
可逆的に不死化された細胞及びそれに関連する方法
本発明は、可逆的に不死化された細胞の作製方法、上記の方法によって入手可能な細胞、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸を含む真核細胞及びこれらの細胞を再分化する方法に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、可逆的に不死化された細胞の作製方法、上記方法によって入手可能な細胞、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸を含む真核細胞及びこれらの細胞を再分化する方法に関する。
【背景技術】
【0002】
早くも19世紀には、摘出された動物心臓の鼓動を体外で維持する方法があった。20世紀の初めには、組織培養の方法論を確立する実験が報告された。細胞培養技術は、1940年代及び1950年代に進歩してウイルス学における研究を支えた。動物細胞培養は、1900年代中頃に一般的な実験室技術になった。
【0003】
一般に、細胞培養の概念には、生体外培養のために組織から細胞、別名初代細胞(primary cells)を単離することが含まれる。このために、コラゲナーゼ、トリプシン又はプロナーゼのような酵素による酵素消化によって初代細胞を遊離してもよい。別法として、組織の断片を適切な増殖培地に置き、そして培養のための増殖用の細胞を入手することができる。
【0004】
いくつかの腫瘍から誘導された細胞を除いて、ほとんどの初代細胞培養物は、限られた寿命を有する。さらにまた、多くの場合、初代細胞は、培養における増殖能が限られており、そして継代延長により複製老化を受けるか(すなわち内皮細胞、平滑筋細胞)又は全く増殖しない(例えば肝細胞、心筋細胞)。
【0005】
従って、細胞に基づく実験は、複数のドナーを必要とし、そしてヒトの場合、細胞により重大な問題:ドナーの遺伝的差異、薬物乱用などが生じる。
【0006】
多くの調査実験は、腫瘍から誘導された細胞株で実施する。しかし、腫瘍由来の細胞は、分化が不正確であることが多く、そして重要な組織特異的遺伝子の発現が遺伝的に不安定である。従って、それらの欠点を回避することによって初代細胞及び腫瘍由来の細胞株の利点を合わせた新たな細胞を提供することが望ましい。
【0007】
初代細胞の増殖能が限られている一つの理由は、細胞分裂ごとに生じる進行性のテロメア短縮化であり、これは最終的に細胞老化に至る。SV40 T抗原を発現させて第2の不可逆的な危機が生じる前にさらなる回数の細胞分裂により細胞を成長させることができる細胞の改変方法が報告されている。生体内で収穫された初代細胞をインビトロで無限に増殖するように誘導する方法を細胞不死化と称する。
【0008】
いくつかの研究は、一連の増殖促進及び抗老化遺伝子を組み合わせて発現することが必要な段階的方法においてヒト細胞を形質転換することができることを示した。従って、適当な遺伝子操作により、培養において制御された細胞増殖を誘導することは可能でありうる。さらに、培養においてヒト不死化細胞株を樹立することは可能であった。ヒトテロメラーゼ触媒ユニットの発現により、増殖停止を阻止し、そしてヒト線維芽細胞及び上皮細胞を不死化することができる。しかし、ヒトテロメラーゼ触媒ユニット単独の発現では、限定数しか不死化することができなかった。ほとんどの細胞タイプは、SV40 T抗原をコードしている遺伝子のような第2の遺伝子の発現を必要とする。培養における不死化細胞の連続増殖は、その初代細胞に相当する物と比較したときに組織特異的遺伝子の発現を下方制御する。2つの癌遺伝子(SV40 T癌タンパク質及びH−rasの癌遺伝子対立遺伝子)と組み合わせたヒトテロメラーゼ触媒サブユニットの発現は、上皮細胞及び線維芽細胞のような正常なヒト細胞に直接的な腫瘍形成転換を生じることが報告されている。さらにまた、ヒトテロメラーゼ触媒ユニットが単独で又は1つ若しくはそれ以上の他の遺伝子と組み合わせて、腎近位尿細管上皮細胞で発現された場合にも初代ヒト細胞の不死化が報告されている。近位尿細管細胞を不死化するこの方法は、これらの細胞により十分に機能したが、HUVEC、肝細胞、平滑筋細胞及び間葉性幹細胞を含むいくつかの他のヒト初代細胞では機能しなかった。
【発明の概要】
【発明が解決しようとする課題】
【0009】
従って、細胞又はその前駆体が種々の細胞タイプから誘導することができ、及び/又は複製老化を示すことなく増殖することができると同時に初代細胞の魅力的な性質、例えば安定な遺伝子型及びその分化レベルを有する細胞及びその作製方法を提供することが本発明の目的である。新規な細胞モデルの開発は、生物学的及び薬学的研究にとって、特に化合物プロファイリング、表現型スクリーニング、標的同定及び/又は標的検証にとって最も重要である。
【課題を解決するための手段】
【0010】
驚くべきことに、ヒト乳頭腫ウイルス(HPV)E6/E7は、上皮細胞、内皮細胞及び肝細胞を含むいくつかの真核細胞中で発現されたときに、細胞を不死化できることが見出された(実施例を参照のこと)。
【0011】
従って、第1の態様において、本発明は、可逆的に不死化された細胞の作製方法であって、
a)真核細胞を準備し、
b)工程a)の細胞を遺伝子改変して細胞中で1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現を可能にし;そして
c)工程b)の細胞中で1つ又はそれ以上の核酸を発現させる
ことを含む方法であって、
その際、1つ又はそれ以上の不死化タンパク質は、ヒト乳頭腫ウイルス(HPV)E6/E7又はその機能的に活性な変異体を含む前記方法に関する。
【0012】
上に詳述したように、ヒト初代細胞の限られた増殖能は、現在、多くの用途におけるその使用を妨げている。細胞を不死化する方法は記載されてきたが、しかしながら、それらは上に詳述された欠点がある。本発明は、細胞を遺伝子改変によって可逆的に不死化された細胞を作製し、そして場合により1つ又はそれ以上の他の遺伝子と併せてHPV E6/E7を発現する新規な方法を開発して代替方法を提供し、そして好ましくはこれらの欠点を克服したことを開示している。
【0013】
本発明の方法の工程a)では、被験者から得た後に直接用いることで真核細胞を準備してもよい。別法として、細胞を培養した後、工程b)の遺伝子改変を実施してもよい。また、細胞は、細胞株から誘導してもよい。しかし、初代細胞、すなわち新たに単離された細胞、又は短期間、すなわち多くとも4週間、好ましくは多くとも3週間、より好ましくは少なくとも2週間、なおより好ましくは多くとも7、6、5、4、3、2又は1日培養された細胞が好ましい。
【0014】
「真核細胞」という用語は、動物、植物、真菌及び原生生物から誘導された細胞を含む。これらの細胞は、膜内に囲まれた複雑な構造に組織化されている。
真核細胞は、核を有する原核細胞とは異なる。多くの真核細胞は、ミトコンドリア、葉緑体及びゴルジ体のような他の膜結合細胞小器官を含む。真核細胞中の細胞分裂は、核のない生物(原核生物)とは異なる。それは、微小管に誘導された運動による重複染色体の分離を含む。2つのタイプの分裂過程がある。有糸分裂では、1つ細胞が分裂して2つの遺伝的に同一の細胞を生じる。減数分裂は、有性生殖に必要である。
【0015】
上に詳述したように、工程a)で得た細胞を工程b)で遺伝子改変して細胞中で1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現を可能にし、その際、1つ又はそれ以上の不死化タンパク質は、ヒト乳頭腫ウイルス(HPV)E6/E7を含む。遺伝子改変の方法、特に遺伝子の導入は、当業者によく知られている。
【0016】
ヒト乳頭腫ウイルス(HPV)E6/E7は、HPVによって発現される重要な発癌性タンパク質に相当する。癌及び前癌性病変に関連する身体的変化を生じることが知られているHPVの菌株の中では、3つの癌タンパク質:E5、E6及びE7が認められている。従って、HPV E6/E7という用語は、個々のもの又は融合タンパク質のずれかとしてのタンパク質の組み合わせのことである。
【0017】
完全長HPV E6遺伝子は、約150〜160個のアミノ酸のタンパク質をコードしており、それは亜鉛結合Cys−X−X−Cysモチーフを含む2つのドメインを含む。E6は、細胞中で複数の役割があり、多くの他のタンパク質と相互作用すると考えられているため、特に興味深い。E6は、主に腫瘍抑制因子として作用するp53又はpRBタンパク質と結合することでそれらを不活性化することによって癌を生じる。腫瘍抑制タンパク質が不活性化されたとき、腫瘍成長は無制限に進行する。E6とp53及びpRBとの相互作用は、ユビキチン化及びユビキチンリガーゼによる分解のためにこれらのタンパク質をマークする。E6は、他の細胞タンパク質上で作用し、そしてテロメラーゼ活性に正の影響を与え、それにより細胞がチェックなしに分裂するのを通常阻止する道筋の1つを不活性化することがわかっている。さらに、E6は、細胞転写因子、E2F1/DP1と相互作用するとき転写補因子、具体的には転写活性化剤として作用することができる。
【0018】
E6は、しばしばシグナル伝達タンパク質中に見出されるPDZドメインに結合することもできる。これらの位置での結合は、DLGタンパク質の形質転換及びその抑制機能の破壊が起こる。また、E6タンパク質は、MAGUK(膜結合性グアニル酸キナーゼファミリー)タンパク質とも相互作用し、通常、構造タンパク質であってDLGの抑制活性に関与していると考えられる。E6がMAGIタンパク質上のPDZドメインと複合体形成するとき、E6はその形状を変形させ、それによってその機能を妨げる。全体として、E6タンパク質は、癌に特有の高められた速度で細胞を増殖及び倍加させるようなやり方で正常なタンパク質活性を妨げる役割を果たす。
【0019】
E7タンパク質は、約100個のアミノ酸のサイズであり、そしてまたE6と同様に2部のCys−X−X−Cysドメインを含む。癌タンパク質として、E7は、網膜芽細胞腫腫瘍抑制遺伝子産物の活性形態、低リン酸化pRBと複合体を形成することにより機能し、そのタンパク質分解を促進する。E7/pRB複合体の形成は、pRBとE2Fとの複合体形成を妨げる。それゆえ、E2FがpRBとの不活性化された組み合わせから解放されると転写活性化剤としてその作用を続けてDNA合成及び細胞増殖を促進する。
【0020】
ハイリスクHPV感染細胞では、E6及びE7は、通常、共発現される。ハイリスクHPV E7癌タンパク質は、pRB並びに関連するpRBファミリーメンバーp107及びp130.17と結合して分解するだけでなく、サイクリン依存性キナーゼ(CDK)阻害剤を不活性化する。その結果、サイクリンE、サイクリンAの発現が異常に増加し、そしてE2Fが介在するCDK2活性が異常になる。また、E2Fは、p14ARFを活性化してp53を安定化することができる。細胞増殖を促進するE7と共同して、ハイリスクHPV E6癌タンパク質は、宿主細胞タンパク質、E6−APと相互作用することによってp53の迅速なプロテアソーム分解を誘導する。E6又はE7の核酸又はアミノ酸配列は、同じものを含むか又は発現するあらゆるHPVタイプから誘導してもよい。100を超える異なるHPVタイプが同定されており、数字で呼ばれている。タイプ16、18、31、33、35、39、45、51、52、56、58、59、68、73及び68は、「ハイリスク」HPV(癌に至ることがあるため)の例であり、そのため上記配列にとっての好ましい供給源である。HPV E6/E7の非常に好ましいcDNAは、SEQ ID NO:4として表記される。
【0021】
また、本発明は、HPV E6/E7の機能的に活性な変異体を包含する。変異体は、E6/E7断片であってもよく、その際、断片は細胞をなお不死化することができる。これは、短いC末端及び/又はN末端欠失(例えば多くとも20、19、18、17、16、15、14、13、12、11、10、9、8、7、6、5、4、3、2又は1個のアミノ酸の欠失)を有するE6/E7タンパク質を含んでもよい。別法として又はさらに、E6/E7タンパク質は、1つ又はそれ以上のアミノ酸置換を含んでもよい。しかし、アミノ酸が化学的に関連するアミノ酸で置換された同類アミノ酸置換(conservative amino acid substitution)は、好ましい。典型的な同類置換は、脂肪族アミノ酸の中からであり、脂肪族ヒドロキシル側鎖を有するアミノ酸の中からであり、酸性残基を有するアミノ酸の中から、アミド誘導体の中から、塩基性残基を有するアミノ酸、又は芳香族残基を有するアミノ酸の中からである。E6/E7変異体は、場合により他の不死化タンパク質と組み合わせて細胞をなお不死化することができる場合、本発明により機能的に活性である。細胞を不死化する能力は、細胞の増殖に関して、そして本発明の実施例において記載されたように、例えば免疫染色により不死化細胞及び対照細胞の量を比較し、全RNAを測定し、細胞形態、細胞生存(場合により不利な条件下)又は細胞数を測定して試験することができる。本発明に関して、変異体の活性が配列変更のないE6/E7タンパク質の活性の少なくとも10%、好ましくは少なくとも25%、より好ましくは少なくとも50%、さらにより好ましくは少なくとも70%、なおより好ましくは少なくとも80%、とりわけ少なくとも90%、特に少なくとも95%、最も好ましくは少なくとも99%に達する場合、変異体は、細胞を不死化する能力を有する。
【0022】
本発明の以下の説明において、E6/E7に関して提供されるすべての詳細は、特に明記しない限り、その機能的に活性な変異体にも関連する。
【0023】
しかし、最も好ましくは、E6/E7は、自然発生E6/E7、なおより好ましくは、HPV E6/E7であり、最も好ましくは、cDNAによってコードされたHPV E6/E7は、SEQ ID NO:4として表記される。
【0024】
「核酸」という用語は、本明細書において詳細に議論されたあらゆる核酸分子、好ましくはDNAのことである。核酸分子は、DNA、cDNA、合成DNA、RNA又はそれらの組み合わせを含むさまざまな供給源から誘導してもよい。このような核酸配列は、ゲノムDNAを含んでもよく、それは自然発生のイントロンを含んでもよいし又は含まなくてもよい。さらに、このようなゲノムDNAは、プロモーター領域、ポリA配列又は他の関連配列と共に入手してもよい。ゲノムDNAは、当分野でよく知られている手段によって適切な細胞から抽出し、そして精製してもよい。別法として、メッセンジャーRNA(mRNA)を細胞から単離して使用し、逆転写又は他の手段によってcDNAを作製することができる。
【0025】
遺伝子改変の前に、通常、興味の遺伝子、ここでは細胞中の1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸を単離する。その後、核酸、好ましくはDNA又はcDNAを、細胞の改変を可能にするやり方で処方する。例えばトランスフェクションによって、すなわち、非ウイルス的方法により細胞に核酸を導入する方法によって、又は形質導入、すなわち、ウイルスによる遺伝物質の挿入により生じる細胞の遺伝的変性によって真核細胞に外来DNAを導入する種々の方法がある。
【0026】
多くの物質がトランスフェクション用の担体として使用されており、それは3種類:(カチオン性)ポリマー、リポソーム及びナノ粒子に分けることができる。これは、トランスフェクションによって、例えばリン酸カルシウム沈殿によって実施してもよい。別の方法は、カチオンポリマー、例えばDEAE−デキストラン又はポリエチレンイミンの使用である。負に荷電されたDNAはポリカチオンに結合し、そして複合体はエンドサイトーシスを介して細胞に取り込まれる。トランスフェクションに対する直接的なアプローチは遺伝子銃であり、ここでは、DNAを不活性固形物(一般に金)のナノ粒子に結合し、次いで、それを標的細胞の核に直接「注射する(shot)」。
【0027】
トランスフェクションでは、遺伝子又はcDNAを適切なベクター中に移してもよい。ベクターは、プラスミド又はウイルスベクターであってもよい。核酸は、当業者に知られている制限酵素及びリガーゼを用いてベクター中に導入してもよい。ベクターを得たら、それを用いて興味の細胞を形質転換することができる。形質転換後、遺伝子改変された細胞を、ベクターの取込みに失敗したものから種々のやり方で選別する。1つの方法は、移植しようとした興味の遺伝子に固着することができるDNAプローブを用いたスクリーニングである。もう1つは、マーカーの使用である。マーカー遺伝子を用いてベクターの存在を検査し、それにより感染及び組込みを確認してもよい。マーカー遺伝子の存在により、挿入物を発現するそれらの宿主細胞のみの選別及び増殖が保証される。典型的な選択遺伝子は、抗生物質並びに他の毒性物質、例えばヒスチジノール、プロマイシン、ハイグロマイシン、ネオマイシン、メトトレキセート及び細胞表面マーカーに対する耐性をもたらすタンパク質をコードしている。別法として、検出可能なタンパク質をコードする遺伝子を、緑色蛍光タンパク質(GFP)又は黄色蛍光タンパク質(YFP)を含む蛍光タンパク質のようなマーカーとして使用してもよい。
【0028】
さらにまた、遺伝子又はcDNAを、核酸の有効な移入又は核酸の発現を可能する適切な要素と組み合わせてもよい。これらは、核酸の転写を制御する要素、例えばプロモーター配列、転写開始配列、エンハンサー配列、選択可能な要素及びレポーター遺伝子であってもよい。さらに、適切な制限酵素認識部位を、興味の核酸に加えてもよい。
【0029】
さらにまた、キャッピング、スプライシング、ポリ(A)末端付加によって翻訳を制御し、そして配列特異的な核外移行速度を変える要素が存在してもよい。
【0030】
ベクターを用いる場合、1つ又はそれ以上の核酸が1つ又はそれ以上のベクターの一部であってもよい。複数のベクターを用いる場合、それらを同時に又は順次、細胞中に導入してもよい。ウイルスベクターは、細胞中に遺伝物質を送達するために一般に用いられる手段である。この方法は、生きている生物内(インビボ)で又は細胞培養(インビトロ)において実施することができる。ウイルスは特殊な分子機構を発達させてそれらが侵入する細胞内部にそれらのゲノムを有効に運ぶ。ウイルスによる遺伝子の送達は、形質導入と称し、そして感染細胞は形質導入されたと記載される。
【0031】
当初、ウイルスベクターは、分子遺伝学実験用の裸のDNAのトランスフェクションに対する代替物として開発された。遺伝子をコードするタンパク質は、一般に特定のタンパク質の機能を研究するためのウイルスベクターを用いて発現することができる。ウイルスベクター、とりわけレトロウイルスは、遺伝物質を細胞に移入するために広く用いられる(また、下記参照)。他のタイプのウイルスベクターは、アデノウイルス及びアデノ随伴ウイルス(AAV)である。AAVは、分裂及び非分裂細胞の両方に感染することができ、そしてそのゲノムを宿主細胞のそれに組み込むことができる。これらの特徴によりAAVは、ウイルスベクターを作成するための非常に魅力的な候補物質となっている。非常に好ましいベクターの配列は、SEQ ID NO:6として表記される。
【0032】
工程b)において細胞を遺伝子改変した後、細胞中で1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸を発現する。細胞は、例えば、発現が構成的に活性なプロモーターによって制御される場合、なんらかのさらなる相互作用なしに、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸を発現することができる。あるいは、例えば、誘導性プロモーターが1つ又はそれ以上の核酸に結合した場合、発現を誘導する必要がありうる。
【0033】
この方法では、真核細胞、好ましくは下記定義された初代細胞を準備し、そして改変された細胞が不死化タンパク質ヒト乳頭腫ウイルス(HPV)E6/E7をコードする1つ又はそれ以上の核酸を発現するように遺伝子改変する。さらに、また、1つ又はそれ以上の核酸は、SV40ラージT抗原(TAG);テロメラーゼ触媒サブユニット(TERT);又はSV40ラージT抗原(TAG)及びテロメラーゼ触媒サブユニット(TERT)を含むさらなる不死化タンパク質をコードしてもよい。従って、HPV E6/E7をコードする核酸は、遺伝子改変によって細胞中に導入された唯一の核酸であってもよい。
【0034】
しかし、本発明の好ましい実施態様において、1つ又はそれ以上の不死化タンパク質は、
・HPV E6/E7及びSV40ラージT抗原(TAG);又は
・HPV E6/E7及びテロメラーゼ触媒サブユニット(TERT);又は
・HPV E6/E7及びSV40ラージT抗原(TAG)及びテロメラーゼ触媒サブユニット(TERT)
を含むか又はそれらからなる。また、これらの機能的に活性な変異体も含まれる。
【0035】
SV40ラージT抗原(Simian Vacuolating Virus 40 Tag、TAGとも称する)は、さまざまな細胞タイプを形質転換することができる、ポリオーマウイルスSV40から誘導された癌遺伝子である六量体タンパク質である。TAGの形質転換活性は、ラージ部分における網膜芽腫(pRB)及びp53腫瘍抑制タンパク質のその摂動(perturbation)による。さらにTAGは、その形質転換機能に寄与しうる転写補活性化因子p300及びCBPを含むいくつかの他の細胞性因子と結合する。
【0036】
TAGは、SV40によるウイルス感染の際に転写される初期遺伝子の産物であり、そしてウイルスのゲノム複製及び宿主細胞周期の制御に関与している。SV40は、二本鎖DNAウイルスであり、パポバウイルスファミリー、ポリオーマウイルス属に属する。ポリオーマウイルスは、多種多様な脊椎動物に感染し、それにより複数の部位で固形腫瘍を生じる。SV40 DNA複製は、ゲノムの起点領域(origin region)にTAGが結合することによって開始される。TAGの機能は、リン酸化によって制御され、それはSV40起点への結合を弱める。TAGとDNAポリメラーゼ−アルファとの間のタンパク質−タンパク質相互作用は、ウイルスゲノムの複製を直接刺激する。また、TAGは、腫瘍抑制タンパク質(p53、p105)と結合して不活性化する。これにより細胞がG1期を離れてS期に入り、DNA複製を促進する。
【0037】
本明細書に使用される「テロメラーゼ触媒サブユニット」又は「TERT」という用語は、テロメラーゼ触媒活性を有するポリペプチド配列のことである。テロメラーゼは、真核染色体の末端に見出される、テロメア領域中DNA鎖の3'(「3'末端」)末端に特異的なDNA反復配列(すべての脊椎動物中の「TTAGGG」)を加えた酵素である。テロメアは、濃縮DNA物質を含み、染色体に安定性を与える。酵素は、それ自体RNA分子を担持する逆転写酵素であり、それはテロメアを延長するときにテンプレートとして用いられ、テロメアは複製周期の後ごとに短縮される。それは、それぞれ2分子のテロメラーゼ触媒サブユニット、別名テロメラーゼ逆転写酵素(TERT);テロメラーゼRNA(hTR又はTERC);及びジスケリン(dyskerin)からなる。TERTは、テンプレートとして一本鎖RNAを用いて一本鎖DNAを作成する逆転写酵素である。
【0038】
また、本発明に関して、TAG及び/又はTERTの機能的に活性な変異体を用いてもよい。「変異体」及び「機能的に活性な」という用語は、E6/E7の変異体に関して上記定義された通りである。本発明の以下の説明において、E6/E7に関して記載されたすべての詳細は、特に明記しない限り、その機能的に活性な変異体にも関連する。
【0039】
一実施例において、第1のベクターは、HPV E6/E7をコードするポリヌクレオチドを含むのに対して、第2のベクターは、SV40ラージT抗原(TAG);テロメラーゼ触媒サブユニット(TERT);又はSV40ラージT抗原(TAG)及びテロメラーゼ触媒サブユニット(TERT)をコードするポリヌクレオチドをコードする核酸を含む。
【0040】
別の態様において、本発明は、可逆的に不死化された細胞の作製方法であって、
a)真核細胞を準備し、
b)工程a)の細胞を遺伝子改変して細胞中で1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現を可能にし;そして
c)工程b)の細胞中で1つ又はそれ以上の核酸を発現する、
ことを含み、
その際、1つ又はそれ以上の不死化タンパク質の少なくとも1つは、ヒト乳頭腫ウイルス(HPV)E6/E7、SV40ラージT抗原及びテロメラーゼ触媒サブユニット(TERT)、又はそれらのいずれかの機能的に活性な変異体からなる群より選ばれ、そして1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現は、EF1アルファプロモーターによって制御されることを含む前記方法に関する。
【0041】
驚くべきことに、プロモーターEF1アルファ(詳細については、下記参照)は、他よりも良好な発現レベルを示すことが見出された。いくつかのプロモーターを異なる初代細胞において試験した。異なるプロモーターの制御下でGFP(緑色蛍光タンパク質)cDNAを発現することによって、CMV(サイトメガロウィルス)プロモーター及びMSCV(マウスサイトメガロウィルス)プロモーターではそれぞれ極めて高い又は極めて低いレベルの発現となったのに対して、EF1アルファプロモーターでは中程度の発現レベルが得られることを示すことができた。本発明のこの態様のすべての用語は、この説明において詳述された通り定義される。
【0042】
本発明のさらなる実施態様において、本発明の方法は、
d)工程c)の細胞を増殖させる
ことをさらに含む。
【0043】
本発明によれば、工程c)の細胞は、細胞生存度、増殖及び完全性を可能にする有効な条件下で細胞を増殖させてもよい。本明細書において、増殖とは、インビトロで培養することによるものである。増殖とは、細胞成長及び分裂の結果として細胞数が増加することであり、それは細胞培養によって達成してもよい。好ましくは、増殖とは、少なくとも1つの細胞の倍加である。増殖をモニタリングする可能な方法としては、増殖に関連する抗原の検出、DNA合成の測定、及び細胞内環境の還元(reduction)の検出が含まれるが、それらに制限されるわけではない。DNA合成は、例えば3H−チミジンの取り込み又は5−ブロモデオキシウリジン(BrdU)の取り込みを定量化することによって測定してもよい。細胞内環境の還元は、テトラゾリウム塩の還元によってモニターしてもよい。生存度は、生存及び複製するための細胞の能力のことである。細胞生存度を測定するために一般に用いられるアッセイには、トリパンブルー色素排除、ニュートラルレッド染色、クリスタルバイオレット包摂、及び51Crリリースが含まれるが、それらに制限されるわけではない。この方法によって産生された安定なクローンは、不死化されたら、培養において少なくとも2ヵ月間培養することができる。好ましい実施態様において、これらの安定なクローンは、少なくとも6ヵ月、好ましくは少なくとも1年、そして最も好ましくは少なくとも2年間、培養において継代することができる。これらの安定なクローンは、それが誘導された細胞タイプに関連する高レベルの機能及び特徴を有し続ける。
【0044】
細胞培養のため、細胞は、細胞産生を助長する適切な条件下で増殖される。温度及びガス混合物を除いて、細胞培養系において最も一般に異なる要因は、増殖培地である。増殖培地の処方は、pH、グルコース濃度、増殖因子及びとりわけ他の栄養成分の存在で変えることができる。補充培地に用いられる増殖因子は、多くの場合、仔ウシ血清のような動物血液に由来する。培養条件は、各細胞タイプで広く変化し、そして特定の細胞タイプについての条件を変えることにより異なる表現型を発現することができる。
【0045】
細胞は、懸濁液又は付着性の培養中で増殖することができる。いくつかの細胞は、血流中に存在する細胞のように表面に付着することなく懸濁液中で自然に生きている。また、可能な付着条件より高密度に増殖することができるように懸濁培養液中で生存することができるように改良された細胞もある。付着細胞は、組織培養プラスチックのような表面を必要とし、これは細胞外マトリックス成分でコートして付着性を高め、そして増殖及び分化に必要な他のシグナルを提供してもよい。固形組織から誘導されたほとんどの細胞は、付着性である。別のタイプの付着培養(adherent culture)は、器官型培養であり、それは二次元の培養皿とは対照的に三次元環境での細胞の増殖を含む。この三次元培養系は、生体内の組織に生化学的及び生理学的により類似しているが、多くの要因(例えば拡散)のため、維持するのが技術的に困難である。適切な方法及びその選択は、当業者に知られている。さらにまた、不死化細胞に適切な方法は、実施例にも記載した。
【0046】
好ましい実施態様において、不死化タンパク質をコードする1つ又はそれ以上の核酸は、1つ又はそれ以上のレトロウイルスベクター、特に1つ又はそれ以上のレンチウイルスベクターを用いることによって導入される。
【0047】
レトロウイルスは、現在の遺伝子移入アプローチの柱の1つである。モロニーマウス白血病ウイルスのような組換え型のレトロウイルスは、安定なやり方で宿主ゲノムに組み込む能力を有する。従って、好ましい実施態様において、不死化タンパク質をコードする1つ又はそれ以上の核酸は、1つ又はそれ以上のレトロウイルスベクター、特に1つ又はそれ以上のレンチウイルスベクターを用いることによって導入される。
【0048】
レトロウイルスは、ウイルス逆転写酵素を用いることによって複製して宿主細胞中でそのRNAをDNAに転写する、RNAゲノムを有するウイルスである。次いで、DNAはインテグラーゼ酵素によって宿主のゲノムに組み込まれる。その後、ウイルスは、宿主細胞のDNAの部分として複製する。レトロウイルスは、レトロウイルス科というウイルスファミリーに属するエンベロープウイルスである。ウイルスそれ自体は、その核酸をmRNA(ビリオン中に5'cap及び3'PolyAを含む)ゲノムの形態で保存しており、そしてそのゲノムの細胞への送達手段として役立ち、偏性寄生生物として目標を定め、そして感染の構成要素となる。宿主の細胞中で、RNA鎖がサイトゾル中で逆転写を受け、そして宿主のゲノムに組み込まれたら、この時点で、レトロウイルスDNAをプロウイルスと称する。一般に、レトロウイルスは、外因性(以下の属、アルファレトロウイルス属、例えばトリ白血病ウイルス;ベータレトロウイルス属、例えばマウス乳癌ウイルス;ガンマレトロウイルス属、例えばマウス白血病ウイルス;デルタレトロウイルス属、例えばウシ白血病ウイルス;ヒトTリンパ球向性ウイルス;イプシロンレトロウイルス属、例えばウォールアイ皮膚肉腫ウイルス;レンチウイルス、例えばヒト免疫不全ウイルス1の他にサル及びネコ免疫不全ウイルス;並びにスプマウイルス属、例えばチンパンジーフォーミーウイルスを含む)及び内因性(ガンマレトロウイルスに最も類似しているクラスI、ベータレトロウイルス及びアルファレトロウイルスに最も類似しているクラスII、及びスプマウイルスに最も類似しているクラスIIIを含む)として特定化されている。
【0049】
レンチウイルスは、一般に長い培養期間を特徴とするレトロウイルス科ファミリーのスローウイルス属である。レンチウイルスは、かなりの量の遺伝情報を宿主細胞のDNAに送達することができるため、遺伝子送達ベクターの最も有効な方法の1つである。レンチウイルスは、主としてインビトロ系又は動物モデルに遺伝子産物を導入するために用いられる研究手段である。レンチウイルス感染は、他の遺伝子治療法よりも分裂及び非分裂細胞の高効率感染、導入遺伝子の長期的に安定な発現、及び低い免疫原性を含む利点を有する。レンチウイルスは、レトロウイルスのサブクラスである。それは、非分裂細胞のゲノム中に組み込む能力のため遺伝子送達ビヒクル(ベクター)として応用されており、他のレトロウイルスは、通常、分裂細胞しか感染させることができないので、これはレンチウイルスの独特な特徴である。RNA形態のウイルスゲノムは、ウイルスが細胞に入ってDNAを作製するときに逆転写され、それは次いで、ウイルスインテグラーゼ酵素によってランダムな位置でゲノム中に挿入される。従って、本発明の好ましい実施態様において、不死化タンパク質をコードする1つ又はそれ以上の核酸は、1つ又はそれ以上のレンチウイルスベクターを用いることによって導入される。レンチウイルスの5つの血清群が知られており、それらが関与する脊椎動物の宿主を反映している(霊長類、ヒツジ及びヤギ、ウマ、ネコ及びウシ)。HIV、SIV及びFIVは、すべてレンチウイルスの例である。伝達は一般に感染粒子を介するが、レンチウイルスは、細胞外粒子を形成する必要なしに宿主細胞と直接接触している隣接細胞を感染させることができる。
【0050】
好ましい実施態様において、本発明に関する真核細胞は、動物細胞、より好ましくは脊索動物細胞、なおより好ましくは脊椎動物、さらにより好ましくは哺乳動物細胞、特にヒト、霊長類又は齧歯類の細胞である。
【0051】
さらに又は別法として、真核細胞は、初代細胞又は非初代細胞であってもよい。しかし、初代細胞、すなわち、本方法の使用直前に被験者から得た細胞を、可逆的に不死化された細胞の作製方法に使用することが好ましい。「初代細胞」という用語は、上記定義された通りである。
【0052】
さらにまた、真核細胞は、あらゆる細胞タイプであってもよい。しかし、生殖系列細胞ではなく、体細胞、すなわち、生物の体を形成しているいずれかの細胞が好ましい。哺乳動物において、生殖系列細胞は、精子及び卵子である。哺乳動物の体中の他のすべての細胞タイプは、体細胞である。体細胞の例としては、内臓、皮膚、骨、血液及び結合組織に由来するものが含まれる。好ましくは、細胞は、分化細胞、すなわちすべての遺伝子の特定のサブセットを発現する特殊化された細胞タイプの細胞である。好ましい分化細胞には、内皮細胞、心筋細胞、平滑筋細胞、肝細胞、有足細胞又は腎臓細胞が含まれる。
【0053】
本発明の別の実施態様において、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現は、EF1アルファプロモーターによって制御される。
【0054】
プロモーターは、特定の遺伝子の転写を促進するDNAの領域である。プロモーターは、典型的には同じ鎖及び上流(センス鎖の5'領域の方)においてそれが制御する遺伝子の近くにある。プロモーターは、遺伝子/DNAの発現を意図する細胞に従って選ばれ、用いることができる。適切なプロモーターの例としては、例えば、EF1アルファ、PGK、モロニーマウス白血病ウイルスプロモーター−エンハンサー要素、ヒトサイトメガロウィルスエンハンサー、ワクシニアP7.5プロモーター又は同様のものが含まれるが、しかし、EF1アルファプロモーターが好ましい。EF1アルファプロモーターは、レンチウイルスベクター及び真核細胞における発現に特に適切である。プロモーターは、真核EF1アルファプロモーター又は好ましくはヒトEF1アルファプロモーターであってもよい。適切なプロモーターの配列は、SEQ ID NO:1として表記される。
【0055】
本発明の別の実施態様において、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸は、loxP(locus of X-over P1)部位の横に位置しており、それはCre又はCre変異体の存在下で1つ又はそれ以上の核酸の除去を可能にする。
【0056】
バクテリオファージP1からの38kDaリコンビナーゼ、Creは、そのエンドヌクレアーゼ活性を用いて2つの同一のloxP部位間の組換えに触媒作用を及ぼす。酵素Creは、補助タンパク質又は補因子を必要とせず、そしてインビトロで及び多種多様な細胞条件下で有効に機能する。Creによって認識される組換え部位は、loxPとして知られている二本鎖DNA配列である。それぞれのloxP部位は、非対称の核領域によって分かれた2つの逆方向反復からなる。Creは、逆方向反復に結合し、そして核領域中でDNAを切断してDNA鎖交換反応を促進する。本来のloxP配列は、34bpからなる。その横に位置する2組の13bpの回文の間に8bpの非対称配列が存在する。詳細な構造を下に示す。
13bp 8bp 13bp
ATAACTTCGTATA−GCATACAT−TATACGAAGTTAT
(SEQ ID NO:7)
好ましいloxP部位は、SEQ IDNO:2及び3として表記される。
【0057】
Cre(環化組換え)タンパク質は、4つのサブユニット及び2つのドメインからなる:大きい方がカルボキシル(C末端)ドメイン、そして小さい方がアミノ(N末端基)ドメインである。総タンパク質は、343個のアミノ酸を有する。Cドメインは、ラムダファージから単離された酵素のインテグラーゼファミリーのドメインに構造が類似している。また、これは、酵素の触媒部位である。Cre変異体ポリペプチドは、特定のアミノ酸配列に関してCreと異なっていてもよいが、Creのリコンビナーゼ活性を維持している。一般に、基準ポリペプチド(reference polypeptide)及び変異体の配列が全体として密接に類似し、そして多くの領域で同一になるように違いは制限される。変異体及び基準ポリペプチドは、あらゆる組み合わせで1つ又はそれ以上の置換、付加又は欠失によってアミノ酸配列が異なっていてもよい。置換された又は挿入されたアミノ酸残基は、遺伝暗号によってコードされたものであってもよいし又はそうでなくてもよい。ポリペプチドの変異体は、自然発生であってもよいし、又は自然発生することが知られていない変異体であってもよい。
【0058】
Cre/lox系は、ゲノムDNA中の部位特異的な組換え事象を制御する遺伝的手段として用いられる。この系により、研究者は、さまざまな遺伝子改変生物を操作して遺伝子発現を制御し、望ましくないDNA配列を欠失させ、そして染色体構造を改変することが可能になる。系はCreを含み、二本鎖DNAは、Creによって両方のloxP部位で切断され、次いで一緒に結合されて戻る。組換えの効率は、loxP部位の方向に左右される。同じ染色体腕上の2つのlox部位について、逆方向loxP部位は、反転を生じるが、loxP部位の同方向反復は、欠失事象が生じる。loxP部位が異なる染色体上にある場合、Creにより誘導された組換えによって転座事象に触媒作用を及ぼすことが可能である。
【0059】
別の態様において、本発明は、本発明の方法によって入手可能な細胞又はその子孫を提供する。
【0060】
なお別の態様において、本発明は、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸を含む真核細胞であって、その際、1つ又はそれ以上の不死化タンパク質の少なくとも1つが、ヒト乳頭腫ウイルス(HPV)E6/E7、SV40ラージT抗原及びテロメラーゼ触媒サブユニット(TERT)からなる群より選ばれ、そして1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現がEF1アルファプロモーターの制御下にある前記真核細胞を提供する。
【0061】
さらに、細胞は、本発明に従って、可逆的に不死化された細胞の作製方法に関して上記詳述された特徴を有しうる。特に、細胞又はその子孫は、以下の性質の1つ又はそれ以上を特徴とすることがある:
i)不死化タンパク質をコードする1つ又はそれ以上の核酸は、1つ又はそれ以上のレトロウイルスベクター、特に1つ又はそれ以上のレンチウイルスベクター上にある;
ii)細胞は、さらに遺伝子改変を含む;
iii)細胞は、哺乳動物細胞、特にヒト、霊長類又は齧歯類の細胞である;
iv)細胞は、初代細胞から誘導される;
v)細胞は、分化細胞、特に内皮細胞、心筋細胞、平滑筋細胞、肝細胞、有足細胞又は腎臓細胞から誘導される;
vi)1つ又はそれ以上の不死化タンパク質をコードする核酸の発現は、EF1アルファプロモーターの制御下にある;及び/又は
vii)1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸は、loxP(locus of X-over P1)部位の横に位置する。
【0062】
別の態様において、本発明は、本発明の細胞、又はその子孫を再分化する方法、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現を阻止することを含む方法を提供する。不死化細胞の使用は、このような方法に利用可能な初代細胞の不足を克服する手段を提供する。不死化が逆転された不死化細胞は、成長及び増殖の低下を示す。2〜3週間後、細胞は、細胞老化に一致する形態を発現し、細胞周期の制御が不死化細胞において無損傷のままであることを示唆している。さらに、不死化が逆転された細胞は、強化された分化機能を示す。不死化が逆転された細胞は、生物学的及び薬学的研究のため、特に化合物プロファイリング、表現型スクリーニング、標的同定及び/又は標的検証のための供給源として用いてもよい。このために、細胞をまず不死化し、そして再分化細胞が必要になるまで培養する(そして増殖させる)ことができる。これにより、多くの実験のための、そして長期間にわたる経済的かつ均一な細胞供給源の使用が確保され、それは望ましいことである。
【0063】
細胞の再分化は、核酸の発現を阻止するなんらかの適切な方法によって実施してもよい。これは、アンチセンス核酸をそれぞれのmRNAに結合してその翻訳を阻止することによって実施してもよい。別法として、制御可能なプロモーター、例えば誘導性プロモーターを用いてもよい。その名称の通り、これらのプロモーターの活性は、生物的又は非生物的因子の有り又は無しによって誘導される。誘導性プロモーターは、それに実施可能に結合された遺伝子の発現を生物発育のある種の段階で又は特定の組織中でオン又はオフにすることができるため、遺伝子工学における非常に強力な手段である。誘導性プロモーターは、アルコール、テトラサイクリン、ステロイド、金属及び他の化合物の有り又は無しによってその転写活性が制御されるプロモーターを含む化学的に制御されるプロモーター;並びに光の有り又は無し及び温度の低い又は高いによってその転写活性が制御されるプロモーターを含む物理的に制御されるプロモーターに分類される。
【0064】
細胞を再分化する方法の好ましい実施態様において、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸は、loxP(locus of X-over P1)部位の横に位置しており、そして阻止は、Creリコンビナーゼ又は機能的に活性なCre変異体によって実施される。
【0065】
好ましくは、loxP部位を用いる場合、不死化過程は、細胞にCre又はCre変異体をコードするポリヌクレオチドを含むベクターを導入することによって逆転させることができる。1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸が、Cre介在性組換えによって細胞から除去されるとき、それぞれの細胞又はクローンの細胞型に特異な活性は、一般に上方制御される。また、細胞不死化に関与する遺伝子を除去すると細胞増殖の速度がかなり低下し、そして最終的に細胞老化に至る。
【0066】
細胞を再分化する方法の別の好ましい実施態様では、レトロウイルスベクター、特にレンチウイルスベクターを用いてCreリコンビナーゼ遺伝子又は機能的に活性なCre変異体遺伝子を細胞に導入する。ベクター及び核酸を移入する方法は、上記詳述した通りである。
【0067】
別の態様において、本発明は、本発明に従って、細胞を再分化する方法によって入手可能な細胞、又はその子孫を提供する。
【0068】
以下の図面及び実施例は、本発明の種々の実施態様を説明することを意図する。このように、議論された特定の改変は、本発明の範囲における制限と解釈すべきではない。本発明の範囲を逸脱することなく種々の同等物、変更及び改変を実施することができることは、当業者に明らかであり、そしてこのような同等の実施態様は、本明細書に含まれるものとすることが理解される。
【図面の簡単な説明】
【0069】
【図1】VECTALYS系のpLV−EF1−Lox1−AgT−Lox2レンチウイルスベクターの略図を示す。cDNAは、両側にlox部位を有するEF1Aプロモーターの制御下でレンチウイルスプラスミド中に作製され(図1)、それは、必要に応じて、「不死化遺伝子(immortogene)」を切除することができる。EF1=EF1アルファプロモーターAgT=SV40ラージT抗原T=SV40スモールt抗原LOX1/LOX2=Creリコンビナーゼ認識部位WPRE=ウッドチャック転写後調節要素
【図2】初代培養された新生ラット有足細胞系統におけるシナプトポジンの発現を示す。A:未分化初代有足細胞の位相コントラスト形態。標準条件下で増殖する未分化有足細胞の丸石形態を示す。細胞は、集密に達するにつれ単層を形成する。B:標準条件下で維持された樹状有足細胞は、非常に大きくかつ扁平状である。C:有足細胞特異的なタンパク質シナプトポジンは、標準培養条件で14日後に発現を開始した(シナプトポジン−薄い灰色、そしてアクチンストレス線維−暗い灰色)。
【図3】分化した有足細胞のマーカーが、「不死化細胞」のCre介在性切除後に再び現れるのを示している。A:(位相コントラスト):LT Ag+E6/E7の過剰発現によって誘導された活性な増殖中に未分化有足細胞の典型的な丸石形態。細胞は、集密に達するにつれ単層を形成する。B:Creリコンビナーゼの形質導入後の樹状有足細胞(大きくかつ扁平状)。C:LT Ag(灰色)の核発現−100%の細胞が陽性である。D:Creリコンビナーゼによる形質導入後、多くの細胞ではSV40 LT Ag(矢印)が失われる。 注:矢印は、失われていると考えられる。E:シナプトポジン及びWT1(F)は、再分化した有足細胞中で発現される。
【図4】初代(対照)及び不死化ヒト肝細胞の免疫細胞化学的な比較を示す。A、B及びC−光学顕微鏡画像;D、E及びF−GFP蛍光(各細胞タイプは、レンチウイルスGFPにより形質導入された)。対照肝細胞は、大きな多角形の細胞であり(A及びD);SV40LTagの発現により極めて小さな卵形細胞に形質転換され(B及びE);HPV E6E7で形質導入した後は、対照細胞と比較してわずかに減少したサイズを有するが、SV40 LTag不死化されたものよりもかなり大きい(FをD及びEと比較する)。G−I:p53(灰色)の免疫染色。対照細胞におけるp53基底レベルは非常に低く(G)、そしてSV40 LTAgでは、極めて高レベルまで核中に蓄積され(H);E6E7不死化細胞において、大半の細胞ではそのレベルは低い(I)。J−L:CDC2、サイクリン依存性キナーゼ1(CDK1)の免疫染色;初代肝細胞では、増殖しないので存在することができなかったが(J)、これに対して、AgT不死化細胞(K)では、そしてまたE6E7不死化細胞(L)では、CDC2発現は、CDC2 mRNAの増加に伴ってかなり増加した(表2参照)。
【図5】内皮細胞特異的マーカーが不死化されたHUVEC中で発現され続けることを示している。新たに単離されたHUVEC(左のパネル)、AgT不死化(中央のパネル)、そしてE6E7不死化HUVEC(右のパネル)。光学顕微鏡画像(A、B及びC)。典型的な内皮マーカー、フォン・ウィルブランド因子(D−F:vWF、薄い灰色)、そしてPECAM(G−I、CD31、暗い灰色)は、対照(D及びG)中、AgT(E及びH)及びE6E7(F及びI)不死化細胞中で等しく発現された。核(暗い灰色)は、ヘキスト422で染色した。
【図6】SV40 LT Ag−及びE6/E7−に誘導された増殖の推定上の機構を示す。非増殖細胞では、cdc2/CDK1、サイクリンA及びB、cdc20の発現は、p53及び「リプレッサーE2F」によって抑制された。SV40 LT Ag及びE6/7不死化細胞では、p53及びRbは不活性化され、そしてE2Fは活性になり、そして、それ自体、cdc2/CDK1、サイクリンA及びB、cdc20の発現を誘導する。
【図7】pLV−EF1−LOX1−E6E7−LOX2(Vectalys)を示す。
【0070】
実施例
物質及び方法
レンチウイルスコンストラクト:すべてのcDNAは、VECTALYS(フランス)に注文した。レンチウイルス粒子は、標準プロトコールに従って生成し、そして1×106〜2×106TU/mlのタイター(titer)を有した。感染効率は、レンチウイルスGFPを用いて測定した。
【0071】
初代細胞の標本及び培養条件:ラット糸球体上皮細胞の初代培養を産生するため、新生仔ラット腎臓から糸球体を単離した。細胞成分のうち95%が糸球体であり、残りが尿細管を含む場合、良好な標本と考えられる。単離した後、コラーゲンコートした24ウェルプレート上の10%FCS及び1%抗生物質入りのDMEM中で糸球体を培養した。24時間後、有足細胞前駆体が、付着した糸球体から外へ増殖し始めた。3日培養した後、増殖している上皮細胞をトリプシン処理し、そして25μm孔径の篩に通過させて残っている糸球体核を除去し、それは主に糸球体間質及び内皮細胞からなる。ヒト由来の初代内皮細胞は、ヒト臍帯からルーチン的に獲得することができる。簡潔に言えば、温かいPBS、pH7.4を用いて臍帯静脈から血液産物をすすぎ、そして静脈に温かいコラゲナーゼA(Roche Diagnostics;PBS中320U/mlの濃度で使用)を充填した。温かいPBS中で20分間インキュベーション後、消化物を熱失活FCS(Gibco/Invitrogen社、カールスルーエ、ドイツによって供給された)を含む遠心管中に集めてコラゲナーゼ活性を無効化し、そして細胞を沈殿させた。上澄みを捨て、そしてFCS、ECGS/H、hEGF、hbFGF及びHC−500を含む内皮細胞増殖培地(PromoCellからのMedium and SupplementPack)中に沈殿物を再懸濁し、そしてコラーゲン−IコートされたT75フラスコ上で培養し、そして37℃加湿インキュベーター中で4〜5日間、集密に増殖させた。細胞を大量に播種して細胞増殖を最小限にし、それによってHUVECがCD14を発現できるようにし、でなければそれは消失した。結果的に、すべての実験では、第1継代HUVECのみを用いた。
【0072】
初代ヒト肝細胞(PHH)は、Cambrex(Weinheim、ドイツ)から入手し、そしてコラーゲン−Iコートされた皿(Becton Dickinson Labware、San Jose、米国)上のHCM Single Quots(Cambrex Bioproducts、ハイデルベルク、ドイツ)で補充されたHBM基礎肝細胞培地(Cambrex Bioproducts、ハイデルベルク、ドイツ)中で培養した。レトロウイルスコンストラクト及びウイルス感染:初代新生仔ラット有足細胞を(下記のように)単離し、そしてloxP部位の横に位置するSV40 LT抗原(LT Ag)遺伝子を含む単一のレンチウイルスコンストラクト又はヒト乳頭腫ウイルス(HPV)E6及びE7を含むベクターを単独で又は組み合わせて感染させた。最初の10日間のうちに、初代有足細胞は、その表現型及び形態を変え、そして増殖し始めた。細胞を3日毎に分け、そして安定なクロzpーンを単離し、そしてさらなる実験のため凍結した。これらの安定なクローンは、分化した有足細胞ではなく、最初に糸球体から増殖した細胞と形態学的に似ていた。Creリコンビナーゼを含むレンチウイルスベクターで感染後、細胞は増殖を終え、そして3〜10日後、その形態を分化した有足細胞にとって特異的なものへと変えた。
【0073】
同じ不死化方法を、HUVEC及びヒト肝細胞に適用した。HUVECは、高発現レベルのLT Ag又はE6/E7にもかかわらずその形態を変えなかったが、活発な増殖を開始した。LT Ag又はE6/E7に感染した後の肝細胞は、その表現型を劇的に変え、そしてCre挿入後、その正常なものを獲得した。
【0074】
抗体:一次抗体:単クローン性WT−1(F−6)、CDC2 p34(B−6)、PCAM−1(CD31)、SV40 T Ag(Pab 108)及び多クローン性vWF(C−20)(Santa Cruz Biotechnology、ハイデルベルク、ドイツ);多クローン性ZO−1(Zymed、サンフランシスコ、CA);単クローン性シナプトポジン(Progen、ハイデルベルク、ドイツ);多クローン性p53(Cell Signalling)。
【0075】
二次抗体:Alexa Fluor(R) 488ヤギ抗マウスIgG(H+L)*2mg/mL*、Alexa Fluor(R) 594ヤギ抗マウスIgG(H+L)*2mg/mL*、Alexa Fluor(R) 488ヤギ抗ウサギIgG(H+L)*2mg/mL*及びAlexa Fluor(R) 594ヤギ抗ウサギIgG(H+L)*2mg/mL*(Invitrogen、カールスルーエ、ドイツ)。
【0076】
免疫染色:コラーゲンタイプIコートされたチャンバースライド(VWR International 社、ダルムシュタット、ドイツ)上で増殖細胞の免疫蛍光染色を実施した。20℃でメタノールを用いて5分間又はリン酸塩緩衝食塩水(PBS)中の4%ホルムアルデヒドを用いて5分間、細胞を固定し、続いてPBS中0.3%トライトンX−100を用いて室温で10〜15分間、透過化処理した。PBSですすいだ後、非特異的結合部位を0.2%Tween 20(Serva、Amstetten、オーストリア)入りのSuperBlock(PBS中のブロッキング緩衝液、Thermo Scientific、ブラウンシュヴァイク、ドイツ)で少なくとも30分間ブロックした。ブロッキング溶液中に予め希釈した一次抗体を室温で60分間又は4℃で一夜適用した。Alexa Fluor(R) 488 又は 594二次抗体(Invitrogen、カールスルーエ、ドイツ)を用いて抗原−抗体複合体を視覚化した。ニコン顕微鏡写真機を用いて画像を得、そしてAdobe Photoshop 6.0ソフトウェアで処理した。
【0077】
全RNAの単離:培養された有足細胞、Huvecs及びPHHから、RNeasy Kit(Qiagen、ヒルデン、ドイツ)を用いて全RNAを単離した。RNase-free DNase-Set(Qiagen、ヒルデン、ドイツ)を用いてゲノムDNA入りの潜在的サンプルコンタミネーションを除去した。質及び純度は、キャピラリー電気泳動(Caliper Lab Chip system Agilent 2100 Bio
analyser、Agilent Technologies社、Waldbronn、ドイツ)を用いて評価した。
【0078】
定量的逆転写−PCR(qRT−PCR):リアルタイム定量的PCRは、QuantiTect
Probe RT-PCR Kit(Qiagen、ヒルデン、ドイツ)を用いて実施した。各サンプルを四つ組で測定した。遺伝子発現の相対的定量のため、対照としてGAPDHを用いてΔΔCt法を使用した。標的及びハウスキーピング遺伝子の増幅は、Applera/Applied Biosystems(Foster City、米国)から入手した異なる蛍光標識Taq Manプローブを用いて同時に検出した。マルチプレックスRT−PCR内の標的及びハウスキーピング遺伝子の増幅直線性は、テンプレートの希釈によりRT−PCR反応を実施することによって評価した。RT−PCR反応及びデータ収集は、iCycler-iQ-Thermocycler(Bio-Rad Laboratories社、ミュンヘン、ドイツ)において実施した。相対的遺伝子発現は、対照サンプルに対する誘導倍率(fold induction)として算出した。負の値は、対照サンプル中の発現レベルと比較した下方制御の倍率(fold down regulation)を示す。
【0079】
スーパーアレイ−技術:PCRアレイは、焦点を絞った遺伝子パネルの発現を分析する手段である。それぞれ96ウェル又は関連の、経路又は疾患に焦点を絞った遺伝子(例えば転写因子、細胞周期、幹細胞)の綿密に調査されたパネル用のSYBR Green最適化プライマーアッセイを含み、そしてSABiosciences(CA、米国)から入手した。
【0080】
PCRアレイ(RT2 Profiler PCR Array System)では、リアルタイムPCR感受性及びマイクロアレイの多重遺伝子のプロファイリング能力を用いて遺伝子発現分析を実施する。実験者のcDNAテンプレート(RT2 First Strand Kit (C-03))と適切なすぐに使用できるPCRマスターミックスとを同じプレートの各ウェルに等体積のアリコートで単純に混合し、次いでリアルタイムPCRサイクリングプログラムを実施する。プレートテンプレート及びリアルタイムPCRサイクリングプログラム(Instrument-Specific Setup Instructions & Protocol Files)は、ファイルhttp://www.sabiosciences.com/pcrarrayprotocolfiles.php.を用いて入手可能である。
【0081】
RT2-First Strand Kitは、有効な第1鎖(first strand)cDNA合成のための迅速かつ便利な方法を提供する。キットは、有効なゲノムDNA除去工程及び組み込まれた外部RNA対照(built-in External RNA Control)も含む。このオールインワンキットは、SABiosciencesのRT2 ProfilerTM PCR Arrays 及び RT2 qPCR Primer AssaysによるリアルタイムPCRベースの遺伝子発現分析のために設計され、そして最適化されている。キットは、逆転写前にRNAサンプルから汚染ゲノムDNAを効果的に除去するための独自の方法を含む。偏りのないやり方(unbiased manner)におけるランダム六量体及びオリゴ−dTプライム逆転写及び逆転写酵素により最適な収量及び長さでcDNA産物を合成する。組み込まれた外部RNA対照は、逆転写効率をモニターし、そしてRT2 RNA QC PCR Array及びRT2 ProfilerTM PCR Arrayと共に用いたときに酵素阻害剤が混入した実験者のRNAサンプルを試験するのに役立つ。マグネシウム及びヌクレオチド濃度及び他の緩衝液成分は、RT2 ProfilerTM PCR Arrays 及びRT2 qPCR Primer Assaysによる遺伝子発現分析に用いたときにRT2 SYBR Green qPCR Master Mixesと最も適合しうる。
【0082】
RT2 SYBR Green/フルオレセインqPCRマスターミックスは、BioRad iCycler(R) 及び MyiQ(R)におけるリアルタイムポリメラーゼ連鎖反応に必要なすべての試薬及び緩衝液:リアルタイムPCR緩衝液、高性能HotStart DNA Taqポリメラーゼ、ヌクレオチド、SYBR(R)Green色素、及び機器の光学的諸特性を規格化するために必要なフルオレセイン参照色素を含む。マスターミックスを実験者のテンプレート及びプライマーと共にPCR管に単に加える。化学修飾及び厳密に制御されたHotStart酵素によりプライマー二量体及び他の非特異的産物の増幅を防ぐことによってより正確なSYBR Greenの結果が独自に得られる。このRT2 qPCRマスターミックスは、BioRad iCycler(R)及びMyiQ(R)におけるSYBR Greenに基づく検出を用いたリアルタイムPCR適用に最も適している。
【0083】
配列:
EF1アルファプロモーター(SEQ ID NO:1)
CGTGAGGCTCCGATGCCCGTCAGTGGGCAGAGCGCACATCGCCCACAGTCCCCGAGAAGTTGGGGGGAGGGGTCGGCAATTGAACCGGTGCCTAGAGAAGGTGGCGCGGGGTAAACTGGGAAAGTGATGTCGTGTACTGGCTCCGCCTTTTTCCCGAGGGTGGGGGAGAACCGTATATAAGTGCAGTAGTCGCCGTGAACGTTCTTTTTCGCAACGGGTTTGCCGCCAGAACACAGG
【0084】
LOX1(SEQ ID NO:2)GATCATAACTTCGTATAGCATACATTATACGAAGTTAT
【0085】
LOX2(SEQ ID NO:3)TCGAATAACTTCGTATAGCATACATTATACGAAGTTAT
【0086】
cDNA E6/E7(SEQ ID NO:4)
ATGTTTCAGGACCCACAGGAGCGACCCGGAAAGTTACCACAGTTATGCACAGAGCTGCAAACAACTATACATGATATAATATTAGAATGTGTGTACTGCAAGCAACAGTTACTGCGACGTGAGGTATATGACTTTGCTTTTCGGGATTTATGCATAGTATATAGAGATGGGAATCCATATGCTGTATGTGATAAATGTTTAAAGTTTTATTCTAAAATTAGTGAGTATAGACATTATTGTTATAGTGTGTATGGAACAACATTAGAACAGCAATACAACAAACCGTTGTGTGATTTGTTAATTAGGTGTATTAACTGTCAAAAGCCACTGTGTCCTGAAGAAAAGCAAAGACATCTGGACAAAAAGCAAAGATTCCATAATATAAGGGGTCGGTGGACCGGTCGATGTATGTCTTGTTGCAGATCATCAAGAACACGTAGAGAAACCCAGCTGTAATCATGCATGGAGATACACCTACATTGCATGAATATATGTTAGATTTGCAACCAGAGACAACTGATCTCTACTGTTATGAGCAATTAAATGACAGCTCAGAGGAGGAGGATGAAATAGATGGTCCAGCTGGACAAGCAGAACCGGACAGAGCCCATTACAATATTGTAACCTTTTGTTGCAAGTGTGACTCTACGCTTCGGTTGTGCGTACAAAGCACACACGTAGACATTCGTACTTTGGAAGACCTGTTAATGGGCACACTAGGAATTGTGTGCCCCATCTGTTCTCAGAAACCATAA
【0087】
WPRE(SEQ ID NO:5)
AATCAACCTCTGGATTACAAAATTTGTGAAAGATTGACTGGTATTCTTAACTATGTTGCTCCTTTTACGCTATGTGGATACGCTGCTTTAATGCCTTTGTATCATGCTATTGCTTCCCGTATGGCTTTCATTTTCTCCTCCTTGTATAAATCCTGGTTGCTGTCTCTTTATGAGGAGTTGTGGCCCGTTGTCAGGCAACGTGGCGTGGTGTGCACTGTGTTTGCTGACGCAACCCCCACTGGTTGGGGCATTGCCACCACCTGTCAGCTCCTTTCCGGGACTTTCGCTTTCCCCCTCCCTATTGCCACGGCGGAACTCATCGCCGCCTGCCTTGCCCGCTGCTGGACAGGGGCTCGGCTGTTGGGCACTGACAATTCCGTGGTGTTGTCGGGGAAGCTGACGTCCTTTCCATGGCTGCTCGCCTGTGTTGCCACCTGGATTCTGCGCGGGACGTCCTTCTGCTACGTCCCTTCGGCCCTCAATCCAGCGGACCTTCCTTCCCGCGGCCTGCTGCCGGCTCTGCGGCCTCTTCCGCGTCTTCGCCTTCGCCCTCAGACGAGTCGGATCTCCCTTTGGGCCGCCTCCCCGCCTG
【0088】
Vector_pLV_EF1a−Lox1−E6E7−Lox2_WPRE
(SEQ ID NO:6)
CGCGTCGTGAGGCTCCGATGCCCGTCAGTGGGCAGAGCGCACATCGCCCACAGTCCCCGAGAAGTTGGGGGGAGGGGTCGGCAATTGAACCGGTGCCTAGAGAAGGTGGCGCGGGGTAAACTGGGAAAGTGATGTCGTGTACTGGCTCCGCCTTTTTCCCGAGGGTGGGGGAGAACCGTATATAAGTGCAGTAGTCGCCGTGAACGTTCTTTTTCGCAACGGGTTTGCCGCCAGAACACAGGATCATAACTTCGTATAGCATACATTATACGAAGTTATGATCCGTTTAAACTTGCTGCGACGCGTACTAGTATGTTTCAGGACCCACAGGAGCGACCCGGAAAGTTACCACAGTTATGCACAGAGCTGCAAACAACTATACATGATATAATATTAGAATGTGTGTACTGCAAGCAACAGTTACTGCGACGTGAGGTATATGACTTTGCTTTTCGGGATTTATGCATAGTATATAGAGATGGGAATCCATATGCTGTATGTGATAAATGTTTAAAGTTTTATTCTAAAATTAGTGAGTATAGACATTATTGTTATAGTGTGTATGGAACAACATTAGAACAGCAATACAACAAACCGTTGTGTGATTTGTTAATTAGGTGTATTAACTGTCAAAAGCCACTGTGTCCTGAAGAAAAGCAAAGACATCTGGACAAAAAGCAAAGATTCCATAATATAAGGGGTCGGTGGACCGGTCGATGTATGTCTTGTTGCAGATCATCAAGAACACGTAGAGAAACCCAGCTGTAATCATGCATGGAGATACACCTACATTGCATGAATATATGTTAGATTTGCAACCAGAGACAACTGATCTCTACTGTTATGAGCAATTAAATGACAGCTCAGAGGAGGAGGATGAAATAGATGGTCCAGCTGGACAAGCAGAACCGGACAGAGCCCATTACAATATTGTAACCTTTTGTTGCAAGTGTGACTCTACGCTTCGGTTGTGCGTACAAAGCACACACGTAGACATTCGTACTTTGGAAGACCTGTTAATGGGCACACTAGGAATTGTGTGCCCCATCTGTTCTCAGAAACCATAATCTAGCGTCGAATAACTTCGTATAGCATACATTATACGAAGTTATTCGACAATCAACCTCTGGATTACAAAATTTGTGAAAGATTGACTGGTATTCTTAACTATGTTGCTCCTTTTACGCTATGTGGATACGCTGCTTTAATGCCTTTGTATCATGCTATTGCTTCCCGTATGGCTTTCATTTTCTCCTCCTTGTATAAATCCTGGTTGCTGTCTCTTTATGAGGAGTTGTGGCCCGTTGTCAGGCAACGTGGCGTGGTGTGCACTGTGTTTGCTGACGCAACCCCCACTGGTTGGGGCATTGCCACCACCTGTCAGCTCCTTTCCGGGACTTTCGCTTTCCCCCTCCCTATTGCCACGGCGGAACTCATCGCCGCCTGCCTTGCCCGCTGCTGGACAGGGGCTCGGCTGTTGGGCACTGACAATTCCGTGGTGTTGTCGGGGAAGCTGACGTCCTTTCCATGGCTGCTCGCCTGTGTTGCCACCTGGATTCTGCGCGGGACGTCCTTCTGCTACGTCCCTTCGGCCCTCAATCCAGCGGACCTTCCTTCCCGCGGCCTGCTGCCGGCTCTGCGGCCTCTTCCGCGTCTTCGCCTTCGCCCTCAGACGAGTCGGATCTCCCTTTGGGCCGCCTCCCCGCCTGGAATTCGAGCT
CGGTACCTTTAAGACCAATGACTTACAAGGCAGCTGTAGATCTTAGCCACTTTTTAAAAGAAAAGGGGGGACTGGAAGGGCTAATTCACTCCCAACGAAGACAAGATCTGCTTTTTGCTTGTACTGGGTCTCTCTGGTTAGACCAGATCTGAGCCTGGGAGCTCTCTGGCTAACTAGGGAACCCACTGCTTAAGCCTCAATAAAGCTTGCCTTGAGTGCTTCAAGTAGTGTGTGCCCGTCTGTTGTGTGACTCTGGTAACTAGAGATCCCTCAGACCCTTTTAGTCAGTGTGGAAAATCTCTAGCAGTAGTAGTTCATGTCATCTTATTATTCAGTATTTATAACTTGCAAAGAAATGAATATCAGAGAGTGAGAGGAACTTGTTTATTGCAGCTTATAATGGTTACAAATAAAGCAATAGCATCACAAATTTCACAAATAAAGCATTTTTTTCACTGCATTCTAGTTGTGGTTTGTCCAAACTCATCAATGTATCTTATCATGTCTGGCTCTAGCTATCCCGCCCCTAACTCCGCCCAGTTCCGCCCATTCTCCGCCCCATGGCTGACTAATTTTTTTTATTTATGCAGAGGCCGAGGCCGCCTCGGCCTCTGAGCTATTCCAGAAGTAGTGAGGAGGCTTTTTTGGAGGCCTAGGCTTTTGCGTCGAGACGTACCCAATTCGCCCTATAGTGAGTCGTATTACGCGCGCTCACTGGCCGTCGTTTTACAACGTCGTGACTGGGAAAACCCTGGCGTTACCCAACTTAATCGCCTTGCAGCACATCCCCCTTTCGCCAGCTGGCGTAATAGCGAAGAGGCCCGCACCGATCGCCCTTCCCAACAGTTGCGCAGCCTGAATGGCGAATGGCGCGACGCGCCCTGTAGCGGCGCATTAAGCGCGGCGGGTGTGGTGGTTACGCGCAGCGTGACCGCTACACTTGCCAGCGCCCTAGCGCCCGCTCCTTTCGCTTTCTTCCCTTCCTTTCTCGCCACGTTCGCCGGCTTTCCCCGTCAAGCTCTAAATCGGGGGCTCCCTTTAGGGTTCCGATTTAGTGCTTTACGGCACCTCGACCCCAAAAAACTTGATTAGGGTGATGGTTCACGTAGTGGGCCATCGCCCTGATAGACGGTTTTTCGCCCTTTGACGTTGGAGTCCACGTTCTTTAATAGTGGACTCTTGTTCCAAACTGGAACAACACTCAACCCTATCTCGGTCTATTCTTTTGATTTATAAGGGATTTTGCCGATTTCGGCCTATTGGTTAAAAAATGAGCTGATTTAACAAAAATTTAACGCGAATTTTAACAAAATATTAACGTTTACAATTTCCCAGGTGGCACTTTTCGGGGAAATGTGCGCGGAACCCCTATTTGTTTATTTTTCTAAATACATTCAAATATGTATCCGCTCATGAGACAATAACCCTGATAAATGCTTCAATAATATTGAAAAAGGAAGAGTATGAGTATTCAACATTTCCGTGTCGCCCTTATTCCCTTTTTTGCGGCATTTTGCCTTCCTGTTTTTGCTCACCCAGAAACGCTGGTGAAAGTAAAAGATGCTGAAGATCAGTTGGGTGCACGAGTGGGTTACATCGAACTGGATCTCAACAGCGGTAAGATCCTTGAGAGTTTTCGCCCCGAAGAACGTTTTCCAATGATGAGCACTTTTAAAGTTCTGCTATGTGGCGCGGTATTATCCCGTATTGACGCCGGGCAAGAGCAACTCGGTCGCCGCATACACTATTCTCAGAATGACTTGGTTGAGTACTCACCAGTCACAGAAAAGCATCTTACGGATGGCATGACAGTAAGAGAATTATGCAGTGCTGCCATAACCATGAGTGATAACACTGCGGCCAACTTACTTCTGACAACGATCGGAGGACCGAAGGAGCTAACCGCTTTTTTGCACAACATGGGGGATCATGTAACTCGCCTTGATCGTTGGGAACCGGAGCTGAATGAAGCCATACCAAACGACGAGCGTGACACCACG
ATGCCTGTAGCAATGGCAACAACGTTGCGCAAACTATTAACTGGCGAACTACTTACTCTAGCTTCCCGGCAACAATTAATAGACTGGATGGAGGCGGATAAAGTTGCAGGACCACTTCTGCGCTCGGCCCTTCCGGCTGGCTGGTTTATTGCTGATAAATCTGGAGCCGGTGAGCGTGGGTCTCGCGGTATCATTGCAGCACTGGGGCCAGATGGTAAGCCCTCCCGTATCGTAGTTATCTACACGACGGGGAGTCAGGCAACTATGGATGAACGAAATAGACAGATCGCTGAGATAGGTGCCTCACTGATTAAGCATTGGTAACTGTCAGACCAAGTTTACTCATATATACTTTAGATTGATTTAAAACTTCATTTTTAATTTAAAAGGATCTAGGTGAAGATCCTTTTTGATAATCTCATGACCAAAATCCCTTAACGTGAGTTTTCGTTCCACTGAGCGTCAGACCCCGTAGAAAAGATCAAAGGATCTTCTTGAGATCCTTTTTTTCTGCGCGTAATCTGCTGCTTGCAAACAAAAAAACCACCGCTACCAGCGGTGGTTTGTTTGCCGGATCAAGAGCTACCAACTCTTTTTCCGAAGGTAACTGGCTTCAGCAGAGCGCAGATACCAAATACTGTCCTTCTAGTGTAGCCGTAGTTAGGCCACCACTTCAAGAACTCTGTAGCACCGCCTACATACCTCGCTCTGCTAATCCTGTTACCAGTGGCTGCTGCCAGTGGCGATAAGTCGTGTCTTACCGGGTTGGACTCAAGACGATAGTTACCGGATAAGGCGCAGCGGTCGGGCTGAACGGGGGGTTCGTGCACACAGCCCAGCTTGGAGCGAACGACCTACACCGAACTGAGATACCTACAGCGTGAGCTATGAGAAAGCGCCACGCTTCCCGAAGGGAGAAAGGCGGACAGGTATCCGGTAAGCGGCAGGGTCGGAACAGGAGAGCGCACGAGGGAGCTTCCAGGGGGAAACGCCTGGTATCTTTATAGTCCTGTCGGGTTTCGCCACCTCTGACTTGAGCGTCGATTTTTGTGATGCTCGTCAGGGGGGCGGAGCCTATGGAAAAACGCCAGCAACGCGGCCTTTTTACGGTTCCTGGCCTTTTGCTGGCCTTTTGCTCACATGTTCTTTCCTGCGTTATCCCCTGATTCTGTGGATAACCGTATTACCGCCTTTGAGTGAGCTGATACCGCTCGCCGCAGCCGAACGACCGAGCGCAGCGAGTCAGTGAGCGAGGAAGCGGAAGAGCGCCCAATACGCAAACCGCCTCTCCCCGCGCGTTGGCCGATTCATTAATGCAGCTGGCACGACAGGTTTCCCGACTGGAAAGCGGGCAGTGAGCGCAACGCAATTAATGTGAGTTAGCTCACTCATTAGGCACCCCAGGCTTTACACTTTATGCTTCCGGCTCGTATGTTGTGTGGAATTGTGAGCGGATAACAATTTCACACAGGAAACAGCTATGACCATGATTACGCCAAGCGCGCAATTAACCCTCACTAAAGGGAACAAAAGCTGGAGCTGCAAGCTTAATGTAGTCTTATGCAATACTCTTGTAGTCTTGCAACATGGTAACGATGAGTTAGCAACATGCCTTACAAGGAGAGAAAAAGCACCGTGCATGCCGATTGGTGGAAGTAAGGTGGTACGATCGTGCCTTATTAGGAAGGCAACAGACGGGTCTGACATGGATTGGACGAACCACTGAATTGCCGCATTGCAGAGATATTGTATTTAAGTGCCTAGCTCGATACAATAAACGGGTCTCTCTGGTTAGACCAGATCTGAGCCTGGGAGCTCTCTGGCTAACTAGGGAACCCACTGCTTAAGCCTCAATAAAGCTTGCCTTGAGTGCTTCAAGTAGTGTGTGCCCGTCTGTTGTGTGACTCTGGTAACTAGAGATCCCTCAGACCCTTTTAGTCAGTGTGGAAAATCTCTAGCAGTGGCGCCCGAACAGGGACCTGAAAGCGAAAGGGAAACCA
GAGCTCTCTCGACGCAGGACTCGGCTTGCTGAAGCGCGCACGGCAAGAGGCGAGGGGCGGCGACTGGTGAGTACGCCAAAAATTTTGACTAGCGGAGGCTAGAAGGAGAGAGATGGGTGCGAGAGCGTCAGTATTAAGCGGGGGAGAATTAGATCGCGATGGGAAAAAATTCGGTTAAGGCCAGGGGGAAAGAAAAAATATAAATTAAAACATATAGTATGGGCAAGCAGGGAGCTAGAACGATTCGCAGTTAATCCTGGCCTGTTAGAAACATCAGAAGGCTGTAGACAAATACTGGGACAGCTACAACCATCCCTTCAGACAGGATCAGAAGAACTTAGATCATTATATAATACAGTAGCAACCCTCTATTGTGTGCATCAAAGGATAGAGATAAAAGACACCAAGGAAGCTTTAGACAAGATAGAGGAAGAGCAAAACAAAAGTAAGACCACCGCACAGCAAGCGGCCGCTGATCTTCAGACCTGGAGGAGGAGATATGAGGGACAATTGGAGAAGTGAATTATATAAATATAAAGTAGTAAAAATTGAACCATTAGGAGTAGCACCCACCAAGGCAAAGAGAAGAGTGGTGCAGAGAGAAAAAAGAGCAGTGGGAATAGGAGCTTTGTTCCTTGGGTTCTTGGGAGCAGCAGGAAGCACTATGGGCGCAGCCTCAATGACGCTGACGGTACAGGCCAGACAATTATTGTCTGGTATAGTGCAGCAGCAGAACAATTTGCTGAGGGCTATTGAGGCGCAACAGCATCTGTTGCAACTCACAGTCTGGGGCATCAAGCAGCTCCAGGCAAGAATCCTGGCTGTGGAAAGATACCTAAAGGATCAACAGCTCCTGGGGATTTGGGGTTGCTCTGGAAAACTCATTTGCACCACTGCTGTGCCTTGGAATGCTAGTTGGAGTAATAAATCTCTGGAACAGATTTGGAATCACACGACCTGGATGGAGTGGGACAGAGAAATTAACAATTACACAAGCTTAATACACTCCTTAATTGAAGAATCGCAAAACCAGCAAGAAAAGAATGAACAAGAATTATTGGAATTAGATAAATGGGCAAGTTTGTGGAATTGGTTTAACATAACAAATTGGCTGTGGTATATAAAATTATTCATAATGATAGTAGGAGGCTTGGTAGGTTTAAGAATAGTTTTTGCTGTACTTTCTATAGTGAATAGAGTTAGGCAGGGATATTCACCATTATCGTTTCAGACCCACCTCCCAACCCCGAGGGGACCCGACAGGCCCGAAGGAATAGAAGAAGAAGGTGGAGAGAGAGACAGAGACAGATCCATTCGATTAGTGAACGGATCTCGACGGTATCGGTTAACTTTTAAAAGAAAAGGGGGGATTGGGGGGTACAGTGCAGGGGAAAGAATAGTAGACATAATAGCAACAGACATACAAACTAAAGAATTACAAAAACAAATTACAAAAATTCAAAATTTTATCGATCACGAGACTAGCCTCGAGAAGCTTGAT
【0089】
結果
A有足細胞
1997年にMundel P 等によって記載されたプロトコールに従って新生仔ラット腎糸球体から有足細胞を単離した。単離した後、コラーゲンコートされた24ウェルプレート上の10%FCS入りのDMEM中で糸球体を培養した。24時間後、有足細胞前駆体は、付着した糸球体から外へ増殖し始めた(図2)。
【0090】
この段階で、細胞に異なるレンチウイルスコンストラクトを形質導入し、そして培養を長期間(2〜4週間)継続した。急速に増殖している細胞の島を単離し、広げ、そしてRT PCR及び免疫染色を用いて特徴づけた。SV40 LTagの発現は、免疫染色によって制御した。細胞の100%は、核中に大量のSV40 LT Agを蓄積し(図3C)、形質導入されてない又はGFP形質導入された細胞では、染色を検出することができなかった(データは示さず)。hTERTによる形質導入は、(定量的RT PCRデータによればhTERTは500倍を超えて過剰発現されたが)増殖速度を高めるには不十分であり、そして数回試行した後、断念した。HPV E6/E7単独による形質導入もまた、有足細胞不死化に十分でなかったが、SV40 LT Agと組み合わせて用いたとき、かなり多くの不死化クローンを増強した。10回を超える継代後、細胞にレンチウイルスCreリコンビナーゼを形質導入した。3日後、特異的な抗SV40 LTag抗体で染色した。多くの細胞は、LT Agの有意な減少又は完全な欠如を示し、これはCreが細胞ゲノムから組込まれたコピーのすべて又は大多数を切除したことを示している(図3D)。LT Agが減少した(すなわち陰性)細胞は、その増殖を停止し、3つはサイズが大きくなり、そして有足細胞の典型的な形態を獲得した(図3A及びB)。切除効率は比較的低く、約30〜50%であった。有足細胞のいくつかの高度に特異的な遺伝子マーカー、例えば、WT1、シナプトポジンは、この細胞中で再発現されたが、ネフリン及びポドシンのような他のものは、極めて低いレベルでしか発現されなかった(図3C〜F及び表1)。
【0091】
【表1】

【0092】
結論
有足細胞は、良好に不死化することができるが、さらなる実験的使用のため「不死化遺伝子」の切除によって非増殖状態に戻らなければならない。目下、アデノウイルスCreを産生しており、そして切除効率を高める目的で試験する。切除しなければ、不死化細胞は、極めて急速に増殖し(倍加時間12〜15時間)そして有足細胞の多くの特異的マーカーを消失するか又は低下したレベルで発現する。
【0093】
B 初代ヒト肝細胞(PHH)
PHHをLonzaから購入し、そしてコラーゲンコートされたプレート上で培養した。細胞は増殖しないが、脱分化の明らかな徴候がなくても培養を10〜14日間持続することができる。PHHにSV40 LT Ag又はE6/E7(hTERTを用いない)を形質導入し、そして数日後、増殖が始まり、そのサイズは、時間と共に縮小し、そしてそれは容易に継代培養することができる。本発明者らは、これらの細胞を、20回を超えて継代培養し、そしてその表現型は変化しなかったが、対照肝細胞と比較すると、それは明らかにその表現型を部分的に失った。特に、CRP及びハプトグロビンのような肝細胞特異的マーカーの発現は、とりわけSV40 LT Ag中で大きく低下しているが、E6/E7不死化細胞中では、ハプトグロビン発現は保たれており、そしてCRP発現は、なお比較的高く、初代細胞と比較して10%である(表2)。重要なことに、肝細胞関連の転写因子HNF4アルファの発現は、E6/E7形質導入PHH中で2倍しか低下しておらず、そしてSV40 LT Ag中ではより大きく8倍低下している。
【0094】
【表2】
【0095】
cDNAアレイ分析は、SV40 LT Ag又はE6/E7ウイルス性初期タンパク質の過剰発現によって生じる不死化が、細胞周期の制御に関与することが知られているいくつかの遺伝子、すなわちE2F1転写因子、サイクリンA2、CDC2/CDK1−サイクリン依存性キナーゼ1の転写を強く誘導することを示している(表2)。トランスクリプトーム変化のより詳細な説明は、後に報告する。
【0096】
SV40 LT Agは、核p53の巨大な蓄積、そしてE6/E7は、p53の分解及び消失をもたらした(図4)。
【0097】
両方の場合、p53は、完全に不活性化され、そしてそのトランス活性化及びトランス抑制性を失う。特に、CDK2の阻害剤、そしてp53の直接標的の1つ、p21WAF1(CIP1又はCDKN1A)の発現は、核中でp53の蓄積にもかかわらず、SV40 LT Ag不死化細胞中で変化しなかった(データは示さず)。一方、別のCDK阻害剤、p16INK4のmRNA発現は、SV40 LT Ag及びE6/E7−不死化細胞中で強く上方制御された(データは示さず)。この複雑な現象の潜在的な機構は、公開されているが、明確な説明はない(Pei 及び Xiong, 2005)。
【0098】
結論:初代ヒト肝細胞は、SV40 LT Ag又はヒト乳頭腫E6/E7初期タンパク質のいずれかのレンチウイルスが介在する送達により非常に有効に不死化することができる。得られた細胞株は、極めて急速に増殖し(倍加時間は、20時間未満である)、そしてとりわけE6/E7に誘導された不死化の場合、肝細胞の多くの重要なマーカーを、なお保っていた。これらの細胞は、細胞分裂しない初代肝細胞の代わりにさまざまな生物学的目的に用いることができる。
C 初代内皮細胞
HUVEC(ヒト臍帯静脈内皮細胞)をPromoCellから購入し、そしてFCS、ECG、hEGF、hbFGF及びHC−500を含む内皮細胞特異的培地を用いて、コラーゲンコートされたプレートにおいて培養した。準集密細胞にSV40 LT Ag又はE6/E7レンチウイルス粒子のいずれかを形質導入した。hTERT単独の形質導入は、有効でなかった(他の試験細胞タイプにおいても同様)。SV40 LT Ag及びE6/E7−形質導入された細胞では、より速い増殖速度が見られたが、hTERTでは見られなかった。FACS分析は、両方の「不死化遺伝子」、LT Ag及びE6/E7により、S−G2−M画分の増加に伴ってG1/G0画分の減少(60から20%まで)が生じることを示した。BrdU取り込み/染色(データは示さず)によりFACSデータを確認した。驚くべきことに、非常により速い増殖速度にもかかわらず、内皮特異的マーカー、CD31(PECAM)及びvWFは、不死化HUVEC中において同じか又はさらにより高レベルで発現された(図5及び表3参照)。
【0099】
【表3】
【0100】
SV40 LT Ag及びE6/E7を過剰発現するHUVECの両方で、内皮特異的なNOS(eNOS)の中程度の減少が検出された。試験した他の初代細胞と同様に、SV40 LT Agは、核p53タンパク質における莫大な増加をもたらした、そしてE6/E7では減少した(データは示さず)。
【0101】
結論:不死化された初代内皮細胞(HUVEC)は、その主要な分化マーカーを維持するが親細胞より3〜5倍急速に増殖する。形態学的に、細胞サイズの縮小のみが示され、そして核多形性により非不死化細胞と区別される。それらは、さまざまな実験タイプに用いることができ、そこでは内皮「バックグラウンド」が必要とされる。
【0102】
不死化の効率。初期ウイルス性タンパク質SV40 LT Ag又はHPV E6/E7を過剰発現するためにレンチウイルス送達系を用いて、本発明者らは、異なる初代ヒト及び齧歯類細胞を不死化することができた。本発明の主要な目標は、初代ラット腎臓有足細胞を不死化することであった。驚くべきことに、レンチウイルスを齧歯類細胞に形質導入することは、初代ヒト細胞と比較してずっと困難であった。これにより、おそらく、なぜ本発明者らがSV40 LT Ag又はE6/E7単独でラット腎臓細胞の不死化を達成することができなかったかを説明することができる。しかし、2つのレンチウイルスコンストラクトの組み合わせ(最初にSV40 LT Ag、そして2日後にE6/E7を細胞に逐次形質導入した)により単離されたラット糸球体からいくつかの永続的な細胞株が得られた。急速に増殖する細胞は、分化マーカーを部分的に失っておりそして形態学的に有足細胞に似ていなかった。従って、Creリコンビナーゼを用いて「不死化遺伝子」を切除する必要があった。レンチウイルスCreにより形質導入すると、細胞は増殖を停止し、サイズが大きくなり、そしてネフリン(NPHS1)、シナプトポジン、ウィルムス腫瘍1(WT1)を含む主な有足細胞特異的マーカーを再発現した。残念なことに、主な有足細胞特異的遺伝子の1つ、ポドシン(NPHS2)は、Cre形質導入された細胞中で再び現れなかった。一般に、本発明者らは、新たに単離されたラット糸球体の有足細胞中でポドシンが短時間(多くとも72時間)しか発現されなかったことを観察した。従って、作製された不死化細胞は、部分的に脱分化したと考えるべきである。ポドシン発現を急速に消失させるもの(多くの他の有足細胞特異的遺伝子が発現され続けるときに)は、今のところ明らかでない。
【0103】
最初に、本発明者らは、2つのさらなる遺伝子、テロメラーゼのヒト触媒サブユニット(hTERT)及び温度感受性(ts)SV40ラージT抗原突然変異体を不死化に使用するつもりであった。これらのcDNAコンストラクトを含むレンチウイルス粒子を作製し、そして形質導入された細胞中でhTERT及びSV40 LT Ag−tsの過剰発現が確認されたが、不死化は達成されなかった。HUVEC、肝細胞、平滑筋細胞、間葉幹細胞を含むいくつかのヒト初代細胞でhTERTを試験したが、全く成功しなかった。従って、少なくとも上記の初代齧歯類及びヒト細胞においてhTERT単独では不死化に不十分であると結論する必要があった。
【0104】
(上記の)初代ヒト細胞の不死化は、SV40 LT Ag及びE6/E7レンチウイルスコンストラクトにより達成に成功した。SV40 LT Agで不死化された細胞(細胞の由来に依存しない)は、E6/E7発現細胞よりもずっとより急速に増殖し、そして分化マーカーの部分的又は完全な喪失を伴って有意な核多形性を示す。E6/E7の発現により増殖が活性化されるが、分化の部分的喪失しか起こらず、従って新規な細胞株の世代にとってより適切な系となる。興味深いことに、HUVECでは、実際に分化状態の喪失は見られず(図6及び表4参照)、そして「不死化遺伝子」を切除する必要なしに不死化細胞を直接用いることができる。
【0105】
不死化の機構。ラージT−抗原(LT Ag)、ウイルス性癌タンパク質は、SV40によってコードされた主要な初期遺伝子産物である。LT Ag及びヘリカーゼファミリーIIIのメンバー(AAA+スーパーファミリーの部分)。このタンパク質は、ウイルスDNA複製にとって重要である(Li 及び Kelly 1984; Neuwald 等 1999)。SV40 LT Agは、休止細胞の増殖を誘導し、いくつかの細胞タイプを形質転換し、そして実験動物における腫瘍形成を誘導する(Fanning 及び Knippers 1992)。LT Agによって生じる不死化/形質転換の正確な機構は、なお明らかではないが、大多数のデータは、p53を含むいくつかの重要な細胞腫瘍抑制因子の機能への干渉を通して細胞周期チェックポイントを乗り越えることができることを示している(Srinivasan 等 1997; Pipas 1998; DeCaprio 1999; Kim 等 2001)。試験したすべての初代ヒト及び齧歯類細胞において、SV40 LT Agをコードするレンチウイルスによる形質導入によりp53の核内蓄積が莫大になった(図3及び4)。反対に、HPV E6/E7で形質導入された細胞では、核p53は消失した(図4)。興味深いことに、いくつかの他のウイルス性癌タンパク質(アデノウイルスE1B、B型肝炎ウイルスXタンパク質、乳頭腫ウイルスE6、エプスタイン−バーウィルスEBNA−5及びヒトTリンパ球向性ウイルスtaxタンパク質)もまたp53を標的とし、強化されたプロテアソーム分解を通してそれを不活性化する。しかし、SV40 LT Agは、主に異なる機構を用い;細胞のp53を安定化する(タンパク質の半減期を高める)と同時にそのトランス活性化活性を阻害する(Deppert 等 1989; Pipas 及び Levine 2001)。p53安定化及び不活性化の機構は、SV40 LT Agとp53との直接結合に左右される。近年では、p53はラージT抗原と共結晶化された。予期せぬことに、構造は、6つのp53モノマーを結合しているLT Agの六量体複合体を示した。結合されたSV40 LTは、p53制御遺伝子のプロモーター中でp53DNA−結合ドメインとp53応答エレメントとの相互作用を阻止する。それは、p53のDNA−結合領域での劇的な立体配置的変化によって生じ、これは、p53内の独特の「メチオニンスイッチ」を通して部分的に達成される(Lilyestrom W 等, 2006)。
【0106】
ヒト乳頭腫ウイルス(HPV)初期タンパク質E6の場合におけるp53の不活性化は、ユビキチン経路を通してp53分解が高められることにより生じる。E6は、細胞ユビキチン−リガーゼを通してp53タンパク質に結合し、E6結合タンパク質(E6−AP)は、酵素のユビキチン複合体を集め、p53上でリシンをユビキチン化し、そしてそのタンパク質分解を開始する。
【0107】
HPV E7は、網膜芽腫腫瘍抑制タンパク質pRbの機能を不活性化する。E7タンパク質は、主に核中に局在化されており、ここで網膜芽腫遺伝子産物pRb(ポケットRb)に結合して細胞周期のS期への進行を促進する(Zur Hausen, 2002)。正常細胞では、pRbは、初期G1期に低リン酸化され、そしてE2F転写因子に結合し、転写リプレッサーとして機能する複合体を形成する。9回リン酸化されると複合体は解離し、E2Fは転写活性化因子として作用することが可能になる。低リン酸化pRbと結合することによって、E7は、それとE2Fとの結合を妨げ、それによって細胞周期進行を促進する。さらに、E7に誘導されたユビキチン介在性pRb分解は、細胞周期停止を有効に克服する際に不可欠であると考えられる(Fehrmann & Laimins, 2003)。
【0108】
従って、SV40及びHPVによる不死化に含まれる一般的な機構は、p53及びpRb不活性化である。非常に最近のデータは、p53が転写活性化だけでなく抑制にも介在していることを示している(Spurgers 等, 2006; Scian 等, 2008)。重要なことに、いくつかのp53に依存して抑制された遺伝子(cdc2、cdk4、サイクリン2A、cdc20、Spurgers 等, 2006参照)は、E2F1−3によってトランス活性化されることが示された(Zhu 等, 2004)。E2F1は、pRbから解離されると(pRbリン酸化又はウイルス性タンパク質によるpRbの機能不活性化のいずれかの結果として)、それ自体の転写を活性化し(正のフィードバック)、そしてG1/S及びG2/M移行にとって重要な標的遺伝子(cdc2、cdk4、サイクリン2A、cdc20)の転写を活性化する。
【0109】
本発明者らは、SV40 LT Ag又はHPV E6/E7によって不死化された異なる細胞タイプにおける増殖及び分化の制御に関与する200を超える遺伝子の発現を分析した。すべての細胞タイプにおいて、E2F1、cdc2/cdk1及びサイクリンA2の増強された発現が見られ、そしていくつかの場合(例えば、初代ヒト肝細胞)、数百倍であった(表2参照)。上記のようなサイクリンA2及びcdc2/cdk1は、p53によって転写的に抑制するか又はE2F1によって転写的に活性化することができる(図6)。従って、本発明者らは、SV40及びHPVが介在する不死化には2つの重要な事象:第1に、p53及びpRbの不活性化(いくつかの群によって以前に示した)、そして第2に、E2Fそれ自体、そしてその標的、サイクリンA2、サイクリンB及びcdc2の転写を刺激することができる転写的に活性なE2Fタンパク質の放出が関与していることを提案する。最近示されたように、cdc2/cdk1は、哺乳動物細胞における唯一の必須な細胞周期Cdkである(Santamaria 等, 2007)。これらのデータは、Cdk1が、不死化、そしておそらくまた二本鎖DNA HPV及びSV40ウイルスによっても生じる発癌の重要な調節因子とみなすことができることを示している。特異的CDK1阻害剤は、確立されたHPV若しくはSV40病因又は突然変異したp53若しくはRbによる腫瘍に対して非常に有効であると期待することができる。
【0110】
文献
DeCaprio JA: The role of the J domain of SV40 large T in cellular transformation. Biologicals. 1999 Mar; 27(1):23-8.
Deppert W, Steinmayer T, Richter W: Cooperation of SV40 large T antigen and the cellular protein p53 in maintenance of cell transformation. Oncogene. 1989 Sep;4(9):1103-10.
Deppert W, Steinmayer T: Metabolic stabilization of p53 in SV40-transformed cells correlates with expression of the transformed phenotype but is independent from complex formation with SV40 large T antigen. Curr Top Microbiol Immunol. 1989; 44:77-83.
Fanning E, Knippers R: Structure and function of simian virus 40 large tumor antigen. Annu Rev Biochem. 1992; 61:55-85. Review.
Fehrmann F, Laimins LA: Human papillomaviruses: targeting differentiating epithelial cells for malignant transformation. Oncogene. 2003 Aug 11;22(33):5201-7. Review.
Li JJ, Kelly TJ: Simian virus 40 DNA replication in vitro. Proc Natl Acad Sci U S A. 1984 Nov; 81(22):6973-7.
Lilyestrom W, Klein MG, Zhang R, Joachimiak A, Chen XS: Crystal structure of SV40 large T-antigen bound to p53: interplay between a viral oncoprotein and a cellular tumor suppressor. Genes Dev. 2006 Sep 1; 20(17):2373-82.
Mundel P, Reiser J, Kriz W: Induction of differentiation in cultured rat and human podocytes. J Am Soc Nephrol 8: 697-705, 1997.
Neuwald AF, Aravind L, Spouge JL, Koonin EV: AAA+: A class of chaperone-like ATPases associated with the assembly, operation, and disassembly of protein complexes. Genome Res. 1999 Jan;9(1):27-43.
Pei XH, Xiong Y: Biochemical and cellular mechanisms of mammalian CDK inhibitors: a few unresolved issues. Oncogene. 2005 Apr 18; 24(17):2787-95. Review.
Pipas JM: Molecular chaperone function of the SV40 large T antigen. Dev Biol Stand.1998; 94:313-9. Review.
Pipas JM, Levine AJ: Role of T antigen interactions with p53 in tumorigenesis. Semin Cancer Biol. 2001 Feb; 11(1):23-30.
Santamaria D, Barriere C, Cerqueira A, Hunt S, Tardy C, Newton K, Caceres JF, Dubus P, Malumbres M, Barbacid M: Cdk1 is sufficient to drive the mammalian cell cycle. Nature. 2007 Aug 16;448(7155):811-5.
Scian MJ, Carchman EH, Mohanraj L, Stagliano KE, Anderson MA, Deb D, Crane BM, Kiyono T, Windle B, Deb SP, Deb S: Wild-type p53 and p73 negatively regulate expression of proliferation related genes. Oncogene. 2008 Apr 17;27(18):2583-93. Epub 2007 Nov 5.
Srinivasan A, McClellan AJ, Vartikar J, Marks I, Cantalupo P, Li Y, Whyte P, Rundell K, Brodsky JL, Pipas JM: The amino-terminal transforming region of simian virus 40 large T and small t antigens functions as a J domain. Mol Cell Biol. 1997 Aug; 17(8):4761-73.
Spurgers KB, Gold DL, Coombes KR, Bohnenstiehl NL, Mullins B, Meyn RE, Logothetis CJ, McDonnell TJ: Identification of cell cycle regulatory genes as principal targets of p53- mediated transcriptional repression. J Biol Chem. 2006 Sep 1;281(35):25134-42. Epub 2006 Jun 23.
Zhu W, Giangrande PH, Nevins JR: E2Fs link the control of G1/S and G2/M transcription. EMBO J. 2004 Nov 24;23(23):4615-26. Epub 2004 Oct 28
Zur Hausen H: Papillomaviruses and cancer: from basic studies to clinical application. Nat Rev Cancer. 2002 May; 2(5):342-50. Review.
【技術分野】
【0001】
本発明は、可逆的に不死化された細胞の作製方法、上記方法によって入手可能な細胞、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸を含む真核細胞及びこれらの細胞を再分化する方法に関する。
【背景技術】
【0002】
早くも19世紀には、摘出された動物心臓の鼓動を体外で維持する方法があった。20世紀の初めには、組織培養の方法論を確立する実験が報告された。細胞培養技術は、1940年代及び1950年代に進歩してウイルス学における研究を支えた。動物細胞培養は、1900年代中頃に一般的な実験室技術になった。
【0003】
一般に、細胞培養の概念には、生体外培養のために組織から細胞、別名初代細胞(primary cells)を単離することが含まれる。このために、コラゲナーゼ、トリプシン又はプロナーゼのような酵素による酵素消化によって初代細胞を遊離してもよい。別法として、組織の断片を適切な増殖培地に置き、そして培養のための増殖用の細胞を入手することができる。
【0004】
いくつかの腫瘍から誘導された細胞を除いて、ほとんどの初代細胞培養物は、限られた寿命を有する。さらにまた、多くの場合、初代細胞は、培養における増殖能が限られており、そして継代延長により複製老化を受けるか(すなわち内皮細胞、平滑筋細胞)又は全く増殖しない(例えば肝細胞、心筋細胞)。
【0005】
従って、細胞に基づく実験は、複数のドナーを必要とし、そしてヒトの場合、細胞により重大な問題:ドナーの遺伝的差異、薬物乱用などが生じる。
【0006】
多くの調査実験は、腫瘍から誘導された細胞株で実施する。しかし、腫瘍由来の細胞は、分化が不正確であることが多く、そして重要な組織特異的遺伝子の発現が遺伝的に不安定である。従って、それらの欠点を回避することによって初代細胞及び腫瘍由来の細胞株の利点を合わせた新たな細胞を提供することが望ましい。
【0007】
初代細胞の増殖能が限られている一つの理由は、細胞分裂ごとに生じる進行性のテロメア短縮化であり、これは最終的に細胞老化に至る。SV40 T抗原を発現させて第2の不可逆的な危機が生じる前にさらなる回数の細胞分裂により細胞を成長させることができる細胞の改変方法が報告されている。生体内で収穫された初代細胞をインビトロで無限に増殖するように誘導する方法を細胞不死化と称する。
【0008】
いくつかの研究は、一連の増殖促進及び抗老化遺伝子を組み合わせて発現することが必要な段階的方法においてヒト細胞を形質転換することができることを示した。従って、適当な遺伝子操作により、培養において制御された細胞増殖を誘導することは可能でありうる。さらに、培養においてヒト不死化細胞株を樹立することは可能であった。ヒトテロメラーゼ触媒ユニットの発現により、増殖停止を阻止し、そしてヒト線維芽細胞及び上皮細胞を不死化することができる。しかし、ヒトテロメラーゼ触媒ユニット単独の発現では、限定数しか不死化することができなかった。ほとんどの細胞タイプは、SV40 T抗原をコードしている遺伝子のような第2の遺伝子の発現を必要とする。培養における不死化細胞の連続増殖は、その初代細胞に相当する物と比較したときに組織特異的遺伝子の発現を下方制御する。2つの癌遺伝子(SV40 T癌タンパク質及びH−rasの癌遺伝子対立遺伝子)と組み合わせたヒトテロメラーゼ触媒サブユニットの発現は、上皮細胞及び線維芽細胞のような正常なヒト細胞に直接的な腫瘍形成転換を生じることが報告されている。さらにまた、ヒトテロメラーゼ触媒ユニットが単独で又は1つ若しくはそれ以上の他の遺伝子と組み合わせて、腎近位尿細管上皮細胞で発現された場合にも初代ヒト細胞の不死化が報告されている。近位尿細管細胞を不死化するこの方法は、これらの細胞により十分に機能したが、HUVEC、肝細胞、平滑筋細胞及び間葉性幹細胞を含むいくつかの他のヒト初代細胞では機能しなかった。
【発明の概要】
【発明が解決しようとする課題】
【0009】
従って、細胞又はその前駆体が種々の細胞タイプから誘導することができ、及び/又は複製老化を示すことなく増殖することができると同時に初代細胞の魅力的な性質、例えば安定な遺伝子型及びその分化レベルを有する細胞及びその作製方法を提供することが本発明の目的である。新規な細胞モデルの開発は、生物学的及び薬学的研究にとって、特に化合物プロファイリング、表現型スクリーニング、標的同定及び/又は標的検証にとって最も重要である。
【課題を解決するための手段】
【0010】
驚くべきことに、ヒト乳頭腫ウイルス(HPV)E6/E7は、上皮細胞、内皮細胞及び肝細胞を含むいくつかの真核細胞中で発現されたときに、細胞を不死化できることが見出された(実施例を参照のこと)。
【0011】
従って、第1の態様において、本発明は、可逆的に不死化された細胞の作製方法であって、
a)真核細胞を準備し、
b)工程a)の細胞を遺伝子改変して細胞中で1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現を可能にし;そして
c)工程b)の細胞中で1つ又はそれ以上の核酸を発現させる
ことを含む方法であって、
その際、1つ又はそれ以上の不死化タンパク質は、ヒト乳頭腫ウイルス(HPV)E6/E7又はその機能的に活性な変異体を含む前記方法に関する。
【0012】
上に詳述したように、ヒト初代細胞の限られた増殖能は、現在、多くの用途におけるその使用を妨げている。細胞を不死化する方法は記載されてきたが、しかしながら、それらは上に詳述された欠点がある。本発明は、細胞を遺伝子改変によって可逆的に不死化された細胞を作製し、そして場合により1つ又はそれ以上の他の遺伝子と併せてHPV E6/E7を発現する新規な方法を開発して代替方法を提供し、そして好ましくはこれらの欠点を克服したことを開示している。
【0013】
本発明の方法の工程a)では、被験者から得た後に直接用いることで真核細胞を準備してもよい。別法として、細胞を培養した後、工程b)の遺伝子改変を実施してもよい。また、細胞は、細胞株から誘導してもよい。しかし、初代細胞、すなわち新たに単離された細胞、又は短期間、すなわち多くとも4週間、好ましくは多くとも3週間、より好ましくは少なくとも2週間、なおより好ましくは多くとも7、6、5、4、3、2又は1日培養された細胞が好ましい。
【0014】
「真核細胞」という用語は、動物、植物、真菌及び原生生物から誘導された細胞を含む。これらの細胞は、膜内に囲まれた複雑な構造に組織化されている。
真核細胞は、核を有する原核細胞とは異なる。多くの真核細胞は、ミトコンドリア、葉緑体及びゴルジ体のような他の膜結合細胞小器官を含む。真核細胞中の細胞分裂は、核のない生物(原核生物)とは異なる。それは、微小管に誘導された運動による重複染色体の分離を含む。2つのタイプの分裂過程がある。有糸分裂では、1つ細胞が分裂して2つの遺伝的に同一の細胞を生じる。減数分裂は、有性生殖に必要である。
【0015】
上に詳述したように、工程a)で得た細胞を工程b)で遺伝子改変して細胞中で1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現を可能にし、その際、1つ又はそれ以上の不死化タンパク質は、ヒト乳頭腫ウイルス(HPV)E6/E7を含む。遺伝子改変の方法、特に遺伝子の導入は、当業者によく知られている。
【0016】
ヒト乳頭腫ウイルス(HPV)E6/E7は、HPVによって発現される重要な発癌性タンパク質に相当する。癌及び前癌性病変に関連する身体的変化を生じることが知られているHPVの菌株の中では、3つの癌タンパク質:E5、E6及びE7が認められている。従って、HPV E6/E7という用語は、個々のもの又は融合タンパク質のずれかとしてのタンパク質の組み合わせのことである。
【0017】
完全長HPV E6遺伝子は、約150〜160個のアミノ酸のタンパク質をコードしており、それは亜鉛結合Cys−X−X−Cysモチーフを含む2つのドメインを含む。E6は、細胞中で複数の役割があり、多くの他のタンパク質と相互作用すると考えられているため、特に興味深い。E6は、主に腫瘍抑制因子として作用するp53又はpRBタンパク質と結合することでそれらを不活性化することによって癌を生じる。腫瘍抑制タンパク質が不活性化されたとき、腫瘍成長は無制限に進行する。E6とp53及びpRBとの相互作用は、ユビキチン化及びユビキチンリガーゼによる分解のためにこれらのタンパク質をマークする。E6は、他の細胞タンパク質上で作用し、そしてテロメラーゼ活性に正の影響を与え、それにより細胞がチェックなしに分裂するのを通常阻止する道筋の1つを不活性化することがわかっている。さらに、E6は、細胞転写因子、E2F1/DP1と相互作用するとき転写補因子、具体的には転写活性化剤として作用することができる。
【0018】
E6は、しばしばシグナル伝達タンパク質中に見出されるPDZドメインに結合することもできる。これらの位置での結合は、DLGタンパク質の形質転換及びその抑制機能の破壊が起こる。また、E6タンパク質は、MAGUK(膜結合性グアニル酸キナーゼファミリー)タンパク質とも相互作用し、通常、構造タンパク質であってDLGの抑制活性に関与していると考えられる。E6がMAGIタンパク質上のPDZドメインと複合体形成するとき、E6はその形状を変形させ、それによってその機能を妨げる。全体として、E6タンパク質は、癌に特有の高められた速度で細胞を増殖及び倍加させるようなやり方で正常なタンパク質活性を妨げる役割を果たす。
【0019】
E7タンパク質は、約100個のアミノ酸のサイズであり、そしてまたE6と同様に2部のCys−X−X−Cysドメインを含む。癌タンパク質として、E7は、網膜芽細胞腫腫瘍抑制遺伝子産物の活性形態、低リン酸化pRBと複合体を形成することにより機能し、そのタンパク質分解を促進する。E7/pRB複合体の形成は、pRBとE2Fとの複合体形成を妨げる。それゆえ、E2FがpRBとの不活性化された組み合わせから解放されると転写活性化剤としてその作用を続けてDNA合成及び細胞増殖を促進する。
【0020】
ハイリスクHPV感染細胞では、E6及びE7は、通常、共発現される。ハイリスクHPV E7癌タンパク質は、pRB並びに関連するpRBファミリーメンバーp107及びp130.17と結合して分解するだけでなく、サイクリン依存性キナーゼ(CDK)阻害剤を不活性化する。その結果、サイクリンE、サイクリンAの発現が異常に増加し、そしてE2Fが介在するCDK2活性が異常になる。また、E2Fは、p14ARFを活性化してp53を安定化することができる。細胞増殖を促進するE7と共同して、ハイリスクHPV E6癌タンパク質は、宿主細胞タンパク質、E6−APと相互作用することによってp53の迅速なプロテアソーム分解を誘導する。E6又はE7の核酸又はアミノ酸配列は、同じものを含むか又は発現するあらゆるHPVタイプから誘導してもよい。100を超える異なるHPVタイプが同定されており、数字で呼ばれている。タイプ16、18、31、33、35、39、45、51、52、56、58、59、68、73及び68は、「ハイリスク」HPV(癌に至ることがあるため)の例であり、そのため上記配列にとっての好ましい供給源である。HPV E6/E7の非常に好ましいcDNAは、SEQ ID NO:4として表記される。
【0021】
また、本発明は、HPV E6/E7の機能的に活性な変異体を包含する。変異体は、E6/E7断片であってもよく、その際、断片は細胞をなお不死化することができる。これは、短いC末端及び/又はN末端欠失(例えば多くとも20、19、18、17、16、15、14、13、12、11、10、9、8、7、6、5、4、3、2又は1個のアミノ酸の欠失)を有するE6/E7タンパク質を含んでもよい。別法として又はさらに、E6/E7タンパク質は、1つ又はそれ以上のアミノ酸置換を含んでもよい。しかし、アミノ酸が化学的に関連するアミノ酸で置換された同類アミノ酸置換(conservative amino acid substitution)は、好ましい。典型的な同類置換は、脂肪族アミノ酸の中からであり、脂肪族ヒドロキシル側鎖を有するアミノ酸の中からであり、酸性残基を有するアミノ酸の中から、アミド誘導体の中から、塩基性残基を有するアミノ酸、又は芳香族残基を有するアミノ酸の中からである。E6/E7変異体は、場合により他の不死化タンパク質と組み合わせて細胞をなお不死化することができる場合、本発明により機能的に活性である。細胞を不死化する能力は、細胞の増殖に関して、そして本発明の実施例において記載されたように、例えば免疫染色により不死化細胞及び対照細胞の量を比較し、全RNAを測定し、細胞形態、細胞生存(場合により不利な条件下)又は細胞数を測定して試験することができる。本発明に関して、変異体の活性が配列変更のないE6/E7タンパク質の活性の少なくとも10%、好ましくは少なくとも25%、より好ましくは少なくとも50%、さらにより好ましくは少なくとも70%、なおより好ましくは少なくとも80%、とりわけ少なくとも90%、特に少なくとも95%、最も好ましくは少なくとも99%に達する場合、変異体は、細胞を不死化する能力を有する。
【0022】
本発明の以下の説明において、E6/E7に関して提供されるすべての詳細は、特に明記しない限り、その機能的に活性な変異体にも関連する。
【0023】
しかし、最も好ましくは、E6/E7は、自然発生E6/E7、なおより好ましくは、HPV E6/E7であり、最も好ましくは、cDNAによってコードされたHPV E6/E7は、SEQ ID NO:4として表記される。
【0024】
「核酸」という用語は、本明細書において詳細に議論されたあらゆる核酸分子、好ましくはDNAのことである。核酸分子は、DNA、cDNA、合成DNA、RNA又はそれらの組み合わせを含むさまざまな供給源から誘導してもよい。このような核酸配列は、ゲノムDNAを含んでもよく、それは自然発生のイントロンを含んでもよいし又は含まなくてもよい。さらに、このようなゲノムDNAは、プロモーター領域、ポリA配列又は他の関連配列と共に入手してもよい。ゲノムDNAは、当分野でよく知られている手段によって適切な細胞から抽出し、そして精製してもよい。別法として、メッセンジャーRNA(mRNA)を細胞から単離して使用し、逆転写又は他の手段によってcDNAを作製することができる。
【0025】
遺伝子改変の前に、通常、興味の遺伝子、ここでは細胞中の1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸を単離する。その後、核酸、好ましくはDNA又はcDNAを、細胞の改変を可能にするやり方で処方する。例えばトランスフェクションによって、すなわち、非ウイルス的方法により細胞に核酸を導入する方法によって、又は形質導入、すなわち、ウイルスによる遺伝物質の挿入により生じる細胞の遺伝的変性によって真核細胞に外来DNAを導入する種々の方法がある。
【0026】
多くの物質がトランスフェクション用の担体として使用されており、それは3種類:(カチオン性)ポリマー、リポソーム及びナノ粒子に分けることができる。これは、トランスフェクションによって、例えばリン酸カルシウム沈殿によって実施してもよい。別の方法は、カチオンポリマー、例えばDEAE−デキストラン又はポリエチレンイミンの使用である。負に荷電されたDNAはポリカチオンに結合し、そして複合体はエンドサイトーシスを介して細胞に取り込まれる。トランスフェクションに対する直接的なアプローチは遺伝子銃であり、ここでは、DNAを不活性固形物(一般に金)のナノ粒子に結合し、次いで、それを標的細胞の核に直接「注射する(shot)」。
【0027】
トランスフェクションでは、遺伝子又はcDNAを適切なベクター中に移してもよい。ベクターは、プラスミド又はウイルスベクターであってもよい。核酸は、当業者に知られている制限酵素及びリガーゼを用いてベクター中に導入してもよい。ベクターを得たら、それを用いて興味の細胞を形質転換することができる。形質転換後、遺伝子改変された細胞を、ベクターの取込みに失敗したものから種々のやり方で選別する。1つの方法は、移植しようとした興味の遺伝子に固着することができるDNAプローブを用いたスクリーニングである。もう1つは、マーカーの使用である。マーカー遺伝子を用いてベクターの存在を検査し、それにより感染及び組込みを確認してもよい。マーカー遺伝子の存在により、挿入物を発現するそれらの宿主細胞のみの選別及び増殖が保証される。典型的な選択遺伝子は、抗生物質並びに他の毒性物質、例えばヒスチジノール、プロマイシン、ハイグロマイシン、ネオマイシン、メトトレキセート及び細胞表面マーカーに対する耐性をもたらすタンパク質をコードしている。別法として、検出可能なタンパク質をコードする遺伝子を、緑色蛍光タンパク質(GFP)又は黄色蛍光タンパク質(YFP)を含む蛍光タンパク質のようなマーカーとして使用してもよい。
【0028】
さらにまた、遺伝子又はcDNAを、核酸の有効な移入又は核酸の発現を可能する適切な要素と組み合わせてもよい。これらは、核酸の転写を制御する要素、例えばプロモーター配列、転写開始配列、エンハンサー配列、選択可能な要素及びレポーター遺伝子であってもよい。さらに、適切な制限酵素認識部位を、興味の核酸に加えてもよい。
【0029】
さらにまた、キャッピング、スプライシング、ポリ(A)末端付加によって翻訳を制御し、そして配列特異的な核外移行速度を変える要素が存在してもよい。
【0030】
ベクターを用いる場合、1つ又はそれ以上の核酸が1つ又はそれ以上のベクターの一部であってもよい。複数のベクターを用いる場合、それらを同時に又は順次、細胞中に導入してもよい。ウイルスベクターは、細胞中に遺伝物質を送達するために一般に用いられる手段である。この方法は、生きている生物内(インビボ)で又は細胞培養(インビトロ)において実施することができる。ウイルスは特殊な分子機構を発達させてそれらが侵入する細胞内部にそれらのゲノムを有効に運ぶ。ウイルスによる遺伝子の送達は、形質導入と称し、そして感染細胞は形質導入されたと記載される。
【0031】
当初、ウイルスベクターは、分子遺伝学実験用の裸のDNAのトランスフェクションに対する代替物として開発された。遺伝子をコードするタンパク質は、一般に特定のタンパク質の機能を研究するためのウイルスベクターを用いて発現することができる。ウイルスベクター、とりわけレトロウイルスは、遺伝物質を細胞に移入するために広く用いられる(また、下記参照)。他のタイプのウイルスベクターは、アデノウイルス及びアデノ随伴ウイルス(AAV)である。AAVは、分裂及び非分裂細胞の両方に感染することができ、そしてそのゲノムを宿主細胞のそれに組み込むことができる。これらの特徴によりAAVは、ウイルスベクターを作成するための非常に魅力的な候補物質となっている。非常に好ましいベクターの配列は、SEQ ID NO:6として表記される。
【0032】
工程b)において細胞を遺伝子改変した後、細胞中で1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸を発現する。細胞は、例えば、発現が構成的に活性なプロモーターによって制御される場合、なんらかのさらなる相互作用なしに、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸を発現することができる。あるいは、例えば、誘導性プロモーターが1つ又はそれ以上の核酸に結合した場合、発現を誘導する必要がありうる。
【0033】
この方法では、真核細胞、好ましくは下記定義された初代細胞を準備し、そして改変された細胞が不死化タンパク質ヒト乳頭腫ウイルス(HPV)E6/E7をコードする1つ又はそれ以上の核酸を発現するように遺伝子改変する。さらに、また、1つ又はそれ以上の核酸は、SV40ラージT抗原(TAG);テロメラーゼ触媒サブユニット(TERT);又はSV40ラージT抗原(TAG)及びテロメラーゼ触媒サブユニット(TERT)を含むさらなる不死化タンパク質をコードしてもよい。従って、HPV E6/E7をコードする核酸は、遺伝子改変によって細胞中に導入された唯一の核酸であってもよい。
【0034】
しかし、本発明の好ましい実施態様において、1つ又はそれ以上の不死化タンパク質は、
・HPV E6/E7及びSV40ラージT抗原(TAG);又は
・HPV E6/E7及びテロメラーゼ触媒サブユニット(TERT);又は
・HPV E6/E7及びSV40ラージT抗原(TAG)及びテロメラーゼ触媒サブユニット(TERT)
を含むか又はそれらからなる。また、これらの機能的に活性な変異体も含まれる。
【0035】
SV40ラージT抗原(Simian Vacuolating Virus 40 Tag、TAGとも称する)は、さまざまな細胞タイプを形質転換することができる、ポリオーマウイルスSV40から誘導された癌遺伝子である六量体タンパク質である。TAGの形質転換活性は、ラージ部分における網膜芽腫(pRB)及びp53腫瘍抑制タンパク質のその摂動(perturbation)による。さらにTAGは、その形質転換機能に寄与しうる転写補活性化因子p300及びCBPを含むいくつかの他の細胞性因子と結合する。
【0036】
TAGは、SV40によるウイルス感染の際に転写される初期遺伝子の産物であり、そしてウイルスのゲノム複製及び宿主細胞周期の制御に関与している。SV40は、二本鎖DNAウイルスであり、パポバウイルスファミリー、ポリオーマウイルス属に属する。ポリオーマウイルスは、多種多様な脊椎動物に感染し、それにより複数の部位で固形腫瘍を生じる。SV40 DNA複製は、ゲノムの起点領域(origin region)にTAGが結合することによって開始される。TAGの機能は、リン酸化によって制御され、それはSV40起点への結合を弱める。TAGとDNAポリメラーゼ−アルファとの間のタンパク質−タンパク質相互作用は、ウイルスゲノムの複製を直接刺激する。また、TAGは、腫瘍抑制タンパク質(p53、p105)と結合して不活性化する。これにより細胞がG1期を離れてS期に入り、DNA複製を促進する。
【0037】
本明細書に使用される「テロメラーゼ触媒サブユニット」又は「TERT」という用語は、テロメラーゼ触媒活性を有するポリペプチド配列のことである。テロメラーゼは、真核染色体の末端に見出される、テロメア領域中DNA鎖の3'(「3'末端」)末端に特異的なDNA反復配列(すべての脊椎動物中の「TTAGGG」)を加えた酵素である。テロメアは、濃縮DNA物質を含み、染色体に安定性を与える。酵素は、それ自体RNA分子を担持する逆転写酵素であり、それはテロメアを延長するときにテンプレートとして用いられ、テロメアは複製周期の後ごとに短縮される。それは、それぞれ2分子のテロメラーゼ触媒サブユニット、別名テロメラーゼ逆転写酵素(TERT);テロメラーゼRNA(hTR又はTERC);及びジスケリン(dyskerin)からなる。TERTは、テンプレートとして一本鎖RNAを用いて一本鎖DNAを作成する逆転写酵素である。
【0038】
また、本発明に関して、TAG及び/又はTERTの機能的に活性な変異体を用いてもよい。「変異体」及び「機能的に活性な」という用語は、E6/E7の変異体に関して上記定義された通りである。本発明の以下の説明において、E6/E7に関して記載されたすべての詳細は、特に明記しない限り、その機能的に活性な変異体にも関連する。
【0039】
一実施例において、第1のベクターは、HPV E6/E7をコードするポリヌクレオチドを含むのに対して、第2のベクターは、SV40ラージT抗原(TAG);テロメラーゼ触媒サブユニット(TERT);又はSV40ラージT抗原(TAG)及びテロメラーゼ触媒サブユニット(TERT)をコードするポリヌクレオチドをコードする核酸を含む。
【0040】
別の態様において、本発明は、可逆的に不死化された細胞の作製方法であって、
a)真核細胞を準備し、
b)工程a)の細胞を遺伝子改変して細胞中で1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現を可能にし;そして
c)工程b)の細胞中で1つ又はそれ以上の核酸を発現する、
ことを含み、
その際、1つ又はそれ以上の不死化タンパク質の少なくとも1つは、ヒト乳頭腫ウイルス(HPV)E6/E7、SV40ラージT抗原及びテロメラーゼ触媒サブユニット(TERT)、又はそれらのいずれかの機能的に活性な変異体からなる群より選ばれ、そして1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現は、EF1アルファプロモーターによって制御されることを含む前記方法に関する。
【0041】
驚くべきことに、プロモーターEF1アルファ(詳細については、下記参照)は、他よりも良好な発現レベルを示すことが見出された。いくつかのプロモーターを異なる初代細胞において試験した。異なるプロモーターの制御下でGFP(緑色蛍光タンパク質)cDNAを発現することによって、CMV(サイトメガロウィルス)プロモーター及びMSCV(マウスサイトメガロウィルス)プロモーターではそれぞれ極めて高い又は極めて低いレベルの発現となったのに対して、EF1アルファプロモーターでは中程度の発現レベルが得られることを示すことができた。本発明のこの態様のすべての用語は、この説明において詳述された通り定義される。
【0042】
本発明のさらなる実施態様において、本発明の方法は、
d)工程c)の細胞を増殖させる
ことをさらに含む。
【0043】
本発明によれば、工程c)の細胞は、細胞生存度、増殖及び完全性を可能にする有効な条件下で細胞を増殖させてもよい。本明細書において、増殖とは、インビトロで培養することによるものである。増殖とは、細胞成長及び分裂の結果として細胞数が増加することであり、それは細胞培養によって達成してもよい。好ましくは、増殖とは、少なくとも1つの細胞の倍加である。増殖をモニタリングする可能な方法としては、増殖に関連する抗原の検出、DNA合成の測定、及び細胞内環境の還元(reduction)の検出が含まれるが、それらに制限されるわけではない。DNA合成は、例えば3H−チミジンの取り込み又は5−ブロモデオキシウリジン(BrdU)の取り込みを定量化することによって測定してもよい。細胞内環境の還元は、テトラゾリウム塩の還元によってモニターしてもよい。生存度は、生存及び複製するための細胞の能力のことである。細胞生存度を測定するために一般に用いられるアッセイには、トリパンブルー色素排除、ニュートラルレッド染色、クリスタルバイオレット包摂、及び51Crリリースが含まれるが、それらに制限されるわけではない。この方法によって産生された安定なクローンは、不死化されたら、培養において少なくとも2ヵ月間培養することができる。好ましい実施態様において、これらの安定なクローンは、少なくとも6ヵ月、好ましくは少なくとも1年、そして最も好ましくは少なくとも2年間、培養において継代することができる。これらの安定なクローンは、それが誘導された細胞タイプに関連する高レベルの機能及び特徴を有し続ける。
【0044】
細胞培養のため、細胞は、細胞産生を助長する適切な条件下で増殖される。温度及びガス混合物を除いて、細胞培養系において最も一般に異なる要因は、増殖培地である。増殖培地の処方は、pH、グルコース濃度、増殖因子及びとりわけ他の栄養成分の存在で変えることができる。補充培地に用いられる増殖因子は、多くの場合、仔ウシ血清のような動物血液に由来する。培養条件は、各細胞タイプで広く変化し、そして特定の細胞タイプについての条件を変えることにより異なる表現型を発現することができる。
【0045】
細胞は、懸濁液又は付着性の培養中で増殖することができる。いくつかの細胞は、血流中に存在する細胞のように表面に付着することなく懸濁液中で自然に生きている。また、可能な付着条件より高密度に増殖することができるように懸濁培養液中で生存することができるように改良された細胞もある。付着細胞は、組織培養プラスチックのような表面を必要とし、これは細胞外マトリックス成分でコートして付着性を高め、そして増殖及び分化に必要な他のシグナルを提供してもよい。固形組織から誘導されたほとんどの細胞は、付着性である。別のタイプの付着培養(adherent culture)は、器官型培養であり、それは二次元の培養皿とは対照的に三次元環境での細胞の増殖を含む。この三次元培養系は、生体内の組織に生化学的及び生理学的により類似しているが、多くの要因(例えば拡散)のため、維持するのが技術的に困難である。適切な方法及びその選択は、当業者に知られている。さらにまた、不死化細胞に適切な方法は、実施例にも記載した。
【0046】
好ましい実施態様において、不死化タンパク質をコードする1つ又はそれ以上の核酸は、1つ又はそれ以上のレトロウイルスベクター、特に1つ又はそれ以上のレンチウイルスベクターを用いることによって導入される。
【0047】
レトロウイルスは、現在の遺伝子移入アプローチの柱の1つである。モロニーマウス白血病ウイルスのような組換え型のレトロウイルスは、安定なやり方で宿主ゲノムに組み込む能力を有する。従って、好ましい実施態様において、不死化タンパク質をコードする1つ又はそれ以上の核酸は、1つ又はそれ以上のレトロウイルスベクター、特に1つ又はそれ以上のレンチウイルスベクターを用いることによって導入される。
【0048】
レトロウイルスは、ウイルス逆転写酵素を用いることによって複製して宿主細胞中でそのRNAをDNAに転写する、RNAゲノムを有するウイルスである。次いで、DNAはインテグラーゼ酵素によって宿主のゲノムに組み込まれる。その後、ウイルスは、宿主細胞のDNAの部分として複製する。レトロウイルスは、レトロウイルス科というウイルスファミリーに属するエンベロープウイルスである。ウイルスそれ自体は、その核酸をmRNA(ビリオン中に5'cap及び3'PolyAを含む)ゲノムの形態で保存しており、そしてそのゲノムの細胞への送達手段として役立ち、偏性寄生生物として目標を定め、そして感染の構成要素となる。宿主の細胞中で、RNA鎖がサイトゾル中で逆転写を受け、そして宿主のゲノムに組み込まれたら、この時点で、レトロウイルスDNAをプロウイルスと称する。一般に、レトロウイルスは、外因性(以下の属、アルファレトロウイルス属、例えばトリ白血病ウイルス;ベータレトロウイルス属、例えばマウス乳癌ウイルス;ガンマレトロウイルス属、例えばマウス白血病ウイルス;デルタレトロウイルス属、例えばウシ白血病ウイルス;ヒトTリンパ球向性ウイルス;イプシロンレトロウイルス属、例えばウォールアイ皮膚肉腫ウイルス;レンチウイルス、例えばヒト免疫不全ウイルス1の他にサル及びネコ免疫不全ウイルス;並びにスプマウイルス属、例えばチンパンジーフォーミーウイルスを含む)及び内因性(ガンマレトロウイルスに最も類似しているクラスI、ベータレトロウイルス及びアルファレトロウイルスに最も類似しているクラスII、及びスプマウイルスに最も類似しているクラスIIIを含む)として特定化されている。
【0049】
レンチウイルスは、一般に長い培養期間を特徴とするレトロウイルス科ファミリーのスローウイルス属である。レンチウイルスは、かなりの量の遺伝情報を宿主細胞のDNAに送達することができるため、遺伝子送達ベクターの最も有効な方法の1つである。レンチウイルスは、主としてインビトロ系又は動物モデルに遺伝子産物を導入するために用いられる研究手段である。レンチウイルス感染は、他の遺伝子治療法よりも分裂及び非分裂細胞の高効率感染、導入遺伝子の長期的に安定な発現、及び低い免疫原性を含む利点を有する。レンチウイルスは、レトロウイルスのサブクラスである。それは、非分裂細胞のゲノム中に組み込む能力のため遺伝子送達ビヒクル(ベクター)として応用されており、他のレトロウイルスは、通常、分裂細胞しか感染させることができないので、これはレンチウイルスの独特な特徴である。RNA形態のウイルスゲノムは、ウイルスが細胞に入ってDNAを作製するときに逆転写され、それは次いで、ウイルスインテグラーゼ酵素によってランダムな位置でゲノム中に挿入される。従って、本発明の好ましい実施態様において、不死化タンパク質をコードする1つ又はそれ以上の核酸は、1つ又はそれ以上のレンチウイルスベクターを用いることによって導入される。レンチウイルスの5つの血清群が知られており、それらが関与する脊椎動物の宿主を反映している(霊長類、ヒツジ及びヤギ、ウマ、ネコ及びウシ)。HIV、SIV及びFIVは、すべてレンチウイルスの例である。伝達は一般に感染粒子を介するが、レンチウイルスは、細胞外粒子を形成する必要なしに宿主細胞と直接接触している隣接細胞を感染させることができる。
【0050】
好ましい実施態様において、本発明に関する真核細胞は、動物細胞、より好ましくは脊索動物細胞、なおより好ましくは脊椎動物、さらにより好ましくは哺乳動物細胞、特にヒト、霊長類又は齧歯類の細胞である。
【0051】
さらに又は別法として、真核細胞は、初代細胞又は非初代細胞であってもよい。しかし、初代細胞、すなわち、本方法の使用直前に被験者から得た細胞を、可逆的に不死化された細胞の作製方法に使用することが好ましい。「初代細胞」という用語は、上記定義された通りである。
【0052】
さらにまた、真核細胞は、あらゆる細胞タイプであってもよい。しかし、生殖系列細胞ではなく、体細胞、すなわち、生物の体を形成しているいずれかの細胞が好ましい。哺乳動物において、生殖系列細胞は、精子及び卵子である。哺乳動物の体中の他のすべての細胞タイプは、体細胞である。体細胞の例としては、内臓、皮膚、骨、血液及び結合組織に由来するものが含まれる。好ましくは、細胞は、分化細胞、すなわちすべての遺伝子の特定のサブセットを発現する特殊化された細胞タイプの細胞である。好ましい分化細胞には、内皮細胞、心筋細胞、平滑筋細胞、肝細胞、有足細胞又は腎臓細胞が含まれる。
【0053】
本発明の別の実施態様において、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現は、EF1アルファプロモーターによって制御される。
【0054】
プロモーターは、特定の遺伝子の転写を促進するDNAの領域である。プロモーターは、典型的には同じ鎖及び上流(センス鎖の5'領域の方)においてそれが制御する遺伝子の近くにある。プロモーターは、遺伝子/DNAの発現を意図する細胞に従って選ばれ、用いることができる。適切なプロモーターの例としては、例えば、EF1アルファ、PGK、モロニーマウス白血病ウイルスプロモーター−エンハンサー要素、ヒトサイトメガロウィルスエンハンサー、ワクシニアP7.5プロモーター又は同様のものが含まれるが、しかし、EF1アルファプロモーターが好ましい。EF1アルファプロモーターは、レンチウイルスベクター及び真核細胞における発現に特に適切である。プロモーターは、真核EF1アルファプロモーター又は好ましくはヒトEF1アルファプロモーターであってもよい。適切なプロモーターの配列は、SEQ ID NO:1として表記される。
【0055】
本発明の別の実施態様において、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸は、loxP(locus of X-over P1)部位の横に位置しており、それはCre又はCre変異体の存在下で1つ又はそれ以上の核酸の除去を可能にする。
【0056】
バクテリオファージP1からの38kDaリコンビナーゼ、Creは、そのエンドヌクレアーゼ活性を用いて2つの同一のloxP部位間の組換えに触媒作用を及ぼす。酵素Creは、補助タンパク質又は補因子を必要とせず、そしてインビトロで及び多種多様な細胞条件下で有効に機能する。Creによって認識される組換え部位は、loxPとして知られている二本鎖DNA配列である。それぞれのloxP部位は、非対称の核領域によって分かれた2つの逆方向反復からなる。Creは、逆方向反復に結合し、そして核領域中でDNAを切断してDNA鎖交換反応を促進する。本来のloxP配列は、34bpからなる。その横に位置する2組の13bpの回文の間に8bpの非対称配列が存在する。詳細な構造を下に示す。
13bp 8bp 13bp
ATAACTTCGTATA−GCATACAT−TATACGAAGTTAT
(SEQ ID NO:7)
好ましいloxP部位は、SEQ IDNO:2及び3として表記される。
【0057】
Cre(環化組換え)タンパク質は、4つのサブユニット及び2つのドメインからなる:大きい方がカルボキシル(C末端)ドメイン、そして小さい方がアミノ(N末端基)ドメインである。総タンパク質は、343個のアミノ酸を有する。Cドメインは、ラムダファージから単離された酵素のインテグラーゼファミリーのドメインに構造が類似している。また、これは、酵素の触媒部位である。Cre変異体ポリペプチドは、特定のアミノ酸配列に関してCreと異なっていてもよいが、Creのリコンビナーゼ活性を維持している。一般に、基準ポリペプチド(reference polypeptide)及び変異体の配列が全体として密接に類似し、そして多くの領域で同一になるように違いは制限される。変異体及び基準ポリペプチドは、あらゆる組み合わせで1つ又はそれ以上の置換、付加又は欠失によってアミノ酸配列が異なっていてもよい。置換された又は挿入されたアミノ酸残基は、遺伝暗号によってコードされたものであってもよいし又はそうでなくてもよい。ポリペプチドの変異体は、自然発生であってもよいし、又は自然発生することが知られていない変異体であってもよい。
【0058】
Cre/lox系は、ゲノムDNA中の部位特異的な組換え事象を制御する遺伝的手段として用いられる。この系により、研究者は、さまざまな遺伝子改変生物を操作して遺伝子発現を制御し、望ましくないDNA配列を欠失させ、そして染色体構造を改変することが可能になる。系はCreを含み、二本鎖DNAは、Creによって両方のloxP部位で切断され、次いで一緒に結合されて戻る。組換えの効率は、loxP部位の方向に左右される。同じ染色体腕上の2つのlox部位について、逆方向loxP部位は、反転を生じるが、loxP部位の同方向反復は、欠失事象が生じる。loxP部位が異なる染色体上にある場合、Creにより誘導された組換えによって転座事象に触媒作用を及ぼすことが可能である。
【0059】
別の態様において、本発明は、本発明の方法によって入手可能な細胞又はその子孫を提供する。
【0060】
なお別の態様において、本発明は、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸を含む真核細胞であって、その際、1つ又はそれ以上の不死化タンパク質の少なくとも1つが、ヒト乳頭腫ウイルス(HPV)E6/E7、SV40ラージT抗原及びテロメラーゼ触媒サブユニット(TERT)からなる群より選ばれ、そして1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現がEF1アルファプロモーターの制御下にある前記真核細胞を提供する。
【0061】
さらに、細胞は、本発明に従って、可逆的に不死化された細胞の作製方法に関して上記詳述された特徴を有しうる。特に、細胞又はその子孫は、以下の性質の1つ又はそれ以上を特徴とすることがある:
i)不死化タンパク質をコードする1つ又はそれ以上の核酸は、1つ又はそれ以上のレトロウイルスベクター、特に1つ又はそれ以上のレンチウイルスベクター上にある;
ii)細胞は、さらに遺伝子改変を含む;
iii)細胞は、哺乳動物細胞、特にヒト、霊長類又は齧歯類の細胞である;
iv)細胞は、初代細胞から誘導される;
v)細胞は、分化細胞、特に内皮細胞、心筋細胞、平滑筋細胞、肝細胞、有足細胞又は腎臓細胞から誘導される;
vi)1つ又はそれ以上の不死化タンパク質をコードする核酸の発現は、EF1アルファプロモーターの制御下にある;及び/又は
vii)1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸は、loxP(locus of X-over P1)部位の横に位置する。
【0062】
別の態様において、本発明は、本発明の細胞、又はその子孫を再分化する方法、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現を阻止することを含む方法を提供する。不死化細胞の使用は、このような方法に利用可能な初代細胞の不足を克服する手段を提供する。不死化が逆転された不死化細胞は、成長及び増殖の低下を示す。2〜3週間後、細胞は、細胞老化に一致する形態を発現し、細胞周期の制御が不死化細胞において無損傷のままであることを示唆している。さらに、不死化が逆転された細胞は、強化された分化機能を示す。不死化が逆転された細胞は、生物学的及び薬学的研究のため、特に化合物プロファイリング、表現型スクリーニング、標的同定及び/又は標的検証のための供給源として用いてもよい。このために、細胞をまず不死化し、そして再分化細胞が必要になるまで培養する(そして増殖させる)ことができる。これにより、多くの実験のための、そして長期間にわたる経済的かつ均一な細胞供給源の使用が確保され、それは望ましいことである。
【0063】
細胞の再分化は、核酸の発現を阻止するなんらかの適切な方法によって実施してもよい。これは、アンチセンス核酸をそれぞれのmRNAに結合してその翻訳を阻止することによって実施してもよい。別法として、制御可能なプロモーター、例えば誘導性プロモーターを用いてもよい。その名称の通り、これらのプロモーターの活性は、生物的又は非生物的因子の有り又は無しによって誘導される。誘導性プロモーターは、それに実施可能に結合された遺伝子の発現を生物発育のある種の段階で又は特定の組織中でオン又はオフにすることができるため、遺伝子工学における非常に強力な手段である。誘導性プロモーターは、アルコール、テトラサイクリン、ステロイド、金属及び他の化合物の有り又は無しによってその転写活性が制御されるプロモーターを含む化学的に制御されるプロモーター;並びに光の有り又は無し及び温度の低い又は高いによってその転写活性が制御されるプロモーターを含む物理的に制御されるプロモーターに分類される。
【0064】
細胞を再分化する方法の好ましい実施態様において、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸は、loxP(locus of X-over P1)部位の横に位置しており、そして阻止は、Creリコンビナーゼ又は機能的に活性なCre変異体によって実施される。
【0065】
好ましくは、loxP部位を用いる場合、不死化過程は、細胞にCre又はCre変異体をコードするポリヌクレオチドを含むベクターを導入することによって逆転させることができる。1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸が、Cre介在性組換えによって細胞から除去されるとき、それぞれの細胞又はクローンの細胞型に特異な活性は、一般に上方制御される。また、細胞不死化に関与する遺伝子を除去すると細胞増殖の速度がかなり低下し、そして最終的に細胞老化に至る。
【0066】
細胞を再分化する方法の別の好ましい実施態様では、レトロウイルスベクター、特にレンチウイルスベクターを用いてCreリコンビナーゼ遺伝子又は機能的に活性なCre変異体遺伝子を細胞に導入する。ベクター及び核酸を移入する方法は、上記詳述した通りである。
【0067】
別の態様において、本発明は、本発明に従って、細胞を再分化する方法によって入手可能な細胞、又はその子孫を提供する。
【0068】
以下の図面及び実施例は、本発明の種々の実施態様を説明することを意図する。このように、議論された特定の改変は、本発明の範囲における制限と解釈すべきではない。本発明の範囲を逸脱することなく種々の同等物、変更及び改変を実施することができることは、当業者に明らかであり、そしてこのような同等の実施態様は、本明細書に含まれるものとすることが理解される。
【図面の簡単な説明】
【0069】
【図1】VECTALYS系のpLV−EF1−Lox1−AgT−Lox2レンチウイルスベクターの略図を示す。cDNAは、両側にlox部位を有するEF1Aプロモーターの制御下でレンチウイルスプラスミド中に作製され(図1)、それは、必要に応じて、「不死化遺伝子(immortogene)」を切除することができる。EF1=EF1アルファプロモーターAgT=SV40ラージT抗原T=SV40スモールt抗原LOX1/LOX2=Creリコンビナーゼ認識部位WPRE=ウッドチャック転写後調節要素
【図2】初代培養された新生ラット有足細胞系統におけるシナプトポジンの発現を示す。A:未分化初代有足細胞の位相コントラスト形態。標準条件下で増殖する未分化有足細胞の丸石形態を示す。細胞は、集密に達するにつれ単層を形成する。B:標準条件下で維持された樹状有足細胞は、非常に大きくかつ扁平状である。C:有足細胞特異的なタンパク質シナプトポジンは、標準培養条件で14日後に発現を開始した(シナプトポジン−薄い灰色、そしてアクチンストレス線維−暗い灰色)。
【図3】分化した有足細胞のマーカーが、「不死化細胞」のCre介在性切除後に再び現れるのを示している。A:(位相コントラスト):LT Ag+E6/E7の過剰発現によって誘導された活性な増殖中に未分化有足細胞の典型的な丸石形態。細胞は、集密に達するにつれ単層を形成する。B:Creリコンビナーゼの形質導入後の樹状有足細胞(大きくかつ扁平状)。C:LT Ag(灰色)の核発現−100%の細胞が陽性である。D:Creリコンビナーゼによる形質導入後、多くの細胞ではSV40 LT Ag(矢印)が失われる。 注:矢印は、失われていると考えられる。E:シナプトポジン及びWT1(F)は、再分化した有足細胞中で発現される。
【図4】初代(対照)及び不死化ヒト肝細胞の免疫細胞化学的な比較を示す。A、B及びC−光学顕微鏡画像;D、E及びF−GFP蛍光(各細胞タイプは、レンチウイルスGFPにより形質導入された)。対照肝細胞は、大きな多角形の細胞であり(A及びD);SV40LTagの発現により極めて小さな卵形細胞に形質転換され(B及びE);HPV E6E7で形質導入した後は、対照細胞と比較してわずかに減少したサイズを有するが、SV40 LTag不死化されたものよりもかなり大きい(FをD及びEと比較する)。G−I:p53(灰色)の免疫染色。対照細胞におけるp53基底レベルは非常に低く(G)、そしてSV40 LTAgでは、極めて高レベルまで核中に蓄積され(H);E6E7不死化細胞において、大半の細胞ではそのレベルは低い(I)。J−L:CDC2、サイクリン依存性キナーゼ1(CDK1)の免疫染色;初代肝細胞では、増殖しないので存在することができなかったが(J)、これに対して、AgT不死化細胞(K)では、そしてまたE6E7不死化細胞(L)では、CDC2発現は、CDC2 mRNAの増加に伴ってかなり増加した(表2参照)。
【図5】内皮細胞特異的マーカーが不死化されたHUVEC中で発現され続けることを示している。新たに単離されたHUVEC(左のパネル)、AgT不死化(中央のパネル)、そしてE6E7不死化HUVEC(右のパネル)。光学顕微鏡画像(A、B及びC)。典型的な内皮マーカー、フォン・ウィルブランド因子(D−F:vWF、薄い灰色)、そしてPECAM(G−I、CD31、暗い灰色)は、対照(D及びG)中、AgT(E及びH)及びE6E7(F及びI)不死化細胞中で等しく発現された。核(暗い灰色)は、ヘキスト422で染色した。
【図6】SV40 LT Ag−及びE6/E7−に誘導された増殖の推定上の機構を示す。非増殖細胞では、cdc2/CDK1、サイクリンA及びB、cdc20の発現は、p53及び「リプレッサーE2F」によって抑制された。SV40 LT Ag及びE6/7不死化細胞では、p53及びRbは不活性化され、そしてE2Fは活性になり、そして、それ自体、cdc2/CDK1、サイクリンA及びB、cdc20の発現を誘導する。
【図7】pLV−EF1−LOX1−E6E7−LOX2(Vectalys)を示す。
【0070】
実施例
物質及び方法
レンチウイルスコンストラクト:すべてのcDNAは、VECTALYS(フランス)に注文した。レンチウイルス粒子は、標準プロトコールに従って生成し、そして1×106〜2×106TU/mlのタイター(titer)を有した。感染効率は、レンチウイルスGFPを用いて測定した。
【0071】
初代細胞の標本及び培養条件:ラット糸球体上皮細胞の初代培養を産生するため、新生仔ラット腎臓から糸球体を単離した。細胞成分のうち95%が糸球体であり、残りが尿細管を含む場合、良好な標本と考えられる。単離した後、コラーゲンコートした24ウェルプレート上の10%FCS及び1%抗生物質入りのDMEM中で糸球体を培養した。24時間後、有足細胞前駆体が、付着した糸球体から外へ増殖し始めた。3日培養した後、増殖している上皮細胞をトリプシン処理し、そして25μm孔径の篩に通過させて残っている糸球体核を除去し、それは主に糸球体間質及び内皮細胞からなる。ヒト由来の初代内皮細胞は、ヒト臍帯からルーチン的に獲得することができる。簡潔に言えば、温かいPBS、pH7.4を用いて臍帯静脈から血液産物をすすぎ、そして静脈に温かいコラゲナーゼA(Roche Diagnostics;PBS中320U/mlの濃度で使用)を充填した。温かいPBS中で20分間インキュベーション後、消化物を熱失活FCS(Gibco/Invitrogen社、カールスルーエ、ドイツによって供給された)を含む遠心管中に集めてコラゲナーゼ活性を無効化し、そして細胞を沈殿させた。上澄みを捨て、そしてFCS、ECGS/H、hEGF、hbFGF及びHC−500を含む内皮細胞増殖培地(PromoCellからのMedium and SupplementPack)中に沈殿物を再懸濁し、そしてコラーゲン−IコートされたT75フラスコ上で培養し、そして37℃加湿インキュベーター中で4〜5日間、集密に増殖させた。細胞を大量に播種して細胞増殖を最小限にし、それによってHUVECがCD14を発現できるようにし、でなければそれは消失した。結果的に、すべての実験では、第1継代HUVECのみを用いた。
【0072】
初代ヒト肝細胞(PHH)は、Cambrex(Weinheim、ドイツ)から入手し、そしてコラーゲン−Iコートされた皿(Becton Dickinson Labware、San Jose、米国)上のHCM Single Quots(Cambrex Bioproducts、ハイデルベルク、ドイツ)で補充されたHBM基礎肝細胞培地(Cambrex Bioproducts、ハイデルベルク、ドイツ)中で培養した。レトロウイルスコンストラクト及びウイルス感染:初代新生仔ラット有足細胞を(下記のように)単離し、そしてloxP部位の横に位置するSV40 LT抗原(LT Ag)遺伝子を含む単一のレンチウイルスコンストラクト又はヒト乳頭腫ウイルス(HPV)E6及びE7を含むベクターを単独で又は組み合わせて感染させた。最初の10日間のうちに、初代有足細胞は、その表現型及び形態を変え、そして増殖し始めた。細胞を3日毎に分け、そして安定なクロzpーンを単離し、そしてさらなる実験のため凍結した。これらの安定なクローンは、分化した有足細胞ではなく、最初に糸球体から増殖した細胞と形態学的に似ていた。Creリコンビナーゼを含むレンチウイルスベクターで感染後、細胞は増殖を終え、そして3〜10日後、その形態を分化した有足細胞にとって特異的なものへと変えた。
【0073】
同じ不死化方法を、HUVEC及びヒト肝細胞に適用した。HUVECは、高発現レベルのLT Ag又はE6/E7にもかかわらずその形態を変えなかったが、活発な増殖を開始した。LT Ag又はE6/E7に感染した後の肝細胞は、その表現型を劇的に変え、そしてCre挿入後、その正常なものを獲得した。
【0074】
抗体:一次抗体:単クローン性WT−1(F−6)、CDC2 p34(B−6)、PCAM−1(CD31)、SV40 T Ag(Pab 108)及び多クローン性vWF(C−20)(Santa Cruz Biotechnology、ハイデルベルク、ドイツ);多クローン性ZO−1(Zymed、サンフランシスコ、CA);単クローン性シナプトポジン(Progen、ハイデルベルク、ドイツ);多クローン性p53(Cell Signalling)。
【0075】
二次抗体:Alexa Fluor(R) 488ヤギ抗マウスIgG(H+L)*2mg/mL*、Alexa Fluor(R) 594ヤギ抗マウスIgG(H+L)*2mg/mL*、Alexa Fluor(R) 488ヤギ抗ウサギIgG(H+L)*2mg/mL*及びAlexa Fluor(R) 594ヤギ抗ウサギIgG(H+L)*2mg/mL*(Invitrogen、カールスルーエ、ドイツ)。
【0076】
免疫染色:コラーゲンタイプIコートされたチャンバースライド(VWR International 社、ダルムシュタット、ドイツ)上で増殖細胞の免疫蛍光染色を実施した。20℃でメタノールを用いて5分間又はリン酸塩緩衝食塩水(PBS)中の4%ホルムアルデヒドを用いて5分間、細胞を固定し、続いてPBS中0.3%トライトンX−100を用いて室温で10〜15分間、透過化処理した。PBSですすいだ後、非特異的結合部位を0.2%Tween 20(Serva、Amstetten、オーストリア)入りのSuperBlock(PBS中のブロッキング緩衝液、Thermo Scientific、ブラウンシュヴァイク、ドイツ)で少なくとも30分間ブロックした。ブロッキング溶液中に予め希釈した一次抗体を室温で60分間又は4℃で一夜適用した。Alexa Fluor(R) 488 又は 594二次抗体(Invitrogen、カールスルーエ、ドイツ)を用いて抗原−抗体複合体を視覚化した。ニコン顕微鏡写真機を用いて画像を得、そしてAdobe Photoshop 6.0ソフトウェアで処理した。
【0077】
全RNAの単離:培養された有足細胞、Huvecs及びPHHから、RNeasy Kit(Qiagen、ヒルデン、ドイツ)を用いて全RNAを単離した。RNase-free DNase-Set(Qiagen、ヒルデン、ドイツ)を用いてゲノムDNA入りの潜在的サンプルコンタミネーションを除去した。質及び純度は、キャピラリー電気泳動(Caliper Lab Chip system Agilent 2100 Bio
analyser、Agilent Technologies社、Waldbronn、ドイツ)を用いて評価した。
【0078】
定量的逆転写−PCR(qRT−PCR):リアルタイム定量的PCRは、QuantiTect
Probe RT-PCR Kit(Qiagen、ヒルデン、ドイツ)を用いて実施した。各サンプルを四つ組で測定した。遺伝子発現の相対的定量のため、対照としてGAPDHを用いてΔΔCt法を使用した。標的及びハウスキーピング遺伝子の増幅は、Applera/Applied Biosystems(Foster City、米国)から入手した異なる蛍光標識Taq Manプローブを用いて同時に検出した。マルチプレックスRT−PCR内の標的及びハウスキーピング遺伝子の増幅直線性は、テンプレートの希釈によりRT−PCR反応を実施することによって評価した。RT−PCR反応及びデータ収集は、iCycler-iQ-Thermocycler(Bio-Rad Laboratories社、ミュンヘン、ドイツ)において実施した。相対的遺伝子発現は、対照サンプルに対する誘導倍率(fold induction)として算出した。負の値は、対照サンプル中の発現レベルと比較した下方制御の倍率(fold down regulation)を示す。
【0079】
スーパーアレイ−技術:PCRアレイは、焦点を絞った遺伝子パネルの発現を分析する手段である。それぞれ96ウェル又は関連の、経路又は疾患に焦点を絞った遺伝子(例えば転写因子、細胞周期、幹細胞)の綿密に調査されたパネル用のSYBR Green最適化プライマーアッセイを含み、そしてSABiosciences(CA、米国)から入手した。
【0080】
PCRアレイ(RT2 Profiler PCR Array System)では、リアルタイムPCR感受性及びマイクロアレイの多重遺伝子のプロファイリング能力を用いて遺伝子発現分析を実施する。実験者のcDNAテンプレート(RT2 First Strand Kit (C-03))と適切なすぐに使用できるPCRマスターミックスとを同じプレートの各ウェルに等体積のアリコートで単純に混合し、次いでリアルタイムPCRサイクリングプログラムを実施する。プレートテンプレート及びリアルタイムPCRサイクリングプログラム(Instrument-Specific Setup Instructions & Protocol Files)は、ファイルhttp://www.sabiosciences.com/pcrarrayprotocolfiles.php.を用いて入手可能である。
【0081】
RT2-First Strand Kitは、有効な第1鎖(first strand)cDNA合成のための迅速かつ便利な方法を提供する。キットは、有効なゲノムDNA除去工程及び組み込まれた外部RNA対照(built-in External RNA Control)も含む。このオールインワンキットは、SABiosciencesのRT2 ProfilerTM PCR Arrays 及び RT2 qPCR Primer AssaysによるリアルタイムPCRベースの遺伝子発現分析のために設計され、そして最適化されている。キットは、逆転写前にRNAサンプルから汚染ゲノムDNAを効果的に除去するための独自の方法を含む。偏りのないやり方(unbiased manner)におけるランダム六量体及びオリゴ−dTプライム逆転写及び逆転写酵素により最適な収量及び長さでcDNA産物を合成する。組み込まれた外部RNA対照は、逆転写効率をモニターし、そしてRT2 RNA QC PCR Array及びRT2 ProfilerTM PCR Arrayと共に用いたときに酵素阻害剤が混入した実験者のRNAサンプルを試験するのに役立つ。マグネシウム及びヌクレオチド濃度及び他の緩衝液成分は、RT2 ProfilerTM PCR Arrays 及びRT2 qPCR Primer Assaysによる遺伝子発現分析に用いたときにRT2 SYBR Green qPCR Master Mixesと最も適合しうる。
【0082】
RT2 SYBR Green/フルオレセインqPCRマスターミックスは、BioRad iCycler(R) 及び MyiQ(R)におけるリアルタイムポリメラーゼ連鎖反応に必要なすべての試薬及び緩衝液:リアルタイムPCR緩衝液、高性能HotStart DNA Taqポリメラーゼ、ヌクレオチド、SYBR(R)Green色素、及び機器の光学的諸特性を規格化するために必要なフルオレセイン参照色素を含む。マスターミックスを実験者のテンプレート及びプライマーと共にPCR管に単に加える。化学修飾及び厳密に制御されたHotStart酵素によりプライマー二量体及び他の非特異的産物の増幅を防ぐことによってより正確なSYBR Greenの結果が独自に得られる。このRT2 qPCRマスターミックスは、BioRad iCycler(R)及びMyiQ(R)におけるSYBR Greenに基づく検出を用いたリアルタイムPCR適用に最も適している。
【0083】
配列:
EF1アルファプロモーター(SEQ ID NO:1)
CGTGAGGCTCCGATGCCCGTCAGTGGGCAGAGCGCACATCGCCCACAGTCCCCGAGAAGTTGGGGGGAGGGGTCGGCAATTGAACCGGTGCCTAGAGAAGGTGGCGCGGGGTAAACTGGGAAAGTGATGTCGTGTACTGGCTCCGCCTTTTTCCCGAGGGTGGGGGAGAACCGTATATAAGTGCAGTAGTCGCCGTGAACGTTCTTTTTCGCAACGGGTTTGCCGCCAGAACACAGG
【0084】
LOX1(SEQ ID NO:2)GATCATAACTTCGTATAGCATACATTATACGAAGTTAT
【0085】
LOX2(SEQ ID NO:3)TCGAATAACTTCGTATAGCATACATTATACGAAGTTAT
【0086】
cDNA E6/E7(SEQ ID NO:4)
ATGTTTCAGGACCCACAGGAGCGACCCGGAAAGTTACCACAGTTATGCACAGAGCTGCAAACAACTATACATGATATAATATTAGAATGTGTGTACTGCAAGCAACAGTTACTGCGACGTGAGGTATATGACTTTGCTTTTCGGGATTTATGCATAGTATATAGAGATGGGAATCCATATGCTGTATGTGATAAATGTTTAAAGTTTTATTCTAAAATTAGTGAGTATAGACATTATTGTTATAGTGTGTATGGAACAACATTAGAACAGCAATACAACAAACCGTTGTGTGATTTGTTAATTAGGTGTATTAACTGTCAAAAGCCACTGTGTCCTGAAGAAAAGCAAAGACATCTGGACAAAAAGCAAAGATTCCATAATATAAGGGGTCGGTGGACCGGTCGATGTATGTCTTGTTGCAGATCATCAAGAACACGTAGAGAAACCCAGCTGTAATCATGCATGGAGATACACCTACATTGCATGAATATATGTTAGATTTGCAACCAGAGACAACTGATCTCTACTGTTATGAGCAATTAAATGACAGCTCAGAGGAGGAGGATGAAATAGATGGTCCAGCTGGACAAGCAGAACCGGACAGAGCCCATTACAATATTGTAACCTTTTGTTGCAAGTGTGACTCTACGCTTCGGTTGTGCGTACAAAGCACACACGTAGACATTCGTACTTTGGAAGACCTGTTAATGGGCACACTAGGAATTGTGTGCCCCATCTGTTCTCAGAAACCATAA
【0087】
WPRE(SEQ ID NO:5)
AATCAACCTCTGGATTACAAAATTTGTGAAAGATTGACTGGTATTCTTAACTATGTTGCTCCTTTTACGCTATGTGGATACGCTGCTTTAATGCCTTTGTATCATGCTATTGCTTCCCGTATGGCTTTCATTTTCTCCTCCTTGTATAAATCCTGGTTGCTGTCTCTTTATGAGGAGTTGTGGCCCGTTGTCAGGCAACGTGGCGTGGTGTGCACTGTGTTTGCTGACGCAACCCCCACTGGTTGGGGCATTGCCACCACCTGTCAGCTCCTTTCCGGGACTTTCGCTTTCCCCCTCCCTATTGCCACGGCGGAACTCATCGCCGCCTGCCTTGCCCGCTGCTGGACAGGGGCTCGGCTGTTGGGCACTGACAATTCCGTGGTGTTGTCGGGGAAGCTGACGTCCTTTCCATGGCTGCTCGCCTGTGTTGCCACCTGGATTCTGCGCGGGACGTCCTTCTGCTACGTCCCTTCGGCCCTCAATCCAGCGGACCTTCCTTCCCGCGGCCTGCTGCCGGCTCTGCGGCCTCTTCCGCGTCTTCGCCTTCGCCCTCAGACGAGTCGGATCTCCCTTTGGGCCGCCTCCCCGCCTG
【0088】
Vector_pLV_EF1a−Lox1−E6E7−Lox2_WPRE
(SEQ ID NO:6)
CGCGTCGTGAGGCTCCGATGCCCGTCAGTGGGCAGAGCGCACATCGCCCACAGTCCCCGAGAAGTTGGGGGGAGGGGTCGGCAATTGAACCGGTGCCTAGAGAAGGTGGCGCGGGGTAAACTGGGAAAGTGATGTCGTGTACTGGCTCCGCCTTTTTCCCGAGGGTGGGGGAGAACCGTATATAAGTGCAGTAGTCGCCGTGAACGTTCTTTTTCGCAACGGGTTTGCCGCCAGAACACAGGATCATAACTTCGTATAGCATACATTATACGAAGTTATGATCCGTTTAAACTTGCTGCGACGCGTACTAGTATGTTTCAGGACCCACAGGAGCGACCCGGAAAGTTACCACAGTTATGCACAGAGCTGCAAACAACTATACATGATATAATATTAGAATGTGTGTACTGCAAGCAACAGTTACTGCGACGTGAGGTATATGACTTTGCTTTTCGGGATTTATGCATAGTATATAGAGATGGGAATCCATATGCTGTATGTGATAAATGTTTAAAGTTTTATTCTAAAATTAGTGAGTATAGACATTATTGTTATAGTGTGTATGGAACAACATTAGAACAGCAATACAACAAACCGTTGTGTGATTTGTTAATTAGGTGTATTAACTGTCAAAAGCCACTGTGTCCTGAAGAAAAGCAAAGACATCTGGACAAAAAGCAAAGATTCCATAATATAAGGGGTCGGTGGACCGGTCGATGTATGTCTTGTTGCAGATCATCAAGAACACGTAGAGAAACCCAGCTGTAATCATGCATGGAGATACACCTACATTGCATGAATATATGTTAGATTTGCAACCAGAGACAACTGATCTCTACTGTTATGAGCAATTAAATGACAGCTCAGAGGAGGAGGATGAAATAGATGGTCCAGCTGGACAAGCAGAACCGGACAGAGCCCATTACAATATTGTAACCTTTTGTTGCAAGTGTGACTCTACGCTTCGGTTGTGCGTACAAAGCACACACGTAGACATTCGTACTTTGGAAGACCTGTTAATGGGCACACTAGGAATTGTGTGCCCCATCTGTTCTCAGAAACCATAATCTAGCGTCGAATAACTTCGTATAGCATACATTATACGAAGTTATTCGACAATCAACCTCTGGATTACAAAATTTGTGAAAGATTGACTGGTATTCTTAACTATGTTGCTCCTTTTACGCTATGTGGATACGCTGCTTTAATGCCTTTGTATCATGCTATTGCTTCCCGTATGGCTTTCATTTTCTCCTCCTTGTATAAATCCTGGTTGCTGTCTCTTTATGAGGAGTTGTGGCCCGTTGTCAGGCAACGTGGCGTGGTGTGCACTGTGTTTGCTGACGCAACCCCCACTGGTTGGGGCATTGCCACCACCTGTCAGCTCCTTTCCGGGACTTTCGCTTTCCCCCTCCCTATTGCCACGGCGGAACTCATCGCCGCCTGCCTTGCCCGCTGCTGGACAGGGGCTCGGCTGTTGGGCACTGACAATTCCGTGGTGTTGTCGGGGAAGCTGACGTCCTTTCCATGGCTGCTCGCCTGTGTTGCCACCTGGATTCTGCGCGGGACGTCCTTCTGCTACGTCCCTTCGGCCCTCAATCCAGCGGACCTTCCTTCCCGCGGCCTGCTGCCGGCTCTGCGGCCTCTTCCGCGTCTTCGCCTTCGCCCTCAGACGAGTCGGATCTCCCTTTGGGCCGCCTCCCCGCCTGGAATTCGAGCT
CGGTACCTTTAAGACCAATGACTTACAAGGCAGCTGTAGATCTTAGCCACTTTTTAAAAGAAAAGGGGGGACTGGAAGGGCTAATTCACTCCCAACGAAGACAAGATCTGCTTTTTGCTTGTACTGGGTCTCTCTGGTTAGACCAGATCTGAGCCTGGGAGCTCTCTGGCTAACTAGGGAACCCACTGCTTAAGCCTCAATAAAGCTTGCCTTGAGTGCTTCAAGTAGTGTGTGCCCGTCTGTTGTGTGACTCTGGTAACTAGAGATCCCTCAGACCCTTTTAGTCAGTGTGGAAAATCTCTAGCAGTAGTAGTTCATGTCATCTTATTATTCAGTATTTATAACTTGCAAAGAAATGAATATCAGAGAGTGAGAGGAACTTGTTTATTGCAGCTTATAATGGTTACAAATAAAGCAATAGCATCACAAATTTCACAAATAAAGCATTTTTTTCACTGCATTCTAGTTGTGGTTTGTCCAAACTCATCAATGTATCTTATCATGTCTGGCTCTAGCTATCCCGCCCCTAACTCCGCCCAGTTCCGCCCATTCTCCGCCCCATGGCTGACTAATTTTTTTTATTTATGCAGAGGCCGAGGCCGCCTCGGCCTCTGAGCTATTCCAGAAGTAGTGAGGAGGCTTTTTTGGAGGCCTAGGCTTTTGCGTCGAGACGTACCCAATTCGCCCTATAGTGAGTCGTATTACGCGCGCTCACTGGCCGTCGTTTTACAACGTCGTGACTGGGAAAACCCTGGCGTTACCCAACTTAATCGCCTTGCAGCACATCCCCCTTTCGCCAGCTGGCGTAATAGCGAAGAGGCCCGCACCGATCGCCCTTCCCAACAGTTGCGCAGCCTGAATGGCGAATGGCGCGACGCGCCCTGTAGCGGCGCATTAAGCGCGGCGGGTGTGGTGGTTACGCGCAGCGTGACCGCTACACTTGCCAGCGCCCTAGCGCCCGCTCCTTTCGCTTTCTTCCCTTCCTTTCTCGCCACGTTCGCCGGCTTTCCCCGTCAAGCTCTAAATCGGGGGCTCCCTTTAGGGTTCCGATTTAGTGCTTTACGGCACCTCGACCCCAAAAAACTTGATTAGGGTGATGGTTCACGTAGTGGGCCATCGCCCTGATAGACGGTTTTTCGCCCTTTGACGTTGGAGTCCACGTTCTTTAATAGTGGACTCTTGTTCCAAACTGGAACAACACTCAACCCTATCTCGGTCTATTCTTTTGATTTATAAGGGATTTTGCCGATTTCGGCCTATTGGTTAAAAAATGAGCTGATTTAACAAAAATTTAACGCGAATTTTAACAAAATATTAACGTTTACAATTTCCCAGGTGGCACTTTTCGGGGAAATGTGCGCGGAACCCCTATTTGTTTATTTTTCTAAATACATTCAAATATGTATCCGCTCATGAGACAATAACCCTGATAAATGCTTCAATAATATTGAAAAAGGAAGAGTATGAGTATTCAACATTTCCGTGTCGCCCTTATTCCCTTTTTTGCGGCATTTTGCCTTCCTGTTTTTGCTCACCCAGAAACGCTGGTGAAAGTAAAAGATGCTGAAGATCAGTTGGGTGCACGAGTGGGTTACATCGAACTGGATCTCAACAGCGGTAAGATCCTTGAGAGTTTTCGCCCCGAAGAACGTTTTCCAATGATGAGCACTTTTAAAGTTCTGCTATGTGGCGCGGTATTATCCCGTATTGACGCCGGGCAAGAGCAACTCGGTCGCCGCATACACTATTCTCAGAATGACTTGGTTGAGTACTCACCAGTCACAGAAAAGCATCTTACGGATGGCATGACAGTAAGAGAATTATGCAGTGCTGCCATAACCATGAGTGATAACACTGCGGCCAACTTACTTCTGACAACGATCGGAGGACCGAAGGAGCTAACCGCTTTTTTGCACAACATGGGGGATCATGTAACTCGCCTTGATCGTTGGGAACCGGAGCTGAATGAAGCCATACCAAACGACGAGCGTGACACCACG
ATGCCTGTAGCAATGGCAACAACGTTGCGCAAACTATTAACTGGCGAACTACTTACTCTAGCTTCCCGGCAACAATTAATAGACTGGATGGAGGCGGATAAAGTTGCAGGACCACTTCTGCGCTCGGCCCTTCCGGCTGGCTGGTTTATTGCTGATAAATCTGGAGCCGGTGAGCGTGGGTCTCGCGGTATCATTGCAGCACTGGGGCCAGATGGTAAGCCCTCCCGTATCGTAGTTATCTACACGACGGGGAGTCAGGCAACTATGGATGAACGAAATAGACAGATCGCTGAGATAGGTGCCTCACTGATTAAGCATTGGTAACTGTCAGACCAAGTTTACTCATATATACTTTAGATTGATTTAAAACTTCATTTTTAATTTAAAAGGATCTAGGTGAAGATCCTTTTTGATAATCTCATGACCAAAATCCCTTAACGTGAGTTTTCGTTCCACTGAGCGTCAGACCCCGTAGAAAAGATCAAAGGATCTTCTTGAGATCCTTTTTTTCTGCGCGTAATCTGCTGCTTGCAAACAAAAAAACCACCGCTACCAGCGGTGGTTTGTTTGCCGGATCAAGAGCTACCAACTCTTTTTCCGAAGGTAACTGGCTTCAGCAGAGCGCAGATACCAAATACTGTCCTTCTAGTGTAGCCGTAGTTAGGCCACCACTTCAAGAACTCTGTAGCACCGCCTACATACCTCGCTCTGCTAATCCTGTTACCAGTGGCTGCTGCCAGTGGCGATAAGTCGTGTCTTACCGGGTTGGACTCAAGACGATAGTTACCGGATAAGGCGCAGCGGTCGGGCTGAACGGGGGGTTCGTGCACACAGCCCAGCTTGGAGCGAACGACCTACACCGAACTGAGATACCTACAGCGTGAGCTATGAGAAAGCGCCACGCTTCCCGAAGGGAGAAAGGCGGACAGGTATCCGGTAAGCGGCAGGGTCGGAACAGGAGAGCGCACGAGGGAGCTTCCAGGGGGAAACGCCTGGTATCTTTATAGTCCTGTCGGGTTTCGCCACCTCTGACTTGAGCGTCGATTTTTGTGATGCTCGTCAGGGGGGCGGAGCCTATGGAAAAACGCCAGCAACGCGGCCTTTTTACGGTTCCTGGCCTTTTGCTGGCCTTTTGCTCACATGTTCTTTCCTGCGTTATCCCCTGATTCTGTGGATAACCGTATTACCGCCTTTGAGTGAGCTGATACCGCTCGCCGCAGCCGAACGACCGAGCGCAGCGAGTCAGTGAGCGAGGAAGCGGAAGAGCGCCCAATACGCAAACCGCCTCTCCCCGCGCGTTGGCCGATTCATTAATGCAGCTGGCACGACAGGTTTCCCGACTGGAAAGCGGGCAGTGAGCGCAACGCAATTAATGTGAGTTAGCTCACTCATTAGGCACCCCAGGCTTTACACTTTATGCTTCCGGCTCGTATGTTGTGTGGAATTGTGAGCGGATAACAATTTCACACAGGAAACAGCTATGACCATGATTACGCCAAGCGCGCAATTAACCCTCACTAAAGGGAACAAAAGCTGGAGCTGCAAGCTTAATGTAGTCTTATGCAATACTCTTGTAGTCTTGCAACATGGTAACGATGAGTTAGCAACATGCCTTACAAGGAGAGAAAAAGCACCGTGCATGCCGATTGGTGGAAGTAAGGTGGTACGATCGTGCCTTATTAGGAAGGCAACAGACGGGTCTGACATGGATTGGACGAACCACTGAATTGCCGCATTGCAGAGATATTGTATTTAAGTGCCTAGCTCGATACAATAAACGGGTCTCTCTGGTTAGACCAGATCTGAGCCTGGGAGCTCTCTGGCTAACTAGGGAACCCACTGCTTAAGCCTCAATAAAGCTTGCCTTGAGTGCTTCAAGTAGTGTGTGCCCGTCTGTTGTGTGACTCTGGTAACTAGAGATCCCTCAGACCCTTTTAGTCAGTGTGGAAAATCTCTAGCAGTGGCGCCCGAACAGGGACCTGAAAGCGAAAGGGAAACCA
GAGCTCTCTCGACGCAGGACTCGGCTTGCTGAAGCGCGCACGGCAAGAGGCGAGGGGCGGCGACTGGTGAGTACGCCAAAAATTTTGACTAGCGGAGGCTAGAAGGAGAGAGATGGGTGCGAGAGCGTCAGTATTAAGCGGGGGAGAATTAGATCGCGATGGGAAAAAATTCGGTTAAGGCCAGGGGGAAAGAAAAAATATAAATTAAAACATATAGTATGGGCAAGCAGGGAGCTAGAACGATTCGCAGTTAATCCTGGCCTGTTAGAAACATCAGAAGGCTGTAGACAAATACTGGGACAGCTACAACCATCCCTTCAGACAGGATCAGAAGAACTTAGATCATTATATAATACAGTAGCAACCCTCTATTGTGTGCATCAAAGGATAGAGATAAAAGACACCAAGGAAGCTTTAGACAAGATAGAGGAAGAGCAAAACAAAAGTAAGACCACCGCACAGCAAGCGGCCGCTGATCTTCAGACCTGGAGGAGGAGATATGAGGGACAATTGGAGAAGTGAATTATATAAATATAAAGTAGTAAAAATTGAACCATTAGGAGTAGCACCCACCAAGGCAAAGAGAAGAGTGGTGCAGAGAGAAAAAAGAGCAGTGGGAATAGGAGCTTTGTTCCTTGGGTTCTTGGGAGCAGCAGGAAGCACTATGGGCGCAGCCTCAATGACGCTGACGGTACAGGCCAGACAATTATTGTCTGGTATAGTGCAGCAGCAGAACAATTTGCTGAGGGCTATTGAGGCGCAACAGCATCTGTTGCAACTCACAGTCTGGGGCATCAAGCAGCTCCAGGCAAGAATCCTGGCTGTGGAAAGATACCTAAAGGATCAACAGCTCCTGGGGATTTGGGGTTGCTCTGGAAAACTCATTTGCACCACTGCTGTGCCTTGGAATGCTAGTTGGAGTAATAAATCTCTGGAACAGATTTGGAATCACACGACCTGGATGGAGTGGGACAGAGAAATTAACAATTACACAAGCTTAATACACTCCTTAATTGAAGAATCGCAAAACCAGCAAGAAAAGAATGAACAAGAATTATTGGAATTAGATAAATGGGCAAGTTTGTGGAATTGGTTTAACATAACAAATTGGCTGTGGTATATAAAATTATTCATAATGATAGTAGGAGGCTTGGTAGGTTTAAGAATAGTTTTTGCTGTACTTTCTATAGTGAATAGAGTTAGGCAGGGATATTCACCATTATCGTTTCAGACCCACCTCCCAACCCCGAGGGGACCCGACAGGCCCGAAGGAATAGAAGAAGAAGGTGGAGAGAGAGACAGAGACAGATCCATTCGATTAGTGAACGGATCTCGACGGTATCGGTTAACTTTTAAAAGAAAAGGGGGGATTGGGGGGTACAGTGCAGGGGAAAGAATAGTAGACATAATAGCAACAGACATACAAACTAAAGAATTACAAAAACAAATTACAAAAATTCAAAATTTTATCGATCACGAGACTAGCCTCGAGAAGCTTGAT
【0089】
結果
A有足細胞
1997年にMundel P 等によって記載されたプロトコールに従って新生仔ラット腎糸球体から有足細胞を単離した。単離した後、コラーゲンコートされた24ウェルプレート上の10%FCS入りのDMEM中で糸球体を培養した。24時間後、有足細胞前駆体は、付着した糸球体から外へ増殖し始めた(図2)。
【0090】
この段階で、細胞に異なるレンチウイルスコンストラクトを形質導入し、そして培養を長期間(2〜4週間)継続した。急速に増殖している細胞の島を単離し、広げ、そしてRT PCR及び免疫染色を用いて特徴づけた。SV40 LTagの発現は、免疫染色によって制御した。細胞の100%は、核中に大量のSV40 LT Agを蓄積し(図3C)、形質導入されてない又はGFP形質導入された細胞では、染色を検出することができなかった(データは示さず)。hTERTによる形質導入は、(定量的RT PCRデータによればhTERTは500倍を超えて過剰発現されたが)増殖速度を高めるには不十分であり、そして数回試行した後、断念した。HPV E6/E7単独による形質導入もまた、有足細胞不死化に十分でなかったが、SV40 LT Agと組み合わせて用いたとき、かなり多くの不死化クローンを増強した。10回を超える継代後、細胞にレンチウイルスCreリコンビナーゼを形質導入した。3日後、特異的な抗SV40 LTag抗体で染色した。多くの細胞は、LT Agの有意な減少又は完全な欠如を示し、これはCreが細胞ゲノムから組込まれたコピーのすべて又は大多数を切除したことを示している(図3D)。LT Agが減少した(すなわち陰性)細胞は、その増殖を停止し、3つはサイズが大きくなり、そして有足細胞の典型的な形態を獲得した(図3A及びB)。切除効率は比較的低く、約30〜50%であった。有足細胞のいくつかの高度に特異的な遺伝子マーカー、例えば、WT1、シナプトポジンは、この細胞中で再発現されたが、ネフリン及びポドシンのような他のものは、極めて低いレベルでしか発現されなかった(図3C〜F及び表1)。
【0091】
【表1】
【0092】
結論
有足細胞は、良好に不死化することができるが、さらなる実験的使用のため「不死化遺伝子」の切除によって非増殖状態に戻らなければならない。目下、アデノウイルスCreを産生しており、そして切除効率を高める目的で試験する。切除しなければ、不死化細胞は、極めて急速に増殖し(倍加時間12〜15時間)そして有足細胞の多くの特異的マーカーを消失するか又は低下したレベルで発現する。
【0093】
B 初代ヒト肝細胞(PHH)
PHHをLonzaから購入し、そしてコラーゲンコートされたプレート上で培養した。細胞は増殖しないが、脱分化の明らかな徴候がなくても培養を10〜14日間持続することができる。PHHにSV40 LT Ag又はE6/E7(hTERTを用いない)を形質導入し、そして数日後、増殖が始まり、そのサイズは、時間と共に縮小し、そしてそれは容易に継代培養することができる。本発明者らは、これらの細胞を、20回を超えて継代培養し、そしてその表現型は変化しなかったが、対照肝細胞と比較すると、それは明らかにその表現型を部分的に失った。特に、CRP及びハプトグロビンのような肝細胞特異的マーカーの発現は、とりわけSV40 LT Ag中で大きく低下しているが、E6/E7不死化細胞中では、ハプトグロビン発現は保たれており、そしてCRP発現は、なお比較的高く、初代細胞と比較して10%である(表2)。重要なことに、肝細胞関連の転写因子HNF4アルファの発現は、E6/E7形質導入PHH中で2倍しか低下しておらず、そしてSV40 LT Ag中ではより大きく8倍低下している。
【0094】
【表2】
【0095】
cDNAアレイ分析は、SV40 LT Ag又はE6/E7ウイルス性初期タンパク質の過剰発現によって生じる不死化が、細胞周期の制御に関与することが知られているいくつかの遺伝子、すなわちE2F1転写因子、サイクリンA2、CDC2/CDK1−サイクリン依存性キナーゼ1の転写を強く誘導することを示している(表2)。トランスクリプトーム変化のより詳細な説明は、後に報告する。
【0096】
SV40 LT Agは、核p53の巨大な蓄積、そしてE6/E7は、p53の分解及び消失をもたらした(図4)。
【0097】
両方の場合、p53は、完全に不活性化され、そしてそのトランス活性化及びトランス抑制性を失う。特に、CDK2の阻害剤、そしてp53の直接標的の1つ、p21WAF1(CIP1又はCDKN1A)の発現は、核中でp53の蓄積にもかかわらず、SV40 LT Ag不死化細胞中で変化しなかった(データは示さず)。一方、別のCDK阻害剤、p16INK4のmRNA発現は、SV40 LT Ag及びE6/E7−不死化細胞中で強く上方制御された(データは示さず)。この複雑な現象の潜在的な機構は、公開されているが、明確な説明はない(Pei 及び Xiong, 2005)。
【0098】
結論:初代ヒト肝細胞は、SV40 LT Ag又はヒト乳頭腫E6/E7初期タンパク質のいずれかのレンチウイルスが介在する送達により非常に有効に不死化することができる。得られた細胞株は、極めて急速に増殖し(倍加時間は、20時間未満である)、そしてとりわけE6/E7に誘導された不死化の場合、肝細胞の多くの重要なマーカーを、なお保っていた。これらの細胞は、細胞分裂しない初代肝細胞の代わりにさまざまな生物学的目的に用いることができる。
C 初代内皮細胞
HUVEC(ヒト臍帯静脈内皮細胞)をPromoCellから購入し、そしてFCS、ECG、hEGF、hbFGF及びHC−500を含む内皮細胞特異的培地を用いて、コラーゲンコートされたプレートにおいて培養した。準集密細胞にSV40 LT Ag又はE6/E7レンチウイルス粒子のいずれかを形質導入した。hTERT単独の形質導入は、有効でなかった(他の試験細胞タイプにおいても同様)。SV40 LT Ag及びE6/E7−形質導入された細胞では、より速い増殖速度が見られたが、hTERTでは見られなかった。FACS分析は、両方の「不死化遺伝子」、LT Ag及びE6/E7により、S−G2−M画分の増加に伴ってG1/G0画分の減少(60から20%まで)が生じることを示した。BrdU取り込み/染色(データは示さず)によりFACSデータを確認した。驚くべきことに、非常により速い増殖速度にもかかわらず、内皮特異的マーカー、CD31(PECAM)及びvWFは、不死化HUVEC中において同じか又はさらにより高レベルで発現された(図5及び表3参照)。
【0099】
【表3】
【0100】
SV40 LT Ag及びE6/E7を過剰発現するHUVECの両方で、内皮特異的なNOS(eNOS)の中程度の減少が検出された。試験した他の初代細胞と同様に、SV40 LT Agは、核p53タンパク質における莫大な増加をもたらした、そしてE6/E7では減少した(データは示さず)。
【0101】
結論:不死化された初代内皮細胞(HUVEC)は、その主要な分化マーカーを維持するが親細胞より3〜5倍急速に増殖する。形態学的に、細胞サイズの縮小のみが示され、そして核多形性により非不死化細胞と区別される。それらは、さまざまな実験タイプに用いることができ、そこでは内皮「バックグラウンド」が必要とされる。
【0102】
不死化の効率。初期ウイルス性タンパク質SV40 LT Ag又はHPV E6/E7を過剰発現するためにレンチウイルス送達系を用いて、本発明者らは、異なる初代ヒト及び齧歯類細胞を不死化することができた。本発明の主要な目標は、初代ラット腎臓有足細胞を不死化することであった。驚くべきことに、レンチウイルスを齧歯類細胞に形質導入することは、初代ヒト細胞と比較してずっと困難であった。これにより、おそらく、なぜ本発明者らがSV40 LT Ag又はE6/E7単独でラット腎臓細胞の不死化を達成することができなかったかを説明することができる。しかし、2つのレンチウイルスコンストラクトの組み合わせ(最初にSV40 LT Ag、そして2日後にE6/E7を細胞に逐次形質導入した)により単離されたラット糸球体からいくつかの永続的な細胞株が得られた。急速に増殖する細胞は、分化マーカーを部分的に失っておりそして形態学的に有足細胞に似ていなかった。従って、Creリコンビナーゼを用いて「不死化遺伝子」を切除する必要があった。レンチウイルスCreにより形質導入すると、細胞は増殖を停止し、サイズが大きくなり、そしてネフリン(NPHS1)、シナプトポジン、ウィルムス腫瘍1(WT1)を含む主な有足細胞特異的マーカーを再発現した。残念なことに、主な有足細胞特異的遺伝子の1つ、ポドシン(NPHS2)は、Cre形質導入された細胞中で再び現れなかった。一般に、本発明者らは、新たに単離されたラット糸球体の有足細胞中でポドシンが短時間(多くとも72時間)しか発現されなかったことを観察した。従って、作製された不死化細胞は、部分的に脱分化したと考えるべきである。ポドシン発現を急速に消失させるもの(多くの他の有足細胞特異的遺伝子が発現され続けるときに)は、今のところ明らかでない。
【0103】
最初に、本発明者らは、2つのさらなる遺伝子、テロメラーゼのヒト触媒サブユニット(hTERT)及び温度感受性(ts)SV40ラージT抗原突然変異体を不死化に使用するつもりであった。これらのcDNAコンストラクトを含むレンチウイルス粒子を作製し、そして形質導入された細胞中でhTERT及びSV40 LT Ag−tsの過剰発現が確認されたが、不死化は達成されなかった。HUVEC、肝細胞、平滑筋細胞、間葉幹細胞を含むいくつかのヒト初代細胞でhTERTを試験したが、全く成功しなかった。従って、少なくとも上記の初代齧歯類及びヒト細胞においてhTERT単独では不死化に不十分であると結論する必要があった。
【0104】
(上記の)初代ヒト細胞の不死化は、SV40 LT Ag及びE6/E7レンチウイルスコンストラクトにより達成に成功した。SV40 LT Agで不死化された細胞(細胞の由来に依存しない)は、E6/E7発現細胞よりもずっとより急速に増殖し、そして分化マーカーの部分的又は完全な喪失を伴って有意な核多形性を示す。E6/E7の発現により増殖が活性化されるが、分化の部分的喪失しか起こらず、従って新規な細胞株の世代にとってより適切な系となる。興味深いことに、HUVECでは、実際に分化状態の喪失は見られず(図6及び表4参照)、そして「不死化遺伝子」を切除する必要なしに不死化細胞を直接用いることができる。
【0105】
不死化の機構。ラージT−抗原(LT Ag)、ウイルス性癌タンパク質は、SV40によってコードされた主要な初期遺伝子産物である。LT Ag及びヘリカーゼファミリーIIIのメンバー(AAA+スーパーファミリーの部分)。このタンパク質は、ウイルスDNA複製にとって重要である(Li 及び Kelly 1984; Neuwald 等 1999)。SV40 LT Agは、休止細胞の増殖を誘導し、いくつかの細胞タイプを形質転換し、そして実験動物における腫瘍形成を誘導する(Fanning 及び Knippers 1992)。LT Agによって生じる不死化/形質転換の正確な機構は、なお明らかではないが、大多数のデータは、p53を含むいくつかの重要な細胞腫瘍抑制因子の機能への干渉を通して細胞周期チェックポイントを乗り越えることができることを示している(Srinivasan 等 1997; Pipas 1998; DeCaprio 1999; Kim 等 2001)。試験したすべての初代ヒト及び齧歯類細胞において、SV40 LT Agをコードするレンチウイルスによる形質導入によりp53の核内蓄積が莫大になった(図3及び4)。反対に、HPV E6/E7で形質導入された細胞では、核p53は消失した(図4)。興味深いことに、いくつかの他のウイルス性癌タンパク質(アデノウイルスE1B、B型肝炎ウイルスXタンパク質、乳頭腫ウイルスE6、エプスタイン−バーウィルスEBNA−5及びヒトTリンパ球向性ウイルスtaxタンパク質)もまたp53を標的とし、強化されたプロテアソーム分解を通してそれを不活性化する。しかし、SV40 LT Agは、主に異なる機構を用い;細胞のp53を安定化する(タンパク質の半減期を高める)と同時にそのトランス活性化活性を阻害する(Deppert 等 1989; Pipas 及び Levine 2001)。p53安定化及び不活性化の機構は、SV40 LT Agとp53との直接結合に左右される。近年では、p53はラージT抗原と共結晶化された。予期せぬことに、構造は、6つのp53モノマーを結合しているLT Agの六量体複合体を示した。結合されたSV40 LTは、p53制御遺伝子のプロモーター中でp53DNA−結合ドメインとp53応答エレメントとの相互作用を阻止する。それは、p53のDNA−結合領域での劇的な立体配置的変化によって生じ、これは、p53内の独特の「メチオニンスイッチ」を通して部分的に達成される(Lilyestrom W 等, 2006)。
【0106】
ヒト乳頭腫ウイルス(HPV)初期タンパク質E6の場合におけるp53の不活性化は、ユビキチン経路を通してp53分解が高められることにより生じる。E6は、細胞ユビキチン−リガーゼを通してp53タンパク質に結合し、E6結合タンパク質(E6−AP)は、酵素のユビキチン複合体を集め、p53上でリシンをユビキチン化し、そしてそのタンパク質分解を開始する。
【0107】
HPV E7は、網膜芽腫腫瘍抑制タンパク質pRbの機能を不活性化する。E7タンパク質は、主に核中に局在化されており、ここで網膜芽腫遺伝子産物pRb(ポケットRb)に結合して細胞周期のS期への進行を促進する(Zur Hausen, 2002)。正常細胞では、pRbは、初期G1期に低リン酸化され、そしてE2F転写因子に結合し、転写リプレッサーとして機能する複合体を形成する。9回リン酸化されると複合体は解離し、E2Fは転写活性化因子として作用することが可能になる。低リン酸化pRbと結合することによって、E7は、それとE2Fとの結合を妨げ、それによって細胞周期進行を促進する。さらに、E7に誘導されたユビキチン介在性pRb分解は、細胞周期停止を有効に克服する際に不可欠であると考えられる(Fehrmann & Laimins, 2003)。
【0108】
従って、SV40及びHPVによる不死化に含まれる一般的な機構は、p53及びpRb不活性化である。非常に最近のデータは、p53が転写活性化だけでなく抑制にも介在していることを示している(Spurgers 等, 2006; Scian 等, 2008)。重要なことに、いくつかのp53に依存して抑制された遺伝子(cdc2、cdk4、サイクリン2A、cdc20、Spurgers 等, 2006参照)は、E2F1−3によってトランス活性化されることが示された(Zhu 等, 2004)。E2F1は、pRbから解離されると(pRbリン酸化又はウイルス性タンパク質によるpRbの機能不活性化のいずれかの結果として)、それ自体の転写を活性化し(正のフィードバック)、そしてG1/S及びG2/M移行にとって重要な標的遺伝子(cdc2、cdk4、サイクリン2A、cdc20)の転写を活性化する。
【0109】
本発明者らは、SV40 LT Ag又はHPV E6/E7によって不死化された異なる細胞タイプにおける増殖及び分化の制御に関与する200を超える遺伝子の発現を分析した。すべての細胞タイプにおいて、E2F1、cdc2/cdk1及びサイクリンA2の増強された発現が見られ、そしていくつかの場合(例えば、初代ヒト肝細胞)、数百倍であった(表2参照)。上記のようなサイクリンA2及びcdc2/cdk1は、p53によって転写的に抑制するか又はE2F1によって転写的に活性化することができる(図6)。従って、本発明者らは、SV40及びHPVが介在する不死化には2つの重要な事象:第1に、p53及びpRbの不活性化(いくつかの群によって以前に示した)、そして第2に、E2Fそれ自体、そしてその標的、サイクリンA2、サイクリンB及びcdc2の転写を刺激することができる転写的に活性なE2Fタンパク質の放出が関与していることを提案する。最近示されたように、cdc2/cdk1は、哺乳動物細胞における唯一の必須な細胞周期Cdkである(Santamaria 等, 2007)。これらのデータは、Cdk1が、不死化、そしておそらくまた二本鎖DNA HPV及びSV40ウイルスによっても生じる発癌の重要な調節因子とみなすことができることを示している。特異的CDK1阻害剤は、確立されたHPV若しくはSV40病因又は突然変異したp53若しくはRbによる腫瘍に対して非常に有効であると期待することができる。
【0110】
文献
DeCaprio JA: The role of the J domain of SV40 large T in cellular transformation. Biologicals. 1999 Mar; 27(1):23-8.
Deppert W, Steinmayer T, Richter W: Cooperation of SV40 large T antigen and the cellular protein p53 in maintenance of cell transformation. Oncogene. 1989 Sep;4(9):1103-10.
Deppert W, Steinmayer T: Metabolic stabilization of p53 in SV40-transformed cells correlates with expression of the transformed phenotype but is independent from complex formation with SV40 large T antigen. Curr Top Microbiol Immunol. 1989; 44:77-83.
Fanning E, Knippers R: Structure and function of simian virus 40 large tumor antigen. Annu Rev Biochem. 1992; 61:55-85. Review.
Fehrmann F, Laimins LA: Human papillomaviruses: targeting differentiating epithelial cells for malignant transformation. Oncogene. 2003 Aug 11;22(33):5201-7. Review.
Li JJ, Kelly TJ: Simian virus 40 DNA replication in vitro. Proc Natl Acad Sci U S A. 1984 Nov; 81(22):6973-7.
Lilyestrom W, Klein MG, Zhang R, Joachimiak A, Chen XS: Crystal structure of SV40 large T-antigen bound to p53: interplay between a viral oncoprotein and a cellular tumor suppressor. Genes Dev. 2006 Sep 1; 20(17):2373-82.
Mundel P, Reiser J, Kriz W: Induction of differentiation in cultured rat and human podocytes. J Am Soc Nephrol 8: 697-705, 1997.
Neuwald AF, Aravind L, Spouge JL, Koonin EV: AAA+: A class of chaperone-like ATPases associated with the assembly, operation, and disassembly of protein complexes. Genome Res. 1999 Jan;9(1):27-43.
Pei XH, Xiong Y: Biochemical and cellular mechanisms of mammalian CDK inhibitors: a few unresolved issues. Oncogene. 2005 Apr 18; 24(17):2787-95. Review.
Pipas JM: Molecular chaperone function of the SV40 large T antigen. Dev Biol Stand.1998; 94:313-9. Review.
Pipas JM, Levine AJ: Role of T antigen interactions with p53 in tumorigenesis. Semin Cancer Biol. 2001 Feb; 11(1):23-30.
Santamaria D, Barriere C, Cerqueira A, Hunt S, Tardy C, Newton K, Caceres JF, Dubus P, Malumbres M, Barbacid M: Cdk1 is sufficient to drive the mammalian cell cycle. Nature. 2007 Aug 16;448(7155):811-5.
Scian MJ, Carchman EH, Mohanraj L, Stagliano KE, Anderson MA, Deb D, Crane BM, Kiyono T, Windle B, Deb SP, Deb S: Wild-type p53 and p73 negatively regulate expression of proliferation related genes. Oncogene. 2008 Apr 17;27(18):2583-93. Epub 2007 Nov 5.
Srinivasan A, McClellan AJ, Vartikar J, Marks I, Cantalupo P, Li Y, Whyte P, Rundell K, Brodsky JL, Pipas JM: The amino-terminal transforming region of simian virus 40 large T and small t antigens functions as a J domain. Mol Cell Biol. 1997 Aug; 17(8):4761-73.
Spurgers KB, Gold DL, Coombes KR, Bohnenstiehl NL, Mullins B, Meyn RE, Logothetis CJ, McDonnell TJ: Identification of cell cycle regulatory genes as principal targets of p53- mediated transcriptional repression. J Biol Chem. 2006 Sep 1;281(35):25134-42. Epub 2006 Jun 23.
Zhu W, Giangrande PH, Nevins JR: E2Fs link the control of G1/S and G2/M transcription. EMBO J. 2004 Nov 24;23(23):4615-26. Epub 2004 Oct 28
Zur Hausen H: Papillomaviruses and cancer: from basic studies to clinical application. Nat Rev Cancer. 2002 May; 2(5):342-50. Review.
【特許請求の範囲】
【請求項1】
可逆的に不死化された細胞の作製方法であって、
a)真核細胞を準備し、
b)工程a)の細胞を遺伝子改変して細胞中で1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現を可能にし;そして
c)工程b)の細胞中で1つ又はそれ以上の核酸を発現する
ことを含み、
その際、1つ又はそれ以上の不死化タンパク質は、場合により(i)SV40ラージT抗原(TAG)又はその機能的に活性な変異体及び/又は(ii)テロメラーゼ触媒サブユニット(TERT)又はその機能的に活性な変異体と組み合わせてヒト乳頭腫ウイルス(HPV)E6/E7又はその機能的に活性な変異体を含む前記方法。
【請求項2】
可逆的に不死化された細胞の作製方法であって、
a)真核細胞を準備し、
b)工程a)の細胞を遺伝子改変して細胞中で1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現を可能にし;そして
c)工程b)の細胞中で1つ又はそれ以上の核酸を発現する
ことを含み、
その際、1つ又はそれ以上の不死化タンパク質の少なくとも1つは、ヒト乳頭腫ウイルス(HPV)E6/E7、SV40ラージT抗原及びテロメラーゼ触媒サブユニット(TERT)、又はその機能的に活性な変異体からなる群より選ばれ、そして1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現がEF1アルファプロモーターによって制御される前記方法。
【請求項3】
方法が、さらに
d)工程c)の細胞を増殖させる
ことを含む、請求項1又は2に記載の方法。
【請求項4】
不死化タンパク質をコードする1つ又はそれ以上の核酸が1つ又はそれ以上のレトロウイルスベクター、特に1つ又はそれ以上のレンチウイルスベクターを用いて導入される、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
工程a)の細胞が、(i)哺乳動物細胞、特にヒト、霊長類又は齧歯類細胞及び/又は(ii)初代細胞である、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
工程a)の細胞が、分化細胞、特に内皮細胞、心筋細胞、平滑筋細胞、肝細胞、有足細胞又は腎臓細胞である、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現がEF1アルファプロモーターによって制御される、請求項1又は3〜6のいずれか1項に記載の方法。
【請求項8】
1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸がloxP(locus of X-over P1)部位に隣接する、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
請求項1〜8のいずれか1項に記載の方法によって入手可能な細胞、又はその子孫。
【請求項10】
1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸を含む真核細胞であって、その際、1つ又はそれ以上の不死化タンパク質の少なくとも1つがヒト乳頭腫ウイルス(HPV)E6/E7、SV40ラージT抗原及びテロメラーゼ触媒サブユニット(TERT)からなる群より選ばれ、そして1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現がEF1アルファプロモーターによって制御される、前記真核細胞。
【請求項11】
i)不死化タンパク質をコードする1つ又はそれ以上の核酸が1つ又はそれ以上のレトロウイルスベクター、特に1つ又はそれ以上のレンチウイルスベクター上にある;
ii)細胞がさらに遺伝子改変を含む;
iii)細胞が哺乳動物細胞、特にヒト、霊長類又は齧歯類細胞である;
iv)細胞が初代細胞から誘導される;
v)細胞が分化細胞、特に内皮細胞、心筋細胞、平滑筋細胞、肝細胞、有足細胞又は腎臓細胞から誘導される;
vi)1つ又はそれ以上の不死化タンパク質をコードする核酸の発現がEF1アルファプロモーターの制御下にある;及び/又は
vii)1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸がloxP(locus of X-over P1)部位の横に位置している、
請求項9若しくは10に記載の細胞又はその子孫。
【請求項12】
請求項9〜11のいずれか1項に記載の細胞、又はその子孫を再分化する方法であって、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現を阻止することを含む前記方法。
【請求項13】
1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸がloxP(locus of X-over P1)部位の横に位置しており、そして阻止がCreリコンビナーゼ又は機能的に活性なCre変異体によって実施される、請求項12に記載の方法。
【請求項14】
Creリコンビナーゼ遺伝子又は機能的に活性なCre変異体遺伝子がレトロウイルスベクター、特にレンチウイルスベクターを用いて細胞に導入される、請求項12又は13に記載の方法。
【請求項15】
請求項12〜14のいずれか1項に記載の方法によって入手可能な細胞、又はその子孫。
【請求項1】
可逆的に不死化された細胞の作製方法であって、
a)真核細胞を準備し、
b)工程a)の細胞を遺伝子改変して細胞中で1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現を可能にし;そして
c)工程b)の細胞中で1つ又はそれ以上の核酸を発現する
ことを含み、
その際、1つ又はそれ以上の不死化タンパク質は、場合により(i)SV40ラージT抗原(TAG)又はその機能的に活性な変異体及び/又は(ii)テロメラーゼ触媒サブユニット(TERT)又はその機能的に活性な変異体と組み合わせてヒト乳頭腫ウイルス(HPV)E6/E7又はその機能的に活性な変異体を含む前記方法。
【請求項2】
可逆的に不死化された細胞の作製方法であって、
a)真核細胞を準備し、
b)工程a)の細胞を遺伝子改変して細胞中で1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現を可能にし;そして
c)工程b)の細胞中で1つ又はそれ以上の核酸を発現する
ことを含み、
その際、1つ又はそれ以上の不死化タンパク質の少なくとも1つは、ヒト乳頭腫ウイルス(HPV)E6/E7、SV40ラージT抗原及びテロメラーゼ触媒サブユニット(TERT)、又はその機能的に活性な変異体からなる群より選ばれ、そして1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現がEF1アルファプロモーターによって制御される前記方法。
【請求項3】
方法が、さらに
d)工程c)の細胞を増殖させる
ことを含む、請求項1又は2に記載の方法。
【請求項4】
不死化タンパク質をコードする1つ又はそれ以上の核酸が1つ又はそれ以上のレトロウイルスベクター、特に1つ又はそれ以上のレンチウイルスベクターを用いて導入される、請求項1〜3のいずれか1項に記載の方法。
【請求項5】
工程a)の細胞が、(i)哺乳動物細胞、特にヒト、霊長類又は齧歯類細胞及び/又は(ii)初代細胞である、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
工程a)の細胞が、分化細胞、特に内皮細胞、心筋細胞、平滑筋細胞、肝細胞、有足細胞又は腎臓細胞である、請求項1〜5のいずれか1項に記載の方法。
【請求項7】
1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現がEF1アルファプロモーターによって制御される、請求項1又は3〜6のいずれか1項に記載の方法。
【請求項8】
1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸がloxP(locus of X-over P1)部位に隣接する、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
請求項1〜8のいずれか1項に記載の方法によって入手可能な細胞、又はその子孫。
【請求項10】
1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸を含む真核細胞であって、その際、1つ又はそれ以上の不死化タンパク質の少なくとも1つがヒト乳頭腫ウイルス(HPV)E6/E7、SV40ラージT抗原及びテロメラーゼ触媒サブユニット(TERT)からなる群より選ばれ、そして1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現がEF1アルファプロモーターによって制御される、前記真核細胞。
【請求項11】
i)不死化タンパク質をコードする1つ又はそれ以上の核酸が1つ又はそれ以上のレトロウイルスベクター、特に1つ又はそれ以上のレンチウイルスベクター上にある;
ii)細胞がさらに遺伝子改変を含む;
iii)細胞が哺乳動物細胞、特にヒト、霊長類又は齧歯類細胞である;
iv)細胞が初代細胞から誘導される;
v)細胞が分化細胞、特に内皮細胞、心筋細胞、平滑筋細胞、肝細胞、有足細胞又は腎臓細胞から誘導される;
vi)1つ又はそれ以上の不死化タンパク質をコードする核酸の発現がEF1アルファプロモーターの制御下にある;及び/又は
vii)1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸がloxP(locus of X-over P1)部位の横に位置している、
請求項9若しくは10に記載の細胞又はその子孫。
【請求項12】
請求項9〜11のいずれか1項に記載の細胞、又はその子孫を再分化する方法であって、1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸の発現を阻止することを含む前記方法。
【請求項13】
1つ又はそれ以上の不死化タンパク質をコードする1つ又はそれ以上の核酸がloxP(locus of X-over P1)部位の横に位置しており、そして阻止がCreリコンビナーゼ又は機能的に活性なCre変異体によって実施される、請求項12に記載の方法。
【請求項14】
Creリコンビナーゼ遺伝子又は機能的に活性なCre変異体遺伝子がレトロウイルスベクター、特にレンチウイルスベクターを用いて細胞に導入される、請求項12又は13に記載の方法。
【請求項15】
請求項12〜14のいずれか1項に記載の方法によって入手可能な細胞、又はその子孫。
【図1】

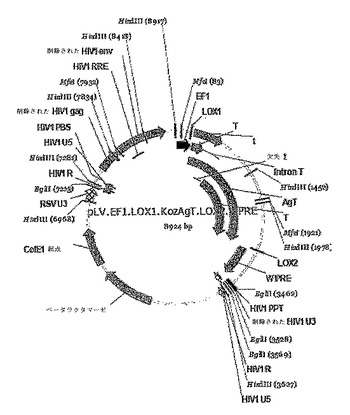
【図2】
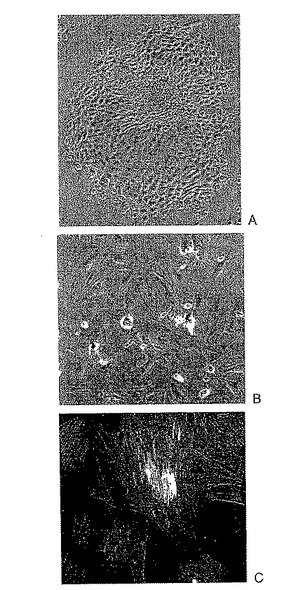
【図3】
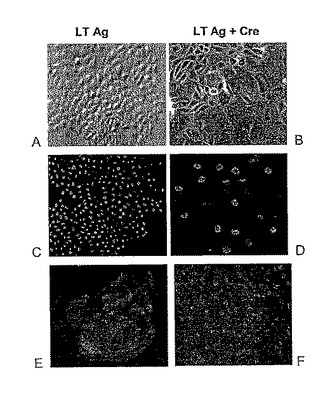
【図4】
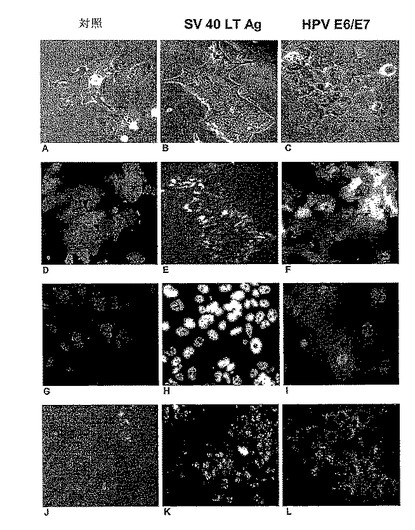
【図5】
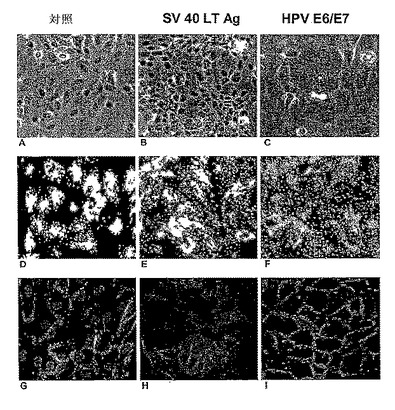
【図6】
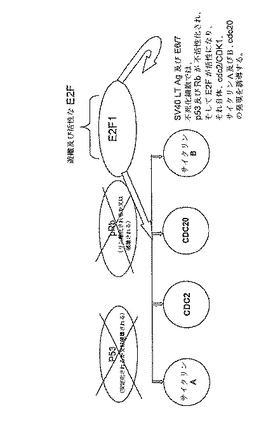
【図7】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公表番号】特表2012−525825(P2012−525825A)
【公表日】平成24年10月25日(2012.10.25)
【国際特許分類】
【出願番号】特願2012−509003(P2012−509003)
【出願日】平成22年5月4日(2010.5.4)
【国際出願番号】PCT/EP2010/055997
【国際公開番号】WO2010/128024
【国際公開日】平成22年11月11日(2010.11.11)
【出願人】(504456798)サノフイ (433)
【Fターム(参考)】
【公表日】平成24年10月25日(2012.10.25)
【国際特許分類】
【出願日】平成22年5月4日(2010.5.4)
【国際出願番号】PCT/EP2010/055997
【国際公開番号】WO2010/128024
【国際公開日】平成22年11月11日(2010.11.11)
【出願人】(504456798)サノフイ (433)
【Fターム(参考)】
[ Back to top ]