
可変部を含むペプチド、当該ペプチドが固定された固相及びその生産方法
【課題】生産効率を向上させた可変部を含むペプチドを提供すること。
【解決手段】抗原結合部位が形成される可変部を含むペプチドであって、重鎖の可変部よりもC末端側又は軽鎖の可変部よりもC末端側に、固相に対して特異的な吸着機能を発現させるアミノ酸配列を有する。
【解決手段】抗原結合部位が形成される可変部を含むペプチドであって、重鎖の可変部よりもC末端側又は軽鎖の可変部よりもC末端側に、固相に対して特異的な吸着機能を発現させるアミノ酸配列を有する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規のペプチド、当該ペプチドの用途及び生産方法に関し、特に、親水性の固相に特異的に結合する機能を発現させるペプチド、当該ペプチドの用途及び生産方法に関する。また、本発明は、固相化法を用いた化合物の検出又は測定に利用可能なペプチド、及び当該ペプチドを利用した化合物の検出又は測定方法に関する。また、本発明は、抗原結合部位が形成される可変部を含むペプチドに関し、免疫グロブリン分子、さらに可変部を含む1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)、定常部融合1本鎖抗体(scFv-Fc)、Fab断片又はF(ab’)2断片及びこれらのペプチドが固定化された固相及びかかる固相の生産方法に関する。
【背景技術】
【0002】
抗体(免疫グロブリン)は、抗原に対して特異的に結合するタンパク質であり、生体内における体液性免疫の主役である。図6(a)は、抗体(免疫グロブリン)の基本構造を示す模式図である。抗体分子61は、2本の分子量5万〜7万の重鎖(H鎖)62と、2本の分子量2.3万の軽鎖(L鎖)63の4本の鎖状ポリペプチドがジスルフィド結合及び非共有結合によって結合して、全体としてY字型を形成している。重鎖(H鎖)62は、4つ又は5つのドメインから構成され、N末端側に可変部(VH)のドメインが配置され、C末端側に向けて3〜4つの定常部(CH1、CH2、CH3、CH4)のドメインが配置される。なお、図6(a)では、4つのドメイン(VH、CH1、CH2、CH3)から構成される重鎖62を示す。軽鎖(L鎖)63は、2つのドメインから構成され、N末端側に可変部(VL)のドメインが配置され、C末端側に定常部(CL)のドメインが配置される。
【0003】
重鎖62及び軽鎖63の可変部(VH及びVL)は、N末端側の2箇所に配置され、両者が一体となって、抗原に対して特異的に結合する立体的な抗原結合部位が形成される。したがって、抗体の特異性は、重鎖及び軽鎖の可変部(VH及びVL)のアミノ酸配列及び組み合わせによって決定され、対応する抗原に応じて、可変部(VH及びVL)のアミノ酸配列や組み合わせが異なる。重鎖及び軽鎖の定常部(CH1、CH2、CH3、CH4及びCL)は、各クラスまたはサブクラス毎にほぼ一定の構造である。
【0004】
抗体分子61は、タンパク質分解酵素であるパパインによって、重鎖のCH1とCH2ドメインの間のヒンジ部64で分解され、図6(b)に示すように、2つのFab断片65と一つのFc断片66とに分解される。なお、別のタンパク質分解酵素であるペプシンによって抗体分子61を分解すると、図6(c)に示すように、2つのFab断片65がヒンジ部64でジスルフィド結合した構造のF(ab’)2断片67を得ることができる。Fab断片65やF(ab’)2断片67は、重鎖及び軽鎖の可変部(VH及びVL)による抗原結合部位を備えているため、抗原に対する特異性を有しており、抗原抗体反応に利用することができる。
【0005】
さらに、重鎖の可変部(VH)と軽鎖の可変部(VL)とをリンカーペプチド69で結合した構造の一本鎖抗体(scFv)68も、重鎖と軽鎖の可変部(VH及びVL)による立体的な抗原結合部位を形成できるため、抗原に対する特異性を有しており、抗原抗体反応に利用することができる。図6(d)は、一本鎖抗体(scFv)68の基本構造を示す模式図である。図6(d)においてが、重鎖の可変部(VH)のC末端が鎖状のリンカーペプチド69によって、軽鎖の可変部(VL)のN末端に連結されている。
【0006】
抗原抗体反応は、抗原と抗体の特異性が高く、抗原結合部位が変われば対応する抗原も変わることから、100万通り以上の重鎖及び軽鎖の可変部(VH及びVL)の組み合わせについて、どのような抗原抗体反応を示すのか検証する基礎的な実験が必要とされている。また、抗原と抗体の特異性が高いことを利用して、抗原抗体反応は、様々な用途及び分野において利用されている。
【0007】
従来から、微量の物質を検出又は測定する方法として、抗原と抗体間の特異的な親和力を利用した免疫測定法(イムノアッセイ)が知られている。免疫測定法は、抗原抗体反応の多様性から、様々な生体成分の分析が可能であり、広い分野において使用されている。さらに、免疫測定法の測定感度を高めるために、放射線化合物、蛍光物質、酵素などの標識を用いた各種の手法が知られており、各標識に応じて、それぞれ放射免疫測定法(ラジオイムノアッセイ:RIA)、免疫蛍光測定法(蛍光イムノアッセイ:FIA)、酵素免疫測定法(ELISA:Enzyme-Linked ImmunoSorbent Assay。エンザイムイムノアッセイとも呼ぶ)などと呼ばれている。特に、ELISAは、高感度で定量性に優れており、精製や前処理といった煩雑なステップを必要としない汎用性の高い検出法であることから、医療診断、環境ホルモン・残留農薬の定量、牛海綿状脳症(BSE)検査、プロテオーム解析等の様々な分析に利用されている。
【0008】
これらの免疫測定法における微量測定では、抗原、抗体又は酵素等のタンパク質を試験管やマイクロプレート等の固相に物理吸着や化学結合を利用して固定化する固相化法が利用されている。固相としては、通常、疎水性のプラスチックが利用される。疎水性の固相は、タンパク質が疎水性相互作用によって強固に結合するので、比較的多くの種類のタンパク質に対して適用できる点で優れている。現在、疎水性のプラスチックとしては、疎水性のポリスチレン(PS)が多用されている。
【0009】
また、免疫測定法は、いずれの方法においても、多数回の吸着・反応処理及び洗浄処理が必要であり、目的物質を検出するまでに長時間を要するものであった。なお、抗原抗体反応を検証する実験においても、試験管やマイクロプレート等の固相に抗体等を固定化して実験するのが一般的であり、固相化法が利用されている。
【0010】
例えば、ELISAは、直接吸着法、サンドイッチ法及び競合法に大別されるが、サンドイッチ法のELISAの場合、特許文献1によれば、第一に、固相に目的物質に対する抗体を結合させた後、固相を複数回洗浄する。第二に、固相上に残った未結合の部位に対して、固相表面にその他の試薬が結合しないように、抗原抗体反応及び酵素反応に関与しない試薬(以下「ブロッキング試薬」という。)を結合させてブロッキングした後、固相を複数回洗浄する。第三に、目的物質を含有する試料と固相上に結合された抗体とを反応させた後、固相を複数回洗浄する。第四に、第2の抗体と目的物質と反応させた後、固相を複数回洗浄する。第五に、第2の抗体と酵素標識とを反応させた後、固相を複数回洗浄する。第六に、酵素標識と基質とを反応させ、吸光度を測定し、試料中の目的物質を検出し、又は濃度を測定する(特許文献1の段落0039参照)。なお、第四の処理で反応させる第2の抗体として、既に酵素標識された抗体を利用すれば、第五の処理は不要となる。
【特許文献1】特表2002-526777号公報
【発明の開示】
【発明が解決しようとする課題】
【0011】
上で述べたとおり、重鎖及び軽鎖の可変部(VH及びVL)の組み合わせは、100万通り以上も存在し、抗原抗体反応を検証するために、膨大な量の実験が必要である。しかし、抗体分子は、分子量が大きく、生産するために動物細胞が必要で、生産コストが高かった。
【0012】
また、検証実験や免疫測定法等の固相化法では、通常、疎水性のプラスチックが利用されるが、疎水性相互作用を利用した結合であるため、抗原、抗体又は酵素等のタンパク質のいずれの部位が固相と結合するのか特定できず、固相と結合する部位によっては、構造変化や不活性化してしまう。例えば、酵素のO−アセチルセリンスルフヒドリラーゼ−A(O-acetyl serine sulfhydryrase-A)は、疎水性ポリスチレンに結合すると、ほぼ完全に失活する。さらに、このようなタンパク質と固相の無秩序な結合により、測定感度が低下し、測定精度も悪くなってしまう。
【0013】
抗体分子61は、図6に示すように、全体としてY字状であり、その上側の2箇所のN末端側に重鎖と軽鎖の可変部(VH及びVL)による立体的な抗原結合部位が形成されている。抗体を固定化する際に、C末端側が固相表面に対して垂直に結合すれば、抗原結合部位が外側を向き、また立体的に正常な配置を取れるため、抗原抗体反応を行うことができる。しかし、N末端側が固相と結合すると、少なくとも結合したN末端側に配置された抗原結合部位は、抗原と反応することができなくなる。また、固相との結合の際に、抗原結合部位の立体構造が変化して、抗体が変性し、抗原抗体反応を示さなくなることもある。
【0014】
Fab断片65やF(ab’)2断片67は、抗体分子61に比べて、分子量が小さくそれら自体を大腸菌や酵母等での生産が可能であるが、抗原結合部位の占める割合が高いので、無秩序に固相表面に結合させた場合、抗原抗体反応を示さなくなる可能性も高くなる。一本鎖抗体(scFv)68に至っては、基本的に抗原結合部位のみによって構成されているので、Fc領域や定常部の影響を排除できるため抗原抗体反応を調べるための抗体側の物質としては好適な形態ではあったが、固定化には極めて不利であり、そのままの状態で疎水性の基板に固定化しても、ほとんど抗原抗体反応を示さなかった。
【0015】
また、一般的に公知のクローン化技術によって製造されたタンパク質は、製造物である目的のタンパク質を回収するための分離、精製する工程に非常に時間と手間がかかっていた。例えば、宿主として大腸菌を使用した場合、大腸菌内には、目的とするタンパク質だけではなく、様々な種類の夾雑物が存在する。このため、大腸菌を培養した後、大腸菌の菌体を破砕して遠心分離することによって、菌体内可溶性画分を採取し、それをカラムによって精製して目的タンパク質を回収する必要があった。
【0016】
また、組換えDNAを導入することによって異種遺伝子を宿主内で大量に発現させた場合、生成されたタンパク質による宿主への悪影響を避けるため、不溶で不活性な凝集体である封入体(inclusion body)として生成されることがある。この封入体(inclusion body)は、遠心分離によって回収した後に、可溶化した上で、そのままでは活性がないので、タンパク質の折りたたみ構造を正常な状態に戻して活性化させるリフォールディングという作業も必要となる。
【0017】
さらに、上述したとおり、免疫測定法は、多数回の吸着・反応処理及び洗浄処理が必要であり、目的物質を検出するまでに長時間を要するものであり、短時間で微量物質を検出又は測定できる手法の開発が望まれている。
【0018】
本発明は、以上の課題及び問題点の一つ若しくは幾つかを解決するためのものであり、抗原抗体反応に利用できる可変部を含むペプチド、これらのペプチドが固定化された固相及びかかる固相の生産方法を提供する。また、生産効率を向上させた可変部を含むペプチド、固定化させても構造変化や不活性化しにくい可変部を含むペプチド、固定化されるタンパク質の構造をより維持し、活性をより保持できる免疫測定法に利用可能な可変部を含むペプチド、測定感度を向上させ、安定した測定精度の免疫測定法に利用可能な可変部を含むペプチド又は従来と比較して短時間で微量物質を検出又は測定できる方法に利用可能な可変部を含むペプチド並びにこれらのペプチドが固定化された固相及びかかる固相の生産方法を提供することを目的とする。
【0019】
なお、本明細書では、新規で有用なペプチド及び当該ペプチドの用途も開示している。また、本発明は、親水性の固相に特異的に結合する機能を発現させるペプチド及び当該ペプチドの用途も開示している。さらに、本発明は、固相化法を用いた化合物の検出又は測定に利用可能なペプチド及び当該ペプチドの用途も開示している。
【課題を解決するための手段】
【0020】
前述した目的を達成するため、抗原結合部位が形成される可変部を含むペプチドであって、重鎖の可変部よりもC末端側又は軽鎖の可変部よりもC末端側に、固相に対して特異的な吸着機能を発現させるアミノ酸配列を有する。また、本発明のペプチドは、抗原結合部位が形成される可変部を含むペプチドであって、重鎖の可変部よりもC末端側又は軽鎖の可変部よりもC末端側に、配列表における配列番号1乃至20に示すアミノ酸配列の何れか一つを有する。さらに、本発明のペプチドは、1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)、定常部融合1本鎖抗体(scFv-Fc)、Fab断片又はF(ab’)2断片であることが好ましい。
【0021】
本発明のペプチドは、前記アミノ酸配列は親水性の樹脂表面に対して特異的な吸着機能を発現させることが好ましく、前記樹脂表面は親水性のポリスチレン表面であってもよい。
【0022】
さらに、前記重鎖及び軽鎖の両方に、前記アミノ酸配列を有する1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)、定常部融合1本鎖抗体(scFv-Fc)、Fab断片又はF(ab’)2断片であることがより好ましい。本発明のペプチドは、前記重鎖可変部と前記軽鎖可変部を結合するリンカーペプチドを有し、前記リンカーペプチドと前記可変部の間又は前記リンカーペプチド内に前記アミノ酸配列を有していてもよい。
【0023】
また、本発明の精製されたペプチドの生産方法では、固相に対して特異的な吸着機能を発現させるアミノ酸配列を有するペプチドと夾雑物とを含む溶液を前記固相表面に接触させることにより、前記ペプチドを前記固相表面に直接吸着させ、前記ペプチドを精製する。
【0024】
さらに、本発明の復元されたペプチドの生産方法では、固相に対して特異的な吸着機能を発現させるアミノ酸配列を有する変性されたペプチドと変性剤とを含む溶液を前記固相表面に接触させることにより、前記ペプチドを前記固相表面に直接吸着させ、前記変性されたペプチドを復元する。ここで、前記変性されたペプチドは、封入体であってもよい。
【0025】
上記ペプチドの生産方法において、前記ペプチドは、抗原結合部位が形成される可変部を含み、重鎖の可変部よりもC末端側又は軽鎖の可変部よりもC末端側に、前記アミノ酸配列を有していてもよい。また、前記溶液は、前記ペプチドを生成した宿主の破砕物、培地または分泌物を含有していてもよい。そして、前記アミノ酸配列としては、配列表における配列番号1乃至20に示すアミノ酸配列を使用することができる。
【0026】
また、本発明では、固相に対して特異的な吸着機能を発現させるアミノ酸配列を有するペプチドをコードする塩基配列を含む遺伝子を宿主に導入して形質転換体とし、その生成物を含む溶液を前記固相表面に接触させることにより、前記ペプチドを前記固相表面に直接吸着させることによって、ペプチドが固定化された固相を生産してもよい。ここで、前記ペプチドは、抗原結合部位が形成される可変部を含み、重鎖の可変部よりもC末端側又は軽鎖の可変部よりもC末端側に有するものでもよく、前記生成物を含む溶液は、前記宿主の破砕物も含有するものであっても、前記宿主の培地を含有するものであっても、酵母から分泌生産された分泌物を含有するものであってもよい。
【0027】
また、本発明の対象としては、抗原結合部位が形成される可変部を含むペプチドであって、重鎖の可変部よりもC末端側又は軽鎖の可変部よりもC末端側に、固相に対して特異的な吸着機能を発現させるアミノ酸配列を有するペプチドをコードする塩基配列を含む遺伝子、当該遺伝子に調製されたベクターまたは当該遺伝子が導入された宿主でもよい。
【0028】
また、本発明のペプチドは、配列表における配列番号1に示すアミノ酸配列を含むペプチドであることが好ましい。また、免疫測定法における固相に固定化されるペプチドであって、配列表における配列番号1に示すアミノ酸配列を含むものでもよい。これらの配列番号1に示すアミノ酸配列は、配列表における配列番号2乃至10に示すアミノ酸配列であってもよい。
【0029】
前記固相は親水性の樹脂表面を有していてもよく、前記固相は親水性のポリスチレン表面を有していてもよい。前記免疫測定法はエンザイムイムノアッセイであってもよい。
【0030】
また、本発明は、配列表における配列番号1に示すアミノ酸配列を含む固定化酵素、配列表における配列番号1に示すアミノ酸配列をコードする塩基配列を含む遺伝子、当該遺伝子に調製されたベクターまたは当該遺伝子が導入された宿主も開示する。そして、これらの配列番号1に示すアミノ酸配列は、配列表における配列番号2乃至10に示すアミノ酸配列であってもよい。
【発明の効果】
【0031】
本発明のペプチドは、特定の固相表面に対して優れた結合性能を示し、特定の固相に直接固定化させることができるので、本ペプチドを容易に分離、精製することができる。すなわち、本ペプチドを生産した後、十分な精製を行わなくても、本ペプチドを含む溶液を特定の固相に接触させるだけで、本ペプチドを固相に直接結合させることができ、本ペプチドの溶液からの分離、精製及び固相への固定化を同時に行うことができる。この結果、従来、長時間必要であった分離、精製工程(例えばアフィニティカラムを使用する精製工程)を省くことができ、本ペプチドの生産から固相への固定化までに要する時間を大幅に短縮させることができるので、本ペプチドでは生産効率が飛躍的に向上する。しかも、この過程において、封入体(inclusion body)の状態で生成されたタンパク質や変性されたタンパク質のリフォールディングも行うことも可能である。
【0032】
また、本ペプチドは、固相化法を用いた化合物の検出又は測定に利用することも可能である。特に免疫測定法において、従来と比較して、特異性のある吸着機能により測定時間を短縮でき、しかも原ペプチドの活性を保ったまま固定化でき、測定感度を向上させ、安定した測定精度を得ることができる。
【図面の簡単な説明】
【0033】
【図1】BSAの存在下における親水性のポリスチレン表面に対する固定化率を示すグラフ。
【図2】(a)〜(d)は、それぞれTween20の添加量が0、0.1、1、10%の条件における親水性のポリスチレンプレートに対する結合性能を示すグラフ。
【図3】(a)〜(d)は、それぞれTween20の添加量が0、0.1、1、10%の条件における疎水性のポリスチレンプレートに対する結合性能を示すグラフ。
【図4】(a)〜(f)は、種々の夾雑物の存在下における親水性及び疎水性のポリスチレンプレートに対する結合性能を示すグラフ。
【図5】(a)〜(f)は、種々の夾雑物の存在下における親水性及び疎水性のポリスチレンプレートに対する活性値を示すグラフ。
【図6】(a)は抗体の基本構造を示す模式図、(b)はFab断片とFc断片の構造を示す模式図、(c)はF(ab’)2断片の構造を示す模式図、(d)は一本鎖抗体(scFv)の基本構造を示す模式図。
【図7】BSAの存在下における親水性のポリスチレン表面に対する固定化率を示すグラフ。
【図8】(a)及び(b)は、種々の夾雑物の存在下における親水性のポリスチレンプレートに対する活性値を示すグラフ。
【図9】BSAの存在下における親水性のポリスチレン表面に対する固定化率を示すグラフ。
【図10】(a)及び(b)は、種々の夾雑物の存在下における親水性のポリスチレンプレートに対する活性値を示すグラフ。
【図11】(a)及び(b)はそれぞれ本ペプチド及び原ペプチドを培養したコロニーにおける菌体濃度、(c)及び(d)はそれぞれ本ペプチド及び原ペプチドの結合性能を示すグラフ。
【図12】夾雑物として各種の培地を加えた状態における親水性のポリスチレン表面に対する結合性能を示すグラフ。
【図13】本アミノ酸配列を一本鎖抗体に導入したときの活性値を示すグラフ。
【図14】一本鎖抗体における本アミノ酸配列の導入位置と活性値の関係を示すグラフ。
【図15】一本鎖抗体における本アミノ酸配列の導入位置と活性値の関係を示すグラフ。
【図16】一本鎖抗体の量と親水性のポリスチレン基板に対する活性値の関係を示すグラフ。
【図17】未精製の変性した状態の本ペプチドのポリスチレン基板に対する結合性能及び活性値を示すグラフ。
【発明を実施するための最良の形態】
【0034】
以下、本発明を詳しく説明する。本発明のペプチドは、固相に対して特異的な吸着機能を発現させるアミノ酸配列(以下「本アミノ酸配列」という)を有する。本アミノ酸配列は、例えば配列表における配列番号1乃至20に示すアミノ酸配列(以下、それぞれ「本アミノ酸配列1」乃至「本アミノ酸配列20」という)である。本アミノ酸配列を有するペプチドを「本ペプチド」という。
【0035】
本アミノ酸配列は、特定の固相(例えば、親水性の樹脂表面を有する固相)に対して特異的な吸着機能を発現させるため、本ペプチドは、特定の固相表面に対して優れた結合性能を示し、特定の固相に直接固定化させることができるので、本ペプチドを容易に分離、精製することができる。すなわち、本ペプチドを生産した後、十分な精製を行わなくても、本ペプチドを含む溶液を特定の固相に接触させるだけで、本ペプチドを固相に直接結合させることができ、本ペプチドの溶液からの分離、精製及び固相への固定化までの工程を同時に行うことができる。この結果、従来、長時間必要であった分離、精製工程(例えばアフィニティカラムを使用する精製工程)を省くことができ、本ペプチドの生産から固相への固定化までに要する時間を大幅に短縮させることができるので、本ペプチドでは生産効率が飛躍的に向上する。しかも、この過程において、封入体(inclusion body)の状態で生成されたタンパク質や変性されたタンパク質のリフォールディングも行うことも可能である。
【0036】
また、本ペプチドは、固相化法を用いた化合物の検出又は測定に利用することも可能である。化合物の検出又は測定に利用される抗原、抗体又は酵素に対し、本アミノ酸配列を導入することにより、抗原、抗体又は酵素を特定の固相に直接固定化することができ、化合物の検出又は測定をすることができる。特に、本ペプチドは、免疫測定法を利用した微量物質の検出又は測定に適用することが好ましい。さらに、本ペプチドは、夾雑物の存在下であっても特異的に、しかも本アミノ酸配列の導入位置を優先して特定の固相上に直接結合させるため、配向及び構造を制御した状態で特定の固相上に固定化できるため、従来と比較して、原ペプチドの活性を保ったまま固定化でき、測定感度を向上させ、安定した測定精度を得ることができる。
【0037】
例えば、サンドイッチ法のELISAに適用した場合、目的物質と特異的に抗原抗体反応を示す抗体に対し、本アミノ酸配列のペプチドを導入し、本ペプチドである抗体を準備する。第一に、かかる抗体を特定の固相に直接結合させた後、固相を複数回洗浄する。第二に、固相上に残った未結合の部位に対して、ブロッキング試薬を結合させてブロッキングした後、固相を複数回洗浄する。第三に、目的物質を含有する試料と固相上に結合された本ペプチドである抗体とを反応させた後、固相を複数回洗浄する。第四に、本ペプチドではない第2の抗体と目的物質と反応させた後、固相を複数回洗浄する。第五に、第2の抗体と酵素標識とを反応させた後、固相を複数回洗浄する。第六に、酵素標識と基質とを反応させ、吸光度を測定し、試料中の目的物質を検出し、又は濃度を測定する。
【0038】
このように、本ペプチドを利用した免疫測定法による化合物の検出又は測定を行うことができる。なお、第四の処理で反応させる第2の抗体として、既に酵素標識された抗体を利用すれば、第五の処理は不要となる。また、本ペプチドが、ブロッキング試薬の存在下においても特異的に結合するので、上記第一の処理において、特定の固相に対し、本ペプチドである抗体とブロッキング試薬とを同時に結合させることで、第二の処理を省略することができ、化合物の検出又は測定時間を短縮できる。
【0039】
さらに、最初に、溶液中において、本ペプチドである抗体と目的物質とを抗原抗体反応させて免疫複合物を形成させ、その後、特定の固相上に免疫複合物を固定化させることにより、化合物の検出又は測定時間を劇的に短縮できる新たな化合物の検出又は測定方法を実現できる。すなわち、上記サンドイッチ法における第一の処理と第三の処理を一回の処理で実行できる。溶液中にブロッキング試薬も添加しておけば、上記第一、第二及び第三の処理を一回の処理で実行できる。より好ましくは、本ペプチドである抗体と目的物質との免疫複合物を形成した後に、溶液中において、さらに酵素標識された第2の抗体とも抗原抗体反応させて、より大きな免疫複合物を形成させ、それを特定の固相上に固定化すれば、上記第一乃至第五の処理を一回の処理で実行できる。
【0040】
本ペプチドは、夾雑物の存在下であっても特異的に、しかも配向及び構造を制御した状態で、特定の固相上に直接固定化できるため、ブロッキング試薬が存在しても、免疫複合物の状態であっても、免疫複合物を固定化して、目的物質の検出又は測定を行うことができる。加えて、溶液中において立体障害なく極めて迅速に抗原抗体反応が行われるため、従来の固定化された状態での抗原抗体反応と比べて、飛躍的に反応速度を増加させることができ、測定感度を向上させ、安定した測定精度を得ることができる。
【0041】
さらに本ペプチドの別の用途として、本ペプチドとして酵素を採用すれば、親水性の樹脂表面における固定化酵素として利用することができる。一般的に、酵素は水溶性であるため、利用する際に使い捨てているが、酵素を固相に結合させて固定化することにより、水溶性の反応生成物と分離することができ、連続的、かつ繰り返し反応させることができる。
【0042】
本アミノ酸配列1は、配列表における配列番号1に示されているように、N末端側からC末端側に向けて順にRXXXRRXRR(R:アルギニン、X:イソロイシン(I)、ロイシン(L)、バリン(V)、アラニン(A)、グリシン(G)、メチオニン(M)、セリン(S)又はトレオニン(T)の一つ又は複数の組合わせ)という配列である。したがって、本アミノ酸配列1には、少なくとも、本アミノ酸配列2乃至10が含まれる。
【0043】
本アミノ酸配列2は、本アミノ酸配列1のXを全てイソロイシン(I)とした配列であり、N末端側からC末端側に向けて順にRIIIRRIRR(R:アルギニン、I:イソロイシン)という配列である。本アミノ酸配列3は、本アミノ酸配列1のXをアラニン(A)とイソロイシン(I)の組合わせとした配列であり、N末端側からC末端側に向けて順にRAIARRIRR(R:アルギニン、A:アラニン、I:イソロイシン)という配列である。本アミノ酸配列4は、本アミノ酸配列1のXを全てロイシン(L)とした配列であり、N末端側からC末端側に向けて順にRLLLRRLRR(R:アルギニン、L:ロイシン)という配列である。本アミノ酸配列5は、本アミノ酸配列1のXを全てバリン(V)とした配列であり、N末端側からC末端側に向けて順にRVVVRRVRR(R:アルギニン、V:バリン)という配列である。本アミノ酸配列6は、本アミノ酸配列1のXを全てアラニン(A)とした配列であり、N末端側からC末端側に向けて順にRAAARRARR(R:アルギニン、A:アラニン)という配列である。本アミノ酸配列7は、本アミノ酸配列1のXを全てグリシン(G)とした配列であり、N末端側からC末端側に向けて順にRGGGRRGRR(R:アルギニン、G:グリシン)という配列である。
【0044】
後述する実施例1乃至6において示すとおり、本アミノ酸配列2乃至7を有するペプチドは、何れも特定の固相表面に対して特異的な吸着機能を発現させることが確認された。本アミノ酸配列2乃至7は、何れもアルギニン(R)の配置は、N末端側から1、5,6、8、9番目と共通であり、その間に配置されるアミノ酸(本アミノ酸配列1のX)を変えたものである。本アミノ酸配列2,4乃至7は、単一のアミノ酸を配置させているが、本アミノ酸配列3では、アラニン(A)とイソロイシン(I)とを組合わせて配置させている。これら全てにおいて共通の性質を示したことから、本アミノ酸配列1のXとして、イソロイシン(I)、ロイシン(L)、バリン(V)、アラニン(A)又はグリシン(G)の一つ又は組み合わせとすることにより特定の固相表面に対して特異的な吸着機能を発現させることが予測される。
【0045】
さらに、本アミノ酸配列8は、本アミノ酸配列1のXを全てメチオニン(M)とした配列であり、N末端側からC末端側に向けて順にRMMMRRMRR(R:アルギニン、M:メチオニン)という配列である。本アミノ酸配列9は、本アミノ酸配列1のXを全てセリン(S)とした配列であり、N末端側からC末端側に向けて順にRSSSRRSRR(R:アルギニン、S:セリン)という配列である。本アミノ酸配列10は、本アミノ酸配列1のXを全てトレオニン(T)とした配列であり、N末端側からC末端側に向けて順にRTTTRRTRR(R:アルギニン、T:トレオニン)という配列である。
【0046】
後述する実施例7乃至9において示すとおり、本アミノ酸配列8乃至10を有するペプチドは、何れも特定の固相表面に対して特異的な吸着機能を発現させることが確認された。メチオニン(M)、セリン(S)又はトレオニン(T)は、アミノ酸(R−CH(NH2)COOH)のR基中に硫黄やヒドロキシ基(OH)が含まれてはいるものの、部分的に鎖状の飽和炭化水素を有し、中性のアミノ酸である点において、R基が鎖状の飽和炭化水素からなるイソロイシン(I)、ロイシン(L)、バリン(V)及びアラニン(A)と共通している。また、メチオニン(M)、セリン(S)及びトレオニン(T)は、R基に環状構造や芳香族化合物を含まず、立体構造の面においても比較的かさの小さいアミノ酸である点においても共通である。このため、本アミノ酸配列1のXとして、メチオニン(M)、セリン(S)又はトレオニン(T)を選択しても、同様の効果が得られることが予測される。
【0047】
さらに、本アミノ酸配列11は、配列表における配列番号11に示されているように、N末端側からC末端側に向けて順にKGLRGWREMISL(リジン(K)、グリシン(G)、ロイシン(L)、アルギニン(R)、グリシン(G)、トリプトファン(W)、アルギニン(R)、グルタミン酸(E)、メチオニン(M)、イソロイシン(I)、セリン(S)、ロイシン(L))という配列である。
【0048】
本アミノ酸配列12は、配列表における配列番号12に示されているように、N末端側からC末端側に向けて順にADYLSRWGSIRN(アラニン(A)、アスパラギン酸(D)、チロシン(Y)、ロイシン(L)、セリン(S)、アルギニン(R)、トリプトファン(W)、グリシン(G)、セリン(S)、イソロイシン(I)、アルギニン(R)、アスパラギン(N))という配列である。
【0049】
本アミノ酸配列13は、配列表における配列番号13に示されているように、N末端側からC末端側に向けて順にSRVHRAVLNGVS(セリン(S)、アルギニン(R)、バリン(V)、ヒスチジン(H)、アルギニン(R)、アラニン(A)、バリン(V)、ロイシン(L)、アスパラギン(N)、グリシン(G)、バリン(V)、セリン(S))という配列である。
【0050】
本アミノ酸配列14は、配列表における配列番号14に示されているように、N末端側からC末端側に向けて順にRPPGVVRRYALG(アルギニン(R)、プロリン(P)、プロリン(P)、グリシン(G)、バリン(V)、バリン(V)、アルギニン(R)、アルギニン(R)、チロシン(Y)、アラニン(A)、ロイシン(L)、グリシン(G))という配列である。
【0051】
本アミノ酸配列15は、配列表における配列番号15に示されているように、N末端側からC末端側に向けて順にVRSWEEQARVTT(バリン(V)、アルギニン(R)、セリン(S)、トリプトファン(W)、グルタミン酸(E)、グルタミン酸(E)、グルタミン(Q)、アラニン(A)、アルギニン(R)、バリン(V)、トレオニン(T)、トレオニン(T))という配列である。
【0052】
本アミノ酸配列16は、配列表における配列番号16に示されているように、N末端側からC末端側に向けて順にRAFIASRRIKRP(アルギニン(R)、アラニン(A)、フェニルアラニン(F)、イソロイシン(I)、アラニン(A)、セリン(S)、アルギニン(R)、アルギニン(R)、イソロイシン(I)、リジン(K)、アルギニン(R)、プロリン(P))という配列である。
【0053】
本アミノ酸配列17は、配列表における配列番号17に示されているように、N末端側からC末端側に向けて順にRESTLKGTSRAV(アルギニン(R)、グルタミン酸(E)、セリン(S)、トレオニン(T)、ロイシン(L)、リジン(K)、グリシン(G)、トレオニン(T)、セリン(S)、アルギニン(R)、アラニン(A)、バリン(V))という配列である。
【0054】
本アミノ酸配列18は、配列表における配列番号18に示されているように、N末端側からC末端側に向けて順にAGLRLKKAAIHR(アラニン(A)、グリシン(G)、ロイシン(L)、アルギニン(R)、ロイシン(L)、リジン(K)、リジン(K)、アラニン(A)、アラニン(A)、イソロイシン(I)、ヒスチジン(H)、アルギニン(R))という配列である。
【0055】
本アミノ酸配列19は、配列表における配列番号19に示されているように、N末端側からC末端側に向けて順にSSLLRAVPEPTG(セリン(S)、セリン(S)、ロイシン(L)、ロイシン(L)、アルギニン(R)、アラニン(A)、バリン(V)、プロリン(P)、グルタミン酸(E)、プロリン(P)、トレオニン(T)、グリシン(G))という配列である。
【0056】
本アミノ酸配列20は、配列表における配列番号20に示されているように、N末端側からC末端側に向けて順にRAFIASRRIRRP(アルギニン(R)、アラニン(A)、フェニルアラニン(F)、イソロイシン(I)、アラニン(A)、セリン(S)、アルギニン(R)、アルギニン(R)、イソロイシン(I)、アルギニン(R)、アルギニン(R)、プロリン(P))という配列である。
【0057】
本アミノ酸配列1乃至20は、特定の固相に対して特異的な吸着機能を発現させることができる。特定の固相とは、現時点で確認されている範囲では、親水性の樹脂表面を有する固相である。すなわち、本ペプチドは、原ペプチドと比較して、親水性の樹脂表面に対する結合性能が向上する。特に、本ペプチドは、親水性のポリスチレン表面に対して優れた結合性能を示す。結合性能は、競合する夾雑物に対して優先的に結合する性能であり、競合する夾雑物、例えば他のペプチド、高分子化合物、界面活性剤等の存在下において、固相に対して結合する本ペプチドの量を比較することで評価できる。
【0058】
親水性の樹脂表面を有する固相としては、ポリスチレン、ポリカーボネート、ポリプロピレン、ポリエチレン、ポリジメチルシロキサン(PDMS)、ポリメタクリル酸メチル(PMMA)等などのプラスチック樹脂表面を変質させて親水化処理したものを用いることができる。通常、製造されたプラスチック樹脂表面は疎水性であるが、この表面に各種の親水化処理を行うことで、親水性の樹脂表面を有する固相とすることができる。例えば、ポリスチレン表面にUV+O3処理やプラズマ酸化処理を行うことにより、親水性のポリスチレン表面を有する固相を準備できた。
【0059】
固相の形態としては、板状(容器やウェルの壁面及び底面を含む)でもよいし、粒状でもよい。特開2007−279018号公報に記載されているように、マイクロウェルプレートの流体取扱部(各ウェル)内に表面が親水化処理された粒状プラスチック基板を充填すれば、ウェル内に本ペプチドを含む溶液を注入するだけで粒状プラスチック基板表面に本ペプチドを固定化することができる。
【0060】
本明細書において「ペプチド」とは、ペプチド結合によって2個以上のアミノ酸が結合したものを指し、オリゴペプチド、ポリペプチド、タンパク質、ホモメリックペプチド及びヘテロメリックペプチドを含む。また、本ペプチドは、電気的に中性の形態、又は塩の形態をとることがあり、配列表における本アミノ酸配列のみからなる形態、特定のタンパク質の末端部または内部に本ペプチドを融合させた融合タンパク質の形態、糖質やポリエチレングリコール、NCS基(isothiocyanate)、NHS基(N-Hydroxy succinimide ester)、マレイミド基、チオール基、ビオチン、蛍光色素等を付加して得られる複合体としての形態、さらには、ペプチドをアセチル化、アミド化及び/又は多官能試験により架橋重合させて得られる誘導体又は重合体としての形態であってもよい。
【0061】
配列表における配列番号21に示す塩基配列は、本アミノ酸配列2をコードする塩基配列の一例である。すなわち、5’末端側から3’末端側に向けて順にcgt atc atc atc cga agg att cga cga(a:アデニン、g:グアニン、c:シトシン、t:チミン)という塩基配列を含むDNAである。配列番号22に示す塩基配列は、本アミノ酸配列3をコードする塩基配列の一例である。すなわち、5’末端側から3’末端側に向けて順にcgt gcg att gcg cga agg att cga cga(a:アデニン、g:グアニン、c:シトシン、t:チミン)という塩基配列を含むDNAである。本アミノ酸配列2又は3をコードする塩基配列としては、配列番号21又は22に示す塩基配列に限定されるものではなく、その他にも、本アミノ酸配列2又は3の各アミノ酸に対応する各コドンの配列を採用できる。本アミノ酸配列をコードする塩基配列を含む遺伝子は、ファージやプラスミド等のベクターを用いて、あるいは宿主のゲノムDNA中に導入し、宿主内において発現させることにより、本ペプチドを生合成できる。
【0062】
本ペプチドとしては、種々のタンパク質を利用することが可能であり、多くの抗原、抗体、レクチン、酵素又は受容体タンパク質等に適用することができる。例えば、グルタチオン転移酵素(GST:Glutathione S-Transferase)、マルトース結合タンパク質(MBP)、アルカリホスファターゼ(ALP)、ペルオキシダーゼ(POD)、ルシフェラーゼ、緑色蛍光タンパク質(GFP)、β−ガラクトシダーゼ(β−Gal)、トリプシン、キモトリプシン、トロンビン、Factor Xa、アンジオテンシン変換酵素、チロシンキナーゼ、インスリンレセプター、EGFレセプター、ストレプトアビジン(SA)、モノクローナル抗体(Mab)、ポリクローナル抗体(Pab)、1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)(例えば2価性1本鎖抗体(sc(Fv)2))、定常部融合1本鎖抗体(scFv-Fc)、Fab断片及びF(ab’)2断片(抗原結合部位を含む抗体の断片)、補体系タンパク質C1q、コンカナバリンA(ConA)、レンチルレクチン(LCA)、抗体結合タンパク質(Protein A、ZZ、Protein G、Protein L等)等に適用することができる。なお、本明細書において、本ペプチドと対比するために、本ペプチドから本アミノ酸配列を除いたペプチド(本アミノ酸配列を導入する前のペプチドを含む)を、以下においては「原ペプチド」と呼ぶ。
【0063】
なお、本ペプチドとして、抗原結合部位が形成される可変部を含むペプチドとする場合、抗体分子それ自体も含むが、大腸菌や酵母での生産が可能である1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)、定常部融合1本鎖抗体(scFv-Fc)、Fab断片、F(ab’)2断片であることが好ましい。多価性1本鎖抗体(sc(Fv)n)は、複数の1本鎖抗体(scFv)をリンカーペプチドで連結したものであり、分子内に複数の抗原結合部位(重鎖可変部(VH)及び軽鎖可変部(VL)ドメイン)を有するペプチドの総称である。複数の抗原結合部位は同じものでもよいし、異なっていてもよい。なお、2価性1本鎖抗体(sc(Fv)2)とは、2つの1本鎖抗体(scFv)をリンカーペプチドで連結したものである。定常部融合1本鎖抗体(scFv-Fc)とは、1本鎖抗体(scFv)のC末端側にFcドメインを有する融合タンパク質の総称である。Fab断片とは、抗体分子をパパイン消化によって得られる断片のうち、VL、CLからなる軽鎖とVH、CH1ドメインからなる重鎖が会合したものを指すが、遺伝子組換え技術で作成することも可能である。F(ab’)2断片とは、抗体分子をペプシン消化後に得られる断片の一つで2分子のFabがヒンジ部のジスルフィド結合で連結されたものを指すが、遺伝子組換え技術で作成することも可能である。
【0064】
本ペプチドにおいて、本アミノ酸配列の導入部位は、原ペプチドが本来有する生理活性を妨げない部位、例えば、抗原の場合は抗原決定基以外、抗体の場合は抗体活性基以外の部位とする。特に、原ペプチドの立体構造を維持したまま固相に固定化させるため、原ペプチドのC末端又はN末端近傍の部位に本アミノ酸配列を導入することが好ましい。なお、本ペプチドには、本アミノ酸配列以外にも、標識用のアフィニティタグ等が別途導入されていてもよい。
【0065】
特に、本ペプチドとして、抗原結合部位が形成される可変部を含むペプチドとする場合は、重鎖の可変部よりもC末端側又は軽鎖の可変部よりもC末端側(両方を含む)に本アミノ酸配列を導入する。可変部よりもC末端側とは、重鎖または軽鎖可変部におけるC末端から本ペプチド全体におけるC末端までの位置であって、可変部の抗原結合を阻害しない位置全てを指す。
【0066】
抗体分子の場合(図6(a)参照)は、重鎖(H鎖)62の可変部(VH)よりもC末端側に位置する定常部(CH1、CH2、CH3、CH4)のドメインにおけるN末端やC末端、軽鎖(L鎖)63の定常部(CL)のドメインにおけるN末端やC末端の一つ若しくは複数箇所に導入することが好ましく、特に、重鎖62のFc領域の定常部(CH2、CH3、CH4)のドメインにおけるN末端やC末端、軽鎖63のC末端の一つ若しくは複数箇所に導入することが好ましい。
【0067】
Fab断片又はF(ab’)2断片の場合(図6(b)及び(c)参照)は、重鎖62又は軽鎖63の可変部のドメイン(VH及びVL)におけるC末端、定常部(CH1及びCL)のドメインにおけるN末端やC末端あるいはの一つ若しくは複数箇所に導入することが好ましく、特に、重鎖62のC末端又は軽鎖63のC末端の一つ若しくは複数箇所に導入することが好ましい。
【0068】
また、1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)、定常部融合1本鎖抗体(scFv-Fc)のように、重鎖と軽鎖を結合するリンカーペプチドを有する場合は、リンカーペプチドと重鎖または軽鎖の間、もしくはリンカーペプチド内部に本アミノ酸配列を導入してもよい。ここで、リンカーペプチドは、重鎖又は軽鎖の一方のC末端と、他方のN末端とを連結しているが、重鎖又は軽鎖のC末端側には、リンカーペプチド全体、すなわち重鎖又は軽鎖のC末端と結合している部分から他方のN末端までの領域も含まれる。なお、リンカーペプチドと重鎖または軽鎖の間、もしくはリンカーペプチド内部に本アミノ酸配列以外の標識用のアフィニティタグ等が別途導入されていてもよい。図6(d)の1本鎖抗体(scFv)の場合、軽鎖の可変部(VL)のC末端側か、重鎖の可変部(VH)のC末端からリンカーペプチド69の内部を含む軽鎖の可変部(VL)のN末端までの間の一方又は両方に本アミノ酸配列を導入する。
【0069】
複数の箇所に本アミノ酸配列を導入することにより、本ペプチドの立体構造をより正常な状態に維持することが可能である。たとえば、Fab断片又はF(ab’)2断片の重鎖及び軽鎖のC末端側にそれぞれ本アミノ酸配列を導入したり、1本鎖抗体(scFv)の重鎖及び軽鎖のC末端側にそれぞれ本アミノ酸配列を導入すれば、その部位が固相表面に結合しやすくなるので、抗原結合部位が外側を向き、抗原抗体反応を起こしやすくなる。
【0070】
本ペプチドは、公知のクローン化技術や化学合成法によって製造することが可能である。例えば、クローン化技術を利用すれば、本アミノ酸配列をコードするDNAを調製し、これを自律複製可能なベクターに挿入して組換えDNAとし、これを大腸菌、枯草菌、放線菌、酵母、糸状菌、植物細胞、昆虫細胞、動物細胞などの適宜宿主に導入して形質転換体とし、その培養物から本アミノ酸配列を含むペプチドを採取することができる。また、本アミノ酸配列をコードするDNAを調製し、小麦胚芽や大腸菌細胞抽出液などを用いて無細胞タンパク質合成系により本ペプチドを合成することができる。また、「固相法」又は「液相法」等の慣用のペプチド化学合成法を利用して、本アミノ酸配列に示されるアミノ酸を逐次、脱水縮合させて伸長させれば、本アミノ酸配列からなる本ペプチドを合成することができる。さらに、かかる本アミノ酸配列からなる本ペプチドを所望の原ペプチドに結合させることにより、本アミノ酸配列を含むより高分子の本ペプチドを合成できる。
【0071】
本ペプチドは、本アミノ酸配列を有しているため、形質転換体からの生成物を含む溶液を固相表面に接触させることにより、本ペプチドを固相表面に直接吸着させて、分離精製することができる。生成物を含む溶液とは、目的となる本ペプチドを含み、且つ宿主に起因する不要な夾雑物が存在する溶液全般を指し、例えば、菌体破砕液、菌体破砕液を遠心分離して得られた可溶性画分、菌体破砕液を遠心分離して得られた不溶性画分を可溶化したもの、細胞膜画分、細胞壁画分、細胞から分泌生産された分泌物、体液、またはこれらの不完全な精製物等を含む。
【0072】
さらに、組換えDNAを導入することによって異種遺伝子を宿主内で大量に発現させた場合、生成されたタンパク質による宿主への悪影響を避けるため、不溶で不活性な凝集体である封入体(inclusion body)として生成されることがあるが、本ペプチドが封入体(inclusion body)として生成されていても、変性剤で封入体(inclusion body)を可溶化後そのまま固相表面に固定化することができ、しかも固定化させた状態で変性剤を除去することによって本ペプチドがリフォールディングされる場合もある。
【0073】
また、封入体に対してだけではなく、何らかの原因によって、立体構造が変性されていても、本ペプチドであれば、そのまま固相表面に固定化することができ、しかも固定化させた状態で適当なリフォールディングバッファを加えることによって本ペプチドをリフォールディングできる場合もある。変性の原因としては、加熱、凍結、高圧、超音波、紫外線、X線、かくはん、吸着、希釈などの物理的な原因と、極端な酸性またはアルカリ性、有機溶媒、重金属塩、変性剤、界面活性剤などの化学的な原因がある。
【0074】
上で述べたとおり、本アミノ酸配列1乃至20は、親水性の樹脂表面に対して優れた結合性能を示すため、本アミノ酸配列1乃至20を有する本ペプチドを親水性の樹脂表面を有する固相に直接固定化することができるので、容易に分離、精製することができる。すなわち、本アミノ酸配列1乃至20を有する本ペプチドを含む溶液を親水性の樹脂表面を有する固相が充填されたカラムを通過させることにより、本アミノ酸配列1乃至20を有する本ペプチドを固相に直接結合させて溶液から分離、精製させることができる。
【0075】
以下に、実施例により本発明を更に具体的に説明するが、本発明は、その要旨を越えない限りこれらの実施例に限定されるものではない。実施例1乃至19は、グルタチオン転移酵素(GST)に本アミノ酸配列を導入した例であり、実施例20は、抗体に本アミノ酸配列を導入した例であり、実施例21乃至28は、一本鎖抗体に本アミノ酸配列を導入した例である。なお、以下の実施例において、本ペプチドの濃度は、ウシ血清由来のアルブミン(BSA)を標準タンパクとして使用したLowry法によって定量した。
【0076】
[実施例1](本アミノ酸配列2を有するGSTの生合成)
まず、グルタチオン転移酵素(GST)をコードする遺伝子を組み込んだプラスミド(GEヘルスケアバイオサイエンス株式会社製「pGEX−3X」、データベース・アクセッション番号:U13852)の制限酵素BamHIの認識する塩基配列部位と制限酵素EcoRIの認識する塩基配列部位の間に、本アミノ酸配列2をコードする遺伝子(配列表の配列番号21に示す塩基配列)を導入したものをベクターとして作製した。
【0077】
次に、宿主として大腸菌(BL21)を使用し、本アミノ酸配列2をコードする遺伝子を導入したベクターを宿主に導入して宿主を形質転換し、アンピシリンを含む寒天培地上で選別した。その後、2xYT培地50ml中で大腸菌を培養し、終濃度が0.1mMになるように、発現誘導物質としてイソプロピル1−チオ−ガラクトシド(IPTG:Isopropyl-1-thio-β-D(-)-galactoside)を加えて、本アミノ酸配列2を含むGST(本ペプチド)を発現させた。
【0078】
そして、菌体破砕後、精製用ゲル(GSH-Sepharose 6B)を用いて菌体内可溶性画分を精製し、リン酸緩衝化生理食塩水(PBS:Phosphate Buffered Saline)中で1晩透析し、滅菌フィルターによって凝集体を除去した。こうして、本ペプチドとして、C末端に本アミノ酸配列2が導入されたグルタチオン転移酵素(GST)を生合成した。以下、実施例1で生合成された本アミノ酸配列2を有するGSTを本ペプチド1という。
【0079】
[実施例2](本アミノ酸配列3を有するGSTの生合成)
プラスミド「pGEX−3X」の制限酵素BamHIの認識する塩基配列部位と制限酵素EcoRIの認識する塩基配列部位の間に、本アミノ酸配列3をコードする遺伝子(配列表の配列番号22に示す塩基配列)を導入した点以外は、実施例1と同様の処理によって、本ペプチドとして、C末端に本アミノ酸配列3が導入されたグルタチオン転移酵素(GST)を生合成した。以下、実施例2で生合成された本アミノ酸配列3を有するGSTを本ペプチド2という。
【0080】
[実施例3〜6](本アミノ酸配列4乃至7を有するGSTの生合成)
プラスミド「pGEX−3X」の制限酵素BamHIの認識する塩基配列部位と制限酵素EcoRIの認識する塩基配列部位の間に、本アミノ酸配列4乃至7をコードする遺伝子を導入した点以外は、実施例1と同様の処理によって、本ペプチドとして、C末端に本アミノ酸配列4乃至7が導入されたグルタチオン転移酵素(GST)をそれぞれ生合成した。なお、実施例3で本アミノ酸配列4を有するGSTを生合成し、実施例4で本アミノ酸配列5を有するGSTを生合成し、実施例5で本アミノ酸配列6を有するGSTを生合成し、実施例6で本アミノ酸配列7を有するGSTを生合成した。以下、実施例3乃至6で生合成された本アミノ酸配列4乃至7を有するGSTを、それぞれ本ペプチド3乃至6という。
【0081】
[実施例7〜9](本アミノ酸配列8乃至10を有するGSTの生合成)
プラスミド「pGEX−3X」の制限酵素BamHIの認識する塩基配列部位と制限酵素EcoRIの認識する塩基配列部位の間に、本アミノ酸配列8乃至10をコードする遺伝子を導入した点以外は、実施例1と同様の処理によって、本ペプチドとして、C末端に本アミノ酸配列8乃至10が導入されたグルタチオン転移酵素(GST)をそれぞれ生合成した。なお、実施例7で本アミノ酸配列8を有するGSTを生合成し、実施例8で本アミノ酸配列9を有するGSTを生合成し、実施例9で本アミノ酸配列10を有するGSTを生合成した。以下、実施例7乃至9で生合成された本アミノ酸配列8乃至10を有するGSTを、それぞれ本ペプチド7乃至9という。
【0082】
[実施例10](本ペプチド1及び2の親水性のポリスチレン表面に対する結合性能評価、夾雑物:BSA)
本ペプチドの親水性のポリスチレン表面に対する結合性能を評価するため、夾雑物としてウシ血清由来のアルブミン(BSA)が存在する状態における実施例1の本ペプチド1、実施例2の本ペプチド2及び原ペプチドであるGSTの親水性のポリスチレン表面に対する固定化率を測定した。
【0083】
まず、BSAを添加しないPBSと、BSA濃度が0.003〜50mg/mlとなるように調製したPBSを用意し、各溶液中に、終濃度が5μg/mlとなるように本ペプチド1を添加して試料を調製した。次に、100μlの各試料を親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、室温で1時間インキュベートして、本ペプチド1を固定化させ、0.1%のTween20を含むPBS(以下「0.1PBST」という)で5回洗浄した。さらに、ブロッキング試薬として、2%のBSAを含む0.1PBSTを300μl加え、室温で1時間インキュベートして、ポリスチレンプレート表面の未結合部位をブロッキングし、0.1PBSTで5回洗浄した。そして、抗体として、0.2%のBSAを含む0.1PBSTで4000倍に希釈された抗GST抗体を100μl加え、室温で1時間インキュベートして、本ペプチド1に含まれるGSTと抗GST抗体とを抗原抗体反応させ、0.1PBSTで5回洗浄した。その後、酵素標識された第2の抗体として、0.2%のBSAを含む0.1PBSTで1000倍に希釈されたHRP-conjugated anti-rabbit IgG antibodyを100μl加え、室温で1時間インキュベートして、さらに抗原抗体反応させ、0.1PBSTで5回洗浄した。最後に、発色基質として、ABTS(2,2'-azinobis(3-ethylbenzthiazoline-6-sulphonic acid))を加え、インキュベートし、マイクロプレートリーダー(TECAN製「SUNRISE Remote」)を用いて、405nmにおける吸光度を測定した。本ペプチド2及びGSTについても同様の処理を行い吸光度を測定した。
【0084】
図1に実施例10の測定結果を示す。図1中の黒丸(●)は本ペプチド1の結果であり、黒三角は本ペプチド2の結果であり、白丸(○)は原ペプチドであるGSTの結果である。なお、図1において、縦軸は固定化率(%)であり、横軸はBSA濃度であるが、固定化率は、BSAを添加しないPBSにおける吸光度(結合した量)を基準(100%)として、その他の吸光度を規格化したものである。
【0085】
図1から、原ペプチドであるGSTでは、夾雑物としてBSAが存在すると、急激に固定化率が低下し、結合する量が減少するが、本ペプチド1及び2では、10mg/mlという高濃度のBSAが存在していても、固定化率は殆ど変わらず、親水性のポリスチレンプレートに特異的に結合することが確認できる。
【0086】
[実施例11](本ペプチド1の親水性のポリスチレン表面に対する結合性能評価、夾雑物:BSA及びTween20)
実施例1の本ペプチド1の結合性能をより詳細に評価するため、夾雑物としてTween20が存在する状態で、疎水性のポリスチレン表面及び親水性のポリスチレン表面に対して結合した量(吸光度)を測定した。
【0087】
まず、Tween20を添加しないPBSと、Tween20濃度が0.1、1,10%となるように調製したPBSを用意し、各溶液中に、終濃度が0〜1μg/mlとなるように本ペプチド1を添加して試料を調製した。次に、100μlの各試料を親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)及び疎水性のポリスチレンプレート(ベクトン・ディッキンソン アンド カンパニー社製「BD Falconマイクロプレート#351172」)に加え、室温で1時間インキュベートして、本ペプチド1を固定化させ、0.1PBSTで5回洗浄した。その後、実施例10と同じ条件で、ブロッキング試薬、抗体、酵素標識された第2の抗体及び発色基質を添加する処理及び洗浄処理を行い吸光度を測定した。原ペプチドであるGSTについても同様の処理を行い吸光度を測定した。
【0088】
図2(a)〜(d)に実施例11の親水性のポリスチレンプレートにおける測定結果を示し、図3(a)〜(d)に実施例11の疎水性のポリスチレンプレートにおける測定結果を示す。図2及び図3の(a)〜(d)は、それぞれTween20の添加量が0、0.1、1、10%の条件の時の測定結果である。図2及び図3中の黒丸(●)は本ペプチド1の結果であり、白丸(○)は原ペプチドであるGSTの結果である。図2及び3において、縦軸は405nmにおける吸光度であり、横軸は本ペプチド1及びGSTの濃度である。なお、図2及び3において、GSTの結果と本ペプチド1の結果が同じだった場合は、GSTの結果である白丸(○)が本ペプチド1の黒丸(●)と重なっており確認できない。例えば、図3(b)〜(d)ではGSTの白丸(○)の全部が本ペプチド1の黒丸(●)と重なっている。
【0089】
図2(a)〜(d)から、本ペプチド1及びGSTは、いずれも添加濃度を増やすと、親水性のポリスチレンプレートに結合する量が増えることが確認できる。また、本ペプチド1では、夾雑物としてTween20を添加しても、親水性のポリスチレンプレートに結合する量は減らず、むしろ若干増加している。GSTでは、夾雑物としてTween20を添加すると、親水性のポリスチレンプレートに結合する量が少し減少する。
【0090】
また、図3(a)によれば、本ペプチド1及びGSTは、いずれも、夾雑物としてTween20が存在しない場合には疎水性のポリスチレンプレートに対し結合する。しかし、図3(b)〜(d)に示すとおり、本ペプチド1及びGSTは、いずれも、夾雑物としてTween20が存在すると、疎水性のポリスチレンプレートには殆ど結合できない。このように、本ペプチドの結合性能は、親水性の樹脂表面を有する固相に特異的なものである。
【0091】
[実施例12](本ペプチド1の親水性のポリスチレン表面に対する結合性能評価、夾雑物:BSA及びTween20)
実施例1の本ペプチド1の結合性能をより詳細に評価するため、夾雑物としてTween20が存在する状態及びTween20及びBSAが存在する状態のそれぞれで、本ペプチド1の添加量を変化させて、疎水性のポリスチレン表面及び親水性のポリスチレン表面に対して結合した量(吸光度)を測定した。
【0092】
まず、PBSと、0.1PBSTと、20mg/mlのBSAを含む0.1PBSTとを用意し、各溶液中に、終濃度が0〜10μg/mlとなるように本ペプチド1を添加して試料を調製した。次に、100μlの各試料を親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)及び疎水性のポリスチレンプレート(ベクトン・ディッキンソン アンド カンパニー社製「BD Falconマイクロプレート#351172」)に加え、室温で2時間インキュベートして、本ペプチド1を固定化させ、0.1PBSTで5回洗浄した。その後、実施例10と同じ条件で、ブロッキング試薬、抗体、酵素標識された第2の抗体及び発色基質を添加する処理及び洗浄処理を行い吸光度を測定した。原ペプチドであるGSTについても同様の処理を行い吸光度を測定した。
【0093】
図4に実施例12の測定結果を示す。図4の(a)〜(c)は、それぞれPBS、0.1PBST及びBSAを含む0.1PBSTにおける親水性のポリスチレンプレートについての測定結果であり、図4の(d)〜(f)は、それぞれPBS、0.1PBST及びBSAを含む0.1PBSTにおける疎水性のポリスチレンプレートについての測定結果である。また、図4中の黒丸(●)は本ペプチド1の結果であり、白丸(○)は原ペプチドであるGSTの結果である。図4において、縦軸は405nmにおける吸光度であり、横軸は本ペプチド1及びGSTの濃度である。なお、図4において、GSTの結果と本ペプチド1の結果が同じだった場合は、GSTの結果である白丸(○)が本ペプチド1の黒丸(●)と重なっており確認できない。例えば、図4(e)及び(f)ではGSTの白丸(○)の全部が本ペプチド1の黒丸(●)と重なっている。
【0094】
図4(a)〜(c)から、本ペプチド1では、夾雑物としてTween20を添加した場合(図4(b))でも、夾雑物としてTween20及びBSAを添加した場合(図4(c))でも、親水性のポリスチレンプレートに結合する量はほとんど変わらないことが確認できる(図4(b)及び(c)の黒丸)。これに対し、GSTでは、夾雑物としてTween20を添加すると、親水性のポリスチレンプレートに結合する量が少し減少し(図4(b)の白丸)、さらにBSAを添加した場合には殆ど結合できない(図4(c)の白丸)。このように、本アミノ酸配列2を導入することにより、親水性のポリスチレンプレートに対する結合性能が向上する。
【0095】
また、図4(d)〜(f)によれば、本ペプチド1及びGSTは、いずれも、夾雑物としてTween20が存在しない場合には疎水性のポリスチレンプレートに対し結合するが、夾雑物としてTween20やBSAが存在すると、疎水性のポリスチレンプレートには殆ど結合できない。
【0096】
[実施例13](本ペプチド1の活性値評価)
実施例1の本ペプチド1の固定化された状態における活性値を評価するため、夾雑物としてTween20が存在する状態及びTween20及びBSAが存在する状態のそれぞれで、疎水性のポリスチレン表面及び親水性のポリスチレン表面に対して結合させ、固定化されたGSTの酵素としての活性値を測定した。
【0097】
まず、PBSと、0.1PBSTと、20mg/mlのBSAを含む0.1PBSTとを用意し、各溶液中に、終濃度が10、20、40μg/mlとなるように本ペプチド1を添加して試料を調製した。次に、100μlの各試料を親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)及び疎水性のポリスチレンプレート(ベクトン・ディッキンソンアンド カンパニー社製「BD Falconマイクロプレート#351172」)に加え、室温で2時間インキュベートして、本ペプチド1を固定化させ、0.1PBSTで5回洗浄した。その後、0.1%Tween20、1mMの還元型グルタチオン及びGSTの基質として1mMのCDNB(1-chloro-2,4-dinitrobenzene)を含む0.1Mリン酸カリウムバッファー(pH6.5)を200μl加え、室温で30分間撹拌しつつ、30秒毎に340nmにおける吸光度を測定した。原ペプチドであるGSTについても同様の処理を行い吸光度を測定した。
【0098】
図5に実施例13の測定結果を示す。図5の(a)〜(c)は、それぞれPBS、0.1PBST及びBSAを含む0.1PBSTにおける親水性のポリスチレンプレートについての測定結果であり、図5の(d)〜(f)は、それぞれPBS、0.1PBST及びBSAを含む0.1PBSTにおける疎水性のポリスチレンプレートについての測定結果である。また、図5中の黒丸(●)は本ペプチド1の結果であり、白丸(○)は原ペプチドであるGSTの結果である。図5において、縦軸は活性値(吸光度の変化率)であり、横軸は本ペプチド1及びGSTの濃度である。なお、図5において、GSTの結果と本ペプチド1の結果が同じだった場合は、GSTの結果である白丸(○)が本ペプチド1の黒丸(●)と重なっており確認できない。例えば、図5(e)及び(f)ではGSTの白丸(○)の全部が本ペプチド1の黒丸(●)と重なっている。
【0099】
図5(a)〜(c)から、親水性のポリスチレンプレートに固定化された本ペプチド1のGSTは、夾雑物としてTween20を添加した場合(図5(b))でも、夾雑物としてTween20及びBSAを添加した場合(図5(c))でも、酵素として非常に活性であることが確認できる。これに対し、GSTは、実施例12における図4(a)及び(b)の白丸で示されるように、夾雑物が添加されていない場合及び夾雑物としてTween20が添加されている場合では親水性のポリスチレンプレートに結合しているが、図5(a)及び(b)に示すとおり、全く酵素の活性を示していない。これは、原ペプチドであるGSTは、親水性のポリスチレンプレートに対し固定化されるが、構造変化や不活性化していることを意味している。なお、GSTは、図5(c)においても全く酵素の活性を示していないが、これは、図4(c)に示すように、そもそもTween20及びBSAの存在下では、親水性のポリスチレンプレートに殆どGSTが結合できないためである。
【0100】
また、図5(d)から、疎水性のポリスチレンプレートに固定化された本ペプチド1のGSTは、殆ど酵素の活性を示していない。実施例12における図4(d)から明らかなように、夾雑物が存在しない状態であれば、本ペプチド1は、疎水性のポリスチレンプレートに対して結合できるが、構造変化や不活性化によって活性が失われることが確認できる。つまり、本アミノ酸配列2の結合性能は、親水性のポリスチレンプレートに対して特異的なものであることが確認され、従来の疎水性のポリスチレンプレートを利用した固定化に比べて、本ペプチド1を親水性のポリスチレンプレートに固定化した方が、活性を維持できることが確認できる。なお、図5(e)及び(f)において、本ペプチド1及びGSTは、いずれも、全く酵素の活性を示していないが、これは、夾雑物としてTween20やBSAが存在すると、疎水性のポリスチレンプレートには殆ど結合できないからである。
【0101】
[実施例14](本ペプチド3乃至6の親水性のポリスチレン表面に対する結合性能評価、夾雑物:BSA及びTween20)
本ペプチド3乃至6の親水性のポリスチレン表面に対する結合性能を評価するため、夾雑物としてBSA及びTween20が存在する状態における実施例3乃至6の本ペプチド3乃至6並びに評価のために本ペプチド1及び原ペプチドであるGSTの親水性のポリスチレン表面に対する固定化率を測定した。
【0102】
まず、BSAを添加しない0.1PBSTと、BSA濃度が0.003〜50mg/mlとなるように調製した0.1PBSTを用意し、各溶液中に、終濃度が5μg/mlとなるように本ペプチド3を添加して試料を調製した。次に、100μlの各試料を親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、25℃で3時間インキュベートして、本ペプチド3を固定化させ、0.1%PBSで6回洗浄した。さらに、ブロッキング試薬として、2%のBSAを含む0.1PBSTを300μl加え、25℃で1時間インキュベートして、ポリスチレンプレート表面の未結合部位をブロッキングし、0.1PBSTで6回洗浄した。そして、抗体として、0.2%のBSAを含む0.1PBSTで5000倍に希釈された抗GST抗体を100μl加え、25℃で1時間インキュベートして、本ペプチド3に含まれるGSTと抗GST抗体とを抗原抗体反応させ、0.1PBSTで6回洗浄した。その後、酵素標識された第2の抗体として、0.2%のBSAを含む0.1PBSTで5000倍に希釈されたHRP-conjugated anti-rabbit IgG antibodyを100μl加え、25℃で1時間インキュベートして、抗原抗体反応させ、さらに0.1PBSTで6回洗浄した。最後に、発色基質として、ABTS(2,2'-azinobis(3-ethylbenzthiazoline-6-sulphonic acid))を加え、インキュベートし、マイクロプレートリーダー(TECAN製「SUNRISE Remote」)を用いて、405nmにおける吸光度を測定した。本ペプチド4乃至6、本ペプチド1及びGSTについても同様の処理を行い吸光度を測定した。
【0103】
図7に実施例14の測定結果を示す。図7中の黒塗りの三角は本ペプチド3の結果であり、白抜き三角(△)は本ペプチド4の結果であり、黒塗りの四角は本ペプチド5の結果であり、白抜き四角(□)は本ペプチド6の結果であり、黒丸(●)は本ペプチド1の結果であり、白丸(○)は原ペプチドであるGSTの結果である。図7において、縦軸は固定化率(%)であり、横軸はBSA濃度であるが、固定化率は、BSAを添加しないPBSTにおける吸光度(結合した量)を基準(100%)として、その他の吸光度を規格化したものである。なお、図7において、本ペプチド1、3乃至6の結果は、類似しており、重畳している部分がある。
【0104】
図7から、原ペプチドであるGSTでは、夾雑物としてBSA及びTween20が存在すると、急激に固定化率が低下し、結合する量が減少するが、本ペプチド3乃至6では、10mg/mlという高濃度のBSAが存在していても、固定化率は殆ど変わらず、本ペプチド1と同じレベルで、親水性のポリスチレンプレートに特異的に結合することが確認できる。
【0105】
[実施例15](本ペプチド3乃至6のポリスチレン表面における活性値評価、夾雑物:BSA及びTween20)
親水性のポリスチレン表面に固定化された状態における本ペプチド3乃至6の活性値を評価するため、夾雑物としてTween20が存在する状態及びTween20及びBSAが存在する状態のそれぞれで、固定化されたGSTの酵素としての活性値を測定した。
【0106】
まず、PBSと、0.1PBSTとを用意し、各溶液中に、終濃度が0.5、10、20、40μg/mlとなるように本ペプチド1、3乃至6をそれぞれ添加して試料を調製した。次に、100μlの各試料を親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、25℃で2時間インキュベートして、本ペプチドを固定化させ、0.1PBSTで6回洗浄し、さらに0.1%Tween20を含む0.1Mリン酸カリウム溶液(pH6.5)で1回洗浄した。その後、0.1%Tween20、1mMの還元型グルタチオン及びGSTの基質として1mMのCDNB(1-chloro-2,4-dinitrobenzene)を含む0.1Mリン酸カリウムバッファー(pH6.5)を200μl加え、室温で30分間撹拌しつつ、30秒毎に340nmにおける吸光度を測定した。原ペプチドであるGSTについても同様の処理を行い吸光度を測定した。
【0107】
図8に実施例15の測定結果を示す。図8の(a)は、PBSにおける測定結果であり、(b)は0.1PBSTにおける測定結果である。また、図8中の黒塗りの三角は本ペプチド3の結果であり、白抜き三角(△)は本ペプチド4の結果であり、黒塗りの四角は本ペプチド5の結果であり、白抜き四角(□)は本ペプチド6の結果であり、黒丸(●)は本ペプチド1の結果であり、白丸(○)は原ペプチドであるGSTの結果である。図8において、縦軸は活性値(吸光度の変化率)であり、横軸は本ペプチド及びGSTの濃度である。なお、図8において、本ペプチド1、3乃至6の結果は、類似しており、重畳している部分がある。
【0108】
図8(a)及び(b)から、親水性のポリスチレンプレートに固定化された本ペプチド1、3乃至6のGSTは、そのままでも(図8(a))、夾雑物としてTween20を添加した場合(図8(b))でも、酵素として非常に活性であることが確認できた。
【0109】
[実施例16](本ペプチド7乃至9の親水性のポリスチレン表面に対する結合性能評価、夾雑物:BSA及びTween20)
実施例7乃至9で生合成した本ペプチド7乃至9の親水性のポリスチレン表面に対する結合性能を評価するため、本ペプチド7乃至9並びに原ペプチドであるGSTについて、実施例14と同様の処理を行い吸光度を測定し、親水性のポリスチレン表面に対する固定化率を評価した。
【0110】
図9に実施例16の測定結果を示す。図9中の黒塗りの丸は本ペプチド7の結果であり、黒塗りの菱形は本ペプチド8の結果であり、白抜き三角(△)は本ペプチド9の結果であり、白丸(○)は原ペプチドであるGSTの結果である。図9において、縦軸は固定化率(%)であり、横軸はBSA濃度であるが、固定化率は、BSAを添加しないPBSTにおける吸光度(結合した量)を基準(100%)として、その他の吸光度を規格化したものである。
【0111】
図9から、原ペプチドであるGSTでは、夾雑物としてBSA及びTween20が存在すると、急激に固定化率が低下し、結合する量が減少するが、本ペプチド7乃至9では、10mg/mlという高濃度のBSAが存在していても、固定化率は殆ど変わらず、親水性のポリスチレンプレートに特異的に結合することが確認できた。
【0112】
[実施例17](本ペプチド7乃至9のポリスチレン表面における活性値評価、夾雑物:BSA及びTween20)
親水性のポリスチレン表面に固定化された状態における本ペプチド7乃至9の活性値を評価するため、本ペプチド7乃至9並びに原ペプチドであるGSTについて、実施例15と同様の処理を行い活性値を測定した。
【0113】
図10に実施例17の測定結果を示す。図10の(a)は、PBSにおける測定結果であり、(b)は0.1PBSTにおける測定結果である。また、図10中の黒塗りの丸は本ペプチド7の結果であり、黒塗りの菱形は本ペプチド8の結果であり、白抜き三角(△)は本ペプチド9の結果であり、白丸(○)は原ペプチドであるGSTの結果である。図10において、縦軸は活性値(吸光度の変化率)であり、横軸は本ペプチド及びGSTの濃度である。
【0114】
図10(a)及び(b)から、親水性のポリスチレンプレートに固定化された本ペプチド7乃至9のGSTは、そのままでも(図10(a))、夾雑物としてTween20を添加した場合(図10(b))でも、酵素として非常に活性であることが確認できた。
【0115】
[実施例18](本ペプチド1の生産時における分離精製工程への適用例、夾雑物:菌体破砕液)
本実施例においては、本ペプチドの特異的な吸着機能を生産時における分離精製工程への適用可能性を評価するため、大腸菌を使用して発現させた本アミノ酸配列2を含むGSTを、特別な精製工程を行うことなく、菌体破砕液をそのまま親水性のポリスチレン表面に固定化させた時の結合した量(吸光度)を測定した。比較用として、原ペプチドであるGSTについても同様に結合した量(吸光度)を測定した。
【0116】
まず、1列につき8個の培養容器(ウェル)が設けられたウェルプレートを5列用いて、39個の培養容器(ウェル)内に200μlの2xYT培地を添加し、各ウェルに、実施例1と同様の本アミノ酸配列2をコードする遺伝子を導入したベクターを導入して形質転換した大腸菌(BL21)を植菌してコロニーを形成し、37℃、200rpmで一晩培養した。原ペプチドであるGSTについても同様に40コロニーを作成し培養した。各コロニーの培養の程度を確認するため、各培養液を100μl回収し、菌体濃度を測定した。図11(a)は本ペプチドを培養した39コロニーにおける菌体濃度(630nmにおける吸光度)であり、図11(b)は原ペプチドを培養した40コロニーにおける菌体濃度である。図11において、横軸はウェルプレートの列番号であり、各列毎に8個のウェルの結果が示されている。図11(a)及び(b)から、全てのコロニーで同程度に培養されていることが確認された。
【0117】
次に、菌体破砕後、菌体破砕液を10μl回収し、10mg/mlのBSAを1%含むPBSを90mlと混合して、親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、室温で1時間インキュベートして、固相上にペプチドを固定化させた。その後、実施例7と同じ条件で、ブロッキング試薬、抗体、酵素標識された第2の抗体及び発色基質を添加する処理及び洗浄処理を行い吸光度を測定した。原ペプチドであるGSTについても同様の処理を行い吸光度を測定した。
【0118】
図11(c)及び(d)に実施例18の測定結果を示す。図11(c)は本ペプチドを培養した39コロニーにおける吸光度(405nm)であり、図11(d)は原ペプチドを培養した40コロニーにおける吸光度(405nm)である。図11(d)に示すように、原ペプチドであるGSTは、吸光度がバックグラウンド程度であり、ほとんど固相表面に固定化されていない。これに対し、図11(c)から、本ペプチドでは、大腸菌の菌体破砕液に含まれた状態のままであっても、固相表面に接触させることで、固相表面に特異的に固定化されることが確認できた。大腸菌の菌体破砕液の中には、夾雑物として、特定されていない大小様々なタンパク質に加えて、タンパク質以外の低分子化合物や脂質なども存在する。これらの夾雑物が存在しても、本ペプチドは親水性のポリスチレンプレートに対し特異的に結合するので、生産時における分離精製工程として利用可能であることが確認できた。
【0119】
[実施例19](本ペプチド1の生産時における分離精製工程への適用例、夾雑物:培地)
本実施例においては、本ペプチドの特異的な吸着機能を生産時における分離精製工程への適用可能性を評価するため、本ペプチド1に夾雑物として各種の培地(YPD、BMMY、2xTY、LB)を加えた状態における親水性のポリスチレン表面に対する結合した量(吸光度)を測定した。比較用として、原ペプチドであるGSTについても同様に結合した量(吸光度)を測定した。YPDは一般的な酵母の培養培地であり、BMMYはP. pastoris専用培地であり、2xTY及びLBは大腸菌専用培地である。
【0120】
まず、YPD、BMMY、2xTY及びLBの4種類の培地を用意し、それぞれ原液〜10000倍に希釈した溶液に対し、5μg/mlとなるように本ペプチド1を添加して試料を調製した。各試料を、親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、室温で1時間インキュベートして、固相上にペプチドを固定化させた。その後、実施例7と同じ条件で、ブロッキング試薬、抗体、酵素標識された第2の抗体及び発色基質を添加する処理及び洗浄処理を行い吸光度を測定した。原ペプチドであるGSTについても同様の処理を行い吸光度を測定した。原ペプチドであるGSTについても同様に吸光度を測定した。
【0121】
図12に実施例19の測定結果を示す。図12中の黒丸(●)は本ペプチド1と培地YPDの混合試料の結果であり、白丸(○)は原ペプチドであるGSTと培地YPDの混合試料の結果であり、黒塗りの三角は本ペプチド1と培地BMMYの混合試料の結果であり、白抜き三角(△)は原ペプチドであるGSTと培地BMMYの混合試料の結果であり、黒塗りの四角は本ペプチド1と培地2xYTの混合試料の結果であり、白抜き四角(□)は原ペプチドであるGSTと培地2xYTの混合試料の結果であり、菱形(◆)は本ペプチド1と培地LBの混合試料の結果であり、白抜き菱形(◇)は原ペプチドであるGSTと培地LBの混合試料の結果でありである。図12において、縦軸は吸光度(405nm)であり、横軸は培地の相対濃度(希釈率)(右端の1が原液)である。図12に示すように、原ペプチドであるGST(白丸、白抜き三角、白抜き四角、白抜き菱形)は、何れも培地が濃くなるにつれて固相表面に結合される量が減っている。これに対し、本ペプチド1(黒丸、黒塗り三角、黒塗り四角、黒塗り菱形)では、培地が濃くなってもかなりの量が固定化されて固相表面に結合されており、培地の濃度にあまり影響されず、固相表面に特異的に固定化されることが確認できた。
【0122】
特に、酵母で使用される培地であるYPD及びBMMYにおいても特異的な吸着性能を発現するので、酵母からの分泌生産に利用可能である。つまり、酵母からの分泌液を直接固相表面に接触させることで、分泌液中の本ペプチドを固定化して分離精製することが可能である。
【0123】
さらに、一本鎖抗体(scFv)の生産に有効な培地であるBMMY及び2xYTでも特異的な吸着性能を発現するので、本ペプチドとして一本鎖抗体(scFv)を生産した場合でも、酵母の分泌液や大腸菌の破砕液から直接一本鎖抗体(scFv)を分離精製することが可能である。
【0124】
[実施例20](本アミノ酸配列2からなる本ペプチドを標識した抗体の生産)
まず、固相法によって、本アミノ酸配列2からなる本ペプチドのN末端側にリジン(K)を有するペプチド(KRIIIRRIRR)を合成した。次に、0.5mgの本アミノ酸配列2からなる本ペプチドを100μlの25%グルタルアルデヒド溶液で溶解し、室温で10分間インキュベートした。そして、9mlのジエチルエーテル及び1mlのエタノールを加えてグルタルアルデヒドで標識された本ペプチドを析出させ、遠心分離により回収した。
【0125】
さらに、0.5mgのマウスモノクローナル抗ヒトCRP抗体(オリエンタル酵母社製、#4715500)を溶解した0.5mlのPBSに、グルタルアルデヒドで標識された本ペプチドを溶解し、25℃で1時間インキュベートした。その後、100μlの2M Tris−HCl(pH8.0)を加え、反応を停止して、限外ろ過法によって、本ペプチドとして、本アミノ酸配列2を含む抗CRP抗体を精製した。以下、実施例20で生成した本アミノ酸配列2を含む抗CRP体を「本ペプチド20」といい、その原ペプチドを「原ペプチド20」という。
【0126】
[実施例21](本アミノ酸配列2をC末端側に有する一本鎖抗体の生成)
本実施例では、配列表の配列番号23、24、25に示す3種類の一本鎖抗体(scFv)のC末端側に本アミノ酸配列2を導入して3種類の一本鎖抗体を生成した。なお、以下、配列表の配列番号23、24及び25に示す3種類の一本鎖抗体(原ペプチド)を、それぞれ「原ペプチド21−1」、「原ペプチド21−2」及び「原ペプチド21−3」といい、本実施例で生成される原ペプチド21−1、原ペプチド21−2及び原ペプチド21−3に本アミノ酸配列2を導入した3種類の一本鎖抗体(本ペプチド)を、それぞれ「本ペプチド21−1」、「本ペプチド21−2」及び「本ペプチド21−3」という。これらの一本鎖抗体は、いずれもC反応性蛋白(C-Reactive Protein:CRP)に対して特異的な抗原抗体反応を示す抗原結合部位を有する。原ペプチド21−1ないし21−3は、重鎖の可変部(VH)のC末端と軽鎖の可変部(VL)のN末端とが4つのグリシン(G)とセリン(S)の組合わせを3組連結した構造 (G4S)3の鎖状のリンカーペプチドで連結した構造である。さらに原ペプチド21−1ないし21−3には軽鎖の可変部(VL)のC末端にヒスタジンタグが導入されている。なお、実施例22及び23の原ペプチドは、配列番号23の一本鎖抗体(原ペプチド21−1)である。
【0127】
まず、5’末端側がリン酸化された本アミノ酸配列2をコードする遺伝子を含むセンス鎖(PStag-sense)及びアンチセンス鎖(PStag-antisense)のオリゴDNAを合成し、アニール後、制限酵素NotI及びXhoIで消化したpET22b(+)ベクター(Novagen製)に導入し、本アミノ酸配列2をコードする遺伝子を含むpET-PStagベクター(pET22b(+)にPStagが導入されたベクター)を調製した。なお、「PStag」とは本アミノ酸配列2を発現させる遺伝子を意味する。以下同じ。
【0128】
次に、重鎖の可変部(VH)をコードする遺伝子を含むセンス鎖(VH-sense)のオリゴDNA及び軽鎖の可変部(VL)をコードする遺伝子を含むアンチセンス鎖(VL-antisense)のオリゴDNAを合成した。これらの合成オリゴDNAをプライマーとし、それぞれ配列番号23,24,25に示す3種類の一本鎖抗体発現ベクターpET-scFv(pET22b(+)にscFvの遺伝子が導入されたベクター)を鋳型としてPCR法によって3種類の一本鎖抗体(scFv)の遺伝子を増幅した。増幅したscFv遺伝子を制限酵素NdeI及びNotIで消化し、同酵素で消化した本アミノ酸配列2をコードする遺伝子を含むpET-PStagベクターに導入し、C末端側に本アミノ酸配列2をコードする遺伝子を含む3種類の一本鎖抗体の発現ベクター(pET-scFv-PStagベクター)を調製した。
【0129】
宿主として大腸菌(BL21(DE3)Rosetta(Novagen))を使用し、上記pET-scFv-PStagベクターを宿主に導入して宿主を形質転換し、アンピシリンを含む寒天培地上で選別した。その後、2xYT培地50ml中で大腸菌を培養し、終濃度が1mMになるように、発現誘導物質としてイソプロピル1−チオ−ガラクトシド(IPTG:Isopropyl-1-thio-β-D(-)-galactoside)を加えて、本アミノ酸配列2をC末端に含む一本鎖抗体(scFv-PStag)を発現させた。
【0130】
そして、菌体破砕後、菌体内不溶性画分を6M塩酸グアニジン(変性剤)及び10mMメルカプトエタノール(還元剤)を含む可溶化液で溶解した。こうして、未精製の変性された本アミノ酸配列2をC末端に含む一本鎖抗体を得ることができた。以下、この状態の実施例21の一本鎖抗体を「未精製の変性本ペプチド21−1」ないし「未精製の変性本ペプチド21−3」という。
【0131】
さらに、未精製の変性本ペプチド21−1ないし21−3を変性状態のままHisTrap HP (GE HealthCare)を用いたNiキレートアフィニティクロマトグラフィ(IMAC)によって精製し、本アミノ酸配列2をC末端側に含む変性した一本鎖抗体を生産した。以下、この状態の実施例21の一本鎖抗体を「精製後の変性本ペプチド21−1」ないし「精製後の変性本ペプチド21−3」という。本ペプチド21−1のアミノ酸配列を配列表の配列番号26に示し、本ペプチド21−2のアミノ酸配列を配列表の配列番号27に示し、本ペプチド21−3のアミノ酸配列を配列表の配列番号28に示す。本ペプチド21−1ないし21−3は、軽鎖の可変部(VL)のC末端側に本アミノ酸配列2を有しており、本アミノ酸配列2のさらにC末端側にヒスタジンタグが連結されている。なお、アミノ酸配列それ自体は、精製の前後でも、変性の前後でも変わらず、配列番号26、27、28に示すとおりである。
【0132】
[実施例22](本アミノ酸配列2を重鎖可変部(VH)とリンカーペプチドの間に有する一本鎖抗体の生成)
本実施例では、本アミノ酸配列2を原ペプチド21−1(配列番号23)及び原ペプチド21−2(配列番号24)の重鎖の可変部(VH)と軽鎖の可変部(VL)を連結するリンカーペプチドに導入した一本鎖抗体(以下で生成されるペプチドを、それぞれ「本ペプチド22−1」及び「本ペプチド22−2」という)を生成する。まず、重鎖の可変部(VH)をコードする遺伝子を含むセンス鎖(VH-sense)のオリゴDNA及びアンチセンス鎖(VH-antisense)のオリゴDNAを合成し、これらの合成オリゴDNAをプライマーとし、配列番号23の一本鎖抗体を発現するpET-scFvベクターを鋳型としてPCR法によって重鎖の可変部(VH)の遺伝子を増幅した。
【0133】
次に、軽鎖の可変部(VL)をコードする遺伝子を含むセンス鎖(VL-sense)のオリゴDNA及びアンチセンス鎖(VL-antisense)のオリゴDNAを合成し、同様に軽鎖の可変部(VL)の遺伝子を増幅した。
【0134】
さらに、重鎖の可変部(VH)のC末端部、本アミノ酸配列2及びリンカーペプチドの順にこれらをコードする遺伝子を持つセンス鎖(VH-Pstag-(G4S)3-sense)のオリゴDNAと本アミノ酸配列2、リンカーペプチド及び軽鎖の可変部(VL)のN末端部分の順にこれらをコードする遺伝子を含むアンチセンス鎖(PStag-(G4S)3-VL-antisense)のオリゴDNAを合成した。
【0135】
重鎖の可変部(VH)の遺伝子、軽鎖の可変部(VL)の遺伝子、センス鎖(VH-PStag-(G4S)3-sense)のオリゴDNA、アンチセンス鎖(PStag-(G4S)3-VL-antisense)のオリゴDNA、センス鎖(VH-sense)のオリゴDNA及びアンチセンス鎖(VL-antisense)のオリゴDNAを用いて、Overlapping PCR法により本アミノ酸配列2を重鎖可変部(VH)とリンカーペプチド(G4S)3との間に有する一本鎖抗体の遺伝子(scFv-(PStag))を調製した。当該遺伝子を制限酵素NdeI及びNotIで消化し、同酵素で消化したpET22b(+)に導入し、リンカーペプチド(G4S)3に本アミノ酸配列2をコードする遺伝子を含む一本鎖抗体の発現ベクター(pET-scFv-(PStag) ベクター)を作製した。なお、「(PStag)」とは一本鎖抗体の途中である重鎖可変部(VH)とリンカーペプチド(G4S)3との間にPStagが導入されることを意味する。以下同じ。
【0136】
後は、実施例21と同様に、当該pET-scFv-(PStag)ベクターを宿主に導入して培養し、菌体破砕後、菌体内不溶性画分を6M塩酸グアニジン(変性剤)及び10mMメルカプトエタノール(還元剤)を含む可溶化液で溶解して、未精製の変性されたリンカーペプチド(G4S)3に本アミノ酸配列2を有する一本鎖抗体を得ることができた。さらに、実施例21と同様の方法で精製し、精製された変性した状態の本ペプチド22−1(以下「精製後の変性本ペプチド22−1」という)を生産した。本ペプチド22−1のアミノ酸配列を配列表の配列番号29に示す。本ペプチド22−1は、重鎖の可変部(VH)のドメインとリンカーペプチド(G4S)3の間に本アミノ酸配列2が配置されている。
【0137】
また、同様の手法によって、原ペプチド21−2(配列番号24)の重鎖の可変部(VH)のドメインとリンカーペプチド(G4S)3の間に本アミノ酸配列2が配置されている精製後の変性本ペプチド22−2を生産した。本ペプチド22−2のアミノ酸配列を配列表の配列番号30に示す。
【0138】
[実施例23](本アミノ酸配列2をC末端及び重鎖可変部(VH)とリンカーペプチドとの間の両方に有する一本鎖抗体の生成)
本実施例では、本アミノ酸配列2を原ペプチド21−1及び原ペプチド21−2の一本鎖抗体のC末端側(すなわち軽鎖の可変部(VL)のC末端側)及び重鎖可変部(VH)とリンカーペプチドの間(すなわち重鎖の可変部(VH)のC末端側)の両方に導入した一本鎖抗体(以下で生成されるペプチドを、それぞれ「本ペプチド23−1」及び「本ペプチド23−2」という)を生成する。まず、実施例22と同様に、本アミノ酸配列2を重鎖可変部(VH)とリンカーペプチド(G4S)3の間に有する一本鎖抗体の遺伝子(scFv-(PStag))を調製した。当該遺伝子を実施例22ではpET22b(+)に導入したが、本実施例では、実施例21で使用した本アミノ酸配列2をコードする遺伝子を含むpET-PStagベクターに導入し、C末端側及び重鎖可変部(VH)とリンカーペプチド(G4S)3との間の両方に本アミノ酸配列2をコードする遺伝子を含む一本鎖抗体の発現ベクター(pET-scFv-(PStag)-PStagベクター)を作製した。
【0139】
後は、実施例21と同様に、当該pET-scFv-(PStag)-PStagベクターを宿主に導入して培養し、菌体破砕後、菌体内不溶性画分を6M塩酸グアニジン(変性剤)及び10mMメルカプトエタノール(還元剤)を含む可溶化液で溶解して、未精製の変性されたC末端側及びリンカーペプチドの両方に本アミノ酸配列2を有する一本鎖抗体(scFv)を得ることができた。さらに、実施例21と同様に精製し、精製された変性した状態の本ペプチド23−1(以下「精製後の変性本ペプチド23−1」という)を生産した。本ペプチド23−1のアミノ酸配列を配列表の配列番号31に示す。本ペプチド23−1は、本ペプチド21−1と同様に軽鎖の可変部(VL)のC末端側に本アミノ酸配列2を有しており、さらに本アミノ酸配列2のC末端側にヒスタジンタグが連結されている。さらに本ペプチド22−1と同様に重鎖の可変部(VH)のドメインとリンカーペプチド(G4S)3との間に本アミノ酸配列2を有している。
【0140】
また、同様の手法によって、原ペプチド21−2(配列番号24)の軽鎖の可変部(VL)のC末端側及び重鎖の可変部(VH)のドメインとリンカーペプチド(G4S)3の間に本アミノ酸配列2が配置されている精製後の変性本ペプチド23−2を生産した。本ペプチド23−2のアミノ酸配列を配列表の配列番号32に示す。
【0141】
[実施例24](本ペプチド21−1ないし21−3の固定化後の活性値評価)
本実施例では、本ペプチド21−1ないし21−3(配列番号26〜28)の固定化された状態における活性値を評価するため、精製後の変性本ペプチド21−1ないし21−3を液相でリフォールディングした後、これらのペプチドを親水性のポリスチレン表面に対して結合させ、抗原抗体反応の活性値を測定した。比較用として、原ペプチド21−1ないし21−3(配列番号23〜25)と、一本鎖抗体ではなく抗体分子である本ペプチド20及び原ペプチド20についても親水性のポリスチレン表面に対して結合させ、活性値を測定した。さらに、従来との比較用として、原ペプチド20(抗体分子)を疎水性のポリスチレン表面に対して結合させ、活性値を測定した。
【0142】
まず、変性剤である尿素が80mMとなるように精製後の変性本ペプチド21−1ないし21−3(配列番号26〜28)を375μM 酸化型グルタチオン(GSSG)を含むPBS中に希釈し、室温で3時間インキュベートしてリフォールディングを行った。遠心分離によって凝集体を除去した後HisTrap HP (GE HealthCare)を用いたNiキレートアフィニティクロマトグラフィ(IMAC)によって精製し、PBSで透析することで可溶性の3種類の本ペプチド21−1ないし21−3を生成した。
【0143】
次に、本ペプチド21−1ないし21−3を5μg/mlの濃度に0.1PBSTで希釈し、100μlを親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、2時間室温でインキュベートして、本ペプチドを固定化させた。0.1PBSTで6回洗浄した後、2%のBSAを含む0.1PBST(ブロッキング試薬)によってポリスチレンプレート表面の未結合部位をブロッキングした。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで1〜5000ng/mlに希釈されたbiotin-CRP (抗原)を100μl加え、室温で1時間インキュベートして、本ペプチドと抗原抗体反応させた。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで5000倍に希釈されたHRP標識ストレプトアビジンを100μl加え、室温で30分インキュベートして、0.1PBSTで6回洗浄した後に、発色基質を加えて30分間発色させた。30分後にマイクロプレートリーダー(TECAN製「SUNRISE Remote」)を用いて、405nmにおける吸光度を測定した。配列番号23〜25に示す原ペプチド21−1ないし21−3である一本鎖抗体、実施例20に記載した本アミノ酸配列2を標識した本ペプチド20、原ペプチド20についても同様の処理を行い吸光度を測定した。さらに、5μg/mlの原ペプチド20をPBS中で疎水性のポリスチレンプレート(ベクトン・ディッキンソン アンド カンパニー社製「BD Falconマイクロプレート#351172」)に固定化して吸光度を測定した。
【0144】
図13に実施例24の測定結果を示す。図13中、黒丸(●)、黒塗りの三角及び黒塗りの四角はそれぞれ配列番号26、27、28に示す本ペプチド21−1、21−2、21−3の結果であり、白丸(○)、白塗りの三角(△)及び白塗りの四角(□)はそれぞれ配列番号23、24、25に示す原ペプチド21−1、21−2、21−3の結果である。さらに、黒塗りの菱形(◆)は、本ペプチド20の結果であり、白塗りの菱形(◇)は、原ペプチド20であるマウスモノクローナル抗ヒトCRP抗体(オリエンタル酵母社製、#4715500)の結果である。×印は原ペプチド20の疎水性のポリスチレンプレートに固定化して実験を行った結果である。図11において、縦軸は、吸光度(405nm)であり、横軸はbiotin-CRPの濃度(ng/ml)である。
【0145】
図13から原ペプチド21−1、21−2、21−3(○、△、□)並びに原ペプチド20(◇)では、親水性のポリスチレン表面においては、ほとんど活性化されていないが、本ペプチド21−1、21−2、21−3(黒丸、黒塗り三角、黒塗りの四角)では、1本鎖抗体のアミノ酸配列の違いによらず何れも高い抗原結合活性が維持されていることが確認できる。また、本アミノ酸配列2を標識した本ペプチド20(◆)においても親水性ポリスチレンプレート上に選択的に固定化され高い抗原結合活性を示していることがわかる。従来ELISAに用いられてきた疎水性ポリスチレンプレート上に固定化されたマウスモノクローナル抗体(原ペプチド20)の結果(×)と比較しても本アミノ酸配列2を有する本ペプチド20並びに21−1、21−2及び21−3は同等もしくはそれ以上に高いシグナルが得られていることが分かる。以上の結果より、様々な1本鎖抗体のC末端側に本アミノ酸配列2を導入した本ペプチド21−1、21−2、21−3を生産することで親水性ポリスチレンプレートに抗原結合活性を高く維持した状態で固定化できることが確認できた。また、本アミノ酸配列2をモノクローナル抗体に標識することで親水性ポリスチレンプレートに抗原結合活性を高く維持した状態で固定化できることが確認できた。
【0146】
[実施例25](精製後の変性本ペプチド21−1,22−1及び23−1の固相リフォールディング及び活性値評価)
本実施例では、精製後の変性本ペプチド21−1,22−1及び23−1の固定化された状態におけるリフォールディング及び活活性値を評価するため、これらのペプチドを親水性のポリスチレン表面に対して結合させ、抗原抗体反応の活性値を測定した。
【0147】
まず、変性剤である尿素の終濃度が4Mとなるように、各精製後の変性本ペプチド21−1(配列番号26),22−1(配列番号29)及び23−1(配列番号31)を調製して試料を作成した。次に、各試料20μlを親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、ミキサーで撹拌後、室温で1時間インキュベートして、本ペプチドを固定化させた(1ウェルあたりのscFvの重量:0.5μg/ウェル)。0.1PBSTで6回洗浄した後、2%のBSAを含む0.1PBST(ブロッキング試薬)によってポリスチレンプレート表面の未結合部位をブロッキングした。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで1〜5000ng/mlに希釈されたbiotin-CRP (抗原)を100μl加え、室温で1時間インキュベートして、本ペプチドと抗原抗体反応させた。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで5000倍に希釈されたHRP標識ストレプトアビジンを100μl加え、室温で30分インキュベートして、0.1PBSTで6回洗浄した後に、発色基質を加えて30分間発色させた。30分後にマイクロプレートリーダー(TECAN製「SUNRISE Remote」)を用いて、405nmにおける吸光度を測定した。原ペプチド21−1(配列番号23)についても同様の処理を行い吸光度を測定した。
【0148】
図14に実施例25の測定結果を示す。図14中、黒丸(●)は精製後の変性本ペプチド21−1の結果であり、黒塗りの三角は精製後の変性本ペプチド22−1の結果であり、黒塗りの四角は精製後の変性本ペプチド23−1の結果であり、白丸(○)は原ペプチド21−1の結果である。図14において、縦軸は、吸光度(405nm)であり、横軸はbiotin-CRPの濃度(ng/ml)である。
【0149】
図14から、原ペプチド21−1である一本鎖抗体(○)では、ほとんど活性化されていないが、精製後の変性本ペプチド21−1,22−1及び23−1は何れも活性化されていることが確認できる。つまり、本アミノ酸配列を用いることにより、変性されている本ペプチド21−1,22−1及び23−1を固相に接触させることによって、固定化させると共に、変性を復元するリフォールディングも行われている。
【0150】
さらに、本アミノ酸配列2をC末端側に導入した本ペプチド21−1も、本アミノ酸配列2を重鎖可変部とリンカーペプチドとの間に導入した本ペプチド22−1も同程度の活性であるのに対し、本アミノ酸配列2をC末端側及び重鎖可変部とリンカーペプチドとの間の両方に導入した本ペプチド23−1ではより高い活性が得られることが確認できた。
【0151】
[実施例26](精製後の変性本ペプチド21−2,22−2及び23−2の固相リフォールディング及び活性値評価)
本実施例では、精製後の変性本ペプチド21−2,22−2及び23−2の固定化された状態におけるリフォールディング及び活活性値を評価するため、これらのペプチドについて、親水性のポリスチレン表面に対して結合させ、抗原抗体反応の活性値を測定した。なお、本実施例は、基本的な操作は実施例25と同様であるが、多少条件を変更しているため、以下念のため記載する。
【0152】
まず、変性剤である尿素の終濃度が4Mとなり、各精製後の変性本ペプチド21−2(配列番号27),22−2(配列番号30)及び23−2(配列番号32)が10μg/mlとなるように、1%のTween20を含むPBSによって溶液を調製して試料を作成した。次に、各試料100μlを親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、ミキサーで撹拌後、室温で3時間インキュベートして、本ペプチドを固定化させた。0.1PBSTで6回洗浄した後、2%のBSAを含む0.1PBST(ブロッキング試薬)を300μl加えて、室温で1時間インキュベートしてポリスチレンプレート表面の未結合部位をブロッキングした。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで1〜5000ng/mlに希釈されたbiotin-CRP (抗原)を100μl加え、室温で1時間インキュベートして、本ペプチドと抗原抗体反応させた。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで5000倍に希釈されたHRP標識ストレプトアビジンを100μl加え、室温で1時間インキュベートした。さらに、0.1PBSTで6回洗浄した後に、発色基質を100μl加えて30分間発色させた。30分後にマイクロプレートリーダー(TECAN製「SUNRISE Remote」)を用いて、405nmにおける吸光度を測定した。原ペプチド21−2(配列番号24)についても同様の処理を行い吸光度を測定した。
【0153】
図15に実施例26の測定結果を示す。図15中、黒丸(●)は精製後の変性本ペプチド21−2の結果であり、黒塗りの三角は精製後の変性本ペプチド22−2の結果であり、黒塗りの四角は精製後の変性本ペプチド23−2の結果であり、白丸(○)は原ペプチド21−2の結果である。図15において、縦軸は、吸光度(405nm)であり、横軸はbiotin-CRPの濃度(ng/ml)である。
【0154】
図15から、原ペプチド21−2である一本鎖抗体(○)では、ほとんど活性化されていないが、精製後の変性本ペプチド21−2,22−2及び23−2は何れも活性化されていることが確認できる。つまり、実施例25と同様に、本実施例においても、本アミノ酸配列を用いることにより、変性されている本ペプチド21−2,22−2及び23−2を固相に接触させることによって、固定化させると共に、変性を復元するリフォールディングも行われることが確認できた。
【0155】
さらに、本アミノ酸配列2をC末端側に導入した本ペプチド21−2の方が、本アミノ酸配列2を重鎖可変部とリンカーペプチドとの間に導入した本ペプチド22−2よりも活性が高いことが確認できるが、それ以上に、本アミノ酸配列2をC末端側及び重鎖可変部とリンカーペプチドとの間の両方に導入した本ペプチド23−2の方がより高い活性が得られることも確認できた。
【0156】
[実施例27](精製後の変性本ペプチド21−1の固相に対する結合性能及び活性値評価)
本実施例では、精製後の変性本ペプチド21−1(配列番号26)の量と親水性のポリスチレン表面に対する結合性能及び活性値との関係を評価した。
【0157】
まず、精製後の変性本ペプチド21−1を3.2μg/ml、32μg/ml及び320μg/mlの各濃度となるように8M尿素で調製して試料を作成した。次に、各10μlの試料をそれぞれ親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、さらに各ウェルに0.1PBSTを90μl加えて10倍に希釈し、4℃で一晩インキュベートして、本ペプチド21−1を固定化させた。0.1PBSTで6回洗浄した後、2%のBSAを含む0.1PBST(ブロッキング試薬)によってポリスチレンプレート表面の未結合部位をブロッキングした。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで1〜5000ng/mlに希釈されたbiotin-CRP (抗原)を100μl加え、室温で1時間インキュベートして、本ペプチドと抗原抗体反応させた。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで1000倍に希釈されたHRP標識ストレプトアビジンを100μl加え、室温で30分インキュベートして、0.1PBSTで6回洗浄した後に、発色基質を加えて30分間発色させた。30分後にマイクロプレートリーダー(TECAN製「SUNRISE Remote」)を用いて、405nmにおける吸光度を測定した。
【0158】
図16に実施例27の測定結果を示す。図16中、黒丸(●)は精製後の変性本ペプチド21−1の量が1ウェルあたり0.032μgの場合、黒塗りの三角は0.32μgの場合、黒塗りの四角は3.2μgの場合の結果である。図16において、縦軸は、吸光度(405nm)であり、横軸はbiotin-CRPの濃度(ng/ml)である。
【0159】
図16から、精製後の変性本ペプチド21−1は、親水性のポリスチレン表面において活性化されていることが確認できる。つまり、本アミノ酸配列を用いることにより、親水性のポリスチレン基板に対して特異的な結合性能を示し、固定化させると共に、変性を復元するリフォールディングもできることが確認された。
【0160】
[実施例28](未精製の変性本ペプチド21−1ないし21−3の固定化による分離精製及び活性値評価)
本実施例では、未精製の変性本ペプチド21−1ないし21−3(配列番号26、27、28)を親水性のポリスチレン表面に固定化することによる分離精製の可能性及び固定化された状態の活性値を評価した。
【0161】
まず、未精製の変性本ペプチド21−1ないし21−3及び3.2mg/mlの精製後の変性本ペプチド21−1を0.1PBSTで10倍に希釈した試料及び100倍に希釈した試料を親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に200μl加え、4℃で一晩インキュベートして、本ペプチドを固定化させた。0.1PBSTで6回洗浄した後、2%のBSAを含む0.1PBST(ブロッキング試薬)によってポリスチレンプレート表面の未結合部位をブロッキングした。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで1〜5000ng/mlに希釈されたbiotin-CRP (抗原)を100μl加え、室温で1時間インキュベートして、本ペプチドと抗原抗体反応させた。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで1000倍に希釈されたHRP標識ストレプトアビジンを100μl加え、室温で30分インキュベートして、0.1PBSTで6回洗浄した後に、発色基質を加えて30分間発色させた。30分後にマイクロプレートリーダー(TECAN製「SUNRISE Remote」)を用いて、405nmにおける吸光度を測定した。
【0162】
図17(a)は10倍に希釈した場合の測定結果であり、図17(b)は100倍に希釈した場合の測定結果である。図17(a)及び(b)中、黒丸(●)は未精製の変性本ペプチド21−1の結果であり、黒塗りの三角は未精製の変性本ペプチド21−2の結果であり、黒塗りの四角は未精製の変性本ペプチド21−3の結果であり、白丸(○)は精製後の変性本ペプチド21−1の結果である。図17(a)及び(b)において、縦軸は、吸光度(405nm)であり、横軸はbiotin-CRPの濃度(ng/ml)である。
【0163】
図17から、未精製の状態であっても、親水性のポリスチレン表面に固定化され、活性化されることが確認できる。つまり、本アミノ酸配列を用いることにより、本ペプチドは親水性のポリスチレンプレートに対し特異的に結合するので、生産時における分離精製工程として利用可能であり、しかも活性化されるのでリフォールディングもされていることが確認できた。
【配列表フリ−テキスト】
【0164】
配列番号1:親水性固体表面に特異的な親和性を有するペプチド。
配列番号2:親水性固体表面に特異的な親和性を有するペプチド。
配列番号3:親水性固体表面に特異的な親和性を有するペプチド。
配列番号4:親水性固体表面に特異的な親和性を有するペプチド。
配列番号5:親水性固体表面に特異的な親和性を有するペプチド。
配列番号6:親水性固体表面に特異的な親和性を有するペプチド。
配列番号7:親水性固体表面に特異的な親和性を有するペプチド。
配列番号8:親水性固体表面に特異的な親和性を有するペプチド。
配列番号9:親水性固体表面に特異的な親和性を有するペプチド。
配列番号10:親水性固体表面に特異的な親和性を有するペプチド。
配列番号11:親水性固体表面に特異的な親和性を有するペプチド。
配列番号12:親水性固体表面に特異的な親和性を有するペプチド。
配列番号13:親水性固体表面に特異的な親和性を有するペプチド。
配列番号14:親水性固体表面に特異的な親和性を有するペプチド。
配列番号15:親水性固体表面に特異的な親和性を有するペプチド。
配列番号16:親水性固体表面に特異的な親和性を有するペプチド。
配列番号17:親水性固体表面に特異的な親和性を有するペプチド。
配列番号18:親水性固体表面に特異的な親和性を有するペプチド。
配列番号19:親水性固体表面に特異的な親和性を有するペプチド。
配列番号20:親水性固体表面に特異的な親和性を有するペプチド。
配列番号21:合成DNA。
配列番号22:合成DNA。
配列番号23:一本鎖抗体。
配列番号24:一本鎖抗体。
配列番号25:一本鎖抗体。
配列番号26:親水性固体表面に特異的な親和性を有する一本鎖抗体。
配列番号27:親水性固体表面に特異的な親和性を有する一本鎖抗体。
配列番号28:親水性固体表面に特異的な親和性を有する一本鎖抗体。
配列番号29:親水性固体表面に特異的な親和性を有する一本鎖抗体。
配列番号30:親水性固体表面に特異的な親和性を有する一本鎖抗体。
配列番号31:親水性固体表面に特異的な親和性を有する一本鎖抗体。
配列番号32:親水性固体表面に特異的な親和性を有する一本鎖抗体。
【技術分野】
【0001】
本発明は、新規のペプチド、当該ペプチドの用途及び生産方法に関し、特に、親水性の固相に特異的に結合する機能を発現させるペプチド、当該ペプチドの用途及び生産方法に関する。また、本発明は、固相化法を用いた化合物の検出又は測定に利用可能なペプチド、及び当該ペプチドを利用した化合物の検出又は測定方法に関する。また、本発明は、抗原結合部位が形成される可変部を含むペプチドに関し、免疫グロブリン分子、さらに可変部を含む1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)、定常部融合1本鎖抗体(scFv-Fc)、Fab断片又はF(ab’)2断片及びこれらのペプチドが固定化された固相及びかかる固相の生産方法に関する。
【背景技術】
【0002】
抗体(免疫グロブリン)は、抗原に対して特異的に結合するタンパク質であり、生体内における体液性免疫の主役である。図6(a)は、抗体(免疫グロブリン)の基本構造を示す模式図である。抗体分子61は、2本の分子量5万〜7万の重鎖(H鎖)62と、2本の分子量2.3万の軽鎖(L鎖)63の4本の鎖状ポリペプチドがジスルフィド結合及び非共有結合によって結合して、全体としてY字型を形成している。重鎖(H鎖)62は、4つ又は5つのドメインから構成され、N末端側に可変部(VH)のドメインが配置され、C末端側に向けて3〜4つの定常部(CH1、CH2、CH3、CH4)のドメインが配置される。なお、図6(a)では、4つのドメイン(VH、CH1、CH2、CH3)から構成される重鎖62を示す。軽鎖(L鎖)63は、2つのドメインから構成され、N末端側に可変部(VL)のドメインが配置され、C末端側に定常部(CL)のドメインが配置される。
【0003】
重鎖62及び軽鎖63の可変部(VH及びVL)は、N末端側の2箇所に配置され、両者が一体となって、抗原に対して特異的に結合する立体的な抗原結合部位が形成される。したがって、抗体の特異性は、重鎖及び軽鎖の可変部(VH及びVL)のアミノ酸配列及び組み合わせによって決定され、対応する抗原に応じて、可変部(VH及びVL)のアミノ酸配列や組み合わせが異なる。重鎖及び軽鎖の定常部(CH1、CH2、CH3、CH4及びCL)は、各クラスまたはサブクラス毎にほぼ一定の構造である。
【0004】
抗体分子61は、タンパク質分解酵素であるパパインによって、重鎖のCH1とCH2ドメインの間のヒンジ部64で分解され、図6(b)に示すように、2つのFab断片65と一つのFc断片66とに分解される。なお、別のタンパク質分解酵素であるペプシンによって抗体分子61を分解すると、図6(c)に示すように、2つのFab断片65がヒンジ部64でジスルフィド結合した構造のF(ab’)2断片67を得ることができる。Fab断片65やF(ab’)2断片67は、重鎖及び軽鎖の可変部(VH及びVL)による抗原結合部位を備えているため、抗原に対する特異性を有しており、抗原抗体反応に利用することができる。
【0005】
さらに、重鎖の可変部(VH)と軽鎖の可変部(VL)とをリンカーペプチド69で結合した構造の一本鎖抗体(scFv)68も、重鎖と軽鎖の可変部(VH及びVL)による立体的な抗原結合部位を形成できるため、抗原に対する特異性を有しており、抗原抗体反応に利用することができる。図6(d)は、一本鎖抗体(scFv)68の基本構造を示す模式図である。図6(d)においてが、重鎖の可変部(VH)のC末端が鎖状のリンカーペプチド69によって、軽鎖の可変部(VL)のN末端に連結されている。
【0006】
抗原抗体反応は、抗原と抗体の特異性が高く、抗原結合部位が変われば対応する抗原も変わることから、100万通り以上の重鎖及び軽鎖の可変部(VH及びVL)の組み合わせについて、どのような抗原抗体反応を示すのか検証する基礎的な実験が必要とされている。また、抗原と抗体の特異性が高いことを利用して、抗原抗体反応は、様々な用途及び分野において利用されている。
【0007】
従来から、微量の物質を検出又は測定する方法として、抗原と抗体間の特異的な親和力を利用した免疫測定法(イムノアッセイ)が知られている。免疫測定法は、抗原抗体反応の多様性から、様々な生体成分の分析が可能であり、広い分野において使用されている。さらに、免疫測定法の測定感度を高めるために、放射線化合物、蛍光物質、酵素などの標識を用いた各種の手法が知られており、各標識に応じて、それぞれ放射免疫測定法(ラジオイムノアッセイ:RIA)、免疫蛍光測定法(蛍光イムノアッセイ:FIA)、酵素免疫測定法(ELISA:Enzyme-Linked ImmunoSorbent Assay。エンザイムイムノアッセイとも呼ぶ)などと呼ばれている。特に、ELISAは、高感度で定量性に優れており、精製や前処理といった煩雑なステップを必要としない汎用性の高い検出法であることから、医療診断、環境ホルモン・残留農薬の定量、牛海綿状脳症(BSE)検査、プロテオーム解析等の様々な分析に利用されている。
【0008】
これらの免疫測定法における微量測定では、抗原、抗体又は酵素等のタンパク質を試験管やマイクロプレート等の固相に物理吸着や化学結合を利用して固定化する固相化法が利用されている。固相としては、通常、疎水性のプラスチックが利用される。疎水性の固相は、タンパク質が疎水性相互作用によって強固に結合するので、比較的多くの種類のタンパク質に対して適用できる点で優れている。現在、疎水性のプラスチックとしては、疎水性のポリスチレン(PS)が多用されている。
【0009】
また、免疫測定法は、いずれの方法においても、多数回の吸着・反応処理及び洗浄処理が必要であり、目的物質を検出するまでに長時間を要するものであった。なお、抗原抗体反応を検証する実験においても、試験管やマイクロプレート等の固相に抗体等を固定化して実験するのが一般的であり、固相化法が利用されている。
【0010】
例えば、ELISAは、直接吸着法、サンドイッチ法及び競合法に大別されるが、サンドイッチ法のELISAの場合、特許文献1によれば、第一に、固相に目的物質に対する抗体を結合させた後、固相を複数回洗浄する。第二に、固相上に残った未結合の部位に対して、固相表面にその他の試薬が結合しないように、抗原抗体反応及び酵素反応に関与しない試薬(以下「ブロッキング試薬」という。)を結合させてブロッキングした後、固相を複数回洗浄する。第三に、目的物質を含有する試料と固相上に結合された抗体とを反応させた後、固相を複数回洗浄する。第四に、第2の抗体と目的物質と反応させた後、固相を複数回洗浄する。第五に、第2の抗体と酵素標識とを反応させた後、固相を複数回洗浄する。第六に、酵素標識と基質とを反応させ、吸光度を測定し、試料中の目的物質を検出し、又は濃度を測定する(特許文献1の段落0039参照)。なお、第四の処理で反応させる第2の抗体として、既に酵素標識された抗体を利用すれば、第五の処理は不要となる。
【特許文献1】特表2002-526777号公報
【発明の開示】
【発明が解決しようとする課題】
【0011】
上で述べたとおり、重鎖及び軽鎖の可変部(VH及びVL)の組み合わせは、100万通り以上も存在し、抗原抗体反応を検証するために、膨大な量の実験が必要である。しかし、抗体分子は、分子量が大きく、生産するために動物細胞が必要で、生産コストが高かった。
【0012】
また、検証実験や免疫測定法等の固相化法では、通常、疎水性のプラスチックが利用されるが、疎水性相互作用を利用した結合であるため、抗原、抗体又は酵素等のタンパク質のいずれの部位が固相と結合するのか特定できず、固相と結合する部位によっては、構造変化や不活性化してしまう。例えば、酵素のO−アセチルセリンスルフヒドリラーゼ−A(O-acetyl serine sulfhydryrase-A)は、疎水性ポリスチレンに結合すると、ほぼ完全に失活する。さらに、このようなタンパク質と固相の無秩序な結合により、測定感度が低下し、測定精度も悪くなってしまう。
【0013】
抗体分子61は、図6に示すように、全体としてY字状であり、その上側の2箇所のN末端側に重鎖と軽鎖の可変部(VH及びVL)による立体的な抗原結合部位が形成されている。抗体を固定化する際に、C末端側が固相表面に対して垂直に結合すれば、抗原結合部位が外側を向き、また立体的に正常な配置を取れるため、抗原抗体反応を行うことができる。しかし、N末端側が固相と結合すると、少なくとも結合したN末端側に配置された抗原結合部位は、抗原と反応することができなくなる。また、固相との結合の際に、抗原結合部位の立体構造が変化して、抗体が変性し、抗原抗体反応を示さなくなることもある。
【0014】
Fab断片65やF(ab’)2断片67は、抗体分子61に比べて、分子量が小さくそれら自体を大腸菌や酵母等での生産が可能であるが、抗原結合部位の占める割合が高いので、無秩序に固相表面に結合させた場合、抗原抗体反応を示さなくなる可能性も高くなる。一本鎖抗体(scFv)68に至っては、基本的に抗原結合部位のみによって構成されているので、Fc領域や定常部の影響を排除できるため抗原抗体反応を調べるための抗体側の物質としては好適な形態ではあったが、固定化には極めて不利であり、そのままの状態で疎水性の基板に固定化しても、ほとんど抗原抗体反応を示さなかった。
【0015】
また、一般的に公知のクローン化技術によって製造されたタンパク質は、製造物である目的のタンパク質を回収するための分離、精製する工程に非常に時間と手間がかかっていた。例えば、宿主として大腸菌を使用した場合、大腸菌内には、目的とするタンパク質だけではなく、様々な種類の夾雑物が存在する。このため、大腸菌を培養した後、大腸菌の菌体を破砕して遠心分離することによって、菌体内可溶性画分を採取し、それをカラムによって精製して目的タンパク質を回収する必要があった。
【0016】
また、組換えDNAを導入することによって異種遺伝子を宿主内で大量に発現させた場合、生成されたタンパク質による宿主への悪影響を避けるため、不溶で不活性な凝集体である封入体(inclusion body)として生成されることがある。この封入体(inclusion body)は、遠心分離によって回収した後に、可溶化した上で、そのままでは活性がないので、タンパク質の折りたたみ構造を正常な状態に戻して活性化させるリフォールディングという作業も必要となる。
【0017】
さらに、上述したとおり、免疫測定法は、多数回の吸着・反応処理及び洗浄処理が必要であり、目的物質を検出するまでに長時間を要するものであり、短時間で微量物質を検出又は測定できる手法の開発が望まれている。
【0018】
本発明は、以上の課題及び問題点の一つ若しくは幾つかを解決するためのものであり、抗原抗体反応に利用できる可変部を含むペプチド、これらのペプチドが固定化された固相及びかかる固相の生産方法を提供する。また、生産効率を向上させた可変部を含むペプチド、固定化させても構造変化や不活性化しにくい可変部を含むペプチド、固定化されるタンパク質の構造をより維持し、活性をより保持できる免疫測定法に利用可能な可変部を含むペプチド、測定感度を向上させ、安定した測定精度の免疫測定法に利用可能な可変部を含むペプチド又は従来と比較して短時間で微量物質を検出又は測定できる方法に利用可能な可変部を含むペプチド並びにこれらのペプチドが固定化された固相及びかかる固相の生産方法を提供することを目的とする。
【0019】
なお、本明細書では、新規で有用なペプチド及び当該ペプチドの用途も開示している。また、本発明は、親水性の固相に特異的に結合する機能を発現させるペプチド及び当該ペプチドの用途も開示している。さらに、本発明は、固相化法を用いた化合物の検出又は測定に利用可能なペプチド及び当該ペプチドの用途も開示している。
【課題を解決するための手段】
【0020】
前述した目的を達成するため、抗原結合部位が形成される可変部を含むペプチドであって、重鎖の可変部よりもC末端側又は軽鎖の可変部よりもC末端側に、固相に対して特異的な吸着機能を発現させるアミノ酸配列を有する。また、本発明のペプチドは、抗原結合部位が形成される可変部を含むペプチドであって、重鎖の可変部よりもC末端側又は軽鎖の可変部よりもC末端側に、配列表における配列番号1乃至20に示すアミノ酸配列の何れか一つを有する。さらに、本発明のペプチドは、1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)、定常部融合1本鎖抗体(scFv-Fc)、Fab断片又はF(ab’)2断片であることが好ましい。
【0021】
本発明のペプチドは、前記アミノ酸配列は親水性の樹脂表面に対して特異的な吸着機能を発現させることが好ましく、前記樹脂表面は親水性のポリスチレン表面であってもよい。
【0022】
さらに、前記重鎖及び軽鎖の両方に、前記アミノ酸配列を有する1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)、定常部融合1本鎖抗体(scFv-Fc)、Fab断片又はF(ab’)2断片であることがより好ましい。本発明のペプチドは、前記重鎖可変部と前記軽鎖可変部を結合するリンカーペプチドを有し、前記リンカーペプチドと前記可変部の間又は前記リンカーペプチド内に前記アミノ酸配列を有していてもよい。
【0023】
また、本発明の精製されたペプチドの生産方法では、固相に対して特異的な吸着機能を発現させるアミノ酸配列を有するペプチドと夾雑物とを含む溶液を前記固相表面に接触させることにより、前記ペプチドを前記固相表面に直接吸着させ、前記ペプチドを精製する。
【0024】
さらに、本発明の復元されたペプチドの生産方法では、固相に対して特異的な吸着機能を発現させるアミノ酸配列を有する変性されたペプチドと変性剤とを含む溶液を前記固相表面に接触させることにより、前記ペプチドを前記固相表面に直接吸着させ、前記変性されたペプチドを復元する。ここで、前記変性されたペプチドは、封入体であってもよい。
【0025】
上記ペプチドの生産方法において、前記ペプチドは、抗原結合部位が形成される可変部を含み、重鎖の可変部よりもC末端側又は軽鎖の可変部よりもC末端側に、前記アミノ酸配列を有していてもよい。また、前記溶液は、前記ペプチドを生成した宿主の破砕物、培地または分泌物を含有していてもよい。そして、前記アミノ酸配列としては、配列表における配列番号1乃至20に示すアミノ酸配列を使用することができる。
【0026】
また、本発明では、固相に対して特異的な吸着機能を発現させるアミノ酸配列を有するペプチドをコードする塩基配列を含む遺伝子を宿主に導入して形質転換体とし、その生成物を含む溶液を前記固相表面に接触させることにより、前記ペプチドを前記固相表面に直接吸着させることによって、ペプチドが固定化された固相を生産してもよい。ここで、前記ペプチドは、抗原結合部位が形成される可変部を含み、重鎖の可変部よりもC末端側又は軽鎖の可変部よりもC末端側に有するものでもよく、前記生成物を含む溶液は、前記宿主の破砕物も含有するものであっても、前記宿主の培地を含有するものであっても、酵母から分泌生産された分泌物を含有するものであってもよい。
【0027】
また、本発明の対象としては、抗原結合部位が形成される可変部を含むペプチドであって、重鎖の可変部よりもC末端側又は軽鎖の可変部よりもC末端側に、固相に対して特異的な吸着機能を発現させるアミノ酸配列を有するペプチドをコードする塩基配列を含む遺伝子、当該遺伝子に調製されたベクターまたは当該遺伝子が導入された宿主でもよい。
【0028】
また、本発明のペプチドは、配列表における配列番号1に示すアミノ酸配列を含むペプチドであることが好ましい。また、免疫測定法における固相に固定化されるペプチドであって、配列表における配列番号1に示すアミノ酸配列を含むものでもよい。これらの配列番号1に示すアミノ酸配列は、配列表における配列番号2乃至10に示すアミノ酸配列であってもよい。
【0029】
前記固相は親水性の樹脂表面を有していてもよく、前記固相は親水性のポリスチレン表面を有していてもよい。前記免疫測定法はエンザイムイムノアッセイであってもよい。
【0030】
また、本発明は、配列表における配列番号1に示すアミノ酸配列を含む固定化酵素、配列表における配列番号1に示すアミノ酸配列をコードする塩基配列を含む遺伝子、当該遺伝子に調製されたベクターまたは当該遺伝子が導入された宿主も開示する。そして、これらの配列番号1に示すアミノ酸配列は、配列表における配列番号2乃至10に示すアミノ酸配列であってもよい。
【発明の効果】
【0031】
本発明のペプチドは、特定の固相表面に対して優れた結合性能を示し、特定の固相に直接固定化させることができるので、本ペプチドを容易に分離、精製することができる。すなわち、本ペプチドを生産した後、十分な精製を行わなくても、本ペプチドを含む溶液を特定の固相に接触させるだけで、本ペプチドを固相に直接結合させることができ、本ペプチドの溶液からの分離、精製及び固相への固定化を同時に行うことができる。この結果、従来、長時間必要であった分離、精製工程(例えばアフィニティカラムを使用する精製工程)を省くことができ、本ペプチドの生産から固相への固定化までに要する時間を大幅に短縮させることができるので、本ペプチドでは生産効率が飛躍的に向上する。しかも、この過程において、封入体(inclusion body)の状態で生成されたタンパク質や変性されたタンパク質のリフォールディングも行うことも可能である。
【0032】
また、本ペプチドは、固相化法を用いた化合物の検出又は測定に利用することも可能である。特に免疫測定法において、従来と比較して、特異性のある吸着機能により測定時間を短縮でき、しかも原ペプチドの活性を保ったまま固定化でき、測定感度を向上させ、安定した測定精度を得ることができる。
【図面の簡単な説明】
【0033】
【図1】BSAの存在下における親水性のポリスチレン表面に対する固定化率を示すグラフ。
【図2】(a)〜(d)は、それぞれTween20の添加量が0、0.1、1、10%の条件における親水性のポリスチレンプレートに対する結合性能を示すグラフ。
【図3】(a)〜(d)は、それぞれTween20の添加量が0、0.1、1、10%の条件における疎水性のポリスチレンプレートに対する結合性能を示すグラフ。
【図4】(a)〜(f)は、種々の夾雑物の存在下における親水性及び疎水性のポリスチレンプレートに対する結合性能を示すグラフ。
【図5】(a)〜(f)は、種々の夾雑物の存在下における親水性及び疎水性のポリスチレンプレートに対する活性値を示すグラフ。
【図6】(a)は抗体の基本構造を示す模式図、(b)はFab断片とFc断片の構造を示す模式図、(c)はF(ab’)2断片の構造を示す模式図、(d)は一本鎖抗体(scFv)の基本構造を示す模式図。
【図7】BSAの存在下における親水性のポリスチレン表面に対する固定化率を示すグラフ。
【図8】(a)及び(b)は、種々の夾雑物の存在下における親水性のポリスチレンプレートに対する活性値を示すグラフ。
【図9】BSAの存在下における親水性のポリスチレン表面に対する固定化率を示すグラフ。
【図10】(a)及び(b)は、種々の夾雑物の存在下における親水性のポリスチレンプレートに対する活性値を示すグラフ。
【図11】(a)及び(b)はそれぞれ本ペプチド及び原ペプチドを培養したコロニーにおける菌体濃度、(c)及び(d)はそれぞれ本ペプチド及び原ペプチドの結合性能を示すグラフ。
【図12】夾雑物として各種の培地を加えた状態における親水性のポリスチレン表面に対する結合性能を示すグラフ。
【図13】本アミノ酸配列を一本鎖抗体に導入したときの活性値を示すグラフ。
【図14】一本鎖抗体における本アミノ酸配列の導入位置と活性値の関係を示すグラフ。
【図15】一本鎖抗体における本アミノ酸配列の導入位置と活性値の関係を示すグラフ。
【図16】一本鎖抗体の量と親水性のポリスチレン基板に対する活性値の関係を示すグラフ。
【図17】未精製の変性した状態の本ペプチドのポリスチレン基板に対する結合性能及び活性値を示すグラフ。
【発明を実施するための最良の形態】
【0034】
以下、本発明を詳しく説明する。本発明のペプチドは、固相に対して特異的な吸着機能を発現させるアミノ酸配列(以下「本アミノ酸配列」という)を有する。本アミノ酸配列は、例えば配列表における配列番号1乃至20に示すアミノ酸配列(以下、それぞれ「本アミノ酸配列1」乃至「本アミノ酸配列20」という)である。本アミノ酸配列を有するペプチドを「本ペプチド」という。
【0035】
本アミノ酸配列は、特定の固相(例えば、親水性の樹脂表面を有する固相)に対して特異的な吸着機能を発現させるため、本ペプチドは、特定の固相表面に対して優れた結合性能を示し、特定の固相に直接固定化させることができるので、本ペプチドを容易に分離、精製することができる。すなわち、本ペプチドを生産した後、十分な精製を行わなくても、本ペプチドを含む溶液を特定の固相に接触させるだけで、本ペプチドを固相に直接結合させることができ、本ペプチドの溶液からの分離、精製及び固相への固定化までの工程を同時に行うことができる。この結果、従来、長時間必要であった分離、精製工程(例えばアフィニティカラムを使用する精製工程)を省くことができ、本ペプチドの生産から固相への固定化までに要する時間を大幅に短縮させることができるので、本ペプチドでは生産効率が飛躍的に向上する。しかも、この過程において、封入体(inclusion body)の状態で生成されたタンパク質や変性されたタンパク質のリフォールディングも行うことも可能である。
【0036】
また、本ペプチドは、固相化法を用いた化合物の検出又は測定に利用することも可能である。化合物の検出又は測定に利用される抗原、抗体又は酵素に対し、本アミノ酸配列を導入することにより、抗原、抗体又は酵素を特定の固相に直接固定化することができ、化合物の検出又は測定をすることができる。特に、本ペプチドは、免疫測定法を利用した微量物質の検出又は測定に適用することが好ましい。さらに、本ペプチドは、夾雑物の存在下であっても特異的に、しかも本アミノ酸配列の導入位置を優先して特定の固相上に直接結合させるため、配向及び構造を制御した状態で特定の固相上に固定化できるため、従来と比較して、原ペプチドの活性を保ったまま固定化でき、測定感度を向上させ、安定した測定精度を得ることができる。
【0037】
例えば、サンドイッチ法のELISAに適用した場合、目的物質と特異的に抗原抗体反応を示す抗体に対し、本アミノ酸配列のペプチドを導入し、本ペプチドである抗体を準備する。第一に、かかる抗体を特定の固相に直接結合させた後、固相を複数回洗浄する。第二に、固相上に残った未結合の部位に対して、ブロッキング試薬を結合させてブロッキングした後、固相を複数回洗浄する。第三に、目的物質を含有する試料と固相上に結合された本ペプチドである抗体とを反応させた後、固相を複数回洗浄する。第四に、本ペプチドではない第2の抗体と目的物質と反応させた後、固相を複数回洗浄する。第五に、第2の抗体と酵素標識とを反応させた後、固相を複数回洗浄する。第六に、酵素標識と基質とを反応させ、吸光度を測定し、試料中の目的物質を検出し、又は濃度を測定する。
【0038】
このように、本ペプチドを利用した免疫測定法による化合物の検出又は測定を行うことができる。なお、第四の処理で反応させる第2の抗体として、既に酵素標識された抗体を利用すれば、第五の処理は不要となる。また、本ペプチドが、ブロッキング試薬の存在下においても特異的に結合するので、上記第一の処理において、特定の固相に対し、本ペプチドである抗体とブロッキング試薬とを同時に結合させることで、第二の処理を省略することができ、化合物の検出又は測定時間を短縮できる。
【0039】
さらに、最初に、溶液中において、本ペプチドである抗体と目的物質とを抗原抗体反応させて免疫複合物を形成させ、その後、特定の固相上に免疫複合物を固定化させることにより、化合物の検出又は測定時間を劇的に短縮できる新たな化合物の検出又は測定方法を実現できる。すなわち、上記サンドイッチ法における第一の処理と第三の処理を一回の処理で実行できる。溶液中にブロッキング試薬も添加しておけば、上記第一、第二及び第三の処理を一回の処理で実行できる。より好ましくは、本ペプチドである抗体と目的物質との免疫複合物を形成した後に、溶液中において、さらに酵素標識された第2の抗体とも抗原抗体反応させて、より大きな免疫複合物を形成させ、それを特定の固相上に固定化すれば、上記第一乃至第五の処理を一回の処理で実行できる。
【0040】
本ペプチドは、夾雑物の存在下であっても特異的に、しかも配向及び構造を制御した状態で、特定の固相上に直接固定化できるため、ブロッキング試薬が存在しても、免疫複合物の状態であっても、免疫複合物を固定化して、目的物質の検出又は測定を行うことができる。加えて、溶液中において立体障害なく極めて迅速に抗原抗体反応が行われるため、従来の固定化された状態での抗原抗体反応と比べて、飛躍的に反応速度を増加させることができ、測定感度を向上させ、安定した測定精度を得ることができる。
【0041】
さらに本ペプチドの別の用途として、本ペプチドとして酵素を採用すれば、親水性の樹脂表面における固定化酵素として利用することができる。一般的に、酵素は水溶性であるため、利用する際に使い捨てているが、酵素を固相に結合させて固定化することにより、水溶性の反応生成物と分離することができ、連続的、かつ繰り返し反応させることができる。
【0042】
本アミノ酸配列1は、配列表における配列番号1に示されているように、N末端側からC末端側に向けて順にRXXXRRXRR(R:アルギニン、X:イソロイシン(I)、ロイシン(L)、バリン(V)、アラニン(A)、グリシン(G)、メチオニン(M)、セリン(S)又はトレオニン(T)の一つ又は複数の組合わせ)という配列である。したがって、本アミノ酸配列1には、少なくとも、本アミノ酸配列2乃至10が含まれる。
【0043】
本アミノ酸配列2は、本アミノ酸配列1のXを全てイソロイシン(I)とした配列であり、N末端側からC末端側に向けて順にRIIIRRIRR(R:アルギニン、I:イソロイシン)という配列である。本アミノ酸配列3は、本アミノ酸配列1のXをアラニン(A)とイソロイシン(I)の組合わせとした配列であり、N末端側からC末端側に向けて順にRAIARRIRR(R:アルギニン、A:アラニン、I:イソロイシン)という配列である。本アミノ酸配列4は、本アミノ酸配列1のXを全てロイシン(L)とした配列であり、N末端側からC末端側に向けて順にRLLLRRLRR(R:アルギニン、L:ロイシン)という配列である。本アミノ酸配列5は、本アミノ酸配列1のXを全てバリン(V)とした配列であり、N末端側からC末端側に向けて順にRVVVRRVRR(R:アルギニン、V:バリン)という配列である。本アミノ酸配列6は、本アミノ酸配列1のXを全てアラニン(A)とした配列であり、N末端側からC末端側に向けて順にRAAARRARR(R:アルギニン、A:アラニン)という配列である。本アミノ酸配列7は、本アミノ酸配列1のXを全てグリシン(G)とした配列であり、N末端側からC末端側に向けて順にRGGGRRGRR(R:アルギニン、G:グリシン)という配列である。
【0044】
後述する実施例1乃至6において示すとおり、本アミノ酸配列2乃至7を有するペプチドは、何れも特定の固相表面に対して特異的な吸着機能を発現させることが確認された。本アミノ酸配列2乃至7は、何れもアルギニン(R)の配置は、N末端側から1、5,6、8、9番目と共通であり、その間に配置されるアミノ酸(本アミノ酸配列1のX)を変えたものである。本アミノ酸配列2,4乃至7は、単一のアミノ酸を配置させているが、本アミノ酸配列3では、アラニン(A)とイソロイシン(I)とを組合わせて配置させている。これら全てにおいて共通の性質を示したことから、本アミノ酸配列1のXとして、イソロイシン(I)、ロイシン(L)、バリン(V)、アラニン(A)又はグリシン(G)の一つ又は組み合わせとすることにより特定の固相表面に対して特異的な吸着機能を発現させることが予測される。
【0045】
さらに、本アミノ酸配列8は、本アミノ酸配列1のXを全てメチオニン(M)とした配列であり、N末端側からC末端側に向けて順にRMMMRRMRR(R:アルギニン、M:メチオニン)という配列である。本アミノ酸配列9は、本アミノ酸配列1のXを全てセリン(S)とした配列であり、N末端側からC末端側に向けて順にRSSSRRSRR(R:アルギニン、S:セリン)という配列である。本アミノ酸配列10は、本アミノ酸配列1のXを全てトレオニン(T)とした配列であり、N末端側からC末端側に向けて順にRTTTRRTRR(R:アルギニン、T:トレオニン)という配列である。
【0046】
後述する実施例7乃至9において示すとおり、本アミノ酸配列8乃至10を有するペプチドは、何れも特定の固相表面に対して特異的な吸着機能を発現させることが確認された。メチオニン(M)、セリン(S)又はトレオニン(T)は、アミノ酸(R−CH(NH2)COOH)のR基中に硫黄やヒドロキシ基(OH)が含まれてはいるものの、部分的に鎖状の飽和炭化水素を有し、中性のアミノ酸である点において、R基が鎖状の飽和炭化水素からなるイソロイシン(I)、ロイシン(L)、バリン(V)及びアラニン(A)と共通している。また、メチオニン(M)、セリン(S)及びトレオニン(T)は、R基に環状構造や芳香族化合物を含まず、立体構造の面においても比較的かさの小さいアミノ酸である点においても共通である。このため、本アミノ酸配列1のXとして、メチオニン(M)、セリン(S)又はトレオニン(T)を選択しても、同様の効果が得られることが予測される。
【0047】
さらに、本アミノ酸配列11は、配列表における配列番号11に示されているように、N末端側からC末端側に向けて順にKGLRGWREMISL(リジン(K)、グリシン(G)、ロイシン(L)、アルギニン(R)、グリシン(G)、トリプトファン(W)、アルギニン(R)、グルタミン酸(E)、メチオニン(M)、イソロイシン(I)、セリン(S)、ロイシン(L))という配列である。
【0048】
本アミノ酸配列12は、配列表における配列番号12に示されているように、N末端側からC末端側に向けて順にADYLSRWGSIRN(アラニン(A)、アスパラギン酸(D)、チロシン(Y)、ロイシン(L)、セリン(S)、アルギニン(R)、トリプトファン(W)、グリシン(G)、セリン(S)、イソロイシン(I)、アルギニン(R)、アスパラギン(N))という配列である。
【0049】
本アミノ酸配列13は、配列表における配列番号13に示されているように、N末端側からC末端側に向けて順にSRVHRAVLNGVS(セリン(S)、アルギニン(R)、バリン(V)、ヒスチジン(H)、アルギニン(R)、アラニン(A)、バリン(V)、ロイシン(L)、アスパラギン(N)、グリシン(G)、バリン(V)、セリン(S))という配列である。
【0050】
本アミノ酸配列14は、配列表における配列番号14に示されているように、N末端側からC末端側に向けて順にRPPGVVRRYALG(アルギニン(R)、プロリン(P)、プロリン(P)、グリシン(G)、バリン(V)、バリン(V)、アルギニン(R)、アルギニン(R)、チロシン(Y)、アラニン(A)、ロイシン(L)、グリシン(G))という配列である。
【0051】
本アミノ酸配列15は、配列表における配列番号15に示されているように、N末端側からC末端側に向けて順にVRSWEEQARVTT(バリン(V)、アルギニン(R)、セリン(S)、トリプトファン(W)、グルタミン酸(E)、グルタミン酸(E)、グルタミン(Q)、アラニン(A)、アルギニン(R)、バリン(V)、トレオニン(T)、トレオニン(T))という配列である。
【0052】
本アミノ酸配列16は、配列表における配列番号16に示されているように、N末端側からC末端側に向けて順にRAFIASRRIKRP(アルギニン(R)、アラニン(A)、フェニルアラニン(F)、イソロイシン(I)、アラニン(A)、セリン(S)、アルギニン(R)、アルギニン(R)、イソロイシン(I)、リジン(K)、アルギニン(R)、プロリン(P))という配列である。
【0053】
本アミノ酸配列17は、配列表における配列番号17に示されているように、N末端側からC末端側に向けて順にRESTLKGTSRAV(アルギニン(R)、グルタミン酸(E)、セリン(S)、トレオニン(T)、ロイシン(L)、リジン(K)、グリシン(G)、トレオニン(T)、セリン(S)、アルギニン(R)、アラニン(A)、バリン(V))という配列である。
【0054】
本アミノ酸配列18は、配列表における配列番号18に示されているように、N末端側からC末端側に向けて順にAGLRLKKAAIHR(アラニン(A)、グリシン(G)、ロイシン(L)、アルギニン(R)、ロイシン(L)、リジン(K)、リジン(K)、アラニン(A)、アラニン(A)、イソロイシン(I)、ヒスチジン(H)、アルギニン(R))という配列である。
【0055】
本アミノ酸配列19は、配列表における配列番号19に示されているように、N末端側からC末端側に向けて順にSSLLRAVPEPTG(セリン(S)、セリン(S)、ロイシン(L)、ロイシン(L)、アルギニン(R)、アラニン(A)、バリン(V)、プロリン(P)、グルタミン酸(E)、プロリン(P)、トレオニン(T)、グリシン(G))という配列である。
【0056】
本アミノ酸配列20は、配列表における配列番号20に示されているように、N末端側からC末端側に向けて順にRAFIASRRIRRP(アルギニン(R)、アラニン(A)、フェニルアラニン(F)、イソロイシン(I)、アラニン(A)、セリン(S)、アルギニン(R)、アルギニン(R)、イソロイシン(I)、アルギニン(R)、アルギニン(R)、プロリン(P))という配列である。
【0057】
本アミノ酸配列1乃至20は、特定の固相に対して特異的な吸着機能を発現させることができる。特定の固相とは、現時点で確認されている範囲では、親水性の樹脂表面を有する固相である。すなわち、本ペプチドは、原ペプチドと比較して、親水性の樹脂表面に対する結合性能が向上する。特に、本ペプチドは、親水性のポリスチレン表面に対して優れた結合性能を示す。結合性能は、競合する夾雑物に対して優先的に結合する性能であり、競合する夾雑物、例えば他のペプチド、高分子化合物、界面活性剤等の存在下において、固相に対して結合する本ペプチドの量を比較することで評価できる。
【0058】
親水性の樹脂表面を有する固相としては、ポリスチレン、ポリカーボネート、ポリプロピレン、ポリエチレン、ポリジメチルシロキサン(PDMS)、ポリメタクリル酸メチル(PMMA)等などのプラスチック樹脂表面を変質させて親水化処理したものを用いることができる。通常、製造されたプラスチック樹脂表面は疎水性であるが、この表面に各種の親水化処理を行うことで、親水性の樹脂表面を有する固相とすることができる。例えば、ポリスチレン表面にUV+O3処理やプラズマ酸化処理を行うことにより、親水性のポリスチレン表面を有する固相を準備できた。
【0059】
固相の形態としては、板状(容器やウェルの壁面及び底面を含む)でもよいし、粒状でもよい。特開2007−279018号公報に記載されているように、マイクロウェルプレートの流体取扱部(各ウェル)内に表面が親水化処理された粒状プラスチック基板を充填すれば、ウェル内に本ペプチドを含む溶液を注入するだけで粒状プラスチック基板表面に本ペプチドを固定化することができる。
【0060】
本明細書において「ペプチド」とは、ペプチド結合によって2個以上のアミノ酸が結合したものを指し、オリゴペプチド、ポリペプチド、タンパク質、ホモメリックペプチド及びヘテロメリックペプチドを含む。また、本ペプチドは、電気的に中性の形態、又は塩の形態をとることがあり、配列表における本アミノ酸配列のみからなる形態、特定のタンパク質の末端部または内部に本ペプチドを融合させた融合タンパク質の形態、糖質やポリエチレングリコール、NCS基(isothiocyanate)、NHS基(N-Hydroxy succinimide ester)、マレイミド基、チオール基、ビオチン、蛍光色素等を付加して得られる複合体としての形態、さらには、ペプチドをアセチル化、アミド化及び/又は多官能試験により架橋重合させて得られる誘導体又は重合体としての形態であってもよい。
【0061】
配列表における配列番号21に示す塩基配列は、本アミノ酸配列2をコードする塩基配列の一例である。すなわち、5’末端側から3’末端側に向けて順にcgt atc atc atc cga agg att cga cga(a:アデニン、g:グアニン、c:シトシン、t:チミン)という塩基配列を含むDNAである。配列番号22に示す塩基配列は、本アミノ酸配列3をコードする塩基配列の一例である。すなわち、5’末端側から3’末端側に向けて順にcgt gcg att gcg cga agg att cga cga(a:アデニン、g:グアニン、c:シトシン、t:チミン)という塩基配列を含むDNAである。本アミノ酸配列2又は3をコードする塩基配列としては、配列番号21又は22に示す塩基配列に限定されるものではなく、その他にも、本アミノ酸配列2又は3の各アミノ酸に対応する各コドンの配列を採用できる。本アミノ酸配列をコードする塩基配列を含む遺伝子は、ファージやプラスミド等のベクターを用いて、あるいは宿主のゲノムDNA中に導入し、宿主内において発現させることにより、本ペプチドを生合成できる。
【0062】
本ペプチドとしては、種々のタンパク質を利用することが可能であり、多くの抗原、抗体、レクチン、酵素又は受容体タンパク質等に適用することができる。例えば、グルタチオン転移酵素(GST:Glutathione S-Transferase)、マルトース結合タンパク質(MBP)、アルカリホスファターゼ(ALP)、ペルオキシダーゼ(POD)、ルシフェラーゼ、緑色蛍光タンパク質(GFP)、β−ガラクトシダーゼ(β−Gal)、トリプシン、キモトリプシン、トロンビン、Factor Xa、アンジオテンシン変換酵素、チロシンキナーゼ、インスリンレセプター、EGFレセプター、ストレプトアビジン(SA)、モノクローナル抗体(Mab)、ポリクローナル抗体(Pab)、1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)(例えば2価性1本鎖抗体(sc(Fv)2))、定常部融合1本鎖抗体(scFv-Fc)、Fab断片及びF(ab’)2断片(抗原結合部位を含む抗体の断片)、補体系タンパク質C1q、コンカナバリンA(ConA)、レンチルレクチン(LCA)、抗体結合タンパク質(Protein A、ZZ、Protein G、Protein L等)等に適用することができる。なお、本明細書において、本ペプチドと対比するために、本ペプチドから本アミノ酸配列を除いたペプチド(本アミノ酸配列を導入する前のペプチドを含む)を、以下においては「原ペプチド」と呼ぶ。
【0063】
なお、本ペプチドとして、抗原結合部位が形成される可変部を含むペプチドとする場合、抗体分子それ自体も含むが、大腸菌や酵母での生産が可能である1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)、定常部融合1本鎖抗体(scFv-Fc)、Fab断片、F(ab’)2断片であることが好ましい。多価性1本鎖抗体(sc(Fv)n)は、複数の1本鎖抗体(scFv)をリンカーペプチドで連結したものであり、分子内に複数の抗原結合部位(重鎖可変部(VH)及び軽鎖可変部(VL)ドメイン)を有するペプチドの総称である。複数の抗原結合部位は同じものでもよいし、異なっていてもよい。なお、2価性1本鎖抗体(sc(Fv)2)とは、2つの1本鎖抗体(scFv)をリンカーペプチドで連結したものである。定常部融合1本鎖抗体(scFv-Fc)とは、1本鎖抗体(scFv)のC末端側にFcドメインを有する融合タンパク質の総称である。Fab断片とは、抗体分子をパパイン消化によって得られる断片のうち、VL、CLからなる軽鎖とVH、CH1ドメインからなる重鎖が会合したものを指すが、遺伝子組換え技術で作成することも可能である。F(ab’)2断片とは、抗体分子をペプシン消化後に得られる断片の一つで2分子のFabがヒンジ部のジスルフィド結合で連結されたものを指すが、遺伝子組換え技術で作成することも可能である。
【0064】
本ペプチドにおいて、本アミノ酸配列の導入部位は、原ペプチドが本来有する生理活性を妨げない部位、例えば、抗原の場合は抗原決定基以外、抗体の場合は抗体活性基以外の部位とする。特に、原ペプチドの立体構造を維持したまま固相に固定化させるため、原ペプチドのC末端又はN末端近傍の部位に本アミノ酸配列を導入することが好ましい。なお、本ペプチドには、本アミノ酸配列以外にも、標識用のアフィニティタグ等が別途導入されていてもよい。
【0065】
特に、本ペプチドとして、抗原結合部位が形成される可変部を含むペプチドとする場合は、重鎖の可変部よりもC末端側又は軽鎖の可変部よりもC末端側(両方を含む)に本アミノ酸配列を導入する。可変部よりもC末端側とは、重鎖または軽鎖可変部におけるC末端から本ペプチド全体におけるC末端までの位置であって、可変部の抗原結合を阻害しない位置全てを指す。
【0066】
抗体分子の場合(図6(a)参照)は、重鎖(H鎖)62の可変部(VH)よりもC末端側に位置する定常部(CH1、CH2、CH3、CH4)のドメインにおけるN末端やC末端、軽鎖(L鎖)63の定常部(CL)のドメインにおけるN末端やC末端の一つ若しくは複数箇所に導入することが好ましく、特に、重鎖62のFc領域の定常部(CH2、CH3、CH4)のドメインにおけるN末端やC末端、軽鎖63のC末端の一つ若しくは複数箇所に導入することが好ましい。
【0067】
Fab断片又はF(ab’)2断片の場合(図6(b)及び(c)参照)は、重鎖62又は軽鎖63の可変部のドメイン(VH及びVL)におけるC末端、定常部(CH1及びCL)のドメインにおけるN末端やC末端あるいはの一つ若しくは複数箇所に導入することが好ましく、特に、重鎖62のC末端又は軽鎖63のC末端の一つ若しくは複数箇所に導入することが好ましい。
【0068】
また、1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)、定常部融合1本鎖抗体(scFv-Fc)のように、重鎖と軽鎖を結合するリンカーペプチドを有する場合は、リンカーペプチドと重鎖または軽鎖の間、もしくはリンカーペプチド内部に本アミノ酸配列を導入してもよい。ここで、リンカーペプチドは、重鎖又は軽鎖の一方のC末端と、他方のN末端とを連結しているが、重鎖又は軽鎖のC末端側には、リンカーペプチド全体、すなわち重鎖又は軽鎖のC末端と結合している部分から他方のN末端までの領域も含まれる。なお、リンカーペプチドと重鎖または軽鎖の間、もしくはリンカーペプチド内部に本アミノ酸配列以外の標識用のアフィニティタグ等が別途導入されていてもよい。図6(d)の1本鎖抗体(scFv)の場合、軽鎖の可変部(VL)のC末端側か、重鎖の可変部(VH)のC末端からリンカーペプチド69の内部を含む軽鎖の可変部(VL)のN末端までの間の一方又は両方に本アミノ酸配列を導入する。
【0069】
複数の箇所に本アミノ酸配列を導入することにより、本ペプチドの立体構造をより正常な状態に維持することが可能である。たとえば、Fab断片又はF(ab’)2断片の重鎖及び軽鎖のC末端側にそれぞれ本アミノ酸配列を導入したり、1本鎖抗体(scFv)の重鎖及び軽鎖のC末端側にそれぞれ本アミノ酸配列を導入すれば、その部位が固相表面に結合しやすくなるので、抗原結合部位が外側を向き、抗原抗体反応を起こしやすくなる。
【0070】
本ペプチドは、公知のクローン化技術や化学合成法によって製造することが可能である。例えば、クローン化技術を利用すれば、本アミノ酸配列をコードするDNAを調製し、これを自律複製可能なベクターに挿入して組換えDNAとし、これを大腸菌、枯草菌、放線菌、酵母、糸状菌、植物細胞、昆虫細胞、動物細胞などの適宜宿主に導入して形質転換体とし、その培養物から本アミノ酸配列を含むペプチドを採取することができる。また、本アミノ酸配列をコードするDNAを調製し、小麦胚芽や大腸菌細胞抽出液などを用いて無細胞タンパク質合成系により本ペプチドを合成することができる。また、「固相法」又は「液相法」等の慣用のペプチド化学合成法を利用して、本アミノ酸配列に示されるアミノ酸を逐次、脱水縮合させて伸長させれば、本アミノ酸配列からなる本ペプチドを合成することができる。さらに、かかる本アミノ酸配列からなる本ペプチドを所望の原ペプチドに結合させることにより、本アミノ酸配列を含むより高分子の本ペプチドを合成できる。
【0071】
本ペプチドは、本アミノ酸配列を有しているため、形質転換体からの生成物を含む溶液を固相表面に接触させることにより、本ペプチドを固相表面に直接吸着させて、分離精製することができる。生成物を含む溶液とは、目的となる本ペプチドを含み、且つ宿主に起因する不要な夾雑物が存在する溶液全般を指し、例えば、菌体破砕液、菌体破砕液を遠心分離して得られた可溶性画分、菌体破砕液を遠心分離して得られた不溶性画分を可溶化したもの、細胞膜画分、細胞壁画分、細胞から分泌生産された分泌物、体液、またはこれらの不完全な精製物等を含む。
【0072】
さらに、組換えDNAを導入することによって異種遺伝子を宿主内で大量に発現させた場合、生成されたタンパク質による宿主への悪影響を避けるため、不溶で不活性な凝集体である封入体(inclusion body)として生成されることがあるが、本ペプチドが封入体(inclusion body)として生成されていても、変性剤で封入体(inclusion body)を可溶化後そのまま固相表面に固定化することができ、しかも固定化させた状態で変性剤を除去することによって本ペプチドがリフォールディングされる場合もある。
【0073】
また、封入体に対してだけではなく、何らかの原因によって、立体構造が変性されていても、本ペプチドであれば、そのまま固相表面に固定化することができ、しかも固定化させた状態で適当なリフォールディングバッファを加えることによって本ペプチドをリフォールディングできる場合もある。変性の原因としては、加熱、凍結、高圧、超音波、紫外線、X線、かくはん、吸着、希釈などの物理的な原因と、極端な酸性またはアルカリ性、有機溶媒、重金属塩、変性剤、界面活性剤などの化学的な原因がある。
【0074】
上で述べたとおり、本アミノ酸配列1乃至20は、親水性の樹脂表面に対して優れた結合性能を示すため、本アミノ酸配列1乃至20を有する本ペプチドを親水性の樹脂表面を有する固相に直接固定化することができるので、容易に分離、精製することができる。すなわち、本アミノ酸配列1乃至20を有する本ペプチドを含む溶液を親水性の樹脂表面を有する固相が充填されたカラムを通過させることにより、本アミノ酸配列1乃至20を有する本ペプチドを固相に直接結合させて溶液から分離、精製させることができる。
【0075】
以下に、実施例により本発明を更に具体的に説明するが、本発明は、その要旨を越えない限りこれらの実施例に限定されるものではない。実施例1乃至19は、グルタチオン転移酵素(GST)に本アミノ酸配列を導入した例であり、実施例20は、抗体に本アミノ酸配列を導入した例であり、実施例21乃至28は、一本鎖抗体に本アミノ酸配列を導入した例である。なお、以下の実施例において、本ペプチドの濃度は、ウシ血清由来のアルブミン(BSA)を標準タンパクとして使用したLowry法によって定量した。
【0076】
[実施例1](本アミノ酸配列2を有するGSTの生合成)
まず、グルタチオン転移酵素(GST)をコードする遺伝子を組み込んだプラスミド(GEヘルスケアバイオサイエンス株式会社製「pGEX−3X」、データベース・アクセッション番号:U13852)の制限酵素BamHIの認識する塩基配列部位と制限酵素EcoRIの認識する塩基配列部位の間に、本アミノ酸配列2をコードする遺伝子(配列表の配列番号21に示す塩基配列)を導入したものをベクターとして作製した。
【0077】
次に、宿主として大腸菌(BL21)を使用し、本アミノ酸配列2をコードする遺伝子を導入したベクターを宿主に導入して宿主を形質転換し、アンピシリンを含む寒天培地上で選別した。その後、2xYT培地50ml中で大腸菌を培養し、終濃度が0.1mMになるように、発現誘導物質としてイソプロピル1−チオ−ガラクトシド(IPTG:Isopropyl-1-thio-β-D(-)-galactoside)を加えて、本アミノ酸配列2を含むGST(本ペプチド)を発現させた。
【0078】
そして、菌体破砕後、精製用ゲル(GSH-Sepharose 6B)を用いて菌体内可溶性画分を精製し、リン酸緩衝化生理食塩水(PBS:Phosphate Buffered Saline)中で1晩透析し、滅菌フィルターによって凝集体を除去した。こうして、本ペプチドとして、C末端に本アミノ酸配列2が導入されたグルタチオン転移酵素(GST)を生合成した。以下、実施例1で生合成された本アミノ酸配列2を有するGSTを本ペプチド1という。
【0079】
[実施例2](本アミノ酸配列3を有するGSTの生合成)
プラスミド「pGEX−3X」の制限酵素BamHIの認識する塩基配列部位と制限酵素EcoRIの認識する塩基配列部位の間に、本アミノ酸配列3をコードする遺伝子(配列表の配列番号22に示す塩基配列)を導入した点以外は、実施例1と同様の処理によって、本ペプチドとして、C末端に本アミノ酸配列3が導入されたグルタチオン転移酵素(GST)を生合成した。以下、実施例2で生合成された本アミノ酸配列3を有するGSTを本ペプチド2という。
【0080】
[実施例3〜6](本アミノ酸配列4乃至7を有するGSTの生合成)
プラスミド「pGEX−3X」の制限酵素BamHIの認識する塩基配列部位と制限酵素EcoRIの認識する塩基配列部位の間に、本アミノ酸配列4乃至7をコードする遺伝子を導入した点以外は、実施例1と同様の処理によって、本ペプチドとして、C末端に本アミノ酸配列4乃至7が導入されたグルタチオン転移酵素(GST)をそれぞれ生合成した。なお、実施例3で本アミノ酸配列4を有するGSTを生合成し、実施例4で本アミノ酸配列5を有するGSTを生合成し、実施例5で本アミノ酸配列6を有するGSTを生合成し、実施例6で本アミノ酸配列7を有するGSTを生合成した。以下、実施例3乃至6で生合成された本アミノ酸配列4乃至7を有するGSTを、それぞれ本ペプチド3乃至6という。
【0081】
[実施例7〜9](本アミノ酸配列8乃至10を有するGSTの生合成)
プラスミド「pGEX−3X」の制限酵素BamHIの認識する塩基配列部位と制限酵素EcoRIの認識する塩基配列部位の間に、本アミノ酸配列8乃至10をコードする遺伝子を導入した点以外は、実施例1と同様の処理によって、本ペプチドとして、C末端に本アミノ酸配列8乃至10が導入されたグルタチオン転移酵素(GST)をそれぞれ生合成した。なお、実施例7で本アミノ酸配列8を有するGSTを生合成し、実施例8で本アミノ酸配列9を有するGSTを生合成し、実施例9で本アミノ酸配列10を有するGSTを生合成した。以下、実施例7乃至9で生合成された本アミノ酸配列8乃至10を有するGSTを、それぞれ本ペプチド7乃至9という。
【0082】
[実施例10](本ペプチド1及び2の親水性のポリスチレン表面に対する結合性能評価、夾雑物:BSA)
本ペプチドの親水性のポリスチレン表面に対する結合性能を評価するため、夾雑物としてウシ血清由来のアルブミン(BSA)が存在する状態における実施例1の本ペプチド1、実施例2の本ペプチド2及び原ペプチドであるGSTの親水性のポリスチレン表面に対する固定化率を測定した。
【0083】
まず、BSAを添加しないPBSと、BSA濃度が0.003〜50mg/mlとなるように調製したPBSを用意し、各溶液中に、終濃度が5μg/mlとなるように本ペプチド1を添加して試料を調製した。次に、100μlの各試料を親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、室温で1時間インキュベートして、本ペプチド1を固定化させ、0.1%のTween20を含むPBS(以下「0.1PBST」という)で5回洗浄した。さらに、ブロッキング試薬として、2%のBSAを含む0.1PBSTを300μl加え、室温で1時間インキュベートして、ポリスチレンプレート表面の未結合部位をブロッキングし、0.1PBSTで5回洗浄した。そして、抗体として、0.2%のBSAを含む0.1PBSTで4000倍に希釈された抗GST抗体を100μl加え、室温で1時間インキュベートして、本ペプチド1に含まれるGSTと抗GST抗体とを抗原抗体反応させ、0.1PBSTで5回洗浄した。その後、酵素標識された第2の抗体として、0.2%のBSAを含む0.1PBSTで1000倍に希釈されたHRP-conjugated anti-rabbit IgG antibodyを100μl加え、室温で1時間インキュベートして、さらに抗原抗体反応させ、0.1PBSTで5回洗浄した。最後に、発色基質として、ABTS(2,2'-azinobis(3-ethylbenzthiazoline-6-sulphonic acid))を加え、インキュベートし、マイクロプレートリーダー(TECAN製「SUNRISE Remote」)を用いて、405nmにおける吸光度を測定した。本ペプチド2及びGSTについても同様の処理を行い吸光度を測定した。
【0084】
図1に実施例10の測定結果を示す。図1中の黒丸(●)は本ペプチド1の結果であり、黒三角は本ペプチド2の結果であり、白丸(○)は原ペプチドであるGSTの結果である。なお、図1において、縦軸は固定化率(%)であり、横軸はBSA濃度であるが、固定化率は、BSAを添加しないPBSにおける吸光度(結合した量)を基準(100%)として、その他の吸光度を規格化したものである。
【0085】
図1から、原ペプチドであるGSTでは、夾雑物としてBSAが存在すると、急激に固定化率が低下し、結合する量が減少するが、本ペプチド1及び2では、10mg/mlという高濃度のBSAが存在していても、固定化率は殆ど変わらず、親水性のポリスチレンプレートに特異的に結合することが確認できる。
【0086】
[実施例11](本ペプチド1の親水性のポリスチレン表面に対する結合性能評価、夾雑物:BSA及びTween20)
実施例1の本ペプチド1の結合性能をより詳細に評価するため、夾雑物としてTween20が存在する状態で、疎水性のポリスチレン表面及び親水性のポリスチレン表面に対して結合した量(吸光度)を測定した。
【0087】
まず、Tween20を添加しないPBSと、Tween20濃度が0.1、1,10%となるように調製したPBSを用意し、各溶液中に、終濃度が0〜1μg/mlとなるように本ペプチド1を添加して試料を調製した。次に、100μlの各試料を親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)及び疎水性のポリスチレンプレート(ベクトン・ディッキンソン アンド カンパニー社製「BD Falconマイクロプレート#351172」)に加え、室温で1時間インキュベートして、本ペプチド1を固定化させ、0.1PBSTで5回洗浄した。その後、実施例10と同じ条件で、ブロッキング試薬、抗体、酵素標識された第2の抗体及び発色基質を添加する処理及び洗浄処理を行い吸光度を測定した。原ペプチドであるGSTについても同様の処理を行い吸光度を測定した。
【0088】
図2(a)〜(d)に実施例11の親水性のポリスチレンプレートにおける測定結果を示し、図3(a)〜(d)に実施例11の疎水性のポリスチレンプレートにおける測定結果を示す。図2及び図3の(a)〜(d)は、それぞれTween20の添加量が0、0.1、1、10%の条件の時の測定結果である。図2及び図3中の黒丸(●)は本ペプチド1の結果であり、白丸(○)は原ペプチドであるGSTの結果である。図2及び3において、縦軸は405nmにおける吸光度であり、横軸は本ペプチド1及びGSTの濃度である。なお、図2及び3において、GSTの結果と本ペプチド1の結果が同じだった場合は、GSTの結果である白丸(○)が本ペプチド1の黒丸(●)と重なっており確認できない。例えば、図3(b)〜(d)ではGSTの白丸(○)の全部が本ペプチド1の黒丸(●)と重なっている。
【0089】
図2(a)〜(d)から、本ペプチド1及びGSTは、いずれも添加濃度を増やすと、親水性のポリスチレンプレートに結合する量が増えることが確認できる。また、本ペプチド1では、夾雑物としてTween20を添加しても、親水性のポリスチレンプレートに結合する量は減らず、むしろ若干増加している。GSTでは、夾雑物としてTween20を添加すると、親水性のポリスチレンプレートに結合する量が少し減少する。
【0090】
また、図3(a)によれば、本ペプチド1及びGSTは、いずれも、夾雑物としてTween20が存在しない場合には疎水性のポリスチレンプレートに対し結合する。しかし、図3(b)〜(d)に示すとおり、本ペプチド1及びGSTは、いずれも、夾雑物としてTween20が存在すると、疎水性のポリスチレンプレートには殆ど結合できない。このように、本ペプチドの結合性能は、親水性の樹脂表面を有する固相に特異的なものである。
【0091】
[実施例12](本ペプチド1の親水性のポリスチレン表面に対する結合性能評価、夾雑物:BSA及びTween20)
実施例1の本ペプチド1の結合性能をより詳細に評価するため、夾雑物としてTween20が存在する状態及びTween20及びBSAが存在する状態のそれぞれで、本ペプチド1の添加量を変化させて、疎水性のポリスチレン表面及び親水性のポリスチレン表面に対して結合した量(吸光度)を測定した。
【0092】
まず、PBSと、0.1PBSTと、20mg/mlのBSAを含む0.1PBSTとを用意し、各溶液中に、終濃度が0〜10μg/mlとなるように本ペプチド1を添加して試料を調製した。次に、100μlの各試料を親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)及び疎水性のポリスチレンプレート(ベクトン・ディッキンソン アンド カンパニー社製「BD Falconマイクロプレート#351172」)に加え、室温で2時間インキュベートして、本ペプチド1を固定化させ、0.1PBSTで5回洗浄した。その後、実施例10と同じ条件で、ブロッキング試薬、抗体、酵素標識された第2の抗体及び発色基質を添加する処理及び洗浄処理を行い吸光度を測定した。原ペプチドであるGSTについても同様の処理を行い吸光度を測定した。
【0093】
図4に実施例12の測定結果を示す。図4の(a)〜(c)は、それぞれPBS、0.1PBST及びBSAを含む0.1PBSTにおける親水性のポリスチレンプレートについての測定結果であり、図4の(d)〜(f)は、それぞれPBS、0.1PBST及びBSAを含む0.1PBSTにおける疎水性のポリスチレンプレートについての測定結果である。また、図4中の黒丸(●)は本ペプチド1の結果であり、白丸(○)は原ペプチドであるGSTの結果である。図4において、縦軸は405nmにおける吸光度であり、横軸は本ペプチド1及びGSTの濃度である。なお、図4において、GSTの結果と本ペプチド1の結果が同じだった場合は、GSTの結果である白丸(○)が本ペプチド1の黒丸(●)と重なっており確認できない。例えば、図4(e)及び(f)ではGSTの白丸(○)の全部が本ペプチド1の黒丸(●)と重なっている。
【0094】
図4(a)〜(c)から、本ペプチド1では、夾雑物としてTween20を添加した場合(図4(b))でも、夾雑物としてTween20及びBSAを添加した場合(図4(c))でも、親水性のポリスチレンプレートに結合する量はほとんど変わらないことが確認できる(図4(b)及び(c)の黒丸)。これに対し、GSTでは、夾雑物としてTween20を添加すると、親水性のポリスチレンプレートに結合する量が少し減少し(図4(b)の白丸)、さらにBSAを添加した場合には殆ど結合できない(図4(c)の白丸)。このように、本アミノ酸配列2を導入することにより、親水性のポリスチレンプレートに対する結合性能が向上する。
【0095】
また、図4(d)〜(f)によれば、本ペプチド1及びGSTは、いずれも、夾雑物としてTween20が存在しない場合には疎水性のポリスチレンプレートに対し結合するが、夾雑物としてTween20やBSAが存在すると、疎水性のポリスチレンプレートには殆ど結合できない。
【0096】
[実施例13](本ペプチド1の活性値評価)
実施例1の本ペプチド1の固定化された状態における活性値を評価するため、夾雑物としてTween20が存在する状態及びTween20及びBSAが存在する状態のそれぞれで、疎水性のポリスチレン表面及び親水性のポリスチレン表面に対して結合させ、固定化されたGSTの酵素としての活性値を測定した。
【0097】
まず、PBSと、0.1PBSTと、20mg/mlのBSAを含む0.1PBSTとを用意し、各溶液中に、終濃度が10、20、40μg/mlとなるように本ペプチド1を添加して試料を調製した。次に、100μlの各試料を親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)及び疎水性のポリスチレンプレート(ベクトン・ディッキンソンアンド カンパニー社製「BD Falconマイクロプレート#351172」)に加え、室温で2時間インキュベートして、本ペプチド1を固定化させ、0.1PBSTで5回洗浄した。その後、0.1%Tween20、1mMの還元型グルタチオン及びGSTの基質として1mMのCDNB(1-chloro-2,4-dinitrobenzene)を含む0.1Mリン酸カリウムバッファー(pH6.5)を200μl加え、室温で30分間撹拌しつつ、30秒毎に340nmにおける吸光度を測定した。原ペプチドであるGSTについても同様の処理を行い吸光度を測定した。
【0098】
図5に実施例13の測定結果を示す。図5の(a)〜(c)は、それぞれPBS、0.1PBST及びBSAを含む0.1PBSTにおける親水性のポリスチレンプレートについての測定結果であり、図5の(d)〜(f)は、それぞれPBS、0.1PBST及びBSAを含む0.1PBSTにおける疎水性のポリスチレンプレートについての測定結果である。また、図5中の黒丸(●)は本ペプチド1の結果であり、白丸(○)は原ペプチドであるGSTの結果である。図5において、縦軸は活性値(吸光度の変化率)であり、横軸は本ペプチド1及びGSTの濃度である。なお、図5において、GSTの結果と本ペプチド1の結果が同じだった場合は、GSTの結果である白丸(○)が本ペプチド1の黒丸(●)と重なっており確認できない。例えば、図5(e)及び(f)ではGSTの白丸(○)の全部が本ペプチド1の黒丸(●)と重なっている。
【0099】
図5(a)〜(c)から、親水性のポリスチレンプレートに固定化された本ペプチド1のGSTは、夾雑物としてTween20を添加した場合(図5(b))でも、夾雑物としてTween20及びBSAを添加した場合(図5(c))でも、酵素として非常に活性であることが確認できる。これに対し、GSTは、実施例12における図4(a)及び(b)の白丸で示されるように、夾雑物が添加されていない場合及び夾雑物としてTween20が添加されている場合では親水性のポリスチレンプレートに結合しているが、図5(a)及び(b)に示すとおり、全く酵素の活性を示していない。これは、原ペプチドであるGSTは、親水性のポリスチレンプレートに対し固定化されるが、構造変化や不活性化していることを意味している。なお、GSTは、図5(c)においても全く酵素の活性を示していないが、これは、図4(c)に示すように、そもそもTween20及びBSAの存在下では、親水性のポリスチレンプレートに殆どGSTが結合できないためである。
【0100】
また、図5(d)から、疎水性のポリスチレンプレートに固定化された本ペプチド1のGSTは、殆ど酵素の活性を示していない。実施例12における図4(d)から明らかなように、夾雑物が存在しない状態であれば、本ペプチド1は、疎水性のポリスチレンプレートに対して結合できるが、構造変化や不活性化によって活性が失われることが確認できる。つまり、本アミノ酸配列2の結合性能は、親水性のポリスチレンプレートに対して特異的なものであることが確認され、従来の疎水性のポリスチレンプレートを利用した固定化に比べて、本ペプチド1を親水性のポリスチレンプレートに固定化した方が、活性を維持できることが確認できる。なお、図5(e)及び(f)において、本ペプチド1及びGSTは、いずれも、全く酵素の活性を示していないが、これは、夾雑物としてTween20やBSAが存在すると、疎水性のポリスチレンプレートには殆ど結合できないからである。
【0101】
[実施例14](本ペプチド3乃至6の親水性のポリスチレン表面に対する結合性能評価、夾雑物:BSA及びTween20)
本ペプチド3乃至6の親水性のポリスチレン表面に対する結合性能を評価するため、夾雑物としてBSA及びTween20が存在する状態における実施例3乃至6の本ペプチド3乃至6並びに評価のために本ペプチド1及び原ペプチドであるGSTの親水性のポリスチレン表面に対する固定化率を測定した。
【0102】
まず、BSAを添加しない0.1PBSTと、BSA濃度が0.003〜50mg/mlとなるように調製した0.1PBSTを用意し、各溶液中に、終濃度が5μg/mlとなるように本ペプチド3を添加して試料を調製した。次に、100μlの各試料を親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、25℃で3時間インキュベートして、本ペプチド3を固定化させ、0.1%PBSで6回洗浄した。さらに、ブロッキング試薬として、2%のBSAを含む0.1PBSTを300μl加え、25℃で1時間インキュベートして、ポリスチレンプレート表面の未結合部位をブロッキングし、0.1PBSTで6回洗浄した。そして、抗体として、0.2%のBSAを含む0.1PBSTで5000倍に希釈された抗GST抗体を100μl加え、25℃で1時間インキュベートして、本ペプチド3に含まれるGSTと抗GST抗体とを抗原抗体反応させ、0.1PBSTで6回洗浄した。その後、酵素標識された第2の抗体として、0.2%のBSAを含む0.1PBSTで5000倍に希釈されたHRP-conjugated anti-rabbit IgG antibodyを100μl加え、25℃で1時間インキュベートして、抗原抗体反応させ、さらに0.1PBSTで6回洗浄した。最後に、発色基質として、ABTS(2,2'-azinobis(3-ethylbenzthiazoline-6-sulphonic acid))を加え、インキュベートし、マイクロプレートリーダー(TECAN製「SUNRISE Remote」)を用いて、405nmにおける吸光度を測定した。本ペプチド4乃至6、本ペプチド1及びGSTについても同様の処理を行い吸光度を測定した。
【0103】
図7に実施例14の測定結果を示す。図7中の黒塗りの三角は本ペプチド3の結果であり、白抜き三角(△)は本ペプチド4の結果であり、黒塗りの四角は本ペプチド5の結果であり、白抜き四角(□)は本ペプチド6の結果であり、黒丸(●)は本ペプチド1の結果であり、白丸(○)は原ペプチドであるGSTの結果である。図7において、縦軸は固定化率(%)であり、横軸はBSA濃度であるが、固定化率は、BSAを添加しないPBSTにおける吸光度(結合した量)を基準(100%)として、その他の吸光度を規格化したものである。なお、図7において、本ペプチド1、3乃至6の結果は、類似しており、重畳している部分がある。
【0104】
図7から、原ペプチドであるGSTでは、夾雑物としてBSA及びTween20が存在すると、急激に固定化率が低下し、結合する量が減少するが、本ペプチド3乃至6では、10mg/mlという高濃度のBSAが存在していても、固定化率は殆ど変わらず、本ペプチド1と同じレベルで、親水性のポリスチレンプレートに特異的に結合することが確認できる。
【0105】
[実施例15](本ペプチド3乃至6のポリスチレン表面における活性値評価、夾雑物:BSA及びTween20)
親水性のポリスチレン表面に固定化された状態における本ペプチド3乃至6の活性値を評価するため、夾雑物としてTween20が存在する状態及びTween20及びBSAが存在する状態のそれぞれで、固定化されたGSTの酵素としての活性値を測定した。
【0106】
まず、PBSと、0.1PBSTとを用意し、各溶液中に、終濃度が0.5、10、20、40μg/mlとなるように本ペプチド1、3乃至6をそれぞれ添加して試料を調製した。次に、100μlの各試料を親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、25℃で2時間インキュベートして、本ペプチドを固定化させ、0.1PBSTで6回洗浄し、さらに0.1%Tween20を含む0.1Mリン酸カリウム溶液(pH6.5)で1回洗浄した。その後、0.1%Tween20、1mMの還元型グルタチオン及びGSTの基質として1mMのCDNB(1-chloro-2,4-dinitrobenzene)を含む0.1Mリン酸カリウムバッファー(pH6.5)を200μl加え、室温で30分間撹拌しつつ、30秒毎に340nmにおける吸光度を測定した。原ペプチドであるGSTについても同様の処理を行い吸光度を測定した。
【0107】
図8に実施例15の測定結果を示す。図8の(a)は、PBSにおける測定結果であり、(b)は0.1PBSTにおける測定結果である。また、図8中の黒塗りの三角は本ペプチド3の結果であり、白抜き三角(△)は本ペプチド4の結果であり、黒塗りの四角は本ペプチド5の結果であり、白抜き四角(□)は本ペプチド6の結果であり、黒丸(●)は本ペプチド1の結果であり、白丸(○)は原ペプチドであるGSTの結果である。図8において、縦軸は活性値(吸光度の変化率)であり、横軸は本ペプチド及びGSTの濃度である。なお、図8において、本ペプチド1、3乃至6の結果は、類似しており、重畳している部分がある。
【0108】
図8(a)及び(b)から、親水性のポリスチレンプレートに固定化された本ペプチド1、3乃至6のGSTは、そのままでも(図8(a))、夾雑物としてTween20を添加した場合(図8(b))でも、酵素として非常に活性であることが確認できた。
【0109】
[実施例16](本ペプチド7乃至9の親水性のポリスチレン表面に対する結合性能評価、夾雑物:BSA及びTween20)
実施例7乃至9で生合成した本ペプチド7乃至9の親水性のポリスチレン表面に対する結合性能を評価するため、本ペプチド7乃至9並びに原ペプチドであるGSTについて、実施例14と同様の処理を行い吸光度を測定し、親水性のポリスチレン表面に対する固定化率を評価した。
【0110】
図9に実施例16の測定結果を示す。図9中の黒塗りの丸は本ペプチド7の結果であり、黒塗りの菱形は本ペプチド8の結果であり、白抜き三角(△)は本ペプチド9の結果であり、白丸(○)は原ペプチドであるGSTの結果である。図9において、縦軸は固定化率(%)であり、横軸はBSA濃度であるが、固定化率は、BSAを添加しないPBSTにおける吸光度(結合した量)を基準(100%)として、その他の吸光度を規格化したものである。
【0111】
図9から、原ペプチドであるGSTでは、夾雑物としてBSA及びTween20が存在すると、急激に固定化率が低下し、結合する量が減少するが、本ペプチド7乃至9では、10mg/mlという高濃度のBSAが存在していても、固定化率は殆ど変わらず、親水性のポリスチレンプレートに特異的に結合することが確認できた。
【0112】
[実施例17](本ペプチド7乃至9のポリスチレン表面における活性値評価、夾雑物:BSA及びTween20)
親水性のポリスチレン表面に固定化された状態における本ペプチド7乃至9の活性値を評価するため、本ペプチド7乃至9並びに原ペプチドであるGSTについて、実施例15と同様の処理を行い活性値を測定した。
【0113】
図10に実施例17の測定結果を示す。図10の(a)は、PBSにおける測定結果であり、(b)は0.1PBSTにおける測定結果である。また、図10中の黒塗りの丸は本ペプチド7の結果であり、黒塗りの菱形は本ペプチド8の結果であり、白抜き三角(△)は本ペプチド9の結果であり、白丸(○)は原ペプチドであるGSTの結果である。図10において、縦軸は活性値(吸光度の変化率)であり、横軸は本ペプチド及びGSTの濃度である。
【0114】
図10(a)及び(b)から、親水性のポリスチレンプレートに固定化された本ペプチド7乃至9のGSTは、そのままでも(図10(a))、夾雑物としてTween20を添加した場合(図10(b))でも、酵素として非常に活性であることが確認できた。
【0115】
[実施例18](本ペプチド1の生産時における分離精製工程への適用例、夾雑物:菌体破砕液)
本実施例においては、本ペプチドの特異的な吸着機能を生産時における分離精製工程への適用可能性を評価するため、大腸菌を使用して発現させた本アミノ酸配列2を含むGSTを、特別な精製工程を行うことなく、菌体破砕液をそのまま親水性のポリスチレン表面に固定化させた時の結合した量(吸光度)を測定した。比較用として、原ペプチドであるGSTについても同様に結合した量(吸光度)を測定した。
【0116】
まず、1列につき8個の培養容器(ウェル)が設けられたウェルプレートを5列用いて、39個の培養容器(ウェル)内に200μlの2xYT培地を添加し、各ウェルに、実施例1と同様の本アミノ酸配列2をコードする遺伝子を導入したベクターを導入して形質転換した大腸菌(BL21)を植菌してコロニーを形成し、37℃、200rpmで一晩培養した。原ペプチドであるGSTについても同様に40コロニーを作成し培養した。各コロニーの培養の程度を確認するため、各培養液を100μl回収し、菌体濃度を測定した。図11(a)は本ペプチドを培養した39コロニーにおける菌体濃度(630nmにおける吸光度)であり、図11(b)は原ペプチドを培養した40コロニーにおける菌体濃度である。図11において、横軸はウェルプレートの列番号であり、各列毎に8個のウェルの結果が示されている。図11(a)及び(b)から、全てのコロニーで同程度に培養されていることが確認された。
【0117】
次に、菌体破砕後、菌体破砕液を10μl回収し、10mg/mlのBSAを1%含むPBSを90mlと混合して、親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、室温で1時間インキュベートして、固相上にペプチドを固定化させた。その後、実施例7と同じ条件で、ブロッキング試薬、抗体、酵素標識された第2の抗体及び発色基質を添加する処理及び洗浄処理を行い吸光度を測定した。原ペプチドであるGSTについても同様の処理を行い吸光度を測定した。
【0118】
図11(c)及び(d)に実施例18の測定結果を示す。図11(c)は本ペプチドを培養した39コロニーにおける吸光度(405nm)であり、図11(d)は原ペプチドを培養した40コロニーにおける吸光度(405nm)である。図11(d)に示すように、原ペプチドであるGSTは、吸光度がバックグラウンド程度であり、ほとんど固相表面に固定化されていない。これに対し、図11(c)から、本ペプチドでは、大腸菌の菌体破砕液に含まれた状態のままであっても、固相表面に接触させることで、固相表面に特異的に固定化されることが確認できた。大腸菌の菌体破砕液の中には、夾雑物として、特定されていない大小様々なタンパク質に加えて、タンパク質以外の低分子化合物や脂質なども存在する。これらの夾雑物が存在しても、本ペプチドは親水性のポリスチレンプレートに対し特異的に結合するので、生産時における分離精製工程として利用可能であることが確認できた。
【0119】
[実施例19](本ペプチド1の生産時における分離精製工程への適用例、夾雑物:培地)
本実施例においては、本ペプチドの特異的な吸着機能を生産時における分離精製工程への適用可能性を評価するため、本ペプチド1に夾雑物として各種の培地(YPD、BMMY、2xTY、LB)を加えた状態における親水性のポリスチレン表面に対する結合した量(吸光度)を測定した。比較用として、原ペプチドであるGSTについても同様に結合した量(吸光度)を測定した。YPDは一般的な酵母の培養培地であり、BMMYはP. pastoris専用培地であり、2xTY及びLBは大腸菌専用培地である。
【0120】
まず、YPD、BMMY、2xTY及びLBの4種類の培地を用意し、それぞれ原液〜10000倍に希釈した溶液に対し、5μg/mlとなるように本ペプチド1を添加して試料を調製した。各試料を、親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、室温で1時間インキュベートして、固相上にペプチドを固定化させた。その後、実施例7と同じ条件で、ブロッキング試薬、抗体、酵素標識された第2の抗体及び発色基質を添加する処理及び洗浄処理を行い吸光度を測定した。原ペプチドであるGSTについても同様の処理を行い吸光度を測定した。原ペプチドであるGSTについても同様に吸光度を測定した。
【0121】
図12に実施例19の測定結果を示す。図12中の黒丸(●)は本ペプチド1と培地YPDの混合試料の結果であり、白丸(○)は原ペプチドであるGSTと培地YPDの混合試料の結果であり、黒塗りの三角は本ペプチド1と培地BMMYの混合試料の結果であり、白抜き三角(△)は原ペプチドであるGSTと培地BMMYの混合試料の結果であり、黒塗りの四角は本ペプチド1と培地2xYTの混合試料の結果であり、白抜き四角(□)は原ペプチドであるGSTと培地2xYTの混合試料の結果であり、菱形(◆)は本ペプチド1と培地LBの混合試料の結果であり、白抜き菱形(◇)は原ペプチドであるGSTと培地LBの混合試料の結果でありである。図12において、縦軸は吸光度(405nm)であり、横軸は培地の相対濃度(希釈率)(右端の1が原液)である。図12に示すように、原ペプチドであるGST(白丸、白抜き三角、白抜き四角、白抜き菱形)は、何れも培地が濃くなるにつれて固相表面に結合される量が減っている。これに対し、本ペプチド1(黒丸、黒塗り三角、黒塗り四角、黒塗り菱形)では、培地が濃くなってもかなりの量が固定化されて固相表面に結合されており、培地の濃度にあまり影響されず、固相表面に特異的に固定化されることが確認できた。
【0122】
特に、酵母で使用される培地であるYPD及びBMMYにおいても特異的な吸着性能を発現するので、酵母からの分泌生産に利用可能である。つまり、酵母からの分泌液を直接固相表面に接触させることで、分泌液中の本ペプチドを固定化して分離精製することが可能である。
【0123】
さらに、一本鎖抗体(scFv)の生産に有効な培地であるBMMY及び2xYTでも特異的な吸着性能を発現するので、本ペプチドとして一本鎖抗体(scFv)を生産した場合でも、酵母の分泌液や大腸菌の破砕液から直接一本鎖抗体(scFv)を分離精製することが可能である。
【0124】
[実施例20](本アミノ酸配列2からなる本ペプチドを標識した抗体の生産)
まず、固相法によって、本アミノ酸配列2からなる本ペプチドのN末端側にリジン(K)を有するペプチド(KRIIIRRIRR)を合成した。次に、0.5mgの本アミノ酸配列2からなる本ペプチドを100μlの25%グルタルアルデヒド溶液で溶解し、室温で10分間インキュベートした。そして、9mlのジエチルエーテル及び1mlのエタノールを加えてグルタルアルデヒドで標識された本ペプチドを析出させ、遠心分離により回収した。
【0125】
さらに、0.5mgのマウスモノクローナル抗ヒトCRP抗体(オリエンタル酵母社製、#4715500)を溶解した0.5mlのPBSに、グルタルアルデヒドで標識された本ペプチドを溶解し、25℃で1時間インキュベートした。その後、100μlの2M Tris−HCl(pH8.0)を加え、反応を停止して、限外ろ過法によって、本ペプチドとして、本アミノ酸配列2を含む抗CRP抗体を精製した。以下、実施例20で生成した本アミノ酸配列2を含む抗CRP体を「本ペプチド20」といい、その原ペプチドを「原ペプチド20」という。
【0126】
[実施例21](本アミノ酸配列2をC末端側に有する一本鎖抗体の生成)
本実施例では、配列表の配列番号23、24、25に示す3種類の一本鎖抗体(scFv)のC末端側に本アミノ酸配列2を導入して3種類の一本鎖抗体を生成した。なお、以下、配列表の配列番号23、24及び25に示す3種類の一本鎖抗体(原ペプチド)を、それぞれ「原ペプチド21−1」、「原ペプチド21−2」及び「原ペプチド21−3」といい、本実施例で生成される原ペプチド21−1、原ペプチド21−2及び原ペプチド21−3に本アミノ酸配列2を導入した3種類の一本鎖抗体(本ペプチド)を、それぞれ「本ペプチド21−1」、「本ペプチド21−2」及び「本ペプチド21−3」という。これらの一本鎖抗体は、いずれもC反応性蛋白(C-Reactive Protein:CRP)に対して特異的な抗原抗体反応を示す抗原結合部位を有する。原ペプチド21−1ないし21−3は、重鎖の可変部(VH)のC末端と軽鎖の可変部(VL)のN末端とが4つのグリシン(G)とセリン(S)の組合わせを3組連結した構造 (G4S)3の鎖状のリンカーペプチドで連結した構造である。さらに原ペプチド21−1ないし21−3には軽鎖の可変部(VL)のC末端にヒスタジンタグが導入されている。なお、実施例22及び23の原ペプチドは、配列番号23の一本鎖抗体(原ペプチド21−1)である。
【0127】
まず、5’末端側がリン酸化された本アミノ酸配列2をコードする遺伝子を含むセンス鎖(PStag-sense)及びアンチセンス鎖(PStag-antisense)のオリゴDNAを合成し、アニール後、制限酵素NotI及びXhoIで消化したpET22b(+)ベクター(Novagen製)に導入し、本アミノ酸配列2をコードする遺伝子を含むpET-PStagベクター(pET22b(+)にPStagが導入されたベクター)を調製した。なお、「PStag」とは本アミノ酸配列2を発現させる遺伝子を意味する。以下同じ。
【0128】
次に、重鎖の可変部(VH)をコードする遺伝子を含むセンス鎖(VH-sense)のオリゴDNA及び軽鎖の可変部(VL)をコードする遺伝子を含むアンチセンス鎖(VL-antisense)のオリゴDNAを合成した。これらの合成オリゴDNAをプライマーとし、それぞれ配列番号23,24,25に示す3種類の一本鎖抗体発現ベクターpET-scFv(pET22b(+)にscFvの遺伝子が導入されたベクター)を鋳型としてPCR法によって3種類の一本鎖抗体(scFv)の遺伝子を増幅した。増幅したscFv遺伝子を制限酵素NdeI及びNotIで消化し、同酵素で消化した本アミノ酸配列2をコードする遺伝子を含むpET-PStagベクターに導入し、C末端側に本アミノ酸配列2をコードする遺伝子を含む3種類の一本鎖抗体の発現ベクター(pET-scFv-PStagベクター)を調製した。
【0129】
宿主として大腸菌(BL21(DE3)Rosetta(Novagen))を使用し、上記pET-scFv-PStagベクターを宿主に導入して宿主を形質転換し、アンピシリンを含む寒天培地上で選別した。その後、2xYT培地50ml中で大腸菌を培養し、終濃度が1mMになるように、発現誘導物質としてイソプロピル1−チオ−ガラクトシド(IPTG:Isopropyl-1-thio-β-D(-)-galactoside)を加えて、本アミノ酸配列2をC末端に含む一本鎖抗体(scFv-PStag)を発現させた。
【0130】
そして、菌体破砕後、菌体内不溶性画分を6M塩酸グアニジン(変性剤)及び10mMメルカプトエタノール(還元剤)を含む可溶化液で溶解した。こうして、未精製の変性された本アミノ酸配列2をC末端に含む一本鎖抗体を得ることができた。以下、この状態の実施例21の一本鎖抗体を「未精製の変性本ペプチド21−1」ないし「未精製の変性本ペプチド21−3」という。
【0131】
さらに、未精製の変性本ペプチド21−1ないし21−3を変性状態のままHisTrap HP (GE HealthCare)を用いたNiキレートアフィニティクロマトグラフィ(IMAC)によって精製し、本アミノ酸配列2をC末端側に含む変性した一本鎖抗体を生産した。以下、この状態の実施例21の一本鎖抗体を「精製後の変性本ペプチド21−1」ないし「精製後の変性本ペプチド21−3」という。本ペプチド21−1のアミノ酸配列を配列表の配列番号26に示し、本ペプチド21−2のアミノ酸配列を配列表の配列番号27に示し、本ペプチド21−3のアミノ酸配列を配列表の配列番号28に示す。本ペプチド21−1ないし21−3は、軽鎖の可変部(VL)のC末端側に本アミノ酸配列2を有しており、本アミノ酸配列2のさらにC末端側にヒスタジンタグが連結されている。なお、アミノ酸配列それ自体は、精製の前後でも、変性の前後でも変わらず、配列番号26、27、28に示すとおりである。
【0132】
[実施例22](本アミノ酸配列2を重鎖可変部(VH)とリンカーペプチドの間に有する一本鎖抗体の生成)
本実施例では、本アミノ酸配列2を原ペプチド21−1(配列番号23)及び原ペプチド21−2(配列番号24)の重鎖の可変部(VH)と軽鎖の可変部(VL)を連結するリンカーペプチドに導入した一本鎖抗体(以下で生成されるペプチドを、それぞれ「本ペプチド22−1」及び「本ペプチド22−2」という)を生成する。まず、重鎖の可変部(VH)をコードする遺伝子を含むセンス鎖(VH-sense)のオリゴDNA及びアンチセンス鎖(VH-antisense)のオリゴDNAを合成し、これらの合成オリゴDNAをプライマーとし、配列番号23の一本鎖抗体を発現するpET-scFvベクターを鋳型としてPCR法によって重鎖の可変部(VH)の遺伝子を増幅した。
【0133】
次に、軽鎖の可変部(VL)をコードする遺伝子を含むセンス鎖(VL-sense)のオリゴDNA及びアンチセンス鎖(VL-antisense)のオリゴDNAを合成し、同様に軽鎖の可変部(VL)の遺伝子を増幅した。
【0134】
さらに、重鎖の可変部(VH)のC末端部、本アミノ酸配列2及びリンカーペプチドの順にこれらをコードする遺伝子を持つセンス鎖(VH-Pstag-(G4S)3-sense)のオリゴDNAと本アミノ酸配列2、リンカーペプチド及び軽鎖の可変部(VL)のN末端部分の順にこれらをコードする遺伝子を含むアンチセンス鎖(PStag-(G4S)3-VL-antisense)のオリゴDNAを合成した。
【0135】
重鎖の可変部(VH)の遺伝子、軽鎖の可変部(VL)の遺伝子、センス鎖(VH-PStag-(G4S)3-sense)のオリゴDNA、アンチセンス鎖(PStag-(G4S)3-VL-antisense)のオリゴDNA、センス鎖(VH-sense)のオリゴDNA及びアンチセンス鎖(VL-antisense)のオリゴDNAを用いて、Overlapping PCR法により本アミノ酸配列2を重鎖可変部(VH)とリンカーペプチド(G4S)3との間に有する一本鎖抗体の遺伝子(scFv-(PStag))を調製した。当該遺伝子を制限酵素NdeI及びNotIで消化し、同酵素で消化したpET22b(+)に導入し、リンカーペプチド(G4S)3に本アミノ酸配列2をコードする遺伝子を含む一本鎖抗体の発現ベクター(pET-scFv-(PStag) ベクター)を作製した。なお、「(PStag)」とは一本鎖抗体の途中である重鎖可変部(VH)とリンカーペプチド(G4S)3との間にPStagが導入されることを意味する。以下同じ。
【0136】
後は、実施例21と同様に、当該pET-scFv-(PStag)ベクターを宿主に導入して培養し、菌体破砕後、菌体内不溶性画分を6M塩酸グアニジン(変性剤)及び10mMメルカプトエタノール(還元剤)を含む可溶化液で溶解して、未精製の変性されたリンカーペプチド(G4S)3に本アミノ酸配列2を有する一本鎖抗体を得ることができた。さらに、実施例21と同様の方法で精製し、精製された変性した状態の本ペプチド22−1(以下「精製後の変性本ペプチド22−1」という)を生産した。本ペプチド22−1のアミノ酸配列を配列表の配列番号29に示す。本ペプチド22−1は、重鎖の可変部(VH)のドメインとリンカーペプチド(G4S)3の間に本アミノ酸配列2が配置されている。
【0137】
また、同様の手法によって、原ペプチド21−2(配列番号24)の重鎖の可変部(VH)のドメインとリンカーペプチド(G4S)3の間に本アミノ酸配列2が配置されている精製後の変性本ペプチド22−2を生産した。本ペプチド22−2のアミノ酸配列を配列表の配列番号30に示す。
【0138】
[実施例23](本アミノ酸配列2をC末端及び重鎖可変部(VH)とリンカーペプチドとの間の両方に有する一本鎖抗体の生成)
本実施例では、本アミノ酸配列2を原ペプチド21−1及び原ペプチド21−2の一本鎖抗体のC末端側(すなわち軽鎖の可変部(VL)のC末端側)及び重鎖可変部(VH)とリンカーペプチドの間(すなわち重鎖の可変部(VH)のC末端側)の両方に導入した一本鎖抗体(以下で生成されるペプチドを、それぞれ「本ペプチド23−1」及び「本ペプチド23−2」という)を生成する。まず、実施例22と同様に、本アミノ酸配列2を重鎖可変部(VH)とリンカーペプチド(G4S)3の間に有する一本鎖抗体の遺伝子(scFv-(PStag))を調製した。当該遺伝子を実施例22ではpET22b(+)に導入したが、本実施例では、実施例21で使用した本アミノ酸配列2をコードする遺伝子を含むpET-PStagベクターに導入し、C末端側及び重鎖可変部(VH)とリンカーペプチド(G4S)3との間の両方に本アミノ酸配列2をコードする遺伝子を含む一本鎖抗体の発現ベクター(pET-scFv-(PStag)-PStagベクター)を作製した。
【0139】
後は、実施例21と同様に、当該pET-scFv-(PStag)-PStagベクターを宿主に導入して培養し、菌体破砕後、菌体内不溶性画分を6M塩酸グアニジン(変性剤)及び10mMメルカプトエタノール(還元剤)を含む可溶化液で溶解して、未精製の変性されたC末端側及びリンカーペプチドの両方に本アミノ酸配列2を有する一本鎖抗体(scFv)を得ることができた。さらに、実施例21と同様に精製し、精製された変性した状態の本ペプチド23−1(以下「精製後の変性本ペプチド23−1」という)を生産した。本ペプチド23−1のアミノ酸配列を配列表の配列番号31に示す。本ペプチド23−1は、本ペプチド21−1と同様に軽鎖の可変部(VL)のC末端側に本アミノ酸配列2を有しており、さらに本アミノ酸配列2のC末端側にヒスタジンタグが連結されている。さらに本ペプチド22−1と同様に重鎖の可変部(VH)のドメインとリンカーペプチド(G4S)3との間に本アミノ酸配列2を有している。
【0140】
また、同様の手法によって、原ペプチド21−2(配列番号24)の軽鎖の可変部(VL)のC末端側及び重鎖の可変部(VH)のドメインとリンカーペプチド(G4S)3の間に本アミノ酸配列2が配置されている精製後の変性本ペプチド23−2を生産した。本ペプチド23−2のアミノ酸配列を配列表の配列番号32に示す。
【0141】
[実施例24](本ペプチド21−1ないし21−3の固定化後の活性値評価)
本実施例では、本ペプチド21−1ないし21−3(配列番号26〜28)の固定化された状態における活性値を評価するため、精製後の変性本ペプチド21−1ないし21−3を液相でリフォールディングした後、これらのペプチドを親水性のポリスチレン表面に対して結合させ、抗原抗体反応の活性値を測定した。比較用として、原ペプチド21−1ないし21−3(配列番号23〜25)と、一本鎖抗体ではなく抗体分子である本ペプチド20及び原ペプチド20についても親水性のポリスチレン表面に対して結合させ、活性値を測定した。さらに、従来との比較用として、原ペプチド20(抗体分子)を疎水性のポリスチレン表面に対して結合させ、活性値を測定した。
【0142】
まず、変性剤である尿素が80mMとなるように精製後の変性本ペプチド21−1ないし21−3(配列番号26〜28)を375μM 酸化型グルタチオン(GSSG)を含むPBS中に希釈し、室温で3時間インキュベートしてリフォールディングを行った。遠心分離によって凝集体を除去した後HisTrap HP (GE HealthCare)を用いたNiキレートアフィニティクロマトグラフィ(IMAC)によって精製し、PBSで透析することで可溶性の3種類の本ペプチド21−1ないし21−3を生成した。
【0143】
次に、本ペプチド21−1ないし21−3を5μg/mlの濃度に0.1PBSTで希釈し、100μlを親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、2時間室温でインキュベートして、本ペプチドを固定化させた。0.1PBSTで6回洗浄した後、2%のBSAを含む0.1PBST(ブロッキング試薬)によってポリスチレンプレート表面の未結合部位をブロッキングした。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで1〜5000ng/mlに希釈されたbiotin-CRP (抗原)を100μl加え、室温で1時間インキュベートして、本ペプチドと抗原抗体反応させた。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで5000倍に希釈されたHRP標識ストレプトアビジンを100μl加え、室温で30分インキュベートして、0.1PBSTで6回洗浄した後に、発色基質を加えて30分間発色させた。30分後にマイクロプレートリーダー(TECAN製「SUNRISE Remote」)を用いて、405nmにおける吸光度を測定した。配列番号23〜25に示す原ペプチド21−1ないし21−3である一本鎖抗体、実施例20に記載した本アミノ酸配列2を標識した本ペプチド20、原ペプチド20についても同様の処理を行い吸光度を測定した。さらに、5μg/mlの原ペプチド20をPBS中で疎水性のポリスチレンプレート(ベクトン・ディッキンソン アンド カンパニー社製「BD Falconマイクロプレート#351172」)に固定化して吸光度を測定した。
【0144】
図13に実施例24の測定結果を示す。図13中、黒丸(●)、黒塗りの三角及び黒塗りの四角はそれぞれ配列番号26、27、28に示す本ペプチド21−1、21−2、21−3の結果であり、白丸(○)、白塗りの三角(△)及び白塗りの四角(□)はそれぞれ配列番号23、24、25に示す原ペプチド21−1、21−2、21−3の結果である。さらに、黒塗りの菱形(◆)は、本ペプチド20の結果であり、白塗りの菱形(◇)は、原ペプチド20であるマウスモノクローナル抗ヒトCRP抗体(オリエンタル酵母社製、#4715500)の結果である。×印は原ペプチド20の疎水性のポリスチレンプレートに固定化して実験を行った結果である。図11において、縦軸は、吸光度(405nm)であり、横軸はbiotin-CRPの濃度(ng/ml)である。
【0145】
図13から原ペプチド21−1、21−2、21−3(○、△、□)並びに原ペプチド20(◇)では、親水性のポリスチレン表面においては、ほとんど活性化されていないが、本ペプチド21−1、21−2、21−3(黒丸、黒塗り三角、黒塗りの四角)では、1本鎖抗体のアミノ酸配列の違いによらず何れも高い抗原結合活性が維持されていることが確認できる。また、本アミノ酸配列2を標識した本ペプチド20(◆)においても親水性ポリスチレンプレート上に選択的に固定化され高い抗原結合活性を示していることがわかる。従来ELISAに用いられてきた疎水性ポリスチレンプレート上に固定化されたマウスモノクローナル抗体(原ペプチド20)の結果(×)と比較しても本アミノ酸配列2を有する本ペプチド20並びに21−1、21−2及び21−3は同等もしくはそれ以上に高いシグナルが得られていることが分かる。以上の結果より、様々な1本鎖抗体のC末端側に本アミノ酸配列2を導入した本ペプチド21−1、21−2、21−3を生産することで親水性ポリスチレンプレートに抗原結合活性を高く維持した状態で固定化できることが確認できた。また、本アミノ酸配列2をモノクローナル抗体に標識することで親水性ポリスチレンプレートに抗原結合活性を高く維持した状態で固定化できることが確認できた。
【0146】
[実施例25](精製後の変性本ペプチド21−1,22−1及び23−1の固相リフォールディング及び活性値評価)
本実施例では、精製後の変性本ペプチド21−1,22−1及び23−1の固定化された状態におけるリフォールディング及び活活性値を評価するため、これらのペプチドを親水性のポリスチレン表面に対して結合させ、抗原抗体反応の活性値を測定した。
【0147】
まず、変性剤である尿素の終濃度が4Mとなるように、各精製後の変性本ペプチド21−1(配列番号26),22−1(配列番号29)及び23−1(配列番号31)を調製して試料を作成した。次に、各試料20μlを親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、ミキサーで撹拌後、室温で1時間インキュベートして、本ペプチドを固定化させた(1ウェルあたりのscFvの重量:0.5μg/ウェル)。0.1PBSTで6回洗浄した後、2%のBSAを含む0.1PBST(ブロッキング試薬)によってポリスチレンプレート表面の未結合部位をブロッキングした。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで1〜5000ng/mlに希釈されたbiotin-CRP (抗原)を100μl加え、室温で1時間インキュベートして、本ペプチドと抗原抗体反応させた。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで5000倍に希釈されたHRP標識ストレプトアビジンを100μl加え、室温で30分インキュベートして、0.1PBSTで6回洗浄した後に、発色基質を加えて30分間発色させた。30分後にマイクロプレートリーダー(TECAN製「SUNRISE Remote」)を用いて、405nmにおける吸光度を測定した。原ペプチド21−1(配列番号23)についても同様の処理を行い吸光度を測定した。
【0148】
図14に実施例25の測定結果を示す。図14中、黒丸(●)は精製後の変性本ペプチド21−1の結果であり、黒塗りの三角は精製後の変性本ペプチド22−1の結果であり、黒塗りの四角は精製後の変性本ペプチド23−1の結果であり、白丸(○)は原ペプチド21−1の結果である。図14において、縦軸は、吸光度(405nm)であり、横軸はbiotin-CRPの濃度(ng/ml)である。
【0149】
図14から、原ペプチド21−1である一本鎖抗体(○)では、ほとんど活性化されていないが、精製後の変性本ペプチド21−1,22−1及び23−1は何れも活性化されていることが確認できる。つまり、本アミノ酸配列を用いることにより、変性されている本ペプチド21−1,22−1及び23−1を固相に接触させることによって、固定化させると共に、変性を復元するリフォールディングも行われている。
【0150】
さらに、本アミノ酸配列2をC末端側に導入した本ペプチド21−1も、本アミノ酸配列2を重鎖可変部とリンカーペプチドとの間に導入した本ペプチド22−1も同程度の活性であるのに対し、本アミノ酸配列2をC末端側及び重鎖可変部とリンカーペプチドとの間の両方に導入した本ペプチド23−1ではより高い活性が得られることが確認できた。
【0151】
[実施例26](精製後の変性本ペプチド21−2,22−2及び23−2の固相リフォールディング及び活性値評価)
本実施例では、精製後の変性本ペプチド21−2,22−2及び23−2の固定化された状態におけるリフォールディング及び活活性値を評価するため、これらのペプチドについて、親水性のポリスチレン表面に対して結合させ、抗原抗体反応の活性値を測定した。なお、本実施例は、基本的な操作は実施例25と同様であるが、多少条件を変更しているため、以下念のため記載する。
【0152】
まず、変性剤である尿素の終濃度が4Mとなり、各精製後の変性本ペプチド21−2(配列番号27),22−2(配列番号30)及び23−2(配列番号32)が10μg/mlとなるように、1%のTween20を含むPBSによって溶液を調製して試料を作成した。次に、各試料100μlを親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、ミキサーで撹拌後、室温で3時間インキュベートして、本ペプチドを固定化させた。0.1PBSTで6回洗浄した後、2%のBSAを含む0.1PBST(ブロッキング試薬)を300μl加えて、室温で1時間インキュベートしてポリスチレンプレート表面の未結合部位をブロッキングした。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで1〜5000ng/mlに希釈されたbiotin-CRP (抗原)を100μl加え、室温で1時間インキュベートして、本ペプチドと抗原抗体反応させた。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで5000倍に希釈されたHRP標識ストレプトアビジンを100μl加え、室温で1時間インキュベートした。さらに、0.1PBSTで6回洗浄した後に、発色基質を100μl加えて30分間発色させた。30分後にマイクロプレートリーダー(TECAN製「SUNRISE Remote」)を用いて、405nmにおける吸光度を測定した。原ペプチド21−2(配列番号24)についても同様の処理を行い吸光度を測定した。
【0153】
図15に実施例26の測定結果を示す。図15中、黒丸(●)は精製後の変性本ペプチド21−2の結果であり、黒塗りの三角は精製後の変性本ペプチド22−2の結果であり、黒塗りの四角は精製後の変性本ペプチド23−2の結果であり、白丸(○)は原ペプチド21−2の結果である。図15において、縦軸は、吸光度(405nm)であり、横軸はbiotin-CRPの濃度(ng/ml)である。
【0154】
図15から、原ペプチド21−2である一本鎖抗体(○)では、ほとんど活性化されていないが、精製後の変性本ペプチド21−2,22−2及び23−2は何れも活性化されていることが確認できる。つまり、実施例25と同様に、本実施例においても、本アミノ酸配列を用いることにより、変性されている本ペプチド21−2,22−2及び23−2を固相に接触させることによって、固定化させると共に、変性を復元するリフォールディングも行われることが確認できた。
【0155】
さらに、本アミノ酸配列2をC末端側に導入した本ペプチド21−2の方が、本アミノ酸配列2を重鎖可変部とリンカーペプチドとの間に導入した本ペプチド22−2よりも活性が高いことが確認できるが、それ以上に、本アミノ酸配列2をC末端側及び重鎖可変部とリンカーペプチドとの間の両方に導入した本ペプチド23−2の方がより高い活性が得られることも確認できた。
【0156】
[実施例27](精製後の変性本ペプチド21−1の固相に対する結合性能及び活性値評価)
本実施例では、精製後の変性本ペプチド21−1(配列番号26)の量と親水性のポリスチレン表面に対する結合性能及び活性値との関係を評価した。
【0157】
まず、精製後の変性本ペプチド21−1を3.2μg/ml、32μg/ml及び320μg/mlの各濃度となるように8M尿素で調製して試料を作成した。次に、各10μlの試料をそれぞれ親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に加え、さらに各ウェルに0.1PBSTを90μl加えて10倍に希釈し、4℃で一晩インキュベートして、本ペプチド21−1を固定化させた。0.1PBSTで6回洗浄した後、2%のBSAを含む0.1PBST(ブロッキング試薬)によってポリスチレンプレート表面の未結合部位をブロッキングした。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで1〜5000ng/mlに希釈されたbiotin-CRP (抗原)を100μl加え、室温で1時間インキュベートして、本ペプチドと抗原抗体反応させた。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで1000倍に希釈されたHRP標識ストレプトアビジンを100μl加え、室温で30分インキュベートして、0.1PBSTで6回洗浄した後に、発色基質を加えて30分間発色させた。30分後にマイクロプレートリーダー(TECAN製「SUNRISE Remote」)を用いて、405nmにおける吸光度を測定した。
【0158】
図16に実施例27の測定結果を示す。図16中、黒丸(●)は精製後の変性本ペプチド21−1の量が1ウェルあたり0.032μgの場合、黒塗りの三角は0.32μgの場合、黒塗りの四角は3.2μgの場合の結果である。図16において、縦軸は、吸光度(405nm)であり、横軸はbiotin-CRPの濃度(ng/ml)である。
【0159】
図16から、精製後の変性本ペプチド21−1は、親水性のポリスチレン表面において活性化されていることが確認できる。つまり、本アミノ酸配列を用いることにより、親水性のポリスチレン基板に対して特異的な結合性能を示し、固定化させると共に、変性を復元するリフォールディングもできることが確認された。
【0160】
[実施例28](未精製の変性本ペプチド21−1ないし21−3の固定化による分離精製及び活性値評価)
本実施例では、未精製の変性本ペプチド21−1ないし21−3(配列番号26、27、28)を親水性のポリスチレン表面に固定化することによる分離精製の可能性及び固定化された状態の活性値を評価した。
【0161】
まず、未精製の変性本ペプチド21−1ないし21−3及び3.2mg/mlの精製後の変性本ペプチド21−1を0.1PBSTで10倍に希釈した試料及び100倍に希釈した試料を親水性のポリスチレンプレート(AGCテクノグラス株式会社製「IWAKIマイクロプレート#3860−096」)に200μl加え、4℃で一晩インキュベートして、本ペプチドを固定化させた。0.1PBSTで6回洗浄した後、2%のBSAを含む0.1PBST(ブロッキング試薬)によってポリスチレンプレート表面の未結合部位をブロッキングした。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで1〜5000ng/mlに希釈されたbiotin-CRP (抗原)を100μl加え、室温で1時間インキュベートして、本ペプチドと抗原抗体反応させた。さらに、0.1PBSTで6回洗浄した後、0.2%のBSAを含む0.1PBSTで1000倍に希釈されたHRP標識ストレプトアビジンを100μl加え、室温で30分インキュベートして、0.1PBSTで6回洗浄した後に、発色基質を加えて30分間発色させた。30分後にマイクロプレートリーダー(TECAN製「SUNRISE Remote」)を用いて、405nmにおける吸光度を測定した。
【0162】
図17(a)は10倍に希釈した場合の測定結果であり、図17(b)は100倍に希釈した場合の測定結果である。図17(a)及び(b)中、黒丸(●)は未精製の変性本ペプチド21−1の結果であり、黒塗りの三角は未精製の変性本ペプチド21−2の結果であり、黒塗りの四角は未精製の変性本ペプチド21−3の結果であり、白丸(○)は精製後の変性本ペプチド21−1の結果である。図17(a)及び(b)において、縦軸は、吸光度(405nm)であり、横軸はbiotin-CRPの濃度(ng/ml)である。
【0163】
図17から、未精製の状態であっても、親水性のポリスチレン表面に固定化され、活性化されることが確認できる。つまり、本アミノ酸配列を用いることにより、本ペプチドは親水性のポリスチレンプレートに対し特異的に結合するので、生産時における分離精製工程として利用可能であり、しかも活性化されるのでリフォールディングもされていることが確認できた。
【配列表フリ−テキスト】
【0164】
配列番号1:親水性固体表面に特異的な親和性を有するペプチド。
配列番号2:親水性固体表面に特異的な親和性を有するペプチド。
配列番号3:親水性固体表面に特異的な親和性を有するペプチド。
配列番号4:親水性固体表面に特異的な親和性を有するペプチド。
配列番号5:親水性固体表面に特異的な親和性を有するペプチド。
配列番号6:親水性固体表面に特異的な親和性を有するペプチド。
配列番号7:親水性固体表面に特異的な親和性を有するペプチド。
配列番号8:親水性固体表面に特異的な親和性を有するペプチド。
配列番号9:親水性固体表面に特異的な親和性を有するペプチド。
配列番号10:親水性固体表面に特異的な親和性を有するペプチド。
配列番号11:親水性固体表面に特異的な親和性を有するペプチド。
配列番号12:親水性固体表面に特異的な親和性を有するペプチド。
配列番号13:親水性固体表面に特異的な親和性を有するペプチド。
配列番号14:親水性固体表面に特異的な親和性を有するペプチド。
配列番号15:親水性固体表面に特異的な親和性を有するペプチド。
配列番号16:親水性固体表面に特異的な親和性を有するペプチド。
配列番号17:親水性固体表面に特異的な親和性を有するペプチド。
配列番号18:親水性固体表面に特異的な親和性を有するペプチド。
配列番号19:親水性固体表面に特異的な親和性を有するペプチド。
配列番号20:親水性固体表面に特異的な親和性を有するペプチド。
配列番号21:合成DNA。
配列番号22:合成DNA。
配列番号23:一本鎖抗体。
配列番号24:一本鎖抗体。
配列番号25:一本鎖抗体。
配列番号26:親水性固体表面に特異的な親和性を有する一本鎖抗体。
配列番号27:親水性固体表面に特異的な親和性を有する一本鎖抗体。
配列番号28:親水性固体表面に特異的な親和性を有する一本鎖抗体。
配列番号29:親水性固体表面に特異的な親和性を有する一本鎖抗体。
配列番号30:親水性固体表面に特異的な親和性を有する一本鎖抗体。
配列番号31:親水性固体表面に特異的な親和性を有する一本鎖抗体。
配列番号32:親水性固体表面に特異的な親和性を有する一本鎖抗体。
【特許請求の範囲】
【請求項1】
重鎖と軽鎖とを結合するリンカーペプチドを有する1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)又は定常部融合1本鎖抗体(scFv-Fc)であって、
前記リンカーペプチドと前記重鎖若しくは前記軽鎖との間、又は前記リンカーペプチド内に、固相に対して特異的な吸着機能を発現させるアミノ酸配列を有するペプチド。
【請求項2】
前記アミノ酸配列は、配列表における配列番号1乃至20に示すアミノ酸配列の何れか一つであることを特徴とする請求項1に記載のペプチド。
【請求項3】
前記アミノ酸配列は親水化処理された樹脂表面の固相に対して特異的な吸着機能を発現させることを特徴とする請求項1又は2に記載のペプチド。
【請求項4】
前記樹脂表面は親水性のポリスチレン表面であることを特徴とする請求項3に記載のペプチド。
【請求項5】
前記リンカーペプチドと前記重鎖との間及び前記リンカーペプチドと前記軽鎖との間の両方に、前記アミノ酸配列を有することを特徴とする請求項1乃至4の何れか1項に記載のペプチド。
【請求項6】
前記アミノ酸配列は免疫測定法における固相に固定化されることを特徴とする請求項1乃至5の何れか1項に記載のペプチド。
【請求項7】
前記免疫測定法はエンザイムイムノアッセイであることを特徴とする請求項6に記載のペプチド。
【請求項8】
固相表面に、請求項1乃至7の何れか1項に記載のペプチドが直接吸着していることを特徴とするペプチドが固定化された固相。
【請求項9】
重鎖と軽鎖とを結合するリンカーペプチドを有する1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)又は定常部融合1本鎖抗体(scFv-Fc)であって、前記リンカーペプチドと前記重鎖若しくは前記軽鎖との間、又は前記リンカーペプチド内に、配列表における配列番号1乃至20に示すアミノ酸配列を有するペプチドをコードする塩基配列を含む遺伝子、当該遺伝子に調製されたベクターまたは当該遺伝子が導入された宿主。
【請求項10】
前記アミノ酸配列は、配列表における配列番号1乃至20に示すアミノ酸配列の何れか一つであることを特徴とする請求項9に記載の遺伝子、ベクターまたは宿主。
【請求項11】
前記アミノ酸配列は親水化処理された樹脂表面の固相に対して特異的な吸着機能を発現させることを特徴とする請求項9又は10に記載の遺伝子、ベクターまたは宿主。
【請求項12】
前記樹脂表面は親水性のポリスチレン表面であることを特徴とする請求項11に記載の遺伝子、ベクターまたは宿主。
【請求項1】
重鎖と軽鎖とを結合するリンカーペプチドを有する1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)又は定常部融合1本鎖抗体(scFv-Fc)であって、
前記リンカーペプチドと前記重鎖若しくは前記軽鎖との間、又は前記リンカーペプチド内に、固相に対して特異的な吸着機能を発現させるアミノ酸配列を有するペプチド。
【請求項2】
前記アミノ酸配列は、配列表における配列番号1乃至20に示すアミノ酸配列の何れか一つであることを特徴とする請求項1に記載のペプチド。
【請求項3】
前記アミノ酸配列は親水化処理された樹脂表面の固相に対して特異的な吸着機能を発現させることを特徴とする請求項1又は2に記載のペプチド。
【請求項4】
前記樹脂表面は親水性のポリスチレン表面であることを特徴とする請求項3に記載のペプチド。
【請求項5】
前記リンカーペプチドと前記重鎖との間及び前記リンカーペプチドと前記軽鎖との間の両方に、前記アミノ酸配列を有することを特徴とする請求項1乃至4の何れか1項に記載のペプチド。
【請求項6】
前記アミノ酸配列は免疫測定法における固相に固定化されることを特徴とする請求項1乃至5の何れか1項に記載のペプチド。
【請求項7】
前記免疫測定法はエンザイムイムノアッセイであることを特徴とする請求項6に記載のペプチド。
【請求項8】
固相表面に、請求項1乃至7の何れか1項に記載のペプチドが直接吸着していることを特徴とするペプチドが固定化された固相。
【請求項9】
重鎖と軽鎖とを結合するリンカーペプチドを有する1本鎖抗体(scFv)、多価性1本鎖抗体(sc(Fv)n)又は定常部融合1本鎖抗体(scFv-Fc)であって、前記リンカーペプチドと前記重鎖若しくは前記軽鎖との間、又は前記リンカーペプチド内に、配列表における配列番号1乃至20に示すアミノ酸配列を有するペプチドをコードする塩基配列を含む遺伝子、当該遺伝子に調製されたベクターまたは当該遺伝子が導入された宿主。
【請求項10】
前記アミノ酸配列は、配列表における配列番号1乃至20に示すアミノ酸配列の何れか一つであることを特徴とする請求項9に記載の遺伝子、ベクターまたは宿主。
【請求項11】
前記アミノ酸配列は親水化処理された樹脂表面の固相に対して特異的な吸着機能を発現させることを特徴とする請求項9又は10に記載の遺伝子、ベクターまたは宿主。
【請求項12】
前記樹脂表面は親水性のポリスチレン表面であることを特徴とする請求項11に記載の遺伝子、ベクターまたは宿主。
【図1】


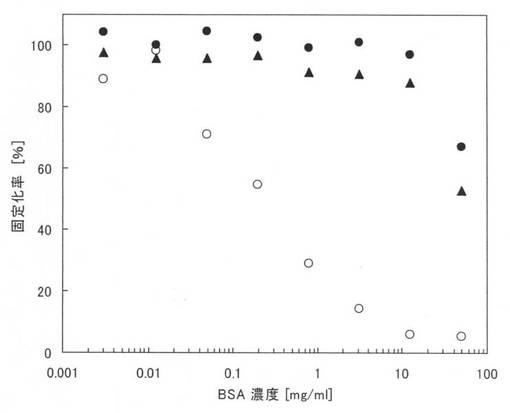
【図2】
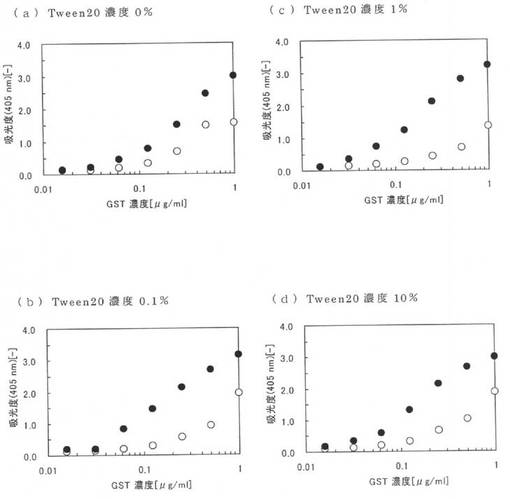
【図3】
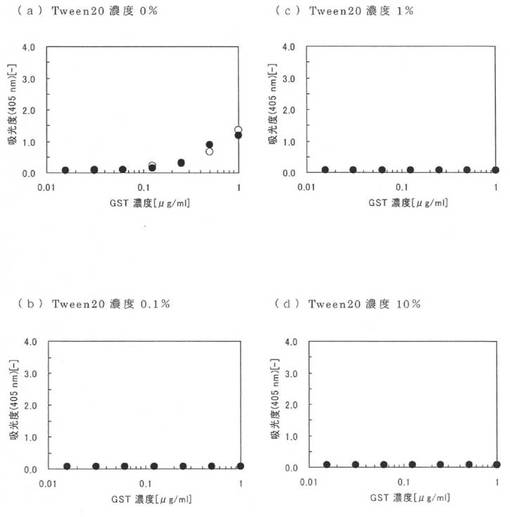
【図4】
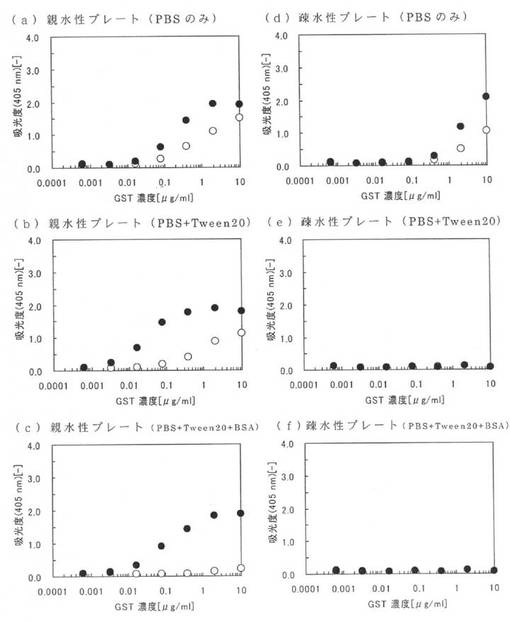
【図5】
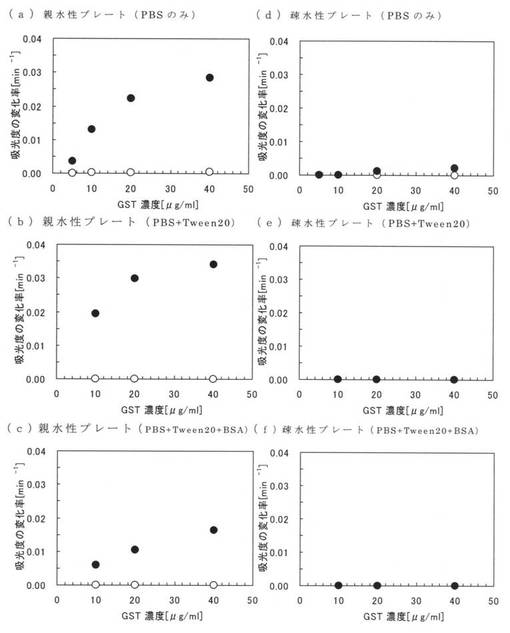
【図6】
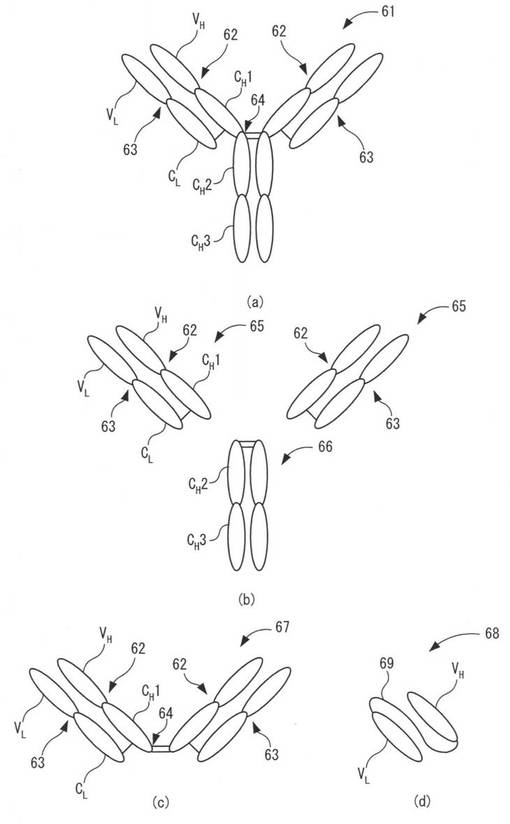
【図7】
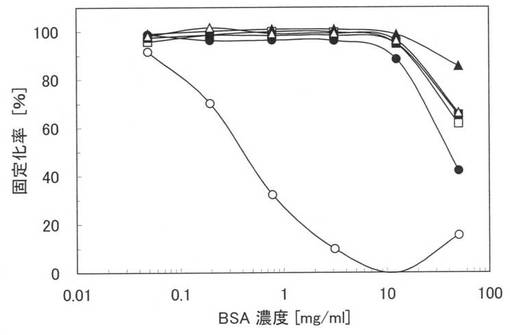
【図8】
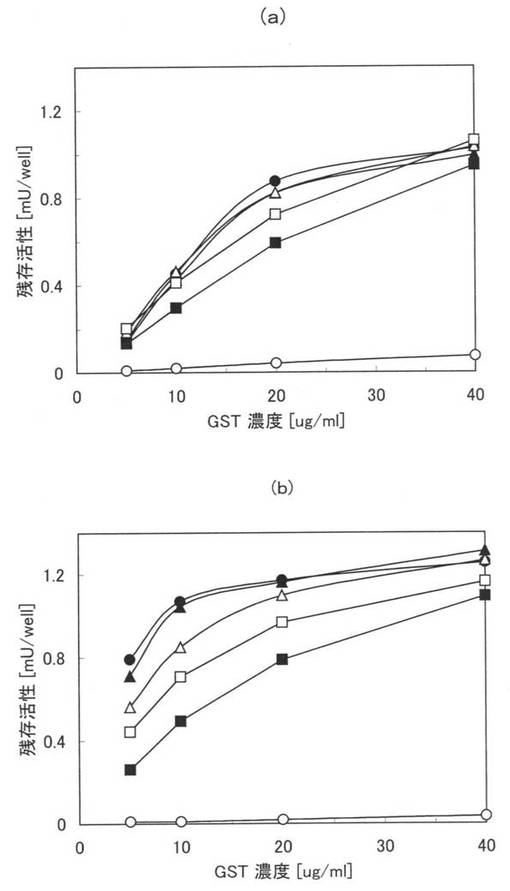
【図9】
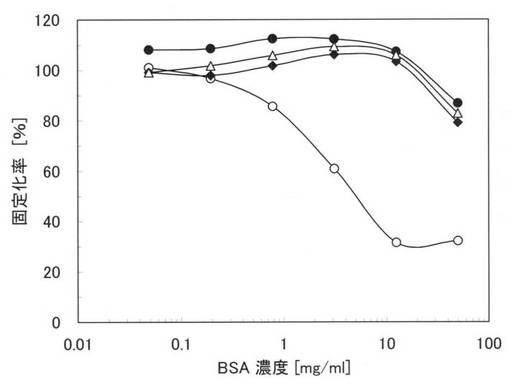
【図10】
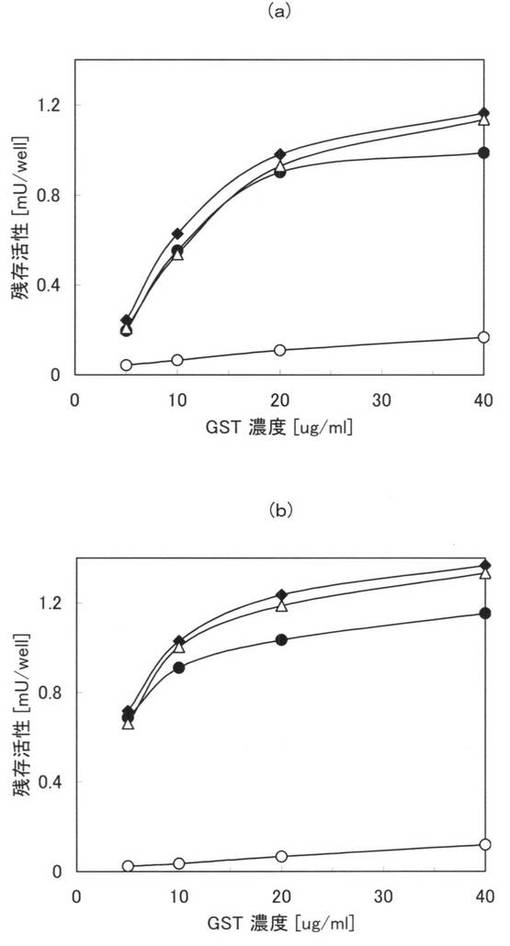
【図11】
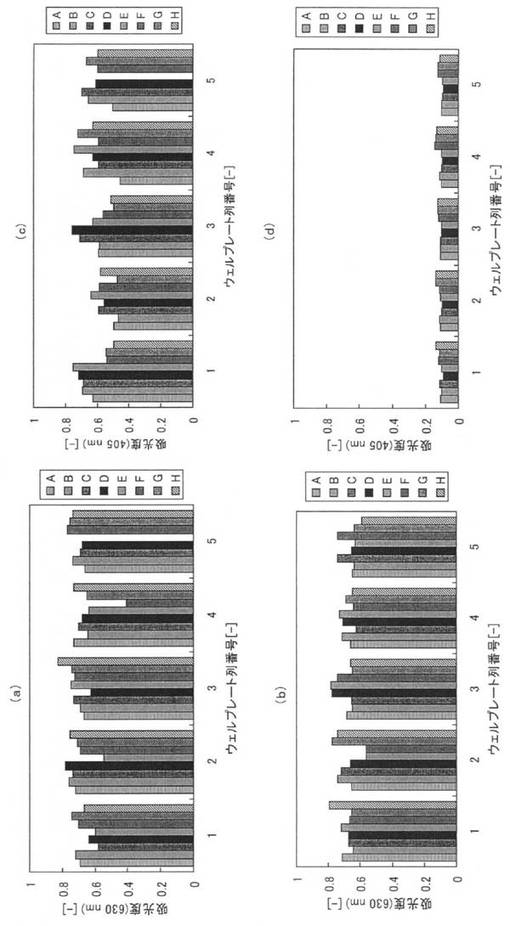
【図12】
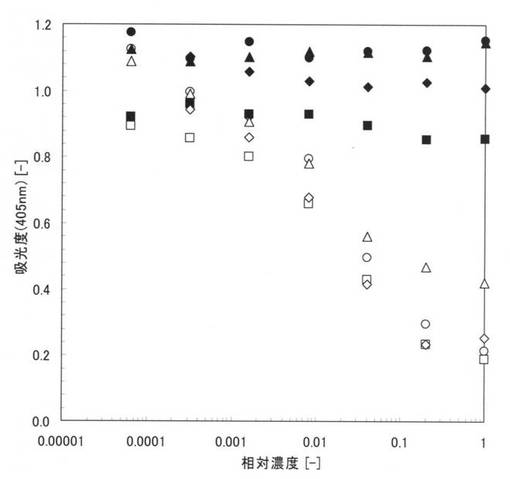
【図13】
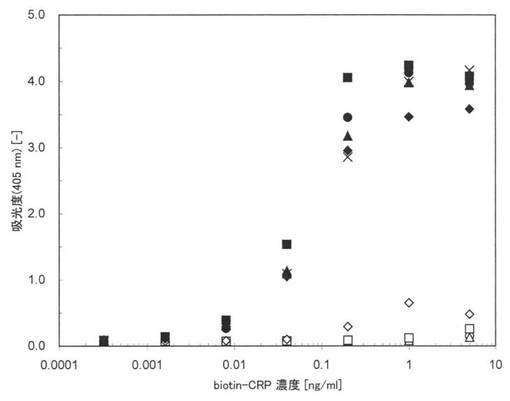
【図14】
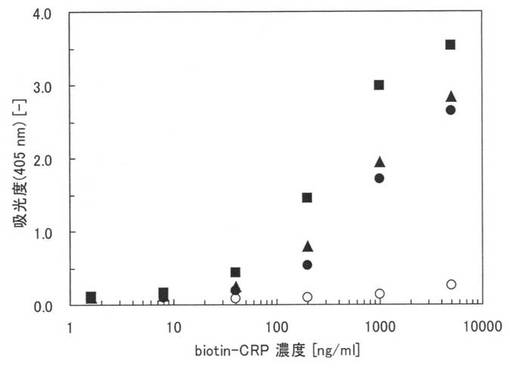
【図15】
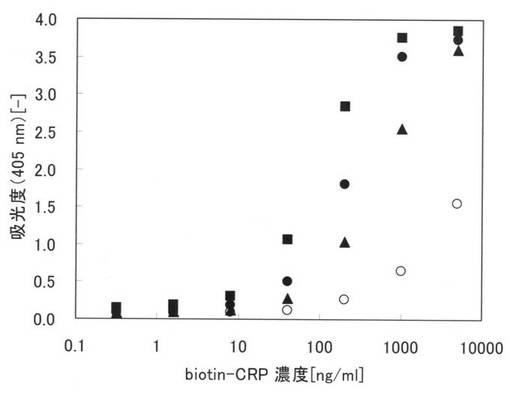
【図16】
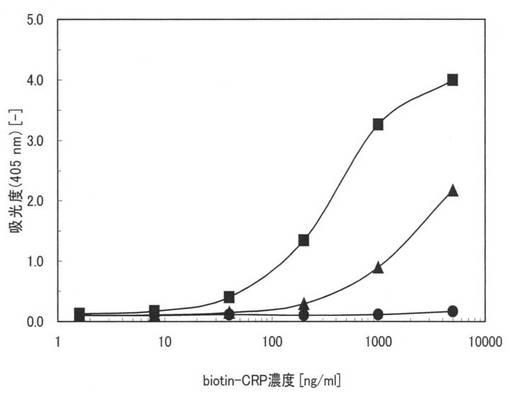
【図17】
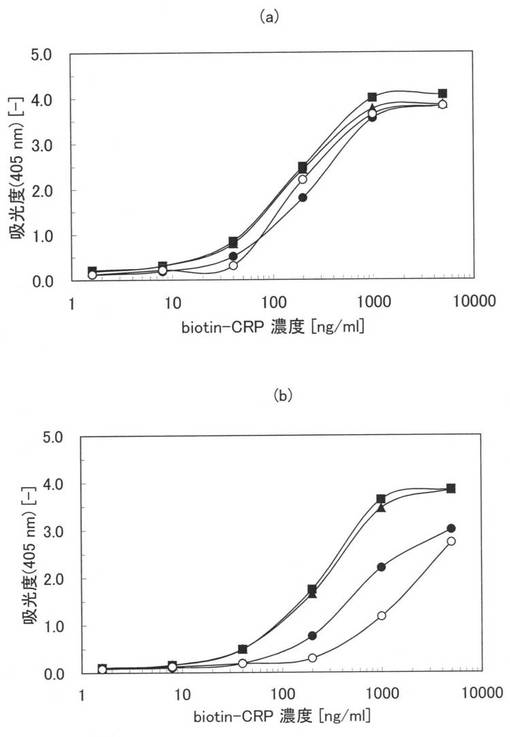
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公開番号】特開2012−250985(P2012−250985A)
【公開日】平成24年12月20日(2012.12.20)
【国際特許分類】
【出願番号】特願2012−177528(P2012−177528)
【出願日】平成24年8月9日(2012.8.9)
【分割の表示】特願2009−553366(P2009−553366)の分割
【原出願日】平成21年2月12日(2009.2.12)
【出願人】(504255685)国立大学法人京都工芸繊維大学 (203)
【出願人】(000208765)株式会社エンプラス (403)
【Fターム(参考)】
【公開日】平成24年12月20日(2012.12.20)
【国際特許分類】
【出願日】平成24年8月9日(2012.8.9)
【分割の表示】特願2009−553366(P2009−553366)の分割
【原出願日】平成21年2月12日(2009.2.12)
【出願人】(504255685)国立大学法人京都工芸繊維大学 (203)
【出願人】(000208765)株式会社エンプラス (403)
【Fターム(参考)】
[ Back to top ]