
可溶性内皮細胞プロテインC/活性化プロテインCレセプターを使用する診断アッセイ
【課題】本発明の目的は、天然に存在する可溶性EPCRについて治療的および診断的使用を同定することである。本発明のさらなる目的は、天然に存在する可溶性EPCRを特徴づけることである。
【解決手段】カルボキシル末端システイン残基が、別のアミノ酸と置換されているか、またはパルミトイル化されていない、改変された内皮プロテインCレセプター。グリコシル化されていない、改変された内皮プロテインCレセプター。単離された、選択的にスプライスされた内皮プロテインCレセプター。膜貫通ドメインの前の天然に存在するタンパク質分解性切断部位で切断された、単離された可溶性内皮プロテインCレセプター。
【解決手段】カルボキシル末端システイン残基が、別のアミノ酸と置換されているか、またはパルミトイル化されていない、改変された内皮プロテインCレセプター。グリコシル化されていない、改変された内皮プロテインCレセプター。単離された、選択的にスプライスされた内皮プロテインCレセプター。膜貫通ドメインの前の天然に存在するタンパク質分解性切断部位で切断された、単離された可溶性内皮プロテインCレセプター。
【発明の詳細な説明】
【背景技術】
【0001】
(発明の背景)
本発明は、一般に、内皮細胞プロテインC/活性化プロテインCレセプターまたはタン
パク質分解もしくは選択的スプライシングのいずれかによって誘導されるその可溶性形態の検出および/または測定を含むアッセイの領域にある。
【0002】
合衆国政府は、Charles T.Esmonに対する国立衛生研究所助成金PO1HL 54804により、本発明に一定の権利を有する。
【0003】
プロテインCのその活性セリンプロテアーゼに対する活性化、活性化プロテインC(APC)は、血液凝固の調節に重要な役割を果たす一連の事象を開始する。プロテインC経路の臨床的重要性は、血栓症を生じる、この経路における多くの機能不全によって示される(非特許文献1;非特許文献2)。プロテインCが欠損した患者は、通常、幼児期に生命を危うくする血栓症合併症を示す(非特許文献3;非特許文献4)。これは、プロテインC投与によって補正される(非特許文献5)。
【0004】
プロテインCおよびAPCはまた、炎症に対する宿主応答の調節に関連している。活性化プロテインC(APC)は、グラム陰性敗血症のヒヒモデルにおいてE.coliの致死的効果を予防し得(非特許文献6;TaylorおよびEsmonに対する特許文献1)、そして予備的な臨床結果は、プロテインCが、ヒト敗血症ショックのある形態を処置することに効果的であることを示唆する(非特許文献7)。プロテインC経路の重要な成分であるプロテインSの阻害は、E.coliの致死量未満のレベルに対する霊長類の応答を悪化させ、そして循環中へのTNFの出現を増大する(非特許文献8)。これらの結果は、プロテインCが、凝固の制御および炎症への影響の両方をなし得ることを示唆する。
【0005】
プロテインCは、凝固系の最終酵素であるトロンビンが、内皮細胞表面タンパク質であるトロンボモジュリンに結合する場合に、活性化される(非特許文献9;非特許文献10;非特許文献11)。細胞培養において、トロンボモジュリン転写は、腫瘍壊死因子(TNF)への内皮細胞の曝露によってブロックされ(非特許文献12そしてトロンボモジュリン活性および抗原は、その後インターナライズされそして分解される(非特許文献13、非特許文献14)。補体系の調節タンパク質であるC4bBPは、プロテインSと結合して、インビトロ(非特許文献15)およびインビボ(非特許文献8)でAPC抗凝固活性を支持することにおいて機能的に不活性である複合体を形成する。C4bBPは、急性期反応物質として行動する(非特許文献16)。したがって、この経路のタンパク質は、炎症を調節するようにみえるだけでなく、これらは炎症を調節する成分と相互作用し、そしてこれら自体は、炎症メディエータによってダウンレギュレーションを受ける。
【0006】
内皮細胞は、効率的なプロテインC活性化を担う公知の2つのレセプター、トロンボモジュリンおよび内皮プロテインC/APCレセプター(EPCR)を発現する点で、プロ
テインC経路において重要な役割を果たす(非特許文献17;非特許文献18)。トロンボモジュリン(CD141)は、高親和性を有する循環トロンビンと結合する膜貫通補因子であり、そして得られる酵素−補因子複合体は、生理学的に関連するプロテインC活性化速度に必要とされる(非特許文献19;非特許文献20)。
【0007】
EPCRは、CD1/MHCクラス1ファミリーとの著しい相同性を有する最近同定さ
れたレセプターである(非特許文献17;非特許文献21;非特許文献22)。プロテインCに対する内皮細胞レセプターのクローニングおよび生物学的役割は、Oklahoma Medical Research Foundationによって「Cloning and Regulation of an Endothelial Cell Protein C/Activated Protein C Receptor」と
題するPCT/US95/09636に記載された。このタンパク質は、238アミノ酸からなると予測され、このアミノ酸は、N末端に15アミノ酸のシグナル配列、およびタイプ1膜貫通タンパク質としてこのレセプターを特徴づける23アミノ酸の膜貫通領域を含む。
【0008】
EPCRは、カルシウムの存在下で類似の親和性(Kdapp約30nM)でプロテイン
CおよびAPCの両方と結合し(非特許文献21)、そして細胞表面上で、トロンビン−トロンボモジュリン活性化複合体に対してプロテインC基質を提示することにより、プロテインC活性化を容易にする(非特許文献18)。内皮細胞レセプターは両方とも、タイプ1膜貫通タンパク質であり、ここでリガンドは、細胞外ドメインに結合し、そして両方とも、短い細胞内細胞質テイルを有する(非特許文献21;非特許文献23;非特許文献24;非特許文献25)。さらに、それらのインビトロ細胞表面発現は、腫瘍壊死因子−αによって同様にダウンレギュレートされる(非特許文献17)。しかし、トロンボモジュリンおよびEPCRの可溶性形態の特徴は、いくつかの点で異なる。組換え可溶性トロンボモジュリンは、膜形態に対して減少した補因子活性を有する(非特許文献26;非特許文献27)。精製した成分および細胞の両方で、トロンボモジュリンによって特異的に誘導されるトロンビンの基質の変化は、トロンビンにおける共有される結合ドメインについての競合、ならびに活性部位ポケットにおけるコンホメーション変化から生じる(非特許文献28;非特許文献29;非特許文献30;非特許文献31;非特許文献32;非特許文献33;非特許文献34)。可溶性トロンボモジュリンもまた、種々のインヒビターによるトロンビンの不活化を加速する(非特許文献35;非特許文献36)。血漿および尿の両方が、検出可能なトロンボモジュリンを含み(非特許文献37;非特許文献38)、そしてトロンボモジュリン遺伝子がイントロンを含まないので(非特許文献23)、これらの可溶性形態は、細胞表面での細胞外ドメインのタンパク質分解による。
【0009】
血漿中のトロンボモジュリンの可溶性分解産物は、種々の疾患状態での内皮細胞損傷の公知のマーカーであり(非特許文献37;非特許文献39;非特許文献40;非特許文献41;非特許文献42;非特許文献43;非特許文献44)、そして種々の減少した親和性を有するトロンビン結合フラグメント、ならびに非結合フラグメントの混合物から構成される(非特許文献37)。
【0010】
対照的に、膜貫通ドメインの直前で短縮された組換え可溶性EPCR(rsEPCR)は、インタクトな細胞表面発現EPCRについて観察される親和性と同様な親和性でプロテインCおよびAPCの両方と結合する(非特許文献21)。おそらくrsEPCRおよび第Va因子の両方が、トロンボモジュリンによって占有されるトロンビン中のアニオン結合外部位(exosite)を暗示する溝に結合決定基を共有するので(非特許文献45)、APC抗凝固活性は、rsEPCRと結合した場合、効果的に阻害される(非特許文献22)。しかし、rsEPCRは、APCによる小さい合成基質のタンパク質分解にも、α1−アンチトリプシンまたはプロテインCインヒビターによるAPCの不活化にも影響を及ぼさないようである(非特許文献22)。プロテインC活性化を増強する膜結合EPCR(非特許文献18)とは異なり、rsEPCRは、可溶性トロンビン−トロンボモジュリン複合体によるプロテインC活性化に対してほとんど効果を有さない(非特許文献22)。このことは、EPCRの任意の可溶性形態が、プロテインCについて膜結合EPCRと競合することによってプロテインC活性化を阻害し得ることを示唆する。
【0011】
免疫組織化学は、EPCRが、主として大きな血管に由来する内皮細胞の表面上に存在し、そしてほとんどの毛細血管内皮細胞上にはないか、または低レベルでしか存在しないことを示す。
【特許文献1】米国特許第5,009,889号明細書
【非特許文献1】EsmonおよびSchwarz,Trends Cardiovasc.Med.,1995,5,p.141−148
【非特許文献2】Reitsmaら,Thromb.Haemost.,1995,73,p.876−879
【非特許文献3】Seligsohnら,N.Engl.J.Med.,1984,310,p.559−562
【非特許文献4】Esmon,Trends Cardiovasc.Med.,1992,2,p.214−220
【非特許文献5】Dreyfusら,N.Engl.J.Med.,1991,325,p.1565−1568
【非特許文献6】Taylorら,J.Clin.Invest.,1987,p.79
【非特許文献7】Gersonら,Pediatrics 91,1993,p.418−422
【非特許文献8】Taylorら,Blood,1991,78,p.357−363
【非特許文献9】Esmon,J.Biol.Chem.,1989,264,p.4743−4746
【非特許文献10】DittmanおよびMajerus,Blood,1990,75,p.329−336
【非特許文献11】Dittman,Trends Cardiovasc.Med.,1991,1,p.331−336
【非特許文献12】ConwayおよびRosenberg,Mol.Cell.Biol.,1988,8,p.5588−5592
【非特許文献13】Lentzら,Blood,1991,77,p.543−550
【非特許文献14】Mooreら,Blood,1989,73,p.159−165
【非特許文献15】Dahlback,J.Biol.Chem.,1986,261,p.12022−12027
【非特許文献16】Dahlback,Thromb.Haemostas.,1991,66,p.49−61
【非特許文献17】FukudomeおよびEsmon,J.Biol.Chem.,1994,269,p.26486−26491
【非特許文献18】Stearns−Kurosawaら,Proc.Natl.Acad.Sci.(USA),1996,93,p.10212−10216
【非特許文献19】EsmonおよびOwen,Proc.Natl.Acad.Sci.(USA),1981,78,p.2249−2252
【非特許文献20】Dittman,W.A.,Trends Cardiovasc.Med.,1991,1,p.331−336
【非特許文献21】Fukudomeら,J.Biol.Chem.,1996,271,p.17491−17498
【非特許文献22】Reganら,J.Biol.Chem.,1996,271,p.17499−17503
【非特許文献23】Jackmanら,Proc.Natl.Acad.Sci.(USA),1987,84,p.6425−6429
【非特許文献24】Wenら,Biochemistory,1987,26,p.4350−4357
【非特許文献25】Suzukiら,EMBO J.,1987,6,p.1891−1897
【非特許文献26】Galvinら,J.Biol.Chem.,1987,262,p.2199−2205
【非特許文献27】Parkinsonら,J.Biol.Chem.,1990,265,p.12602−12610
【非特許文献28】Yeら,J.Biol.Chem.,1991,266,p.23016−23021
【非特許文献29】Luら,J.Biol.Chem.,1989,264,p.12956−12962
【非特許文献30】Yeら,J.Biol.Chem.,1992,267,p.11023−11028
【非特許文献31】Hofsteengeら,Biochem.J.,1986,237,p.243−251
【非特許文献32】Mathews,Biochemistry,1994,33,p.13547−13552
【非特許文献33】Esmonら,J.Biol.Chem.,1982,257,p.7944−7947
【非特許文献34】Sadlerら,Haemostasis,1993,23,p.183−193
【非特許文献35】BourinおよびLindahl,Biochem.J.,1993,289,p.313−330
【非特許文献36】Rezaie,J.Biol.Chem.,1995,270,p.25336−25339
【非特許文献37】Takanoら,Blood,1990,76,p.2024−2029
【非特許文献38】IshiiおよびMajerus,J.Clin.Invest.,1985,76,p.2178−2181
【非特許文献39】Tanakaら,Clin.Chem.,1991,37,p.269−272
【非特許文献40】Takahashiら,Am.J.Hematol.,1991,38,p.174−177
【非特許文献41】Asakuraら,Am.J.Hematol.,1991,38,p.281−287
【非特許文献42】Wadaら,Am.J.Hematol.,1992,39,p.20−24
【非特許文献43】Takahashiら,Am.J.Hematol.,1992,41:32−39
【非特許文献44】Ohdamaら,Chest,1994,106,p.666−671
【非特許文献45】Matherら,EMBO J.,1996,15,p.6822−6831
【発明の開示】
【発明が解決しようとする課題】
【0012】
したがって、本発明の目的は、天然に存在する可溶性EPCRについて治療的および診断的使用を同定することである。
【0013】
本発明のさらなる目的は、天然に存在する可溶性EPCRを特徴づけることである。
【課題を解決するための手段】
【0014】
(発明の要旨)
血漿EPCRは、単離され、特徴づけられ、そして細胞性プロテインC活性化およびAPC抗凝固活性をブロックすることが示されている。血漿EPCRは、約43,000ダ
ルトンであるようであり、そして約100ng/mlで循環する(98.4±27.8ng/ml、n=22)。血漿EPCRを、イオン交換、イムノアフィニティー、およびプロテインCアフィニティークロマトグラフィーを使用して、ヒトのクエン酸処理血漿から精製した。フローサイトメトリー実験は、血漿EPCRが、膜貫通および細胞質ドメインを含まないEPCRとして定義される組換え短縮型EPCRから予め決定された親和性(Kdapp約30nM)と同様の親和性で、活性化プロテインCと結合することを示した。さら
に、血漿EPCRは、内皮細胞株におけるプロテインC活性化および1段階第Xa因子凝血アッセイにおけるAPC抗凝固活性の両方を阻害した。可溶性EPCRはまた、ヒト尿において検出されている。EPCRをコードする遺伝子のクローニングは、少なくともヒトEPCRが選択的にスプライスされ得、選択的にスプライスされた形態(sEPCR)に特有のインサートを含む短縮型可溶性EPCRを生じることを示す。これらの結果は、血漿EPCRが、細胞表面でのタンパク質分解または選択的スプライシングのいずれかによって誘導され得ることを示す。
【0015】
血漿EPCRの局所濃度が、特に疾患状態において、十分に高い場合、そのデータは、短縮型可溶性血漿EPCRが、プロテインC活性化の膜結合EPCR増大および活性化プロテインCの抗凝固機能を減弱させ得ることを示す。自己免疫疾患(全身性エリテマトーデス、SLE)および敗血症(炎症および凝固異常の両方を含む障害)の患者からのEPCRのレベルと正常EPCRを比較する実施例によって示されるように、可溶性EPCRのレベルは、炎症および異常凝固に関連する疾患状態と相関するようである。凝固、炎症、および大血管疾患を含む疾患状態を示す、可溶性EPCRの測定に基づくアッセイが記載される。単離精製された可溶性EPCR、組換え短縮型可溶性EPCR、および可溶性EPCRに対する抗体を含む、アッセイ試薬が記載される。
【0016】
本発明はさらに、以下を提供する。
(1)可溶性内皮プロテインCレセプターについてのアッセイであって、
テストされる患者からサンプルを得る工程、および
可溶性内皮プロテインCレセプターの量を測定する工程、
を包含する、アッセイ。
(2)上記サンプルが、尿、血漿、血清、組織サンプル、および間質液からなる群より選択される、項目1に記載のアッセイ。
(3)可溶性内皮プロテインCレセプターの量を較正標準と相関させる工程をさらに包含する、項目1に記載のアッセイ。
(4)上記患者が、凝固および炎症状態および障害、内皮に対する損傷に関連する障害または疾患、ならびに大血管疾患からなる群より選択される障害または疾患を有する、項目1に記載のアッセイ。
(5)上記障害または疾患が、自己免疫疾患、移植、敗血症、ショック、子癇前症、糖尿病、血管疾患、腎臓疾患、および肝臓疾患からなる群より選択される、項目4に記載のアッセイ。
(6)上記血管疾患が、心肺バイパス、不安定性アンギナ、再狭窄、および血管形成からなる群より選択される、項目5に記載のアッセイ。
(7)大血管に対する損傷を検出するための、項目1に記載のアッセイ。
(8)内皮プロテインCレセプターの検出および測定のためのキットであって、
内皮プロテインCレセプターと免疫反応性である抗体、
患者からのサンプル中で上記抗体と内皮プロテインCレセプターとの間の反応を検出するための試薬、および
上記反応量を、内皮プロテインCレセプターの正常および異常レベルと相関させるための標準
を含む、キット。
(9)上記抗体が、膜貫通ドメインを含まない内皮プロテインCレセプターについてより、膜貫通ドメインを含む内皮プロテインCレセプターについて、より高い親和性を有する、項目8に記載のキット。
(10)上記抗体が、選択的にスプライスされた内皮プロテインCレセプターにおけるインサートと免疫反応性である、項目8に記載のキット。
(11)上記抗体が、内皮プロテインCレセプターと、活性化プロテインCまたはプロテインCとの結合をブロックする、項目8に記載のキット。
(12)カルボキシル末端システイン残基が、別のアミノ酸と置換されているか、またはパルミトイル化されていない、改変された内皮プロテインCレセプター。
(13)グリコシル化されていない、改変された内皮プロテインCレセプター。
(14)単離された、選択的にスプライスされた内皮プロテインCレセプター。
(15)膜貫通ドメインの前の天然に存在するタンパク質分解性切断部位で切断された、単離された可溶性内皮プロテインCレセプター。
【発明の効果】
【0017】
自己免疫疾患(全身性エリテマトーデス、SLE)および敗血症(炎症および凝固異常の両方を含む障害)の患者からのEPCRのレベルと正常EPCRを比較する実施例によって示されるように、可溶性EPCRのレベルは、炎症および異常凝固に関連する疾患状態と相関するようである。凝固、炎症、および大血管疾患を含む疾患状態を示す、可溶性EPCRの測定に基づくアッセイが提供される。単離精製された可溶性EPCR、組換え短縮型可溶性EPCR、および可溶性EPCRに対する抗体を含む、アッセイ試薬が提供される。これらは従来技術からは予測不可能であった格別の効果である。
【発明を実施するための最良の形態】
【0018】
(発明の詳細な説明)
(定義)
内皮プロテインCレセプター、EPCR。
【0019】
可溶性、溶液中にあり、そして細胞表面に結合していない。
【0020】
短縮型、膜貫通ドメインおよび細胞質ドメインを含まない;タンパク質分解または選択的スプライシングのいずれかの結果であり得る。
【0021】
(可溶性EPCRの検出および特徴付け;生理学的役割およびマーカーとしての有用性)
EPCRの機能のこれまでの研究は、膜形態のEPCRに結合するプロテインCが、細胞表面上のトロンビン−トロンボモジュリン複合体によりプロテインC活性化の促進を生じる(Stearns−Kurosawaら,1996)が、可溶性組換えEPCRはAPC抗凝固活性を阻害する(Reganら,1996)ことを見出した。これらの観察は、血漿中の可溶性トロンボモジュリン分解産物が、種々の疾患状態における内皮損傷のマーカーであるという知識と共に、可溶性循環形態のEPCRが存在するかどうか、そして存在するのであれば、プロテインC経路においてどういう役割を有するかという疑問に至った。
【0022】
実施例は、可溶性形態のEPCRが血漿中で循環し、そして尿中に存在することを示す
。健常ドナー集団において、血漿EPCRレベルは、約100ng/mlであり、そして
約43,000ダルトンの単一抗原種であるようであった。血漿からの可溶性EPCRの
その後の精製および機能的研究は、血漿EPCRが、インタクトな膜結合EPCRと同様な親和性で、プロテインCおよびAPCの両方と結合し得ることを決定した。内皮細胞株を使用するインビトロ研究は、血漿EPCRが、生理学的濃度に近い濃度のプロテインCおよびトロンビンで、プロテインCを阻害することを示した。さらに、血漿への精製血漿EPCRの直接添加は、rsEPCRに対するモノクローナル抗体で逆転されるAPC抗凝固活性の阻害を生じた。
【0023】
この精製血漿タンパク質のEPCRとの同定は、rsEPCRの特性との比較に基づいた。これらのタンパク質は両方とも、同じ群のモノクローナル抗体およびポリクローナル抗体と反応し、同じアミノ末端配列を有し、Ca2+依存的様式で固定されたプロテインCに結合し、そして類似の用量応答曲線でプロテインC活性化およびAPC抗凝固活性をブロックした。さらに、rsEPCRおよび血漿EPCRに対するプロテインCおよびAPCの親和性は共に、インタクトな膜結合EPCRの親和性に類似する。これらの特性は、EPCRに特有であるようである。
【0024】
これまでの研究は、内皮細胞で発現した膜結合EPCRが、3〜5倍の間のファクターでプロテインC活性化を増大することを示したが、実施例は、血漿から精製された可溶性形態のEPCRが、内皮細胞でのプロテインC活性化およびAPC抗凝固活性を阻害することを示す。これは、EPCRが、いくつかの方法で、プロテインC経路を調節し得ることを予測する。第1に、より大きな血管(ここでは、トロンボモジュリン濃度が微小循環に対して低い)において、EPCR発現は、対応して増加する(Laszikら、Circulation 1997)。免疫組織化学は、ほとんどの器官において、EPCR発現は、大血管で最も高密度であり、そして血管サイズが減少するにつれて次第に減少し、最も豊富な内皮細胞タイプである毛細管内皮ではほとんどまたは全く発現しない。EPCR発現は、循環からプロテインC基質を捕獲し、そして活性化のためにトロンビン−トロンボモジュリン複合体に提示することにおいて、重要な役割を果たし得る。このことは、EA.hy926内皮細胞株およびヒト臍静脈内皮細胞の両方ともが、トロンボモジュリ
ンより、少なくとも6倍多い表面発現EPCR抗原を有するというインビトロ観察によって支持される。トロンボモジュリン濃度が高くそしてEPCRが低い微小循環では、プロテインC活性化にほとんど影響がないと予測される。最後に、循環可溶性EPCRは、APCの生成およびAPCが第Va因子を不活化する能力を減少させ得る。
【0025】
健常個体において、可溶性EPCRレベルは、約2.5nMであり、この濃度は、Kdapp(約30nM)および循環にける80nMのプロテインC濃度の両方よりも十分に低い。可溶性血漿EPCRの効果(APC抗凝固活性およびプロテインC活性化の阻害)の両方ともが、正常血漿に存在する濃度よりかなり高い濃度を必要とする。このことが、血漿EPCRの生理学的役割の疑問を、不確定のままにしている。40nMを越える可溶性EPCRレベルを有する患者が、実施例3(狼瘡)に記載のように同定された。したがって、内皮細胞表面に近い局所濃度が、全身濃度を超えるならば、可溶性EPCR濃度は、APCの生成および活性の両方を減弱するレベルに達し、血栓症の危険性に寄与する。
【0026】
可溶性形態のレセプターは、膜結合レセプターのタンパク質分解的切断、または選択的スプライシングメカニズムによって産生され得る。膜表面でのタンパク質分解は、可溶性トロンボモジュリン、ならびにTNF、IL−1、IL−2、M−CSF、PDGF、およびNGFのレセプターを遊離する(Heaneyら,1996,Blood 87:847−857)。可溶性レセプターは、膜レセプターのアンタゴニストとして作用すること、リガンドを安定化すること、リガンド媒介されたシグナリングを開始すること、膜形態のダウンモジュレーション、およびレセプターインヒビターへ結合してレセプター−リ
ガンド活性を間接的に容易にすることを含む、多くの可能性のある機能を有する。後者のメカニズムは、両方のIL−1レセプターの可溶性イソ形態が、タンパク質分解的切断によって生成され、そしてIL−1αおよびIL−1βへの応答性を厳格に調節する、IL−1レセプター系によって使用される(Arendら,1994,J.Immunol.
153:4766−4774)。以下に議論するように、膜貫通ドメインの直前で短縮された可溶性タンパク質をコードするEPCRゲノム構造は、選択的スプライシング部位を含む(FukudomeおよびEsmon,1995,J.Biol.Chem. 270:5571−5577)。可溶性IL−6レセプターは、タンパク質分解メカニズムおよび選択的スプライシングメカニズムの両方によって生成されるようである(Mullbergら,1994,J.Immunol. 152:4958−4968;Lustら,1992,Cytokine 4:96−100;Horiuchiら,1994,Eur.J.Immunol. 24:1945−1948)。この切断部位はまた、MorrisseyおよびEsmonへの米国特許第5,298,599号(その教示は本明細書に援用される)に記載のように、短縮型EPCR、続いて直ぐペプチド配列(抗体はこの配列に対して特異的に指向する)をコードする発現ベクターを構築することによって、大量の可溶性EPCRを回収することにおいて有用であり得る。次いで、エピトープは、患者への投与の前または後に、タンパク質分解によって切断される。Hoppらへの米国特許第4,782,137号もまた参照。
【0027】
免疫組織化学研究は、EPCRが、主として大血管の内皮に位置し、そして毛細血管ではかろうじて検出可能であることを示した。したがって、膜結合EPCRに由来する血漿EPCRは、大血管疾患プロセスのマーカーとして用い得る。血漿EPCRは、血漿トロンボモジュリンレベルとの有用な比較として用い得る。血漿トロンボモジュリンレベルは、種々の疾患状態において調整されることが示されているが、大血管および小血管の両方の疾患プロセスを反映し、ほとんどの内皮が微小血管性であるのでおそらく小血管の寄与に支配される。
【0028】
(EPCRのヌクレオチドおよび予測タンパク質構造分析)
EPCRのcDNAは、238アミノ酸(配列番号2)のタンパク質をコードすると予測され、これは、N末端に15アミノ酸のシグナル配列を含む(vonHeijne,(1986)Nucleic Acids Res. 14,4683−4690)。したがって、成熟タンパク質は、223アミノ酸を含むと予測される。組換えタンパク質の直接配列決定は、成熟タンパク質がSer18で始まることを示した。配列番号2は、EPCRの予測アミノ酸配列である。配列番号2のアミノ酸1〜15(MLTTLLPILLLSGWA)は、von Heijne(von Heijne,1986)の方法によって決定される推定シグナル配列である。配列番号2のアミノ酸211〜236(LVLGVLVGGFIIAGVAVGIFLCTGGR)は、推定膜貫通ドメインである。可能性のあるN−グリコシル化部位は、配列番号2のアミノ酸47〜49、64〜66、136〜138、および172〜174に存在する。細胞外システイン残基は、配列番号2のアミノ酸17(血漿EPCRでは除去されている)、114、118、および186に存在する。23アミノ酸からなる可能性のある膜貫通領域(Engelmanら,(1986)Annu.Rev.Biophys.Chem. 15,321−53)を、C末端で同定した(配列番号2のアミノ酸211で始まる)。
【0029】
このタンパク質はタイプ1膜貫通タンパク質である。細胞外ドメインは、4つの可能性のあるN−グリコシル化部位および3つのCys残基を含む。グリコシル化は、N−グリカナーゼ消化によって示されるように、活性に必須ではない。細胞質領域は、3アミノ酸のみを含み、そしてCysで終結し、パルミトイル化されている。末端システインが適切にパルミトイル化されてない場合、タンパク質は分泌され得る。EPCRの配列を変化させて、このシステインを別のアミノ酸と置換することにより、膜結合する代わりに分泌さ
れる、本質的に完全長のEPCRを作成する手段が提供される。
【0030】
本明細書で使用される場合、レセプターをコードするヌクレオチド配列は、配列番号1に示される配列、およびストリンジェントな条件下で配列番号1にハイブリダイズする、その保存的置換、付加、または欠失を有する配列を含む。本明細書で使用される場合、レセプターを構成するアミノ酸配列は、配列番号2に示される配列、および機能的に等価な生物学的活性を有するレセプターを形成する、その保存的置換、付加、または欠失を有する配列を含む。何が、保存的置換、付加、または欠失を構成し、そしてどれが、本明細書に記載の機能的アッセイを使用して機能的に等価なレセプター分子をコードまたは形成すると容易に確認され得るかは、当業者に周知である。これは、さらに、図3を参照することによって例示され、以下に議論される。
【0031】
(選択的スプライシング)
レセプターは、特異的リガンドに結合して生理学的応答を生じることを担う細胞の、外部に曝された部分を有する、細胞膜に繋がれたタンパク質として最も頻繁に思い描かれる。多くの場合、もはや細胞に制限されないという事実にもかかわらず、頻繁にそのリガンドを確かに結合し得る、可溶性形態のレセプターが存在する。可溶性レセプターイソ形態へのリガンド結合はまた、多くの形態をとる応答を生じ得、これには、膜結合レセプター相互作用のアップモジュレーションまたはダウンモジュレーション、あるいは正常には応答性ではない細胞へリガンドを輸送することによって応答を伝達させることを含む(Heaney,MLおよびDW Golde. Soluble cytokine receptors. Blood 87:847−857, 1996)。
【0032】
可溶性レセプターを産生するための2つの公知のメカニズムが存在する:膜結合レセプターをタンパク質分解して、細胞外ドメインを遊離し、そして膜アンカーを残すことによるもの、およびmRNAの選択的スプライシングによるもの(図1)。後者のメカニズムは多くの形態を取り得るが、リーディングフレームがエキソン−イントロン境界により連続し、そして膜貫通アンカーをコードする配列に達する前に停止コドンで終結する場合、最も単純である。これは、膜形態に類似するが、重要な差異を有するタンパク質を生成する。これは、可溶性タンパク質として、作製され、そして分泌され、そして特有のカルボキシル末端テイルを有する。このテイルを、膜結合レセプターの形成において無視されるイントロンmRNA配列の一部を読むことによって形成した。選択的スプライシングによる可溶性レセプターの生成はまた、共に同じmRNAテンプレートを起源にするという事実にもかかわらず、膜結合レセプターとは独立して調整され得る(Heaneyら,Proc.Natl.Acad.Sci.U.S.A. 92:2365−2369,1995)。
【0033】
可溶性レセプターが選択的スプライシングメカニズムによって生成されることを証明するためには、ゲノム配列および関連領域のイントロン−エキソン境界を知らなければならない。生理学的応答と可溶性レセプターを関連付けて、プロテインC遺伝子およびプロテインS遺伝子の発現の間にかなり頻繁に含まれることが見られる異常なmRNAスプライシングと区別することもまた役立つ(Bergら,Blood Coag Fibrinol. 7:625−631,1996)。
【0034】
以下の研究および結果の詳細は、実施例に記載される。ヒト血漿は、約100ng/m
lの可溶性EPCRを含む(表1)。これを、2つのモノクローナル抗体(1494mAbおよび1495mAb)および標準的技法を使用する酵素結合イムノアッセイ(ELISA)によって測定した。著しく上昇した可溶性EPCRレベルを、全身性エリテマトーデスおよび敗血症の患者で見いだした。これらのレベルは、ほとんど例外なく、大血管の表面上のみに存在する膜結合レセプターについてかなり高いようであった。これを真相正
しく認識するために、トロンボモジュリン(TM)は、すべての内皮、ならびにいくつかの非血管細胞上で発現されるが、正常可溶性TMレベルは、わずかに約10〜40ng/
mlである(Takanoら,Blood 76:2024−2029,1990)。可溶性TMレベルは、狼瘡の患者で上昇していたが、敗血症では上昇していなかった。重要なことには、これらの患者群において、血漿EPCRとTMレベルとの間に相関がなかったことである(それぞれ、r2=0.028および0.034)。
【0035】
【表1】

【0036】
TMゲノム構造は、イントロンを含まず(Jackmanら,Proc.Natl.Acad.Sci.(USA)84:6425−6429,1987)、そのため可溶性TMイソ形態を生成するための唯一の方法は、膜結合レセプターのタンパク質分解による。好中球エラスターゼおよびカテプシンGによる内皮TMのタンパク質分解は、インビトロで示されており、これは、種々の疾患状態で見出される上昇した可溶性TMレベルが、内皮表面で、活性化された炎症性細胞の産物によって媒介されるタンパク質分解から生じることを示唆する(Boehmeら,Immunology 87:134−140,1996、およびAbeら,J.Lab.Clin.Med. 123:874−881,1994)。
【0037】
血漿EPCRおよびTMレベルと高い血漿EPCR濃度との間の相関の欠如は、血漿EPCRがタンパク質分解および選択的スプライシングの両方のメカニズムを起源にするという概念に一致する。ヒトEPCRのゲノム構造は、イントロンによって分離される、4つのエキソンを含む。この配列を概観することにより、7527位にTAA停止コドンを含むエキソンIII−イントロンIII境界(5'GTで)の後のインフレームリーディ
ング配列が明らかになる。この停止コドンは、膜貫通ドメインをコードするエキソンIVの上流にあるので、予測されるタンパク質は、特有な48残基のカルボキシル末端テイル(イントロン配列によってコードされる)を含み、そして膜貫通アンカーを含まない。
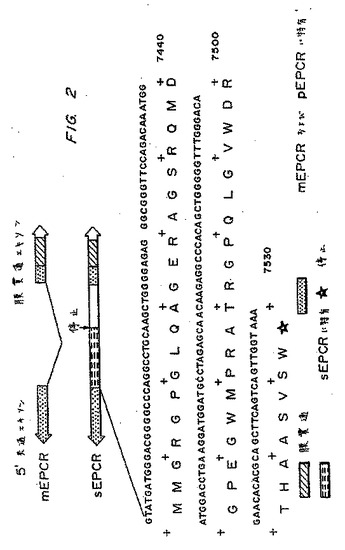
【0038】
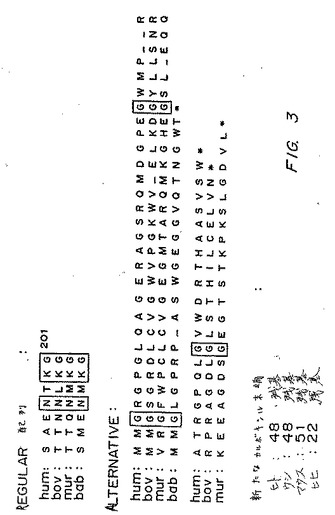
図1は、短縮型EPCRが誘導され得る2つの可能性のある方法の図である:膜貫通ドメインの直前でのタンパク質分解によるか、または選択的スプライシングによる。図2に示されるように、選択的スプライシングは、選択的にスプライスされた短縮型EPCR中にあるペプチド配列が含まれることを生じる。図3に示されるように、この配列は、種間で高度に保存されるが、わずかな差が存在し、ヒトおよびウシのEPCRについては48残基、マウスEPCRについては51残基、そしてヒヒEPCRについては22残基の新しいカルボキシル末端テイルを生じる。
【0039】
(レセプタータンパク質の発現についての患者サンプルのスクリーニング)
患者サンプルは、EPCR、EPCR中の選択的にスプライスされたインサートに存在する特有のインサートのいずれかに対する抗体、またはコンホメーションの差に起因してEPCRまたはsEPCRのいずれかに対してより大きな親和性で結合する抗体を使用して、sEPCRまたはEPCRの存在および量について、スクリーニングされ得る。サンプルはまた、存在するタンパク質を特異的に定量するための他の標準的技法を使用して、スクリーニングされ得る。
【0040】
(診断的または治療的使用のための抗体の生成)
EPCR、および特に、可溶性EPCR(「sEPCR」)、および組換え可溶性EPCR(「rsEPCR」)に対する抗体が生成され得、これらは、レセプタータンパク質の検出、特徴付け、または単離に有用であり、そしてほとんどの場合、リガンド結合の阻害によって、レセプタータンパク質活性を調節するために有用である。抗体は、免疫原として、ヒトまたは動物の精製または組換えレセプタータンパク質もしくはそのフラグメントを使用して、標準的技法によって生成される。
【0041】
EPCRに対するモノクローナル抗体を、Esmonら,1993,Methods Enzymol. 222:359−385によって他のタンパク質について記載されたように得た。1494、1495、および1496mAbと呼ばれる抗体は、組換え可溶性EPCRおよび細胞表面発現EPCRに結合するIgG1κ抗体である。1494および1496mAbは、EPCRへのプロテインCおよびAPCの結合をブロックし、そして細胞性EPCRがトロンビン−トロンボモジュリン複合体によるプロテインC活性化を容易にする能力を阻害する。1495mAbは、EPCRへのリガンド結合をブロックせず、細胞表面プロテインCを変化させず、そして1494または1486mAbのエピトープとは異なる結合エピトープを有する。抗体は、標準的技法、例えば、放射標識、酵素標識、フルオレセインのような蛍光標識、金粒子、染料、および抗体の検出のための他の手段を使用して標識され得る。例えば、抗体は、標準的な手順を使用して、ビオチンアミドカプロン酸N−ヒドロキシスクシンイミドエステルでビオチン化され得る。抗体は、イムノアッセイにおける使用のための固体支持体、例えば、AffiGel−10TM、ニトロセルロース、またはマイクロタイターウェル、あるいは溶液相イムノアッセイにおける使用のための固体支持体に固定され得る。
【0042】
好ましい実施態様では、EPCRは、50マイクロリットルの、15mM Na2CO3、35mM NaHCO3(pH9.6)中4マイクログラム/mlの1495mAbで4
℃にて一晩コーティングしたマイクロタイタープレート(MaxisorpTM,NUNC
NS,Roskilde,Denmark)を使用して測定される。室温で、次いでプレートを20mM Tris−HCl、0.1M NaCl、0.05% Tween 20(pH7.5)(アッセイ緩衝液)で3回洗浄し、そして0.1%(wt/vol)ゼラ
チンを含むアッセイ緩衝液で少なくとも1時間ブロックする。次いで、ウェルを洗浄し、50マイクロリットルサンプルを3連のウェルに添加し、そしてプレートを1時間インキュベートする。ウェルを吸引し、アッセイ緩衝液で3回洗浄し、そして50マイクロリットル、2マイクログラム/mlのビオチン−1494mAbを添加する。プレートを1時
間インキュベートし、3回洗浄し、そして50マイクロリットルの、0.25マイクログ
ラム/mlのストレプトアビジン−アルカリホスファターゼ結合体(GIBCO BRL
)を添加し、そしてさらに1時間インキュベートする。ウェルを5回洗浄し、そしてELISA増幅キット(GIBCO BRL)の基質および増幅試薬を、製造業者の指示に従って15分間隔で連続して添加する。発色を0.3M H2SO4で停止し、そして終点吸
光度を、Vmaxマイクロプレートリーダーにて490mmで読み取る。3連のウェルでの
標準は、20mM Tris−HCl、0.1M NaCl、および1mM EDTA、
0.1%ゼラチン(pH7.5)中の1.5〜100ng rsEPCR/mlである。標
準曲線は、1.5〜12.5ng/mlで直線であり、そしてサンプルを、直線範囲に入る
ように同じ緩衝液で希釈する。研究は、1〜2パーセントの間で、血漿が、アッセイの直線性にも標準曲線の感度にも影響を及ぼさないことを示す。健常ボランティアからの血漿サンプルを、1mM EDTAを含むアッセイ緩衝液で最終的に2%血漿まで希釈した。そしてEPCR抗原レベルを、同じプレート上で決定された標準曲線を参照して、3連ウェルの平均から算出する。
【0043】
(障害)
可溶性EPCRについてのアッセイは、本明細書で議論されるような凝固および炎症状態および障害、例えば、狼瘡のような自己免疫疾患、移植モニタリング、敗血症、ショック、子癇前症、糖尿病、心肺バイパス、不安定性アンギナ、再狭窄、血管形成(すなわち、血管疾患)、腎臓または肝臓疾患の検出および分析に有用である。例えば、EPCRは、大血管についてのマーカー、したがって大血管に対する損傷についてのマーカーである。可溶性EPCRの量の増加は、大血管傷害を示し、EPCRのタンパク質分解またはsEPCR合成の刺激のいずれかを生じる。EPCR対トロンボモジュリンの比はまた、血液または尿のいずれかのサンプルに基づいて決定され得、これは微小血管対大血管の相対的な程度を示す。サイトカイン、白血球活性化マーカー、および補体因子または活性化マーカーに対するEPCRの相対量もまた、疾患状態を示すために使用され得る。
【0044】
EPCRは内皮細胞上に存在するので、内皮細胞損傷のマーカーとして有用である。これは、薬物の効果(毒性および効力の両方)の指標として使用され得る。例えば、狼瘡患者では、炎症性/凝固媒介性大血管傷害を効果的に最小にする薬物は、EPCRレベルの
減少を生じる。
【0045】
本発明は、以下の非限定的な実施例を参照することによってさらに理解される。
【実施例】
【0046】
(実施例1:ヒト血漿における機能的内皮プロテインCレセプターの同定)
以下の略号を使用する:rsEPCR、膜貫通ドメインおよびサイトゾルテイルの代わりに挿入されたHPC4エピトープを有する組換え可溶性EPCR;mAb、モノクローナル抗体;SDS−PAGE、ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動。
【0047】
(方法)
材料。以下の試薬を示された供給業者から購入した:
ブタ腸粘膜ヘパリン、ジイソプロピルフルオロリン酸、ビオチンアミドカプロン酸N−ヒドロキシスクシンイミドエステル、ウシ血清アルブミン、Sigma(St.Lois,MO);Spectrozyme PCa、American
Diagnostica(Greenwich,CT);ELISA増幅キット、GibcoBRL(Gaithersburg,MD);AffiGel−10、BioRad(Hercules,CA);ハンクス平衡化塩溶液、3−(N−モルホリン)プロパン硫酸(MOPS)、Fisher Scientific(Fair Lawn,NJ)。すべての他の試薬は、市販の最高品質なものであった。
【0048】
タンパク質。ヒトプロテインC(Esmonら,1993,Methods Enzymol. 222:359−385)、ウシトロンビン(Owenら,1974,J.BIol.Chem. 249:594−605)、およびウシアンチトロンビン(Esmon 1977「Factors regulating the inhibition of thrombin by antithrombin III. In Chemistry and Biology of Thrombin」R.L.Lund
blad,J.W.Fenton,II,およびK.G.Mann編 Ann Arbor Science,Ann Arbor.403−411)を、記載のように精製した。組換え可溶性EPCR、rsEPCRは、膜貫通ドメインの直前の残基210で短縮されたEPCRの細胞外ドメイン、続くHPC4モノクローナル抗体におけるカルシウム依存的イムノアフィニティー精製を可能にする12残基配列からなる(Takahashiら,1992;Stearnsら,1988,J.Biol.Chem. 263:826−832)。rsEPCRの構築、精製、およびプロテインC/APC結合特性(Fu
kudomeら,1996)。ヤギ免疫前血清およびrsEPCRに対するヤギポリクローナル抗血清を調製し、そしてIgGを精製した(Fukudomeら,1996)。ヤギ抗rsEPCRポリクローナル抗体を、標準的手順を使用して、ビオチンアミドカプロン酸N−ヒドロキシスクシンイミドエステルでビオチン化した。
【0049】
モノクローナル抗体。rsEPCRに対するモノクローナル抗体(mAb)を、他のタンパク質について記載されたように得た(Esmonら,1993)。1494、1495、および1496mAbは、rsEPCRおよび細胞表面発現EPCRに結合するIgG1κ抗体である。1494および1496mAbは、EPCRへのプロテインCおよびAPCの結合をブロックし、そして細胞性EPCRがトロンビン−トロンボモジュリン複合体によるプロテインC活性化を容易にする能力を阻害する(Stearns−Kurosawaら,1996)。1495mAbは、EPCRへのリガンド結合をブロックせず、細胞表面プロテインC活性化を変化させず、そして1494または1496mAbのエピトープとは異なる結合エピトープを有する。1494および1495mAbを、標準的手順を使用して、ビオチンアミドカプロン酸N−ヒドロキシスクシンイミドエステルでビオチン化した。1494mAbを、血漿EPCRのイムノアフィニティー精製のために、製造業者の指示に従って、AffiGel−10にカップリングした。抗EPCR mAbのスクリーニングを、Stearns−Kurosawaら(1996);Fukudomeら(1996)に記載の方法を使用して行った。
【0050】
凝血アッセイ。1段階第Xa因子凝血アッセイにおけるAPC(25nM)抗凝固活性に対するrsEPCRまたは精製血漿EPCRの効果を、83μg/mlの1496mA
b(APC−EPCR相互作用をブロックする抗体)の存在または非存在下で行った(Reganら,1996)。可溶性EPCRおよび1496mAbを、アッセイ前15分間プレインキュベートした。
【0051】
細胞培養。すべてのヒト細胞株を、以前に記載されたように維持した(Fukudomeら,1996)。EA.hy926細胞(形質転換されたヒト内皮細胞株;Edgel
lら,1983,Proc.Natl.Acad.Sci.(USA)80:3734−3737)は、Cora−Jean Edgell(University of North Carolina at Chapel Hill)が好意により提供した。
【0052】
フローサイトメトリー分析。蛍光プローブとして用いるために、APCを、記載されるように(FukudomeおよびEsmon、1994;Bock,P.E.1988.Biochemistry 27:6633−6639)、フルオレセインで活性部位(fl−APC)において標識した。EA.hy926細胞へのAPC結合に対するrsE
PCRまたは血漿EPCRの効果を、フローサイトメトリーによって研究した(Fukudomeら、1996)。手短には、収集された細胞を、漸増する濃度のいずれかの可溶性EPCR調製物の非存在下または存在下において、60nMのfl−APCとともに、氷上で30分間インキュベートし、洗浄し、そして細胞に結合した蛍光を、フローサイトメトリーによって1サンプルごとに10,000事象を計数して測定した。全てのアッセ
イを、1%のウシ血清アルブミン、3mM CaCl2、0.6mM MgCl2、および
0.02%アジ化ナトリウムを補充したハンクス平衡化塩溶液において行った。
【0053】
細胞表面プロテインC活性化。EA.hy926細胞を96ウェル組織培養ディッシュ
において培養した(Stearns−Kurosawaら、1996)。コンフルエントな単層を、1%(w/v)のウシ血清アルブミン、3mM CaCl2、0.6mM Mg
Cl2、および0.02%アジ化ナトリウムを補充したハンクス平衡化塩溶液を用いて、3回洗浄した。全てのアッセイを、室温において、同一の緩衝液中で、60μlの最終容量にて行った。そして、全てのタンパク質濃度は、アッセイにおける最終濃度を示す。プロテインC(0.1μM)を、示された濃度のrsEPCR、血漿EPCR、または149
4mAbの非存在下または存在下で添加し、細胞とともに15分間プレインキュベートした。トロンビンを混合液に添加して(2nM)、活性化反応を開始した。示された時間に、50μlのアリコートを取り出し、そして96ウェルマイクロタイタープレート中の10μlのアンチトロンビン(0.7μM最終)およびヘパリン(5U/ml最終)に添加した。APCアミド分解活性をSpectrozyme PCa基質(0.2mM)の添加
によって測定し、405nmでの吸光度の変化速度(mOD/分)をVmaxキネティッ
クマイクロプレートリーダー(Molecular Devices,Menlo Park,CA)で測定した。すべてのアッセイ点は、3連ウェルにおいて行われ、そして十分に活性化されたプロテインC対mOD/分の標準曲線を参照することにより決定される
場合、プロテインC基質の10%未満が活性化された。
【0054】
血漿および血清の収集。健常な成人ボランティア(女性12人および男性10人)から、3.8%緩衝化クエン酸溶液中かまたは抗凝固剤を含まないチューブ中に、静脈穿刺に
よって全血を収集した(Vacutainer tube;Becton Dickinson,Franklin Lakes,NJ)。年齢、食餌、または他の変数に関するドナーのスクリーニングは試みなかった。全てのボランティアに研究についての情報を開示し、彼らから書面による同意を得た。血液を、1160×gで10分間遠心分離した。血漿および血清を等分し、アッセイまで−80℃で冷凍保存した。
【0055】
血漿EPCRの定量のためのELISA。血漿におけるEPCR抗原の検出のための酵素結合免疫吸着アッセイを開発した。マイクロタイタープレート(Maxisorp;Nunc,Roskilde,Denmark)を、50μlの、15mM Na2CO3、35mM NaHCO3(pH9.6)中4μg/mlの1495mAbを用いて、4℃にて一晩コートした。以下の工程を室温で行った。ウェルを、20mM Tris−HCl、0.1M NaCl、0.05% Tween20(pH7.5)(アッセイ緩衝液)で
3回洗浄し、0.1%(w/v)ゼラチンを含有するアッセイ緩衝液で少なくとも1時間ブロックした。ウェルを洗浄し、50μlのサンプルを3連のウェルに添加し、そしてプレートを1時間インキュベートした。ウェルを吸引して、アッセイ緩衝液で3回洗浄し、50μlの2μg/mlビオチン−1494mAbを添加した。プレートを1時間インキュ
ベートし、3回洗浄し、50μlの0.25μg/mlストレプトアビジン−アルカリホスファターゼ結合体(GibcoBRL)を添加し、さらに1時間インキュベートした。ウェルを5回洗浄し、ELISA増幅キット(GibcoBRL)の基質および増幅試薬を、製造業者の指示に従い、15分の間隔で連続的に添加した。発色を0.3M H2SO4
を用いて停止し、490nmの終点吸光度をVmaxマイクロプレートリーダーで読み取った。各プレートは、20mM Tris−HCl、0.1M NaCl、1mM ED
TA、0.1%ゼラチン(pH7.5)中1.5〜100ng/mlのrsEPCRの標準を3連のウェルに含んだ。標準曲線は、1.5〜12.5ng/mlで直線であり(r=0.99)、そして血漿サンプルを直線範囲に入るように同じ緩衝液で希釈した。予備的な実験により、再終濃度1〜2%のヒト血漿は、標準曲線の直線性にも感度にも影響しないことを実証した。健常ボランティア由来の血漿サンプルを、1mMEDTAを含有するアッセイ緩衝液で、最終的に2%血漿に希釈し、そしてEPCR抗原レベルを、同一プレート上で決定された標準曲線を参照して、3連ウェルの平均から算出した。
【0056】
コーティング抗体と検出抗体を逆にし(1494mAbをコーティングに;ビオチン−1495mAbを検出に)、そして抗体結合を、Blue Phos基質(KPL Laboratories;Gaithersburg,MD)で検出する、別のアッセイを開発した。この方法を使用して、敗血症患者における血漿EPCRをアッセイした。このアッセイは、おそらく親和性の差異のために、より高感度であったが、両アッセイとも、定性的には、同様の結果を与えた。
【0057】
ウエスタンブロット。血漿または血清サンプルのドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動(SDS−PAGE)を、10%アクリルアミドゲルを、Laemmli緩衝液系(Nature 227:680−685)と共に用いて、非還元条件下で標準的な手順を用いて行った。ゲルをポリビニリデン膜(PVDF;Millipore,Bedford,MA)に転写し、膜をブロックし、次に免疫前ヤギIgG(50μg/ml)か、またはヤギ抗rsEPCRポリクローナルIgG(50μg/ml)のいずれかと30分間インキュベートした。洗浄後、膜を1:20,000希釈のマウス抗ヤギI
gG−西洋わさびペルオキシダーゼ結合体(Pierce,Rockford,IL)と30分間インキュベートした。膜を洗浄し、結合した抗体−酵素結合体を増強化学発光基質(Pierce)で製造業者の指示に従って検出した。
【0058】
免疫吸着。健常ボランティアからの血清またはクエン酸処理血漿サンプル(400μl)を、AffiGel−10に結合させた50μlの1495mAb(5mg IgG/
ml樹脂)とともに、混合しながら4℃にて一晩インキュベートした。サンプルを遠心分離し、上清を除去し、そして樹脂を1mlの20mM
Tris−HCl、0.1M NaCl、0.02%アジ化ナトリウム(pH7.5)で
3回洗浄した。最終20mM ジチオスレイトールを含有するSDS−PAGEサンプル緩衝液を、洗浄した樹脂に添加し、サンプルを3分間沸騰し、そしてSDS−PAGEおよびウエスタンブロッティングのために処理した。膜を4μg/mlのビオチン化ヤギ抗
rsEPCRポリクローナル抗体でプローブし、そして結合抗体をストレプトアビジン−西洋わさびペルオキシダーゼ結合体(Pierce)および増強化学発光検出系で検出した。予備的実験により、室温にて1〜4時間の、100μlのTris不活化AffiGel−10樹脂でのサンプルの事前吸着、続く1495mAb−AffiGel−10での一晩の免疫吸着が、同一のウエスタンブロットの結果を与えることが決定された。
【0059】
血漿EPCRの精製。血漿EPCRを、イオン交換クロマトグラフィー、抗rsEPCR mAbイムノアフィニティークロマトグラフィー、およびプロテインC親和性カラムでのクロマトグラフィーの組み合わせを用いて、ヒトクエン酸処理血漿(Oklahoma Blood Institute)から精製した。わずかに異なる方法で2つの調製を行った。
【0060】
第1の調製において、血漿(1L)を等量の20mM Tris−HCl(pH7.5
)、10mM ベンズアミド、400単位のヘパリンナトリウムで希釈し、そして1gの膨潤前のQAE樹脂でバッチ吸着した。沈澱後、樹脂をプロテインCの精製のために処理した(Esmonら、1993)。固体硫酸アンモニウムを上清に4℃にて40%飽和まで添加し、遠心分離し、そしてさらなる硫酸アンモニウムをその上清に70%飽和に達するまで添加した。遠心分離後、柔らかなペレットを透析バッグに入れ、12Lの20mM
Tris−HCl、0.02%アジ化ナトリウム(pH7.4)に対し一晩透析した。透析物を、20mM Tris−HCl、0.1M NaCl、0.02%アジ化ナトリウム(pH7.4)で平衡化した、1496mAb−AffiGel−10イムノアフィニテ
ィーカラム(6ml樹脂;5mg IgG/ml樹脂)にかけた。カラムを、12mlを
超える同じ緩衝液で洗浄し、20mM Tris−HCl(pH7.4)中の50%(v/
v)エチレングリコールで溶出した(Jun Xu、未発表の観察)。溶出物からのピーク画分をプールし(0.37の全OD280)、濃縮し(Centriprep30、Millipore)、そして緩衝液を20mM Tris−HCl、0.1M NaCl、3
mM CaCl2、0.6mMMgCl2、0.02%アジ化ナトリウム(pH7.4)に交
換した。この物質をプロテインCアフィニティーカラムにかけた。このカラムは、同じ緩衝液においてHPC4−AffiGel−10カラム(5mg IgG/ml樹脂;0.9×8cm)に精製プロテインC(3mg)をかけることにより事前に調製した。HPC4mAbは、カルシウム依存的様式でプロテインC活性化領域に結合し(Esmonら、1993;Stearnsら、1988)、その後のプロテインCへのEPCRの結合を妨げない。血漿EPCRを含有するサンプルをカラムにかけた後、カラムを約12mlの緩衝液で洗浄し、20mM Tris−HCl、0.1M NaCl、5mM EDTA、
10mM MOPS、0.02%アジ化ナトリウム(pH7.5)で溶出した。画分を、280nmでの吸光度について、そして上記のELISAを用いてEPCR抗原についてモニターした。プロテインCおよび血漿EPCRの両方を含有する溶出物を、FPLC(Pharmacia−LKB、Uppsala、Sweden)MonoQカラムにかけ、カラムを20mM Tris−HCl(pH7.5)中の0.1〜1M NaClの直線勾配で展開した。約半分の血漿EPCRは、MonoQカラムに結合せず、半分は、約0.
2M NaClに溶出し、そしてプロテインCは、約0.5M NaClに溶出した。血
漿EPCRの両方のイオン種は、還元条件下または非還元条件下での銀染色、クマシーBB染色、またはPVDF膜への転写後の金染色(Pierce)を用いるSDS−PAGE、およびビオチン−ポリクローナル抗rsEPCR抗体プローブでのウエスタンブロットで、同一のようであった。
【0061】
血漿EPCRの第2の調製は、4Lの血漿を用いて開始し、機能研究に十分なタンパク
質を精製した。この場合、1496−AffiGel−10樹脂(20mlの5mg IgG/ml樹脂)を、10mMベンズアミジン、1mMジイソプロピルフルオロホスフェ
ートおよび0.5単位/mlのヘパリンナトリウム(最終濃度)と共に、クエン酸処理血漿に、直接添加した。血漿を、穏やかに混合しながら、4℃にて一晩バッチ吸着した。樹脂の沈澱後、上清をプロテインC精製のために処理した(Esmonら、1993)。樹脂を2.5×30cmのカラムに充填し、20mM Tris−HCl、0.1M NaCl、0.02%アジ化ナトリウム(pH7.4)で、十分に洗浄し、そして20mM Tr
is−HCl(pH7.4)中50%のエチレングリコールで溶出した。溶出物をプール
し、濃縮し(5.5の全OD280)、MonoQカラムにかけ、2つのイオン種(素通り、および0.2M NaCl溶出ピーク)を、1496−AffiGel−10樹脂(1.5×11cm)に再びかけた。カラムを前のとおりに、50%エチレングリコールで溶出した。溶出物(0.71 OD)を濃縮し、緩衝液を、Cetriprep30を含む、2
0mM Tris−HCl、0.1MNaCl、3mM CaCl2、0.6mM MgC
l2、0.02%アジ化ナトリウムに交換した。次に、この物質をアフィニティーカラムにかけた。ここで、プロテインC(2.9mg)を、同じ緩衝液中で、HPC2−Affi
Gel−10カラム(0.6×17cm)に最初にかけた。HPC2mAbは、プロテイ
ンCのセリンプロテアーゼドメインに結合し、EPCR結合を妨げない(Fukudomeら、1996)。結合したEPCRを、5mM EDTAを含有する緩衝液で溶出した。混入している血清アミロイドP(プロテインCサンプル由来)を、FPLC MonoQカラムでのイオン交換クロマトグラフィーによって除去した。サンプルを0.2M N
aCl中でカラムにかけ、その結果、血漿EPCRは結合せず、そして0.4〜0.5M NaClにおいて溶出する混入物から分離された。得られる精製血漿EPCR(0.19
3 OD280)は、銀染色を用いるSDS−PAGE、およびポリクローナル抗rsEP
CRを用いるウエスタンブロットにより、均質のようであった。この物質を機能研究およびアミノ末端配列分析に使用した。
【0062】
タンパク質配列決定。可溶性血漿EPCRのアミノ末端配列分析を、Molecular Biology Research Facility、W.K.Warren Medical Research Institute(Oklahoma City)のKenneth Jackson博士の研究室において行った。アミノ酸を標準の1文字コードによって表す。
【0063】
(結果)
第1のアプローチとして、3人の健常ボランティア由来の血漿および血清サンプルを希釈し(4%v/v)、非還元条件下で10%SDS−PAGEゲルで泳動し、そしてrs
EPCRに対して惹起したヤギポリクローナル抗体を用いるウエスタンブロットのために処理した。健常ボランティア由来の血漿および血清サンプル(4%v/v)を非還元条件
下の10%ゲルでのSDS−PAGEのために処理し、膜に転写し、そして膜をヤギ抗rsEPCRポリクローナル抗体でプローブした。結果をrsEPCR(0.2ng)と比
較した。結合した抗体を、マウス抗ヤギIgGおよび増強化学発光検出系で検出した。2人の健常ボランティア由来の血漿サンプルを1495 AffiGel−10樹脂で免疫吸着した。洗浄した樹脂を溶出させ、そして還元条件下でのSDS−PAGEのために処理した。ウエスタンブロッティングを、ビオチン−ヤギ抗rsEPCRをプローブとして使用して行った。
【0064】
血漿EPCRの純度を、銀染色したSDS−PAGE 10%ゲル、およびビオチン−ヤギ抗rsEPCRでプローブした膜のウエスタンブロットから決定した(還元および非還元)。膜をポリクローナル抗体でプローブした後、血清および血漿サンプルの両方で、約43,000Daの単一のバンドが現れる。検出されたタンパク質のサイズは、rsE
PCRよりわずかに大きいようである。検出された他のバンドは、免疫前IgGおよびより長い暴露時間でのプロービングより判断すると、IgGのバックグラウンド結合であった。AffiGel−10樹脂と結合した抗EPCR 1495mAbと共に血漿サンプルを一晩インキュベートすること、続く洗浄および還元条件下での結合抗原の溶出は、ビオチン−ヤギ抗rsEPCRポリクローナル抗体でのウエスタンブロットにより検出される単一バンドを生じた。
【0065】
mAb 1495をコーティング抗体として使用するELISAによる、健常ボランティア由来の血漿における可溶性EPCR抗原の測定は、女性(n=12)では91.1+/−24.5ng/mlの抗原レベル、そして男性(n=10)では107.2+/−30.2
ng/mlの抗原レベルを見出した。ともに計算する場合、平均血漿EPCR抗原レベル
は、98.4+/−27.8ng/mlであった。男性の値は、トロンボモジュリンと同様に(Quehenbergerら、Thromb.Haemost. 76:792−734)、女性の値よりわずかに高いようであるが、研究した集団は、統計学的分析にはあまりに限定されており、そしてこの研究は、性別、年齢、食餌または他の変数に起因する差異を評価するためには設計されていなかった。
【0066】
血漿EPCRは、約100ng/mlの単一種であるようであるので、血漿EPCRは
、循環EPCRが、プロテインCおよびAPCに結合し得るか否かを決定するために重要になった。可溶性EPCRを、イオン交換クロマトグラフィー、硫酸アンモニウムでの沈降、および実験手順において記載されるような抗EPCR 1496mAb−AffiGel−10カラムクロマトグラフィーによる免疫吸着の組み合わせによって、ヒト血漿から精製した。
【0067】
3mM CaCl2、0.6mM MgCl2を含む緩衝液中で抗プロテインCHPC4
mAb−AffiGel−10カラムにプロテインC(3mg)を適用することによって調製したプロテインCアフィニティーカラムに、この血漿EPCR(約110μg)を適
用した。カラムを洗浄し、そして血漿EPCRを画分19に適用した。カラムを洗浄し、そして画分35で開始する5mM EDTAを含む緩衝液で溶出した。それらの画分について、280nmでの吸光度およびEPCR抗原を決定した。EPCR抗原を、ELISAによって決定した。
【0068】
適用した血漿EPCR抗原の98%以上が、プロテインCアフィニティーカラムに結合した。吸光度プロフィールは、この抗体へのプロテインC結合のカルシウム依存性と一致して、抗体カラムからのEPCRおよびプロテインCの同時溶出を示す(Stearnsら、1988)。
【0069】
機能研究および構造研究のために十分なタンパク質を精製するために、類似しているがわずかに改変した手順を用いて、4Lの血漿からEPCRを精製した。プロテインC−抗体アフィニティーカラムからの溶出後、タンパク質を夾雑する残渣を、FPLC Mono Qカラムでのイオン交換クロマトグラフィーによって取り出した。得られた血漿EPCRの調製物は、銀染色を用いるSDS−PAGE 10%ゲルで均質のようであり、そして同一の結果が、ビオチン−ヤギ抗rsEPCRポリクローナル抗体でプローブしたウェスタンブロットに関して、還元条件下および非還元条件下の両方で得られた。精製されたタンパク質のアミノ末端配列分析は、唯一の配列S−Q−D−A−S−D(この配列は、組換え可溶性EPCRのアミノ末端配列に同一である:配列番号2)を生じた。これは、天然の供給源からのEPCRの最初のアミノ末端配列決定である。
【0070】
血漿EPCRの、APCに結合する能力を、競合研究によって評価した。この競合研究において、血漿EPCRを、APCについて細胞性EPCRと競合させ、そして細胞性EPCRに結合し得る得られた遊離のAPCを、フローサイトメトリーによって評価した(図4a)。活性部位においてフルオレセインで標識したAPC(fl−APC)を、血漿EPCRまたはrsEPCRのいずれかの、存在下または非存在下で、EA.hy926
細胞とともにインキュベートした。細胞へのAPC結合の阻害についてのEPCR濃度依存性は、EPCRの両方の可溶性形態について類似であった。この観察は、APCに結合することについての血漿EPCRの親和力が、rsEPCR−APC結合相互作用について以前に決定された親和力(Kdapp約30nM)に類似することを示す。
【0071】
rsEPCRは、可溶性系においてプロテインC活性化に対してほとんど効果を有さないが(Reganら、1996)、膜結合EPCRは、細胞表面上での活性化を促進する、非常に強力な能力を有する(Stearns−Kurosawaら、1996)。プロテインCおよびAPCに結合し得るEPCRの循環形態の存在を示す最新のデータは、血漿EPCRが、プロテインCの細胞表面活性化を変更する能力を有することを示唆した。EA.hy926細胞上でのほぼ生理学的レベルのプロテインC(0.1μM)のトロンビン依存性活性化は、過剰なrsEPCRによって、図4bによって示されるように、ほぼ、EPCR−プロテインC結合相互作用をブロックする抗rsEPCR 1494mAbを用いて観察されたプロテインCのレベルにまで阻害された。以前の研究は、rsEPCRが小さな合成基質を使用するAPCアミド分解活性(amidolyticactivity)に対する効果を有さないことを実証した(Reganら、1996)。血漿EPCRは、図4cに示されるように、rsEPCRと比較してEA.hy926細胞上の細
胞表面プロテインC活性化を阻害する能力においてわずかにより有効であった。
【0072】
1段Xa因子凝固アッセイにおいて、精製された血漿および可溶性組換えEPCRは、同様に凝固時間のAPC延長を阻害した(図4d)。rsEPCRによるAPC抗凝固活性の阻害が、以前に観察されている(Reganら、1996)。予測されるように、1496mAbは、APC−血漿EPCR結合相互作用をブロックすることによって、この効果を逆転した。
【0073】
(実施例2:尿における可溶性EPCRの検出)
可溶性EPCRが尿中に存在するかどうかという疑問を検討するために、4つの尿サンプル(朝一番の排尿)を回収し、そしてウェスタンブロッティングおよびELISAによって可溶性EPCRの存在について分析した。
【0074】
希釈していない小児の尿サンプルを、4%の正常な血漿および組換え可溶性EPCR(1ng)と比較した。サンプルを、ビオチン−ヤギ−抗rsEPCRおよびストレプトアビジン−アルカリホスファターゼ検出系とともにインキュベートした。
【0075】
ウェスタンブロットは、a)可溶性EPCRが尿中に存在すること、およびb)可溶性EPCR抗原が、血漿中で観察されるサイズと類似のサイズで存在することを示す。明らかな分解は観察されない。ELISAによって定量される、4つのサンプル中の可溶性EPCRの量は、40.3、6.1、35.6、および90.1ng/mlであった。
【0076】
(実施例3:狼瘡患者由来の血漿EPCRの測定)
正常なヒト血漿EPCR濃度は、上記のように約100ng/ml(98.4±27.8
ng/ml;2.5nM)である。エリテマトーデスに罹患した患者由来のサンプルのパネル(n=54)をアッセイし、そして可溶性EPCRレベルが、検出不能レベルから1,
700ng/ml以上までの範囲であることを見出した。15人の患者は、200ng/ml以上の可溶性EPCRレベルを有した。
【0077】
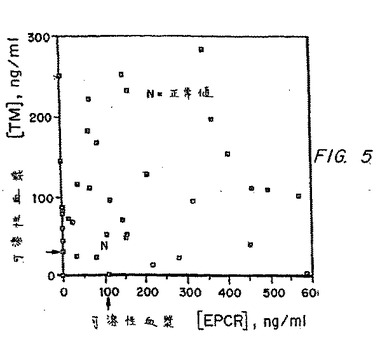
以前の研究は、内皮損傷に起因する、狼瘡患者における上昇した可溶性血漿TMレベルを示し、そして最新の狼瘡患者サンプルを、参照として血漿TMについてアッセイした。それらの可溶性TMレベルが、図5によって示されるように、それらの可溶性EPCRレベルと完全に相関を有さないことが見出された。このことは、可溶性血漿EPCRの供給源が、単に、ランダムに損傷を受けた内皮由来ではないことを示唆する重要な観察である。TMとは対照的に、ヒトおよび霊長類における膜結合EPCR発現は大きな脈管の内皮に主に制限され、毛細管はほとんどEPCRを発現しない。EPCRの特有の局在化は、大きな脈管の血栓症を予防するために、局所的にプロテインC活性化を増大すると予測される。大きな脈管への膜結合EPCRの主な局在化は、可溶性血漿EPCR濃度によって予測され得る大きな脈管における標的化された血栓症の危険を示す。
【0078】
(実施例4:敗血症性ショック患者における血漿可溶性EPCR)
敗血症(accp/sccm consensus conference,ches
t 1992;101:1644−1655)は、以下の臨床的発現のうちの1つより多くを含むが、それらに限定されない、感染に対する全身性炎症応答として定義される:
1)38℃以上または36℃未満の体温;
2)1分あたり90拍以上の心拍数;
3)以下によって示される頻呼吸:
a)1分あたり20呼吸以上の呼吸率;
b)32mmHg未満のPaCO2の過度呼吸;
4)12,000/mm3以上もしくは4,000/mm3未満のWBCカウント、または10%以上の未熟好中球(バンド)の存在。
【0079】
サンプルを、器官機能不全、低灌流(hypoperfusion)、または低血圧と関連する敗血症によって規定されるような、重篤な敗血症を伴うか、または伴わない、手術後の合併症を有する患者から得た。灌流異常は、乳酸アシドーシス、尿量過少、または精神状態における急性変化を含み得る。敗血症性ショックは、十分な流体(fluid)蘇生、および心臓性ショックの非存在にもかかわらず、24時間以上の間、血管作用薬を
必要とする低血圧を伴う敗血症をいう。
【0080】
本研究において含まれるすべての患者は、以下の診断基準を満たした:
a)呼吸の支持(24時間以上の制御された換気)およびまたは血流力学的支持(5マイクログラム/Kg/分以上の強心薬、ドパミン、もしくはドブタミン、および/または血
管作用性アミン、エピネフリン、もしくはノルエピネフリンの要求)を必要とする、敗血症および/または手術後の合併症に起因する、集中治療室への入室;
b)18歳〜75歳の間の年齢;
c)70%未満の抗トロンビン活性(局所にて試験される)。
患者が、多くの外傷、肝硬変、または急性肝不全、末期ガン、免疫不全、白血病、妊娠、またはヘパリン治療を有する場合、その患者を除外した。
【0081】
患者の血液サンプルを、時間0(集中治療室ICUへの入室)の時点および抗トロンビンII(ATIII)または偽薬での処置後2日目および6日目に採取した。血漿可溶性EPCRおよび可溶性トロンボモジュリン(TM)を、時間0のサンプルに関してのみアッセイした。
sEPCR:
正常:133.4±53.4ng/ml(平均±標準偏差)
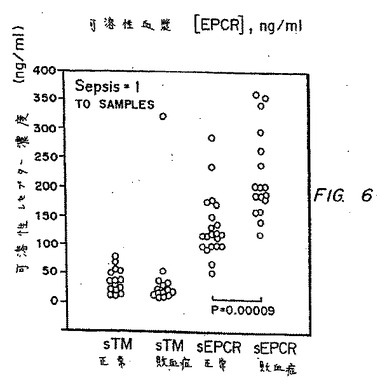
敗血症:224.9±74.5ng/ml
平均間の有意な差あり、P=0.00009
sTM:
正常:35.5±20.4ng/ml(平均±標準偏差)
敗血症:39.9±73.1ng/ml
平均間の有意な差なし、P=0.81
血漿中のsEPCRレベルとsTMレベルとの相関関係なし、r2=0.34。
【0082】
これらの結果を、図6にグラフで示す。狼瘡患者と同様に、敗血症に罹患した患者は、可溶性TMレベルと相関せず、血漿EPCRレベルにおいて非常に顕著な上昇を示す。
【0083】
可溶性血漿EPCRが、プロテインC活性化および活性化したプロテインC抗凝固活性の両方を阻害するという観察によって、これらの患者における上昇した血漿EPCRレベルが、さらなる血栓の危険性を引き起こし、そして脈管損傷/応答の証拠を表すことが示
される。これらが示す状態の例は、内皮細胞刺激、アテローム発生、白血球接着、およびプラークの破裂に関連する障害を含む。
【0084】
(実施例5:ヒヒ組織およびヒト組織におけるEPCRの選択的スプライシング形態の同定)
可溶性EPCRイソ型が、選択的スプライシング機構によって生成され得るか否かを決定するための最初のアプローチとして、RNAを、ヒト組織およびヒヒ組織から単離し、そして逆転写酵素−PCR(RT−PCR)を遺伝子特異的プライマーを用いて実施した。ヒヒEPCRゲノム配列は公知ではないが、ヒヒとヒトとは進化のスケールにおいて密接に関連しているという論理に基づいて、ヒト配列に基づくプライマーを使用した。
【0085】
一般にRT−PCR手順において、総RNAをホモジナイズした組織から単離する。RNAを、特定のアンチセンスプライマー、ヌクレオチド、および逆転写酵素と混合する。混合において、RNAは、逆転写酵素についての鋳型として作用し、第1鎖cDNAを作製する。次いで、この新しいcDNA鋳型を、特定のプライマーおよびTaqポリメラーゼを用いて従来のPCRによって増幅する。EPCRの膜形態(424bp)および推定の選択的スプライシング産物(674bp)の両方を増幅するプライマーを選択した。EPCRの両方の形態に対応する産物を、種々のヒヒ組織(図4)ならびにヒトの肺および
胎盤から増幅した。逆転写酵素を含まないコントロール、および組織との反応における1,885bpのバンドの欠失によって判断されるように、ゲノムDNAに関する可能性の
ある夾雑は存在しないようであった。
【0086】
ヒヒゲノムDNA配列が、選択的スプライシングのための適切なエキソン−イントロン境界およびイントロン−インフレームリーディング配列を有することを確認するために、ヒヒ腎臓ゲノムDNA由来のイントロン配列を、従来のPCRによって増幅した。ゲノム構造は種間で保持されるという前提を立て、そしてイントロンIIIに隣接するプライマー(ヒト配列)を使用した。これは、選択的スプライシング配列を含むと考えられるイントロン(ヒト配列)である。ヒヒ腎臓組織をホモジナイズし、そしてDNAを抽出した。DNAを、特定のプライマーと混合し、そして産物をPCRによって増幅した。DNA産物を精製し、そしてアガロースゲルで電気泳動した。
【0087】
(手順の詳細)
A.EPCR ELISA:被覆する抗体は、EPCRのリガンド結合ドメインに結合する1494mAbである。検出する抗体は、ビオチン化1495mAbであり、これはプロテインC/APC結合をブロックせず、そして1494mAbと交差反応しない。検
出系は、ストレプトアジビン−アルカリホスファターゼおよびBluePhos基質(KPL製)である。
【0088】
B.組織のRT−PCR:組織(50〜100mg)をTrizol(Gibco BRL)中にホモジナイズした。RNAを含む上層を、クロロホルムで抽出し、イソプロパノールで沈殿させ、洗浄し、そしてDEPC−水中に溶解した。RNA(1〜5μg)を、適切な緩衝液中で、製造業者の指示書に従って、ヌクレオチド、CREAアンチセンスプライマー、および逆転写酵素と混合した(第1鎖cDNA合成のためのSuperscriptTM Preamplification system,Gibco BRL)。cDNA産物を、CRESおよびCREAプライマーを用いて、従来のPCRによって、30サイクル増幅した。cDNA産物を、標準的な手順を用いて、クロロホルム抽出およびアルコール沈殿によって精製し、水中に溶解し、そして2%アガロースゲルで電気泳動した。ゲルを、Vistra Green(Amersham)で染色し、そしてリン光画像機(phosphoimager)(StormTM scanner,Molecular Dynamics,Inc.)で画像化した。
【0089】
C.ヒヒゲノムDNAのPCR:ヒヒ腎臓DNA(82mg)を、Trizol試薬中にホモジナイズした。DNAを含む下層を、抽出し、沈殿させ、そして滅菌水中に溶解した。DNAを、緩衝液、ヌクレオチド、ならびにHRT−1およびHRT−2プライマーとの混合液中で、従来のPCRによって、30サイクル増幅した。増幅したDNAを、標準的な手順を用いて、抽出し、沈殿させ、滅菌水中に溶解し、そして2%アガロースゲルで電気泳動した。単一のバンド(465bp)を、エチジウムブロミドで可視化し、切り出し、そしてPCR産物を、製造業者の指示書(Qiagen)に従ってスピンカラムで精製した。PCR産物を、同じプライマーを用いて配列決定した。
【0090】
(D.プライマー配列)
CRES:5'−TCGTGCGCCTGGTGCACCAGGAGC−3'
(エキソンIIの末端近くの5'センスプライマー)
CREA:5'−CGCCGTCCACCTGTGCACAGGAAG−3'
(エキソンIV内の3'アンチセンスプライマー)
HRT−1:5'−AGCAGCTCAATGCCTACAACCG−3'
(エキソンIIIの末端近くの5'センスプライマー)
HRT−2:5'−CCGTAGAAGGACACGTGTCCACCTGCCGC−
3'
(エキソンIV内の3'アンチセンスプライマー)
(ヒヒ組織に関する結果)
ゲルから切り出され、精製され、そして配列決定された腎臓ゲノムDNAから増幅された単一のバンドが存在した。その配列は、ヒト配列に対して92%同一であり、そしてエキソン−イントロン境界は保存された。このイントロン配列における高レベルの類似性は重要である。なぜなら、イントロン配列は、代表的には種間において十分には保存されないからである。終止コドンを有するイントロン内のインフレームリーディング配列もまた存在し、このことは、ヒヒの選択的にスプライシングした可溶性タンパク質中の特有の22残基のカルボキシル末端テイルを予測する。
【0091】
推定の可溶性EPCRイソ型が、特有のカルボキシル末端テイルを有するという観察は、イソ型特異的抗体を使用するイソ型間の区別のための構造的差異を提供する。実用モデルとしては、タンパク質分解された可溶性EPCRの血漿レベルが内皮損傷を報告する一方で、選択的スプライシングを受けた可溶性EPCRのレベルが刺激に対する内皮応答を報告するというものである。可溶性EPCRイソ型の相対的血漿レベルが、特定の病理学における大きな脈管の内皮機能不全および損傷に関する情報を提供することが期待される。
【0092】
(ヒト組織に関する結果)
ヒト組織(胎盤、肺、および舌)由来のRT−PCR産物を、EPCRに特異的なCRES/CREAプライマーを用いて電気泳動した。手順は、ヒヒ組織について使用された
手順と同じであった。EPCRの膜イソ型(mEPCR)および選択的スプライシングを受けた可溶性EPCRイソ型(sEPCR)に対応する産物を観察した。産物は、ヒヒ組織を用いて観察された産物と本質的に同じようである。唯一の差異は、胎盤組織がさらなる産物を有するようであることである。
【0093】
(配列表)
【0094】
【表2】
【0095】
【表3】
【0096】
【表4】
【0097】
【表5】
【0098】
【表6】
【0099】
【表7】
【0100】
【表8】
【図面の簡単な説明】
【0101】
【図1】図1は、膜結合レセプターをタンパク質分解して細胞外ドメインを遊離しそして膜アンカーを残すこと、およびmRNAの選択的スプライシングによって、EPCRに適用されるような可溶性レセプターを産生するための2つの既知のメカニズムの概略図であり、膜結合EPCR(mEPCR)およびタンパク質分解血漿EPCR(pEPCR)に特有の配列、ならびに可溶性EPCR(sEPCR)に特有の配列を示す。
【図2】図2は、mEPCRおよびsEPCRを比較する概略図であり、ヌクレオチドインサートおよびsEPCRに特有のコードされたアミノ酸配列を示す。
【図3】図3は、選択的スプライシングによってヒト、ウシ、マウス、およびヒヒのEPCRに挿入された配列を示す。
【図4】図4aは、可溶性血漿EPCRがヒトプロテインCおよびAPCに結合することを示すグラフである。EA.hy926細胞を、氷上で30分間0〜500nMrsEPCR(黒丸)または血漿EPCR(白丸)の存在下で、60nM fl−APCと共にインキュベートした。細胞を洗浄し、そして細胞結合した蛍光を、記載のようにフローサイトメトリーによって決定した。fl−APCを添加しない場合での固有の細胞蛍光を、矢印で示す。プロットした平均細胞蛍光(MCF)は、2連のMCF測定の平均を表す。 図4bおよび4cは、可溶性血漿EPCRおよびrsEPCRが、細胞表面上でプロテインC活性化を阻害することを示すグラフである。図4bでは、EA.hy926細胞単層を、0.1μMプロテインC単独(白四角)、または1μM rsEPCR(黒丸)、または2μg/ml 1494mAb(白丸)と共に、室温にて15分間プレインキュベートした。プロテインC活性化を、トロンビン(2nM最終)の添加によって開始し、そして反応を、所定の時間で停止した。活性化プロテインCを、アミド分解アッセイで決定し、そして活性速度をmOD/分で各時点についてプロットした。トロンビンを添加しないコントロールウェルを含ませた(黒四角)。各データ点は、3連のウェル測定の平均を表す。図4cでは、EA.hy926細胞単層を、0.1μMのプロテインCおよび示された濃度の血漿EPCR(白丸)またはrsEPCR(黒丸)と共に、室温にて15分間プレインキュベートした。トロンビン(最終2nM)を添加し、そして活性化を室温にて60分間行った。上清を、アンチトロンビンおよびヘパリンの混合物に添加し、そして活性化プロテインC活性(mOD/分)を、アミド分解アッセイで決定した。各データ点は、3連のウェル測定の平均を表す。 図4dは、可溶性血漿EPCRがAPC抗凝固活性を阻害することを示すグラフである。APC(25nM)の抗凝固活性を、460nMの血漿EPCRまたはrsEPCRの存在下で、1段階Xa凝血アッセイを用いて決定した。この効果は、いずれかの可溶性EPCRを、EPCRへのAPCの結合をブロックする、42μg/mlの1496mAbと共に、5分間プレインキュベートした場合には逆転した。データは、4〜6測定の平均±S.D.を表す。
【図5】図5は、狼瘡患者において、可溶性血漿TMのレベルを可溶性血漿PCRと比較するグラフであり、TM値とEPCR値との間に相関はないが、狼瘡患者の大多数が、可溶性血漿EPCRの非常に上昇したレベルを示すことを証明する。
【図6】図6は、sTM(正常)、sTM(敗血症)、sEPCR(正常)、およびsEPCR(敗血症)についての可溶性レセプター濃度(ng/ml)のグラフである。sTMとsEPCRとの間に相関はない(r2=0.034)。
【背景技術】
【0001】
(発明の背景)
本発明は、一般に、内皮細胞プロテインC/活性化プロテインCレセプターまたはタン
パク質分解もしくは選択的スプライシングのいずれかによって誘導されるその可溶性形態の検出および/または測定を含むアッセイの領域にある。
【0002】
合衆国政府は、Charles T.Esmonに対する国立衛生研究所助成金PO1HL 54804により、本発明に一定の権利を有する。
【0003】
プロテインCのその活性セリンプロテアーゼに対する活性化、活性化プロテインC(APC)は、血液凝固の調節に重要な役割を果たす一連の事象を開始する。プロテインC経路の臨床的重要性は、血栓症を生じる、この経路における多くの機能不全によって示される(非特許文献1;非特許文献2)。プロテインCが欠損した患者は、通常、幼児期に生命を危うくする血栓症合併症を示す(非特許文献3;非特許文献4)。これは、プロテインC投与によって補正される(非特許文献5)。
【0004】
プロテインCおよびAPCはまた、炎症に対する宿主応答の調節に関連している。活性化プロテインC(APC)は、グラム陰性敗血症のヒヒモデルにおいてE.coliの致死的効果を予防し得(非特許文献6;TaylorおよびEsmonに対する特許文献1)、そして予備的な臨床結果は、プロテインCが、ヒト敗血症ショックのある形態を処置することに効果的であることを示唆する(非特許文献7)。プロテインC経路の重要な成分であるプロテインSの阻害は、E.coliの致死量未満のレベルに対する霊長類の応答を悪化させ、そして循環中へのTNFの出現を増大する(非特許文献8)。これらの結果は、プロテインCが、凝固の制御および炎症への影響の両方をなし得ることを示唆する。
【0005】
プロテインCは、凝固系の最終酵素であるトロンビンが、内皮細胞表面タンパク質であるトロンボモジュリンに結合する場合に、活性化される(非特許文献9;非特許文献10;非特許文献11)。細胞培養において、トロンボモジュリン転写は、腫瘍壊死因子(TNF)への内皮細胞の曝露によってブロックされ(非特許文献12そしてトロンボモジュリン活性および抗原は、その後インターナライズされそして分解される(非特許文献13、非特許文献14)。補体系の調節タンパク質であるC4bBPは、プロテインSと結合して、インビトロ(非特許文献15)およびインビボ(非特許文献8)でAPC抗凝固活性を支持することにおいて機能的に不活性である複合体を形成する。C4bBPは、急性期反応物質として行動する(非特許文献16)。したがって、この経路のタンパク質は、炎症を調節するようにみえるだけでなく、これらは炎症を調節する成分と相互作用し、そしてこれら自体は、炎症メディエータによってダウンレギュレーションを受ける。
【0006】
内皮細胞は、効率的なプロテインC活性化を担う公知の2つのレセプター、トロンボモジュリンおよび内皮プロテインC/APCレセプター(EPCR)を発現する点で、プロ
テインC経路において重要な役割を果たす(非特許文献17;非特許文献18)。トロンボモジュリン(CD141)は、高親和性を有する循環トロンビンと結合する膜貫通補因子であり、そして得られる酵素−補因子複合体は、生理学的に関連するプロテインC活性化速度に必要とされる(非特許文献19;非特許文献20)。
【0007】
EPCRは、CD1/MHCクラス1ファミリーとの著しい相同性を有する最近同定さ
れたレセプターである(非特許文献17;非特許文献21;非特許文献22)。プロテインCに対する内皮細胞レセプターのクローニングおよび生物学的役割は、Oklahoma Medical Research Foundationによって「Cloning and Regulation of an Endothelial Cell Protein C/Activated Protein C Receptor」と
題するPCT/US95/09636に記載された。このタンパク質は、238アミノ酸からなると予測され、このアミノ酸は、N末端に15アミノ酸のシグナル配列、およびタイプ1膜貫通タンパク質としてこのレセプターを特徴づける23アミノ酸の膜貫通領域を含む。
【0008】
EPCRは、カルシウムの存在下で類似の親和性(Kdapp約30nM)でプロテイン
CおよびAPCの両方と結合し(非特許文献21)、そして細胞表面上で、トロンビン−トロンボモジュリン活性化複合体に対してプロテインC基質を提示することにより、プロテインC活性化を容易にする(非特許文献18)。内皮細胞レセプターは両方とも、タイプ1膜貫通タンパク質であり、ここでリガンドは、細胞外ドメインに結合し、そして両方とも、短い細胞内細胞質テイルを有する(非特許文献21;非特許文献23;非特許文献24;非特許文献25)。さらに、それらのインビトロ細胞表面発現は、腫瘍壊死因子−αによって同様にダウンレギュレートされる(非特許文献17)。しかし、トロンボモジュリンおよびEPCRの可溶性形態の特徴は、いくつかの点で異なる。組換え可溶性トロンボモジュリンは、膜形態に対して減少した補因子活性を有する(非特許文献26;非特許文献27)。精製した成分および細胞の両方で、トロンボモジュリンによって特異的に誘導されるトロンビンの基質の変化は、トロンビンにおける共有される結合ドメインについての競合、ならびに活性部位ポケットにおけるコンホメーション変化から生じる(非特許文献28;非特許文献29;非特許文献30;非特許文献31;非特許文献32;非特許文献33;非特許文献34)。可溶性トロンボモジュリンもまた、種々のインヒビターによるトロンビンの不活化を加速する(非特許文献35;非特許文献36)。血漿および尿の両方が、検出可能なトロンボモジュリンを含み(非特許文献37;非特許文献38)、そしてトロンボモジュリン遺伝子がイントロンを含まないので(非特許文献23)、これらの可溶性形態は、細胞表面での細胞外ドメインのタンパク質分解による。
【0009】
血漿中のトロンボモジュリンの可溶性分解産物は、種々の疾患状態での内皮細胞損傷の公知のマーカーであり(非特許文献37;非特許文献39;非特許文献40;非特許文献41;非特許文献42;非特許文献43;非特許文献44)、そして種々の減少した親和性を有するトロンビン結合フラグメント、ならびに非結合フラグメントの混合物から構成される(非特許文献37)。
【0010】
対照的に、膜貫通ドメインの直前で短縮された組換え可溶性EPCR(rsEPCR)は、インタクトな細胞表面発現EPCRについて観察される親和性と同様な親和性でプロテインCおよびAPCの両方と結合する(非特許文献21)。おそらくrsEPCRおよび第Va因子の両方が、トロンボモジュリンによって占有されるトロンビン中のアニオン結合外部位(exosite)を暗示する溝に結合決定基を共有するので(非特許文献45)、APC抗凝固活性は、rsEPCRと結合した場合、効果的に阻害される(非特許文献22)。しかし、rsEPCRは、APCによる小さい合成基質のタンパク質分解にも、α1−アンチトリプシンまたはプロテインCインヒビターによるAPCの不活化にも影響を及ぼさないようである(非特許文献22)。プロテインC活性化を増強する膜結合EPCR(非特許文献18)とは異なり、rsEPCRは、可溶性トロンビン−トロンボモジュリン複合体によるプロテインC活性化に対してほとんど効果を有さない(非特許文献22)。このことは、EPCRの任意の可溶性形態が、プロテインCについて膜結合EPCRと競合することによってプロテインC活性化を阻害し得ることを示唆する。
【0011】
免疫組織化学は、EPCRが、主として大きな血管に由来する内皮細胞の表面上に存在し、そしてほとんどの毛細血管内皮細胞上にはないか、または低レベルでしか存在しないことを示す。
【特許文献1】米国特許第5,009,889号明細書
【非特許文献1】EsmonおよびSchwarz,Trends Cardiovasc.Med.,1995,5,p.141−148
【非特許文献2】Reitsmaら,Thromb.Haemost.,1995,73,p.876−879
【非特許文献3】Seligsohnら,N.Engl.J.Med.,1984,310,p.559−562
【非特許文献4】Esmon,Trends Cardiovasc.Med.,1992,2,p.214−220
【非特許文献5】Dreyfusら,N.Engl.J.Med.,1991,325,p.1565−1568
【非特許文献6】Taylorら,J.Clin.Invest.,1987,p.79
【非特許文献7】Gersonら,Pediatrics 91,1993,p.418−422
【非特許文献8】Taylorら,Blood,1991,78,p.357−363
【非特許文献9】Esmon,J.Biol.Chem.,1989,264,p.4743−4746
【非特許文献10】DittmanおよびMajerus,Blood,1990,75,p.329−336
【非特許文献11】Dittman,Trends Cardiovasc.Med.,1991,1,p.331−336
【非特許文献12】ConwayおよびRosenberg,Mol.Cell.Biol.,1988,8,p.5588−5592
【非特許文献13】Lentzら,Blood,1991,77,p.543−550
【非特許文献14】Mooreら,Blood,1989,73,p.159−165
【非特許文献15】Dahlback,J.Biol.Chem.,1986,261,p.12022−12027
【非特許文献16】Dahlback,Thromb.Haemostas.,1991,66,p.49−61
【非特許文献17】FukudomeおよびEsmon,J.Biol.Chem.,1994,269,p.26486−26491
【非特許文献18】Stearns−Kurosawaら,Proc.Natl.Acad.Sci.(USA),1996,93,p.10212−10216
【非特許文献19】EsmonおよびOwen,Proc.Natl.Acad.Sci.(USA),1981,78,p.2249−2252
【非特許文献20】Dittman,W.A.,Trends Cardiovasc.Med.,1991,1,p.331−336
【非特許文献21】Fukudomeら,J.Biol.Chem.,1996,271,p.17491−17498
【非特許文献22】Reganら,J.Biol.Chem.,1996,271,p.17499−17503
【非特許文献23】Jackmanら,Proc.Natl.Acad.Sci.(USA),1987,84,p.6425−6429
【非特許文献24】Wenら,Biochemistory,1987,26,p.4350−4357
【非特許文献25】Suzukiら,EMBO J.,1987,6,p.1891−1897
【非特許文献26】Galvinら,J.Biol.Chem.,1987,262,p.2199−2205
【非特許文献27】Parkinsonら,J.Biol.Chem.,1990,265,p.12602−12610
【非特許文献28】Yeら,J.Biol.Chem.,1991,266,p.23016−23021
【非特許文献29】Luら,J.Biol.Chem.,1989,264,p.12956−12962
【非特許文献30】Yeら,J.Biol.Chem.,1992,267,p.11023−11028
【非特許文献31】Hofsteengeら,Biochem.J.,1986,237,p.243−251
【非特許文献32】Mathews,Biochemistry,1994,33,p.13547−13552
【非特許文献33】Esmonら,J.Biol.Chem.,1982,257,p.7944−7947
【非特許文献34】Sadlerら,Haemostasis,1993,23,p.183−193
【非特許文献35】BourinおよびLindahl,Biochem.J.,1993,289,p.313−330
【非特許文献36】Rezaie,J.Biol.Chem.,1995,270,p.25336−25339
【非特許文献37】Takanoら,Blood,1990,76,p.2024−2029
【非特許文献38】IshiiおよびMajerus,J.Clin.Invest.,1985,76,p.2178−2181
【非特許文献39】Tanakaら,Clin.Chem.,1991,37,p.269−272
【非特許文献40】Takahashiら,Am.J.Hematol.,1991,38,p.174−177
【非特許文献41】Asakuraら,Am.J.Hematol.,1991,38,p.281−287
【非特許文献42】Wadaら,Am.J.Hematol.,1992,39,p.20−24
【非特許文献43】Takahashiら,Am.J.Hematol.,1992,41:32−39
【非特許文献44】Ohdamaら,Chest,1994,106,p.666−671
【非特許文献45】Matherら,EMBO J.,1996,15,p.6822−6831
【発明の開示】
【発明が解決しようとする課題】
【0012】
したがって、本発明の目的は、天然に存在する可溶性EPCRについて治療的および診断的使用を同定することである。
【0013】
本発明のさらなる目的は、天然に存在する可溶性EPCRを特徴づけることである。
【課題を解決するための手段】
【0014】
(発明の要旨)
血漿EPCRは、単離され、特徴づけられ、そして細胞性プロテインC活性化およびAPC抗凝固活性をブロックすることが示されている。血漿EPCRは、約43,000ダ
ルトンであるようであり、そして約100ng/mlで循環する(98.4±27.8ng/ml、n=22)。血漿EPCRを、イオン交換、イムノアフィニティー、およびプロテインCアフィニティークロマトグラフィーを使用して、ヒトのクエン酸処理血漿から精製した。フローサイトメトリー実験は、血漿EPCRが、膜貫通および細胞質ドメインを含まないEPCRとして定義される組換え短縮型EPCRから予め決定された親和性(Kdapp約30nM)と同様の親和性で、活性化プロテインCと結合することを示した。さら
に、血漿EPCRは、内皮細胞株におけるプロテインC活性化および1段階第Xa因子凝血アッセイにおけるAPC抗凝固活性の両方を阻害した。可溶性EPCRはまた、ヒト尿において検出されている。EPCRをコードする遺伝子のクローニングは、少なくともヒトEPCRが選択的にスプライスされ得、選択的にスプライスされた形態(sEPCR)に特有のインサートを含む短縮型可溶性EPCRを生じることを示す。これらの結果は、血漿EPCRが、細胞表面でのタンパク質分解または選択的スプライシングのいずれかによって誘導され得ることを示す。
【0015】
血漿EPCRの局所濃度が、特に疾患状態において、十分に高い場合、そのデータは、短縮型可溶性血漿EPCRが、プロテインC活性化の膜結合EPCR増大および活性化プロテインCの抗凝固機能を減弱させ得ることを示す。自己免疫疾患(全身性エリテマトーデス、SLE)および敗血症(炎症および凝固異常の両方を含む障害)の患者からのEPCRのレベルと正常EPCRを比較する実施例によって示されるように、可溶性EPCRのレベルは、炎症および異常凝固に関連する疾患状態と相関するようである。凝固、炎症、および大血管疾患を含む疾患状態を示す、可溶性EPCRの測定に基づくアッセイが記載される。単離精製された可溶性EPCR、組換え短縮型可溶性EPCR、および可溶性EPCRに対する抗体を含む、アッセイ試薬が記載される。
【0016】
本発明はさらに、以下を提供する。
(1)可溶性内皮プロテインCレセプターについてのアッセイであって、
テストされる患者からサンプルを得る工程、および
可溶性内皮プロテインCレセプターの量を測定する工程、
を包含する、アッセイ。
(2)上記サンプルが、尿、血漿、血清、組織サンプル、および間質液からなる群より選択される、項目1に記載のアッセイ。
(3)可溶性内皮プロテインCレセプターの量を較正標準と相関させる工程をさらに包含する、項目1に記載のアッセイ。
(4)上記患者が、凝固および炎症状態および障害、内皮に対する損傷に関連する障害または疾患、ならびに大血管疾患からなる群より選択される障害または疾患を有する、項目1に記載のアッセイ。
(5)上記障害または疾患が、自己免疫疾患、移植、敗血症、ショック、子癇前症、糖尿病、血管疾患、腎臓疾患、および肝臓疾患からなる群より選択される、項目4に記載のアッセイ。
(6)上記血管疾患が、心肺バイパス、不安定性アンギナ、再狭窄、および血管形成からなる群より選択される、項目5に記載のアッセイ。
(7)大血管に対する損傷を検出するための、項目1に記載のアッセイ。
(8)内皮プロテインCレセプターの検出および測定のためのキットであって、
内皮プロテインCレセプターと免疫反応性である抗体、
患者からのサンプル中で上記抗体と内皮プロテインCレセプターとの間の反応を検出するための試薬、および
上記反応量を、内皮プロテインCレセプターの正常および異常レベルと相関させるための標準
を含む、キット。
(9)上記抗体が、膜貫通ドメインを含まない内皮プロテインCレセプターについてより、膜貫通ドメインを含む内皮プロテインCレセプターについて、より高い親和性を有する、項目8に記載のキット。
(10)上記抗体が、選択的にスプライスされた内皮プロテインCレセプターにおけるインサートと免疫反応性である、項目8に記載のキット。
(11)上記抗体が、内皮プロテインCレセプターと、活性化プロテインCまたはプロテインCとの結合をブロックする、項目8に記載のキット。
(12)カルボキシル末端システイン残基が、別のアミノ酸と置換されているか、またはパルミトイル化されていない、改変された内皮プロテインCレセプター。
(13)グリコシル化されていない、改変された内皮プロテインCレセプター。
(14)単離された、選択的にスプライスされた内皮プロテインCレセプター。
(15)膜貫通ドメインの前の天然に存在するタンパク質分解性切断部位で切断された、単離された可溶性内皮プロテインCレセプター。
【発明の効果】
【0017】
自己免疫疾患(全身性エリテマトーデス、SLE)および敗血症(炎症および凝固異常の両方を含む障害)の患者からのEPCRのレベルと正常EPCRを比較する実施例によって示されるように、可溶性EPCRのレベルは、炎症および異常凝固に関連する疾患状態と相関するようである。凝固、炎症、および大血管疾患を含む疾患状態を示す、可溶性EPCRの測定に基づくアッセイが提供される。単離精製された可溶性EPCR、組換え短縮型可溶性EPCR、および可溶性EPCRに対する抗体を含む、アッセイ試薬が提供される。これらは従来技術からは予測不可能であった格別の効果である。
【発明を実施するための最良の形態】
【0018】
(発明の詳細な説明)
(定義)
内皮プロテインCレセプター、EPCR。
【0019】
可溶性、溶液中にあり、そして細胞表面に結合していない。
【0020】
短縮型、膜貫通ドメインおよび細胞質ドメインを含まない;タンパク質分解または選択的スプライシングのいずれかの結果であり得る。
【0021】
(可溶性EPCRの検出および特徴付け;生理学的役割およびマーカーとしての有用性)
EPCRの機能のこれまでの研究は、膜形態のEPCRに結合するプロテインCが、細胞表面上のトロンビン−トロンボモジュリン複合体によりプロテインC活性化の促進を生じる(Stearns−Kurosawaら,1996)が、可溶性組換えEPCRはAPC抗凝固活性を阻害する(Reganら,1996)ことを見出した。これらの観察は、血漿中の可溶性トロンボモジュリン分解産物が、種々の疾患状態における内皮損傷のマーカーであるという知識と共に、可溶性循環形態のEPCRが存在するかどうか、そして存在するのであれば、プロテインC経路においてどういう役割を有するかという疑問に至った。
【0022】
実施例は、可溶性形態のEPCRが血漿中で循環し、そして尿中に存在することを示す
。健常ドナー集団において、血漿EPCRレベルは、約100ng/mlであり、そして
約43,000ダルトンの単一抗原種であるようであった。血漿からの可溶性EPCRの
その後の精製および機能的研究は、血漿EPCRが、インタクトな膜結合EPCRと同様な親和性で、プロテインCおよびAPCの両方と結合し得ることを決定した。内皮細胞株を使用するインビトロ研究は、血漿EPCRが、生理学的濃度に近い濃度のプロテインCおよびトロンビンで、プロテインCを阻害することを示した。さらに、血漿への精製血漿EPCRの直接添加は、rsEPCRに対するモノクローナル抗体で逆転されるAPC抗凝固活性の阻害を生じた。
【0023】
この精製血漿タンパク質のEPCRとの同定は、rsEPCRの特性との比較に基づいた。これらのタンパク質は両方とも、同じ群のモノクローナル抗体およびポリクローナル抗体と反応し、同じアミノ末端配列を有し、Ca2+依存的様式で固定されたプロテインCに結合し、そして類似の用量応答曲線でプロテインC活性化およびAPC抗凝固活性をブロックした。さらに、rsEPCRおよび血漿EPCRに対するプロテインCおよびAPCの親和性は共に、インタクトな膜結合EPCRの親和性に類似する。これらの特性は、EPCRに特有であるようである。
【0024】
これまでの研究は、内皮細胞で発現した膜結合EPCRが、3〜5倍の間のファクターでプロテインC活性化を増大することを示したが、実施例は、血漿から精製された可溶性形態のEPCRが、内皮細胞でのプロテインC活性化およびAPC抗凝固活性を阻害することを示す。これは、EPCRが、いくつかの方法で、プロテインC経路を調節し得ることを予測する。第1に、より大きな血管(ここでは、トロンボモジュリン濃度が微小循環に対して低い)において、EPCR発現は、対応して増加する(Laszikら、Circulation 1997)。免疫組織化学は、ほとんどの器官において、EPCR発現は、大血管で最も高密度であり、そして血管サイズが減少するにつれて次第に減少し、最も豊富な内皮細胞タイプである毛細管内皮ではほとんどまたは全く発現しない。EPCR発現は、循環からプロテインC基質を捕獲し、そして活性化のためにトロンビン−トロンボモジュリン複合体に提示することにおいて、重要な役割を果たし得る。このことは、EA.hy926内皮細胞株およびヒト臍静脈内皮細胞の両方ともが、トロンボモジュリ
ンより、少なくとも6倍多い表面発現EPCR抗原を有するというインビトロ観察によって支持される。トロンボモジュリン濃度が高くそしてEPCRが低い微小循環では、プロテインC活性化にほとんど影響がないと予測される。最後に、循環可溶性EPCRは、APCの生成およびAPCが第Va因子を不活化する能力を減少させ得る。
【0025】
健常個体において、可溶性EPCRレベルは、約2.5nMであり、この濃度は、Kdapp(約30nM)および循環にける80nMのプロテインC濃度の両方よりも十分に低い。可溶性血漿EPCRの効果(APC抗凝固活性およびプロテインC活性化の阻害)の両方ともが、正常血漿に存在する濃度よりかなり高い濃度を必要とする。このことが、血漿EPCRの生理学的役割の疑問を、不確定のままにしている。40nMを越える可溶性EPCRレベルを有する患者が、実施例3(狼瘡)に記載のように同定された。したがって、内皮細胞表面に近い局所濃度が、全身濃度を超えるならば、可溶性EPCR濃度は、APCの生成および活性の両方を減弱するレベルに達し、血栓症の危険性に寄与する。
【0026】
可溶性形態のレセプターは、膜結合レセプターのタンパク質分解的切断、または選択的スプライシングメカニズムによって産生され得る。膜表面でのタンパク質分解は、可溶性トロンボモジュリン、ならびにTNF、IL−1、IL−2、M−CSF、PDGF、およびNGFのレセプターを遊離する(Heaneyら,1996,Blood 87:847−857)。可溶性レセプターは、膜レセプターのアンタゴニストとして作用すること、リガンドを安定化すること、リガンド媒介されたシグナリングを開始すること、膜形態のダウンモジュレーション、およびレセプターインヒビターへ結合してレセプター−リ
ガンド活性を間接的に容易にすることを含む、多くの可能性のある機能を有する。後者のメカニズムは、両方のIL−1レセプターの可溶性イソ形態が、タンパク質分解的切断によって生成され、そしてIL−1αおよびIL−1βへの応答性を厳格に調節する、IL−1レセプター系によって使用される(Arendら,1994,J.Immunol.
153:4766−4774)。以下に議論するように、膜貫通ドメインの直前で短縮された可溶性タンパク質をコードするEPCRゲノム構造は、選択的スプライシング部位を含む(FukudomeおよびEsmon,1995,J.Biol.Chem. 270:5571−5577)。可溶性IL−6レセプターは、タンパク質分解メカニズムおよび選択的スプライシングメカニズムの両方によって生成されるようである(Mullbergら,1994,J.Immunol. 152:4958−4968;Lustら,1992,Cytokine 4:96−100;Horiuchiら,1994,Eur.J.Immunol. 24:1945−1948)。この切断部位はまた、MorrisseyおよびEsmonへの米国特許第5,298,599号(その教示は本明細書に援用される)に記載のように、短縮型EPCR、続いて直ぐペプチド配列(抗体はこの配列に対して特異的に指向する)をコードする発現ベクターを構築することによって、大量の可溶性EPCRを回収することにおいて有用であり得る。次いで、エピトープは、患者への投与の前または後に、タンパク質分解によって切断される。Hoppらへの米国特許第4,782,137号もまた参照。
【0027】
免疫組織化学研究は、EPCRが、主として大血管の内皮に位置し、そして毛細血管ではかろうじて検出可能であることを示した。したがって、膜結合EPCRに由来する血漿EPCRは、大血管疾患プロセスのマーカーとして用い得る。血漿EPCRは、血漿トロンボモジュリンレベルとの有用な比較として用い得る。血漿トロンボモジュリンレベルは、種々の疾患状態において調整されることが示されているが、大血管および小血管の両方の疾患プロセスを反映し、ほとんどの内皮が微小血管性であるのでおそらく小血管の寄与に支配される。
【0028】
(EPCRのヌクレオチドおよび予測タンパク質構造分析)
EPCRのcDNAは、238アミノ酸(配列番号2)のタンパク質をコードすると予測され、これは、N末端に15アミノ酸のシグナル配列を含む(vonHeijne,(1986)Nucleic Acids Res. 14,4683−4690)。したがって、成熟タンパク質は、223アミノ酸を含むと予測される。組換えタンパク質の直接配列決定は、成熟タンパク質がSer18で始まることを示した。配列番号2は、EPCRの予測アミノ酸配列である。配列番号2のアミノ酸1〜15(MLTTLLPILLLSGWA)は、von Heijne(von Heijne,1986)の方法によって決定される推定シグナル配列である。配列番号2のアミノ酸211〜236(LVLGVLVGGFIIAGVAVGIFLCTGGR)は、推定膜貫通ドメインである。可能性のあるN−グリコシル化部位は、配列番号2のアミノ酸47〜49、64〜66、136〜138、および172〜174に存在する。細胞外システイン残基は、配列番号2のアミノ酸17(血漿EPCRでは除去されている)、114、118、および186に存在する。23アミノ酸からなる可能性のある膜貫通領域(Engelmanら,(1986)Annu.Rev.Biophys.Chem. 15,321−53)を、C末端で同定した(配列番号2のアミノ酸211で始まる)。
【0029】
このタンパク質はタイプ1膜貫通タンパク質である。細胞外ドメインは、4つの可能性のあるN−グリコシル化部位および3つのCys残基を含む。グリコシル化は、N−グリカナーゼ消化によって示されるように、活性に必須ではない。細胞質領域は、3アミノ酸のみを含み、そしてCysで終結し、パルミトイル化されている。末端システインが適切にパルミトイル化されてない場合、タンパク質は分泌され得る。EPCRの配列を変化させて、このシステインを別のアミノ酸と置換することにより、膜結合する代わりに分泌さ
れる、本質的に完全長のEPCRを作成する手段が提供される。
【0030】
本明細書で使用される場合、レセプターをコードするヌクレオチド配列は、配列番号1に示される配列、およびストリンジェントな条件下で配列番号1にハイブリダイズする、その保存的置換、付加、または欠失を有する配列を含む。本明細書で使用される場合、レセプターを構成するアミノ酸配列は、配列番号2に示される配列、および機能的に等価な生物学的活性を有するレセプターを形成する、その保存的置換、付加、または欠失を有する配列を含む。何が、保存的置換、付加、または欠失を構成し、そしてどれが、本明細書に記載の機能的アッセイを使用して機能的に等価なレセプター分子をコードまたは形成すると容易に確認され得るかは、当業者に周知である。これは、さらに、図3を参照することによって例示され、以下に議論される。
【0031】
(選択的スプライシング)
レセプターは、特異的リガンドに結合して生理学的応答を生じることを担う細胞の、外部に曝された部分を有する、細胞膜に繋がれたタンパク質として最も頻繁に思い描かれる。多くの場合、もはや細胞に制限されないという事実にもかかわらず、頻繁にそのリガンドを確かに結合し得る、可溶性形態のレセプターが存在する。可溶性レセプターイソ形態へのリガンド結合はまた、多くの形態をとる応答を生じ得、これには、膜結合レセプター相互作用のアップモジュレーションまたはダウンモジュレーション、あるいは正常には応答性ではない細胞へリガンドを輸送することによって応答を伝達させることを含む(Heaney,MLおよびDW Golde. Soluble cytokine receptors. Blood 87:847−857, 1996)。
【0032】
可溶性レセプターを産生するための2つの公知のメカニズムが存在する:膜結合レセプターをタンパク質分解して、細胞外ドメインを遊離し、そして膜アンカーを残すことによるもの、およびmRNAの選択的スプライシングによるもの(図1)。後者のメカニズムは多くの形態を取り得るが、リーディングフレームがエキソン−イントロン境界により連続し、そして膜貫通アンカーをコードする配列に達する前に停止コドンで終結する場合、最も単純である。これは、膜形態に類似するが、重要な差異を有するタンパク質を生成する。これは、可溶性タンパク質として、作製され、そして分泌され、そして特有のカルボキシル末端テイルを有する。このテイルを、膜結合レセプターの形成において無視されるイントロンmRNA配列の一部を読むことによって形成した。選択的スプライシングによる可溶性レセプターの生成はまた、共に同じmRNAテンプレートを起源にするという事実にもかかわらず、膜結合レセプターとは独立して調整され得る(Heaneyら,Proc.Natl.Acad.Sci.U.S.A. 92:2365−2369,1995)。
【0033】
可溶性レセプターが選択的スプライシングメカニズムによって生成されることを証明するためには、ゲノム配列および関連領域のイントロン−エキソン境界を知らなければならない。生理学的応答と可溶性レセプターを関連付けて、プロテインC遺伝子およびプロテインS遺伝子の発現の間にかなり頻繁に含まれることが見られる異常なmRNAスプライシングと区別することもまた役立つ(Bergら,Blood Coag Fibrinol. 7:625−631,1996)。
【0034】
以下の研究および結果の詳細は、実施例に記載される。ヒト血漿は、約100ng/m
lの可溶性EPCRを含む(表1)。これを、2つのモノクローナル抗体(1494mAbおよび1495mAb)および標準的技法を使用する酵素結合イムノアッセイ(ELISA)によって測定した。著しく上昇した可溶性EPCRレベルを、全身性エリテマトーデスおよび敗血症の患者で見いだした。これらのレベルは、ほとんど例外なく、大血管の表面上のみに存在する膜結合レセプターについてかなり高いようであった。これを真相正
しく認識するために、トロンボモジュリン(TM)は、すべての内皮、ならびにいくつかの非血管細胞上で発現されるが、正常可溶性TMレベルは、わずかに約10〜40ng/
mlである(Takanoら,Blood 76:2024−2029,1990)。可溶性TMレベルは、狼瘡の患者で上昇していたが、敗血症では上昇していなかった。重要なことには、これらの患者群において、血漿EPCRとTMレベルとの間に相関がなかったことである(それぞれ、r2=0.028および0.034)。
【0035】
【表1】
【0036】
TMゲノム構造は、イントロンを含まず(Jackmanら,Proc.Natl.Acad.Sci.(USA)84:6425−6429,1987)、そのため可溶性TMイソ形態を生成するための唯一の方法は、膜結合レセプターのタンパク質分解による。好中球エラスターゼおよびカテプシンGによる内皮TMのタンパク質分解は、インビトロで示されており、これは、種々の疾患状態で見出される上昇した可溶性TMレベルが、内皮表面で、活性化された炎症性細胞の産物によって媒介されるタンパク質分解から生じることを示唆する(Boehmeら,Immunology 87:134−140,1996、およびAbeら,J.Lab.Clin.Med. 123:874−881,1994)。
【0037】
血漿EPCRおよびTMレベルと高い血漿EPCR濃度との間の相関の欠如は、血漿EPCRがタンパク質分解および選択的スプライシングの両方のメカニズムを起源にするという概念に一致する。ヒトEPCRのゲノム構造は、イントロンによって分離される、4つのエキソンを含む。この配列を概観することにより、7527位にTAA停止コドンを含むエキソンIII−イントロンIII境界(5'GTで)の後のインフレームリーディ
ング配列が明らかになる。この停止コドンは、膜貫通ドメインをコードするエキソンIVの上流にあるので、予測されるタンパク質は、特有な48残基のカルボキシル末端テイル(イントロン配列によってコードされる)を含み、そして膜貫通アンカーを含まない。
【0038】
図1は、短縮型EPCRが誘導され得る2つの可能性のある方法の図である:膜貫通ドメインの直前でのタンパク質分解によるか、または選択的スプライシングによる。図2に示されるように、選択的スプライシングは、選択的にスプライスされた短縮型EPCR中にあるペプチド配列が含まれることを生じる。図3に示されるように、この配列は、種間で高度に保存されるが、わずかな差が存在し、ヒトおよびウシのEPCRについては48残基、マウスEPCRについては51残基、そしてヒヒEPCRについては22残基の新しいカルボキシル末端テイルを生じる。
【0039】
(レセプタータンパク質の発現についての患者サンプルのスクリーニング)
患者サンプルは、EPCR、EPCR中の選択的にスプライスされたインサートに存在する特有のインサートのいずれかに対する抗体、またはコンホメーションの差に起因してEPCRまたはsEPCRのいずれかに対してより大きな親和性で結合する抗体を使用して、sEPCRまたはEPCRの存在および量について、スクリーニングされ得る。サンプルはまた、存在するタンパク質を特異的に定量するための他の標準的技法を使用して、スクリーニングされ得る。
【0040】
(診断的または治療的使用のための抗体の生成)
EPCR、および特に、可溶性EPCR(「sEPCR」)、および組換え可溶性EPCR(「rsEPCR」)に対する抗体が生成され得、これらは、レセプタータンパク質の検出、特徴付け、または単離に有用であり、そしてほとんどの場合、リガンド結合の阻害によって、レセプタータンパク質活性を調節するために有用である。抗体は、免疫原として、ヒトまたは動物の精製または組換えレセプタータンパク質もしくはそのフラグメントを使用して、標準的技法によって生成される。
【0041】
EPCRに対するモノクローナル抗体を、Esmonら,1993,Methods Enzymol. 222:359−385によって他のタンパク質について記載されたように得た。1494、1495、および1496mAbと呼ばれる抗体は、組換え可溶性EPCRおよび細胞表面発現EPCRに結合するIgG1κ抗体である。1494および1496mAbは、EPCRへのプロテインCおよびAPCの結合をブロックし、そして細胞性EPCRがトロンビン−トロンボモジュリン複合体によるプロテインC活性化を容易にする能力を阻害する。1495mAbは、EPCRへのリガンド結合をブロックせず、細胞表面プロテインCを変化させず、そして1494または1486mAbのエピトープとは異なる結合エピトープを有する。抗体は、標準的技法、例えば、放射標識、酵素標識、フルオレセインのような蛍光標識、金粒子、染料、および抗体の検出のための他の手段を使用して標識され得る。例えば、抗体は、標準的な手順を使用して、ビオチンアミドカプロン酸N−ヒドロキシスクシンイミドエステルでビオチン化され得る。抗体は、イムノアッセイにおける使用のための固体支持体、例えば、AffiGel−10TM、ニトロセルロース、またはマイクロタイターウェル、あるいは溶液相イムノアッセイにおける使用のための固体支持体に固定され得る。
【0042】
好ましい実施態様では、EPCRは、50マイクロリットルの、15mM Na2CO3、35mM NaHCO3(pH9.6)中4マイクログラム/mlの1495mAbで4
℃にて一晩コーティングしたマイクロタイタープレート(MaxisorpTM,NUNC
NS,Roskilde,Denmark)を使用して測定される。室温で、次いでプレートを20mM Tris−HCl、0.1M NaCl、0.05% Tween 20(pH7.5)(アッセイ緩衝液)で3回洗浄し、そして0.1%(wt/vol)ゼラ
チンを含むアッセイ緩衝液で少なくとも1時間ブロックする。次いで、ウェルを洗浄し、50マイクロリットルサンプルを3連のウェルに添加し、そしてプレートを1時間インキュベートする。ウェルを吸引し、アッセイ緩衝液で3回洗浄し、そして50マイクロリットル、2マイクログラム/mlのビオチン−1494mAbを添加する。プレートを1時
間インキュベートし、3回洗浄し、そして50マイクロリットルの、0.25マイクログ
ラム/mlのストレプトアビジン−アルカリホスファターゼ結合体(GIBCO BRL
)を添加し、そしてさらに1時間インキュベートする。ウェルを5回洗浄し、そしてELISA増幅キット(GIBCO BRL)の基質および増幅試薬を、製造業者の指示に従って15分間隔で連続して添加する。発色を0.3M H2SO4で停止し、そして終点吸
光度を、Vmaxマイクロプレートリーダーにて490mmで読み取る。3連のウェルでの
標準は、20mM Tris−HCl、0.1M NaCl、および1mM EDTA、
0.1%ゼラチン(pH7.5)中の1.5〜100ng rsEPCR/mlである。標
準曲線は、1.5〜12.5ng/mlで直線であり、そしてサンプルを、直線範囲に入る
ように同じ緩衝液で希釈する。研究は、1〜2パーセントの間で、血漿が、アッセイの直線性にも標準曲線の感度にも影響を及ぼさないことを示す。健常ボランティアからの血漿サンプルを、1mM EDTAを含むアッセイ緩衝液で最終的に2%血漿まで希釈した。そしてEPCR抗原レベルを、同じプレート上で決定された標準曲線を参照して、3連ウェルの平均から算出する。
【0043】
(障害)
可溶性EPCRについてのアッセイは、本明細書で議論されるような凝固および炎症状態および障害、例えば、狼瘡のような自己免疫疾患、移植モニタリング、敗血症、ショック、子癇前症、糖尿病、心肺バイパス、不安定性アンギナ、再狭窄、血管形成(すなわち、血管疾患)、腎臓または肝臓疾患の検出および分析に有用である。例えば、EPCRは、大血管についてのマーカー、したがって大血管に対する損傷についてのマーカーである。可溶性EPCRの量の増加は、大血管傷害を示し、EPCRのタンパク質分解またはsEPCR合成の刺激のいずれかを生じる。EPCR対トロンボモジュリンの比はまた、血液または尿のいずれかのサンプルに基づいて決定され得、これは微小血管対大血管の相対的な程度を示す。サイトカイン、白血球活性化マーカー、および補体因子または活性化マーカーに対するEPCRの相対量もまた、疾患状態を示すために使用され得る。
【0044】
EPCRは内皮細胞上に存在するので、内皮細胞損傷のマーカーとして有用である。これは、薬物の効果(毒性および効力の両方)の指標として使用され得る。例えば、狼瘡患者では、炎症性/凝固媒介性大血管傷害を効果的に最小にする薬物は、EPCRレベルの
減少を生じる。
【0045】
本発明は、以下の非限定的な実施例を参照することによってさらに理解される。
【実施例】
【0046】
(実施例1:ヒト血漿における機能的内皮プロテインCレセプターの同定)
以下の略号を使用する:rsEPCR、膜貫通ドメインおよびサイトゾルテイルの代わりに挿入されたHPC4エピトープを有する組換え可溶性EPCR;mAb、モノクローナル抗体;SDS−PAGE、ドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動。
【0047】
(方法)
材料。以下の試薬を示された供給業者から購入した:
ブタ腸粘膜ヘパリン、ジイソプロピルフルオロリン酸、ビオチンアミドカプロン酸N−ヒドロキシスクシンイミドエステル、ウシ血清アルブミン、Sigma(St.Lois,MO);Spectrozyme PCa、American
Diagnostica(Greenwich,CT);ELISA増幅キット、GibcoBRL(Gaithersburg,MD);AffiGel−10、BioRad(Hercules,CA);ハンクス平衡化塩溶液、3−(N−モルホリン)プロパン硫酸(MOPS)、Fisher Scientific(Fair Lawn,NJ)。すべての他の試薬は、市販の最高品質なものであった。
【0048】
タンパク質。ヒトプロテインC(Esmonら,1993,Methods Enzymol. 222:359−385)、ウシトロンビン(Owenら,1974,J.BIol.Chem. 249:594−605)、およびウシアンチトロンビン(Esmon 1977「Factors regulating the inhibition of thrombin by antithrombin III. In Chemistry and Biology of Thrombin」R.L.Lund
blad,J.W.Fenton,II,およびK.G.Mann編 Ann Arbor Science,Ann Arbor.403−411)を、記載のように精製した。組換え可溶性EPCR、rsEPCRは、膜貫通ドメインの直前の残基210で短縮されたEPCRの細胞外ドメイン、続くHPC4モノクローナル抗体におけるカルシウム依存的イムノアフィニティー精製を可能にする12残基配列からなる(Takahashiら,1992;Stearnsら,1988,J.Biol.Chem. 263:826−832)。rsEPCRの構築、精製、およびプロテインC/APC結合特性(Fu
kudomeら,1996)。ヤギ免疫前血清およびrsEPCRに対するヤギポリクローナル抗血清を調製し、そしてIgGを精製した(Fukudomeら,1996)。ヤギ抗rsEPCRポリクローナル抗体を、標準的手順を使用して、ビオチンアミドカプロン酸N−ヒドロキシスクシンイミドエステルでビオチン化した。
【0049】
モノクローナル抗体。rsEPCRに対するモノクローナル抗体(mAb)を、他のタンパク質について記載されたように得た(Esmonら,1993)。1494、1495、および1496mAbは、rsEPCRおよび細胞表面発現EPCRに結合するIgG1κ抗体である。1494および1496mAbは、EPCRへのプロテインCおよびAPCの結合をブロックし、そして細胞性EPCRがトロンビン−トロンボモジュリン複合体によるプロテインC活性化を容易にする能力を阻害する(Stearns−Kurosawaら,1996)。1495mAbは、EPCRへのリガンド結合をブロックせず、細胞表面プロテインC活性化を変化させず、そして1494または1496mAbのエピトープとは異なる結合エピトープを有する。1494および1495mAbを、標準的手順を使用して、ビオチンアミドカプロン酸N−ヒドロキシスクシンイミドエステルでビオチン化した。1494mAbを、血漿EPCRのイムノアフィニティー精製のために、製造業者の指示に従って、AffiGel−10にカップリングした。抗EPCR mAbのスクリーニングを、Stearns−Kurosawaら(1996);Fukudomeら(1996)に記載の方法を使用して行った。
【0050】
凝血アッセイ。1段階第Xa因子凝血アッセイにおけるAPC(25nM)抗凝固活性に対するrsEPCRまたは精製血漿EPCRの効果を、83μg/mlの1496mA
b(APC−EPCR相互作用をブロックする抗体)の存在または非存在下で行った(Reganら,1996)。可溶性EPCRおよび1496mAbを、アッセイ前15分間プレインキュベートした。
【0051】
細胞培養。すべてのヒト細胞株を、以前に記載されたように維持した(Fukudomeら,1996)。EA.hy926細胞(形質転換されたヒト内皮細胞株;Edgel
lら,1983,Proc.Natl.Acad.Sci.(USA)80:3734−3737)は、Cora−Jean Edgell(University of North Carolina at Chapel Hill)が好意により提供した。
【0052】
フローサイトメトリー分析。蛍光プローブとして用いるために、APCを、記載されるように(FukudomeおよびEsmon、1994;Bock,P.E.1988.Biochemistry 27:6633−6639)、フルオレセインで活性部位(fl−APC)において標識した。EA.hy926細胞へのAPC結合に対するrsE
PCRまたは血漿EPCRの効果を、フローサイトメトリーによって研究した(Fukudomeら、1996)。手短には、収集された細胞を、漸増する濃度のいずれかの可溶性EPCR調製物の非存在下または存在下において、60nMのfl−APCとともに、氷上で30分間インキュベートし、洗浄し、そして細胞に結合した蛍光を、フローサイトメトリーによって1サンプルごとに10,000事象を計数して測定した。全てのアッセ
イを、1%のウシ血清アルブミン、3mM CaCl2、0.6mM MgCl2、および
0.02%アジ化ナトリウムを補充したハンクス平衡化塩溶液において行った。
【0053】
細胞表面プロテインC活性化。EA.hy926細胞を96ウェル組織培養ディッシュ
において培養した(Stearns−Kurosawaら、1996)。コンフルエントな単層を、1%(w/v)のウシ血清アルブミン、3mM CaCl2、0.6mM Mg
Cl2、および0.02%アジ化ナトリウムを補充したハンクス平衡化塩溶液を用いて、3回洗浄した。全てのアッセイを、室温において、同一の緩衝液中で、60μlの最終容量にて行った。そして、全てのタンパク質濃度は、アッセイにおける最終濃度を示す。プロテインC(0.1μM)を、示された濃度のrsEPCR、血漿EPCR、または149
4mAbの非存在下または存在下で添加し、細胞とともに15分間プレインキュベートした。トロンビンを混合液に添加して(2nM)、活性化反応を開始した。示された時間に、50μlのアリコートを取り出し、そして96ウェルマイクロタイタープレート中の10μlのアンチトロンビン(0.7μM最終)およびヘパリン(5U/ml最終)に添加した。APCアミド分解活性をSpectrozyme PCa基質(0.2mM)の添加
によって測定し、405nmでの吸光度の変化速度(mOD/分)をVmaxキネティッ
クマイクロプレートリーダー(Molecular Devices,Menlo Park,CA)で測定した。すべてのアッセイ点は、3連ウェルにおいて行われ、そして十分に活性化されたプロテインC対mOD/分の標準曲線を参照することにより決定される
場合、プロテインC基質の10%未満が活性化された。
【0054】
血漿および血清の収集。健常な成人ボランティア(女性12人および男性10人)から、3.8%緩衝化クエン酸溶液中かまたは抗凝固剤を含まないチューブ中に、静脈穿刺に
よって全血を収集した(Vacutainer tube;Becton Dickinson,Franklin Lakes,NJ)。年齢、食餌、または他の変数に関するドナーのスクリーニングは試みなかった。全てのボランティアに研究についての情報を開示し、彼らから書面による同意を得た。血液を、1160×gで10分間遠心分離した。血漿および血清を等分し、アッセイまで−80℃で冷凍保存した。
【0055】
血漿EPCRの定量のためのELISA。血漿におけるEPCR抗原の検出のための酵素結合免疫吸着アッセイを開発した。マイクロタイタープレート(Maxisorp;Nunc,Roskilde,Denmark)を、50μlの、15mM Na2CO3、35mM NaHCO3(pH9.6)中4μg/mlの1495mAbを用いて、4℃にて一晩コートした。以下の工程を室温で行った。ウェルを、20mM Tris−HCl、0.1M NaCl、0.05% Tween20(pH7.5)(アッセイ緩衝液)で
3回洗浄し、0.1%(w/v)ゼラチンを含有するアッセイ緩衝液で少なくとも1時間ブロックした。ウェルを洗浄し、50μlのサンプルを3連のウェルに添加し、そしてプレートを1時間インキュベートした。ウェルを吸引して、アッセイ緩衝液で3回洗浄し、50μlの2μg/mlビオチン−1494mAbを添加した。プレートを1時間インキュ
ベートし、3回洗浄し、50μlの0.25μg/mlストレプトアビジン−アルカリホスファターゼ結合体(GibcoBRL)を添加し、さらに1時間インキュベートした。ウェルを5回洗浄し、ELISA増幅キット(GibcoBRL)の基質および増幅試薬を、製造業者の指示に従い、15分の間隔で連続的に添加した。発色を0.3M H2SO4
を用いて停止し、490nmの終点吸光度をVmaxマイクロプレートリーダーで読み取った。各プレートは、20mM Tris−HCl、0.1M NaCl、1mM ED
TA、0.1%ゼラチン(pH7.5)中1.5〜100ng/mlのrsEPCRの標準を3連のウェルに含んだ。標準曲線は、1.5〜12.5ng/mlで直線であり(r=0.99)、そして血漿サンプルを直線範囲に入るように同じ緩衝液で希釈した。予備的な実験により、再終濃度1〜2%のヒト血漿は、標準曲線の直線性にも感度にも影響しないことを実証した。健常ボランティア由来の血漿サンプルを、1mMEDTAを含有するアッセイ緩衝液で、最終的に2%血漿に希釈し、そしてEPCR抗原レベルを、同一プレート上で決定された標準曲線を参照して、3連ウェルの平均から算出した。
【0056】
コーティング抗体と検出抗体を逆にし(1494mAbをコーティングに;ビオチン−1495mAbを検出に)、そして抗体結合を、Blue Phos基質(KPL Laboratories;Gaithersburg,MD)で検出する、別のアッセイを開発した。この方法を使用して、敗血症患者における血漿EPCRをアッセイした。このアッセイは、おそらく親和性の差異のために、より高感度であったが、両アッセイとも、定性的には、同様の結果を与えた。
【0057】
ウエスタンブロット。血漿または血清サンプルのドデシル硫酸ナトリウムポリアクリルアミドゲル電気泳動(SDS−PAGE)を、10%アクリルアミドゲルを、Laemmli緩衝液系(Nature 227:680−685)と共に用いて、非還元条件下で標準的な手順を用いて行った。ゲルをポリビニリデン膜(PVDF;Millipore,Bedford,MA)に転写し、膜をブロックし、次に免疫前ヤギIgG(50μg/ml)か、またはヤギ抗rsEPCRポリクローナルIgG(50μg/ml)のいずれかと30分間インキュベートした。洗浄後、膜を1:20,000希釈のマウス抗ヤギI
gG−西洋わさびペルオキシダーゼ結合体(Pierce,Rockford,IL)と30分間インキュベートした。膜を洗浄し、結合した抗体−酵素結合体を増強化学発光基質(Pierce)で製造業者の指示に従って検出した。
【0058】
免疫吸着。健常ボランティアからの血清またはクエン酸処理血漿サンプル(400μl)を、AffiGel−10に結合させた50μlの1495mAb(5mg IgG/
ml樹脂)とともに、混合しながら4℃にて一晩インキュベートした。サンプルを遠心分離し、上清を除去し、そして樹脂を1mlの20mM
Tris−HCl、0.1M NaCl、0.02%アジ化ナトリウム(pH7.5)で
3回洗浄した。最終20mM ジチオスレイトールを含有するSDS−PAGEサンプル緩衝液を、洗浄した樹脂に添加し、サンプルを3分間沸騰し、そしてSDS−PAGEおよびウエスタンブロッティングのために処理した。膜を4μg/mlのビオチン化ヤギ抗
rsEPCRポリクローナル抗体でプローブし、そして結合抗体をストレプトアビジン−西洋わさびペルオキシダーゼ結合体(Pierce)および増強化学発光検出系で検出した。予備的実験により、室温にて1〜4時間の、100μlのTris不活化AffiGel−10樹脂でのサンプルの事前吸着、続く1495mAb−AffiGel−10での一晩の免疫吸着が、同一のウエスタンブロットの結果を与えることが決定された。
【0059】
血漿EPCRの精製。血漿EPCRを、イオン交換クロマトグラフィー、抗rsEPCR mAbイムノアフィニティークロマトグラフィー、およびプロテインC親和性カラムでのクロマトグラフィーの組み合わせを用いて、ヒトクエン酸処理血漿(Oklahoma Blood Institute)から精製した。わずかに異なる方法で2つの調製を行った。
【0060】
第1の調製において、血漿(1L)を等量の20mM Tris−HCl(pH7.5
)、10mM ベンズアミド、400単位のヘパリンナトリウムで希釈し、そして1gの膨潤前のQAE樹脂でバッチ吸着した。沈澱後、樹脂をプロテインCの精製のために処理した(Esmonら、1993)。固体硫酸アンモニウムを上清に4℃にて40%飽和まで添加し、遠心分離し、そしてさらなる硫酸アンモニウムをその上清に70%飽和に達するまで添加した。遠心分離後、柔らかなペレットを透析バッグに入れ、12Lの20mM
Tris−HCl、0.02%アジ化ナトリウム(pH7.4)に対し一晩透析した。透析物を、20mM Tris−HCl、0.1M NaCl、0.02%アジ化ナトリウム(pH7.4)で平衡化した、1496mAb−AffiGel−10イムノアフィニテ
ィーカラム(6ml樹脂;5mg IgG/ml樹脂)にかけた。カラムを、12mlを
超える同じ緩衝液で洗浄し、20mM Tris−HCl(pH7.4)中の50%(v/
v)エチレングリコールで溶出した(Jun Xu、未発表の観察)。溶出物からのピーク画分をプールし(0.37の全OD280)、濃縮し(Centriprep30、Millipore)、そして緩衝液を20mM Tris−HCl、0.1M NaCl、3
mM CaCl2、0.6mMMgCl2、0.02%アジ化ナトリウム(pH7.4)に交
換した。この物質をプロテインCアフィニティーカラムにかけた。このカラムは、同じ緩衝液においてHPC4−AffiGel−10カラム(5mg IgG/ml樹脂;0.9×8cm)に精製プロテインC(3mg)をかけることにより事前に調製した。HPC4mAbは、カルシウム依存的様式でプロテインC活性化領域に結合し(Esmonら、1993;Stearnsら、1988)、その後のプロテインCへのEPCRの結合を妨げない。血漿EPCRを含有するサンプルをカラムにかけた後、カラムを約12mlの緩衝液で洗浄し、20mM Tris−HCl、0.1M NaCl、5mM EDTA、
10mM MOPS、0.02%アジ化ナトリウム(pH7.5)で溶出した。画分を、280nmでの吸光度について、そして上記のELISAを用いてEPCR抗原についてモニターした。プロテインCおよび血漿EPCRの両方を含有する溶出物を、FPLC(Pharmacia−LKB、Uppsala、Sweden)MonoQカラムにかけ、カラムを20mM Tris−HCl(pH7.5)中の0.1〜1M NaClの直線勾配で展開した。約半分の血漿EPCRは、MonoQカラムに結合せず、半分は、約0.
2M NaClに溶出し、そしてプロテインCは、約0.5M NaClに溶出した。血
漿EPCRの両方のイオン種は、還元条件下または非還元条件下での銀染色、クマシーBB染色、またはPVDF膜への転写後の金染色(Pierce)を用いるSDS−PAGE、およびビオチン−ポリクローナル抗rsEPCR抗体プローブでのウエスタンブロットで、同一のようであった。
【0061】
血漿EPCRの第2の調製は、4Lの血漿を用いて開始し、機能研究に十分なタンパク
質を精製した。この場合、1496−AffiGel−10樹脂(20mlの5mg IgG/ml樹脂)を、10mMベンズアミジン、1mMジイソプロピルフルオロホスフェ
ートおよび0.5単位/mlのヘパリンナトリウム(最終濃度)と共に、クエン酸処理血漿に、直接添加した。血漿を、穏やかに混合しながら、4℃にて一晩バッチ吸着した。樹脂の沈澱後、上清をプロテインC精製のために処理した(Esmonら、1993)。樹脂を2.5×30cmのカラムに充填し、20mM Tris−HCl、0.1M NaCl、0.02%アジ化ナトリウム(pH7.4)で、十分に洗浄し、そして20mM Tr
is−HCl(pH7.4)中50%のエチレングリコールで溶出した。溶出物をプール
し、濃縮し(5.5の全OD280)、MonoQカラムにかけ、2つのイオン種(素通り、および0.2M NaCl溶出ピーク)を、1496−AffiGel−10樹脂(1.5×11cm)に再びかけた。カラムを前のとおりに、50%エチレングリコールで溶出した。溶出物(0.71 OD)を濃縮し、緩衝液を、Cetriprep30を含む、2
0mM Tris−HCl、0.1MNaCl、3mM CaCl2、0.6mM MgC
l2、0.02%アジ化ナトリウムに交換した。次に、この物質をアフィニティーカラムにかけた。ここで、プロテインC(2.9mg)を、同じ緩衝液中で、HPC2−Affi
Gel−10カラム(0.6×17cm)に最初にかけた。HPC2mAbは、プロテイ
ンCのセリンプロテアーゼドメインに結合し、EPCR結合を妨げない(Fukudomeら、1996)。結合したEPCRを、5mM EDTAを含有する緩衝液で溶出した。混入している血清アミロイドP(プロテインCサンプル由来)を、FPLC MonoQカラムでのイオン交換クロマトグラフィーによって除去した。サンプルを0.2M N
aCl中でカラムにかけ、その結果、血漿EPCRは結合せず、そして0.4〜0.5M NaClにおいて溶出する混入物から分離された。得られる精製血漿EPCR(0.19
3 OD280)は、銀染色を用いるSDS−PAGE、およびポリクローナル抗rsEP
CRを用いるウエスタンブロットにより、均質のようであった。この物質を機能研究およびアミノ末端配列分析に使用した。
【0062】
タンパク質配列決定。可溶性血漿EPCRのアミノ末端配列分析を、Molecular Biology Research Facility、W.K.Warren Medical Research Institute(Oklahoma City)のKenneth Jackson博士の研究室において行った。アミノ酸を標準の1文字コードによって表す。
【0063】
(結果)
第1のアプローチとして、3人の健常ボランティア由来の血漿および血清サンプルを希釈し(4%v/v)、非還元条件下で10%SDS−PAGEゲルで泳動し、そしてrs
EPCRに対して惹起したヤギポリクローナル抗体を用いるウエスタンブロットのために処理した。健常ボランティア由来の血漿および血清サンプル(4%v/v)を非還元条件
下の10%ゲルでのSDS−PAGEのために処理し、膜に転写し、そして膜をヤギ抗rsEPCRポリクローナル抗体でプローブした。結果をrsEPCR(0.2ng)と比
較した。結合した抗体を、マウス抗ヤギIgGおよび増強化学発光検出系で検出した。2人の健常ボランティア由来の血漿サンプルを1495 AffiGel−10樹脂で免疫吸着した。洗浄した樹脂を溶出させ、そして還元条件下でのSDS−PAGEのために処理した。ウエスタンブロッティングを、ビオチン−ヤギ抗rsEPCRをプローブとして使用して行った。
【0064】
血漿EPCRの純度を、銀染色したSDS−PAGE 10%ゲル、およびビオチン−ヤギ抗rsEPCRでプローブした膜のウエスタンブロットから決定した(還元および非還元)。膜をポリクローナル抗体でプローブした後、血清および血漿サンプルの両方で、約43,000Daの単一のバンドが現れる。検出されたタンパク質のサイズは、rsE
PCRよりわずかに大きいようである。検出された他のバンドは、免疫前IgGおよびより長い暴露時間でのプロービングより判断すると、IgGのバックグラウンド結合であった。AffiGel−10樹脂と結合した抗EPCR 1495mAbと共に血漿サンプルを一晩インキュベートすること、続く洗浄および還元条件下での結合抗原の溶出は、ビオチン−ヤギ抗rsEPCRポリクローナル抗体でのウエスタンブロットにより検出される単一バンドを生じた。
【0065】
mAb 1495をコーティング抗体として使用するELISAによる、健常ボランティア由来の血漿における可溶性EPCR抗原の測定は、女性(n=12)では91.1+/−24.5ng/mlの抗原レベル、そして男性(n=10)では107.2+/−30.2
ng/mlの抗原レベルを見出した。ともに計算する場合、平均血漿EPCR抗原レベル
は、98.4+/−27.8ng/mlであった。男性の値は、トロンボモジュリンと同様に(Quehenbergerら、Thromb.Haemost. 76:792−734)、女性の値よりわずかに高いようであるが、研究した集団は、統計学的分析にはあまりに限定されており、そしてこの研究は、性別、年齢、食餌または他の変数に起因する差異を評価するためには設計されていなかった。
【0066】
血漿EPCRは、約100ng/mlの単一種であるようであるので、血漿EPCRは
、循環EPCRが、プロテインCおよびAPCに結合し得るか否かを決定するために重要になった。可溶性EPCRを、イオン交換クロマトグラフィー、硫酸アンモニウムでの沈降、および実験手順において記載されるような抗EPCR 1496mAb−AffiGel−10カラムクロマトグラフィーによる免疫吸着の組み合わせによって、ヒト血漿から精製した。
【0067】
3mM CaCl2、0.6mM MgCl2を含む緩衝液中で抗プロテインCHPC4
mAb−AffiGel−10カラムにプロテインC(3mg)を適用することによって調製したプロテインCアフィニティーカラムに、この血漿EPCR(約110μg)を適
用した。カラムを洗浄し、そして血漿EPCRを画分19に適用した。カラムを洗浄し、そして画分35で開始する5mM EDTAを含む緩衝液で溶出した。それらの画分について、280nmでの吸光度およびEPCR抗原を決定した。EPCR抗原を、ELISAによって決定した。
【0068】
適用した血漿EPCR抗原の98%以上が、プロテインCアフィニティーカラムに結合した。吸光度プロフィールは、この抗体へのプロテインC結合のカルシウム依存性と一致して、抗体カラムからのEPCRおよびプロテインCの同時溶出を示す(Stearnsら、1988)。
【0069】
機能研究および構造研究のために十分なタンパク質を精製するために、類似しているがわずかに改変した手順を用いて、4Lの血漿からEPCRを精製した。プロテインC−抗体アフィニティーカラムからの溶出後、タンパク質を夾雑する残渣を、FPLC Mono Qカラムでのイオン交換クロマトグラフィーによって取り出した。得られた血漿EPCRの調製物は、銀染色を用いるSDS−PAGE 10%ゲルで均質のようであり、そして同一の結果が、ビオチン−ヤギ抗rsEPCRポリクローナル抗体でプローブしたウェスタンブロットに関して、還元条件下および非還元条件下の両方で得られた。精製されたタンパク質のアミノ末端配列分析は、唯一の配列S−Q−D−A−S−D(この配列は、組換え可溶性EPCRのアミノ末端配列に同一である:配列番号2)を生じた。これは、天然の供給源からのEPCRの最初のアミノ末端配列決定である。
【0070】
血漿EPCRの、APCに結合する能力を、競合研究によって評価した。この競合研究において、血漿EPCRを、APCについて細胞性EPCRと競合させ、そして細胞性EPCRに結合し得る得られた遊離のAPCを、フローサイトメトリーによって評価した(図4a)。活性部位においてフルオレセインで標識したAPC(fl−APC)を、血漿EPCRまたはrsEPCRのいずれかの、存在下または非存在下で、EA.hy926
細胞とともにインキュベートした。細胞へのAPC結合の阻害についてのEPCR濃度依存性は、EPCRの両方の可溶性形態について類似であった。この観察は、APCに結合することについての血漿EPCRの親和力が、rsEPCR−APC結合相互作用について以前に決定された親和力(Kdapp約30nM)に類似することを示す。
【0071】
rsEPCRは、可溶性系においてプロテインC活性化に対してほとんど効果を有さないが(Reganら、1996)、膜結合EPCRは、細胞表面上での活性化を促進する、非常に強力な能力を有する(Stearns−Kurosawaら、1996)。プロテインCおよびAPCに結合し得るEPCRの循環形態の存在を示す最新のデータは、血漿EPCRが、プロテインCの細胞表面活性化を変更する能力を有することを示唆した。EA.hy926細胞上でのほぼ生理学的レベルのプロテインC(0.1μM)のトロンビン依存性活性化は、過剰なrsEPCRによって、図4bによって示されるように、ほぼ、EPCR−プロテインC結合相互作用をブロックする抗rsEPCR 1494mAbを用いて観察されたプロテインCのレベルにまで阻害された。以前の研究は、rsEPCRが小さな合成基質を使用するAPCアミド分解活性(amidolyticactivity)に対する効果を有さないことを実証した(Reganら、1996)。血漿EPCRは、図4cに示されるように、rsEPCRと比較してEA.hy926細胞上の細
胞表面プロテインC活性化を阻害する能力においてわずかにより有効であった。
【0072】
1段Xa因子凝固アッセイにおいて、精製された血漿および可溶性組換えEPCRは、同様に凝固時間のAPC延長を阻害した(図4d)。rsEPCRによるAPC抗凝固活性の阻害が、以前に観察されている(Reganら、1996)。予測されるように、1496mAbは、APC−血漿EPCR結合相互作用をブロックすることによって、この効果を逆転した。
【0073】
(実施例2:尿における可溶性EPCRの検出)
可溶性EPCRが尿中に存在するかどうかという疑問を検討するために、4つの尿サンプル(朝一番の排尿)を回収し、そしてウェスタンブロッティングおよびELISAによって可溶性EPCRの存在について分析した。
【0074】
希釈していない小児の尿サンプルを、4%の正常な血漿および組換え可溶性EPCR(1ng)と比較した。サンプルを、ビオチン−ヤギ−抗rsEPCRおよびストレプトアビジン−アルカリホスファターゼ検出系とともにインキュベートした。
【0075】
ウェスタンブロットは、a)可溶性EPCRが尿中に存在すること、およびb)可溶性EPCR抗原が、血漿中で観察されるサイズと類似のサイズで存在することを示す。明らかな分解は観察されない。ELISAによって定量される、4つのサンプル中の可溶性EPCRの量は、40.3、6.1、35.6、および90.1ng/mlであった。
【0076】
(実施例3:狼瘡患者由来の血漿EPCRの測定)
正常なヒト血漿EPCR濃度は、上記のように約100ng/ml(98.4±27.8
ng/ml;2.5nM)である。エリテマトーデスに罹患した患者由来のサンプルのパネル(n=54)をアッセイし、そして可溶性EPCRレベルが、検出不能レベルから1,
700ng/ml以上までの範囲であることを見出した。15人の患者は、200ng/ml以上の可溶性EPCRレベルを有した。
【0077】
以前の研究は、内皮損傷に起因する、狼瘡患者における上昇した可溶性血漿TMレベルを示し、そして最新の狼瘡患者サンプルを、参照として血漿TMについてアッセイした。それらの可溶性TMレベルが、図5によって示されるように、それらの可溶性EPCRレベルと完全に相関を有さないことが見出された。このことは、可溶性血漿EPCRの供給源が、単に、ランダムに損傷を受けた内皮由来ではないことを示唆する重要な観察である。TMとは対照的に、ヒトおよび霊長類における膜結合EPCR発現は大きな脈管の内皮に主に制限され、毛細管はほとんどEPCRを発現しない。EPCRの特有の局在化は、大きな脈管の血栓症を予防するために、局所的にプロテインC活性化を増大すると予測される。大きな脈管への膜結合EPCRの主な局在化は、可溶性血漿EPCR濃度によって予測され得る大きな脈管における標的化された血栓症の危険を示す。
【0078】
(実施例4:敗血症性ショック患者における血漿可溶性EPCR)
敗血症(accp/sccm consensus conference,ches
t 1992;101:1644−1655)は、以下の臨床的発現のうちの1つより多くを含むが、それらに限定されない、感染に対する全身性炎症応答として定義される:
1)38℃以上または36℃未満の体温;
2)1分あたり90拍以上の心拍数;
3)以下によって示される頻呼吸:
a)1分あたり20呼吸以上の呼吸率;
b)32mmHg未満のPaCO2の過度呼吸;
4)12,000/mm3以上もしくは4,000/mm3未満のWBCカウント、または10%以上の未熟好中球(バンド)の存在。
【0079】
サンプルを、器官機能不全、低灌流(hypoperfusion)、または低血圧と関連する敗血症によって規定されるような、重篤な敗血症を伴うか、または伴わない、手術後の合併症を有する患者から得た。灌流異常は、乳酸アシドーシス、尿量過少、または精神状態における急性変化を含み得る。敗血症性ショックは、十分な流体(fluid)蘇生、および心臓性ショックの非存在にもかかわらず、24時間以上の間、血管作用薬を
必要とする低血圧を伴う敗血症をいう。
【0080】
本研究において含まれるすべての患者は、以下の診断基準を満たした:
a)呼吸の支持(24時間以上の制御された換気)およびまたは血流力学的支持(5マイクログラム/Kg/分以上の強心薬、ドパミン、もしくはドブタミン、および/または血
管作用性アミン、エピネフリン、もしくはノルエピネフリンの要求)を必要とする、敗血症および/または手術後の合併症に起因する、集中治療室への入室;
b)18歳〜75歳の間の年齢;
c)70%未満の抗トロンビン活性(局所にて試験される)。
患者が、多くの外傷、肝硬変、または急性肝不全、末期ガン、免疫不全、白血病、妊娠、またはヘパリン治療を有する場合、その患者を除外した。
【0081】
患者の血液サンプルを、時間0(集中治療室ICUへの入室)の時点および抗トロンビンII(ATIII)または偽薬での処置後2日目および6日目に採取した。血漿可溶性EPCRおよび可溶性トロンボモジュリン(TM)を、時間0のサンプルに関してのみアッセイした。
sEPCR:
正常:133.4±53.4ng/ml(平均±標準偏差)
敗血症:224.9±74.5ng/ml
平均間の有意な差あり、P=0.00009
sTM:
正常:35.5±20.4ng/ml(平均±標準偏差)
敗血症:39.9±73.1ng/ml
平均間の有意な差なし、P=0.81
血漿中のsEPCRレベルとsTMレベルとの相関関係なし、r2=0.34。
【0082】
これらの結果を、図6にグラフで示す。狼瘡患者と同様に、敗血症に罹患した患者は、可溶性TMレベルと相関せず、血漿EPCRレベルにおいて非常に顕著な上昇を示す。
【0083】
可溶性血漿EPCRが、プロテインC活性化および活性化したプロテインC抗凝固活性の両方を阻害するという観察によって、これらの患者における上昇した血漿EPCRレベルが、さらなる血栓の危険性を引き起こし、そして脈管損傷/応答の証拠を表すことが示
される。これらが示す状態の例は、内皮細胞刺激、アテローム発生、白血球接着、およびプラークの破裂に関連する障害を含む。
【0084】
(実施例5:ヒヒ組織およびヒト組織におけるEPCRの選択的スプライシング形態の同定)
可溶性EPCRイソ型が、選択的スプライシング機構によって生成され得るか否かを決定するための最初のアプローチとして、RNAを、ヒト組織およびヒヒ組織から単離し、そして逆転写酵素−PCR(RT−PCR)を遺伝子特異的プライマーを用いて実施した。ヒヒEPCRゲノム配列は公知ではないが、ヒヒとヒトとは進化のスケールにおいて密接に関連しているという論理に基づいて、ヒト配列に基づくプライマーを使用した。
【0085】
一般にRT−PCR手順において、総RNAをホモジナイズした組織から単離する。RNAを、特定のアンチセンスプライマー、ヌクレオチド、および逆転写酵素と混合する。混合において、RNAは、逆転写酵素についての鋳型として作用し、第1鎖cDNAを作製する。次いで、この新しいcDNA鋳型を、特定のプライマーおよびTaqポリメラーゼを用いて従来のPCRによって増幅する。EPCRの膜形態(424bp)および推定の選択的スプライシング産物(674bp)の両方を増幅するプライマーを選択した。EPCRの両方の形態に対応する産物を、種々のヒヒ組織(図4)ならびにヒトの肺および
胎盤から増幅した。逆転写酵素を含まないコントロール、および組織との反応における1,885bpのバンドの欠失によって判断されるように、ゲノムDNAに関する可能性の
ある夾雑は存在しないようであった。
【0086】
ヒヒゲノムDNA配列が、選択的スプライシングのための適切なエキソン−イントロン境界およびイントロン−インフレームリーディング配列を有することを確認するために、ヒヒ腎臓ゲノムDNA由来のイントロン配列を、従来のPCRによって増幅した。ゲノム構造は種間で保持されるという前提を立て、そしてイントロンIIIに隣接するプライマー(ヒト配列)を使用した。これは、選択的スプライシング配列を含むと考えられるイントロン(ヒト配列)である。ヒヒ腎臓組織をホモジナイズし、そしてDNAを抽出した。DNAを、特定のプライマーと混合し、そして産物をPCRによって増幅した。DNA産物を精製し、そしてアガロースゲルで電気泳動した。
【0087】
(手順の詳細)
A.EPCR ELISA:被覆する抗体は、EPCRのリガンド結合ドメインに結合する1494mAbである。検出する抗体は、ビオチン化1495mAbであり、これはプロテインC/APC結合をブロックせず、そして1494mAbと交差反応しない。検
出系は、ストレプトアジビン−アルカリホスファターゼおよびBluePhos基質(KPL製)である。
【0088】
B.組織のRT−PCR:組織(50〜100mg)をTrizol(Gibco BRL)中にホモジナイズした。RNAを含む上層を、クロロホルムで抽出し、イソプロパノールで沈殿させ、洗浄し、そしてDEPC−水中に溶解した。RNA(1〜5μg)を、適切な緩衝液中で、製造業者の指示書に従って、ヌクレオチド、CREAアンチセンスプライマー、および逆転写酵素と混合した(第1鎖cDNA合成のためのSuperscriptTM Preamplification system,Gibco BRL)。cDNA産物を、CRESおよびCREAプライマーを用いて、従来のPCRによって、30サイクル増幅した。cDNA産物を、標準的な手順を用いて、クロロホルム抽出およびアルコール沈殿によって精製し、水中に溶解し、そして2%アガロースゲルで電気泳動した。ゲルを、Vistra Green(Amersham)で染色し、そしてリン光画像機(phosphoimager)(StormTM scanner,Molecular Dynamics,Inc.)で画像化した。
【0089】
C.ヒヒゲノムDNAのPCR:ヒヒ腎臓DNA(82mg)を、Trizol試薬中にホモジナイズした。DNAを含む下層を、抽出し、沈殿させ、そして滅菌水中に溶解した。DNAを、緩衝液、ヌクレオチド、ならびにHRT−1およびHRT−2プライマーとの混合液中で、従来のPCRによって、30サイクル増幅した。増幅したDNAを、標準的な手順を用いて、抽出し、沈殿させ、滅菌水中に溶解し、そして2%アガロースゲルで電気泳動した。単一のバンド(465bp)を、エチジウムブロミドで可視化し、切り出し、そしてPCR産物を、製造業者の指示書(Qiagen)に従ってスピンカラムで精製した。PCR産物を、同じプライマーを用いて配列決定した。
【0090】
(D.プライマー配列)
CRES:5'−TCGTGCGCCTGGTGCACCAGGAGC−3'
(エキソンIIの末端近くの5'センスプライマー)
CREA:5'−CGCCGTCCACCTGTGCACAGGAAG−3'
(エキソンIV内の3'アンチセンスプライマー)
HRT−1:5'−AGCAGCTCAATGCCTACAACCG−3'
(エキソンIIIの末端近くの5'センスプライマー)
HRT−2:5'−CCGTAGAAGGACACGTGTCCACCTGCCGC−
3'
(エキソンIV内の3'アンチセンスプライマー)
(ヒヒ組織に関する結果)
ゲルから切り出され、精製され、そして配列決定された腎臓ゲノムDNAから増幅された単一のバンドが存在した。その配列は、ヒト配列に対して92%同一であり、そしてエキソン−イントロン境界は保存された。このイントロン配列における高レベルの類似性は重要である。なぜなら、イントロン配列は、代表的には種間において十分には保存されないからである。終止コドンを有するイントロン内のインフレームリーディング配列もまた存在し、このことは、ヒヒの選択的にスプライシングした可溶性タンパク質中の特有の22残基のカルボキシル末端テイルを予測する。
【0091】
推定の可溶性EPCRイソ型が、特有のカルボキシル末端テイルを有するという観察は、イソ型特異的抗体を使用するイソ型間の区別のための構造的差異を提供する。実用モデルとしては、タンパク質分解された可溶性EPCRの血漿レベルが内皮損傷を報告する一方で、選択的スプライシングを受けた可溶性EPCRのレベルが刺激に対する内皮応答を報告するというものである。可溶性EPCRイソ型の相対的血漿レベルが、特定の病理学における大きな脈管の内皮機能不全および損傷に関する情報を提供することが期待される。
【0092】
(ヒト組織に関する結果)
ヒト組織(胎盤、肺、および舌)由来のRT−PCR産物を、EPCRに特異的なCRES/CREAプライマーを用いて電気泳動した。手順は、ヒヒ組織について使用された
手順と同じであった。EPCRの膜イソ型(mEPCR)および選択的スプライシングを受けた可溶性EPCRイソ型(sEPCR)に対応する産物を観察した。産物は、ヒヒ組織を用いて観察された産物と本質的に同じようである。唯一の差異は、胎盤組織がさらなる産物を有するようであることである。
【0093】
(配列表)
【0094】
【表2】
【0095】
【表3】
【0096】
【表4】
【0097】
【表5】
【0098】
【表6】
【0099】
【表7】
【0100】
【表8】
【図面の簡単な説明】
【0101】
【図1】図1は、膜結合レセプターをタンパク質分解して細胞外ドメインを遊離しそして膜アンカーを残すこと、およびmRNAの選択的スプライシングによって、EPCRに適用されるような可溶性レセプターを産生するための2つの既知のメカニズムの概略図であり、膜結合EPCR(mEPCR)およびタンパク質分解血漿EPCR(pEPCR)に特有の配列、ならびに可溶性EPCR(sEPCR)に特有の配列を示す。
【図2】図2は、mEPCRおよびsEPCRを比較する概略図であり、ヌクレオチドインサートおよびsEPCRに特有のコードされたアミノ酸配列を示す。
【図3】図3は、選択的スプライシングによってヒト、ウシ、マウス、およびヒヒのEPCRに挿入された配列を示す。
【図4】図4aは、可溶性血漿EPCRがヒトプロテインCおよびAPCに結合することを示すグラフである。EA.hy926細胞を、氷上で30分間0〜500nMrsEPCR(黒丸)または血漿EPCR(白丸)の存在下で、60nM fl−APCと共にインキュベートした。細胞を洗浄し、そして細胞結合した蛍光を、記載のようにフローサイトメトリーによって決定した。fl−APCを添加しない場合での固有の細胞蛍光を、矢印で示す。プロットした平均細胞蛍光(MCF)は、2連のMCF測定の平均を表す。 図4bおよび4cは、可溶性血漿EPCRおよびrsEPCRが、細胞表面上でプロテインC活性化を阻害することを示すグラフである。図4bでは、EA.hy926細胞単層を、0.1μMプロテインC単独(白四角)、または1μM rsEPCR(黒丸)、または2μg/ml 1494mAb(白丸)と共に、室温にて15分間プレインキュベートした。プロテインC活性化を、トロンビン(2nM最終)の添加によって開始し、そして反応を、所定の時間で停止した。活性化プロテインCを、アミド分解アッセイで決定し、そして活性速度をmOD/分で各時点についてプロットした。トロンビンを添加しないコントロールウェルを含ませた(黒四角)。各データ点は、3連のウェル測定の平均を表す。図4cでは、EA.hy926細胞単層を、0.1μMのプロテインCおよび示された濃度の血漿EPCR(白丸)またはrsEPCR(黒丸)と共に、室温にて15分間プレインキュベートした。トロンビン(最終2nM)を添加し、そして活性化を室温にて60分間行った。上清を、アンチトロンビンおよびヘパリンの混合物に添加し、そして活性化プロテインC活性(mOD/分)を、アミド分解アッセイで決定した。各データ点は、3連のウェル測定の平均を表す。 図4dは、可溶性血漿EPCRがAPC抗凝固活性を阻害することを示すグラフである。APC(25nM)の抗凝固活性を、460nMの血漿EPCRまたはrsEPCRの存在下で、1段階Xa凝血アッセイを用いて決定した。この効果は、いずれかの可溶性EPCRを、EPCRへのAPCの結合をブロックする、42μg/mlの1496mAbと共に、5分間プレインキュベートした場合には逆転した。データは、4〜6測定の平均±S.D.を表す。
【図5】図5は、狼瘡患者において、可溶性血漿TMのレベルを可溶性血漿PCRと比較するグラフであり、TM値とEPCR値との間に相関はないが、狼瘡患者の大多数が、可溶性血漿EPCRの非常に上昇したレベルを示すことを証明する。
【図6】図6は、sTM(正常)、sTM(敗血症)、sEPCR(正常)、およびsEPCR(敗血症)についての可溶性レセプター濃度(ng/ml)のグラフである。sTMとsEPCRとの間に相関はない(r2=0.034)。
【特許請求の範囲】
【請求項1】
明細書に記載の発明。
【請求項1】
明細書に記載の発明。
【図1】


【図2】
【図3】
【図4】

【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2008−164635(P2008−164635A)
【公開日】平成20年7月17日(2008.7.17)
【国際特許分類】
【出願番号】特願2008−66863(P2008−66863)
【出願日】平成20年3月14日(2008.3.14)
【分割の表示】特願2006−200128(P2006−200128)の分割
【原出願日】平成10年6月26日(1998.6.26)
【出願人】(594003676)オクラホマ メディカル リサーチ ファウンデーション (14)
【氏名又は名称原語表記】OKLAHOMA MEDICAL RESEARCH FOUNDATION
【住所又は居所原語表記】825 N.E. 13th Street,Oklahoma City,Oklahoma 73104,United States of America
【Fターム(参考)】
【公開日】平成20年7月17日(2008.7.17)
【国際特許分類】
【出願日】平成20年3月14日(2008.3.14)
【分割の表示】特願2006−200128(P2006−200128)の分割
【原出願日】平成10年6月26日(1998.6.26)
【出願人】(594003676)オクラホマ メディカル リサーチ ファウンデーション (14)
【氏名又は名称原語表記】OKLAHOMA MEDICAL RESEARCH FOUNDATION
【住所又は居所原語表記】825 N.E. 13th Street,Oklahoma City,Oklahoma 73104,United States of America
【Fターム(参考)】
[ Back to top ]