1セットの識別可能に標識されたマイクロスフェアまたはビーズを意味する。好ましくは、マイクロスフェアは、ポリスチレン・ジビニルベンゼンビーズである。特異的蛍光色素でマークされ、特異的蛍光プロフィールを有するマイクロスフェアのセットは、例えばLuminex Corporation(テキサス州オースティン)から市販で入手できる。
【0058】
本発明は、さらに複数の本発明のポリペプチドを提供する。任意で、ポリペプチドのLRRは、ポリペプチド全体にわたり高度に可変性である。そこで、大多数は、内部LRRの変動性に基づいて、相違する結合特異性を備えるポリペプチドを含むことができる。
【0059】
さらにまた、本発明のポリペプチドを備える容器または本発明のポリペプチドを備える固定性固体支持体もしくは可動性固体支持体を含むキットもまた提供される。任意で、ポリペプチドは固体支持体またはキットへ結合させられる。任意で、キットは、ポリペプチド、固体支持体、およびポリペプチドを固体支持体へ結合させるための連結手段を含有している。
【0060】
本発明は、本発明のポリペプチドをコードする単離された核酸を提供する。そのような核酸の1つの例は、配列番号156のヌクレオチド配列(代表的VLRのORF)を含んでいる。VLRまたはそのフラグメントをコードする核酸の他の例には、配列番号44、配列番号53〜55、配列番号57〜59、配列番号66〜67、配列番号73〜74、配列番号76、配列番号79〜80、および配列番号172〜302が挙げられる。VLR遺伝子に関連する種々の配列には、Genbank受託番号AY57791〜AY578059、AY964719〜AY964931、AY965520〜AY965612、AY965658〜AY965681、およびCK988414〜CK988652を有する様々な配列がある。これらの配列は、全体が、ならびにその中に含まれる個別のサブ配列(領域またはフラグメント)が参照として本明細書に援用される。
【0061】
そのような核酸配列は、一例として核酸の属によって提供されるが、限定することは意図されていない。さらに、これらの核酸を含む発現ベクターも提供されるが、このとき核酸は発現制御配列へ機能的に連結している。さらに発現ベクターを含む培養細胞も提供される。そのような発現ベクターおよび培養細胞を使用すると、本発明のポリペプチドを作製できる。
【0062】
本明細書では、例えばVLRまたはそのフラグメントもしくは変異体をコードする核酸を含む、核酸に基づく様々な分子が開示されている。開示された核酸は、ヌクレオチド、ヌクレオチドアナログ、またはヌクレオチド置換基から作製される。
【0063】
ヌクレオチドアナログは、塩基、糖、またはリン酸塩成分のいずれかへの一部のタイプの修飾を含有するヌクレオチドである。塩基成分に対する修飾は、A、C、G、およびT/Uの天然および合成修飾、ならびにウラシル−5−イル(.psi.)、ヒポキサンチン−9−イル(I)、および2−アミノアデニン−9−イルなどの様々なプリンもしくはピリミジン塩基の天然および合成修飾を含む。修飾された塩基には、5−メチルシトシン(5−me−C)、5−ヒドロキシメチルシトシン、キサンチン、ヒポキサンチン、2−アミノアデニン、アデニンおよびグアニンの6−メチルおよびその他のアルキル誘導体、アデニンおよびグアニンの2−プロピルおよびその他のアルキル誘導体、2−チオウラシル、2−チオチミンおよび2−チオシトシン、5−ハロウラシルおよび5−ハロシトシン、5−プロピニルウラシルおよび5−プロピニルシトシン、6−アゾウラシル、6−アゾシトシンおよび6−アゾチミン、5−ウラシル(プソイドウラシル)、4−チオウラシル、8−ハロ、8−アミノ、8−チオール、8−チオアルキル、8−ヒドロキシルならびに他の8−置換のアデニンおよびグアニン、5−ハロ、特に5−ブロモ、5−トリフルオロメチルおよび他の5−置換のウラシルおよびシトシン、7−メチルグアニンおよび7−メチルアデニン、8−アザグアニンおよび8−アザアデニン、7−デアザグアニンおよび7−デアザアデニン、ならびに3−デアザグアニンおよび3−デアザアデニンが挙げられるが、それらに限定されない。追加の塩基修飾は、例えば米国特許第3,687,808号、Englisch et al.,Angewandte Chemie,International Edition,1991,30,613、およびSanghvi,Y.S.,Chapter 15,Antisense Research and Applications,pages 289−302,Crooke,S.T.およびLebleu,B.編,CRC Press,1993に見いだすことができる。5−置換ピリミジン、6−アザピリミジンなどの所定のヌクレオチドアナログならびに2−アミノプロピルアデニン、5−プロピニルウラシルおよび5−プロピニルシトシン、5−メチルシトシンを含むN−2、N−6およびO−6置換プリンは、二本鎖形成の安定性を増加させることができる。多くの場合、塩基修飾は例えば、二本鎖安定性上昇などの独特の特性を達成するために、2’−O−メトキシエチルなどの糖修飾と組み合わせることができる。広範囲の塩基修飾について詳述している第4,845,205号;第5,130,302号;第5,134,066号;第5,175,273号;第5,367,066号;第5,432,272号;第5,457,187号;第5,459,255号;第5,484,908号;第5,502,177号;第5,525,711号;第5,552,540号;第5,587,469号;第5,594,121号;第5,596,091号;第5,614,617号;および第5,681,941号などの多数の米国特許がある。これらの各特許は、参照して本明細書に組み込まれる。
【0064】
ヌクレオチドアナログは、さらに糖成分の修飾を含むことができる。糖部分の修飾は、リボースおよびデオキシリボースの天然修飾ならびに合成修飾を含むであろう。糖修飾は、2’位での以下の修飾を含むがそれらに限定されない:OH;F;O−、S−、もしくはN−アルキル;O−、S−、もしくはN−アルケニル;O−、S−もしくはN−アルキニル;またはO−アルキル−O−アルキル。このときアルキル、アルケニルおよびアルキニルは置換または未置換C1〜C10アルキルまたはC2〜C10アルケニルおよびアルキニルであってよい。2’糖修飾には−O[(CH2)nO]mCH3、−O(CH2)nOCH3、−O(CH2)nNH2、−O(CH2)nCH3、−O(CH2)n−ONH2、および−O(CH2)nON[(CH2)nCH3)]2(式中、nおよびmは1〜約10である)が含まれるがそれらに限定されない。
【0065】
2’位でのその他の糖修飾には:C1〜C10低級アルキル、置換低級アルキル、アルカリル、アラルキル、O−アルカリルもしくはO−アラルキル、SH、SCH3、OCN、Cl、Br、CN、CF3、OCF3、SOCH3、SO2CH3、ONO2、NO2、N3、NH2、ヘテロシクロアルキル、ヘテロシクロアルカリル、アミノアルキルアミノ、ポリアルキルアミノ、置換シリル、RNA開裂基、レポーター基、インターカレーター、オリゴヌクレオチドの薬物動態特性を改良するための基、またはオリゴヌクレオチドの薬力学的特性を改良するための基、および類似の特性を有する他の置換基が含まれるがそれらに限定されない。類似の修飾は、糖の上の他の位置、特に3’末端ヌクレオチド上または2’〜5’結合オリゴヌクレオチド内の糖の3’位、および5’末端ヌクレオチドの5’位で行なうことができる。修飾された糖は、例えばCH2およびSなどの架橋環状酸素での修飾を含有する糖も含むであろう。ヌクレオチド糖アナログは、ペントフラノシル糖の代わりにシクロブチル成分などの糖ミメティックもまた有する可能性がある。そのような修飾された糖構造の調製を教示する第4,981,957号;第5,118,800号;第5,319,080号;第5,359,044号;第5,393,878号;第5,446,137号;第5,466,786号;第5,514,785号;第5,519,134号;第5,567,811号;第5,576,427号;第5,591,722号;第5,597,909号;第5,610,300号;第5,627,053号;第5,639,873号;第5,646,265号;第5,658,873号;第5,670,633号;および第5,700,920号などの多数の米国特許が存在しており、それらは各々全体として参照して本明細書に組み込まれる。
【0066】
ヌクレオチドアナログは、さらにリン酸塩成分で修飾することもできる。修飾されたリン酸塩成分には、2つのヌクレオチド間の連結がホスホロチオエート、キラルホスホロチオエート、ホスホロジチオエート、ホスホトリエステル、アミノアルキルホスホトリエステル、メチルならびに3’−アルキレンホスホネートおよびキラルホスホネートを含む他のアルキルホスホネート、ホスフィネート、3’−アミノホスホルアミデートおよびアミノアルキルホスホルアミデート、チオホスホルアミデートを含むホスホルアミデート、チオノアルキルホスホネート、チオノアルキルホスホトリエステル、およびボラノホスフェートを含むように修飾できる成分が含まれるがそれらに限定されない。これらのホスフェートまたは2つのヌクレオチド間の修飾ホスフェート結合は3’−5’結合または2’−5’結合を通して行なうことができ、この結合は3’−5’から5’−3’または2’−5’から5’−2’などの逆極性を含有できることを理解されたい。様々な塩、混合塩および遊離酸形もまた含まれる。多数の米国特許が修飾リン酸エステルを含有するヌクレオチドの作製方法および使用方法を教示しており、それらにはその各々が参照して本明細書に組み込まれる第3,687,808号;第4,469,863号;第4,476,301号;第5,023,243号;第5,177,196号;第5,188,897号;第5,264,423号;第5,276,019号;第5,278,302号;第5,286,717号;第5,321,131号;第5,399,676号;第5,405,939号;第5,453,496号;第5,455,233号;第5,466,677号;第5,476,925号;第5,519,126号;第5,536,821号;第5,541,306号;第5,550,111号;第5,563,253号;第5,571,799号;第5,587,361号;および第5,625,050号が含まれるがそれらに限定されない。
【0067】
ヌクレオチドアナログは必ずしも単一修飾を含有している必要はなく、成分の1つの中または相違する成分間に複数の修飾を含有していてよいことも理解されたい。
【0068】
ヌクレオチド置換基は、ヌクレオチドと類似の機能的特性を有する分子であるが、ペプチド核酸(PNA)などはリン酸エステル成分を含有していない。ヌクレオチド置換基は、Watson−Crick様式またはHoogsteen様式で核酸を認識するであろうが、リン酸エステル成分以外の成分を通して連結される分子である。ヌクレオチド置換基は、適切な標的核酸と相互作用すると二本鎖型構造に適合することができる。
【0069】
ヌクレオチド置換基は、リン酸エステル成分および/または糖成分が置換されているヌクレオチドまたはヌクレオチドアナログである。ヌクレオチド置換基は、標準的なリン原子を含有していない。リン酸エステルに対する置換基は、例えば、短鎖アルキルもしくはシクロアルキルヌクレオシド間結合、混合ヘテロ原子およびアルキルもしくはシクロアルキルヌクレオシド間結合、または1つまたは複数の短鎖ヘテロ原子もしくは複素環ヌクレオシド間結合であってよい。これらには、モルホリノ結合(一部にはヌクレオシドの糖部分から形成される);シロキサン骨格;硫化物、スルホキシドおよびスルホン骨格;ホルムアセチルおよびチオホルムアセチル骨格;メチレンホルムアセチルおよびチオホルムアセチル骨格;アルケン含有骨格;スルファメート骨格;メチレンイミノおよびメチレンヒドラジノ骨格;スルホン酸エステル骨格およびスルホンアミド骨格;アミド骨格;ならびに混合N、O、SおよびCH2成分部分を有するその他が含まれる。多数の米国特許がリン酸エステル置換基の作製方法および使用方法を教示しており、それらにはその各々が参照して本明細書に組み込まれる第5,034,506号;第5,166,315号;第5,185,444号;第5,214,134号;第5,216,141号;第5,235,033号;第5,264,562号;第5,264,564号;第5,405,938号;第5,434,257号;第5,466,677号;第5,470,967号;第5,489,677号;第5,541,307号;第5,561,225号;第5,596,086号;第5,602,240号;第5,610,289号;第5,602,240号;第5,608,046号;第5,610,289号;第5,618,704号;第5,623,070号;第5,663,312号;第5,633,360号;第5,677,437号;および第5,677,439号が含まれるがそれらに限定されない。
【0070】
さらにヌクレオチド置換基ではヌクレオチドの糖およびリン酸部分の両方の成分が、例えばアミド型結合(アミノエチルグリシン)(PNA)によって置換されてよいことを理解されたい。各々が参照して本明細書に組み込まれる米国特許第5,539,082号;第5,714,331号;および第5,719,262号は、PNA分子の製造方法および使用方法を教示している。(さらに、Nielsen et al.,Science,1991,254,1497−1500も参照されたい)。
【0071】
例えば細胞取り込みを強化させるために、他のタイプの分子(コンジュゲート)をヌクレオチドまたはヌクレオチドアナログへ結合させることも可能である。コンジュゲートは、ヌクレオチドまたはヌクレオチドアナログへ化学的に結合させることができる。そのようなコンジュゲートには、コレステロール成分などの脂質成分(Letsinger et al.,Proc.Natl.Acad.Sci.USA,1989,86,6553−6556)、コール酸(Manoharan et al.,Bioorg.Med.Chem.Let.,1994,4,1053−1060)、チオエーテル、例えばヘキシル−S−トリチルチオール(Manoharan et al.,Ann.N.Y.Acad.Sci.,1992,660,306−309;Manoharan et al.,Bioorg.Med.Chem.Let.,1993,3,2765−2770)、チオコレステロール(Oberhauser et al.,Nucl.Acids Res.,1992,20,533−538)、脂肪族鎖、例えばドデカンジオールもしくはウンデシル残基(Saison−Behmoaras et al.,EMBO
J,1991,10,1111−1118;Kabanov et al,FEBS Lett.,1990,259,327−330;Svinarchuk et al.,Biochimie,1993,75,49−54)、リン脂質、例えばジ−ヘキサデシル−rac−グリセロールもしくはトリエチルアンモニウム1,2−ジ−O−ヘキサデシル−rac−グリセロ−3−H−ホスホン酸塩(Manoharan et al.,Tetrahedron Lett.,1995,36,3651−3654;Shea
et al.,Nucl.Acids Res.,1990,18,3777−3783)、ポリアミンもしくはポリエチレングリコール鎖(Manoharan et al.,Nucleosides & Nucleotides,1995,14,969−973)、またはアダマンタン酢酸(Manoharan et al.,Tetrahedron Lett.,1995,36,3651−3654)、パルミチル成分(Mishra et al.,Biochem.Biophys.Acta,1995,1264,229−237)、またはオクタデシルアミンもしくはヘキシルアミノ−カルボニル−オキシコレステロール成分(Crooke et al.,J.Pharmacol.Exp.Ther.,1996,277,923−937が含まれるがそれらに限定されない。多数の米国特許がそのようなコンジュゲートの調製を教示しており、それらには各々が参照して本明細書に組み込まれる米国特許第4,828,979号;第4,948,882号;第5,218,105号;第5,525,465号;第5,541,313号;第5,545,730号;第5,552,538号;第5,578,717,第5,580,731号;第5,580,731号;第5,591,584号;第5,109,124号;第5,118,802号;第5,138,045号;第5,414,077号;第5,486,603号;第5,512,439号;第5,578,718号;第5,608,046号;第4,587,044号;第4,605,735号;第4,667,025号;第4,762,779号;第4,789,737号;第4,824,941号;第4,835,263号;第4,876,335号;第4,904,582号;第4,958,013号;第5,082,830号;第5,112,963号;第5,214,136号;第5,082,830号;第5,112,963号;第5,214,136号;第5,245,022号;第5,254,469号;第5,258,506号;第5,262,536号;第5,272,250号;第5,292,873号;第5,317,098号;第5,371,241号、第5,391,723号;第5,416,203号、第5,451,463号;第5,510,475号;第5,512,667号;第5,514,785号;第5,565,552号;第5,567,810号;第5,574,142号;第5,585,481号;第5,587,371号;第5,595,726号;第5,597,696号;第5,599,923号;第5,599,928号;および第5,688,941号が含まれるがそれらに限定されない。
【0072】
VLR遺伝子、または匹敵する遺伝子と相互作用できるプライマーおよびプローブを含む組成物が開示される。所定の実施形態では、プライマーはDNA増幅反応を支援するために使用される。典型的には、プライマーは配列特異的方法で延長させることができるであろう。配列特異的方法でのプライマーの伸長には、それにプライマーがハイブリダイズされる、さもなければ関連付けられる核酸分子の配列および/または組成がプライマーの伸長によって生成する生成物の組成または配列を指示する、または影響を及ぼす任意の方法が含まれる。このため配列特異的方法でのプライマーの伸長には、PCR、DNAシーケンシング、DNA伸長、DNA重合、RNA転写、または逆転写が含まれるがそれらに限定されない。配列特異的方法でプライマーを増幅させる技術および条件が好ましい。所定の実施形態では、プライマーはPCRまたは直接的シーケンシングなどのDNA増幅反応のために使用される。所定の実施形態では、プライマーはさらにまた非酵素的技術を用いて伸長させることもできることを理解されたいが、このとき例えばプライマーを伸長させるために使用されるヌクレオチドもしくはオリゴヌクレオチドは、それらが配列特異的方法でプライマーを伸長させるように化学的に反応するように修飾させられる。
【0073】
所定の実施形態においてVLR遺伝子と相互作用するためのプライマーもしくはプローブのサイズは、DNA増幅などのプライマーの所望の酵素的操作またはプローブもしくはプライマーの単純なハイブリダイゼーションを支援する任意のサイズであってよい。典型的なVLRプライマーもしくはプローブは、少なくとも6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、125、150、175、200、225、250、275、300、325、350、375、400、425、450、475、500、550、600、650、700、750、800、850、900、950、1000、1250、1500、1750、2000、2250、2500、2750、3000、3500、または4000ヌクレオチド長であろう。
【0074】
本発明のポリペプチドおよび核酸は、様々な技術において使用できる。例えば、本ポリペプチドは選択された作用因子を検出するため、選択された作用因子の活性を遮断するため、作用因子を精製するために、イメージングツールとして、および治療薬として使用できる。
【0075】
本明細書では、サンプル中の作用因子を検出する方法であって、ポリペプチドがサンプル中の作用因子へ結合できる条件下でサンプルをポリペプチドと接触させる工程、およびサンプル中の作用因子へ結合したポリペプチドを検出する工程、を含む方法が提供される。結合したポリペプチドは、サンプル中に作用因子が含まれることを示す。検出方法は、当技術分野においてよく知られている。例えば、ポリペプチドは、上述したように検出可能なタグを用いて標識できる。検出方法を使用すると、サンプル中の作用因子の存在または不在を記録することができる。しかし、検出方法はさらに定量方法と結合することができる。インビトロアッセイ法には、標準物質に比較して量を確定するために使用できる知られている作用因子量のコントロールサンプルとの比較に基づいて作用因子の定量を可能にするELISAなどの比色アッセイ法が含まれる。この方法は、さらに、放出された放射線に基づいて定量を可能にする放射分析アッセイおよび蛍光アッセイまたは上述した可視化および定量の任意の手段を含むことができる。
【0076】
サンプルは、任意の生物学的サンプルを含む、試験される任意のサンプルであってよい。サンプルには、液体サンプル(水、血液、尿など)、組織サンプル、培養サンプル、細胞サンプルなどを含むことができる。
【0077】
本発明のポリペプチドは、さらにまた遮断抗体に匹敵して、それへ結合する作用因子の活性を遮断するために使用することもできる。そこで、作用因子の活性を遮断する方法であって、作用因子にポリペプチドが結合できる条件下でその作用因子を本発明のポリペプチドと接触させる工程を含む方法もまた開示される。作用因子へのポリペプチドの結合は、その作用因子の活性を遮断する。接触させるステップは、インビボまたはインビトロのいずれであってもよい。そこで、例えば、サンプルの汚染を減少させるために、毒素へ結合するポリペプチドをサンプルに添加して、毒素活性を遮断することができる。
【0078】
本発明のポリペプチドは、さらにまた拮抗抗体に匹敵して、それへ結合する任意の作用因子の活性を促進するために使用することもできる。そこで、作用因子の活性を促進する方法であって、作用因子にポリペプチドが結合できる条件下でその作用因子を本発明のポリペプチドと接触させる工程を含む方法もまた開示される。作用因子へのポリペプチドの結合は、その作用因子の活性を促進する。
【0079】
本明細書に開示したポリペプチドは、知られていない機能を備える遺伝子の機能を決定するために使用することができる。そこで、開示されたポリペプチドをタンパク質ノックダウンアッセイにおいて使用する方法が開示される。例えば、開示されたポリペプチドは、機能が知られていない遺伝子を含む細胞の細胞質内で発現させることができる。RNA転写産物が細胞の細胞質内で翻訳されている場合は、開示されたポリペプチドは問題の遺伝子のタンパク質産物へ結合することができる。タンパク質発現の消失が細胞に及ぼす作用を監視することによって、タンパク質機能を決定することができる。そこで、機能が知られていない遺伝子産物に対して特異的なポリペプチドが特別に開示される。さらに、遺伝子の機能を決定する方法であって、遺伝子のタンパク質産物に対して特異的なポリペプチドを、その遺伝子を発現する細胞の細胞質内へ導入する工程、および機能が知られていない遺伝子のタンパク質産物の消失に起因する作用を監視する工程、を含む方法もまた開示される。
【0080】
本発明のポリペプチドは、イメージング法においても使用できる。例えば、本発明は、被験者に有効量のポリペプチドを投与する工程、および被験者において結合したポリペプチドの所在位置を検出する工程、を含むイメージング法を提供する。イメージング法の例は、上述されている。
【0081】
本発明は、さらに精製方法を提供する。本明細書では、サンプルから作用因子を精製する方法であって、ポリペプチドがそのサンプルへ結合してポリペプチド/作用因子複合体を形成する条件下でサンプルをポリペプチドと接触させる工程;およびその作用因子をポリペプチド/作用因子複合体から単離する工程、を含む方法が開示される。例えば、ポリペプチドはカラムへ結合させ、サンプルは、サンプル内の作用因子が結合したポリペプチドへ結合できる条件下でカラムを通過させることができる。作用因子は、引き続いて所望の溶離剤中でカラムから溶出させることができる。精製方法は、研究方法として、および商業的方法として有用であろう。例えば、そのような方法は、薬理学的化合物から汚染物質を除去する際に有用であろう。
【0082】
本ポリペプチドは、治療方法においても使用できる。例えば、本明細書では、被験者における病原性作用を減少させる、または予防する方法であって、病原体へ結合する有効量のポリペプチドを被験者に投与する工程を含む方法が提供される。さらにまた、有害作用を減少させられる、または有効な作用を促進できるように作用因子の活性を遮断または促進する方法も提供される。
【0083】
本明細書では、本発明のポリペプチドまたは核酸および医薬上許容される担体を含む組成物が提供される。本発明の組成物は、インビボで投与することもできる。組成物は、経口、非経口(例、静脈内)、筋肉内注射、腹腔内注射、経皮的、体外、局所的などによって投与されてよいが、典型的には局所的鼻腔内投与または吸入剤による投与が好ましい。本明細書で使用する「局所的鼻腔内投与」は、鼻孔の一方または両方を通して鼻および鼻腔内への組成物の送達を意味しており、スプレー機構もしくは滴下機構、または核酸もしくはベクターのエーロゾル化を通しての送達を含むことができる。後者は、大量の動物を同時に治療しなければならない場合には有効なことがある。吸入剤による組成物の投与は、スプレーまたは滴下機構による送達を介して鼻または口を通して行なうことができる。送達は、挿管を通して呼吸器系の任意の領域(例、肺)へ直接的に行なうこともできる。必要とされる組成物の正確な量は、被験者の種、年齢、体重および全身状態、治療されるアレルギー性障害の重症度、使用される特定の核酸またはベクター、投与様式などに依存して、被験者毎に相違するであろう。そこで、各組成物について正確な量を規定するのは不可能である。しかし、適切な量は、当業者が本明細書の教示を前提に日常的な実験器具を使用さえすれば決定することができる。
【0084】
使用する場合、組成物の非経口投与は一般に注射を特徴とする。注射剤は、液剤もしくは懸濁剤、注射前に液体中の懸濁液にするために適合する固体形、またはエマルジョンとしてのいずれかの便宜的形態で調製できる。非経口投与のために最近改良されたアプローチは、一定用量が維持されるような徐放系または持続放出系の使用が含まれる。例えば、参照して本明細書に組み込まれる米国特許第3,610,795号を参照されたい。
【0085】
物質は、(例えば、微粒子、リポソーム、または細胞内に組み込まれて)溶液、懸濁液中にあってよい。これらは、抗体、受容体、または受容体リガンドを介して特定細胞タイプを標的とすることができる。下記の参考文献は、腫瘍組織へ特異的タンパク質をターゲッティングするためのこのテクノロジーの使用例である(Senter,et al.,Bioconjugate Chem.,2:447−451,(1991);Bagshawe,K.D.,Br.J.Cancer,60:275−281,(1989);Bagshawe,et al.,Br.J.Cancer,58:700−703,(1988);Senter,et al.,Bioconjugate Chem.,4:3−9,(1993);Battelli,et al.,Cancer Immunol.Immunother.,35:421−425,(1992);PieterszおよびMcKenzie,Immunolog.Reviews,129:57−80,(1992);ならびにRoffler,et al.,Biochem.Pharmacol,42:2062−2065,(1991))。「ステルス」などのビヒクルおよびその他の抗体結合リポソーム(大腸癌を標的とする脂質媒介性薬物を含む)、細胞特異的リガンドを通してのDNAの受容体媒介性ターゲッティング、リンパ球指向性腫瘍ターゲッティング、およびインビボのマウス神経膠腫細胞に対する高度に特異的な治療的レトロウイルスターゲッティング。下記の参考文献は、腫瘍組織へ特異的タンパク質をターゲッティングするためのこのテクノロジーの使用例である(Hughes et al.,Cancer Research,49:6214−6220,(1989);およびLitzingerおよびHuang,Biochimica et Biophysica Acta,1104:179−187,(1992))。一般に、受容体は、構成的またはリガンド誘導性のいずれかで、エンドサイトーシスの経路に含まれている。これらの受容体は、クラスリン被覆ピット内でクラスターを形成し、クラスリン被覆小胞を介して細胞内に進入し、その中で受容体が分類される酸性化エンドソームを通過し、そして次に細胞表面へ再循環する、細胞内に蓄積される、またはリソソーム内で分解される、のいずれかとなる。内在化経路は、栄養素取り込み、活性化タンパク質の除去、高分子のクリアランス、ウイルスおよび毒素の日和見進入、リガンドの解離および分解、ならびに受容体レベル調節などの様々な機能に役立つ。多数の受容体は、細胞タイプ、受容体濃度、リガンドのタイプ、リガンドの価数、およびリガンドの濃度に依存して、2つ以上の細胞内経路にしたがう。受容体媒介性エンドサイトーシスの分子および細胞機序は精査されている(BrownおよびGreene,DNA and Cell Biology 10:6,399−409 (1991))。
【0086】
「医薬上許容される」は、生物学的またはその他の点で不都合ではない物質を意味しており、すなわちその物質は、何の不都合な生物学的作用を誘発することなく、またはそれが含有される医薬組成物の他の成分のいずれかと有害な方法で相互作用することなく本発明のポリペプチドと一緒に被験者に投与することができる。担体は、当業者にはよく知られているように、当然ながら有効成分の分解を最小限に抑えるように、そして被験者における有害な副作用を最小限に抑えるように選択されるであろう。適切な担体およびそれらの調製物は、Remington:The Science and Practice
of Pharmacy(第19版)編者A.R.Gennaro,Mack Publishing Company,Easton,PA 1995に記載されている。典型的には、調製物を等張性にするために、調製物中では適切な量の医薬上許容される塩が使用される。医薬上許容される担体の例には、食塩液、リンゲル液およびデキストロース液が含まれるがそれらに限定されない。溶液のpHは、好ましくは約5〜約8、より好ましくは約7〜約7.5である。その他の担体には、可変性リンパ球受容体を含有する固体疎水性ポリマーの半透性マトリックスなどの徐放性調製物が含まれるが、そのマトリックスは成形製品、例えばフィルム、リポソームまたは微粒子の形状にある。当業者には、例えば投与経路、投与される可変性リンパ球受容体の濃度に依存して所定の担体がより好ましいことは明白であろう。
【0087】
医薬担体は、当業者には知られている。これらは、最も典型的には無菌水、食塩液、および生理的pHにある緩衝液などの溶液を含むヒトに薬物を投与するための標準的担体であろう。本組成物は、例えば筋肉内または皮下投与することができる。その他の化合物は、当業者によって使用される標準方法にしたがって投与されるであろう。
【0088】
本医薬組成物は、精選分子に加えて、担体、増粘剤、希釈剤、バッファー、保存剤、界面活性剤などを含むことができる。本医薬組成物は、さらに抗菌物質、抗炎症薬、麻酔薬などの1つまたは複数の有効成分を含むことができる。
【0089】
非経口投与のための本調製物には、無菌の水性または非水性液剤、懸濁剤、およびエマルジョンが含まれる。非水性溶媒の例は、プロピレングリコール、ポリエチレングリコール、オリーブ油などの植物油、およびオレイン酸エチルなどの注射用有機エステルである。水性担体には、水、食塩液および緩衝溶媒を含むアルコール性/水性の溶液、エマルジョンもしくは懸濁液が含まれる。非経口ビヒクルには、塩化ナトリウム溶液、リンガーデキストロース、デキストロースおよび塩化ナトリウム、乳酸加リンガー液、または固定油が含まれる。静注用ビヒクルには、液体および栄養補充剤、電解質補充剤(リンガーデキストロースをベースとする補充剤など)などが含まれる。例えば抗菌物質、抗酸化剤、キレート剤、および不活性ガスなどの保存剤およびその他の添加剤もまた存在していてよい。
【0090】
局所適用のための本調製物は、軟膏、ローション剤、クリーム剤、ゲル剤、滴剤、坐剤、スプレー剤、液剤および散剤を含むことができる。従来型医薬担体、水性、粉末状または油性基剤、増粘剤などが必要になる、または望ましいことがある。
【0091】
経口投与のための本組成物には、散剤もしくは顆粒剤、水もしくは非水性溶媒中の懸濁剤もしくは液剤、カプセル剤、サシェ剤、または錠剤が含まれる。増粘剤、矯味剤、希釈剤、乳化剤、分散助剤または結合剤が望ましいことがある。
【0092】
本組成物の一部は、もしかすると塩酸塩、臭化水素酸、過塩素酸、硝酸、チオシアン酸、硫酸、およびリン酸などの無機酸と、ギ酸、酢酸、プロピオン酸、グリコール酸、乳酸、ピルビン酸、シュウ酸、マロン酸、コハク酸、マレイン酸、およびフマル酸などの有機酸との反応によって、または水酸化ナトリウム、水酸化アンモニウム、水酸化カリウムなどの無機塩基、およびモノ−、ジ−、トリアルキルおよびアリールエミンならびに置換されたエタノールアミンなどの有機塩基との反応によって生成される医薬上許容される酸付加塩もしくは塩基付加塩として投与できる。
【0093】
組成物を投与するための用量範囲は、症状や障害に有効である所望の作用を生じさせるために十分な量である。用量は、望ましくない交差反応、アナフィラキシー反応などの有害な副作用を惹起するほど大量であってはならない。一般に、用量は、患者における年齢、状態、性別および疾患の程度に伴って変動するが、当業者であれば決定することができる。用量は、万一何らかの禁忌がある場合に個々の医師によって調整することができる。用量は変動することがあり、1日もしくは数日にわたって、1回または複数回の投与で投与することができる。
【0094】
本発明の可変性リンパ球受容体および可変性リンパ球受容体フラグメントおよび変異体は、さらにまた患者もしくは被験者に、その患者もしくは被験者自身の細胞が核酸を取り込んで、コードされた可変性リンパ球受容体もしくは可変性リンパ球受容体フラグメントを生成かつ分泌するように、可変性リンパ球受容体または可変性リンパ球受容体フラグメントもしくは変異体をコードする核酸調製物(例、DNAもしくはRNA)として投与することもできる。
【0095】
インビトロまたはインビボのいずれかで、核酸を細胞に送達するために使用できる多数の組成物および方法がある。これらの方法および組成物は、大きく分けると次の2つのクラスに分類できる:ウイルスベースの送達系および非ウイルスベースの送達系。例えば、核酸は、エレクトロポレーション、リポフェクション、リン酸カルシウム沈降法、プラスミド、ウイルスベクター、ウイルス核酸、ファージ核酸、ファージ、コスミドなどの多数の直接送達系を通して、またはカチオン性リポソームなどの細胞もしくは担体中の遺伝物質の輸送を介して送達できる。ウイルスベクター、化学的トランスフェクタントを含むトランスフェクションのための適切な手段、またはエレクトロポレーションおよびDNAの直接拡散法などの物理−機械的方法は、例えばWolff,J.A.,et al.,Science,247,1465−1468,(1990);およびWolff,J.A.Nature,352,815−818,(1991)によって記載されている。そのような方法は当技術分野においてよく知られており、本明細書に記載した組成物および方法と一緒に使用するために容易に適合させることができる。所定の場合には、これらの方法は大きなDNA分子とともに特異的に機能するように修飾されるであろう。さらに、これらの方法を使用すると、担体のターゲッティング特性を使用することによって、所定の疾患および細胞集団を標的とすることができる。
【0096】
トランスファーベクターは、核酸を細胞内へ送達するために、または遺伝子を送達するための一般的戦略の一部として、例えば組み換えレトロウイルスまたはアデノウイルスの一部として使用される任意のヌクレオチド構造(例、プラスミド)であってよい(Ram
et al.Cancer Res.53:83−88,(1993))。本明細書で使用するプラスミドまたはウイルスベクターは、VLRなどの開示された核酸を細胞内へ分解させずに輸送する作用因子であり、その中にそれが送達される細胞内での遺伝子の発現を生じさせるプロモーターが含まれる。ウイルスベクターは、例えば、アデノウイルス、アデノ関連ウイルス、ヘルペスウイルス、ワクシニアウイルス、ポリオウイルス、AIDSウイルス、ニューロン栄養ウイルス、シンドビスウイルス、およびHIV骨格を備えるウイルスを含むその他のRNAウイルスである。さらに好ましいのは、それらをベクターとして使用するために適合させるこれらのウイルスの特性を共有する任意のウイルスファミリーである。レトロウイルスには、マウスマロネー白血病ウイルス(MMLV)、およびベクターとしてのMMLVの所望の特性を発現するレトロウイルスが含まれる。レトロウイルスベクターは、他のウイルスベクターより大きな遺伝的ペイロード、すなわち導入遺伝子またはマーカー遺伝子を運ぶことができ、このために一般的に使用されるベクターである。しかし、それらは非増殖細胞中では有用ではない。アデノウイルスベクターは比較的安定性であり、作業するのが容易であり、高い力価を有し、エーロゾル調製物中で送達することができ、非分割細胞をトランスフェクトすることができる。ポックスウイルスベクターは大きく、遺伝子を挿入するための幾つかの部位を有しており、それらは熱安定性で、室温で保管することができる。好ましい実施形態は、ウイルス抗原によって惹起される、宿主生体の免疫応答を抑制できるように組み換えられているウイルスベクターである。このタイプの好ましいベクターは、インターロイキン8または10のためのコーディング領域を有するであろう。
【0097】
ウイルスベクターは、細胞内へ遺伝子を導入するために化学的または物理的方法より高度のトランスアクション能力(遺伝子を導入する能力)を有する可能性がある。典型的には、ウイルスベクターは、非構造的初期遺伝子、構造的後期遺伝子、RNAポリメラーゼIII転写産物、複製およびエンカプシデーションのために必要な逆方向末端反復、およびウイルスゲノムの転写および複製を制御するためのプロモーターを含有する。ベクターとして組み換えられると、典型的にはウイルスから1つまたは複数の初期遺伝子が除去され、遺伝子もしくは遺伝子/プロモーターカセットが除去されたウイルスDNAの代わりにウイルスゲノム内に挿入される。このタイプの構築体は、約8kbまでの外来遺伝物質を運ぶことができる。除去された初期遺伝子の必要な機能は、典型的には初期遺伝子の遺伝子産物をトランスで発現するように組み換えられている細胞系によって補充される。
【0098】
レトロウイルスは、任意のタイプ、サブファミリー、属、または向性を含むレトロウイルスのウイルスファミリーに属する動物ウイルスである。レトロウイルスベクターは、一般に、参照して本明細書組み込まれるVerma,I.M.,Retroviral vectors for gene transfer.In Microbiology−1985,American Society for Microbiology,pp.229−232,Washington,(1985)の中に記載されている。遺伝子療法のためにレトロウイルスベクターを使用する方法の例は、それらの教示が参照して本明細書に組み込まれる米国特許第4,868,116号および第4,980,286号;PCT国際特許公開WO90/02806およびWO89/07136;ならびにMulligan,(Science 260:926−932(1993))に記載されている。
【0099】
レトロウイルスは、本質的に核酸カーゴ内にパッキングされているパッケージである。核酸カーゴは、それとともにパッケージングシグナルを運ぶが、これは複製された娘分子がパッケージコート内に効率的にパッケージングされることを保証する。パッケージングシグナルに加えて、複製、および複製されたウイルスのパッケージングのためにシス形に必要とされる多数の分子が存在する。典型的には、レトロウイルスゲノムは、タンパク質コートを作製する際に関係しているgag、pol、およびenv遺伝子を含有している。標的細胞へ輸送されるべき異質DNAによって典型的に置換されるのはgag、pol、およびenv遺伝子である。レトロウイルスベクターは、典型的にはパッケージコート内に組み込むためのパッケージングシグナル、gag転写ユニットの開始をシグナルする配列、逆転写のtRNAプライマーへ結合するためのプライマー結合部位を含む逆転写のために必要なエレメント、DNA合成中のRNAストランドのスイッチを誘導する末端反復配列、DNA合成の第2ストランドの合成のための開始部位として機能する3’LTRに対して5’のプリンリッチ配列、および宿主ゲノム内に挿入するためにDNA状態のレトロウイルスの挿入を可能にするLTRの両末端近くの特異的配列を含有する。gag、pol、およびenv遺伝子の除去は、約8kbの異質配列をウイルスゲノム内に挿入し、逆転写させ、複製後には新規なレトロウイルス粒子内にパッケージングすることを可能にする。この核酸の量は、各転写産物のサイズに依存して、1つから多数までの遺伝子を送達するために十分である。挿入物内に他の遺伝子と一緒に陽性または陰性いずれかの選択可能なマーカーを含めることが好ましい。
【0100】
大多数のレトロウイルスベクター内の複製機構およびパッケージングタンパク質は除去されているので(gag、pol、およびenv)、これらのベクターは典型的にはそれらをパッケージング細胞系内へ配置することによって生成される。パッケージング細胞系は、複製およびパッケージング機構を含有するが、任意のパッケージングシグナルが欠けているレトロウイルスを用いてトランスフェクトまたは形質転換されている細胞系である。精選DNAを運ぶベクターがこれらの細胞系内にトランスフェクトされると、当該遺伝子を含有するベクターが複製され、ヘルパー細胞によってシス形で提供される機構によって新規なレトロウイルス粒子内にパッケージングされる。この機構のためのゲノムは、それらに必要なシグナルが欠如しているためにパッケージングされない。
【0101】
複製欠損アデノウイルスの構築については記載されている(Berkner et al.,J.Virology 61:1213−1220(1987);Massie et al.,Mol.Cell.Biol.6:2872−2883(1986);Haj−Ahmad et al.,J.Virology 57:267−274(1986);Davidson et al.,J.Virology 61:1226−1239(1987);Zhang “Generation and identification of recombinant adenovirus by liposome−mediated transfection and PCR analysis” BioTechniques 15:868−872(1993))。これらのウイルスをベクターとして使用する有益性は、それらが初期感染細胞内で複製できるが、新規な感染性ウイルス粒子を形成することはできないために、それらが他の細胞タイプへ拡大できる程度が限定されることにある。組み換えアデノウイルスは、気道上皮、肝細胞、血管内皮、CNS実質、および多数の他の組織部位へ直接的なインビボ送達後に高い効率の遺伝子導入を達成することが証明されている(Morsy,J.Clin.Invest.92:1580−1586(1993);Kirshenbaum,J Clin.Invest.92:381−387(1993);Roessler,J.Clin.Invest.92:1085−1092(1993);Moullier,Nature Genetics 4:154−159(1993);La Salle,Science 259:988−990(1993);Gomez−Foix,J.Biol.Chem.267:25129−25134(1992);Rich,Human Gene Therapy 4:461−476(1993);Zabner,Nature Genetics 6:75−83(1994);Guzman,Circulation Research 73:1201−1207(1993);Bout,Human Gene Therapy 5:3−10(1994);Zabner,Cell 75:207−216(1993);Caillaud,Eur.J.Neuroscience 5:1287−1291(1993);およびRagot,J.Gen.Virology 74:501−507(1993))。組み換えアデノウイルスは、特異的細胞表面受容体へ結合することによって遺伝子形質導入を達成するが、その後にウイルスは、野生型または複製欠損アデノウイルスと同一方法で受容体媒介性エンドサイトーシスによって内在化される(ChardonnetおよびDales,Virolog
y 40:462−477(1970);BrownおよびBurlingham,J Virology 12:386−396(1973);SvenssonおよびPersson,J.Virology 55:442−449(1985);Seth,et
al.,J.Virol.51:650−655(1984);Seth,et al.,Mol.Cell.Biol.4:1528−1533(1984);Varga et al.,J Virology 65:6061−6070(1991);Wickham et al.,Cell 73:309−319(1993))。
【0102】
ウイルスベクターは、E1遺伝子が除去されているアデノウイルスをベースとするウイルスベクターであってよく、これらのバイロン(viron)はヒト293細胞系などの細胞系において生成される。また別の好ましい実施形態では、E1およびE3遺伝子の両方がアデノウイルスゲノムから除去される。
【0103】
また別のタイプのウイルスベクターは、アデノ関連ウイルス(AAV)をベースとしている。この欠陥のあるパルボウイルスは、多数の細胞タイプを感染させることができ、ヒトにとって非病原性であるので好ましい。AAV型ベクターは約4〜5kbを輸送することができ、野生型AAVは19番染色体中へ安定性で挿入できることが知られている。この部位特異的組み込み特性を含有するベクターが好ましい。このタイプのベクターの特に好ましい実施形態は、Avigen(カリフォルニア州サンフランシスコ)によって製造された、単純ヘルペスウイルスチミジンキナーゼ遺伝子、HSV−tk、および/または緑色蛍光タンパク質(GFP)をコードする遺伝子などのマーカー遺伝子を含有することのできるP4.1Cベクターである。
【0104】
また別のタイプのAAVウイルスでは、AAVは、異種遺伝子に機能的に連結した細胞特異的発現を指令するプロモーターを含有する少なくとも1つのカセットを挟んでいる1対の逆方向末端反復(ITR)を含有している。この状況における異種とは、AAVまたはB19パルボウイルスにとって自然ではない任意のヌクレオチド配列または遺伝子を意味する。
【0105】
典型的には、AAVおよびB19コーディング領域は、安全な非細胞毒性ベクターを生じさせるように欠失させられている。AAV ITR、またはそれらの修飾は、感染性および部位特異的組み込みを付与するが、細胞毒性は付与せず、そしてこのプロモーターは細胞特異的発現を指令する。AAVベクターに関連する物質については、本明細書に参照して組み込まれる米国特許第6,261,834号に記載されている。
【0106】
そこで本発明のベクターは、実質的毒性を伴わずに哺乳動物染色体内へ組み込むことのできるDNA分子を提供する。
【0107】
ウイルスおよびレトロウイルス内に挿入された遺伝子は、通常は、所望の遺伝子産物の発現を制御するのに役立つプロモーター、および/またはエンハンサーを含有している。プロモーターは、一般に、転写開始部位に関して相当に固定された場所にある場合に機能するDNAの配列である。プロモーターは、RNAポリメラーゼと転写因子との基本的相互作用のために必要とされるコアエレメントを含有しており、上流エレメントおよび応答エレメントを含有していてよい。
【0108】
大きなヒトヘルペスウイルスを用いた分子的遺伝学実験は、それによって大きな異種DNAフラグメントをクローン化し、増殖させ、そしてヘルペスウイルスを用いた感染のために許容される細胞内で確立することができる手段を提供してきた(Sun et al.,Nature genetics 8:33−41,1994;CotterおよびRobertson,.Curr Opin Mol Ther 5:633−644,1999)。これらの大きなDNAウイルス(単純ヘルペスウイルス(HSV)およびエプスタイン・バーウイルス(EBV))は、特定細胞へ150kbを超えるヒト異種DNAのフラグメントを送達する能力を有している。EBV組み換え体は、エピソームDNAとしての感染したB細胞内に大きなDNAの塊を維持することができる。330kbまでのヒトゲノム挿入物を運ぶ個別クローンは、遺伝的に安定性であると思われた。これらのエピソームを維持するためには、EBVによる感染中に構成的に発現した特異的EBV核タンパク質EBNA1を必要とする。さらに、これらのベクターは、大量のタンパク質をインビトロで一時的に生成できるトランスフェクションのために使用できる。ヘルペスウイルスアンプリコン系もまた、220kbを超えるDNA破片をパッケージングし、DNAをエピソームとして安定性で維持できる細胞を感染させるために使用されている。
【0109】
その他の有用な系には、例えば、複製および宿主限定非複製ワクシニアウイルスベクターが含まれる。
【0110】
開示された組成物は、様々な方法で標的細胞へ送達することができる。例えば、本組成物はエレクトロポレーション、またはリポフェクション、またはリン酸カルシウム沈降法を通して送達できる。選択される送達機構は、一部には標的とされる細胞のタイプ、およびその送達が例えばインビボまたはインビトロのどちらで発生するのかに依存するであろう。
【0111】
そこで、本組成物は、開示されたベクターに加えて、例えばカチオン性リポソーム(例、DOTMA、DOPE、DC−コレステロール)またはアニオン性リポソームなどのリポソームのような脂質を含むことができる。リポソームは、所望であれば特定細胞をターゲッティングすることを促進するためのタンパク質をさらに含むことができる。化合物およびカチオン性リポソームを含む組成物の投与は、標的器官へ輸入性の血液に投与する、または気道の細胞へターゲッティングするために気道内へ吸入させることができる。リポソームに関しては、例えば、Brigham et al.Am.J.Resp.Cell.Mol.Biol.1:95−100 (1989);Felgner et al.Proc.Natl.Acad.Sci USA 84:7413−7417 (1987);米国特許第4,897,355号を参照されたい。さらに、化合物はマクロファージなどの特定細胞タイプへターゲッティングできる、またはマイクロカプセルからの化合物の拡散もしくは化合物の送達が特定の速度もしくは用量のために設計されているマイクロカプセルの構成成分として投与できる。
【0112】
投与および被験者の細胞内への外因性DNAの取り込み(すなわち、遺伝子形質導入またはトランスフェクション)を含む上述した方法では、細胞への組成物の送達は様々な機構を介して行なうことができる。1つの例として、送達は、市販で入手できるLIPOFECTIN、LIPOFECTAMINE(GIBCO−BRL,Inc.、メリーランド州ゲーサーズバーグ)、SUPERFECT(Qiagen,Inc.、独国ヒルデン)およびTRANSFECTAM(Promega Biotec,Inc.,、ワイオミング州マディソン)などのリポソーム調製物、ならびに当技術分野において標準的な方法にしたがって開発された他のリポソームを使用して、リポソームを介して行なうことができる。さらに、本発明の核酸またはベクターは、エレクトロポレーションによってインビボで送達することができ、そのためのテクノロジーはGenetronics,Inc.(カリフォルニア州サンディエゴ)から、ならびにSONOPORATION装置(ImaRx Pharmaceutical Corp.、アリゾナ州トゥーソン)によって入手できる。
【0113】
物質は、(例えば、微粒子、リポソーム、または細胞内に組み込まれて)溶液や懸濁液中に含まれていてよい。これらは、VLR、抗体、受容体、または受容体リガンドを介して特定細胞タイプを標的とすることができる。下記の参考文献は、腫瘍組織へ特異的タンパク質をターゲッティングするためのこのテクノロジーの使用例である(Senter,et al.,Bioconjugate Chem.,2:447−451,(1991);Bagshawe,K.D.,Br.J.Cancer,60:275−281,(1989);Bagshawe,et al.,Br.J.Cancer,58:700−703,(1988);Senter,et al.,Bioconjugate Chem.,4:3−9,(1993);Battelli,et al.,Cancer Immunol.Immunother.,35:421−425,(1992);PieterszおよびMcKenzie,Immunolog.Reviews,129:57−80,(1992);およびRoffler,et al.,Biochem.Pharmacol,42:2062−2065,(1991))。これらの技術は、様々な他の特定細胞タイプに対して使用できる。「ステルス」などのビヒクルおよびその他の抗体またはVLR結合リポソーム(大腸癌を標的とする脂質媒介性薬物を含む)、細胞特異的リガンドを通してのDNAの受容体媒介性ターゲッティング、リンパ球指向性腫瘍ターゲッティング、およびインビボのマウス神経膠腫細胞に対する高度に特異的な治療的レトロウイルスターゲッティング。下記の参考文献は、腫瘍組織へ特異的タンパク質をターゲッティングするためのこのテクノロジーの使用例である(Hughes et al.,Cancer Research,49:6214−6220,(1989);およびLitzingerおよびHuang,Biochimica et Biophysica Acta,1104:179−187,(1992))。一般に、受容体は、構成的またはリガンド誘導性のいずれかで、エンドサイトーシスの経路に含まれている。これらの受容体は、クラスリン被覆ピット内でクラスターを形成し、クラスリン被覆小胞を介して細胞内に進入し、その中で受容体が分類される酸性化エンドソームを通過し、そして次に細胞表面へ再循環する、細胞内に蓄積される、またはリソソーム内で分解される、のいずれかとなる。内在化経路は、栄養素取り込み、活性化タンパク質の除去、高分子のクリアランス、ウイルスおよび毒素の日和見進入、リガンドの解離および分解、ならびに受容体レベル調節などの様々な機能に役立つ。多数の受容体は、細胞タイプ、受容体濃度、リガンドのタイプ、リガンドの価数、およびリガンドの濃度に依存して、2つ以上の細胞内経路にしたがう。受容体媒介性エンドサイトーシスの分子および細胞機序は精査されている(BrownおよびGreene,DNA and Cell Biology
10:6,399−409 (1991))。
【0114】
宿主細胞ゲノム内に組み込まれるべき細胞へ送達される核酸は、典型的には組み込み配列を含有している。これらの配列は、特にウイルスをベースとする系が使用される場合は、ウイルス関連配列であることが多い。これらのウイルス組み込み系は、送達系内に含有される核酸を宿主ゲノム内に組み込むことができるように、リポソームなどの非核酸をベースとする送達系を用いて送達されるべき核酸内に組み込むこともできる。
【0115】
その他の宿主ゲノム内に組み込むための一般的技術には、例えば、宿主ゲノムとの同種組み換えを促進するように設計された系が含まれる。これらの系は、典型的には、送達された核酸が宿主ゲノム内に組み込まれるのを誘発するように、ベクター核酸と標的核酸との組み換えが発生する宿主細胞ゲノム内で標的配列と十分な相同性を有する、発現する核酸に隣接する配列に依存している。同種組み換えを促進するために必要なこれらの系および方法は、当業者には知られている。
【0116】
上述したように、本組成物は医薬上許容される担体中で投与することができ、当技術分野においてよく知られている様々な機構(例、裸のDNAの取り込み、リポソーム融合、遺伝子銃を介してのDNAの筋肉内注射、エンドサイトーシスなど)によって、被験者の細胞へインビボおよび/またはエックスビボで送達することができる。
【0117】
エクソビボ法を使用する場合は、細胞または組織を取り出し、当技術分野においてよく知られている標準プロトコールにしたがって体外で維持することができる。本組成物は、例えばリン酸カルシウム媒介性遺伝子送達、エレクトロポレーション、マイクロインジェクションまたはプロテオリポソームなどの任意の遺伝子輸送機構を介して細胞内へ導入することができる。形質導入された細胞は、次に(例えば医薬上許容される担体中で)注入する、または細胞もしくは組織タイプにとって標準的方法によって被験者に同位置へ逆移植することができる。被験者に様々な細胞を移植または注入するための標準方法は知られている。
【0118】
細胞へ送達される核酸は、典型的には発現制御系を含有している。例えば、ウイルスおよびレトロウイルス内に挿入された遺伝子は、通常は、所望の遺伝子産物の発現を制御するのに役立つようにプロモーター、および/またはエンハンサーを含有している。プロモーターは、一般に、転写開始部位に関して比較的固定された場所にある場合に機能するDNAの配列である。プロモーターは、RNAポリメラーゼと転写因子との基本的相互作用のために必要とされるコアエレメントを含有しており、上流エレメントおよび応答エレメントを含有していてよい。
【0119】
哺乳動物宿主細胞中のベクターからの転写を制御する好ましいプロモーターは、例えば、ポリオーマウイルス、サルウイルス40(SV40)、アデノウイルス、レトロウイルス、B型肝炎ウイルスおよび最も好ましくはサイトメガロウイルスなどのウイルスゲノムの様々な起源から、または異種哺乳動物プロモーター、例えばβアクチンプロモーターから入手できる。SV40ウイルスの初期および後期プロモーターは、便宜的には、SV40ウイルス複製起点も含有するSV40制限フラグメントとして入手される(Fiers
et al.,Nature,273:113(1978))。ヒトサイトメガロウイルスの前初期プロモーターは、便宜的にはHindIII E制限フラグメントとして入手される(Greenway,P.J.et al.,Gene 18:355−360(1982))。当然ながら、宿主細胞または関連種由来のプロモーターもまたこの場合に有用である。
【0120】
エンハンサーは、一般に転写開始部位から固定されない距離で機能し、転写ユニットに対して5’(Laimins,L.et al.,Proc.Natl.Acad.Sci.78:993(1981))または3’(Lusky,M.L.,et al.,Mol.Cell Bio.3:1108(1983))のいずれかであってよいDNAの配列を意味する。さらに、エンハンサーはイントロン内(Banerji,J.L.et
al.,Cell 33:729(1983))ならびにコーディング配列自体の内側(Osborne,T.F.,et al.,Mol.Cell Bio.4:1293(1984))にあってよい。それらは通常は10〜300塩基対長であり、それらはシス形で機能する。エンハンサーは、隣接プロモーターからの転写を増加させるように機能する。エンハンサーは、さらにまた転写の調節を媒介する応答エレメントを含有することが多い。プロモーターは、さらにまた転写の調節を媒介する応答エレメントを含有することができる。エンハンサーは、しばしば遺伝子の発現の調節を決定する。多くのエンハンサー配列は哺乳動物遺伝子から現在では知られているが(グロビン、エラスターゼ、アルブミン、−フェトプロテインおよびインスリン)、典型的には一般的発現のためには真核細胞ウイルス由来のエンハンサーが使用されるであろう。好ましい例は、複製起点の後期側(100〜270塩基対)上のSV40エンハンサー、サイトメガロウイルス初期プロモーターエンハンサー、複製起点の後期側上のポリオーマエンハンサー、およびアデノウイルスエンハンサーである。
【0121】
プロモーターおよび/またはエンハンサーは、それらの機能を始動させる光線または特異的化学事象のいずれかによって特異的に活性化することができる。系は、テトラサイクリンおよびデキサメタゾンなどの試薬によって調節できる。ガンマ線照射などの照射への曝露、またはアルキル化化学療法薬によってウイルスベクター遺伝子発現を強化する方法もある。
【0122】
所定の実施形態では、プロモーターおよび/またはエンハンサー領域は、転写されるべき転写ユニットの領域の発現を最大限にするための構成的プロモーターおよび/またはエンハンサーとして機能することができる。所定の構築物では、プロモーターおよび/またはエンハンサー領域は、特定時間に特定タイプの細胞中でしか発現しないとはいえ、全真核細胞タイプで活性な可能性がある。このタイプの好ましいプロモーターは、CMVプロモーター(650塩基)である。その他の好ましいプロモーターは、SV40プロモーター、サイトメガロウイルス(全長プロモーター)、およびレトロウイルスベクターLTFである。
【0123】
真核宿主細胞(酵母、真菌、昆虫、植物、動物、ヒトまたは有核細胞)中で使用される発現ベクターは、さらにまたmRNA発現に影響を及ぼすことのできる転写の終結のために必要な配列を含有することができる。これらの領域は、組織因子タンパク質をコードするmRNAの非翻訳部分におけるポリアデニル化セグメントとして転写される。3’非翻訳領域には、転写終結部位も含まれる。転写ユニットがさらにポリアデニル化領域も含有することが好ましい。この領域の1つの利点は、転写されたユニットがプロセッシングされてmRNAと同様に輸送される可能性をそれが増加させることにある。発現構築物中のポリアデニル化シグナルの同定および使用は、明白に確立されている。導入遺伝子構築物中で同種ポリアデニル化シグナルが使用されるのが好ましい。所定の転写ユニットでは、ポリアデニル化領域はSV40初期ポリアデニル化シグナルから引き出され、約400塩基からなる。さらにまた、転写されたユニットは他の標準配列を単独で、または上記の配列と組み合わせて含有し、その構築物からの発現、またはその構築物の安定性を改良するのが好ましい。
【0124】
ウイルスベクターは、マーカー産物をコードする核酸配列を含有することができる。このマーカー産物は、遺伝子が細胞に送達されているかどうか、送達されている場合は発現しているかどうかを決定するために使用される。好ましいマーカー遺伝子は、β−ガラクトシダーゼをコードする大腸菌lacZ遺伝子、および緑色蛍光タンパク質である。
【0125】
一部の実施形態では、マーカーは選択可能なマーカーであってよい。哺乳動物細胞のために適切な選択可能なマーカーの例は、ジヒドロ葉酸還元酵素(DHFR)、チミジンキナーゼ、ネオマイシン、ネオマイシンアナログG418、ハイドロマイシン、およびピューロマイシンである。そのような選択可能なマーカーを哺乳動物宿主細胞内へ移すのに成功すると、形質転換された哺乳動物宿主細胞は、選択的圧力下に置かれたときに生存することができる。選択的レジメンには広く使用されている2つの明確なカテゴリーが存在する。第1カテゴリーは細胞の代謝、および補給された培地とは無関係に増殖する能力が欠如する突然変異細胞系の使用に基づいている。2つの例は、CHO DHFR細胞およびマウスLTK細胞である。これらの細胞は、チミジンまたはヒポキサンチンなどの栄養素を添加せずに増殖する能力が欠如している。これらの細胞は完全なヌクレオチド合成経路のために必要な所定の遺伝子が欠如しているので、それらは欠けているヌクレオチドが補給培地中に提供されない限り生存することはできない。培地への補給の代替策は、各遺伝子が欠けている細胞中へ無傷DHFRまたはTK遺伝子を導入すること、これによりそれらの増殖要件を変化させることである。DHFRまたはTK遺伝子を用いて形質転換されなかった個々の細胞は、非補給培地中では生存することができないであろう。
【0126】
第2カテゴリーは、任意の細胞タイプで使用される選択スキームに関する、突然変異細胞系の使用を必要としない優性選択である。これらのスキームは、典型的には宿主細胞の増殖を停止させるための薬剤を使用する。新規な遺伝子を有するそれらの細胞は、薬物耐性を備えるタンパク質を発現し、その選択を生き延びるであろう。そのような優性選択の例は、薬物のネオマイシン(Southern P.and Berg,P.,J.Molec.Appl.Genet.1:327 (1982))、ミコフェノール酸(Mulligan,R.C.and Berg,P.Science 209:1422 (1980))、またはハイグロマイシン(Sugden,B.et al.,Mol.Cell.Biol.5:410−413 (1985))を使用する。これら3つの例は、適切な薬剤G418もしくはネオマイシン(ゲネシチン)、xgpt(ミコフェノール酸)またはハイグロマイシン各々に対する耐性を備える真核生物制御下の細菌遺伝子を使用する。その他には、ネオマイシンアナログG418およびpuramycinが含まれる。
【0127】
被験者の細胞内への外因性DNAの投与および取り込み(すなわち遺伝子形質導入またはトランスフェクション)を含んでいる上述した方法では、本発明の核酸は裸のDNAもしくはRNAの形状にあってよい、または、当業者には明白であるように、それによって抗体をコードするDNAフラグメントがプロモーターの転写調節下となるように、これらの核酸は核酸を細胞へ送達するためのベクター内に含まれていてよい。ベクターは、アデノウイルスベクター(Quantum Biotechnologies,Inc.(カナダ国ケベック州ラバル)などの市販で入手できる調製物であってよい。細胞への核酸またはベクターの送達は、様々な機構を介して行なわれる。1つの例として、送達は、市販で入手できるLIPOFECTIN、LIPOFECTAMINE(GIBCO−BRL,Inc.、メリーランド州ゲーサーズバーグ)、SUPERFECT(Qiagen,Inc.、独国ヒルデン)およびTRANSFECTAM(Promega Biotec,Inc.,、ワイオミング州マディソン)などのリポソーム調製物、ならびに当技術分野において標準的な方法にしたがって開発された他のリポソームを使用して、リポソームを介して行なうことができる。さらに、本発明の核酸またはベクターは、エレクトロポレーションによってインビボで送達することができ、そのためのテクノロジーはGenetronics,Inc.(San Diego,CA)から、ならびにSONOPORATION装置(ImaRx Pharmaceutical Corp.、アリゾナ州トゥーソン)によって入手できる。
【0128】
1つの例として、ベクターの送達は、組み換えレトロウイルスゲノムをパッケージできるレトロウイルスベクター系などのウイルス系によって行なうことができる(例えば、Pastan et al.,Proc.Natl.Acad.Sci.U.S.A.85:4486,1988;Miller et al.,Mol.Cell.Biol.6:2895,1986を参照されたい)。組み換えレトロウイルスは、感染させ、それによって感染した細胞へ本発明の広範な中和抗体をコードする核酸(またはその活性フラグメント)を送達するために使用できる。変化した核酸を哺乳動物細胞中へ導入する正確な方法は、当然ながらレトロウイルスベクターの使用に限定されない。この方法のためには、アデノウイルスベクター(Mitani et al.,Hum.Gene Ther.5:941−948,1994)、アデノ関連ウイルス(AAV)ベクター(Goodman et al.,Blood 84:1492−1500,1994)、レンチウイルスベクター(Naidini et al.,Science 272:263−267,1996)、シュードタイプ化レトロウイルスベクター(Agrawal et al.,Exper.Hematol.24:738−747,1996)の使用を含む他の技術を広汎に利用できる。リポソーム送達および受容体媒介性ならびにその他のエンドサイトーシス機構などの物理的形質導入技術もまた使用できる(例えば、Schwartzenberger et al.,Blood 87:472−478,1996を参照されたい)。本発明は、これらやその他の一般に使用される遺伝子導入法のいずれかと結び付けて使用できる。
【0129】
1つの例として、本発明の抗体をコードする核酸がアデノウイルスベクター内で被験者の細胞へ送達される場合は、ヒトへアデノウイルスを投与するための用量は注射1回当たり約107〜109プラーク形成単位(pfu)の範囲に及んでよいが、注射1回当たり1012pfuという高用量でもよい(Crystal,Hum.Gene Ther.8:985−1001,1997;AlvarezおよびCuriel,Hum.Gene Ther.8:597−613,1997)。被験者は単回注射を受けることができる。または追加の注射が必要な場合は、6カ月間隔(または熟練の医師によって決定される他の適切な時間間隔)で無期限の期間にわたり、および/または治療の有効性が確定されるまで繰り返すことができる。
使用する場合、本発明の核酸またはベクターの非経口投与は一般に注射を特徴とする。注射剤は、液剤もしくは懸濁剤、注射前に液体中の懸濁液にするために適合する固体形、またはエマルジョンとしてのいずれかの従来の形態で調製できる。非経口投与のために最近改訂されたアプローチは、一定用量が維持されるような徐放系または持続放出系の使用が含まれる。例えば、参照して本明細書に組み込まれる米国特許第3,610,795号を参照されたい。適切な調製物および治療用化合物の様々な投与経路についてのこれ以上の考察については、例えば、Remington:The Science and Practice of Pharmacy(第19版)編者A.R.Gennaro,Mack Publishing Company,Easton,PA 1995を参照されたい。
【0130】
本発明は、さらに本発明のポリペプチドの製造方法であって、ポリペプチドをコードする核酸を含むベクターを含む細胞を培養する工程と、およびそのポリペプチドを細胞から、または培地から精製する工程と、を含む方法を提供する。さらにまた、タンパク質合成技術を使用した本発明のポリペプチドの製造方法が提供される。
【0131】
さらに、被験者における1つまたは複数の可変性リンパ球受容体をスクリーニングする方法であって、被験者においてN末端ロイシンリッチリピート(LRRNT)、1つまたは複数のロイシンリッチリピート(LRR)、C末端ロイシンリッチリピート(LRRCT)、および連結ペプチドを含む1つまたは複数のポリペプチドを同定する工程を含む方法が開示されるが、このとき連結ペプチドはαヘリックスを含んでいる。
【実施例】
【0132】
下記の実施例は、当業者に、本明細書で特許請求した化合物、組成物、製品、装置および/または方法の製造方法および評価方法に関する完全な開示および説明を提供できるように提示するものであり、純粋に本発明の例示となることを意図しており、本発明者らが本発明であると見なす本発明の範囲を限定することは意図されていない。数字(例、量、温度など)に関して正確さを保証できるように努力はしてきたが、いくらかの誤差および偏差が含まれる可能性がある。他に特に記載しない限り、部は重量部であり、温度は℃または周囲温度であり、そして圧力は大気圧またはほぼ大気圧である。
【0133】
(実施例1:ウミヤツメにおける可変性リンパ球受容体)
(免疫賦活された血液リンパ球由来の転写産物の分析)
活性化リンパ球のトランスクリプトームを調査するために、生きている大腸菌、ヒツジ赤血球、フィトヘマグルチニンおよびアメリカヤマゴボウマイトジェンを含有する抗原/マイトジェンカクテルの腹腔内注射によって、ヤツメウナギの幼生を1週間間隔で2〜4回免疫賦活した。第2回追加免疫賦活から3日後の末梢白血球中の大リンパ球の分画は、非免疫賦活個体に比して13倍大きく、骨髄細胞の分画もまた6倍大きかった(図1a)。小リンパ球に比較して、大リンパ球はサイズがほぼ2倍であり、広範囲のアズール親和性細胞質を有し、顕著な核小体を特徴とした(図1b)。これらの細胞を選別し、ヤツメウナギ活性化骨髄細胞または赤血球由来のcDNAに対する差引きによって活性化リンパ球のメッセージが富裕なcDNAライブラリーを構築するために使用した。
【0134】
差引きされたライブラリーからの1,507クローン中で同定された最も豊富な配列群から、非免疫賦活リンパ球転写産物の調査からの1セットの52個のLRRを含有する発現配列タグ(EST)とともにクラスター化した、可変数の様々なロイシンリッチリピート(LRR)モチーフを備える319種のタンパク質が予測された。3’末端配列をパージングした後、1セットの239種の固有に多様なLRRタンパク質が同定され、それらのうちの22種が239〜304アミノ酸のオープンリーディングフレーム(ORF)の大多数または全部をコードしていた(図6)。これらのヤツメウナギタンパク質は、これらの239の配列各々が固有であり、それらの転写産物は主としてリンパ球によって発現することが見いだされたために、暫定的に可変性リンパ球受容体(VLR)と命名された(図1c)。造血細胞組織由来のリンパ球は非免疫賦活動物において最高VLRレベルを示し、そして免疫賦活は大リンパ球によるVLR転写の増強を生じさせた。これらのVLRの基本的組成は、保存されたシグナルペプチド、N末端LRR(LRRNT)、可変数の多様なLRR、および連結ペプチドを含んでおり、その後にC末端LRR(LRRCT)ならびにトレオニンリッチおよびプラリンリッチ柄部、全体的グリコシル−ホスファチジル−イノシトール(GPI)−アンカー部位および疎水性尾部から構成される保存されたC末端が続いた(図1dおよび図7)。エピトープタグ付きVLRをコードするレトロウイルス構築物が哺乳動物細胞系内にトランスフェクトされると、免疫蛍光分析により、タンパク質の細胞表面局在化が確証され、細菌性GPI特異的ホスホリパーゼCを用いた処置は、細胞表面発現のレベルを有意に減少させ(図1e)、上清中へVLRタンパク質を放出した。3次元構造モデルを生成するために、11個のLRRからなる最長VLR配列を関連LRRタンパク質の結晶構造座標上に通した(Schwede et al.,2000)。このモデルは、Toll様受容体(TLR)エクトドメインのために予測されたモデル(Bell et al.,2003)に類似して、9つのβシートがLRRNTおよびLRRCTによって両端上でキャップされる凹状ソレノイド構造(図1f)を提供する。
【0135】
(VLRレパートリーは個々のヤツメウナギにおいて高度に多様である)
個々のヤツメウナギにおいてRT−PCRによってVLR多様性について調査した。3匹の免疫賦活幼生および4匹の非免疫賦活幼生由来の白血球mRNAを、VLR多様性領域に隣接するプライマーを用いて増幅させた。動物1匹に付き約10クローンのシーケンシングにより69個の固有のVLRが生じ、1匹の個体から同一のクローンは2個しか生じなかった。2匹の動物由来の20個のVLRの可変性配列は、タンパク質多様性を示している(図2;図3および図8に含まれた全セット)。134〜214アミノ酸のサイズ変動は、主としてLRRモジュール数における差に起因した。各配列は、LRRNT、18アミノ酸LRR1、ほぼ変わりなく24アミノ酸長の1〜9個のLRR、13アミノ酸の連結ペプチドおよびC末端LRRCTを含有する;LRRNTは、30〜38アミノ酸を有し、LRRCTは48〜58アミノ酸を有する。顕著な配列多様性の領域は各LRRモチーフから明白であるが、LRRNTにおける最初から7個の残基およびLRRCTにおける最終20個の残基はほぼ不変である。
【0136】
個々のリンパ球のレベルでのVLR多様性を評価するために、全ORFに隣接するプライマーを用いたRT−PCRを使用した。単細胞単離体を免疫賦活および非免疫賦活幼生の血液から選別した。非免疫賦活動物からの6回の単細胞反応および免疫賦活幼生からの7回の反応から入手したPCR産物の分析は、リンパ球13個中12個が単一VLRを発現し(図3)、10個の非免疫賦活細胞のコントロールプールからのVLRクローン6個中5個が固有であることを証明した。1個の細胞単離体は2個のVLR(9.16S、9.16L)を生じたが、この単離体が2個のリンパ球を含有していた可能性を排除することはできない。3個のVLRは、短縮されたタンパク質を予測させるインフレーム終止コドンを有していた。興味深いことに、免疫賦活幼生由来の5個のリンパ球(9.1=9.16S;9.2=9.16L;9.7=9.9)間では、同一VLRの組み合わせが同定された。追加の3匹の免疫賦活幼生(#5〜7)由来の血液サンプルの分析では固有のVLRしか明らかにならなかった(N=27)。これらの所見は、多様なVLRの単一対立遺伝子発現を示唆しており、VLRを有するリンパ球のクローン性増殖についての予備的証拠を提供する。
【0137】
(VLR遺伝子座の複雑性)
保存されたC末端プローブを用いたゲノムブロット・ハイブリダイゼーションにより単一バンドが明らかになった(図4a)。保存された5’UTRおよびシグナルペプチドからなるN末端プローブは、そのブロットが2つの追加のBamHIハンドを示した個体を除いて、使用した制限酵素に依存して、2〜3つのバンドと反応した。さらに、ゲノムパルスフィールドCHEFブロットにより、全6個の消化物中でC末端プローブとの単一ハイブリダイゼーションバンドが明らかになったが、他方N末端プローブは1つの追加の350kbのNotIバンドとのマッチングパターンを生じた(図4b)。これらの所見は、100〜150kbのゲノム内に含有される生殖細胞系VLR遺伝子(gVLR)のN末端およびC末端を備える単一VLR遺伝子座を示している(図4b;PacI消化物)。この遺伝子座を詳細に特性付けるために、これらのプローブを使用して、成体の赤血球DNAから構築された大きな挿入物ウミヤツメP1細菌人工染色体(PAC)ライブラリーをスクリーニングした。両方のプローブを用いてハイブリダイズした5つのPACの分析では、単一の14kbのVLR遺伝子(gVLR)アンプリコンが、ORFフランキングプライマーを用いたロングレンジPCR(LR−PCR)によって同定された。PCR産物の制限酵素分析では、同一のEcoRIバンドおよび2つの対立遺伝子BamHIパターンが明らかになった。2つのgVLR対立遺伝子を表しているPACクローンは、各々33および44kb挿入物を備えるPAC3およびPAC16であるとシーケンシングされた。それらの配列はgVLRを含有する20kbの領域にわたりオーバーラップしていた;PAC16は、gVLRから25kb上流に伸長し、PAC3は18kb下流に伸長した。PAC3および16の間のオーバーラップ領域は、PAC16のgVLRにおける短い欠失を除いて、ほぼ同一であった(24、43および78塩基対)。これらの配列は、このためPAC3のわずかに長い配列を保存しているgVLRコンティーグ内へ融合させられた(図5a)。
【0138】
PAC3/16コンティーグ内のgVLRは4つのエクソンからなる。第1は5’UTRの一部を含有する;エクソン2は5’UTRの残り、シグナルペプチドおよびLRRNTの5’側半分を含有する;エクソン3はLRRCTの5’側半分を含有し、そしてエクソン4はLRRCTの3’側半分、C末端および3’UTRをコードする。標準的な真核細胞スプライス部位は5’UTRイントロンでしか同定されなかったが、他方gVLR内の他のエクソン/イントロン境界はcDNA配列とのアライメントによって決定された。注目すべきことに、gVLR配列は、3’LRRNT、LRR1または24アミノ酸LRRのいずれも含有していなかった。このgVLRから上流では、順方向または逆方向のいずれかに配置されたLRRNT、LRR1およびLRRを含む、一重または二重の可変性LRRモジュールの6つのカセットが同定された。これらのLRRカセットはコンティーグの最初の6kbに及んだが、他方2つの多様な5’LRRCTカセットはgVLRから7kb下流に位置した。
【0139】
また別のクローンのPAC4は、N末端プローブを用いた場合しかハイブリダイズしなかったが、LRRNTおよびLRR1コンセンサスプライマーを用いるPCRによって同定された複数のLRRをコードすることが見いだされた。全挿入物は58kb長であり(図5b)、この配列は小さなギャップ(PAC4内の210〜738塩基対の4つのギャップおよびPAC3/16コンティーグ内の25〜55塩基対の8つのギャップ)を備える11.7kbのgVLRコンティーグとオーバーラップした。このオーバーラップはgVLRエクソン2と3の間の介在配列内に伸長したが、PAC4の553塩基対の末端配列は固有であった。総計30の、1〜3個の多様なLRRモジュールの17のカセットは、部分gVLRから15kb上流に位置したPAC4内の31kb領域内でコードされた。PAC3/16gVLRコンティーグとPAC4配列との比較により、追加の1〜5kbの領域が90%を超える同一性を備えることが明らかになったが、これらは非関連配列によって中断された。PAC4は、5’フランクおよび約半分のgVLRを含む約12kbの重複、または高度に相違するVLR対立遺伝子のいずれかを表すことができよう。これらの可能性を区別するために、これらのPAC挿入物からのgVLR内の制限部位のマップに対するN末端プローブ(図4a)とゲノムハイブリダイゼーションのパターンを比較した。ブロットパターンおよび制限マップは、ブロット内の2kbバンドとは相違していたPAC4からの5.7kbのHindIIIフラグメントを除いて、全フラグメントについて匹敵していた(図4a)。3つのブロットされたゲノムおよびPACライブラリーからのゲノム間のそのような限定された変動性を考えると、PAC4は多型性gVLR対立遺伝子を表している可能性はないと思われる。限定されたVLR対立遺伝子変動は、マイクロサテライト遺伝子座においてさえ低対立遺伝子多様性の他の証拠と一致しており(Bryan et al.,2003)、北米五大湖内のウミヤツメ集団およびその他の淡水に生息する集団が高度に近親交配されていることを示している。そこでこの分析は、単一ヤツメウナギgVLR遺伝子座がgVLRのN末端半分の追加のコピーを有していることを示している。
【0140】
(体細胞性gVLR再配列は多様な成熟VLRを生成する)
VLR多様性領域に隣接するプライマーを用いたPCR増幅によって幼生DNAサンプルを分析すると、6個の固有のイントロンレスVLR ORFが得られた(図3、動物番号#10、12)。この興味のある所見にしたがって、ORFフランキングプライマーを用いた幼生DNAサンプルのPCR増幅は、5’UTRイントロンを含む1.5〜2kbのVLRクローンを生成し、これはクローン14個中13個の固有の配列を明らかにした(#10、11)。これらのゲノムPCRクローンは連続したVLR ORFを含有していたので、それらは「不完全」生殖細胞系VLRからそれらを区別するために暫定的に成熟VLRと名付けられた。配列分析は、これらの成熟VLRがN末端プローブとハイブリダイズする1〜1.3kbの多型性EcoRIバンドを生成することを示唆したが、これらのバンドはリンパ球DNAブロット内でしか観察されなかった(図4cが含まれる)。これらの観察は、ペレット化された赤血球または幼生全身から抽出されたヤツメウナギDNAサンプルが成熟VLRを含有しているが、これらのサンプルからのDNAブロットで検出されるほど十分に豊富なのは生殖細胞系VLRのコピーだけであることを指摘している。
【0141】
この不可解な謎を解決するために、ヤツメウナギリンパ球内での体細胞性遺伝子再配列が、生殖細胞系gVLRからの非コーディングDNAを上流および下流カセットからの多様なLRRと置換して、小さな成熟VLRを生成すると理論付けた。この仮説を試験するために、生殖細胞系gVLRにわたるPCR増幅のための、上流の約3kbおよび下流フランクの約3kbを含むプライマーを設計した(図5a)。幼生DNAサンプルからのLR−PCR増幅は、PAC16プラスミドからのgVLRアンプリコンに類似する約20kbの小さなバンドにプラスして約8kbの追加の顕著なバンドを生じた(図5c)。2匹の幼生サンプルからの8kbアンプリコンの配列分析は、クローン10個中9個が固有の成熟VLRをコードすることを明らかにし(図3)、それらのフランクはgVLRのフランクと同一であった(図5d)。全28個の固有の成熟VLRは、4つの幼生DNAサンプルからのPCR産物間で同定された。リンパ球DNAは、おそらくこれらのDNAサンプルを抽出するために使用されたペレット化赤血球または幼生全身の小さな分画としての、これらの成熟VLRにとってのテンプレートであった。明らかに、LR−PCR中にはリンパ球成熟VLRのより短いテンプレートが優先的に増幅された。gVLR ORFに隣接するプライマーを用いて増幅した場合に、1.5〜2kbの成熟VLRおよび14kbのgVLRの2つのアンプリコンを生じさせる類似のPCRバイアスが観察された。
【0142】
そこでヤツメウナギにおける適応免疫応答を惹起するであろうリンパ球受容体についての探索は、有顎類脊椎動物のIgおよびTCRとは完全に相違する可変性リンパ球受容体の系を同定する。VLRは、複数のLRRモジュールおよびGPIアンカーを介してリンパ球血漿膜へ結合させられている不変柄領域からなる。N末端およびC末端LRRのフランキングチップは不変であり、介在LRRの数および配列における変化は顕著なVLR多様性の一因となる。潜在的VLR多様性は354の固有配列中345を備えて膨大であり、免疫賦活リンパ球由来の同一VLRは3対に過ぎず、ほぼ同一のVLRは3個であった。したがってVLRは、この代表的無顎類に多様なリンパ球受容体レバートリーを与えている。
【0143】
これらの高度に多様なVLRは、病原体の認識において役割を果たす。多様なLRRモジュールを特徴とするタンパク質は、それらが並外れた広範囲のリガンドアレイと相互作用する傾向を示すために、動物および植物の主要な本質的免疫受容体である。動物TLRは、ウイルス、細菌、真菌および原生動物上で保存されたエピトープの認識に関係しており、炎症反応において最高点に達するシグナル伝達カスケードを活性化する(Beutler,2004)。可溶形でも見いだされるGPI固定LRRタンパク質であるCD14は、細菌リポ多糖およびリン脂質へ結合して、TLR4受容体とのシグナリング複合体を形成する(Landmann,2000)。さらにまた別のサイトゾルLRRタンパク質の哺乳動物ファミリーであるNBS−LRRは、細胞内病原体を認識する(Chamaillard et al.,2003)。植物疾患耐性遺伝子は、数百種ものNBS−LRRタンパク質、LRR受容体様キナーゼおよびLRR受容体様タンパク質を含む大きな多重遺伝子ファミリーのメンバーであり、それらの多くは抗病原体反応の特異的活性化に関係することが証明されている(Jones et al.,2004)。顕著な多様性を備える抗原結合VLRは、ヤツメウナギにおいて観察される適応免疫応答を媒介する。リンパ球の表面へのVLRのGPI固定は、これらの受容体のGPI特異的ホスホリパーゼ放出を許容し(Ikezawa 2002)、VLRに予想免疫系における表面受容体および体液性アグルチニンの両方としての二重機能性を付与する。
【0144】
ゲノムPACクローンのシーケンシングにより、シグナルペプチド、5’LRRNT、5’LRRCT、3’LRRCTおよびC末端だけをコードする4つのエクソンからなる生殖細胞系gVLRが同定された。gVLRは5’LRRCTを除いて多様性LRRモジュールを欠如しており、これは修飾しないと高度に多様なVLRメッセージをコードできないであろうことを示唆していた。しかし、複数の多様なLRRカセットはgVLRの上流および下流で見いだされ、これらは成熟VLR遺伝子を組み立てるためにgVLR内へ挿入するために利用できよう。成熟VLRは上流および下流LRRカセットを備える生殖細胞系gVLR内の非コーディングDNAの体細胞性置換を通して生成されるという仮説を試験するために、LR−PCRを使用して生殖細胞系および成熟VLR遺伝子両方の存在を検出した。gVLRからの約20kbの予想された産物は2匹のヤツメウナギのゲノムDNA、そしてさらに、多様なセットの成熟VLRをコードすることが見いだされた成熟VLRからの予測8kbのアンプリコンから入手された。さらに、少数の例では、候補LRRドナーはVLR配列に対する同一性に基づいてgVLR隣接カセット間で同定することができ、そしてgVLR 5’LRRNTおよび3’LRRCT内の高度に保存された配列は遺伝子変換プロセスのための固定領域として潜在的に機能できるであろう。VLRは、体細胞性DNA再配列の機構によって生成される。
【0145】
非減数分裂性DNA再配列は、他の系から知られている。例えば、表面構成成分をコードする遺伝子の再配列は、慢性感染中の免疫認識から逃れるために数種の病原体によって使用される戦略である。淋菌(Neisseria gonorrhoeae)のピリンにおける抗原性変化は、pilE遺伝子座と複数のサイレントなpilSコピーとの間の非可逆的組み換えを含んでおり(Hamrick,2001)、ライム病ボレリア属スピロヘータ類(Borrelia spirochaetes)における抗原性変化は1アレイの15のサイレントカセットとvlsE発現部位間の遺伝子変換によって生成される(Wang et al.,2003)。同様に原生動物ブルーストリパノソーマ(Trypanosoma brucei)は反復されたDNA再配列によるそれらの変異体表面コート糖タンパク質の発現を交互に繰り返し(Donelson,2003)、ならびにマラリア原虫熱帯熱マラリア原虫(Plasmodium falciparum)および腸常在菌であるランブル鞭毛虫(Giardia lamblia)は複数の表面抗原遺伝子間で頻回に切り替わる。宿主と寄生虫との間の進化上の武器競争(arm race)において、脊椎動物は生殖細胞系受容体の体細胞性再配列によって感染性疾患に対抗するために類似の戦略を採用した。多様なリンパ球抗原受容体は、有顎類では対のトランスポザーゼ様RAG1およびRAG2のカットアンドペースト活性によって(Schluter et al.,1999)および無顎類ではいまだ特性付けられていない機構によって組み立てられる。
【0146】
ヤツメウナギVLR系の特徴は、2つの顕著な相違とともに、有顎類脊椎動物リンパ球のIgおよびTCRとの類似性を有する。第1に、ヤツメウナギVLRはLRRモジュールからなるが、他方有顎類抗原受容体はIgドメインからなる。ヤツメウナギの免疫は、祖先の生殖細胞系をコードするLRR受容体の多様性を、その生殖細胞系VLR遺伝子を体細胞性で多様化したものである可変性リンパ球LRR受容体の系と置換する、段階的進化プロセスを受ける。これとは対照的に、有顎類脊椎動物の組み換えリンパ球受容体のコア成分としてのIgドメインは、Igスーパーファミリーメンバーが有顎類脊椎動物以外の動物における病原体または同種移植片の任意のタイプの免疫認識において何らかの役割を果たすことは証明されていないので、それらの祖先からの興味深い追跡不能な進化的推進力である。(Kaufman,2002)。第2に、ヤツメウナギ中のMHC分子の存在についての証拠は見いだされていない。有顎類脊椎動物においては、抗原ペプチドをT細胞へ効率的に提示させるためには多型性MHC分子が必須であるが、他方近親交配されたMHCホモ接合体は疾患耐性の損傷に苦しむと思われる(Penn et al.,2002;Grimholt et al.,2003)。ヤツメウナギは五大湖内の近親交配集団として成長するので、これはそれらのVLR系が多型性構成成分とは無関係に機能するように進化してきた可能性を示している。
【0147】
(動物)
ウミヤツメの幼生(体長8〜13cm)はミシガン湖の支流(Lamprey Services、ミシガン州ラジントン)、またはヒューロン湖の支流(Hammond Bay Biological Station、ミシガン州ミラーズバーグ)からであった。免疫賦活のための幼生は鎮静させ(100mg/L MS222;Sigma)、107大腸菌BL21(DE3)、107ヒツジ赤血球、50μgのフィトヘマグルチニンおよび25μgのヤマゴボウマイトジェン(Sigma)を含有する75μLの0.67×PBSを腹腔内注射した。免疫賦活は1週間間隔で2または4回繰返し、細胞は最終免疫賦活の3〜4日後に採取した。尾を切断した幼生から採血し、0.57×PBSおよび30mM EDTAを用いて1:1に希釈した。バフィコート白血球を50gでの5分間の遠心によって収集した。MoFlo血球計算器を用いて(Mayer et al.,2002a)に記載されたように細胞を選別した。
【0148】
(差引きした免疫賦活白血球cDNAライブラリー)
Super SMART PCR cDNA合成法(BD Biosciences)を1週間間隔で4回免疫賦活した幼生から選別した大リンパ球、骨髄細胞および赤血球からのmRNAとともに使用した。2回の反応において活性化リンパ球cDNAは、骨髄細胞または赤血球のcDNAに対して差引きした(PCR−Select,BD Biosciences)。差引きした産物をpGEM−T Easy(Promega)内でクローン化し、1,507配列について分析した。
【0149】
【表3】
(VLR RT−PCR)
非免疫賦活幼生(#1〜4)、または1週間間隔で2回免疫賦活した幼生(#5〜7)からのバフィコート白血球を300gで5分間かけてペレット化した。第1鎖cDNAは50ngのランダムヘキサマー(Superscript III;Invitrogen)を用いてプライミングした。VLR多様性領域は、LRR.F1+LRR.R1を用いるExpand High Fidelity(Roche)を使用して増幅させた(表3)。熱サイクルは次のとおりであった:94℃で1分間、次に94℃で30秒間、59℃で30秒間、72℃で1分間の35サイクル。動物1匹当たり10〜12クローンをシーケンシングした。
【0150】
(VLR単細胞RT−PCR)
非免疫賦活幼生(#8)、および1週間間隔で2回免疫賦活した幼生(#9)のバフィコート由来の単一リンパ球、または10−細胞プールを0.2mLのTRIzol(Invitrogen)内へ選別した。第1鎖cDNAはLRR_C.R2を用いてプライミングした。VLRは、最初はAdvantage II(BD Biosciences)を用いるSlit.F+LRR_C.R2および次にExpand High Fidelityを用いるLRR_N.F1+LRR_C.R1の2ラウンドのネステッドPCRによって増幅させた。サイクリングパラメータは次のとおりであった:94℃で1分間、次に94℃で30秒間、60℃で30秒間、72℃で1分間の40サイクル。ベクタープライマーを用いたコロニーPCRによって、12個の細胞各々からの6コロニー内で単一サイズ挿入物が明らかになり、それらのうちの3つをシーケンシングした。細胞9.16からのコロニーは2サイズであることが明らかになり、3つの短い挿入物および3つの長い挿入物をシーケンシングした。10個の非免疫賦活細胞プールからは、6個のクローンをシーケンシングした。
【0151】
(ゲノムDNAおよびゲノムPCR)
ゲノムDNAは、幼生全身の1/3、50gで5分間かけてペレット化した0.25mLの血液からの赤血球、または107の選別されたリンパ球から単離した。PCRは、Expand Long Template(Roche)を用いて400ngのgDNAから実施した。VLR多様性領域は、LRR.F1+LRR.R1を用いて幼生#10および12から増幅させた。成熟VLRは、Slit.F+LRR_C.R2またはLRR_N.F1+LRR_C.R1を用いて、動物#10および11から増幅させた。gVLRを超える増幅は、gVLR.F1+gVLR.R1を用いて動物#10および13からであった。8kbバンドは、pCR−XL(Invitrogen)内でクローン化し、M13.Forward、M13.Reverse、Slit.FおよびLRR_C.R2を用いてシーケンシングした。
【0152】
(実際のノーザンブロットおよびDNAブロット)
実際のノーザンは、推奨されたように調製した(Super SMARTマニュアル)。20サイクルで増幅させたcDNAは、幼生尾部、肝臓ならびに非免疫賦活動物の血液、盲樋および腎臓から選別したリンパ球から、または1週間間隔4回免疫賦活した幼生の血液から選別した小および大リンパ球、骨髄細胞および赤血球からであった。
【0153】
1レーン当たり10μgの幼生#10、12および13からのゲノムDNAは、BamHI、EcoRIまたはHindIII(Roche)を用いて消化した;5μgのリンパ球DNAはEcoRIを用いて消化した。パルスフィールドCHEFブロットのために、10匹の幼生からの赤血球をアガロース中に包埋し、1レーン当たり20μgのDNAはAscI、FseI、NotI、PacI、PmeI、またはSfiIを用いて消化した。
【0154】
次の32P標識プローブを使用した:Slit.F+Slit.Rを用いてクローンLRR−2913からPCR増幅させたVLR N末端プローブ(196塩基対)、およびLRR_C.F1+LRR_C.R1を用いて増幅させたC末端プローブ(208塩基対);GAPDH.F+GAPDH.Rを用いてクローンPmGAPDHから増幅させたGAPDHプローブ(314塩基対)。
【0155】
(PACライブラリーおよびクローン)
pCYPAC6(AF133437)中のアレイ化ウミヤツメPACライブラリーは、部分MboI消化物を用いて1匹のミシガン湖成体の赤血球DNAから構築した。6×104クローンは、1〜2倍のゲノム被度を有する65kbの平均挿入物を有していた。ライブラリーは、N末端およびC末端プローブ両方を用いてスクリーニングした。陽性クローンのプラスミドは、EcoRI消化し、ブロッティングし、そしてN末端またはC末端プローブのどちらかとハイブリダイズした。5種のPACは両方のプローブとハイブリダイズし(2、3、15、16、17)、5種のPACはN末端プローブとのみハイブリダイズした(4、9、14、35、42、43)。
【0156】
gVLRは、Slit.F+LRR_C.R2を用いてPAC2、3、15、16および17のプラスミドからのExpand Long Templateを用いて増幅させた。全PCR産物は、2セットのBamHIパターン(PAC2、3および15〜17)を含む14kbであった。PAC3、4および16はMcGill University(カナダ国ケベック州)でシーケンシングされた。
【0157】
(VLR GPIアンカー)
LRRNTから終止コドンまでのVLR挿入物は、Dis_LRR.F+Dis_LRR.R1を用いるExpand High Fidelityを使用してクローンLRR−2913から増幅させ、pDisplay(Invitrogen)内のIgκシグナルペプチドおよびヘマグルチニンエピトープへ融合させた。表面局在化およびVLR GPIアンカーは、BW1547細胞、またはmFcγRIIbを発現するコントロール中で分析した。細胞は、30℃で45分間にわたり1単位/mLの細菌性GPI特異的ホスホリパーゼC(Sigma)を用いて処理した。エピトープタグ付けタンパク質は、抗HA−タグmAb 12CA5を用いて表面染色した。
【0158】
(配列分析)
配列変動性は、MEGA2.1 UPGMA(Kumar et al.,2001)を用いて推定した。GPIアンカー部位は:http://129.194.185.165/dgpi/によって同定した。SWISS−MODEL VLR 3D構造は:http://cubic.bioc.columbia.edu/predictprotein/submit_meta.htmlによって同定した。クローン12.26からの残基22〜319は、CD42a(1m10.pdb)およびNOGO−66受容体(lp8t.pdb)の結晶座標上に進ませた。
【0159】
(実施例2:メクラウナギにおける可変性リンパ球受容体)
(円口類VLRホモログ)
VLRの2つの明確なタイプであるVLR−AおよびVLR−Bは、沿岸メクラウナギであるEptatretus burgeriの12,000個の白血球cDNAクローンから発現した配列タグ間で同定された(Suzuki et al,2004B)。適合するVLRは次に、太平洋メクラウナギE.stoutiiのリンパ球様細胞の転写産物からRT−PCRによってクローニングした。図9は、メクラウナギVLR−AおよびVLR−B、ウミヤツメVLR(Petromyzon marinus)および2つの非寄生性ヤツメウナギのVLR、アメリカカワヤツメ(Lampetra appendix)および北部カワヤツメ(Ichthyomyzon fossor)のアミノ酸配列のアライメントを描出している。これらのVLRは類似の構造的ドメインを共有している:シグナルペプチド(SP)、N末端LRR(LRRNT)、18残基LRR1、その後に可変数の24残基LRR、13残基連結ペプチド(CP)およびC末端LRR(LRRCT)が続く。C末端の最初では、ヤツメウナギVLRおよびメクラウナギVLR−Bはトレオニン/プロリンリッチ領域を有しているが、この領域はメクラウナギVLR−A内では明確には保存されていない。全VLRタンパク質は、グリコシル−ホスファチジル−イノシトール(GPI)細胞表面膜アンカーを加えるためにタンパク質を修飾するために必要とされる疎水性尾部領域で終了する。ウミヤツメVLRと同様に、メクラウナギVLR−AはGPI固定タンパク質であると予測されたが、ω開裂部位は同定されなかった(DGPI http://129.194.185.165/dgpi/);VLR−BについてのC末端疎水性プロファイルもまたGPI修飾の予測因子である。
【0160】
メクラウナギVLRの転写産物はリンパ球様細胞中では豊富であるが、それらの光散乱特性によって選別された骨髄細胞または赤血球中では豊富ではない。VLR−A転写産物レベルは、白血球サンプル中のVLR−Bレベルより約3倍高かった。太平洋メクラウナギの両方のVLRタイプは高度に異質であり(図10AおよびB)、これは可変数の24残基のLRRモジュールおよび顕著なLRR配列多様性を示していた。匹敵する多様性は沿岸メクラウナギからのVLR−A配列(N=66)およびVLR−B配列(N=18)について観察された(図13)。興味深いことに、同一であった、または1〜2個の残基しか相違していなかった2〜4個のVLR−Aクローンの5つのクラスターは、抗原とマイトジェンとのカクテルの4回にわたる週1回の注射が投与されたメクラウナギ#5(図10Aにおいて星印で表示した)からの40の転写産物間で見いだされた。このメクラウナギからのVLR−A転写産物の30%が関連配列のクラスターから構成されるという所見は、リンパ球を有するVLR−Aのクローン性増殖を示唆している。1〜2のアミノ酸置換を備えるクローンは、体細胞性超変異を通しての追加のVLR多様化を反映している。
【0161】
固有の配列である太平洋メクラウナギVLR−Aのデータセット(N=130)から、24残基LRRの転写産物による2〜6コピーが明らかになる(N=527;平均4)。VLR−Bデータセット(N=69)内には、1〜6コピーの24残基LRRが存在したが(N=195;平均2.8)、他方129のウミヤツメVLR(19;GenBank受託番号AY577943〜AY578059)のデータセット内には1〜9コピーの24残基LRRが存在した(N=325;平均2.5)。そして、これらのVLRの個別構成成分を、確実なアライメントのために種間で相違し過ぎるLRRNTおよびLRRCTを除いて(表4;328個のLRR1ドメイン、328個のCPドメイン、および24残基LRRの1,047個の単一ドメイン)、近隣結合系統樹において個別に分析した。
【0162】
【表4】
これらのクラスターは、ほぼ排他的に同一タイプおよび種起源、すなわち太平洋メクラウナギVLR−A、VLR−B、またはウミヤツメVLRクラスタリングであった。相違するVLRタイプ間に同一LRRドメインの例はなかった。しかし、メクラウナギVLR−Aクラスター内およびヤツメウナギVLRクラスター内のLRR1およびCPドメインの大部分は同一であった(表4)。これとは対照的に、メクラウナギVLR−B内のLRR1ドメインは98%固有であった;24残基LRRのセットは、さらに主として固有配列から構成された:97%はメクラウナギVLR−B中で、90%はVLR−A中で、および83%はウミヤツメVLR中で固有であった。この顕著に高度の多様性は、24残基LRRタイプ各々について引き出されたコンセンサス配列が少なくとも10フレームワーク残基を共有することを前提にすると特に注目に値する。
【0163】
(メクラウナギVLR遺伝子)
太平洋メクラウナギおよび沿岸メクラウナギのVLR遺伝子座のゲノム組織は、細菌性人工染色体(BAC)ライブラリーから単離した大きな挿入物のゲノムクローンの配列から決定したが、BACは各VLRタイプに対して1つであった(図11)。メクラウナギゲノム内では、各gVLRについて1コピーしか同定されなかった。遺伝子座の配列および組織は、両方の種でほぼ同一であり、gVLR−AおよびgVLR−B間で相当に保存されている。メクラウナギgVLRは、5’非翻訳領域(UTR)で始まり、2つのコーディング領域が続く(図12A)。ウミヤツメgVLRにおけると同様に、5’UTRは、太平洋メクラウナギgVLR−A内では6.4kb長およびgVLR−B中では220塩基対であるイントロンによって分割されている。メクラウナギgVLR内の第1コーディング領域は、gVLR−A内のシグナルペプチドおよびLRRNTドメインならびにgVLR−B内の23残基シグナルペプチド中の残基1〜13だけをコードする。次に、gVLR−AおよびgVLR−Bに対して各々171および211塩基対の短い介在配列が存在する。第2コーディング領域は、LRRCTの5’末端をコードするヤツメウナギ領域が欠けていること以外は、ウミヤツメgVLR中と同様に、LRRCTの3’末端およびC末端からなる。メクラウナギgVLRは、開始コドンから終止コドンまでがgVLR−A内では671塩基対そしてgVLR−B内では410塩基対とコンパクトである。
【0164】
メクラウナギgVLR遺伝子座は、生殖細胞系遺伝子から約20〜40kb下流に位置する多様なLRRモチーフをコードするカセットを有している(図11)。VLR−A遺伝子座では、多様なCPドメインの6〜8末端残基およびgVLR−A 3’LRRCTとの4残基の同一オーバーラップを含む5’LRRCTをコードするカセットが存在する。さらに下流には、gVLR−Aに比較して逆方向に配置された2つの多様なLRRおよび次に逆方向不完全5’LRRCTを含むカセットが存在する。gVLR−B遺伝子座には、シグナルペプチドの残基7〜23および5’LRRNT、次に多様なCPドメインおよび5’LRRCT、1つの逆方向LRRをコードするカセットがあり、さらにより下流では、LRRの12末端残基および8個の近位残基からなる別の逆方向LRRカセットが存在する。gVLRから約50kb上流および約70kb下流にわたるフランキングDNA内では他の多様なLRRモジュールは同定されなかった。しかし、リンパ球様細胞からのゲノムPCRアンプリコンのサンプル:2匹の動物からの35個の固有の成熟VLR−Aおよび38個のVLR−B配列中で同定された成熟VLR遺伝子の欠如している構成成分を提供するために多様なLRRエレメントがゲノム内のどこかに存在すると思われる(図10)。そこで、メクラウナギ成熟VLR遺伝子は、ヤツメウナギのための場合と同様に、体細胞性組み換えを通して組み立てられなければならない。
【0165】
メクラウナギリンパ球様細胞中の生殖細胞系VLR遺伝子は、遺伝子再配列の前に能動的に転写される。VLR−A生殖細胞系転写産物のPCRアンプリコンは、約0.7kb長およびVLR−Bについては約0.5kbであるが(図12B、RT−PCR;PCRプライマーの位置は図12Aに示されている)、他方より大きなアンプリコンは再配列された成熟VLR遺伝子からの転写産物に対応して、VLR−AおよびVLR−Bに対して各々約1.1および約0.8kbであった。血中ゲノムDNAからの対応するPCRアンプリコンは、生殖細胞系遺伝子に対しては約0.7kbならびに成熟VLR−A遺伝子および成熟VLR−B遺伝子に対して約1.1kbである(図12B、ゲノムPCR)。生殖細胞系および成熟VLR遺伝子からの転写産物中では、5’イントロンはスプライシングされて出され、対応するゲノムPCRアンプリコンより短いRT−PCR産物が産生する(図12BにおけるVLR−Bを参照;gVLR−Aアンプリコンは6.4kbイントロンを含んでいない)。しかし、コーディングエクソン間の介在配列は、それらにコンセンサスな真核細胞のスプライス部位が欠けているために、生殖細胞系転写産物中に保持されている。それに対して生殖細胞系スイッチ領域転写が必須である哺乳動物抗体クラススイッチ組み換えにおける場合と同様に、gVLR再配列のために生殖細胞系転写が必要とされることがある(Bottaro et al,1994;Hein et al.,1998)。
【0166】
(VLR系統発生学)
無顎類VLRタンパク質の系統発生学的分析により、ヤツメウナギVLR配列、メクラウナギVLR−A配列およびVLR−B配列によって各々構成される3つの別個のクラスターが明らかになる(図12C)。メクラウナギVLR−BおよびヤツメウナギVLRは、メクラウナギVLR−Aとの分科とは別個の分科でクラスターを形成している。VLR多様性領域であるLRRNT〜LRRCTまたはLRR1〜CPだけがアライメントされた場合は同一樹形が見られた。そこでメクラウナギVLR−Aは祖先遺伝子の重複によって発生した(図12D)か、あるいは4億9,900万年±3,800万年前のカンブリア紀においてメクラウナギとヤツメウナギ系統に分裂した後にVLR−Aオルソログを消失した(Hedges et al.,2001)。ヤツメウナギVLR−Aオルソログが存在するが、それは極めて低レベルで発現する、または非リンパ系細胞中では発現しないために、ヤツメウナギリンパ球様細胞由来の18,000を超えるcDNA配列中では検出されなかった可能性もある(Pancer et al.,2004)。
【0167】
現存する円口類の両方種類におけるVLRの存在は、脊椎動物が先行の分子認識系を発達させるための強力な進化圧力の指標である。この分析は、カンブリア紀における4,000万年未満以内に、LRRまたはIgいずれかの遺伝子フラグメントが組み換えアッセンブリーを受けてリンパ球受容体の多様なレパートリーを生成する2つの根本的に相違する系が無顎類および有顎類において進化したことを示唆している。この進化のシナリオは、多くの興味深い疑問を引き起こすが、それらの1つは2つの適応免疫戦略が収束進化を意味するかどうか、または一方が他方の先祖であるかどうかという問題に関する。VLRが先駆脊椎動物免疫受容体であったのか、または個別に進化したVLRおよびIgの再配列であったのかは、現存および絶滅無顎類および有顎類脊椎動物の群間での系統発生的関係が明確に解決された場合にしか明らかにならないであろう(Mallatt et al.,2003;Meyer et al.,2003)。しかしこの点について、現代の無顎類の両方の種類におけるVLRの存在は、円口類の単一系統起源に好都合な追加の分子的証拠を提供している。
【0168】
(動物)
太平洋メクラウナギEptatretus stoutii(体長30〜60cm)の生きている標本は、Marinus Scientific(カリフォルニア州ロングビーチ)から購入し、12℃の人工海水(Oceanic System、テキサス州ダラス)中で2カ月間維持した。アメリカカワヤツメ(Lampetra appendix)および北部カワヤツメ(Ichthyomyzon fossor)の幼生(体長:15〜20cm)は、五大湖の支流から入手した(Lamprey Services、ミシガン州ラジントン)。
【0169】
メクラウナギはpH=7に緩衝した0.5g/LのMS222(Sigma、ミズーリ州セントルイス)中へ15分間浸漬することにより鎮静し、その後に0.5mLのメクラウナギPBS中の抗原/マイトジェンカクテル(1Lに付き:28gのNaCl、0.2gのKCL、1.44gのNa2HPO4、0.24gのKH2PO4、pH=7.4、1オスモル)を用いて腹腔内注射した。このカクテルは、109の生きている大腸菌TG1細菌、109のヒツジ赤血球(Colorado Serum Company、コロラド州デンバー)ならびに各100μgのフィトヘマグルチニンおよびヤマゴボウマイトジェン(Sigma)を含有していた。免疫賦活は1週間間隔で繰返し、第4回目の免疫賦活から4日後にシリンジを用いて尾血洞から血液を採取し、30mM EDTAを含有するメクラウナギPBSを用いて1:1に希釈した。50×gでの5分間の遠心後に採取したバフィコート白血球は報告されたように(Newton et al.,1994;Raison et al.,1994)、MoFlo血球計算器(Cytomation,コロラド州フォートコリンズ)を用いて光散乱特性によって選別した。
【0170】
(メクラウナギVLR)
沿岸メクラウナギEptatretus burgeriのVLRホモログは、非免疫賦活動物#7、8の白血球RNAから発現した配列タグのデータベースに対するBLASTクエリーとしてヤツメウナギVLRを用いて同定した(Suzuki et al.,2004B)。両方のストランド上の有意な適合を有するクローンを:64個のVLR−A cDNAクローンおよび15個のVLR−B cDNAクローン上でシーケンシングした。太平洋メクラウナギに対しては、3匹の非免疫賦活個体(#1〜3、6)からの分離されていない血球およびバフィコート白血球、および2匹の免疫賦活動物(#4、5)からのバフィコート白血球を血液ゲノムDNAおよび白血球RNAを抽出するために使用した。RNAの抽出はTRIzol試薬(Invitrogen、カリフォルニア州カールズバッド)を用いて行い、ポリA RNAはDynabeads mRNA精製キット(Dynal Biotech、ニューヨーク州レークサクセス)を用いて選択した。第1ストランドcDNA合成は、Superscript III First Strand cDNA合成キット(Invitrogen)を用いて、VLR−Aについては20pmoleのHgVLRA.F1(表5)、またはVLR−BについてはHgVLRB.F1を用いてプライムし、生成物はカラム精製した(QIAquick PCR精製;QIAGEN、カリフォルニア州バレンシア)。
【0171】
【表5】
VLRは次に、10pmole/μLでフォワードプライマーおよびリバースプライマーのセット(F1またはF2およびR1またはR2)各1μL、5μLの10×バッファー、36.25μLのDDW、5μLのcDNAもしくはゲノムDNA(250ng)および0.75μLの伸長酵素を含有する50μL反応液中で、Expand High Fidelity PCR(Roche Applied Science、インディアナ州インディアナポリス)を用いて、cDNAまたはゲノムDNAからPCR増幅させた。反応液は、94℃で1分間の1サイクル、次に94℃で30秒間、58℃で30秒間および72℃で1分間の35サイクル、および最後に72℃で7分間の伸長を用いて増幅させた。生成物をカラム精製し、pCRII−TOPO(Invitrogen)中でクローン化し、挿入物をシーケンシングした。太平洋メクラウナギについては、109個のVLR−A RT−PCRクローンをシーケンシングし(4個はインフレーム終止コドンを含有していた)、そして36個のゲノム成熟VLR−Aアンプリコンをシーケンシングした(2個がインフレーム終止コドンを含有していた)。VLR−Bについては、37個のRT−PCRクローンをシーケンシングし(1個はインフレーム終止コドンを含有していた)、そして38個のゲノム成熟VLR−Bアンプリコンをシーケンシングした(4個がインフレーム終止コドンを含有していた)。沿岸メクラウナギ#9からの肝臓ゲノムDNA(Suzuki et al.,2004B)をPCRクローニングのために使用し、成熟VLRについてシーケンシングした:4個の成熟VLR−Aアンプリコン(2個はインフレーム終止コドンを含有していた)および3個の成熟VLR−Bアンプリコン。
【0172】
(非寄生性ヤツメウナギVLR)
第1鎖cDNAは、リバースプライマーVLR_3UT.R(ウミヤツメ3’UTRプライマー、表5)を用いて上述したように合成した。アメリカカワヤツメについてはフォワードプライマーおよびリバースプライマーはSlit.F(ウミヤツメ5’UTRプライマー)であり、北部カワヤツメについてはLRR_N.F1(また別のウミヤツメ5’UTRプライマー)であった。アメリカカワヤツメの総計13個および北部カワヤツメの7個の固有のVLRクローンをシーケンシングした。
【0173】
(BACライブラリーおよびクローン)
沿岸メクラウナギBACライブラリー(Suzuki et al.,2004A)は、上述したようにVLRプライマーを用いるPCRによってスクリーニングした(F1またはF2およびR1またはR2)。太平洋メクラウナギBACライブラリー(VMRC23)は、ベクターpCCBACE1(Epicentre Technologies、ワイオミング州マディソン)内の単一標本からの赤血球DNAのEcoRI部分消化物から構築した。このライブラリーは、約184,000個の組み換え体からなり、メクラウナギゲノムの約5×の有効範囲を含んでいる。全ライブラリーは5’および3’のVLR−AプローブおよびVLR−Bプローブを用いたハイブリダイゼーションによってスクリーニングし、陽性クローンはPCRによって正しいことを確証された。太平洋および沿岸メクラウナギ由来の各VLRタイプについて1つのBACを約10×有効範囲でシーケンシングし、コンティーグに組み立てた(Macrogen、韓国ソウル)。挿入物の不完全配列の場合は、コンティーグ内にギャップを占めないgVLRおよびLRRカセットを含有する部分だけを含めた:Eb_gVLR−A、43,362bp;Eb_gVLR−B、92,072bp;Es_gVLR−A、81,648bp;Es_gVLR−B,76,730bp。
【0174】
(配列分析)
隣接結合およびUPGMA樹は、MEGA3 Molecular Evolutionary Genetics Analysisからのプログラムを用いてペアワイズ欠失選択を用いて構築した(Kumar et al.,2004)。BAC挿入物中の遺伝子の予測は、ftp://ftp.ncbi.nlm.nih.gov/blast/executables/およびGenScanサーバー:genes.mit.edu/GENSCAN.htmlからダウンロードしたローカルBLASTを使用することによって遂行した。
【0175】
本出願を通して、様々な刊行物が参照されている。これらの刊行物の開示は、本発明が関係する技術の現在の状態をより十分に記載するために、これにより全体として本出願に組み込まれる。開示した参考文献は、その参考文献が依拠している文章の中で考察されている、その中に含有される物質については、本明細書に個別かつ詳細に参照して組み込まれる。
【0176】
当業者には、本発明において、本発明の範囲または精神から逸脱せずに様々な修飾および変化を加えられることは明白であろう。本発明のその他の実施形態は、当業者には本明細書に開示した本明細書の説明および実践の考察から明白であろう。本明細書および実施例は例示に過ぎないと見なされ、本発明の真の範囲および精神は添付の特許請求の範囲によって指示されることが意図されている。
【0177】
【表6】
【0178】
【表7】
【0179】
【表8】
【0180】
【表9】
【0181】
【表10】
【数1】
【数2】
【数3】
【数4】
【数5】
【数6】
【数7】
【数8】
【数9】
【数10】
【数11】
【数12】
【数13】
【数14】
【数15】
【数16】
【数17】
【数18】
【数19】
【数20】
【数21】
【数22】
【数23】
【数24】
【数25】
【数26】
【数27】
【数28】
【数29】
【数30】
【数31】
【数32】
【数33】
【数34】
【数35】
【数36】
【数37】
【数38】
【数39】
【数40】
【数41】
【数42】
【数43】
【数44】
【数45】
【数46】
【数47】
【数48】
【数49】
【数50】
【数51】
【数52】
【数53】
【数54】
【数55】
【数56】
【数57】
【数58】
【数59】
【数60】
【数61】
【数62】
【数63】
【数64】
【数65】
【数66】
【数67】
【数68】
【数69】
【数70】
【数71】
【数72】
【数73】
【数74】
【数75】
【数76】
【数77】
【数78】
【数79】
【数80】
【数81】
【数82】
【数83】
【数84】
【数85】
【数86】
【数87】
【数88】
【数89】
【数90】
【数91】
【数92】
【数93】
【数94】
【数95】
【数96】
【数97】
【数98】
【数99】
【数100】
【数101】
【数102】
【数103】
【数104】
【数105】
【数106】
【数107】
【数108】
【数109】
【数110】
【数111】
【数112】
【数113】
【数114】
【数115】
【数116】
【数117】
【数118】
【数119】
【数120】
【数121】
【数122】
【数123】
【数124】
【数125】
【数126】
【数127】
【数128】
【数129】
【数130】
【数131】
【数132】
【数133】
【数134】
【数135】
【数136】
【数137】
【数138】
【数139】
【数140】
【数141】
【数142】
【数143】
【数144】
【数145】
【数146】
【数147】
【数148】
【数149】
【数150】
【数151】
【数152】
【数153】
【数154】
【数155】
【数156】
【数157】
【数158】
【特許請求の範囲】
【請求項1】
本願明細書に記載された発明。
【請求項1】
本願明細書に記載された発明。
【図1】

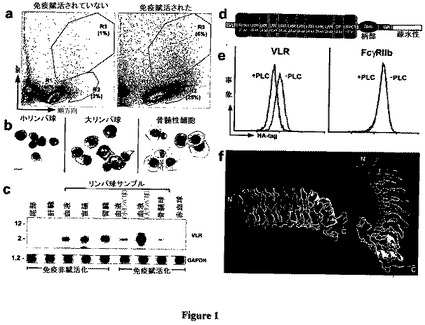
【図2】

【図3】
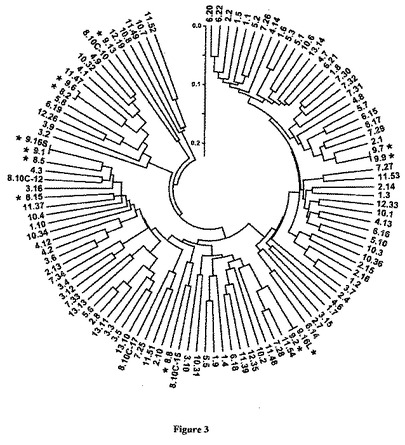
【図4】
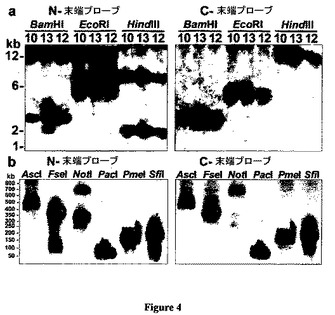
【図5】
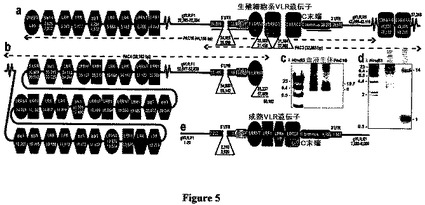
【図6】

【図7】

【図8−1】

【図8−2】

【図9】
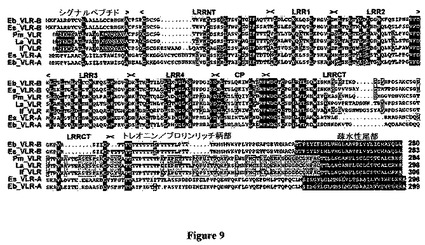
【図10】
【図11】
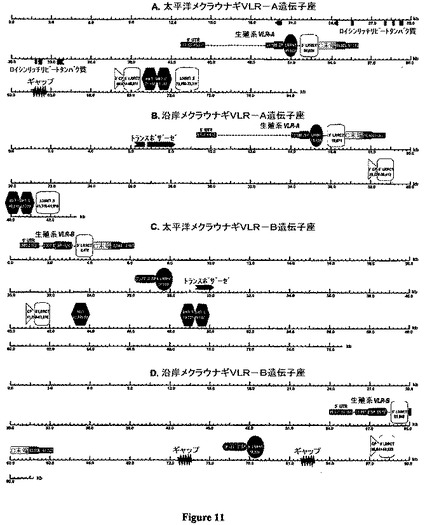
【図12】
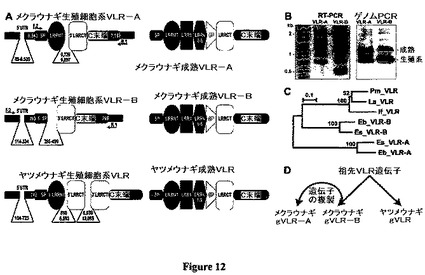
【図13】
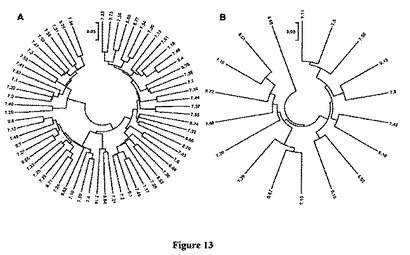
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8−1】
【図8−2】
【図9】
【図10】
【図11】
【図12】
【図13】
【公開番号】特開2011−206060(P2011−206060A)
【公開日】平成23年10月20日(2011.10.20)
【国際特許分類】
【外国語出願】
【出願番号】特願2011−158483(P2011−158483)
【出願日】平成23年7月19日(2011.7.19)
【分割の表示】特願2007−527508(P2007−527508)の分割
【原出願日】平成17年5月23日(2005.5.23)
【出願人】(506385335)ザ ユーエービー リサーチ ファウンデーション (10)
【出願人】(506386284)ベナロヤ リサーチ インスティテュート (2)
【Fターム(参考)】
【公開日】平成23年10月20日(2011.10.20)
【国際特許分類】
【出願番号】特願2011−158483(P2011−158483)
【出願日】平成23年7月19日(2011.7.19)
【分割の表示】特願2007−527508(P2007−527508)の分割
【原出願日】平成17年5月23日(2005.5.23)
【出願人】(506385335)ザ ユーエービー リサーチ ファウンデーション (10)
【出願人】(506386284)ベナロヤ リサーチ インスティテュート (2)
【Fターム(参考)】
[ Back to top ]