
可溶性CEAに対する抵抗性を有する医薬組成物
本発明は、ヒトにおける上皮性腫瘍の治療のための医薬組成物に関し、前記医薬組成物は、ヒトCD3と特異的に結合する第1の結合ドメインとヒトCEAと特異的に結合する第2の結合ドメインを有する二重特異性一本鎖抗体を含み、前記第2の結合ドメインは、少なくともCDR−H3の一部分またはマウスモノクローナル抗体A5B7の完全なCDR−H3を含む。さらに、前記医薬組成物の製造のためのプロセスならびにヒトCD3抗原およびヒトCEA抗原に特異性を有する特異的な二重特異性一本鎖抗体分子の医学的/製薬的使用が開示される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、可溶性CEAに対する抵抗性を有する医薬組成物に関する。
【背景技術】
【0002】
ゴールドおよびフリードマンが最初にヒト結腸癌組織抽出物における腫瘍関連癌胎児性抗原(CEA)を記載してから30年以上が経過した(Gold and Freedman; J. Exp. Med. 122 (1965); 467-481)。
その間、CEA遺伝子ファミリーに関連する28のその他の遺伝子/偽遺伝子が見出された。CEA遺伝子ファミリーのメンバーに用いる命名を簡略化するために、該ファミリーは最近「CEA関連細胞接着分子」(CEACAM)と改名され、そのメンバーの命名が統一された(Beauchemin, Exp. Cell Res. 252 (1999), 243-249)。例えば、この命名に従って、ヒトCEA(CD66e)はCEACAM5と名付けられる。
【0003】
ヒトCEA遺伝子ファミリーは、染色体19q13.2の上に集まる(Olsen et al., Genomics 23 (1994); 659-668)。その29遺伝子および偽遺伝子は3つのサブグループ、すなわち、7つの発現遺伝子を含有するCEAサブグループ、11の発現遺伝子を含有する妊娠特異的糖タンパク質(PSG)サブグループおよび偽遺伝子のみを含有する第3のサブグループに分割することができる(Hammarstroem, Sem. Cancer Biol. 9 (1999), 67-81; Beauchemin, Exp. Cell Res. 252 (1999), 243-249)。CEAのアミノ酸配列および他のファミリーメンバーの解析により、それらが免疫グロブリン(Ig)スーパーファミリーに属することが明らかとなった(Williams and Barclay, Annul. Rev. Immunol. 6 (1988), 381-405)。CEAサブグループの全てのメンバーは細胞表面膜に付着している。胆汁糖タンパク質(CEACAM1;BGP1;TM−CEA;CD66a)、CEA遺伝子ファミリーメンバー1(CEACAM3;CGM1;CD66d)およびCEA遺伝子ファミリーメンバー7(CEACAM4;CGM7)は疎水性の膜貫通ドメインを有するのに対し、癌胎児性抗原(癌胎児性抗原関連細胞接着分子5;CEACAM5;CEA;CD66e)、非特異的交差反応性抗原(CEACAM6;NCA;NCA−50/90;CD66c)、CEA遺伝子ファミリーメンバー2(CEACAM7;CGM2)およびCEA遺伝子ファミリーメンバー6(CEACAM8;CGM6;CD66b)はグリコシルホスファチジルイノシトール(GPI)脂質部分によって血漿膜と繋がれている。CEAタンパク質は、Igドメインの数に応じて約300kDaまでの分子量で高度にグリコシル化されている。
【0004】
CEAタンパク質の生物活性に関して、腫瘍細胞株を用いるインビトロ研究から、胆汁糖タンパク質、CEAおよび非特異的交差反応性抗原を含むいくつかのCEAサブファミリーは、腫瘍細胞表面に発現すると、同種親和性の異型細胞接着分子として作用することができることが示唆された(Oikawa et al., Biochem. Biophys. Res. Commun. 186 (1992), 881-887;Zhou et al., Cancer Res. 53 (1993), 3817-3822)。より最近には、微生物による攻撃から結腸を守る自然免疫応答におけるCEAおよび非特異的交差反応抗原の可能性のある役割が考察されている(Hammarstroem and Baranov, Trends Microbiol. 9 (2001), p. 119-125)。特に、これらのタンパク質は微生物と結合および捕獲することにより、それらが微絨毛の上皮細胞に到達し、侵入することを防ぐことが提唱されている。
【0005】
CEAは、胎児期の間に発現し、健常成人には存在せず、再び癌に発現する癌胎児性抗原であると仮定された。しかし、CEAは、正常成人組織にも発現する。例えば、胆汁糖タンパク質、CEA、非特異的交差反応性抗原およびCEA遺伝子ファミリーメンバー2は、正常ヒト結腸に、特に腸管内腔に面している成熟した円柱上皮細胞におよび陰窩口(crypt mouth)で高度に分化した細胞に発現する(Fraengsmyr et al., Cancer Res. 55 (1995), 2963-2967;Fraengsmyr et al., Tumor Biol. 20 (1999), 277-292)。より具体的には、これらのタンパク質は、フリーな管腔表面の内側を覆う成熟した結腸細胞の刷子縁多糖外被に局在する。胆汁糖タンパク質、CEAおよび非特異的交差反応性抗原はまた、多数の上皮性起源の腫瘍にも発現する(Hammarstriem, Sem. Cancer Biol. 9 (1999), 67-81; Shively and Beatty CRC Crit. Rev. Oncol. Hematol. 2 (1985), 355-399)。
【0006】
既に1970年代後半および1980年代前半に、CEAは結腸直腸およびその他の上皮腫瘍の放射免疫学的局在に好まれる標的抗原となった。これは、CEAが胃腸癌および膵臓癌の95%に、ならびに大部分の小細胞および非小細胞肺癌腫に過剰発現するという事実に起因する。また、それは乳癌ならびに頭部および頚部の扁平上皮癌にも発現する(Primus et al., Cancer 42 (1978), 1540-1545)。実際に、CEAは最も広く用いられている臨床腫瘍マーカーの1つである。それは、その安定性、正常成人組織での相当に制限された発現、ならびに上皮性起源の腫瘍での高度な発現のために、結腸直腸癌およびその他のいくつかの癌の血清腫瘍マーカーとして用いられている。健康な個体においてCEAの大半は結腸で産生される。そこでそれは成熟した円柱細胞の頂端膜側から腸管内腔に放出され、糞便の中で消失する。従って、健康な個体からは、血液中に極めて低いレベルしか通常見出されない。例えば、健康な個体の血液中のCEAレベルは2μg/l未満である。それに対して、結腸直腸癌およびその他の癌腫の患者からの血清中のCEAレベルは2000μg/lを超える範囲まで増加している(Thomson et al., PNAS 64 (1969), 161-167)。特に、進行性、悪性、または後期の上皮腫瘍は、しばしば高い血清濃度の可溶性CEAを伴う(Fletcher; Ann. Intern. Med. 104 (1986), 66-73)。CEAを含む血漿膜からの成分は、血漿膜由来小胞のように表面から継続的に剥脱することが知られ(Taylor and Black, J. Natl. Cancer Inst. 74 (1985), 859-866;Sack et al., J Clin Invest. 82 (1988), 586-93)、それはリンパ管および血管を流れて最後に血液に行き着き得る。腫瘍サイズが増大するにつれて、より多くのCEAが血液に蓄積する。腫瘍マーカーとしての血清CEA決定の主な用途は、結腸癌の手術後のサーベイランスにある。増加したCEAレベルは、患者のそれぞれ81%(Minton et al., Cancer 55 (1985), 1284-1290)および89%(Wanebo et al., Surg. Gynecol. Obstet. 169 (1989), 479-487)において、再発性疾患の最初の指標であった。血清CEAレベルはまた、予後指標としても用いることができる(Mulcahy and Benson, Curr. Oncol. Rep. 1 (1999), 168-172)。
【0007】
多くの上皮癌でのその過剰発現に起因して、CEAは腫瘍マーカーとして用いられるだけでなく、抗腫瘍療法の標的としても用いられる。例えば、胃腸癌は高い割合のヒト上皮腫瘍の原因であり、米国において2001年の胃癌の新規症例は21,700、結腸直腸癌の新規症例は135,400と推定される(Greenlee; CA Cancer J Clin 51 (2001), 15-36)。結腸直腸癌は3番目に頻度の高い悪性腫瘍であり、男性および女性の両方において癌死の3番目の主な原因である(Ries; Cancer 88 (2000), 2398-2424)。これらの腫瘍に対する新規な治療法を見出すために、抗CEAモノクローナル抗体がCEA陽性癌の可能性のある治療法として調査されてきた(Murakami et al., Immunol. Invest. 25 (1996), 23-35)。
【0008】
腫瘍量の少ない患者(低い血清CEAレベルに相当する)が首尾よく治療を受けるアプローチの一例は、ベアらによって行われた研究である。このアプローチでは、ラベツズマブ(labetuzumab)(ラベツズマブは、ヒト化形態の抗CEAモノクローナル抗体MN−14である;Behr et al., Cancer, 94: 1373-1381, (2002), 1559-64)の131I標識変異体が、5−フルオロウラシルおよびフォリン酸に、または肝転移後のアジュバント・セッティングに対して化学療法抵抗性の、少量の転移性疾患を有する30名のCRC患者が登録された第II相試験において分析された。131I標識ラベツズマブの単回注射が行われた。評価可能な19名の患者のうち、3名が部分的緩解を有し、8名がわずかな応答を15ヶ月まで持続して示した。アジュバント・セッティングにおいて、9名のうち7名の患者が3年間まで疾患に罹らなかったのに対して、対照群の再発率は同じ期間で67%であった。患者の血清CEAレベルは3.9〜45ng/mlの範囲であった(Behr et al., Cancer, 94: 1373-1381, 2002)。CEA血清レベルの低い(<5ng/ml)患者を特徴とする別の研究では、131I−ラベツズマブ(上記引用文中)を用いるCEAの放射免疫療法は、肝臓における結腸直腸癌転移の救済切除(salvage resection)後の生存を向上したことが示された。23名の患者に40〜60mCi/m2 131I−ラベツズマブの用量が投与された。5年生存率は、それぞれ処置群について51.3%、対照群について7.4%であった(Liersch et al., JCO, 2005, ASCO Proc, Vol 23, No 16S: 3627)。
【0009】
けれども、高い血清CEA濃度に対応する治療的アプローチは、しばしば低いかまたは全くない抗腫瘍応答という結果をもたらした。例えば、ヒト化抗CEAモノクローナル抗体を臨床で評価するために行われた臨床研究では、MN−14のCDR移植形(version)(hMN−14;Sharkey, Cancer Res. 55 (23 Suppl) (1995) 5935s- 5945s)が131Iで標識された。19名の進行したCEA産生腫瘍患者に131I標識したhMN−14を与えた。hMN−14の体内分布、腫瘍ターゲッティング、および薬物動態挙動は、マウスMN−14に見られるものに類似していた。しかし、血漿中CEAの上昇した(>200ng/ml)患者は、注射後1時間以内に30%を上回る標識抗体が複合体化した。これらの患者の一部では、合併症の増加の結果、血漿中CEAの低い患者において見られるものよりも、血液からのクリアランスが急速な抗体の代謝の促進がもたらされた(Sharkey、前掲)。Yuらにより実施された別の第1相試験では、CEAに対する131I標識した高親和性マウスモノクローナル抗体(mAb)、COL−1(Muraro, Cancer Res. 45 (1985), 5769-80)が胃腸悪性腫瘍の患者において調査された。特に、血清CEAおよび腫瘍体積の薬物動態学への影響が分析された。本目的のため、18名の進行した胃腸悪性腫瘍患者に、20mgの131I標識したCOL−1を10mCi/m2〜75mCi/m2の用量で投与した。血清CEAレベルは6〜2739ng/mLの範囲であった(平均+/−SD、500+/−639)。すべての腫瘍の浸潤した(tumor-involved)器官のうち82%が陽性であり、全ての病変の58%であった。しかし、今回も上昇した血清CEA(>500ng/mL)および腫瘍体積が血清放射能のクリアランスと直接相互に関連があることが見出された。著者らは、高度に上昇した循環CEAレベルおよび/または増大した腫瘍体積を有する患者は、131I標識したCOL−1をより急速に循環から排除すると結論付けた(Yu et al., J. Cli. Oncol. 14 (1996), 1798-1809)。Hajjarらによる、転移性胃腸および大腸直腸癌患者におけるヨウ素131標識ヒト化MN−14抗CEAモノクローナル抗体での研究において同様の結果が得られた。この第1相試験では、21名の患者が事前外部照射の後かまたは標準的な化学療法の後に抗体で処置された。21名のうち7名の患者がヒト抗ヒト抗体(HAHA)を有したが、有害作用はなかった。抗腫瘍応答は認められなかった。この場合も、上昇した血漿CEAレベルが血液および身体全体からの抗体のクリアランスを増大させることが見出され(Hajjar et al., Clin Colorectal Cancer, 2 (2002), 31-42)、そのことは、少なくとも一部分、この研究において見出された抗腫瘍応答の欠如の説明をもたらし得る。血液および身体からの治療用抗体の急速なクリアランスという現象は、器官の損傷を防ぐために身体から急速に取り除かれる必要のある免疫複合体の形成増加により説明され得る。最も可能性が高いと思われるモノクローナル抗体の急速なクリアランスに起因して、これらの研究に登録した腫瘍患者に治療効果を見出すことはできなかった。
【0010】
免疫複合体形成および、大部分が免疫細胞のFc受容体により認識される抗体のFc部分に媒介される、治療用モノクローナル抗体の急速なクリアランスにより生じる問題を回避するために、Fc部分を含まない抗体誘導体(例えばscFv構築物)または断片(例えばFabおよびFab2断片)を産生し、臨床で分析した。これらの研究の大部分は、腫瘍画像形成および検出/局在化を対象とする(例えば、Chester et al., Cancer Chemother Pharmacol, 46 (2000) Suppl: S8-12;Mayer et al., Clin Cancer Res, 6: (2000) 1711-1719; Begent et al., Nat Med, 2 (1996): 979-984参照)。そのような抗体誘導体/断片の治療効力を臨床で調査したものはほんのわずかの研究しかなかった。例えば、Francisらによる臨床アプローチでは(Francis, Br. J. Cancer 87(6) (2002), 600-607)、scFv−カルボキシペプチダーゼ構築物の抗腫瘍活性が調査された。この第1相試験では、酵素プロドラッグ療法(ADEPT)に対する抗体が、進行した結腸直腸癌またはその他のCEA産生腫瘍を有する患者に使用された。本目的のため、抗体酵素標的化薬剤として細菌酵素カルボキシペプチダーゼ(CPG2)と結合した、CEAに対するマウスモノクローナル抗体(A5B7)のF(ab)2断片からなるA5CP、およびZD2767P、ビス−ヨードフェノールマスタードプロドラッグが用いられた。結果として、この研究において臨床応答および放射性応答は見られなかった。処置前の血清CEAレベルは1000ng/mlまでの範囲であった。これらの処置した腫瘍患者の高い血清中CEA濃度は、少なくとも一部分、この研究で観察された抗腫瘍応答の欠如の原因であり得る。
【発明の開示】
【0011】
上に述べた課題を考慮すると、進行性、悪性、または後期上皮腫瘍のための効果的な治療法のための手段および方法の提供が大いに望まれる。
【0012】
従って、本発明の一態様は医薬組成物に関し、前記医薬組成物は、
(a)ヒトCD3と特異的に結合する第1の結合ドメイン、および
(b)ヒトCEAと特異的に結合する第2の結合ドメイン
を有する二重特異性一本鎖抗体を含み、
前記第2の結合ドメインが少なくともアミノ酸配列「DX1X2X3X4FYFDY」(配列番号65)を含み、「X1」、「X2」、「X3」または「X4」は、任意のアミノ酸残基を表し、該アミノ酸残基「D」はマウスモノクローナル抗体A5B7のCDR−H3のKabat位置95に相当し、該アミノ酸残基「FYFDY」は、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置100、100a、100b、101、および102にそれぞれ相当する。一実施形態では、「X1」は、「R」(アルギニン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;「X2」は、「G」(グリシン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表し;「X3」は、「L」(ロイシン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;かつ、「X4」は、「R」(アルギニン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表す。
【0013】
本発明の医薬組成物の一実施形態では、本明細書に定義される二重特異性一本鎖抗体のヒトCEAに特異的な前記第2の結合ドメインは、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102に相当するアミノ酸配列「DRGLRFYFDY」(配列番号66)を少なくとも含む。
【0014】
本発明は、その血漿中の可溶性CEA濃度の高い上皮性腫瘍患者の治療に特に適した手段および方法を提供する。そのような高い可溶性CEA濃度は、進行性腫瘍、再発性、転移性、後期腫瘍を有する上皮性腫瘍患者および高腫瘍量の患者の血清/血漿に見出される。アミノ酸配列「DRGLRFYFDY」(配列番号66)を含むCEA結合ドメインを含む二重特異性一本鎖抗体はCEA陽性標的細胞と結合するだけでなく、可溶性CEAとも結合することが見出された;本発明の実施例3および図2;ならびにEP B1 491031号参照。示されるアミノ酸配列「DRGLRFYFDY」は、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102(配列番号66)に相当する(Harwood, Br J Cancer. 54 (1986), 75-82)。驚くことに、可溶性CEAと結合しているにもかかわらず、前記二重特異性一本鎖抗体は、高い濃度の可溶性CEAの存在下でさえも(最大1μg/mlの可溶性CEAを試験)、CEAを有する腫瘍細胞を死滅させる。言い換えれば、前記二重特異性構築物は、可溶性CEAによって、CEA陽性の腫瘍細胞に対するそれらの細胞障害活性を阻害されない。
【発明を実施するための最良の形態】
【0015】
以下の実施例5および8(併せて図5、6、8、10、19、20、22および27)に示されるように、アミノ酸配列「DRGLRFYFDY」を含むCEA結合ドメインを含む二重特異性一本鎖抗体は、高い濃度の可溶性CEAの存在下でさえもCEA陽性腫瘍細胞に対する細胞障害性を媒介した。例えば、図10は、漸増量の可溶性CEA抗原の存在下で、KatoIII細胞(CEA陽性のヒト胃癌細胞株)に再指向されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイを示す。刺激を受けたヒトCD8陽性細胞障害性T細胞(CTL)をエフェクター細胞として用いた。CEAI VHVLx配列番号77VHVLに媒介される細胞障害性は、可溶性CEAに対して抵抗性である。対照的に、CEAII VHVLx配列番号77VHVLに媒介される細胞障害活性は、漸増量の可溶性CEAにより阻害される。CEAIは、マウスmAb A5B7に由来する可変領域であるのに対して、CEAII VHVLは、mAb T84.66に由来する。
【0016】
重要なことに、抗CEAx抗CD3二重特異性一本鎖抗体のヒトCEA結合ドメイン(すなわち、ヒトCEAと特異的に結合するヒト結合ドメイン)で使用される場合、アミノ酸配列「DRGLRFYFDY」が可溶性CEAに対する抵抗性を媒介するために十分であることが見出された;例えば、図19、20、22および27参照。下記において、本明細書に定義される二重特異性一本鎖抗体は、従って可溶性CEA抗原に対して抵抗性であると称される。本明細書において、用語「可溶性CEA抗原に対する抵抗性」、「可溶性CEAに対する抵抗性」または関連する用語は、前記二重特異性一本鎖抗体により媒介されるCEA陽性標的または腫瘍細胞に対する細胞障害性が漸増濃度の可溶性CEAにより影響を受けないという事実を指す。特に、細胞障害活性は、高い濃度の可溶性CEA(最大1μg/mlまでを試験)にさえ阻害されない。上に述べたように、健康な個体の血液中のCEAレベルは、2ng/ml未満である。腫瘍患者の血清/血漿中の高い可溶性CEA濃度は、進行性、再発性、転移性、または後期腫瘍および高腫瘍量の患者に特有である。従って、本発明は、その血漿中の可溶性CEA濃度がそのように高い上皮性腫瘍患者の治療に特に適した手段および方法を提供する。本明細書において、用語「高い可溶性CEA濃度」は、10、20、50、70、80、90または100ng/mlよりも高い可溶性血清/血漿CEA濃度を示す。この血清/血漿CEA濃度は、特に、ELISAにより測定することができる。好ましくは、前記可溶性血清/血漿CEA濃度は、例えばELISAにより測定して、100ng/mlよりも高い。
【0017】
可溶性CEA抗原に抵抗性を有する前記二重特異性一本鎖抗体の作製は、以下の実施例から明らかなように平凡な作業ではなかった。例えば、膜結合CEAと結合するが可溶性CEAとは結合しないことの知られているモノクローナル抗体(mAb)に由来するCEA結合ドメインを有する二重特異性一本鎖抗体、すなわち、mAb PR1A3(Durbin, Proc Natl Acad Sci U S A. 91 (1994), 4313-7)は、産生させることができなかった。二重特異性一本鎖抗体形式中で用いた場合、抗CD3x抗CEA二重特異性一本鎖構築物の発現/分泌は達成できなかった。PR1A3のヒト化形(version)(Durbin、前掲)を作製に利用した場合、二重特異性一本鎖抗体構築物は発現し、宿主細胞から分泌された。しかし、抗CEA結合ドメインと膜結合CEAの結合は得られなかった。
【0018】
十分に説明されているモノクローナル抗体T84.66(Neumaier, M. et al., Cancer Res 50 (1990), 2128-34)またはMFE−23(Boehm, M. K. Biochem J 2 (2000), 519-28)に由来する二重特異性一本鎖抗体を作製した場合、これらの二重特異性抗体は可溶性CEA抗原に対し高い感受性があった、すなわち、CEA陽性標的または腫瘍細胞に対するそれらの細胞障害活性は、可溶性CEA抗原の存在下で阻害された。前記構築物は可溶性CEAと結合することが見出されていることから、可溶性CEA抗原は抗体が膜結合CEAと結合することを妨げ、それにより抗体に媒介される細胞障害活性を阻害するとの結論が下された。例えば、図7は、可溶性ヒトCEAの存在下で、CEAをトランスフェクトしたCHO細胞に再指向されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイを示す。刺激を受けたヒトCD8陽性細胞障害性T細胞(CTL)をエフェクター細胞として用いた。CEAII VHVLx配列番号77VHVLの細胞障害活性は、漸増量の可溶性CEAにより明らかに阻害される。CEAII VHVLはmAb T84.66に由来する;配列番号77は抗CD3VH−VLドメインである。
【0019】
可溶性CEA抗原に対する抵抗性は二重特異性一本鎖抗体にのみ見出すことができ、そのCEA結合ドメインはマウスモノクローナル抗体A5B7のCDR−H3のアミノ酸配列「DRGLRFYFDY」を含んだ(Harwood, Br J Cancer. 54 (1986), 75-82)。MFE−23由来およびT84.66由来の二重特異性一本鎖構築物に関して、A5B7由来の二重特異性一本鎖抗体は可溶性CEAと結合する。MFE−23由来およびT84.66由来の二重特異性一本鎖構築物に関して得られた結果を鑑みると、可溶性CEAは、A5B7由来一本鎖二重特異性抗体構築物において細胞障害活性に影響を及ぼさないことが予期され得る。
【0020】
上に述べたように、ヒトにおけるCEAを有する上皮性腫瘍に対して向けられる多くの治療的アプローチは、癌患者の血漿中に高いレベルの可溶性CEA抗原が存在することにより著しく妨害される。例えば、血漿中の高いCEA濃度の存在下での、増加した免疫複合体形成および治療用抗CEAモノクローナル抗体のクリアランスは、いくつかの臨床研究で観察されている。さらに、可溶性CEA抗原(しばしば、進行性腫瘍、再発性癌、転移性腫瘍、高腫瘍量、または後期腫瘍の癌患者の血清中に高い濃度で存在する)は、CEA陽性腫瘍細胞に対して向けられる治療薬を阻害し、従って腫瘍細胞の認識および破壊を妨げる。従って、腫瘍に到達する治療薬の実際の量は減少し、結果として抗腫瘍活性が低下し、低いかさらに全くなくなる。この制限は、これまでのところ、例えば、治療薬と腫瘍細胞の相互作用を妨げる可能性が低い、極めて低い量の可溶性CEA抗原を有する患者に対する抗体に基づくアプローチを制限する。
【0021】
本発明において、ヒトCD3およびヒトCEAに特異的な二重特異性一本鎖抗体治療薬を作製することが可能であることが見出され、その腫瘍細胞に対する細胞障害活性は、高濃度の可溶性CEA抗原(最大1μg/mlの可溶性CEAを試験)に対してさえも抵抗性である。この知見は、本発明の二重特異性一本鎖抗体が可溶性CEA抗原と結合するという事実を考えると、全く予期せぬものである(本発明の実施例3および図2;ならびにEP B1 491031号も参照)。それにもかかわらず、本明細書に定義される二重特異性一本鎖抗体は、腫瘍細胞に対するその細胞障害活性において、高レベルの可溶性CEAの存在に対してさえも完全に抵抗性である。従って、本発明は、例えば、腫瘍進行中に観察される、血漿中の可溶性CEA濃度の高い腫瘍患者の治療に、再発性癌に、転移に、高腫瘍量の患者に、または後期腫瘍に特に適した手段および方法を提供する。
【0022】
本発明によれば、用語「医薬組成物」は、ヒト患者への投与のための組成物に関する。好ましくは、医薬組成物は、担体、安定剤および/または賦形剤からなる適した製剤を含む。好ましい実施形態では、医薬組成物は、非経口、経皮、管腔内、動脈内、くも膜下腔内および/または鼻腔内投与のための、または組織への直接注射による組成物を含む。前記組成物は、注入または注射によって患者へ投与されることが特に想定される。適した組成物の投与は、様々な方法によって、例えば、静脈内、腹腔内、皮下、筋肉内、局所または皮内投与によって達成されてよい。本発明の組成物は、製薬上許容される担体をさらに含んでよい。適した医薬担体の例は当分野で周知であり、それにはリン酸緩衝生理食塩水溶液、水、様々な種類の湿潤剤、滅菌溶液、リポソームなどが含まれる。このような担体を含む組成物は、周知の従来法によって処方されてよい。これらの組成物は、被験体に適した用量で投与することができ、それは、本明細書に記載される可溶性血清CEA抗原に対して抵抗性を示す二重特異性一本鎖抗体の漸増用量の投与による用量漸増試験により決定することができる。上記に示したように、本明細書に記載される、可溶性血清CEA抗原に対する抵抗性を有する二重特異性一本鎖抗体は、高いCEA血清中濃度を有する癌患者、例えば、進行性腫瘍、再発性癌、転移性腫瘍、腫瘍量の大きな腫瘍、または後期腫瘍の治療に有利に用いることができる。これらの組成物はまた、その他のタンパク質性および非タンパク質性薬剤と組み合わせて、例えば併用療法(co-therapy)の形で投与することができる。これらの薬剤は、本明細書に定義される二重特異性一本鎖抗体を含む組成物と同時に投与してもよいし、前記二重特異性一本鎖抗体の投与の前後に適時に定められた間隔および用量で別々に投与してもよい。投与計画は主治医および臨床学的要因により決定される。医学分野で周知のように、任意の患者一人への投薬量は多くの要因によって決まり、それには患者の大きさ、体表面積、年齢、投与される特定の化合物、性別、投与時間および経路、全体的な健康状態、および同時に投与されるその他の薬剤が含まれる。非経口投与のための製剤としては、滅菌水溶液または非水溶液、および懸濁液が挙げられる。非水溶媒の例は、プロピレングリコール、ポリエチレングリコール、植物油、および注射用の有機エステル類、例えばオレイン酸エチルである。水性担体としては、水、水溶液、または生理食塩水および緩衝媒体を含む懸濁液が挙げられる。非経口媒体としては、塩化ナトリウム溶液、リンゲルデキストロース、デキストロースおよび塩化ナトリウム、または乳酸化リンゲル液が挙げられる。静脈内媒体としては、液体および栄養素補充液、電解質補充液(リンゲルデキストロースに基づくものなど)などが挙げられる。防腐剤およびその他の添加剤も存在してよく、例えば、抗菌剤、抗酸化剤、キレート剤、不活性ガスなどが挙げられる。さらに、本発明の組成物は、例えば、好ましくはヒト起源の血清アルブミンまたは免疫グロブリンなどのタンパク質性担体を含んでもよい。併用療法は、本明細書に定義される二重特異性一本鎖抗体に加えて、組成物の意図される使用によって、さらなる生物活性物質を含むことが想定される。このような物質は、胃腸管系に作用する薬剤、抗悪性腫瘍薬として作用する薬剤、化学療法薬、細胞増殖抑制剤(cytostatica)、高尿酸血症を予防する薬剤、免疫反応を阻害する薬剤(例えばコルチコステロイド)、炎症応答を調節する薬剤、循環系に作用する薬剤および/またはサイトカインなどの当分野で公知の薬剤であってよい。
【0023】
好ましくは、本明細書に定義される二重特異性一本鎖抗体は、緩衝液、安定剤および界面活性剤中に処方される。緩衝液は、リン酸緩衝液、クエン酸緩衝液、コハク酸緩衝液または酢酸緩衝液であってよい。安定剤は、アミノ酸および/または糖であってよい。界面活性剤は、合成洗剤、PEGなどであってよい。より好ましくは、本明細書に定義される二重特異性一本鎖抗体は、クエン酸塩、リシン、トレハロースおよびTween80中に処方される。本発明の医薬組成物の希釈液として、等張生理食塩水およびTween80が好ましい。
【0024】
本明細書において、「二重特異性一本鎖抗体」は、2つの結合ドメインを含む単一ポリペプチド鎖を意味する。各「結合ドメイン」は、本明細書において、抗体重鎖由来の1つの可変領域(「VH領域」)を含み、第1の結合ドメインのVH領域は、前記第1の分子、すなわちヒトCD3分子と特異的に結合し、第2の結合ドメインのVH領域は、下により詳細に定義されるヒトCEAと特異的に結合する。2つの結合ドメインは、場合により、一般に約5アミノ酸を含む短いポリペプチドスペーサーにより相互に連結されている。各結合ドメインは、抗体軽鎖(「VL領域」)由来の1つの可変領域をさらに含み、第1および第2の結合ドメインのそれぞれの中のVH領域およびVL領域は、ポリペプチドリンカー(例えば、EP B1 623679に開示およびクレームされる種類のもの)を介して相互に連結されているが、いずれの場合も、第1の結合ドメインのVH領域およびVL領域と、第2の結合ドメインのVH領域およびVL領域を、ともにそれぞれの第1および第2の分子と特異的に結合することが可能であるように、相互に2つで一組とさせるのに十分なほど長い。第1または第2の結合ドメインのV領域の配置は、VH−VLであっても、VL−VHであってもよい。好ましくは、ヒトCD3と特異的に結合する第1の結合ドメインの配置は、以下の実施例に示されるように、VH−VLである。第1の結合ドメインは、第2の結合ドメインに対してN末端に位置してもC末端に位置してもよいことが想定されている。従って、本明細書において定義される二重特異性一本鎖抗体の結合ドメインの配置は、VHCEA−VLCEA−VHCD3−VLCD3、VLCEA−VHCEA−VHCD3−VLCD3、VHCD3−VLCD3−VHCEA−VLCEAまたはVHCD3−VLCD3−VLCEA−VHCEAであってよい。好ましくは、CD3に特異的な前記第1の結合ドメインは、第2の結合ドメインに対してC末端に位置している。より好ましくは、本明細書において定義される二重特異性一本鎖抗体の結合ドメインは、VHCEA−VLCEA−VHCD3−VLCD3またはVLCEA−VHCEA−VHCD3−VLCD3の順序で配置される。さらにより好ましくは、配置はVLCEA−VHCEA−VHCD3−VLCD3である。最も好ましいのは、配列番号34に定義される二重特異性一本鎖抗体構築物A240VL−B9VHx配列番号77VHVLである。本明細書において定義される二重特異性一本鎖抗体の前記第1および/または第2の結合ドメインは、非ヒト起源(すなわち、非ヒト配列に由来)であってよいことが想定されている。例えば、前記第1および/または第2の結合ドメインは、マウスモノクローナル抗体に由来してよい。しかし、マウス抗体に由来する二重特異性一本鎖抗体は、ヒト患者に投与される場合、異物と認識される可能性がある。従って、本明細書において定義される二重特異性一本鎖抗体の前記第1および/または第2の結合ドメインは、ヒト起源(すなわち、ヒト配列に由来)であることが好ましい。そのようなCEAまたはCD3と特異的に結合するヒト結合ドメインは、例えば、ファージディスプレイに基づく技法により同定することができる。例えば、第1(または第2)の結合ドメインのVH領域はヒトVH領域であるのに対して、第1(または第2)の結合ドメインの対応するVL領域は非ヒト起源であってよいことも想定される。そのような結合ドメインは、キメラ結合ドメインとも称され得る。あるいは、前記結合ドメインの1つが非ヒト起源であるのに対して、他方はヒト起源であり、その結果キメラ二重特異性一本鎖抗体がもたらされる。前記第1および/または第2の結合ドメインは、ヒト患者に投与される際に、本明細書に記載される二重特異性一本鎖抗体の免疫原性を低下させるために、さらに修飾してもよい。例えば、本明細書において定義される二重特異性一本鎖抗体の前記第1または第2の結合ドメインのうちの少なくとも1つは、下により詳細に述べられるように、ヒト化、CDR移植、キメラおよび/または脱免疫化またはヒトであってよい。第1および/または第2の結合ドメイン中でVHおよびVL領域を連結するポリペプチドリンカーが脱免疫化されていることも想定される。好ましくは、脱免疫化された第1の結合ドメイン(CD3に特異的)内のVHおよびVL領域を連結するポリペプチドリンカーは、配列「GEGTSTGS(G2S)2GGAD」(配列番号141)を有する脱免疫化されたポリペプチドリンカーである。本明細書において定義される二重特異性一本鎖抗体の前記結合ドメインの一方または両方が、例えば、タンパク質の発現、精製、検出または富化に用いることのできる、いわゆる「タグ」(Flagタグ、c−mycタグ、GSTタグまたはHisタグなど)を有することもさらに想定されている。例えば、Flagタグに最も広く用いられている親水性のオクタペプチドは、現在DYKDDDDKであるが(Chubet and Brizzard, Biotechniques 20 (1996): 136- 141)、最近の研究から、より短いペプチド、DYKDが、M1モノクローナル抗体にほぼ同じ親和性で認識され得ることが示唆されている(Knappik A, Pluckthun A; Biotechniques 17 (1994): 754-761)。Flagタグ、c−mycタグ、GSTタグ、Hisタグなどは、例えば、タグ−VHCEA−VLCEA−VHCD3−VLCD3またはVLCEA−VHCEA−VHCD3−VLCD3−タグなど、二重特異性一本鎖抗体のN末端かまたはC末端のいずれかに位置してよい。発現、検出または精製目的のそのようなタグの起源および特性は当分野で十分説明されている;例えば、Lichty, Protein Expr Purif. 41 (2005), 98-105参照。
【0025】
本明細書において、用語「一本鎖Fv」または「scFv」とは、抗体のVHドメインおよびVLドメインを含む抗体フラグメントを指し、これらのドメインは単一ポリペプチド鎖中に存在する。可変ドメインはVH−VLまたはVL−VHの順序で配置することができる。一般に、Fvポリペプチドは、scFvが抗原結合のための所望の構造を形成することを可能にするポリペプチドリンカーをVHドメインおよびVLドメインの間にさらに含む。scFvの概説には、Pluckthun in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds. Springer-Verlag, New York, pp. 269-315 (1994)参照。特定の実施形態では、本発明は、本明細書において定義される二重特異性一本鎖抗体に由来する抗CEA scFvに関する。
【0026】
本発明によれば、Ig由来抗原相互作用に関する文脈中で用いられる用語「結合ドメイン」または「可変領域」は、抗体、抗体断片またはその誘導体に由来する少なくとも1つのCDRを含む、ポリペプチドの断片および誘導体を含む。本明細書において定義される二重特異性一本鎖抗体のヒトCEAと特異的に結合する第2の結合ドメインは、少なくとも1つのCDR、好ましくはCDR−H3、より好ましくはマウスモノクローナル抗体A5B7のCDR−H3のKabatの位置100、100a、100b、101、および102にそれぞれ相当するアミノ酸配列「FYFDY」(配列番号112)を含むマウスモノクローナル抗体A5B7のCDR−H3の一部分;さらにより好ましくはアミノ酸配列「DX1X2X3X4FYFDY」(配列番号65)(「X1」、「X2」、「X3」または「X4」は、任意のアミノ酸残基を表し、該アミノ酸残基「D」は、マウスモノクローナル抗体A5B7のCDR−H3のKabat位置95に相当し、アミノ酸残基「FYFDY」は、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置100、100a、100b、101、および102にそれぞれ相当する)を含むマウスモノクローナル抗体A5B7のCDR−H3の一部分を含むことは、本発明により想定されている。マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置96(「X1」)、97(「X2」)、98(「X3」)および99(「X4」)にそれぞれ相当する「X1」、「X2」、「X3」または「X4」は、アミノ酸残基「R」(アルギニン)、「G」(グリシン)、「L」(ロイシン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、「S」(セリン)、「W」(トリプトファン)、「F」(フェニルアラニン)または「T」(トレオニン)を表すことが想定されている。本明細書において、「X1」、「X2」、「X3」および「X4」が同じアミノ酸を表す、例えば「X1」、「X2」、「X3」および「X4」がすべて「F」(フェニルアラニン)である場合は、本発明の範囲から除外される。好ましくは、「X1」が、「R」(アルギニン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;「X2」が、「G」(グリシン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表し;「X3」が、「L」(ロイシン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;かつ、「X4」が、「R」(アルギニン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表す。あるいは最も好ましいものは、本明細書において定義される二重特異性一本鎖抗体のヒトCEAと特異的に結合する第2の結合ドメインは、A5B7のCDR−H3のKabatの位置95〜102に相当するアミノ酸配列「DRGLRFYFDY」(配列番号66)を含むA5B7の完全なCDR−H3を含む。以下の実施例に示されるように、前記mAb A5B7由来CDR−H3「DRGLRFYFDY」(配列番号66)アミノ酸配列をCEAと相互作用する第2の結合ドメイン中に含む、本明細書において定義される二重特異性一本鎖抗体の腫瘍細胞に対する細胞障害活性は、可溶性CEA抗原に対して抵抗性であり、それによりその血漿中の血清CEA濃度の高い腫瘍患者の治療を可能にする。CDRの決定は、当業者に公知である;例えば、http://www.bioinf.org.uk/abs/#cdrid参照。抗体中のアミノ酸配列の番号付けは、例えば、当分野に記載されるKabatの番号付けスキームに従って行ってよい;例えば、Kabat, E. A., T. T. Wu, H. M. Perry, K. S. Gottesman, and C. Foeller. 1991. Sequences of Proteins of Immunological Interest, 5th ed. Bethesda, Md.: National Center for Biotechnology Information, National Library of Medicine参照。
【0027】
最も好ましくは、そして添付の実施例に記録されるように、本発明の医薬組成物に用いられる「二重特異性一本鎖抗体」は、脱免疫化された抗CD3結合ドメイン(WO 2005/040220号)と、少なくともマウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102(配列番号66)に相当するアミノ酸配列「DRGLRFYFDY」を含むヒト抗CEA結合ドメインを含む、二重特異性一本鎖Fv(scFv)である。二重特異性一本鎖分子は当分野で公知であり、例えば、WO 99/54440号またはMack, PNAS, (1995), 92, 7021-7025に記載されている。
【0028】
本発明に従って用いられる用語「一本鎖」は、好ましくは、単一の核酸分子によりコードすることのできる共直線性(co-linear)アミノ酸配列の形態で、二重特異性一本鎖構築物の前記第1および第2のドメインが共有結合していることを意味する。
【0029】
本明細書において、「ヒト」とは、種Homo sapiensを指す。「ヒト」分子、例えばヒトCEAまたはヒトCD3(CD3ε)は、従って、Homo sapiensにおいて天然に発現するその分子の変異体である。
【0030】
用語「上皮腫瘍」とは、本明細書において、CEA陽性である上皮性起源の腫瘍を意味する(Cancer Medicine;6th ed.; Kufe, Donald W.; Pollock, Raphael E.; Weichselbaum, Ralph R.; Bast, Robert C., Jr.; Gansler, Ted S.; Holland, James F.; Frei III, Emil., editors. Hamilton (Canada): BC Decker Inc. 2003;http://www.dkfz.de: http://www.krebsinformationsdienst.de/Krebsarten/index.html)。治療し得る上皮腫瘍は、胃腸腺癌、乳腺腺癌または肺腺癌である。前記胃腸腺癌は、好ましくは結腸直腸腺癌、膵臓腺癌、食道腺癌または胃腺癌である。本明細書に示されるように、本発明の医薬組成物は、腫瘍患者の血漿中の高レベルの可溶性CEA抗原を特徴とする、進行性腫瘍、転移、再発性癌、後期上皮腫瘍、上皮腫瘍量の多い患者、またはCEA血清中濃度が100ng/mlより高い腫瘍患者(例えばELISAにより測定)の治療に特に有利である。前記医薬組成物を原発腫瘍の外科切除の後に使用することも本発明の範囲内である。従って例えば、CEA産生上皮腫瘍に由来する散在性の(disseminated)残存腫瘍細胞も、CEAをそれらの微小環境に流し、その周囲では可溶性CEAのレベルも高い。従って、本発明の医薬組成物の細胞障害活性の可溶性CEAに対する抵抗性は、最小残存病変の治療にも有利である。前記医薬組成物は、残りの腫瘍細胞を死滅させるために、(CEA源、すなわち原発腫瘍の除去によって)血清CEAレベルの低下する期間に投与し得ることが想定される。また、前記医薬組成物は、二次性腫瘍または転移の形成によって血清CEAレベルが増大する場合に、原発腫瘍の除去後に有用であり得る。CEA血清中濃度は、例えばCEA ELISAアッセイ(例えばIBL CEA EIA、IBL Hamburg,Germany参照)により決定することができる。上記に示したように、多くの抗体に基づく治療的アプローチにおいて、前記血清CEAは、抗体と腫瘍細胞上の膜結合CEAの結合を阻害し、抗体の活性を遮断し、それにより抗腫瘍療法の成功を悪化させる。
【0031】
本明細書において、用語「特異的に結合する」または「特異的結合」または「〜との/〜に対する特異的反応性」などの関連表現は、本明細書に定義される二重特異性一本鎖抗体の第1および/または第2の結合ドメインの、可能な結合パートナーとしての複数の異なる分子のプールから、前記それぞれの第1および/または第2の分子にのみ結合する、あるいは有意に結合する程度までそれぞれの第1および/または第2の分子を識別する能力を指す。このような結合測定は、例えばBiacore装置で、ELISA、FACS分析などにより日常的に行うことができる。より具体的には、本明細書において定義される二重特異性一本鎖抗体の第1の結合ドメインは、ヒトCD3、好ましくは、ヒトCD3εと結合する。本明細書において定義される二重特異性一本鎖抗体の第2の結合ドメインは、下に述べるように、上皮性腫瘍抗原、すなわち、ヒトCEA(癌胎児性抗原、癌胎児性抗原関連細胞接着分子5;CEACAM5;CD66e)と結合する。用語「特異的結合」は、本発明によれば、二重特異性一本鎖抗体分子が本明細書に定義されるヒト標的分子それぞれの少なくとも2、3、4、5、6、7、8、またはさらにそれより多くのアミノ酸と特異的に相互作用かつ/または結合できることを意味する。前記用語は、抗体分子の特異性、すなわち、本明細書に定義されるヒト標的分子の特定の領域を識別するその能力に関する。抗原相互作用部位とその特異的抗原との特異的相互作用は、例えば抗原の立体構造の変化の誘導、抗原のオリゴマー化などに起因する、シグナルの開始をもたらし得る。さらに、前記結合は、「キーロックの原則(key-lock-principle)」という特異性に例証され得る。従って、抗原相互作用部位および抗原のアミノ酸配列中の特異的モチーフは、それらの一次、二次または三次構造の結果ならびに前記構造の二次修飾の結果として相互に結合する。抗原相互作用部位のその特異的抗原との特異的相互作用は、さらに前記部位と抗原との結合をもたらし得る。
【0032】
抗体の「特異的結合」は、主に2つのパラメータ:定性的パラメータ(結合エピトープ、すなわち抗体が結合する場所)および定量的パラメータ(結合親和性、すなわちそれが結合する場所でどの程度強く結合するか)に特徴付けられる。どのエピトープが抗体によって結合されるかは、例えば公知のFACS方法論、ペプチドスポットエピトープマッピング、質量分析またはペプチドELISAにより有利に決定することができる。特定のエピトープと結合している抗体の強度は、例えば公知のBiacoreおよび/またはELISA法により有利に決定することができる。このような技法を組み合わせることにより、結合特異性の代表的な尺度としてのシグナル対ノイズ比の算出が可能となる。そのようなシグナル対ノイズ比では、シグナルは注目されるエピトープと結合する抗体の強度を表すのに対して、ノイズは注目されるエピトープとは異なる他の非関連エピトープと結合する抗体の強度を表す。好ましくは、注目されるエピトープに対するシグナル:ノイズ比(注目されるエピトープと異なるその他のエピトープに対するものより約50倍高い)は、評価される抗体が注目されるエピトープと特異的な方法で結合する指標として考えることができる、言い換えれば「特異的結合剤」である。
【0033】
本発明に従って用いられる用語「特異的結合」または「特異的相互作用」は、二重特異性一本鎖抗体構築物が類似構造のポリペプチドと交差反応しないか、または本質的に交差反応しないことを意味する。研究中の二重特異性一本鎖のパネルの交差反応は、例えば、注目されるポリペプチドならびに程度の差はあるが(構造的かつ/または機能的に)密接に関連した多数のポリペプチドとの、従来条件下で二重特異性一本鎖抗体構築物の前記パネルの結合(例えば、Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, 1988 ならびに Using Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, 1999参照)を評価することにより試験することができる。例えば、本発明の二重特異性一本鎖抗体の最初の結合ドメインが、ヒトCEA(癌胎児性抗原;CEACAM5;CEA;CD66e)、すなわち可溶性CEA抗原及び膜結合CEAの双方と結合することは本発明の範囲内であるが、それに対して、その他のCEAファミリーメンバー、例えば胆汁糖タンパク質(CEACAM1;BGP1;TM−CEA;CD66a)と結合している二重特異性抗体は、前記範囲から除外される。
【0034】
抗原相互作用部位と特異的抗原との特異的相互作用の例は、その受容体のリガンドの特異性を含む。前記定義は、特にその特異的な受容体との結合の際にシグナルを誘導するリガンドの相互作用を含む。対応するリガンドの例は、その特異的なサイトカイン受容体と相互作用/結合するサイトカインを含む。また、前記定義により特に含まれるものは、抗原相互作用部位と、セレクチンファミリー、インテグリンおよびEGFなどの成長因子のファミリーの抗原などの抗原との結合である。前記相互作用の別の例(それも前記定義により特に含まれる)は、抗原決定基(エピトープ)と抗体の抗原結合部位の相互作用である。
【0035】
用語「〜と結合/相互作用する」もまた、立体構造エピトープ、構造エピトープまたはヒト標的分子の2つの領域からなる不連続エピトープまたはその部分に関連する。本発明の文脈において、立体構造エピトープは、ポリペプチドが折りたたまれて未変性タンパク質となる際に分子の表面上に集まる一次配列において分離した2またはそれ以上の別個のアミノ酸配列により定義される(Sela, (1969) Science 166, 1365 およびLaver, (1990) Cell 61, 553-6)。
【0036】
用語「不連続エピトープ」とは、本発明の文脈において、ポリペプチド鎖の離れた部分の残基から構築される非直線状エピトープを意味する。これらの残基は、ポリペプチド鎖が三次元構造に折りたたまれて立体構造/構造エピトープを構成する際に分子の表面上に集まる。
【0037】
本明細書において「CD3」は、多分子T細胞受容体複合体の一部としてT細胞で発現し、少なくとも5つの異なる鎖、CD3−γ、−δ、−ε、−ζ、および−ηからなる抗原を意味する。T細胞でのCD3のクラスター形成(例えば、固定化された抗CD3抗体による)は、T細胞受容体の関与に似たT細胞の活性化を引き起こすが、そのクローンに典型的な特異性とは無関係である。実際に、大部分の抗CD3抗体はCD3ε鎖を認識する。ヒトCD3εのアミノ酸配列はGenBank受託番号NM_000733に表され、配列番号111を含む。
【0038】
「CEA」とは、上皮性起源の多数の腫瘍に発現した抗原である、癌胎児性抗原(癌胎児性抗原関連細胞接着分子5;CEACAM5;CEA;CD66e)を意味する(Hammarstroem, Sem. Cancer Biol. 9 (1999), 67-81; Shively and Beatty CRC Crit. Rev. Oncol. Hematol. 2 (1985), 355-399)。ヒトCEAのアミノ酸配列は、GenBank受託番号NM_004363に表され、配列番号76を含む。
【0039】
本発明において、驚くことに、ヒトCD3およびヒトCEAに対して特異性を有する抗体に基づく治療薬を生成することが可能であり、腫瘍細胞に対して向けられる細胞障害活性がさらに高い濃度の可溶性CEA抗原に対して抵抗性であることが見出された。この知見は、本発明の二重特異性一本鎖抗体が可溶性CEA抗原と結合するという事実を考えると全く予期されないものである。例えば、モノクローナル抗体T84.66またはMFE−23に由来する二重特異性一本鎖抗体構築物が作製される場合、これらの抗体は可溶性CEA抗原に感受性が高かった、すなわちそれらの細胞障害活性は可溶性CEA抗原の存在下で遮断されていた。可溶性CEAによる前記構築物の細胞障害活性の阻害は、抗体の量を増加させることによっても克服できなかった。これらの構築物はまた、可溶性CEAとの結合が可能であることが見出されている。このことを考えて、可溶性CEA抗原は、抗体がその細胞障害活性を発揮することを妨げると結論付けられた。その一方、本明細書に定義される二重特異性一本鎖抗体は、腫瘍細胞に対するそれらの細胞障害活性においてさらに高いレベルの可溶性CEAの存在に対して完全に抵抗性である。さらに、その高い細胞障害活性に起因して、本明細書において定義される前記二重特異性構築物は、低い濃度であってさえもそれらの生物活性を誘起する。故に、本明細書において定義される二重特異性一本鎖抗体を含む少量の医薬組成物は、血清/血漿中の可溶性CEA濃度が高いことを特徴とする上皮性腫瘍患者において治療効果を達成するために十分である。上皮性腫瘍患者の血清/血漿中の可溶性CEA濃度が高いことは、進行性、再発性、転移性、または後期腫瘍および高腫瘍量の患者の特徴である。さらに一層驚くことに、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102に相当するアミノ酸配列「DRGLRFYFDY」(配列番号66)は、抗CEAx抗CD3二重特異性一本鎖抗体のヒトCEA結合ドメイン(すなわち、ヒトCEAと特異的に結合するヒト結合ドメイン)に使用した場合、可溶性CEA抗原に対する抵抗性を媒介するのに十分であることが見出された。それがヒト起源であることに起因して、前記構築物はヒト腫瘍患者に投与される場合、免疫原性が低いか免疫原性がない。要約すると、本明細書において定義される二重特異性一本鎖抗体を含む医薬組成物は、例えば、腫瘍進行中に観察される、血漿中の可溶性CEA濃度の高い上皮性腫瘍患者の治療に、再発性癌に、転移に、高腫瘍量の患者に、または後期腫瘍に特に有用である。
【0040】
本発明の医薬組成物のもう1つの好ましい実施形態では、本明細書において定義される二重特異性一本鎖抗体のCD3に特異的な前記第1の結合ドメインは、第2の結合ドメインに対してC末端に位置している。
【0041】
本発明のおよび全ての実施形態の範囲内で、単一ポリペプチド鎖上の、すなわち、本明細書において定義される二重特異性一本鎖抗体の中の、第1および第2の結合ドメインの配置の順序は関連性がある。本明細書において定義される二重特異性一本鎖抗体の結合ドメインの配置は、VHCEA−VLCEA−VHCD3−VLCD3、VLCEA−VHCEA−VHCD3−VLCD3、VHCD3−VLCD3−VHCEA−VLCEAまたはVHCD3−VLCD3−VLCEA−VHCEAであり得ることが想定されている。以下の実施例に示されるように、上述の利点は、第1の結合ドメイン(CD3と特異的に結合する)が第2の結合ドメインに対してC末端に位置している、すなわち、第2の結合ドメインよりも二重特異性一本鎖抗体のC末端に近い場合、特に実現可能である。ヒトCD3と特異的に結合する第1の結合ドメインは、VH−VLの配向に配置されていることが好ましい。例えば、本明細書において定義される二重特異性一本鎖抗体の結合ドメインは、VHCEA−VLCEA−VHCD3−VLCD3またはVLCEA−VHCEA−VHCD3−VLCD3の順序で配置されてよい。本明細書において、「〜に対してN末端に」または「〜に対してC末端に」およびそれらの文法上の変化形は、二重特異性一本鎖抗体の絶対的なNまたはC末端での配置よりもむしろ一次アミノ酸配列内の相対的な位置を意味する。故に、限定されない例として、「第2の結合ドメインに対してC末端に位置する」第1の結合ドメインとは、単に第1の結合ドメインが二重特異性一本鎖抗体内の第2の結合ドメインのカルボキシル側に位置することを意味し、さらなる配列、例えば上に述べたタグ、または別のタンパク質性もしくは非タンパク質性化合物(放射性同位元素など)が二重特異性一本鎖抗体の最終的なC末端に位置する可能性を排除するものではない。
【0042】
好ましくは、本明細書において定義される二重特異性一本鎖抗体の前記結合ドメインは、VHCEA−VLCEA−VHCD3−VLCD3またはVLCEA−VHCEA−VHCD3−VLCD3の順序で配置される。さらにより好ましくは、配置は、VLCEA−VHCEA−VHCD3−VLCD3である。最も好ましいのは、配列番号34に定義される二重特異性一本鎖抗体構築物A240VL−B9VHx配列番号77VHVLである。
【0043】
本明細書において定義される二重特異性一本鎖抗体のヒトCEAと特異的に結合する第2の結合ドメインは、少なくとも1つのCDR、好ましくはCDR−H3、より好ましくはマウスモノクローナル抗体A5B7のCDR−H3のKabatの位置100、100a、100b、101、および102にそれぞれ相当するアミノ酸配列「FYFDY」(配列番号112)を含むマウスモノクローナル抗体A5B7のCDR−H3の一部分;さらにより好ましくはアミノ酸配列「DX1X2X3X4FYFDY」(配列番号65)(「X1」、「X2」、「X3」または「X4」は、任意のアミノ酸残基を表し、該アミノ酸残基「D」は、マウスモノクローナル抗体A5B7のCDR−H3のKabat位置95に相当し、アミノ酸残基「FYFDY」は、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置100、100a、100b、101、および102にそれぞれ相当する)を含むマウスモノクローナル抗体A5B7のCDR−H3の一部分を含むことが好ましい。本明細書において、「X1」、「X2」、「X3」および「X4」は、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置96(「X1」)、97(「X2」)、98(「X3」)および99(「X4」)にそれぞれ相当する。「X1」、「X2」、「X3」または「X4」は、アミノ酸残基「R」(アルギニン)、「G」(グリシン)、「L」(ロイシン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、「S」(セリン)、「W」(トリプトファン)、「F」(フェニルアラニン)または「T」(トレオニン)を表すことが想定されている。本明細書において、「X1」、「X2」、「X3」および「X4」が同じアミノ酸を表す、例えば「X1」、「X2」、「X3」および「X4」がすべて「F」(フェニルアラニン)である場合は、本発明の範囲から除外される。好ましくは、「X1」が、「R」(アルギニン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;「X2」が、「G」(グリシン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表し;「X3」が、「L」(ロイシン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;かつ、「X4」が、「R」(アルギニン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表す。さらにより好ましくは、ヒトCEAに特異的な第2の結合ドメインは、モノクローナル抗体A5B7のCDR−H3のアミノ酸配列「RFYFDY」(配列番号113)、「LRFYFDY」(配列番号114)、「GLRFYFDY」(配列番号115)、または「RGLRFYFDY」(配列番号116)を少なくとも含む。最も好ましいのは、Kabatの位置95(「D」、アスパラギン酸)、96(「R」;アルギニン)、97(「G」;グリシン)、98(「L」;ロイシン)、99(「R」;アルギニン)、100(「F」;フェニルアラニン)、100a(「Y」;チロシン)、100b(「F」;フェニルアラニン)、101(「D」;アスパラギン酸)、および102(「Y」;チロシン)にそれぞれ相当する、アミノ酸配列「DRGLRFYFDY」(配列番号66)を含むA5B7の完全なCDR−H3である。Kabatの方式に従う番号付けは、例えば、Kabat, E. A., T. T. Wu, H. M. Perry, K. S. Gottesman, and C. Foeller. 1991. Sequences of Proteins of Immunological Interest, 5th ed. Bethesda, Md.: National Center for Biotechnology Information, National Library of Medicineに説明されている。
【0044】
以下の実施例に示されるように、CEAと相互作用する第二の結合ドメインの前記mAb A5B7由来のCDR−H3「DRGLRFYFDY」(配列番号66)アミノ酸配列を含む、本明細書に定義される二重特異性一本鎖抗体の腫瘍細胞に対する細胞障害活性は、可溶性CEA抗原に対して抵抗性であり、それにより血漿中の血清CEA濃度の高い腫瘍患者の治療が可能となる。
【0045】
このA5B7由来「DRGLRFYFDY」CDR−H3アミノ酸配列は、例えば、(上皮腫瘍細胞上の)CEA標的抗原に対する親和性を改良するため、かつ/または本明細書に定義される二重特異性一本鎖抗体の「微細な(fine)特異性」を最適化するためにさらに修飾することが望ましい。本目的のため、アミノ酸配列「DX1X2X3X4FYFDY」(配列番号65)において、改良された親和性および/または微細な特異性を有する、修飾されたCDR−H3を同定するために、様々なアミノ酸残基を位置「X1」、「X2」、「X3」および/または「X4」(それぞれ、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置96(「X1」)、97(「X2」)、98(「X3」)および99(「X4」)に対応)で試験してよい。例えば、「X1」、「X2」、「X3」または「X4」は、アミノ酸残基「R」(アルギニン)、「G」(グリシン)、「L」(ロイシン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、「S」(セリン)、「W」(トリプトファン)、「F」(フェニルアラニン)または「T」(トレオニン)で表され得る。ここで、表示される「X」の位置の1、2、3または4つ全ては、CDR−H3「DRGLRFYFDY」(配列番号66)アミノ酸配列中のKabatの位置96〜99にて本来の「RGLR」アミノ酸配列と比較して交換されてよい。しかし、「X1」、「X2」、「X3」および「X4」が同じアミノ酸を表す場合、例えば「X1」、「X2」、「X3」および「X4」が全て「F」(フェニルアラニン)である場合は本発明の特許請求の範囲から除外される。上記のA5B7由来の「DRGLRFYFDY」CDR−H3アミノ酸配列の修飾は当分野で公知の方法、例えば、CEA結合ドメイン中に、このような修飾されたCDR−H3領域を含む二重特異性一本鎖抗体の作製を可能にするランダム化されたプライマーを用いるPCRにより達成することができる。これらの修飾された二重特異性一本鎖抗体の親和性または微細な特異性は、当分野で記載される方法、例えばELISA、BiacoreまたはFACS分析により試験することができる。そのような修飾されたCDR−H3を含む二重特異性一本鎖抗体の可溶性CEA抗原に対する抵抗性は、以下の実施例に記載されるように、漸増量の可溶性CEAの存在下で、細胞障害活性アッセイで試験することができる。
【0046】
より好ましくは、本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な前記第2の結合ドメインは、配列番号65もしくは66および/またはアミノ酸配列「SYWMH」(配列番号68)を有するCDR−H1および/またはアミノ酸配列「FIRNKANGGTTEYAASVKG」(配列番号67)もしくは「FILNKANGGTTEYAASVKG」(配列番号145)を有するCDR−H2を含む。従って、本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な前記第2の結合ドメインは、上に定義される1、2または3のCDR−H領域を含んでよい。あるいは、本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な前記第2の結合ドメインは、配列番号65もしくは66および/またはアミノ酸配列「TYAMH」(配列番号70)を有するCDR−H1および/またはアミノ酸配列「LISNDGSNKYYADSVKG」(配列番号69)を有するCDR−H2を含む。従って、その代わりとして、本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な前記第2の結合ドメインは、上に定義される1、2または3のCDR−H領域を含んでよい。さらにより好ましくは、本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な前記第2の結合ドメインは、上に示される1、2または3のCDR−H領域に加えて、アミノ酸配列「TLRRGINVGAYSIY」(配列番号73)を有するCDR−L1および/またはアミノ酸配列「YKSDSDKQQGS」(配列番号72)を有するCDR−L2および/またはアミノ酸配列「MIWHSGASAV」(配列番号71)を有するCDR−L3を含む。
【0047】
本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な第2の結合ドメインのVH領域のアミノ酸配列は、好ましくは、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102に相当する「DRGLRFYFDY」(配列番号66)、およびアミノ酸配列「SYWMH」(配列番号68)を有するCDR−H1およびアミノ酸配列「FIRNKANGGTTEYAASVKG」(配列番号67)を有するCDR−H2を含む、配列番号60である。
【0048】
本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な第2の結合ドメインのVH領域のアミノ酸配列は、好ましくは、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102に相当する「DRGLRFYFDY」(配列番号66)、およびアミノ酸配列「SYWMH」(配列番号68)を有するCDR−H1、およびアミノ酸配列「FILNKANGGTTEYAASVKG」(配列番号145)を有するCDR−H2を含む、配列番号146である。
【0049】
本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な第2の結合ドメインのVH領域のアミノ酸配列は、好ましくは、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102に相当する「DRGLRFYFDY」(配列番号66)、およびアミノ酸配列「TYAMH」(配列番号70)を有するCDR−H1、およびアミノ酸配列「LISNDGSNKYYADSVKG」(配列番号69)を有するCDR−H2を含む、配列番号58または配列番号62である。
【0050】
本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な第2の結合ドメインのVL領域は、好ましくは、アミノ酸配列「TLRRGINVGAYSIY」(配列番号73)を有するCDR−L1、およびアミノ酸配列「YKSDSDKQQGS」(配列番号72)を有するCDR−L2、およびアミノ酸配列「MIWHSGASAV」(配列番号71)を有するCDR−L3を含む、配列番号64である。
【0051】
上に述べたように、CEAと特異的に結合する第2の結合ドメインの可変領域の順序または配置は、VH−VLまたはVL−VHであってよい。両方の配置が本発明の範囲内である。配列番号60のVHと配列番号64のVLを含む第2の結合ドメインについて、VH−VL配置は配列番号52に示され、一方、VL−VH配置は配列番号122に示される。配列番号146のVHと配列番号64のVLを含む第2の結合ドメインについて、VH−VL配置は配列番号147に示される。
【0052】
配列番号58のVHと配列番号64のVLを含む第2の結合ドメインについて、VH−VL配置は配列番号50に示され、一方、VL−VH配置は配列番号120に示される。配列番号62のVHと配列番号64のVLを含む第2の結合ドメインについて、VH−VL配置は配列番号54に示され、一方、VL−VH配置は配列番号124に示される。配列番号56のVHと配列番号64のVLを含む第2の結合ドメインについて、VH−VL配置は配列番号48に示され、一方、VL−VH配置は配列番号118に示される。
【0053】
さらにより好ましくは、本明細書において定義される二重特異性一本鎖抗体のCEAに特異的な第2の結合ドメインのV領域は、以下、
(a)配列番号60に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;
(b)配列番号146に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;
(c)配列番号58に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;
(d)配列番号62に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;ならびに
(e)配列番号56に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域
からなる群より選択される。
【0054】
最も好ましくは、前記二重特異性一本鎖抗体は、以下、
(a)配列番号6、8、16、18、24、26、32、34、40、42、126、130、134または143のいずれかに表されるアミノ酸配列;
(b)配列番号5、7、15、17、23、25、31、33、39、41、125、129、133または142に示される核酸配列によりコードされるアミノ酸配列;
(c)ストリンジェントな条件下で(b)の相補的な核酸配列とハイブリダイズする核酸配列によりコードされるアミノ酸配列;
(d)(b)の核酸配列に対する遺伝暗号の結果として縮重した核酸配列によりコードされるアミノ酸配列;ならびに
(e)(a)または(b)のアミノ酸配列と少なくとも85%同一、より好ましくは少なくとも90%同一、最も好ましくは少なくとも95%同一であるアミノ酸配列
からなる群より選択されるアミノ酸配列を含む。
【0055】
本発明の医薬組成物のもう1つの好ましい実施形態では、治療されるべき前記上皮腫瘍は、胃腸腺癌、乳腺腺癌または肺腺癌である。前記胃腸腺癌は、好ましくは結腸直腸腺癌、膵臓腺癌、食道腺癌または胃腺癌である。
【0056】
より好ましくは、本発明の前記医薬組成物は、進行性腫瘍、後期腫瘍、腫瘍量の多い腫瘍患者、転移性腫瘍、またはCEA血清中濃度が100ng/mlより高い腫瘍患者の治療のためのものである。前記CEA血清中濃度は、例えばELISAにより測定され得る。
【0057】
本発明の医薬組成物のさらに好ましい実施形態では、本明細書において定義される二重特異性一本鎖抗体の前記第1または第2の結合ドメインの少なくとも1つは、キメラ、ヒト化、CDR移植、および/または脱免疫化またはヒトである。
【0058】
本明細書において、用語「キメラ」は、上に定義されている。本明細書において、用語「ヒト」結合ドメイン、例えば、ヒトCEAと特異的に結合するヒト結合ドメインは、本明細書において定義される二重特異性一本鎖抗体が、マウスモノクローナル抗体A5B7のCDR−H3または上記定義のA5B7由来CDR−H3のKabatの位置100、100a、100b、101、および102(配列番号112)に相当するアミノ酸配列「FYFDY」を少なくとも有する、ヒト生殖細胞系列抗体レパートリーまたは抗体レパートリーに含有されているアミノ酸配列を含むことを意味するものと理解される。本明細書において定義される二重特異性一本鎖抗体はまた、それが、体細胞突然変異のインプリントに起因して予測され得る程度に、その(それらの)最も近いヒト生殖細胞系列配列に由来する配列からなる場合にヒトとみなされ得る。さらに、多くの非ヒト哺乳類、例えばマウスおよびラットなどのげっ歯類の抗体は、発現したヒト抗体レパートリー中にも存在することが予期され得るVH CDRアミノ酸配列を含む。発現したヒト抗体レパートリー中に存在することが予期され得る、ヒトまたは非ヒト起源の任意のそのような配列も本発明の目的において「ヒト」とみなされ得る。
【0059】
本明細書において、用語「ヒト化した」、「ヒト化」または「ヒト様」は、同義的に用いられて、その結合ドメインの少なくとも1つにおいて、非ヒト抗体またはその断片由来の少なくとも1つの相補性決定領域(「CDR」)を含む二重特異性一本鎖抗体を指す。ヒト化アプローチは、例えばWO 91/09968号および米国特許第6,407,213号に記載されている。限定されない例として、この用語は、少なくとも1つの結合ドメインの可変領域が別の非ヒト動物(例えばげっ歯類)由来の単一のCDR領域(例えばVHの3番目のCDR領域)を含む場合、ならびに、一方または両方の可変領域がそれらのそれぞれの1番目、2番目および3番目のCDRの各々に前記非ヒト動物由来のCDRを含む場合を包含する。二重特異性一本鎖抗体の結合ドメインの全てのCDRが、例えばげっ歯類由来のそれらの対応する同等物で置換されている場合、通常「CDR移植(CDR-grafting)」といい、この用語は本明細書において用語「ヒト化」またはその文法上の変化形も包含していると理解される。用語「ヒト化」または文法上の変化形はまた、第一および/または第二の結合ドメインのVHおよび/またはVL内での1またはそれ以上のCDR領域の置換に加えて、CDR間のフレームワーク(「FR」)領域内の少なくとも単一のアミノ酸残基のさらなる変異(例えば置換)が、その位置のアミノ酸を、置換に用いたCDR領域の由来する動物におけるその位置のアミノ酸に合致させている場合も包含する。当分野で公知のように、このような個々の変異は、その標的分子のCDR供与体として用いられる非ヒト抗体の本来の結合親和性を回復させるために、CDR移植の後にフレームワーク領域に作製される場合が多い。用語「ヒト化」は、上記のフレームワーク領域中のアミノ酸置換に加えて、非ヒト動物由来のCDR領域における、ヒト抗体由来の相当するCDR領域のアミノ酸へのアミノ酸置換(1または複数個所)をさらに包含する。
【0060】
本明細書において、用語「脱免疫化された」または「脱免疫化」は、元の野生型構築物と向かい合う第一および/または第二の結合ドメインを、ヒトにおいて前記野生型構築物を非免疫原性または低免疫原性にすることにより修飾することを意味する。脱免疫化アプローチは、例えばWO 00/34317号、WO 98/52976号、WO 02/079415号またはWO 92/10755号に示されている。用語「脱免疫化された」はまた、T細胞エピトープを形成する傾向の低下を示す構築物に関する。本発明に従って、用語「T細胞エピトープを形成する傾向の低下」とは、特異的なT細胞活性化をもたらすT細胞エピトープの除去に関する。さらに、「T細胞エピトープを形成する傾向の低下」とは、T細胞エピトープの形成に寄与するアミノ酸の置換、すなわち、T細胞エピトープの形成に必須のアミノ酸の置換を意味する。言い換えれば、「T細胞エピトープを形成する傾向の低下」は、免疫原性の低下または抗原非依存性のT細胞増殖を誘導する能力の低下に関する。用語「T細胞エピトープ」は、細胞内でペプチド、ポリペプチドまたはタンパク質の分解中に放出され、その後に、T細胞の活性化を引き起こすために主要組織適合抗原複合体(MHC)の分子より提示される短いペプチド配列に関する;特に、WO 02/066514号参照。MHCクラスIIで表されるペプチドに関して、このようなT細胞の活性化は、次に、T細胞の直接刺激による抗体応答を引き起こし、前記抗体を産生することができる。「T細胞エピトープを形成する傾向の低下」および/または「脱免疫化」は、当分野で公知の技法により測定することができる。T細胞増殖アッセイによりインビトロでタンパク質の脱免疫化を試験することが好ましい。このアッセイでは、世界で80%を超えるHLA−DR対立遺伝子を提示するドナー由来のPBMCを、野生型かまたは脱免疫化されたペプチドのいずれかに応答する増殖についてスクリーニングする。理想的には、細胞増殖は野生型ペプチドに抗原提示細胞を負荷する(loading)際にのみ検出される。あるいは、全てのハプロタイプを提示するHLA−DR四量体を発現させることにより、脱免疫化を試験することができる。これらの四量体は、ペプチド結合について試験するか、または増殖アッセイにおいて抗原提示細胞の代わりをするペプチドを負荷してよい。脱免疫化されたペプチドがHLA−DRハプロタイプに提示されるかどうかを試験するため、例えばPBMC上の蛍光標識ペプチドの結合を測定することができる。さらに、脱免疫化は、患者への投与後に脱免疫化分子に対する抗体が形成されたかどうかを判定することにより証明することができる。好ましくは、CDR領域の結合親和性が影響を受けないように、T細胞エピトープを誘導する傾向の低下をもたらすために、抗体由来分子をフレームワーク領域で脱免疫化し、大部分のCDR領域は修飾されない。たとえ1つのT細胞エピトープの除去であっても、結果として免疫原性が低下する。
【0061】
要約すると、上記のアプローチは、上皮性腫瘍患者に投与された場合、本明細書において定義される治療用二重特異性一本鎖抗体の免疫原性を低下させるのに役立つ。例えば、配列番号77に示されるCD3と特異的に結合している第1の結合ドメインは脱免疫化されている;WO 2005/040220号も参照のこと。好ましくは、このCD3結合ドメインにおけるV領域の配置は、VH−VLである。
【0062】
別の態様では、本発明は、
(a)配列番号6、8、16、18、24、26、32、34、40、42、126、130、134または143のいずれか1つに表されるアミノ酸配列;
(b)配列番号5、7、15、17、23、25、31、33、39、41、125、129、133または142に示される核酸配列によりコードされるアミノ酸配列;
(c)ストリンジェントな条件下で(b)の相補的な核酸配列とハイブリダイズする核酸配列によりコードされるアミノ酸配列;
(d)(b)の核酸配列に対する遺伝暗号の結果として縮重した核酸配列によりコードされるアミノ酸配列;および
(e)(a)または(b)のアミノ酸配列と少なくとも85%同一、より好ましくは少なくとも90%同一、最も好ましくは少なくとも95%同一であるアミノ酸配列、
からなる群より選択されるアミノ酸配列を含む、二重特異性一本鎖抗体に関する。
【0063】
一実施形態では、本発明は、上記に定義される二重特異性一本鎖抗体を含む組成物に関する。好ましくは、上記に定義される前記二重特異性一本鎖抗体は、ヒトにおける上皮性腫瘍の治療のための医薬組成物として使用される。前記上皮性腫瘍は、CEA陽性である。これらの医薬組成物のCEA陽性上皮性腫瘍細胞に対する細胞障害活性は、腫瘍患者の血漿中の高い濃度の可溶性CEA抗原にさえも抵抗性である。さらに、上記に定義される前記二重特異性一本鎖抗体またはそれに由来する抗CEA scFvは、下にさらに詳細に述べられるようにヒトにおける上皮性腫瘍の検出のための診断用組成物として使用することができる。
【0064】
本明細書において、用語「ストリンジェントな条件下でハイブリダイズする」とは、ストリンジェントなハイブリダイゼーション条件下で、配列番号5、7、15、17、23、25、31、33、39、41、125、129、133または142に示される配列、またはその相補物とハイブリダイズすることができ、CEA陽性腫瘍細胞に対して細胞障害活性を有する二重特異性一本鎖抗体をコードする核酸配列を指す。「ストリンジェントなハイブリダイゼーション条件」とは、50% ホルムアミド、5xSSC(750mM NaCl、75mM クエン酸三ナトリウム)、50mM リン酸ナトリウム(pH7.6)、5x デンハート液、10% 硫酸デキストラン、および20μg/mlの変性断片化サケ精子DNAを含む溶液中、42℃にて一晩のインキュベーション、その後、約65℃にて0.1x SSC中でのフィルターの洗浄を指す。
【0065】
任意の特定の核酸分子またはポリペプチドが本明細書に定義されるヌクレオチドまたはアミノ酸配列と少なくとも80%、85%、90%、95%、96%、97%、98%または99%同一であるかどうかは、公知のコンピュータープログラムを用いて従来法で決定してよい。クエリー配列(本明細書に定義される配列)と対象配列(subject sequence)との間で最も全体的に適合するものを決定するための好ましい方法(グローバル配列アラインメントとも称される)は、Bultragらのアルゴリズム(Comp. App. Biosci. 6:237-245 (1990))に基づくFASTDBコンピュータープログラムを用いて決定してよい。配列アラインメントにおいてクエリー配列と対象配列は、双方ともDNA配列である。RNA配列はUをTに変換することにより比較され得る。
【0066】
本発明はまた、本明細書に定義される二重特異性一本鎖抗体をコードする核酸配列を含む医薬組成物を提供する。前記核酸は、以下により詳細に示されるように、ヒトにおいて上皮腫瘍を治療するために、例えば遺伝子治療アプローチに利用することができる。
【0067】
本発明はさらに上記に定義される核酸配列を含むベクターを含む医薬組成物に関する。好ましくは、前記ベクターは、上記に定義される前記核酸配列と作動可能なように連結されている調節配列をさらに含む。より好ましくは、前記ベクターは発現ベクターである。
【0068】
さらに、本発明のベクターはまた、遺伝子導入または標的遺伝子組換えベクターであってよい。エキソビボ(ex-vivo)またはインビボ技術により治療遺伝子または核酸を細胞に導入することに基づく遺伝子治療は、遺伝子導入の最も重要な適用の一つである。インビトロまたはインビボでの遺伝子治療に適したベクター、方法または遺伝子送達系は、文献に記載され、当業者に公知である;例えば、Giordano, Nature Medicine 2 (1996), 534-539; Schaper, Circ. Res. 79 (1996), 911-919; Anderson, Science 256 (1992), 808-813, Isner, Lancet 348 (1996), 370-374; Muhlhauser, Circ. Res. 77 (1995), 1077-1086;Onodua, Blood 91 (1998), 30-36; Verzeletti, Hum. Gene Ther. 9 (1998), 2243-2251; Verma, Nature 389 (1997), 239-242; Anderson, Nature 392 (Supp. 1998), 25-30; Wang, Gene Therapy 4 (1997), 393-400;Wang, Nature Medicine 2 (1996), 714-716;WO 94/29469;WO 97/00957;US5,580,859;US5,589,466;US4,394,448またはSchaper, Current Opinion in Biotechnology 7 (1996), 635-640、ならびにそれに引用される参照文献を参照。本明細書に定義される核酸分子およびベクターは、細胞への直接導入用に、またはリポソーム、ウイルス(例えばアデノウイルス、レトロウイルス)ベクター、エレクトロポレーション、またはその他の送達系を介する細胞への導入用に設計することができる。さらに、バキュロウイルス系を本明細書に定義される核酸分子のための真核生物発現系として用いてもよい。導入および遺伝子治療アプローチは、好ましくは、本明細書に定義される機能的二重特異性一本鎖抗体構築物の発現をもたらすべきであり、それによって、前記発現した二重特異性一本鎖抗体構築物はヒトにおける上皮腫瘍の治療、寛解および/または予防に特に有用である。
【0069】
さらなる態様では、本発明は、上記に定義されるベクターまたは核酸で形質転換またはトランスフェクトされた宿主を含む医薬組成物に関する。
【0070】
本発明のさらなる態様は、免疫エフェクター細胞の活性化シグナルを提供することのできるタンパク質性化合物をさらに含む、上記に定義される医薬組成物に関する。
【0071】
好ましくは、医薬組成物は、担体、安定剤および/または賦形剤からなる適した製剤をさらに含む。
【0072】
別の態様では、本発明は、上記に定義される医薬組成物の製造のためのプロセスに関し、前記プロセスは上記に定義される宿主を上記に定義される二重特異性一本鎖抗体の発現を可能にする条件下で培養することと、産生された二重特異性一本鎖抗体を培養物から回収することを含む。
【0073】
本発明のさらなる態様は、ヒトにおける上皮腫瘍の予防、治療または寛解のための医薬組成物の調製のための、上記に定義される二重特異性一本鎖抗体または上記に定義されるプロセスにより産生された二重特異性一本鎖抗体、上記に定義される核酸分子、上記に定義されるベクターまたは上記に定義される宿主の使用に関する。本発明の別の態様は、ヒトにおける上皮腫瘍の予防、治療または寛解のための方法に関し、前記方法は本発明の、または上に示されるプロセスによって製造された医薬組成物の有効量を投与する段階を含む。当業者、特に主治医であれば、本発明の二重特異的分子/二重特異的一本鎖抗体の投与を必要とする患者の治療の成功を評価することができる。従って、投与計画ならびに投薬量および投与回数は前記当業者により判断され得る。つまり、判断される、対応する「寛解」および/または「治療」は下に定義される。
【0074】
本明細書において、上皮性腫瘍との関連で、本発明の医薬組成物の「有効量」または「治療上有効な量」とは、一次、局所または転移性腫瘍組織を破壊、改変、制御または除去するために十分な治療薬の量を指す。治療上有効な量とは、上皮性腫瘍の広がりを遅延または最小化するために十分な治療薬の量を指し得る。また、治療上有効な量とは、上皮性腫瘍の治療または管理において治療上の利益をもたらす治療薬または医薬品の量も指し得る。さらに、本発明の治療薬または医薬品に関して治療上有効な量とは、上皮性腫瘍の治療または管理において治療上の利益をもたらす治療薬または医薬品の、単独の、または他の治療法と組み合わせた量を意味する。本明細書において定義される二重特異性一本鎖抗体の量に関連して用いられる場合、この用語は、全体的な治療法を改善するか、望ましくない効果を低下または回避するか、または別の治療薬の治療効力または別の治療薬との相乗効果(本明細書において定義される通り)を促進する量を包含し得る。好ましくは、治療薬の治療上有効な量は、上皮性腫瘍の治療において、全体的な治療法を改善するか、望ましくない効果を低下または回避するか、または別の治療薬の治療効力または別の治療薬との相乗効果を促進する。例えば、本明細書において定義される二重特異性一本鎖抗体は、単一療法として患者に投与される場合、上皮性腫瘍の直径を20%縮小させることができる。これに対し、第2の治療薬、例えば、下に定義される抗癌剤は、腫瘍を10%縮小させることができる。しかし、本明細書において定義される二重特異性一本鎖抗体と前記第2の治療薬の両方を併用療法の形態で組み合わせて投与する場合、50%の腫瘍の縮小を観察することができる。このような効果は本明細書において相乗効果として理解される。
【0075】
本明細書において言及されるように、用語「治療法」とは、上皮性腫瘍の予防、治療または寛解に用いられ得る任意の投与計画、方法および/または薬剤を指す。用語「上皮性腫瘍の予防、治療または寛解」は、下に詳細に説明されている。用語「治療法(therapies)」および「治療法(therapy)」は、上皮性腫瘍、あるいはその1またはそれ以上の症状の治療、予防、または寛解に有用な生物学的療法、支持療法、化学療法、放射線療法および/またはその他の治療法を称し得る。
【0076】
本明細書において、患者に1または複数の治療法を投与することとの関連で、用語「治療する」、「治療」および「治療すること」は、上皮性腫瘍の進行、重篤度、および/または持続の低下または寛解を指す。前記上皮性腫瘍は、異常な発現(例えば、CEAの過剰発現もしくは過剰活性)および/または、1もしくはそれ以上の治療法(1またはそれ以上の医薬品または治療薬の投与を含む)の投与の結果として生じるその1またはそれ以上の症状の寛解を伴い得る。
【0077】
最も好ましい投与方法は、所与の時間/期間の静脈内投与である。本明細書に定義される二重特異性一本鎖抗体は1回につき単独で投与されてよいが、好ましいものは、製薬上許容される担体中の投与である。適した医薬担体の例は当分野で周知であり、リン酸緩衝生理食塩水溶液、水、リポソーム、様々な種類の湿潤剤、滅菌溶液などが含まれる。このような担体を含む組成物は、周知の従来法により処方されてよい。これらの医薬組成物は被験体に適した用量で投与することができる。投与計画は主治医および臨床学的因子により決定される。医学分野で周知のように、任意の患者一人への投薬量は多くの要因によって決まり、それには患者の大きさ、体表面積、年齢、投与される特定の化合物、性別、投与時間および経路、全体的な健康状態、および同時に投与されるその他の薬剤が含まれる。非経口投与のための製剤としては、滅菌水溶液または非水溶液、および懸濁液が挙げられる。非水溶媒の例は、プロピレングリコール、ポリエチレングリコール、および注射用の有機エステル類、例えばオレイン酸エチルである。水性担体としては、水、水溶液、または生理食塩水および緩衝媒体を含む懸濁液が挙げられる。非経口媒体としては、塩化ナトリウム溶液、リンゲルデキストロース、デキストロースおよび塩化ナトリウム、乳酸化リンゲル液、または硬化油が挙げられる。静脈内媒体としては、液体および栄養素補充液、電解質補充液(リンゲルデキストロースに基づくものなど)などが挙げられる。防腐剤およびその他の添加剤も存在してよく、例えば、抗菌剤、抗酸化剤、キレート剤、不活性ガスなどが挙げられる。さらに、組成物は、例えば、好ましくはヒト起源の血清アルブミンまたは免疫グロブリンなどのタンパク質性担体を含んでもよい。併用療法は、タンパク質性二重特異性一本鎖抗体に加えて、医薬組成物の意図される使用に応じて、さらなる生物活性物質を含むことが想定される。このような物質は、胃腸管系に作用する物質、細胞増殖抑制剤として作用する物質、高尿酸血症を予防する物質、免疫反応を阻害する物質(例えばコルチコステロイド、FK506)、循環系に作用する薬剤および/またはT細胞共刺激分子またはサイトカインなどの当分野で公知の薬剤であってよい。好ましくは、本明細書に定義される二重特異性一本鎖抗体は、緩衝液、安定剤および界面活性剤中に処方される。緩衝液は、リン酸緩衝液、クエン酸緩衝液、コハク酸緩衝液または酢酸緩衝液であってよい。安定剤は、アミノ酸および/または糖であってよい。界面活性剤は、合成洗剤、PEGなどであってよい。より好ましくは、本明細書に定義される二重特異性一本鎖抗体は、クエン酸塩、リシン、トレハロースおよびTween80中に処方される。前記医薬組成物の希釈液として、等張生理食塩水およびTween80が好ましい。
【0078】
本明細書において用語「寛解」とは、疾患、すなわち上皮腫瘍の重篤度の改善または緩和を指す。例えば、そのような寛解は、本発明の医薬組成物の投与に起因する、安定な疾患の達成、またさらにより好ましくは、上皮腫瘍の縮小、すなわち、微小、部分的応答または完全寛解であり得る。「安定な疾患」とは、臨床および/または組織学的診断方法により腫瘍進行/増殖が全くまたはほとんど観察されないまたは検出されない疾患状態を指す。例えば、指標病変の断面積の合計の50%より多くの縮小である、腫瘍の縮小は、「部分応答」と考えられる。「完全寛解」は、治療後に病変がこれ以上全く検出されない状態を意味する。安定疾患と部分応答との間の腫瘍縮小を伴う応答は、微小応答と考えられ得る。例えば、指標病変の断面積の合計の20%、25%または30%縮小は、微小応答と称され得る。
【0079】
用語「寛解」は、本明細書において、上皮腫瘍の数の減少も包含する。それは、さらに腫瘍進行の阻止/減速を意味する。さらに、治療しなかった腫瘍患者と比較して、治療した腫瘍患者の全体的な生存の改善は、本明細書において「寛解」と考えられる。これは、必要な変更を加えて、治療しなかった腫瘍患者と比較して、治療した腫瘍患者の進行のない生存または再発のない生存の改善にも当てはまる。さらに、用語「寛解」はまた、上皮腫瘍の症状の強度の減少も指し、その結果得られる、例えば治療した腫瘍患者の生活の質の改善も指す。
【0080】
用語「上皮腫瘍の予防」は、本明細書において、次のように理解される。原発上皮腫瘍のヒト患者からの外科切除後、および/または原発上皮腫瘍の化学療法または放射線治療後に、腫瘍細胞全てが身体から排除されない場合もあり得る。しかし、これらの残存腫瘍細胞は患者において再発性癌、すなわち、局所再発および/または転移を引き起こす可能性がある。転移はよく見られる癌の合併症であるが、癌細胞が原発腫瘍から播種されて遠位のコロニーを形成する過程ははあまり分かっていない。転移性癌は、ほぼ例外なく難治性で新しい治療方法の必要性が生じる。本発明の医薬組成物を用いると、これらの播種された腫瘍細胞を死滅させて二次性腫瘍(一次治療後に身体に残存する腫瘍細胞から生じる)の形成を予防することができる。このように、本医薬組成物は、腫瘍患者において局所再発および/または転移の形成を予防するために役立つ。
【0081】
抗腫瘍治療の成功は、それぞれの疾病について、例えばコンピュータ断層撮影、X線、核磁気共鳴断層撮影(例えば、米国国立癌研究所の判定基準に基づく応答評価(Cheson (1999), J. Clin. Oncol.; 17 (4):1244))、陽電子放射断層撮影走査、内視鏡検査、蛍光活性化セルソーター、骨髄の吸引、胸水または腹水、組織/組織学、および様々な上皮腫瘍特異的臨床化学パラメータ(例えば血清中の可溶性CEA濃度)により確立された標準的な方法でモニターすることができ、さらに、その他の確立された標準的な方法を用いてよい。さらに、T細胞の活性化を測定するアッセイを用いてよい;例えば、WO 99/054440号を参照。治療しなかった腫瘍患者と比較して、治療した腫瘍患者の全体的な生存、進行のない生存または再発のない生存の決定のために統計学を用いてもよい。
【0082】
好ましくは、前記上皮腫瘍は、胃腸腺癌、乳腺腺癌または肺腺癌である。前記胃腸腺癌は、より好ましくは、結腸直腸腺癌、膵臓腺癌、食道腺癌または胃腺癌である。
【0083】
さらにより好ましくは、前記本発明の医薬組成物は、進行性腫瘍、後期腫瘍、腫瘍量の多い腫瘍患者、転移性腫瘍、またはCEA血清中濃度が100ng/mlより高い腫瘍患者の治療のためのものである(例えばELISAにより測定され得る)。
【0084】
本発明の使用または方法のもう1つの好ましい実施形態では、上記に定義される前記医薬組成物は、さらなる薬剤と組み合わせて、すなわち、併用療法の一部として投与されるのに適している。
【0085】
特定の実施形態では、本明細書において定義される二重特異性一本鎖抗体または医薬組成物は、1またはそれ以上のその他の治療法と組み合わせて投与される。特定の実施形態では、本明細書において定義される二重特異性一本鎖抗体または医薬組成物は、1またはそれ以上のその他の治療法と同時に患者に投与される。そのような治療法は上皮性腫瘍の治療に有用であることが好ましい。用語「同時に」は、医薬組成物または治療薬を全く同時に投与することに限定するのではなく、むしろ、本明細書において定義される二重特異性一本鎖抗体または医薬組成物とその他の薬剤が、患者に順番に投与されること、及び本明細書において定義される二重特異性一本鎖抗体または医薬組成物が他の薬剤と一緒に作用して、別の方法で投与された場合よりも増大した利益をもたらすことのできるような間隔で投与されることを意味する。例えば、それぞれの治療薬は同時にまたは任意の順序で異なる時点に順次投与されてよい;しかし、同時に投与されない場合、それらは所望の治療効果を得るために時間的に十分に接近して投与されるべきである。
【0086】
それぞれの治療薬は、任意の適当な形態で、任意の適した経路から別々に投与することができる。他の実施形態では、本明細書において定義される二重特異性一本鎖抗体または医薬組成物は、外科的手術の前に、同時に、または後に投与される。該外科的手術は局在した上皮性腫瘍を完全に除去するか、または大きな上皮性腫瘍のサイズを縮小させることが好ましい。外科的手術はまた、予防的手段としてなされるか、または疼痛を軽減するためになされてよい。
【0087】
本明細書において提供される投薬量および投与の頻度は、上に定義される「治療上有効な」という用語に包含される。投薬量および頻度は、一般に、投与される特定の治療薬または予防薬、上皮性腫瘍の重篤度および種類、投与経路、ならびに患者の年齢、体重、応答、および過去の病歴に依存する各患者についての特異的な因子に従ってさらに変動する。当業者は、そのような因子を考慮すること、および、例えば、文献に報告され、Physicians' Desk Reference (59th ed., 2005) で推奨されている投薬量に従うことにより、適した投与計画を選択することができる。
【0088】
一部の実施形態では、本明細書において定義される二重特異性一本鎖抗体または医薬組成物の投与による治療法は、1またはそれ以上の治療法、例えば化学療法、放射線療法、ホルモン療法、および/または生物学的療法/免疫療法の投与と組み合わされる。治療薬としては、限定されるものではないが、タンパク質性分子(限定されるものではないが、ペプチド、ポリペプチド、タンパク質(翻訳後修飾されたタンパク質、抗体などを含む)を含む);または小分子(1000ダルトン未満)、無機もしくは有機化合物;または核酸分子(二本鎖もしくは一本鎖DNA、または二本鎖もしくは一本鎖RNA、ならびに三重らせん核酸分子を含む)が挙げられる。治療薬は、任意の既知生物(限定されるものではないが、動物、植物、細菌、真菌、および原生生物、またはウイルスを含む)に由来するか、または合成分子ライブラリーに由来してよい。
【0089】
特定の実施形態では、本発明の方法および使用は、キナーゼ、例えば当分野に記載されるゲフィチニブ(イレッサ)、エルロチニブ(タルセバ)、抗EGFR抗体(例えば、セツキシマブ;エルビタックス)、または抗Her2/neu抗体(例えば、トラスツズマブ;ハーセプチン)の阻害剤である1またはそれ以上の治療薬の投与と組み合わせた、本明細書において定義される二重特異性一本鎖抗体または医薬組成物の投与を包含する;例えば、Hardie and Hanks (1995) The Protein Kinase Facts Book, I and II, Academic Press, San Diego, California 参照。
【0090】
別の特定の実施形態では、本発明の方法および使用は、血管形成阻害剤、例えば当分野に記載される、抗VEGF抗体(例えば、ベバシズマブ;アバスチン)、小分子化合物(例えば、バタラニブもしくはソラフェニブ)またはCOX阻害剤である、1またはそれ以上の治療薬の投与と組み合わせた、本明細書において定義される二重特異性一本鎖抗体または医薬組成物の投与を包含する。
【0091】
別の特定の実施形態では、本発明の方法および使用は、抗癌剤、例えば当分野に記載される、5−フルオロウラシル、ロイコボリン、カペシタビン、オキサリプラチン、イリノテカン、ゲムシタビン、ドキソルビシン、エピルビシン、エトポシド、シスプラチン、カルボプラチン、タキサン類(例えば、ドセタキセル、パクリタキセル)である、1またはそれ以上の治療薬の投与と組み合わせた、本明細書において定義される二重特異性一本鎖抗体または医薬組成物の投与を包含する。
【0092】
本明細書において定義される二重特異性一本鎖抗体または医薬組成物をさらなる治療薬と組み合わせて用いる上皮性腫瘍患者の併用療法は、相乗作用をもたらすことが好ましい。本明細書において、用語「相乗的な」とは、任意の2またはそれ以上の単一の治療法(例えば、1またはそれ以上の治療薬)の付加的効果よりも効果的な、治療法の組み合わせ(例えば、上に説明される、本明細書において定義される二重特異性一本鎖抗体とさらなる治療薬の組み合わせ)を指す。例えば、本明細書において定義される二重特異性一本鎖抗体は、単一療法として患者に投与される場合、20%の上皮性腫瘍の直径の縮小をもたらすことができる。その一方、第2の治療薬、例えば、下に定義される抗癌剤は、腫瘍を10%縮小させることができる。しかし、本明細書において定義される二重特異性一本鎖抗体と前記第2の治療薬の両方を併用療法の形態で組み合わせて投与する場合、50%の腫瘍の縮小を観察することができる。
【0093】
治療法の組み合わせ(例えば、上に説明される、本明細書において定義される二重特異性一本鎖抗体と、さらなる治療薬の組み合わせ)の相乗作用により、低投薬量の1またはそれ以上の治療法(例えば、1またはそれ以上の治療薬)の使用および/または疾患、例えば、上皮性腫瘍のある患者へ前記治療法の低頻度の投与が可能となる。低い投薬量の治療法(例えば、治療薬)を用いることができ、かつ/または前記治療法を低頻度で投与することができると、疾患(例えば上皮性腫瘍)の予防または治療において前記治療法の効力を低下させることなく、前記治療法の被験体への投与に関連する毒性が低減される。さらに、相乗作用は、上皮性腫瘍(CEAの異常な発現(例えば、過剰発現)または活性を伴う可能性がある)の予防、管理、治療および/または寛解において、治療法(例えば、治療薬)の改善された有効性をもたらし得る。最後に、治療法(例えば、治療薬)の組み合わせの相乗作用は、任意の単一療法の使用に関連する、有害なまたは望ましくない副作用を回避または減少させることができる。
【0094】
前記併用療法において、活性薬剤は場合により本明細書において定義される二重特異性一本鎖抗体と同じ医薬組成物に含まれても、別個の医薬組成物に含まれてもよい。この後者の場合、前記別個の医薬組成物は、本明細書において定義される二重特異性一本鎖抗体を含む前記医薬組成物投与よりも前に、同時に、または後に、投与されるのに適している。さらなる薬剤または医薬組成物は、非タンパク質性化合物もしくはタンパク質性化合物であり得る。さらなる薬剤がタンパク質性化合物である場合、該タンパク質性化合物は免疫エフェクター細胞の活性化シグナルを提供できることが有利である。
【0095】
好ましくは、前記タンパク質性化合物または非タンパク質性化合物は、同時にまたは非同時に、上記において定義される二重特異性一本鎖抗体、上記において定義される核酸分子、上記において定義されるベクター、または上記において定義される宿主とともに投与されてよい。好ましくは、処置されるべき前記被験体は、ヒトである。
【0096】
さらなる実施形態では、本明細書において定義される一本鎖二重特異性抗体または抗CEA scFvは、診断用または検出可能な薬剤と複合体化され得る。そのような診断および検出は、抗体またはscFvを、検出可能な物質、例えば様々な酵素類、例えば西洋ワサビペルオキシダーゼ、アルカリ性ホスファターゼ、β−ガラクトシダーゼ、またはアセチルコリンエステラーゼ;補欠分子族、例えばストレプトアビジン/ビオチンおよびアビジン/ビオチン;蛍光物質、例えば、ウンベリフェロン、フルオレセイン、フルオレセインイソチオシアネート、ローダミン、ジクロロトリアジニルアミンフルオレセイン、ダンシルクロリドまたはフィコエリトリン;発光物質、例えば、ルミノール;生物発光物質、例えば、ルシフェラーゼ、ルシフェリン、およびエクオリン;放射性物質および同位元素、例えばコバルト(57Co)、インジウム(115In、113In、112In、111In)、ヨウ素(131I、125I、123I、121I)、またはイットリウム(90Y)、様々な陽電子放出断層撮影を用いる陽電子放出金属、および非放射性常磁性金属イオンと結合させることにより達成することができる。
【0097】
部分と抗体を複合体化するための技法は周知である。部分は、当分野で公知の任意の方法により抗体と複合体化させることができ、それには、限定されるものではないが、アルデヒド/シッフ結合、スルフヒドリル結合、酸に不安定な結合、シス−アコニチル結合、ヒドラゾン結合、酵素分解可能な結合が含まれる;一般に、 Garnett, 2002, Adv. Drug Deliv. Rev. 53:171-216 参照。部分と抗体を複合体化するためのさらなる技法は当分野で周知である、例えば、Arnon et al., Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy. In Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), pp. 243-56 (Alan R. Liss, Inc. 1985)参照。抗体とポリペプチド部分を融合または複合体化させるための方法は当分野で公知である;例えば、Ashkenazi et al., 1991, PNAS 88: 10535-10539 参照。抗体と部分の融合は必ずしも直接的である必要はなく、リンカー配列によって生じるものであってよい。そのようなリンカー分子は一般に当分野で公知であり、Denardo et al., 1998, Clin Cancer Res. 4:2483-90; Peterson et al., 1999, Bioconjug. Chem. 10:553 に記載されている。
【0098】
さらなる態様において、本発明は、上記に定義される二重特異性一本鎖抗体、上記に定義される核酸分子、上記に定義されるベクター、または上記に定義される宿主を含むキットに関する。
【0099】
これらのおよびその他の実施形態は、本発明の説明および実施例に開示され、包含される。免疫学分野の組換え技術および方法は、例えば、Sambrook et al., Molecular Cloning: A Laboratory Manual;Cold Spring Harbor Laboratory Press, 3rd edition 2001; Lefkovits; Immunology Methods Manual;The Comprehensive Sourcebook of Techniques;Academic Press, 1997; Golemis; Protein-Protein Interactions: A Molecular Cloning Manual;Cold Spring Laboratory Press, 2002に記載されている。本発明に従って用いられ得る抗体、方法、用途および化合物のいずれかに関するさらなる参照文献は、例えば電子装置を用いて公開されているライブラリーおよびデータベースから得ることができる。例えば、インターネットで利用可能な公開データベース「Medline」を、例えばhttp://www.ncbi.nlm.nih.qov/PubMed/medline.htmlで利用してよい。さらなるデータベースおよびアドレス、例えばhttp://www.ncbi.nim.nih.qov/、http://www.infobioaen.fr/、http://www.fmi.ch/bioloqv/research tools.html、http://www.tiqr.orQ/が当業者に公知であり、例えば、http://www.lvcos.comを用いて得ることもできる。腫瘍関連の話題は、例えば、http://www.nih.govまたはhttp://www.dkfz.deを参照のこと。
【実施例】
【0100】
以下の実施例は本発明を説明する。
【0101】
実施例1:ヒトCEA(癌胎児性抗原関連細胞接着分子5;CEACAM5)がトランスフェクトされたCHO細胞の作製
CEA陽性KatoIII細胞(ヒト胃癌腫細胞株;ATCC HTB−103)を用いて全RNAを得、それをキットマニュアルの説明書(Qiagen、RNeasy Mini Kit)に従って単離した。得られたRNAを、ランダムに開始される逆転写によるcDNA合成に用いた。CEA抗原の全長配列のクローニングのため、以下のオリゴヌクレオチドを用いた。5’CEACAM5 EcoRI GAATTCGCCACCATGGAGTCTCCCTCGGCCCC(配列番号74)および3’CEACAM5 Sal I GTCGACCTATATCAGAGCAACCCC(配列番号75)。PCR(第1サイクルに関して、93℃にて5分間の変性、58℃にて1分間のアニーリング、72℃にて1分間の伸張;93℃にて1分間の変性、58℃にて1分間のアニーリング、72℃にて1分間の伸張を30サイクル;72℃にて5分間の末端伸張)を用いてコード配列を増幅した。その後、PCR産物をEcoRIおよびSalIで消化し、適切に消化した発現ベクターpEF−DHFRに連結し、大腸菌に形質転換した。単離したプラスミドDNAを配列決定し、確立されたCEACAM5の核酸配列(米国国立バイオテクノロジー情報センター(National Center for biotechnology information)のNM_004363、http://www.ncbi.nlm.nih.gov/)と比較した。標準的なプロトコールに従って、上述の手順を実行した(Sambrook, Molecular Cloning; A Laboratory Manual, Cold Spring Harbour Laboratory Press, Cold Spring Harbour, New York (1989;2001)。確認された核酸配列を含むクローンを、構築物の真核生物における発現のためにDHFR欠損CHO細胞にトランスフェクトした。DHFR欠損CHO細胞における真核生物タンパク質発現を、Kaufmannに記載のとおり行った(Kaufmann R.J., Methods Enzymol. 185 (1990), 537-566)。構築物の遺伝子増幅を、最大20nM MTXの終濃度までの、漸増濃度のMTXにより誘導した。次に、トランスフェクトした細胞を、FACSアッセイを用いてCEA抗原の発現について試験した。その目的のため、2.5×105のトランスフェクト細胞を、濃度5μg/mlのマウスモノクローナル抗体COL−1(No.MS−613−P1ABX,Neomakers;Fremont,CA,USA)とともにインキュベートした。50μl PBS中2% FCS(Dianova,Hamburg,Germanyより入手)を用いて1:100で希釈した、R−フィコエリトリン結合アフィニティー精製F(ab’)2断片、ヤギ抗マウスIgG、Fc−γ断片特異的抗体を用いて、抗体の結合を検出した。細胞を、FACS−Calibur(Becton Dickinson,Heidelberg)でフローサイトメトリーにより分析した。FACS染色および蛍光強度の測定を、Current Protocols in Immunology (Coligan, Kruisbeek, Margulies, Shevach and Strober, Wiley-Interscience, 2002)に記載のとおり行った。結果として、形質転換体は、ヒトCEA抗原について明らかな陽性染色を実証した。
【0102】
実施例2:CEAxCD3二重特異性一本鎖抗体の生成
一般に、それぞれヒトCEA抗原に対して結合特異性を有するドメインを含む二重特異性一本鎖抗体分子、ならびに配列番号77に示されるヒトCD3抗原に対して結合特異性を持つ脱免疫化されたドメインを、表1に提示されるように設計した。このCD3結合ドメイン中のV領域の配置は常にVH−VLである。本明細書において定義される二重特異性一本鎖抗体において用いられる、この脱免疫化された抗CD3結合ドメインは、例えば、WO 2005/040220号において既に記載されている。
【0103】
1.抗CEA特異性および抗CD3特異性を含む二重特異性一本鎖抗体分子の形式(表1)
【表1】

【0104】
モノクローナル抗体若しくはハイブリドーマに由来するか、またはファージディスプレイ誘導選択(PDGS)により得た、ヒトCEA抗原に特異的な可変軽鎖(VL)および可変重鎖(VH)領域を含有する前述の構築物は、遺伝子合成およびその後の、CD3に特異的なVHとVLの組み合わせを含む発現ベクターへのクローニングにより得た。前記二重特異性一本鎖構築物の作製はまた、例えば、Sambrook(前掲)に記載の組換え技術に従っても行うことができる。そのための詳細な指示は、例えば、WO 99/054440号に規定されている。
【0105】
抗CD3結合ドメインは、ヒトCD3抗原に対する結合特異性を有する脱免疫化されたドメインに相当する。本明細書に記載される二重特異性一本鎖構築物中の脱免疫化された抗CD3結合ドメインのV領域の配置は、常にVH−VLである。対応する前記VH−VLドメインのアミノ酸配列は配列番号77に示される。ヒト癌胎児性抗原に特異的なCEAI、CEAIIおよびCEAIIIは、それぞれ、mAb A5B7(Chester, K. A. et al., Int J Cancer 57 (1994), 67-72)、T84.66(Neumaier, M. et al., Cancer Res 50 (1990), 2128-34)およびMFE−23(Boehm, M. K. Biochem J 2 (2000), 519-28)に由来する可変軽鎖(VL)および可変重鎖(VH)領域を含む。A5、B9、D8、およびE12は、ヒトCEAに特異的なヒトVH領域であり、一方、A240は同じ特異性を有するヒトVL領域である。ヒトA5、B9、D8、E12およびA240V領域の作製は、実施例6および7に詳細に記載されている。本明細書に記載されるすべての二重特異性一本鎖抗体の対応するヌクレオチドおよびアミノ酸配列はリストする配列に示される。
【0106】
以下では、CEAI VLVHx配列番号77VHVL(配列番号6)構築物の作製がさらに詳細に説明される。上に言及されるその他の構築物の作製は、十分当業者の範囲内である方法に必要な改変を施して適宜に行うことができる。前述のCEAI特異性および脱免疫化された抗CD3(配列番号77)特異性を含む二重特異性一本鎖抗体分子を作製するため、最初に、標準的なプロトコールに従う遺伝子合成により得たCEAIの可変領域をPCRにより改変して相当する一本鎖Fv抗体フラグメントを得る必要があった。本目的のため、2段階の融合PCRを用いて可変領域をコードする配列を増幅させた。一組の適切なプライマーを設計してPCRに基づくクローニング段階を行い、最後に、その結果として、15アミノ酸リンカー([Gly4Ser]3)をVL−リンカー−VHの順序で含む2つの可変ドメインを連結している一本鎖抗体がもたらされた。
【0107】
要するに、以下のプライマーの組み合わせを用いた。
【表2】
【0108】
オリゴヌクレオチドプライマーの核酸配列を下に示す。
5’CEAI LH:5'AGGTGTACACTCCGACATTGAGCTCACCCAG3’(配列番号137)
3’CEAI VLリンカー:5'GGAGCCGCCGCCGCCAGAACCACCACCACCTTTGATCTCGAGCTTGG3'(配列番号138)
5’CEAI VHリンカー:5'GGCGGCGGCGGCTCCGGTGGTGGTGGTTCTCAGGTCCAACTGCAGGAG3'(配列番号139)
3’CEAI LH:5’AATCCGGAGGAGACGGTGACCG3'(配列番号140)
【0109】
一本鎖抗体を作製するため、上記のそれぞれのプライマーの組み合わせで2回のPCRをPCR段階1および2として行った。このPCRの間、重複する相補的配列を(それぞれのリンカープライマーから生じる)PCR生成物に導入し、それは一体となって(combined)その後の融合PCRの間に15アミノ酸リンカーのコード配列を形成した。その後、増幅したVHおよびVLドメインを、外側のプライマーのみと両方のPCR生成物を必要とする、この融合PCR中で結合させた。得られるscFv抗体は、5’末端でBsrGIの制限酵素認識部位に隣接し、3’末端でBspEIの制限酵素認識部位に隣接している。WO 2005/040220号に記載されるようにマウス免疫グロブリンリーダーペプチドのコード配列とのインフレーム融合を可能にするために、BsrGI部位の付加を行った。二重特異性一本鎖抗体を作製するためのCD3特異的一本鎖抗体をコードする配列とのインフレーム融合を可能にするために、BspEI部位を作製した。一本鎖Fv抗体の融合を達成するため、かつ真核生物の発現を可能にするため、CEA特異的一本鎖Fv抗体のコード配列は、BsrGIおよびBspEIを介して、WO 2005/040220号に記載されるように脱免疫化された抗CD3一本鎖Fv抗体(本発明において配列番号77と称される)を含有するpEFDHFR発現ベクターにクローン化された(pEFDHFRは、Mack et al. Proc. Natl. Acad. Sci. USA 92 (1995) 7021-7025 に記載されている)。構築物の単一のクローンを単離し、標準的なプロトコールに従って発現ベクター中で隣接領域と相補的なプライマーを用いて配列決定した(Sambrock, Molecular Cloning; A Laboratory Manual, 2nd edition, Cold Spring Harbour laboratory Press, Cold Spring Harbour, New York (1989))。さらなる実験のため、検証された核酸配列を含む構築物のクローンが選択された。
【0110】
2.CEAxCD3二重特異性一本鎖抗体の発現および精製
二重特異性一本鎖抗体をチャイニーズハムスター卵巣(CHO)細胞で発現させた。DHFR欠損CHO細胞における真核生物タンパク質発現を、Kaufmann(前掲)に記載のとおり行った。構築物の遺伝子増幅は漸増濃度のMTXにより20nM MTXまでの終濃度まで誘導した。静置培養の二代継代後、細胞をCHO改変DMEM培地(HiQ(登録商標)、HiClone)を含むローラーボトル中で7日間増殖させた後、回収した。細胞を遠心分離により取り除き、発現タンパク質を含有する上清を−20℃で保存した。
【0111】
Akta(登録商標)FPLCシステム(Pharmacia)およびUnicorn(登録商標)ソフトウェアをクロマトグラフィーに使用した。全ての化学物質は研究等級のものであり、Sigma社(Deisenhofen)またはMerck社(Darmstadt)より購入した。固定化金属アフィニティークロマトグラフィー(「IMAC」)を、製造業者の提供するプロトコールに従って、ZnCl2を装填したFractogel(登録商標)カラム(Merck)を用いて行った。カラムをバッファーA2(20mM リン酸ナトリウムバッファー、pH7.5、0.4M NaCl)で平衡化し、細胞培養上清(500ml)を3ml/分の流速にてカラム(10ml)に適用した。カラムをバッファーA2で洗浄して結合しなかったサンプルを取り除いた。結合したタンパク質は、2段階勾配のバッファーB2(20mM リン酸ナトリウムバッファー、pH7.5、0.4M NaCl、0.5M イミダゾール)を用いて、以下に従って溶出した。
段階1:6カラム容積中バッファーB2 20%;
段階2:6カラム容積中バッファーB2 100%。
【0112】
段階2からの溶出したタンパク質画分を、さらなる精製のためにプールした。ゲルろ過クロマトグラフィーを、PBS(Gibco)で平衡化したSephadex S200 HiPrepカラム(Pharmacia)で行った。溶出したタンパク質サンプル(流速1ml/分)を、検出のため、標準的なSDS−PAGEおよびウエスタンブロットに付した。精製より前に、カラムを分子量決定のために較正した(分子量マーカーキット、Sigma MW GF−200)。プロテインアッセイ染料(MicroBCA、Pierce)およびIgG(Biorad)を標準タンパク質として用いて、タンパク質濃度を測定した。
【0113】
CEAxCD3二重特異性一本鎖抗体を、IMACの2段階精製プロセスおよびゲルろ過で単離した。主生成物の分子量は、未変性条件下でPBSでのゲルろ過により測定して約52kDaであった。この分子量は二重特異性一本鎖抗体に一致する。全ての構築物がこの方法に従って精製された。
【0114】
精製された二重特異性一本鎖抗体タンパク質を、還元条件下で事前成形された4〜12% Bis Trisゲル(Invitrogen)で行われたSDS PAGEで分析した。製造業者の提供するプロトコールに従ってサンプルの調製および適用を行った。MultiMarkタンパク質標準品(Invitrogen)を用いて分子量を測定した。コロイドクマシーでゲルを染色した(Invitrogenプロトコール)。単離したタンパク質の純度は、SDS−PAGEで測定して>95%であった。Optitran(登録商標)BA−S83膜およびInvitrogenブロットモジュールを製造業者の提供するプロトコールに従って用いてウエスタンブロットを行った。使用した抗体は、基質としてHisタグ(Penta His、Qiagen)およびアルカリ性ホスファターゼ(AP)で標識したヤギ抗マウスIg(Sigma)、およびBCIP/NBT(Sigma)に対して指向された。二重特異性一本鎖抗体は、ウエスタンブロットにより特異的に検出することができた。精製した二重特異性一本鎖抗体分子に対応する52kDで単一のバンドが検出された。
【0115】
3.CEAxCD3二重特異性一本鎖抗体のフローサイトメトリー結合分析
膜結合ヒトCEAおよびヒトCD3との結合能に関して構築物の官能性を試験するため、FACS分析を行った。本目的のため、ヒトCEAトランスフェクトCHO細胞とCD3陽性ヒトT細胞白血病細胞株HPB−AII(DSMZ,Braunschweig,ACC483)を使用した。200,000個のCEA陽性CHO細胞または200,000個のHPB−AII細胞を、氷上で30分間、異なる配置のCEAおよびCD3のVHおよびVL領域(上記の通り)を含む二重特異性抗体をそれぞれ発現しているCHO細胞培養の純粋な細胞上清50μlとともにインキュベートした。細胞をPBSで2回洗浄し、構築物の結合を、構築物のC末端のヒスチジンタグを介して細胞と結合した構築物と特異的に結合する、標識されていないマウスPenta His抗体(2% FCSを含む50μl PBSに1:20で希釈;Qiagen)を用いて検出した。洗浄段階の後に、結合していないマウスPenta His抗体を取り除いた。結合した抗His抗体を、フィコエリトリンと複合体化し、2% FCSを含む50μl PBSに1:100で希釈されたFcγ特異的抗体(Dianova)を用いて検出した。ヒトCEAとの結合の陽性対照として、モノクローナル抗体Col−1(実施例3参照)を用いた。ヒトCD3との結合の対照には、WO 99/054440号に記載のCD19xCD3二重特異性一本鎖構築物を使用した。陰性対照として、培養上清の代わりに新鮮な培地を用いた。
【0116】
細胞をフローサイトメトリーで分析した(FACS−Calibur;Becton Dickinson,Heidelberg)。FACS染色および蛍光強度の測定を、Current Protocols in Immunology(Coligan, Kruisbeek, Margulies, Shevach and Strober, Wiley-lnterscience, 2002)に記載のとおり行った。
【0117】
図1に示されるように、二重特異性一本鎖抗体の数個のドメイン配置、すなわち、CEAI VHVLx配列番号77VHVL(配列番号8)、CEAI VLVHx配列番号77VHVL(配列番号6)、CEAII VHVLx配列番号77VHVL(配列番号10)、CEAIII VLVHx配列番号77VHVL(配列番号12)およびCEAIII VHVLx配列番号77VHVL(配列番号14)は、ヒト膜結合CEAおよびヒトCD3と結合した。陰性対照として、培地ならびに1.および2.検出抗体を使用した。
【0118】
実施例3:CEAxCD3二重特異性一本鎖抗体と可溶性ヒトCEAの結合
可溶性ヒトCEAに対する特異性を決定するため、様々なCEAxCD3二重特異性一本鎖抗体をELISAで試験した。
【0119】
本目的のため、可溶性ヒトCEA抗原を最初にビオチン化した。ビオチン化は、5% DMSO(Sigma)を含有するPBS中、15倍モル過剰のEZ−Link Sulfo NHS−LC−LC−ビオチン(Pierce)とともに、1時間室温にてサンプルミキサー(Dynal)中で達成した。遊離ビオチンとビオチン化CEA抗原を分離するため、アッセイを標準的なプロトコールに従ってPBSに対して過剰に透析した。保持された、ビオチン標識CEAの生物活性をELISA結合実験で確認した。
【0120】
可溶性ヒトCEAに対するCEAxCD3二重特異性一本鎖抗体の特異性を測定するための直接ELISAを、標準的な手順に従って行った。要するに、50μl/ウェルのPBSまたは可溶性ビオチン化ヒトCEA(Abcam;1xPBS中5μg/ml)を、4℃にて約16時間インキュベートすることにより96ウェルストレプトアビジンコートELISAプレート(Nunc)上に固定化した。ウェルあたり200μlの水で洗浄した後、200μlのブロッキング溶液(PBS/3% BSA)を添加した。室温にて1時間ブロッキングした後、ブロッキング溶液を除去した。その後の洗浄段階(200μl/ウェルの1xPBS/0.05%(v/v) Tween20)およびインキュベーション段階はすべて室温にて行った。1回洗浄した後、CEAI VHVLx配列番号77VHVL(配列番号8;mAb A5B7に由来する抗CEA部分)、CEAII VHVLx配列番号77VHVL(配列番号10;mAb T84.66に由来する抗CEA部分)、およびCEAIII VHVLx配列番号77VHVL(配列番号14;mAb MFE−23に由来する抗CEA部分)二重特異性一本鎖抗体と、マウスモノクローナル抗体CEA/CD66 Ab−3(Col−1;Dunn)を異なる濃度(1xPBS中0.5μg/mlおよび5μg/ml;50μl/ウェル)中で1時間インキュベートした。1xPBS(50μl/ウェル)を非特異的結合の対照として添加した。3回の洗浄段階の後、50μl/ウェルのPenta−His IgG(Qiagen;1xPBS中2μg/ml)をHis−タグ二重特異性一本鎖抗体の検出のために添加した。その後、ウェルを3回洗浄し、50μlの西洋ワサビペルオキシダーゼ標識されたヤギ抗マウスFcγ特異的抗体(Jackson ImmuneResearch;1xPBS中1:1000)とともに1時間インキュベートした。3回洗浄した後、ELISAはABTS基質溶液(Roche)を添加することにより発色(develop)させ、405nmの波長で吸光度を測定した。
【0121】
図2は、ELISAで検出された様々な二重特異性一本鎖抗体の吸光度を示す。CEAI VHVLx配列番号77VHVL(配列番号8;mAb A5B7に由来する抗CEA部分)、CEAII VHVLx配列番号77VHVL(配列番号10;mAb T84.66に由来する抗CEA部分)、およびCEAIII VHVLx配列番号77VHVL(配列番号14;mAb MFE−23に由来する抗CEA部分)二重特異性一本鎖抗体と、マウスモノクローナル抗体Col−1は、可溶性ヒトCEAと特異的に結合した。CEA抗原の不在下(PBS対照)で、結合シグナルは観察されなかった。要約すると、mAb A5B7、T84.66およびMFE−23に由来する抗CEA結合ドメインは、可溶性および膜結合CEAの両方と結合する。
【0122】
実施例4:CEAxCD3二重特異性一本鎖抗体の生物活性
作製したCEAxCD3二重特異性一本鎖抗体の生物活性を、標的細胞としてヒト胃癌細胞株KatoIIIまたはヒトCEAトランスフェクトCHO細胞、およびエフェクター細胞として刺激を受けたヒトCD8陽性T細胞または未変性のPBMCをそれぞれ用いるインビトロクロム放出細胞障害性アッセイにより分析した。
【0123】
刺激を受けたCD8+T細胞の作製は、以下の通り行った。
ペトリ皿(直径145mm、greiner bio−one)を抗CD3抗体(OKT3 Janssen−Cilag GmbH,Orthoclone1mg/ml;終濃度1μg/ml)および抗CD28抗体(BD、1mg/ml;終濃度1μg/ml)で37℃にて1時間プレコートした。インキュベーション時間の後、結合しなかったタンパク質をPBSでの1回の洗浄段階により除去した。新鮮なPBMCを、標準的なプロトコールに従ってフィコール勾配遠心分離により末梢血(30〜50ml)から単離した。3〜5×107PBMCを、150ml RPMI1640/10% FCS/IL−2 20U/ml(Proleukin,Chiron)中のプレコートしたペトリ皿に添加し、2日間刺激した。3日目に細胞を回収し、RPMI1640で1回洗浄し、大きなTフラスコに移した。IL−2を20U/mlの終濃度に添加し、再び1日間培養した。マニュアルの指示に従ってCD8陰性単離キット(Dynal Biotech)を使用してCD8+CTLを単離した。未変性のPBMCは、フィコール勾配遠心分離の後に刺激手順を踏まずに直接用いた。標的細胞をPBSで2回洗浄し、50%FCSを含む最終容量100μlのRPMI中で37℃にて45分間、11.1MBq51Crで標識した。その後、標識した標的細胞を5mlのRPMIで3回洗浄し、次に細胞障害性アッセイで使用した。アッセイは、E:T比が10:1(1ウェルあたり5000個の標的細胞および50000個のエフェクター細胞に相当)のRPMI(上記の通り)を補給した96丸底プレートにおいて総量250μlで行った。構築物の評価のため、アッセイ容量中出発濃度1μg/mlの二重特異性一本鎖分子および12個のその三倍希釈液を用いた。アッセイ時間は18時間であり、細胞障害性は、最大溶解(Triton−Xの添加)および自発的溶解(エフェクター細胞なし)の差に関連した、上清中に放出されたクロムの相対値として測定された。全ての測定は三つ組みで行われた。上清中のクロム活性の測定は、Wizard3ガンマカウンター(Perkin Elmer Life Sciences GmbH,Koln,Germany)で行った。実験データの分析は、Prism4 for Windows(登録商標)(バージョン4.02、GraphPad Software Inc.,San Diego,California,USA)を用いて行った。シグモイド型用量応答曲線は、一般に、ソフトウェアで測定してR2値が>0.90である。分析プログラムにより計算されるEC50値を生物活性の比較に用いた。
【0124】
図3は、様々なドメイン配置、すなわち、配列番号77VHVLxCEAI VHVL(配列番号4)および配列番号77VHVLxCEAI VLVH(配列番号2)(両構築物はN末端に抗CD3部分を含む)、ならびにCEAI VLVHx配列番号77VHVL(配列番号6)およびCEAI VHVLx配列番号77VHVL(配列番号8)(C末端に抗CD3を含む)についてのヒトCEAトランスフェクト標的細胞(CHO−CEA+細胞)に対する細胞障害活性を示す。非トランスフェクトCHO細胞(ヒトCEAを欠く)を陰性対照として用いた。
【0125】
図4では、CEAI VHVLx配列番号77VHVL(配列番号8)、CEAIII VLVHx配列番号77VHVL(配列番号12)、CEAIII VHVLx配列番号77VHVL(配列番号14)、およびCEAII VHVLx配列番号77VHVL(配列番号10)は、ヒトCEAトランスフェクトCHO細胞に対する細胞障害活性を提示した。非トランスフェクトCHO細胞(ヒトCEAを欠く)を陰性対照として用いた。上に述べたように、CEAIは、マウスmAb A5B7に由来する可変領域を意味し、CEAIIは、マウスmAb T84.66に由来する可変領域であり、CEAIIIは、マウスmAb MFE−23由来の可変領域を指す。
【0126】
要約すると、試験した構築物はすべて、可溶性ヒトCEAの不在下で、(ヒト)CEA発現KatoIIIおよびヒトCEAトランスフェクトCHO細胞に対する細胞障害活性を示した。
【0127】
実施例5:可溶性CEA抗原の存在下で特異的なCEAxCD3二重特異性一本鎖抗体の生物活性
競合アッセイを、次の例外:CEAxCD3二重特異性一本鎖抗体の生物活性を様々な濃度の可溶性ヒトCEA抗原の存在下で試験する、を除いて実施例4に記載のとおり行う。使用した可溶性ヒトCEA抗原(AbCAM Ltd.Cambridge UK)を単独の患者の肝臓からの転移性結腸癌から単離した。細胞を添加する前に、所定量の二重特異性一本鎖抗体を漸増量の可溶性ヒトCEA抗原(0μg/ml;0.1μg/ml;1μg/ml、または0μg/ml;0.004μg/ml;0.02μg/ml;0.1μg/ml;0.5μg/ml;1μg/mlのいずれか)で37℃にて30分間プレインキュベーションすることにより実験的に競合アッセイを行った。残りのアッセイは、実施例4に記載されるとおり行った。これらの競合実験の結果を図5〜11に示す。
【0128】
ヒトCEA陽性標的細胞に対する二重特異性一本鎖抗体、mAb T84.66に由来する抗CEA部分を含むCEAII VHVLx配列番号77VHVL(配列番号10;図7および9〜11)、およびmAb MFE−23に由来する抗CEA領域を含むCEAIII VLVHx配列番号77VHVL(配列番号12;図8参照)の細胞障害活性は、漸増量の可溶性ヒトCEA抗原により大幅に阻害された。上に述べたように、前記構築物は膜結合と可溶性ヒトCEA抗原の両方と結合する;例えば、実施例2および3ならびに図1および2参照。従って、最も確かなことには、可溶性ヒトCEAは、前記二重特異性一本鎖抗体の抗CEA部分が標的細胞、例えば、CHO−CEA+またはKatoIII腫瘍細胞上の膜結合ヒトCEAと結合することを阻害し、それにより前記抗体構築物により媒介される細胞障害活性を阻害する。
【0129】
それに対して、驚くことに、mAb A5B7に由来する抗CEA部分を有する二重特異性一本鎖抗体が可溶性ヒトCEAに対して抵抗性であることが見出された。例えば、CEAI VLVHx配列番号77VHVL(配列番号6;図5参照)、CEAI VHVLx配列番号77VHVL(配列番号8;図5、8および10参照)、配列番号77VHVLxCEAI VHVL(配列番号4;図6参照)および配列番号77VHVLxCEAI VLVH(配列番号2;図6参照)は、漸増量の可溶性ヒトCEAにより阻害されず、高い濃度(1μg/ml)によってさえも阻害されない。これは、前記二重特異性一本鎖抗体の抗CEA部分が可溶性ヒトCEAと結合するという事実を考慮すると予測することができなかった(図2)。むしろ、漸増量の可溶性ヒトCEAによるCEA陽性標的細胞に対する細胞障害活性の阻害を、T84.66由来構築物およびMFE−23由来構築物の場合と同じように、予測することができた;上記参照。
【0130】
従って、本発明は、高いレベルの可溶性CEA抗原の存在下でさえ細胞障害性抗腫瘍活性を有する医薬組成物を提供する。そのため、これらの医薬組成物はその血漿中の可溶性CEA抗原濃度の高い腫瘍患者、例えば、進行性上皮性腫瘍、転移性上皮性腫瘍、高腫瘍量、後期上皮性腫瘍の患者またはELISAで測定してCEA血清濃度が100ng/mlよりも高い腫瘍患者の治療に特に適している。
【0131】
実施例6:ヒトVL領域の選択
癌患者に投与される際に免疫原性の低下した医薬組成物を提供するため、可溶性CEA抗原に対する抵抗性を有するヒト二重特異性一本鎖抗体を作製した。第1段階で、可溶性CEAに対する抵抗性を有するヒトVL領域が単離された。従って、この実験の目的は、モノクローナル抗体(mAb)A5B7の母親由来のマウスVHと対となり得るヒトVL領域の選択である。
【0132】
1.可溶性ヒトCEA抗原のビオチン化
ファージライブラリー選択のため、可溶性CEA抗原をビオチン化した。ビオチン化は、5% DMSO(Sigma)を含有するPBS中で、サンプルミキサー(Dynal)中で室温にて1時間15倍モル過剰のEZ−Link Sulfo NHS−LC−LC−ビオチン(Pierce)を用いて達成した。遊離ビオチンとビオチン化CEA抗原の分離のため、標準的なプロトコールに従って、アッセイをPBSに対して過剰に透析した。ビオチン標識したCEAの保持された生物活性をELISA結合アッセイで確認した。
【0133】
2.RNAのヒトB細胞からの単離
100mLの血液を5名の健康なヒトドナーから採取した。末梢血単核細胞(PBMC)を標準的な方法に従ってフィコール勾配により単離した。RNeasy(登録商標)Midi Kit(QIAGEN)を製造業者の説明書に従って用いて、全RNAを単離細胞から単離した。標準的な方法に従ってcDNAを合成した(Sambrook, Cold Spring Harbor Laboratory Press 1989, 2001)。
【0134】
3.可変軽鎖領域(VL−領域)のPCR増幅
軽鎖V領域DNAの単離のため、RT−PCRを、V−κ−(5’−hu−VK1−SacI−2001(5’−GAGCCGCACG AGCCCGAGCT CCAGATGACC CAGTCTCC−3’)(配列番号78)、5’−hu−VK2/4−SacI−2001(5’−GAGCCGCACG AGCCCGAGCT CGTGATGACY CAGTCTCC−3’)(配列番号79)、5’−hu−VK3−SacI−2001(5’−GAGCCGCACG AGCCCGAGCT CGTGWTGACR CAGTCTCC−3’)(配列番号80)、5’−hu−VK5−SacI−2001(5’−GAGCCGCACG AGCCCGAGCT CACACTCACG CAGTCTCC−3’)(配列番号81)、5’−hu−VK6−SacI−2001(5’−GAGCCGCACG AGCCCGAGCT CGTGCTGACT CAGTCTCC−3’)(配列番号82)、3’−hu−Vk−J1−SpeI−BsiWI(5’−GACGACACTA GTTGCAGCCA CCGTACGTTT GATTTCCACC TTGGTCC−3’)(配列番号83)、3’−hu−Vk−J2/4−SpeI−BsiWI(5’−GACGACACTA GTTGCAGCCA CCGTACGTTT GATCTCCASC TTGGTCC−3’)(配列番号84)、3’−hu−Vk−J3−SpeI−BsiWI(5’−GACGACACTA GTTGCAGCCA CCGTACGTTT GATATCCACT TTGGTCC−3’)(配列番号85)、3’−hu−Vk−J5−SpeI−BsiWI(5’−GACGACACTA GTTGCAGCCA CCGTACGTTT AATCTCCAGT CGTGTCC−3’)(配列番号86))およびVλ(5’−huVL1a−SacI−2001(GAG CCG CAC GAG CCC GAG CTC GTG TTG ACG CAG CCG CCC TC)(配列番号87)、5’−huVL1b−SacI−2001(GAG CCG CAC GAG CCC GAG CTC GTG CTG ACT CAG CCA CCC TC)(配列番号88)、5’−huVL2−SacI−2001(GAG CCG CAG GAG CCC GAG CTC GCC CTG ACT CAG CCT SCC TCC GT)(配列番号89)、5’−huVL4−SacI−2001(ACC TGC GAG CTC GTG CTG ACT CAR YCM YCC TCT GC)(配列番号90)、5’−huVL5−SacI−2001(ACC TGC GAG CTC GTG CTG ACT CAG CCR SCT TCC)(配列番号91)、5’−huVL6−SacI−2001(ACC TGC GAG CTC ATG CTG ACT CAG CCC CAC TC)(配列番号92)、5’−huVL3/9−SacI−2001(GAG CCG CAC GAG CCC GAG CTC GWG CTG ACT CAG CCA CCY TC)(配列番号93)、5’−huVL7/8−SacI−2001(GAG CCG CAC GAG CCC GAG CTC GTG GTG ACY CAG GAG CCM TC)(配列番号94)、3’−hu−Vlam−BlnI−SpeI−2001(CGT GGG ACT AGT CTT GGG CTG ACC TAG GAC GGT)(配列番号95)、3’−hu−Vlam2−BlnI−SpeI−2002:CGT GGG ACT AGT CTT GGG CTG ACC GAG GAC GGT)(配列番号96)プライマーセットを用いて行った。
【0135】
ヒトB細胞由来のRNAをcDNAに転写し(上記の通り)、PCR反応の鋳型DNAとして用いた。1回のPCR反応につき、1個の5’−プライマーを1個の3’−プライマーと組み合わせた。異なるPCR反応の数を、5’−および3’−プライマーの可能な組み合わせの数により決定した。増幅には次のPCRプログラムを用いた。すなわち94℃にて15秒間の変性、52℃にて50秒間のプライマーアニーリングおよび72℃にて90秒間のプライマー伸張を40サイクル以上行い、それに続いて72℃にて10分間の最終伸張を行った。次に、軽鎖DNA V断片を標準的なプロトコールに従って単離した。
【0136】
4.ライブラリー構築−ヒトVLプールのクローニング
ファージディスプレイライブラリーは、概して、例えば「Phage Display: A Laboratory Manual」; Ed. Barbas, Burton, Scott & Silverman; Cold Spring Harbor Laboratory Press, 2001に開示されるような標準的な手順に基づいて構築された。
【0137】
PCR増幅のために選択されるプライマーは、軽鎖V断片の5’−SacIおよび3’−SpeI認識部位を生じた。4回の連結反応をセットアップし、各々400ngの軽鎖断片(SacI−SpeI消化、2×κおよび2×λ)および1400ngのファージミドpComb3H5BHis(SacI−SpeI消化;大きな断片;このベクターは、Ralf Lutterbuese博士の論文に記載されている)で構成された。次に、得られる4つの抗体V軽鎖プールを、エレクトロポレーション(2.5kV、0.2cmギャップキュベット、25mF、200 Ohm、Biorad gene−pulser)により300μlのエレクトロコンピテント(electrocompetent)大腸菌(Escherichia coli)XL1 Blue株に各々形質転換して、
κ1:2×108
κ2:6×107
λ1:9×107
λ2:6×107
の独立したクローンからなるライブラリーサイズを得た。
【0138】
様々なPCR増幅から得たκ(軽鎖)DNA断片を、各連結について次のように秤量した。それぞれの5’−プライマーは特定のグループを定義する。これらのグループの中で、3’−プライマーはサブグループを定義する。κサブグループを、1:2:1:1(プライマー3’−hu−Vk−J1−SpeI−BsiWI:3’−hu−Vk−J2/4−SpeI−BsiWI:3’−hu−Vk−J3−SpeI−BsiWI:3’−hu−Vk−J5−SpeI−BsiWIに対応)に秤量した。グループを、それらの生殖系列分布1:1:1:0.2:0.2(プライマー5’−huVK1−Sac−2001:5’−huVK3−Sac−2001:5’−huVK2/4−Sac−2001:5’−huVK5−Sac−2001:5’−huVK6−Sac−2001に対応)に従って秤量した。
【0139】
異なるPCR増幅から得たλ(軽鎖)DNA断片を、各連結について次のように秤量した。それぞれの5’−プライマーは、特定のグループを定義する。これらのグループの中で、3’−プライマーはサブグループを定義する。λサブグループを、3:1(プライマー3’−hu−Vlam−BlnI−SpeI−2001:3’−hu−Vlam2−BlnI−SpeI−2002に対応)に秤量した。
【0140】
グループを、それらの生殖系列分布1:1:2:2:2:3(プライマー5’−huVL1a−SacI−2001:5’−huVL1b−SacI−2001:5’−huVL2−SacI−2001:5’−huVL4−SacI−2001+5’−huVL5−SacI−2001:5’−huVL6−SacI−2001+5’−huVL7/8−SacI−2001:5’−huVL3/9−SacI−2001に対応)に従って秤量した。
【0141】
エレクトロポレーション後、それぞれの反応液を表現型発現のためにSOC培養液(Fluka)中でインキュベートした。2つのκ培養物を組み合わせ、同様に2つのλ培養物も組み合わせた。得られるκ培養物および得られるλ培養物を、次にそれぞれ500mLの、50μg/ml カルベニシリンおよび2% w/v グルコースを含有するSB選択培地中で一晩インキュベートした。翌日、細胞を遠心分離により回収し、市販のプラスミド調製キット(Qiagen)を用いてプラスミドの調製を行った。
【0142】
5.抗体ライブラリーの構築−ヒトVL−母親由来VH
PCRを行ってmAb A5B7の母親由来のVHを前記母親由来のVHを含有するベクターから増幅した。増幅のために、5’−プライマー5’−AVH−Xho I(5’−GTC ACA CTC GAG TCA GGA GGA GGC TTG GTA C−3’)(配列番号97)および3’−プライマー3’−AVH−BstEII(5’−GTC ACA GGT GAC CGT GGT CCC TTG GCC CCA G−3’(配列番号98)を用いて標準的手順に従うPCRプロトコールに従った。分析用アガロースゲルからの約350bp増幅産物の精製後、DNA断片を制限酵素BstEIIおよびXho Iで切り出した。ファージミドpComb3H5BHis(このベクターはRalf Lutterbuese博士の論文に記載されている。)をしかるべく消化させ、大きな断片を上述の断片と連結させた。大腸菌XL1 blueへの形質転換後、単一のクローンを100mL SB培地(50μg/ml カルベニシリン含有)で培養し、プラスミドを標準的なプロトコールに従って調製した。挿入物を配列決定することにより首尾よいクローニングを確認した(Sequiserve,Munich)。
【0143】
このベクターpComb3H5BHis/mAb A5B7の母親由来VHを制限酵素SacIおよびSpeIで制限した。大きなベクター断片を単離した。プラスミド−DNAを含有するVκライブラリーおよびVλライブラリーを制限酵素SacIおよびSpeIで制限した。小さなVκ断片およびそれぞれのVλ断片(各およそ350bp)を標準的なプロトコールに従って単離した。1200ngのベクター断片を各200ngのVκおよびVλ断片両方の混合と連結した。連結反応物をエレクトロポレーション(2.5kV、0.2cmギャップキュベット、25mF、200 Ohm)により300μLのエレクトロコンピテント(electrocompetent)大腸菌XL1 Blue株に形質転換して、1.2×108の独立したクローンからなる全scFvライブラリーサイズを得た。
【0144】
表現型発現およびカルベニシリンへの緩やかな適応の後、抗体ライブラリーをSB−カルベニシリン(50μg/ml)選択培地へ移した。次に、抗体ライブラリーを感染量の1×1012粒子のヘルパーファージVCSM13で感染させ、その結果、繊維状のM13ファージの産生および分泌が得られ、この各ファージ粒子は半ヒトscFv−断片をコードする一本鎖pComb3H5BHis−DNAを含み、ファージコートタンパク質IIIへの翻訳融合として対応するscFv−タンパク質を提示した。
【0145】
6.ヒトVLのファージディスプレイ選択
scFv−レパートリーを有するファージ粒子をPEG8000/NaCl沈殿および遠心分離により培養上清から回収した。次に、約1×1011〜1×1012scFvファージ粒子を0.5mLのTBS/1% BSAに再懸濁し、ストレプトアビジンコートされたELISAプレート(Nunc)のウェル中で1時間固定されたビオチン化可溶性CEAとともにインキュベートした。抗原10μg/mlのPBS溶液(50μl)をストレプトアビジンコートされたウェル中で4℃にて一晩インキュベートし、水で1回洗浄し、それに続いてTBS中200μlの3% BSAで37℃にて1時間ブロッキングし、それをインキュベーション後に取り除いた。
【0146】
標的抗原と特異的結合しなかったscFvファージをTBS/0.05% Tweenでの洗浄段階により除去した。この洗浄手順は、さらなるラウンドで10回まで繰り返された。
【0147】
洗浄後、HCl−グリシン、pH 2.2を用いることにより結合体を溶出した。2M Tris、pH 12での中和後、溶出液を新鮮な非感染大腸菌XL1 Blue培養株の感染に用いた。
【0148】
残っている高結合体を溶出するため、50μlの新鮮な大腸菌XL1 blue培養株(OD600≧0.5)をウェルに添加し、15分間インキュベートした。次に、両方の培養物を混合し、ヒトscFv−断片をコードするファージミドコピーで首尾よく形質導入した細胞を再びカルベニシリン抵抗性について選択し、その後にVCMS13ヘルパーファージで感染させて、抗体提示およびインビトロでの選択の第2のラウンドを開始した。
【0149】
4ラウンドのパニングに対応するプラスミドDNAを大腸菌培養物から単離した。可溶性scFv−タンパク質の産生のため、VH−VL−DNA断片をプラスミドから切り出し(Xho I−SpeI)、プラスミドpComb3H5BFIag/His中の同じ制限部位を介してクローニングし、その発現構築物(例えばscFv)は、scFvとHis6タグとの間にFlagタグ(TGDYKDDDDK)(配列番号99)を含み、さらなるファージタンパク質が欠失している。
【0150】
連結後、各プール(異なるラウンドのパニング)のプラスミドDNAを100μL熱ショックコンピテント大腸菌TG1に形質転換し、カルベニシリンLB寒天上にプレーティングした。単一のコロニーを選定し、96ウェルプレート(Greiner)中の120μLのLBカルベニシリン(50μg/ml)1% グルコースに播種した。ウェルを半透膜(Greiner)で密封し、振盪インキュベータでプレートを37℃にて一晩インキュベートした(マスタープレート)。次に、10μlのマスタープレート培養物を、ウェルあたり90μL LBカルベニシリン(50μg/ml)0.1% グルコースを含有する第2の96ウェルプレート(ワーキングプレート)に移した。37℃の振盪インキュベータ中で4時間のインキュベーションの後、20μL LBカルベニシリン、6mM IPTGを各ウェルに添加することによりscFv生成を誘導した。振盪しながら30℃にて一晩の別のインキュベーション段階の後、細胞を、40μL 溶解緩衝液(400mM ホウ酸、320mM NaCl、4mM EDTA pH8、2.5mg/ml リゾチーム)での室温にて1時間のインキュベーションで溶解した。残りの細胞および細胞片を1.900×gで12分間の遠心分離により分離した(Hettich)。
【0151】
次に、scFv分子を含有する上清を、フローサイトメトリー結合アッセイで結合について試験した。ヒトCEAでトランスフェクトしたCHO細胞をCEA陽性細胞株として用いた。最初に100,000〜200,000個の細胞をヒトscFvまたは関連する対照を含有する周辺質調製物とともにインキュベートすることにより細胞結合アッセイを行った。インキュベーション後、細胞をPBS/1% FCS(ウシ胎児血清)中で洗浄し、さらに5〜10μg/mlの抗FLAG M2抗体(Sigma)とともにインキュベートした。細胞を再び洗浄した後、それらをポリクローナル、PE標識抗マウス抗体(Dianova)とともにインキュベートし、その後フローサイトメトリーにより分析した。およそ600個のクローンをCEA陽性のCHO細胞で結合シグナルについて試験した。27個の陽性クローンが得られた。それぞれのscFv DNAを配列決定した後、合計9個の異なる配列が得られた。
【0152】
図12は、フローサイトメトリー分析により測定される、9個の様々な半ヒトscFv(すなわち、マウスA5B7 VH−ヒトVL)構築物と多様な細胞株の結合を示す。前記図は複数の図表を含み、1つが試験された各構築物を表す。任意の所与図表中、黒色の分布は、いずれの構築物も不在下であるが、scFvの検出に用いられるあらゆる適切な検出剤を含むPBS単独でインキュベートした細胞の蛍光強度を示す。このようにして、観察されるいずれの蛍光シフトも間違いなく検出剤または緩衝液よりもscFv構築物に起因し得る。構築物とそれぞれの細胞株との結合の指標である蛍光のシフトは、各図表中灰色の線で示される。一般に、(黒色の)対照から離れてより高い強度のシフト、すなわち対照よりもはるかに遠いシフトはより強い結合を示し、一方、(黒色の)対照から離れてより低い強度のシフト、すなわち対照により近いシフトはより弱い結合を示す。
【0153】
図12から、構築物A−121、A−183、A−240、A−313、A−290、A−315、A4−35、A4−52、MP2−A5は、それぞれの対照と比較して、蛍光強度において明らかに識別できるシフトを示し、scFvとCHO標的細胞上の膜結合CEAとの結合を示すことが分かる。以下で、scFv A−240(配列番号48)のヒトVL領域を選択し、ヒトVH領域の単離に用いた。前記ヒトVL領域は、配列番号63(核酸配列)および配列番号64(アミノ酸配列)に示される。
【0154】
実施例7:可溶性CEA抗原に対して抵抗性のあるヒトVH領域の抗体ライブラリーの構築およびファージディスプレイ選択
以下の実験の目的は、実施例6に記載されるように選択される、scFv A−240のヒトVL領域と対となる可溶性CEA抗原に対して抵抗性のある一連のヒトVH領域の選択である。前記ヒトVL領域は、配列番号63(核酸配列)および配列番号64(アミノ酸配列)に示される。
【0155】
1.末梢血単核細胞(PBMC)からのRNAの単離
100mLの血液を5名の健康なヒトドナーから採取した。末梢血単核細胞(PBMC)を、標準的な方法に従ってフィコール勾配により単離した。全RNAを、RNeasy(登録商標)Midi Kit(QIAGEN)を製造業者の説明書に従って用いてPBMCから単離した。cDNAを標準的な方法に従って合成した(Sambrook, Cold Spring Harbor Laboratory Press 1989, 2001)。
【0156】
2.可変重鎖領域(VH領域)のPCR増幅
VHライブラリーを構築し、Lib 134−VHと名付けた。このVH−ライブラリーは、その後にヒトFR4生殖系列配列が続く母親由来の抗体のVHCDR3と作動可能なように連結された、PCR増幅された上記のPBMCプールのVH領域由来のFR1−CDR1−FR2−CDR2−FR3のヒトレパートリーで構成される。
【0157】
ヒト鋳型VH領域の単離のため、5’−VH特異的プライマーセット(5’−huVH1,3,5−Xho I−2001(5’−AGG TGC AGC TGC TCG AGT CTG G−3’)(配列番号100)、5’−huVH4−Xho I−2001(5’−CAG GTG CAG CTG CTC GAG TCG GG−3’)(配列番号101)、5’−huVH4B−Xho I−2001(5’−CAG GTG CAG CTA CTC GAG TGG GG−3’)(配列番号102))および2つの3’−VH特異的プライマーセット(3’−hu−VH−BstEII−2001(5’−CTG AGG AGA CGG TGA CC−3’)(配列番号103)、3’−hu−VH−J3−BstEII−2001(5’−CTG AAG AGA CGG TGA CC−3’)(配列番号104))を用いてRT−PCRを行った。1回のPCR反応につき、1個の5’−プライマーを1個の3’−プライマーと組み合わせた;異なるPCR反応の数は、5’−および3’−プライマーの可能な組み合わせの数により決定した。5つのドナーのPBMC cDNAを、VH遺伝子の供給源として用いた。次のPCRプログラム:94℃にて15秒間の変性、52℃にて50秒間のプライマーアニーリングおよび72℃にて60秒間のプライマー伸張を40サイクル以上行い、それに続いて72℃にて10分間の最終伸張、を増幅に用いた。約350bpのサイズの増幅産物を標準的な方法に従って単離した。
【0158】
Lib 134−VH領域の単離のため、RT−PCRを二段階で行った。最初に、ヒト重鎖VH−セグメント(FR1−CDR1−FR2−CDR2−FR3)を、上記と同じ5’−VH特異的プライマーセット(5’−huVH1,3,5−Xho I−2001、5’−huVH4−Xho I−2001、5’−huVH4B−Xho I−2001;配列番号100〜102)および3’−特異的プライマーセット(3’−A134−VH1A(5’−GTA GTC AAA GTA GAA CCG TAG CCC CCT ATC TCT YGC ACA GTA ATA CAC GGC−3’)(配列番号105)、3’−A134−VH1B(5’−GTA GTC AAA GTA GAA CCG TAG CCC CCT ATC TCT YGC ACA GTA ATA CAY RGC−3’)(配列番号106)、3’−A134−VH3A(5’−GTA GTC AAA GTA GAA CCG TAG CCC CCT ATC TCT TGY ACA GTA ATA CAC RGC−3’)(配列番号107)(示される「T」は「A」、「C」または「G」にも置き換えられる)、3’−A134−VH3B(5’−GTA GTC AAA GTA GAA CCG TAG CCC CCT ATC TCT TGC ACA GTA ATA CAA RGC−3’)(配列番号108)(示される「T」は「A」、「C」または「G」にも置き換えられる)、3’−A134−VH4(5’−GTA GTC AAA GTA GAA CCG TAG CCC CCT ATC TCT SGC ACA GTA ATA CAC RGC−3’)(配列番号109))を、FR3の非常に末端の領域で一致するヒトVHサブファミリー1、3および4に対して用いて、単離された鋳型VH断片からPCR増幅した。
【0159】
次のプライマーの組み合わせを用いた。
a)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH1A
b)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH1B
c)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH3A
d)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH3B
e)5’−huVH4−Xho I−2001 × 3’−A134−VH4
f)5’−huVH4B−Xho I−2001 × 3’−A134−VH4
【0160】
1回のPCR反応につき、1個の5’−プライマーを1個の3’−プライマーと組み合わせた;異なるPCR反応の数は、5’−および3’−プライマーの可能な組み合わせの数により決定した。次のPCRプログラム:94℃にて15秒間の変性、52℃にて50秒間のプライマーアニーリングおよび72℃にて90秒間のプライマー伸張を40サイクル以上行い、それに続いて72℃にて10分間の最終伸張、を増幅に用いた。このPCR段階およびそれぞれの3’−プライマー配列によって、ヒトVHセグメントが母親由来のVH CDR3の一部分について延長された。それは言い換えると次の2番目の段階のPCR3’−プライマーのプライミングサイトである。
【0161】
これらのVH−(FR1−CDR1−FR2−CDR2−FR3)DNA断片を、次に、それぞれの5’VH特異的プライマーと、増幅されたDNA断片の普遍的な3’−末端に一致する普遍的な3’プライマー(3’A134−JH6−BstEII、5’−CGA GAC GGT GAC CGT GGT CCC TTG GCC CCA GTA GTC AAA GTA GAA CCG TAG CC−3’)(配列番号110)を再び用いる2番目のPCR反応の鋳型として用いた。
【0162】
次のPCRプログラム:
94℃にて15秒間の変性、52℃にて50秒間のプライマーアニーリングおよび72℃にて60秒間のプライマー伸張を40サイクル以上行い、それに続いて72℃にて10分間の最終伸張、を増幅に用いた。DNA V断片を標準的なプロトコールに従って単離した。
【0163】
3.ライブラリー構築−ヒトVHプールのクローニング
前述の方法の第2ラウンドでは、1回目の前の選択(実施例6参照)で同定されたscFv A−240のヒトVLを選択し、それに続いてヒトscFvを作製する目的でヒトVH断片のライブラリーと合成した。ファージディスプレイライブラリーは、概して、例えば「Phage Display: A Laboratory Manual」;Ed. Barbas, Burton, Scott & Silverman; Cold Spring Harbor Laboratory Press, 2001に開示されるような標準的な手順に基づいて構築された。
【0164】
異なるPCR増幅からの重鎖DNA断片を各連結について次のように秤量した。
a:b:c:d:e:f=3:1:3:1:1:1、ここでa〜fは次の意味:
a)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH1A
b)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH1B
c)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH3A
d)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH3B
e)5’−huVH4−Xho I−2001 × 3’−A134−VH4
f)5’−huVH4B−Xho I−2001 × 3’−A134−VH4
を有する。
【0165】
400ngのヒトLib 134−VH断片プール(Xho I-BstEII消化)および1200ngのプラスミドpComb3H5BHis/A−240 VL(scFv A−240のVL領域をコードするDNAを、制限部位SacIおよびSpeIを介して標準手順に従ってpComb3H5BHisにクローニングした)で構成される、1回の連結反応を設定した。得られる抗体ヒトVHプールを、次に、300μLのエレクトロコンピテント大腸菌XL1 Blue株にエレクトロポレーション(2.5kV、0.2cmギャップキュベット、25mF、200 Ohm、Biorad gene−pulser)により形質転換し、合計で0.8×108の独立したクローンからなるライブラリーサイズを得た。
【0166】
エレクトロポレーション後、アッセイをSOC培養液(Fluka)中で表現型発現のためにインキュベートした。次に培養物を、50μg/ml カルベニシリンおよび2% v/v グルコースを含有する500mLのSB選択培地中で一晩それぞれインキュベートした。翌日、培養物の細胞を遠心分離により回収し、市販のプラスミド調製キット(QIAGEN)を用いてプラスミド調製を行ってDNAライブラリーを保存した。
【0167】
それぞれのscFvプールをコードする、1.5μgのこのプラスミドプールを、次に、大腸菌XL1 blue(2.5kV、0.2cmギャップキュベット、25mF、200 Ohm、Biorad gene−pulser)に電気穿孔して合計で2.4×109の独立したクローンからなるライブラリーサイズを得た。表現型発現およびカルベニシリンへの緩やかな適応の後、抗体ライブラリーをSB−カルベニシリン(50μg/ml)選択培地へ移した。次に、抗体ライブラリーを感染量の1×1012粒子のヘルパーファージVCSM13で感染させ、その結果、繊維状のM13ファージの産生および分泌が得られ、この各ファージ粒子はヒトscFv−断片をコードする一本鎖pComb3H5BHis−DNAを含み、ファージコートタンパク質IIIへの翻訳融合として対応するscFv−タンパク質を提示した。
【0168】
4.ヒトVHのファージディスプレイ選択
ヒトscFv−レパートリーを有するファージ粒子をPEG8000/NaCl沈殿および遠心分離により培養上清から回収した。次に、約1×1011〜1×1012scFvファージ粒子を0.5mLのTBS/1% BSAに再懸濁し、ストレプトアビジンコートされたELISAプレート(Nunc)のウェル中で1時間固定されたビオチン化可溶性CEAとともにインキュベートした。抗原10μg/mlのPBS溶液(50μl)をストレプトアビジンコートされたウェル中で4℃にて一晩インキュベートし、水で1回洗浄し、それに続いてTBS中200μlの3% BSAで37℃にて1時間ブロッキングし、それをインキュベーション後に取り除いた。
【0169】
標的抗原と特異的結合しなかったscFvファージをTBS/0.05% Tweenでの洗浄段階により除去した。この洗浄手順は、さらなるラウンドで10回まで繰り返された。
【0170】
洗浄後、HCl−グリシン、pH 2.2を用いることにより結合体を溶出した。2M Tris、pH 12での中和後、溶出液を新鮮な非感染大腸菌XL1 Blue培養株の感染に用いた。
【0171】
残っている高結合体を溶出するため、50μlの新鮮な大腸菌XL1 blue培養株(OD600≧0.5)をウェルに添加し、15分間インキュベートした。次に、両方の培養物を混合し、ヒトscFv−断片をコードするファージミドコピーで首尾よく形質導入した細胞を再びカルベニシリン抵抗性について選択し、その後にVCMS13ヘルパーファージで感染させて、抗体提示およびインビトロでの選択の第2のラウンドを開始した。
【0172】
4ラウンドのパニングに対応するプラスミドDNAを大腸菌培養物から単離した。可溶性scFv−タンパク質の産生のため、VH−VL−DNA断片をプラスミドから切り出し(XhoI−SpeI)、プラスミドpComb3H5BFlag/His中の同じ制限部位を介してクローニングし、その発現構築物(例えばscFv)は、scFvとHis6タグとの間にFlagタグ(TGDYKDDDDK)(配列番号99)を含み、さらなるファージタンパク質が欠失している。
【0173】
連結後、各プール(異なるラウンドのパニング)のプラスミドDNAを100μL熱ショックコンピテント大腸菌TG1に形質転換し、カルベニシリンLB寒天上にプレーティングした。単一のコロニーを選定し、96ウェルプレート(Greiner)中の120μLのLBカルベニシリン(50μg/ml)1% グルコースに播種した。ウェルに半透膜(Greiner)で密封し、インキュベータでプレートを37℃にて一晩インキュベートした(マスタープレート)。次に、10μlのマスタープレート培養物を、ウェルあたり90μL LBカルベニシリン(50μg/ml)0.1% グルコースを含有する第2の96ウェルプレート(ワーキングプレート)に移した。37℃の振盪インキュベータ中で4時間のインキュベーションの後、20μL LBカルベニシリン、6mM IPTGを各ウェルに添加することによりscFv生成を誘導した。振盪しながら30℃にて一晩の別のインキュベーション段階の後、細胞を、40μL 溶解緩衝液(400mM ホウ酸、320mM NaCl、4mM EDTA pH8、2.5mg/ml リゾチーム)での室温にて1時間のインキュベーションで溶解した。残りの細胞および細胞片は1,900×gで12分間の遠心分離により分離した(Hettich)。
【0174】
次に、scFv分子を含有する上清を、フローサイトメトリー結合アッセイで結合について試験した。
【0175】
ヒトCEAをトランスフェクトしたCHO細胞をCEA陽性細胞株として用いた。最初に100,000〜200,000個の細胞をヒトscFvまたは関連する対照を含有する周辺質調製物とともにインキュベートすることにより細胞結合アッセイを行った。インキュベーション後、細胞をPBS/1% FCS(ウシ胎児血清)中で洗浄し、さらに5〜10μg/mlの抗FLAG M2抗体とともにインキュベートした。細胞を再び洗浄した後、それらをポリクローナル、PE標識抗マウス抗体(Dianova)とともにインキュベートし、その後フローサイトメトリーにより分析した。46個のクローンをCEA陽性のCHO細胞で結合シグナルについて試験した。それらの全てが陽性のシグナルを示した。それぞれのscFv DNAを配列決定した後、合計9個の異なる配列が得られ、そのうちの8個が高度の相同性を示した。ヒト構築物MP510_3A−5.3(MP510−A5;配列番号50)、MP510_3−B9.1(MP511−B9;配列番号52)、MP510_3−D8.1(MP511−D8;配列番号54)が、さらなる特性決定のために選択された。対応するアミノ酸配列を配列表に示す。
【0176】
前記ヒト構築物MP510−A5、MP511−B9、MP511−D8ならびに半ヒト構築物A−240 Vλ.3(マウスVH A5B7/ヒトVLA240;配列番号48)の周辺質抽出物を、CEA陽性および陰性の細胞株でのフローサイトメトリー実験でさらに分析した。図13から、ヒト構築物MP510−A5(配列番号50)、MP511−B9(配列番号52)、MP511−D8(配列番号54)は、それぞれの半ヒト対照A−240 Vλ.3(マウスVH A5B7/ヒトVLA240;配列番号48)と比較して、明らかに識別できるシフトを蛍光強度で示すことが分かる。従って、ヒトscFv構築物は、膜結合ヒトCEAに対して半ヒト構築物A−240 Vλ.3よりも強い結合活性を示す。さらに、全てのヒト構築物は、CEA陽性のヒトKATO III細胞(ヒト胃癌細胞株)との明確な結合を示したのに対し、それらのどれもがCEA陰性の非トランスフェクトCHO細胞ならびにCEA陰性のヒトNALM6細胞(ヒトB細胞株)との結合を示さなかった(データは示さず)。
【0177】
実施例8:ヒトCEAxCD3二重特異性一本鎖抗体の作製および細胞障害活性
1.配置
次の段階では、ヒト二重特異性一本鎖抗体分子の様々なドメイン配置が作製された。これらの分子はヒト抗CEA結合ドメイン(その作製は上記実施例7に記載されている)を含み、配列番号77VHVLに示されるヒトCD3抗原に対して特異性を有する脱免疫化結合ドメインは表1に示されるように設計された;上記実施例2も参照。本明細書に記載されるヒト抗CEA結合ドメインを含む二重特異性一本鎖抗体構築物はELISAプレート上に固定化された可溶性CEA抗原で4ラウンドのファージディスプレイ選択後に単離されているので、それらはすべて(明白に)可溶性ヒトCEA抗原と結合する。
【0178】
特に、次の配置が作製された。
(a)N末端に位置するヒト抗CEA部分:
(i)VH−VL配向の抗CEA:
A5VH−A240VLx配列番号77VHVL(配列番号24)、
A5VH−A240VL#x配列番号77VHVL(配列番号126)、
B9VH−A240VLx配列番号77VHVL(配列番号32)、
B9VH−A240VL#x配列番号77VHVL(配列番号130)、
D8VH−A240VLx配列番号77VHVL(配列番号40)
D8VH−A240VL#x配列番号77VHVL(配列番号134)、および
CEAI VH−A240VLx配列番号77VHVL(配列番号16)。
(ii)VL−VH配向の抗CEA:
A240VL−A5VHx配列番号77VHVL(配列番号26)、
A240VL−B9VHx配列番号77VHVL(配列番号34)、
A240VL−D8VHx配列番号77VHVL(配列番号42)、
およびA240VL−CEAI VHx配列番号77VHVL(配列番号18)。
【0179】
(b)C末端に位置するヒト抗CEA部分:
(i)VH−VL配向の抗CEA:
配列番号77VHVLxA5VH−A240VL(配列番号30)、
配列番号77VHVLxA5VH−A240VL#(配列番号128)、
配列番号77VHVLxB9VH−A240VL(配列番号36)、
配列番号77VHVLxE12VH−A240VL(配列番号143)、
配列番号77VHVLxB9VH−A240VL#(配列番号132)、配列番号77VHVLxD8VH−A240VL(配列番号44)、
配列番号77VHVLxD8VH−A240VL#(配列番号136)、
および配列番号77VHVLxCEAI VH−A240VL(配列番号20)。
(ii)VL−VH配向の抗CEA:
配列番号77VHVLxA240VL−A5VH(配列番号28)、
配列番号77VHVLxA240VL−B9VH(配列番号38)、
配列番号77VHVLxA240VL−D8VH(配列番号46)、および
配列番号77VHVLxA240VL−CEAI VH(配列番号22)。
【0180】
CEAI VH(配列番号56)は、マウスmAb A5B7に由来するVH領域を意味し、一方、CEAI VHVLまたはCEAI VLVHは、mAb A5B7に由来するVH−VLドメインおよびVL−VHドメインをそれぞれ指す。A240はヒトVL領域に相当する(配列番号64および実施例7参照)。従って、例えば、CEAI VH−A240VLx配列番号77VHVL(配列番号16)は、mAb7由来のマウスVH領域およびヒトVL領域A240を有する半ヒトCEA結合ドメインを含む二重特異性構築物に相当する。ヒト抗体ライブラリーのクローニングのため、A240VL領域の元のN末端をコードする核酸配列は制限部位へ変換する必要があった。A5VH−A240VL#x配列番号77VHVL(配列番号126)、配列番号77VHVLxA5VH−A240VL#(配列番号128)、B9VH−A240VL#x配列番号77VHVL(配列番号130)、配列番号77VHVLxB9VH−A240VL#(配列番号132)、D8VH−A240VL#x配列番号77VHVL(配列番号134)、および配列番号77VHVLxD8VH−A240VL#(配列番号136)において、元のN末端が再導入された。
【0181】
二重特異性一本鎖抗体構築物である配列番号77VHVLxE12VH−A240VL(配列番号143)は、配列番号77VHVLxB9VH−A240VL(配列番号36)構築物とただ1つのアミノ酸残基が異なる。E12VHではCDR−H2配列は「FILNKANGGTTEYAASVKG」(配列番号145)と読まれるが、B9VHではそれは「FIRNKANGGTTEYAASVKG」(配列番号67)と読まれる。ヒトE12VH領域を実施例7に示すように単離した。
【0182】
2.発現、精製およびフローサイトメトリー分析
これらのヒトCEAxCD3二重特異性一本鎖抗体の発現、精製およびフローサイトメトリー分析は、上記実施例2に記載される方法により実行された。
【0183】
3.結合活性
図14に例示されるように、ヒト二重特異性一本鎖抗体構築物A5VH−A240VLx配列番号77VHVL(配列番号24)、B9VH−A240VLx配列番号77VHVL(配列番号32)、D8VH−A240VLx配列番号77VHVL(配列番号40)およびCEAI VH−A240VLx配列番号77VHVL(配列番号16)は、CEA陽性ヒトKatoIII細胞と結合し、かつCD3陽性HPB−ALL細胞と結合する。
【0184】
4.可溶性CEA抗原の不在下での細胞障害活性
CHO−CEA+標的細胞に対する細胞障害活性を、ヒト二重特異性一本鎖抗体の次のドメイン配置について実証した。
【0185】
N末端に位置する抗CEA部分を含む構築物と抗CEA部分のVH−VL配向について、図15は、A5VH−A240VLx配列番号77VHVL(配列番号24)、B9VH−A240VLx配列番号77VHVL(配列番号32)、D8VH−A240VLx配列番号77VHVL(配列番号40)およびCEAI VH−A240VLx配列番号77VHVL(配列番号16)の細胞障害活性を示す。図16は、A240VL−A5VHx配列番号77VHVL(配列番号26)、A240VL−B9VHx配列番号77VHVL(配列番号34)、A240VL−D8VHx配列番号77VHVL(配列番号42)、およびA240VL−CEAI VHx配列番号77VHVL(配列番号18)に関する抗CEAドメイン(N末端に位置する)のVL−VH配向に対する細胞障害活性を示す。
【0186】
C末端に位置する抗CEA部分を含む構築物と抗CEA部分のVH−VL配向について、図17は、配列番号77VHVLxA5VH−A240VL(配列番号30)、配列番号77VHVLxB9VH−A240VL(配列番号36)、配列番号77VHVLxD8VH−A240VL(配列番号44)、および配列番号77VHVLxCEAI VH−A240VL(配列番号20)に関するCEA+標的細胞に対する細胞障害性を提示する。配列番号77VHVLxCEAI LHを陽性対照として用いた。図18は、抗CEAドメイン(C末端に位置する)のVL−VH配向を含む構築物の配列番号77VHVLxA240VL−A5VH(配列番号28)、配列番号77VHVLxA240VL−B9VH(配列番号38)、配列番号77VHVLxA240VL−D8VH(配列番号46)、および配列番号77VHVLxA240VL−CEAI VH(配列番号22)に関する細胞障害活性を示す。配列番号77VHVLxCEAI LH(配列番号2)を陽性対照として用いた。
【0187】
図21は、ヒトB9VH領域とヒトA240VL領域を異なる配置で含む二重特異性一本鎖抗体構築物の細胞障害活性を示す。A240VL−B9VHx配列番号77VHVL(配列番号34)、配列番号77VHVLxA240VL−B9VH(配列番号38)、配列番号77VHVLxB9VH−A240VL(配列番号36)、およびB9VH−A240VLx配列番号77VHVL(配列番号32)は、ヒトCEAトランスフェクトCHO細胞に対する細胞障害活性を示した。
【0188】
5.ヒトCEAxCD3二重特異性一本鎖抗体の可溶性CEA抗原に対する抵抗性
可溶性ヒトCEA抗原の存在下、ヒト二重特異性一本鎖抗体についての競合アッセイを上記実施例5に示すように行った。図19および20は、構築物A5VH−A240VLx配列番号77VHVL(配列番号24)、B9VH−A240VLx配列番号77VHVL(配列番号32)、D8VH−A240VLx配列番号77VHVL(配列番号40)およびCEAI VH−A240VLx配列番号77VHVL(配列番号16)に例示されるように、ヒト構築物についても、細胞障害活性の可溶性CEA抗原に対する抵抗性を実証する;マウス由来二重特異性一本鎖抗体の可溶性CEAに対する抵抗性については、実施例5参照。さらに、図22は、CEA反応性の二重特異性一本鎖構築物A240VL−B9VHx配列番号77VHVL(配列番号34)が、漸増量の可溶性CEA抗原の存在下でT細胞を再指向してCHO−CEA+細胞を溶解させたことを示す。この実験は、A240VL−B9VHx配列番号77VHVLに媒介される細胞障害活性も可溶性CEAに対して抵抗性であることを実証する。最後に、図27から導かれるように、二重特異性一本鎖構築物である配列番号77VHVLxE12VH−A240VL(配列番号143)に媒介される細胞障害活性は、可溶性CEAに対して抵抗性である。
【0189】
従って、本発明は、高いレベルの可溶性CEA抗原の存在下でさえ細胞障害性抗腫瘍活性を有する医薬組成物を提供する。上に述べたように、上皮性腫瘍患者において血清CEA濃度が高いと、抗CEA抗体に基づく治療法の成功が低下する。従って、本発明の医薬組成物は、患者の血清/血漿中の高い可溶性CEA抗原濃度を特徴とする、進行性上皮性腫瘍(例えば、原発腫瘍の外科切除後の続発性、転移性腫瘍)、悪性上皮性腫瘍、高い(上皮性)腫瘍量および後期上皮性腫瘍の患者の治療に特に適している。さらに、本発明の医薬組成物はまた、低い投薬量で投与されることが予期される。それに加えて、前記医薬組成物は、本発明の医薬組成物中の二重特異性一本鎖抗体のヒト起源の抗CEA部分と脱免疫化された抗CD3部分に起因して、患者に投与された場合に免疫原性となる可能性は低い。さらに、本発明の医薬組成物は、低分子量および小型の二重特異性一本鎖構築物に起因して、高い腫瘍浸透をもたらすことが予期される。その上、腫瘍治療のためには、該分子の高い細胞障害活性のために少量の二重特異性一本鎖抗体が用いられる。患者には少量が投与されると予期されるため、患者に対する有害作用は低減されるとも予期される。最後に、可溶性CEAに対する抵抗性のない二重特異性一本鎖抗体は、上に説明したように、それらは低い濃度で投与されるため、腫瘍患者の血漿中の低い濃度の可溶性CEA抗原にさえも非常に感受性が高い。この問題もまた、本発明の医薬組成物により回避される。
【0190】
実施例9:高分解能陽イオン交換クロマトグラフィーによるA240VL−B9VHx配列番号77VHVL(配列番号34)の精製モノマーのタンパク質の同質性評価
精製されたA240VL−B9VHx配列番号77VHVL(配列番号34)構築物の同質性をさらに特性決定するため、単離モノマー画分を高分解能陽イオン交換クロマトグラフィーに付した。クロマトグラフィーは、20mM MESバッファーpH5.5で平衡化したMiniSカラム(Mini S PE 4.6/50 CatX 0.8ml;GE Healthcare 17−5005−01)で行われた。カラムに充填する前にサンプルを同じバッファーで1:3希釈した。結合したタンパク質を、60カラム量中に1M NaClを0〜30%含有する平衡バッファー勾配を用いて溶出させた。残りのタンパク質を3カラム量の1M NaClで溶出した。結果として得られるクロマトグラムは図23に示され、単一の主ピークを有する同質のタンパク質画分を提示する。
【0191】
実施例10:A240VL−B9VHx配列番号77VHVL(配列番号34)の血漿安定性
A240VL−B9VHx配列番号77VHVL(配列番号34)構築物の血漿安定性を、様々なインキュベーション条件下で、続いて実施例4に記載の標準的な51クロム放出に基づく細胞障害性アッセイにより試験した。
【0192】
5名の健康なドナーの血液を含むヒト血漿プールを、EDTAコートされたシリンジに血液を採取することにより作製した。細胞成分を遠心分離により取り除き、上部の血漿相を回収し、その後プールした。A240VL−B9VHx配列番号77VHVL(配列番号34)構築物を、血漿の存在下または不在下、37℃または4℃にてインキュベートした。対照として、構築物を細胞障害性アッセイの直前に血漿またはRPMI1640培地にそれぞれ希釈した。CHO−CEA+は標的細胞の役割を果たし;刺激を受けたCD8+T細胞をエフェクター細胞として用いた。効果:標的(E:T)比は10:1であった。このアッセイの持続時間は18時間であった。
【0193】
図24に示されるように、CEA反応性の二重特異性一本鎖構築物A240VL−B9VHx配列番号77VHVL(配列番号34)は、極めて安定性であることが実証された;37℃にて24時間ヒト血漿中でのインキュベーションの後、細胞障害活性の損失は検出することができなかった。
【0194】
実施例11:高分解能陽イオン交換クロマトグラフィーによる配列番号77VHVLxE12VH−A240VL(配列番号143)の精製モノマーのタンパク質同質性評価
この実験は、配列番号77VHVLxE12VH−A240VL(配列番号143)を分析することを除いて実施例9と同様に行った。結果として得られるクロマトグラムは図25に示され、単一の主ピークを有する同質のタンパク質画分を提示する。
【0195】
実施例12:配列番号77VHVLxE12VH−A240VL(配列番号143)の血漿安定性
この実験は、配列番号77VHVLxE12VH−A240VL(配列番号143)を分析することを除いて実施例10と同様に行った。図26に示されるように、CEA反応性の二重特異性一本鎖構築物である配列番号77VHVLxE12VH−A240VL(配列番号143)は極めて安定性であることが実証された;37℃にて24時間ヒト血漿中でのインキュベーションの後、細胞障害活性の損失は検出することができなかった。
【図面の簡単な説明】
【0196】
【図1】様々なヒトCEA反応性の二重特異性一本鎖構築物とヒトCEAトランスフェクトCHO細胞およびCD3陽性HPB−AII細胞とのそれぞれのFACS結合分析。CEA結合の陽性対照として、モノクローナル抗体CoI−1を使用した。ヒトCD3結合の陽性対照には、WO 99/054440号に記載のCD19xCD3二重特異性一本鎖構築物を使用した。この陽性対照では、太い線は、10μg/mlの精製CD19xCD3二重特異性一本鎖抗体とともにインキュベートし、その後に抗His抗体および検出抗体とともにインキュベートした細胞を表す。細い柱状グラフ線は、陰性対照:抗His抗体および検出抗体とともにインキュベートした細胞を反映する。ヒト(膜結合)CEAおよびヒトCD3の結合活性は、CEAI VHVLx配列番号77VHVL、CEAI VLVHx配列番号77VHVL、CEAII VHVLx配列番号77VHVL、CEAIII VLVHx配列番号77VHVLおよびCEAIII VHVLx配列番号77VHVLについて検出可能であった。本発明に記載される二重特異性一本鎖抗体に対応するそれぞれの柱状グラフにおいて、細い線は陰性対照を表し、太い輝線は培養上清とともにインキュベートした細胞を表し、一方、太い暗線(大部分が右側)は、10μg/mlの精製二重特異性一本鎖抗体とともにインキュベートした細胞を表す。
【図2】直接ELISAにより検出された、可溶性CEAに対する二重特異性一本鎖抗CEA/抗CD3抗体CEAI VHVLx配列番号77VHVL、CEAII VHVLx配列番号77VHVLおよびCEAIII VHVLx配列番号77VHVLならびに抗CEA抗体Col−1の結合シグナル。CEAI VHVLx配列番号77VHVL(mAb A5B7に由来する抗CEA結合ドメイン)、CEAII VHVLx配列番号77VHVL(mAb T84.66に由来する抗CEA結合ドメイン)、およびCEAIII VHVLx配列番号77VHVL(mAb MFE−23に由来する抗CEA結合ドメイン)二重特異性一本鎖抗体ならびにマウスモノクローナル抗体Col−1は、固定化された可溶性ヒトCEAと特異的に結合した。可溶性CEA抗原(PBS対照)の不在下で結合シグナルは観察されなかった。
【図3】示されるCEA反応性の二重特異性一本鎖構築物は、可溶性CEAの不在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。再指向による溶解(redirected lysis)の特異性を実証するため、非CEA反応性の二重特異性一本鎖構築物を陰性対照として含めた。ヒトCEAトランスフェクト標的細胞(CHO−CEA+細胞)に対する細胞障害活性は、様々なドメイン配置、すなわち、配列番号77VHVLxCEAI VHVLおよび配列番号77VHVLxCEAI VLVH(両構築物は抗CD3結合ドメインをN末端に含む)、ならびにCEAI VLVHx配列番号77VHVLおよびCEAI VHVLx配列番号77VHVL(抗CD3結合ドメインをC末端に含む)について示すことができた。非トランスフェクトCHO細胞(ヒトCEAを欠く)を陰性対照として用いた。
【図4】示されるCEA反応性の二重特異性一本鎖構築物は、可溶性CEAの不在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。再指向による溶解の特異性を実証するため、非トランスフェクトCHO細胞を陰性対照として含めた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。CEAI−HL(CEAI VHVLx配列番号77VHVL)、CEAIII−LH(CEAIII VLVHx配列番号77VHVL)、CEAIII−HL(CEAIII VHVLx配列番号77VHVL)、およびCEAII HL(CEAII VHVLx配列番号77VHVL)は、ヒトCEAトランスフェクトCHO細胞に対する細胞障害活性を示した。非トランスフェクトCHO細胞(ヒトCEAを欠く)をCEAI−LH(CEAI VLVHx配列番号77VHVL)、CEAIII−HL(CEAIII VHVLx配列番号77VHVL)およびCEAII−HL(CEAII VHVLx配列番号77VHVL)の陰性対照として用いた。CEAIは、マウスmAb A5B7に由来する可変領域を意味し、CEAIIは、マウスmAb T84.66に由来する可変領域であり、CEAIIIは、マウスmAb MFE−23由来の可変領域を指す。
【図5】示されるCEA反応性の二重特異性一本鎖構築物は、可溶性ヒトCEAの存在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。CEAI VLVHx配列番号77VHVLおよびCEAI VHVLx配列番号77VHVLに媒介される細胞障害活性は、1μg/mlまでの漸増量の可溶性ヒトCEAにより阻害されない。CEAIは、マウスmAb A5B7に由来する可変領域である。
【図6】示されるCEA反応性の二重特異性一本鎖構築物は、可溶性ヒトCEAの存在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。配列番号77VHVLxCEAI VHVLおよび配列番号77VHVLxCEAI VLVHに媒介される細胞障害活性は、1μg/mlまでの漸増量の可溶性ヒトCEAにより阻害されない。CEAIは、マウスmAb A5B7に由来する可変領域である。
【図7】示されるCEA反応性の二重特異性一本鎖構築物は、可溶性ヒトCEAの存在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。CEAII VHVLx配列番号77VHVLの細胞障害活性は、漸増量の可溶性CEAにより阻害された。CEAII VHVLは、mAb T84.66に由来する。
【図8】示されるCEA反応性の二重特異性一本鎖構築物は、可溶性ヒトCEAの存在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。CEAI VHVLx配列番号77VHVLに媒介される細胞障害性が可溶性CEA抗原による阻害に抵抗性であるのに対して、CEAIII VHVLx配列番号77VHVLに媒介される細胞障害活性は、少量の可溶性CEAにさえも阻害される。CEAIII VHVLは、mAb MFE−23に由来するのに対し、CEAIは、マウスmAb A5B7に由来する可変領域である。
【図9】示されるCEA反応性の二重特異性一本鎖構築物は、漸増量の可溶性CEA抗原の存在下で、T細胞を再指向してKatoIII細胞を溶解させた。未変性のヒトPBMCをエフェクター細胞として用いた。CEAII VHVLx配列番号77に媒介される細胞障害活性は、可溶性CEAに対して抵抗性でない。CEAII VHVLは、mAb T84.66に由来する。
【図10】示されるCEA反応性の二重特異性一本鎖構築物は、漸増量の可溶性CEA抗原の存在下で、T細胞を再指向してKatoIII細胞を溶解させた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。CEAI VHVLx配列番号77VHVLに媒介される細胞障害性は、可溶性CEAに対して抵抗性である。対照的に、CEAII VHVLx配列番号77VHVLに媒介される細胞障害活性は、漸増量の可溶性CEAにより阻害される。CEAII VHVLは、mAb T84.66に由来するのに対し、CEAIは、マウスmAb A5B7に由来する可変領域である。
【図11】示されるCEA反応性の二重特異性一本鎖構築物は、漸増量の可溶性CEA抗原の存在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。CEAII VHVLx配列番号77VHVLに媒介される細胞障害活性は、漸増量の可溶性CEAにより阻害される。CEAII VHVLは、mAb T84.66に由来する。
【図12】Flagがタグされた、選択されたクローン由来のscFvタンパク質断片を含有する周辺質調製物のフローサイトメトリー分析を示す。可溶性scFvタンパク質断片の周辺質調製物を100,000〜200,000個のCEAをトランスフェクトしたCHO細胞に添加した。検出のため、モノクローナル抗Flag抗体を用い、その後にPE標識ポリクローナル抗マウス抗体を用いた。細胞と結合しているScFvを、蛍光強度を増加させて、PBSのみでインキュベートした細胞と比較して測定した。蛍光強度をX軸にブロットし、イベントの数をY軸にブロットした。陰性対照(PBSおよび検出試薬)は黒く塗られた曲線で示し、個別のscFvは灰色の線で示す。右への移動は細胞との正の結合を示す。全てのscFv、すなわち、A−121、A−183、A240、A−313、A−290、A−315、A4−35、A4−52およびMP2−A5は、CHO細胞上で膜と結合したCEAと結合する。scFvの各々は、実施例6に記載されるように、マウスA5B7VH領域とヒトVL領域からなる。
【図13】Flagがタグされた、選択されたクローン由来のscFvタンパク質断片を含有する周辺質調製物のフローサイトメトリー分析を示す。可溶性scFvタンパク質断片の周辺質調製物を100,000〜200,000個のCEAをトランスフェクトしたCHO細胞に添加した。モノクローナル抗Flag抗体、続いてPE標識ポリクローナル抗マウス抗体で検出を行った。細胞と結合しているScFvを、蛍光強度を増加させて、PBSのみでインキュベートした細胞と比較して測定した。蛍光強度をX軸にブロットし、イベントの数をY軸にブロットした。陰性対照(PBSおよび検出試薬)は黒く塗られた曲線で示し、個別のscFvは灰色の線で示す。右への移動は細胞との正の結合を示す。完全ヒト化scFv構築物MP510_3A5.3、MP510_3−B9.1、およびMP510_3−D8.1はCHO細胞上で膜と結合したCEAと結合する。これらのscFvの各々は、実施例7に記載されるように、ヒトVH領域とヒトVL領域A240からなる。240Vλ.3は、マウスA5B7 VH領域とヒトVL A−240領域からなるscFvである。この構築物はまた、CEA結合活性を示す。
【図14】様々なヒトCEA反応性の二重特異性一本鎖構築物とKatoIII細胞およびHPB−AII細胞とのそれぞれのFACS結合分析。太い線は、抗His抗体および検出抗体とともにインキュベートした、トランスフェクトCHO細胞の細胞培養上清でインキュベートした細胞を表す。細い柱状グラフ線は、陰性対照:抗His抗体および検出抗体とともにインキュベートした細胞を反映する。ヒト二重特異性一本鎖抗体構築物A5VH−A240VLx配列番号77VHVL、B9VH−A240VLx配列番号77VHVL、およびD8VH−A240VLx配列番号77VHVLは、Kato細胞上でヒトCEAと、HPB−AII細胞上でヒトCD3と結合する。mAb A5B7に由来するCEA結合ドメインのVH領域を含むCEAI VH−A240VLx配列番号77VHVLは、同じ結合活性を示す。
【図15】可溶性CEAの不在下で、CEAでトランスフェクトしたCHO細胞に再指向された、示されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイ。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。細胞障害活性は、構築物A5VH−A240VLx配列番号77VHVL、B9VH−A240VLx配列番号77VHVL、D8VH−A240VLx配列番号77VHVLおよびCEAI VH−A240VLx配列番号77VHVLについて検出することができた。CEAI VHはmAb A5B7に由来するVH領域である。
【図16】可溶性CEAの不在下で、CEAをトランスフェクトしたCHO細胞に再指向された、示されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイ。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。この図は、A240VL−A5VHx配列番号77VHVL、A240VL−B9VHx配列番号77VHVL、A240VL−D8VHx配列番号77VHVL、およびA240VL−CEAI VHx配列番号77VHVLの細胞障害活性を実証する。
【図17】可溶性CEAの不在下で、CEAをトランスフェクトしたCHO細胞に再指向された、示されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイ。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。CEA+標的細胞に対する細胞障害性は、配列番号77VHVLxA5VH−A240VL、配列番号77VHVLxB9VH−A240VL、配列番号77VHVLxD8VH−A240VL、配列番号77VHVLxCEAI VH−A240VLおよび配列番号77VHVLxCEAI VLVHについて示される。
【図18】可溶性CEAの不在下で、CEAをトランスフェクトしたCHO細胞に再指向された、示されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイ。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。細胞障害活性は、配列番号77VHVLxA240VL−A5VH、配列番号77VHVLxA240VL−B9VH、配列番号77VHVLxA240VL−D8VH、および配列番号77VHVLxA240VL−CEAI VHおよび配列番号77VHVLxCEAI VLVHついて示される。
【図19】漸増量の可溶性CEA抗原の存在下で、CEAをトランスフェクトしたCHO細胞に再指向された、示されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイ。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。この図は、A5VH−A240VLx配列番号77VHVLおよびB9VH−A240VLx配列番号77VHVLについて例証されるように、可溶性CEA抗原に対するヒト二重特異性一本鎖抗体構築物の細胞障害活性の抵抗性を実証する。
【図20】漸増量の可溶性CEA抗原の存在下で、CEAをトランスフェクトしたCHO細胞に再指向された、示されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイ。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。ヒト二重特異性一本鎖抗体構築物D8VH−A240VLx配列番号77VHVLおよびCEAI VH−A240VLx配列番号77VHVLもまた、可溶性CEA抗原に対する抵抗性を示す。
【図21】示されるCEA反応性の二重特異性一本鎖構築物は、可溶性CEAの不在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。再指向による溶解の特異性を実証するため、非トランスフェクトCHO細胞を陰性対照として含めた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。A240VL−B9VHx配列番号77VHVL、配列番号77VHVLxA240VL−B9VH、配列番号77VHVLxB9VH−A240VL、B9VH−A240VLx配列番号77VHVL、および配列番号77VHVLxCEAI VHVLは、ヒトCEAトランスフェクトCHO細胞に対する細胞障害活性を示した。
【図22】示されるCEA反応性の二重特異性一本鎖構築物は、漸増量の可溶性CEA抗原の存在下で、T細胞を再指向してCHO−CEA+細胞を溶解させた。刺激を受けたヒトCD8+T細胞をエフェクター細胞として用いた。A240VL−B9VHx配列番号77VHVLに媒介される細胞障害活性は、可溶性CEAに対して抵抗性である。
【図23】二重特異性一本鎖構築物A240VL−B9VHx配列番号77VHVLの高分解能陽イオン交換クロマトグラム、青色の線(上側の曲線)は、タンパク質の全体的な電荷イソ型を示す。構築物の高同質性を示す、1つの単一ピークが検出された。
【図24】ヒト血漿における24時間のインキュベーション後の細胞障害性の評価に基づくタンパク質安定性アッセイ。CEA反応性の二重特異性一本鎖構築物は、可溶性CEAの不在下で、CEAをトランスフェクトしたCHO細胞に再指向された。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。この図は、ヒト血漿中の二重特異性一本鎖構築物A240VL−B9VHx配列番号77VHVLの血漿安定性を実証する。構築物の細胞障害活性は、生理的条件下で、血漿タンパク質の影響を受けない。
【図25】二重特異性一本鎖構築物である配列番号77VHVLxE12VH−A240VLの高分解能陽イオン交換クロマトグラム、青色の線(上側の曲線)は、タンパク質の全体的な電荷イソ型を示す。この図は、二重特異性一本鎖構築物である配列番号77VHVLxE12VH−A240VLの同質性を実証する。
【図26】ヒト血漿における24時間のインキュベーション後の細胞障害性の評価に基づくタンパク質安定性アッセイ。CEA反応性の二重特異性一本鎖構築物は、可溶性CEAの不在下で、CEAをトランスフェクトしたCHO細胞に再指向された。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。この図は、ヒト血漿中の二重特異性一本鎖構築物である配列番号77VHVLxE12VH−A240VLの血漿安定性を実証する。構築物の細胞障害活性は、生理的条件下で、血漿タンパク質の影響を受けない。
【図27】示されるCEA反応性の二重特異性一本鎖構築物は、漸増量の可溶性CEA抗原の存在下で、T細胞を再指向してCHO−CEA+細胞を溶解させた。刺激を受けたヒトCD8+T細胞をエフェクター細胞として用いた。二重特異性一本鎖構築物である配列番号77VHVLxE12VH−A240VLは、可溶性CEAに対して抵抗性である細胞障害活性を媒介した。
【技術分野】
【0001】
本発明は、可溶性CEAに対する抵抗性を有する医薬組成物に関する。
【背景技術】
【0002】
ゴールドおよびフリードマンが最初にヒト結腸癌組織抽出物における腫瘍関連癌胎児性抗原(CEA)を記載してから30年以上が経過した(Gold and Freedman; J. Exp. Med. 122 (1965); 467-481)。
その間、CEA遺伝子ファミリーに関連する28のその他の遺伝子/偽遺伝子が見出された。CEA遺伝子ファミリーのメンバーに用いる命名を簡略化するために、該ファミリーは最近「CEA関連細胞接着分子」(CEACAM)と改名され、そのメンバーの命名が統一された(Beauchemin, Exp. Cell Res. 252 (1999), 243-249)。例えば、この命名に従って、ヒトCEA(CD66e)はCEACAM5と名付けられる。
【0003】
ヒトCEA遺伝子ファミリーは、染色体19q13.2の上に集まる(Olsen et al., Genomics 23 (1994); 659-668)。その29遺伝子および偽遺伝子は3つのサブグループ、すなわち、7つの発現遺伝子を含有するCEAサブグループ、11の発現遺伝子を含有する妊娠特異的糖タンパク質(PSG)サブグループおよび偽遺伝子のみを含有する第3のサブグループに分割することができる(Hammarstroem, Sem. Cancer Biol. 9 (1999), 67-81; Beauchemin, Exp. Cell Res. 252 (1999), 243-249)。CEAのアミノ酸配列および他のファミリーメンバーの解析により、それらが免疫グロブリン(Ig)スーパーファミリーに属することが明らかとなった(Williams and Barclay, Annul. Rev. Immunol. 6 (1988), 381-405)。CEAサブグループの全てのメンバーは細胞表面膜に付着している。胆汁糖タンパク質(CEACAM1;BGP1;TM−CEA;CD66a)、CEA遺伝子ファミリーメンバー1(CEACAM3;CGM1;CD66d)およびCEA遺伝子ファミリーメンバー7(CEACAM4;CGM7)は疎水性の膜貫通ドメインを有するのに対し、癌胎児性抗原(癌胎児性抗原関連細胞接着分子5;CEACAM5;CEA;CD66e)、非特異的交差反応性抗原(CEACAM6;NCA;NCA−50/90;CD66c)、CEA遺伝子ファミリーメンバー2(CEACAM7;CGM2)およびCEA遺伝子ファミリーメンバー6(CEACAM8;CGM6;CD66b)はグリコシルホスファチジルイノシトール(GPI)脂質部分によって血漿膜と繋がれている。CEAタンパク質は、Igドメインの数に応じて約300kDaまでの分子量で高度にグリコシル化されている。
【0004】
CEAタンパク質の生物活性に関して、腫瘍細胞株を用いるインビトロ研究から、胆汁糖タンパク質、CEAおよび非特異的交差反応性抗原を含むいくつかのCEAサブファミリーは、腫瘍細胞表面に発現すると、同種親和性の異型細胞接着分子として作用することができることが示唆された(Oikawa et al., Biochem. Biophys. Res. Commun. 186 (1992), 881-887;Zhou et al., Cancer Res. 53 (1993), 3817-3822)。より最近には、微生物による攻撃から結腸を守る自然免疫応答におけるCEAおよび非特異的交差反応抗原の可能性のある役割が考察されている(Hammarstroem and Baranov, Trends Microbiol. 9 (2001), p. 119-125)。特に、これらのタンパク質は微生物と結合および捕獲することにより、それらが微絨毛の上皮細胞に到達し、侵入することを防ぐことが提唱されている。
【0005】
CEAは、胎児期の間に発現し、健常成人には存在せず、再び癌に発現する癌胎児性抗原であると仮定された。しかし、CEAは、正常成人組織にも発現する。例えば、胆汁糖タンパク質、CEA、非特異的交差反応性抗原およびCEA遺伝子ファミリーメンバー2は、正常ヒト結腸に、特に腸管内腔に面している成熟した円柱上皮細胞におよび陰窩口(crypt mouth)で高度に分化した細胞に発現する(Fraengsmyr et al., Cancer Res. 55 (1995), 2963-2967;Fraengsmyr et al., Tumor Biol. 20 (1999), 277-292)。より具体的には、これらのタンパク質は、フリーな管腔表面の内側を覆う成熟した結腸細胞の刷子縁多糖外被に局在する。胆汁糖タンパク質、CEAおよび非特異的交差反応性抗原はまた、多数の上皮性起源の腫瘍にも発現する(Hammarstriem, Sem. Cancer Biol. 9 (1999), 67-81; Shively and Beatty CRC Crit. Rev. Oncol. Hematol. 2 (1985), 355-399)。
【0006】
既に1970年代後半および1980年代前半に、CEAは結腸直腸およびその他の上皮腫瘍の放射免疫学的局在に好まれる標的抗原となった。これは、CEAが胃腸癌および膵臓癌の95%に、ならびに大部分の小細胞および非小細胞肺癌腫に過剰発現するという事実に起因する。また、それは乳癌ならびに頭部および頚部の扁平上皮癌にも発現する(Primus et al., Cancer 42 (1978), 1540-1545)。実際に、CEAは最も広く用いられている臨床腫瘍マーカーの1つである。それは、その安定性、正常成人組織での相当に制限された発現、ならびに上皮性起源の腫瘍での高度な発現のために、結腸直腸癌およびその他のいくつかの癌の血清腫瘍マーカーとして用いられている。健康な個体においてCEAの大半は結腸で産生される。そこでそれは成熟した円柱細胞の頂端膜側から腸管内腔に放出され、糞便の中で消失する。従って、健康な個体からは、血液中に極めて低いレベルしか通常見出されない。例えば、健康な個体の血液中のCEAレベルは2μg/l未満である。それに対して、結腸直腸癌およびその他の癌腫の患者からの血清中のCEAレベルは2000μg/lを超える範囲まで増加している(Thomson et al., PNAS 64 (1969), 161-167)。特に、進行性、悪性、または後期の上皮腫瘍は、しばしば高い血清濃度の可溶性CEAを伴う(Fletcher; Ann. Intern. Med. 104 (1986), 66-73)。CEAを含む血漿膜からの成分は、血漿膜由来小胞のように表面から継続的に剥脱することが知られ(Taylor and Black, J. Natl. Cancer Inst. 74 (1985), 859-866;Sack et al., J Clin Invest. 82 (1988), 586-93)、それはリンパ管および血管を流れて最後に血液に行き着き得る。腫瘍サイズが増大するにつれて、より多くのCEAが血液に蓄積する。腫瘍マーカーとしての血清CEA決定の主な用途は、結腸癌の手術後のサーベイランスにある。増加したCEAレベルは、患者のそれぞれ81%(Minton et al., Cancer 55 (1985), 1284-1290)および89%(Wanebo et al., Surg. Gynecol. Obstet. 169 (1989), 479-487)において、再発性疾患の最初の指標であった。血清CEAレベルはまた、予後指標としても用いることができる(Mulcahy and Benson, Curr. Oncol. Rep. 1 (1999), 168-172)。
【0007】
多くの上皮癌でのその過剰発現に起因して、CEAは腫瘍マーカーとして用いられるだけでなく、抗腫瘍療法の標的としても用いられる。例えば、胃腸癌は高い割合のヒト上皮腫瘍の原因であり、米国において2001年の胃癌の新規症例は21,700、結腸直腸癌の新規症例は135,400と推定される(Greenlee; CA Cancer J Clin 51 (2001), 15-36)。結腸直腸癌は3番目に頻度の高い悪性腫瘍であり、男性および女性の両方において癌死の3番目の主な原因である(Ries; Cancer 88 (2000), 2398-2424)。これらの腫瘍に対する新規な治療法を見出すために、抗CEAモノクローナル抗体がCEA陽性癌の可能性のある治療法として調査されてきた(Murakami et al., Immunol. Invest. 25 (1996), 23-35)。
【0008】
腫瘍量の少ない患者(低い血清CEAレベルに相当する)が首尾よく治療を受けるアプローチの一例は、ベアらによって行われた研究である。このアプローチでは、ラベツズマブ(labetuzumab)(ラベツズマブは、ヒト化形態の抗CEAモノクローナル抗体MN−14である;Behr et al., Cancer, 94: 1373-1381, (2002), 1559-64)の131I標識変異体が、5−フルオロウラシルおよびフォリン酸に、または肝転移後のアジュバント・セッティングに対して化学療法抵抗性の、少量の転移性疾患を有する30名のCRC患者が登録された第II相試験において分析された。131I標識ラベツズマブの単回注射が行われた。評価可能な19名の患者のうち、3名が部分的緩解を有し、8名がわずかな応答を15ヶ月まで持続して示した。アジュバント・セッティングにおいて、9名のうち7名の患者が3年間まで疾患に罹らなかったのに対して、対照群の再発率は同じ期間で67%であった。患者の血清CEAレベルは3.9〜45ng/mlの範囲であった(Behr et al., Cancer, 94: 1373-1381, 2002)。CEA血清レベルの低い(<5ng/ml)患者を特徴とする別の研究では、131I−ラベツズマブ(上記引用文中)を用いるCEAの放射免疫療法は、肝臓における結腸直腸癌転移の救済切除(salvage resection)後の生存を向上したことが示された。23名の患者に40〜60mCi/m2 131I−ラベツズマブの用量が投与された。5年生存率は、それぞれ処置群について51.3%、対照群について7.4%であった(Liersch et al., JCO, 2005, ASCO Proc, Vol 23, No 16S: 3627)。
【0009】
けれども、高い血清CEA濃度に対応する治療的アプローチは、しばしば低いかまたは全くない抗腫瘍応答という結果をもたらした。例えば、ヒト化抗CEAモノクローナル抗体を臨床で評価するために行われた臨床研究では、MN−14のCDR移植形(version)(hMN−14;Sharkey, Cancer Res. 55 (23 Suppl) (1995) 5935s- 5945s)が131Iで標識された。19名の進行したCEA産生腫瘍患者に131I標識したhMN−14を与えた。hMN−14の体内分布、腫瘍ターゲッティング、および薬物動態挙動は、マウスMN−14に見られるものに類似していた。しかし、血漿中CEAの上昇した(>200ng/ml)患者は、注射後1時間以内に30%を上回る標識抗体が複合体化した。これらの患者の一部では、合併症の増加の結果、血漿中CEAの低い患者において見られるものよりも、血液からのクリアランスが急速な抗体の代謝の促進がもたらされた(Sharkey、前掲)。Yuらにより実施された別の第1相試験では、CEAに対する131I標識した高親和性マウスモノクローナル抗体(mAb)、COL−1(Muraro, Cancer Res. 45 (1985), 5769-80)が胃腸悪性腫瘍の患者において調査された。特に、血清CEAおよび腫瘍体積の薬物動態学への影響が分析された。本目的のため、18名の進行した胃腸悪性腫瘍患者に、20mgの131I標識したCOL−1を10mCi/m2〜75mCi/m2の用量で投与した。血清CEAレベルは6〜2739ng/mLの範囲であった(平均+/−SD、500+/−639)。すべての腫瘍の浸潤した(tumor-involved)器官のうち82%が陽性であり、全ての病変の58%であった。しかし、今回も上昇した血清CEA(>500ng/mL)および腫瘍体積が血清放射能のクリアランスと直接相互に関連があることが見出された。著者らは、高度に上昇した循環CEAレベルおよび/または増大した腫瘍体積を有する患者は、131I標識したCOL−1をより急速に循環から排除すると結論付けた(Yu et al., J. Cli. Oncol. 14 (1996), 1798-1809)。Hajjarらによる、転移性胃腸および大腸直腸癌患者におけるヨウ素131標識ヒト化MN−14抗CEAモノクローナル抗体での研究において同様の結果が得られた。この第1相試験では、21名の患者が事前外部照射の後かまたは標準的な化学療法の後に抗体で処置された。21名のうち7名の患者がヒト抗ヒト抗体(HAHA)を有したが、有害作用はなかった。抗腫瘍応答は認められなかった。この場合も、上昇した血漿CEAレベルが血液および身体全体からの抗体のクリアランスを増大させることが見出され(Hajjar et al., Clin Colorectal Cancer, 2 (2002), 31-42)、そのことは、少なくとも一部分、この研究において見出された抗腫瘍応答の欠如の説明をもたらし得る。血液および身体からの治療用抗体の急速なクリアランスという現象は、器官の損傷を防ぐために身体から急速に取り除かれる必要のある免疫複合体の形成増加により説明され得る。最も可能性が高いと思われるモノクローナル抗体の急速なクリアランスに起因して、これらの研究に登録した腫瘍患者に治療効果を見出すことはできなかった。
【0010】
免疫複合体形成および、大部分が免疫細胞のFc受容体により認識される抗体のFc部分に媒介される、治療用モノクローナル抗体の急速なクリアランスにより生じる問題を回避するために、Fc部分を含まない抗体誘導体(例えばscFv構築物)または断片(例えばFabおよびFab2断片)を産生し、臨床で分析した。これらの研究の大部分は、腫瘍画像形成および検出/局在化を対象とする(例えば、Chester et al., Cancer Chemother Pharmacol, 46 (2000) Suppl: S8-12;Mayer et al., Clin Cancer Res, 6: (2000) 1711-1719; Begent et al., Nat Med, 2 (1996): 979-984参照)。そのような抗体誘導体/断片の治療効力を臨床で調査したものはほんのわずかの研究しかなかった。例えば、Francisらによる臨床アプローチでは(Francis, Br. J. Cancer 87(6) (2002), 600-607)、scFv−カルボキシペプチダーゼ構築物の抗腫瘍活性が調査された。この第1相試験では、酵素プロドラッグ療法(ADEPT)に対する抗体が、進行した結腸直腸癌またはその他のCEA産生腫瘍を有する患者に使用された。本目的のため、抗体酵素標的化薬剤として細菌酵素カルボキシペプチダーゼ(CPG2)と結合した、CEAに対するマウスモノクローナル抗体(A5B7)のF(ab)2断片からなるA5CP、およびZD2767P、ビス−ヨードフェノールマスタードプロドラッグが用いられた。結果として、この研究において臨床応答および放射性応答は見られなかった。処置前の血清CEAレベルは1000ng/mlまでの範囲であった。これらの処置した腫瘍患者の高い血清中CEA濃度は、少なくとも一部分、この研究で観察された抗腫瘍応答の欠如の原因であり得る。
【発明の開示】
【0011】
上に述べた課題を考慮すると、進行性、悪性、または後期上皮腫瘍のための効果的な治療法のための手段および方法の提供が大いに望まれる。
【0012】
従って、本発明の一態様は医薬組成物に関し、前記医薬組成物は、
(a)ヒトCD3と特異的に結合する第1の結合ドメイン、および
(b)ヒトCEAと特異的に結合する第2の結合ドメイン
を有する二重特異性一本鎖抗体を含み、
前記第2の結合ドメインが少なくともアミノ酸配列「DX1X2X3X4FYFDY」(配列番号65)を含み、「X1」、「X2」、「X3」または「X4」は、任意のアミノ酸残基を表し、該アミノ酸残基「D」はマウスモノクローナル抗体A5B7のCDR−H3のKabat位置95に相当し、該アミノ酸残基「FYFDY」は、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置100、100a、100b、101、および102にそれぞれ相当する。一実施形態では、「X1」は、「R」(アルギニン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;「X2」は、「G」(グリシン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表し;「X3」は、「L」(ロイシン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;かつ、「X4」は、「R」(アルギニン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表す。
【0013】
本発明の医薬組成物の一実施形態では、本明細書に定義される二重特異性一本鎖抗体のヒトCEAに特異的な前記第2の結合ドメインは、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102に相当するアミノ酸配列「DRGLRFYFDY」(配列番号66)を少なくとも含む。
【0014】
本発明は、その血漿中の可溶性CEA濃度の高い上皮性腫瘍患者の治療に特に適した手段および方法を提供する。そのような高い可溶性CEA濃度は、進行性腫瘍、再発性、転移性、後期腫瘍を有する上皮性腫瘍患者および高腫瘍量の患者の血清/血漿に見出される。アミノ酸配列「DRGLRFYFDY」(配列番号66)を含むCEA結合ドメインを含む二重特異性一本鎖抗体はCEA陽性標的細胞と結合するだけでなく、可溶性CEAとも結合することが見出された;本発明の実施例3および図2;ならびにEP B1 491031号参照。示されるアミノ酸配列「DRGLRFYFDY」は、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102(配列番号66)に相当する(Harwood, Br J Cancer. 54 (1986), 75-82)。驚くことに、可溶性CEAと結合しているにもかかわらず、前記二重特異性一本鎖抗体は、高い濃度の可溶性CEAの存在下でさえも(最大1μg/mlの可溶性CEAを試験)、CEAを有する腫瘍細胞を死滅させる。言い換えれば、前記二重特異性構築物は、可溶性CEAによって、CEA陽性の腫瘍細胞に対するそれらの細胞障害活性を阻害されない。
【発明を実施するための最良の形態】
【0015】
以下の実施例5および8(併せて図5、6、8、10、19、20、22および27)に示されるように、アミノ酸配列「DRGLRFYFDY」を含むCEA結合ドメインを含む二重特異性一本鎖抗体は、高い濃度の可溶性CEAの存在下でさえもCEA陽性腫瘍細胞に対する細胞障害性を媒介した。例えば、図10は、漸増量の可溶性CEA抗原の存在下で、KatoIII細胞(CEA陽性のヒト胃癌細胞株)に再指向されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイを示す。刺激を受けたヒトCD8陽性細胞障害性T細胞(CTL)をエフェクター細胞として用いた。CEAI VHVLx配列番号77VHVLに媒介される細胞障害性は、可溶性CEAに対して抵抗性である。対照的に、CEAII VHVLx配列番号77VHVLに媒介される細胞障害活性は、漸増量の可溶性CEAにより阻害される。CEAIは、マウスmAb A5B7に由来する可変領域であるのに対して、CEAII VHVLは、mAb T84.66に由来する。
【0016】
重要なことに、抗CEAx抗CD3二重特異性一本鎖抗体のヒトCEA結合ドメイン(すなわち、ヒトCEAと特異的に結合するヒト結合ドメイン)で使用される場合、アミノ酸配列「DRGLRFYFDY」が可溶性CEAに対する抵抗性を媒介するために十分であることが見出された;例えば、図19、20、22および27参照。下記において、本明細書に定義される二重特異性一本鎖抗体は、従って可溶性CEA抗原に対して抵抗性であると称される。本明細書において、用語「可溶性CEA抗原に対する抵抗性」、「可溶性CEAに対する抵抗性」または関連する用語は、前記二重特異性一本鎖抗体により媒介されるCEA陽性標的または腫瘍細胞に対する細胞障害性が漸増濃度の可溶性CEAにより影響を受けないという事実を指す。特に、細胞障害活性は、高い濃度の可溶性CEA(最大1μg/mlまでを試験)にさえ阻害されない。上に述べたように、健康な個体の血液中のCEAレベルは、2ng/ml未満である。腫瘍患者の血清/血漿中の高い可溶性CEA濃度は、進行性、再発性、転移性、または後期腫瘍および高腫瘍量の患者に特有である。従って、本発明は、その血漿中の可溶性CEA濃度がそのように高い上皮性腫瘍患者の治療に特に適した手段および方法を提供する。本明細書において、用語「高い可溶性CEA濃度」は、10、20、50、70、80、90または100ng/mlよりも高い可溶性血清/血漿CEA濃度を示す。この血清/血漿CEA濃度は、特に、ELISAにより測定することができる。好ましくは、前記可溶性血清/血漿CEA濃度は、例えばELISAにより測定して、100ng/mlよりも高い。
【0017】
可溶性CEA抗原に抵抗性を有する前記二重特異性一本鎖抗体の作製は、以下の実施例から明らかなように平凡な作業ではなかった。例えば、膜結合CEAと結合するが可溶性CEAとは結合しないことの知られているモノクローナル抗体(mAb)に由来するCEA結合ドメインを有する二重特異性一本鎖抗体、すなわち、mAb PR1A3(Durbin, Proc Natl Acad Sci U S A. 91 (1994), 4313-7)は、産生させることができなかった。二重特異性一本鎖抗体形式中で用いた場合、抗CD3x抗CEA二重特異性一本鎖構築物の発現/分泌は達成できなかった。PR1A3のヒト化形(version)(Durbin、前掲)を作製に利用した場合、二重特異性一本鎖抗体構築物は発現し、宿主細胞から分泌された。しかし、抗CEA結合ドメインと膜結合CEAの結合は得られなかった。
【0018】
十分に説明されているモノクローナル抗体T84.66(Neumaier, M. et al., Cancer Res 50 (1990), 2128-34)またはMFE−23(Boehm, M. K. Biochem J 2 (2000), 519-28)に由来する二重特異性一本鎖抗体を作製した場合、これらの二重特異性抗体は可溶性CEA抗原に対し高い感受性があった、すなわち、CEA陽性標的または腫瘍細胞に対するそれらの細胞障害活性は、可溶性CEA抗原の存在下で阻害された。前記構築物は可溶性CEAと結合することが見出されていることから、可溶性CEA抗原は抗体が膜結合CEAと結合することを妨げ、それにより抗体に媒介される細胞障害活性を阻害するとの結論が下された。例えば、図7は、可溶性ヒトCEAの存在下で、CEAをトランスフェクトしたCHO細胞に再指向されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイを示す。刺激を受けたヒトCD8陽性細胞障害性T細胞(CTL)をエフェクター細胞として用いた。CEAII VHVLx配列番号77VHVLの細胞障害活性は、漸増量の可溶性CEAにより明らかに阻害される。CEAII VHVLはmAb T84.66に由来する;配列番号77は抗CD3VH−VLドメインである。
【0019】
可溶性CEA抗原に対する抵抗性は二重特異性一本鎖抗体にのみ見出すことができ、そのCEA結合ドメインはマウスモノクローナル抗体A5B7のCDR−H3のアミノ酸配列「DRGLRFYFDY」を含んだ(Harwood, Br J Cancer. 54 (1986), 75-82)。MFE−23由来およびT84.66由来の二重特異性一本鎖構築物に関して、A5B7由来の二重特異性一本鎖抗体は可溶性CEAと結合する。MFE−23由来およびT84.66由来の二重特異性一本鎖構築物に関して得られた結果を鑑みると、可溶性CEAは、A5B7由来一本鎖二重特異性抗体構築物において細胞障害活性に影響を及ぼさないことが予期され得る。
【0020】
上に述べたように、ヒトにおけるCEAを有する上皮性腫瘍に対して向けられる多くの治療的アプローチは、癌患者の血漿中に高いレベルの可溶性CEA抗原が存在することにより著しく妨害される。例えば、血漿中の高いCEA濃度の存在下での、増加した免疫複合体形成および治療用抗CEAモノクローナル抗体のクリアランスは、いくつかの臨床研究で観察されている。さらに、可溶性CEA抗原(しばしば、進行性腫瘍、再発性癌、転移性腫瘍、高腫瘍量、または後期腫瘍の癌患者の血清中に高い濃度で存在する)は、CEA陽性腫瘍細胞に対して向けられる治療薬を阻害し、従って腫瘍細胞の認識および破壊を妨げる。従って、腫瘍に到達する治療薬の実際の量は減少し、結果として抗腫瘍活性が低下し、低いかさらに全くなくなる。この制限は、これまでのところ、例えば、治療薬と腫瘍細胞の相互作用を妨げる可能性が低い、極めて低い量の可溶性CEA抗原を有する患者に対する抗体に基づくアプローチを制限する。
【0021】
本発明において、ヒトCD3およびヒトCEAに特異的な二重特異性一本鎖抗体治療薬を作製することが可能であることが見出され、その腫瘍細胞に対する細胞障害活性は、高濃度の可溶性CEA抗原(最大1μg/mlの可溶性CEAを試験)に対してさえも抵抗性である。この知見は、本発明の二重特異性一本鎖抗体が可溶性CEA抗原と結合するという事実を考えると、全く予期せぬものである(本発明の実施例3および図2;ならびにEP B1 491031号も参照)。それにもかかわらず、本明細書に定義される二重特異性一本鎖抗体は、腫瘍細胞に対するその細胞障害活性において、高レベルの可溶性CEAの存在に対してさえも完全に抵抗性である。従って、本発明は、例えば、腫瘍進行中に観察される、血漿中の可溶性CEA濃度の高い腫瘍患者の治療に、再発性癌に、転移に、高腫瘍量の患者に、または後期腫瘍に特に適した手段および方法を提供する。
【0022】
本発明によれば、用語「医薬組成物」は、ヒト患者への投与のための組成物に関する。好ましくは、医薬組成物は、担体、安定剤および/または賦形剤からなる適した製剤を含む。好ましい実施形態では、医薬組成物は、非経口、経皮、管腔内、動脈内、くも膜下腔内および/または鼻腔内投与のための、または組織への直接注射による組成物を含む。前記組成物は、注入または注射によって患者へ投与されることが特に想定される。適した組成物の投与は、様々な方法によって、例えば、静脈内、腹腔内、皮下、筋肉内、局所または皮内投与によって達成されてよい。本発明の組成物は、製薬上許容される担体をさらに含んでよい。適した医薬担体の例は当分野で周知であり、それにはリン酸緩衝生理食塩水溶液、水、様々な種類の湿潤剤、滅菌溶液、リポソームなどが含まれる。このような担体を含む組成物は、周知の従来法によって処方されてよい。これらの組成物は、被験体に適した用量で投与することができ、それは、本明細書に記載される可溶性血清CEA抗原に対して抵抗性を示す二重特異性一本鎖抗体の漸増用量の投与による用量漸増試験により決定することができる。上記に示したように、本明細書に記載される、可溶性血清CEA抗原に対する抵抗性を有する二重特異性一本鎖抗体は、高いCEA血清中濃度を有する癌患者、例えば、進行性腫瘍、再発性癌、転移性腫瘍、腫瘍量の大きな腫瘍、または後期腫瘍の治療に有利に用いることができる。これらの組成物はまた、その他のタンパク質性および非タンパク質性薬剤と組み合わせて、例えば併用療法(co-therapy)の形で投与することができる。これらの薬剤は、本明細書に定義される二重特異性一本鎖抗体を含む組成物と同時に投与してもよいし、前記二重特異性一本鎖抗体の投与の前後に適時に定められた間隔および用量で別々に投与してもよい。投与計画は主治医および臨床学的要因により決定される。医学分野で周知のように、任意の患者一人への投薬量は多くの要因によって決まり、それには患者の大きさ、体表面積、年齢、投与される特定の化合物、性別、投与時間および経路、全体的な健康状態、および同時に投与されるその他の薬剤が含まれる。非経口投与のための製剤としては、滅菌水溶液または非水溶液、および懸濁液が挙げられる。非水溶媒の例は、プロピレングリコール、ポリエチレングリコール、植物油、および注射用の有機エステル類、例えばオレイン酸エチルである。水性担体としては、水、水溶液、または生理食塩水および緩衝媒体を含む懸濁液が挙げられる。非経口媒体としては、塩化ナトリウム溶液、リンゲルデキストロース、デキストロースおよび塩化ナトリウム、または乳酸化リンゲル液が挙げられる。静脈内媒体としては、液体および栄養素補充液、電解質補充液(リンゲルデキストロースに基づくものなど)などが挙げられる。防腐剤およびその他の添加剤も存在してよく、例えば、抗菌剤、抗酸化剤、キレート剤、不活性ガスなどが挙げられる。さらに、本発明の組成物は、例えば、好ましくはヒト起源の血清アルブミンまたは免疫グロブリンなどのタンパク質性担体を含んでもよい。併用療法は、本明細書に定義される二重特異性一本鎖抗体に加えて、組成物の意図される使用によって、さらなる生物活性物質を含むことが想定される。このような物質は、胃腸管系に作用する薬剤、抗悪性腫瘍薬として作用する薬剤、化学療法薬、細胞増殖抑制剤(cytostatica)、高尿酸血症を予防する薬剤、免疫反応を阻害する薬剤(例えばコルチコステロイド)、炎症応答を調節する薬剤、循環系に作用する薬剤および/またはサイトカインなどの当分野で公知の薬剤であってよい。
【0023】
好ましくは、本明細書に定義される二重特異性一本鎖抗体は、緩衝液、安定剤および界面活性剤中に処方される。緩衝液は、リン酸緩衝液、クエン酸緩衝液、コハク酸緩衝液または酢酸緩衝液であってよい。安定剤は、アミノ酸および/または糖であってよい。界面活性剤は、合成洗剤、PEGなどであってよい。より好ましくは、本明細書に定義される二重特異性一本鎖抗体は、クエン酸塩、リシン、トレハロースおよびTween80中に処方される。本発明の医薬組成物の希釈液として、等張生理食塩水およびTween80が好ましい。
【0024】
本明細書において、「二重特異性一本鎖抗体」は、2つの結合ドメインを含む単一ポリペプチド鎖を意味する。各「結合ドメイン」は、本明細書において、抗体重鎖由来の1つの可変領域(「VH領域」)を含み、第1の結合ドメインのVH領域は、前記第1の分子、すなわちヒトCD3分子と特異的に結合し、第2の結合ドメインのVH領域は、下により詳細に定義されるヒトCEAと特異的に結合する。2つの結合ドメインは、場合により、一般に約5アミノ酸を含む短いポリペプチドスペーサーにより相互に連結されている。各結合ドメインは、抗体軽鎖(「VL領域」)由来の1つの可変領域をさらに含み、第1および第2の結合ドメインのそれぞれの中のVH領域およびVL領域は、ポリペプチドリンカー(例えば、EP B1 623679に開示およびクレームされる種類のもの)を介して相互に連結されているが、いずれの場合も、第1の結合ドメインのVH領域およびVL領域と、第2の結合ドメインのVH領域およびVL領域を、ともにそれぞれの第1および第2の分子と特異的に結合することが可能であるように、相互に2つで一組とさせるのに十分なほど長い。第1または第2の結合ドメインのV領域の配置は、VH−VLであっても、VL−VHであってもよい。好ましくは、ヒトCD3と特異的に結合する第1の結合ドメインの配置は、以下の実施例に示されるように、VH−VLである。第1の結合ドメインは、第2の結合ドメインに対してN末端に位置してもC末端に位置してもよいことが想定されている。従って、本明細書において定義される二重特異性一本鎖抗体の結合ドメインの配置は、VHCEA−VLCEA−VHCD3−VLCD3、VLCEA−VHCEA−VHCD3−VLCD3、VHCD3−VLCD3−VHCEA−VLCEAまたはVHCD3−VLCD3−VLCEA−VHCEAであってよい。好ましくは、CD3に特異的な前記第1の結合ドメインは、第2の結合ドメインに対してC末端に位置している。より好ましくは、本明細書において定義される二重特異性一本鎖抗体の結合ドメインは、VHCEA−VLCEA−VHCD3−VLCD3またはVLCEA−VHCEA−VHCD3−VLCD3の順序で配置される。さらにより好ましくは、配置はVLCEA−VHCEA−VHCD3−VLCD3である。最も好ましいのは、配列番号34に定義される二重特異性一本鎖抗体構築物A240VL−B9VHx配列番号77VHVLである。本明細書において定義される二重特異性一本鎖抗体の前記第1および/または第2の結合ドメインは、非ヒト起源(すなわち、非ヒト配列に由来)であってよいことが想定されている。例えば、前記第1および/または第2の結合ドメインは、マウスモノクローナル抗体に由来してよい。しかし、マウス抗体に由来する二重特異性一本鎖抗体は、ヒト患者に投与される場合、異物と認識される可能性がある。従って、本明細書において定義される二重特異性一本鎖抗体の前記第1および/または第2の結合ドメインは、ヒト起源(すなわち、ヒト配列に由来)であることが好ましい。そのようなCEAまたはCD3と特異的に結合するヒト結合ドメインは、例えば、ファージディスプレイに基づく技法により同定することができる。例えば、第1(または第2)の結合ドメインのVH領域はヒトVH領域であるのに対して、第1(または第2)の結合ドメインの対応するVL領域は非ヒト起源であってよいことも想定される。そのような結合ドメインは、キメラ結合ドメインとも称され得る。あるいは、前記結合ドメインの1つが非ヒト起源であるのに対して、他方はヒト起源であり、その結果キメラ二重特異性一本鎖抗体がもたらされる。前記第1および/または第2の結合ドメインは、ヒト患者に投与される際に、本明細書に記載される二重特異性一本鎖抗体の免疫原性を低下させるために、さらに修飾してもよい。例えば、本明細書において定義される二重特異性一本鎖抗体の前記第1または第2の結合ドメインのうちの少なくとも1つは、下により詳細に述べられるように、ヒト化、CDR移植、キメラおよび/または脱免疫化またはヒトであってよい。第1および/または第2の結合ドメイン中でVHおよびVL領域を連結するポリペプチドリンカーが脱免疫化されていることも想定される。好ましくは、脱免疫化された第1の結合ドメイン(CD3に特異的)内のVHおよびVL領域を連結するポリペプチドリンカーは、配列「GEGTSTGS(G2S)2GGAD」(配列番号141)を有する脱免疫化されたポリペプチドリンカーである。本明細書において定義される二重特異性一本鎖抗体の前記結合ドメインの一方または両方が、例えば、タンパク質の発現、精製、検出または富化に用いることのできる、いわゆる「タグ」(Flagタグ、c−mycタグ、GSTタグまたはHisタグなど)を有することもさらに想定されている。例えば、Flagタグに最も広く用いられている親水性のオクタペプチドは、現在DYKDDDDKであるが(Chubet and Brizzard, Biotechniques 20 (1996): 136- 141)、最近の研究から、より短いペプチド、DYKDが、M1モノクローナル抗体にほぼ同じ親和性で認識され得ることが示唆されている(Knappik A, Pluckthun A; Biotechniques 17 (1994): 754-761)。Flagタグ、c−mycタグ、GSTタグ、Hisタグなどは、例えば、タグ−VHCEA−VLCEA−VHCD3−VLCD3またはVLCEA−VHCEA−VHCD3−VLCD3−タグなど、二重特異性一本鎖抗体のN末端かまたはC末端のいずれかに位置してよい。発現、検出または精製目的のそのようなタグの起源および特性は当分野で十分説明されている;例えば、Lichty, Protein Expr Purif. 41 (2005), 98-105参照。
【0025】
本明細書において、用語「一本鎖Fv」または「scFv」とは、抗体のVHドメインおよびVLドメインを含む抗体フラグメントを指し、これらのドメインは単一ポリペプチド鎖中に存在する。可変ドメインはVH−VLまたはVL−VHの順序で配置することができる。一般に、Fvポリペプチドは、scFvが抗原結合のための所望の構造を形成することを可能にするポリペプチドリンカーをVHドメインおよびVLドメインの間にさらに含む。scFvの概説には、Pluckthun in The Pharmacology of Monoclonal Antibodies, vol. 113, Rosenburg and Moore eds. Springer-Verlag, New York, pp. 269-315 (1994)参照。特定の実施形態では、本発明は、本明細書において定義される二重特異性一本鎖抗体に由来する抗CEA scFvに関する。
【0026】
本発明によれば、Ig由来抗原相互作用に関する文脈中で用いられる用語「結合ドメイン」または「可変領域」は、抗体、抗体断片またはその誘導体に由来する少なくとも1つのCDRを含む、ポリペプチドの断片および誘導体を含む。本明細書において定義される二重特異性一本鎖抗体のヒトCEAと特異的に結合する第2の結合ドメインは、少なくとも1つのCDR、好ましくはCDR−H3、より好ましくはマウスモノクローナル抗体A5B7のCDR−H3のKabatの位置100、100a、100b、101、および102にそれぞれ相当するアミノ酸配列「FYFDY」(配列番号112)を含むマウスモノクローナル抗体A5B7のCDR−H3の一部分;さらにより好ましくはアミノ酸配列「DX1X2X3X4FYFDY」(配列番号65)(「X1」、「X2」、「X3」または「X4」は、任意のアミノ酸残基を表し、該アミノ酸残基「D」は、マウスモノクローナル抗体A5B7のCDR−H3のKabat位置95に相当し、アミノ酸残基「FYFDY」は、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置100、100a、100b、101、および102にそれぞれ相当する)を含むマウスモノクローナル抗体A5B7のCDR−H3の一部分を含むことは、本発明により想定されている。マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置96(「X1」)、97(「X2」)、98(「X3」)および99(「X4」)にそれぞれ相当する「X1」、「X2」、「X3」または「X4」は、アミノ酸残基「R」(アルギニン)、「G」(グリシン)、「L」(ロイシン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、「S」(セリン)、「W」(トリプトファン)、「F」(フェニルアラニン)または「T」(トレオニン)を表すことが想定されている。本明細書において、「X1」、「X2」、「X3」および「X4」が同じアミノ酸を表す、例えば「X1」、「X2」、「X3」および「X4」がすべて「F」(フェニルアラニン)である場合は、本発明の範囲から除外される。好ましくは、「X1」が、「R」(アルギニン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;「X2」が、「G」(グリシン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表し;「X3」が、「L」(ロイシン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;かつ、「X4」が、「R」(アルギニン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表す。あるいは最も好ましいものは、本明細書において定義される二重特異性一本鎖抗体のヒトCEAと特異的に結合する第2の結合ドメインは、A5B7のCDR−H3のKabatの位置95〜102に相当するアミノ酸配列「DRGLRFYFDY」(配列番号66)を含むA5B7の完全なCDR−H3を含む。以下の実施例に示されるように、前記mAb A5B7由来CDR−H3「DRGLRFYFDY」(配列番号66)アミノ酸配列をCEAと相互作用する第2の結合ドメイン中に含む、本明細書において定義される二重特異性一本鎖抗体の腫瘍細胞に対する細胞障害活性は、可溶性CEA抗原に対して抵抗性であり、それによりその血漿中の血清CEA濃度の高い腫瘍患者の治療を可能にする。CDRの決定は、当業者に公知である;例えば、http://www.bioinf.org.uk/abs/#cdrid参照。抗体中のアミノ酸配列の番号付けは、例えば、当分野に記載されるKabatの番号付けスキームに従って行ってよい;例えば、Kabat, E. A., T. T. Wu, H. M. Perry, K. S. Gottesman, and C. Foeller. 1991. Sequences of Proteins of Immunological Interest, 5th ed. Bethesda, Md.: National Center for Biotechnology Information, National Library of Medicine参照。
【0027】
最も好ましくは、そして添付の実施例に記録されるように、本発明の医薬組成物に用いられる「二重特異性一本鎖抗体」は、脱免疫化された抗CD3結合ドメイン(WO 2005/040220号)と、少なくともマウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102(配列番号66)に相当するアミノ酸配列「DRGLRFYFDY」を含むヒト抗CEA結合ドメインを含む、二重特異性一本鎖Fv(scFv)である。二重特異性一本鎖分子は当分野で公知であり、例えば、WO 99/54440号またはMack, PNAS, (1995), 92, 7021-7025に記載されている。
【0028】
本発明に従って用いられる用語「一本鎖」は、好ましくは、単一の核酸分子によりコードすることのできる共直線性(co-linear)アミノ酸配列の形態で、二重特異性一本鎖構築物の前記第1および第2のドメインが共有結合していることを意味する。
【0029】
本明細書において、「ヒト」とは、種Homo sapiensを指す。「ヒト」分子、例えばヒトCEAまたはヒトCD3(CD3ε)は、従って、Homo sapiensにおいて天然に発現するその分子の変異体である。
【0030】
用語「上皮腫瘍」とは、本明細書において、CEA陽性である上皮性起源の腫瘍を意味する(Cancer Medicine;6th ed.; Kufe, Donald W.; Pollock, Raphael E.; Weichselbaum, Ralph R.; Bast, Robert C., Jr.; Gansler, Ted S.; Holland, James F.; Frei III, Emil., editors. Hamilton (Canada): BC Decker Inc. 2003;http://www.dkfz.de: http://www.krebsinformationsdienst.de/Krebsarten/index.html)。治療し得る上皮腫瘍は、胃腸腺癌、乳腺腺癌または肺腺癌である。前記胃腸腺癌は、好ましくは結腸直腸腺癌、膵臓腺癌、食道腺癌または胃腺癌である。本明細書に示されるように、本発明の医薬組成物は、腫瘍患者の血漿中の高レベルの可溶性CEA抗原を特徴とする、進行性腫瘍、転移、再発性癌、後期上皮腫瘍、上皮腫瘍量の多い患者、またはCEA血清中濃度が100ng/mlより高い腫瘍患者(例えばELISAにより測定)の治療に特に有利である。前記医薬組成物を原発腫瘍の外科切除の後に使用することも本発明の範囲内である。従って例えば、CEA産生上皮腫瘍に由来する散在性の(disseminated)残存腫瘍細胞も、CEAをそれらの微小環境に流し、その周囲では可溶性CEAのレベルも高い。従って、本発明の医薬組成物の細胞障害活性の可溶性CEAに対する抵抗性は、最小残存病変の治療にも有利である。前記医薬組成物は、残りの腫瘍細胞を死滅させるために、(CEA源、すなわち原発腫瘍の除去によって)血清CEAレベルの低下する期間に投与し得ることが想定される。また、前記医薬組成物は、二次性腫瘍または転移の形成によって血清CEAレベルが増大する場合に、原発腫瘍の除去後に有用であり得る。CEA血清中濃度は、例えばCEA ELISAアッセイ(例えばIBL CEA EIA、IBL Hamburg,Germany参照)により決定することができる。上記に示したように、多くの抗体に基づく治療的アプローチにおいて、前記血清CEAは、抗体と腫瘍細胞上の膜結合CEAの結合を阻害し、抗体の活性を遮断し、それにより抗腫瘍療法の成功を悪化させる。
【0031】
本明細書において、用語「特異的に結合する」または「特異的結合」または「〜との/〜に対する特異的反応性」などの関連表現は、本明細書に定義される二重特異性一本鎖抗体の第1および/または第2の結合ドメインの、可能な結合パートナーとしての複数の異なる分子のプールから、前記それぞれの第1および/または第2の分子にのみ結合する、あるいは有意に結合する程度までそれぞれの第1および/または第2の分子を識別する能力を指す。このような結合測定は、例えばBiacore装置で、ELISA、FACS分析などにより日常的に行うことができる。より具体的には、本明細書において定義される二重特異性一本鎖抗体の第1の結合ドメインは、ヒトCD3、好ましくは、ヒトCD3εと結合する。本明細書において定義される二重特異性一本鎖抗体の第2の結合ドメインは、下に述べるように、上皮性腫瘍抗原、すなわち、ヒトCEA(癌胎児性抗原、癌胎児性抗原関連細胞接着分子5;CEACAM5;CD66e)と結合する。用語「特異的結合」は、本発明によれば、二重特異性一本鎖抗体分子が本明細書に定義されるヒト標的分子それぞれの少なくとも2、3、4、5、6、7、8、またはさらにそれより多くのアミノ酸と特異的に相互作用かつ/または結合できることを意味する。前記用語は、抗体分子の特異性、すなわち、本明細書に定義されるヒト標的分子の特定の領域を識別するその能力に関する。抗原相互作用部位とその特異的抗原との特異的相互作用は、例えば抗原の立体構造の変化の誘導、抗原のオリゴマー化などに起因する、シグナルの開始をもたらし得る。さらに、前記結合は、「キーロックの原則(key-lock-principle)」という特異性に例証され得る。従って、抗原相互作用部位および抗原のアミノ酸配列中の特異的モチーフは、それらの一次、二次または三次構造の結果ならびに前記構造の二次修飾の結果として相互に結合する。抗原相互作用部位のその特異的抗原との特異的相互作用は、さらに前記部位と抗原との結合をもたらし得る。
【0032】
抗体の「特異的結合」は、主に2つのパラメータ:定性的パラメータ(結合エピトープ、すなわち抗体が結合する場所)および定量的パラメータ(結合親和性、すなわちそれが結合する場所でどの程度強く結合するか)に特徴付けられる。どのエピトープが抗体によって結合されるかは、例えば公知のFACS方法論、ペプチドスポットエピトープマッピング、質量分析またはペプチドELISAにより有利に決定することができる。特定のエピトープと結合している抗体の強度は、例えば公知のBiacoreおよび/またはELISA法により有利に決定することができる。このような技法を組み合わせることにより、結合特異性の代表的な尺度としてのシグナル対ノイズ比の算出が可能となる。そのようなシグナル対ノイズ比では、シグナルは注目されるエピトープと結合する抗体の強度を表すのに対して、ノイズは注目されるエピトープとは異なる他の非関連エピトープと結合する抗体の強度を表す。好ましくは、注目されるエピトープに対するシグナル:ノイズ比(注目されるエピトープと異なるその他のエピトープに対するものより約50倍高い)は、評価される抗体が注目されるエピトープと特異的な方法で結合する指標として考えることができる、言い換えれば「特異的結合剤」である。
【0033】
本発明に従って用いられる用語「特異的結合」または「特異的相互作用」は、二重特異性一本鎖抗体構築物が類似構造のポリペプチドと交差反応しないか、または本質的に交差反応しないことを意味する。研究中の二重特異性一本鎖のパネルの交差反応は、例えば、注目されるポリペプチドならびに程度の差はあるが(構造的かつ/または機能的に)密接に関連した多数のポリペプチドとの、従来条件下で二重特異性一本鎖抗体構築物の前記パネルの結合(例えば、Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, 1988 ならびに Using Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, 1999参照)を評価することにより試験することができる。例えば、本発明の二重特異性一本鎖抗体の最初の結合ドメインが、ヒトCEA(癌胎児性抗原;CEACAM5;CEA;CD66e)、すなわち可溶性CEA抗原及び膜結合CEAの双方と結合することは本発明の範囲内であるが、それに対して、その他のCEAファミリーメンバー、例えば胆汁糖タンパク質(CEACAM1;BGP1;TM−CEA;CD66a)と結合している二重特異性抗体は、前記範囲から除外される。
【0034】
抗原相互作用部位と特異的抗原との特異的相互作用の例は、その受容体のリガンドの特異性を含む。前記定義は、特にその特異的な受容体との結合の際にシグナルを誘導するリガンドの相互作用を含む。対応するリガンドの例は、その特異的なサイトカイン受容体と相互作用/結合するサイトカインを含む。また、前記定義により特に含まれるものは、抗原相互作用部位と、セレクチンファミリー、インテグリンおよびEGFなどの成長因子のファミリーの抗原などの抗原との結合である。前記相互作用の別の例(それも前記定義により特に含まれる)は、抗原決定基(エピトープ)と抗体の抗原結合部位の相互作用である。
【0035】
用語「〜と結合/相互作用する」もまた、立体構造エピトープ、構造エピトープまたはヒト標的分子の2つの領域からなる不連続エピトープまたはその部分に関連する。本発明の文脈において、立体構造エピトープは、ポリペプチドが折りたたまれて未変性タンパク質となる際に分子の表面上に集まる一次配列において分離した2またはそれ以上の別個のアミノ酸配列により定義される(Sela, (1969) Science 166, 1365 およびLaver, (1990) Cell 61, 553-6)。
【0036】
用語「不連続エピトープ」とは、本発明の文脈において、ポリペプチド鎖の離れた部分の残基から構築される非直線状エピトープを意味する。これらの残基は、ポリペプチド鎖が三次元構造に折りたたまれて立体構造/構造エピトープを構成する際に分子の表面上に集まる。
【0037】
本明細書において「CD3」は、多分子T細胞受容体複合体の一部としてT細胞で発現し、少なくとも5つの異なる鎖、CD3−γ、−δ、−ε、−ζ、および−ηからなる抗原を意味する。T細胞でのCD3のクラスター形成(例えば、固定化された抗CD3抗体による)は、T細胞受容体の関与に似たT細胞の活性化を引き起こすが、そのクローンに典型的な特異性とは無関係である。実際に、大部分の抗CD3抗体はCD3ε鎖を認識する。ヒトCD3εのアミノ酸配列はGenBank受託番号NM_000733に表され、配列番号111を含む。
【0038】
「CEA」とは、上皮性起源の多数の腫瘍に発現した抗原である、癌胎児性抗原(癌胎児性抗原関連細胞接着分子5;CEACAM5;CEA;CD66e)を意味する(Hammarstroem, Sem. Cancer Biol. 9 (1999), 67-81; Shively and Beatty CRC Crit. Rev. Oncol. Hematol. 2 (1985), 355-399)。ヒトCEAのアミノ酸配列は、GenBank受託番号NM_004363に表され、配列番号76を含む。
【0039】
本発明において、驚くことに、ヒトCD3およびヒトCEAに対して特異性を有する抗体に基づく治療薬を生成することが可能であり、腫瘍細胞に対して向けられる細胞障害活性がさらに高い濃度の可溶性CEA抗原に対して抵抗性であることが見出された。この知見は、本発明の二重特異性一本鎖抗体が可溶性CEA抗原と結合するという事実を考えると全く予期されないものである。例えば、モノクローナル抗体T84.66またはMFE−23に由来する二重特異性一本鎖抗体構築物が作製される場合、これらの抗体は可溶性CEA抗原に感受性が高かった、すなわちそれらの細胞障害活性は可溶性CEA抗原の存在下で遮断されていた。可溶性CEAによる前記構築物の細胞障害活性の阻害は、抗体の量を増加させることによっても克服できなかった。これらの構築物はまた、可溶性CEAとの結合が可能であることが見出されている。このことを考えて、可溶性CEA抗原は、抗体がその細胞障害活性を発揮することを妨げると結論付けられた。その一方、本明細書に定義される二重特異性一本鎖抗体は、腫瘍細胞に対するそれらの細胞障害活性においてさらに高いレベルの可溶性CEAの存在に対して完全に抵抗性である。さらに、その高い細胞障害活性に起因して、本明細書において定義される前記二重特異性構築物は、低い濃度であってさえもそれらの生物活性を誘起する。故に、本明細書において定義される二重特異性一本鎖抗体を含む少量の医薬組成物は、血清/血漿中の可溶性CEA濃度が高いことを特徴とする上皮性腫瘍患者において治療効果を達成するために十分である。上皮性腫瘍患者の血清/血漿中の可溶性CEA濃度が高いことは、進行性、再発性、転移性、または後期腫瘍および高腫瘍量の患者の特徴である。さらに一層驚くことに、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102に相当するアミノ酸配列「DRGLRFYFDY」(配列番号66)は、抗CEAx抗CD3二重特異性一本鎖抗体のヒトCEA結合ドメイン(すなわち、ヒトCEAと特異的に結合するヒト結合ドメイン)に使用した場合、可溶性CEA抗原に対する抵抗性を媒介するのに十分であることが見出された。それがヒト起源であることに起因して、前記構築物はヒト腫瘍患者に投与される場合、免疫原性が低いか免疫原性がない。要約すると、本明細書において定義される二重特異性一本鎖抗体を含む医薬組成物は、例えば、腫瘍進行中に観察される、血漿中の可溶性CEA濃度の高い上皮性腫瘍患者の治療に、再発性癌に、転移に、高腫瘍量の患者に、または後期腫瘍に特に有用である。
【0040】
本発明の医薬組成物のもう1つの好ましい実施形態では、本明細書において定義される二重特異性一本鎖抗体のCD3に特異的な前記第1の結合ドメインは、第2の結合ドメインに対してC末端に位置している。
【0041】
本発明のおよび全ての実施形態の範囲内で、単一ポリペプチド鎖上の、すなわち、本明細書において定義される二重特異性一本鎖抗体の中の、第1および第2の結合ドメインの配置の順序は関連性がある。本明細書において定義される二重特異性一本鎖抗体の結合ドメインの配置は、VHCEA−VLCEA−VHCD3−VLCD3、VLCEA−VHCEA−VHCD3−VLCD3、VHCD3−VLCD3−VHCEA−VLCEAまたはVHCD3−VLCD3−VLCEA−VHCEAであり得ることが想定されている。以下の実施例に示されるように、上述の利点は、第1の結合ドメイン(CD3と特異的に結合する)が第2の結合ドメインに対してC末端に位置している、すなわち、第2の結合ドメインよりも二重特異性一本鎖抗体のC末端に近い場合、特に実現可能である。ヒトCD3と特異的に結合する第1の結合ドメインは、VH−VLの配向に配置されていることが好ましい。例えば、本明細書において定義される二重特異性一本鎖抗体の結合ドメインは、VHCEA−VLCEA−VHCD3−VLCD3またはVLCEA−VHCEA−VHCD3−VLCD3の順序で配置されてよい。本明細書において、「〜に対してN末端に」または「〜に対してC末端に」およびそれらの文法上の変化形は、二重特異性一本鎖抗体の絶対的なNまたはC末端での配置よりもむしろ一次アミノ酸配列内の相対的な位置を意味する。故に、限定されない例として、「第2の結合ドメインに対してC末端に位置する」第1の結合ドメインとは、単に第1の結合ドメインが二重特異性一本鎖抗体内の第2の結合ドメインのカルボキシル側に位置することを意味し、さらなる配列、例えば上に述べたタグ、または別のタンパク質性もしくは非タンパク質性化合物(放射性同位元素など)が二重特異性一本鎖抗体の最終的なC末端に位置する可能性を排除するものではない。
【0042】
好ましくは、本明細書において定義される二重特異性一本鎖抗体の前記結合ドメインは、VHCEA−VLCEA−VHCD3−VLCD3またはVLCEA−VHCEA−VHCD3−VLCD3の順序で配置される。さらにより好ましくは、配置は、VLCEA−VHCEA−VHCD3−VLCD3である。最も好ましいのは、配列番号34に定義される二重特異性一本鎖抗体構築物A240VL−B9VHx配列番号77VHVLである。
【0043】
本明細書において定義される二重特異性一本鎖抗体のヒトCEAと特異的に結合する第2の結合ドメインは、少なくとも1つのCDR、好ましくはCDR−H3、より好ましくはマウスモノクローナル抗体A5B7のCDR−H3のKabatの位置100、100a、100b、101、および102にそれぞれ相当するアミノ酸配列「FYFDY」(配列番号112)を含むマウスモノクローナル抗体A5B7のCDR−H3の一部分;さらにより好ましくはアミノ酸配列「DX1X2X3X4FYFDY」(配列番号65)(「X1」、「X2」、「X3」または「X4」は、任意のアミノ酸残基を表し、該アミノ酸残基「D」は、マウスモノクローナル抗体A5B7のCDR−H3のKabat位置95に相当し、アミノ酸残基「FYFDY」は、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置100、100a、100b、101、および102にそれぞれ相当する)を含むマウスモノクローナル抗体A5B7のCDR−H3の一部分を含むことが好ましい。本明細書において、「X1」、「X2」、「X3」および「X4」は、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置96(「X1」)、97(「X2」)、98(「X3」)および99(「X4」)にそれぞれ相当する。「X1」、「X2」、「X3」または「X4」は、アミノ酸残基「R」(アルギニン)、「G」(グリシン)、「L」(ロイシン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、「S」(セリン)、「W」(トリプトファン)、「F」(フェニルアラニン)または「T」(トレオニン)を表すことが想定されている。本明細書において、「X1」、「X2」、「X3」および「X4」が同じアミノ酸を表す、例えば「X1」、「X2」、「X3」および「X4」がすべて「F」(フェニルアラニン)である場合は、本発明の範囲から除外される。好ましくは、「X1」が、「R」(アルギニン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;「X2」が、「G」(グリシン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表し;「X3」が、「L」(ロイシン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;かつ、「X4」が、「R」(アルギニン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表す。さらにより好ましくは、ヒトCEAに特異的な第2の結合ドメインは、モノクローナル抗体A5B7のCDR−H3のアミノ酸配列「RFYFDY」(配列番号113)、「LRFYFDY」(配列番号114)、「GLRFYFDY」(配列番号115)、または「RGLRFYFDY」(配列番号116)を少なくとも含む。最も好ましいのは、Kabatの位置95(「D」、アスパラギン酸)、96(「R」;アルギニン)、97(「G」;グリシン)、98(「L」;ロイシン)、99(「R」;アルギニン)、100(「F」;フェニルアラニン)、100a(「Y」;チロシン)、100b(「F」;フェニルアラニン)、101(「D」;アスパラギン酸)、および102(「Y」;チロシン)にそれぞれ相当する、アミノ酸配列「DRGLRFYFDY」(配列番号66)を含むA5B7の完全なCDR−H3である。Kabatの方式に従う番号付けは、例えば、Kabat, E. A., T. T. Wu, H. M. Perry, K. S. Gottesman, and C. Foeller. 1991. Sequences of Proteins of Immunological Interest, 5th ed. Bethesda, Md.: National Center for Biotechnology Information, National Library of Medicineに説明されている。
【0044】
以下の実施例に示されるように、CEAと相互作用する第二の結合ドメインの前記mAb A5B7由来のCDR−H3「DRGLRFYFDY」(配列番号66)アミノ酸配列を含む、本明細書に定義される二重特異性一本鎖抗体の腫瘍細胞に対する細胞障害活性は、可溶性CEA抗原に対して抵抗性であり、それにより血漿中の血清CEA濃度の高い腫瘍患者の治療が可能となる。
【0045】
このA5B7由来「DRGLRFYFDY」CDR−H3アミノ酸配列は、例えば、(上皮腫瘍細胞上の)CEA標的抗原に対する親和性を改良するため、かつ/または本明細書に定義される二重特異性一本鎖抗体の「微細な(fine)特異性」を最適化するためにさらに修飾することが望ましい。本目的のため、アミノ酸配列「DX1X2X3X4FYFDY」(配列番号65)において、改良された親和性および/または微細な特異性を有する、修飾されたCDR−H3を同定するために、様々なアミノ酸残基を位置「X1」、「X2」、「X3」および/または「X4」(それぞれ、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置96(「X1」)、97(「X2」)、98(「X3」)および99(「X4」)に対応)で試験してよい。例えば、「X1」、「X2」、「X3」または「X4」は、アミノ酸残基「R」(アルギニン)、「G」(グリシン)、「L」(ロイシン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、「S」(セリン)、「W」(トリプトファン)、「F」(フェニルアラニン)または「T」(トレオニン)で表され得る。ここで、表示される「X」の位置の1、2、3または4つ全ては、CDR−H3「DRGLRFYFDY」(配列番号66)アミノ酸配列中のKabatの位置96〜99にて本来の「RGLR」アミノ酸配列と比較して交換されてよい。しかし、「X1」、「X2」、「X3」および「X4」が同じアミノ酸を表す場合、例えば「X1」、「X2」、「X3」および「X4」が全て「F」(フェニルアラニン)である場合は本発明の特許請求の範囲から除外される。上記のA5B7由来の「DRGLRFYFDY」CDR−H3アミノ酸配列の修飾は当分野で公知の方法、例えば、CEA結合ドメイン中に、このような修飾されたCDR−H3領域を含む二重特異性一本鎖抗体の作製を可能にするランダム化されたプライマーを用いるPCRにより達成することができる。これらの修飾された二重特異性一本鎖抗体の親和性または微細な特異性は、当分野で記載される方法、例えばELISA、BiacoreまたはFACS分析により試験することができる。そのような修飾されたCDR−H3を含む二重特異性一本鎖抗体の可溶性CEA抗原に対する抵抗性は、以下の実施例に記載されるように、漸増量の可溶性CEAの存在下で、細胞障害活性アッセイで試験することができる。
【0046】
より好ましくは、本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な前記第2の結合ドメインは、配列番号65もしくは66および/またはアミノ酸配列「SYWMH」(配列番号68)を有するCDR−H1および/またはアミノ酸配列「FIRNKANGGTTEYAASVKG」(配列番号67)もしくは「FILNKANGGTTEYAASVKG」(配列番号145)を有するCDR−H2を含む。従って、本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な前記第2の結合ドメインは、上に定義される1、2または3のCDR−H領域を含んでよい。あるいは、本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な前記第2の結合ドメインは、配列番号65もしくは66および/またはアミノ酸配列「TYAMH」(配列番号70)を有するCDR−H1および/またはアミノ酸配列「LISNDGSNKYYADSVKG」(配列番号69)を有するCDR−H2を含む。従って、その代わりとして、本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な前記第2の結合ドメインは、上に定義される1、2または3のCDR−H領域を含んでよい。さらにより好ましくは、本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な前記第2の結合ドメインは、上に示される1、2または3のCDR−H領域に加えて、アミノ酸配列「TLRRGINVGAYSIY」(配列番号73)を有するCDR−L1および/またはアミノ酸配列「YKSDSDKQQGS」(配列番号72)を有するCDR−L2および/またはアミノ酸配列「MIWHSGASAV」(配列番号71)を有するCDR−L3を含む。
【0047】
本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な第2の結合ドメインのVH領域のアミノ酸配列は、好ましくは、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102に相当する「DRGLRFYFDY」(配列番号66)、およびアミノ酸配列「SYWMH」(配列番号68)を有するCDR−H1およびアミノ酸配列「FIRNKANGGTTEYAASVKG」(配列番号67)を有するCDR−H2を含む、配列番号60である。
【0048】
本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な第2の結合ドメインのVH領域のアミノ酸配列は、好ましくは、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102に相当する「DRGLRFYFDY」(配列番号66)、およびアミノ酸配列「SYWMH」(配列番号68)を有するCDR−H1、およびアミノ酸配列「FILNKANGGTTEYAASVKG」(配列番号145)を有するCDR−H2を含む、配列番号146である。
【0049】
本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な第2の結合ドメインのVH領域のアミノ酸配列は、好ましくは、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102に相当する「DRGLRFYFDY」(配列番号66)、およびアミノ酸配列「TYAMH」(配列番号70)を有するCDR−H1、およびアミノ酸配列「LISNDGSNKYYADSVKG」(配列番号69)を有するCDR−H2を含む、配列番号58または配列番号62である。
【0050】
本明細書において定義される二重特異性一本鎖抗体のヒトCEAに特異的な第2の結合ドメインのVL領域は、好ましくは、アミノ酸配列「TLRRGINVGAYSIY」(配列番号73)を有するCDR−L1、およびアミノ酸配列「YKSDSDKQQGS」(配列番号72)を有するCDR−L2、およびアミノ酸配列「MIWHSGASAV」(配列番号71)を有するCDR−L3を含む、配列番号64である。
【0051】
上に述べたように、CEAと特異的に結合する第2の結合ドメインの可変領域の順序または配置は、VH−VLまたはVL−VHであってよい。両方の配置が本発明の範囲内である。配列番号60のVHと配列番号64のVLを含む第2の結合ドメインについて、VH−VL配置は配列番号52に示され、一方、VL−VH配置は配列番号122に示される。配列番号146のVHと配列番号64のVLを含む第2の結合ドメインについて、VH−VL配置は配列番号147に示される。
【0052】
配列番号58のVHと配列番号64のVLを含む第2の結合ドメインについて、VH−VL配置は配列番号50に示され、一方、VL−VH配置は配列番号120に示される。配列番号62のVHと配列番号64のVLを含む第2の結合ドメインについて、VH−VL配置は配列番号54に示され、一方、VL−VH配置は配列番号124に示される。配列番号56のVHと配列番号64のVLを含む第2の結合ドメインについて、VH−VL配置は配列番号48に示され、一方、VL−VH配置は配列番号118に示される。
【0053】
さらにより好ましくは、本明細書において定義される二重特異性一本鎖抗体のCEAに特異的な第2の結合ドメインのV領域は、以下、
(a)配列番号60に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;
(b)配列番号146に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;
(c)配列番号58に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;
(d)配列番号62に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;ならびに
(e)配列番号56に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域
からなる群より選択される。
【0054】
最も好ましくは、前記二重特異性一本鎖抗体は、以下、
(a)配列番号6、8、16、18、24、26、32、34、40、42、126、130、134または143のいずれかに表されるアミノ酸配列;
(b)配列番号5、7、15、17、23、25、31、33、39、41、125、129、133または142に示される核酸配列によりコードされるアミノ酸配列;
(c)ストリンジェントな条件下で(b)の相補的な核酸配列とハイブリダイズする核酸配列によりコードされるアミノ酸配列;
(d)(b)の核酸配列に対する遺伝暗号の結果として縮重した核酸配列によりコードされるアミノ酸配列;ならびに
(e)(a)または(b)のアミノ酸配列と少なくとも85%同一、より好ましくは少なくとも90%同一、最も好ましくは少なくとも95%同一であるアミノ酸配列
からなる群より選択されるアミノ酸配列を含む。
【0055】
本発明の医薬組成物のもう1つの好ましい実施形態では、治療されるべき前記上皮腫瘍は、胃腸腺癌、乳腺腺癌または肺腺癌である。前記胃腸腺癌は、好ましくは結腸直腸腺癌、膵臓腺癌、食道腺癌または胃腺癌である。
【0056】
より好ましくは、本発明の前記医薬組成物は、進行性腫瘍、後期腫瘍、腫瘍量の多い腫瘍患者、転移性腫瘍、またはCEA血清中濃度が100ng/mlより高い腫瘍患者の治療のためのものである。前記CEA血清中濃度は、例えばELISAにより測定され得る。
【0057】
本発明の医薬組成物のさらに好ましい実施形態では、本明細書において定義される二重特異性一本鎖抗体の前記第1または第2の結合ドメインの少なくとも1つは、キメラ、ヒト化、CDR移植、および/または脱免疫化またはヒトである。
【0058】
本明細書において、用語「キメラ」は、上に定義されている。本明細書において、用語「ヒト」結合ドメイン、例えば、ヒトCEAと特異的に結合するヒト結合ドメインは、本明細書において定義される二重特異性一本鎖抗体が、マウスモノクローナル抗体A5B7のCDR−H3または上記定義のA5B7由来CDR−H3のKabatの位置100、100a、100b、101、および102(配列番号112)に相当するアミノ酸配列「FYFDY」を少なくとも有する、ヒト生殖細胞系列抗体レパートリーまたは抗体レパートリーに含有されているアミノ酸配列を含むことを意味するものと理解される。本明細書において定義される二重特異性一本鎖抗体はまた、それが、体細胞突然変異のインプリントに起因して予測され得る程度に、その(それらの)最も近いヒト生殖細胞系列配列に由来する配列からなる場合にヒトとみなされ得る。さらに、多くの非ヒト哺乳類、例えばマウスおよびラットなどのげっ歯類の抗体は、発現したヒト抗体レパートリー中にも存在することが予期され得るVH CDRアミノ酸配列を含む。発現したヒト抗体レパートリー中に存在することが予期され得る、ヒトまたは非ヒト起源の任意のそのような配列も本発明の目的において「ヒト」とみなされ得る。
【0059】
本明細書において、用語「ヒト化した」、「ヒト化」または「ヒト様」は、同義的に用いられて、その結合ドメインの少なくとも1つにおいて、非ヒト抗体またはその断片由来の少なくとも1つの相補性決定領域(「CDR」)を含む二重特異性一本鎖抗体を指す。ヒト化アプローチは、例えばWO 91/09968号および米国特許第6,407,213号に記載されている。限定されない例として、この用語は、少なくとも1つの結合ドメインの可変領域が別の非ヒト動物(例えばげっ歯類)由来の単一のCDR領域(例えばVHの3番目のCDR領域)を含む場合、ならびに、一方または両方の可変領域がそれらのそれぞれの1番目、2番目および3番目のCDRの各々に前記非ヒト動物由来のCDRを含む場合を包含する。二重特異性一本鎖抗体の結合ドメインの全てのCDRが、例えばげっ歯類由来のそれらの対応する同等物で置換されている場合、通常「CDR移植(CDR-grafting)」といい、この用語は本明細書において用語「ヒト化」またはその文法上の変化形も包含していると理解される。用語「ヒト化」または文法上の変化形はまた、第一および/または第二の結合ドメインのVHおよび/またはVL内での1またはそれ以上のCDR領域の置換に加えて、CDR間のフレームワーク(「FR」)領域内の少なくとも単一のアミノ酸残基のさらなる変異(例えば置換)が、その位置のアミノ酸を、置換に用いたCDR領域の由来する動物におけるその位置のアミノ酸に合致させている場合も包含する。当分野で公知のように、このような個々の変異は、その標的分子のCDR供与体として用いられる非ヒト抗体の本来の結合親和性を回復させるために、CDR移植の後にフレームワーク領域に作製される場合が多い。用語「ヒト化」は、上記のフレームワーク領域中のアミノ酸置換に加えて、非ヒト動物由来のCDR領域における、ヒト抗体由来の相当するCDR領域のアミノ酸へのアミノ酸置換(1または複数個所)をさらに包含する。
【0060】
本明細書において、用語「脱免疫化された」または「脱免疫化」は、元の野生型構築物と向かい合う第一および/または第二の結合ドメインを、ヒトにおいて前記野生型構築物を非免疫原性または低免疫原性にすることにより修飾することを意味する。脱免疫化アプローチは、例えばWO 00/34317号、WO 98/52976号、WO 02/079415号またはWO 92/10755号に示されている。用語「脱免疫化された」はまた、T細胞エピトープを形成する傾向の低下を示す構築物に関する。本発明に従って、用語「T細胞エピトープを形成する傾向の低下」とは、特異的なT細胞活性化をもたらすT細胞エピトープの除去に関する。さらに、「T細胞エピトープを形成する傾向の低下」とは、T細胞エピトープの形成に寄与するアミノ酸の置換、すなわち、T細胞エピトープの形成に必須のアミノ酸の置換を意味する。言い換えれば、「T細胞エピトープを形成する傾向の低下」は、免疫原性の低下または抗原非依存性のT細胞増殖を誘導する能力の低下に関する。用語「T細胞エピトープ」は、細胞内でペプチド、ポリペプチドまたはタンパク質の分解中に放出され、その後に、T細胞の活性化を引き起こすために主要組織適合抗原複合体(MHC)の分子より提示される短いペプチド配列に関する;特に、WO 02/066514号参照。MHCクラスIIで表されるペプチドに関して、このようなT細胞の活性化は、次に、T細胞の直接刺激による抗体応答を引き起こし、前記抗体を産生することができる。「T細胞エピトープを形成する傾向の低下」および/または「脱免疫化」は、当分野で公知の技法により測定することができる。T細胞増殖アッセイによりインビトロでタンパク質の脱免疫化を試験することが好ましい。このアッセイでは、世界で80%を超えるHLA−DR対立遺伝子を提示するドナー由来のPBMCを、野生型かまたは脱免疫化されたペプチドのいずれかに応答する増殖についてスクリーニングする。理想的には、細胞増殖は野生型ペプチドに抗原提示細胞を負荷する(loading)際にのみ検出される。あるいは、全てのハプロタイプを提示するHLA−DR四量体を発現させることにより、脱免疫化を試験することができる。これらの四量体は、ペプチド結合について試験するか、または増殖アッセイにおいて抗原提示細胞の代わりをするペプチドを負荷してよい。脱免疫化されたペプチドがHLA−DRハプロタイプに提示されるかどうかを試験するため、例えばPBMC上の蛍光標識ペプチドの結合を測定することができる。さらに、脱免疫化は、患者への投与後に脱免疫化分子に対する抗体が形成されたかどうかを判定することにより証明することができる。好ましくは、CDR領域の結合親和性が影響を受けないように、T細胞エピトープを誘導する傾向の低下をもたらすために、抗体由来分子をフレームワーク領域で脱免疫化し、大部分のCDR領域は修飾されない。たとえ1つのT細胞エピトープの除去であっても、結果として免疫原性が低下する。
【0061】
要約すると、上記のアプローチは、上皮性腫瘍患者に投与された場合、本明細書において定義される治療用二重特異性一本鎖抗体の免疫原性を低下させるのに役立つ。例えば、配列番号77に示されるCD3と特異的に結合している第1の結合ドメインは脱免疫化されている;WO 2005/040220号も参照のこと。好ましくは、このCD3結合ドメインにおけるV領域の配置は、VH−VLである。
【0062】
別の態様では、本発明は、
(a)配列番号6、8、16、18、24、26、32、34、40、42、126、130、134または143のいずれか1つに表されるアミノ酸配列;
(b)配列番号5、7、15、17、23、25、31、33、39、41、125、129、133または142に示される核酸配列によりコードされるアミノ酸配列;
(c)ストリンジェントな条件下で(b)の相補的な核酸配列とハイブリダイズする核酸配列によりコードされるアミノ酸配列;
(d)(b)の核酸配列に対する遺伝暗号の結果として縮重した核酸配列によりコードされるアミノ酸配列;および
(e)(a)または(b)のアミノ酸配列と少なくとも85%同一、より好ましくは少なくとも90%同一、最も好ましくは少なくとも95%同一であるアミノ酸配列、
からなる群より選択されるアミノ酸配列を含む、二重特異性一本鎖抗体に関する。
【0063】
一実施形態では、本発明は、上記に定義される二重特異性一本鎖抗体を含む組成物に関する。好ましくは、上記に定義される前記二重特異性一本鎖抗体は、ヒトにおける上皮性腫瘍の治療のための医薬組成物として使用される。前記上皮性腫瘍は、CEA陽性である。これらの医薬組成物のCEA陽性上皮性腫瘍細胞に対する細胞障害活性は、腫瘍患者の血漿中の高い濃度の可溶性CEA抗原にさえも抵抗性である。さらに、上記に定義される前記二重特異性一本鎖抗体またはそれに由来する抗CEA scFvは、下にさらに詳細に述べられるようにヒトにおける上皮性腫瘍の検出のための診断用組成物として使用することができる。
【0064】
本明細書において、用語「ストリンジェントな条件下でハイブリダイズする」とは、ストリンジェントなハイブリダイゼーション条件下で、配列番号5、7、15、17、23、25、31、33、39、41、125、129、133または142に示される配列、またはその相補物とハイブリダイズすることができ、CEA陽性腫瘍細胞に対して細胞障害活性を有する二重特異性一本鎖抗体をコードする核酸配列を指す。「ストリンジェントなハイブリダイゼーション条件」とは、50% ホルムアミド、5xSSC(750mM NaCl、75mM クエン酸三ナトリウム)、50mM リン酸ナトリウム(pH7.6)、5x デンハート液、10% 硫酸デキストラン、および20μg/mlの変性断片化サケ精子DNAを含む溶液中、42℃にて一晩のインキュベーション、その後、約65℃にて0.1x SSC中でのフィルターの洗浄を指す。
【0065】
任意の特定の核酸分子またはポリペプチドが本明細書に定義されるヌクレオチドまたはアミノ酸配列と少なくとも80%、85%、90%、95%、96%、97%、98%または99%同一であるかどうかは、公知のコンピュータープログラムを用いて従来法で決定してよい。クエリー配列(本明細書に定義される配列)と対象配列(subject sequence)との間で最も全体的に適合するものを決定するための好ましい方法(グローバル配列アラインメントとも称される)は、Bultragらのアルゴリズム(Comp. App. Biosci. 6:237-245 (1990))に基づくFASTDBコンピュータープログラムを用いて決定してよい。配列アラインメントにおいてクエリー配列と対象配列は、双方ともDNA配列である。RNA配列はUをTに変換することにより比較され得る。
【0066】
本発明はまた、本明細書に定義される二重特異性一本鎖抗体をコードする核酸配列を含む医薬組成物を提供する。前記核酸は、以下により詳細に示されるように、ヒトにおいて上皮腫瘍を治療するために、例えば遺伝子治療アプローチに利用することができる。
【0067】
本発明はさらに上記に定義される核酸配列を含むベクターを含む医薬組成物に関する。好ましくは、前記ベクターは、上記に定義される前記核酸配列と作動可能なように連結されている調節配列をさらに含む。より好ましくは、前記ベクターは発現ベクターである。
【0068】
さらに、本発明のベクターはまた、遺伝子導入または標的遺伝子組換えベクターであってよい。エキソビボ(ex-vivo)またはインビボ技術により治療遺伝子または核酸を細胞に導入することに基づく遺伝子治療は、遺伝子導入の最も重要な適用の一つである。インビトロまたはインビボでの遺伝子治療に適したベクター、方法または遺伝子送達系は、文献に記載され、当業者に公知である;例えば、Giordano, Nature Medicine 2 (1996), 534-539; Schaper, Circ. Res. 79 (1996), 911-919; Anderson, Science 256 (1992), 808-813, Isner, Lancet 348 (1996), 370-374; Muhlhauser, Circ. Res. 77 (1995), 1077-1086;Onodua, Blood 91 (1998), 30-36; Verzeletti, Hum. Gene Ther. 9 (1998), 2243-2251; Verma, Nature 389 (1997), 239-242; Anderson, Nature 392 (Supp. 1998), 25-30; Wang, Gene Therapy 4 (1997), 393-400;Wang, Nature Medicine 2 (1996), 714-716;WO 94/29469;WO 97/00957;US5,580,859;US5,589,466;US4,394,448またはSchaper, Current Opinion in Biotechnology 7 (1996), 635-640、ならびにそれに引用される参照文献を参照。本明細書に定義される核酸分子およびベクターは、細胞への直接導入用に、またはリポソーム、ウイルス(例えばアデノウイルス、レトロウイルス)ベクター、エレクトロポレーション、またはその他の送達系を介する細胞への導入用に設計することができる。さらに、バキュロウイルス系を本明細書に定義される核酸分子のための真核生物発現系として用いてもよい。導入および遺伝子治療アプローチは、好ましくは、本明細書に定義される機能的二重特異性一本鎖抗体構築物の発現をもたらすべきであり、それによって、前記発現した二重特異性一本鎖抗体構築物はヒトにおける上皮腫瘍の治療、寛解および/または予防に特に有用である。
【0069】
さらなる態様では、本発明は、上記に定義されるベクターまたは核酸で形質転換またはトランスフェクトされた宿主を含む医薬組成物に関する。
【0070】
本発明のさらなる態様は、免疫エフェクター細胞の活性化シグナルを提供することのできるタンパク質性化合物をさらに含む、上記に定義される医薬組成物に関する。
【0071】
好ましくは、医薬組成物は、担体、安定剤および/または賦形剤からなる適した製剤をさらに含む。
【0072】
別の態様では、本発明は、上記に定義される医薬組成物の製造のためのプロセスに関し、前記プロセスは上記に定義される宿主を上記に定義される二重特異性一本鎖抗体の発現を可能にする条件下で培養することと、産生された二重特異性一本鎖抗体を培養物から回収することを含む。
【0073】
本発明のさらなる態様は、ヒトにおける上皮腫瘍の予防、治療または寛解のための医薬組成物の調製のための、上記に定義される二重特異性一本鎖抗体または上記に定義されるプロセスにより産生された二重特異性一本鎖抗体、上記に定義される核酸分子、上記に定義されるベクターまたは上記に定義される宿主の使用に関する。本発明の別の態様は、ヒトにおける上皮腫瘍の予防、治療または寛解のための方法に関し、前記方法は本発明の、または上に示されるプロセスによって製造された医薬組成物の有効量を投与する段階を含む。当業者、特に主治医であれば、本発明の二重特異的分子/二重特異的一本鎖抗体の投与を必要とする患者の治療の成功を評価することができる。従って、投与計画ならびに投薬量および投与回数は前記当業者により判断され得る。つまり、判断される、対応する「寛解」および/または「治療」は下に定義される。
【0074】
本明細書において、上皮性腫瘍との関連で、本発明の医薬組成物の「有効量」または「治療上有効な量」とは、一次、局所または転移性腫瘍組織を破壊、改変、制御または除去するために十分な治療薬の量を指す。治療上有効な量とは、上皮性腫瘍の広がりを遅延または最小化するために十分な治療薬の量を指し得る。また、治療上有効な量とは、上皮性腫瘍の治療または管理において治療上の利益をもたらす治療薬または医薬品の量も指し得る。さらに、本発明の治療薬または医薬品に関して治療上有効な量とは、上皮性腫瘍の治療または管理において治療上の利益をもたらす治療薬または医薬品の、単独の、または他の治療法と組み合わせた量を意味する。本明細書において定義される二重特異性一本鎖抗体の量に関連して用いられる場合、この用語は、全体的な治療法を改善するか、望ましくない効果を低下または回避するか、または別の治療薬の治療効力または別の治療薬との相乗効果(本明細書において定義される通り)を促進する量を包含し得る。好ましくは、治療薬の治療上有効な量は、上皮性腫瘍の治療において、全体的な治療法を改善するか、望ましくない効果を低下または回避するか、または別の治療薬の治療効力または別の治療薬との相乗効果を促進する。例えば、本明細書において定義される二重特異性一本鎖抗体は、単一療法として患者に投与される場合、上皮性腫瘍の直径を20%縮小させることができる。これに対し、第2の治療薬、例えば、下に定義される抗癌剤は、腫瘍を10%縮小させることができる。しかし、本明細書において定義される二重特異性一本鎖抗体と前記第2の治療薬の両方を併用療法の形態で組み合わせて投与する場合、50%の腫瘍の縮小を観察することができる。このような効果は本明細書において相乗効果として理解される。
【0075】
本明細書において言及されるように、用語「治療法」とは、上皮性腫瘍の予防、治療または寛解に用いられ得る任意の投与計画、方法および/または薬剤を指す。用語「上皮性腫瘍の予防、治療または寛解」は、下に詳細に説明されている。用語「治療法(therapies)」および「治療法(therapy)」は、上皮性腫瘍、あるいはその1またはそれ以上の症状の治療、予防、または寛解に有用な生物学的療法、支持療法、化学療法、放射線療法および/またはその他の治療法を称し得る。
【0076】
本明細書において、患者に1または複数の治療法を投与することとの関連で、用語「治療する」、「治療」および「治療すること」は、上皮性腫瘍の進行、重篤度、および/または持続の低下または寛解を指す。前記上皮性腫瘍は、異常な発現(例えば、CEAの過剰発現もしくは過剰活性)および/または、1もしくはそれ以上の治療法(1またはそれ以上の医薬品または治療薬の投与を含む)の投与の結果として生じるその1またはそれ以上の症状の寛解を伴い得る。
【0077】
最も好ましい投与方法は、所与の時間/期間の静脈内投与である。本明細書に定義される二重特異性一本鎖抗体は1回につき単独で投与されてよいが、好ましいものは、製薬上許容される担体中の投与である。適した医薬担体の例は当分野で周知であり、リン酸緩衝生理食塩水溶液、水、リポソーム、様々な種類の湿潤剤、滅菌溶液などが含まれる。このような担体を含む組成物は、周知の従来法により処方されてよい。これらの医薬組成物は被験体に適した用量で投与することができる。投与計画は主治医および臨床学的因子により決定される。医学分野で周知のように、任意の患者一人への投薬量は多くの要因によって決まり、それには患者の大きさ、体表面積、年齢、投与される特定の化合物、性別、投与時間および経路、全体的な健康状態、および同時に投与されるその他の薬剤が含まれる。非経口投与のための製剤としては、滅菌水溶液または非水溶液、および懸濁液が挙げられる。非水溶媒の例は、プロピレングリコール、ポリエチレングリコール、および注射用の有機エステル類、例えばオレイン酸エチルである。水性担体としては、水、水溶液、または生理食塩水および緩衝媒体を含む懸濁液が挙げられる。非経口媒体としては、塩化ナトリウム溶液、リンゲルデキストロース、デキストロースおよび塩化ナトリウム、乳酸化リンゲル液、または硬化油が挙げられる。静脈内媒体としては、液体および栄養素補充液、電解質補充液(リンゲルデキストロースに基づくものなど)などが挙げられる。防腐剤およびその他の添加剤も存在してよく、例えば、抗菌剤、抗酸化剤、キレート剤、不活性ガスなどが挙げられる。さらに、組成物は、例えば、好ましくはヒト起源の血清アルブミンまたは免疫グロブリンなどのタンパク質性担体を含んでもよい。併用療法は、タンパク質性二重特異性一本鎖抗体に加えて、医薬組成物の意図される使用に応じて、さらなる生物活性物質を含むことが想定される。このような物質は、胃腸管系に作用する物質、細胞増殖抑制剤として作用する物質、高尿酸血症を予防する物質、免疫反応を阻害する物質(例えばコルチコステロイド、FK506)、循環系に作用する薬剤および/またはT細胞共刺激分子またはサイトカインなどの当分野で公知の薬剤であってよい。好ましくは、本明細書に定義される二重特異性一本鎖抗体は、緩衝液、安定剤および界面活性剤中に処方される。緩衝液は、リン酸緩衝液、クエン酸緩衝液、コハク酸緩衝液または酢酸緩衝液であってよい。安定剤は、アミノ酸および/または糖であってよい。界面活性剤は、合成洗剤、PEGなどであってよい。より好ましくは、本明細書に定義される二重特異性一本鎖抗体は、クエン酸塩、リシン、トレハロースおよびTween80中に処方される。前記医薬組成物の希釈液として、等張生理食塩水およびTween80が好ましい。
【0078】
本明細書において用語「寛解」とは、疾患、すなわち上皮腫瘍の重篤度の改善または緩和を指す。例えば、そのような寛解は、本発明の医薬組成物の投与に起因する、安定な疾患の達成、またさらにより好ましくは、上皮腫瘍の縮小、すなわち、微小、部分的応答または完全寛解であり得る。「安定な疾患」とは、臨床および/または組織学的診断方法により腫瘍進行/増殖が全くまたはほとんど観察されないまたは検出されない疾患状態を指す。例えば、指標病変の断面積の合計の50%より多くの縮小である、腫瘍の縮小は、「部分応答」と考えられる。「完全寛解」は、治療後に病変がこれ以上全く検出されない状態を意味する。安定疾患と部分応答との間の腫瘍縮小を伴う応答は、微小応答と考えられ得る。例えば、指標病変の断面積の合計の20%、25%または30%縮小は、微小応答と称され得る。
【0079】
用語「寛解」は、本明細書において、上皮腫瘍の数の減少も包含する。それは、さらに腫瘍進行の阻止/減速を意味する。さらに、治療しなかった腫瘍患者と比較して、治療した腫瘍患者の全体的な生存の改善は、本明細書において「寛解」と考えられる。これは、必要な変更を加えて、治療しなかった腫瘍患者と比較して、治療した腫瘍患者の進行のない生存または再発のない生存の改善にも当てはまる。さらに、用語「寛解」はまた、上皮腫瘍の症状の強度の減少も指し、その結果得られる、例えば治療した腫瘍患者の生活の質の改善も指す。
【0080】
用語「上皮腫瘍の予防」は、本明細書において、次のように理解される。原発上皮腫瘍のヒト患者からの外科切除後、および/または原発上皮腫瘍の化学療法または放射線治療後に、腫瘍細胞全てが身体から排除されない場合もあり得る。しかし、これらの残存腫瘍細胞は患者において再発性癌、すなわち、局所再発および/または転移を引き起こす可能性がある。転移はよく見られる癌の合併症であるが、癌細胞が原発腫瘍から播種されて遠位のコロニーを形成する過程ははあまり分かっていない。転移性癌は、ほぼ例外なく難治性で新しい治療方法の必要性が生じる。本発明の医薬組成物を用いると、これらの播種された腫瘍細胞を死滅させて二次性腫瘍(一次治療後に身体に残存する腫瘍細胞から生じる)の形成を予防することができる。このように、本医薬組成物は、腫瘍患者において局所再発および/または転移の形成を予防するために役立つ。
【0081】
抗腫瘍治療の成功は、それぞれの疾病について、例えばコンピュータ断層撮影、X線、核磁気共鳴断層撮影(例えば、米国国立癌研究所の判定基準に基づく応答評価(Cheson (1999), J. Clin. Oncol.; 17 (4):1244))、陽電子放射断層撮影走査、内視鏡検査、蛍光活性化セルソーター、骨髄の吸引、胸水または腹水、組織/組織学、および様々な上皮腫瘍特異的臨床化学パラメータ(例えば血清中の可溶性CEA濃度)により確立された標準的な方法でモニターすることができ、さらに、その他の確立された標準的な方法を用いてよい。さらに、T細胞の活性化を測定するアッセイを用いてよい;例えば、WO 99/054440号を参照。治療しなかった腫瘍患者と比較して、治療した腫瘍患者の全体的な生存、進行のない生存または再発のない生存の決定のために統計学を用いてもよい。
【0082】
好ましくは、前記上皮腫瘍は、胃腸腺癌、乳腺腺癌または肺腺癌である。前記胃腸腺癌は、より好ましくは、結腸直腸腺癌、膵臓腺癌、食道腺癌または胃腺癌である。
【0083】
さらにより好ましくは、前記本発明の医薬組成物は、進行性腫瘍、後期腫瘍、腫瘍量の多い腫瘍患者、転移性腫瘍、またはCEA血清中濃度が100ng/mlより高い腫瘍患者の治療のためのものである(例えばELISAにより測定され得る)。
【0084】
本発明の使用または方法のもう1つの好ましい実施形態では、上記に定義される前記医薬組成物は、さらなる薬剤と組み合わせて、すなわち、併用療法の一部として投与されるのに適している。
【0085】
特定の実施形態では、本明細書において定義される二重特異性一本鎖抗体または医薬組成物は、1またはそれ以上のその他の治療法と組み合わせて投与される。特定の実施形態では、本明細書において定義される二重特異性一本鎖抗体または医薬組成物は、1またはそれ以上のその他の治療法と同時に患者に投与される。そのような治療法は上皮性腫瘍の治療に有用であることが好ましい。用語「同時に」は、医薬組成物または治療薬を全く同時に投与することに限定するのではなく、むしろ、本明細書において定義される二重特異性一本鎖抗体または医薬組成物とその他の薬剤が、患者に順番に投与されること、及び本明細書において定義される二重特異性一本鎖抗体または医薬組成物が他の薬剤と一緒に作用して、別の方法で投与された場合よりも増大した利益をもたらすことのできるような間隔で投与されることを意味する。例えば、それぞれの治療薬は同時にまたは任意の順序で異なる時点に順次投与されてよい;しかし、同時に投与されない場合、それらは所望の治療効果を得るために時間的に十分に接近して投与されるべきである。
【0086】
それぞれの治療薬は、任意の適当な形態で、任意の適した経路から別々に投与することができる。他の実施形態では、本明細書において定義される二重特異性一本鎖抗体または医薬組成物は、外科的手術の前に、同時に、または後に投与される。該外科的手術は局在した上皮性腫瘍を完全に除去するか、または大きな上皮性腫瘍のサイズを縮小させることが好ましい。外科的手術はまた、予防的手段としてなされるか、または疼痛を軽減するためになされてよい。
【0087】
本明細書において提供される投薬量および投与の頻度は、上に定義される「治療上有効な」という用語に包含される。投薬量および頻度は、一般に、投与される特定の治療薬または予防薬、上皮性腫瘍の重篤度および種類、投与経路、ならびに患者の年齢、体重、応答、および過去の病歴に依存する各患者についての特異的な因子に従ってさらに変動する。当業者は、そのような因子を考慮すること、および、例えば、文献に報告され、Physicians' Desk Reference (59th ed., 2005) で推奨されている投薬量に従うことにより、適した投与計画を選択することができる。
【0088】
一部の実施形態では、本明細書において定義される二重特異性一本鎖抗体または医薬組成物の投与による治療法は、1またはそれ以上の治療法、例えば化学療法、放射線療法、ホルモン療法、および/または生物学的療法/免疫療法の投与と組み合わされる。治療薬としては、限定されるものではないが、タンパク質性分子(限定されるものではないが、ペプチド、ポリペプチド、タンパク質(翻訳後修飾されたタンパク質、抗体などを含む)を含む);または小分子(1000ダルトン未満)、無機もしくは有機化合物;または核酸分子(二本鎖もしくは一本鎖DNA、または二本鎖もしくは一本鎖RNA、ならびに三重らせん核酸分子を含む)が挙げられる。治療薬は、任意の既知生物(限定されるものではないが、動物、植物、細菌、真菌、および原生生物、またはウイルスを含む)に由来するか、または合成分子ライブラリーに由来してよい。
【0089】
特定の実施形態では、本発明の方法および使用は、キナーゼ、例えば当分野に記載されるゲフィチニブ(イレッサ)、エルロチニブ(タルセバ)、抗EGFR抗体(例えば、セツキシマブ;エルビタックス)、または抗Her2/neu抗体(例えば、トラスツズマブ;ハーセプチン)の阻害剤である1またはそれ以上の治療薬の投与と組み合わせた、本明細書において定義される二重特異性一本鎖抗体または医薬組成物の投与を包含する;例えば、Hardie and Hanks (1995) The Protein Kinase Facts Book, I and II, Academic Press, San Diego, California 参照。
【0090】
別の特定の実施形態では、本発明の方法および使用は、血管形成阻害剤、例えば当分野に記載される、抗VEGF抗体(例えば、ベバシズマブ;アバスチン)、小分子化合物(例えば、バタラニブもしくはソラフェニブ)またはCOX阻害剤である、1またはそれ以上の治療薬の投与と組み合わせた、本明細書において定義される二重特異性一本鎖抗体または医薬組成物の投与を包含する。
【0091】
別の特定の実施形態では、本発明の方法および使用は、抗癌剤、例えば当分野に記載される、5−フルオロウラシル、ロイコボリン、カペシタビン、オキサリプラチン、イリノテカン、ゲムシタビン、ドキソルビシン、エピルビシン、エトポシド、シスプラチン、カルボプラチン、タキサン類(例えば、ドセタキセル、パクリタキセル)である、1またはそれ以上の治療薬の投与と組み合わせた、本明細書において定義される二重特異性一本鎖抗体または医薬組成物の投与を包含する。
【0092】
本明細書において定義される二重特異性一本鎖抗体または医薬組成物をさらなる治療薬と組み合わせて用いる上皮性腫瘍患者の併用療法は、相乗作用をもたらすことが好ましい。本明細書において、用語「相乗的な」とは、任意の2またはそれ以上の単一の治療法(例えば、1またはそれ以上の治療薬)の付加的効果よりも効果的な、治療法の組み合わせ(例えば、上に説明される、本明細書において定義される二重特異性一本鎖抗体とさらなる治療薬の組み合わせ)を指す。例えば、本明細書において定義される二重特異性一本鎖抗体は、単一療法として患者に投与される場合、20%の上皮性腫瘍の直径の縮小をもたらすことができる。その一方、第2の治療薬、例えば、下に定義される抗癌剤は、腫瘍を10%縮小させることができる。しかし、本明細書において定義される二重特異性一本鎖抗体と前記第2の治療薬の両方を併用療法の形態で組み合わせて投与する場合、50%の腫瘍の縮小を観察することができる。
【0093】
治療法の組み合わせ(例えば、上に説明される、本明細書において定義される二重特異性一本鎖抗体と、さらなる治療薬の組み合わせ)の相乗作用により、低投薬量の1またはそれ以上の治療法(例えば、1またはそれ以上の治療薬)の使用および/または疾患、例えば、上皮性腫瘍のある患者へ前記治療法の低頻度の投与が可能となる。低い投薬量の治療法(例えば、治療薬)を用いることができ、かつ/または前記治療法を低頻度で投与することができると、疾患(例えば上皮性腫瘍)の予防または治療において前記治療法の効力を低下させることなく、前記治療法の被験体への投与に関連する毒性が低減される。さらに、相乗作用は、上皮性腫瘍(CEAの異常な発現(例えば、過剰発現)または活性を伴う可能性がある)の予防、管理、治療および/または寛解において、治療法(例えば、治療薬)の改善された有効性をもたらし得る。最後に、治療法(例えば、治療薬)の組み合わせの相乗作用は、任意の単一療法の使用に関連する、有害なまたは望ましくない副作用を回避または減少させることができる。
【0094】
前記併用療法において、活性薬剤は場合により本明細書において定義される二重特異性一本鎖抗体と同じ医薬組成物に含まれても、別個の医薬組成物に含まれてもよい。この後者の場合、前記別個の医薬組成物は、本明細書において定義される二重特異性一本鎖抗体を含む前記医薬組成物投与よりも前に、同時に、または後に、投与されるのに適している。さらなる薬剤または医薬組成物は、非タンパク質性化合物もしくはタンパク質性化合物であり得る。さらなる薬剤がタンパク質性化合物である場合、該タンパク質性化合物は免疫エフェクター細胞の活性化シグナルを提供できることが有利である。
【0095】
好ましくは、前記タンパク質性化合物または非タンパク質性化合物は、同時にまたは非同時に、上記において定義される二重特異性一本鎖抗体、上記において定義される核酸分子、上記において定義されるベクター、または上記において定義される宿主とともに投与されてよい。好ましくは、処置されるべき前記被験体は、ヒトである。
【0096】
さらなる実施形態では、本明細書において定義される一本鎖二重特異性抗体または抗CEA scFvは、診断用または検出可能な薬剤と複合体化され得る。そのような診断および検出は、抗体またはscFvを、検出可能な物質、例えば様々な酵素類、例えば西洋ワサビペルオキシダーゼ、アルカリ性ホスファターゼ、β−ガラクトシダーゼ、またはアセチルコリンエステラーゼ;補欠分子族、例えばストレプトアビジン/ビオチンおよびアビジン/ビオチン;蛍光物質、例えば、ウンベリフェロン、フルオレセイン、フルオレセインイソチオシアネート、ローダミン、ジクロロトリアジニルアミンフルオレセイン、ダンシルクロリドまたはフィコエリトリン;発光物質、例えば、ルミノール;生物発光物質、例えば、ルシフェラーゼ、ルシフェリン、およびエクオリン;放射性物質および同位元素、例えばコバルト(57Co)、インジウム(115In、113In、112In、111In)、ヨウ素(131I、125I、123I、121I)、またはイットリウム(90Y)、様々な陽電子放出断層撮影を用いる陽電子放出金属、および非放射性常磁性金属イオンと結合させることにより達成することができる。
【0097】
部分と抗体を複合体化するための技法は周知である。部分は、当分野で公知の任意の方法により抗体と複合体化させることができ、それには、限定されるものではないが、アルデヒド/シッフ結合、スルフヒドリル結合、酸に不安定な結合、シス−アコニチル結合、ヒドラゾン結合、酵素分解可能な結合が含まれる;一般に、 Garnett, 2002, Adv. Drug Deliv. Rev. 53:171-216 参照。部分と抗体を複合体化するためのさらなる技法は当分野で周知である、例えば、Arnon et al., Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy. In Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), pp. 243-56 (Alan R. Liss, Inc. 1985)参照。抗体とポリペプチド部分を融合または複合体化させるための方法は当分野で公知である;例えば、Ashkenazi et al., 1991, PNAS 88: 10535-10539 参照。抗体と部分の融合は必ずしも直接的である必要はなく、リンカー配列によって生じるものであってよい。そのようなリンカー分子は一般に当分野で公知であり、Denardo et al., 1998, Clin Cancer Res. 4:2483-90; Peterson et al., 1999, Bioconjug. Chem. 10:553 に記載されている。
【0098】
さらなる態様において、本発明は、上記に定義される二重特異性一本鎖抗体、上記に定義される核酸分子、上記に定義されるベクター、または上記に定義される宿主を含むキットに関する。
【0099】
これらのおよびその他の実施形態は、本発明の説明および実施例に開示され、包含される。免疫学分野の組換え技術および方法は、例えば、Sambrook et al., Molecular Cloning: A Laboratory Manual;Cold Spring Harbor Laboratory Press, 3rd edition 2001; Lefkovits; Immunology Methods Manual;The Comprehensive Sourcebook of Techniques;Academic Press, 1997; Golemis; Protein-Protein Interactions: A Molecular Cloning Manual;Cold Spring Laboratory Press, 2002に記載されている。本発明に従って用いられ得る抗体、方法、用途および化合物のいずれかに関するさらなる参照文献は、例えば電子装置を用いて公開されているライブラリーおよびデータベースから得ることができる。例えば、インターネットで利用可能な公開データベース「Medline」を、例えばhttp://www.ncbi.nlm.nih.qov/PubMed/medline.htmlで利用してよい。さらなるデータベースおよびアドレス、例えばhttp://www.ncbi.nim.nih.qov/、http://www.infobioaen.fr/、http://www.fmi.ch/bioloqv/research tools.html、http://www.tiqr.orQ/が当業者に公知であり、例えば、http://www.lvcos.comを用いて得ることもできる。腫瘍関連の話題は、例えば、http://www.nih.govまたはhttp://www.dkfz.deを参照のこと。
【実施例】
【0100】
以下の実施例は本発明を説明する。
【0101】
実施例1:ヒトCEA(癌胎児性抗原関連細胞接着分子5;CEACAM5)がトランスフェクトされたCHO細胞の作製
CEA陽性KatoIII細胞(ヒト胃癌腫細胞株;ATCC HTB−103)を用いて全RNAを得、それをキットマニュアルの説明書(Qiagen、RNeasy Mini Kit)に従って単離した。得られたRNAを、ランダムに開始される逆転写によるcDNA合成に用いた。CEA抗原の全長配列のクローニングのため、以下のオリゴヌクレオチドを用いた。5’CEACAM5 EcoRI GAATTCGCCACCATGGAGTCTCCCTCGGCCCC(配列番号74)および3’CEACAM5 Sal I GTCGACCTATATCAGAGCAACCCC(配列番号75)。PCR(第1サイクルに関して、93℃にて5分間の変性、58℃にて1分間のアニーリング、72℃にて1分間の伸張;93℃にて1分間の変性、58℃にて1分間のアニーリング、72℃にて1分間の伸張を30サイクル;72℃にて5分間の末端伸張)を用いてコード配列を増幅した。その後、PCR産物をEcoRIおよびSalIで消化し、適切に消化した発現ベクターpEF−DHFRに連結し、大腸菌に形質転換した。単離したプラスミドDNAを配列決定し、確立されたCEACAM5の核酸配列(米国国立バイオテクノロジー情報センター(National Center for biotechnology information)のNM_004363、http://www.ncbi.nlm.nih.gov/)と比較した。標準的なプロトコールに従って、上述の手順を実行した(Sambrook, Molecular Cloning; A Laboratory Manual, Cold Spring Harbour Laboratory Press, Cold Spring Harbour, New York (1989;2001)。確認された核酸配列を含むクローンを、構築物の真核生物における発現のためにDHFR欠損CHO細胞にトランスフェクトした。DHFR欠損CHO細胞における真核生物タンパク質発現を、Kaufmannに記載のとおり行った(Kaufmann R.J., Methods Enzymol. 185 (1990), 537-566)。構築物の遺伝子増幅を、最大20nM MTXの終濃度までの、漸増濃度のMTXにより誘導した。次に、トランスフェクトした細胞を、FACSアッセイを用いてCEA抗原の発現について試験した。その目的のため、2.5×105のトランスフェクト細胞を、濃度5μg/mlのマウスモノクローナル抗体COL−1(No.MS−613−P1ABX,Neomakers;Fremont,CA,USA)とともにインキュベートした。50μl PBS中2% FCS(Dianova,Hamburg,Germanyより入手)を用いて1:100で希釈した、R−フィコエリトリン結合アフィニティー精製F(ab’)2断片、ヤギ抗マウスIgG、Fc−γ断片特異的抗体を用いて、抗体の結合を検出した。細胞を、FACS−Calibur(Becton Dickinson,Heidelberg)でフローサイトメトリーにより分析した。FACS染色および蛍光強度の測定を、Current Protocols in Immunology (Coligan, Kruisbeek, Margulies, Shevach and Strober, Wiley-Interscience, 2002)に記載のとおり行った。結果として、形質転換体は、ヒトCEA抗原について明らかな陽性染色を実証した。
【0102】
実施例2:CEAxCD3二重特異性一本鎖抗体の生成
一般に、それぞれヒトCEA抗原に対して結合特異性を有するドメインを含む二重特異性一本鎖抗体分子、ならびに配列番号77に示されるヒトCD3抗原に対して結合特異性を持つ脱免疫化されたドメインを、表1に提示されるように設計した。このCD3結合ドメイン中のV領域の配置は常にVH−VLである。本明細書において定義される二重特異性一本鎖抗体において用いられる、この脱免疫化された抗CD3結合ドメインは、例えば、WO 2005/040220号において既に記載されている。
【0103】
1.抗CEA特異性および抗CD3特異性を含む二重特異性一本鎖抗体分子の形式(表1)
【表1】
【0104】
モノクローナル抗体若しくはハイブリドーマに由来するか、またはファージディスプレイ誘導選択(PDGS)により得た、ヒトCEA抗原に特異的な可変軽鎖(VL)および可変重鎖(VH)領域を含有する前述の構築物は、遺伝子合成およびその後の、CD3に特異的なVHとVLの組み合わせを含む発現ベクターへのクローニングにより得た。前記二重特異性一本鎖構築物の作製はまた、例えば、Sambrook(前掲)に記載の組換え技術に従っても行うことができる。そのための詳細な指示は、例えば、WO 99/054440号に規定されている。
【0105】
抗CD3結合ドメインは、ヒトCD3抗原に対する結合特異性を有する脱免疫化されたドメインに相当する。本明細書に記載される二重特異性一本鎖構築物中の脱免疫化された抗CD3結合ドメインのV領域の配置は、常にVH−VLである。対応する前記VH−VLドメインのアミノ酸配列は配列番号77に示される。ヒト癌胎児性抗原に特異的なCEAI、CEAIIおよびCEAIIIは、それぞれ、mAb A5B7(Chester, K. A. et al., Int J Cancer 57 (1994), 67-72)、T84.66(Neumaier, M. et al., Cancer Res 50 (1990), 2128-34)およびMFE−23(Boehm, M. K. Biochem J 2 (2000), 519-28)に由来する可変軽鎖(VL)および可変重鎖(VH)領域を含む。A5、B9、D8、およびE12は、ヒトCEAに特異的なヒトVH領域であり、一方、A240は同じ特異性を有するヒトVL領域である。ヒトA5、B9、D8、E12およびA240V領域の作製は、実施例6および7に詳細に記載されている。本明細書に記載されるすべての二重特異性一本鎖抗体の対応するヌクレオチドおよびアミノ酸配列はリストする配列に示される。
【0106】
以下では、CEAI VLVHx配列番号77VHVL(配列番号6)構築物の作製がさらに詳細に説明される。上に言及されるその他の構築物の作製は、十分当業者の範囲内である方法に必要な改変を施して適宜に行うことができる。前述のCEAI特異性および脱免疫化された抗CD3(配列番号77)特異性を含む二重特異性一本鎖抗体分子を作製するため、最初に、標準的なプロトコールに従う遺伝子合成により得たCEAIの可変領域をPCRにより改変して相当する一本鎖Fv抗体フラグメントを得る必要があった。本目的のため、2段階の融合PCRを用いて可変領域をコードする配列を増幅させた。一組の適切なプライマーを設計してPCRに基づくクローニング段階を行い、最後に、その結果として、15アミノ酸リンカー([Gly4Ser]3)をVL−リンカー−VHの順序で含む2つの可変ドメインを連結している一本鎖抗体がもたらされた。
【0107】
要するに、以下のプライマーの組み合わせを用いた。
【表2】
【0108】
オリゴヌクレオチドプライマーの核酸配列を下に示す。
5’CEAI LH:5'AGGTGTACACTCCGACATTGAGCTCACCCAG3’(配列番号137)
3’CEAI VLリンカー:5'GGAGCCGCCGCCGCCAGAACCACCACCACCTTTGATCTCGAGCTTGG3'(配列番号138)
5’CEAI VHリンカー:5'GGCGGCGGCGGCTCCGGTGGTGGTGGTTCTCAGGTCCAACTGCAGGAG3'(配列番号139)
3’CEAI LH:5’AATCCGGAGGAGACGGTGACCG3'(配列番号140)
【0109】
一本鎖抗体を作製するため、上記のそれぞれのプライマーの組み合わせで2回のPCRをPCR段階1および2として行った。このPCRの間、重複する相補的配列を(それぞれのリンカープライマーから生じる)PCR生成物に導入し、それは一体となって(combined)その後の融合PCRの間に15アミノ酸リンカーのコード配列を形成した。その後、増幅したVHおよびVLドメインを、外側のプライマーのみと両方のPCR生成物を必要とする、この融合PCR中で結合させた。得られるscFv抗体は、5’末端でBsrGIの制限酵素認識部位に隣接し、3’末端でBspEIの制限酵素認識部位に隣接している。WO 2005/040220号に記載されるようにマウス免疫グロブリンリーダーペプチドのコード配列とのインフレーム融合を可能にするために、BsrGI部位の付加を行った。二重特異性一本鎖抗体を作製するためのCD3特異的一本鎖抗体をコードする配列とのインフレーム融合を可能にするために、BspEI部位を作製した。一本鎖Fv抗体の融合を達成するため、かつ真核生物の発現を可能にするため、CEA特異的一本鎖Fv抗体のコード配列は、BsrGIおよびBspEIを介して、WO 2005/040220号に記載されるように脱免疫化された抗CD3一本鎖Fv抗体(本発明において配列番号77と称される)を含有するpEFDHFR発現ベクターにクローン化された(pEFDHFRは、Mack et al. Proc. Natl. Acad. Sci. USA 92 (1995) 7021-7025 に記載されている)。構築物の単一のクローンを単離し、標準的なプロトコールに従って発現ベクター中で隣接領域と相補的なプライマーを用いて配列決定した(Sambrock, Molecular Cloning; A Laboratory Manual, 2nd edition, Cold Spring Harbour laboratory Press, Cold Spring Harbour, New York (1989))。さらなる実験のため、検証された核酸配列を含む構築物のクローンが選択された。
【0110】
2.CEAxCD3二重特異性一本鎖抗体の発現および精製
二重特異性一本鎖抗体をチャイニーズハムスター卵巣(CHO)細胞で発現させた。DHFR欠損CHO細胞における真核生物タンパク質発現を、Kaufmann(前掲)に記載のとおり行った。構築物の遺伝子増幅は漸増濃度のMTXにより20nM MTXまでの終濃度まで誘導した。静置培養の二代継代後、細胞をCHO改変DMEM培地(HiQ(登録商標)、HiClone)を含むローラーボトル中で7日間増殖させた後、回収した。細胞を遠心分離により取り除き、発現タンパク質を含有する上清を−20℃で保存した。
【0111】
Akta(登録商標)FPLCシステム(Pharmacia)およびUnicorn(登録商標)ソフトウェアをクロマトグラフィーに使用した。全ての化学物質は研究等級のものであり、Sigma社(Deisenhofen)またはMerck社(Darmstadt)より購入した。固定化金属アフィニティークロマトグラフィー(「IMAC」)を、製造業者の提供するプロトコールに従って、ZnCl2を装填したFractogel(登録商標)カラム(Merck)を用いて行った。カラムをバッファーA2(20mM リン酸ナトリウムバッファー、pH7.5、0.4M NaCl)で平衡化し、細胞培養上清(500ml)を3ml/分の流速にてカラム(10ml)に適用した。カラムをバッファーA2で洗浄して結合しなかったサンプルを取り除いた。結合したタンパク質は、2段階勾配のバッファーB2(20mM リン酸ナトリウムバッファー、pH7.5、0.4M NaCl、0.5M イミダゾール)を用いて、以下に従って溶出した。
段階1:6カラム容積中バッファーB2 20%;
段階2:6カラム容積中バッファーB2 100%。
【0112】
段階2からの溶出したタンパク質画分を、さらなる精製のためにプールした。ゲルろ過クロマトグラフィーを、PBS(Gibco)で平衡化したSephadex S200 HiPrepカラム(Pharmacia)で行った。溶出したタンパク質サンプル(流速1ml/分)を、検出のため、標準的なSDS−PAGEおよびウエスタンブロットに付した。精製より前に、カラムを分子量決定のために較正した(分子量マーカーキット、Sigma MW GF−200)。プロテインアッセイ染料(MicroBCA、Pierce)およびIgG(Biorad)を標準タンパク質として用いて、タンパク質濃度を測定した。
【0113】
CEAxCD3二重特異性一本鎖抗体を、IMACの2段階精製プロセスおよびゲルろ過で単離した。主生成物の分子量は、未変性条件下でPBSでのゲルろ過により測定して約52kDaであった。この分子量は二重特異性一本鎖抗体に一致する。全ての構築物がこの方法に従って精製された。
【0114】
精製された二重特異性一本鎖抗体タンパク質を、還元条件下で事前成形された4〜12% Bis Trisゲル(Invitrogen)で行われたSDS PAGEで分析した。製造業者の提供するプロトコールに従ってサンプルの調製および適用を行った。MultiMarkタンパク質標準品(Invitrogen)を用いて分子量を測定した。コロイドクマシーでゲルを染色した(Invitrogenプロトコール)。単離したタンパク質の純度は、SDS−PAGEで測定して>95%であった。Optitran(登録商標)BA−S83膜およびInvitrogenブロットモジュールを製造業者の提供するプロトコールに従って用いてウエスタンブロットを行った。使用した抗体は、基質としてHisタグ(Penta His、Qiagen)およびアルカリ性ホスファターゼ(AP)で標識したヤギ抗マウスIg(Sigma)、およびBCIP/NBT(Sigma)に対して指向された。二重特異性一本鎖抗体は、ウエスタンブロットにより特異的に検出することができた。精製した二重特異性一本鎖抗体分子に対応する52kDで単一のバンドが検出された。
【0115】
3.CEAxCD3二重特異性一本鎖抗体のフローサイトメトリー結合分析
膜結合ヒトCEAおよびヒトCD3との結合能に関して構築物の官能性を試験するため、FACS分析を行った。本目的のため、ヒトCEAトランスフェクトCHO細胞とCD3陽性ヒトT細胞白血病細胞株HPB−AII(DSMZ,Braunschweig,ACC483)を使用した。200,000個のCEA陽性CHO細胞または200,000個のHPB−AII細胞を、氷上で30分間、異なる配置のCEAおよびCD3のVHおよびVL領域(上記の通り)を含む二重特異性抗体をそれぞれ発現しているCHO細胞培養の純粋な細胞上清50μlとともにインキュベートした。細胞をPBSで2回洗浄し、構築物の結合を、構築物のC末端のヒスチジンタグを介して細胞と結合した構築物と特異的に結合する、標識されていないマウスPenta His抗体(2% FCSを含む50μl PBSに1:20で希釈;Qiagen)を用いて検出した。洗浄段階の後に、結合していないマウスPenta His抗体を取り除いた。結合した抗His抗体を、フィコエリトリンと複合体化し、2% FCSを含む50μl PBSに1:100で希釈されたFcγ特異的抗体(Dianova)を用いて検出した。ヒトCEAとの結合の陽性対照として、モノクローナル抗体Col−1(実施例3参照)を用いた。ヒトCD3との結合の対照には、WO 99/054440号に記載のCD19xCD3二重特異性一本鎖構築物を使用した。陰性対照として、培養上清の代わりに新鮮な培地を用いた。
【0116】
細胞をフローサイトメトリーで分析した(FACS−Calibur;Becton Dickinson,Heidelberg)。FACS染色および蛍光強度の測定を、Current Protocols in Immunology(Coligan, Kruisbeek, Margulies, Shevach and Strober, Wiley-lnterscience, 2002)に記載のとおり行った。
【0117】
図1に示されるように、二重特異性一本鎖抗体の数個のドメイン配置、すなわち、CEAI VHVLx配列番号77VHVL(配列番号8)、CEAI VLVHx配列番号77VHVL(配列番号6)、CEAII VHVLx配列番号77VHVL(配列番号10)、CEAIII VLVHx配列番号77VHVL(配列番号12)およびCEAIII VHVLx配列番号77VHVL(配列番号14)は、ヒト膜結合CEAおよびヒトCD3と結合した。陰性対照として、培地ならびに1.および2.検出抗体を使用した。
【0118】
実施例3:CEAxCD3二重特異性一本鎖抗体と可溶性ヒトCEAの結合
可溶性ヒトCEAに対する特異性を決定するため、様々なCEAxCD3二重特異性一本鎖抗体をELISAで試験した。
【0119】
本目的のため、可溶性ヒトCEA抗原を最初にビオチン化した。ビオチン化は、5% DMSO(Sigma)を含有するPBS中、15倍モル過剰のEZ−Link Sulfo NHS−LC−LC−ビオチン(Pierce)とともに、1時間室温にてサンプルミキサー(Dynal)中で達成した。遊離ビオチンとビオチン化CEA抗原を分離するため、アッセイを標準的なプロトコールに従ってPBSに対して過剰に透析した。保持された、ビオチン標識CEAの生物活性をELISA結合実験で確認した。
【0120】
可溶性ヒトCEAに対するCEAxCD3二重特異性一本鎖抗体の特異性を測定するための直接ELISAを、標準的な手順に従って行った。要するに、50μl/ウェルのPBSまたは可溶性ビオチン化ヒトCEA(Abcam;1xPBS中5μg/ml)を、4℃にて約16時間インキュベートすることにより96ウェルストレプトアビジンコートELISAプレート(Nunc)上に固定化した。ウェルあたり200μlの水で洗浄した後、200μlのブロッキング溶液(PBS/3% BSA)を添加した。室温にて1時間ブロッキングした後、ブロッキング溶液を除去した。その後の洗浄段階(200μl/ウェルの1xPBS/0.05%(v/v) Tween20)およびインキュベーション段階はすべて室温にて行った。1回洗浄した後、CEAI VHVLx配列番号77VHVL(配列番号8;mAb A5B7に由来する抗CEA部分)、CEAII VHVLx配列番号77VHVL(配列番号10;mAb T84.66に由来する抗CEA部分)、およびCEAIII VHVLx配列番号77VHVL(配列番号14;mAb MFE−23に由来する抗CEA部分)二重特異性一本鎖抗体と、マウスモノクローナル抗体CEA/CD66 Ab−3(Col−1;Dunn)を異なる濃度(1xPBS中0.5μg/mlおよび5μg/ml;50μl/ウェル)中で1時間インキュベートした。1xPBS(50μl/ウェル)を非特異的結合の対照として添加した。3回の洗浄段階の後、50μl/ウェルのPenta−His IgG(Qiagen;1xPBS中2μg/ml)をHis−タグ二重特異性一本鎖抗体の検出のために添加した。その後、ウェルを3回洗浄し、50μlの西洋ワサビペルオキシダーゼ標識されたヤギ抗マウスFcγ特異的抗体(Jackson ImmuneResearch;1xPBS中1:1000)とともに1時間インキュベートした。3回洗浄した後、ELISAはABTS基質溶液(Roche)を添加することにより発色(develop)させ、405nmの波長で吸光度を測定した。
【0121】
図2は、ELISAで検出された様々な二重特異性一本鎖抗体の吸光度を示す。CEAI VHVLx配列番号77VHVL(配列番号8;mAb A5B7に由来する抗CEA部分)、CEAII VHVLx配列番号77VHVL(配列番号10;mAb T84.66に由来する抗CEA部分)、およびCEAIII VHVLx配列番号77VHVL(配列番号14;mAb MFE−23に由来する抗CEA部分)二重特異性一本鎖抗体と、マウスモノクローナル抗体Col−1は、可溶性ヒトCEAと特異的に結合した。CEA抗原の不在下(PBS対照)で、結合シグナルは観察されなかった。要約すると、mAb A5B7、T84.66およびMFE−23に由来する抗CEA結合ドメインは、可溶性および膜結合CEAの両方と結合する。
【0122】
実施例4:CEAxCD3二重特異性一本鎖抗体の生物活性
作製したCEAxCD3二重特異性一本鎖抗体の生物活性を、標的細胞としてヒト胃癌細胞株KatoIIIまたはヒトCEAトランスフェクトCHO細胞、およびエフェクター細胞として刺激を受けたヒトCD8陽性T細胞または未変性のPBMCをそれぞれ用いるインビトロクロム放出細胞障害性アッセイにより分析した。
【0123】
刺激を受けたCD8+T細胞の作製は、以下の通り行った。
ペトリ皿(直径145mm、greiner bio−one)を抗CD3抗体(OKT3 Janssen−Cilag GmbH,Orthoclone1mg/ml;終濃度1μg/ml)および抗CD28抗体(BD、1mg/ml;終濃度1μg/ml)で37℃にて1時間プレコートした。インキュベーション時間の後、結合しなかったタンパク質をPBSでの1回の洗浄段階により除去した。新鮮なPBMCを、標準的なプロトコールに従ってフィコール勾配遠心分離により末梢血(30〜50ml)から単離した。3〜5×107PBMCを、150ml RPMI1640/10% FCS/IL−2 20U/ml(Proleukin,Chiron)中のプレコートしたペトリ皿に添加し、2日間刺激した。3日目に細胞を回収し、RPMI1640で1回洗浄し、大きなTフラスコに移した。IL−2を20U/mlの終濃度に添加し、再び1日間培養した。マニュアルの指示に従ってCD8陰性単離キット(Dynal Biotech)を使用してCD8+CTLを単離した。未変性のPBMCは、フィコール勾配遠心分離の後に刺激手順を踏まずに直接用いた。標的細胞をPBSで2回洗浄し、50%FCSを含む最終容量100μlのRPMI中で37℃にて45分間、11.1MBq51Crで標識した。その後、標識した標的細胞を5mlのRPMIで3回洗浄し、次に細胞障害性アッセイで使用した。アッセイは、E:T比が10:1(1ウェルあたり5000個の標的細胞および50000個のエフェクター細胞に相当)のRPMI(上記の通り)を補給した96丸底プレートにおいて総量250μlで行った。構築物の評価のため、アッセイ容量中出発濃度1μg/mlの二重特異性一本鎖分子および12個のその三倍希釈液を用いた。アッセイ時間は18時間であり、細胞障害性は、最大溶解(Triton−Xの添加)および自発的溶解(エフェクター細胞なし)の差に関連した、上清中に放出されたクロムの相対値として測定された。全ての測定は三つ組みで行われた。上清中のクロム活性の測定は、Wizard3ガンマカウンター(Perkin Elmer Life Sciences GmbH,Koln,Germany)で行った。実験データの分析は、Prism4 for Windows(登録商標)(バージョン4.02、GraphPad Software Inc.,San Diego,California,USA)を用いて行った。シグモイド型用量応答曲線は、一般に、ソフトウェアで測定してR2値が>0.90である。分析プログラムにより計算されるEC50値を生物活性の比較に用いた。
【0124】
図3は、様々なドメイン配置、すなわち、配列番号77VHVLxCEAI VHVL(配列番号4)および配列番号77VHVLxCEAI VLVH(配列番号2)(両構築物はN末端に抗CD3部分を含む)、ならびにCEAI VLVHx配列番号77VHVL(配列番号6)およびCEAI VHVLx配列番号77VHVL(配列番号8)(C末端に抗CD3を含む)についてのヒトCEAトランスフェクト標的細胞(CHO−CEA+細胞)に対する細胞障害活性を示す。非トランスフェクトCHO細胞(ヒトCEAを欠く)を陰性対照として用いた。
【0125】
図4では、CEAI VHVLx配列番号77VHVL(配列番号8)、CEAIII VLVHx配列番号77VHVL(配列番号12)、CEAIII VHVLx配列番号77VHVL(配列番号14)、およびCEAII VHVLx配列番号77VHVL(配列番号10)は、ヒトCEAトランスフェクトCHO細胞に対する細胞障害活性を提示した。非トランスフェクトCHO細胞(ヒトCEAを欠く)を陰性対照として用いた。上に述べたように、CEAIは、マウスmAb A5B7に由来する可変領域を意味し、CEAIIは、マウスmAb T84.66に由来する可変領域であり、CEAIIIは、マウスmAb MFE−23由来の可変領域を指す。
【0126】
要約すると、試験した構築物はすべて、可溶性ヒトCEAの不在下で、(ヒト)CEA発現KatoIIIおよびヒトCEAトランスフェクトCHO細胞に対する細胞障害活性を示した。
【0127】
実施例5:可溶性CEA抗原の存在下で特異的なCEAxCD3二重特異性一本鎖抗体の生物活性
競合アッセイを、次の例外:CEAxCD3二重特異性一本鎖抗体の生物活性を様々な濃度の可溶性ヒトCEA抗原の存在下で試験する、を除いて実施例4に記載のとおり行う。使用した可溶性ヒトCEA抗原(AbCAM Ltd.Cambridge UK)を単独の患者の肝臓からの転移性結腸癌から単離した。細胞を添加する前に、所定量の二重特異性一本鎖抗体を漸増量の可溶性ヒトCEA抗原(0μg/ml;0.1μg/ml;1μg/ml、または0μg/ml;0.004μg/ml;0.02μg/ml;0.1μg/ml;0.5μg/ml;1μg/mlのいずれか)で37℃にて30分間プレインキュベーションすることにより実験的に競合アッセイを行った。残りのアッセイは、実施例4に記載されるとおり行った。これらの競合実験の結果を図5〜11に示す。
【0128】
ヒトCEA陽性標的細胞に対する二重特異性一本鎖抗体、mAb T84.66に由来する抗CEA部分を含むCEAII VHVLx配列番号77VHVL(配列番号10;図7および9〜11)、およびmAb MFE−23に由来する抗CEA領域を含むCEAIII VLVHx配列番号77VHVL(配列番号12;図8参照)の細胞障害活性は、漸増量の可溶性ヒトCEA抗原により大幅に阻害された。上に述べたように、前記構築物は膜結合と可溶性ヒトCEA抗原の両方と結合する;例えば、実施例2および3ならびに図1および2参照。従って、最も確かなことには、可溶性ヒトCEAは、前記二重特異性一本鎖抗体の抗CEA部分が標的細胞、例えば、CHO−CEA+またはKatoIII腫瘍細胞上の膜結合ヒトCEAと結合することを阻害し、それにより前記抗体構築物により媒介される細胞障害活性を阻害する。
【0129】
それに対して、驚くことに、mAb A5B7に由来する抗CEA部分を有する二重特異性一本鎖抗体が可溶性ヒトCEAに対して抵抗性であることが見出された。例えば、CEAI VLVHx配列番号77VHVL(配列番号6;図5参照)、CEAI VHVLx配列番号77VHVL(配列番号8;図5、8および10参照)、配列番号77VHVLxCEAI VHVL(配列番号4;図6参照)および配列番号77VHVLxCEAI VLVH(配列番号2;図6参照)は、漸増量の可溶性ヒトCEAにより阻害されず、高い濃度(1μg/ml)によってさえも阻害されない。これは、前記二重特異性一本鎖抗体の抗CEA部分が可溶性ヒトCEAと結合するという事実を考慮すると予測することができなかった(図2)。むしろ、漸増量の可溶性ヒトCEAによるCEA陽性標的細胞に対する細胞障害活性の阻害を、T84.66由来構築物およびMFE−23由来構築物の場合と同じように、予測することができた;上記参照。
【0130】
従って、本発明は、高いレベルの可溶性CEA抗原の存在下でさえ細胞障害性抗腫瘍活性を有する医薬組成物を提供する。そのため、これらの医薬組成物はその血漿中の可溶性CEA抗原濃度の高い腫瘍患者、例えば、進行性上皮性腫瘍、転移性上皮性腫瘍、高腫瘍量、後期上皮性腫瘍の患者またはELISAで測定してCEA血清濃度が100ng/mlよりも高い腫瘍患者の治療に特に適している。
【0131】
実施例6:ヒトVL領域の選択
癌患者に投与される際に免疫原性の低下した医薬組成物を提供するため、可溶性CEA抗原に対する抵抗性を有するヒト二重特異性一本鎖抗体を作製した。第1段階で、可溶性CEAに対する抵抗性を有するヒトVL領域が単離された。従って、この実験の目的は、モノクローナル抗体(mAb)A5B7の母親由来のマウスVHと対となり得るヒトVL領域の選択である。
【0132】
1.可溶性ヒトCEA抗原のビオチン化
ファージライブラリー選択のため、可溶性CEA抗原をビオチン化した。ビオチン化は、5% DMSO(Sigma)を含有するPBS中で、サンプルミキサー(Dynal)中で室温にて1時間15倍モル過剰のEZ−Link Sulfo NHS−LC−LC−ビオチン(Pierce)を用いて達成した。遊離ビオチンとビオチン化CEA抗原の分離のため、標準的なプロトコールに従って、アッセイをPBSに対して過剰に透析した。ビオチン標識したCEAの保持された生物活性をELISA結合アッセイで確認した。
【0133】
2.RNAのヒトB細胞からの単離
100mLの血液を5名の健康なヒトドナーから採取した。末梢血単核細胞(PBMC)を標準的な方法に従ってフィコール勾配により単離した。RNeasy(登録商標)Midi Kit(QIAGEN)を製造業者の説明書に従って用いて、全RNAを単離細胞から単離した。標準的な方法に従ってcDNAを合成した(Sambrook, Cold Spring Harbor Laboratory Press 1989, 2001)。
【0134】
3.可変軽鎖領域(VL−領域)のPCR増幅
軽鎖V領域DNAの単離のため、RT−PCRを、V−κ−(5’−hu−VK1−SacI−2001(5’−GAGCCGCACG AGCCCGAGCT CCAGATGACC CAGTCTCC−3’)(配列番号78)、5’−hu−VK2/4−SacI−2001(5’−GAGCCGCACG AGCCCGAGCT CGTGATGACY CAGTCTCC−3’)(配列番号79)、5’−hu−VK3−SacI−2001(5’−GAGCCGCACG AGCCCGAGCT CGTGWTGACR CAGTCTCC−3’)(配列番号80)、5’−hu−VK5−SacI−2001(5’−GAGCCGCACG AGCCCGAGCT CACACTCACG CAGTCTCC−3’)(配列番号81)、5’−hu−VK6−SacI−2001(5’−GAGCCGCACG AGCCCGAGCT CGTGCTGACT CAGTCTCC−3’)(配列番号82)、3’−hu−Vk−J1−SpeI−BsiWI(5’−GACGACACTA GTTGCAGCCA CCGTACGTTT GATTTCCACC TTGGTCC−3’)(配列番号83)、3’−hu−Vk−J2/4−SpeI−BsiWI(5’−GACGACACTA GTTGCAGCCA CCGTACGTTT GATCTCCASC TTGGTCC−3’)(配列番号84)、3’−hu−Vk−J3−SpeI−BsiWI(5’−GACGACACTA GTTGCAGCCA CCGTACGTTT GATATCCACT TTGGTCC−3’)(配列番号85)、3’−hu−Vk−J5−SpeI−BsiWI(5’−GACGACACTA GTTGCAGCCA CCGTACGTTT AATCTCCAGT CGTGTCC−3’)(配列番号86))およびVλ(5’−huVL1a−SacI−2001(GAG CCG CAC GAG CCC GAG CTC GTG TTG ACG CAG CCG CCC TC)(配列番号87)、5’−huVL1b−SacI−2001(GAG CCG CAC GAG CCC GAG CTC GTG CTG ACT CAG CCA CCC TC)(配列番号88)、5’−huVL2−SacI−2001(GAG CCG CAG GAG CCC GAG CTC GCC CTG ACT CAG CCT SCC TCC GT)(配列番号89)、5’−huVL4−SacI−2001(ACC TGC GAG CTC GTG CTG ACT CAR YCM YCC TCT GC)(配列番号90)、5’−huVL5−SacI−2001(ACC TGC GAG CTC GTG CTG ACT CAG CCR SCT TCC)(配列番号91)、5’−huVL6−SacI−2001(ACC TGC GAG CTC ATG CTG ACT CAG CCC CAC TC)(配列番号92)、5’−huVL3/9−SacI−2001(GAG CCG CAC GAG CCC GAG CTC GWG CTG ACT CAG CCA CCY TC)(配列番号93)、5’−huVL7/8−SacI−2001(GAG CCG CAC GAG CCC GAG CTC GTG GTG ACY CAG GAG CCM TC)(配列番号94)、3’−hu−Vlam−BlnI−SpeI−2001(CGT GGG ACT AGT CTT GGG CTG ACC TAG GAC GGT)(配列番号95)、3’−hu−Vlam2−BlnI−SpeI−2002:CGT GGG ACT AGT CTT GGG CTG ACC GAG GAC GGT)(配列番号96)プライマーセットを用いて行った。
【0135】
ヒトB細胞由来のRNAをcDNAに転写し(上記の通り)、PCR反応の鋳型DNAとして用いた。1回のPCR反応につき、1個の5’−プライマーを1個の3’−プライマーと組み合わせた。異なるPCR反応の数を、5’−および3’−プライマーの可能な組み合わせの数により決定した。増幅には次のPCRプログラムを用いた。すなわち94℃にて15秒間の変性、52℃にて50秒間のプライマーアニーリングおよび72℃にて90秒間のプライマー伸張を40サイクル以上行い、それに続いて72℃にて10分間の最終伸張を行った。次に、軽鎖DNA V断片を標準的なプロトコールに従って単離した。
【0136】
4.ライブラリー構築−ヒトVLプールのクローニング
ファージディスプレイライブラリーは、概して、例えば「Phage Display: A Laboratory Manual」; Ed. Barbas, Burton, Scott & Silverman; Cold Spring Harbor Laboratory Press, 2001に開示されるような標準的な手順に基づいて構築された。
【0137】
PCR増幅のために選択されるプライマーは、軽鎖V断片の5’−SacIおよび3’−SpeI認識部位を生じた。4回の連結反応をセットアップし、各々400ngの軽鎖断片(SacI−SpeI消化、2×κおよび2×λ)および1400ngのファージミドpComb3H5BHis(SacI−SpeI消化;大きな断片;このベクターは、Ralf Lutterbuese博士の論文に記載されている)で構成された。次に、得られる4つの抗体V軽鎖プールを、エレクトロポレーション(2.5kV、0.2cmギャップキュベット、25mF、200 Ohm、Biorad gene−pulser)により300μlのエレクトロコンピテント(electrocompetent)大腸菌(Escherichia coli)XL1 Blue株に各々形質転換して、
κ1:2×108
κ2:6×107
λ1:9×107
λ2:6×107
の独立したクローンからなるライブラリーサイズを得た。
【0138】
様々なPCR増幅から得たκ(軽鎖)DNA断片を、各連結について次のように秤量した。それぞれの5’−プライマーは特定のグループを定義する。これらのグループの中で、3’−プライマーはサブグループを定義する。κサブグループを、1:2:1:1(プライマー3’−hu−Vk−J1−SpeI−BsiWI:3’−hu−Vk−J2/4−SpeI−BsiWI:3’−hu−Vk−J3−SpeI−BsiWI:3’−hu−Vk−J5−SpeI−BsiWIに対応)に秤量した。グループを、それらの生殖系列分布1:1:1:0.2:0.2(プライマー5’−huVK1−Sac−2001:5’−huVK3−Sac−2001:5’−huVK2/4−Sac−2001:5’−huVK5−Sac−2001:5’−huVK6−Sac−2001に対応)に従って秤量した。
【0139】
異なるPCR増幅から得たλ(軽鎖)DNA断片を、各連結について次のように秤量した。それぞれの5’−プライマーは、特定のグループを定義する。これらのグループの中で、3’−プライマーはサブグループを定義する。λサブグループを、3:1(プライマー3’−hu−Vlam−BlnI−SpeI−2001:3’−hu−Vlam2−BlnI−SpeI−2002に対応)に秤量した。
【0140】
グループを、それらの生殖系列分布1:1:2:2:2:3(プライマー5’−huVL1a−SacI−2001:5’−huVL1b−SacI−2001:5’−huVL2−SacI−2001:5’−huVL4−SacI−2001+5’−huVL5−SacI−2001:5’−huVL6−SacI−2001+5’−huVL7/8−SacI−2001:5’−huVL3/9−SacI−2001に対応)に従って秤量した。
【0141】
エレクトロポレーション後、それぞれの反応液を表現型発現のためにSOC培養液(Fluka)中でインキュベートした。2つのκ培養物を組み合わせ、同様に2つのλ培養物も組み合わせた。得られるκ培養物および得られるλ培養物を、次にそれぞれ500mLの、50μg/ml カルベニシリンおよび2% w/v グルコースを含有するSB選択培地中で一晩インキュベートした。翌日、細胞を遠心分離により回収し、市販のプラスミド調製キット(Qiagen)を用いてプラスミドの調製を行った。
【0142】
5.抗体ライブラリーの構築−ヒトVL−母親由来VH
PCRを行ってmAb A5B7の母親由来のVHを前記母親由来のVHを含有するベクターから増幅した。増幅のために、5’−プライマー5’−AVH−Xho I(5’−GTC ACA CTC GAG TCA GGA GGA GGC TTG GTA C−3’)(配列番号97)および3’−プライマー3’−AVH−BstEII(5’−GTC ACA GGT GAC CGT GGT CCC TTG GCC CCA G−3’(配列番号98)を用いて標準的手順に従うPCRプロトコールに従った。分析用アガロースゲルからの約350bp増幅産物の精製後、DNA断片を制限酵素BstEIIおよびXho Iで切り出した。ファージミドpComb3H5BHis(このベクターはRalf Lutterbuese博士の論文に記載されている。)をしかるべく消化させ、大きな断片を上述の断片と連結させた。大腸菌XL1 blueへの形質転換後、単一のクローンを100mL SB培地(50μg/ml カルベニシリン含有)で培養し、プラスミドを標準的なプロトコールに従って調製した。挿入物を配列決定することにより首尾よいクローニングを確認した(Sequiserve,Munich)。
【0143】
このベクターpComb3H5BHis/mAb A5B7の母親由来VHを制限酵素SacIおよびSpeIで制限した。大きなベクター断片を単離した。プラスミド−DNAを含有するVκライブラリーおよびVλライブラリーを制限酵素SacIおよびSpeIで制限した。小さなVκ断片およびそれぞれのVλ断片(各およそ350bp)を標準的なプロトコールに従って単離した。1200ngのベクター断片を各200ngのVκおよびVλ断片両方の混合と連結した。連結反応物をエレクトロポレーション(2.5kV、0.2cmギャップキュベット、25mF、200 Ohm)により300μLのエレクトロコンピテント(electrocompetent)大腸菌XL1 Blue株に形質転換して、1.2×108の独立したクローンからなる全scFvライブラリーサイズを得た。
【0144】
表現型発現およびカルベニシリンへの緩やかな適応の後、抗体ライブラリーをSB−カルベニシリン(50μg/ml)選択培地へ移した。次に、抗体ライブラリーを感染量の1×1012粒子のヘルパーファージVCSM13で感染させ、その結果、繊維状のM13ファージの産生および分泌が得られ、この各ファージ粒子は半ヒトscFv−断片をコードする一本鎖pComb3H5BHis−DNAを含み、ファージコートタンパク質IIIへの翻訳融合として対応するscFv−タンパク質を提示した。
【0145】
6.ヒトVLのファージディスプレイ選択
scFv−レパートリーを有するファージ粒子をPEG8000/NaCl沈殿および遠心分離により培養上清から回収した。次に、約1×1011〜1×1012scFvファージ粒子を0.5mLのTBS/1% BSAに再懸濁し、ストレプトアビジンコートされたELISAプレート(Nunc)のウェル中で1時間固定されたビオチン化可溶性CEAとともにインキュベートした。抗原10μg/mlのPBS溶液(50μl)をストレプトアビジンコートされたウェル中で4℃にて一晩インキュベートし、水で1回洗浄し、それに続いてTBS中200μlの3% BSAで37℃にて1時間ブロッキングし、それをインキュベーション後に取り除いた。
【0146】
標的抗原と特異的結合しなかったscFvファージをTBS/0.05% Tweenでの洗浄段階により除去した。この洗浄手順は、さらなるラウンドで10回まで繰り返された。
【0147】
洗浄後、HCl−グリシン、pH 2.2を用いることにより結合体を溶出した。2M Tris、pH 12での中和後、溶出液を新鮮な非感染大腸菌XL1 Blue培養株の感染に用いた。
【0148】
残っている高結合体を溶出するため、50μlの新鮮な大腸菌XL1 blue培養株(OD600≧0.5)をウェルに添加し、15分間インキュベートした。次に、両方の培養物を混合し、ヒトscFv−断片をコードするファージミドコピーで首尾よく形質導入した細胞を再びカルベニシリン抵抗性について選択し、その後にVCMS13ヘルパーファージで感染させて、抗体提示およびインビトロでの選択の第2のラウンドを開始した。
【0149】
4ラウンドのパニングに対応するプラスミドDNAを大腸菌培養物から単離した。可溶性scFv−タンパク質の産生のため、VH−VL−DNA断片をプラスミドから切り出し(Xho I−SpeI)、プラスミドpComb3H5BFIag/His中の同じ制限部位を介してクローニングし、その発現構築物(例えばscFv)は、scFvとHis6タグとの間にFlagタグ(TGDYKDDDDK)(配列番号99)を含み、さらなるファージタンパク質が欠失している。
【0150】
連結後、各プール(異なるラウンドのパニング)のプラスミドDNAを100μL熱ショックコンピテント大腸菌TG1に形質転換し、カルベニシリンLB寒天上にプレーティングした。単一のコロニーを選定し、96ウェルプレート(Greiner)中の120μLのLBカルベニシリン(50μg/ml)1% グルコースに播種した。ウェルを半透膜(Greiner)で密封し、振盪インキュベータでプレートを37℃にて一晩インキュベートした(マスタープレート)。次に、10μlのマスタープレート培養物を、ウェルあたり90μL LBカルベニシリン(50μg/ml)0.1% グルコースを含有する第2の96ウェルプレート(ワーキングプレート)に移した。37℃の振盪インキュベータ中で4時間のインキュベーションの後、20μL LBカルベニシリン、6mM IPTGを各ウェルに添加することによりscFv生成を誘導した。振盪しながら30℃にて一晩の別のインキュベーション段階の後、細胞を、40μL 溶解緩衝液(400mM ホウ酸、320mM NaCl、4mM EDTA pH8、2.5mg/ml リゾチーム)での室温にて1時間のインキュベーションで溶解した。残りの細胞および細胞片を1.900×gで12分間の遠心分離により分離した(Hettich)。
【0151】
次に、scFv分子を含有する上清を、フローサイトメトリー結合アッセイで結合について試験した。ヒトCEAでトランスフェクトしたCHO細胞をCEA陽性細胞株として用いた。最初に100,000〜200,000個の細胞をヒトscFvまたは関連する対照を含有する周辺質調製物とともにインキュベートすることにより細胞結合アッセイを行った。インキュベーション後、細胞をPBS/1% FCS(ウシ胎児血清)中で洗浄し、さらに5〜10μg/mlの抗FLAG M2抗体(Sigma)とともにインキュベートした。細胞を再び洗浄した後、それらをポリクローナル、PE標識抗マウス抗体(Dianova)とともにインキュベートし、その後フローサイトメトリーにより分析した。およそ600個のクローンをCEA陽性のCHO細胞で結合シグナルについて試験した。27個の陽性クローンが得られた。それぞれのscFv DNAを配列決定した後、合計9個の異なる配列が得られた。
【0152】
図12は、フローサイトメトリー分析により測定される、9個の様々な半ヒトscFv(すなわち、マウスA5B7 VH−ヒトVL)構築物と多様な細胞株の結合を示す。前記図は複数の図表を含み、1つが試験された各構築物を表す。任意の所与図表中、黒色の分布は、いずれの構築物も不在下であるが、scFvの検出に用いられるあらゆる適切な検出剤を含むPBS単独でインキュベートした細胞の蛍光強度を示す。このようにして、観察されるいずれの蛍光シフトも間違いなく検出剤または緩衝液よりもscFv構築物に起因し得る。構築物とそれぞれの細胞株との結合の指標である蛍光のシフトは、各図表中灰色の線で示される。一般に、(黒色の)対照から離れてより高い強度のシフト、すなわち対照よりもはるかに遠いシフトはより強い結合を示し、一方、(黒色の)対照から離れてより低い強度のシフト、すなわち対照により近いシフトはより弱い結合を示す。
【0153】
図12から、構築物A−121、A−183、A−240、A−313、A−290、A−315、A4−35、A4−52、MP2−A5は、それぞれの対照と比較して、蛍光強度において明らかに識別できるシフトを示し、scFvとCHO標的細胞上の膜結合CEAとの結合を示すことが分かる。以下で、scFv A−240(配列番号48)のヒトVL領域を選択し、ヒトVH領域の単離に用いた。前記ヒトVL領域は、配列番号63(核酸配列)および配列番号64(アミノ酸配列)に示される。
【0154】
実施例7:可溶性CEA抗原に対して抵抗性のあるヒトVH領域の抗体ライブラリーの構築およびファージディスプレイ選択
以下の実験の目的は、実施例6に記載されるように選択される、scFv A−240のヒトVL領域と対となる可溶性CEA抗原に対して抵抗性のある一連のヒトVH領域の選択である。前記ヒトVL領域は、配列番号63(核酸配列)および配列番号64(アミノ酸配列)に示される。
【0155】
1.末梢血単核細胞(PBMC)からのRNAの単離
100mLの血液を5名の健康なヒトドナーから採取した。末梢血単核細胞(PBMC)を、標準的な方法に従ってフィコール勾配により単離した。全RNAを、RNeasy(登録商標)Midi Kit(QIAGEN)を製造業者の説明書に従って用いてPBMCから単離した。cDNAを標準的な方法に従って合成した(Sambrook, Cold Spring Harbor Laboratory Press 1989, 2001)。
【0156】
2.可変重鎖領域(VH領域)のPCR増幅
VHライブラリーを構築し、Lib 134−VHと名付けた。このVH−ライブラリーは、その後にヒトFR4生殖系列配列が続く母親由来の抗体のVHCDR3と作動可能なように連結された、PCR増幅された上記のPBMCプールのVH領域由来のFR1−CDR1−FR2−CDR2−FR3のヒトレパートリーで構成される。
【0157】
ヒト鋳型VH領域の単離のため、5’−VH特異的プライマーセット(5’−huVH1,3,5−Xho I−2001(5’−AGG TGC AGC TGC TCG AGT CTG G−3’)(配列番号100)、5’−huVH4−Xho I−2001(5’−CAG GTG CAG CTG CTC GAG TCG GG−3’)(配列番号101)、5’−huVH4B−Xho I−2001(5’−CAG GTG CAG CTA CTC GAG TGG GG−3’)(配列番号102))および2つの3’−VH特異的プライマーセット(3’−hu−VH−BstEII−2001(5’−CTG AGG AGA CGG TGA CC−3’)(配列番号103)、3’−hu−VH−J3−BstEII−2001(5’−CTG AAG AGA CGG TGA CC−3’)(配列番号104))を用いてRT−PCRを行った。1回のPCR反応につき、1個の5’−プライマーを1個の3’−プライマーと組み合わせた;異なるPCR反応の数は、5’−および3’−プライマーの可能な組み合わせの数により決定した。5つのドナーのPBMC cDNAを、VH遺伝子の供給源として用いた。次のPCRプログラム:94℃にて15秒間の変性、52℃にて50秒間のプライマーアニーリングおよび72℃にて60秒間のプライマー伸張を40サイクル以上行い、それに続いて72℃にて10分間の最終伸張、を増幅に用いた。約350bpのサイズの増幅産物を標準的な方法に従って単離した。
【0158】
Lib 134−VH領域の単離のため、RT−PCRを二段階で行った。最初に、ヒト重鎖VH−セグメント(FR1−CDR1−FR2−CDR2−FR3)を、上記と同じ5’−VH特異的プライマーセット(5’−huVH1,3,5−Xho I−2001、5’−huVH4−Xho I−2001、5’−huVH4B−Xho I−2001;配列番号100〜102)および3’−特異的プライマーセット(3’−A134−VH1A(5’−GTA GTC AAA GTA GAA CCG TAG CCC CCT ATC TCT YGC ACA GTA ATA CAC GGC−3’)(配列番号105)、3’−A134−VH1B(5’−GTA GTC AAA GTA GAA CCG TAG CCC CCT ATC TCT YGC ACA GTA ATA CAY RGC−3’)(配列番号106)、3’−A134−VH3A(5’−GTA GTC AAA GTA GAA CCG TAG CCC CCT ATC TCT TGY ACA GTA ATA CAC RGC−3’)(配列番号107)(示される「T」は「A」、「C」または「G」にも置き換えられる)、3’−A134−VH3B(5’−GTA GTC AAA GTA GAA CCG TAG CCC CCT ATC TCT TGC ACA GTA ATA CAA RGC−3’)(配列番号108)(示される「T」は「A」、「C」または「G」にも置き換えられる)、3’−A134−VH4(5’−GTA GTC AAA GTA GAA CCG TAG CCC CCT ATC TCT SGC ACA GTA ATA CAC RGC−3’)(配列番号109))を、FR3の非常に末端の領域で一致するヒトVHサブファミリー1、3および4に対して用いて、単離された鋳型VH断片からPCR増幅した。
【0159】
次のプライマーの組み合わせを用いた。
a)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH1A
b)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH1B
c)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH3A
d)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH3B
e)5’−huVH4−Xho I−2001 × 3’−A134−VH4
f)5’−huVH4B−Xho I−2001 × 3’−A134−VH4
【0160】
1回のPCR反応につき、1個の5’−プライマーを1個の3’−プライマーと組み合わせた;異なるPCR反応の数は、5’−および3’−プライマーの可能な組み合わせの数により決定した。次のPCRプログラム:94℃にて15秒間の変性、52℃にて50秒間のプライマーアニーリングおよび72℃にて90秒間のプライマー伸張を40サイクル以上行い、それに続いて72℃にて10分間の最終伸張、を増幅に用いた。このPCR段階およびそれぞれの3’−プライマー配列によって、ヒトVHセグメントが母親由来のVH CDR3の一部分について延長された。それは言い換えると次の2番目の段階のPCR3’−プライマーのプライミングサイトである。
【0161】
これらのVH−(FR1−CDR1−FR2−CDR2−FR3)DNA断片を、次に、それぞれの5’VH特異的プライマーと、増幅されたDNA断片の普遍的な3’−末端に一致する普遍的な3’プライマー(3’A134−JH6−BstEII、5’−CGA GAC GGT GAC CGT GGT CCC TTG GCC CCA GTA GTC AAA GTA GAA CCG TAG CC−3’)(配列番号110)を再び用いる2番目のPCR反応の鋳型として用いた。
【0162】
次のPCRプログラム:
94℃にて15秒間の変性、52℃にて50秒間のプライマーアニーリングおよび72℃にて60秒間のプライマー伸張を40サイクル以上行い、それに続いて72℃にて10分間の最終伸張、を増幅に用いた。DNA V断片を標準的なプロトコールに従って単離した。
【0163】
3.ライブラリー構築−ヒトVHプールのクローニング
前述の方法の第2ラウンドでは、1回目の前の選択(実施例6参照)で同定されたscFv A−240のヒトVLを選択し、それに続いてヒトscFvを作製する目的でヒトVH断片のライブラリーと合成した。ファージディスプレイライブラリーは、概して、例えば「Phage Display: A Laboratory Manual」;Ed. Barbas, Burton, Scott & Silverman; Cold Spring Harbor Laboratory Press, 2001に開示されるような標準的な手順に基づいて構築された。
【0164】
異なるPCR増幅からの重鎖DNA断片を各連結について次のように秤量した。
a:b:c:d:e:f=3:1:3:1:1:1、ここでa〜fは次の意味:
a)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH1A
b)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH1B
c)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH3A
d)5’−huVH1,3,5−Xho I−2001 × 3’−A134−VH3B
e)5’−huVH4−Xho I−2001 × 3’−A134−VH4
f)5’−huVH4B−Xho I−2001 × 3’−A134−VH4
を有する。
【0165】
400ngのヒトLib 134−VH断片プール(Xho I-BstEII消化)および1200ngのプラスミドpComb3H5BHis/A−240 VL(scFv A−240のVL領域をコードするDNAを、制限部位SacIおよびSpeIを介して標準手順に従ってpComb3H5BHisにクローニングした)で構成される、1回の連結反応を設定した。得られる抗体ヒトVHプールを、次に、300μLのエレクトロコンピテント大腸菌XL1 Blue株にエレクトロポレーション(2.5kV、0.2cmギャップキュベット、25mF、200 Ohm、Biorad gene−pulser)により形質転換し、合計で0.8×108の独立したクローンからなるライブラリーサイズを得た。
【0166】
エレクトロポレーション後、アッセイをSOC培養液(Fluka)中で表現型発現のためにインキュベートした。次に培養物を、50μg/ml カルベニシリンおよび2% v/v グルコースを含有する500mLのSB選択培地中で一晩それぞれインキュベートした。翌日、培養物の細胞を遠心分離により回収し、市販のプラスミド調製キット(QIAGEN)を用いてプラスミド調製を行ってDNAライブラリーを保存した。
【0167】
それぞれのscFvプールをコードする、1.5μgのこのプラスミドプールを、次に、大腸菌XL1 blue(2.5kV、0.2cmギャップキュベット、25mF、200 Ohm、Biorad gene−pulser)に電気穿孔して合計で2.4×109の独立したクローンからなるライブラリーサイズを得た。表現型発現およびカルベニシリンへの緩やかな適応の後、抗体ライブラリーをSB−カルベニシリン(50μg/ml)選択培地へ移した。次に、抗体ライブラリーを感染量の1×1012粒子のヘルパーファージVCSM13で感染させ、その結果、繊維状のM13ファージの産生および分泌が得られ、この各ファージ粒子はヒトscFv−断片をコードする一本鎖pComb3H5BHis−DNAを含み、ファージコートタンパク質IIIへの翻訳融合として対応するscFv−タンパク質を提示した。
【0168】
4.ヒトVHのファージディスプレイ選択
ヒトscFv−レパートリーを有するファージ粒子をPEG8000/NaCl沈殿および遠心分離により培養上清から回収した。次に、約1×1011〜1×1012scFvファージ粒子を0.5mLのTBS/1% BSAに再懸濁し、ストレプトアビジンコートされたELISAプレート(Nunc)のウェル中で1時間固定されたビオチン化可溶性CEAとともにインキュベートした。抗原10μg/mlのPBS溶液(50μl)をストレプトアビジンコートされたウェル中で4℃にて一晩インキュベートし、水で1回洗浄し、それに続いてTBS中200μlの3% BSAで37℃にて1時間ブロッキングし、それをインキュベーション後に取り除いた。
【0169】
標的抗原と特異的結合しなかったscFvファージをTBS/0.05% Tweenでの洗浄段階により除去した。この洗浄手順は、さらなるラウンドで10回まで繰り返された。
【0170】
洗浄後、HCl−グリシン、pH 2.2を用いることにより結合体を溶出した。2M Tris、pH 12での中和後、溶出液を新鮮な非感染大腸菌XL1 Blue培養株の感染に用いた。
【0171】
残っている高結合体を溶出するため、50μlの新鮮な大腸菌XL1 blue培養株(OD600≧0.5)をウェルに添加し、15分間インキュベートした。次に、両方の培養物を混合し、ヒトscFv−断片をコードするファージミドコピーで首尾よく形質導入した細胞を再びカルベニシリン抵抗性について選択し、その後にVCMS13ヘルパーファージで感染させて、抗体提示およびインビトロでの選択の第2のラウンドを開始した。
【0172】
4ラウンドのパニングに対応するプラスミドDNAを大腸菌培養物から単離した。可溶性scFv−タンパク質の産生のため、VH−VL−DNA断片をプラスミドから切り出し(XhoI−SpeI)、プラスミドpComb3H5BFlag/His中の同じ制限部位を介してクローニングし、その発現構築物(例えばscFv)は、scFvとHis6タグとの間にFlagタグ(TGDYKDDDDK)(配列番号99)を含み、さらなるファージタンパク質が欠失している。
【0173】
連結後、各プール(異なるラウンドのパニング)のプラスミドDNAを100μL熱ショックコンピテント大腸菌TG1に形質転換し、カルベニシリンLB寒天上にプレーティングした。単一のコロニーを選定し、96ウェルプレート(Greiner)中の120μLのLBカルベニシリン(50μg/ml)1% グルコースに播種した。ウェルに半透膜(Greiner)で密封し、インキュベータでプレートを37℃にて一晩インキュベートした(マスタープレート)。次に、10μlのマスタープレート培養物を、ウェルあたり90μL LBカルベニシリン(50μg/ml)0.1% グルコースを含有する第2の96ウェルプレート(ワーキングプレート)に移した。37℃の振盪インキュベータ中で4時間のインキュベーションの後、20μL LBカルベニシリン、6mM IPTGを各ウェルに添加することによりscFv生成を誘導した。振盪しながら30℃にて一晩の別のインキュベーション段階の後、細胞を、40μL 溶解緩衝液(400mM ホウ酸、320mM NaCl、4mM EDTA pH8、2.5mg/ml リゾチーム)での室温にて1時間のインキュベーションで溶解した。残りの細胞および細胞片は1,900×gで12分間の遠心分離により分離した(Hettich)。
【0174】
次に、scFv分子を含有する上清を、フローサイトメトリー結合アッセイで結合について試験した。
【0175】
ヒトCEAをトランスフェクトしたCHO細胞をCEA陽性細胞株として用いた。最初に100,000〜200,000個の細胞をヒトscFvまたは関連する対照を含有する周辺質調製物とともにインキュベートすることにより細胞結合アッセイを行った。インキュベーション後、細胞をPBS/1% FCS(ウシ胎児血清)中で洗浄し、さらに5〜10μg/mlの抗FLAG M2抗体とともにインキュベートした。細胞を再び洗浄した後、それらをポリクローナル、PE標識抗マウス抗体(Dianova)とともにインキュベートし、その後フローサイトメトリーにより分析した。46個のクローンをCEA陽性のCHO細胞で結合シグナルについて試験した。それらの全てが陽性のシグナルを示した。それぞれのscFv DNAを配列決定した後、合計9個の異なる配列が得られ、そのうちの8個が高度の相同性を示した。ヒト構築物MP510_3A−5.3(MP510−A5;配列番号50)、MP510_3−B9.1(MP511−B9;配列番号52)、MP510_3−D8.1(MP511−D8;配列番号54)が、さらなる特性決定のために選択された。対応するアミノ酸配列を配列表に示す。
【0176】
前記ヒト構築物MP510−A5、MP511−B9、MP511−D8ならびに半ヒト構築物A−240 Vλ.3(マウスVH A5B7/ヒトVLA240;配列番号48)の周辺質抽出物を、CEA陽性および陰性の細胞株でのフローサイトメトリー実験でさらに分析した。図13から、ヒト構築物MP510−A5(配列番号50)、MP511−B9(配列番号52)、MP511−D8(配列番号54)は、それぞれの半ヒト対照A−240 Vλ.3(マウスVH A5B7/ヒトVLA240;配列番号48)と比較して、明らかに識別できるシフトを蛍光強度で示すことが分かる。従って、ヒトscFv構築物は、膜結合ヒトCEAに対して半ヒト構築物A−240 Vλ.3よりも強い結合活性を示す。さらに、全てのヒト構築物は、CEA陽性のヒトKATO III細胞(ヒト胃癌細胞株)との明確な結合を示したのに対し、それらのどれもがCEA陰性の非トランスフェクトCHO細胞ならびにCEA陰性のヒトNALM6細胞(ヒトB細胞株)との結合を示さなかった(データは示さず)。
【0177】
実施例8:ヒトCEAxCD3二重特異性一本鎖抗体の作製および細胞障害活性
1.配置
次の段階では、ヒト二重特異性一本鎖抗体分子の様々なドメイン配置が作製された。これらの分子はヒト抗CEA結合ドメイン(その作製は上記実施例7に記載されている)を含み、配列番号77VHVLに示されるヒトCD3抗原に対して特異性を有する脱免疫化結合ドメインは表1に示されるように設計された;上記実施例2も参照。本明細書に記載されるヒト抗CEA結合ドメインを含む二重特異性一本鎖抗体構築物はELISAプレート上に固定化された可溶性CEA抗原で4ラウンドのファージディスプレイ選択後に単離されているので、それらはすべて(明白に)可溶性ヒトCEA抗原と結合する。
【0178】
特に、次の配置が作製された。
(a)N末端に位置するヒト抗CEA部分:
(i)VH−VL配向の抗CEA:
A5VH−A240VLx配列番号77VHVL(配列番号24)、
A5VH−A240VL#x配列番号77VHVL(配列番号126)、
B9VH−A240VLx配列番号77VHVL(配列番号32)、
B9VH−A240VL#x配列番号77VHVL(配列番号130)、
D8VH−A240VLx配列番号77VHVL(配列番号40)
D8VH−A240VL#x配列番号77VHVL(配列番号134)、および
CEAI VH−A240VLx配列番号77VHVL(配列番号16)。
(ii)VL−VH配向の抗CEA:
A240VL−A5VHx配列番号77VHVL(配列番号26)、
A240VL−B9VHx配列番号77VHVL(配列番号34)、
A240VL−D8VHx配列番号77VHVL(配列番号42)、
およびA240VL−CEAI VHx配列番号77VHVL(配列番号18)。
【0179】
(b)C末端に位置するヒト抗CEA部分:
(i)VH−VL配向の抗CEA:
配列番号77VHVLxA5VH−A240VL(配列番号30)、
配列番号77VHVLxA5VH−A240VL#(配列番号128)、
配列番号77VHVLxB9VH−A240VL(配列番号36)、
配列番号77VHVLxE12VH−A240VL(配列番号143)、
配列番号77VHVLxB9VH−A240VL#(配列番号132)、配列番号77VHVLxD8VH−A240VL(配列番号44)、
配列番号77VHVLxD8VH−A240VL#(配列番号136)、
および配列番号77VHVLxCEAI VH−A240VL(配列番号20)。
(ii)VL−VH配向の抗CEA:
配列番号77VHVLxA240VL−A5VH(配列番号28)、
配列番号77VHVLxA240VL−B9VH(配列番号38)、
配列番号77VHVLxA240VL−D8VH(配列番号46)、および
配列番号77VHVLxA240VL−CEAI VH(配列番号22)。
【0180】
CEAI VH(配列番号56)は、マウスmAb A5B7に由来するVH領域を意味し、一方、CEAI VHVLまたはCEAI VLVHは、mAb A5B7に由来するVH−VLドメインおよびVL−VHドメインをそれぞれ指す。A240はヒトVL領域に相当する(配列番号64および実施例7参照)。従って、例えば、CEAI VH−A240VLx配列番号77VHVL(配列番号16)は、mAb7由来のマウスVH領域およびヒトVL領域A240を有する半ヒトCEA結合ドメインを含む二重特異性構築物に相当する。ヒト抗体ライブラリーのクローニングのため、A240VL領域の元のN末端をコードする核酸配列は制限部位へ変換する必要があった。A5VH−A240VL#x配列番号77VHVL(配列番号126)、配列番号77VHVLxA5VH−A240VL#(配列番号128)、B9VH−A240VL#x配列番号77VHVL(配列番号130)、配列番号77VHVLxB9VH−A240VL#(配列番号132)、D8VH−A240VL#x配列番号77VHVL(配列番号134)、および配列番号77VHVLxD8VH−A240VL#(配列番号136)において、元のN末端が再導入された。
【0181】
二重特異性一本鎖抗体構築物である配列番号77VHVLxE12VH−A240VL(配列番号143)は、配列番号77VHVLxB9VH−A240VL(配列番号36)構築物とただ1つのアミノ酸残基が異なる。E12VHではCDR−H2配列は「FILNKANGGTTEYAASVKG」(配列番号145)と読まれるが、B9VHではそれは「FIRNKANGGTTEYAASVKG」(配列番号67)と読まれる。ヒトE12VH領域を実施例7に示すように単離した。
【0182】
2.発現、精製およびフローサイトメトリー分析
これらのヒトCEAxCD3二重特異性一本鎖抗体の発現、精製およびフローサイトメトリー分析は、上記実施例2に記載される方法により実行された。
【0183】
3.結合活性
図14に例示されるように、ヒト二重特異性一本鎖抗体構築物A5VH−A240VLx配列番号77VHVL(配列番号24)、B9VH−A240VLx配列番号77VHVL(配列番号32)、D8VH−A240VLx配列番号77VHVL(配列番号40)およびCEAI VH−A240VLx配列番号77VHVL(配列番号16)は、CEA陽性ヒトKatoIII細胞と結合し、かつCD3陽性HPB−ALL細胞と結合する。
【0184】
4.可溶性CEA抗原の不在下での細胞障害活性
CHO−CEA+標的細胞に対する細胞障害活性を、ヒト二重特異性一本鎖抗体の次のドメイン配置について実証した。
【0185】
N末端に位置する抗CEA部分を含む構築物と抗CEA部分のVH−VL配向について、図15は、A5VH−A240VLx配列番号77VHVL(配列番号24)、B9VH−A240VLx配列番号77VHVL(配列番号32)、D8VH−A240VLx配列番号77VHVL(配列番号40)およびCEAI VH−A240VLx配列番号77VHVL(配列番号16)の細胞障害活性を示す。図16は、A240VL−A5VHx配列番号77VHVL(配列番号26)、A240VL−B9VHx配列番号77VHVL(配列番号34)、A240VL−D8VHx配列番号77VHVL(配列番号42)、およびA240VL−CEAI VHx配列番号77VHVL(配列番号18)に関する抗CEAドメイン(N末端に位置する)のVL−VH配向に対する細胞障害活性を示す。
【0186】
C末端に位置する抗CEA部分を含む構築物と抗CEA部分のVH−VL配向について、図17は、配列番号77VHVLxA5VH−A240VL(配列番号30)、配列番号77VHVLxB9VH−A240VL(配列番号36)、配列番号77VHVLxD8VH−A240VL(配列番号44)、および配列番号77VHVLxCEAI VH−A240VL(配列番号20)に関するCEA+標的細胞に対する細胞障害性を提示する。配列番号77VHVLxCEAI LHを陽性対照として用いた。図18は、抗CEAドメイン(C末端に位置する)のVL−VH配向を含む構築物の配列番号77VHVLxA240VL−A5VH(配列番号28)、配列番号77VHVLxA240VL−B9VH(配列番号38)、配列番号77VHVLxA240VL−D8VH(配列番号46)、および配列番号77VHVLxA240VL−CEAI VH(配列番号22)に関する細胞障害活性を示す。配列番号77VHVLxCEAI LH(配列番号2)を陽性対照として用いた。
【0187】
図21は、ヒトB9VH領域とヒトA240VL領域を異なる配置で含む二重特異性一本鎖抗体構築物の細胞障害活性を示す。A240VL−B9VHx配列番号77VHVL(配列番号34)、配列番号77VHVLxA240VL−B9VH(配列番号38)、配列番号77VHVLxB9VH−A240VL(配列番号36)、およびB9VH−A240VLx配列番号77VHVL(配列番号32)は、ヒトCEAトランスフェクトCHO細胞に対する細胞障害活性を示した。
【0188】
5.ヒトCEAxCD3二重特異性一本鎖抗体の可溶性CEA抗原に対する抵抗性
可溶性ヒトCEA抗原の存在下、ヒト二重特異性一本鎖抗体についての競合アッセイを上記実施例5に示すように行った。図19および20は、構築物A5VH−A240VLx配列番号77VHVL(配列番号24)、B9VH−A240VLx配列番号77VHVL(配列番号32)、D8VH−A240VLx配列番号77VHVL(配列番号40)およびCEAI VH−A240VLx配列番号77VHVL(配列番号16)に例示されるように、ヒト構築物についても、細胞障害活性の可溶性CEA抗原に対する抵抗性を実証する;マウス由来二重特異性一本鎖抗体の可溶性CEAに対する抵抗性については、実施例5参照。さらに、図22は、CEA反応性の二重特異性一本鎖構築物A240VL−B9VHx配列番号77VHVL(配列番号34)が、漸増量の可溶性CEA抗原の存在下でT細胞を再指向してCHO−CEA+細胞を溶解させたことを示す。この実験は、A240VL−B9VHx配列番号77VHVLに媒介される細胞障害活性も可溶性CEAに対して抵抗性であることを実証する。最後に、図27から導かれるように、二重特異性一本鎖構築物である配列番号77VHVLxE12VH−A240VL(配列番号143)に媒介される細胞障害活性は、可溶性CEAに対して抵抗性である。
【0189】
従って、本発明は、高いレベルの可溶性CEA抗原の存在下でさえ細胞障害性抗腫瘍活性を有する医薬組成物を提供する。上に述べたように、上皮性腫瘍患者において血清CEA濃度が高いと、抗CEA抗体に基づく治療法の成功が低下する。従って、本発明の医薬組成物は、患者の血清/血漿中の高い可溶性CEA抗原濃度を特徴とする、進行性上皮性腫瘍(例えば、原発腫瘍の外科切除後の続発性、転移性腫瘍)、悪性上皮性腫瘍、高い(上皮性)腫瘍量および後期上皮性腫瘍の患者の治療に特に適している。さらに、本発明の医薬組成物はまた、低い投薬量で投与されることが予期される。それに加えて、前記医薬組成物は、本発明の医薬組成物中の二重特異性一本鎖抗体のヒト起源の抗CEA部分と脱免疫化された抗CD3部分に起因して、患者に投与された場合に免疫原性となる可能性は低い。さらに、本発明の医薬組成物は、低分子量および小型の二重特異性一本鎖構築物に起因して、高い腫瘍浸透をもたらすことが予期される。その上、腫瘍治療のためには、該分子の高い細胞障害活性のために少量の二重特異性一本鎖抗体が用いられる。患者には少量が投与されると予期されるため、患者に対する有害作用は低減されるとも予期される。最後に、可溶性CEAに対する抵抗性のない二重特異性一本鎖抗体は、上に説明したように、それらは低い濃度で投与されるため、腫瘍患者の血漿中の低い濃度の可溶性CEA抗原にさえも非常に感受性が高い。この問題もまた、本発明の医薬組成物により回避される。
【0190】
実施例9:高分解能陽イオン交換クロマトグラフィーによるA240VL−B9VHx配列番号77VHVL(配列番号34)の精製モノマーのタンパク質の同質性評価
精製されたA240VL−B9VHx配列番号77VHVL(配列番号34)構築物の同質性をさらに特性決定するため、単離モノマー画分を高分解能陽イオン交換クロマトグラフィーに付した。クロマトグラフィーは、20mM MESバッファーpH5.5で平衡化したMiniSカラム(Mini S PE 4.6/50 CatX 0.8ml;GE Healthcare 17−5005−01)で行われた。カラムに充填する前にサンプルを同じバッファーで1:3希釈した。結合したタンパク質を、60カラム量中に1M NaClを0〜30%含有する平衡バッファー勾配を用いて溶出させた。残りのタンパク質を3カラム量の1M NaClで溶出した。結果として得られるクロマトグラムは図23に示され、単一の主ピークを有する同質のタンパク質画分を提示する。
【0191】
実施例10:A240VL−B9VHx配列番号77VHVL(配列番号34)の血漿安定性
A240VL−B9VHx配列番号77VHVL(配列番号34)構築物の血漿安定性を、様々なインキュベーション条件下で、続いて実施例4に記載の標準的な51クロム放出に基づく細胞障害性アッセイにより試験した。
【0192】
5名の健康なドナーの血液を含むヒト血漿プールを、EDTAコートされたシリンジに血液を採取することにより作製した。細胞成分を遠心分離により取り除き、上部の血漿相を回収し、その後プールした。A240VL−B9VHx配列番号77VHVL(配列番号34)構築物を、血漿の存在下または不在下、37℃または4℃にてインキュベートした。対照として、構築物を細胞障害性アッセイの直前に血漿またはRPMI1640培地にそれぞれ希釈した。CHO−CEA+は標的細胞の役割を果たし;刺激を受けたCD8+T細胞をエフェクター細胞として用いた。効果:標的(E:T)比は10:1であった。このアッセイの持続時間は18時間であった。
【0193】
図24に示されるように、CEA反応性の二重特異性一本鎖構築物A240VL−B9VHx配列番号77VHVL(配列番号34)は、極めて安定性であることが実証された;37℃にて24時間ヒト血漿中でのインキュベーションの後、細胞障害活性の損失は検出することができなかった。
【0194】
実施例11:高分解能陽イオン交換クロマトグラフィーによる配列番号77VHVLxE12VH−A240VL(配列番号143)の精製モノマーのタンパク質同質性評価
この実験は、配列番号77VHVLxE12VH−A240VL(配列番号143)を分析することを除いて実施例9と同様に行った。結果として得られるクロマトグラムは図25に示され、単一の主ピークを有する同質のタンパク質画分を提示する。
【0195】
実施例12:配列番号77VHVLxE12VH−A240VL(配列番号143)の血漿安定性
この実験は、配列番号77VHVLxE12VH−A240VL(配列番号143)を分析することを除いて実施例10と同様に行った。図26に示されるように、CEA反応性の二重特異性一本鎖構築物である配列番号77VHVLxE12VH−A240VL(配列番号143)は極めて安定性であることが実証された;37℃にて24時間ヒト血漿中でのインキュベーションの後、細胞障害活性の損失は検出することができなかった。
【図面の簡単な説明】
【0196】
【図1】様々なヒトCEA反応性の二重特異性一本鎖構築物とヒトCEAトランスフェクトCHO細胞およびCD3陽性HPB−AII細胞とのそれぞれのFACS結合分析。CEA結合の陽性対照として、モノクローナル抗体CoI−1を使用した。ヒトCD3結合の陽性対照には、WO 99/054440号に記載のCD19xCD3二重特異性一本鎖構築物を使用した。この陽性対照では、太い線は、10μg/mlの精製CD19xCD3二重特異性一本鎖抗体とともにインキュベートし、その後に抗His抗体および検出抗体とともにインキュベートした細胞を表す。細い柱状グラフ線は、陰性対照:抗His抗体および検出抗体とともにインキュベートした細胞を反映する。ヒト(膜結合)CEAおよびヒトCD3の結合活性は、CEAI VHVLx配列番号77VHVL、CEAI VLVHx配列番号77VHVL、CEAII VHVLx配列番号77VHVL、CEAIII VLVHx配列番号77VHVLおよびCEAIII VHVLx配列番号77VHVLについて検出可能であった。本発明に記載される二重特異性一本鎖抗体に対応するそれぞれの柱状グラフにおいて、細い線は陰性対照を表し、太い輝線は培養上清とともにインキュベートした細胞を表し、一方、太い暗線(大部分が右側)は、10μg/mlの精製二重特異性一本鎖抗体とともにインキュベートした細胞を表す。
【図2】直接ELISAにより検出された、可溶性CEAに対する二重特異性一本鎖抗CEA/抗CD3抗体CEAI VHVLx配列番号77VHVL、CEAII VHVLx配列番号77VHVLおよびCEAIII VHVLx配列番号77VHVLならびに抗CEA抗体Col−1の結合シグナル。CEAI VHVLx配列番号77VHVL(mAb A5B7に由来する抗CEA結合ドメイン)、CEAII VHVLx配列番号77VHVL(mAb T84.66に由来する抗CEA結合ドメイン)、およびCEAIII VHVLx配列番号77VHVL(mAb MFE−23に由来する抗CEA結合ドメイン)二重特異性一本鎖抗体ならびにマウスモノクローナル抗体Col−1は、固定化された可溶性ヒトCEAと特異的に結合した。可溶性CEA抗原(PBS対照)の不在下で結合シグナルは観察されなかった。
【図3】示されるCEA反応性の二重特異性一本鎖構築物は、可溶性CEAの不在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。再指向による溶解(redirected lysis)の特異性を実証するため、非CEA反応性の二重特異性一本鎖構築物を陰性対照として含めた。ヒトCEAトランスフェクト標的細胞(CHO−CEA+細胞)に対する細胞障害活性は、様々なドメイン配置、すなわち、配列番号77VHVLxCEAI VHVLおよび配列番号77VHVLxCEAI VLVH(両構築物は抗CD3結合ドメインをN末端に含む)、ならびにCEAI VLVHx配列番号77VHVLおよびCEAI VHVLx配列番号77VHVL(抗CD3結合ドメインをC末端に含む)について示すことができた。非トランスフェクトCHO細胞(ヒトCEAを欠く)を陰性対照として用いた。
【図4】示されるCEA反応性の二重特異性一本鎖構築物は、可溶性CEAの不在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。再指向による溶解の特異性を実証するため、非トランスフェクトCHO細胞を陰性対照として含めた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。CEAI−HL(CEAI VHVLx配列番号77VHVL)、CEAIII−LH(CEAIII VLVHx配列番号77VHVL)、CEAIII−HL(CEAIII VHVLx配列番号77VHVL)、およびCEAII HL(CEAII VHVLx配列番号77VHVL)は、ヒトCEAトランスフェクトCHO細胞に対する細胞障害活性を示した。非トランスフェクトCHO細胞(ヒトCEAを欠く)をCEAI−LH(CEAI VLVHx配列番号77VHVL)、CEAIII−HL(CEAIII VHVLx配列番号77VHVL)およびCEAII−HL(CEAII VHVLx配列番号77VHVL)の陰性対照として用いた。CEAIは、マウスmAb A5B7に由来する可変領域を意味し、CEAIIは、マウスmAb T84.66に由来する可変領域であり、CEAIIIは、マウスmAb MFE−23由来の可変領域を指す。
【図5】示されるCEA反応性の二重特異性一本鎖構築物は、可溶性ヒトCEAの存在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。CEAI VLVHx配列番号77VHVLおよびCEAI VHVLx配列番号77VHVLに媒介される細胞障害活性は、1μg/mlまでの漸増量の可溶性ヒトCEAにより阻害されない。CEAIは、マウスmAb A5B7に由来する可変領域である。
【図6】示されるCEA反応性の二重特異性一本鎖構築物は、可溶性ヒトCEAの存在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。配列番号77VHVLxCEAI VHVLおよび配列番号77VHVLxCEAI VLVHに媒介される細胞障害活性は、1μg/mlまでの漸増量の可溶性ヒトCEAにより阻害されない。CEAIは、マウスmAb A5B7に由来する可変領域である。
【図7】示されるCEA反応性の二重特異性一本鎖構築物は、可溶性ヒトCEAの存在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。CEAII VHVLx配列番号77VHVLの細胞障害活性は、漸増量の可溶性CEAにより阻害された。CEAII VHVLは、mAb T84.66に由来する。
【図8】示されるCEA反応性の二重特異性一本鎖構築物は、可溶性ヒトCEAの存在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。CEAI VHVLx配列番号77VHVLに媒介される細胞障害性が可溶性CEA抗原による阻害に抵抗性であるのに対して、CEAIII VHVLx配列番号77VHVLに媒介される細胞障害活性は、少量の可溶性CEAにさえも阻害される。CEAIII VHVLは、mAb MFE−23に由来するのに対し、CEAIは、マウスmAb A5B7に由来する可変領域である。
【図9】示されるCEA反応性の二重特異性一本鎖構築物は、漸増量の可溶性CEA抗原の存在下で、T細胞を再指向してKatoIII細胞を溶解させた。未変性のヒトPBMCをエフェクター細胞として用いた。CEAII VHVLx配列番号77に媒介される細胞障害活性は、可溶性CEAに対して抵抗性でない。CEAII VHVLは、mAb T84.66に由来する。
【図10】示されるCEA反応性の二重特異性一本鎖構築物は、漸増量の可溶性CEA抗原の存在下で、T細胞を再指向してKatoIII細胞を溶解させた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。CEAI VHVLx配列番号77VHVLに媒介される細胞障害性は、可溶性CEAに対して抵抗性である。対照的に、CEAII VHVLx配列番号77VHVLに媒介される細胞障害活性は、漸増量の可溶性CEAにより阻害される。CEAII VHVLは、mAb T84.66に由来するのに対し、CEAIは、マウスmAb A5B7に由来する可変領域である。
【図11】示されるCEA反応性の二重特異性一本鎖構築物は、漸増量の可溶性CEA抗原の存在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。CEAII VHVLx配列番号77VHVLに媒介される細胞障害活性は、漸増量の可溶性CEAにより阻害される。CEAII VHVLは、mAb T84.66に由来する。
【図12】Flagがタグされた、選択されたクローン由来のscFvタンパク質断片を含有する周辺質調製物のフローサイトメトリー分析を示す。可溶性scFvタンパク質断片の周辺質調製物を100,000〜200,000個のCEAをトランスフェクトしたCHO細胞に添加した。検出のため、モノクローナル抗Flag抗体を用い、その後にPE標識ポリクローナル抗マウス抗体を用いた。細胞と結合しているScFvを、蛍光強度を増加させて、PBSのみでインキュベートした細胞と比較して測定した。蛍光強度をX軸にブロットし、イベントの数をY軸にブロットした。陰性対照(PBSおよび検出試薬)は黒く塗られた曲線で示し、個別のscFvは灰色の線で示す。右への移動は細胞との正の結合を示す。全てのscFv、すなわち、A−121、A−183、A240、A−313、A−290、A−315、A4−35、A4−52およびMP2−A5は、CHO細胞上で膜と結合したCEAと結合する。scFvの各々は、実施例6に記載されるように、マウスA5B7VH領域とヒトVL領域からなる。
【図13】Flagがタグされた、選択されたクローン由来のscFvタンパク質断片を含有する周辺質調製物のフローサイトメトリー分析を示す。可溶性scFvタンパク質断片の周辺質調製物を100,000〜200,000個のCEAをトランスフェクトしたCHO細胞に添加した。モノクローナル抗Flag抗体、続いてPE標識ポリクローナル抗マウス抗体で検出を行った。細胞と結合しているScFvを、蛍光強度を増加させて、PBSのみでインキュベートした細胞と比較して測定した。蛍光強度をX軸にブロットし、イベントの数をY軸にブロットした。陰性対照(PBSおよび検出試薬)は黒く塗られた曲線で示し、個別のscFvは灰色の線で示す。右への移動は細胞との正の結合を示す。完全ヒト化scFv構築物MP510_3A5.3、MP510_3−B9.1、およびMP510_3−D8.1はCHO細胞上で膜と結合したCEAと結合する。これらのscFvの各々は、実施例7に記載されるように、ヒトVH領域とヒトVL領域A240からなる。240Vλ.3は、マウスA5B7 VH領域とヒトVL A−240領域からなるscFvである。この構築物はまた、CEA結合活性を示す。
【図14】様々なヒトCEA反応性の二重特異性一本鎖構築物とKatoIII細胞およびHPB−AII細胞とのそれぞれのFACS結合分析。太い線は、抗His抗体および検出抗体とともにインキュベートした、トランスフェクトCHO細胞の細胞培養上清でインキュベートした細胞を表す。細い柱状グラフ線は、陰性対照:抗His抗体および検出抗体とともにインキュベートした細胞を反映する。ヒト二重特異性一本鎖抗体構築物A5VH−A240VLx配列番号77VHVL、B9VH−A240VLx配列番号77VHVL、およびD8VH−A240VLx配列番号77VHVLは、Kato細胞上でヒトCEAと、HPB−AII細胞上でヒトCD3と結合する。mAb A5B7に由来するCEA結合ドメインのVH領域を含むCEAI VH−A240VLx配列番号77VHVLは、同じ結合活性を示す。
【図15】可溶性CEAの不在下で、CEAでトランスフェクトしたCHO細胞に再指向された、示されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイ。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。細胞障害活性は、構築物A5VH−A240VLx配列番号77VHVL、B9VH−A240VLx配列番号77VHVL、D8VH−A240VLx配列番号77VHVLおよびCEAI VH−A240VLx配列番号77VHVLについて検出することができた。CEAI VHはmAb A5B7に由来するVH領域である。
【図16】可溶性CEAの不在下で、CEAをトランスフェクトしたCHO細胞に再指向された、示されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイ。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。この図は、A240VL−A5VHx配列番号77VHVL、A240VL−B9VHx配列番号77VHVL、A240VL−D8VHx配列番号77VHVL、およびA240VL−CEAI VHx配列番号77VHVLの細胞障害活性を実証する。
【図17】可溶性CEAの不在下で、CEAをトランスフェクトしたCHO細胞に再指向された、示されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイ。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。CEA+標的細胞に対する細胞障害性は、配列番号77VHVLxA5VH−A240VL、配列番号77VHVLxB9VH−A240VL、配列番号77VHVLxD8VH−A240VL、配列番号77VHVLxCEAI VH−A240VLおよび配列番号77VHVLxCEAI VLVHについて示される。
【図18】可溶性CEAの不在下で、CEAをトランスフェクトしたCHO細胞に再指向された、示されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイ。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。細胞障害活性は、配列番号77VHVLxA240VL−A5VH、配列番号77VHVLxA240VL−B9VH、配列番号77VHVLxA240VL−D8VH、および配列番号77VHVLxA240VL−CEAI VHおよび配列番号77VHVLxCEAI VLVHついて示される。
【図19】漸増量の可溶性CEA抗原の存在下で、CEAをトランスフェクトしたCHO細胞に再指向された、示されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイ。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。この図は、A5VH−A240VLx配列番号77VHVLおよびB9VH−A240VLx配列番号77VHVLについて例証されるように、可溶性CEA抗原に対するヒト二重特異性一本鎖抗体構築物の細胞障害活性の抵抗性を実証する。
【図20】漸増量の可溶性CEA抗原の存在下で、CEAをトランスフェクトしたCHO細胞に再指向された、示されたCEA反応性の二重特異性一本鎖構築物の細胞障害性アッセイ。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。ヒト二重特異性一本鎖抗体構築物D8VH−A240VLx配列番号77VHVLおよびCEAI VH−A240VLx配列番号77VHVLもまた、可溶性CEA抗原に対する抵抗性を示す。
【図21】示されるCEA反応性の二重特異性一本鎖構築物は、可溶性CEAの不在下で、T細胞を再指向してCEAをトランスフェクトしたCHO細胞を溶解させた。再指向による溶解の特異性を実証するため、非トランスフェクトCHO細胞を陰性対照として含めた。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。A240VL−B9VHx配列番号77VHVL、配列番号77VHVLxA240VL−B9VH、配列番号77VHVLxB9VH−A240VL、B9VH−A240VLx配列番号77VHVL、および配列番号77VHVLxCEAI VHVLは、ヒトCEAトランスフェクトCHO細胞に対する細胞障害活性を示した。
【図22】示されるCEA反応性の二重特異性一本鎖構築物は、漸増量の可溶性CEA抗原の存在下で、T細胞を再指向してCHO−CEA+細胞を溶解させた。刺激を受けたヒトCD8+T細胞をエフェクター細胞として用いた。A240VL−B9VHx配列番号77VHVLに媒介される細胞障害活性は、可溶性CEAに対して抵抗性である。
【図23】二重特異性一本鎖構築物A240VL−B9VHx配列番号77VHVLの高分解能陽イオン交換クロマトグラム、青色の線(上側の曲線)は、タンパク質の全体的な電荷イソ型を示す。構築物の高同質性を示す、1つの単一ピークが検出された。
【図24】ヒト血漿における24時間のインキュベーション後の細胞障害性の評価に基づくタンパク質安定性アッセイ。CEA反応性の二重特異性一本鎖構築物は、可溶性CEAの不在下で、CEAをトランスフェクトしたCHO細胞に再指向された。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。この図は、ヒト血漿中の二重特異性一本鎖構築物A240VL−B9VHx配列番号77VHVLの血漿安定性を実証する。構築物の細胞障害活性は、生理的条件下で、血漿タンパク質の影響を受けない。
【図25】二重特異性一本鎖構築物である配列番号77VHVLxE12VH−A240VLの高分解能陽イオン交換クロマトグラム、青色の線(上側の曲線)は、タンパク質の全体的な電荷イソ型を示す。この図は、二重特異性一本鎖構築物である配列番号77VHVLxE12VH−A240VLの同質性を実証する。
【図26】ヒト血漿における24時間のインキュベーション後の細胞障害性の評価に基づくタンパク質安定性アッセイ。CEA反応性の二重特異性一本鎖構築物は、可溶性CEAの不在下で、CEAをトランスフェクトしたCHO細胞に再指向された。刺激を受けたヒトCD8陽性CTLをエフェクター細胞として用いた。この図は、ヒト血漿中の二重特異性一本鎖構築物である配列番号77VHVLxE12VH−A240VLの血漿安定性を実証する。構築物の細胞障害活性は、生理的条件下で、血漿タンパク質の影響を受けない。
【図27】示されるCEA反応性の二重特異性一本鎖構築物は、漸増量の可溶性CEA抗原の存在下で、T細胞を再指向してCHO−CEA+細胞を溶解させた。刺激を受けたヒトCD8+T細胞をエフェクター細胞として用いた。二重特異性一本鎖構築物である配列番号77VHVLxE12VH−A240VLは、可溶性CEAに対して抵抗性である細胞障害活性を媒介した。
【特許請求の範囲】
【請求項1】
ヒトにおける上皮性腫瘍の治療のための医薬組成物であって、前記医薬組成物が、
(a)ヒトCD3と特異的に結合する第1の結合ドメイン、および
(b)ヒトCEAと特異的に結合する第2の結合ドメインを有する二重特異性一本鎖抗体を含み、
前記第2の結合ドメインが少なくともアミノ酸配列「DX1X2X3X4FYFDY」(配列番号65)を含み、「X1」、「X2」、「X3」または「X4」は任意のアミノ酸残基を表し、アミノ酸残基「D」はマウスモノクローナル抗体A5B7のCDR−H3のKabat位置95に相当し、アミノ酸残基「FYFDY」は、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置100、100a、100b、101、および102にそれぞれ相当する、医薬組成物。
【請求項2】
「X1」が、「R」(アルギニン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;
「X2」が、「G」(グリシン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表し;
「X3」が、「L」(ロイシン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;かつ
「X4」が、「R」(アルギニン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表す、
請求項1に記載の医薬組成物。
【請求項3】
ヒトCEAに特異的な前記第2の結合ドメインが、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102に相当するアミノ酸配列「DRGLRFYFDY」(配列番号66)を少なくとも含む、請求項1または2に記載の医薬組成物。
【請求項4】
CD3に特異的な前記第1の結合ドメインがC末端に位置している、請求項1から3のいずれか一項に記載の医薬組成物。
【請求項5】
前記結合ドメインが、VHCEA−VLCEA−VHCD3−VLCD3またはVLCEA−VHCEA−VHCD3−VLCD3の順序で配置されている、請求項1から4のいずれかに記載の医薬組成物。
【請求項6】
ヒトCEAに特異的な前記第2の結合ドメインが、アミノ酸配列「SYWMH」(配列番号68)を有するCDR−H1および/またはアミノ酸配列「FIRNKANGGTTEYAASVKG」(配列番号67)もしくは「FILNKANGGTTEYAASVKG」(配列番号145)を有するCDR−H2を含む、請求項1から5のいずれかに記載の医薬組成物。
【請求項7】
ヒトCEAに特異的な前記第2の結合ドメインが、アミノ酸配列「TYAMH」(配列番号70)を有するCDR−H1および/またはアミノ酸配列「LISNDGSNKYYADSVKG」(配列番号69)を有するCDR−H2を含む、請求項1から5のいずれかに記載の医薬組成物。
【請求項8】
ヒトCEAに特異的な前記第2の結合ドメインが、アミノ酸配列「TLRRGINVGAYSIY」(配列番号73)を有するCDR−L1および/またはアミノ酸配列「YKSDSDKQQGS」(配列番号72)を有するCDR−L2および/またはアミノ酸配列「MIWHSGASAV」(配列番号71)を有するCDR−L3を含む、請求項1から7のいずれかに記載の医薬組成物。
【請求項9】
ヒトCEAに特異的な第2の結合ドメインのVH領域のアミノ酸配列が、配列番号60または146である、請求項6に記載の医薬組成物。
【請求項10】
ヒトCEAに特異的な第2の結合ドメインのVH領域のアミノ酸配列が配列番号58または配列番号62である、請求項7に記載の医薬組成物。
【請求項11】
ヒトCEAに特異的な第2の結合ドメインのVL領域のアミノ酸配列が、配列番号64である、請求項8に記載の医薬組成物。
【請求項12】
CEAに特異的な第2の結合ドメインのV領域が、以下、
(a)配列番号60に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;
(b)配列番号146に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;
(c)配列番号58に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;
(d)配列番号62に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;ならびに
(e)配列番号56に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域
からなる群より選択される、請求項1から11のいずれか一項に記載の医薬組成物。
【請求項13】
前記二重特異性一本鎖抗体が、
(a)配列番号6、8、16、18、24、26、32、34、40、42、126、130、134または143のいずれか1つに表されるアミノ酸配列;
(b)配列番号5、7、15、17、23、25、31、33、39、41、125、129、133または142に示される核酸配列によりコードされるアミノ酸配列;
(c)ストリンジェントな条件下で(b)の相補的な核酸配列とハイブリダイズする核酸配列によりコードされるアミノ酸配列;
(d)(b)の核酸配列に対する遺伝暗号の結果として縮重した核酸配列によりコードされるアミノ酸配列;および
(e)(a)または(b)のアミノ酸配列と少なくとも85%同一、より好ましくは少なくとも90%同一、最も好ましくは少なくとも95%同一であるアミノ酸配列
からなる群より選択されるアミノ酸配列を含む、請求項1から12のいずれか一項に記載の医薬組成物。
【請求項14】
前記上皮性腫瘍が、胃腸腺癌、乳腺癌または肺腺癌である、請求項1〜13のいずれかに記載の医薬組成物。
【請求項15】
前記胃腸腺癌が結腸直腸腺癌、膵臓腺癌、食道腺癌または胃腺癌である、請求項14に記載の医薬組成物。
【請求項16】
前記医薬組成物が進行性腫瘍、後期腫瘍、腫瘍量の多い腫瘍患者、転移性腫瘍、またはCEA血清中濃度が100ng/mlより高い腫瘍患者の治療のためのものである、請求項1から15のいずれかに記載の医薬組成物。
【請求項17】
前記第1または第2の結合ドメインの少なくとも1つが、キメラ、ヒト化、CDR移植、および/または脱免疫化またはヒトである、請求項1〜16のいずれかに記載の医薬組成物。
【請求項18】
請求項1から17のいずれかに記載の二重特異性一本鎖抗体をコードする核酸配列を含む、医薬組成物。
【請求項19】
請求項18に記載の核酸配列を含むベクターを含む、医薬組成物。
【請求項20】
前記ベクターが、請求項18に記載の前記核酸配列と作動可能なように連結されている調節配列をさらに含む、請求項19に記載の医薬組成物。
【請求項21】
前記ベクターが発現ベクターである、請求項19または20に記載の医薬組成物。
【請求項22】
請求項18に記載の核酸または請求項19から21のいずれかに記載のベクターで形質転換またはトランスフェクトされた宿主を含む、医薬組成物。
【請求項23】
免疫エフェクター細胞の活性化シグナルを提供することのできるタンパク質性化合物をさらに含む、請求項1から22のいずれかに記載の医薬組成物。
【請求項24】
請求項1から23のいずれかに記載の医薬組成物の製造のためのプロセスであって、前記プロセスが、請求項22に記載の宿主を請求項1から17のいずれかに記載の二重特異性一本鎖抗体の発現を可能にする条件下で培養する段階と、産生された二重特異性一本鎖抗体を培養物から回収する段階とを含む、プロセス。
【請求項25】
担体、安定剤および/または賦形剤の適した製剤をさらに含む、請求項1から24のいずれかに記載の医薬組成物。
【請求項26】
請求項1から17のいずれかに記載の二重特異性一本鎖抗体、請求項18に記載の核酸分子、請求項19から21のいずれかに記載のベクター、または請求項22に記載の宿主の、ヒトにおける上皮腫瘍の予防、治療または寛解のための医薬組成物の調製のための使用。
【請求項27】
前記上皮腫瘍が、胃腸腺癌、乳腺腺癌または肺腺癌である、請求項26に記載の使用。
【請求項28】
前記胃腸腺癌が、結腸直腸腺癌、膵臓腺癌、食道腺癌または胃腺癌である、請求項27に記載の使用。
【請求項29】
医薬組成物が、進行性腫瘍、後期腫瘍、高腫瘍量の腫瘍患者、転移性腫瘍、またはCEA血清濃度が100ng/mlよりも高い腫瘍患者の治療のための、請求項1から28のいずれかに記載の使用。
【請求項30】
前記医薬組成物が、さらなる薬剤と組み合わせて投与されることに適している、請求項1から29のいずれかに記載の使用。
【請求項31】
上皮性腫瘍の予防、治療または寛解を必要とする被験体における、上皮性腫瘍の予防、治療または寛解のための方法であって、前記方法が、請求項1から17のいずれか一項に記載の医薬組成物または請求項24に記載のプロセスに従って製造された医薬組成物の有効量を十分な時間をかけて投与する段階を含む、方法。
【請求項32】
前記上皮性腫瘍が、胃腸腺癌、乳腺癌または肺腺癌である、請求項31に記載の方法。
【請求項33】
前記胃腸腺癌が、結腸直腸腺癌、膵臓腺癌、食道腺癌または胃腺癌である、請求項32に記載の方法。
【請求項34】
医薬組成物が、進行性腫瘍、後期腫瘍、高腫瘍量の腫瘍患者、転移性腫瘍、またはCEA血清濃度が100ng/mlよりも高い腫瘍患者の治療のためのものである、請求項31から33のいずれかに記載の方法。
【請求項35】
前記医薬組成物が、さらなる薬剤と組み合わせて投与される、請求項31から34のいずれかに記載の方法。
【請求項36】
前記薬剤が、非タンパク質性化合物またはタンパク質性化合物である、請求項35に記載の方法。
【請求項37】
免疫エフェクター細胞の活性化シグナルを提供することのできるタンパク質性化合物の投与を含む、請求項36に記載の方法。
【請求項38】
前記タンパク質性化合物または非タンパク質性化合物が、請求項1から17のいずれかに記載の二重特異性一本鎖抗体、請求項18に記載の核酸分子、請求項19から21のいずれかに記載のベクター、または請求項22に記載の宿主と同時または非同時に投与される、請求項36または37に記載の方法。
【請求項39】
前記被験体がヒトである、請求項31から38のいずれか一項に記載の方法。
【請求項40】
請求項1から17のいずれかに記載の二重特異性一本鎖抗体、請求項18に記載の核酸分子、請求項19から21のいずれかに記載のベクター、または請求項22に記載の宿主を含む、キット。
【請求項41】
前記CEA血清中濃度がELISAにより測定される、請求項16に記載の医薬組成物、請求項29に記載の使用、または請求項34に記載の方法。
【請求項1】
ヒトにおける上皮性腫瘍の治療のための医薬組成物であって、前記医薬組成物が、
(a)ヒトCD3と特異的に結合する第1の結合ドメイン、および
(b)ヒトCEAと特異的に結合する第2の結合ドメインを有する二重特異性一本鎖抗体を含み、
前記第2の結合ドメインが少なくともアミノ酸配列「DX1X2X3X4FYFDY」(配列番号65)を含み、「X1」、「X2」、「X3」または「X4」は任意のアミノ酸残基を表し、アミノ酸残基「D」はマウスモノクローナル抗体A5B7のCDR−H3のKabat位置95に相当し、アミノ酸残基「FYFDY」は、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置100、100a、100b、101、および102にそれぞれ相当する、医薬組成物。
【請求項2】
「X1」が、「R」(アルギニン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;
「X2」が、「G」(グリシン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表し;
「X3」が、「L」(ロイシン)、「F」(フェニルアラニン)、「M」(メチオニン)、「E」(グルタミン酸)、または「T」(トレオニン)を表し;かつ
「X4」が、「R」(アルギニン)、「Y」(チロシン)、「A」(アラニン)、「D」(アスパラギン酸)、または「S」(セリン)を表す、
請求項1に記載の医薬組成物。
【請求項3】
ヒトCEAに特異的な前記第2の結合ドメインが、マウスモノクローナル抗体A5B7のCDR−H3のKabatの位置95〜102に相当するアミノ酸配列「DRGLRFYFDY」(配列番号66)を少なくとも含む、請求項1または2に記載の医薬組成物。
【請求項4】
CD3に特異的な前記第1の結合ドメインがC末端に位置している、請求項1から3のいずれか一項に記載の医薬組成物。
【請求項5】
前記結合ドメインが、VHCEA−VLCEA−VHCD3−VLCD3またはVLCEA−VHCEA−VHCD3−VLCD3の順序で配置されている、請求項1から4のいずれかに記載の医薬組成物。
【請求項6】
ヒトCEAに特異的な前記第2の結合ドメインが、アミノ酸配列「SYWMH」(配列番号68)を有するCDR−H1および/またはアミノ酸配列「FIRNKANGGTTEYAASVKG」(配列番号67)もしくは「FILNKANGGTTEYAASVKG」(配列番号145)を有するCDR−H2を含む、請求項1から5のいずれかに記載の医薬組成物。
【請求項7】
ヒトCEAに特異的な前記第2の結合ドメインが、アミノ酸配列「TYAMH」(配列番号70)を有するCDR−H1および/またはアミノ酸配列「LISNDGSNKYYADSVKG」(配列番号69)を有するCDR−H2を含む、請求項1から5のいずれかに記載の医薬組成物。
【請求項8】
ヒトCEAに特異的な前記第2の結合ドメインが、アミノ酸配列「TLRRGINVGAYSIY」(配列番号73)を有するCDR−L1および/またはアミノ酸配列「YKSDSDKQQGS」(配列番号72)を有するCDR−L2および/またはアミノ酸配列「MIWHSGASAV」(配列番号71)を有するCDR−L3を含む、請求項1から7のいずれかに記載の医薬組成物。
【請求項9】
ヒトCEAに特異的な第2の結合ドメインのVH領域のアミノ酸配列が、配列番号60または146である、請求項6に記載の医薬組成物。
【請求項10】
ヒトCEAに特異的な第2の結合ドメインのVH領域のアミノ酸配列が配列番号58または配列番号62である、請求項7に記載の医薬組成物。
【請求項11】
ヒトCEAに特異的な第2の結合ドメインのVL領域のアミノ酸配列が、配列番号64である、請求項8に記載の医薬組成物。
【請求項12】
CEAに特異的な第2の結合ドメインのV領域が、以下、
(a)配列番号60に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;
(b)配列番号146に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;
(c)配列番号58に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;
(d)配列番号62に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域;ならびに
(e)配列番号56に示されるアミノ酸配列からなるVH領域および配列番号64に示されるアミノ酸配列からなるVL領域
からなる群より選択される、請求項1から11のいずれか一項に記載の医薬組成物。
【請求項13】
前記二重特異性一本鎖抗体が、
(a)配列番号6、8、16、18、24、26、32、34、40、42、126、130、134または143のいずれか1つに表されるアミノ酸配列;
(b)配列番号5、7、15、17、23、25、31、33、39、41、125、129、133または142に示される核酸配列によりコードされるアミノ酸配列;
(c)ストリンジェントな条件下で(b)の相補的な核酸配列とハイブリダイズする核酸配列によりコードされるアミノ酸配列;
(d)(b)の核酸配列に対する遺伝暗号の結果として縮重した核酸配列によりコードされるアミノ酸配列;および
(e)(a)または(b)のアミノ酸配列と少なくとも85%同一、より好ましくは少なくとも90%同一、最も好ましくは少なくとも95%同一であるアミノ酸配列
からなる群より選択されるアミノ酸配列を含む、請求項1から12のいずれか一項に記載の医薬組成物。
【請求項14】
前記上皮性腫瘍が、胃腸腺癌、乳腺癌または肺腺癌である、請求項1〜13のいずれかに記載の医薬組成物。
【請求項15】
前記胃腸腺癌が結腸直腸腺癌、膵臓腺癌、食道腺癌または胃腺癌である、請求項14に記載の医薬組成物。
【請求項16】
前記医薬組成物が進行性腫瘍、後期腫瘍、腫瘍量の多い腫瘍患者、転移性腫瘍、またはCEA血清中濃度が100ng/mlより高い腫瘍患者の治療のためのものである、請求項1から15のいずれかに記載の医薬組成物。
【請求項17】
前記第1または第2の結合ドメインの少なくとも1つが、キメラ、ヒト化、CDR移植、および/または脱免疫化またはヒトである、請求項1〜16のいずれかに記載の医薬組成物。
【請求項18】
請求項1から17のいずれかに記載の二重特異性一本鎖抗体をコードする核酸配列を含む、医薬組成物。
【請求項19】
請求項18に記載の核酸配列を含むベクターを含む、医薬組成物。
【請求項20】
前記ベクターが、請求項18に記載の前記核酸配列と作動可能なように連結されている調節配列をさらに含む、請求項19に記載の医薬組成物。
【請求項21】
前記ベクターが発現ベクターである、請求項19または20に記載の医薬組成物。
【請求項22】
請求項18に記載の核酸または請求項19から21のいずれかに記載のベクターで形質転換またはトランスフェクトされた宿主を含む、医薬組成物。
【請求項23】
免疫エフェクター細胞の活性化シグナルを提供することのできるタンパク質性化合物をさらに含む、請求項1から22のいずれかに記載の医薬組成物。
【請求項24】
請求項1から23のいずれかに記載の医薬組成物の製造のためのプロセスであって、前記プロセスが、請求項22に記載の宿主を請求項1から17のいずれかに記載の二重特異性一本鎖抗体の発現を可能にする条件下で培養する段階と、産生された二重特異性一本鎖抗体を培養物から回収する段階とを含む、プロセス。
【請求項25】
担体、安定剤および/または賦形剤の適した製剤をさらに含む、請求項1から24のいずれかに記載の医薬組成物。
【請求項26】
請求項1から17のいずれかに記載の二重特異性一本鎖抗体、請求項18に記載の核酸分子、請求項19から21のいずれかに記載のベクター、または請求項22に記載の宿主の、ヒトにおける上皮腫瘍の予防、治療または寛解のための医薬組成物の調製のための使用。
【請求項27】
前記上皮腫瘍が、胃腸腺癌、乳腺腺癌または肺腺癌である、請求項26に記載の使用。
【請求項28】
前記胃腸腺癌が、結腸直腸腺癌、膵臓腺癌、食道腺癌または胃腺癌である、請求項27に記載の使用。
【請求項29】
医薬組成物が、進行性腫瘍、後期腫瘍、高腫瘍量の腫瘍患者、転移性腫瘍、またはCEA血清濃度が100ng/mlよりも高い腫瘍患者の治療のための、請求項1から28のいずれかに記載の使用。
【請求項30】
前記医薬組成物が、さらなる薬剤と組み合わせて投与されることに適している、請求項1から29のいずれかに記載の使用。
【請求項31】
上皮性腫瘍の予防、治療または寛解を必要とする被験体における、上皮性腫瘍の予防、治療または寛解のための方法であって、前記方法が、請求項1から17のいずれか一項に記載の医薬組成物または請求項24に記載のプロセスに従って製造された医薬組成物の有効量を十分な時間をかけて投与する段階を含む、方法。
【請求項32】
前記上皮性腫瘍が、胃腸腺癌、乳腺癌または肺腺癌である、請求項31に記載の方法。
【請求項33】
前記胃腸腺癌が、結腸直腸腺癌、膵臓腺癌、食道腺癌または胃腺癌である、請求項32に記載の方法。
【請求項34】
医薬組成物が、進行性腫瘍、後期腫瘍、高腫瘍量の腫瘍患者、転移性腫瘍、またはCEA血清濃度が100ng/mlよりも高い腫瘍患者の治療のためのものである、請求項31から33のいずれかに記載の方法。
【請求項35】
前記医薬組成物が、さらなる薬剤と組み合わせて投与される、請求項31から34のいずれかに記載の方法。
【請求項36】
前記薬剤が、非タンパク質性化合物またはタンパク質性化合物である、請求項35に記載の方法。
【請求項37】
免疫エフェクター細胞の活性化シグナルを提供することのできるタンパク質性化合物の投与を含む、請求項36に記載の方法。
【請求項38】
前記タンパク質性化合物または非タンパク質性化合物が、請求項1から17のいずれかに記載の二重特異性一本鎖抗体、請求項18に記載の核酸分子、請求項19から21のいずれかに記載のベクター、または請求項22に記載の宿主と同時または非同時に投与される、請求項36または37に記載の方法。
【請求項39】
前記被験体がヒトである、請求項31から38のいずれか一項に記載の方法。
【請求項40】
請求項1から17のいずれかに記載の二重特異性一本鎖抗体、請求項18に記載の核酸分子、請求項19から21のいずれかに記載のベクター、または請求項22に記載の宿主を含む、キット。
【請求項41】
前記CEA血清中濃度がELISAにより測定される、請求項16に記載の医薬組成物、請求項29に記載の使用、または請求項34に記載の方法。
【図1】

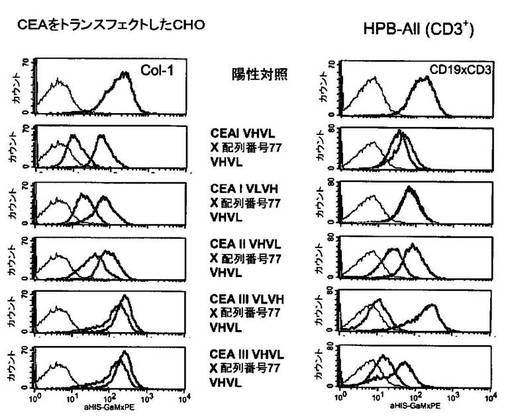
【図2】
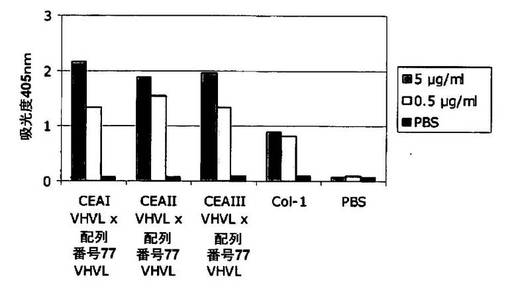
【図3】
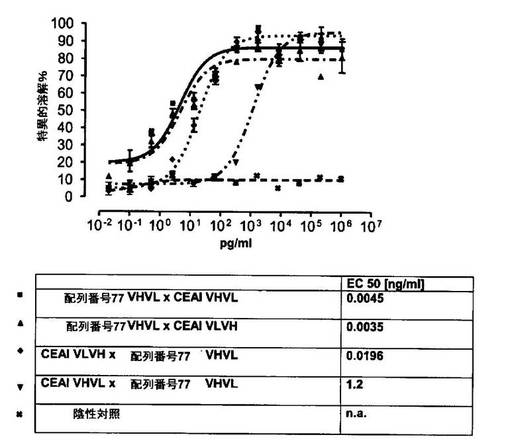
【図4】
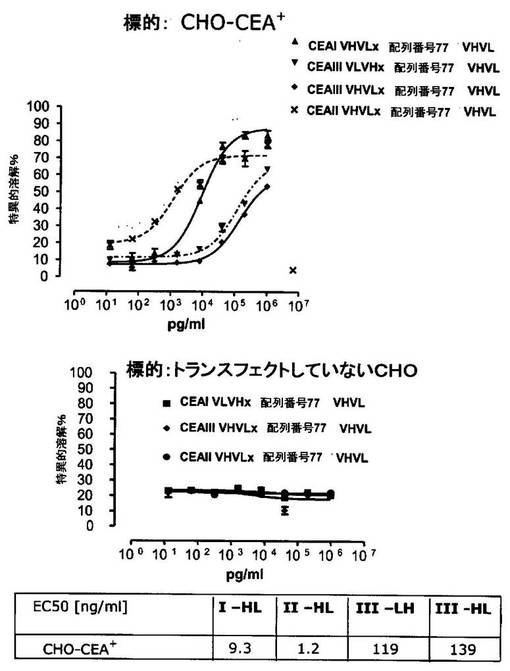
【図5】
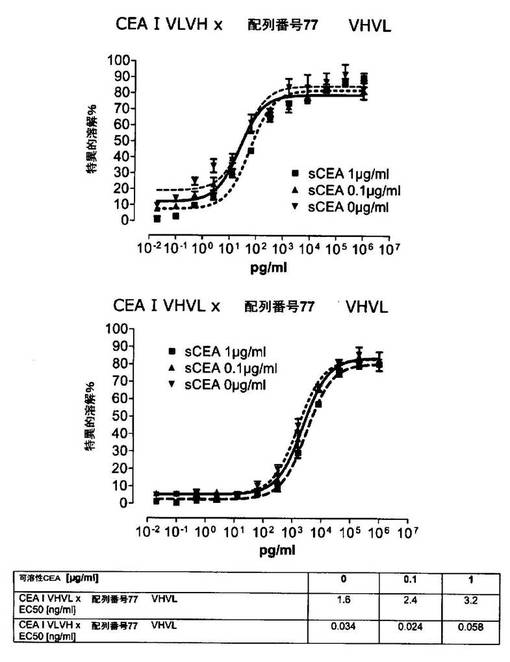
【図6】
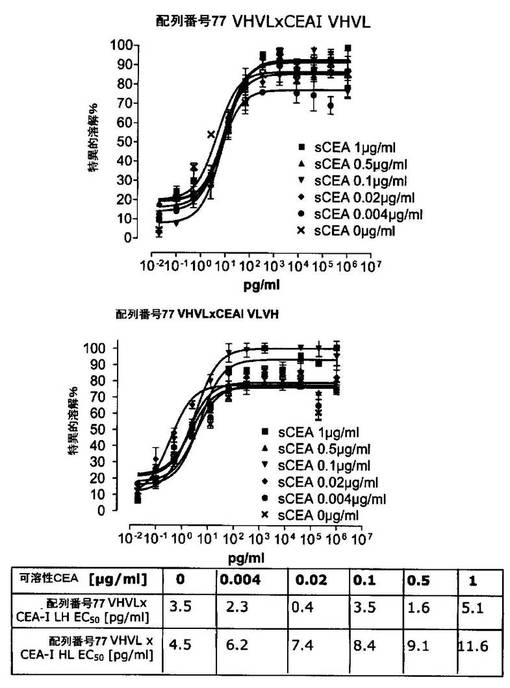
【図7】
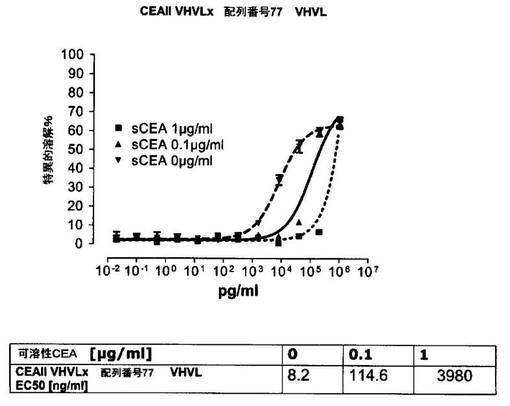
【図8】
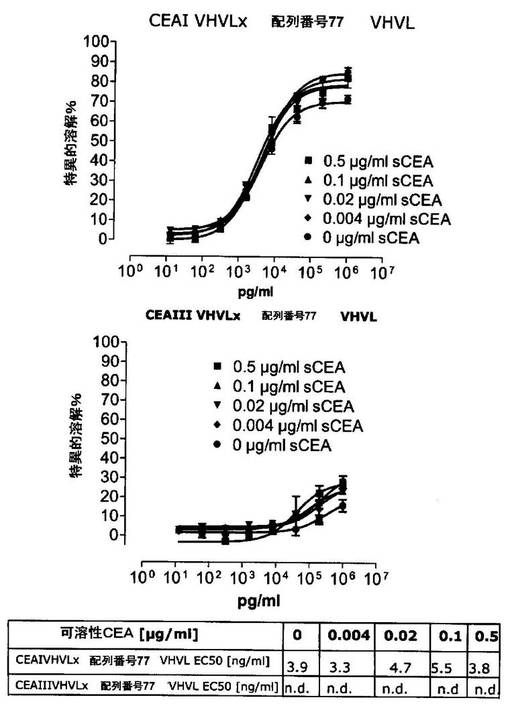
【図9】
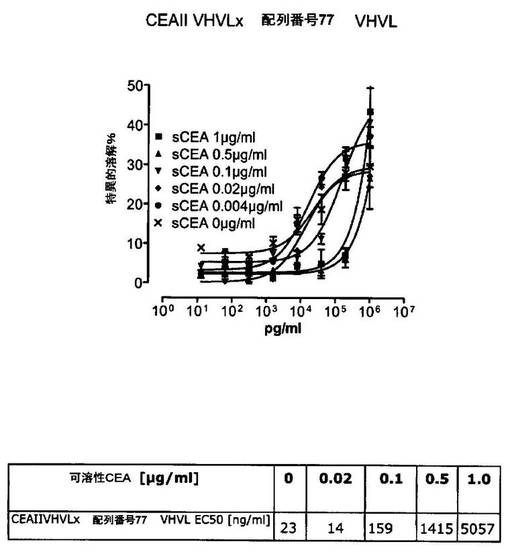
【図10】
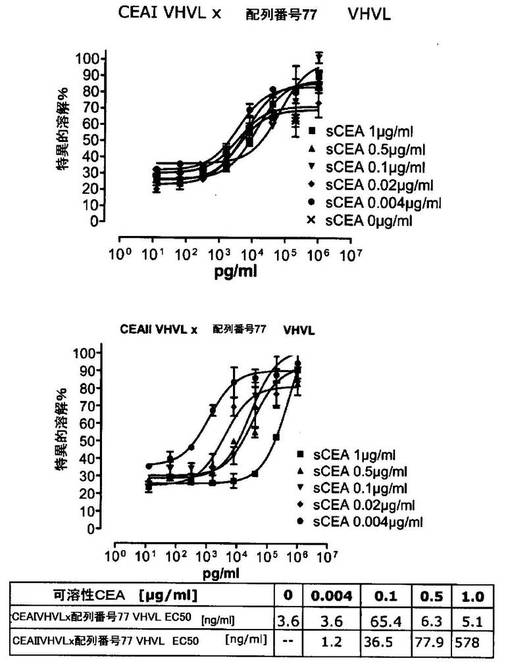
【図11】
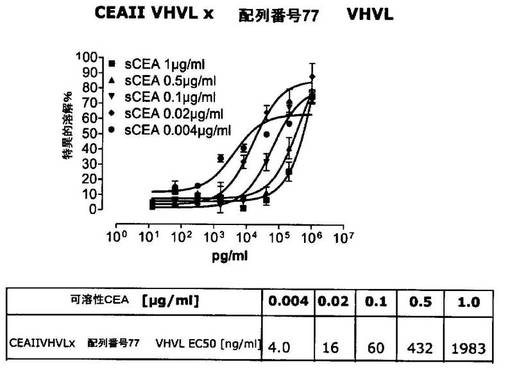
【図12】
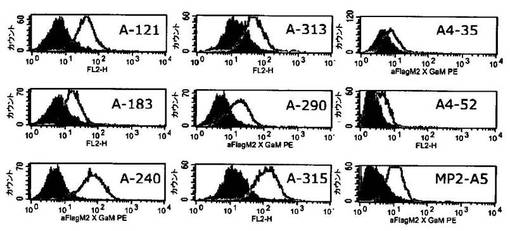
【図13】
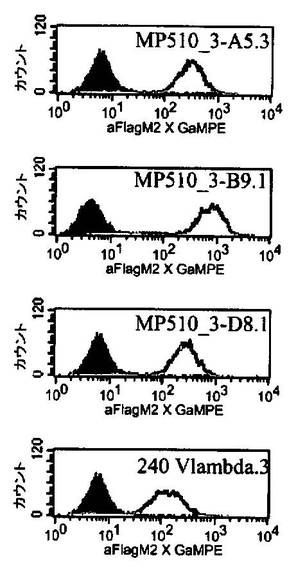
【図14】
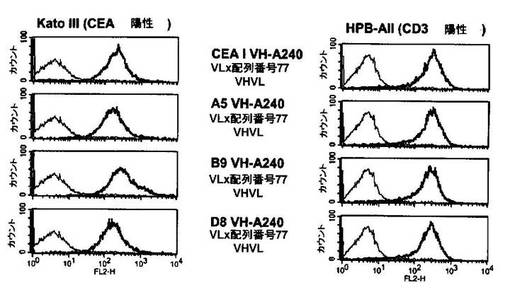
【図15】
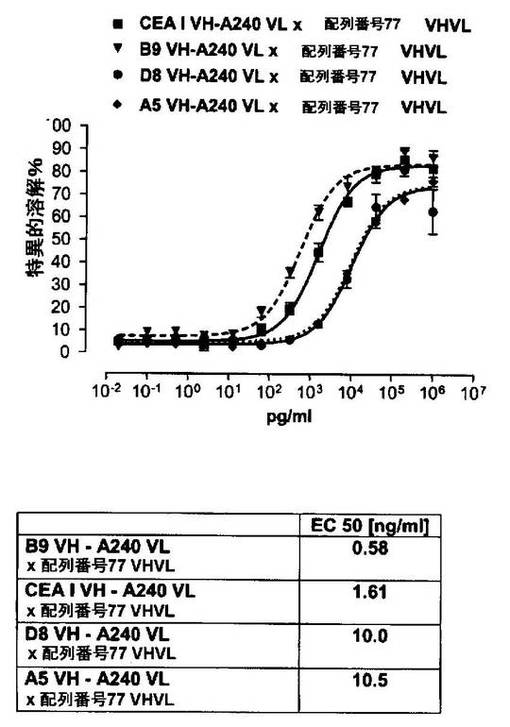
【図16】
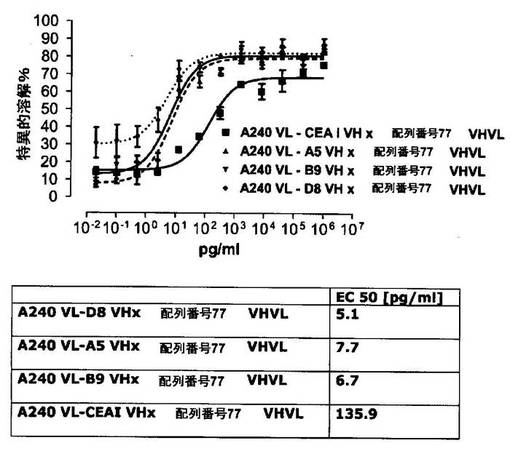
【図17】
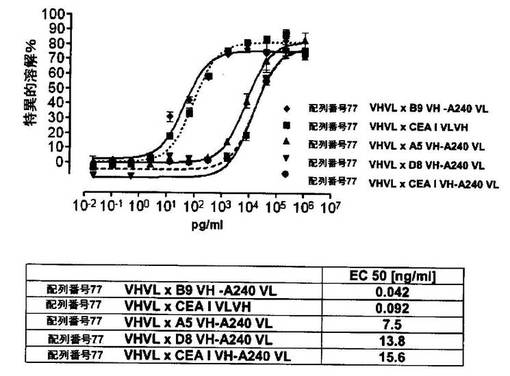
【図18】
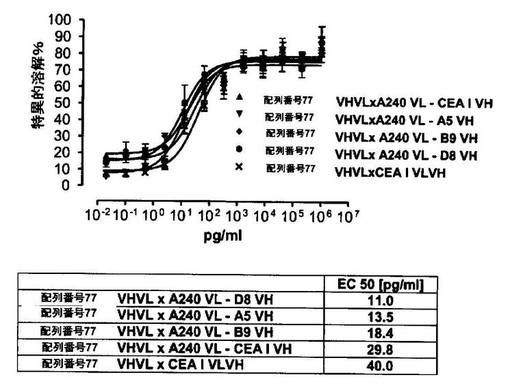
【図19】
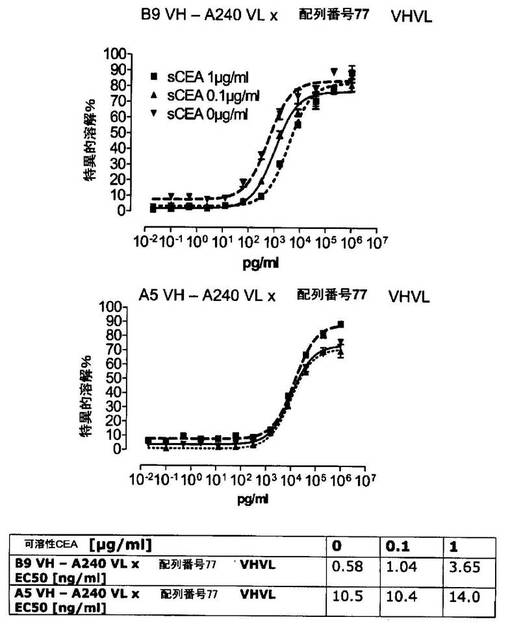
【図20】
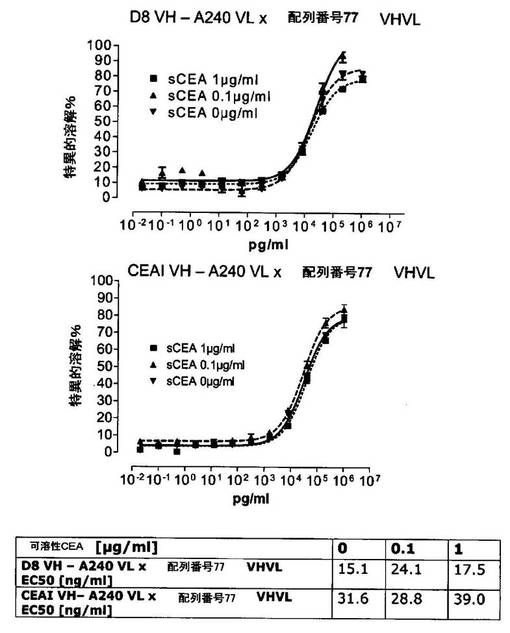
【図21】
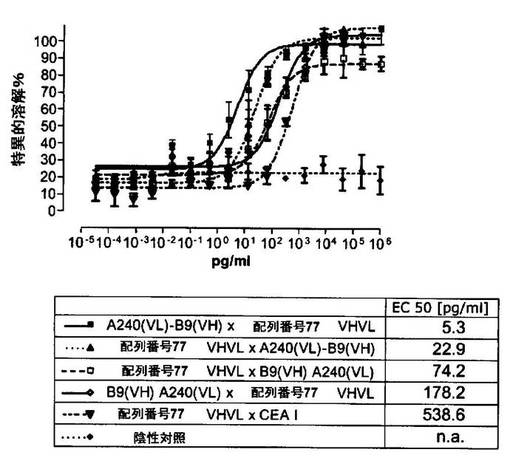
【図22】
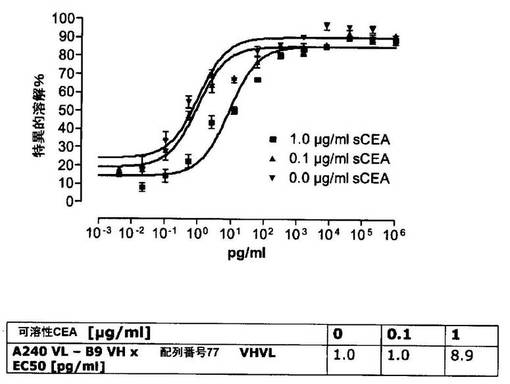
【図23】
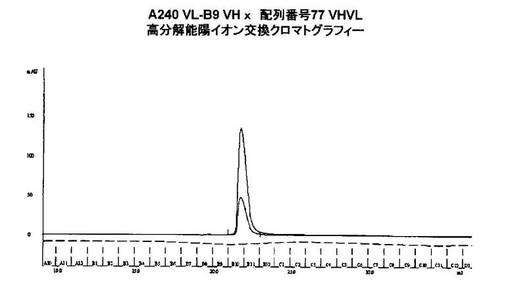
【図24】
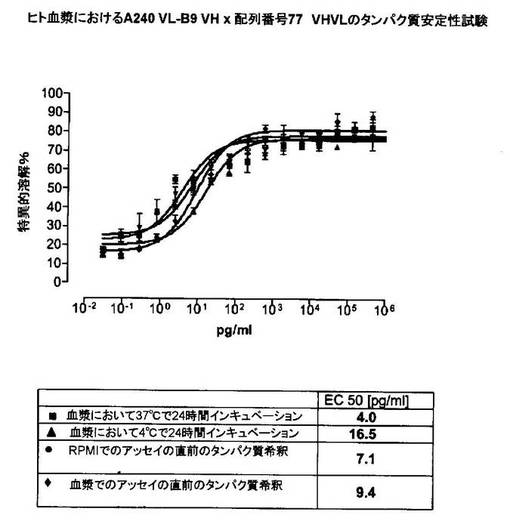
【図25】
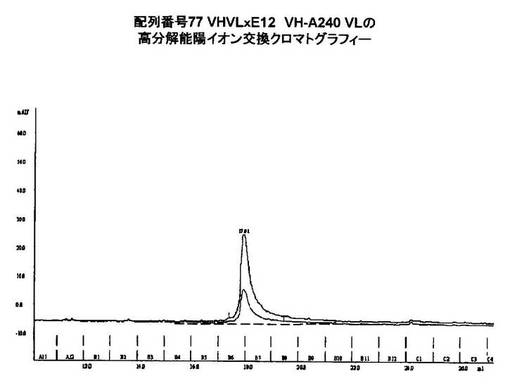
【図26】
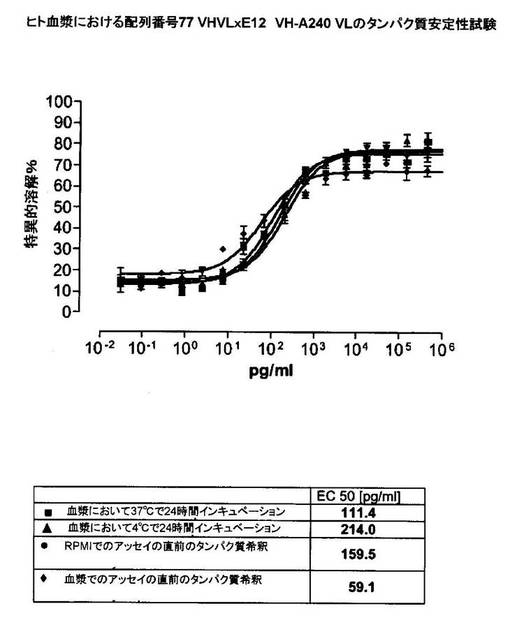
【図27】
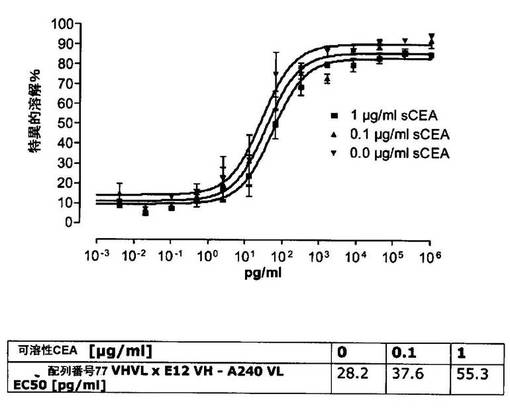
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【公表番号】特表2009−520734(P2009−520734A)
【公表日】平成21年5月28日(2009.5.28)
【国際特許分類】
【出願番号】特願2008−546272(P2008−546272)
【出願日】平成18年12月21日(2006.12.21)
【国際出願番号】PCT/EP2006/012425
【国際公開番号】WO2007/071426
【国際公開日】平成19年6月28日(2007.6.28)
【出願人】(502343861)マイクロメット アーゲー (6)
【Fターム(参考)】
【公表日】平成21年5月28日(2009.5.28)
【国際特許分類】
【出願日】平成18年12月21日(2006.12.21)
【国際出願番号】PCT/EP2006/012425
【国際公開番号】WO2007/071426
【国際公開日】平成19年6月28日(2007.6.28)
【出願人】(502343861)マイクロメット アーゲー (6)
【Fターム(参考)】
[ Back to top ]