
含硫化合物含量が増加した酵母、そのスクリーニング方法、及び培養方法
γ−グルタミルシステインやグルタチオンなどの含硫化合物の細胞内の含量が増加した酵母をセレン酸感受性と赤色呈色の性質を組み合わせることによって得る。赤色呈色の方法は、アデニン要求性の付与およびメチオニンを添加した最小培地での培養を含む。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、含硫化合物含有酵母、そのスクリーニング方法、及び該酵母を培養する方法に関するものである。本発明によって得られる含硫化合物高含有酵母は、食品、医薬品、化成品、飼料等の分野で有用である。
【背景技術】
【0002】
現在、含硫化合物は、食品、医薬品、化成品などの幅広い分野で使用されている。例えば、グルタミン酸、システインおよびグリシンからなるトリペプチドであるグルタチオン(GSH)は、医薬品としての薬効を有することが知られている。現在、医薬品としてのGSH製剤は、解毒剤及び眼科用剤として位置づけられており、各種の中毒、慢性肝臓疾患、抗癌剤の副作用や放射線療法による障害の防止、皮膚疾患および白内障や角膜損傷の治療に用いられている(非特許文献1)。
【0003】
一方、食品用途として、グルタチオンは食品にコク味を付与する物質であることが知られており(非特許文献2)、グルタチオンを高含有する酵母エキスが調味料用途に用いられている。また、グルタミン酸とシステインからなるジペプチドであるγ−グルタミルシステインは食品用途で有用であると報告されつつある。
【0004】
前述のように、含硫化合物は幅広い産業上の有用性を有しているため、これらを効率的に生産する微生物を取得しようと様々な検討が行われてきた。例えば、含硫化合物の生合成経路においてターゲットとする酵素を予測し、その機能を改変することにより細胞内の含硫化合物含量を増加させる方法が開示されている。例えば、大腸菌のγ−グルタミルシステイン合成酵素(非特許文献3)やグルタチオン合成酵素(非特許文献4)を酵母に導入することによって細胞内のグルタチオン含量が増加することが報告されている。
【0005】
これら以外にも、グルタチオン合成に関与する酵素を酵母に導入することによって含硫化合物であるグルタチオン含量を増加させる方法が報告されている(特許文献1、特許文献2、特許文献3、特許文献4)。
【0006】
一方、親株を突然変異処理することにより無作為な遺伝子改変を施した微生物を、各種薬剤を含む培地にスプレッドし生育可能な菌株(薬剤耐性株)を選択すること、又はレプリカ法により各種薬剤を含む培地で生育不可能な菌株(薬剤感受性株)を選択することにより含硫化合物の細胞内含量が増加した微生物を選抜する方法が報告されている。例えば、キャンディダ属酵母を変異処理し、エチオニン及び亜硫酸塩含有培地上で生育可能な菌株を選抜する方法(特許文献5、特許文献6、特許文献7)、サッカロミセス属酵母を変異処理し、亜鉛耐性度が向上した菌株を選抜する方法(特許文献8)などがある。これらの方法以外にも、含硫化合物の細胞内含量を増加させるための各種薬剤が検討されてきた(特許文献9、特許文献10)。
【0007】
ところが、近年、菌体内にグルタチオンを5重量%以上含有するサッカロミセス・セレビシエに属する酵母菌株をランダム変異により取得することに成功したと報告された(特許文献11)。同文献では変異株のグルタチオン含有量を一株一株測定することによって高生産株を取得していた。同文献の方法によって大量の菌株を評価可能にしたと報告していたが、同文献に記載されている方法でどれだけの速度で菌株が測定可能か検討したところ、1人が1日に評価できる菌株数は、精々100株程度であった。それゆえ、同文献に記載の方法は操作が極めて煩雑かつ高コストで経済的に負担の大きいことは明らかである。
【0008】
ところで、アデニン要求性を有する酵母、特に、ADE1遺伝子又はADE2遺伝子に変異を有する酵母は赤く発色することが古くから知られている。近年、遺伝子レベルでこのメカニズムが解析された。即ち、プリン合成系の第6ステップを触媒する酵素およびAIR中間体をコードするADE2遺伝子又はプリン合成系の第7ステップを触媒する酵素およびCAIR中間体をコードするADE1遺伝子の変異によって蓄積するアデニン生合成の中間体であるAIR(phosphoribosylaminoimidazole)やCIAR(phosphoribosylaminoimidazole carboxylate)がグルタチオンと結合し液胞に運ばれることによって赤く発色することが明らかとなった(非特許文献5)。しかし、同文献では、GSHの生合成の律速酵素をコードするGSH1遺伝子を過剰発現させたが、赤色レベルは変わらなかったと報告している。このことから、当業者であっても、アデニン要求性をGSH高含有酵母のスクリーニングに利用しようとの発想は持ち得ない状況であった。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】特開昭61−52299号公報
【特許文献2】特開昭62−275685号公報
【特許文献3】特開昭63−129985号公報
【特許文献4】特開平4−179484号公報
【特許文献5】特開昭59−151894号公報
【特許文献6】特開平03−18872号公報
【特許文献7】特開平10−191963号公報
【特許文献8】特開平02−295480号公報
【特許文献9】特開平06−70752号公報
【特許文献10】特開平08−70884号公報
【特許文献11】特開2004−180509号公報
【非特許文献】
【0010】
【非特許文献1】蛋白質核酸酵素 1988-7 VOL.33 NO.9 ISSN 003909450臨時増刊 「グルタチオン研究のエポック」p1626
【非特許文献2】Y.Ueda et al, Biosci. Biotech. Biochem. ,61,1977-1980(1997)
【非特許文献3】Yasuyuki OHTAKE et al, Agric. Biol. Chem., 52(11), 2753〜2762, 1988
【非特許文献4】Yasuyuki OHTAKE et al, Journal of FERMENTATION AND BIOENGINEERING, Vol.68, No.6, 390-394, 1989
【非特許文献5】K.G. Sharma et al, Arch Microbiol (2003) 180:108-117
【発明の概要】
【発明が解決しようとする課題】
【0011】
本発明は、細胞内の含硫化合物含有量が増加した酵母、酵母を効率よくスクリーニングする方法、及びそれら酵母に細胞内の含硫化合物を高含有させる培養方法を提供することを目的とする。
【課題を解決するための手段】
【0012】
本発明者らは、K.G. Sharmaらの報告(非特許文献5)を詳細に検討し、アデニン生合成は生物に不可欠なATP生合成に関与していることから、ADE1遺伝子破壊株又はADE2遺伝子破壊株のようなアデニン要求性株ではAIRやCAIRといった中間体は細胞内に十分に蓄積しているとの仮説を持った。本発明者らは、この仮説が正しいとすれば、細胞内のグルタチオン含量が酵母の赤色発色をコントロールしていると考え、より赤く発色する酵母はよりグルタチオン含量が高くなっていると考えるにいたった。鋭意検討を行った結果、K.G.
Sharmaらの仮説とは異なり、本発明者らは、酵母の赤色を指標にすれば、グルタチオンを高含有する酵母がスクリーニング可能であることを見いだし、また、グルタチオンを含有する含硫化合物の細胞内含有量が、セレン酸感受性と、アデニン要求性の付与又はメチオニンを添加した最少培地での培養に起因する赤色呈色の特性との組み合わせにより増加可能であることを見出し、それにより本発明を完成させた。
【0013】
本発明の一態様は、細胞内の含硫化合物含量が増加した酵母であって、セレン酸感受性を有し、かつ培地上で培養されたときに該培地上で赤色を呈する酵母を提供することである。
本発明の別の態様は、前記培地はメチオニンが添加された培地である、上記の酵母を提供することである。
本発明の別の態様は、アデニン要求性の付与により赤色を呈色する、上記の酵母を提供することである。
本発明の別の態様は、アデニン要求性がADE1遺伝子又はADE2遺伝子の改変により付された、上記の酵母を提供することである。
本発明の別の態様は、さらに、ADE4遺伝子またはADE8遺伝子に変異が導入されたことにより、前記培地上で白色を呈するようになった、上記の酵母を提供することである。
本発明の別の態様は、セレン酸感受性がMET25遺伝子の発現を増強する改変により付された、上記の酵母を提供することである。
本発明の別の態様は、含硫化合物がシステイン、γ−グルタミルシステイン、グルタチオン、システニルグリシンからなる群から選択される少なくとも1つ以上の化合物である上記の酵母を提供することである。
本発明の別の態様は、菌体量を増加させるために細胞内のアデニンが充足する状態で酵母を培養する工程と、細胞内の含硫化合物含量を増加させるために細胞内のアデニンが不足する状態で酵母を培養する工程を含む、上記の酵母の培養方法を提供することである。
本発明の別の態様は、アデニン要求性およびセレン酸感受性を有する酵母に遺伝子改変処理を施す工程、アデニンが不足した時に酵母が赤く発色する培地に改変酵母をスプレッドし、酵母コロニーを形成する工程、及び改変前よりも赤く発色する酵母を選択する工程を含む、細胞内の含硫化合物含量が増加した酵母のスクリーニング方法を提供することである。
本発明の別の態様は、セレン酸感受性を有する酵母に遺伝子改変処理を行う工程、最小培地にメチオニンを追加した培地に改変酵母をスプレッドし、コロニーを形成する工程、及び改変前よりも赤く発色する酵母を選択する工程を含む、細胞内の含硫化合物含量が増加した酵母のスクリーニング方法を提供することである。
本発明の別の態様は、前記含硫化合物がシステイン、γ−グルタミルシステイン、グルタチオン、システニルグリシンからなる群から選択される少なくとも1つ以上の化合物である、上記方法を提供することである。
【図面の簡単な説明】
【0014】
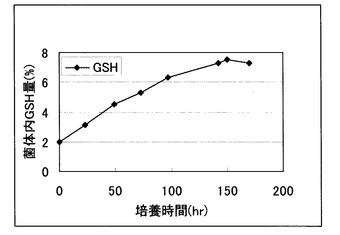
【図1】Y-3256株のGSH含量の経時変化を示す図。
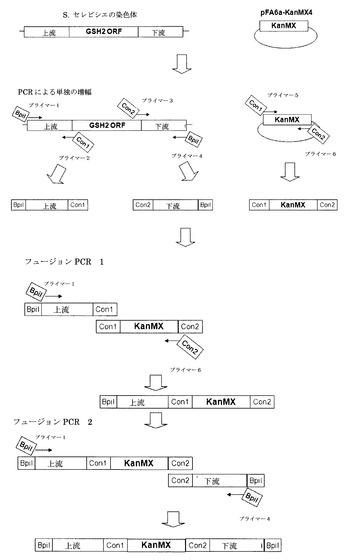
【図2】GSH2遺伝子破壊カセットの構築手順を示す図。
【発明を実施するための形態】
【0015】
本発明の酵母は、セレン酸感受性を有し、かつ培地上で培養されたときに該培地上で赤色を呈することによって細胞内の含硫化合物含量が増加した酵母であれば特に制限されない。サッカロミセス・セレビシエ等のサッカロミセス属、キャンディダ・ユティリス等のキャンディダ属、ピヒア・パストリス等のピヒア属、シゾサッカロミセス・ポンベ等のシゾサッカロミセス属等に属する酵母を例示することができる。中でも、含硫化合物の製造によく用いられているサッカロミセス・セレビシエやキャンディダ・ユティリスが好ましい。本発明の酵母は、1倍体でもよいし、2倍性またはそれ以上の倍数性を有するもので
あってもよい。
ここで、赤色性を評価する培地としては、酵母細胞内のアデニンが不足した時に酵母が赤く発色することができる培地であれば特に制限されない。アデニン含量が25mg/L以下の固体培地を挙げることができ、具体的にはYPD培地(組成:Bacto-yeast extract 1%、Bacto-peptone 2%、Glucose 2%:METHODS IN YEAST GENETICS 2000 Edition p171参照。ISBN 0-87969-588-9)PGC培地(組成:Casamine acid(vitamin free) 0.5%、Bacto-peptone 1%、Glucose 2%。)等を例示することができる。微量成分との影響でより赤く発色しやすいPGC培地の方が好ましい。培地中にカドミウムを添加すれば、より鮮明に赤く発色する為、カドミウムを添加することがより好ましい。培地は、メチオニンを追加することが好ましい。なお、株はコロニーを形成し、コロニーの色で赤色性を評価できるため、液体培地よりも寒天培地を用いることが好ましい。培地上で赤色を呈するようにするための方法は特に制限されないが、アデニン要求性を付与することやメチオニンが添加された最小培地で培養することが例示される。
【0016】
本発明において、アデニン要求性を有するとは、アデニンを含まない固体培地上では株がコロニーを形成できないか、あるいは後述するY-3219株と同程度の小さいコロニーを形成することを意味する。アデニン要求性の原因となる遺伝子は特定されていても特定されていなくてもよいが、特定されている方が好ましい。アデニン要求性の原因となる遺伝子は、得られたアデニン要求性の株を相補性分析または配列分析によって特定してもよい。本発明の酵母は、アデニン生合成経路の遺伝子のいずれかが不活性化するように改変されたことにより、アデニン要求性を有することが好ましい。アデニン生合成経路の遺伝子としては、GSHと結合する中間体AIRやCAIRの蓄積にダイレクトに影響するADE1遺伝子、ADE2遺伝子などが挙げられる。本発明において、「不活性化するように改変する」とは、遺伝子産物の活性が失われるような変異を遺伝子に導入すること、および遺伝子が発現しなくなるようにすることを含む。
【0017】
本発明において、遺伝子改変処理とは、親株の塩基配列が変異することを意味する。従来法による変異技術を用いても良いし、遺伝子組み換え技術を用いてもよい。従来法による変異技術の例としては、紫外線、レーザーなどの照射によって変異をもたらす方法、EMS(メタンスルホン酸エチル)、MNNG(N-メチル-N'-ニトロ-N-ニトロソグアニジン)、DAPA(4-ジメチルアミノベンゼンジアゾスルホン酸ナトリウム)等の変異誘発剤を用いる方法などがある。また、酵母を培養している際に生じる自然変異を利用してもよい。酵母に変異処理を施し、アデニンを含む培地では生育し、アデニンを含まない培地では生育しないか、あるいはほとんど生育できない株を選択することにより、アデニン要求性株を得ることができる。また、ADE1遺伝子又はADE2遺伝子に変異を有する酵母はアデニン要求性であり、赤く発色する。
【0018】
遺伝子組み換え技術を用いる方法としては、相同組み換えを利用して染色体上の野生型遺伝子を変異型遺伝子(不活性型遺伝子または破壊型遺伝子)で置換する方法などが挙げられる。
【0019】
なお、上述の遺伝子置換は、以下のようにして行うことができる。まず、変異型ADE1遺伝子を含む組換えDNAで酵母を形質転換し、変異型ADE1遺伝子と染色体上のADE1遺伝子との間で組換えを起こさせる。その際、組換えDNAには、宿主の栄養要求性等の形質にしたがって、マーカー遺伝子を含ませておくと操作がしやすい。また、組換えDNAは、制限酵素で切断する等により直鎖状にし、さらに、酵母で機能する複製制御領域を除いておくと、染色体に組換えDNAが組み込まれた株を効率よく取得することができる。
酵母の形質転換は、プロトプラスト法、KU法、KUR法、エレクトロポレーション法等、通常酵母の形質転換に用いられる方法を採用することができる。上記のようにして染色体に組換えDNAが組み込まれた株は、変異型ADE1遺伝子と染色体上にもともと存在す
るADE1遺伝子との組換えを起こし、野生型ADE1遺伝子と変異型ADE1遺伝子との融合遺伝子2個が組換えDNAの他の部分(ベクター部分及びマーカー遺伝子)を挟んだ状態で染色体に挿入されている。
融合遺伝子2個のうち、変異型ADE1遺伝子のみを残すために、2個のADE1遺伝子の組換えにより1コピーのADE1遺伝子を、ベクター部分(マーカー遺伝子を含む)とともに染色体DNAから脱落させる。その際、2つのケースが起こる。1つのケースでは、野生型ADE1遺伝子が染色体DNA上に残され、変異型ADE1遺伝子が切り出される。もう一つのケースでは、反対に変異型ADE1遺伝子が染色体DNA上に残され、野生型ADE1遺伝子が切り出される。いずれの場合もマーカー遺伝子が脱落するので、2回目の組換えが生じたことは、マーカー遺伝子に対応する表現形質によって確認することができる。また、目的とする遺伝子置換株は、PCRによりADE1遺伝子を増幅し、その構造を調べることによって、選択することができる。
以上、ADE1遺伝子を例に説明したが、ADE2遺伝子やその他の遺伝子も同様にして改変することができる。
【0020】
なお、ADE1遺伝子及び/またはADE2遺伝子に加えて、ADE4遺伝子またはADE8遺伝子に変異が入った場合、酵母は赤色ではなく通常の白色となる為更に好ましい。なお、ADE4遺伝子またはADE8遺伝子に変異を導入する場合のスクリーニングは、まずADE1遺伝子やADE2遺伝子に変異を導入し、赤色の発色度合いを指標に含硫化合物含量が増加した酵母を取得した後、ADE4遺伝子またはADE8遺伝子を変異させれば効率的に白色の含硫化合物高含有酵母を取得できる。
ADE4遺伝子またはADE8遺伝子の変異もADE1遺伝子、ADE2遺伝子と同様に導入することができる。
【0021】
ADE1遺伝子、ADE2遺伝子、ADE4遺伝子、ADE8遺伝子は改変対象の酵母自身の遺伝子を用いることができ、例えば、Saccharomyces cerevisiaeのADE1遺伝子、ADE2遺伝子、ADE4遺伝子、ADE8遺伝子としては、それぞれ、配列番号1、3、5、15の塩基配列を有するDNAが挙げられる。また、ADE1遺伝子、ADE2遺伝子、ADE4遺伝子、ADE8遺伝子は、遺伝子改変株の創製に用いる酵母の各遺伝子と相同組換えを起こす程度の同一性を有していれば特に制限されない。それゆえ、ADE1遺伝子、ADE2遺伝子、ADE4遺伝子、ADE8遺伝子は、それぞれ配列番号2、4、6、16に示すアミノ酸配列と、80%以上、好ましくは90%以上、より好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列をコードするDNAであればよい。また、ADE1遺伝子、ADE2遺伝子、ADE4遺伝子、ADE8遺伝子は、配列番号1、3、5、15に示す塩基配列を有するDNAと、ストリンジェントな条件下でハイブリダイズするDNAであってよい。ストリンジェントな条件としては、65℃、0.1×SSC、0.1%SDSに相当する塩濃度で1回の洗浄、好ましくは2回または3回の洗浄が行われる条件が挙げられる。
【0022】
本発明において、セレン酸感受性を有するとは、セレン酸とメチオニンを含有する培地(例:5mMのセレン酸と1mMのメチオニンを含有する培地)上ではコロニーを形成できないことをいう。セレン酸は硫酸のアナログで細胞毒性を示す。通常の酵母は硫酸の取り込み系がメチオニンなどの有機硫黄化合物によって抑制されているのでセレン酸は細胞内に取り込まれず、セレン酸含有培地でも生育する。一方、セレン酸感受性株は硫酸の取り込み系がメチオニンなどの有機硫黄化合物によって抑制されず、セレン酸を取り込んでしまう。硫酸の取り込み系が抑制されないため、セレン酸感受性株では含硫化合物の生合成が野生株より向上している(MOLLECULAR AND CELLUAR BIOLOGY Dec, 1995, p6526-6534)。
【0023】
セレン酸感受性は変異処理により付与されてもよいし、遺伝子組み換えによって付与されてもよい。例えば、酵母に変異処理を施し、セレン酸とメチオニンを含む培地では生育
できない株を選択することにより、セレン酸感受性株を得ることができる。
【0024】
一方、セレン酸感受性を付与するための遺伝子組み換えとしては、O-acetylhomoserine
sulfhydrylaseをコードするMET25遺伝子の発現を野生株と比べて増強するように改変することが挙げられる。MET25遺伝子の発現を増強する方法としては、細胞内へのプラスミドの導入や、染色体上への導入などによりMET25遺伝子のコピー数を増大させることや、MET25遺伝子のプロモーターを強力なプロモーターに置換する方法が例示される。
【0025】
また、MET25遺伝子の転写調節因子を改変することによってMET25遺伝子の発現を増強することもできる。MET25遺伝子の発現については、次のように考えられている。MET4産物は、MET25遺伝子の発現に正の因子として働く。通常は、MET4産物は、MET30産物及び他の数種のタンパク質とともにSCFMET30複合体を形成し、ユビキチン化を受け、26Sプロテアソームによるタンパク分解系によりMET30産物とともに分解されるため、MET25遺伝子の発現は抑制されている。一方、SCFMET30複合体の機能が低下すると、MET4産物及びMET30産物は分解を受けず、MET25遺伝子が発現する。(Patton et al.,Genes Dev.12: 692-705, 1998、Rouillon et al., EMBO Journal 19: 282-294, 2000)したがって、MET4遺伝子やMET30遺伝子に変異を導入し、MET25遺伝子の発現を増強することもできる。
MET25遺伝子の発現量を増強することができる変異型MET4遺伝子としては、215番目のセリンがプロリンに、または、156番目のイソロイシンがセリンに変異したMET4遺伝子が報告されている(大村ら FEBS Letters 387(1996) 179-183、特開平10-33161号公報)。
また、MET25遺伝子の発現量を増大させる変異型MET30遺伝子としては、WD40リピートに変異を導入することにより得られる変異型MET30遺伝子が報告されている。また、特開2004-201677には、569位のセリンをフェニルアラニン等の他のアミノ酸に置換する変異を有する変異型MET30遺伝子が報告されている。
このような変異型遺伝子で染色体上の野生型遺伝子を置換することにより、MET25遺伝子を発現増強させることができ、セレン酸感受性を付与できる。
【0026】
MET25遺伝子、MET30遺伝子、MET4遺伝子としては、改変された酵母のように、何らかの酵母の遺伝子であってよい。例えば、Saccharomyces cerevisiaeのMET25遺伝子、MET30遺伝子、MET4遺伝子として、それぞれ、配列番号9、11、7の塩基配列を有するDNAが挙げられる。また、MET25遺伝子はO-acetylhomoserine sulfhydrylase活性を有するタンパク質をコードする限り、また、MET30遺伝子、MET4遺伝子は、Met25遺伝子の発現を抑制するタンパク質をコードする限り特に制限されない。それゆえ、MET25遺伝子、MET30遺伝子、MET4遺伝子は、それぞれ、配列番号10、12、8に示すアミノ酸配列と、80%以上、好ましくは90%以上、より好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列をコードするDNAであってもよい。MET25遺伝子、MET30遺伝子、MET4遺伝子は配列番号9、11、7に示す塩基配列を有するDNAと、ストリンジェントな条件下でハイブリダイズするDNAであってもよい。ストリンジェントな条件としては前述のものが挙げられる。
【0027】
本発明の酵母はセレン酸感受性にアデニン要求性を組み合わせることによって、対応する酵母のセレン酸感受性かつアデニン非要求性の株よりも細胞内の含硫化合物含量が増加している。
本発明において含硫化合物とは、化学式内に-SH基を有するものをいい、タンパク質、ペプチド、アミノ酸、或いはその他の物質であってもよい。タンパク質としては、構成アミノ酸の30%がシステイン残基からなるメタロチオネイン、ペプチドとしてはグルタチオン、γ−グルタミルシステイン、システニルグリシンなどを、アミノ酸としてはシステインを、その他の物質としてはホモシステインなどを例示することができるが特にこれらに限定されるものでもない。グルタチオン、γ−グルタミルシステイン、システインは、
現在幅広く産業上で利用されているので、対象とする含硫化合物として特に好ましい。
【0028】
含硫化合物としてグルタチオンを酵母内に蓄積させる場合は、さらに、酵母細胞内でグルタチオン合成酵素とγ−グルタミルシステイン合成酵素の一方又は両方の活性を増強させてもよい。酵素活性の増強は、これらの酵素をコードする遺伝子のコピー数を増加させるか、あるいはプロモーターを強力なプロモーターに置換することによって行うことができる。Saccharomyces cerevisiaeのγ−グルタミルシステイン合成酵素遺伝子の配列と、Candida utilisのグルタチオン合成酵素遺伝子およびγ−グルタミルシステイン合成酵素遺伝子の配列は特開2005−073638に開示されている。また、Saccharomyces cerevisiaeのグルタチオン合成酵素遺伝子の塩基配列とコードされるアミノ酸配列を配列番号13、14に示した。
【0029】
酵母の細胞内に、含硫化合物として、γ−グルタミルシステインを蓄積させる場合には、酵母をグルタチオン合成酵素活性が低下するように改変することが好ましく、グルタチオン合成酵素活性が低下し、かつ、γ−グルタミルシステイン合成酵素活性が増大するように改変することがより好ましい。グルタチオン合成酵素活性が低下するような改変は、グルタチオン合成酵素遺伝子を破壊するか、あるいは、グルタチオン合成酵素活性が低下するような変異を導入することが挙げられる。
グルタチオン合成酵素活性が低下するような変異としては、例えば、配列番号14のアミノ酸配列の370位のアルギニン残基を終止コドンに変更する変異が挙げられる。
グルタチオン合成酵素活性が低下するような変異の他の例として下記の変異が挙げられる(国際公開03/046155号パンフレット)
(1)配列番号14のアミノ酸配列の47位のスレオニンをイソロイシンに置換する変異。
(2)配列番号14のアミノ酸配列の387位のグリシンをアスパラギン酸に置換する変異。
(3)配列番号14のアミノ酸配列の54位のプロリンをロイシンに置換する変異。
上記変異は、(1)または(2)の単独変異でも、(1)〜(3)の任意の組合せでもよいが、(1)と(3)の組合せ、及び(2)と(3)の組合せが好ましい。
なお、システインはγ−グルタミルシステインを熱処理することによって生成するため、γ−グルタミルシステインを蓄積した酵母を熱処理すればシステインが蓄積した酵母が得られる。
また、含硫化合物として、システニルグリシンを蓄積させる場合には、酵母をECM38活性が増強するように改変することが好ましい(Yeast. 2003 Jul 30;20(10):857-63)。
グルタチオンやγ−グルタミルシステイン等の含硫化合物の細胞内含量が増加した酵母は、ラパマイシン耐性を示す酵母から得ることもできる(WO2006/013736)。
【0030】
上記のアデニン要求性およびセレン酸感受性を有する酵母については、さらに遺伝子改変処理、好ましくは変異処理を施して、アデニンが不足した時に酵母が赤く発色する培地にスプレッドし、改変前よりもより赤く発色する酵母を選択することにより、さらに含硫化合物の含量を増加させることができる。
本発明において、スクリーニングに用いる培地は、酵母細胞内のアデニンが不足した時に酵母が赤く発色することができる培地であれば特に制限されない。アデニン含量が25mg/L以下の培地を挙げることができ、具体的にはYPD培地(組成:Bacto-yeast extract 1%、Bacto-peptone 2%、Glucose 2%。METHODS IN YEAST GENETICS 2000 Edition p171参照。ISBN 0-87969-588-9)PGC培地(組成:Casamine acid(vitamin free) 0.5%、Bacto-peptone 1%、Glucose 2%。)等を例示することができる。微量成分との影響でより赤く発色しやすいPGC培地の方が好ましい。培地中にカドミウムを添加すれば、より鮮明に赤く発色する為、カドミウムを添加することがより好ましい。なお、菌株の単離を考えれば液体培地よりも寒天培地を用いることが好ましい。発色度合いを識別するためには、固体
培地上で20〜30℃で約1週間培養することが好ましい。
【0031】
また、セレン酸感受性を有する酵母について、遺伝子改変処理、好ましくは変異剤処理を行い、最小培地にメチオニンを追加した培地に該処理された酵母をスプレッドし、培地中に出現した改変酵母の発色度合いを識別し改変前よりも赤く発色する酵母を選択することにより、細胞内の含硫化合物含量が増加した酵母をスクリーニングすることもできる。ここで使用する培地としては、擬似的にアデニン要求性と同じ状態を作り出すことのできるメチオニン含有培地であればよいが、ビオチンの非存在下でメチオニンが添加された最小培地が好ましく、具体的には、例えば、後述のmin-met(+)-biotin(-)プレートなどが挙げられる。
【0032】
本発明の酵母を培養することにより含硫化合物が蓄積した酵母を製造することができる。培養方法として好ましくは、本発明の酵母を、十分な量のアデニンを含む培地(アデニン充足状態)で培養して同酵母を増殖させた後、アデニン量が制限された培地で培養し、菌体内の含硫化合物含量を増加させる方法が挙げられる。これにより、含硫化合物が蓄積した酵母を効率よく製造することができる。
好ましくは、前記アデニンの「十分な量」は、予め一定量の酵母菌体を得るのに必要なアデニン要求量を実験的に測定し、目的の量の菌体を得るために必要なアデニン量を算出することによって、決定することができる。例えば、100mg/L以上が挙げられる。
アデニン量以外の培地及び培養条件は、通常の酵母培養に用いられる培地及び条件を採用することができる。尚、必要に応じて、用いる酵母の形質にしたがって必要な栄養素を培地に添加する。
【0033】
十分な量の酵母菌体が得られたら、アデニン量が制限された培地で培養する。アデニン量が制限された培地としては、アデニン含量が25mg/L以下の培地が挙げられる。アデニン量が制限された培地で培養する間に、酵母菌体内の含硫化合物の蓄積量は経時的に上昇する。
好ましくは、目的とする含硫化合物の蓄積量に達したら、培養を終了する。通常は、培養時間は10〜30時間、好ましくは15〜27時間である。
【0034】
上記のようにして得られる培養物又はその分画物は、含硫化合物を含有する。培養物は、酵母菌体を含む培養培地であってもよいし、それから採取された酵母菌体であってもよい。含硫化合物を含む分画物は、細胞破砕物または酵母抽出物であっても良い。酵母エキス等の調製は、通常の酵母エキスの調製と同様にして行えばよい。酵母エキスは、酵母菌体を熱水抽出したものを処理したものでもよいし、酵母菌体を酵素消化したものを処理したものでもよい。
【0035】
含硫化合物は上記の酵母細胞から単離することができる。上記含硫化合物、酵母培養物又はその分画物は、食品、医薬品、化成品、飼料等の製造に用いることができる。食品としては、アルコール飲料、パン食品、又は発酵食品調味料が挙げられる。食品は、含硫化合物、培養細胞又はその分画物を、食品原料に混合し、食品に加工することによって製造される。
【実施例】
【0036】
以下、本発明を実施例に基づき説明する。なお、本発明は以下の実施例になんら限定されるものではない。
【0037】
<実施例1>
(アデニン要求性酵母の取得)
変異型MET30遺伝子を保有するサッカロミセス・セレビシエ1倍体AJ14819株(MATα型
))とサッカロミセス・セレビシエ1倍体AJ14810株(MATa型)を接合させることにより、サッカロミセス・セレビシエ2倍体株を取得した。AJ14819株はセレン酸感受性株であり、2003年10月1日にブダペスト条約に基づき独立行政法人 産業技術総合研究所 特許生物寄託センター(日本国、305-8566 茨城県つくば市東1丁目1-1つくばセンター 中央第6)にFERM BP-8502の受託番号で寄託されている。AJ14180株は、2002年11月1日にブダペスト条約に基づき、独立行政法人 産業技術総合研究所 特許生物寄託センターにFERM BP-8229の受託番号で寄託されている。
同2倍体株を胞子形成させ、4分子解析することにより以下の性質を有する菌株を取得した。
A:MATa型1倍体、変異型MET30遺伝子
B:MATα型1倍体、変異型MET30遺伝子
C:MATa型1倍体、野生型MET30遺伝子
D:MATα型1倍体、野生型MET30遺伝子
【0038】
次に、取得した4菌株のMET25遺伝子の発現量を比較した。特開2004-201677実施例1記載の方法に基づき、これら4株のMET25遺伝子発現量を以下のようにして測定した。菌株を各々YPD培地に植菌し(坂口フラスコ500ml容、50ml張り込み)、30℃で振とう培養した。その対数増殖期に集菌し、菌体内に含まれているRNAを回収し、MET25遺伝子の発現を内部標準としてACT1遺伝子を用いて定量した。定量は、PCR5700(Applied Biosystems社)を用い、TaqMan One-Step RT-PCRキット(Applied Biosystems社)を用いて行なった。TaqMan Probe(Applied Biosystems社)に、特開2004-201677実施例1記載のACT1-986T及びMET25-1077Tを、ACT1遺伝子の増幅用に特開2004-201677実施例1記載のACT1-963FとACT1-1039Rを、そして、MET25遺伝子の増幅用にMET25-1056FとMET25-1134Rをプライマーとして用いた。その結果、変異型MET30遺伝子およびセレン酸感受性を有するA株及びB株のMET25遺伝子の発現量は、変異型MET30遺伝子を有しないC株及びD株よりも高いことを確認した。B株にプライベート番号AJ14889及びA株にプライベート番号AJ14890を付した。
【0039】
次に、常法に従いAJ14889株を変異剤MNNG(1-methyl-3-nitro-1-nitrosoguanidine)で生存率5〜10%になる条件で処理した後、YPDプレートにスプレッドし30℃で約1週間培養した。出現したコロニーの中から赤色に発色する菌株を選択した。次に、得られた菌株につき、常法に従い、アデニン要求性の相補テストを行った。その結果、ADE1遺伝子変異によるアデニン要求性酵母N1株及びADE2遺伝子変異によるアデニンリーキー変異酵母N2株を取得した。(N1株およびN2株は、2008年12月23日、ブダペスト条約に基づきRussian
National Collection of Industrial Microorganisms(VKPM)(1 Dorozhny proezd., 1 モスクワ 117545, ロシア)にそれぞれVKPM Y-3218、VKPM Y-3219で国際寄託されている。)
【0040】
<実施例2>
(強い赤色発色を指標とした高GSH蓄積変異体のスクリーニング)
次に、常法に基づきADE1遺伝子に変異を有するY-3218株を変異剤MNNGで生存率5〜10%になる条件で処理した後、PGC又はYPDプレートにスプレッドし30℃で約1週間培養した。出現したコロニーの中から、Y-3218株よりもより赤く発色する菌株を選択し、GSH含量をAJ14889株(親株)およびY-3218株と比較した。選択した株、AJ14889株およびY-3218株をそれぞれ5mlのYPD液体培地に植菌し、30℃、250rpmの振とう数で24時間培養した。50mlのYPD液体培地にそれぞれの株の得られた培養物を植菌し、30℃、250rpmの振とう数で24時間培養した。それぞれの株のGSH含有量を承保に従って測定した結果、AJ14889株に比べY-3218株のGSH含量は高かった(表1)。選択した株(計180株)のうち7株がY-3218株よりGSH含量が高いことがわかった(表1)。
【0041】
【表1】
【0042】
ADE2遺伝子変異株Y-3219株を用いて同様の検討を行った。常法に基づきY-3219株を変異剤MNNGで生存率5〜10%になる条件で処理した後、PGC又はYPDプレートにスプレッドし30℃で約1週間培養した。出現したコロニーの中から、別途スポットしていたY-3219株よりもより赤く発色する菌株を選択し、GSH含量をAJ14889株(親株)およびY-3219株と比較した。YPD寒天培地よりコロニーをストリークし、選択した株、AJ14889株およびY-3218株をそれぞれ5mlのYPD液体培地に植菌し、30℃、250rpmの振とう数で24時間培養した。50mlのYPD液体培地にそれぞれの株の得られた培養物を植菌し、30℃、250rpmの振とう数で24時間培養した。それぞれの株のGSH含有量を承保に従って測定した結果、AJ14889株よりもY-3219株のGSH含量が高いことがわかった。選択した株(計118株)のうち10株がY-3218株よりGSH含量が高いことがわかった(表2)。
【0043】
【表2】
【0044】
表1、表2に示すようにアデニン要求性による酵母の赤色の発色を指標としてGSH含量
が向上した酵母を単離することが可能であった。
【0045】
<実施例3>
(GSH高含量の白色コロニーの単離)
野生型の酵母は赤色ではなく、白色〜クリーム色である。そこで、発明者らは赤色の発色度合いを指標にスクリーニングした酵母から通常の色のコロニーを形成する株の取得を検討した。まず、実施例2と同様にY-3219株をMNNGで変異処理し、PGC培地またはYPD培地に播いてより赤色の強い株を得た。その結果、実施例2と同様に表3のY-3219-20株が得られた。次に、Y-3219-20株をMNNGで、生存率5〜10%になるように変異処理を行い、PGCプレートにスプレッドし、30℃で約1週間培養した。出現したコロニーの中から、3つの白コロニー(Y-3219-20-52、Y-3219-20-53、Y-3219-20-56)と1つの赤コロニー(Y-3219-20-1)を選抜し、それらのGSH含量を測定し、Y-3219株、Y-3219-20株と比較した。すなわち、得られた株と、Y-3219株及びY-3219-20株を5mLのYPD液体培地に植菌し、30℃、250rpmの振とう数で24時間培養した。次に、50mlのYPD液体培地にそれぞれの株の得られた培養物を植菌し、30℃、250rpmの振とう数で24時間培養した。それぞれの菌体内のGSH量を常法に従って測定した。結果を表3に示す。
【0046】
【表3】
【0047】
白色の菌株を試験株を用いて遺伝解析した。その結果、GSH含量が減少したY-3219-20-52株ではアデニン要求性を失っていた。一方、高いGSH含量を維持したY-3219-20-56株では、ADE2遺伝子変異に加えて、ADE4遺伝子も変異していた。Y-3219-20-53株では、ADE2遺伝子変異に加えて、ADE8遺伝子も変異していた。この結果より、赤色コロニーを形成するアデニン要求性の株はADE4遺伝子またはADE8遺伝子が変異することで、通常の白色コロニーを形成する株になり得ることがわかった。
Y-3219-20-56株は、2008年12月23日、ブダペスト条約に基づきRussian National Collection of Industrial Microorganisms(VKPM)(1 Dorozhny proezd., 1 モスクワ 117545, ロシア)にVKPM Y-3256で国際寄託されている。
【0048】
<実施例4>
(アデニン不充足による高GSH蓄積)
アデニン充足度合いとGSH含量との関係について検討した。Y-3256株を500ml容坂口フラスコ中の50mlのYPD培地(D-glucose 20g/L、Bact Peptone 20g/L、Yeast Extract 10g/L)で30℃、120rpmで24時間振とう培養し、得られた培養物を、500ml容坂口フラスコ中の異なる濃度のアデニン(最終アデニン濃度0mg/L、10mg/Lまたは20 mg/L)を加えた50mlのYPD培地に、初期吸光度660nm=0.1になるように植菌し、30℃、120rpmで振とう培養した。その結果、GSH量経時変化は表4の通りであった。
【0049】
【表4】
【0050】
これらの結果から、アデニン要求性Y-3256株のGSH含量は培地中のアデニン量に反比例していることが明らかとなった。なお、YPD培地には微量のアデニンが含まれているため、アデニン添加量なし(Ade 0mg/L)の場合でもY-3256株は生育可能である(YPD培地のアデニン含量は約10mg/Lであった)。
【0051】
(Jar Fermentor評価)
Y-3256株をJar Fermentorで培養し、GSH含量の経時変化を評価した。YPD寒天培地より菌株細胞を拾い、50mlのYPD培地を含む750ml容三角フラスコ3つに植菌し、30℃、250rpmで20時間振とう培養した。得られたシード培養液(120ml)を3L容Jar-Fermentor中の1.2Lのメイン培地(YPD培地)に植菌し、30℃、攪拌速度1,100rpmで培養した。この時の通気量は1/1vvmに設定し、pHはアンモニア水により6.0に制御した。培養0hr〜24hrの間は1.5ml/hrの速度で、24hr以降は1.8ml/hrの速度で流加培地を連続フィードした。なお、このとき使用した流加培地の組成は、1LあたりGlucose600g、Bacto-yeast extract 10g、corn
extract 10g、Bacto-peptone 10g、(NH4)2SO4 0.274g、KH2PO4 0.11g、KCl 0.732g、MgSO4 0.466g、CuSO40.0012g、ZnSO4 0.014g、MnSO4 0.00334g、NaMoO40.00012g、KCl 0.002g、H3BO30.00004g、CoSO4 0.0001g、CaCl2 0.28g、FeSO4 0.2g、Biotine 0.05mg、riboflavin 0.2mg、thiamine 0.5mgである。
この時のY-3256株のGSH含量変化は図1の通りであった。
【0052】
<実施例6>
(2倍体株の取得及び培地組成の検討)
常法に基づき、Y-3219-20-53株の自己2倍体化を試みた。実験の条件は以下のとおりである。Y-3219-20-53株を常法により、生存率5〜10%になるような条件で変異剤MNNGで処理し、YPD寒天培地に100〜200個のコロニーが出現するようにスプレッドした。30℃で5日間培養後、アデニンを添加したSD寒天培地及びYPD寒天培地にレプリカした。YPD寒天培地では生育可能であるが、アデニンを添加したSD寒天培地では生育できない菌株を選抜した。アデニン以外の栄養要求性を調べ、栄養要求性が異なるY-3219-20-53-aux1株及びY-3219-20-53-aux2株を取得した。Y-3219-20-53-aux1株をアデニンを添加したSD寒天培地に縦方向の直線になるようにストリークし、Y-3219-20-53-aux2株を同じアデニンを添加したSD寒天培地に横方向の直線になるようにストリークした。但し、この時縦方向の直線と横方向の直線が1点で交差するように行った。このアデニンを添加したSD寒天培地を30℃で20日間培養し、交差点に出現するコロニーを選択した。この様にしてアデニン要求性の2倍体株であるD1-3株を取得した。本菌株は、2008年12月23日、ブダペスト条約に基づきRussian National Collection of Industrial Microorganisms(VKPM)(1 Dorozhny proezd., 1 モスクワ 117545, ロシア)にVKPM Y-3309で国際寄託されている。なお、D1-3株をアデニンを添加したSD寒天培地にストリークし、30℃で2日間培養したところ、コロニーが出現したことから、D1-3株はアデニン以外の栄養要求性が付与されていないことが確認できた。
また、アデニン非要求性の対象区として、AJ14889株とAJ14890株を接合させることによ
り2倍体Dip株を取得した。
D1-3株及びDip株を、5mlのYPD液体培地を入れた試験管に植菌し、30℃、250rpmの振とう数で24時間培養した。得られた培養物を、表5に示す組成の各培地50mlに植菌し、30℃、250rpmの振とう数で24時間培養した。各菌体内のGSH含量を測定した。その結果は表6に示すとおりである。
【0053】
【表5】
【0054】
【表6】
【0055】
このようにいずれの培地においてもアデニン要求性によってGSH含量が増加していた。
【0056】
<実施例7>
(アデニン要求性のγ−グルタミルシステイン高含有酵母の育種及びγ−グルタミルシステイン含量向上効果の評価)
実施例3にて取得したY-3256株のグルタチオン合成酵素をコードするGSH2遺伝子を破壊することによりγ−グルタミルシステインを蓄積する1倍体株を取得した。GSH2遺伝子破壊カセットを構築するために、プライマー17〜22を用い、S. cerevisiae(野生株)のゲノムDNAおよびプラスミドpFA6a-KanMX6(Chiara et al., Yeast 2000, 16:1089-1097)を鋳型にしてPCRを行った(図2)。詳細な条件は以下のとおりである。
【0057】
まず、GSH2のORFから約400bp上流の領域をX2180-1B(S. cerevisiae 野生型株:ATCCにて受託番号ATCC204505で入手可能)のゲノムDNAを鋳型に、GSH2-up-Fプライマー(配列番号17)およびGSH2-up-Rプライマー(配列番号18)を用いて増幅し、GSH2上流フラグメントを得た。GSH2上流フラグメントは、GSH2-up-FプライマーおよびGSH2-up-Rプライマーの配列に基づき、その一方の末端側にBpiI制限部位を有し、もう一方の末端側に後述するフュージョンPCR用のcon1部位を有する。
また、GSH2のORFから約300bp下流の領域をX2180-1BのゲノムDNAを鋳型に、GSH2-down-Fプライマー(配列番号19)およびGSH2-down-Rプライマー(配列番号20)を用いて増幅しGSH2下流フラグメントを得た。GSH2下流フラグメントは、GSH2-down-FプライマーおよびGSH2-down-Rプライマーの配列に基づき、その一方の末端側にBpiI制限部位を有し、もう一方の末端側にフュージョンPCR用のcon2部位を有する。
KanMX遺伝子をpFA6a-KanMX6プラスミドを鋳型に、マーカーFプライマー(配列番号21
)とマーカーRプライマー(配列番号22)を用いてPCR増幅した。このPCRで増幅されたKanMX遺伝子は、マーカーFプライマーおよびマーカーRプライマーの配列に基づき、その一方の末端側にcon1部位を有し、もう一方の末端側にcon2部位を有する。
これらのPCRの条件は次のとおりである。DNAポリメラーゼの混合物(Pfu:Taq=1:10;Fermentas, Lithuaniaより入手可能)をそれぞれの反応に用いた。PCRは94℃30秒、50℃30秒、68℃3分のサイクルを30回繰り返した。
【0058】
得られた3フラグメントをフュージョンPCRで結合させてGSH破壊カセットを構築した。GSH2上流フラグメントとKanMX遺伝子フラグメントを鋳型に用い、GSH2-up-FプライマーとマーカーRプライマーを用いてフュージョンPCRを行った。GSH2上流フラグメントとKanMX遺伝子フラグメントはいずれも末端にcon1部位を有するため、フュージョンPCRを行うことにより、これらが連結されたGSH2上流-KanMXフラグメントが得られる。このフュージョンPCRは前記と同様のDNAポリメラーゼの混合物を用いた。PCRは、94℃30秒、61℃30秒、68℃4.5分のサイクルを5回、次いで94℃30秒、50℃30秒、68℃4.5分のサイクルを25回繰り返した。
次に、GSH2上流-KanMXフラグメントとGSH2下流フラグメントを鋳型に用い、GSH2-up-FプライマーとGSH2-down-Rプライマーを用いてフュージョンPCRを行った。GSH2上流-KanMXフラグメントとGSH2下流フラグメントはいずれも末端にcon2部位を有するため、GSH2上流-KanMXフラグメントとGSH2下流フラグメントが連結され、GSH2破壊カセットが得られる。このフュージョンPCRは前記と同様のDNAポリメラーゼの混合物を用いた。PCRは、94℃30秒、61℃30秒、68℃5.3分のサイクルを5回、次いで94℃30秒、50℃30秒、68℃5.3分のサイクルを25回繰り返した。
【0059】
プライマーの配列は以下のとおりである。
(1) GSH2-up-F, CCGAAGACCTTCGTTTGGTGTTATGGT (配列番号17)
(2) GSH2-up-R, GAGAGGGGGGGGGTGGGGGGAAGGTGGATAGTGTGCC (配列番号18)
(3) GSH2-down-F, CCTCCTCCCCCCGCCCACGGCAGGATTCGGATGTTTG (配列番号19)
(4) GSH2-down-R, CGAAGACTCAGTACGAGCATTACGCAA (配列番号20)
(5) Marker-F, 5’-CCCCACCCCCCCCCTCTCTACCGTTCGTATAATGTATGCTATACGAAGTTATACTGGATGGCGGCGTTAG-3’ (配列番号21)
(6) Marker-R, 5’-GTGGGCGGGGGGAGGAGGTACCGTTCGTATAGCATACATTATACGAAGTTATGTTTAGCTTGCCTCGTCC-3’ (配列番号22)
【0060】
GSH2破壊カセットを用いてY-3256株を形質転換し相同組み換えを起こさせた、得られた形質転換体をG418(50μg/ml)を含有するYPD寒天培地にスプレッドした。出現したコロニーから、GSH2遺伝子が破壊された菌株であるN8△GSH2株を取得した。
同菌株を500ml容坂口フラスコ中の50mlのYPD培地(D-glucose 20g/L、Bact Peptone 20g/L、Yeast Extract 10g/L)に植菌し、30℃、120rpmで24時間振とう培養後、得られた培養物を、異なる濃度のアデニン(最終アデニン濃度0mg/L、10mg/L、20mg/L)を添加した500ml容坂口フラスコ中のYPD培地50mlに初期吸光度660nm=0.1になるように植菌し、30℃、120rpmで振とう培養した。γ−グルタミルシステイン濃度を常法に従い測定した。表7にγ−グルタミルシステインの経時変化を示す。γ−グルタミルシステイン濃度はアデニン要求性の付与により高められ得ることがわかった。
【0061】
【表7】
【0062】
<実施例8>
(擬似的なアデニン要求性条件を作り出す培地での評価)
常法に従い、AJ14889株を変異剤MNNGで生存率が約10%になる条件で処理した後、表8に記載の成分および150mg/Lのメチオニンを含むmin-met(+)-biotin(-)プレートに、1プレートあたりのコロニー数が350個以下になるようにスプレッドした。30℃で8〜10日間培養した後、30枚のプレートに出現したコロニーの中から、薄いピンク色から赤色に発色しているコロニーを106株ピックアップした。
次に、AJ14889株(親株)及び選抜した菌株を各々YPD培地に植菌し菌体内のGSH含量を測定した。具体的には、YPD培地で培養した選抜した菌株とAJ14889株をそれぞれ、5mlのYPD液体培地を入れた試験管に植菌し、30℃、250rpmの振とう数で24時間培養した。そして、750ml容の三角フラスコに50mlのYPD培地を入れたメイン培地に前培養液を、600nmでの吸光度が0.1になるように植菌し、30℃、250rpmの振とう数で24時間及び48時間培養した。その結果、選択された106株中8株は、24hr時又は48hr時にAJ14889株よりもGSH含量が上回っていた(表9)。選択した108株中8株がGSH含量が向上した菌株であったことから、本方法は非常に効率的なスクリーニング方法であることが示された。
【0063】
【表8】
【0064】
【表9】
【0065】
次に、上記8株より選択した89-6株、89-28株、89-31株について、生育とGSH含量をAJ14889株と比較した。これらの株は、600nmでの吸光度が0.3となるように植菌したことを除いては、前述のように培養した。生育は乾燥菌体重量(DCW)の変化に基づいて測定した。表10に示すように、選択した3株は、24hr時及び48hr時において生育が親株とほぼ同等で、GSH含量は親株を上回っていた。
【0066】
【表10】
【0067】
また、この3株に予期しない栄養要求性が付与されていないか検証するために、AJ14889株及びこの3株をSD培地に植菌し、その生育の有無を調べた。具体的には、YPD寒天培地で培養しておいた菌株を、5mlのSD培地を入れた試験管に植菌し、30℃、250rpmの振とう数で、それぞれ24時間および48時間培養した。その結果、選択した3株はSD培地でAJ14889株とほぼ同等の生育を示し(表11)、これら3株に予期しない栄養要求性は付与されていないことが示された。
【0068】
【表11】
【産業上の利用可能性】
【0069】
本発明によって、細胞内の含硫化合物含有量が増加したアデニン要求性酵母、該酵母のスクリーニング方法、及び該酵母の培養方法が提供される。本発明は、食品、医薬品、化成品、飼料等の幅広い産業で使用することが可能である。
【技術分野】
【0001】
本発明は、含硫化合物含有酵母、そのスクリーニング方法、及び該酵母を培養する方法に関するものである。本発明によって得られる含硫化合物高含有酵母は、食品、医薬品、化成品、飼料等の分野で有用である。
【背景技術】
【0002】
現在、含硫化合物は、食品、医薬品、化成品などの幅広い分野で使用されている。例えば、グルタミン酸、システインおよびグリシンからなるトリペプチドであるグルタチオン(GSH)は、医薬品としての薬効を有することが知られている。現在、医薬品としてのGSH製剤は、解毒剤及び眼科用剤として位置づけられており、各種の中毒、慢性肝臓疾患、抗癌剤の副作用や放射線療法による障害の防止、皮膚疾患および白内障や角膜損傷の治療に用いられている(非特許文献1)。
【0003】
一方、食品用途として、グルタチオンは食品にコク味を付与する物質であることが知られており(非特許文献2)、グルタチオンを高含有する酵母エキスが調味料用途に用いられている。また、グルタミン酸とシステインからなるジペプチドであるγ−グルタミルシステインは食品用途で有用であると報告されつつある。
【0004】
前述のように、含硫化合物は幅広い産業上の有用性を有しているため、これらを効率的に生産する微生物を取得しようと様々な検討が行われてきた。例えば、含硫化合物の生合成経路においてターゲットとする酵素を予測し、その機能を改変することにより細胞内の含硫化合物含量を増加させる方法が開示されている。例えば、大腸菌のγ−グルタミルシステイン合成酵素(非特許文献3)やグルタチオン合成酵素(非特許文献4)を酵母に導入することによって細胞内のグルタチオン含量が増加することが報告されている。
【0005】
これら以外にも、グルタチオン合成に関与する酵素を酵母に導入することによって含硫化合物であるグルタチオン含量を増加させる方法が報告されている(特許文献1、特許文献2、特許文献3、特許文献4)。
【0006】
一方、親株を突然変異処理することにより無作為な遺伝子改変を施した微生物を、各種薬剤を含む培地にスプレッドし生育可能な菌株(薬剤耐性株)を選択すること、又はレプリカ法により各種薬剤を含む培地で生育不可能な菌株(薬剤感受性株)を選択することにより含硫化合物の細胞内含量が増加した微生物を選抜する方法が報告されている。例えば、キャンディダ属酵母を変異処理し、エチオニン及び亜硫酸塩含有培地上で生育可能な菌株を選抜する方法(特許文献5、特許文献6、特許文献7)、サッカロミセス属酵母を変異処理し、亜鉛耐性度が向上した菌株を選抜する方法(特許文献8)などがある。これらの方法以外にも、含硫化合物の細胞内含量を増加させるための各種薬剤が検討されてきた(特許文献9、特許文献10)。
【0007】
ところが、近年、菌体内にグルタチオンを5重量%以上含有するサッカロミセス・セレビシエに属する酵母菌株をランダム変異により取得することに成功したと報告された(特許文献11)。同文献では変異株のグルタチオン含有量を一株一株測定することによって高生産株を取得していた。同文献の方法によって大量の菌株を評価可能にしたと報告していたが、同文献に記載されている方法でどれだけの速度で菌株が測定可能か検討したところ、1人が1日に評価できる菌株数は、精々100株程度であった。それゆえ、同文献に記載の方法は操作が極めて煩雑かつ高コストで経済的に負担の大きいことは明らかである。
【0008】
ところで、アデニン要求性を有する酵母、特に、ADE1遺伝子又はADE2遺伝子に変異を有する酵母は赤く発色することが古くから知られている。近年、遺伝子レベルでこのメカニズムが解析された。即ち、プリン合成系の第6ステップを触媒する酵素およびAIR中間体をコードするADE2遺伝子又はプリン合成系の第7ステップを触媒する酵素およびCAIR中間体をコードするADE1遺伝子の変異によって蓄積するアデニン生合成の中間体であるAIR(phosphoribosylaminoimidazole)やCIAR(phosphoribosylaminoimidazole carboxylate)がグルタチオンと結合し液胞に運ばれることによって赤く発色することが明らかとなった(非特許文献5)。しかし、同文献では、GSHの生合成の律速酵素をコードするGSH1遺伝子を過剰発現させたが、赤色レベルは変わらなかったと報告している。このことから、当業者であっても、アデニン要求性をGSH高含有酵母のスクリーニングに利用しようとの発想は持ち得ない状況であった。
【先行技術文献】
【特許文献】
【0009】
【特許文献1】特開昭61−52299号公報
【特許文献2】特開昭62−275685号公報
【特許文献3】特開昭63−129985号公報
【特許文献4】特開平4−179484号公報
【特許文献5】特開昭59−151894号公報
【特許文献6】特開平03−18872号公報
【特許文献7】特開平10−191963号公報
【特許文献8】特開平02−295480号公報
【特許文献9】特開平06−70752号公報
【特許文献10】特開平08−70884号公報
【特許文献11】特開2004−180509号公報
【非特許文献】
【0010】
【非特許文献1】蛋白質核酸酵素 1988-7 VOL.33 NO.9 ISSN 003909450臨時増刊 「グルタチオン研究のエポック」p1626
【非特許文献2】Y.Ueda et al, Biosci. Biotech. Biochem. ,61,1977-1980(1997)
【非特許文献3】Yasuyuki OHTAKE et al, Agric. Biol. Chem., 52(11), 2753〜2762, 1988
【非特許文献4】Yasuyuki OHTAKE et al, Journal of FERMENTATION AND BIOENGINEERING, Vol.68, No.6, 390-394, 1989
【非特許文献5】K.G. Sharma et al, Arch Microbiol (2003) 180:108-117
【発明の概要】
【発明が解決しようとする課題】
【0011】
本発明は、細胞内の含硫化合物含有量が増加した酵母、酵母を効率よくスクリーニングする方法、及びそれら酵母に細胞内の含硫化合物を高含有させる培養方法を提供することを目的とする。
【課題を解決するための手段】
【0012】
本発明者らは、K.G. Sharmaらの報告(非特許文献5)を詳細に検討し、アデニン生合成は生物に不可欠なATP生合成に関与していることから、ADE1遺伝子破壊株又はADE2遺伝子破壊株のようなアデニン要求性株ではAIRやCAIRといった中間体は細胞内に十分に蓄積しているとの仮説を持った。本発明者らは、この仮説が正しいとすれば、細胞内のグルタチオン含量が酵母の赤色発色をコントロールしていると考え、より赤く発色する酵母はよりグルタチオン含量が高くなっていると考えるにいたった。鋭意検討を行った結果、K.G.
Sharmaらの仮説とは異なり、本発明者らは、酵母の赤色を指標にすれば、グルタチオンを高含有する酵母がスクリーニング可能であることを見いだし、また、グルタチオンを含有する含硫化合物の細胞内含有量が、セレン酸感受性と、アデニン要求性の付与又はメチオニンを添加した最少培地での培養に起因する赤色呈色の特性との組み合わせにより増加可能であることを見出し、それにより本発明を完成させた。
【0013】
本発明の一態様は、細胞内の含硫化合物含量が増加した酵母であって、セレン酸感受性を有し、かつ培地上で培養されたときに該培地上で赤色を呈する酵母を提供することである。
本発明の別の態様は、前記培地はメチオニンが添加された培地である、上記の酵母を提供することである。
本発明の別の態様は、アデニン要求性の付与により赤色を呈色する、上記の酵母を提供することである。
本発明の別の態様は、アデニン要求性がADE1遺伝子又はADE2遺伝子の改変により付された、上記の酵母を提供することである。
本発明の別の態様は、さらに、ADE4遺伝子またはADE8遺伝子に変異が導入されたことにより、前記培地上で白色を呈するようになった、上記の酵母を提供することである。
本発明の別の態様は、セレン酸感受性がMET25遺伝子の発現を増強する改変により付された、上記の酵母を提供することである。
本発明の別の態様は、含硫化合物がシステイン、γ−グルタミルシステイン、グルタチオン、システニルグリシンからなる群から選択される少なくとも1つ以上の化合物である上記の酵母を提供することである。
本発明の別の態様は、菌体量を増加させるために細胞内のアデニンが充足する状態で酵母を培養する工程と、細胞内の含硫化合物含量を増加させるために細胞内のアデニンが不足する状態で酵母を培養する工程を含む、上記の酵母の培養方法を提供することである。
本発明の別の態様は、アデニン要求性およびセレン酸感受性を有する酵母に遺伝子改変処理を施す工程、アデニンが不足した時に酵母が赤く発色する培地に改変酵母をスプレッドし、酵母コロニーを形成する工程、及び改変前よりも赤く発色する酵母を選択する工程を含む、細胞内の含硫化合物含量が増加した酵母のスクリーニング方法を提供することである。
本発明の別の態様は、セレン酸感受性を有する酵母に遺伝子改変処理を行う工程、最小培地にメチオニンを追加した培地に改変酵母をスプレッドし、コロニーを形成する工程、及び改変前よりも赤く発色する酵母を選択する工程を含む、細胞内の含硫化合物含量が増加した酵母のスクリーニング方法を提供することである。
本発明の別の態様は、前記含硫化合物がシステイン、γ−グルタミルシステイン、グルタチオン、システニルグリシンからなる群から選択される少なくとも1つ以上の化合物である、上記方法を提供することである。
【図面の簡単な説明】
【0014】
【図1】Y-3256株のGSH含量の経時変化を示す図。
【図2】GSH2遺伝子破壊カセットの構築手順を示す図。
【発明を実施するための形態】
【0015】
本発明の酵母は、セレン酸感受性を有し、かつ培地上で培養されたときに該培地上で赤色を呈することによって細胞内の含硫化合物含量が増加した酵母であれば特に制限されない。サッカロミセス・セレビシエ等のサッカロミセス属、キャンディダ・ユティリス等のキャンディダ属、ピヒア・パストリス等のピヒア属、シゾサッカロミセス・ポンベ等のシゾサッカロミセス属等に属する酵母を例示することができる。中でも、含硫化合物の製造によく用いられているサッカロミセス・セレビシエやキャンディダ・ユティリスが好ましい。本発明の酵母は、1倍体でもよいし、2倍性またはそれ以上の倍数性を有するもので
あってもよい。
ここで、赤色性を評価する培地としては、酵母細胞内のアデニンが不足した時に酵母が赤く発色することができる培地であれば特に制限されない。アデニン含量が25mg/L以下の固体培地を挙げることができ、具体的にはYPD培地(組成:Bacto-yeast extract 1%、Bacto-peptone 2%、Glucose 2%:METHODS IN YEAST GENETICS 2000 Edition p171参照。ISBN 0-87969-588-9)PGC培地(組成:Casamine acid(vitamin free) 0.5%、Bacto-peptone 1%、Glucose 2%。)等を例示することができる。微量成分との影響でより赤く発色しやすいPGC培地の方が好ましい。培地中にカドミウムを添加すれば、より鮮明に赤く発色する為、カドミウムを添加することがより好ましい。培地は、メチオニンを追加することが好ましい。なお、株はコロニーを形成し、コロニーの色で赤色性を評価できるため、液体培地よりも寒天培地を用いることが好ましい。培地上で赤色を呈するようにするための方法は特に制限されないが、アデニン要求性を付与することやメチオニンが添加された最小培地で培養することが例示される。
【0016】
本発明において、アデニン要求性を有するとは、アデニンを含まない固体培地上では株がコロニーを形成できないか、あるいは後述するY-3219株と同程度の小さいコロニーを形成することを意味する。アデニン要求性の原因となる遺伝子は特定されていても特定されていなくてもよいが、特定されている方が好ましい。アデニン要求性の原因となる遺伝子は、得られたアデニン要求性の株を相補性分析または配列分析によって特定してもよい。本発明の酵母は、アデニン生合成経路の遺伝子のいずれかが不活性化するように改変されたことにより、アデニン要求性を有することが好ましい。アデニン生合成経路の遺伝子としては、GSHと結合する中間体AIRやCAIRの蓄積にダイレクトに影響するADE1遺伝子、ADE2遺伝子などが挙げられる。本発明において、「不活性化するように改変する」とは、遺伝子産物の活性が失われるような変異を遺伝子に導入すること、および遺伝子が発現しなくなるようにすることを含む。
【0017】
本発明において、遺伝子改変処理とは、親株の塩基配列が変異することを意味する。従来法による変異技術を用いても良いし、遺伝子組み換え技術を用いてもよい。従来法による変異技術の例としては、紫外線、レーザーなどの照射によって変異をもたらす方法、EMS(メタンスルホン酸エチル)、MNNG(N-メチル-N'-ニトロ-N-ニトロソグアニジン)、DAPA(4-ジメチルアミノベンゼンジアゾスルホン酸ナトリウム)等の変異誘発剤を用いる方法などがある。また、酵母を培養している際に生じる自然変異を利用してもよい。酵母に変異処理を施し、アデニンを含む培地では生育し、アデニンを含まない培地では生育しないか、あるいはほとんど生育できない株を選択することにより、アデニン要求性株を得ることができる。また、ADE1遺伝子又はADE2遺伝子に変異を有する酵母はアデニン要求性であり、赤く発色する。
【0018】
遺伝子組み換え技術を用いる方法としては、相同組み換えを利用して染色体上の野生型遺伝子を変異型遺伝子(不活性型遺伝子または破壊型遺伝子)で置換する方法などが挙げられる。
【0019】
なお、上述の遺伝子置換は、以下のようにして行うことができる。まず、変異型ADE1遺伝子を含む組換えDNAで酵母を形質転換し、変異型ADE1遺伝子と染色体上のADE1遺伝子との間で組換えを起こさせる。その際、組換えDNAには、宿主の栄養要求性等の形質にしたがって、マーカー遺伝子を含ませておくと操作がしやすい。また、組換えDNAは、制限酵素で切断する等により直鎖状にし、さらに、酵母で機能する複製制御領域を除いておくと、染色体に組換えDNAが組み込まれた株を効率よく取得することができる。
酵母の形質転換は、プロトプラスト法、KU法、KUR法、エレクトロポレーション法等、通常酵母の形質転換に用いられる方法を採用することができる。上記のようにして染色体に組換えDNAが組み込まれた株は、変異型ADE1遺伝子と染色体上にもともと存在す
るADE1遺伝子との組換えを起こし、野生型ADE1遺伝子と変異型ADE1遺伝子との融合遺伝子2個が組換えDNAの他の部分(ベクター部分及びマーカー遺伝子)を挟んだ状態で染色体に挿入されている。
融合遺伝子2個のうち、変異型ADE1遺伝子のみを残すために、2個のADE1遺伝子の組換えにより1コピーのADE1遺伝子を、ベクター部分(マーカー遺伝子を含む)とともに染色体DNAから脱落させる。その際、2つのケースが起こる。1つのケースでは、野生型ADE1遺伝子が染色体DNA上に残され、変異型ADE1遺伝子が切り出される。もう一つのケースでは、反対に変異型ADE1遺伝子が染色体DNA上に残され、野生型ADE1遺伝子が切り出される。いずれの場合もマーカー遺伝子が脱落するので、2回目の組換えが生じたことは、マーカー遺伝子に対応する表現形質によって確認することができる。また、目的とする遺伝子置換株は、PCRによりADE1遺伝子を増幅し、その構造を調べることによって、選択することができる。
以上、ADE1遺伝子を例に説明したが、ADE2遺伝子やその他の遺伝子も同様にして改変することができる。
【0020】
なお、ADE1遺伝子及び/またはADE2遺伝子に加えて、ADE4遺伝子またはADE8遺伝子に変異が入った場合、酵母は赤色ではなく通常の白色となる為更に好ましい。なお、ADE4遺伝子またはADE8遺伝子に変異を導入する場合のスクリーニングは、まずADE1遺伝子やADE2遺伝子に変異を導入し、赤色の発色度合いを指標に含硫化合物含量が増加した酵母を取得した後、ADE4遺伝子またはADE8遺伝子を変異させれば効率的に白色の含硫化合物高含有酵母を取得できる。
ADE4遺伝子またはADE8遺伝子の変異もADE1遺伝子、ADE2遺伝子と同様に導入することができる。
【0021】
ADE1遺伝子、ADE2遺伝子、ADE4遺伝子、ADE8遺伝子は改変対象の酵母自身の遺伝子を用いることができ、例えば、Saccharomyces cerevisiaeのADE1遺伝子、ADE2遺伝子、ADE4遺伝子、ADE8遺伝子としては、それぞれ、配列番号1、3、5、15の塩基配列を有するDNAが挙げられる。また、ADE1遺伝子、ADE2遺伝子、ADE4遺伝子、ADE8遺伝子は、遺伝子改変株の創製に用いる酵母の各遺伝子と相同組換えを起こす程度の同一性を有していれば特に制限されない。それゆえ、ADE1遺伝子、ADE2遺伝子、ADE4遺伝子、ADE8遺伝子は、それぞれ配列番号2、4、6、16に示すアミノ酸配列と、80%以上、好ましくは90%以上、より好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列をコードするDNAであればよい。また、ADE1遺伝子、ADE2遺伝子、ADE4遺伝子、ADE8遺伝子は、配列番号1、3、5、15に示す塩基配列を有するDNAと、ストリンジェントな条件下でハイブリダイズするDNAであってよい。ストリンジェントな条件としては、65℃、0.1×SSC、0.1%SDSに相当する塩濃度で1回の洗浄、好ましくは2回または3回の洗浄が行われる条件が挙げられる。
【0022】
本発明において、セレン酸感受性を有するとは、セレン酸とメチオニンを含有する培地(例:5mMのセレン酸と1mMのメチオニンを含有する培地)上ではコロニーを形成できないことをいう。セレン酸は硫酸のアナログで細胞毒性を示す。通常の酵母は硫酸の取り込み系がメチオニンなどの有機硫黄化合物によって抑制されているのでセレン酸は細胞内に取り込まれず、セレン酸含有培地でも生育する。一方、セレン酸感受性株は硫酸の取り込み系がメチオニンなどの有機硫黄化合物によって抑制されず、セレン酸を取り込んでしまう。硫酸の取り込み系が抑制されないため、セレン酸感受性株では含硫化合物の生合成が野生株より向上している(MOLLECULAR AND CELLUAR BIOLOGY Dec, 1995, p6526-6534)。
【0023】
セレン酸感受性は変異処理により付与されてもよいし、遺伝子組み換えによって付与されてもよい。例えば、酵母に変異処理を施し、セレン酸とメチオニンを含む培地では生育
できない株を選択することにより、セレン酸感受性株を得ることができる。
【0024】
一方、セレン酸感受性を付与するための遺伝子組み換えとしては、O-acetylhomoserine
sulfhydrylaseをコードするMET25遺伝子の発現を野生株と比べて増強するように改変することが挙げられる。MET25遺伝子の発現を増強する方法としては、細胞内へのプラスミドの導入や、染色体上への導入などによりMET25遺伝子のコピー数を増大させることや、MET25遺伝子のプロモーターを強力なプロモーターに置換する方法が例示される。
【0025】
また、MET25遺伝子の転写調節因子を改変することによってMET25遺伝子の発現を増強することもできる。MET25遺伝子の発現については、次のように考えられている。MET4産物は、MET25遺伝子の発現に正の因子として働く。通常は、MET4産物は、MET30産物及び他の数種のタンパク質とともにSCFMET30複合体を形成し、ユビキチン化を受け、26Sプロテアソームによるタンパク分解系によりMET30産物とともに分解されるため、MET25遺伝子の発現は抑制されている。一方、SCFMET30複合体の機能が低下すると、MET4産物及びMET30産物は分解を受けず、MET25遺伝子が発現する。(Patton et al.,Genes Dev.12: 692-705, 1998、Rouillon et al., EMBO Journal 19: 282-294, 2000)したがって、MET4遺伝子やMET30遺伝子に変異を導入し、MET25遺伝子の発現を増強することもできる。
MET25遺伝子の発現量を増強することができる変異型MET4遺伝子としては、215番目のセリンがプロリンに、または、156番目のイソロイシンがセリンに変異したMET4遺伝子が報告されている(大村ら FEBS Letters 387(1996) 179-183、特開平10-33161号公報)。
また、MET25遺伝子の発現量を増大させる変異型MET30遺伝子としては、WD40リピートに変異を導入することにより得られる変異型MET30遺伝子が報告されている。また、特開2004-201677には、569位のセリンをフェニルアラニン等の他のアミノ酸に置換する変異を有する変異型MET30遺伝子が報告されている。
このような変異型遺伝子で染色体上の野生型遺伝子を置換することにより、MET25遺伝子を発現増強させることができ、セレン酸感受性を付与できる。
【0026】
MET25遺伝子、MET30遺伝子、MET4遺伝子としては、改変された酵母のように、何らかの酵母の遺伝子であってよい。例えば、Saccharomyces cerevisiaeのMET25遺伝子、MET30遺伝子、MET4遺伝子として、それぞれ、配列番号9、11、7の塩基配列を有するDNAが挙げられる。また、MET25遺伝子はO-acetylhomoserine sulfhydrylase活性を有するタンパク質をコードする限り、また、MET30遺伝子、MET4遺伝子は、Met25遺伝子の発現を抑制するタンパク質をコードする限り特に制限されない。それゆえ、MET25遺伝子、MET30遺伝子、MET4遺伝子は、それぞれ、配列番号10、12、8に示すアミノ酸配列と、80%以上、好ましくは90%以上、より好ましくは95%以上、特に好ましくは98%以上の同一性を有するアミノ酸配列をコードするDNAであってもよい。MET25遺伝子、MET30遺伝子、MET4遺伝子は配列番号9、11、7に示す塩基配列を有するDNAと、ストリンジェントな条件下でハイブリダイズするDNAであってもよい。ストリンジェントな条件としては前述のものが挙げられる。
【0027】
本発明の酵母はセレン酸感受性にアデニン要求性を組み合わせることによって、対応する酵母のセレン酸感受性かつアデニン非要求性の株よりも細胞内の含硫化合物含量が増加している。
本発明において含硫化合物とは、化学式内に-SH基を有するものをいい、タンパク質、ペプチド、アミノ酸、或いはその他の物質であってもよい。タンパク質としては、構成アミノ酸の30%がシステイン残基からなるメタロチオネイン、ペプチドとしてはグルタチオン、γ−グルタミルシステイン、システニルグリシンなどを、アミノ酸としてはシステインを、その他の物質としてはホモシステインなどを例示することができるが特にこれらに限定されるものでもない。グルタチオン、γ−グルタミルシステイン、システインは、
現在幅広く産業上で利用されているので、対象とする含硫化合物として特に好ましい。
【0028】
含硫化合物としてグルタチオンを酵母内に蓄積させる場合は、さらに、酵母細胞内でグルタチオン合成酵素とγ−グルタミルシステイン合成酵素の一方又は両方の活性を増強させてもよい。酵素活性の増強は、これらの酵素をコードする遺伝子のコピー数を増加させるか、あるいはプロモーターを強力なプロモーターに置換することによって行うことができる。Saccharomyces cerevisiaeのγ−グルタミルシステイン合成酵素遺伝子の配列と、Candida utilisのグルタチオン合成酵素遺伝子およびγ−グルタミルシステイン合成酵素遺伝子の配列は特開2005−073638に開示されている。また、Saccharomyces cerevisiaeのグルタチオン合成酵素遺伝子の塩基配列とコードされるアミノ酸配列を配列番号13、14に示した。
【0029】
酵母の細胞内に、含硫化合物として、γ−グルタミルシステインを蓄積させる場合には、酵母をグルタチオン合成酵素活性が低下するように改変することが好ましく、グルタチオン合成酵素活性が低下し、かつ、γ−グルタミルシステイン合成酵素活性が増大するように改変することがより好ましい。グルタチオン合成酵素活性が低下するような改変は、グルタチオン合成酵素遺伝子を破壊するか、あるいは、グルタチオン合成酵素活性が低下するような変異を導入することが挙げられる。
グルタチオン合成酵素活性が低下するような変異としては、例えば、配列番号14のアミノ酸配列の370位のアルギニン残基を終止コドンに変更する変異が挙げられる。
グルタチオン合成酵素活性が低下するような変異の他の例として下記の変異が挙げられる(国際公開03/046155号パンフレット)
(1)配列番号14のアミノ酸配列の47位のスレオニンをイソロイシンに置換する変異。
(2)配列番号14のアミノ酸配列の387位のグリシンをアスパラギン酸に置換する変異。
(3)配列番号14のアミノ酸配列の54位のプロリンをロイシンに置換する変異。
上記変異は、(1)または(2)の単独変異でも、(1)〜(3)の任意の組合せでもよいが、(1)と(3)の組合せ、及び(2)と(3)の組合せが好ましい。
なお、システインはγ−グルタミルシステインを熱処理することによって生成するため、γ−グルタミルシステインを蓄積した酵母を熱処理すればシステインが蓄積した酵母が得られる。
また、含硫化合物として、システニルグリシンを蓄積させる場合には、酵母をECM38活性が増強するように改変することが好ましい(Yeast. 2003 Jul 30;20(10):857-63)。
グルタチオンやγ−グルタミルシステイン等の含硫化合物の細胞内含量が増加した酵母は、ラパマイシン耐性を示す酵母から得ることもできる(WO2006/013736)。
【0030】
上記のアデニン要求性およびセレン酸感受性を有する酵母については、さらに遺伝子改変処理、好ましくは変異処理を施して、アデニンが不足した時に酵母が赤く発色する培地にスプレッドし、改変前よりもより赤く発色する酵母を選択することにより、さらに含硫化合物の含量を増加させることができる。
本発明において、スクリーニングに用いる培地は、酵母細胞内のアデニンが不足した時に酵母が赤く発色することができる培地であれば特に制限されない。アデニン含量が25mg/L以下の培地を挙げることができ、具体的にはYPD培地(組成:Bacto-yeast extract 1%、Bacto-peptone 2%、Glucose 2%。METHODS IN YEAST GENETICS 2000 Edition p171参照。ISBN 0-87969-588-9)PGC培地(組成:Casamine acid(vitamin free) 0.5%、Bacto-peptone 1%、Glucose 2%。)等を例示することができる。微量成分との影響でより赤く発色しやすいPGC培地の方が好ましい。培地中にカドミウムを添加すれば、より鮮明に赤く発色する為、カドミウムを添加することがより好ましい。なお、菌株の単離を考えれば液体培地よりも寒天培地を用いることが好ましい。発色度合いを識別するためには、固体
培地上で20〜30℃で約1週間培養することが好ましい。
【0031】
また、セレン酸感受性を有する酵母について、遺伝子改変処理、好ましくは変異剤処理を行い、最小培地にメチオニンを追加した培地に該処理された酵母をスプレッドし、培地中に出現した改変酵母の発色度合いを識別し改変前よりも赤く発色する酵母を選択することにより、細胞内の含硫化合物含量が増加した酵母をスクリーニングすることもできる。ここで使用する培地としては、擬似的にアデニン要求性と同じ状態を作り出すことのできるメチオニン含有培地であればよいが、ビオチンの非存在下でメチオニンが添加された最小培地が好ましく、具体的には、例えば、後述のmin-met(+)-biotin(-)プレートなどが挙げられる。
【0032】
本発明の酵母を培養することにより含硫化合物が蓄積した酵母を製造することができる。培養方法として好ましくは、本発明の酵母を、十分な量のアデニンを含む培地(アデニン充足状態)で培養して同酵母を増殖させた後、アデニン量が制限された培地で培養し、菌体内の含硫化合物含量を増加させる方法が挙げられる。これにより、含硫化合物が蓄積した酵母を効率よく製造することができる。
好ましくは、前記アデニンの「十分な量」は、予め一定量の酵母菌体を得るのに必要なアデニン要求量を実験的に測定し、目的の量の菌体を得るために必要なアデニン量を算出することによって、決定することができる。例えば、100mg/L以上が挙げられる。
アデニン量以外の培地及び培養条件は、通常の酵母培養に用いられる培地及び条件を採用することができる。尚、必要に応じて、用いる酵母の形質にしたがって必要な栄養素を培地に添加する。
【0033】
十分な量の酵母菌体が得られたら、アデニン量が制限された培地で培養する。アデニン量が制限された培地としては、アデニン含量が25mg/L以下の培地が挙げられる。アデニン量が制限された培地で培養する間に、酵母菌体内の含硫化合物の蓄積量は経時的に上昇する。
好ましくは、目的とする含硫化合物の蓄積量に達したら、培養を終了する。通常は、培養時間は10〜30時間、好ましくは15〜27時間である。
【0034】
上記のようにして得られる培養物又はその分画物は、含硫化合物を含有する。培養物は、酵母菌体を含む培養培地であってもよいし、それから採取された酵母菌体であってもよい。含硫化合物を含む分画物は、細胞破砕物または酵母抽出物であっても良い。酵母エキス等の調製は、通常の酵母エキスの調製と同様にして行えばよい。酵母エキスは、酵母菌体を熱水抽出したものを処理したものでもよいし、酵母菌体を酵素消化したものを処理したものでもよい。
【0035】
含硫化合物は上記の酵母細胞から単離することができる。上記含硫化合物、酵母培養物又はその分画物は、食品、医薬品、化成品、飼料等の製造に用いることができる。食品としては、アルコール飲料、パン食品、又は発酵食品調味料が挙げられる。食品は、含硫化合物、培養細胞又はその分画物を、食品原料に混合し、食品に加工することによって製造される。
【実施例】
【0036】
以下、本発明を実施例に基づき説明する。なお、本発明は以下の実施例になんら限定されるものではない。
【0037】
<実施例1>
(アデニン要求性酵母の取得)
変異型MET30遺伝子を保有するサッカロミセス・セレビシエ1倍体AJ14819株(MATα型
))とサッカロミセス・セレビシエ1倍体AJ14810株(MATa型)を接合させることにより、サッカロミセス・セレビシエ2倍体株を取得した。AJ14819株はセレン酸感受性株であり、2003年10月1日にブダペスト条約に基づき独立行政法人 産業技術総合研究所 特許生物寄託センター(日本国、305-8566 茨城県つくば市東1丁目1-1つくばセンター 中央第6)にFERM BP-8502の受託番号で寄託されている。AJ14180株は、2002年11月1日にブダペスト条約に基づき、独立行政法人 産業技術総合研究所 特許生物寄託センターにFERM BP-8229の受託番号で寄託されている。
同2倍体株を胞子形成させ、4分子解析することにより以下の性質を有する菌株を取得した。
A:MATa型1倍体、変異型MET30遺伝子
B:MATα型1倍体、変異型MET30遺伝子
C:MATa型1倍体、野生型MET30遺伝子
D:MATα型1倍体、野生型MET30遺伝子
【0038】
次に、取得した4菌株のMET25遺伝子の発現量を比較した。特開2004-201677実施例1記載の方法に基づき、これら4株のMET25遺伝子発現量を以下のようにして測定した。菌株を各々YPD培地に植菌し(坂口フラスコ500ml容、50ml張り込み)、30℃で振とう培養した。その対数増殖期に集菌し、菌体内に含まれているRNAを回収し、MET25遺伝子の発現を内部標準としてACT1遺伝子を用いて定量した。定量は、PCR5700(Applied Biosystems社)を用い、TaqMan One-Step RT-PCRキット(Applied Biosystems社)を用いて行なった。TaqMan Probe(Applied Biosystems社)に、特開2004-201677実施例1記載のACT1-986T及びMET25-1077Tを、ACT1遺伝子の増幅用に特開2004-201677実施例1記載のACT1-963FとACT1-1039Rを、そして、MET25遺伝子の増幅用にMET25-1056FとMET25-1134Rをプライマーとして用いた。その結果、変異型MET30遺伝子およびセレン酸感受性を有するA株及びB株のMET25遺伝子の発現量は、変異型MET30遺伝子を有しないC株及びD株よりも高いことを確認した。B株にプライベート番号AJ14889及びA株にプライベート番号AJ14890を付した。
【0039】
次に、常法に従いAJ14889株を変異剤MNNG(1-methyl-3-nitro-1-nitrosoguanidine)で生存率5〜10%になる条件で処理した後、YPDプレートにスプレッドし30℃で約1週間培養した。出現したコロニーの中から赤色に発色する菌株を選択した。次に、得られた菌株につき、常法に従い、アデニン要求性の相補テストを行った。その結果、ADE1遺伝子変異によるアデニン要求性酵母N1株及びADE2遺伝子変異によるアデニンリーキー変異酵母N2株を取得した。(N1株およびN2株は、2008年12月23日、ブダペスト条約に基づきRussian
National Collection of Industrial Microorganisms(VKPM)(1 Dorozhny proezd., 1 モスクワ 117545, ロシア)にそれぞれVKPM Y-3218、VKPM Y-3219で国際寄託されている。)
【0040】
<実施例2>
(強い赤色発色を指標とした高GSH蓄積変異体のスクリーニング)
次に、常法に基づきADE1遺伝子に変異を有するY-3218株を変異剤MNNGで生存率5〜10%になる条件で処理した後、PGC又はYPDプレートにスプレッドし30℃で約1週間培養した。出現したコロニーの中から、Y-3218株よりもより赤く発色する菌株を選択し、GSH含量をAJ14889株(親株)およびY-3218株と比較した。選択した株、AJ14889株およびY-3218株をそれぞれ5mlのYPD液体培地に植菌し、30℃、250rpmの振とう数で24時間培養した。50mlのYPD液体培地にそれぞれの株の得られた培養物を植菌し、30℃、250rpmの振とう数で24時間培養した。それぞれの株のGSH含有量を承保に従って測定した結果、AJ14889株に比べY-3218株のGSH含量は高かった(表1)。選択した株(計180株)のうち7株がY-3218株よりGSH含量が高いことがわかった(表1)。
【0041】
【表1】
【0042】
ADE2遺伝子変異株Y-3219株を用いて同様の検討を行った。常法に基づきY-3219株を変異剤MNNGで生存率5〜10%になる条件で処理した後、PGC又はYPDプレートにスプレッドし30℃で約1週間培養した。出現したコロニーの中から、別途スポットしていたY-3219株よりもより赤く発色する菌株を選択し、GSH含量をAJ14889株(親株)およびY-3219株と比較した。YPD寒天培地よりコロニーをストリークし、選択した株、AJ14889株およびY-3218株をそれぞれ5mlのYPD液体培地に植菌し、30℃、250rpmの振とう数で24時間培養した。50mlのYPD液体培地にそれぞれの株の得られた培養物を植菌し、30℃、250rpmの振とう数で24時間培養した。それぞれの株のGSH含有量を承保に従って測定した結果、AJ14889株よりもY-3219株のGSH含量が高いことがわかった。選択した株(計118株)のうち10株がY-3218株よりGSH含量が高いことがわかった(表2)。
【0043】
【表2】
【0044】
表1、表2に示すようにアデニン要求性による酵母の赤色の発色を指標としてGSH含量
が向上した酵母を単離することが可能であった。
【0045】
<実施例3>
(GSH高含量の白色コロニーの単離)
野生型の酵母は赤色ではなく、白色〜クリーム色である。そこで、発明者らは赤色の発色度合いを指標にスクリーニングした酵母から通常の色のコロニーを形成する株の取得を検討した。まず、実施例2と同様にY-3219株をMNNGで変異処理し、PGC培地またはYPD培地に播いてより赤色の強い株を得た。その結果、実施例2と同様に表3のY-3219-20株が得られた。次に、Y-3219-20株をMNNGで、生存率5〜10%になるように変異処理を行い、PGCプレートにスプレッドし、30℃で約1週間培養した。出現したコロニーの中から、3つの白コロニー(Y-3219-20-52、Y-3219-20-53、Y-3219-20-56)と1つの赤コロニー(Y-3219-20-1)を選抜し、それらのGSH含量を測定し、Y-3219株、Y-3219-20株と比較した。すなわち、得られた株と、Y-3219株及びY-3219-20株を5mLのYPD液体培地に植菌し、30℃、250rpmの振とう数で24時間培養した。次に、50mlのYPD液体培地にそれぞれの株の得られた培養物を植菌し、30℃、250rpmの振とう数で24時間培養した。それぞれの菌体内のGSH量を常法に従って測定した。結果を表3に示す。
【0046】
【表3】
【0047】
白色の菌株を試験株を用いて遺伝解析した。その結果、GSH含量が減少したY-3219-20-52株ではアデニン要求性を失っていた。一方、高いGSH含量を維持したY-3219-20-56株では、ADE2遺伝子変異に加えて、ADE4遺伝子も変異していた。Y-3219-20-53株では、ADE2遺伝子変異に加えて、ADE8遺伝子も変異していた。この結果より、赤色コロニーを形成するアデニン要求性の株はADE4遺伝子またはADE8遺伝子が変異することで、通常の白色コロニーを形成する株になり得ることがわかった。
Y-3219-20-56株は、2008年12月23日、ブダペスト条約に基づきRussian National Collection of Industrial Microorganisms(VKPM)(1 Dorozhny proezd., 1 モスクワ 117545, ロシア)にVKPM Y-3256で国際寄託されている。
【0048】
<実施例4>
(アデニン不充足による高GSH蓄積)
アデニン充足度合いとGSH含量との関係について検討した。Y-3256株を500ml容坂口フラスコ中の50mlのYPD培地(D-glucose 20g/L、Bact Peptone 20g/L、Yeast Extract 10g/L)で30℃、120rpmで24時間振とう培養し、得られた培養物を、500ml容坂口フラスコ中の異なる濃度のアデニン(最終アデニン濃度0mg/L、10mg/Lまたは20 mg/L)を加えた50mlのYPD培地に、初期吸光度660nm=0.1になるように植菌し、30℃、120rpmで振とう培養した。その結果、GSH量経時変化は表4の通りであった。
【0049】
【表4】
【0050】
これらの結果から、アデニン要求性Y-3256株のGSH含量は培地中のアデニン量に反比例していることが明らかとなった。なお、YPD培地には微量のアデニンが含まれているため、アデニン添加量なし(Ade 0mg/L)の場合でもY-3256株は生育可能である(YPD培地のアデニン含量は約10mg/Lであった)。
【0051】
(Jar Fermentor評価)
Y-3256株をJar Fermentorで培養し、GSH含量の経時変化を評価した。YPD寒天培地より菌株細胞を拾い、50mlのYPD培地を含む750ml容三角フラスコ3つに植菌し、30℃、250rpmで20時間振とう培養した。得られたシード培養液(120ml)を3L容Jar-Fermentor中の1.2Lのメイン培地(YPD培地)に植菌し、30℃、攪拌速度1,100rpmで培養した。この時の通気量は1/1vvmに設定し、pHはアンモニア水により6.0に制御した。培養0hr〜24hrの間は1.5ml/hrの速度で、24hr以降は1.8ml/hrの速度で流加培地を連続フィードした。なお、このとき使用した流加培地の組成は、1LあたりGlucose600g、Bacto-yeast extract 10g、corn
extract 10g、Bacto-peptone 10g、(NH4)2SO4 0.274g、KH2PO4 0.11g、KCl 0.732g、MgSO4 0.466g、CuSO40.0012g、ZnSO4 0.014g、MnSO4 0.00334g、NaMoO40.00012g、KCl 0.002g、H3BO30.00004g、CoSO4 0.0001g、CaCl2 0.28g、FeSO4 0.2g、Biotine 0.05mg、riboflavin 0.2mg、thiamine 0.5mgである。
この時のY-3256株のGSH含量変化は図1の通りであった。
【0052】
<実施例6>
(2倍体株の取得及び培地組成の検討)
常法に基づき、Y-3219-20-53株の自己2倍体化を試みた。実験の条件は以下のとおりである。Y-3219-20-53株を常法により、生存率5〜10%になるような条件で変異剤MNNGで処理し、YPD寒天培地に100〜200個のコロニーが出現するようにスプレッドした。30℃で5日間培養後、アデニンを添加したSD寒天培地及びYPD寒天培地にレプリカした。YPD寒天培地では生育可能であるが、アデニンを添加したSD寒天培地では生育できない菌株を選抜した。アデニン以外の栄養要求性を調べ、栄養要求性が異なるY-3219-20-53-aux1株及びY-3219-20-53-aux2株を取得した。Y-3219-20-53-aux1株をアデニンを添加したSD寒天培地に縦方向の直線になるようにストリークし、Y-3219-20-53-aux2株を同じアデニンを添加したSD寒天培地に横方向の直線になるようにストリークした。但し、この時縦方向の直線と横方向の直線が1点で交差するように行った。このアデニンを添加したSD寒天培地を30℃で20日間培養し、交差点に出現するコロニーを選択した。この様にしてアデニン要求性の2倍体株であるD1-3株を取得した。本菌株は、2008年12月23日、ブダペスト条約に基づきRussian National Collection of Industrial Microorganisms(VKPM)(1 Dorozhny proezd., 1 モスクワ 117545, ロシア)にVKPM Y-3309で国際寄託されている。なお、D1-3株をアデニンを添加したSD寒天培地にストリークし、30℃で2日間培養したところ、コロニーが出現したことから、D1-3株はアデニン以外の栄養要求性が付与されていないことが確認できた。
また、アデニン非要求性の対象区として、AJ14889株とAJ14890株を接合させることによ
り2倍体Dip株を取得した。
D1-3株及びDip株を、5mlのYPD液体培地を入れた試験管に植菌し、30℃、250rpmの振とう数で24時間培養した。得られた培養物を、表5に示す組成の各培地50mlに植菌し、30℃、250rpmの振とう数で24時間培養した。各菌体内のGSH含量を測定した。その結果は表6に示すとおりである。
【0053】
【表5】
【0054】
【表6】
【0055】
このようにいずれの培地においてもアデニン要求性によってGSH含量が増加していた。
【0056】
<実施例7>
(アデニン要求性のγ−グルタミルシステイン高含有酵母の育種及びγ−グルタミルシステイン含量向上効果の評価)
実施例3にて取得したY-3256株のグルタチオン合成酵素をコードするGSH2遺伝子を破壊することによりγ−グルタミルシステインを蓄積する1倍体株を取得した。GSH2遺伝子破壊カセットを構築するために、プライマー17〜22を用い、S. cerevisiae(野生株)のゲノムDNAおよびプラスミドpFA6a-KanMX6(Chiara et al., Yeast 2000, 16:1089-1097)を鋳型にしてPCRを行った(図2)。詳細な条件は以下のとおりである。
【0057】
まず、GSH2のORFから約400bp上流の領域をX2180-1B(S. cerevisiae 野生型株:ATCCにて受託番号ATCC204505で入手可能)のゲノムDNAを鋳型に、GSH2-up-Fプライマー(配列番号17)およびGSH2-up-Rプライマー(配列番号18)を用いて増幅し、GSH2上流フラグメントを得た。GSH2上流フラグメントは、GSH2-up-FプライマーおよびGSH2-up-Rプライマーの配列に基づき、その一方の末端側にBpiI制限部位を有し、もう一方の末端側に後述するフュージョンPCR用のcon1部位を有する。
また、GSH2のORFから約300bp下流の領域をX2180-1BのゲノムDNAを鋳型に、GSH2-down-Fプライマー(配列番号19)およびGSH2-down-Rプライマー(配列番号20)を用いて増幅しGSH2下流フラグメントを得た。GSH2下流フラグメントは、GSH2-down-FプライマーおよびGSH2-down-Rプライマーの配列に基づき、その一方の末端側にBpiI制限部位を有し、もう一方の末端側にフュージョンPCR用のcon2部位を有する。
KanMX遺伝子をpFA6a-KanMX6プラスミドを鋳型に、マーカーFプライマー(配列番号21
)とマーカーRプライマー(配列番号22)を用いてPCR増幅した。このPCRで増幅されたKanMX遺伝子は、マーカーFプライマーおよびマーカーRプライマーの配列に基づき、その一方の末端側にcon1部位を有し、もう一方の末端側にcon2部位を有する。
これらのPCRの条件は次のとおりである。DNAポリメラーゼの混合物(Pfu:Taq=1:10;Fermentas, Lithuaniaより入手可能)をそれぞれの反応に用いた。PCRは94℃30秒、50℃30秒、68℃3分のサイクルを30回繰り返した。
【0058】
得られた3フラグメントをフュージョンPCRで結合させてGSH破壊カセットを構築した。GSH2上流フラグメントとKanMX遺伝子フラグメントを鋳型に用い、GSH2-up-FプライマーとマーカーRプライマーを用いてフュージョンPCRを行った。GSH2上流フラグメントとKanMX遺伝子フラグメントはいずれも末端にcon1部位を有するため、フュージョンPCRを行うことにより、これらが連結されたGSH2上流-KanMXフラグメントが得られる。このフュージョンPCRは前記と同様のDNAポリメラーゼの混合物を用いた。PCRは、94℃30秒、61℃30秒、68℃4.5分のサイクルを5回、次いで94℃30秒、50℃30秒、68℃4.5分のサイクルを25回繰り返した。
次に、GSH2上流-KanMXフラグメントとGSH2下流フラグメントを鋳型に用い、GSH2-up-FプライマーとGSH2-down-Rプライマーを用いてフュージョンPCRを行った。GSH2上流-KanMXフラグメントとGSH2下流フラグメントはいずれも末端にcon2部位を有するため、GSH2上流-KanMXフラグメントとGSH2下流フラグメントが連結され、GSH2破壊カセットが得られる。このフュージョンPCRは前記と同様のDNAポリメラーゼの混合物を用いた。PCRは、94℃30秒、61℃30秒、68℃5.3分のサイクルを5回、次いで94℃30秒、50℃30秒、68℃5.3分のサイクルを25回繰り返した。
【0059】
プライマーの配列は以下のとおりである。
(1) GSH2-up-F, CCGAAGACCTTCGTTTGGTGTTATGGT (配列番号17)
(2) GSH2-up-R, GAGAGGGGGGGGGTGGGGGGAAGGTGGATAGTGTGCC (配列番号18)
(3) GSH2-down-F, CCTCCTCCCCCCGCCCACGGCAGGATTCGGATGTTTG (配列番号19)
(4) GSH2-down-R, CGAAGACTCAGTACGAGCATTACGCAA (配列番号20)
(5) Marker-F, 5’-CCCCACCCCCCCCCTCTCTACCGTTCGTATAATGTATGCTATACGAAGTTATACTGGATGGCGGCGTTAG-3’ (配列番号21)
(6) Marker-R, 5’-GTGGGCGGGGGGAGGAGGTACCGTTCGTATAGCATACATTATACGAAGTTATGTTTAGCTTGCCTCGTCC-3’ (配列番号22)
【0060】
GSH2破壊カセットを用いてY-3256株を形質転換し相同組み換えを起こさせた、得られた形質転換体をG418(50μg/ml)を含有するYPD寒天培地にスプレッドした。出現したコロニーから、GSH2遺伝子が破壊された菌株であるN8△GSH2株を取得した。
同菌株を500ml容坂口フラスコ中の50mlのYPD培地(D-glucose 20g/L、Bact Peptone 20g/L、Yeast Extract 10g/L)に植菌し、30℃、120rpmで24時間振とう培養後、得られた培養物を、異なる濃度のアデニン(最終アデニン濃度0mg/L、10mg/L、20mg/L)を添加した500ml容坂口フラスコ中のYPD培地50mlに初期吸光度660nm=0.1になるように植菌し、30℃、120rpmで振とう培養した。γ−グルタミルシステイン濃度を常法に従い測定した。表7にγ−グルタミルシステインの経時変化を示す。γ−グルタミルシステイン濃度はアデニン要求性の付与により高められ得ることがわかった。
【0061】
【表7】
【0062】
<実施例8>
(擬似的なアデニン要求性条件を作り出す培地での評価)
常法に従い、AJ14889株を変異剤MNNGで生存率が約10%になる条件で処理した後、表8に記載の成分および150mg/Lのメチオニンを含むmin-met(+)-biotin(-)プレートに、1プレートあたりのコロニー数が350個以下になるようにスプレッドした。30℃で8〜10日間培養した後、30枚のプレートに出現したコロニーの中から、薄いピンク色から赤色に発色しているコロニーを106株ピックアップした。
次に、AJ14889株(親株)及び選抜した菌株を各々YPD培地に植菌し菌体内のGSH含量を測定した。具体的には、YPD培地で培養した選抜した菌株とAJ14889株をそれぞれ、5mlのYPD液体培地を入れた試験管に植菌し、30℃、250rpmの振とう数で24時間培養した。そして、750ml容の三角フラスコに50mlのYPD培地を入れたメイン培地に前培養液を、600nmでの吸光度が0.1になるように植菌し、30℃、250rpmの振とう数で24時間及び48時間培養した。その結果、選択された106株中8株は、24hr時又は48hr時にAJ14889株よりもGSH含量が上回っていた(表9)。選択した108株中8株がGSH含量が向上した菌株であったことから、本方法は非常に効率的なスクリーニング方法であることが示された。
【0063】
【表8】
【0064】
【表9】
【0065】
次に、上記8株より選択した89-6株、89-28株、89-31株について、生育とGSH含量をAJ14889株と比較した。これらの株は、600nmでの吸光度が0.3となるように植菌したことを除いては、前述のように培養した。生育は乾燥菌体重量(DCW)の変化に基づいて測定した。表10に示すように、選択した3株は、24hr時及び48hr時において生育が親株とほぼ同等で、GSH含量は親株を上回っていた。
【0066】
【表10】
【0067】
また、この3株に予期しない栄養要求性が付与されていないか検証するために、AJ14889株及びこの3株をSD培地に植菌し、その生育の有無を調べた。具体的には、YPD寒天培地で培養しておいた菌株を、5mlのSD培地を入れた試験管に植菌し、30℃、250rpmの振とう数で、それぞれ24時間および48時間培養した。その結果、選択した3株はSD培地でAJ14889株とほぼ同等の生育を示し(表11)、これら3株に予期しない栄養要求性は付与されていないことが示された。
【0068】
【表11】
【産業上の利用可能性】
【0069】
本発明によって、細胞内の含硫化合物含有量が増加したアデニン要求性酵母、該酵母のスクリーニング方法、及び該酵母の培養方法が提供される。本発明は、食品、医薬品、化成品、飼料等の幅広い産業で使用することが可能である。
【特許請求の範囲】
【請求項1】
細胞内の含硫化合物含量が増加した酵母であって、セレン酸感受性を有し、かつ培地上で培養されたときに該培地上で赤色を呈する酵母。
【請求項2】
前記培地はメチオニンが添加された培地である、請求項1に記載の酵母。
【請求項3】
アデニン要求性の付与により培地上で赤色を呈する、請求項1または2に記載の酵母。
【請求項4】
アデニン要求性がADE1遺伝子又はADE2遺伝子の改変により付された、請求項3に記載の酵母。
【請求項5】
さらに、ADE4遺伝子またはADE8遺伝子に変異が導入されたことにより、前記培地上で白色を呈するようになった、請求項3または4に記載の酵母。
【請求項6】
セレン酸感受性がMET25遺伝子の発現を増強する改変により付された、請求項1〜5のいずれか一項に記載の酵母。
【請求項7】
含硫化合物がシステイン、γ−グルタミルシステイン、グルタチオン、システニルグリシンからなる群から選択される少なくとも1つ以上の化合物である請求項1〜6のいずれか一項に記載の酵母。
【請求項8】
菌体量を増加させるために細胞内のアデニンが充足する状態で酵母を培養する工程と、細胞内の含硫化合物含量を増加させるために細胞内のアデニンが不足する状態で酵母を培養する工程を含む、請求項1〜7のいずれか一項に記載の酵母の培養方法。
【請求項9】
アデニン要求性およびセレン酸感受性を有する酵母に遺伝子改変処理を施す工程、アデニンが不足した時に酵母が赤く発色する培地に改変酵母をスプレッドし酵母コロニーを形成する工程、及び改変前よりも赤く発色する酵母コロニーを選択する工程を含む、細胞内の含硫化合物含量が増加した酵母のスクリーニング方法。
【請求項10】
セレン酸感受性を有する酵母に遺伝子改変処理を行う工程、最小培地にメチオニンを追加した培地に改変酵母をスプレッドし酵母コロニーを形成する工程、及び改変前よりも赤く発色する酵母コロニーを選択する工程を含む、細胞内の含硫化合物含量が増加した酵母のスクリーニング方法。
【請求項11】
含硫化合物がシステイン、γ−グルタミルシステイン、グルタチオン、システニルグリシンからなる群から選択される少なくとも1つ以上の化合物である、請求項9または10に記載のスクリーニング方法。
【請求項1】
細胞内の含硫化合物含量が増加した酵母であって、セレン酸感受性を有し、かつ培地上で培養されたときに該培地上で赤色を呈する酵母。
【請求項2】
前記培地はメチオニンが添加された培地である、請求項1に記載の酵母。
【請求項3】
アデニン要求性の付与により培地上で赤色を呈する、請求項1または2に記載の酵母。
【請求項4】
アデニン要求性がADE1遺伝子又はADE2遺伝子の改変により付された、請求項3に記載の酵母。
【請求項5】
さらに、ADE4遺伝子またはADE8遺伝子に変異が導入されたことにより、前記培地上で白色を呈するようになった、請求項3または4に記載の酵母。
【請求項6】
セレン酸感受性がMET25遺伝子の発現を増強する改変により付された、請求項1〜5のいずれか一項に記載の酵母。
【請求項7】
含硫化合物がシステイン、γ−グルタミルシステイン、グルタチオン、システニルグリシンからなる群から選択される少なくとも1つ以上の化合物である請求項1〜6のいずれか一項に記載の酵母。
【請求項8】
菌体量を増加させるために細胞内のアデニンが充足する状態で酵母を培養する工程と、細胞内の含硫化合物含量を増加させるために細胞内のアデニンが不足する状態で酵母を培養する工程を含む、請求項1〜7のいずれか一項に記載の酵母の培養方法。
【請求項9】
アデニン要求性およびセレン酸感受性を有する酵母に遺伝子改変処理を施す工程、アデニンが不足した時に酵母が赤く発色する培地に改変酵母をスプレッドし酵母コロニーを形成する工程、及び改変前よりも赤く発色する酵母コロニーを選択する工程を含む、細胞内の含硫化合物含量が増加した酵母のスクリーニング方法。
【請求項10】
セレン酸感受性を有する酵母に遺伝子改変処理を行う工程、最小培地にメチオニンを追加した培地に改変酵母をスプレッドし酵母コロニーを形成する工程、及び改変前よりも赤く発色する酵母コロニーを選択する工程を含む、細胞内の含硫化合物含量が増加した酵母のスクリーニング方法。
【請求項11】
含硫化合物がシステイン、γ−グルタミルシステイン、グルタチオン、システニルグリシンからなる群から選択される少なくとも1つ以上の化合物である、請求項9または10に記載のスクリーニング方法。
【図1】

【図2】
【図2】
【公表番号】特表2012−523230(P2012−523230A)
【公表日】平成24年10月4日(2012.10.4)
【国際特許分類】
【出願番号】特願2012−504171(P2012−504171)
【出願日】平成22年2月26日(2010.2.26)
【国際出願番号】PCT/JP2010/053695
【国際公開番号】WO2010/116833
【国際公開日】平成22年10月14日(2010.10.14)
【出願人】(000000066)味の素株式会社 (887)
【Fターム(参考)】
【公表日】平成24年10月4日(2012.10.4)
【国際特許分類】
【出願日】平成22年2月26日(2010.2.26)
【国際出願番号】PCT/JP2010/053695
【国際公開番号】WO2010/116833
【国際公開日】平成22年10月14日(2010.10.14)
【出願人】(000000066)味の素株式会社 (887)
【Fターム(参考)】
[ Back to top ]