
呼吸性ウイルス感染を処置するための組成物およびその使用
本発明は、RSウイルスおよびトリインフルエンザA(H5N1株を含む)を含む、呼吸性ウイルス感染におけるウイルス複製に干渉するsiRNA組成物を、提供する。本発明は、呼吸性ウイルス感染細胞におけるウイルス遺伝子の発現を阻害するためのsiRNA組成物の使用、および被験体における呼吸性ウイルス感染の処置における使用のためのsiRNA組成物の使用を、さらに提供する。一般に本発明は、RSウイルスもしくはインフルエンザAウイルスのゲノムを標的化する15〜30塩基の第1のヌクレオチド配列、その相補体、二本鎖ポリヌクレオチドもしくはヘアピンポリヌクレオチドを含むポリヌクレオチドを提供する。さらに、本発明は、siRNA配列を含むベクター、細胞および薬学的組成物を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明は、呼吸性ウイルス感染(特にRSウイルスおよびトリインフルエンザA(H5N1株を含む))におけるウイルス複製に干渉するsiRNA組成物に関連する。本発明は、呼吸性ウイルス感染細胞におけるウイルス遺伝子の発現を阻害するためのsiRNA組成物の使用、および被験体における呼吸性ウイルス感染の処置における使用のためのsiRNA組成物の使用に、さらに関連する。
【背景技術】
【0002】
(発明の背景)
呼吸性ウイルス感染は、何世紀にもわたって、ヒトの健康および生命に対する重大な脅威であった。悪名高いエピソードとしては、インフルエンザ株、RSウイルス、および重症急性呼吸性症候群(SARS)によって引き起こされる感染が挙げられる。これらは、世界中で約2〜4千万人の人々を殺傷した1918年の世界的なインフルエンザ汎流行を含む。より近年の年代に、他のインフルエンザ汎流行もまた、存在した。SARSは、2002年に発生し、およそ800人を犠牲にした(2)。
【0003】
(RSウイルス)
RSウイルス(RSV)感染は、深刻な汎流行性の気道疾患の主要な原因である。約3分の2の乳児が、人生の最初の1年間にRSVに感染し、そして約100%が2歳までに感染している。現在、RSV感染を処置するために利用可能である、特定かつ有効な治療薬は存在しない。
【0004】
RSウイルス(RSV)は、エンベロープを有する(enveloped)、非セグメント化一本鎖ネガティブRNAウイルス(NNR)であり、パラミクソウイルス科(Paramyxoviradae)モノネガウイルス(mononegaviruses)属に属する(14、29)。パラミクソウイルスは、以下の特徴を共有する。1)これらは、ウイルスヌクレオカプシドタンパク質(N)によってしっかりと被包されている一本鎖RNAゲノムを有する(29、30)。2)サブゲノムmRNAは、RdRPによって正常なゲノムから転写される。3)ウイルス複製は、宿主細胞の細胞質において起こる。感染から子孫ビリオンの放出までのRSV生活環の詳細は、よく研究されている(15)。
【0005】
RSVゲノムは、3’−NS1、NS2、N、P、M、SH、G、F、M2、L−5’の遺伝子を含む15.5kb長の、ネガティブ鎖である(図1を参照).これらの7つの遺伝子の産物(すなわち、N、P、M、SH、G、F、およびL)は、他のパラミクソウイルス3と共通している。いくつかのウイルス因子もしくは宿主因子は、RNA転写および複製の調節に関与する(20)。さらに、mRNAの転写およびRNA複製において調節的な役割を果たすウイルスのシス作動性シグナルが存在する(3)。
【0006】
RSV感染の潜伏期は、約4〜5日間である。RSVは、まず鼻咽頭に影響を及ぼし、次いで、数日のうちに、気管支および細気管支に達し、感染は呼吸器上皮の表面層に限られる。
【0007】
(インフルエンザA)
1997年の初めに、トリインフルエンザAの新型株であるH5N1が出現した。ほぼトリ(野生集団および家禽の両方)に限定されていたが、このウイルスは、感染したトリに直接的に接触したヒトにのみ感染するようであった。ヒトにおいて、感染は深刻な疾患を引き起こし、ヒトにおいて重症の呼吸性疾病および死をもたらす(3〜12)。多くの症例および発生が、東南アジアの種々の国において起こった。このトリウイルスのヒトに感染する能力を考慮すると、接触感染性ヒト改変体への突然変異の増大した危険性が存在し、この突然変異は、効率的かつ継続的なヒトからヒトへの伝播による新型インフルエンザ汎流行の出現ならびに大勢の死の危険をもたらす。
【0008】
トリインフルエンザH5N1は、肺炎に関連する新しく出現した感染性因子であるので、その病理学およびメカニズムがあまり明らかになっておらず、ヒト疾患症例におけるH5N1トリインフルエンザのための特定かつ有効な処置は存在しない。現在、インフルエンザ感染は、2種の(ノイラミニダーゼインヒビターのクラスの)薬物のような抗ウイルス薬:オセルタミビル(市場ではタミフルとして知られる)およびザナミビル(市場ではリレンザとして知られる)、またはより旧式なM2インヒビターであるアマンタジンおよびリマンタジンによって処置される。
【0009】
H5N1は、インフルエンザウイルスA型のサブタイプである。このウイルスは、エンベロープを有し、フラグメント化された、ネガティブ一本鎖RNAウイルスであるので、オルトミクソウイルス科に属する。インフルエンザAウイルス(H5N1を含む)の生活環の間、ウイルスゲノムRNA(vRNA)は、相補的RNA(cRNA)産生のためのテンプレートとして働き、このcRNAはまた、伝令RNA(mRNA)産生のためのテンプレートとして働く。ウイルス複製の間に生じるRNA分子のこれらの3つの形態の各々は、全て、センスsiRNAもしくはアンチセンスsiRNAのいずれかを用いて、siRNA媒介型分解のために標的化され得る。インフルエンザAゲノム(8個の分離したRNAセグメントからなり、少なくとも10個のオープンリーディングフレーム(ORF)を含む)は、ウイルスゲノム複製およびサブゲノムのmRNA合成もしくは遺伝子指向型mRNA合成の両方のためのテンプレートとして働く。図2は、インフルエンザAビリオンの構造を表す概略図を示す。ポリメラーゼPB2、PB1(ポリメラーゼ塩基性タンパク質1およびポリメラーゼ塩基性タンパク質2)ならびにPA(ポリメラーゼ酸性タンパク質)は、それぞれRNA1、RNA2およびRNA3によってコードされていた。4種のウイルス構造タンパク質である、H(血球凝集素)、N(ノイラミニダーゼ)、M1およびM2(マトリックスタンパク質1およびマトリックスタンパク質2)は、それぞれ、RNAセグメント4、RNAセグメント6およびRNAセグメント7によってコードされ、一方、RNA5は、NP(ヌクレオカプシドタンパク質)をコードし、そしてRNA8は、NS1およびNS2(非構造タンパク質1および非構造タンパク質2)をコードする。
【0010】
(RNA干渉)
RNA干渉(RNAi)は、配列特異的RNA分解プロセスであり、理論的に任意の遺伝子を、ノックダウンするかまたはサイレンシングするための、比較的容易かつ直接的な方法を提供する(17、18、19)。天然に存在するRNA干渉において、二本鎖RNAは、RNase III/ヘリカーゼタンパク質であるDicerによって、低分子干渉(small interfering)RNA(siRNA)分子へと切断される。このsiRNA分子は、2ヌクレオチド(nt)オーバーハング(overhang)を3’末端において有する、19〜23ヌクレオチドのdsRNAである。これらのsiRNAは、RNA誘導型サイレンシング複合体(RNA−induced−silencing−complex:RISC)と呼ばれる多成分リボヌクレアーゼ内に組み込まれる。siRNAの一本の鎖は、RISCに結合したままであり、この複合体を同族の(cognate)RNAへと導き、この同族のRNAは、RISC中のこのガイド因子(guider)ss−siRNAに対して相補的である配列を有する。このsiRNA指向型エンドヌクレアーゼは、このRNAを消化し、それによってこのRNAを不活性化する。近年の研究は、化学的に合成された21〜25−nt siRNAの使用は、哺乳動物細胞においてRNAiの作用を示すことを明らかにし(20)、そしてsiRNAハイブリダイゼーションの(末端におけるかもしくは中央部における)熱力学的安定性は、分子の機能を決定する際に中枢的な役割を果たすことを明らかにしている(21、22)。これらならびにRISC、siRNA分子およびRNAiの他の特徴は、記載されている(23〜28)。
【0011】
哺乳動物細胞におけるRNAiの、研究室における用途、または潜在的に、治療用途は、化学的に合成されたsiRNAもしくは内因的に発現された分子のいずれかを使用する(非特許文献1(2)、非特許文献2(21))。内因性siRNAは、最初に低分子ヘアピンRNA(shRNA)として、発現ベクター(プラスミドもしくはウイルスベクター)によって発現され、次いで、DicerによってsiRNAへとプロセシングされる。siRNAは、特にウイルス感染によって引き起こされるヒト疾患のための治療薬であるという非常な有望性を有すると考えられる(非特許文献3(19)、非特許文献4(20)、27〜30)。
【0012】
重要なことには、現在、ウイルスゲノム配列(例えば、約16〜30塩基対のオリゴヌクレオチド)を標的化する可能性のある多くの潜在的候補siRNA配列のうちのどれが、実際に有効なsiRNA活性を示すかは、いかなる程度の確信をもって予測することも不可能である。その代わり、個々の特異的候補siRNAポリヌクレオチド配列もしくは特異的候補siRNAオリゴヌクレオチド配列は、標的化された遺伝子の発現の意図される干渉が起こるか否かを決定するために、作製されそして試験されなければならない。したがって、所定のmRNAの発現の特異的な改変を可能にするsiRNAポリヌクレオチドを、確信を持って設計するための慣用的方法は、当該分野に存在しない。
【非特許文献1】Abbot,A.Nature(2003)424:121〜123
【非特許文献2】Reynolds,A.ら、Nat.Biotechnology(2004)22:326〜330
【非特許文献3】Schiwarz,D.S.ら、Cell(2003)115:199〜208
【非特許文献4】Khvorova,A.ら、Cell(2003)115:209〜216
【発明の開示】
【発明が解決しようとする課題】
【0013】
ウイルス病原体遺伝子およびその同族タンパク質産物の発現を阻害する組成物および方法についての、非常な需要が存在し続けている。特に、ウイルス感染した細胞における病原性呼吸性ウイルス遺伝子の発現を阻害するための組成物および方法、ならびに被験体における呼吸性ウイルス感染を処置するための組成物および方法についての、差し迫った需要が存在する。さらに、RSVおよびトリインフルエンザA(特にH5N1株)による感染に取り組む組成物および方法についての需要が、さらに存在する。非常に有効で、かつ公知の抗ウイルス剤の使用もしくは公知の抗ウイルス剤の改変に依存しない処置のための組成物および方法についての需要が、さらに存在する。本発明は、これらおよびこれらと関連する需要に取り組む。
【課題を解決するための手段】
【0014】
(発明の要旨)
本発明は、ウイルス病原体(例えば、インフルエンザA H5N1およびRSVを含む、呼吸性ウイルス感染を引き起こすウイルス病原体)のウイルスRNA分子の破壊を通して、ウイルス感染および複製を阻害するための、RNA干渉の使用に関連する組成物および方法を提供する。これらのウイルスは、ヒトおよび他の哺乳動物において重症の呼吸性疾患を引き起こす病原体である。ウイルス複製の阻害は、このウイルスに感染した培養細胞ならびにこのウイルスに感染した被験体において、ウイルス感染と戦う。このウイルス複製の阻害は、その症状の軽減を含む。
【0015】
第1の局面において、本発明は、長さが200以下かつ15以上の任意の数のヌクレオチドであり得る単離されたポリヌクレオチドを提供する。このポリヌクレオチドは、吸性合胞体ウイルスもしくはインフルエンザAウイルスのゲノムを標的化する第1のヌクレオチド配列を含む。このポリヌクレオチドにおいて、任意のT(チミジン)もしくは任意のU(ウリジン)は、必要に応じて他と置換され得る。さらに、このポリヌクレオチドにおいて、第1のヌクレオチド配列は、(a)長さが15〜30の任意の数のヌクレオチドである配列、または(b)(a)において与えられた配列の相補体からなる。このようなポリヌクレオチドは、本明細書中で、直鎖状ポリヌクレオチドと呼ばれ得る。
【0016】
本発明の関連の局面において、上記のポリヌクレオチドは、ループ配列によって第1のヌクレオチド配列から分離される第2のヌクレオチド配列をさらに含み、この第2のヌクレオチド配列は、
(a)第1のヌクレオチド配列と実質的に同一の長さを有し、かつ
(b)第1のヌクレオチド配列に対し実質的に相補的である。この後者の構造(ヘアピンポリヌクレオチドと呼ばれる)において、上記第1のヌクレオチド配列は、上記第2のヌクレオチド配列とハイブリダイズして、その相補的配列が、上記ループ配列によって連結されている、ヘアピンを形成する。
【0017】
上記直鎖状ポリヌクレオチドの多くの実施形態およびこのヘアピンポリヌクレオチドの多くの実施形態において、上記第1のヌクレオチド配列は、以下のいずれかである:
(a)配列番号1〜263から選択される配列、
(b)項目(a)において与えられた配列より長い標的性(targeting)配列であって、この標的性配列は、呼吸性ウイルスのゲノムを標的化し、かつ配列番号1〜263から選択される配列を含む、
(c)配列番号1〜263から選択される配列のフラグメントであって、このフラグメントは、少なくとも15ヌクレオチドの長さでありかつこの選択される配列より長くとも1塩基短い、連続する塩基の配列からなる、フラグメント、
(d)配列番号1〜263から選択される配列と5ヌクレオチドまで異なる配列、または
(e)(a)〜(d)において与えられた任意の配列の相補体。
【0018】
直鎖状ポリヌクレオチドもしくはヘアピンポリヌクレオチドの種々の実施形態において、上記第1のヌクレオチド配列の長さは、21〜25の任意の数のヌクレオチドである。多くの実施形態において、直鎖状ポリヌクレオチドもしくはヘアピンポリヌクレオチドは、配列番号1〜263から選択される配列からなり、そして必要に応じて、この選択される配列の3’に結合するジヌクレオチドオーバーハングを含む。直鎖状ポリヌクレオチドもしくはヘアピンポリヌクレオチドのなおさらなる実施形態において、この第1のヌクレオチド配列の3’末端におけるジヌクレオチド配列は、TT、TU、UT、もしくはUUであり、そしてリボヌクレオチドもしくはデオキシリボヌクレオチドのいずれか、または両方を含む。種々のさらなる実施形態において、直鎖状ポリヌクレオチドもしくはヘアピンポリヌクレオチドは、DNAであってもよく、またはこれはRNAであってもよく、あるいはこれはデオキシリボヌクレオチドおよびリボヌクレオチドの両方から構成されてもよい。
【0019】
さらなる局面において、本発明は、二本鎖ポリヌクレオチドを提供する。この二本鎖ポリヌクレオチドは、請求項1に記載の第1のポリヌクレオチド鎖および少なくともこの第1の鎖のこの第1のヌクレオチド配列に対して相補的でありかつこれにハイブリダイズして二本鎖組成物を形成する第2のポリヌクレオチド鎖を含む。これらのポリヌクレオチド構造もまた、直鎖状ポリヌクレオチドと呼ばれ得る。
【0020】
なおさらなる局面において、本発明は、請求項1、請求項2もしくは両方に記載の2つもしくはそれより多くの標的性ポリヌクレオチドを含む組み合わせであって、この組み合わせの各ポリヌクレオチドは、標的ウイルスのゲノムにおける異なる配列を標的化する組み合わせを提供する。
【0021】
呼吸性ウイルス病原体標的に対する高い程度の類似性および同一性に起因して、そして理論に縛られることを望まないが、ウイルスに感染した細胞内への導入の際に、このポリヌクレオチドは、RNA干渉を誘導し、病原体ゲノムRNA、相補的RNAおよび伝令RNAの消化をもたらす。特に、本発明のこれらの局面の重要な実施形態において、これらのポリヌクレオチドにおける第1のヌクレオチド配列もしくはその相補体は、RNA誘導型サイレンシング複合体(RISC)を形成し、このRISCは、上記ポリヌクレオチドsiRNA配列を病原体ゲノムRNA配列に誘導し、それによって病原体ゲノムRNAの切断を促進すると考えられる。
【0022】
さらなる局面において、本発明は、本発明の直鎖状ポリヌクレオチドもしくはヘアピンポリヌクレオチドによって与えられる配列を有するベクターを提供する。種々の実施形態において、これらのベクターのいずれかは、プラスミド、組換えウイルス、トランスポゾン、もしくは微小染色体であり得る。なおさらなる局面は、本発明の1種もしくはそれより多くの直鎖状ポリヌクレオチド、または本発明の1種もしくはそれより多くのヘアピンポリヌクレオチドによってトランスフェクトされた細胞を提供する。
【0023】
なおさらなる局面において、本発明は、1種もしくはそれより多くの直鎖状ポリヌクレオチドもしくはヘアピンポリヌクレオチドまたはそれらの混合物、ならびに薬学的に受容可能なキャリアを含有する薬学的組成物を提供し、ここで、各ポリヌクレオチドは、標的ウイルスのゲノムにおける異なる配列を標的化する。
【0024】
なおさらなる局面において、本発明は、直鎖状ポリヌクレオチドを有する1種もしくはそれより多くのベクター、またはヘアピンポリヌクレオチドを有するベクター、あるいはそれらの混合物、ならびに薬学的に受容可能なキャリアを含有する薬学的組成物を提供し、ここで、各ベクターは、標的ウイルスのゲノムにおける異なる配列を標的化するポリヌクレオチドを有する。
【0025】
薬学的組成物の種々の実施形態において、このキャリアとしては、合成カチオン性ポリマー、リポソーム、デキストロース、界面活性剤、またはこれらの任意の2種もしくはそれより多くの組み合わせが挙げられる。
【0026】
なおさらなる局面において、本発明は、RSウイルスもしくはインフルエンザAウイルスのゲノムを標的化する配列を有する直鎖状ポリヌクレオチドもしくはヘアピンポリヌクレオチドを合成する方法を提供する。この方法は、以下の工程を包含する:
(a)反応性末端(live reactive end)を含みかつこの配列の第1の末端におけるヌクレオチドに対応するヌクレオチド試薬を提供する工程;
(b)反応性末端を含みかつ該配列の後に続く位置に対応する、さらなるヌクレオチド試薬を添加して、上記工程(a)の反応性末端と反応させ、そして伸長中のポリヌクレオチド配列の長さを1ヌクレオチド増加させる工程、ならびに所望されない産物および過剰な試薬を除去する工程、ならびに
(c)上記配列の第2の末端におけるヌクレオチドに対応する上記ヌクレオチド試薬が添加され、それによって完全なポリヌクレオチドが提供されるまで、工程(b)を繰り返す工程。
【0027】
なおさらなる局面において、本発明は、RNAインヒビターによって細胞をトランスフェクトするための方法を提供し、ここで、この方法は、この細胞を、1種もしくはそれより多くの直鎖状ポリヌクレオチド、または1種もしくはそれより多くのヘアピンポリヌクレオチドを含有する組成物と接触させる工程を包含する。多くの実施形態において、このようにトランスフェクトされる細胞としては、1種もしくはそれより多くのポリヌクレオチドによって標的化される呼吸性ウイルスが挙げられる。
【0028】
なおさらなる局面において、本発明は、呼吸性ウイルスに感染した細胞におけるこのウイルスの複製を阻害する方法を提供し、この方法は、この細胞を、1種もしくはそれより多くの直鎖状ポリヌクレオチド、または1種もしくはそれより多くのヘアピンポリヌクレオチドを含有する組成物と接触させる工程を包含し、ここで、この1種もしくはそれより多くのポリヌクレオチドは、このウイルスを標的化する。
【0029】
なおさらなる局面において、本発明は、被験体における呼吸性ウイルスに起因する感染を処置するために有効な薬学的組成物の製造における、この呼吸性ウイルスを標的化する直鎖状ポリヌクレオチドもしくはこれらの2種もしくはそれより多くの混合物の使用、またはこの呼吸性ウイルスを標的化するヘアピンポリヌクレオチドもしくはこれらの2種もしくはそれより多くの混合物の使用を提供する。この使用の種々の実施形態において、この被験体は、ヒトである。
【0030】
なおさらなる局面において、本発明は、被験体における呼吸性ウイルスに起因する感染を処置する方法を提供する。この方法は、呼吸性ウイルスを標的化する直鎖状ポリヌクレオチドもしくはこれらの2種もしくはそれより多くの混合物の有効用量、または呼吸性ウイルスを標的化するヘアピンポリヌクレオチドもしくはこれらの2種もしくはそれより多くの混合物の有効用量を、この被験体に投与する工程を包含する。この方法の種々の実施形態において、この被験体は、ヒトである。さらなる実施形態において、直鎖状ポリヌクレオチドおよびヘアピンポリヌクレオチドの両方が、この被験体に投与される。
【発明を実施するための最良の形態】
【0031】
(発明の詳細な説明)
本明細書中で記載された全ての特許、特許出願公開、および特許公報は、その全体が、本明細書中で逐語的に示されたのと同様に参考として援用される。本明細書中で記載された全ての技術的刊行物もまた、参考として援用される。
【0032】
本明細書中で、冠詞「a」、「an」および「the」は、単数形もしくは複数形として、同等の意味を表す。これらの冠詞についての特定の意味は、これらが使用される文脈から明らかである。
【0033】
本明細書中で使用される場合、用語「標的」配列ならびに類似の用語および語句は、本発明のポリヌクレオチドが指向される病原体のゲノムにおいて存在するヌクレオチド配列をいう。ポリヌクレオチドは、病原体配列を、(a)この病原体のゲノム内に含まれる特定の下位配列(標的配列と呼ばれる)に対し相同であるかもしくは同一である配列を含むこと、または(b)相補体が標的配列と相同であるかもしくは同一である配列を含むことのいずれかによって、標的化する。病原体配列を標的化する任意のポリヌクレオチドは、RNA干渉現象にしたがって標的配列とハイブリダイズする能力を有し、それによってRNA干渉を開始する。
【0034】
本明細書中で使用される場合、用語「相補体」、「相補性」、ならびに類似の用語および語句は、塩基が、塩基対塩基で、相補的塩基対を形成する2つの配列に関連し、生物化学、分子生物学、遺伝学および本発明の分野に関する類似の分野のような分野における当業者によって従来理解される通りである。
【0035】
本明細書中で使用される場合、第1の配列もしくは下位配列が、第2の配列もしくは下位配列と、この配列もしくは下位配列の全ての位置において同じ塩基を有する場合、第1の配列もしくは下位配列は、第2の配列もしくは下位配列と「同一である」か、「100%の同一性」を有するか、または100%同一性の概念を伝達する用語もしくは語句によって記載される。同一性の決定において、T(チミジン)もしくはその任意の誘導体またはU(ウリジン)もしくはその任意の誘導体を含む任意の特定の塩基位置は、互いに同等であり、したがって同一であるとみなされる。
【0036】
本明細書中で記載される場合、標的性ポリヌクレオチドの配列、またはその相補体は、標的配列に対して完全に同一であってもよく、またはこれは、標的配列における特定の位置において不一致(mismatched)な塩基を含んでもよい。ミスマッチ(mismatch)の組み込みは、本明細書中で完全に記載される。理論に縛られることを望まないが、ミスマッチの組み込みは、目的の特定の標的配列のためのRNA干渉現象を最適化する生理学的条件下での、意図される程度のハイブリダイゼーションの安定性を提供する。同一性の程度は、2つの配列における塩基が互いに同一である位置の百分率を決定する。「配列同一性の百分率」は、最適に並べられた2つの配列を、比較の領域にわたって比較し、両配列において同一な核酸塩基(例えば、核酸の場合、A、TもしくはU、C、G、またはI)が存在する位置の数を決定して一致した位置の数を得、この一致した位置の数を比較の領域内の位置の総数(すなわち、ウインドウサイズ)で割り、そしてその結果に100を掛けて、配列同一性の百分率を得ることによって、計算される。互いに100%未満同一である配列は、互いに「類似する」かもしくは「相同である」;相同性の程度もしくは同一性百分率は、以下の節において決定されるような2つの配列もしくは下位配列の間の同一性の百分率を意味する、類義語である。例えば、互いに、少なくとも60%の同一性、もしくは好ましくは少なくとも65%の同一性、もしくは好ましくは少なくとも70%の同一性、もしくは好ましくは少なくとも75%の同一性、もしくは好ましくは少なくとも80%の同一性、もしくはより好ましくは少なくとも85%の同一性、もしくはより好ましくは少なくとも90%の同一性、もしくはなおより好ましくは少なくとも95%の同一性を示す、2つの配列は、互いに「類似である」かもしくは「相同である」。あるいは、siRNA分子のオリゴヌクレオチド配列に関して、5もしくはそれ未満の塩基、または4もしくはそれ未満の塩基、または3もしくはそれ未満の塩基、または2もしくはそれ未満の塩基、または1塩基が異なる2つの配列は、互いに「類似である」かもしくは「相同である」。
【0037】
「同一性」は、当該分野で公知であるように、2つもしくはそれより多くのポリペプチド配列もしくは2つもしくはそれより多くのポリヌクレオチド配列の間の関連であり、配列を比較することによって決定される。当該分野では、「同一性」はまた、場合によって、ポリペプチド配列もしくはポリヌクレオチド配列の間の配列の、このような配列の鎖(string)の間のマッチ(match)によって決定される関連性の程度をも意味する。「同一性」および「類似性」は、公知の方法によって容易に計算され得る。これらの方法としては、以下に記載される方法が挙げられるが、これらに限定されない:Computational Molecular Biology,Lesk.A.M.,編,Oxford University Press,New York,1988;Biocomputing:Informatics and Genome Projects,Smith,D.W.,編,Academic Press,New York,1993;Computer Analysis of Sequence Data,Part I.Griffin,A.M.,およびGriffin,H.G.,編Humana Press,New Jersey,1994;Sequence Analysis in Molecular Biology,von Heinje,G.,Academic Press,1987;ならびにSequence Analysis Primer,Gribskov,M.およびDevereux,J.,編,M Stockton Press.New York,1991;ならびにCarillo,H.,およびLipman,D.,SIAM J.Applied Math.(1988)48:1073。同一性を決定する好ましい方法は、試験される配列の間の最大のマッチを与えるように設計される。同一性および類似性を決定する方法は、公に利用可能なコンピュータープログラムにおいてまとめられている。2つの配列間の同一性および類似性を決定するための好ましいコンピュータープログラム方法としては、以下が挙げられるが、これらに限定されない:GCGプログラムパッケージ(Devercux,J.,ら(1984)Nucleic Acids Research 12(1):387)、BLASTP、BLASTN、およびFASTA(Atschul,S.F.ら(1990)J.Molec.Biol.215:403−410)。BLAST Xプログラムは、NCBIおよび他の供給源から、公に利用可能である(BLAST Manual,Altschul,S.,ら,NCBI NLM NIH Bethesda,Md.20894;Altschul,S.,ら(1990)J.Mol.Biol.215:403−410。周知のSmith Watermanアルゴリズムもまた、同一性を決定するために使用され得る。
【0038】
配列比較のためのパラメータとしては、以下が挙げられる:アルゴリズム:NeedlemanおよびWunsch、J.Mol Biol.48:443−453(1970)。
【0039】
比較マトリックス:HentikoffおよびHentikoff,(1992)Proc.Natl.Acad.Sci.USA.89:10915−10919からのBLOSSUM62。
【0040】
本明細書中で使用される場合、用語「単離された」、および類似の語は、核酸、ポリヌクレオチド、もしくはオリゴヌクレオチドを記載するために使用される場合、その天然の状態もしくは元の状態から取り出されることをいう。したがって、これらが天然において存在する場合、これらは、その元の環境から取り出されている。これらが合成的に調製された場合、これらは、合成から得られた元の産物混合物から取り出されている。例えば、この用語が本明細書中で使用される場合、天然の状態において生きている生物中に天然に存在している、天然に存在するポリヌクレオチドは、「単離され」ていないが、その天然の状態において共に存在する物質から分離されている同じポリヌクレオチドは、「単離されて」いる。一般に、少なくとも1種の有意な共に存在する物質の除去は、核酸、ポリヌクレオチド、オリゴヌクレオチドの「単離」を構成する。多くの場合、いくつかの、多くの、もしくはほとんどの、共に存在する物質が、核酸、ポリヌクレオチド、オリゴヌクレオチド、タンパク質、ポリペプチド、もしくはオリゴペプチドを単離するために除去され得る。非限定の例として、ポリヌクレオチドに関して、用語「単離された」は、天然に存在する染色体および細胞からこのポリヌクレオチドが分離されることを意味し得る。さらに、例として、タンパク質もしくはポリペプチドの「単離」は、細胞溶解物もしくは細胞ホモジネートにおける別の成分からの分離を意味し得る。
【0041】
インビトロ合成プロセスもしくは化学合成プロセスの産物である核酸、ポリヌクレオチド、もしくはオリゴヌクレオチドは、合成プロセスの結果として、本質的に単離されている。重要な実施形態において、このような合成産物は、使用される試薬および前駆体、ならびにそのプロセスによる副産物を除去するために処理される。
【0042】
同様に、ポリヌクレオチドおよびポリペプチドは、例えば、処方物、ポリヌクレオチドの細胞への導入のための組成物、化学反応もしくは酵素的反応のための組成物もしくは溶液のような組成物中に存在し得る。これらの組成物は、天然に存在する組成物ではなく、その中に、本明細書中で使用される用語の意味の範囲内である単離されたポリヌクレオチドもしくはポリペプチドを維持する。
【0043】
本明細書中および特許請求の範囲において使用される場合、「核酸」もしくは「ポリヌクレオチド」、およびこれらに基づく類似の用語は、天然に存在するヌクレオチドからなるポリマーおよび合成ヌクレオチドもしくは改変ヌクレオチドからなるポリマーをいう。したがって、本明細書中で使用される場合、RNAであるポリヌクレオチド、もしくはDNAであるポリヌクレオチドは、天然に存在する部分(例えば、天然に存在する塩基およびリボース環もしくはデオキシリボース環)を含み得るか、またはこれらは、以下で記載されるような合成部分もしくは改変部分から構成され得る。ヌクレオチド間の結合は、一般に、3’−5’リン酸結合であり、これは、天然のホスホジエステル結合であっても、ホスホチオエステル結合であっても、なお他の合成的結合であってもよい。改変骨格の例としては、ホスホロチオエート、キラルホスホロチオエート、ホスホロジチオエート、ホスホトリエステル、アミノアルキルホスホトリエステル、メチルホスホネートおよび他のアルキルホスホネート(3’−アルキレンホスホネート、5’−アルキレンホスホネートおよびキラルホスホネートを含む)、ホスフィネート、ホスホルアミデート(3’−アミノホスホルアミデートおよびアミノアルキルホスホルアミデートを含む)、チオノホスホルアミデート、チオノアルキルホスホネート、チオノアルキルホスホトリエステル、セレノホスフェートおよびボラノホスフェートが挙げられる。さらなる結合としては、ホスホトリエステル、シロキサン、カーボネート、カルボキシメチルエステル、アセトアミデート、カルバメート、チオエーテル、架橋されたホスホルアミデート、架橋されたメチレンホスホネート、架橋されたホスホロチオエートおよびスルホンのヌクレオチド内結合が挙げられる。他のポリマー性結合としては、これらの2’−5’結合アナログが挙げられる。米国特許第6,503,754号および同第6,506,735号ならびにこれらの中に引用されている参考文献(本明細書中で参考として援用される)を参照のこと。
【0044】
核酸およびポリヌクレオチドは、20もしくはそれより多くのヌクレオチドの長さ、または30もしくはそれより多くのヌクレオチドの長さ、または50もしくはそれより多くのヌクレオチドの長さ、または100もしくはそれより多いか、または1000もしくはそれより多いか、または10,000もしくはそれより多いか、または100,000もしくはそれより多い長さである。siRNAは、本明細書中で規定されるポリヌクレオチドであり得る。本明細書中で使用される場合、「オリゴヌクレオチド」およびこれに基づく類似の用語は、直前の段落で記載されるように、天然に存在するヌクレオチドからなる短いポリマー、ならびに合成ヌクレオチドもしくは改変ヌクレオチドからなるポリマーをいう。オリゴヌクレオチドは、10もしくはそれより多くのヌクレオチドの長さ、または15、もしくは16、もしくは17、もしくは18、もしくは19、もしくは20またはそれより多くのヌクレオチドの長さ、または21、もしくは22、もしくは23、もしくは24またはそれより多くのヌクレオチドの長さ、または25、もしくは26、もしくは27、もしくは28もしくは29、もしくは30またはそれより多くのヌクレオチドの長さ、35もしくはそれより多く、40もしくはそれより多く、45もしくはそれより多く、約50までのヌクレオチドの長さであり得る。siRNAであるオリゴヌクレオチドは、15と30との間の任意の数のヌクレオチドを有し得る。多くの実施形態において、siRNAは、21と25との間の任意の数のヌクレオチドを有し得る。
【0045】
大きさの範囲の重複ゆえに、用語「ポリヌクレオチド」および「オリゴヌクレオチド」は、本発明のsiRNAをいうために本明細書中で同じように使用され得ることが、上で示された定義から理解される。
【0046】
本明細書中および特許請求の範囲において使用される場合、「ヌクレオチド配列」、「オリゴヌクレオチド配列」もしくは「ポリヌクレオチド配列」、および類似の用語は、オリゴヌクレオチドもしくはポリヌクレオチドが有する塩基の配列と、この配列を有するオリゴヌクレオチドもしくはポリヌクレオチドの構造との両方に、相互交換可能に関連する。ヌクレオチド配列もしくはポリヌクレオチド配列は、さらに、任意の天然のポリヌクレオチドもしくはオリゴヌクレオチド、または任意の合成のポリヌクレオチドもしくはオリゴヌクレオチドに関連し、これらのポリヌクレオチドもしくはオリゴヌクレオチドにおいて、塩基の配列は、当該分野で従来使用されるように、塩基を示す文字の特定の配列の記載もしくは言及によって規定される。
【0047】
オリゴヌクレオチドおよびポリヌクレオチドにおける塩基は、プリン塩基(アデニン(A)およびグアニン(G))ならびにピリミジン塩基(チミジン(T)、シトシン(C)およびウラシル(U))を含む、「未改変」塩基もしくは「天然」塩基であり得る。さらに、これらは、改変もしくは置換を有する塩基であり得る。本明細書中で使用される場合、改変塩基の非限定の例は、他の合成塩基および天然塩基を含む。これらの塩基は、例えば、5−メチルシトシン(5−me−C)、5−ヒドロキシメチルシトシン、キサンチン、ヒポキサンチン、2−アミノアデニン、アデニンおよびグアニンの6−メチル誘導体および他のアルキル誘導体、アデニンおよびグアニンの2−プロピル誘導体および他のアルキル誘導体、2−チオウラシル、2−チオチミジンおよび2−チオシトシン、5−ハロウラシルおよび5−ハロシトシン、5−プロピニルウラシルおよび5−プロピニルシトシンおよびピリミジン塩基の他のアルキニル誘導体、6−アゾウラシル、6−アゾシトシンおよび6−アゾチミジン、5−ウラシル(プソイドウラシル)、4−チオウラシル、8−ハロ、8−アミノ、8−チオール、8−チオアルキル、8−ヒドロキシルおよび他の8−置換アデニンおよびグアニン、5−ハロ(特に5−ブロモ)、5−トリフルオロメチルおよび他の5−置換ウラシルおよびシトシン、7−メチルグアニンおよび7−メチルアデニン、2−フルオロ−アデニン、2−アミノアデニン、8−アザグアニンおよび8−アザアデニン、7−デアザグアニンおよび7−デアザアデニンおよび3−デアザグアニンおよび3−デアザアデニンである。さらなる改変塩基としては、三環式ピリミジン(例えば、フェノキサジンシチジン(1H−ピリミド[5,4−b][1,4]ベンゾキサジン−2(3H)−オン)、フェノチアジンシチジン(1−ピリミド[5,4−b][1,4]ベンゾチアジン−2(3H)−オン))、G−クランプ(clamp)(例えば、置換フェノキサジンシチジン(例えば、9−(2−アミノエトキシ)−H−ピリミド[5,4−b][1,4]ベンゾキサジン−2(3H)−オン)、カルバゾールシチジン(2H−ピリミド[4,5−b]インドール−2−オン)、ピリドインドールシチジン(H−ピリド[3’、2’:4,5]ピロロ[2,3−d]ピリミジン−2−オン))が挙げられる。改変塩基はまた、プリン塩基もしくはピリミジン塩基が他の複素環と置換されている改変塩基(例えば、7−デアザ−アデニン、7−デアザグアノシン、2−アミノピリジンおよび2−ピリドン)を含む。さらなる塩基としては、米国特許第3,687,808号に開示される塩基、The Concise Encyclopedia Of Polymer Science And Engineering,858−859頁,Kroschwitz,J.L編,John Wiley & Sons,1990によって開示される塩基、Englischら,Angewandte Chemie,International Edition(1991)30,613によって開示される塩基、ならびにSanghvi,Y.S.,第15章,Antisense Research and Applications,289−302頁、Crooke,S.T.およびLebleu,B.,編,CRC Press,1993によって開示される塩基が挙げられる。特定のこれらの塩基は、本発明のオリゴマー化合物の結合親和性を増大させるために、特に有用である。これらとしては、5−置換ピリミジン、6−アザピリミジンおよびN−2置換プリン、N−6置換プリンおよびO−6置換プリン(2−アミノプロピルアデニン、5−プロピニルウラシルおよび5−プロピニルシトシンを含む)が挙げられる。5−メチルシトシン置換は、核酸二本鎖の安定性を0.6〜1.2℃増大することが示されており(Sanghvi,Y.S.,Crooke,S.T.およびLebleu,B.,編,Antisense Research and Applications,CRC Press,Boca Raton,1993,pp.276−278)、ここで好ましい塩基置換であり、なおより好ましくは、2’−O−メトキシエチル糖修飾と結合される場合である。米国特許第6,503,754号および同第6,506,735号ならびにこれらにおいて引用された参考文献(本明細書中で参考として援用される)を参照のこと。任意の改変塩基の使用は、当業者に理解されるとおり、同じ塩基対特性を有する天然に存在する塩基の使用と同等である。
【0048】
本明細書中および特許請求の範囲において使用される場合、用語「相補的」およびこれに基づく類似の語は、核酸、ポリヌクレオチドもしくはオリゴヌクレオチドの1つの鎖における第1の核酸塩基の、核酸、ポリヌクレオチドもしくはオリゴヌクレオチドの1つの鎖における特定の第2の核酸塩基とのみ特異的に相互作用する能力をいう。非限定の例として、天然に存在する塩基が考慮される場合、AとTもしくはUとは、互いに相互作用し、そしてGとCとは互いに相互作用する。本発明および特許請求の範囲において使用される場合、「相補的」は、「完全に相補的」であることを意味し、すなわち、2つのポリヌクレオチド鎖が互いに並列される場合、1本の鎖における連続する塩基の配列における各塩基が、対向する鎖における同じ長さの連続する塩基の配列における相互作用する塩基に対して相補的である、少なくとも一部分の鎖が存在する。
【0049】
本明細書中で使用される場合、「ハイブリダイズ」、「ハイブリダイゼーション」および類似の語は、鎖と相補的な配列とを互いに相互作用させることによって、核酸、ポリヌクレオチド、もしくはオリゴヌクレオチドの2本鎖を形成するプロセスをいう。この相互作用は、各々の鎖における相補的な塩基が、特異的に相互作用して対を形成することによって起こる。鎖が互いにハイブリダイズする能力は、以下で示すような種々の条件に依存する。核酸鎖は、各鎖における十分な数の対応する位置が、互いに相互作用し得るヌクレオチドによって占められている場合、互いにハイブリダイズする。二本鎖を形成する鎖の配列は、特異的にハイブリダイズすることができるために互いに100%の相補性を必要とはしないことが、本発明の分野の当業者(非限定の例として、分子生物学者および細胞生物学者が挙げられる)によって理解される。
【0050】
本明細書中で使用される場合、「フラグメント」およびこれに基づく類似の語は、参照の全体配列よりも短い、核酸、ポリヌクレオチドもしくはオリゴヌクレオチドの一部分に関連する。フラグメントにおける塩基の配列は、このフラグメントが生じた分子の対応する部分の配列から改変されない;フラグメントにおいて、このフラグメントが生じた分子の対応する部分の配列と比較して、挿入も欠失も存在しない。本明細書中で企図されるように、核酸もしくはポリヌクレオチドのフラグメント(例えば、オリゴヌクレオチド)は、15もしくはそれより多くの塩基の長さ、または16もしくはそれより多く、17もしくはそれより多く、18もしくはそれより多く、または19もしくはそれより多く、または20もしくはそれより多く、または21もしくはそれより多く、または22もしくはそれより多く、または23もしくはそれより多く、または24もしくはそれより多く、または25もしくはそれより多く、または26もしくはそれより多く、または27もしくはそれより多く、または28もしくはそれより多く、または29もしくはそれより多く、30もしくはそれより多く、50もしくはそれより多く、75もしくはそれより多く、100もしくはそれより多くの塩基の長さであり、全長配列より1塩基短い長さまでの長さである。オリゴヌクレオチドは、化学的に合成され得、そしてsiRNA、PCRプライマー、またはプローブとして使用され得る。
【0051】
検出および標識。標的性ポリヌクレオチド、例えばsiRNA配列を含むポリヌクレオチド、ならびにウイルスポリヌクレオチド標的は、多くの手段によって検出され得る。検出としては、標的性ポリヌクレオチドの存在もしくは量を観察する能力をもたらす、1種もしくはそれより多くの任意のプロセスが挙げられ得る。1つの実施形態において、標的性ポリヌクレオチドもしくはウイルス標的を含むサンプル核酸は、拡張(expansion)の前に検出され得る。代替の実施形態において、サンプル中の標的性ポリヌクレオチドは、拡張されて、拡張された標的性ポリヌクレオチドもしくは拡張されたウイルス標的を提供し得、そしてこの拡張されたポリヌクレオチドが、検出されるかもしくは定量され得る。物理的方法、化学的方法もしくは生物学的方法が、標的性ポリヌクレオチドを検出しそして定量するために使用され得る。物理的方法としては、非限定の例として、表面プラスモン共鳴(SPR)検出(例えば、プローブの表面への結合)および標的性ポリヌクレオチドの固定化プローブへの結合を検出するためのSPRの使用、またはクロマトグラフィー媒体中にプローブを有し、そして標的性ポリヌクレオチドの結合をクロマトグラフィー媒体中で検出することをが挙げられる。物理的方法は、標的性ポリヌクレオチドが他のポリヌクレオチドから分離され、そしてこの分離されたポリヌクレオチドが検出される、ゲル電気泳動形式もしくはキャピラリー電気泳動形式をさらに含む。化学的方法としては、ポリメラーゼ連鎖反応(PCR)方法、および一般に標的性ポリヌクレオチドがプローブにハイブリダイズする、ハイブリダイゼーション方法が挙げられる。生物学的方法としては、標的性ポリヌクレオチドもしくは標的ポリヌクレオチドに、細胞において生物学的効果を発揮させ、そしてこの効果を検出することが挙げられる。本発明は、生物学的アッセイにおいて使用され得る生物学的効果の例を開示する。これらとしては、粒子を数えることによるビリオンの計数、プラークアッセイ、感染細胞における細胞変性効果の評価などが挙げられる。多くの実施形態において、ポリヌクレオチドは、以下で記載されるように標識されて、検出および定量を補助する。例えば、拡張を包含しない実施形態において、サンプル核酸は、標識化ヌクレオチドもしくは標識化オリゴヌクレオチドリンカーのような標識化部分の化学的付加もしくは酵素的付加によって標識され得る。
【0052】
拡張されたポリヌクレオチドが、直接的に検出され得、そして/または定量され得る。例えば、拡張されたポリヌクレオチドは、ゲル中の電気泳動(大きさによって分離する)に供され、そして色素によって染色されて、その存在および量を明らかにし得る。あるいは、拡張された標的性ポリヌクレオチドは、プローブ核酸のハイブリダイゼーション条件(以下を参照)下での曝露の際に検出され得、そしてハイブリダイゼーションによる結合が、検出されそして/または定量される。検出は、標的性ポリヌクレオチドがプローブに結合していることの決定を可能にすることを何らかの手段で達成する。このことは、フラグメントをハイブリダイズすることによってもたらされるプローブの物理的特性における変化を検出することによって達成され得る。このような物理的検出方法の非限定の例は、SPRである。
【0053】
検出を達成する代替の方法は、この拡張されたポリヌクレオチドの標識化形態を使用すること、および結合した標識を検出することである。標識は、例えば、125I、35S、32P、14C、もしくは3Hのような放射性同位体標識であり得、これはその放射活性によって検出され得る。あるいは、標識は、分光学的方法を用いて(例えば、蛍光、リン光もしくは化学発光によって)検出され得るように選択され得る。したがって、蛍光標識もしくはリン光標識または化学発光反応を誘導する標識が、使用され得る。標識は、なおさらに、特定のリガンド−レセプター対におけるリガンドであり得る;このリガンドの存在は、次いで、通常はそれ自体が検出のために標識されている特異的レセプターの二次結合によって検出される。
【0054】
(干渉性RNA)
本発明に従って、呼吸性ウイルス標的の遺伝子発現は、RNA干渉によって低減される。ウイルス遺伝子の発現産物は、このウイルス標的の少なくとも1つのセグメントに対して相補的である特異的二本鎖siRNAヌクレオチド配列によって標的化され、このセグメントは、15と30との間の任意の数のヌクレオチドを含むか、または多くの場合、21と25との間の任意の数のヌクレオチドを含む。この標的は、5’非翻訳(UT)領域中、コード配列中、または3’UT領域中に存在し得る。例えば、PCT出願WO00/44895、WO99/32619、WO01/75164、WO01/92513、WO01/29058、WO01/89304、WO02/16620、およびWO02/29858(各々は、本明細書中でその全体が参考として援用される)を参照のこと。
【0055】
本発明の方法に従い、呼吸性ウイルス遺伝子発現およびそれによる呼吸性ウイルス複製は、siRNAを用いて抑制される。本発明に従う標的性ポリヌクレオチドは、siRNAオリゴヌクレオチドを含む。このようなsiRNAはまた、ウイルス配列と同一もしくは類似であるヌクレオチド配列の化学的合成によって調製され得る。例えば、Tuschl,Zamore,Lehmann,BartelおよびSharp(1999),Genes & Dev.13:3191−3197(本明細書中でその全体が参考として援用される)を参照のこと。あるいは、標的性siRNAは、標的性ポリヌクレオチド配列を用いて、例えば、無細胞系(例えばDrosophila抽出物であるがこれに限定されない)において呼吸性ウイルスリボポリヌクレオチド配列を消化することによって、または組換え二本鎖ウイルスcRNAの転写によって、得られ得る。
【0056】
効率的なサイレンシングは、一般に、同じ長さの16〜30ntのセンス鎖と16〜30ntのアンチセンス鎖とから構成されるsiRNA二本鎖によって観察される。多くの実施形態において、siRNA対形成二本鎖の各鎖は、3’末端において2−ntのオーバーハングの付加を有する。2−ntの3’オーバーハングの配列は、siRNA標的認識の特異性に対し、さらに僅かに寄与する。1つの実施形態において、3’オーバーハング中のヌクレオチドは、リボヌクレオチドである。代替の実施形態において、3’オーバーハング中のヌクレオチドは、デオキシリボヌクレオチドである。3’デオキシヌクレオチドの使用は、細胞内安定性の増強を提供する。
【0057】
本発明の組換え発現ベクターは、細胞内に導入された場合、呼吸性ウイルスを標的化するsiRNA配列を含むRNAを提供するようにプロセシングされる。このようなベクターは、発現を可能にする様式でウイルス標的化配列に隣接(flanking)する作動可能に連結された制御配列を含む発現ベクター内にクローニングされたDNA分子である。このベクターからは、ウイルスRNAに対するアンチセンスであるRNA分子は、第1のプロモーター(例えば、クローニングされたDNAの3’のプロモーター配列)によって転写され、そしてウイルスRNA標的についてのセンス鎖であるRNA分子は、第2のプロモーター(例えば、クローニングされたDNAの5’のプロモーター配列)によって転写される。このセンス鎖およびアンチセンス鎖は、次いで、インビボでハイブリダイズして、呼吸性ウイルス遺伝子のサイレンシングのためにこのウイルスを標的化するsiRNA構築物を産生する。あるいは、2つの構築物が、siRNA構築物のセンス鎖およびアンチセンス鎖を作製するために使用され得る。さらに、クローニングされたDNAは、二次構造を有する転写物をコードし得、ここで、1つの転写物は、標的遺伝子からのセンス配列および相補的アンチセンス配列の両方を有する。この実施形態の例において、ヘアピンRNAi産物は、標的遺伝子の全てもしくは一部分に類似する。別の例において、ヘアピンRNAi産物は、siRNAである。ウイルス配列に隣接(flanking)する調節配列は、同一であっても異なっていてもよく、それによって、その発現は、独立して調節されてもよく、または時間的様式もしくは空間的様式で調節されてもよい。
【0058】
特定の実施形態において、siRNAは、例えば、より低分子の核RNA(snRNA)U6もしくはヒトRNase P RNA H1由来のRNApol III転写単位を含むベクター内に、ウイルス遺伝子テンプレートをクローニングすることによって細胞内で転写される。ベクター系の1つの例は、GeneSuppressorTM RNA干渉キット(Imgenexから市販される)である。U6プロモーターおよびH1プロモーターは、Pol IIIプロモーターのIII型クラスのメンバーである。U6様プロモーターの+1ヌクレオチドは、常にグアノシンであり、一方、H1プロモーターについての+1は、アデノシンである。これらのプロモーターについての終止シグナルは、5つの連続するチミジンによって規定される。この転写物は、代表的に、第2のウリジンの後で切断される。この位置における切断は、発現されたsiRNAにおいて3’UUオーバーハングを産生し、これは、合成siRNAの3’オーバーハングと類似する。400ヌクレオチド未満の長さの任意の配列が、これらのプロモーターによって転写され得るので、これらは、例えば、約50ヌクレオチドのRNAステム−ループ転写物における約21ヌクレオチドのsiRNAの発現に理想的に適合する。RNAiの特徴およびsiRNAの効力に影響を及ぼす要因が、研究されている(例えば、Elbashir,LendeckelおよびTuschl(2001)Genes & Dev.15:188−200)。
【0059】
選択されたsiRNA配列についての最初のBLAST相同性検索は、入手可能なヌクレオチド配列ライブラリーに対して行われ、ウイルス遺伝子のみが標的化され、宿主遺伝子が標的化されないことを確認する。Elbashirら,2001 EMBO J.20(23):6877−88を参照。
【0060】
(ポリヌクレオチドの合成)
標的性ヌクレオチド配列に対応するオリゴヌクレオチドおよび標的性配列を含むポリヌクレオチドは、例えば、自動化DNA合成装置を用いて、標準的合成技術によって調製され得る。オリゴヌクレオチドを合成するための方法は、周知の化学的プロセスを含み、これらとしては、表面誘導体化粒子へのヌクレオチドホスホルアミダイトの連続的付加(T,BrownおよびDorcas J.S.Brown,Oligonucleotides and Analogues A Practical Approach,F.Eckstein,編,Oxford University Press,Oxford,pp.1−24(1991)によって記載され、本明細書中で参考として援用される)が挙げられるが、これらに限定されない。
【0061】
合成手順の一例は、Expedite RNAホスホルアミダイトおよびチミジンホスホルアミダイト(Proligo,Germany)を使用する。合成オリゴヌクレオチドは、脱保護されそしてゲル精製されて(Elbashirら(2001)Genes & Dev.15,188−200)、その後、Sep−PakC18カートリッジ(Waters,Milford,Mass.,USA)精製される(Tuschlら(1993)Biochemistry,32:11658−11668)。相補的ssRNAは、アニーリング緩衝液(100mM酢酸カリウム、30mM HEPES−KOH(pH7.4)、2mM酢酸マグネシウム)中で90℃で、1分間インキュベーションされ、その後、37℃で1時間インキュベーションされて、対応するds−siRNAとハイブリダイズする。
【0062】
オリゴヌクレオチド合成の他の方法としては、以下が挙げられるが、これらに限定されない:ホスホトリエステル方法およびホスホジエステル方法(Narangら,(1979)Meth.Enzymol.68:90)ならびにH−ホスホネート方法(とりわけGaregg,P.J.ら,(1985)「Formation of internucleotidic bonds via phosphonate intermediates」,Chem.Scripta 25,280−282;およびFroehler,B.C,ら,(1986a)「Synthesis of DNA via deoxynucleoside H−phosphonate intermediates」,Nucleic Acid Res.,14,5399−5407)に従う固相オリゴヌクレオチド合成、ならびに支持体上の合成(Beaucageら(1981)Tetrahedron Letters 22:1859−1862)、ならびにホスホルアミデート技術(Caruthers,M.H.,ら,「Methods in Enzymology」,第154巻,pp.287−314(1988))、米国特許第5,153,319号、同第5,132,418号、同第4,500,707号、同第4,458,066号、同第4,973,679号、同第4,668,777号、および同第4,415,732号ならびに「Synthesis and Applications of DNA and RNA」,S.A.Narang,編,Academic Press,New York,1987およびその中に含まれる参考文献に記載される技術、ならびに非ホスホルアミダイト技術。固相合成は、オリゴヌクレオチドを不純物および過剰な試薬から単離することを助ける。一旦固体支持体から切断されると、このオリゴヌクレオチドは、公知の技術によってさらに単離され得る。
【0063】
(本発明の阻害性ポリヌクレオチド)
本発明は、呼吸性ウイルス病原体に感染した細胞内に侵入する際にRNA干渉現象を引き起こすように意図されるオリゴヌクレオチドを、広く提供する。本発明は、呼吸性ウイルス標的の性質に制限されないが、RSVおよびインフルエンザAのいくつかの株を標的化するオリゴヌクレオチドを強調する。RNA干渉は、適切な二本鎖RNAによって細胞内で起こり、この二本鎖RNAの鎖のうちの1本は、ウイルスの標的遺伝子における配列と同一であるかこれに高度に類似する。一般に、呼吸性ウイルスを標的化するオリゴヌクレオチドは、DNAもしくはRNAであり得るか、または、リボヌクレオチドおよびデオキシリボヌクレオチドの混合物を含み得る。後者の一例は、デオキシジヌクレオチド配列(例えば、d(TT)、d(UU)、d(TU)、d(UT)および他の可能性のあるジヌクレオチド)で3’末端において終止するRNA配列である。さらなる実施形態において、3’オーバーハングは、上で特定された塩基を有するリボヌクレオチドなどから構成され得る。さらに、このオリゴヌクレオチド薬剤は、一本鎖もしくは二本鎖であり得る。本発明の治療用オリゴヌクレオチドのいくつかの実施形態は、オリゴリボヌクレオチドであるか、もしくは3’d(TT)末端を有するオリゴリボヌクレオチドであることが予測される。一本鎖標的性ポリヌクレオチドは、哺乳動物細胞への侵入の際、細胞内に存在する内因性酵素活性によって、容易に二本鎖分子に変換される。
【0064】
最も一般的には、本発明は、オリゴヌクレオチドもしくはポリヌクレオチドを提供し、これは、15ヌクレオチドから200ヌクレオチドの長さまでのいずれかの長さの範囲であり得る。このポリヌクレオチドは、RSウイルスもしくはインフルエンザAウイルスのゲノムを標的化する第1のヌクレオチド配列を含む。この第1のヌクレオチド配列は、(a)15〜30の任意の数のヌクレオチドの長さである配列、または(b)その相補体のいずれかから構成される。このようなポリヌクレオチドは、本明細書中で、直鎖状ポリヌクレオチドと呼ばれる。
【0065】
図3は、本発明のポリヌクレオチドの特定の実施形態の概略図を提供する。本発明は、RSVもしくはインフルエンザAの種々の株における標的配列を開示するか、または特定の場合には、標的配列と僅かに不適合であるsiRNA配列を開示し、これらの全ては、配列番号1〜263に提供される。配列番号1〜263に提供される配列は、19ヌクレオチド〜25ヌクレオチドの長さの範囲である。この標的性配列は、図3において薄い影をつけたブロックによって表される。図3、パネルAのa)は、「SEQ」として示される開示された配列が、必要に応じてより大きなポリヌクレオチド内に含まれ得、このポリヌクレオチドの全長が200ヌクレオチドまでの範囲に及び得る、実施形態を図示する。
【0066】
本発明は、標的性ポリヌクレオチドにおいて、配列番号1〜263から選択される配列が、SEQによって表される第1のヌクレオチド配列よりも長いウイルスゲノム中の配列を標的化するようなより長い標的性配列の一部分であり得ることを、さらに提供する。これは、図3、パネルAのb)に図示され、ここでは、完全な標的性配列が、ポリヌクレオチドの上の横線およびSEQブロックの周囲の濃い影によって示される。ポリヌクレオチドの全ての実施形態において、このより長い配列は、必要に応じて、なおより長い(200もしくはより少ない塩基の長さである)ポリヌクレオチドに含まれ得る(図3、パネルA、b))。
【0067】
本発明は、配列番号1〜263のいずれかのフラグメントであって、少なくとも15ヌクレオチドの長さである(そして長くとも参照配列番号よりも1塩基短い;図3、パネルA、c)において図示される)配列、ならびに5ヌクレオチドまでが配列番号1〜263において与えられる標的配列から異なり得る(図3、パネルA、d)において図示される)配列(この例において、3つの改変塩基が、3本の濃い縦棒によって表される)をさらに提供する。
【0068】
なおさらに、本発明は、上記の配列のいずれかの相補体である配列を提供する(図3、パネルAのe)に示され、「COMPL」と名付けられる)。任意のこれらの配列が、本発明のオリゴヌクレオチドもしくはポリヌクレオチドに含まれる。本発明の任意の直鎖状ポリヌクレオチドは、上記のa)〜e)に記載される配列のみによって構成されてもよく、または必要に応じて、上限200ヌクレオチドまでのさらなる塩基を含んでもよい。RNA干渉は、二本鎖RNAを必要とするので、標的性ポリヌクレオチド自身は、少なくとも配列番号1〜263において与えられる配列に対して相補的でありかつこれにハイブリダイズする第2の鎖を含む二本鎖であり得るか、または細胞内プロセシングに依存して相補鎖を産生し得る。
【0069】
従って、上記のポリヌクレオチドは、一本鎖であってもよく、または二本鎖であってもよい。なおさらなる実施形態において、このポリヌクレオチドは、デオキシリボヌクレオチドのみを含むか、またはリボヌクレオチドのみを含むか、あるいはデオキシリボヌクレオチドとリボヌクレオチドとの両方を含む。本明細書中で記載されるポリヌクレオチドの重要な実施形態において、標的配列は、15ヌクレオチド(nt)、もしくは16nt、もしくは17nt、もしくは18nt、もしくは19nt、もしくは20nt、もしくは21nt、もしくは22nt、もしくは23nt、もしくは24nt、もしくは25nt、もしくは26nt、もしくは27nt、もしくは28nt、もしくは29、もしくは30ntのいずれかの長さであり得る配列からなる。なおさなる有利な実施形態において、この標的性配列は、ウイルス病原体ゲノムにおける標的配列と5塩基まで異なり得る。
【0070】
本発明のいくつかの実施形態において、上記ポリヌクレオチドは、標的性配列からなり、そして必要に応じて、本明細書中に記載される3’ジヌクレオチドオーバーハングを含む、siRNAである。
【0071】
あるいは、RNA干渉における二本鎖RNAの必要を認識して、上記オリゴヌクレオチドもしくはポリヌクレオチドは、分子内ヘアピンループ二本鎖分子を形成するように調製され得る。このような分子は、上の段落の実施形態のいずれかにおいて記載される第1の配列、その後ろの短いループ配列、そのさらに後ろの第2の配列であって第1の配列に対して相補的な配列から形成される。このような構造は、所望の分子内ヘアピンを形成する。その上さらに、また、挙げられた3つの所望の構造が、200ヌクレオチドまでの任意の全体の長さを有する任意のオリゴヌクレオチドもしくはポリヌクレオチドにおいて構成され得るような、最大200ヌクレオチドの長さを有するポリヌクレオチドが開示される。ヘアピンループポリヌクレオチドは、図3、パネルBにおいて図示される。
【0072】
(標的としてのRSV株)
RSVについて、ウイルスゲノムの任意の部分が標的化され得るが、理屈からすれば、ウイルス特異的酵素的機能をコードする標的性遺伝子が潜在的siRNA候補を提供するはずである。これらの遺伝子としては、L遺伝子、F遺伝子、G遺伝子およびP遺伝子が挙げられる。ウイルスゲノムの5’末端に位置するL遺伝子は、少量でしか発現されず、従って、有効なRNAiサイレンシングは、少量のRNAiのみを必要とし得る。構造遺伝子もまた、標的化され得る。
【0073】
以下のRSVサブグループAおよびBの代表的な株の配列は、サブグループAとサブグループBとの間で共通である(表1)か、またはサブグループA(表2)もしくはサブグループB(表3)に特異的である標的配列を選択するために使用された。
【0074】
サブグループA:株A2(GenBank M74568)および株Long(P−mRNA、GenBank M22644、およびF−mRNA、GenBank M22643のみ)。
【0075】
サブグループB:株B1(GenBank NC_001781)および株9320(GenBank AY353550)。
【0076】
ウイルス遺伝子配列は、共通領域もしくは独特の領域を探索するために並列された。各標的化遺伝子もしくは標的化領域について、少なくとも2つの標的を、適用不可能(NA)でない限り選択した。両サブグループに共通の標的配列を見出すことは、これらのどちらかに独特の標的を見出すよりも難しかった。何故なら、利用可能な、相同であるかもしくは同一である配列は、非常に少ないからである。この場合(表1)、時々、RNA二本鎖の1端の5’末端にミスマッチを導入する必要があった。
【0077】
【表1】
(表2.サブグループA(株A2 & Long株のF/P)に特異的な遺伝子標的)
【0078】
【表2】
(表3.サブグループB(株B1および株9320)に特異的な遺伝子標的)
【0079】
【表3】
ウイルスゲノムの5’末端に対して指向されるsiRNA因子を提供するために、株B1もしくは株9320のいずれか個々に独特の尾部(trail)標的が、以下に示される(表4)。(+)鎖(アンチ−ゲノムRNA)の5’末端が標的化され、すなわち、ポジティブRNA鎖(アンチゲノムRNA)をテンプレートとして用いてウイルスゲノムが複製を開始する際、新生リーダー配列が産生される。
【0080】
(表4.RSVサブグループBの2つの株についての尾部標的)
【0081】
【表4】
上の表1〜表4に列挙した標的に基づき、siRNAは、以下のように作製され得る:a)二本鎖siRNAの各鎖に、3’−dTdTオーバーハングが提供される、b)「ミスマッチ」が必要な場合、G:Cは、G:TもしくはG:Aに変えられ、A:Tは、A:CもしくはA:Gに変えられ、そしてC:GはC:TもしくはC:Aに変えられる。
【0082】
(標的としてのインフルエンザA株)
(+)鎖mRNA配列内の25ヌクレオチドおよび19ヌクレオチドの長さのsiRNA標的配列が、全ての8つのセグメントで同定されている。この配列は、表5〜表12に示される。siRNA配列は、この8つのセグメントの各々に由来する。各配列についての記載(entry)はまた、5’末端からのmRNAにおける開始ヌクレオチド位置を示す。
【0083】
(表5.インフルエンザAウイルスのヘマグルチニン遺伝子(A/chicken/Thailand/CH−2/2004(H5N1);GenBank AY649382)を標的化するsiRNA配列。)
【0084】
【表5】
(表6.インフルエンザAウイルスのマトリックスタンパク質2およびマトリックスタンパク質1(M)遺伝子(A/Thailand/1(KAN−1)/2004(H5N1);GenBank AY626144)を標的化するsiRNA配列。)
【0085】
【表6】
(表7.インフルエンザAウイルスのノイラミニダーゼ遺伝子(A/chicken/Thailand/CH−2/2004(H5N1);GenBank AY649383)を標的化するsiRNA配列。)
【0086】
【表7】
(表8.インフルエンザAウイルスのヌクレオカプシドタンパク質(NP)遺伝子(A/Thailand/1(KAN−1)/2004(H5N1);GenBank AY626145)を標的化するsiRNA配列。)
【0087】
【表8】
(表9.インフルエンザAウイルスの非構造タンパク質1および非構造タンパク質2(NS)の遺伝子(A/Thailand/1(KAN−1)/2004(H5N1);GenBank AY626146)を標的化するsiRNA配列。)
【0088】
【表9】
(表10.インフルエンザAウイルスのポリメラーゼ酸性タンパク質(PA)遺伝子(A/Thailand/1(KAN−1)/2004(H5N1);GenBank AY626147)を標的化するsiRNA配列。)
【0089】
【表10】
(表11.インフルエンザAウイルスのポリメラーゼ塩基性タンパク質1(PB1)遺伝子(A/Thailand/1(KAN−1)/2004(H5N1);GenBank AY626148)を標的化するsiRNA配列。)
【0090】
【表11】
(表12.インフルエンザAウイルスのポリメラーゼ塩基性タンパク質2(PB2)遺伝子(A/Thailand/1(KAN−1)/2004(H5N1);GenBank AY626149)を標的化するsiRNA配列。)
【0091】
【表12】
インフルエンザAを標的化する潜在的siRNA候補配列の選択のための別のアプローチは、2つの主要なウイルス表面糖タンパク質であるヘマグルチニン(HA)およびノイラミニダーゼ(NA)のサブタイプ特異性に着目することである。ヒト感染を引き起こすことが報告されているサブタイプは、H5N1、H7N7、およびH9N2である。以下は、サブタイプ特異的なHA標的およびNA標的の例である。
【0092】
(表13.H5N1ヘマグルチニン(HA)のsiRNA標的化配列(GenBank DQ023145,GI:66775624に基づく):
【0093】
【表13】
表13における配列は、中国において2000年と2003年との間にHPAIウイルスによって感染したニワトリから単離された6種のH5N1株によって共有される。これらのH5N1株は、表14に列挙される。
【0094】
(表14)
【0095】
【表14】
(表15.H5N1ノイラミニダーゼ(NA)のsiRNA標的化配列(GenBank DQ023147,GI:66775628に基づく):
【0096】
【表15】
上の配列は、中国において2001年と2003年との間に感染したニワトリもしくはブタから単離された6種のH5N1株によって共有される。これらのH5N1株は、以下の表16に列挙される。
【0097】
(表16)
【0098】
【表16】
(表17.H7N7ヘマグルチニン(HA)のsiRNA標的化配列(GenBank AY999986,GI:66394837に基づく):
【0099】
【表17】
上の全ての配列は、スウェーデンにおいて2002年にマガモから単離された6種のH7N7株によって共有される。これらのH7N7株は、以下の表18に列挙される。
【0100】
6種のHA配列を比較して、HA1関連ドメインがより高い頻度の点変異を有することが見出されており、従って、10個の標的がHA2ドメインから選択される。
【0101】
(表18)
【0102】
【表18】
いくつかのインフルエンザA H5N1 mRNA配列に対するさらなるsiRNA二本鎖が、同定された。これらを表19に示す。これらの位置は図4に図示される。
【0103】
(表19.H5N1トリインフルエンザを標的化するRNAi配列)
【0104】
【表19】
(siRNAの組み合わせ)
本発明のいくつかの実施形態は、2種もしくはそれより多くのオリゴヌクレオチドまたはポリヌクレオチドを含有する薬学的組成物を提供し、これらのオリゴヌクレオチドまたはポリヌクレオチドの各々は、呼吸性ウイルスのゲノムにおける遺伝子を標的化する配列を含む。関連の実施形態は、この組み合わせを用いて細胞を処置する方法、および呼吸性ウイルス感染を処置する方法を提供し、そしてこのような組み合わせ組成物の、呼吸性ウイルス感染を処置することを意図される薬学的組成物の製造における使用を提供する。この組み合わせの個々のポリヌクレオチド成分は、ウイルス病原体のゲノムにおける同じ遺伝子の異なる部分、または異なる遺伝子、あるいは1つの遺伝子および1つより多くの遺伝子のいくつかの部分を、標的化し得る。オリゴヌクレオチドもしくはポリヌクレオチドの組み合わせを用いる利点は、所定の遺伝子の発現を阻害する利益が、この組み合わせにおいて大きくなることである。多数の標的性配列の使用により、遺伝子のノックダウンもしくはウイルスゲノムのサイレンシングにおいて、より大きな効力が、達成される。ウイルス複製の阻害における増大した効力は、ウイルスゲノムにおける1種より多くの遺伝子を標的化することによって達成される。
【0105】
(薬学的組成物)
本発明の標的性ポリヌクレオチドは、本明細書中で「活性化合物」もしくは「治療薬」と記載される。これらの治療薬は、被験体への投与のために適する薬学的組成物中に組み込まれ得る。
【0106】
本明細書中で使用される場合、「薬学的に受容可能なキャリア」は、薬学的投与に適合性の、任意のそして全ての溶媒、分散媒体、コーティング、抗細菌剤および抗真菌剤、等張剤および吸収遅延剤などを含むことを意図される。適切なキャリアは、Remington’s Pharmaceutical Sciences,Gennaro AR(編)第20版(2000)Williams & Wilkins PA,USAならびにWilson and Gisvold’s Textbook of Organic Medicinal and Pharmaceutical Chemistry(DelgadoおよびRemers,Lippincott−Raven.による)のような教科書(本明細書中で参考として援用される)に記載される。このようなキャリアもしくは希釈剤において使用され得る成分の好ましい例としては、以下が挙げられるが、これらに限定されない:水、生理食塩水、リン酸塩、カルボン酸塩、アミノ酸溶液、リンガー液、デキストロース(グルコースについての異名)溶液、および5%ヒト血清アルブミン。非限定の例として、デキストロースは、5%もしくは10%の水溶液として使用され得る。リポソームおよび非水性ビヒクル(例えば、不揮発性油)もまた、使用され得る。このような媒体および薬剤の薬学的に活性な物質のための使用は、当該分野で周知である。追加の活性化合物もまた、組成物中に組み込まれ得る。
【0107】
本発明の薬学的組成物は、意図される投与の経路に適合するように処方される。投与の経路の例としては、非経口投与(例えば、静脈内投与、皮内投与、皮下投与)、経口投与、鼻投与、吸入投与、経皮投与(局所投与)、経粘膜投与および直腸投与が挙げられる。非経口適用、静脈内適用、皮内適用もしくは皮下適用のために使用される溶液もしくは懸濁液は、以下の成分を含み得る:注射用水、生理食塩水溶液、不揮発性油、ポリエチレングリコール、グリセリン、プロピレングリコールもしくは他の合成溶媒のような無菌希釈剤;ベンジルアルコールもしくはメチルパラベンのような抗細菌剤;アスコルビン酸もしくは亜硫酸水素ナトリウムのような抗酸化剤;エチレンジアミンテトラ酢酸のようなキレート剤;酢酸塩、クエン酸塩もしくはリン酸塩のような緩衝剤、ならびに張度を調整するための薬剤(例えば、塩化ナトリウムまたはデキストロース)。
【0108】
吸入による投与のために、上記化合物は、適切な噴霧体(例えば、二酸化炭素のような気体)を含む加圧式容器もしくはディスペンサーまたは噴霧器からエアロゾルスプレーの形態で送達される。
【0109】
1つの実施形態において、活性化合物は、制御放出処方物のように、この化合物を身体からの迅速な除去から保護するキャリアと共に調製され、移植物およびマイクロカプセル化送達系を含む。持続放出調製物の適切な例としては、抗体を含む固体疎水性ポリマーの半透性マトリックスが挙げられ、このマトリックスは、成形された製品の形態(例えば、フィルムもしくはマイクロカプセル)である。持続放出マトリックスの例としては、ポリエステル、ヒドロゲル(例えばポリ(2−ヒドロキシエチル−メタクリレート)もしくはポリ(ビニルアルコール))、ポリラクチド(米国特許第3,773,919号)、L−グルタミン酸とγエチル−L−グルタメートとのコポリマー、非分解性エチレン−酢酸ビニル、分解性乳酸−グリコール酸コポリマー(例えばLUPRON DEPOTTM(乳酸−グリコール酸コポリマーおよび酢酸ロイプロリドから構成される注射可能ミクロスフェア))、ならびにポリ−D−(−)−3−ヒドロキシ酪酸が挙げられる。エチレン−酢酸ビニルおよび乳酸−グリコール酸のようなポリマーは、100日間にわたって分子を放出可能であり、特定のヒドロゲルは、より短い期間にわたって薬学的活性薬剤を放出する。有益なポリマーは、生分解性もしくは生物適合性である。リポソーム懸濁液(ウイルス抗原に対するモノクローナル抗体によって感染細胞を標的化するリポソームを含む)もまた、薬学的に受容可能なキャリアとして使用され得る。これらは、例えば米国特許第4,522,811号において記載されるような当業者に公知の方法に従って調製され得る。有益な形態を有する持続放出調製物(例えばミクロスフェア)は、上記のような材料から調製され得る。
【0110】
本発明のsiRNAポリヌクレオチドは、ベクターに挿入され得、そして遺伝子治療ベクターとして使用され得る。遺伝子治療ベクターは、多くの経路(例えば、米国特許第5,703,055号に記載される)のいずれかによって被験体に送達され得る。従ってまた、送達としては、例えば、静脈内注射、局所投与(米国特許第5,328,470号を参照)または定位注射(stereotactic injection)(例えば、Chenら(1994)Proc.Natl.Acad.Sci.USA 91:3054−3057を参照)が挙げられる。遺伝子治療ベクターの薬学的調製物は、受容可能な希釈剤中に治療ベクターを含み得るか、または遺伝子送達ビヒクルが埋め込まれた徐放性マトリックスを含み得る。あるいは、完全な遺伝子送達ベクターが組換え細胞からインタクトに産生され得る(例えば、レトロウイルスベクター)場合、この薬学的調製物は、この遺伝子送達系を産生する1つもしくはそれより多くの細胞を含み得る。
【0111】
薬学的組成物は、例えば容器、パックもしくはディスペンサー中で、投与の指示書と共にキットに含まれ得る。
【0112】
被験体における呼吸性ウイルス感染を処置するための薬学的組成物もしくは医薬の製造における治療薬の使用もまた、本発明の範囲内である。
【0113】
(送達)
いくつかの実施形態において、本発明のsiRNAポリヌクレオチドは、リポソーム媒介型トランスフェクションによって、例えば市販の試薬もしくは技術(例えば、OligofectamineTM、LipofectAmineTM試薬、LipofectAmine 2000TM(Invitrogen)、ならびにエレクトロポレーションおよび同様の技術)を使用して、培養物中の細胞に送達される。さらに、siRNAポリヌクレオチドは、吸入もしくは気道への点滴注入を介して、動物モデル(例えばげっ歯類もしくは非ヒト霊長類)に送達される。動物モデルでの使用のためのさらなる経路としては、静脈内(IV)、皮下(SC)、および関連の投与の経路が挙げられる。siRNAを含有する薬学的組成物は、siRNAの安定性を保護するか、siRNAの持続時間を延長させるか、siRNA機能を助長するか、または特異的な組織/細胞にsiRNAを標的化させる、さらなる成分を含む。これらの成分としては、種々の生分解性ポリマー、カチオン性ポリマー(例えばポリエチレンイミン)、ヒスチジン−リジン(HK)ポリペプチドのようなカチオン性コポリペプチド(例えば、Mixsonらに対するPCT公開番号WO01/47496、Biomerieuxに対するWO02/096941およびMassachusetts Institute of Technologyに対するWO99/42091を参照)、ペグ化カチオン性ポリペプチド、およびリガンドを組み込んだポリマーなど、正に荷電したポリペプチド、PolyTranポリマー(天然の多糖類、スクレログルカンとしても知られる)、標的性リガンドと結合体化したポリマーからなるナノ粒子(TargeTran改変体)、界面活性剤(Infasurf;Forest Laboratories,Inc.;ONY Inc.)、ならびにカチオン性ポリマー(例えばポリエチレンイミン)が挙げられる。Infasurf(登録商標)(カルファクタント(calfactant))は、気管内点滴注入における使用のための、仔ウシ肺から単離された天然の肺界面活性剤である;これは、リン脂質、中性脂質、ならびに疎水性界面活性剤関連タンパク質Bおよび疎水性界面活性剤関連タンパク質Cを含む。このポリマーは、一次元ポリマーもしくは多次元ポリマーのいずれかであり得、そしてまた、直径20ミクロン未満、20ミクロンと100ミクロンとの間もしくは100ミクロンを超える、ミクロ粒子もしくはナノ粒子であり得る。このポリマーは、特定の組織もしくは細胞のレセプターまたは分子に特異的なリガンド分子を有し得、従って、siRNAの送達を標的化するために使用され得る。siRNAポリヌクレオチドはまた、カチオン性リポソームベースのキャリア(例えば、DOTAP、DOTAP/コレステロール(Qbiogene,Inc.))および他の型の脂質水溶液によって送達される。さらに、低パーセント(5〜10%)のグルコース水溶液およびInfasurfは、siRNAの気道送達のための有効なキャリアである30。
【0114】
5%グルコースおよびInfasurfの経口気道送達溶液に懸濁した蛍光標識化siRNAを用い、蛍光顕微鏡で試験して、siRNAが外鼻孔を介してもしくは経口−気道経路を介してマウスに送達されそして肺組織を洗浄した後に、このsiRNAは肺において広汎に分布していることが示されている(共有に係るWO2005/01940(本明細書中でその全体が参考として援用される)を参照)。マウスの鼻道および肺(上気道および気道深部(deeper respiratory tract))へのsiRNAの送達は、siRNAと同時にプラスミド中で送達した指示遺伝子(GFPもしくはルシフェラーゼ)を、首尾よくサイレンシングすることを示した(このプラスミドは、この指示遺伝子とsiRNA標的との融合物を有した)(共有に係るWO2005/01940を参照)。さらに、本発明者ら(他と共に研究した)によって報告された実験は、siRNA種がSARSコロナウイルスの複製を阻害し、それによって肺の病理学を緩和することを、SARS感染したアカゲザルにおいて示した。
【0115】
(siRNA組換えベクター)
本発明の別の局面は、本発明のsiRNAポリヌクレオチドを有するベクター、好ましくは発現ベクターに関する。本明細書中で使用される場合、用語「ベクター」は、連結した別の核酸を輸送可能な核酸分子をいう。ベクターの1つの型は、「プラスミド」であり、これは、さらなるDNAセグメントを連結し得る、環状二本鎖DNAループをいう。ベクターの別の型は、ウイルスベクターであり、ここで、さらなるDNAセグメントが、ウイルスゲノム中に連結され得る。特定のベクターは、作動可能に連結した遺伝子の発現を指示し得る。このようなベクターは、本明細書中で「発現ベクター」と呼ばれる。一般に、組換えDNA技術において使用される発現ベクターは、多くの場合、プラスミドの形態である。プラスミドが最も一般的に使用されるベクターの形態であるので、本明細書において、「プラスミド」および「ベクター」は、相互交換可能に使用され得る。しかし、本発明は、ウイルスベクター(例えば、複製欠損型のレトロウイルス、アデノウイルスおよびアデノ関連ウイルス)のような発現ベクターの他の形態を含むことを意図し、これらは、同等の機能をもたらす。
【0116】
本発明の組換え発現ベクターは、本発明の核酸を、この核酸の宿主細胞における発現のために適する形態で含み、これは、この組換え発現ベクターが、発現のために使用される宿主細胞に基づいて選択される1つもしくはそれより多くの調節配列を含むことを意味し、これらの調節配列は、発現される核酸配列に作動可能に連結される。組換え発現ベクター内で、「作動可能に連結される」は、目的のヌクレオチド配列が、このヌクレオチド配列の(例えば、インビトロ転写/翻訳系において、もしくはこのベクターが宿主細胞に導入される場合には宿主細胞においての)発現を可能にする様式で、調節配列に連結されることを意味する。用語「調節配列」は、プロモーター、エンハンサーおよび他の発現制御エレメント(例えば、ポリアデニル化シグナル)を含むことが意図される。このような調節配列は、例えば、Goeddel(1990)GENE EXPRESSION TECHNOLOGY:METHODS IN ENZYMOLOGY 185,Academic Press,San Diego,Calif.において記載される。調節配列としては、多くの型の宿主細胞におけるヌクレオチド配列の構成的発現を指示する調節配列、ならびに特定の宿主細胞においてのみヌクレオチド配列の構成的発現を指示する調節配列(例えば、組織特異的調節配列)が挙げられる。なお別の実施形態において、本発明の核酸は、哺乳動物発現ベクターを用いて哺乳動物細胞において発現される。哺乳動物発現ベクターの例としては、pCDM8(Seed(1987)Nature329:840)およびpMT2PC(Kaufmanら(1987)EMBO J 6:187−195)が挙げられる。哺乳動物細胞において使用される場合、発現ベクターの制御機能は、多くの場合、ウイルス調節エレメントによって提供される。例えば、一般に使用されるプロモーターは、ポリオーマ、アデノウイルス2、サイトメガロウイルスおよびシミアンウイルス40に由来する。さらなるベクターは、細菌人工染色体、酵母人工染色体もしくは哺乳動物人工染色体のような微小染色体を含む。他の適切な発現系は、原核生物細胞および真核細胞の両方についての発現系である。例えば、Sambrookら,MOLECULAR CLONING:A LABORATORY MANUAL.第2版,Cold Spring Harbor Laboratory,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.,1989の第16章および第17章を参照のこと。
【0117】
別の実施形態において、組換え哺乳動物発現ベクターは、特定の細胞型(例えば、気道の細胞)において選択的に核酸の発現を指示し得る。組織特異的調節エレメントは、当該分野で公知である。本発明は、組換え発現ベクター内にクローニングされた本発明のDNA分子を含む発現ベクターを、さらに提供する。このDNA分子は、(このDNA分子の転写による)ウイルスRNAを標的化するsiRNAを含むRNA分子の発現を可能にする様式で、調節配列に作動可能に連結される。種々の細胞型において上記RNA分子の連続的な発現を指示する、核酸に作動可能に連結される調節配列(例えば、ウイルスプロモーターおよび/もしくはエンハンサー)が選択され得るか、またはアンチセンスRNAの、構成的発現、組織特異的発現もしくは細胞型特異的発現を指示する調節配列が選択され得る。
【0118】
ベクターDNAは、従来的な形質転換技術もしくはトランスフェクション技術を介して、原核生物細胞もしくは真核細胞内に導入され得る。本明細書中で使用される場合、用語「形質転換」および「トランスフェクション」は、外来性の核酸(例えば、DNA)を宿主細胞内に導入するための、種々の当該分野で認識されている技術をいうことが意図され、これらとしては、リン酸カルシウムもしくは塩化カルシウムの共沈殿、DEAE−デキストラン−媒介型トランスフェクション、リポフェクション、もしくはエレクトロポレーションが挙げられる。宿主細胞を形質転換するかもしくはトランスフェクションするための適切な方法は、Sambrook,ら(2001),Ausubelら(2002),および他の研究室マニュアルにおいて見出され得る。
【0119】
(ウイルスの力価もしくは量についてのアッセイの方法)
本発明の実施において、ウイルスについてのアッセイは、いくつかの手順によって実施され得る。これらの中でも、非限定の例として、免疫ブロット(ウエスタンブロット)、免疫沈降(I.P.)、および複合型逆転写ポリメラーゼ連鎖反応(RT−PCR)アッセイなどが挙げられる。このような手順は、トランスフェクトされた組織培養物中の溶解物もしくは上清に存在し得る、標的化したmRNAもしくはそのタンパク質産物の合成の減少を測定する。同様に、これらのアッセイは、診断的手順として、感染した動物被験体もしくはヒト被験体から得られた、ホモジナイズした組織サンプル、鼻咽頭洗浄液、分泌液中に存在し得る、標的化したmRNAもしくはそのタンパク質産物の合成の減少を測定するために用いられ得る。
【0120】
RT−PCRアッセイ方法において、ウイルスゲノムRNAの合成は、NS1 ORFとNS2 ORFとの間の結合部(joint)にまたがった配列に相補的であるプライマーを用いて検出される。NS1/NS2プライマーを用いたこのRT−PCRの結果は、全ゲノムRNAの合成を反映する。
【0121】
この方法は、組織培養物におけるsiRNAによって誘導されたウイルス複製の干渉が、本発明の分野における当業者に公知であるように、定量的リアルタイムPCR(RTQ−PCR)、TCID50、ウイルスプラークアッセイ、免疫蛍光アッセイおよび免疫組織化学などによって測定されるアッセイをさらに含む。さらに、この方法は、上記TCID50法を用いて組織培養物中のウイルス複製の阻害をモニタリングする、ウイルスの存在についてのアッセイをさらに含む。ここで、ウイルス力価は、任意の種類の細胞病原性終末点(非限定の例として、細胞融合、細胞変性効果(CPE)、細胞吸着などが挙げられる)によって測定される。
【0122】
上記の方法は、試験動物における上記siRNA媒介型ウイルス複製の干渉が、RTQ−PCR、病理学、免疫組織化学およびウイルスの再単離などによって測定されるアッセイを、さらに含む。
【0123】
プライマーが、RNAiによるmRNA合成の減少を測定するために使用されるRT−PCR検出のために設計される。このRT反応は、ヘキサマーもしくはポリ−dTプライマーによって開始される;そしてPCRは、siRNAによって標的化される各遺伝子に特異的な上流もしくは下流のプライマーを用いて実施される。
【0124】
ゲノムRNA合成の検出について、一対のプライマー(上および下)が、NS1 ORFおよびNS2 ORFの中の配列に対応して設計される。ウイルスゲノムのどちらかの端に対して相補的なプライマーではなく「結合部にまたがる(joint−crossing)」プライマーを使用する目的は、取り扱いにくい大型の全体のRNA転写物(約15K−ntの長さ)を避けるためである。このRT−PCRの産物は、この設計により、ゲノムRNA全体の合成を反映する。
【0125】
同定されているプライマーおよびRT−PCRの産物が、以下の表20、表21および表22に列挙される。
【0126】
(表20.RSV株A2(サブグループA)についてのプライマー)
【0127】
【表20】
(表21.RSV株B1(サブグループB)についてのプライマー)
【0128】
【表21】
(表22.PCR産物の大きさ)
【0129】
【表22】
【実施例】
【0130】
(実施例1.細胞培養物における抗H5N1インフルエンザA siRNA分子の効果。)
本実施例は、培養物におけるH5N1感染細胞に対するsiRNAの阻害性効果の評価を報告する。
【0131】
(siRNAの設計および合成)
各々、H5N1のNP遺伝子(NP−1およびNP−2;表19、配列番号)遺伝子およびM2遺伝子(M2−1およびM2−2;表19、配列番号)を標的化する2つのsiRNAを、開発した。これらのsiRNA標的配列は、H5N1ウイルスのいくつかの株において保存されている(例えば、gi|47834945_10−1506、gi|8452827_1−1497、gi|13925158_46−1542、gi|61927237_46−1542、gi|59940391_44−>1535、gi|9802277_46−1542)。5’センス鎖においてフルオレセイン標識されているds−siRNAを、Proligo BioTech Ltd(Paris,France)によって化学的に合成した。その抗H5N1効果を、siRNAでトランスフェクトした、H5N1ウイルスに感染したMDCK細胞において検出した(Madin−Darbyイヌ腎臓;ATCC登録番号:CCL−34,Manassas,VA)。H5N1の阻害を、以下に記載されるように、細胞変性効果(CPE)、標準的TCID50(50%組織培養感染用量)プロトコールによる培養培地中に放出されたウイルスの逆滴定、およびリアルタイムRT−PCRを用いる細胞内ウイルスRNAの定量によって、決定した。
【0132】
(細胞培養、siRNAトランスフェクションおよびH5N1ウイルス感染)
MDCK細胞を、10%ウシ胎仔血清(FBS,Invitrogen)含有MEM培地中で培養し、そして維持した。約5000個の細胞を、ウイルス感染および複製アッセイのために、96ウェルプレートの各ウェル中に入れた。細胞を、0.5μlのLipofectamine 2000(Invitrogen,CA)と混合した100nM、50nM、25nMおよび12.5nMのsiRNAでトランスフェクションした。ルシフェラーゼを標的化する無関係のsiRNA(GL2i)、もしくはSARSコロナウイルス(C−1)を標的化するsiRNA、およびトランスフェクト体(transfectant)Lipofectamine 2000のみ(C−2)を、本実験においてネガティブコントロールとして含めた。トランスフェクションの6時間後、この培養培地を除去し、そしてこの細胞を、H5N1ウイルス感染の前にPBSで2回洗浄した。1% FBS含有MEM中に希釈した100μlの100 TCID50 H5N1ウイルス(株:A/HongKong/486/97)7を、トランスフェクトされた細胞に加えた。培養上清および細胞を、それぞれ感染の12時間後、16時間後および24時間後に、放出されたウイルス力価および細胞内ウイルスRNAコピー数の検出のために収集した。感染の24時間後、位相差顕微鏡下でCPEを観察し、そして記録した。この実験を、3連で行い、そして少なくとも3回繰り返した。
【0133】
(定量的RT−PCR)
Q−RT−PCRを、以下のように実行する。全細胞内RNAを、RNeasy Miniキット(Qiagen,Germany)を製造業者の指示に従って用いて単離した。逆転写実験を、ThermoScript RT−PCRシステム(Invitrogen,CA)を用いるオリゴ−dTプライミングによって実施した。次いで、リアルタイムPCRを、表23に示すプライマーを用いて実施した。
【0134】
(表23.Q−RT−PCRのためのH5N1プライマー)
【0135】
【表23】
5μlのRT産物(テンプレート)、各1μlの順(forward)プライマーおよび逆(reverse)プライマー(最終濃度500nM)、2μlの25mM MgCl2、ならびに9μlのH2Oを、2μlのSYBR Green I Master Mix(Roche,USA)と混合し、そしてリアルタイム定量を、ABI7900 Sequence Detection Systemを用いて実施した。PCR条件は、以下であった:50℃を5分間、95℃を10分間、次いで95℃を10秒間、61℃を5秒間および72℃を5秒間を40サイクル。β−アクチンのコピー数もまた、内部コントロールとして測定した。1000コピーのβ−アクチンあたりの細胞内ウイルスRNAのコピー数を、計算し、そして未処理コントロールと比較した相対的ウイルスRNAコピー数として表した(処理したコピー数/未処理のコピー数×100%)。
【0136】
(ウイルス力価の測定)
siRNA処理を行ったかまたは行わない培養物中のウイルス力価を、以下のように試験した。簡潔には、感染細胞からの馴化培地を、1% FBS含有MEM中に、10倍の連続工程で希釈した。各希釈物を、標準的TCID50プロトコールに従って細胞を感染させるために使用した。簡潔には、細胞を、96ウェル皿に感染の16時間前に入れた。感染の72時間後、CPEを、位相差顕微鏡下で観察し、そしてCPE陽性(CPE+)細胞を決定した。次いで、TCID50を、以下のように評価した:
【0137】
【化1】
ここで、h=希釈工程の補間されたlog10値、これを50%を超える値の工程のlog10に加えた。感染性ウイルス力価を、計算しそして未処理コントロールと比較した相対的ウイルス収率として表した(処理した力価/未処理の力価×100%)。
【0138】
(結果)
siRNA NP−1(表19)処理は、培養培地中のH5N1ウイルス産生(図5、パネルA)および細胞におけるウイルスRNAコピーの複製(図5、パネルB)を、12.5〜100nMの濃度において99%を超えて低下させた。一方、NP−2を標的化するsiRNA(表19)による処理は、100nMの濃度において、細胞培養物中のウイルス増殖の約60%しか阻害しなかった。対照的に、無関係のsiRNA処理コントロール(C−1)およびキャリア処理コントロール(C−2)は、未処理コントロールと比較して、ウイルス増殖を有意に阻害しなかった。
【0139】
M2−1(表19)に対して指向されるsiRNA(表19)による処理もまた、種々の濃度で、ウイルス増殖(図5、パネルC)およびウイルスRNAの複製(図5、パネルD)に対して、約80%〜94%の阻害性効果を示した。ここで、M2−2は、100nMの濃度で約60%の阻害性効果を示したが、より低い濃度は試験しなかった。
【0140】
第2の実験において、培養物中の細胞を、50nMの種々のsiRNAで処理した。培養上清培地中に産生されるウイルスを、3回の時点で収集した。サンプルを、培養物中の細胞の50%に感染するために必要な組織培養感染用量(TCID50)を決定することによって滴定した(図6、パネルA;これは対数座標スケール上に示されていることに留意されたい)。ウイルスRNAのコピーの数/1000コピーのβ−アクチンを、リアルタイムRT−PCRによって決定した(図6、パネルB)。無関係のsiRNA(C−1)およびトランスフェクト体(C−2)を、コントロールとして実験に適用した。NP−1 siRNAおよびNP−2 siRNAが、ウイルス粒子の増殖およびウイルスRNAの増大の阻害において非常に有効であることが示される。M1−1 siRNAおよびM2−2 siRNAの使用は、ウイルス複製の妨害において部分的に有効である。
【0141】
この実施例における結果は、H5N1ゲノムにおける異なる遺伝子に対して指向される種々のsiRNAが、培養中の感染細胞増殖におけるウイルス複製の阻害において、非常に有効であることを実証する。
【0142】
(実施例2.H5N1インフルエンザAに対して指向されるsiRNAの組み合わせ。)
H5N1ゲノム中の種々の遺伝子に対して指向されるsiRNAの組み合わせを、有効な抗H5N1活性を提供するために同定した。この実施例は、このような2つの組み合わせを示す。
【0143】
【化2】
(実施例3.インビトロの細胞中のsiRNAによる標的遺伝子およびウイルス複製の阻害)
この実施例において、種々の呼吸性ウイルス遺伝子を標的化する、同族(cognate)の標的遺伝子をサイレンシングする能力もしくはウイルス複製を阻害する能力において最適に有効なsiRNAおよび最適に有効なsiRNAの組み合わせが、細胞培養物中の実験によって同定される。インビトロで有効なsiRNAは、インビボでさらに試験されそして使用される候補である。
【0144】
培養した許容細胞株(例えば、非限定の例として、A549(ATCC登録番号:CCL−185TM、II型肺胞上皮肺癌腫細胞株))を、RSVウイルス株もしくはインフルエンザA株(例えば、H5N1)で感染させ、次いで種々のsiRNAにより、個々にもしくは組み合わせて、トランスフェクトする。組み合わせの場合、1つのシグナル遺伝子を標的化する2種のsiRNA、もしくは2つもしくはそれより多くの遺伝子を標的化するsiRNAが、使用される。単独のsiRNAもしくは組み合わせのsiRNAの総siRNA用量を一定にする。異なる実験プロトコールにおいて、siRNAトランスフェクションは、RSV感染もしくはインフルエンザA感染の数時間前に実施されるか、ウイルス感染と同時に実施されるか、または感染の数時間後に実施される。これらの異なる手順は、試験したsiRNAが、細胞レベルにおいて予防的効果および/もしくは治療的効果を示すか否かについての情報を提供する。
【0145】
標的遺伝子の阻害の程度、もしくはウイルス複製の阻害を、種々の方法によってアッセイする。アッセイの非限定の例としては、以下の手順が挙げられる:
(1)免疫ブロット(ウエスタン)を、細胞溶解物および所定のウイルス抗原に対するRSV特異的抗体もしくはインフルエンザA特異的抗体を用いて実施する。
【0146】
(2)免疫沈降(IP)を、細胞溶解物および上記の1つの遺伝子産物に対するRSV特異的抗体もしくはインフルエンザA特異的抗体を用いて実施する。
【0147】
(3)rvtr−PCRを行って、mRNA転写もしくはウイルスRNA複製の阻害を実証する。標的化遺伝子の転写物を検出するため、もしくは試験したsiRNAオリゴがゲノムRNAをもまた標的化し得るか否かを検出するための、特別なプライマーを設計する。
【0148】
(4)細胞融合体ベースのTCID50の測定を使用して、無関係のコントロールsiRNAの特異的siRNAで処理した細胞培養物と比較して、ウイルス複製の阻害をモニタリングし得る。
【0149】
(5)免疫蛍光もしくは免疫組織化学もまた、ウイルス力価を滴定するために使用され得、それによって、ウイルス複製のsiRNA媒介型阻害をモニタリングし得る。
【0150】
(実施例4.小動物モデルにおける、siRNAによる標的遺伝子の阻害)
この実施例において、種々のsiRNA(個々にもしくは組み合わせ)が、動物モデルにおけるRSVもしくはトリインフルエンザH5N1の処置におけるその効力を決定するために試験される。RSV株もしくはインフルエンザH5N1株を、吸入もしくは点滴注入により気道を通して試験動物に感染させるために使用する。siRNAは、同じ経路を通って送達され、そしてRSV感染もしくはインフルエンザH5N1感染の前に、感染と同時に、もしくは感染後に適用される。siRNA送達時間を変えることにより、試験動物におけるRSVもしくはインフルエンザH5N1の複製の阻害、RSVもしくはインフルエンザH5N1によって誘導される病理学の低減、およびRSVもしくはインフルエンザH5N1様の症状の軽減における、siRNAの効力の情報を得ることが可能であり、そしてRSVもしくはインフルエンザH5N1に特異的なsiRNAが、動物における実験的なRSV感染もしくはインフルエンザH5N1感染に対する予防効果もしくは治療効果を示すか否かの情報を得ることが可能である。
【0151】
RSVおよびインフルエンザH5N1は広範な範囲の動物種に感染し得るが、RSVもしくはインフルエンザH5N1の動物モデル研究において、マウスおよびコットンラット(cotton rat)が、従来使用されている。げっ歯類の感染は、通常、低レベル〜中レベルのウイルス複製をもたらし、これは、4日目にピークを迎え、そして迅速に除去される。これらの動物は、明らかな気道疾患を示さないが、肺の病理学を示す。高用量のウイルス感染において、これらは、体重減少、肺機能の変化、および毛皮の乱れ(ruffled fur)を示し、これらは疾患の指標である。
【0152】
上の実施例3に記載したものと同じ診断アッセイを、動物から採取したサンプル(例えば、鼻、頬および咽頭のスワブ)におけるsiRNA媒介型遺伝子サイレンシングおよびRSV感染もしくはインフルエンザH5N1感染の阻害をモニタリングするために使用する。さらに、肺の病理学、肺の免疫組織化学および症状観察を実施し、インビボでのRSV感染もしくはインフルエンザH5N1感染におけるsiRNAの効力を決定する。
【0153】
(実施例5.非ヒト霊長類モデルにおける、siRNAによる標的遺伝子の阻害)
本実施例は、非ヒト霊長類における、RSVもしくはインフルエンザH5N1のウイルス複製のsiRNA媒介型阻害の研究を記載する。用量投与の効力および安全性が、本研究の主要な目的である。
【0154】
本発明者らのプロトコールに基づき、別の呼吸性疾患であるSARSについてのアカゲザルモデルを用いて、30mg/kg体重までのsiRNAの用量は耐容可能であり、動物に毒性の徴候を示さない。(22)非ヒト霊長類哺乳動物によって事前にスクリーニングされているsiRNAの呼吸器送達(吸入もしくは点滴注入による)は、ウイルス複製の阻害ならびにウイルス誘導型病理学および疾患様症状の軽減に対して実質的に影響することを示した。RSVもしくはインフルエンザH5N1の本研究のために、同じ送達経路を使用し、そして類似の投薬量のsiRNAを送達する。
【0155】
小動物研究について記載された実験(実施例4)に加え、以下のアッセイが、霊長類において容易に実施され、そして治療効力のより直接的な評価に寄与する。
【0156】
(1)ウイルス散布性:ウイルス散布性を、感染したサルの鼻咽頭洗浄サンプルを用いて測定する。ウイルス収量を、TCID50(細胞融合体ベース)および/または免疫蛍光アッセイのいずれかによって滴定する。
【0157】
(2)鼻咽頭洗浄サンプルのRTQ−PCR:ウイルスゲノムコピー数の変化をモニタリングする。
【0158】
(3)症状モニタリング:ヒト乳児患者において観察されるような細気管支炎症状を見出しやすい。
【0159】
本発明は、本発明の種々の例示的実施形態に関して記載され、そして説明されているが、均等な実施形態および種々の他の改変、付加および削除が、これらの実施形態において、そしてこれらの実施形態に対して、本発明の精神および範囲から逸脱することなく、なされ得る。
【0160】
(参考文献)
【0161】
【化3】
【図面の簡単な説明】
【0162】
【図1】ヒトRSウイルス(hRSV、RSV)(−)ssRNAゲノムの概略図。GenBank登録番号NC_001781に基づく。
【図2】インフルエンザA型ウイルス構造の概略図。この図は、ウイルスゲノムの8つのセグメントが、ビリオン内に組み込まれていることを示す。
【図3】本発明のポリヌクレオチドの種々の実施形態の概略図。パネルAは、直鎖状ポリヌクレオチドの実施形態。長さは、200ヌクレオチド以下かつ15ヌクレオチド以上である。(b)において、記載の標的性配列は、より大きな標的性配列内に含まれる。(d)において、暗色の垂直の棒は、置換されたヌクレオチドを図解する。パネルBは、全体の長さが200ヌクレオチド以下のヘアピンポリヌクレオチドの実施形態である。
【図4】siRNA配列のH5N1ゲノム内の配置を示す概略図である。
【図5−1】MDCK細胞培養物におけるH5N1ウイルス増殖の阻害。
【図5−2】MDCK細胞培養物におけるH5N1ウイルス増殖の阻害。
【図6】異なる時点で決定されたsiRNAの阻害性効果。
【技術分野】
【0001】
(発明の分野)
本発明は、呼吸性ウイルス感染(特にRSウイルスおよびトリインフルエンザA(H5N1株を含む))におけるウイルス複製に干渉するsiRNA組成物に関連する。本発明は、呼吸性ウイルス感染細胞におけるウイルス遺伝子の発現を阻害するためのsiRNA組成物の使用、および被験体における呼吸性ウイルス感染の処置における使用のためのsiRNA組成物の使用に、さらに関連する。
【背景技術】
【0002】
(発明の背景)
呼吸性ウイルス感染は、何世紀にもわたって、ヒトの健康および生命に対する重大な脅威であった。悪名高いエピソードとしては、インフルエンザ株、RSウイルス、および重症急性呼吸性症候群(SARS)によって引き起こされる感染が挙げられる。これらは、世界中で約2〜4千万人の人々を殺傷した1918年の世界的なインフルエンザ汎流行を含む。より近年の年代に、他のインフルエンザ汎流行もまた、存在した。SARSは、2002年に発生し、およそ800人を犠牲にした(2)。
【0003】
(RSウイルス)
RSウイルス(RSV)感染は、深刻な汎流行性の気道疾患の主要な原因である。約3分の2の乳児が、人生の最初の1年間にRSVに感染し、そして約100%が2歳までに感染している。現在、RSV感染を処置するために利用可能である、特定かつ有効な治療薬は存在しない。
【0004】
RSウイルス(RSV)は、エンベロープを有する(enveloped)、非セグメント化一本鎖ネガティブRNAウイルス(NNR)であり、パラミクソウイルス科(Paramyxoviradae)モノネガウイルス(mononegaviruses)属に属する(14、29)。パラミクソウイルスは、以下の特徴を共有する。1)これらは、ウイルスヌクレオカプシドタンパク質(N)によってしっかりと被包されている一本鎖RNAゲノムを有する(29、30)。2)サブゲノムmRNAは、RdRPによって正常なゲノムから転写される。3)ウイルス複製は、宿主細胞の細胞質において起こる。感染から子孫ビリオンの放出までのRSV生活環の詳細は、よく研究されている(15)。
【0005】
RSVゲノムは、3’−NS1、NS2、N、P、M、SH、G、F、M2、L−5’の遺伝子を含む15.5kb長の、ネガティブ鎖である(図1を参照).これらの7つの遺伝子の産物(すなわち、N、P、M、SH、G、F、およびL)は、他のパラミクソウイルス3と共通している。いくつかのウイルス因子もしくは宿主因子は、RNA転写および複製の調節に関与する(20)。さらに、mRNAの転写およびRNA複製において調節的な役割を果たすウイルスのシス作動性シグナルが存在する(3)。
【0006】
RSV感染の潜伏期は、約4〜5日間である。RSVは、まず鼻咽頭に影響を及ぼし、次いで、数日のうちに、気管支および細気管支に達し、感染は呼吸器上皮の表面層に限られる。
【0007】
(インフルエンザA)
1997年の初めに、トリインフルエンザAの新型株であるH5N1が出現した。ほぼトリ(野生集団および家禽の両方)に限定されていたが、このウイルスは、感染したトリに直接的に接触したヒトにのみ感染するようであった。ヒトにおいて、感染は深刻な疾患を引き起こし、ヒトにおいて重症の呼吸性疾病および死をもたらす(3〜12)。多くの症例および発生が、東南アジアの種々の国において起こった。このトリウイルスのヒトに感染する能力を考慮すると、接触感染性ヒト改変体への突然変異の増大した危険性が存在し、この突然変異は、効率的かつ継続的なヒトからヒトへの伝播による新型インフルエンザ汎流行の出現ならびに大勢の死の危険をもたらす。
【0008】
トリインフルエンザH5N1は、肺炎に関連する新しく出現した感染性因子であるので、その病理学およびメカニズムがあまり明らかになっておらず、ヒト疾患症例におけるH5N1トリインフルエンザのための特定かつ有効な処置は存在しない。現在、インフルエンザ感染は、2種の(ノイラミニダーゼインヒビターのクラスの)薬物のような抗ウイルス薬:オセルタミビル(市場ではタミフルとして知られる)およびザナミビル(市場ではリレンザとして知られる)、またはより旧式なM2インヒビターであるアマンタジンおよびリマンタジンによって処置される。
【0009】
H5N1は、インフルエンザウイルスA型のサブタイプである。このウイルスは、エンベロープを有し、フラグメント化された、ネガティブ一本鎖RNAウイルスであるので、オルトミクソウイルス科に属する。インフルエンザAウイルス(H5N1を含む)の生活環の間、ウイルスゲノムRNA(vRNA)は、相補的RNA(cRNA)産生のためのテンプレートとして働き、このcRNAはまた、伝令RNA(mRNA)産生のためのテンプレートとして働く。ウイルス複製の間に生じるRNA分子のこれらの3つの形態の各々は、全て、センスsiRNAもしくはアンチセンスsiRNAのいずれかを用いて、siRNA媒介型分解のために標的化され得る。インフルエンザAゲノム(8個の分離したRNAセグメントからなり、少なくとも10個のオープンリーディングフレーム(ORF)を含む)は、ウイルスゲノム複製およびサブゲノムのmRNA合成もしくは遺伝子指向型mRNA合成の両方のためのテンプレートとして働く。図2は、インフルエンザAビリオンの構造を表す概略図を示す。ポリメラーゼPB2、PB1(ポリメラーゼ塩基性タンパク質1およびポリメラーゼ塩基性タンパク質2)ならびにPA(ポリメラーゼ酸性タンパク質)は、それぞれRNA1、RNA2およびRNA3によってコードされていた。4種のウイルス構造タンパク質である、H(血球凝集素)、N(ノイラミニダーゼ)、M1およびM2(マトリックスタンパク質1およびマトリックスタンパク質2)は、それぞれ、RNAセグメント4、RNAセグメント6およびRNAセグメント7によってコードされ、一方、RNA5は、NP(ヌクレオカプシドタンパク質)をコードし、そしてRNA8は、NS1およびNS2(非構造タンパク質1および非構造タンパク質2)をコードする。
【0010】
(RNA干渉)
RNA干渉(RNAi)は、配列特異的RNA分解プロセスであり、理論的に任意の遺伝子を、ノックダウンするかまたはサイレンシングするための、比較的容易かつ直接的な方法を提供する(17、18、19)。天然に存在するRNA干渉において、二本鎖RNAは、RNase III/ヘリカーゼタンパク質であるDicerによって、低分子干渉(small interfering)RNA(siRNA)分子へと切断される。このsiRNA分子は、2ヌクレオチド(nt)オーバーハング(overhang)を3’末端において有する、19〜23ヌクレオチドのdsRNAである。これらのsiRNAは、RNA誘導型サイレンシング複合体(RNA−induced−silencing−complex:RISC)と呼ばれる多成分リボヌクレアーゼ内に組み込まれる。siRNAの一本の鎖は、RISCに結合したままであり、この複合体を同族の(cognate)RNAへと導き、この同族のRNAは、RISC中のこのガイド因子(guider)ss−siRNAに対して相補的である配列を有する。このsiRNA指向型エンドヌクレアーゼは、このRNAを消化し、それによってこのRNAを不活性化する。近年の研究は、化学的に合成された21〜25−nt siRNAの使用は、哺乳動物細胞においてRNAiの作用を示すことを明らかにし(20)、そしてsiRNAハイブリダイゼーションの(末端におけるかもしくは中央部における)熱力学的安定性は、分子の機能を決定する際に中枢的な役割を果たすことを明らかにしている(21、22)。これらならびにRISC、siRNA分子およびRNAiの他の特徴は、記載されている(23〜28)。
【0011】
哺乳動物細胞におけるRNAiの、研究室における用途、または潜在的に、治療用途は、化学的に合成されたsiRNAもしくは内因的に発現された分子のいずれかを使用する(非特許文献1(2)、非特許文献2(21))。内因性siRNAは、最初に低分子ヘアピンRNA(shRNA)として、発現ベクター(プラスミドもしくはウイルスベクター)によって発現され、次いで、DicerによってsiRNAへとプロセシングされる。siRNAは、特にウイルス感染によって引き起こされるヒト疾患のための治療薬であるという非常な有望性を有すると考えられる(非特許文献3(19)、非特許文献4(20)、27〜30)。
【0012】
重要なことには、現在、ウイルスゲノム配列(例えば、約16〜30塩基対のオリゴヌクレオチド)を標的化する可能性のある多くの潜在的候補siRNA配列のうちのどれが、実際に有効なsiRNA活性を示すかは、いかなる程度の確信をもって予測することも不可能である。その代わり、個々の特異的候補siRNAポリヌクレオチド配列もしくは特異的候補siRNAオリゴヌクレオチド配列は、標的化された遺伝子の発現の意図される干渉が起こるか否かを決定するために、作製されそして試験されなければならない。したがって、所定のmRNAの発現の特異的な改変を可能にするsiRNAポリヌクレオチドを、確信を持って設計するための慣用的方法は、当該分野に存在しない。
【非特許文献1】Abbot,A.Nature(2003)424:121〜123
【非特許文献2】Reynolds,A.ら、Nat.Biotechnology(2004)22:326〜330
【非特許文献3】Schiwarz,D.S.ら、Cell(2003)115:199〜208
【非特許文献4】Khvorova,A.ら、Cell(2003)115:209〜216
【発明の開示】
【発明が解決しようとする課題】
【0013】
ウイルス病原体遺伝子およびその同族タンパク質産物の発現を阻害する組成物および方法についての、非常な需要が存在し続けている。特に、ウイルス感染した細胞における病原性呼吸性ウイルス遺伝子の発現を阻害するための組成物および方法、ならびに被験体における呼吸性ウイルス感染を処置するための組成物および方法についての、差し迫った需要が存在する。さらに、RSVおよびトリインフルエンザA(特にH5N1株)による感染に取り組む組成物および方法についての需要が、さらに存在する。非常に有効で、かつ公知の抗ウイルス剤の使用もしくは公知の抗ウイルス剤の改変に依存しない処置のための組成物および方法についての需要が、さらに存在する。本発明は、これらおよびこれらと関連する需要に取り組む。
【課題を解決するための手段】
【0014】
(発明の要旨)
本発明は、ウイルス病原体(例えば、インフルエンザA H5N1およびRSVを含む、呼吸性ウイルス感染を引き起こすウイルス病原体)のウイルスRNA分子の破壊を通して、ウイルス感染および複製を阻害するための、RNA干渉の使用に関連する組成物および方法を提供する。これらのウイルスは、ヒトおよび他の哺乳動物において重症の呼吸性疾患を引き起こす病原体である。ウイルス複製の阻害は、このウイルスに感染した培養細胞ならびにこのウイルスに感染した被験体において、ウイルス感染と戦う。このウイルス複製の阻害は、その症状の軽減を含む。
【0015】
第1の局面において、本発明は、長さが200以下かつ15以上の任意の数のヌクレオチドであり得る単離されたポリヌクレオチドを提供する。このポリヌクレオチドは、吸性合胞体ウイルスもしくはインフルエンザAウイルスのゲノムを標的化する第1のヌクレオチド配列を含む。このポリヌクレオチドにおいて、任意のT(チミジン)もしくは任意のU(ウリジン)は、必要に応じて他と置換され得る。さらに、このポリヌクレオチドにおいて、第1のヌクレオチド配列は、(a)長さが15〜30の任意の数のヌクレオチドである配列、または(b)(a)において与えられた配列の相補体からなる。このようなポリヌクレオチドは、本明細書中で、直鎖状ポリヌクレオチドと呼ばれ得る。
【0016】
本発明の関連の局面において、上記のポリヌクレオチドは、ループ配列によって第1のヌクレオチド配列から分離される第2のヌクレオチド配列をさらに含み、この第2のヌクレオチド配列は、
(a)第1のヌクレオチド配列と実質的に同一の長さを有し、かつ
(b)第1のヌクレオチド配列に対し実質的に相補的である。この後者の構造(ヘアピンポリヌクレオチドと呼ばれる)において、上記第1のヌクレオチド配列は、上記第2のヌクレオチド配列とハイブリダイズして、その相補的配列が、上記ループ配列によって連結されている、ヘアピンを形成する。
【0017】
上記直鎖状ポリヌクレオチドの多くの実施形態およびこのヘアピンポリヌクレオチドの多くの実施形態において、上記第1のヌクレオチド配列は、以下のいずれかである:
(a)配列番号1〜263から選択される配列、
(b)項目(a)において与えられた配列より長い標的性(targeting)配列であって、この標的性配列は、呼吸性ウイルスのゲノムを標的化し、かつ配列番号1〜263から選択される配列を含む、
(c)配列番号1〜263から選択される配列のフラグメントであって、このフラグメントは、少なくとも15ヌクレオチドの長さでありかつこの選択される配列より長くとも1塩基短い、連続する塩基の配列からなる、フラグメント、
(d)配列番号1〜263から選択される配列と5ヌクレオチドまで異なる配列、または
(e)(a)〜(d)において与えられた任意の配列の相補体。
【0018】
直鎖状ポリヌクレオチドもしくはヘアピンポリヌクレオチドの種々の実施形態において、上記第1のヌクレオチド配列の長さは、21〜25の任意の数のヌクレオチドである。多くの実施形態において、直鎖状ポリヌクレオチドもしくはヘアピンポリヌクレオチドは、配列番号1〜263から選択される配列からなり、そして必要に応じて、この選択される配列の3’に結合するジヌクレオチドオーバーハングを含む。直鎖状ポリヌクレオチドもしくはヘアピンポリヌクレオチドのなおさらなる実施形態において、この第1のヌクレオチド配列の3’末端におけるジヌクレオチド配列は、TT、TU、UT、もしくはUUであり、そしてリボヌクレオチドもしくはデオキシリボヌクレオチドのいずれか、または両方を含む。種々のさらなる実施形態において、直鎖状ポリヌクレオチドもしくはヘアピンポリヌクレオチドは、DNAであってもよく、またはこれはRNAであってもよく、あるいはこれはデオキシリボヌクレオチドおよびリボヌクレオチドの両方から構成されてもよい。
【0019】
さらなる局面において、本発明は、二本鎖ポリヌクレオチドを提供する。この二本鎖ポリヌクレオチドは、請求項1に記載の第1のポリヌクレオチド鎖および少なくともこの第1の鎖のこの第1のヌクレオチド配列に対して相補的でありかつこれにハイブリダイズして二本鎖組成物を形成する第2のポリヌクレオチド鎖を含む。これらのポリヌクレオチド構造もまた、直鎖状ポリヌクレオチドと呼ばれ得る。
【0020】
なおさらなる局面において、本発明は、請求項1、請求項2もしくは両方に記載の2つもしくはそれより多くの標的性ポリヌクレオチドを含む組み合わせであって、この組み合わせの各ポリヌクレオチドは、標的ウイルスのゲノムにおける異なる配列を標的化する組み合わせを提供する。
【0021】
呼吸性ウイルス病原体標的に対する高い程度の類似性および同一性に起因して、そして理論に縛られることを望まないが、ウイルスに感染した細胞内への導入の際に、このポリヌクレオチドは、RNA干渉を誘導し、病原体ゲノムRNA、相補的RNAおよび伝令RNAの消化をもたらす。特に、本発明のこれらの局面の重要な実施形態において、これらのポリヌクレオチドにおける第1のヌクレオチド配列もしくはその相補体は、RNA誘導型サイレンシング複合体(RISC)を形成し、このRISCは、上記ポリヌクレオチドsiRNA配列を病原体ゲノムRNA配列に誘導し、それによって病原体ゲノムRNAの切断を促進すると考えられる。
【0022】
さらなる局面において、本発明は、本発明の直鎖状ポリヌクレオチドもしくはヘアピンポリヌクレオチドによって与えられる配列を有するベクターを提供する。種々の実施形態において、これらのベクターのいずれかは、プラスミド、組換えウイルス、トランスポゾン、もしくは微小染色体であり得る。なおさらなる局面は、本発明の1種もしくはそれより多くの直鎖状ポリヌクレオチド、または本発明の1種もしくはそれより多くのヘアピンポリヌクレオチドによってトランスフェクトされた細胞を提供する。
【0023】
なおさらなる局面において、本発明は、1種もしくはそれより多くの直鎖状ポリヌクレオチドもしくはヘアピンポリヌクレオチドまたはそれらの混合物、ならびに薬学的に受容可能なキャリアを含有する薬学的組成物を提供し、ここで、各ポリヌクレオチドは、標的ウイルスのゲノムにおける異なる配列を標的化する。
【0024】
なおさらなる局面において、本発明は、直鎖状ポリヌクレオチドを有する1種もしくはそれより多くのベクター、またはヘアピンポリヌクレオチドを有するベクター、あるいはそれらの混合物、ならびに薬学的に受容可能なキャリアを含有する薬学的組成物を提供し、ここで、各ベクターは、標的ウイルスのゲノムにおける異なる配列を標的化するポリヌクレオチドを有する。
【0025】
薬学的組成物の種々の実施形態において、このキャリアとしては、合成カチオン性ポリマー、リポソーム、デキストロース、界面活性剤、またはこれらの任意の2種もしくはそれより多くの組み合わせが挙げられる。
【0026】
なおさらなる局面において、本発明は、RSウイルスもしくはインフルエンザAウイルスのゲノムを標的化する配列を有する直鎖状ポリヌクレオチドもしくはヘアピンポリヌクレオチドを合成する方法を提供する。この方法は、以下の工程を包含する:
(a)反応性末端(live reactive end)を含みかつこの配列の第1の末端におけるヌクレオチドに対応するヌクレオチド試薬を提供する工程;
(b)反応性末端を含みかつ該配列の後に続く位置に対応する、さらなるヌクレオチド試薬を添加して、上記工程(a)の反応性末端と反応させ、そして伸長中のポリヌクレオチド配列の長さを1ヌクレオチド増加させる工程、ならびに所望されない産物および過剰な試薬を除去する工程、ならびに
(c)上記配列の第2の末端におけるヌクレオチドに対応する上記ヌクレオチド試薬が添加され、それによって完全なポリヌクレオチドが提供されるまで、工程(b)を繰り返す工程。
【0027】
なおさらなる局面において、本発明は、RNAインヒビターによって細胞をトランスフェクトするための方法を提供し、ここで、この方法は、この細胞を、1種もしくはそれより多くの直鎖状ポリヌクレオチド、または1種もしくはそれより多くのヘアピンポリヌクレオチドを含有する組成物と接触させる工程を包含する。多くの実施形態において、このようにトランスフェクトされる細胞としては、1種もしくはそれより多くのポリヌクレオチドによって標的化される呼吸性ウイルスが挙げられる。
【0028】
なおさらなる局面において、本発明は、呼吸性ウイルスに感染した細胞におけるこのウイルスの複製を阻害する方法を提供し、この方法は、この細胞を、1種もしくはそれより多くの直鎖状ポリヌクレオチド、または1種もしくはそれより多くのヘアピンポリヌクレオチドを含有する組成物と接触させる工程を包含し、ここで、この1種もしくはそれより多くのポリヌクレオチドは、このウイルスを標的化する。
【0029】
なおさらなる局面において、本発明は、被験体における呼吸性ウイルスに起因する感染を処置するために有効な薬学的組成物の製造における、この呼吸性ウイルスを標的化する直鎖状ポリヌクレオチドもしくはこれらの2種もしくはそれより多くの混合物の使用、またはこの呼吸性ウイルスを標的化するヘアピンポリヌクレオチドもしくはこれらの2種もしくはそれより多くの混合物の使用を提供する。この使用の種々の実施形態において、この被験体は、ヒトである。
【0030】
なおさらなる局面において、本発明は、被験体における呼吸性ウイルスに起因する感染を処置する方法を提供する。この方法は、呼吸性ウイルスを標的化する直鎖状ポリヌクレオチドもしくはこれらの2種もしくはそれより多くの混合物の有効用量、または呼吸性ウイルスを標的化するヘアピンポリヌクレオチドもしくはこれらの2種もしくはそれより多くの混合物の有効用量を、この被験体に投与する工程を包含する。この方法の種々の実施形態において、この被験体は、ヒトである。さらなる実施形態において、直鎖状ポリヌクレオチドおよびヘアピンポリヌクレオチドの両方が、この被験体に投与される。
【発明を実施するための最良の形態】
【0031】
(発明の詳細な説明)
本明細書中で記載された全ての特許、特許出願公開、および特許公報は、その全体が、本明細書中で逐語的に示されたのと同様に参考として援用される。本明細書中で記載された全ての技術的刊行物もまた、参考として援用される。
【0032】
本明細書中で、冠詞「a」、「an」および「the」は、単数形もしくは複数形として、同等の意味を表す。これらの冠詞についての特定の意味は、これらが使用される文脈から明らかである。
【0033】
本明細書中で使用される場合、用語「標的」配列ならびに類似の用語および語句は、本発明のポリヌクレオチドが指向される病原体のゲノムにおいて存在するヌクレオチド配列をいう。ポリヌクレオチドは、病原体配列を、(a)この病原体のゲノム内に含まれる特定の下位配列(標的配列と呼ばれる)に対し相同であるかもしくは同一である配列を含むこと、または(b)相補体が標的配列と相同であるかもしくは同一である配列を含むことのいずれかによって、標的化する。病原体配列を標的化する任意のポリヌクレオチドは、RNA干渉現象にしたがって標的配列とハイブリダイズする能力を有し、それによってRNA干渉を開始する。
【0034】
本明細書中で使用される場合、用語「相補体」、「相補性」、ならびに類似の用語および語句は、塩基が、塩基対塩基で、相補的塩基対を形成する2つの配列に関連し、生物化学、分子生物学、遺伝学および本発明の分野に関する類似の分野のような分野における当業者によって従来理解される通りである。
【0035】
本明細書中で使用される場合、第1の配列もしくは下位配列が、第2の配列もしくは下位配列と、この配列もしくは下位配列の全ての位置において同じ塩基を有する場合、第1の配列もしくは下位配列は、第2の配列もしくは下位配列と「同一である」か、「100%の同一性」を有するか、または100%同一性の概念を伝達する用語もしくは語句によって記載される。同一性の決定において、T(チミジン)もしくはその任意の誘導体またはU(ウリジン)もしくはその任意の誘導体を含む任意の特定の塩基位置は、互いに同等であり、したがって同一であるとみなされる。
【0036】
本明細書中で記載される場合、標的性ポリヌクレオチドの配列、またはその相補体は、標的配列に対して完全に同一であってもよく、またはこれは、標的配列における特定の位置において不一致(mismatched)な塩基を含んでもよい。ミスマッチ(mismatch)の組み込みは、本明細書中で完全に記載される。理論に縛られることを望まないが、ミスマッチの組み込みは、目的の特定の標的配列のためのRNA干渉現象を最適化する生理学的条件下での、意図される程度のハイブリダイゼーションの安定性を提供する。同一性の程度は、2つの配列における塩基が互いに同一である位置の百分率を決定する。「配列同一性の百分率」は、最適に並べられた2つの配列を、比較の領域にわたって比較し、両配列において同一な核酸塩基(例えば、核酸の場合、A、TもしくはU、C、G、またはI)が存在する位置の数を決定して一致した位置の数を得、この一致した位置の数を比較の領域内の位置の総数(すなわち、ウインドウサイズ)で割り、そしてその結果に100を掛けて、配列同一性の百分率を得ることによって、計算される。互いに100%未満同一である配列は、互いに「類似する」かもしくは「相同である」;相同性の程度もしくは同一性百分率は、以下の節において決定されるような2つの配列もしくは下位配列の間の同一性の百分率を意味する、類義語である。例えば、互いに、少なくとも60%の同一性、もしくは好ましくは少なくとも65%の同一性、もしくは好ましくは少なくとも70%の同一性、もしくは好ましくは少なくとも75%の同一性、もしくは好ましくは少なくとも80%の同一性、もしくはより好ましくは少なくとも85%の同一性、もしくはより好ましくは少なくとも90%の同一性、もしくはなおより好ましくは少なくとも95%の同一性を示す、2つの配列は、互いに「類似である」かもしくは「相同である」。あるいは、siRNA分子のオリゴヌクレオチド配列に関して、5もしくはそれ未満の塩基、または4もしくはそれ未満の塩基、または3もしくはそれ未満の塩基、または2もしくはそれ未満の塩基、または1塩基が異なる2つの配列は、互いに「類似である」かもしくは「相同である」。
【0037】
「同一性」は、当該分野で公知であるように、2つもしくはそれより多くのポリペプチド配列もしくは2つもしくはそれより多くのポリヌクレオチド配列の間の関連であり、配列を比較することによって決定される。当該分野では、「同一性」はまた、場合によって、ポリペプチド配列もしくはポリヌクレオチド配列の間の配列の、このような配列の鎖(string)の間のマッチ(match)によって決定される関連性の程度をも意味する。「同一性」および「類似性」は、公知の方法によって容易に計算され得る。これらの方法としては、以下に記載される方法が挙げられるが、これらに限定されない:Computational Molecular Biology,Lesk.A.M.,編,Oxford University Press,New York,1988;Biocomputing:Informatics and Genome Projects,Smith,D.W.,編,Academic Press,New York,1993;Computer Analysis of Sequence Data,Part I.Griffin,A.M.,およびGriffin,H.G.,編Humana Press,New Jersey,1994;Sequence Analysis in Molecular Biology,von Heinje,G.,Academic Press,1987;ならびにSequence Analysis Primer,Gribskov,M.およびDevereux,J.,編,M Stockton Press.New York,1991;ならびにCarillo,H.,およびLipman,D.,SIAM J.Applied Math.(1988)48:1073。同一性を決定する好ましい方法は、試験される配列の間の最大のマッチを与えるように設計される。同一性および類似性を決定する方法は、公に利用可能なコンピュータープログラムにおいてまとめられている。2つの配列間の同一性および類似性を決定するための好ましいコンピュータープログラム方法としては、以下が挙げられるが、これらに限定されない:GCGプログラムパッケージ(Devercux,J.,ら(1984)Nucleic Acids Research 12(1):387)、BLASTP、BLASTN、およびFASTA(Atschul,S.F.ら(1990)J.Molec.Biol.215:403−410)。BLAST Xプログラムは、NCBIおよび他の供給源から、公に利用可能である(BLAST Manual,Altschul,S.,ら,NCBI NLM NIH Bethesda,Md.20894;Altschul,S.,ら(1990)J.Mol.Biol.215:403−410。周知のSmith Watermanアルゴリズムもまた、同一性を決定するために使用され得る。
【0038】
配列比較のためのパラメータとしては、以下が挙げられる:アルゴリズム:NeedlemanおよびWunsch、J.Mol Biol.48:443−453(1970)。
【0039】
比較マトリックス:HentikoffおよびHentikoff,(1992)Proc.Natl.Acad.Sci.USA.89:10915−10919からのBLOSSUM62。
【0040】
本明細書中で使用される場合、用語「単離された」、および類似の語は、核酸、ポリヌクレオチド、もしくはオリゴヌクレオチドを記載するために使用される場合、その天然の状態もしくは元の状態から取り出されることをいう。したがって、これらが天然において存在する場合、これらは、その元の環境から取り出されている。これらが合成的に調製された場合、これらは、合成から得られた元の産物混合物から取り出されている。例えば、この用語が本明細書中で使用される場合、天然の状態において生きている生物中に天然に存在している、天然に存在するポリヌクレオチドは、「単離され」ていないが、その天然の状態において共に存在する物質から分離されている同じポリヌクレオチドは、「単離されて」いる。一般に、少なくとも1種の有意な共に存在する物質の除去は、核酸、ポリヌクレオチド、オリゴヌクレオチドの「単離」を構成する。多くの場合、いくつかの、多くの、もしくはほとんどの、共に存在する物質が、核酸、ポリヌクレオチド、オリゴヌクレオチド、タンパク質、ポリペプチド、もしくはオリゴペプチドを単離するために除去され得る。非限定の例として、ポリヌクレオチドに関して、用語「単離された」は、天然に存在する染色体および細胞からこのポリヌクレオチドが分離されることを意味し得る。さらに、例として、タンパク質もしくはポリペプチドの「単離」は、細胞溶解物もしくは細胞ホモジネートにおける別の成分からの分離を意味し得る。
【0041】
インビトロ合成プロセスもしくは化学合成プロセスの産物である核酸、ポリヌクレオチド、もしくはオリゴヌクレオチドは、合成プロセスの結果として、本質的に単離されている。重要な実施形態において、このような合成産物は、使用される試薬および前駆体、ならびにそのプロセスによる副産物を除去するために処理される。
【0042】
同様に、ポリヌクレオチドおよびポリペプチドは、例えば、処方物、ポリヌクレオチドの細胞への導入のための組成物、化学反応もしくは酵素的反応のための組成物もしくは溶液のような組成物中に存在し得る。これらの組成物は、天然に存在する組成物ではなく、その中に、本明細書中で使用される用語の意味の範囲内である単離されたポリヌクレオチドもしくはポリペプチドを維持する。
【0043】
本明細書中および特許請求の範囲において使用される場合、「核酸」もしくは「ポリヌクレオチド」、およびこれらに基づく類似の用語は、天然に存在するヌクレオチドからなるポリマーおよび合成ヌクレオチドもしくは改変ヌクレオチドからなるポリマーをいう。したがって、本明細書中で使用される場合、RNAであるポリヌクレオチド、もしくはDNAであるポリヌクレオチドは、天然に存在する部分(例えば、天然に存在する塩基およびリボース環もしくはデオキシリボース環)を含み得るか、またはこれらは、以下で記載されるような合成部分もしくは改変部分から構成され得る。ヌクレオチド間の結合は、一般に、3’−5’リン酸結合であり、これは、天然のホスホジエステル結合であっても、ホスホチオエステル結合であっても、なお他の合成的結合であってもよい。改変骨格の例としては、ホスホロチオエート、キラルホスホロチオエート、ホスホロジチオエート、ホスホトリエステル、アミノアルキルホスホトリエステル、メチルホスホネートおよび他のアルキルホスホネート(3’−アルキレンホスホネート、5’−アルキレンホスホネートおよびキラルホスホネートを含む)、ホスフィネート、ホスホルアミデート(3’−アミノホスホルアミデートおよびアミノアルキルホスホルアミデートを含む)、チオノホスホルアミデート、チオノアルキルホスホネート、チオノアルキルホスホトリエステル、セレノホスフェートおよびボラノホスフェートが挙げられる。さらなる結合としては、ホスホトリエステル、シロキサン、カーボネート、カルボキシメチルエステル、アセトアミデート、カルバメート、チオエーテル、架橋されたホスホルアミデート、架橋されたメチレンホスホネート、架橋されたホスホロチオエートおよびスルホンのヌクレオチド内結合が挙げられる。他のポリマー性結合としては、これらの2’−5’結合アナログが挙げられる。米国特許第6,503,754号および同第6,506,735号ならびにこれらの中に引用されている参考文献(本明細書中で参考として援用される)を参照のこと。
【0044】
核酸およびポリヌクレオチドは、20もしくはそれより多くのヌクレオチドの長さ、または30もしくはそれより多くのヌクレオチドの長さ、または50もしくはそれより多くのヌクレオチドの長さ、または100もしくはそれより多いか、または1000もしくはそれより多いか、または10,000もしくはそれより多いか、または100,000もしくはそれより多い長さである。siRNAは、本明細書中で規定されるポリヌクレオチドであり得る。本明細書中で使用される場合、「オリゴヌクレオチド」およびこれに基づく類似の用語は、直前の段落で記載されるように、天然に存在するヌクレオチドからなる短いポリマー、ならびに合成ヌクレオチドもしくは改変ヌクレオチドからなるポリマーをいう。オリゴヌクレオチドは、10もしくはそれより多くのヌクレオチドの長さ、または15、もしくは16、もしくは17、もしくは18、もしくは19、もしくは20またはそれより多くのヌクレオチドの長さ、または21、もしくは22、もしくは23、もしくは24またはそれより多くのヌクレオチドの長さ、または25、もしくは26、もしくは27、もしくは28もしくは29、もしくは30またはそれより多くのヌクレオチドの長さ、35もしくはそれより多く、40もしくはそれより多く、45もしくはそれより多く、約50までのヌクレオチドの長さであり得る。siRNAであるオリゴヌクレオチドは、15と30との間の任意の数のヌクレオチドを有し得る。多くの実施形態において、siRNAは、21と25との間の任意の数のヌクレオチドを有し得る。
【0045】
大きさの範囲の重複ゆえに、用語「ポリヌクレオチド」および「オリゴヌクレオチド」は、本発明のsiRNAをいうために本明細書中で同じように使用され得ることが、上で示された定義から理解される。
【0046】
本明細書中および特許請求の範囲において使用される場合、「ヌクレオチド配列」、「オリゴヌクレオチド配列」もしくは「ポリヌクレオチド配列」、および類似の用語は、オリゴヌクレオチドもしくはポリヌクレオチドが有する塩基の配列と、この配列を有するオリゴヌクレオチドもしくはポリヌクレオチドの構造との両方に、相互交換可能に関連する。ヌクレオチド配列もしくはポリヌクレオチド配列は、さらに、任意の天然のポリヌクレオチドもしくはオリゴヌクレオチド、または任意の合成のポリヌクレオチドもしくはオリゴヌクレオチドに関連し、これらのポリヌクレオチドもしくはオリゴヌクレオチドにおいて、塩基の配列は、当該分野で従来使用されるように、塩基を示す文字の特定の配列の記載もしくは言及によって規定される。
【0047】
オリゴヌクレオチドおよびポリヌクレオチドにおける塩基は、プリン塩基(アデニン(A)およびグアニン(G))ならびにピリミジン塩基(チミジン(T)、シトシン(C)およびウラシル(U))を含む、「未改変」塩基もしくは「天然」塩基であり得る。さらに、これらは、改変もしくは置換を有する塩基であり得る。本明細書中で使用される場合、改変塩基の非限定の例は、他の合成塩基および天然塩基を含む。これらの塩基は、例えば、5−メチルシトシン(5−me−C)、5−ヒドロキシメチルシトシン、キサンチン、ヒポキサンチン、2−アミノアデニン、アデニンおよびグアニンの6−メチル誘導体および他のアルキル誘導体、アデニンおよびグアニンの2−プロピル誘導体および他のアルキル誘導体、2−チオウラシル、2−チオチミジンおよび2−チオシトシン、5−ハロウラシルおよび5−ハロシトシン、5−プロピニルウラシルおよび5−プロピニルシトシンおよびピリミジン塩基の他のアルキニル誘導体、6−アゾウラシル、6−アゾシトシンおよび6−アゾチミジン、5−ウラシル(プソイドウラシル)、4−チオウラシル、8−ハロ、8−アミノ、8−チオール、8−チオアルキル、8−ヒドロキシルおよび他の8−置換アデニンおよびグアニン、5−ハロ(特に5−ブロモ)、5−トリフルオロメチルおよび他の5−置換ウラシルおよびシトシン、7−メチルグアニンおよび7−メチルアデニン、2−フルオロ−アデニン、2−アミノアデニン、8−アザグアニンおよび8−アザアデニン、7−デアザグアニンおよび7−デアザアデニンおよび3−デアザグアニンおよび3−デアザアデニンである。さらなる改変塩基としては、三環式ピリミジン(例えば、フェノキサジンシチジン(1H−ピリミド[5,4−b][1,4]ベンゾキサジン−2(3H)−オン)、フェノチアジンシチジン(1−ピリミド[5,4−b][1,4]ベンゾチアジン−2(3H)−オン))、G−クランプ(clamp)(例えば、置換フェノキサジンシチジン(例えば、9−(2−アミノエトキシ)−H−ピリミド[5,4−b][1,4]ベンゾキサジン−2(3H)−オン)、カルバゾールシチジン(2H−ピリミド[4,5−b]インドール−2−オン)、ピリドインドールシチジン(H−ピリド[3’、2’:4,5]ピロロ[2,3−d]ピリミジン−2−オン))が挙げられる。改変塩基はまた、プリン塩基もしくはピリミジン塩基が他の複素環と置換されている改変塩基(例えば、7−デアザ−アデニン、7−デアザグアノシン、2−アミノピリジンおよび2−ピリドン)を含む。さらなる塩基としては、米国特許第3,687,808号に開示される塩基、The Concise Encyclopedia Of Polymer Science And Engineering,858−859頁,Kroschwitz,J.L編,John Wiley & Sons,1990によって開示される塩基、Englischら,Angewandte Chemie,International Edition(1991)30,613によって開示される塩基、ならびにSanghvi,Y.S.,第15章,Antisense Research and Applications,289−302頁、Crooke,S.T.およびLebleu,B.,編,CRC Press,1993によって開示される塩基が挙げられる。特定のこれらの塩基は、本発明のオリゴマー化合物の結合親和性を増大させるために、特に有用である。これらとしては、5−置換ピリミジン、6−アザピリミジンおよびN−2置換プリン、N−6置換プリンおよびO−6置換プリン(2−アミノプロピルアデニン、5−プロピニルウラシルおよび5−プロピニルシトシンを含む)が挙げられる。5−メチルシトシン置換は、核酸二本鎖の安定性を0.6〜1.2℃増大することが示されており(Sanghvi,Y.S.,Crooke,S.T.およびLebleu,B.,編,Antisense Research and Applications,CRC Press,Boca Raton,1993,pp.276−278)、ここで好ましい塩基置換であり、なおより好ましくは、2’−O−メトキシエチル糖修飾と結合される場合である。米国特許第6,503,754号および同第6,506,735号ならびにこれらにおいて引用された参考文献(本明細書中で参考として援用される)を参照のこと。任意の改変塩基の使用は、当業者に理解されるとおり、同じ塩基対特性を有する天然に存在する塩基の使用と同等である。
【0048】
本明細書中および特許請求の範囲において使用される場合、用語「相補的」およびこれに基づく類似の語は、核酸、ポリヌクレオチドもしくはオリゴヌクレオチドの1つの鎖における第1の核酸塩基の、核酸、ポリヌクレオチドもしくはオリゴヌクレオチドの1つの鎖における特定の第2の核酸塩基とのみ特異的に相互作用する能力をいう。非限定の例として、天然に存在する塩基が考慮される場合、AとTもしくはUとは、互いに相互作用し、そしてGとCとは互いに相互作用する。本発明および特許請求の範囲において使用される場合、「相補的」は、「完全に相補的」であることを意味し、すなわち、2つのポリヌクレオチド鎖が互いに並列される場合、1本の鎖における連続する塩基の配列における各塩基が、対向する鎖における同じ長さの連続する塩基の配列における相互作用する塩基に対して相補的である、少なくとも一部分の鎖が存在する。
【0049】
本明細書中で使用される場合、「ハイブリダイズ」、「ハイブリダイゼーション」および類似の語は、鎖と相補的な配列とを互いに相互作用させることによって、核酸、ポリヌクレオチド、もしくはオリゴヌクレオチドの2本鎖を形成するプロセスをいう。この相互作用は、各々の鎖における相補的な塩基が、特異的に相互作用して対を形成することによって起こる。鎖が互いにハイブリダイズする能力は、以下で示すような種々の条件に依存する。核酸鎖は、各鎖における十分な数の対応する位置が、互いに相互作用し得るヌクレオチドによって占められている場合、互いにハイブリダイズする。二本鎖を形成する鎖の配列は、特異的にハイブリダイズすることができるために互いに100%の相補性を必要とはしないことが、本発明の分野の当業者(非限定の例として、分子生物学者および細胞生物学者が挙げられる)によって理解される。
【0050】
本明細書中で使用される場合、「フラグメント」およびこれに基づく類似の語は、参照の全体配列よりも短い、核酸、ポリヌクレオチドもしくはオリゴヌクレオチドの一部分に関連する。フラグメントにおける塩基の配列は、このフラグメントが生じた分子の対応する部分の配列から改変されない;フラグメントにおいて、このフラグメントが生じた分子の対応する部分の配列と比較して、挿入も欠失も存在しない。本明細書中で企図されるように、核酸もしくはポリヌクレオチドのフラグメント(例えば、オリゴヌクレオチド)は、15もしくはそれより多くの塩基の長さ、または16もしくはそれより多く、17もしくはそれより多く、18もしくはそれより多く、または19もしくはそれより多く、または20もしくはそれより多く、または21もしくはそれより多く、または22もしくはそれより多く、または23もしくはそれより多く、または24もしくはそれより多く、または25もしくはそれより多く、または26もしくはそれより多く、または27もしくはそれより多く、または28もしくはそれより多く、または29もしくはそれより多く、30もしくはそれより多く、50もしくはそれより多く、75もしくはそれより多く、100もしくはそれより多くの塩基の長さであり、全長配列より1塩基短い長さまでの長さである。オリゴヌクレオチドは、化学的に合成され得、そしてsiRNA、PCRプライマー、またはプローブとして使用され得る。
【0051】
検出および標識。標的性ポリヌクレオチド、例えばsiRNA配列を含むポリヌクレオチド、ならびにウイルスポリヌクレオチド標的は、多くの手段によって検出され得る。検出としては、標的性ポリヌクレオチドの存在もしくは量を観察する能力をもたらす、1種もしくはそれより多くの任意のプロセスが挙げられ得る。1つの実施形態において、標的性ポリヌクレオチドもしくはウイルス標的を含むサンプル核酸は、拡張(expansion)の前に検出され得る。代替の実施形態において、サンプル中の標的性ポリヌクレオチドは、拡張されて、拡張された標的性ポリヌクレオチドもしくは拡張されたウイルス標的を提供し得、そしてこの拡張されたポリヌクレオチドが、検出されるかもしくは定量され得る。物理的方法、化学的方法もしくは生物学的方法が、標的性ポリヌクレオチドを検出しそして定量するために使用され得る。物理的方法としては、非限定の例として、表面プラスモン共鳴(SPR)検出(例えば、プローブの表面への結合)および標的性ポリヌクレオチドの固定化プローブへの結合を検出するためのSPRの使用、またはクロマトグラフィー媒体中にプローブを有し、そして標的性ポリヌクレオチドの結合をクロマトグラフィー媒体中で検出することをが挙げられる。物理的方法は、標的性ポリヌクレオチドが他のポリヌクレオチドから分離され、そしてこの分離されたポリヌクレオチドが検出される、ゲル電気泳動形式もしくはキャピラリー電気泳動形式をさらに含む。化学的方法としては、ポリメラーゼ連鎖反応(PCR)方法、および一般に標的性ポリヌクレオチドがプローブにハイブリダイズする、ハイブリダイゼーション方法が挙げられる。生物学的方法としては、標的性ポリヌクレオチドもしくは標的ポリヌクレオチドに、細胞において生物学的効果を発揮させ、そしてこの効果を検出することが挙げられる。本発明は、生物学的アッセイにおいて使用され得る生物学的効果の例を開示する。これらとしては、粒子を数えることによるビリオンの計数、プラークアッセイ、感染細胞における細胞変性効果の評価などが挙げられる。多くの実施形態において、ポリヌクレオチドは、以下で記載されるように標識されて、検出および定量を補助する。例えば、拡張を包含しない実施形態において、サンプル核酸は、標識化ヌクレオチドもしくは標識化オリゴヌクレオチドリンカーのような標識化部分の化学的付加もしくは酵素的付加によって標識され得る。
【0052】
拡張されたポリヌクレオチドが、直接的に検出され得、そして/または定量され得る。例えば、拡張されたポリヌクレオチドは、ゲル中の電気泳動(大きさによって分離する)に供され、そして色素によって染色されて、その存在および量を明らかにし得る。あるいは、拡張された標的性ポリヌクレオチドは、プローブ核酸のハイブリダイゼーション条件(以下を参照)下での曝露の際に検出され得、そしてハイブリダイゼーションによる結合が、検出されそして/または定量される。検出は、標的性ポリヌクレオチドがプローブに結合していることの決定を可能にすることを何らかの手段で達成する。このことは、フラグメントをハイブリダイズすることによってもたらされるプローブの物理的特性における変化を検出することによって達成され得る。このような物理的検出方法の非限定の例は、SPRである。
【0053】
検出を達成する代替の方法は、この拡張されたポリヌクレオチドの標識化形態を使用すること、および結合した標識を検出することである。標識は、例えば、125I、35S、32P、14C、もしくは3Hのような放射性同位体標識であり得、これはその放射活性によって検出され得る。あるいは、標識は、分光学的方法を用いて(例えば、蛍光、リン光もしくは化学発光によって)検出され得るように選択され得る。したがって、蛍光標識もしくはリン光標識または化学発光反応を誘導する標識が、使用され得る。標識は、なおさらに、特定のリガンド−レセプター対におけるリガンドであり得る;このリガンドの存在は、次いで、通常はそれ自体が検出のために標識されている特異的レセプターの二次結合によって検出される。
【0054】
(干渉性RNA)
本発明に従って、呼吸性ウイルス標的の遺伝子発現は、RNA干渉によって低減される。ウイルス遺伝子の発現産物は、このウイルス標的の少なくとも1つのセグメントに対して相補的である特異的二本鎖siRNAヌクレオチド配列によって標的化され、このセグメントは、15と30との間の任意の数のヌクレオチドを含むか、または多くの場合、21と25との間の任意の数のヌクレオチドを含む。この標的は、5’非翻訳(UT)領域中、コード配列中、または3’UT領域中に存在し得る。例えば、PCT出願WO00/44895、WO99/32619、WO01/75164、WO01/92513、WO01/29058、WO01/89304、WO02/16620、およびWO02/29858(各々は、本明細書中でその全体が参考として援用される)を参照のこと。
【0055】
本発明の方法に従い、呼吸性ウイルス遺伝子発現およびそれによる呼吸性ウイルス複製は、siRNAを用いて抑制される。本発明に従う標的性ポリヌクレオチドは、siRNAオリゴヌクレオチドを含む。このようなsiRNAはまた、ウイルス配列と同一もしくは類似であるヌクレオチド配列の化学的合成によって調製され得る。例えば、Tuschl,Zamore,Lehmann,BartelおよびSharp(1999),Genes & Dev.13:3191−3197(本明細書中でその全体が参考として援用される)を参照のこと。あるいは、標的性siRNAは、標的性ポリヌクレオチド配列を用いて、例えば、無細胞系(例えばDrosophila抽出物であるがこれに限定されない)において呼吸性ウイルスリボポリヌクレオチド配列を消化することによって、または組換え二本鎖ウイルスcRNAの転写によって、得られ得る。
【0056】
効率的なサイレンシングは、一般に、同じ長さの16〜30ntのセンス鎖と16〜30ntのアンチセンス鎖とから構成されるsiRNA二本鎖によって観察される。多くの実施形態において、siRNA対形成二本鎖の各鎖は、3’末端において2−ntのオーバーハングの付加を有する。2−ntの3’オーバーハングの配列は、siRNA標的認識の特異性に対し、さらに僅かに寄与する。1つの実施形態において、3’オーバーハング中のヌクレオチドは、リボヌクレオチドである。代替の実施形態において、3’オーバーハング中のヌクレオチドは、デオキシリボヌクレオチドである。3’デオキシヌクレオチドの使用は、細胞内安定性の増強を提供する。
【0057】
本発明の組換え発現ベクターは、細胞内に導入された場合、呼吸性ウイルスを標的化するsiRNA配列を含むRNAを提供するようにプロセシングされる。このようなベクターは、発現を可能にする様式でウイルス標的化配列に隣接(flanking)する作動可能に連結された制御配列を含む発現ベクター内にクローニングされたDNA分子である。このベクターからは、ウイルスRNAに対するアンチセンスであるRNA分子は、第1のプロモーター(例えば、クローニングされたDNAの3’のプロモーター配列)によって転写され、そしてウイルスRNA標的についてのセンス鎖であるRNA分子は、第2のプロモーター(例えば、クローニングされたDNAの5’のプロモーター配列)によって転写される。このセンス鎖およびアンチセンス鎖は、次いで、インビボでハイブリダイズして、呼吸性ウイルス遺伝子のサイレンシングのためにこのウイルスを標的化するsiRNA構築物を産生する。あるいは、2つの構築物が、siRNA構築物のセンス鎖およびアンチセンス鎖を作製するために使用され得る。さらに、クローニングされたDNAは、二次構造を有する転写物をコードし得、ここで、1つの転写物は、標的遺伝子からのセンス配列および相補的アンチセンス配列の両方を有する。この実施形態の例において、ヘアピンRNAi産物は、標的遺伝子の全てもしくは一部分に類似する。別の例において、ヘアピンRNAi産物は、siRNAである。ウイルス配列に隣接(flanking)する調節配列は、同一であっても異なっていてもよく、それによって、その発現は、独立して調節されてもよく、または時間的様式もしくは空間的様式で調節されてもよい。
【0058】
特定の実施形態において、siRNAは、例えば、より低分子の核RNA(snRNA)U6もしくはヒトRNase P RNA H1由来のRNApol III転写単位を含むベクター内に、ウイルス遺伝子テンプレートをクローニングすることによって細胞内で転写される。ベクター系の1つの例は、GeneSuppressorTM RNA干渉キット(Imgenexから市販される)である。U6プロモーターおよびH1プロモーターは、Pol IIIプロモーターのIII型クラスのメンバーである。U6様プロモーターの+1ヌクレオチドは、常にグアノシンであり、一方、H1プロモーターについての+1は、アデノシンである。これらのプロモーターについての終止シグナルは、5つの連続するチミジンによって規定される。この転写物は、代表的に、第2のウリジンの後で切断される。この位置における切断は、発現されたsiRNAにおいて3’UUオーバーハングを産生し、これは、合成siRNAの3’オーバーハングと類似する。400ヌクレオチド未満の長さの任意の配列が、これらのプロモーターによって転写され得るので、これらは、例えば、約50ヌクレオチドのRNAステム−ループ転写物における約21ヌクレオチドのsiRNAの発現に理想的に適合する。RNAiの特徴およびsiRNAの効力に影響を及ぼす要因が、研究されている(例えば、Elbashir,LendeckelおよびTuschl(2001)Genes & Dev.15:188−200)。
【0059】
選択されたsiRNA配列についての最初のBLAST相同性検索は、入手可能なヌクレオチド配列ライブラリーに対して行われ、ウイルス遺伝子のみが標的化され、宿主遺伝子が標的化されないことを確認する。Elbashirら,2001 EMBO J.20(23):6877−88を参照。
【0060】
(ポリヌクレオチドの合成)
標的性ヌクレオチド配列に対応するオリゴヌクレオチドおよび標的性配列を含むポリヌクレオチドは、例えば、自動化DNA合成装置を用いて、標準的合成技術によって調製され得る。オリゴヌクレオチドを合成するための方法は、周知の化学的プロセスを含み、これらとしては、表面誘導体化粒子へのヌクレオチドホスホルアミダイトの連続的付加(T,BrownおよびDorcas J.S.Brown,Oligonucleotides and Analogues A Practical Approach,F.Eckstein,編,Oxford University Press,Oxford,pp.1−24(1991)によって記載され、本明細書中で参考として援用される)が挙げられるが、これらに限定されない。
【0061】
合成手順の一例は、Expedite RNAホスホルアミダイトおよびチミジンホスホルアミダイト(Proligo,Germany)を使用する。合成オリゴヌクレオチドは、脱保護されそしてゲル精製されて(Elbashirら(2001)Genes & Dev.15,188−200)、その後、Sep−PakC18カートリッジ(Waters,Milford,Mass.,USA)精製される(Tuschlら(1993)Biochemistry,32:11658−11668)。相補的ssRNAは、アニーリング緩衝液(100mM酢酸カリウム、30mM HEPES−KOH(pH7.4)、2mM酢酸マグネシウム)中で90℃で、1分間インキュベーションされ、その後、37℃で1時間インキュベーションされて、対応するds−siRNAとハイブリダイズする。
【0062】
オリゴヌクレオチド合成の他の方法としては、以下が挙げられるが、これらに限定されない:ホスホトリエステル方法およびホスホジエステル方法(Narangら,(1979)Meth.Enzymol.68:90)ならびにH−ホスホネート方法(とりわけGaregg,P.J.ら,(1985)「Formation of internucleotidic bonds via phosphonate intermediates」,Chem.Scripta 25,280−282;およびFroehler,B.C,ら,(1986a)「Synthesis of DNA via deoxynucleoside H−phosphonate intermediates」,Nucleic Acid Res.,14,5399−5407)に従う固相オリゴヌクレオチド合成、ならびに支持体上の合成(Beaucageら(1981)Tetrahedron Letters 22:1859−1862)、ならびにホスホルアミデート技術(Caruthers,M.H.,ら,「Methods in Enzymology」,第154巻,pp.287−314(1988))、米国特許第5,153,319号、同第5,132,418号、同第4,500,707号、同第4,458,066号、同第4,973,679号、同第4,668,777号、および同第4,415,732号ならびに「Synthesis and Applications of DNA and RNA」,S.A.Narang,編,Academic Press,New York,1987およびその中に含まれる参考文献に記載される技術、ならびに非ホスホルアミダイト技術。固相合成は、オリゴヌクレオチドを不純物および過剰な試薬から単離することを助ける。一旦固体支持体から切断されると、このオリゴヌクレオチドは、公知の技術によってさらに単離され得る。
【0063】
(本発明の阻害性ポリヌクレオチド)
本発明は、呼吸性ウイルス病原体に感染した細胞内に侵入する際にRNA干渉現象を引き起こすように意図されるオリゴヌクレオチドを、広く提供する。本発明は、呼吸性ウイルス標的の性質に制限されないが、RSVおよびインフルエンザAのいくつかの株を標的化するオリゴヌクレオチドを強調する。RNA干渉は、適切な二本鎖RNAによって細胞内で起こり、この二本鎖RNAの鎖のうちの1本は、ウイルスの標的遺伝子における配列と同一であるかこれに高度に類似する。一般に、呼吸性ウイルスを標的化するオリゴヌクレオチドは、DNAもしくはRNAであり得るか、または、リボヌクレオチドおよびデオキシリボヌクレオチドの混合物を含み得る。後者の一例は、デオキシジヌクレオチド配列(例えば、d(TT)、d(UU)、d(TU)、d(UT)および他の可能性のあるジヌクレオチド)で3’末端において終止するRNA配列である。さらなる実施形態において、3’オーバーハングは、上で特定された塩基を有するリボヌクレオチドなどから構成され得る。さらに、このオリゴヌクレオチド薬剤は、一本鎖もしくは二本鎖であり得る。本発明の治療用オリゴヌクレオチドのいくつかの実施形態は、オリゴリボヌクレオチドであるか、もしくは3’d(TT)末端を有するオリゴリボヌクレオチドであることが予測される。一本鎖標的性ポリヌクレオチドは、哺乳動物細胞への侵入の際、細胞内に存在する内因性酵素活性によって、容易に二本鎖分子に変換される。
【0064】
最も一般的には、本発明は、オリゴヌクレオチドもしくはポリヌクレオチドを提供し、これは、15ヌクレオチドから200ヌクレオチドの長さまでのいずれかの長さの範囲であり得る。このポリヌクレオチドは、RSウイルスもしくはインフルエンザAウイルスのゲノムを標的化する第1のヌクレオチド配列を含む。この第1のヌクレオチド配列は、(a)15〜30の任意の数のヌクレオチドの長さである配列、または(b)その相補体のいずれかから構成される。このようなポリヌクレオチドは、本明細書中で、直鎖状ポリヌクレオチドと呼ばれる。
【0065】
図3は、本発明のポリヌクレオチドの特定の実施形態の概略図を提供する。本発明は、RSVもしくはインフルエンザAの種々の株における標的配列を開示するか、または特定の場合には、標的配列と僅かに不適合であるsiRNA配列を開示し、これらの全ては、配列番号1〜263に提供される。配列番号1〜263に提供される配列は、19ヌクレオチド〜25ヌクレオチドの長さの範囲である。この標的性配列は、図3において薄い影をつけたブロックによって表される。図3、パネルAのa)は、「SEQ」として示される開示された配列が、必要に応じてより大きなポリヌクレオチド内に含まれ得、このポリヌクレオチドの全長が200ヌクレオチドまでの範囲に及び得る、実施形態を図示する。
【0066】
本発明は、標的性ポリヌクレオチドにおいて、配列番号1〜263から選択される配列が、SEQによって表される第1のヌクレオチド配列よりも長いウイルスゲノム中の配列を標的化するようなより長い標的性配列の一部分であり得ることを、さらに提供する。これは、図3、パネルAのb)に図示され、ここでは、完全な標的性配列が、ポリヌクレオチドの上の横線およびSEQブロックの周囲の濃い影によって示される。ポリヌクレオチドの全ての実施形態において、このより長い配列は、必要に応じて、なおより長い(200もしくはより少ない塩基の長さである)ポリヌクレオチドに含まれ得る(図3、パネルA、b))。
【0067】
本発明は、配列番号1〜263のいずれかのフラグメントであって、少なくとも15ヌクレオチドの長さである(そして長くとも参照配列番号よりも1塩基短い;図3、パネルA、c)において図示される)配列、ならびに5ヌクレオチドまでが配列番号1〜263において与えられる標的配列から異なり得る(図3、パネルA、d)において図示される)配列(この例において、3つの改変塩基が、3本の濃い縦棒によって表される)をさらに提供する。
【0068】
なおさらに、本発明は、上記の配列のいずれかの相補体である配列を提供する(図3、パネルAのe)に示され、「COMPL」と名付けられる)。任意のこれらの配列が、本発明のオリゴヌクレオチドもしくはポリヌクレオチドに含まれる。本発明の任意の直鎖状ポリヌクレオチドは、上記のa)〜e)に記載される配列のみによって構成されてもよく、または必要に応じて、上限200ヌクレオチドまでのさらなる塩基を含んでもよい。RNA干渉は、二本鎖RNAを必要とするので、標的性ポリヌクレオチド自身は、少なくとも配列番号1〜263において与えられる配列に対して相補的でありかつこれにハイブリダイズする第2の鎖を含む二本鎖であり得るか、または細胞内プロセシングに依存して相補鎖を産生し得る。
【0069】
従って、上記のポリヌクレオチドは、一本鎖であってもよく、または二本鎖であってもよい。なおさらなる実施形態において、このポリヌクレオチドは、デオキシリボヌクレオチドのみを含むか、またはリボヌクレオチドのみを含むか、あるいはデオキシリボヌクレオチドとリボヌクレオチドとの両方を含む。本明細書中で記載されるポリヌクレオチドの重要な実施形態において、標的配列は、15ヌクレオチド(nt)、もしくは16nt、もしくは17nt、もしくは18nt、もしくは19nt、もしくは20nt、もしくは21nt、もしくは22nt、もしくは23nt、もしくは24nt、もしくは25nt、もしくは26nt、もしくは27nt、もしくは28nt、もしくは29、もしくは30ntのいずれかの長さであり得る配列からなる。なおさなる有利な実施形態において、この標的性配列は、ウイルス病原体ゲノムにおける標的配列と5塩基まで異なり得る。
【0070】
本発明のいくつかの実施形態において、上記ポリヌクレオチドは、標的性配列からなり、そして必要に応じて、本明細書中に記載される3’ジヌクレオチドオーバーハングを含む、siRNAである。
【0071】
あるいは、RNA干渉における二本鎖RNAの必要を認識して、上記オリゴヌクレオチドもしくはポリヌクレオチドは、分子内ヘアピンループ二本鎖分子を形成するように調製され得る。このような分子は、上の段落の実施形態のいずれかにおいて記載される第1の配列、その後ろの短いループ配列、そのさらに後ろの第2の配列であって第1の配列に対して相補的な配列から形成される。このような構造は、所望の分子内ヘアピンを形成する。その上さらに、また、挙げられた3つの所望の構造が、200ヌクレオチドまでの任意の全体の長さを有する任意のオリゴヌクレオチドもしくはポリヌクレオチドにおいて構成され得るような、最大200ヌクレオチドの長さを有するポリヌクレオチドが開示される。ヘアピンループポリヌクレオチドは、図3、パネルBにおいて図示される。
【0072】
(標的としてのRSV株)
RSVについて、ウイルスゲノムの任意の部分が標的化され得るが、理屈からすれば、ウイルス特異的酵素的機能をコードする標的性遺伝子が潜在的siRNA候補を提供するはずである。これらの遺伝子としては、L遺伝子、F遺伝子、G遺伝子およびP遺伝子が挙げられる。ウイルスゲノムの5’末端に位置するL遺伝子は、少量でしか発現されず、従って、有効なRNAiサイレンシングは、少量のRNAiのみを必要とし得る。構造遺伝子もまた、標的化され得る。
【0073】
以下のRSVサブグループAおよびBの代表的な株の配列は、サブグループAとサブグループBとの間で共通である(表1)か、またはサブグループA(表2)もしくはサブグループB(表3)に特異的である標的配列を選択するために使用された。
【0074】
サブグループA:株A2(GenBank M74568)および株Long(P−mRNA、GenBank M22644、およびF−mRNA、GenBank M22643のみ)。
【0075】
サブグループB:株B1(GenBank NC_001781)および株9320(GenBank AY353550)。
【0076】
ウイルス遺伝子配列は、共通領域もしくは独特の領域を探索するために並列された。各標的化遺伝子もしくは標的化領域について、少なくとも2つの標的を、適用不可能(NA)でない限り選択した。両サブグループに共通の標的配列を見出すことは、これらのどちらかに独特の標的を見出すよりも難しかった。何故なら、利用可能な、相同であるかもしくは同一である配列は、非常に少ないからである。この場合(表1)、時々、RNA二本鎖の1端の5’末端にミスマッチを導入する必要があった。
【0077】
【表1】
(表2.サブグループA(株A2 & Long株のF/P)に特異的な遺伝子標的)
【0078】
【表2】
(表3.サブグループB(株B1および株9320)に特異的な遺伝子標的)
【0079】
【表3】
ウイルスゲノムの5’末端に対して指向されるsiRNA因子を提供するために、株B1もしくは株9320のいずれか個々に独特の尾部(trail)標的が、以下に示される(表4)。(+)鎖(アンチ−ゲノムRNA)の5’末端が標的化され、すなわち、ポジティブRNA鎖(アンチゲノムRNA)をテンプレートとして用いてウイルスゲノムが複製を開始する際、新生リーダー配列が産生される。
【0080】
(表4.RSVサブグループBの2つの株についての尾部標的)
【0081】
【表4】
上の表1〜表4に列挙した標的に基づき、siRNAは、以下のように作製され得る:a)二本鎖siRNAの各鎖に、3’−dTdTオーバーハングが提供される、b)「ミスマッチ」が必要な場合、G:Cは、G:TもしくはG:Aに変えられ、A:Tは、A:CもしくはA:Gに変えられ、そしてC:GはC:TもしくはC:Aに変えられる。
【0082】
(標的としてのインフルエンザA株)
(+)鎖mRNA配列内の25ヌクレオチドおよび19ヌクレオチドの長さのsiRNA標的配列が、全ての8つのセグメントで同定されている。この配列は、表5〜表12に示される。siRNA配列は、この8つのセグメントの各々に由来する。各配列についての記載(entry)はまた、5’末端からのmRNAにおける開始ヌクレオチド位置を示す。
【0083】
(表5.インフルエンザAウイルスのヘマグルチニン遺伝子(A/chicken/Thailand/CH−2/2004(H5N1);GenBank AY649382)を標的化するsiRNA配列。)
【0084】
【表5】
(表6.インフルエンザAウイルスのマトリックスタンパク質2およびマトリックスタンパク質1(M)遺伝子(A/Thailand/1(KAN−1)/2004(H5N1);GenBank AY626144)を標的化するsiRNA配列。)
【0085】
【表6】
(表7.インフルエンザAウイルスのノイラミニダーゼ遺伝子(A/chicken/Thailand/CH−2/2004(H5N1);GenBank AY649383)を標的化するsiRNA配列。)
【0086】
【表7】
(表8.インフルエンザAウイルスのヌクレオカプシドタンパク質(NP)遺伝子(A/Thailand/1(KAN−1)/2004(H5N1);GenBank AY626145)を標的化するsiRNA配列。)
【0087】
【表8】
(表9.インフルエンザAウイルスの非構造タンパク質1および非構造タンパク質2(NS)の遺伝子(A/Thailand/1(KAN−1)/2004(H5N1);GenBank AY626146)を標的化するsiRNA配列。)
【0088】
【表9】
(表10.インフルエンザAウイルスのポリメラーゼ酸性タンパク質(PA)遺伝子(A/Thailand/1(KAN−1)/2004(H5N1);GenBank AY626147)を標的化するsiRNA配列。)
【0089】
【表10】
(表11.インフルエンザAウイルスのポリメラーゼ塩基性タンパク質1(PB1)遺伝子(A/Thailand/1(KAN−1)/2004(H5N1);GenBank AY626148)を標的化するsiRNA配列。)
【0090】
【表11】
(表12.インフルエンザAウイルスのポリメラーゼ塩基性タンパク質2(PB2)遺伝子(A/Thailand/1(KAN−1)/2004(H5N1);GenBank AY626149)を標的化するsiRNA配列。)
【0091】
【表12】
インフルエンザAを標的化する潜在的siRNA候補配列の選択のための別のアプローチは、2つの主要なウイルス表面糖タンパク質であるヘマグルチニン(HA)およびノイラミニダーゼ(NA)のサブタイプ特異性に着目することである。ヒト感染を引き起こすことが報告されているサブタイプは、H5N1、H7N7、およびH9N2である。以下は、サブタイプ特異的なHA標的およびNA標的の例である。
【0092】
(表13.H5N1ヘマグルチニン(HA)のsiRNA標的化配列(GenBank DQ023145,GI:66775624に基づく):
【0093】
【表13】
表13における配列は、中国において2000年と2003年との間にHPAIウイルスによって感染したニワトリから単離された6種のH5N1株によって共有される。これらのH5N1株は、表14に列挙される。
【0094】
(表14)
【0095】
【表14】
(表15.H5N1ノイラミニダーゼ(NA)のsiRNA標的化配列(GenBank DQ023147,GI:66775628に基づく):
【0096】
【表15】
上の配列は、中国において2001年と2003年との間に感染したニワトリもしくはブタから単離された6種のH5N1株によって共有される。これらのH5N1株は、以下の表16に列挙される。
【0097】
(表16)
【0098】
【表16】
(表17.H7N7ヘマグルチニン(HA)のsiRNA標的化配列(GenBank AY999986,GI:66394837に基づく):
【0099】
【表17】
上の全ての配列は、スウェーデンにおいて2002年にマガモから単離された6種のH7N7株によって共有される。これらのH7N7株は、以下の表18に列挙される。
【0100】
6種のHA配列を比較して、HA1関連ドメインがより高い頻度の点変異を有することが見出されており、従って、10個の標的がHA2ドメインから選択される。
【0101】
(表18)
【0102】
【表18】
いくつかのインフルエンザA H5N1 mRNA配列に対するさらなるsiRNA二本鎖が、同定された。これらを表19に示す。これらの位置は図4に図示される。
【0103】
(表19.H5N1トリインフルエンザを標的化するRNAi配列)
【0104】
【表19】
(siRNAの組み合わせ)
本発明のいくつかの実施形態は、2種もしくはそれより多くのオリゴヌクレオチドまたはポリヌクレオチドを含有する薬学的組成物を提供し、これらのオリゴヌクレオチドまたはポリヌクレオチドの各々は、呼吸性ウイルスのゲノムにおける遺伝子を標的化する配列を含む。関連の実施形態は、この組み合わせを用いて細胞を処置する方法、および呼吸性ウイルス感染を処置する方法を提供し、そしてこのような組み合わせ組成物の、呼吸性ウイルス感染を処置することを意図される薬学的組成物の製造における使用を提供する。この組み合わせの個々のポリヌクレオチド成分は、ウイルス病原体のゲノムにおける同じ遺伝子の異なる部分、または異なる遺伝子、あるいは1つの遺伝子および1つより多くの遺伝子のいくつかの部分を、標的化し得る。オリゴヌクレオチドもしくはポリヌクレオチドの組み合わせを用いる利点は、所定の遺伝子の発現を阻害する利益が、この組み合わせにおいて大きくなることである。多数の標的性配列の使用により、遺伝子のノックダウンもしくはウイルスゲノムのサイレンシングにおいて、より大きな効力が、達成される。ウイルス複製の阻害における増大した効力は、ウイルスゲノムにおける1種より多くの遺伝子を標的化することによって達成される。
【0105】
(薬学的組成物)
本発明の標的性ポリヌクレオチドは、本明細書中で「活性化合物」もしくは「治療薬」と記載される。これらの治療薬は、被験体への投与のために適する薬学的組成物中に組み込まれ得る。
【0106】
本明細書中で使用される場合、「薬学的に受容可能なキャリア」は、薬学的投与に適合性の、任意のそして全ての溶媒、分散媒体、コーティング、抗細菌剤および抗真菌剤、等張剤および吸収遅延剤などを含むことを意図される。適切なキャリアは、Remington’s Pharmaceutical Sciences,Gennaro AR(編)第20版(2000)Williams & Wilkins PA,USAならびにWilson and Gisvold’s Textbook of Organic Medicinal and Pharmaceutical Chemistry(DelgadoおよびRemers,Lippincott−Raven.による)のような教科書(本明細書中で参考として援用される)に記載される。このようなキャリアもしくは希釈剤において使用され得る成分の好ましい例としては、以下が挙げられるが、これらに限定されない:水、生理食塩水、リン酸塩、カルボン酸塩、アミノ酸溶液、リンガー液、デキストロース(グルコースについての異名)溶液、および5%ヒト血清アルブミン。非限定の例として、デキストロースは、5%もしくは10%の水溶液として使用され得る。リポソームおよび非水性ビヒクル(例えば、不揮発性油)もまた、使用され得る。このような媒体および薬剤の薬学的に活性な物質のための使用は、当該分野で周知である。追加の活性化合物もまた、組成物中に組み込まれ得る。
【0107】
本発明の薬学的組成物は、意図される投与の経路に適合するように処方される。投与の経路の例としては、非経口投与(例えば、静脈内投与、皮内投与、皮下投与)、経口投与、鼻投与、吸入投与、経皮投与(局所投与)、経粘膜投与および直腸投与が挙げられる。非経口適用、静脈内適用、皮内適用もしくは皮下適用のために使用される溶液もしくは懸濁液は、以下の成分を含み得る:注射用水、生理食塩水溶液、不揮発性油、ポリエチレングリコール、グリセリン、プロピレングリコールもしくは他の合成溶媒のような無菌希釈剤;ベンジルアルコールもしくはメチルパラベンのような抗細菌剤;アスコルビン酸もしくは亜硫酸水素ナトリウムのような抗酸化剤;エチレンジアミンテトラ酢酸のようなキレート剤;酢酸塩、クエン酸塩もしくはリン酸塩のような緩衝剤、ならびに張度を調整するための薬剤(例えば、塩化ナトリウムまたはデキストロース)。
【0108】
吸入による投与のために、上記化合物は、適切な噴霧体(例えば、二酸化炭素のような気体)を含む加圧式容器もしくはディスペンサーまたは噴霧器からエアロゾルスプレーの形態で送達される。
【0109】
1つの実施形態において、活性化合物は、制御放出処方物のように、この化合物を身体からの迅速な除去から保護するキャリアと共に調製され、移植物およびマイクロカプセル化送達系を含む。持続放出調製物の適切な例としては、抗体を含む固体疎水性ポリマーの半透性マトリックスが挙げられ、このマトリックスは、成形された製品の形態(例えば、フィルムもしくはマイクロカプセル)である。持続放出マトリックスの例としては、ポリエステル、ヒドロゲル(例えばポリ(2−ヒドロキシエチル−メタクリレート)もしくはポリ(ビニルアルコール))、ポリラクチド(米国特許第3,773,919号)、L−グルタミン酸とγエチル−L−グルタメートとのコポリマー、非分解性エチレン−酢酸ビニル、分解性乳酸−グリコール酸コポリマー(例えばLUPRON DEPOTTM(乳酸−グリコール酸コポリマーおよび酢酸ロイプロリドから構成される注射可能ミクロスフェア))、ならびにポリ−D−(−)−3−ヒドロキシ酪酸が挙げられる。エチレン−酢酸ビニルおよび乳酸−グリコール酸のようなポリマーは、100日間にわたって分子を放出可能であり、特定のヒドロゲルは、より短い期間にわたって薬学的活性薬剤を放出する。有益なポリマーは、生分解性もしくは生物適合性である。リポソーム懸濁液(ウイルス抗原に対するモノクローナル抗体によって感染細胞を標的化するリポソームを含む)もまた、薬学的に受容可能なキャリアとして使用され得る。これらは、例えば米国特許第4,522,811号において記載されるような当業者に公知の方法に従って調製され得る。有益な形態を有する持続放出調製物(例えばミクロスフェア)は、上記のような材料から調製され得る。
【0110】
本発明のsiRNAポリヌクレオチドは、ベクターに挿入され得、そして遺伝子治療ベクターとして使用され得る。遺伝子治療ベクターは、多くの経路(例えば、米国特許第5,703,055号に記載される)のいずれかによって被験体に送達され得る。従ってまた、送達としては、例えば、静脈内注射、局所投与(米国特許第5,328,470号を参照)または定位注射(stereotactic injection)(例えば、Chenら(1994)Proc.Natl.Acad.Sci.USA 91:3054−3057を参照)が挙げられる。遺伝子治療ベクターの薬学的調製物は、受容可能な希釈剤中に治療ベクターを含み得るか、または遺伝子送達ビヒクルが埋め込まれた徐放性マトリックスを含み得る。あるいは、完全な遺伝子送達ベクターが組換え細胞からインタクトに産生され得る(例えば、レトロウイルスベクター)場合、この薬学的調製物は、この遺伝子送達系を産生する1つもしくはそれより多くの細胞を含み得る。
【0111】
薬学的組成物は、例えば容器、パックもしくはディスペンサー中で、投与の指示書と共にキットに含まれ得る。
【0112】
被験体における呼吸性ウイルス感染を処置するための薬学的組成物もしくは医薬の製造における治療薬の使用もまた、本発明の範囲内である。
【0113】
(送達)
いくつかの実施形態において、本発明のsiRNAポリヌクレオチドは、リポソーム媒介型トランスフェクションによって、例えば市販の試薬もしくは技術(例えば、OligofectamineTM、LipofectAmineTM試薬、LipofectAmine 2000TM(Invitrogen)、ならびにエレクトロポレーションおよび同様の技術)を使用して、培養物中の細胞に送達される。さらに、siRNAポリヌクレオチドは、吸入もしくは気道への点滴注入を介して、動物モデル(例えばげっ歯類もしくは非ヒト霊長類)に送達される。動物モデルでの使用のためのさらなる経路としては、静脈内(IV)、皮下(SC)、および関連の投与の経路が挙げられる。siRNAを含有する薬学的組成物は、siRNAの安定性を保護するか、siRNAの持続時間を延長させるか、siRNA機能を助長するか、または特異的な組織/細胞にsiRNAを標的化させる、さらなる成分を含む。これらの成分としては、種々の生分解性ポリマー、カチオン性ポリマー(例えばポリエチレンイミン)、ヒスチジン−リジン(HK)ポリペプチドのようなカチオン性コポリペプチド(例えば、Mixsonらに対するPCT公開番号WO01/47496、Biomerieuxに対するWO02/096941およびMassachusetts Institute of Technologyに対するWO99/42091を参照)、ペグ化カチオン性ポリペプチド、およびリガンドを組み込んだポリマーなど、正に荷電したポリペプチド、PolyTranポリマー(天然の多糖類、スクレログルカンとしても知られる)、標的性リガンドと結合体化したポリマーからなるナノ粒子(TargeTran改変体)、界面活性剤(Infasurf;Forest Laboratories,Inc.;ONY Inc.)、ならびにカチオン性ポリマー(例えばポリエチレンイミン)が挙げられる。Infasurf(登録商標)(カルファクタント(calfactant))は、気管内点滴注入における使用のための、仔ウシ肺から単離された天然の肺界面活性剤である;これは、リン脂質、中性脂質、ならびに疎水性界面活性剤関連タンパク質Bおよび疎水性界面活性剤関連タンパク質Cを含む。このポリマーは、一次元ポリマーもしくは多次元ポリマーのいずれかであり得、そしてまた、直径20ミクロン未満、20ミクロンと100ミクロンとの間もしくは100ミクロンを超える、ミクロ粒子もしくはナノ粒子であり得る。このポリマーは、特定の組織もしくは細胞のレセプターまたは分子に特異的なリガンド分子を有し得、従って、siRNAの送達を標的化するために使用され得る。siRNAポリヌクレオチドはまた、カチオン性リポソームベースのキャリア(例えば、DOTAP、DOTAP/コレステロール(Qbiogene,Inc.))および他の型の脂質水溶液によって送達される。さらに、低パーセント(5〜10%)のグルコース水溶液およびInfasurfは、siRNAの気道送達のための有効なキャリアである30。
【0114】
5%グルコースおよびInfasurfの経口気道送達溶液に懸濁した蛍光標識化siRNAを用い、蛍光顕微鏡で試験して、siRNAが外鼻孔を介してもしくは経口−気道経路を介してマウスに送達されそして肺組織を洗浄した後に、このsiRNAは肺において広汎に分布していることが示されている(共有に係るWO2005/01940(本明細書中でその全体が参考として援用される)を参照)。マウスの鼻道および肺(上気道および気道深部(deeper respiratory tract))へのsiRNAの送達は、siRNAと同時にプラスミド中で送達した指示遺伝子(GFPもしくはルシフェラーゼ)を、首尾よくサイレンシングすることを示した(このプラスミドは、この指示遺伝子とsiRNA標的との融合物を有した)(共有に係るWO2005/01940を参照)。さらに、本発明者ら(他と共に研究した)によって報告された実験は、siRNA種がSARSコロナウイルスの複製を阻害し、それによって肺の病理学を緩和することを、SARS感染したアカゲザルにおいて示した。
【0115】
(siRNA組換えベクター)
本発明の別の局面は、本発明のsiRNAポリヌクレオチドを有するベクター、好ましくは発現ベクターに関する。本明細書中で使用される場合、用語「ベクター」は、連結した別の核酸を輸送可能な核酸分子をいう。ベクターの1つの型は、「プラスミド」であり、これは、さらなるDNAセグメントを連結し得る、環状二本鎖DNAループをいう。ベクターの別の型は、ウイルスベクターであり、ここで、さらなるDNAセグメントが、ウイルスゲノム中に連結され得る。特定のベクターは、作動可能に連結した遺伝子の発現を指示し得る。このようなベクターは、本明細書中で「発現ベクター」と呼ばれる。一般に、組換えDNA技術において使用される発現ベクターは、多くの場合、プラスミドの形態である。プラスミドが最も一般的に使用されるベクターの形態であるので、本明細書において、「プラスミド」および「ベクター」は、相互交換可能に使用され得る。しかし、本発明は、ウイルスベクター(例えば、複製欠損型のレトロウイルス、アデノウイルスおよびアデノ関連ウイルス)のような発現ベクターの他の形態を含むことを意図し、これらは、同等の機能をもたらす。
【0116】
本発明の組換え発現ベクターは、本発明の核酸を、この核酸の宿主細胞における発現のために適する形態で含み、これは、この組換え発現ベクターが、発現のために使用される宿主細胞に基づいて選択される1つもしくはそれより多くの調節配列を含むことを意味し、これらの調節配列は、発現される核酸配列に作動可能に連結される。組換え発現ベクター内で、「作動可能に連結される」は、目的のヌクレオチド配列が、このヌクレオチド配列の(例えば、インビトロ転写/翻訳系において、もしくはこのベクターが宿主細胞に導入される場合には宿主細胞においての)発現を可能にする様式で、調節配列に連結されることを意味する。用語「調節配列」は、プロモーター、エンハンサーおよび他の発現制御エレメント(例えば、ポリアデニル化シグナル)を含むことが意図される。このような調節配列は、例えば、Goeddel(1990)GENE EXPRESSION TECHNOLOGY:METHODS IN ENZYMOLOGY 185,Academic Press,San Diego,Calif.において記載される。調節配列としては、多くの型の宿主細胞におけるヌクレオチド配列の構成的発現を指示する調節配列、ならびに特定の宿主細胞においてのみヌクレオチド配列の構成的発現を指示する調節配列(例えば、組織特異的調節配列)が挙げられる。なお別の実施形態において、本発明の核酸は、哺乳動物発現ベクターを用いて哺乳動物細胞において発現される。哺乳動物発現ベクターの例としては、pCDM8(Seed(1987)Nature329:840)およびpMT2PC(Kaufmanら(1987)EMBO J 6:187−195)が挙げられる。哺乳動物細胞において使用される場合、発現ベクターの制御機能は、多くの場合、ウイルス調節エレメントによって提供される。例えば、一般に使用されるプロモーターは、ポリオーマ、アデノウイルス2、サイトメガロウイルスおよびシミアンウイルス40に由来する。さらなるベクターは、細菌人工染色体、酵母人工染色体もしくは哺乳動物人工染色体のような微小染色体を含む。他の適切な発現系は、原核生物細胞および真核細胞の両方についての発現系である。例えば、Sambrookら,MOLECULAR CLONING:A LABORATORY MANUAL.第2版,Cold Spring Harbor Laboratory,Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.,1989の第16章および第17章を参照のこと。
【0117】
別の実施形態において、組換え哺乳動物発現ベクターは、特定の細胞型(例えば、気道の細胞)において選択的に核酸の発現を指示し得る。組織特異的調節エレメントは、当該分野で公知である。本発明は、組換え発現ベクター内にクローニングされた本発明のDNA分子を含む発現ベクターを、さらに提供する。このDNA分子は、(このDNA分子の転写による)ウイルスRNAを標的化するsiRNAを含むRNA分子の発現を可能にする様式で、調節配列に作動可能に連結される。種々の細胞型において上記RNA分子の連続的な発現を指示する、核酸に作動可能に連結される調節配列(例えば、ウイルスプロモーターおよび/もしくはエンハンサー)が選択され得るか、またはアンチセンスRNAの、構成的発現、組織特異的発現もしくは細胞型特異的発現を指示する調節配列が選択され得る。
【0118】
ベクターDNAは、従来的な形質転換技術もしくはトランスフェクション技術を介して、原核生物細胞もしくは真核細胞内に導入され得る。本明細書中で使用される場合、用語「形質転換」および「トランスフェクション」は、外来性の核酸(例えば、DNA)を宿主細胞内に導入するための、種々の当該分野で認識されている技術をいうことが意図され、これらとしては、リン酸カルシウムもしくは塩化カルシウムの共沈殿、DEAE−デキストラン−媒介型トランスフェクション、リポフェクション、もしくはエレクトロポレーションが挙げられる。宿主細胞を形質転換するかもしくはトランスフェクションするための適切な方法は、Sambrook,ら(2001),Ausubelら(2002),および他の研究室マニュアルにおいて見出され得る。
【0119】
(ウイルスの力価もしくは量についてのアッセイの方法)
本発明の実施において、ウイルスについてのアッセイは、いくつかの手順によって実施され得る。これらの中でも、非限定の例として、免疫ブロット(ウエスタンブロット)、免疫沈降(I.P.)、および複合型逆転写ポリメラーゼ連鎖反応(RT−PCR)アッセイなどが挙げられる。このような手順は、トランスフェクトされた組織培養物中の溶解物もしくは上清に存在し得る、標的化したmRNAもしくはそのタンパク質産物の合成の減少を測定する。同様に、これらのアッセイは、診断的手順として、感染した動物被験体もしくはヒト被験体から得られた、ホモジナイズした組織サンプル、鼻咽頭洗浄液、分泌液中に存在し得る、標的化したmRNAもしくはそのタンパク質産物の合成の減少を測定するために用いられ得る。
【0120】
RT−PCRアッセイ方法において、ウイルスゲノムRNAの合成は、NS1 ORFとNS2 ORFとの間の結合部(joint)にまたがった配列に相補的であるプライマーを用いて検出される。NS1/NS2プライマーを用いたこのRT−PCRの結果は、全ゲノムRNAの合成を反映する。
【0121】
この方法は、組織培養物におけるsiRNAによって誘導されたウイルス複製の干渉が、本発明の分野における当業者に公知であるように、定量的リアルタイムPCR(RTQ−PCR)、TCID50、ウイルスプラークアッセイ、免疫蛍光アッセイおよび免疫組織化学などによって測定されるアッセイをさらに含む。さらに、この方法は、上記TCID50法を用いて組織培養物中のウイルス複製の阻害をモニタリングする、ウイルスの存在についてのアッセイをさらに含む。ここで、ウイルス力価は、任意の種類の細胞病原性終末点(非限定の例として、細胞融合、細胞変性効果(CPE)、細胞吸着などが挙げられる)によって測定される。
【0122】
上記の方法は、試験動物における上記siRNA媒介型ウイルス複製の干渉が、RTQ−PCR、病理学、免疫組織化学およびウイルスの再単離などによって測定されるアッセイを、さらに含む。
【0123】
プライマーが、RNAiによるmRNA合成の減少を測定するために使用されるRT−PCR検出のために設計される。このRT反応は、ヘキサマーもしくはポリ−dTプライマーによって開始される;そしてPCRは、siRNAによって標的化される各遺伝子に特異的な上流もしくは下流のプライマーを用いて実施される。
【0124】
ゲノムRNA合成の検出について、一対のプライマー(上および下)が、NS1 ORFおよびNS2 ORFの中の配列に対応して設計される。ウイルスゲノムのどちらかの端に対して相補的なプライマーではなく「結合部にまたがる(joint−crossing)」プライマーを使用する目的は、取り扱いにくい大型の全体のRNA転写物(約15K−ntの長さ)を避けるためである。このRT−PCRの産物は、この設計により、ゲノムRNA全体の合成を反映する。
【0125】
同定されているプライマーおよびRT−PCRの産物が、以下の表20、表21および表22に列挙される。
【0126】
(表20.RSV株A2(サブグループA)についてのプライマー)
【0127】
【表20】
(表21.RSV株B1(サブグループB)についてのプライマー)
【0128】
【表21】
(表22.PCR産物の大きさ)
【0129】
【表22】
【実施例】
【0130】
(実施例1.細胞培養物における抗H5N1インフルエンザA siRNA分子の効果。)
本実施例は、培養物におけるH5N1感染細胞に対するsiRNAの阻害性効果の評価を報告する。
【0131】
(siRNAの設計および合成)
各々、H5N1のNP遺伝子(NP−1およびNP−2;表19、配列番号)遺伝子およびM2遺伝子(M2−1およびM2−2;表19、配列番号)を標的化する2つのsiRNAを、開発した。これらのsiRNA標的配列は、H5N1ウイルスのいくつかの株において保存されている(例えば、gi|47834945_10−1506、gi|8452827_1−1497、gi|13925158_46−1542、gi|61927237_46−1542、gi|59940391_44−>1535、gi|9802277_46−1542)。5’センス鎖においてフルオレセイン標識されているds−siRNAを、Proligo BioTech Ltd(Paris,France)によって化学的に合成した。その抗H5N1効果を、siRNAでトランスフェクトした、H5N1ウイルスに感染したMDCK細胞において検出した(Madin−Darbyイヌ腎臓;ATCC登録番号:CCL−34,Manassas,VA)。H5N1の阻害を、以下に記載されるように、細胞変性効果(CPE)、標準的TCID50(50%組織培養感染用量)プロトコールによる培養培地中に放出されたウイルスの逆滴定、およびリアルタイムRT−PCRを用いる細胞内ウイルスRNAの定量によって、決定した。
【0132】
(細胞培養、siRNAトランスフェクションおよびH5N1ウイルス感染)
MDCK細胞を、10%ウシ胎仔血清(FBS,Invitrogen)含有MEM培地中で培養し、そして維持した。約5000個の細胞を、ウイルス感染および複製アッセイのために、96ウェルプレートの各ウェル中に入れた。細胞を、0.5μlのLipofectamine 2000(Invitrogen,CA)と混合した100nM、50nM、25nMおよび12.5nMのsiRNAでトランスフェクションした。ルシフェラーゼを標的化する無関係のsiRNA(GL2i)、もしくはSARSコロナウイルス(C−1)を標的化するsiRNA、およびトランスフェクト体(transfectant)Lipofectamine 2000のみ(C−2)を、本実験においてネガティブコントロールとして含めた。トランスフェクションの6時間後、この培養培地を除去し、そしてこの細胞を、H5N1ウイルス感染の前にPBSで2回洗浄した。1% FBS含有MEM中に希釈した100μlの100 TCID50 H5N1ウイルス(株:A/HongKong/486/97)7を、トランスフェクトされた細胞に加えた。培養上清および細胞を、それぞれ感染の12時間後、16時間後および24時間後に、放出されたウイルス力価および細胞内ウイルスRNAコピー数の検出のために収集した。感染の24時間後、位相差顕微鏡下でCPEを観察し、そして記録した。この実験を、3連で行い、そして少なくとも3回繰り返した。
【0133】
(定量的RT−PCR)
Q−RT−PCRを、以下のように実行する。全細胞内RNAを、RNeasy Miniキット(Qiagen,Germany)を製造業者の指示に従って用いて単離した。逆転写実験を、ThermoScript RT−PCRシステム(Invitrogen,CA)を用いるオリゴ−dTプライミングによって実施した。次いで、リアルタイムPCRを、表23に示すプライマーを用いて実施した。
【0134】
(表23.Q−RT−PCRのためのH5N1プライマー)
【0135】
【表23】
5μlのRT産物(テンプレート)、各1μlの順(forward)プライマーおよび逆(reverse)プライマー(最終濃度500nM)、2μlの25mM MgCl2、ならびに9μlのH2Oを、2μlのSYBR Green I Master Mix(Roche,USA)と混合し、そしてリアルタイム定量を、ABI7900 Sequence Detection Systemを用いて実施した。PCR条件は、以下であった:50℃を5分間、95℃を10分間、次いで95℃を10秒間、61℃を5秒間および72℃を5秒間を40サイクル。β−アクチンのコピー数もまた、内部コントロールとして測定した。1000コピーのβ−アクチンあたりの細胞内ウイルスRNAのコピー数を、計算し、そして未処理コントロールと比較した相対的ウイルスRNAコピー数として表した(処理したコピー数/未処理のコピー数×100%)。
【0136】
(ウイルス力価の測定)
siRNA処理を行ったかまたは行わない培養物中のウイルス力価を、以下のように試験した。簡潔には、感染細胞からの馴化培地を、1% FBS含有MEM中に、10倍の連続工程で希釈した。各希釈物を、標準的TCID50プロトコールに従って細胞を感染させるために使用した。簡潔には、細胞を、96ウェル皿に感染の16時間前に入れた。感染の72時間後、CPEを、位相差顕微鏡下で観察し、そしてCPE陽性(CPE+)細胞を決定した。次いで、TCID50を、以下のように評価した:
【0137】
【化1】
ここで、h=希釈工程の補間されたlog10値、これを50%を超える値の工程のlog10に加えた。感染性ウイルス力価を、計算しそして未処理コントロールと比較した相対的ウイルス収率として表した(処理した力価/未処理の力価×100%)。
【0138】
(結果)
siRNA NP−1(表19)処理は、培養培地中のH5N1ウイルス産生(図5、パネルA)および細胞におけるウイルスRNAコピーの複製(図5、パネルB)を、12.5〜100nMの濃度において99%を超えて低下させた。一方、NP−2を標的化するsiRNA(表19)による処理は、100nMの濃度において、細胞培養物中のウイルス増殖の約60%しか阻害しなかった。対照的に、無関係のsiRNA処理コントロール(C−1)およびキャリア処理コントロール(C−2)は、未処理コントロールと比較して、ウイルス増殖を有意に阻害しなかった。
【0139】
M2−1(表19)に対して指向されるsiRNA(表19)による処理もまた、種々の濃度で、ウイルス増殖(図5、パネルC)およびウイルスRNAの複製(図5、パネルD)に対して、約80%〜94%の阻害性効果を示した。ここで、M2−2は、100nMの濃度で約60%の阻害性効果を示したが、より低い濃度は試験しなかった。
【0140】
第2の実験において、培養物中の細胞を、50nMの種々のsiRNAで処理した。培養上清培地中に産生されるウイルスを、3回の時点で収集した。サンプルを、培養物中の細胞の50%に感染するために必要な組織培養感染用量(TCID50)を決定することによって滴定した(図6、パネルA;これは対数座標スケール上に示されていることに留意されたい)。ウイルスRNAのコピーの数/1000コピーのβ−アクチンを、リアルタイムRT−PCRによって決定した(図6、パネルB)。無関係のsiRNA(C−1)およびトランスフェクト体(C−2)を、コントロールとして実験に適用した。NP−1 siRNAおよびNP−2 siRNAが、ウイルス粒子の増殖およびウイルスRNAの増大の阻害において非常に有効であることが示される。M1−1 siRNAおよびM2−2 siRNAの使用は、ウイルス複製の妨害において部分的に有効である。
【0141】
この実施例における結果は、H5N1ゲノムにおける異なる遺伝子に対して指向される種々のsiRNAが、培養中の感染細胞増殖におけるウイルス複製の阻害において、非常に有効であることを実証する。
【0142】
(実施例2.H5N1インフルエンザAに対して指向されるsiRNAの組み合わせ。)
H5N1ゲノム中の種々の遺伝子に対して指向されるsiRNAの組み合わせを、有効な抗H5N1活性を提供するために同定した。この実施例は、このような2つの組み合わせを示す。
【0143】
【化2】
(実施例3.インビトロの細胞中のsiRNAによる標的遺伝子およびウイルス複製の阻害)
この実施例において、種々の呼吸性ウイルス遺伝子を標的化する、同族(cognate)の標的遺伝子をサイレンシングする能力もしくはウイルス複製を阻害する能力において最適に有効なsiRNAおよび最適に有効なsiRNAの組み合わせが、細胞培養物中の実験によって同定される。インビトロで有効なsiRNAは、インビボでさらに試験されそして使用される候補である。
【0144】
培養した許容細胞株(例えば、非限定の例として、A549(ATCC登録番号:CCL−185TM、II型肺胞上皮肺癌腫細胞株))を、RSVウイルス株もしくはインフルエンザA株(例えば、H5N1)で感染させ、次いで種々のsiRNAにより、個々にもしくは組み合わせて、トランスフェクトする。組み合わせの場合、1つのシグナル遺伝子を標的化する2種のsiRNA、もしくは2つもしくはそれより多くの遺伝子を標的化するsiRNAが、使用される。単独のsiRNAもしくは組み合わせのsiRNAの総siRNA用量を一定にする。異なる実験プロトコールにおいて、siRNAトランスフェクションは、RSV感染もしくはインフルエンザA感染の数時間前に実施されるか、ウイルス感染と同時に実施されるか、または感染の数時間後に実施される。これらの異なる手順は、試験したsiRNAが、細胞レベルにおいて予防的効果および/もしくは治療的効果を示すか否かについての情報を提供する。
【0145】
標的遺伝子の阻害の程度、もしくはウイルス複製の阻害を、種々の方法によってアッセイする。アッセイの非限定の例としては、以下の手順が挙げられる:
(1)免疫ブロット(ウエスタン)を、細胞溶解物および所定のウイルス抗原に対するRSV特異的抗体もしくはインフルエンザA特異的抗体を用いて実施する。
【0146】
(2)免疫沈降(IP)を、細胞溶解物および上記の1つの遺伝子産物に対するRSV特異的抗体もしくはインフルエンザA特異的抗体を用いて実施する。
【0147】
(3)rvtr−PCRを行って、mRNA転写もしくはウイルスRNA複製の阻害を実証する。標的化遺伝子の転写物を検出するため、もしくは試験したsiRNAオリゴがゲノムRNAをもまた標的化し得るか否かを検出するための、特別なプライマーを設計する。
【0148】
(4)細胞融合体ベースのTCID50の測定を使用して、無関係のコントロールsiRNAの特異的siRNAで処理した細胞培養物と比較して、ウイルス複製の阻害をモニタリングし得る。
【0149】
(5)免疫蛍光もしくは免疫組織化学もまた、ウイルス力価を滴定するために使用され得、それによって、ウイルス複製のsiRNA媒介型阻害をモニタリングし得る。
【0150】
(実施例4.小動物モデルにおける、siRNAによる標的遺伝子の阻害)
この実施例において、種々のsiRNA(個々にもしくは組み合わせ)が、動物モデルにおけるRSVもしくはトリインフルエンザH5N1の処置におけるその効力を決定するために試験される。RSV株もしくはインフルエンザH5N1株を、吸入もしくは点滴注入により気道を通して試験動物に感染させるために使用する。siRNAは、同じ経路を通って送達され、そしてRSV感染もしくはインフルエンザH5N1感染の前に、感染と同時に、もしくは感染後に適用される。siRNA送達時間を変えることにより、試験動物におけるRSVもしくはインフルエンザH5N1の複製の阻害、RSVもしくはインフルエンザH5N1によって誘導される病理学の低減、およびRSVもしくはインフルエンザH5N1様の症状の軽減における、siRNAの効力の情報を得ることが可能であり、そしてRSVもしくはインフルエンザH5N1に特異的なsiRNAが、動物における実験的なRSV感染もしくはインフルエンザH5N1感染に対する予防効果もしくは治療効果を示すか否かの情報を得ることが可能である。
【0151】
RSVおよびインフルエンザH5N1は広範な範囲の動物種に感染し得るが、RSVもしくはインフルエンザH5N1の動物モデル研究において、マウスおよびコットンラット(cotton rat)が、従来使用されている。げっ歯類の感染は、通常、低レベル〜中レベルのウイルス複製をもたらし、これは、4日目にピークを迎え、そして迅速に除去される。これらの動物は、明らかな気道疾患を示さないが、肺の病理学を示す。高用量のウイルス感染において、これらは、体重減少、肺機能の変化、および毛皮の乱れ(ruffled fur)を示し、これらは疾患の指標である。
【0152】
上の実施例3に記載したものと同じ診断アッセイを、動物から採取したサンプル(例えば、鼻、頬および咽頭のスワブ)におけるsiRNA媒介型遺伝子サイレンシングおよびRSV感染もしくはインフルエンザH5N1感染の阻害をモニタリングするために使用する。さらに、肺の病理学、肺の免疫組織化学および症状観察を実施し、インビボでのRSV感染もしくはインフルエンザH5N1感染におけるsiRNAの効力を決定する。
【0153】
(実施例5.非ヒト霊長類モデルにおける、siRNAによる標的遺伝子の阻害)
本実施例は、非ヒト霊長類における、RSVもしくはインフルエンザH5N1のウイルス複製のsiRNA媒介型阻害の研究を記載する。用量投与の効力および安全性が、本研究の主要な目的である。
【0154】
本発明者らのプロトコールに基づき、別の呼吸性疾患であるSARSについてのアカゲザルモデルを用いて、30mg/kg体重までのsiRNAの用量は耐容可能であり、動物に毒性の徴候を示さない。(22)非ヒト霊長類哺乳動物によって事前にスクリーニングされているsiRNAの呼吸器送達(吸入もしくは点滴注入による)は、ウイルス複製の阻害ならびにウイルス誘導型病理学および疾患様症状の軽減に対して実質的に影響することを示した。RSVもしくはインフルエンザH5N1の本研究のために、同じ送達経路を使用し、そして類似の投薬量のsiRNAを送達する。
【0155】
小動物研究について記載された実験(実施例4)に加え、以下のアッセイが、霊長類において容易に実施され、そして治療効力のより直接的な評価に寄与する。
【0156】
(1)ウイルス散布性:ウイルス散布性を、感染したサルの鼻咽頭洗浄サンプルを用いて測定する。ウイルス収量を、TCID50(細胞融合体ベース)および/または免疫蛍光アッセイのいずれかによって滴定する。
【0157】
(2)鼻咽頭洗浄サンプルのRTQ−PCR:ウイルスゲノムコピー数の変化をモニタリングする。
【0158】
(3)症状モニタリング:ヒト乳児患者において観察されるような細気管支炎症状を見出しやすい。
【0159】
本発明は、本発明の種々の例示的実施形態に関して記載され、そして説明されているが、均等な実施形態および種々の他の改変、付加および削除が、これらの実施形態において、そしてこれらの実施形態に対して、本発明の精神および範囲から逸脱することなく、なされ得る。
【0160】
(参考文献)
【0161】
【化3】
【図面の簡単な説明】
【0162】
【図1】ヒトRSウイルス(hRSV、RSV)(−)ssRNAゲノムの概略図。GenBank登録番号NC_001781に基づく。
【図2】インフルエンザA型ウイルス構造の概略図。この図は、ウイルスゲノムの8つのセグメントが、ビリオン内に組み込まれていることを示す。
【図3】本発明のポリヌクレオチドの種々の実施形態の概略図。パネルAは、直鎖状ポリヌクレオチドの実施形態。長さは、200ヌクレオチド以下かつ15ヌクレオチド以上である。(b)において、記載の標的性配列は、より大きな標的性配列内に含まれる。(d)において、暗色の垂直の棒は、置換されたヌクレオチドを図解する。パネルBは、全体の長さが200ヌクレオチド以下のヘアピンポリヌクレオチドの実施形態である。
【図4】siRNA配列のH5N1ゲノム内の配置を示す概略図である。
【図5−1】MDCK細胞培養物におけるH5N1ウイルス増殖の阻害。
【図5−2】MDCK細胞培養物におけるH5N1ウイルス増殖の阻害。
【図6】異なる時点で決定されたsiRNAの阻害性効果。
【特許請求の範囲】
【請求項1】
長さが200ヌクレオチドもしくはそれより少ないヌクレオチドである、単離されたポリヌクレオチドであって、該ポリヌクレオチドは、第1のヌクレオチド配列を含み、該第1のヌクレオチド配列は、RSウイルスもしくはインフルエンザAウイルスのゲノムを標的化し、ここで任意のT(チミジン)もしくは任意のU(ウリジン)は、必要に応じて他と置換され得、そしてここで、該第1のヌクレオチド配列は、
(a)長さが15〜30個の任意の数のヌクレオチドである配列、または
(b)(a)に記載の配列の相補体
からなる、ポリヌクレオチド。
【請求項2】
前記第1のヌクレオチド配列からループ配列によって分離される第2のヌクレオチド配列をさらに含む、請求項1に記載のポリヌクレオチドであって、該第2のヌクレオチド配列は、
(a)該第1のヌクレオチド配列と実質的に同一な長さを有し、かつ
(b)該第1のヌクレオチド配列に対して実質的に相補的である、
ポリヌクレオチド。
【請求項3】
請求項1または請求項2に記載のポリヌクレオチドであって、前記第1のヌクレオチド配列は、
(a)配列番号1〜263から選択される配列、
(b)項目(a)に記載の配列より長い標的性配列であって、ここで該標的性配列は、呼吸性ウイルスのゲノムを標的化し、そして配列番号1〜263から選択される配列を含む、標的性配列、
(c)配列番号1〜263から選択される配列のフラグメントであって、該フラグメントは、少なくとも15ヌクレオチド長でありかつ該選択される配列より長くとも1塩基短い長さである、連続する塩基の配列からなる、フラグメント
(d)5ヌクレオチドまでが、配列番号1〜263から選択される配列とは異なる配列、または
(e)(a)〜(d)に記載の配列の相補体
からなる、ポリヌクレオチド。
【請求項4】
請求項1もしくは請求項2に記載のポリヌクレオチドであって、前記第1のヌクレオチド配列の長さが、21〜25個の任意の数のヌクレオチドである、ポリヌクレオチド。
【請求項5】
配列番号1〜263から選択される配列からなる、請求項1もしくは請求項2に記載のポリヌクレオチドであって、必要に応じて、該選択される配列の3’に結合するジヌクレオチドオーバーハングを含む、ポリヌクレオチド。
【請求項6】
請求項1もしくは請求項2に記載のポリヌクレオチドであって、ここで前記第1のヌクレオチド配列の3’末端におけるジヌクレオチド配列は、TT、TU、UT、もしくはUUであり、そしてここで、該ジヌクレオチドは、リボヌクレオチドもしくはデオキシリボヌクレオチドまたはその両方を含む、ポリヌクレオチド。
【請求項7】
請求項1もしくは請求項2に記載のポリヌクレオチドであって、該ポリヌクレオチドは、DNAである、ポリヌクレオチド。
【請求項8】
請求項1もしくは請求項2に記載のポリヌクレオチドであって、該ポリヌクレオチドは、RNAである、ポリヌクレオチド。
【請求項9】
請求項1もしくは請求項2に記載のポリヌクレオチドであって、該ポリヌクレオチドは、デオキシリボヌクレオチドおよびリボヌクレオチドの両方を含む、ポリヌクレオチド。
【請求項10】
請求項1に記載の第1のポリヌクレオチド鎖、および少なくとも該第1のポリヌクレオチド鎖の該第1のヌクレオチド配列に対して相補的でありかつ該配列にハイブリダイズされる第2のポリヌクレオチド鎖を含む、二本鎖ポリヌクレオチド。
【請求項11】
請求項1、請求項2またはその両方に記載の複数の標的性ポリヌクレオチドを含む組み合わせであって、ここで、各ポリヌクレオチドは、前記標的ウイルスのゲノムにおける異なった配列を標的化する、組み合わせ。
【請求項12】
請求項1に記載のポリヌクレオチドを含むベクター。
【請求項13】
請求項2に記載のポリヌクレオチドを含む、ベクター。
【請求項14】
請求項12もしくは請求項13に記載のベクターであって、該ベクターは、プラスミド、組換えウイルス、トランスポゾン、もしくは微小染色体である、ベクター。
【請求項15】
1種もしくはそれより多くの請求項1に記載のポリヌクレオチドによってトランスフェクトされた細胞。
【請求項16】
1種もしくはそれより多くの請求項2に記載のポリヌクレオチドによってトランスフェクトされた細胞。
【請求項17】
1種もしくはそれより多くの請求項1もしくは請求項2に記載のポリヌクレオチド、またはこれらの混合物、ならびに薬学的に受容可能なキャリアを含有する薬学的組成物であって、各ポリヌクレオチドは、前記標的ウイルスのゲノムにおける異なった配列を標的化する、薬学的組成物。
【請求項18】
1種もしくはそれより多くの請求項12もしくは請求項13に記載のベクターまたはその混合物、ならびに薬学的に受容可能なキャリアを含有する薬学的組成物であって、各ベクターは、前記標的ウイルスのゲノムにおける異なった配列を標的化するポリヌクレオチドを保有する、薬学的組成物。
【請求項19】
請求項17もしくは請求項18に記載の薬学的組成物であって、前記キャリアは、合成ポリマー、リポソーム、デキストロース、界面活性剤、またはこれらの任意の2種またはそれより多くの組み合わせを含む、薬学的組成物。
【請求項20】
RSウイルスもしくはインフルエンザAウイルスのゲノムを標的化する配列を有する請求項1もしくは請求項2に記載のポリヌクレオチドを合成する方法であって、該方法は、以下:
(a)反応性末端を含み、かつ該配列の第1の末端におけるヌクレオチドに対応する、ヌクレオチド試薬を提供する工程、
(b)反応性末端を含みかつ該配列の後に続く位置に対応する、さらなるヌクレオチド試薬を添加して、該工程(a)の該反応性末端と反応させ、そして伸長中の該ポリヌクレオチド配列の長さを1ヌクレオチド増加させる工程、ならびに所望されない産物および過剰な試薬を除去する工程、ならびに
(c)該配列の第2の末端におけるヌクレオチドに対応する該ヌクレオチド試薬が添加され、それによって完全なポリヌクレオチドが提供されるまで、工程(b)を繰り返す工程
を、包含する、方法。
【請求項21】
細胞を、RNAインヒビターによってトランスフェクトする方法であって、該方法は、該細胞を、1種もしくはそれより多くの請求項1に記載のポリヌクレオチドを含有する組成物と接触させる工程を包含する、方法。
【請求項22】
前記細胞は、前記1種もしくはそれより多くのポリヌクレオチドによって標的化される呼吸性ウイルスを含む、請求項21に記載の方法。
【請求項23】
細胞を、RNAインヒビターによってトランスフェクトする方法であって、該方法は、該細胞を、1種もしくはそれより多くの請求項2に記載のポリヌクレオチドを含有する組成物と接触させる工程を包含する、方法。
【請求項24】
前記細胞は、前記1種もしくはそれより多くのポリヌクレオチドによって標的化される呼吸性ウイルスを含む、請求項23に記載の方法。
【請求項25】
呼吸性ウイルスに感染した細胞における該ウイルスの複製を阻害する方法であって、該方法は、該細胞を、1種もしくはそれより多くの請求項1に記載のポリヌクレオチドを含有する組成物と接触させる工程を包含し、ここで、該1種もしくはそれより多くのポリヌクレオチドは、該ウイルスを標的化する、方法。
【請求項26】
呼吸性ウイルスに感染した細胞における該ウイルスの複製を阻害する方法であって、該細胞を、1種もしくはそれより多くの請求項2に記載のポリヌクレオチドを含有する組成物と接触させる工程を包含し、ここで、該1種もしくはそれより多くのポリヌクレオチドは、該ウイルスを標的化する、方法。
【請求項27】
被験体における呼吸性ウイルスに起因する感染を処置するために有効な薬学的組成物の製造における、該呼吸性ウイルスを標的化する、請求項1に記載のポリヌクレオチドの使用、または2種もしくはそれより多くの該ポリヌクレオチドの混合物の使用。
【請求項28】
前記被験体は、ヒトである、請求項27に記載の使用。
【請求項29】
被験体における呼吸性ウイルスに起因する感染を処置するために有効な薬学的組成物の製造における、該呼吸性ウイルスを標的化する、請求項2に記載のポリヌクレオチドの使用、または2種もしくはそれより多くの該ポリヌクレオチドの混合物の使用。
【請求項30】
前記被験体は、ヒトである、請求項29に記載の使用。
【請求項1】
長さが200ヌクレオチドもしくはそれより少ないヌクレオチドである、単離されたポリヌクレオチドであって、該ポリヌクレオチドは、第1のヌクレオチド配列を含み、該第1のヌクレオチド配列は、RSウイルスもしくはインフルエンザAウイルスのゲノムを標的化し、ここで任意のT(チミジン)もしくは任意のU(ウリジン)は、必要に応じて他と置換され得、そしてここで、該第1のヌクレオチド配列は、
(a)長さが15〜30個の任意の数のヌクレオチドである配列、または
(b)(a)に記載の配列の相補体
からなる、ポリヌクレオチド。
【請求項2】
前記第1のヌクレオチド配列からループ配列によって分離される第2のヌクレオチド配列をさらに含む、請求項1に記載のポリヌクレオチドであって、該第2のヌクレオチド配列は、
(a)該第1のヌクレオチド配列と実質的に同一な長さを有し、かつ
(b)該第1のヌクレオチド配列に対して実質的に相補的である、
ポリヌクレオチド。
【請求項3】
請求項1または請求項2に記載のポリヌクレオチドであって、前記第1のヌクレオチド配列は、
(a)配列番号1〜263から選択される配列、
(b)項目(a)に記載の配列より長い標的性配列であって、ここで該標的性配列は、呼吸性ウイルスのゲノムを標的化し、そして配列番号1〜263から選択される配列を含む、標的性配列、
(c)配列番号1〜263から選択される配列のフラグメントであって、該フラグメントは、少なくとも15ヌクレオチド長でありかつ該選択される配列より長くとも1塩基短い長さである、連続する塩基の配列からなる、フラグメント
(d)5ヌクレオチドまでが、配列番号1〜263から選択される配列とは異なる配列、または
(e)(a)〜(d)に記載の配列の相補体
からなる、ポリヌクレオチド。
【請求項4】
請求項1もしくは請求項2に記載のポリヌクレオチドであって、前記第1のヌクレオチド配列の長さが、21〜25個の任意の数のヌクレオチドである、ポリヌクレオチド。
【請求項5】
配列番号1〜263から選択される配列からなる、請求項1もしくは請求項2に記載のポリヌクレオチドであって、必要に応じて、該選択される配列の3’に結合するジヌクレオチドオーバーハングを含む、ポリヌクレオチド。
【請求項6】
請求項1もしくは請求項2に記載のポリヌクレオチドであって、ここで前記第1のヌクレオチド配列の3’末端におけるジヌクレオチド配列は、TT、TU、UT、もしくはUUであり、そしてここで、該ジヌクレオチドは、リボヌクレオチドもしくはデオキシリボヌクレオチドまたはその両方を含む、ポリヌクレオチド。
【請求項7】
請求項1もしくは請求項2に記載のポリヌクレオチドであって、該ポリヌクレオチドは、DNAである、ポリヌクレオチド。
【請求項8】
請求項1もしくは請求項2に記載のポリヌクレオチドであって、該ポリヌクレオチドは、RNAである、ポリヌクレオチド。
【請求項9】
請求項1もしくは請求項2に記載のポリヌクレオチドであって、該ポリヌクレオチドは、デオキシリボヌクレオチドおよびリボヌクレオチドの両方を含む、ポリヌクレオチド。
【請求項10】
請求項1に記載の第1のポリヌクレオチド鎖、および少なくとも該第1のポリヌクレオチド鎖の該第1のヌクレオチド配列に対して相補的でありかつ該配列にハイブリダイズされる第2のポリヌクレオチド鎖を含む、二本鎖ポリヌクレオチド。
【請求項11】
請求項1、請求項2またはその両方に記載の複数の標的性ポリヌクレオチドを含む組み合わせであって、ここで、各ポリヌクレオチドは、前記標的ウイルスのゲノムにおける異なった配列を標的化する、組み合わせ。
【請求項12】
請求項1に記載のポリヌクレオチドを含むベクター。
【請求項13】
請求項2に記載のポリヌクレオチドを含む、ベクター。
【請求項14】
請求項12もしくは請求項13に記載のベクターであって、該ベクターは、プラスミド、組換えウイルス、トランスポゾン、もしくは微小染色体である、ベクター。
【請求項15】
1種もしくはそれより多くの請求項1に記載のポリヌクレオチドによってトランスフェクトされた細胞。
【請求項16】
1種もしくはそれより多くの請求項2に記載のポリヌクレオチドによってトランスフェクトされた細胞。
【請求項17】
1種もしくはそれより多くの請求項1もしくは請求項2に記載のポリヌクレオチド、またはこれらの混合物、ならびに薬学的に受容可能なキャリアを含有する薬学的組成物であって、各ポリヌクレオチドは、前記標的ウイルスのゲノムにおける異なった配列を標的化する、薬学的組成物。
【請求項18】
1種もしくはそれより多くの請求項12もしくは請求項13に記載のベクターまたはその混合物、ならびに薬学的に受容可能なキャリアを含有する薬学的組成物であって、各ベクターは、前記標的ウイルスのゲノムにおける異なった配列を標的化するポリヌクレオチドを保有する、薬学的組成物。
【請求項19】
請求項17もしくは請求項18に記載の薬学的組成物であって、前記キャリアは、合成ポリマー、リポソーム、デキストロース、界面活性剤、またはこれらの任意の2種またはそれより多くの組み合わせを含む、薬学的組成物。
【請求項20】
RSウイルスもしくはインフルエンザAウイルスのゲノムを標的化する配列を有する請求項1もしくは請求項2に記載のポリヌクレオチドを合成する方法であって、該方法は、以下:
(a)反応性末端を含み、かつ該配列の第1の末端におけるヌクレオチドに対応する、ヌクレオチド試薬を提供する工程、
(b)反応性末端を含みかつ該配列の後に続く位置に対応する、さらなるヌクレオチド試薬を添加して、該工程(a)の該反応性末端と反応させ、そして伸長中の該ポリヌクレオチド配列の長さを1ヌクレオチド増加させる工程、ならびに所望されない産物および過剰な試薬を除去する工程、ならびに
(c)該配列の第2の末端におけるヌクレオチドに対応する該ヌクレオチド試薬が添加され、それによって完全なポリヌクレオチドが提供されるまで、工程(b)を繰り返す工程
を、包含する、方法。
【請求項21】
細胞を、RNAインヒビターによってトランスフェクトする方法であって、該方法は、該細胞を、1種もしくはそれより多くの請求項1に記載のポリヌクレオチドを含有する組成物と接触させる工程を包含する、方法。
【請求項22】
前記細胞は、前記1種もしくはそれより多くのポリヌクレオチドによって標的化される呼吸性ウイルスを含む、請求項21に記載の方法。
【請求項23】
細胞を、RNAインヒビターによってトランスフェクトする方法であって、該方法は、該細胞を、1種もしくはそれより多くの請求項2に記載のポリヌクレオチドを含有する組成物と接触させる工程を包含する、方法。
【請求項24】
前記細胞は、前記1種もしくはそれより多くのポリヌクレオチドによって標的化される呼吸性ウイルスを含む、請求項23に記載の方法。
【請求項25】
呼吸性ウイルスに感染した細胞における該ウイルスの複製を阻害する方法であって、該方法は、該細胞を、1種もしくはそれより多くの請求項1に記載のポリヌクレオチドを含有する組成物と接触させる工程を包含し、ここで、該1種もしくはそれより多くのポリヌクレオチドは、該ウイルスを標的化する、方法。
【請求項26】
呼吸性ウイルスに感染した細胞における該ウイルスの複製を阻害する方法であって、該細胞を、1種もしくはそれより多くの請求項2に記載のポリヌクレオチドを含有する組成物と接触させる工程を包含し、ここで、該1種もしくはそれより多くのポリヌクレオチドは、該ウイルスを標的化する、方法。
【請求項27】
被験体における呼吸性ウイルスに起因する感染を処置するために有効な薬学的組成物の製造における、該呼吸性ウイルスを標的化する、請求項1に記載のポリヌクレオチドの使用、または2種もしくはそれより多くの該ポリヌクレオチドの混合物の使用。
【請求項28】
前記被験体は、ヒトである、請求項27に記載の使用。
【請求項29】
被験体における呼吸性ウイルスに起因する感染を処置するために有効な薬学的組成物の製造における、該呼吸性ウイルスを標的化する、請求項2に記載のポリヌクレオチドの使用、または2種もしくはそれより多くの該ポリヌクレオチドの混合物の使用。
【請求項30】
前記被験体は、ヒトである、請求項29に記載の使用。
【図1】

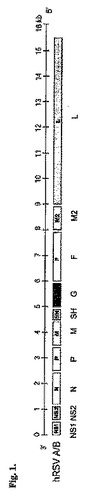
【図2】
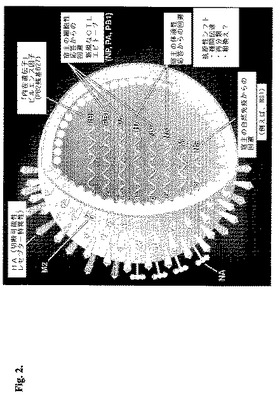
【図3】
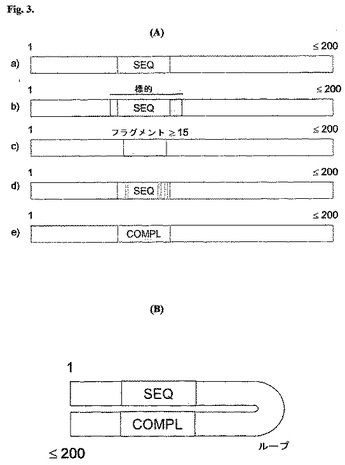
【図4】
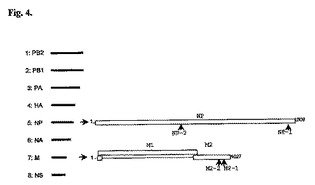
【図5−1】

【図5−2】
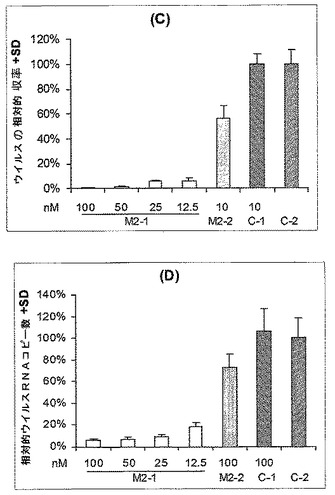
【図6】
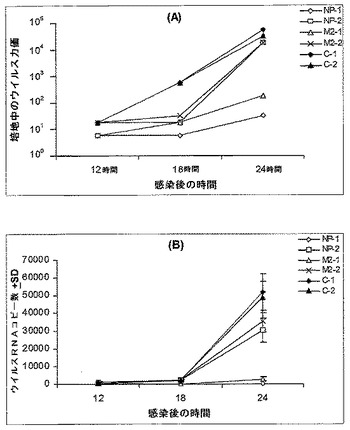
【図2】
【図3】
【図4】
【図5−1】
【図5−2】
【図6】
【公表番号】特表2009−526516(P2009−526516A)
【公表日】平成21年7月23日(2009.7.23)
【国際特許分類】
【出願番号】特願2007−539358(P2007−539358)
【出願日】平成17年11月4日(2005.11.4)
【国際出願番号】PCT/US2005/040048
【国際公開番号】WO2006/121464
【国際公開日】平成18年11月16日(2006.11.16)
【出願人】(509127310)サーナオミクス インコーポレイテッド (1)
【Fターム(参考)】
【公表日】平成21年7月23日(2009.7.23)
【国際特許分類】
【出願日】平成17年11月4日(2005.11.4)
【国際出願番号】PCT/US2005/040048
【国際公開番号】WO2006/121464
【国際公開日】平成18年11月16日(2006.11.16)
【出願人】(509127310)サーナオミクス インコーポレイテッド (1)
【Fターム(参考)】
[ Back to top ]