
味覚受容体の潜在的モジュレーター化合物をスクリーニングする改善された方法
BRET技術を使用する、味覚受容体の潜在的モジュレーター化合物をスクリーニングする方法。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、味覚受容体の潜在的モジュレーター化合物をスクリーニングする方法であって、BRET技術を使用する方法に関する。
【背景技術】
【0002】
風味は、我々が快適な食物(即ち栄養食物)を消費し、不快な食物(潜在的毒素を含有する。)を回避するように食物摂取1を制御する一次知覚系の一部である。風味は、視覚、味覚、香りおよび食感の入力から形成される感覚である。しかしながら、食物の選択および我々が消費する量は、5種の基本味の3種(塩味、甘味および旨味)によってより動かされ、他の風味性によってはあまり影響を受けないと考えられる。これらの特質を含有する食物は、ヒトおよび多くの哺乳動物により好まれる食物である傾向があり;これに関して、旨味はタンパク質についてのマーカーとして、甘味は炭水化物についてのマーカーとして機能する。
【0003】
近年、これらの味覚様式の検出に関与する受容体が同定され、クローニングされ2−4、こうして味覚受容体の活性化をインビトロで調査することが可能となっている。甘味、旨味および苦味についての受容体は、Gタンパク質共役受容体(GPCR)のクラスに属する一方、塩味および酸味はイオンチャネルにより検出される可能性が最も高い。
【0004】
甘味は異種受容体二量体T1R2/T1R3により感知される一方、旨味は主としてT1R1/T1R3受容体により検出されるが2、他の受容体も同様に旨味に関与することが示されている5。
【0005】
インビトロ受容体活性化をインビボ知覚に良好な相関で計測するために、HEK293細胞などの哺乳動物細胞系内での味覚受容体の異種発現を含む種々の細胞系を使用することができる2,6−9。現在利用可能な機能的インビトロスクリーニング系は、通常、プロミスカスなGタンパク質、例えばGα15、Gα16またはこれらのGタンパク質のC末端ドメインの種々の適応部とのキメラを使用し;このことは、対象の受容体のPLC(ホスホリパーゼC)へのシグナリングカスケードおよび細胞内カルシウムの放出を指向する。このアプローチは純粋化合物の調査に極めて成功したが、抽出物または複合体試料の試験がより困難であることが証明されており:Gタンパク質の普遍的性質に起因してGタンパク質はスクリーニング細胞系内で(過剰)発現された組換え受容体に結合し得るだけでなく、低レベルにおいて内因的に存在する多くの受容体にも結合し得る。このことは、これらの内因性受容体を活性化する天然混合物中に存在するアゴニストにより誘導される非特異的カルシウムシグナルをもたらし得る。さらに、抽出物または複合試験試料は、GPCRを介する手段以外の手段により細胞内カルシウムを上昇させる物質も含有することが多く、これらのシグナルは受容体により誘導されたカルシウム放出と区別することができない。多くの天然混合物について観察される高い非特異的バックグラウンドシグナルは、大規模な分別手順を行わないこれらの試料の直接的なスクリーニングを妨げる。このような抽出物または複合試料の使用は、例えば食物材料を評価する場合にとてもありふれていることに留意すべきである。
【発明の概要】
【発明が解決しようとする課題】
【0006】
従って、味覚受容体の潜在的モジュレーター化合物のための改善されたスクリーニング方法であって、既存の方法の欠点のそれぞれを有しない方法が依然として必要とされている。
【課題を解決するための手段】
【0007】
第1の態様において、味覚受容体の潜在的モジュレーター化合物をスクリーニングする方法であって、BRET(生物発光共鳴エネルギー転移)(10,11)技術を使用する方法が提供される。本方法の各特徴を以下に詳細に定義する。
【0008】
好ましい方法は、以下の工程:
a)発光タンパク質、例えばルシフェラーゼタンパク質に融合している味覚受容体およびβ−アレスチンに融合している蛍光タンパク質を発現し、またはこれらの発現を誘導する細胞を提供する工程、
b)工程a)において得られた細胞を潜在的モジュレーター化合物によりチャレンジする工程ならびに
c)工程b)において得られた細胞のBRETシグナルを、該潜在的モジュレーターの不存在下でb)において得られた細胞のBRETシグナルと比較する工程
を含む。
【0009】
代替的に、第1の態様において、本発明は、味覚受容体をモジュレートする化合物を同定する方法であって、(a)(i)発光タンパク質に融合している味覚受容体および(ii)β−アレスチンに融合している蛍光タンパク質を発現する細胞を提供する工程;(b)細胞を潜在的モジュレーター化合物と接触させ、およびBRETシグナルを測定する工程;ならびに(c)工程(b)において得られたBRETシグナルを、該潜在的モジュレーター化合物の不存在下で細胞から得られたBRETシグナルと比較する工程を含み、(b)において得られたBRETシグナルと潜在的モジュレーター化合物の不存在下で得られたBRETシグナルとの差が、該潜在的モジュレーター化合物が味覚受容体をモジュレートする化合物であることを示す方法を提供する。
【0010】
本発明者らの発明は、本発明の方法により高い特異性を付与するBRET技術またはアッセイを使用し:対象の味覚受容体はドナー発光タンパク質、例えばルシフェラーゼタンパク質と融合しており、前記受容体に由来するシグナルに影響を与え、BRETシグナルを引き起こし得る細胞構成成分はない。このことは、味覚受容体の潜在的モジュレーター化合物を同定するためにスクリーニングに利用可能であることが多い天然混合物に関して特に興味深く:内因性受容体を活性化する構成成分も他の経路を介して細胞内カルシウムの非特異的上昇を事前に引き起こす物質も、BRETシグナルを引き起こすことはできない。読取り窓は受容体−発光融合タンパク質に唯一フォーカスされ、こうしてこの方法が、未精製の粗製抽出物を使用する高度に特異的な受容体活性化の直接調査に例外的に有用となる。
【0011】
本発明の方法は、GPCR(Gタンパク質共役受容体)である味覚受容体の受容体刺激時にβ−アレスチンをトランスロケートする能力を基礎とし、エネルギードナーを含有する融合タンパク質(発光タンパク質、例えばルシフェラーゼ)と、所与の波長において光を吸収し、より長い波長において再発光するエネルギーアクセプタータンパク質(フルオロフォア、典型的には蛍光タンパク質)との間のエネルギー転移を計測することにより受容体−β−アレスチン相互作用を計測するためにBRETアッセイを利用する10。GPCR活性化アッセイの場合、発光タンパク質、例えばルシフェラーゼは受容体のC末端に融合しており、蛍光タンパク質はβ−アレスチンに融合している。受容体が活性化される場合、細胞質β−アレスチンは細胞膜にリクルートされ、受容体を内在化について標的化する。β−アレスチン/蛍光タンパク質と発光タンパク質融合受容体との相互作用の間、ドナーおよびアクセプタータンパク質は近接し、BRETシグナルを誘導する。
【0012】
従って、BRET技術はシグナルまたはBRETシグナルを生成することができる技術またはアッセイであり、前記シグナルは、発光タンパク質に融合している味覚受容体と蛍光タンパク質との間のエネルギー転移であり、前記シグナルは、潜在的モジュレーター化合物の存在に起因して前記味覚受容体の活性化を反映する。
【0013】
本発明の方法の工程a)は、発光タンパク質、例えばルシフェラーゼタンパク質に融合している味覚受容体およびβ−アレスチンに融合している蛍光タンパク質を発現し、またはこれらの発現を誘導する細胞を提供する。本発明の方法の工程a)は、(i)発光タンパク質に融合している味覚受容体および(ii)β−アレスチンに融合している蛍光タンパク質を発現する細胞を提供することもできる。
【0014】
味覚受容体は、ヒトの口内において味覚に関連することが公知の任意の受容体であり得る。味覚受容体は、味覚認識に関与するものとして後に発見される任意の受容体でもあり得る。味覚受容体は、舌内で発現され得る:MSG(グルタミン酸ナトリウム)または旨味受容体、甘味受容体、苦味受容体または脂肪受容体。甘味認識に関与することが公知の受容体は、2種のサブユニットT1R2(Taste1 Receptor2)およびT1R3(Taste1 Receptor3)を含むヘテロ二量体である。旨味認識に関与することが公知の受容体は、2種のサブユニットT1R1(Taste1 Receptor1)およびT1R3を含む別のヘテロ二量体である。別のMSGまたは旨味受容体は、mGluR4(aまたはc)(代謝型グルタミン酸受容体4(aまたはc))の1種以上のサブユニットから構成される。苦味受容体は、TAS2(Taste2)受容体の1種以上のサブユニットから構成される。脂肪酸受容体は、GPR120(Gタンパク質共役受容体120)の1種以上のサブユニットから構成される。ヒトT1R1を表す好ましい核酸配列は、配列番号1である。ヒトT1R1タンパク質を表す対応する好ましいアミノ酸配列は、配列番号2により表される。ヒトT1R3を表す好ましい核酸配列は、配列番号3である。ヒトT1R3タンパク質を表す対応する好ましいアミノ酸配列は、配列番号4により表される。
【0015】
本発明に関して、味覚受容体は、ヒトの腸管内で栄養/脂肪酸感知に関与する受容体でもあり得る。このような受容体としては:カルシウム感知受容体、Gタンパク質共役受容体ファミリーCグループ6サブタイプA(GPRC6A)、L−アルファ−アミノ酸を感知する味覚受容体二量体T1R1/T1R3、炭水化物感知T1R2/T1R3受容体二量体、タンパク質分解産物センサGPR93(GPR92とも称される。)ならびに遊離脂肪酸(FFA)感知受容体FFA1、FFA2、FFA3、GPR84およびGPR120が挙げられる5。表3に同定された受容体のそれぞれを本発明の方法において使用することができる。これらの受容体のそれぞれをコードする好ましい核酸分子を配列表に挙げる。対応する好ましいコードされる受容体も配列表に挙げる(表3も参照。)。
【0016】
本発明の方法は、T1R1およびT1R3サブユニットを含む味覚受容体を使用して、および発光タンパク質としてのルシフェラーゼを使用して例示される。しかしながら、当業者は、本発明が前記ヘテロ二量体およびこの発光タンパク質を使用する方法に限定されるものではないことを理解する。本発明は、味覚受容体、好ましくはT1R1、T1R3ヘテロ二量体を発現する細胞を提供する。前記味覚受容体は、好ましくは機能的である。このことは、潜在的モジュレーターを添加することなく実施される本発明のスクリーニング方法において、この味覚受容体を活性化することが公知の物質が前記細胞に添加される場合に検出可能なBRETシグナルが存在することを意味する。各味覚受容体について、このような物質は公知である。このような物質、即ちアゴニストの例を表1に同定する。
【0017】
本発明はまた、蛍光タンパク質に融合している味覚受容体およびβ−アレスチンに融合している発光タンパク質、例えばルシフェラーゼタンパク質を発現し、またはこれらの発現を誘導する細胞を提供する工程a)を提供する。味覚受容体に融合している発光タンパク質、例えばルシフェラーゼタンパク質についての本明細書に定義の各特徴は、β−アレスチンに融合している発光タンパク質、例えばルシフェラーゼタンパク質についても保持される。β−アレスチンに融合している蛍光タンパク質についての本明細書に定義の各特徴は、発光タンパク質、例えばルシフェラーゼタンパク質に融合している蛍光タンパク質についても保持される。従って、本発明はまた、味覚受容体が蛍光タンパク質に融合しており、β−アレスチンが発光タンパク質に融合している方法を提供する。より一般的には、当業者は、発光タンパク質が味覚受容体に融合しており、蛍光タンパク質がβ−アレスチンに融合している本発明の任意の実施形態を、蛍光タンパク質が味覚受容体に融合しており、発光タンパク質がβ−アレスチンに融合している、他の点では同一の実施形態により置き換えることができることを理解する。
【0018】
本発明は、本発明の方法において使用される細胞を得るために使用すべき、核酸配列により表される好ましい核酸分子またはアミノ酸配列により表されるコードされるタンパク質を同定する。しかしながら、本明細書において同定される核酸配列のそれぞれは、天然に生じる形態、SNP(単一ヌクレオチド多型)を含有するバリアント、選択的スプライスによる形態、形態の組合せまたは当分野において公知の任意の機能的バリアントにより置き換えることができる。本明細書に定義の核酸分子は、本明細書に前述の細胞内で発現される場合、機能的であるべきである。核酸配列のバリアントは、この核酸配列の断片であり得る。好ましいバリアントは、サイレント突然変異を含有する。代替的にまたは組合せにおいて、核酸配列バリアントは、別のアミノ酸配列を生じさせないが、前記核酸配列が発現される宿主細胞のコドン使用頻度に対応する核酸置換の導入により得ることもできる。好ましくは、核酸配列バリアントは、本明細書に上記定義の核酸配列の任意の1種から出発して5’および/または3’末端からの1個以上のヌクレオチドが欠失されたものである。代替的にまたは組合せにおいて、核酸配列バリアントは、好ましくは別の生物および/または別の本明細書に上記定義の核酸配列のファミリーメンバーから単離された核酸配列である。これらのバリアントは全て、選択種、例えば哺乳動物種、例えばヒトからのcDNAまたはゲノミックライブラリーを使用する典型的なアプローチにおいて得ることができる。続いて、ライブラリーは厳しい条件下でハイブリダイゼーションにより本明細書に上記定義の核酸配列の1種またはこの一部を用いてスクリーニングすることができる。厳しい条件は、5×SSPE、0.3%SDS、200pg/mlの剪断および変性されたサケ精子DNAならびに50%ホルムアミド中での42℃におけるプレハイブリダイゼーションおよびハイブリダイゼーションを意味する。続いて、2×SSC、0.2%SDSおよび75℃をそれぞれ使用してハイブリダイゼーション反応物を30分間3回洗浄する。代替的にまたは組合せにおいて、核酸配列バリアントは、データベースにおけるアミノ酸同一性および/または類似性の探索ならびに検索により同定された好適なアミノ酸配列をコードする核酸配列の合成により得ることができる。
【0019】
ヒトは好ましい種である。別の好ましい実施形態によれば、核酸配列バリアントは、対立遺伝子バリアントである。対立遺伝子バリアントは、同一染色体座を占有する遺伝子の2種以上の別の代わりの形態のいずれかを意味する。対立遺伝子変異は突然変異を介して天然に生じ、個体群内の表現型多型をもたらし得る。別の好ましい実施形態によれば、核酸配列バリアントは、遺伝コードの縮退に関して本明細書に上記定義の核酸配列のいずれとも異なる。
【0020】
使用される核酸分子に関するさらなる説明を、標題「配列番号により定義される核酸分子および配列同一性」セクションに挙げる。好ましい実施形態において、使用される核酸分子はヒトに由来する。より好ましくは、この好ましい実施形態に定義される核酸分子は、哺乳動物、よりいっそう好ましくはヒト細胞における機能的発現のためのものである。ヒト配列と高度に相同性(少なくとも85%の同一性)である配列の使用は、魅力的である。これというのも、この核酸分子が哺乳動物細胞、好ましくはヒト細胞内で発現され、機能的であることを予期することができるからである。さらに、この配列は、ヒト配列と高度に相同性であるので、こうして調製された細胞型がヒト配列に対してより低い同一性を有する配列を用いて調製された細胞型よりも効率的にヒト味覚を模倣することが予期される。よりいっそう好ましくは、本明細書に上記定義の同一性は、85%以上、よりいっそう好ましくは90%以上、よりいっそう好ましくは91%以上、よりいっそう好ましくは92%以上、よりいっそう好ましくは93%以上、よりいっそう好ましくは94%以上、よりいっそう好ましくは95%以上、よりいっそう好ましくは96%以上、よりいっそう好ましくは97%以上、よりいっそう好ましくは98%以上、よりいっそう好ましくは99%以上、最も好ましくは100%である。
【0021】
本発明において、味覚受容体またはこのサブユニットをコードする核酸分子は、発光タンパク質、例えばルシフェラーゼタンパク質に融合している。好ましい実施形態において、発光タンパク質はルシフェラーゼタンパク質である。発光タンパク質、例えばルシフェラーゼタンパク質は、好ましくは細胞内部分である受容体のC末端部において融合している。当業者は、発光タンパク質、例えばルシフェラーゼタンパク質を味覚受容体の細胞内部分のいずれにも融合していてよいことを理解する。しかしながら、こうして得られたタンパク質は依然として機能的であるべきであり;即ち、活性化可能であるべきである。従って、発光タンパク質、例えばルシフェラーゼタンパク質を味覚受容体の細胞内部分におけるある場所に融合させた場合、好ましくは組換えタンパク質であるこうして得られたこのようなタンパク質をこの機能性に関して試験することが好ましい。好ましい実施形態において、発光タンパク質、例えばルシフェラーゼタンパク質は、味覚受容体のC末端部分の端部において、または前記味覚受容体のC末端部分の端部に可能な限り近くで融合している。前記味覚受容体のC末端部分の端部に可能な限り近くとは、好ましくは、発光タンパク質、例えばルシフェラーゼタンパク質の最初のアミノ酸が味覚受容体のC末端部分の最後のアミノ酸に対応する場所または味覚受容体のC末端部分の最後のアミノ酸から1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20個前のアミノ酸において存在することを意味する。ルシフェラーゼは、ホタルルシフェラーゼであり得、または任意のレニラ(Renilla)種からのものであり得る。ルシフェラーゼをコードする好ましい核酸分子は、哺乳動物細胞内でのこの発現レベルを改善するためにヒト化コドンを用いて改善されてきた。好ましいルシフェラーゼをコードするより好ましい核酸分子を配列番号77として挙げる。好ましいコードされるルシフェラーゼタンパク質を配列番号78として同定する。当業者は、2種の核酸分子をインフレームでいかに融合させるかを認識する。旨味受容体の好ましいT1R1およびT1R3サブユニットとしての1種超の区別されるサブユニットを有する味覚受容体の場合、各サブユニットを発光タンパク質、例えばルシフェラーゼタンパク質またはサブユニットの1種のみと融合させることができる。このことは、各味覚受容体について保持され、即ち、当業者は、このことが他の(ヘテロ)多量体味覚受容体についても保持されることを理解する。味覚受容体が1種超の区別されるサブユニットを有する場合、各サブユニットは、発光タンパク質、例えばルシフェラーゼタンパク質に融合されていてよい。代替的に、サブユニットの1種以上のタイプのみが発光タンパク質、例えばルシフェラーゼに融合されている。
【0022】
本発明において、蛍光タンパク質はβ−アレスチンに融合している。好ましい蛍光タンパク質は緑色蛍光タンパク質(GFP)である。GFP2がより好ましい。
【0023】
好ましいβ−アレスチンは、ヒトまたは哺乳動物β−アレスチンである。より好ましくは、β−アレスチンは非視覚β−アレスチン、例えばβ−アレスチン2または3などである。より好ましくは、β−アレスチンは配列番号79により表されるβ−アレスチン2である。前記β−アレスチン2をコードする好ましい核酸分子は、配列番号80により表される。よりいっそう好ましいβ−アレスチン2は、WO2004/065963またはWO2004/034054に記載されている。蛍光タンパク質とβ−アレスチンとの融合体も当業者に公知である。本発明の方法において、各核酸分子(即ち、ルシフェラーゼに融合している味覚受容体をコードするものおよびβ−アレスチンに融合している蛍光タンパク質をコードするもの)は、核酸構築物中に存在する。各構築物を細胞内に導入する。
【0024】
従って、本発明の細胞は、本明細書に定義の核酸構築物を含む。当業者は、細胞の選択が味覚受容体をコードする核酸配列の起源に大いに依存することを認識する。発現される味覚受容体が機能的である限り、任意の細胞を選択することができる。好ましくは、味覚受容体および/またはβ−アレスチンの発現は安定性であり、場合により誘導性である。代替的に、味覚受容体および/またはβ−アレスチンの発現は一過性である。誘導性発現は、「味覚受容体の発現」セクションに詳細に説明する。好ましくは、細胞は原核または真核細胞である。より好ましくは、細胞は昆虫または哺乳動物細胞である。よりいっそう好ましくは、哺乳動物細胞はヒト細胞である。哺乳動物細胞の例は、HEK293、HEK293T、MDCK、CHO、COS、NIH3T3、Swiss3T3、BHKおよびA549である。よりいっそう好ましくは、細胞は哺乳動物細胞、例えばHEK293である。本発明の細胞は、組換え細胞として把握することができる。本発明の細胞は、有利には、本発明の方法において使用される。
【0025】
選択される発現系のタイプに応じて、当業者は、味覚受容体およびβ−アレスチンの最も好ましい発現レベルを得るために培養条件を適合させることができると考えられる。誘導性発現系の場合、当業者は、誘導条件を最適化することもできると考えられる。発現の誘導の期間および発現の誘導の間の温度を最適化することもできると考えられる。好ましい実施形態によれば、味覚受容体の発現の誘導の開始時、サブコンフルエント細胞を、好適な培養培地を有する96ウェルプレート中に装入することができる。サブコンフルエントは、好ましくは70%コンフルエント、より好ましくは80%コンフルエントを意味する。好ましい実施形態において、添加される誘導剤は、誘導性発現系、好ましくはテトラサイクリン調節性プロモーターを使用する場合、テトラサイクリンまたはドキシサイクリンである。
【0026】
一実施形態において、細胞に、発光タンパク質、例えばルシフェラーゼタンパク質に融合している味覚受容体をコードする核酸分子およびβ−アレスチンに融合している蛍光タンパク質をコードする核酸分子を一過的に形質移入することができる。味覚受容体が1種超の区別されるサブユニットを有する場合、サブユニット当たり1種の核酸分子を使用することができる。代替的に、サブユニットの1種超のタイプを含む1種の単一核酸分子を使用することができる。一過性形質移入は、Lipofectamine2000を製造業者のプロトコルに従って使用して実施することができる。手短に述べると、細胞は、ウェル(12ウェルプレート、1ml培地/ウェル)当たり2×105個の細胞の密度において播種することができ、翌日に約80−90%のコンフルエンシーを目的とする。24時間後、発光タンパク質、例えばルシフェラーゼに融合している味覚受容体をコードする核酸分子を、β−アレスチンに融合している蛍光タンパク質をコードする核酸分子とともに同時形質移入することができる。ウェル当たり合計15gのトータルDNAを使用することができる。前記DNAを含む混合物を室温において30分間インキュベートし、各ウェルに添加し、細胞を48時間増殖させることができる。BRET計測は、形質移入の48−52時間後に実施することができる。代替的に、上記と同一のプロトコルを使用して発光タンパク質、例えばルシフェラーゼタンパク質に融合している味覚受容体をコードする核酸分子を、β−アレスチンに融合している蛍光タンパク質を安定的に発現する細胞内に形質移入することができる。好ましい形質移入プロトコルは、HEK293細胞についての実験部に記載する。
【0027】
好ましい実施形態において、味覚受容体はT1R1およびT1R3サブユニットを含み、サブユニットの少なくとも1種:T1R1もしくはT1R3または両サブユニットは発光タンパク質、例えばルシフェラーゼに融合している。好ましくは、ルシフェラーゼタンパク質はレニラルシフェラーゼである。レニラルシフェラーゼに融合している好ましいT1R1サブユニットをコードする好ましい核酸配列は、配列番号5により表される。対応する好ましいコードされるアミノ酸配列は、配列番号6により表される。レニラルシフェラーゼに融合している好ましいT1R3サブユニットをコードする好ましい核酸配列は、配列番号7により表される。対応する好ましいコードされるアミノ酸配列は、配列番号8により表される。
【0028】
さらに好ましい実施形態において、β−アレスチンに融合している蛍光タンパク質はGFPタンパク質、好ましくはGFP2であり、β−アレスチンはβ−アレスチン2である。
【0029】
よりいっそう好ましくは、味覚受容体は、T1R1およびT1R3サブユニットを含み、各サブユニットは、発光タンパク質、例えばルシフェラーゼタンパク質、好ましくはルシフェラーゼ、より好ましくはレニラルシフェラーゼに融合しており、β−アレスチンに融合している蛍光タンパク質はGFPタンパク質、好ましくはGFP2であり、β−アレスチンはβ−アレスチン2である。
【0030】
発光タンパク質がルシフェラーゼタンパク質であり得ることが、本発明によりさらに包含される。代替的に、発光タンパク質は、別の好適なエネルギードナーであり得る。GFPを別の好適なエネルギーアクセプターにより置き換えることができることも、本発明により包含される。理論的には、これ自体のポリペプチド配列内で可視波長発色団を形成することができるタンパク質の構造的に相同性のクラスのメンバーであると定義される、任意の蛍光タンパク質または分子を使用することができる。幾つかの蛍光タンパク質が、BRET技術において既に使用されている(Bacart J.Et al,(2008)Biotechnol.J.3:311−324およびPfleger K.D.,et al,(2006),Nature Methods,3:165−174参照。)。
【0031】
発光タンパク質、例えばルシフェラーゼおよびGFPは、それぞれ好ましいエネルギードナーおよびエネルギーアクセプターとして本明細書に提示される。発光タンパク質、例えばルシフェラーゼまたはGFPについて定義される特徴のそれぞれは、それぞれ任意の他のエネルギードナーまたはエネルギーアクセプターについても保持される。
【0032】
従って、本発明の方法において:
(a)味覚受容体のサブユニットの少なくとも1種、好ましくはT1R1およびT1R3の少なくとも1種は、発光タンパク質、例えばルシフェラーゼタンパク質に融合しており、もしくは味覚受容体の各サブユニット、好ましくはT1R1およびT1R3は、それぞれ発光タンパク質、例えばルシフェラーゼタンパク質に融合しており、
ならびに/または
(b)β−アレスチンに融合している蛍光タンパク質はGFPタンパク質であり、好ましくはGFPはGFP2であり、および/もしくはβ−アレスチンはβ−アレスチン2である。
【0033】
代替的に、本発明の方法において:
(a)味覚受容体のサブユニットの少なくとも1種、好ましくはT1R1およびT1R3の少なくとも1種は蛍光タンパク質に融合しており、もしくは味覚受容体の各サブユニット、好ましくはT1R1およびT1R3は蛍光タンパク質に融合しており、
ならびに/または
(b)発光タンパク質はβ−アレスチンに融合している。
【0034】
代替的に、本発明の方法において:
(a)味覚受容体はT1R1、T1R3ヘテロ二量体であり、サブユニットT1R1およびT1R3の少なくとも1種が発光タンパク質に融合しており、および蛍光タンパク質がβ−アレスチンに融合しており;または
(b)味覚受容体はT1R1、T1R3ヘテロ二量体であり、サブユニットT1R1およびT1R3の少なくとも1種が蛍光タンパク質に融合しており、および発光タンパク質がβ−アレスチンに融合している。
【0035】
好ましくは、本発明の方法において、(a)発光タンパク質の少なくとも1種がルシフェラーゼであり;(b)蛍光タンパク質の少なくとも1種がGFPであり;および(c)β−アレスチンの少なくとも1種が非視覚β−アレスチンである。より好ましくは、(a)ルシフェラーゼの少なくとも1種がレニラルシフェラーゼであり;および(b)非視覚β−アレスチンの少なくとも1種がβ−アレスチン2である。
【0036】
工程b)は、工程a)において得られた細胞を潜在的モジュレーター化合物によりチャレンジすることを含む。
【0037】
この工程に関して、チャレンジすることは、工程a)において得られた細胞を潜在的モジュレーター化合物と接触させることを意味することができる。
【0038】
味覚受容体の潜在的モジュレーター化合物は、味覚受容体の活性化される能力を遮断し、阻害し、モジュレートし、または向上させることにより味覚認識を遮断し、阻害し、モジュレートし、または向上させ、こうして細胞内シグナルを細胞内にトランスデュースすることができる化合物として本明細書に定義される。任意の分子、例えば天然または合成の任意の有機分子、例えばタンパク質、オリゴペプチド、有機小分子、多糖、脂質(例えばスフィンゴ脂質)、脂肪酸、ポリヌクレオチド、オリゴヌクレオチドなどを本発明において試験することができる。潜在的モジュレーター化合物は、化合物のライブラリー、例えば十分な範囲の多様性を提供するコンビナトリアルまたはランダム化ライブラリーの形態であり得る。
【0039】
潜在的モジュレーター化合物は、通常、水性試料溶液中に存在する。このような水性試料溶液は、(a)食品;(b)食品の抽出物;および(c)バイオマスの抽出物、好ましくは可食性バイオマスの抽出物の少なくとも1種を含むことができる。従って、試料溶液は、液体食品またはこの希釈物を含むことができる。液体食品は、例えば飲料または醤油などのようなソースであり得る。水性試料溶液は、食品の抽出物、例えば固体食品(例えばチーズ)または脂肪食品の抽出物を含むこともできる。または、水性試料溶液は、植物、動物および微生物の少なくとも1種からのバイオマスの抽出物を含むことができる。従って、水性試料溶液は、植物、動物および微生物の少なくとも1種からの可溶性分子を含むことができる。従って、水性試料溶液は、発酵食品からの抽出物を含むことができる。好ましい実施形態において、試料溶液は、トマト抽出物を含む。抽出物はさらに以下に本明細書に定義されるとおりであり、以下に本明細書に定義されるとおり調製することができる。
【0040】
本発明の方法の利点は、本発明の方法により複合試料中の潜在的モジュレーターの同定が可能となることである。従って、好ましくは、水性試料溶液は、複合試料溶液である。水性試料溶液は、少なくとも2種の区別される有機分子を含む、好ましくは少なくとも3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20種または50種超または100種超の区別される有機分子を含む混合物であり得る。化合物の数は、好ましくは以下に同定されるとおり評価される。水性試料溶液は、この溶液が10種超の区別される有機分子、好ましくは10種超の区別される潜在的モジュレーターを含む場合、複合試料溶液と定義することができる。水性試料溶液は、通常、例えば、所定量の1種以上の特定されたモジュレーターを含む参照溶液とは対照的に未特定組成物の溶液であることがさらに理解される。従って、未特定組成物の溶液は、アイデンティティおよび/または濃度が未知であり、または定義されていない化合物を含む溶液である。
【0041】
水性試料溶液は、好ましくは可溶物のみを含む溶液である。不溶物は、当分野において公知であり、抽出物について以下に本明細書に記載されている手段により試料溶液から除去することができる。水性試料溶液は、溶液を生理学的に許容されるpHおよび浸透圧値に調整することにより、工程a)において提供された細胞と生理学的に適合性を示すまでさらに調整することができる。
【0042】
好ましい実施形態において、このようなモジュレーターは抽出物中に存在する。抽出物は、少なくとも2種の区別される分子、少なくとも3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20種または50種超または100種超の区別される分子を含む混合物である。化合物の数は、好ましくは、以下に同定されるとおり評価される。抽出物は、この抽出物が10種超の区別される分子を含む場合、複合抽出物と定義することができる。抽出物は、少なくとも2種の区別される有機分子、少なくとも3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20種または50種超または100種超の区別される有機分子を含む混合物である。化合物の数は、好ましくは、以下に同定されるとおり評価される。抽出物は、この抽出物が10種超の区別される有機分子を含む場合、複合抽出物と定義することができる。抽出物は精製しなくてよく、植物ベースの抽出物、好ましくはトマト抽出物または動物ベースの抽出物および/または食物ベースの抽出物であり得る。抽出物は、食物ベースまたは食物由来の抽出物であり得る。食物ベースまたは食物由来の抽出物は、食物組成物もしくは食品として、または食物組成物もしくは食品をベースとし、もしくはこれらに由来するものとして把握することができる。抽出物のアイデンティティ、この組成および抽出物が調製された手法は、本発明について重要ではない。対照的に、本発明の利点は、本発明の方法を使用して潜在的に各抽出物から味覚受容体の潜在的モジュレーターを同定することができることである。
【0043】
抽出物は、植物、動物および/または微生物バイオマスの抽出のために溶媒(例えば水)または溶媒混合物(例えば水/アルコール)を使用することにより得られた未精製混合物、例えば前記材料からの全ての可溶性化合物を好ましくは元の材料中に存在するのと同様の比、好ましくはこれらの溶解度に従って補正された比で含有する生の、加工された、発酵されたおよび/または乾燥された植物または動物または食物材料であり得る。
【0044】
抽出物中の化合物の数および濃度は、元の植物または動物または食物材料中のこれらの存在、使用される溶媒系におけるこれらの溶解度を認識し、当業者に公知の技術、例えばクロマトグラフィー(HPLC、GC)および/または分光技術(NMR、質量分析)を使用して評価することができる。
【0045】
本発明において使用すべき抽出物の調製は、通常、少なくとも2種の工程:1)可溶化および/または均一化工程;ならびに2)不溶物および場合により非水相もしくは溶媒を除去する工程を含む。好適な抽出物を生成する考えられる手法は、慣用の加工条件(例えば加熱、撹拌、粉砕)下で好適な溶媒(例えば水、アルコール)または溶媒混合物(例えば水/アルコール)により(例えば切断、温浸、乾燥、粉化などにより)予備加工し、下流で加工(例えば濾過、遠心分離、乾燥)することができる植物または動物または食物材料の処理であり得る。
【0046】
好ましい方法において、味覚受容体の公知のアクチベーターを工程b)においてさらに添加する。味覚受容体の好ましい公知のアクチベーターは、表1に既に同定されている。アクチベーターは、阻害剤についての探索において使用することができ:味覚受容体を最初にアクチベーターと接触させてBRETシグナル(即ち、前記受容体の定常活性よりも高いシグナル)を生成する。続いて、潜在的阻害化合物を前記アクチベーターと一緒に接触させる。生成されたシグナルが低減すれば、前記潜在的阻害化合物は阻害剤である。アクチベーターは、好ましくは、これに関連して次いで阻害剤により低下される受容体活性化のレベルを増加させるために使用し;このことにより、読取り窓およびひいては感度が増加する。
【0047】
工程c)は、工程b)において得られた細胞のBRETシグナルを、潜在的モジュレーター化合物の不存在下でb)において得られた細胞のBRETシグナルと比較することを含む。代替的に、工程c)は、工程b)において得られたBRETシグナルを、潜在的モジュレーター化合物の不存在下で細胞から得られたBRETシグナルと比較することを含み、b)において得られたBRETシグナルと潜在的モジュレーター化合物の不存在下で得られたBRETシグナルとの差は、潜在的モジュレーター化合物が味覚受容体をモジュレートする化合物であることを示す。
【0048】
BRETシグナルの生成を導くBRET技術またはアッセイは、当業者に公知の任意のBRET技術であり得る。味覚受容体に融合している発光ドナー発光タンパク質、例えばルシフェラーゼタンパク質の任意の公知のバリアントを使用することができる。同じことが、フルオロフォアもしくは蛍光タンパク質とも称される発光アクセプターまたは蛍光タンパク質に融合しているエネルギーアクセプタータンパク質、例えばβ−アレスチンについて保持される。発光ドナーおよび発光アクセプターのアイデンティティに応じて、BRET1またはBRET2またはBRET3技術を使用する。幾つかのBRETアッセイについて使用されるタンパク質のアイデンティティ、条件および波長は当業者に公知であり、例えばBacart J.et al(2008),Biotechnol.J.3:311−324を参照。しかしながら、BRET技術において使用されるタンパク質のそれぞれは改変してこれらのスペクトル特性を改善することができ、これらをBRET技術により好適とする。好ましくは、BRET2技術を使用する。これというのも、この技術はより良好な感度および効率:BRETシグナルのより早期の検出を導く関与する波長のより良好な分離を提供することが予期されるからである。
【0049】
好ましくは抽出物中に存在する潜在的モジュレーター化合物を用いる受容体刺激後、蛍光タンパク質に融合しているβ−アレスチンタンパク質は、活性化された味覚受容体と相互作用し、こうして蛍光タンパク質を前記活性化された味覚受容体に融合している発光タンパク質、例えばルシフェラーゼと近接して接触させ、考えられるこれらの2種のタンパク質間のエネルギー転移をもたらし、BRETシグナルを生成する。BRET2の場合、アッセイは好ましくは以下のとおりである:酸素の存在下、ルシフェラーゼは基質DeepBlueCの395−410nmにおいて計測することができるコエレンテルアミドへの転換を触媒する。蛍光タンパク質が発光タンパク質、例えばルシフェラーゼに近接しており、エネルギー転移が生じる場合、発光は510nmにシフトし;このことはBRETシグナルと称され、アクセプター(蛍光タンパク質)とドナー(発光タンパク質、例えばルシフェラーゼ)との比として表現される。BRETシグナルは、本明細書において比が0よりも大きい検出可能なBRETシグナルとして定義される。BRETシグナルがもっぱら味覚受容体の特異的活性化により引き起こされることを確保するため、好ましくは発光タンパク質、例えばルシフェラーゼタンパク質に融合している味覚受容体およびβ−アレスチンに融合している蛍光タンパク質を発現する細胞を用いて得られたBRETシグナルを、前記受容体を発現しない細胞および/または発光タンパク質、例えばルシフェラーゼタンパク質に融合している味覚受容体を発現しない細胞および/またはβ−アレスチンに融合している蛍光タンパク質を発現しない細胞を用いて得られたBRETシグナルと比較する。当業者は、BRETアッセイをいかに実施するかを認識する。蛍光は、典型的には、蛍光プレートリーダーにより計測する。一実施形態において、Mithras LB 940プレートリーダー(Berthold Technologies)を使用することができる。使用することができるプロトコルは、本質的には、Packard BioOneに記載されているものと同一であり、またはわずかな改変を有する刊行されたプロトコル13であり:細胞は、本明細書に上記のとおり形質移入することができる。形質移入48時間後、細胞を回収し、BRET緩衝液(2μg/mlアプロチニンを含有するD−PBS)中に2×106個の細胞/mlの密度において溶解させることができる。細胞を平衡化のために室温において1時間放置した後、約1×105個の細胞を含有する30μlを白色96ウェルプレートの各ウェルに移すことができる。10μlの潜在的モジュレ−ティング化合物または抽出物(または緩衝液)および10μlの基質コエレンテラジン(最終濃度5μM)を、インジェクターを使用して同時に添加することができる。
【0050】
最終インジェクション直後、発光シグナルの生成に使用される基質を考慮して繰り返し連続読取りをドナーの規定の発光波長およびアクセプターの規定の発光波長において行う。BRETシグナルは、アクセプタータンパク質について計測された光シグナルと、ドナータンパク質について計測された光シグナルとの比として測定することができる。
【0051】
BRET技術がBRET1技術である場合、ドナーはレニラルシフェラーゼ[Rluc]および基質としてのコエレンテラジンhであり;アクセプターは強化黄色蛍光タンパク質[強化YFP]、YFPトパーズ(topaz)、YFPシトリン(citrine)、YFPビーナス(venus)、YPet.)である。
【0052】
最終インジェクション直後、繰り返し連続読取りを、Rluc/コエレンテラジンhの規定の発光波長のピーク(480nm)およびアクセプターの規定の発光波長のピーク(530nm)をカバーする波長において行う。BRETシグナルは、アクセプタータンパク質について計測された光シグナルとドナータンパク質について計測された光シグナルとの比として決定することができる。
【0053】
技術がBRET2技術である場合、ドナーはレニラルシフェラーゼ[Rluc]またはレニラルシフェラーゼ突然変異体8[Rluc8](Loening AM,et al,Protein Eng Des Sel 2006 September;19(9):391−400およびBacart J,et al,Biotechnol J 2008 March;3(3):311−24.)および基質としてのDeepBlueC(商標)またはコエレンテラジン400aであり;アクセプターは、緑色蛍光タンパク質−2[GFP2]または緑色蛍光タンパク質10[GFP10])である。
【0054】
最終インジェクション直後、繰り返し連続読取りを、Rluc/コエレンテラジン400aの規定の発光波長のピーク(395nm)およびアクセプターの規定の発光波長のピーク(510nm)をカバーする波長において行う。BRETシグナルは、アクセプタータンパク質について計測された光シグナルとドナータンパク質について計測された光シグナルとの比として決定することができる。
【0055】
工程c)において得られた細胞のBRETシグナルを、潜在的モジュレーター化合物の不存在下でc)において得られた細胞の対応する不存在のBRETシグナルと比較する。代替的に、工程c)において得られた細胞のBRETシグナルの変化を、潜在的モジュレーター化合物の不存在下でc)において得られた細胞の対応する元のBRETシグナルと比較する。
【0056】
代替的に、工程b)において得られたBRETシグナルを、潜在的モジュレーター化合物の不存在下で細胞から得られたバックグラウンド(または不存在の)BRETシグナルと比較する。b)において得られたBRETシグナルと、潜在的モジュレーター化合物の不存在下で得られたBRETシグナルとの差は、潜在的モジュレーター化合物が味覚受容体をモジュレートする化合物であることを示す。代替的に、工程b)において得られたBRETシグナルを、潜在的モジュレーター化合物の不存在下で工程b)において得られた細胞の対応する元のBRETシグナルと比較することができる。b)において得られたBRETシグナルと、潜在的モジュレーター化合物の不存在下で得られたBRETシグナルとの差は、潜在的モジュレーター化合物が味覚受容体をモジュレートする化合物であることを示す。同様に、2種以上の異なる試料溶液を用いて得られたBRETシグナルを比較することができ、このことにより異なる試料溶液は、上記の食品、食品の抽出物および/またはバイオマスの抽出物の異なるフラクションを含有することができる。
【0057】
BRETシグナルを増加させ、または検出可能なBRETシグナルを誘導する化合物は、味覚の潜在的エンハンサーである。対照的に、BRETシグナルを減少させる化合物は味覚の潜在的マスカーである。
【0058】
好ましい実施形態によれば、味覚の潜在的エンハンサーは、工程c)において実施された比較がBRETシグナルもしくは検出可能なBRETシグナルの存在または少なくとも2%のBRETシグナルの検出可能な増加を示す場合に同定された。より好ましくは、味覚の潜在的エンハンサーは、工程c)において実施された比較がBRETシグナルの少なくとも4%、少なくとも6%、少なくとも8%、少なくとも10%、少なくとも12%、少なくとも14%、少なくとも16%、少なくとも18%、少なくとも20%、少なくとも22%、少なくとも24%、少なくとも26%、少なくとも28%、少なくとも30%、少なくとも32%、少なくとも34%、少なくとも36%、少なくとも38%、少なくとも40%、少なくとも42%以上の増加を示す場合に同定された。
【0059】
別の好ましい実施形態によれば、味覚の潜在的マスカーは、工程c)において実施される比較がBRETシグナルの少なくとも2%の減少を示す場合に同定された。より好ましくは、味覚の潜在的マスカーは、工程c)において実施される比較がBRETシグナルの少なくとも4%、少なくとも6%、少なくとも8%、少なくとも10%、少なくとも12%、少なくとも14%、少なくとも16%、少なくとも18%、少なくとも20%、少なくとも22%、少なくとも24%、少なくとも26%、少なくとも28%、少なくとも30%、少なくとも32%、少なくとも34%、少なくとも36%、少なくとも38%、少なくとも40%、少なくとも42%以上の減少を示す場合に同定された。
【0060】
味覚受容体の公知の阻害剤またはアクチベーターが存在する場合、BRETシグナルのそれぞれの増加または減少を、前記潜在的モジュレーター化合物の不存在下で得られたBRETシグナルと比較する。
【0061】
さらなる態様において、本明細書に同定される方法により同定された味覚受容体の潜在的モジュレーター化合物が提供される。
【0062】
別の態様において、本発明は、味覚受容体をモジュレートする化合物を生成する方法に関する。本方法は、本明細書に定義の方法において味覚受容体をモジュレートする化合物を同定し、該化合物を回収する工程を含む。味覚をモジュレートする化合物の回収および/または(部分)精製の方法は、当業者に自体公知である。
【0063】
さらに別の態様において、本発明は、味覚受容体をモジュレートする化合物の同定のための、BRET技術またはアッセイの使用に関する。BRET技術またはアッセイは、本明細書に記載の本発明の方法に従って使用することができる。
【0064】
一般的技術情報
配列番号により定義される核酸分子および配列同一性
所与の配列同一性番号(例えば配列番号1)により本明細書に同定される各遺伝子または核酸分子は、開示されているこの規定の配列に限定されるものではないことを理解すべきである。本明細書に同定されている各遺伝子配列またはヌクレオチド配列は、本明細書に同定されている所与のタンパク質またはポリペプチドをコードする。本願全体にわたり、規定のヌクレオチド配列の配列番号(配列番号1を例にとる。)を指すごとに、これを
i.配列番号1と少なくとも60%の配列同一性を有するヌクレオチド配列を含むヌクレオチド配列、
ii.相補的ストランドが(i)または(ii)の配列の核酸分子にハイブリダイズするヌクレオチド配列;
iii.配列が遺伝コードの縮退に起因して(i)の核酸分子の配列とは異なるヌクレオチド配列、
iv.ヌクレオチド配列配列番号1によりコードされるアミノ酸配列と少なくとも60%のアミノ酸同一性を有するアミノ酸配列をコードするヌクレオチド配列
により置き換えることができる。
【0065】
本明細書に記載の各ヌクレオチド配列またはアミノ酸配列は、それぞれ所与のヌクレオチド配列またはアミノ酸配列とのこの同一性の割合(少なくとも60%)に関して、さらに好ましい実施形態において、それぞれ所与のヌクレオチドまたはアミノ酸配列との少なくとも65%、70%、75%、80%、85%、90%、95%、97%、98%、99%以上の同一性を有する。好ましい実施形態において、配列同一性は、本明細書に同定されている配列の全長を比較することにより決定する。
【0066】
「配列同一性」は、本明細書において、配列を比較することにより決定される、2種以上のアミノ酸(ポリペプチドまたはタンパク質)配列間または2種以上の核酸(ポリヌクレオチド)配列間の関係と定義される。当分野において「同一性」も、場合によってはこのような配列のストリング間の一致により決定されるアミノ酸または核酸配列間の配列関連性の程度を意味する。2種のアミノ酸配列間の「類似性」は、アミノ酸配列およびあるポリペプチドの第2のポリペプチドの配列に対してこの保存されているアミノ酸置換物を比較することにより決定する。「同一性」および「類似性」は、限定されるものではないが、(Computational Molecular Biology,Lesk,A.M.,ed.,Oxford University Press,New York,1988;Biocomputing:Informatics and Genome Projects,Smith,D.W.,ed.,Academic Press,New York,1993;Computer Analysis of Sequence Data,Part I,Griffin,A.M.,and Griffin,H.G.,eds.,Humana Press,New Jersey,1994;Sequence Analysis in Molecular Biology,von Heine,G.,Academic Press,1987;およびSequence Analysis Primer,Gribskov,M.and Devereux,J.,eds.,M Stockton Press,New York,1991;およびCarillo,H.,and Lipman,D.,SIAM J.Applied Math.,48:1073(1988)に記載のものを含む公知の方法により容易に計算することができる。
【0067】
同一性を決定する好ましい方法は、試験される配列間の最大の一致を与えるように設計する。同一性および類似性を決定する方法は、公的に入手可能なコンピュータプログラムにおいてコード化されている。2種の配列間の同一性および類似性を決定する好ましいコンピュータプログラム方法としては、例えば、GCGプログラムパッケージ(Devereux,J.,et al.,Nucleic Acids Research 12(1):387(1984)),BestFit,BLASTP,BLASTNおよびFASTA(Altschul,S.F.et al.,J.Mol.Biol.215:403−410(1990)が挙げられる。BLAST Xプログラムは、NCBIおよび他の供給源(BLAST Manual,Altschul,S.,et al.,NCBI NLM NIH Bethesda,MD 20894;Altschul,S.,et al.,J.Mol.Biol.215:403−410(1990)から公的に入手可能である。周知のSmith Watermanアルゴリズムを使用して同一性を決定することもできる。
【0068】
ポリペプチド配列比較についての好ましいパラメーターとしては、以下のもの:アルゴリズム:Needleman and Wunsch,J.Mol.Biol.48:443−453(1970);比較マトリックス:Hentikoff and Hentikoff,Proc.Natl.Acad.Sci.USA.89:10915−10919(1992)からのBLOSSUM62;ギャップペナルティ:12;およびギャップ長さペナルティ:4が挙げられる。これらのパラメーターを用いる有用なプログラムは、Genetics Computer Group,located in Madison,Wlからの「Ogap」プログラムとして公的に入手可能である。上記パラメーターは、(末端基についてペナルティを伴わない)アミノ酸比較についてのデフォルトパラメーターである。
【0069】
核酸比較についての好ましいパラメーターとしては、以下のもの:アルゴリズム:Needleman and Wunsch,J.Mol.Biol.48:443−453(1970);比較マトリックス:一致=+10、不一致=0;ギャップペナルティ:50;ギャップ長さペナルティ:3が挙げられる。Madison,Wisに位置するGenetics Computer GroupからのGapプログラムとして入手可能。核酸比較についてのデフォルトパラメーターは上記に挙げる。
【0070】
場合により、アミノ酸類似性の程度の決定において、当業者は、当業者に明確であるいわゆる「保存」アミノ酸置換を考慮することもできる。保存アミノ酸置換は、類似側鎖を有する残基の交換可能性を指す。例えば、脂肪族側鎖を有するアミノ酸のグループはグリシン、アラニン、バリン、ロイシンおよびイソロイシンであり;脂肪族−ヒドロキシル側鎖を有するアミノ酸のグループはセリンおよびトレオニンであり;アミド含有側鎖を有するアミノ酸のグループはアスパラギンおよびグルタミンであり;芳香族側鎖を有するアミノ酸のグループはフェニルアラニン、チロシンおよびトリプトファンであり;塩基性側鎖を有するアミノ酸のグループはリジン、アルギニンおよびヒスチジンであり;硫黄含有側鎖を有するアミノ酸のグループはシステインおよびメチオニンである。好ましい保存アミノ酸置換基は:バリン−ロイシン−イソロイシン、フェニルアラニン−チロシン、リジン−アルギニン、アラニン−バリンおよびアスパラギン−グルタミンである。本明細書に開示のアミノ酸配列の置換バリアントは、開示配列中の少なくとも1個の残基が除去され、異なる残基がこの場所に挿入されたものである。好ましくは、アミノ酸変化は保存的である。天然に生じるアミノ酸のそれぞれについて好ましい保存的置換は、以下のとおりである:Alaからser;Argからlys;Asnからglnまたはhis;Aspからglu;Cysからserまたはala;Glnからasn;Gluからasp;Glyからpro;Hisからasnまたはgln;Ileからleuまたはval;Leuからileまたはval;Lysからarg;glnまたはglu;Metからleuまたはile;Pheからmet、leuまたはtyr;Serからthr;Thrからser;Trpからtyr;Tyrからtrpまたはphe;およびValからileまたはleu。
【0071】
細胞内での味覚受容体の発現
本発明の方法において使用される味覚受容体およびβ−アレスチンは、ルシフェラーゼタンパク質に融合している味覚受容体をコードするヌクレオチド配列および蛍光タンパク質に融合しているβ−アレスチンをコードする別の配列を好適な細胞内で発現させる組換え技術を使用して調製することができる。従って本発明はまた、上記定義の前記ヌクレオチド配列により表される前記核酸分子を含むベクターの使用に関する。好ましくは、ベクターは、ベクターに好適な宿主内でのベクターの増幅を確保する複製起源(または自律複製配列)を含む複製ベクターである。代替的に、ベクターは、例えば相同組換えまたは別の手段を介して細胞のゲノム中に統合することができる。特に好ましいベクターは、上記定義のポリペプチドをコードするヌクレオチド配列が、ベクターのための細胞内でコード配列の発現を指向することができるプロモーターに作動可能に連結している発現ベクターである。
【0072】
本明細書において使用される用語「プロモーター」は、1種以上の遺伝子の転写を制御するように機能し、遺伝子の転写開始部位の転写の方向に関して上流に局在しており、DNA依存性RNAポリメラーゼについての結合部位、転写開始部位ならびに限定されるものではないが、転写因子結合部位、レプレッサーおよびアクチベータータンパク質結合部位を含む任意の他のDNA配列ならびにプロモーターからの転写の量を調節するように直接的または間接的に作用することが当業者に公知のヌクレオチドの任意の他の配列の存在により構造的に同定されている核酸断片を指す。「構成性」プロモーターは、ほとんどの生理学的および発育条件下で活性であるプロモーターである。「誘導性」プロモーターは、生理学的または発育条件に応じて調節されるプロモーターである。発現ベクターにより、上記定義のポリペプチドを、前記ポリペプチドをコードするヌクレオチド配列を好適な細胞、例えば培養細胞内で発現させる組換え技術を使用して調製することが可能となる。
【0073】
好ましくは、上記定義の本発明の核酸配列のそれぞれの発現は、誘導性である。各核酸配列の発現の誘導性は、当業者に公知の任意の手法により実現することができる。例えば、Invitrogen T−Rex系(テトラサイクリン調節性発現、tetオペロンに基づき、培地に誘導物質が添加されるまで挿入遺伝子の発現が抑制される。)、Invitrogen Gene−Switch系(GaW−Elbプロモーターの活性化に基づく)、Stratagene Complete Control誘導性哺乳動物発現系(ドロソフィア・メラノガステル(Drosophila melanogaster)エクジソン受容体についての遺伝子およびエクジソン受容体についての結合部位を含有するプロモーターの両方を内部に持つ哺乳動物細胞内での昆虫ホルモンのエクジソンまたはこの類似体であるポナステロンA(ponA)による転写の活性化に基づく)、New England Biolabs RheoSwitch(登録商標)哺乳動物誘導性発現系(合成誘導物質であるRheoSwitch Ligand RSLlおよび2つの領域からなるキメラ核受容体との高度に特異的な相互作用に基づく)、Qbiogene Q−mate(商標)誘導性発現系またはQ−mate(商標)CymR系(誘導物質分子であるクマートの不存在下、オペレーター部位に結合しているクマートリプレッサータンパク質CymRによる遺伝子発現の抑制に基づく。クマートの存在下、CymRはクマートに結合し、立体配置の変化を受け、オペレーター部位からのこの放出をもたらす。)、Stratagene製LacSwitch(登録商標)II誘導性哺乳動物発現系(lacオペロンに基づき、誘導物質が培地に添加されるまで挿入遺伝子の発現が抑制される。)がある。よりいっそう好ましくは、各核酸部分配列の発現は、本発明の核酸配列中に存在する核酸部分配列のそれぞれに作動可能に連結している誘導性プロモーターの存在により誘導性となる。本発明に関して、「作動可能に連結している」は、制御配列、本明細書においてはプロモーター配列が、制御配列が核酸部分配列の発現を指向するように、核酸部分配列に対する位置に適切に配置される立体配置と定義される。
【0074】
誘導性プロモーターは、上記定義の所与の核酸部分配列について、および所与の細胞内で機能的である任意のプロモーターまたはこの部分であり得、プロモーターの転写開始活性は、細胞を培養する間に所与の誘導剤を添加したときに細胞内で誘導され得る。より好ましくは、誘導性プロモーターはテトラサイクリン調節性プロモーターである。よりいっそう好ましくは、誘導性プロモーターは、Yao et al(Yao,F.et al.(1998)Hum.Gene Therapy 9:1939−1950およびYao,F and Eriksson,E.(1999)Hum.Gene Therapy 10:419−427)に記載のテトラサイクリン調節性ハイブリッドヒトサイトメガロウイルスプロモーターである。この系は、InvitrogenからT−Rex発現系として購入することができる。)。誘導性発現系の使用により、安定発現系を使用して記載された毒性の問題を回避することができる。
【0075】
典型的には、上記定義のポリペプチドをコードする核酸を発現ベクター中で使用する。語句「発現ベクター」は、一般に、このような配列と適合性の宿主内で遺伝子の発現を行うことができるヌクレオチド配列を指す。これらの発現ベクターは、典型的には、少なくとも好適なプロモーター配列および場合により転写終結シグナルを含む。発現の実施に必要な、または一助となる追加の因子を本明細書に記載のとおり使用することもできる。ポリペプチドをコードする核酸またはDNAをインビトロ細胞培養物中に導入し、発現させることができるDNA構築物中に取り込む。具体的には、DNA構築物は、原核宿主、例えば細菌、例えばE.コリ(E.coli)内での複製に好適であり、または培養された哺乳動物、植物、昆虫、例えばSf9、酵母、真菌もしくは別の真核細胞系中に導入することができる。
【0076】
特定の細胞内への導入のために調製されたDNA構築物は、典型的には、宿主により認識される複製系、所望のポリペプチドをコードする目的DNAセグメントならびにポリペプチドコードセグメントに作動可能に連結している転写および翻訳開始および終結調節配列を含む。DNAセグメントは、このセグメントが別のDNAセグメントとの機能的関係に置かれている場合、「作動可能に連結」している。例えば、プロモーターまたはエンハンサーは、これが配列の転写を刺激する場合、コード配列に作動可能に連結している。しかしながら、エンハンサーは、エンハンサーが転写を制御するコード配列と隣接している必要はない。連結は、簡便な制限部位またはこの代わりに挿入されたアダプターもしくはリンカーにおけるライゲーションにより達成される。
【0077】
適切なプロモーター配列の選択は、一般に、DNAセグメントの発現について選択された細胞タイプに依存する。好適なプロモーター配列の例としては、当分野において周知の原核および真核プロモーターが挙げられる(例えばSambrook and Russell,2001,上掲参照。)。転写調節配列としては、典型的には、宿主により認識される異種エンハンサーまたはプロモーターが挙げられる。適切なプロモーターの選択は細胞に依存するが、プロモーター、例えばtrp、lacおよびファージプロモーター、tRNAプロモーターおよび糖分解酵素プロモーターが公知であり、入手可能である(例えばSambrook and Russell,2001,上掲参照。)。複製系ならびに転写および翻訳調節配列をポリペプチドコードセグメントについての挿入部位と一緒に含む発現ベクターを用いることができる。細胞系および発現ベクターの作用可能な組合せの例は、Sambrook and Russell(2001,上掲)およびMetzger et al.(1988)Nature 334:31−36に記載されている。例えば、好適な発現ベクターは、酵母、例えばS.セレビシエ(S.cerevisiae)、例えば昆虫細胞、例えばSf9細胞、哺乳動物細胞、例えばCHO細胞および細菌細胞、例えばE.コリ内で発現させることができる。従って、細胞は原核または真核宿主細胞であり得る。細胞は、液体または固体培地中での培養に好適な細胞であり得る。細胞は、好ましくは、上記定義の本発明の方法において使用する。
【0078】
本明細書およびこの特許請求の範囲において、動詞「含むこと」およびこの活用形は、この非限定的な意味において、語の後の項目が含まれることを意味するように使用されるが、特に言及されなかった項目が除外されるわけではない。さらに動詞「からなる」は、本明細書に定義の方法が具体的に同定されているものよりも追加の工程を含むことができ、前記追加の工程は本発明の特有の特性を変化させないことを意味する「から本質的になる」により置き換えることができる。さらに、不定冠詞「a」または「an」による要素に対する言及は、文脈が唯一の要素の存在を明確に要求しない限り、1つ超の要素が存在する可能性を除外するものではない。従って、不定冠詞「a」または「an」は、通常「少なくとも1つ」を意味する。
【0079】
本明細書に引用されている全ての特許および参考文献は、参照により全体として本明細書に組み込まれる。
【0080】
以下の実施例を説明の目的のためにのみ提供するが、以下の実施例は本発明の範囲を決して限定するものではない。
【図面の簡単な説明】
【0081】
【図1】実施例2に記載のカルシウム蛍光マーカーFluo−4−AMを使用して細胞内カルシウムの放出を追跡することにより計測されたHEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を示す。旨味受容体の活性は、試験溶液添加後の最大蛍光を、試験溶液添加前に計測されたベースライン蛍光から差し引くことにより計算された蛍光の変化として計測した。試験溶液は示されている抽出物を含み、実施例2に記載のとおり調製した。MSG=10mM;MSG+IMP=1mMのMSG+500μMのIMP。
【図2a】実施例2に記載のBRETアッセイにより計測されたHEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を示す。示されている試験溶液(または緩衝液)および基質を細胞に添加した。BRETシグナルは、ドナー(400nm)について測定されたシグナルにより割ったアクセプター波長(515nm)において取られた読取り値の比として測定した。アクセプター波長におけるドナー発光の重複に起因するバックグラウンドシグナルについて補正するため、BRET比を、ドナー単独(T1R1/T1R3−Rluc)を発現する細胞についてパラレルで測定した。このBRETバックグラウンド値を、両方のBRETパートナーを発現する細胞について得られたBRET値から差し引いた(BRET=BRET比−バックグラウンド比)。緩衝液単独について達成された値を上回るBRETシグナルを、旨味受容体特異的として定義した(表2Bも参照。)。
【図2b】実施例2に記載のBRETアッセイにより計測されたHEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を示す。示されている試験溶液(または緩衝液)および基質を細胞に添加した。BRETシグナルは、ドナー(400nm)について測定されたシグナルにより割ったアクセプター波長(515nm)において取られた読取り値の比として測定した。アクセプター波長におけるドナー発光の重複に起因するバックグラウンドシグナルについて補正するため、BRET比を、ドナー単独(T1R1/T1R3−Rluc)を発現する細胞についてパラレルで測定した。このBRETバックグラウンド値を、両方のBRETパートナーを発現する細胞について得られたBRET値から差し引いた(BRET=BRET比−バックグラウンド比)。緩衝液単独について達成された値を上回るBRETシグナルを、旨味受容体特異的として定義した(表2Bも参照。)。
【図2c】実施例2に記載のBRETアッセイにより計測されたHEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を示す。示されている試験溶液(または緩衝液)および基質を細胞に添加した。BRETシグナルは、ドナー(400nm)について測定されたシグナルにより割ったアクセプター波長(515nm)において取られた読取り値の比として測定した。アクセプター波長におけるドナー発光の重複に起因するバックグラウンドシグナルについて補正するため、BRET比を、ドナー単独(T1R1/T1R3−Rluc)を発現する細胞についてパラレルで測定した。このBRETバックグラウンド値を、両方のBRETパートナーを発現する細胞について得られたBRET値から差し引いた(BRET=BRET比−バックグラウンド比)。緩衝液単独について達成された値を上回るBRETシグナルを、旨味受容体特異的として定義した(表2Bも参照。)。
【図2d】実施例2に記載のBRETアッセイにより計測されたHEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を示す。示されている試験溶液(または緩衝液)および基質を細胞に添加した。BRETシグナルは、ドナー(400nm)について測定されたシグナルにより割ったアクセプター波長(515nm)において取られた読取り値の比として測定した。アクセプター波長におけるドナー発光の重複に起因するバックグラウンドシグナルについて補正するため、BRET比を、ドナー単独(T1R1/T1R3−Rluc)を発現する細胞についてパラレルで測定した。このBRETバックグラウンド値を、両方のBRETパートナーを発現する細胞について得られたBRET値から差し引いた(BRET=BRET比−バックグラウンド比)。緩衝液単独について達成された値を上回るBRETシグナルを、旨味受容体特異的として定義した(表2Bも参照。)。
【図2e】実施例2に記載のBRETアッセイにより計測されたHEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を示す。示されている試験溶液(または緩衝液)および基質を細胞に添加した。BRETシグナルは、ドナー(400nm)について測定されたシグナルにより割ったアクセプター波長(515nm)において取られた読取り値の比として測定した。アクセプター波長におけるドナー発光の重複に起因するバックグラウンドシグナルについて補正するため、BRET比を、ドナー単独(T1R1/T1R3−Rluc)を発現する細胞についてパラレルで測定した。このBRETバックグラウンド値を、両方のBRETパートナーを発現する細胞について得られたBRET値から差し引いた(BRET=BRET比−バックグラウンド比)。緩衝液単独について達成された値を上回るBRETシグナルを、旨味受容体特異的として定義した(表2Bも参照。)。
【図2f】実施例2に記載のBRETアッセイにより計測されたHEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を示す。示されている試験溶液(または緩衝液)および基質を細胞に添加した。BRETシグナルは、ドナー(400nm)について測定されたシグナルにより割ったアクセプター波長(515nm)において取られた読取り値の比として測定した。アクセプター波長におけるドナー発光の重複に起因するバックグラウンドシグナルについて補正するため、BRET比を、ドナー単独(T1R1/T1R3−Rluc)を発現する細胞についてパラレルで測定した。このBRETバックグラウンド値を、両方のBRETパートナーを発現する細胞について得られたBRET値から差し引いた(BRET=BRET比−バックグラウンド比)。緩衝液単独について達成された値を上回るBRETシグナルを、旨味受容体特異的として定義した(表2Bも参照。)。
【発明を実施するための形態】
【実施例】
【0082】
[実施例1]
ヒト旨味受容体T1R1/T1R3を使用する原理の証明:
本発明者らは、BRET技術を使用してトマトおよび他の粗製植物または食物抽出物中の旨味を評価および定量する特有の機能的旨味受容体アッセイを開発し:原理を試験するため、本発明者らはBRET−2に関するドナーおよびアクセプタータンパク質を使用した。要するに、ヒトT1R1およびT1R3味覚受容体をクローニングし、ドナータンパク質としてのレニラルシフェラーゼ(Rluc)と融合させ;緑色蛍光タンパク質−2(GFP2)をアクセプタータンパク質として機能させ、β−アレスチン−2に融合させた。両方の構築物をタンパク質の異種発現のためにHEK293細胞系細胞)内に形質移入した。種々の哺乳動物細胞系が、受容体または他のタンパク質の発現に高度に好適であると報告されており、HEK293(ヒト胚腎臓)細胞およびCHO(チャイニーズハムスター卵巣)細胞は最も多用途で好適な細胞の一部である12。同様にGPCRは、実験の性質、各受容体または利用可能な時間に応じて、複数の刊行物に記載の種々の手法、例えば一過性6,8,13−15または安定性(構成性または誘導性)6,9,16,17発現において発現させることができる。
【0083】
アゴニストまたは粗製抽出物を用いる受容体刺激後、β−アレスチン−2/GFP−2タンパク質は活性化された受容体と相互作用し、こうしてGFP−2をルシフェラーゼと近接させ、考えられるこれらの2種のタンパク質間のエネルギー転移をもたらし、BRETシグナルを生成する。酸素の存在下、ルシフェラーゼは基質DeepBlueCの、395−410nmにおいて計測することができるコエレンテルアミドへの転換を触媒する。GFP−2がルシフェラーゼに近接し、エネルギー転移が生じる場合、発光は510nmにシフトし;このことをBRETシグナルと称し、アクセプター(GFP−2)とドナー(レニラルシフェラーゼ)との比として表現する。
【0084】
以下の実験は、慣用のカルシウムベースの機能的受容体アッセイが粗製抽出物の効果の計測に好適でなく;対照的に、BRETアッセイ(本明細書においては旨味について示される。)が粗製天然抽出物を使用して特異的受容体応答を明確に計測することができることを説明するために設計する。さらに、本アッセイは、異なるトマト試料間の受容体活性化の差の検出についても十分に感受性である。
【0085】
材料および方法:
化学薬品および培地:
Fluo−4 AMはMolecular Probes製(#F−14202、DMSO中5mM原液として調製)、DMEM(4.5g/lグルコースおよびウルトラグルタミンを有する、#BE12−604F)はLonza製、トリプシン−EDTA、Lipofectamine 2000、OptiMemおよびFCSは、Life Technologies Invitrogenから入手した。グルタミン酸ナトリウム(MSG)、イノシン5’−一リン酸および全ての他の化学薬品はSigma−Aldrichから入手した。
【0086】
GFP2−β−アレスチン−2をコードするプラスミドは、BioSignal Packard(#6310176)から購入した。
【0087】
コエレンテラジン400A(DeepBlueC誘導体)およびコエレンテラジン−Hは、VWR International(それぞれ#BTIU10125−1および233903−50)製であった。
【0088】
全ての他の細胞培養供給物はGreiner BioOne製であった。
【0089】
受容体融合構築物、細胞系および培地:
標準的分子クローニング技術およびPerkinElmer(#6310220)製コドンヒト化pRluc−N3ベクターを使用してヒトT1R1およびヒトT1R3旨味受容体を、これらのC末端においてインフレームでレニラルシフェラーゼ(Rluc)に融合させた。旨味受容体およびGFP−2/β−アレスチン−2をコードする構築物の発現のため、慣習的な形質移入法(以下も参照。)を使用してHEK293細胞(ヒト胚腎臓細胞、ATCC)を使用した。
【0090】
HEK293細胞をDMEMおよび10%FBS中で37℃/5%CO2において維持した。
【0091】
トマト抽出物:
代表的な天然複合試験材料として機能するトマト抽出物を以下のとおり調製した:冷凍トマト試料(−80℃)を秤量し、等量の水中に溶解させた。種子および小室組織(軟塊)を除去し取り分けた後、乳鉢および乳棒を使用してトマト果皮(果肉)を粉砕した。次いで、種子および軟塊を混合物に添加して戻し、これらが十分に混合されることを確保し、種子は破砕しなかった。混合物を4000rpmにおいて15分間遠心分離し、上澄み(漿液)を除去し、10mlアリコート中で凍結乾燥させた。計測前、等容量の水を添加して試料を溶解させた。
【0092】
T1R1/T1R3およびRluc融合構築物の生成:
上記のとおり、標準的分子クローニング技術を使用してヒトT1R1およびヒトT1R3旨味受容体のC末端Rluc受容体融合構築物を生成した。このことは、2種の異なる構築物:T1R1−RlucおよびT1R3−Rlucをもたらした。これらは、一緒に形質移入することができ、2種のRluc部分を含有する機能的受容体ヘテロ二量体をもたらし;代替的にいずれかの構築物を野性型受容体との組合せにおいて形質移入することができ、こうしてT1R3と組み合わせたT1R1−RlucまたはT1R3−Rlucと組み合わせたT1R1をもたらす。
【0093】
野性型受容体ならびに融合構築物およびレニラルシフェラーゼについての配列(cDNAおよびタンパク質)を配列表に挙げる(配列番号5−8)。
【0094】
形質移入:
HEK293細胞に、Lipofectamine2000を製造業者のプロトコルに従って使用してT1R1/T1R3受容体(Rluc融合タンパク質を含有)およびGFP−2−β−アレスチン−2をコードするプラスミドを一過的に形質移入する。要するに、HEK293細胞をウェル(12ウェルプレート、1ml培地/ウェル)当たり2×105個の細胞の密度において播種し、翌日に約80−90%のコンフルエンシーを目的とする。24時間後、ウェル当たり15gのトータルDNAを使用して旨味受容体構築物(T1R1−Rluc、T1R3−Rlucまたは野性型受容体)を、GFP−2−β−アレスチン−2をコードするプラスミドと同時形質移入する。DNAを100μlのOptiMem中に溶解させ、これを、4μlのlipofectamine2000を含有する100μlのOptiMemと合わせる。次いで、混合物を室温において30分間インキュベートし、各ウェルに添加し、細胞を48時間増殖させる。BRET計測は、形質移入48−52時間後に実施する。代替的に、上記と同一のプロトコルを使用して旨味受容体をコードするプラスミドを、GFP−2−β−アレスチン−2を安定的に発現するHEK293細胞内に形質移入することができる。
【0095】
カルシウムベースの受容体アッセイについては、Gα15を安定的に発現するHEK293細胞にT1R1およびT1R3受容体を上記の手順に従って形質移入するが、96ウェルプレートフォーマットについてのプロトコルをスケールダウンする(ポリリジン被覆μClear96ウェルプレートを使用して10倍低減)。
【0096】
カルシウムベースの受容体アッセイ:
HEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を、細胞内カルシウムの放出を追跡することにより計測した。増殖培地を除去し、2.5μMのカルシウムの蛍光マーカーFluo−4−AMを含有し、細胞からのFluo−4の漏れを防止するために0.5mMプロベニシドおよび0.5%FCSを補給した50μlのTyrode緩衝液(140mM NaCl、5mM KCl、10mMグルコース、1mM CaCl2.2H2O、1mM MgCl2.6H2O、10mMピルビン酸Naおよび50mM HEPES pH7.4)により細胞を1時間負荷し、次いで37℃において1時間インキュベートした。混合物を取り出し、0.5mMプロベニシドを含有する150μlのTyrode緩衝液を添加した。Flexstation II 384(Molecular Devices)を使用して細胞内カルシウムレベルを追跡した。蛍光計測は、485nmの励起波長および520nmの発光波長において37℃において実施した。50μlの化合物またはトマト抽出物を、20秒のベースライン計測後に毎秒104μlのピペッター速度において細胞に添加した。旨味受容体の活性は、試験溶液添加後の最大蛍光を試験溶液添加前に計測されたベースライン蛍光から差し引くことにより計算される蛍光変化(ΔF)として計測した。
【0097】
表2Aに示すデータは、純粋な旨味化合物の使用が旨味受容体特異的応答(受容体を発現しない対照細胞は増加を付与しない。)をもたらし;対照的に、種々のトマト抽出物の使用が対照細胞において高い非特異的応答をもたらし、旨味を示すシグナルを同定することができないことを極めて明確に示す。
【0098】
BRETアッセイ:
BRETアッセイには、Mithras LB 940プレートリーダー(Berthold Technologies)を使用する。使用するプロトコルは、本質的には、Packard BioOneにより記載されているものと同一であり、またはわずかな改変を有する刊行されたプロトコル13であり:HEK293細胞を上記のとおり形質移入する。形質移入48時間後、細胞を回収し、BRET緩衝液(2μg/mlアプロチニンを含有するD−PBS)中に2×106個の細胞/mlの密度において溶解させた。細胞を平衡化のために室温において1時間放置した後、1×105個の細胞を含有する30μlを白色96ウェルプレートの各ウェルに移した。10μlの試験化合物(または緩衝液)および10μlの基質コエレンテラジン(最終濃度5μM)を、インジェクターを使用して細胞に同時に添加する。最終インジェクション直後、連続読取りを410nmおよび515nmにおいて行う。BRETシグナルは、GFP−2発光(515nm)およびRluc/コエレンテラジン発光(410nm)の比として測定する。
【0099】
[実施例2]
複合試料中の味覚モジュレーターの検出
試験試料(抽出物)の調製および選択
下記の抽出物または粗製混合物は、これらの高い非特異的バックグラウンドシグナルに起因してカルシウムベースの受容体アッセイを用いる試験に不適な天然試料についての例である。これらの抽出物または粗製混合物は、異なる起源からのものであり、広範な潜在的試験試料を説明するために選択される。選択試料はグルタマート、リボチドまたはこれら両方を含有することも公知であり、従って旨味受容体T1R1/T1R3を例として使用するBRETアッセイにおいて試験するための良好な候補である。
トマト:トマト抽出物の調製は実施例1に既に記載した。
酵母抽出物:5’−リボチドが濃縮された市販の酵母抽出物(DSM,Maxarome Select酵母抽出物)。酵母抽出物を水中に溶解させた(10×酵母抽出物の重量)。
醤油:市販の醤油を使用した(Conimex ketjap manis)。
チキンブイヨン:市販のチキンブイヨンキューブ(Knorr)を製造業者により示されるとおりに温水中に溶解させた。粒子を試験試料から除去するため、ブイヨンを5000rpmにおいて10分間遠心分離し、上澄みを新たなチューブに移した。
チーズ抽出物:市販のロクフォールチーズを秤量し、等量の水中に溶解させ、乳鉢および乳棒を使用してホモジナイズした。混合物を5000rpmにおいて10分間遠心分離して粒子を除去し、上澄みを新たなチューブに移した。
【0100】
実験手順
形質移入:
HEK293細胞に、Lipofectamine2000を製造業者のプロトコルに従って使用してT1R1およびT1R3受容体(本実験においてT1R3受容体はRluc融合タンパク質を含有する。)ならびにGFP−2−β−アレスチン−2をコードするプラスミドを、一過的に形質移入した。要するに、HEK293細胞をウェル(12ウェルプレート、1ml培地/ウェル)当たり2×105個の細胞の密度において播種し、翌日に約80−90%のコンフルエンシーを目的とした。24時間後、ウェル当たり15μgのトータルDNAを使用して旨味受容体構築物(T1R1およびT1R3−Rluc受容体)を、GFP−2−β−アレスチン−2をコードするプラスミドと同時形質移入した。DNAを100μlのOptiMem中に溶解させ、これを、4μlのlipofectamine2000を含有する100μlのOptiMemと合わせた。次いで、混合物を室温において30分間インキュベートし、各ウェルに添加し、細胞を48時間増殖させた。BRET計測は、形質移入48−52時間後に実施した。
【0101】
代替的に、上記と同一のプロトコルを使用して旨味受容体をコードするプラスミドを、GFP−2−β−アレスチン−2を安定的に発現するHEK293細胞内に形質移入した。
【0102】
カルシウムベースの受容体アッセイについては、Gα15を安定的に発現するHEK293細胞にT1R1およびT1R3受容体を上記の手順に従って形質移入するが、96ウェルプレートフォーマットについてのプロトコルをスケールダウンする(ポリリジン被覆μClear96ウェルプレートを使用して10倍低減)。
【0103】
カルシウムベースの受容体アッセイ:
HEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を、細胞内カルシウムの放出を追跡することにより計測した。増殖培地を除去し、2.5μMのカルシウムの蛍光マーカーFluo−4−AMを含有し、細胞からのFluo−4の漏れを防止するために0.5mMプロベニシドおよび0.5%FCSを補給した50μlのTyrode緩衝液(140mM NaCl、5mM KCl、10mMグルコース、1mM CaCl2.2H2O、1mM MgCl2.6H2O、10mMピルビン酸Naおよび50mM HEPES pH7.4)により細胞を1時間負荷し、次いで37℃において1時間インキュベートした。混合物を取り出し、0.5mMプロベニシドを含有する150μlのTyrode緩衝液を添加した。Flexstation II 384(Molecular Devices)を使用して細胞内カルシウムレベルを追跡した。蛍光計測は、485nmの励起波長および520nmの発光波長において37℃において実施した。50μlの化合物または抽出物を20秒のベースライン計測後に毎秒104μlのピペッター速度において細胞に添加した。旨味受容体の活性は、試験溶液添加後の最大蛍光を試験溶液添加前に計測されたベースライン蛍光から差し引くことにより計算される蛍光変化(ΔF)として計測した。
【0104】
表2Bおよび図1は、純粋な旨味化合物(MSGおよびMSG+IMP)の使用が旨味受容体特異的応答をもたらし;対照的に、種々の抽出物および天然混合物の使用が旨味受容体を同様に発現しない対照細胞において高い非特異的応答をもたらし、このことにより旨味を示す受容体特異的シグナルを計算することができないことを説明する。
【0105】
BRETアッセイ:
BRETアッセイには、Mithras LB 940プレートリーダー(Berthold Technologies)を使用した。アッセイプロトコルは、本質的には、Packard BioOneにより記載されているものと同一であり、またはわずかな改変を有する他の刊行されたプロトコル10,13であり:HEK293細胞を上記のとおり形質移入した。形質移入40時間後、細胞を回収し、BRET緩衝液(1g/l D−グルコースおよび2μg/mlアプロチニンを含有するD−PBS)中に2×106個の細胞/mlの密度において溶解させる。細胞を平衡化のために室温において1時間放置した後、約1×105個の細胞を含有する30μlを白色96ウェルプレートの各ウェルに形質移入する。10μlの試験化合物(または緩衝液)および10μlの基質コエレンテラジン400(DeepBlueC、最終濃度5μM)を、インジェクターを使用して細胞に同時に添加した。最終インジェクション直後、連続読取りを400nmおよび515nmにおいて行った。代替的に、化合物を細胞に手作業で添加し、基質のインジェクション前に5分間インキュベートした。BRETシグナルは、ドナー(Rluc/コエレンテラジン400;400nm)について測定されたシグナルにより割ったアクセプター波長(GFP−2;515nm)において取られた読取り値の比として測定した。アクセプター波長におけるドナー発光の重複に起因するバックグラウンドシグナルについて補正するため、BRET比を、ドナー単独(T1R1/T1R3−Rluc)を発現する細胞についてパラレルで測定した。このBRETバックグラウンド値を、両方のBRETパートナーを発現する細胞について得られたBRET値から差し引いた(BRET=BRET比−バックグラウンド比)。緩衝液単独について達成された値を上回るBRETシグナルを、旨味受容体特異的として定義した(表2B参照。)。
【0106】
結果:
表2Bおよび図1に示されるとおり、カルシウムベースの受容体アッセイを使用して代表的抽出物および他の複合混合物を使用して旨味受容体特異的シグナルを達成することができなかった。対照的に、図2は、記載のBRETアッセイを使用してこれらの複合試料についての旨味受容体活性化を特異的に計測することができたことを示す。
【0107】
受容体特異的シグナルの定義は、技術的理由についてわずかに異なる:カルシウムアッセイについては、旨味受容体およびGα15を発現する細胞をGα15のみを発現する細胞と比較し;これらのシグナル間の差を旨味受容体特異的シグナルとして定義する。BRETアッセイについては、ドナー(T1R1/T1R3−Rluc)のみを発現する細胞を使用してバックグラウンドを測定し;陽性値が特異的BRETシグナルを示す。
【0108】
【表1】
【0109】
【表2】
【0110】
【表3】
【0111】
【表4】
【表5】
【技術分野】
【0001】
本発明は、味覚受容体の潜在的モジュレーター化合物をスクリーニングする方法であって、BRET技術を使用する方法に関する。
【背景技術】
【0002】
風味は、我々が快適な食物(即ち栄養食物)を消費し、不快な食物(潜在的毒素を含有する。)を回避するように食物摂取1を制御する一次知覚系の一部である。風味は、視覚、味覚、香りおよび食感の入力から形成される感覚である。しかしながら、食物の選択および我々が消費する量は、5種の基本味の3種(塩味、甘味および旨味)によってより動かされ、他の風味性によってはあまり影響を受けないと考えられる。これらの特質を含有する食物は、ヒトおよび多くの哺乳動物により好まれる食物である傾向があり;これに関して、旨味はタンパク質についてのマーカーとして、甘味は炭水化物についてのマーカーとして機能する。
【0003】
近年、これらの味覚様式の検出に関与する受容体が同定され、クローニングされ2−4、こうして味覚受容体の活性化をインビトロで調査することが可能となっている。甘味、旨味および苦味についての受容体は、Gタンパク質共役受容体(GPCR)のクラスに属する一方、塩味および酸味はイオンチャネルにより検出される可能性が最も高い。
【0004】
甘味は異種受容体二量体T1R2/T1R3により感知される一方、旨味は主としてT1R1/T1R3受容体により検出されるが2、他の受容体も同様に旨味に関与することが示されている5。
【0005】
インビトロ受容体活性化をインビボ知覚に良好な相関で計測するために、HEK293細胞などの哺乳動物細胞系内での味覚受容体の異種発現を含む種々の細胞系を使用することができる2,6−9。現在利用可能な機能的インビトロスクリーニング系は、通常、プロミスカスなGタンパク質、例えばGα15、Gα16またはこれらのGタンパク質のC末端ドメインの種々の適応部とのキメラを使用し;このことは、対象の受容体のPLC(ホスホリパーゼC)へのシグナリングカスケードおよび細胞内カルシウムの放出を指向する。このアプローチは純粋化合物の調査に極めて成功したが、抽出物または複合体試料の試験がより困難であることが証明されており:Gタンパク質の普遍的性質に起因してGタンパク質はスクリーニング細胞系内で(過剰)発現された組換え受容体に結合し得るだけでなく、低レベルにおいて内因的に存在する多くの受容体にも結合し得る。このことは、これらの内因性受容体を活性化する天然混合物中に存在するアゴニストにより誘導される非特異的カルシウムシグナルをもたらし得る。さらに、抽出物または複合試験試料は、GPCRを介する手段以外の手段により細胞内カルシウムを上昇させる物質も含有することが多く、これらのシグナルは受容体により誘導されたカルシウム放出と区別することができない。多くの天然混合物について観察される高い非特異的バックグラウンドシグナルは、大規模な分別手順を行わないこれらの試料の直接的なスクリーニングを妨げる。このような抽出物または複合試料の使用は、例えば食物材料を評価する場合にとてもありふれていることに留意すべきである。
【発明の概要】
【発明が解決しようとする課題】
【0006】
従って、味覚受容体の潜在的モジュレーター化合物のための改善されたスクリーニング方法であって、既存の方法の欠点のそれぞれを有しない方法が依然として必要とされている。
【課題を解決するための手段】
【0007】
第1の態様において、味覚受容体の潜在的モジュレーター化合物をスクリーニングする方法であって、BRET(生物発光共鳴エネルギー転移)(10,11)技術を使用する方法が提供される。本方法の各特徴を以下に詳細に定義する。
【0008】
好ましい方法は、以下の工程:
a)発光タンパク質、例えばルシフェラーゼタンパク質に融合している味覚受容体およびβ−アレスチンに融合している蛍光タンパク質を発現し、またはこれらの発現を誘導する細胞を提供する工程、
b)工程a)において得られた細胞を潜在的モジュレーター化合物によりチャレンジする工程ならびに
c)工程b)において得られた細胞のBRETシグナルを、該潜在的モジュレーターの不存在下でb)において得られた細胞のBRETシグナルと比較する工程
を含む。
【0009】
代替的に、第1の態様において、本発明は、味覚受容体をモジュレートする化合物を同定する方法であって、(a)(i)発光タンパク質に融合している味覚受容体および(ii)β−アレスチンに融合している蛍光タンパク質を発現する細胞を提供する工程;(b)細胞を潜在的モジュレーター化合物と接触させ、およびBRETシグナルを測定する工程;ならびに(c)工程(b)において得られたBRETシグナルを、該潜在的モジュレーター化合物の不存在下で細胞から得られたBRETシグナルと比較する工程を含み、(b)において得られたBRETシグナルと潜在的モジュレーター化合物の不存在下で得られたBRETシグナルとの差が、該潜在的モジュレーター化合物が味覚受容体をモジュレートする化合物であることを示す方法を提供する。
【0010】
本発明者らの発明は、本発明の方法により高い特異性を付与するBRET技術またはアッセイを使用し:対象の味覚受容体はドナー発光タンパク質、例えばルシフェラーゼタンパク質と融合しており、前記受容体に由来するシグナルに影響を与え、BRETシグナルを引き起こし得る細胞構成成分はない。このことは、味覚受容体の潜在的モジュレーター化合物を同定するためにスクリーニングに利用可能であることが多い天然混合物に関して特に興味深く:内因性受容体を活性化する構成成分も他の経路を介して細胞内カルシウムの非特異的上昇を事前に引き起こす物質も、BRETシグナルを引き起こすことはできない。読取り窓は受容体−発光融合タンパク質に唯一フォーカスされ、こうしてこの方法が、未精製の粗製抽出物を使用する高度に特異的な受容体活性化の直接調査に例外的に有用となる。
【0011】
本発明の方法は、GPCR(Gタンパク質共役受容体)である味覚受容体の受容体刺激時にβ−アレスチンをトランスロケートする能力を基礎とし、エネルギードナーを含有する融合タンパク質(発光タンパク質、例えばルシフェラーゼ)と、所与の波長において光を吸収し、より長い波長において再発光するエネルギーアクセプタータンパク質(フルオロフォア、典型的には蛍光タンパク質)との間のエネルギー転移を計測することにより受容体−β−アレスチン相互作用を計測するためにBRETアッセイを利用する10。GPCR活性化アッセイの場合、発光タンパク質、例えばルシフェラーゼは受容体のC末端に融合しており、蛍光タンパク質はβ−アレスチンに融合している。受容体が活性化される場合、細胞質β−アレスチンは細胞膜にリクルートされ、受容体を内在化について標的化する。β−アレスチン/蛍光タンパク質と発光タンパク質融合受容体との相互作用の間、ドナーおよびアクセプタータンパク質は近接し、BRETシグナルを誘導する。
【0012】
従って、BRET技術はシグナルまたはBRETシグナルを生成することができる技術またはアッセイであり、前記シグナルは、発光タンパク質に融合している味覚受容体と蛍光タンパク質との間のエネルギー転移であり、前記シグナルは、潜在的モジュレーター化合物の存在に起因して前記味覚受容体の活性化を反映する。
【0013】
本発明の方法の工程a)は、発光タンパク質、例えばルシフェラーゼタンパク質に融合している味覚受容体およびβ−アレスチンに融合している蛍光タンパク質を発現し、またはこれらの発現を誘導する細胞を提供する。本発明の方法の工程a)は、(i)発光タンパク質に融合している味覚受容体および(ii)β−アレスチンに融合している蛍光タンパク質を発現する細胞を提供することもできる。
【0014】
味覚受容体は、ヒトの口内において味覚に関連することが公知の任意の受容体であり得る。味覚受容体は、味覚認識に関与するものとして後に発見される任意の受容体でもあり得る。味覚受容体は、舌内で発現され得る:MSG(グルタミン酸ナトリウム)または旨味受容体、甘味受容体、苦味受容体または脂肪受容体。甘味認識に関与することが公知の受容体は、2種のサブユニットT1R2(Taste1 Receptor2)およびT1R3(Taste1 Receptor3)を含むヘテロ二量体である。旨味認識に関与することが公知の受容体は、2種のサブユニットT1R1(Taste1 Receptor1)およびT1R3を含む別のヘテロ二量体である。別のMSGまたは旨味受容体は、mGluR4(aまたはc)(代謝型グルタミン酸受容体4(aまたはc))の1種以上のサブユニットから構成される。苦味受容体は、TAS2(Taste2)受容体の1種以上のサブユニットから構成される。脂肪酸受容体は、GPR120(Gタンパク質共役受容体120)の1種以上のサブユニットから構成される。ヒトT1R1を表す好ましい核酸配列は、配列番号1である。ヒトT1R1タンパク質を表す対応する好ましいアミノ酸配列は、配列番号2により表される。ヒトT1R3を表す好ましい核酸配列は、配列番号3である。ヒトT1R3タンパク質を表す対応する好ましいアミノ酸配列は、配列番号4により表される。
【0015】
本発明に関して、味覚受容体は、ヒトの腸管内で栄養/脂肪酸感知に関与する受容体でもあり得る。このような受容体としては:カルシウム感知受容体、Gタンパク質共役受容体ファミリーCグループ6サブタイプA(GPRC6A)、L−アルファ−アミノ酸を感知する味覚受容体二量体T1R1/T1R3、炭水化物感知T1R2/T1R3受容体二量体、タンパク質分解産物センサGPR93(GPR92とも称される。)ならびに遊離脂肪酸(FFA)感知受容体FFA1、FFA2、FFA3、GPR84およびGPR120が挙げられる5。表3に同定された受容体のそれぞれを本発明の方法において使用することができる。これらの受容体のそれぞれをコードする好ましい核酸分子を配列表に挙げる。対応する好ましいコードされる受容体も配列表に挙げる(表3も参照。)。
【0016】
本発明の方法は、T1R1およびT1R3サブユニットを含む味覚受容体を使用して、および発光タンパク質としてのルシフェラーゼを使用して例示される。しかしながら、当業者は、本発明が前記ヘテロ二量体およびこの発光タンパク質を使用する方法に限定されるものではないことを理解する。本発明は、味覚受容体、好ましくはT1R1、T1R3ヘテロ二量体を発現する細胞を提供する。前記味覚受容体は、好ましくは機能的である。このことは、潜在的モジュレーターを添加することなく実施される本発明のスクリーニング方法において、この味覚受容体を活性化することが公知の物質が前記細胞に添加される場合に検出可能なBRETシグナルが存在することを意味する。各味覚受容体について、このような物質は公知である。このような物質、即ちアゴニストの例を表1に同定する。
【0017】
本発明はまた、蛍光タンパク質に融合している味覚受容体およびβ−アレスチンに融合している発光タンパク質、例えばルシフェラーゼタンパク質を発現し、またはこれらの発現を誘導する細胞を提供する工程a)を提供する。味覚受容体に融合している発光タンパク質、例えばルシフェラーゼタンパク質についての本明細書に定義の各特徴は、β−アレスチンに融合している発光タンパク質、例えばルシフェラーゼタンパク質についても保持される。β−アレスチンに融合している蛍光タンパク質についての本明細書に定義の各特徴は、発光タンパク質、例えばルシフェラーゼタンパク質に融合している蛍光タンパク質についても保持される。従って、本発明はまた、味覚受容体が蛍光タンパク質に融合しており、β−アレスチンが発光タンパク質に融合している方法を提供する。より一般的には、当業者は、発光タンパク質が味覚受容体に融合しており、蛍光タンパク質がβ−アレスチンに融合している本発明の任意の実施形態を、蛍光タンパク質が味覚受容体に融合しており、発光タンパク質がβ−アレスチンに融合している、他の点では同一の実施形態により置き換えることができることを理解する。
【0018】
本発明は、本発明の方法において使用される細胞を得るために使用すべき、核酸配列により表される好ましい核酸分子またはアミノ酸配列により表されるコードされるタンパク質を同定する。しかしながら、本明細書において同定される核酸配列のそれぞれは、天然に生じる形態、SNP(単一ヌクレオチド多型)を含有するバリアント、選択的スプライスによる形態、形態の組合せまたは当分野において公知の任意の機能的バリアントにより置き換えることができる。本明細書に定義の核酸分子は、本明細書に前述の細胞内で発現される場合、機能的であるべきである。核酸配列のバリアントは、この核酸配列の断片であり得る。好ましいバリアントは、サイレント突然変異を含有する。代替的にまたは組合せにおいて、核酸配列バリアントは、別のアミノ酸配列を生じさせないが、前記核酸配列が発現される宿主細胞のコドン使用頻度に対応する核酸置換の導入により得ることもできる。好ましくは、核酸配列バリアントは、本明細書に上記定義の核酸配列の任意の1種から出発して5’および/または3’末端からの1個以上のヌクレオチドが欠失されたものである。代替的にまたは組合せにおいて、核酸配列バリアントは、好ましくは別の生物および/または別の本明細書に上記定義の核酸配列のファミリーメンバーから単離された核酸配列である。これらのバリアントは全て、選択種、例えば哺乳動物種、例えばヒトからのcDNAまたはゲノミックライブラリーを使用する典型的なアプローチにおいて得ることができる。続いて、ライブラリーは厳しい条件下でハイブリダイゼーションにより本明細書に上記定義の核酸配列の1種またはこの一部を用いてスクリーニングすることができる。厳しい条件は、5×SSPE、0.3%SDS、200pg/mlの剪断および変性されたサケ精子DNAならびに50%ホルムアミド中での42℃におけるプレハイブリダイゼーションおよびハイブリダイゼーションを意味する。続いて、2×SSC、0.2%SDSおよび75℃をそれぞれ使用してハイブリダイゼーション反応物を30分間3回洗浄する。代替的にまたは組合せにおいて、核酸配列バリアントは、データベースにおけるアミノ酸同一性および/または類似性の探索ならびに検索により同定された好適なアミノ酸配列をコードする核酸配列の合成により得ることができる。
【0019】
ヒトは好ましい種である。別の好ましい実施形態によれば、核酸配列バリアントは、対立遺伝子バリアントである。対立遺伝子バリアントは、同一染色体座を占有する遺伝子の2種以上の別の代わりの形態のいずれかを意味する。対立遺伝子変異は突然変異を介して天然に生じ、個体群内の表現型多型をもたらし得る。別の好ましい実施形態によれば、核酸配列バリアントは、遺伝コードの縮退に関して本明細書に上記定義の核酸配列のいずれとも異なる。
【0020】
使用される核酸分子に関するさらなる説明を、標題「配列番号により定義される核酸分子および配列同一性」セクションに挙げる。好ましい実施形態において、使用される核酸分子はヒトに由来する。より好ましくは、この好ましい実施形態に定義される核酸分子は、哺乳動物、よりいっそう好ましくはヒト細胞における機能的発現のためのものである。ヒト配列と高度に相同性(少なくとも85%の同一性)である配列の使用は、魅力的である。これというのも、この核酸分子が哺乳動物細胞、好ましくはヒト細胞内で発現され、機能的であることを予期することができるからである。さらに、この配列は、ヒト配列と高度に相同性であるので、こうして調製された細胞型がヒト配列に対してより低い同一性を有する配列を用いて調製された細胞型よりも効率的にヒト味覚を模倣することが予期される。よりいっそう好ましくは、本明細書に上記定義の同一性は、85%以上、よりいっそう好ましくは90%以上、よりいっそう好ましくは91%以上、よりいっそう好ましくは92%以上、よりいっそう好ましくは93%以上、よりいっそう好ましくは94%以上、よりいっそう好ましくは95%以上、よりいっそう好ましくは96%以上、よりいっそう好ましくは97%以上、よりいっそう好ましくは98%以上、よりいっそう好ましくは99%以上、最も好ましくは100%である。
【0021】
本発明において、味覚受容体またはこのサブユニットをコードする核酸分子は、発光タンパク質、例えばルシフェラーゼタンパク質に融合している。好ましい実施形態において、発光タンパク質はルシフェラーゼタンパク質である。発光タンパク質、例えばルシフェラーゼタンパク質は、好ましくは細胞内部分である受容体のC末端部において融合している。当業者は、発光タンパク質、例えばルシフェラーゼタンパク質を味覚受容体の細胞内部分のいずれにも融合していてよいことを理解する。しかしながら、こうして得られたタンパク質は依然として機能的であるべきであり;即ち、活性化可能であるべきである。従って、発光タンパク質、例えばルシフェラーゼタンパク質を味覚受容体の細胞内部分におけるある場所に融合させた場合、好ましくは組換えタンパク質であるこうして得られたこのようなタンパク質をこの機能性に関して試験することが好ましい。好ましい実施形態において、発光タンパク質、例えばルシフェラーゼタンパク質は、味覚受容体のC末端部分の端部において、または前記味覚受容体のC末端部分の端部に可能な限り近くで融合している。前記味覚受容体のC末端部分の端部に可能な限り近くとは、好ましくは、発光タンパク質、例えばルシフェラーゼタンパク質の最初のアミノ酸が味覚受容体のC末端部分の最後のアミノ酸に対応する場所または味覚受容体のC末端部分の最後のアミノ酸から1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20個前のアミノ酸において存在することを意味する。ルシフェラーゼは、ホタルルシフェラーゼであり得、または任意のレニラ(Renilla)種からのものであり得る。ルシフェラーゼをコードする好ましい核酸分子は、哺乳動物細胞内でのこの発現レベルを改善するためにヒト化コドンを用いて改善されてきた。好ましいルシフェラーゼをコードするより好ましい核酸分子を配列番号77として挙げる。好ましいコードされるルシフェラーゼタンパク質を配列番号78として同定する。当業者は、2種の核酸分子をインフレームでいかに融合させるかを認識する。旨味受容体の好ましいT1R1およびT1R3サブユニットとしての1種超の区別されるサブユニットを有する味覚受容体の場合、各サブユニットを発光タンパク質、例えばルシフェラーゼタンパク質またはサブユニットの1種のみと融合させることができる。このことは、各味覚受容体について保持され、即ち、当業者は、このことが他の(ヘテロ)多量体味覚受容体についても保持されることを理解する。味覚受容体が1種超の区別されるサブユニットを有する場合、各サブユニットは、発光タンパク質、例えばルシフェラーゼタンパク質に融合されていてよい。代替的に、サブユニットの1種以上のタイプのみが発光タンパク質、例えばルシフェラーゼに融合されている。
【0022】
本発明において、蛍光タンパク質はβ−アレスチンに融合している。好ましい蛍光タンパク質は緑色蛍光タンパク質(GFP)である。GFP2がより好ましい。
【0023】
好ましいβ−アレスチンは、ヒトまたは哺乳動物β−アレスチンである。より好ましくは、β−アレスチンは非視覚β−アレスチン、例えばβ−アレスチン2または3などである。より好ましくは、β−アレスチンは配列番号79により表されるβ−アレスチン2である。前記β−アレスチン2をコードする好ましい核酸分子は、配列番号80により表される。よりいっそう好ましいβ−アレスチン2は、WO2004/065963またはWO2004/034054に記載されている。蛍光タンパク質とβ−アレスチンとの融合体も当業者に公知である。本発明の方法において、各核酸分子(即ち、ルシフェラーゼに融合している味覚受容体をコードするものおよびβ−アレスチンに融合している蛍光タンパク質をコードするもの)は、核酸構築物中に存在する。各構築物を細胞内に導入する。
【0024】
従って、本発明の細胞は、本明細書に定義の核酸構築物を含む。当業者は、細胞の選択が味覚受容体をコードする核酸配列の起源に大いに依存することを認識する。発現される味覚受容体が機能的である限り、任意の細胞を選択することができる。好ましくは、味覚受容体および/またはβ−アレスチンの発現は安定性であり、場合により誘導性である。代替的に、味覚受容体および/またはβ−アレスチンの発現は一過性である。誘導性発現は、「味覚受容体の発現」セクションに詳細に説明する。好ましくは、細胞は原核または真核細胞である。より好ましくは、細胞は昆虫または哺乳動物細胞である。よりいっそう好ましくは、哺乳動物細胞はヒト細胞である。哺乳動物細胞の例は、HEK293、HEK293T、MDCK、CHO、COS、NIH3T3、Swiss3T3、BHKおよびA549である。よりいっそう好ましくは、細胞は哺乳動物細胞、例えばHEK293である。本発明の細胞は、組換え細胞として把握することができる。本発明の細胞は、有利には、本発明の方法において使用される。
【0025】
選択される発現系のタイプに応じて、当業者は、味覚受容体およびβ−アレスチンの最も好ましい発現レベルを得るために培養条件を適合させることができると考えられる。誘導性発現系の場合、当業者は、誘導条件を最適化することもできると考えられる。発現の誘導の期間および発現の誘導の間の温度を最適化することもできると考えられる。好ましい実施形態によれば、味覚受容体の発現の誘導の開始時、サブコンフルエント細胞を、好適な培養培地を有する96ウェルプレート中に装入することができる。サブコンフルエントは、好ましくは70%コンフルエント、より好ましくは80%コンフルエントを意味する。好ましい実施形態において、添加される誘導剤は、誘導性発現系、好ましくはテトラサイクリン調節性プロモーターを使用する場合、テトラサイクリンまたはドキシサイクリンである。
【0026】
一実施形態において、細胞に、発光タンパク質、例えばルシフェラーゼタンパク質に融合している味覚受容体をコードする核酸分子およびβ−アレスチンに融合している蛍光タンパク質をコードする核酸分子を一過的に形質移入することができる。味覚受容体が1種超の区別されるサブユニットを有する場合、サブユニット当たり1種の核酸分子を使用することができる。代替的に、サブユニットの1種超のタイプを含む1種の単一核酸分子を使用することができる。一過性形質移入は、Lipofectamine2000を製造業者のプロトコルに従って使用して実施することができる。手短に述べると、細胞は、ウェル(12ウェルプレート、1ml培地/ウェル)当たり2×105個の細胞の密度において播種することができ、翌日に約80−90%のコンフルエンシーを目的とする。24時間後、発光タンパク質、例えばルシフェラーゼに融合している味覚受容体をコードする核酸分子を、β−アレスチンに融合している蛍光タンパク質をコードする核酸分子とともに同時形質移入することができる。ウェル当たり合計15gのトータルDNAを使用することができる。前記DNAを含む混合物を室温において30分間インキュベートし、各ウェルに添加し、細胞を48時間増殖させることができる。BRET計測は、形質移入の48−52時間後に実施することができる。代替的に、上記と同一のプロトコルを使用して発光タンパク質、例えばルシフェラーゼタンパク質に融合している味覚受容体をコードする核酸分子を、β−アレスチンに融合している蛍光タンパク質を安定的に発現する細胞内に形質移入することができる。好ましい形質移入プロトコルは、HEK293細胞についての実験部に記載する。
【0027】
好ましい実施形態において、味覚受容体はT1R1およびT1R3サブユニットを含み、サブユニットの少なくとも1種:T1R1もしくはT1R3または両サブユニットは発光タンパク質、例えばルシフェラーゼに融合している。好ましくは、ルシフェラーゼタンパク質はレニラルシフェラーゼである。レニラルシフェラーゼに融合している好ましいT1R1サブユニットをコードする好ましい核酸配列は、配列番号5により表される。対応する好ましいコードされるアミノ酸配列は、配列番号6により表される。レニラルシフェラーゼに融合している好ましいT1R3サブユニットをコードする好ましい核酸配列は、配列番号7により表される。対応する好ましいコードされるアミノ酸配列は、配列番号8により表される。
【0028】
さらに好ましい実施形態において、β−アレスチンに融合している蛍光タンパク質はGFPタンパク質、好ましくはGFP2であり、β−アレスチンはβ−アレスチン2である。
【0029】
よりいっそう好ましくは、味覚受容体は、T1R1およびT1R3サブユニットを含み、各サブユニットは、発光タンパク質、例えばルシフェラーゼタンパク質、好ましくはルシフェラーゼ、より好ましくはレニラルシフェラーゼに融合しており、β−アレスチンに融合している蛍光タンパク質はGFPタンパク質、好ましくはGFP2であり、β−アレスチンはβ−アレスチン2である。
【0030】
発光タンパク質がルシフェラーゼタンパク質であり得ることが、本発明によりさらに包含される。代替的に、発光タンパク質は、別の好適なエネルギードナーであり得る。GFPを別の好適なエネルギーアクセプターにより置き換えることができることも、本発明により包含される。理論的には、これ自体のポリペプチド配列内で可視波長発色団を形成することができるタンパク質の構造的に相同性のクラスのメンバーであると定義される、任意の蛍光タンパク質または分子を使用することができる。幾つかの蛍光タンパク質が、BRET技術において既に使用されている(Bacart J.Et al,(2008)Biotechnol.J.3:311−324およびPfleger K.D.,et al,(2006),Nature Methods,3:165−174参照。)。
【0031】
発光タンパク質、例えばルシフェラーゼおよびGFPは、それぞれ好ましいエネルギードナーおよびエネルギーアクセプターとして本明細書に提示される。発光タンパク質、例えばルシフェラーゼまたはGFPについて定義される特徴のそれぞれは、それぞれ任意の他のエネルギードナーまたはエネルギーアクセプターについても保持される。
【0032】
従って、本発明の方法において:
(a)味覚受容体のサブユニットの少なくとも1種、好ましくはT1R1およびT1R3の少なくとも1種は、発光タンパク質、例えばルシフェラーゼタンパク質に融合しており、もしくは味覚受容体の各サブユニット、好ましくはT1R1およびT1R3は、それぞれ発光タンパク質、例えばルシフェラーゼタンパク質に融合しており、
ならびに/または
(b)β−アレスチンに融合している蛍光タンパク質はGFPタンパク質であり、好ましくはGFPはGFP2であり、および/もしくはβ−アレスチンはβ−アレスチン2である。
【0033】
代替的に、本発明の方法において:
(a)味覚受容体のサブユニットの少なくとも1種、好ましくはT1R1およびT1R3の少なくとも1種は蛍光タンパク質に融合しており、もしくは味覚受容体の各サブユニット、好ましくはT1R1およびT1R3は蛍光タンパク質に融合しており、
ならびに/または
(b)発光タンパク質はβ−アレスチンに融合している。
【0034】
代替的に、本発明の方法において:
(a)味覚受容体はT1R1、T1R3ヘテロ二量体であり、サブユニットT1R1およびT1R3の少なくとも1種が発光タンパク質に融合しており、および蛍光タンパク質がβ−アレスチンに融合しており;または
(b)味覚受容体はT1R1、T1R3ヘテロ二量体であり、サブユニットT1R1およびT1R3の少なくとも1種が蛍光タンパク質に融合しており、および発光タンパク質がβ−アレスチンに融合している。
【0035】
好ましくは、本発明の方法において、(a)発光タンパク質の少なくとも1種がルシフェラーゼであり;(b)蛍光タンパク質の少なくとも1種がGFPであり;および(c)β−アレスチンの少なくとも1種が非視覚β−アレスチンである。より好ましくは、(a)ルシフェラーゼの少なくとも1種がレニラルシフェラーゼであり;および(b)非視覚β−アレスチンの少なくとも1種がβ−アレスチン2である。
【0036】
工程b)は、工程a)において得られた細胞を潜在的モジュレーター化合物によりチャレンジすることを含む。
【0037】
この工程に関して、チャレンジすることは、工程a)において得られた細胞を潜在的モジュレーター化合物と接触させることを意味することができる。
【0038】
味覚受容体の潜在的モジュレーター化合物は、味覚受容体の活性化される能力を遮断し、阻害し、モジュレートし、または向上させることにより味覚認識を遮断し、阻害し、モジュレートし、または向上させ、こうして細胞内シグナルを細胞内にトランスデュースすることができる化合物として本明細書に定義される。任意の分子、例えば天然または合成の任意の有機分子、例えばタンパク質、オリゴペプチド、有機小分子、多糖、脂質(例えばスフィンゴ脂質)、脂肪酸、ポリヌクレオチド、オリゴヌクレオチドなどを本発明において試験することができる。潜在的モジュレーター化合物は、化合物のライブラリー、例えば十分な範囲の多様性を提供するコンビナトリアルまたはランダム化ライブラリーの形態であり得る。
【0039】
潜在的モジュレーター化合物は、通常、水性試料溶液中に存在する。このような水性試料溶液は、(a)食品;(b)食品の抽出物;および(c)バイオマスの抽出物、好ましくは可食性バイオマスの抽出物の少なくとも1種を含むことができる。従って、試料溶液は、液体食品またはこの希釈物を含むことができる。液体食品は、例えば飲料または醤油などのようなソースであり得る。水性試料溶液は、食品の抽出物、例えば固体食品(例えばチーズ)または脂肪食品の抽出物を含むこともできる。または、水性試料溶液は、植物、動物および微生物の少なくとも1種からのバイオマスの抽出物を含むことができる。従って、水性試料溶液は、植物、動物および微生物の少なくとも1種からの可溶性分子を含むことができる。従って、水性試料溶液は、発酵食品からの抽出物を含むことができる。好ましい実施形態において、試料溶液は、トマト抽出物を含む。抽出物はさらに以下に本明細書に定義されるとおりであり、以下に本明細書に定義されるとおり調製することができる。
【0040】
本発明の方法の利点は、本発明の方法により複合試料中の潜在的モジュレーターの同定が可能となることである。従って、好ましくは、水性試料溶液は、複合試料溶液である。水性試料溶液は、少なくとも2種の区別される有機分子を含む、好ましくは少なくとも3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20種または50種超または100種超の区別される有機分子を含む混合物であり得る。化合物の数は、好ましくは以下に同定されるとおり評価される。水性試料溶液は、この溶液が10種超の区別される有機分子、好ましくは10種超の区別される潜在的モジュレーターを含む場合、複合試料溶液と定義することができる。水性試料溶液は、通常、例えば、所定量の1種以上の特定されたモジュレーターを含む参照溶液とは対照的に未特定組成物の溶液であることがさらに理解される。従って、未特定組成物の溶液は、アイデンティティおよび/または濃度が未知であり、または定義されていない化合物を含む溶液である。
【0041】
水性試料溶液は、好ましくは可溶物のみを含む溶液である。不溶物は、当分野において公知であり、抽出物について以下に本明細書に記載されている手段により試料溶液から除去することができる。水性試料溶液は、溶液を生理学的に許容されるpHおよび浸透圧値に調整することにより、工程a)において提供された細胞と生理学的に適合性を示すまでさらに調整することができる。
【0042】
好ましい実施形態において、このようなモジュレーターは抽出物中に存在する。抽出物は、少なくとも2種の区別される分子、少なくとも3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20種または50種超または100種超の区別される分子を含む混合物である。化合物の数は、好ましくは、以下に同定されるとおり評価される。抽出物は、この抽出物が10種超の区別される分子を含む場合、複合抽出物と定義することができる。抽出物は、少なくとも2種の区別される有機分子、少なくとも3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20種または50種超または100種超の区別される有機分子を含む混合物である。化合物の数は、好ましくは、以下に同定されるとおり評価される。抽出物は、この抽出物が10種超の区別される有機分子を含む場合、複合抽出物と定義することができる。抽出物は精製しなくてよく、植物ベースの抽出物、好ましくはトマト抽出物または動物ベースの抽出物および/または食物ベースの抽出物であり得る。抽出物は、食物ベースまたは食物由来の抽出物であり得る。食物ベースまたは食物由来の抽出物は、食物組成物もしくは食品として、または食物組成物もしくは食品をベースとし、もしくはこれらに由来するものとして把握することができる。抽出物のアイデンティティ、この組成および抽出物が調製された手法は、本発明について重要ではない。対照的に、本発明の利点は、本発明の方法を使用して潜在的に各抽出物から味覚受容体の潜在的モジュレーターを同定することができることである。
【0043】
抽出物は、植物、動物および/または微生物バイオマスの抽出のために溶媒(例えば水)または溶媒混合物(例えば水/アルコール)を使用することにより得られた未精製混合物、例えば前記材料からの全ての可溶性化合物を好ましくは元の材料中に存在するのと同様の比、好ましくはこれらの溶解度に従って補正された比で含有する生の、加工された、発酵されたおよび/または乾燥された植物または動物または食物材料であり得る。
【0044】
抽出物中の化合物の数および濃度は、元の植物または動物または食物材料中のこれらの存在、使用される溶媒系におけるこれらの溶解度を認識し、当業者に公知の技術、例えばクロマトグラフィー(HPLC、GC)および/または分光技術(NMR、質量分析)を使用して評価することができる。
【0045】
本発明において使用すべき抽出物の調製は、通常、少なくとも2種の工程:1)可溶化および/または均一化工程;ならびに2)不溶物および場合により非水相もしくは溶媒を除去する工程を含む。好適な抽出物を生成する考えられる手法は、慣用の加工条件(例えば加熱、撹拌、粉砕)下で好適な溶媒(例えば水、アルコール)または溶媒混合物(例えば水/アルコール)により(例えば切断、温浸、乾燥、粉化などにより)予備加工し、下流で加工(例えば濾過、遠心分離、乾燥)することができる植物または動物または食物材料の処理であり得る。
【0046】
好ましい方法において、味覚受容体の公知のアクチベーターを工程b)においてさらに添加する。味覚受容体の好ましい公知のアクチベーターは、表1に既に同定されている。アクチベーターは、阻害剤についての探索において使用することができ:味覚受容体を最初にアクチベーターと接触させてBRETシグナル(即ち、前記受容体の定常活性よりも高いシグナル)を生成する。続いて、潜在的阻害化合物を前記アクチベーターと一緒に接触させる。生成されたシグナルが低減すれば、前記潜在的阻害化合物は阻害剤である。アクチベーターは、好ましくは、これに関連して次いで阻害剤により低下される受容体活性化のレベルを増加させるために使用し;このことにより、読取り窓およびひいては感度が増加する。
【0047】
工程c)は、工程b)において得られた細胞のBRETシグナルを、潜在的モジュレーター化合物の不存在下でb)において得られた細胞のBRETシグナルと比較することを含む。代替的に、工程c)は、工程b)において得られたBRETシグナルを、潜在的モジュレーター化合物の不存在下で細胞から得られたBRETシグナルと比較することを含み、b)において得られたBRETシグナルと潜在的モジュレーター化合物の不存在下で得られたBRETシグナルとの差は、潜在的モジュレーター化合物が味覚受容体をモジュレートする化合物であることを示す。
【0048】
BRETシグナルの生成を導くBRET技術またはアッセイは、当業者に公知の任意のBRET技術であり得る。味覚受容体に融合している発光ドナー発光タンパク質、例えばルシフェラーゼタンパク質の任意の公知のバリアントを使用することができる。同じことが、フルオロフォアもしくは蛍光タンパク質とも称される発光アクセプターまたは蛍光タンパク質に融合しているエネルギーアクセプタータンパク質、例えばβ−アレスチンについて保持される。発光ドナーおよび発光アクセプターのアイデンティティに応じて、BRET1またはBRET2またはBRET3技術を使用する。幾つかのBRETアッセイについて使用されるタンパク質のアイデンティティ、条件および波長は当業者に公知であり、例えばBacart J.et al(2008),Biotechnol.J.3:311−324を参照。しかしながら、BRET技術において使用されるタンパク質のそれぞれは改変してこれらのスペクトル特性を改善することができ、これらをBRET技術により好適とする。好ましくは、BRET2技術を使用する。これというのも、この技術はより良好な感度および効率:BRETシグナルのより早期の検出を導く関与する波長のより良好な分離を提供することが予期されるからである。
【0049】
好ましくは抽出物中に存在する潜在的モジュレーター化合物を用いる受容体刺激後、蛍光タンパク質に融合しているβ−アレスチンタンパク質は、活性化された味覚受容体と相互作用し、こうして蛍光タンパク質を前記活性化された味覚受容体に融合している発光タンパク質、例えばルシフェラーゼと近接して接触させ、考えられるこれらの2種のタンパク質間のエネルギー転移をもたらし、BRETシグナルを生成する。BRET2の場合、アッセイは好ましくは以下のとおりである:酸素の存在下、ルシフェラーゼは基質DeepBlueCの395−410nmにおいて計測することができるコエレンテルアミドへの転換を触媒する。蛍光タンパク質が発光タンパク質、例えばルシフェラーゼに近接しており、エネルギー転移が生じる場合、発光は510nmにシフトし;このことはBRETシグナルと称され、アクセプター(蛍光タンパク質)とドナー(発光タンパク質、例えばルシフェラーゼ)との比として表現される。BRETシグナルは、本明細書において比が0よりも大きい検出可能なBRETシグナルとして定義される。BRETシグナルがもっぱら味覚受容体の特異的活性化により引き起こされることを確保するため、好ましくは発光タンパク質、例えばルシフェラーゼタンパク質に融合している味覚受容体およびβ−アレスチンに融合している蛍光タンパク質を発現する細胞を用いて得られたBRETシグナルを、前記受容体を発現しない細胞および/または発光タンパク質、例えばルシフェラーゼタンパク質に融合している味覚受容体を発現しない細胞および/またはβ−アレスチンに融合している蛍光タンパク質を発現しない細胞を用いて得られたBRETシグナルと比較する。当業者は、BRETアッセイをいかに実施するかを認識する。蛍光は、典型的には、蛍光プレートリーダーにより計測する。一実施形態において、Mithras LB 940プレートリーダー(Berthold Technologies)を使用することができる。使用することができるプロトコルは、本質的には、Packard BioOneに記載されているものと同一であり、またはわずかな改変を有する刊行されたプロトコル13であり:細胞は、本明細書に上記のとおり形質移入することができる。形質移入48時間後、細胞を回収し、BRET緩衝液(2μg/mlアプロチニンを含有するD−PBS)中に2×106個の細胞/mlの密度において溶解させることができる。細胞を平衡化のために室温において1時間放置した後、約1×105個の細胞を含有する30μlを白色96ウェルプレートの各ウェルに移すことができる。10μlの潜在的モジュレ−ティング化合物または抽出物(または緩衝液)および10μlの基質コエレンテラジン(最終濃度5μM)を、インジェクターを使用して同時に添加することができる。
【0050】
最終インジェクション直後、発光シグナルの生成に使用される基質を考慮して繰り返し連続読取りをドナーの規定の発光波長およびアクセプターの規定の発光波長において行う。BRETシグナルは、アクセプタータンパク質について計測された光シグナルと、ドナータンパク質について計測された光シグナルとの比として測定することができる。
【0051】
BRET技術がBRET1技術である場合、ドナーはレニラルシフェラーゼ[Rluc]および基質としてのコエレンテラジンhであり;アクセプターは強化黄色蛍光タンパク質[強化YFP]、YFPトパーズ(topaz)、YFPシトリン(citrine)、YFPビーナス(venus)、YPet.)である。
【0052】
最終インジェクション直後、繰り返し連続読取りを、Rluc/コエレンテラジンhの規定の発光波長のピーク(480nm)およびアクセプターの規定の発光波長のピーク(530nm)をカバーする波長において行う。BRETシグナルは、アクセプタータンパク質について計測された光シグナルとドナータンパク質について計測された光シグナルとの比として決定することができる。
【0053】
技術がBRET2技術である場合、ドナーはレニラルシフェラーゼ[Rluc]またはレニラルシフェラーゼ突然変異体8[Rluc8](Loening AM,et al,Protein Eng Des Sel 2006 September;19(9):391−400およびBacart J,et al,Biotechnol J 2008 March;3(3):311−24.)および基質としてのDeepBlueC(商標)またはコエレンテラジン400aであり;アクセプターは、緑色蛍光タンパク質−2[GFP2]または緑色蛍光タンパク質10[GFP10])である。
【0054】
最終インジェクション直後、繰り返し連続読取りを、Rluc/コエレンテラジン400aの規定の発光波長のピーク(395nm)およびアクセプターの規定の発光波長のピーク(510nm)をカバーする波長において行う。BRETシグナルは、アクセプタータンパク質について計測された光シグナルとドナータンパク質について計測された光シグナルとの比として決定することができる。
【0055】
工程c)において得られた細胞のBRETシグナルを、潜在的モジュレーター化合物の不存在下でc)において得られた細胞の対応する不存在のBRETシグナルと比較する。代替的に、工程c)において得られた細胞のBRETシグナルの変化を、潜在的モジュレーター化合物の不存在下でc)において得られた細胞の対応する元のBRETシグナルと比較する。
【0056】
代替的に、工程b)において得られたBRETシグナルを、潜在的モジュレーター化合物の不存在下で細胞から得られたバックグラウンド(または不存在の)BRETシグナルと比較する。b)において得られたBRETシグナルと、潜在的モジュレーター化合物の不存在下で得られたBRETシグナルとの差は、潜在的モジュレーター化合物が味覚受容体をモジュレートする化合物であることを示す。代替的に、工程b)において得られたBRETシグナルを、潜在的モジュレーター化合物の不存在下で工程b)において得られた細胞の対応する元のBRETシグナルと比較することができる。b)において得られたBRETシグナルと、潜在的モジュレーター化合物の不存在下で得られたBRETシグナルとの差は、潜在的モジュレーター化合物が味覚受容体をモジュレートする化合物であることを示す。同様に、2種以上の異なる試料溶液を用いて得られたBRETシグナルを比較することができ、このことにより異なる試料溶液は、上記の食品、食品の抽出物および/またはバイオマスの抽出物の異なるフラクションを含有することができる。
【0057】
BRETシグナルを増加させ、または検出可能なBRETシグナルを誘導する化合物は、味覚の潜在的エンハンサーである。対照的に、BRETシグナルを減少させる化合物は味覚の潜在的マスカーである。
【0058】
好ましい実施形態によれば、味覚の潜在的エンハンサーは、工程c)において実施された比較がBRETシグナルもしくは検出可能なBRETシグナルの存在または少なくとも2%のBRETシグナルの検出可能な増加を示す場合に同定された。より好ましくは、味覚の潜在的エンハンサーは、工程c)において実施された比較がBRETシグナルの少なくとも4%、少なくとも6%、少なくとも8%、少なくとも10%、少なくとも12%、少なくとも14%、少なくとも16%、少なくとも18%、少なくとも20%、少なくとも22%、少なくとも24%、少なくとも26%、少なくとも28%、少なくとも30%、少なくとも32%、少なくとも34%、少なくとも36%、少なくとも38%、少なくとも40%、少なくとも42%以上の増加を示す場合に同定された。
【0059】
別の好ましい実施形態によれば、味覚の潜在的マスカーは、工程c)において実施される比較がBRETシグナルの少なくとも2%の減少を示す場合に同定された。より好ましくは、味覚の潜在的マスカーは、工程c)において実施される比較がBRETシグナルの少なくとも4%、少なくとも6%、少なくとも8%、少なくとも10%、少なくとも12%、少なくとも14%、少なくとも16%、少なくとも18%、少なくとも20%、少なくとも22%、少なくとも24%、少なくとも26%、少なくとも28%、少なくとも30%、少なくとも32%、少なくとも34%、少なくとも36%、少なくとも38%、少なくとも40%、少なくとも42%以上の減少を示す場合に同定された。
【0060】
味覚受容体の公知の阻害剤またはアクチベーターが存在する場合、BRETシグナルのそれぞれの増加または減少を、前記潜在的モジュレーター化合物の不存在下で得られたBRETシグナルと比較する。
【0061】
さらなる態様において、本明細書に同定される方法により同定された味覚受容体の潜在的モジュレーター化合物が提供される。
【0062】
別の態様において、本発明は、味覚受容体をモジュレートする化合物を生成する方法に関する。本方法は、本明細書に定義の方法において味覚受容体をモジュレートする化合物を同定し、該化合物を回収する工程を含む。味覚をモジュレートする化合物の回収および/または(部分)精製の方法は、当業者に自体公知である。
【0063】
さらに別の態様において、本発明は、味覚受容体をモジュレートする化合物の同定のための、BRET技術またはアッセイの使用に関する。BRET技術またはアッセイは、本明細書に記載の本発明の方法に従って使用することができる。
【0064】
一般的技術情報
配列番号により定義される核酸分子および配列同一性
所与の配列同一性番号(例えば配列番号1)により本明細書に同定される各遺伝子または核酸分子は、開示されているこの規定の配列に限定されるものではないことを理解すべきである。本明細書に同定されている各遺伝子配列またはヌクレオチド配列は、本明細書に同定されている所与のタンパク質またはポリペプチドをコードする。本願全体にわたり、規定のヌクレオチド配列の配列番号(配列番号1を例にとる。)を指すごとに、これを
i.配列番号1と少なくとも60%の配列同一性を有するヌクレオチド配列を含むヌクレオチド配列、
ii.相補的ストランドが(i)または(ii)の配列の核酸分子にハイブリダイズするヌクレオチド配列;
iii.配列が遺伝コードの縮退に起因して(i)の核酸分子の配列とは異なるヌクレオチド配列、
iv.ヌクレオチド配列配列番号1によりコードされるアミノ酸配列と少なくとも60%のアミノ酸同一性を有するアミノ酸配列をコードするヌクレオチド配列
により置き換えることができる。
【0065】
本明細書に記載の各ヌクレオチド配列またはアミノ酸配列は、それぞれ所与のヌクレオチド配列またはアミノ酸配列とのこの同一性の割合(少なくとも60%)に関して、さらに好ましい実施形態において、それぞれ所与のヌクレオチドまたはアミノ酸配列との少なくとも65%、70%、75%、80%、85%、90%、95%、97%、98%、99%以上の同一性を有する。好ましい実施形態において、配列同一性は、本明細書に同定されている配列の全長を比較することにより決定する。
【0066】
「配列同一性」は、本明細書において、配列を比較することにより決定される、2種以上のアミノ酸(ポリペプチドまたはタンパク質)配列間または2種以上の核酸(ポリヌクレオチド)配列間の関係と定義される。当分野において「同一性」も、場合によってはこのような配列のストリング間の一致により決定されるアミノ酸または核酸配列間の配列関連性の程度を意味する。2種のアミノ酸配列間の「類似性」は、アミノ酸配列およびあるポリペプチドの第2のポリペプチドの配列に対してこの保存されているアミノ酸置換物を比較することにより決定する。「同一性」および「類似性」は、限定されるものではないが、(Computational Molecular Biology,Lesk,A.M.,ed.,Oxford University Press,New York,1988;Biocomputing:Informatics and Genome Projects,Smith,D.W.,ed.,Academic Press,New York,1993;Computer Analysis of Sequence Data,Part I,Griffin,A.M.,and Griffin,H.G.,eds.,Humana Press,New Jersey,1994;Sequence Analysis in Molecular Biology,von Heine,G.,Academic Press,1987;およびSequence Analysis Primer,Gribskov,M.and Devereux,J.,eds.,M Stockton Press,New York,1991;およびCarillo,H.,and Lipman,D.,SIAM J.Applied Math.,48:1073(1988)に記載のものを含む公知の方法により容易に計算することができる。
【0067】
同一性を決定する好ましい方法は、試験される配列間の最大の一致を与えるように設計する。同一性および類似性を決定する方法は、公的に入手可能なコンピュータプログラムにおいてコード化されている。2種の配列間の同一性および類似性を決定する好ましいコンピュータプログラム方法としては、例えば、GCGプログラムパッケージ(Devereux,J.,et al.,Nucleic Acids Research 12(1):387(1984)),BestFit,BLASTP,BLASTNおよびFASTA(Altschul,S.F.et al.,J.Mol.Biol.215:403−410(1990)が挙げられる。BLAST Xプログラムは、NCBIおよび他の供給源(BLAST Manual,Altschul,S.,et al.,NCBI NLM NIH Bethesda,MD 20894;Altschul,S.,et al.,J.Mol.Biol.215:403−410(1990)から公的に入手可能である。周知のSmith Watermanアルゴリズムを使用して同一性を決定することもできる。
【0068】
ポリペプチド配列比較についての好ましいパラメーターとしては、以下のもの:アルゴリズム:Needleman and Wunsch,J.Mol.Biol.48:443−453(1970);比較マトリックス:Hentikoff and Hentikoff,Proc.Natl.Acad.Sci.USA.89:10915−10919(1992)からのBLOSSUM62;ギャップペナルティ:12;およびギャップ長さペナルティ:4が挙げられる。これらのパラメーターを用いる有用なプログラムは、Genetics Computer Group,located in Madison,Wlからの「Ogap」プログラムとして公的に入手可能である。上記パラメーターは、(末端基についてペナルティを伴わない)アミノ酸比較についてのデフォルトパラメーターである。
【0069】
核酸比較についての好ましいパラメーターとしては、以下のもの:アルゴリズム:Needleman and Wunsch,J.Mol.Biol.48:443−453(1970);比較マトリックス:一致=+10、不一致=0;ギャップペナルティ:50;ギャップ長さペナルティ:3が挙げられる。Madison,Wisに位置するGenetics Computer GroupからのGapプログラムとして入手可能。核酸比較についてのデフォルトパラメーターは上記に挙げる。
【0070】
場合により、アミノ酸類似性の程度の決定において、当業者は、当業者に明確であるいわゆる「保存」アミノ酸置換を考慮することもできる。保存アミノ酸置換は、類似側鎖を有する残基の交換可能性を指す。例えば、脂肪族側鎖を有するアミノ酸のグループはグリシン、アラニン、バリン、ロイシンおよびイソロイシンであり;脂肪族−ヒドロキシル側鎖を有するアミノ酸のグループはセリンおよびトレオニンであり;アミド含有側鎖を有するアミノ酸のグループはアスパラギンおよびグルタミンであり;芳香族側鎖を有するアミノ酸のグループはフェニルアラニン、チロシンおよびトリプトファンであり;塩基性側鎖を有するアミノ酸のグループはリジン、アルギニンおよびヒスチジンであり;硫黄含有側鎖を有するアミノ酸のグループはシステインおよびメチオニンである。好ましい保存アミノ酸置換基は:バリン−ロイシン−イソロイシン、フェニルアラニン−チロシン、リジン−アルギニン、アラニン−バリンおよびアスパラギン−グルタミンである。本明細書に開示のアミノ酸配列の置換バリアントは、開示配列中の少なくとも1個の残基が除去され、異なる残基がこの場所に挿入されたものである。好ましくは、アミノ酸変化は保存的である。天然に生じるアミノ酸のそれぞれについて好ましい保存的置換は、以下のとおりである:Alaからser;Argからlys;Asnからglnまたはhis;Aspからglu;Cysからserまたはala;Glnからasn;Gluからasp;Glyからpro;Hisからasnまたはgln;Ileからleuまたはval;Leuからileまたはval;Lysからarg;glnまたはglu;Metからleuまたはile;Pheからmet、leuまたはtyr;Serからthr;Thrからser;Trpからtyr;Tyrからtrpまたはphe;およびValからileまたはleu。
【0071】
細胞内での味覚受容体の発現
本発明の方法において使用される味覚受容体およびβ−アレスチンは、ルシフェラーゼタンパク質に融合している味覚受容体をコードするヌクレオチド配列および蛍光タンパク質に融合しているβ−アレスチンをコードする別の配列を好適な細胞内で発現させる組換え技術を使用して調製することができる。従って本発明はまた、上記定義の前記ヌクレオチド配列により表される前記核酸分子を含むベクターの使用に関する。好ましくは、ベクターは、ベクターに好適な宿主内でのベクターの増幅を確保する複製起源(または自律複製配列)を含む複製ベクターである。代替的に、ベクターは、例えば相同組換えまたは別の手段を介して細胞のゲノム中に統合することができる。特に好ましいベクターは、上記定義のポリペプチドをコードするヌクレオチド配列が、ベクターのための細胞内でコード配列の発現を指向することができるプロモーターに作動可能に連結している発現ベクターである。
【0072】
本明細書において使用される用語「プロモーター」は、1種以上の遺伝子の転写を制御するように機能し、遺伝子の転写開始部位の転写の方向に関して上流に局在しており、DNA依存性RNAポリメラーゼについての結合部位、転写開始部位ならびに限定されるものではないが、転写因子結合部位、レプレッサーおよびアクチベータータンパク質結合部位を含む任意の他のDNA配列ならびにプロモーターからの転写の量を調節するように直接的または間接的に作用することが当業者に公知のヌクレオチドの任意の他の配列の存在により構造的に同定されている核酸断片を指す。「構成性」プロモーターは、ほとんどの生理学的および発育条件下で活性であるプロモーターである。「誘導性」プロモーターは、生理学的または発育条件に応じて調節されるプロモーターである。発現ベクターにより、上記定義のポリペプチドを、前記ポリペプチドをコードするヌクレオチド配列を好適な細胞、例えば培養細胞内で発現させる組換え技術を使用して調製することが可能となる。
【0073】
好ましくは、上記定義の本発明の核酸配列のそれぞれの発現は、誘導性である。各核酸配列の発現の誘導性は、当業者に公知の任意の手法により実現することができる。例えば、Invitrogen T−Rex系(テトラサイクリン調節性発現、tetオペロンに基づき、培地に誘導物質が添加されるまで挿入遺伝子の発現が抑制される。)、Invitrogen Gene−Switch系(GaW−Elbプロモーターの活性化に基づく)、Stratagene Complete Control誘導性哺乳動物発現系(ドロソフィア・メラノガステル(Drosophila melanogaster)エクジソン受容体についての遺伝子およびエクジソン受容体についての結合部位を含有するプロモーターの両方を内部に持つ哺乳動物細胞内での昆虫ホルモンのエクジソンまたはこの類似体であるポナステロンA(ponA)による転写の活性化に基づく)、New England Biolabs RheoSwitch(登録商標)哺乳動物誘導性発現系(合成誘導物質であるRheoSwitch Ligand RSLlおよび2つの領域からなるキメラ核受容体との高度に特異的な相互作用に基づく)、Qbiogene Q−mate(商標)誘導性発現系またはQ−mate(商標)CymR系(誘導物質分子であるクマートの不存在下、オペレーター部位に結合しているクマートリプレッサータンパク質CymRによる遺伝子発現の抑制に基づく。クマートの存在下、CymRはクマートに結合し、立体配置の変化を受け、オペレーター部位からのこの放出をもたらす。)、Stratagene製LacSwitch(登録商標)II誘導性哺乳動物発現系(lacオペロンに基づき、誘導物質が培地に添加されるまで挿入遺伝子の発現が抑制される。)がある。よりいっそう好ましくは、各核酸部分配列の発現は、本発明の核酸配列中に存在する核酸部分配列のそれぞれに作動可能に連結している誘導性プロモーターの存在により誘導性となる。本発明に関して、「作動可能に連結している」は、制御配列、本明細書においてはプロモーター配列が、制御配列が核酸部分配列の発現を指向するように、核酸部分配列に対する位置に適切に配置される立体配置と定義される。
【0074】
誘導性プロモーターは、上記定義の所与の核酸部分配列について、および所与の細胞内で機能的である任意のプロモーターまたはこの部分であり得、プロモーターの転写開始活性は、細胞を培養する間に所与の誘導剤を添加したときに細胞内で誘導され得る。より好ましくは、誘導性プロモーターはテトラサイクリン調節性プロモーターである。よりいっそう好ましくは、誘導性プロモーターは、Yao et al(Yao,F.et al.(1998)Hum.Gene Therapy 9:1939−1950およびYao,F and Eriksson,E.(1999)Hum.Gene Therapy 10:419−427)に記載のテトラサイクリン調節性ハイブリッドヒトサイトメガロウイルスプロモーターである。この系は、InvitrogenからT−Rex発現系として購入することができる。)。誘導性発現系の使用により、安定発現系を使用して記載された毒性の問題を回避することができる。
【0075】
典型的には、上記定義のポリペプチドをコードする核酸を発現ベクター中で使用する。語句「発現ベクター」は、一般に、このような配列と適合性の宿主内で遺伝子の発現を行うことができるヌクレオチド配列を指す。これらの発現ベクターは、典型的には、少なくとも好適なプロモーター配列および場合により転写終結シグナルを含む。発現の実施に必要な、または一助となる追加の因子を本明細書に記載のとおり使用することもできる。ポリペプチドをコードする核酸またはDNAをインビトロ細胞培養物中に導入し、発現させることができるDNA構築物中に取り込む。具体的には、DNA構築物は、原核宿主、例えば細菌、例えばE.コリ(E.coli)内での複製に好適であり、または培養された哺乳動物、植物、昆虫、例えばSf9、酵母、真菌もしくは別の真核細胞系中に導入することができる。
【0076】
特定の細胞内への導入のために調製されたDNA構築物は、典型的には、宿主により認識される複製系、所望のポリペプチドをコードする目的DNAセグメントならびにポリペプチドコードセグメントに作動可能に連結している転写および翻訳開始および終結調節配列を含む。DNAセグメントは、このセグメントが別のDNAセグメントとの機能的関係に置かれている場合、「作動可能に連結」している。例えば、プロモーターまたはエンハンサーは、これが配列の転写を刺激する場合、コード配列に作動可能に連結している。しかしながら、エンハンサーは、エンハンサーが転写を制御するコード配列と隣接している必要はない。連結は、簡便な制限部位またはこの代わりに挿入されたアダプターもしくはリンカーにおけるライゲーションにより達成される。
【0077】
適切なプロモーター配列の選択は、一般に、DNAセグメントの発現について選択された細胞タイプに依存する。好適なプロモーター配列の例としては、当分野において周知の原核および真核プロモーターが挙げられる(例えばSambrook and Russell,2001,上掲参照。)。転写調節配列としては、典型的には、宿主により認識される異種エンハンサーまたはプロモーターが挙げられる。適切なプロモーターの選択は細胞に依存するが、プロモーター、例えばtrp、lacおよびファージプロモーター、tRNAプロモーターおよび糖分解酵素プロモーターが公知であり、入手可能である(例えばSambrook and Russell,2001,上掲参照。)。複製系ならびに転写および翻訳調節配列をポリペプチドコードセグメントについての挿入部位と一緒に含む発現ベクターを用いることができる。細胞系および発現ベクターの作用可能な組合せの例は、Sambrook and Russell(2001,上掲)およびMetzger et al.(1988)Nature 334:31−36に記載されている。例えば、好適な発現ベクターは、酵母、例えばS.セレビシエ(S.cerevisiae)、例えば昆虫細胞、例えばSf9細胞、哺乳動物細胞、例えばCHO細胞および細菌細胞、例えばE.コリ内で発現させることができる。従って、細胞は原核または真核宿主細胞であり得る。細胞は、液体または固体培地中での培養に好適な細胞であり得る。細胞は、好ましくは、上記定義の本発明の方法において使用する。
【0078】
本明細書およびこの特許請求の範囲において、動詞「含むこと」およびこの活用形は、この非限定的な意味において、語の後の項目が含まれることを意味するように使用されるが、特に言及されなかった項目が除外されるわけではない。さらに動詞「からなる」は、本明細書に定義の方法が具体的に同定されているものよりも追加の工程を含むことができ、前記追加の工程は本発明の特有の特性を変化させないことを意味する「から本質的になる」により置き換えることができる。さらに、不定冠詞「a」または「an」による要素に対する言及は、文脈が唯一の要素の存在を明確に要求しない限り、1つ超の要素が存在する可能性を除外するものではない。従って、不定冠詞「a」または「an」は、通常「少なくとも1つ」を意味する。
【0079】
本明細書に引用されている全ての特許および参考文献は、参照により全体として本明細書に組み込まれる。
【0080】
以下の実施例を説明の目的のためにのみ提供するが、以下の実施例は本発明の範囲を決して限定するものではない。
【図面の簡単な説明】
【0081】
【図1】実施例2に記載のカルシウム蛍光マーカーFluo−4−AMを使用して細胞内カルシウムの放出を追跡することにより計測されたHEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を示す。旨味受容体の活性は、試験溶液添加後の最大蛍光を、試験溶液添加前に計測されたベースライン蛍光から差し引くことにより計算された蛍光の変化として計測した。試験溶液は示されている抽出物を含み、実施例2に記載のとおり調製した。MSG=10mM;MSG+IMP=1mMのMSG+500μMのIMP。
【図2a】実施例2に記載のBRETアッセイにより計測されたHEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を示す。示されている試験溶液(または緩衝液)および基質を細胞に添加した。BRETシグナルは、ドナー(400nm)について測定されたシグナルにより割ったアクセプター波長(515nm)において取られた読取り値の比として測定した。アクセプター波長におけるドナー発光の重複に起因するバックグラウンドシグナルについて補正するため、BRET比を、ドナー単独(T1R1/T1R3−Rluc)を発現する細胞についてパラレルで測定した。このBRETバックグラウンド値を、両方のBRETパートナーを発現する細胞について得られたBRET値から差し引いた(BRET=BRET比−バックグラウンド比)。緩衝液単独について達成された値を上回るBRETシグナルを、旨味受容体特異的として定義した(表2Bも参照。)。
【図2b】実施例2に記載のBRETアッセイにより計測されたHEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を示す。示されている試験溶液(または緩衝液)および基質を細胞に添加した。BRETシグナルは、ドナー(400nm)について測定されたシグナルにより割ったアクセプター波長(515nm)において取られた読取り値の比として測定した。アクセプター波長におけるドナー発光の重複に起因するバックグラウンドシグナルについて補正するため、BRET比を、ドナー単独(T1R1/T1R3−Rluc)を発現する細胞についてパラレルで測定した。このBRETバックグラウンド値を、両方のBRETパートナーを発現する細胞について得られたBRET値から差し引いた(BRET=BRET比−バックグラウンド比)。緩衝液単独について達成された値を上回るBRETシグナルを、旨味受容体特異的として定義した(表2Bも参照。)。
【図2c】実施例2に記載のBRETアッセイにより計測されたHEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を示す。示されている試験溶液(または緩衝液)および基質を細胞に添加した。BRETシグナルは、ドナー(400nm)について測定されたシグナルにより割ったアクセプター波長(515nm)において取られた読取り値の比として測定した。アクセプター波長におけるドナー発光の重複に起因するバックグラウンドシグナルについて補正するため、BRET比を、ドナー単独(T1R1/T1R3−Rluc)を発現する細胞についてパラレルで測定した。このBRETバックグラウンド値を、両方のBRETパートナーを発現する細胞について得られたBRET値から差し引いた(BRET=BRET比−バックグラウンド比)。緩衝液単独について達成された値を上回るBRETシグナルを、旨味受容体特異的として定義した(表2Bも参照。)。
【図2d】実施例2に記載のBRETアッセイにより計測されたHEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を示す。示されている試験溶液(または緩衝液)および基質を細胞に添加した。BRETシグナルは、ドナー(400nm)について測定されたシグナルにより割ったアクセプター波長(515nm)において取られた読取り値の比として測定した。アクセプター波長におけるドナー発光の重複に起因するバックグラウンドシグナルについて補正するため、BRET比を、ドナー単独(T1R1/T1R3−Rluc)を発現する細胞についてパラレルで測定した。このBRETバックグラウンド値を、両方のBRETパートナーを発現する細胞について得られたBRET値から差し引いた(BRET=BRET比−バックグラウンド比)。緩衝液単独について達成された値を上回るBRETシグナルを、旨味受容体特異的として定義した(表2Bも参照。)。
【図2e】実施例2に記載のBRETアッセイにより計測されたHEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を示す。示されている試験溶液(または緩衝液)および基質を細胞に添加した。BRETシグナルは、ドナー(400nm)について測定されたシグナルにより割ったアクセプター波長(515nm)において取られた読取り値の比として測定した。アクセプター波長におけるドナー発光の重複に起因するバックグラウンドシグナルについて補正するため、BRET比を、ドナー単独(T1R1/T1R3−Rluc)を発現する細胞についてパラレルで測定した。このBRETバックグラウンド値を、両方のBRETパートナーを発現する細胞について得られたBRET値から差し引いた(BRET=BRET比−バックグラウンド比)。緩衝液単独について達成された値を上回るBRETシグナルを、旨味受容体特異的として定義した(表2Bも参照。)。
【図2f】実施例2に記載のBRETアッセイにより計測されたHEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を示す。示されている試験溶液(または緩衝液)および基質を細胞に添加した。BRETシグナルは、ドナー(400nm)について測定されたシグナルにより割ったアクセプター波長(515nm)において取られた読取り値の比として測定した。アクセプター波長におけるドナー発光の重複に起因するバックグラウンドシグナルについて補正するため、BRET比を、ドナー単独(T1R1/T1R3−Rluc)を発現する細胞についてパラレルで測定した。このBRETバックグラウンド値を、両方のBRETパートナーを発現する細胞について得られたBRET値から差し引いた(BRET=BRET比−バックグラウンド比)。緩衝液単独について達成された値を上回るBRETシグナルを、旨味受容体特異的として定義した(表2Bも参照。)。
【発明を実施するための形態】
【実施例】
【0082】
[実施例1]
ヒト旨味受容体T1R1/T1R3を使用する原理の証明:
本発明者らは、BRET技術を使用してトマトおよび他の粗製植物または食物抽出物中の旨味を評価および定量する特有の機能的旨味受容体アッセイを開発し:原理を試験するため、本発明者らはBRET−2に関するドナーおよびアクセプタータンパク質を使用した。要するに、ヒトT1R1およびT1R3味覚受容体をクローニングし、ドナータンパク質としてのレニラルシフェラーゼ(Rluc)と融合させ;緑色蛍光タンパク質−2(GFP2)をアクセプタータンパク質として機能させ、β−アレスチン−2に融合させた。両方の構築物をタンパク質の異種発現のためにHEK293細胞系細胞)内に形質移入した。種々の哺乳動物細胞系が、受容体または他のタンパク質の発現に高度に好適であると報告されており、HEK293(ヒト胚腎臓)細胞およびCHO(チャイニーズハムスター卵巣)細胞は最も多用途で好適な細胞の一部である12。同様にGPCRは、実験の性質、各受容体または利用可能な時間に応じて、複数の刊行物に記載の種々の手法、例えば一過性6,8,13−15または安定性(構成性または誘導性)6,9,16,17発現において発現させることができる。
【0083】
アゴニストまたは粗製抽出物を用いる受容体刺激後、β−アレスチン−2/GFP−2タンパク質は活性化された受容体と相互作用し、こうしてGFP−2をルシフェラーゼと近接させ、考えられるこれらの2種のタンパク質間のエネルギー転移をもたらし、BRETシグナルを生成する。酸素の存在下、ルシフェラーゼは基質DeepBlueCの、395−410nmにおいて計測することができるコエレンテルアミドへの転換を触媒する。GFP−2がルシフェラーゼに近接し、エネルギー転移が生じる場合、発光は510nmにシフトし;このことをBRETシグナルと称し、アクセプター(GFP−2)とドナー(レニラルシフェラーゼ)との比として表現する。
【0084】
以下の実験は、慣用のカルシウムベースの機能的受容体アッセイが粗製抽出物の効果の計測に好適でなく;対照的に、BRETアッセイ(本明細書においては旨味について示される。)が粗製天然抽出物を使用して特異的受容体応答を明確に計測することができることを説明するために設計する。さらに、本アッセイは、異なるトマト試料間の受容体活性化の差の検出についても十分に感受性である。
【0085】
材料および方法:
化学薬品および培地:
Fluo−4 AMはMolecular Probes製(#F−14202、DMSO中5mM原液として調製)、DMEM(4.5g/lグルコースおよびウルトラグルタミンを有する、#BE12−604F)はLonza製、トリプシン−EDTA、Lipofectamine 2000、OptiMemおよびFCSは、Life Technologies Invitrogenから入手した。グルタミン酸ナトリウム(MSG)、イノシン5’−一リン酸および全ての他の化学薬品はSigma−Aldrichから入手した。
【0086】
GFP2−β−アレスチン−2をコードするプラスミドは、BioSignal Packard(#6310176)から購入した。
【0087】
コエレンテラジン400A(DeepBlueC誘導体)およびコエレンテラジン−Hは、VWR International(それぞれ#BTIU10125−1および233903−50)製であった。
【0088】
全ての他の細胞培養供給物はGreiner BioOne製であった。
【0089】
受容体融合構築物、細胞系および培地:
標準的分子クローニング技術およびPerkinElmer(#6310220)製コドンヒト化pRluc−N3ベクターを使用してヒトT1R1およびヒトT1R3旨味受容体を、これらのC末端においてインフレームでレニラルシフェラーゼ(Rluc)に融合させた。旨味受容体およびGFP−2/β−アレスチン−2をコードする構築物の発現のため、慣習的な形質移入法(以下も参照。)を使用してHEK293細胞(ヒト胚腎臓細胞、ATCC)を使用した。
【0090】
HEK293細胞をDMEMおよび10%FBS中で37℃/5%CO2において維持した。
【0091】
トマト抽出物:
代表的な天然複合試験材料として機能するトマト抽出物を以下のとおり調製した:冷凍トマト試料(−80℃)を秤量し、等量の水中に溶解させた。種子および小室組織(軟塊)を除去し取り分けた後、乳鉢および乳棒を使用してトマト果皮(果肉)を粉砕した。次いで、種子および軟塊を混合物に添加して戻し、これらが十分に混合されることを確保し、種子は破砕しなかった。混合物を4000rpmにおいて15分間遠心分離し、上澄み(漿液)を除去し、10mlアリコート中で凍結乾燥させた。計測前、等容量の水を添加して試料を溶解させた。
【0092】
T1R1/T1R3およびRluc融合構築物の生成:
上記のとおり、標準的分子クローニング技術を使用してヒトT1R1およびヒトT1R3旨味受容体のC末端Rluc受容体融合構築物を生成した。このことは、2種の異なる構築物:T1R1−RlucおよびT1R3−Rlucをもたらした。これらは、一緒に形質移入することができ、2種のRluc部分を含有する機能的受容体ヘテロ二量体をもたらし;代替的にいずれかの構築物を野性型受容体との組合せにおいて形質移入することができ、こうしてT1R3と組み合わせたT1R1−RlucまたはT1R3−Rlucと組み合わせたT1R1をもたらす。
【0093】
野性型受容体ならびに融合構築物およびレニラルシフェラーゼについての配列(cDNAおよびタンパク質)を配列表に挙げる(配列番号5−8)。
【0094】
形質移入:
HEK293細胞に、Lipofectamine2000を製造業者のプロトコルに従って使用してT1R1/T1R3受容体(Rluc融合タンパク質を含有)およびGFP−2−β−アレスチン−2をコードするプラスミドを一過的に形質移入する。要するに、HEK293細胞をウェル(12ウェルプレート、1ml培地/ウェル)当たり2×105個の細胞の密度において播種し、翌日に約80−90%のコンフルエンシーを目的とする。24時間後、ウェル当たり15gのトータルDNAを使用して旨味受容体構築物(T1R1−Rluc、T1R3−Rlucまたは野性型受容体)を、GFP−2−β−アレスチン−2をコードするプラスミドと同時形質移入する。DNAを100μlのOptiMem中に溶解させ、これを、4μlのlipofectamine2000を含有する100μlのOptiMemと合わせる。次いで、混合物を室温において30分間インキュベートし、各ウェルに添加し、細胞を48時間増殖させる。BRET計測は、形質移入48−52時間後に実施する。代替的に、上記と同一のプロトコルを使用して旨味受容体をコードするプラスミドを、GFP−2−β−アレスチン−2を安定的に発現するHEK293細胞内に形質移入することができる。
【0095】
カルシウムベースの受容体アッセイについては、Gα15を安定的に発現するHEK293細胞にT1R1およびT1R3受容体を上記の手順に従って形質移入するが、96ウェルプレートフォーマットについてのプロトコルをスケールダウンする(ポリリジン被覆μClear96ウェルプレートを使用して10倍低減)。
【0096】
カルシウムベースの受容体アッセイ:
HEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を、細胞内カルシウムの放出を追跡することにより計測した。増殖培地を除去し、2.5μMのカルシウムの蛍光マーカーFluo−4−AMを含有し、細胞からのFluo−4の漏れを防止するために0.5mMプロベニシドおよび0.5%FCSを補給した50μlのTyrode緩衝液(140mM NaCl、5mM KCl、10mMグルコース、1mM CaCl2.2H2O、1mM MgCl2.6H2O、10mMピルビン酸Naおよび50mM HEPES pH7.4)により細胞を1時間負荷し、次いで37℃において1時間インキュベートした。混合物を取り出し、0.5mMプロベニシドを含有する150μlのTyrode緩衝液を添加した。Flexstation II 384(Molecular Devices)を使用して細胞内カルシウムレベルを追跡した。蛍光計測は、485nmの励起波長および520nmの発光波長において37℃において実施した。50μlの化合物またはトマト抽出物を、20秒のベースライン計測後に毎秒104μlのピペッター速度において細胞に添加した。旨味受容体の活性は、試験溶液添加後の最大蛍光を試験溶液添加前に計測されたベースライン蛍光から差し引くことにより計算される蛍光変化(ΔF)として計測した。
【0097】
表2Aに示すデータは、純粋な旨味化合物の使用が旨味受容体特異的応答(受容体を発現しない対照細胞は増加を付与しない。)をもたらし;対照的に、種々のトマト抽出物の使用が対照細胞において高い非特異的応答をもたらし、旨味を示すシグナルを同定することができないことを極めて明確に示す。
【0098】
BRETアッセイ:
BRETアッセイには、Mithras LB 940プレートリーダー(Berthold Technologies)を使用する。使用するプロトコルは、本質的には、Packard BioOneにより記載されているものと同一であり、またはわずかな改変を有する刊行されたプロトコル13であり:HEK293細胞を上記のとおり形質移入する。形質移入48時間後、細胞を回収し、BRET緩衝液(2μg/mlアプロチニンを含有するD−PBS)中に2×106個の細胞/mlの密度において溶解させた。細胞を平衡化のために室温において1時間放置した後、1×105個の細胞を含有する30μlを白色96ウェルプレートの各ウェルに移した。10μlの試験化合物(または緩衝液)および10μlの基質コエレンテラジン(最終濃度5μM)を、インジェクターを使用して細胞に同時に添加する。最終インジェクション直後、連続読取りを410nmおよび515nmにおいて行う。BRETシグナルは、GFP−2発光(515nm)およびRluc/コエレンテラジン発光(410nm)の比として測定する。
【0099】
[実施例2]
複合試料中の味覚モジュレーターの検出
試験試料(抽出物)の調製および選択
下記の抽出物または粗製混合物は、これらの高い非特異的バックグラウンドシグナルに起因してカルシウムベースの受容体アッセイを用いる試験に不適な天然試料についての例である。これらの抽出物または粗製混合物は、異なる起源からのものであり、広範な潜在的試験試料を説明するために選択される。選択試料はグルタマート、リボチドまたはこれら両方を含有することも公知であり、従って旨味受容体T1R1/T1R3を例として使用するBRETアッセイにおいて試験するための良好な候補である。
トマト:トマト抽出物の調製は実施例1に既に記載した。
酵母抽出物:5’−リボチドが濃縮された市販の酵母抽出物(DSM,Maxarome Select酵母抽出物)。酵母抽出物を水中に溶解させた(10×酵母抽出物の重量)。
醤油:市販の醤油を使用した(Conimex ketjap manis)。
チキンブイヨン:市販のチキンブイヨンキューブ(Knorr)を製造業者により示されるとおりに温水中に溶解させた。粒子を試験試料から除去するため、ブイヨンを5000rpmにおいて10分間遠心分離し、上澄みを新たなチューブに移した。
チーズ抽出物:市販のロクフォールチーズを秤量し、等量の水中に溶解させ、乳鉢および乳棒を使用してホモジナイズした。混合物を5000rpmにおいて10分間遠心分離して粒子を除去し、上澄みを新たなチューブに移した。
【0100】
実験手順
形質移入:
HEK293細胞に、Lipofectamine2000を製造業者のプロトコルに従って使用してT1R1およびT1R3受容体(本実験においてT1R3受容体はRluc融合タンパク質を含有する。)ならびにGFP−2−β−アレスチン−2をコードするプラスミドを、一過的に形質移入した。要するに、HEK293細胞をウェル(12ウェルプレート、1ml培地/ウェル)当たり2×105個の細胞の密度において播種し、翌日に約80−90%のコンフルエンシーを目的とした。24時間後、ウェル当たり15μgのトータルDNAを使用して旨味受容体構築物(T1R1およびT1R3−Rluc受容体)を、GFP−2−β−アレスチン−2をコードするプラスミドと同時形質移入した。DNAを100μlのOptiMem中に溶解させ、これを、4μlのlipofectamine2000を含有する100μlのOptiMemと合わせた。次いで、混合物を室温において30分間インキュベートし、各ウェルに添加し、細胞を48時間増殖させた。BRET計測は、形質移入48−52時間後に実施した。
【0101】
代替的に、上記と同一のプロトコルを使用して旨味受容体をコードするプラスミドを、GFP−2−β−アレスチン−2を安定的に発現するHEK293細胞内に形質移入した。
【0102】
カルシウムベースの受容体アッセイについては、Gα15を安定的に発現するHEK293細胞にT1R1およびT1R3受容体を上記の手順に従って形質移入するが、96ウェルプレートフォーマットについてのプロトコルをスケールダウンする(ポリリジン被覆μClear96ウェルプレートを使用して10倍低減)。
【0103】
カルシウムベースの受容体アッセイ:
HEK293−Gα15細胞内で発現されたT1R1/T1R3旨味受容体の活性化を、細胞内カルシウムの放出を追跡することにより計測した。増殖培地を除去し、2.5μMのカルシウムの蛍光マーカーFluo−4−AMを含有し、細胞からのFluo−4の漏れを防止するために0.5mMプロベニシドおよび0.5%FCSを補給した50μlのTyrode緩衝液(140mM NaCl、5mM KCl、10mMグルコース、1mM CaCl2.2H2O、1mM MgCl2.6H2O、10mMピルビン酸Naおよび50mM HEPES pH7.4)により細胞を1時間負荷し、次いで37℃において1時間インキュベートした。混合物を取り出し、0.5mMプロベニシドを含有する150μlのTyrode緩衝液を添加した。Flexstation II 384(Molecular Devices)を使用して細胞内カルシウムレベルを追跡した。蛍光計測は、485nmの励起波長および520nmの発光波長において37℃において実施した。50μlの化合物または抽出物を20秒のベースライン計測後に毎秒104μlのピペッター速度において細胞に添加した。旨味受容体の活性は、試験溶液添加後の最大蛍光を試験溶液添加前に計測されたベースライン蛍光から差し引くことにより計算される蛍光変化(ΔF)として計測した。
【0104】
表2Bおよび図1は、純粋な旨味化合物(MSGおよびMSG+IMP)の使用が旨味受容体特異的応答をもたらし;対照的に、種々の抽出物および天然混合物の使用が旨味受容体を同様に発現しない対照細胞において高い非特異的応答をもたらし、このことにより旨味を示す受容体特異的シグナルを計算することができないことを説明する。
【0105】
BRETアッセイ:
BRETアッセイには、Mithras LB 940プレートリーダー(Berthold Technologies)を使用した。アッセイプロトコルは、本質的には、Packard BioOneにより記載されているものと同一であり、またはわずかな改変を有する他の刊行されたプロトコル10,13であり:HEK293細胞を上記のとおり形質移入した。形質移入40時間後、細胞を回収し、BRET緩衝液(1g/l D−グルコースおよび2μg/mlアプロチニンを含有するD−PBS)中に2×106個の細胞/mlの密度において溶解させる。細胞を平衡化のために室温において1時間放置した後、約1×105個の細胞を含有する30μlを白色96ウェルプレートの各ウェルに形質移入する。10μlの試験化合物(または緩衝液)および10μlの基質コエレンテラジン400(DeepBlueC、最終濃度5μM)を、インジェクターを使用して細胞に同時に添加した。最終インジェクション直後、連続読取りを400nmおよび515nmにおいて行った。代替的に、化合物を細胞に手作業で添加し、基質のインジェクション前に5分間インキュベートした。BRETシグナルは、ドナー(Rluc/コエレンテラジン400;400nm)について測定されたシグナルにより割ったアクセプター波長(GFP−2;515nm)において取られた読取り値の比として測定した。アクセプター波長におけるドナー発光の重複に起因するバックグラウンドシグナルについて補正するため、BRET比を、ドナー単独(T1R1/T1R3−Rluc)を発現する細胞についてパラレルで測定した。このBRETバックグラウンド値を、両方のBRETパートナーを発現する細胞について得られたBRET値から差し引いた(BRET=BRET比−バックグラウンド比)。緩衝液単独について達成された値を上回るBRETシグナルを、旨味受容体特異的として定義した(表2B参照。)。
【0106】
結果:
表2Bおよび図1に示されるとおり、カルシウムベースの受容体アッセイを使用して代表的抽出物および他の複合混合物を使用して旨味受容体特異的シグナルを達成することができなかった。対照的に、図2は、記載のBRETアッセイを使用してこれらの複合試料についての旨味受容体活性化を特異的に計測することができたことを示す。
【0107】
受容体特異的シグナルの定義は、技術的理由についてわずかに異なる:カルシウムアッセイについては、旨味受容体およびGα15を発現する細胞をGα15のみを発現する細胞と比較し;これらのシグナル間の差を旨味受容体特異的シグナルとして定義する。BRETアッセイについては、ドナー(T1R1/T1R3−Rluc)のみを発現する細胞を使用してバックグラウンドを測定し;陽性値が特異的BRETシグナルを示す。
【0108】
【表1】
【0109】
【表2】
【0110】
【表3】
【0111】
【表4】
【表5】
【特許請求の範囲】
【請求項1】
味覚受容体をモジュレートする化合物を同定する方法であって:
a)(i)発光タンパク質に融合している味覚受容体および(ii)β−アレスチンに融合している蛍光タンパク質を発現する細胞を提供する工程;
b)細胞を潜在的モジュレーター化合物と接触させ、およびBRETシグナルを測定する工程;
c)工程b)において得られたBRETシグナルを、該潜在的モジュレーター化合物の不存在下で細胞から得られたBRETシグナルと比較する工程
を含み、
b)において得られたBRETシグナルと潜在的モジュレーター化合物の不存在下で得られたBRETシグナルとの差が、該潜在的モジュレーター化合物が味覚受容体をモジュレートする化合物であることを示す方法。
【請求項2】
BRETシグナルがBRET1またはBRET2技術において得られる、請求項1に記載の方法。
【請求項3】
味覚受容体が蛍光タンパク質に、およびβ−アレスチンが発光タンパク質に融合している、請求項1または2に記載の方法。
【請求項4】
潜在的モジュレーター化合物が水性試料溶液中に存在する、請求項1から3のいずれか一項に記載の方法。
【請求項5】
水性試料溶液が、(a)食品;(b)食品の抽出物:および(c)(可食性)バイオマスの抽出物の少なくとも1種を含む、請求項4に記載の方法。
【請求項6】
バイオマスの抽出物が、植物、動物および微生物の少なくとも1種からの抽出物(分子を含む。)である、請求項5に記載の方法。
【請求項7】
抽出物がトマト抽出物である、請求項6に記載の方法。
【請求項8】
水性試料溶液が、少なくとも10種の区別される有機分子を含む、請求項3から7のいずれか一項に記載の方法。
【請求項9】
水性試料溶液が未特定組成物の溶液である、請求項3から8のいずれか一項に記載の方法。
【請求項10】
味覚受容体が、MSGまたは旨味受容体、甘味受容体、苦味受容体、脂肪受容体および栄養/脂肪酸感知腸管受容体からなる群から選択される、請求項1から9のいずれか一項に記載の方法。
【請求項11】
(a)味覚受容体がT1R1、T1R3ヘテロ二量体であり、サブユニットT1R1およびT1R3の少なくとも1種が発光タンパク質に融合しており、および蛍光タンパク質がβ−アレスチンに融合しており;または
(b)味覚受容体がT1R1、T1R3ヘテロ二量体であり、サブユニットT1R1およびT1R3の少なくとも1種が蛍光タンパク質に融合しており、および発光タンパク質がβ−アレスチンに融合している、
請求項12に記載の方法。
【請求項12】
(a)発光タンパク質の少なくとも1種がルシフェラーゼであり:
(b)蛍光タンパク質の少なくとも1種がGFPであり;および
(c)β−アレスチンの少なくとも1種が非視覚β−アレスチンである、
請求項1から11のいずれか一項に記載の方法。
【請求項13】
(a)ルシフェラーゼの少なくとも1種がレニラルシフェラーゼであり;および
(b)非視覚β−アレスチンの少なくとも1種がβ−アレスチン2である、
請求項12に記載の方法。
【請求項14】
味覚受容体をモジュレートする化合物を生成する方法であって、請求項1から13のいずれか一項に記載の方法により味覚受容体をモジュレートする化合物を同定する工程および該化合物を回収する工程を含む方法。
【請求項15】
味覚受容体をモジュレートする化合物の同定のための、BRET技術の使用。
【請求項1】
味覚受容体をモジュレートする化合物を同定する方法であって:
a)(i)発光タンパク質に融合している味覚受容体および(ii)β−アレスチンに融合している蛍光タンパク質を発現する細胞を提供する工程;
b)細胞を潜在的モジュレーター化合物と接触させ、およびBRETシグナルを測定する工程;
c)工程b)において得られたBRETシグナルを、該潜在的モジュレーター化合物の不存在下で細胞から得られたBRETシグナルと比較する工程
を含み、
b)において得られたBRETシグナルと潜在的モジュレーター化合物の不存在下で得られたBRETシグナルとの差が、該潜在的モジュレーター化合物が味覚受容体をモジュレートする化合物であることを示す方法。
【請求項2】
BRETシグナルがBRET1またはBRET2技術において得られる、請求項1に記載の方法。
【請求項3】
味覚受容体が蛍光タンパク質に、およびβ−アレスチンが発光タンパク質に融合している、請求項1または2に記載の方法。
【請求項4】
潜在的モジュレーター化合物が水性試料溶液中に存在する、請求項1から3のいずれか一項に記載の方法。
【請求項5】
水性試料溶液が、(a)食品;(b)食品の抽出物:および(c)(可食性)バイオマスの抽出物の少なくとも1種を含む、請求項4に記載の方法。
【請求項6】
バイオマスの抽出物が、植物、動物および微生物の少なくとも1種からの抽出物(分子を含む。)である、請求項5に記載の方法。
【請求項7】
抽出物がトマト抽出物である、請求項6に記載の方法。
【請求項8】
水性試料溶液が、少なくとも10種の区別される有機分子を含む、請求項3から7のいずれか一項に記載の方法。
【請求項9】
水性試料溶液が未特定組成物の溶液である、請求項3から8のいずれか一項に記載の方法。
【請求項10】
味覚受容体が、MSGまたは旨味受容体、甘味受容体、苦味受容体、脂肪受容体および栄養/脂肪酸感知腸管受容体からなる群から選択される、請求項1から9のいずれか一項に記載の方法。
【請求項11】
(a)味覚受容体がT1R1、T1R3ヘテロ二量体であり、サブユニットT1R1およびT1R3の少なくとも1種が発光タンパク質に融合しており、および蛍光タンパク質がβ−アレスチンに融合しており;または
(b)味覚受容体がT1R1、T1R3ヘテロ二量体であり、サブユニットT1R1およびT1R3の少なくとも1種が蛍光タンパク質に融合しており、および発光タンパク質がβ−アレスチンに融合している、
請求項12に記載の方法。
【請求項12】
(a)発光タンパク質の少なくとも1種がルシフェラーゼであり:
(b)蛍光タンパク質の少なくとも1種がGFPであり;および
(c)β−アレスチンの少なくとも1種が非視覚β−アレスチンである、
請求項1から11のいずれか一項に記載の方法。
【請求項13】
(a)ルシフェラーゼの少なくとも1種がレニラルシフェラーゼであり;および
(b)非視覚β−アレスチンの少なくとも1種がβ−アレスチン2である、
請求項12に記載の方法。
【請求項14】
味覚受容体をモジュレートする化合物を生成する方法であって、請求項1から13のいずれか一項に記載の方法により味覚受容体をモジュレートする化合物を同定する工程および該化合物を回収する工程を含む方法。
【請求項15】
味覚受容体をモジュレートする化合物の同定のための、BRET技術の使用。
【図1】

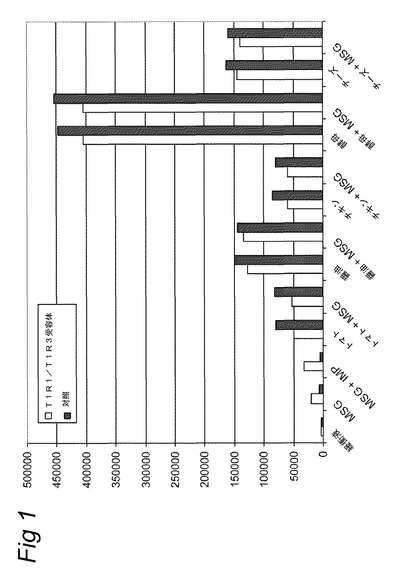
【図2a】
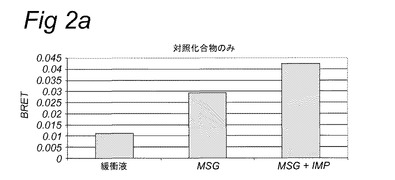
【図2b】
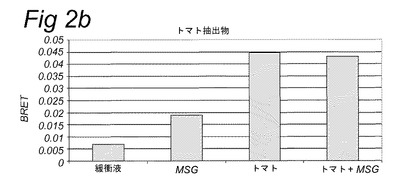
【図2c】
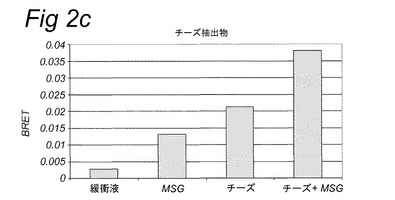
【図2d】
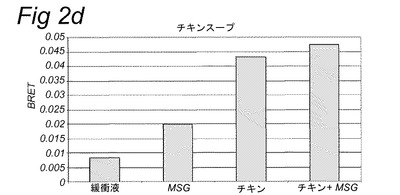
【図2e】
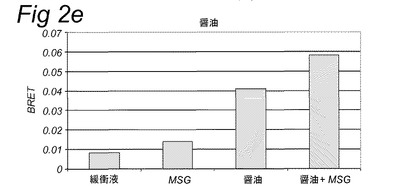
【図2f】
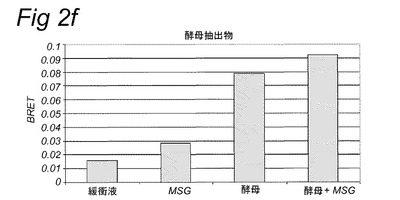
【図2a】
【図2b】
【図2c】
【図2d】
【図2e】
【図2f】
【公表番号】特表2013−512664(P2013−512664A)
【公表日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願番号】特願2012−541437(P2012−541437)
【出願日】平成22年11月29日(2010.11.29)
【国際出願番号】PCT/EP2010/068399
【国際公開番号】WO2011/067202
【国際公開日】平成23年6月9日(2011.6.9)
【出願人】(590003065)ユニリーバー・ナームローゼ・ベンノートシヤープ (494)
【Fターム(参考)】
【公表日】平成25年4月18日(2013.4.18)
【国際特許分類】
【出願日】平成22年11月29日(2010.11.29)
【国際出願番号】PCT/EP2010/068399
【国際公開番号】WO2011/067202
【国際公開日】平成23年6月9日(2011.6.9)
【出願人】(590003065)ユニリーバー・ナームローゼ・ベンノートシヤープ (494)
【Fターム(参考)】
[ Back to top ]