
呼吸障害に関連する方法
哺乳動物被験体において誘導されたPGE2経路を阻害することにより呼吸障害を処置する方法、被験体由来のサンプルにおけるPGE2またはその代謝産物レベルが対照レベルよりも上昇していることを検出することにより無呼吸、低酸素性虚血性脳症または周産期仮死を評価する方法、ならびに呼吸障害を処置するための医薬のインビトロおよびインビボスクリーニング方法が開示されている。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、呼吸障害(breathing disorder)、例えば無呼吸の処置方法、診断およびスクリーニング方法、ならびにこのような方法において使用する組成物に関する。
【背景技術】
【0002】
無呼吸および乳幼児突然死症候群(SIDS)は、新生児の大きな医学的懸案事項であり、これらの病因には感染が重要な役割を果たしている可能性がある。無呼吸は、新生児における感染の際に一般的にみられる徴候であり、SIDS患者の大部分において死の前に軽度のウイルスまたは細菌感染が確認される(1、2、111)。
【0003】
脳幹の呼吸調節に非最適性または遅延がみられる小児、例えば未熟児(すべてがその生後初年度に、一部は小児初期以降も)、先天性中枢性低換気症候群(CCHS)(79)、レット症候群およびプラダー・ウィリー症候群(PWS)(80)を発症した小児は、無呼吸を伴う周期性の不規則な呼吸を行い、それは睡眠中および感染エピソード時に増え、その際、その無呼吸は、外的蘇生または自然蘇生(autoresuscitation)がなされない場合、致命的となる可能性があり、時に致命的となる。
【0004】
SIDSで死亡する小児においてしばしば死の前に軽度の感染が確認され、最新の研究成果は、脳幹機能障害および低酸素イベントからの自然蘇生不全がこれらの原因不明の死の大部分と関連していることを示している(81、82)。
【0005】
呼吸調節に後天的または先天的な欠陥がある、例えばレット症候群およびPWSを発症した小児および成人は、潜在的に致死性の呼吸機能障害(respiratory dysfunction)の危険が高いが、睡眠時無呼吸症候群の小児および成人ならびにパーキンソン病の成人もまた、呼吸調節を故障し、しばしば感染を伴い死亡する(83)。呼吸系の障害(呼吸不全または感染)は、PWS児における最大の死因であることが明らかにされた(107)。さらに、小児期のいびきおよび閉塞性睡眠時無呼吸症候群(OSAS)は、睡眠を妨害し、神経認知学的発達を妨げ、それによって長期的な機能障害を引き起こす可能性がある。これは、呼吸器感染および後発的な危険因子、例えば社会生活における喫煙および喘息の出現によって悪化する(108−111)。
【0006】
潜在的に危険であり生命の脅威となる呼吸障害は、成人でもみられる。したがって、低酸素に対する換気応答の故障は、成人期のパーキンソン病、睡眠に関係する呼吸障害、例えば睡眠時無呼吸症候群およびOSASで重要な役割を果たしている可能性がある。
【0007】
炎症性サイトカイン、例えばインターロイキン1β(IL−1β)は、これらのイベントのカギとなるメディエーターの可能性がある(3)。IL−1βは、感染および炎症に対する急性免疫反応の際に産生され、様々な病的挙動を誘起する(レビューには(4)を参照されたい)。これまでの研究から、この免疫調節因子は呼吸および自然蘇生をも変化させることが示されている(5〜10)。IL−1βは、脳幹の呼吸関連領域、例えば弧束核(NTS)および吻側延髄腹外側(RVLM)において前初期遺伝子c−fosの発現を誘導する(11)。しかし、IL−1βは巨大な疎油性タンパク質であり、血液脳関門をまたいで容易に拡散しない。さらに、NTSおよびRVLMはIL−1受容体のmRNAを発現していないようであり(12)、かつIL−1βはインビトロで脳幹の呼吸関連ニューロン活動を変化させない(5)。
【0008】
我々はこれまでに、非特異的COX阻害剤であるインドメタシンがIL−1βにより誘導される呼吸抑制を軽減することを示している(5)。PGE2それ自体は、インビボで、ヒツジの胎仔および新生仔において呼吸を抑制し(17〜19)、かつインビトロで、呼吸関連ニューロンを阻害する(5)。早産児および満期産児において新生児尿中プロスタノイド排出が調査され(112)、早産児においてPGE−Mと無呼吸の関係が確認されている(113)。
【0009】
インドメタシンは、これまで、未熟児の無呼吸を処置するのに使用されている(45)。しかし、インドメタシンは、新生児において多くの副作用を引き起こす(46)。新生児におけるインドメタシンの利用に関連する副作用には、腎臓、腸および脳における薬物誘発性の血流減少が含まれる(46)。呼吸機能障害の処置にはカフェインが使用され、持続的気道陽圧法(CPAP)および酸素補給も同様である。さらに、ナロキソン(オピオイド受容体アンタゴニスト)による急性期処置もまた利用されている。それでもなお、呼吸障害の処置方法、特に無呼吸の処置に関する要望は確実に存在する。
【発明の概要】
【課題を解決するための手段】
【0010】
本発明者らは今回、誘導されたPGE2経路が感染および低酸素に対する呼吸応答のカギとなる調節因子であることを発見した(合わせて114も参照されたい)。誘導されたPGE2経路は図6に示されている。
【0011】
IL−1βは、血液脳関門の血管内皮細胞のILー1受容体に結合して、シクロオキシゲナーゼ2(COX−2)およびミクロソームプロスタグランジンEシンターゼ1(mPGES−1)の活性を誘導する(レビューには(13)を参照されたい)。COX−2はアラキドン酸からのプロスタグランジンH2(PGH2)の生成を触媒し、mPGES−1は、引き続く、PGH2からのプロスタグランジンE2(PGE2)の生成を触媒する。その後PGE2は脳実質に放出され、そこでIL−1βのいくつかの中枢的作用、例えば熱誘導(14)、行動反応(15)および神経内分泌変化(16)を媒介することが最近報告されている。本願明細書にさらに記載されるように、プロスタグランジンは、IL−1βの換気作用も媒介する(54)。さらに、PGE2受容体であるEプロスタノイド受容体サブタイプ3(EP3R)が、脳幹の呼吸関連領域:NTSおよびRVLMに局在する(20、21)。
【0012】
本願明細書に記載されるように、IL−1βは、mPGES−1の活性化およびPGE2の脳幹EP3Rへの結合を通じて中枢呼吸に悪影響を及ぼし、無呼吸頻度の増加および低酸素イベント後の自然蘇生不全を引き起こす。したがって、誘導されたPGE2経路に関連する呼吸障害は、一またはそれ以上の場所でこの経路を狙い撃ちすることによって、例えばCOX−2を阻害することによって、mPGES−1を阻害することによって、および/またはEP3Rを阻害することによって軽減され得る。
【0013】
したがって、一つの局面において、本発明は、哺乳動物対象における呼吸障害の処置方法であって、Eプロスタノイド受容体サブタイプ3(EP3R)阻害剤、ミクロソームプロスタグランジンEシンターゼ1(mPGES−1)阻害剤および/またはシクロオキシゲナーゼ2(COX−2)選択的阻害剤を含む治療有効量の組成物を処置が必要な対象に投与することを含む、方法、を提供する。
【0014】
本願明細書に記載される誘導性のPGE2経路における一つまたはそれ以上のステップを標的とする組成物を用いて呼吸障害、例えば無呼吸の誘導に関与する適当な経路を遮断することにより、選択性の低い治療法に付随する有害な影響を最小限に抑えることができると考えられる。例えば、COX−2を選択的に、mPGES−1を、および/またはEP3Rを標的化することによって、本願明細書中に記載されるような呼吸障害は、有害な副作用、例えば非選択的COX阻害剤インドメタシンの使用時にみられる副作用を最小限に抑えつつ軽減することができる。
【0015】
さらなる局面において、本発明は、EP3R阻害剤、mPGES−1阻害剤および/またはCOX−2選択的阻害剤を含む、哺乳動物被験体における呼吸障害の処置に使用できる組成物を提供する。
【0016】
さらなる局面において、本発明は、哺乳動物被験体における呼吸障害の処置するための医薬の製造における、EP3R阻害剤、mPGES−1阻害剤および/またはCOX−2選択的阻害剤を含む組成物の使用を提供する。
【0017】
さらなる局面において、本発明は、哺乳動物対象における呼吸障害に対する感受性またはその存在の評価方法であって、当該哺乳動物由来のサンプル中のプロスタグランジンE2(PGE2)またはその代謝産物のレベルを検出すること、およびPGE2またはその代謝産物の対照レベルと当該サンプル中のレベルを比較することを含み、PGE2またはその代謝産物の対照レベルと比較して当該サンプル中のPGE2またはその代謝産物レベルが高いことが、当該対象における呼吸障害に対する感受性またはその存在を指示する、方法、を提供する。
【0018】
本発明者らは、本願において、呼吸障害、例えば無呼吸および低酸素後の脆弱な自然蘇生におけるPGE2の中枢的役割に関する証拠を提供する。具体的には、脳脊髄液(CSF)および/または尿中のPGE2および/またはその代謝産物レベルの上昇が、無呼吸頻度の増加および低酸素後に自然蘇生する能力の低下に関連している。C反応性タンパク質(CRP)レベル、PGE2レベルと無呼吸の間の相関関係は、PGE2および/もしくはその代謝産物のレベルを単独でまたは感染マーカー、例えばCRPと組み合わせてモニタリングすることで、呼吸障害およびそれに対する感受性に関する診断的利益を得ることができることを示している。PGE2はサイトカインおよび低酸素刺激に反応して迅速に生成されるため、そのモニタリングは、感染および仮死が疑われる哺乳動物における呼吸障害、例えば乳幼児における無呼吸の増加の診断および監視に特に有用である。
【0019】
驚くべきことに、本発明者らは、尿プロスタグランジン代謝産物(u−PGEM)のレベルが、感染の進行および随伴性の無呼吸がみられる乳幼児、PWS児および睡眠時無呼吸がみられる一部の成人(高い無呼吸インデックスを有する成人を含む)において上昇することを発見した。特異的かつ高感度のアッセイを用いて尿中のPGE2レベルの測定することで、呼吸障害(特に無呼吸)の予見および評価のための非侵襲的方法が実現し、これは驚くほど広い患者年齢層に適用できると考えられる。感染および随伴性の無呼吸がみられる乳幼児において、u−PGEMレベルの上昇は、CRPレベルの上昇よりも早い段階で生じるようである。したがって、生物学的サンプル(例えば、尿、血液またはCSF)中のPGE2および/またはその代謝産物レベルの評価は、感染随伴性の炎症および呼吸機能障害患者の診断、処置および管理の面で、CRPレベルの評価よりも優れている。
【0020】
したがって、本発明は、ヒト対象における無呼吸の存在および/またはその重症度の評価方法であって、当該対象から採取した尿サンプル中の一つまたはそれ以上のPGE2代謝産物のレベルを検出すること、および当該一つまたはそれ以上のPGE2代謝産物の対照レベルと当該サンプル中のレベルを比較することを含み、当該サンプル中の一つまたはそれ以上のPGE2代謝産物のレベルが当該一つまたはそれ以上のPGE2代謝産物の対照レベルと比較して少なくとも20%、少なくとも50%、少なくとも100%または少なくとも200%高いことが、当該対象における無呼吸の存在および/またはその重症度の高さを指示する、方法、を提供する。特定の例において、ヒト対象は、閉塞性睡眠時無呼吸症候群(OSAS)、プラダー・ウィリー症候群、先天性低換気症候群および/またはレット症候群に罹患している。特定の例において、ヒト対象は、16歳以上、1〜16歳、または0〜1歳である。
【0021】
本発明者らは、本願において、出生時仮死後の被験体におけるPGE2の上昇およびPGE2と低酸素性虚血性脳症(HIE)の相関関係について記載している。これらの結果は、PGE2およびその代謝産物が、周産期の脳における酸素供給の欠陥に起因する神経学的損傷の強力な診断マーカーとなることを示している。さらに、これらの結果は、被験体が罹患した低酸素の程度が、サンプル(例えばCSF、尿または血液サンプル)中で検出されるPGE2およびその代謝産物のレベルに反映されることを示している。
【0022】
したがって、さらなる局面において、本発明は、哺乳動物対象における低酸素性虚血性脳症(HIE)に対する感受性またはその存在の評価方法であって、当該対象由来のサンプル中のプロスタグランジンE2(PGE2)またはその代謝産物のレベルを検出すること、およびPGE2またはその代謝産物の対照レベルと当該サンプル中のレベルを比較することを含み、PGE2の対照レベルと比較して当該サンプル中のPGE2またはその代謝産物レベルが高いことが、当該対象におけるHIEに対する感受性またはその存在を指示する、方法、を提供する。
【0023】
さらなる局面において、本発明は、哺乳動物被験体が罹患した低酸素または重度の低酸素仮死(例えば周産期仮死)の評価方法であって、当該対象由来のサンプル中のプロスタグランジンE2(PGE2)またはその代謝産物のレベルを検出すること、およびPGE2またはその代謝産物の対照レベルと当該サンプル中のレベルを比較することを含み、PGE2の対照レベルと比較して当該サンプル中のPGE2またはその代謝産物レベルが高いことが、当該対象が低酸素または重度の低酸素仮死(例えば周産期仮死)を罹患したことを指示する、方法、を提供する。
【0024】
さらなる局面において、本発明は、哺乳動物における呼吸障害の処置に使用できる物質の同定方法であって、誘導されたPGE2経路を阻害する能力について試験物質をアッセイすること、例えば、
(a)COX−2により媒介されるPGH2の生成、
(b)mPGES−1により媒介される、mPGES−1の環状エンドペルオキシド基質から当該基質の9−ケト,11αヒドロキシ型である生産物への変換、および
(c)EP3Rアゴニストにより媒介されるEP3Rの活性化、
の一つまたはそれ以上を阻害する能力について試験物質をアッセイすることを含み、誘導されたPGE2経路の阻害、例えば、(a)、(b)および(c)の一つまたはそれ以上の阻害は、当該試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを指示する、方法、を提供する。
【0025】
誘導されたPGE2経路を阻害する能力を有することが見出された試験物質は、一つまたはそれ以上のさらなる成分、例えば薬学的に許容できる補助成分(excipient)を含む組成物に配合され得る。このような組成物は、哺乳動物における呼吸障害の処置方法において使用され得る。
【0026】
誘導されたPGE2経路の中枢的な重要性およびその呼吸障害、例えば無呼吸に対する寄与(図6参照)の認識は、呼吸障害の処置において治療的有用性を有し得る物質を同定する上での基礎となる。特に、
(a)COX−2により媒介されるPGH2の生成、
(b)mPGES−1により媒介される、mPGES−1の環状エンドペルオキシド基質から当該基質の9−ケト,11αヒドロキシ型である生産物への変換、および
(c)EP3Rアゴニストにより媒介されるEP3Rの活性化、
の一つまたはそれ以上を阻害する能力について試験物質をスクリーニングする方法が、一つまたはそれ以上のインビトロアッセイを用いて実行され得る。阻害活性についての試験物質のスクリーニングは、呼吸障害の動物モデルに対する試験物質の効果の測定に基づくスクリーニング法よりも容易にスケールアップすることができる。このことは、呼吸障害の動物モデルを用いて試験物質をスクリーニングする前にイニシャルインビトロスクリーニングが行われる場合に有益である。このようにして、適当なインビトロ薬理活性を有する見込みのある物質が、その後のインビボ試験のために選択され得る。
【0027】
さらなる局面において、本発明は、哺乳動物における呼吸障害の処置に使用できる物質の同定方法であって、試験物質を試験哺乳動物に投与すること、ここで、当該試験物質は、誘導されたPGE2経路の阻害剤、例えば、EP3R阻害剤、mPGES−1阻害剤および/またはCOX−2選択的阻害剤である、および当該試験物質を投与しなかった対照の哺乳動物における徴候または症状との比較で、当該試験哺乳動物における呼吸障害の徴候または症状の重症度を決定することを含み、当該試験哺乳動物における呼吸障害の徴候または症状の重症度が対照哺乳動物のそれらよりも低いことは、当該試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを指示する、方法、を提供する。
【0028】
本発明のこの局面の方法はさらに、試験物質が誘導されたPGE2経路を阻害する能力を有するかどうか、例えばEP3R阻害剤、mPGES−1阻害剤および/またはCOX−2選択的阻害剤として機能する能力を有するかどうかを決定する前段階を含み得る。
【0029】
呼吸障害の徴候または症状の重症度を低下させ、それによって呼吸障害を処置する能力を有することが見出された試験化合物は、一つまたはそれ以上の別の成分、例えば薬学的に許容できる補助成分を含む組成物に配合され得る。このような組成物は、哺乳動物における呼吸障害の処置方法において使用され得る。
【0030】
さらなる局面において、本発明は、PGE2以外のEプロスタノイド受容体サブタイプ3(EP3R)アゴニスト、ミクロソームプロスタグランジンEシンターゼ1(mPGES−1)活性化剤および/またはシクロオキシゲナーゼ2(COX−2)選択的活性化剤を含む有効量の組成物を哺乳動物に投与することを含む、哺乳動物における呼吸抑制の誘導方法を提供する。
【0031】
哺乳動物における呼吸抑制の誘導は、呼吸障害の研究に特に有用であり得る。例えば、哺乳動物における呼吸抑制の誘導は、呼吸障害、例えば無呼吸、低酸素および/または脆弱な自然蘇生の動物モデルの作製に有用であり得る。このようなモデルは、EP3RまたはmPGES−1の活性化が無呼吸、例えば睡眠時無呼吸およびパーキンソン病、例えばパーキンソン病に付随する呼吸機能障害の動物モデルにおいて見られるかどうかを試験するのに有用であり得る。
【0032】
低酸素下で放出されるPGE2は、例えば、EP3R−Gi活性化の刺激ならびにそれに続くcAMPの減少およびニューロン活動の低下による脳の急性低酸素抵に対する耐性の強化を通じて急性的な神経保護効果を有し得る。
【0033】
本発明は、明らかに不可能な場合および明示的に回避される旨の説明がある場合を除いて、記載される局面および好ましい特徴の組み合わせを包含する。本発明のこれらおよびさらなる局面および実施態様については、実施例および添付図面を参照しつつ以下でより詳細に説明されている。
【図面の簡単な説明】
【0034】
【図1】IL−1βおよび無酸素によって迅速に誘導される脳幹のmPGES−1を示す。皮質および脳幹のミクロソームフラクション(血液脳関門(BBB)の内皮細胞を含む)におけるmPGES−1活性は、IL−1βまたはビヒクルで処置した9日齢マウス(n=33)および酸素正常状態または正常酸素および無酸素(100%N2、5分間)に曝した同じマウスにおいて分析した。A)野生型マウスにおいて、mPGES−1活性は、NaCl(対照)処置の90分後またはIL−1β処置の90分後および180分後に測定した。対照mPGES−1+/+マウスにおける内因性mPGES−1活性は、皮質よりも脳幹において高いことを観察した。加えて、IL−1βは、時間依存的な様式でmPGES−1活性を誘導した。B)90分の時点で、IL−1β処置マウスは、脳幹において、生理食塩水処置マウスよりもおよそ2倍高い活性を示した。無酸素もまた、mPGES−1活性を有意に誘導した。さらに、IL−1βの効果と一過的な無酸素暴露の効果は加算的なものであった。IL−1β処置マウスを無酸素に曝すと、脳幹において、対照マウスよりも4倍高い活性を観察した。しかし、mPGES−1遺伝子を欠失したマウスは、IL−1βおよび無酸素に対して無視できる程度の活性しか示さなかった。データは、平均±SEMで示されている。**P<0.01、***P<0.001
【図2】IL−1βによる、mPGES−1の活性化を通じた呼吸の抑制を示す。全身フロープレチスモグラフィーを使用し、IL−1β(n=52)またはNaCl(n=48)のいずれかのi.p.投与後の9日齢mPGES−1 WTマウス(n=66)およびmPGES−1 KOマウス(n=34)において、基準呼吸および過酸素に対する換気反応を試験した。A)プレスチモグラフ記録は、NaClまたはIL−1βを与えた野生型マウスにおける正常酸素および過酸素下での呼吸を示している(5秒間、呼吸の振幅1μl/s)。B、C)全てのマウスが過酸素に反応し、呼吸頻度(fR、呼吸/分)を減らした。IL−1βは、mPGES−1+/+マウスにおいて、NaClよりも大きくfRを減少させたが、mPGES−1-/-マウスにおいては、正常酸素または過酸素下での呼吸を変化させなかった。mPGES−1+/+マウスは、mPGES−1-/-マウスよりも大きな、過酸素下での呼吸抑制を示した。データは、平均±SEMで示されている。*NaClを与えたmPGES−1+/+マウスとの比較でP<0.05。
【図3】IL−1βによる、mPGES−1を通じた無酸素下生存率の低下を示す。9日齢のmPGES−1+/+マウス(n=37)およびmPGES−1-/-マウス(n=20)を、IL−1β(n=29)またはビヒクル(n=28)の末梢投与から80分後に、5分間の無酸素(100% N2)に曝した。A)NaClを与えたmPGES−1+/+マウスのプレスチモグラフ記録は、無酸素に対する初期の過呼吸およびその後のあえぎ呼吸を示している。このマウスは、100% O2を与えた後に自然蘇生した。B)IL−1βを与えたmPGES−1+/+マウスのプレスチモグラフ記録は、無酸素に対する短い過呼吸期間およびその後のあえぎ呼吸を示している。このマウスは、100% O2を与えた後の自然蘇生に失敗した。あえぎ回数(C)は、グループ間で相違する傾向があった(ウィルコクソンX2、P=0.06)。各遺伝子型内で処置の効果を比較すると、IL−1βは野生型マウスにおいてあえぎ回数を減らしたが、その効果はmPGES−1を欠くマウスにおいては観察されなかった。D)IL−1βは、mPGES−1+/+マウスにおいて、NaClよりも、無酸素下での生存率を低下させたが、mPGES−1-/-マウスにおいてはそうならなかった。データは、平均±SEMで示されている。*P<0.05、**P<0.01
【図4】PGE2による脳幹呼吸活動の抑制および脳幹EP3受容体を通じた無呼吸の誘導を示す。呼吸は、遺伝子型がEP3R+/+(n=13)およびEP3R-/-(n=25)の新生仔マウスにおいて、PGE2(n=19)またはNaCl(n=19)投与後に試験した。A)新生仔EP3R+/+(■)およびEP3R-/-(□)マウスにおいて、0分時にPGE2を注射(icv)し、その後、正常酸素および1分間の過酸素チャレンジを行った。EP3R+/+マウスは、正常酸素および過酸素下で、無呼吸に起因する少ない呼吸頻度(fR、呼吸/分)および変動係数(C.V.)の高い不規則な呼吸律動を示した。EP3R-/-マウスにおいては、麻酔期間後も基準fRが変化せず、呼吸パターンの変動が小さかった。PGE2のicv投与後の最初の20分間の間に、温度変化または温度依存性は見られなかった。B)プレスチモグラフ記録(10秒間、呼吸の振幅1μl/s)は、EP3R+/+マウスにおいて正常酸素下でPGE2により無呼吸エピソードが誘導されたが、EP3R-/-マウスにおいては誘導されなかったことが実証されている。C)EP3R+/+マウスにおいて、PGE2は、正常酸素および過酸素下で、ビヒクルよりも多くの無呼吸を誘導した。このPGE2の効果は、EP3R-/-マウスにおいては観察されなかった。D)2〜3日齢EP3R+/+仔(■、n=5)由来の脳幹脊髄「全(en bloc)」調製物において、PGE2(20μg/l)は、呼吸律動の形成を、対照頻度(fR)の64±5%まで可逆的に抑制した(ANOVA反復測定法、P<0.01)。PGE2は、EP3R-/-マウス由来の調製物(□、n=6)の呼吸活動に影響しなかった。E)横延髄切片において、疑核(NA)の腹側の吻側延髄腹外側(RVLM)内に存在し、プレベッツィンガー複合体を含む呼吸関連ニューロンが、NK1RおよびEP3Rを共発現している。NK1RおよびEP3Rの両方の発現が示されている。矢印は、いくつかのRVLM呼吸関連ニューロンにおけるEP3RおよびNK1Rの共存を示している。F)EP3R-/-マウスにおいて、NK1Rは発現するがEP3Rは発現しないことが確認された。スケールバーは100μmを表す。データは、平均±SEMで示されている。*NaClを与えたEP3R+/+マウスとの比較でP<0.05。
【図5】新生児における脳脊髄液中のPGE2と無呼吸インデックスの相関関係を示す。脳脊髄液(CSF)は、腰椎穿刺の臨床評価を行った新生児集中治療室の乳幼児(n=12、生後平均16±4日、在胎齢平均32±2週)から得た。次いで、乳幼児の心拍呼吸の記録を行った(9.2±2.4時間)。CSF中のPGE2濃度は、標準化された酵素イムノアッセイ(EIA)プロトコルを用いて分析し、感染マーカーであるC反応性タンパク質(CRP)および無呼吸インデックス(#無呼吸/時間)との相関関係を得た。中枢PGE2濃度は、血中CRPレベルと正の相関関係があった(P=0.01)。さらに、中枢PGE2濃度と無呼吸インデックスとの間で決定的な関係が観察された(P<0.05)。図5において、我々は、検出不可能なレベルのPGE2(0±0pg/ml)と高レベルのPGE2(52±22pg/ml)を区別した。データは、平均±SEMで示されている。
【図6】IL−1βにより誘導される、プロスタグランジンE2を経由する経路を通じた呼吸抑制および自然蘇生不全の模式図を示す。全身免疫反応の際、炎症性サイトカインであるインターロイキン1β(IL−1β)は末梢血流中に放出される。IL−1βは、血液脳関門(BBB)の内皮細胞上に存在するその受容体(IL−1R)に結合する。IL−1Rの活性化は、シクロオキシゲナーゼ2(COX−2)を通じたアラキドン酸(AA)からのプロスタグランジンH2(PGH2)の合成および律速酵素ミクロソームプロスタグランジンEシンターゼ1(mPGES−1)を通じたPGH2からのプロスタグランジンE2(PGE2)の合成を誘導する。PGE2は脳の実質に放出され、脳幹の呼吸調節領域、例えば例えば弧束核(NTS)および吻側延髄腹外側(RVLM)に存在するそのEP3受容体(EP3R)に結合する。その結果、中枢呼吸関連ニューロンおよび呼吸活動が抑制され、これによって、低酸素イベント下でのあえぎ呼吸および自然蘇生能力が致死的に低下し得る。
【図7】A)は、ヒト乳幼児における、CSF中のPGE2代謝産物濃度と仮死の程度および有害な結果との相関関係を示す。CSF中のPGE2代謝産物は、生後24時間以内に行った腰椎穿刺の中で獲得し、これは低酸素性虚血性脳症(HIE)と相関する。B)は、ヒト乳幼児の出生5分後のアプガールスコアとCSF中のPGE2代謝産物濃度の相関関係を示す。
【図8】対照の健常な成人vs閉塞性睡眠時無呼吸症候群を患っている成人の尿プロスタグランジン代謝産物(u−PGEM)レベルを示す。測定は、三重項四重極質量分析・テトラノルPGEM法により行った(PGE代謝産物は、pmolPGEM/μgクレアチンで表される)。無呼吸群は、対照よりも数値のばらつきがずっと大きく、その中にはPGEMレベルが極めて高い部分集団(点線の円)が含まれる。
【図9】対照の健常な小児vsプラダー・ウィリー症候群(PWS)を患っている小児(3〜16歳)の尿プロスタグランジン(u−PGEM)レベルを示す。測定は、三重項四重極質量分析・テトラノルPGEM法により行った(PGE代謝産物は、pmolPGEM/μgクレアチンで表される)。PWS群は、対照よりも有意に高いu−PGEMレベルを示す。
【図10】対照の健常な乳幼児(1月〜1歳)vs進行中の炎症、ウイルス性細気管支炎および随伴性の無呼吸がみられる乳幼児の尿プロスタグランジン(u−PGEM)レベルを示す。測定は、三重項四重極質量分析・テトラノルPGEM法により行った(PGE代謝産物は、pmolPGEM/μgクレアチンで表される)。無呼吸および炎症群は、対照よりも有意に高いu−PGEMレベルを示す。
【発明を実施するための形態】
【0035】
発明の詳細な説明
呼吸障害
本発明は、呼吸および/または換気の中枢調節の異常が関与する一群の呼吸障害を想定している。特に、呼吸障害は、異常な、例えば不規則または少ない呼吸頻度、少ないおよび/もしくは短いあえぎ呼吸、少ない1回換気量ならびに/または低酸素に対する呼吸反応の弱体化であり得る。呼吸障害は周期性呼吸であり得る。
【0036】
無呼吸
呼吸障害は無呼吸であり得る。無呼吸は呼吸の停止を意味し、これは一時的または恒久的なものであり得る。無呼吸は、例えば、インピーダンスニューモグラフィーによって確認され、本明細書中に記載されるようなイベントモニタリングシステムを通じて記録され得る。無呼吸頻度は、規定の無呼吸閾値を超えるイベント数と定義され得る。定義は、対象となる対象の年齢に応じて異なることが知られている。いくつかの実施態様において、例えば、哺乳動物が5歳未満のヒト乳幼児の場合、無呼吸は、直前の0.5秒間における平均インピーダンス信号振幅の≧10秒の減少から直前の25秒間に測定される平均振幅の16%未満の減少と定義され得る。他の実施態様において、例えば、哺乳動物がヒト成人の場合、無呼吸は、>10秒の呼吸停止と定義され得る。特定の実施態様においては、無呼吸は、2回の呼吸サイクルを上回る呼吸停止と定義され得る。
【0037】
睡眠関連呼吸障害
呼吸障害は睡眠中に起こる障害であり得る。乳幼児における睡眠時無呼吸は、重症例では、乳幼児突然死症候群(SIDS)の危険度上昇と関連する。成人の睡眠時無呼吸もまた本発明において想定されており、これにはいびきが含まれ得る。
【0038】
周期性呼吸
睡眠時の呼吸障害は、周期性呼吸、低酸素のエピソードおよび反復的な睡眠からの覚醒によって特徴付けられ、その症状には、過剰な日中の眠気、記憶力、学習力および注意力の低下が含まれる。間欠的な低酸素および睡眠の断片化は、それぞれ独自に、海馬および前頭前皮質、記憶および実行機能の神経処理と関係が深い領域におけるニューロンの故障の原因となり得る。
【0039】
周期性呼吸または過呼吸と無呼吸の交互出現は、未熟児に共通する呼吸パターンである。臨床的に重要な未熟児の無呼吸はほとんど常に周期性呼吸に関連するものである。呼吸低下期にはPaO2が減少し、これは、初期の小児または過去に脳幹呼吸中枢を損傷している患者において呼吸を減少させ得る。これは、低酸素により誘導される脳幹呼吸中枢の抑制により起こり、部分的にはアデノシンおよびPGE2の放出により媒介される(54、85)。過呼吸期または過換気期にはPaCO2が減少して呼吸刺激が減少し、それによって無呼吸が引き起こされ得る。
【0040】
後期早産児(late preterm infant)は、CO2に対して比較的鈍い換気応答を続け、睡眠時間の50%超をREMに費やし、かつ無呼吸および周期性呼吸を示し続け、その有病率は、1500g未満で誕生した乳幼児における60%に対して10%である。
【0041】
真の周期性呼吸または無呼吸は、呼吸サイクルにおいて呼吸深度が最低の部分が実際に休止点となった場合に出現する(無呼吸の場合)。
【0042】
新生児、小児、および成人において、睡眠障害性周期呼吸および間欠的低酸素は神経の故障と関連し、そのような損傷は認知機能障害を引き起こし得る(92、93)。
【0043】
自然蘇生不全
呼吸障害は、低酸素イベント後の自然蘇生不全であり得る。自然蘇生は、睡眠または重度の低酸素による呼吸動作の抑制から自己を呼び戻す脳の能力であり、長期の低酸素下では強制的な規則性・吸息性のあえぎ呼吸がみられる。これにより体および血液がその酸素供給を取り戻し飽和することができる。
【0044】
哺乳動物は典型的に、無酸素に対して二相反応を示し、まず換気を増加させ(すなわち過呼吸)、その後に低酸素型の換気抑制を行う(すなわち一次無呼吸、あえぎ呼吸、二次無呼吸)。次いで、低酸素後の酸素供給により自然蘇生が導かれる。低酸素後の自然蘇生不全は、介入がないと死に至る場合もある。
【0045】
SIDS
呼吸障害は、乳幼児突然死症候群(SIDS)を引き起こす障害であり得る。SIDS(「ゆりかご死」とも呼ばれる)は、概ね2歳未満で起きる乳幼児の予期せぬ突然死である。睡眠中に起こり得る呼吸停止および自然蘇生不全が、SIDSと呼ばれる死をもたらし得る。したがって、特に重度の呼吸障害は、予期せぬ突然死を導く可能性がある。特定の実施態様において、本発明は特に、予期せぬ突然死を十分にもたらし得る重度の呼吸障害を想定している。
【0046】
感染性呼吸障害
呼吸障害はウイルスおよび/または細菌感染に関係し得る。様々な感染マーカー、例えば、CRP、白血球数およびIL−1βを含む炎症性サイトカインが、感染時に増大し、これらは、呼吸障害が感染要因を有することを示唆する。
【0047】
本発明の特定の実施態様において、呼吸障害は、IL−1β関連呼吸障害であり得る。IL−1βは、感染および炎症に対する急性免疫反応の際に産生される。本明細書中に開示されているように、IL−1βは血液脳関門の血管内皮細胞上のIL−1受容体に対して作用し、COX−2を誘導し、誘導されたPGE2経路を刺激し、そして最終的には無呼吸頻度の増加および低酸素イベント後の自然蘇生不全の原因となる中枢呼吸の抑制をもたらす。IL−1βの血中レベルが対照レベルよりも上昇していることは、呼吸障害がIL−1β関連呼吸障害であることを示唆する。
【0048】
特定の実施態様において、哺乳動物または哺乳動物対象は、自律神経機能障害(autonomic dysfunction)、例えばプラダー・ウィリー症候群(PWS)、先天性中枢性低換気症候群(「CCHS」、「オンディーヌの呪い」とも呼ばれる)および/またはレット症候群を含む獲得性または先天性の呼吸調節障害に罹患したヒトであり得る。PWS、CCHSまたはレット症候群を患った乳幼児は、感染イベントの際、呼吸機能障害に起因する死亡の危険性が高くなる。
【0049】
低酸素性虚血性脳症
低酸素性虚血性脳症(HIE)は、周産期の脳における酸素供給に故障が生じ、脳のエネルギー代謝が妨害された正期産児の状態を表すのに使用される用語である(97)。この状態は、死亡または重度の神経学的後遺症を引き起こし得る。
【0050】
磁気共鳴分光法を用いた脳エネルギー代謝の研究は、脳細胞への酸素供給の一次妨害の後に、数時間または数日遅れることがある二次段階として神経脱落が起こるとの仮説を打ち立て(98、99)、これは動物研究においても示されている(100)。この神経損傷の遅延は、部分的には、炎症メディエーターがその傷害に反応して近辺の環境に放出されることが原因であると考えられている。
【0051】
神経系と免疫系の間の相互作用は、疾患の多くの局面において重要である。HIEは、病態生理も病因も十分に理解されていない。最近、低酸素虚血以外の原因、例えば子宮内または新生児期の炎症(101、102)が脚光を浴びており、傷害のメディエーターとしてサイトカインが注目されている(103)。虚血性脳傷害の病因における炎症カスケードの関与を示す証拠も存在する(104)。星状細胞および小グリア細胞により分泌されるサイトカインはこの炎症反応のメディエーターとして特に重要な役割を果たしており、これらは周産期仮死後の脳においてアポトーシスを誘発し神経細胞死に寄与し得る多様なシグナルの一種であると考えられている。しかし、体内の別の場所と同様、CNSにおいても特定のサイトカインは、当初は疾患プロセスを増幅し、後にそれを軽減するよう機能し得る。PGE2は、サイトカインおよび低酸素刺激に応じて迅速に生成されることから、出生児仮死を罹患した乳幼児の診断および調査において特に有用となり得る。
【0052】
本明細書でさらに記載されるように(特に以下の実施例7を参照されたい)、PGE2が周産期仮死の結果として脳で放出されることが見出された。このことは、mPGES−1が、迅速に活性化され、哺乳動物、例えばヒトおよびマウスにおける重度の低酸素に対する応答に関与することを示唆している。低酸素、例えば周産期仮死に対する応答における誘導されたPGE2経路の役割の発見は、治療的介入における標的および診断ツール、特に周産期仮死を罹患した新生児に対する治療標的および診断ツールを提供する。
【0053】
哺乳動物
本発明の任意の局面において、哺乳動物または哺乳動物対象は、成人(成体)、小児(仔)または乳幼児(仔)、例えば新生児(仔)であり得る。哺乳動物または哺乳動物対象は、好ましくはヒトである。特定の実施態様において、ヒトは、任意の年齢または一定範囲の年齢、例えば16歳未満、10歳未満、0〜5歳および0〜24月であり得る。特定の例において、対象は、自律神機能経障害、例えばPWS、CCHSまたはレット症候群を患ったヒトの小児である。したがって、本発明の任意の局面において、対象は家族性自律神経失調症を患ったヒト(乳幼児、小児もしくは成人)または脳幹から生じる原因未知の呼吸系およびそれに関連する自律神経障害を患ったヒト(乳幼児、小児もしくは成人)であり得る。
【0054】
特定の例において、対象は、OSASに罹患したヒト小児(0〜18歳)である。対象は、産後齢0〜25週および在胎齢28〜36週のヒト乳幼児であり得る。特定の実施態様において、ヒトは成人、例えば18歳以上であり得る。哺乳動物は、睡眠時無呼吸(例えばOSAS、いびき)および/またはパーキンソン病に罹患したヒト成人であり得る。本明細書中に記載される研究の結果として、u−PGEMの増加が、OSASを患いボディマスインデックス(BMI)が30未満の成人からなる部分集団における無呼吸(睡眠時無呼吸を含む)の感受性および/または重症度の上昇に特に重要となり得る、ということが示唆される。BMIは、対象のkg体重を彼または彼女のメートル身長の2乗で割ることによって算出される。したがって、BMI>30の対象は、典型的には肥満体とみなされる。本発明の任意の局面に従う特定の実施態様において、対象は、BMI>30のヒト成人であり得る。
【0055】
誘導されたPGE2経路
本発明は、本明細書で定義されるような呼吸障害の治療的処置のために、誘導されたPGE2経路を操作することを想定している。本発明者らは、誘導されたPGE2経路が、無呼吸頻度の増加および低酸素イベント後の自然蘇生不全を引き起こす要因となることを発見した。誘導されたPGE2経路は図6で説明されている。全身免疫反応の際、炎症性サイトカインIL−1βは末梢血流中に放出される。IL−1βは、血液脳関門の内皮細胞上に存在するその受容体(IL−1R)に結合する。IL−1Rの活性化は、COX−2を通じたアラキドン酸からのPGH2の生成および律速酵素mPGES−1を通じたPGH2からのPGE2の生成を誘導する。PGE2は脳の実質に放出され、脳幹の呼吸調節領域、例えば例えば弧束核(NTS)および吻側延髄腹外側(RVLM)に存在するEP3Rに結合する。
【0056】
本発明は、脳幹呼吸調節領域への波及効果を遮断または軽減するために、一つまたはそれ以上の場所で誘導されたPGE2経路を操作、例えば薬理学的操作を行うことを想定している。誘導されたPGE2経路は、脳幹の呼吸調節領域への波及効果を遮断または軽減する効果を有する任意の点で阻害され得る。特に、誘導されたPGE2経路は、本明細書にさらに記載されるように、COX−2、mPGES−1および/またはEP3Rを阻害することによって遮断され得る。
【0057】
誘導されたPGE2経路の阻害剤
誘導されたPGE2経路の阻害剤は、脳幹の呼吸調節領域への波及効果を遮断または軽減する能力を有する。この阻害剤は、誘導されたPGE2経路の任意の点で、直接的または間接的に作用し得る。例えば、この阻害剤は、
(a)この経路に関与するポリペプチド(「誘導されたPGE2経路ポリペプチド」)、例えばCOX−2ポリペプチド、mPGES−1ポリペプチドおよび/もしくはEP3Rポリペプチドと直接的に相互作用し得る;
(b)この経路に関与するポリペプチドと間接的に相互作用し得る、例えばCOX−2ポリペプチド、mPGES−1ポリペプチドおよび/もしくはEP3Rポリペプチドの活性化因子に結合しそれを阻害することによって相互作用し得る;ならびに/または
(c)誘導されたPGE2経路ポリペプチドをコードする遺伝子の発現を妨害し得る、例えばCOX−2コード遺伝子、mPGES−1コード遺伝子および/もしくはEP3Rコード遺伝子の発現(例えば転写および/もしくは翻訳)をダウンレギュレートし得る。
【0058】
EP3R
Eプロスタノイド受容体サブタイプ3(EP3R)ポリペプチドは、EP3Rアゴニスト、例えばPGE2に結合し、下流へシグナルを伝達する、例えばGタンパク質を通じてシグナル伝達する能力を有する。ヒトおよびマウスのEP3Rアミノ酸配列は、すでに報告されている(84、その開示内容を参照により本明細書で援用することを明記する)。ヒトEP3Rヌクレオチド配列は、GenBankデータベースに寄託されている(アクセッション番号L26976、その開示内容を参照により本明細書で援用することを明記する)。EP3Rポリペプチドは、好ましくは配列番号2のヒトEP3Rアミノ酸配列を含むかまたはこれからなる。しかし、EP3Rポリペプチドは、非ヒト哺乳動物、例えばマウスまたは他のげっ歯類由来のホモログであってもよい。EP3Rポリプチドは、一つまたはそれ以上のアミノ酸が挿入、欠失または置換によって変更されたヒトEP3Rタンパク質の変異体または誘導体であってもよい。好ましくは、EP3Rポリペプチドは、配列番号2の全長アミノ酸配列と少なくとも70%、より好ましくは80%、さらにより好ましくは90%、さらにより好ましくは95%、最も好ましくは99%のアミノ酸同一性を有し、かつEP3Rアゴニスト、例えばPGE2に結合し、下流へシグナルを伝達する能力を有するアミノ酸配列を含む。いくつかの実施態様において、EP3Rポリペプチドは単離されている。
【0059】
ヒトEP3Rの活性化は、[cAMP]iを低下させ、[Ca++]iをやや上昇させる(84)。cAMPの減少は、呼吸に関連する脳幹ニューロンにおける発火度および発火頻度を減少させる、従って呼吸活動を低下させることが示されている(85)。ニューロンにおいて、EP3Rの活性化はプロテインキナーゼC非依存性Rho活性化経路を通じた神経突起の伸展を妨げ得る(86、87)。さらに、EP3Rは腎臓で高度に発現され、腎臓におけるEP3Rの活性化は血管収縮作用を示す(88)。
【0060】
EP3Rポリペプチドは、配列番号2のアミノ酸配列を有する全長EP3Rポリペプチドより短いが、その本来の生物学的活性を保持している活性部分であってもよい。特に、この活性部分は、EP3Rアゴニスト、例えばPGE2に結合し、下流へシグナルを伝達する、例えばGタンパク質を通じてシグナル伝達することができるものである。
【0061】
EP3Rコード遺伝子は、本明細書で定義されるEP3Rポリペプチドをコードするヌクレオチド配列を含み得る。EP3Rコード遺伝子は、配列番号1の全長ヌクレオチド配列と少なくとも70%、より好ましくは80%、さらにより好ましくは90%、さらにより好ましくは95%、最も好ましくは99%のヌクレオチド配列同一性を有するヌクレオチド配列を含み得る。
【0062】
EP3R阻害剤
EP3R阻害剤は、脳幹の呼吸調節領域に対するEP3R媒介性の効果を阻止または減少、例えばEP3R媒介性の無呼吸、呼吸抑制および/または自然蘇生不全を阻止または減少させる。
【0063】
本発明は、多くの異なるタイプのEP3R阻害剤の使用を想定している。例えば、EP3R阻害剤は、本明細書で定義されるEP3Rポリペプチドに結合し、アゴニストによる誘導される(例えばPGE2誘導性の)下流へのシグナル伝達(Gタンパク質共役シグナル伝達を含む)を阻止または減少させるアンタゴニストであり得る。さらに、この阻害剤は、EP3Rポリペプチドの活性化因子に結合しこれを阻害することによって間接的に作用するものであり得る。本明細書で定義されるEP3Rコード遺伝子の発現を(例えば、EP3Rコード遺伝子の転写および/または翻訳を阻害することによって)ダウンレギュレートするEP3R阻害剤もまた想定されている。
【0064】
EP3Rポリペプチドに結合する阻害剤の例には、特異的結合物質、例えば抗体分子、およびEP3Rポリペプチドへの結合においてPGE2と競合する低分子が含まれる。EP3Rコード遺伝子の発現をダウンレギュレートする阻害剤の例には、EP3Rコード遺伝子またはその一部と相補的な核酸分子およびEP3Rをコードする遺伝子またはそのフラグメントの配列に対応する二本鎖RNAが含まれる。EP3Rコード遺伝子の発現をダウンレギュレートする阻害剤には、リボザイムおよび/または三重らせん因子も含まれる。低分子、特異的結合物質および核酸を含む、多くの異なるクラスの阻害剤のさらなる詳細は本明細書中に記載されている。
【0065】
EP3Rの低分子阻害剤
本発明は、EP3Rポリペプチドに結合しアゴニストにより誘導される(例えばPGE2誘導性の)下流へのシグナル伝達、例えばGタンパク質系シグナル伝達を阻止または減少させる、約2000ダルトン以下、例えば50〜1000ダルトンの有機化合物または無機化合物の使用を想定している。EP3Rの低分子阻害剤は、EP3Rポリペプチドに競合的に結合し、PGE2と同じ部位への結合において競合する、または非競合的に結合するアンタゴニストであり得る。この低分子性EP3Rアンタゴニストは、好ましくは、中枢作用性の(すなわち血液脳関門を通過することができる)ものである。しかし、血液脳関門を通過できない低分子性EP3Rアンタゴニストもまた想定されており、これらは、例えば脳室内(i.c.v.)投与により中枢に送達され得る。
【0066】
低分子性EP3Rアンタゴニストは、(2E)−N−[(5−ブロモ−2−メトキシフェニル)スルホニル]−3−[5−クロロ−2−(2−ナフチルメチル)フェニル]アクリルアミド(L826266)または薬学的に許容できるその塩を含み得る。
【0067】
さらなる低分子性EP3Rアンタゴニストは、本明細書に記載されるスクリーニング法を用いて同定され得る。
【0068】
EP3Rの特異的結合阻害剤
いくつかの実施態様において、EP3R阻害剤は、本明細書で定義されるEP3Rポリペプチドに結合しアゴニストにより誘導される(例えばPGE2誘導性の)下流へのシグナル伝達、例えばGタンパク質系シグナル伝達を阻止または減少させる特異的結合物質であり得る。
【0069】
いくつかの実施態様において、特異的結合物質は抗体分子であり得る。他の実施態様において、特異的結合物質は、非抗体分子内に抗原結合部位、例えば非抗体タンパク質スキャホールド上にCDRのセットを含み得る。
【0070】
「抗体分子」は、天然由来であるか部分合成または完全合成であるかに関係なく、免疫グロブリンを意味する。全長抗体のフラグメントがその抗原結合機能を発揮できることが示されている。従って、抗体分子への言及は、全長抗体に及ぶし、抗体結合フラグメントを含む任意のポリペプチドまたはタンパク質にも及ぶ。
【0071】
結合フラグメントの例は、(i)VL、VH、CLおよびCH1ドメインからなるFabフラグメント、(ii)VHおよびCH1ドメインからなるFdフラグメント、(iii)単一抗体のVLおよびVHドメインからなるFvフラグメント、(iv)VHドメインからなるdAbフラグメント(55)、(v)単離されたCDR領域、(vi)Fabフラグメントが二つ連結された二価フラグメントである、F(ab’)2フラグメント、(vii)VHドメインおよびVLドメインがペプチドリンカーによって連結されこの二つのドメインが連携して抗原結合部位を形成している、短鎖Fv分子(scFv)(56〜57)、(viii)二特異性短鎖Fv二量体(WO93/11161)および(ix)遺伝子融合によって構築された多価または多特異性フラグメントである「ダイアボディ(diabody)」(WO94/13804;58)である。Fv、scFvまたはダイアボディ分子は、VHドメインとVLドメインをつなぐジスルフィド架橋を導入することによって安定化され得る(59)。scFvにCH3ドメインが連結されたミニボディも作製することができる(60)。
【0072】
EP3Rの核酸阻害剤
本発明はまた、EP3R遺伝子の発現のダウンレギュレーションに関する当該分野で公知の技術を使用する。これらには、RNA干渉(RNAi)の使用が含まれる。
【0073】
ヒトにおいて、EP3Rは、配列番号1のヌクレオチド配列を有する遺伝子によってコードされている。ヒトEP3Rのアミノ酸配列は配列番号2に示されている。このヌクレオチド配列は、本明細書にさらに記載されるように、EP3Rコード遺伝子の発現をダウンレギュレートすることができる核酸分子を設計するのに利用され得る。
【0074】
低分子RNAも、遺伝子発現の調節に利用され得る。これらには、低分子干渉RNA(siRNA)によるmRNAの特異的分解、転写後遺伝子サイレンシング(PTG)、マイクロmRNA(miRNA)によるmRNAの発生学的に調節された配列特異的翻訳抑制および特異的転写遺伝子サイレンシングが含まれる。
【0075】
ヘテロクロマチン複合体の標的化および特定の染色体部位での後成的な遺伝子サイレンシングにおけるRNAi機構および低分子RNAの役割についても実証されている。RNA干渉(RNAi)としても知られている、二本鎖RNA(dsRNA)依存的な転写後サイレンシングは、dsRNA複合体が短期的なサイレンシングのために特定の相同遺伝子を狙い撃ちする現象である。RNA干渉は、配列同一性を有するmRNAの分解を促進するシグナルとして機能する。一般的に、20nt siRNAは、遺伝子特異的なサイレンシングを誘導するのに十分長く、宿主反応を回避するのに十分短い。標的化された遺伝子産物の発現減少は顕著で、少数のsiRNA分子によって90%サイレンシングを誘導することができる。
【0076】
当該分野において、これらのRNA配列は、それらの起源によって、「短鎖または低分子干渉RNA」(siRNA)または「マイクロRNA」(miRNA)と称される。両タイプの配列は、相補的なRNAに結合し、mRNA分解を誘導する(RNAi)かまたはmRNAからタンパク質への翻訳を停止させることによって遺伝子発現をダウンレギュレートするのに使用され得る。siRNAは、長い二本鎖RNAのプロセシングによって生じ、自然環境で見出される場合典型的には外因性である。マイクロRNA(miRNA)は、内因的にコードされる小さな非コーディングRNAであり、ショートヘアピンのプロセシングにより生じる。siRNAとmiRNAは両方とも、部分的に相補的な標的配列を有するmRNAの翻訳をRNAの切断を行わずに阻害することができ、かつ完全に相補的な配列を有するmRNAを分解することができる。
【0077】
siRNAリガンドは典型的には二本鎖であり、RNA媒介性の標的遺伝子機能のダウンレギュレーションの効果を最適化するためには、siRNAによるmRNA標的の認識を媒介するRISC複合体によるsiRNAの正確な認識を確実にし、かつ宿主反応を減らせる程度にsiRNAが短くなるよう、siRNA分子の長さを選択するのが好ましい。
【0078】
miRNAリガンドは典型的には一本鎖であり、ヘアピンを形成できる部分的に相補的な領域を有する。miRNAはDNAから転写されるがタンパク質に翻訳されないRNA遺伝子である。miRNA遺伝子をコードするDNA配列はmiRNAよりも長い。このDNA配列は、miRNA配列およびほぼ逆の相補鎖を含む。このDNA配列が一本鎖RNA分子に転写されると、miRNA配列およびその逆相補性塩基は対をなして部分的に二本鎖のRNAセグメントを形成する。マイクロRNA配列の設計については(61)で説明されている。
【0079】
典型的には、siRNAまたはmiRNAの効果を模倣するためのRNAリガンドは、10〜40リボヌクレオチド(またはその合成アナログ)、より好ましくは17〜30リボヌクレオチド、より好ましくは19〜25リボヌクレオチドおよび最も好ましくは21〜23リボヌクレオチドを有する。二本鎖siRNAを利用する本発明のいくつかの実施態様において、この分子は、対称な3’オーバーハング、例えば一つまたは二つの(リボ)ヌクレオチドのオーバーハング、典型的にはUUまたはdTdTの3’オーバーハングを有し得る。本明細書の開示に基づき、当業者は、適当なsiRNAおよびmiRNA配列を容易に、例えばAmbion社のsiRNAファインダー(http://www.ambion.com/techlib/misc/siRNA_finder.htmlを参照されたい)等のリソースを用いて容易に設計することができる。siRNAおよびmiRNA配列は、合成的に作製しこれを外的に導入することにより遺伝子をダウンレギュレートすることもできるし、発現系(例えばベクター)を用いて生成させることもできる。好ましい実施態様において、siRNAは合成される。
【0080】
siRNAを生成するために長鎖の二本鎖RNAを細胞中でプロセシングさせることができる(例えば(62)を参照されたい)。この長鎖dsRNA分子は、対称な3’または5’オーバーハング、例えば一つまたは二つの(リボ)ヌクレオチドからなるオーバーハングを有していてもよいし、平滑末端であってもよい。この長鎖dsRNA分子は25ヌクレオチドまたはそれ以上であり得る。好ましくは、長鎖dsRNA分子は25〜30ヌクレオチド長である。より好ましくは、長鎖dsRNA分子は25〜27ヌクレオチド長である。最も好ましくは、長鎖dsRNA分子は27ヌクレオチド長である。30ヌクレオチドまたはそれ以上の長さのdsRNAを、ベクターpDECAPを用いて発現される場合もある(63)。
【0081】
代替手段は、細胞内でのショートヘアピンRNA分子(shRNA)の発現である。shRNAは合成性のsiRNAよりも安定である。shRNAは、小さなループ配列によって隔てられた短い逆位反復配列からなる。逆位反復配列の一つは遺伝子標的に相補的である。shRNAは、細胞内で、DICERによってsiRNAへのプロセシングを受け、このsiRNAが標的遺伝子であるmRNAを分解し、発現を抑制する。好ましい実施態様において、shRNAは、ベクター、例えば本発明のアデノウイルスベクターからの転写によって内因的に(細胞内で)生成される。shRNAは、RNAポリメラーゼIIIプロモーター、例えばヒトH1または7SKプロモーター、またはRNAポリメラーゼIIプロモーターの制御下にshRNA配列をコードするベクターを細胞にトランスフェクトすることによって細胞内で生成される場合もある。あるいは、shRNAは、ベクターからの転写によって外因的に(インビトロで)生成され得る。shRNAは次いで、細胞に直接的に導入され得る。好ましくは、shRNA分子は、EP3Rコード遺伝子の部分配列を含む。好ましくは、shRNA配列は、40〜100塩基長、より好ましくは40〜70塩基長である。ヘアピンの幹部分は、好ましくは19〜30塩基対である。この幹部分は、ヘアピン構造を安定化させるGU対を含み得る。
【0082】
siRNA分子、長鎖dsRNA分子またはmiRNA分子は、核酸配列、好ましくはベクターに含まれる核酸配列の転写により組換え生成されることができる。好ましくは、siRNA分子、長鎖dsRNA分子またはmiRNA分子は、EP3Rコード遺伝子の部分配列を含む。
【0083】
一つの実施態様において、siRNA、長鎖dsRNAまたはmiRNAは、ベクターからの転写により内因的に(細胞内で)生成される。ベクターは、当該分野で公知の任意の方法で細胞に導入され得る。場合により、RNA配列の発現は、組織特異的プロモーターを用いて調節され得る。さらなる実施態様において、siRNA、長鎖dsRNAまたはmiRNAは、ベクターからの転写により外因的に(インビトロで)生成される。
【0084】
一つの実施態様において、ベクターは、センス方向およびアンチセンス方向の両方に、EP3Rコード遺伝子の全長または部分核酸配列を含み得、それらのセンスおよびアンチセンス部分は、RNAとして発現されると集合して二本鎖RNAを形成する。好ましくは、ベクターは配列番号1の核酸配列またはその変異体もしくはフラグメントを含む。別の実施態様において、センスおよびアンチセンス配列は、異なるベクターにより提供される。好ましくは、ベクターは、配列番号1の核酸配列またはその変異体もしくはフラグメントを含む。
【0085】
あるいは、siRNA分子は、当該分野で公知の標準的な固相または液相合成技術を用いて合成することができる。ヌクレオチド間の結合は、ホスホジエステル結合またはその代替手段として、例えば、式P(O)S、(チオエート);P(S)S、(ジチオエート);P(O)NR’2;P(O)R’;P(O)OR6;CO;またはCONR’2の連結基であり得る(式中、RはH(もしくは塩)またはアルキル(1〜12C)であり、R6はアルキル(1〜9C)であり隣接するヌクレオチドに−O−または−S−を通じて接続される)。天然の塩基に加えて修飾ヌクレオチド塩基を使用することができ、これらを含むsiRNA分子に有益な特性を付与することができる。
【0086】
例えば、修飾塩基は、siRNA分子の安定性を向上し、それによってサイレンシングに必要となる量を減らすことができる。修飾塩基の導入はまた、非修飾siRNAよりも安定性の高いまたは低いsiRNA分子を実現し得る。
【0087】
用語「修飾ヌクレオチド塩基」には、共有結合性の修飾がなされた塩基および/または糖を有するヌクレオチドが含まれる。例えば、修飾ヌクレオチドには、3’位にヒドロキシル基以外および5’位にリン酸基以外の低分子量有機基が共有結合により付加された糖を有するヌクレオチドが含まれる。この種の修飾ヌクレオチドには、2’置換型の糖、例えば2’−O−メチル−;2−O−アルキル;2−O−アリル;2’−S−アルキル;2’−S−アリル;2’−フルオロ−;2’−ハロまたは2’−アジド−リボース、炭素環系糖アナログα−アノマー糖、エピマー糖、例えばアラビノース、キシロースまたはリキソース、ピラノース糖、フラノース糖、およびセドヘプツロースも含み得る。
【0088】
修飾ヌクレオチドは当該分野で公知であり、アルキル化プリンおよびピリミジン、アシル化プリンおよびピリミジン、ならびにその他の複素環系が含まれる。これらのクラスのピリミジンおよびプリンは当該分野で公知であり、プソイドイソシトシン、N4,N4−エタノシトシン、8−ヒドロキシ−N6−メチルアデニン、4−アセチルシトシン、5−(カルボキシヒドロキシルメチル)ウラシル、5−フルオロウラシル、5−ブロモウラシル、5−カルボキシメチルアミノメチル−2−チオウラシル、5−カルボキシメチルアミノメチルウラシル、ジヒドロウラシル、イノシン、N6−イソペンチル−アデニン、1−メチルアデニン、1−メチルプソイドウラシル、1−メチルグアニン、2,2−ジメチルグアニン、2−メチルアデニン、2−メチルグアニン、3−メチルシトシン、5−メチルシトシン、N6−メチルアデニン、7−メチルグアニン、5−メチルアミノメチルウラシル、5−メトキシアミノメチル−2−チオウラシル、−D−マンノシルケオシン、5−メトキシカルボニルメチルウラシル、5−メトキシウラシル、2−メチルチオ−N6−イソペンテニルアデニン、ウラシル−5−オキシ酢酸メチルエステル、プソイドウラシル(psueouracil)、2−チオシトシン、5−メチル−2−チオウラシル、2−チオウラシル、4−チオウラシル、5−メチルウラシル、N−ウラシル−5−オキシ酢酸メチルエステル、ウラシル−5−オキシ酢酸、ケオシン(queosine)、2−チオシトシン、5−プロピルウラシル、5−プロピルシトシン、5−エチルウラシル、5−エチルシトシン、5−ブチルウラシル、5−ペンチルウラシル、5−ペンチルシトシン、および2,6−ジアミノプリン、メチルプソイドウラシル、1−メチルグアニン、1−メチルシトシンが含まれる。
【0089】
RNAiを使用して線虫(C.elegans)、ショウジョウバエ(Drosophila)、植物および哺乳動物において遺伝子を抑制する方法は当該分野で公知である(WO01/29058;WO99/32619;64〜74、これら全てを参照により本明細書で援用することを明記する)。
【0090】
EP3Rコード遺伝子の発現をダウンレギュレートするリボザイムは、好ましくは、EP3Rコード遺伝子、例えば配列番号1のDNA配列を有するEP3Rコード遺伝子、のRNA配列に特異的なものである。リボザイムは、一本鎖RNA、例えばmRNAを規定の配列で特異的に切断する核酸分子、実際はRNA、であり、それらの特異性は操作することができる。ハンマーヘッド型リボザイムは、約11〜18塩基長の塩基配列を認識し、したがって約4塩基長の配列を認識するテトラヒメナ型のリボザイムよりも高い特異性を有する点で好ましい場合があるが、後者のタイプのリボザイムも特定の状況で有用である。リボザイムの利用に関する参考文献には、Marschall,et al.1994;Hasselhoff,1988およびCech,1988が含まれる。
mPGES−1 ミクロソームプロスタグランジンEシンターゼ1(mPGES−1)ポリペプチドは、グルタチオンの存在下でPGH2からのPGE2の合成を触媒する能力を有する。mPGES−1ポリペプチドは、好ましくは配列番号4のヒトmPGES−1アミノ酸配列を含むまたはそれからなる。しかし、mPGES−1ポリペプチドは、非ヒト動物、例えばマウスまたは他のげっ歯類由来のホモログであってもよい。mPGES−1ポリプチドは、一つまたはそれ以上のアミノ酸が挿入、欠失または置換によって変更されたヒトmPGES−1タンパク質の変異体または誘導体であってもよい。好ましくは、mPGES−1ポリペプチドは、配列番号4の全長アミノ酸配列と少なくとも70%、より好ましくは80%、さらにより好ましくは90%、さらにより好ましくは95%、最も好ましくは99%のアミノ酸同一性を有し、かつグルタチオンの存在下でPGH2からのPGE2の合成を触媒する能力を有するアミノ酸配列を含む。いくつかの実施態様において、mPGES−1ポリペプチドは単離されている。
【0091】
ヒトmPGES−1 cDNAのコード配列は、以下では配列番号3として示される。5’および3’非翻訳末端を含む全長cDNAはGenBankアクセッション番号NM_004878.3の下で入F手可能である。
【0092】
mPGES−1ポリペプチドは、配列番号4のアミノ酸配列を有する全長mPGES−1ポリペプチドより短いが、その本来の生物学的活性を保持する活性部分であってもよい。特にこの活性部分は、グルタチオンの存在下でPGH2からのPGE2の合成を触媒する能力を有するものである。
【0093】
mPGES−1コード遺伝子は、本明細書で定義されるmPGES−1ポリペプチドをコードするヌクレオチド配列を含み得る。mPGES−1コード遺伝子は、配列番号3の全長ヌクレオチド配列と少なくとも70%、より好ましくは80%、さらにより好ましくは90%、さらにより好ましくは95%、最も好ましくは99%のヌクレオチド配列同一性を有するヌクレオチド配列を含み得る。
【0094】
mPGES−1阻害剤
mPGES−1阻害剤は、mPGES−1により媒介されるPGE2の合成を阻止または減少させる。mPGES−1阻害剤は、mPGES−1により媒介されるPGE2レベル、特に血液脳関門の内皮細胞および/または脳の実質におけるPGE2レベルの上昇を阻止または減少させる。PGE2合成を阻止または減少させることにより、mPGES−1阻害剤は、誘導されたPGE2経路により媒介される無呼吸、呼吸抑制および/または自然蘇生不全を軽減し得る。
【0095】
本発明は、多くの異なるタイプのmPGES−1阻害剤の使用を想定している。例えば、阻害剤は、本明細書で定義されるmPGES−1ポリペプチドに結合してその触媒機能を妨害し得るものであり、そのような阻害剤には、mPGES−1ポリペプチドの触媒活性部位に結合する競合的阻害剤および触媒活性部位から離れた部位でmPGES−1ポリペプチドと結合するアロステリック阻害剤が含まれる。さらに、この阻害剤は、mPGES−1ポリペプチドの活性化因子に結合しこれを阻害することによって間接的に作用するものであり得る。mPGES−1コード遺伝子の発現を(例えば、mPGES−1コード遺伝子の転写および/または翻訳を阻害することによって)ダウンレギュレートするmPGES−1阻害剤もまた想定されている。
【0096】
mPGES−1ポリペプチドに結合する阻害剤の例には、特異的結合物質、例えば抗体分子、およびmPGES−1ポリペプチドと競合的または非競合的に結合する低分子が含まれる。mPGES−1コード遺伝子の発現をダウンレギュレートする阻害剤の例には、mPGES−1コード遺伝子またはその一部と相補的な核酸分子およびmPGES−1をコードする遺伝子またはそのフラグメントの配列に対応する二本鎖RNAが含まれる。mPGES−1コード遺伝子の発現をダウンレギュレートする阻害剤には、リボザイムおよび/または三重らせん因子も含まれる。低分子、特異的結合物質および核酸を含む、多くの異なるクラスの阻害剤のさらなる詳細は本明細書中に記載されている。
【0097】
mPGES−1の低分子阻害剤
低分子性のmPGES−1阻害剤は、mPGES−1ポリペプチドと結合して、mPGES−1ポリペプチドによる環状エンドペルオキシド基質から当該基質の9−ケト,11αヒドロキシル型である生産物への変換を阻止または制限し得る。この低分子は、mPGES−1ポリペプチドの活性部位に結合するものでもその遠位部位に結合するものでもよく、可逆的に結合するものでも不可逆的に結合するものでもよい。
【0098】
ロイコトリエンC4、NS−398、スリンダク硫化物を含む多くの化合物がmPGES−1酵素を阻害することが見出されており、これらのIC50値はそれぞれ5、20および80μMである(75、その開示を参照により本明細書で援用することを明記する)。また、15−デオキシ−Δ12,14−PGJ2、アラキドン酸、ドコサヘキサエン酸、エイコサペンタエン酸および3−[tert−ブチルチオ−1−(4−クロロベンジル)−5−イソプロピル−1H−インドール−2−イル]−2,2−ジメチルプロピオン酸(MK−886)はすべて、0.3μMという同様のIC50値でmPGESを阻害することが報告されている(76−77)。
【0099】
さらなる低分子性mPGES−1阻害剤は、本明細書に記載されるスクリーニング法を用いて同定され得る。
【0100】
mPGES−1の特異的結合阻害剤
いくつかの実施態様において、mPGES−1阻害剤は、本明細書で定義されるmPGES−1ポリペプチドに結合し、mPGES−1により媒介される環状エンドペルオキシド基質から当該基質の9−ケト,11αヒドロキシル型である生産物への変換を阻止または減少させる特異的結合物質であり得る。
【0101】
mPGES−1の特異的結合阻害剤は抗体分子であり得る。様々なタイプの抗体分子が、EP3Rの特異的結合阻害剤に関連して記載されている。抗体分子は、それがEP3RポリペプチドではなくmPGES−1ポリペプチドに結合することを除いては上述の通りである。
【0102】
mPGES−1の核酸阻害剤
本発明はまた、mPGES−1コード遺伝子の発現をダウンレギュレートする阻害剤を想定している。
【0103】
ヒトにおいて、mPGES−1は、配列番号3のヌクレオチド配列を有する遺伝子によってコードされる。ヒトmPGES−1のアミノ酸配列は配列番号4に示されている。このヌクレオチド配列は、mPGES−1コード遺伝子の発現をダウンレギュレートすることができる核酸分子を設計するのに利用され得るが、これについては、その核酸分子がEP3Rコード遺伝子ではなくmPGES−1コード遺伝子の発現をダウンレギュレートすることを除いてEP3R阻害剤に関連して説明した通りである。したがって、EP3Rコード遺伝子の配列、部分配列または相補配列に関する言及は、必要な修正を加えた上で、mPGES−1コード遺伝子の配列、部分配列または相補配列にも適用される。
【0104】
COX−2
シクロオキシゲナーゼ2(COX−2)ポリペプチドは、アラキドン酸からのPGH2の合成を触媒する能力を有する。ヒトCOX−2のアミノ酸配列は、GenBankアクセッション番号NP_000954の下で寄託されており(これを参照により本明細書で援用することを明記する)、また、以下では配列番号6として示されている。COX−2ポリペプチドは、好ましくは配列番号6のヒトCOX−2アミノ酸配列を含むかまたはこれからなる。しかし、COX−2ポリペプチドは、非ヒト動物、例えばマウスまたは他のげっ歯類由来のホモログであってもよい。COX−2ポリプチドは、一つまたはそれ以上のアミノ酸が挿入、欠失または置換によって変更されたヒトCOX−2タンパク質の変異体または誘導体であってもよい。好ましくは、COX−2ポリペプチドは、配列番号6の全長アミノ酸配列と少なくとも70%、より好ましくは80%、さらにより好ましくは90%、さらにより好ましくは95%、最も好ましくは99%のアミノ酸同一性を有し、かつアラキドン酸からのPGH2合成を触媒する能力を有するアミノ酸配列を含む。いくつかの実施態様において、COX−2ポリペプチドは単離されている。
【0105】
COX−2ポリペプチドは、配列番号6のアミノ酸配列を有する全長COX−2ポリペプチドより短いが、その本来の生物学的活性を保持する活性部分であってもよい。特にこの活性部分は、アラキドン酸からのPGH2合成を触媒する能力を有するものである。
【0106】
ヒトCOX−2のcDNA配列は、GenBankに寄託されており(アクセッション番号NM_000963、これを参照により本明細書で援用することを明記する)、以下では配列番号5として示される。そのコード配列は、太字で示されたヌクレオチド135〜1949である。
【0107】
COX−2コード遺伝子は、本明細書で定義されるCOX−2ポリペプチドをコードするヌクレオチド配列を含み得る。COX−2コード遺伝子は、配列番号5またはそのコード領域(配列番号5のヌクレオチド135〜1949)のヌクレオチド配列と少なくとも70%、より好ましくは80%、さらにより好ましくは90%、さらにより好ましくは95%、最も好ましくは99%のヌクレオチド配列同一性を有するヌクレオチド配列を含み得る。
【0108】
COX−2の選択的阻害剤
COX−2の選択的阻害剤は、COX−2によって媒介されるPGH2合成を阻止または減少させる。COX−2選択的阻害剤は、COX−2によって媒介されるPGH2レベルの上昇を阻止または減少させ、それによって誘導されたPGE2経路により媒介される無呼吸、呼吸抑制および/または自然蘇生不全を軽減し得る。
【0109】
さらに、COX−2選択的阻害剤は、COX−1に対する阻害活性よりも大きな阻害活性をCOX−2に対して有する。COX−2阻害剤の選択性は、一般に、非選択的なCOX阻害に伴う有害な影響、例えば重要な構成性のCOX−1活性の阻害によって引き起こされる影響を軽減する。COX−2選択的阻害剤は、COX−1の2倍またはそれ以上、例えば5倍または10倍高い阻害活性をCOX−2に対して有し得る。したがって、COX−2選択的阻害剤のIC50値は、同じ阻害剤のCOX−1に対するIC50値よりも2倍低い、好ましくは5倍または10倍低い場合がある。
【0110】
本発明は、多くの異なるタイプのCOX−2選択的阻害剤の使用を想定している。例えば、阻害剤は、本明細書で定義されるCOX−2ポリペプチドに結合してその触媒機能を妨害するものであり、そのような阻害剤には、COX−2ポリペプチドの触媒活性部位に結合する競合的阻害剤および触媒活性部位から離れた部位でCOX−2ポリペプチドと結合するアロステリック阻害剤が含まれる。さらに、さらに、この阻害剤は、COX−2ポリペプチドの活性化因子に結合しこれを阻害することによって間接的に作用するものであり得る。COX−2コード遺伝子の発現を(例えば、COX−2コード遺伝子の転写および/または翻訳を阻害することによって)ダウンレギュレートするCOX−2阻害剤もまた想定されている。
【0111】
COX−2ポリペプチドに結合する阻害剤の例には、特異的結合物質、例えば抗体分子、およびCOX−2ポリペプチドと競合的または非競合的に結合する低分子が含まれる。COX−2コード遺伝子の発現をダウンレギュレートする阻害剤の例には、COX−2コード遺伝子またはその一部と相補的な核酸分子およびCOX−2ポリペプチドをコードする遺伝子またはそのフラグメントの配列に対応する二本鎖RNAが含まれる。COX−2コード遺伝子の発現をダウンレギュレートする阻害剤には、リボザイムおよび/または三重らせん因子も含まれる。低分子、特異的結合物質および核酸を含む、多くの異なるクラスの阻害剤のさらなる詳細は本明細書中に記載されている。
【0112】
COX−2の低分子阻害剤
COX−2の低分子性選択的阻害剤は、COX−2ポリペプチドに結合し、COX−2により媒介されるアラキドン酸からPGH2への変換を阻止または減少させ得る。この低分子は、COX−2ポリペプチドの触媒活性部位に結合するものでもその遠位部位に結合するものでもよく、可逆的に結合するものでも不可逆的に結合するものでもよい。
【0113】
COX−2選択的阻害剤として作用する多くの化合物が記載されている。COX−2選択的阻害剤の一例は、「コキシブ類(coxibs)」として知られている薬物である。
【0114】
いくつかの実施態様において、COX−2の低分子性選択的阻害剤は、4−(5−メチル−3−フェニルイソキサゾール−4−イル)ベンゼンスルホンアミド(バルデコキシブ)もしくは薬学的に許容できるその塩、4−[5−(4−メチルフェニル)−3−(トリフルオロメチル)ピラゾール−1−イル]ベンゼンスルホンアミド(セレコキシブ)もしくは薬学的に許容できるその塩、および/または4−(4−メチルスルホニルフェニル)−3−フェニル−5H−フラン−2−オン(ロフェコキシブ)もしくは薬学的に許容できるその塩を含み得る。
【0115】
本発明において有用な多くのCOX−2阻害剤はこれまでに報告されている(COX阻害、特にCOX−2阻害の薬理のレビューについては94を参照されたい。この文献の開示内容を参照により本明細書で援用することを明記する)。
【0116】
さらなるCOX−2の低分子性選択的阻害剤は、本明細書に記載されるスクリーニング法を用いて同定され得る。
【0117】
COX−2の特異的結合阻害剤
いくつかの実施態様において、COX−2の選択的阻害剤は、本明細書で定義されるCOX−2ポリペプチドに結合し、COX−2により媒介されるアラキドン酸からPGH2への変換を阻止または減少させる特異的結合物質であり得る。
【0118】
COX−2の特異的結合阻害剤は抗体分子であり得る。様々なタイプの抗体分子が、EP3Rの特異的結合阻害剤に関連して記載されている。抗体分子は、それがEP3RポリペプチドではなくCOX−2ポリペプチドに結合することを除いては上述の通りである。好ましくは、COX−2の特異的結合阻害剤はCOX−1ポリペプチドと交差反応しないものである。
【0119】
COX−2の核酸阻害剤
本発明はまた、COX−2コード遺伝子の発現をダウンレギュレートする阻害剤を想定している。
ヒトにおいて、COX−2は、配列番号5のヌクレオチド配列を有する遺伝子によってコードされる。ヒトCOX−2のアミノ酸配列は配列番号6に示されている。このヌクレオチド配列は、COX−2コード遺伝子の発現をダウンレギュレートすることができる核酸分子を設計するのに利用され得るが、これについては、その核酸分子がEP3Rコード遺伝子ではなくCOX−2コード遺伝子の発現をダウンレギュレートすることを除いてEP3R阻害剤に関連して説明した通りである。したがって、EP3Rコード遺伝子の配列、部分配列または相補配列に関する言及は、必要な修正を加えた上で、COX−2コード遺伝子の配列、部分配列または相補配列にも適用される。
【0120】
治療
本発明は、本明細書で定義される呼吸障害の治療的処置および予防的処置の両方を想定している。この処置は、呼吸障害に対する哺乳動物の感受性を低下させる、および/または哺乳動物における呼吸障害の一つまたはそれ以上の臨床的側面を完全にもしくは部分的に好転させるものであり得る。例えば、本発明は、無呼吸を罹患した患者の呼吸を正常化させることを想定している。低酸素イベント後の自然蘇生の改善もまた想定されている。
好ましい実施態様において、哺乳動物は、本明細書で定義される呼吸障害の危険があると判断された患者であり得る。例えば、感染、特にIL−1βレベルを上昇させる感染がみられるヒト乳幼児が、無呼吸の可能性およびその重症度を低下させるために、EP3R阻害剤、mPGES−1阻害剤、および/またはCOX−2選択的阻害剤を含む薬剤で処置され得る。
【0121】
処方
本発明は、本明細書で定義される阻害剤の様々な薬学的組成物を想定している。薬学的組成物は、一般に、一つ又はそれ以上の薬学的に許容できる塩、担体または補助成分を含む。さらに、本明細書で定義される阻害剤を一つ以上含む薬学的組成物が想定される。例えば、組成物は、EP3R阻害剤、mPGES−1阻害剤およびCOX−2選択的阻害剤から選択される二つまたはそれ以上の薬剤を含み得る。あるいは、一つ以上の阻害剤が用いられる場合、その薬剤は、同時または連続投与用の別々の組成物に配合され得る。
【0122】
投与方法
本発明にしたがい適当な投与経路が利用され得る。典型的には、本明細書で定義される阻害剤を含む組成物は、口腔から、直腸から、鼻腔内に、静脈内注射、筋内注射、皮下注射、腹腔内注射もしくは脳室内注射によって、経皮パッチまたは小型ポンプによって投与され得る。血液脳関門を通過することができないEP3R阻害剤を含む組成物の場合、脳室内注射が好ましい。
【0123】
評価および診断
本発明は、哺乳動物由来のサンプル中の一つまたはそれ以上の誘導されたPGE2経路のマーカーを検出することによって哺乳動物における呼吸障害の感受性またはその存在を評価する方法を想定している。呼吸障害に罹患していることまたは呼吸障害の危険が高いことが確認された対象は、次いで、本明細書で定義される阻害剤により処置され得る。
【0124】
患者において誘導されたPGE2経路の活性が上昇しているかどうかを評価する多くの方法が想定されている。いくつかの実施態様において、PGE2またはその代謝産物のレベルが対象由来のサンプルにおいて検出され、対照レベルと比較される。対照レベルは、好ましくは既定の「正常」範囲である。例えば、対照レベルは、健常な対照由来の類似サンプルにおいて見られるPGE2またはその代謝産物のレベルであり得る。対照レベルは、過去に健常な対照対象に関して測定または報告された値であり得るし、ある集団から得られた平均値でもあり得る。
【0125】
PGE2および/または一つもしくはそれ以上のその代謝産物は、本明細書にさらに記載されるような生物学的サンプルにおいて測定され得る。PGE2代謝産物は数が多いが、その大部分はLC−MS/MS(液体クロマトグラフィー三重項四重極質量分析器)によって検出することができる(105、その開示内容全体を参照により本明細書で援用することを明記する)。
【0126】
本発明におけるPGE2代謝産物の例には、7α−ヒドロキシ−5,11−ジケト−2,3,4,5,20−ペンタ−19−カルボキシプロスタン酸およびE系よびF系の13,14−ジヒドロ−15−ケト代謝産物が含まれる。PGE2および/または一つもしくはそれ以上のその代謝産物(7α−ヒドロキシ−5,11−ジケト−2,3,4,5,20−ペンタ−19−カルボキシプロスタン酸およびE系よびF系の13,14−ジヒドロ−15−ケト代謝産物を含む)は、対象サンプルに適した任意の技術によって測定され得る。本発明におけるPGE2代謝産物ならびにその検出および測定技術については(106、その開示内容全体を参照により本明細書で援用することを明記する)にも記載されている。
【0127】
PGE2およびその代謝産物の測定アッセイの具体例には酵素免疫アッセイ(EIA)が含まれ、これについては以下の実施例の節でさらに詳細に記載されている。EIAキットは商業的に入手可能であり、個々の化合物の高感度検出を実現する。
【0128】
さらなる例として、PGE2および/または一つもしくはそれ以上のその代謝産物(7α−ヒドロキシ−5,11−ジケト−2,3,4,5,20−ペンタ−19−カルボキシプロスタン酸およびE系よびF系の13,14−ジヒドロ−15−ケト代謝産物を含む)の測定または検出は、LC.MS/MSおよび/または三重項四重極質量分析(三重項四重極法(QQQ)としても知られている)を用いるものであり得る。三重項四重極質量分析の使用は、この分析法がフェムト/ピコモル濃度の化合物を検出する能力を有することから、特定状況下で好ましい。タンデム四重極(三重項四重極)機器は、既知の代謝産物およびペプチド(例えばPGE2および/または一つもしくはそれ以上のその代謝産物)の定量に利用される。この機器は、アラキドン酸カスケードの量的経路分析に使用することができる。さらに、この機器は、臨床材料におけるペプチドの量的検証および臨床材料間での相違が代謝学的に確認された代謝産物の量的検証に使用され得る。推奨される機器は、電子スプレーイオン化インターフェース(ESI)を経由して超高速液体クロマトグラフィー(UPLC)に接続され得る。液体クロマトグラフィーにおいてサイズの小さい粒子(<1.8μm)を使用することで、クロマトグラフピーク幅を劇的に、典型的には30〜60秒(従来型LC)から3〜5秒(UPLC)に狭めることができる。これにより分離能が向上し、したがってより多くの化合物をより短時間で分離することができる。三重項四重極質量分析器において、特定の代謝産物の分子イオンが第一の四重極で選択され、代謝産物の断片化がコリージョンガスを含むコリージョンセル中で誘導される。第二の四重極では電子遷移痕を生じる特定の「娘イオン(daughter ion)」が選択される(反応モニタリング)。代謝産物/ペプチドは個別に断片化するため、この娘イオンは極めて化合物特異的なトレーサーとなる。典型的には、約100のトレースを同時にモニタリング(多重反応モニタリング、MRM)、一つの分析物中の多くの代謝産物を特異的かつ高感度で定量することができる。好ましくは、本発明の方法は、尿サンプル中の一つまたはそれ以上のPGE2代謝産物の測定を含み、三重項四重極質量分析を利用するものである。特に好ましい尿PGE2代謝産物(u−PGEM)測定アッセイは実施例8に記載されている。いくつかの例において、本発明の方法は、尿サンプル中の一つまたはそれ以上のPGE2代謝産物の測定を含み、この方法はさらに、PGE2の尿中レベルが尿クレアチニンレベルに対する相対レベルである場合、尿サンプル中のクレアチニン濃度の測定を含む。
【0129】
サンプル中のPGE2またはその代謝産物のレベルと対照レベルとの比較は、既定の対照値または対照値の範囲を記録した図表、データベースまたは文献を調べることによって行える。いくつかの例において、例えば既定の対照値が入手できない場合、サンプルレベルと対照レベルの比較は、調査対象の対象由来のサンプル中のPGE2またはその代謝産物レベルの検出に続いてまたはそれと平行して、健常な対象由来の対照サンプル中のPGE2またはその代謝産物レベルを検出することを含むことになる。
【0130】
PGE2レベルまたはPGE2代謝産物レベルが対照レベルと比較して上昇していることは、呼吸障害の存在またはその危険が高いこと、例えば無呼吸頻度が増えていることを示すと考えられる。
【0131】
以下に記載されるデータは、PGE2代謝産物が、乳幼児が出生時に罹患した仮死(「周産期仮死」)の程度および/または哺乳動物対象における低酸素性虚血性脳症(HIE)の存在もしくは重症度を推測する上で有用な指標として使用できる証拠となる。PGE2レベルまたはPGE2代謝産物レベルが対照レベルと比較して上昇、特に対象の出生から数日以内、例えば96、48、24、12、6、4、3もしくは2時間以内または60、30、20、10または5分以内に対象から採取したサンプルにおいて上昇していることは、哺乳動物対象におけるHIEの存在を示唆すること、および/または対象が周産期仮死を罹患したことを示すことが見出された。対照レベルとの比較の上でのPGE2またはPGE2代謝産物の増加率は、周産期仮死の程度および/またはHIEの重症度の程度と相関し、したがって対象の可能性のある神経学的結果と相関することが見出された。
【0132】
したがって本発明の方法は、予後および長期的な神経学的結果の推測に有用であり、したがって処置に関する迅速な決断を補助できる点で価値がある。
【0133】
実験結果は、PGE2の半減期が、いくつかの例において、約12〜18時間であることを示している。PGE2およびその代謝産物は、72時間以上経過した後でさえ存在し測定できる場合もある。PGE2分解のハーフタイムはその細胞環境に依存して大きく変化する。PGE2の半減期は、数分から数時間まで変化し得る。体内で産生されたPGE2および尿またはその他の体液中に分泌されたPGE2を評価する場合、その代謝産物も合わせて測定するのが重要である。
【0134】
いくつかの実施態様において、サンプル中のPGE2またはその代謝産物レベルは、PGE2またはその代謝産物の参考レベルと比較される。参考レベルは対照レベルとは別も
のであり得る。例えば、参考レベルは、本明細書で定義される呼吸障害または哺乳動物被験体における周産期仮死もしくはHIEを示す値または値の範囲であり得る。このような例において、PGE2またはその代謝産物レベルがほぼ参考レベルまたは参考値の範囲内であることは、本明細書で定義される呼吸障害の存在もしくはその危険が高いこと、乳幼児が出生時に罹患した仮死の程度、および/または被験体におけるHIEの存在もしくはその重症度を示す。参考レベルは、呼吸障害、乳幼児が出生時に罹患した仮死および/または被験体におけるHIEの特定の重症度または段階に関連する値または一定範囲の値であり得る。
【0135】
いくつかの実施態様において、本発明の方法は、患者において誘導されたPGE2経路の活性が高まっているかどうかを、mPGES−1コード遺伝子の発現を検出することによって評価することを含む。これには、mPGES−1コード遺伝子のmRNAのレベルを測定、例えば定量的、準定量的またはリアルタイムPCRベースの方法を用いて測定することが含まれる。mPGES−1コード遺伝子の発現の上昇は、呼吸障害の危険の上昇を示唆する。患者において誘導されたPGE2経路の活性が高まっているかどうかを評価するその他の方法には、PGH2レベルの上昇、COX−2遺伝子発現の増大および/またはIL−1βレベルの上昇を検出することが含まれる。本発明は、一つまたはそれ以上の、誘導されたPGE2経路の活性の増加に関するマーカーを検出することを想定している。例えば、PGE2レベルの検出は、PGH2レベル、mPGES−1発現、COX−2発現および/またはIL−1βレベルの検出と組み合わされ得る。
【0136】
いくつかの実施態様において、本発明の方法は、mPGES−1、COX−2および/またはEP3Rをコードする遺伝子における一つまたはそれ以上の変異を同定することを含み得る。例えば、mPGES−1、COX−2および/またはEP3Rをコードする遺伝子における単一ヌクレオチド多型(SNP)は、本明細書で定義される呼吸障害に対する感受性の上昇と関連し得る。
【0137】
サンプル
サンプルは、液体サンプル、例えばCSFサンプル、血液サンプル、尿サンプル、または非液体サンプル、例えば組織検体サンプルであり得る。好ましくは、サンプルはCSF、尿または血液サンプルである。特定の実施態様において、尿サンプルが特に好ましい。
サンプルは、本明細書で詳述される状態の実際の原因もしくは疑われる原因またはその状態の発症の後の既定の時点で哺乳動物被験体、例えばヒト被験体から採取され得る。例えば、サンプルは、被験体の出生後または入院後もしくは医師との面会後96、48、24、12、6、4、3もしくは2時間以内または60、30、20、10もしくは5分以内にヒト乳幼児から採取され得る。いくつかの例において、サンプルは、低温で(例えば4℃付近でまたは−80℃〜−20℃で)保存されたヒト尿サンプルであり得る。
【0138】
感染マーカー
本発明者らは、PGE2レベル、CRPおよび無呼吸インデックスが相関していることを発見した(図5を参照されたい)。いくつかの実施態様において、診断方法は、追加的に、感染に関連するマーカーのレベルを検出することを含み得る。例えば、CRPレベルが、患者由来のサンプル、好ましくは血液または尿サンプルにおいて評価され得る。感染マーカーのレベルが対照レベルと比較して上昇していることは、特にPGE2レベルまたは誘導されたPGE2経路の活性の増加に関する他のマーカーのレベルの上昇と組み合わせた場合に、呼吸障害の危険が高いことを示唆する。
【0139】
対照レベルは、好ましくは既定の「正常」範囲である。例えば、対照レベルは、健常な対照由来の類似のサンプルにおいて見出されるCRPのレベルであり得る。対照レベルは、過去に健常な対照対象に関して測定または報告された値であり得るし、ある集団から得られた平均値でもあり得る。
【0140】
サンプル中のCRPレベルと対照レベルとの比較は、既定の対照値または対照値の範囲を記録した図表、データベースまたは文献を調べることによって行える。いくつかの例において、例えば既定の対照値が入手できない場合、サンプルレベルと対照レベルの比較は、調査対象の対象由来のサンプル中のCRPレベルの検出に続いてまたはそれと平行して、健常な対象由来の対照サンプル中のCRPレベルを検出することを含むことになる。
【0141】
CRPレベルが対照レベルと比較して上昇していることは、呼吸障害の存在またはその危険が高いこと、例えば無呼吸頻度が増えていることを示すと考えられる。
【0142】
いくつかの実施態様において、サンプル中のCRPレベルは、CRPの参考レベルと比較される。参考レベルは対照レベルとは別ものであり得る。例えば、参考レベルは、本明細書で定義される呼吸障害を示す値または値の範囲であり得る。このような例において、CRPレベルがほぼ参考レベルまたは参考値の範囲内であることは、本明細書で定義される呼吸障害の存在またはその危険が高いことを示す。参考レベルは、本明細書で定義される呼吸障害の特定の重症度または段階に関連する値または値の範囲であり得る。
【0143】
さらに、PGE2またはその代謝産物の測定が、炎症マーカーとしてのCRPまたは高感受性CRP(hsCRP)の測定の補足としてまたはその代わりに使用され得る。
【0144】
スクリーニング法
本発明は、哺乳動物における呼吸障害の処置に使用できる物質の同定を想定している。したがって、哺乳動物における呼吸障害の処置に使用できる物質の同定方法は、誘導されたPGE2経路を阻害する能力について試験物質をアッセイすること、例えばEP3R阻害剤、mPGES−1阻害剤および/またはCOX−2選択的阻害剤として機能するかどうかについて試験物質をアッセイすることを含み得る。誘導されたPGE2経路の阻害は、試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを示す。試験物質、これは候補化合物または候補組成物であり得る、は、
(a)この経路に関係するポリペプチド(「誘導されたPGE2経路ポリペプチド」)、例えばCOX−2ポリペプチド、mPGES−1ポリペプチドおよび/もしくはEP3Rポリペプチドと直接的に相互作用することによって、
(b)この経路に関係するポリペプチドと、例えばCOX−2ポリペプチド、mPGES−1ポリペプチドおよび/もしくはEP3Rポリペプチドの活性化因子に結合しこれを阻害することにより、間接的に相互作用することによって、ならびに/または
(c)誘導されたPGE2経路ポリペプチドをコードする遺伝子の発現をダウンレギュレートすることによって、例えば、COX−2コード遺伝子、mPGES−1コード遺伝子および/もしくはEP3Rコード遺伝子の発現(例えば転写および/もしくは翻訳)をダウンレギュレートすることによって、
誘導されたPGE2経路を阻害し得る。
【0145】
ポリペプチドの阻害剤のスクリーニング
誘導されたPGE2経路ポリペプチドと相互作用および/または結合する試験物質の能力の測定は、その試験物質が、誘導されたPGE2経路の潜在的阻害剤であることを同定するのに使用され得る。この方法は、相互作用または結合を検出または観察すること、次いで、その試験物質を、それが誘導されたPGE2経路ポリペプチドの活性、例えば酵素活性または受容体を媒介するシグナル伝達を阻害するかどうかを決定するさらなるアッセイ法において使用することを含み得る。
【0146】
本発明アッセイの細部は慣用的な技術および知識を用いて当業者によって変更され得る。例えば、ポリペプチドまたはペプチド間の相互作用は、その一方を検出標識で標識してこれを固相に固定したもう一方と接触させることによって、インビトロで試験され得る。適当な検出標識には35Sメチオニンが含まれ、これは組み換え法で作製されたペプチドおよびポリペプチドに組み込まれ得る。組み換え法で作製されたペプチドおよびポリペプチドはまた、抗体で標識することができるエピトープを含む融合タンパク質として発現され得る。
【0147】
固相に固定されるタンパク質またはペプチドは、固相に結合されたそのタンパク質に対する抗体を用いて、または公知の他の技術を通じて固定され得る。好ましいインビトロ相互作用は、グルタチオン−S−トランスフェラーゼ(GST)を含む融合タンパク質を利用するものである。これは、グルタチオンアガロースビーズ上に固定され得る。上記タイプのインビトロアッセイフォーマットにおいて、試験化合物は、固定されたGST融合ポリペプチドに結合する標識ペプチドまたはポリペプチドの量を減少させる能力を測定することによってアッセイされ得る。これは、SDSポリアクリルアミドゲル電気泳動によってグルタチオンアガロースビーズを分画することによって測定され得る。あるいは、ビーズは未結合タンパク質を除去するために洗浄され、結合したタンパク質の量は、例えば適当なシンチレーションカウンター内に存在する標識の量を計測することによって測定することができる。
【0148】
一般的に、誘導されたPGE2経路ポリペプチドと結合または相互作用する試験物質の能力の同定およびその潜在的なPGE2経路阻害剤としての同定に続いて、試験物質が誘導されたPGE2経路ポリペプチドの活性を阻害できるかできないかの測定を含む一つまたはそれ以上のアッセイが行われる。通常、誘導されたPGE2経路ポリペプチドを阻害する試験物質の能力の測定を含むアッセイは、その試験物質が誘導されたPGE2経路ポリペプチドと結合または相互作用することができるかどうかが不明な状況下で行われ得るが、多数の試験化合物を試験するためのスクリーンとして以前の結合/相互作用アッセイを使用し、誘導されたPGE2経路ポリペプチドの活性を阻害する能力の測定を含む機能アッセイを実行可能なレベルまで、潜在的阻害剤の数を削減することができる。
【0149】
試験物質が誘導されたPGE2経路ポリペプチドの阻害剤として機能するかどうかを測定するアッセイ法、特にCOX−2、mPGES−1およびEP3Rアッセイが本明細書中でさらに記載されている。
【0150】
コンビナトリアルライブラリ技術(78)は、ポリペプチドの活性を調節する能力について潜在的に膨大な数の異なる物質を試験するのに効果的な手法を提供する。
【0151】
本発明のアッセイに投入され得る試験物質または試験化合物の量は、通常、使用する化合物の種類に基づく試行錯誤により決定され得る。典型的には、約0.1nM〜10μM濃度の試験化合物(例えば推定阻害剤)が使用され得る。ペプチドが試験物質である場合は、それよりも高い濃度が使用され得る。使用され得る化合物は、天然化合物であってもドラッグスクリーニングプログラムで使用される合成化合物であってもよい。複数の既知または未知成分を含有する植物抽出物もまた使用され得る。他の阻害剤または阻害剤候補は、ポリペプチドまたはペプチドフラグメントの3次元構造のモデリングに基づくもの、ならびに特定の分子形状、サイズおよび電荷特性を有する潜在的阻害化合物を生み出す合理的なドラッグデザインの使用に基づくものであり得る。
【0152】
遺伝子発現の阻害剤のスクリーニング
誘導されたPGE2経路の阻害剤は、誘導されたPGE2経路ポリペプチドをコードする遺伝子、例えばCOX−2コード遺伝子、mPGES−1コード遺伝子および/またはEP3Rコード遺伝子の発現を妨げることによってこの経路を阻害し得る。したがって、本発明のアッセイ法は、試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを同定することを含み得、この方法は、誘導されたPGE2経路ポリペプチドをコードする遺伝子の発現を減少または阻害することができる物質をスクリーニングすることを含み、このスクリーニングは、
(a)上記遺伝子のプロモーターを含むDNAと試験物質を接触させ(プロモーターは遺伝子に機能的に連結されている)、
(b)このプロモーターからの遺伝子発現のレベルを決定し、そして
(c)試験物質の存在下での遺伝子発現レベルと、同条件かつ試験物質の非存在下での遺伝子発現レベルを比較する、
ことを含み、試験物質の存在下で遺伝子発現レベルが低下することは、その試験物質が、誘導されたPGE2経路ポリペプチドをコードする遺伝子の発現を阻害できることを示す。
【0153】
この方法はさらに、試験物質が、誘導されたPGE2経路ポリペプチドをコードする遺伝子の発現の阻害剤であること、すなわち哺乳動物における呼吸障害の処置に使用できる物質であることを同定することを含み得る。
【0154】
したがって工程(c)は、試験物質の存在下での遺伝子発現レベルが同条件かつ試験物質の非存在下での遺伝子発現レベルと比較して低下していることを検出し、それによってその試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを同定することを含み得る。
【0155】
この方法は、試験物質と発現系、例えば遺伝子に機能的に連結された遺伝子プロモーターを含む宿主細胞を接触させること、および遺伝子発現を測定することを含み得る。この遺伝子は、誘導されたPGE2経路ポリペプチドをコードする遺伝子であってもよいし、異種遺伝子、例えばレポーター遺伝子であってもよい。「レポーター遺伝子」は、自身のコードする生産物が発現された後にアッセイされ得る遺伝子、すなわちプロモーター活性に関する「レポート」を行う遺伝子である。
【0156】
「プロモーター」は、機能的に連結されたDNAの下流に(すなわち、二本鎖DNAのセンス鎖における3’方向に)向かう転写の開始点となるヌクレオチド配列を意味する。遺伝子のプロモーターは、ヒト染色体においてその遺伝子の5’側に存在するヌクレオチド配列または別の種、例えばラットまたはマウスにおける等価な配列を含むかまたは本質的にそれからなるものでよい。
【0157】
プロモーター活性のレベルは、例えば、そのプロモーターからの転写によって生成されるmRNAの量の評価またはそのプロモーターからの転写によって生成されたmRNAの翻訳によって生成されるタンパク質産物の量の評価によって定量可能である。発現系内に存在する特定のmRNAの量は、例えば、そのmRNAとハイブリダイズすることができかつ標識されているかまたは特定の増幅反応、例えばポリメラーゼ連鎖反応(PCR)において使用できる特定のオリゴヌクレオチドを用いて測定され得る。
【0158】
レポーター遺伝子の使用は、タンパク質産物に基づくプロモーター活性の測定を容易にする。レポーター遺伝子は、好ましくは、検出シグナル、好ましくは視覚的に検出可能なシグナル、例えば有色生成物を生じる反応を触媒する酵素をコードする。βガラクトシダーゼおよびルシフェラーゼを含む多くの例が知られている。βガラクトシダーゼ活性は、基質の青色化によってアッセイされ得、このアッセイは肉眼により、または分光光度計を使用して吸光度を測定することにより行われる。蛍光、例えばルシフェラーゼ活性の結果として生じる蛍光は、分光光度計を用いて定量され得る。放射能アッセイは、例えばクロラムフェニコールアセチルトランスフェラーゼを用いて行われ得る。ただしこの化合物は、非放射能アッセイにおいても使用され得る。レポーター遺伝子からの発現により生じる遺伝子産物の存在および/または量は、その生成物に結合できる分子、例えば抗体またはそのフラグメントを用いて測定され得る。結合性分子は、任意の標準技術を用いて直接的または間接的に標識され得る。
【0159】
プロモーター構築物は任意の適当な技術を用いて細胞株に導入され、そのゲノムにレポーター構築物が統合された安定な細胞株が作製され得る。その細胞は、様々な時間、試験化合物と共に培養およびインキュベートされ得る。その細胞は、多数の化合物の分析を容易にするため、96ウェルプレート上で培養され得る。次いでその細胞は洗浄され、レポーター遺伝子の発現が分析され得る。一部のレポーター、例えばルシフェラーゼについては、細胞は溶解された後に分析される。
【0160】
当業者は、多くの利用可能なレポーター遺伝子およびアッセイ技術が遺伝子の活性を測定するのに使用できることを認識している。さらなる例については、Sambrook and Russell, Molecular Cloning: a Laboratory Manual: 3rd edition, 2001, Cold Spring Harbor Laboratory Pressを参照されたい。
【0161】
COX−2アッセイ
本発明は、試験物質、これは候補化合物または候補組成物であり得る、がCOX−2選択的阻害活性を有するかどうかを測定し、それによってCOX−2選択的阻害活性を有することが確認された試験物質を、呼吸障害の処置に使用できる物質として同定するアッセイ法を想定している。
【0162】
いくつかの実施態様において、このアッセイ法は、COX−2ポリペプチドと試験物質およびアラキドン酸を、アラキドン酸が当該試験物質の非存在下でCOX−2によってPGH2に変換される条件下で接触させること、および当該試験物質の非存在下でのPGH2合成の対照レベルとの比較で当該試験物質の存在下でのPGH2合成レベルを決定することを含み、当該試験物質の存在下でのPGH2合成レベルが上述の対照レベルよりも低いことは、当該試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを示す。
【0163】
COX−2阻害剤の同定方法には、これまでに報告されている方法が含まれる(89、90、91、その開示内容を参照により本明細書で援用することを明記する)。COX−2を阻害することが見出された候補化合物または候補組成物は、その化合物または組成物が哺乳動物における呼吸障害を処置する能力を有するかどうかを確認するため、本明細書に記載されるさらなる試験、例えばインビボ試験に供され得る。
【0164】
多くのCOX−2阻害剤用スクリーニングキットが市販されている。例えば、Cayman Chemicals社の製品番号560131「COX Inhibitor Screening Assay」は、必要なヒトCOX−2補因子を提供し、その検出はSnCl2によるPGH2から主としてPGF2αへの還元に基づくものである(http://www.caymanchem.com/app/template/Product.vm/catalog/560131/a/zを参照されたい)。
【0165】
これ以外にも、生成したPGH2を検出するための代替法がいくつか存在する。例えば、PGH2は、塩化鉄処理後に12−HHTおよびマロンジアルデヒドに変換でき、両者は高スループットで測定できる。また、COX−2のペルオキシダーゼ活性は、例えば、これもCayman chemicals社製のキットに記載されるようにして使用することができる(http://www.caymanchem.com/app/template/Product.vm/catalog/760111/a/zを参照されたい)。
【0166】
この方法は、通常、試験物質を、または酵素および酵素基質と共に試験物質をインキュベートすることを含む。基質は、生理学的基質、例えばアラキドン酸であってもよいし、修飾されているかまたは非生理学的な基質、例えばその酵素反応の中で検出可能な(例えば有色の)生成物を生じるよう設計された基質であってもよい。
【0167】
COX−2ポリペプチドを試験物質および基質、例えばアラキドン酸と接触させる順序は変更され得る。例えば、COX−2ポリペプチドは最初に試験物質と共にインキュベートされ、次いで基質と接触され得るし、その逆でもよい。
【0168】
このようにして、試験物質の存在下での生成物の生成は、試験物質の非存在下での生成物の生成と比較され得る。生成物レベルが低いまたは生成物の形成速度が遅いことは、試験物質が酵素活性を阻害していることを示す。
【0169】
阻害剤のアッセイのさらなる可能性は、COX−2を(生来的にまたは組み換え的に)発現する適当な細胞株によるPGH2生成に影響を及ぼす物質の能力の試験である。本発明のアッセイは、導入された一つまたはそれ以上のベクターから関連ポリペプチドまたはペプチドを発現する細胞株、例えば酵母株において実施され得る。
【0170】
それ以外のさらなるアッセイの可能性は、COX−2を含む不均一タンパク質調製物(ヒトでもそれ以外の哺乳動物でもよい)によるPGH2生成に影響を及ぼす物質の能力の試験である。本発明の好ましいアッセイは、単離された/精製されたCOX−2ポリペプチド(全長COX−2またはその活性部分を含む)のCOX−2活性を阻害する試験物質の能力を測定することを含む。
【0171】
本発明のアッセイ法において、生成物の生成は、基質レベルを定量することにより、および/または生成物レベルを定量することにより測定することができる。残存する基質レベルが高ければ、生成物の生成レベルは低いということである。
【0172】
いくつかの実施態様において、アッセイ法は、COX−2阻害についての試験物質の選択性を他のポリペプチド、例えばCOX−1と比較の上で測定することを含み得る。例えば、このアッセイ法は、試験物質のCOX−1に対する阻害活性、例えばIC50、および試験物質のCOX−2に対する阻害活性、例えばIC50を測定することを含み得る。好ましくは、COX−2選択的阻害剤として同定される試験物質は、COX−1に対する阻害活性の2倍またはそれ以上、例えば5倍または10倍高い阻害活性をCOX−2に対して有する。したがって、COX−2阻害に関する試験物質のIC50値は、COX−1阻害に関する同じ試験物質のIC50値よりも2倍またはそれ以上、例えば5倍または10倍低い。
【0173】
生成物の測定には、HPLC、UV分析、放射能測定、またはRIA(例えば、市販のPGE検出用RIAキット)を利用することができる。生成物の形成は、ガスクロマトグラフィー(GC)または質量分析(MS)、またはTLCおよび放射能走査により分析することができる。
【0174】
COX−2タンパク質を使用する本発明の方法において、完全(全長)COX−2タンパク質配列を使用する必要はない。二分子間の結合を試験するまたはCOX−2酵素活性を試験する本発明のアッセイは、フラグメントまたは変異体を使用することができる。フラグメントは、当業者に公知の任意の適当な方法によって作製および使用され得る。適当なフラグメント作製法には、コードDNAからのフラグメントの組み換え発現が含まれるがこれに限定されない。このようなフラグメントは、コードDNAを入手し、発現される部分のいずれかの側にある適当な制限酵素認識部位を確認し、DNAからその部分を切り出すことによって作製され得る。この部分は、次いで、標準的な市販の発現系内の適当なプロモーターに機能的に連結され得る。別の組み換えアプローチは、DNAの関連部分を適当なPCRプライマーを用いて増幅することである。小フラグメント(例えば、約20または30アミノ酸未満)はまた、当該分野で周知のペプチド合成法を用いて作製され得る。COX−2の活性部分は、アッセイ法に使用され得る。
【0175】
COX−2ポリペプチドの「活性部分」は、本発明の方法において使用することができる。活性部分は、全長ポリペプチドより短いがその本来の生物学的活性を保持しているペプチドを意味する。特に、活性部分は、適当な条件下でアラキドン酸からのPGH2合成を触媒する能力を保持しているものである。
【0176】
mPGES−1アッセイ
本発明は、試験物質、これは候補化合物または候補組成物であり得る、がmPGES−1阻害活性を有するかどうかを測定し、mPGES−1阻害活性を有することが確認された試験物質を、哺乳動物における呼吸障害の処置に使用できる物質として同定するアッセイ法を想定している。
【0177】
いくつかの実施態様において、このアッセイ法は、mPGES−1ポリペプチドと試験物質およびmPGES−1の環状エンドペルオキシド基質を、当該mPGES−1の環状エンドペルオキシド基質が当該試験物質の非存在下でmPGES−1によって当該基質の9−ケト,11αヒドロキシ型である生産物に変換される条件下で接触させること、および当該試験物質の非存在下での当該生産物の対照生成レベルとの比較で当該試験物質の存在下でのPGH2またはその非酵素的分解産物(PGE2、PGD2またはPGF2α)のレベルを測定することを含み、当該試験物質の存在下での当該生産物の生成レベルが上述の対照レベルよりも低いことは、当該試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを示す。
【0178】
この方法は、通常、試験物質を、または酵素および酵素基質と共に試験物質をインキュベートすることを含む。基質は、生理学的基質、例えばPGH2であってもよいし、修飾されているかまたは非生理学的な基質、例えばその酵素反応の中で検出可能な(例えば有色の)生成物を生じるよう設計された基質であってもよい。
【0179】
mPGES−1ポリペプチドを試験物質および基質、例えばPGH2と接触させる順序は変更され得る。例えば、mPGES−1ポリペプチドは最初に試験物質と共にインキュベートされ、次いで基質と接触され得るし、その逆であってもよい。
【0180】
このようにして、試験物質の存在下での生成物の生成は、試験物質の非存在下での生成物の生成と比較され得る。生成物レベルが低いまたは生成物の形成速度が遅いことは、試験物質が酵素活性を阻害していることを示す。
【0181】
阻害剤アッセイのさらなる可能性は、mPGES−1を(生来的にまたは組み換え的に)発現する適当な細胞株によるPGE2生成に影響を及ぼす物質の能力の試験である。本発明のアッセイは、導入された一つまたはそれ以上のベクターから関連ポリペプチドまたはペプチドを発現する細胞株、例えば酵母株において実施され得る。
【0182】
それ以外のさらなるアッセイの可能性は、mPGES−1を含む不均一タンパク質調製物(ヒトでもそれ以外の哺乳動物でもよい)によるPGE2生成に影響を及ぼす物質の能力の試験である。本発明の好ましいアッセイは、単離された/精製されたmPGES−1ポリペプチド(全長mPGES−1またはその活性部分を含む)のmPGES−1活性を阻害する試験物質の能力を測定することを含む。
【0183】
mPGES−1ポリペプチドの活性を阻害する物質(すなわちmPGES−1阻害剤)のスクリーニング方法は、適当な反応媒体中で一つまたはそれ以上の試験物質と当該ポリペプチドを接触し、処理されたポリペプチドの活性を試験し、その活性を試験物質で処理していない同等の反応媒体中での当該ポリペプチドの活性と比較することを含み得る。処理されたポリペプチドと未処理のポリペプチドとの間の活性の違いは、関係する試験物質の調節効果を示唆する。
【0184】
このアッセイ法は、
(a)PGE2が正常に生成される条件の下、還元型グルタチオンおよびPGH2の存在下でmPGES−1ポリペプチドおよび試験化合物をインキュベートし、そして
(b)PGE2の生成を測定すること、
を含み得る。
【0185】
mPGES−1のPGH2基質は、COX−2およびAAのインキュベーションによって提供され得、したがってこれらがPGH2を提供するためにアッセイ媒体中に提供されてもよい。さらに、mPGES−1は環状エンドペルオキシドからの9−ケト,11αヒドロキシプロスタグランジンの立体特異的な形成を触媒するので、適当な生成物を測定することにより、他のmPGES−1基質を、mPGES−1活性およびその活性に対する試験化合物の効果の測定に使用することもできる。
【0186】
基質 生成物
PGH2 PGE2
PGH1 PGE1
PGH3 PGE3
PGG2 15(S)ヒドロペルオキシPGE2
PGG1 15(S)ヒドロペルオキシPGE1
PGG3 15(S)ヒドロペルオキシPGE3
【0187】
上記の通り、基質は、これまでに議論してきた基質のいずれかであってもよいし、任意の他の適当な基質でもよく、これは当業者の自由である。基質はPGH2であり得、その場合生成物はPGE2である。
【0188】
本発明のアッセイ法において、生成物の生成は、基質レベルを定量することにより、および/または生成物レベルを定量することにより測定することができる。アッセイ終了時にまたはアッセイ反応が終了した時点で残存する基質は、塩化鉄または塩化スズを添加することによって、それぞれ、12−ヒドロキシヘプタデカトリエン酸およびマロンジアルデヒドまたはPGF2αに変換することができる。したがって、これらの化合物の量は、PGE2の形成を間接的に反映するものである。これらの化合物の定量は、残存する基質量の定量により生成物の生成を測定する手段である。残存基質のレベルが高ければ、生成物の生成レベルが低いということである。
【0189】
mPGES−1阻害剤は、PGE2または他の生成物(使用する基質に依存する)の生成量が、試験物質を用いない対照実験と比較して減少していることを測定することによって同定され得る(またはmPGES−1阻害剤であると推定される候補物質が実際にそうであると確認され得る)。したがって、試験物質の存在下での生成物の生成は、試験物質の非存在下での生成物の生成と比較され得る。生成物レベルが低いことまたは生成物の形成速度が遅いことは、試験物質がmPGES−1活性を阻害していることを示す。このようにして、試験物質は、哺乳動物における呼吸障害の処置に使用できる薬剤であることが確認され得る。
【0190】
生成物の測定には、HPLC、UV分析、放射能測定、またはRIA(例えば、市販のPGE検出用RIAキット)を利用することができる。生成物の形成は、ガスクロマトグラフィー(GC)または質量分析(MS)、またはTLCおよび放射能走査により分析することができる。
【0191】
mPGES−1タンパク質を使用する本発明の方法において、完全(全長)mPGES−1タンパク質配列を使用する必要はない。二分子間の結合を試験するまたはPGEシンターゼ活性を試験する本発明のアッセイは、フラグメントまたは変異体を使用することができる。フラグメントは、当業者に公知の任意の適当な方法によって作製および使用され得る。適当なフラグメント作製法には、コードDNAからのフラグメントの組み換え発現が含まれるがこれに限定されない。このようなフラグメントは、コードDNAを入手し、発現される部分のいずれかの側にある適当な制限酵素認識部位を確認し、DNAからその部分を切り出すことによって作製され得る。この部分は、次いで、標準的な市販の発現系内の適当なプロモーターに機能的に連結され得る。別の組み換えアプローチは、DNAの関連部分を適当なPCRプライマーを用いて増幅することである。小フラグメント(例えば、約20または30アミノ酸未満)はまた、当該分野で周知のペプチド合成法を用いて作製され得る。mPGES−1の活性部分は、アッセイ法に使用され得る。
【0192】
mPGES−1ポリペプチドの「活性部分」は、本発明の方法において使用することができる。活性部分は、全長ポリペプチドより短いがその本来の生物学的活性を保持しているペプチドを意味する。特に、活性部分は、グルタチオンの存在下でのPGH2からのPGE2合成を触媒する能力を保持しているものである。
【0193】
EP3Rアッセイ
本発明は、試験物質、これは候補化合物または候補組成物であり得る、がEP3R阻害活性を有するかどうかを測定し、EP3R阻害活性を有することが確認された試験物質を、呼吸障害の処置に使用できる物質として同定するアッセイ法を想定している。
【0194】
いくつかの実施態様において、このアッセイ法は、EP3Rポリペプチドと試験物質およびEP3Rアゴニストを、EP3Rアゴニストが当該試験物質の非存在下でEP3Rポリペプチドを活性化する条件下で接触させること、および当該試験物質の非存在下でのEP3Rポリペプチドの活性化の対照レベルとの比較で当該試験物質の存在下でのEP3Rポリペプチドの活性化レベルを測定することを含み、当該試験物質の存在下でのEP3Rポリペプチドの活性化レベルが上述の対照レベルよりも低いことは、当該試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを示す。
【0195】
EP3Rアゴニストは、天然アゴニスト、例えばPGE2であってもよいし、合成アゴニストでもよい。多くのEP3Rアゴニストが市販、例えばBiomol社から販売されている。一つの十分に特徴付けられている例は、Sulprostoneである(http://www.caymanchem.com/app/template/Product.vm/catalog/14765を参照されたい)。EP3Rポリペプチドの活性化は、受容体タンパク質におけるコンホメーション変化であり得、それによってGタンパク質との結合がもたらされる。EP3Rポリペプチドの活性化は、アデニリルシクラーゼ活性に対する効果をモニタリングすることによって検出され得る。例えば、細胞ベースのアッセイにおいて、細胞表面に存在するEP3Rポリペプチドの活性化は、その細胞内のcAMP濃度の増加または減少をモニタリングすることによって検出され得る。
【0196】
いくつかの実施態様において、EP3Rポリペプチドは細胞表面に存在し、そこでEP3Rはレポート手段と結合する。このレポート手段は、受容体活性化の指標となる。例えば、レポート手段は、EP3Rにより媒介されるシグナル伝達経路においてEP3Rより下流の物質を含み得る。そのような下流物質のレベルにおける何らかの変化をモニタリングすることによって、EP3Rの活性化をモニタリングすることができる。レポート手段は、蛍光または放射能標識の検出を含む多くの技術のいずれかによってモニタリングされ得る。特定の実施態様において、EP3RはGタンパク質を通じてアデニリルシクラーゼに結合し、それによってcAMP生成を調節し得る。試験化合物の存在下および非存在下でEP3Rアゴニストに反応するcAMPレベルをモニタリングすることによって、EP3Rポリペプチドのアンタゴニストとして機能する試験化合物の能力が測定され得る。ヒトEP3Rの活性化は、[cAMP]iを低下させ、[Ca++]iをやや上昇させ得る。したがって、EP3Rアゴニストは、細胞内[cAMP]の低下および/または細胞内[Ca++]の上昇を誘導し得る。これは、例えば、FLIPRベースのアッセイを用いてモニタリングされ得る。EP3Rのアンタゴニストは、任意のEP3Rアゴニストにより誘導される細胞内[cAMP]の低下および/または細胞内[Ca++]の上昇を阻止または制限し得る。
【0197】
インビボスクリーニング
本発明は、哺乳動物における呼吸障害の処置に使用できる物質の同定方法を想定している。この方法は、誘導されたPGE2経路を阻害することが知られているまたは阻害すると期待される一つまたはそれ以上の物質を使用し得る。
【0198】
したがって、本発明は、哺乳動物における呼吸障害の処置に使用できる物質の同定方法であって、試験物質を試験哺乳動物に投与すること、ここで、当該試験物質は、EP3R阻害剤、mPGES−1阻害剤および/またはCOX−2選択的阻害剤である、および当該試験物質を投与しなかった対照の哺乳動物における徴候または症状との比較で、当該試験哺乳動物における呼吸障害の徴候または症状の重症度を決定することを含み、当該試験哺乳動物における呼吸障害の徴候または症状の重症度が当該対照哺乳動物のそれらよりも低いことは、当該試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを示す、方法、を想定している。
【0199】
例えば、試験物質は、
(a)COX−2により媒介されるPGH2の合成、
(b)mPGES−1により媒介される、mPGES−1の環状エンドペルオキシド基質から当該基質の9−ケト,11αヒドロキシ型である生産物への変換、および
(c)EP3Rアゴニストにより媒介されるEP3Rの活性化、
の一つまたはそれ以上を阻害する能力を有することが見出された物質であり得る。
【0200】
EP3R阻害剤、mPGES−1阻害剤またはCOX−2選択的阻害剤である試験物質の同定方法については、本明細書にさらに記載されている。試験物質をEP3R阻害剤、mPGES−1阻害剤またはCOX−2選択的阻害剤として同定することは、インビボスクリーニングよりも前の初期段階で行われ得る。このようにして、複数の化合物が、所望の薬理学的活性についてインビトロでスクリーニングされ、次いでその所望の薬理学的活性を有することが見出された化合物がインビボでスクリーニングされ得る。EP3R阻害剤、mPGES−1阻害剤およびCOX−2選択的阻害剤については本明細書にさらに詳細に記載されている。
【0201】
呼吸障害の徴候または症状には、呼吸抑制、無呼吸頻度、低酸素後の自然蘇生の脆弱さ、呼吸頻度の減少、1回換気量の減少および/または低酸素に対するあえぎ呼吸の減少が含まれ得る。徴候または症状の重症度の決定は、試験/対照哺乳動物を低酸素圧、低酸素に曝した後、および/または試験/対照哺乳動物にIL−1β、リポ多糖(LPS)またはPGE2を投与した後に、その徴候または症状を決定することを含み得る。
【0202】
本明細書で使用される場合、呼吸障害の徴候または症状の重症度が低いことは、その徴候または症状が哺乳動物に害を及ぼす可能性が低いことを意味する。例えば、本発明の方法が、IL−1β投与後の無呼吸頻度を決定することを含む場合、無呼吸頻度の少なさおよび/または無呼吸エピソードの短さが、徴候または症状の重症度の低さとみなされるであろう。
【0203】
呼吸障害の徴候または症状をモニタリングするのに適した技術については本明細書にさらに記載されている。例えば、本発明の方法は、プレチスモグラフィーまたはインピーダンスニューモグラフィーを利用し得る。この方法は、内部酸素圧を変えることができる空気管理チャンバーを利用し得る。好ましくは、このチャンバーは温度管理がなされているものである。
【0204】
あるいは、呼吸障害の徴候または症状の決定は、脳幹の呼吸活動を、例えば試験/対照哺乳動物から単離された脳幹・脊髄調製物を用いてモニタリングすることを含み得る。脳幹呼吸活動は電極を用いてモニタリングされ得るが、これについては本明細書にさらに記載されている。本発明の方法が試験/対照哺乳動物から単離された脳幹・脊髄調製物を用いて脳幹呼吸活動をモニタリングすることを含む場合、試験物質は、脳幹・脊髄の単離前に投与されてもよいし、試験/対照哺乳動物からの単離後に脳幹・脊髄調製物に対して直接投与されてもよい。
【0205】
本発明の方法は、エクスビボ脳幹脊髄全調製物または脳幹スライス調製物を利用し得る。このような調製物は、アゴニストおよび/またはアンタゴニスト、例えば誘導されたPGE2経路のアゴニストおよび/またはアンタゴニスト、ならびに環境変化の細胞、ネットワークおよび行動への効果の平行モニタリングを実現する。この方法は、本明細書で定義されるようなインサイチュおよびインビボ法と組み合わせることができる。無呼吸の誘導は、環境変化、例えばO2濃度の低下、例えば低酸素によって達成され得る。あるいはまたは加えて、無呼吸の誘導は、薬学的または麻酔処置、例えばオピオイド受容体アゴニストおよび/またはフォースコリンを含むcAMP濃度上昇薬によって達成され得る。
【0206】
試験哺乳動物および対照哺乳動物はげっ歯類であり得、各々好ましくはマウスまたはラットである。本発明の方法は、好ましくは、ヒトにおける呼吸障害の処置に使用できる薬剤を同定する方法である。
【0207】
本発明の方法は、バロメーター技術またはフロープレチスモグラフィー技術を用いて呼吸障害の徴候または症状の重症度を決定することを含み得る。このような技術は、試験哺乳動物および対照哺乳動物がげっ歯類、例えばマウスまたはラットである場合に好ましい。特定の実施態様において、試験哺乳動物はヒトである。このような場合、呼吸障害の徴候または症状の重症度の決定は、睡眠ポリグラフ記録法の使用を含み得る。
【0208】
試験哺乳動物および対照哺乳動物は、好ましくは、対照動物に試験物質を与えない点を除いて同一条件下に置かれる。好ましくは、対照投与、例えば生理食塩水が対照哺乳動物に与えられ、そしてそれは、好ましくは、試験物質を試験哺乳動物に投与するのと同一経路によって対照哺乳動物に投与される。
【0209】
特定の実施態様において、試験哺乳動物および対照哺乳動物は、同じ動物であり得る。この場合、試験物質を投与されなかった対照哺乳動物における徴候または症状との比較で試験哺乳動物における呼吸障害の徴候または症状の重症度を決定することは、まず、試験物質の投与前に哺乳動物における呼吸障害の徴候または症状の重症度を決定し(「対照記録」)、次に、試験物質の投与後に哺乳動物における呼吸障害の徴候または症状の重症度を決定する(「試験記録」)ことにより実行され得る。次いで、対照記録および試験記録は比較され、対照記録の重症度よりも試験記録の重症度が低いことが、試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを示す。同じ動物を試験哺乳動物および対照哺乳動物として使用することは、哺乳動物がヒト、例えば臨床研究下にあるヒトである場合に好ましい。
【0210】
以下は実施例であり、特許請求の範囲を制限するものと解釈されるべきではない。
【実施例】
【0211】
材料および方法
動物
DBA/1lacJ純系(n=158)(Jackson Laboratory, Bar Harbor, ME)およびC57BL/6系(n=75)(Beverly Koller博士、University of North Carolina, Chapel Hill, NCの厚意により寄贈された)の新生仔を使用した。ミクロソームプロスタグランジンEシンターゼ1(mPGES−1)およびEP3受容体(EP3R)の遺伝子を以前に記載されたようにしてノックアウトマウスにおいて選択的に欠失させた(47、48、両方の開示内容を参照により本明細書で援用することを明記する)。全ての動物を実験後直ちに断頭により屠殺し、PCRおよびサザンブロット分析を用いてジェノタイピングを行った。野生型DBA/1lacJマウスの一部から得たデータを、新生仔DBA/1lacJマウスにおける呼吸活動の特徴付けに利用した(6)。全てのマウスを、12時間明:12時間暗サイクルという標準化された条件下で飼育した。餌および水分は自由にとらせた。
【0212】
ヒト被験者
カロリンスカ大学病院の新生児集中治療室の乳児(平均在胎齢32±2週)に協力していただいた(生後年齢平均16±4日)(n=12)。その乳児のうち、臨床適応のための腰椎穿刺を受け、書面上のインフォームドコンセントが得られたものを適格者とした。これらの研究は、欧州共同体のガイドラインに従って行ない、内部の倫理委員会による承認を受けた。乳児のうち、臨床適応、例えば感染の疑い、神経学的変化および心拍呼吸上の問題について腰椎穿刺を受けたものを実験適格者とした。脳室内出血(等級≧2)、白質病変(PVL、脳室周囲白質軟化症)、発作、出血後水頭症または先天性異常が見られた乳児は除外した。新生児分娩データ、医学的状態、感染マーカー、呼吸治療および投薬を含む関連医学情報(pertinent medical information)を記録した。心拍呼吸の記録は、腰椎穿刺後12時間以内に行った(平均4.8±1.7時間)。
【0213】
薬物
組換えマウスインターロイキン1β(IL−1β)(Nordic Biosite AB, Taby, Sweden)を滅菌したNaClで再構成し、1μg/ml溶液とした。プロスタグランジンE2(PGE2)(Cayman Chemicals, Ann Arbor, MI,USA)は、合成CSF(aCSF)に希釈して、インビボ実験用に2nmol/μl、インビトロ実験用に20μg/l(60nM)の濃度とした。
【0214】
非拘束全身フロープレチスモグラフィー
プレシキグラスチャンバー(35ml)を高感度ダイレクトエアフローセンサー(0〜200ml/分;TRN3100、 Kent Scientific Corporation, Litchfield, CT, USA)に接続した。フローシグナルは4チャンネル増幅器(P/N 770 S/N 5; SENSElab, Somedic Sales, Horby, Sweden)によって増幅し、デジタルデータに変換し、DasyLabソフトウェア(Datalog GmbH & Co.KG, Monchengladbach, Germany)を用いてオンラインコンピュータにより100Hzで記録した。呼吸頻度(fR、呼吸/分)1回換気量(VT、μl/呼吸)および毎分換気量(VE、μl/分)を算出した。チャンバー温度は、チャンバーをサーモスタット付水浴に沈めることによって、記録されている新生仔マウスの温熱中間範囲に従い30.1±0.1℃で維持した(49)。以前に記述されているように、プリセット高精度シリンジ(Hamilton Bonaduz AG, Switzerland)を用いて標準化された量の空気(5〜200μl)を繰り返し注入することによってチャンバーをキャリブレートした(6)。ガス交換の95%が投与の35秒以内に行われたことを、CO2濃度分析(Metek CD-3AおよびS-3A, PA, USA)によって確認した。
【0215】
インピーダンスニューモグラフィー
乳児の心拍呼吸活動を、インピーダンスニューモグラフィーを用いて非侵襲的に測定し、イベントモニタリングシステム(KIDS, Hoffrichter GmbH, Schwerin, Germany)を介して記録した。ベースライン呼吸速度および無呼吸閾値を超えるイベントを記録するようモニターをプログラムした。無呼吸は、直前の0.5秒間における平均インピーダンス信号振幅の≧10秒の減少から直前の25秒間に測定される平均振幅の16%未満の減少と定義した。このイベント前後の60秒間もモニターのメモリに保存した。
【0216】
IL−1βまたはNaClのi.p.注射後のプレチスモグラフィー
mPGES−1およびEP3Rを様々に発現する、それぞれ、9日齢DBA/1lacJマウス(n=143)およびC57BL/6マウス(n=16)における呼吸を、フロープレチスモグラフィーを用いて試験した。各マウスに、IL−1β(10μg/kg)またはビヒクルの腹腔内注射(0.01ml/g)を行った。70分後に、マウスを、プレスチモグラフチャンバーに誘導した。4分間の正常酸素(21% O2)およびその後の1分間の過酸素(100% O2)の間の呼吸を評価した。正常酸素下での5分間の回復期間の後、無酸素(100% N2)に対する呼吸応答を試驗した。最後に、100% O2を8分間与え、自然蘇生の能力を評価した。ベースライン、70分、およびチャンバーから出た後の皮膚温度を記録した。直腸温度は、直腸プローブの設置が呼吸行動を変化せる可能性を考慮して測定しなかった。性別を推定するために肛門生殖突起間距離を測定した。
【0217】
PGE2またはビヒクルのicv注射後のプレチスモグラフィー
EP3Rを様々に発現する、9日齢C57BL/6マウス(n=38)における呼吸を、フロープレチスモグラフィーを用いて試験した。約60秒間のセボフルラン麻酔投与を行った後、PGE2(4nmol、2〜4μl aCSF)またはビヒクルを、ポリエチレンチューブに取り付けた先細ガラスピペットを用いて側脳室にゆっくりと注入した。次いでマウスを直ちにプレスチモグラフチャンバーに入れた。10分間の正常酸素下での回復期間の後、マウスを前記のような過酸素および無酸素チャレンジに暴露した。ベースラインおよびその後の各時点での動物の皮膚温度を、サーミスタ温度プローブを用いて記録した。
【0218】
脳幹呼吸活動
脳幹脊髄標本は、以前に記載されるようにして(50、51、両方の開示内容を参照により本明細書で援用することを明記する)、EP3R+/+およびEP3R-/-の遺伝型を有する2日齢C57BL/6マウス(n=11)から素早く単離した。C4前根における吸息律動に対応する呼吸関連活動を、ガラス吸引電極を用いて追跡し、記録し(5kHz)、オフラインで分析した。対照の記録は、PGE2を含有するaCSFの灌流およびその後のaCSF洗浄前少なくとも20分間行った。
【0219】
mPGES−1活性の測定
新生仔マウスの脳(n=33)を、0.25Mスクロース、1×コンプリートプロテアーゼインヒビター(Roche Diagnostics)および1mM還元型グルタチオンを含有する0.1M KPi(無機リン酸カリウム)緩衝液中でホモジナイズし、その後に超音波処置した。膜画分は、細胞成分分画法によって単離した。膜画分におけるmPGES−1活性は、以前に記載されたようにして測定した(52、その開示内容を参照により本明細書で援用することを明記する)。
【0220】
免疫組織化学
9日齢野生型およびEP3Rノックアウト仔から脳幹を断頭後に素早く切り出し、4%パラホルモアルデヒドで固定し、15%スクロース含有リン酸緩衝化生理食塩水(PBS)、pH7.4中で一晩凍結保護した。次いでこの脳幹を素早く凍結し、14μmの横断切片をクリオスタット(Leica CM3050 S, Leica Microsystems Nussloch GmbH)に逐次集めた。切片を風乾し、PBSで再度水和させ、そして0.3%過酸化水素を使用して内因性ペルオキシダーゼを10分間阻害した。その後のPBS洗浄の後、この切片を5%ヤギ血清(Jackson Immunoresearch Laboratories, West Grove, PA)、1%ウシ血清アルブミン(Sigma-Aldrich)および0.3% Triton X−100(Sigma-Aldrich)を含有するPBS中、45分間ブロックおよび透過処理を行い、ウサギNK−1R抗体(1:20,000希釈;Sigma-Aldrich)と共に一晩インキュベートした。次いでこの切片をPBS中で洗浄し、1:50希釈のビオチニル化二次抗体(ヤギ抗ウサギ;Vector Laboratories, Burlingame, CA)と共にインキュベートした。1時間のインキュベーション後、切片をリンスし、ペルオキシダーゼ結合型Vectastain ABC(1:100希釈;Vector Laboratories)と30分間、次いでCy3結合型Tyramideシグナル増幅液(TSA、1:50、PerkinElmer, Boston, MA)と2分間インキュベートした。反応をPBS中停止させ、5%ロバ血清(Jackson)、1%ウシ血清アルブミン(Sigma-Aldrich)および0.3% Triton X−100(Sigma-Aldrich)を含有するPBSで45分間ブロックした。次いで切片を、1:50希釈のウサギEP3R抗体(Cayman Chemical, MI)と共に4℃(C)で一晩インキュベートした。次の日、切片をPBS中でリンスし、Alexa488結合型二次抗体(ロバ抗ウサギ;Molecular probes)と共に1時間インキュベートした。その後のPBS洗浄の後、切片をVectashield Hard Setマウンティングメディウム(Vector Laboratories)にマウントした。潜在的な交差反応の危険性を除去するため、一次抗体を滴定して最適な希釈を決め、各一次抗体が除かれた対照スライドをインキュベートした。さらに、EP3Rノックアウトマウス(n=4)由来の脳幹スライスを、通常のNK1R染色による上記のプロトコルを用いて研究したが、EP3Rは検出されなかった。画像は、ImageJソフトウェア(NIH, Bethesda, MD)で処理した。
【0221】
CSF分析および心拍呼吸の記録
脳脊髄液サンプルを、標準化された酵素免疫アッセイ(EIA)プロトコル(Cayman Chemicals, Ann Arbor, MI, USA)を用いて、PGE2およびPGE2代謝産物について分析した。腰椎穿刺の後できるだけ早く乳児の心拍呼吸の記録を行った(平均記録時間:9.2±2.4時間)。腰椎穿刺前12時間以内に測定した感染マーカー(例えば、C反応性タンパク質、白血球)の血中濃度も記録した。
【0222】
プレチスモグラフィーデータの分析
動きアーチファクトを伴わない平穏状態下呼吸の期間を分析に使用した。正常酸素および過酸素ならびに無酸素応答(すなわち、過呼吸、一次無呼吸、あえぎ呼吸、二次無呼吸および自然蘇生)の平均fR、VTおよびVE値を以前に記載されているようにして分析した(6、その開示内容を参照により本明細書で援用することを明記する)。全ての動物について生存を記録した。無呼吸は≧3回の呼吸サイクル分の呼吸停止と定義した。呼吸の規則性は、変動係数(C.V.)(すなわち、SDを、60秒の間の呼吸と呼吸の間隔の平均で割ったもの)を用いて定量した。
【0223】
乳児の心拍呼吸データの分析
モニタリングソフトウェアを使用してベースライン呼吸速度を記録し、全ての心拍呼吸イベントを可視化した。無呼吸インデックス(A.I.、無呼吸数/記録時間)を決定した。心拍呼吸活動、感染状況およびCSF中のPGE2レベルの間の相関関係を評価した。全ての動きアーチファクトを分析から除外した。
【0224】
脳幹脊髄標本
脳幹は、橋が除去されるように第VI脳神経根と台形体の下境界部の間で吻側から取り出した。この標本を、合成脳脊髄液(aCSF):130mM NaCl、3.3mM KCl、0.8mM KH2PO4、0.8mM CaCl2、1.0mM MgCl2、26mM NaHCO3および30mM D−グルコースを入れた1.5mlチャンバー中、28℃で連続的に灌流させた(流速3〜4ml/分)。この溶液を95%O2および5%CO2で連続的に平衡化してpH7.4とした(50、51)。
【0225】
プレスチモグラフデータの分析
無酸素に対する応答は年齢によって変わるので(53)、我々はP9齢の時にすべての記録をとるようにした;しかし、問題となる年齢による影響を最小限に抑えるため、年齢の関連値として体重を使用し、集団平均体重の1SD以内の体重を有する動物のみ無酸素および生存分析に含めた(6)。
【0226】
動物の特徴
IL−1βまたはNaClのi.p.注射後のプレチスモグラフィー実験において、mPGES−1+/+マウスはmPGES−1-/-マウスよりも体重が軽かった(それぞれ、4.4±0.1g vs 4.9±0.1g)。動物の性別に違いはなかった。ベースラインにおける動物の皮膚温度(34.7±0.1℃)および注射70分後の温度(34.8±0.1℃)はグループ間で類似していた。無酸素後、mPGES−1+/+マウスはmPGES−1-/-マウスよりも皮膚温度が高かった(それぞれ、32.2±0.1℃ vs 31.4±0.2℃)。C57BL/6マウスにおいて、動物の体重(4.5±0.1g)、動物の性別、ベースライン温度(34.4±0.2℃)、70分の温度(34.5±0.5℃)または無酸素後の温度(30.4±0.1℃)に違いはなかった。PGE2またはビヒクルのicv注射後のプレチスモグラフィー実験において、C57BL/6マウスは、動物の性別および麻酔後温度(31.0±0.2℃)において違いを示さなかった。しかし、EP3R+/+マウスは、EP3R-/-マウスよりも体重が重かった(それぞれ、4.9±0.1g vs 4.1±0.1g)。9日齢のEP3R+/+マウス(n=13)およびEP3R-/-マウス(n=26)の皮膚温度を、ベースラインならびにPGE2またはビヒクルのicv注射後の正常酸素、過酸素および無酸素下での各時点で測定した。注射23分後の無酸素暴露まで温度の違いはみられなかった。この時点で、EP3R-/-マウスは、EP3R+/+マウスよりも皮膚温度が低かった(それぞれ、30.9±0.3℃ vs 31.8±0.3℃)。温度類似性は、30〜31分の無酸素後期間で異なった(それぞれ、29.8±0.2℃ vs 30.4±0.1℃)。
【0227】
統計
1要因のANOVAにより、正規分布および等分散のパラメータを比較した。スチューデントt post−hoc検定を用いて多重比較を行った。ノンパラメトリックな測定および非ガウス分布型のデータについては、ウィルコクソンχ2テストを使用した。変数の経時的変化は、MANOVA反復測定デザインを使用して試験した。スピアマンのRho相関テストにより変数間の相関関係を決定した。データは平均±SEMで示されている。P<0.05の値を統計的に有意であるとした。
【0228】
実施例1:内因性の脳幹mPGES−1活性および呼吸強壮効果
我々はまず、9日齢のmPGES−1+/+マウスおよびmPGES−1-/-マウスにおけ
る内因的なPGE2産生およびその換気に対する効果を試験した。野生型マウスの示した基準となるミクロソームプロスタグランジンEシンターゼ1(mPGES−1)活性は、ホモジナイズした皮質よりもホモジナイズした脳幹において高かった(図1)。正常酸素下での呼吸は遺伝子型間で類似したが、fRはmPGES−1-/-マウスよりもmPGES−1+/+マウスの方が低い傾向があった(クラスカル・ウォリス、P=0.03;スチューデントt post−hoc検定、P=0.18)(表1)。中枢呼吸の駆動を、1分間の過酸素チャレンジ(100% O2、1分間)によって試験した。両方の遺伝子型のマウスは、呼吸頻度(fR)を減らすことで、過酸素に対して応答した(図2)。しかし、呼吸抑制は、mPGES−1+/+マウスの方が、mPGES−1-/-マウスよりも大きかった(それぞれ、27±2% vs 19±3%)。
【0229】
【表1】

【0230】
正常酸素および過酸素(100% O2)下での呼吸頻度(fR、呼吸/分)、1回換気量(VT、μl/呼吸/g)および毎分換気量(VE、μl/分/g)を、IL−1βまたはビヒクルの腹腔内注射後の9日齢mPGES−1+/+マウスおよびmPGES−1-/-マウスにおいて試験した。各遺伝子型内での処置の効果を比較すると、IL−1βは、mPGES−1+/+マウスにおいて基準fRを減らす傾向があり(ウィルコクソンχ2、P=0.17)、それはmPGES−1-/-マウスではみられなかった。全てのマウスは、fRを減らすことで過酸素に応答した。IL−1βは、mPGES−1+/+マウスにおける過酸素下でfRを抑制したが、この効果は、mPGES−1-/-マウスではみられなかった。mPGES−1+/+マウスは、mPGES−1-/-マウスと比較して、過酸素下での呼吸抑制の程度が大きかった。データは平均±SEMで示されている。*P<0.05、#P<0.05(体重で正規化した場合)。
【0231】
この結果は、mPGES−1活性の内因的な発現、特に脳幹における発現を実証している。mPGES−1は、主として、血液脳関門(BBB)付近の内皮細胞によって発現される(25)。重要な呼吸中枢付近の、脳幹を覆う内皮細胞におけるmPGES−1の構成的および急速誘導性発現は、呼吸の調節におけるPGE2の重要な役割を示唆している。mPGES−1を欠くマウスよりも野生型マウスにおいて過酸素下で有意な呼吸抑制がみられたこともまた、内因的なPGE2が周産期における呼吸律動形成に対して強壮効果を有することの証拠を与える。
【0232】
これまでの研究は、内因的なプロスタグランジン産生を遮断するプロスタグランジン合成阻害剤が、産後初期に胎児の呼吸動作および中枢呼吸を増加させることを報告している(26〜28)。プロスタグランジンの調節効果には、周産期の換気の初期阻害を含む発生学的変化が見られるが(18、26、27、29)、その後年齢の増加と共に呼吸における変化は小さくなる(19)。しかし、PGE2は、加齢後にも無呼吸の誘導により通常の呼吸を妨害する可能性がある(19)。EP3Rの遺伝子およびタンパク質はげっ歯類成体のRVLMにおいても発現されるが(20、21、30)、発生学的変化は、周産期にとどまらない脳幹でのPGE2受容体発現の変化の副産物的なものである可能性がある。さらに、プロスタグランジンの結合密度は減少するものの、これは全年齢にわたって同じ脳幹領域に局在する(31)。EP3R発現の個体発生(ontogenesis)およびPGE2の呼吸効果における潜在的な発生学的変化の根底にあるメカニズム、例えばEP3Rの翻訳後修飾、橋上側(suprapontine)への影響、のさらなる調査が必要とされる。
【0233】
実施例2:IL−1βおよび無酸素はマウス脳幹におけるmPGES−1活性を誘導する
我々は、9日齢mPGES−1+/+、mPGES−1-/-およびEP3R+/+マウスのホモジナイズした脳幹および皮質におけるmPGES−1活性に対する、IL−1βおよび短期間の無酸素暴露(100% N2、5分間)の効果も測定した(図1)。IL−1βは、時間依存的なmPGES−1活性の増加を、特に脳幹において誘導した。特に、脳幹のmPGES−1活性は、IL−1β投与90分後および180分後にそれぞれ2倍および4倍増加したが、皮質における活性は90分と180分の間で変化しなかった。無酸素暴露もまた、脳幹および皮質の両方においてmPGES−1活性を誘導した。注目すべきは、mPGES−1活性に対するIL−1βおよび短期間の無酸素暴露の相加的効果であり、これは脳幹においてより顕著であった。EP3R野生型マウスは、IL−1β投与90分後に、mPGES−1野生型マウスと類似のmPGES−1活性を示した。さらに、EP3Rマウスはまた、脳幹において、皮質よりも高いmPGES−1活性を有していた(それぞれ、PGE2:1111±49および710±44pmol/分/mpタンパク質)。
【0234】
PGE2は、無酸素に応答する呼吸においても重要な役割を果たすようである。短期間の無酸素暴露は、ホモジナイズしたマウスの脳においてmPGES−1活性を増加させた。このインビボでのmPGES−1活性の迅速な増加は新しい知見である。これまでの研究は、無酸素が、エクスビボでのマウス皮質片におけるPGE2産生およびブタの脳におけるプロスタグランジンHシンターゼ2 mRNAの発現を誘導することを示していた(32、33)。一過性の仮死も同様に、新生仔モルモットの脳においてPGE2濃度を増加させ、この効果はインドメタシン処置によって阻害される(34)。
【0235】
既知のmPGES−1酵素調節メカニズムはいずれも、本願において実証されたmPGES−1活性の迅速な変化を説明できるものではない。遺伝子発現の誘導は、このような短期間の無酸素イベントの間に起こりそうにない。しかし、構成的に発現されるmPGES−1の転写後調節、例えばリン酸化は、考えられる原因であろう。それ以外の可能性は、以前にヒト細胞系においてCOX−2 mRNAを用いて示され(35)、最近では心筋細胞において示されたような(36)、mPGES−1 mRNAの安定化である。基礎をなすメカニズムを明らかにするためさらなる研究が必要である。
【0236】
実施例3:IL−1βは、mPGES−1+/+マウスにおいて呼吸を抑制するが、この効果はmPGES−1-/-またはEP3R-/-マウスにおいてはみられない
IL−1βの換気効果を仲介するPGE2の役割を試験するため、我々は、9日齢のmPGES−1+/+、mPGES−1-/-およびEP3R-/-マウスにおいて、IL−1βまたはビヒクルをi.p.投与した後の正常酸素および過酸素(100% O2、1分間)下での呼吸を、フロープレチスモグラフィーを用いて分析した(図2、表1)。全てのマ
ウスが、処置に関係なく、fRを減らすことで過酸素チャレンジに対して応答したが、IL−1β処置した野生型マウスは、ビヒクル処置した野生型マウスよりも大きな呼吸抑制を示した。IL−1βはまた、mPGES−1+/+マウスにおいて基準fRを減少させる傾向があった(クラスカル・ウォリス、P=0.03;スチューデントt post−hoc検定、P=0.17)。逆に、IL−1βは、mPGES−1-/-またはEP3R-/-マウスにおいて正常酸素または過酸素下で換気を変化させなかった。
【0237】
この結果は、IL−1βが中枢呼吸を抑制するのにmPGES−1の活性化が必要とされることを指示している。第一に、IL−1βは、時間依存的な様式で脳幹のmPGES−1活性を増加させた。第二に、IL−1βは、mPGES−1+/+マウスにおいて呼吸を抑制したが、mPGES−1-/-マウスにおいては抑制しなかった。インドメタシンは、プロスタグランジン合成を遮断することによって、基準呼吸に対するIL−1βの効果を同じように軽減することが示されている(5)。
【0238】
実施例4:IL−1βは野生型マウスの無酸素下生存率を悪化させるが、この効果はmPGES−1またはEP3Rを欠くマウスにおいてはみられない
次に、我々は、IL−1βが、PGE2を仲介するメカニズムを通じて、低酸素換気応答および低酸素性無呼吸後の自然蘇生に影響するかどうかを調査した。フロープレチスモグラフィーを用いて、無酸素(100% N2、5分間)およびその後の過酸素(100% O2、8分間)下での呼吸を、mPGES−1+/+、mPGES−1-/-およびEP3R-/-マウスへのIL−1βまたはビヒクルのi.p.注射の80分後から試験した(図3、表2)。全てのマウスは、初期の換気亢進(すなわち、過呼吸)およびそれに続く低酸素型換気抑制(すなわち、一次無呼吸、あえぎ呼吸、二次無呼吸)という、無酸素に対する二相性の応答を示した。IL−1βは、mPGES−1+/+マウスにおいてあえぎ数を減少させたが、この効果はmPGES−1-/-マウスにおいてはみられなかった。IL−1β処置したmPGES−1+/+マウスはまた、IL−1β処置したmPGES−1-/-マウスよりもあえぎの持続時間が短い傾向があった(クラスカル・ウォリス、P=0.19;スチューデントt post−hoc検定、P=0.003)。あえぎ数の少なさおよびあえぎ持続時間の短さは、無酸素下生存率の低下と相関していた。IL−1βは、mPGES−1+/+マウスにおいて無酸素下生存率を有意に低下させたが、mPGES−1またはEP3R遺伝子を欠くマウスの生存率は低下しなかった。IL−1βは、EP3R-/-マウスの低酸素型換気応答に影響しなかった。
【0239】
【表2】
【0240】
ミクロソームプロスタグランジンEシンターゼ1(mPGES−1)の発現が様々な新生仔マウスにIL−1βまたはビヒクルを末梢投与してから80分後に無酸素に暴露した。マウスは、過呼吸期にfR、VTおよびVEの初期増加およびおよびそれに続く低酸素型換気抑制期にあえぎ応答を示した。各遺伝子型内で処置効果を比較すると、IL−1βは野生型マウスにおいてあえぎ数を減少させたが、この効果はmPGES−1の発現が少ないマウスにおいては観察されなかった。データは平均±S.E.M.で示されている。**P<0.01
【0241】
この研究は、PGE2が、IL−1βの無酸素下換気効果を仲介する上でも重要な役割を果たしていることを実証している。IL−1βは、野生型マウスにおいて低酸素無呼吸後の自然蘇生を阻害したが、この効果はmPGES−1またはEP3Rを欠くマウスにおいてはみられなかった。これまでの研究は、インドメタシンが、新生仔ラットにおける低酸素下でのあえぎ呼吸および無酸素下生存率に対するIL−1βの有害な効果を軽減することを示している(5)。
【0242】
実施例5:PGE2は脳幹の呼吸関連活動を低下させ、EP3Rを通じて無呼吸を誘発する
PGE2が脳幹EP3受容体に特異的に結合することにより呼吸を抑制するかどうかをより正確に把握するために、人工脳脊髄液またはPGE2を投与した2〜3日齢のEP3R+/+およびEP3R-/-マウスの脳幹脊髄のひとかたまりの標本を用いて中枢呼吸活動を測定した。対照条件下では、同様の呼吸活動をEP3R+/+およびEP3R-/-マウス由来の標本において記録した。しかし、PGE2はEP3R+/+標本における呼吸頻度を可逆的に阻害したが、EP3R-/-標本に対しては影響しなかった(図4)。
【0243】
EP3Rを通じて呼吸を変化させるPGE2の能力を、フロープレチスモグラフィーを用いてさらに評価した。EP3R+/+およびEP3R-/-マウスにおけるPGE2またはビヒクルのicv注射後、正常酸素および過酸素下での呼吸を分析した(図4および表3)。PGE2は、EP3R+/+マウスにおいて、正常酸素および過酸素下で有意に多い無呼吸頻度および不規則な呼吸パターンを誘発したが、この効果はEP3R-/-マウスにおいてはみられなかった。次に、このマウスを、無酸素およびそれに続いき過酸素に曝し、自然蘇生を試みさせた。全てのマウスは5分間の無酸素暴露期間を超えてあえぎ呼吸を継続し、38匹のうちの一匹のみが自然蘇生に失敗した(PGE2処置したEP3R-/-マウス)。PGE2は、ビヒクルと比較して、EP3R+/+またはEP3R-/-マウスのあえぎ応答または無酸素下生存率を変化させなかった。最後に、我々は、吻側延髄腹外側(RVLM)の呼吸関連ニューロンがEP3Rを発現しているかどうかを調査した。特に、疑核の腹側に位置しプレベッツィンガー複合体を含むRVLMに局在する呼吸関連ニューロンを同定するツールとしてNK1Rの免疫標識を使用した(22〜24)。我々は、これらのニューロンが、NK1RおよびEP3Rを共発現していることを証明した(図4)。
【0244】
【表3】
【0245】
正常酸素、過酸素(100% O2)および無酸素(100% N2)下での呼吸頻度(fR、呼吸/分)、1回換気量(VT、μl/呼吸/g)および毎分換気量(VE、μl/呼吸/g)を、PGE2またはビヒクルの脳室内(icv)注射後の9日齢EP3R+/+マウス(n=13)およびEP3R-/-マウス(n=25)において試験した。各遺伝子型内で処置効果を比較すると、PGE2は、EP3R+/+マウスにおいて正常酸素および過酸素下でのfRを有意に抑制したが、この効果はEP3R-/-マウスにおいてはみられなかった。PGE2はまた、EP3R+/+マウスにおいて過酸素下でのfRを減少させる傾向があった(ANOVA、p=0.11)が、この傾向はEP3R-/-マウスにおいてはみられなかった。データは平均±SEMで示されている。*p<0.05、**p<0.01
【0246】
先の実施例において示された結果は、mPGES−1の活性化後、新たに合成されるPGE2がIL−1βの中枢呼吸作用を発揮させることの証拠を与えている。我々は、本願において、PGE2が野生型マウスにおける呼吸を妨げることを証明しているが、これは、PGE2が動物胎仔および新生仔における呼吸を抑制することを実証する研究と矛盾するものではない(18、29、37)。さらに、PGE2はインビボで末梢での化学的感受性を変化させず、インビトロで脳幹呼吸活動を直接的に阻害することから、これらの効果は中枢において生じていると言える。これまでの研究は、PGE2が、新生仔ラットにおける呼吸関連ニューロンを阻害し(5)、かつ頸動脈洞および迷走神経の疑似手術または除神経後のヒツジにおける胎仔の呼吸動作も同様に阻害することを示している(38)。
【0247】
さらに、このPGE2の調節効果は、脳幹EP3受容体への結合を通じて起こる。IL−1βは、EP3R-/-マウスにおける呼吸を変化させることができなかった。PGE2は、EP3R+/+マウスにおいてインビボで無呼吸および不規則な呼吸を誘発したが、この効果はEP3R-/-マウスにおいてはみられなかった。最後に、EP3受容体の存在は、インビトロでの脳幹の呼吸関連律動活動を阻害するのに必要であった。特定のプロスタグランジン受容体であるサブタイプEP3RはNTSおよびRVLMに局在するが(20、21)、プロスタグランジンの呼吸への効果がこれらの受容体における挙動を通じて生じることおよびこれらが呼吸関連ニューロンにおいて発現されることを示す先行技術は存在しない。
【0248】
この実施例の結果は、IL−1βにより誘導されるPGE2および低酸素が、プレベッツィンガー複合体(preBotC)を含むRVLMにおける呼吸関連ニューロンを、EP3Rを通じて選択的に調節することを示唆している。PGE1を含む他の神経モジュレーターが、preBotCニューロンを阻害し、呼吸関連律動を弛緩させることが示されており(22、23)、preBotCの病変は、無酸素下でのあえぎ呼吸を妨げ、中枢性無呼吸および運動失調呼吸を惹起する可能性がある(39、40)。さらに、これらの呼吸関連ニューロンは最近、低酸素に対する十分な応答、あえぎ呼吸および自然蘇生による脳幹のホメオスタシスの維持、したがって酸素レベルの回復に重要であることが示されている(41)。この生きている脳幹の神経網のPGE2誘導性の抑制、例えば感染反応時抑制は、あえぎ呼吸および自然蘇生の不全ならびに究極的には死を引き起こし得る。
【0249】
実施例6:中枢PGE2濃度はヒト乳児における無呼吸頻度と相関する
ヒト新生児における感染と無呼吸の間の関係の根底にあるメカニズムをさらに解明するため、我々は、新生児において、感染マーカーC反応性タンパク質(CRP)、脳脊髄液PGE2レベルおよび無呼吸イベントの間の関係を試験した。CRPは、中枢PGE2と正の相関をし、CSF中のPGE2濃度と無呼吸頻度の間には正の関係があった(図5)。
【0250】
無呼吸は、新生児における敗血症の一般的にみられる徴候であるが(1)、この関係の基礎となるメカニズムは未だ解明されていない。我々は、本願において、感染マーカーCRPがヒト新生児のCSFにおけるPGE2レベルの上昇と相関関係にあることを証明する。重要なことは、我々は、PGE2が無呼吸頻度の増加と関連することも実証していることである。これらの発見は、ヒト新生児において、感染が、サイトカインの全身放出ならびにその後のPGE2の生合成および中枢作用を通じて呼吸を抑制することを示唆している。本明細書に記載されるメカニズムは、睡眠時無呼吸を患った小児におけるCRPレベルと無呼吸/呼吸低下インデックスの間の独立的関係(42)およびヒト乳児の咽頭分泌物中のIL−1β濃度と無呼吸の臨床的重症度との間の正の相関(8)を示す以前の報告を説明することができる。一過性の無呼吸もまた、ヒト新生児におけるプロスタグランジン処置の一般的な副作用であり(43)、これは脳幹の呼吸中枢におけるEP3受容体の活性化に起因するものであり得る。さらに、我々のデータは、新生児における中枢性無呼吸と尿中PGE代謝産物の間の正の相関(44)を説明するものである。
【0251】
炎症メディエーターは、新生児において感染および仮死を検出する上で重要なマーカーとして提案されている。サイトカインおよび低酸素刺激に対するPGE2の迅速な合成は、感染または仮死に起因する可能性のある無呼吸が増加している乳児の診断およびサーベイランスに利用できる。PGE2モニタリングの潜在的な診断的利益を、他の感染マーカー、例えばCRPとの比較で評価する研究が必要とされる。
【0252】
この結果は、IL−1βの有害な効果がmPGES−1およびEP3R遺伝子の選択的欠失によって軽減されたというものであり、感染に関連する新生児無呼吸の処置に関する重要な示唆を含んでいる。これまで、未熟児の無呼吸を処置するのにインドメタシンが使用されていた(45)。しかし、インドメタシンは新生児において多数の副作用を引き起こし(46)、したがって、mPGES−1またはEP3受容体を選択的に標的にする処置様式は有用性が高いと考えられる。
【0253】
先の実施例は、全身性のインターロイキン1βが、脳幹の呼吸関連領域におけるmPGES−1の活性化およびEP3受容体へのPGE2の結合を介して呼吸および自然蘇生を抑制することを実証している(図6)。さらに、重度の低酸素はmPGES−1活性を迅速に誘導し、これにより内因的なPGE2が新生児期の低酸素条件下で脳幹の呼吸ニューロンを調節し得ることが示唆されている。最後に、ヒト新生児における感染、中枢PGE2および無呼吸イベントの間の相関が明らかにされた。
【0254】
実施例7:PGE2の代謝産物と出生時仮死およびHIEの程度に対する相関
本発明者らは、ヒト乳児における周産期仮死がPGE2の迅速な放出および神経学的損傷を引き起こすという仮説を調査した。
患者
ストックホルムのカロリンスカ病院で処置をうけている63名の乳児(妊娠>37週)を、1999年10月から2004年9月の間、親の同意を得て本研究に登録した。43名の乳児が出生時仮死に関する以下の基準を充足した:1)後期の減速というカルディオトコグラフィーパターン、変動の不存在または徐脈、羊水の胎便染色、頭皮のpH<7.2または乳酸>4.8mmolによって示される胎児仮死の徴候、2)5分後のアプガールスコア<6および分娩室で>3分間の蘇生を行う必要性または生後60分以内に採取した患者由来の臍帯血または静脈血におけるpH<7.1、BE<−15(または乳酸>4.8mm/L)により示される生後ストレス、3)生後6時間以内の脳症の神経学的徴候。
除外基準は、先天性奇形、染色体異常および仮死に関係のない脳症、代謝病、髄膜炎が確認された子宮内/周産期感染であった。
対照群は、感染の疑いがあるが血液およびCSFからの細菌およびウイルス培養が陰性であった、CSF中に白血球がみられずかつ正常のタンパク量である、およびCNS病を示唆する所見がない、20名の新生児からなるものであった。
【0255】
臨床評価
最初にこれらの患者を本研究に登録する数時間前、次いで出産約12、36および72時間後ならびに新生児集中治療室の患者については第7日目に、神経学的評価(95、その開示内容を参照により本明細書で援用することを明記する)を行った。低酸素性虚血性脳症(HIE)は、サルナットおよびサルナットの基準(96、その開示内容を参照により本明細書で援用することを明記する)に従い、軽度(mild)、中等度(moderate)または重度(severe)に分類した。全ての患者を出産直後の数日の間に評価するために連続振幅表示型EEG(continuous amplitude-integrated EEG)を使用した。生後3日目および最初の週におけるEEG登録日に、中等度および重度のHIEを有する全ての患者に対して、脳のCTおよびMRIスキャンを行った。
生存患者の神経学的評価は、生後3、6および18月に、小児神経科医が行った。その評価に基づいて、小児を(1)正常アウトカム(normal outcome)、(2)軽度の運動障害(mild motor impairment);軽度の異常な筋緊張もしくは運動発達遅延の症状、または(3)有害事象(adverse outcome);脳性麻痺(両麻痺、片麻痺、四肢麻痺)、精神遅滞、痙攣もしくは死、に分類した。
【0256】
アプガールスコア
アプガールスコアは、出産直後の新生児の身体的状態を評価するための実用的方法である。アプガールスコアは、心拍数、呼吸努力、筋緊張、皮膚の色および刺激(例えば外鼻孔へのカテーテル挿入または足裏の摩擦)に対する反応を採点することによって得られる数値である。これらの対象となる各徴候には、0、1または2点が付与される。アプガールスコア最高点である10点は、乳児が考えうる限り最高の状態にあることを意味する。アプガールスコア0〜3点の乳児は、直ちに蘇生が必要である。アプガールスコアは、通常、乳児の出産60秒後に行われ(アプガー1分)、次いで、一般的には、出産5分後に繰り返される(アプガー5分)。困難な蘇生の場合、アプガールスコアは、10、15および20分後に再度行われることもある。生後20分時のアプガールスコア0〜3点は、高い罹患率(病気)および死亡率(死)を示唆するものである。
【0257】
CSFのサンプリング
生後最初の24時間(13.9+/−5.8)および/または30〜80時間の間(57.8+/−9.9)にCSFスピナルタブ(Spinal tab)を行った。各スピナルタブにより1〜2mlの量のCSFを集めた。このサンプルを3000rpm、4℃で10分間遠心し、その上清を、分析するまで0.5mlずつ−80℃で保存した。
【0258】
PGE2アッセイ
脳脊髄液サンプル中のPGE2およびPGE2代謝産物は、標準化された酵素免疫アッセイ(EIA)プロトコル(Cayman Chemicals, Ann Arbor, MI, USA)を用いて分析した。
【0259】
タンパク質の分析(BCAアッセイ)
サンプル中のタンパク質レベルを決定するためにBCAアッセイを行った。
【0260】
統計学的分析
臨床データは、そうでないことが明記されない限り、記載される目的についての中央値および四分位範囲で示されている。患者と対照の間の差を分析するのにマン・ホイットニー検定トを用いた。PGE2代謝産物またはサイトカインレベルとHIEまたは臨床評価の程度の間の関係を決定するのにクラスカル・ウォリス検定を使用した。
【0261】
結果
患者群(n=43)を、HIEのサルナットおよびサルナットの分類に従い3つの下位グループに分けた。この分類によれば、13名の乳児が軽度のHIE(HIE I)であり、その全員が正常アウトカムであった。16名の乳児は中等度のHIE(HIE II)であり、そのうちの8名の乳児が脳性麻痺、精神運動遅滞および痙攣の問題がある有害の神経学的アウトカムであり、さらに、2名の乳児は軽度の運動障害、6名は正常アウトカムであった。14名の乳児は重度のHIE(HIE III)であり、そのうちの8名は出産直後〜12日目に死亡し、6名の患者は生存した上で、有害の神経学的アウトカム;痙性四肢麻痺性脳性麻痺、精神運動遅滞、小頭症および複雑型痙攣を有した。
【0262】
患者群および対照群の臨床データは以下の表4に示されている。在胎齢および出産時体重に関しては患者と対照の間に差がなかったが、5分アプガールスコアおよび臍動脈または初期患者pHに関して差がみられた(p<0.001)。アプガールスコアを、刺激(例えば乳児の鼻へのカテーテル挿入または乳児の足裏の摩擦)に対して得た。いずれの臨床データについても、患者群の間で差がみられなかった。血中CRPレベルは、対照および患者の両方において有意ではなかった。
【0263】
図7Aに示されるように、出生時仮死の程度(5分および10分アプガールスコア)および神経学的評価は、正期産児におけるCSF PGE2代謝産物レベルに相関する。
【0264】
同様に、図7Bに示されるように、PGE2代謝産物はまた、新生児の状態の指標である出産5分後のアプガールスコアに相関し、したがって出生時の仮死の程度に相関する可能性がある。
【0265】
これらの結果は、PGE2がヒト乳児における重度の低酸素(仮死)時に迅速に放出されること、したがって、新生児仮死を罹患した赤ん坊における診断ツールおよび/または治療的介入の標的として使用できることを示唆している。
【0266】
【表4】
【0267】
実施例8−尿プロスタグランジン代謝産物、炎症および呼吸機能障害との相関
本発明者らは、三連四重極質量分析・テトラノルPGEMプロトコルを用いる尿プロスタグランジンE代謝産物(u−PGEM)の高感度かつ特異的な検出方法を開発した。
バリデーション研究は、この三連四重極質量分析・テトラノルPGEM法が、同じ被験者から採取したサンプル間で<5%の実験内変動を示すことを示している。室温で保存した尿サンプルではPGE代謝産物が分解されることが見出され、そのt1/2をおよそ2時間と概算した。対照的に、4℃での直接保存は、サンプルの分解を有意に減少させた。−20℃〜−80℃で保存したサンプルは、サンプル間で比較した場合、見かけ上、PGE代謝産物の分解を実質的に示さなかった。
【0268】
サンプルの調製
尿サンプルを、2%(v/v)の1Mクエン酸の添加によりpH3.0付近まで酸性化した。次いで、145μlの酸性尿アリコートを、エタノール中に9pmol/μlテトラノルPGEM−d6および0.45pmol/μl 11β−PGF2α−d4を含有する5μlの内部標準溶液でスパイクした。100μlをLC−MS/MS機器に注入した。標準曲線用および品質管理用のサンプルを、2%(v/v)の1Mクエン酸で酸性にしたPBSを用いて準備した。次いで、140μlの酸性PBSアリコートを、5μlの内部標準溶液(上記参照)および5μlの標準溶液(30〜900pmol/μlテトラノルPGEMおよび3〜90pmol/μl 11β−PGF2α)を用いてスパイクした。100μlをLC−MS/MS機器に注入し、100〜3000pmolテトラノルPGEMおよび10〜300pmol 11β−PGF2αの標準曲線を得た。LC−MS/MSの条件:分析物は、0.0005%FAを含むH2Oおよび移動相として0.0
005%FAを含むACNを用いて、Phenomenex Synergi Hydro RPカラム(100mm×2mm i.d.、2.5μm粒子径および100Å孔径)上で分離した。サンプル注入の直後に、15〜60% ACN、0.0005%FAの直線勾配を15分間適用し、その後、95%ACN、0.0005% FAで洗浄し、再平衡化した。総泳動時間は21分間であった。質量分析器は、陰イオンモード、電気スプレー電圧−3000V、350℃で作動させた。プロスタグランジン代謝産物の検出および定量のために、多重反応モニタリング(MRM)を使用して、テトラノルPGEMについての327.1>255.3およびテトラノルPGEM−d6の333.1>263.3(分裂エネルギー70V、衝突エネルギー−20V、滞留時間100m秒)ならびに11β−PGF2α、−PGF2αの353.3>309.3および11β−PGF2α−d4の357.3>313.3(分裂エネルギー150V、衝突エネルギー−15V、滞留時間100m秒)の遷移を記録した。全ての四重極は、最高感度を得る単一解像度で作動させた。
【0269】
本明細書に記載される結果は、成人、小児(1〜16歳)および乳児(0〜1歳)から得られたu−PGEMレベルの上昇が炎症の指標として信頼でき、呼吸機能障害(無呼吸を含む)と有意に関連することを示している。
【0270】
健常な成人対照由来の尿サンプル(n=10)を、「閉塞性」睡眠時無呼吸症候群(OSAS)患者から得た尿サンプル(n=24、22〜55歳)と比較した。睡眠に関係する無呼吸症候群(「閉塞性睡眠時無呼吸症候群」(OSAS)−いびき)は、成人女性の3%程度および成人男性の5%程度でみられると見積もられる。この結果は図8に示されており、y軸は尿中のPGE代謝産物をピコモルPGEM/μgクレアチニンの単位で示している。閉塞性睡眠時無呼吸症候群と診断された全ての患者に対して、夜間睡眠ポリグラフ記録実験(night-time sleep polysomnographic recording Laboratory test)を行い、この中で、尿サンプルを睡眠ポリグラフ記録(呼吸および飽和を含む)後の朝に採取した。
【0271】
睡眠時無呼吸(いびき)群は、対照群よりもu−PGEMレベルの多様性が有意に大きかった(大きな値の分散を観察した)。本発明者らは、u−PGEMレベルの上昇が無呼吸インデックス、すなわち無呼吸数/時間と相関する明確な傾向を観察した。さらに、重度のOSAS患者は、無呼吸インデックスとCRP(炎症およびPGE2の間接的マーカー)の間で有意な相関を有した。
【0272】
睡眠時無呼吸を示した3名の成人のおよそ1名でu−PGEMが上昇しており、これが無呼吸の重症度と相関していた。図8に示されるグループ間の比較はp=0.12を示した。しかし、重度の無呼吸問題を抱える患者のみを含め、閉塞性の問題(BMI値>過体重)を抱える患者を排除した場合、無呼吸とu−PGEMレベルの間に有意な関連性がみられる。
【0273】
本発明者らは、無呼吸インデックスが高い個体を、u−PGEMが高い被験者(すなわち、対照レベルよりも高い対象、図8の点線の囲みを参照されたい)と重ねることができることを発見した。
【0274】
本発明者らは、プラダー・ウィリー症候群(PWS)の小児(3〜16歳)におけるu−PGEMレベルについても調査した。プラダー・ウィリー症候群(15q11〜q13の欠失)の患者は、呼吸および心血管調節系に故障があり、特に睡眠中に無呼吸がみられる(115)。心肺の障害に起因する死は通常睡眠中に起こり、原因となる要因は特定されていないものの、3つの死亡例のうつ2つで軽度の感染エピソードが関係している(107)。
【0275】
我々は、mPGES−1経路の活性化が感染時に起こる潜在的に致死性の過度の呼吸の故障に関与しているという仮説をたてた(合わせてNature Medicine 2007, Vol.13, No.7, p.789, Research Highlights:“Baby's breath”も参照されたい)。
【0276】
既知の感染および炎症マーカーであるhs−CRP、CRP、WBCおよびサイトカイン(IL−1β)ならびにPGE2の尿中代謝産物を、循環器疾患登録(cardiovascular registration)と平行して試験した。これを、プラダー・ウィリー症候群を患った乳児および成人に対して、1)定期年次身体検査時、2)感染の徴候(体温>38.5℃)から24時間後および3)臨床的感染からの回復から少なくとも1週間後に行った。分析は主たる臨床研究室およびカロリンスカ・コアプロテオミクス施設の研究室において行い、三連四重極質量分析機器を用いて既知の代謝産物およびペプチドの定量を行った。
【0277】
PWS小児は呼吸パターンおよび自律調節に故障があり、しばしば軽度の上気道感染を伴い突然死することが知られている(年間罹患率2〜3%)。図9に示されるように、PWS小児(n=6)における尿中PGEMレベルは、健常な対照小児と比較して有意に上昇することが見出された。図9において、y軸は尿中のPGE代謝産物をピコモルPGEM/μgクレアチニンの単位で示している。この患者群(PWS)におけるu−PGEMレベルの上昇は、呼吸障害(特に無呼吸)、炎症およびPGE2(例えばu−PGEMレベル)の間の関連性のさらなる証拠となる。これにより、(PWSを患ったまたは患っていない)小児から得たサンプル中のプロスタグランジン代謝産物(例えばu−PGEM)の増加が、呼吸障害、例えば無呼吸、OSAS、SIDSおよび/もしくは炎症関連呼吸障害を患っているまたはそれらを発症する可能性が高いことを示すものであると考えることができる。さらに、感染に関連する呼吸機能障害を患った小児の部分集団は特に、呼吸障害とサンプル中のプロスタグランジン代謝産物(例えばu−PGEM)の増加の間で有意な相関を示す。この部分集団は、a)OSASおよび/またはb)自律神経機能障害の徴候、例えばPWS、レット症候群またはCCHS(先天性中枢性低換気症候群、「オンディーヌの呪い」とも呼ばれる)と関連する徴候を有する小児を含む。
【0278】
さらに、本発明者らは、進行中の炎症、ウイルス性細気管支炎および随伴性の無呼吸がみられる乳児(n=10)におけるu−PGEMレベルを調査した。その結果は図10に示されており、y軸は尿中のPGE代謝産物をピコモルPGEM/μgクレアチニンの単位で示している。進行中の炎症および随伴性の無呼吸がみられる乳児群は、対照(n=10、進行中の炎症または無呼吸がみられない乳児および小児)と比較して非常に高いu−PGEMレベルを示した。さらに、日常的な臨床ケアにおいて感染の評価に広く使用されているCRP(C反応性タンパク質)レベルはわずかに上昇したにすぎなかった。したがって、u−PGEMレベルの測定は、進行中の炎症を評価する上で、CRPの測定よりも有用であり、一部の乳児に見られる失調性の呼吸調節の潜在的なメカニズムを明らかにする。生後1〜6月の過敏な小児における炎症は、主として睡眠中の不規則な呼吸および無呼吸に関係しているようである。
【0279】
ウイルス感染(例えば、ウイルス性細気管支炎)は、重度の呼吸妨害および脳幹における「呼吸ペースメーカー」の中枢的抑制を引き起こす。しかし、このような感染は、典型的には、進行中の炎症障害の存在についての従来的なマーカーであるCRPのわずかな増加しか引き起こさない。したがって、プロスタグランジン代謝産物(例えば、u−PGEMレベル)の測定は、感染の初期段階において潜在的な炎症および/または呼吸障害の示唆を提供すると考えられる。したがって、プロスタグランジン代謝産物レベルのアッセイは臨床の場で有益であり、医師が炎症の重症度、予後および可能な治療的介入を「ベッドの上で」決定することを可能にする。
【0280】
本明細書において引用した全ての参考文献は、個々の刊行物または特許または特許出願について特別かつ個別にその全体が参照により援用されることが示されているのと同程度に、それら全体をすべての目的のために参照により援用する。
【0281】
本明細書に記載される具体的な実施態様は例示の目的で提供されるものであり、限定を意図していない。本明細書中のあらゆる副題は利便性のためのみに含まれるのであり、いかなる様式によってもその開示内容を限定するものとみなすべきではない。
【0282】
参考文献
1. Fanaroff, A. A., et al. (1998) Pediatr Infect Dis J 17, 593-598.
2. Prandota, J. (2004) Am J Ther 11, 517-546.
3. Guntheroth, W. (1989) Med Hypotheses 28, 121-123.
4. Dantzer, R. (2001) Ann N Y Acad Sci 933, 222-234.
5. Olsson, A., et al. (2003) Pediatr Res 54, 326-331.
6. Hofstetter, A. O. & Herlenius, E. (2005) Respir Physiol Neurobiol 146, 135-146.
7. Froen, J. F., et al. (2000) Pediatrics 105, E52.
8. Lindgren, C., Grogaard, J (1996) Acta Paediatr 85, 798-803.
9. Stoltenberg, L., et al. (1994) J Perinat Med 22, 421-432.
10. Vege, A., et al. (1998) Acta Paediatr 87, 819-824.
11. Ericsson, A., et al. (1994) Journal of Neuroscience 14, 897-913.
12. Ericsson, A., et al. (1995) J Comp Neurol 361, 681-698.
13. Engblom, D., Ek, M, Saha, S, Ericsson-Dahlstrand, A, Jakobsson, PJ, Blomqvist, A (2002) J Mol Med 80, 5-15.
14. Coceani, F. & Akarsu, E. S. (1998) Ann N Y Acad Sci 856, 76-82.
15. Crestani, F., Seguy, F, Dantzer, R (1991) Brain Res 542, 330-335.
16. Ericsson, A., Arias, C, Sawchenko, PE (1997) J Neurosci 17, 7166-7179.
17. Guerra, F., Savich, RD, Wallen, LD, Lee, CH, Clyman, RI, Mauray, FE, Kitterman, JA (1988) Journal of Applied Physiology 64, 2160-2166.
18. Kitterman, J., Liggins, GC, Fewell, JE, Tooley, WH (1983) J Appl Physiol 54,
687-692.
19. Tai, T. C. & Adamson, S. L. (2000) Am J Physiol 278, 1460-1473.
20. Ek, M., Arias, C., Sawchenko, P., & Ericsson-Dahlstrand, A. (2000) J Comp Neurol 428, 5-20.
21. Nakamura, K., Kaneko, T., Yamashita, Y., Hasegawa, H., Katoh, H., & Negishi,
M. (2000) J Comp Neurol 421, 543-569.
22. Gray, P. A., Rekling, J. C., Bocchiaro, C. M., & Feldman, J. L. (1999) Science 286, 1566-1568.
23. Ballanyi, K., Lalley, P. M., Hoch, B., & Richter, D. W. (1997) J Physiol 504, 127-134.
24. Pagliardini, S., Ren, J., & Greer, J. J. (2003) J Neurosci 23, 9575-9584.
25. Yamagata, K., Matsumura, K., Inoue, W., Shiraki, T., Suzuki, K., Yasuda, S.,
Sugiura, H., Cao, C., Watanabe, Y., & Kobayashi, S. (2001) J Neurosci 21, 2669-2677.
26. Kitterman, J., Liggins, GC, Clements, JA, Tooley, WH (1979) J Dev Physiol 1,
453-466.
27. Guerra, F. A., Savich, R. D., Clyman, R. I., & Kitterman, J. A. (1989) J Dev
Physiol 11, 1-6.
28. Long, W. (1988) J Appl Physiol 64, 409-418.
29. Guerra, F. A., Savich, R. D., Wallen, L. D., Lee, C. H., Clyman, R. I., Maur
ay, F. E., & Kitterman, J. A. (1988) J Appl Physiol 64, 2160-2166.
30. Lein, E. S., Hawrylycz, M. J., Ao, N., Ayres, M., Bensinger, A., Bernard, A., Boe, A. F., Boguski, M. S., Brockway, K. S., Byrnes, E. J., et al. (2007) Nature 445, 168-176.
【0283】
31. Tai, T., MacLusky, NJ, Adamson, SL (1994) Brain Res 652, 28-39.
32. Degi, R., Bari, F., Thore, C., Beasley, T., Thrikawala, N., & Busija, D. W. (1998) Neurobiology (Bp) 6, 467-468.
33. Shohami, E. & Gross, J. (1986) J Neurochem 47, 1678-1681.
34. Allen, L. G., Louis, T. M., & Kopelman, A. E. (1982) Biol Neonate 42, 8-14.
35. Ristimaki, A., Garfinkel, S., Wessendorf, J., Maciag, T., & Hla, T. (1994) J
Biol Chem 269, 11769-11775.
36. Degousee, N., Angoulvant, D., Fazel, S., Stefanski, E., Saha, S., Iliescu, K., Lindsay, T. F., Fish, J. E., Marsden, P. A., Li, R. K., et al. (2006) J Biol Chem 281, 16443-16452.
37. Savich, R. D., Guerra, F. A., Lee, C. C., & Kitterman, J. A. (1995) J Appl Physiol 78, 1477-1484.
38. Murai, D. T., Wallen, L. D., Lee, C. C., Clyman, R. I., Mauray, F., & Kitterman, J. A. (1987) J Appl Physiol 62, 271-277.
39. Feldman, J. L. & Del Negro, C. A. (2006) 7, 232-241.
40. McKay, L. C., Janczewski, W. A., & Feldman, J. L. (2005) Nature Neuroscience
8, 1142-1144.
41. Paton, J. F., Abdala, A. P., Koizumi, H., Smith, J. C., & St-John, W. M. (2006) Nat Neurosci 9, 311-313.
42. Tauman, R., Ivanenko, A., O'Brien, L. M., & Gozal, D. (2004) Pediatrics 113,
e564-569.
43. Singh, G., Fong, LV, Salmon, AP, Keeton, BR (1994) Eur Heart J 15, 377-381.
44. Hoch, B. & Bernhard, M. (2000) Acta Paediatr 89, 1364-1368.
45. Hammerman, C. & Zangen, D. (1993) Crit Care Med 21, 154-155.
46. Schmidt, B., Davis, P., Moddemann, D., Ohlsson, A., Roberts, R. S., Saigal, S., Solimano, A., Vincer, M., & Wright, L. L. (2001) N Engl J Med 344, 1966-1972.
47. Trebino, C. E., Stock, J. L., Gibbons, C. P., Naiman, B. M., Wachtmann, T. S., Umland, J. P., Pandher, K., Lapointe, J. M., Saha, S., Roach, M. L., et al. (2003) Proc Natl Acad Sci U S A 100, 9044-9049.
48. Fleming, E. F., Athirakul, K., Oliverio, M. I., Key, M., Goulet, J., Koller,
B. H., & Coffman, T. M. (1998) Am J Physiol 275, F955-961.
49. Jacobi, M. S. & Thach, B. T. (1989) J Appl Physiol 66, 2384-2390.
50. Herlenius, E., Lagercrantz, H, Yamamoto, Y (1997) Pediatr Res 42, 46-53.
51. Suzue, T. (1984) J Physiol 354, 173-183.
52. Engblom, D., Saha, S., Engstrom, L., Westman, M., Audoly, L. P., Jakobsson, P. J., & Blomqvist, A. (2003) Nat Neurosci 6, 1137-1138.
53. Fewell, J. E., Smith, F. G., Ng, V. K., Wong, V. H., & Wang, Y. (2000) Am J Physiol Regul Integr Comp Physiol 279, R39-46.
54. Hofstetter, A. O., et al. (2007) PNAS USA 104(23), 9894-9899.
55. Ward, E.S., et al. (1989) Nature 341, 544-546.
56. Bird et al. (1988) Science, 242, 423-426.
57. Huston et al. (1988) PNAS USA, 85, 5879-5883.
58. P. Holliger et al. (1993) PNAS USA, 90, 6444-6448.
59. Y. Reiter et al. (1996) Nature Biotech, 14, 1239-1245.
60. S. Hu et al. (1996) Cancer Res., 56, 3055-3061.
【0284】
61. John et al. (2004) PLoS Biology, 11(2), 1862-1879.
62. Myers (2003) Nature Biotechnology, 21, 324-328.
63. Shinagawa et al. (2003) Genes and Dev., 17, 1340-1345.
64. Fire, A., et al. (1998) Nature 391, 806-811.
65. Fire, A., (1999) Trends Genet. 15, 358-363.
66. Sharp, P. A. (2001) Genes Dev. 15, 485-490.
67. Hammond, S. M., et al. (2001) Nature Rev. Genet. 2, 1110-1119.
68. Tuschl, T. (2001) Chem. Biochem. 2, 239-245.
69. Hamilton, A., et al. (1999) Science 286, 950-952.
70. Hammond, S. M., et al. (2000) Nature 404, 293-296.
71. Zamore, P. D., et al. (2000) Cell 101, 25-33.
72. Bernstein, E., et al. (2001) Nature 409 363-366.
73. Elbashir, S. M., et al. (2001) Genes Dev. 15, 188-200.
74. Elbashir, S. M., et al. (2001) Nature 411, 494-498.
75. Thoren et al. (2000) Eur. J. Biochem. 267, 6428.
76. Mancini et al. (2001) J. Biol. Chem. 276, 4469.
77. Quraishi et al. (2002) Biochem. Pharmacol. 63, 1183.
78. Schultz, J. S., (1996) Biotechnol. Prog. 12, 729-743.
79. Cutz, E. et al. (1997) Am. J. Respir. Crit. Care Med. 155(1), 358-363.
80. Festen, D.A. et al. (2006) J. Clin. Endocrinol. Metab. 91(12), 4911-4915.
81. Debra, E. et al. (2007) Am. J. Med. Gen. Part A, 771-788.
82. Paterson, D.S. et al. (2006) JAMA 296(17), 2124-2132.
83. Onodera, H. et al. (2000) Lancet 356(9231), 739-740.
84. Yang, J. et al. (1994) Biochem. Biophys. Res. Commun. 198(3), 999-1006.
85. Herlenius, E. and Lagercrantz, H. (1999), J. Physiol. 518 (Pt 1), 159-172.
86. Ballanyi, K. et al. (1997) J. Physiol. 504(1), 127-134.
87. Katoh, H. et al. (1996) Journal of Biological Chemistry 271(47), 29780-29784.
88. van Rodijnen, W.F. et al. (2007) Am. J. Physiol. Renal Physiol. 292(3), F1094-1101.
89. Rowland, S.E. et al. (2007) Eur. J. Pharmacol. 560(2-3), 216-224.
90. Habib, A. et al. (2007) Faseb. J. 21(8), 1665-1674.
【0285】
91. Yao, J.C. et al. (2007) Yakugaku Zasshi 127(3), 527-532.
92. Morrell, M.J., and Twigg, G. (2006) Adv. Exp. Med. Biol. 588, 75-88.
93. Rigatto, H., (2003) “Periodic Breathing”. In: Mathew OP, editor. Respiratory control and disorders in the newborn, Vol 173. New York: Marcel Dekker, Inc;
237-72.
94. Simmons, D.L. et al. (2004) Pharmacol Rev. 56(3), 387-437.
95. Dubowitz, L. and Dubowitz, V. (1981) Clinical Developmental Medicine 79, London, England: Mac Keith Press.
96. Sarnat, H.B. and Sarnat, M.S. (1976) Arch Neurol. 33, 696-705.
97. Volpe, J.J. (1994) Hypoxic-ischaemic encephalopathy: Neuropathology and pathogenesis In Neurology of the Newborn, pp. 279-314. Philadelphia: WB Saunders Co.98. Roth, S.C., et al. (1992) Dev Med and Child Neurol. 34:285-295.
99. Hanrahan, D., et al. (1999) Dev Med and Child Neurol. 41:76-82.
100. Mehmet, H., et al. (1994) Neurosci Lett. 181:121-125.
101. Greher, J.K. and Nelson, K.B. (1997). J. Am Med Association. 278:207-211.
102. Nelson, K.B., et al. (1998) Ann Neurol. 44(4):665-75.
103. Foster-Barber, A., and Ferriero, D.M. (2002) Mental retard and dev disabilities. 8:20-24.
104. Silveira, R.C., and Procianoy, R.S. (2003). J. Pediatr. 625-629.
105. Deems et al. (2007) Methods Enzymol. 432, 59-82.
106. Welsh, T. N. et al. (2007) Prostaglandins & other Lipid Mediators 83, 304-310.
107. Tauber, M. et al. (2008) Am J Med Genet A. 146(7):881-887.
108. Perez, I.A. and Ward, S.L. (2008) Pediatr Ann. 37(7):465-470.
109. Wildhaber, J.H. and Moeller, A. (2007) Swiss Med Wkly. 137(49-50):689-694.
110. Tarasiuk, A. (2007) Am J Respir Crit Care Med. 175(1):55-61.
111. Weber, M.A. (2008) Lancet 371:1848-1853.
112. Hoch, B. et al. (2000) Prostaglandins Other Lipid Mediat. 60(1-3):9-14.
113. Hoch, B. and Bernhard, M. (2000) Acta Paediatr. 89(11):1364-1368.
114. Hofstetter, A.O. et al. (2007) PNAS 104(23):9894-9899.
115. Festen, D.A. et al. (2006) J Clin Endocrinol Metab. 91(12):4911-4915.
【0286】
本明細書中で参照された配列
【化1】
【0287】
【化2】
【0288】
【化3】
【0289】
【化4】
【0290】
【化5】
【技術分野】
【0001】
本発明は、呼吸障害(breathing disorder)、例えば無呼吸の処置方法、診断およびスクリーニング方法、ならびにこのような方法において使用する組成物に関する。
【背景技術】
【0002】
無呼吸および乳幼児突然死症候群(SIDS)は、新生児の大きな医学的懸案事項であり、これらの病因には感染が重要な役割を果たしている可能性がある。無呼吸は、新生児における感染の際に一般的にみられる徴候であり、SIDS患者の大部分において死の前に軽度のウイルスまたは細菌感染が確認される(1、2、111)。
【0003】
脳幹の呼吸調節に非最適性または遅延がみられる小児、例えば未熟児(すべてがその生後初年度に、一部は小児初期以降も)、先天性中枢性低換気症候群(CCHS)(79)、レット症候群およびプラダー・ウィリー症候群(PWS)(80)を発症した小児は、無呼吸を伴う周期性の不規則な呼吸を行い、それは睡眠中および感染エピソード時に増え、その際、その無呼吸は、外的蘇生または自然蘇生(autoresuscitation)がなされない場合、致命的となる可能性があり、時に致命的となる。
【0004】
SIDSで死亡する小児においてしばしば死の前に軽度の感染が確認され、最新の研究成果は、脳幹機能障害および低酸素イベントからの自然蘇生不全がこれらの原因不明の死の大部分と関連していることを示している(81、82)。
【0005】
呼吸調節に後天的または先天的な欠陥がある、例えばレット症候群およびPWSを発症した小児および成人は、潜在的に致死性の呼吸機能障害(respiratory dysfunction)の危険が高いが、睡眠時無呼吸症候群の小児および成人ならびにパーキンソン病の成人もまた、呼吸調節を故障し、しばしば感染を伴い死亡する(83)。呼吸系の障害(呼吸不全または感染)は、PWS児における最大の死因であることが明らかにされた(107)。さらに、小児期のいびきおよび閉塞性睡眠時無呼吸症候群(OSAS)は、睡眠を妨害し、神経認知学的発達を妨げ、それによって長期的な機能障害を引き起こす可能性がある。これは、呼吸器感染および後発的な危険因子、例えば社会生活における喫煙および喘息の出現によって悪化する(108−111)。
【0006】
潜在的に危険であり生命の脅威となる呼吸障害は、成人でもみられる。したがって、低酸素に対する換気応答の故障は、成人期のパーキンソン病、睡眠に関係する呼吸障害、例えば睡眠時無呼吸症候群およびOSASで重要な役割を果たしている可能性がある。
【0007】
炎症性サイトカイン、例えばインターロイキン1β(IL−1β)は、これらのイベントのカギとなるメディエーターの可能性がある(3)。IL−1βは、感染および炎症に対する急性免疫反応の際に産生され、様々な病的挙動を誘起する(レビューには(4)を参照されたい)。これまでの研究から、この免疫調節因子は呼吸および自然蘇生をも変化させることが示されている(5〜10)。IL−1βは、脳幹の呼吸関連領域、例えば弧束核(NTS)および吻側延髄腹外側(RVLM)において前初期遺伝子c−fosの発現を誘導する(11)。しかし、IL−1βは巨大な疎油性タンパク質であり、血液脳関門をまたいで容易に拡散しない。さらに、NTSおよびRVLMはIL−1受容体のmRNAを発現していないようであり(12)、かつIL−1βはインビトロで脳幹の呼吸関連ニューロン活動を変化させない(5)。
【0008】
我々はこれまでに、非特異的COX阻害剤であるインドメタシンがIL−1βにより誘導される呼吸抑制を軽減することを示している(5)。PGE2それ自体は、インビボで、ヒツジの胎仔および新生仔において呼吸を抑制し(17〜19)、かつインビトロで、呼吸関連ニューロンを阻害する(5)。早産児および満期産児において新生児尿中プロスタノイド排出が調査され(112)、早産児においてPGE−Mと無呼吸の関係が確認されている(113)。
【0009】
インドメタシンは、これまで、未熟児の無呼吸を処置するのに使用されている(45)。しかし、インドメタシンは、新生児において多くの副作用を引き起こす(46)。新生児におけるインドメタシンの利用に関連する副作用には、腎臓、腸および脳における薬物誘発性の血流減少が含まれる(46)。呼吸機能障害の処置にはカフェインが使用され、持続的気道陽圧法(CPAP)および酸素補給も同様である。さらに、ナロキソン(オピオイド受容体アンタゴニスト)による急性期処置もまた利用されている。それでもなお、呼吸障害の処置方法、特に無呼吸の処置に関する要望は確実に存在する。
【発明の概要】
【課題を解決するための手段】
【0010】
本発明者らは今回、誘導されたPGE2経路が感染および低酸素に対する呼吸応答のカギとなる調節因子であることを発見した(合わせて114も参照されたい)。誘導されたPGE2経路は図6に示されている。
【0011】
IL−1βは、血液脳関門の血管内皮細胞のILー1受容体に結合して、シクロオキシゲナーゼ2(COX−2)およびミクロソームプロスタグランジンEシンターゼ1(mPGES−1)の活性を誘導する(レビューには(13)を参照されたい)。COX−2はアラキドン酸からのプロスタグランジンH2(PGH2)の生成を触媒し、mPGES−1は、引き続く、PGH2からのプロスタグランジンE2(PGE2)の生成を触媒する。その後PGE2は脳実質に放出され、そこでIL−1βのいくつかの中枢的作用、例えば熱誘導(14)、行動反応(15)および神経内分泌変化(16)を媒介することが最近報告されている。本願明細書にさらに記載されるように、プロスタグランジンは、IL−1βの換気作用も媒介する(54)。さらに、PGE2受容体であるEプロスタノイド受容体サブタイプ3(EP3R)が、脳幹の呼吸関連領域:NTSおよびRVLMに局在する(20、21)。
【0012】
本願明細書に記載されるように、IL−1βは、mPGES−1の活性化およびPGE2の脳幹EP3Rへの結合を通じて中枢呼吸に悪影響を及ぼし、無呼吸頻度の増加および低酸素イベント後の自然蘇生不全を引き起こす。したがって、誘導されたPGE2経路に関連する呼吸障害は、一またはそれ以上の場所でこの経路を狙い撃ちすることによって、例えばCOX−2を阻害することによって、mPGES−1を阻害することによって、および/またはEP3Rを阻害することによって軽減され得る。
【0013】
したがって、一つの局面において、本発明は、哺乳動物対象における呼吸障害の処置方法であって、Eプロスタノイド受容体サブタイプ3(EP3R)阻害剤、ミクロソームプロスタグランジンEシンターゼ1(mPGES−1)阻害剤および/またはシクロオキシゲナーゼ2(COX−2)選択的阻害剤を含む治療有効量の組成物を処置が必要な対象に投与することを含む、方法、を提供する。
【0014】
本願明細書に記載される誘導性のPGE2経路における一つまたはそれ以上のステップを標的とする組成物を用いて呼吸障害、例えば無呼吸の誘導に関与する適当な経路を遮断することにより、選択性の低い治療法に付随する有害な影響を最小限に抑えることができると考えられる。例えば、COX−2を選択的に、mPGES−1を、および/またはEP3Rを標的化することによって、本願明細書中に記載されるような呼吸障害は、有害な副作用、例えば非選択的COX阻害剤インドメタシンの使用時にみられる副作用を最小限に抑えつつ軽減することができる。
【0015】
さらなる局面において、本発明は、EP3R阻害剤、mPGES−1阻害剤および/またはCOX−2選択的阻害剤を含む、哺乳動物被験体における呼吸障害の処置に使用できる組成物を提供する。
【0016】
さらなる局面において、本発明は、哺乳動物被験体における呼吸障害の処置するための医薬の製造における、EP3R阻害剤、mPGES−1阻害剤および/またはCOX−2選択的阻害剤を含む組成物の使用を提供する。
【0017】
さらなる局面において、本発明は、哺乳動物対象における呼吸障害に対する感受性またはその存在の評価方法であって、当該哺乳動物由来のサンプル中のプロスタグランジンE2(PGE2)またはその代謝産物のレベルを検出すること、およびPGE2またはその代謝産物の対照レベルと当該サンプル中のレベルを比較することを含み、PGE2またはその代謝産物の対照レベルと比較して当該サンプル中のPGE2またはその代謝産物レベルが高いことが、当該対象における呼吸障害に対する感受性またはその存在を指示する、方法、を提供する。
【0018】
本発明者らは、本願において、呼吸障害、例えば無呼吸および低酸素後の脆弱な自然蘇生におけるPGE2の中枢的役割に関する証拠を提供する。具体的には、脳脊髄液(CSF)および/または尿中のPGE2および/またはその代謝産物レベルの上昇が、無呼吸頻度の増加および低酸素後に自然蘇生する能力の低下に関連している。C反応性タンパク質(CRP)レベル、PGE2レベルと無呼吸の間の相関関係は、PGE2および/もしくはその代謝産物のレベルを単独でまたは感染マーカー、例えばCRPと組み合わせてモニタリングすることで、呼吸障害およびそれに対する感受性に関する診断的利益を得ることができることを示している。PGE2はサイトカインおよび低酸素刺激に反応して迅速に生成されるため、そのモニタリングは、感染および仮死が疑われる哺乳動物における呼吸障害、例えば乳幼児における無呼吸の増加の診断および監視に特に有用である。
【0019】
驚くべきことに、本発明者らは、尿プロスタグランジン代謝産物(u−PGEM)のレベルが、感染の進行および随伴性の無呼吸がみられる乳幼児、PWS児および睡眠時無呼吸がみられる一部の成人(高い無呼吸インデックスを有する成人を含む)において上昇することを発見した。特異的かつ高感度のアッセイを用いて尿中のPGE2レベルの測定することで、呼吸障害(特に無呼吸)の予見および評価のための非侵襲的方法が実現し、これは驚くほど広い患者年齢層に適用できると考えられる。感染および随伴性の無呼吸がみられる乳幼児において、u−PGEMレベルの上昇は、CRPレベルの上昇よりも早い段階で生じるようである。したがって、生物学的サンプル(例えば、尿、血液またはCSF)中のPGE2および/またはその代謝産物レベルの評価は、感染随伴性の炎症および呼吸機能障害患者の診断、処置および管理の面で、CRPレベルの評価よりも優れている。
【0020】
したがって、本発明は、ヒト対象における無呼吸の存在および/またはその重症度の評価方法であって、当該対象から採取した尿サンプル中の一つまたはそれ以上のPGE2代謝産物のレベルを検出すること、および当該一つまたはそれ以上のPGE2代謝産物の対照レベルと当該サンプル中のレベルを比較することを含み、当該サンプル中の一つまたはそれ以上のPGE2代謝産物のレベルが当該一つまたはそれ以上のPGE2代謝産物の対照レベルと比較して少なくとも20%、少なくとも50%、少なくとも100%または少なくとも200%高いことが、当該対象における無呼吸の存在および/またはその重症度の高さを指示する、方法、を提供する。特定の例において、ヒト対象は、閉塞性睡眠時無呼吸症候群(OSAS)、プラダー・ウィリー症候群、先天性低換気症候群および/またはレット症候群に罹患している。特定の例において、ヒト対象は、16歳以上、1〜16歳、または0〜1歳である。
【0021】
本発明者らは、本願において、出生時仮死後の被験体におけるPGE2の上昇およびPGE2と低酸素性虚血性脳症(HIE)の相関関係について記載している。これらの結果は、PGE2およびその代謝産物が、周産期の脳における酸素供給の欠陥に起因する神経学的損傷の強力な診断マーカーとなることを示している。さらに、これらの結果は、被験体が罹患した低酸素の程度が、サンプル(例えばCSF、尿または血液サンプル)中で検出されるPGE2およびその代謝産物のレベルに反映されることを示している。
【0022】
したがって、さらなる局面において、本発明は、哺乳動物対象における低酸素性虚血性脳症(HIE)に対する感受性またはその存在の評価方法であって、当該対象由来のサンプル中のプロスタグランジンE2(PGE2)またはその代謝産物のレベルを検出すること、およびPGE2またはその代謝産物の対照レベルと当該サンプル中のレベルを比較することを含み、PGE2の対照レベルと比較して当該サンプル中のPGE2またはその代謝産物レベルが高いことが、当該対象におけるHIEに対する感受性またはその存在を指示する、方法、を提供する。
【0023】
さらなる局面において、本発明は、哺乳動物被験体が罹患した低酸素または重度の低酸素仮死(例えば周産期仮死)の評価方法であって、当該対象由来のサンプル中のプロスタグランジンE2(PGE2)またはその代謝産物のレベルを検出すること、およびPGE2またはその代謝産物の対照レベルと当該サンプル中のレベルを比較することを含み、PGE2の対照レベルと比較して当該サンプル中のPGE2またはその代謝産物レベルが高いことが、当該対象が低酸素または重度の低酸素仮死(例えば周産期仮死)を罹患したことを指示する、方法、を提供する。
【0024】
さらなる局面において、本発明は、哺乳動物における呼吸障害の処置に使用できる物質の同定方法であって、誘導されたPGE2経路を阻害する能力について試験物質をアッセイすること、例えば、
(a)COX−2により媒介されるPGH2の生成、
(b)mPGES−1により媒介される、mPGES−1の環状エンドペルオキシド基質から当該基質の9−ケト,11αヒドロキシ型である生産物への変換、および
(c)EP3Rアゴニストにより媒介されるEP3Rの活性化、
の一つまたはそれ以上を阻害する能力について試験物質をアッセイすることを含み、誘導されたPGE2経路の阻害、例えば、(a)、(b)および(c)の一つまたはそれ以上の阻害は、当該試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを指示する、方法、を提供する。
【0025】
誘導されたPGE2経路を阻害する能力を有することが見出された試験物質は、一つまたはそれ以上のさらなる成分、例えば薬学的に許容できる補助成分(excipient)を含む組成物に配合され得る。このような組成物は、哺乳動物における呼吸障害の処置方法において使用され得る。
【0026】
誘導されたPGE2経路の中枢的な重要性およびその呼吸障害、例えば無呼吸に対する寄与(図6参照)の認識は、呼吸障害の処置において治療的有用性を有し得る物質を同定する上での基礎となる。特に、
(a)COX−2により媒介されるPGH2の生成、
(b)mPGES−1により媒介される、mPGES−1の環状エンドペルオキシド基質から当該基質の9−ケト,11αヒドロキシ型である生産物への変換、および
(c)EP3Rアゴニストにより媒介されるEP3Rの活性化、
の一つまたはそれ以上を阻害する能力について試験物質をスクリーニングする方法が、一つまたはそれ以上のインビトロアッセイを用いて実行され得る。阻害活性についての試験物質のスクリーニングは、呼吸障害の動物モデルに対する試験物質の効果の測定に基づくスクリーニング法よりも容易にスケールアップすることができる。このことは、呼吸障害の動物モデルを用いて試験物質をスクリーニングする前にイニシャルインビトロスクリーニングが行われる場合に有益である。このようにして、適当なインビトロ薬理活性を有する見込みのある物質が、その後のインビボ試験のために選択され得る。
【0027】
さらなる局面において、本発明は、哺乳動物における呼吸障害の処置に使用できる物質の同定方法であって、試験物質を試験哺乳動物に投与すること、ここで、当該試験物質は、誘導されたPGE2経路の阻害剤、例えば、EP3R阻害剤、mPGES−1阻害剤および/またはCOX−2選択的阻害剤である、および当該試験物質を投与しなかった対照の哺乳動物における徴候または症状との比較で、当該試験哺乳動物における呼吸障害の徴候または症状の重症度を決定することを含み、当該試験哺乳動物における呼吸障害の徴候または症状の重症度が対照哺乳動物のそれらよりも低いことは、当該試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを指示する、方法、を提供する。
【0028】
本発明のこの局面の方法はさらに、試験物質が誘導されたPGE2経路を阻害する能力を有するかどうか、例えばEP3R阻害剤、mPGES−1阻害剤および/またはCOX−2選択的阻害剤として機能する能力を有するかどうかを決定する前段階を含み得る。
【0029】
呼吸障害の徴候または症状の重症度を低下させ、それによって呼吸障害を処置する能力を有することが見出された試験化合物は、一つまたはそれ以上の別の成分、例えば薬学的に許容できる補助成分を含む組成物に配合され得る。このような組成物は、哺乳動物における呼吸障害の処置方法において使用され得る。
【0030】
さらなる局面において、本発明は、PGE2以外のEプロスタノイド受容体サブタイプ3(EP3R)アゴニスト、ミクロソームプロスタグランジンEシンターゼ1(mPGES−1)活性化剤および/またはシクロオキシゲナーゼ2(COX−2)選択的活性化剤を含む有効量の組成物を哺乳動物に投与することを含む、哺乳動物における呼吸抑制の誘導方法を提供する。
【0031】
哺乳動物における呼吸抑制の誘導は、呼吸障害の研究に特に有用であり得る。例えば、哺乳動物における呼吸抑制の誘導は、呼吸障害、例えば無呼吸、低酸素および/または脆弱な自然蘇生の動物モデルの作製に有用であり得る。このようなモデルは、EP3RまたはmPGES−1の活性化が無呼吸、例えば睡眠時無呼吸およびパーキンソン病、例えばパーキンソン病に付随する呼吸機能障害の動物モデルにおいて見られるかどうかを試験するのに有用であり得る。
【0032】
低酸素下で放出されるPGE2は、例えば、EP3R−Gi活性化の刺激ならびにそれに続くcAMPの減少およびニューロン活動の低下による脳の急性低酸素抵に対する耐性の強化を通じて急性的な神経保護効果を有し得る。
【0033】
本発明は、明らかに不可能な場合および明示的に回避される旨の説明がある場合を除いて、記載される局面および好ましい特徴の組み合わせを包含する。本発明のこれらおよびさらなる局面および実施態様については、実施例および添付図面を参照しつつ以下でより詳細に説明されている。
【図面の簡単な説明】
【0034】
【図1】IL−1βおよび無酸素によって迅速に誘導される脳幹のmPGES−1を示す。皮質および脳幹のミクロソームフラクション(血液脳関門(BBB)の内皮細胞を含む)におけるmPGES−1活性は、IL−1βまたはビヒクルで処置した9日齢マウス(n=33)および酸素正常状態または正常酸素および無酸素(100%N2、5分間)に曝した同じマウスにおいて分析した。A)野生型マウスにおいて、mPGES−1活性は、NaCl(対照)処置の90分後またはIL−1β処置の90分後および180分後に測定した。対照mPGES−1+/+マウスにおける内因性mPGES−1活性は、皮質よりも脳幹において高いことを観察した。加えて、IL−1βは、時間依存的な様式でmPGES−1活性を誘導した。B)90分の時点で、IL−1β処置マウスは、脳幹において、生理食塩水処置マウスよりもおよそ2倍高い活性を示した。無酸素もまた、mPGES−1活性を有意に誘導した。さらに、IL−1βの効果と一過的な無酸素暴露の効果は加算的なものであった。IL−1β処置マウスを無酸素に曝すと、脳幹において、対照マウスよりも4倍高い活性を観察した。しかし、mPGES−1遺伝子を欠失したマウスは、IL−1βおよび無酸素に対して無視できる程度の活性しか示さなかった。データは、平均±SEMで示されている。**P<0.01、***P<0.001
【図2】IL−1βによる、mPGES−1の活性化を通じた呼吸の抑制を示す。全身フロープレチスモグラフィーを使用し、IL−1β(n=52)またはNaCl(n=48)のいずれかのi.p.投与後の9日齢mPGES−1 WTマウス(n=66)およびmPGES−1 KOマウス(n=34)において、基準呼吸および過酸素に対する換気反応を試験した。A)プレスチモグラフ記録は、NaClまたはIL−1βを与えた野生型マウスにおける正常酸素および過酸素下での呼吸を示している(5秒間、呼吸の振幅1μl/s)。B、C)全てのマウスが過酸素に反応し、呼吸頻度(fR、呼吸/分)を減らした。IL−1βは、mPGES−1+/+マウスにおいて、NaClよりも大きくfRを減少させたが、mPGES−1-/-マウスにおいては、正常酸素または過酸素下での呼吸を変化させなかった。mPGES−1+/+マウスは、mPGES−1-/-マウスよりも大きな、過酸素下での呼吸抑制を示した。データは、平均±SEMで示されている。*NaClを与えたmPGES−1+/+マウスとの比較でP<0.05。
【図3】IL−1βによる、mPGES−1を通じた無酸素下生存率の低下を示す。9日齢のmPGES−1+/+マウス(n=37)およびmPGES−1-/-マウス(n=20)を、IL−1β(n=29)またはビヒクル(n=28)の末梢投与から80分後に、5分間の無酸素(100% N2)に曝した。A)NaClを与えたmPGES−1+/+マウスのプレスチモグラフ記録は、無酸素に対する初期の過呼吸およびその後のあえぎ呼吸を示している。このマウスは、100% O2を与えた後に自然蘇生した。B)IL−1βを与えたmPGES−1+/+マウスのプレスチモグラフ記録は、無酸素に対する短い過呼吸期間およびその後のあえぎ呼吸を示している。このマウスは、100% O2を与えた後の自然蘇生に失敗した。あえぎ回数(C)は、グループ間で相違する傾向があった(ウィルコクソンX2、P=0.06)。各遺伝子型内で処置の効果を比較すると、IL−1βは野生型マウスにおいてあえぎ回数を減らしたが、その効果はmPGES−1を欠くマウスにおいては観察されなかった。D)IL−1βは、mPGES−1+/+マウスにおいて、NaClよりも、無酸素下での生存率を低下させたが、mPGES−1-/-マウスにおいてはそうならなかった。データは、平均±SEMで示されている。*P<0.05、**P<0.01
【図4】PGE2による脳幹呼吸活動の抑制および脳幹EP3受容体を通じた無呼吸の誘導を示す。呼吸は、遺伝子型がEP3R+/+(n=13)およびEP3R-/-(n=25)の新生仔マウスにおいて、PGE2(n=19)またはNaCl(n=19)投与後に試験した。A)新生仔EP3R+/+(■)およびEP3R-/-(□)マウスにおいて、0分時にPGE2を注射(icv)し、その後、正常酸素および1分間の過酸素チャレンジを行った。EP3R+/+マウスは、正常酸素および過酸素下で、無呼吸に起因する少ない呼吸頻度(fR、呼吸/分)および変動係数(C.V.)の高い不規則な呼吸律動を示した。EP3R-/-マウスにおいては、麻酔期間後も基準fRが変化せず、呼吸パターンの変動が小さかった。PGE2のicv投与後の最初の20分間の間に、温度変化または温度依存性は見られなかった。B)プレスチモグラフ記録(10秒間、呼吸の振幅1μl/s)は、EP3R+/+マウスにおいて正常酸素下でPGE2により無呼吸エピソードが誘導されたが、EP3R-/-マウスにおいては誘導されなかったことが実証されている。C)EP3R+/+マウスにおいて、PGE2は、正常酸素および過酸素下で、ビヒクルよりも多くの無呼吸を誘導した。このPGE2の効果は、EP3R-/-マウスにおいては観察されなかった。D)2〜3日齢EP3R+/+仔(■、n=5)由来の脳幹脊髄「全(en bloc)」調製物において、PGE2(20μg/l)は、呼吸律動の形成を、対照頻度(fR)の64±5%まで可逆的に抑制した(ANOVA反復測定法、P<0.01)。PGE2は、EP3R-/-マウス由来の調製物(□、n=6)の呼吸活動に影響しなかった。E)横延髄切片において、疑核(NA)の腹側の吻側延髄腹外側(RVLM)内に存在し、プレベッツィンガー複合体を含む呼吸関連ニューロンが、NK1RおよびEP3Rを共発現している。NK1RおよびEP3Rの両方の発現が示されている。矢印は、いくつかのRVLM呼吸関連ニューロンにおけるEP3RおよびNK1Rの共存を示している。F)EP3R-/-マウスにおいて、NK1Rは発現するがEP3Rは発現しないことが確認された。スケールバーは100μmを表す。データは、平均±SEMで示されている。*NaClを与えたEP3R+/+マウスとの比較でP<0.05。
【図5】新生児における脳脊髄液中のPGE2と無呼吸インデックスの相関関係を示す。脳脊髄液(CSF)は、腰椎穿刺の臨床評価を行った新生児集中治療室の乳幼児(n=12、生後平均16±4日、在胎齢平均32±2週)から得た。次いで、乳幼児の心拍呼吸の記録を行った(9.2±2.4時間)。CSF中のPGE2濃度は、標準化された酵素イムノアッセイ(EIA)プロトコルを用いて分析し、感染マーカーであるC反応性タンパク質(CRP)および無呼吸インデックス(#無呼吸/時間)との相関関係を得た。中枢PGE2濃度は、血中CRPレベルと正の相関関係があった(P=0.01)。さらに、中枢PGE2濃度と無呼吸インデックスとの間で決定的な関係が観察された(P<0.05)。図5において、我々は、検出不可能なレベルのPGE2(0±0pg/ml)と高レベルのPGE2(52±22pg/ml)を区別した。データは、平均±SEMで示されている。
【図6】IL−1βにより誘導される、プロスタグランジンE2を経由する経路を通じた呼吸抑制および自然蘇生不全の模式図を示す。全身免疫反応の際、炎症性サイトカインであるインターロイキン1β(IL−1β)は末梢血流中に放出される。IL−1βは、血液脳関門(BBB)の内皮細胞上に存在するその受容体(IL−1R)に結合する。IL−1Rの活性化は、シクロオキシゲナーゼ2(COX−2)を通じたアラキドン酸(AA)からのプロスタグランジンH2(PGH2)の合成および律速酵素ミクロソームプロスタグランジンEシンターゼ1(mPGES−1)を通じたPGH2からのプロスタグランジンE2(PGE2)の合成を誘導する。PGE2は脳の実質に放出され、脳幹の呼吸調節領域、例えば例えば弧束核(NTS)および吻側延髄腹外側(RVLM)に存在するそのEP3受容体(EP3R)に結合する。その結果、中枢呼吸関連ニューロンおよび呼吸活動が抑制され、これによって、低酸素イベント下でのあえぎ呼吸および自然蘇生能力が致死的に低下し得る。
【図7】A)は、ヒト乳幼児における、CSF中のPGE2代謝産物濃度と仮死の程度および有害な結果との相関関係を示す。CSF中のPGE2代謝産物は、生後24時間以内に行った腰椎穿刺の中で獲得し、これは低酸素性虚血性脳症(HIE)と相関する。B)は、ヒト乳幼児の出生5分後のアプガールスコアとCSF中のPGE2代謝産物濃度の相関関係を示す。
【図8】対照の健常な成人vs閉塞性睡眠時無呼吸症候群を患っている成人の尿プロスタグランジン代謝産物(u−PGEM)レベルを示す。測定は、三重項四重極質量分析・テトラノルPGEM法により行った(PGE代謝産物は、pmolPGEM/μgクレアチンで表される)。無呼吸群は、対照よりも数値のばらつきがずっと大きく、その中にはPGEMレベルが極めて高い部分集団(点線の円)が含まれる。
【図9】対照の健常な小児vsプラダー・ウィリー症候群(PWS)を患っている小児(3〜16歳)の尿プロスタグランジン(u−PGEM)レベルを示す。測定は、三重項四重極質量分析・テトラノルPGEM法により行った(PGE代謝産物は、pmolPGEM/μgクレアチンで表される)。PWS群は、対照よりも有意に高いu−PGEMレベルを示す。
【図10】対照の健常な乳幼児(1月〜1歳)vs進行中の炎症、ウイルス性細気管支炎および随伴性の無呼吸がみられる乳幼児の尿プロスタグランジン(u−PGEM)レベルを示す。測定は、三重項四重極質量分析・テトラノルPGEM法により行った(PGE代謝産物は、pmolPGEM/μgクレアチンで表される)。無呼吸および炎症群は、対照よりも有意に高いu−PGEMレベルを示す。
【発明を実施するための形態】
【0035】
発明の詳細な説明
呼吸障害
本発明は、呼吸および/または換気の中枢調節の異常が関与する一群の呼吸障害を想定している。特に、呼吸障害は、異常な、例えば不規則または少ない呼吸頻度、少ないおよび/もしくは短いあえぎ呼吸、少ない1回換気量ならびに/または低酸素に対する呼吸反応の弱体化であり得る。呼吸障害は周期性呼吸であり得る。
【0036】
無呼吸
呼吸障害は無呼吸であり得る。無呼吸は呼吸の停止を意味し、これは一時的または恒久的なものであり得る。無呼吸は、例えば、インピーダンスニューモグラフィーによって確認され、本明細書中に記載されるようなイベントモニタリングシステムを通じて記録され得る。無呼吸頻度は、規定の無呼吸閾値を超えるイベント数と定義され得る。定義は、対象となる対象の年齢に応じて異なることが知られている。いくつかの実施態様において、例えば、哺乳動物が5歳未満のヒト乳幼児の場合、無呼吸は、直前の0.5秒間における平均インピーダンス信号振幅の≧10秒の減少から直前の25秒間に測定される平均振幅の16%未満の減少と定義され得る。他の実施態様において、例えば、哺乳動物がヒト成人の場合、無呼吸は、>10秒の呼吸停止と定義され得る。特定の実施態様においては、無呼吸は、2回の呼吸サイクルを上回る呼吸停止と定義され得る。
【0037】
睡眠関連呼吸障害
呼吸障害は睡眠中に起こる障害であり得る。乳幼児における睡眠時無呼吸は、重症例では、乳幼児突然死症候群(SIDS)の危険度上昇と関連する。成人の睡眠時無呼吸もまた本発明において想定されており、これにはいびきが含まれ得る。
【0038】
周期性呼吸
睡眠時の呼吸障害は、周期性呼吸、低酸素のエピソードおよび反復的な睡眠からの覚醒によって特徴付けられ、その症状には、過剰な日中の眠気、記憶力、学習力および注意力の低下が含まれる。間欠的な低酸素および睡眠の断片化は、それぞれ独自に、海馬および前頭前皮質、記憶および実行機能の神経処理と関係が深い領域におけるニューロンの故障の原因となり得る。
【0039】
周期性呼吸または過呼吸と無呼吸の交互出現は、未熟児に共通する呼吸パターンである。臨床的に重要な未熟児の無呼吸はほとんど常に周期性呼吸に関連するものである。呼吸低下期にはPaO2が減少し、これは、初期の小児または過去に脳幹呼吸中枢を損傷している患者において呼吸を減少させ得る。これは、低酸素により誘導される脳幹呼吸中枢の抑制により起こり、部分的にはアデノシンおよびPGE2の放出により媒介される(54、85)。過呼吸期または過換気期にはPaCO2が減少して呼吸刺激が減少し、それによって無呼吸が引き起こされ得る。
【0040】
後期早産児(late preterm infant)は、CO2に対して比較的鈍い換気応答を続け、睡眠時間の50%超をREMに費やし、かつ無呼吸および周期性呼吸を示し続け、その有病率は、1500g未満で誕生した乳幼児における60%に対して10%である。
【0041】
真の周期性呼吸または無呼吸は、呼吸サイクルにおいて呼吸深度が最低の部分が実際に休止点となった場合に出現する(無呼吸の場合)。
【0042】
新生児、小児、および成人において、睡眠障害性周期呼吸および間欠的低酸素は神経の故障と関連し、そのような損傷は認知機能障害を引き起こし得る(92、93)。
【0043】
自然蘇生不全
呼吸障害は、低酸素イベント後の自然蘇生不全であり得る。自然蘇生は、睡眠または重度の低酸素による呼吸動作の抑制から自己を呼び戻す脳の能力であり、長期の低酸素下では強制的な規則性・吸息性のあえぎ呼吸がみられる。これにより体および血液がその酸素供給を取り戻し飽和することができる。
【0044】
哺乳動物は典型的に、無酸素に対して二相反応を示し、まず換気を増加させ(すなわち過呼吸)、その後に低酸素型の換気抑制を行う(すなわち一次無呼吸、あえぎ呼吸、二次無呼吸)。次いで、低酸素後の酸素供給により自然蘇生が導かれる。低酸素後の自然蘇生不全は、介入がないと死に至る場合もある。
【0045】
SIDS
呼吸障害は、乳幼児突然死症候群(SIDS)を引き起こす障害であり得る。SIDS(「ゆりかご死」とも呼ばれる)は、概ね2歳未満で起きる乳幼児の予期せぬ突然死である。睡眠中に起こり得る呼吸停止および自然蘇生不全が、SIDSと呼ばれる死をもたらし得る。したがって、特に重度の呼吸障害は、予期せぬ突然死を導く可能性がある。特定の実施態様において、本発明は特に、予期せぬ突然死を十分にもたらし得る重度の呼吸障害を想定している。
【0046】
感染性呼吸障害
呼吸障害はウイルスおよび/または細菌感染に関係し得る。様々な感染マーカー、例えば、CRP、白血球数およびIL−1βを含む炎症性サイトカインが、感染時に増大し、これらは、呼吸障害が感染要因を有することを示唆する。
【0047】
本発明の特定の実施態様において、呼吸障害は、IL−1β関連呼吸障害であり得る。IL−1βは、感染および炎症に対する急性免疫反応の際に産生される。本明細書中に開示されているように、IL−1βは血液脳関門の血管内皮細胞上のIL−1受容体に対して作用し、COX−2を誘導し、誘導されたPGE2経路を刺激し、そして最終的には無呼吸頻度の増加および低酸素イベント後の自然蘇生不全の原因となる中枢呼吸の抑制をもたらす。IL−1βの血中レベルが対照レベルよりも上昇していることは、呼吸障害がIL−1β関連呼吸障害であることを示唆する。
【0048】
特定の実施態様において、哺乳動物または哺乳動物対象は、自律神経機能障害(autonomic dysfunction)、例えばプラダー・ウィリー症候群(PWS)、先天性中枢性低換気症候群(「CCHS」、「オンディーヌの呪い」とも呼ばれる)および/またはレット症候群を含む獲得性または先天性の呼吸調節障害に罹患したヒトであり得る。PWS、CCHSまたはレット症候群を患った乳幼児は、感染イベントの際、呼吸機能障害に起因する死亡の危険性が高くなる。
【0049】
低酸素性虚血性脳症
低酸素性虚血性脳症(HIE)は、周産期の脳における酸素供給に故障が生じ、脳のエネルギー代謝が妨害された正期産児の状態を表すのに使用される用語である(97)。この状態は、死亡または重度の神経学的後遺症を引き起こし得る。
【0050】
磁気共鳴分光法を用いた脳エネルギー代謝の研究は、脳細胞への酸素供給の一次妨害の後に、数時間または数日遅れることがある二次段階として神経脱落が起こるとの仮説を打ち立て(98、99)、これは動物研究においても示されている(100)。この神経損傷の遅延は、部分的には、炎症メディエーターがその傷害に反応して近辺の環境に放出されることが原因であると考えられている。
【0051】
神経系と免疫系の間の相互作用は、疾患の多くの局面において重要である。HIEは、病態生理も病因も十分に理解されていない。最近、低酸素虚血以外の原因、例えば子宮内または新生児期の炎症(101、102)が脚光を浴びており、傷害のメディエーターとしてサイトカインが注目されている(103)。虚血性脳傷害の病因における炎症カスケードの関与を示す証拠も存在する(104)。星状細胞および小グリア細胞により分泌されるサイトカインはこの炎症反応のメディエーターとして特に重要な役割を果たしており、これらは周産期仮死後の脳においてアポトーシスを誘発し神経細胞死に寄与し得る多様なシグナルの一種であると考えられている。しかし、体内の別の場所と同様、CNSにおいても特定のサイトカインは、当初は疾患プロセスを増幅し、後にそれを軽減するよう機能し得る。PGE2は、サイトカインおよび低酸素刺激に応じて迅速に生成されることから、出生児仮死を罹患した乳幼児の診断および調査において特に有用となり得る。
【0052】
本明細書でさらに記載されるように(特に以下の実施例7を参照されたい)、PGE2が周産期仮死の結果として脳で放出されることが見出された。このことは、mPGES−1が、迅速に活性化され、哺乳動物、例えばヒトおよびマウスにおける重度の低酸素に対する応答に関与することを示唆している。低酸素、例えば周産期仮死に対する応答における誘導されたPGE2経路の役割の発見は、治療的介入における標的および診断ツール、特に周産期仮死を罹患した新生児に対する治療標的および診断ツールを提供する。
【0053】
哺乳動物
本発明の任意の局面において、哺乳動物または哺乳動物対象は、成人(成体)、小児(仔)または乳幼児(仔)、例えば新生児(仔)であり得る。哺乳動物または哺乳動物対象は、好ましくはヒトである。特定の実施態様において、ヒトは、任意の年齢または一定範囲の年齢、例えば16歳未満、10歳未満、0〜5歳および0〜24月であり得る。特定の例において、対象は、自律神機能経障害、例えばPWS、CCHSまたはレット症候群を患ったヒトの小児である。したがって、本発明の任意の局面において、対象は家族性自律神経失調症を患ったヒト(乳幼児、小児もしくは成人)または脳幹から生じる原因未知の呼吸系およびそれに関連する自律神経障害を患ったヒト(乳幼児、小児もしくは成人)であり得る。
【0054】
特定の例において、対象は、OSASに罹患したヒト小児(0〜18歳)である。対象は、産後齢0〜25週および在胎齢28〜36週のヒト乳幼児であり得る。特定の実施態様において、ヒトは成人、例えば18歳以上であり得る。哺乳動物は、睡眠時無呼吸(例えばOSAS、いびき)および/またはパーキンソン病に罹患したヒト成人であり得る。本明細書中に記載される研究の結果として、u−PGEMの増加が、OSASを患いボディマスインデックス(BMI)が30未満の成人からなる部分集団における無呼吸(睡眠時無呼吸を含む)の感受性および/または重症度の上昇に特に重要となり得る、ということが示唆される。BMIは、対象のkg体重を彼または彼女のメートル身長の2乗で割ることによって算出される。したがって、BMI>30の対象は、典型的には肥満体とみなされる。本発明の任意の局面に従う特定の実施態様において、対象は、BMI>30のヒト成人であり得る。
【0055】
誘導されたPGE2経路
本発明は、本明細書で定義されるような呼吸障害の治療的処置のために、誘導されたPGE2経路を操作することを想定している。本発明者らは、誘導されたPGE2経路が、無呼吸頻度の増加および低酸素イベント後の自然蘇生不全を引き起こす要因となることを発見した。誘導されたPGE2経路は図6で説明されている。全身免疫反応の際、炎症性サイトカインIL−1βは末梢血流中に放出される。IL−1βは、血液脳関門の内皮細胞上に存在するその受容体(IL−1R)に結合する。IL−1Rの活性化は、COX−2を通じたアラキドン酸からのPGH2の生成および律速酵素mPGES−1を通じたPGH2からのPGE2の生成を誘導する。PGE2は脳の実質に放出され、脳幹の呼吸調節領域、例えば例えば弧束核(NTS)および吻側延髄腹外側(RVLM)に存在するEP3Rに結合する。
【0056】
本発明は、脳幹呼吸調節領域への波及効果を遮断または軽減するために、一つまたはそれ以上の場所で誘導されたPGE2経路を操作、例えば薬理学的操作を行うことを想定している。誘導されたPGE2経路は、脳幹の呼吸調節領域への波及効果を遮断または軽減する効果を有する任意の点で阻害され得る。特に、誘導されたPGE2経路は、本明細書にさらに記載されるように、COX−2、mPGES−1および/またはEP3Rを阻害することによって遮断され得る。
【0057】
誘導されたPGE2経路の阻害剤
誘導されたPGE2経路の阻害剤は、脳幹の呼吸調節領域への波及効果を遮断または軽減する能力を有する。この阻害剤は、誘導されたPGE2経路の任意の点で、直接的または間接的に作用し得る。例えば、この阻害剤は、
(a)この経路に関与するポリペプチド(「誘導されたPGE2経路ポリペプチド」)、例えばCOX−2ポリペプチド、mPGES−1ポリペプチドおよび/もしくはEP3Rポリペプチドと直接的に相互作用し得る;
(b)この経路に関与するポリペプチドと間接的に相互作用し得る、例えばCOX−2ポリペプチド、mPGES−1ポリペプチドおよび/もしくはEP3Rポリペプチドの活性化因子に結合しそれを阻害することによって相互作用し得る;ならびに/または
(c)誘導されたPGE2経路ポリペプチドをコードする遺伝子の発現を妨害し得る、例えばCOX−2コード遺伝子、mPGES−1コード遺伝子および/もしくはEP3Rコード遺伝子の発現(例えば転写および/もしくは翻訳)をダウンレギュレートし得る。
【0058】
EP3R
Eプロスタノイド受容体サブタイプ3(EP3R)ポリペプチドは、EP3Rアゴニスト、例えばPGE2に結合し、下流へシグナルを伝達する、例えばGタンパク質を通じてシグナル伝達する能力を有する。ヒトおよびマウスのEP3Rアミノ酸配列は、すでに報告されている(84、その開示内容を参照により本明細書で援用することを明記する)。ヒトEP3Rヌクレオチド配列は、GenBankデータベースに寄託されている(アクセッション番号L26976、その開示内容を参照により本明細書で援用することを明記する)。EP3Rポリペプチドは、好ましくは配列番号2のヒトEP3Rアミノ酸配列を含むかまたはこれからなる。しかし、EP3Rポリペプチドは、非ヒト哺乳動物、例えばマウスまたは他のげっ歯類由来のホモログであってもよい。EP3Rポリプチドは、一つまたはそれ以上のアミノ酸が挿入、欠失または置換によって変更されたヒトEP3Rタンパク質の変異体または誘導体であってもよい。好ましくは、EP3Rポリペプチドは、配列番号2の全長アミノ酸配列と少なくとも70%、より好ましくは80%、さらにより好ましくは90%、さらにより好ましくは95%、最も好ましくは99%のアミノ酸同一性を有し、かつEP3Rアゴニスト、例えばPGE2に結合し、下流へシグナルを伝達する能力を有するアミノ酸配列を含む。いくつかの実施態様において、EP3Rポリペプチドは単離されている。
【0059】
ヒトEP3Rの活性化は、[cAMP]iを低下させ、[Ca++]iをやや上昇させる(84)。cAMPの減少は、呼吸に関連する脳幹ニューロンにおける発火度および発火頻度を減少させる、従って呼吸活動を低下させることが示されている(85)。ニューロンにおいて、EP3Rの活性化はプロテインキナーゼC非依存性Rho活性化経路を通じた神経突起の伸展を妨げ得る(86、87)。さらに、EP3Rは腎臓で高度に発現され、腎臓におけるEP3Rの活性化は血管収縮作用を示す(88)。
【0060】
EP3Rポリペプチドは、配列番号2のアミノ酸配列を有する全長EP3Rポリペプチドより短いが、その本来の生物学的活性を保持している活性部分であってもよい。特に、この活性部分は、EP3Rアゴニスト、例えばPGE2に結合し、下流へシグナルを伝達する、例えばGタンパク質を通じてシグナル伝達することができるものである。
【0061】
EP3Rコード遺伝子は、本明細書で定義されるEP3Rポリペプチドをコードするヌクレオチド配列を含み得る。EP3Rコード遺伝子は、配列番号1の全長ヌクレオチド配列と少なくとも70%、より好ましくは80%、さらにより好ましくは90%、さらにより好ましくは95%、最も好ましくは99%のヌクレオチド配列同一性を有するヌクレオチド配列を含み得る。
【0062】
EP3R阻害剤
EP3R阻害剤は、脳幹の呼吸調節領域に対するEP3R媒介性の効果を阻止または減少、例えばEP3R媒介性の無呼吸、呼吸抑制および/または自然蘇生不全を阻止または減少させる。
【0063】
本発明は、多くの異なるタイプのEP3R阻害剤の使用を想定している。例えば、EP3R阻害剤は、本明細書で定義されるEP3Rポリペプチドに結合し、アゴニストによる誘導される(例えばPGE2誘導性の)下流へのシグナル伝達(Gタンパク質共役シグナル伝達を含む)を阻止または減少させるアンタゴニストであり得る。さらに、この阻害剤は、EP3Rポリペプチドの活性化因子に結合しこれを阻害することによって間接的に作用するものであり得る。本明細書で定義されるEP3Rコード遺伝子の発現を(例えば、EP3Rコード遺伝子の転写および/または翻訳を阻害することによって)ダウンレギュレートするEP3R阻害剤もまた想定されている。
【0064】
EP3Rポリペプチドに結合する阻害剤の例には、特異的結合物質、例えば抗体分子、およびEP3Rポリペプチドへの結合においてPGE2と競合する低分子が含まれる。EP3Rコード遺伝子の発現をダウンレギュレートする阻害剤の例には、EP3Rコード遺伝子またはその一部と相補的な核酸分子およびEP3Rをコードする遺伝子またはそのフラグメントの配列に対応する二本鎖RNAが含まれる。EP3Rコード遺伝子の発現をダウンレギュレートする阻害剤には、リボザイムおよび/または三重らせん因子も含まれる。低分子、特異的結合物質および核酸を含む、多くの異なるクラスの阻害剤のさらなる詳細は本明細書中に記載されている。
【0065】
EP3Rの低分子阻害剤
本発明は、EP3Rポリペプチドに結合しアゴニストにより誘導される(例えばPGE2誘導性の)下流へのシグナル伝達、例えばGタンパク質系シグナル伝達を阻止または減少させる、約2000ダルトン以下、例えば50〜1000ダルトンの有機化合物または無機化合物の使用を想定している。EP3Rの低分子阻害剤は、EP3Rポリペプチドに競合的に結合し、PGE2と同じ部位への結合において競合する、または非競合的に結合するアンタゴニストであり得る。この低分子性EP3Rアンタゴニストは、好ましくは、中枢作用性の(すなわち血液脳関門を通過することができる)ものである。しかし、血液脳関門を通過できない低分子性EP3Rアンタゴニストもまた想定されており、これらは、例えば脳室内(i.c.v.)投与により中枢に送達され得る。
【0066】
低分子性EP3Rアンタゴニストは、(2E)−N−[(5−ブロモ−2−メトキシフェニル)スルホニル]−3−[5−クロロ−2−(2−ナフチルメチル)フェニル]アクリルアミド(L826266)または薬学的に許容できるその塩を含み得る。
【0067】
さらなる低分子性EP3Rアンタゴニストは、本明細書に記載されるスクリーニング法を用いて同定され得る。
【0068】
EP3Rの特異的結合阻害剤
いくつかの実施態様において、EP3R阻害剤は、本明細書で定義されるEP3Rポリペプチドに結合しアゴニストにより誘導される(例えばPGE2誘導性の)下流へのシグナル伝達、例えばGタンパク質系シグナル伝達を阻止または減少させる特異的結合物質であり得る。
【0069】
いくつかの実施態様において、特異的結合物質は抗体分子であり得る。他の実施態様において、特異的結合物質は、非抗体分子内に抗原結合部位、例えば非抗体タンパク質スキャホールド上にCDRのセットを含み得る。
【0070】
「抗体分子」は、天然由来であるか部分合成または完全合成であるかに関係なく、免疫グロブリンを意味する。全長抗体のフラグメントがその抗原結合機能を発揮できることが示されている。従って、抗体分子への言及は、全長抗体に及ぶし、抗体結合フラグメントを含む任意のポリペプチドまたはタンパク質にも及ぶ。
【0071】
結合フラグメントの例は、(i)VL、VH、CLおよびCH1ドメインからなるFabフラグメント、(ii)VHおよびCH1ドメインからなるFdフラグメント、(iii)単一抗体のVLおよびVHドメインからなるFvフラグメント、(iv)VHドメインからなるdAbフラグメント(55)、(v)単離されたCDR領域、(vi)Fabフラグメントが二つ連結された二価フラグメントである、F(ab’)2フラグメント、(vii)VHドメインおよびVLドメインがペプチドリンカーによって連結されこの二つのドメインが連携して抗原結合部位を形成している、短鎖Fv分子(scFv)(56〜57)、(viii)二特異性短鎖Fv二量体(WO93/11161)および(ix)遺伝子融合によって構築された多価または多特異性フラグメントである「ダイアボディ(diabody)」(WO94/13804;58)である。Fv、scFvまたはダイアボディ分子は、VHドメインとVLドメインをつなぐジスルフィド架橋を導入することによって安定化され得る(59)。scFvにCH3ドメインが連結されたミニボディも作製することができる(60)。
【0072】
EP3Rの核酸阻害剤
本発明はまた、EP3R遺伝子の発現のダウンレギュレーションに関する当該分野で公知の技術を使用する。これらには、RNA干渉(RNAi)の使用が含まれる。
【0073】
ヒトにおいて、EP3Rは、配列番号1のヌクレオチド配列を有する遺伝子によってコードされている。ヒトEP3Rのアミノ酸配列は配列番号2に示されている。このヌクレオチド配列は、本明細書にさらに記載されるように、EP3Rコード遺伝子の発現をダウンレギュレートすることができる核酸分子を設計するのに利用され得る。
【0074】
低分子RNAも、遺伝子発現の調節に利用され得る。これらには、低分子干渉RNA(siRNA)によるmRNAの特異的分解、転写後遺伝子サイレンシング(PTG)、マイクロmRNA(miRNA)によるmRNAの発生学的に調節された配列特異的翻訳抑制および特異的転写遺伝子サイレンシングが含まれる。
【0075】
ヘテロクロマチン複合体の標的化および特定の染色体部位での後成的な遺伝子サイレンシングにおけるRNAi機構および低分子RNAの役割についても実証されている。RNA干渉(RNAi)としても知られている、二本鎖RNA(dsRNA)依存的な転写後サイレンシングは、dsRNA複合体が短期的なサイレンシングのために特定の相同遺伝子を狙い撃ちする現象である。RNA干渉は、配列同一性を有するmRNAの分解を促進するシグナルとして機能する。一般的に、20nt siRNAは、遺伝子特異的なサイレンシングを誘導するのに十分長く、宿主反応を回避するのに十分短い。標的化された遺伝子産物の発現減少は顕著で、少数のsiRNA分子によって90%サイレンシングを誘導することができる。
【0076】
当該分野において、これらのRNA配列は、それらの起源によって、「短鎖または低分子干渉RNA」(siRNA)または「マイクロRNA」(miRNA)と称される。両タイプの配列は、相補的なRNAに結合し、mRNA分解を誘導する(RNAi)かまたはmRNAからタンパク質への翻訳を停止させることによって遺伝子発現をダウンレギュレートするのに使用され得る。siRNAは、長い二本鎖RNAのプロセシングによって生じ、自然環境で見出される場合典型的には外因性である。マイクロRNA(miRNA)は、内因的にコードされる小さな非コーディングRNAであり、ショートヘアピンのプロセシングにより生じる。siRNAとmiRNAは両方とも、部分的に相補的な標的配列を有するmRNAの翻訳をRNAの切断を行わずに阻害することができ、かつ完全に相補的な配列を有するmRNAを分解することができる。
【0077】
siRNAリガンドは典型的には二本鎖であり、RNA媒介性の標的遺伝子機能のダウンレギュレーションの効果を最適化するためには、siRNAによるmRNA標的の認識を媒介するRISC複合体によるsiRNAの正確な認識を確実にし、かつ宿主反応を減らせる程度にsiRNAが短くなるよう、siRNA分子の長さを選択するのが好ましい。
【0078】
miRNAリガンドは典型的には一本鎖であり、ヘアピンを形成できる部分的に相補的な領域を有する。miRNAはDNAから転写されるがタンパク質に翻訳されないRNA遺伝子である。miRNA遺伝子をコードするDNA配列はmiRNAよりも長い。このDNA配列は、miRNA配列およびほぼ逆の相補鎖を含む。このDNA配列が一本鎖RNA分子に転写されると、miRNA配列およびその逆相補性塩基は対をなして部分的に二本鎖のRNAセグメントを形成する。マイクロRNA配列の設計については(61)で説明されている。
【0079】
典型的には、siRNAまたはmiRNAの効果を模倣するためのRNAリガンドは、10〜40リボヌクレオチド(またはその合成アナログ)、より好ましくは17〜30リボヌクレオチド、より好ましくは19〜25リボヌクレオチドおよび最も好ましくは21〜23リボヌクレオチドを有する。二本鎖siRNAを利用する本発明のいくつかの実施態様において、この分子は、対称な3’オーバーハング、例えば一つまたは二つの(リボ)ヌクレオチドのオーバーハング、典型的にはUUまたはdTdTの3’オーバーハングを有し得る。本明細書の開示に基づき、当業者は、適当なsiRNAおよびmiRNA配列を容易に、例えばAmbion社のsiRNAファインダー(http://www.ambion.com/techlib/misc/siRNA_finder.htmlを参照されたい)等のリソースを用いて容易に設計することができる。siRNAおよびmiRNA配列は、合成的に作製しこれを外的に導入することにより遺伝子をダウンレギュレートすることもできるし、発現系(例えばベクター)を用いて生成させることもできる。好ましい実施態様において、siRNAは合成される。
【0080】
siRNAを生成するために長鎖の二本鎖RNAを細胞中でプロセシングさせることができる(例えば(62)を参照されたい)。この長鎖dsRNA分子は、対称な3’または5’オーバーハング、例えば一つまたは二つの(リボ)ヌクレオチドからなるオーバーハングを有していてもよいし、平滑末端であってもよい。この長鎖dsRNA分子は25ヌクレオチドまたはそれ以上であり得る。好ましくは、長鎖dsRNA分子は25〜30ヌクレオチド長である。より好ましくは、長鎖dsRNA分子は25〜27ヌクレオチド長である。最も好ましくは、長鎖dsRNA分子は27ヌクレオチド長である。30ヌクレオチドまたはそれ以上の長さのdsRNAを、ベクターpDECAPを用いて発現される場合もある(63)。
【0081】
代替手段は、細胞内でのショートヘアピンRNA分子(shRNA)の発現である。shRNAは合成性のsiRNAよりも安定である。shRNAは、小さなループ配列によって隔てられた短い逆位反復配列からなる。逆位反復配列の一つは遺伝子標的に相補的である。shRNAは、細胞内で、DICERによってsiRNAへのプロセシングを受け、このsiRNAが標的遺伝子であるmRNAを分解し、発現を抑制する。好ましい実施態様において、shRNAは、ベクター、例えば本発明のアデノウイルスベクターからの転写によって内因的に(細胞内で)生成される。shRNAは、RNAポリメラーゼIIIプロモーター、例えばヒトH1または7SKプロモーター、またはRNAポリメラーゼIIプロモーターの制御下にshRNA配列をコードするベクターを細胞にトランスフェクトすることによって細胞内で生成される場合もある。あるいは、shRNAは、ベクターからの転写によって外因的に(インビトロで)生成され得る。shRNAは次いで、細胞に直接的に導入され得る。好ましくは、shRNA分子は、EP3Rコード遺伝子の部分配列を含む。好ましくは、shRNA配列は、40〜100塩基長、より好ましくは40〜70塩基長である。ヘアピンの幹部分は、好ましくは19〜30塩基対である。この幹部分は、ヘアピン構造を安定化させるGU対を含み得る。
【0082】
siRNA分子、長鎖dsRNA分子またはmiRNA分子は、核酸配列、好ましくはベクターに含まれる核酸配列の転写により組換え生成されることができる。好ましくは、siRNA分子、長鎖dsRNA分子またはmiRNA分子は、EP3Rコード遺伝子の部分配列を含む。
【0083】
一つの実施態様において、siRNA、長鎖dsRNAまたはmiRNAは、ベクターからの転写により内因的に(細胞内で)生成される。ベクターは、当該分野で公知の任意の方法で細胞に導入され得る。場合により、RNA配列の発現は、組織特異的プロモーターを用いて調節され得る。さらなる実施態様において、siRNA、長鎖dsRNAまたはmiRNAは、ベクターからの転写により外因的に(インビトロで)生成される。
【0084】
一つの実施態様において、ベクターは、センス方向およびアンチセンス方向の両方に、EP3Rコード遺伝子の全長または部分核酸配列を含み得、それらのセンスおよびアンチセンス部分は、RNAとして発現されると集合して二本鎖RNAを形成する。好ましくは、ベクターは配列番号1の核酸配列またはその変異体もしくはフラグメントを含む。別の実施態様において、センスおよびアンチセンス配列は、異なるベクターにより提供される。好ましくは、ベクターは、配列番号1の核酸配列またはその変異体もしくはフラグメントを含む。
【0085】
あるいは、siRNA分子は、当該分野で公知の標準的な固相または液相合成技術を用いて合成することができる。ヌクレオチド間の結合は、ホスホジエステル結合またはその代替手段として、例えば、式P(O)S、(チオエート);P(S)S、(ジチオエート);P(O)NR’2;P(O)R’;P(O)OR6;CO;またはCONR’2の連結基であり得る(式中、RはH(もしくは塩)またはアルキル(1〜12C)であり、R6はアルキル(1〜9C)であり隣接するヌクレオチドに−O−または−S−を通じて接続される)。天然の塩基に加えて修飾ヌクレオチド塩基を使用することができ、これらを含むsiRNA分子に有益な特性を付与することができる。
【0086】
例えば、修飾塩基は、siRNA分子の安定性を向上し、それによってサイレンシングに必要となる量を減らすことができる。修飾塩基の導入はまた、非修飾siRNAよりも安定性の高いまたは低いsiRNA分子を実現し得る。
【0087】
用語「修飾ヌクレオチド塩基」には、共有結合性の修飾がなされた塩基および/または糖を有するヌクレオチドが含まれる。例えば、修飾ヌクレオチドには、3’位にヒドロキシル基以外および5’位にリン酸基以外の低分子量有機基が共有結合により付加された糖を有するヌクレオチドが含まれる。この種の修飾ヌクレオチドには、2’置換型の糖、例えば2’−O−メチル−;2−O−アルキル;2−O−アリル;2’−S−アルキル;2’−S−アリル;2’−フルオロ−;2’−ハロまたは2’−アジド−リボース、炭素環系糖アナログα−アノマー糖、エピマー糖、例えばアラビノース、キシロースまたはリキソース、ピラノース糖、フラノース糖、およびセドヘプツロースも含み得る。
【0088】
修飾ヌクレオチドは当該分野で公知であり、アルキル化プリンおよびピリミジン、アシル化プリンおよびピリミジン、ならびにその他の複素環系が含まれる。これらのクラスのピリミジンおよびプリンは当該分野で公知であり、プソイドイソシトシン、N4,N4−エタノシトシン、8−ヒドロキシ−N6−メチルアデニン、4−アセチルシトシン、5−(カルボキシヒドロキシルメチル)ウラシル、5−フルオロウラシル、5−ブロモウラシル、5−カルボキシメチルアミノメチル−2−チオウラシル、5−カルボキシメチルアミノメチルウラシル、ジヒドロウラシル、イノシン、N6−イソペンチル−アデニン、1−メチルアデニン、1−メチルプソイドウラシル、1−メチルグアニン、2,2−ジメチルグアニン、2−メチルアデニン、2−メチルグアニン、3−メチルシトシン、5−メチルシトシン、N6−メチルアデニン、7−メチルグアニン、5−メチルアミノメチルウラシル、5−メトキシアミノメチル−2−チオウラシル、−D−マンノシルケオシン、5−メトキシカルボニルメチルウラシル、5−メトキシウラシル、2−メチルチオ−N6−イソペンテニルアデニン、ウラシル−5−オキシ酢酸メチルエステル、プソイドウラシル(psueouracil)、2−チオシトシン、5−メチル−2−チオウラシル、2−チオウラシル、4−チオウラシル、5−メチルウラシル、N−ウラシル−5−オキシ酢酸メチルエステル、ウラシル−5−オキシ酢酸、ケオシン(queosine)、2−チオシトシン、5−プロピルウラシル、5−プロピルシトシン、5−エチルウラシル、5−エチルシトシン、5−ブチルウラシル、5−ペンチルウラシル、5−ペンチルシトシン、および2,6−ジアミノプリン、メチルプソイドウラシル、1−メチルグアニン、1−メチルシトシンが含まれる。
【0089】
RNAiを使用して線虫(C.elegans)、ショウジョウバエ(Drosophila)、植物および哺乳動物において遺伝子を抑制する方法は当該分野で公知である(WO01/29058;WO99/32619;64〜74、これら全てを参照により本明細書で援用することを明記する)。
【0090】
EP3Rコード遺伝子の発現をダウンレギュレートするリボザイムは、好ましくは、EP3Rコード遺伝子、例えば配列番号1のDNA配列を有するEP3Rコード遺伝子、のRNA配列に特異的なものである。リボザイムは、一本鎖RNA、例えばmRNAを規定の配列で特異的に切断する核酸分子、実際はRNA、であり、それらの特異性は操作することができる。ハンマーヘッド型リボザイムは、約11〜18塩基長の塩基配列を認識し、したがって約4塩基長の配列を認識するテトラヒメナ型のリボザイムよりも高い特異性を有する点で好ましい場合があるが、後者のタイプのリボザイムも特定の状況で有用である。リボザイムの利用に関する参考文献には、Marschall,et al.1994;Hasselhoff,1988およびCech,1988が含まれる。
mPGES−1 ミクロソームプロスタグランジンEシンターゼ1(mPGES−1)ポリペプチドは、グルタチオンの存在下でPGH2からのPGE2の合成を触媒する能力を有する。mPGES−1ポリペプチドは、好ましくは配列番号4のヒトmPGES−1アミノ酸配列を含むまたはそれからなる。しかし、mPGES−1ポリペプチドは、非ヒト動物、例えばマウスまたは他のげっ歯類由来のホモログであってもよい。mPGES−1ポリプチドは、一つまたはそれ以上のアミノ酸が挿入、欠失または置換によって変更されたヒトmPGES−1タンパク質の変異体または誘導体であってもよい。好ましくは、mPGES−1ポリペプチドは、配列番号4の全長アミノ酸配列と少なくとも70%、より好ましくは80%、さらにより好ましくは90%、さらにより好ましくは95%、最も好ましくは99%のアミノ酸同一性を有し、かつグルタチオンの存在下でPGH2からのPGE2の合成を触媒する能力を有するアミノ酸配列を含む。いくつかの実施態様において、mPGES−1ポリペプチドは単離されている。
【0091】
ヒトmPGES−1 cDNAのコード配列は、以下では配列番号3として示される。5’および3’非翻訳末端を含む全長cDNAはGenBankアクセッション番号NM_004878.3の下で入F手可能である。
【0092】
mPGES−1ポリペプチドは、配列番号4のアミノ酸配列を有する全長mPGES−1ポリペプチドより短いが、その本来の生物学的活性を保持する活性部分であってもよい。特にこの活性部分は、グルタチオンの存在下でPGH2からのPGE2の合成を触媒する能力を有するものである。
【0093】
mPGES−1コード遺伝子は、本明細書で定義されるmPGES−1ポリペプチドをコードするヌクレオチド配列を含み得る。mPGES−1コード遺伝子は、配列番号3の全長ヌクレオチド配列と少なくとも70%、より好ましくは80%、さらにより好ましくは90%、さらにより好ましくは95%、最も好ましくは99%のヌクレオチド配列同一性を有するヌクレオチド配列を含み得る。
【0094】
mPGES−1阻害剤
mPGES−1阻害剤は、mPGES−1により媒介されるPGE2の合成を阻止または減少させる。mPGES−1阻害剤は、mPGES−1により媒介されるPGE2レベル、特に血液脳関門の内皮細胞および/または脳の実質におけるPGE2レベルの上昇を阻止または減少させる。PGE2合成を阻止または減少させることにより、mPGES−1阻害剤は、誘導されたPGE2経路により媒介される無呼吸、呼吸抑制および/または自然蘇生不全を軽減し得る。
【0095】
本発明は、多くの異なるタイプのmPGES−1阻害剤の使用を想定している。例えば、阻害剤は、本明細書で定義されるmPGES−1ポリペプチドに結合してその触媒機能を妨害し得るものであり、そのような阻害剤には、mPGES−1ポリペプチドの触媒活性部位に結合する競合的阻害剤および触媒活性部位から離れた部位でmPGES−1ポリペプチドと結合するアロステリック阻害剤が含まれる。さらに、この阻害剤は、mPGES−1ポリペプチドの活性化因子に結合しこれを阻害することによって間接的に作用するものであり得る。mPGES−1コード遺伝子の発現を(例えば、mPGES−1コード遺伝子の転写および/または翻訳を阻害することによって)ダウンレギュレートするmPGES−1阻害剤もまた想定されている。
【0096】
mPGES−1ポリペプチドに結合する阻害剤の例には、特異的結合物質、例えば抗体分子、およびmPGES−1ポリペプチドと競合的または非競合的に結合する低分子が含まれる。mPGES−1コード遺伝子の発現をダウンレギュレートする阻害剤の例には、mPGES−1コード遺伝子またはその一部と相補的な核酸分子およびmPGES−1をコードする遺伝子またはそのフラグメントの配列に対応する二本鎖RNAが含まれる。mPGES−1コード遺伝子の発現をダウンレギュレートする阻害剤には、リボザイムおよび/または三重らせん因子も含まれる。低分子、特異的結合物質および核酸を含む、多くの異なるクラスの阻害剤のさらなる詳細は本明細書中に記載されている。
【0097】
mPGES−1の低分子阻害剤
低分子性のmPGES−1阻害剤は、mPGES−1ポリペプチドと結合して、mPGES−1ポリペプチドによる環状エンドペルオキシド基質から当該基質の9−ケト,11αヒドロキシル型である生産物への変換を阻止または制限し得る。この低分子は、mPGES−1ポリペプチドの活性部位に結合するものでもその遠位部位に結合するものでもよく、可逆的に結合するものでも不可逆的に結合するものでもよい。
【0098】
ロイコトリエンC4、NS−398、スリンダク硫化物を含む多くの化合物がmPGES−1酵素を阻害することが見出されており、これらのIC50値はそれぞれ5、20および80μMである(75、その開示を参照により本明細書で援用することを明記する)。また、15−デオキシ−Δ12,14−PGJ2、アラキドン酸、ドコサヘキサエン酸、エイコサペンタエン酸および3−[tert−ブチルチオ−1−(4−クロロベンジル)−5−イソプロピル−1H−インドール−2−イル]−2,2−ジメチルプロピオン酸(MK−886)はすべて、0.3μMという同様のIC50値でmPGESを阻害することが報告されている(76−77)。
【0099】
さらなる低分子性mPGES−1阻害剤は、本明細書に記載されるスクリーニング法を用いて同定され得る。
【0100】
mPGES−1の特異的結合阻害剤
いくつかの実施態様において、mPGES−1阻害剤は、本明細書で定義されるmPGES−1ポリペプチドに結合し、mPGES−1により媒介される環状エンドペルオキシド基質から当該基質の9−ケト,11αヒドロキシル型である生産物への変換を阻止または減少させる特異的結合物質であり得る。
【0101】
mPGES−1の特異的結合阻害剤は抗体分子であり得る。様々なタイプの抗体分子が、EP3Rの特異的結合阻害剤に関連して記載されている。抗体分子は、それがEP3RポリペプチドではなくmPGES−1ポリペプチドに結合することを除いては上述の通りである。
【0102】
mPGES−1の核酸阻害剤
本発明はまた、mPGES−1コード遺伝子の発現をダウンレギュレートする阻害剤を想定している。
【0103】
ヒトにおいて、mPGES−1は、配列番号3のヌクレオチド配列を有する遺伝子によってコードされる。ヒトmPGES−1のアミノ酸配列は配列番号4に示されている。このヌクレオチド配列は、mPGES−1コード遺伝子の発現をダウンレギュレートすることができる核酸分子を設計するのに利用され得るが、これについては、その核酸分子がEP3Rコード遺伝子ではなくmPGES−1コード遺伝子の発現をダウンレギュレートすることを除いてEP3R阻害剤に関連して説明した通りである。したがって、EP3Rコード遺伝子の配列、部分配列または相補配列に関する言及は、必要な修正を加えた上で、mPGES−1コード遺伝子の配列、部分配列または相補配列にも適用される。
【0104】
COX−2
シクロオキシゲナーゼ2(COX−2)ポリペプチドは、アラキドン酸からのPGH2の合成を触媒する能力を有する。ヒトCOX−2のアミノ酸配列は、GenBankアクセッション番号NP_000954の下で寄託されており(これを参照により本明細書で援用することを明記する)、また、以下では配列番号6として示されている。COX−2ポリペプチドは、好ましくは配列番号6のヒトCOX−2アミノ酸配列を含むかまたはこれからなる。しかし、COX−2ポリペプチドは、非ヒト動物、例えばマウスまたは他のげっ歯類由来のホモログであってもよい。COX−2ポリプチドは、一つまたはそれ以上のアミノ酸が挿入、欠失または置換によって変更されたヒトCOX−2タンパク質の変異体または誘導体であってもよい。好ましくは、COX−2ポリペプチドは、配列番号6の全長アミノ酸配列と少なくとも70%、より好ましくは80%、さらにより好ましくは90%、さらにより好ましくは95%、最も好ましくは99%のアミノ酸同一性を有し、かつアラキドン酸からのPGH2合成を触媒する能力を有するアミノ酸配列を含む。いくつかの実施態様において、COX−2ポリペプチドは単離されている。
【0105】
COX−2ポリペプチドは、配列番号6のアミノ酸配列を有する全長COX−2ポリペプチドより短いが、その本来の生物学的活性を保持する活性部分であってもよい。特にこの活性部分は、アラキドン酸からのPGH2合成を触媒する能力を有するものである。
【0106】
ヒトCOX−2のcDNA配列は、GenBankに寄託されており(アクセッション番号NM_000963、これを参照により本明細書で援用することを明記する)、以下では配列番号5として示される。そのコード配列は、太字で示されたヌクレオチド135〜1949である。
【0107】
COX−2コード遺伝子は、本明細書で定義されるCOX−2ポリペプチドをコードするヌクレオチド配列を含み得る。COX−2コード遺伝子は、配列番号5またはそのコード領域(配列番号5のヌクレオチド135〜1949)のヌクレオチド配列と少なくとも70%、より好ましくは80%、さらにより好ましくは90%、さらにより好ましくは95%、最も好ましくは99%のヌクレオチド配列同一性を有するヌクレオチド配列を含み得る。
【0108】
COX−2の選択的阻害剤
COX−2の選択的阻害剤は、COX−2によって媒介されるPGH2合成を阻止または減少させる。COX−2選択的阻害剤は、COX−2によって媒介されるPGH2レベルの上昇を阻止または減少させ、それによって誘導されたPGE2経路により媒介される無呼吸、呼吸抑制および/または自然蘇生不全を軽減し得る。
【0109】
さらに、COX−2選択的阻害剤は、COX−1に対する阻害活性よりも大きな阻害活性をCOX−2に対して有する。COX−2阻害剤の選択性は、一般に、非選択的なCOX阻害に伴う有害な影響、例えば重要な構成性のCOX−1活性の阻害によって引き起こされる影響を軽減する。COX−2選択的阻害剤は、COX−1の2倍またはそれ以上、例えば5倍または10倍高い阻害活性をCOX−2に対して有し得る。したがって、COX−2選択的阻害剤のIC50値は、同じ阻害剤のCOX−1に対するIC50値よりも2倍低い、好ましくは5倍または10倍低い場合がある。
【0110】
本発明は、多くの異なるタイプのCOX−2選択的阻害剤の使用を想定している。例えば、阻害剤は、本明細書で定義されるCOX−2ポリペプチドに結合してその触媒機能を妨害するものであり、そのような阻害剤には、COX−2ポリペプチドの触媒活性部位に結合する競合的阻害剤および触媒活性部位から離れた部位でCOX−2ポリペプチドと結合するアロステリック阻害剤が含まれる。さらに、さらに、この阻害剤は、COX−2ポリペプチドの活性化因子に結合しこれを阻害することによって間接的に作用するものであり得る。COX−2コード遺伝子の発現を(例えば、COX−2コード遺伝子の転写および/または翻訳を阻害することによって)ダウンレギュレートするCOX−2阻害剤もまた想定されている。
【0111】
COX−2ポリペプチドに結合する阻害剤の例には、特異的結合物質、例えば抗体分子、およびCOX−2ポリペプチドと競合的または非競合的に結合する低分子が含まれる。COX−2コード遺伝子の発現をダウンレギュレートする阻害剤の例には、COX−2コード遺伝子またはその一部と相補的な核酸分子およびCOX−2ポリペプチドをコードする遺伝子またはそのフラグメントの配列に対応する二本鎖RNAが含まれる。COX−2コード遺伝子の発現をダウンレギュレートする阻害剤には、リボザイムおよび/または三重らせん因子も含まれる。低分子、特異的結合物質および核酸を含む、多くの異なるクラスの阻害剤のさらなる詳細は本明細書中に記載されている。
【0112】
COX−2の低分子阻害剤
COX−2の低分子性選択的阻害剤は、COX−2ポリペプチドに結合し、COX−2により媒介されるアラキドン酸からPGH2への変換を阻止または減少させ得る。この低分子は、COX−2ポリペプチドの触媒活性部位に結合するものでもその遠位部位に結合するものでもよく、可逆的に結合するものでも不可逆的に結合するものでもよい。
【0113】
COX−2選択的阻害剤として作用する多くの化合物が記載されている。COX−2選択的阻害剤の一例は、「コキシブ類(coxibs)」として知られている薬物である。
【0114】
いくつかの実施態様において、COX−2の低分子性選択的阻害剤は、4−(5−メチル−3−フェニルイソキサゾール−4−イル)ベンゼンスルホンアミド(バルデコキシブ)もしくは薬学的に許容できるその塩、4−[5−(4−メチルフェニル)−3−(トリフルオロメチル)ピラゾール−1−イル]ベンゼンスルホンアミド(セレコキシブ)もしくは薬学的に許容できるその塩、および/または4−(4−メチルスルホニルフェニル)−3−フェニル−5H−フラン−2−オン(ロフェコキシブ)もしくは薬学的に許容できるその塩を含み得る。
【0115】
本発明において有用な多くのCOX−2阻害剤はこれまでに報告されている(COX阻害、特にCOX−2阻害の薬理のレビューについては94を参照されたい。この文献の開示内容を参照により本明細書で援用することを明記する)。
【0116】
さらなるCOX−2の低分子性選択的阻害剤は、本明細書に記載されるスクリーニング法を用いて同定され得る。
【0117】
COX−2の特異的結合阻害剤
いくつかの実施態様において、COX−2の選択的阻害剤は、本明細書で定義されるCOX−2ポリペプチドに結合し、COX−2により媒介されるアラキドン酸からPGH2への変換を阻止または減少させる特異的結合物質であり得る。
【0118】
COX−2の特異的結合阻害剤は抗体分子であり得る。様々なタイプの抗体分子が、EP3Rの特異的結合阻害剤に関連して記載されている。抗体分子は、それがEP3RポリペプチドではなくCOX−2ポリペプチドに結合することを除いては上述の通りである。好ましくは、COX−2の特異的結合阻害剤はCOX−1ポリペプチドと交差反応しないものである。
【0119】
COX−2の核酸阻害剤
本発明はまた、COX−2コード遺伝子の発現をダウンレギュレートする阻害剤を想定している。
ヒトにおいて、COX−2は、配列番号5のヌクレオチド配列を有する遺伝子によってコードされる。ヒトCOX−2のアミノ酸配列は配列番号6に示されている。このヌクレオチド配列は、COX−2コード遺伝子の発現をダウンレギュレートすることができる核酸分子を設計するのに利用され得るが、これについては、その核酸分子がEP3Rコード遺伝子ではなくCOX−2コード遺伝子の発現をダウンレギュレートすることを除いてEP3R阻害剤に関連して説明した通りである。したがって、EP3Rコード遺伝子の配列、部分配列または相補配列に関する言及は、必要な修正を加えた上で、COX−2コード遺伝子の配列、部分配列または相補配列にも適用される。
【0120】
治療
本発明は、本明細書で定義される呼吸障害の治療的処置および予防的処置の両方を想定している。この処置は、呼吸障害に対する哺乳動物の感受性を低下させる、および/または哺乳動物における呼吸障害の一つまたはそれ以上の臨床的側面を完全にもしくは部分的に好転させるものであり得る。例えば、本発明は、無呼吸を罹患した患者の呼吸を正常化させることを想定している。低酸素イベント後の自然蘇生の改善もまた想定されている。
好ましい実施態様において、哺乳動物は、本明細書で定義される呼吸障害の危険があると判断された患者であり得る。例えば、感染、特にIL−1βレベルを上昇させる感染がみられるヒト乳幼児が、無呼吸の可能性およびその重症度を低下させるために、EP3R阻害剤、mPGES−1阻害剤、および/またはCOX−2選択的阻害剤を含む薬剤で処置され得る。
【0121】
処方
本発明は、本明細書で定義される阻害剤の様々な薬学的組成物を想定している。薬学的組成物は、一般に、一つ又はそれ以上の薬学的に許容できる塩、担体または補助成分を含む。さらに、本明細書で定義される阻害剤を一つ以上含む薬学的組成物が想定される。例えば、組成物は、EP3R阻害剤、mPGES−1阻害剤およびCOX−2選択的阻害剤から選択される二つまたはそれ以上の薬剤を含み得る。あるいは、一つ以上の阻害剤が用いられる場合、その薬剤は、同時または連続投与用の別々の組成物に配合され得る。
【0122】
投与方法
本発明にしたがい適当な投与経路が利用され得る。典型的には、本明細書で定義される阻害剤を含む組成物は、口腔から、直腸から、鼻腔内に、静脈内注射、筋内注射、皮下注射、腹腔内注射もしくは脳室内注射によって、経皮パッチまたは小型ポンプによって投与され得る。血液脳関門を通過することができないEP3R阻害剤を含む組成物の場合、脳室内注射が好ましい。
【0123】
評価および診断
本発明は、哺乳動物由来のサンプル中の一つまたはそれ以上の誘導されたPGE2経路のマーカーを検出することによって哺乳動物における呼吸障害の感受性またはその存在を評価する方法を想定している。呼吸障害に罹患していることまたは呼吸障害の危険が高いことが確認された対象は、次いで、本明細書で定義される阻害剤により処置され得る。
【0124】
患者において誘導されたPGE2経路の活性が上昇しているかどうかを評価する多くの方法が想定されている。いくつかの実施態様において、PGE2またはその代謝産物のレベルが対象由来のサンプルにおいて検出され、対照レベルと比較される。対照レベルは、好ましくは既定の「正常」範囲である。例えば、対照レベルは、健常な対照由来の類似サンプルにおいて見られるPGE2またはその代謝産物のレベルであり得る。対照レベルは、過去に健常な対照対象に関して測定または報告された値であり得るし、ある集団から得られた平均値でもあり得る。
【0125】
PGE2および/または一つもしくはそれ以上のその代謝産物は、本明細書にさらに記載されるような生物学的サンプルにおいて測定され得る。PGE2代謝産物は数が多いが、その大部分はLC−MS/MS(液体クロマトグラフィー三重項四重極質量分析器)によって検出することができる(105、その開示内容全体を参照により本明細書で援用することを明記する)。
【0126】
本発明におけるPGE2代謝産物の例には、7α−ヒドロキシ−5,11−ジケト−2,3,4,5,20−ペンタ−19−カルボキシプロスタン酸およびE系よびF系の13,14−ジヒドロ−15−ケト代謝産物が含まれる。PGE2および/または一つもしくはそれ以上のその代謝産物(7α−ヒドロキシ−5,11−ジケト−2,3,4,5,20−ペンタ−19−カルボキシプロスタン酸およびE系よびF系の13,14−ジヒドロ−15−ケト代謝産物を含む)は、対象サンプルに適した任意の技術によって測定され得る。本発明におけるPGE2代謝産物ならびにその検出および測定技術については(106、その開示内容全体を参照により本明細書で援用することを明記する)にも記載されている。
【0127】
PGE2およびその代謝産物の測定アッセイの具体例には酵素免疫アッセイ(EIA)が含まれ、これについては以下の実施例の節でさらに詳細に記載されている。EIAキットは商業的に入手可能であり、個々の化合物の高感度検出を実現する。
【0128】
さらなる例として、PGE2および/または一つもしくはそれ以上のその代謝産物(7α−ヒドロキシ−5,11−ジケト−2,3,4,5,20−ペンタ−19−カルボキシプロスタン酸およびE系よびF系の13,14−ジヒドロ−15−ケト代謝産物を含む)の測定または検出は、LC.MS/MSおよび/または三重項四重極質量分析(三重項四重極法(QQQ)としても知られている)を用いるものであり得る。三重項四重極質量分析の使用は、この分析法がフェムト/ピコモル濃度の化合物を検出する能力を有することから、特定状況下で好ましい。タンデム四重極(三重項四重極)機器は、既知の代謝産物およびペプチド(例えばPGE2および/または一つもしくはそれ以上のその代謝産物)の定量に利用される。この機器は、アラキドン酸カスケードの量的経路分析に使用することができる。さらに、この機器は、臨床材料におけるペプチドの量的検証および臨床材料間での相違が代謝学的に確認された代謝産物の量的検証に使用され得る。推奨される機器は、電子スプレーイオン化インターフェース(ESI)を経由して超高速液体クロマトグラフィー(UPLC)に接続され得る。液体クロマトグラフィーにおいてサイズの小さい粒子(<1.8μm)を使用することで、クロマトグラフピーク幅を劇的に、典型的には30〜60秒(従来型LC)から3〜5秒(UPLC)に狭めることができる。これにより分離能が向上し、したがってより多くの化合物をより短時間で分離することができる。三重項四重極質量分析器において、特定の代謝産物の分子イオンが第一の四重極で選択され、代謝産物の断片化がコリージョンガスを含むコリージョンセル中で誘導される。第二の四重極では電子遷移痕を生じる特定の「娘イオン(daughter ion)」が選択される(反応モニタリング)。代謝産物/ペプチドは個別に断片化するため、この娘イオンは極めて化合物特異的なトレーサーとなる。典型的には、約100のトレースを同時にモニタリング(多重反応モニタリング、MRM)、一つの分析物中の多くの代謝産物を特異的かつ高感度で定量することができる。好ましくは、本発明の方法は、尿サンプル中の一つまたはそれ以上のPGE2代謝産物の測定を含み、三重項四重極質量分析を利用するものである。特に好ましい尿PGE2代謝産物(u−PGEM)測定アッセイは実施例8に記載されている。いくつかの例において、本発明の方法は、尿サンプル中の一つまたはそれ以上のPGE2代謝産物の測定を含み、この方法はさらに、PGE2の尿中レベルが尿クレアチニンレベルに対する相対レベルである場合、尿サンプル中のクレアチニン濃度の測定を含む。
【0129】
サンプル中のPGE2またはその代謝産物のレベルと対照レベルとの比較は、既定の対照値または対照値の範囲を記録した図表、データベースまたは文献を調べることによって行える。いくつかの例において、例えば既定の対照値が入手できない場合、サンプルレベルと対照レベルの比較は、調査対象の対象由来のサンプル中のPGE2またはその代謝産物レベルの検出に続いてまたはそれと平行して、健常な対象由来の対照サンプル中のPGE2またはその代謝産物レベルを検出することを含むことになる。
【0130】
PGE2レベルまたはPGE2代謝産物レベルが対照レベルと比較して上昇していることは、呼吸障害の存在またはその危険が高いこと、例えば無呼吸頻度が増えていることを示すと考えられる。
【0131】
以下に記載されるデータは、PGE2代謝産物が、乳幼児が出生時に罹患した仮死(「周産期仮死」)の程度および/または哺乳動物対象における低酸素性虚血性脳症(HIE)の存在もしくは重症度を推測する上で有用な指標として使用できる証拠となる。PGE2レベルまたはPGE2代謝産物レベルが対照レベルと比較して上昇、特に対象の出生から数日以内、例えば96、48、24、12、6、4、3もしくは2時間以内または60、30、20、10または5分以内に対象から採取したサンプルにおいて上昇していることは、哺乳動物対象におけるHIEの存在を示唆すること、および/または対象が周産期仮死を罹患したことを示すことが見出された。対照レベルとの比較の上でのPGE2またはPGE2代謝産物の増加率は、周産期仮死の程度および/またはHIEの重症度の程度と相関し、したがって対象の可能性のある神経学的結果と相関することが見出された。
【0132】
したがって本発明の方法は、予後および長期的な神経学的結果の推測に有用であり、したがって処置に関する迅速な決断を補助できる点で価値がある。
【0133】
実験結果は、PGE2の半減期が、いくつかの例において、約12〜18時間であることを示している。PGE2およびその代謝産物は、72時間以上経過した後でさえ存在し測定できる場合もある。PGE2分解のハーフタイムはその細胞環境に依存して大きく変化する。PGE2の半減期は、数分から数時間まで変化し得る。体内で産生されたPGE2および尿またはその他の体液中に分泌されたPGE2を評価する場合、その代謝産物も合わせて測定するのが重要である。
【0134】
いくつかの実施態様において、サンプル中のPGE2またはその代謝産物レベルは、PGE2またはその代謝産物の参考レベルと比較される。参考レベルは対照レベルとは別も
のであり得る。例えば、参考レベルは、本明細書で定義される呼吸障害または哺乳動物被験体における周産期仮死もしくはHIEを示す値または値の範囲であり得る。このような例において、PGE2またはその代謝産物レベルがほぼ参考レベルまたは参考値の範囲内であることは、本明細書で定義される呼吸障害の存在もしくはその危険が高いこと、乳幼児が出生時に罹患した仮死の程度、および/または被験体におけるHIEの存在もしくはその重症度を示す。参考レベルは、呼吸障害、乳幼児が出生時に罹患した仮死および/または被験体におけるHIEの特定の重症度または段階に関連する値または一定範囲の値であり得る。
【0135】
いくつかの実施態様において、本発明の方法は、患者において誘導されたPGE2経路の活性が高まっているかどうかを、mPGES−1コード遺伝子の発現を検出することによって評価することを含む。これには、mPGES−1コード遺伝子のmRNAのレベルを測定、例えば定量的、準定量的またはリアルタイムPCRベースの方法を用いて測定することが含まれる。mPGES−1コード遺伝子の発現の上昇は、呼吸障害の危険の上昇を示唆する。患者において誘導されたPGE2経路の活性が高まっているかどうかを評価するその他の方法には、PGH2レベルの上昇、COX−2遺伝子発現の増大および/またはIL−1βレベルの上昇を検出することが含まれる。本発明は、一つまたはそれ以上の、誘導されたPGE2経路の活性の増加に関するマーカーを検出することを想定している。例えば、PGE2レベルの検出は、PGH2レベル、mPGES−1発現、COX−2発現および/またはIL−1βレベルの検出と組み合わされ得る。
【0136】
いくつかの実施態様において、本発明の方法は、mPGES−1、COX−2および/またはEP3Rをコードする遺伝子における一つまたはそれ以上の変異を同定することを含み得る。例えば、mPGES−1、COX−2および/またはEP3Rをコードする遺伝子における単一ヌクレオチド多型(SNP)は、本明細書で定義される呼吸障害に対する感受性の上昇と関連し得る。
【0137】
サンプル
サンプルは、液体サンプル、例えばCSFサンプル、血液サンプル、尿サンプル、または非液体サンプル、例えば組織検体サンプルであり得る。好ましくは、サンプルはCSF、尿または血液サンプルである。特定の実施態様において、尿サンプルが特に好ましい。
サンプルは、本明細書で詳述される状態の実際の原因もしくは疑われる原因またはその状態の発症の後の既定の時点で哺乳動物被験体、例えばヒト被験体から採取され得る。例えば、サンプルは、被験体の出生後または入院後もしくは医師との面会後96、48、24、12、6、4、3もしくは2時間以内または60、30、20、10もしくは5分以内にヒト乳幼児から採取され得る。いくつかの例において、サンプルは、低温で(例えば4℃付近でまたは−80℃〜−20℃で)保存されたヒト尿サンプルであり得る。
【0138】
感染マーカー
本発明者らは、PGE2レベル、CRPおよび無呼吸インデックスが相関していることを発見した(図5を参照されたい)。いくつかの実施態様において、診断方法は、追加的に、感染に関連するマーカーのレベルを検出することを含み得る。例えば、CRPレベルが、患者由来のサンプル、好ましくは血液または尿サンプルにおいて評価され得る。感染マーカーのレベルが対照レベルと比較して上昇していることは、特にPGE2レベルまたは誘導されたPGE2経路の活性の増加に関する他のマーカーのレベルの上昇と組み合わせた場合に、呼吸障害の危険が高いことを示唆する。
【0139】
対照レベルは、好ましくは既定の「正常」範囲である。例えば、対照レベルは、健常な対照由来の類似のサンプルにおいて見出されるCRPのレベルであり得る。対照レベルは、過去に健常な対照対象に関して測定または報告された値であり得るし、ある集団から得られた平均値でもあり得る。
【0140】
サンプル中のCRPレベルと対照レベルとの比較は、既定の対照値または対照値の範囲を記録した図表、データベースまたは文献を調べることによって行える。いくつかの例において、例えば既定の対照値が入手できない場合、サンプルレベルと対照レベルの比較は、調査対象の対象由来のサンプル中のCRPレベルの検出に続いてまたはそれと平行して、健常な対象由来の対照サンプル中のCRPレベルを検出することを含むことになる。
【0141】
CRPレベルが対照レベルと比較して上昇していることは、呼吸障害の存在またはその危険が高いこと、例えば無呼吸頻度が増えていることを示すと考えられる。
【0142】
いくつかの実施態様において、サンプル中のCRPレベルは、CRPの参考レベルと比較される。参考レベルは対照レベルとは別ものであり得る。例えば、参考レベルは、本明細書で定義される呼吸障害を示す値または値の範囲であり得る。このような例において、CRPレベルがほぼ参考レベルまたは参考値の範囲内であることは、本明細書で定義される呼吸障害の存在またはその危険が高いことを示す。参考レベルは、本明細書で定義される呼吸障害の特定の重症度または段階に関連する値または値の範囲であり得る。
【0143】
さらに、PGE2またはその代謝産物の測定が、炎症マーカーとしてのCRPまたは高感受性CRP(hsCRP)の測定の補足としてまたはその代わりに使用され得る。
【0144】
スクリーニング法
本発明は、哺乳動物における呼吸障害の処置に使用できる物質の同定を想定している。したがって、哺乳動物における呼吸障害の処置に使用できる物質の同定方法は、誘導されたPGE2経路を阻害する能力について試験物質をアッセイすること、例えばEP3R阻害剤、mPGES−1阻害剤および/またはCOX−2選択的阻害剤として機能するかどうかについて試験物質をアッセイすることを含み得る。誘導されたPGE2経路の阻害は、試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを示す。試験物質、これは候補化合物または候補組成物であり得る、は、
(a)この経路に関係するポリペプチド(「誘導されたPGE2経路ポリペプチド」)、例えばCOX−2ポリペプチド、mPGES−1ポリペプチドおよび/もしくはEP3Rポリペプチドと直接的に相互作用することによって、
(b)この経路に関係するポリペプチドと、例えばCOX−2ポリペプチド、mPGES−1ポリペプチドおよび/もしくはEP3Rポリペプチドの活性化因子に結合しこれを阻害することにより、間接的に相互作用することによって、ならびに/または
(c)誘導されたPGE2経路ポリペプチドをコードする遺伝子の発現をダウンレギュレートすることによって、例えば、COX−2コード遺伝子、mPGES−1コード遺伝子および/もしくはEP3Rコード遺伝子の発現(例えば転写および/もしくは翻訳)をダウンレギュレートすることによって、
誘導されたPGE2経路を阻害し得る。
【0145】
ポリペプチドの阻害剤のスクリーニング
誘導されたPGE2経路ポリペプチドと相互作用および/または結合する試験物質の能力の測定は、その試験物質が、誘導されたPGE2経路の潜在的阻害剤であることを同定するのに使用され得る。この方法は、相互作用または結合を検出または観察すること、次いで、その試験物質を、それが誘導されたPGE2経路ポリペプチドの活性、例えば酵素活性または受容体を媒介するシグナル伝達を阻害するかどうかを決定するさらなるアッセイ法において使用することを含み得る。
【0146】
本発明アッセイの細部は慣用的な技術および知識を用いて当業者によって変更され得る。例えば、ポリペプチドまたはペプチド間の相互作用は、その一方を検出標識で標識してこれを固相に固定したもう一方と接触させることによって、インビトロで試験され得る。適当な検出標識には35Sメチオニンが含まれ、これは組み換え法で作製されたペプチドおよびポリペプチドに組み込まれ得る。組み換え法で作製されたペプチドおよびポリペプチドはまた、抗体で標識することができるエピトープを含む融合タンパク質として発現され得る。
【0147】
固相に固定されるタンパク質またはペプチドは、固相に結合されたそのタンパク質に対する抗体を用いて、または公知の他の技術を通じて固定され得る。好ましいインビトロ相互作用は、グルタチオン−S−トランスフェラーゼ(GST)を含む融合タンパク質を利用するものである。これは、グルタチオンアガロースビーズ上に固定され得る。上記タイプのインビトロアッセイフォーマットにおいて、試験化合物は、固定されたGST融合ポリペプチドに結合する標識ペプチドまたはポリペプチドの量を減少させる能力を測定することによってアッセイされ得る。これは、SDSポリアクリルアミドゲル電気泳動によってグルタチオンアガロースビーズを分画することによって測定され得る。あるいは、ビーズは未結合タンパク質を除去するために洗浄され、結合したタンパク質の量は、例えば適当なシンチレーションカウンター内に存在する標識の量を計測することによって測定することができる。
【0148】
一般的に、誘導されたPGE2経路ポリペプチドと結合または相互作用する試験物質の能力の同定およびその潜在的なPGE2経路阻害剤としての同定に続いて、試験物質が誘導されたPGE2経路ポリペプチドの活性を阻害できるかできないかの測定を含む一つまたはそれ以上のアッセイが行われる。通常、誘導されたPGE2経路ポリペプチドを阻害する試験物質の能力の測定を含むアッセイは、その試験物質が誘導されたPGE2経路ポリペプチドと結合または相互作用することができるかどうかが不明な状況下で行われ得るが、多数の試験化合物を試験するためのスクリーンとして以前の結合/相互作用アッセイを使用し、誘導されたPGE2経路ポリペプチドの活性を阻害する能力の測定を含む機能アッセイを実行可能なレベルまで、潜在的阻害剤の数を削減することができる。
【0149】
試験物質が誘導されたPGE2経路ポリペプチドの阻害剤として機能するかどうかを測定するアッセイ法、特にCOX−2、mPGES−1およびEP3Rアッセイが本明細書中でさらに記載されている。
【0150】
コンビナトリアルライブラリ技術(78)は、ポリペプチドの活性を調節する能力について潜在的に膨大な数の異なる物質を試験するのに効果的な手法を提供する。
【0151】
本発明のアッセイに投入され得る試験物質または試験化合物の量は、通常、使用する化合物の種類に基づく試行錯誤により決定され得る。典型的には、約0.1nM〜10μM濃度の試験化合物(例えば推定阻害剤)が使用され得る。ペプチドが試験物質である場合は、それよりも高い濃度が使用され得る。使用され得る化合物は、天然化合物であってもドラッグスクリーニングプログラムで使用される合成化合物であってもよい。複数の既知または未知成分を含有する植物抽出物もまた使用され得る。他の阻害剤または阻害剤候補は、ポリペプチドまたはペプチドフラグメントの3次元構造のモデリングに基づくもの、ならびに特定の分子形状、サイズおよび電荷特性を有する潜在的阻害化合物を生み出す合理的なドラッグデザインの使用に基づくものであり得る。
【0152】
遺伝子発現の阻害剤のスクリーニング
誘導されたPGE2経路の阻害剤は、誘導されたPGE2経路ポリペプチドをコードする遺伝子、例えばCOX−2コード遺伝子、mPGES−1コード遺伝子および/またはEP3Rコード遺伝子の発現を妨げることによってこの経路を阻害し得る。したがって、本発明のアッセイ法は、試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを同定することを含み得、この方法は、誘導されたPGE2経路ポリペプチドをコードする遺伝子の発現を減少または阻害することができる物質をスクリーニングすることを含み、このスクリーニングは、
(a)上記遺伝子のプロモーターを含むDNAと試験物質を接触させ(プロモーターは遺伝子に機能的に連結されている)、
(b)このプロモーターからの遺伝子発現のレベルを決定し、そして
(c)試験物質の存在下での遺伝子発現レベルと、同条件かつ試験物質の非存在下での遺伝子発現レベルを比較する、
ことを含み、試験物質の存在下で遺伝子発現レベルが低下することは、その試験物質が、誘導されたPGE2経路ポリペプチドをコードする遺伝子の発現を阻害できることを示す。
【0153】
この方法はさらに、試験物質が、誘導されたPGE2経路ポリペプチドをコードする遺伝子の発現の阻害剤であること、すなわち哺乳動物における呼吸障害の処置に使用できる物質であることを同定することを含み得る。
【0154】
したがって工程(c)は、試験物質の存在下での遺伝子発現レベルが同条件かつ試験物質の非存在下での遺伝子発現レベルと比較して低下していることを検出し、それによってその試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを同定することを含み得る。
【0155】
この方法は、試験物質と発現系、例えば遺伝子に機能的に連結された遺伝子プロモーターを含む宿主細胞を接触させること、および遺伝子発現を測定することを含み得る。この遺伝子は、誘導されたPGE2経路ポリペプチドをコードする遺伝子であってもよいし、異種遺伝子、例えばレポーター遺伝子であってもよい。「レポーター遺伝子」は、自身のコードする生産物が発現された後にアッセイされ得る遺伝子、すなわちプロモーター活性に関する「レポート」を行う遺伝子である。
【0156】
「プロモーター」は、機能的に連結されたDNAの下流に(すなわち、二本鎖DNAのセンス鎖における3’方向に)向かう転写の開始点となるヌクレオチド配列を意味する。遺伝子のプロモーターは、ヒト染色体においてその遺伝子の5’側に存在するヌクレオチド配列または別の種、例えばラットまたはマウスにおける等価な配列を含むかまたは本質的にそれからなるものでよい。
【0157】
プロモーター活性のレベルは、例えば、そのプロモーターからの転写によって生成されるmRNAの量の評価またはそのプロモーターからの転写によって生成されたmRNAの翻訳によって生成されるタンパク質産物の量の評価によって定量可能である。発現系内に存在する特定のmRNAの量は、例えば、そのmRNAとハイブリダイズすることができかつ標識されているかまたは特定の増幅反応、例えばポリメラーゼ連鎖反応(PCR)において使用できる特定のオリゴヌクレオチドを用いて測定され得る。
【0158】
レポーター遺伝子の使用は、タンパク質産物に基づくプロモーター活性の測定を容易にする。レポーター遺伝子は、好ましくは、検出シグナル、好ましくは視覚的に検出可能なシグナル、例えば有色生成物を生じる反応を触媒する酵素をコードする。βガラクトシダーゼおよびルシフェラーゼを含む多くの例が知られている。βガラクトシダーゼ活性は、基質の青色化によってアッセイされ得、このアッセイは肉眼により、または分光光度計を使用して吸光度を測定することにより行われる。蛍光、例えばルシフェラーゼ活性の結果として生じる蛍光は、分光光度計を用いて定量され得る。放射能アッセイは、例えばクロラムフェニコールアセチルトランスフェラーゼを用いて行われ得る。ただしこの化合物は、非放射能アッセイにおいても使用され得る。レポーター遺伝子からの発現により生じる遺伝子産物の存在および/または量は、その生成物に結合できる分子、例えば抗体またはそのフラグメントを用いて測定され得る。結合性分子は、任意の標準技術を用いて直接的または間接的に標識され得る。
【0159】
プロモーター構築物は任意の適当な技術を用いて細胞株に導入され、そのゲノムにレポーター構築物が統合された安定な細胞株が作製され得る。その細胞は、様々な時間、試験化合物と共に培養およびインキュベートされ得る。その細胞は、多数の化合物の分析を容易にするため、96ウェルプレート上で培養され得る。次いでその細胞は洗浄され、レポーター遺伝子の発現が分析され得る。一部のレポーター、例えばルシフェラーゼについては、細胞は溶解された後に分析される。
【0160】
当業者は、多くの利用可能なレポーター遺伝子およびアッセイ技術が遺伝子の活性を測定するのに使用できることを認識している。さらなる例については、Sambrook and Russell, Molecular Cloning: a Laboratory Manual: 3rd edition, 2001, Cold Spring Harbor Laboratory Pressを参照されたい。
【0161】
COX−2アッセイ
本発明は、試験物質、これは候補化合物または候補組成物であり得る、がCOX−2選択的阻害活性を有するかどうかを測定し、それによってCOX−2選択的阻害活性を有することが確認された試験物質を、呼吸障害の処置に使用できる物質として同定するアッセイ法を想定している。
【0162】
いくつかの実施態様において、このアッセイ法は、COX−2ポリペプチドと試験物質およびアラキドン酸を、アラキドン酸が当該試験物質の非存在下でCOX−2によってPGH2に変換される条件下で接触させること、および当該試験物質の非存在下でのPGH2合成の対照レベルとの比較で当該試験物質の存在下でのPGH2合成レベルを決定することを含み、当該試験物質の存在下でのPGH2合成レベルが上述の対照レベルよりも低いことは、当該試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを示す。
【0163】
COX−2阻害剤の同定方法には、これまでに報告されている方法が含まれる(89、90、91、その開示内容を参照により本明細書で援用することを明記する)。COX−2を阻害することが見出された候補化合物または候補組成物は、その化合物または組成物が哺乳動物における呼吸障害を処置する能力を有するかどうかを確認するため、本明細書に記載されるさらなる試験、例えばインビボ試験に供され得る。
【0164】
多くのCOX−2阻害剤用スクリーニングキットが市販されている。例えば、Cayman Chemicals社の製品番号560131「COX Inhibitor Screening Assay」は、必要なヒトCOX−2補因子を提供し、その検出はSnCl2によるPGH2から主としてPGF2αへの還元に基づくものである(http://www.caymanchem.com/app/template/Product.vm/catalog/560131/a/zを参照されたい)。
【0165】
これ以外にも、生成したPGH2を検出するための代替法がいくつか存在する。例えば、PGH2は、塩化鉄処理後に12−HHTおよびマロンジアルデヒドに変換でき、両者は高スループットで測定できる。また、COX−2のペルオキシダーゼ活性は、例えば、これもCayman chemicals社製のキットに記載されるようにして使用することができる(http://www.caymanchem.com/app/template/Product.vm/catalog/760111/a/zを参照されたい)。
【0166】
この方法は、通常、試験物質を、または酵素および酵素基質と共に試験物質をインキュベートすることを含む。基質は、生理学的基質、例えばアラキドン酸であってもよいし、修飾されているかまたは非生理学的な基質、例えばその酵素反応の中で検出可能な(例えば有色の)生成物を生じるよう設計された基質であってもよい。
【0167】
COX−2ポリペプチドを試験物質および基質、例えばアラキドン酸と接触させる順序は変更され得る。例えば、COX−2ポリペプチドは最初に試験物質と共にインキュベートされ、次いで基質と接触され得るし、その逆でもよい。
【0168】
このようにして、試験物質の存在下での生成物の生成は、試験物質の非存在下での生成物の生成と比較され得る。生成物レベルが低いまたは生成物の形成速度が遅いことは、試験物質が酵素活性を阻害していることを示す。
【0169】
阻害剤のアッセイのさらなる可能性は、COX−2を(生来的にまたは組み換え的に)発現する適当な細胞株によるPGH2生成に影響を及ぼす物質の能力の試験である。本発明のアッセイは、導入された一つまたはそれ以上のベクターから関連ポリペプチドまたはペプチドを発現する細胞株、例えば酵母株において実施され得る。
【0170】
それ以外のさらなるアッセイの可能性は、COX−2を含む不均一タンパク質調製物(ヒトでもそれ以外の哺乳動物でもよい)によるPGH2生成に影響を及ぼす物質の能力の試験である。本発明の好ましいアッセイは、単離された/精製されたCOX−2ポリペプチド(全長COX−2またはその活性部分を含む)のCOX−2活性を阻害する試験物質の能力を測定することを含む。
【0171】
本発明のアッセイ法において、生成物の生成は、基質レベルを定量することにより、および/または生成物レベルを定量することにより測定することができる。残存する基質レベルが高ければ、生成物の生成レベルは低いということである。
【0172】
いくつかの実施態様において、アッセイ法は、COX−2阻害についての試験物質の選択性を他のポリペプチド、例えばCOX−1と比較の上で測定することを含み得る。例えば、このアッセイ法は、試験物質のCOX−1に対する阻害活性、例えばIC50、および試験物質のCOX−2に対する阻害活性、例えばIC50を測定することを含み得る。好ましくは、COX−2選択的阻害剤として同定される試験物質は、COX−1に対する阻害活性の2倍またはそれ以上、例えば5倍または10倍高い阻害活性をCOX−2に対して有する。したがって、COX−2阻害に関する試験物質のIC50値は、COX−1阻害に関する同じ試験物質のIC50値よりも2倍またはそれ以上、例えば5倍または10倍低い。
【0173】
生成物の測定には、HPLC、UV分析、放射能測定、またはRIA(例えば、市販のPGE検出用RIAキット)を利用することができる。生成物の形成は、ガスクロマトグラフィー(GC)または質量分析(MS)、またはTLCおよび放射能走査により分析することができる。
【0174】
COX−2タンパク質を使用する本発明の方法において、完全(全長)COX−2タンパク質配列を使用する必要はない。二分子間の結合を試験するまたはCOX−2酵素活性を試験する本発明のアッセイは、フラグメントまたは変異体を使用することができる。フラグメントは、当業者に公知の任意の適当な方法によって作製および使用され得る。適当なフラグメント作製法には、コードDNAからのフラグメントの組み換え発現が含まれるがこれに限定されない。このようなフラグメントは、コードDNAを入手し、発現される部分のいずれかの側にある適当な制限酵素認識部位を確認し、DNAからその部分を切り出すことによって作製され得る。この部分は、次いで、標準的な市販の発現系内の適当なプロモーターに機能的に連結され得る。別の組み換えアプローチは、DNAの関連部分を適当なPCRプライマーを用いて増幅することである。小フラグメント(例えば、約20または30アミノ酸未満)はまた、当該分野で周知のペプチド合成法を用いて作製され得る。COX−2の活性部分は、アッセイ法に使用され得る。
【0175】
COX−2ポリペプチドの「活性部分」は、本発明の方法において使用することができる。活性部分は、全長ポリペプチドより短いがその本来の生物学的活性を保持しているペプチドを意味する。特に、活性部分は、適当な条件下でアラキドン酸からのPGH2合成を触媒する能力を保持しているものである。
【0176】
mPGES−1アッセイ
本発明は、試験物質、これは候補化合物または候補組成物であり得る、がmPGES−1阻害活性を有するかどうかを測定し、mPGES−1阻害活性を有することが確認された試験物質を、哺乳動物における呼吸障害の処置に使用できる物質として同定するアッセイ法を想定している。
【0177】
いくつかの実施態様において、このアッセイ法は、mPGES−1ポリペプチドと試験物質およびmPGES−1の環状エンドペルオキシド基質を、当該mPGES−1の環状エンドペルオキシド基質が当該試験物質の非存在下でmPGES−1によって当該基質の9−ケト,11αヒドロキシ型である生産物に変換される条件下で接触させること、および当該試験物質の非存在下での当該生産物の対照生成レベルとの比較で当該試験物質の存在下でのPGH2またはその非酵素的分解産物(PGE2、PGD2またはPGF2α)のレベルを測定することを含み、当該試験物質の存在下での当該生産物の生成レベルが上述の対照レベルよりも低いことは、当該試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを示す。
【0178】
この方法は、通常、試験物質を、または酵素および酵素基質と共に試験物質をインキュベートすることを含む。基質は、生理学的基質、例えばPGH2であってもよいし、修飾されているかまたは非生理学的な基質、例えばその酵素反応の中で検出可能な(例えば有色の)生成物を生じるよう設計された基質であってもよい。
【0179】
mPGES−1ポリペプチドを試験物質および基質、例えばPGH2と接触させる順序は変更され得る。例えば、mPGES−1ポリペプチドは最初に試験物質と共にインキュベートされ、次いで基質と接触され得るし、その逆であってもよい。
【0180】
このようにして、試験物質の存在下での生成物の生成は、試験物質の非存在下での生成物の生成と比較され得る。生成物レベルが低いまたは生成物の形成速度が遅いことは、試験物質が酵素活性を阻害していることを示す。
【0181】
阻害剤アッセイのさらなる可能性は、mPGES−1を(生来的にまたは組み換え的に)発現する適当な細胞株によるPGE2生成に影響を及ぼす物質の能力の試験である。本発明のアッセイは、導入された一つまたはそれ以上のベクターから関連ポリペプチドまたはペプチドを発現する細胞株、例えば酵母株において実施され得る。
【0182】
それ以外のさらなるアッセイの可能性は、mPGES−1を含む不均一タンパク質調製物(ヒトでもそれ以外の哺乳動物でもよい)によるPGE2生成に影響を及ぼす物質の能力の試験である。本発明の好ましいアッセイは、単離された/精製されたmPGES−1ポリペプチド(全長mPGES−1またはその活性部分を含む)のmPGES−1活性を阻害する試験物質の能力を測定することを含む。
【0183】
mPGES−1ポリペプチドの活性を阻害する物質(すなわちmPGES−1阻害剤)のスクリーニング方法は、適当な反応媒体中で一つまたはそれ以上の試験物質と当該ポリペプチドを接触し、処理されたポリペプチドの活性を試験し、その活性を試験物質で処理していない同等の反応媒体中での当該ポリペプチドの活性と比較することを含み得る。処理されたポリペプチドと未処理のポリペプチドとの間の活性の違いは、関係する試験物質の調節効果を示唆する。
【0184】
このアッセイ法は、
(a)PGE2が正常に生成される条件の下、還元型グルタチオンおよびPGH2の存在下でmPGES−1ポリペプチドおよび試験化合物をインキュベートし、そして
(b)PGE2の生成を測定すること、
を含み得る。
【0185】
mPGES−1のPGH2基質は、COX−2およびAAのインキュベーションによって提供され得、したがってこれらがPGH2を提供するためにアッセイ媒体中に提供されてもよい。さらに、mPGES−1は環状エンドペルオキシドからの9−ケト,11αヒドロキシプロスタグランジンの立体特異的な形成を触媒するので、適当な生成物を測定することにより、他のmPGES−1基質を、mPGES−1活性およびその活性に対する試験化合物の効果の測定に使用することもできる。
【0186】
基質 生成物
PGH2 PGE2
PGH1 PGE1
PGH3 PGE3
PGG2 15(S)ヒドロペルオキシPGE2
PGG1 15(S)ヒドロペルオキシPGE1
PGG3 15(S)ヒドロペルオキシPGE3
【0187】
上記の通り、基質は、これまでに議論してきた基質のいずれかであってもよいし、任意の他の適当な基質でもよく、これは当業者の自由である。基質はPGH2であり得、その場合生成物はPGE2である。
【0188】
本発明のアッセイ法において、生成物の生成は、基質レベルを定量することにより、および/または生成物レベルを定量することにより測定することができる。アッセイ終了時にまたはアッセイ反応が終了した時点で残存する基質は、塩化鉄または塩化スズを添加することによって、それぞれ、12−ヒドロキシヘプタデカトリエン酸およびマロンジアルデヒドまたはPGF2αに変換することができる。したがって、これらの化合物の量は、PGE2の形成を間接的に反映するものである。これらの化合物の定量は、残存する基質量の定量により生成物の生成を測定する手段である。残存基質のレベルが高ければ、生成物の生成レベルが低いということである。
【0189】
mPGES−1阻害剤は、PGE2または他の生成物(使用する基質に依存する)の生成量が、試験物質を用いない対照実験と比較して減少していることを測定することによって同定され得る(またはmPGES−1阻害剤であると推定される候補物質が実際にそうであると確認され得る)。したがって、試験物質の存在下での生成物の生成は、試験物質の非存在下での生成物の生成と比較され得る。生成物レベルが低いことまたは生成物の形成速度が遅いことは、試験物質がmPGES−1活性を阻害していることを示す。このようにして、試験物質は、哺乳動物における呼吸障害の処置に使用できる薬剤であることが確認され得る。
【0190】
生成物の測定には、HPLC、UV分析、放射能測定、またはRIA(例えば、市販のPGE検出用RIAキット)を利用することができる。生成物の形成は、ガスクロマトグラフィー(GC)または質量分析(MS)、またはTLCおよび放射能走査により分析することができる。
【0191】
mPGES−1タンパク質を使用する本発明の方法において、完全(全長)mPGES−1タンパク質配列を使用する必要はない。二分子間の結合を試験するまたはPGEシンターゼ活性を試験する本発明のアッセイは、フラグメントまたは変異体を使用することができる。フラグメントは、当業者に公知の任意の適当な方法によって作製および使用され得る。適当なフラグメント作製法には、コードDNAからのフラグメントの組み換え発現が含まれるがこれに限定されない。このようなフラグメントは、コードDNAを入手し、発現される部分のいずれかの側にある適当な制限酵素認識部位を確認し、DNAからその部分を切り出すことによって作製され得る。この部分は、次いで、標準的な市販の発現系内の適当なプロモーターに機能的に連結され得る。別の組み換えアプローチは、DNAの関連部分を適当なPCRプライマーを用いて増幅することである。小フラグメント(例えば、約20または30アミノ酸未満)はまた、当該分野で周知のペプチド合成法を用いて作製され得る。mPGES−1の活性部分は、アッセイ法に使用され得る。
【0192】
mPGES−1ポリペプチドの「活性部分」は、本発明の方法において使用することができる。活性部分は、全長ポリペプチドより短いがその本来の生物学的活性を保持しているペプチドを意味する。特に、活性部分は、グルタチオンの存在下でのPGH2からのPGE2合成を触媒する能力を保持しているものである。
【0193】
EP3Rアッセイ
本発明は、試験物質、これは候補化合物または候補組成物であり得る、がEP3R阻害活性を有するかどうかを測定し、EP3R阻害活性を有することが確認された試験物質を、呼吸障害の処置に使用できる物質として同定するアッセイ法を想定している。
【0194】
いくつかの実施態様において、このアッセイ法は、EP3Rポリペプチドと試験物質およびEP3Rアゴニストを、EP3Rアゴニストが当該試験物質の非存在下でEP3Rポリペプチドを活性化する条件下で接触させること、および当該試験物質の非存在下でのEP3Rポリペプチドの活性化の対照レベルとの比較で当該試験物質の存在下でのEP3Rポリペプチドの活性化レベルを測定することを含み、当該試験物質の存在下でのEP3Rポリペプチドの活性化レベルが上述の対照レベルよりも低いことは、当該試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを示す。
【0195】
EP3Rアゴニストは、天然アゴニスト、例えばPGE2であってもよいし、合成アゴニストでもよい。多くのEP3Rアゴニストが市販、例えばBiomol社から販売されている。一つの十分に特徴付けられている例は、Sulprostoneである(http://www.caymanchem.com/app/template/Product.vm/catalog/14765を参照されたい)。EP3Rポリペプチドの活性化は、受容体タンパク質におけるコンホメーション変化であり得、それによってGタンパク質との結合がもたらされる。EP3Rポリペプチドの活性化は、アデニリルシクラーゼ活性に対する効果をモニタリングすることによって検出され得る。例えば、細胞ベースのアッセイにおいて、細胞表面に存在するEP3Rポリペプチドの活性化は、その細胞内のcAMP濃度の増加または減少をモニタリングすることによって検出され得る。
【0196】
いくつかの実施態様において、EP3Rポリペプチドは細胞表面に存在し、そこでEP3Rはレポート手段と結合する。このレポート手段は、受容体活性化の指標となる。例えば、レポート手段は、EP3Rにより媒介されるシグナル伝達経路においてEP3Rより下流の物質を含み得る。そのような下流物質のレベルにおける何らかの変化をモニタリングすることによって、EP3Rの活性化をモニタリングすることができる。レポート手段は、蛍光または放射能標識の検出を含む多くの技術のいずれかによってモニタリングされ得る。特定の実施態様において、EP3RはGタンパク質を通じてアデニリルシクラーゼに結合し、それによってcAMP生成を調節し得る。試験化合物の存在下および非存在下でEP3Rアゴニストに反応するcAMPレベルをモニタリングすることによって、EP3Rポリペプチドのアンタゴニストとして機能する試験化合物の能力が測定され得る。ヒトEP3Rの活性化は、[cAMP]iを低下させ、[Ca++]iをやや上昇させ得る。したがって、EP3Rアゴニストは、細胞内[cAMP]の低下および/または細胞内[Ca++]の上昇を誘導し得る。これは、例えば、FLIPRベースのアッセイを用いてモニタリングされ得る。EP3Rのアンタゴニストは、任意のEP3Rアゴニストにより誘導される細胞内[cAMP]の低下および/または細胞内[Ca++]の上昇を阻止または制限し得る。
【0197】
インビボスクリーニング
本発明は、哺乳動物における呼吸障害の処置に使用できる物質の同定方法を想定している。この方法は、誘導されたPGE2経路を阻害することが知られているまたは阻害すると期待される一つまたはそれ以上の物質を使用し得る。
【0198】
したがって、本発明は、哺乳動物における呼吸障害の処置に使用できる物質の同定方法であって、試験物質を試験哺乳動物に投与すること、ここで、当該試験物質は、EP3R阻害剤、mPGES−1阻害剤および/またはCOX−2選択的阻害剤である、および当該試験物質を投与しなかった対照の哺乳動物における徴候または症状との比較で、当該試験哺乳動物における呼吸障害の徴候または症状の重症度を決定することを含み、当該試験哺乳動物における呼吸障害の徴候または症状の重症度が当該対照哺乳動物のそれらよりも低いことは、当該試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを示す、方法、を想定している。
【0199】
例えば、試験物質は、
(a)COX−2により媒介されるPGH2の合成、
(b)mPGES−1により媒介される、mPGES−1の環状エンドペルオキシド基質から当該基質の9−ケト,11αヒドロキシ型である生産物への変換、および
(c)EP3Rアゴニストにより媒介されるEP3Rの活性化、
の一つまたはそれ以上を阻害する能力を有することが見出された物質であり得る。
【0200】
EP3R阻害剤、mPGES−1阻害剤またはCOX−2選択的阻害剤である試験物質の同定方法については、本明細書にさらに記載されている。試験物質をEP3R阻害剤、mPGES−1阻害剤またはCOX−2選択的阻害剤として同定することは、インビボスクリーニングよりも前の初期段階で行われ得る。このようにして、複数の化合物が、所望の薬理学的活性についてインビトロでスクリーニングされ、次いでその所望の薬理学的活性を有することが見出された化合物がインビボでスクリーニングされ得る。EP3R阻害剤、mPGES−1阻害剤およびCOX−2選択的阻害剤については本明細書にさらに詳細に記載されている。
【0201】
呼吸障害の徴候または症状には、呼吸抑制、無呼吸頻度、低酸素後の自然蘇生の脆弱さ、呼吸頻度の減少、1回換気量の減少および/または低酸素に対するあえぎ呼吸の減少が含まれ得る。徴候または症状の重症度の決定は、試験/対照哺乳動物を低酸素圧、低酸素に曝した後、および/または試験/対照哺乳動物にIL−1β、リポ多糖(LPS)またはPGE2を投与した後に、その徴候または症状を決定することを含み得る。
【0202】
本明細書で使用される場合、呼吸障害の徴候または症状の重症度が低いことは、その徴候または症状が哺乳動物に害を及ぼす可能性が低いことを意味する。例えば、本発明の方法が、IL−1β投与後の無呼吸頻度を決定することを含む場合、無呼吸頻度の少なさおよび/または無呼吸エピソードの短さが、徴候または症状の重症度の低さとみなされるであろう。
【0203】
呼吸障害の徴候または症状をモニタリングするのに適した技術については本明細書にさらに記載されている。例えば、本発明の方法は、プレチスモグラフィーまたはインピーダンスニューモグラフィーを利用し得る。この方法は、内部酸素圧を変えることができる空気管理チャンバーを利用し得る。好ましくは、このチャンバーは温度管理がなされているものである。
【0204】
あるいは、呼吸障害の徴候または症状の決定は、脳幹の呼吸活動を、例えば試験/対照哺乳動物から単離された脳幹・脊髄調製物を用いてモニタリングすることを含み得る。脳幹呼吸活動は電極を用いてモニタリングされ得るが、これについては本明細書にさらに記載されている。本発明の方法が試験/対照哺乳動物から単離された脳幹・脊髄調製物を用いて脳幹呼吸活動をモニタリングすることを含む場合、試験物質は、脳幹・脊髄の単離前に投与されてもよいし、試験/対照哺乳動物からの単離後に脳幹・脊髄調製物に対して直接投与されてもよい。
【0205】
本発明の方法は、エクスビボ脳幹脊髄全調製物または脳幹スライス調製物を利用し得る。このような調製物は、アゴニストおよび/またはアンタゴニスト、例えば誘導されたPGE2経路のアゴニストおよび/またはアンタゴニスト、ならびに環境変化の細胞、ネットワークおよび行動への効果の平行モニタリングを実現する。この方法は、本明細書で定義されるようなインサイチュおよびインビボ法と組み合わせることができる。無呼吸の誘導は、環境変化、例えばO2濃度の低下、例えば低酸素によって達成され得る。あるいはまたは加えて、無呼吸の誘導は、薬学的または麻酔処置、例えばオピオイド受容体アゴニストおよび/またはフォースコリンを含むcAMP濃度上昇薬によって達成され得る。
【0206】
試験哺乳動物および対照哺乳動物はげっ歯類であり得、各々好ましくはマウスまたはラットである。本発明の方法は、好ましくは、ヒトにおける呼吸障害の処置に使用できる薬剤を同定する方法である。
【0207】
本発明の方法は、バロメーター技術またはフロープレチスモグラフィー技術を用いて呼吸障害の徴候または症状の重症度を決定することを含み得る。このような技術は、試験哺乳動物および対照哺乳動物がげっ歯類、例えばマウスまたはラットである場合に好ましい。特定の実施態様において、試験哺乳動物はヒトである。このような場合、呼吸障害の徴候または症状の重症度の決定は、睡眠ポリグラフ記録法の使用を含み得る。
【0208】
試験哺乳動物および対照哺乳動物は、好ましくは、対照動物に試験物質を与えない点を除いて同一条件下に置かれる。好ましくは、対照投与、例えば生理食塩水が対照哺乳動物に与えられ、そしてそれは、好ましくは、試験物質を試験哺乳動物に投与するのと同一経路によって対照哺乳動物に投与される。
【0209】
特定の実施態様において、試験哺乳動物および対照哺乳動物は、同じ動物であり得る。この場合、試験物質を投与されなかった対照哺乳動物における徴候または症状との比較で試験哺乳動物における呼吸障害の徴候または症状の重症度を決定することは、まず、試験物質の投与前に哺乳動物における呼吸障害の徴候または症状の重症度を決定し(「対照記録」)、次に、試験物質の投与後に哺乳動物における呼吸障害の徴候または症状の重症度を決定する(「試験記録」)ことにより実行され得る。次いで、対照記録および試験記録は比較され、対照記録の重症度よりも試験記録の重症度が低いことが、試験物質が哺乳動物における呼吸障害の処置に使用できる物質であることを示す。同じ動物を試験哺乳動物および対照哺乳動物として使用することは、哺乳動物がヒト、例えば臨床研究下にあるヒトである場合に好ましい。
【0210】
以下は実施例であり、特許請求の範囲を制限するものと解釈されるべきではない。
【実施例】
【0211】
材料および方法
動物
DBA/1lacJ純系(n=158)(Jackson Laboratory, Bar Harbor, ME)およびC57BL/6系(n=75)(Beverly Koller博士、University of North Carolina, Chapel Hill, NCの厚意により寄贈された)の新生仔を使用した。ミクロソームプロスタグランジンEシンターゼ1(mPGES−1)およびEP3受容体(EP3R)の遺伝子を以前に記載されたようにしてノックアウトマウスにおいて選択的に欠失させた(47、48、両方の開示内容を参照により本明細書で援用することを明記する)。全ての動物を実験後直ちに断頭により屠殺し、PCRおよびサザンブロット分析を用いてジェノタイピングを行った。野生型DBA/1lacJマウスの一部から得たデータを、新生仔DBA/1lacJマウスにおける呼吸活動の特徴付けに利用した(6)。全てのマウスを、12時間明:12時間暗サイクルという標準化された条件下で飼育した。餌および水分は自由にとらせた。
【0212】
ヒト被験者
カロリンスカ大学病院の新生児集中治療室の乳児(平均在胎齢32±2週)に協力していただいた(生後年齢平均16±4日)(n=12)。その乳児のうち、臨床適応のための腰椎穿刺を受け、書面上のインフォームドコンセントが得られたものを適格者とした。これらの研究は、欧州共同体のガイドラインに従って行ない、内部の倫理委員会による承認を受けた。乳児のうち、臨床適応、例えば感染の疑い、神経学的変化および心拍呼吸上の問題について腰椎穿刺を受けたものを実験適格者とした。脳室内出血(等級≧2)、白質病変(PVL、脳室周囲白質軟化症)、発作、出血後水頭症または先天性異常が見られた乳児は除外した。新生児分娩データ、医学的状態、感染マーカー、呼吸治療および投薬を含む関連医学情報(pertinent medical information)を記録した。心拍呼吸の記録は、腰椎穿刺後12時間以内に行った(平均4.8±1.7時間)。
【0213】
薬物
組換えマウスインターロイキン1β(IL−1β)(Nordic Biosite AB, Taby, Sweden)を滅菌したNaClで再構成し、1μg/ml溶液とした。プロスタグランジンE2(PGE2)(Cayman Chemicals, Ann Arbor, MI,USA)は、合成CSF(aCSF)に希釈して、インビボ実験用に2nmol/μl、インビトロ実験用に20μg/l(60nM)の濃度とした。
【0214】
非拘束全身フロープレチスモグラフィー
プレシキグラスチャンバー(35ml)を高感度ダイレクトエアフローセンサー(0〜200ml/分;TRN3100、 Kent Scientific Corporation, Litchfield, CT, USA)に接続した。フローシグナルは4チャンネル増幅器(P/N 770 S/N 5; SENSElab, Somedic Sales, Horby, Sweden)によって増幅し、デジタルデータに変換し、DasyLabソフトウェア(Datalog GmbH & Co.KG, Monchengladbach, Germany)を用いてオンラインコンピュータにより100Hzで記録した。呼吸頻度(fR、呼吸/分)1回換気量(VT、μl/呼吸)および毎分換気量(VE、μl/分)を算出した。チャンバー温度は、チャンバーをサーモスタット付水浴に沈めることによって、記録されている新生仔マウスの温熱中間範囲に従い30.1±0.1℃で維持した(49)。以前に記述されているように、プリセット高精度シリンジ(Hamilton Bonaduz AG, Switzerland)を用いて標準化された量の空気(5〜200μl)を繰り返し注入することによってチャンバーをキャリブレートした(6)。ガス交換の95%が投与の35秒以内に行われたことを、CO2濃度分析(Metek CD-3AおよびS-3A, PA, USA)によって確認した。
【0215】
インピーダンスニューモグラフィー
乳児の心拍呼吸活動を、インピーダンスニューモグラフィーを用いて非侵襲的に測定し、イベントモニタリングシステム(KIDS, Hoffrichter GmbH, Schwerin, Germany)を介して記録した。ベースライン呼吸速度および無呼吸閾値を超えるイベントを記録するようモニターをプログラムした。無呼吸は、直前の0.5秒間における平均インピーダンス信号振幅の≧10秒の減少から直前の25秒間に測定される平均振幅の16%未満の減少と定義した。このイベント前後の60秒間もモニターのメモリに保存した。
【0216】
IL−1βまたはNaClのi.p.注射後のプレチスモグラフィー
mPGES−1およびEP3Rを様々に発現する、それぞれ、9日齢DBA/1lacJマウス(n=143)およびC57BL/6マウス(n=16)における呼吸を、フロープレチスモグラフィーを用いて試験した。各マウスに、IL−1β(10μg/kg)またはビヒクルの腹腔内注射(0.01ml/g)を行った。70分後に、マウスを、プレスチモグラフチャンバーに誘導した。4分間の正常酸素(21% O2)およびその後の1分間の過酸素(100% O2)の間の呼吸を評価した。正常酸素下での5分間の回復期間の後、無酸素(100% N2)に対する呼吸応答を試驗した。最後に、100% O2を8分間与え、自然蘇生の能力を評価した。ベースライン、70分、およびチャンバーから出た後の皮膚温度を記録した。直腸温度は、直腸プローブの設置が呼吸行動を変化せる可能性を考慮して測定しなかった。性別を推定するために肛門生殖突起間距離を測定した。
【0217】
PGE2またはビヒクルのicv注射後のプレチスモグラフィー
EP3Rを様々に発現する、9日齢C57BL/6マウス(n=38)における呼吸を、フロープレチスモグラフィーを用いて試験した。約60秒間のセボフルラン麻酔投与を行った後、PGE2(4nmol、2〜4μl aCSF)またはビヒクルを、ポリエチレンチューブに取り付けた先細ガラスピペットを用いて側脳室にゆっくりと注入した。次いでマウスを直ちにプレスチモグラフチャンバーに入れた。10分間の正常酸素下での回復期間の後、マウスを前記のような過酸素および無酸素チャレンジに暴露した。ベースラインおよびその後の各時点での動物の皮膚温度を、サーミスタ温度プローブを用いて記録した。
【0218】
脳幹呼吸活動
脳幹脊髄標本は、以前に記載されるようにして(50、51、両方の開示内容を参照により本明細書で援用することを明記する)、EP3R+/+およびEP3R-/-の遺伝型を有する2日齢C57BL/6マウス(n=11)から素早く単離した。C4前根における吸息律動に対応する呼吸関連活動を、ガラス吸引電極を用いて追跡し、記録し(5kHz)、オフラインで分析した。対照の記録は、PGE2を含有するaCSFの灌流およびその後のaCSF洗浄前少なくとも20分間行った。
【0219】
mPGES−1活性の測定
新生仔マウスの脳(n=33)を、0.25Mスクロース、1×コンプリートプロテアーゼインヒビター(Roche Diagnostics)および1mM還元型グルタチオンを含有する0.1M KPi(無機リン酸カリウム)緩衝液中でホモジナイズし、その後に超音波処置した。膜画分は、細胞成分分画法によって単離した。膜画分におけるmPGES−1活性は、以前に記載されたようにして測定した(52、その開示内容を参照により本明細書で援用することを明記する)。
【0220】
免疫組織化学
9日齢野生型およびEP3Rノックアウト仔から脳幹を断頭後に素早く切り出し、4%パラホルモアルデヒドで固定し、15%スクロース含有リン酸緩衝化生理食塩水(PBS)、pH7.4中で一晩凍結保護した。次いでこの脳幹を素早く凍結し、14μmの横断切片をクリオスタット(Leica CM3050 S, Leica Microsystems Nussloch GmbH)に逐次集めた。切片を風乾し、PBSで再度水和させ、そして0.3%過酸化水素を使用して内因性ペルオキシダーゼを10分間阻害した。その後のPBS洗浄の後、この切片を5%ヤギ血清(Jackson Immunoresearch Laboratories, West Grove, PA)、1%ウシ血清アルブミン(Sigma-Aldrich)および0.3% Triton X−100(Sigma-Aldrich)を含有するPBS中、45分間ブロックおよび透過処理を行い、ウサギNK−1R抗体(1:20,000希釈;Sigma-Aldrich)と共に一晩インキュベートした。次いでこの切片をPBS中で洗浄し、1:50希釈のビオチニル化二次抗体(ヤギ抗ウサギ;Vector Laboratories, Burlingame, CA)と共にインキュベートした。1時間のインキュベーション後、切片をリンスし、ペルオキシダーゼ結合型Vectastain ABC(1:100希釈;Vector Laboratories)と30分間、次いでCy3結合型Tyramideシグナル増幅液(TSA、1:50、PerkinElmer, Boston, MA)と2分間インキュベートした。反応をPBS中停止させ、5%ロバ血清(Jackson)、1%ウシ血清アルブミン(Sigma-Aldrich)および0.3% Triton X−100(Sigma-Aldrich)を含有するPBSで45分間ブロックした。次いで切片を、1:50希釈のウサギEP3R抗体(Cayman Chemical, MI)と共に4℃(C)で一晩インキュベートした。次の日、切片をPBS中でリンスし、Alexa488結合型二次抗体(ロバ抗ウサギ;Molecular probes)と共に1時間インキュベートした。その後のPBS洗浄の後、切片をVectashield Hard Setマウンティングメディウム(Vector Laboratories)にマウントした。潜在的な交差反応の危険性を除去するため、一次抗体を滴定して最適な希釈を決め、各一次抗体が除かれた対照スライドをインキュベートした。さらに、EP3Rノックアウトマウス(n=4)由来の脳幹スライスを、通常のNK1R染色による上記のプロトコルを用いて研究したが、EP3Rは検出されなかった。画像は、ImageJソフトウェア(NIH, Bethesda, MD)で処理した。
【0221】
CSF分析および心拍呼吸の記録
脳脊髄液サンプルを、標準化された酵素免疫アッセイ(EIA)プロトコル(Cayman Chemicals, Ann Arbor, MI, USA)を用いて、PGE2およびPGE2代謝産物について分析した。腰椎穿刺の後できるだけ早く乳児の心拍呼吸の記録を行った(平均記録時間:9.2±2.4時間)。腰椎穿刺前12時間以内に測定した感染マーカー(例えば、C反応性タンパク質、白血球)の血中濃度も記録した。
【0222】
プレチスモグラフィーデータの分析
動きアーチファクトを伴わない平穏状態下呼吸の期間を分析に使用した。正常酸素および過酸素ならびに無酸素応答(すなわち、過呼吸、一次無呼吸、あえぎ呼吸、二次無呼吸および自然蘇生)の平均fR、VTおよびVE値を以前に記載されているようにして分析した(6、その開示内容を参照により本明細書で援用することを明記する)。全ての動物について生存を記録した。無呼吸は≧3回の呼吸サイクル分の呼吸停止と定義した。呼吸の規則性は、変動係数(C.V.)(すなわち、SDを、60秒の間の呼吸と呼吸の間隔の平均で割ったもの)を用いて定量した。
【0223】
乳児の心拍呼吸データの分析
モニタリングソフトウェアを使用してベースライン呼吸速度を記録し、全ての心拍呼吸イベントを可視化した。無呼吸インデックス(A.I.、無呼吸数/記録時間)を決定した。心拍呼吸活動、感染状況およびCSF中のPGE2レベルの間の相関関係を評価した。全ての動きアーチファクトを分析から除外した。
【0224】
脳幹脊髄標本
脳幹は、橋が除去されるように第VI脳神経根と台形体の下境界部の間で吻側から取り出した。この標本を、合成脳脊髄液(aCSF):130mM NaCl、3.3mM KCl、0.8mM KH2PO4、0.8mM CaCl2、1.0mM MgCl2、26mM NaHCO3および30mM D−グルコースを入れた1.5mlチャンバー中、28℃で連続的に灌流させた(流速3〜4ml/分)。この溶液を95%O2および5%CO2で連続的に平衡化してpH7.4とした(50、51)。
【0225】
プレスチモグラフデータの分析
無酸素に対する応答は年齢によって変わるので(53)、我々はP9齢の時にすべての記録をとるようにした;しかし、問題となる年齢による影響を最小限に抑えるため、年齢の関連値として体重を使用し、集団平均体重の1SD以内の体重を有する動物のみ無酸素および生存分析に含めた(6)。
【0226】
動物の特徴
IL−1βまたはNaClのi.p.注射後のプレチスモグラフィー実験において、mPGES−1+/+マウスはmPGES−1-/-マウスよりも体重が軽かった(それぞれ、4.4±0.1g vs 4.9±0.1g)。動物の性別に違いはなかった。ベースラインにおける動物の皮膚温度(34.7±0.1℃)および注射70分後の温度(34.8±0.1℃)はグループ間で類似していた。無酸素後、mPGES−1+/+マウスはmPGES−1-/-マウスよりも皮膚温度が高かった(それぞれ、32.2±0.1℃ vs 31.4±0.2℃)。C57BL/6マウスにおいて、動物の体重(4.5±0.1g)、動物の性別、ベースライン温度(34.4±0.2℃)、70分の温度(34.5±0.5℃)または無酸素後の温度(30.4±0.1℃)に違いはなかった。PGE2またはビヒクルのicv注射後のプレチスモグラフィー実験において、C57BL/6マウスは、動物の性別および麻酔後温度(31.0±0.2℃)において違いを示さなかった。しかし、EP3R+/+マウスは、EP3R-/-マウスよりも体重が重かった(それぞれ、4.9±0.1g vs 4.1±0.1g)。9日齢のEP3R+/+マウス(n=13)およびEP3R-/-マウス(n=26)の皮膚温度を、ベースラインならびにPGE2またはビヒクルのicv注射後の正常酸素、過酸素および無酸素下での各時点で測定した。注射23分後の無酸素暴露まで温度の違いはみられなかった。この時点で、EP3R-/-マウスは、EP3R+/+マウスよりも皮膚温度が低かった(それぞれ、30.9±0.3℃ vs 31.8±0.3℃)。温度類似性は、30〜31分の無酸素後期間で異なった(それぞれ、29.8±0.2℃ vs 30.4±0.1℃)。
【0227】
統計
1要因のANOVAにより、正規分布および等分散のパラメータを比較した。スチューデントt post−hoc検定を用いて多重比較を行った。ノンパラメトリックな測定および非ガウス分布型のデータについては、ウィルコクソンχ2テストを使用した。変数の経時的変化は、MANOVA反復測定デザインを使用して試験した。スピアマンのRho相関テストにより変数間の相関関係を決定した。データは平均±SEMで示されている。P<0.05の値を統計的に有意であるとした。
【0228】
実施例1:内因性の脳幹mPGES−1活性および呼吸強壮効果
我々はまず、9日齢のmPGES−1+/+マウスおよびmPGES−1-/-マウスにおけ
る内因的なPGE2産生およびその換気に対する効果を試験した。野生型マウスの示した基準となるミクロソームプロスタグランジンEシンターゼ1(mPGES−1)活性は、ホモジナイズした皮質よりもホモジナイズした脳幹において高かった(図1)。正常酸素下での呼吸は遺伝子型間で類似したが、fRはmPGES−1-/-マウスよりもmPGES−1+/+マウスの方が低い傾向があった(クラスカル・ウォリス、P=0.03;スチューデントt post−hoc検定、P=0.18)(表1)。中枢呼吸の駆動を、1分間の過酸素チャレンジ(100% O2、1分間)によって試験した。両方の遺伝子型のマウスは、呼吸頻度(fR)を減らすことで、過酸素に対して応答した(図2)。しかし、呼吸抑制は、mPGES−1+/+マウスの方が、mPGES−1-/-マウスよりも大きかった(それぞれ、27±2% vs 19±3%)。
【0229】
【表1】
【0230】
正常酸素および過酸素(100% O2)下での呼吸頻度(fR、呼吸/分)、1回換気量(VT、μl/呼吸/g)および毎分換気量(VE、μl/分/g)を、IL−1βまたはビヒクルの腹腔内注射後の9日齢mPGES−1+/+マウスおよびmPGES−1-/-マウスにおいて試験した。各遺伝子型内での処置の効果を比較すると、IL−1βは、mPGES−1+/+マウスにおいて基準fRを減らす傾向があり(ウィルコクソンχ2、P=0.17)、それはmPGES−1-/-マウスではみられなかった。全てのマウスは、fRを減らすことで過酸素に応答した。IL−1βは、mPGES−1+/+マウスにおける過酸素下でfRを抑制したが、この効果は、mPGES−1-/-マウスではみられなかった。mPGES−1+/+マウスは、mPGES−1-/-マウスと比較して、過酸素下での呼吸抑制の程度が大きかった。データは平均±SEMで示されている。*P<0.05、#P<0.05(体重で正規化した場合)。
【0231】
この結果は、mPGES−1活性の内因的な発現、特に脳幹における発現を実証している。mPGES−1は、主として、血液脳関門(BBB)付近の内皮細胞によって発現される(25)。重要な呼吸中枢付近の、脳幹を覆う内皮細胞におけるmPGES−1の構成的および急速誘導性発現は、呼吸の調節におけるPGE2の重要な役割を示唆している。mPGES−1を欠くマウスよりも野生型マウスにおいて過酸素下で有意な呼吸抑制がみられたこともまた、内因的なPGE2が周産期における呼吸律動形成に対して強壮効果を有することの証拠を与える。
【0232】
これまでの研究は、内因的なプロスタグランジン産生を遮断するプロスタグランジン合成阻害剤が、産後初期に胎児の呼吸動作および中枢呼吸を増加させることを報告している(26〜28)。プロスタグランジンの調節効果には、周産期の換気の初期阻害を含む発生学的変化が見られるが(18、26、27、29)、その後年齢の増加と共に呼吸における変化は小さくなる(19)。しかし、PGE2は、加齢後にも無呼吸の誘導により通常の呼吸を妨害する可能性がある(19)。EP3Rの遺伝子およびタンパク質はげっ歯類成体のRVLMにおいても発現されるが(20、21、30)、発生学的変化は、周産期にとどまらない脳幹でのPGE2受容体発現の変化の副産物的なものである可能性がある。さらに、プロスタグランジンの結合密度は減少するものの、これは全年齢にわたって同じ脳幹領域に局在する(31)。EP3R発現の個体発生(ontogenesis)およびPGE2の呼吸効果における潜在的な発生学的変化の根底にあるメカニズム、例えばEP3Rの翻訳後修飾、橋上側(suprapontine)への影響、のさらなる調査が必要とされる。
【0233】
実施例2:IL−1βおよび無酸素はマウス脳幹におけるmPGES−1活性を誘導する
我々は、9日齢mPGES−1+/+、mPGES−1-/-およびEP3R+/+マウスのホモジナイズした脳幹および皮質におけるmPGES−1活性に対する、IL−1βおよび短期間の無酸素暴露(100% N2、5分間)の効果も測定した(図1)。IL−1βは、時間依存的なmPGES−1活性の増加を、特に脳幹において誘導した。特に、脳幹のmPGES−1活性は、IL−1β投与90分後および180分後にそれぞれ2倍および4倍増加したが、皮質における活性は90分と180分の間で変化しなかった。無酸素暴露もまた、脳幹および皮質の両方においてmPGES−1活性を誘導した。注目すべきは、mPGES−1活性に対するIL−1βおよび短期間の無酸素暴露の相加的効果であり、これは脳幹においてより顕著であった。EP3R野生型マウスは、IL−1β投与90分後に、mPGES−1野生型マウスと類似のmPGES−1活性を示した。さらに、EP3Rマウスはまた、脳幹において、皮質よりも高いmPGES−1活性を有していた(それぞれ、PGE2:1111±49および710±44pmol/分/mpタンパク質)。
【0234】
PGE2は、無酸素に応答する呼吸においても重要な役割を果たすようである。短期間の無酸素暴露は、ホモジナイズしたマウスの脳においてmPGES−1活性を増加させた。このインビボでのmPGES−1活性の迅速な増加は新しい知見である。これまでの研究は、無酸素が、エクスビボでのマウス皮質片におけるPGE2産生およびブタの脳におけるプロスタグランジンHシンターゼ2 mRNAの発現を誘導することを示していた(32、33)。一過性の仮死も同様に、新生仔モルモットの脳においてPGE2濃度を増加させ、この効果はインドメタシン処置によって阻害される(34)。
【0235】
既知のmPGES−1酵素調節メカニズムはいずれも、本願において実証されたmPGES−1活性の迅速な変化を説明できるものではない。遺伝子発現の誘導は、このような短期間の無酸素イベントの間に起こりそうにない。しかし、構成的に発現されるmPGES−1の転写後調節、例えばリン酸化は、考えられる原因であろう。それ以外の可能性は、以前にヒト細胞系においてCOX−2 mRNAを用いて示され(35)、最近では心筋細胞において示されたような(36)、mPGES−1 mRNAの安定化である。基礎をなすメカニズムを明らかにするためさらなる研究が必要である。
【0236】
実施例3:IL−1βは、mPGES−1+/+マウスにおいて呼吸を抑制するが、この効果はmPGES−1-/-またはEP3R-/-マウスにおいてはみられない
IL−1βの換気効果を仲介するPGE2の役割を試験するため、我々は、9日齢のmPGES−1+/+、mPGES−1-/-およびEP3R-/-マウスにおいて、IL−1βまたはビヒクルをi.p.投与した後の正常酸素および過酸素(100% O2、1分間)下での呼吸を、フロープレチスモグラフィーを用いて分析した(図2、表1)。全てのマ
ウスが、処置に関係なく、fRを減らすことで過酸素チャレンジに対して応答したが、IL−1β処置した野生型マウスは、ビヒクル処置した野生型マウスよりも大きな呼吸抑制を示した。IL−1βはまた、mPGES−1+/+マウスにおいて基準fRを減少させる傾向があった(クラスカル・ウォリス、P=0.03;スチューデントt post−hoc検定、P=0.17)。逆に、IL−1βは、mPGES−1-/-またはEP3R-/-マウスにおいて正常酸素または過酸素下で換気を変化させなかった。
【0237】
この結果は、IL−1βが中枢呼吸を抑制するのにmPGES−1の活性化が必要とされることを指示している。第一に、IL−1βは、時間依存的な様式で脳幹のmPGES−1活性を増加させた。第二に、IL−1βは、mPGES−1+/+マウスにおいて呼吸を抑制したが、mPGES−1-/-マウスにおいては抑制しなかった。インドメタシンは、プロスタグランジン合成を遮断することによって、基準呼吸に対するIL−1βの効果を同じように軽減することが示されている(5)。
【0238】
実施例4:IL−1βは野生型マウスの無酸素下生存率を悪化させるが、この効果はmPGES−1またはEP3Rを欠くマウスにおいてはみられない
次に、我々は、IL−1βが、PGE2を仲介するメカニズムを通じて、低酸素換気応答および低酸素性無呼吸後の自然蘇生に影響するかどうかを調査した。フロープレチスモグラフィーを用いて、無酸素(100% N2、5分間)およびその後の過酸素(100% O2、8分間)下での呼吸を、mPGES−1+/+、mPGES−1-/-およびEP3R-/-マウスへのIL−1βまたはビヒクルのi.p.注射の80分後から試験した(図3、表2)。全てのマウスは、初期の換気亢進(すなわち、過呼吸)およびそれに続く低酸素型換気抑制(すなわち、一次無呼吸、あえぎ呼吸、二次無呼吸)という、無酸素に対する二相性の応答を示した。IL−1βは、mPGES−1+/+マウスにおいてあえぎ数を減少させたが、この効果はmPGES−1-/-マウスにおいてはみられなかった。IL−1β処置したmPGES−1+/+マウスはまた、IL−1β処置したmPGES−1-/-マウスよりもあえぎの持続時間が短い傾向があった(クラスカル・ウォリス、P=0.19;スチューデントt post−hoc検定、P=0.003)。あえぎ数の少なさおよびあえぎ持続時間の短さは、無酸素下生存率の低下と相関していた。IL−1βは、mPGES−1+/+マウスにおいて無酸素下生存率を有意に低下させたが、mPGES−1またはEP3R遺伝子を欠くマウスの生存率は低下しなかった。IL−1βは、EP3R-/-マウスの低酸素型換気応答に影響しなかった。
【0239】
【表2】
【0240】
ミクロソームプロスタグランジンEシンターゼ1(mPGES−1)の発現が様々な新生仔マウスにIL−1βまたはビヒクルを末梢投与してから80分後に無酸素に暴露した。マウスは、過呼吸期にfR、VTおよびVEの初期増加およびおよびそれに続く低酸素型換気抑制期にあえぎ応答を示した。各遺伝子型内で処置効果を比較すると、IL−1βは野生型マウスにおいてあえぎ数を減少させたが、この効果はmPGES−1の発現が少ないマウスにおいては観察されなかった。データは平均±S.E.M.で示されている。**P<0.01
【0241】
この研究は、PGE2が、IL−1βの無酸素下換気効果を仲介する上でも重要な役割を果たしていることを実証している。IL−1βは、野生型マウスにおいて低酸素無呼吸後の自然蘇生を阻害したが、この効果はmPGES−1またはEP3Rを欠くマウスにおいてはみられなかった。これまでの研究は、インドメタシンが、新生仔ラットにおける低酸素下でのあえぎ呼吸および無酸素下生存率に対するIL−1βの有害な効果を軽減することを示している(5)。
【0242】
実施例5:PGE2は脳幹の呼吸関連活動を低下させ、EP3Rを通じて無呼吸を誘発する
PGE2が脳幹EP3受容体に特異的に結合することにより呼吸を抑制するかどうかをより正確に把握するために、人工脳脊髄液またはPGE2を投与した2〜3日齢のEP3R+/+およびEP3R-/-マウスの脳幹脊髄のひとかたまりの標本を用いて中枢呼吸活動を測定した。対照条件下では、同様の呼吸活動をEP3R+/+およびEP3R-/-マウス由来の標本において記録した。しかし、PGE2はEP3R+/+標本における呼吸頻度を可逆的に阻害したが、EP3R-/-標本に対しては影響しなかった(図4)。
【0243】
EP3Rを通じて呼吸を変化させるPGE2の能力を、フロープレチスモグラフィーを用いてさらに評価した。EP3R+/+およびEP3R-/-マウスにおけるPGE2またはビヒクルのicv注射後、正常酸素および過酸素下での呼吸を分析した(図4および表3)。PGE2は、EP3R+/+マウスにおいて、正常酸素および過酸素下で有意に多い無呼吸頻度および不規則な呼吸パターンを誘発したが、この効果はEP3R-/-マウスにおいてはみられなかった。次に、このマウスを、無酸素およびそれに続いき過酸素に曝し、自然蘇生を試みさせた。全てのマウスは5分間の無酸素暴露期間を超えてあえぎ呼吸を継続し、38匹のうちの一匹のみが自然蘇生に失敗した(PGE2処置したEP3R-/-マウス)。PGE2は、ビヒクルと比較して、EP3R+/+またはEP3R-/-マウスのあえぎ応答または無酸素下生存率を変化させなかった。最後に、我々は、吻側延髄腹外側(RVLM)の呼吸関連ニューロンがEP3Rを発現しているかどうかを調査した。特に、疑核の腹側に位置しプレベッツィンガー複合体を含むRVLMに局在する呼吸関連ニューロンを同定するツールとしてNK1Rの免疫標識を使用した(22〜24)。我々は、これらのニューロンが、NK1RおよびEP3Rを共発現していることを証明した(図4)。
【0244】
【表3】
【0245】
正常酸素、過酸素(100% O2)および無酸素(100% N2)下での呼吸頻度(fR、呼吸/分)、1回換気量(VT、μl/呼吸/g)および毎分換気量(VE、μl/呼吸/g)を、PGE2またはビヒクルの脳室内(icv)注射後の9日齢EP3R+/+マウス(n=13)およびEP3R-/-マウス(n=25)において試験した。各遺伝子型内で処置効果を比較すると、PGE2は、EP3R+/+マウスにおいて正常酸素および過酸素下でのfRを有意に抑制したが、この効果はEP3R-/-マウスにおいてはみられなかった。PGE2はまた、EP3R+/+マウスにおいて過酸素下でのfRを減少させる傾向があった(ANOVA、p=0.11)が、この傾向はEP3R-/-マウスにおいてはみられなかった。データは平均±SEMで示されている。*p<0.05、**p<0.01
【0246】
先の実施例において示された結果は、mPGES−1の活性化後、新たに合成されるPGE2がIL−1βの中枢呼吸作用を発揮させることの証拠を与えている。我々は、本願において、PGE2が野生型マウスにおける呼吸を妨げることを証明しているが、これは、PGE2が動物胎仔および新生仔における呼吸を抑制することを実証する研究と矛盾するものではない(18、29、37)。さらに、PGE2はインビボで末梢での化学的感受性を変化させず、インビトロで脳幹呼吸活動を直接的に阻害することから、これらの効果は中枢において生じていると言える。これまでの研究は、PGE2が、新生仔ラットにおける呼吸関連ニューロンを阻害し(5)、かつ頸動脈洞および迷走神経の疑似手術または除神経後のヒツジにおける胎仔の呼吸動作も同様に阻害することを示している(38)。
【0247】
さらに、このPGE2の調節効果は、脳幹EP3受容体への結合を通じて起こる。IL−1βは、EP3R-/-マウスにおける呼吸を変化させることができなかった。PGE2は、EP3R+/+マウスにおいてインビボで無呼吸および不規則な呼吸を誘発したが、この効果はEP3R-/-マウスにおいてはみられなかった。最後に、EP3受容体の存在は、インビトロでの脳幹の呼吸関連律動活動を阻害するのに必要であった。特定のプロスタグランジン受容体であるサブタイプEP3RはNTSおよびRVLMに局在するが(20、21)、プロスタグランジンの呼吸への効果がこれらの受容体における挙動を通じて生じることおよびこれらが呼吸関連ニューロンにおいて発現されることを示す先行技術は存在しない。
【0248】
この実施例の結果は、IL−1βにより誘導されるPGE2および低酸素が、プレベッツィンガー複合体(preBotC)を含むRVLMにおける呼吸関連ニューロンを、EP3Rを通じて選択的に調節することを示唆している。PGE1を含む他の神経モジュレーターが、preBotCニューロンを阻害し、呼吸関連律動を弛緩させることが示されており(22、23)、preBotCの病変は、無酸素下でのあえぎ呼吸を妨げ、中枢性無呼吸および運動失調呼吸を惹起する可能性がある(39、40)。さらに、これらの呼吸関連ニューロンは最近、低酸素に対する十分な応答、あえぎ呼吸および自然蘇生による脳幹のホメオスタシスの維持、したがって酸素レベルの回復に重要であることが示されている(41)。この生きている脳幹の神経網のPGE2誘導性の抑制、例えば感染反応時抑制は、あえぎ呼吸および自然蘇生の不全ならびに究極的には死を引き起こし得る。
【0249】
実施例6:中枢PGE2濃度はヒト乳児における無呼吸頻度と相関する
ヒト新生児における感染と無呼吸の間の関係の根底にあるメカニズムをさらに解明するため、我々は、新生児において、感染マーカーC反応性タンパク質(CRP)、脳脊髄液PGE2レベルおよび無呼吸イベントの間の関係を試験した。CRPは、中枢PGE2と正の相関をし、CSF中のPGE2濃度と無呼吸頻度の間には正の関係があった(図5)。
【0250】
無呼吸は、新生児における敗血症の一般的にみられる徴候であるが(1)、この関係の基礎となるメカニズムは未だ解明されていない。我々は、本願において、感染マーカーCRPがヒト新生児のCSFにおけるPGE2レベルの上昇と相関関係にあることを証明する。重要なことは、我々は、PGE2が無呼吸頻度の増加と関連することも実証していることである。これらの発見は、ヒト新生児において、感染が、サイトカインの全身放出ならびにその後のPGE2の生合成および中枢作用を通じて呼吸を抑制することを示唆している。本明細書に記載されるメカニズムは、睡眠時無呼吸を患った小児におけるCRPレベルと無呼吸/呼吸低下インデックスの間の独立的関係(42)およびヒト乳児の咽頭分泌物中のIL−1β濃度と無呼吸の臨床的重症度との間の正の相関(8)を示す以前の報告を説明することができる。一過性の無呼吸もまた、ヒト新生児におけるプロスタグランジン処置の一般的な副作用であり(43)、これは脳幹の呼吸中枢におけるEP3受容体の活性化に起因するものであり得る。さらに、我々のデータは、新生児における中枢性無呼吸と尿中PGE代謝産物の間の正の相関(44)を説明するものである。
【0251】
炎症メディエーターは、新生児において感染および仮死を検出する上で重要なマーカーとして提案されている。サイトカインおよび低酸素刺激に対するPGE2の迅速な合成は、感染または仮死に起因する可能性のある無呼吸が増加している乳児の診断およびサーベイランスに利用できる。PGE2モニタリングの潜在的な診断的利益を、他の感染マーカー、例えばCRPとの比較で評価する研究が必要とされる。
【0252】
この結果は、IL−1βの有害な効果がmPGES−1およびEP3R遺伝子の選択的欠失によって軽減されたというものであり、感染に関連する新生児無呼吸の処置に関する重要な示唆を含んでいる。これまで、未熟児の無呼吸を処置するのにインドメタシンが使用されていた(45)。しかし、インドメタシンは新生児において多数の副作用を引き起こし(46)、したがって、mPGES−1またはEP3受容体を選択的に標的にする処置様式は有用性が高いと考えられる。
【0253】
先の実施例は、全身性のインターロイキン1βが、脳幹の呼吸関連領域におけるmPGES−1の活性化およびEP3受容体へのPGE2の結合を介して呼吸および自然蘇生を抑制することを実証している(図6)。さらに、重度の低酸素はmPGES−1活性を迅速に誘導し、これにより内因的なPGE2が新生児期の低酸素条件下で脳幹の呼吸ニューロンを調節し得ることが示唆されている。最後に、ヒト新生児における感染、中枢PGE2および無呼吸イベントの間の相関が明らかにされた。
【0254】
実施例7:PGE2の代謝産物と出生時仮死およびHIEの程度に対する相関
本発明者らは、ヒト乳児における周産期仮死がPGE2の迅速な放出および神経学的損傷を引き起こすという仮説を調査した。
患者
ストックホルムのカロリンスカ病院で処置をうけている63名の乳児(妊娠>37週)を、1999年10月から2004年9月の間、親の同意を得て本研究に登録した。43名の乳児が出生時仮死に関する以下の基準を充足した:1)後期の減速というカルディオトコグラフィーパターン、変動の不存在または徐脈、羊水の胎便染色、頭皮のpH<7.2または乳酸>4.8mmolによって示される胎児仮死の徴候、2)5分後のアプガールスコア<6および分娩室で>3分間の蘇生を行う必要性または生後60分以内に採取した患者由来の臍帯血または静脈血におけるpH<7.1、BE<−15(または乳酸>4.8mm/L)により示される生後ストレス、3)生後6時間以内の脳症の神経学的徴候。
除外基準は、先天性奇形、染色体異常および仮死に関係のない脳症、代謝病、髄膜炎が確認された子宮内/周産期感染であった。
対照群は、感染の疑いがあるが血液およびCSFからの細菌およびウイルス培養が陰性であった、CSF中に白血球がみられずかつ正常のタンパク量である、およびCNS病を示唆する所見がない、20名の新生児からなるものであった。
【0255】
臨床評価
最初にこれらの患者を本研究に登録する数時間前、次いで出産約12、36および72時間後ならびに新生児集中治療室の患者については第7日目に、神経学的評価(95、その開示内容を参照により本明細書で援用することを明記する)を行った。低酸素性虚血性脳症(HIE)は、サルナットおよびサルナットの基準(96、その開示内容を参照により本明細書で援用することを明記する)に従い、軽度(mild)、中等度(moderate)または重度(severe)に分類した。全ての患者を出産直後の数日の間に評価するために連続振幅表示型EEG(continuous amplitude-integrated EEG)を使用した。生後3日目および最初の週におけるEEG登録日に、中等度および重度のHIEを有する全ての患者に対して、脳のCTおよびMRIスキャンを行った。
生存患者の神経学的評価は、生後3、6および18月に、小児神経科医が行った。その評価に基づいて、小児を(1)正常アウトカム(normal outcome)、(2)軽度の運動障害(mild motor impairment);軽度の異常な筋緊張もしくは運動発達遅延の症状、または(3)有害事象(adverse outcome);脳性麻痺(両麻痺、片麻痺、四肢麻痺)、精神遅滞、痙攣もしくは死、に分類した。
【0256】
アプガールスコア
アプガールスコアは、出産直後の新生児の身体的状態を評価するための実用的方法である。アプガールスコアは、心拍数、呼吸努力、筋緊張、皮膚の色および刺激(例えば外鼻孔へのカテーテル挿入または足裏の摩擦)に対する反応を採点することによって得られる数値である。これらの対象となる各徴候には、0、1または2点が付与される。アプガールスコア最高点である10点は、乳児が考えうる限り最高の状態にあることを意味する。アプガールスコア0〜3点の乳児は、直ちに蘇生が必要である。アプガールスコアは、通常、乳児の出産60秒後に行われ(アプガー1分)、次いで、一般的には、出産5分後に繰り返される(アプガー5分)。困難な蘇生の場合、アプガールスコアは、10、15および20分後に再度行われることもある。生後20分時のアプガールスコア0〜3点は、高い罹患率(病気)および死亡率(死)を示唆するものである。
【0257】
CSFのサンプリング
生後最初の24時間(13.9+/−5.8)および/または30〜80時間の間(57.8+/−9.9)にCSFスピナルタブ(Spinal tab)を行った。各スピナルタブにより1〜2mlの量のCSFを集めた。このサンプルを3000rpm、4℃で10分間遠心し、その上清を、分析するまで0.5mlずつ−80℃で保存した。
【0258】
PGE2アッセイ
脳脊髄液サンプル中のPGE2およびPGE2代謝産物は、標準化された酵素免疫アッセイ(EIA)プロトコル(Cayman Chemicals, Ann Arbor, MI, USA)を用いて分析した。
【0259】
タンパク質の分析(BCAアッセイ)
サンプル中のタンパク質レベルを決定するためにBCAアッセイを行った。
【0260】
統計学的分析
臨床データは、そうでないことが明記されない限り、記載される目的についての中央値および四分位範囲で示されている。患者と対照の間の差を分析するのにマン・ホイットニー検定トを用いた。PGE2代謝産物またはサイトカインレベルとHIEまたは臨床評価の程度の間の関係を決定するのにクラスカル・ウォリス検定を使用した。
【0261】
結果
患者群(n=43)を、HIEのサルナットおよびサルナットの分類に従い3つの下位グループに分けた。この分類によれば、13名の乳児が軽度のHIE(HIE I)であり、その全員が正常アウトカムであった。16名の乳児は中等度のHIE(HIE II)であり、そのうちの8名の乳児が脳性麻痺、精神運動遅滞および痙攣の問題がある有害の神経学的アウトカムであり、さらに、2名の乳児は軽度の運動障害、6名は正常アウトカムであった。14名の乳児は重度のHIE(HIE III)であり、そのうちの8名は出産直後〜12日目に死亡し、6名の患者は生存した上で、有害の神経学的アウトカム;痙性四肢麻痺性脳性麻痺、精神運動遅滞、小頭症および複雑型痙攣を有した。
【0262】
患者群および対照群の臨床データは以下の表4に示されている。在胎齢および出産時体重に関しては患者と対照の間に差がなかったが、5分アプガールスコアおよび臍動脈または初期患者pHに関して差がみられた(p<0.001)。アプガールスコアを、刺激(例えば乳児の鼻へのカテーテル挿入または乳児の足裏の摩擦)に対して得た。いずれの臨床データについても、患者群の間で差がみられなかった。血中CRPレベルは、対照および患者の両方において有意ではなかった。
【0263】
図7Aに示されるように、出生時仮死の程度(5分および10分アプガールスコア)および神経学的評価は、正期産児におけるCSF PGE2代謝産物レベルに相関する。
【0264】
同様に、図7Bに示されるように、PGE2代謝産物はまた、新生児の状態の指標である出産5分後のアプガールスコアに相関し、したがって出生時の仮死の程度に相関する可能性がある。
【0265】
これらの結果は、PGE2がヒト乳児における重度の低酸素(仮死)時に迅速に放出されること、したがって、新生児仮死を罹患した赤ん坊における診断ツールおよび/または治療的介入の標的として使用できることを示唆している。
【0266】
【表4】
【0267】
実施例8−尿プロスタグランジン代謝産物、炎症および呼吸機能障害との相関
本発明者らは、三連四重極質量分析・テトラノルPGEMプロトコルを用いる尿プロスタグランジンE代謝産物(u−PGEM)の高感度かつ特異的な検出方法を開発した。
バリデーション研究は、この三連四重極質量分析・テトラノルPGEM法が、同じ被験者から採取したサンプル間で<5%の実験内変動を示すことを示している。室温で保存した尿サンプルではPGE代謝産物が分解されることが見出され、そのt1/2をおよそ2時間と概算した。対照的に、4℃での直接保存は、サンプルの分解を有意に減少させた。−20℃〜−80℃で保存したサンプルは、サンプル間で比較した場合、見かけ上、PGE代謝産物の分解を実質的に示さなかった。
【0268】
サンプルの調製
尿サンプルを、2%(v/v)の1Mクエン酸の添加によりpH3.0付近まで酸性化した。次いで、145μlの酸性尿アリコートを、エタノール中に9pmol/μlテトラノルPGEM−d6および0.45pmol/μl 11β−PGF2α−d4を含有する5μlの内部標準溶液でスパイクした。100μlをLC−MS/MS機器に注入した。標準曲線用および品質管理用のサンプルを、2%(v/v)の1Mクエン酸で酸性にしたPBSを用いて準備した。次いで、140μlの酸性PBSアリコートを、5μlの内部標準溶液(上記参照)および5μlの標準溶液(30〜900pmol/μlテトラノルPGEMおよび3〜90pmol/μl 11β−PGF2α)を用いてスパイクした。100μlをLC−MS/MS機器に注入し、100〜3000pmolテトラノルPGEMおよび10〜300pmol 11β−PGF2αの標準曲線を得た。LC−MS/MSの条件:分析物は、0.0005%FAを含むH2Oおよび移動相として0.0
005%FAを含むACNを用いて、Phenomenex Synergi Hydro RPカラム(100mm×2mm i.d.、2.5μm粒子径および100Å孔径)上で分離した。サンプル注入の直後に、15〜60% ACN、0.0005%FAの直線勾配を15分間適用し、その後、95%ACN、0.0005% FAで洗浄し、再平衡化した。総泳動時間は21分間であった。質量分析器は、陰イオンモード、電気スプレー電圧−3000V、350℃で作動させた。プロスタグランジン代謝産物の検出および定量のために、多重反応モニタリング(MRM)を使用して、テトラノルPGEMについての327.1>255.3およびテトラノルPGEM−d6の333.1>263.3(分裂エネルギー70V、衝突エネルギー−20V、滞留時間100m秒)ならびに11β−PGF2α、−PGF2αの353.3>309.3および11β−PGF2α−d4の357.3>313.3(分裂エネルギー150V、衝突エネルギー−15V、滞留時間100m秒)の遷移を記録した。全ての四重極は、最高感度を得る単一解像度で作動させた。
【0269】
本明細書に記載される結果は、成人、小児(1〜16歳)および乳児(0〜1歳)から得られたu−PGEMレベルの上昇が炎症の指標として信頼でき、呼吸機能障害(無呼吸を含む)と有意に関連することを示している。
【0270】
健常な成人対照由来の尿サンプル(n=10)を、「閉塞性」睡眠時無呼吸症候群(OSAS)患者から得た尿サンプル(n=24、22〜55歳)と比較した。睡眠に関係する無呼吸症候群(「閉塞性睡眠時無呼吸症候群」(OSAS)−いびき)は、成人女性の3%程度および成人男性の5%程度でみられると見積もられる。この結果は図8に示されており、y軸は尿中のPGE代謝産物をピコモルPGEM/μgクレアチニンの単位で示している。閉塞性睡眠時無呼吸症候群と診断された全ての患者に対して、夜間睡眠ポリグラフ記録実験(night-time sleep polysomnographic recording Laboratory test)を行い、この中で、尿サンプルを睡眠ポリグラフ記録(呼吸および飽和を含む)後の朝に採取した。
【0271】
睡眠時無呼吸(いびき)群は、対照群よりもu−PGEMレベルの多様性が有意に大きかった(大きな値の分散を観察した)。本発明者らは、u−PGEMレベルの上昇が無呼吸インデックス、すなわち無呼吸数/時間と相関する明確な傾向を観察した。さらに、重度のOSAS患者は、無呼吸インデックスとCRP(炎症およびPGE2の間接的マーカー)の間で有意な相関を有した。
【0272】
睡眠時無呼吸を示した3名の成人のおよそ1名でu−PGEMが上昇しており、これが無呼吸の重症度と相関していた。図8に示されるグループ間の比較はp=0.12を示した。しかし、重度の無呼吸問題を抱える患者のみを含め、閉塞性の問題(BMI値>過体重)を抱える患者を排除した場合、無呼吸とu−PGEMレベルの間に有意な関連性がみられる。
【0273】
本発明者らは、無呼吸インデックスが高い個体を、u−PGEMが高い被験者(すなわち、対照レベルよりも高い対象、図8の点線の囲みを参照されたい)と重ねることができることを発見した。
【0274】
本発明者らは、プラダー・ウィリー症候群(PWS)の小児(3〜16歳)におけるu−PGEMレベルについても調査した。プラダー・ウィリー症候群(15q11〜q13の欠失)の患者は、呼吸および心血管調節系に故障があり、特に睡眠中に無呼吸がみられる(115)。心肺の障害に起因する死は通常睡眠中に起こり、原因となる要因は特定されていないものの、3つの死亡例のうつ2つで軽度の感染エピソードが関係している(107)。
【0275】
我々は、mPGES−1経路の活性化が感染時に起こる潜在的に致死性の過度の呼吸の故障に関与しているという仮説をたてた(合わせてNature Medicine 2007, Vol.13, No.7, p.789, Research Highlights:“Baby's breath”も参照されたい)。
【0276】
既知の感染および炎症マーカーであるhs−CRP、CRP、WBCおよびサイトカイン(IL−1β)ならびにPGE2の尿中代謝産物を、循環器疾患登録(cardiovascular registration)と平行して試験した。これを、プラダー・ウィリー症候群を患った乳児および成人に対して、1)定期年次身体検査時、2)感染の徴候(体温>38.5℃)から24時間後および3)臨床的感染からの回復から少なくとも1週間後に行った。分析は主たる臨床研究室およびカロリンスカ・コアプロテオミクス施設の研究室において行い、三連四重極質量分析機器を用いて既知の代謝産物およびペプチドの定量を行った。
【0277】
PWS小児は呼吸パターンおよび自律調節に故障があり、しばしば軽度の上気道感染を伴い突然死することが知られている(年間罹患率2〜3%)。図9に示されるように、PWS小児(n=6)における尿中PGEMレベルは、健常な対照小児と比較して有意に上昇することが見出された。図9において、y軸は尿中のPGE代謝産物をピコモルPGEM/μgクレアチニンの単位で示している。この患者群(PWS)におけるu−PGEMレベルの上昇は、呼吸障害(特に無呼吸)、炎症およびPGE2(例えばu−PGEMレベル)の間の関連性のさらなる証拠となる。これにより、(PWSを患ったまたは患っていない)小児から得たサンプル中のプロスタグランジン代謝産物(例えばu−PGEM)の増加が、呼吸障害、例えば無呼吸、OSAS、SIDSおよび/もしくは炎症関連呼吸障害を患っているまたはそれらを発症する可能性が高いことを示すものであると考えることができる。さらに、感染に関連する呼吸機能障害を患った小児の部分集団は特に、呼吸障害とサンプル中のプロスタグランジン代謝産物(例えばu−PGEM)の増加の間で有意な相関を示す。この部分集団は、a)OSASおよび/またはb)自律神経機能障害の徴候、例えばPWS、レット症候群またはCCHS(先天性中枢性低換気症候群、「オンディーヌの呪い」とも呼ばれる)と関連する徴候を有する小児を含む。
【0278】
さらに、本発明者らは、進行中の炎症、ウイルス性細気管支炎および随伴性の無呼吸がみられる乳児(n=10)におけるu−PGEMレベルを調査した。その結果は図10に示されており、y軸は尿中のPGE代謝産物をピコモルPGEM/μgクレアチニンの単位で示している。進行中の炎症および随伴性の無呼吸がみられる乳児群は、対照(n=10、進行中の炎症または無呼吸がみられない乳児および小児)と比較して非常に高いu−PGEMレベルを示した。さらに、日常的な臨床ケアにおいて感染の評価に広く使用されているCRP(C反応性タンパク質)レベルはわずかに上昇したにすぎなかった。したがって、u−PGEMレベルの測定は、進行中の炎症を評価する上で、CRPの測定よりも有用であり、一部の乳児に見られる失調性の呼吸調節の潜在的なメカニズムを明らかにする。生後1〜6月の過敏な小児における炎症は、主として睡眠中の不規則な呼吸および無呼吸に関係しているようである。
【0279】
ウイルス感染(例えば、ウイルス性細気管支炎)は、重度の呼吸妨害および脳幹における「呼吸ペースメーカー」の中枢的抑制を引き起こす。しかし、このような感染は、典型的には、進行中の炎症障害の存在についての従来的なマーカーであるCRPのわずかな増加しか引き起こさない。したがって、プロスタグランジン代謝産物(例えば、u−PGEMレベル)の測定は、感染の初期段階において潜在的な炎症および/または呼吸障害の示唆を提供すると考えられる。したがって、プロスタグランジン代謝産物レベルのアッセイは臨床の場で有益であり、医師が炎症の重症度、予後および可能な治療的介入を「ベッドの上で」決定することを可能にする。
【0280】
本明細書において引用した全ての参考文献は、個々の刊行物または特許または特許出願について特別かつ個別にその全体が参照により援用されることが示されているのと同程度に、それら全体をすべての目的のために参照により援用する。
【0281】
本明細書に記載される具体的な実施態様は例示の目的で提供されるものであり、限定を意図していない。本明細書中のあらゆる副題は利便性のためのみに含まれるのであり、いかなる様式によってもその開示内容を限定するものとみなすべきではない。
【0282】
参考文献
1. Fanaroff, A. A., et al. (1998) Pediatr Infect Dis J 17, 593-598.
2. Prandota, J. (2004) Am J Ther 11, 517-546.
3. Guntheroth, W. (1989) Med Hypotheses 28, 121-123.
4. Dantzer, R. (2001) Ann N Y Acad Sci 933, 222-234.
5. Olsson, A., et al. (2003) Pediatr Res 54, 326-331.
6. Hofstetter, A. O. & Herlenius, E. (2005) Respir Physiol Neurobiol 146, 135-146.
7. Froen, J. F., et al. (2000) Pediatrics 105, E52.
8. Lindgren, C., Grogaard, J (1996) Acta Paediatr 85, 798-803.
9. Stoltenberg, L., et al. (1994) J Perinat Med 22, 421-432.
10. Vege, A., et al. (1998) Acta Paediatr 87, 819-824.
11. Ericsson, A., et al. (1994) Journal of Neuroscience 14, 897-913.
12. Ericsson, A., et al. (1995) J Comp Neurol 361, 681-698.
13. Engblom, D., Ek, M, Saha, S, Ericsson-Dahlstrand, A, Jakobsson, PJ, Blomqvist, A (2002) J Mol Med 80, 5-15.
14. Coceani, F. & Akarsu, E. S. (1998) Ann N Y Acad Sci 856, 76-82.
15. Crestani, F., Seguy, F, Dantzer, R (1991) Brain Res 542, 330-335.
16. Ericsson, A., Arias, C, Sawchenko, PE (1997) J Neurosci 17, 7166-7179.
17. Guerra, F., Savich, RD, Wallen, LD, Lee, CH, Clyman, RI, Mauray, FE, Kitterman, JA (1988) Journal of Applied Physiology 64, 2160-2166.
18. Kitterman, J., Liggins, GC, Fewell, JE, Tooley, WH (1983) J Appl Physiol 54,
687-692.
19. Tai, T. C. & Adamson, S. L. (2000) Am J Physiol 278, 1460-1473.
20. Ek, M., Arias, C., Sawchenko, P., & Ericsson-Dahlstrand, A. (2000) J Comp Neurol 428, 5-20.
21. Nakamura, K., Kaneko, T., Yamashita, Y., Hasegawa, H., Katoh, H., & Negishi,
M. (2000) J Comp Neurol 421, 543-569.
22. Gray, P. A., Rekling, J. C., Bocchiaro, C. M., & Feldman, J. L. (1999) Science 286, 1566-1568.
23. Ballanyi, K., Lalley, P. M., Hoch, B., & Richter, D. W. (1997) J Physiol 504, 127-134.
24. Pagliardini, S., Ren, J., & Greer, J. J. (2003) J Neurosci 23, 9575-9584.
25. Yamagata, K., Matsumura, K., Inoue, W., Shiraki, T., Suzuki, K., Yasuda, S.,
Sugiura, H., Cao, C., Watanabe, Y., & Kobayashi, S. (2001) J Neurosci 21, 2669-2677.
26. Kitterman, J., Liggins, GC, Clements, JA, Tooley, WH (1979) J Dev Physiol 1,
453-466.
27. Guerra, F. A., Savich, R. D., Clyman, R. I., & Kitterman, J. A. (1989) J Dev
Physiol 11, 1-6.
28. Long, W. (1988) J Appl Physiol 64, 409-418.
29. Guerra, F. A., Savich, R. D., Wallen, L. D., Lee, C. H., Clyman, R. I., Maur
ay, F. E., & Kitterman, J. A. (1988) J Appl Physiol 64, 2160-2166.
30. Lein, E. S., Hawrylycz, M. J., Ao, N., Ayres, M., Bensinger, A., Bernard, A., Boe, A. F., Boguski, M. S., Brockway, K. S., Byrnes, E. J., et al. (2007) Nature 445, 168-176.
【0283】
31. Tai, T., MacLusky, NJ, Adamson, SL (1994) Brain Res 652, 28-39.
32. Degi, R., Bari, F., Thore, C., Beasley, T., Thrikawala, N., & Busija, D. W. (1998) Neurobiology (Bp) 6, 467-468.
33. Shohami, E. & Gross, J. (1986) J Neurochem 47, 1678-1681.
34. Allen, L. G., Louis, T. M., & Kopelman, A. E. (1982) Biol Neonate 42, 8-14.
35. Ristimaki, A., Garfinkel, S., Wessendorf, J., Maciag, T., & Hla, T. (1994) J
Biol Chem 269, 11769-11775.
36. Degousee, N., Angoulvant, D., Fazel, S., Stefanski, E., Saha, S., Iliescu, K., Lindsay, T. F., Fish, J. E., Marsden, P. A., Li, R. K., et al. (2006) J Biol Chem 281, 16443-16452.
37. Savich, R. D., Guerra, F. A., Lee, C. C., & Kitterman, J. A. (1995) J Appl Physiol 78, 1477-1484.
38. Murai, D. T., Wallen, L. D., Lee, C. C., Clyman, R. I., Mauray, F., & Kitterman, J. A. (1987) J Appl Physiol 62, 271-277.
39. Feldman, J. L. & Del Negro, C. A. (2006) 7, 232-241.
40. McKay, L. C., Janczewski, W. A., & Feldman, J. L. (2005) Nature Neuroscience
8, 1142-1144.
41. Paton, J. F., Abdala, A. P., Koizumi, H., Smith, J. C., & St-John, W. M. (2006) Nat Neurosci 9, 311-313.
42. Tauman, R., Ivanenko, A., O'Brien, L. M., & Gozal, D. (2004) Pediatrics 113,
e564-569.
43. Singh, G., Fong, LV, Salmon, AP, Keeton, BR (1994) Eur Heart J 15, 377-381.
44. Hoch, B. & Bernhard, M. (2000) Acta Paediatr 89, 1364-1368.
45. Hammerman, C. & Zangen, D. (1993) Crit Care Med 21, 154-155.
46. Schmidt, B., Davis, P., Moddemann, D., Ohlsson, A., Roberts, R. S., Saigal, S., Solimano, A., Vincer, M., & Wright, L. L. (2001) N Engl J Med 344, 1966-1972.
47. Trebino, C. E., Stock, J. L., Gibbons, C. P., Naiman, B. M., Wachtmann, T. S., Umland, J. P., Pandher, K., Lapointe, J. M., Saha, S., Roach, M. L., et al. (2003) Proc Natl Acad Sci U S A 100, 9044-9049.
48. Fleming, E. F., Athirakul, K., Oliverio, M. I., Key, M., Goulet, J., Koller,
B. H., & Coffman, T. M. (1998) Am J Physiol 275, F955-961.
49. Jacobi, M. S. & Thach, B. T. (1989) J Appl Physiol 66, 2384-2390.
50. Herlenius, E., Lagercrantz, H, Yamamoto, Y (1997) Pediatr Res 42, 46-53.
51. Suzue, T. (1984) J Physiol 354, 173-183.
52. Engblom, D., Saha, S., Engstrom, L., Westman, M., Audoly, L. P., Jakobsson, P. J., & Blomqvist, A. (2003) Nat Neurosci 6, 1137-1138.
53. Fewell, J. E., Smith, F. G., Ng, V. K., Wong, V. H., & Wang, Y. (2000) Am J Physiol Regul Integr Comp Physiol 279, R39-46.
54. Hofstetter, A. O., et al. (2007) PNAS USA 104(23), 9894-9899.
55. Ward, E.S., et al. (1989) Nature 341, 544-546.
56. Bird et al. (1988) Science, 242, 423-426.
57. Huston et al. (1988) PNAS USA, 85, 5879-5883.
58. P. Holliger et al. (1993) PNAS USA, 90, 6444-6448.
59. Y. Reiter et al. (1996) Nature Biotech, 14, 1239-1245.
60. S. Hu et al. (1996) Cancer Res., 56, 3055-3061.
【0284】
61. John et al. (2004) PLoS Biology, 11(2), 1862-1879.
62. Myers (2003) Nature Biotechnology, 21, 324-328.
63. Shinagawa et al. (2003) Genes and Dev., 17, 1340-1345.
64. Fire, A., et al. (1998) Nature 391, 806-811.
65. Fire, A., (1999) Trends Genet. 15, 358-363.
66. Sharp, P. A. (2001) Genes Dev. 15, 485-490.
67. Hammond, S. M., et al. (2001) Nature Rev. Genet. 2, 1110-1119.
68. Tuschl, T. (2001) Chem. Biochem. 2, 239-245.
69. Hamilton, A., et al. (1999) Science 286, 950-952.
70. Hammond, S. M., et al. (2000) Nature 404, 293-296.
71. Zamore, P. D., et al. (2000) Cell 101, 25-33.
72. Bernstein, E., et al. (2001) Nature 409 363-366.
73. Elbashir, S. M., et al. (2001) Genes Dev. 15, 188-200.
74. Elbashir, S. M., et al. (2001) Nature 411, 494-498.
75. Thoren et al. (2000) Eur. J. Biochem. 267, 6428.
76. Mancini et al. (2001) J. Biol. Chem. 276, 4469.
77. Quraishi et al. (2002) Biochem. Pharmacol. 63, 1183.
78. Schultz, J. S., (1996) Biotechnol. Prog. 12, 729-743.
79. Cutz, E. et al. (1997) Am. J. Respir. Crit. Care Med. 155(1), 358-363.
80. Festen, D.A. et al. (2006) J. Clin. Endocrinol. Metab. 91(12), 4911-4915.
81. Debra, E. et al. (2007) Am. J. Med. Gen. Part A, 771-788.
82. Paterson, D.S. et al. (2006) JAMA 296(17), 2124-2132.
83. Onodera, H. et al. (2000) Lancet 356(9231), 739-740.
84. Yang, J. et al. (1994) Biochem. Biophys. Res. Commun. 198(3), 999-1006.
85. Herlenius, E. and Lagercrantz, H. (1999), J. Physiol. 518 (Pt 1), 159-172.
86. Ballanyi, K. et al. (1997) J. Physiol. 504(1), 127-134.
87. Katoh, H. et al. (1996) Journal of Biological Chemistry 271(47), 29780-29784.
88. van Rodijnen, W.F. et al. (2007) Am. J. Physiol. Renal Physiol. 292(3), F1094-1101.
89. Rowland, S.E. et al. (2007) Eur. J. Pharmacol. 560(2-3), 216-224.
90. Habib, A. et al. (2007) Faseb. J. 21(8), 1665-1674.
【0285】
91. Yao, J.C. et al. (2007) Yakugaku Zasshi 127(3), 527-532.
92. Morrell, M.J., and Twigg, G. (2006) Adv. Exp. Med. Biol. 588, 75-88.
93. Rigatto, H., (2003) “Periodic Breathing”. In: Mathew OP, editor. Respiratory control and disorders in the newborn, Vol 173. New York: Marcel Dekker, Inc;
237-72.
94. Simmons, D.L. et al. (2004) Pharmacol Rev. 56(3), 387-437.
95. Dubowitz, L. and Dubowitz, V. (1981) Clinical Developmental Medicine 79, London, England: Mac Keith Press.
96. Sarnat, H.B. and Sarnat, M.S. (1976) Arch Neurol. 33, 696-705.
97. Volpe, J.J. (1994) Hypoxic-ischaemic encephalopathy: Neuropathology and pathogenesis In Neurology of the Newborn, pp. 279-314. Philadelphia: WB Saunders Co.98. Roth, S.C., et al. (1992) Dev Med and Child Neurol. 34:285-295.
99. Hanrahan, D., et al. (1999) Dev Med and Child Neurol. 41:76-82.
100. Mehmet, H., et al. (1994) Neurosci Lett. 181:121-125.
101. Greher, J.K. and Nelson, K.B. (1997). J. Am Med Association. 278:207-211.
102. Nelson, K.B., et al. (1998) Ann Neurol. 44(4):665-75.
103. Foster-Barber, A., and Ferriero, D.M. (2002) Mental retard and dev disabilities. 8:20-24.
104. Silveira, R.C., and Procianoy, R.S. (2003). J. Pediatr. 625-629.
105. Deems et al. (2007) Methods Enzymol. 432, 59-82.
106. Welsh, T. N. et al. (2007) Prostaglandins & other Lipid Mediators 83, 304-310.
107. Tauber, M. et al. (2008) Am J Med Genet A. 146(7):881-887.
108. Perez, I.A. and Ward, S.L. (2008) Pediatr Ann. 37(7):465-470.
109. Wildhaber, J.H. and Moeller, A. (2007) Swiss Med Wkly. 137(49-50):689-694.
110. Tarasiuk, A. (2007) Am J Respir Crit Care Med. 175(1):55-61.
111. Weber, M.A. (2008) Lancet 371:1848-1853.
112. Hoch, B. et al. (2000) Prostaglandins Other Lipid Mediat. 60(1-3):9-14.
113. Hoch, B. and Bernhard, M. (2000) Acta Paediatr. 89(11):1364-1368.
114. Hofstetter, A.O. et al. (2007) PNAS 104(23):9894-9899.
115. Festen, D.A. et al. (2006) J Clin Endocrinol Metab. 91(12):4911-4915.
【0286】
本明細書中で参照された配列
【化1】
【0287】
【化2】
【0288】
【化3】
【0289】
【化4】
【0290】
【化5】
【特許請求の範囲】
【請求項1】
哺乳動物対象における呼吸障害の処置方法であって、Eプロスタノイド受容体サブタイプ3(EP3R)阻害剤、ミクロソームプロスタグランジンEシンターゼ1(mPGES−1)阻害剤および/またはシクロオキシゲナーゼ2(COX−2)選択的阻害剤を含む組成物を対象に投与することを含む、上記方法。
【請求項2】
組成物がEP3R阻害剤を含む、請求項1に記載の方法。
【請求項3】
EP3R阻害剤が、EP3Rポリペプチドに結合する特異的結合物質またはEP3Rコード遺伝子の発現をダウンレギュレートする核酸である、請求項2に記載の方法。
【請求項4】
EP3R阻害剤が、(2E)−N−[(5−ブロモ−2−メトキシフェニル)スルホニル]−3−[5−クロロ−2−(2−ナフチルメチル)フェニル]アクリルアミド(L826266)または薬学的に許容できるその塩である、請求項2に記載の方法。
【請求項5】
組成物がmPGES−1阻害剤を含む、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
mPGES−1阻害剤が、mPGES−1ポリペプチドに結合する特異的結合物質またはmPGES−1コード遺伝子の発現をダウンレギュレートする核酸である、請求項5に記載の方法。
【請求項7】
mPGES−1阻害剤が、3−[tert−ブチルチオ−1−(4−クロロベンジル)−5−イソプロピル−1H−インドール−2−イル]−2,2−ジメチルプロピオン酸(MK−886)または薬学的に許容できるその塩である、請求項5に記載の方法。
【請求項8】
組成物がCOX−2選択的阻害剤を含む、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
COX−2選択的阻害剤が、COX−2ポリペプチドに結合する特異的結合物質またはCOX−2コード遺伝子の発現をダウンレギュレートする核酸である、請求項8に記載の方法。
【請求項10】
COX−2選択的阻害剤が、4−(5−メチル−3−フェニルイソキサゾール−4−イル)ベンゼンスルホンアミド(バルデコキシブ)もしくは薬学的に許容できるその塩、4−[5−(4−メチルフェニル)−3−(トリフルオロメチル)ピラゾール−1−イル]ベンゼンスルホンアミド(セレコキシブ)もしくは薬学的に許容できるその塩、または4−(4−メチルスルホニルフェニル)−3−フェニル−5H−フラン−2−オン(ロフェコキシブ)もしくは薬学的に許容できるその塩である、請求項8に記載の方法。
【請求項11】
哺乳動物対象における呼吸障害に対する感受性またはその存在の評価方法であって、対象由来のサンプル中のプロスタグランジンE2(PGE2)またはその代謝産物のレベルを検出すること、およびPGE2またはその代謝産物の対照レベルとサンプル中のレベルを比較することを含み、ここでPGE2またはその代謝産物の対照レベルと比較してサンプル中のPGE2またはその代謝産物の高レベルが、対象における呼吸障害に対する感受性またはその存在を指示する、上記方法。
【請求項12】
サンプルが尿サンプルまたは脳脊髄液(CSF)サンプルを含む、請求項11に記載の方法。
【請求項13】
対象由来のサンプル中のC反応性タンパク質(CRP)のレベルを検出すること、およびCRPの対照レベルとサンプル中のレベルを比較することをさらに含み、ここでCRPの対照レベルと比較して高レベルのサンプル中のCRPレベルの高レベルが、対象における呼吸障害に対する感受性またはその存在を指示する、請求項11または12に記載の方法。
【請求項14】
呼吸障害が、無呼吸、周期性呼吸または低酸素イベント後の自然蘇生不全である、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
呼吸障害が、睡眠中に起きる呼吸障害、特に閉塞性睡眠時無呼吸症候群である、請求項1〜14のいずれか1項に記載の方法。
【請求項16】
呼吸障害が感染性呼吸障害である、請求項1〜15のいずれか1項に記載の方法。
【請求項17】
感染性呼吸障害がIL−1β関連呼吸障害である、請求項16に記載の方法。
【請求項18】
呼吸障害が低酸素イベント後の無呼吸である、請求項14に記載の方法。
【請求項19】
無呼吸が低酸素イベントにより誘発される、請求項15に記載の方法。
【請求項20】
哺乳動物対象がヒト対象である、請求項1〜19のいずれか1項に記載の方法。
【請求項21】
ヒト対象が5歳未満である、請求項20に記載の方法。
【請求項22】
呼吸障害が、乳児突然死症候群(SIDS)を引き起こすまたはその尤度を高める障害である、請求項21に記載の方法。
【請求項23】
低酸素イベントが周産期仮死である、請求項18または19に記載の方法。
【請求項24】
ヒト対象が18歳以上である、請求項20に記載の方法。
【請求項25】
呼吸障害が成人の睡眠時無呼吸である、請求項24に記載の方法。
【請求項26】
哺乳動物対象における低酸素性虚血性脳症(HIE)に対する感受性またはその存在の評価方法であって、対象由来のサンプル中のプロスタグランジンE2(PGE2)またはその代謝産物のレベルを検出すること、およびPGE2またはその代謝産物の対照レベルとサンプル中のレベルを比較することを含み、ここでPGE2の対照レベルと比較してサンプル中のPGE2またはその代謝産物の高レベルが、対象におけるHIEに対する感受性またはその存在を指示する、上記方法。
【請求項27】
PGE2またはその代謝産物の対照レベルと比較してサンプル中のPGE2またはその代謝産物レベルの上昇度を測定することによって対象におけるHIEの重症度を類別することを含む、請求項26に記載の方法。
【請求項28】
哺乳動物対象が周産期仮死に罹患していることを評価する方法であって、対象由来のサンプル中のプロスタグランジンE2(PGE2)またはその代謝産物のレベルを検出すること、およびPGE2またはその代謝産物の対照レベルとサンプル中のレベルを比較することを含み、ここでPGE2の対照レベルと比較してサンプル中のPGE2またはその代謝産物の高レベルが、対象が周産期仮死に罹患していることを指示する、上記方法。
【請求項29】
PGE2またはその代謝産物の対照レベルと比較してサンプル中のPGE2またはその代謝産物レベルの上昇度を測定することによって対象が罹患している周産期仮死の重症度を類別することを含む、請求項28に記載の方法。
【請求項30】
サンプルが対象の出生7日以内に採取された脳脊髄液(CSF)、尿または血液サンプルである、請求項26〜29のいずれか1項に記載の方法。
【請求項31】
サンプルが対象の出生24時間以内に採取される、請求項30に記載の方法。
【請求項32】
哺乳動物対象がヒト対象である、請求項26〜31のいずれか1項に記載の方法。
【請求項33】
ヒト対象のアプガールスコアを出生30分以内に測定することをさらに含む、請求項32に記載の方法。
【請求項34】
アプガールスコアが出生から約1、5、10、15および/または20分後に測定される、請求項33に記載の方法。
【請求項35】
哺乳動物における呼吸障害の処置に使用できる物質を同定する方法であって、
(a)COX−2により仲介されるPGH2の合成、
(b)mPGES−1により仲介される、mPGES−1の環状エンドペルオキシド基質から基質の9−ケト,11αヒドロキシ型である生産物への変換、および
(c)EP3Rアゴニストにより仲介されるEP3Rの活性化、
の一つまたはそれ以上を阻害する能力について試験物質をアッセイすることを含み、
ここで(a)、(b)および(c)の一つまたはそれ以上の阻害は、試験物質が哺乳動物における呼吸障害の処置に使用するための物質であることを指示する、上記方法。
【請求項36】
COX−2ポリペプチドと試験物質およびアラキドン酸を、アラキドン酸が試験物質の非存在下でCOX−2によってPGH2に変換される条件下で接触させること、および
試験物質の非存在下でのPGH2生産の対照レベルと比較される試験物質の存在下でのPGH2生産レベルを決定することを含み、
ここで試験物質の存在下でのPGH2生産レベルが上記対照レベルとの比較でより低いことは、試験物質が哺乳動物における呼吸障害の処置に使用するための物質であることを指示する、請求項35に記載の方法。
【請求項37】
試験物質の存在下でのPGH2生産レベルが対照レベルとの比較でより低いことを検出すること、およびそれによって試験物質を哺乳動物における呼吸障害の処置に使用するための物質であると同定することを含む、請求項36に記載の方法。
【請求項38】
mPGES−1ポリペプチドと試験物質およびmPGES−1の環状エンドペルオキシド基質を、mPGES−1の環状エンドペルオキシド基質が試験物質の非存在下でmPGES−1によって基質の9−ケト,11αヒドロキシ型である生産物に変換される条件下で接触させること、および
試験物質の非存在下での生産物生産の対照レベルと比較される試験物質の存在下での生産物の生産レベルを決定することを含み、
ここで試験物質の存在下での生産物の生産レベルが上記対照レベルとの比較でより低いことは、試験物質が哺乳動物における呼吸障害の処置に使用するための物質であることを指示する、請求項35に記載の方法。
【請求項39】
試験物質の存在下での生産物の生産レベルが対照レベルより低いことを検出すること、およびそれによって試験物質を、哺乳動物における呼吸障害の処置に使用するための物質であると同定することを含む、請求項38に記載の方法。
【請求項40】
EP3Rポリペプチドと試験物質およびEP3Rアゴニストを、EP3Rアゴニストが試験物質の非存在下でEP3Rポリペプチドを活性化する条件下で接触させること、および
試験物質の非存在下でのEP3Rポリペプチドの活性化の対照レベルと比較される当該試験物質の存在下でのEP3Rポリペプチドの活性化レベルを決定することを含み、
ここで試験物質の存在下でのEP3Rポリペプチドの活性化レベルが対照レベルとの比較でより低いことは、試験物質が哺乳動物における呼吸障害の処置に使用するための物質であることを指示する、請求項35に記載の方法。
【請求項41】
試験物質の存在下でのEP3Rポリペプチドの活性化レベルが対照レベルとの比較でより低いことを検出すること、およびそれによって試験物質を、哺乳動物における呼吸障害の処置に使用するための物質であると同定することを含む、請求項40に記載の方法。
【請求項42】
哺乳動物における呼吸障害の処置に使用できる物質の同定方法であって、試験物質を試験哺乳動物に投与すること、ここで試験物質はEP3R阻害剤、mPGES−1阻害剤および/またはCOX−2選択的阻害剤である、および
試験物質を投与していない対照の哺乳動物における徴候または症状と比較される試験哺乳動物における呼吸障害の徴候または症状の重症度を決定することを含み、
ここで試験哺乳動物における呼吸障害の徴候または症状の重症度が上記対照哺乳動物のそれらよりも低いことは、試験物質が哺乳動物における呼吸障害の処置に使用するための物質であることを指示する、上記方法。
【請求項43】
徴候または症状が、呼吸抑制、呼吸頻度の減少、1回換気量の減少および低酸素に応答するあえぎ呼吸の減少から選択される、請求項42に記載の方法。
【請求項44】
徴候または症状の重症度を決定する前にIL−1βまたはLPSを投与することを含む、請求項42または43に記載の方法。
【請求項45】
試験物質が哺乳動物における呼吸障害の処置に使用するための物質であると同定され、そしてここで試験物質を、薬学的に許容できる賦形剤を含む組成物に製剤化することをさらに含む、請求項35〜44のいずれか1項に記載の方法。
【請求項46】
ヒト対象における低酸素および/または無呼吸の存在および/またはそれらの重症度の評価方法であって、対象から採取した尿サンプル中の一つまたはそれ以上のPGE2代謝産物のレベルを検出すること、および
該一つまたはそれ以上のPGE2代謝産物の対照レベルとサンプル中のレベルを比較することを含み、
ここでサンプル中の該一つまたはそれ以上のPGE2代謝産物のレベルが該一つまたはそれ以上のPGE2代謝産物の対照レベルと比較して少なくとも20%、少なくとも50%、少なくとも100%または少なくとも200%高いことが、対象における低酸素および/または無呼吸の存在および/またはそれらのより高い重症度を指示する、上記方法。
【請求項47】
ヒト対象が、閉塞性睡眠時無呼吸症候群(OSAS)、自律神経機能障害、例えばプラダー・ウィリー症候群、先天性低換気症候群またはレット症候群を有する、請求項46に記載の方法。
【請求項48】
ヒト対象が16歳以上である、請求項46または47に記載の方法。
【請求項49】
ヒト対象が1〜16歳である、請求項46または47に記載の方法。
【請求項50】
ヒト対象が0〜1歳である、請求項46または47に記載の方法。
【請求項1】
哺乳動物対象における呼吸障害の処置方法であって、Eプロスタノイド受容体サブタイプ3(EP3R)阻害剤、ミクロソームプロスタグランジンEシンターゼ1(mPGES−1)阻害剤および/またはシクロオキシゲナーゼ2(COX−2)選択的阻害剤を含む組成物を対象に投与することを含む、上記方法。
【請求項2】
組成物がEP3R阻害剤を含む、請求項1に記載の方法。
【請求項3】
EP3R阻害剤が、EP3Rポリペプチドに結合する特異的結合物質またはEP3Rコード遺伝子の発現をダウンレギュレートする核酸である、請求項2に記載の方法。
【請求項4】
EP3R阻害剤が、(2E)−N−[(5−ブロモ−2−メトキシフェニル)スルホニル]−3−[5−クロロ−2−(2−ナフチルメチル)フェニル]アクリルアミド(L826266)または薬学的に許容できるその塩である、請求項2に記載の方法。
【請求項5】
組成物がmPGES−1阻害剤を含む、請求項1〜4のいずれか1項に記載の方法。
【請求項6】
mPGES−1阻害剤が、mPGES−1ポリペプチドに結合する特異的結合物質またはmPGES−1コード遺伝子の発現をダウンレギュレートする核酸である、請求項5に記載の方法。
【請求項7】
mPGES−1阻害剤が、3−[tert−ブチルチオ−1−(4−クロロベンジル)−5−イソプロピル−1H−インドール−2−イル]−2,2−ジメチルプロピオン酸(MK−886)または薬学的に許容できるその塩である、請求項5に記載の方法。
【請求項8】
組成物がCOX−2選択的阻害剤を含む、請求項1〜7のいずれか1項に記載の方法。
【請求項9】
COX−2選択的阻害剤が、COX−2ポリペプチドに結合する特異的結合物質またはCOX−2コード遺伝子の発現をダウンレギュレートする核酸である、請求項8に記載の方法。
【請求項10】
COX−2選択的阻害剤が、4−(5−メチル−3−フェニルイソキサゾール−4−イル)ベンゼンスルホンアミド(バルデコキシブ)もしくは薬学的に許容できるその塩、4−[5−(4−メチルフェニル)−3−(トリフルオロメチル)ピラゾール−1−イル]ベンゼンスルホンアミド(セレコキシブ)もしくは薬学的に許容できるその塩、または4−(4−メチルスルホニルフェニル)−3−フェニル−5H−フラン−2−オン(ロフェコキシブ)もしくは薬学的に許容できるその塩である、請求項8に記載の方法。
【請求項11】
哺乳動物対象における呼吸障害に対する感受性またはその存在の評価方法であって、対象由来のサンプル中のプロスタグランジンE2(PGE2)またはその代謝産物のレベルを検出すること、およびPGE2またはその代謝産物の対照レベルとサンプル中のレベルを比較することを含み、ここでPGE2またはその代謝産物の対照レベルと比較してサンプル中のPGE2またはその代謝産物の高レベルが、対象における呼吸障害に対する感受性またはその存在を指示する、上記方法。
【請求項12】
サンプルが尿サンプルまたは脳脊髄液(CSF)サンプルを含む、請求項11に記載の方法。
【請求項13】
対象由来のサンプル中のC反応性タンパク質(CRP)のレベルを検出すること、およびCRPの対照レベルとサンプル中のレベルを比較することをさらに含み、ここでCRPの対照レベルと比較して高レベルのサンプル中のCRPレベルの高レベルが、対象における呼吸障害に対する感受性またはその存在を指示する、請求項11または12に記載の方法。
【請求項14】
呼吸障害が、無呼吸、周期性呼吸または低酸素イベント後の自然蘇生不全である、請求項1〜13のいずれか1項に記載の方法。
【請求項15】
呼吸障害が、睡眠中に起きる呼吸障害、特に閉塞性睡眠時無呼吸症候群である、請求項1〜14のいずれか1項に記載の方法。
【請求項16】
呼吸障害が感染性呼吸障害である、請求項1〜15のいずれか1項に記載の方法。
【請求項17】
感染性呼吸障害がIL−1β関連呼吸障害である、請求項16に記載の方法。
【請求項18】
呼吸障害が低酸素イベント後の無呼吸である、請求項14に記載の方法。
【請求項19】
無呼吸が低酸素イベントにより誘発される、請求項15に記載の方法。
【請求項20】
哺乳動物対象がヒト対象である、請求項1〜19のいずれか1項に記載の方法。
【請求項21】
ヒト対象が5歳未満である、請求項20に記載の方法。
【請求項22】
呼吸障害が、乳児突然死症候群(SIDS)を引き起こすまたはその尤度を高める障害である、請求項21に記載の方法。
【請求項23】
低酸素イベントが周産期仮死である、請求項18または19に記載の方法。
【請求項24】
ヒト対象が18歳以上である、請求項20に記載の方法。
【請求項25】
呼吸障害が成人の睡眠時無呼吸である、請求項24に記載の方法。
【請求項26】
哺乳動物対象における低酸素性虚血性脳症(HIE)に対する感受性またはその存在の評価方法であって、対象由来のサンプル中のプロスタグランジンE2(PGE2)またはその代謝産物のレベルを検出すること、およびPGE2またはその代謝産物の対照レベルとサンプル中のレベルを比較することを含み、ここでPGE2の対照レベルと比較してサンプル中のPGE2またはその代謝産物の高レベルが、対象におけるHIEに対する感受性またはその存在を指示する、上記方法。
【請求項27】
PGE2またはその代謝産物の対照レベルと比較してサンプル中のPGE2またはその代謝産物レベルの上昇度を測定することによって対象におけるHIEの重症度を類別することを含む、請求項26に記載の方法。
【請求項28】
哺乳動物対象が周産期仮死に罹患していることを評価する方法であって、対象由来のサンプル中のプロスタグランジンE2(PGE2)またはその代謝産物のレベルを検出すること、およびPGE2またはその代謝産物の対照レベルとサンプル中のレベルを比較することを含み、ここでPGE2の対照レベルと比較してサンプル中のPGE2またはその代謝産物の高レベルが、対象が周産期仮死に罹患していることを指示する、上記方法。
【請求項29】
PGE2またはその代謝産物の対照レベルと比較してサンプル中のPGE2またはその代謝産物レベルの上昇度を測定することによって対象が罹患している周産期仮死の重症度を類別することを含む、請求項28に記載の方法。
【請求項30】
サンプルが対象の出生7日以内に採取された脳脊髄液(CSF)、尿または血液サンプルである、請求項26〜29のいずれか1項に記載の方法。
【請求項31】
サンプルが対象の出生24時間以内に採取される、請求項30に記載の方法。
【請求項32】
哺乳動物対象がヒト対象である、請求項26〜31のいずれか1項に記載の方法。
【請求項33】
ヒト対象のアプガールスコアを出生30分以内に測定することをさらに含む、請求項32に記載の方法。
【請求項34】
アプガールスコアが出生から約1、5、10、15および/または20分後に測定される、請求項33に記載の方法。
【請求項35】
哺乳動物における呼吸障害の処置に使用できる物質を同定する方法であって、
(a)COX−2により仲介されるPGH2の合成、
(b)mPGES−1により仲介される、mPGES−1の環状エンドペルオキシド基質から基質の9−ケト,11αヒドロキシ型である生産物への変換、および
(c)EP3Rアゴニストにより仲介されるEP3Rの活性化、
の一つまたはそれ以上を阻害する能力について試験物質をアッセイすることを含み、
ここで(a)、(b)および(c)の一つまたはそれ以上の阻害は、試験物質が哺乳動物における呼吸障害の処置に使用するための物質であることを指示する、上記方法。
【請求項36】
COX−2ポリペプチドと試験物質およびアラキドン酸を、アラキドン酸が試験物質の非存在下でCOX−2によってPGH2に変換される条件下で接触させること、および
試験物質の非存在下でのPGH2生産の対照レベルと比較される試験物質の存在下でのPGH2生産レベルを決定することを含み、
ここで試験物質の存在下でのPGH2生産レベルが上記対照レベルとの比較でより低いことは、試験物質が哺乳動物における呼吸障害の処置に使用するための物質であることを指示する、請求項35に記載の方法。
【請求項37】
試験物質の存在下でのPGH2生産レベルが対照レベルとの比較でより低いことを検出すること、およびそれによって試験物質を哺乳動物における呼吸障害の処置に使用するための物質であると同定することを含む、請求項36に記載の方法。
【請求項38】
mPGES−1ポリペプチドと試験物質およびmPGES−1の環状エンドペルオキシド基質を、mPGES−1の環状エンドペルオキシド基質が試験物質の非存在下でmPGES−1によって基質の9−ケト,11αヒドロキシ型である生産物に変換される条件下で接触させること、および
試験物質の非存在下での生産物生産の対照レベルと比較される試験物質の存在下での生産物の生産レベルを決定することを含み、
ここで試験物質の存在下での生産物の生産レベルが上記対照レベルとの比較でより低いことは、試験物質が哺乳動物における呼吸障害の処置に使用するための物質であることを指示する、請求項35に記載の方法。
【請求項39】
試験物質の存在下での生産物の生産レベルが対照レベルより低いことを検出すること、およびそれによって試験物質を、哺乳動物における呼吸障害の処置に使用するための物質であると同定することを含む、請求項38に記載の方法。
【請求項40】
EP3Rポリペプチドと試験物質およびEP3Rアゴニストを、EP3Rアゴニストが試験物質の非存在下でEP3Rポリペプチドを活性化する条件下で接触させること、および
試験物質の非存在下でのEP3Rポリペプチドの活性化の対照レベルと比較される当該試験物質の存在下でのEP3Rポリペプチドの活性化レベルを決定することを含み、
ここで試験物質の存在下でのEP3Rポリペプチドの活性化レベルが対照レベルとの比較でより低いことは、試験物質が哺乳動物における呼吸障害の処置に使用するための物質であることを指示する、請求項35に記載の方法。
【請求項41】
試験物質の存在下でのEP3Rポリペプチドの活性化レベルが対照レベルとの比較でより低いことを検出すること、およびそれによって試験物質を、哺乳動物における呼吸障害の処置に使用するための物質であると同定することを含む、請求項40に記載の方法。
【請求項42】
哺乳動物における呼吸障害の処置に使用できる物質の同定方法であって、試験物質を試験哺乳動物に投与すること、ここで試験物質はEP3R阻害剤、mPGES−1阻害剤および/またはCOX−2選択的阻害剤である、および
試験物質を投与していない対照の哺乳動物における徴候または症状と比較される試験哺乳動物における呼吸障害の徴候または症状の重症度を決定することを含み、
ここで試験哺乳動物における呼吸障害の徴候または症状の重症度が上記対照哺乳動物のそれらよりも低いことは、試験物質が哺乳動物における呼吸障害の処置に使用するための物質であることを指示する、上記方法。
【請求項43】
徴候または症状が、呼吸抑制、呼吸頻度の減少、1回換気量の減少および低酸素に応答するあえぎ呼吸の減少から選択される、請求項42に記載の方法。
【請求項44】
徴候または症状の重症度を決定する前にIL−1βまたはLPSを投与することを含む、請求項42または43に記載の方法。
【請求項45】
試験物質が哺乳動物における呼吸障害の処置に使用するための物質であると同定され、そしてここで試験物質を、薬学的に許容できる賦形剤を含む組成物に製剤化することをさらに含む、請求項35〜44のいずれか1項に記載の方法。
【請求項46】
ヒト対象における低酸素および/または無呼吸の存在および/またはそれらの重症度の評価方法であって、対象から採取した尿サンプル中の一つまたはそれ以上のPGE2代謝産物のレベルを検出すること、および
該一つまたはそれ以上のPGE2代謝産物の対照レベルとサンプル中のレベルを比較することを含み、
ここでサンプル中の該一つまたはそれ以上のPGE2代謝産物のレベルが該一つまたはそれ以上のPGE2代謝産物の対照レベルと比較して少なくとも20%、少なくとも50%、少なくとも100%または少なくとも200%高いことが、対象における低酸素および/または無呼吸の存在および/またはそれらのより高い重症度を指示する、上記方法。
【請求項47】
ヒト対象が、閉塞性睡眠時無呼吸症候群(OSAS)、自律神経機能障害、例えばプラダー・ウィリー症候群、先天性低換気症候群またはレット症候群を有する、請求項46に記載の方法。
【請求項48】
ヒト対象が16歳以上である、請求項46または47に記載の方法。
【請求項49】
ヒト対象が1〜16歳である、請求項46または47に記載の方法。
【請求項50】
ヒト対象が0〜1歳である、請求項46または47に記載の方法。
【図1】

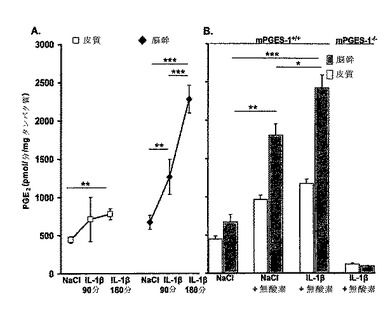
【図2】
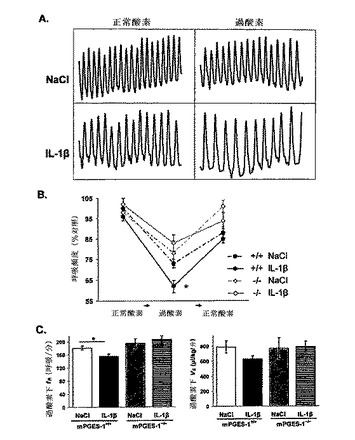
【図3】
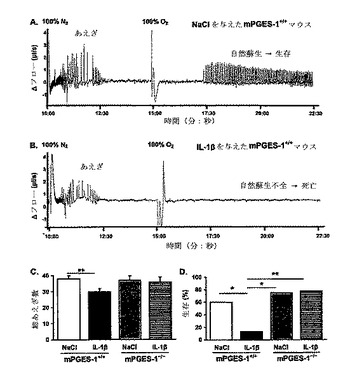
【図4】
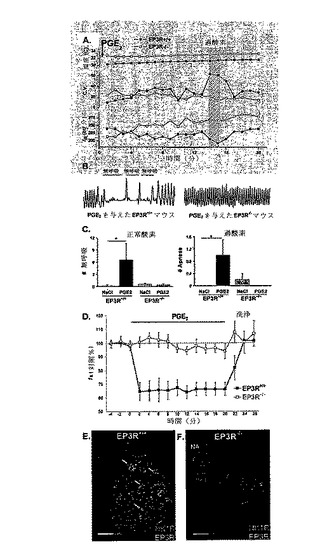
【図5】
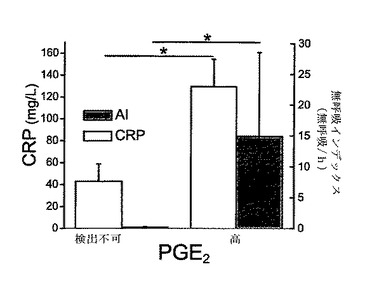
【図6】
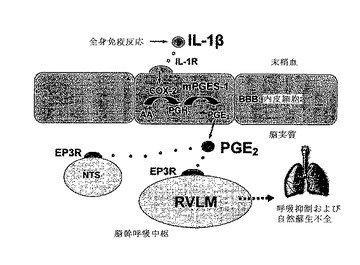
【図7】
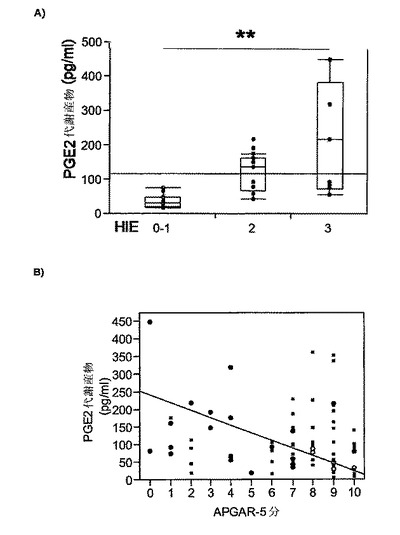
【図8】
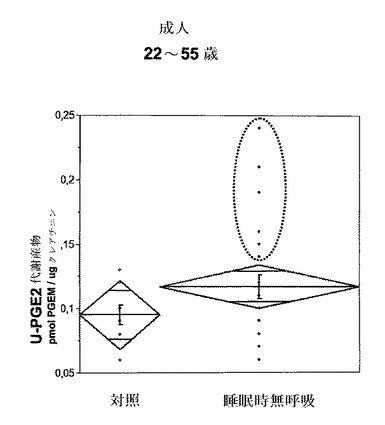
【図9】
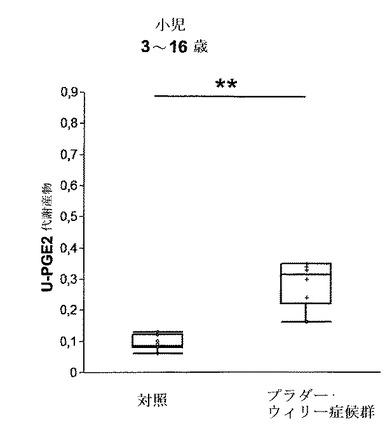
【図10】
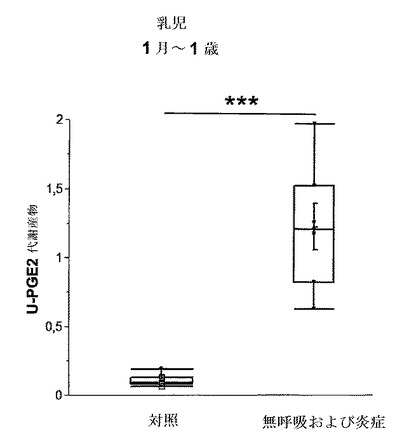
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公表番号】特表2011−503164(P2011−503164A)
【公表日】平成23年1月27日(2011.1.27)
【国際特許分類】
【出願番号】特願2010−533659(P2010−533659)
【出願日】平成20年11月12日(2008.11.12)
【国際出願番号】PCT/GB2008/003856
【国際公開番号】WO2009/063226
【国際公開日】平成21年5月22日(2009.5.22)
【出願人】(510130583)サムサラ・メディシン・アクチエボラーグ (1)
【Fターム(参考)】
【公表日】平成23年1月27日(2011.1.27)
【国際特許分類】
【出願日】平成20年11月12日(2008.11.12)
【国際出願番号】PCT/GB2008/003856
【国際公開番号】WO2009/063226
【国際公開日】平成21年5月22日(2009.5.22)
【出願人】(510130583)サムサラ・メディシン・アクチエボラーグ (1)
【Fターム(参考)】
[ Back to top ]