
哺乳動物細胞の培養において高生細胞密度を得るための方法
真核細胞の流加培養における生存率を高める方法を開示する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、高濃度のグルコース添加により、流加細胞培養中で高い生細胞密度及び長い培養寿命を実現するための方法に関する。本方法は、対象とする分泌タンパク質の産生量を高めるうえで有用である。
【背景技術】
【0002】
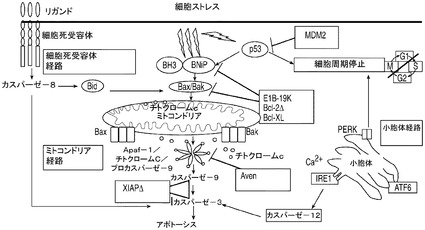
哺乳動物細胞の培養は、適当な翻訳後修飾がなされたタンパク質を生産することが可能であることから、多くの組換えタンパク質製造プロセスにおいて選択されている系である。製造需要が高まるにつれ、産物の収率を高めることによってプロセス効率を改善するとう強い動機づけがある。商業的な製造プロセスにおいて生物学的治療を目的としたタンパク質又は他のタンパク質の1L当たり数グラムの製造レベルを実現するには、哺乳動物細胞の培養及び操作方法を最適化することが不可欠である。現在の高密度、無タンパク質の哺乳動物細胞培養に内在する問題として細胞死の問題があり、一般的な流加バイオリアクターではアポトーシスによるものが最大でその80%を占める。こうした細胞死は、栄養及び増殖因子の欠乏、酸素欠乏、毒素の蓄積、及び剪断応力といった諸条件に応じて誘導されるものである(Goswami et al.,Biotechnol Bioeng 62:632〜640(1999))。アポトーシスは最大の生細胞密度を制限し、死滅期の開始を早め、異種由来タンパク質の収率を潜在的に低下させる(Chiang and Sisk,Biotechnol Bioeng 91:779〜792(2005);Figueroa et al.,Biotechnol Bioeng.73:211〜222(2001),Metab Eng 5:230〜245(2003),Biotechnol Bioeng 85:589〜600(2004);Mercille and Massie,Biotechnol Bioeng 44:1140〜1154(1994))。
【0003】
アポトーシスは細胞の内外で開始されるシグナル伝達経路の複雑なネットワークによって引き起こされるものであり、細胞死の最終ステージを媒介するシステインアスパラギン酸プロテアーゼ(カスパーゼ)が最終的に活性化される。これについては図1を参照されたい。哺乳動物の細胞培養における長期の産生培養の間に細胞生存率を維持する目的でアポトーシスを防止する様々な方法が用いられている(Arden and Betenbaugh,Trends Biotechnol 22:174〜180(2004);Vives et al.,Metab Eng 5:124〜132(2003))。増殖因子、加水分解物、及び律速栄養素を培地に添加して細胞外環境を変化させることによってタンパク質の産生量が高まり、アポトーシスが減少する(Burteau et al.,In Vitro Cell Dev Biol Anim.39:291〜296(2003);Zhang and Robinson,Cytotechnology 48:59〜74(2005))。細胞内からのアポトーシスのシグナル伝達カスケードを阻害する化学的及び遺伝子的手法を模索する研究者もいる(Sauerwald et al.,Biotechnol Bioeng 77:704〜716(2002),Biotechnol Bioeng 81:329〜340(2003))。
【0004】
ガン細胞において発現量が増大している遺伝子の過剰発現が、カスパーゼ活性化の上流でアポトーシスを防止することによってバイオリアクター中で増殖させた細胞の生存期間を延ばしうることが研究者によって見出されている(Goswami et al.,supra;Mastrangelo et al.,Trends Biotechnol 16:88〜95(1998);Meents et al.,Biotechnol Bioeng 80:706〜716(2002);Tey et al.,J Biotechnol 79:147〜159(2000)、及び、Biotechnol Bioeng 68:31〜43(2000))。生産用細胞株におけるこれらのタンパク質の発現量の増大は、アポトーシスを引き起こす細胞内のシグナル伝達を効果的に抑制することによって細胞死を制限することにより、特定の場合において生存率を維持し生物治療物質の産生量を増大させた。
【0005】
高密度、無タンパク質の哺乳動物細胞培養にやはり内在する問題点として、老廃物の蓄積及びそれが細胞増殖にもたらす悪影響がある。細胞培養における最も一般的な2つの老廃物は、乳酸及びアンモニアである。過剰な乳酸の蓄積の問題に対処するために多くの手法が考案されており、1)培地のグルコース濃度を低く維持する(Kurokawa et al.,BiotechnolBioeng 44:95〜103(1994);Xie and Wang,Biotechnol Bioeng 43:1175〜1189(1993);Zhang et al.,J Chem Technol Biotechnol 79:171〜181(2004);Zhou et al.,Biotechnol Bioeng 46:579〜587(1995))、2)フルクトースなどの代替的な糖を添加する(Martinelle et al.,Biotechnol Bioeng 60:508〜517(1998),Altamirano et al.,J Biotechnol 110:171〜179(2004)and J Biotechnol 125:547〜556(2006),Walschin and Wu,J Biotechnol 131:168〜176(2007))、3)相同組み換え又はSiRNA技術により乳酸脱水素酵素(LDH)の発現を部分的にノックアウトする、4)ピルビン酸カルボキシラーゼの過剰発現、5)ピルビン酸脱水素酵素(PDH)の活性化因子(PDHキナーゼ阻害を介した)であるジクロロ酢酸塩(DCA)の使用、6)LDH競合阻害剤であるオキサミン酸、及び7)灌流による除去(米国特許出願公開第2009/0042253A1号)などが挙げられる。
【0006】
当初は、多くのアポトーシス経路のタンパク質の固有の機能は、ミトコンドリア膜に結合してミトコンドリアの透過性を調節することによりアポトーシスを調節することと考えられていた。最近の研究により、アポトーシスを引き起こすシグナル伝達に関与する主要なタンパク質は、細胞の代謝及びエネルギー恒常性を制御するタンパク質と相互作用してこれらのタンパク質に影響を及ぼすことが示された。Majors et al.,Metab Eng 9:317〜326(2007)(概論)、及びWhite et al.,Nat Cell Biol 7:1021〜1028(2005)を参照されたい。最近の研究では、BcL−XLを過剰発現しているCHO細胞のマイクロアレイ分析により、糖新生の主要な酵素である乳酸脱水素酵素の発現量が増大していることが示された。
【0007】
特定の細胞及びウイルスは、ミトコンドリアのアポトーシス経路において機能するアポトーシス耐性遺伝子を生産する。これらの遺伝子は、以下の3つのグループ、すなわち、1)経路の早期に働くもの(例えば、Bcl−2ファミリーメンバーのタンパク質)、2)経路の中期に働いてアポトソーム複合体を妨害又は阻害するもの(例えば、Aven)、及び3)経路の後期に働くもの(例えば、XIAPなどのカスパーゼ阻害因子)に分類することができる。これらの遺伝子の大半のものの機能は、哺乳動物の発現系においてこれらを過剰発現させることによって研究されており、特定の場合では、それぞれが経路の異なる部分に由来する2以上の遺伝子の過剰発現の組み合わせによる影響が調べられている。その例としては、1)CHO細胞におけるBcL−XLとXIAPの欠失突然変異(XIAPΔ)との相加的効果(Figueroa et al.,Metab.Eng.5:230〜245(2003))、2)BHK細胞におけるE1B−19KとAven(Nivitchanyong et al.,Biotechnol Bioeng 98:825〜841(2007))、及び3)BcL−XL、Aven、及びXIAPΔ(Sauerwald et al.,supra,(2003);Sauerwald et al,Biotechnol Bioeng 94:362〜369(2006))が挙げられる。
【発明の概要】
【発明が解決しようとする課題】
【0008】
しかしながら、これらの研究では、細胞の細胞代謝状態に対するアポトーシス耐性遺伝子の影響については調べられていない。したがって、高い生細胞密度、寿命、及び生産性を実現するために栄養消費及び代謝産物の蓄積状態を最適化するための哺乳動物細胞の培養システムが求められている。
【課題を解決するための手段】
【0009】
本発明の一態様は、真核細胞の流加培養において高い生細胞密度を得るための方法であって、
a)1種以上の異種由来のアポトーシス耐性(アポトーシスR)遺伝子及び1種以上の対象遺伝子を発現する真核細胞株を培養する工程と、
b)前記細胞培養の指数増殖期及び定常期に高グルコース添加培地を維持する工程と、を含む方法である。
【0010】
本発明の別の態様は、真核細胞の流加培養において分泌タンパク質の産生量を高めるための方法であって、
a)1種以上の異種由来のアポトーシス耐性(アポトーシスR)遺伝子及び1種以上の対象遺伝子を発現する真核細胞株を培養する工程と、
b)前記細胞培養の指数増殖期及び定常期に高グルコース添加培地を維持する工程と、を含む方法である。
【図面の簡単な説明】
【0011】
【図1】ミトコンドリアのアポトーシス経路の構成要素を示す図。
【図2A】アポトーシス耐性細胞株の特性評価を示す図。
【図2B】アポトーシス耐性細胞株の特性評価を示す図。
【図2C】アポトーシス耐性細胞株の特性評価を示す図。
【図3】フローサイトメトリー分析によりEA167及びEAX197のアポトーシス耐性を証明した図。
【図4A】E1B19K、E1B19K+AVEN、及び、E1B19K+AVEN+XIAPΔをトランスフェクトすることによって生成されたアポトーシス耐性細胞株の増殖プロファイルである。コントロール(●)、E64(◆)、EA167(■)及びEAX197(▼)の震盪フラスコ回分培養を、生細胞密度及び生存率について観測した。
【図4B】E1B19K、E1B19K+AVEN、及び、E1B19K+AVEN+XIAPΔをトランスフェクトすることによって生成されたアポトーシス耐性細胞株の増殖プロファイルである。コントロール(●)、E64(◆)、EA167(■)及びEAX197(▼)の震盪フラスコ回分培養を、生細胞密度及び生存率について観測した。
【図4C】E1B19K、E1B19K+AVEN、及び、E1B19K+AVEN+XIAPΔをトランスフェクトすることによって生成されたアポトーシス耐性細胞株の増殖プロファイルである。コントロール(●)、E64(◆)、EA167(■)及びEAX197(▼)の震盪フラスコ回分培養を、生細胞密度及び生存率について観測した。
【図5A】E1B19K+AVEN±XIAPΔでトランスフェクトすることによって生成されたアポトーシス耐性細胞株の代謝産物のプロファイルである。コントロール(●)、EA167(■)及びEAX197(▼)の震盪フラスコ回分培養を、異なる代謝産物について観測した。
【図5B】E1B19K+AVEN±XIAPΔでトランスフェクトすることによって生成されたアポトーシス耐性細胞株の代謝産物のプロファイルである。コントロール(●)、EA167(■)及びEAX197(▼)の震盪フラスコ回分培養を、異なる代謝産物について観測した。
【図5C】E1B19K+AVEN±XIAPΔでトランスフェクトすることによって生成されたアポトーシス耐性細胞株の代謝産物のプロファイルである。コントロール(●)、EA167(■)及びEAX197(▼)の震盪フラスコ回分培養を、異なる代謝産物について観測した。
【図6A】アポトーシス耐性細胞株に連日乳酸を補充した結果を示す図である。A)コントロール(●);EA167(無添加□、乳酸添加■)、及びB)EAX197(無添加△、乳酸添加▼)。
【図6B】アポトーシス耐性細胞株に連日乳酸を補充した結果を示す図である。A)コントロール(●);EA167(無添加□、乳酸添加■)、及びB)EAX167(添加△、乳酸無添加▼)。
【図7A】添加乳酸の存在下又は不在下における、コントロール並びにアポトーシス耐性細胞株EA167及びEAX197の増殖及び代謝産物のプロファイルである。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図7B】添加乳酸の存在下又は不在下における、コントロール並びにアポトーシス耐性細胞株EA167及びEAX197の増殖及び代謝産物のプロファイルである。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図7C】添加乳酸の存在下又は不在下における、コントロール並びにアポトーシス耐性細胞株EA167及びEAX197の増殖及び代謝産物のプロファイルである。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図7D】添加乳酸の存在下又は不在下における、コントロール並びにアポトーシス耐性細胞株EA167及びEAX197の増殖及び代謝産物のプロファイルである。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図7E】添加乳酸の存在下又は不在下における、コントロール並びにアポトーシス耐性細胞株EA167及びEAX197の増殖及び代謝産物のプロファイルである。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図7F】添加乳酸の存在下又は不在下における、コントロール並びにアポトーシス耐性細胞株EA167及びEAX197の増殖及び代謝産物のプロファイルである。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図7G】添加乳酸の存在下又は不在下における、コントロール並びにアポトーシス耐性細胞株EA167及びEAX197の増殖及び代謝産物のプロファイルである。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図8A】乳酸添加時にアポトーシス耐性細胞株によって減少するアミノ酸のプロファイルを示す図である。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図8B】乳酸添加時にアポトーシス耐性細胞株によって減少するアミノ酸のプロファイルを示す図である。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図8C】乳酸添加時にアポトーシス耐性細胞株によって減少するアミノ酸のプロファイルを示す図である。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図8D】乳酸添加時にアポトーシス耐性細胞株によって減少するアミノ酸のプロファイルを示す図である。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図8E】乳酸添加時にアポトーシス耐性細胞株によって減少するアミノ酸のプロファイルを示す図である。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図8F】乳酸添加時にアポトーシス耐性細胞株によって減少するアミノ酸のプロファイルを示す図である。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図8G】乳酸添加時にアポトーシス耐性細胞株によって減少するアミノ酸のプロファイルを示す図である。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図9A】特別配合培地中、高濃度のグルコース中で培養されたコントロール及びアポトーシス耐性細胞株EAX197の増殖プロファイル及び乳酸蓄積プロファイルである。アポトーシス耐性細胞株EAX197(■)、コントロール細胞株(◇)。
【図9B】特別配合培地中、高濃度のグルコース中で培養されたコントロール及びアポトーシス耐性細胞株EAX197の増殖プロファイル及び乳酸蓄積プロファイルである。アポトーシス耐性細胞株EAX197(■)、コントロール細胞株(◇)。
【図9C】特別配合培地中、高濃度のグルコース中で培養されたコントロール及びアポトーシス耐性細胞株EAX197の増殖プロファイル及び乳酸蓄積プロファイルである。アポトーシス耐性細胞株EAX197(■)、コントロール細胞株(◇)。
【図9D】特別配合培地中、高濃度のグルコース中で培養されたコントロール及びアポトーシス耐性細胞株EAX197の増殖プロファイル及び乳酸蓄積プロファイルである。アポトーシス耐性細胞株EAX197(■)、コントロール細胞株(◇)。
【図10A】30mM又は60mMのグルコースを含む特別配合培地中で培養されたEAX197及びコントロールホスト細胞株から生成された細胞株の抗体力価を示す図である。アポトーシス耐性細胞株EAX197(■)、コントロール細胞株(□)。
【図10B】30mM又は60mMのグルコースを含む特別配合培地中で培養されたEAX197及びコントロールホスト細胞株から生成された細胞株の抗体力価を示す図である。アポトーシス耐性細胞株EAX197(■)、コントロール細胞株(□)。
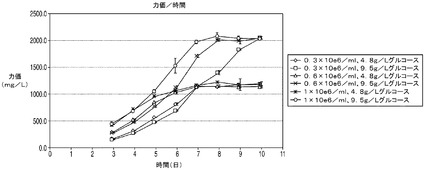
【図11A】30mM又は60mMのグルコースを含む培地中に高細胞播種密度及び低細胞播種密度で播種した、アポトーシス耐性遺伝子Bcl−2を有する細胞株のVCD及び力価を示す図。
【図11B】30mM又は60mMのグルコースを含む培地中に高細胞播種密度及び低細胞播種密度で播種した、アポトーシス耐性遺伝子Bcl−2を有する細胞株のVCD及び力価を示す図。
【発明を実施するための形態】
【0012】
本明細書に引用される特許及び特許出願など(ただしこれらに限定されない)のすべての刊行物は、本明細書にそれらの全体が記載されているものと同様にして本願に援用するものである。
【0013】
説明文中で使用する用語「流加細胞培養」とは、増殖を律速する栄養基質を培養に添加することに基づいた細胞培養プロセスを意味する。流加法は、バイオリアクター中で高い細胞密度に達するためにバイオ産業のプロセスで一般的に使用されている方法である。 HYPERLINK "http://en.wikipedia.org/wiki/Bioreactor" \o "Bioreactor" しかしながら、グルコースなどの栄養素を添加すると乳酸やアンモニアなどの代謝老廃物が形成される。濃度18mMの乳酸(Kurano et al.,1990)及び8mMのアンモニア(Hansen and Emborg,1994)は真核細胞の増殖を阻害することが報告されている。
【0014】
流加細胞培養における過剰な乳酸の蓄積の問題に対処するためにこれまでに多くの手法が考案されており、それらの手法について上記に述べた。本発明では、乳酸濃度を低下させて生細胞密度を高めることに代わる手法を提供する。この方法では、1種以上のアポトーシス耐性遺伝子をホスト細胞株で過剰発現させる。得られたアポトーシス耐性細胞株ではミトコンドリアでの呼吸が刺激され、乳酸の蓄積量が減少することになる。したがって、これらの細胞株は、高濃度のグルコースを添加しても生存することが可能である。その結果、これらのアポトーシス耐性細胞株の培養では高い生細胞密度に達することが可能である。これらの細胞株はまた、そのアポトーシス耐性のために長い寿命を有する。
【0015】
本発明の方法は、チャイニーズハムスター子宮細胞(CHO)、ミエローマ又はハイブリドーマ細胞培養などの流加細胞培養における生細胞の密度及び生存率を高めるうえで有用である。詳細には、本発明の方法はCHO細胞培養の積分生細胞数(IVCC)を高めるうえで有用である。本発明の方法は、1種以上の異種由来のアポトーシス耐性遺伝子(アポトーシスR)遺伝子及び1種以上の対象遺伝子を発現する細胞株を培養する工程と、細胞培養の指数増殖期及び定常期の間に高濃度のグルコースの培地への添加を維持する工程とを含む。これらの細胞系は、ペプチド、ペプチド融合体、増殖因子、ホルモン抗体、設計アンキリン反復タンパク質(DARPins)、及び治療、診断又は研究目的に有用な他のタンパク質などの対象タンパク質を発現する生産用細胞株の作出のめの優れたホストとなる。本発明の方法において有用なCHO細胞株としては、CHO−K1(インビトロジェン社(Invitrogen)カリフォルニア州カールスバッド)及びCHOK1SV(ロンザ・バイオロジックス社(Lonza Biologics)英国、スラウ)が挙げられる。本発明の方法において有用なミエローマ株としては、NS0及びSp2/0が挙げられる。
【0016】
本発明の方法において有用な細胞株は、1種以上の異種由来のアポトーシス耐性遺伝子を発現する。詳細には、E1B19K(配列番号1及び2)及びAven(配列番号3及び4)が有用である。XIAPΔ(配列番号:5及び6)をコードした遺伝子を使用することもできる。これらのアポトーシス耐性遺伝子は、それぞれアポトーシスを引き起こすシグナル伝達経路の早期、中期、及び後期に働く遺伝子を代表するものである。更に、Bcl−2Δをコードする遺伝子を使用することもできる(配列番号7及び8)。アポトーシス耐性遺伝子の発現は、当業者には周知の形質移入(トランスフェクション)技術によって実現させることができる。MDM2(配列番号9及び10)及びBcl−XL(配列番号11及び12)などの、アポトーシスを引き起こすシグナル伝達経路の上記のステージからの他のアポトーシス耐性遺伝子も本発明の方法において有用である。
【0017】
本発明においては、異種由来のアポトーシス耐性遺伝子を発現する細胞株の使用により、これらの細胞株の乳酸の分泌量が低減されるかあるいは蓄積された乳酸が消費されることによって、高濃度のグルコースの培養中への添加を維持しつつ、積分生細胞数(IVCC)の値がコントロール細胞株と比較して約2倍高い値に達する。培養中の生産用細胞株の生細胞密度を高めることにより、バイオリアクターの使用によって得られる産物の収率が高まる。このように生産性が高められることにより、複雑な生物製剤の製造コストが低減されると同時に、非生細胞の細胞溶解がないために優れた品質の産物が生産される(溶解した細胞は産物を分解するプロテアーゼを放出するため)。したがってこれらの細胞株は、対象タンパク質(単数又は複数)を発現する生産用細胞株を作出するための優れたホストとなる。
【0018】
本発明の方法では、老廃物として蓄積される乳酸を消費する、又はその産生量が低減したアポトーシス耐性細胞株の能力によって、高濃度グルコース条件下、及びグルコース欠乏後の増殖を可能とする培養手法が与えられる。異なる実施形態において、本発明の方法は、流加細胞培養における高いピーク生細胞密度、長い寿命、高い分泌タンパク質力価、高い積分生細胞数、低い細胞カルシウム流束、及び高いミトコンドリア膜電位を与えるものである。更に、高濃度の有毒な乳酸及びアンモニアを制限するために用いられてきた流加培養法は、これらの細胞株の高い生産性にとって必要ではない可能性がある。
【0019】
トランスフェクトした後、アポトーシス耐性導入遺伝子の染色体への組み込み部位によってその発現レベルが決まる。更に、複数のアポトーシス耐性遺伝子をトランスフェクトする場合、これらのタンパク質の多くは直接的又は間接的に相互作用することによって細胞の生理学的変化を引き起こすことから、特定のアポトーシス耐性導入遺伝子の発現レベルがその細胞が有する他のアポトーシス耐性タンパク質の活性に影響を及ぼしうる。したがって、複数の遺伝子を有する細胞株の全体のアポトーシス耐性に対する各アポトーシス耐性遺伝子の定量的寄与は、各アポトーシス耐性遺伝子の相乗効果のために決定することが困難である。更に、すべてのクローンが同じ導入遺伝子の集合をトランスフェクトされていたとしてもアポトーシス耐性に関しては各クローン間で大きなばらつきがある。後述する実施例に示される結果からは、試験した各アポトーシス耐性遺伝子は、各遺伝子をトランスフェクトしたホスト細胞株のアポトーシス耐性を高める方向に漸増する正の値を与えることが明らかである。上記の束縛条件内では、3重形質導入体は一般に2重形質導入体よりも優れており、2重形質導入体は1個のみのアポトーシス耐性遺伝子を過剰発現している形質導入体よりも優れていた。
【0020】
したがって、適当な発現ベクターをトランスフェクトすることでE1B−19K、Aven及びXIAPΔを異なるレベルで発現するクローンが得られた。E1B−19Kの発現レベルは比較的低く、E1B−19Kを高い発現レベルで発現する稀なクローンは不安定であるか増殖性が低かった(データは示さず)。下記実施例において、E1B−19Kを単独で発現しているCHO細胞では細胞死に対する保護は限定的なものに過ぎなかった(図4)。
【0021】
E1B−19Kを単独で発現している細胞株で得られた結果に対し、下記実施例において示されるデータは、Aven(EA167)、又はAven及びXIAPΔ(EAX197)を同時に発現しているCHO K1細胞株では、生存率、細胞密度及びIVCCに向上が見られたことを示している。下記実施例に示される結果は、CHO K1細胞株における2個のアポトーシス耐性遺伝子の発現により、スタウロスポリン及び長期の回分培養といった障害の存在下でカスパーゼ活性が大幅に低下し、ミトコンドリアの膜電位が高くなったことを更に示している。
【0022】
下記実施例では、Bcl−XLの機能的ホモログであるE1B−19KとAven及びXIAPΔとの組み合わせの発現が細胞死を遅らせる効果について調べた。これらのトランスフェクト細胞におけるXIAPΔの発現レベルは、細胞内に既に存在する内因性の野生型XIAPタンパク質に対して高くはなかったが、これはこのシステムにおいて2個のプラスミドをトランスフェクトするために必要な条件によるものと恐らく考えられる。しかしながら、EAX197におけるXIAPΔの追加(Aven及びE1B−19Kに加えて)により、無添加(図5及び7)及び乳酸を添加した培養(図7)の両方で最大生細胞密度が一貫して向上した。実際、乳酸を添加したEAX197細胞株は、60%以上の高い生存率を14日間にわたって維持したが、これはEA167細胞よりも2日長く、コントロール培養よりも4日間長かった。XIAPΔを有することによりカスパーゼ活性に対する細胞の耐性が高まると考えられ、このことは図3に、スタウロスポリンによる処理後においてもEAX197のカスパーゼ活性が極めて低かったことにより実証されている。更に、XIAPの突然変異(XIAPΔ)の使用により、野生型XIAPタンパク質と比較して高いアポトーシス耐性が与えられる。したがって、XIAPの欠失突然変異(XIAPΔ)の保護能力は、これらのCHO株においてこのタンパク質の発現レベルが高くなくとも充分なものである可能性がある。
【0023】
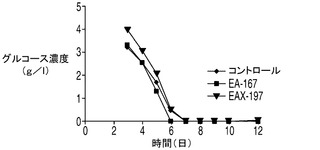
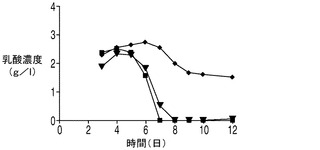
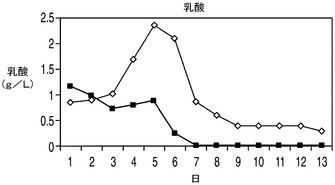
下記実施例では、細胞の代謝における1種以上のアポトーシス耐性遺伝子の役割を調べた。Aven及び/又はXIAPΔとともにE1B−19Kを発現しているアポトーシス耐性細胞株において栄養素の消費量及び代謝産物の産生量を調べた。詳細には、2つの最も一般的な細胞培養の老廃物(すなわち乳酸とアンモニア)の蓄積量についてアポトーシス耐性細胞株とコントロール細胞株との間で比較を行った。下記に示すデータでは、CD−CHO培地中の震盪フラスコ回分培養において、播種後6日目〜7日目にコントロール細胞株は2.8g/L(31mM)の乳酸及び12mMのアンモニア(図5)を蓄積したが、これらは細胞増殖を阻害することが報告されている濃度18mMの乳酸(Kuranoら、前出)及び8mMのアンモニア(Hansen及びEmborg、前出)を大幅に上回る濃度であった。これらの濃度は、CHOコントロール細胞株において播種後8日目に見られた生存率の低下の理由の一部となっていることが考えられる。
【0024】
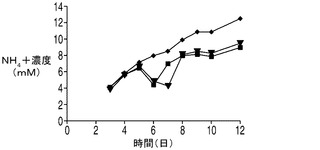
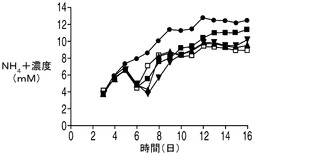
下記に示すデータは、アポトーシス抵抗細胞株がある程度の乳酸を初期に蓄積することを示しているが、これらの細胞株は指数増殖期に乳酸を消費しはじめ、VCDはコントロール培養を大幅に上回って増大し続けた。グルコースの枯渇後に直ちに乳酸を消費する能力、又はアポトーシス阻害性の増大、又は両方の因子が細胞増殖の継続に寄与していた。更に、アポトーシス耐性細胞は内因性の乳酸が枯渇した場合にのみ定常期に入った。乳酸の消費がこれらの操作された細胞株の一般的な特性であることを調べるため、アポトーシス耐性培養に外因性の乳酸を加えて欠乏した乳酸を補充した(図6)。下記実施例に示される結果は、アポトーシス耐性培養は添加された乳酸を消費し、コントロール培養と比較して高いVCD及び生存率を更に4〜7日間にわたって維持した(無添加及び乳酸添加培養を図7A及びBにそれぞれ示す)。このプロセスの更なる利点としては、アポトーシス耐性細胞株の培養中の乳酸が低濃度であることによって、浸透圧がより低く、pHがより高くなることがあり、これらはいずれも産物の高い品質にとって極めて望ましい。
【0025】
研究者によりこれまでに、ハイブリドーマ、NS0ミエローマ、及びCHOの培養中において乳酸の消費が検出されている(deZengotita et al.,2000;Zhou et al.,1995 and 1997;Burky et al.,2007;Pascoe et al.,2007)。しかしながら、これらの細胞は通常、指数増殖期では乳酸を産生し、定常期に消費へと移行するが、このことは、乳酸が中間の副産物及び炭水化物エネルギー源の両方でありうることを示唆するものである(Burky et al.,2007;Brooks et al.,1985)。下記に示す結果では、コントロールのCHO培養は、上記の実験において見られるように定常期において特に「高濃度」グルコース培地で(図9)一定量の乳酸を消費したようであった。これに対し、アポトーシス耐性細胞では、指数増殖期に乳酸が消費され、外因性の乳酸が欠乏した場合にのみ細胞が定常期に入るという大きく異なったプロファイルを示した。
【0026】
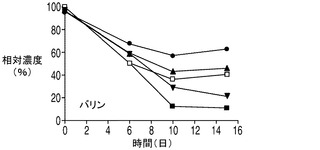
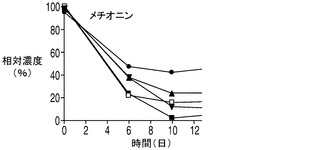
乳酸脱水素酵素(LDH)によって触媒されるピルビン酸の乳酸への変換は可逆反応であるが、平衡定数が3.6×104/Mと乳酸を生成する方向に大きく傾いている。しかしながら、解糖系がトリカルボン酸(TCA)の需要を満たすことができない場合、乳酸はピルビン酸へとその反応を進めるLDHの複数のアイソザイムの1つによって逆に変換される。アポトーシス耐性細胞株によって消費された乳酸は、TCA回路に入って細胞のエネルギー及びアミノ酸の産生に使われるものと考えられる。興味深い点として、アポトーシス耐性培養のアンモニアのプロファイルは、無添加及び乳酸添加細胞株の両方においてアンモニアの産生量が繰り返し一時的に低下するとともに全体的にアミノ酸の産生量が減少し、コントロール培養のアンモニアプロファイルと大きく異なった。これは、アミノ酸がTCA回路でエネルギー源として使用されることによってアンモニアが蓄積したものと考えられる。したがって、アンモニアの産生量の減少は、アポトーシス耐性細胞では、コントロール細胞と比較してTCA回路におけるエネルギーの必要量の内、相対的に小さな割合がアミノ酸から得られている可能性を示唆しており、このことはアポトーシス耐性細胞が炭素源として乳酸もより多く消費していることを考えると妥当な結果である。しかしながらアンモニア産生量の低下は一時的なものであり、アポトーシス耐性細胞が依然、アミノ酸をエネルギー源として利用している可能性がある点は注意を要する。実際、いずれも分枝鎖アミノ酸であるイソロイシン、ロイシン、及びバリンの3種類のアミノ酸は、アポトーシス耐性細胞によってより急速に消費され、その消費量は乳酸添加培養で大きかった(図8A〜C)。アポトーシス耐性細胞株はコントロール細胞株と比較して継続的な増殖を示したことから、これら3種類のアミノ酸が脂肪酸及びアポトーシス耐性細胞によって必要とされる他のタンパク質の構成単位を構成するとともに、TCA回路のエネルギー源として異化反応に用いられている可能性がある。メチオニンについても幾分似た消費プロファイルが観察された(図8D)。
【0027】
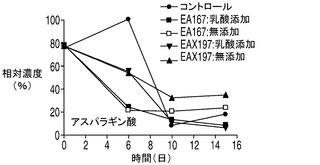
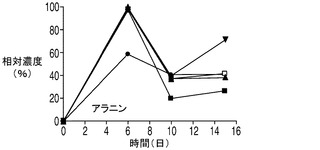
減少することが観察された上記のアミノ酸に対し、アラニン及びアスパラギン酸などの幾つかのアミノ酸がコントロール培養中に一時的に分泌された。ピルビン酸の一部が、アラニンアミノトランスフェラーゼによってアラニンに変換されている可能性がある。この経路はアポトーシス耐性細胞でより活性が高くなる場合があるので、コントロール細胞株に対してアラニン濃度がより大きな上昇につながる(図8F)。一方、アスパラギン酸は、アスパラギン酸アミノ酸ファミリーの第1のアミノ酸であり、アスパラギン酸トランスアミナーゼを用いてTCA回路の中間体であるオキザロ酢酸とグルタミン酸の変換によってα−ケトグルタル酸とともに生成する。その濃度は、コントロール細胞においてのみ最初の6日間上昇したのに対して、アポトーシス耐性細胞株では連続的に減少した。この反応は可逆反応であるため、増殖期の初期におけるアポトーシス耐性細胞株でのアスパラギン酸の欠乏は、アポトーシス耐性細胞株ではTCA回路における可逆的なオキザロ酢酸産物の産生量が制限されている可能性を示している。オキザロ酢酸がTCA回路の最初の段階においてアセチルCoAとも反応することを考慮すると、アポトーシス耐性細胞株における乳酸からアセチルCoAへのピルビン酸の流入量の増加は、オキザロ酢酸の需要を大きくしている可能性がある。これに対して、コントロール細胞株は乳酸を消費しておらず、したがってオキザロ酢酸からアスパラギン酸を生成しているため、コントロール細胞株ではアセチルCoAに対する需要は低いものと考えられる。実際、こうしたアセチルCoAに対する需要の不足は、細胞培養の最初の7日間にわたって見られた、アセチルCoAに代わる副生成物としての乳酸のコントロール細胞内への蓄積を引き起こした可能性が考えられる。また、コントロール細胞における乳酸消費の不足がアセチルCoAの供給を制限し、これにともなうアスパラギン酸の生成量の増加を引き起こした可能性も考えられる。
【0028】
これらの結果は、アポトーシス耐性遺伝子を発現する細胞株ではTCA回路のエネルギー効率が高くなっていることを示すものであるが、こうした細胞反応の正確な解釈にはより詳細な代謝フラックスの分析が待たれるところである。消費される乳酸の運命には関係なく、本発明ではアポトーシス耐性細胞株は蓄積した乳酸を消費し、これにより細胞株の長い寿命及び高いIVCCに寄与するものである。
【0029】
アポトーシス耐性細胞株が蓄積された乳酸を消費することによって寿命が延びるという点で独特であるならば、これらの細胞株は乳酸を含む培地又は標準よりも高い濃度のグルコースを含む培地中で培養できる可能性がある。乳酸が唯一の炭素源として与えられた場合には増殖及び生存率はいずれのタイプの細胞株においても支持されなかったが、乳酸がグルコースに加えて(データは示さず)一成分として与えられた場合、又は乳酸が老廃物として蓄積された場合には乳酸の消費が認められた。このことは、アポトーシス耐性細胞はピルビン酸から糖新生を行うことはできず、乳酸からエネルギーを得ることもできないが、グルコースが乳酸とともに与えられた場合には、細胞はグルコース及び培養に蓄積又は添加された乳酸の両方を消費することができることを示唆している。更に、高濃度(60mM)のグルコースを含む培地では、アポトーシス耐性細胞株のIVCC(並びにVCD及び生存率、図9A〜C)はコントロール細胞株のIVCC(並びにVCD及び生存率)よりも大幅に高かった。これは、高グルコース培地では、コントロール細胞株が20mMよりも高濃度の乳酸を蓄積したことによるものであり、高濃度の乳酸が毒性を示した可能性がある。これに対して、アポトーシス耐性細胞株では増殖期の初期に乳酸を消費することにより、より多くの生細胞を維持することが可能であったため、コントロールと比較して170%というIVCCの増大につながった(図9C)。
【0030】
アポトーシス耐性細胞株は乳酸を消費して高いIVCCが実現されることから、治療用抗体などの対象とするタンパク質を発現するアポトーシス耐性ホスト細胞株由来の生産用細胞株では、コントロール細胞株に由来する生産用細胞株と比較して大幅に高い力価が実現された。このように高い力価は、治療用タンパク質又は他の対象とするタンパク質を生産する哺乳動物細胞株においてアポトーシス耐性遺伝子を使用することの潜在的な商業的利益を例証するものである。
【0031】
次に、本発明を以下の具体的かつ非限定的な実施例を参照しながら説明する。
【実施例】
【0032】
以下の実施例では、アポトーシス耐性遺伝子を過剰発現するCHO細胞株を震盪フラスコ培養中で、ピーク生細胞密度、寿命、カスパーゼ−3活性化、及びミトコンドリア膜電位(MMP)について分析を行った。更に、これらの細胞株における栄養素の消費量及び代謝産物の産生量をコントロール細胞と比較することによって、アポトーシス耐性遺伝子の発現が哺乳動物細胞の培養システム中の栄養素の消費及び代謝産物の蓄積に与える影響について調べた。
【0033】
材料及び方法
細胞培養:
コントロール細胞株C1013Aとして指定したCHOK1SV細胞株(ロンザ・バイオロジックス社(Lonza Biologics)英国、スラウ)を、30mMのグルコースを含み、6mMのL−グルタミン(インビトロジェン社(Invitrogen)カタログ番号10313〜021)を添加したCD−CHO培地(カタログ番号10743〜011、インビトロジェン社(Invitrogen)カリフォルニア州カールスバッド)中で培養した。場合に応じて、60mM(高グルコース培地として定義される)など異なる濃度のグルコースを含んだ別の無動物タンパク質培地を使用した。ウシ胎児血清はハイクローン・ラボ社(Hyclone Labs)(ユタ州ローガン)(カタログ番号SH30071.03)より購入した。細胞培養をCedex自動細胞計数装置(イノバティス社(Innovatis)ドイツ)により観測した。積分生細胞数(IVCC、細胞・日/ml)を下式:
IVCC(d1)=[VCD(d0)+VCD(d1)]/2+VCD(d0)
(式中、VCD=生細胞密度)により計算した。
【0034】
プラスミドの構築:
E1B−19K(EF−1aプロモーター)を単独で、又はAven(CMVプロモーター)と組み合わせて構成的に発現するように設計した、これまでに述べられている(Nivitchanyong et.al.,2007)pBUDCE4.1ベクターを使用した。XIAPΔ(CMVプロモーター)を発現するベクターは、サワーウォルド(Sauerwald et al.,2002)により述べられるようなものを使用した。ブランクベクターとは最初のpBUDCE4.1ベクターのことを指す。
【0035】
グルタミンシンターゼ(GS)発現ベクター(ロンザ・バイオロジックス社(Lonza Biologics)英国、スラウより研究用ライセンスにて取得)に重鎖及び軽鎖cDNAをクローニングすることによってモデル抗体(抗体1)発現ベクターを構築した。
【0036】
アポトーシス耐性細胞株の生成:
CHOK1SV細胞株の指数増殖期の培養に、1)pBUDCE4.1、2)pBUDCE4.1−E1B−19K、3)pBUDCE4.1−E1B−19K−Aven、並びに4)pBUDCE4.1−E1B−19K−Aven及びpCMV−XIAPΔをトランスフェクトした。トランスフェクションは製造者の推奨するところにしたがってFugene(ロシュ社()カタログ番号1815075、スイス、バーゼル)を使用して行った。トランスフェクトした2日後に、300μg/mlのゼオシン(上記のトランスフェクション1、2及び3に対し)、300μg/mlのゼオシン及び400μg/mlのハイグロマイシン(トランスフェクション4に対し)を含む増殖培地中、96穴プレートに播いた(表1を参照)。各トランスフェクションから約200個の抗生物質耐性クローンを増殖させて下記に述べるようにしてカスパーゼ3/7の活性について分析した。有望なクローンは、各抗生物質の不在下で10代継代して安定性試験を行った。
【0037】
【表1】

【0038】
最初に、CHOK1SV細胞株にブランクベクターをトランスフェクトし、ゼオシン耐性コロニーを選択することによってコントロール細胞株を作出した。これらの細胞株は、トランスフェクトしていないコントロールCHOK1SVと比較してIVCCが平均で約20%低かったため、CHOK1SVを後のすべての実験においてコントロール細胞株として用いた。次いで、最も有望なアポトーシス耐性細胞株であるEAX197及びコントロール細胞株を用い、GS発現ベクターを製造者の推奨するところにしたがって使用してモデル抗体を発現する生産用細胞株を作出した。細胞培養中の抗体をNephelometry(ベックマン・アレイ・システム社(Beckman Array System))によって測定した。
【0039】
アポトーシス耐性細胞株の震盪フラスコ培養:
選択されたアポトーシス耐性細胞株を、6mMのグルタミン及び必要な選択用抗生物質を添加したCD−CHO培地中で無添加の回分方式で培養した。一部の実験では、アポトーシス耐性細胞株の回分式培養に乳酸を添加して細胞株が消費した乳酸を補充した。更に、選択された抗体発現細胞株を、6mMのグルタミン及び60mMのグルコースを添加した特別に配合した無動物タンパク質培地中で培養した。
【0040】
カスパーゼ3/7活性のアッセイ:
約3×105個の各クローンの細胞を1mlの増殖培地中、24穴プレートに播種した。播種4日目後(d4)に、約1×105個の細胞を3重に96穴プレートに移した。スタウロスポリン(2μM fc)を添加し、細胞を16時間インキュベートした後、APO−ONEキット(ビー・ディー・ラボ社(BD Labs))によりカスパーゼ3/7の活性についてアッセイした。この手順をスタウロスポリンを加えない点を除き、10日目において繰り返した。4日目及び10日目の両方においてカスパーゼ3/7の活性が有意に低かったクローンをSF培地中で増殖させた。選択されたクローンのアポトーシス耐性を、フローサイトメトリー分析(下記を参照)及び場合によりミトコンドリア膜電位を測定することによって確認した。
【0041】
フローサイトメトリーによるアポトーシス耐性クローンの分析:
指数増殖期の約1×106個の細胞を各震盪フラスコから抜き取って24穴プレートに移し、スタウロスポリン(2μM fc)と16時間インキュベートし、収穫してPBS中で1回洗った。次いで細胞をCytoPerm(カタログ番号2075KK、ビー・ディー・バイオサイエンス社(BD BioScience))とインキュベートして固定するとともに細胞を透過性とした。PBSによる洗浄後、細胞をFITC標識抗カスパーゼ3(カタログ番号68654、ビー・ディー・バイオサイエンス社(BD BioScience))抗体とインキュベートした後、フローサイトメトリーにより分析した。
【0042】
ミトコンドリア膜電位(MMP)アッセイ:
細胞を震盪フラスコ培養から播種後6日目に抜き取り、スタウロスポリン(5μM fc)と2時間インキュベートした。この後、細胞を洗い、製造者の推奨するところにしたがって親油性カチオン性色素であるJC−1(ケイマン・ラボ社(Cayman Labs)ミシガン州アナーバー)で処理した。プレートをFL535及びFL595で読み取り、その比を計算した。
【0043】
ウェスタンブロッティング:
示される細胞株から約1×107個の細胞を収穫し、PBS中で洗浄し、1% NP−40、120mMのTris−Hcl、150mMのNaCl、0.2mMのPMSF、及び1mMのEDTAを含むRIPAバッファー中で溶解した。各試料からの細胞質タンパク質50μgを4〜12% NuPAGE勾配ゲルにロードし、Novexシステム(インビトロジェン社(Invitrogen))でSDS−PAGEによって分離した。分離したタンパク質のバンドをニトロセルロース上に転写して、1)ヒトE1B−19Kに対する抗マウス抗体(希釈率1:40)(カタログ番号DP−17、カルバイオケム社(CalBiochem)メリーランド州ギブスタウン)、2)ヒトAvenに対する抗ウサギ抗体(希釈率1:1000)(カタログ番号612521、ビー・ディー・バイオサイエンス社(BD BioScience)カリフォルニア州サンホセ)、及び3)ヒトXIAPに対する抗マウス抗体(カタログ番号616713、ビー・ディー・バイオサイエンス社)を用いて分析した。使用した抗体を含め、上記のプロトコールは、販売業者から購入した既知量の精製したE1B−19K、Aven、及びXIAP(それぞれ約10ng)を用いて最適化かつ標準化した。タンパク質のバンドはECL検出キット(ジー・イー・ヘルスケア社(GE Healthcare)ニュージャージー州ピスカタウェイ)を使用して可視化した。
【0044】
培地中の代謝産物濃度の測定:
すべての重要な代謝産物及び老廃物の濃度をYSI2700自動分析装置を使用して測定した。アンモニアイオン濃度はフローインジェクション分析(Campmajo et al,1994)により測定した。アミノ酸は逆相カラム(ウォーターズ社(Waters)マサチューセッツ州ミルフォード)を用い、HPLC(ウォーターズ社)によって測定した。
【0045】
(実施例1)
アポトーシス耐性細胞株の生成及び特性評価
本実験で使用した細胞株、各細胞株を生成するためにそれぞれトランスフェクトした発現プラスミド、及び形質移入細胞(トランスフェクトーマ(Transfectoma))を単離するために使用した選択用物質のリストを上記表1に示す。表に示した細胞株は、各トランスフェクションから生成された多数のクローンの内の代表的なものである。各細胞株の名称は、ホスト細胞株にトランスフェクトされたアポトーシス耐性遺伝子に由来している。例えばEAX197は、E1B−19K(E)、Aven(A)、及びXIAPΔ(X)をトランスフェクトした細胞株である。アポトーシス耐性(アポトーシスR)という表記は、これらのアポトーシス耐性遺伝子の1以上がトランスフェクトされた細胞株を指して用いる。
【0046】
本実験で用いたすべてのアポトーシス耐性細胞株について、ウェスタンブロッティング、カスパーゼ3/7、及びミトコンドリア膜電位による特性評価を行った(データは示さず)。1重形質移入体E64、又は2重形質移入体EA63、EA112、EA167及びEA190はいずれも、コントロールであるトランスフェクトされていない細胞株C1013Aで見られたバックグラウンドレベルよりも高い発現レベルでE1B−19Kを発現した。4つの2重形質移入体の内、細胞株EA112及びEA167は、E1B−19K(約19kDa)及びAven(約55kDa)の両方を発現した。細胞株EA63及びEA190は、E1B−19Kを高レベルで発現したが、Avenの発現レベルは極めて低かった。2重ベクターpBUDCE4.1−E1B−19K−AvenとpCMV−XIAPΔをトランスフェクトすることによって生成された4つの細胞株、すなわちEAX64、EAX99、EAX148及びEAX197の内、EAX197のみが3種類のタンパク質のすべてを検出可能なレベルで発現した。
【0047】
トランスフェクトしていないコントロールのレーンC1013Aを含むすべてのレーンにおいて組換えAvenの上側に観察された約55kDaのバンドは、免疫後の血清では可視化されたが免疫前の血清では可視化されなかったことから内因性のAvenタンパク質であると考えられた(Sauerwald et al.,2002,Chau et al.,2000)。トランスフェクトされたAven遺伝子は最初の6個のアミノ酸が欠失しているが、これらのアミノ酸の欠失はタンパク質の生物学的活性に影響しないことが示されている。このため、トランスフェクトされたAvenは内因性のAvenよりも若干小さい。Avenの分解産物である可能性のある40kDaのバンドがこのタンパク質をE1B−19Kとともに過剰発現したすべての細胞株において観察された。同様のサイズの分解生成物が、AvenがBcl−XL(Sauerwald et al.,2005)又はBcl−2(Figueroa、未発表の観察)とともに過剰発現された場合に観察されている。更に、Avenの発現は、E1B−19K、Aven及びXIAPΔを過剰発現するEAX197を含む3重形質移入体において顕著に低かった。
【0048】
興味深い点として、トランスフェクトしたXIAPΔの発現レベルは内因性の完全なXIAPタンパク質の発現レベルと比較して顕著に低かった。XIAPとXIAPΔとの間の発現レベルの明瞭な差は、アポトーシス耐性タンパク質のこれら2つの形態の抗体結合効率の差によって一部説明することができる。内因性のXIAPと形質移入されたXIAPΔとの間の明瞭な発現レベルの差は、アポトーシスの防止におけるこのトランスフェクトタンパク質の寄与に疑問を投げかけるものである。実際、この実験は、所定のアポトーシス耐性遺伝子の発現レベルと、そのタンパク質が細胞株に与えることのできるアポトーシス阻害のレベルとの間には必ずしも直接的な相関が認められないことを示唆している。
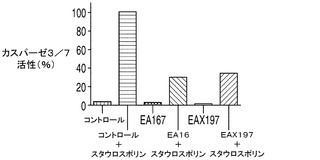
【0049】
カスパーゼ3/7の活性はアポトーシスに対する細胞株の耐性の大きさの指標を与えることから、カスパーゼ3/7活性についてAPO−ONEによりそれぞれのアポトーシス耐性細胞株を調べた。簡単に述べると、このアッセイでは、蛍光前駆基質であるZ−DEVD−R110がカスパーゼ3/7によって用量依存的に開裂される。EA167及びEAX197並びにコントロール細胞株の指数増殖期の培養をスタウロスポリンで16時間処理することによってアポトーシスを誘導した後、カスパーゼ3/7を測定した。スタウロスポリン処理したコントロール細胞のカスパーゼ3/7活性を任意に100%に設定した。これらの条件下で、EA167及びEAX197細胞のカスパーゼ活性はそれぞれコントロールの30%及び35%であったが、このことはこれらの細胞株がアポトーシス誘導に対して少なくとも部分的に耐性を有することを示唆している(図2A)。EA167及びEAX197のアポトーシス耐性の更なる根拠として、これら2つの細胞株とコントロール細胞株のスタウロスポリン処理培養をFITC結合抗カスパーゼ3抗体で標識した後、フローサイトメトリーで分析した。図3に見られるように、非処理又は溶媒(DMSO)処理によらず、3つの細胞株のすべてにおいてカスパーゼ3陽性細胞は集団の1%以下であった。しかしながら、スタウロスポリンによる処理後にはコントロール細胞集団の91%がカスパーゼ3について陽性であった。これに対し、細胞株EAX197に由来する集団のわずか9%、及び細胞株EA167に由来する集団の30%がカスパーゼ3について陽性であり、これら2つの細胞株におけるアポトーシス耐性の表現型が証明された。
【0050】
アポトーシスに対する耐性が寿命の増大につながるか否かを調べるため、複数のEA細胞株を震盪フラスコ(回分方式)中で増殖させ、生細胞密度(VCD)及び積分生細胞数(IVCC)の両方を観測した。細胞の比生産性(specific productivity)が変化しないものと仮定すると、積分生細胞数が大きくなると培養の体積生産性が向上する。したがってIVCCを、生産用細胞株の作出のためのホストとして機能しうる優れたアポトーシス耐性細胞株をスクリーニングするためのパラメータとして用いることができる。図2Bに示されるように、相対カスパーゼ3/7活性が低い細胞株は、IVCCの高い細胞株を生成する傾向が見られた。EA細胞株のそれぞれはコントロールに対して低いカスパーゼ3/7活性を示したが、IVCCとそれぞれの細胞株が有するカスパーゼ3/7のレベルとの間には厳密な相関は認められなかった。更に、特定のアポトーシス耐性細胞株はアポトーシス耐性の表現型の程度に関してその姉妹クローンよりも安定していた。EA167は強力な抗アポトーシス活性及び震盪フラスコ培養中での安定した増殖性の両方を有することから、この細胞株を更なる実験用に選択した。更に、EA167はこのパネルでは最も安定した細胞株であり、ウェスタンブロットによって検出されるようにAvenを高レベルで発現する。
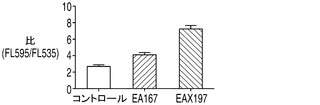
【0051】
アポトーシス経路のシグナル伝達分子の多くがミトコンドリアと相互作用するかあるいはミトコンドリア内部に存在している。ミトコンドリアでこれらのタンパク質はミトコンドリアの膜と相互作用してミトコンドリアの透過性を調節することによりアポトーシスを制御する。したがって、ミトコンドリア膜電位(MMP)は、コントロールに対するEA167及びEAX197のアポトーシス耐性の特性を比較するための細胞生理学の別のモニターとして機能する。図2Cのアッセイの結果に見られるように、EA167及びEAX197はいずれもコントロール細胞株よりも高いMMPを有していた。
【0052】
(実施例2)
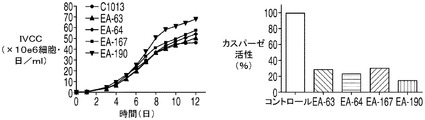
CHOK1SVの生細胞密度に対するE1B−19Kの影響
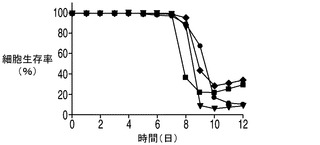
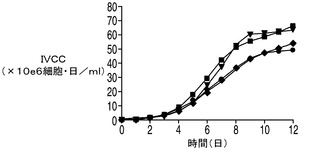
E1B−19KをAvenとともに過剰発現する2重形質移入体(EA167)、E1B−19KをAven及びXIAPΔとともに過剰発現する3重形質形質移入体(EAX197)、並びにE1B−19Kのみを過剰発現する形質移入体(E64)を図4でコントロール(ホスト)細胞株と比較した。コントロール細胞株のピークVCDは7.7×106細胞/mlに達し、E64(E1B−19Kのみを発現)のピークVCDは8.9×106細胞/mlであったのに対して、EA167のピークVCDは1.4×107細胞/mlであり、EAX197のピークVCDは1.5×107細胞/mlよりも高く、コントロール細胞株と比べて190%以上の増加を示した。EA167及びEAX197の生存率は、恐らく栄養素の枯渇のために播種後8日目又は9日目に急激に低下したが、コントロール及びE64細胞株の生存率の低下はより緩慢であった(図4B)。IVCCに対する正味の影響を図4Cに示す。コントロール細胞株のIVCCは4.9×107細胞・日/mlであり、次がE64 のIVCC(5.2×107細胞・日/ml)であった。明らかに2つのアポトーシス耐性細胞株EA167(6.3×107細胞・日/ml)及びEAX197(6.6×107細胞・日/ml)のIVCCは、コントロール細胞株のIVCCよりも有意に高かった(EA167及びEAX197でそれぞれ136%及び140%)。
【0053】
EA167及びEAX197以外にも、幾つかの他のアポトーシス耐性細胞株が同様に低い乳酸産生表現型を示した(データは示さず)。更に、低い乳酸産生特性を示す野生型細胞株も稀ではあるが報告されている(Pascoe et al,2007)。
【0054】
(実施例3)
アポトーシス細胞株の代謝プロファイル
アポトーシスからの保護を与えるアポトーシス耐性細胞株EA167及びEAX197の生理学的性質を回分培養中でコントロール細胞と比較するため、回分培養の開始時に与えられる主要な栄養素及びアミノ酸、すなわちグルコース、グルタミン酸及びグルタミンの濃度を観測した。更に、老廃物であるアンモニア及び乳酸の蓄積についても観測を行った。図5に、EAX197及びEA167並びにコントロール細胞株を市販のCD−CHO培地中、震盪フラスコ中で回分方式で培養した場合のこれらの代謝物質の濃度を示す。図5Aに示されるように、グルコースは播種後6日目又は7日目までに3つの細胞株のすべての培地から急速に枯渇した。グルタミンの枯渇についてはアポトーシス耐性細胞株とコントロール細胞株との間に有意差は認められず、グルタミン酸濃度は全日数にわたって3つの細胞株で大きく変化しなかった。3つの細胞株のすべてにおいて、4日目までの細胞増殖期の初期において乳酸の蓄積が認められた(図5B)。コントロール細胞株では、乳酸は6日目まで増加し続けた(その時点で2.8g/Lに達した)後、乳酸濃度に若干の低下が認められ、最終的に1.8g/Lに達した。こうした量の乳酸は、培養のpHを低下させることが予想され、重炭酸塩を添加することによってpHが調節されているバイオリアクター中では浸透圧が上昇することになる。これらの特性のすべて及び他のストレス因子によって動物細胞にとって有毒な環境がもたらされうる。これに対して、アポトーシス耐性細胞株中の乳酸濃度は短期間増大した後、5日目以降は急速に低下し、7日目(EA167)又は8日目(EAX197)までには乳酸は完全に消失した。興味深い点として、図5Aにおいてこれら2つの細胞株では乳酸が0にまで低下した直後にVCDが急激に低下しており、細胞生存率の低下が培地中に利用可能なグルコース又は乳酸がなくなったことによるものであることを示唆している。
【0055】
アポトーシス耐性細胞株とコントロール細胞株との間に認められた別の差異はアンモニアのプロファイルである(図5C)。EA167及びEAX197中のアンモニア濃度は、乳酸濃度の低下が始まった6日目に若干低下し、その後やがて再び増大した。しかしながら、アポトーシス耐性細胞株の全体のアンモニア濃度はコントロール細胞株よりも低く、このことはアポトーシス耐性細胞株がコントロール細胞株と比較してより高いIVCCに達し、したがって全体の細胞増殖率がより高かったことから予想外の結果である(図4C)。
【0056】
上記のデータから、アポトーシス耐性細胞株の特性がコントロール細胞株とは大きく異なっていることが明らかである。しかしながら、複数のクローンについて幾分異なる実験条件下で調べたところ、各アポトーシス耐性形質移入体間で幅広い表現型が観察された。表現型の変異はコントロール細胞株の単離クローンにおいても見られることから、適当なスクリーニング法を用いることによってコントロール細胞株から稀なアポトーシス耐性クローンを選択することが可能である。この点に関しては、マタノビッチ(Mattanovich)及びボース(Borth)(2006)によって細胞分別を応用して所望の性質を有する稀な変異体を野生型(コントロール)細胞株から単離することについて概説されている。
【0057】
(実施例4)
乳酸添加の影響
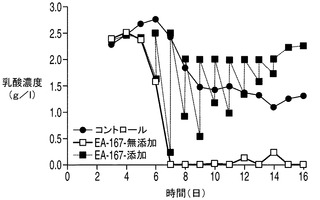
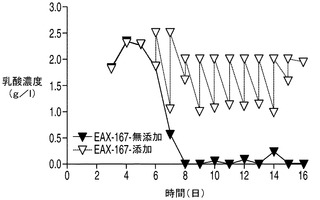
任意の時点においてある細胞培養中に存在する乳酸の量は、その細胞株によって分泌される乳酸の量から細胞株によって消費される乳酸の量を差し引いた量によって決まる。アポトーシス耐性細胞株が実際に乳酸を消費しているか否か、すなわちこれらの細胞株において正味の乳酸産生量が低下しているか否かを調べるため、3つの細胞株すべての培養を毎日観察し、アポトーシス耐性細胞株とコントロール細胞株との間で乳酸濃度に目立った差があれば外因性の乳酸を添加することによって埋めた。上記で観察したように、無添加のアポトーシス耐性細胞株EA167及びEAX197の乳酸濃度は5日目に低下しはじめ(図6A及び6B)、7日目(EA167、無添加)又は8日目(EAX197、無添加)までに乳酸は枯渇した。コントロール細胞株の乳酸プロファイルは6日目まで増加しつづけ、そこから次の3日間濃度は若干低下したが0にまで低下することはなかった。アポトーシス耐性細胞株を毎日観察し、個別の流加培養に7日目に2.5g/L(EA167に対して、図6A)及び8日目に2g/L(EAX197に対して、図6B)となるように乳酸を加えてコントロール細胞株の濃度と同様の濃度とした。補充分の乳酸の添加にも関わらず、アポトーシス耐性細胞株ではコントロールと比較して6日目〜12日目にかけて乳酸濃度は大幅に急速に低下した。実際、EAX197の流加培養では6日目〜7日目にかけて乳酸濃度が2.5〜0.2g/Lにまで低下したのに対して、コントロール培養では同じ期間に乳酸濃度は2.7〜2.4g/Lに低下したに過ぎなかった。したがって、乳酸はアポトーシス耐性細胞株ではコントロール細胞株と比較して補充分の乳酸の添加後少なくとも7日間にわたって加速した速度で消費されたものと考えられる。
【0058】
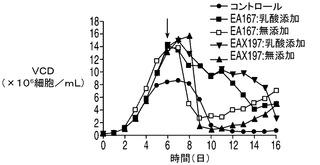
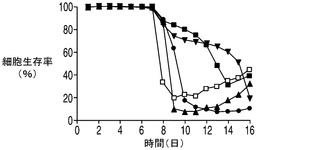
VCD及び細胞生存率に対する乳酸補充の影響をそれぞれ図7A及び7Bに示す。上記で観察したように(図5A)、無添加のアポトーシス耐性細胞株のVCDは8日目(EA167)及び9日目(EAX197)から急速に低下した。これは恐らく、グルコース及び/又は乳酸を含む1以上の不可欠な培地成分がこれらの培養中で枯渇したことによるものと考えられる。コントロール細胞株では7日目から緩慢な低下を示した。しかしながら、乳酸を補充した培養(EA167(乳酸添加)及びEAX197(乳酸添加))ではVCDの低下はより緩やかであり、一方の細胞株(EAX197(乳酸添加))では、VCDは、14日目にこの培養が最終的に死滅するまで8×106〜10×106細胞/mlの間で安定した。図7Bに示される細胞生存率は、培地に乳酸を補充した2つのアポトーシス耐性培養において同様のより緩やかな低下を示した。これらの結果は、アポトーシス耐性細胞株が内因性及び補充された乳酸の両方を効率的に利用して正細胞密度及び生存率を維持することが可能であるのに対して、コントロール細胞株は同様の効率的な乳酸消費を行うことができないことを示唆している。この差は、無添加のアポトーシス耐性細胞株よりも約180〜220%高く、コントロール培養よりも約235%〜250%高かった乳酸添加アポトーシス耐性細胞株のIVCCに反映されている。
【0059】
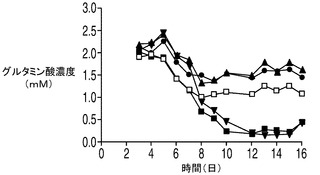
図7D〜Fに示される他の栄養素及び代謝産物の多くは、図5の無添加培養で上記に観察されたものと同様のプロファイルを示した。グルコース及びグルタミンは培養の初期の時点で急速に消費された。アポトーシス耐性細胞株のアンモニアのプロファイルもやはり、無補充及び乳酸補充培養の両方において乳酸の消費が始まった時点でわずかに低下した。上記の各実験と同様、各アポトーシス耐性細胞株の最終的なアンモニア濃度は、IVCCが4つのアポトーシス耐性細胞株のすべてにおいて大幅に高かったにも関わらずコントロールの濃度よりも低かった。グルタミン酸の濃度は乳酸添加培養と無添加培養との間で異なり、無添加及びコントロール細胞株と比較して添加培養でグルタミン酸の枯渇は大幅に大きかった。
【0060】
(実施例5)
アポトーシス耐性細胞株及びコントロール細胞株のアミノ酸プロファイル
20種類のアミノ酸のそれぞれの濃度を、コントロール細胞株、並びに無添加及び乳酸添加アポトーシス耐性細胞株の培養において観測した。グルタミン及びグルタミン酸以外のアミノ酸の消費又は産生に関しては、表2に示されるように9種類の必須アミノ酸(フェニルアラニン、トレオニン、メチオニン、リジン、トリプトファン、ロイシン、イソロイシン、バリン及びヒスチジン)のいずれも枯渇しなかった点を最初に述べておく必要がある。アポトーシス耐性細胞株(無添加)及びコントロール細胞株の培養では、アスパラギン、セリン、トリプトファン及びシステイン(斜体で示す)はほぼ完全に枯渇した。興味深い点として、3種類の分枝鎖アミノ酸、すなわちイソロイシン、ロイシン及びバリン(表2に灰色の網かけで示す)はいずれも、コントロールと比較してアポトーシス耐性細胞株においてより速やかに培地から欠乏し、この欠乏は乳酸添加培養においてより顕著であった(図8A〜D)。グルタミン酸に加え、これら3つのアミノ酸の最終濃度は乳酸添加培養において培地から大幅に低下した。消費率が高かった他のアミノ酸(最終濃度が培地中の最初の濃度の50%以下)としては、チロシン、フェニルアラニン、メチオニン(図8D)及びアスパラギン酸(図8E)がある。
【0061】
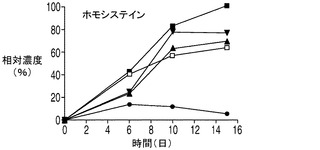
培地中に分泌されたアミノ酸についてコントロール細胞株とアポトーシス耐性細胞株との間で比較を行うのも考慮の対象として興味深い。図8Eに示されるように、コントロール細胞株が最初の6日間にわたって培地中にアスパラギン酸を蓄積したのに対して、4つのアポトーシス耐性細胞株培養のすべてにおいて同じ期間でアスパラギン酸濃度は低下した。更に、各アポトーシス耐性細胞株はコントロール細胞株と比較して最初の6日間にわたって大幅に多量のアラニンを分泌した(図8F)のに対して、これよりも遅い期間ではすべての細胞株がアラニンを消費した。更に、各アポトーシス耐性細胞株はコントロール細胞株と比較して細胞培養の全体を通じてより多くのホモシスチンを産生した(図8G)。
【0062】
(実施例6)
高濃度グルコースの存在下での回分培養
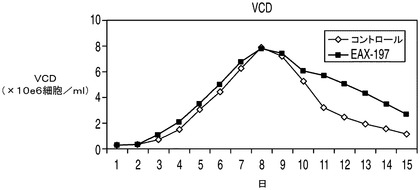
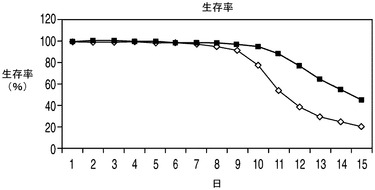
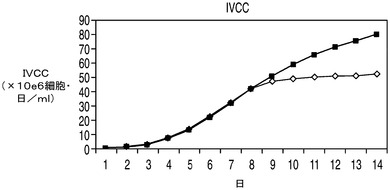
アポトーシス耐性細胞株が乳酸を利用できるならば、有毒なレベルの乳酸が蓄積するためにコントロール細胞にとっては阻害性を示すようなグルコース濃度においてもこれらの細胞株は生存できる可能性がある。更に、これらの細胞株の増殖は乳酸を唯一の炭素源として含む培地によって支持されうる可能性がある。高濃度のグルコースなどのため浸透圧が維持できなくなるために上記の実験で使用したCD−CHO培地を使用することは不可能であり、そのためこれらの実験では新しい培地を使用する必要があった。したがって、上記の実験で最も高い持続的増殖性を示したEAX197の増殖速度論を、60mMのグルコース(高濃度グルコース)又は30mMの乳酸を含む特別に配合した無動物タンパク質培地中でコントロール細胞株と比較した。アポトーシス耐性細胞株及びコントロール細胞株のいずれにおいても、グルコースがまったく存在しない場合、すなわち乳酸のみが存在する場合には持続的増殖が見られなかった(データは示さず)。両方の細胞株を「高濃度」(60mM)のグルコースを含む同じ特別配合培地中で培養したところ、EAX197は、特に細胞培養の後期においてコントロール細胞株よりも高いVCD、生存率及びIVCCを示した(図9A〜C)。EAX197細胞株は、上記のCD−CHO培地中におけるよりもコントロールと比較した場合の全体のIVCCの増加率は低かった(130%)が、後期の高い生存率のためにその差は依然有意であった。高濃度グルコース中で培養したこれら2つの細胞株の乳酸プロファイルを図9Dに示す。IVCCと同様、両細胞株の乳酸プロファイルは、CD−CHO培地で観察されたものとは幾分異なっていた(図9D)。CD−CHO培地を用いた上記の結果と同様、特別配合培地中のアポトーシス耐性細胞株ではコントロールよりも早期に正味の乳酸消費が開始され(2日目まで、図9Dに示される)、最大乳酸濃度は大幅に低かった(コントロール細胞株の2.4g/Lに対して1.2g/L)。コントロール細胞は、培地中の乳酸の消費も開始した5日目までは乳酸の正味の産生を示した。CD−CHO培地は適当な組成を有する市販の配合物であるため、これらの異なる配合物中において乳酸消費プロファイルが異なる原因を理解する目的で2つの配合物中の培地成分の正確な比較を行うことはできなかった。
【0063】
(実施例7)
高グルコース培地中のアポトーシス耐性細胞株の生産性
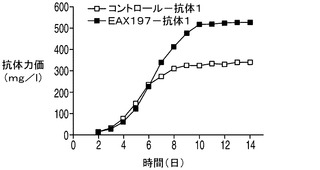
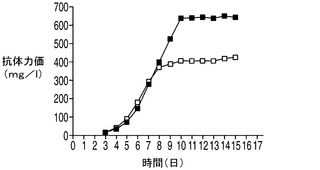
続いて、アポトーシス耐性細胞株であるEAX197及びコントロール細胞株をホストとして使用して、モデル抗体を発現する生産用細胞株を作出した。各細胞株について同数のクローンを調べ、各ホストから生成した最も優れたクローンを抗体の生産性について震盪フラスコ回分方式で比較した。高グルコース環境下における各細胞株の性能を比較するため、60mM(高濃度)グルコースを含む特別配合培地を炭素源として使用した。標準培地(図10A)中では、EAX197から誘導された生産用細胞株では力価が489mg/Lであり、337mg/Lの力価を示したコントロール細胞株と比較して145%の増加率であった。高グルコース培地(図10B)中では同じ細胞株が678mg/Lの力価を示し、コントロール細胞株(348mg/L)の力価と比較して178%の増加率であった。産生量の向上は、EAX197ではIVCCがコントロール細胞株に対して188%に増大していることに一部よるものである(データは示さず)。
【0064】
(実施例8)
E1B19K+AVEN±XIAPΔの過剰発現のアポトーシスの阻害に対する影響
ミトコンドリア内のアポトーシスカスケードの主要な活性化因子を図1に示した。アポトーシス耐性CHO細胞株を作出するため、アポトーシスの早期、中期及び後期に働く3つのアポトーシス耐性遺伝子を選択した。これらの遺伝子は、E1B19K(E)、AVEN(A)及びXIAPΔ(X)である。広く用いられている形質移入(トランスフェクション)用ホスト細胞株であるCHOK1SVに、E1B19K及びAVENを、XIAPΔとともに、又はXIAPΔ抜きでトランスフェクトした。多数の形質移入細胞をスクリーニングして各トランスフェクションから数個のクローンを更なる実験用に選択した。これらの細胞株の増殖プロファイルの実験により、これらの細胞株は極めて高いVCDに達し、一般にそのIVCCはコントロールのIVCCの約2倍高いことが示された。アポトーシス耐性細胞株では播種後8日目までに生存率が低下したが、乳酸を消費することによって10日目以降は生存率が回復した。E1B19K+AVEN±XIAPΔを発現しているアポトーシス耐性細胞株の培養液は、アンモニア及び乳酸の濃度が低下しているか検出不能なレベルであり、グルタミン酸は培地から枯渇していた。
【0065】
(実施例9)
抗体力価を高めるためのBcl−2dの使用
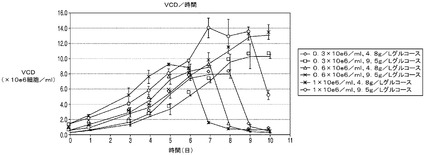
上記の結果は、アポトーシス耐性細胞株はコントロール細胞と比較して乳酸の蓄積量が少なく、アポトーシス耐性培養の生存率を高グルコース培地中で維持することが可能であることを示している。以下の震盪フラスコ回分培養実験では、Bcl−2d並びに抗体の重鎖及び軽鎖を発現するアポトーシス耐性細胞株(C2088B)を標準的濃度(4.8g/L)及び高濃度(9.5g/L)のグルコース中、異なる播種密度で播種した。この結果は、アポトーシス耐性細胞株は9.5g/Lのグルコースを含む培地中に1e6細胞/mlの密度で播種した場合、播種後7日目に14e6細胞/mlのピークVCD(図11A)、及び2g/Lの抗体力価に達したことを示す。これに対して、標準条件下(接種密度0.3e5細胞/ml及び4.8g/L)で培養したC2088BはビークVCDが7e6細胞/ml、抗体力価は1g/Lであった(図11B)。これは、高グルコース条件では標準条件(すなわち、4.8g/Lのグルコースを含む培地中、0.3e5細胞/mlの播種密度)と比較して抗体力価が200%に増大することを意味する。
【0066】
以上に本発明の全容を説明したが、本発明には、多くの変更及び改良を、添付の特許請求の範囲の趣旨及び範囲から逸脱することなく行いうることは当業者には明白であろう。
【技術分野】
【0001】
本発明は、高濃度のグルコース添加により、流加細胞培養中で高い生細胞密度及び長い培養寿命を実現するための方法に関する。本方法は、対象とする分泌タンパク質の産生量を高めるうえで有用である。
【背景技術】
【0002】
哺乳動物細胞の培養は、適当な翻訳後修飾がなされたタンパク質を生産することが可能であることから、多くの組換えタンパク質製造プロセスにおいて選択されている系である。製造需要が高まるにつれ、産物の収率を高めることによってプロセス効率を改善するとう強い動機づけがある。商業的な製造プロセスにおいて生物学的治療を目的としたタンパク質又は他のタンパク質の1L当たり数グラムの製造レベルを実現するには、哺乳動物細胞の培養及び操作方法を最適化することが不可欠である。現在の高密度、無タンパク質の哺乳動物細胞培養に内在する問題として細胞死の問題があり、一般的な流加バイオリアクターではアポトーシスによるものが最大でその80%を占める。こうした細胞死は、栄養及び増殖因子の欠乏、酸素欠乏、毒素の蓄積、及び剪断応力といった諸条件に応じて誘導されるものである(Goswami et al.,Biotechnol Bioeng 62:632〜640(1999))。アポトーシスは最大の生細胞密度を制限し、死滅期の開始を早め、異種由来タンパク質の収率を潜在的に低下させる(Chiang and Sisk,Biotechnol Bioeng 91:779〜792(2005);Figueroa et al.,Biotechnol Bioeng.73:211〜222(2001),Metab Eng 5:230〜245(2003),Biotechnol Bioeng 85:589〜600(2004);Mercille and Massie,Biotechnol Bioeng 44:1140〜1154(1994))。
【0003】
アポトーシスは細胞の内外で開始されるシグナル伝達経路の複雑なネットワークによって引き起こされるものであり、細胞死の最終ステージを媒介するシステインアスパラギン酸プロテアーゼ(カスパーゼ)が最終的に活性化される。これについては図1を参照されたい。哺乳動物の細胞培養における長期の産生培養の間に細胞生存率を維持する目的でアポトーシスを防止する様々な方法が用いられている(Arden and Betenbaugh,Trends Biotechnol 22:174〜180(2004);Vives et al.,Metab Eng 5:124〜132(2003))。増殖因子、加水分解物、及び律速栄養素を培地に添加して細胞外環境を変化させることによってタンパク質の産生量が高まり、アポトーシスが減少する(Burteau et al.,In Vitro Cell Dev Biol Anim.39:291〜296(2003);Zhang and Robinson,Cytotechnology 48:59〜74(2005))。細胞内からのアポトーシスのシグナル伝達カスケードを阻害する化学的及び遺伝子的手法を模索する研究者もいる(Sauerwald et al.,Biotechnol Bioeng 77:704〜716(2002),Biotechnol Bioeng 81:329〜340(2003))。
【0004】
ガン細胞において発現量が増大している遺伝子の過剰発現が、カスパーゼ活性化の上流でアポトーシスを防止することによってバイオリアクター中で増殖させた細胞の生存期間を延ばしうることが研究者によって見出されている(Goswami et al.,supra;Mastrangelo et al.,Trends Biotechnol 16:88〜95(1998);Meents et al.,Biotechnol Bioeng 80:706〜716(2002);Tey et al.,J Biotechnol 79:147〜159(2000)、及び、Biotechnol Bioeng 68:31〜43(2000))。生産用細胞株におけるこれらのタンパク質の発現量の増大は、アポトーシスを引き起こす細胞内のシグナル伝達を効果的に抑制することによって細胞死を制限することにより、特定の場合において生存率を維持し生物治療物質の産生量を増大させた。
【0005】
高密度、無タンパク質の哺乳動物細胞培養にやはり内在する問題点として、老廃物の蓄積及びそれが細胞増殖にもたらす悪影響がある。細胞培養における最も一般的な2つの老廃物は、乳酸及びアンモニアである。過剰な乳酸の蓄積の問題に対処するために多くの手法が考案されており、1)培地のグルコース濃度を低く維持する(Kurokawa et al.,BiotechnolBioeng 44:95〜103(1994);Xie and Wang,Biotechnol Bioeng 43:1175〜1189(1993);Zhang et al.,J Chem Technol Biotechnol 79:171〜181(2004);Zhou et al.,Biotechnol Bioeng 46:579〜587(1995))、2)フルクトースなどの代替的な糖を添加する(Martinelle et al.,Biotechnol Bioeng 60:508〜517(1998),Altamirano et al.,J Biotechnol 110:171〜179(2004)and J Biotechnol 125:547〜556(2006),Walschin and Wu,J Biotechnol 131:168〜176(2007))、3)相同組み換え又はSiRNA技術により乳酸脱水素酵素(LDH)の発現を部分的にノックアウトする、4)ピルビン酸カルボキシラーゼの過剰発現、5)ピルビン酸脱水素酵素(PDH)の活性化因子(PDHキナーゼ阻害を介した)であるジクロロ酢酸塩(DCA)の使用、6)LDH競合阻害剤であるオキサミン酸、及び7)灌流による除去(米国特許出願公開第2009/0042253A1号)などが挙げられる。
【0006】
当初は、多くのアポトーシス経路のタンパク質の固有の機能は、ミトコンドリア膜に結合してミトコンドリアの透過性を調節することによりアポトーシスを調節することと考えられていた。最近の研究により、アポトーシスを引き起こすシグナル伝達に関与する主要なタンパク質は、細胞の代謝及びエネルギー恒常性を制御するタンパク質と相互作用してこれらのタンパク質に影響を及ぼすことが示された。Majors et al.,Metab Eng 9:317〜326(2007)(概論)、及びWhite et al.,Nat Cell Biol 7:1021〜1028(2005)を参照されたい。最近の研究では、BcL−XLを過剰発現しているCHO細胞のマイクロアレイ分析により、糖新生の主要な酵素である乳酸脱水素酵素の発現量が増大していることが示された。
【0007】
特定の細胞及びウイルスは、ミトコンドリアのアポトーシス経路において機能するアポトーシス耐性遺伝子を生産する。これらの遺伝子は、以下の3つのグループ、すなわち、1)経路の早期に働くもの(例えば、Bcl−2ファミリーメンバーのタンパク質)、2)経路の中期に働いてアポトソーム複合体を妨害又は阻害するもの(例えば、Aven)、及び3)経路の後期に働くもの(例えば、XIAPなどのカスパーゼ阻害因子)に分類することができる。これらの遺伝子の大半のものの機能は、哺乳動物の発現系においてこれらを過剰発現させることによって研究されており、特定の場合では、それぞれが経路の異なる部分に由来する2以上の遺伝子の過剰発現の組み合わせによる影響が調べられている。その例としては、1)CHO細胞におけるBcL−XLとXIAPの欠失突然変異(XIAPΔ)との相加的効果(Figueroa et al.,Metab.Eng.5:230〜245(2003))、2)BHK細胞におけるE1B−19KとAven(Nivitchanyong et al.,Biotechnol Bioeng 98:825〜841(2007))、及び3)BcL−XL、Aven、及びXIAPΔ(Sauerwald et al.,supra,(2003);Sauerwald et al,Biotechnol Bioeng 94:362〜369(2006))が挙げられる。
【発明の概要】
【発明が解決しようとする課題】
【0008】
しかしながら、これらの研究では、細胞の細胞代謝状態に対するアポトーシス耐性遺伝子の影響については調べられていない。したがって、高い生細胞密度、寿命、及び生産性を実現するために栄養消費及び代謝産物の蓄積状態を最適化するための哺乳動物細胞の培養システムが求められている。
【課題を解決するための手段】
【0009】
本発明の一態様は、真核細胞の流加培養において高い生細胞密度を得るための方法であって、
a)1種以上の異種由来のアポトーシス耐性(アポトーシスR)遺伝子及び1種以上の対象遺伝子を発現する真核細胞株を培養する工程と、
b)前記細胞培養の指数増殖期及び定常期に高グルコース添加培地を維持する工程と、を含む方法である。
【0010】
本発明の別の態様は、真核細胞の流加培養において分泌タンパク質の産生量を高めるための方法であって、
a)1種以上の異種由来のアポトーシス耐性(アポトーシスR)遺伝子及び1種以上の対象遺伝子を発現する真核細胞株を培養する工程と、
b)前記細胞培養の指数増殖期及び定常期に高グルコース添加培地を維持する工程と、を含む方法である。
【図面の簡単な説明】
【0011】
【図1】ミトコンドリアのアポトーシス経路の構成要素を示す図。
【図2A】アポトーシス耐性細胞株の特性評価を示す図。
【図2B】アポトーシス耐性細胞株の特性評価を示す図。
【図2C】アポトーシス耐性細胞株の特性評価を示す図。
【図3】フローサイトメトリー分析によりEA167及びEAX197のアポトーシス耐性を証明した図。
【図4A】E1B19K、E1B19K+AVEN、及び、E1B19K+AVEN+XIAPΔをトランスフェクトすることによって生成されたアポトーシス耐性細胞株の増殖プロファイルである。コントロール(●)、E64(◆)、EA167(■)及びEAX197(▼)の震盪フラスコ回分培養を、生細胞密度及び生存率について観測した。
【図4B】E1B19K、E1B19K+AVEN、及び、E1B19K+AVEN+XIAPΔをトランスフェクトすることによって生成されたアポトーシス耐性細胞株の増殖プロファイルである。コントロール(●)、E64(◆)、EA167(■)及びEAX197(▼)の震盪フラスコ回分培養を、生細胞密度及び生存率について観測した。
【図4C】E1B19K、E1B19K+AVEN、及び、E1B19K+AVEN+XIAPΔをトランスフェクトすることによって生成されたアポトーシス耐性細胞株の増殖プロファイルである。コントロール(●)、E64(◆)、EA167(■)及びEAX197(▼)の震盪フラスコ回分培養を、生細胞密度及び生存率について観測した。
【図5A】E1B19K+AVEN±XIAPΔでトランスフェクトすることによって生成されたアポトーシス耐性細胞株の代謝産物のプロファイルである。コントロール(●)、EA167(■)及びEAX197(▼)の震盪フラスコ回分培養を、異なる代謝産物について観測した。
【図5B】E1B19K+AVEN±XIAPΔでトランスフェクトすることによって生成されたアポトーシス耐性細胞株の代謝産物のプロファイルである。コントロール(●)、EA167(■)及びEAX197(▼)の震盪フラスコ回分培養を、異なる代謝産物について観測した。
【図5C】E1B19K+AVEN±XIAPΔでトランスフェクトすることによって生成されたアポトーシス耐性細胞株の代謝産物のプロファイルである。コントロール(●)、EA167(■)及びEAX197(▼)の震盪フラスコ回分培養を、異なる代謝産物について観測した。
【図6A】アポトーシス耐性細胞株に連日乳酸を補充した結果を示す図である。A)コントロール(●);EA167(無添加□、乳酸添加■)、及びB)EAX197(無添加△、乳酸添加▼)。
【図6B】アポトーシス耐性細胞株に連日乳酸を補充した結果を示す図である。A)コントロール(●);EA167(無添加□、乳酸添加■)、及びB)EAX167(添加△、乳酸無添加▼)。
【図7A】添加乳酸の存在下又は不在下における、コントロール並びにアポトーシス耐性細胞株EA167及びEAX197の増殖及び代謝産物のプロファイルである。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図7B】添加乳酸の存在下又は不在下における、コントロール並びにアポトーシス耐性細胞株EA167及びEAX197の増殖及び代謝産物のプロファイルである。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図7C】添加乳酸の存在下又は不在下における、コントロール並びにアポトーシス耐性細胞株EA167及びEAX197の増殖及び代謝産物のプロファイルである。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図7D】添加乳酸の存在下又は不在下における、コントロール並びにアポトーシス耐性細胞株EA167及びEAX197の増殖及び代謝産物のプロファイルである。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図7E】添加乳酸の存在下又は不在下における、コントロール並びにアポトーシス耐性細胞株EA167及びEAX197の増殖及び代謝産物のプロファイルである。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図7F】添加乳酸の存在下又は不在下における、コントロール並びにアポトーシス耐性細胞株EA167及びEAX197の増殖及び代謝産物のプロファイルである。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図7G】添加乳酸の存在下又は不在下における、コントロール並びにアポトーシス耐性細胞株EA167及びEAX197の増殖及び代謝産物のプロファイルである。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図8A】乳酸添加時にアポトーシス耐性細胞株によって減少するアミノ酸のプロファイルを示す図である。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図8B】乳酸添加時にアポトーシス耐性細胞株によって減少するアミノ酸のプロファイルを示す図である。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図8C】乳酸添加時にアポトーシス耐性細胞株によって減少するアミノ酸のプロファイルを示す図である。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図8D】乳酸添加時にアポトーシス耐性細胞株によって減少するアミノ酸のプロファイルを示す図である。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図8E】乳酸添加時にアポトーシス耐性細胞株によって減少するアミノ酸のプロファイルを示す図である。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図8F】乳酸添加時にアポトーシス耐性細胞株によって減少するアミノ酸のプロファイルを示す図である。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図8G】乳酸添加時にアポトーシス耐性細胞株によって減少するアミノ酸のプロファイルを示す図である。コントロール(●)、EA167(無添加□、乳酸添加■)、EAX197(無添加▲、乳酸添加▼)。
【図9A】特別配合培地中、高濃度のグルコース中で培養されたコントロール及びアポトーシス耐性細胞株EAX197の増殖プロファイル及び乳酸蓄積プロファイルである。アポトーシス耐性細胞株EAX197(■)、コントロール細胞株(◇)。
【図9B】特別配合培地中、高濃度のグルコース中で培養されたコントロール及びアポトーシス耐性細胞株EAX197の増殖プロファイル及び乳酸蓄積プロファイルである。アポトーシス耐性細胞株EAX197(■)、コントロール細胞株(◇)。
【図9C】特別配合培地中、高濃度のグルコース中で培養されたコントロール及びアポトーシス耐性細胞株EAX197の増殖プロファイル及び乳酸蓄積プロファイルである。アポトーシス耐性細胞株EAX197(■)、コントロール細胞株(◇)。
【図9D】特別配合培地中、高濃度のグルコース中で培養されたコントロール及びアポトーシス耐性細胞株EAX197の増殖プロファイル及び乳酸蓄積プロファイルである。アポトーシス耐性細胞株EAX197(■)、コントロール細胞株(◇)。
【図10A】30mM又は60mMのグルコースを含む特別配合培地中で培養されたEAX197及びコントロールホスト細胞株から生成された細胞株の抗体力価を示す図である。アポトーシス耐性細胞株EAX197(■)、コントロール細胞株(□)。
【図10B】30mM又は60mMのグルコースを含む特別配合培地中で培養されたEAX197及びコントロールホスト細胞株から生成された細胞株の抗体力価を示す図である。アポトーシス耐性細胞株EAX197(■)、コントロール細胞株(□)。
【図11A】30mM又は60mMのグルコースを含む培地中に高細胞播種密度及び低細胞播種密度で播種した、アポトーシス耐性遺伝子Bcl−2を有する細胞株のVCD及び力価を示す図。
【図11B】30mM又は60mMのグルコースを含む培地中に高細胞播種密度及び低細胞播種密度で播種した、アポトーシス耐性遺伝子Bcl−2を有する細胞株のVCD及び力価を示す図。
【発明を実施するための形態】
【0012】
本明細書に引用される特許及び特許出願など(ただしこれらに限定されない)のすべての刊行物は、本明細書にそれらの全体が記載されているものと同様にして本願に援用するものである。
【0013】
説明文中で使用する用語「流加細胞培養」とは、増殖を律速する栄養基質を培養に添加することに基づいた細胞培養プロセスを意味する。流加法は、バイオリアクター中で高い細胞密度に達するためにバイオ産業のプロセスで一般的に使用されている方法である。 HYPERLINK "http://en.wikipedia.org/wiki/Bioreactor" \o "Bioreactor" しかしながら、グルコースなどの栄養素を添加すると乳酸やアンモニアなどの代謝老廃物が形成される。濃度18mMの乳酸(Kurano et al.,1990)及び8mMのアンモニア(Hansen and Emborg,1994)は真核細胞の増殖を阻害することが報告されている。
【0014】
流加細胞培養における過剰な乳酸の蓄積の問題に対処するためにこれまでに多くの手法が考案されており、それらの手法について上記に述べた。本発明では、乳酸濃度を低下させて生細胞密度を高めることに代わる手法を提供する。この方法では、1種以上のアポトーシス耐性遺伝子をホスト細胞株で過剰発現させる。得られたアポトーシス耐性細胞株ではミトコンドリアでの呼吸が刺激され、乳酸の蓄積量が減少することになる。したがって、これらの細胞株は、高濃度のグルコースを添加しても生存することが可能である。その結果、これらのアポトーシス耐性細胞株の培養では高い生細胞密度に達することが可能である。これらの細胞株はまた、そのアポトーシス耐性のために長い寿命を有する。
【0015】
本発明の方法は、チャイニーズハムスター子宮細胞(CHO)、ミエローマ又はハイブリドーマ細胞培養などの流加細胞培養における生細胞の密度及び生存率を高めるうえで有用である。詳細には、本発明の方法はCHO細胞培養の積分生細胞数(IVCC)を高めるうえで有用である。本発明の方法は、1種以上の異種由来のアポトーシス耐性遺伝子(アポトーシスR)遺伝子及び1種以上の対象遺伝子を発現する細胞株を培養する工程と、細胞培養の指数増殖期及び定常期の間に高濃度のグルコースの培地への添加を維持する工程とを含む。これらの細胞系は、ペプチド、ペプチド融合体、増殖因子、ホルモン抗体、設計アンキリン反復タンパク質(DARPins)、及び治療、診断又は研究目的に有用な他のタンパク質などの対象タンパク質を発現する生産用細胞株の作出のめの優れたホストとなる。本発明の方法において有用なCHO細胞株としては、CHO−K1(インビトロジェン社(Invitrogen)カリフォルニア州カールスバッド)及びCHOK1SV(ロンザ・バイオロジックス社(Lonza Biologics)英国、スラウ)が挙げられる。本発明の方法において有用なミエローマ株としては、NS0及びSp2/0が挙げられる。
【0016】
本発明の方法において有用な細胞株は、1種以上の異種由来のアポトーシス耐性遺伝子を発現する。詳細には、E1B19K(配列番号1及び2)及びAven(配列番号3及び4)が有用である。XIAPΔ(配列番号:5及び6)をコードした遺伝子を使用することもできる。これらのアポトーシス耐性遺伝子は、それぞれアポトーシスを引き起こすシグナル伝達経路の早期、中期、及び後期に働く遺伝子を代表するものである。更に、Bcl−2Δをコードする遺伝子を使用することもできる(配列番号7及び8)。アポトーシス耐性遺伝子の発現は、当業者には周知の形質移入(トランスフェクション)技術によって実現させることができる。MDM2(配列番号9及び10)及びBcl−XL(配列番号11及び12)などの、アポトーシスを引き起こすシグナル伝達経路の上記のステージからの他のアポトーシス耐性遺伝子も本発明の方法において有用である。
【0017】
本発明においては、異種由来のアポトーシス耐性遺伝子を発現する細胞株の使用により、これらの細胞株の乳酸の分泌量が低減されるかあるいは蓄積された乳酸が消費されることによって、高濃度のグルコースの培養中への添加を維持しつつ、積分生細胞数(IVCC)の値がコントロール細胞株と比較して約2倍高い値に達する。培養中の生産用細胞株の生細胞密度を高めることにより、バイオリアクターの使用によって得られる産物の収率が高まる。このように生産性が高められることにより、複雑な生物製剤の製造コストが低減されると同時に、非生細胞の細胞溶解がないために優れた品質の産物が生産される(溶解した細胞は産物を分解するプロテアーゼを放出するため)。したがってこれらの細胞株は、対象タンパク質(単数又は複数)を発現する生産用細胞株を作出するための優れたホストとなる。
【0018】
本発明の方法では、老廃物として蓄積される乳酸を消費する、又はその産生量が低減したアポトーシス耐性細胞株の能力によって、高濃度グルコース条件下、及びグルコース欠乏後の増殖を可能とする培養手法が与えられる。異なる実施形態において、本発明の方法は、流加細胞培養における高いピーク生細胞密度、長い寿命、高い分泌タンパク質力価、高い積分生細胞数、低い細胞カルシウム流束、及び高いミトコンドリア膜電位を与えるものである。更に、高濃度の有毒な乳酸及びアンモニアを制限するために用いられてきた流加培養法は、これらの細胞株の高い生産性にとって必要ではない可能性がある。
【0019】
トランスフェクトした後、アポトーシス耐性導入遺伝子の染色体への組み込み部位によってその発現レベルが決まる。更に、複数のアポトーシス耐性遺伝子をトランスフェクトする場合、これらのタンパク質の多くは直接的又は間接的に相互作用することによって細胞の生理学的変化を引き起こすことから、特定のアポトーシス耐性導入遺伝子の発現レベルがその細胞が有する他のアポトーシス耐性タンパク質の活性に影響を及ぼしうる。したがって、複数の遺伝子を有する細胞株の全体のアポトーシス耐性に対する各アポトーシス耐性遺伝子の定量的寄与は、各アポトーシス耐性遺伝子の相乗効果のために決定することが困難である。更に、すべてのクローンが同じ導入遺伝子の集合をトランスフェクトされていたとしてもアポトーシス耐性に関しては各クローン間で大きなばらつきがある。後述する実施例に示される結果からは、試験した各アポトーシス耐性遺伝子は、各遺伝子をトランスフェクトしたホスト細胞株のアポトーシス耐性を高める方向に漸増する正の値を与えることが明らかである。上記の束縛条件内では、3重形質導入体は一般に2重形質導入体よりも優れており、2重形質導入体は1個のみのアポトーシス耐性遺伝子を過剰発現している形質導入体よりも優れていた。
【0020】
したがって、適当な発現ベクターをトランスフェクトすることでE1B−19K、Aven及びXIAPΔを異なるレベルで発現するクローンが得られた。E1B−19Kの発現レベルは比較的低く、E1B−19Kを高い発現レベルで発現する稀なクローンは不安定であるか増殖性が低かった(データは示さず)。下記実施例において、E1B−19Kを単独で発現しているCHO細胞では細胞死に対する保護は限定的なものに過ぎなかった(図4)。
【0021】
E1B−19Kを単独で発現している細胞株で得られた結果に対し、下記実施例において示されるデータは、Aven(EA167)、又はAven及びXIAPΔ(EAX197)を同時に発現しているCHO K1細胞株では、生存率、細胞密度及びIVCCに向上が見られたことを示している。下記実施例に示される結果は、CHO K1細胞株における2個のアポトーシス耐性遺伝子の発現により、スタウロスポリン及び長期の回分培養といった障害の存在下でカスパーゼ活性が大幅に低下し、ミトコンドリアの膜電位が高くなったことを更に示している。
【0022】
下記実施例では、Bcl−XLの機能的ホモログであるE1B−19KとAven及びXIAPΔとの組み合わせの発現が細胞死を遅らせる効果について調べた。これらのトランスフェクト細胞におけるXIAPΔの発現レベルは、細胞内に既に存在する内因性の野生型XIAPタンパク質に対して高くはなかったが、これはこのシステムにおいて2個のプラスミドをトランスフェクトするために必要な条件によるものと恐らく考えられる。しかしながら、EAX197におけるXIAPΔの追加(Aven及びE1B−19Kに加えて)により、無添加(図5及び7)及び乳酸を添加した培養(図7)の両方で最大生細胞密度が一貫して向上した。実際、乳酸を添加したEAX197細胞株は、60%以上の高い生存率を14日間にわたって維持したが、これはEA167細胞よりも2日長く、コントロール培養よりも4日間長かった。XIAPΔを有することによりカスパーゼ活性に対する細胞の耐性が高まると考えられ、このことは図3に、スタウロスポリンによる処理後においてもEAX197のカスパーゼ活性が極めて低かったことにより実証されている。更に、XIAPの突然変異(XIAPΔ)の使用により、野生型XIAPタンパク質と比較して高いアポトーシス耐性が与えられる。したがって、XIAPの欠失突然変異(XIAPΔ)の保護能力は、これらのCHO株においてこのタンパク質の発現レベルが高くなくとも充分なものである可能性がある。
【0023】
下記実施例では、細胞の代謝における1種以上のアポトーシス耐性遺伝子の役割を調べた。Aven及び/又はXIAPΔとともにE1B−19Kを発現しているアポトーシス耐性細胞株において栄養素の消費量及び代謝産物の産生量を調べた。詳細には、2つの最も一般的な細胞培養の老廃物(すなわち乳酸とアンモニア)の蓄積量についてアポトーシス耐性細胞株とコントロール細胞株との間で比較を行った。下記に示すデータでは、CD−CHO培地中の震盪フラスコ回分培養において、播種後6日目〜7日目にコントロール細胞株は2.8g/L(31mM)の乳酸及び12mMのアンモニア(図5)を蓄積したが、これらは細胞増殖を阻害することが報告されている濃度18mMの乳酸(Kuranoら、前出)及び8mMのアンモニア(Hansen及びEmborg、前出)を大幅に上回る濃度であった。これらの濃度は、CHOコントロール細胞株において播種後8日目に見られた生存率の低下の理由の一部となっていることが考えられる。
【0024】
下記に示すデータは、アポトーシス抵抗細胞株がある程度の乳酸を初期に蓄積することを示しているが、これらの細胞株は指数増殖期に乳酸を消費しはじめ、VCDはコントロール培養を大幅に上回って増大し続けた。グルコースの枯渇後に直ちに乳酸を消費する能力、又はアポトーシス阻害性の増大、又は両方の因子が細胞増殖の継続に寄与していた。更に、アポトーシス耐性細胞は内因性の乳酸が枯渇した場合にのみ定常期に入った。乳酸の消費がこれらの操作された細胞株の一般的な特性であることを調べるため、アポトーシス耐性培養に外因性の乳酸を加えて欠乏した乳酸を補充した(図6)。下記実施例に示される結果は、アポトーシス耐性培養は添加された乳酸を消費し、コントロール培養と比較して高いVCD及び生存率を更に4〜7日間にわたって維持した(無添加及び乳酸添加培養を図7A及びBにそれぞれ示す)。このプロセスの更なる利点としては、アポトーシス耐性細胞株の培養中の乳酸が低濃度であることによって、浸透圧がより低く、pHがより高くなることがあり、これらはいずれも産物の高い品質にとって極めて望ましい。
【0025】
研究者によりこれまでに、ハイブリドーマ、NS0ミエローマ、及びCHOの培養中において乳酸の消費が検出されている(deZengotita et al.,2000;Zhou et al.,1995 and 1997;Burky et al.,2007;Pascoe et al.,2007)。しかしながら、これらの細胞は通常、指数増殖期では乳酸を産生し、定常期に消費へと移行するが、このことは、乳酸が中間の副産物及び炭水化物エネルギー源の両方でありうることを示唆するものである(Burky et al.,2007;Brooks et al.,1985)。下記に示す結果では、コントロールのCHO培養は、上記の実験において見られるように定常期において特に「高濃度」グルコース培地で(図9)一定量の乳酸を消費したようであった。これに対し、アポトーシス耐性細胞では、指数増殖期に乳酸が消費され、外因性の乳酸が欠乏した場合にのみ細胞が定常期に入るという大きく異なったプロファイルを示した。
【0026】
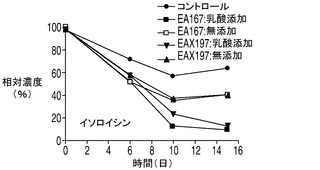
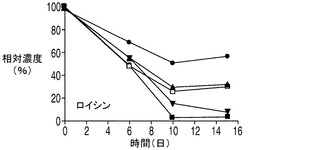
乳酸脱水素酵素(LDH)によって触媒されるピルビン酸の乳酸への変換は可逆反応であるが、平衡定数が3.6×104/Mと乳酸を生成する方向に大きく傾いている。しかしながら、解糖系がトリカルボン酸(TCA)の需要を満たすことができない場合、乳酸はピルビン酸へとその反応を進めるLDHの複数のアイソザイムの1つによって逆に変換される。アポトーシス耐性細胞株によって消費された乳酸は、TCA回路に入って細胞のエネルギー及びアミノ酸の産生に使われるものと考えられる。興味深い点として、アポトーシス耐性培養のアンモニアのプロファイルは、無添加及び乳酸添加細胞株の両方においてアンモニアの産生量が繰り返し一時的に低下するとともに全体的にアミノ酸の産生量が減少し、コントロール培養のアンモニアプロファイルと大きく異なった。これは、アミノ酸がTCA回路でエネルギー源として使用されることによってアンモニアが蓄積したものと考えられる。したがって、アンモニアの産生量の減少は、アポトーシス耐性細胞では、コントロール細胞と比較してTCA回路におけるエネルギーの必要量の内、相対的に小さな割合がアミノ酸から得られている可能性を示唆しており、このことはアポトーシス耐性細胞が炭素源として乳酸もより多く消費していることを考えると妥当な結果である。しかしながらアンモニア産生量の低下は一時的なものであり、アポトーシス耐性細胞が依然、アミノ酸をエネルギー源として利用している可能性がある点は注意を要する。実際、いずれも分枝鎖アミノ酸であるイソロイシン、ロイシン、及びバリンの3種類のアミノ酸は、アポトーシス耐性細胞によってより急速に消費され、その消費量は乳酸添加培養で大きかった(図8A〜C)。アポトーシス耐性細胞株はコントロール細胞株と比較して継続的な増殖を示したことから、これら3種類のアミノ酸が脂肪酸及びアポトーシス耐性細胞によって必要とされる他のタンパク質の構成単位を構成するとともに、TCA回路のエネルギー源として異化反応に用いられている可能性がある。メチオニンについても幾分似た消費プロファイルが観察された(図8D)。
【0027】
減少することが観察された上記のアミノ酸に対し、アラニン及びアスパラギン酸などの幾つかのアミノ酸がコントロール培養中に一時的に分泌された。ピルビン酸の一部が、アラニンアミノトランスフェラーゼによってアラニンに変換されている可能性がある。この経路はアポトーシス耐性細胞でより活性が高くなる場合があるので、コントロール細胞株に対してアラニン濃度がより大きな上昇につながる(図8F)。一方、アスパラギン酸は、アスパラギン酸アミノ酸ファミリーの第1のアミノ酸であり、アスパラギン酸トランスアミナーゼを用いてTCA回路の中間体であるオキザロ酢酸とグルタミン酸の変換によってα−ケトグルタル酸とともに生成する。その濃度は、コントロール細胞においてのみ最初の6日間上昇したのに対して、アポトーシス耐性細胞株では連続的に減少した。この反応は可逆反応であるため、増殖期の初期におけるアポトーシス耐性細胞株でのアスパラギン酸の欠乏は、アポトーシス耐性細胞株ではTCA回路における可逆的なオキザロ酢酸産物の産生量が制限されている可能性を示している。オキザロ酢酸がTCA回路の最初の段階においてアセチルCoAとも反応することを考慮すると、アポトーシス耐性細胞株における乳酸からアセチルCoAへのピルビン酸の流入量の増加は、オキザロ酢酸の需要を大きくしている可能性がある。これに対して、コントロール細胞株は乳酸を消費しておらず、したがってオキザロ酢酸からアスパラギン酸を生成しているため、コントロール細胞株ではアセチルCoAに対する需要は低いものと考えられる。実際、こうしたアセチルCoAに対する需要の不足は、細胞培養の最初の7日間にわたって見られた、アセチルCoAに代わる副生成物としての乳酸のコントロール細胞内への蓄積を引き起こした可能性が考えられる。また、コントロール細胞における乳酸消費の不足がアセチルCoAの供給を制限し、これにともなうアスパラギン酸の生成量の増加を引き起こした可能性も考えられる。
【0028】
これらの結果は、アポトーシス耐性遺伝子を発現する細胞株ではTCA回路のエネルギー効率が高くなっていることを示すものであるが、こうした細胞反応の正確な解釈にはより詳細な代謝フラックスの分析が待たれるところである。消費される乳酸の運命には関係なく、本発明ではアポトーシス耐性細胞株は蓄積した乳酸を消費し、これにより細胞株の長い寿命及び高いIVCCに寄与するものである。
【0029】
アポトーシス耐性細胞株が蓄積された乳酸を消費することによって寿命が延びるという点で独特であるならば、これらの細胞株は乳酸を含む培地又は標準よりも高い濃度のグルコースを含む培地中で培養できる可能性がある。乳酸が唯一の炭素源として与えられた場合には増殖及び生存率はいずれのタイプの細胞株においても支持されなかったが、乳酸がグルコースに加えて(データは示さず)一成分として与えられた場合、又は乳酸が老廃物として蓄積された場合には乳酸の消費が認められた。このことは、アポトーシス耐性細胞はピルビン酸から糖新生を行うことはできず、乳酸からエネルギーを得ることもできないが、グルコースが乳酸とともに与えられた場合には、細胞はグルコース及び培養に蓄積又は添加された乳酸の両方を消費することができることを示唆している。更に、高濃度(60mM)のグルコースを含む培地では、アポトーシス耐性細胞株のIVCC(並びにVCD及び生存率、図9A〜C)はコントロール細胞株のIVCC(並びにVCD及び生存率)よりも大幅に高かった。これは、高グルコース培地では、コントロール細胞株が20mMよりも高濃度の乳酸を蓄積したことによるものであり、高濃度の乳酸が毒性を示した可能性がある。これに対して、アポトーシス耐性細胞株では増殖期の初期に乳酸を消費することにより、より多くの生細胞を維持することが可能であったため、コントロールと比較して170%というIVCCの増大につながった(図9C)。
【0030】
アポトーシス耐性細胞株は乳酸を消費して高いIVCCが実現されることから、治療用抗体などの対象とするタンパク質を発現するアポトーシス耐性ホスト細胞株由来の生産用細胞株では、コントロール細胞株に由来する生産用細胞株と比較して大幅に高い力価が実現された。このように高い力価は、治療用タンパク質又は他の対象とするタンパク質を生産する哺乳動物細胞株においてアポトーシス耐性遺伝子を使用することの潜在的な商業的利益を例証するものである。
【0031】
次に、本発明を以下の具体的かつ非限定的な実施例を参照しながら説明する。
【実施例】
【0032】
以下の実施例では、アポトーシス耐性遺伝子を過剰発現するCHO細胞株を震盪フラスコ培養中で、ピーク生細胞密度、寿命、カスパーゼ−3活性化、及びミトコンドリア膜電位(MMP)について分析を行った。更に、これらの細胞株における栄養素の消費量及び代謝産物の産生量をコントロール細胞と比較することによって、アポトーシス耐性遺伝子の発現が哺乳動物細胞の培養システム中の栄養素の消費及び代謝産物の蓄積に与える影響について調べた。
【0033】
材料及び方法
細胞培養:
コントロール細胞株C1013Aとして指定したCHOK1SV細胞株(ロンザ・バイオロジックス社(Lonza Biologics)英国、スラウ)を、30mMのグルコースを含み、6mMのL−グルタミン(インビトロジェン社(Invitrogen)カタログ番号10313〜021)を添加したCD−CHO培地(カタログ番号10743〜011、インビトロジェン社(Invitrogen)カリフォルニア州カールスバッド)中で培養した。場合に応じて、60mM(高グルコース培地として定義される)など異なる濃度のグルコースを含んだ別の無動物タンパク質培地を使用した。ウシ胎児血清はハイクローン・ラボ社(Hyclone Labs)(ユタ州ローガン)(カタログ番号SH30071.03)より購入した。細胞培養をCedex自動細胞計数装置(イノバティス社(Innovatis)ドイツ)により観測した。積分生細胞数(IVCC、細胞・日/ml)を下式:
IVCC(d1)=[VCD(d0)+VCD(d1)]/2+VCD(d0)
(式中、VCD=生細胞密度)により計算した。
【0034】
プラスミドの構築:
E1B−19K(EF−1aプロモーター)を単独で、又はAven(CMVプロモーター)と組み合わせて構成的に発現するように設計した、これまでに述べられている(Nivitchanyong et.al.,2007)pBUDCE4.1ベクターを使用した。XIAPΔ(CMVプロモーター)を発現するベクターは、サワーウォルド(Sauerwald et al.,2002)により述べられるようなものを使用した。ブランクベクターとは最初のpBUDCE4.1ベクターのことを指す。
【0035】
グルタミンシンターゼ(GS)発現ベクター(ロンザ・バイオロジックス社(Lonza Biologics)英国、スラウより研究用ライセンスにて取得)に重鎖及び軽鎖cDNAをクローニングすることによってモデル抗体(抗体1)発現ベクターを構築した。
【0036】
アポトーシス耐性細胞株の生成:
CHOK1SV細胞株の指数増殖期の培養に、1)pBUDCE4.1、2)pBUDCE4.1−E1B−19K、3)pBUDCE4.1−E1B−19K−Aven、並びに4)pBUDCE4.1−E1B−19K−Aven及びpCMV−XIAPΔをトランスフェクトした。トランスフェクションは製造者の推奨するところにしたがってFugene(ロシュ社()カタログ番号1815075、スイス、バーゼル)を使用して行った。トランスフェクトした2日後に、300μg/mlのゼオシン(上記のトランスフェクション1、2及び3に対し)、300μg/mlのゼオシン及び400μg/mlのハイグロマイシン(トランスフェクション4に対し)を含む増殖培地中、96穴プレートに播いた(表1を参照)。各トランスフェクションから約200個の抗生物質耐性クローンを増殖させて下記に述べるようにしてカスパーゼ3/7の活性について分析した。有望なクローンは、各抗生物質の不在下で10代継代して安定性試験を行った。
【0037】
【表1】
【0038】
最初に、CHOK1SV細胞株にブランクベクターをトランスフェクトし、ゼオシン耐性コロニーを選択することによってコントロール細胞株を作出した。これらの細胞株は、トランスフェクトしていないコントロールCHOK1SVと比較してIVCCが平均で約20%低かったため、CHOK1SVを後のすべての実験においてコントロール細胞株として用いた。次いで、最も有望なアポトーシス耐性細胞株であるEAX197及びコントロール細胞株を用い、GS発現ベクターを製造者の推奨するところにしたがって使用してモデル抗体を発現する生産用細胞株を作出した。細胞培養中の抗体をNephelometry(ベックマン・アレイ・システム社(Beckman Array System))によって測定した。
【0039】
アポトーシス耐性細胞株の震盪フラスコ培養:
選択されたアポトーシス耐性細胞株を、6mMのグルタミン及び必要な選択用抗生物質を添加したCD−CHO培地中で無添加の回分方式で培養した。一部の実験では、アポトーシス耐性細胞株の回分式培養に乳酸を添加して細胞株が消費した乳酸を補充した。更に、選択された抗体発現細胞株を、6mMのグルタミン及び60mMのグルコースを添加した特別に配合した無動物タンパク質培地中で培養した。
【0040】
カスパーゼ3/7活性のアッセイ:
約3×105個の各クローンの細胞を1mlの増殖培地中、24穴プレートに播種した。播種4日目後(d4)に、約1×105個の細胞を3重に96穴プレートに移した。スタウロスポリン(2μM fc)を添加し、細胞を16時間インキュベートした後、APO−ONEキット(ビー・ディー・ラボ社(BD Labs))によりカスパーゼ3/7の活性についてアッセイした。この手順をスタウロスポリンを加えない点を除き、10日目において繰り返した。4日目及び10日目の両方においてカスパーゼ3/7の活性が有意に低かったクローンをSF培地中で増殖させた。選択されたクローンのアポトーシス耐性を、フローサイトメトリー分析(下記を参照)及び場合によりミトコンドリア膜電位を測定することによって確認した。
【0041】
フローサイトメトリーによるアポトーシス耐性クローンの分析:
指数増殖期の約1×106個の細胞を各震盪フラスコから抜き取って24穴プレートに移し、スタウロスポリン(2μM fc)と16時間インキュベートし、収穫してPBS中で1回洗った。次いで細胞をCytoPerm(カタログ番号2075KK、ビー・ディー・バイオサイエンス社(BD BioScience))とインキュベートして固定するとともに細胞を透過性とした。PBSによる洗浄後、細胞をFITC標識抗カスパーゼ3(カタログ番号68654、ビー・ディー・バイオサイエンス社(BD BioScience))抗体とインキュベートした後、フローサイトメトリーにより分析した。
【0042】
ミトコンドリア膜電位(MMP)アッセイ:
細胞を震盪フラスコ培養から播種後6日目に抜き取り、スタウロスポリン(5μM fc)と2時間インキュベートした。この後、細胞を洗い、製造者の推奨するところにしたがって親油性カチオン性色素であるJC−1(ケイマン・ラボ社(Cayman Labs)ミシガン州アナーバー)で処理した。プレートをFL535及びFL595で読み取り、その比を計算した。
【0043】
ウェスタンブロッティング:
示される細胞株から約1×107個の細胞を収穫し、PBS中で洗浄し、1% NP−40、120mMのTris−Hcl、150mMのNaCl、0.2mMのPMSF、及び1mMのEDTAを含むRIPAバッファー中で溶解した。各試料からの細胞質タンパク質50μgを4〜12% NuPAGE勾配ゲルにロードし、Novexシステム(インビトロジェン社(Invitrogen))でSDS−PAGEによって分離した。分離したタンパク質のバンドをニトロセルロース上に転写して、1)ヒトE1B−19Kに対する抗マウス抗体(希釈率1:40)(カタログ番号DP−17、カルバイオケム社(CalBiochem)メリーランド州ギブスタウン)、2)ヒトAvenに対する抗ウサギ抗体(希釈率1:1000)(カタログ番号612521、ビー・ディー・バイオサイエンス社(BD BioScience)カリフォルニア州サンホセ)、及び3)ヒトXIAPに対する抗マウス抗体(カタログ番号616713、ビー・ディー・バイオサイエンス社)を用いて分析した。使用した抗体を含め、上記のプロトコールは、販売業者から購入した既知量の精製したE1B−19K、Aven、及びXIAP(それぞれ約10ng)を用いて最適化かつ標準化した。タンパク質のバンドはECL検出キット(ジー・イー・ヘルスケア社(GE Healthcare)ニュージャージー州ピスカタウェイ)を使用して可視化した。
【0044】
培地中の代謝産物濃度の測定:
すべての重要な代謝産物及び老廃物の濃度をYSI2700自動分析装置を使用して測定した。アンモニアイオン濃度はフローインジェクション分析(Campmajo et al,1994)により測定した。アミノ酸は逆相カラム(ウォーターズ社(Waters)マサチューセッツ州ミルフォード)を用い、HPLC(ウォーターズ社)によって測定した。
【0045】
(実施例1)
アポトーシス耐性細胞株の生成及び特性評価
本実験で使用した細胞株、各細胞株を生成するためにそれぞれトランスフェクトした発現プラスミド、及び形質移入細胞(トランスフェクトーマ(Transfectoma))を単離するために使用した選択用物質のリストを上記表1に示す。表に示した細胞株は、各トランスフェクションから生成された多数のクローンの内の代表的なものである。各細胞株の名称は、ホスト細胞株にトランスフェクトされたアポトーシス耐性遺伝子に由来している。例えばEAX197は、E1B−19K(E)、Aven(A)、及びXIAPΔ(X)をトランスフェクトした細胞株である。アポトーシス耐性(アポトーシスR)という表記は、これらのアポトーシス耐性遺伝子の1以上がトランスフェクトされた細胞株を指して用いる。
【0046】
本実験で用いたすべてのアポトーシス耐性細胞株について、ウェスタンブロッティング、カスパーゼ3/7、及びミトコンドリア膜電位による特性評価を行った(データは示さず)。1重形質移入体E64、又は2重形質移入体EA63、EA112、EA167及びEA190はいずれも、コントロールであるトランスフェクトされていない細胞株C1013Aで見られたバックグラウンドレベルよりも高い発現レベルでE1B−19Kを発現した。4つの2重形質移入体の内、細胞株EA112及びEA167は、E1B−19K(約19kDa)及びAven(約55kDa)の両方を発現した。細胞株EA63及びEA190は、E1B−19Kを高レベルで発現したが、Avenの発現レベルは極めて低かった。2重ベクターpBUDCE4.1−E1B−19K−AvenとpCMV−XIAPΔをトランスフェクトすることによって生成された4つの細胞株、すなわちEAX64、EAX99、EAX148及びEAX197の内、EAX197のみが3種類のタンパク質のすべてを検出可能なレベルで発現した。
【0047】
トランスフェクトしていないコントロールのレーンC1013Aを含むすべてのレーンにおいて組換えAvenの上側に観察された約55kDaのバンドは、免疫後の血清では可視化されたが免疫前の血清では可視化されなかったことから内因性のAvenタンパク質であると考えられた(Sauerwald et al.,2002,Chau et al.,2000)。トランスフェクトされたAven遺伝子は最初の6個のアミノ酸が欠失しているが、これらのアミノ酸の欠失はタンパク質の生物学的活性に影響しないことが示されている。このため、トランスフェクトされたAvenは内因性のAvenよりも若干小さい。Avenの分解産物である可能性のある40kDaのバンドがこのタンパク質をE1B−19Kとともに過剰発現したすべての細胞株において観察された。同様のサイズの分解生成物が、AvenがBcl−XL(Sauerwald et al.,2005)又はBcl−2(Figueroa、未発表の観察)とともに過剰発現された場合に観察されている。更に、Avenの発現は、E1B−19K、Aven及びXIAPΔを過剰発現するEAX197を含む3重形質移入体において顕著に低かった。
【0048】
興味深い点として、トランスフェクトしたXIAPΔの発現レベルは内因性の完全なXIAPタンパク質の発現レベルと比較して顕著に低かった。XIAPとXIAPΔとの間の発現レベルの明瞭な差は、アポトーシス耐性タンパク質のこれら2つの形態の抗体結合効率の差によって一部説明することができる。内因性のXIAPと形質移入されたXIAPΔとの間の明瞭な発現レベルの差は、アポトーシスの防止におけるこのトランスフェクトタンパク質の寄与に疑問を投げかけるものである。実際、この実験は、所定のアポトーシス耐性遺伝子の発現レベルと、そのタンパク質が細胞株に与えることのできるアポトーシス阻害のレベルとの間には必ずしも直接的な相関が認められないことを示唆している。
【0049】
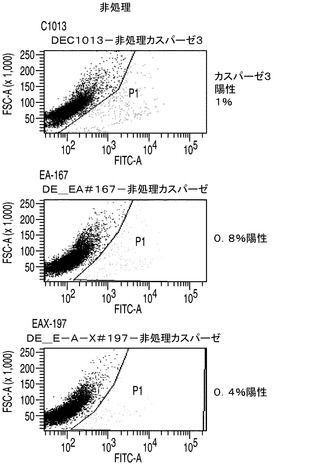
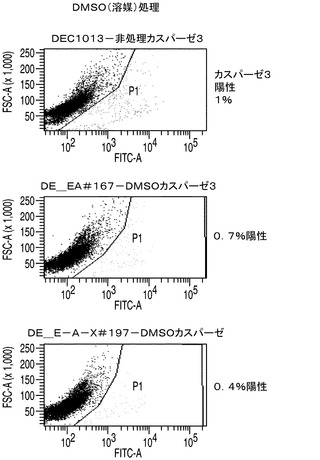
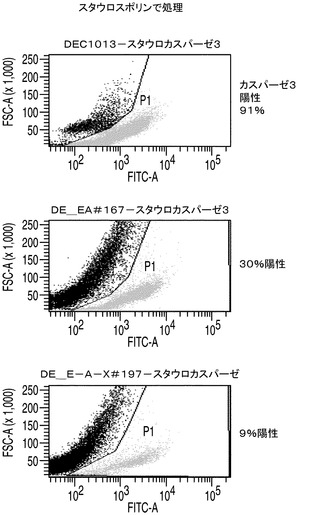
カスパーゼ3/7の活性はアポトーシスに対する細胞株の耐性の大きさの指標を与えることから、カスパーゼ3/7活性についてAPO−ONEによりそれぞれのアポトーシス耐性細胞株を調べた。簡単に述べると、このアッセイでは、蛍光前駆基質であるZ−DEVD−R110がカスパーゼ3/7によって用量依存的に開裂される。EA167及びEAX197並びにコントロール細胞株の指数増殖期の培養をスタウロスポリンで16時間処理することによってアポトーシスを誘導した後、カスパーゼ3/7を測定した。スタウロスポリン処理したコントロール細胞のカスパーゼ3/7活性を任意に100%に設定した。これらの条件下で、EA167及びEAX197細胞のカスパーゼ活性はそれぞれコントロールの30%及び35%であったが、このことはこれらの細胞株がアポトーシス誘導に対して少なくとも部分的に耐性を有することを示唆している(図2A)。EA167及びEAX197のアポトーシス耐性の更なる根拠として、これら2つの細胞株とコントロール細胞株のスタウロスポリン処理培養をFITC結合抗カスパーゼ3抗体で標識した後、フローサイトメトリーで分析した。図3に見られるように、非処理又は溶媒(DMSO)処理によらず、3つの細胞株のすべてにおいてカスパーゼ3陽性細胞は集団の1%以下であった。しかしながら、スタウロスポリンによる処理後にはコントロール細胞集団の91%がカスパーゼ3について陽性であった。これに対し、細胞株EAX197に由来する集団のわずか9%、及び細胞株EA167に由来する集団の30%がカスパーゼ3について陽性であり、これら2つの細胞株におけるアポトーシス耐性の表現型が証明された。
【0050】
アポトーシスに対する耐性が寿命の増大につながるか否かを調べるため、複数のEA細胞株を震盪フラスコ(回分方式)中で増殖させ、生細胞密度(VCD)及び積分生細胞数(IVCC)の両方を観測した。細胞の比生産性(specific productivity)が変化しないものと仮定すると、積分生細胞数が大きくなると培養の体積生産性が向上する。したがってIVCCを、生産用細胞株の作出のためのホストとして機能しうる優れたアポトーシス耐性細胞株をスクリーニングするためのパラメータとして用いることができる。図2Bに示されるように、相対カスパーゼ3/7活性が低い細胞株は、IVCCの高い細胞株を生成する傾向が見られた。EA細胞株のそれぞれはコントロールに対して低いカスパーゼ3/7活性を示したが、IVCCとそれぞれの細胞株が有するカスパーゼ3/7のレベルとの間には厳密な相関は認められなかった。更に、特定のアポトーシス耐性細胞株はアポトーシス耐性の表現型の程度に関してその姉妹クローンよりも安定していた。EA167は強力な抗アポトーシス活性及び震盪フラスコ培養中での安定した増殖性の両方を有することから、この細胞株を更なる実験用に選択した。更に、EA167はこのパネルでは最も安定した細胞株であり、ウェスタンブロットによって検出されるようにAvenを高レベルで発現する。
【0051】
アポトーシス経路のシグナル伝達分子の多くがミトコンドリアと相互作用するかあるいはミトコンドリア内部に存在している。ミトコンドリアでこれらのタンパク質はミトコンドリアの膜と相互作用してミトコンドリアの透過性を調節することによりアポトーシスを制御する。したがって、ミトコンドリア膜電位(MMP)は、コントロールに対するEA167及びEAX197のアポトーシス耐性の特性を比較するための細胞生理学の別のモニターとして機能する。図2Cのアッセイの結果に見られるように、EA167及びEAX197はいずれもコントロール細胞株よりも高いMMPを有していた。
【0052】
(実施例2)
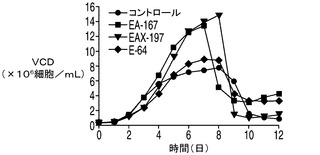
CHOK1SVの生細胞密度に対するE1B−19Kの影響
E1B−19KをAvenとともに過剰発現する2重形質移入体(EA167)、E1B−19KをAven及びXIAPΔとともに過剰発現する3重形質形質移入体(EAX197)、並びにE1B−19Kのみを過剰発現する形質移入体(E64)を図4でコントロール(ホスト)細胞株と比較した。コントロール細胞株のピークVCDは7.7×106細胞/mlに達し、E64(E1B−19Kのみを発現)のピークVCDは8.9×106細胞/mlであったのに対して、EA167のピークVCDは1.4×107細胞/mlであり、EAX197のピークVCDは1.5×107細胞/mlよりも高く、コントロール細胞株と比べて190%以上の増加を示した。EA167及びEAX197の生存率は、恐らく栄養素の枯渇のために播種後8日目又は9日目に急激に低下したが、コントロール及びE64細胞株の生存率の低下はより緩慢であった(図4B)。IVCCに対する正味の影響を図4Cに示す。コントロール細胞株のIVCCは4.9×107細胞・日/mlであり、次がE64 のIVCC(5.2×107細胞・日/ml)であった。明らかに2つのアポトーシス耐性細胞株EA167(6.3×107細胞・日/ml)及びEAX197(6.6×107細胞・日/ml)のIVCCは、コントロール細胞株のIVCCよりも有意に高かった(EA167及びEAX197でそれぞれ136%及び140%)。
【0053】
EA167及びEAX197以外にも、幾つかの他のアポトーシス耐性細胞株が同様に低い乳酸産生表現型を示した(データは示さず)。更に、低い乳酸産生特性を示す野生型細胞株も稀ではあるが報告されている(Pascoe et al,2007)。
【0054】
(実施例3)
アポトーシス細胞株の代謝プロファイル
アポトーシスからの保護を与えるアポトーシス耐性細胞株EA167及びEAX197の生理学的性質を回分培養中でコントロール細胞と比較するため、回分培養の開始時に与えられる主要な栄養素及びアミノ酸、すなわちグルコース、グルタミン酸及びグルタミンの濃度を観測した。更に、老廃物であるアンモニア及び乳酸の蓄積についても観測を行った。図5に、EAX197及びEA167並びにコントロール細胞株を市販のCD−CHO培地中、震盪フラスコ中で回分方式で培養した場合のこれらの代謝物質の濃度を示す。図5Aに示されるように、グルコースは播種後6日目又は7日目までに3つの細胞株のすべての培地から急速に枯渇した。グルタミンの枯渇についてはアポトーシス耐性細胞株とコントロール細胞株との間に有意差は認められず、グルタミン酸濃度は全日数にわたって3つの細胞株で大きく変化しなかった。3つの細胞株のすべてにおいて、4日目までの細胞増殖期の初期において乳酸の蓄積が認められた(図5B)。コントロール細胞株では、乳酸は6日目まで増加し続けた(その時点で2.8g/Lに達した)後、乳酸濃度に若干の低下が認められ、最終的に1.8g/Lに達した。こうした量の乳酸は、培養のpHを低下させることが予想され、重炭酸塩を添加することによってpHが調節されているバイオリアクター中では浸透圧が上昇することになる。これらの特性のすべて及び他のストレス因子によって動物細胞にとって有毒な環境がもたらされうる。これに対して、アポトーシス耐性細胞株中の乳酸濃度は短期間増大した後、5日目以降は急速に低下し、7日目(EA167)又は8日目(EAX197)までには乳酸は完全に消失した。興味深い点として、図5Aにおいてこれら2つの細胞株では乳酸が0にまで低下した直後にVCDが急激に低下しており、細胞生存率の低下が培地中に利用可能なグルコース又は乳酸がなくなったことによるものであることを示唆している。
【0055】
アポトーシス耐性細胞株とコントロール細胞株との間に認められた別の差異はアンモニアのプロファイルである(図5C)。EA167及びEAX197中のアンモニア濃度は、乳酸濃度の低下が始まった6日目に若干低下し、その後やがて再び増大した。しかしながら、アポトーシス耐性細胞株の全体のアンモニア濃度はコントロール細胞株よりも低く、このことはアポトーシス耐性細胞株がコントロール細胞株と比較してより高いIVCCに達し、したがって全体の細胞増殖率がより高かったことから予想外の結果である(図4C)。
【0056】
上記のデータから、アポトーシス耐性細胞株の特性がコントロール細胞株とは大きく異なっていることが明らかである。しかしながら、複数のクローンについて幾分異なる実験条件下で調べたところ、各アポトーシス耐性形質移入体間で幅広い表現型が観察された。表現型の変異はコントロール細胞株の単離クローンにおいても見られることから、適当なスクリーニング法を用いることによってコントロール細胞株から稀なアポトーシス耐性クローンを選択することが可能である。この点に関しては、マタノビッチ(Mattanovich)及びボース(Borth)(2006)によって細胞分別を応用して所望の性質を有する稀な変異体を野生型(コントロール)細胞株から単離することについて概説されている。
【0057】
(実施例4)
乳酸添加の影響
任意の時点においてある細胞培養中に存在する乳酸の量は、その細胞株によって分泌される乳酸の量から細胞株によって消費される乳酸の量を差し引いた量によって決まる。アポトーシス耐性細胞株が実際に乳酸を消費しているか否か、すなわちこれらの細胞株において正味の乳酸産生量が低下しているか否かを調べるため、3つの細胞株すべての培養を毎日観察し、アポトーシス耐性細胞株とコントロール細胞株との間で乳酸濃度に目立った差があれば外因性の乳酸を添加することによって埋めた。上記で観察したように、無添加のアポトーシス耐性細胞株EA167及びEAX197の乳酸濃度は5日目に低下しはじめ(図6A及び6B)、7日目(EA167、無添加)又は8日目(EAX197、無添加)までに乳酸は枯渇した。コントロール細胞株の乳酸プロファイルは6日目まで増加しつづけ、そこから次の3日間濃度は若干低下したが0にまで低下することはなかった。アポトーシス耐性細胞株を毎日観察し、個別の流加培養に7日目に2.5g/L(EA167に対して、図6A)及び8日目に2g/L(EAX197に対して、図6B)となるように乳酸を加えてコントロール細胞株の濃度と同様の濃度とした。補充分の乳酸の添加にも関わらず、アポトーシス耐性細胞株ではコントロールと比較して6日目〜12日目にかけて乳酸濃度は大幅に急速に低下した。実際、EAX197の流加培養では6日目〜7日目にかけて乳酸濃度が2.5〜0.2g/Lにまで低下したのに対して、コントロール培養では同じ期間に乳酸濃度は2.7〜2.4g/Lに低下したに過ぎなかった。したがって、乳酸はアポトーシス耐性細胞株ではコントロール細胞株と比較して補充分の乳酸の添加後少なくとも7日間にわたって加速した速度で消費されたものと考えられる。
【0058】
VCD及び細胞生存率に対する乳酸補充の影響をそれぞれ図7A及び7Bに示す。上記で観察したように(図5A)、無添加のアポトーシス耐性細胞株のVCDは8日目(EA167)及び9日目(EAX197)から急速に低下した。これは恐らく、グルコース及び/又は乳酸を含む1以上の不可欠な培地成分がこれらの培養中で枯渇したことによるものと考えられる。コントロール細胞株では7日目から緩慢な低下を示した。しかしながら、乳酸を補充した培養(EA167(乳酸添加)及びEAX197(乳酸添加))ではVCDの低下はより緩やかであり、一方の細胞株(EAX197(乳酸添加))では、VCDは、14日目にこの培養が最終的に死滅するまで8×106〜10×106細胞/mlの間で安定した。図7Bに示される細胞生存率は、培地に乳酸を補充した2つのアポトーシス耐性培養において同様のより緩やかな低下を示した。これらの結果は、アポトーシス耐性細胞株が内因性及び補充された乳酸の両方を効率的に利用して正細胞密度及び生存率を維持することが可能であるのに対して、コントロール細胞株は同様の効率的な乳酸消費を行うことができないことを示唆している。この差は、無添加のアポトーシス耐性細胞株よりも約180〜220%高く、コントロール培養よりも約235%〜250%高かった乳酸添加アポトーシス耐性細胞株のIVCCに反映されている。
【0059】
図7D〜Fに示される他の栄養素及び代謝産物の多くは、図5の無添加培養で上記に観察されたものと同様のプロファイルを示した。グルコース及びグルタミンは培養の初期の時点で急速に消費された。アポトーシス耐性細胞株のアンモニアのプロファイルもやはり、無補充及び乳酸補充培養の両方において乳酸の消費が始まった時点でわずかに低下した。上記の各実験と同様、各アポトーシス耐性細胞株の最終的なアンモニア濃度は、IVCCが4つのアポトーシス耐性細胞株のすべてにおいて大幅に高かったにも関わらずコントロールの濃度よりも低かった。グルタミン酸の濃度は乳酸添加培養と無添加培養との間で異なり、無添加及びコントロール細胞株と比較して添加培養でグルタミン酸の枯渇は大幅に大きかった。
【0060】
(実施例5)
アポトーシス耐性細胞株及びコントロール細胞株のアミノ酸プロファイル
20種類のアミノ酸のそれぞれの濃度を、コントロール細胞株、並びに無添加及び乳酸添加アポトーシス耐性細胞株の培養において観測した。グルタミン及びグルタミン酸以外のアミノ酸の消費又は産生に関しては、表2に示されるように9種類の必須アミノ酸(フェニルアラニン、トレオニン、メチオニン、リジン、トリプトファン、ロイシン、イソロイシン、バリン及びヒスチジン)のいずれも枯渇しなかった点を最初に述べておく必要がある。アポトーシス耐性細胞株(無添加)及びコントロール細胞株の培養では、アスパラギン、セリン、トリプトファン及びシステイン(斜体で示す)はほぼ完全に枯渇した。興味深い点として、3種類の分枝鎖アミノ酸、すなわちイソロイシン、ロイシン及びバリン(表2に灰色の網かけで示す)はいずれも、コントロールと比較してアポトーシス耐性細胞株においてより速やかに培地から欠乏し、この欠乏は乳酸添加培養においてより顕著であった(図8A〜D)。グルタミン酸に加え、これら3つのアミノ酸の最終濃度は乳酸添加培養において培地から大幅に低下した。消費率が高かった他のアミノ酸(最終濃度が培地中の最初の濃度の50%以下)としては、チロシン、フェニルアラニン、メチオニン(図8D)及びアスパラギン酸(図8E)がある。
【0061】
培地中に分泌されたアミノ酸についてコントロール細胞株とアポトーシス耐性細胞株との間で比較を行うのも考慮の対象として興味深い。図8Eに示されるように、コントロール細胞株が最初の6日間にわたって培地中にアスパラギン酸を蓄積したのに対して、4つのアポトーシス耐性細胞株培養のすべてにおいて同じ期間でアスパラギン酸濃度は低下した。更に、各アポトーシス耐性細胞株はコントロール細胞株と比較して最初の6日間にわたって大幅に多量のアラニンを分泌した(図8F)のに対して、これよりも遅い期間ではすべての細胞株がアラニンを消費した。更に、各アポトーシス耐性細胞株はコントロール細胞株と比較して細胞培養の全体を通じてより多くのホモシスチンを産生した(図8G)。
【0062】
(実施例6)
高濃度グルコースの存在下での回分培養
アポトーシス耐性細胞株が乳酸を利用できるならば、有毒なレベルの乳酸が蓄積するためにコントロール細胞にとっては阻害性を示すようなグルコース濃度においてもこれらの細胞株は生存できる可能性がある。更に、これらの細胞株の増殖は乳酸を唯一の炭素源として含む培地によって支持されうる可能性がある。高濃度のグルコースなどのため浸透圧が維持できなくなるために上記の実験で使用したCD−CHO培地を使用することは不可能であり、そのためこれらの実験では新しい培地を使用する必要があった。したがって、上記の実験で最も高い持続的増殖性を示したEAX197の増殖速度論を、60mMのグルコース(高濃度グルコース)又は30mMの乳酸を含む特別に配合した無動物タンパク質培地中でコントロール細胞株と比較した。アポトーシス耐性細胞株及びコントロール細胞株のいずれにおいても、グルコースがまったく存在しない場合、すなわち乳酸のみが存在する場合には持続的増殖が見られなかった(データは示さず)。両方の細胞株を「高濃度」(60mM)のグルコースを含む同じ特別配合培地中で培養したところ、EAX197は、特に細胞培養の後期においてコントロール細胞株よりも高いVCD、生存率及びIVCCを示した(図9A〜C)。EAX197細胞株は、上記のCD−CHO培地中におけるよりもコントロールと比較した場合の全体のIVCCの増加率は低かった(130%)が、後期の高い生存率のためにその差は依然有意であった。高濃度グルコース中で培養したこれら2つの細胞株の乳酸プロファイルを図9Dに示す。IVCCと同様、両細胞株の乳酸プロファイルは、CD−CHO培地で観察されたものとは幾分異なっていた(図9D)。CD−CHO培地を用いた上記の結果と同様、特別配合培地中のアポトーシス耐性細胞株ではコントロールよりも早期に正味の乳酸消費が開始され(2日目まで、図9Dに示される)、最大乳酸濃度は大幅に低かった(コントロール細胞株の2.4g/Lに対して1.2g/L)。コントロール細胞は、培地中の乳酸の消費も開始した5日目までは乳酸の正味の産生を示した。CD−CHO培地は適当な組成を有する市販の配合物であるため、これらの異なる配合物中において乳酸消費プロファイルが異なる原因を理解する目的で2つの配合物中の培地成分の正確な比較を行うことはできなかった。
【0063】
(実施例7)
高グルコース培地中のアポトーシス耐性細胞株の生産性
続いて、アポトーシス耐性細胞株であるEAX197及びコントロール細胞株をホストとして使用して、モデル抗体を発現する生産用細胞株を作出した。各細胞株について同数のクローンを調べ、各ホストから生成した最も優れたクローンを抗体の生産性について震盪フラスコ回分方式で比較した。高グルコース環境下における各細胞株の性能を比較するため、60mM(高濃度)グルコースを含む特別配合培地を炭素源として使用した。標準培地(図10A)中では、EAX197から誘導された生産用細胞株では力価が489mg/Lであり、337mg/Lの力価を示したコントロール細胞株と比較して145%の増加率であった。高グルコース培地(図10B)中では同じ細胞株が678mg/Lの力価を示し、コントロール細胞株(348mg/L)の力価と比較して178%の増加率であった。産生量の向上は、EAX197ではIVCCがコントロール細胞株に対して188%に増大していることに一部よるものである(データは示さず)。
【0064】
(実施例8)
E1B19K+AVEN±XIAPΔの過剰発現のアポトーシスの阻害に対する影響
ミトコンドリア内のアポトーシスカスケードの主要な活性化因子を図1に示した。アポトーシス耐性CHO細胞株を作出するため、アポトーシスの早期、中期及び後期に働く3つのアポトーシス耐性遺伝子を選択した。これらの遺伝子は、E1B19K(E)、AVEN(A)及びXIAPΔ(X)である。広く用いられている形質移入(トランスフェクション)用ホスト細胞株であるCHOK1SVに、E1B19K及びAVENを、XIAPΔとともに、又はXIAPΔ抜きでトランスフェクトした。多数の形質移入細胞をスクリーニングして各トランスフェクションから数個のクローンを更なる実験用に選択した。これらの細胞株の増殖プロファイルの実験により、これらの細胞株は極めて高いVCDに達し、一般にそのIVCCはコントロールのIVCCの約2倍高いことが示された。アポトーシス耐性細胞株では播種後8日目までに生存率が低下したが、乳酸を消費することによって10日目以降は生存率が回復した。E1B19K+AVEN±XIAPΔを発現しているアポトーシス耐性細胞株の培養液は、アンモニア及び乳酸の濃度が低下しているか検出不能なレベルであり、グルタミン酸は培地から枯渇していた。
【0065】
(実施例9)
抗体力価を高めるためのBcl−2dの使用
上記の結果は、アポトーシス耐性細胞株はコントロール細胞と比較して乳酸の蓄積量が少なく、アポトーシス耐性培養の生存率を高グルコース培地中で維持することが可能であることを示している。以下の震盪フラスコ回分培養実験では、Bcl−2d並びに抗体の重鎖及び軽鎖を発現するアポトーシス耐性細胞株(C2088B)を標準的濃度(4.8g/L)及び高濃度(9.5g/L)のグルコース中、異なる播種密度で播種した。この結果は、アポトーシス耐性細胞株は9.5g/Lのグルコースを含む培地中に1e6細胞/mlの密度で播種した場合、播種後7日目に14e6細胞/mlのピークVCD(図11A)、及び2g/Lの抗体力価に達したことを示す。これに対して、標準条件下(接種密度0.3e5細胞/ml及び4.8g/L)で培養したC2088BはビークVCDが7e6細胞/ml、抗体力価は1g/Lであった(図11B)。これは、高グルコース条件では標準条件(すなわち、4.8g/Lのグルコースを含む培地中、0.3e5細胞/mlの播種密度)と比較して抗体力価が200%に増大することを意味する。
【0066】
以上に本発明の全容を説明したが、本発明には、多くの変更及び改良を、添付の特許請求の範囲の趣旨及び範囲から逸脱することなく行いうることは当業者には明白であろう。
【特許請求の範囲】
【請求項1】
真核細胞の流加培養における生細胞密度を高める方法であって、
a)1種以上の異種由来のアポトーシス耐性(アポトーシスR)遺伝子及び1種以上の対象遺伝子を発現する真核細胞株を培養する工程と、
b)前記細胞培養の指数増殖期及び定常期に高グルコース添加培地を維持する工程と、を含む方法。
【請求項2】
前記真核細胞株が、チャイニーズハムスター子宮細胞(CHO)株である、請求項1に記載の方法。
【請求項3】
前記CHO細胞株がCHO−K1である、請求項2に記載の方法。
【請求項4】
前記CHO細胞株がCHO−K1SVである、請求項2に記載の方法。
【請求項5】
前記真核細胞株がミエローマ細胞株である、請求項1に記載の方法。
【請求項6】
前記ミエローマ細胞株がNS0である、請求項5に記載の方法。
【請求項7】
前記ミエローマ細胞株がSp2/0である、請求項5に記載の方法。
【請求項8】
前記真核細胞株がハイブリドーマである、請求項1に記載の方法。
【請求項9】
前記高グルコース添加培地が約60mMのグルコースを含む、請求項1に記載の方法。
【請求項10】
前記アポトーシス耐性遺伝子がE1B19K及びAVENを含む、請求項1に記載の方法。
【請求項11】
前記アポトーシスR耐性細胞遺伝子がXIAPをさらに含む、請求項10に記載の方法。
【請求項12】
前記アポトーシスR耐性遺伝子がBcl−2Δを含む、請求項1に記載の方法。
【請求項13】
前記CHO細胞株が、前記細胞培養の指数増殖期に蓄積した乳酸を消費する、請求項1に記載の方法。
【請求項14】
前記細胞培養の指数増殖期における前記CHO細胞株の乳酸分泌量が、1種以上のアポトーシス耐性細胞株を含有していないCHO細胞株よりも小さい、請求項1に記載の方法。
【請求項15】
ピーク生細胞密度(VCD)が高められている、請求項1に記載の方法。
【請求項16】
前記細胞培養の寿命が延びている、請求項1に記載の方法。
【請求項17】
前記細胞培養の力価が高められている、請求項1に記載の方法。
【請求項18】
前記細胞培養の積分生細胞数(IVCC)が高められている、請求項1に記載の方法。
【請求項19】
細胞のカルシウム流束が低下している、請求項1に記載の方法。
【請求項20】
ミトコンドリア膜電位が高められている、請求項1に記載の方法。
【請求項21】
前記対象遺伝子が抗体の重鎖及び抗体の軽鎖をコードしている、請求項1に記載の方法。
【請求項1】
真核細胞の流加培養における生細胞密度を高める方法であって、
a)1種以上の異種由来のアポトーシス耐性(アポトーシスR)遺伝子及び1種以上の対象遺伝子を発現する真核細胞株を培養する工程と、
b)前記細胞培養の指数増殖期及び定常期に高グルコース添加培地を維持する工程と、を含む方法。
【請求項2】
前記真核細胞株が、チャイニーズハムスター子宮細胞(CHO)株である、請求項1に記載の方法。
【請求項3】
前記CHO細胞株がCHO−K1である、請求項2に記載の方法。
【請求項4】
前記CHO細胞株がCHO−K1SVである、請求項2に記載の方法。
【請求項5】
前記真核細胞株がミエローマ細胞株である、請求項1に記載の方法。
【請求項6】
前記ミエローマ細胞株がNS0である、請求項5に記載の方法。
【請求項7】
前記ミエローマ細胞株がSp2/0である、請求項5に記載の方法。
【請求項8】
前記真核細胞株がハイブリドーマである、請求項1に記載の方法。
【請求項9】
前記高グルコース添加培地が約60mMのグルコースを含む、請求項1に記載の方法。
【請求項10】
前記アポトーシス耐性遺伝子がE1B19K及びAVENを含む、請求項1に記載の方法。
【請求項11】
前記アポトーシスR耐性細胞遺伝子がXIAPをさらに含む、請求項10に記載の方法。
【請求項12】
前記アポトーシスR耐性遺伝子がBcl−2Δを含む、請求項1に記載の方法。
【請求項13】
前記CHO細胞株が、前記細胞培養の指数増殖期に蓄積した乳酸を消費する、請求項1に記載の方法。
【請求項14】
前記細胞培養の指数増殖期における前記CHO細胞株の乳酸分泌量が、1種以上のアポトーシス耐性細胞株を含有していないCHO細胞株よりも小さい、請求項1に記載の方法。
【請求項15】
ピーク生細胞密度(VCD)が高められている、請求項1に記載の方法。
【請求項16】
前記細胞培養の寿命が延びている、請求項1に記載の方法。
【請求項17】
前記細胞培養の力価が高められている、請求項1に記載の方法。
【請求項18】
前記細胞培養の積分生細胞数(IVCC)が高められている、請求項1に記載の方法。
【請求項19】
細胞のカルシウム流束が低下している、請求項1に記載の方法。
【請求項20】
ミトコンドリア膜電位が高められている、請求項1に記載の方法。
【請求項21】
前記対象遺伝子が抗体の重鎖及び抗体の軽鎖をコードしている、請求項1に記載の方法。
【図1】

【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図3C】
【図4A】
【図4B】
【図4C】
【図5A】
【図5B】
【図5C】
【図6A】
【図6B】
【図7A】
【図7B】
【図7C】
【図7D】
【図7E】
【図7F】
【図7G】
【図8A】
【図8B】
【図8C】
【図8D】
【図8E】
【図8F】
【図8G】
【図9A】
【図9B】
【図9C】
【図9D】
【図10A】
【図10B】
【図11A】
【図11B】
【図2A】
【図2B】
【図2C】
【図3A】
【図3B】
【図3C】
【図4A】
【図4B】
【図4C】
【図5A】
【図5B】
【図5C】
【図6A】
【図6B】
【図7A】
【図7B】
【図7C】
【図7D】
【図7E】
【図7F】
【図7G】
【図8A】
【図8B】
【図8C】
【図8D】
【図8E】
【図8F】
【図8G】
【図9A】
【図9B】
【図9C】
【図9D】
【図10A】
【図10B】
【図11A】
【図11B】
【公表番号】特表2011−524172(P2011−524172A)
【公表日】平成23年9月1日(2011.9.1)
【国際特許分類】
【出願番号】特願2011−513722(P2011−513722)
【出願日】平成21年6月12日(2009.6.12)
【国際出願番号】PCT/US2009/047178
【国際公開番号】WO2009/152413
【国際公開日】平成21年12月17日(2009.12.17)
【出願人】(509087759)セントコア・オーソ・バイオテツク・インコーポレーテツド (77)
【Fターム(参考)】
【公表日】平成23年9月1日(2011.9.1)
【国際特許分類】
【出願日】平成21年6月12日(2009.6.12)
【国際出願番号】PCT/US2009/047178
【国際公開番号】WO2009/152413
【国際公開日】平成21年12月17日(2009.12.17)
【出願人】(509087759)セントコア・オーソ・バイオテツク・インコーポレーテツド (77)
【Fターム(参考)】
[ Back to top ]