
同定方法
本発明は、広域スペクトルケモカイン阻害剤(BSCI)活性を有し、且つ抗炎症剤としての用途を有する、新規化学物質、及び医薬組成物の発見方法に関する。ある態様において、BSCI活性を有する化合物又は物質の同定方法は、以下の工程:(a)1又は2以上の候補化合物又は物質が、白血球上で発現されることが知られているGPCR神経受容体への結合についてスクリーニングされ;その後(b)選択されたGPCR神経受容体に対して結合を示す化合物又は物質が、機能アッセイでBSCI活性について試験され;そして(c)選択されたGPCR神経受容体に対して結合を示す化合物又は物質が、機能アッセイで古典的アゴニスト活性について試験される、を含み、ここで工程(a)は最初に行われるが、工程(b)及び(c)は任意の順番で、又は同時に行われる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、広域スペクトルケモカイン阻害剤(BSCI)活性を有し、且つ抗炎症剤としての用途を有する、新規化学物質、及び医薬組成物の発見方法に関する。
【0002】
ケモカインは、哺乳動物の免疫系の制御において重要な役割を果たす約50個の、構造が関連している、大部分が小さい(8〜12kDa)タンパク質の群である。ケモカインリガンドは、約20個の密接に関連した受容体を介してシグナルを出し、そしてそれらはGタンパク質結合受容体(GPCR)の1型クラスのメンバーである。重要なことは、リガンドの多くは2種以上の受容体に結合し、かつこれを介して生産的にシグナルを出すことである。同様に、多くの受容体が2種以上のリガンドに結合する。
【0003】
特定の種類の白血球上に、これらのケモカイン受容体の種々のサブセットが発現されている。この複雑な縮重の結果として、白血球動員の厳密な時間的及び空間的制御が可能である。ケモカインは免疫系の「交通警官」として中心的な役割を果たし、種々の白血球が、骨髄から血液を介して免疫応答の2次的リンパ組織又は免疫応答部位に取る複雑な経路をコードしている可能性がある。
【0004】
このため、ケモカインシグナル伝達を調節する化合物は長い間、炎症性要素を有する非常に広範囲の疾患(例えば、アテローム性動脈硬化症、自己免疫疾患、アレルギー、湿疹、乾癬、又は神経変性障害)に有用な、新規抗炎症剤開発のための有望な候補と考えられている。これらの疾患のための新しい薬に対して大きなニーズがある:既存の小分子治療薬は一般に、有効であるが重篤な副作用がある(例えばコルチコステロイド)か、又は有効性は小さいが比較的安全である(例えばジクロフェナックのようなNSAID)。生物学的治療薬(例えば、TNF−αに対する抗体)はより特異的であるが、製造コストが高く、単純な経口投与には適していない。小分子ケモカイン阻害剤は、既存の生物学的抗炎症剤と比較して、他の群の小分子抗炎症剤より有効性と安全性の兼ね合いが優れている可能性があり、改良された薬剤学的性質(コストを含む)を与える。
【0005】
残念ながら、特異的ケモカイン受容体アンタゴニストはこの可能性を実現するには進歩が遅い。まずこれらは、従来の大量処理分子スクリーニング法(例えば、リガンド:受容体相互作用を阻止することができる物質についての受容体結合アッセイで、ランダムなライブラリーがスクリーニングされる)を使用して見つけることが難しい。ケモカイン受容体間の類似性は、化合物がしばしばケモカイン受容体のサブセット(単一の受容体についての特異性を有する代わりに)と反応することを意味し、さらに悪いことに多くの初期ケモカイン受容体調節物質は、セロトニンやカテコールアミン系の関連受容体に結合し、挙動、血圧、及び他の表現型に好ましくない副作用を引き起こす。
【0006】
第2に、あるクラス内の単一の受容体に対する優れた特異性(これは、従来の医薬開発思想の最大の目的である)は、これが最終的に達成されても特に有効ではないことが明らかとなった。同じ受容体に結合する多くの関連リガンド、及び多くのリガンドのためにシグナルを変換できるいくつかの受容体がある、ケモカインシステムの縮重のために、単一の受容体を阻止する物質は通常、一般的抗炎症性は無く、免疫系に対して失望するほどに弱い作用を有する。これは音楽の増幅器に例えることができる:抗炎症作用は「ボリュームコントロール」の調節を必要とするが、特異的ケモカイン受容体アンタゴニストは「グラフィックエコライザー」に対して作用する。すなわちこのような物質は、免疫応答の種類をわずかに調節する(おそらく、存在する白血球サブセットにより浸入物の細胞組成を変化させる)が、炎症の程度を定量的に低下させることはない。
【0007】
その結果、ケモカインシステムが最初に記載されてから20年以上経過し、その後主要な免疫調節標的として出現している2009年でさえ、ヒトの治療薬として認可されているケモカイン阻害剤は無い(しかし、幾つかは特定の適応症で臨床治験の後期に入っており、炎症応答の種類の調節がある程度の効果を有している、例えばリウマチ様関節炎におけるCCR1ケモカイン受容体の特異的アンタゴニストであるBX471がある(Horuk (2005) Mini Rev. Med. Chem. 5:791-804))。従って別のアプローチが必要である:多くのケモカインによるシグナルを同時に阻害する化合物(我々が広域スペクトルケモカイン阻害剤(broad-spectrum chemokine inhibitor)、又はBSCIと名付けた物質;Grainger & Reckless (2003) Biochem. Pharmacol. 65:1027-34; Grainger et al. (2005) Mini Rev. Med. Chem. 5:825-32)。
【0008】
広域スペクトルケモカイン阻害剤(BSCI)活性を有する化合物は、広範囲の疾患を治療または予防するための抗炎症性薬物として有用な新しいクラスの物質(agent)である。ヒト疾患の動物モデルでは、アテローム性動脈硬化症、卒中、喘息、術的接着形成、子宮内膜症、皮膚炎、内毒素血症、及びリウマチ様関節炎において、BSCIは白血球蓄積や疾患重症度の他のマーカーを低下させるのに有効であることが証明されている(Reckless et al. (2001) Immunology 103:244-54; Beech et al. (2001) J. Cereb. Blood Flow Metabol. 21:683-9; Grainger & Reckless (2003) Biochem. Pharmacol. 65:1027-34; Naidu et al. (2003) Ann. Thorac. Surg. 78:1118-22; Naidu et al. (2004) J. Heart Lung Transplant. 23:128-34; Reckless et al. (2005) J. Vasc. Res. 42:492-502; Berkkanoglu et al. (2005) Hum. Reprod. 20:3047-52; Kayisli et al. (2007) Reprod. Sci. 14:825-35; Hildebrandt et al. (2009)、印刷中)。BSCIはまた、インビトロでHIV複製を抑制することが証明されている(HIVの細胞侵入のケモカイン依存性により生じる、抗炎症性に無関係の活性;Grainger & Lever (2005) Retrovirology 2:23)。
【0009】
このような広範囲の疾患モデルで強力な抗炎症活性を有するにもかかわらず、クラスとしてのBSCIは、注目すべきことに毒性が無いか又は副作用が小さい(Schroff et al. (2005) Mini Rev Med Chem. 5:849-55)。例えばアミノラクタムBSCIは、広範囲の急性及び慢性毒性試験で多くの投与経路で評価されているが、作用の無いレベルが最大治療用量より少なくとも1,000倍高い。これらのアミノラクタムBSCIはまた、優れた安全性薬理学的プロフィールを示す(hERGテール電流に対する作用がないか、又は遺伝毒性のアッセイにおいて作用がない;例えばWO2009/016390を参照)。
【0010】
これらの知見はまとめると、BSCIがクラスとして既存の抗炎症薬物(コルチコステロイドやNSAIDを含む)と比較して、有効性と副作用との優れた兼ね合いを与えることができることを示唆する。従って、BSCI活性を有する組成物を同定する方法に対するニーズがある。
【0011】
残念ながら、現在までに記載されているBSCIは運良く偶然同定されたものであり、新規の、潜在的に構造的関連性がないBSCIを同定する簡単な方法は無い。BSCI活性を有する最初の化合物は、ペプチド3(Peptide 3)と呼ぶペプチドであり、これはMCP−1の配列から選択され、複数のケモカイン受容体に結合する能力について試験された(Reckless & Grainger (1999) Biochem. J. 340:803-11)。ペプチド3は複数のケモカイン受容体に結合しないことがわかった(しかし実際は、細胞表面上野単一のまだ同定されていない高親和性部位に結合する;Grainger & Reckless (2003) Biochem. Pharmacol. 65:1027-34)が、少なくとも10個の異なる受容体に結合することができるそれらの間で、少なくとも5個のケモカインに対するBSCI活性を有した。
【0012】
このペプチド3の構造鋳型から、BSCIの一連の異なる構造群(ペプチド、ペプトイド、及び非ペプチド構造体を含む)が設計され、合成され、特徴づけされた。重要なことは、ペプチドのBSCI活性の原因である構造モチーフは、WxQトリペプチド(ここで、Wはアミノ酸トリプトファンであり、xは任意のタンパク新生アミノ酸であり、Qはグルタミンである)まで絞られたことである。これは、ペプチドではないWxQ模倣物の設計を可能にし、その1つ(N−アシルグルタリミドNR58,4)はBSCIとしてナノモル活性を有する(Fox et al. (2002) J. Med. Chem. 45:360-70)。
【0013】
NR58,4はBSCIとしてインビボで活性である(内毒素血症のような急性炎症のモデルで強力な抗炎症作用を有する)が、薬剤開発のための理想的な化合物ではないことが明らかになった。その必須のイミド環が血漿中で未特定のアミダーゼによる加水分解を受けやすく、その結果化合物はインビボで急速に不活性化された(Fox et al. (2005) J. Med. Chem. 48:867-74)。
【0014】
その後、さらなる医学化学的研究により、BSCIとしての効力(potency)を保持するがインビボでは安定である、関連性を有する種類の置換アミノラクタムが同定された。このクラスのBSCIの例には、N−オレオイル−3−アミノカプロラクタム(BN83253; Fox et al. (2005) J. Med. Chem. 48:867-74)がある。広範囲の、構造的関連性がある他のアミノラクタムBSCIも記載されている(例えば、WO2005/053702、WO2006/016152、WO2006/018609、WO2006/085096;WO2006/134385、WO2006/134384、及びWO2009/016390を参照)が、その少なくとも一部は、ヒトに対する医薬として開発するのに適した性質を有する。
【0015】
必須であるWxQファーマコフォアの構造に基づいて、別の群の非ペプチドBSCIも同定されている。アルカロイドであるヨヒンビン(yohimbine)(インドジャボク(Rauwolfia Serpentina)の根から得られるアルファ2−アドレナリン受容体ゴニスト)は、WVQトリペプチドの特定のコンフォメーションに似た炭素骨格を有し、ヨヒンビンの17−アミド誘導体はBSCIのように高活性である(WO99/12968; Grainger et al. (2005) Mini Rev Med Chem. 5:825-32)。
【0016】
非ペプチドBSCIの第3の群も記載されており、これも再度WVQトリペプチドの炭素骨格とのリセルグアミド核の類似性に基づく。クラスA禁止物質であるLSD(リセルグ酸のN,N−ジエチルアミド)のようなすでに報告されたリセルグアミド類は、複数のGPCRでアゴニストとして作用する(従って、幻覚誘発性を含む広範囲の生理学的作用を有する)が、我々は、化合物L−II(ジヒドロレセルグ酸のN−アスパラギンアミド)及びリセルグ酸の他のグルタミンとアスパラギンアミドが、他のリセルグ酸アミドの予測される副作用無しでBSCIのように高活性であることを証明した(WO99/12968)。
【0017】
現在までに記載されているすべての群のBSCI間の構造的関係を図1に例示する。この図から、BSCIのすべての群は互いの化学構造の類似性により特定されていることが明らかである。
【0018】
既知の化合物と構造が関連していないBSCIを同定するための一般的方法は、現在存在しない。この限界に関する理由は、BSCIの分子標的についての情報が欠如しているためである。これらのBSCIがケモカイン受容体アンタゴニストとして作用せず、試験したケモカイン受容体のいずれにも結合しないが、その代わりに細胞表面上の明確な(しかし同定されていない)高親和性部位に結合することがいったん明らかになると(Grainger & Reckless (2003) Biochem. Pharmacol. 65:1027-34)、従来の分子スクリーニング手段(例えば受容体結合アッセイ)を使用して、新規の、構造的に固有のBSCI群を捜すことは不可能であった。機能スクリーニングツール(すなわち、ライブラリー要素が、いくつかの異なるケモカイン(非ケモカインではない)に応答して細胞遊走を阻止するかどうかを調べる)を使用して、BSCI活性について非常に小さいライブラリーを直接スクリーニングすることができるかも知れないが、このようなスクリーニング方策は、機能スクリーニングアッセイの労働集約的性質のために直ちに非現実的となる(細胞遊走の阻害剤を同定するための機能スクリーニングアッセイを実施するための技術困難さの詳細な総説については、Frow et al. (2004) Med. Res. Rev. 24:276-98を参照)。さらにこのような機能スクリーニングは、生物学的適合性がある緩衝液に可溶性でありかつ低細胞毒性(大量処理薬物スクリーニングについて使用される典型的なランダムライブラリーで大部分(確実に90%を超える)の化合物を除外するパラメータ)を示す化合物にのみ適用することができる。簡単に説明すると、このようなアプローチは、ランダム化合物ライブラリーから完全に新しいクラスのBSCIを見つけるには全く現実的ではない。
【0019】
新規の、構造的に固有であるBSCIを同定できる能力は、この薬剤群の臨床開発のために重要である。既存のBSCIは、臨床開発を通して進歩するため、最適ではない性質が特定される可能性は低い。ヒトに投与されたBSCIは現在まで無く、現在までに同定されていない副作用又は毒性のリスクは大きいままである。さらに悪いことに、明らかとされている副作用又は毒性の大部分は、関連構造を有する化合物クラスによって共有される、クラス効果の可能性がある。従って残念ながら、BSCIについて現在まで記載されているすべてのクラスの化合物は、その構造類似性の結果として同定されたため、共有される副作用が見つかるリスクは高くなっている。
【発明の概要】
【0020】
ここで我々は、異なる群のBSCIにより使用される3つの異なる受容体を説明する。3つの受容体はすべてGPCR群のメンバーであり、その全ては、白血球上及び脳中に高発現される。驚くべきことに我々はまた、報告されるBSCIが、標的である特定の受容体に無関係に、機能的選択性をもって作用することを証明する。これらの結果は、本発明に従って、既知の化合物と同じ構造モチーフを含まない、構造的に多様なBSCI群を同定するのに有用な分子スクリーニング例を提供する。本発明は、一部においてこれらの結果に基づく。
【0021】
本発明は、添付の特許請求の範囲及び以下で定義される。ある態様において、1又は2以上の白血球サブセット上で発現されるGPCR神経受容体との相互作用について、1又は2以上の化合物(compound)若しくは物質(agent)がスクリーニングされ、その後の第2工程で、標的受容体と相互作用するこれらの化合物若しくは物質が、ケモカインにより誘導される白血球遊走を阻害する能力についてスクリーニングされるという方法が提供される。ケモカイン誘導性の白血球遊走を阻害する化合物若しくは物質は、その後インビトロでBSCI活性について試験される。重要なことは、標的受容体に対してアゴニストである化合物又は物質必ずしもすべてが、BSCI活性を持ってはいないことである。従って本発明の2つの工程は、BSCI活性を有する化合物を同定するのに必要かつ十分である。
【0022】
本発明の具体的な非限定例や追加の特徴は、以下の図面を参照して説明される。
【図面の簡単な説明】
【0023】
【図1】本明細書に記載の主要なBSCIの構造を示し、そのすべてが共有する相互関係(及び共通の構造モチーフ)を例示する。
【図2】単一の受容体(灰色の形)での機能的選択性の概念を示す図である。
【図3】タンパク質コード枠中の単一のイントロン(より暗い四角)の選択的スプライシングの結果として、sstr2遺伝子座から得られる2つの公知の代替転写体(T#1とT#2)を示す図である。
【図4】BSCIBN83253の用量応答(x軸、nM)と、MCP−1誘導性THP−1細胞遊走(y軸、阻害%)とに対する、sstr2アンタゴニストBIM23627(「I」)の濃度上昇の影響を示すグラフである。
【図5】BSCI(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)ーテトラヒドロピリジン−2−オンの用量応答(「バレロラクタム(valerolactam)」;x軸、nM)と、MCP−1誘導性THP−1細胞遊走(y軸、阻害%)に対する、sstr2アンタゴニストBIM23627(「I」)の濃度上昇の影響を示すグラフである。
【図6】細菌内毒素の単回腹腔内投与に応答する、TNF−アルファ産生(y軸、阻害%)に対する、sstr2アンタゴニストBIM23627とBSCIBN83250の同時投与(x軸、mg/kg用量)の影響を示すグラフである。
【図7】細菌内毒素の単回腹腔内投与に応答する、TNF−アルファ産生(y軸、阻害%)に対する、sstr2アンタゴニストBIM23627とBSCI(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オンの同時投与(「バレロラクタム」;x軸、mg/kg用量)の影響を示すグラフである。
【図8】細菌内毒素の単回腹腔内投与に応答する、TNF−アルファ産生(y軸、阻害%)に対する、sstr2アンタゴニストBIM23627と合成コルチコステロイドであるデキサメタゾンの同時投与(x軸、mg/kg用量)の影響を示すグラフである。
【図9】THP−1細胞(左のパネル)と陽性対照として使用したPanc−1細胞(右のパネル)から調製したmRNA試料からの、逆転写(「RT」)有り(+)及び無し(−)のPCR反応からのアンプリコンを明らかとする、臭化エチジウムで染色しUV光透過により観察したアガロースゲルを示す。
【図10】BN83250の標識類縁体である4’,5’−3[H]−(S)−3−(2’,2’−ジメチルドデカノイル)アミノ−カプロラクタム(4,5−3[H]−BN83250)の合成スキームを示す。化合物(S,E)−3−(2’,2’−ジメチルドデク−4’−エノイル)アミノ−カプロラクタム(上)はPd触媒の存在下でトリチウムガスと反応して、4,5−3[H]−BN83250(下)を形成する。
【図11】THP−1細胞への4,5−3[H]−BN83250(x軸、nM)の特異的結合(y軸、cpm)を示すグラフである。値は各BN83250濃度での3回測定の平均(±SD)であり、非特異結合(過剰の非標識BN83250の存在下)を引いてある。中心線は、単純な結合モデルを使用した時の最適フィットであり、外の線は95%信頼限界を示す。
【図12】THP−1細胞への10nMの4,5−[3H]−BN83250の結合(y軸、結合した標識[cpm])に対する、種々の濃度の非標識(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オン(「バレロラクタム」)の影響を示すグラフとヒストグラムである。
【図13】ポリリンカー中のEcoRI(5’)部位とXhoI(3’)部位との間でクローン化されたヒトsstr2Aをコードする1,100bp cDNA(「sstr2a」として示す)を有するpcDNA3.1+(Invitrogen)のプラスミド地図を示す。
【図14】親Jurkat T細胞(白丸、実線)、sstr2を発現するJurkat D3B細胞(黒丸、実線)、及びTHP−1細胞(点線)のSDF−1α誘導性遊走に対するBSCI(BN83250)の影響を比較する。(A)では、BN83250の濃度をx軸(nM)に示し、阻害%をy軸に示す。各細胞株の染色の分布を示す頻度ヒストグラムを、(B)THP−1細胞(*=特異的結合の375中央値単位)、(C)Jurkat D3B細胞(*=特異的結合の40中央値単位)、及び(C)Jurkat T細胞(*=特異的結合の1中央値単位)に示す。各頻度ヒストグラムでは、x軸はFl1−Hを示し、y軸はカウントを示す。頻度ヒストグラム(B)〜(D)の黒い線は、PBSのみによる染色を示す;薄い灰色の線は一次抗体対照を示し、中程度の灰色の線は、完全な染色反応を示す。
【図15】THP−1細胞のMCP−1誘導性遊走(y軸、阻害%)に対する種々の濃度のソマトスタチン14(x軸、nM)の影響を示す。点線は完全な阻害を示し、これはBSCIにより達成される。
【図16】広範囲の「古典的」sstr2アゴニスト(A側)と「BSCI型」sstr2アゴニスト(又は「ソマトタキシン類」、B側)の両方について、sstr2結合の見かけの親和性(x軸;sstr2での「Ka」、nM)に対するケモカイン誘導性遊走の阻害の効力(y軸;ED50対ケモカイン誘導性遊走、nM)をプロットしたグラフである。
【図17】本発明の方法に従って試験するための候補BSCIとして設計された、種々の新規ベータ−メチオニントリプトファンベースのsstr2アゴニスト(L−799,976と化合物A〜F)の構造を示す。
【図18】本発明の1つの実施態様に従う化合物P(x軸、nM)によるケモカインMCP−1誘導性THP−1細胞遊走の阻害(y軸、パーセント値として示す)を示すグラフである。
【図19】化合物P(x軸、nM)による成長ホルモン(GH)放出の阻害(y軸、パーセント値として示す)を示すグラフである。
【図20】Medrol(登録商標)(A)又は化合物P(B)で治療した患者からの血清試料中の、炎症の代替マーカーとしてのC−反応性タンパク質(CRP;y軸、ng/ml)のレベルを比較するヒストグラムである。カラムは、投与前(a)と最後の投与日(b)のCRPレベルを示す。星印はp=0.02であることを示す。
【発明を実施するための形態】
【0024】
BSCIのペプチド及びアミノラクタム群の両方の標的受容体がソマトスタチンの2型受容体(sstr2)として開示(後述の実施例参照)され、ヨヒンバミド(例えばヨヒンバン−16−アミド)の標的受容体がα2−アドレナリン受容体であり、リセルグアミド類(例えばL−II)の標的受容体が5−HT2aであり、すべてが機能的選択性をもって作用しているため、これらのすべての受容体について、ケモカイン誘導性遊走の阻害を促進する機能的選択性リガンドが存在することが明らかである。これらの受容体のすべては、白血球上で発現されるGPCR神経受容体のより大きなクラスのメンバーであるため、白血球上に発現されるすべてではなくても多くのGPCR神経受容体について、同様の機能的選択性を有するリガンドが見つかると推定することは妥当である。従って本発明の方法はある態様において、白血球上で発現されるGPCR神経受容体との相互作用について、1又は2以上の候補化合物又は物質をスクリーニングすることを含む。
【0025】
本発明の方法のある応用において、スクリーニングすべき1又は2以上の化合物はまず、選択された受容体を発現する細胞を、受容体に結合することができる放射性リガンドに曝露することにより、白血球上に発現されたGPCR神経受容体と相互作用する能力について試験される。例えば、ソマトスタチン受容体sstr2を発現する細胞は、試験化合物の存在下及び非存在下で、sstr2のリガンド(例えば標識ソマトスタチン)に曝露される。試験化合物がsstr2受容体と相互作用する場合、受容体に特異的に結合するソマトスタチンの量は変化するであろう。このような受容体結合アッセイは、同じ目的を達成するために多くの異なる方法で行うことができる:異なるリガンドが使用され得る(例えば、sstr2特異的アンタゴニストBIM23627;Tulipano et al. (2002) 143:1218-24);リガンド結合を視覚化するために、異なるレポーター系が使用され得る(シンチレーション近傍(scintillation proximity)アッセイと蛍光分極は、このような2つの例である);組換え系とは異なる細胞(細胞膜調製物、又はsstr2発現細胞を含む)が使用され得る;細胞を試験化合物に曝露するのに、異なるビヒクルが使用され得る;リガンドの特異的に結合を観察するのに、異なる緩衝液条件が使用され得る、など。
【0026】
上記例の第2の工程で、sstr2受容体と相互作用を示した化合物は、ケモカイン誘導性白血球遊走の阻害についてスクリーニングされる。例えば白血球が、ケモカインMCP−1の勾配の存在下でTranswell(登録商標)遊走アッセイプレートに入れられ、試験化合物の存在下又は非存在下において、37℃で2時間インキュベートされる(Frow et al. (2004) Med. Res. Rev. 24:276-98)。次に、この実験の最後に、遊走を行った細胞の数を、例えば生体染色色素MTTと標準曲線への内挿を用いて測定する。試験化合物がケモカイン誘導性遊走を阻害する場合、化合物を受けるウェル中で遊走するものは少ないであろう。このような遊走アッセイは、同じ目的を達成するために多くの異なる方法で行い得る:異なる遊走チャンバーを使用し得る(マルチウェルフォーマットTranswell(登録商標)遊走チャンバー、従来のボデンチャンバー、ゲル内遊走ウェル、ダンチャンバーなど);異なるケモカインを使用し得る(SDF−1α、IL−8、MIP−1α、RANTESなど);異なる白血球をチャンバーに適用し得る(新たに調製された末梢血好中球若しくはリンパ球、腹膜マクロファージ、又はTHP−1若しくはJurkatのような細胞株を含む);遊走した細胞の数を定量するために異なる検出系を使用し得る、など。
【0027】
同様に本発明の方法は、白血球上で発現される任意の他のGPCR神経受容体に適用され得る。そのような受容体の例は、特に限定されないが、アドレナリン受容体(α2−アドレナリン受容体を含む)、セロトニン受容体(5HT2a受容体を含む)、ソマトスタチン受容体(sstr2を含む)、ドーパミン受容体、ムスカリン性アセチルコリン受容体、物質P/NPY受容体、バソプレシン受容体、甲状腺ホルモン受容体、カンナビノイド受容体、コレシストキニン受容体、オピオイド受容体、アデノシン受容体、グルカゴン受容体、PACAP/VIP受容体、メラトニン受容体、ブラジキニン受容体、カルシトニン受容体、及びメラノコルチン受容体を含み得る。
【0028】
好ましくは本発明の方法は、アドレナリン受容体、セロトニン受容体、又はソマトスタチン受容体に適用される。さらに好ましくは本発明の方法は、α2−アドレナリン受容体、5HT2a受容体、又はsstr2受容体に適用される。
【0029】
この方法がソマトスタチン受容体(sstr2を含む)に適用される場合、候補化合物又は物質はソマトスタチンを除外し得る。
【0030】
本発明の方法は、BSCI活性を有する可能性が最も高いものを同定するために、化合物のライブラリーをスクリーニングするのに容易に適用し得る。このようなライブラリーは、ランダムライブラリー(例えば、大量処理スクリーニング用の多くの薬品会社により使用される構造の多様な化合物の大きなライブラリー)であるか、又は指向性(directed)ライブラリー(例えば、限定された数の前駆体化合物に適用されるコンビナトリアルケミストリーにより合成されるライブラリー)であり得る。いくつかの指向性ライブラリーは、BSCI化合物(又は、選択された受容体に対する従来のアゴニスト)を含有する可能性が高く、従ってこのようなライブラリーは本発明の方法を用いてスクリーニングする時、特に重要となり得る。例えば、我々がすでに開示した(WO2006/085096参照)Gタンパク質アゴニスト濃縮ライブラリー(GAEL)ライブラリーは、BSCI化合物が特定のGPCRで特殊なクラスのアゴニストとして作用していることが明らかなため、本発明の方法に従って生産的にスクリーニングされ得る。指向性ライブラリー(例えばGAEL)の使用は、本明細書に開示した基準に従って候補BSCI活性を有する化合物を同定するために、スクリーニングされる必要のある化合物の数を減少させ得る。しかし本発明の方法は、充分にハイスループットであり、これはランダムライブラリーへうまく適用され、このような応用は本発明に包含され得る。
【0031】
場合により、選択された受容体に対する「古典的(classical)」アゴニストとしてのBSCIの活性も測定される。好ましくは、本発明の方法に従って同定される化合物又は物質は、両方の種類の機能アッセイ(functional assay)(すなわち、BSCI活性についての細胞遊走アッセイ、及び選択された受容体に対する古典的アゴニスト活性についての機能アッセイ)で活性であるが、古典的受容体アゴニストとしてのその効力より強力な(すなわち、より低濃度で作用する)、ケモカイン誘導性白血球遊走の阻害剤である。
【0032】
すなわち、最適なBSCIは本発明の方法に従って、以下の3つの性質のすべてを有する化合物として同定され得る:
【0033】
(i)化合物は、白血球上で発現された選択されたGPCR神経受容体と、少なくとも10μMの親和性で相互作用する(すなわち、化合物が10μMで存在する時、存在するsstr2受容体の半分以上はそれらに結合された化合物を有する);
(ii)化合物は、ケモカインにより誘導された白血球遊走を、少なくとも1μMの効力で阻害する(すなわち、化合物が1μMで存在する時、細胞遊走の程度は少なくとも50%阻害される);及び
(iii)選択された受容体における古典的sstr2アゴニスト活性の、1又は2以上のアッセイ中の化合物の効力(potency)は、白血球遊走の阻害についての効力より低い。
【0034】
さらに好ましくは、化合物は、選択された受容体と少なくとも1μMの親和性で相互作用し得る;さらに好ましくは、化合物はケモカイン誘導性白血球遊走を、少なくとも100nMの効力で阻害し得る;さらに好ましくは白血球遊走に対する化合物の効力は、選択された受容体との相互作用の親和性又は古典的アゴニスト活性の効力より、10倍以上高いものであり得る。
【0035】
さらに好ましくは、化合物は、ケモカイン誘導性白血球遊走を少なくとも10nMの効力で阻害し得る;さらに好ましくは白血球遊走に対するか化合物の効力は、選択された受容体との相互作用の親和性又は古典的アゴニスト活性の効力より、50倍以上高いものであり得る。
【0036】
本発明の方法により、これらの神経受容体に対する従来のアゴニスト(特に限定されないが、天然のリガンドを含む)と、BSCI活性を有する化合物とを区別可能であることは明らかである。従来のアゴニストである化合物は、ケモカイン誘導性白血球遊走を阻害し得るが、これらは、特定の神経受容体との相互作用の親和性と、同様の効力又はより低い効力で阻害する。
【0037】
同様に、これらの受容体に対する従来のアゴニストは、標的受容体を介して従来の作用(例えば、成長ホルモン放出の阻害、又は血圧調節、インスリン放出の調節、例えば試験中の特定の受容体に依存して)を示すのと同様の濃度で、ケモカイン誘導性白血球遊走に対する作用を示すのみである。従って、これらの神経受容体に対する従来のアゴニストは、ある投与量で抗炎症活性を有するが、このような化合物はまた天然のリガンドの作用を示し、同じ化合物投与量で標的受容体を介して作用する。本発明の方法の重要な利点は、抗炎症性を有する物質を単に同定するのみでなく、その抗炎症性が副作用を超える、優れたサブクラスの物質を同定することであることを、強調すべきである。具体的には同定される物質は、白血球上で発現されるGPCR神経受容体に対する古典的アゴニストより大きな、抗炎症剤としての効力を有する(白血球上で発現されるGPCR神経受容体に対する大部分の既知のアゴニストにより共有されない性質)。
【0038】
ペプチドBSCI及びアミノラクタムBSCIは、sstr2アゴニストの特別な(及び、まだ同定されていない)サブセットである。これらのBSCIと従来のsstr2アゴニストは共通の分子標的(sstr2受容体)を共有するが、この2群の物質は受容体への結合に対して異なる結果を引き起こす。この概念は、他の受容体について別のところで詳細に記載(Urban et al (2007) J. Pharmacol. Exp. Ther. 320:1-13)されており、機能的選択性(functional selectivity)と呼ばれている。有用なたとえは、受容体(この場合sstr2)をピアノ鍵盤と見なすことである:多くの異なる手の位置(異なるリガンドに相当)は鍵盤(生産性結合に相当)と相互作用することができるが、それぞれ異なる和音の音を発生する(受容体からの異なる機能的結果に相当、例えば異なる第2のメッセンジャーが産生される)。この機能的選択性の概念を図2に示す。
【0039】
ヨヒンバミドBSCI(ヨヒンバン−16−アミドを含む)は、α2−アドレナリン受容体アゴニストの特別な(及び、まだ同定されていない)サブセットである。これらのBSCIと従来のα2−アドレナリン受容体ゴニスト(例えばレセルピン)は分子標的を共有するが、この2群の物質は受容体への結合に対して異なる結果(Urban et al (2007) J. Pharmacol. Exp. Ther. 320:1-13)(機能的選択性と呼ばれる現象)を引き起こす。ヨヒンバミドは、ケモカイン誘導性白血球遊走を強力に阻害するが、血圧には影響を与えない;レセルピンは血圧を強力に低下させるが、ケモカイン誘導性白血球遊走には影響を与えない。
【0040】
リセルグアミドBSCI(L−IIを含む)は、5HT2aアゴニストの特別な(及び、まだ同定されていない)サブセットである。これらのBSCIと従来の5HT2aアゴニスト(例えばTCB2;McLean et al. (2006) J. Med. Chem. 49:5793-803)は共通の分子標的(5HT2a)を共有するが、この2群の物質は受容体への結合に対して異なる結果(Urban et al (2007) J. Pharmacol. Exp. Ther. 320:1-13)(機能的選択性と呼ばれる現象)を引き起こす。リセルグアミドは、ケモカイン誘導性白血球遊走を強力に阻害するが、幻覚誘発性(げっ歯類における特徴的な首ひねり)には影響を与えない;TCB2は幻覚誘発活性の特徴的な首ひねり(head twitch)を強力に誘発するが、ケモカイン誘導性白血球遊走には影響を与えない。
【0041】
BSCIが、白血球上で発現されることが公知である種々のGPCR神経受容体に対する特別なクラスの機能選択的アゴニストであるという、本明細書に開示された知識は、BSCI活性を有する化合物を予め構造情報無しで(すなわち、すでに記載されているBSCI化合物群に共通のファーマコフォアの情報無しで)同定することを可能にする。
【0042】
このような化合物は、本発明の方法に従って、例えば以下のように同定され得る。最初の工程では、白血球上で発現されることが公知の選択されたGPCR神経受容体との相互作用についてスクリーニングされる。次にBSCIは、従来の標的又はその受容体に対する古典的アゴニスト活性より強力な効力で、ケモカイン誘導性白血球遊走を阻害する能力により、その受容体に対する従来のアゴニストから区別される。場合により第4の工程で、これらのすでに開示されているものと構造的に無関係の新しいクラスのBSCIを定義する構造モチーフが同定され得る。
【0043】
第一の工程は、白血球上で発現されることが公知の選択されたGPCR神経受容体との相互作用についてのスクリーニングを含む。この定義を満足するために受容体は、(a)当該分野で公知の構造的及び操作的基準に従う、GPCRスーパーファミリーのメンバー;(b)機能的に有意なレベルで中枢神経系において発現される受容体;及び(c)白血球又は白血球のサブセット上で機能的に有意なレベルで発現されることが公知、でなければならない。本発明の実施のために選択される受容体がGPCRスーパーファミリーのメンバーであり、中枢神経系中で発現されるという要件は、BSCIが、この定義を満足するすべての3つの異なる受容体で機能的選択性リガンドであるという知見に基づく。この受容体が白血球上で発現されるという要件は、選択された物質がケモカイン誘導性白血球遊走を抑制してほしいという要求に基づく。
【0044】
本発明の方法の具体的な実施のために使用される1又は2以上の受容体の選択は、主に異なる白血球サブセットにわたる受容体の発現パターンに基づくことが企図される。例えばケモカインに応答して好中球遊走を選択的に抑制する物質が必要なら、(他の白血球と比較して)好中球上に選択的に発現されるGPCR神経受容体が選択され得る。同様に、肥満細胞遊走を選択的に抑制する物質が必要なら、肥満細胞上で発現されるGPCR神経受容体が選択され得る。あるいは、疾患中におけるGPCR神経受容体の発現パターンの変化は、特定の受容体を選択するための刺激となり得る;喘息の肺で好酸球上で選択的に発現される物質は、喘息で使用される抗炎症剤を同定するための理想的な標的となり得る。従って本発明の方法を適用する固体又は群の要求は、容易に、本発明の応用のための適切なGPCR神経受容体を選択するための基準を提供し得るが、本発明の使用は、使用されるGPCR神経受容体を選択するための特定のアプローチに決して限定されない。
【0045】
第2及び第3の工程は、(1)ケモカイン誘導性白血球遊走の阻害に関する、及び(2)選択される受容体に対する古典的アゴニスト活性に関する機能アッセイを行うことを含む。これらの2つの工程は両方とも、GPCR神経受容体により仲介されるBSCI活性について機能的選択性を有する物質を明確に定義するのに必要である。しかしこれらの2つの工程は、順番に又は基本的に同時に行うことができる。第1の工程で選択された特定の標的であるGPCR神経受容体を発現することが公知であるBSCI活性の機能アッセイのために、白血球集団を使用することは重要である。これができないと、この方法から擬陰性結果が得られるが、それは、BSCI活性に対する機能的選択性を有し得る全ての物質は、標的受容体を発現せず、且つこれと相互作用しない細胞のケモカイン誘導性遊走に対して作用を有しないためである。
【0046】
機能的選択性BSCI活性に関連する共通の構造モチーフを同定するために、場合により第4の工程が適用される。典型的にはこのような工程は、反復する構造要素が、BSCI活性を付与するのに必要なモチーフに一致する可能性が高くなるように、本発明の方法に従ってBSCIとして新たに同定された化合物間で構造要素を比較することを含み得る。次に、例えば関連する構造を有する種々の分子を合成して、モチーフの存在がBSCI活性の存在に関連していることを証明することにより、そのようなモチーフの重要性を実験的に証明することができる。その結果、たとえクラスの単一のメンバーのみが、白血球上で発現されるGPCR神経受容体との相互作用についてのスクリーニングを介して独自に同定されたとしても、本発明の方法はBSCI化合物のクラス全体を定義するために使用され得る(各クラスのメンバーは、構造が互いに関連している)。
【0047】
すなわち、本発明の方法に従って同定されるBSCI化合物は、新規であって構造が関連したBSCI群の合成と特徴づけのための構造的リードとして働くことが企図される。その結果、本発明の方法により同定される最初のヒットのそのような構造誘導体は、本発明の方法の適用により直接同定はされないが、それらの同定における重要な工程は本発明の方法に依存し、従ってこの方法での、関連するBSCI群の間接的同定が、本発明のこの態様に従って企図される。
【0048】
本発明の方法に従ってライブラリーがスクリーニングされる場合、ライブラリーの要素は個々の化合物(又は、少なくとも実質的に純粋な化合物、ここで標的化合物は存在する物質の少なくとも25%を占め、それより高レベルでは他の化合物は存在しない)でもよく、又はライブラリーの要素は化合物の混合物でもよい。要素が化合物の混合物である場合、混合物の成分は公知であり得るか又は(多くのコンビナトリアルライブラリー中のように)予測され得るか、又は未知(例えば、天然の生成物の単離体)であり得る。
【0049】
本発明の方法に従ってスクリーニングされるライブラリー要素の一部又はすべてが(組成が既知であるか又は未知である)混合物である場合、本発明の方法の第1工程後、白血球上で発現される選択されたGPCR神経受容体との相互作用の原因である混合物から特定の化合物が単離され、同定された後、第2工程(ケモカイン誘導性白血球遊走に対する化合物の作用の評価)が適用されることが企図される。そのような単離工程がその方法に含まれていない場合、その方法は混合物中の単一の化合物が(必要に応じて)BSCI活性を有する場合を、混合物中の1つの化合物が選択される神経受容体と相互作用し、第2の無関係の化合物がBSCI活性と関係の無い機構により白血球遊走を阻害した場合から区別できないであろう。
【0050】
例えば、ソマトスタチン−14(従来のsstr2アゴニストであるがBSCIではない)とアジ化ナトリウム(細胞の遊走を阻止するがBSCIではない細胞毒素)との混合物からなるライブラリー要素に本発明の方法を適用した場合、この混合物はsstr2と強い相互作用を示し、(存在するアジ化物の相対的濃度に依存して)sstr2での親和性から予測される効力より高い効力でケモカイン誘導性白血球遊走を阻害し得る。そのような混合物は、間違ってBSCIとして同定され得る。しかし、もし上記したようにsstr2(本例におけるソマトスタチン)との相互作用の原因である混合物の成分が、ケモカイン誘導性遊走に対する作用をアッセイする第2の工程を適用する前に混合物から単離された場合、この方法は混合物をBSCI活性が欠如しているとして正しく同定し得る。
【0051】
原則的には、本発明の方法の工程は任意の順序で適用できる(例えば、試験中の化合物の、ケモカイン誘導性遊走の阻害剤としての効力を測定し、次に白血球上で発現されたGPCR神経受容体との化合物の相互作用(もしあれば)の親和性を測定することが、原則的に可能である)が、そのような方法は本発明の好適な態様の利点が欠如している。これらの状況では、機能アッセイを使用してランダムライブラリーをスクリーニングすることができない(毒性で不溶性の化合物の存在のために、ならびにそのようなスクリーニングの資源の関係で)。従って好適な本発明では、白血球上で発現されたGPCR神経受容体との相互作用についてのスクリーニングが最初の工程として行われ、次にケモカイン誘導性白血球遊走の阻害のために機能アッセイが行われる。白血球上で発現されるGPCR神経受容体との相互作用を評価するためのスクリーニング(分子スクリーニングの例)は、ケモカイン誘導性白血球遊走に対する活性の測定(機能性スクリーニングの例)よりも、スクリーニングされる化合物当たりの必要な資源量がはるかに少ないため、この工程順序は、入手可能な資源の最適使用を反映する。
【0052】
本発明の方法の適用は、仮想のスクリーニング、ならびに実際の若しくは物理的スクリーニングを含むことが企図される。すなわち、本発明の方法に従って候補BSCIとして化合物を同定するのに必要な情報の一部(又はすべて)は、すでに利用可能である。例えば化合物のライブラリーは、本開示前に白血球上で発現された種々のGPCR神経受容体との相互作用についてすでにスクリーニングされており、結果が(例えば、コンピューターのハードディスク中に)保存されているかも知れない。従って、ライブラリーからの化合物のサブセットが、白血球中のケモカイン誘導性遊走の阻害剤として選択され、次にその効力が試験され、本発明の方法に従って候補BSCIとして同定され得る。そのような手順が、本明細書に開示された方法の適用であり、候補BSCIを同定するために開示された基準に依存することは明らかである。同様に、一連の化合物について、白血球上で発現される1又は2以上のGPCR神経受容体とのそれぞれの相互作用の親和性がすでに公知であり、かつ別にケモカイン誘導性白血球動員の阻害のための化合物の効力がすでに公知である場合、どの化合物が候補BSCIであるかを同定するための既存の情報の組合せは、新規の実験が必要ではない場合でも、本発明の方法の適用であり得る。
【0053】
特に、本発明の1又は2以上の工程が新規の実験を必要しない場合、その工程は任意の順序で実施できる(すなわち、本明細書に記載の好適な順序である必要は無い)。
【0054】
白血球上で発現されるGPCR神経受容体に対するアゴニストに帰因される多くの抗炎症活性が記載されている(例えば、Paran & Paran (2003) Curr. Opin. Invetig. Drugs 4:578-82; Sener et al. (2005) Peptides 26:493-9; Gonzales-Rey et al. (2007) Ann. Rheum. Dis. 66:582-8、及びその中の文献; Armani et al. (2007) J. Leukoc. Biol. 81:845-55; Cury et al. (2008) Dig. Dis. Sci. 53:2516-20を参照、これらは、白血球上で発現される単一のGPCR神経受容体であるsstr2についての報告に限定される;多くの他のGPCR神経受容体について同様の例が存在する)。従って本発明の新規性と進歩性は、白血球上で発現されるGPCR神経受容体を介して作用するリガンドが、臨床的に有用な抗炎症作用を有し得るという発見(単一の工程のみを有する発明を教示する発見、すなわち白血球上で発現されるGPCR神経受容体との相互作用についてスクリーニングして、有用となり得る抗炎症剤を同定すること)には無い。その代わりに本発明は、白血球上で発現されるGPCR神経受容体に対するすべてのアゴニストの一部(及び、おそらく非常に小さい比率)が、実質的に古典的アゴニスト活性(例えば、成長ホルモン分泌の抑制、血圧又は幻覚誘発性の調節)の代わりに、抗炎症活性を有するという予想外の知見に関する。
【0055】
この種の機能的選択性(異なるリガンドが、同じ受容体に結合しこれを活性化するが、顕著に異なる機能的結果を誘発する)は薬学の文献で公知である(総説については、Urban et al (2007) J. Pharmacol. Exp. Ther. 320:1-13を参照)が、確固たる文献は少ない。従って、ある受容体について又はある対の機能について、機能的選択性リガンドが存在するかどうかを推定することは不可能であろう。そのため我々の開示以前には、当業者は、GPCR神経受容体の活性化に関連する抗炎症活性は必ず古典的アゴニスト応答が伴うと妥当に結論付けており、抗炎症活性を有する新規薬剤の発見用の標的としてのGPCR神経受容体の有用性を限定していた。我々の開示は、機能的選択性は1つのGPCR神経受容体にのみ存在するのではなく、3つの異なるGPCR神経受容体で存在し、さらに、これらの各場合にほとんどまたは全くアゴニスト様活性の無い実質的な抗炎症活性を示すことがあり得ることを、明確に示す。我々の開示前に利用可能な情報では、当業者(本発明者らを含む)でさえ、この知見は予測されてはおらず、また可能であることも、もちろん確率が高いことも推定されなかった。
【0056】
本発明は、機能的選択性を示し、選択されたGPCR神経受容体に対する古典的アゴニスト活性よりもBSCI活性を促進する物質の明確な同定のための方法を提供する。本発明を有効に実施するために、この機能的選択性が如何に達成されるかの分子的詳細も、所望の結果を規定するリガンド間の構造特性も理解する必要は無い。本開示において提供される情報は、本来は等しく有望な候補のプールから有用と思われる機能的選択性物質を選択することを、当業者に教示するのに必要かつ十分である。しかし、本知見が予測できない程度を例示し、如何に機能的選択性を達成しなければならないかを例示することが有用かも知れない。
【0057】
リガンドが受容体に結合する時、リガンド分子と受容体タンパク質との間には複数の非共有結合的相互作用がある(特に限定されないが、ファンデアワールス力、水素結合、双極子相互作用、及び塩ブリッジを含む;図2参照)。これらの相互作用は、リガンドと受容体との相互作用の親和性(従って、他のタンパク質ではなく標的受容体への結合の選択性)と、リガンド結合に対するシグナル伝達応答の両方に関与する。天然のリガンドでは、これらの相互作用は受容体タンパク質のコンフォメーションの変化を引き起こす(受容体:リガンド複合体の最小エネルギーコンフォメーションは、分離した時の受容体とリガンドの最小エネルギーコンフォメーションとは顕著に異なる)。このコンフォメーションの変化は、シグナルを細胞の外へのリガンド結合から細胞内シグナル伝達カスケードに変換する。この変換は、直接的(内因性キナーゼ活性を有する受容体の場合のように)又は間接的(Gタンパク質結合受容体、例えばsstr2の場合のように)であり、コンフォメーションの変化は、1又は2以上の追加のタンパク質の会合を促進し、これはシグナルを変換する。受容体に結合し、天然のリガンドで見られる受容体中のコンフォメーション変化を再現する物質は、受容体に対するアゴニストであり得る(図2A)。これらの変化を誘導することなく受容体に結合する物質はアンタゴニストであり得る(これが天然のリガンドへの結合を阻止すると仮定して;図2B)。しかし、異常な状況下では第3の可能性が出てくる:物質が結合し、非常に異なるセットのコンフォメーション変化を誘導し、これはシグナルの性質に定量的ではなく定性的変化を引き起こし、次にこれが変換される(図2C及びD)。機能的選択性(及びある程度は、スーパーアゴニスト及びインバースアゴニスト)の薬理学的現象を指示するのは、この第3の異常なシナリオである。
【0058】
この分子的シナリオは、音楽に例えるとより理解し易いかも知れない。このモデルでは、ピアノ鍵盤は受容体の例えであり、ピアニストの手はリガンドである。通常の条件下では、手の作用(特定のコンフォメーション)は特定の和音を打つ。この「シグナル」は受容体に対するアゴニスト応答である。ピアノの蓋を閉じることは、鍵盤の位置でコンフォメーション変化を誘導することなくリガンドと受容体との相互作用を防止し、従って和音(アゴニストシグナル)は聞こえない。ピアノの蓋はアンタゴニストの類縁体である。ピアニストを鍵盤の弾き方を知らない小児(ライブラリーからランダムに選択される化合物の例え)に交換すると、ほとんどすべての場合に音楽の和音ではなく不可解な応答を出すであろう。しかしまれに、天才児に遭遇してピアニストとはまったく異なるきれいな和音を演奏するかも知れない。同じ受容体が機能的選択性で応答するようになった:異なるリガンドが同じ受容体から定性的に異なる応答を誘発する。
【0059】
上記より、同じ受容体に結合する異なるリガンドから、機能的結果を予測する方法が存在しないことは明らかであろう。実際、ある受容体にとって、どのシグナルが可能であるかを予測することさえ不可能である(どのように鍵盤を押しても、ピアノの鍵盤からバイオリンの音を出すことはできないであろうという意味で)。偶然の発見後に、開示された情報に基づいて実験的スクリーニングを行うことが、所望の機能的選択性を有する物質(例えば、本明細書に記載のBSCI)を発見する方法である。
【0060】
本発明が、単独のそれぞれの工程が当該分野で公知の方法である、3又は4以上の工程からなることは明らかである。例えば、天然のリガンドの既知の作用を再現するために、又は天然のリガンドの既知の作用を阻止するために、白血球上で発現されるGPCR神経受容体と相互作用する物質を捜すために、多くの場合に第1の工程が使用されていることは明らかである。同様に我々と他の研究者は、BSCI活性のために物質を特徴づけすることを目的として、ケモカイン誘導性白血球遊走の機能アッセイの使用を開示した。しかし本開示前には、全く異なる結果(ランダムライブラリー中のBSCI活性を有する化合物についてスクリーニングする能力;ほとんどのソマトスタチン受容体相互作用性物質はBSCIではなく、ランダムライブラリー中のほとんどの要素は、BSCI活性に関する機能アッセイに適合しないため、当該能力は、いずれかの工程を単独で使用して達成することはできない)を達成するために、これらの2つの方法を結びつけ、これらを連続して行うための可能な理論的根拠はなかった。しかし、BSCIが、白血球上で発現されるGPCR神経受容体と相互作用し、機能的選択性により作用するという情報(これは、本文書において初めて開示される)は、これらの2つの無関係のプロセスを組合せて、予想外の有用性を有する新規方法とすることにより、本発明を可能にする。本発明の方法のうまいたとえは、新しいペプチドの組成物である:ペプチドは(ほとんどすべて)同じアミノ酸構成単位からなり、同じ方法で結合される。しかし、非常に多くの組合せがあるため、特定の機能を有する特定の組合せ(新しいペプチド配列)の同定は進歩性があると見なされる。同様に、BSCIが作用できる非常に多くの可能な受容体があり(検索が、インビボで抗炎症活性を仲介できることが公知のものに限定されていても)、それぞれの受容体に対する分子スクリーニングを、BSCI活性についての機能スクリーニングと組合せようとすることは実現不可能であった。本発明は、ランダムライブラリーからBSCIを同定するための方法を提供することにより、最初は等しく可能なはずであった多くの他の組合せより予想外かつ特に優れた有用な組合せについての、この実現不可能な検索を克服する。
【0061】
本明細書に記載の方法は、他の種(特に限定されないが、マウス、ラット、イヌ、及び霊長類、ならびに非哺乳動物種、例えば鳥)のGPCR神経受容体のホモログに等しく適用し得る。GPCR神経受容体の非ヒトホモログが、ヒトでの使用を目的(例えば、抗炎症性薬物として)とした本発明に従って、化合物を同定するために使用されるなら、非ヒト受容体を使用して同定される物質をヒト受容体に対する機能的選択性についてチェックする追加の工程が必要とされ得る。しかし好ましくは、ヒト系でのBSCI活性がスクリーニング操作の所望の結果であるなら、本発明の方法は白血球上で発現されるヒトGPCR神経受容体に適用されるはずである。もちろん、所望の結果が方法の非ヒト系でのBSCI活性(例えば、獣医学的薬剤)であるなら、1又は2以上の標的種からのGPCR神経受容体類縁体を使用することが最も好適であり得る。
【0062】
本発明の方法は、受容体の断片が完全長受容体の天然のリガンド又は別のリガンドに結合する能力を保持するなら、受容体の不完全な断片を使用して適用することができることが企図される。典型的にはそのような機能性断片は、タンパク質配列の大部分を含有するが、リガンド結合に関与しない部分は(例えば、発現又は安定性を改善するために)、本発明の適用に影響を与えることなく欠失し得る。
【0063】
GPCR神経受容体への試験物質の結合が、間接的(受容体に対する、あらかじめ標識されていた別のリガンドとの結合の競合により)に又は直接的(試験物質自体が標識物を有する)に測定できることは明らかであろう。本発明の範囲は、試験物質と受容体との相互作用を検出するために使用される方法には決して限定されない。
【0064】
試験物質のBSCI活性を検出するために使用される機能アッセイが、インビトロ(例えば、Transwell(登録商標)遊走アッセイを使用して)又はインビボ(例えば、ケモカインの腹腔内注入後、応答する白血球動員の評価を使用して)でも実施できることは明らかであろう。さらに、BSCI活性の評価のために白血球動員(インビトロとインビボの両方)の代替マーカーを使用することができる。例えば白血球は、(活性化されている時は特に)サイトカインTNF−アルファを発現することが知られている。その結果、インビトロ又はインビボでのTNF−アルファの測定レベルは、白血球動員を評価するための代替法であり、従ってBSCI活性を測定するための方法である。本発明の範囲は決して、特定の試験化合物に関連するBSCI活性を検出するために使用される方法に限定されるものではない。
【0065】
試験物質の古典的アゴニスト活性を検出するために使用される機能アッセイが、インビトロ(例えば、ソマトスタチン受容体アゴニストについて下垂体細胞からのGH放出を測定することにより)又はインビボ(例えば、ソマトスタチン受容体アゴニストについて血清中のGHレベルを測定することにより)でも実施できることは明らかであろう。さらに、古典的sstr2アゴニスト活性を推定するために、代替マーカーを使用することができる。GHレベルの指標として、従って古典的ソマトスタチン受容体アゴニスト活性の推定値として、IGF−Iレベルが測定されるように、再度ソマトスタチン受容体アゴニストの例を使用して、GHはインスリン様増殖因子I(IGF−I)のレベルを制御する。本発明の範囲は決して、特定の試験化合物に関連する古典的アゴニスト活性を検出するために使用される方法に限定されるものではない。
【0066】
場合により、本発明の方法の前工程中に選択された化合物の1又は2以上の構造的特徴の存在に基づいて、新規な構造クラスのBSCIの同定を可能にするように、追加の工程を本発明の方法に付加してもよい。そのような構造モチーフの同定は、本発明の方法の適用を通じて、ライブラリー(ランダムライブラリーでも、選択される候補からなるものでも)からのヒットを同定するために、その性質が実験的又は理論的に探索された化合物のみよりはるかに広範囲のBSCIのアレイを可能とする。例えば、白血球上で発現されるGPCR神経受容体への結合についてのランダムライブラリーのスクリーニング(本発明の第1工程)後に、ケモカイン誘導性白血球遊走の阻害の測定(第2工程)、そして選択された受容体の「古典的」標的に対する効力の随時測定により、本発明の方法に従っていくつかの新しい化合物を選択し得る。2又は3以上のこれらの選択された化合物は、共通の構造モチーフ(例えば、3−置換シス−デカリン環系の存在)を共有し得る。従って任意の追加の工程では、3−置換シス−デカリン環系の存在が化合物に所望の性質を付与し、適切な予測によりさらなる3−置換シス−デカリン含有分子を合成でき、その少なくとも一部が所望の性質を有し得ることは明らかである。そのような状況下では、BSCI活性又は「ソマトタキシン」活性に関連する新しい構造モチーフの定義は、本発明の方法の適用を構成し得る。
【0067】
したがって本発明の方法は、少なくとも3つの工程を含む(白血球上で発現されるGPCR神経受容体との相互作用の測定、次にBSCI活性についての機能アッセイ、及び選択された受容体に対する「古典的」アゴニスト活性について機能アッセイ)。場合により最終工程を加えてもよい(BSCI活性に関連する構造モチーフを定義する)。この最終工程の利点は、アッセイベースのスクリーニングではなく構造相同性により、所望の性質を有する化合物を選択することを可能にすることである。そのような「ヒット・トゥー・リード(hit to lead)」最適化プログラムは当該分野で公知であり、ヒトの薬剤として使用するのに適した新しい物質の発見への本発明の方法の適用を促進する。しかし特定の適用に依存して、この任意の最終工程を実施する資源の問題が、同定される化合物の範囲が拡張される利点より大きいかも知れない。
【0068】
本明細書において用語「化合物(compound)」及び「物質(agent)」は一般に同義であると考えられ、従って文脈が特に明記しない場合は、相互に変換可能である。
【0069】
用語「広域スペクトル(broad-spectrum)ケモカイン阻害剤」又は「BSCI」は、通常は異なる受容体を介して遊走を誘導するシグナルを出すが、非ケモカイン化学誘引物質により誘導される細胞遊走にはほとんど又は全く影響を与えない広範囲のケモカインに応答して、細胞遊走を阻害できる化合物又は物質を意味する。これは操作的な定義であり、完全に異なる分子機構の結果として異なる化合物若しくは物質がBSCIとなり得ることは明らかである。しかし本発明は、重要でかつ広範に研究されたBSCIの下位区分である、本発明で同定された、sstr2又は関連する受容体を介して作用するBSCIの同定にのみ関連する。
【0070】
用語「BSCI活性」は、白血球上で発現されるGPCR神経受容体で作用するいくつかのアゴニストが、インビトロ若しくはインビボでケモカイン誘導性細胞遊走に、ならびに関連するインビボでの免疫学的変化(例えば、Tヘルパー(CD4+)細胞サイトカイン分泌プロフィールの再調整)に対して有する作用を示す。
【0071】
用語「古典的(classical)アゴニスト活性」は、あるGPCR神経受容体を充分なレベルで発現する細胞が、その受容体についての任意の公知のアゴニスト化合物(当該受容体の天然の1又は2以上のリガンドを含む)に曝露された時、おそらく異なる細胞種で、起きることが公知である一群の細胞応答を示す。例えば、sstr2受容体について古典的アゴニスト活性の例は、下垂体細胞によるGH放出の抑制、膵臓細胞によるインスリン放出の抑制であろう(Olias et al. (2004) J. Neurochem. 89:1057)。しかし種々の細胞に作用するsstr2アゴニストの他の広範囲の作用も記載されており、これら(上記した「BSCI活性」を除いて)もまた、古典的sstr2アゴニスト活性を含む(Olias et al. (2004) J. Neurochem. 89:1057)。すなわち古典的アゴニスト活性は、BSCI活性を除く選択されたGPCR神経受容体に対するアゴニストのすべての公知の活性として定義され、従ってアゴニストのすべての公知の活性は本定義により包含される。
【0072】
用語「sstr2」は、ヒト染色体17q24上のsstr2遺伝子(HGNC ID:11331;配列番号1に示される完全な遺伝子配列)によりコードされ、後述の配列表中の配列番号2で示されるアミノ酸配列をコードする、ヒトの2型ソマトスタチン受容体を示す。
【0073】
特に明記しない場合、用語sstr2は、配列番号1に示す遺伝子座の転写から得られる、オルタナティブ・スプライシングされたか又は示差的プロセシングされた任意のmRNAの生成物を包含することを企図する。疑問を避けるため、用語sstr2は、タンパク質コード枠中に位置する単一のイントロンでsstr2 mRNAの選択的スプライシングから得られるsstr2aタンパク質とsstr2bタンパク質の片方又は両方を包含する。他の種の類縁体(analog)又は相同体(homolog)もまた、特定の文脈でナンセンスでない限りは、この用語に包含される。
【0074】
用語「ソマトスタチン受容体」は、ソマトスタチンが高親和性(典型的には1μM又はそれ以下)で結合する任意の受容体(現在公知であっても公知でなくても)を示す。疑問を避けるため、これは、sstr1、sstr2、sstr3、sstr4、及びsstr5を含む。他の種の類縁体又は相同体もまた、特定の文脈でナンセンスでない限りは、この用語に包含される。
【0075】
用語「ソマトスタチン受容体」はさらに、ヒトsstr2(上記した)のアミノ酸配列と、少なくとも50%の同一性、例えば少なくとも55%、60%、65%、70%、75%、80%、85%、90%、95%の同一性、又は少なくとも96%、97%、98%、さらには99%の同一性を有するアミノ酸配列を有するポリペプチドを包含する。ポリペプチドは好ましくは、上記のソマトスタチン受容体活性を有する。
【0076】
アミノ酸配列間の配列同一性は、配列の整列を比較することにより決定することができる。比較される配列中の同等の位置が同じアミノ酸で占められる時、分子はその位置で同一である。整列を同一性のパーセントとしてスコア化することは、比較される配列が共有する位置での同一のアミノ酸又は塩基の数の関数である。配列を比較する時、最適な整列は1又は2以上の配列へギャップを導入して、配列中の可能な挿入や欠失を考慮することが必要な場合がある。配列比較法は、比較される配列中の同一分子の同じ数について、2つの比較配列間のより高い関連を反映するできるだけ少ないギャップの配列整列が、多くのギャップを有するものより、高いスコアを有するように、ギャップペナルティを使用する。最大同一性パーセントの計算は、ギャップペナルティを考慮して、最適整列を作成することを含む。
【0077】
配列比較を行うための適切なコンピュータープログラムは、商業部門や公共部門で入手できる。例には、MatGat (Campanella et al., 2003, BMC Bioinformatics 4: 29; http://bitincka.com/ledion/matgatから入手できるプログラム)、Gap (Needleman & Wunsch, 1970, J. Mol. Biol. 48: 443-453)、FASTA (Altschul et al., 1990, J. Mol. Biol. 215: 403-410; http://www.ebi.ac.uk/fastaから入手できるプログラム)、Clustal W 2.0 and X 2.0 (Larkin et al., 2007, Bioinformatics 23: 2947-2948; http://www.ebi.ac.uk/tools/clustalw2から入手できるプログラム)、及び EMBOSS Pairwise Alignment Algorithms (Needleman & Wunsch, 1970, supra; Kruskal, 1983, In: Time warps, string edits and macromolecules: the theory and practice of sequence comparison, Sankoff & Kruskal (eds), pp 1-44, Addison Wesley; http://www.ebi.ac.uk/tools/emboss/alignから入手できるプログラム)がある。すべてのプログラムは、デフォールト・パラメータを使用して使用される。
【0078】
例えば、配列比較は、EMBOSS Pairwise Alignment Algorithmsの「ニードル」法を使用して行われ、これは、長さ全体にわたって考慮される時の2つの配列の最適整列(ギャップを含む)を決定し、同一性パーセントスコアを提供する。アミノ酸配列比較のデフォールト・パラメータ(「タンパク質分子」オプション)は、ギャップエクステンションペナルティ:0.5、ギャップオープニングペナルティ:10.0、マトリックス:Blosum 62である。ヌクレオチド配列比較のデフォールト・パラメータ(「DNA分子」オプション)は、ギャップエクステンションペナルティ:0.5、ギャップオープニングペナルティ:10.0、マトリックス:DNAfullである。
【0079】
用語「ソマトスタチン」は、ソマトスタチンをコードする遺伝子の任意の産物(ヒトにおいては染色体3q28上のsst遺伝子(HGNC ID:11329)によりコードされる)を示し、そしてそれは当該遺伝子産物の制御されたタンパク質分解により生成される断片を含む。特に明記しない場合、用語ソマトスタチン(又はsst)は、生物学的機能が割り当てられたタンパク質分解により生成される任意の断片(特に限定されないが、sst−14及びsst−28を含む)を包含することが意図される。他の種の類縁体又は相同体もまた、特定の文脈でナンセンスでない限りは、この用語に包含される。
【0080】
用語「白血球」は、血液中の任意の又はすべての白血球集団、ならびに骨髄又は2次的リンパ系臓器(例えば脾臓)中のこれらの細胞の前駆体、及び組織中のこれらの細胞の後代(例えば、破骨細胞、クプファー細胞、小神経膠細胞、肥満細胞、樹状細胞など)を示す。白血球の例には、単球、リンパ球(Bリンパ球とTリンパ球の両方、キラーT細胞、ヘルパーT細胞、制御T細胞、及びナチュラルキラー細胞を含む)、及び顆粒球(好中球、好塩基球、好酸球)がある。
【0081】
用語「GPCR神経受容体」は、中枢神経系の細胞(ニューロン、星状細胞、及び小神経膠細胞を含む)上で機能的に関連するレベルで発現されるGタンパク質結合受容体(GPCR)のスーパーファミリーの任意のメンバーを包含する。Gタンパク質結合受容体は、充分定義されたクラスであり、受容体が、その配列と構造に基づいて及びそのシグナル伝達機構に基づいて、GPCRスーパーファミリーのメンバーであるかどうかを明確に決定するための方法が当該分野に存在する。例えば、GPCRは典型的には、7つの膜貫通ドメイン(個々には12〜20個の疎水性アミノ酸残基の区域で、いずれかの端に1又は2以上の荷電残基を有する、同定可能な特徴を有する)の存在により特徴づけされる。GPCRはまた、GTP結合タンパク質(Gタンパク質を呼ぶ)の群のメンバーと相互作用する能力、及びこれを活性化する能力により特徴づけされる。GPCR神経受容体の例には、特に限定されないが以下の群が含まれる:アドレナリン受容体(α2−アドレナリン受容体を含む)、セロトニン受容体(5HT2a受容体を含む)、ソマトスタチン受容体(sstr2を含む)、ドーパミン受容体、ムスカリン性アセチルコリン受容体、物質P/NPY受容体、バソプレシン受容体、甲状腺ホルモン受容体、カンナビノイド受容体、コレシストキニン受容体、オピオイド受容体、アデノシン受容体、グルカゴン受容体、PACAP/VIP受容体、メラトニン受容体、ブラジキニン受容体、カルシトニン受容体、及びメラノコルチン受容体を含む。
【0082】
用語「白血球上で発現されるGPCR神経受容体」は、白血球又は白血球の少なくとも1つのサブセットにより機能的に有意なレベルで受容体が発現されている、すべてのGPCR神経受容体のサブセットを示す。すべての白血球サブセット上で発現されるGPCRの完全な相補体は現在未知であるが、この用語は、これらが白血球の少なくとも1つのサブセットで発現されることが公知であるため、本発明での使用のために選択されるすべてのGPCR神経受容体、ならびに白血球中の発現パターンのスクリーニングに基づいて選択されたGPCR神経受容体(これらは、以前は白血球上で発現されることは知られていなかった)を明確に包含するために使用される。白血球上に存在すると記載されておらず、白血球に基づく発現を調べて選択されたのではないGPCR神経受容体は、本定義に包含され得ない。
【0083】
用語「効力(potency)」は、特定の作用を有するのに必要な特定の物質の濃度を説明するために使用される。典型的には効力は、試験中のエンドポイントを50%阻害するのに必要な化合物の濃度とされる(ED50又はIC50と呼ぶ)。従って低濃度でのED50又はIC50は、高い効力を有する化合物を示す。効力は、試験した濃度で試験中のエンドポイントに対する化合物の最大作用を示す「出力(power)」(後述)とは完全に異なる。化合物は、高効力であるが低い出力を有し(例えば、ED50が0.05nMでケモカイン誘導性遊走に対して作用するが、非常に高濃度でも遊走を40%しか阻害しない)、又は低効力で高い出力を有する(例えば、ED50が10mMであるが、100μMより高い濃度でエンドポイントを100%阻害する)ことがある。
【0084】
用語「出力(power)」は薬理学的アッセイに適用される時、そのアッセイでその物質の最適濃度を選択することにより得られる試験中のエンドポイントに対する、特定の物質の最大作用を説明するために使用される。最適濃度は、典型的には試験される最も高い濃度であるが、ある例(例えば、ベル型の用量応答が一般的に観察される場合、例えばケモカインに対する化学走化性応答)では、低濃度が試験中のエンドポイントに対して最大作用を有する場合がある。出力は効力とは関係無く、低効力を有する化合物が高い出力を有していたり、逆の場合もある。
【0085】
用語「約」は、考慮される値の周りの間隔を示す。本特許出願において使用されるように、「約X]は、(X−Xの10%)〜(X+Xの10%)、好ましくは、(X−Xの5%)〜(X+Xの5%)の間隔である。
【0086】
本説明中の数値範囲の使用は、その範囲内のすべての個々の整数と、ある範囲の最も広い範囲内の上限数と下限数とのすべての組合せを、本発明の範囲内に含めることを明確に企図する。
【0087】
本明細書において、用語「含んでなる」は、含むことと、からなることの両方を意味するとして読まれるべきである。逆に発明が、ある化合物を「活性成分として含む医薬組成物」という用語は、この用語が、他の活性成分が存在してもよい組成物と、定義した1つの活性成分のみからなる組成物との両方を包含することを意味する。
【0088】
以下の略語は、使用される場合、文脈から適切であるとしてポリペプチド又は他のアミド含有化合物の一部として単離されるものであっても、20の天然に存在するタンパク質新生アミノ酸を意味する。
【0089】
【表1】

【0090】
特に明記しない場合は、ここで使用されるすべての技術的・科学的用語は、本発明が関連する分野の当業者により通常理解されるものと同じ意味を有する。
【0091】
本発明の具体的な非限定例や追加の特徴は、以下の図面を参照して説明される。
【0092】
いくつかの図をさらに詳細に説明する。
図1は、ペプチド3(示していない)から得られる配列の環状レトロインベルソ(retroinverso)類縁体であるNR58−3.14.3(「A」)を示す。トリペプチドモチーフWxQは主要なファーマコフォアとして同定され、WVQ(「B」)のようなトリペプチドはBSCI活性を有する(低効力であるが)。ヨヒンバン(Yohinban)−16−アミド(「C」)とNR58,4(「D」)は、WxQトリペプチドの構造類縁体として同定された。次にBN83253(「E」)、BN83250(「F」)、及び(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オン(「G」)のようなラクタムが、NR58,4のようなイミドの安定な類縁体として同定された。リセルグ酸のアスパラギン及びグルタミンアミド(例えばL−II)は、ヨヒンバン−16−アミドの構造相同体として同定された。
【0093】
図2ではパネルAは、天然のリガンドが受容体と相互作用し、複数の応答(矢印)を発生することを示し、これは細胞内シグナル又は機能的結果であると解釈される。パネルBでは、アンタゴニストは受容体に結合し、天然のリガンドが結合することを妨害するが、活性なシグナル伝達を誘導するのに必要な受容体部分とは反応しない。パネルCでは、機能選択性アゴニストが、ある応答(太い矢印)を他の応答(細い矢印)より優先的に活性化するように、受容体に結合する。パネルDでは、反対の機能的選択性を有する異なるアゴニストが例示される。
【0094】
図3を参照して、転写体A(「T#1」)ではイントロンはスプライシングにより除去されておらず、翻訳停止コドンは、末端切断型タンパク質(sstr2aと呼ぶ)を与える。転写体B(「T#2」)では、イントロンはスプライシングにより除去されており、翻訳は第2エクソン及び代替翻訳停止コードに進み、sstr2bタンパク質(これは短いC末端テイルのみでsstr2aとは異なる)を与える。sstr2転写体を明確に同定するために使用されるPCRプライマーの位置を、sstr2プライマーセットについては(a)として、sstr2Aプライマーセットについては(b)として示す。この2つの異なる転写体から得られるアンプリコンのサイズは、398bp(T#1、sstr2プライマーセット)、87bp(T#1、sstr2Aプライマーセット)、57bp(T#2、sstr2プライマーセット)、及びバンド無し(T#2 sstr2Aプライマーセット)である。
【0095】
図4では、アンタゴニスト(黒い線、小さい四角)BN83253の不在が、MCP−1誘導性細胞遊走を用量依存的に阻害することがわかる。アンタゴニストの濃度が上昇すると(薄い灰色、大きい記号)、BN832653のBSCI活性が次第に失われる。
【0096】
図5では、アンタゴニスト(黒い線、小さい四角)の非存在下では、バレロラクタムはMCP−1誘導性細胞遊走を用量依存的に阻害する。アンタゴニストの濃度が上昇すると(薄い灰色、大きい記号)、BN832653のBSCI活性が次第に失われる。このバレロラクタムBSCIはBIM83253より30〜50倍強力であり、その結果、NIM83253からBSCI活性を排除するには、より高濃度の競合的アンタゴニスト(BIM23627)が必要であることに注意されたい(このグラフを図4と比較)。
【0097】
図6と7の両方では、BSCIは、内毒素に応答したTNF−アルファ産生を用量依存的に阻害した(実線、四角記号)。BSCIの見かけの効力は、BIM23627の同時投与により低下した(点線、三角記号)。
【0098】
図8ではデキサメタゾンは、内毒素に応答したTNF−アルファ産生を用量依存的に阻害した(実線、四角記号)。BIM23627(点線、三角記号)の同時投与は、デキサメタゾンの見かけの効力に対して影響はなかった。
【0099】
図9では、両方のタイプの細胞について、2つの異なるプライマーセットを使用し(図3参照)、1つはsstr2a転写体からアンプリコン(「A」)のみを産生し、他はsstr2aとsstr2b(ここで「sstr2」と記載する)から異なるサイズのアンプリコン(「A*」と「B」)を生成する。図9では、M1は398bpであり、M2は87bpであり、M3は57bpである。このデータは、THP−1細胞中のsstr2aの発現を確認し、sstr2bの存在については何の証拠も示さない。
【0100】
図12では,値は各薬剤濃度での三回測定値(±SD)である。過剰の非標識BN83250(「A」、10nM 3H−BN83250)との競合を、非特異結合の対照として示し、特異的結合を示す上の点線を決定する。陰性対照(「B」、過剰のコールド)が提供され、これは非特異結合を示す下の点線を決定し、一方、BN83250の結合に対するsstr2アンタゴニストBIM23627(「C」;100μM)の影響を比較のために示す。
【0101】
図13では、ポリリンカー中の「*」は、XbaI部位の上流にATGがあることを示す。細菌とJurkat細胞中のプラスミドの存在について選択するために使用される、それぞれ「アンピシリン」と「ネオマイシン」抵抗性コーディング読み取り枠に注目されたい。cDNA挿入体は真核生物系ではCMVプロモーター(「P CMV」)下で発現され、標準的なポリアデニル化配列(「BGH pA」)を有する。
【0102】
図14を参照して(A)は、BN83250がJurkat 3DBのSDF−1α誘導性遊走を阻害(THP−1細胞で観察されるものより低効力であるが)するが、親Jurkat T細胞株のSDF−1α誘導性遊走にはほとんどまたは全く影響が無いことを示す。各細胞株について、フローサイトメトリーにより測定されるsstr2Aの細胞表面染色のレベルを、頻度ヒストグラム(B)〜(D)に示す。
【0103】
図16では、遊走に影響を与えない化合物は効力10μMでプロットされ、従ってこれらがグラフに現れる。BN83250(白三角)、(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オン(白四角)、ソマトスタチン(白菱形)、及びL−779,976(白丸)は、グラフから特定することができる。すべての既知の「BSCI型」sstr2アゴニストが右下に有り、すべての既知の「古典的」sstr2アゴニストが左上にあることを強調するために、ED50とKaが等しい同一性の線が示される。
【実施例】
【0104】
実施例1:sstr2アンタゴニストはインビトロでBSCI活性を阻止する
既知のペプチドとアミノラクタムBSCI化合物がsstr2受容体を介して作用することを証明するために、我々は一連の特異的sstr2アンタゴニストを使用した。種々の構造が異なる化合物のBSCI作用が、特定の受容体に対する一連のアンタゴニストにより阻止されるなら、BSCI活性が、使用したアンタゴニストに阻止される受容体への結合に依存すると結論する。
【0105】
ここで我々は、ケモカインに応答して遊走を阻止する種々のBSCIの能力(BSCIの標準活性)が、2型ソマトスタチン受容体(sstr2)に特異的ないくつかのアンタゴニストによりインビトロで阻害されることを証明する。
【0106】
方法:
これらの実験ではヒト骨髄単球性細胞株THP−1を使用したが、ヒト末梢血単球でも同様の結果が得られている。白血球遊走は、Neuroprobe(Gaithersburg, MD, USA)の96ウェルフォーマットのTranswell(登録商標)アッセイシステムを使用して測定した。原理的にこのアッセイは、多孔性膜により分離された2つのチャンバーからなる。化学誘引物質を下の区画に入れ、細胞を上の区画に入れる。37℃である時間インキュベーション後、細胞を化学誘引物質の方に動かすと、下の区画の細胞数は化学誘引物質の活性に比例する(一連の対照に対して)。
【0107】
種々のアッセイ条件が、ケモカイン誘導性白血球遊走(ならびに、sstr2アンタゴニストによるBSCIの又はBSCI活性の影響)を実証するために許容されるが、ここでは具体例が提供される。この方法は、基本的に既に記載されているように行った(Reckless & Grainger (1999) Biochem. J. 340:803-11)。
【0108】
材料
Transwell(登録商標)アッセイシステムは、Neuroprobe(Gaithersburg, MD, USA)により製造された。
【0109】
使用したプレートは、ChemoTx(商標)プレート(Neuroprobe 101-8)と30μlの透明プレート(Neuroprobe MP30)である。
【0110】
ゲイの平衡塩類溶液(Geys' Balanced Salt Solution)を、Sigma(Sigma G-9779)から購入した。
【0111】
脂肪酸を含まないBSAを、Sigma(Sigma A-8806)から購入した。
【0112】
MTT、すなわち3−(4,5−ジメチルチアゾール−2−イル)−2,5−ジフェニルテトラゾリウムブロミドを、Sigma(Sigma M-5655)から購入した。
【0113】
フェノールレッドを含まないRPMI1640を、Sigma(Sigma R-8755)から購入した。
【0114】
THP−1細胞株(欧州セルカルチャーコレクション(European Cell culture Collection))を、白血球細胞集団として使用した。
【0115】
試験プロトコール
まず、上の区画に入れる細胞懸濁物を調製した。THP−1細胞を遠心分離(770×g、4分)によりペレット化し、1mg/ml BSA含有ゲイの平衡塩類溶液(GBSS+BSA)で洗浄した。次にこの洗浄を繰り返し、細胞を再度ペレット化した後、例えば標準的血球計算器を使用して計数するために少量のGBSS+BSAに再懸濁した。
【0116】
次に存在する細胞の数に応じてGBSS+BSAの容量を調整し、細胞の最終密度が1mlのGBSS+BSA当たり4.45×106細胞になるようにした。これにより、プレートの上部チャンバーに入れる各25μlの溶液当たり100,000個のTHP−1細胞が確保された。
【0117】
MCP−1誘導性遊走に対するBSCI活性の逆転を証明するために、3ロットの細胞を調製する必要があった。4.45×106細胞/mlのTHP−1細胞の懸濁物を、3つの瓶に分割した。1つの瓶にBSCIを適切な最終濃度で、適切なビヒクル中に加えた(例えば、(S)−3−(2’,2’−ジメチルプロパノイルアミノ)−テトラヒドロピリジン−2−オンを、GBSS中のストック溶液から1nMで加えた)。2つ目の瓶にBSCIを最初の瓶と同じ濃度で加えたが、さらにsstr2アンタゴニストも加えた(例えば、BIM23454[Tulipano et al. (2002) 143:1218-24]又はBIM23627をGBSS中のストック溶液から66nMで加えた)。3番目の瓶に、適宜GBSS+BSA+ビヒクルを加え、3つの瓶の中の容量が等しく対照として作用するようにした。
【0118】
次に、下の区画に入れる化学誘引物質溶液を調製した。MCP−1をGBSS+BSAで希釈して、最終濃度を25ng/ml(すべて添加後)とした。Frow et al (Med. Res. Rev. 24:276-98) が推奨するように、これはこのシステムで最大遊走を誘導するMCP−1の濃度である。この溶液を、細胞懸濁物と同様に3つの瓶に分割した。1つの瓶に、BSCIを細胞懸濁物に加えたものと同じ最終濃度になるように加え、2番目の瓶に、BSCIとsstr2アンタゴニストを両方とも細胞懸濁物に加えたものと同じ濃度になるように加え、残りの瓶にはGBSS+BSA+ビヒクルを適宜加えて、3つの瓶の中の容量を等しくした。下の区画の溶液中のMCP−1の最終濃度と上の区画の細胞の最終濃度とを確立する時、BSCIとsstr2アンタゴニストを添加するために、液体の容量を考慮する必要があることに注意されたい。
【0119】
下のウェル用の化学誘引物質溶液と上のチャンバー用の細胞溶液をいったん調製した後、遊走チャンバーを組み立てた。29μlの適切な化学誘引物質溶液をチャンバーの下のウェルに入れた。各条件を少なくとも3回測定で行った。すべての下のチャンバーを充填した後、多孔性膜を製造業者の説明書に従ってチャンバーに適用した。最後に25μlの適切な細胞溶液を上の各チャンバーに加えた。蒸発を防ぐために、装置全体の上にプラスチックの蓋をかぶせた。
【0120】
組み立てたチャンバーを37℃、5%CO2で2時間インキュベートした。GBSS+BSA中の細胞懸濁物も、同じ条件下にて試験管中でインキュベートした:これらの細胞は、各条件下で下のチャンバーに遊走した細胞の数を測定するための標準曲線を構築するために使用した。
【0121】
インキュベーションの最後に、上のチャンバーから液体の細胞懸濁物を静かに取り出し、20μlの氷冷した20mM EDTAのPBS液を加え、装置を4℃で15分インキュベートした。この操作により、膜の下側に付着していたすべての細胞が下のチャンバーに落ちた。
【0122】
インキュベーション後、フィルターを注意深くGBSS+BSAでフラッシュしてEDTAを洗い流し、次にフィルターを取り出した。
【0123】
次に、各条件下で下のチャンバーに遊走した細胞の数は、多くの方法(直接計数、蛍光性若しくは放射活性マーカー又は生体染色色素の使用による標識を含む)により測定することができる。典型的には我々は、生体染色色素MTTを使用した。3μlのストックMTTを各ウェルに加え、次にプレートを37℃で1〜2時間インキュベートし、その間、細胞内の脱水素酵素は可溶性MTTを不溶性の青いホルマザン生成物に変換し、これは分光光学的に定量することができる。
【0124】
並行して、8点標準曲線を準備した。上の各チャンバーに加えられた細胞の数(100,000)で開始して、GBSS+BSAで2倍連続希釈を行い、プレートに細胞を25μl加え、3μlのMTTストック溶液を加えた。標準曲線プレートを遊走プレートとともにインキュベートした。このインキュベーションの最後に、沈殿したホルマザン生成物を乱さないように、下のチャンバーから注意深く液体を取り出した。短時間風乾後、下の各チャンバーに20μlのDMSOを加えて青い色素を可溶化し、96ウェルプレートリーダーを使用して595nmの吸光度を測定した。次に各ウェルの吸光度を標準曲線に内挿して、下の各チャンバー中の細胞数を推定した。
【0125】
MCP−1刺激遊走は、MCP−1が25ng/mlで存在した下の区画に到達した平均細胞数から、MCP−1を加えなかったウェル中の下の区画に到達した平均細胞数を引いて、計算した。
【0126】
種々の濃度のBSCIの存在下又は非存在下で起きたMCP−1誘導性遊走を比較することにより、BSCIの影響を計算した。典型的には遊走の阻害を、BSCIの存在下により阻止された総MCP−1誘導性遊走のパーセントとして表わした。ほとんどの化合物について、種々の異なる化合物濃度(典型的には、1nM〜1μM、又はあまり活性ではない化合物の場合それ以上)で起きるMCP−1誘導性遊走の阻害を測定することにより、用量応答グラフを構築した。次に各化合物の阻害活性を、MCP−1誘導性遊走を50%だけ低下させるのに必要な化合物の濃度として表わした(ED50濃度)。sstr2アンタゴニストとBSCIとの存在下で起きるMCP−1誘導性遊走を、BSCI単独(同じ濃度のBSCI化合物)の存在下で起きるMCP−1誘導性遊走と比較することにより、sstr2アンタゴニストの影響を計算した。sstr2アンタゴニストがBSCI化合物のBSCI活性を阻害したなら、BSCI単独で処理したウェルより、sstr2アンタゴニストとBSCIの両方で処理したウェルで、より多くの遊走が起きていることが予測された。
【0127】
結果:MCP−1(25ng/ml)は、THP−1細胞の実質的な遊走を誘導した(MCP−1の無い対照ウェル中よりMCP−1の存在下で、25,000〜40,000個多い細胞が下のチャンバーに遊走する)。この遊走は、広範囲のBSCIにより用量依存的に阻害された。図4では、MCP−1の阻害が、上昇する濃度のBSCI BN83253(N−オレオイル−3−アミノカプロラクタム;Fox et al. (2005) J. Med. Chem. 48:867-74)に応答して、THP−1細胞遊走を誘導したことを示す(小さい四角)。図5では、上昇する濃度のバレロラクタムBSCI (S)−3−(2’,2’−ジメチルプロピオニルアミノ)−テトラヒドロピリジン−2−オンに応答したMCP−1誘導性THP−1細胞遊走の阻害を示す(小さい四角)。両方の場合に、600nMのsstr2アンタゴニストBIM23627の存在(sstr2でこの化合物のKiより約100倍高い;Tulipano et al. (2002) 143:1218-24)は、BSCI介在阻害を完全に防止した(すなわち、加えたBSCIの濃度に無関係に、高用量のBIM23627の存在により、MCP−1誘導性遊走は対照レベルまで回復される;図4と図5の三角)。
【0128】
低用量のBIM23627(例えば、この受容体で作用するこの化合物についてのKiに近い6nM;図4と図5の中程度の大きさの菱形記号)では、BSCI作用は低用量のBSCIで阻害されたが、高用量THP−1では、誘導性細胞遊走はなお阻害された。これは、同じ部位への結合に対するBSCIとsstr2アンタゴニストとの競合に一致する。低BSCI濃度では、sstr2アンタゴニストが結合し、遊走の阻害は見られない。BSCI濃度が上昇すると、徐々にsstr2がBSCIにより占められ、sstr2により占められることはなく、BSCI活性が現れる。
【0129】
非常に低用量のBIM23627(例えば、sstr2で作用するこの化合物についてのKiより約100倍低い0.06nM;図4と図5の小さい菱形)では、BSCI作用はすべての濃度のBSCI化合物で見られたが、低濃度BSCIでは程度の小さい逆転が見られ、同じ部位へ結合するこれらの物質間の競合と再度一致する。
【0130】
これらの結果は、異なるBSCI((S)−3−(アダマンタン−1−カルボニルアミノ)−カプロラクタム(WO2006/016152参照)とペプチド3(Reckless & Grainger (1999) Biochem. J. 340:803-11)を含む)を使用して、何度も再現されている。さらに、異なるsstr2アンタゴニスト(BIM 23454; Tulipano et al. (2002) 143:1218-24)を使用しても、同じ結果が得られている。従って我々は、ペプチドからアミノカプロラクタム、そして第3世代アミノバレロラクタムにわたる非常に異なる構造クラスの広範囲のBSCI化合物について、BSCI活性はsstr2(又は、使用した両方のアンタゴニストにより阻害される密接に関連する受容体)により仲介されると結論付けた。
【0131】
他のソマトスタチン受容体について選択性のアンタゴニストを使用して、同様の実験も行われている。例えば、sstr3とsstr5について選択性のアンタゴニストであるBIM23627(Ipsen(Paris, France)から供与された)は、標的受容体のKiより10倍超高い濃度のアンタゴニストでさえ、BSCI活性の逆転をほとんど起こさない(<10%)。
【0132】
実施例2:sstr2アンタゴニストはインビボでBSCI活性を阻止する
インビトロのBSCI活性はsstr2(又は、BIM23627とBIM23454に阻害される密接に関連した受容体)に仲介される。次に我々は、インビボのBSCIの抗炎症活性もsstr2依存性であることの証明を試みた。幸いにBIM23627はインビボで活性であることが証明されており、急性の高用量投与後の少なくとも短時間で、古典的sstr2介在シグナルを阻止することができる(Tulipano et al. (2002) 143:1218-24)。従って、急性炎症のモデルを採用するなら、BSCIの抗炎症作用が、同時のsstr2阻止により阻害されたかどうかを決定することができるであろう。
【0133】
マウスの致死的ではない内毒素血症モデルが我々の実験室で詳細に特徴づけされているため、我々はこのモデルを選択した(例えば、Fox et al. (2002) J. Med. Chem. 45:360-70; Fox et al. (2005) J. Med. Chem. 48:867-74を参照)。細菌性リポ多糖(LPS)の単回腹腔内注射に応答した全身性炎症の程度は、抗原刺激投与後2時間目に血清TNF−アルファを測定することにより推定することができる。さらにこのモデルでは、BSCIが炎症を強力に阻害することが知られている(Fox et al. (2005) J. Med. Chem. 48:867-74)。
【0134】
方法:マウス(CD1系統;成体のオス;1群6匹)を、適切な経路でBSCI(適切なビヒクル中)又はビヒクル単独で処理した。例えば皮下注射(これは経口バイオアベイラビリティが悪いため)によりBN83250(図1参照)を投与し、一方(S)−3−(2’,2’−ジメチルプロピオニルアミノ)−テトラヒドロピリジン−2−オンを強制経口投与した。同時に各BSCI濃度で1セットのマウスに、4mg/動物(120mg/kg)のBIM23627を皮下注射により投与した(ラットで、古典的sstr2介在シグナル伝達(例えばGH放出)を抑制することが知られている処理条件;Tulipano et al. (2002) 143:1218-24)。投与の30分〜1時間後(BSCIの最大血漿濃度が達成される時間)に、750μgのLPS(Sigma; L4130 lot#024K4077; 900,000 EU/mg)の腹腔内注射により炎症性抗原刺激が誘発された。
【0135】
さらに2時間後、CO2窒息により動物を屠殺し、心臓穿刺により血液を採取した。従来法で血清を調製し、血清中のTNF−アルファのレベルを製造業者の説明書に従ってELISA(R&D Systems; Murine Quantikine code# MTA00)を使用して測定した。
【0136】
結果:解析後、各群の動物の平均を報告した(残りの点から3標準偏差より高いか又は低いデータ点は除外した)。予想されるようにLPS抗原刺激は、3,000ng/mlを超える平均に達するTNF−アルファのレベルで大規模な全身性炎症を誘導した(LPS抗原刺激を受けなかった対照動物からの血清中の<10ng/mlと比較)。この炎症は、使用したBSCIに関係なく、BSCIを用いる治療により用量依存的に劇的に阻害された(図6はBN82350を示す[図1];図7は、(S)−3−(2’,2’−ジメチルプロピオニルアミノ)−テトラヒドロピリジン−2−オン[バレロラクタム]を示す)。
【0137】
インビボで活性のあるsstr2アンタゴニストであるBIM23627の同時投与は、試験したすべての用量でBSCIの抗炎症性を顕著に低下させた(点線、図6と7の三角記号)。この観察結果は、別々の実験で各BSCIについて2回繰り返された。我々は、BIM23627の同時投与により、BSCIの抗炎症性効力が(採用されるBSCIに関係無く)10〜100倍低下すると予想している。これらの結果は、丁度我々がインビトロで観察したように(実施例1参照)、インビボのBSCI活性がsstr2(又はBIM23627により阻害される密接に関連した受容体)に依存することを明確に証明している。
【0138】
別のシリーズの対照実験で、BSCIとは完全に無関係な機構により抗炎症活性を示す合成コルチコステロイドであるデキサメタゾンの抗炎症活性に対するBIM23627の作用を調べた。デキサメタゾンは血清TNF−アルファレベル(全身性炎症のマーカーとして)をBSCIと同程度に低下させた(図8を図6及び7と比較)が、非常に高用量ではデキサメタゾンはわずかにより強力(BSCIの最大用量での約80%の阻害と比較して、90%を超えるTNF−アルファの阻害)であり、BSCIはわずかにより強力であった(デキサメタゾンより3〜10倍低い用量で、TNF−アルファの統計的に有意な低下)。しかし、このモデルでの量の活性にもかかわらず、BIM23627の同時投与はデキサメタゾンの抗炎症性に全く影響を与えなかった(図8中の点線と三角記号)。従って我々は、BIM23627は炎症経路に対して特異的な作用は無く、BSCI(異なる機構で作用する他の抗炎症薬剤を除く)は、インビボでsstr2に高度に依存すると結論付けた。
【0139】
実施例3:sstr2発現はBSCI感受性に相関する
複数のsstr2特異的アンタゴニストの使用は、BSCI活性がsstr2との相互作用に、又は同じアンタゴニストにより阻害される異なる受容体(sstr2に密接に関連している可能性がある)に依存することを示す。sstr2がBSCIの分子標的であることをさらに示すために、我々は、BSCIに対して異なる感受性を有する異なるタイプの一連の白血球中で、フローサイトメトリーにより及び定性的PCRにより、sstr2レベルを測定した。sstr2の存在とBSCIに対する感受性との相関は、BSCI活性がsstr2への結合又はその活性化を必要とするという結論を強固にした。
【0140】
方法:sstr2に特異的な抗体を用いるフローサイトメトリーを使用して、ヒト末梢血白血球上のsstr2発現の相対レベルを推定した。全血を、製造業者の説明書に従ってErythrolyse (Serotec cat number: BUF04B)で処理した。得られた総白血球集団をウサギポリクローナル抗ヒトsstr2抗体 (Sigma, Cat code S0694; 1μg/ml) で染色し、次にFITCで標識した抗ウサギ2次抗体 (Sigma F9887; 50μg/ml) により、具体的な白血球サブセットを明確に同定するために使用できる種々の白血球CDマーカーに対するrPEで標識した一群のモノクローナル抗体を用いて並行して染色した。そのcDNAマーカーに基づいて細胞集団をゲート制御した後、sstr2染色の平均レベル(イソタイプ対照を引いて)を、3回測定値について平均±SEMとして報告した。THP−1とJurkat細胞の培養物を、細胞をErythrolyseで処理しなかった以外は同じプロトコールを使用して分析した。
【0141】
図3に示したプライマーセットを使用してPCRを行い、sstr2aと2b転写体とを区別した。製造業者の説明書に従ってRNAqueous 4PCR キット (Ambion, Cat code #1914) を使用して、THP−1とJurkat細胞からmRNAを調製した。次に、得られたmRNA調製物を、逆転写酵素(RT)を使用してcDNAに変換し、次にこのcDNAを使用して、sstr2プライマー、dNTP(Ambion)、PCR緩衝液、及びTaqポリメラーゼを用いて、総量45μlで以下のPCR反応を構成した。プライマー配列は以下の通りである:sstr2前進プライマー 5'GCCAAGATGAAGACCATCAC 3'(配列番号3)、及び逆進プライマー 5'GATGAACCCTGTGTACCAAGC 3'(配列番号4)。PCRは、Peltier PTC-200サーマルサイクラーを製造業者の説明書に従って使用して、以下のサイクル時間と条件を使用して行った:94℃で2分後、94℃で30秒、55℃で30秒、72℃で1分を30サイクル、そして最後に72℃で10分。増幅の最後に、生成物を臭化エチジウムの存在下で2%アガロースゲルで分離し、UVトランスイルミネータ(Stratagene Eagle Eye II cabinet)で視覚化した。sstr2を高レベルで発現することが知られているpanc−1細胞(Li et al. (2004) J. Surg Res. 119:130-137)を陽性対照として使用した。逆転写酵素の無い反応物を陰性対照として使用した(mRNA調製物に混入しているゲノムDNAの結果として偽陽性シグナルが得られないことを確認する)。さらに、ゲノムアンプリコンが大きさから容易に区別できるように、プライマーをエクソンの境界をまたぐように設計した。
【0142】
結果:sstr2がいくつかの白血球集団で検出され、以前の報告と一致した(Hiruma et al. (1990) Immunology 71:480-5; Tsutsumi et al. (1997) Cell. Immunol. 181:41-9; ten Bokum et al. (1999) J. Endocrinol. 161:167-75 and the references therein; Elliot et al. (1999) Eur. J. Immunol. 29:2454-63; Talme et al. (2001) Clin. Exp. Immunol. 125:71-9; Dalm et al. (2003) Am. J. Physiol. Endocrinol. Metab. 285:E344-53; Lichtenauer-Kaligis et al. (2004) Eur. J. Endocrinol. 150:565-77)。任意の蛍光単位の相対レベルを以下の表1に示す。興味深いことに、THP−1細胞(これはBSCIに対して感受性が高い)は、sstr2について高レベルの表面染色を示し、一方Jurkat T細胞(これは、BSCIに対してほとんど完全に非感受性である)は、実質的に検出できない染色レベルを示す。
【0143】
ヒト末梢血白血球は、一般に2つの細胞株間の中間のsstr2の染色を示す。顆粒球(主に好中球)と単球は高レベルのsstr2を発現し、両方のタイプのヒト末梢血白血球はBSCIに対して感受性であることが知られている。Tリンパ球(CD3+)は、より低いが検出できるレベルを発現し、全CD3+集団が均一に染色されたため、これはCD4+(ヘルパー)とCD8+(キラー)T細胞集団の両方で同様の可能性がある。これに対してsstr2は、BSCIに対して非感受性の細胞であるBリンパ球集団上で検出されなかった(インビボで新抗原に応答した抗体生成に対するBSCIの作用の欠如に一致する)。
【0144】
【表2】
【0145】
表1.種々の白血球集団でフローサイトメトリーにより推定したsstr2細胞表面染色の相対レベル。aSSCは、サイドスキャッターに基づくゲート制御を示す。顆粒球が、その細胞質顆粒の結果として、特徴的に高い光分散性を有するためである。
【0146】
sstr2のmRNAの存在はTHP−1細胞で確認され(図9)、定性的PCRによりJurkat T細胞中に存在しないことが証明された。これは、これらの試験で抗体試薬の予測された特異性を確認している。さらに我々は、THP−1細胞中に存在する大部分のsstr2がsstr2aアイソフォームであることを証明した(これらの細胞中のmRNAのスプライシングパターンに基づく;図3と9)。フローサイトメトリー実験で使用した抗体がsstr2aとsstr2bアイソフォームを区別するかどうかは不明であるが、示された結果は、抗体がsstr2aアイソフォームを検出できることを強く示唆する。
【0147】
我々は、白血球の原形質膜上のsstr2の存在がBSCIに対する感受性と相関すると結論付けた。特に、検出可能なsstr2発現の無い細胞が、ケモカイン誘導性細胞遊走のBSCI介在阻害に対して非感受性であることを注目している。
【0148】
実施例4:BSCIはsstr2に結合する
我々は、標識BSCIリガンドを使用する従来の平衡結合試験を使用して、BSCIとsstr2受容体との分子相互作用を調べた。これらの試験について我々は、不飽和前駆体の接触水素化によりトリチウム(すなわち[3H])で標識した型で容易に合成できるアミノラクタムBSCI BN83250を使用した(図10)。不飽和前駆体である(S,E)−3−(2’,2’−ジメチルドデク−4’−エノイルアミノ)−カプロラクタムを、すでに報告されているように合成した(WO2005/053702)。
【0149】
方法:(S,E)−3−(2’,2’−ジメチルドデク−4’−エノイルアミノ)−カプロラクタムから、パラジウム触媒(CRB Ltd, Cleveland, UK)上でトリチウムガスを使用した接触水素化により、[3H]−BN83250を合成し、トリチウム化生成物をエタノール中270μMの濃度で比活性220Ci/mmolで得た。(S,E)−3−(2’,2’−ジメチルドデク−4’−エノイルアミノ)−カプロラクタムは以下のように製造した:水(30ml)中の(S,S)−3−アミノカプロラクタムヒドロ−ピロリジン−5−カルボキシレート(10mmol)とNa2CO3(30mmol)を、2,2−ジメチル−ドデカ−4−エノイルクロリド(10mmol)のジクロロメタン(30ml)溶液に周囲温度で加え、反応物を12時間攪拌した。次に有機層を分離し、水層を追加のジクロロメタンで抽出した(2×25ml)。一緒にした有機層をNa2CO3上で乾燥し、真空下で濃縮した。残渣をシリカカラムクロマトグラフィー(1:1 EtOAc:ヘキサンからEtOAc)で精製して、(S,E)−3−(2’,2’−ジメチル−ドデク−4’−エノイル)アミノ−カプロラクタムを無色の油状物として得た(2.12g,63%)。放射能標識化合物を暗所で4℃で、必要な時まで保存した。使用前に、非標識BN83250を添加して比活性を調整した。
【0150】
結合の解析のために、ゲイの平衡塩類溶液中の1×106のTHP−1細胞、10mM [3H]−BN83250(50 Ci/mmol)、及び種々の濃度の(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オン(10pM〜10μM)を用いて、個々の反応物を構成した(各条件について3回測定)。過剰(100μM)の非標識BN83250を使用して、非特異結合のレベルを測定した。4℃で2時間後(平衡結合を達成することがすでに証明されている条件、かつこれは結合の過剰推定を引き起こす放射能標識の取り込みを防ぐ)、過剰の標識物を遠心分離(5mlのゲイの平衡塩類溶液を用いて3回洗浄)して洗い流し、可溶化した細胞ペレットのシンチレーション計測により、結合した放射活性を測定した。
【0151】
結果:[3H]−BN83250は、平衡条件下でTHP−1細胞への特異的で競合性の結合を示す。結合のScatchard解析は、約80nMの親和性を有する、細胞当たり100,000〜200,000部位で存在する1つの結合部位に一致する(図11)。BN83250のTHP−1細胞への特異的結合は、過剰(100μM)のsstr2アンタゴニストBN23627により60〜80%(実験に依存する)阻害される(図12)。sstr2が欠如したJurkat T細胞では特異的結合はほとんど検出されなかった(<20,000部位/細胞)が、sstr2を発現するベクターを用いた安定なトランスフェクション後に特異的結合が回復された(sstr2を発現する安定なJurkat T細胞株の調製については実施例5を参照)。まとめるとこれらの結果は、この結合部位がsstr2であり、BN83250の観察されたBSCI活性の原因である結合相互作用であることを証明する。
【0152】
我々が以前最も高効力のBSCIの1つであるとして記載した(WO2009/016390)(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オンが、この結合部位で競合する作用を図12に示す。(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オンは、[3H]−BN83250と、sstr2での結合に対して用量依存的に競合し、ED50が約50pMであり、Hillスロープが約−1.0であり、それらは単純モードの競合結合を示唆する。高レベルの(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オン(1nM以上)は、BN83250結合を80〜90%阻害し、2つの分子が基本的に同一の結合部位を共有することを示唆する。ペプチド3(Reckless & Grainger (1999) Biochem. J. 340:803-11)、NR58−3.14.3(Reckless et al. (2001) Immunology 103:244-54; WO99/12968)、(S)−3−(アダマンタン−1’−カルボニルアミノ)−カプロラクタム(WO2006/016152)、及び(S)−3−(1’−メチオニンシクロヘキシル−1’−カルボニルアミノ)−カプロラクタム(WO2006/134384)を含む、試験した他のBSCI化合物もまた、sstr2への結合に対して、BN83250と同程度に競合する。
【0153】
THP−1細胞への(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オンの結合の見かけの親和性定数は、インビトロアッセイ(実施例1参照)でのBSCI活性のピコモーラーED50に一致するが、古典的sstr2アゴニスト活性についてのED50より数オーダー低い(実施例7参照)。我々は、現在まで記載されているBSCIのすべての群がsstr2に結合し、この相互作用が化合物のBSCI活性の原因であると結論付けた。
【0154】
実施例5:sstr2の発現は細胞株にBSCI感受性を付与する
ここで我々は、sstr2受容体の(強制)発現がBSCIに感受性を付与するのに必要かつ十分であることを証明する。我々はすでに、フローサイトメトリーにより測定されるsstr2の細胞表面発現がBSCIに対する感受性と相関し、従ってsstr2をほとんどまたは全く含まない細胞はBSCIに対して非感受性であるが、高レベルのsstr2を有する細胞は、ケモカイン誘導性遊走のBSCI介在阻害に対して感受性であることを既に示している。より高レベルのsstr2は、BSCI作用の効力の上昇に関連している。しかしこの相関は、sstr2がBSCI活性の原因であることを確証的に実証していない。
【0155】
我々は組換えDNA技術を使用して、白血球細胞株であるJurkat T細胞株(これは通常sstr2を発現しない)中のsstr2の異所性発現を強制的に行った。達成された発現レベルをフローサイトメトリーにより測定した。次に、安定にトランスフェクトされた細胞株のBSCIに対する感受性を測定し、親Jurkat T細胞株と比較した。
【0156】
方法:RNAqueous 4PCR キット(Ambion, Cat code #1914)を使用して製造業者の説明書に従って、Panc−1細胞(高レベルのsstr2を発現することが知られているヒト膵臓細胞株)から総mRNAを調製した。次にmRNAを逆転写(RETROscriptキットを使用; Ambio)して、cDNA調製物を得た。次にPCR(Peltier PTC-200サーマルサイクラーを製造業者の説明書に従って使用して以下のサイクル時間と条件を使用して行った:94℃で2分後、94℃で30秒、55℃で30秒、72℃で1分を30サイクル、そして最後に72℃で10分)により、sstr2遺伝子の完全長cDNAを得て、分取ゲル電気泳動により単離した。sstr2 cDNAの配列を配列決定により確認した。sstr2 cDNAをEcoRIとXhoIにより処理して粘性末端を得た。これはポリリンカーへのプラスミドpcDNA3.1+(Invitrogen)の指向性クローン化と次の連結を可能にした(10μlの連結反応を400 T4 DNAリガーゼ(New England Biolabs)を用いて行い、リガーゼ緩衝液中挿入体:プラスミド比3:1を有する最大量のDNAを13℃で一晩インキュベートした)。得られたベクター(図13参照)を使用して、標準的方法によりXL−10 Ultracompetent 細胞(Stratagene)を形質転換した。MiniPrep キット((Wizard Plus SV Minipreps DNA purification System; Promega)を使用して、Jurkat T細胞へのトランスフェクションについて製造業者の説明書に従って、プラスミドDNAを調製した。
【0157】
Jurkat T細胞はRPMI+10%胎児牛血清(FCS)で培養し、4日毎に新鮮な培地で1:3希釈して継代培養した。継代培養の48時間後の対数増殖期のJurkat T細胞を遠心分離して洗浄し、無血清RPMI培地に7.5×105細胞/mlで再懸濁した。次に、DNAとトランスフェクション試薬の混合物(2μgのDNA;3μlのFugene 6 トランスフェクション試薬; Roche)を加え、細胞を37℃で48時間インキュベートした。次に細胞をRPMI+10%FCSで遠心分離して3回洗浄し、次に新鮮な培地に1×105細胞/mlで懸濁した。次に、生じた細胞懸濁液の68mlを96ウェルプレート(Corning)中の全部で226ウェルに蒔いた。48時間後、培地にG418(0.9mg/ml Active G418)を加えてトランスフェクション体を選択し、継代培養を繰り返して細胞を拡張し、個々のウェルからの集団(クローン性又はオリゴクローン性と見なされた)を別々に維持した。細胞集団を25cm2のフラスコ(全部で約5×106細胞)に拡張した後、少量の試料を取りだし、フローサイトメトリーによりsstr2の発現を試験した。
【0158】
こうして、ヒトsstr2抗体(Sigma、S0694;使用したフローサイトメトリープロトコールの詳細については実施例3を参照)を使用してフローサイトメトリーにより検出された、高レベルのsstr2を細胞表面に発現する多くの個々の細胞株が得られた。これらのうち1つの株(Jurkat−D3Bと命名)は、末梢血の単球(健常なヒト末梢血中の最大発現細胞集団;実施例3を参照)に匹敵する高レベルのsstr2を発現することがわかったが、Jurkat−D3B中のsstr2のレベルはTHP−1細胞中よりはまだかなり低かった(図14)。
【0159】
結果:SDF−1α受容体であるCXCR4のレベルは、Jurkat−D3B細胞株と親Jurkat T細胞株とで同じであった。さらにJurkat−D3B細胞は、Transwell(登録商標)化学走化性アッセイ(使用した方法については実施例1を参照)で親細胞株と同様の濃度範囲にわたって、SDF−1αに応答した。我々は、Jurkat T細胞とJurkat−D3B細胞がこの実験の条件下で同じケモカイン誘導性遊走を示し、従ってBSCIに対するその感受性の比較に適していると結論付けた。
【0160】
Jurkat T細胞及びJurkat−D3B細胞は、100ng/ml SDF−1α(R&D systems;両方の細胞株で最大の細胞遊走を誘導したSDF−1αの濃度)に応答して誘導され遊走した。次にBSCI BN83250を0nM〜1000nM(それぞれ3回測定)の種々の濃度で上の区画と下の区画に加えた(BSCI濃度の勾配を生じないように、各区画で同じ濃度で)。DMSO(1%最終濃度)をビヒクルとして使用し、すべてのウェルで等しい濃度で存在させた。
【0161】
以前の結果(実施例3参照)に一致して、低レベル又は検出できないレベルのsstr2を発現する(図14)親Jurkat T細胞は、BN83250に対して実質的に非感受性であった(THP−1細胞株のような感受性細胞上でこのBSCIについてのED50<1nMと比較して、ED50は1μMよりかなり高かった)。これに対して、中程度のレベルのsstr2を発現する(図14)Jurkat−D3B細胞は、BN83250に対して実質的により感受性であった(図14)。Jurkat−D3B上のBN83250のED50は約10nMであり、sstr2の異所性発現の結果として、BSCI感受性の100倍を超える上昇であった。挿入体の無いpcDNA3.1のトランスフェクションと選択により、親Jurkat T細胞と同様にBN83250に対して非感受性であるJurkat T細胞株が得られた。
【0162】
Jurkat−D3Bはまた、ペプチドNR58−3.14.3やバレロラクタム(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オンのような、構造が異なる他のBSCIによる阻害に対しても感受性であったが、各場合に、その効力はTHP−1細胞上より有意に低かった。これはおそらく、THP−1細胞と比較してJurkat−D3B中で達成される低レベルのsstr2発現を反映している(図14)。薬理学的物質に対する応答の効力は、(いわゆる「スペア受容体」の結果として)受容体発現のレベルと相関することは公知である。我々のデータは、BSCIに対する感受性が細胞表面のsstr2発現のレベルに依存するという提唱と一致する。
【0163】
我々は、白血球細胞にBSCIに対する感受性を付与するのに、sstr2発現が必要かつ十分であると結論付けた。
【0164】
実施例6:従来のsstr2アゴニストはインビトロでBSCI活性が無い
BSCI活性を有する種々の構造が異なる化合物が、sstr2を介して作用することが証明されている。(sstr2を、これらのBSCIの活性の分子標的として確認している。実施例1〜5を参照。)sstr2ですべてのアゴニストがBSCI活性を有するか、又はBSCIとして機能的に同定された化合物がsstr2アゴニストの特殊なクラスであるかどうかを調べるために、我々は一連の古典的ソマトスタチン受容体を、インビトロでのBSCI活性について試験した。
【0165】
方法:実施例1に記載されたように正確に白血球遊走アッセイを行った。簡単に説明するとTHP−1細胞は、ChemoTx(商標)96ウェルプレート遊走系(Neuroprobe Inc., Gaithersburg, MD)中で25ng/mlのMCP−1に応答して、遊走が誘導された。種々の古典的sstr2アゴニストを異なるウェル(3回測定)に、等しい濃度で上の区画と下の区画の両方に加えた(正確に、前述のアシルアミノラクタムのBSCI活性を証明する実験と同じように;Reckless & Grainger (1999) Biochem. J. 340:803-11; Fox et al. (2002) J. Med. Chem. 45:360-70; Fox et al. (2005) J. Med. Chem. 48:867-74)。37℃で2時間後、生体染色色素であるMTT及び標準曲線の内挿を使用して、すべて実施例1と同じように正確に、下の区画に遊走した細胞の数を調べた。
【0166】
結果:sstr2で天然のペプチドリガンドであるソマトスタチン(sst14)は、BSCI活性をほんのわずかに再現するのみであった(図15)。注目すべきは、sst14の最大用量(1μM)でさえ、達成される白血球遊走の最大阻害は、具体的な実験に依存して40〜60%であった。10nMより高いsst14濃度では、細胞上のすべてのsstr2受容体が占有され、従って遊走を完全に阻止できないことは、BSCIと比較してsst14に応答してsstr2がシグナルを出す方法の定性的差を表す。
【0167】
さらにsst14は、sstr2結合の結合定数(約0.05nM)より高い濃度で白血球遊走を阻害するのみであるが、現在まで試験したすべてのBSCIは、その結合親和性より低い濃度で白血球遊走を阻害する。これらの観察結果は、BSCIが古典的sstr2アゴニスト(例えば天然のリガンドであるsst14)より強力なシグナル(白血球遊走に関して)を誘導することを示唆する。その結果BSCIは、白血球遊走を最大阻害するのに細胞表面受容体のほんのわずかを占有すればよいが、従来のsstr2アゴニストは基本的にすべての受容体を占有しなければならず、それでも最大阻害は達成されない。
【0168】
別にプロセシングされたsstr28天然のリガンドを使用して同様の実験を行い、同様の結果が得られた。白血球遊走はsstr2での結合親和性より高い濃度でのみ阻害され、最大阻害は非常に高用量でもわずかに40〜60%であった。臨床的に先端巨大症で成長ホルモン放出を抑制するのに使用される、合成ソマトスタチン類縁体ナンレオチド(lanreotide)(Prevost et al. (1991) Endcrinology 129:323-9)を使用して、再度同様の結果が得られた。
【0169】
我々はまた、その受容体結合特性が完全には決定されていない、最近記載されたソマトスタチン類縁体であるコルチスタチン(de Lecea et al. (1996) Nature 381:242-5)を試験した。コルチスタチンは、sstr2での異なるクラスの天然のリガンドである可能性がある。しかし我々は、コルチスタチンが、白血球遊走に対してその作用がソマトスタチンと非常によく似ている(非常に高用量で使用しても、約50%の遊走を阻害するのみ)が、はるかに低効力である(ED50=約75nM;表2)ことを見いだした。従って現在まで、sstr2受容体に対する天然のリガンドは、同じ受容体で作用するアシルアミノラクタムの強力なBSCI活性を再現できていない。
【0170】
最後に我々は、Ipsen(Paris, France)により提供され、後述の表2と3で「BIM」と呼ぶ、一連の合成sstr2選択的及び特異的アゴニストを試験した。ソマトスタチンは広く同様の親和性ですべてのソマトスタチン受容体に結合し、弱いBSCI様活性は、異なる受容体を介して生成された反対のシグナルによる可能性がある。しかし、10μMで白血球遊走を阻害した(しかし、これより低用量では阻害しなかった)BIM23065を除いて、これらの選択的又は特異的な従来のsstr2アゴニストはいずれも、白血球遊走阻害アッセイで大きな活性を示さなかった(表2)。
【0171】
【表3】
【0172】
表2.種々の公知の又は疑いのあるsstr2アゴニスト化合物のsstr2での結合定数(「Ka」)、及びMCP−1誘導性THP−1細胞の遊走の阻害のためのED50。試験した最高用量(1〜10μM)で観察された最大阻害(「最大作用」)も示される。
【0173】
sstr2でのコルチスタチンの親和性は明確に報告されていない。BIM23065は、試験した最大の可能な濃度(10μM)で細胞遊走を85%阻害し、従ってこれが達成できる最大作用かどうかは不明である。BIM23A387は、Pawlikowsky et al. (2007) J. Physiol. Pharmacol. 58:179-88に記載され、BIM23120はFerrante et al. (2006) Endocrine-Related Cancer 13:955-62に記載されている。
【0174】
これらの結果はまとめると、BSCIが特殊なクラスのsstr2アゴニストであり、ほとんどの化合物(天然のリガンドを含む)は、BSCIとして知られている物質より、白血球遊走を阻害する(及び従ってインビボでの抗炎症活性を示す)能力がはるかに低いことを示す。BSCIが古典的アゴニストとは異なる程度を図16(この図はケモカイン誘導性白血球遊走の阻害についてのsstr2での親和性対ED50をプロットする)に示す。BSCIは一緒に集まり、従来のアゴニストにより顕著に分離される。従って、この試験の組合せを使用して、新規であって、構造が特有のBSCI群(既知のBSCIと構造類似性が無い)を明確に同定することができる。
【0175】
さらに、ここに示した方法は、天然のリガンドsst14とその臨床的類縁体ランレオチド(lanreotide)(Paran & Paran (2003) Curr. Opin. Invetig. Drugs 4:578-82; Sener et al. (2005) Peptides 26:493-9; Gonzales-Rey et al. (2007) Ann. Rheum. Dis. 66:582-8 and the references therein; Armani et al. (2007) J. Leukoc. Biol. 81:845-55; Cury et al. (2008) Dig. Dis. Sci. 53:2516-20)を含むいくつかのソマトスタチン受容体アゴニストについて提唱されている、弱くて臨床的にあまり有用ではない抗炎症活性から、BSCIを区別することは明らかである。
【0176】
BSCIを同定するための既存の方法は基本的に、構造活性相関を規定することに基づいて元々のヒット構造周りのさらなるリードを得るための従来の医化学法とともに、ヒットを同定する機能アッセイを使用するランダムスクリーニングに依存した。本明細書に記載の方法は、資源集約的な低〜中処理能力の機能アッセイを、大量処理の低コスト分子スクリーニングで置き換えることを可能にするため、はるかに有用である。本発明のこの実施において、sstr2との相互作用に基づいてまず化合物が選択され、次に比較的少数の陽性ヒットが、白血球阻害活性についてスクリーニングされて、新しいBSCIが同定される。この適用は、後述の実施例8と9でさらに例示される。
【0177】
実施例7:BSCIは白血球遊走を他の従来のsstr2標的より強力に阻害する
BSCIはsstr2受容体に対するアゴニストであるが、必ずしもすべてのsstr2受容体アゴニストがBSCI活性を示さない。ここで我々は、その逆が真かどうかを調べた:BSCIは、他の機能アッセイにより評価される強力なsstr2アゴニスト様活性を有するか?
【0178】
「古典的」sstr2機能のための2つの異なるアッセイを採用した:(1)sstr2アゴニストにより用量依存的に阻害される成長ホルモンを分泌した胎児ラットの下垂体細胞を培養し、及び(2)sstr2アゴニストにより用量依存的に阻害されるインスリンを分泌したヒトPanc−1細胞株のグルコース感受性サブクローンを選択した。
【0179】
方法:胎児ラットの下垂体細胞培養物を、既に記載されているように、酵素分散により正確に樹立した(Murray et al. (2004) J. Clin. Endocrinol. Metab. 89:3027-32)。低グルコースDMEM+10%FCSで48時間後、48ウェルプレート中の細胞(5×104細胞/ウェル)を無血清DMEMで4時間洗浄し、次に無血清DMEM中でさらに24時間、種々の濃度の試験物質とともにインキュベートした。この時間の最後に、調整培地を採取し、細胞破片を遠心分離して除去し、市販のELISA(Linco; cat# EZRMGH-45K)を使用して、製造業者の説明書に従ってGHについて測定した。
【0180】
ヒトPanc−1(外分泌膵臓の多型類上皮癌から得られる細胞株)のグルコース感受性サブクローンを、基本的にHamil et al. (2008) Biomed. Sci. Instrum. 44:441-6が記載したように得た。簡単に説明するとPanc−1細胞を24ウェルプレートで継代培養し、培地に5%グルコースを補足した。72時間後、グルコース感受性の丸い表現型への最も顕著な変化を示したウェルを選択し、通常の培地で拡張した。次に得られたサブ株を、96ウェルプレートのウェル(5×104細胞/ウェル)を調製することによりグルコース感受性について試験し、これを次に一連のグルコース濃度を含有する培養培地(低グルコースDMEM、DMEM、DMEM+1%グルコース、DMEM+2.5%グルコース、及びDMEM+5%グルコース)に曝露した。24時間後、調整培地を採取し、ELISA(Linco; cat# EZHI-14K)により製造業者の説明書に従ってインスリン分泌を測定した。
【0181】
最も高いグルコース感受性(低グルコースDMEMと比較して5%グルコース中でインスリン分泌が3.4倍増加した)を有し、実施例3に記載した方法を使用してフローサイトメトリーによりsstr2を発現することが証明されたクローン(Panc−2H4)を使用して、我々はグルコース依存性インスリン分泌を調節する種々のsstr2アゴニストの能力を試験した。DMEM中のPanc−2H4(5×104細胞/ウェル)を使用して、追加の96ウェルプレートを調製した。24時間後、培地を種々の濃度の試験物質を含有するDMEM+5%グルコースに切り替えた(1つの条件について3つのウェル)。さらに24時間後、調整培地を採取し、インスリン濃度を上記したようにELISAにより測定した。
【0182】
両方のアッセイで、各試験化合物の存在下で2番目に高い濃度の標準物質に曝露したELISA中の追加の対照ウェルを含めることにより、ELISAに対する試験化合物(試験した最も高い濃度)の作用の欠如を証明した。結果を、ビヒクルのみの存在下で細胞からの変化倍数として表した平均±SDとして報告した。
【0183】
結果:予想したように、天然のリガンドであるソマトスタチン−14を含む古典的sstr2アゴニストは、ラット下垂体細胞からGH分泌を用量依存的に低下させ、10nMで最大で64±9%阻害であった(表3)。古典的アゴニストはPanc−2H4によるインスリン分泌に対して同様の作用を有したが、最大作用はより小さかった(43±7%阻害)。両方のアッセイで、GH又はインスリン分泌を抑制するのに必要な濃度は、sstr2への結合についての種々の物質の公知の親和性と一致した。
【0184】
これとは顕著に異なり、これらの古典的sstr2アゴニストは、ケモカインに応答してTHP−1細胞や末梢血白血球の遊走を阻害することはないか、又はGH及び/又はインスリン分泌を阻害するのに必要な濃度よりかなり高い濃度でのみ阻害した(表3)。
【0185】
ペプチドであるペプチド3とNR58−3.14.3、ならびにアミノカプロラクタム類BN83250とBN83470、及びバレロラクタム(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オンを含む、構造が異なる種々のBSCIは、はるかに高濃度でのみGH放出を阻害し(表3)、最大作用は小さかった。これらの物質のいずれも、試験した最大濃度でもPanc−2H4によるインスリン分泌を阻害しなかった。しかし予想したように、これらの物質はケモカインに応答してTHP−1細胞とラット末梢血白血球の遊走を強力に阻害した(表3)。
【0186】
表3の結果をまとめると、sstr2に結合し、この受容体でアゴニストとして作用する化合物は、容易に2つの群に分類されることを示す。1つの群の物質(本適用の目的のために「古典的」sstr2アゴニストと呼ぶ)は、ケモカイン誘導性白血球遊走を阻害するよりも高効力でかつ強力にGH分泌とインスリン分泌とを阻害する。第2の新規の群の物質(本適用の目的のために「BSCI型」sstr2アゴニスト又はソマトタキシン類と呼ぶ)は、ケモカイン誘導性白血球遊走を阻害するよりも高効力でかつ強力にケモカイン誘導性白血球遊走を阻害する。
【0187】
【表4】
【0188】
表3.「古典的」対「BSCI」標的に対する種々のsstr2アゴニストの効力の比較。4つの別々のアッセイにおける12の構造の異なるsstr2アゴニストの作用の効力(ED50、nMで)と出力(ビヒクルと比較した抑制パーセントとしての最大作用)を示す。2つのアッセイは、古典的sstr2依存性応答(GH抑制とインスリン抑制)を測定し、2つのアッセイはsstr2を介するBSCI活性(ケモカインに応答する白血球遊走の阻害)を測定する。これらの2つの分類の応答の相対的効力は、本発明の方法に従ってsstr2アゴニストを機能的に分類することを可能にする:最初の7つの物質は「古典的」sstr2アゴニストと分類され、後の5つの物質は「BSCI」sstr2アゴニストと分類される。略語:SST=ソマトスタチン;n.d.=測定されない;「バレロラクタム」=(3)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オン。BN83470=(S)−3−(アダマンタン−1’−カルボニルアミノ)−カプロラクタム。
【0189】
これらの機能的に異なるクラスのsstr2アゴニストが存在することを当業者に認めさせたであろう情報はまだ開示されていないが、この現象のいくつかの他の例があり、ここでは同じ受容体に対する異なるアゴニストが、「機能的選択性」として知られている異なる細胞応答を引き起こすことができる(他の受容体におけるこの現象の優れた最近の総説については、Kenakin (2007) Mol. Pharmacol. 72:1393-401を参照)。多くの分子機構がこの現象(例えば、活性状態ベースの選択性、偏りのある作動性、刺激の移動など)の基礎となっているが、基礎となっている分子機構の性質は、本明細書に記載したような単純な機能アッセイにより、群のメンバーを明確に区別する能力にはあまり関連していない。
【0190】
ここでsstr2アゴニストの2つのクラスを例示するのに使用される化合物群は、非常に明確であり、容易に区別される(すなわち、いくつかの異なるアッセイで生成する機能的応答の点で、各クラスのすべてのメンバーは、他のクラスのメンバーとよりも、互いによく似ている)。しかし、すべての化合物スペースにわたって、「ハイブリッド挙動」を示す物質が存在する可能性がある。すなわちこれらの物質は、GH/インスリン分泌及びケモカイン誘導性白血球遊走を、ほぼ等しい効力と出力で阻害する。そのような物質は、ここに提供した方法に基づいて、いずれかのクラスに容易に割り当てることはできないであろう。従って我々は、各クラスのメンバーを明確に規定し、いずれかの分類から「ハイブリッド挙動」化合物を除外する定義を提供した。ケモカイン誘導性白血球遊走アッセイ(すなわち、表3の右側のカラム中で2より大きい比率)よりも、GH又はインスリン分泌アッセイ中で少なくとも2倍高い効力(すなわち、半最大効力に必要な濃度)を有する分子は、「古典的」sstr2アゴニストと定義される。逆に、GH又はインスリン分泌アッセイ(すなわち、表3の右側のカラム中で0.5より小さい比率)よりも、ケモカイン誘導性白血球遊走アッセイ中の少なくとも2倍高い効力又は出力を有する分子は、本発明の方法に従って「BSCI」sstr2アゴニスト(ソマトタキシン類)と定義される。
【0191】
本発明の方法の実施において、同じ種からの細胞を使用してアッセイを比較することが好ましいことに注意されたい。古典的sstr2標的に対する物質の作用を測定するためにラットの下垂体細胞が使用される本例では、結果は、ラット末梢血白血球のケモカイン誘導性遊走に対する、同じ物質の作用と比較される。同様に、古典的sstr2標的に対する物質の作用を測定するためにヒトPanc−2H4細胞が使用される場合、結果は、ヒトTHP−1細胞のケモカイン誘導性遊走に対する、同じ物質の作用と比較される。本発明の方法を、種を超えて適用することは、純粋に2つの種からのsstr2に対する親和性が異なることの結果として、化合物の分類を間違える(両方の方向に)危険がある。例えば、ヒトsstr2よりラットsstr2で10倍低い親和性を有する化合物は、もしその物質が、ヒトTHP−1ケモカイン誘導性白血球遊走アッセイにおけるより、ラット下垂体Gh抑制アッセイで3倍低い効力であった場合、その物質は本発明の方法により「BSCI」sstr2アゴニストとして間違って分類されるであろう(なぜなら、受容体に対する結合親和性の10倍の差を補正すると、「古典的」活性は実際には「BSCI」活性より高かったため)。このため、表3中の「古典的」活性と「BSCI」活性との比は、ラットとヒトについて別々に計算される。
【0192】
実施例8:ヨヒンバン−16−アミドはα2−アドレナリン受容体で作用する機能的選択性リガンドである
ヨヒンビンは、トウワタ(milkweed)植物(インドジャボク(Rauwolfia Serpentina))の根からから得られる古典的α−アドレナリン作用模倣物のクラスのメンバーであり、これはまたレセルピンとレシンナミンとを含む。これらの物質は強力な降圧薬(すなわち、血圧を下げる)であるが、これらは好ましくない副作用を伴う(これは、記憶と多くの他の神経機能に重要なα2−アドレナリン受容体調節の中心的作用に関連する催眠作用や他の精神的作用を含む)。その結果ヨヒンビンとレセルピンは、もはや医薬として使用されていないが、米国ではまだ催淫薬として広範に使用される漢方薬として入手できる(この血管拡張活性は、Viagra(商標)で見られるような陰茎勃起を引き起こすが、降圧危機の結果として入院リスクが高く、ヨヒンビンを含む薬草抽出物の規制違反的使用が、米国において毎年10,000例の入院と何人かの死亡を引き起こしていると推定されている)。
【0193】
ここで我々は、既に記載されているヨヒンビンの16−アミド誘導体(WO99/12968参照)もまたα2−アドレナリン受容体と相互作用するが、実質的に古典的α2−アドレナリン受容体アゴニスト活性の代わりにケモカイン誘導性白血球遊走の阻害(すなわち、BSCI活性)を引き起こすことを示す。すなわちヨヒンビンの16−アミド誘導体は、α2−アドレナリン受容体で機能的選択性リガンドである(アミノラクタムBSCIがsstr2受容体で機能的選択性リガンドであるのとほとんど同じように)。
【0194】
方法:α2−アドレナリン受容体を発現する細胞からの市販の膜調製物(Sigma)を用いて製造業者の説明書に従って、放射性リガンド結合実験を行った。[3H]−ジヒドロエルゴクリプチン([3H]−DHE)を5nM濃度(反応当たり0.05μCi)で放射性リガンドプローブ(Hoffman et al. (1980) J. Biol. Chem. 255:4645-52の方法に従って)として使用した。実験は3回測定で行い、非特異結合は過剰(10μM)のオキシメタゾリンの存在下で測定した。
【0195】
MCP−1誘導性THP−1細胞遊走の阻害は、正確に上記したように測定した(実施例1)。すべての実験でDMSO(1%最終濃度)をビヒクルとして使用した。
【0196】
結果:ヨヒンバン−16−アミドはα2−アドレナリン受容体への[3H]−DHEの結合を、用量依存的に見かけのKa=89±13nMで阻害した。これは、同じ条件下で塩酸ヨヒンビンの結合の見かけのKaより約10倍低い(6.8±1.2nM)が、これは、ヨヒンバン−16−アミドが妥当な親和性でまだα2−アドレナリン受容体に結合することを裏付けている。
【0197】
異なるα2−アドレナリン受容体リガンドであるRS−79948−197(Grainger et al. (2005) Mini Rev. Med. Chem. 5:825-32)を使用して同様の結果がすでに得られているが、16−アミド基の導入によるα2−アドレナリン受容体に対する見かけの親和性の低下は、この放射性リガンドを使用すると、より大きかった(塩酸ヨヒンビンの3.8nMと比較してヨヒンバン−16−アミドでは770nM)。これは、ヨヒンバン−16−アミドの結合部位がヨヒンビンの結合部位とはわずかに異なり、RS−79948−197結合部位よりはるかにDHE結合部位と重複していることを示唆する。結合部位のこの小さなシフトは、我々は観察した機能的選択性の根拠となり得る。しかし我々は、ヨヒンバン−16−アミドがα2−アドレナリン受容体に結合すると結論付けた。
【0198】
ヨヒンバン−16−アミドは、THP−1細胞(これは、相当レベルのα2−アドレナリン受容体を発現する)のMCP−1誘導性遊走をED50が0.5nMで阻害し、一方ヨヒンビンはこのアッセイでは極めて不活性である(ED50=30nM)。しかしいずれの物質も、α2−アドレナリン受容体を発現しないJurkat T細胞のSDF−1a誘導性遊走を阻害しない。これらの結果をまとめると、ヨヒンバン−16−アミドと、程度ははるかに小さいがヨヒンビンとの両方は、これらが結合するα2−アドレナリン受容体への作用を介してBSCI活性を示す。
【0199】
しかし重要なことは、ヨヒンバン−16−アミドは古典的α−アドレナリン作用活性を有しないことである。例えばヨヒンバン−16−アミドは、ADPへの応答で血小板の活性化に効果を及ぼさないが、古典的α2−アドレナリンリガンド(ヨヒンビン、クロニジン、及びジヒドロエルゴクリプチンを含む)はすべて、顕著なADP介在血小板凝集を強化する。
【0200】
まとめると、これらの結果は、ヨヒンバン−16−アミドがα2−アドレナリン受容体で作用する機能的選択性リガンドであることを明確に証明している。この化合物は受容体への結合がヨヒンビンより弱いけれども(競合実験の見かけの親和性は、使用される放射性リガンドプローブに依存して大きく異なるため、標識ヨヒンバン−16−アミド誘導体が合成され特徴付けされるまで、相互作用の実際の親和性は不明であるけれども)、それでもBSCIよりほとんど100倍高効力である。したがって、ヨヒンバン−16−アミドの結合は、BSCI活性に必要なシグナル伝達出力の活性化を促進する。これに対してヨヒンバン−16−アミドは、ADP介在血小板活性化を強化しない(古典的α2−アドレナリン受容体活性)。その結果、我々は、ヨヒンバン−16−アミドが白血球(BSCI活性は受容体で古典的活性を実質的に置換する)上で発現されるGPCR神経受容体に対する機能的選択性リガンドの例であると結論付けた。そのような化合物は、抗炎症剤として潜在的に非常に有用であり、標的受容体の活性化に関連すると予測される副作用が無く、本発明の方法に従って容易に同定される。
【0201】
実施例9:L−IIは5HT2a受容体で作用する機能的選択性リガンドである
リセルグ酸のアミド誘導体(N,N−ジエチルアミド、LSDを含む)は、セロトニン受容体5HT2aでアゴニスト活性を介して(医学的に)所望ではない幻覚誘発活性を有し、その結果、その精神活性作用のための潜在的に有害なレクリエーション的使用の結果として多くの国で禁止されている物質である。
【0202】
ここで我々は、既に記載されているリセルグ酸のグルタミンとアスパラギンアミド結合体(WO99/12968参照)もまた5HT2aと相互作用するが、実質的に古典的5HT2aアゴニスト活性の代わりにケモカイン誘導性白血球遊走の阻害(すなわち、BSCI活性)を引き起こすことを示す。すなわちリセルグ酸のグルタミンとアスパラギンアミド誘導体は、5HT2a受容体で機能的選択性リガンドである(アミノラクタムBSCIがsstr2受容体で機能的選択性リガンドであり、ヨヒンバン−16−アミドがα2−アドレナリン受容体で機能的選択性リガンドであるのとほとんど同じように)。
【0203】
方法:5HT2a受容体を発現する細胞からの市販の膜調製物(Sigma)を用いて製造業者の説明書に従って、放射性リガンド結合実験を行った。[3H]−ケタンセリンを放射性リガンドプローブとして(Leysen et al. (1988) Pharmacol. Exp. Therap. 244:310-21の方法に従って)1nM濃度(反応当たり0.05μCi)で放射性リガンドプローブ(Hoffman et al. (1980) J. Biol. Chem. 255:4645-52の方法に従って)として使用した。実験は3回測定で行い、非特異結合は過剰(10μM)のメテルゴリン(比較的非特異的な麦角由来セロトニン受容体アンタゴニスト)の存在下で測定した。
【0204】
MCP−1誘導性ヒト末梢血単核球細胞遊走の阻害は、正確に上記したように測定された(実施例1)。すべての実験でDMSO(1%最終濃度)をビヒクルとして使用した。
【0205】
結果:L−IIは5HT2a受容体受容体への[3H]−ケタンセリンの結合を、用量依存的に見かけのKa=23±5nMで、同じ条件下でリセルグ酸の結合の見かけのKaと同様に阻害した。従ってL−IIは、リセルグ酸の他のアミド誘導体のように、妥当な親和性で5HT2a受容体に結合する。
【0206】
L−IIは、ヒト末梢血単球細胞(これは、高レベルの5HT2a受容体を発現する)のMCP−1誘導性遊走を、ED50が2nMで阻害し、一方リセルグ酸はこのアッセイでは極めて不活性である(ED50>1nM)。しかしいずれの物質も、Jurkat T細胞のSDF−1α誘導性遊走もTHP−1細胞のMCP−1誘導性遊走も阻害しない(両方の細胞株とも、高レベルの5HT2aを発現しない)。これらの結果をまとめると、L−IIは5HT2a受容体への作用を介してBSCI活性を示す。
【0207】
しかし重要なことは、L−IIは古典的5HT2aアゴニスト活性を有さないことである。例えばL−IIは、げっ歯類で幻覚誘発剤のマーカーとして従来の理解されている首ひねりを誘導しない。5HT2aのほとんどの古典的アゴニスト(LSD、メスカリン、シロシン、TCB−2、及びBromo-DrangonFLY)は、この古典的作用を誘導する。
【0208】
これらの結果はまとめると、L−IIが5HT2a受容体で作用する機能的選択性リガンドであることを明確に証明している。L−IIは5HT2aに多くの古典的アゴニストと同様の親和性で結合するが、それでもこれは、BSCIより少なくとも500倍の効力を有する。従ってL−II結合は、BSCI活性に必要なシグナル伝達出力の活性化を促進する。これに対してL−IIは、首ひねりを誘導しない(古典的5HT2aアゴニスト活性)。従って我々は、L−IIが、白血球(ここで、BSCI活性は受容体で古典的活性を実質的に置換する)上で発現されるGPCR神経受容体に対する機能的選択性リガンドの例であると結論付けた。そのような化合物は、抗炎症剤として潜在的に非常に有用であり、標的受容体の活性化に関連すると予測される副作用が無く、本発明の方法に従って容易に同定される。
【0209】
少なくとも細胞内シグナル伝達のレベルで機能的選択性は、5HT2a受容体について既に記載されており(例えば、Moya et al. (2007) J. Pharmacol. Exp. Ther. 321:1054-61を参照)、ここで2,5−DMAは、ホスホリパーゼA2(PLA2)を活性化することなくホスホリパーゼC(PLC)活性化を介してリン酸イノシトールの放出を誘導するが、関連するリガンド2C−Nは、PLCを活性化することなくPLA2によるアラキドン酸の放出を活性化する。これは我々の知見と一致するが、この機能的選択性シグナル伝達の結果としてどの機能的応答が異なるかの教示は無く、白血球上に発現される広範囲のGPCR神経受容体に対する機能的選択性が、有用な抗炎症活性を有する一連の化合物を立証できるという我々の発見を予測してはいない。
【0210】
実施例10:本発明の方法に従う、候補BSCIの同定(1)
本発明の方法をさらに例示するために、BSCI候補についてL−779,976に関連するsstr2アゴニストを含有する一連のベータ−メチルトリプトファンをスクリーニングするために、それは適用された。L−779,976は、記載されている(Rohrer et al. (1998) Science 282:737-40)初期の非ペプチドsstr2選択的アゴニストの1つであり、これは、従来のスクリーニング法、分子設計、及び医化学的アプローチにより発見された。これは、天然のソマトスタチンリガンド中の主要なQFWKテトラペプチド(配列番号5)に弱く基づくため、古典的sstr2アゴニスト性を示す(実施例7を参照)。
【0211】
興味深いことに、QxWK(配列番号6)もまた、初期ペプチドBSCI中の主要なモチーフとして同定された(Fox et al. (2002) J. Med. Chem. 45:360-70)が、現在までに記載されている非ペプチドBSCIであるsstr2アゴニストはQxW末端を形成し(図1参照)、一方古典的sstr2アゴニストはWK末端を形成している。これは、グルタミン模倣物グループと融合したL−779,976の変種が新しいクラスのBSCIであるという考えにつながる。
【0212】
ここで我々は、本発明の方法に従って半ダースの単純な新規L−779,976誘導体を試験し、その1つ(「化合物C」と呼ぶ)が新規構造クラスのBSCI化合物の最初のメンバーであることを示す。
【0213】
図17に示す6つの化合物を、DMSOビヒクル(1%最終濃度)中0.01nM〜1,000nMの種々の濃度で、Panc−2H4インスリン抑制アッセイ(正確に実施例7に記載されたように)とケモカイン誘導性THP−1遊走抑制アッセイ(正確に実施例1に記載されたように)とに付した。結果を表4に示す。
【0214】
【表5】
【0215】
表4.L−779,976の6つの類縁体の古典的sstr2アゴニストとBSCI sstr2アゴニスト活性。Panc−2H4細胞中のグルコース依存性インスリン分泌の抑制を、正確に実施例7に記載されたように測定した。ケモカイン誘導性THP−1細胞遊走の抑制を、正確に実施例1に記載されたように測定した。n.c.=比率は計算していない(いずれかのアッセイで不充分な活性を示す化合物について)。
【0216】
親L−779,976及び化合物Aは、古典的sstr2アゴニストである。化合物D、E及びFは、この実験で本発明の方法に従って評価するには、いずれかのアッセイで不充分な活性を有する。化合物Bは、古典的sstr2アゴニスト活性とBSCI sstr2アゴニスト活性の両方を有する。しかしケモカイン誘導性白血球遊走の抑制効力は、インスリン分泌の抑制効力より2倍以上高いため、本発明の方法は、化合物Cを、新規であって構造が固有のBSCI化合物(ソマトタキシン)として同定する。
【0217】
ここで我々は、抗炎症剤としてさらに開発するのに適した、BSCI活性を有する化合物を明確に同定するために、既知のsstr2アゴニスト化合物の一連の類縁体に本発明の方法が如何に適用されるかを例示する。機能アッセイを使用してスクリーニングされる少数の分子を選択するために、sstr2結合の知識が第1工程で使用されたため、sstr2への結合の実験的スクリーニングは行なわなかったが、これは本発明の方法の適用であることは明らかである。
【0218】
実施例11:本発明の方法に従う、候補BSCIの同定(2)
前の例ではBSCIの分子標的の知識を使用して、既知のsstr2受容体アゴニストの修飾に基づく、機能的スクリーニングのための候補BSCI化合物を設計した。しかし本発明の方法はまた、非常に大きなライブラリーのランダムスクリーニングにも適用することができる。この適用では、まずsstr2を標的として使用する従来の分子スクリーニングが、まずランダムライブラリーからのヒットを同定するために使用され、次に第2段階において、ヒット(又はその類縁体)は、「古典的」sstr2アゴニスト活性、及び「BSCI」sstr2アゴニスト(ソマトタキシン)活性のについての一対の機能アッセイを使用してスクリーニングされる。
【0219】
方法:sstr2を発現するように安定にトランスフェクトしたJurkat T細胞(Jurkat D3B;実施例5参照)から調製した膜を使用して、sstr2と相互作用する化合物の分子スクリーニングを確立した。簡単に説明するとJurkat D3B細胞(調製物当たり5×107個の細胞)をダルベッコーPBSで遠心分離して洗浄し、次にプロテアーゼ阻害剤(1mM PMSF、100mM ロイペプチン)を含有する1mlの氷冷ダルベッコーPBSに再懸濁した。次に細胞懸濁液を、Dounceホモジナイザーを使用して溶解し、得られたホモジネートを700×gで4℃にて1分遠心分離して、壊れていない細胞と大きな細胞断片をペレット化した。上清を取り出し、超遠心分離管に移し、60,000×gで4℃にて2時間遠心分離して、膜断片のペレットを得た。ペレットを、プロテアーゼ阻害剤を含有する氷冷ダルベッコ(Dulbecco)−PBSで静かに洗浄し、次にプロテアーゼ阻害剤を含有する100μlのダルベッコ−PBSに再懸濁した。次にクマシープラス(Coomassie Plus)キット(Pierce)を使用して製造業者の説明書に従ってタンパク質濃度を測定し、プロテアーゼ阻害剤を含有するダルベッコ−PBSのさらなる添加により最終濃度を2mg/mlに調整した。次に凍結防止剤として等量のグリセロールを加え(50%グリセロール中1mg/mlの総タンパク質濃度の膜のストック調製物を与える)、使用するまでストックを−80℃で最大1ヶ月間保存した。
【0220】
スクリーニングは384ウェルマイクロタイタープレートで行った。各ウェルで、結合緩衝液(ゲイの平衡塩類溶液)中1μgのJurkat D3B膜画分タンパク質、1nMの[125I]標識ソマトスタチン−14(400 mCi/mmol)、及びDMSO中100mg/mlからの最終濃度1mg/mlのライブラリーからの試験化合物からなる1つの反応を、最終反応容量25μlで構成した。37℃で2時間(平衡結合に至ることがすでに確立された条件)結合させ、次に反応物を384ウェルのマニホールド中GFCろ過シートでろ過し、0.5%トリトンX−100と300mM NaClをさらに含む10mlの結合緩衝液を用いて連続流で洗浄した。結合したカウントは定量的シンチグラフィにより評価した。各プレートで、結合の陽性対照としてBIM23627(10μM)を使用し、非特異結合を測定するために100nMの非標識ソマトスタチンを使用した。結合は、ある化合物の存在下での特異的カウントの抑制パーセントとして表した。試験化合物を加えずに全384ウェルプレートをアッセイして評価すると、特異的カウントの38%までの抑制は陰性対照(ビヒクルのみ)レベルより4標準偏差低いレベルであった。その結果、特異的カウントの76%又はそれ以上の抑制を与える試験物質を陽性ヒットとした。
【0221】
このアッセイからのヒットを、正確に実施例4に記載したように、従来のScatchard解析を使用して確認した。sstr2への結合の見かけのKaが1μM又はそれ以上の場合は、化合物をさらに調べた。
【0222】
次に、sstr2で相互作用が確認された化合物を対の機能アッセイで評価した:panc−2H4インスリン抑制アッセイ(「古典的」sstr2アゴニスト活性の推定として、正確に実施例7に記載のように行った)及びケモカイン誘導性THP−1細胞遊走抑制アッセイ(「BSCI」sstr2アゴニスト活性の推定として、正確に実施例1に記載のように行った)。本発明の方法に従って、ケモカイン誘導性THP−1細胞遊走抑制アッセイで少なくとも2倍高い効力を有する物質が、抗炎症剤としてさらなる研究と開発に適した候補BSCIとして選択される。
【0223】
結果:26枚のアッセイプレートを流して(全部で9,880化合物をスクリーニングした)、標識ソマトスタチン−14とsstr2の結合を、76%を超えて抑制した28のヒット(0.28%)が得られた。
【0224】
これらの1次ヒットのうち11個(1次ヒットの39%;ライブラリーの0.11%)はScatchard解析でうまく再現され、9個(繰り返しヒットの81%;ライブラリーの0.09%)は、sstr2に対する見かけの結合親和性が1μMを超えた。
【0225】
次に、再現されたヒット(sstr2についての見かけの結合親和性の順位でASX.A〜ASX.Iと呼び、ASX.Aが最も高い見かけの親和性を有する)を、正確に実施例7に記載のように対の機能アッセイを使用して評価し、その結果を表5に示す。
【0226】
【表6】
【0227】
表5.9,880の化合物のスクリーニングからの再現された中程度の親和性(1μM又はそれ以上)のヒットの古典的sstr2アゴニスト及びBSCI sstr2アゴニスト活性。Panc−2H4細胞中のグルコース依存性インスリン分泌の抑制を、正確に実施例7に記載のように測定した。ケモカイン誘導性THP−1細胞遊走の抑制は、正確に実施例1に記載のように測定した。n.c.=比率は計算していない(いずれかのアッセイで不充分な活性を示す化合物について)。L−779,976とBN83250のデータを、比較のために古典的タイプのsstr2アゴニストとBSCIタイプ(ソマトタキシン)のsstr2アゴニストの例として示す。
【0228】
驚くべきことに、11個の化合物のうちの9個はいずれかの機能アッセイで検出可能なアゴニスト活性を有した。ほとんどのGPCRについて、大量処理スクリーニングにより同定されたヒットの中で、10%(10個のうちの1個)未満はアゴニスト活性を有すると予測され得る。この珍しい知見は、sstr2でのアゴニスト:受容体相互作用の相対的複雑さを反映しているか、又は我々は確立(天然のリガンドの結合について競合)するために選択した具体的なスクリーニングの結果かも知れない。さらにスクリーニングしたライブラリーはランダムではなく、既知のGPCRアゴニストと構造類似性を有する鋳型を用いてすでに濃縮されていた。
【0229】
これらの9個のうち3個は、本発明の方法に従って「古典的」タイプのsstr2アゴニスト活性を有するとして明確に特定できる高いアゴニスト活性を有し(ASX.A、ASX.C、及びASX.D)、細胞遊走抑制アッセイよりインスリン抑制アッセイで少なくとも2倍高い効力を有した。これに対して、再現されたヒットのうちで最も低い見かけの結合親和性であった1個のみ(ASX.H)は、インスリン抑制アッセイより細胞遊走抑制アッセイで少なくとも2倍高い効力を有して、明確に「BSCI」タイプのsstr2アゴニスト(1つのソタトタキシン)として特定された。これらの知見は、1個の化合物(ASX.D)がこの細胞遊走抑制アッセイでASX.Hに匹敵する効力を有したが、「古典的」sstr2アゴニスト活性についてのインスリン抑制アッセイでも高い効力を有したため、細胞遊走の抑制の機能アッセイでさえ、「BSCI」タイプのsstr2アゴニスト(ソマトタキシン類)を明確に特定するのに、単独では使用できないことを例示する。本発明の方法は、完全に新規な「BSCI」タイプのsstr2アゴニスト(ソマトタキシン類)を特定するのに必要かつ十分である。
【0230】
本発明の方法を使用して我々は9,880個の化合物をスクリーニングして、BSCIとして充分な性質(もちろんED50が45nMでは、それは、我々がすでに記載した最適化リード候補よりインビトロで約1,000倍低い効力である;例えばWO2009/016390参照)を有し、従来の医化学リード最適化プログラムのひな型として機能できるであろう1つの化合物を見つけた。機能アッセイによる9,880個の化合物すべての直接スクリーニングは、何カ月もかかったであろう(通常の1週間労働で約100個の化合物をスクリーニングすることができる)。BSCIの分子標的の知識を利用する本発明の方法の適用は、この全モデルスクリーニングをたった1週間で完了することを可能にした。これは、消費される資源の約99%の削減である。典型的には、医薬業界で行われる大量処理スクリーニングは、本例で我々が使用したものより10倍〜100倍多い化合物をスクリーニングできるであろう。そのようなスクリーニングは、本発明の方法を使用すると容易であるが、機能的スクリーニングアッセイを使用すると実現不可能であり、同じスクリーニング量を達成するのにほとんど10年必要であろうと我々は推定する。
【0231】
実施例12:本発明の方法で同定される候補BSCIのインビボ評価
抗炎症特性を有するが、古典的ソマトスタチン受容体アゴニストの内分泌調節性を持たない薬剤として有用な物質を同定するために、本発明の方法に従って化合物Pが同定された。
【0232】
化合物Pはsstr2に結合し、ここでアゴニストとして作用する。図18に示すように化合物Pは、本発明の方法に従うアッセイでケモカインMCP−1に応答してTHP−1細胞の遊走を、ED50が約0.05nMで阻害する。
【0233】
しかし、図19に示すように化合物Pは、本発明の方法に従って行われたアッセイで、培養下垂体細胞からの成長ホルモン(GH)の放出を、ED50が約2000nMで阻害したのみである。図19において、点線は、例えば10nMのソマトスタチン−14(古典的sstr2アゴニスト)を用いて、このアッセイで達成される最大作用(成長ホルモン分泌の約75%の低下)を示す。その結果、ED50用量は、GH分泌を約37%低下させた化合物Pの用量である。
【0234】
こうして化合物Pの古典的タイプの活性とBSCIタイプの活性の比は、約0.000025であると計算される。
【0235】
インビトロで行われたアッセイがインビボの化合物の活性を予測するものであることを証明するために、36人の正常で健常なヒト被験者(男性又は女性)を、12人の被験者の3つの群の臨床試験に入れた。各群12人の被験者のうち、9人は経口的に化合物Pを60mLの水中の液体薬剤として投与され、残りの3人はプラセボとして60mLの水のみを投与された。第1群は150mgの化合物Pを7日間毎日投与され、第2群は500mgの化合物Pを7日間毎日投与され、第3群は1500mgの化合物Pを10日間毎日投与された。化合物Pの投与前、投与期間中、及び投与後の様々な時間に血液試料を採取した。
【0236】
化合物Pは、約25時間の半減期で、ヒトでほぼ100%の経口バイオアベイラビリティを有することがわかった。化合物Pへの被験者の予測された曝露は、被験者の血清中の化合物Pについて評価済みLC−MSアッセイを使用して確認した。
【0237】
予測されたように化合物Pは、ヒトで抗炎症性であった。C−反応性タンパク質(CRP)はヒトの炎症レベルの充分認められたマーカーであり、急性及び慢性の炎症反応中に上昇する(Coventry et al. (2009) J. Translational Med. 7: 102を参照)。その結果、血清CRPレベルは炎症の代替マーカーとして広く受け入れられている。CRPレベルは、投与の前(投与前)と投与の最後の日(投与後)の血清試料で、市販のELISAアッセイを使用して測定された。すべての3つの用量の化合物Pで処理すると、処理された被験者中のCRPレベルが低下したが、プラセボを投与された被験者では低下しなかった。化合物P処理の効果(用量に無関係に)を図21に示す。
【0238】
図20では、充分に確立された臨床的な抗炎症治療薬Medrol(登録商標)(メチルプレドニソロン、コルチコステロイド)と化合物Pの作用を比較した。両方の化合物がCRPを同程度に低下させたが、作用は化合物Pについてのみ統計的に有意であった(p=0.02)。
【0239】
図20の結果は、化合物Pが合成コルチコステロイドMedrol(登録商標)と少なくとも同様に有効な抗炎症剤であることを裏付けている。
【0240】
しかし、また本発明の方法に従って予測されるように、我々は化合物Pが2型ソマトスタチン受容体に結合するが、視床下部下垂体副腎軸(HPA軸)のマーカーのパネルに対して効果が無く、従ってヒトで古典的タイプの活性が完全に欠如していることを見いだした。
【0241】
化合物Pは、非常に高用量であっても、7日間治療したヒトでGHレベルに対して何の作用もなかった。さらに化合物Pは、非常に高用量であっても、7日間治療したヒトでACTHレベルに対して何の作用もなかった。ACTHは、ヒトのHPA軸の乱れの最も感受性の高いマーカーであることが確立されており(Denef (2008) J. Neuroendocrinol. 20: 1-70を参照)、これらの研究をまとめると、化合物Pはヒトで臨床的に有用な抗炎症作用を有するが、同じ用量で古典的タイプのソマトスタチン模倣作用を持たないことを裏付けている。
【0242】
本例は、2型ソマトスタチン受容体に結合する、本発明の方法に従って同定された化合物が、ヒトで臨床的に有用な抗炎症活性を有し、(これに関連して)好ましくない内分泌調節作用(又は「古典的タイプ」作用)を全く有さないことを裏付けている。
【0243】
実施例の要約
実施例をまとめると我々は、4つの異なる群のBSCIが、白血球上で発現される3個の異なるGPCR神経受容体でアゴニストとして作用することを証明した。ペプチド3関連のポリアミドとアミノラクタムは、2型ソマトスタチン受容体(sstr2;実施例1〜5)のアゴニストとして作用する。ヨヒンビンの16アミド誘導体は、α2−アドレナリン受容体を介して作用する(実施例8)。リセルグ酸のグルタミン及びアスパラギンアミド結合体は、5HT2aセロトニン受容体のアゴニストとして作用する(実施例9)。しかし、各場合にこれらの受容体に対する他のアゴニストの大部分はBSCIとして作用することができない(実施例6、8、及び9)か、又は実質的に低い効力及び/又は出力で作用する。その結果我々は、BSCIが、これらの受容体に対するアゴニストの、より多くの一般的クラスの、特別な、使用可能に定義されたサブクラスであると結論付けた。
【0244】
さらに、これらの群は互いの構造的関係によってのみ同定されたため、このような機能的選択性リガンドがすべてのGPCR神経受容体について存在すると結論することは妥当である。白血球上で発現された3つの異なるGPCR神経受容体で、我々が機能的選択性リガンドを発見したことは驚くべきことである(これらの化合物を同定するために使用した方法は、これらの分子標的の情報に全く依存しなかったため)。これらの物質の作用機構が開示された(本明細書)ため、この情報を使用して、白血球上で発現される他のGPCR神経受容体で同様に作用するリガンドを見つけることができる。このような探索を行う方法は、本発明の一態様である。
【0245】
古典的アゴニスト活性に対するBSCI活性の機能的選択性に基づいて、白血球上で発現されるGPCR神経受容体でアゴニストを下位分類することは、有用(これは、既知のクラスとの構造的関係に無関係に新しいBSCIの同定を可能にするため)であり、新規(これまでは記載されていないため)であり、充分定義され(実施例10と11に例示される、サブクラスのメンバーを明確に区別する方法が示されるため)、且つ驚くべきことである(既存の科学文献又は特許文献はいずれも、古典的活性に対して抗炎症活性を促進する白血球上で発現されるGPCR神経受容体の機能的に明確なサブクラスが存在し得ることを示唆していない)。
【0246】
実施例10と実施例11の両方において、新規であって構造が固有のBSCI化合物を同定するために、本発明の主題であるBSCIの分子標的の先行情報を使用した。実施例10では、類縁体作成ための適切な骨格としてL−779,976を選択するために、標的の情報が使用され、そして抗炎症剤としての以後の開発のための最も有望な候補を同定するために、これらの類縁体を、その後に本発明の方法に従う一対のアッセイ(「古典的」sstr2アゴニスト活性のアッセイと「BSCI」sstr2アゴニスト活性のアッセイ)を使用してスクリーニングした。実施例11では、sstr2相互作用性構造について直接的分子スクリーニングを計画及び展開するために、標的の情報が使用された。これは次に、本発明の方法に従って一つのアッセイ(「古典的」sstr2アゴニスト活性のアッセイと「BSCI」sstr2アゴニスト活性のアッセイ)の適用によりさらに洗練された。こうして、抗炎症剤としてさらなる開発に最も有望なライブラリー要素が同定された。
【0247】
最後に実施例12において、2型ソマトスタチン受容体に結合する、本発明で同定された化合物が、好ましくない内分泌調節(又は「古典的タイプの」)作用無しで、ヒトで臨床的に有用な抗炎症活性を有することを証明することにより、本発明の方法は有効であることが確認された。
【0248】
BSCIが、白血球上で発現されるGPCR神経受容体において機能的選択性により作用するという驚くべきこと知見は、非常に広範囲の疾患を治療するための新規抗炎症剤として有望なBSCI活性を有する物質を同定するためのユニークで有用な新しい方法を提供する。現在まで、新規であって構造が特有であるBSCI化合物を同定する唯一の方法は、既存のBSCI化合物の構造をさらに修飾する(既存の構造クラス内で既知のBSCI化合物の数を有効に拡大する)か、又はBSCI活性の機能アッセイで選択されない化合物ライブラリーをスクリーニングする(例えば、本発明の実施例で広く使用されるTranswell(登録商標)細胞遊走アッセイ)ことであった。同じ期間に従来の分子スクリーニングにより数万の化合物が処理されるのと比較すると、現在までに入手できる最良の方法でさえ、機能アッセイによりわずかに数百の新しい化合物しかスクリーニングできない。分子スクリーニングと機能的スクリーニングによる処理速度の2〜3オーダーの差は、本発明の有用性にとって重要である:ランダムな機能的スクリーニングにより新しい構造クラスのBSCIを見つけることにおいて、資源問題は非現実的なほど大きい。しかし本発明の方法は、まず白血球上で発現されるGPCR神経受容体を使用して従来の分子スクリーニングを行い、次に第2の工程で、分子スクリーニングからのヒットを機能アッセイによりさらにスクリーニングして、BSCIの機能的特性(すなわち、GH又はインスリン放出の抑制より、ケモカイン誘導性白血球遊走の抑制に対してより高効力を有する物質)を同定する。第1工程の分子スクリーニングは、ランダムライブラリー中の化合物の数を2〜3オーダー減少させるため(妥当な効力ヒットは典型的には、ランダムライブラリーのわずかに0.1〜1%であるため)、以後の機能的スクリーニングの資源の要求性は、スクリーニングされる化合物当たりの利用はより大きくなるが、現実的になる。
【0249】
本発明を好適な実施態様を参照して説明してきたが、この種々の修飾態様や変更態様は本発明の精神と範囲を逸脱することなく実施可能であり、このような修飾が本発明に明確に包含されることを当業者は認識し得る。本明細書に開示し、特許請求の範囲に記載した具体例は、全く制限的ではなく、また制限的であるべきではない。
【0250】
本明細書に記載したすべての文書は参照のためその全体が本明細書に組み込まれる。
【0251】
以下の配列表に記載された配列は本発明の一部を構成する。
【配列表フリーテキスト】
【0252】
配列表のフリーテキスト
合成オリゴヌクレオチド(配列番号3〜4);
天然のソマトスタチンリガンドの主要なBSCIモチーフ(配列番号5);
初期ペプチドBSCIの主要なモチーフ(配列番号6);
Xaaは任意の天然に存在するアミノ酸でよい(配列番号6)。
【技術分野】
【0001】
本発明は、広域スペクトルケモカイン阻害剤(BSCI)活性を有し、且つ抗炎症剤としての用途を有する、新規化学物質、及び医薬組成物の発見方法に関する。
【0002】
ケモカインは、哺乳動物の免疫系の制御において重要な役割を果たす約50個の、構造が関連している、大部分が小さい(8〜12kDa)タンパク質の群である。ケモカインリガンドは、約20個の密接に関連した受容体を介してシグナルを出し、そしてそれらはGタンパク質結合受容体(GPCR)の1型クラスのメンバーである。重要なことは、リガンドの多くは2種以上の受容体に結合し、かつこれを介して生産的にシグナルを出すことである。同様に、多くの受容体が2種以上のリガンドに結合する。
【0003】
特定の種類の白血球上に、これらのケモカイン受容体の種々のサブセットが発現されている。この複雑な縮重の結果として、白血球動員の厳密な時間的及び空間的制御が可能である。ケモカインは免疫系の「交通警官」として中心的な役割を果たし、種々の白血球が、骨髄から血液を介して免疫応答の2次的リンパ組織又は免疫応答部位に取る複雑な経路をコードしている可能性がある。
【0004】
このため、ケモカインシグナル伝達を調節する化合物は長い間、炎症性要素を有する非常に広範囲の疾患(例えば、アテローム性動脈硬化症、自己免疫疾患、アレルギー、湿疹、乾癬、又は神経変性障害)に有用な、新規抗炎症剤開発のための有望な候補と考えられている。これらの疾患のための新しい薬に対して大きなニーズがある:既存の小分子治療薬は一般に、有効であるが重篤な副作用がある(例えばコルチコステロイド)か、又は有効性は小さいが比較的安全である(例えばジクロフェナックのようなNSAID)。生物学的治療薬(例えば、TNF−αに対する抗体)はより特異的であるが、製造コストが高く、単純な経口投与には適していない。小分子ケモカイン阻害剤は、既存の生物学的抗炎症剤と比較して、他の群の小分子抗炎症剤より有効性と安全性の兼ね合いが優れている可能性があり、改良された薬剤学的性質(コストを含む)を与える。
【0005】
残念ながら、特異的ケモカイン受容体アンタゴニストはこの可能性を実現するには進歩が遅い。まずこれらは、従来の大量処理分子スクリーニング法(例えば、リガンド:受容体相互作用を阻止することができる物質についての受容体結合アッセイで、ランダムなライブラリーがスクリーニングされる)を使用して見つけることが難しい。ケモカイン受容体間の類似性は、化合物がしばしばケモカイン受容体のサブセット(単一の受容体についての特異性を有する代わりに)と反応することを意味し、さらに悪いことに多くの初期ケモカイン受容体調節物質は、セロトニンやカテコールアミン系の関連受容体に結合し、挙動、血圧、及び他の表現型に好ましくない副作用を引き起こす。
【0006】
第2に、あるクラス内の単一の受容体に対する優れた特異性(これは、従来の医薬開発思想の最大の目的である)は、これが最終的に達成されても特に有効ではないことが明らかとなった。同じ受容体に結合する多くの関連リガンド、及び多くのリガンドのためにシグナルを変換できるいくつかの受容体がある、ケモカインシステムの縮重のために、単一の受容体を阻止する物質は通常、一般的抗炎症性は無く、免疫系に対して失望するほどに弱い作用を有する。これは音楽の増幅器に例えることができる:抗炎症作用は「ボリュームコントロール」の調節を必要とするが、特異的ケモカイン受容体アンタゴニストは「グラフィックエコライザー」に対して作用する。すなわちこのような物質は、免疫応答の種類をわずかに調節する(おそらく、存在する白血球サブセットにより浸入物の細胞組成を変化させる)が、炎症の程度を定量的に低下させることはない。
【0007】
その結果、ケモカインシステムが最初に記載されてから20年以上経過し、その後主要な免疫調節標的として出現している2009年でさえ、ヒトの治療薬として認可されているケモカイン阻害剤は無い(しかし、幾つかは特定の適応症で臨床治験の後期に入っており、炎症応答の種類の調節がある程度の効果を有している、例えばリウマチ様関節炎におけるCCR1ケモカイン受容体の特異的アンタゴニストであるBX471がある(Horuk (2005) Mini Rev. Med. Chem. 5:791-804))。従って別のアプローチが必要である:多くのケモカインによるシグナルを同時に阻害する化合物(我々が広域スペクトルケモカイン阻害剤(broad-spectrum chemokine inhibitor)、又はBSCIと名付けた物質;Grainger & Reckless (2003) Biochem. Pharmacol. 65:1027-34; Grainger et al. (2005) Mini Rev. Med. Chem. 5:825-32)。
【0008】
広域スペクトルケモカイン阻害剤(BSCI)活性を有する化合物は、広範囲の疾患を治療または予防するための抗炎症性薬物として有用な新しいクラスの物質(agent)である。ヒト疾患の動物モデルでは、アテローム性動脈硬化症、卒中、喘息、術的接着形成、子宮内膜症、皮膚炎、内毒素血症、及びリウマチ様関節炎において、BSCIは白血球蓄積や疾患重症度の他のマーカーを低下させるのに有効であることが証明されている(Reckless et al. (2001) Immunology 103:244-54; Beech et al. (2001) J. Cereb. Blood Flow Metabol. 21:683-9; Grainger & Reckless (2003) Biochem. Pharmacol. 65:1027-34; Naidu et al. (2003) Ann. Thorac. Surg. 78:1118-22; Naidu et al. (2004) J. Heart Lung Transplant. 23:128-34; Reckless et al. (2005) J. Vasc. Res. 42:492-502; Berkkanoglu et al. (2005) Hum. Reprod. 20:3047-52; Kayisli et al. (2007) Reprod. Sci. 14:825-35; Hildebrandt et al. (2009)、印刷中)。BSCIはまた、インビトロでHIV複製を抑制することが証明されている(HIVの細胞侵入のケモカイン依存性により生じる、抗炎症性に無関係の活性;Grainger & Lever (2005) Retrovirology 2:23)。
【0009】
このような広範囲の疾患モデルで強力な抗炎症活性を有するにもかかわらず、クラスとしてのBSCIは、注目すべきことに毒性が無いか又は副作用が小さい(Schroff et al. (2005) Mini Rev Med Chem. 5:849-55)。例えばアミノラクタムBSCIは、広範囲の急性及び慢性毒性試験で多くの投与経路で評価されているが、作用の無いレベルが最大治療用量より少なくとも1,000倍高い。これらのアミノラクタムBSCIはまた、優れた安全性薬理学的プロフィールを示す(hERGテール電流に対する作用がないか、又は遺伝毒性のアッセイにおいて作用がない;例えばWO2009/016390を参照)。
【0010】
これらの知見はまとめると、BSCIがクラスとして既存の抗炎症薬物(コルチコステロイドやNSAIDを含む)と比較して、有効性と副作用との優れた兼ね合いを与えることができることを示唆する。従って、BSCI活性を有する組成物を同定する方法に対するニーズがある。
【0011】
残念ながら、現在までに記載されているBSCIは運良く偶然同定されたものであり、新規の、潜在的に構造的関連性がないBSCIを同定する簡単な方法は無い。BSCI活性を有する最初の化合物は、ペプチド3(Peptide 3)と呼ぶペプチドであり、これはMCP−1の配列から選択され、複数のケモカイン受容体に結合する能力について試験された(Reckless & Grainger (1999) Biochem. J. 340:803-11)。ペプチド3は複数のケモカイン受容体に結合しないことがわかった(しかし実際は、細胞表面上野単一のまだ同定されていない高親和性部位に結合する;Grainger & Reckless (2003) Biochem. Pharmacol. 65:1027-34)が、少なくとも10個の異なる受容体に結合することができるそれらの間で、少なくとも5個のケモカインに対するBSCI活性を有した。
【0012】
このペプチド3の構造鋳型から、BSCIの一連の異なる構造群(ペプチド、ペプトイド、及び非ペプチド構造体を含む)が設計され、合成され、特徴づけされた。重要なことは、ペプチドのBSCI活性の原因である構造モチーフは、WxQトリペプチド(ここで、Wはアミノ酸トリプトファンであり、xは任意のタンパク新生アミノ酸であり、Qはグルタミンである)まで絞られたことである。これは、ペプチドではないWxQ模倣物の設計を可能にし、その1つ(N−アシルグルタリミドNR58,4)はBSCIとしてナノモル活性を有する(Fox et al. (2002) J. Med. Chem. 45:360-70)。
【0013】
NR58,4はBSCIとしてインビボで活性である(内毒素血症のような急性炎症のモデルで強力な抗炎症作用を有する)が、薬剤開発のための理想的な化合物ではないことが明らかになった。その必須のイミド環が血漿中で未特定のアミダーゼによる加水分解を受けやすく、その結果化合物はインビボで急速に不活性化された(Fox et al. (2005) J. Med. Chem. 48:867-74)。
【0014】
その後、さらなる医学化学的研究により、BSCIとしての効力(potency)を保持するがインビボでは安定である、関連性を有する種類の置換アミノラクタムが同定された。このクラスのBSCIの例には、N−オレオイル−3−アミノカプロラクタム(BN83253; Fox et al. (2005) J. Med. Chem. 48:867-74)がある。広範囲の、構造的関連性がある他のアミノラクタムBSCIも記載されている(例えば、WO2005/053702、WO2006/016152、WO2006/018609、WO2006/085096;WO2006/134385、WO2006/134384、及びWO2009/016390を参照)が、その少なくとも一部は、ヒトに対する医薬として開発するのに適した性質を有する。
【0015】
必須であるWxQファーマコフォアの構造に基づいて、別の群の非ペプチドBSCIも同定されている。アルカロイドであるヨヒンビン(yohimbine)(インドジャボク(Rauwolfia Serpentina)の根から得られるアルファ2−アドレナリン受容体ゴニスト)は、WVQトリペプチドの特定のコンフォメーションに似た炭素骨格を有し、ヨヒンビンの17−アミド誘導体はBSCIのように高活性である(WO99/12968; Grainger et al. (2005) Mini Rev Med Chem. 5:825-32)。
【0016】
非ペプチドBSCIの第3の群も記載されており、これも再度WVQトリペプチドの炭素骨格とのリセルグアミド核の類似性に基づく。クラスA禁止物質であるLSD(リセルグ酸のN,N−ジエチルアミド)のようなすでに報告されたリセルグアミド類は、複数のGPCRでアゴニストとして作用する(従って、幻覚誘発性を含む広範囲の生理学的作用を有する)が、我々は、化合物L−II(ジヒドロレセルグ酸のN−アスパラギンアミド)及びリセルグ酸の他のグルタミンとアスパラギンアミドが、他のリセルグ酸アミドの予測される副作用無しでBSCIのように高活性であることを証明した(WO99/12968)。
【0017】
現在までに記載されているすべての群のBSCI間の構造的関係を図1に例示する。この図から、BSCIのすべての群は互いの化学構造の類似性により特定されていることが明らかである。
【0018】
既知の化合物と構造が関連していないBSCIを同定するための一般的方法は、現在存在しない。この限界に関する理由は、BSCIの分子標的についての情報が欠如しているためである。これらのBSCIがケモカイン受容体アンタゴニストとして作用せず、試験したケモカイン受容体のいずれにも結合しないが、その代わりに細胞表面上の明確な(しかし同定されていない)高親和性部位に結合することがいったん明らかになると(Grainger & Reckless (2003) Biochem. Pharmacol. 65:1027-34)、従来の分子スクリーニング手段(例えば受容体結合アッセイ)を使用して、新規の、構造的に固有のBSCI群を捜すことは不可能であった。機能スクリーニングツール(すなわち、ライブラリー要素が、いくつかの異なるケモカイン(非ケモカインではない)に応答して細胞遊走を阻止するかどうかを調べる)を使用して、BSCI活性について非常に小さいライブラリーを直接スクリーニングすることができるかも知れないが、このようなスクリーニング方策は、機能スクリーニングアッセイの労働集約的性質のために直ちに非現実的となる(細胞遊走の阻害剤を同定するための機能スクリーニングアッセイを実施するための技術困難さの詳細な総説については、Frow et al. (2004) Med. Res. Rev. 24:276-98を参照)。さらにこのような機能スクリーニングは、生物学的適合性がある緩衝液に可溶性でありかつ低細胞毒性(大量処理薬物スクリーニングについて使用される典型的なランダムライブラリーで大部分(確実に90%を超える)の化合物を除外するパラメータ)を示す化合物にのみ適用することができる。簡単に説明すると、このようなアプローチは、ランダム化合物ライブラリーから完全に新しいクラスのBSCIを見つけるには全く現実的ではない。
【0019】
新規の、構造的に固有であるBSCIを同定できる能力は、この薬剤群の臨床開発のために重要である。既存のBSCIは、臨床開発を通して進歩するため、最適ではない性質が特定される可能性は低い。ヒトに投与されたBSCIは現在まで無く、現在までに同定されていない副作用又は毒性のリスクは大きいままである。さらに悪いことに、明らかとされている副作用又は毒性の大部分は、関連構造を有する化合物クラスによって共有される、クラス効果の可能性がある。従って残念ながら、BSCIについて現在まで記載されているすべてのクラスの化合物は、その構造類似性の結果として同定されたため、共有される副作用が見つかるリスクは高くなっている。
【発明の概要】
【0020】
ここで我々は、異なる群のBSCIにより使用される3つの異なる受容体を説明する。3つの受容体はすべてGPCR群のメンバーであり、その全ては、白血球上及び脳中に高発現される。驚くべきことに我々はまた、報告されるBSCIが、標的である特定の受容体に無関係に、機能的選択性をもって作用することを証明する。これらの結果は、本発明に従って、既知の化合物と同じ構造モチーフを含まない、構造的に多様なBSCI群を同定するのに有用な分子スクリーニング例を提供する。本発明は、一部においてこれらの結果に基づく。
【0021】
本発明は、添付の特許請求の範囲及び以下で定義される。ある態様において、1又は2以上の白血球サブセット上で発現されるGPCR神経受容体との相互作用について、1又は2以上の化合物(compound)若しくは物質(agent)がスクリーニングされ、その後の第2工程で、標的受容体と相互作用するこれらの化合物若しくは物質が、ケモカインにより誘導される白血球遊走を阻害する能力についてスクリーニングされるという方法が提供される。ケモカイン誘導性の白血球遊走を阻害する化合物若しくは物質は、その後インビトロでBSCI活性について試験される。重要なことは、標的受容体に対してアゴニストである化合物又は物質必ずしもすべてが、BSCI活性を持ってはいないことである。従って本発明の2つの工程は、BSCI活性を有する化合物を同定するのに必要かつ十分である。
【0022】
本発明の具体的な非限定例や追加の特徴は、以下の図面を参照して説明される。
【図面の簡単な説明】
【0023】
【図1】本明細書に記載の主要なBSCIの構造を示し、そのすべてが共有する相互関係(及び共通の構造モチーフ)を例示する。
【図2】単一の受容体(灰色の形)での機能的選択性の概念を示す図である。
【図3】タンパク質コード枠中の単一のイントロン(より暗い四角)の選択的スプライシングの結果として、sstr2遺伝子座から得られる2つの公知の代替転写体(T#1とT#2)を示す図である。
【図4】BSCIBN83253の用量応答(x軸、nM)と、MCP−1誘導性THP−1細胞遊走(y軸、阻害%)とに対する、sstr2アンタゴニストBIM23627(「I」)の濃度上昇の影響を示すグラフである。
【図5】BSCI(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)ーテトラヒドロピリジン−2−オンの用量応答(「バレロラクタム(valerolactam)」;x軸、nM)と、MCP−1誘導性THP−1細胞遊走(y軸、阻害%)に対する、sstr2アンタゴニストBIM23627(「I」)の濃度上昇の影響を示すグラフである。
【図6】細菌内毒素の単回腹腔内投与に応答する、TNF−アルファ産生(y軸、阻害%)に対する、sstr2アンタゴニストBIM23627とBSCIBN83250の同時投与(x軸、mg/kg用量)の影響を示すグラフである。
【図7】細菌内毒素の単回腹腔内投与に応答する、TNF−アルファ産生(y軸、阻害%)に対する、sstr2アンタゴニストBIM23627とBSCI(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オンの同時投与(「バレロラクタム」;x軸、mg/kg用量)の影響を示すグラフである。
【図8】細菌内毒素の単回腹腔内投与に応答する、TNF−アルファ産生(y軸、阻害%)に対する、sstr2アンタゴニストBIM23627と合成コルチコステロイドであるデキサメタゾンの同時投与(x軸、mg/kg用量)の影響を示すグラフである。
【図9】THP−1細胞(左のパネル)と陽性対照として使用したPanc−1細胞(右のパネル)から調製したmRNA試料からの、逆転写(「RT」)有り(+)及び無し(−)のPCR反応からのアンプリコンを明らかとする、臭化エチジウムで染色しUV光透過により観察したアガロースゲルを示す。
【図10】BN83250の標識類縁体である4’,5’−3[H]−(S)−3−(2’,2’−ジメチルドデカノイル)アミノ−カプロラクタム(4,5−3[H]−BN83250)の合成スキームを示す。化合物(S,E)−3−(2’,2’−ジメチルドデク−4’−エノイル)アミノ−カプロラクタム(上)はPd触媒の存在下でトリチウムガスと反応して、4,5−3[H]−BN83250(下)を形成する。
【図11】THP−1細胞への4,5−3[H]−BN83250(x軸、nM)の特異的結合(y軸、cpm)を示すグラフである。値は各BN83250濃度での3回測定の平均(±SD)であり、非特異結合(過剰の非標識BN83250の存在下)を引いてある。中心線は、単純な結合モデルを使用した時の最適フィットであり、外の線は95%信頼限界を示す。
【図12】THP−1細胞への10nMの4,5−[3H]−BN83250の結合(y軸、結合した標識[cpm])に対する、種々の濃度の非標識(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オン(「バレロラクタム」)の影響を示すグラフとヒストグラムである。
【図13】ポリリンカー中のEcoRI(5’)部位とXhoI(3’)部位との間でクローン化されたヒトsstr2Aをコードする1,100bp cDNA(「sstr2a」として示す)を有するpcDNA3.1+(Invitrogen)のプラスミド地図を示す。
【図14】親Jurkat T細胞(白丸、実線)、sstr2を発現するJurkat D3B細胞(黒丸、実線)、及びTHP−1細胞(点線)のSDF−1α誘導性遊走に対するBSCI(BN83250)の影響を比較する。(A)では、BN83250の濃度をx軸(nM)に示し、阻害%をy軸に示す。各細胞株の染色の分布を示す頻度ヒストグラムを、(B)THP−1細胞(*=特異的結合の375中央値単位)、(C)Jurkat D3B細胞(*=特異的結合の40中央値単位)、及び(C)Jurkat T細胞(*=特異的結合の1中央値単位)に示す。各頻度ヒストグラムでは、x軸はFl1−Hを示し、y軸はカウントを示す。頻度ヒストグラム(B)〜(D)の黒い線は、PBSのみによる染色を示す;薄い灰色の線は一次抗体対照を示し、中程度の灰色の線は、完全な染色反応を示す。
【図15】THP−1細胞のMCP−1誘導性遊走(y軸、阻害%)に対する種々の濃度のソマトスタチン14(x軸、nM)の影響を示す。点線は完全な阻害を示し、これはBSCIにより達成される。
【図16】広範囲の「古典的」sstr2アゴニスト(A側)と「BSCI型」sstr2アゴニスト(又は「ソマトタキシン類」、B側)の両方について、sstr2結合の見かけの親和性(x軸;sstr2での「Ka」、nM)に対するケモカイン誘導性遊走の阻害の効力(y軸;ED50対ケモカイン誘導性遊走、nM)をプロットしたグラフである。
【図17】本発明の方法に従って試験するための候補BSCIとして設計された、種々の新規ベータ−メチオニントリプトファンベースのsstr2アゴニスト(L−799,976と化合物A〜F)の構造を示す。
【図18】本発明の1つの実施態様に従う化合物P(x軸、nM)によるケモカインMCP−1誘導性THP−1細胞遊走の阻害(y軸、パーセント値として示す)を示すグラフである。
【図19】化合物P(x軸、nM)による成長ホルモン(GH)放出の阻害(y軸、パーセント値として示す)を示すグラフである。
【図20】Medrol(登録商標)(A)又は化合物P(B)で治療した患者からの血清試料中の、炎症の代替マーカーとしてのC−反応性タンパク質(CRP;y軸、ng/ml)のレベルを比較するヒストグラムである。カラムは、投与前(a)と最後の投与日(b)のCRPレベルを示す。星印はp=0.02であることを示す。
【発明を実施するための形態】
【0024】
BSCIのペプチド及びアミノラクタム群の両方の標的受容体がソマトスタチンの2型受容体(sstr2)として開示(後述の実施例参照)され、ヨヒンバミド(例えばヨヒンバン−16−アミド)の標的受容体がα2−アドレナリン受容体であり、リセルグアミド類(例えばL−II)の標的受容体が5−HT2aであり、すべてが機能的選択性をもって作用しているため、これらのすべての受容体について、ケモカイン誘導性遊走の阻害を促進する機能的選択性リガンドが存在することが明らかである。これらの受容体のすべては、白血球上で発現されるGPCR神経受容体のより大きなクラスのメンバーであるため、白血球上に発現されるすべてではなくても多くのGPCR神経受容体について、同様の機能的選択性を有するリガンドが見つかると推定することは妥当である。従って本発明の方法はある態様において、白血球上で発現されるGPCR神経受容体との相互作用について、1又は2以上の候補化合物又は物質をスクリーニングすることを含む。
【0025】
本発明の方法のある応用において、スクリーニングすべき1又は2以上の化合物はまず、選択された受容体を発現する細胞を、受容体に結合することができる放射性リガンドに曝露することにより、白血球上に発現されたGPCR神経受容体と相互作用する能力について試験される。例えば、ソマトスタチン受容体sstr2を発現する細胞は、試験化合物の存在下及び非存在下で、sstr2のリガンド(例えば標識ソマトスタチン)に曝露される。試験化合物がsstr2受容体と相互作用する場合、受容体に特異的に結合するソマトスタチンの量は変化するであろう。このような受容体結合アッセイは、同じ目的を達成するために多くの異なる方法で行うことができる:異なるリガンドが使用され得る(例えば、sstr2特異的アンタゴニストBIM23627;Tulipano et al. (2002) 143:1218-24);リガンド結合を視覚化するために、異なるレポーター系が使用され得る(シンチレーション近傍(scintillation proximity)アッセイと蛍光分極は、このような2つの例である);組換え系とは異なる細胞(細胞膜調製物、又はsstr2発現細胞を含む)が使用され得る;細胞を試験化合物に曝露するのに、異なるビヒクルが使用され得る;リガンドの特異的に結合を観察するのに、異なる緩衝液条件が使用され得る、など。
【0026】
上記例の第2の工程で、sstr2受容体と相互作用を示した化合物は、ケモカイン誘導性白血球遊走の阻害についてスクリーニングされる。例えば白血球が、ケモカインMCP−1の勾配の存在下でTranswell(登録商標)遊走アッセイプレートに入れられ、試験化合物の存在下又は非存在下において、37℃で2時間インキュベートされる(Frow et al. (2004) Med. Res. Rev. 24:276-98)。次に、この実験の最後に、遊走を行った細胞の数を、例えば生体染色色素MTTと標準曲線への内挿を用いて測定する。試験化合物がケモカイン誘導性遊走を阻害する場合、化合物を受けるウェル中で遊走するものは少ないであろう。このような遊走アッセイは、同じ目的を達成するために多くの異なる方法で行い得る:異なる遊走チャンバーを使用し得る(マルチウェルフォーマットTranswell(登録商標)遊走チャンバー、従来のボデンチャンバー、ゲル内遊走ウェル、ダンチャンバーなど);異なるケモカインを使用し得る(SDF−1α、IL−8、MIP−1α、RANTESなど);異なる白血球をチャンバーに適用し得る(新たに調製された末梢血好中球若しくはリンパ球、腹膜マクロファージ、又はTHP−1若しくはJurkatのような細胞株を含む);遊走した細胞の数を定量するために異なる検出系を使用し得る、など。
【0027】
同様に本発明の方法は、白血球上で発現される任意の他のGPCR神経受容体に適用され得る。そのような受容体の例は、特に限定されないが、アドレナリン受容体(α2−アドレナリン受容体を含む)、セロトニン受容体(5HT2a受容体を含む)、ソマトスタチン受容体(sstr2を含む)、ドーパミン受容体、ムスカリン性アセチルコリン受容体、物質P/NPY受容体、バソプレシン受容体、甲状腺ホルモン受容体、カンナビノイド受容体、コレシストキニン受容体、オピオイド受容体、アデノシン受容体、グルカゴン受容体、PACAP/VIP受容体、メラトニン受容体、ブラジキニン受容体、カルシトニン受容体、及びメラノコルチン受容体を含み得る。
【0028】
好ましくは本発明の方法は、アドレナリン受容体、セロトニン受容体、又はソマトスタチン受容体に適用される。さらに好ましくは本発明の方法は、α2−アドレナリン受容体、5HT2a受容体、又はsstr2受容体に適用される。
【0029】
この方法がソマトスタチン受容体(sstr2を含む)に適用される場合、候補化合物又は物質はソマトスタチンを除外し得る。
【0030】
本発明の方法は、BSCI活性を有する可能性が最も高いものを同定するために、化合物のライブラリーをスクリーニングするのに容易に適用し得る。このようなライブラリーは、ランダムライブラリー(例えば、大量処理スクリーニング用の多くの薬品会社により使用される構造の多様な化合物の大きなライブラリー)であるか、又は指向性(directed)ライブラリー(例えば、限定された数の前駆体化合物に適用されるコンビナトリアルケミストリーにより合成されるライブラリー)であり得る。いくつかの指向性ライブラリーは、BSCI化合物(又は、選択された受容体に対する従来のアゴニスト)を含有する可能性が高く、従ってこのようなライブラリーは本発明の方法を用いてスクリーニングする時、特に重要となり得る。例えば、我々がすでに開示した(WO2006/085096参照)Gタンパク質アゴニスト濃縮ライブラリー(GAEL)ライブラリーは、BSCI化合物が特定のGPCRで特殊なクラスのアゴニストとして作用していることが明らかなため、本発明の方法に従って生産的にスクリーニングされ得る。指向性ライブラリー(例えばGAEL)の使用は、本明細書に開示した基準に従って候補BSCI活性を有する化合物を同定するために、スクリーニングされる必要のある化合物の数を減少させ得る。しかし本発明の方法は、充分にハイスループットであり、これはランダムライブラリーへうまく適用され、このような応用は本発明に包含され得る。
【0031】
場合により、選択された受容体に対する「古典的(classical)」アゴニストとしてのBSCIの活性も測定される。好ましくは、本発明の方法に従って同定される化合物又は物質は、両方の種類の機能アッセイ(functional assay)(すなわち、BSCI活性についての細胞遊走アッセイ、及び選択された受容体に対する古典的アゴニスト活性についての機能アッセイ)で活性であるが、古典的受容体アゴニストとしてのその効力より強力な(すなわち、より低濃度で作用する)、ケモカイン誘導性白血球遊走の阻害剤である。
【0032】
すなわち、最適なBSCIは本発明の方法に従って、以下の3つの性質のすべてを有する化合物として同定され得る:
【0033】
(i)化合物は、白血球上で発現された選択されたGPCR神経受容体と、少なくとも10μMの親和性で相互作用する(すなわち、化合物が10μMで存在する時、存在するsstr2受容体の半分以上はそれらに結合された化合物を有する);
(ii)化合物は、ケモカインにより誘導された白血球遊走を、少なくとも1μMの効力で阻害する(すなわち、化合物が1μMで存在する時、細胞遊走の程度は少なくとも50%阻害される);及び
(iii)選択された受容体における古典的sstr2アゴニスト活性の、1又は2以上のアッセイ中の化合物の効力(potency)は、白血球遊走の阻害についての効力より低い。
【0034】
さらに好ましくは、化合物は、選択された受容体と少なくとも1μMの親和性で相互作用し得る;さらに好ましくは、化合物はケモカイン誘導性白血球遊走を、少なくとも100nMの効力で阻害し得る;さらに好ましくは白血球遊走に対する化合物の効力は、選択された受容体との相互作用の親和性又は古典的アゴニスト活性の効力より、10倍以上高いものであり得る。
【0035】
さらに好ましくは、化合物は、ケモカイン誘導性白血球遊走を少なくとも10nMの効力で阻害し得る;さらに好ましくは白血球遊走に対するか化合物の効力は、選択された受容体との相互作用の親和性又は古典的アゴニスト活性の効力より、50倍以上高いものであり得る。
【0036】
本発明の方法により、これらの神経受容体に対する従来のアゴニスト(特に限定されないが、天然のリガンドを含む)と、BSCI活性を有する化合物とを区別可能であることは明らかである。従来のアゴニストである化合物は、ケモカイン誘導性白血球遊走を阻害し得るが、これらは、特定の神経受容体との相互作用の親和性と、同様の効力又はより低い効力で阻害する。
【0037】
同様に、これらの受容体に対する従来のアゴニストは、標的受容体を介して従来の作用(例えば、成長ホルモン放出の阻害、又は血圧調節、インスリン放出の調節、例えば試験中の特定の受容体に依存して)を示すのと同様の濃度で、ケモカイン誘導性白血球遊走に対する作用を示すのみである。従って、これらの神経受容体に対する従来のアゴニストは、ある投与量で抗炎症活性を有するが、このような化合物はまた天然のリガンドの作用を示し、同じ化合物投与量で標的受容体を介して作用する。本発明の方法の重要な利点は、抗炎症性を有する物質を単に同定するのみでなく、その抗炎症性が副作用を超える、優れたサブクラスの物質を同定することであることを、強調すべきである。具体的には同定される物質は、白血球上で発現されるGPCR神経受容体に対する古典的アゴニストより大きな、抗炎症剤としての効力を有する(白血球上で発現されるGPCR神経受容体に対する大部分の既知のアゴニストにより共有されない性質)。
【0038】
ペプチドBSCI及びアミノラクタムBSCIは、sstr2アゴニストの特別な(及び、まだ同定されていない)サブセットである。これらのBSCIと従来のsstr2アゴニストは共通の分子標的(sstr2受容体)を共有するが、この2群の物質は受容体への結合に対して異なる結果を引き起こす。この概念は、他の受容体について別のところで詳細に記載(Urban et al (2007) J. Pharmacol. Exp. Ther. 320:1-13)されており、機能的選択性(functional selectivity)と呼ばれている。有用なたとえは、受容体(この場合sstr2)をピアノ鍵盤と見なすことである:多くの異なる手の位置(異なるリガンドに相当)は鍵盤(生産性結合に相当)と相互作用することができるが、それぞれ異なる和音の音を発生する(受容体からの異なる機能的結果に相当、例えば異なる第2のメッセンジャーが産生される)。この機能的選択性の概念を図2に示す。
【0039】
ヨヒンバミドBSCI(ヨヒンバン−16−アミドを含む)は、α2−アドレナリン受容体アゴニストの特別な(及び、まだ同定されていない)サブセットである。これらのBSCIと従来のα2−アドレナリン受容体ゴニスト(例えばレセルピン)は分子標的を共有するが、この2群の物質は受容体への結合に対して異なる結果(Urban et al (2007) J. Pharmacol. Exp. Ther. 320:1-13)(機能的選択性と呼ばれる現象)を引き起こす。ヨヒンバミドは、ケモカイン誘導性白血球遊走を強力に阻害するが、血圧には影響を与えない;レセルピンは血圧を強力に低下させるが、ケモカイン誘導性白血球遊走には影響を与えない。
【0040】
リセルグアミドBSCI(L−IIを含む)は、5HT2aアゴニストの特別な(及び、まだ同定されていない)サブセットである。これらのBSCIと従来の5HT2aアゴニスト(例えばTCB2;McLean et al. (2006) J. Med. Chem. 49:5793-803)は共通の分子標的(5HT2a)を共有するが、この2群の物質は受容体への結合に対して異なる結果(Urban et al (2007) J. Pharmacol. Exp. Ther. 320:1-13)(機能的選択性と呼ばれる現象)を引き起こす。リセルグアミドは、ケモカイン誘導性白血球遊走を強力に阻害するが、幻覚誘発性(げっ歯類における特徴的な首ひねり)には影響を与えない;TCB2は幻覚誘発活性の特徴的な首ひねり(head twitch)を強力に誘発するが、ケモカイン誘導性白血球遊走には影響を与えない。
【0041】
BSCIが、白血球上で発現されることが公知である種々のGPCR神経受容体に対する特別なクラスの機能選択的アゴニストであるという、本明細書に開示された知識は、BSCI活性を有する化合物を予め構造情報無しで(すなわち、すでに記載されているBSCI化合物群に共通のファーマコフォアの情報無しで)同定することを可能にする。
【0042】
このような化合物は、本発明の方法に従って、例えば以下のように同定され得る。最初の工程では、白血球上で発現されることが公知の選択されたGPCR神経受容体との相互作用についてスクリーニングされる。次にBSCIは、従来の標的又はその受容体に対する古典的アゴニスト活性より強力な効力で、ケモカイン誘導性白血球遊走を阻害する能力により、その受容体に対する従来のアゴニストから区別される。場合により第4の工程で、これらのすでに開示されているものと構造的に無関係の新しいクラスのBSCIを定義する構造モチーフが同定され得る。
【0043】
第一の工程は、白血球上で発現されることが公知の選択されたGPCR神経受容体との相互作用についてのスクリーニングを含む。この定義を満足するために受容体は、(a)当該分野で公知の構造的及び操作的基準に従う、GPCRスーパーファミリーのメンバー;(b)機能的に有意なレベルで中枢神経系において発現される受容体;及び(c)白血球又は白血球のサブセット上で機能的に有意なレベルで発現されることが公知、でなければならない。本発明の実施のために選択される受容体がGPCRスーパーファミリーのメンバーであり、中枢神経系中で発現されるという要件は、BSCIが、この定義を満足するすべての3つの異なる受容体で機能的選択性リガンドであるという知見に基づく。この受容体が白血球上で発現されるという要件は、選択された物質がケモカイン誘導性白血球遊走を抑制してほしいという要求に基づく。
【0044】
本発明の方法の具体的な実施のために使用される1又は2以上の受容体の選択は、主に異なる白血球サブセットにわたる受容体の発現パターンに基づくことが企図される。例えばケモカインに応答して好中球遊走を選択的に抑制する物質が必要なら、(他の白血球と比較して)好中球上に選択的に発現されるGPCR神経受容体が選択され得る。同様に、肥満細胞遊走を選択的に抑制する物質が必要なら、肥満細胞上で発現されるGPCR神経受容体が選択され得る。あるいは、疾患中におけるGPCR神経受容体の発現パターンの変化は、特定の受容体を選択するための刺激となり得る;喘息の肺で好酸球上で選択的に発現される物質は、喘息で使用される抗炎症剤を同定するための理想的な標的となり得る。従って本発明の方法を適用する固体又は群の要求は、容易に、本発明の応用のための適切なGPCR神経受容体を選択するための基準を提供し得るが、本発明の使用は、使用されるGPCR神経受容体を選択するための特定のアプローチに決して限定されない。
【0045】
第2及び第3の工程は、(1)ケモカイン誘導性白血球遊走の阻害に関する、及び(2)選択される受容体に対する古典的アゴニスト活性に関する機能アッセイを行うことを含む。これらの2つの工程は両方とも、GPCR神経受容体により仲介されるBSCI活性について機能的選択性を有する物質を明確に定義するのに必要である。しかしこれらの2つの工程は、順番に又は基本的に同時に行うことができる。第1の工程で選択された特定の標的であるGPCR神経受容体を発現することが公知であるBSCI活性の機能アッセイのために、白血球集団を使用することは重要である。これができないと、この方法から擬陰性結果が得られるが、それは、BSCI活性に対する機能的選択性を有し得る全ての物質は、標的受容体を発現せず、且つこれと相互作用しない細胞のケモカイン誘導性遊走に対して作用を有しないためである。
【0046】
機能的選択性BSCI活性に関連する共通の構造モチーフを同定するために、場合により第4の工程が適用される。典型的にはこのような工程は、反復する構造要素が、BSCI活性を付与するのに必要なモチーフに一致する可能性が高くなるように、本発明の方法に従ってBSCIとして新たに同定された化合物間で構造要素を比較することを含み得る。次に、例えば関連する構造を有する種々の分子を合成して、モチーフの存在がBSCI活性の存在に関連していることを証明することにより、そのようなモチーフの重要性を実験的に証明することができる。その結果、たとえクラスの単一のメンバーのみが、白血球上で発現されるGPCR神経受容体との相互作用についてのスクリーニングを介して独自に同定されたとしても、本発明の方法はBSCI化合物のクラス全体を定義するために使用され得る(各クラスのメンバーは、構造が互いに関連している)。
【0047】
すなわち、本発明の方法に従って同定されるBSCI化合物は、新規であって構造が関連したBSCI群の合成と特徴づけのための構造的リードとして働くことが企図される。その結果、本発明の方法により同定される最初のヒットのそのような構造誘導体は、本発明の方法の適用により直接同定はされないが、それらの同定における重要な工程は本発明の方法に依存し、従ってこの方法での、関連するBSCI群の間接的同定が、本発明のこの態様に従って企図される。
【0048】
本発明の方法に従ってライブラリーがスクリーニングされる場合、ライブラリーの要素は個々の化合物(又は、少なくとも実質的に純粋な化合物、ここで標的化合物は存在する物質の少なくとも25%を占め、それより高レベルでは他の化合物は存在しない)でもよく、又はライブラリーの要素は化合物の混合物でもよい。要素が化合物の混合物である場合、混合物の成分は公知であり得るか又は(多くのコンビナトリアルライブラリー中のように)予測され得るか、又は未知(例えば、天然の生成物の単離体)であり得る。
【0049】
本発明の方法に従ってスクリーニングされるライブラリー要素の一部又はすべてが(組成が既知であるか又は未知である)混合物である場合、本発明の方法の第1工程後、白血球上で発現される選択されたGPCR神経受容体との相互作用の原因である混合物から特定の化合物が単離され、同定された後、第2工程(ケモカイン誘導性白血球遊走に対する化合物の作用の評価)が適用されることが企図される。そのような単離工程がその方法に含まれていない場合、その方法は混合物中の単一の化合物が(必要に応じて)BSCI活性を有する場合を、混合物中の1つの化合物が選択される神経受容体と相互作用し、第2の無関係の化合物がBSCI活性と関係の無い機構により白血球遊走を阻害した場合から区別できないであろう。
【0050】
例えば、ソマトスタチン−14(従来のsstr2アゴニストであるがBSCIではない)とアジ化ナトリウム(細胞の遊走を阻止するがBSCIではない細胞毒素)との混合物からなるライブラリー要素に本発明の方法を適用した場合、この混合物はsstr2と強い相互作用を示し、(存在するアジ化物の相対的濃度に依存して)sstr2での親和性から予測される効力より高い効力でケモカイン誘導性白血球遊走を阻害し得る。そのような混合物は、間違ってBSCIとして同定され得る。しかし、もし上記したようにsstr2(本例におけるソマトスタチン)との相互作用の原因である混合物の成分が、ケモカイン誘導性遊走に対する作用をアッセイする第2の工程を適用する前に混合物から単離された場合、この方法は混合物をBSCI活性が欠如しているとして正しく同定し得る。
【0051】
原則的には、本発明の方法の工程は任意の順序で適用できる(例えば、試験中の化合物の、ケモカイン誘導性遊走の阻害剤としての効力を測定し、次に白血球上で発現されたGPCR神経受容体との化合物の相互作用(もしあれば)の親和性を測定することが、原則的に可能である)が、そのような方法は本発明の好適な態様の利点が欠如している。これらの状況では、機能アッセイを使用してランダムライブラリーをスクリーニングすることができない(毒性で不溶性の化合物の存在のために、ならびにそのようなスクリーニングの資源の関係で)。従って好適な本発明では、白血球上で発現されたGPCR神経受容体との相互作用についてのスクリーニングが最初の工程として行われ、次にケモカイン誘導性白血球遊走の阻害のために機能アッセイが行われる。白血球上で発現されるGPCR神経受容体との相互作用を評価するためのスクリーニング(分子スクリーニングの例)は、ケモカイン誘導性白血球遊走に対する活性の測定(機能性スクリーニングの例)よりも、スクリーニングされる化合物当たりの必要な資源量がはるかに少ないため、この工程順序は、入手可能な資源の最適使用を反映する。
【0052】
本発明の方法の適用は、仮想のスクリーニング、ならびに実際の若しくは物理的スクリーニングを含むことが企図される。すなわち、本発明の方法に従って候補BSCIとして化合物を同定するのに必要な情報の一部(又はすべて)は、すでに利用可能である。例えば化合物のライブラリーは、本開示前に白血球上で発現された種々のGPCR神経受容体との相互作用についてすでにスクリーニングされており、結果が(例えば、コンピューターのハードディスク中に)保存されているかも知れない。従って、ライブラリーからの化合物のサブセットが、白血球中のケモカイン誘導性遊走の阻害剤として選択され、次にその効力が試験され、本発明の方法に従って候補BSCIとして同定され得る。そのような手順が、本明細書に開示された方法の適用であり、候補BSCIを同定するために開示された基準に依存することは明らかである。同様に、一連の化合物について、白血球上で発現される1又は2以上のGPCR神経受容体とのそれぞれの相互作用の親和性がすでに公知であり、かつ別にケモカイン誘導性白血球動員の阻害のための化合物の効力がすでに公知である場合、どの化合物が候補BSCIであるかを同定するための既存の情報の組合せは、新規の実験が必要ではない場合でも、本発明の方法の適用であり得る。
【0053】
特に、本発明の1又は2以上の工程が新規の実験を必要しない場合、その工程は任意の順序で実施できる(すなわち、本明細書に記載の好適な順序である必要は無い)。
【0054】
白血球上で発現されるGPCR神経受容体に対するアゴニストに帰因される多くの抗炎症活性が記載されている(例えば、Paran & Paran (2003) Curr. Opin. Invetig. Drugs 4:578-82; Sener et al. (2005) Peptides 26:493-9; Gonzales-Rey et al. (2007) Ann. Rheum. Dis. 66:582-8、及びその中の文献; Armani et al. (2007) J. Leukoc. Biol. 81:845-55; Cury et al. (2008) Dig. Dis. Sci. 53:2516-20を参照、これらは、白血球上で発現される単一のGPCR神経受容体であるsstr2についての報告に限定される;多くの他のGPCR神経受容体について同様の例が存在する)。従って本発明の新規性と進歩性は、白血球上で発現されるGPCR神経受容体を介して作用するリガンドが、臨床的に有用な抗炎症作用を有し得るという発見(単一の工程のみを有する発明を教示する発見、すなわち白血球上で発現されるGPCR神経受容体との相互作用についてスクリーニングして、有用となり得る抗炎症剤を同定すること)には無い。その代わりに本発明は、白血球上で発現されるGPCR神経受容体に対するすべてのアゴニストの一部(及び、おそらく非常に小さい比率)が、実質的に古典的アゴニスト活性(例えば、成長ホルモン分泌の抑制、血圧又は幻覚誘発性の調節)の代わりに、抗炎症活性を有するという予想外の知見に関する。
【0055】
この種の機能的選択性(異なるリガンドが、同じ受容体に結合しこれを活性化するが、顕著に異なる機能的結果を誘発する)は薬学の文献で公知である(総説については、Urban et al (2007) J. Pharmacol. Exp. Ther. 320:1-13を参照)が、確固たる文献は少ない。従って、ある受容体について又はある対の機能について、機能的選択性リガンドが存在するかどうかを推定することは不可能であろう。そのため我々の開示以前には、当業者は、GPCR神経受容体の活性化に関連する抗炎症活性は必ず古典的アゴニスト応答が伴うと妥当に結論付けており、抗炎症活性を有する新規薬剤の発見用の標的としてのGPCR神経受容体の有用性を限定していた。我々の開示は、機能的選択性は1つのGPCR神経受容体にのみ存在するのではなく、3つの異なるGPCR神経受容体で存在し、さらに、これらの各場合にほとんどまたは全くアゴニスト様活性の無い実質的な抗炎症活性を示すことがあり得ることを、明確に示す。我々の開示前に利用可能な情報では、当業者(本発明者らを含む)でさえ、この知見は予測されてはおらず、また可能であることも、もちろん確率が高いことも推定されなかった。
【0056】
本発明は、機能的選択性を示し、選択されたGPCR神経受容体に対する古典的アゴニスト活性よりもBSCI活性を促進する物質の明確な同定のための方法を提供する。本発明を有効に実施するために、この機能的選択性が如何に達成されるかの分子的詳細も、所望の結果を規定するリガンド間の構造特性も理解する必要は無い。本開示において提供される情報は、本来は等しく有望な候補のプールから有用と思われる機能的選択性物質を選択することを、当業者に教示するのに必要かつ十分である。しかし、本知見が予測できない程度を例示し、如何に機能的選択性を達成しなければならないかを例示することが有用かも知れない。
【0057】
リガンドが受容体に結合する時、リガンド分子と受容体タンパク質との間には複数の非共有結合的相互作用がある(特に限定されないが、ファンデアワールス力、水素結合、双極子相互作用、及び塩ブリッジを含む;図2参照)。これらの相互作用は、リガンドと受容体との相互作用の親和性(従って、他のタンパク質ではなく標的受容体への結合の選択性)と、リガンド結合に対するシグナル伝達応答の両方に関与する。天然のリガンドでは、これらの相互作用は受容体タンパク質のコンフォメーションの変化を引き起こす(受容体:リガンド複合体の最小エネルギーコンフォメーションは、分離した時の受容体とリガンドの最小エネルギーコンフォメーションとは顕著に異なる)。このコンフォメーションの変化は、シグナルを細胞の外へのリガンド結合から細胞内シグナル伝達カスケードに変換する。この変換は、直接的(内因性キナーゼ活性を有する受容体の場合のように)又は間接的(Gタンパク質結合受容体、例えばsstr2の場合のように)であり、コンフォメーションの変化は、1又は2以上の追加のタンパク質の会合を促進し、これはシグナルを変換する。受容体に結合し、天然のリガンドで見られる受容体中のコンフォメーション変化を再現する物質は、受容体に対するアゴニストであり得る(図2A)。これらの変化を誘導することなく受容体に結合する物質はアンタゴニストであり得る(これが天然のリガンドへの結合を阻止すると仮定して;図2B)。しかし、異常な状況下では第3の可能性が出てくる:物質が結合し、非常に異なるセットのコンフォメーション変化を誘導し、これはシグナルの性質に定量的ではなく定性的変化を引き起こし、次にこれが変換される(図2C及びD)。機能的選択性(及びある程度は、スーパーアゴニスト及びインバースアゴニスト)の薬理学的現象を指示するのは、この第3の異常なシナリオである。
【0058】
この分子的シナリオは、音楽に例えるとより理解し易いかも知れない。このモデルでは、ピアノ鍵盤は受容体の例えであり、ピアニストの手はリガンドである。通常の条件下では、手の作用(特定のコンフォメーション)は特定の和音を打つ。この「シグナル」は受容体に対するアゴニスト応答である。ピアノの蓋を閉じることは、鍵盤の位置でコンフォメーション変化を誘導することなくリガンドと受容体との相互作用を防止し、従って和音(アゴニストシグナル)は聞こえない。ピアノの蓋はアンタゴニストの類縁体である。ピアニストを鍵盤の弾き方を知らない小児(ライブラリーからランダムに選択される化合物の例え)に交換すると、ほとんどすべての場合に音楽の和音ではなく不可解な応答を出すであろう。しかしまれに、天才児に遭遇してピアニストとはまったく異なるきれいな和音を演奏するかも知れない。同じ受容体が機能的選択性で応答するようになった:異なるリガンドが同じ受容体から定性的に異なる応答を誘発する。
【0059】
上記より、同じ受容体に結合する異なるリガンドから、機能的結果を予測する方法が存在しないことは明らかであろう。実際、ある受容体にとって、どのシグナルが可能であるかを予測することさえ不可能である(どのように鍵盤を押しても、ピアノの鍵盤からバイオリンの音を出すことはできないであろうという意味で)。偶然の発見後に、開示された情報に基づいて実験的スクリーニングを行うことが、所望の機能的選択性を有する物質(例えば、本明細書に記載のBSCI)を発見する方法である。
【0060】
本発明が、単独のそれぞれの工程が当該分野で公知の方法である、3又は4以上の工程からなることは明らかである。例えば、天然のリガンドの既知の作用を再現するために、又は天然のリガンドの既知の作用を阻止するために、白血球上で発現されるGPCR神経受容体と相互作用する物質を捜すために、多くの場合に第1の工程が使用されていることは明らかである。同様に我々と他の研究者は、BSCI活性のために物質を特徴づけすることを目的として、ケモカイン誘導性白血球遊走の機能アッセイの使用を開示した。しかし本開示前には、全く異なる結果(ランダムライブラリー中のBSCI活性を有する化合物についてスクリーニングする能力;ほとんどのソマトスタチン受容体相互作用性物質はBSCIではなく、ランダムライブラリー中のほとんどの要素は、BSCI活性に関する機能アッセイに適合しないため、当該能力は、いずれかの工程を単独で使用して達成することはできない)を達成するために、これらの2つの方法を結びつけ、これらを連続して行うための可能な理論的根拠はなかった。しかし、BSCIが、白血球上で発現されるGPCR神経受容体と相互作用し、機能的選択性により作用するという情報(これは、本文書において初めて開示される)は、これらの2つの無関係のプロセスを組合せて、予想外の有用性を有する新規方法とすることにより、本発明を可能にする。本発明の方法のうまいたとえは、新しいペプチドの組成物である:ペプチドは(ほとんどすべて)同じアミノ酸構成単位からなり、同じ方法で結合される。しかし、非常に多くの組合せがあるため、特定の機能を有する特定の組合せ(新しいペプチド配列)の同定は進歩性があると見なされる。同様に、BSCIが作用できる非常に多くの可能な受容体があり(検索が、インビボで抗炎症活性を仲介できることが公知のものに限定されていても)、それぞれの受容体に対する分子スクリーニングを、BSCI活性についての機能スクリーニングと組合せようとすることは実現不可能であった。本発明は、ランダムライブラリーからBSCIを同定するための方法を提供することにより、最初は等しく可能なはずであった多くの他の組合せより予想外かつ特に優れた有用な組合せについての、この実現不可能な検索を克服する。
【0061】
本明細書に記載の方法は、他の種(特に限定されないが、マウス、ラット、イヌ、及び霊長類、ならびに非哺乳動物種、例えば鳥)のGPCR神経受容体のホモログに等しく適用し得る。GPCR神経受容体の非ヒトホモログが、ヒトでの使用を目的(例えば、抗炎症性薬物として)とした本発明に従って、化合物を同定するために使用されるなら、非ヒト受容体を使用して同定される物質をヒト受容体に対する機能的選択性についてチェックする追加の工程が必要とされ得る。しかし好ましくは、ヒト系でのBSCI活性がスクリーニング操作の所望の結果であるなら、本発明の方法は白血球上で発現されるヒトGPCR神経受容体に適用されるはずである。もちろん、所望の結果が方法の非ヒト系でのBSCI活性(例えば、獣医学的薬剤)であるなら、1又は2以上の標的種からのGPCR神経受容体類縁体を使用することが最も好適であり得る。
【0062】
本発明の方法は、受容体の断片が完全長受容体の天然のリガンド又は別のリガンドに結合する能力を保持するなら、受容体の不完全な断片を使用して適用することができることが企図される。典型的にはそのような機能性断片は、タンパク質配列の大部分を含有するが、リガンド結合に関与しない部分は(例えば、発現又は安定性を改善するために)、本発明の適用に影響を与えることなく欠失し得る。
【0063】
GPCR神経受容体への試験物質の結合が、間接的(受容体に対する、あらかじめ標識されていた別のリガンドとの結合の競合により)に又は直接的(試験物質自体が標識物を有する)に測定できることは明らかであろう。本発明の範囲は、試験物質と受容体との相互作用を検出するために使用される方法には決して限定されない。
【0064】
試験物質のBSCI活性を検出するために使用される機能アッセイが、インビトロ(例えば、Transwell(登録商標)遊走アッセイを使用して)又はインビボ(例えば、ケモカインの腹腔内注入後、応答する白血球動員の評価を使用して)でも実施できることは明らかであろう。さらに、BSCI活性の評価のために白血球動員(インビトロとインビボの両方)の代替マーカーを使用することができる。例えば白血球は、(活性化されている時は特に)サイトカインTNF−アルファを発現することが知られている。その結果、インビトロ又はインビボでのTNF−アルファの測定レベルは、白血球動員を評価するための代替法であり、従ってBSCI活性を測定するための方法である。本発明の範囲は決して、特定の試験化合物に関連するBSCI活性を検出するために使用される方法に限定されるものではない。
【0065】
試験物質の古典的アゴニスト活性を検出するために使用される機能アッセイが、インビトロ(例えば、ソマトスタチン受容体アゴニストについて下垂体細胞からのGH放出を測定することにより)又はインビボ(例えば、ソマトスタチン受容体アゴニストについて血清中のGHレベルを測定することにより)でも実施できることは明らかであろう。さらに、古典的sstr2アゴニスト活性を推定するために、代替マーカーを使用することができる。GHレベルの指標として、従って古典的ソマトスタチン受容体アゴニスト活性の推定値として、IGF−Iレベルが測定されるように、再度ソマトスタチン受容体アゴニストの例を使用して、GHはインスリン様増殖因子I(IGF−I)のレベルを制御する。本発明の範囲は決して、特定の試験化合物に関連する古典的アゴニスト活性を検出するために使用される方法に限定されるものではない。
【0066】
場合により、本発明の方法の前工程中に選択された化合物の1又は2以上の構造的特徴の存在に基づいて、新規な構造クラスのBSCIの同定を可能にするように、追加の工程を本発明の方法に付加してもよい。そのような構造モチーフの同定は、本発明の方法の適用を通じて、ライブラリー(ランダムライブラリーでも、選択される候補からなるものでも)からのヒットを同定するために、その性質が実験的又は理論的に探索された化合物のみよりはるかに広範囲のBSCIのアレイを可能とする。例えば、白血球上で発現されるGPCR神経受容体への結合についてのランダムライブラリーのスクリーニング(本発明の第1工程)後に、ケモカイン誘導性白血球遊走の阻害の測定(第2工程)、そして選択された受容体の「古典的」標的に対する効力の随時測定により、本発明の方法に従っていくつかの新しい化合物を選択し得る。2又は3以上のこれらの選択された化合物は、共通の構造モチーフ(例えば、3−置換シス−デカリン環系の存在)を共有し得る。従って任意の追加の工程では、3−置換シス−デカリン環系の存在が化合物に所望の性質を付与し、適切な予測によりさらなる3−置換シス−デカリン含有分子を合成でき、その少なくとも一部が所望の性質を有し得ることは明らかである。そのような状況下では、BSCI活性又は「ソマトタキシン」活性に関連する新しい構造モチーフの定義は、本発明の方法の適用を構成し得る。
【0067】
したがって本発明の方法は、少なくとも3つの工程を含む(白血球上で発現されるGPCR神経受容体との相互作用の測定、次にBSCI活性についての機能アッセイ、及び選択された受容体に対する「古典的」アゴニスト活性について機能アッセイ)。場合により最終工程を加えてもよい(BSCI活性に関連する構造モチーフを定義する)。この最終工程の利点は、アッセイベースのスクリーニングではなく構造相同性により、所望の性質を有する化合物を選択することを可能にすることである。そのような「ヒット・トゥー・リード(hit to lead)」最適化プログラムは当該分野で公知であり、ヒトの薬剤として使用するのに適した新しい物質の発見への本発明の方法の適用を促進する。しかし特定の適用に依存して、この任意の最終工程を実施する資源の問題が、同定される化合物の範囲が拡張される利点より大きいかも知れない。
【0068】
本明細書において用語「化合物(compound)」及び「物質(agent)」は一般に同義であると考えられ、従って文脈が特に明記しない場合は、相互に変換可能である。
【0069】
用語「広域スペクトル(broad-spectrum)ケモカイン阻害剤」又は「BSCI」は、通常は異なる受容体を介して遊走を誘導するシグナルを出すが、非ケモカイン化学誘引物質により誘導される細胞遊走にはほとんど又は全く影響を与えない広範囲のケモカインに応答して、細胞遊走を阻害できる化合物又は物質を意味する。これは操作的な定義であり、完全に異なる分子機構の結果として異なる化合物若しくは物質がBSCIとなり得ることは明らかである。しかし本発明は、重要でかつ広範に研究されたBSCIの下位区分である、本発明で同定された、sstr2又は関連する受容体を介して作用するBSCIの同定にのみ関連する。
【0070】
用語「BSCI活性」は、白血球上で発現されるGPCR神経受容体で作用するいくつかのアゴニストが、インビトロ若しくはインビボでケモカイン誘導性細胞遊走に、ならびに関連するインビボでの免疫学的変化(例えば、Tヘルパー(CD4+)細胞サイトカイン分泌プロフィールの再調整)に対して有する作用を示す。
【0071】
用語「古典的(classical)アゴニスト活性」は、あるGPCR神経受容体を充分なレベルで発現する細胞が、その受容体についての任意の公知のアゴニスト化合物(当該受容体の天然の1又は2以上のリガンドを含む)に曝露された時、おそらく異なる細胞種で、起きることが公知である一群の細胞応答を示す。例えば、sstr2受容体について古典的アゴニスト活性の例は、下垂体細胞によるGH放出の抑制、膵臓細胞によるインスリン放出の抑制であろう(Olias et al. (2004) J. Neurochem. 89:1057)。しかし種々の細胞に作用するsstr2アゴニストの他の広範囲の作用も記載されており、これら(上記した「BSCI活性」を除いて)もまた、古典的sstr2アゴニスト活性を含む(Olias et al. (2004) J. Neurochem. 89:1057)。すなわち古典的アゴニスト活性は、BSCI活性を除く選択されたGPCR神経受容体に対するアゴニストのすべての公知の活性として定義され、従ってアゴニストのすべての公知の活性は本定義により包含される。
【0072】
用語「sstr2」は、ヒト染色体17q24上のsstr2遺伝子(HGNC ID:11331;配列番号1に示される完全な遺伝子配列)によりコードされ、後述の配列表中の配列番号2で示されるアミノ酸配列をコードする、ヒトの2型ソマトスタチン受容体を示す。
【0073】
特に明記しない場合、用語sstr2は、配列番号1に示す遺伝子座の転写から得られる、オルタナティブ・スプライシングされたか又は示差的プロセシングされた任意のmRNAの生成物を包含することを企図する。疑問を避けるため、用語sstr2は、タンパク質コード枠中に位置する単一のイントロンでsstr2 mRNAの選択的スプライシングから得られるsstr2aタンパク質とsstr2bタンパク質の片方又は両方を包含する。他の種の類縁体(analog)又は相同体(homolog)もまた、特定の文脈でナンセンスでない限りは、この用語に包含される。
【0074】
用語「ソマトスタチン受容体」は、ソマトスタチンが高親和性(典型的には1μM又はそれ以下)で結合する任意の受容体(現在公知であっても公知でなくても)を示す。疑問を避けるため、これは、sstr1、sstr2、sstr3、sstr4、及びsstr5を含む。他の種の類縁体又は相同体もまた、特定の文脈でナンセンスでない限りは、この用語に包含される。
【0075】
用語「ソマトスタチン受容体」はさらに、ヒトsstr2(上記した)のアミノ酸配列と、少なくとも50%の同一性、例えば少なくとも55%、60%、65%、70%、75%、80%、85%、90%、95%の同一性、又は少なくとも96%、97%、98%、さらには99%の同一性を有するアミノ酸配列を有するポリペプチドを包含する。ポリペプチドは好ましくは、上記のソマトスタチン受容体活性を有する。
【0076】
アミノ酸配列間の配列同一性は、配列の整列を比較することにより決定することができる。比較される配列中の同等の位置が同じアミノ酸で占められる時、分子はその位置で同一である。整列を同一性のパーセントとしてスコア化することは、比較される配列が共有する位置での同一のアミノ酸又は塩基の数の関数である。配列を比較する時、最適な整列は1又は2以上の配列へギャップを導入して、配列中の可能な挿入や欠失を考慮することが必要な場合がある。配列比較法は、比較される配列中の同一分子の同じ数について、2つの比較配列間のより高い関連を反映するできるだけ少ないギャップの配列整列が、多くのギャップを有するものより、高いスコアを有するように、ギャップペナルティを使用する。最大同一性パーセントの計算は、ギャップペナルティを考慮して、最適整列を作成することを含む。
【0077】
配列比較を行うための適切なコンピュータープログラムは、商業部門や公共部門で入手できる。例には、MatGat (Campanella et al., 2003, BMC Bioinformatics 4: 29; http://bitincka.com/ledion/matgatから入手できるプログラム)、Gap (Needleman & Wunsch, 1970, J. Mol. Biol. 48: 443-453)、FASTA (Altschul et al., 1990, J. Mol. Biol. 215: 403-410; http://www.ebi.ac.uk/fastaから入手できるプログラム)、Clustal W 2.0 and X 2.0 (Larkin et al., 2007, Bioinformatics 23: 2947-2948; http://www.ebi.ac.uk/tools/clustalw2から入手できるプログラム)、及び EMBOSS Pairwise Alignment Algorithms (Needleman & Wunsch, 1970, supra; Kruskal, 1983, In: Time warps, string edits and macromolecules: the theory and practice of sequence comparison, Sankoff & Kruskal (eds), pp 1-44, Addison Wesley; http://www.ebi.ac.uk/tools/emboss/alignから入手できるプログラム)がある。すべてのプログラムは、デフォールト・パラメータを使用して使用される。
【0078】
例えば、配列比較は、EMBOSS Pairwise Alignment Algorithmsの「ニードル」法を使用して行われ、これは、長さ全体にわたって考慮される時の2つの配列の最適整列(ギャップを含む)を決定し、同一性パーセントスコアを提供する。アミノ酸配列比較のデフォールト・パラメータ(「タンパク質分子」オプション)は、ギャップエクステンションペナルティ:0.5、ギャップオープニングペナルティ:10.0、マトリックス:Blosum 62である。ヌクレオチド配列比較のデフォールト・パラメータ(「DNA分子」オプション)は、ギャップエクステンションペナルティ:0.5、ギャップオープニングペナルティ:10.0、マトリックス:DNAfullである。
【0079】
用語「ソマトスタチン」は、ソマトスタチンをコードする遺伝子の任意の産物(ヒトにおいては染色体3q28上のsst遺伝子(HGNC ID:11329)によりコードされる)を示し、そしてそれは当該遺伝子産物の制御されたタンパク質分解により生成される断片を含む。特に明記しない場合、用語ソマトスタチン(又はsst)は、生物学的機能が割り当てられたタンパク質分解により生成される任意の断片(特に限定されないが、sst−14及びsst−28を含む)を包含することが意図される。他の種の類縁体又は相同体もまた、特定の文脈でナンセンスでない限りは、この用語に包含される。
【0080】
用語「白血球」は、血液中の任意の又はすべての白血球集団、ならびに骨髄又は2次的リンパ系臓器(例えば脾臓)中のこれらの細胞の前駆体、及び組織中のこれらの細胞の後代(例えば、破骨細胞、クプファー細胞、小神経膠細胞、肥満細胞、樹状細胞など)を示す。白血球の例には、単球、リンパ球(Bリンパ球とTリンパ球の両方、キラーT細胞、ヘルパーT細胞、制御T細胞、及びナチュラルキラー細胞を含む)、及び顆粒球(好中球、好塩基球、好酸球)がある。
【0081】
用語「GPCR神経受容体」は、中枢神経系の細胞(ニューロン、星状細胞、及び小神経膠細胞を含む)上で機能的に関連するレベルで発現されるGタンパク質結合受容体(GPCR)のスーパーファミリーの任意のメンバーを包含する。Gタンパク質結合受容体は、充分定義されたクラスであり、受容体が、その配列と構造に基づいて及びそのシグナル伝達機構に基づいて、GPCRスーパーファミリーのメンバーであるかどうかを明確に決定するための方法が当該分野に存在する。例えば、GPCRは典型的には、7つの膜貫通ドメイン(個々には12〜20個の疎水性アミノ酸残基の区域で、いずれかの端に1又は2以上の荷電残基を有する、同定可能な特徴を有する)の存在により特徴づけされる。GPCRはまた、GTP結合タンパク質(Gタンパク質を呼ぶ)の群のメンバーと相互作用する能力、及びこれを活性化する能力により特徴づけされる。GPCR神経受容体の例には、特に限定されないが以下の群が含まれる:アドレナリン受容体(α2−アドレナリン受容体を含む)、セロトニン受容体(5HT2a受容体を含む)、ソマトスタチン受容体(sstr2を含む)、ドーパミン受容体、ムスカリン性アセチルコリン受容体、物質P/NPY受容体、バソプレシン受容体、甲状腺ホルモン受容体、カンナビノイド受容体、コレシストキニン受容体、オピオイド受容体、アデノシン受容体、グルカゴン受容体、PACAP/VIP受容体、メラトニン受容体、ブラジキニン受容体、カルシトニン受容体、及びメラノコルチン受容体を含む。
【0082】
用語「白血球上で発現されるGPCR神経受容体」は、白血球又は白血球の少なくとも1つのサブセットにより機能的に有意なレベルで受容体が発現されている、すべてのGPCR神経受容体のサブセットを示す。すべての白血球サブセット上で発現されるGPCRの完全な相補体は現在未知であるが、この用語は、これらが白血球の少なくとも1つのサブセットで発現されることが公知であるため、本発明での使用のために選択されるすべてのGPCR神経受容体、ならびに白血球中の発現パターンのスクリーニングに基づいて選択されたGPCR神経受容体(これらは、以前は白血球上で発現されることは知られていなかった)を明確に包含するために使用される。白血球上に存在すると記載されておらず、白血球に基づく発現を調べて選択されたのではないGPCR神経受容体は、本定義に包含され得ない。
【0083】
用語「効力(potency)」は、特定の作用を有するのに必要な特定の物質の濃度を説明するために使用される。典型的には効力は、試験中のエンドポイントを50%阻害するのに必要な化合物の濃度とされる(ED50又はIC50と呼ぶ)。従って低濃度でのED50又はIC50は、高い効力を有する化合物を示す。効力は、試験した濃度で試験中のエンドポイントに対する化合物の最大作用を示す「出力(power)」(後述)とは完全に異なる。化合物は、高効力であるが低い出力を有し(例えば、ED50が0.05nMでケモカイン誘導性遊走に対して作用するが、非常に高濃度でも遊走を40%しか阻害しない)、又は低効力で高い出力を有する(例えば、ED50が10mMであるが、100μMより高い濃度でエンドポイントを100%阻害する)ことがある。
【0084】
用語「出力(power)」は薬理学的アッセイに適用される時、そのアッセイでその物質の最適濃度を選択することにより得られる試験中のエンドポイントに対する、特定の物質の最大作用を説明するために使用される。最適濃度は、典型的には試験される最も高い濃度であるが、ある例(例えば、ベル型の用量応答が一般的に観察される場合、例えばケモカインに対する化学走化性応答)では、低濃度が試験中のエンドポイントに対して最大作用を有する場合がある。出力は効力とは関係無く、低効力を有する化合物が高い出力を有していたり、逆の場合もある。
【0085】
用語「約」は、考慮される値の周りの間隔を示す。本特許出願において使用されるように、「約X]は、(X−Xの10%)〜(X+Xの10%)、好ましくは、(X−Xの5%)〜(X+Xの5%)の間隔である。
【0086】
本説明中の数値範囲の使用は、その範囲内のすべての個々の整数と、ある範囲の最も広い範囲内の上限数と下限数とのすべての組合せを、本発明の範囲内に含めることを明確に企図する。
【0087】
本明細書において、用語「含んでなる」は、含むことと、からなることの両方を意味するとして読まれるべきである。逆に発明が、ある化合物を「活性成分として含む医薬組成物」という用語は、この用語が、他の活性成分が存在してもよい組成物と、定義した1つの活性成分のみからなる組成物との両方を包含することを意味する。
【0088】
以下の略語は、使用される場合、文脈から適切であるとしてポリペプチド又は他のアミド含有化合物の一部として単離されるものであっても、20の天然に存在するタンパク質新生アミノ酸を意味する。
【0089】
【表1】
【0090】
特に明記しない場合は、ここで使用されるすべての技術的・科学的用語は、本発明が関連する分野の当業者により通常理解されるものと同じ意味を有する。
【0091】
本発明の具体的な非限定例や追加の特徴は、以下の図面を参照して説明される。
【0092】
いくつかの図をさらに詳細に説明する。
図1は、ペプチド3(示していない)から得られる配列の環状レトロインベルソ(retroinverso)類縁体であるNR58−3.14.3(「A」)を示す。トリペプチドモチーフWxQは主要なファーマコフォアとして同定され、WVQ(「B」)のようなトリペプチドはBSCI活性を有する(低効力であるが)。ヨヒンバン(Yohinban)−16−アミド(「C」)とNR58,4(「D」)は、WxQトリペプチドの構造類縁体として同定された。次にBN83253(「E」)、BN83250(「F」)、及び(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オン(「G」)のようなラクタムが、NR58,4のようなイミドの安定な類縁体として同定された。リセルグ酸のアスパラギン及びグルタミンアミド(例えばL−II)は、ヨヒンバン−16−アミドの構造相同体として同定された。
【0093】
図2ではパネルAは、天然のリガンドが受容体と相互作用し、複数の応答(矢印)を発生することを示し、これは細胞内シグナル又は機能的結果であると解釈される。パネルBでは、アンタゴニストは受容体に結合し、天然のリガンドが結合することを妨害するが、活性なシグナル伝達を誘導するのに必要な受容体部分とは反応しない。パネルCでは、機能選択性アゴニストが、ある応答(太い矢印)を他の応答(細い矢印)より優先的に活性化するように、受容体に結合する。パネルDでは、反対の機能的選択性を有する異なるアゴニストが例示される。
【0094】
図3を参照して、転写体A(「T#1」)ではイントロンはスプライシングにより除去されておらず、翻訳停止コドンは、末端切断型タンパク質(sstr2aと呼ぶ)を与える。転写体B(「T#2」)では、イントロンはスプライシングにより除去されており、翻訳は第2エクソン及び代替翻訳停止コードに進み、sstr2bタンパク質(これは短いC末端テイルのみでsstr2aとは異なる)を与える。sstr2転写体を明確に同定するために使用されるPCRプライマーの位置を、sstr2プライマーセットについては(a)として、sstr2Aプライマーセットについては(b)として示す。この2つの異なる転写体から得られるアンプリコンのサイズは、398bp(T#1、sstr2プライマーセット)、87bp(T#1、sstr2Aプライマーセット)、57bp(T#2、sstr2プライマーセット)、及びバンド無し(T#2 sstr2Aプライマーセット)である。
【0095】
図4では、アンタゴニスト(黒い線、小さい四角)BN83253の不在が、MCP−1誘導性細胞遊走を用量依存的に阻害することがわかる。アンタゴニストの濃度が上昇すると(薄い灰色、大きい記号)、BN832653のBSCI活性が次第に失われる。
【0096】
図5では、アンタゴニスト(黒い線、小さい四角)の非存在下では、バレロラクタムはMCP−1誘導性細胞遊走を用量依存的に阻害する。アンタゴニストの濃度が上昇すると(薄い灰色、大きい記号)、BN832653のBSCI活性が次第に失われる。このバレロラクタムBSCIはBIM83253より30〜50倍強力であり、その結果、NIM83253からBSCI活性を排除するには、より高濃度の競合的アンタゴニスト(BIM23627)が必要であることに注意されたい(このグラフを図4と比較)。
【0097】
図6と7の両方では、BSCIは、内毒素に応答したTNF−アルファ産生を用量依存的に阻害した(実線、四角記号)。BSCIの見かけの効力は、BIM23627の同時投与により低下した(点線、三角記号)。
【0098】
図8ではデキサメタゾンは、内毒素に応答したTNF−アルファ産生を用量依存的に阻害した(実線、四角記号)。BIM23627(点線、三角記号)の同時投与は、デキサメタゾンの見かけの効力に対して影響はなかった。
【0099】
図9では、両方のタイプの細胞について、2つの異なるプライマーセットを使用し(図3参照)、1つはsstr2a転写体からアンプリコン(「A」)のみを産生し、他はsstr2aとsstr2b(ここで「sstr2」と記載する)から異なるサイズのアンプリコン(「A*」と「B」)を生成する。図9では、M1は398bpであり、M2は87bpであり、M3は57bpである。このデータは、THP−1細胞中のsstr2aの発現を確認し、sstr2bの存在については何の証拠も示さない。
【0100】
図12では,値は各薬剤濃度での三回測定値(±SD)である。過剰の非標識BN83250(「A」、10nM 3H−BN83250)との競合を、非特異結合の対照として示し、特異的結合を示す上の点線を決定する。陰性対照(「B」、過剰のコールド)が提供され、これは非特異結合を示す下の点線を決定し、一方、BN83250の結合に対するsstr2アンタゴニストBIM23627(「C」;100μM)の影響を比較のために示す。
【0101】
図13では、ポリリンカー中の「*」は、XbaI部位の上流にATGがあることを示す。細菌とJurkat細胞中のプラスミドの存在について選択するために使用される、それぞれ「アンピシリン」と「ネオマイシン」抵抗性コーディング読み取り枠に注目されたい。cDNA挿入体は真核生物系ではCMVプロモーター(「P CMV」)下で発現され、標準的なポリアデニル化配列(「BGH pA」)を有する。
【0102】
図14を参照して(A)は、BN83250がJurkat 3DBのSDF−1α誘導性遊走を阻害(THP−1細胞で観察されるものより低効力であるが)するが、親Jurkat T細胞株のSDF−1α誘導性遊走にはほとんどまたは全く影響が無いことを示す。各細胞株について、フローサイトメトリーにより測定されるsstr2Aの細胞表面染色のレベルを、頻度ヒストグラム(B)〜(D)に示す。
【0103】
図16では、遊走に影響を与えない化合物は効力10μMでプロットされ、従ってこれらがグラフに現れる。BN83250(白三角)、(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オン(白四角)、ソマトスタチン(白菱形)、及びL−779,976(白丸)は、グラフから特定することができる。すべての既知の「BSCI型」sstr2アゴニストが右下に有り、すべての既知の「古典的」sstr2アゴニストが左上にあることを強調するために、ED50とKaが等しい同一性の線が示される。
【実施例】
【0104】
実施例1:sstr2アンタゴニストはインビトロでBSCI活性を阻止する
既知のペプチドとアミノラクタムBSCI化合物がsstr2受容体を介して作用することを証明するために、我々は一連の特異的sstr2アンタゴニストを使用した。種々の構造が異なる化合物のBSCI作用が、特定の受容体に対する一連のアンタゴニストにより阻止されるなら、BSCI活性が、使用したアンタゴニストに阻止される受容体への結合に依存すると結論する。
【0105】
ここで我々は、ケモカインに応答して遊走を阻止する種々のBSCIの能力(BSCIの標準活性)が、2型ソマトスタチン受容体(sstr2)に特異的ないくつかのアンタゴニストによりインビトロで阻害されることを証明する。
【0106】
方法:
これらの実験ではヒト骨髄単球性細胞株THP−1を使用したが、ヒト末梢血単球でも同様の結果が得られている。白血球遊走は、Neuroprobe(Gaithersburg, MD, USA)の96ウェルフォーマットのTranswell(登録商標)アッセイシステムを使用して測定した。原理的にこのアッセイは、多孔性膜により分離された2つのチャンバーからなる。化学誘引物質を下の区画に入れ、細胞を上の区画に入れる。37℃である時間インキュベーション後、細胞を化学誘引物質の方に動かすと、下の区画の細胞数は化学誘引物質の活性に比例する(一連の対照に対して)。
【0107】
種々のアッセイ条件が、ケモカイン誘導性白血球遊走(ならびに、sstr2アンタゴニストによるBSCIの又はBSCI活性の影響)を実証するために許容されるが、ここでは具体例が提供される。この方法は、基本的に既に記載されているように行った(Reckless & Grainger (1999) Biochem. J. 340:803-11)。
【0108】
材料
Transwell(登録商標)アッセイシステムは、Neuroprobe(Gaithersburg, MD, USA)により製造された。
【0109】
使用したプレートは、ChemoTx(商標)プレート(Neuroprobe 101-8)と30μlの透明プレート(Neuroprobe MP30)である。
【0110】
ゲイの平衡塩類溶液(Geys' Balanced Salt Solution)を、Sigma(Sigma G-9779)から購入した。
【0111】
脂肪酸を含まないBSAを、Sigma(Sigma A-8806)から購入した。
【0112】
MTT、すなわち3−(4,5−ジメチルチアゾール−2−イル)−2,5−ジフェニルテトラゾリウムブロミドを、Sigma(Sigma M-5655)から購入した。
【0113】
フェノールレッドを含まないRPMI1640を、Sigma(Sigma R-8755)から購入した。
【0114】
THP−1細胞株(欧州セルカルチャーコレクション(European Cell culture Collection))を、白血球細胞集団として使用した。
【0115】
試験プロトコール
まず、上の区画に入れる細胞懸濁物を調製した。THP−1細胞を遠心分離(770×g、4分)によりペレット化し、1mg/ml BSA含有ゲイの平衡塩類溶液(GBSS+BSA)で洗浄した。次にこの洗浄を繰り返し、細胞を再度ペレット化した後、例えば標準的血球計算器を使用して計数するために少量のGBSS+BSAに再懸濁した。
【0116】
次に存在する細胞の数に応じてGBSS+BSAの容量を調整し、細胞の最終密度が1mlのGBSS+BSA当たり4.45×106細胞になるようにした。これにより、プレートの上部チャンバーに入れる各25μlの溶液当たり100,000個のTHP−1細胞が確保された。
【0117】
MCP−1誘導性遊走に対するBSCI活性の逆転を証明するために、3ロットの細胞を調製する必要があった。4.45×106細胞/mlのTHP−1細胞の懸濁物を、3つの瓶に分割した。1つの瓶にBSCIを適切な最終濃度で、適切なビヒクル中に加えた(例えば、(S)−3−(2’,2’−ジメチルプロパノイルアミノ)−テトラヒドロピリジン−2−オンを、GBSS中のストック溶液から1nMで加えた)。2つ目の瓶にBSCIを最初の瓶と同じ濃度で加えたが、さらにsstr2アンタゴニストも加えた(例えば、BIM23454[Tulipano et al. (2002) 143:1218-24]又はBIM23627をGBSS中のストック溶液から66nMで加えた)。3番目の瓶に、適宜GBSS+BSA+ビヒクルを加え、3つの瓶の中の容量が等しく対照として作用するようにした。
【0118】
次に、下の区画に入れる化学誘引物質溶液を調製した。MCP−1をGBSS+BSAで希釈して、最終濃度を25ng/ml(すべて添加後)とした。Frow et al (Med. Res. Rev. 24:276-98) が推奨するように、これはこのシステムで最大遊走を誘導するMCP−1の濃度である。この溶液を、細胞懸濁物と同様に3つの瓶に分割した。1つの瓶に、BSCIを細胞懸濁物に加えたものと同じ最終濃度になるように加え、2番目の瓶に、BSCIとsstr2アンタゴニストを両方とも細胞懸濁物に加えたものと同じ濃度になるように加え、残りの瓶にはGBSS+BSA+ビヒクルを適宜加えて、3つの瓶の中の容量を等しくした。下の区画の溶液中のMCP−1の最終濃度と上の区画の細胞の最終濃度とを確立する時、BSCIとsstr2アンタゴニストを添加するために、液体の容量を考慮する必要があることに注意されたい。
【0119】
下のウェル用の化学誘引物質溶液と上のチャンバー用の細胞溶液をいったん調製した後、遊走チャンバーを組み立てた。29μlの適切な化学誘引物質溶液をチャンバーの下のウェルに入れた。各条件を少なくとも3回測定で行った。すべての下のチャンバーを充填した後、多孔性膜を製造業者の説明書に従ってチャンバーに適用した。最後に25μlの適切な細胞溶液を上の各チャンバーに加えた。蒸発を防ぐために、装置全体の上にプラスチックの蓋をかぶせた。
【0120】
組み立てたチャンバーを37℃、5%CO2で2時間インキュベートした。GBSS+BSA中の細胞懸濁物も、同じ条件下にて試験管中でインキュベートした:これらの細胞は、各条件下で下のチャンバーに遊走した細胞の数を測定するための標準曲線を構築するために使用した。
【0121】
インキュベーションの最後に、上のチャンバーから液体の細胞懸濁物を静かに取り出し、20μlの氷冷した20mM EDTAのPBS液を加え、装置を4℃で15分インキュベートした。この操作により、膜の下側に付着していたすべての細胞が下のチャンバーに落ちた。
【0122】
インキュベーション後、フィルターを注意深くGBSS+BSAでフラッシュしてEDTAを洗い流し、次にフィルターを取り出した。
【0123】
次に、各条件下で下のチャンバーに遊走した細胞の数は、多くの方法(直接計数、蛍光性若しくは放射活性マーカー又は生体染色色素の使用による標識を含む)により測定することができる。典型的には我々は、生体染色色素MTTを使用した。3μlのストックMTTを各ウェルに加え、次にプレートを37℃で1〜2時間インキュベートし、その間、細胞内の脱水素酵素は可溶性MTTを不溶性の青いホルマザン生成物に変換し、これは分光光学的に定量することができる。
【0124】
並行して、8点標準曲線を準備した。上の各チャンバーに加えられた細胞の数(100,000)で開始して、GBSS+BSAで2倍連続希釈を行い、プレートに細胞を25μl加え、3μlのMTTストック溶液を加えた。標準曲線プレートを遊走プレートとともにインキュベートした。このインキュベーションの最後に、沈殿したホルマザン生成物を乱さないように、下のチャンバーから注意深く液体を取り出した。短時間風乾後、下の各チャンバーに20μlのDMSOを加えて青い色素を可溶化し、96ウェルプレートリーダーを使用して595nmの吸光度を測定した。次に各ウェルの吸光度を標準曲線に内挿して、下の各チャンバー中の細胞数を推定した。
【0125】
MCP−1刺激遊走は、MCP−1が25ng/mlで存在した下の区画に到達した平均細胞数から、MCP−1を加えなかったウェル中の下の区画に到達した平均細胞数を引いて、計算した。
【0126】
種々の濃度のBSCIの存在下又は非存在下で起きたMCP−1誘導性遊走を比較することにより、BSCIの影響を計算した。典型的には遊走の阻害を、BSCIの存在下により阻止された総MCP−1誘導性遊走のパーセントとして表わした。ほとんどの化合物について、種々の異なる化合物濃度(典型的には、1nM〜1μM、又はあまり活性ではない化合物の場合それ以上)で起きるMCP−1誘導性遊走の阻害を測定することにより、用量応答グラフを構築した。次に各化合物の阻害活性を、MCP−1誘導性遊走を50%だけ低下させるのに必要な化合物の濃度として表わした(ED50濃度)。sstr2アンタゴニストとBSCIとの存在下で起きるMCP−1誘導性遊走を、BSCI単独(同じ濃度のBSCI化合物)の存在下で起きるMCP−1誘導性遊走と比較することにより、sstr2アンタゴニストの影響を計算した。sstr2アンタゴニストがBSCI化合物のBSCI活性を阻害したなら、BSCI単独で処理したウェルより、sstr2アンタゴニストとBSCIの両方で処理したウェルで、より多くの遊走が起きていることが予測された。
【0127】
結果:MCP−1(25ng/ml)は、THP−1細胞の実質的な遊走を誘導した(MCP−1の無い対照ウェル中よりMCP−1の存在下で、25,000〜40,000個多い細胞が下のチャンバーに遊走する)。この遊走は、広範囲のBSCIにより用量依存的に阻害された。図4では、MCP−1の阻害が、上昇する濃度のBSCI BN83253(N−オレオイル−3−アミノカプロラクタム;Fox et al. (2005) J. Med. Chem. 48:867-74)に応答して、THP−1細胞遊走を誘導したことを示す(小さい四角)。図5では、上昇する濃度のバレロラクタムBSCI (S)−3−(2’,2’−ジメチルプロピオニルアミノ)−テトラヒドロピリジン−2−オンに応答したMCP−1誘導性THP−1細胞遊走の阻害を示す(小さい四角)。両方の場合に、600nMのsstr2アンタゴニストBIM23627の存在(sstr2でこの化合物のKiより約100倍高い;Tulipano et al. (2002) 143:1218-24)は、BSCI介在阻害を完全に防止した(すなわち、加えたBSCIの濃度に無関係に、高用量のBIM23627の存在により、MCP−1誘導性遊走は対照レベルまで回復される;図4と図5の三角)。
【0128】
低用量のBIM23627(例えば、この受容体で作用するこの化合物についてのKiに近い6nM;図4と図5の中程度の大きさの菱形記号)では、BSCI作用は低用量のBSCIで阻害されたが、高用量THP−1では、誘導性細胞遊走はなお阻害された。これは、同じ部位への結合に対するBSCIとsstr2アンタゴニストとの競合に一致する。低BSCI濃度では、sstr2アンタゴニストが結合し、遊走の阻害は見られない。BSCI濃度が上昇すると、徐々にsstr2がBSCIにより占められ、sstr2により占められることはなく、BSCI活性が現れる。
【0129】
非常に低用量のBIM23627(例えば、sstr2で作用するこの化合物についてのKiより約100倍低い0.06nM;図4と図5の小さい菱形)では、BSCI作用はすべての濃度のBSCI化合物で見られたが、低濃度BSCIでは程度の小さい逆転が見られ、同じ部位へ結合するこれらの物質間の競合と再度一致する。
【0130】
これらの結果は、異なるBSCI((S)−3−(アダマンタン−1−カルボニルアミノ)−カプロラクタム(WO2006/016152参照)とペプチド3(Reckless & Grainger (1999) Biochem. J. 340:803-11)を含む)を使用して、何度も再現されている。さらに、異なるsstr2アンタゴニスト(BIM 23454; Tulipano et al. (2002) 143:1218-24)を使用しても、同じ結果が得られている。従って我々は、ペプチドからアミノカプロラクタム、そして第3世代アミノバレロラクタムにわたる非常に異なる構造クラスの広範囲のBSCI化合物について、BSCI活性はsstr2(又は、使用した両方のアンタゴニストにより阻害される密接に関連する受容体)により仲介されると結論付けた。
【0131】
他のソマトスタチン受容体について選択性のアンタゴニストを使用して、同様の実験も行われている。例えば、sstr3とsstr5について選択性のアンタゴニストであるBIM23627(Ipsen(Paris, France)から供与された)は、標的受容体のKiより10倍超高い濃度のアンタゴニストでさえ、BSCI活性の逆転をほとんど起こさない(<10%)。
【0132】
実施例2:sstr2アンタゴニストはインビボでBSCI活性を阻止する
インビトロのBSCI活性はsstr2(又は、BIM23627とBIM23454に阻害される密接に関連した受容体)に仲介される。次に我々は、インビボのBSCIの抗炎症活性もsstr2依存性であることの証明を試みた。幸いにBIM23627はインビボで活性であることが証明されており、急性の高用量投与後の少なくとも短時間で、古典的sstr2介在シグナルを阻止することができる(Tulipano et al. (2002) 143:1218-24)。従って、急性炎症のモデルを採用するなら、BSCIの抗炎症作用が、同時のsstr2阻止により阻害されたかどうかを決定することができるであろう。
【0133】
マウスの致死的ではない内毒素血症モデルが我々の実験室で詳細に特徴づけされているため、我々はこのモデルを選択した(例えば、Fox et al. (2002) J. Med. Chem. 45:360-70; Fox et al. (2005) J. Med. Chem. 48:867-74を参照)。細菌性リポ多糖(LPS)の単回腹腔内注射に応答した全身性炎症の程度は、抗原刺激投与後2時間目に血清TNF−アルファを測定することにより推定することができる。さらにこのモデルでは、BSCIが炎症を強力に阻害することが知られている(Fox et al. (2005) J. Med. Chem. 48:867-74)。
【0134】
方法:マウス(CD1系統;成体のオス;1群6匹)を、適切な経路でBSCI(適切なビヒクル中)又はビヒクル単独で処理した。例えば皮下注射(これは経口バイオアベイラビリティが悪いため)によりBN83250(図1参照)を投与し、一方(S)−3−(2’,2’−ジメチルプロピオニルアミノ)−テトラヒドロピリジン−2−オンを強制経口投与した。同時に各BSCI濃度で1セットのマウスに、4mg/動物(120mg/kg)のBIM23627を皮下注射により投与した(ラットで、古典的sstr2介在シグナル伝達(例えばGH放出)を抑制することが知られている処理条件;Tulipano et al. (2002) 143:1218-24)。投与の30分〜1時間後(BSCIの最大血漿濃度が達成される時間)に、750μgのLPS(Sigma; L4130 lot#024K4077; 900,000 EU/mg)の腹腔内注射により炎症性抗原刺激が誘発された。
【0135】
さらに2時間後、CO2窒息により動物を屠殺し、心臓穿刺により血液を採取した。従来法で血清を調製し、血清中のTNF−アルファのレベルを製造業者の説明書に従ってELISA(R&D Systems; Murine Quantikine code# MTA00)を使用して測定した。
【0136】
結果:解析後、各群の動物の平均を報告した(残りの点から3標準偏差より高いか又は低いデータ点は除外した)。予想されるようにLPS抗原刺激は、3,000ng/mlを超える平均に達するTNF−アルファのレベルで大規模な全身性炎症を誘導した(LPS抗原刺激を受けなかった対照動物からの血清中の<10ng/mlと比較)。この炎症は、使用したBSCIに関係なく、BSCIを用いる治療により用量依存的に劇的に阻害された(図6はBN82350を示す[図1];図7は、(S)−3−(2’,2’−ジメチルプロピオニルアミノ)−テトラヒドロピリジン−2−オン[バレロラクタム]を示す)。
【0137】
インビボで活性のあるsstr2アンタゴニストであるBIM23627の同時投与は、試験したすべての用量でBSCIの抗炎症性を顕著に低下させた(点線、図6と7の三角記号)。この観察結果は、別々の実験で各BSCIについて2回繰り返された。我々は、BIM23627の同時投与により、BSCIの抗炎症性効力が(採用されるBSCIに関係無く)10〜100倍低下すると予想している。これらの結果は、丁度我々がインビトロで観察したように(実施例1参照)、インビボのBSCI活性がsstr2(又はBIM23627により阻害される密接に関連した受容体)に依存することを明確に証明している。
【0138】
別のシリーズの対照実験で、BSCIとは完全に無関係な機構により抗炎症活性を示す合成コルチコステロイドであるデキサメタゾンの抗炎症活性に対するBIM23627の作用を調べた。デキサメタゾンは血清TNF−アルファレベル(全身性炎症のマーカーとして)をBSCIと同程度に低下させた(図8を図6及び7と比較)が、非常に高用量ではデキサメタゾンはわずかにより強力(BSCIの最大用量での約80%の阻害と比較して、90%を超えるTNF−アルファの阻害)であり、BSCIはわずかにより強力であった(デキサメタゾンより3〜10倍低い用量で、TNF−アルファの統計的に有意な低下)。しかし、このモデルでの量の活性にもかかわらず、BIM23627の同時投与はデキサメタゾンの抗炎症性に全く影響を与えなかった(図8中の点線と三角記号)。従って我々は、BIM23627は炎症経路に対して特異的な作用は無く、BSCI(異なる機構で作用する他の抗炎症薬剤を除く)は、インビボでsstr2に高度に依存すると結論付けた。
【0139】
実施例3:sstr2発現はBSCI感受性に相関する
複数のsstr2特異的アンタゴニストの使用は、BSCI活性がsstr2との相互作用に、又は同じアンタゴニストにより阻害される異なる受容体(sstr2に密接に関連している可能性がある)に依存することを示す。sstr2がBSCIの分子標的であることをさらに示すために、我々は、BSCIに対して異なる感受性を有する異なるタイプの一連の白血球中で、フローサイトメトリーにより及び定性的PCRにより、sstr2レベルを測定した。sstr2の存在とBSCIに対する感受性との相関は、BSCI活性がsstr2への結合又はその活性化を必要とするという結論を強固にした。
【0140】
方法:sstr2に特異的な抗体を用いるフローサイトメトリーを使用して、ヒト末梢血白血球上のsstr2発現の相対レベルを推定した。全血を、製造業者の説明書に従ってErythrolyse (Serotec cat number: BUF04B)で処理した。得られた総白血球集団をウサギポリクローナル抗ヒトsstr2抗体 (Sigma, Cat code S0694; 1μg/ml) で染色し、次にFITCで標識した抗ウサギ2次抗体 (Sigma F9887; 50μg/ml) により、具体的な白血球サブセットを明確に同定するために使用できる種々の白血球CDマーカーに対するrPEで標識した一群のモノクローナル抗体を用いて並行して染色した。そのcDNAマーカーに基づいて細胞集団をゲート制御した後、sstr2染色の平均レベル(イソタイプ対照を引いて)を、3回測定値について平均±SEMとして報告した。THP−1とJurkat細胞の培養物を、細胞をErythrolyseで処理しなかった以外は同じプロトコールを使用して分析した。
【0141】
図3に示したプライマーセットを使用してPCRを行い、sstr2aと2b転写体とを区別した。製造業者の説明書に従ってRNAqueous 4PCR キット (Ambion, Cat code #1914) を使用して、THP−1とJurkat細胞からmRNAを調製した。次に、得られたmRNA調製物を、逆転写酵素(RT)を使用してcDNAに変換し、次にこのcDNAを使用して、sstr2プライマー、dNTP(Ambion)、PCR緩衝液、及びTaqポリメラーゼを用いて、総量45μlで以下のPCR反応を構成した。プライマー配列は以下の通りである:sstr2前進プライマー 5'GCCAAGATGAAGACCATCAC 3'(配列番号3)、及び逆進プライマー 5'GATGAACCCTGTGTACCAAGC 3'(配列番号4)。PCRは、Peltier PTC-200サーマルサイクラーを製造業者の説明書に従って使用して、以下のサイクル時間と条件を使用して行った:94℃で2分後、94℃で30秒、55℃で30秒、72℃で1分を30サイクル、そして最後に72℃で10分。増幅の最後に、生成物を臭化エチジウムの存在下で2%アガロースゲルで分離し、UVトランスイルミネータ(Stratagene Eagle Eye II cabinet)で視覚化した。sstr2を高レベルで発現することが知られているpanc−1細胞(Li et al. (2004) J. Surg Res. 119:130-137)を陽性対照として使用した。逆転写酵素の無い反応物を陰性対照として使用した(mRNA調製物に混入しているゲノムDNAの結果として偽陽性シグナルが得られないことを確認する)。さらに、ゲノムアンプリコンが大きさから容易に区別できるように、プライマーをエクソンの境界をまたぐように設計した。
【0142】
結果:sstr2がいくつかの白血球集団で検出され、以前の報告と一致した(Hiruma et al. (1990) Immunology 71:480-5; Tsutsumi et al. (1997) Cell. Immunol. 181:41-9; ten Bokum et al. (1999) J. Endocrinol. 161:167-75 and the references therein; Elliot et al. (1999) Eur. J. Immunol. 29:2454-63; Talme et al. (2001) Clin. Exp. Immunol. 125:71-9; Dalm et al. (2003) Am. J. Physiol. Endocrinol. Metab. 285:E344-53; Lichtenauer-Kaligis et al. (2004) Eur. J. Endocrinol. 150:565-77)。任意の蛍光単位の相対レベルを以下の表1に示す。興味深いことに、THP−1細胞(これはBSCIに対して感受性が高い)は、sstr2について高レベルの表面染色を示し、一方Jurkat T細胞(これは、BSCIに対してほとんど完全に非感受性である)は、実質的に検出できない染色レベルを示す。
【0143】
ヒト末梢血白血球は、一般に2つの細胞株間の中間のsstr2の染色を示す。顆粒球(主に好中球)と単球は高レベルのsstr2を発現し、両方のタイプのヒト末梢血白血球はBSCIに対して感受性であることが知られている。Tリンパ球(CD3+)は、より低いが検出できるレベルを発現し、全CD3+集団が均一に染色されたため、これはCD4+(ヘルパー)とCD8+(キラー)T細胞集団の両方で同様の可能性がある。これに対してsstr2は、BSCIに対して非感受性の細胞であるBリンパ球集団上で検出されなかった(インビボで新抗原に応答した抗体生成に対するBSCIの作用の欠如に一致する)。
【0144】
【表2】
【0145】
表1.種々の白血球集団でフローサイトメトリーにより推定したsstr2細胞表面染色の相対レベル。aSSCは、サイドスキャッターに基づくゲート制御を示す。顆粒球が、その細胞質顆粒の結果として、特徴的に高い光分散性を有するためである。
【0146】
sstr2のmRNAの存在はTHP−1細胞で確認され(図9)、定性的PCRによりJurkat T細胞中に存在しないことが証明された。これは、これらの試験で抗体試薬の予測された特異性を確認している。さらに我々は、THP−1細胞中に存在する大部分のsstr2がsstr2aアイソフォームであることを証明した(これらの細胞中のmRNAのスプライシングパターンに基づく;図3と9)。フローサイトメトリー実験で使用した抗体がsstr2aとsstr2bアイソフォームを区別するかどうかは不明であるが、示された結果は、抗体がsstr2aアイソフォームを検出できることを強く示唆する。
【0147】
我々は、白血球の原形質膜上のsstr2の存在がBSCIに対する感受性と相関すると結論付けた。特に、検出可能なsstr2発現の無い細胞が、ケモカイン誘導性細胞遊走のBSCI介在阻害に対して非感受性であることを注目している。
【0148】
実施例4:BSCIはsstr2に結合する
我々は、標識BSCIリガンドを使用する従来の平衡結合試験を使用して、BSCIとsstr2受容体との分子相互作用を調べた。これらの試験について我々は、不飽和前駆体の接触水素化によりトリチウム(すなわち[3H])で標識した型で容易に合成できるアミノラクタムBSCI BN83250を使用した(図10)。不飽和前駆体である(S,E)−3−(2’,2’−ジメチルドデク−4’−エノイルアミノ)−カプロラクタムを、すでに報告されているように合成した(WO2005/053702)。
【0149】
方法:(S,E)−3−(2’,2’−ジメチルドデク−4’−エノイルアミノ)−カプロラクタムから、パラジウム触媒(CRB Ltd, Cleveland, UK)上でトリチウムガスを使用した接触水素化により、[3H]−BN83250を合成し、トリチウム化生成物をエタノール中270μMの濃度で比活性220Ci/mmolで得た。(S,E)−3−(2’,2’−ジメチルドデク−4’−エノイルアミノ)−カプロラクタムは以下のように製造した:水(30ml)中の(S,S)−3−アミノカプロラクタムヒドロ−ピロリジン−5−カルボキシレート(10mmol)とNa2CO3(30mmol)を、2,2−ジメチル−ドデカ−4−エノイルクロリド(10mmol)のジクロロメタン(30ml)溶液に周囲温度で加え、反応物を12時間攪拌した。次に有機層を分離し、水層を追加のジクロロメタンで抽出した(2×25ml)。一緒にした有機層をNa2CO3上で乾燥し、真空下で濃縮した。残渣をシリカカラムクロマトグラフィー(1:1 EtOAc:ヘキサンからEtOAc)で精製して、(S,E)−3−(2’,2’−ジメチル−ドデク−4’−エノイル)アミノ−カプロラクタムを無色の油状物として得た(2.12g,63%)。放射能標識化合物を暗所で4℃で、必要な時まで保存した。使用前に、非標識BN83250を添加して比活性を調整した。
【0150】
結合の解析のために、ゲイの平衡塩類溶液中の1×106のTHP−1細胞、10mM [3H]−BN83250(50 Ci/mmol)、及び種々の濃度の(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オン(10pM〜10μM)を用いて、個々の反応物を構成した(各条件について3回測定)。過剰(100μM)の非標識BN83250を使用して、非特異結合のレベルを測定した。4℃で2時間後(平衡結合を達成することがすでに証明されている条件、かつこれは結合の過剰推定を引き起こす放射能標識の取り込みを防ぐ)、過剰の標識物を遠心分離(5mlのゲイの平衡塩類溶液を用いて3回洗浄)して洗い流し、可溶化した細胞ペレットのシンチレーション計測により、結合した放射活性を測定した。
【0151】
結果:[3H]−BN83250は、平衡条件下でTHP−1細胞への特異的で競合性の結合を示す。結合のScatchard解析は、約80nMの親和性を有する、細胞当たり100,000〜200,000部位で存在する1つの結合部位に一致する(図11)。BN83250のTHP−1細胞への特異的結合は、過剰(100μM)のsstr2アンタゴニストBN23627により60〜80%(実験に依存する)阻害される(図12)。sstr2が欠如したJurkat T細胞では特異的結合はほとんど検出されなかった(<20,000部位/細胞)が、sstr2を発現するベクターを用いた安定なトランスフェクション後に特異的結合が回復された(sstr2を発現する安定なJurkat T細胞株の調製については実施例5を参照)。まとめるとこれらの結果は、この結合部位がsstr2であり、BN83250の観察されたBSCI活性の原因である結合相互作用であることを証明する。
【0152】
我々が以前最も高効力のBSCIの1つであるとして記載した(WO2009/016390)(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オンが、この結合部位で競合する作用を図12に示す。(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オンは、[3H]−BN83250と、sstr2での結合に対して用量依存的に競合し、ED50が約50pMであり、Hillスロープが約−1.0であり、それらは単純モードの競合結合を示唆する。高レベルの(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オン(1nM以上)は、BN83250結合を80〜90%阻害し、2つの分子が基本的に同一の結合部位を共有することを示唆する。ペプチド3(Reckless & Grainger (1999) Biochem. J. 340:803-11)、NR58−3.14.3(Reckless et al. (2001) Immunology 103:244-54; WO99/12968)、(S)−3−(アダマンタン−1’−カルボニルアミノ)−カプロラクタム(WO2006/016152)、及び(S)−3−(1’−メチオニンシクロヘキシル−1’−カルボニルアミノ)−カプロラクタム(WO2006/134384)を含む、試験した他のBSCI化合物もまた、sstr2への結合に対して、BN83250と同程度に競合する。
【0153】
THP−1細胞への(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オンの結合の見かけの親和性定数は、インビトロアッセイ(実施例1参照)でのBSCI活性のピコモーラーED50に一致するが、古典的sstr2アゴニスト活性についてのED50より数オーダー低い(実施例7参照)。我々は、現在まで記載されているBSCIのすべての群がsstr2に結合し、この相互作用が化合物のBSCI活性の原因であると結論付けた。
【0154】
実施例5:sstr2の発現は細胞株にBSCI感受性を付与する
ここで我々は、sstr2受容体の(強制)発現がBSCIに感受性を付与するのに必要かつ十分であることを証明する。我々はすでに、フローサイトメトリーにより測定されるsstr2の細胞表面発現がBSCIに対する感受性と相関し、従ってsstr2をほとんどまたは全く含まない細胞はBSCIに対して非感受性であるが、高レベルのsstr2を有する細胞は、ケモカイン誘導性遊走のBSCI介在阻害に対して感受性であることを既に示している。より高レベルのsstr2は、BSCI作用の効力の上昇に関連している。しかしこの相関は、sstr2がBSCI活性の原因であることを確証的に実証していない。
【0155】
我々は組換えDNA技術を使用して、白血球細胞株であるJurkat T細胞株(これは通常sstr2を発現しない)中のsstr2の異所性発現を強制的に行った。達成された発現レベルをフローサイトメトリーにより測定した。次に、安定にトランスフェクトされた細胞株のBSCIに対する感受性を測定し、親Jurkat T細胞株と比較した。
【0156】
方法:RNAqueous 4PCR キット(Ambion, Cat code #1914)を使用して製造業者の説明書に従って、Panc−1細胞(高レベルのsstr2を発現することが知られているヒト膵臓細胞株)から総mRNAを調製した。次にmRNAを逆転写(RETROscriptキットを使用; Ambio)して、cDNA調製物を得た。次にPCR(Peltier PTC-200サーマルサイクラーを製造業者の説明書に従って使用して以下のサイクル時間と条件を使用して行った:94℃で2分後、94℃で30秒、55℃で30秒、72℃で1分を30サイクル、そして最後に72℃で10分)により、sstr2遺伝子の完全長cDNAを得て、分取ゲル電気泳動により単離した。sstr2 cDNAの配列を配列決定により確認した。sstr2 cDNAをEcoRIとXhoIにより処理して粘性末端を得た。これはポリリンカーへのプラスミドpcDNA3.1+(Invitrogen)の指向性クローン化と次の連結を可能にした(10μlの連結反応を400 T4 DNAリガーゼ(New England Biolabs)を用いて行い、リガーゼ緩衝液中挿入体:プラスミド比3:1を有する最大量のDNAを13℃で一晩インキュベートした)。得られたベクター(図13参照)を使用して、標準的方法によりXL−10 Ultracompetent 細胞(Stratagene)を形質転換した。MiniPrep キット((Wizard Plus SV Minipreps DNA purification System; Promega)を使用して、Jurkat T細胞へのトランスフェクションについて製造業者の説明書に従って、プラスミドDNAを調製した。
【0157】
Jurkat T細胞はRPMI+10%胎児牛血清(FCS)で培養し、4日毎に新鮮な培地で1:3希釈して継代培養した。継代培養の48時間後の対数増殖期のJurkat T細胞を遠心分離して洗浄し、無血清RPMI培地に7.5×105細胞/mlで再懸濁した。次に、DNAとトランスフェクション試薬の混合物(2μgのDNA;3μlのFugene 6 トランスフェクション試薬; Roche)を加え、細胞を37℃で48時間インキュベートした。次に細胞をRPMI+10%FCSで遠心分離して3回洗浄し、次に新鮮な培地に1×105細胞/mlで懸濁した。次に、生じた細胞懸濁液の68mlを96ウェルプレート(Corning)中の全部で226ウェルに蒔いた。48時間後、培地にG418(0.9mg/ml Active G418)を加えてトランスフェクション体を選択し、継代培養を繰り返して細胞を拡張し、個々のウェルからの集団(クローン性又はオリゴクローン性と見なされた)を別々に維持した。細胞集団を25cm2のフラスコ(全部で約5×106細胞)に拡張した後、少量の試料を取りだし、フローサイトメトリーによりsstr2の発現を試験した。
【0158】
こうして、ヒトsstr2抗体(Sigma、S0694;使用したフローサイトメトリープロトコールの詳細については実施例3を参照)を使用してフローサイトメトリーにより検出された、高レベルのsstr2を細胞表面に発現する多くの個々の細胞株が得られた。これらのうち1つの株(Jurkat−D3Bと命名)は、末梢血の単球(健常なヒト末梢血中の最大発現細胞集団;実施例3を参照)に匹敵する高レベルのsstr2を発現することがわかったが、Jurkat−D3B中のsstr2のレベルはTHP−1細胞中よりはまだかなり低かった(図14)。
【0159】
結果:SDF−1α受容体であるCXCR4のレベルは、Jurkat−D3B細胞株と親Jurkat T細胞株とで同じであった。さらにJurkat−D3B細胞は、Transwell(登録商標)化学走化性アッセイ(使用した方法については実施例1を参照)で親細胞株と同様の濃度範囲にわたって、SDF−1αに応答した。我々は、Jurkat T細胞とJurkat−D3B細胞がこの実験の条件下で同じケモカイン誘導性遊走を示し、従ってBSCIに対するその感受性の比較に適していると結論付けた。
【0160】
Jurkat T細胞及びJurkat−D3B細胞は、100ng/ml SDF−1α(R&D systems;両方の細胞株で最大の細胞遊走を誘導したSDF−1αの濃度)に応答して誘導され遊走した。次にBSCI BN83250を0nM〜1000nM(それぞれ3回測定)の種々の濃度で上の区画と下の区画に加えた(BSCI濃度の勾配を生じないように、各区画で同じ濃度で)。DMSO(1%最終濃度)をビヒクルとして使用し、すべてのウェルで等しい濃度で存在させた。
【0161】
以前の結果(実施例3参照)に一致して、低レベル又は検出できないレベルのsstr2を発現する(図14)親Jurkat T細胞は、BN83250に対して実質的に非感受性であった(THP−1細胞株のような感受性細胞上でこのBSCIについてのED50<1nMと比較して、ED50は1μMよりかなり高かった)。これに対して、中程度のレベルのsstr2を発現する(図14)Jurkat−D3B細胞は、BN83250に対して実質的により感受性であった(図14)。Jurkat−D3B上のBN83250のED50は約10nMであり、sstr2の異所性発現の結果として、BSCI感受性の100倍を超える上昇であった。挿入体の無いpcDNA3.1のトランスフェクションと選択により、親Jurkat T細胞と同様にBN83250に対して非感受性であるJurkat T細胞株が得られた。
【0162】
Jurkat−D3Bはまた、ペプチドNR58−3.14.3やバレロラクタム(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オンのような、構造が異なる他のBSCIによる阻害に対しても感受性であったが、各場合に、その効力はTHP−1細胞上より有意に低かった。これはおそらく、THP−1細胞と比較してJurkat−D3B中で達成される低レベルのsstr2発現を反映している(図14)。薬理学的物質に対する応答の効力は、(いわゆる「スペア受容体」の結果として)受容体発現のレベルと相関することは公知である。我々のデータは、BSCIに対する感受性が細胞表面のsstr2発現のレベルに依存するという提唱と一致する。
【0163】
我々は、白血球細胞にBSCIに対する感受性を付与するのに、sstr2発現が必要かつ十分であると結論付けた。
【0164】
実施例6:従来のsstr2アゴニストはインビトロでBSCI活性が無い
BSCI活性を有する種々の構造が異なる化合物が、sstr2を介して作用することが証明されている。(sstr2を、これらのBSCIの活性の分子標的として確認している。実施例1〜5を参照。)sstr2ですべてのアゴニストがBSCI活性を有するか、又はBSCIとして機能的に同定された化合物がsstr2アゴニストの特殊なクラスであるかどうかを調べるために、我々は一連の古典的ソマトスタチン受容体を、インビトロでのBSCI活性について試験した。
【0165】
方法:実施例1に記載されたように正確に白血球遊走アッセイを行った。簡単に説明するとTHP−1細胞は、ChemoTx(商標)96ウェルプレート遊走系(Neuroprobe Inc., Gaithersburg, MD)中で25ng/mlのMCP−1に応答して、遊走が誘導された。種々の古典的sstr2アゴニストを異なるウェル(3回測定)に、等しい濃度で上の区画と下の区画の両方に加えた(正確に、前述のアシルアミノラクタムのBSCI活性を証明する実験と同じように;Reckless & Grainger (1999) Biochem. J. 340:803-11; Fox et al. (2002) J. Med. Chem. 45:360-70; Fox et al. (2005) J. Med. Chem. 48:867-74)。37℃で2時間後、生体染色色素であるMTT及び標準曲線の内挿を使用して、すべて実施例1と同じように正確に、下の区画に遊走した細胞の数を調べた。
【0166】
結果:sstr2で天然のペプチドリガンドであるソマトスタチン(sst14)は、BSCI活性をほんのわずかに再現するのみであった(図15)。注目すべきは、sst14の最大用量(1μM)でさえ、達成される白血球遊走の最大阻害は、具体的な実験に依存して40〜60%であった。10nMより高いsst14濃度では、細胞上のすべてのsstr2受容体が占有され、従って遊走を完全に阻止できないことは、BSCIと比較してsst14に応答してsstr2がシグナルを出す方法の定性的差を表す。
【0167】
さらにsst14は、sstr2結合の結合定数(約0.05nM)より高い濃度で白血球遊走を阻害するのみであるが、現在まで試験したすべてのBSCIは、その結合親和性より低い濃度で白血球遊走を阻害する。これらの観察結果は、BSCIが古典的sstr2アゴニスト(例えば天然のリガンドであるsst14)より強力なシグナル(白血球遊走に関して)を誘導することを示唆する。その結果BSCIは、白血球遊走を最大阻害するのに細胞表面受容体のほんのわずかを占有すればよいが、従来のsstr2アゴニストは基本的にすべての受容体を占有しなければならず、それでも最大阻害は達成されない。
【0168】
別にプロセシングされたsstr28天然のリガンドを使用して同様の実験を行い、同様の結果が得られた。白血球遊走はsstr2での結合親和性より高い濃度でのみ阻害され、最大阻害は非常に高用量でもわずかに40〜60%であった。臨床的に先端巨大症で成長ホルモン放出を抑制するのに使用される、合成ソマトスタチン類縁体ナンレオチド(lanreotide)(Prevost et al. (1991) Endcrinology 129:323-9)を使用して、再度同様の結果が得られた。
【0169】
我々はまた、その受容体結合特性が完全には決定されていない、最近記載されたソマトスタチン類縁体であるコルチスタチン(de Lecea et al. (1996) Nature 381:242-5)を試験した。コルチスタチンは、sstr2での異なるクラスの天然のリガンドである可能性がある。しかし我々は、コルチスタチンが、白血球遊走に対してその作用がソマトスタチンと非常によく似ている(非常に高用量で使用しても、約50%の遊走を阻害するのみ)が、はるかに低効力である(ED50=約75nM;表2)ことを見いだした。従って現在まで、sstr2受容体に対する天然のリガンドは、同じ受容体で作用するアシルアミノラクタムの強力なBSCI活性を再現できていない。
【0170】
最後に我々は、Ipsen(Paris, France)により提供され、後述の表2と3で「BIM」と呼ぶ、一連の合成sstr2選択的及び特異的アゴニストを試験した。ソマトスタチンは広く同様の親和性ですべてのソマトスタチン受容体に結合し、弱いBSCI様活性は、異なる受容体を介して生成された反対のシグナルによる可能性がある。しかし、10μMで白血球遊走を阻害した(しかし、これより低用量では阻害しなかった)BIM23065を除いて、これらの選択的又は特異的な従来のsstr2アゴニストはいずれも、白血球遊走阻害アッセイで大きな活性を示さなかった(表2)。
【0171】
【表3】
【0172】
表2.種々の公知の又は疑いのあるsstr2アゴニスト化合物のsstr2での結合定数(「Ka」)、及びMCP−1誘導性THP−1細胞の遊走の阻害のためのED50。試験した最高用量(1〜10μM)で観察された最大阻害(「最大作用」)も示される。
【0173】
sstr2でのコルチスタチンの親和性は明確に報告されていない。BIM23065は、試験した最大の可能な濃度(10μM)で細胞遊走を85%阻害し、従ってこれが達成できる最大作用かどうかは不明である。BIM23A387は、Pawlikowsky et al. (2007) J. Physiol. Pharmacol. 58:179-88に記載され、BIM23120はFerrante et al. (2006) Endocrine-Related Cancer 13:955-62に記載されている。
【0174】
これらの結果はまとめると、BSCIが特殊なクラスのsstr2アゴニストであり、ほとんどの化合物(天然のリガンドを含む)は、BSCIとして知られている物質より、白血球遊走を阻害する(及び従ってインビボでの抗炎症活性を示す)能力がはるかに低いことを示す。BSCIが古典的アゴニストとは異なる程度を図16(この図はケモカイン誘導性白血球遊走の阻害についてのsstr2での親和性対ED50をプロットする)に示す。BSCIは一緒に集まり、従来のアゴニストにより顕著に分離される。従って、この試験の組合せを使用して、新規であって、構造が特有のBSCI群(既知のBSCIと構造類似性が無い)を明確に同定することができる。
【0175】
さらに、ここに示した方法は、天然のリガンドsst14とその臨床的類縁体ランレオチド(lanreotide)(Paran & Paran (2003) Curr. Opin. Invetig. Drugs 4:578-82; Sener et al. (2005) Peptides 26:493-9; Gonzales-Rey et al. (2007) Ann. Rheum. Dis. 66:582-8 and the references therein; Armani et al. (2007) J. Leukoc. Biol. 81:845-55; Cury et al. (2008) Dig. Dis. Sci. 53:2516-20)を含むいくつかのソマトスタチン受容体アゴニストについて提唱されている、弱くて臨床的にあまり有用ではない抗炎症活性から、BSCIを区別することは明らかである。
【0176】
BSCIを同定するための既存の方法は基本的に、構造活性相関を規定することに基づいて元々のヒット構造周りのさらなるリードを得るための従来の医化学法とともに、ヒットを同定する機能アッセイを使用するランダムスクリーニングに依存した。本明細書に記載の方法は、資源集約的な低〜中処理能力の機能アッセイを、大量処理の低コスト分子スクリーニングで置き換えることを可能にするため、はるかに有用である。本発明のこの実施において、sstr2との相互作用に基づいてまず化合物が選択され、次に比較的少数の陽性ヒットが、白血球阻害活性についてスクリーニングされて、新しいBSCIが同定される。この適用は、後述の実施例8と9でさらに例示される。
【0177】
実施例7:BSCIは白血球遊走を他の従来のsstr2標的より強力に阻害する
BSCIはsstr2受容体に対するアゴニストであるが、必ずしもすべてのsstr2受容体アゴニストがBSCI活性を示さない。ここで我々は、その逆が真かどうかを調べた:BSCIは、他の機能アッセイにより評価される強力なsstr2アゴニスト様活性を有するか?
【0178】
「古典的」sstr2機能のための2つの異なるアッセイを採用した:(1)sstr2アゴニストにより用量依存的に阻害される成長ホルモンを分泌した胎児ラットの下垂体細胞を培養し、及び(2)sstr2アゴニストにより用量依存的に阻害されるインスリンを分泌したヒトPanc−1細胞株のグルコース感受性サブクローンを選択した。
【0179】
方法:胎児ラットの下垂体細胞培養物を、既に記載されているように、酵素分散により正確に樹立した(Murray et al. (2004) J. Clin. Endocrinol. Metab. 89:3027-32)。低グルコースDMEM+10%FCSで48時間後、48ウェルプレート中の細胞(5×104細胞/ウェル)を無血清DMEMで4時間洗浄し、次に無血清DMEM中でさらに24時間、種々の濃度の試験物質とともにインキュベートした。この時間の最後に、調整培地を採取し、細胞破片を遠心分離して除去し、市販のELISA(Linco; cat# EZRMGH-45K)を使用して、製造業者の説明書に従ってGHについて測定した。
【0180】
ヒトPanc−1(外分泌膵臓の多型類上皮癌から得られる細胞株)のグルコース感受性サブクローンを、基本的にHamil et al. (2008) Biomed. Sci. Instrum. 44:441-6が記載したように得た。簡単に説明するとPanc−1細胞を24ウェルプレートで継代培養し、培地に5%グルコースを補足した。72時間後、グルコース感受性の丸い表現型への最も顕著な変化を示したウェルを選択し、通常の培地で拡張した。次に得られたサブ株を、96ウェルプレートのウェル(5×104細胞/ウェル)を調製することによりグルコース感受性について試験し、これを次に一連のグルコース濃度を含有する培養培地(低グルコースDMEM、DMEM、DMEM+1%グルコース、DMEM+2.5%グルコース、及びDMEM+5%グルコース)に曝露した。24時間後、調整培地を採取し、ELISA(Linco; cat# EZHI-14K)により製造業者の説明書に従ってインスリン分泌を測定した。
【0181】
最も高いグルコース感受性(低グルコースDMEMと比較して5%グルコース中でインスリン分泌が3.4倍増加した)を有し、実施例3に記載した方法を使用してフローサイトメトリーによりsstr2を発現することが証明されたクローン(Panc−2H4)を使用して、我々はグルコース依存性インスリン分泌を調節する種々のsstr2アゴニストの能力を試験した。DMEM中のPanc−2H4(5×104細胞/ウェル)を使用して、追加の96ウェルプレートを調製した。24時間後、培地を種々の濃度の試験物質を含有するDMEM+5%グルコースに切り替えた(1つの条件について3つのウェル)。さらに24時間後、調整培地を採取し、インスリン濃度を上記したようにELISAにより測定した。
【0182】
両方のアッセイで、各試験化合物の存在下で2番目に高い濃度の標準物質に曝露したELISA中の追加の対照ウェルを含めることにより、ELISAに対する試験化合物(試験した最も高い濃度)の作用の欠如を証明した。結果を、ビヒクルのみの存在下で細胞からの変化倍数として表した平均±SDとして報告した。
【0183】
結果:予想したように、天然のリガンドであるソマトスタチン−14を含む古典的sstr2アゴニストは、ラット下垂体細胞からGH分泌を用量依存的に低下させ、10nMで最大で64±9%阻害であった(表3)。古典的アゴニストはPanc−2H4によるインスリン分泌に対して同様の作用を有したが、最大作用はより小さかった(43±7%阻害)。両方のアッセイで、GH又はインスリン分泌を抑制するのに必要な濃度は、sstr2への結合についての種々の物質の公知の親和性と一致した。
【0184】
これとは顕著に異なり、これらの古典的sstr2アゴニストは、ケモカインに応答してTHP−1細胞や末梢血白血球の遊走を阻害することはないか、又はGH及び/又はインスリン分泌を阻害するのに必要な濃度よりかなり高い濃度でのみ阻害した(表3)。
【0185】
ペプチドであるペプチド3とNR58−3.14.3、ならびにアミノカプロラクタム類BN83250とBN83470、及びバレロラクタム(S)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オンを含む、構造が異なる種々のBSCIは、はるかに高濃度でのみGH放出を阻害し(表3)、最大作用は小さかった。これらの物質のいずれも、試験した最大濃度でもPanc−2H4によるインスリン分泌を阻害しなかった。しかし予想したように、これらの物質はケモカインに応答してTHP−1細胞とラット末梢血白血球の遊走を強力に阻害した(表3)。
【0186】
表3の結果をまとめると、sstr2に結合し、この受容体でアゴニストとして作用する化合物は、容易に2つの群に分類されることを示す。1つの群の物質(本適用の目的のために「古典的」sstr2アゴニストと呼ぶ)は、ケモカイン誘導性白血球遊走を阻害するよりも高効力でかつ強力にGH分泌とインスリン分泌とを阻害する。第2の新規の群の物質(本適用の目的のために「BSCI型」sstr2アゴニスト又はソマトタキシン類と呼ぶ)は、ケモカイン誘導性白血球遊走を阻害するよりも高効力でかつ強力にケモカイン誘導性白血球遊走を阻害する。
【0187】
【表4】
【0188】
表3.「古典的」対「BSCI」標的に対する種々のsstr2アゴニストの効力の比較。4つの別々のアッセイにおける12の構造の異なるsstr2アゴニストの作用の効力(ED50、nMで)と出力(ビヒクルと比較した抑制パーセントとしての最大作用)を示す。2つのアッセイは、古典的sstr2依存性応答(GH抑制とインスリン抑制)を測定し、2つのアッセイはsstr2を介するBSCI活性(ケモカインに応答する白血球遊走の阻害)を測定する。これらの2つの分類の応答の相対的効力は、本発明の方法に従ってsstr2アゴニストを機能的に分類することを可能にする:最初の7つの物質は「古典的」sstr2アゴニストと分類され、後の5つの物質は「BSCI」sstr2アゴニストと分類される。略語:SST=ソマトスタチン;n.d.=測定されない;「バレロラクタム」=(3)−3−(2’,2’−ジメチルプロピオノイルアミノ)−テトラヒドロピリジン−2−オン。BN83470=(S)−3−(アダマンタン−1’−カルボニルアミノ)−カプロラクタム。
【0189】
これらの機能的に異なるクラスのsstr2アゴニストが存在することを当業者に認めさせたであろう情報はまだ開示されていないが、この現象のいくつかの他の例があり、ここでは同じ受容体に対する異なるアゴニストが、「機能的選択性」として知られている異なる細胞応答を引き起こすことができる(他の受容体におけるこの現象の優れた最近の総説については、Kenakin (2007) Mol. Pharmacol. 72:1393-401を参照)。多くの分子機構がこの現象(例えば、活性状態ベースの選択性、偏りのある作動性、刺激の移動など)の基礎となっているが、基礎となっている分子機構の性質は、本明細書に記載したような単純な機能アッセイにより、群のメンバーを明確に区別する能力にはあまり関連していない。
【0190】
ここでsstr2アゴニストの2つのクラスを例示するのに使用される化合物群は、非常に明確であり、容易に区別される(すなわち、いくつかの異なるアッセイで生成する機能的応答の点で、各クラスのすべてのメンバーは、他のクラスのメンバーとよりも、互いによく似ている)。しかし、すべての化合物スペースにわたって、「ハイブリッド挙動」を示す物質が存在する可能性がある。すなわちこれらの物質は、GH/インスリン分泌及びケモカイン誘導性白血球遊走を、ほぼ等しい効力と出力で阻害する。そのような物質は、ここに提供した方法に基づいて、いずれかのクラスに容易に割り当てることはできないであろう。従って我々は、各クラスのメンバーを明確に規定し、いずれかの分類から「ハイブリッド挙動」化合物を除外する定義を提供した。ケモカイン誘導性白血球遊走アッセイ(すなわち、表3の右側のカラム中で2より大きい比率)よりも、GH又はインスリン分泌アッセイ中で少なくとも2倍高い効力(すなわち、半最大効力に必要な濃度)を有する分子は、「古典的」sstr2アゴニストと定義される。逆に、GH又はインスリン分泌アッセイ(すなわち、表3の右側のカラム中で0.5より小さい比率)よりも、ケモカイン誘導性白血球遊走アッセイ中の少なくとも2倍高い効力又は出力を有する分子は、本発明の方法に従って「BSCI」sstr2アゴニスト(ソマトタキシン類)と定義される。
【0191】
本発明の方法の実施において、同じ種からの細胞を使用してアッセイを比較することが好ましいことに注意されたい。古典的sstr2標的に対する物質の作用を測定するためにラットの下垂体細胞が使用される本例では、結果は、ラット末梢血白血球のケモカイン誘導性遊走に対する、同じ物質の作用と比較される。同様に、古典的sstr2標的に対する物質の作用を測定するためにヒトPanc−2H4細胞が使用される場合、結果は、ヒトTHP−1細胞のケモカイン誘導性遊走に対する、同じ物質の作用と比較される。本発明の方法を、種を超えて適用することは、純粋に2つの種からのsstr2に対する親和性が異なることの結果として、化合物の分類を間違える(両方の方向に)危険がある。例えば、ヒトsstr2よりラットsstr2で10倍低い親和性を有する化合物は、もしその物質が、ヒトTHP−1ケモカイン誘導性白血球遊走アッセイにおけるより、ラット下垂体Gh抑制アッセイで3倍低い効力であった場合、その物質は本発明の方法により「BSCI」sstr2アゴニストとして間違って分類されるであろう(なぜなら、受容体に対する結合親和性の10倍の差を補正すると、「古典的」活性は実際には「BSCI」活性より高かったため)。このため、表3中の「古典的」活性と「BSCI」活性との比は、ラットとヒトについて別々に計算される。
【0192】
実施例8:ヨヒンバン−16−アミドはα2−アドレナリン受容体で作用する機能的選択性リガンドである
ヨヒンビンは、トウワタ(milkweed)植物(インドジャボク(Rauwolfia Serpentina))の根からから得られる古典的α−アドレナリン作用模倣物のクラスのメンバーであり、これはまたレセルピンとレシンナミンとを含む。これらの物質は強力な降圧薬(すなわち、血圧を下げる)であるが、これらは好ましくない副作用を伴う(これは、記憶と多くの他の神経機能に重要なα2−アドレナリン受容体調節の中心的作用に関連する催眠作用や他の精神的作用を含む)。その結果ヨヒンビンとレセルピンは、もはや医薬として使用されていないが、米国ではまだ催淫薬として広範に使用される漢方薬として入手できる(この血管拡張活性は、Viagra(商標)で見られるような陰茎勃起を引き起こすが、降圧危機の結果として入院リスクが高く、ヨヒンビンを含む薬草抽出物の規制違反的使用が、米国において毎年10,000例の入院と何人かの死亡を引き起こしていると推定されている)。
【0193】
ここで我々は、既に記載されているヨヒンビンの16−アミド誘導体(WO99/12968参照)もまたα2−アドレナリン受容体と相互作用するが、実質的に古典的α2−アドレナリン受容体アゴニスト活性の代わりにケモカイン誘導性白血球遊走の阻害(すなわち、BSCI活性)を引き起こすことを示す。すなわちヨヒンビンの16−アミド誘導体は、α2−アドレナリン受容体で機能的選択性リガンドである(アミノラクタムBSCIがsstr2受容体で機能的選択性リガンドであるのとほとんど同じように)。
【0194】
方法:α2−アドレナリン受容体を発現する細胞からの市販の膜調製物(Sigma)を用いて製造業者の説明書に従って、放射性リガンド結合実験を行った。[3H]−ジヒドロエルゴクリプチン([3H]−DHE)を5nM濃度(反応当たり0.05μCi)で放射性リガンドプローブ(Hoffman et al. (1980) J. Biol. Chem. 255:4645-52の方法に従って)として使用した。実験は3回測定で行い、非特異結合は過剰(10μM)のオキシメタゾリンの存在下で測定した。
【0195】
MCP−1誘導性THP−1細胞遊走の阻害は、正確に上記したように測定した(実施例1)。すべての実験でDMSO(1%最終濃度)をビヒクルとして使用した。
【0196】
結果:ヨヒンバン−16−アミドはα2−アドレナリン受容体への[3H]−DHEの結合を、用量依存的に見かけのKa=89±13nMで阻害した。これは、同じ条件下で塩酸ヨヒンビンの結合の見かけのKaより約10倍低い(6.8±1.2nM)が、これは、ヨヒンバン−16−アミドが妥当な親和性でまだα2−アドレナリン受容体に結合することを裏付けている。
【0197】
異なるα2−アドレナリン受容体リガンドであるRS−79948−197(Grainger et al. (2005) Mini Rev. Med. Chem. 5:825-32)を使用して同様の結果がすでに得られているが、16−アミド基の導入によるα2−アドレナリン受容体に対する見かけの親和性の低下は、この放射性リガンドを使用すると、より大きかった(塩酸ヨヒンビンの3.8nMと比較してヨヒンバン−16−アミドでは770nM)。これは、ヨヒンバン−16−アミドの結合部位がヨヒンビンの結合部位とはわずかに異なり、RS−79948−197結合部位よりはるかにDHE結合部位と重複していることを示唆する。結合部位のこの小さなシフトは、我々は観察した機能的選択性の根拠となり得る。しかし我々は、ヨヒンバン−16−アミドがα2−アドレナリン受容体に結合すると結論付けた。
【0198】
ヨヒンバン−16−アミドは、THP−1細胞(これは、相当レベルのα2−アドレナリン受容体を発現する)のMCP−1誘導性遊走をED50が0.5nMで阻害し、一方ヨヒンビンはこのアッセイでは極めて不活性である(ED50=30nM)。しかしいずれの物質も、α2−アドレナリン受容体を発現しないJurkat T細胞のSDF−1a誘導性遊走を阻害しない。これらの結果をまとめると、ヨヒンバン−16−アミドと、程度ははるかに小さいがヨヒンビンとの両方は、これらが結合するα2−アドレナリン受容体への作用を介してBSCI活性を示す。
【0199】
しかし重要なことは、ヨヒンバン−16−アミドは古典的α−アドレナリン作用活性を有しないことである。例えばヨヒンバン−16−アミドは、ADPへの応答で血小板の活性化に効果を及ぼさないが、古典的α2−アドレナリンリガンド(ヨヒンビン、クロニジン、及びジヒドロエルゴクリプチンを含む)はすべて、顕著なADP介在血小板凝集を強化する。
【0200】
まとめると、これらの結果は、ヨヒンバン−16−アミドがα2−アドレナリン受容体で作用する機能的選択性リガンドであることを明確に証明している。この化合物は受容体への結合がヨヒンビンより弱いけれども(競合実験の見かけの親和性は、使用される放射性リガンドプローブに依存して大きく異なるため、標識ヨヒンバン−16−アミド誘導体が合成され特徴付けされるまで、相互作用の実際の親和性は不明であるけれども)、それでもBSCIよりほとんど100倍高効力である。したがって、ヨヒンバン−16−アミドの結合は、BSCI活性に必要なシグナル伝達出力の活性化を促進する。これに対してヨヒンバン−16−アミドは、ADP介在血小板活性化を強化しない(古典的α2−アドレナリン受容体活性)。その結果、我々は、ヨヒンバン−16−アミドが白血球(BSCI活性は受容体で古典的活性を実質的に置換する)上で発現されるGPCR神経受容体に対する機能的選択性リガンドの例であると結論付けた。そのような化合物は、抗炎症剤として潜在的に非常に有用であり、標的受容体の活性化に関連すると予測される副作用が無く、本発明の方法に従って容易に同定される。
【0201】
実施例9:L−IIは5HT2a受容体で作用する機能的選択性リガンドである
リセルグ酸のアミド誘導体(N,N−ジエチルアミド、LSDを含む)は、セロトニン受容体5HT2aでアゴニスト活性を介して(医学的に)所望ではない幻覚誘発活性を有し、その結果、その精神活性作用のための潜在的に有害なレクリエーション的使用の結果として多くの国で禁止されている物質である。
【0202】
ここで我々は、既に記載されているリセルグ酸のグルタミンとアスパラギンアミド結合体(WO99/12968参照)もまた5HT2aと相互作用するが、実質的に古典的5HT2aアゴニスト活性の代わりにケモカイン誘導性白血球遊走の阻害(すなわち、BSCI活性)を引き起こすことを示す。すなわちリセルグ酸のグルタミンとアスパラギンアミド誘導体は、5HT2a受容体で機能的選択性リガンドである(アミノラクタムBSCIがsstr2受容体で機能的選択性リガンドであり、ヨヒンバン−16−アミドがα2−アドレナリン受容体で機能的選択性リガンドであるのとほとんど同じように)。
【0203】
方法:5HT2a受容体を発現する細胞からの市販の膜調製物(Sigma)を用いて製造業者の説明書に従って、放射性リガンド結合実験を行った。[3H]−ケタンセリンを放射性リガンドプローブとして(Leysen et al. (1988) Pharmacol. Exp. Therap. 244:310-21の方法に従って)1nM濃度(反応当たり0.05μCi)で放射性リガンドプローブ(Hoffman et al. (1980) J. Biol. Chem. 255:4645-52の方法に従って)として使用した。実験は3回測定で行い、非特異結合は過剰(10μM)のメテルゴリン(比較的非特異的な麦角由来セロトニン受容体アンタゴニスト)の存在下で測定した。
【0204】
MCP−1誘導性ヒト末梢血単核球細胞遊走の阻害は、正確に上記したように測定された(実施例1)。すべての実験でDMSO(1%最終濃度)をビヒクルとして使用した。
【0205】
結果:L−IIは5HT2a受容体受容体への[3H]−ケタンセリンの結合を、用量依存的に見かけのKa=23±5nMで、同じ条件下でリセルグ酸の結合の見かけのKaと同様に阻害した。従ってL−IIは、リセルグ酸の他のアミド誘導体のように、妥当な親和性で5HT2a受容体に結合する。
【0206】
L−IIは、ヒト末梢血単球細胞(これは、高レベルの5HT2a受容体を発現する)のMCP−1誘導性遊走を、ED50が2nMで阻害し、一方リセルグ酸はこのアッセイでは極めて不活性である(ED50>1nM)。しかしいずれの物質も、Jurkat T細胞のSDF−1α誘導性遊走もTHP−1細胞のMCP−1誘導性遊走も阻害しない(両方の細胞株とも、高レベルの5HT2aを発現しない)。これらの結果をまとめると、L−IIは5HT2a受容体への作用を介してBSCI活性を示す。
【0207】
しかし重要なことは、L−IIは古典的5HT2aアゴニスト活性を有さないことである。例えばL−IIは、げっ歯類で幻覚誘発剤のマーカーとして従来の理解されている首ひねりを誘導しない。5HT2aのほとんどの古典的アゴニスト(LSD、メスカリン、シロシン、TCB−2、及びBromo-DrangonFLY)は、この古典的作用を誘導する。
【0208】
これらの結果はまとめると、L−IIが5HT2a受容体で作用する機能的選択性リガンドであることを明確に証明している。L−IIは5HT2aに多くの古典的アゴニストと同様の親和性で結合するが、それでもこれは、BSCIより少なくとも500倍の効力を有する。従ってL−II結合は、BSCI活性に必要なシグナル伝達出力の活性化を促進する。これに対してL−IIは、首ひねりを誘導しない(古典的5HT2aアゴニスト活性)。従って我々は、L−IIが、白血球(ここで、BSCI活性は受容体で古典的活性を実質的に置換する)上で発現されるGPCR神経受容体に対する機能的選択性リガンドの例であると結論付けた。そのような化合物は、抗炎症剤として潜在的に非常に有用であり、標的受容体の活性化に関連すると予測される副作用が無く、本発明の方法に従って容易に同定される。
【0209】
少なくとも細胞内シグナル伝達のレベルで機能的選択性は、5HT2a受容体について既に記載されており(例えば、Moya et al. (2007) J. Pharmacol. Exp. Ther. 321:1054-61を参照)、ここで2,5−DMAは、ホスホリパーゼA2(PLA2)を活性化することなくホスホリパーゼC(PLC)活性化を介してリン酸イノシトールの放出を誘導するが、関連するリガンド2C−Nは、PLCを活性化することなくPLA2によるアラキドン酸の放出を活性化する。これは我々の知見と一致するが、この機能的選択性シグナル伝達の結果としてどの機能的応答が異なるかの教示は無く、白血球上に発現される広範囲のGPCR神経受容体に対する機能的選択性が、有用な抗炎症活性を有する一連の化合物を立証できるという我々の発見を予測してはいない。
【0210】
実施例10:本発明の方法に従う、候補BSCIの同定(1)
本発明の方法をさらに例示するために、BSCI候補についてL−779,976に関連するsstr2アゴニストを含有する一連のベータ−メチルトリプトファンをスクリーニングするために、それは適用された。L−779,976は、記載されている(Rohrer et al. (1998) Science 282:737-40)初期の非ペプチドsstr2選択的アゴニストの1つであり、これは、従来のスクリーニング法、分子設計、及び医化学的アプローチにより発見された。これは、天然のソマトスタチンリガンド中の主要なQFWKテトラペプチド(配列番号5)に弱く基づくため、古典的sstr2アゴニスト性を示す(実施例7を参照)。
【0211】
興味深いことに、QxWK(配列番号6)もまた、初期ペプチドBSCI中の主要なモチーフとして同定された(Fox et al. (2002) J. Med. Chem. 45:360-70)が、現在までに記載されている非ペプチドBSCIであるsstr2アゴニストはQxW末端を形成し(図1参照)、一方古典的sstr2アゴニストはWK末端を形成している。これは、グルタミン模倣物グループと融合したL−779,976の変種が新しいクラスのBSCIであるという考えにつながる。
【0212】
ここで我々は、本発明の方法に従って半ダースの単純な新規L−779,976誘導体を試験し、その1つ(「化合物C」と呼ぶ)が新規構造クラスのBSCI化合物の最初のメンバーであることを示す。
【0213】
図17に示す6つの化合物を、DMSOビヒクル(1%最終濃度)中0.01nM〜1,000nMの種々の濃度で、Panc−2H4インスリン抑制アッセイ(正確に実施例7に記載されたように)とケモカイン誘導性THP−1遊走抑制アッセイ(正確に実施例1に記載されたように)とに付した。結果を表4に示す。
【0214】
【表5】
【0215】
表4.L−779,976の6つの類縁体の古典的sstr2アゴニストとBSCI sstr2アゴニスト活性。Panc−2H4細胞中のグルコース依存性インスリン分泌の抑制を、正確に実施例7に記載されたように測定した。ケモカイン誘導性THP−1細胞遊走の抑制を、正確に実施例1に記載されたように測定した。n.c.=比率は計算していない(いずれかのアッセイで不充分な活性を示す化合物について)。
【0216】
親L−779,976及び化合物Aは、古典的sstr2アゴニストである。化合物D、E及びFは、この実験で本発明の方法に従って評価するには、いずれかのアッセイで不充分な活性を有する。化合物Bは、古典的sstr2アゴニスト活性とBSCI sstr2アゴニスト活性の両方を有する。しかしケモカイン誘導性白血球遊走の抑制効力は、インスリン分泌の抑制効力より2倍以上高いため、本発明の方法は、化合物Cを、新規であって構造が固有のBSCI化合物(ソマトタキシン)として同定する。
【0217】
ここで我々は、抗炎症剤としてさらに開発するのに適した、BSCI活性を有する化合物を明確に同定するために、既知のsstr2アゴニスト化合物の一連の類縁体に本発明の方法が如何に適用されるかを例示する。機能アッセイを使用してスクリーニングされる少数の分子を選択するために、sstr2結合の知識が第1工程で使用されたため、sstr2への結合の実験的スクリーニングは行なわなかったが、これは本発明の方法の適用であることは明らかである。
【0218】
実施例11:本発明の方法に従う、候補BSCIの同定(2)
前の例ではBSCIの分子標的の知識を使用して、既知のsstr2受容体アゴニストの修飾に基づく、機能的スクリーニングのための候補BSCI化合物を設計した。しかし本発明の方法はまた、非常に大きなライブラリーのランダムスクリーニングにも適用することができる。この適用では、まずsstr2を標的として使用する従来の分子スクリーニングが、まずランダムライブラリーからのヒットを同定するために使用され、次に第2段階において、ヒット(又はその類縁体)は、「古典的」sstr2アゴニスト活性、及び「BSCI」sstr2アゴニスト(ソマトタキシン)活性のについての一対の機能アッセイを使用してスクリーニングされる。
【0219】
方法:sstr2を発現するように安定にトランスフェクトしたJurkat T細胞(Jurkat D3B;実施例5参照)から調製した膜を使用して、sstr2と相互作用する化合物の分子スクリーニングを確立した。簡単に説明するとJurkat D3B細胞(調製物当たり5×107個の細胞)をダルベッコーPBSで遠心分離して洗浄し、次にプロテアーゼ阻害剤(1mM PMSF、100mM ロイペプチン)を含有する1mlの氷冷ダルベッコーPBSに再懸濁した。次に細胞懸濁液を、Dounceホモジナイザーを使用して溶解し、得られたホモジネートを700×gで4℃にて1分遠心分離して、壊れていない細胞と大きな細胞断片をペレット化した。上清を取り出し、超遠心分離管に移し、60,000×gで4℃にて2時間遠心分離して、膜断片のペレットを得た。ペレットを、プロテアーゼ阻害剤を含有する氷冷ダルベッコ(Dulbecco)−PBSで静かに洗浄し、次にプロテアーゼ阻害剤を含有する100μlのダルベッコ−PBSに再懸濁した。次にクマシープラス(Coomassie Plus)キット(Pierce)を使用して製造業者の説明書に従ってタンパク質濃度を測定し、プロテアーゼ阻害剤を含有するダルベッコ−PBSのさらなる添加により最終濃度を2mg/mlに調整した。次に凍結防止剤として等量のグリセロールを加え(50%グリセロール中1mg/mlの総タンパク質濃度の膜のストック調製物を与える)、使用するまでストックを−80℃で最大1ヶ月間保存した。
【0220】
スクリーニングは384ウェルマイクロタイタープレートで行った。各ウェルで、結合緩衝液(ゲイの平衡塩類溶液)中1μgのJurkat D3B膜画分タンパク質、1nMの[125I]標識ソマトスタチン−14(400 mCi/mmol)、及びDMSO中100mg/mlからの最終濃度1mg/mlのライブラリーからの試験化合物からなる1つの反応を、最終反応容量25μlで構成した。37℃で2時間(平衡結合に至ることがすでに確立された条件)結合させ、次に反応物を384ウェルのマニホールド中GFCろ過シートでろ過し、0.5%トリトンX−100と300mM NaClをさらに含む10mlの結合緩衝液を用いて連続流で洗浄した。結合したカウントは定量的シンチグラフィにより評価した。各プレートで、結合の陽性対照としてBIM23627(10μM)を使用し、非特異結合を測定するために100nMの非標識ソマトスタチンを使用した。結合は、ある化合物の存在下での特異的カウントの抑制パーセントとして表した。試験化合物を加えずに全384ウェルプレートをアッセイして評価すると、特異的カウントの38%までの抑制は陰性対照(ビヒクルのみ)レベルより4標準偏差低いレベルであった。その結果、特異的カウントの76%又はそれ以上の抑制を与える試験物質を陽性ヒットとした。
【0221】
このアッセイからのヒットを、正確に実施例4に記載したように、従来のScatchard解析を使用して確認した。sstr2への結合の見かけのKaが1μM又はそれ以上の場合は、化合物をさらに調べた。
【0222】
次に、sstr2で相互作用が確認された化合物を対の機能アッセイで評価した:panc−2H4インスリン抑制アッセイ(「古典的」sstr2アゴニスト活性の推定として、正確に実施例7に記載のように行った)及びケモカイン誘導性THP−1細胞遊走抑制アッセイ(「BSCI」sstr2アゴニスト活性の推定として、正確に実施例1に記載のように行った)。本発明の方法に従って、ケモカイン誘導性THP−1細胞遊走抑制アッセイで少なくとも2倍高い効力を有する物質が、抗炎症剤としてさらなる研究と開発に適した候補BSCIとして選択される。
【0223】
結果:26枚のアッセイプレートを流して(全部で9,880化合物をスクリーニングした)、標識ソマトスタチン−14とsstr2の結合を、76%を超えて抑制した28のヒット(0.28%)が得られた。
【0224】
これらの1次ヒットのうち11個(1次ヒットの39%;ライブラリーの0.11%)はScatchard解析でうまく再現され、9個(繰り返しヒットの81%;ライブラリーの0.09%)は、sstr2に対する見かけの結合親和性が1μMを超えた。
【0225】
次に、再現されたヒット(sstr2についての見かけの結合親和性の順位でASX.A〜ASX.Iと呼び、ASX.Aが最も高い見かけの親和性を有する)を、正確に実施例7に記載のように対の機能アッセイを使用して評価し、その結果を表5に示す。
【0226】
【表6】
【0227】
表5.9,880の化合物のスクリーニングからの再現された中程度の親和性(1μM又はそれ以上)のヒットの古典的sstr2アゴニスト及びBSCI sstr2アゴニスト活性。Panc−2H4細胞中のグルコース依存性インスリン分泌の抑制を、正確に実施例7に記載のように測定した。ケモカイン誘導性THP−1細胞遊走の抑制は、正確に実施例1に記載のように測定した。n.c.=比率は計算していない(いずれかのアッセイで不充分な活性を示す化合物について)。L−779,976とBN83250のデータを、比較のために古典的タイプのsstr2アゴニストとBSCIタイプ(ソマトタキシン)のsstr2アゴニストの例として示す。
【0228】
驚くべきことに、11個の化合物のうちの9個はいずれかの機能アッセイで検出可能なアゴニスト活性を有した。ほとんどのGPCRについて、大量処理スクリーニングにより同定されたヒットの中で、10%(10個のうちの1個)未満はアゴニスト活性を有すると予測され得る。この珍しい知見は、sstr2でのアゴニスト:受容体相互作用の相対的複雑さを反映しているか、又は我々は確立(天然のリガンドの結合について競合)するために選択した具体的なスクリーニングの結果かも知れない。さらにスクリーニングしたライブラリーはランダムではなく、既知のGPCRアゴニストと構造類似性を有する鋳型を用いてすでに濃縮されていた。
【0229】
これらの9個のうち3個は、本発明の方法に従って「古典的」タイプのsstr2アゴニスト活性を有するとして明確に特定できる高いアゴニスト活性を有し(ASX.A、ASX.C、及びASX.D)、細胞遊走抑制アッセイよりインスリン抑制アッセイで少なくとも2倍高い効力を有した。これに対して、再現されたヒットのうちで最も低い見かけの結合親和性であった1個のみ(ASX.H)は、インスリン抑制アッセイより細胞遊走抑制アッセイで少なくとも2倍高い効力を有して、明確に「BSCI」タイプのsstr2アゴニスト(1つのソタトタキシン)として特定された。これらの知見は、1個の化合物(ASX.D)がこの細胞遊走抑制アッセイでASX.Hに匹敵する効力を有したが、「古典的」sstr2アゴニスト活性についてのインスリン抑制アッセイでも高い効力を有したため、細胞遊走の抑制の機能アッセイでさえ、「BSCI」タイプのsstr2アゴニスト(ソマトタキシン類)を明確に特定するのに、単独では使用できないことを例示する。本発明の方法は、完全に新規な「BSCI」タイプのsstr2アゴニスト(ソマトタキシン類)を特定するのに必要かつ十分である。
【0230】
本発明の方法を使用して我々は9,880個の化合物をスクリーニングして、BSCIとして充分な性質(もちろんED50が45nMでは、それは、我々がすでに記載した最適化リード候補よりインビトロで約1,000倍低い効力である;例えばWO2009/016390参照)を有し、従来の医化学リード最適化プログラムのひな型として機能できるであろう1つの化合物を見つけた。機能アッセイによる9,880個の化合物すべての直接スクリーニングは、何カ月もかかったであろう(通常の1週間労働で約100個の化合物をスクリーニングすることができる)。BSCIの分子標的の知識を利用する本発明の方法の適用は、この全モデルスクリーニングをたった1週間で完了することを可能にした。これは、消費される資源の約99%の削減である。典型的には、医薬業界で行われる大量処理スクリーニングは、本例で我々が使用したものより10倍〜100倍多い化合物をスクリーニングできるであろう。そのようなスクリーニングは、本発明の方法を使用すると容易であるが、機能的スクリーニングアッセイを使用すると実現不可能であり、同じスクリーニング量を達成するのにほとんど10年必要であろうと我々は推定する。
【0231】
実施例12:本発明の方法で同定される候補BSCIのインビボ評価
抗炎症特性を有するが、古典的ソマトスタチン受容体アゴニストの内分泌調節性を持たない薬剤として有用な物質を同定するために、本発明の方法に従って化合物Pが同定された。
【0232】
化合物Pはsstr2に結合し、ここでアゴニストとして作用する。図18に示すように化合物Pは、本発明の方法に従うアッセイでケモカインMCP−1に応答してTHP−1細胞の遊走を、ED50が約0.05nMで阻害する。
【0233】
しかし、図19に示すように化合物Pは、本発明の方法に従って行われたアッセイで、培養下垂体細胞からの成長ホルモン(GH)の放出を、ED50が約2000nMで阻害したのみである。図19において、点線は、例えば10nMのソマトスタチン−14(古典的sstr2アゴニスト)を用いて、このアッセイで達成される最大作用(成長ホルモン分泌の約75%の低下)を示す。その結果、ED50用量は、GH分泌を約37%低下させた化合物Pの用量である。
【0234】
こうして化合物Pの古典的タイプの活性とBSCIタイプの活性の比は、約0.000025であると計算される。
【0235】
インビトロで行われたアッセイがインビボの化合物の活性を予測するものであることを証明するために、36人の正常で健常なヒト被験者(男性又は女性)を、12人の被験者の3つの群の臨床試験に入れた。各群12人の被験者のうち、9人は経口的に化合物Pを60mLの水中の液体薬剤として投与され、残りの3人はプラセボとして60mLの水のみを投与された。第1群は150mgの化合物Pを7日間毎日投与され、第2群は500mgの化合物Pを7日間毎日投与され、第3群は1500mgの化合物Pを10日間毎日投与された。化合物Pの投与前、投与期間中、及び投与後の様々な時間に血液試料を採取した。
【0236】
化合物Pは、約25時間の半減期で、ヒトでほぼ100%の経口バイオアベイラビリティを有することがわかった。化合物Pへの被験者の予測された曝露は、被験者の血清中の化合物Pについて評価済みLC−MSアッセイを使用して確認した。
【0237】
予測されたように化合物Pは、ヒトで抗炎症性であった。C−反応性タンパク質(CRP)はヒトの炎症レベルの充分認められたマーカーであり、急性及び慢性の炎症反応中に上昇する(Coventry et al. (2009) J. Translational Med. 7: 102を参照)。その結果、血清CRPレベルは炎症の代替マーカーとして広く受け入れられている。CRPレベルは、投与の前(投与前)と投与の最後の日(投与後)の血清試料で、市販のELISAアッセイを使用して測定された。すべての3つの用量の化合物Pで処理すると、処理された被験者中のCRPレベルが低下したが、プラセボを投与された被験者では低下しなかった。化合物P処理の効果(用量に無関係に)を図21に示す。
【0238】
図20では、充分に確立された臨床的な抗炎症治療薬Medrol(登録商標)(メチルプレドニソロン、コルチコステロイド)と化合物Pの作用を比較した。両方の化合物がCRPを同程度に低下させたが、作用は化合物Pについてのみ統計的に有意であった(p=0.02)。
【0239】
図20の結果は、化合物Pが合成コルチコステロイドMedrol(登録商標)と少なくとも同様に有効な抗炎症剤であることを裏付けている。
【0240】
しかし、また本発明の方法に従って予測されるように、我々は化合物Pが2型ソマトスタチン受容体に結合するが、視床下部下垂体副腎軸(HPA軸)のマーカーのパネルに対して効果が無く、従ってヒトで古典的タイプの活性が完全に欠如していることを見いだした。
【0241】
化合物Pは、非常に高用量であっても、7日間治療したヒトでGHレベルに対して何の作用もなかった。さらに化合物Pは、非常に高用量であっても、7日間治療したヒトでACTHレベルに対して何の作用もなかった。ACTHは、ヒトのHPA軸の乱れの最も感受性の高いマーカーであることが確立されており(Denef (2008) J. Neuroendocrinol. 20: 1-70を参照)、これらの研究をまとめると、化合物Pはヒトで臨床的に有用な抗炎症作用を有するが、同じ用量で古典的タイプのソマトスタチン模倣作用を持たないことを裏付けている。
【0242】
本例は、2型ソマトスタチン受容体に結合する、本発明の方法に従って同定された化合物が、ヒトで臨床的に有用な抗炎症活性を有し、(これに関連して)好ましくない内分泌調節作用(又は「古典的タイプ」作用)を全く有さないことを裏付けている。
【0243】
実施例の要約
実施例をまとめると我々は、4つの異なる群のBSCIが、白血球上で発現される3個の異なるGPCR神経受容体でアゴニストとして作用することを証明した。ペプチド3関連のポリアミドとアミノラクタムは、2型ソマトスタチン受容体(sstr2;実施例1〜5)のアゴニストとして作用する。ヨヒンビンの16アミド誘導体は、α2−アドレナリン受容体を介して作用する(実施例8)。リセルグ酸のグルタミン及びアスパラギンアミド結合体は、5HT2aセロトニン受容体のアゴニストとして作用する(実施例9)。しかし、各場合にこれらの受容体に対する他のアゴニストの大部分はBSCIとして作用することができない(実施例6、8、及び9)か、又は実質的に低い効力及び/又は出力で作用する。その結果我々は、BSCIが、これらの受容体に対するアゴニストの、より多くの一般的クラスの、特別な、使用可能に定義されたサブクラスであると結論付けた。
【0244】
さらに、これらの群は互いの構造的関係によってのみ同定されたため、このような機能的選択性リガンドがすべてのGPCR神経受容体について存在すると結論することは妥当である。白血球上で発現された3つの異なるGPCR神経受容体で、我々が機能的選択性リガンドを発見したことは驚くべきことである(これらの化合物を同定するために使用した方法は、これらの分子標的の情報に全く依存しなかったため)。これらの物質の作用機構が開示された(本明細書)ため、この情報を使用して、白血球上で発現される他のGPCR神経受容体で同様に作用するリガンドを見つけることができる。このような探索を行う方法は、本発明の一態様である。
【0245】
古典的アゴニスト活性に対するBSCI活性の機能的選択性に基づいて、白血球上で発現されるGPCR神経受容体でアゴニストを下位分類することは、有用(これは、既知のクラスとの構造的関係に無関係に新しいBSCIの同定を可能にするため)であり、新規(これまでは記載されていないため)であり、充分定義され(実施例10と11に例示される、サブクラスのメンバーを明確に区別する方法が示されるため)、且つ驚くべきことである(既存の科学文献又は特許文献はいずれも、古典的活性に対して抗炎症活性を促進する白血球上で発現されるGPCR神経受容体の機能的に明確なサブクラスが存在し得ることを示唆していない)。
【0246】
実施例10と実施例11の両方において、新規であって構造が固有のBSCI化合物を同定するために、本発明の主題であるBSCIの分子標的の先行情報を使用した。実施例10では、類縁体作成ための適切な骨格としてL−779,976を選択するために、標的の情報が使用され、そして抗炎症剤としての以後の開発のための最も有望な候補を同定するために、これらの類縁体を、その後に本発明の方法に従う一対のアッセイ(「古典的」sstr2アゴニスト活性のアッセイと「BSCI」sstr2アゴニスト活性のアッセイ)を使用してスクリーニングした。実施例11では、sstr2相互作用性構造について直接的分子スクリーニングを計画及び展開するために、標的の情報が使用された。これは次に、本発明の方法に従って一つのアッセイ(「古典的」sstr2アゴニスト活性のアッセイと「BSCI」sstr2アゴニスト活性のアッセイ)の適用によりさらに洗練された。こうして、抗炎症剤としてさらなる開発に最も有望なライブラリー要素が同定された。
【0247】
最後に実施例12において、2型ソマトスタチン受容体に結合する、本発明で同定された化合物が、好ましくない内分泌調節(又は「古典的タイプの」)作用無しで、ヒトで臨床的に有用な抗炎症活性を有することを証明することにより、本発明の方法は有効であることが確認された。
【0248】
BSCIが、白血球上で発現されるGPCR神経受容体において機能的選択性により作用するという驚くべきこと知見は、非常に広範囲の疾患を治療するための新規抗炎症剤として有望なBSCI活性を有する物質を同定するためのユニークで有用な新しい方法を提供する。現在まで、新規であって構造が特有であるBSCI化合物を同定する唯一の方法は、既存のBSCI化合物の構造をさらに修飾する(既存の構造クラス内で既知のBSCI化合物の数を有効に拡大する)か、又はBSCI活性の機能アッセイで選択されない化合物ライブラリーをスクリーニングする(例えば、本発明の実施例で広く使用されるTranswell(登録商標)細胞遊走アッセイ)ことであった。同じ期間に従来の分子スクリーニングにより数万の化合物が処理されるのと比較すると、現在までに入手できる最良の方法でさえ、機能アッセイによりわずかに数百の新しい化合物しかスクリーニングできない。分子スクリーニングと機能的スクリーニングによる処理速度の2〜3オーダーの差は、本発明の有用性にとって重要である:ランダムな機能的スクリーニングにより新しい構造クラスのBSCIを見つけることにおいて、資源問題は非現実的なほど大きい。しかし本発明の方法は、まず白血球上で発現されるGPCR神経受容体を使用して従来の分子スクリーニングを行い、次に第2の工程で、分子スクリーニングからのヒットを機能アッセイによりさらにスクリーニングして、BSCIの機能的特性(すなわち、GH又はインスリン放出の抑制より、ケモカイン誘導性白血球遊走の抑制に対してより高効力を有する物質)を同定する。第1工程の分子スクリーニングは、ランダムライブラリー中の化合物の数を2〜3オーダー減少させるため(妥当な効力ヒットは典型的には、ランダムライブラリーのわずかに0.1〜1%であるため)、以後の機能的スクリーニングの資源の要求性は、スクリーニングされる化合物当たりの利用はより大きくなるが、現実的になる。
【0249】
本発明を好適な実施態様を参照して説明してきたが、この種々の修飾態様や変更態様は本発明の精神と範囲を逸脱することなく実施可能であり、このような修飾が本発明に明確に包含されることを当業者は認識し得る。本明細書に開示し、特許請求の範囲に記載した具体例は、全く制限的ではなく、また制限的であるべきではない。
【0250】
本明細書に記載したすべての文書は参照のためその全体が本明細書に組み込まれる。
【0251】
以下の配列表に記載された配列は本発明の一部を構成する。
【配列表フリーテキスト】
【0252】
配列表のフリーテキスト
合成オリゴヌクレオチド(配列番号3〜4);
天然のソマトスタチンリガンドの主要なBSCIモチーフ(配列番号5);
初期ペプチドBSCIの主要なモチーフ(配列番号6);
Xaaは任意の天然に存在するアミノ酸でよい(配列番号6)。
【特許請求の範囲】
【請求項1】
広域スペクトルケモカイン阻害剤(BSCI)活性を有する化合物又は物質(agent)の同定方法であって、以下の工程:
(a)第一に、1又は2以上の候補化合物又は物質が、白血球上で発現されることが知られているGPCR神経受容体への結合についてスクリーニングされ;その後、
(b)選択された前記GPCR神経受容体に対して結合を示す化合物又は物質が、機能アッセイでBSCI活性について試験され;そして、
(c)選択された前記GPCR神経受容体に対して結合を示す化合物又は物質が、機能アッセイで古典的アゴニスト活性(classical agonist activity)について試験される、
を含み、ここで、工程(a)は最初に行われるが、工程(b)及び(c)は任意の順番で、又は同時に行われる、前記方法。
【請求項2】
BSCI活性についての前記機能アッセイが、インビトロでのケモカイン誘導性細胞遊走の抑制についてのアッセイである、請求項1に記載の方法。
【請求項3】
BSCI活性についての前記機能アッセイが、インビボでのケモカイン誘導性細胞遊走の抑制についてのアッセイである、請求項1に記載の方法。
【請求項4】
BSCI活性についての前記機能アッセイが、Tヘルパー細胞のサイトカイン分泌についてのアッセイである、請求項1に記載の方法。
【請求項5】
古典的アゴニスト活性についての前記機能アッセイがインビトロで行われる、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
古典的アゴニスト活性についての前記機能アッセイがインビボで行われる、請求項1〜4のいずれか1項に記載の方法。
【請求項7】
BSCI活性の結果を検出するために代替バイオマーカーが使用される、請求項1〜6のいずれか1項に記載の方法。
【請求項8】
古典的アゴニスト活性の結果を検出するために代替バイオマーカーが使用される、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
スクリーニングされる候補化合物又は物質が、ライブラリーから選択されるか又は一緒になってライブラリーを構成する、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
スクリーニングされる候補化合物又は物質が、コンビナトリアルケミストリーにより製造される、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
試験される候補化合物又は物質が、化合物の混合物である、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
前記GPCR神経受容体との分子相互作用が、選択された前記GPCR神経受容体を発現する細胞全体、又は選択された前記GPCR神経受容体を発現する細胞由来の膜を使用して行われる、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
前記細胞が、選択された前記GPCR神経受容体を過剰発現するように遺伝子操作されている、請求項12に記載の方法。
【請求項14】
前記細胞が、選択された前記GPCR神経受容体の断片を発現するか、又は発現するように遺伝子操作されている、請求項12又は13に記載の方法。
【請求項15】
選択された前記GPCR神経受容体との分子相互作用が、単離されたか又は精製された受容体を使用して行われる、請求項1〜14のいずれか1項に記載の方法。
【請求項16】
単離されたか又は精製されたソマトスタチン受容体の任意の断片が使用される、請求項15に記載の方法。
【請求項17】
候補化合物又は物質と選択された前記GPCR神経受容体との間の相互作用が、前記選択されたGPCR神経受容体に結合することができる別の標識リガンドとの競合により測定される、請求項1〜16のいずれか1項に記載の方法。
【請求項18】
候補化合物又は物質間の相互作用が直接測定される、請求項1〜17のいずれか1項に記載の方法。
【請求項19】
前記GPCR神経受容体がソマトスタチン受容体である、請求項1〜18のいずれか1項に記載の方法。
【請求項20】
前記GPCR神経受容体がsstr2受容体である、請求項19の方法。
【請求項21】
前記ソマトスタチン受容体がsstr2Aである、請求項19の方法。
【請求項22】
前記ソマトスタチン受容体がsstr2Bである、請求項19の方法。
【請求項23】
前記候補化合物又は物質は、ソマトスタチンを除外する、請求項19〜22のいずれか1項に記載の方法。
【請求項24】
前記GPCR神経受容体がα2−アドレナリン受容体である、請求項1〜18のいずれか1項に記載の方法。
【請求項25】
前記α2−アドレナリン受容体が、ADRA2A、ADRA2B、又はADRA2Cである、請求項24の方法。
【請求項26】
GPCR神経受容体がセロトニン受容体である、請求項1〜18のいずれか1項に記載の方法。
【請求項27】
セロトニン受容体が5HT2aである、請求項26に記載の方法。
【請求項28】
1又は2以上の前記試験物質と前記GPCR神経受容体との相互作用が、測定されるか、又は任意の種類の科学文献又は既存のデータベースの調査により推測される、請求項1〜27のいずれか1項に記載の方法。
【請求項29】
以前の工程の適用により選択された化合物内に存在する1又は2以上の構造モチーフに基づく、新規構造クラスのBSCIを定義するための追加の工程を有する、請求項1〜28のいずれか1項に記載の方法。
【請求項1】
広域スペクトルケモカイン阻害剤(BSCI)活性を有する化合物又は物質(agent)の同定方法であって、以下の工程:
(a)第一に、1又は2以上の候補化合物又は物質が、白血球上で発現されることが知られているGPCR神経受容体への結合についてスクリーニングされ;その後、
(b)選択された前記GPCR神経受容体に対して結合を示す化合物又は物質が、機能アッセイでBSCI活性について試験され;そして、
(c)選択された前記GPCR神経受容体に対して結合を示す化合物又は物質が、機能アッセイで古典的アゴニスト活性(classical agonist activity)について試験される、
を含み、ここで、工程(a)は最初に行われるが、工程(b)及び(c)は任意の順番で、又は同時に行われる、前記方法。
【請求項2】
BSCI活性についての前記機能アッセイが、インビトロでのケモカイン誘導性細胞遊走の抑制についてのアッセイである、請求項1に記載の方法。
【請求項3】
BSCI活性についての前記機能アッセイが、インビボでのケモカイン誘導性細胞遊走の抑制についてのアッセイである、請求項1に記載の方法。
【請求項4】
BSCI活性についての前記機能アッセイが、Tヘルパー細胞のサイトカイン分泌についてのアッセイである、請求項1に記載の方法。
【請求項5】
古典的アゴニスト活性についての前記機能アッセイがインビトロで行われる、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
古典的アゴニスト活性についての前記機能アッセイがインビボで行われる、請求項1〜4のいずれか1項に記載の方法。
【請求項7】
BSCI活性の結果を検出するために代替バイオマーカーが使用される、請求項1〜6のいずれか1項に記載の方法。
【請求項8】
古典的アゴニスト活性の結果を検出するために代替バイオマーカーが使用される、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
スクリーニングされる候補化合物又は物質が、ライブラリーから選択されるか又は一緒になってライブラリーを構成する、請求項1〜8のいずれか1項に記載の方法。
【請求項10】
スクリーニングされる候補化合物又は物質が、コンビナトリアルケミストリーにより製造される、請求項1〜9のいずれか1項に記載の方法。
【請求項11】
試験される候補化合物又は物質が、化合物の混合物である、請求項1〜10のいずれか1項に記載の方法。
【請求項12】
前記GPCR神経受容体との分子相互作用が、選択された前記GPCR神経受容体を発現する細胞全体、又は選択された前記GPCR神経受容体を発現する細胞由来の膜を使用して行われる、請求項1〜11のいずれか1項に記載の方法。
【請求項13】
前記細胞が、選択された前記GPCR神経受容体を過剰発現するように遺伝子操作されている、請求項12に記載の方法。
【請求項14】
前記細胞が、選択された前記GPCR神経受容体の断片を発現するか、又は発現するように遺伝子操作されている、請求項12又は13に記載の方法。
【請求項15】
選択された前記GPCR神経受容体との分子相互作用が、単離されたか又は精製された受容体を使用して行われる、請求項1〜14のいずれか1項に記載の方法。
【請求項16】
単離されたか又は精製されたソマトスタチン受容体の任意の断片が使用される、請求項15に記載の方法。
【請求項17】
候補化合物又は物質と選択された前記GPCR神経受容体との間の相互作用が、前記選択されたGPCR神経受容体に結合することができる別の標識リガンドとの競合により測定される、請求項1〜16のいずれか1項に記載の方法。
【請求項18】
候補化合物又は物質間の相互作用が直接測定される、請求項1〜17のいずれか1項に記載の方法。
【請求項19】
前記GPCR神経受容体がソマトスタチン受容体である、請求項1〜18のいずれか1項に記載の方法。
【請求項20】
前記GPCR神経受容体がsstr2受容体である、請求項19の方法。
【請求項21】
前記ソマトスタチン受容体がsstr2Aである、請求項19の方法。
【請求項22】
前記ソマトスタチン受容体がsstr2Bである、請求項19の方法。
【請求項23】
前記候補化合物又は物質は、ソマトスタチンを除外する、請求項19〜22のいずれか1項に記載の方法。
【請求項24】
前記GPCR神経受容体がα2−アドレナリン受容体である、請求項1〜18のいずれか1項に記載の方法。
【請求項25】
前記α2−アドレナリン受容体が、ADRA2A、ADRA2B、又はADRA2Cである、請求項24の方法。
【請求項26】
GPCR神経受容体がセロトニン受容体である、請求項1〜18のいずれか1項に記載の方法。
【請求項27】
セロトニン受容体が5HT2aである、請求項26に記載の方法。
【請求項28】
1又は2以上の前記試験物質と前記GPCR神経受容体との相互作用が、測定されるか、又は任意の種類の科学文献又は既存のデータベースの調査により推測される、請求項1〜27のいずれか1項に記載の方法。
【請求項29】
以前の工程の適用により選択された化合物内に存在する1又は2以上の構造モチーフに基づく、新規構造クラスのBSCIを定義するための追加の工程を有する、請求項1〜28のいずれか1項に記載の方法。
【図1】

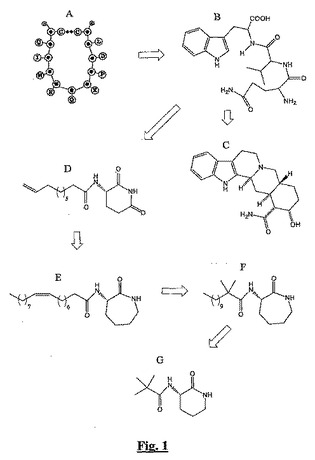
【図2】
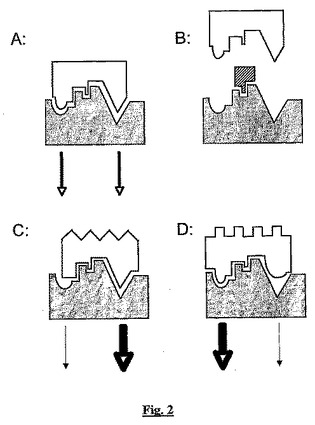
【図3】
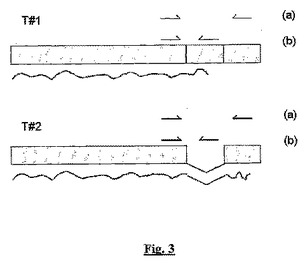
【図4】
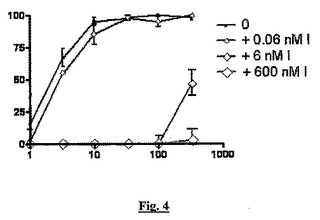
【図5】
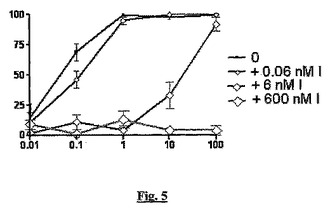
【図6】
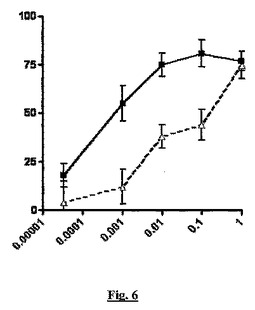
【図7】
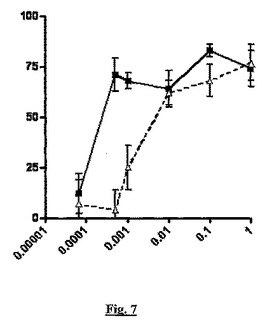
【図8】
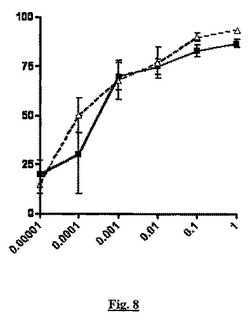
【図9】
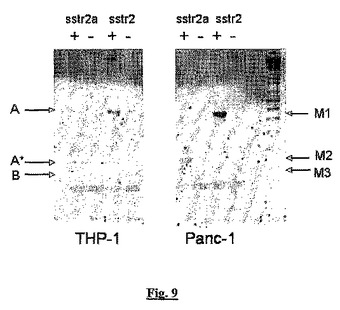
【図10】
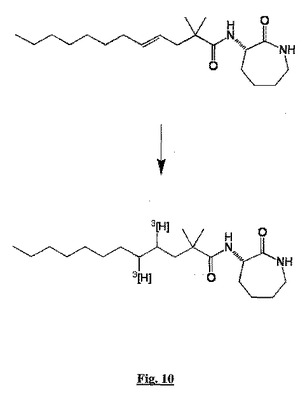
【図11】
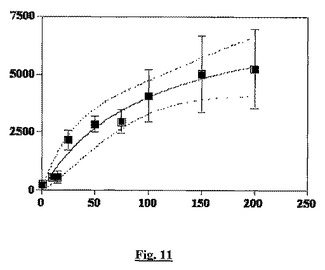
【図12】
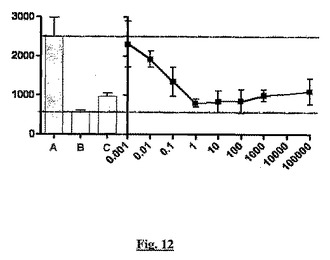
【図13】
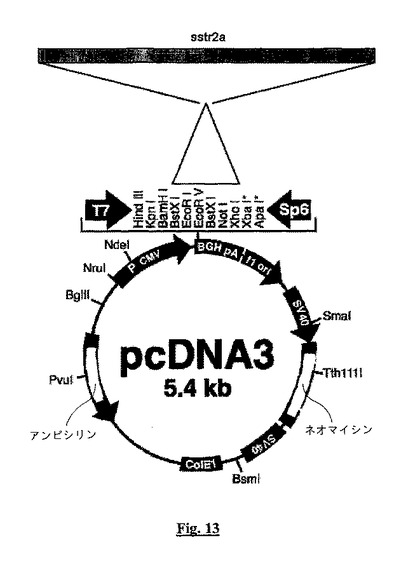
【図14】
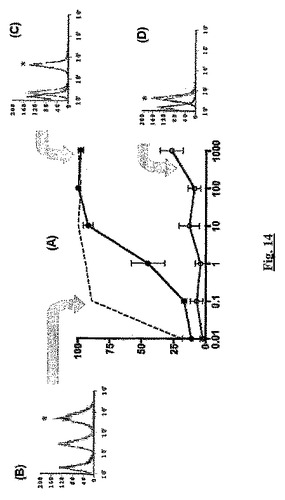
【図15】
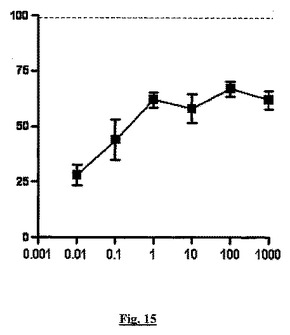
【図16】
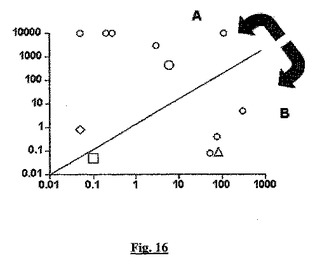
【図17】
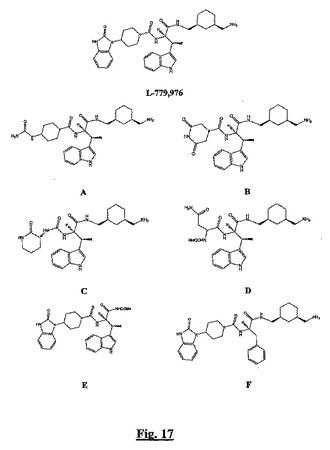
【図18】
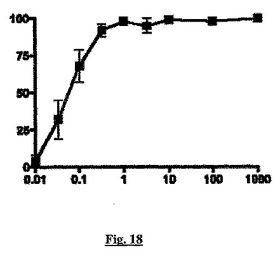
【図19】
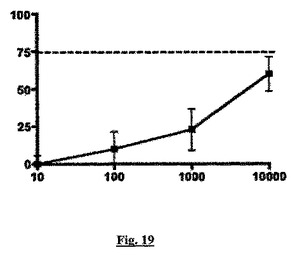
【図20】
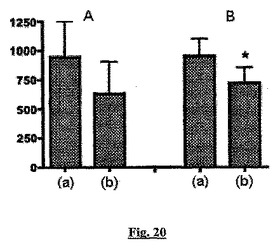
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【公表番号】特表2012−518994(P2012−518994A)
【公表日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願番号】特願2011−551520(P2011−551520)
【出願日】平成22年2月26日(2010.2.26)
【国際出願番号】PCT/GB2010/000342
【国際公開番号】WO2010/097591
【国際公開日】平成22年9月2日(2010.9.2)
【出願人】(503372842)ケンブリッジ エンタープライズ リミティド (32)
【Fターム(参考)】
【公表日】平成24年8月23日(2012.8.23)
【国際特許分類】
【出願日】平成22年2月26日(2010.2.26)
【国際出願番号】PCT/GB2010/000342
【国際公開番号】WO2010/097591
【国際公開日】平成22年9月2日(2010.9.2)
【出願人】(503372842)ケンブリッジ エンタープライズ リミティド (32)
【Fターム(参考)】
[ Back to top ]