
哺乳動物T1R3甘味レセプター
本発明は、甘味レセプターの単離された核酸配列およびアミノ酸配列、T1R3 Gタンパク質共役型レセプターポリペプチドのモノマーまたはホモダイマーを含むか、それらからなるレセプター(これらは、T1R3が細胞中に発現された場合、甘味リガンドに応答してシグナルを変換する)、そのようなレセプターに対する抗体、そのような核酸およびレセプターを検出する方法、ならびに甘味レセプターおよびアミノ酸味レセプターの調節因子をスクリーニングする方法を提供する。

【発明の詳細な説明】
【技術分野】
【0001】
(関連出願に対する相互参照)
本出願は、米国特許出願第10/679,102号(2003年10月2日出願)(その全体が参考として本明細書において援用)の優先権を主張する。
【0002】
(連邦政府により支援される研究および開発の下になされた発明の権利に関する声明書)
該当なし。
【0003】
(発明の分野)
本発明は、単離された甘味レセプターの核酸配列およびアミノ酸配列、T1R3 Gタンパク質共役型レセプターポリペプチドのモノマーまたはホモダイマーを含むか、それからなる上記レセプター、そのようなレセプターに対する抗体、そのような核酸およびレセプターを検出する方法、ならびに甘味レセプターの調節因子をスクリーニングする方法を提供する。
【背景技術】
【0004】
(発明の背景)
味覚は、甘味、苦味、酸味、塩味および旨味(アミノ酸)の刺激の検出ならびに応答を担っている。それはまた、これらの種々の味の様式を区別し、固有の行動の応答を生じ得る。例えば、動物は、苦味のする化合物を極度に嫌うが、甘味および旨味の刺激には引き付けられる。味シグナルの検出および情報処理を調査するために、本発明者らは、甘味レセプター、旨味レセプターおよび苦味レセプターの単離および特徴付けに焦点を定めた。これらのレセプターは、味覚系の機構を表し、そして味コーディング(taste coding)の論理を規定することを助ける、強力な分子的手段を提供する。
【0005】
候補哺乳動物味レセプターの2つのファミリー(T1RおよびT2R)が、甘味、旨味および苦味の検出において関連付けられてきた。T2Rは、約30の味特異的GPCRのファミリーであり、オプシンと遠縁であり、そしてヒトおよびマウスにおいて苦味と遺伝的に関連付けられたゲノムの領域中でクラスター化されている(非特許文献1;非特許文献2)。いくつかのT2Rは、異種発現アッセイにおいて苦味レセプターとして機能することが示されており、苦味センサーとしてのそれらの役割を実証している(非特許文献3;非特許文献4)。ほとんどのT2Rは、味レセプター細胞の同じサブセット中に同時発現され(非特許文献1)、これらの細胞が、一般的苦味検出器(detector)として機能することを示唆している。
【0006】
T1Rは、舌上皮および口蓋上皮の味細胞において発現される3つのGPCRの小ファミリーであり、代謝生成物産性グルタメートレセプター、カルシウム感知レセプター、および鋤鼻レセプターと遠縁である(非特許文献5;非特許文献6;非特許文献7;非特許文献8;非特許文献9;非特許文献10)。T1Rは、少なくとも2つのヘテロマーレセプターを産生するように組み合わされる:T1R1およびT1R3は、L−アミノ酸センサーを形成し、このセンサーは、げっ歯類においてほとんどのアミノ酸を認識し、そしてT1R1およびT1R3は、広範囲にわたるように調節された甘味レセプターとして機能するように結合する(非特許文献9;非特許文献11;非特許文献12;ならびに特許文献1、特許文献2、および特許文献3も参照のこと)。
【0007】
動物は、広範囲の化学的に別個の甘味分子(天然の糖、人工甘味料、D−アミノ酸、および強烈に甘いタンパク質が挙げられる)を検出し得る。全ての甘み(sweet universe)甘さを味わうのに何種類の異なるレセプターが必要であろうか?ヒトおよびげっ歯類のT1R2+T1R3ヘテロマー甘味レセプターが、細胞ベースのアッセイにおいて、全てのクラスの甘味化合物に応答し、そしてインビボでのそれぞれの心理物理学的閾値および/または行動学的閾値に相当する親和性を伴って、そのように応答する(非特許文献9;非特許文献12)。根本的なレベルにおいて、甘味の進化は、潜在的な食料の供給源中の糖含有量を検出しそして測定する必要性を反映している可能性が高い。従って、天然の糖に対して広範囲にわたるように調節された1つのレセプターが、必要とされる全てであり得る。他方、種々の糖および人工甘味料の多くの研究は、1より多い甘味レセプターの可能性を暗示する(非特許文献13;非特許文献14)。
【0008】
ヒトにおいて、L−グルタメート一ナトリウム(MSG)およびL−アスパルテートは、明白な良い味の感覚(いわゆる、旨味(Maga、1983))を誘発する(しかし他のアミノ酸は誘発しない)。特に、げっ歯類のT1R1+T1R3と違って、ヒトT1R1+T1R3アミノ酸味レセプターは、他のL−アミノ酸よりも、L−グルタメートおよびL−アスパルテートに実質的に敏感である(非特許文献12)。これらの発見は、T1R1+T1R3が、哺乳動物旨味レセプターであり得るという提唱に結び付いた(非特許文献11;非特許文献12)。しかし、多数の研究(T1R3 KOマウスの最近の分析(非特許文献15)を含む)は、旨味は、mGluR4t(代謝生成物産性グルタメートレセプターの短縮型改変体)によって媒介されることを示唆している(非特許文献16;非特許文献17)。
【0009】
異なる味の特質は、どのように味細胞レベルにおいてコードされるのであろうか?哺乳動物において、味レセプター細胞は、味蕾に構築されており、この味蕾は、舌上皮の異なる乳頭に分布されている。各味蕾は、50〜150の細胞(前駆体細胞、支持細胞、および味レセプター細胞が挙げられる)を含んでいる(非特許文献18)。このレセプター細胞は、脳幹および視床のシナプスを通じて皮質の味覚中枢に情報を伝える、求心性神経繊維によって神経支配される。末梢における味コーディングの最も単純なモデルにおいて、各味様式は、特異的なレセプターを発現する細胞の特有の集団(例えば、甘味細胞、苦味細胞、塩感知細胞など)によってコードされる。このシナリオにおいて、任意の1つの味の特質についての我々の知覚は、舌における異なる細胞の型の活性化から生ずる(専有回線モデル(labeled line model))。あるいは、個々の味細胞は、複数の味覚様式を認識し得、そして多くのそのような広範囲にわたるように調節されたレセプター細胞の集団発火パターンが、味の特質をコードする(総ニューロンモデル(across fiber model))。
【0010】
最近、本発明者らは、T1RおよびT2Rが、舌上皮のレセプター細胞の完全に非重複の集団において発現されることを示し(非特許文献9)、そして苦味レセプター発現細胞は、苦味への応答を媒介するが、甘味またはアミノ酸味覚物質への応答を媒介しないことを示した(非特許文献19)。合わせて、これらの結果は、味レセプター細胞は、全ての様式にわたって広範囲に調節されるのではないことを主張し、そして末梢における味コーディングの専有回線モデルを強く支持した。ここで本発明者らが取り組む根本的な問題は、何種類の細胞およびレセプターが甘味および旨味(2つの主要な誘引性の味覚)を媒介するのに必要であるかということである。本発明者らはここで、甘味および旨味が、T1Rによって排他的に媒介されることを示し、そして個々のT1Rサブユニットの遺伝的除去が、これら2つの誘引性の味覚様式に選択的に影響することを示す。甘味および旨味感知のための細胞およびレセプターの同定はまた、本発明者らが、細胞刺激から、レセプター活性の味覚応答をコードする役割を分離する戦略を考案することを可能にした。本発明者らは、T1R2+T1R3発現細胞において改変k−オピオイドレセプターを発現するよう操作された動物が、k−オピオイドアゴニストに対して特異的に引き付けられることを示し、(T1Rレセプター自身というよりも)甘味レセプター発現細胞の活性化が、甘味物質への行動的誘引性の重要な決定要素であることを証明する。最後に、本発明者らはここで、T1R1は単独で(モノマーとしてかまたはホモダイマーとしてのいずれかで)、天然に存在する糖のためのレセプターとして作用することを示す。
【特許文献1】国際公開第00/06592号パンフレット
【特許文献2】国際公開第00/06593号パンフレット
【特許文献3】国際公開第03/004992号パンフレット
【非特許文献1】Adlerら、Cell、2000年、100号、693−702頁
【非特許文献2】Matsunamiら、Nature、2000年、404号、601−604頁
【非特許文献3】Chandrashekarら、Cell、2000年、100号、703−711頁
【非特許文献4】Bufeら、Nat Genet、2002年、32号、397−401頁
【非特許文献5】Hoonら、Cell.1999年、96号、541−551頁
【非特許文献6】Kitagawaら、Biochem Biophys Res Commun、2001年、283号、236−242頁
【非特許文献7】Maxら、Sac.Nat Genet、2001年、28号、58−63頁
【非特許文献8】Montmayeurら、Nat Neurosci、2001年、4号、492−498頁
【非特許文献9】Nelsonら、Cell、2001年、106号、381−390頁
【非特許文献10】Sainzら、J Neurochem、2001年、77号、896−903頁
【非特許文献11】Nelson,G.ら、Nature、2002年、416号、199−202頁
【非特許文献12】Li,X.ら、Proc Natl Acad Sci U S A、2002年、99号、4692−4696頁
【非特許文献13】Schiffmanら、Pharmacol Biochem Behav、1981年、15号、377−388頁
【非特許文献14】Ninomiyaら、J Neurophysiol、1999年、81号、3087−3091頁
【非特許文献15】Damakら、Science、2003年、301号、850−853頁
【非特許文献16】Chaudhariら、Neurosci、1996年、16号、3817−3826頁
【非特許文献17】Chaudhariら、Nat Neurosci、2000年、3号、113−119頁
【非特許文献18】Lindermann、Physiol Rev、1996年、76号、718−766頁
【非特許文献19】Zhangら、Cell、2003年、112号、293−301頁
【発明の開示】
【課題を解決するための手段】
【0011】
(発明の簡単な要旨)
従って、本発明は、最初に、ホモダイマーの甘味レセプターを提供する。このレセプターは、2つのT1R3ポリペプチドを含むかまたは2つのT1R3ポリペプチドからなる。本発明はまた、1つのT1R3ポリペプチドを含むかまたは1つのT1R3ポリペプチドからなる、モノマーの甘味レセプターを提供する。これらのレセプターは、T1R3が細胞中に発現された場合、甘味リガンドに応答してシグナルを変換する。一実施形態において、この甘味リガンドは、天然に存在する甘味分子である。別の実施形態において、この甘味リガンドは、人工的であり、そして天然に存在する甘味分子を模倣する。一実施形態において、ホモダイマーのT1R3ポリペプチドは、非共有結合性に結合される。
【0012】
一局面において、本発明は、T1R3ポリペプチドを含む甘味レセプターを提供し、このT1R3ポリペプチドは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列に対して、約80%を超えるアミノ酸配列の同一性を有するか、あるいは、中程度もしくは高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる。
【0013】
一実施形態において、T1R3を含むレセプターは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31に対して生じた、ポリクローナル抗体に特異的に結合する。別の実施形態において、このレセプターは、Gタンパク質共役型レセプター活性を有する。別の実施形態において、T1R3ポリペプチドは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列を有する。別の実施形態において、このレセプターは、ヒト、ラット、またはマウス由来である。
【0014】
別の実施形態において、甘味レセプターは、T1R3ポリペプチドを含み、そして天然の糖(例えば、グルコース、ガラクトース、フルクトース、マルトース、ラクトース、およびスクロース)を認識する。
【0015】
一局面において、本発明は、単離されたポリペプチドを提供し、このポリペプチドは、T1R3を含むホモダイマーもしくはモノマーの甘味レセプターの細胞外、膜貫通ドメイン、または細胞質ドメインを含み、この細胞外、膜貫通ドメイン、または細胞質ドメインは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31の細胞外、膜貫通ドメイン、または細胞質ドメインとの、約80%を超えるアミノ酸配列の同一性を有する。別の実施形態において、細胞外ドメイン、膜貫通ドメイン、または細胞質ドメインは、高度にストリンジェントな条件の下、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列の細胞外ドメイン、膜貫通ドメイン、または細胞質ドメインにハイブリダイズする。
【0016】
一実施形態において、ポリペプチドは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31の細胞外、膜貫通ドメイン、または細胞質ドメインをコードする。別の実施形態において、細胞外、膜貫通ドメイン、または細胞質ドメインは、異種ポリペプチドに共有結合され、キメラポリペプチドを形成する。別の実施形態において、キメラポリペプチドは、Gタンパク質共役型レセプター活性を有する。
【0017】
一局面において、本発明は、ホモダイマーのまたはモノマーの甘味レセプターと選択的に結合する抗体を提供し、このレセプターは、1つかもしくは2つのT1R3ポリペプチドを含み(しかし、T1R1もしくはT1R2ポリペプチドを含まない)、この抗体は、配列番号15、配列番号20、配列番号23、配列番号25、配列番号31のアミノ酸配列に対して、約80%を超えるアミノ酸配列の同一性を含むか、あるいは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる、T1R3ポリペプチドを含むレセプターに対し生産される。
【0018】
別の局面において、本発明は、味細胞において甘味シグナルを調節する化合物を同定するための方法を提供し、この方法は、(i)T1R3ポリペプチドを含む(しかし、T1R1もしくはT1R2ポリペプチドを含まない)ホモダイマーまたはモノマーのレセプターとこの化合物を接触させる工程であって、このポリペプチドは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31に対して、約80%を超えるアミノ酸配列の同一性を含むか;あるいは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる、工程;ならびに(ii)上記レセプター上の化合物の機能的効果を決定する工程を包含する。
【0019】
一実施形態において、上記機能的効果は、インビトロで決定される。一実施形態において、上記ポリペプチドは、細胞かまたは細胞膜において発現される。別の実施形態において、上記レセプターは、固相に共有結合性かまたは非共有結合性のいずれかで結合される。
【0020】
別の局面において、本発明は、味細胞において甘味シグナルを調節する化合物を同定するための方法を提供し、この方法は、(i)この化合物とT1R3ポリペプチドを含むが、T1R1もしくはT1R2ポリペプチドを含まないホモダイマーかまたはモノマーのレセプターを発現する、細胞を接触させる工程であって、このT1R3ポリペプチドは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31に対して、約80%を超えるアミノ酸配列の同一性を含むか;あるいは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる、工程;ならびに(ii)上記レセプター上の化合物の機能的効果を決定する工程を包含する。
【0021】
一実施形態において、上記機能的効果は、細胞内のcAMP、IP3、またはCa2+の変化を測定することによって決定される。別の実施形態において、上記機能的効果は、化学的効果または表現型効果である。別の実施形態において、上記機能的効果は、物理的効果である。別の実施形態において、上記機能的効果は、上記レセプターの細胞外ドメインへの上記化合物の結合を測定することによって決定される。別の実施形態において、ポリペプチドは、組換え型である。別の実施形態において、上記細胞は、真核生物細胞(例えば、哺乳動物細胞(例えば、ヒト細胞))である。別の実施形態において、上記細胞は、Gタンパク質(Gα15)を発現する。
【発明を実施するための最良の形態】
【0022】
(発明の詳細な説明)
(序論)
T1RおよびT2Rは、味レセプター細胞のサブセットに選択的に発現されるGタンパク質共役型レセプター(GPCR)の2つのファミリーである(Hoonら、Cell 96:541−551(1999);Adlerら、Cell 100:693−702(2000);Chandrashekarら、Cell 100:703−711(2000);Matsunamiら、Nature 404:601−604(2000);Nelsonら、Cell 106:381−390(2001);Kitagawaら、Biochem.Biophys.Res.Cummun.283:236−242(2001);Montmayeurら、Nature Neurosci.4:492−498(2001);Maxら、Nature Genet.28:58−63(2001);Sainzら、J.Neurochem.77:896−903(2001))。T2Rは、苦味の検出に関与する(Adlerら、Cell 100:693−702(2000);Chandrashekarら、Cell、100:703−711(2000));T1R2およびT1R3は、組み合わさって甘味レセプターとして機能する(Nelsonら、Cell 106:381−390(2001)を参照のこと;そしてT1R1およびT1R3は、本明細書中に記載されるように、組み合わさってアミノ酸味レセプターとして機能する(Nelsonら、Nature 2002年2月24日およびWO 03/004992もまた参照のこと)))。本発明者らは、ここでホモダイマーの味レセプターを同定し、この味レセプター中で、2つのT1R3ポリペプチドは、組み合わさって甘味レセプターとして機能する。T1R3のモノマー形態はまた、甘味レセプターとして作用する。
【0023】
異種発現系を使用して、本発明者らは、T1R3が、それ自身と組み合わさり、そしてまたモノマーとして作用して甘味レセプターとして機能し、甘味分子(例えば、スクロース、ガラクトース、フルクトース、グルコース、マルトース、およびラクトース)を認識することを示す。候補レセプターは、Gα16−GαzおよびGα15乱交雑Gタンパク質を含むヒト胎児由来腎臓(HEK)細胞内に発現され(Offermannsら、J.Biol.Chem.270:15175−15180(1995);Modyら、Mol.Pharmacol.57:13−23(2000))、そして刺激によって引き出された細胞内カルシウムの変化ついてアッセイされる。この系において、レセプターの活性化は、ホスホリパーゼCβ(PLC-βの活性化およびカルシウム指示染料(indicator dye)を使用して、単一の細胞レベルでモニタリングされ得る内部貯蔵からのカルシウムの放出に結び付く(Chandrashekarら、Cell 100:703−711(2000);Nelsonら、Cell 106:381−390(2001);Tsienら、Cell Calcium 6:145−157(1985))。
【0024】
上記レセプターをコードするこれらの核酸およびタンパク質は、その核酸が、味細胞中で特異的に発現される場合、味細胞の同定のための有用なプローブを提供する。このレセプターは、新規の味覚物質(例えば、人工甘味料分子)の検定について有用である。例えば、GPCRポリペプチドおよびタンパク質のためのプローブは、味細胞(例えば、葉状細胞、口蓋細胞、および有郭細胞)のサブセット、または特異的な味レセプター細胞(例えば、甘味レセプター細胞)を同定するために使用され得る。これらはまた、舌の味細胞と脳の味中枢につながっている味感覚ニューロンとの間の関係を明らかにする味地形図(topographic map)の作製のための手段として役立つ。さらに、これらがコードする核酸およびタンパク質は、味誘導行動を分析するためのプローブとして使用され得る。
【0025】
本発明はまた、これら新規のT1R3を含むモノマーまたはホモダイマーの甘味レセプターの調節因子(例えば、アクチベーター、インヒビター、刺激物質、エンハンサー、アゴニスト、およびアンタゴニスト)をスクリーニングする方法を提供する。一実施形態において、本発明のモノマーまたはホモダイマーのT1R3を含むレセプターは、天然に存在するかもしくは人工の甘味分子もしくは甘味変換の調節因子(例えば、小有機分子、アミノ酸、ペプチド、炭水化物、脂質、多糖類など)をスクリーニングために使用され得る。例えば、本発明のホモダイマーまたはモノマーのT1R3を含むレセプターは、以下の実施例のセクションに記載されるように、天然に存在する甘味味覚物質を認識する。そのようなレセプターは、人工甘味料または変更された天然に存在する甘味料(これらの甘味料は、ホモダイマーまたはモノマーのT1R3を含むレセプターの天然に存在する糖のリガンドを模倣する)をスクリーニングするために使用され得る。そのような甘味変換の調節因子は、甘味シグナル伝達経路の薬理学的および遺伝的調節のために、そして新規の甘味リガンドの発見のために有用である。これらのスクリーニングの方法は、甘味細胞活性のアゴニストおよびアンタゴニストを同定するために使用され得る。次いで、これらの調節化合物は、食品産業および薬品産業において味を作るために使用され得る。従って、本発明は、T1R3を含むレセプターが、甘味変換に対する調節因子の効果についての直接的または間接的なレポーター分子として作用する場合、味変換のためのアッセイを提供する。GPCRは、例えば、リガンド結合、Gタンパク質結合、調節分子結合、イオン濃度、膜電位、電流の流れ、イオン流入、転写、シグナル変換、レセプター−リガンド相互作用、神経伝達物質およびホルモン放出;ならびにインビトロ、インビボ、およびエクスビボでのセカンドメッセンジャーの濃度の変化を測定するために、アッセイにおいて使用され得る。一実施形態において、T1R3を含むレセプターは、第二のレポーター分子(例えば、緑蛍光タンパク質)への付着を介した間接的なレポーターとして使用され得る(例えば、MistiliおよびSpector,Nature Biotechnology 15:961−964(1997)を参照のこと)。別の実施形態において、T1R3を含むレセプターは、T1R1またはT1R2のいずれも発現しない細胞において組換えによって発現され、そしてGPCR活性を介した味変換の調節は、Ca2+レベルの変化を測定することによって検定される。
【0026】
味変換の調節因子を検定する方法としては、T1R3、その一部を(例えば、細胞外ドメイン)、または1つ以上のT1R3のドメインを含むキメラタンパク質を含むレセプターを使用したインビトロリガンド結合アッセイ、ならびにインビボ(細胞ベースおよび動物)アッセイ(例えば、卵母細胞T1R3レセプター発現;組織培養細胞T1R3レセプター発現;T1R3の転写活性化;GPCRのリン酸化および脱リン酸化;GPCRへのGタンパク質結合;リガンド結合アッセイ;電圧、膜電位および伝導度の変化;イオン流入アッセイ;細胞内セカンドメッセンジャー(例えば、cAMPおよびイノシトール三リン酸)の変化;細胞内カルシウムレベルの変化;および神経伝達物質の放出)が挙げられる。
【0027】
(定義)
「T1Rファミリー味レセプター」とは、ホモダイマーのレセプター、ヘテロダイマーのレセプターまたはモノマーのレセプターとしての、Gタンパク質共役型レセプター(例えば、T1R1、T1R2、およびT1R3、またはこれらの任意の組み合わせ)のT1Rファミリーのメンバーを含むレセプターをいう。一実施形態において、このT1Rファミリーレセプターは、T1R3(「T1R3を含む味レセプター」または「T1R3を含む甘味レセプター」)を含む。一実施形態において、T1Rファミリーレセプターは、共有結合性にまたは非共有結合性のいずれかで結合されるホモダイマーのレセプターを形成する、第一のT1R3ポリペプチドおよび第二のT1R3ポリペプチドを含む。別の実施形態において、T1Rファミリーレセプターは、1つのT1R3ポリペプチドを含むが、他のT1Rポリペプチドを含まず、そしてモノマーのレセプターを形成する。別の実施形態において、T1Rファミリーレセプターは、T1R3およびT1Rファミリーの異種のポリペプチドを含む。一実施形態において、レセプターは、T1R1およびT1R3を含む。別の実施形態において、レセプターは、T1R2およびT1R3を含む。一実施形態において、T1R3を含むレセプターは、そのレセプターの2つのメンバーが同じ細胞において同時発現される場合(例えば、T1R3およびT1R3、またはT1R1およびT1R3、またはT1R2およびT1R3)、活性である。別の実施形態において、T1Rポリペプチドは、同じ細胞において同時発現され、そしてヘテロダイマーかまたはホモダイマーのレセプターを形成し、ここで、レセプターのT1Rポリペプチドは、非共有結合性に結合されるかまたは共有結合性に結合される。レセプターは、例えば、天然に存在する甘味分子および/または人工甘味分子(例えば、スクロース、フルクトース、ガラクトース、マンノース、グルコース、ラクトース、サッカリン、ズルチン、アセスルファーム−K)ならびに(甘いおよび甘くない)他の分子を認識する能力を有する。これらの分子は、T1R3を含み、味を変換するGタンパク質共役型レセプターのためのリガンドとして作用することによって、「甘味シグナル変換を調節する」化合物の例である。
【0028】
用語「GPCR−B3またはT1R1」、「GPCR−B4またはT1R2」および「T1R3」あるいは「GPCR−B3またはT1R1」、「GPCR−B4またはT1R2」および「T1R3」をコードする核酸とは、Gタンパク質共役型レセプターのT1Rファミリーのメンバーである、核酸およびポリペプチド多型改変体、対立遺伝子、変異体、および種間のホモログをいい、そして(1)配列番号1、2、3、7、8、9、15、18、20、23、25、27、または31によってコードされるアミノ酸配列に対して、約60%を超えるアミノ酸配列の同一性、65%、70%、75%、80%、85%、90%、好ましくは、91%、92%、93%、94%、95%、96%、97%、98%または99%以上のアミノ酸配列の同一性を有する、好ましくは少なくとも約25、50、100、200、500、1000またはそれより多くのアミノ酸にわたる領域にわたるアミノ酸配列を有し;(2)配列番号1、2、3、7、8、9、15、18、20、23、25、27、または31によってコードされたアミノ酸配列を含む免疫原に対して惹起された抗体(例えば、ポリクローナル抗体)およびこれらの保存的に改変された改変体に結合し;(3)ストリンジェントなハイブリダイゼーション条件下で、T1Rタンパク質をコードする核酸配列(例えば、配列番号4、5、6、10、11、12、13、14、16、17、19、21、22、24、26、28、および30)に対応するアンチセンス鎖およびこれらの保存的に改変された改変体に、特異的にハイブリダイズし、(4)配列番号4、5、6、10、11、12、13、14、16、17、19、21、22、24、26、または28、または30に対して、約60%を超える配列の同一性、65%、70%、75%、80%、85%、90%、好ましくは91%、92%、93%、94%、95%、96%、97%、98%または99%以上のヌクレオチド配列の同一性を有する、好ましくは少なくとも25、50、100、200、500、1000以上のヌクレオチドの領域にわたる核酸配列を有する。さらに、本発明のT1Rファミリーポリペプチド(例えば、T1R1、T1R2、もしくはT1R3)またはT1R3を含むレセプター(例えば、T1R3、T1R3+T1R3、T1R1+T1R3もしくはT1R2+T1R3)は、単独でもしくは同じ細胞で同時発現した場合、または別のT1Rファミリーメンバーとモノマー、ホモダイマー、またはヘテロダイマーとして同時発現した場合のいずれかにおいて、Gタンパク質共役型レセプター活性を有する。ヒト、ラット、およびマウスのT1R1、T1R2、およびT1R3の、アミノ酸配列およびヌクレオチド配列の登録番号は、GenBankにおいて見出され得る(ヒトT1R1アミノ酸配列については、例えば、登録番号DAA00012およびNP_619642を参照のこと;ヒトT1R1ヌクレオチド配列については、例えば、登録番号BK000153を参照のこと;ヒトT1R2アミノ酸配列については、例えば、登録番号DAA00019、AAM12239、およびNP_619642.1を参照のこと;ヒトT1R2ヌクレオチド配列については、例えば、登録番号BK000151、NM_138697.1、AF458149S1−6を参照のこと;ヒトT1R3アミノ酸配列については、例えば、登録番号DAA00013を参照のこと;ヒトT1R3ヌクレオチド配列については、例えば、登録番号BK000152を参照のこと)。T1R1、T1R2、およびT1R3のアミノ酸配列およびヌクレオチド配列については、WO 00/06592、WO 00/06593、WO 01/66563、WO 03/001876、WO 02/064631、WO 03/004992、WO 03/025137、WO 02/086079、およびWO 01/83749もまた参照のこと(各々の全体は、本明細書において参考として援用されている)。
【0029】
T1Rタンパク質は、「Gタンパク質共役型レセプター活性」を有する。例えば、このタンパク質は、細胞外の刺激(例えば、リガンド結合(例えば、甘味リガンド))に応答してGタンパク質に結合し、そして酵素(例えば、ホスホリパーゼCおよびアデニレートシクラーゼ)の刺激を介してセカンドメッセンジャー(例えば、IP3、cAMP、およびCa2+)の産生を促進する。そのような活性は、Gタンパク質または乱交雑Gタンパク質(例えば、Gα15もしくはGα16−Gαz)のいずれかと酵素(例えば、PLC)とに、GPCR(またはキメラGPCR)を結合することによって、ならびに細胞内カルシウム利用の増加を測定することによって、異種細胞中で測定され得る(OffermansおよびSimon,J.Biol.Chem.270:15175−15180(1995))。レセプターの活性は、例えば、蛍光Ca2+指示染料および蛍光イメージングを使用して、[Ca2+]iにおけるリガンドによって誘導された変化を記録することによって効果的に測定され得る。
【0030】
そのようなGPCRは、当業者に公知の方法(例えば、疎水性ドメインおよび親水性ドメインを同定する配列分析プログラム(例えば、KyteおよびDoolittle,J.Mol.Biol.157:105−132(1982)を参照のこと))を使用して、構造的に同定され得る、膜貫通ドメイン、細胞外ドメインおよび細胞質ドメインを有する。そのようなドメインは、キメラタンパク質を作製するために、および本発明のインビトロアッセイのために有用である(例えば、WO 94/05695および米国特許第5,508,384号を参照のこと)。
【0031】
本発明の甘味レセプターまたはタンパク質の活性(例えば、シグナル変換)を調節する化合物をテストするためのアッセイの状況における語句「機能的効果」は、間接的にまたは直接的にGPCRまたは甘味レセプターの影響下にあるパラメータ(例えば、物理的効果、表現型効果、または化学的効果(例えば、外部の刺激(例えば、リガンド結合)に応じて細胞のシグナルを変換する能力、またはリガンドに結合する能力))の決定を含む。それは、結合活性およびシグナル変換を含む。「機能的効果」は、インビトロ、インビボ、およびエクスビボでの活性を含む。
【0032】
「機能的効果を決定すること」によってとは、間接的にまたは直接的にT1R GPCRタンパク質または1つ以上のT1R GPCRタンパク質を含む甘味レセプターの影響下にある、パラメータ(例えば、物理的効果および化学的効果、または表現型効果)を増加または減少する化合物をアッセイすることを意味する。そのような機能的効果は、当業者に公知の任意の手段によって測定され得る(例えば、分光特性(例えば、蛍光、吸収度、屈折率)の変化;流体力学的特性(例えば、形状);クロマトグラフィー特性;またはタンパク質の溶解特性;タンパク質の誘導マーカーまたは転写活性化の測定;結合活性または結合アッセイ(例えば、抗体への結合)の測定;リガンド結合活性または天然に存在するかもしくは合成のいずれかのそのアナログの変化の測定;細胞増殖の測定;細胞表面マーカー表現の測定;T1R関連配列のタンパク質レベルの変化の測定;RNA安定性の測定;Gタンパク質結合;GPCRリン酸化または脱リン酸化;シグナル変換(例えば、レセプター−リガンド相互作用、セカンドメッセンジャー濃度(例えば、cAMP、cGMP、IP3、PI、または細胞内Ca2+));神経伝達物質放出;ホルモン放出;電圧、膜電位およびコンダクタンスの変化;イオン流入;調節分子結合;例えば、化学発光、蛍光、比色の反応、抗体結合、および誘導マーカーを介した下流またはレポーター遺伝子発現(CAT、ルシフェラーゼ、β−gal、GFPなど)の同定)。
【0033】
T1Rファミリーポリヌクレオチド配列およびポリペプチド配列およびT1Rファミリー味レセプターの「インヒビター」、「アクチベーター」、および「調節因子」は、T1Rポリヌクレオチド配列およびポリペプチド配列ならびにT1Rファミリー味レセプター(モノマーレセプター、ホモダイマーレセプターおよびヘテロダイマーレセプターを含む)のインビトロアッセイおよびインビボアッセイを使用して同定される、活性化分子、阻害分子、または調節分子に言及するために使用される。インヒビターは、例えば、T1Rファミリーの味レセプター(例えば、T1R3ポリペプチドを含むレセプター)に結合し、部分的にもしくは完全に活性を遮断するか、活性を減少、予防、遅延するか、の活性もしくは発現を不活性化、脱感作、またはダウンレギュレートする化合物(例えば、アンタゴニスト)である。「アクチベーター」は、T1Rファミリーの味レセプター(例えば、T1R3ポリペプチドを含むレセプター)を増加、開放、活性化、促進、活性化を増強、感作、アゴニスト化、またはアップレギュレートする化合物である(例えば、アゴニスト)。インヒビター、アクチベーター、または調節因子としてはまた、遺伝的に改変されたバージョンのT1Rファミリー味レセプター(例えば、変更された活性を有するバージョン)ならびに天然に存在するリガンドおよび合成のリガンド、アンタゴニスト、アゴニスト、抗体、アンチセンス分子、リボザイム、小化学分子などが挙げられる。インヒビターおよびアクチベーターのためのそのようなアッセイは、例えば、インビトロで、細胞で、もしくは細胞膜でT1Rファミリー味レセプターを発現する工程、推定される調節因子化合物を適用する工程、次いで上述のように、活性に対する機能的効果を決定する工程を包含する。一実施形態において、T1R3ポリペプチドを含む味レセプターは、甘味分子(例えば、スクロース、グルコース、フルクトース、ラクトース、マンノース、ガラクトース、サッカリン、ズルチン、アセスルファーム−K)を認識する能力を有する。別の実施形態において、T1R3ポリペプチドを含む味レセプターは、他の分子(例えば、潜在的人工甘味料)を認識する能力を有する。これらの分子は、Gタンパク質共役型レセプターの細胞外リガンドとして作用することによって、およびそのレセプターを活性化することによって、味シグナル変換を調節する化合物の例である。他の実施形態において、味シグナル変換を調節する化合物は、レセプターの細胞内リガンドとして作用するか、あるいは細胞外リガンドの結合を阻害もしくは活性化するか、あるいはレセプターの細胞内リガンドの結合を阻害もしくは活性化する分子である。
【0034】
味レセプターのT1Rファミリーを含むサンプルまたはアッセイは、潜在的なアクチベーター、インヒビター、または調節因子で処理され、そして阻害の程度を検査するために、上記インヒビター、アクチベーター、または調節因子を含まないコントロールサンプルと比較される。(インヒビターで処理されていない)コントロールサンプルは、100%の相対タンパク質活性値を割り当てられる。T1Rファミリーレセプターの阻害は、コントロールと比較した活性値が、約80%(好ましくは50%、より好ましくは25〜0%)である場合に、達成される。T1Rファミリーレセプターの活性化は、(アクチベーターで処理されていない)コントロールと比較した活性値が、約110%、より好ましくは150%、より好ましくは200〜500%(すなわち、コントロールと比較して2倍から5倍高い)、より好ましくは1000〜3000%高い場合に、達成される。
【0035】
本明細書において使用される場合、用語「テスト化合物」または「薬物候補」または「調節因子」または文法上の等価物は、直接的にまたは間接的に味を調節する能力を試験される任意の分子(例えば、天然に存在するかもしくは合成の、人工甘味料または天然に存在する糖(例えば、タンパク質、オリゴペプチド(例えば、約5から約25アミノ酸長、好ましくは約10から20アミノ酸長、または12から18アミノ酸長、好ましくは12アミノ酸長、15アミノ酸長、または18アミノ酸長)、小有機分子、多糖類、脂質、脂肪酸、ポリヌクレオチド、オリゴヌクレオチドなど))を説明する。テスト化合物は、テスト化合物のライブラリー(例えば、十分な範囲の多様性を提供するコンビナトリアルライブラリーまたはランダム化されたライブラリー)の形態であり得る。テスト化合物は、必要に応じて、融合パートナー(例えば、標的化合物、救助(rescue)化合物、ダイマー化合物、安定化合物、取り組み可能な化合物(addressable compound)、および他の機能的部分)に結合される。従来、有用な特性を有する新規の化学実体は、ある所望の特性もしくは活性(例えば、阻害活性)を有するテスト化合物(「リード化合物」といわれる)を同定し、このリード化合物の改変体を作製し、そしてこれらの改変体化合物の特性および活性を評価することによって生成される。多くの場合、ハイスループットスクリーニング(HTS)方法が、そのような分析のために使用される。
【0036】
「小有機分子」とは、約50ダルトンを超え、そして約2500ダルトン未満(好ましくは約2000ダルトン未満、好ましくは約100から約1000ダルトンの間、より好ましくは約200から約500ダルトンの間)の分子量を有する、天然に存在するかまたは合成のいずれかの有機化合物をいう。
【0037】
「生物学的サンプル」としては、組織の切片(例えば、生検サンプルおよび剖検サンプル)および組織学的目的のために取られた凍結切片が挙げられる。そのようなサンプルとしては、血液、痰、組織、培養細胞(例えば、初代培養)、外移植片、および形質転換細胞、糞便、尿などが挙げられる。生物学的サンプルは、代表的に、真核生物体(最も好ましくは、哺乳動物(例えば、霊長類(例えば、チンパンジーもしくはヒト);ウシ;イヌ;ネコ;げっ歯類(例えば、モルモット、ラット、マウス);ウサギ);または鳥類;爬虫類;あるいは魚類)から得られる。
【0038】
「ヘテロダイマー」とは、2つの異なるポリペプチドサブユニット(例えば、分子が、共有結合性(例えば、リンカーもしくは化学結合を通じて)の連結か、または非共有結合性(例えば、イオン結合、ファンデルワールス結合、静電結合、もしくは水素結合)の連結のいずれかを介して結合される、2つの異なるポリペプチド)を含む、ダイマーのレセプターである。同じ細胞内で同時発現された場合(好ましくは、このレセプターが、共有結合性または非共有結合性のいずれかで連結されたヘテロダイマーを形成するように同時発現された場合)、本発明のT1R3を含むレセプターは機能する。例えば、T1R1およびT1R3は、ヘテロマーのレセプターを形成し、そしてT1R2およびT1R3は、ヘテロマーのレセプターを形成する。
【0039】
「ホモダイマー」とは、2つの同じポリペプチドサブユニット(例えば、分子が、共有結合性(例えば、リンカーもしくは化学結合を通じて)の連結か、または非共有結合性(例えば、イオン結合、ファンデルワールス結合、静電結合、もしくは水素結合)の連結のいずれかを介して結合される、2つのT1R3ポリペプチド)を含むダイマーのレセプターである。本発明のT1R3を含むレセプターは、同じ細胞内で同時発現された場合(好ましくは、このレセプターが、共有結合性または非共有結合性のいずれかで連結されたホモダイマーを形成するように同時発現された場合)、機能する。
【0040】
「モノマー」は、1つのポリペプチドサブユニット(例えば、1つのT1R3ポリペプチド)を含むレセプターである。
【0041】
2つ以上の核酸またはポリペプチド配列の状況において、用語「同一の」または「同一性」パーセントとは、以下に記載されるデフォルトパラメータ(default parameter)を有するBLASTまたはBLAST 2.0配列比較アルゴリズムを使用して、または手動のアラインメントおよび視覚的検査(例えば、NCBIウェブサイトなどを参照のこと)によって測定される場合、同じであるか、あるいは特定のパーセンテージの同じであるアミノ酸残基またはヌクレオチド(すなわち、比較ウィンドウ(comparison window)または指定された領域にわたる最大限の一致について比較されそしてアラインメントされた場合の、特定された領域(例えば、ヌクレオチド配列、配列番号1〜25)にわたる、約60%の同一性(好ましくは、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%以上の同一性))を有する、2つ以上の配列もしくは部分配列をいう。そして、そのような配列は、「実質的に同一」であると言われる。この定義はまた、テスト配列の相補性(compliment)をいうか、または相補性に対して適用され得る。この定義はまた、欠失および/または付加を有する配列、ならびに置換を有する配列を含む。以下に記載されるように、好ましいアルゴリズムは、ギャップなどを説明し得る。好ましくは、同一性は、少なくとも約25アミノ酸長もしくは約25ヌクレオチド長である領域にわたって(またはより好ましくは50〜100アミノ酸長もしくは50〜100ヌクレオチド長である領域にわたって)存在する。
【0042】
配列の比較について、代表的には、1つの配列は、テスト配列が比較される参照配列として作用する。配列比較アルゴリズムを使用する場合、テスト配列および参照配列がコンピューターに入力され、必要な場合、部分配列座標が指定され、そして配列アルゴリズムプログラムパラメータが指定される。好ましくは、デフォルトプログラムパラメータが使用され得るか、または代替的パラメータが指定され得る。次いで、この配列比較アルゴリズムは、上記プログラムパラメータに基づいて参照配列と比較したテスト配列のパーセント配列同一性を計算する。
【0043】
本明細書において使用される場合、「比較ウィンドウ」とは、20から600(通常約50〜約200、より通常的に約100〜約150)からなる群より選択される連続する位置の数のうちのいずれか1つのセグメントの参照を含み、この中で、配列は、同じ数の連続する位置の参照配列と、2つの配列が最適にアラインメントされた後、比較され得る。比較のために配列をアラインメントする方法は、当該分野において周知である。比較のために配列の最適なアラインメントは、例えば、SmithおよびWaterman,Adv.Appl.Math.2:482(1981)の局所相同性アルゴリズムによって、NeedlemanおよびWunsch,J.Mol.Biol.48:443(1970)の相同性アラインメントアルゴリズムによって、PearsonおよびLipman,Proc.Nat’l.Acad.Sci.USA 85:2444(1988)の類似性方法(similarity method)の検索によって、これらのアルゴリズムのコンピューター化された(Wisconsin Genetics Software Package,Genetics Computer Group,575 Science Dr.,Madison,WI中のGAP、BESTFIT、FASTA、およびTFASTA)実施によって、または手動の配列および視覚的検査(例えば、Current Protocols in Molecular Biology(Ausubelら(編)、1995年 補遺)を参照のこと)によって実行され得る。
【0044】
パーセント配列同一性および配列類似性の決定に適しているアルゴリズムの好ましい例は、BLASTおよびBLAST 2.0アルゴリズムであり、これらは、それぞれ、Altschulら、Nuc.Acids Res.25:3389−3402(1977)およびAltschulら、J.Mol.Biol.215:403−410(1990)に記載されている。BLASTおよびBLAST 2.0は、本発明の核酸およびタンパク質のパーセント配列同一性を決定するために本明細書において記載されるパラメータを用いて使用される。BLAST分析を実行するためのソフトウェアは、National Center for Biotechnology Information(http://www.ncbi.nlm.nih.gov/)を通して公的に入手可能である。このアルゴリズムは、最初にクエリー配列の長さWの短い語(この語は、データベース配列の同じ長さの語とアラインメントされる場合、ある正の値の閾値スコアTにマッチ(match)するかまたはTを満たすかのいずれかである)を同定することによって、高スコア配列ペア(HSP)を同定することに関与する。Tは、近隣語スコア閾値(score threshold)といわれる(Altschulら、前出)。これらの最初の近隣語のヒット(hit)は、それらを含むより長いHSPを見付けるために検索を開始するための源として作用する。語のヒットは、累積アラインメントスコアが増加され得る限り、各配列に沿って両方向に拡張される。累積スコアは、ヌクレオチド配列については、パラメータM(マッチ残基のペアに対しての報酬スコア(reward score);常に>0)およびパラメータN(ミスマッチ残基に対してのペナルティスコア;常に<0)を使用して計算される。アミノ酸配列については、スコアマトリックスが、累積スコアを計算するために使用される。各方向の語のヒットの拡張は、累積アラインメントスコアが最大限に達成された値から量Xが減少する場合;累積スコアが、1つ以上の負のスコア残基アラインメントの蓄積に起因して0以下になる場合;あるいは、どちらかの配列の末端に到達する場合に、停止される。BLASTアルゴリズムパラメータW、T、およびXは、配列の感度およびスピードを決定する。(ヌクレオチド配列のための)BLASTNプログラムは、デフォルトとして、語の長さ(W)11、期待値(E)10、M=5、N=−4および両方の鎖の比較を、使用する。アミノ酸配列のために、BLASTPプログラムは、デフォルトとして、語の長さ3、期待値(E)10、BLOSUM62スコアマトリックス(HenikoffおよびHenikoff、Proc.Natl.Acad.Sci.USA 89:10915(1989)を参照のこと)アラインメント(B)50、期待値(E)10、M=5、N=−4、および両方の鎖の比較、を使用する。
【0045】
用語「ポリペプチド」、「ペプチド」、および「タンパク質」とは、本明細書において、アミノ酸残基のポリマーをいうために相互交換可能に使用される。これらの用語は、1つ以上のアミノ酸残基が対応する天然に存在するアミノ酸の人工的な化学的模倣体であるアミノ酸ポリマー、ならびに天然に存在するアミノ酸ポリマーおよび天然に存在しないアミノ酸ポリマーに当てはめられる。
【0046】
用語「アミノ酸」とは、天然に存在するアミノ酸および合成のアミノ酸、鏡像異性体(D−およびL−形態)、およびアキラルアミノ酸、ならびに天然に存在するアミノ酸と類似の様式で機能するアミノ酸アナログおよびアミノ酸模倣体をいう。天然に存在するアミノ酸は、遺伝子コードによってコードされるもの、ならびに、その後改変されるアミノ酸である(例えば、ヒドロキシプロリン、γ−カルボキシグルタメート、およびO−ホスホセリン)。アミノ酸アナログとは、天然に存在するアミノ酸と同じ基本的化学構造を有する化合物をいう(すなわち、水素、カルボキシル基、アミノ基、およびR基(例えば、ホモセリン、ノルロイシン、メチオニン、スルホキシド、メチオニンメチルスルホニウム)に結合されるα−炭素)。そのようなアナログは、改変R基(例えば、ノルロイシン)または改変ペプチド骨格を有するが、天然に存在するアミノ酸と同じ基本的化学構造を保持する。アミノ酸模倣体は、アミノ酸の一般の化学構造と異なる構造を有するが、天然に存在するアミノ酸と類似の様式で機能する化学化合物をいう。
【0047】
本明細書において、アミノ酸は、一般に公知の3文字記号、またはIUPAC−IUB Biochemical Nomenclature Commissionによって推奨される1文字記号のいずれかによって言及され得る。同様に、ヌクレオチドは、その一般に容認されている1文字コードによって言及され得る。
【0048】
「保存的に改変された改変体」は、アミノ酸配列と核酸配列の両方に適用される。特定の核酸配列に関して、保存的に改変された改変体とは、同一のまたは本質的に同一のアミン酸配列をコードする核酸をいうか、あるいは核酸がアミノ酸配列をコードしない場合は、本質的に同一の配列をいう。遺伝子コードの縮重に起因して、多数の機能的に同一の核酸が、任意の所与のタンパク質をコードする。例えば、コドンGCA、GCC、GCGおよびGCUは全て、アミノ酸のアラニンをコードする。従って、アラニンがコドンによって特定される全ての位置において、そのコドンは、コードされたポリペプチドを変更することなしに、記載の任意の対応するコドンに変更され得る。そのような核酸の多様性は、「サイレント多様性(silent variation)」であり、これは、1種の保存的に改変された改変体(variation)である。本明細書において、ポリペプチドをコードする全ての核酸配列はまた、核酸の全ての可能なサイレント多様性を説明する。当業者は、核酸の各コドン(通常メチオニンのための唯一のコドンであるAUG、および通常トリプトファンのための唯一のコドンであるTGGを除く)が、機能的に同一の分子を産生するために改変され得ることを認識する。従って、ポリペプチドをコードする核酸の各々のサイレント多様性は、実際のプローブの配列に関してではなく、発現産物に関して、各々の記載された配列について暗示する。
【0049】
アミノ酸配列に関して、当業者は、コードされた配列中の1つのアミノ酸もしくはわずかな割合のアミノ酸を変更、付加または欠失させる核酸配列、ペプチド配列、ポリペプチド配列、またはタンパク質配列への個々の置換、欠失または付加が、その変更が化学的に類似のアミノ酸でのアミノ酸の置換を生じる「保存的に改変された改変体」であることを認識する。機能的に類似のアミノ酸を提供する保存的置換表は、当該分野において周知である。そのような保存的に改変された改変体は、本発明の多型改変体、種間のホモログ、および対立遺伝子に加えられ、そしてこれらを排除しない。
【0050】
以下の8つのグループは、それぞれ、相互に保存的置換であるアミノ酸を含む:1)アラニン(A)、グリシン(G);2)アスパラギン酸(D)、グルタミン酸(E);3)アスパラギン(N)、グルタミン(Q);4)アルギニン(R)、リジン(K);5)イソロイシン(I)、ロイシン(L)、メチオニン(M)、バリン(V);6)フェニルアラニン(F)、チロシン(Y)、トリプトファン(W);7)セリン(S)、スレオニン(T);ならびに8)システイン(C)、メチオニン(M)(例えば、Creighton,Proteins(1984)を参照のこと)。
【0051】
マクロ分子構造(例えば、ポリペプチド構造)は、種々のレベルの構成の点から考察され得る。この構成の一般的考察については、例えば、Albertsら、Molecular Biology of the Cell(第3版、1994年)ならびにCantorおよびSchimmel、Biophysical Chemistry Part I:The Conformation of Biological Macromolecules(1980)を参照のこと。「一次構造」とは、特定のペプチドのアミノ酸配列をいう。「二次構造」とは、ポリペプチド内の局所的に整列した、3次元構造をいう。これらの構造は、一般には、ドメイン(例えば、細胞外ドメイン、膜貫通ドメイン、および細胞質ドメイン)として公知である。ドメインは、ポリペプチドの圧縮したユニットを形成するポリペプチドの部分であり、そして代表的に、15〜350アミノ酸長である。代表的なドメインは、より小さい構成(例えば、βシートおよびαヘリックスの伸張)の部分からなりたっている。「三次構造」とは、ポリペプチドモノマーの完全な三次元構造をいう。「四次構造」とは、独立の三次ユニットの非共有結合性の会合によって形成された3次元構造をいう。異方性の用語はまた、エネルギー用語としても公知である。
【0052】
「核酸」とは、デオキシリボヌクレオチドまたはリボヌクレオチド、および単鎖または二本鎖のいずれかの形態のそれらのポリマー、ならびに任意のそのような配列の相補体をいう。核酸にはまた、DNA、cDNA、RNA、ポリヌクレオチド、ヌクレオチドなどが含まれる。この用語は、公知のヌクレオチドアナログまたは改変された骨格の残基または連結を含む核酸を包含し、この核酸は、合成されるもの、天然に存在するもの、および天然に存在しないものであり、参照核酸と類似の結合特性を有し、そして参照ヌクレオチドと類似の様式で代謝される。そのようなアナログの例としては、限定されることなく、ホスホロチオエート、ホスホルアミデート、メチルホスホネート、キラル−メチルホスホネート、2−O−メチルリボヌクレオチド、ペプチド−核酸(PNA)が挙げられる。
【0053】
特定の核酸配列はまた、暗黙のうちに、「スプライス改変体」を含む。同様に、核酸によってコードされる特定のタンパク質は、その核酸のスプライス改変体によってコードされる任意のタンパク質を暗黙のうちに含む。その名称が示唆するように、「スプライス改変体」は、遺伝子の代替的なスプライシングの産物である。転写後、最初の核酸転写産物が、スプライシングされ得、それゆえ、異なる(代替的な)核酸スプライス産物が、異なるポリペプチドをコードし得る。スプライス改変体の産物の機構は様々であるが、エキソンの代替的なスプライシングを含む。読み過し転写による同じ核酸に由来する代替的なポリペプチドもまた、この定義に含まれる。スプライシング反応の任意の産物(スプライス産物の組換え型形態を含む)が、この定義の中に含まれる。
【0054】
「標識」または「検出可能部分」とは、分光的手段、光化学的手段、生化学的手段、免疫化学的手段、化学的手段、または他の物理的手段によって検出可能な組成物である。例えば、有用な標識としては、32P、蛍光染料、電子密集試薬(electron−dense reagent)、酵素(例えば、ELISA中で一般に使用されるようなもの)、ビオチン、ジゴキシゲニン、またはハプテンおよびタンパク質が挙げられ、これらは、例えば、ペプチドの中へ放射標識を取り込ませることによって検出可能とされ得るか、またはこのペプチドと特異的に反応する抗体を検出するために使用され得る。
【0055】
例えば、細胞、または核酸、タンパク質、またはベクターを参照して使用する場合、用語「組換え型」とは、細胞、核酸、タンパク質またはベクターが、異種の核酸もしくはタンパク質の導入、またはネイティブの核酸もしくはタンパク質の変更によって改変されたこと、あるいは、細胞が、そのように改変された細胞に由来することを示す。従って、例えば、組換え型細胞は、細胞のネイティブ(非組換え型)形態内に見出されない遺伝子を発現するか、あるいは発現される条件下で別のやり方で異常発現されるネイティブの遺伝子を発現するか、または全く発現されない。
【0056】
核酸の一部への参照として使用される場合、用語「異種」とは、核酸が天然において相互に同じ関係で見出されない2つ以上の部分配列を含むことを示す。例えば、この核酸は、代表的に、組換えで産生され、新規の機能的核酸を産生するように整列された関連しない遺伝子由来の2つ以上の配列を有する(例えば、1つの供給源に由来するプロモーターおよび別の供給源に由来するコーディング領域)。同様に、異種のタンパク質は、このタンパク質が、天然において相互に同じ関係で見出されない2つ以上の部分配列を含むことを示す(例えば、融合タンパク質)。
【0057】
語句「ストリンジェントなハイブリダイゼーション条件」とは、プローブが、代表的に核酸の複合混合物中でその標的の部分配列にハイブリダイズするが、他の配列にはハイブリダイズしない条件をいう。ストリンジェントな条件は、配列に依存し、そして異なる環境においては、その条件は異なる。より長い配列は、より高温で特異的にハイブリダイズする。核酸のハイブリダイゼーションに関する広範なガイドは、Tijssen,Techniques in Biochemistry and Molecular Biology−−Hybridization with Nucleic Probes,「Overview of principles of hybridization and the strategy of nucleic acid assays」(1993)中に見出される。一般に、ストリンジェントな条件は、規定のイオン強度、pHにおいて特異的配列の熱融点(Tm)よりも約5〜10℃低くなるように選択される。このTmは、標的に相補的な50%のプローブが、平衡状態にある標的配列にハイブリダイズする(標的配列が過剰に存在する場合、Tmにおいて50%のプローブが平行状態において占有される)(規定のイオン強度、pH、および核濃度の下での)温度である。ストリンジェントな条件はまた、非安定化因子(例えば、ホルムアミド)の添加によって達成され得る。選択的または特異的なハイブリダイゼーションについて、正のシグナルは、少なくともバックグラウンドの2倍(好ましくは、バックグラウンドのハイブリダイゼーションの10倍)である。例示的なストリンジェントなハイブリダイゼーション条件は、以下のようなものであり得る:50%ホルムアミド、5×SSC、および1% SDS、42℃にてインキュベートされるか、または5×SSC、1% SDS、65℃にてインキュベートされ、0.2×SSC中で洗浄、そして0.1% SDS、65℃。
【0058】
ストリンジェントな条件下で相互にハイブリダイズしない核酸は、それらがコードするポリペプチドが実質的に同一である場合、それでも実質的に同一である。このことは、例えば、核酸のコピーが、遺伝子コードによって可能とされた最大限のコドンの縮重を使用して生成される場合に、発生する。そのような場合、この核酸は、代表的には、中程度にストリンジェントなハイブリダイゼーション条件下で、ハイブリダイズする。例示的な「中程度にストリンジェントなハイブリダイゼーション条件」は、40%ホルムアミド、1M NaCl、1% SDS、37℃の緩衝液中のハイブリダイゼーションおよび1×SSC、45℃の洗浄を含む。正のハイブリダイゼーションは、バックグラウンドの少なくとも2倍である。技術者は、代替的なハイブリダイゼーションおよび洗浄の条件が、類似のストリンジェントな条件を提供するために利用され得ることを容易に認識する。ハイブリダイゼーションパラメータを決定するためのさらなるガイドラインが、多数の参考文献(例えば、Current Protocols in Molecular Biology,Ausubelら(編))において提供されている。
【0059】
PCRについて、アニーリング温度は、プライマーの長さに依存して、約32℃と48℃との間で変動し得るが、約36℃の温度が低ストリンジェントな増幅のために典型的である。プライマーの長さおよび特異性に依存して、高ストリンジェントなアニーリング温度は、約50℃から約65℃までの範囲であり得るが、高ストリンジェントなPCR増幅について、約62℃の温度が典型的である。高ストリンジェントな増幅および低ストリンジェントな増幅の両方についての典型的なサイクルの条件は、以下の条件の30〜40サイクルを含む:30秒〜2分間の90℃〜95℃の変性期、30秒〜2分続くアニーリング期、および1分〜2分間の約72℃の伸張期。低ストリンジェントな増幅反応および高ストリンジェントな増幅反応のためのプロトコルおよびガイドラインは、例えば、Innisら、PCR Protocols,A Guide to Methods and Applications(1990)中に提供される。
【0060】
「抗体」とは、特異的に抗原に結合し、そして認識する免疫グロブリン遺伝子またはそのフラグメント由来のフレームワーク領域を含むポリペプチドをいう。この認識された免疫グロブリン遺伝子としては、κ、λ、α、γ、δ、ε、およびμ定常領域遺伝子、ならびに無数の免疫グロブリン可変領域遺伝子が挙げられる。軽鎖は、κまたはλのいずれかとして分類される。重鎖は、γ、μ、α、δ、またはεとして分類され、これらは、順番に、それぞれ、免疫グロブリンクラスのIgG、IgM、IgA、IgDおよびIgEを規定する。代表的に、抗体の抗原結合領域は、結合の特異性および親和性において最も重要である。
【0061】
例示的な免疫グロブリン(抗体)構造単位は、テトラマーを含む。各テトラマーは、ポリペプチド鎖の2つの同一のペアからなり、各ペアは、1つの「軽」鎖(約25kD)および1つの「重」鎖(約50〜70kD)を有する。各鎖のN末端は、主に抗原認識を担う約100から110以上のアミノ酸の可変領域を規定する。用語、可変軽鎖(VL)および可変重鎖(VH)とは、それぞれ、これらの軽鎖および重鎖をいう。
【0062】
抗体は、例えば、インタクトな免疫グロブリンとしてか、あるいは種々のぺプチダーゼによる消化によって産生される複数のよく特徴付けされたフラグメントとして存在する。従って、例えば、ペプシンは、ヒンジ領域のジスルフィド結合下で抗体を消化し、F(ab)’2(それ自身が、ジスルフィド結合によってVH−CH1に結合される軽鎖である、Fabのダイマー)を産生する。このF(ab)’2は、緩やかな条件下で還元され、ヒンジ領域のジスルフィド結合を分解し得、それによって、F(ab)’2ダイマーはFab’モノマーに変換する。このFab’モノマーは、本質的にヒンジ領域の一部を有するFabである(Fundamental Immunology(Paul(編)、第3版、1993年)参照のこと)。種々の抗体フラグメントが、インタクトな抗体の消化に関して規定されるが、当業者は、そのようなフラグメントが、化学的にかまたは組換えDNA技術を使用することによって、新規に合成され得ることを理解する。従って、本明細書において使用される場合、用語、抗体はまた、抗体全体の改変によって産生された抗体フラグメントか、または組換えDNA方法論を使用して新規に合成された抗体フラグメント(例えば、単鎖Fv)か、またはファージディスプレイライブラリー(例えば、McCaffertyら、Nature 348:552−554(1990)を参照のこと)を使用して同定された抗体フラグメントを含む。
【0063】
抗体(例えば、組換え型抗体、モノクローナル抗体、またはポリクローナル抗体)の調製について、当該分野において公知の多くの技術が使用され得る(例えば、KohlerおよびMilstein,Nature 256:495−497(1975);Kozborら、Immunology Today 4:72(1983);Coleら、Monoclonal Antibodies and Cancer Therapy,77−96頁、Alan R.Liss,Inc.(1985);Coligan,Current Protocols in Immunology(1991);HarlowおよびLane,Antibodies,A Laboratory Manual(1988);およびGoding,Monoclonal Antibodies:Principles and Practice(第2版、1986年)を参照のこと)。単鎖抗体の産生のための技術(米国特許第4,946,778号)が、本発明のポリペプチドに対する抗体を産生するために採用され得る。さらに、トランスジェニックマウス、または他の生物体(例えば、他の哺乳動物)は、ヒト化抗体を発現するために使用され得る。あるいは、ファージディスプレイ技術が、選択された抗原に特異的に結合する抗体およびヘテロマーのFabフラグメントを同定するために使用され得る(例えば、McCaffertyら、Nature 348:552−554(1990);Marksら、Biotechnology 10:779−783(1992)を参照のこと)。
【0064】
「キメラ抗体」とは、(a)定常領域、もしくはその一部が、変更、置換、もしくは交換され、それゆえ抗原結合部位(可変領域)が、異なるかもしくは変更された、クラス、エフェクター機能および/もしくは種の定常領域に結合されるか、またはキメラ抗体に新規の特性を与える完全に異なる分子(例えば、酵素、トキシン、ホルモン、成長因子、薬物など)に結合される抗体分子か;あるいは、(b)可変領域、もしくはその一部が、異なるかもしくは変更された抗原特異性を有する可変領域と変更、置換、もしくは交換される抗体分子である。
【0065】
一実施形態において、抗体は、「エフェクター」部分に接合される。このエフェクター部分は、任意の数の分子(標識化部分(例えば、放射性標識または蛍光標識)を含む)であり得るか、または治療部分であり得る。一局面において、抗体は、タンパク質の活性を調節する。
【0066】
タンパク質またはペプチドを参照する場合、語句、抗体に「特異的に(もしくは選択的に)結合する」または抗体と「特異的に(もしくは選択的に)免疫応答する」とは、多くの場合異種集団のタンパク質および他の生物学的製剤中において、タンパク質の存在を測定する結合反応をいう。従って、指定されたイムノアッセイ条件下で、特異化された抗体は、バックグラウンドの少なくとも2倍、そしてより代表的にはバックグラウンドの10倍から100倍を超えて特定のタンパク質に結合する。そのような条件下での抗体への特異的な結合は、特定のタンパク質についての特異性のために選択される抗体を必要とする。例えば、T1Rタンパク質、あるいは、配列番号1〜25によってコードされる配列を含み、ホモダイマーかもしくはヘテロダイマーのT1R3を含む味レセプター、多型改変体、対立遺伝子、オルソログ、および保存的に改変された改変体、もしくはスプライス改変体、またはそれらの部分に対して惹起されたポリクローナル抗体は、T1Rタンパク質および/あるいはホモダイマーかもしくはヘテロダイマーのT1R3を含む味レセプターと特異的に免疫応答し、そして他のタンパク質と特異的に免疫応答しないポリクローナル抗体のみを得るために選択され得る。一実施形態において、抗体は、ホモダイマーのT1R3を含む味レセプターに反応するが、T1Rファミリーの個々のタンパク質メンバーには反応しない。この選択は、他の分子と交差反応する抗体を削除することによって達成され得る。種々のイムノアッセイ形式が、特定のタンパク質と特異的に免疫応答する抗体を選択するために使用され得る。例えば、固相ELISAイムノアッセイが、タンパク質と特異的に免疫応答する抗体を選択するために慣用的に使用される(特異的な免疫反応性を測定するために使用され得るイムノアッセイの形式および条件の説明については、例えば、HarlowおよびLane,Antibodies,A Laboratory Manual(1988)を参照のこと)。
【0067】
(T1Rファミリーメンバーをコードする核酸の単離)
本発明は、組換え遺伝学の分野の慣例的な技術に依存する。本発明における使用の一般的な方法を開示する基本的テキストとしては、Sambrookら、Molecular Cloning,A Laboratory Manual(第2版、1989年);Kriegler,Gene Transfer and Expression:A Laboratory Manual(1990);およびCurrent Protocols in Molecular Biology(Ausubelら(編)、1994年)が挙げられる。
【0068】
本明細書において開示されるアミノ酸配列に実質的に同一である、T1R核酸、多型改変体、オルソログ、および対立遺伝子は、T1R核酸プローブ、およびオリゴヌクレオチドを使用して、ストリンジェントなハイブリダイゼーション条件下でライブラリーをスクリーニングすることによって単離し得る。あるいは、発現ライブラリーは、ヒトT1Rまたはその一部分に対して産生された抗血清または精製された抗体を用いて、免疫学的に発現されたホモログを検出することによって、T1Rタンパク質、多型改変体、オルソログ、および対立遺伝子をクローニングするために使用され得る。
【0069】
cDNAライブラリーを作製するために、当業者は、T1R RNAが豊富な供給源(例えば、味蕾(例えば、有郭、葉状、茸状、および口蓋))を選択するべきである。このmRNAは、次いで、逆転写酵素を使用してcDNAにされ、組換えベクターに連結され、そして増殖、スクリーニング、およびクローニングのために組換え型宿主へトランスフェクトされる。cDNAライブラリーを作製およびスクリーニングするための方法は、周知である(例えば、GublerおよびHoffman,Gene 25:263−269(1983);Sambrookら、前出;Ausubelら、前出、を参照のこと)。
【0070】
ゲノムライブラリーのために、上記DNAは、組織から抽出され、そして約12〜20kbのフラグメントを生成するために、機械的に剪断されるか、または酵素的に消化される。次いで、このフラグメントは、勾配遠心分離によって所望されない大きさのものから分離され、そしてバクテリオファージラムダベクター中に構築される。これらのベクターおよびファージは、インビトロでパッケージされる。組換え型ファージは、BentonおよびDavis,Science 196:180−182(1977)中に記載されるようにプラークハイブリダイゼーションによって分析される。コロニーハイブリダイゼーションが、Grunsteinら、Proc.Natl.Acad.Sci.USA.,72:3961−3965(1975)中に一般的に記載されるように実行される。
【0071】
T1R核酸およびそのオルソログ、対立遺伝子、変異体、多型改変体、および保存的に改変された改変体を単離する代替的な方法は、合成オリゴヌクレオチドプライマーの使用とRNAもしくはDNAテンプレートの増幅とを組み合わせる(米国特許第4,683,195号および同第4,683,202号;PCR Protocols:A Guide to Methods and Applications(Innisら(編)、1990年)を参照のこと)。ポリメラーゼ連鎖反応(PCR)およびリガーゼ連鎖反応(LCR)のような方法を、直接的に、mRNAから、cDNAから、ゲノムライブラリーからまたはcDNAライブラリーからヒトT1Rの核酸配列を増幅するために使用し得る。変質したオリゴヌクレオチドを、本明細書において提供される配列を使用してT1Rホモログを増幅するために、設計し得る。制限エンドヌクレアーゼ部位は、プライマーの中へ取り込まれ得る。ポリメラーゼ連鎖反応または他のインビトロ増幅方法はまた、例えば、核酸配列決定または他の目的のために、発現されるタンパク質をコードする核酸配列をクローニングするのに、生理学的サンプルにおいてT1RをコードするmRNAの存在を検出するためのプローブとして使用する核酸を作製するのに、有用であり得る。PCR反応によって増幅される遺伝子は、アガロースゲルから精製され得、そして適切なベクター中へクローニングされ得る。
【0072】
T1Rの遺伝子発現はまた、当該分野において公知の技術(例えば、mRNAの逆転写および増幅、全RNAまたはポリA+RNAの単離、ノザンブロット法、ドットブロット法、インサイチュハイブリダイゼーション、RNase防御(protection)、高密度ポリヌクレオチドアレイ技術など)によって分析され得る。
【0073】
T1Rタンパク質をコードする核酸は、本発明のT1Rタンパク質、オルソログ、対立遺伝子、保存的に改変された改変体、および多型改変体を同定するために高密度オリゴヌクレオチドアレイ技術(例えば、GeneChipTM)とともに使用され得る(例えば、Gunthandら、AIDS Res.Hum.Retroviruses 14:869−876(1998);Kozalら、Nat.Med.2:753−759(1996);Matsonら、Anal.Biochem.224:110−106(1995);Lockhartら、Nat.Biotechnol.14:1675−1680(1996);Gingerasら、Genome Res.8:435−448(1998);Haciaら、Nucleic Acids Res.26:3865−3866(1998)を参照のこと)。
【0074】
T1Rの遺伝子は、代表的に、複製および/または発現のための原核生物細胞または真核生物細胞の中への変換の前に、中間ベクター中にクローニングされる。これらの中間ベクターは、代表的に、原核生物ベクター(例えば、プラスミド)またはシャトルベクターである。
【0075】
(原核生物および真核生物の発現)
クローニングされた遺伝子の高レベルの発現(例えば、T1Rタンパク質をコードするcDNAの高レベルの発現)を得るために、当業者は、代表的に、T1Rを発現ベクター中にサブクローニングする。この発現ベクターは、転写、転写/翻訳ターミネーターに指向するための強力なプロモーターを、そして核酸がタンパク質をコードする場合、翻訳開始のためのリボソーム結合部位を含む。T1R核酸は、同じかまたは異なるベクターに、同時発現され得るか、または分離して発現され得る(好ましくは同時発現され得る)。適切な細菌プロモーターは、当該分野において周知であり、そして例えば、Sambrookら、およびAusubelら(前出)に記載される。T1Rタンパク質を発現するための細菌発現系は、例えば、E.coli、Bacillus種、およびSalmonella(Palvaら、Gene 22:229−235(1983);Mosbachら、Nature 302:543−545(1983))において入手可能である。そのような発現系のためのキットは、市販されている。哺乳動物細胞、酵母、および昆虫細胞のための真核生物発現系は、当該分野において周知であり、そしてまた市販されている。好ましい一実施形態において、レトロウイルス発現系が、本発明中で使用される。
【0076】
異種の核酸の発現を導くために使用されるプロモーターの選択は、特定の用途に依存している。プロモーターは、好ましくは、異種の転写開始部位から、その天然の設定における、転写開始部位からの距離とほぼ同じ距離に位置付けされる。しかし、当該分野において公知のように、この距離のいくらかの多様性は、プロモーター機能の損失なしに順応され得る。
【0077】
プロモーターに加えて、発現ベクターは、代表的に、宿主細胞中でT1Rをコードする核酸の発現に必要とされる全てのさらなる要素を含む、転写ユニットまたは発現カセットを含む。従って、代表的な発現カセットは、T1Rをコードする核酸配列に作動可能に結合するプロモーター、ならびに、転写物の効率的なポリアデニル化、リボソーム結合部位、および翻訳終止のために必要とされるシグナルを含む。このカセットのさらなる要素は、エンハンサー、ならびに、ゲノムDNAが構造遺伝子として使用される場合、機能的スプライスドナーおよびアクセプター部位を伴うイントロンを含み得る。
【0078】
プロモーター配列に加えて、上記発現カセットはまた、効率的な終止を提供するために構造遺伝子の下流に転写終止領域を含むはずである。この終止領域は、プロモーター配列と同じ遺伝子から得られ得るか、または異なる遺伝子から得られ得る。
【0079】
細胞に遺伝情報を輸送するために使用される特定の発現ベクターは、特に重要ではない。真核生物細胞または原核生物細胞における発現のために使用される任意の従来のベクターが、使用され得る。標準的な細菌発現ベクターは、プラスミド(例えば、pBR322ベースのプラスミド、pSKF、pET23D)および融合発現系(例えば、MBP、GST、およびLacZ)を含む。エピトープのタグがまた、組換えタンパク質に付加され、単離のための便利な方法(例えば、c−myc)を提供し得る。配列のタグが、核酸回収(rescue)のために発現カセットに含まれ得る。マーカー(例えば、蛍光タンパク質、緑蛍光タンパク質、または赤蛍光タンパク質、β−gal、CATなど)が、ベクター変換のためのマーカーとしてベクター中に含まれ得る。
【0080】
真核生物のウイルス由来の調節エレメントを含む発現ベクターは、代表的に、真核生物発現ベクター(例えば、SV40ベクター、乳頭腫ウイルスベクター、レトロウイルスベクター、およびエプスタイン−バーウイルス由来のベクター)において使用される。他の例示的な真核生物ベクターとしては、pMSG、pAV009/A+、pMTO10/A+、pMAMneo−5、バキュロウイルスpDSVE、および任意の他のベクター(CMVプロモーター、SV40早期プロモーター、SV40後期プロモーター、メタロチオネインプロモーター、マウス乳腺癌ウイルスプロモーター、ラウス肉腫ウイルスプロモーター、ポリへドリンプロモーター、または真核生物細胞の発現について有効性が示された他のプロモーターの指示の下、タンパク質の発現を可能にするベクター)が挙げられる。
【0081】
真核生物ベクター由来のタンパク質の発現はまた、誘導プロモーターを使用して調節され得る。誘導プロモーターを用いて、発現レベルは、プロモーターへの誘導因子(例えば、テトラサイクリンまたはエクジソン)のための応答エレメントの取り込みによってこれらの因子の濃度に関連する。概して、高レベルの発現が、誘導因子の存在下のみで誘導プロモーターから得られる;基本の発現レベルは、最小限である。
【0082】
一実施形態において、本発明のベクターは、調節可能なプロモーター(例えば、tet−調節系およびRU−486系)を有する(例えば、GossenおよびBujard,Proc.Nat’l Acad.Sci.USA 89:5547(1992);Oliginoら、Gene Ther.5:491−496(1998);Wangら、Gene Ther.4:432−441(1997);Neeringら、Blood 88:1147−1155(1996);およびRendahlら、Nat.Biotechnol.16:757−761(1998)を参照のこと)。これらは、候補標的核酸の発現における小分子の制御を与える。この有益な特徴は、所望の表現型が、体細胞の変異よりも、トランスフェクトされたcDNAによってもたらされることを決定するために使用され得る。
【0083】
いくつかの発現系は、遺伝子増幅を提供するマーカーを有する(例えば、チミジンキナーゼおよびジヒドロ葉酸還元酵素)。あるいは、遺伝子増幅に関与しない高収量発現系もまた適切である(例えば、昆虫細胞のバキュロウイルスベクターをポリへドリンプロモーターまたは他の強力なバキュロウイルスプロモーターの指示の下、T1Rをコードする配列とともに使用する)。
【0084】
発現ベクターに代表的に含まれるエレメントはまた、E.coliにおいて機能するレプリコン、組換え型プラスミドを有する細菌の選択を可能にさせる抗生物質耐性をコードする遺伝子、および真核生物の配列の挿入を可能にするプラスミドの本質的ではない領域における特有の制限部位を含む。選択された特定の抗生物質耐性遺伝子は重要ではなく、当該分野において公知の任意の多くの耐性遺伝子が適している。好ましくは、原核生物配列は、必要な場合、それが、真核生物細胞のDNAの複製を干渉しないように、選ばれる。
【0085】
標準的なトランスフェクション方法が、大量のT1Rタンパク質を発現する細菌細胞株、哺乳動物細胞株、酵母細胞株または昆虫細胞株を生成するために使用され、次いで、このタンパク質は、標準的な技術を使用して精製される(例えば、Colleyら、J.Biol.Chem.264:17619−17622(1989);Methods in Enzyrrcology,182巻(Deutscher,(編),1990年)中の、Guide to Protein Purificationを参照のこと)。真核生物細胞および原核生物細胞の変換は、標準的な技術に従って実行される(例えば、Morrison,J Bact.132:349−351(1977);Clark−CurtissおよびCurtiss,Methods in Enzymology 101:347−362(Wuら(編)、1983年)を参照のこと)。
【0086】
宿主細胞に外来のヌクレオチド配列を導入するための任意の周知の手順が、使用され得る。これらは、リン酸カルシウムトランスフェクション、ポリブレン、プロトプラスト融合、エレクトロポレーション、微粒子銃、リポソーム、マイクロインジェクション、細胞質ベクター(plasma vector)、ウイルスベクターならびに宿主細胞中へクローニングされたゲノムDNA、cDNA、合成DNAまたは他の外来遺伝物質を導入するための他の周知の任意の方法の使用を含む(例えば、Sambrookら(前出)を参照のこと)。使用された特定の遺伝子工学手順が、T1Rを発現し得る宿主細胞に、少なくとも1つの遺伝子を首尾よく導入し得ることのみが重要である。
【0087】
細胞に発現ベクターが導入された後、トランスフェクトされた細胞は、T1Rの発現を有利にする条件下で培養され、このT1Rは、以下に同定される標準的な技術を使用して、培養物から回収される。
【0088】
(T1Rポリペプチドの精製)
天然に存在するT1RポリペプチドもしくはT1R3を含むレセプターか、または組換え型T1RポリペプチドまたはT1R3を含むレセプターは、機能的アッセイにおける使用のために精製され得る。天然に存在するT1Rタンパク質またはT1R3を含むレセプターは、例えば、ヒト組織から精製され得る。組換え型T1RポリペプチドまたはT1R3を含むレセプターは、任意の適切な発現系から精製され得る。T1Rポリペプチドは、代表的には、T1R3を含むレセプターを形成するために同じ細胞中で同時発現される。
【0089】
上記T1Rタンパク質またはT1R3を含むレセプターは、標準的な技術(硫酸アンモニアのような物質を用いた選択的沈降反応;カラムクロマトグラフィー;免疫精製方法、および他の方法(例えば、Scopes,Protein Purification:Principles and Practice(1982);米国特許第4,673,641号;Ausubelら、前出;およびSambrookら、前出、を参照のこと)を含む)によって相当の純度まで精製され得る。
【0090】
複数の手順が、組換え型T1Rタンパク質またはT1R3を含むレセプターを精製する場合に、使用され得る。例えば、確立した分子付着特性を有するタンパク質は、可逆的にあり、T1Rタンパク質またはT1R3を含むレセプターに融合され得る。適切なリガンドを使用して、T1Rタンパク質またはT1R3を含むレセプターは、選択的に精製カラムに吸着され、次いで、比較的純粋な形態でそのカラムから遊離され得る。次いで、この融合されたタンパク質は、酵素の活性によって除去される。最終的に、T1Rタンパク質またはT1R3を含むレセプターは、イムノアフィニティーカラムを使用して精製され得る。
【0091】
(A.組換え型細菌からのT1Rの精製)
組換え型タンパク質が、代表的に、プロモーターの誘導後、変換された細菌によって大量に発現される。しかし、発現は、構成的であり得る。IPTGを用いたプロモーターの誘導は、誘導プロモーター系の一例である。細菌は、当該分野において標準的な手順によって増殖される。新鮮なもしくは冷凍された細菌細胞が、タンパク質の分離のために使用される。
【0092】
細菌中に発現されたタンパク質は、不溶性の凝集体(「封入体」)を形成し得る。いくつかのプロトコルが、T1Rタンパク質またはT1R3を含むレセプターの封入体の精製に適している。例えば、封入体の精製は、代表的に、細菌細胞の分裂による(例えば、50mM TRIS/HCL pH 7.5、50mM NaCl、5mM MgCl2、1mM DTT、0.1mM ATP、および1mM PMSFの緩衝液中のインキュベーションによる)封入体の抽出、分離および/または精製を含む。この細胞の懸濁液は、French Pressを2〜3回通すことを使用して溶解され得るか、Polytron(Brinkman Instruments)を使用して均質化され得るか、あるいは氷上で超音波をかけられ得る。細菌を溶解する代替的な方法は、当業者に明白である(例えば、Sambrookら、前出;Ausubelら、前出、を参照のこと)。
【0093】
必要な場合、上記封入体は、可溶化され、そしてその溶解した細胞懸濁液は、代表的に、所望されない不溶性物質を除去するために遠心分離される。封入体を形成したタンパク質は、適合する緩衝液を用いた希釈または透析によって再生され得る。適切な溶媒としては、尿素(約4Mから約8Mまで)、ホルムアミド(少なくとも約80%体積(体積/体積ベース))、および塩酸グアニジン(約4Mから約8Mまで)が挙げられるが、これらに限定されない。凝集体を形成するタンパク質を可溶化し得るいくつかの溶媒(例えば、SDS(ドデシル硫酸ナトリウム)、70%ギ酸)は、免疫原性および/または活性の欠如を伴うタンパク質の不可逆な変性の可能性のため、この手順における使用に不適切である。塩酸グアニジンおよび類似の因子は変性剤であるが、この変性は不可逆性ではなく、そして再生が変性剤の除去(透析による、例えば)または希釈の際に起こり得、免疫学的におよび/または生物学的に活性なタンパク質の再形成を可能にする。他の適切な緩衝液は、当業者に公知である。ヒトT1Rタンパク質またはT1Rを含むレセプターは、標準的な分離技術(例えば、Ni−NTAアガロース樹脂を用いる)によって細菌の他のタンパク質から分離される。
【0094】
あるいは、細菌のペリプラズムからT1Rタンパク質またはT1Rを含むレセプターを精製することが可能である。細菌の溶解後、このT1Rタンパク質またはT1Rを含むレセプターが細菌のペリプラズムに放出される場合、この細菌のペリプラズム画分は、低温浸透圧性ショックおよび当業者に公知の他の方法によって単離され得る。このペリプラズムから組換え型タンパク質を単離するために、細菌細胞は遠心分離され、ペレットを形成する。このペレットは、20%スクロースを含む緩衝液中で再懸濁される。この細胞を溶解するために、細菌を遠心分離し、そしてそのペレットを氷冷した5mM MgSO4中で再懸濁し、そして約10分間氷浴中器に保存する。この細胞懸濁液を遠心分離し、そしてその上清をデカントし、そして保存する。この上清に存在する組換え型タンパク質は、当業者に周知の標準的な分離技術によって宿主タンパク質から分離され得る。
【0095】
(B.T1Rタンパク質を精製するための標準的なタンパク質分離技術)
(溶解度による分別)
初めの工程として,多くの場合(特にタンパク質混合物が複雑である場合に)、初めの塩分別は、目的の組換え型タンパク質から多くの所望されない宿主細胞タンパク質(もしくは細胞培養培地由来のタンパク質)を分離し得る。好ましい塩は、硫酸アンモニウムである。硫酸アンモニウムは、タンパク質混合物中の水の量を効果的に減少させることによってタンパク質を沈殿させる。次いで、タンパク質は、その溶解度に基づいて沈殿する。タンパク質がより疎水性であるほど、タンパク質が低い硫酸アンモニウムの濃度で沈殿する可能性がより高い。代表的なプロトコルは、結果として生じる硫酸アンモニウム濃度を20〜30%の間にするように、タンパク質溶液に飽和硫酸アンモニウムを添加する工程を包含する。この濃度は、最も疎水性であるタンパク質を沈殿させる。次いで、この沈殿物は廃棄され(目的のタンパク質が疎水性でない限り)、そして硫酸アンモニウムが、目的のタンパク質を沈殿することが公知である濃度まで上清に添加される。次いで、この沈殿物は、緩衝液中で可溶化され、そして必要な場合、過剰の塩が、透析もしくはダイアフィルトレーション(diafiltration)のいずれかを通じて除去される。タンパク質の溶解度に依存する他の方法(例えば、低温エタノール沈殿)は、当業者に周知であり、そして複雑なタンパク質混合物を分別するために使用され得る。
【0096】
(サイズ差別的ろ過)
T1Rタンパク質またはT1R3を含むレセプターの分子量を、異なる孔のサイズの膜(例えば、Amicon membraneもしくはMillipore membrane)を通した限外ろ過を使用して、より大きいサイズのタンパク質およびより小さいサイズのタンパク質からその分子量を単離するために使用し得る。第一の工程として、タンパク質混合物が、目的のタンパク質の分子量よりも低いカットオフ分子量を有する孔のサイズの膜を通して限外ろ過される。次いで、この限外ろ過の未透過物は、目的のタンパク質の分子量よりも大きい分子のカットオフを有する膜に対して限外ろ過される。上記組換え型タンパク質は、この膜を通してろ液へと通過する。次いで、このろ液で、以下に記載されるようにクロマトグラフィーを行い得る。
【0097】
(カラムクロマトグラフィー)
T1Rタンパク質またはT1R3を含むレセプターはまた、そのサイズ、正味の表面電荷(net surface charge)、疎水性、およびリガンドに対する親和性に基づき、他のタンパク質から分離され得る。さらに、タンパク質に対して惹起された抗体は、カラムマトリックスに接合され、そしてこのタンパク質は、免疫精製され得る。これらの方法の全てが、当該分野において周知である。クロマトグラフィー技術が、任意のスケールで、そして多くの異なる製造者(例えば、Pharmacia Biotech)からの器具を使用して実行され得ることが、当業者に明白である。
【0098】
(T1Rタンパク質の調節因子のためのアッセイ)
(A.アッセイ)
T1R3を含む味レセプターの調整、および対応する味の調整は、種々のインビトロおよびインビボアッセイを使用して評価され得る。そのようなアッセイは、T1R3を含む味レセプターのインヒビターおよびアクチベーター、および結果的に味のインヒビターおよびアクチベーターについてテストするために使用され得る。T1R3を含む甘味レセプターのそのような調節因子は、味シグナル変換に関与する。T1R3を含む味レセプターの調節因子は、組換え型もしくは天然に存在するT1R3を含む味レセプター(好ましくは、ヒトレセプター)を使用してテストされる。
【0099】
一実施形態において、本発明のモノマーのもしくはホモダイマーのT1R3を含むレセプターは、天然に存在するかもしくは人工の甘味分子(例えば、小有機分子、アミノ酸、ペプチド、炭水化物、脂質、多糖類など)についてスクリーニングするために使用され得る。例えば、本発明のホモダイマーのもしくはモノマーのT1R3を含むレセプターは、以下の実施例のセクションに記載されるように、天然に存在する甘味味覚物質を認識する。そのようなレセプターは、人工甘味料または変更された天然に存在する甘味料(これらの甘味料は、ホモダイマーもしくはモノマーのT1R3を含むレセプターの天然に存在する糖のリガンドを模倣する)についてスクリーニングするために使用され得る。
【0100】
好ましくは、そのT1R3を含む味レセプターは、本明細書において提供される配列かまたはその保存的に改変された改変体によってコードされるような配列を有する。あるいは、このアッセイのT1R3を含む味レセプターは、真核生物に由来し、そして本明細書において提供されるアミノ酸配列に対して、実質的なアミノ酸配列同一性を有するアミノ酸配列を含むか、あるいは、(中程度もしくは高度に)ストリンジェントな条件下で、本明細書において記載されるようなヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる。概して、アミノ酸配列の同一性は、少なくとも60%(好ましくは、少なくとも65%、70%、75%、80%、85%、または90%、最も好ましくは少なくとも95%)である。
【0101】
甘味シグナル変換の測定、あるいはT1R3を含む味レセプターもしくはT1R3を含む味レセプターを発現する細胞(組換え型かもしくは天然に存在するもののいずれか)の甘味シグナル変換表現型の損失の測定が、本明細書において記載されるように、種々のアッセイ(インビトロ、インビボ、およびエクスビボ)を使用して実行され得る。活性または結合に影響を及ぼす適切な物理的変化、化学的変化もしくは表現型変化が、本発明のポリペプチドに対するテスト化合物の影響を評価するために使用され得る。これらの機能的効果がインタクトな細胞または動物を使用して決定される場合、実施者はまた、種々の効果(例えば、シグナル変換の場合、例えば、リガンド結合、ホルモン放出、公知の遺伝マーカーおよび特徴付けされていない遺伝マーカー(例えば、ノザンブロット)の両方に対する転写の変化、細胞代謝の変化(例えば、pHの変化)、ならびに、細胞内セカンドメッセンジャー(Ca2+、IP3、cGMP、もしくはcAMP)の変化)も測定し得る。
【0102】
(インビトロアッセイ)
活性を調整するT1R3を含む味レセプターを有する化合物を同定するためのアッセイが、インビトロで実行され得る。そのようなアッセイは、全長のT1R3を含む味レセプターもしくはその改変体、またはキメラを形成するために異種のタンパク質に融合される、T1R3を含む味レセプターのフラグメント(例えば、細胞外ドメイン)を使用し得る(例えば、WO 01/66563、WO 03/001876、WO 02/064631、およびWO 03/004992を参照のこと)。精製された組換え型もしくは天然に存在するT1R3を含む味レセプターは、本発明のインビトロ方法において使用され得る。精製されたT1R3を含む味レセプターに加えて、その組換え型もしくは天然に存在するT1R3を含む味レセプターは、細胞の溶解物または細胞膜の一部であり得る。以下に記載されるように、結合アッセイは、固体状態かもしくは可溶性状態のいずれかであり得る。好ましくは、そのタンパク質または膜は、固体の支持体に(共有結合性または非共有結合性に)結合される。多くの場合、本発明のインビトロアッセイは、リガンド結合アッセイまたはリガンド親和性アッセイ(本明細書において記載されるような、公知の細胞外リガンドと、または公知の細胞内リガンドGTPのいずれかと、非競合または競合するいずれかのアッセイ)である。他のインビトロアッセイは、このタンパク質についての分光特性(例えば、蛍光、吸収度、屈折率)、流体力学的特性(例えば、形状)、クロマトグラフィー特性、または溶解特性の変化を測定する工程を包含する。
【0103】
一実施形態において、ハイスループット結合アッセイが実行され、その中で、T1R3を含む味レセプターまたはそのフラグメントを含むキメラが、潜在的な調節因子と接触され、そして適切な期間、インキュベートされる。一実施形態において、この潜在的な調節因子は固体の支持体に結合され、そしてT1R3を含む味レセプターが添加される。別の実施形態において、このT1R3を含む味レセプターは、固体の支持体に結合される。広範な種類の調節因子(小有機分子、ペプチド、抗体、およびT1R3を含む味レセプターリガンドアナログを含む)が、以下に記載されるように、使用され得る。広範な種類のアッセイ(標識化タンパク質−タンパク質結合アッセイ、電気泳動移動度シフト、イムノアッセイ、酵素アッセイ(例えば、リン酸化アッセイ)などを含む)が、T1R3を含む味レセプター−調節因子結合を同定するために使用され得る。いくつかの場合、候補調節因子の結合は、競合結合アッセイの使用を通じて決定され、ここで、公知のリガンドの結合との干渉が、潜在的な調節因子の存在下で測定される。T1R3を含む味レセプターリガンドが、本明細書において提供される。この調節因子または公知のリガンドのいずれかがまず結合し、次いで、その競合相手が添加される。このT1R3を含む味レセプターが洗浄された後、潜在的な調節因子または公知のリガンドのいずれかの結合との干渉が、決定される。多くの場合、この潜在的な調節因子または公知のリガンドのいずれかが標識される。
【0104】
(細胞ベースのインビボアッセイ)
別の実施形態において、T1R3を含む味レセプターが細胞中に発現され(例えば、T1Rファミリー(例えば、T1R1およびT1R3またはT1R2およびT1R3)の1つかもしくは2つのメンバーの発現または同時発現による、好ましくは、任意の他のT1Rファミリーメンバーの発現を伴わないT1R3単独の発現による)、そして機能的(例えば、物理的および化学的または表現型)変化が、T1R3を含む味レセプター味調節因子を同定するためにアッセイされる。T1R3を含む味レセプターを発現する細胞もまた、結合アッセイにおいて使用され得る。任意の適切な機能的効果が、本明細書において記載されるように測定され得る。例えば、リガンド結合、Gタンパク質結合、およびGPCRシグナル変換(例えば、細胞内Ca2+レベルの変化)は全て、細胞ベースの系を使用して潜在的な調節因子を同定するための適切なアッセイである。そのような細胞ベースのアッセイのために適切な細胞は、本明細書において記載されるように、初代細胞と細胞株との両方を含む。T1R3を含む味レセプターは、天然に存在するかまたは組換え型であり得る。さらに、上述のように、GPCR活性を有するキメラT1R3を含む味レセプターが、細胞ベースのアッセイにおいて使用され得る。例えば、T1Rタンパク質の細胞外ドメインは、異種のタンパク質(好ましくは、異種のGPCR)の膜貫通ドメインおよび/または細胞質ドメインに融合し得る。そのようなキメラGPCRは、GPCR活性を有し、そして本発明の細胞ベースのアッセイにおいて使用され得る。
【0105】
別の実施形態において、細胞のT1Rポリペプチドレベルは、タンパク質またはmRNAのレベルを測定することによって決定される。T1Rタンパク質またはT1Rシグナル変換に関連するタンパク質のレベルは、T1R3を含む味レセプターもしくはそのフラグメントに選択的に結合する抗体を用いて、イムノアッセイ(例えば、ウェスタンブロット法、ELISAなど)を使用して測定される。mRNAの測定については、増幅(例えば、PCR、LCRを用いる)、またはハイブリダイゼーションアッセイ(例えば、ノザンハイブリダイゼイーション)、RNase防御、ドットブロット法)を使用して)が好ましい。タンパク質またはmRNAのレベルは、本明細書において記載されるように、直接的にまたは間接的に標識された検出因子(例えば、蛍光で標識された核酸または放射性標識された核酸、放射性標識された抗体または酵素で標識された抗体など)を使用して検出される。
【0106】
あるいは、T1R3を含むレセプター発現は、レポーター遺伝子系を使用して測定され得る。そのような系は、レポーター遺伝子(例えば、クロラムフェニコールアセチルトランスフェラーゼ、ホタルルシフェラーゼ、細菌のルシフェラーゼ、β−ガラクトシダーゼおよびアルカリホスファターゼ)に作動可能に結合したT1Rタンパク質プロモーターを使用して考案され得る。さらに、目的のタンパク質は、第二のレポーター(例えば、赤蛍光タンパク質または緑蛍光タンパク質への付着を介した間接的なレポーターとして使用され得る(例えば、MistiliおよびSpector,Nature Biotechnology 15:961−964(1997)を参照のこと)。レポーター構築物は、代表的に、細胞へトランスフェクトされる。潜在的な調節因子での処理後、レポーター遺伝子の転写、翻訳、または活性の量が、当業者に公知の標準的な技術に従って測定される。
【0107】
別の実施形態において、GPCRシグナル変換に関連する機能的効果が、測定され得る。活性化もしくは阻害された、T1R3を含むGタンパク質共役型レセプターは、標的酵素、セカンドメッセンジャー、チャネル、および他のエフェクタータンパク質の特性を変更する。その例としては、cGMPホスホジエステラーゼ、アデニル酸シクラーゼ、ホスホリパーゼC、IP3の活性、ならびにGタンパク質による種々のチャネルの調節が挙げられる。下流での結果がまた調査され得る(例えば、ホスホリパーゼCによるジアシルグリセロールおよびIP3の産生、これに次ぐ、IP3によるカルシウム可動化について)。活性化されたGPCRレセプターは、このレセプターのC末端尾部(およびおそらく他の部位も同様に)をリン酸化するキナーゼのための基質となる。従って、アクチベーターは、γ標識化GTPから上記レセプターへの32Pの転移を促進し、このことは、シンチレーション計数器を用いてアッセイされ得る。C末端尾部のリン酸化は、アレスチンのようなタンパク質の結合を促進し、そしてGタンパク質の結合に干渉する。GPCRシグナル変換、およびシグナル変換をアッセイする方法の一般的な概説については、例えば、Methods in Enzymology,第237巻および第238巻(1994)および第96巻(1983);Bourneら、Nature 10:349:117−27(1991);Bourneら、Nature 348:125−32(1990);Pitcherら、Annu.Rev.Biochem.67:653−92(1998)を参照のこと。
【0108】
上述のように、いくつかのGタンパク質共役型レセプターの活性化は、ホスファチジルイノシトールのホスホリパーゼCを媒介した加水分解を通じたイノシトール三リン酸(IP3)の形成を刺激する(BerridgeおよびIrvine,Nature 312:315−21(1984))。IP3は、次いで、細胞内カルシウムイオンの貯蔵の放出を刺激する。従って、細胞質のカルシウムイオンのレベルの変化、またはセカンドメッセンジャー(例えば、IP3)のレベルの変化を、Gタンパク質共役型レセプター機能を評価するために使用し得る。そのようなGタンパク質共役型レセプターを発現する細胞は、細胞内貯蔵とイオンチャネルの活性化を介することの両方からの寄与の結果として増加した細胞質カルシウムレベルを示し得、この場合、必要ではないが、内部貯蔵からのカルシウム放出から生じる蛍光応答を区別するために、カルシウムを含まない緩衝液中でそのようなアッセイを実行すること(必要に応じてキレート化剤(例えば、EGTA)を補充される)が所望され得る。
【0109】
一例において、T1R3を含む味レセプターGPCR活性は、このレセプターをホスホリパーゼCシグナル変換経路に結合する乱交雑したGタンパク質を有する異種の細胞中のT1R3を含む味レセプターを発現することによって測定される(OffermannsおよびSimon,J.Biol.Chem.270:15175−15180(1995)を参照のこと)。シグナル変換の調節は、細胞内Ca2+レベルの変化を測定することによってアッセイされ、この細胞内Ca2+レベルは、T1R3を含む味レセプターと結合する分子の投与を介するGPCRシグナル変換経路の調節に応じて変化する。Ca2+レベルの変化は、必要に応じて、蛍光Ca2+指示染料および蛍光イメージングを使用して測定される。
【0110】
別の例において、ホスファチジルイノシトール(PI)の加水分解は、本明細書において参考として援用されている米国特許第5,436,128号に従って分析され得る。簡潔にいうと、そのアッセイは、48時間以上、3H−ミオイノシトールを用いて細胞の標識する工程を包含する。この標識された細胞は、1時間、テスト化合物で処理される。この処理された細胞は、クロロホルム−メタノール−水中で溶解および抽出され、その後、イノシトールリン酸はイオン交換クロマトグラフィーによって分離され、そしてシンチレーション計数(scintillation counting)によって定量化された。刺激倍率(fold stimulation)は、緩衝液コントロール存在下のcpmに対するアゴニスト存在下のcpmの比を計算することによって決定される。同様に、阻害倍率(fold inhibition)は、緩衝液コントロール(アゴニストを含んでいても、含んでいなくてもよい)存在下のcpmに対するアンタゴニスト存在下のcpmの比を計算することによって決定される。
【0111】
他のアッセイは、活性化された場合、酵素(例えば、アデニル酸シクラーゼ)を活性化または阻害することによって、細胞内のサイクリックヌクレオチド(例えば、cAMPまたはcGMP)のレベルの変化をもたらすレセプターの活性を決定する工程を包含し得る。レセプターの活性化がサイクリックヌクレオチドレベルの減少をもたらす場合、アッセイ中の細胞にレセプターを活性化する化合物を添加する前に、細胞内のサイクリックヌクレオチドレベルを増加する因子(例えば、フォルスコリン)に細胞を暴露することが好ましくあり得る。
【0112】
一例において、細胞内cAMPまたはcGMPの変化が、イムノアッセイを使用して測定され得る。OffermannsおよびSimon,J.Biol.Chem.270:15175−15180(1995)に記載される方法が、cAMPのレベルを決定するために使用され得る。さらに、Felley−Boscoら、Am.J.Resp.Cell and Mol.Biol.11:159−164(1994)に記載される方法が、cGMPのレベルを決定するために使用され得る。さらに、cAMPおよび/またはGMPを測定するためのアッセイキットが、米国特許第4,115,538号(本明細書において参考として援用されている)に記載されている。
【0113】
一例において、Gタンパク質共役型レセプター活性のためのアッセイは、レセプター活性を報告するために、イオン感受性色素または電位(voltage)感受性色素で負荷される細胞を含む。そのようなレセプターの活性を決定するためのアッセイはまた、テストされた化合物の活性を評価するための陰性のコントロールまたは陽性のコントロールとして、他のGタンパク質共役型レセプターの公知のアゴニストおよびアンタゴニストを使用し得る。調節化合物(例えば、アゴニスト、アンタゴニスト)を同定するためのアッセイにおいて、細胞質中のイオンのレベルまたは膜電位の変化が、それぞれ、イオン感受性指示薬(ion sensitive indicator)または膜電位蛍光指示薬(membrane voltage fluorescent indicator)を使用してモニタリングされる。とりわけ、使用され得るイオン感受性指示薬および電位プローブは、Molecular Probes 1997 Catalog中で開示される。Gタンパク質共役型レセプターについて、乱交雑Gタンパク質(例えば、Gα15およびGα16)が、最適なアッセイにおいて使用され得る(Wilkieら、Proc.Nat’l Acad.Sci.USA 88:10049−10053(1991))。そのような乱交雑Gタンパク質は、広範囲のレセプターの共役を可能にする。
【0114】
(動物モデル)
味覚の動物モデルはまた、味覚の調節因子のためのスクリーニングにおける使用を見出す(例えば、本明細書において記載されるようなT1Rノックアウトマウス系統)。トランスジェニック動物技術(遺伝子ノックアウト技術(例えば、適切な遺伝子標的ベクターを用いる異種の組換えの結果として)、または遺伝子過剰発現を含む)は、T1R3を含むレセプターもしくはその構成要素の欠如もしくは発現の増加をもたらす。所望される場合、T1R3を含むレセプターもしくはその構成要素の組織特異的な発現もしくはノックアウトが必要であり得る。そのような方法によって作製されたトランスジェニック動物は、味調節の動物モデルとしての用途を見出し、そしてさらに、味調節の調節因子をスクリーニングすることに有用である。
【0115】
(B.調節因子)
T1R3を含む味レセプターの調節因子としてテストされる化合物は、任意の小有機分子、または生物学的実体(例えば、タンパク質(例えば、抗体またはペプチド)、アミノ酸、脂質、脂肪、糖(例えば、単糖類、二糖類、または多糖類)、核酸(例えば、アンチセンスオリゴヌクレオチドまたはリボザイム)、または小有機分子)であり得る。あるいは、調節因子は、T1R3を含む味レセプターの遺伝的に変更されたバージョンであり得る。代表的に、テスト化合物は、小有機分子、アミノ酸、ペプチド、脂質、および単糖類、二糖類および多糖類である。
【0116】
本質的に、任意の化学化合物は、本発明のアッセイにおいて潜在的な調節因子またはリガンドとして使用され得るが、非常に多くの場合、水溶液中あるいは、有機(特にDMSOベースの)溶液中に溶解され得る化合物が使用される。このアッセイは、アッセイ工程を自動化することによって、そして代表的には平行して(例えば、ロボットアッセイにおけるマイクロタイタープレート上のマイクロタイター形式において)実行される、任意のアッセイに便利な供給源由来の化合物を提供することによって、多くの化学ライブラリーをスクリーニングするように設計される。多くの化学化合物の供給元(Sigma(St.Louis,MO),Aldrich(St.Louis,MO),Sigma−Aldrich(St.Louis,MO),Fluka Chemika−Biochemica Analytika(Buchs Switzerland)などが挙げられる)が存在することが理解される。
【0117】
好ましい一実施形態において、ハイスループットスクリーニング方法は、多数の潜在的治療用化合物(潜在的調節因子化合物または潜在的リガンド化合物)を含むコンビナトリアル小有機分子ライブラリーまたはペプチドライブラリーを提供する工程を包含する。次いで、そのような「コンビナトリアルケミカルライブラリー」または「リガンドライブラリー」は、本明細書において記載されるように1つ以上のアッセイにおいてスクリーニングされ、所望の特徴的活性を示すそれらのライブラリーメンバー(特定の化学的種またはサブクラス)を同定する。従って、同定された化合物は、従来の「リード化合物」として役立ち得るか、またはそれら自身、潜在的な治療法または実際の治療法として使用され得る。
【0118】
コンビナトリアルケミカルライブラリーは、化学的合成または生物学的合成のいずれかによって、複数の化学的「ビルディングブロック(building block)」(例えば、試薬)を組み合わせることによって、生成された種々の化学化合物の集合である。例えば、線状のコンビナトリアルケミカルライブラリー(例えば、ポリペプチドライブラリー)が、所与の化合物の長さ(すなわち、ポリペプチド化合物中のアミノ酸の数)に対する全ての可能な方法において、一組の化学的ビルディングブロック(アミノ酸)を組み合わせることによって形成される。何百万の化学化合物が、そのような化学的ビルディングブロックのコンビナトリアル混合を通して合成され得る。
【0119】
コンビナトリアルケミカルライブラリーの調製およびスクリーニングは、当業者に周知である。そのようなコンビナトリアルケミカルライブラリーとしては、ペプチドライブラリー(例えば、米国特許第5,010,175号、Furka,Int.J.Pept.Prot.Res.37:487−493(1991)およびHoughtonら、Nature 354:84−88(1991)を参照のこと)が挙げられるが、これらに限定されない。化学的多様性ライブラリーを生成するための他の化学物質もまた、使用され得る。そのような化学物質としては、ペプトイド(例えば、PCT公開番号WO 91/19735)、コードされたペプチド(例えば、PCT公開番号WO 93/20242)、ランダムバイオオリゴマー(例えば、PCT公開番号WO 92/00091)、ベンゾジアゼピン(例えば、米国特許第5,288,514号)、ダイバーソマー(diversomer)(例えば、ヒダントイン、ベンゾジアゼピンおよびジペプチド)(Hobbsら、Proc.Nat.Acad.Sci.USA 90:6909−6913(1993))、ビニル性ポリペプチド(Hagiharaら、J.Amer.Chem.Soc.114:6568(1992))、グルコース骨格を有する非ペプチドペプチド模倣物(nonpeptidal peptidomimetic)(Hirschmannら、J.Amer.Chem.Soc.114:9217−9218(1992))、低化合物ライブラリーのアナログの有機合成(Chenら、J.Amer.Chem.Soc.116:2661(1994)、オリゴカルバメート(Choら、Science 261:1303(1993))、および/またはペプチジルホスホネート(Campbellら、J.Org.Chem.59:658(1994))、核酸ライブラリー(Ausubel、BergerおよびSambrook、全て前出、を参照のこと)、ペプチド核酸ライブラリー(例えば、米国特許5,539,083号を参照のこと)、抗体ライブラリー(例えば、Vaughnら、Nature Biotechnology、14(3):309−314(1996)およびPCT/US96/10287を参照のこと)、炭水化物ライブラリー(例えば、Liangら、Science、274:1520−1522(1996)および米国特許第5,593,853号を参照のこと)、低有機分子ライブラリー(例えば、ベンゾジアゼピン、Baum CおよびEN、1月18日、33頁(1993年);イソプレノイド、米国特許第5,569,588号;チアゾリジノンおよびメタチアザノン(metathiazanone)、米国特許第5,549,974号;ピロリジン、米国特許第5,525,735号および同第5,519,134号;モルホリノ化合物、米国特許第5,506,337号;ベンゾジアゼピン、同第5,288,514号など)が挙げられるが、これらに限定されない。
【0120】
コンビナトリアルライブラリーの調製のための装置は、市販されている(例えば、357 MPS,390 MPS,Advanced Chem Tech,Louisville KY,Symphony,Rainin,Woburn,MA,433A Applied Biosystems,Foster City,CA,9050 Plus,Millipore,Bedford,MAを参照のこと)。さらに、多数のコンビナトリアルライブラリー自体が、市販されている(例えば、ComGenex,Princeton,N.J.,Asinex,Moscow,Ru,Tripos,Inc.,St.Louis,MO,ChemStar,Ltd,Moscow,RU,3D Pharmaceuticals,Exton,PA,Martek Biosciences,Columbia,MD,などを参照のこと)。
【0121】
(C.固体状態ハイスループットアッセイおよび可溶性ハイスループットアッセイ)
一実施形態において、本発明は、T1R3を含む味レセプター、またはT1R3を含む味レセプターを発現する細胞もしくは組織(天然に存在するかまたは組換え型かのいずれか)を使用して、可溶性アッセイを提供する。別の実施形態において、本発明は、ハイスループット形式における固相ベースのインビトロアッセイを提供し、ここで、上記T1R3を含む味レセプターは、固相に付着される。本明細書において記載される任意のアッセイは、ハイスループットスクリーニングのために適合され得る(例えば、リガンド結合、細胞増殖、細胞表面マーカー流動(例えば、スクリーニング、放射性標識化GTP結合)、セカンドメッセンジャー流動(例えば、Ca2+、IP3、cGMP、またはcAMP、サイトカイン産生)など)。
【0122】
本発明のハイスループットアッセイ(可溶性状態または固体状態のいずれか)において、一日で、数千までの異なる調節因子またはリガンドをスクリーニングすることが可能である。この方法論は、インビトロでT1R3を含む味レセプターのために、またはT1R3を含む味レセプターを含む細胞ベースのアッセイもしくは膜ベースのアッセイのために使用され得る。特に、マイクロタイタープレートの各ウェルは、選択された潜在的な調節因子に対する分離アッセイを実行するために使用され得るか、あるいは、濃度もしくはインキュベーション時間効果が観察される場合、5〜10ウェルごとに、1つの調節因子をテストし得る。従って、1つの標準マイクロタイタープレートは、約100(例えば、96)の調節因子をアッセイし得る。1536のウェルプレートが使用される場合、1つのプレートは容易に約100〜約1500の異なる化合物をアッセイし得る。一日につき多くのプレートをアッセイすることが、可能である;約6,000までの、20,000までの、50,000までの、または100,000を超える、異なる化合物についてスクリーニングするアッセイは、本発明の統合的なシステムを使用して可能である。
【0123】
固体状態の反応について、目的のタンパク質もしくはそのフラグメント(例えば、細胞外ドメイン)、または融合タンパク質の一部として目的のタンパク質もしくはそのフラグメントを含む細胞もしくは膜が、共有結合性または非共有結合性の結合を介して(例えば、タグを介して)、直接的にかまたは間接的に、固体状態の構成要素に結合され得る。このタグは、任意の種々の構成要素であり得る。一般的に、タグ(タグ結合因子)に結合する分子は、固体の支持体に固定され、そしてタグ化された目的の分子は、そのタグとそのタグ結合因子との相互作用によって固体の支持体に付着される。
【0124】
多数のタグおよびタグ結合因子が、文献に十分に記載されている公知の分子相互作用に基づいて使用され得る。例えば、タグが天然の結合剤(例えば、ビオチン、タンパク質A、またはタンパク質G)を有する場合、それは適切なタグ結合因子(アビジン、ストレプトアビジン、ニュートラアビジン(neutravidin)、免疫グロブリンのFc領域など)と共に使用され得る。天然の結合剤(例えば、ビオチン)を有する分子に対する抗体もまた、広く入手可能であり、そしてその抗体も、適切なタグ結合因子である;SIGMA Immunochemicals 1998 catalogue SIGMA,St.Louis MOを参照のこと。
【0125】
同様に、任意のハプテン化合物または抗原性化合物が、タグ/タグ結合因子のペアを形成するために、適切な抗体と共に使用され得る。数千の特異的抗体が市販されており、そして多くのさらなる抗体が文献に記載されている。例えば、1つの一般的構造において、タグは第一の抗体であり、そしてタグ結合因子は、第一の抗体を認識する第二の抗体である。抗体−抗原相互作用に加えて、レセプター−リガンド相互作用もまた、タグおよびタグ結合因子のペアとして適切である。例えば、細胞膜レセプターのアゴニストおよびアンタゴニスト(例えば、細胞レセプター−リガンド相互作用(例えば、トランスフェリン、c−キット、ウイルスレセプターリガンド、サイトカインレセプター、ケモカインレセプター、インターロイキンレセプター、免疫グロブリンレセプターおよび抗体、カドヘリンファミリー、インテグリンファミリー、セレクチンファミリーなど));例えば、PigottおよびPower,The Adhesion Molecule Facts Book I(1993)を参照のこと。同様に、トキシンおよび毒物(venom)、ウイルスエピトープ、ホルモン(例えば、オピエート、ステロイドなど)、細胞内レセプター(例えば、種々の小リガンド(ステロイド、甲状腺ホルモン、レチノイド、およびビタミンDを含む);ペプチドの効果を媒介するもの)、薬物、レクチン、糖、核酸(線状ポリマー構造と環状ポリマー構造との両方)、オリゴ糖、タンパク質、リン脂質および抗体は全て、種々の細胞レセプターと相互作用し得る。
【0126】
合成ポリマー(例えば、ポリウレタン、ポリエステル、ポリカーボネート、ポリ尿素、ポリアミド、ポリエチレンイミン、ポリアリーレンスルフィド、ポリシロキサン、ポリイミド、およびポリアセテート)もまた、適切なタグまたはタグ結合因子を形成し得る。多くの他のタグ/タグ結合因子ペアもまた、本開示の概説の際に当業者に理解されるように、本明細書において記載されたアッセイ系において有用である。
【0127】
一般的なリンカー(例えば、ペプチド、ポリエーテルなど)もまたタグとして役立ち得、そしてこれはポリペプチド配列(例えば、約5アミノ酸と200アミノ酸との間のポリグリシン配列)を含み得る。そのような柔軟なリンカーは、当業者に公知である。例えば、ポリ(エチレングリコール)リンカーは、Shearwater Polymers,Inc.Huntsville,Alabamaから入手可能である。これらのリンカーは、必要に応じて、アミド結合、スルフヒドリル結合、またはヘテロ官能基結合(heterofunctional linkage)を有する。
【0128】
タグ結合因子は、現在利用可能な任意の種々の方法を使用して固体の基質に固定される。固体の基質は、一般に、タグ結合因子の一部と反応する表面へ化学基を固定する化学試薬に、その基質の全てまたは一部を暴露することによって誘導体化されるかまたは官能基化される。例えば、より長い鎖の部分へ付着するために適している基としては、アミン基、ヒドロキシル基、チオール基、およびカルボキシル基が挙げられる。アミノアルキルシランおよびヒドロキシアルキルシランは、種々の表面(例えば、ガラス表面)を官能基化するために使用され得る。そのような固相のバイオポリマーアレイの構築物は、文献に十分に記載されている。例えば、Merrifield,J.Am.Chem.Soc.85:2149−2154(1963)(例えば、ペプチドの固相合成を記載している);Geysenら、J.Immun.Meth.102:259−274(1987)(ピン(pin)上での固相構成要素の合成を記載している);FrankおよびDoring,Tetrahedron 44:60316040(1988)(セルロースディスク上での種々のペプチド配列の合成を記載している);Fodorら、Science,251:767−777(1991);Sheldonら、Clinical Chemistry 39(4):718−719(1993);およびKozalら、Nature Medicine 2(7):753759(1996)(いずれも、固体の基質へ固定されるバイオポリマーのアレイを記載している)を参照のこと。基質にタグ結合因子を固定するための非化学的アプローチは、他の一般的方法(例えば、加熱、紫外線放射による架橋など)を含む。
【0129】
(T1R3を含むレセプターの免疫学的検出)
核酸ハイブリダイゼーション技術を使用するT1R遺伝子および遺伝子発現の検出に加えて、当業者はまた、本発明のT1R3を含む味レセプターを検出するためにイムノアッセイを使用し得る。そのようなアッセイは、T1R3を含む味レセプターの調節因子についてスクリーニングするため、ならびに治療上の用途および診断上の用途のために有用である。イムノアッセイは、T1R3を含む味レセプターを定性的にまたは定量的に分析するために使用され得る。この適用可能な技術の一般的概要は、HarlowおよびLane,Antibodies:A Laboratory Manual(1988)に見出され得る。
【0130】
(A.抗体の産生)
T1Rタンパク質およびT1R3を含む味レセプターと特異的に反応するポリクローナル抗体およびモノクローナル抗体を産生する方法は、当業者に公知である(例えば、Coligan,Current Protocols in Immunology(1991);HarlowおよびLane(前出);Goding,Monoclonal Antibodies:Principles and Practice(第2版、1986年);ならびに、KohlerおよびMilstein,Nature 256:495−497(1975)を参照のこと)。そのような技術としては、ファージまたは類似のベクター中の組換え型抗体のライブラリー由来の抗体の選択による抗体の調製、ならびに、ウサギまたはマウスを免疫化することによるポリクローナル抗体およびモノクローナル抗体の調製が挙げられる(例えば、Huseら、Science 246:1275−1281(1989);Wardら、Nature 341:544−546(1989)を参照のこと)。
【0131】
T1Rタンパク質またはT1R3を含む味レセプターの一部を含む多数の免疫原が、T1Rタンパク質に特異的に反応する抗体を産生するために使用され得る。例えば、組換え型タンパク質またはその抗原性のフラグメントが、本明細書において記載されるように単離され得る。組換え型タンパク質は、上述のように真核生物細胞または原核生物細胞中に発現され得、そして一般に上述のように精製され得る。組換え型タンパク質は、モノクローナル抗体またはポリクローナル抗体の産生のための好ましい免疫原である。あるいは、本明細書において開示されるような配列に由来し、そしてキャリアタンパク質に接合される合成ペプチドは、免疫原として使用され得る。天然に存在するタンパク質はまた、純粋な形態かまたは純粋ではない形態かのいずれかで使用され得る。次いで、この生成物は、抗体を産生し得る動物に注入される。モノクローナル抗体またはポリクローナル抗体のいずれかが、上記タンパク質を測定するためのイムノアッセイにおけるその後の使用のために産生され得る。
【0132】
ポリクローナル抗体を産生する方法は、当業者に公知である。マウス(例えば、BALB/Cマウス)またはウサギの純系が、標準的なアジュバント(例えば、フロイントアジュバント)および標準的な免疫化プロトコルを使用して上記タンパク質で免疫化される。この動物の免疫原調製物への免疫応答が、テスト採血を採取しそしてβサブユニットへの反応性の力価を決定することによって、モニタリングされる。抗体の免疫原への適切に高い力価が得られる場合、この動物から血液が収集され、そして抗血清が調製される。上記タンパク質に反応する抗体を濃縮するための抗血清のさらなる分画化は、所望される場合に、行われる(HarlowおよびLane(前出)を参照のこと)。
【0133】
モノクローナル抗体は、当業者によく知られた種々の技術によって得られ得る。簡潔にいうと、所望の抗原で免疫化された動物由来の脾臓細胞が、一般に骨髄腫細胞との融合によって不死化される(KohlerおよびMilstein,Eur.J.Immunol.6:511−519(1976)を参照のこと)。不死化の代替的方法としては、エプスタイン−バーウイルス、癌遺伝子、またはレトロウイルスを用いた形質転換、あるいは当該分野において周知の他の方法が挙げられる。1つの不死化細胞から生じたコロニーは、抗原に対する所望の特異性および親和性を有する抗体の産生のためにスクリーニングされ、そしてそのような細胞によって産生についてモノクローナル抗体の収量は、種々の技術(脊椎動物宿主の腹腔内への注射を含む)によって増大され得る。あるいは、当業者は、Huseら、Science 246:1275−1281(1989)によって概説される一般的プロトコルに従ってヒトB細胞由来のDNAライブラリーをスクリーニングすることにより、モノクローナル抗体またはその結合フラグメントをコードするDNA配列を単離し得る。
【0134】
モノクローナル抗体およびポリクローナル血清は、収集されそしてイムノアッセイ(例えば、固体の支持体上に固定された免疫原を用いた固相イムノアッセイ)において免疫原タンパク質に対して力価を測定(titered)される。代表的に、104以上の力価を有するポリクローナル抗血清が、競合結合イムノアッセイを使用して、T1Rではないタンパク質、またはT1R3も含む味レセプターではないタンパク質に対する交差反応性について選択されそしてテストされる。特異的ポリクローナル抗血清およびモノクローナル抗体は、一般的に少なくとも約0.1mM(より一般的には少なくとも約1μM、好ましくは少なくとも約0.1μM以下、そして最も好ましくは0.01μM以下)のKdで結合する。特定のT1R3を含む味レセプターのオルソログ(例えば、ヒトT1R3を含む味レセプター)にのみ特異的な抗体は、1つの種(例えば、非ヒト哺乳動物)から他の交差反応性オルソログを除去することによっても産生され得る。さらに、個々のT1Rタンパク質が、レセプターと個々のT1Rタンパク質との両方へ結合する抗体を除去するために使用され得る。この様式において、特定のレセプターにのみ結合する抗体が得られ得る。
【0135】
一度T1R3を含む味レセプターに対する特異的な抗体が利用可能となった場合、上記タンパク質は、種々のイムノアッセイ方法によって検出され得る。さらに、この抗体は、T1R3を含む味レセプター調節因子として治療用に使用され得る。免疫学的手順およびイムノアッセイ手順の概説については、Basic and Clinical Immunology(StitesおよびTerr(編)、第7版、1991年)を参照のこと。さらに、本発明のイムノアッセイは、Enzyme Immunoassay(Maggio(編)、1980年);およびHarlowおよびLane(前出)中に広範に論評される任意のいくつかの構成において実行され得る。
【0136】
(B.免疫結合アッセイ)
T1R3を含む味レセプターが、任意の多数のよく認識されている免疫結合アッセイ(例えば、米国特許第4,366,241号、同4,376,110号、同4,517,288号、および同4,837,168号を参照のこと)を使用して検出され得、そして定量化され得る。一般のイムノアッセイの概説については、Methods in Cell Biology:Antibodies in Cell Biology,第37巻(Asai(編)、1993年);Basic and Clinical Immunology(StitesおよびTerr(編)、第7版、1991年)もまた参照のこと。免疫結合アッセイ(またはイムノアッセイ)は、代表的に、最適なタンパク質または抗原(この場合、T1R3を含む味レセプターまたはその抗原部分配列)に特異的に結合する抗体を使用する。抗体(例えば、抗T1R3を含む味レセプター)が、任意の多数の当業者に周知の手段のいずれかおよび上述のような手段によって産生され得る。
【0137】
イムノアッセイはまた、多くの場合、上記抗体および抗原によって形成される複合体に特異的に結合しそしてその複合体を標識するために、標識性因子を使用する。この標識性因子は、それ自身、抗体/抗原複合体を含む部分の1つであり得る。従って、この標識性因子は、標識されたT1R3を含む味レセプターであり得るか、または標識された抗T1R3を含む味レセプター抗体であり得る。あるいは、この標識性因子は、抗体/T1R3を含む味レセプター複合体に特異的に結合する第三の部分(例えば、二次抗体)であり得る(二次抗体は、代表的に、一次抗体が由来する種の抗体に特異的である)。免疫グロブリン定常領域(例えば、タンパク質Aまたはタンパク質G)に特異的に結合し得る他のタンパク質もまた、標識性因子として使用され得る。これらのタンパク質は、種々の種に由来する免疫グロブリン定常領域に、強い非免疫原性の反応を示す(例えば、Kronvalら、J.Immunol.111:1401−1406(1973);Akerstromら、J.Immunol.135:2589−2542(1985)を参照のこと)。標識性因子は、別の分子(例えば、ストレプトアビジン)が特異的に結合し得る検出可能部分(例えば、ビオチン)を用いて改変され得る。種々の検出可能部分は、当業者に周知である。
【0138】
このアッセイの全体を通して、インキュベーション工程および/または洗浄工程が、試薬の各組み合わせの後に必要とされ得る。インキュベーション工程は、約5秒から数時間まで(必要に応じて、約5分から約24時間まで)変動し得る。しかし、インキュベーション時間は、このアッセイの構成、抗原、溶液の体積、濃度などに依存する。通常、アッセイは周囲温度で実行されるが、例えば、10℃から40℃の温度の範囲にわたって実行され得る。
【0139】
(非競合アッセイ構成)
サンプル中の、T1R3を含む味レセプターを検出するためのイムノアッセイは、競合アッセイかまたは非競合アッセイのいずれかであり得る。非競合イムノアッセイは、抗原の量が直接測定されるアッセイである。1つの好ましい「サンドイッチ」アッセイにおいて、例えば、抗T1R3を含む味レセプターの抗体は、それらが固定化される固体の基質に直接的に結合され得る。次いで、これらの固定化された抗体は、このテストサンプルに存在するT1R3を含む味レセプターを捕捉する。従って、固定化された、T1R3を含む味レセプターは、次いで、標識性因子(例えば、標識を保有する第二のT1R3を含む味レセプター抗体)によって結合される。あるいは、この第二の抗体は、標識を欠いていてもよいが、それは代わりに、第二の抗体が由来する種の抗体に特異的な、標識された第三の抗体によって結合され得る。第二または第三の抗体は、代表的に、検出可能部分を提供するために、別の分子(例えば、ストレプトアビジン)が特異的に結合する検出可能部分(例えば、ビオチン)によって改変され得る。
【0140】
(競合アッセイ形式)
競合アッセイにおいて、このサンプルに存在するT1R3を含む味レセプターの量は、サンプルに存在する未知のT1R3を含む味レセプターによって抗T1R3を含む味レセプター抗体から置換された(追いやられた)、既知の添加された(外因性)T1R3を含む味レセプターの量を測定することによって間接的に測定される。一競合アッセイにおいて、既知の量のT1R3を含む味レセプターがサンプルに添加され、次いで、このサンプルはT1R3を含む味レセプターに特異的に結合する抗体に接触される。抗体に結合する外因性T1R3を含む味レセプターの量は、このサンプルに存在するT1R3を含む味レセプターの濃度に反比例する。特に好ましい実施形態において、この抗体は、固体の基質上に固定化される。抗体に結合するT1R3を含む味レセプターの量は、T1R3を含む味レセプター/抗体複合体に存在するT1R3を含む味レセプターの量を測定することによってか、または代替的に複合体化されていない残りのタンパク質の量を測定することによってかのいずれかで決定され得る。T1R3を含む味レセプターの量は、標識されたT1R3を含む味レセプター分子を提供することによって検出され得る。
【0141】
ハプテン阻害アッセイは、別の好ましい競合アッセイである。このアッセイにおいて、既知のT1R3を含む味レセプターは、固体の基質上に固定化される。既知の量の抗T1R3を含む味レセプターの抗体が、このサンプルに添加され、次いで、このサンプルは、固定化されたT1R3を含む味レセプターに接触される。既知の固定化されたT1R3を含む味レセプターに結合された抗T1R3を含む味レセプターの抗体の量は、このサンプルに存在するT1R3を含む味レセプターの量に反比例する。ふたたび、固定化された抗体の量は、抗体の固定化された分画かもしくは溶液中に残存する抗体の分画のいずれかを検出することによって検出され得る。検出は、抗体が標識化される直接的なものであるか、または上述のように抗体に特異的に結合する標識化部分のその後の添加による間接的なものであり得る。
【0142】
(交差反応性決定)
競合結合形式におけるイムノアッセイはまた、交差反応性を決定するために使用され得る。例えば、T1R3を含む味レセプターは、固体の支持体に固定化され得る。固定化された抗原への抗血清の結合について競合する、タンパク質(例えば、T1R3を含む味レセプターおよびホモログ)が、アッセイに添加される。この固定化されたタンパク質への抗血清の結合について競合する添加されたタンパク質の能力は、それ自身と競合するT1R3を含む味レセプターの能力と比較される。上記タンパク質のパーセント交差反応性は、標準的な計算を使用して計算される。上に列挙された添加されたタンパク質のそれぞれと10%未満の交差反応性を有する抗血清が、選択されそして集められる。この交差反応する抗体は、添加された考えられたタンパク質(added considered protein)(例えば、遠縁のホモログ)との免疫吸収によって、集められた抗血清から必要に応じて除去される。
【0143】
次いで、上記免疫吸収されそして集められた抗血清を、第二のタンパク質(おそらくT1R3を含む味レセプターの対立遺伝子または多型改変体と考えられる)をその免疫原タンパク質と比較するために、上述のような競合結合イムノアッセイにおいて使用する。この比較を行うために、この2つのタンパク質は、それぞれ、広範な濃度でアッセイされ、そして固定化されたタンパク質へのこの抗血清の結合の50%を阻害するために必要とされる各タンパク質の量が決定される。結合の50%を阻害するために必要とされる第二のタンパク質の量が、結合の50%を阻害するために必要とされるT1R3を含む味レセプターの量の10倍未満の場合には、この第二のタンパク質は、T1R3を含む味レセプター免疫原に対して産生されたポリクローナル抗体に特異的に結合するといわれる。
【0144】
(他のアッセイ形式)
ウェスタンブロット(イムノブロット)分析が、このサンプル中のT1R3を含む味レセプターの存在を検出および定量化するために使用される。この技術は、概して、分子量に基づくゲル電気泳動によるサンプルタンパク質を分離する工程、この分離されたタンパク質を適切な固体の支持体(例えば、ニトロセルロースフィルター、ナイロンフィルター、または誘導化ナイロンフィルター)へ移動する工程、およびT1R3を含む味レセプターに特異的に結合する抗体とともにそのサンプルをインキュベートする工程を包含する。抗T1R3を含む味レセプターの抗体は、固体の支持体上のT1R3を含む味レセプターに特異的に結合する。これらの抗体は、直接的に標識化され得るか、または代替的に、抗T1R3を含む味レセプターの抗体に特異的に結合する、標識化抗体(例えば、標識化ヒツジ抗マウス抗体)を使用して、その後検出され得る。
【0145】
他のアッセイ形式は、特定の分子(例えば、抗体)に結合しそしてカプセル化された試薬またはマーカーを放出するように設計されたリポソームを使用する、リポソームイムノアッセイ(LIA)を含む。次いで、放出された化学物質は、標準的な技術に従って検出される(Monroeら、Amer.Clin.Prod.Rev.5:34−41(1986)を参照のこと)。
【0146】
(非特異的結合の減少)
当業者は、多くの場合、イムノアッセイにおいて非特異的な結合を最小限にすることが望ましいことを理解する。特に、このアッセイが、固体の基質上で固定された抗原または抗体に関連する場合、その基質への非特異的な結合の量を最小限にすることが望ましい。そのような非特異的な結合を減少する手段は、当業者に周知である。代表的に、この技術は、タンパク様の組成物でその基質をコーティングする工程を包含する。特に、タンパク質組成物(例えば、ウシ血清アルブミン(BSA)、無脂粉乳、およびゼラチン)が、広く使用され、粉乳が最も好まれている。
【0147】
(標識)
このアッセイで使用される特定の標識または検出可能なグループは、このアッセイにおいて使用された抗体の特異的な結合を著しく干渉しない限り、本発明の重要な局面ではない。検出可能なグループは、検出可能な物理的もしくは化学的特性を有する任意の物質であり得る。そのような検出可能な標識は、イムノアッセイの分野においてよく開発され、そして概して、そのような方法において有用な任意の標識のほとんどが、本発明に適用され得る。従って、標識は、分光的手段、光化学的手段、生化学的手段、免疫化学的手段、電気的手段、光学的手段、または化学的手段によって検出可能な任意の組成物である。本発明において有用な標識としては、磁気ビーズ(例えば、DYNABEADSTM)、蛍光染料(例えば、フルオレセインイソチオシアネート、テキサスレッド、ローダミンなど)、放射性標識(例えば、3H、125I、35S、14C、または32P)、酵素(例えば、西洋ワサビペルオキシダーゼ、アルカリホスファターゼおよびELISAにおいて一般に使用される他の酵素)、および比色標識(例えば、コロイド金または着色ガラスまたはプラスチックビーズ(例えば、ポリスチレン、ポリプロピレン、ラテックスなど))が挙げられる。
【0148】
その標識は、当該分野に周知の方法によるアッセイの所望の構成要素に直接的にまたは間接的に結合され得る。上に示したように、広範な標識が使用され得、その標識の選択は、必要とされる感度、化合物との接合の容易さ、安定性の必要条件、利用可能な機器、および処理設備に依存する。
【0149】
非放射性標識は、多くの場合、間接的な手段によって付着される。概して、リガンド分子(例えば、ビオチン)は、分子に共有結合性に結合される。次いで、このリガンドは、別の分子(例えば、ストレプトアビジン)に結合し、この分子は、生来に検出可能であるかまたはシグナル系(例えば、検出可能な酵素、蛍光化合物、または化学発光化合物)に共有結合性に結合される。このリガンドおよびこれらの標的は、T1R3を含む味レセプターを認識する抗体、または抗T1R3を含む味レセプターを認識する二次抗体との任意の適切な組み合わせにおいて使用され得る。
【0150】
この分子はまた、シグナルを生成する化合物に直接的に接合され得る(例えば、酵素または発蛍光団との接合によって)。標識として目的の酵素は、主に、ヒドロラーゼ(特に、ホスファターゼ、エステラーゼ、およびグリコシダーゼ)、またはオキシドターゼ(oxidotase)(特に、ペルオキシダーゼ)である。蛍光化合物としては、フルオレセインおよびその誘導体、ローダミンおよびその誘導体、ダンシル、ウンベリフェロンなどが挙げられる。化学発光化合物としては、ルシフェリン、および2,3−ジヒドロフタラジンジオン(dihydrophthalazinedione)(例えば、ルミノール)が挙げられる。使用され得る種々の標識化またはシグナル生成系の概説については、米国特許第4,391,904号を参照のこと。
【0151】
標識を検出する手段は、当業者に周知である。従って、例えば、標識が放射性標識である場合、検出のための手段としては、シンチレーション計数器あるいはオートラジオグラフィー中のような写真フィルムが挙げられる。標識が蛍光標識である場合、それは、適切な光の波長で蛍光色素を励起させ、そして結果として生じる蛍光を検出することによって検出され得る。この蛍光は、写真フィルムによって、電子検出器(例えば、電荷結合デバイス(CCD)または光電子増倍管など)の使用によって、視覚的に検出され得る。同様に、酵素標識は、酵素のための適切な基質を提供し、そして結果として生じた反応産物を検出することによって検出され得る。最終的に、単純な比色標識が、単純に、標識に関連する色を観察することによって、検出され得る。従って、種々のディップスティックアッセイ(dipstick assay)において、接合された金は、多くの場合、ピンク色に見えるが、種々の接合されたビーズは、そのビーズの色に見える。
【0152】
いくつかのアッセイ形式は、標識化構成要素の使用を必要としない。例えば、凝集アッセイ(agglutination assay)は、標的抗体の存在を検出するために使用され得る。この場合において、抗原をコーティングした粒子は、標的抗体を含むサンプルによって凝集される。この形式において、この構成要素は、標識化されることを必要とせず、そして標的抗体の存在は、単純な視覚的検査によって検出される。
【0153】
(薬学的組成物および投与)
薬学的に受容可能なキャリアは、投与される特定の組成物(例えば、核酸、オリゴヌクレオチド、アミノ酸、タンパク質、ペプチド、小有機分子、脂質、炭水化物、単糖類、二糖類、または多糖類、粒子、または変換された細胞)によって、ならびに、その組成物を投与するために使用される特定の方法によって部分的に決定される。従って、本発明の薬学的組成物の多岐にわたる適切な処方物が存在する(例えば、Remington’s Pharmaceutical Sciences,第17版、1989年、を参照のこと)。投与は、任意の都合のよい様式で行われ得る(例えば、注射、経口投与、吸引、経皮的適用、または直腸投与)。
【0154】
経口投与に適している処方物は、(a)溶液(例えば、希釈液(例えば、水、生理食塩水、またはPEG 400)に懸濁している有効量のパッケージ化された核酸);(b)カプセル、小袋(sachet)または錠剤、それぞれが所定の量の活性成分(液体、固体、顆粒またはゼラチンとして)を含んでいる;(c)適切な液体中の懸濁剤;ならびに(d)適切なエマルジョンからなり得る。錠剤形態としては、1つ以上のラクトース、スクロース、マンニトール、ソルビトール、リン酸カルシウム、コーンスターチ、ポテトデンプン、微晶質セルロース、ゼラチン、コロイド二酸化ケイ素、滑石、ステアリン酸マグネシウム、ステアリン酸、および他の賦形剤、着色料、充填剤、結合剤、希釈液、緩衝剤、湿潤剤、保存薬、香料、染料、崩壊剤、および薬学的に適合するキャリアが挙げられ得る。ロゼンジ形態は、香料(例えば、スクロース)中に活性成分を、ならびに不活性塩基中に活性成分を含む香錠(例えば、活性成分に加えて当該分野において公知のキャリアを含んでいる、ゼラチンおよびグリセリンまたはスクロースおよびアカシアエマルジョン、ゲルなど)を含み得る。
【0155】
最適な化合物は、単独でまたは他の適切な構成要素との組み合わせにおいて、吸引を介して投与される、エーロゾル処方物中に作製され得る(すなわち、それらは「噴霧」され得る)。エーロゾル処方物は、加圧された受容可能な推進薬(例えば、ジクロロジフルオロメタン、プロパン、窒素など)中に配置され得る。
【0156】
非経口投与(例えば、関節内(関節(joint)内の)経路、静脈内経路、筋肉経路、皮内経路、腹腔内経路、および皮下経路)に適している処方物は、水溶性および非水溶性の等張無菌注入溶液を含み、この溶液は、抗酸化剤、緩衝液、静菌薬、ならびに意図されるレシピエントの血液とその処方物を等張にする溶質、ならびに水溶性および非水溶性の無菌懸濁液を含み得、この懸濁液は、懸濁剤、溶解剤、シックニング剤(thickening agent)、安定剤、および保存剤を含み得る。本発明の実施において、組成物は、例えば、経口的に、局所的に、腹腔内に、嚢内に(intravesically)、または髄膜内に、静脈内注入によって投与され得る。非経口的投与および静脈内投与が、好ましい投与の方法である。推薦される処方物は、単位用量または複数用量の密封された容器(例えば、アンプルおよびバイアル)中に提供され得る。
【0157】
注入溶液および懸濁液は、既に記載された種類の無菌の粉末、顆粒、および錠剤から調製され得る。エクスビボ治療用の核酸によって変換された細胞もまた、上述のように静脈内にまたは非経口的に投与され得る。
【0158】
患者に投与される用量は、本発明の状況において、長期にわたって患者の有益な治療上の応答をもたらすのに十分であるべきである。この用量は、特定の使用されたベクターの有効性および患者の状態、ならびに処置される患者の体重または表面積によって決定される。用量の大きさはまた、特定のベクターの投与に伴う任意の有害な副作用の存在、性質、および程度、あるいは特定の患者中の変換された細胞型によって決定される。
【0159】
T1R3を含む味レセプターの減少した発現もしくは異常な発現のための状態の処置または予防において投与されるベクターの有効量を決定する際に、医師は、ベクターの循環している血しょうレベル、ベクターの毒性、疾患の進行、および抗ベクター抗体の産生を評価する。概して、ベクター由来の裸の核酸に相当する用量は、代表的な70キログラムの患者について約1μgから100μgまでであり、そしてレトロウイルス粒子を含むベクターの用量は、等量の治療用核酸を産生するために計算される。
【0160】
投与について、本発明の化合物および変換された細胞は、集団のおよび総合的に健康な患者に適用された場合の、インヒビター、ベクター、または変換された細胞型のLD−50、ならびに、種々の濃度のインヒビター、ベクターまたは細胞型の副作用によって決定された頻度で投与され得る。投与は、一回かまたは分割した投与を介して遂行され得る。
【0161】
(細胞トランスフェクションおよび遺伝子治療)
本発明は、インビトロおよびインビボでの細胞のトランスフェクションのためのT1R3を含む味レセプターの核酸を提供する。これらの核酸は、以下に記載されるような標的細胞および生物体のトランスフェクションのための複数の周知のベクターのいずれかへ挿入され得る。この核酸は、そのベクターと標的細胞との相互作用を通じてエクスビボまたはインビボで細胞にトランスフェクトされる。次いで、この核酸は、プロモーターの制御下で、T1Rファミリーの2つのメンバーを同時発現させ、それにより、T1R3を含む味レセプターの欠如、部分的不活性化、または異常な発現の効果を緩和することによって、本発明のT1R3を含む味レセプターを発現する。この組成物は、患者に治療に対する応答を引き起こさせるために十分な量において、患者に投与される。このことを達成するのに適切な量は、「治療有効用量または治療有効量」として規定される。
【0162】
そのような遺伝子治療手順は、複数の状況において、後天的および先天的な遺伝子欠損および他の疾患を矯正するように使用されてきた。ヒトにおいて合成遺伝子を発現する能力は、多くの重要なヒト疾患(他の治療による処置を受けられない多くの疾患を含む)の予防および/または治癒を促進する(遺伝子治療手順の概説については、Anderson,Science 256:808−813(1992);NabelおよびFelgner,TIBTECH 11:211−217(1993);MitaniおよびCaskey,TIBTECH 11:162−166(1993);Mulligan,Science 926−932 (1993);Dillon,TIBTECH 11:167−175(1993);Miller,Nature 357:455−460(1992);Van Brunt,Biotechnology 6(10):1149−1154 (1998);Vigne,Restorative Neurology and Neuroscience 8:35−36(1995);KremerおよびPerricaudet,British Medical Bulletin 51(1):31−44 (1995);Current Topics in Microbiology and Immunology(DoerflerおよびBohm(編)、1995年)中のHaddadaら;およびYuら、Gene Therapy 1:13−26(1994)を参照のこと)。
【0163】
(参考文献)
【0164】
【化001】
【0165】
【化002】
【0166】
【化003】
【0167】
【化004】
【0168】
【化005】
【0169】
【化006】
【実施例】
【0170】
以下の実施例は、説明のために提供されるのであり、本発明の特許請求の範囲を制限するためのものではない。
【0171】
(実施例1)
(結果)
(T1R1 KOマウス、T1R2 KOマウスおよびT1R3 KOマウスの作製)
T1Rレセプターの発現は、舌および口蓋中の味細胞の重複していない3つの主な集団を規定する:T1R1およびT1R3を同時発現する細胞(T1R1+T1R3)、T1R2およびT1R3を同時発現する細胞(T1R2+T1R3)、ならびにT1R3のみを発現する細胞(Nelson,G.ら、Cell,106,381−390(2001))。HEK細胞のT1Rの異種発現の研究は、T1R1およびT1R3は組み合わさって、広範囲にわたるように調節されたL−アミノ酸レセプターを形成し、他方で、T1R2およびT1R3の同時発現は、甘味化合物の全てのクラスに応答する甘味レセプターを生成することを示した(Nelson,G.ら、Cell,106,381−390(2001)、Nelson,G.ら、Nature,416,199−202(2002);Li,X.ら、Proc Natl Acad Sci U S A,99,4692−4696(2002))。T1R3がインビボで甘味レセプターおよびアミノ酸味レセプターの共通の構成要素として機能する場合、このGPCRのノックアウトは、甘味受容能力およびアミノ酸味受容能力を欠くマウスを生成するはずである。対照的に、T1R1またはT1R2のノックアウトは、1つの味様式を選択的に影響すると予測される。
【0172】
インビボでのT1Rの役割を規定するために、本発明者らは、レセプター機能にとって本質的なドメインをコードするエキソンを欠失することによってそれぞれのT1Rを欠くノックアウトマウスを生成した。図1は、KO戦略を示し、そして対応しているホモ接合体KO動物において特異的T1R染色の完全な欠如を示すインサイチュハイブリダイゼーション実験を示す。任意の1つのT1Rの損失が、味細胞の生存率または一体性を影響しないことを確実にするために、本発明者らはまた、コントロールおよびKO動物中の他のT1R、T2R、PLCb2(Rossler,P.ら、Eur J Cell Biol,77,253−261(1998);Zhang,Y.ら、Cell,112,293−301(2003))およびTRPM5(Perez,C.A.ら、Nat Neurosci,5,1169−1176(2002);Zhang,Y.ら、Cell,112,293−301(2003))の発現を比較した。野生型とKO味組織との間のT1R−、T2R、PLCb2およびTRPM5陽性細胞の数においてまたはそれらの分布において、有意差は観察されなかった(図1およびデータは示されていない)。
【0173】
2つの相補的戦略を、遺伝子的に改変されたマウスの味応答をアッセイするために使用した。第一に、本発明者らは、舌の味レセプター細胞を神経支配する主要な神経(鼓索神経)の1つから味覚物質誘導性活動電位を記録した。この生理学的アッセイは、末梢における味覚系の活性をモニタリングし、そして味レセプター細胞の機能の測定を提供する。第二に、本発明者らは、標準的な長期二瓶採取嗜好性アッセイにおける味の選択を測定することによって、または多チャンネル味覚計における即時なめ応答の直接計数によって味に対する行動を調査した(Glendinning,J.I.ら、Chem Senses,27,461−474(2002);Zhang,Y.ら、Cell,112,293−301(2003);実験手順を参照のこと)。この第二の方法は、味覚物質への非常に短い暴露(全体で30分にわたる5秒の事象 対 二瓶嗜好性アッセイについては48時間)により、それゆえ、他の感覚入力の衝撃、ならびにこのアッセイからの摂取後および学習効果を最小限にするという大きな利点を有する。
【0174】
図2は、T1Rのノックアウトは、クエン酸、塩化ナトリウム、および種々の苦味味覚物質に対する生理学的応答または行動学的応答のいずれについても有意な効果を有さないことを示す。これらの結果は、苦味、塩味、および酸味の受容または知覚が、T1Rレセプターと独立した経路を通って作動することを示し、そしてさらに個々の様式が相互に独立して作動する、末梢におけるコーディングモデルを実証する。
【0175】
(T1R1+3は旨味レセプターである)
以前に、Chaudhariらは、代謝生成物産性グルタミン酸レセプター4(mGluR4t)の短縮型改変体を記載し、そしてそれが旨味レセプターとして機能することを示唆した(Chaudhari,N.ら、Nat Neurosci,3,113−119(2000))。本発明者らは,この提唱を多くの理由から満足のいくものではないと見出す。(1)このmGluR4t改変体は、表面標的に必要なmGluR4シグナル配列を欠いている。(2)この推定レセプターはまた、mGluRのグルタメート結合ドメインの結晶構造によって解明されるように、グルタメート認識のために重要なドメインの大きな分画を欠く(Kunishima,N.ら、Nature,407,971−977(2000))。(3)mGluR4t旨味シグナルは、cAMP経路を介して作動すると提唱されてきた(Abaffy,T.ら、Am J Physiol Cell Physiol,284,C1420−1428(2003);Chaudhari,N.ら、Nat Neurosci,3,113−119(2000))。しかし、アミノ酸/旨味は、PLCβ2/TRPM5依存性プロセスである(Zhang,Y.ら、Cell,112,293−301(2003))。(4)旨味は、旨味エンハンサー(IMPおよびGMP)によって強く影響されるが、mGluR4活性は影響されない。(5)最後に、mGluR4 KO動物は、旨味刺激への応答を保持する(Chaudhari,N.およびRoper,S.D.,Ann N Y Acad Sci,855,398−406(1998))。対照的に、最近の証拠は、このT1R1+3アミノ酸レセプターが、哺乳動物旨味(グルタメート)味センサーとして機能し得ることを示唆する:第一に、ヒトおよびげっ歯類T1R1+3レセプターは、げっ歯類とヒトとの間のアミノ酸味の差異を模倣する選択性および感受性の差異を示す(Nelson,G.ら、Nature,416,199−202(2002);Yoshiiら、1986)。第二に、T1R1+3活性は、IMPおよびGMP(2つの最も公知なインビボでの旨味の増強剤)によって依存的に増強される(Nelson,G.ら、Nature,416,199−202(2002);Li,X.ら、Proc Natl Acad Sci U S A,99,4692−4696(2002))。第三に,T1R1+3は、旨味アゴニストL−AspおよびL−AP4の心理物理学的に適切な濃度によって活性化される(Nelson,G.ら、Nature,416,199−202(2002);Li,X.ら、Proc Natl Acad Sci U S A,99,4692−4696(2002))。旨味におけるT1R1+3の役割を厳しく評価するために、本発明者らは、T1R1 KO動物およびT1R3 KO動物を調査した(図1を参照のこと)。
【0176】
そのNa+含有量のため、グルタミン酸一ナトリウム(MSG)は、塩味と旨味の両方を引き出す。それゆえ、本発明者らは、行動学的および電気生理学的研究において、グルタメートの塩味から塩味を単離することを可能にしたいくつかの計画を使用して旨味応答をアッセイした。これらは、MPG(グルタメートのカリウム塩)を使用してナトリウムチャネルブロッカーアミロライドの存在下でMSGをテストする工程、ならびに、旨味エンハンサーIMPの存在下または非存在下で旨味アゴニストAP4およびアスパラギン酸の全てをテストする工程を包含した。図3は、塩効果が最小限にされる場合、T1R3 KOマウスは行動学的誘引性の劇的な損失、およびそれに対応する、全ての旨味味覚物質(グルタメート、アスパルテート、グルタメート+IMP、およびIMPのみを含む)に対する生理学的応答の対応している重大な不足を有することを示す。ごく最近、Damakらは、独立して、T1R3 KO動物を作製したが、複数の旨味レセプターが、彼らのKOマウスの研究において残る顕著なMSG応答として存在するはずであると結論付けた(Damak,S.ら、Science,301,850−853(2003))。特に、KO動物のMSG応答は、IMPと完全に独立であり、IMPの増大は、旨味様式の特徴であることと矛盾する。塩効果は考慮されなかったので、本発明者らは、それらの残存する応答の多くは、旨味よりもむしろ、MSG中のNa+含有量を反映すると疑っている(図3E〜図3FのMSG+IMP対MPG+IMPまたはMSG+IMP+アミロライドに対して応答を比較すること)。
【0177】
T1R1がT1R3と組み合わさって(T1R1+3)哺乳動物旨味レセプターを生成する場合、T1R1のノックアウトもまた、全ての旨味応答を除去するはずである。図3は、まさにこの場合を示す。対照的に、これらの非常に同じ味覚物質は、コントロールとT1R2 KO動物において、正常で、強力な応答を引き起こす。合わせて、これらの結果は、T1R1+3が哺乳動物の旨味レセプターであることを証明する。
【0178】
以前に、本発明者らは、代表的な旨味味覚物質に加えて、マウスT1R1+3レセプターがまた、他のL−アミノ酸によって活性化され、そしてIMPの存在下で広範囲にわたるように調節されたL−アミノ酸センサーとして機能することを示した(Nelson,Gら、Nature,416,199−202(2002))。それゆえ、本発明者らは、IMPの存在下または非存在下におけるL−アミノ酸に対するT1R1 KO動物およびT1R3 KO動物の応答を試験した。実際、アミノ酸味覚物質に対する応答は、T1R1 KO系統およびT1R3 KO系統において重度に欠陥があるが、T1R2 KO系統においては欠陥がなく(図3)、広範なL−アミノ酸およびIMPのための味レセプターとしてのT1R1+3ヘテロマーGPCR複合体を堅固に確立している。興味深いことに、本発明者らがヒトにとって甘く感じる過剰に高濃度のL−アミノ酸(例えば、>300mM Ala、Ser、およびThr)をアッセイした場合、T1R1 KO動物は、わずかな残りの誘引性を保持したが、T1R3 KOマウスでは保持しなかった(図3のパネルDを参照のこと);これらの微量の行動学的応答は、T1R2+3甘味レセプターの活性化を反映する可能性が高い(Nelson,G.ら、Cell,106,381−390(2001);以下を参照のこと)。
【0179】
(T1R2+3およびT1R3は、甘味の受容および知覚のために必要とされる)
T1R2+3は、細胞ベースのアッセイにおいて、天然の糖、人工甘味料、D−アミノ酸および甘味タンパク質を含む種々の化学物質のクラスのためのヘテロマーレセプターとして機能する(Nelson,G.ら、Cell,106,381−390(2001);Li,X.ら、Proc Natl Acad Sci U S A,99,4692−4696(2002))。しかし、複数の研究は、動物が甘味レセプターの別個の型を発現し得ることを示唆している(Schiffman,S.S.ら、Pharmacol Biochem Behav,15,377−388(1981);Ninomiya,Y.ら、J Neurophysiol,81,3087−3091(1999))。インビボでのT1R2+3の役割を規定するために、本発明者らは、機能的T1R2タンパク質およびT1R3タンパク質を欠くノックアウトマウスの甘味応答を調査した。図4および図5は、全てのクラスの甘味味覚物質への応答が、T1R2ノックアウト系統+T1R3ノックアウト系統において劇的に弱められていることを示す。本発明者らは、広範なパネルの糖、人工甘味料およびD−アミノ酸を試験し、そして全ての場合において、応答が重度に障害されていた:行動学的誘引性がほとんど消失し、そして神経応答も大いに減少する。これらの結果は、T1R2+T1R3がインビボでの主要な甘味センサーであることを確証する。
【0180】
特に、非常に高濃度(>300mM)の天然の糖は、T1R2とT1R3との両方の系統において中程度だが検出可能な誘引的応答を誘発したが人工甘味料またはD−アミノ酸では誘発しなかった。従って、さらなる甘味レセプター(すなわち、T1R非依存経路)が存在するか、またはT1R2およびT1R3はまた、それらのヘテロマーのパートナーの存在しない状態で天然の糖に対する低親和性レセプターとしてそれら自身で機能し得る。実際、残存する応答がT1R2またはT1R3に起因する場合、これらのGPCRのダブルノックアウトは、全ての甘味応答を除去するはずである。T1R2およびT1R3遺伝子座は、第4染色体の遠位末端において連鎖されるので(Nelson,G.ら、Cell,106,381−390(2001))、本発明者らは、組み換え型T1R2 KO、T1R3 KOマウスを作製し、次いで、これらを生理学的および行動学的に試験した。図4および図5(赤線)は、T1R2、T1R3ダブルKOマウスが高濃度の糖への全ての応答を失ったことを示す。合わせて、これらの結果は、T1Rのコンビナトリアル集合物のインビボでの重要性を示し、そして全ての甘味受容はT1R2およびT1R3のGPCRを介して作動することを示す。
【0181】
T1R2またはT1R3ホモダイマーレセプターは、野生型マウスの甘味感知において重要な役割を果たすのであろうか?T1R2は、常にT1R3を含む細胞中で発現される(T1R2+3細胞;Nelson,G.ら、Cell,106,381−390(2001))。それゆえ、たとえいくつかのT1R2がこれらの細胞中でT1R3と会合しない場合でも、甘味味覚物質のためのT1R2+3のヘテロマーのレセプターのより高い親和性は、細胞応答を支配しやすい。対照的に、本発明者らは、既に、T1R3がまた、T1R1およびT1R2とは独立して、舌および口蓋上皮の有意な割合の細胞において見出されることを報告した(T1R3単独の細胞;Nelson,G.ら、Cell,106,381−390(2001))。このクラスの細胞は、高濃度の糖を検出し、そしてそれに応答するさらなる手段を動物に提供し得る。T1R3単独で天然の糖のための低い親和性のレセプターとして機能し得ることを示すために、本発明者らは、安定的にT1R3およびT1Rと共役するように操作された最適なGタンパク質キメラを発現するHEK細胞を生成した(実験手順を参照のこと)。図6は、実際、T1R3は単独で非常に高濃度の天然の糖に応答するが、低濃度(<300mM)または人工甘味料には応答しないことを示す。これらの結果は、T1R3が低い親和性の糖レセプターであることを確証し、そしてT1R3単独の細胞はインビボでさらなる甘味センサーとして機能するという仮定を支持する(Nelson,G.ら、Cell,106,381−390(2001))。天然甘味料および人工甘味料を感知するこの部分的な細胞の分離が、なぜ人工甘味料が、高濃度の天然の糖によってもたらされる甘さのレベルを達成しないかを説明することを補助し得る(すなわち、T1R2+3細胞の活性化 対 T1R2+3細胞およびT1R3単独の細胞の活性化)。
【0182】
(T1R2は種特異的甘味嗜好性の範囲を定める)
ヒトは、げっ歯類が味わい得ない複数の天然甘味料および人工甘味料を味わい得る。例えば、モネリン、タウマチン、アスパルテーム、およびネオへスペリジンジヒドロカルコンは、ヒトにはmM以下の濃度で甘いと感じさせるが、他方で、げっ歯類は100倍高い濃度でさえも嗜好性を示さない(Danilova,V.ら、J Neurophysiol,80,2102−2112(1998))。以前に、本発明者らは、げっ歯類T1RおよびヒトT1Rがそれらのアミノ酸配列において30%を超えて類似していないことを報告し、そしてそのような違いは甘味検出における種に特異的な選択性の基礎になっていると仮定した(Nelson,G.ら、Cell,106,381−390(2001);Nelson,G.ら、Nature,416,199−202(2002))。T1R2は排他的に甘味検出に関わるが、他方で、T1R3は甘味認識とアミノ酸認識との両方に関与しているので、本発明者らは、T1R2がインビボでの甘味選択性の特に重要な決定要素であると理由付けした。それゆえ、本発明者らは、T1R2 KOマウス中にヒトT1R2遺伝子を導入することが甘味応答を回復し、そして甘味応答を「ヒト化する」はずであると予測した。
【0183】
本発明者らは、T1R2 KO対立遺伝子であるが、代わりにネイティブ「T1R2細胞」においてヒトT1R2トランスジーンを発現させたホモ接合であるマウスを生成した。T1R2調節配列を含む12kbのゲノムクローンを、hT1R2全長cDNAに融合し、そしてT1R2 KOマウスに導入した。複数の独立した系統を、甘味味覚物質に対するそれらの選択性および感受性についてアッセイした。hTlR2の発現を調査するために、本発明者らは、野生型mT1R2対立遺伝子を保有するトランスジェニック動物で二色蛍光インサイチュハイブリダイゼーション実験を実行した。図7(パネルA〜D)は、ヒトT1R2がT1R2発現細胞中に選択的に発現され、そして甘味機能を効果的に回復することを示す。より重要なことに、ヒトトランスジーンは、ここで、ヒトに甘いと感じさせるが、通常げっ歯類には誘引性ではないいくつかの化合物を検出し、そして応答する能力をこれらのマウスに与える;これらの化合物としては、アスパルテーム、グリチルリジン酸および甘味タンパク質(タウマチンおよびモネリン)が挙げられる。興味深いことに、ヒト化T1R2マウスは、強度の甘味化合物(ネオへスペリジンジヒドロカルコン)になお応答せず、ヒトT1R2およびマウスT1R3 GPCRでトランスフェクトされたHEK細胞も応答しない。しかし、細胞をヒトT1R2およびヒトT1R3でトランスフェクトする場合、この細胞は、ネオへスペリジンジヒドロカルコンに強く応答する。まとめると、これらの実験は、げっ歯類とヒトとの間の甘味選択性および甘味特異性の違いの重要な決定要素がT1Rであることを証明し、そしてさらに、T1R2+3およびT1R2発現細胞をインビボでの甘味の主要なメディエーターとして実証する。最後に、本発明者らは、T1R2とT1R3との両方の多型性が、ヒトの個々の甘味嗜好性の重要な決定要素であることを提案する。
【0184】
(T1R2を発現する細胞は、インビボでの行動学的誘引性をコードする)
味レセプターの活性化は、動物の個々の行動学的応答を誘発する。例えば、T1R2+3レセプターの興奮は、マウスの糖および甘味化合物に対する行動学的誘引性を刺激する。この応答は、これらが発現されるレセプターまたは細胞の特性であろうか?この質問に答える1つの手段は、T1R2+3細胞において味覚系に関連のない新規のレセプターを発現することであり、そしてその選択的刺激が誘引性応答を誘発するか否かを調査することである(Troemel,E.R.ら、Cell 91,161−169(1997))。
【0185】
本発明者らのアプローチは、T1R2+3細胞の内因性シグナル伝達経路へ共役し得るが、非天然のリガンドによって活性化し得るのみであるGPCRの発現を標的にすることであった。レセプター発現の前後にまさにその同じ動物の味応答を調査するために、本発明者らは誘導系を利用した。これを達成するために、本発明者らはトランスジェニックマウスを使用し、このマウスの中で、合成リガンドによってのみ活性化される改変k−オピオイドレセプター(RASSL;Redfern,C.H.ら、Nat Biotechnol,17,165−169(1999))を、Tet−on誘導系の制御の下、T1R2発現細胞を標的にした(実験手順を参照のこと)。
【0186】
図7Eは、非誘導動物、またはドキシサイクリンで処理された野生型コントロールが、完全にk−オピオイドアゴニスト(スピラドリン)に非感受性であることを示す。注目すべきことに、T1R2細胞中のRASSL発現の誘導が、ここで、ナノモルの濃度のスピラドリンに強力に誘引される動物を生成する(図7、赤線)。従って、本発明者らは、T1R2発現細胞の活性化は(それらが発現するレセプターよりも)、マウスにおいて行動学的誘引性を決定することを結論付ける。さらに、これらの結果は、1つの細胞型を活性化することが、特異的味応答を誘発するのに十分であることを明白に示す;それゆえ、活性のコンビナトリアルパターンまたは一時的コーディングを必要とするモデルは、T1R2発現細胞によって媒介される誘引性を説明するために必要とされない。拡張することによって、本発明者らは、上記レセプターの性質に関わらず、ヒトT1R2+3細胞におけるこれらの味シグナル伝達経路の活性が甘味を誘発することを示唆する。
【0187】
複数のレセプターが、哺乳動物の甘味および旨味を媒介するものと提案されている。特に、これらの2つの様式の各々の範囲内でさえも、細胞を透過する味覚物質によって直接的に活性化される細胞内標的を引き起こす数個のGPCR、イオンチャネル、およびモデルが仮定されてきた(Kinnamon,S.C.Neuron,25,507−510(2000);Margolskee,R.F.,J Biol Chem,277,1−4(2002))。本発明者らは、哺乳動物の甘味および旨味のためのレセプターがT1Rであることを証明するために、細胞ベースのアッセイ、遺伝学的アプローチ、生理学的アプローチ、および行動学的アプローチの組み合わせを使用してきた;旨味はT1R1+3のヘテロマーのGPCRによって媒介され、そして甘味は2つのT1Rベースのレセプター(T1R2およびT1R3)(T1R2+3、そして最も可能性の高いものとしては、ホモダイマーのT1R3)によって媒介される。それゆえ、甘味およびアミノ酸味(旨味)(行動学的誘引性を誘発する2つの化学感覚性入力)は、共通のレセプターレパートリーおよび進化の起源を共有する。
【0188】
ヒトT1R1+3レセプターは、他のアミノ酸よりもはるかに効果的にグルタメートおよびアスパラテートによって活性化される(Li,X.ら、Proc Natl Acad Sci U S A,99,4692−4696(2002))。対照的に、マウスT1R1+3レセプターは、細胞ベースのアッセイ(Nelson,G.ら、Nature,416,199−202(2002))とインビボ(本紙)との両方において非常に広範囲のL−アミノ酸を認識する。このT1R1+3レセプターの進化上の役割が、タンパク質の豊富な食料に対する誘引性応答を媒介することであった場合、当業者は、霊長類における2つのアミノ酸のみに対するレセプターの選択性の調整が、実質的にこれらの栄養が豊富な食物を検出する能力を変更したか否かということを問題とし得る。そのようなアミノ酸は、通常、複合体の混合物として見出され、いずれのものも概して適切であり、従って、この味覚物質の選択性の「限定」は食事に影響を顕著に有さないはずである。同じ細胞および同じレセプターが、グルタメート、他のアミノ酸およびIMPを認識すると仮定すると、本発明者らは、げっ歯類において、この旨味様式が、ほとんどのL−アミノ酸を含むように一般化されなければならないこと、そしてげっ歯類における明白なグルタメート味のまさにその概念(Chaudhari,N.ら、Nat Neurosci,3,113−119(2000);Lin,W.ら、J Neurophysiol,89,1434−1439(2003))が再評価される必要があることを示唆する
スプーン一杯の糖または数錠の人工甘味料?本発明者らの日々の経験が、天然の甘味料および人工甘味料は同じ味がしないことを本発明者らに伝えている。この原稿において、本発明者らは、T1R2およびT1R3が、全ての甘味感知の原因であることを示した。それらは、どのように甘味味覚物質の間の知覚された味の違いを明らかにするのであろうか?多くの甘味料は、他の味様式(のためのレセプターを活性化する可能性がある例えば、サッカリンの苦い後味を説明するT2R苦味感知細胞(データは示さず))。それゆえ、1つの甘味分子種の「味」さえも、異なる味様式に調節される細胞の組み合わさった活性を反映し得、そして甘味感知細胞の活性のみを反映し得るわけではない。本発明者らはまた、より高いが、なお生理学的に関連のある濃度の糖(>300mM)において、天然甘味料および人工甘味料が部分的に重複するが、なお別個の甘味レセプター型(T1R2+3およびT1R3のみ)を活性化することを示してきた。
【0189】
本発明者らは、T1Rが2つの主要な誘引性味様式のメディエーターであることを示し、そして、RASSLオピオイドレセプターを発現するマウスがスピラドリン(通常、味がなくそして栄養的に関連のない化合物)に強力に誘引されるようになったことを示し、味わうことが信じることであることを証明した。ここで、苦味、甘味、および旨味のための細胞およびレセプターの発見および機能的特徴付けは、どのように味が末梢においてコードされるかという説得力のある視点を提供する、専用の味レセプター細胞が誘引される行動および回避的な行動を媒介する(例えば、Zhang,Y.ら、Cell,112,293−301(2003)を参照のこと)。
【0190】
(実験手順)
(T1R1、T1R2およびT1R3の遺伝子標的)
T1Rノックアウト動物を作製するために使用される戦略を、図1に示す。T1R1について、7つの予測されたレセプターの膜貫通ドメインをコードするエキソン6を、PGK−neorカセットによって置換した。R1 ES細胞におけるホモログの組み換えを、標的構築物の外でのプローブとの、診断サザンハイブリゼーションによって検出した。2つの標的ESクローンを、C57BL/6胚盤胞中へ注入した。キメラマウスを、C57BL/6マウスと交配させ、そしてホモ接合のノックアウトコロニーを確立する前に、2世代にわたってC57BL/6マウスと後代戻し交雑を行った。
【0191】
T1R2について、類似のアプローチが、エキソン5およびエキソン6を除去した(図1を参照のこと)。キメラ動物を、C57BL/6マウスと交配させ、そして4世代にわたってC57BL/6マウスと後代戻し交雑を行った。T1R3味覚因子(taster)(C57)および非味覚因子(129)対立遺伝子(Nelson,G.ら、Cell,106,381−390(2001))を、EcoRI多型性(T1R3の開始ATGの約12kb上流)に基づき同定した。この研究において使用されたT1R2ノックアウト動物の全てが、T1R3の味覚因子対立遺伝子を保有していた。しかし、非味覚因子T1R3対立遺伝子とホモ接合のT1R2 KOマウスの研究は、定性的に類似の結果を生じた(データは示さず)。T1R3 KOノックアウト動物を作製するために、本発明者らは、エキソン1をエキソン5に置換し、PGK−neorカセットを有するN末端細胞外ドメインをコードした(図1を参照のこと)。キメラマウスを、C57BL/6マウスと交配させ、2世代にわたってC57BL/6マウスと後代戻し交雑を行った。
【0192】
T1Rノックアウトは、正常な生存率、体重、総合的な解剖学的構造および一般的行動を有する。同様に、味レセプター細胞は、全てのノックアウトバックグラウンドにおいて形態学的におよび数的に正常に見える。
【0193】
(インサイチュハイブリダイゼーション)
新鮮凍結切片(16μm/切片)を、シラン化スライドに付着させ、そして既に記載したようにインサイチュハイブリダイゼーションまたは免疫組織化学のために調製した(Hoon,M.A.ら、Cell.,96,541−551(1999))。単一標識インサイチュハイブリダイゼーション(Single label in situ hybridization)を、ジゴキシゲニンで標識されたプローブを使用して実行した;T1R1およびT1R2プローブは、予測された膜貫通ドメインに対してだが、他方で、T1R3およびRASSL(Redfern,C.H.ら、Nat Biotechnol,17,165−169(1999))プローブは完全なコーディング配列を利用した。二重標識蛍光検出は、高いストリンジェンシーにおいて(ハイブリダイゼーション、5×SSC、50%ホルムアミド、65〜72℃;洗浄、0.2×SSC、72℃)、フルオレセイン(全長hT1R2)プローブおよびジゴキシゲニン(全長mT1R2)プローブを使用した。ハイブリダイゼーションを、明白な蛍光基質を用いて検出し(Adler,E.ら、Cell,100,693−702(2000))、そして標識化の特異性をT1R2ノックアウトおよび非トランスジェニックコントロールを使用して確認した。
【0194】
(ヒトT1R2およびRASSLを発現するトランスジェニックマウスの作製)
マウスT1R2の上流の約12kbのゲノムフラグメントを、ヒトT1R2 cDNAに、ならびに、逆テトラサイクリン依存性トランスアクチベーター(rtTA)構築物に融合した(Gossen,M.ら、Curr Opin Biotechnol,5,516−520(1994))。トランスジェニック系統を、FVB/Nマウス由来の接合体の前核注入によって生成した。3つの独立したヒトT1R2トランスジェニック系統は、アスパルテーム(10mM)に対する行動学的誘引性を示した。1つの系統をT1R2ノックアウトバックグラウンド中へ交配し、そして味応答およびトランスジーン発現についてアッセイした。TlR2細胞の外では発現を検出しなかった。TlR2−rtTAトランスジェニック系統をtetO−Ro1/tetO−lacZトランスジェニック動物と交配した(Redfern,C.H.ら、Nat Biotechnol,17,165−169(1999))。ダブルヘテロ接合の子孫を、3日間、ドキシサイクリン処理(6gm/kg)(Bio−Serv)によって誘導し(Gogos,J.A.ら、Cell,103,609−620(2000))、そしてガラクトシダーゼ活性(Zack,D.J.ら、Neuron 6,187−199(1991))および舌および口蓋のRASSL発現について調査した。適切なβガラクトシダーゼ染色およびRASSL発現パターンを示す系統を、行動学的アッセイのために選択した。
【0195】
(行動学的アッセイ)
味覚行動を、マルチチャネル味覚計(Davis MS160−Mouse gustometer;DiLog Instruments,Tallahassee,FL)において即時なめ応答を数えることによって味嗜好性を直接的に測定する短期アッセイを使用してアッセイした。マウスを、既に記載したように訓練し、そして試験した(Zhang,Y.ら、Cell,112,293−301(2003))。個々のマウスを30分間、味覚計に配置し、そして刺激を、その刺激が噴出したマウスのなめる行動により開始される5秒間の試行の間、ランダムな順で提示した。サッカリンナトリウム、グルタメートおよびアスパルテートについて、100μMのアミロライドを全ての溶液(コントロールを含む)に添加し、塩味の効果を最小限にした。データポイントは、適切なコントロール味覚物質のサンプリングに対する、マウスが味覚物質をなめる平均速度(コントロールに対するなめる速さとして規定した比)を示す;なめの抑制を、1からコントロールに対するなめる速さを引いた値、として規定した。ほとんどの場合、上記コントロール味覚物質は、水であったが、アミノ酸+1mM IMP、200mM MSGおよび10mM IMPについては、コントロールは、それぞれ、1mM IMP、200mMグルコン酸ナトリウムおよび10mM CMPであった。
【0196】
標準的な二瓶嗜好性アッセイを、既に記載されたように実行した(Nelson,G.ら、Cell,106,381−390(2001))。TlR2−rtTAおよびtetO−Ro1/tetO−lacZトランスジーンを保有するマウスについて、発現を、行動学的テストの前に3日間およびその間、ドキシサイクリン処理によって誘導した。コントロールは、誘導されていない同じマウス、ならびに、ドキシサイクリンで処理されたT1R2−rtTAトランスジーンのみを保有するマウスをテストする工程を包含した。3つの全てのグループは、スクロースへの正常な応答を示した。
【0197】
本発明者らは、二瓶アッセイにおいてT1R−KO動物が非常に高濃度の天然の糖(>500mM)を含む溶液を同定することを「学習」するように見えることに気付いた;連続した暴露は、検出閾値の減少および嗜好性比の増加をもたらした。マウスは、標準的な二瓶アッセイにおいて、48時間、反復してテスト化合物に暴露されるので、マウスは、味覚物質を水と区別するために、他の感覚の入力(例えば、テクスチャーまたはにおい)を使用し得る。適切に制御されない場合、このことは、味経路を介した行動学的誘引性によるものと容易に誤解され得る。この問題を避けるために、本発明者らは、短期の即時なめ応答アッセイ(上を参照のこと)またはナイーブなノックアウトマウス(すなわち、訓練またはテストのいずれかの間にそのような味覚物質に暴露されないかったマウス)での二瓶アッセイのいずれかを使用した。
【0198】
(神経記録)
舌の刺激および記録手順を、既に記載されているように実行した(Dahl,M.ら、Brain Res,756,22−34(1997);Nelson,G.ら、Nature,416,199−202(2002))。ニューロンのシグナルを、Grass P511 AC増幅器(Astro−Med)を用いて、増幅し(5,000×)、Digidata 1200B A/D変換器(Axon Instruments)でデジタル化し、そして0.5秒の時定数で取り込んだ(r.m.s.電圧)。味刺激を、20秒間隔の4ml/分の一定の流動速度にて提示し、提示の間に人工だ液(Danilova,V.,およびHellekant,G.,BMC Neurosci,4,5.(2003))を用いて2分間リンスすることによって散在させた。全てのデータ分析は、刺激の適用の直後の25秒間にわたって取り込まれた応答を使用した。各実験シリーズは、記録の安定性を確実にするために0.1Mのクエン酸の提示によって分けられる6個の味覚物質の適用からなっていた。0.1Mのクエン酸への平均応答を、各実験シリーズへの応答を標準化するために使用した。
【0199】
神経記録のために使用された味覚物質(最大濃度)は:スクロース、グルコース、マルトース(600mM);サッカリンナトリウム(40mM);AceK(60mM);クエン酸(100mM);NaCl(100mM);NH4Cl(100mM);6−n−プロピル−チオウラシル(10mM)、キニーネ(10mM);シクロヘキシミド(1mM);L−Ser、L−Ala、(30mM、0.5mM IMPが添加されている)、MSGおよびMPG(300mM、0.5mM IMPありまたはなし);D−Ala、D−Phe、およびD−Trp(l00mM)であった。アミロライド(50μM)を、図の説明文中に示されるようにナトリウム応答を減少するために添加した。
【0200】
(T1R異種発現およびカルシウムイメージング)
改変HEK−293細胞(PEAKrapid細胞;Edge BioSystems,MD)を増殖し、T1Rおよび乱交雑Gタンパク質でトランスフェクトし、そして本質的に以前に記載されたように(Nelson,G.ら、Cell,106,381−390(2001))Caイメージングによって味覚物質への機能的応答についてアッセイした。FURA−2負荷およびCa−イメージングにおける小さな違いは、アッセイ緩衝液として0.1% BSA,100μM EGTAおよび200μM CaCl2を含む199(H)Medium(Biosource)を使用する工程、ならびに、FURA−AMエステル切断を可能にする時間を10分まで減少する工程を包含した。このイメージングシステムは,l0×/0.5N.A.蛍光(fluor)対物レンズ(Zeiss)を備えるOlympus IX50顕微鏡、TILLイメージングシステム(TILL Photonics GmbH)、および冷却CCDカメラであった。蛍光イメージの取得および分析は、TILL−Visionソフトウェアを使用した。
【0201】
[Ca2+]iの変化に対するT1R応答の共役を最適化するために、ヒトGα16のC末端残基(Offermanns,S.,およびSimon,M.I.,J Biol Chem,270,15175−15180(1995))を、Gz(Mody,S.M.ら、Mol Pharmacol,57,13−23(2000))、ガストデューシン(McLaughlin,S.K.ら、Nature,357,563−569(1992))またはGαi2に由来する対応する残基で置換した。ガストデューシン(Ggust−25)のC末端の25残基を含むキメラは、一過性トランスフェクションアッセイにおいて、マウスT1R2+3およびT1R1+3の応答を媒介することに特に効果的であると証明し、そしてさらなる研究のために使用された。T1R3およびGgust−25を安定的に発現する細胞系を、ピューロマイシンおよびゼオシン(Invitrogen)選択を使用して確立した。T1R3およびGgust−25を発現する3つの独立の系を、T1R3レセプターの特異性および用量応答を調査するために使用された。スクロースおよびマルトース(>300mM)は、T1R3およびGgust−25に依存する用量依存応答を惹起したが、高濃度の他のいくつかの糖(グルコース、フルクトース、トレハロースおよびガラクトース)を使用する試みは、それらが[Ca2+]iにおける有意なレセプター非依存性の増加を誘発するので、非実用的であると判明した。
【0202】
本明細書において記載される実施例および実施形態は、説明する目的のみのためであり、それを考慮した種々の改変または変化は当業者に示唆され、そして本出願の精神および範囲ならびに添付された特許請求の範囲の範囲内に包含されていることが理解される。本明細書において引用されている全ての刊行物、特許、および特許出願は、あらゆる目的のためにその全体が本明細書において参考として援用されている。
【0203】
(配列表)
(T1R1配列)
(ラットT1R1アミノ酸配列−−配列番号1)
【0204】
【化007】
【0205】
(マウスT1R1アミノ酸配列−−配列番号2)
【0206】
【化008】
【0207】
(ヒトT1R1アミノ酸配列−−配列番号3)
【0208】
【化009】
【0209】
【化010】
【0210】
(ラットT1R1ヌクレオチド配列−−配列番号4)
【0211】
【化011】
【0212】
(マウスT1R1ヌクレオチド配列−−配列番号5)
【0213】
【化012】
【0214】
(ヒトT1R1ヌクレオチド配列−−配列番号6)
【0215】
【化013】
【0216】
【化014】
【0217】
(T1R2配列)
(ラットT1R2アミノ酸配列−−配列番号7)
【0218】
【化015】
【0219】
(マウスT1R2アミノ酸配列−−配列番号8)
【0220】
【化016】
【0221】
(ヒトT1R2アミノ酸配列−−配列番号9)
【0222】
【化017】
【0223】
(ラットT1R2ヌクレオチド配列−−配列番号10)
【0224】
【化018】
【0225】
【化019】
【0226】
(マウスT1R2ヌクレオチド配列−−配列番号11)
【0227】
【化020】
【0228】
【化021】
【0229】
(ヒトT1R2ヌクレオチド配列−−配列番号12)
【0230】
【化022】
【0231】
【化023】
【0232】
(T1R3配列)
(ヒトT1R3ゲノムヌクレオチド配列−−配列番号13)
【0233】
【化024】
【0234】
【化025】
【0235】
(ヒトT1R3 cdsヌクレオチド配列−−配列番号14)
【0236】
【化026】
【0237】
【化027】
【0238】
(ヒトT1R3アミノ酸配列−−配列番号15)
【0239】
【化028】
【0240】
(マウスT1R3 Sac非味覚因子129ゲノムヌクレオチド配列−−配列番号16)
【0241】
【化029】
【0242】
【化030】
【0243】
(マウスT1R3 Sac非味覚因子129 cdsヌクレオチド配列−−配列番号17)
【0244】
【化031】
【0245】
【化032】
【0246】
(マウスT1R3 Sac非味覚因子129アミノ酸配列−−配列番号18)
【0247】
【化033】
【0248】
(マウスT1R3 Sac味覚因子SWR cdsヌクレオチド配列−−配列番号19)
【0249】
【化034】
【0250】
【化035】
【0251】
(マウスT1R3 Sac味覚因子SWRアミノ酸配列−−配列番号20)
【0252】
【化036】
【0253】
(マウスT1R3 Sac味覚因子c57ゲノムヌクレオチド配列−−配列番号21)
【0254】
【化037】
【0255】
【化038】
【0256】
(マウスT1R3 Sac味覚因子C57 cdsヌクレオチド配列−−配列番号22)
【0257】
【化039】
【0258】
(マウスT1R3 Sac味覚因子C57アミノ酸配列−−配列番号23)
【0259】
【化040】
【0260】
【化041】
【0261】
(ラットT1R3 CDSヌクレオチド配列−−配列番号24)
【0262】
【化042】
【0263】
【化043】
【0264】
(ラットT1R3アミノ酸配列−−配列番号25)
【0265】
【化044】
【図面の簡単な説明】
【0266】
【図1A】T1R1、T1R2およびT1R3の標的KO。(a)3つのT1R遺伝子の構造およびノックアウト動物を作製するための戦略を示す模式図。標的構築物は、T1R1およびT1R2の7つの予測された膜貫通へリックスの全て、およびT1R3の細胞外リガンド結合ドメイン全体を除去した。(b)T1Rの野生型動物の味蕾における強い発現、そして対応するノックアウトマウスにおける完全な欠如、を示しているインサイチュハイブリダイゼーション標識。
【図1B】T1R1、T1R2およびT1R3の標的KO。(a)3つのT1R遺伝子の構造およびノックアウト動物を作製するための戦略を示す模式図。標的構築物は、T1R1およびT1R2の7つの予測された膜貫通へリックスの全て、およびT1R3の細胞外リガンド結合ドメイン全体を除去した。(b)T1Rの野生型動物の味蕾における強い発現、そして対応するノックアウトマウスにおける完全な欠如、を示しているインサイチュハイブリダイゼーション標識。
【図2A】T1R変異体は、通常、酸味、塩味および苦味の刺激に応答する。(a)野生型マウス(WT)、T1R1ノックアウトマウス(1−KO)、T1R2ノックアウトマウス(2−KO)およびT1R3ノックアウトマウス(3−KO)は、酸味(100mMクエン酸)、塩味(100mM NaCl)および苦味(10mM PROP)味覚物質に強い神経応答を示す。(b)統合神経応答(例えば、(a)に示されたもの)が、100mMクエン酸によって誘発された応答に対して標準化された;コントロールおよびKO動物は、相互に区別不能である。値は、平均±標準誤差である(n=4)。データは、鼓索神経の応答を示す(詳細については実験手順を参照のこと)。(c)野生型動物およびT1Rノックアウト動物の、味覚嗜好性が、短期接近味試験(brief access taste test)(Zhang,Y.ら、Cell,112,293−301(2003))を使用して水に対して測定された。4つの全ての系統が、酸味、塩味および苦味の刺激に対する正常な応答を示した。値は、平均±標準誤差である(n=7)。類似の結果が、標準的な二瓶嗜好性アッセイ(two bottle preference assay)を使用して得られた(データは示さず)。Cyx、シクロヘキシミド;Den、安息香酸デナトニウム;PROP、6−n−プロピル−チオウラシル;Qui、キニーネ。
【図2BC】T1R変異体は、通常、酸味、塩味および苦味の刺激に応答する。(a)野生型マウス(WT)、T1R1ノックアウトマウス(1−KO)、T1R2ノックアウトマウス(2−KO)およびT1R3ノックアウトマウス(3−KO)は、酸味(100mMクエン酸)、塩味(100mM NaCl)および苦味(10mM PROP)味覚物質に強い神経応答を示す。(b)統合神経応答(例えば、(a)に示されたもの)が、100mMクエン酸によって誘発された応答に対して標準化された;コントロールおよびKO動物は、相互に区別不能である。値は、平均±標準誤差である(n=4)。データは、鼓索神経の応答を示す(詳細については実験手順を参照のこと)。(c)野生型動物およびT1Rノックアウト動物の、味覚嗜好性が、短期接近味試験(Zhang,Y.ら、Cell,112,293−301(2003))を使用して水に対して測定された。4つの全ての系統が、酸味、塩味および苦味の刺激に対する正常な応答を示した。値は、平均±標準誤差である(n=7)。類似の結果が、標準的な二瓶嗜好性アッセイを使用して得られた(データは示さず)。Cyx、シクロヘキシミド;Den、安息香酸デナトニウム;PROP、6−n−プロピル−チオウラシル;Qui、キニーネ。
【図3AD】T1R1+T1R3は、哺乳動物旨味レセプターとして機能する。(a−d)野生型マウス(白丸、点線)、T1R1 KOマウス(青丸およびバー)、T1R2 KOマウス(グレーの丸およびバー)およびT1R3 KOマウス(茶丸およびバー)の味覚嗜好性が、水に対して、短期接近味試験を使用して測定された。T1R2 KOマウスは、野生型コントロールと同等である。対照的に、T1R1およびT1R3ノックアウト動物は、旨味味覚物質(a)MSG+1mM IMP、(b)MSG、(c)IMP、および(d)L−Asp(100mM)、およびAP4(30mM)に対する嗜好性の完全な消失を示す。さらに、両方のノックアウトは、他のアミノ酸応答において顕著な障害を有する。L−Asn(100mM)およびL−Argが、各100mMにて使用された。(e−f)旨味味覚物質およびアミノ酸に対する統合された鼓索神経応答。アミロライドまたはMSGのカリウム塩(MPG)のいずれかを使用することによって、塩効果が回避される場合、T1R1およびT1R3ノックアウトは、(e)旨味アゴニストおよびL−アミノ酸に対する応答を完全に消失されている。対照的に、(f)高濃度の塩が使用される場合(例えば、100mM MSG)、残存応答が検出される。
【図3EF】T1R1+T1R3は、哺乳動物旨味レセプターとして機能する。(a−d)野生型マウス(白丸、点線)、T1R1 KOマウス(青丸およびバー)、T1R2 KOマウス(グレーの丸およびバー)およびT1R3 KOマウス(茶丸およびバー)の味覚嗜好性が、水に対して、短期接近味試験を使用して測定された。T1R2 KOマウスは、野生型コントロールと同等である。対照的に、T1R1およびT1R3ノックアウト動物は、旨味味覚物質(a)MSG+1mM IMP、(b)MSG、(c)IMP、および(d)L−Asp(100mM)、およびAP4(30mM)に対する嗜好性の完全な消失を示す。さらに、両方のノックアウトは、他のアミノ酸応答において顕著な障害を有する。L−Asn(100mM)およびL−Argが、各100mMにて使用された。(e−f)旨味味覚物質およびアミノ酸に対する統合された鼓索神経応答。アミロライドまたはMSGのカリウム塩(MPG)のいずれかを使用することによって、塩効果が回避される場合、T1R1およびT1R3ノックアウトは、(e)旨味アゴニストおよびL−アミノ酸に対する応答を完全に消失されている。対照的に、(f)高濃度の塩が使用される場合(例えば、100mM MSG)、残存応答が検出される。
【図4】T1R2およびT1R3は、甘味の知覚において重要である。(a)野生型マウス(白丸、点線)、T1R1 KOマウス(グレーの丸およびバー)、T1R2 KOマウス(緑の丸およびバー)およびT1R3 KOマウス(茶丸およびバー)の味覚嗜好性が、水に対して、短期接近味試験を使用して測定された。T1R1 KOマウスは、野生型コントロールと同等である。対照的に、T1R2およびT1R3ノックアウト動物は、人工甘味量およびD−アミノ酸に対する嗜好性の完全な消失を示すが、高濃度の天然の糖に対する残存応答を保持する。これらは、マルトース、スクロースおよびグルコースについて、拡大スケールの用量反応として、(b)において強調されている。しかし、T1R2/T1R3ダブルノックアウト動物(赤丸)は、全ての甘味応答の完全な消失を示す。値は、平均±標準誤差である(n=7)。D−AsnおよびD−Pheは、各々100mMであり、そしてD−Trpは、30mMで使用された。
【図5】T1R2およびT1R3は、哺乳動物甘味レセプターをコードする。パネル(a)は、野生型(WT)およびT1Rノックアウト動物(1−KO、2−KO、3−KO)における天然の糖、人工甘味料およびD−アミノ酸への統合された鼓索神経の応答を示す。T1R2およびT1R3ノックアウトは、人工甘味料およびD−アミノ酸に対する応答を完全に消失させているが(赤線)、高濃度の天然の糖への小さな神経応答を示す。しかし、これらは、T1R2/T1R3ダブルノックアウトマウスにおいて完全に消滅した(下部の青線)。パネル(b)は、増大したパネルの味覚物質に対する平均神経応答を示す;野生型、白いバー;T1R2 KO、緑のバー;T1R3KO、茶色のバー;T1R2/T1R3ダブルKO、赤いバー。値は、標準化された鼓索神経応答の平均±標準誤差である(n=4)。
【図6】hT1R3は、高濃度の天然の糖に応答する。乱交雑したGタンパク質Ggust−25(実験手順を参照のこと)およびマウスT1R3 GPCRを同時発現するHEK−293細胞、またはT1R2プラスT1R3を両方同時トランスフェクトされたHEK−293細胞を、種々の甘味化合物を用いて刺激した。上部のパネルは、500mMスクロースによるT1R3発現細胞の刺激の際の、[Ca2+]iの増加(しかし、300mMスクロースでは増加がないこと)を示す。人工甘味料(300mMサッカリン、右のパネル)を用いた場合、またはレセプターもしくはGgust−25のない細胞中において、応答は検出されなかった;スケールは、FURA−2 F340/F380比より決定された[Ca2+]i(nM)を示す。予期されたように、T1R2+T1R3を発現するコントロール細胞(下部のパネル)は、低濃度の天然の甘味料(300mMスクロース)および人工甘味料(30mMサッカリン)に強く応答する。
【図7】T1R2発現細胞の活性は、行動の誘引を誘発する。
【0267】
(a)野生型マウスおよび強力なT1R2プロモーターの制御下でヒトT1R2遺伝子を発現するT1R2 KOマウスが、(b−d)種々のヒト甘味味覚物質への行動の応答について試験された:(b)Ace−K、アセスルファーム−K、(c)アスパルテーム、および(d)MON、モネリン(約10μM);THAU、タウマチン(約5μM);ASP、アスパルテーム(10mM);GA、グリチルリジン酸(500μM);NH、ネオへスペリジンジヒドロカルコン(400μM)。ヒトT1R2味レセプターは、(a)T1R2細胞において選択的に発現され、そして(b)T1R2 KOマウスの甘味応答を効果的に救助する。重要なことに、トランスジーンの存在は(c−d)、トランスジェニック動物の甘味の嗜好性をヒトのようにする。詳細についてはテキストを参照のこと。(e)T1R2細胞中のRASSL(Redfern,C.H.ら、Nat Biotechnol,17,165−169(1999))の発現は、スピラドリンへの特異的な行動の誘引を示す動物を作製する。RASSL発現動物における強い応答を誘発するために必要な濃度の100×においてでさえも、非誘導動物またはコントロールマウスにおいては応答が見られないことに注意すること。値は、平均±標準誤差である(n=7)。
【図8】図8は、hT1R1のヌクレオチド配列(配列番号26)を提供する。
【図9】図9は、hT1R1のアミノ酸配列(配列番号27)を提供する。
【図10】図10は、hT1R2のヌクレオチド配列(配列番号28)を提供する。
【図11】図11は、hT1R2のアミノ酸配列(配列番号29)を提供する。
【図12】図12は、hT1R3のヌクレオチド配列(配列番号30)を提供する。
【図13】図13は、hT1R3のアミノ酸配列(配列番号31)を提供する。
【技術分野】
【0001】
(関連出願に対する相互参照)
本出願は、米国特許出願第10/679,102号(2003年10月2日出願)(その全体が参考として本明細書において援用)の優先権を主張する。
【0002】
(連邦政府により支援される研究および開発の下になされた発明の権利に関する声明書)
該当なし。
【0003】
(発明の分野)
本発明は、単離された甘味レセプターの核酸配列およびアミノ酸配列、T1R3 Gタンパク質共役型レセプターポリペプチドのモノマーまたはホモダイマーを含むか、それからなる上記レセプター、そのようなレセプターに対する抗体、そのような核酸およびレセプターを検出する方法、ならびに甘味レセプターの調節因子をスクリーニングする方法を提供する。
【背景技術】
【0004】
(発明の背景)
味覚は、甘味、苦味、酸味、塩味および旨味(アミノ酸)の刺激の検出ならびに応答を担っている。それはまた、これらの種々の味の様式を区別し、固有の行動の応答を生じ得る。例えば、動物は、苦味のする化合物を極度に嫌うが、甘味および旨味の刺激には引き付けられる。味シグナルの検出および情報処理を調査するために、本発明者らは、甘味レセプター、旨味レセプターおよび苦味レセプターの単離および特徴付けに焦点を定めた。これらのレセプターは、味覚系の機構を表し、そして味コーディング(taste coding)の論理を規定することを助ける、強力な分子的手段を提供する。
【0005】
候補哺乳動物味レセプターの2つのファミリー(T1RおよびT2R)が、甘味、旨味および苦味の検出において関連付けられてきた。T2Rは、約30の味特異的GPCRのファミリーであり、オプシンと遠縁であり、そしてヒトおよびマウスにおいて苦味と遺伝的に関連付けられたゲノムの領域中でクラスター化されている(非特許文献1;非特許文献2)。いくつかのT2Rは、異種発現アッセイにおいて苦味レセプターとして機能することが示されており、苦味センサーとしてのそれらの役割を実証している(非特許文献3;非特許文献4)。ほとんどのT2Rは、味レセプター細胞の同じサブセット中に同時発現され(非特許文献1)、これらの細胞が、一般的苦味検出器(detector)として機能することを示唆している。
【0006】
T1Rは、舌上皮および口蓋上皮の味細胞において発現される3つのGPCRの小ファミリーであり、代謝生成物産性グルタメートレセプター、カルシウム感知レセプター、および鋤鼻レセプターと遠縁である(非特許文献5;非特許文献6;非特許文献7;非特許文献8;非特許文献9;非特許文献10)。T1Rは、少なくとも2つのヘテロマーレセプターを産生するように組み合わされる:T1R1およびT1R3は、L−アミノ酸センサーを形成し、このセンサーは、げっ歯類においてほとんどのアミノ酸を認識し、そしてT1R1およびT1R3は、広範囲にわたるように調節された甘味レセプターとして機能するように結合する(非特許文献9;非特許文献11;非特許文献12;ならびに特許文献1、特許文献2、および特許文献3も参照のこと)。
【0007】
動物は、広範囲の化学的に別個の甘味分子(天然の糖、人工甘味料、D−アミノ酸、および強烈に甘いタンパク質が挙げられる)を検出し得る。全ての甘み(sweet universe)甘さを味わうのに何種類の異なるレセプターが必要であろうか?ヒトおよびげっ歯類のT1R2+T1R3ヘテロマー甘味レセプターが、細胞ベースのアッセイにおいて、全てのクラスの甘味化合物に応答し、そしてインビボでのそれぞれの心理物理学的閾値および/または行動学的閾値に相当する親和性を伴って、そのように応答する(非特許文献9;非特許文献12)。根本的なレベルにおいて、甘味の進化は、潜在的な食料の供給源中の糖含有量を検出しそして測定する必要性を反映している可能性が高い。従って、天然の糖に対して広範囲にわたるように調節された1つのレセプターが、必要とされる全てであり得る。他方、種々の糖および人工甘味料の多くの研究は、1より多い甘味レセプターの可能性を暗示する(非特許文献13;非特許文献14)。
【0008】
ヒトにおいて、L−グルタメート一ナトリウム(MSG)およびL−アスパルテートは、明白な良い味の感覚(いわゆる、旨味(Maga、1983))を誘発する(しかし他のアミノ酸は誘発しない)。特に、げっ歯類のT1R1+T1R3と違って、ヒトT1R1+T1R3アミノ酸味レセプターは、他のL−アミノ酸よりも、L−グルタメートおよびL−アスパルテートに実質的に敏感である(非特許文献12)。これらの発見は、T1R1+T1R3が、哺乳動物旨味レセプターであり得るという提唱に結び付いた(非特許文献11;非特許文献12)。しかし、多数の研究(T1R3 KOマウスの最近の分析(非特許文献15)を含む)は、旨味は、mGluR4t(代謝生成物産性グルタメートレセプターの短縮型改変体)によって媒介されることを示唆している(非特許文献16;非特許文献17)。
【0009】
異なる味の特質は、どのように味細胞レベルにおいてコードされるのであろうか?哺乳動物において、味レセプター細胞は、味蕾に構築されており、この味蕾は、舌上皮の異なる乳頭に分布されている。各味蕾は、50〜150の細胞(前駆体細胞、支持細胞、および味レセプター細胞が挙げられる)を含んでいる(非特許文献18)。このレセプター細胞は、脳幹および視床のシナプスを通じて皮質の味覚中枢に情報を伝える、求心性神経繊維によって神経支配される。末梢における味コーディングの最も単純なモデルにおいて、各味様式は、特異的なレセプターを発現する細胞の特有の集団(例えば、甘味細胞、苦味細胞、塩感知細胞など)によってコードされる。このシナリオにおいて、任意の1つの味の特質についての我々の知覚は、舌における異なる細胞の型の活性化から生ずる(専有回線モデル(labeled line model))。あるいは、個々の味細胞は、複数の味覚様式を認識し得、そして多くのそのような広範囲にわたるように調節されたレセプター細胞の集団発火パターンが、味の特質をコードする(総ニューロンモデル(across fiber model))。
【0010】
最近、本発明者らは、T1RおよびT2Rが、舌上皮のレセプター細胞の完全に非重複の集団において発現されることを示し(非特許文献9)、そして苦味レセプター発現細胞は、苦味への応答を媒介するが、甘味またはアミノ酸味覚物質への応答を媒介しないことを示した(非特許文献19)。合わせて、これらの結果は、味レセプター細胞は、全ての様式にわたって広範囲に調節されるのではないことを主張し、そして末梢における味コーディングの専有回線モデルを強く支持した。ここで本発明者らが取り組む根本的な問題は、何種類の細胞およびレセプターが甘味および旨味(2つの主要な誘引性の味覚)を媒介するのに必要であるかということである。本発明者らはここで、甘味および旨味が、T1Rによって排他的に媒介されることを示し、そして個々のT1Rサブユニットの遺伝的除去が、これら2つの誘引性の味覚様式に選択的に影響することを示す。甘味および旨味感知のための細胞およびレセプターの同定はまた、本発明者らが、細胞刺激から、レセプター活性の味覚応答をコードする役割を分離する戦略を考案することを可能にした。本発明者らは、T1R2+T1R3発現細胞において改変k−オピオイドレセプターを発現するよう操作された動物が、k−オピオイドアゴニストに対して特異的に引き付けられることを示し、(T1Rレセプター自身というよりも)甘味レセプター発現細胞の活性化が、甘味物質への行動的誘引性の重要な決定要素であることを証明する。最後に、本発明者らはここで、T1R1は単独で(モノマーとしてかまたはホモダイマーとしてのいずれかで)、天然に存在する糖のためのレセプターとして作用することを示す。
【特許文献1】国際公開第00/06592号パンフレット
【特許文献2】国際公開第00/06593号パンフレット
【特許文献3】国際公開第03/004992号パンフレット
【非特許文献1】Adlerら、Cell、2000年、100号、693−702頁
【非特許文献2】Matsunamiら、Nature、2000年、404号、601−604頁
【非特許文献3】Chandrashekarら、Cell、2000年、100号、703−711頁
【非特許文献4】Bufeら、Nat Genet、2002年、32号、397−401頁
【非特許文献5】Hoonら、Cell.1999年、96号、541−551頁
【非特許文献6】Kitagawaら、Biochem Biophys Res Commun、2001年、283号、236−242頁
【非特許文献7】Maxら、Sac.Nat Genet、2001年、28号、58−63頁
【非特許文献8】Montmayeurら、Nat Neurosci、2001年、4号、492−498頁
【非特許文献9】Nelsonら、Cell、2001年、106号、381−390頁
【非特許文献10】Sainzら、J Neurochem、2001年、77号、896−903頁
【非特許文献11】Nelson,G.ら、Nature、2002年、416号、199−202頁
【非特許文献12】Li,X.ら、Proc Natl Acad Sci U S A、2002年、99号、4692−4696頁
【非特許文献13】Schiffmanら、Pharmacol Biochem Behav、1981年、15号、377−388頁
【非特許文献14】Ninomiyaら、J Neurophysiol、1999年、81号、3087−3091頁
【非特許文献15】Damakら、Science、2003年、301号、850−853頁
【非特許文献16】Chaudhariら、Neurosci、1996年、16号、3817−3826頁
【非特許文献17】Chaudhariら、Nat Neurosci、2000年、3号、113−119頁
【非特許文献18】Lindermann、Physiol Rev、1996年、76号、718−766頁
【非特許文献19】Zhangら、Cell、2003年、112号、293−301頁
【発明の開示】
【課題を解決するための手段】
【0011】
(発明の簡単な要旨)
従って、本発明は、最初に、ホモダイマーの甘味レセプターを提供する。このレセプターは、2つのT1R3ポリペプチドを含むかまたは2つのT1R3ポリペプチドからなる。本発明はまた、1つのT1R3ポリペプチドを含むかまたは1つのT1R3ポリペプチドからなる、モノマーの甘味レセプターを提供する。これらのレセプターは、T1R3が細胞中に発現された場合、甘味リガンドに応答してシグナルを変換する。一実施形態において、この甘味リガンドは、天然に存在する甘味分子である。別の実施形態において、この甘味リガンドは、人工的であり、そして天然に存在する甘味分子を模倣する。一実施形態において、ホモダイマーのT1R3ポリペプチドは、非共有結合性に結合される。
【0012】
一局面において、本発明は、T1R3ポリペプチドを含む甘味レセプターを提供し、このT1R3ポリペプチドは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列に対して、約80%を超えるアミノ酸配列の同一性を有するか、あるいは、中程度もしくは高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる。
【0013】
一実施形態において、T1R3を含むレセプターは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31に対して生じた、ポリクローナル抗体に特異的に結合する。別の実施形態において、このレセプターは、Gタンパク質共役型レセプター活性を有する。別の実施形態において、T1R3ポリペプチドは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列を有する。別の実施形態において、このレセプターは、ヒト、ラット、またはマウス由来である。
【0014】
別の実施形態において、甘味レセプターは、T1R3ポリペプチドを含み、そして天然の糖(例えば、グルコース、ガラクトース、フルクトース、マルトース、ラクトース、およびスクロース)を認識する。
【0015】
一局面において、本発明は、単離されたポリペプチドを提供し、このポリペプチドは、T1R3を含むホモダイマーもしくはモノマーの甘味レセプターの細胞外、膜貫通ドメイン、または細胞質ドメインを含み、この細胞外、膜貫通ドメイン、または細胞質ドメインは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31の細胞外、膜貫通ドメイン、または細胞質ドメインとの、約80%を超えるアミノ酸配列の同一性を有する。別の実施形態において、細胞外ドメイン、膜貫通ドメイン、または細胞質ドメインは、高度にストリンジェントな条件の下、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列の細胞外ドメイン、膜貫通ドメイン、または細胞質ドメインにハイブリダイズする。
【0016】
一実施形態において、ポリペプチドは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31の細胞外、膜貫通ドメイン、または細胞質ドメインをコードする。別の実施形態において、細胞外、膜貫通ドメイン、または細胞質ドメインは、異種ポリペプチドに共有結合され、キメラポリペプチドを形成する。別の実施形態において、キメラポリペプチドは、Gタンパク質共役型レセプター活性を有する。
【0017】
一局面において、本発明は、ホモダイマーのまたはモノマーの甘味レセプターと選択的に結合する抗体を提供し、このレセプターは、1つかもしくは2つのT1R3ポリペプチドを含み(しかし、T1R1もしくはT1R2ポリペプチドを含まない)、この抗体は、配列番号15、配列番号20、配列番号23、配列番号25、配列番号31のアミノ酸配列に対して、約80%を超えるアミノ酸配列の同一性を含むか、あるいは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる、T1R3ポリペプチドを含むレセプターに対し生産される。
【0018】
別の局面において、本発明は、味細胞において甘味シグナルを調節する化合物を同定するための方法を提供し、この方法は、(i)T1R3ポリペプチドを含む(しかし、T1R1もしくはT1R2ポリペプチドを含まない)ホモダイマーまたはモノマーのレセプターとこの化合物を接触させる工程であって、このポリペプチドは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31に対して、約80%を超えるアミノ酸配列の同一性を含むか;あるいは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる、工程;ならびに(ii)上記レセプター上の化合物の機能的効果を決定する工程を包含する。
【0019】
一実施形態において、上記機能的効果は、インビトロで決定される。一実施形態において、上記ポリペプチドは、細胞かまたは細胞膜において発現される。別の実施形態において、上記レセプターは、固相に共有結合性かまたは非共有結合性のいずれかで結合される。
【0020】
別の局面において、本発明は、味細胞において甘味シグナルを調節する化合物を同定するための方法を提供し、この方法は、(i)この化合物とT1R3ポリペプチドを含むが、T1R1もしくはT1R2ポリペプチドを含まないホモダイマーかまたはモノマーのレセプターを発現する、細胞を接触させる工程であって、このT1R3ポリペプチドは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31に対して、約80%を超えるアミノ酸配列の同一性を含むか;あるいは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる、工程;ならびに(ii)上記レセプター上の化合物の機能的効果を決定する工程を包含する。
【0021】
一実施形態において、上記機能的効果は、細胞内のcAMP、IP3、またはCa2+の変化を測定することによって決定される。別の実施形態において、上記機能的効果は、化学的効果または表現型効果である。別の実施形態において、上記機能的効果は、物理的効果である。別の実施形態において、上記機能的効果は、上記レセプターの細胞外ドメインへの上記化合物の結合を測定することによって決定される。別の実施形態において、ポリペプチドは、組換え型である。別の実施形態において、上記細胞は、真核生物細胞(例えば、哺乳動物細胞(例えば、ヒト細胞))である。別の実施形態において、上記細胞は、Gタンパク質(Gα15)を発現する。
【発明を実施するための最良の形態】
【0022】
(発明の詳細な説明)
(序論)
T1RおよびT2Rは、味レセプター細胞のサブセットに選択的に発現されるGタンパク質共役型レセプター(GPCR)の2つのファミリーである(Hoonら、Cell 96:541−551(1999);Adlerら、Cell 100:693−702(2000);Chandrashekarら、Cell 100:703−711(2000);Matsunamiら、Nature 404:601−604(2000);Nelsonら、Cell 106:381−390(2001);Kitagawaら、Biochem.Biophys.Res.Cummun.283:236−242(2001);Montmayeurら、Nature Neurosci.4:492−498(2001);Maxら、Nature Genet.28:58−63(2001);Sainzら、J.Neurochem.77:896−903(2001))。T2Rは、苦味の検出に関与する(Adlerら、Cell 100:693−702(2000);Chandrashekarら、Cell、100:703−711(2000));T1R2およびT1R3は、組み合わさって甘味レセプターとして機能する(Nelsonら、Cell 106:381−390(2001)を参照のこと;そしてT1R1およびT1R3は、本明細書中に記載されるように、組み合わさってアミノ酸味レセプターとして機能する(Nelsonら、Nature 2002年2月24日およびWO 03/004992もまた参照のこと)))。本発明者らは、ここでホモダイマーの味レセプターを同定し、この味レセプター中で、2つのT1R3ポリペプチドは、組み合わさって甘味レセプターとして機能する。T1R3のモノマー形態はまた、甘味レセプターとして作用する。
【0023】
異種発現系を使用して、本発明者らは、T1R3が、それ自身と組み合わさり、そしてまたモノマーとして作用して甘味レセプターとして機能し、甘味分子(例えば、スクロース、ガラクトース、フルクトース、グルコース、マルトース、およびラクトース)を認識することを示す。候補レセプターは、Gα16−GαzおよびGα15乱交雑Gタンパク質を含むヒト胎児由来腎臓(HEK)細胞内に発現され(Offermannsら、J.Biol.Chem.270:15175−15180(1995);Modyら、Mol.Pharmacol.57:13−23(2000))、そして刺激によって引き出された細胞内カルシウムの変化ついてアッセイされる。この系において、レセプターの活性化は、ホスホリパーゼCβ(PLC-βの活性化およびカルシウム指示染料(indicator dye)を使用して、単一の細胞レベルでモニタリングされ得る内部貯蔵からのカルシウムの放出に結び付く(Chandrashekarら、Cell 100:703−711(2000);Nelsonら、Cell 106:381−390(2001);Tsienら、Cell Calcium 6:145−157(1985))。
【0024】
上記レセプターをコードするこれらの核酸およびタンパク質は、その核酸が、味細胞中で特異的に発現される場合、味細胞の同定のための有用なプローブを提供する。このレセプターは、新規の味覚物質(例えば、人工甘味料分子)の検定について有用である。例えば、GPCRポリペプチドおよびタンパク質のためのプローブは、味細胞(例えば、葉状細胞、口蓋細胞、および有郭細胞)のサブセット、または特異的な味レセプター細胞(例えば、甘味レセプター細胞)を同定するために使用され得る。これらはまた、舌の味細胞と脳の味中枢につながっている味感覚ニューロンとの間の関係を明らかにする味地形図(topographic map)の作製のための手段として役立つ。さらに、これらがコードする核酸およびタンパク質は、味誘導行動を分析するためのプローブとして使用され得る。
【0025】
本発明はまた、これら新規のT1R3を含むモノマーまたはホモダイマーの甘味レセプターの調節因子(例えば、アクチベーター、インヒビター、刺激物質、エンハンサー、アゴニスト、およびアンタゴニスト)をスクリーニングする方法を提供する。一実施形態において、本発明のモノマーまたはホモダイマーのT1R3を含むレセプターは、天然に存在するかもしくは人工の甘味分子もしくは甘味変換の調節因子(例えば、小有機分子、アミノ酸、ペプチド、炭水化物、脂質、多糖類など)をスクリーニングために使用され得る。例えば、本発明のホモダイマーまたはモノマーのT1R3を含むレセプターは、以下の実施例のセクションに記載されるように、天然に存在する甘味味覚物質を認識する。そのようなレセプターは、人工甘味料または変更された天然に存在する甘味料(これらの甘味料は、ホモダイマーまたはモノマーのT1R3を含むレセプターの天然に存在する糖のリガンドを模倣する)をスクリーニングするために使用され得る。そのような甘味変換の調節因子は、甘味シグナル伝達経路の薬理学的および遺伝的調節のために、そして新規の甘味リガンドの発見のために有用である。これらのスクリーニングの方法は、甘味細胞活性のアゴニストおよびアンタゴニストを同定するために使用され得る。次いで、これらの調節化合物は、食品産業および薬品産業において味を作るために使用され得る。従って、本発明は、T1R3を含むレセプターが、甘味変換に対する調節因子の効果についての直接的または間接的なレポーター分子として作用する場合、味変換のためのアッセイを提供する。GPCRは、例えば、リガンド結合、Gタンパク質結合、調節分子結合、イオン濃度、膜電位、電流の流れ、イオン流入、転写、シグナル変換、レセプター−リガンド相互作用、神経伝達物質およびホルモン放出;ならびにインビトロ、インビボ、およびエクスビボでのセカンドメッセンジャーの濃度の変化を測定するために、アッセイにおいて使用され得る。一実施形態において、T1R3を含むレセプターは、第二のレポーター分子(例えば、緑蛍光タンパク質)への付着を介した間接的なレポーターとして使用され得る(例えば、MistiliおよびSpector,Nature Biotechnology 15:961−964(1997)を参照のこと)。別の実施形態において、T1R3を含むレセプターは、T1R1またはT1R2のいずれも発現しない細胞において組換えによって発現され、そしてGPCR活性を介した味変換の調節は、Ca2+レベルの変化を測定することによって検定される。
【0026】
味変換の調節因子を検定する方法としては、T1R3、その一部を(例えば、細胞外ドメイン)、または1つ以上のT1R3のドメインを含むキメラタンパク質を含むレセプターを使用したインビトロリガンド結合アッセイ、ならびにインビボ(細胞ベースおよび動物)アッセイ(例えば、卵母細胞T1R3レセプター発現;組織培養細胞T1R3レセプター発現;T1R3の転写活性化;GPCRのリン酸化および脱リン酸化;GPCRへのGタンパク質結合;リガンド結合アッセイ;電圧、膜電位および伝導度の変化;イオン流入アッセイ;細胞内セカンドメッセンジャー(例えば、cAMPおよびイノシトール三リン酸)の変化;細胞内カルシウムレベルの変化;および神経伝達物質の放出)が挙げられる。
【0027】
(定義)
「T1Rファミリー味レセプター」とは、ホモダイマーのレセプター、ヘテロダイマーのレセプターまたはモノマーのレセプターとしての、Gタンパク質共役型レセプター(例えば、T1R1、T1R2、およびT1R3、またはこれらの任意の組み合わせ)のT1Rファミリーのメンバーを含むレセプターをいう。一実施形態において、このT1Rファミリーレセプターは、T1R3(「T1R3を含む味レセプター」または「T1R3を含む甘味レセプター」)を含む。一実施形態において、T1Rファミリーレセプターは、共有結合性にまたは非共有結合性のいずれかで結合されるホモダイマーのレセプターを形成する、第一のT1R3ポリペプチドおよび第二のT1R3ポリペプチドを含む。別の実施形態において、T1Rファミリーレセプターは、1つのT1R3ポリペプチドを含むが、他のT1Rポリペプチドを含まず、そしてモノマーのレセプターを形成する。別の実施形態において、T1Rファミリーレセプターは、T1R3およびT1Rファミリーの異種のポリペプチドを含む。一実施形態において、レセプターは、T1R1およびT1R3を含む。別の実施形態において、レセプターは、T1R2およびT1R3を含む。一実施形態において、T1R3を含むレセプターは、そのレセプターの2つのメンバーが同じ細胞において同時発現される場合(例えば、T1R3およびT1R3、またはT1R1およびT1R3、またはT1R2およびT1R3)、活性である。別の実施形態において、T1Rポリペプチドは、同じ細胞において同時発現され、そしてヘテロダイマーかまたはホモダイマーのレセプターを形成し、ここで、レセプターのT1Rポリペプチドは、非共有結合性に結合されるかまたは共有結合性に結合される。レセプターは、例えば、天然に存在する甘味分子および/または人工甘味分子(例えば、スクロース、フルクトース、ガラクトース、マンノース、グルコース、ラクトース、サッカリン、ズルチン、アセスルファーム−K)ならびに(甘いおよび甘くない)他の分子を認識する能力を有する。これらの分子は、T1R3を含み、味を変換するGタンパク質共役型レセプターのためのリガンドとして作用することによって、「甘味シグナル変換を調節する」化合物の例である。
【0028】
用語「GPCR−B3またはT1R1」、「GPCR−B4またはT1R2」および「T1R3」あるいは「GPCR−B3またはT1R1」、「GPCR−B4またはT1R2」および「T1R3」をコードする核酸とは、Gタンパク質共役型レセプターのT1Rファミリーのメンバーである、核酸およびポリペプチド多型改変体、対立遺伝子、変異体、および種間のホモログをいい、そして(1)配列番号1、2、3、7、8、9、15、18、20、23、25、27、または31によってコードされるアミノ酸配列に対して、約60%を超えるアミノ酸配列の同一性、65%、70%、75%、80%、85%、90%、好ましくは、91%、92%、93%、94%、95%、96%、97%、98%または99%以上のアミノ酸配列の同一性を有する、好ましくは少なくとも約25、50、100、200、500、1000またはそれより多くのアミノ酸にわたる領域にわたるアミノ酸配列を有し;(2)配列番号1、2、3、7、8、9、15、18、20、23、25、27、または31によってコードされたアミノ酸配列を含む免疫原に対して惹起された抗体(例えば、ポリクローナル抗体)およびこれらの保存的に改変された改変体に結合し;(3)ストリンジェントなハイブリダイゼーション条件下で、T1Rタンパク質をコードする核酸配列(例えば、配列番号4、5、6、10、11、12、13、14、16、17、19、21、22、24、26、28、および30)に対応するアンチセンス鎖およびこれらの保存的に改変された改変体に、特異的にハイブリダイズし、(4)配列番号4、5、6、10、11、12、13、14、16、17、19、21、22、24、26、または28、または30に対して、約60%を超える配列の同一性、65%、70%、75%、80%、85%、90%、好ましくは91%、92%、93%、94%、95%、96%、97%、98%または99%以上のヌクレオチド配列の同一性を有する、好ましくは少なくとも25、50、100、200、500、1000以上のヌクレオチドの領域にわたる核酸配列を有する。さらに、本発明のT1Rファミリーポリペプチド(例えば、T1R1、T1R2、もしくはT1R3)またはT1R3を含むレセプター(例えば、T1R3、T1R3+T1R3、T1R1+T1R3もしくはT1R2+T1R3)は、単独でもしくは同じ細胞で同時発現した場合、または別のT1Rファミリーメンバーとモノマー、ホモダイマー、またはヘテロダイマーとして同時発現した場合のいずれかにおいて、Gタンパク質共役型レセプター活性を有する。ヒト、ラット、およびマウスのT1R1、T1R2、およびT1R3の、アミノ酸配列およびヌクレオチド配列の登録番号は、GenBankにおいて見出され得る(ヒトT1R1アミノ酸配列については、例えば、登録番号DAA00012およびNP_619642を参照のこと;ヒトT1R1ヌクレオチド配列については、例えば、登録番号BK000153を参照のこと;ヒトT1R2アミノ酸配列については、例えば、登録番号DAA00019、AAM12239、およびNP_619642.1を参照のこと;ヒトT1R2ヌクレオチド配列については、例えば、登録番号BK000151、NM_138697.1、AF458149S1−6を参照のこと;ヒトT1R3アミノ酸配列については、例えば、登録番号DAA00013を参照のこと;ヒトT1R3ヌクレオチド配列については、例えば、登録番号BK000152を参照のこと)。T1R1、T1R2、およびT1R3のアミノ酸配列およびヌクレオチド配列については、WO 00/06592、WO 00/06593、WO 01/66563、WO 03/001876、WO 02/064631、WO 03/004992、WO 03/025137、WO 02/086079、およびWO 01/83749もまた参照のこと(各々の全体は、本明細書において参考として援用されている)。
【0029】
T1Rタンパク質は、「Gタンパク質共役型レセプター活性」を有する。例えば、このタンパク質は、細胞外の刺激(例えば、リガンド結合(例えば、甘味リガンド))に応答してGタンパク質に結合し、そして酵素(例えば、ホスホリパーゼCおよびアデニレートシクラーゼ)の刺激を介してセカンドメッセンジャー(例えば、IP3、cAMP、およびCa2+)の産生を促進する。そのような活性は、Gタンパク質または乱交雑Gタンパク質(例えば、Gα15もしくはGα16−Gαz)のいずれかと酵素(例えば、PLC)とに、GPCR(またはキメラGPCR)を結合することによって、ならびに細胞内カルシウム利用の増加を測定することによって、異種細胞中で測定され得る(OffermansおよびSimon,J.Biol.Chem.270:15175−15180(1995))。レセプターの活性は、例えば、蛍光Ca2+指示染料および蛍光イメージングを使用して、[Ca2+]iにおけるリガンドによって誘導された変化を記録することによって効果的に測定され得る。
【0030】
そのようなGPCRは、当業者に公知の方法(例えば、疎水性ドメインおよび親水性ドメインを同定する配列分析プログラム(例えば、KyteおよびDoolittle,J.Mol.Biol.157:105−132(1982)を参照のこと))を使用して、構造的に同定され得る、膜貫通ドメイン、細胞外ドメインおよび細胞質ドメインを有する。そのようなドメインは、キメラタンパク質を作製するために、および本発明のインビトロアッセイのために有用である(例えば、WO 94/05695および米国特許第5,508,384号を参照のこと)。
【0031】
本発明の甘味レセプターまたはタンパク質の活性(例えば、シグナル変換)を調節する化合物をテストするためのアッセイの状況における語句「機能的効果」は、間接的にまたは直接的にGPCRまたは甘味レセプターの影響下にあるパラメータ(例えば、物理的効果、表現型効果、または化学的効果(例えば、外部の刺激(例えば、リガンド結合)に応じて細胞のシグナルを変換する能力、またはリガンドに結合する能力))の決定を含む。それは、結合活性およびシグナル変換を含む。「機能的効果」は、インビトロ、インビボ、およびエクスビボでの活性を含む。
【0032】
「機能的効果を決定すること」によってとは、間接的にまたは直接的にT1R GPCRタンパク質または1つ以上のT1R GPCRタンパク質を含む甘味レセプターの影響下にある、パラメータ(例えば、物理的効果および化学的効果、または表現型効果)を増加または減少する化合物をアッセイすることを意味する。そのような機能的効果は、当業者に公知の任意の手段によって測定され得る(例えば、分光特性(例えば、蛍光、吸収度、屈折率)の変化;流体力学的特性(例えば、形状);クロマトグラフィー特性;またはタンパク質の溶解特性;タンパク質の誘導マーカーまたは転写活性化の測定;結合活性または結合アッセイ(例えば、抗体への結合)の測定;リガンド結合活性または天然に存在するかもしくは合成のいずれかのそのアナログの変化の測定;細胞増殖の測定;細胞表面マーカー表現の測定;T1R関連配列のタンパク質レベルの変化の測定;RNA安定性の測定;Gタンパク質結合;GPCRリン酸化または脱リン酸化;シグナル変換(例えば、レセプター−リガンド相互作用、セカンドメッセンジャー濃度(例えば、cAMP、cGMP、IP3、PI、または細胞内Ca2+));神経伝達物質放出;ホルモン放出;電圧、膜電位およびコンダクタンスの変化;イオン流入;調節分子結合;例えば、化学発光、蛍光、比色の反応、抗体結合、および誘導マーカーを介した下流またはレポーター遺伝子発現(CAT、ルシフェラーゼ、β−gal、GFPなど)の同定)。
【0033】
T1Rファミリーポリヌクレオチド配列およびポリペプチド配列およびT1Rファミリー味レセプターの「インヒビター」、「アクチベーター」、および「調節因子」は、T1Rポリヌクレオチド配列およびポリペプチド配列ならびにT1Rファミリー味レセプター(モノマーレセプター、ホモダイマーレセプターおよびヘテロダイマーレセプターを含む)のインビトロアッセイおよびインビボアッセイを使用して同定される、活性化分子、阻害分子、または調節分子に言及するために使用される。インヒビターは、例えば、T1Rファミリーの味レセプター(例えば、T1R3ポリペプチドを含むレセプター)に結合し、部分的にもしくは完全に活性を遮断するか、活性を減少、予防、遅延するか、の活性もしくは発現を不活性化、脱感作、またはダウンレギュレートする化合物(例えば、アンタゴニスト)である。「アクチベーター」は、T1Rファミリーの味レセプター(例えば、T1R3ポリペプチドを含むレセプター)を増加、開放、活性化、促進、活性化を増強、感作、アゴニスト化、またはアップレギュレートする化合物である(例えば、アゴニスト)。インヒビター、アクチベーター、または調節因子としてはまた、遺伝的に改変されたバージョンのT1Rファミリー味レセプター(例えば、変更された活性を有するバージョン)ならびに天然に存在するリガンドおよび合成のリガンド、アンタゴニスト、アゴニスト、抗体、アンチセンス分子、リボザイム、小化学分子などが挙げられる。インヒビターおよびアクチベーターのためのそのようなアッセイは、例えば、インビトロで、細胞で、もしくは細胞膜でT1Rファミリー味レセプターを発現する工程、推定される調節因子化合物を適用する工程、次いで上述のように、活性に対する機能的効果を決定する工程を包含する。一実施形態において、T1R3ポリペプチドを含む味レセプターは、甘味分子(例えば、スクロース、グルコース、フルクトース、ラクトース、マンノース、ガラクトース、サッカリン、ズルチン、アセスルファーム−K)を認識する能力を有する。別の実施形態において、T1R3ポリペプチドを含む味レセプターは、他の分子(例えば、潜在的人工甘味料)を認識する能力を有する。これらの分子は、Gタンパク質共役型レセプターの細胞外リガンドとして作用することによって、およびそのレセプターを活性化することによって、味シグナル変換を調節する化合物の例である。他の実施形態において、味シグナル変換を調節する化合物は、レセプターの細胞内リガンドとして作用するか、あるいは細胞外リガンドの結合を阻害もしくは活性化するか、あるいはレセプターの細胞内リガンドの結合を阻害もしくは活性化する分子である。
【0034】
味レセプターのT1Rファミリーを含むサンプルまたはアッセイは、潜在的なアクチベーター、インヒビター、または調節因子で処理され、そして阻害の程度を検査するために、上記インヒビター、アクチベーター、または調節因子を含まないコントロールサンプルと比較される。(インヒビターで処理されていない)コントロールサンプルは、100%の相対タンパク質活性値を割り当てられる。T1Rファミリーレセプターの阻害は、コントロールと比較した活性値が、約80%(好ましくは50%、より好ましくは25〜0%)である場合に、達成される。T1Rファミリーレセプターの活性化は、(アクチベーターで処理されていない)コントロールと比較した活性値が、約110%、より好ましくは150%、より好ましくは200〜500%(すなわち、コントロールと比較して2倍から5倍高い)、より好ましくは1000〜3000%高い場合に、達成される。
【0035】
本明細書において使用される場合、用語「テスト化合物」または「薬物候補」または「調節因子」または文法上の等価物は、直接的にまたは間接的に味を調節する能力を試験される任意の分子(例えば、天然に存在するかもしくは合成の、人工甘味料または天然に存在する糖(例えば、タンパク質、オリゴペプチド(例えば、約5から約25アミノ酸長、好ましくは約10から20アミノ酸長、または12から18アミノ酸長、好ましくは12アミノ酸長、15アミノ酸長、または18アミノ酸長)、小有機分子、多糖類、脂質、脂肪酸、ポリヌクレオチド、オリゴヌクレオチドなど))を説明する。テスト化合物は、テスト化合物のライブラリー(例えば、十分な範囲の多様性を提供するコンビナトリアルライブラリーまたはランダム化されたライブラリー)の形態であり得る。テスト化合物は、必要に応じて、融合パートナー(例えば、標的化合物、救助(rescue)化合物、ダイマー化合物、安定化合物、取り組み可能な化合物(addressable compound)、および他の機能的部分)に結合される。従来、有用な特性を有する新規の化学実体は、ある所望の特性もしくは活性(例えば、阻害活性)を有するテスト化合物(「リード化合物」といわれる)を同定し、このリード化合物の改変体を作製し、そしてこれらの改変体化合物の特性および活性を評価することによって生成される。多くの場合、ハイスループットスクリーニング(HTS)方法が、そのような分析のために使用される。
【0036】
「小有機分子」とは、約50ダルトンを超え、そして約2500ダルトン未満(好ましくは約2000ダルトン未満、好ましくは約100から約1000ダルトンの間、より好ましくは約200から約500ダルトンの間)の分子量を有する、天然に存在するかまたは合成のいずれかの有機化合物をいう。
【0037】
「生物学的サンプル」としては、組織の切片(例えば、生検サンプルおよび剖検サンプル)および組織学的目的のために取られた凍結切片が挙げられる。そのようなサンプルとしては、血液、痰、組織、培養細胞(例えば、初代培養)、外移植片、および形質転換細胞、糞便、尿などが挙げられる。生物学的サンプルは、代表的に、真核生物体(最も好ましくは、哺乳動物(例えば、霊長類(例えば、チンパンジーもしくはヒト);ウシ;イヌ;ネコ;げっ歯類(例えば、モルモット、ラット、マウス);ウサギ);または鳥類;爬虫類;あるいは魚類)から得られる。
【0038】
「ヘテロダイマー」とは、2つの異なるポリペプチドサブユニット(例えば、分子が、共有結合性(例えば、リンカーもしくは化学結合を通じて)の連結か、または非共有結合性(例えば、イオン結合、ファンデルワールス結合、静電結合、もしくは水素結合)の連結のいずれかを介して結合される、2つの異なるポリペプチド)を含む、ダイマーのレセプターである。同じ細胞内で同時発現された場合(好ましくは、このレセプターが、共有結合性または非共有結合性のいずれかで連結されたヘテロダイマーを形成するように同時発現された場合)、本発明のT1R3を含むレセプターは機能する。例えば、T1R1およびT1R3は、ヘテロマーのレセプターを形成し、そしてT1R2およびT1R3は、ヘテロマーのレセプターを形成する。
【0039】
「ホモダイマー」とは、2つの同じポリペプチドサブユニット(例えば、分子が、共有結合性(例えば、リンカーもしくは化学結合を通じて)の連結か、または非共有結合性(例えば、イオン結合、ファンデルワールス結合、静電結合、もしくは水素結合)の連結のいずれかを介して結合される、2つのT1R3ポリペプチド)を含むダイマーのレセプターである。本発明のT1R3を含むレセプターは、同じ細胞内で同時発現された場合(好ましくは、このレセプターが、共有結合性または非共有結合性のいずれかで連結されたホモダイマーを形成するように同時発現された場合)、機能する。
【0040】
「モノマー」は、1つのポリペプチドサブユニット(例えば、1つのT1R3ポリペプチド)を含むレセプターである。
【0041】
2つ以上の核酸またはポリペプチド配列の状況において、用語「同一の」または「同一性」パーセントとは、以下に記載されるデフォルトパラメータ(default parameter)を有するBLASTまたはBLAST 2.0配列比較アルゴリズムを使用して、または手動のアラインメントおよび視覚的検査(例えば、NCBIウェブサイトなどを参照のこと)によって測定される場合、同じであるか、あるいは特定のパーセンテージの同じであるアミノ酸残基またはヌクレオチド(すなわち、比較ウィンドウ(comparison window)または指定された領域にわたる最大限の一致について比較されそしてアラインメントされた場合の、特定された領域(例えば、ヌクレオチド配列、配列番号1〜25)にわたる、約60%の同一性(好ましくは、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%以上の同一性))を有する、2つ以上の配列もしくは部分配列をいう。そして、そのような配列は、「実質的に同一」であると言われる。この定義はまた、テスト配列の相補性(compliment)をいうか、または相補性に対して適用され得る。この定義はまた、欠失および/または付加を有する配列、ならびに置換を有する配列を含む。以下に記載されるように、好ましいアルゴリズムは、ギャップなどを説明し得る。好ましくは、同一性は、少なくとも約25アミノ酸長もしくは約25ヌクレオチド長である領域にわたって(またはより好ましくは50〜100アミノ酸長もしくは50〜100ヌクレオチド長である領域にわたって)存在する。
【0042】
配列の比較について、代表的には、1つの配列は、テスト配列が比較される参照配列として作用する。配列比較アルゴリズムを使用する場合、テスト配列および参照配列がコンピューターに入力され、必要な場合、部分配列座標が指定され、そして配列アルゴリズムプログラムパラメータが指定される。好ましくは、デフォルトプログラムパラメータが使用され得るか、または代替的パラメータが指定され得る。次いで、この配列比較アルゴリズムは、上記プログラムパラメータに基づいて参照配列と比較したテスト配列のパーセント配列同一性を計算する。
【0043】
本明細書において使用される場合、「比較ウィンドウ」とは、20から600(通常約50〜約200、より通常的に約100〜約150)からなる群より選択される連続する位置の数のうちのいずれか1つのセグメントの参照を含み、この中で、配列は、同じ数の連続する位置の参照配列と、2つの配列が最適にアラインメントされた後、比較され得る。比較のために配列をアラインメントする方法は、当該分野において周知である。比較のために配列の最適なアラインメントは、例えば、SmithおよびWaterman,Adv.Appl.Math.2:482(1981)の局所相同性アルゴリズムによって、NeedlemanおよびWunsch,J.Mol.Biol.48:443(1970)の相同性アラインメントアルゴリズムによって、PearsonおよびLipman,Proc.Nat’l.Acad.Sci.USA 85:2444(1988)の類似性方法(similarity method)の検索によって、これらのアルゴリズムのコンピューター化された(Wisconsin Genetics Software Package,Genetics Computer Group,575 Science Dr.,Madison,WI中のGAP、BESTFIT、FASTA、およびTFASTA)実施によって、または手動の配列および視覚的検査(例えば、Current Protocols in Molecular Biology(Ausubelら(編)、1995年 補遺)を参照のこと)によって実行され得る。
【0044】
パーセント配列同一性および配列類似性の決定に適しているアルゴリズムの好ましい例は、BLASTおよびBLAST 2.0アルゴリズムであり、これらは、それぞれ、Altschulら、Nuc.Acids Res.25:3389−3402(1977)およびAltschulら、J.Mol.Biol.215:403−410(1990)に記載されている。BLASTおよびBLAST 2.0は、本発明の核酸およびタンパク質のパーセント配列同一性を決定するために本明細書において記載されるパラメータを用いて使用される。BLAST分析を実行するためのソフトウェアは、National Center for Biotechnology Information(http://www.ncbi.nlm.nih.gov/)を通して公的に入手可能である。このアルゴリズムは、最初にクエリー配列の長さWの短い語(この語は、データベース配列の同じ長さの語とアラインメントされる場合、ある正の値の閾値スコアTにマッチ(match)するかまたはTを満たすかのいずれかである)を同定することによって、高スコア配列ペア(HSP)を同定することに関与する。Tは、近隣語スコア閾値(score threshold)といわれる(Altschulら、前出)。これらの最初の近隣語のヒット(hit)は、それらを含むより長いHSPを見付けるために検索を開始するための源として作用する。語のヒットは、累積アラインメントスコアが増加され得る限り、各配列に沿って両方向に拡張される。累積スコアは、ヌクレオチド配列については、パラメータM(マッチ残基のペアに対しての報酬スコア(reward score);常に>0)およびパラメータN(ミスマッチ残基に対してのペナルティスコア;常に<0)を使用して計算される。アミノ酸配列については、スコアマトリックスが、累積スコアを計算するために使用される。各方向の語のヒットの拡張は、累積アラインメントスコアが最大限に達成された値から量Xが減少する場合;累積スコアが、1つ以上の負のスコア残基アラインメントの蓄積に起因して0以下になる場合;あるいは、どちらかの配列の末端に到達する場合に、停止される。BLASTアルゴリズムパラメータW、T、およびXは、配列の感度およびスピードを決定する。(ヌクレオチド配列のための)BLASTNプログラムは、デフォルトとして、語の長さ(W)11、期待値(E)10、M=5、N=−4および両方の鎖の比較を、使用する。アミノ酸配列のために、BLASTPプログラムは、デフォルトとして、語の長さ3、期待値(E)10、BLOSUM62スコアマトリックス(HenikoffおよびHenikoff、Proc.Natl.Acad.Sci.USA 89:10915(1989)を参照のこと)アラインメント(B)50、期待値(E)10、M=5、N=−4、および両方の鎖の比較、を使用する。
【0045】
用語「ポリペプチド」、「ペプチド」、および「タンパク質」とは、本明細書において、アミノ酸残基のポリマーをいうために相互交換可能に使用される。これらの用語は、1つ以上のアミノ酸残基が対応する天然に存在するアミノ酸の人工的な化学的模倣体であるアミノ酸ポリマー、ならびに天然に存在するアミノ酸ポリマーおよび天然に存在しないアミノ酸ポリマーに当てはめられる。
【0046】
用語「アミノ酸」とは、天然に存在するアミノ酸および合成のアミノ酸、鏡像異性体(D−およびL−形態)、およびアキラルアミノ酸、ならびに天然に存在するアミノ酸と類似の様式で機能するアミノ酸アナログおよびアミノ酸模倣体をいう。天然に存在するアミノ酸は、遺伝子コードによってコードされるもの、ならびに、その後改変されるアミノ酸である(例えば、ヒドロキシプロリン、γ−カルボキシグルタメート、およびO−ホスホセリン)。アミノ酸アナログとは、天然に存在するアミノ酸と同じ基本的化学構造を有する化合物をいう(すなわち、水素、カルボキシル基、アミノ基、およびR基(例えば、ホモセリン、ノルロイシン、メチオニン、スルホキシド、メチオニンメチルスルホニウム)に結合されるα−炭素)。そのようなアナログは、改変R基(例えば、ノルロイシン)または改変ペプチド骨格を有するが、天然に存在するアミノ酸と同じ基本的化学構造を保持する。アミノ酸模倣体は、アミノ酸の一般の化学構造と異なる構造を有するが、天然に存在するアミノ酸と類似の様式で機能する化学化合物をいう。
【0047】
本明細書において、アミノ酸は、一般に公知の3文字記号、またはIUPAC−IUB Biochemical Nomenclature Commissionによって推奨される1文字記号のいずれかによって言及され得る。同様に、ヌクレオチドは、その一般に容認されている1文字コードによって言及され得る。
【0048】
「保存的に改変された改変体」は、アミノ酸配列と核酸配列の両方に適用される。特定の核酸配列に関して、保存的に改変された改変体とは、同一のまたは本質的に同一のアミン酸配列をコードする核酸をいうか、あるいは核酸がアミノ酸配列をコードしない場合は、本質的に同一の配列をいう。遺伝子コードの縮重に起因して、多数の機能的に同一の核酸が、任意の所与のタンパク質をコードする。例えば、コドンGCA、GCC、GCGおよびGCUは全て、アミノ酸のアラニンをコードする。従って、アラニンがコドンによって特定される全ての位置において、そのコドンは、コードされたポリペプチドを変更することなしに、記載の任意の対応するコドンに変更され得る。そのような核酸の多様性は、「サイレント多様性(silent variation)」であり、これは、1種の保存的に改変された改変体(variation)である。本明細書において、ポリペプチドをコードする全ての核酸配列はまた、核酸の全ての可能なサイレント多様性を説明する。当業者は、核酸の各コドン(通常メチオニンのための唯一のコドンであるAUG、および通常トリプトファンのための唯一のコドンであるTGGを除く)が、機能的に同一の分子を産生するために改変され得ることを認識する。従って、ポリペプチドをコードする核酸の各々のサイレント多様性は、実際のプローブの配列に関してではなく、発現産物に関して、各々の記載された配列について暗示する。
【0049】
アミノ酸配列に関して、当業者は、コードされた配列中の1つのアミノ酸もしくはわずかな割合のアミノ酸を変更、付加または欠失させる核酸配列、ペプチド配列、ポリペプチド配列、またはタンパク質配列への個々の置換、欠失または付加が、その変更が化学的に類似のアミノ酸でのアミノ酸の置換を生じる「保存的に改変された改変体」であることを認識する。機能的に類似のアミノ酸を提供する保存的置換表は、当該分野において周知である。そのような保存的に改変された改変体は、本発明の多型改変体、種間のホモログ、および対立遺伝子に加えられ、そしてこれらを排除しない。
【0050】
以下の8つのグループは、それぞれ、相互に保存的置換であるアミノ酸を含む:1)アラニン(A)、グリシン(G);2)アスパラギン酸(D)、グルタミン酸(E);3)アスパラギン(N)、グルタミン(Q);4)アルギニン(R)、リジン(K);5)イソロイシン(I)、ロイシン(L)、メチオニン(M)、バリン(V);6)フェニルアラニン(F)、チロシン(Y)、トリプトファン(W);7)セリン(S)、スレオニン(T);ならびに8)システイン(C)、メチオニン(M)(例えば、Creighton,Proteins(1984)を参照のこと)。
【0051】
マクロ分子構造(例えば、ポリペプチド構造)は、種々のレベルの構成の点から考察され得る。この構成の一般的考察については、例えば、Albertsら、Molecular Biology of the Cell(第3版、1994年)ならびにCantorおよびSchimmel、Biophysical Chemistry Part I:The Conformation of Biological Macromolecules(1980)を参照のこと。「一次構造」とは、特定のペプチドのアミノ酸配列をいう。「二次構造」とは、ポリペプチド内の局所的に整列した、3次元構造をいう。これらの構造は、一般には、ドメイン(例えば、細胞外ドメイン、膜貫通ドメイン、および細胞質ドメイン)として公知である。ドメインは、ポリペプチドの圧縮したユニットを形成するポリペプチドの部分であり、そして代表的に、15〜350アミノ酸長である。代表的なドメインは、より小さい構成(例えば、βシートおよびαヘリックスの伸張)の部分からなりたっている。「三次構造」とは、ポリペプチドモノマーの完全な三次元構造をいう。「四次構造」とは、独立の三次ユニットの非共有結合性の会合によって形成された3次元構造をいう。異方性の用語はまた、エネルギー用語としても公知である。
【0052】
「核酸」とは、デオキシリボヌクレオチドまたはリボヌクレオチド、および単鎖または二本鎖のいずれかの形態のそれらのポリマー、ならびに任意のそのような配列の相補体をいう。核酸にはまた、DNA、cDNA、RNA、ポリヌクレオチド、ヌクレオチドなどが含まれる。この用語は、公知のヌクレオチドアナログまたは改変された骨格の残基または連結を含む核酸を包含し、この核酸は、合成されるもの、天然に存在するもの、および天然に存在しないものであり、参照核酸と類似の結合特性を有し、そして参照ヌクレオチドと類似の様式で代謝される。そのようなアナログの例としては、限定されることなく、ホスホロチオエート、ホスホルアミデート、メチルホスホネート、キラル−メチルホスホネート、2−O−メチルリボヌクレオチド、ペプチド−核酸(PNA)が挙げられる。
【0053】
特定の核酸配列はまた、暗黙のうちに、「スプライス改変体」を含む。同様に、核酸によってコードされる特定のタンパク質は、その核酸のスプライス改変体によってコードされる任意のタンパク質を暗黙のうちに含む。その名称が示唆するように、「スプライス改変体」は、遺伝子の代替的なスプライシングの産物である。転写後、最初の核酸転写産物が、スプライシングされ得、それゆえ、異なる(代替的な)核酸スプライス産物が、異なるポリペプチドをコードし得る。スプライス改変体の産物の機構は様々であるが、エキソンの代替的なスプライシングを含む。読み過し転写による同じ核酸に由来する代替的なポリペプチドもまた、この定義に含まれる。スプライシング反応の任意の産物(スプライス産物の組換え型形態を含む)が、この定義の中に含まれる。
【0054】
「標識」または「検出可能部分」とは、分光的手段、光化学的手段、生化学的手段、免疫化学的手段、化学的手段、または他の物理的手段によって検出可能な組成物である。例えば、有用な標識としては、32P、蛍光染料、電子密集試薬(electron−dense reagent)、酵素(例えば、ELISA中で一般に使用されるようなもの)、ビオチン、ジゴキシゲニン、またはハプテンおよびタンパク質が挙げられ、これらは、例えば、ペプチドの中へ放射標識を取り込ませることによって検出可能とされ得るか、またはこのペプチドと特異的に反応する抗体を検出するために使用され得る。
【0055】
例えば、細胞、または核酸、タンパク質、またはベクターを参照して使用する場合、用語「組換え型」とは、細胞、核酸、タンパク質またはベクターが、異種の核酸もしくはタンパク質の導入、またはネイティブの核酸もしくはタンパク質の変更によって改変されたこと、あるいは、細胞が、そのように改変された細胞に由来することを示す。従って、例えば、組換え型細胞は、細胞のネイティブ(非組換え型)形態内に見出されない遺伝子を発現するか、あるいは発現される条件下で別のやり方で異常発現されるネイティブの遺伝子を発現するか、または全く発現されない。
【0056】
核酸の一部への参照として使用される場合、用語「異種」とは、核酸が天然において相互に同じ関係で見出されない2つ以上の部分配列を含むことを示す。例えば、この核酸は、代表的に、組換えで産生され、新規の機能的核酸を産生するように整列された関連しない遺伝子由来の2つ以上の配列を有する(例えば、1つの供給源に由来するプロモーターおよび別の供給源に由来するコーディング領域)。同様に、異種のタンパク質は、このタンパク質が、天然において相互に同じ関係で見出されない2つ以上の部分配列を含むことを示す(例えば、融合タンパク質)。
【0057】
語句「ストリンジェントなハイブリダイゼーション条件」とは、プローブが、代表的に核酸の複合混合物中でその標的の部分配列にハイブリダイズするが、他の配列にはハイブリダイズしない条件をいう。ストリンジェントな条件は、配列に依存し、そして異なる環境においては、その条件は異なる。より長い配列は、より高温で特異的にハイブリダイズする。核酸のハイブリダイゼーションに関する広範なガイドは、Tijssen,Techniques in Biochemistry and Molecular Biology−−Hybridization with Nucleic Probes,「Overview of principles of hybridization and the strategy of nucleic acid assays」(1993)中に見出される。一般に、ストリンジェントな条件は、規定のイオン強度、pHにおいて特異的配列の熱融点(Tm)よりも約5〜10℃低くなるように選択される。このTmは、標的に相補的な50%のプローブが、平衡状態にある標的配列にハイブリダイズする(標的配列が過剰に存在する場合、Tmにおいて50%のプローブが平行状態において占有される)(規定のイオン強度、pH、および核濃度の下での)温度である。ストリンジェントな条件はまた、非安定化因子(例えば、ホルムアミド)の添加によって達成され得る。選択的または特異的なハイブリダイゼーションについて、正のシグナルは、少なくともバックグラウンドの2倍(好ましくは、バックグラウンドのハイブリダイゼーションの10倍)である。例示的なストリンジェントなハイブリダイゼーション条件は、以下のようなものであり得る:50%ホルムアミド、5×SSC、および1% SDS、42℃にてインキュベートされるか、または5×SSC、1% SDS、65℃にてインキュベートされ、0.2×SSC中で洗浄、そして0.1% SDS、65℃。
【0058】
ストリンジェントな条件下で相互にハイブリダイズしない核酸は、それらがコードするポリペプチドが実質的に同一である場合、それでも実質的に同一である。このことは、例えば、核酸のコピーが、遺伝子コードによって可能とされた最大限のコドンの縮重を使用して生成される場合に、発生する。そのような場合、この核酸は、代表的には、中程度にストリンジェントなハイブリダイゼーション条件下で、ハイブリダイズする。例示的な「中程度にストリンジェントなハイブリダイゼーション条件」は、40%ホルムアミド、1M NaCl、1% SDS、37℃の緩衝液中のハイブリダイゼーションおよび1×SSC、45℃の洗浄を含む。正のハイブリダイゼーションは、バックグラウンドの少なくとも2倍である。技術者は、代替的なハイブリダイゼーションおよび洗浄の条件が、類似のストリンジェントな条件を提供するために利用され得ることを容易に認識する。ハイブリダイゼーションパラメータを決定するためのさらなるガイドラインが、多数の参考文献(例えば、Current Protocols in Molecular Biology,Ausubelら(編))において提供されている。
【0059】
PCRについて、アニーリング温度は、プライマーの長さに依存して、約32℃と48℃との間で変動し得るが、約36℃の温度が低ストリンジェントな増幅のために典型的である。プライマーの長さおよび特異性に依存して、高ストリンジェントなアニーリング温度は、約50℃から約65℃までの範囲であり得るが、高ストリンジェントなPCR増幅について、約62℃の温度が典型的である。高ストリンジェントな増幅および低ストリンジェントな増幅の両方についての典型的なサイクルの条件は、以下の条件の30〜40サイクルを含む:30秒〜2分間の90℃〜95℃の変性期、30秒〜2分続くアニーリング期、および1分〜2分間の約72℃の伸張期。低ストリンジェントな増幅反応および高ストリンジェントな増幅反応のためのプロトコルおよびガイドラインは、例えば、Innisら、PCR Protocols,A Guide to Methods and Applications(1990)中に提供される。
【0060】
「抗体」とは、特異的に抗原に結合し、そして認識する免疫グロブリン遺伝子またはそのフラグメント由来のフレームワーク領域を含むポリペプチドをいう。この認識された免疫グロブリン遺伝子としては、κ、λ、α、γ、δ、ε、およびμ定常領域遺伝子、ならびに無数の免疫グロブリン可変領域遺伝子が挙げられる。軽鎖は、κまたはλのいずれかとして分類される。重鎖は、γ、μ、α、δ、またはεとして分類され、これらは、順番に、それぞれ、免疫グロブリンクラスのIgG、IgM、IgA、IgDおよびIgEを規定する。代表的に、抗体の抗原結合領域は、結合の特異性および親和性において最も重要である。
【0061】
例示的な免疫グロブリン(抗体)構造単位は、テトラマーを含む。各テトラマーは、ポリペプチド鎖の2つの同一のペアからなり、各ペアは、1つの「軽」鎖(約25kD)および1つの「重」鎖(約50〜70kD)を有する。各鎖のN末端は、主に抗原認識を担う約100から110以上のアミノ酸の可変領域を規定する。用語、可変軽鎖(VL)および可変重鎖(VH)とは、それぞれ、これらの軽鎖および重鎖をいう。
【0062】
抗体は、例えば、インタクトな免疫グロブリンとしてか、あるいは種々のぺプチダーゼによる消化によって産生される複数のよく特徴付けされたフラグメントとして存在する。従って、例えば、ペプシンは、ヒンジ領域のジスルフィド結合下で抗体を消化し、F(ab)’2(それ自身が、ジスルフィド結合によってVH−CH1に結合される軽鎖である、Fabのダイマー)を産生する。このF(ab)’2は、緩やかな条件下で還元され、ヒンジ領域のジスルフィド結合を分解し得、それによって、F(ab)’2ダイマーはFab’モノマーに変換する。このFab’モノマーは、本質的にヒンジ領域の一部を有するFabである(Fundamental Immunology(Paul(編)、第3版、1993年)参照のこと)。種々の抗体フラグメントが、インタクトな抗体の消化に関して規定されるが、当業者は、そのようなフラグメントが、化学的にかまたは組換えDNA技術を使用することによって、新規に合成され得ることを理解する。従って、本明細書において使用される場合、用語、抗体はまた、抗体全体の改変によって産生された抗体フラグメントか、または組換えDNA方法論を使用して新規に合成された抗体フラグメント(例えば、単鎖Fv)か、またはファージディスプレイライブラリー(例えば、McCaffertyら、Nature 348:552−554(1990)を参照のこと)を使用して同定された抗体フラグメントを含む。
【0063】
抗体(例えば、組換え型抗体、モノクローナル抗体、またはポリクローナル抗体)の調製について、当該分野において公知の多くの技術が使用され得る(例えば、KohlerおよびMilstein,Nature 256:495−497(1975);Kozborら、Immunology Today 4:72(1983);Coleら、Monoclonal Antibodies and Cancer Therapy,77−96頁、Alan R.Liss,Inc.(1985);Coligan,Current Protocols in Immunology(1991);HarlowおよびLane,Antibodies,A Laboratory Manual(1988);およびGoding,Monoclonal Antibodies:Principles and Practice(第2版、1986年)を参照のこと)。単鎖抗体の産生のための技術(米国特許第4,946,778号)が、本発明のポリペプチドに対する抗体を産生するために採用され得る。さらに、トランスジェニックマウス、または他の生物体(例えば、他の哺乳動物)は、ヒト化抗体を発現するために使用され得る。あるいは、ファージディスプレイ技術が、選択された抗原に特異的に結合する抗体およびヘテロマーのFabフラグメントを同定するために使用され得る(例えば、McCaffertyら、Nature 348:552−554(1990);Marksら、Biotechnology 10:779−783(1992)を参照のこと)。
【0064】
「キメラ抗体」とは、(a)定常領域、もしくはその一部が、変更、置換、もしくは交換され、それゆえ抗原結合部位(可変領域)が、異なるかもしくは変更された、クラス、エフェクター機能および/もしくは種の定常領域に結合されるか、またはキメラ抗体に新規の特性を与える完全に異なる分子(例えば、酵素、トキシン、ホルモン、成長因子、薬物など)に結合される抗体分子か;あるいは、(b)可変領域、もしくはその一部が、異なるかもしくは変更された抗原特異性を有する可変領域と変更、置換、もしくは交換される抗体分子である。
【0065】
一実施形態において、抗体は、「エフェクター」部分に接合される。このエフェクター部分は、任意の数の分子(標識化部分(例えば、放射性標識または蛍光標識)を含む)であり得るか、または治療部分であり得る。一局面において、抗体は、タンパク質の活性を調節する。
【0066】
タンパク質またはペプチドを参照する場合、語句、抗体に「特異的に(もしくは選択的に)結合する」または抗体と「特異的に(もしくは選択的に)免疫応答する」とは、多くの場合異種集団のタンパク質および他の生物学的製剤中において、タンパク質の存在を測定する結合反応をいう。従って、指定されたイムノアッセイ条件下で、特異化された抗体は、バックグラウンドの少なくとも2倍、そしてより代表的にはバックグラウンドの10倍から100倍を超えて特定のタンパク質に結合する。そのような条件下での抗体への特異的な結合は、特定のタンパク質についての特異性のために選択される抗体を必要とする。例えば、T1Rタンパク質、あるいは、配列番号1〜25によってコードされる配列を含み、ホモダイマーかもしくはヘテロダイマーのT1R3を含む味レセプター、多型改変体、対立遺伝子、オルソログ、および保存的に改変された改変体、もしくはスプライス改変体、またはそれらの部分に対して惹起されたポリクローナル抗体は、T1Rタンパク質および/あるいはホモダイマーかもしくはヘテロダイマーのT1R3を含む味レセプターと特異的に免疫応答し、そして他のタンパク質と特異的に免疫応答しないポリクローナル抗体のみを得るために選択され得る。一実施形態において、抗体は、ホモダイマーのT1R3を含む味レセプターに反応するが、T1Rファミリーの個々のタンパク質メンバーには反応しない。この選択は、他の分子と交差反応する抗体を削除することによって達成され得る。種々のイムノアッセイ形式が、特定のタンパク質と特異的に免疫応答する抗体を選択するために使用され得る。例えば、固相ELISAイムノアッセイが、タンパク質と特異的に免疫応答する抗体を選択するために慣用的に使用される(特異的な免疫反応性を測定するために使用され得るイムノアッセイの形式および条件の説明については、例えば、HarlowおよびLane,Antibodies,A Laboratory Manual(1988)を参照のこと)。
【0067】
(T1Rファミリーメンバーをコードする核酸の単離)
本発明は、組換え遺伝学の分野の慣例的な技術に依存する。本発明における使用の一般的な方法を開示する基本的テキストとしては、Sambrookら、Molecular Cloning,A Laboratory Manual(第2版、1989年);Kriegler,Gene Transfer and Expression:A Laboratory Manual(1990);およびCurrent Protocols in Molecular Biology(Ausubelら(編)、1994年)が挙げられる。
【0068】
本明細書において開示されるアミノ酸配列に実質的に同一である、T1R核酸、多型改変体、オルソログ、および対立遺伝子は、T1R核酸プローブ、およびオリゴヌクレオチドを使用して、ストリンジェントなハイブリダイゼーション条件下でライブラリーをスクリーニングすることによって単離し得る。あるいは、発現ライブラリーは、ヒトT1Rまたはその一部分に対して産生された抗血清または精製された抗体を用いて、免疫学的に発現されたホモログを検出することによって、T1Rタンパク質、多型改変体、オルソログ、および対立遺伝子をクローニングするために使用され得る。
【0069】
cDNAライブラリーを作製するために、当業者は、T1R RNAが豊富な供給源(例えば、味蕾(例えば、有郭、葉状、茸状、および口蓋))を選択するべきである。このmRNAは、次いで、逆転写酵素を使用してcDNAにされ、組換えベクターに連結され、そして増殖、スクリーニング、およびクローニングのために組換え型宿主へトランスフェクトされる。cDNAライブラリーを作製およびスクリーニングするための方法は、周知である(例えば、GublerおよびHoffman,Gene 25:263−269(1983);Sambrookら、前出;Ausubelら、前出、を参照のこと)。
【0070】
ゲノムライブラリーのために、上記DNAは、組織から抽出され、そして約12〜20kbのフラグメントを生成するために、機械的に剪断されるか、または酵素的に消化される。次いで、このフラグメントは、勾配遠心分離によって所望されない大きさのものから分離され、そしてバクテリオファージラムダベクター中に構築される。これらのベクターおよびファージは、インビトロでパッケージされる。組換え型ファージは、BentonおよびDavis,Science 196:180−182(1977)中に記載されるようにプラークハイブリダイゼーションによって分析される。コロニーハイブリダイゼーションが、Grunsteinら、Proc.Natl.Acad.Sci.USA.,72:3961−3965(1975)中に一般的に記載されるように実行される。
【0071】
T1R核酸およびそのオルソログ、対立遺伝子、変異体、多型改変体、および保存的に改変された改変体を単離する代替的な方法は、合成オリゴヌクレオチドプライマーの使用とRNAもしくはDNAテンプレートの増幅とを組み合わせる(米国特許第4,683,195号および同第4,683,202号;PCR Protocols:A Guide to Methods and Applications(Innisら(編)、1990年)を参照のこと)。ポリメラーゼ連鎖反応(PCR)およびリガーゼ連鎖反応(LCR)のような方法を、直接的に、mRNAから、cDNAから、ゲノムライブラリーからまたはcDNAライブラリーからヒトT1Rの核酸配列を増幅するために使用し得る。変質したオリゴヌクレオチドを、本明細書において提供される配列を使用してT1Rホモログを増幅するために、設計し得る。制限エンドヌクレアーゼ部位は、プライマーの中へ取り込まれ得る。ポリメラーゼ連鎖反応または他のインビトロ増幅方法はまた、例えば、核酸配列決定または他の目的のために、発現されるタンパク質をコードする核酸配列をクローニングするのに、生理学的サンプルにおいてT1RをコードするmRNAの存在を検出するためのプローブとして使用する核酸を作製するのに、有用であり得る。PCR反応によって増幅される遺伝子は、アガロースゲルから精製され得、そして適切なベクター中へクローニングされ得る。
【0072】
T1Rの遺伝子発現はまた、当該分野において公知の技術(例えば、mRNAの逆転写および増幅、全RNAまたはポリA+RNAの単離、ノザンブロット法、ドットブロット法、インサイチュハイブリダイゼーション、RNase防御(protection)、高密度ポリヌクレオチドアレイ技術など)によって分析され得る。
【0073】
T1Rタンパク質をコードする核酸は、本発明のT1Rタンパク質、オルソログ、対立遺伝子、保存的に改変された改変体、および多型改変体を同定するために高密度オリゴヌクレオチドアレイ技術(例えば、GeneChipTM)とともに使用され得る(例えば、Gunthandら、AIDS Res.Hum.Retroviruses 14:869−876(1998);Kozalら、Nat.Med.2:753−759(1996);Matsonら、Anal.Biochem.224:110−106(1995);Lockhartら、Nat.Biotechnol.14:1675−1680(1996);Gingerasら、Genome Res.8:435−448(1998);Haciaら、Nucleic Acids Res.26:3865−3866(1998)を参照のこと)。
【0074】
T1Rの遺伝子は、代表的に、複製および/または発現のための原核生物細胞または真核生物細胞の中への変換の前に、中間ベクター中にクローニングされる。これらの中間ベクターは、代表的に、原核生物ベクター(例えば、プラスミド)またはシャトルベクターである。
【0075】
(原核生物および真核生物の発現)
クローニングされた遺伝子の高レベルの発現(例えば、T1Rタンパク質をコードするcDNAの高レベルの発現)を得るために、当業者は、代表的に、T1Rを発現ベクター中にサブクローニングする。この発現ベクターは、転写、転写/翻訳ターミネーターに指向するための強力なプロモーターを、そして核酸がタンパク質をコードする場合、翻訳開始のためのリボソーム結合部位を含む。T1R核酸は、同じかまたは異なるベクターに、同時発現され得るか、または分離して発現され得る(好ましくは同時発現され得る)。適切な細菌プロモーターは、当該分野において周知であり、そして例えば、Sambrookら、およびAusubelら(前出)に記載される。T1Rタンパク質を発現するための細菌発現系は、例えば、E.coli、Bacillus種、およびSalmonella(Palvaら、Gene 22:229−235(1983);Mosbachら、Nature 302:543−545(1983))において入手可能である。そのような発現系のためのキットは、市販されている。哺乳動物細胞、酵母、および昆虫細胞のための真核生物発現系は、当該分野において周知であり、そしてまた市販されている。好ましい一実施形態において、レトロウイルス発現系が、本発明中で使用される。
【0076】
異種の核酸の発現を導くために使用されるプロモーターの選択は、特定の用途に依存している。プロモーターは、好ましくは、異種の転写開始部位から、その天然の設定における、転写開始部位からの距離とほぼ同じ距離に位置付けされる。しかし、当該分野において公知のように、この距離のいくらかの多様性は、プロモーター機能の損失なしに順応され得る。
【0077】
プロモーターに加えて、発現ベクターは、代表的に、宿主細胞中でT1Rをコードする核酸の発現に必要とされる全てのさらなる要素を含む、転写ユニットまたは発現カセットを含む。従って、代表的な発現カセットは、T1Rをコードする核酸配列に作動可能に結合するプロモーター、ならびに、転写物の効率的なポリアデニル化、リボソーム結合部位、および翻訳終止のために必要とされるシグナルを含む。このカセットのさらなる要素は、エンハンサー、ならびに、ゲノムDNAが構造遺伝子として使用される場合、機能的スプライスドナーおよびアクセプター部位を伴うイントロンを含み得る。
【0078】
プロモーター配列に加えて、上記発現カセットはまた、効率的な終止を提供するために構造遺伝子の下流に転写終止領域を含むはずである。この終止領域は、プロモーター配列と同じ遺伝子から得られ得るか、または異なる遺伝子から得られ得る。
【0079】
細胞に遺伝情報を輸送するために使用される特定の発現ベクターは、特に重要ではない。真核生物細胞または原核生物細胞における発現のために使用される任意の従来のベクターが、使用され得る。標準的な細菌発現ベクターは、プラスミド(例えば、pBR322ベースのプラスミド、pSKF、pET23D)および融合発現系(例えば、MBP、GST、およびLacZ)を含む。エピトープのタグがまた、組換えタンパク質に付加され、単離のための便利な方法(例えば、c−myc)を提供し得る。配列のタグが、核酸回収(rescue)のために発現カセットに含まれ得る。マーカー(例えば、蛍光タンパク質、緑蛍光タンパク質、または赤蛍光タンパク質、β−gal、CATなど)が、ベクター変換のためのマーカーとしてベクター中に含まれ得る。
【0080】
真核生物のウイルス由来の調節エレメントを含む発現ベクターは、代表的に、真核生物発現ベクター(例えば、SV40ベクター、乳頭腫ウイルスベクター、レトロウイルスベクター、およびエプスタイン−バーウイルス由来のベクター)において使用される。他の例示的な真核生物ベクターとしては、pMSG、pAV009/A+、pMTO10/A+、pMAMneo−5、バキュロウイルスpDSVE、および任意の他のベクター(CMVプロモーター、SV40早期プロモーター、SV40後期プロモーター、メタロチオネインプロモーター、マウス乳腺癌ウイルスプロモーター、ラウス肉腫ウイルスプロモーター、ポリへドリンプロモーター、または真核生物細胞の発現について有効性が示された他のプロモーターの指示の下、タンパク質の発現を可能にするベクター)が挙げられる。
【0081】
真核生物ベクター由来のタンパク質の発現はまた、誘導プロモーターを使用して調節され得る。誘導プロモーターを用いて、発現レベルは、プロモーターへの誘導因子(例えば、テトラサイクリンまたはエクジソン)のための応答エレメントの取り込みによってこれらの因子の濃度に関連する。概して、高レベルの発現が、誘導因子の存在下のみで誘導プロモーターから得られる;基本の発現レベルは、最小限である。
【0082】
一実施形態において、本発明のベクターは、調節可能なプロモーター(例えば、tet−調節系およびRU−486系)を有する(例えば、GossenおよびBujard,Proc.Nat’l Acad.Sci.USA 89:5547(1992);Oliginoら、Gene Ther.5:491−496(1998);Wangら、Gene Ther.4:432−441(1997);Neeringら、Blood 88:1147−1155(1996);およびRendahlら、Nat.Biotechnol.16:757−761(1998)を参照のこと)。これらは、候補標的核酸の発現における小分子の制御を与える。この有益な特徴は、所望の表現型が、体細胞の変異よりも、トランスフェクトされたcDNAによってもたらされることを決定するために使用され得る。
【0083】
いくつかの発現系は、遺伝子増幅を提供するマーカーを有する(例えば、チミジンキナーゼおよびジヒドロ葉酸還元酵素)。あるいは、遺伝子増幅に関与しない高収量発現系もまた適切である(例えば、昆虫細胞のバキュロウイルスベクターをポリへドリンプロモーターまたは他の強力なバキュロウイルスプロモーターの指示の下、T1Rをコードする配列とともに使用する)。
【0084】
発現ベクターに代表的に含まれるエレメントはまた、E.coliにおいて機能するレプリコン、組換え型プラスミドを有する細菌の選択を可能にさせる抗生物質耐性をコードする遺伝子、および真核生物の配列の挿入を可能にするプラスミドの本質的ではない領域における特有の制限部位を含む。選択された特定の抗生物質耐性遺伝子は重要ではなく、当該分野において公知の任意の多くの耐性遺伝子が適している。好ましくは、原核生物配列は、必要な場合、それが、真核生物細胞のDNAの複製を干渉しないように、選ばれる。
【0085】
標準的なトランスフェクション方法が、大量のT1Rタンパク質を発現する細菌細胞株、哺乳動物細胞株、酵母細胞株または昆虫細胞株を生成するために使用され、次いで、このタンパク質は、標準的な技術を使用して精製される(例えば、Colleyら、J.Biol.Chem.264:17619−17622(1989);Methods in Enzyrrcology,182巻(Deutscher,(編),1990年)中の、Guide to Protein Purificationを参照のこと)。真核生物細胞および原核生物細胞の変換は、標準的な技術に従って実行される(例えば、Morrison,J Bact.132:349−351(1977);Clark−CurtissおよびCurtiss,Methods in Enzymology 101:347−362(Wuら(編)、1983年)を参照のこと)。
【0086】
宿主細胞に外来のヌクレオチド配列を導入するための任意の周知の手順が、使用され得る。これらは、リン酸カルシウムトランスフェクション、ポリブレン、プロトプラスト融合、エレクトロポレーション、微粒子銃、リポソーム、マイクロインジェクション、細胞質ベクター(plasma vector)、ウイルスベクターならびに宿主細胞中へクローニングされたゲノムDNA、cDNA、合成DNAまたは他の外来遺伝物質を導入するための他の周知の任意の方法の使用を含む(例えば、Sambrookら(前出)を参照のこと)。使用された特定の遺伝子工学手順が、T1Rを発現し得る宿主細胞に、少なくとも1つの遺伝子を首尾よく導入し得ることのみが重要である。
【0087】
細胞に発現ベクターが導入された後、トランスフェクトされた細胞は、T1Rの発現を有利にする条件下で培養され、このT1Rは、以下に同定される標準的な技術を使用して、培養物から回収される。
【0088】
(T1Rポリペプチドの精製)
天然に存在するT1RポリペプチドもしくはT1R3を含むレセプターか、または組換え型T1RポリペプチドまたはT1R3を含むレセプターは、機能的アッセイにおける使用のために精製され得る。天然に存在するT1Rタンパク質またはT1R3を含むレセプターは、例えば、ヒト組織から精製され得る。組換え型T1RポリペプチドまたはT1R3を含むレセプターは、任意の適切な発現系から精製され得る。T1Rポリペプチドは、代表的には、T1R3を含むレセプターを形成するために同じ細胞中で同時発現される。
【0089】
上記T1Rタンパク質またはT1R3を含むレセプターは、標準的な技術(硫酸アンモニアのような物質を用いた選択的沈降反応;カラムクロマトグラフィー;免疫精製方法、および他の方法(例えば、Scopes,Protein Purification:Principles and Practice(1982);米国特許第4,673,641号;Ausubelら、前出;およびSambrookら、前出、を参照のこと)を含む)によって相当の純度まで精製され得る。
【0090】
複数の手順が、組換え型T1Rタンパク質またはT1R3を含むレセプターを精製する場合に、使用され得る。例えば、確立した分子付着特性を有するタンパク質は、可逆的にあり、T1Rタンパク質またはT1R3を含むレセプターに融合され得る。適切なリガンドを使用して、T1Rタンパク質またはT1R3を含むレセプターは、選択的に精製カラムに吸着され、次いで、比較的純粋な形態でそのカラムから遊離され得る。次いで、この融合されたタンパク質は、酵素の活性によって除去される。最終的に、T1Rタンパク質またはT1R3を含むレセプターは、イムノアフィニティーカラムを使用して精製され得る。
【0091】
(A.組換え型細菌からのT1Rの精製)
組換え型タンパク質が、代表的に、プロモーターの誘導後、変換された細菌によって大量に発現される。しかし、発現は、構成的であり得る。IPTGを用いたプロモーターの誘導は、誘導プロモーター系の一例である。細菌は、当該分野において標準的な手順によって増殖される。新鮮なもしくは冷凍された細菌細胞が、タンパク質の分離のために使用される。
【0092】
細菌中に発現されたタンパク質は、不溶性の凝集体(「封入体」)を形成し得る。いくつかのプロトコルが、T1Rタンパク質またはT1R3を含むレセプターの封入体の精製に適している。例えば、封入体の精製は、代表的に、細菌細胞の分裂による(例えば、50mM TRIS/HCL pH 7.5、50mM NaCl、5mM MgCl2、1mM DTT、0.1mM ATP、および1mM PMSFの緩衝液中のインキュベーションによる)封入体の抽出、分離および/または精製を含む。この細胞の懸濁液は、French Pressを2〜3回通すことを使用して溶解され得るか、Polytron(Brinkman Instruments)を使用して均質化され得るか、あるいは氷上で超音波をかけられ得る。細菌を溶解する代替的な方法は、当業者に明白である(例えば、Sambrookら、前出;Ausubelら、前出、を参照のこと)。
【0093】
必要な場合、上記封入体は、可溶化され、そしてその溶解した細胞懸濁液は、代表的に、所望されない不溶性物質を除去するために遠心分離される。封入体を形成したタンパク質は、適合する緩衝液を用いた希釈または透析によって再生され得る。適切な溶媒としては、尿素(約4Mから約8Mまで)、ホルムアミド(少なくとも約80%体積(体積/体積ベース))、および塩酸グアニジン(約4Mから約8Mまで)が挙げられるが、これらに限定されない。凝集体を形成するタンパク質を可溶化し得るいくつかの溶媒(例えば、SDS(ドデシル硫酸ナトリウム)、70%ギ酸)は、免疫原性および/または活性の欠如を伴うタンパク質の不可逆な変性の可能性のため、この手順における使用に不適切である。塩酸グアニジンおよび類似の因子は変性剤であるが、この変性は不可逆性ではなく、そして再生が変性剤の除去(透析による、例えば)または希釈の際に起こり得、免疫学的におよび/または生物学的に活性なタンパク質の再形成を可能にする。他の適切な緩衝液は、当業者に公知である。ヒトT1Rタンパク質またはT1Rを含むレセプターは、標準的な分離技術(例えば、Ni−NTAアガロース樹脂を用いる)によって細菌の他のタンパク質から分離される。
【0094】
あるいは、細菌のペリプラズムからT1Rタンパク質またはT1Rを含むレセプターを精製することが可能である。細菌の溶解後、このT1Rタンパク質またはT1Rを含むレセプターが細菌のペリプラズムに放出される場合、この細菌のペリプラズム画分は、低温浸透圧性ショックおよび当業者に公知の他の方法によって単離され得る。このペリプラズムから組換え型タンパク質を単離するために、細菌細胞は遠心分離され、ペレットを形成する。このペレットは、20%スクロースを含む緩衝液中で再懸濁される。この細胞を溶解するために、細菌を遠心分離し、そしてそのペレットを氷冷した5mM MgSO4中で再懸濁し、そして約10分間氷浴中器に保存する。この細胞懸濁液を遠心分離し、そしてその上清をデカントし、そして保存する。この上清に存在する組換え型タンパク質は、当業者に周知の標準的な分離技術によって宿主タンパク質から分離され得る。
【0095】
(B.T1Rタンパク質を精製するための標準的なタンパク質分離技術)
(溶解度による分別)
初めの工程として,多くの場合(特にタンパク質混合物が複雑である場合に)、初めの塩分別は、目的の組換え型タンパク質から多くの所望されない宿主細胞タンパク質(もしくは細胞培養培地由来のタンパク質)を分離し得る。好ましい塩は、硫酸アンモニウムである。硫酸アンモニウムは、タンパク質混合物中の水の量を効果的に減少させることによってタンパク質を沈殿させる。次いで、タンパク質は、その溶解度に基づいて沈殿する。タンパク質がより疎水性であるほど、タンパク質が低い硫酸アンモニウムの濃度で沈殿する可能性がより高い。代表的なプロトコルは、結果として生じる硫酸アンモニウム濃度を20〜30%の間にするように、タンパク質溶液に飽和硫酸アンモニウムを添加する工程を包含する。この濃度は、最も疎水性であるタンパク質を沈殿させる。次いで、この沈殿物は廃棄され(目的のタンパク質が疎水性でない限り)、そして硫酸アンモニウムが、目的のタンパク質を沈殿することが公知である濃度まで上清に添加される。次いで、この沈殿物は、緩衝液中で可溶化され、そして必要な場合、過剰の塩が、透析もしくはダイアフィルトレーション(diafiltration)のいずれかを通じて除去される。タンパク質の溶解度に依存する他の方法(例えば、低温エタノール沈殿)は、当業者に周知であり、そして複雑なタンパク質混合物を分別するために使用され得る。
【0096】
(サイズ差別的ろ過)
T1Rタンパク質またはT1R3を含むレセプターの分子量を、異なる孔のサイズの膜(例えば、Amicon membraneもしくはMillipore membrane)を通した限外ろ過を使用して、より大きいサイズのタンパク質およびより小さいサイズのタンパク質からその分子量を単離するために使用し得る。第一の工程として、タンパク質混合物が、目的のタンパク質の分子量よりも低いカットオフ分子量を有する孔のサイズの膜を通して限外ろ過される。次いで、この限外ろ過の未透過物は、目的のタンパク質の分子量よりも大きい分子のカットオフを有する膜に対して限外ろ過される。上記組換え型タンパク質は、この膜を通してろ液へと通過する。次いで、このろ液で、以下に記載されるようにクロマトグラフィーを行い得る。
【0097】
(カラムクロマトグラフィー)
T1Rタンパク質またはT1R3を含むレセプターはまた、そのサイズ、正味の表面電荷(net surface charge)、疎水性、およびリガンドに対する親和性に基づき、他のタンパク質から分離され得る。さらに、タンパク質に対して惹起された抗体は、カラムマトリックスに接合され、そしてこのタンパク質は、免疫精製され得る。これらの方法の全てが、当該分野において周知である。クロマトグラフィー技術が、任意のスケールで、そして多くの異なる製造者(例えば、Pharmacia Biotech)からの器具を使用して実行され得ることが、当業者に明白である。
【0098】
(T1Rタンパク質の調節因子のためのアッセイ)
(A.アッセイ)
T1R3を含む味レセプターの調整、および対応する味の調整は、種々のインビトロおよびインビボアッセイを使用して評価され得る。そのようなアッセイは、T1R3を含む味レセプターのインヒビターおよびアクチベーター、および結果的に味のインヒビターおよびアクチベーターについてテストするために使用され得る。T1R3を含む甘味レセプターのそのような調節因子は、味シグナル変換に関与する。T1R3を含む味レセプターの調節因子は、組換え型もしくは天然に存在するT1R3を含む味レセプター(好ましくは、ヒトレセプター)を使用してテストされる。
【0099】
一実施形態において、本発明のモノマーのもしくはホモダイマーのT1R3を含むレセプターは、天然に存在するかもしくは人工の甘味分子(例えば、小有機分子、アミノ酸、ペプチド、炭水化物、脂質、多糖類など)についてスクリーニングするために使用され得る。例えば、本発明のホモダイマーのもしくはモノマーのT1R3を含むレセプターは、以下の実施例のセクションに記載されるように、天然に存在する甘味味覚物質を認識する。そのようなレセプターは、人工甘味料または変更された天然に存在する甘味料(これらの甘味料は、ホモダイマーもしくはモノマーのT1R3を含むレセプターの天然に存在する糖のリガンドを模倣する)についてスクリーニングするために使用され得る。
【0100】
好ましくは、そのT1R3を含む味レセプターは、本明細書において提供される配列かまたはその保存的に改変された改変体によってコードされるような配列を有する。あるいは、このアッセイのT1R3を含む味レセプターは、真核生物に由来し、そして本明細書において提供されるアミノ酸配列に対して、実質的なアミノ酸配列同一性を有するアミノ酸配列を含むか、あるいは、(中程度もしくは高度に)ストリンジェントな条件下で、本明細書において記載されるようなヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる。概して、アミノ酸配列の同一性は、少なくとも60%(好ましくは、少なくとも65%、70%、75%、80%、85%、または90%、最も好ましくは少なくとも95%)である。
【0101】
甘味シグナル変換の測定、あるいはT1R3を含む味レセプターもしくはT1R3を含む味レセプターを発現する細胞(組換え型かもしくは天然に存在するもののいずれか)の甘味シグナル変換表現型の損失の測定が、本明細書において記載されるように、種々のアッセイ(インビトロ、インビボ、およびエクスビボ)を使用して実行され得る。活性または結合に影響を及ぼす適切な物理的変化、化学的変化もしくは表現型変化が、本発明のポリペプチドに対するテスト化合物の影響を評価するために使用され得る。これらの機能的効果がインタクトな細胞または動物を使用して決定される場合、実施者はまた、種々の効果(例えば、シグナル変換の場合、例えば、リガンド結合、ホルモン放出、公知の遺伝マーカーおよび特徴付けされていない遺伝マーカー(例えば、ノザンブロット)の両方に対する転写の変化、細胞代謝の変化(例えば、pHの変化)、ならびに、細胞内セカンドメッセンジャー(Ca2+、IP3、cGMP、もしくはcAMP)の変化)も測定し得る。
【0102】
(インビトロアッセイ)
活性を調整するT1R3を含む味レセプターを有する化合物を同定するためのアッセイが、インビトロで実行され得る。そのようなアッセイは、全長のT1R3を含む味レセプターもしくはその改変体、またはキメラを形成するために異種のタンパク質に融合される、T1R3を含む味レセプターのフラグメント(例えば、細胞外ドメイン)を使用し得る(例えば、WO 01/66563、WO 03/001876、WO 02/064631、およびWO 03/004992を参照のこと)。精製された組換え型もしくは天然に存在するT1R3を含む味レセプターは、本発明のインビトロ方法において使用され得る。精製されたT1R3を含む味レセプターに加えて、その組換え型もしくは天然に存在するT1R3を含む味レセプターは、細胞の溶解物または細胞膜の一部であり得る。以下に記載されるように、結合アッセイは、固体状態かもしくは可溶性状態のいずれかであり得る。好ましくは、そのタンパク質または膜は、固体の支持体に(共有結合性または非共有結合性に)結合される。多くの場合、本発明のインビトロアッセイは、リガンド結合アッセイまたはリガンド親和性アッセイ(本明細書において記載されるような、公知の細胞外リガンドと、または公知の細胞内リガンドGTPのいずれかと、非競合または競合するいずれかのアッセイ)である。他のインビトロアッセイは、このタンパク質についての分光特性(例えば、蛍光、吸収度、屈折率)、流体力学的特性(例えば、形状)、クロマトグラフィー特性、または溶解特性の変化を測定する工程を包含する。
【0103】
一実施形態において、ハイスループット結合アッセイが実行され、その中で、T1R3を含む味レセプターまたはそのフラグメントを含むキメラが、潜在的な調節因子と接触され、そして適切な期間、インキュベートされる。一実施形態において、この潜在的な調節因子は固体の支持体に結合され、そしてT1R3を含む味レセプターが添加される。別の実施形態において、このT1R3を含む味レセプターは、固体の支持体に結合される。広範な種類の調節因子(小有機分子、ペプチド、抗体、およびT1R3を含む味レセプターリガンドアナログを含む)が、以下に記載されるように、使用され得る。広範な種類のアッセイ(標識化タンパク質−タンパク質結合アッセイ、電気泳動移動度シフト、イムノアッセイ、酵素アッセイ(例えば、リン酸化アッセイ)などを含む)が、T1R3を含む味レセプター−調節因子結合を同定するために使用され得る。いくつかの場合、候補調節因子の結合は、競合結合アッセイの使用を通じて決定され、ここで、公知のリガンドの結合との干渉が、潜在的な調節因子の存在下で測定される。T1R3を含む味レセプターリガンドが、本明細書において提供される。この調節因子または公知のリガンドのいずれかがまず結合し、次いで、その競合相手が添加される。このT1R3を含む味レセプターが洗浄された後、潜在的な調節因子または公知のリガンドのいずれかの結合との干渉が、決定される。多くの場合、この潜在的な調節因子または公知のリガンドのいずれかが標識される。
【0104】
(細胞ベースのインビボアッセイ)
別の実施形態において、T1R3を含む味レセプターが細胞中に発現され(例えば、T1Rファミリー(例えば、T1R1およびT1R3またはT1R2およびT1R3)の1つかもしくは2つのメンバーの発現または同時発現による、好ましくは、任意の他のT1Rファミリーメンバーの発現を伴わないT1R3単独の発現による)、そして機能的(例えば、物理的および化学的または表現型)変化が、T1R3を含む味レセプター味調節因子を同定するためにアッセイされる。T1R3を含む味レセプターを発現する細胞もまた、結合アッセイにおいて使用され得る。任意の適切な機能的効果が、本明細書において記載されるように測定され得る。例えば、リガンド結合、Gタンパク質結合、およびGPCRシグナル変換(例えば、細胞内Ca2+レベルの変化)は全て、細胞ベースの系を使用して潜在的な調節因子を同定するための適切なアッセイである。そのような細胞ベースのアッセイのために適切な細胞は、本明細書において記載されるように、初代細胞と細胞株との両方を含む。T1R3を含む味レセプターは、天然に存在するかまたは組換え型であり得る。さらに、上述のように、GPCR活性を有するキメラT1R3を含む味レセプターが、細胞ベースのアッセイにおいて使用され得る。例えば、T1Rタンパク質の細胞外ドメインは、異種のタンパク質(好ましくは、異種のGPCR)の膜貫通ドメインおよび/または細胞質ドメインに融合し得る。そのようなキメラGPCRは、GPCR活性を有し、そして本発明の細胞ベースのアッセイにおいて使用され得る。
【0105】
別の実施形態において、細胞のT1Rポリペプチドレベルは、タンパク質またはmRNAのレベルを測定することによって決定される。T1Rタンパク質またはT1Rシグナル変換に関連するタンパク質のレベルは、T1R3を含む味レセプターもしくはそのフラグメントに選択的に結合する抗体を用いて、イムノアッセイ(例えば、ウェスタンブロット法、ELISAなど)を使用して測定される。mRNAの測定については、増幅(例えば、PCR、LCRを用いる)、またはハイブリダイゼーションアッセイ(例えば、ノザンハイブリダイゼイーション)、RNase防御、ドットブロット法)を使用して)が好ましい。タンパク質またはmRNAのレベルは、本明細書において記載されるように、直接的にまたは間接的に標識された検出因子(例えば、蛍光で標識された核酸または放射性標識された核酸、放射性標識された抗体または酵素で標識された抗体など)を使用して検出される。
【0106】
あるいは、T1R3を含むレセプター発現は、レポーター遺伝子系を使用して測定され得る。そのような系は、レポーター遺伝子(例えば、クロラムフェニコールアセチルトランスフェラーゼ、ホタルルシフェラーゼ、細菌のルシフェラーゼ、β−ガラクトシダーゼおよびアルカリホスファターゼ)に作動可能に結合したT1Rタンパク質プロモーターを使用して考案され得る。さらに、目的のタンパク質は、第二のレポーター(例えば、赤蛍光タンパク質または緑蛍光タンパク質への付着を介した間接的なレポーターとして使用され得る(例えば、MistiliおよびSpector,Nature Biotechnology 15:961−964(1997)を参照のこと)。レポーター構築物は、代表的に、細胞へトランスフェクトされる。潜在的な調節因子での処理後、レポーター遺伝子の転写、翻訳、または活性の量が、当業者に公知の標準的な技術に従って測定される。
【0107】
別の実施形態において、GPCRシグナル変換に関連する機能的効果が、測定され得る。活性化もしくは阻害された、T1R3を含むGタンパク質共役型レセプターは、標的酵素、セカンドメッセンジャー、チャネル、および他のエフェクタータンパク質の特性を変更する。その例としては、cGMPホスホジエステラーゼ、アデニル酸シクラーゼ、ホスホリパーゼC、IP3の活性、ならびにGタンパク質による種々のチャネルの調節が挙げられる。下流での結果がまた調査され得る(例えば、ホスホリパーゼCによるジアシルグリセロールおよびIP3の産生、これに次ぐ、IP3によるカルシウム可動化について)。活性化されたGPCRレセプターは、このレセプターのC末端尾部(およびおそらく他の部位も同様に)をリン酸化するキナーゼのための基質となる。従って、アクチベーターは、γ標識化GTPから上記レセプターへの32Pの転移を促進し、このことは、シンチレーション計数器を用いてアッセイされ得る。C末端尾部のリン酸化は、アレスチンのようなタンパク質の結合を促進し、そしてGタンパク質の結合に干渉する。GPCRシグナル変換、およびシグナル変換をアッセイする方法の一般的な概説については、例えば、Methods in Enzymology,第237巻および第238巻(1994)および第96巻(1983);Bourneら、Nature 10:349:117−27(1991);Bourneら、Nature 348:125−32(1990);Pitcherら、Annu.Rev.Biochem.67:653−92(1998)を参照のこと。
【0108】
上述のように、いくつかのGタンパク質共役型レセプターの活性化は、ホスファチジルイノシトールのホスホリパーゼCを媒介した加水分解を通じたイノシトール三リン酸(IP3)の形成を刺激する(BerridgeおよびIrvine,Nature 312:315−21(1984))。IP3は、次いで、細胞内カルシウムイオンの貯蔵の放出を刺激する。従って、細胞質のカルシウムイオンのレベルの変化、またはセカンドメッセンジャー(例えば、IP3)のレベルの変化を、Gタンパク質共役型レセプター機能を評価するために使用し得る。そのようなGタンパク質共役型レセプターを発現する細胞は、細胞内貯蔵とイオンチャネルの活性化を介することの両方からの寄与の結果として増加した細胞質カルシウムレベルを示し得、この場合、必要ではないが、内部貯蔵からのカルシウム放出から生じる蛍光応答を区別するために、カルシウムを含まない緩衝液中でそのようなアッセイを実行すること(必要に応じてキレート化剤(例えば、EGTA)を補充される)が所望され得る。
【0109】
一例において、T1R3を含む味レセプターGPCR活性は、このレセプターをホスホリパーゼCシグナル変換経路に結合する乱交雑したGタンパク質を有する異種の細胞中のT1R3を含む味レセプターを発現することによって測定される(OffermannsおよびSimon,J.Biol.Chem.270:15175−15180(1995)を参照のこと)。シグナル変換の調節は、細胞内Ca2+レベルの変化を測定することによってアッセイされ、この細胞内Ca2+レベルは、T1R3を含む味レセプターと結合する分子の投与を介するGPCRシグナル変換経路の調節に応じて変化する。Ca2+レベルの変化は、必要に応じて、蛍光Ca2+指示染料および蛍光イメージングを使用して測定される。
【0110】
別の例において、ホスファチジルイノシトール(PI)の加水分解は、本明細書において参考として援用されている米国特許第5,436,128号に従って分析され得る。簡潔にいうと、そのアッセイは、48時間以上、3H−ミオイノシトールを用いて細胞の標識する工程を包含する。この標識された細胞は、1時間、テスト化合物で処理される。この処理された細胞は、クロロホルム−メタノール−水中で溶解および抽出され、その後、イノシトールリン酸はイオン交換クロマトグラフィーによって分離され、そしてシンチレーション計数(scintillation counting)によって定量化された。刺激倍率(fold stimulation)は、緩衝液コントロール存在下のcpmに対するアゴニスト存在下のcpmの比を計算することによって決定される。同様に、阻害倍率(fold inhibition)は、緩衝液コントロール(アゴニストを含んでいても、含んでいなくてもよい)存在下のcpmに対するアンタゴニスト存在下のcpmの比を計算することによって決定される。
【0111】
他のアッセイは、活性化された場合、酵素(例えば、アデニル酸シクラーゼ)を活性化または阻害することによって、細胞内のサイクリックヌクレオチド(例えば、cAMPまたはcGMP)のレベルの変化をもたらすレセプターの活性を決定する工程を包含し得る。レセプターの活性化がサイクリックヌクレオチドレベルの減少をもたらす場合、アッセイ中の細胞にレセプターを活性化する化合物を添加する前に、細胞内のサイクリックヌクレオチドレベルを増加する因子(例えば、フォルスコリン)に細胞を暴露することが好ましくあり得る。
【0112】
一例において、細胞内cAMPまたはcGMPの変化が、イムノアッセイを使用して測定され得る。OffermannsおよびSimon,J.Biol.Chem.270:15175−15180(1995)に記載される方法が、cAMPのレベルを決定するために使用され得る。さらに、Felley−Boscoら、Am.J.Resp.Cell and Mol.Biol.11:159−164(1994)に記載される方法が、cGMPのレベルを決定するために使用され得る。さらに、cAMPおよび/またはGMPを測定するためのアッセイキットが、米国特許第4,115,538号(本明細書において参考として援用されている)に記載されている。
【0113】
一例において、Gタンパク質共役型レセプター活性のためのアッセイは、レセプター活性を報告するために、イオン感受性色素または電位(voltage)感受性色素で負荷される細胞を含む。そのようなレセプターの活性を決定するためのアッセイはまた、テストされた化合物の活性を評価するための陰性のコントロールまたは陽性のコントロールとして、他のGタンパク質共役型レセプターの公知のアゴニストおよびアンタゴニストを使用し得る。調節化合物(例えば、アゴニスト、アンタゴニスト)を同定するためのアッセイにおいて、細胞質中のイオンのレベルまたは膜電位の変化が、それぞれ、イオン感受性指示薬(ion sensitive indicator)または膜電位蛍光指示薬(membrane voltage fluorescent indicator)を使用してモニタリングされる。とりわけ、使用され得るイオン感受性指示薬および電位プローブは、Molecular Probes 1997 Catalog中で開示される。Gタンパク質共役型レセプターについて、乱交雑Gタンパク質(例えば、Gα15およびGα16)が、最適なアッセイにおいて使用され得る(Wilkieら、Proc.Nat’l Acad.Sci.USA 88:10049−10053(1991))。そのような乱交雑Gタンパク質は、広範囲のレセプターの共役を可能にする。
【0114】
(動物モデル)
味覚の動物モデルはまた、味覚の調節因子のためのスクリーニングにおける使用を見出す(例えば、本明細書において記載されるようなT1Rノックアウトマウス系統)。トランスジェニック動物技術(遺伝子ノックアウト技術(例えば、適切な遺伝子標的ベクターを用いる異種の組換えの結果として)、または遺伝子過剰発現を含む)は、T1R3を含むレセプターもしくはその構成要素の欠如もしくは発現の増加をもたらす。所望される場合、T1R3を含むレセプターもしくはその構成要素の組織特異的な発現もしくはノックアウトが必要であり得る。そのような方法によって作製されたトランスジェニック動物は、味調節の動物モデルとしての用途を見出し、そしてさらに、味調節の調節因子をスクリーニングすることに有用である。
【0115】
(B.調節因子)
T1R3を含む味レセプターの調節因子としてテストされる化合物は、任意の小有機分子、または生物学的実体(例えば、タンパク質(例えば、抗体またはペプチド)、アミノ酸、脂質、脂肪、糖(例えば、単糖類、二糖類、または多糖類)、核酸(例えば、アンチセンスオリゴヌクレオチドまたはリボザイム)、または小有機分子)であり得る。あるいは、調節因子は、T1R3を含む味レセプターの遺伝的に変更されたバージョンであり得る。代表的に、テスト化合物は、小有機分子、アミノ酸、ペプチド、脂質、および単糖類、二糖類および多糖類である。
【0116】
本質的に、任意の化学化合物は、本発明のアッセイにおいて潜在的な調節因子またはリガンドとして使用され得るが、非常に多くの場合、水溶液中あるいは、有機(特にDMSOベースの)溶液中に溶解され得る化合物が使用される。このアッセイは、アッセイ工程を自動化することによって、そして代表的には平行して(例えば、ロボットアッセイにおけるマイクロタイタープレート上のマイクロタイター形式において)実行される、任意のアッセイに便利な供給源由来の化合物を提供することによって、多くの化学ライブラリーをスクリーニングするように設計される。多くの化学化合物の供給元(Sigma(St.Louis,MO),Aldrich(St.Louis,MO),Sigma−Aldrich(St.Louis,MO),Fluka Chemika−Biochemica Analytika(Buchs Switzerland)などが挙げられる)が存在することが理解される。
【0117】
好ましい一実施形態において、ハイスループットスクリーニング方法は、多数の潜在的治療用化合物(潜在的調節因子化合物または潜在的リガンド化合物)を含むコンビナトリアル小有機分子ライブラリーまたはペプチドライブラリーを提供する工程を包含する。次いで、そのような「コンビナトリアルケミカルライブラリー」または「リガンドライブラリー」は、本明細書において記載されるように1つ以上のアッセイにおいてスクリーニングされ、所望の特徴的活性を示すそれらのライブラリーメンバー(特定の化学的種またはサブクラス)を同定する。従って、同定された化合物は、従来の「リード化合物」として役立ち得るか、またはそれら自身、潜在的な治療法または実際の治療法として使用され得る。
【0118】
コンビナトリアルケミカルライブラリーは、化学的合成または生物学的合成のいずれかによって、複数の化学的「ビルディングブロック(building block)」(例えば、試薬)を組み合わせることによって、生成された種々の化学化合物の集合である。例えば、線状のコンビナトリアルケミカルライブラリー(例えば、ポリペプチドライブラリー)が、所与の化合物の長さ(すなわち、ポリペプチド化合物中のアミノ酸の数)に対する全ての可能な方法において、一組の化学的ビルディングブロック(アミノ酸)を組み合わせることによって形成される。何百万の化学化合物が、そのような化学的ビルディングブロックのコンビナトリアル混合を通して合成され得る。
【0119】
コンビナトリアルケミカルライブラリーの調製およびスクリーニングは、当業者に周知である。そのようなコンビナトリアルケミカルライブラリーとしては、ペプチドライブラリー(例えば、米国特許第5,010,175号、Furka,Int.J.Pept.Prot.Res.37:487−493(1991)およびHoughtonら、Nature 354:84−88(1991)を参照のこと)が挙げられるが、これらに限定されない。化学的多様性ライブラリーを生成するための他の化学物質もまた、使用され得る。そのような化学物質としては、ペプトイド(例えば、PCT公開番号WO 91/19735)、コードされたペプチド(例えば、PCT公開番号WO 93/20242)、ランダムバイオオリゴマー(例えば、PCT公開番号WO 92/00091)、ベンゾジアゼピン(例えば、米国特許第5,288,514号)、ダイバーソマー(diversomer)(例えば、ヒダントイン、ベンゾジアゼピンおよびジペプチド)(Hobbsら、Proc.Nat.Acad.Sci.USA 90:6909−6913(1993))、ビニル性ポリペプチド(Hagiharaら、J.Amer.Chem.Soc.114:6568(1992))、グルコース骨格を有する非ペプチドペプチド模倣物(nonpeptidal peptidomimetic)(Hirschmannら、J.Amer.Chem.Soc.114:9217−9218(1992))、低化合物ライブラリーのアナログの有機合成(Chenら、J.Amer.Chem.Soc.116:2661(1994)、オリゴカルバメート(Choら、Science 261:1303(1993))、および/またはペプチジルホスホネート(Campbellら、J.Org.Chem.59:658(1994))、核酸ライブラリー(Ausubel、BergerおよびSambrook、全て前出、を参照のこと)、ペプチド核酸ライブラリー(例えば、米国特許5,539,083号を参照のこと)、抗体ライブラリー(例えば、Vaughnら、Nature Biotechnology、14(3):309−314(1996)およびPCT/US96/10287を参照のこと)、炭水化物ライブラリー(例えば、Liangら、Science、274:1520−1522(1996)および米国特許第5,593,853号を参照のこと)、低有機分子ライブラリー(例えば、ベンゾジアゼピン、Baum CおよびEN、1月18日、33頁(1993年);イソプレノイド、米国特許第5,569,588号;チアゾリジノンおよびメタチアザノン(metathiazanone)、米国特許第5,549,974号;ピロリジン、米国特許第5,525,735号および同第5,519,134号;モルホリノ化合物、米国特許第5,506,337号;ベンゾジアゼピン、同第5,288,514号など)が挙げられるが、これらに限定されない。
【0120】
コンビナトリアルライブラリーの調製のための装置は、市販されている(例えば、357 MPS,390 MPS,Advanced Chem Tech,Louisville KY,Symphony,Rainin,Woburn,MA,433A Applied Biosystems,Foster City,CA,9050 Plus,Millipore,Bedford,MAを参照のこと)。さらに、多数のコンビナトリアルライブラリー自体が、市販されている(例えば、ComGenex,Princeton,N.J.,Asinex,Moscow,Ru,Tripos,Inc.,St.Louis,MO,ChemStar,Ltd,Moscow,RU,3D Pharmaceuticals,Exton,PA,Martek Biosciences,Columbia,MD,などを参照のこと)。
【0121】
(C.固体状態ハイスループットアッセイおよび可溶性ハイスループットアッセイ)
一実施形態において、本発明は、T1R3を含む味レセプター、またはT1R3を含む味レセプターを発現する細胞もしくは組織(天然に存在するかまたは組換え型かのいずれか)を使用して、可溶性アッセイを提供する。別の実施形態において、本発明は、ハイスループット形式における固相ベースのインビトロアッセイを提供し、ここで、上記T1R3を含む味レセプターは、固相に付着される。本明細書において記載される任意のアッセイは、ハイスループットスクリーニングのために適合され得る(例えば、リガンド結合、細胞増殖、細胞表面マーカー流動(例えば、スクリーニング、放射性標識化GTP結合)、セカンドメッセンジャー流動(例えば、Ca2+、IP3、cGMP、またはcAMP、サイトカイン産生)など)。
【0122】
本発明のハイスループットアッセイ(可溶性状態または固体状態のいずれか)において、一日で、数千までの異なる調節因子またはリガンドをスクリーニングすることが可能である。この方法論は、インビトロでT1R3を含む味レセプターのために、またはT1R3を含む味レセプターを含む細胞ベースのアッセイもしくは膜ベースのアッセイのために使用され得る。特に、マイクロタイタープレートの各ウェルは、選択された潜在的な調節因子に対する分離アッセイを実行するために使用され得るか、あるいは、濃度もしくはインキュベーション時間効果が観察される場合、5〜10ウェルごとに、1つの調節因子をテストし得る。従って、1つの標準マイクロタイタープレートは、約100(例えば、96)の調節因子をアッセイし得る。1536のウェルプレートが使用される場合、1つのプレートは容易に約100〜約1500の異なる化合物をアッセイし得る。一日につき多くのプレートをアッセイすることが、可能である;約6,000までの、20,000までの、50,000までの、または100,000を超える、異なる化合物についてスクリーニングするアッセイは、本発明の統合的なシステムを使用して可能である。
【0123】
固体状態の反応について、目的のタンパク質もしくはそのフラグメント(例えば、細胞外ドメイン)、または融合タンパク質の一部として目的のタンパク質もしくはそのフラグメントを含む細胞もしくは膜が、共有結合性または非共有結合性の結合を介して(例えば、タグを介して)、直接的にかまたは間接的に、固体状態の構成要素に結合され得る。このタグは、任意の種々の構成要素であり得る。一般的に、タグ(タグ結合因子)に結合する分子は、固体の支持体に固定され、そしてタグ化された目的の分子は、そのタグとそのタグ結合因子との相互作用によって固体の支持体に付着される。
【0124】
多数のタグおよびタグ結合因子が、文献に十分に記載されている公知の分子相互作用に基づいて使用され得る。例えば、タグが天然の結合剤(例えば、ビオチン、タンパク質A、またはタンパク質G)を有する場合、それは適切なタグ結合因子(アビジン、ストレプトアビジン、ニュートラアビジン(neutravidin)、免疫グロブリンのFc領域など)と共に使用され得る。天然の結合剤(例えば、ビオチン)を有する分子に対する抗体もまた、広く入手可能であり、そしてその抗体も、適切なタグ結合因子である;SIGMA Immunochemicals 1998 catalogue SIGMA,St.Louis MOを参照のこと。
【0125】
同様に、任意のハプテン化合物または抗原性化合物が、タグ/タグ結合因子のペアを形成するために、適切な抗体と共に使用され得る。数千の特異的抗体が市販されており、そして多くのさらなる抗体が文献に記載されている。例えば、1つの一般的構造において、タグは第一の抗体であり、そしてタグ結合因子は、第一の抗体を認識する第二の抗体である。抗体−抗原相互作用に加えて、レセプター−リガンド相互作用もまた、タグおよびタグ結合因子のペアとして適切である。例えば、細胞膜レセプターのアゴニストおよびアンタゴニスト(例えば、細胞レセプター−リガンド相互作用(例えば、トランスフェリン、c−キット、ウイルスレセプターリガンド、サイトカインレセプター、ケモカインレセプター、インターロイキンレセプター、免疫グロブリンレセプターおよび抗体、カドヘリンファミリー、インテグリンファミリー、セレクチンファミリーなど));例えば、PigottおよびPower,The Adhesion Molecule Facts Book I(1993)を参照のこと。同様に、トキシンおよび毒物(venom)、ウイルスエピトープ、ホルモン(例えば、オピエート、ステロイドなど)、細胞内レセプター(例えば、種々の小リガンド(ステロイド、甲状腺ホルモン、レチノイド、およびビタミンDを含む);ペプチドの効果を媒介するもの)、薬物、レクチン、糖、核酸(線状ポリマー構造と環状ポリマー構造との両方)、オリゴ糖、タンパク質、リン脂質および抗体は全て、種々の細胞レセプターと相互作用し得る。
【0126】
合成ポリマー(例えば、ポリウレタン、ポリエステル、ポリカーボネート、ポリ尿素、ポリアミド、ポリエチレンイミン、ポリアリーレンスルフィド、ポリシロキサン、ポリイミド、およびポリアセテート)もまた、適切なタグまたはタグ結合因子を形成し得る。多くの他のタグ/タグ結合因子ペアもまた、本開示の概説の際に当業者に理解されるように、本明細書において記載されたアッセイ系において有用である。
【0127】
一般的なリンカー(例えば、ペプチド、ポリエーテルなど)もまたタグとして役立ち得、そしてこれはポリペプチド配列(例えば、約5アミノ酸と200アミノ酸との間のポリグリシン配列)を含み得る。そのような柔軟なリンカーは、当業者に公知である。例えば、ポリ(エチレングリコール)リンカーは、Shearwater Polymers,Inc.Huntsville,Alabamaから入手可能である。これらのリンカーは、必要に応じて、アミド結合、スルフヒドリル結合、またはヘテロ官能基結合(heterofunctional linkage)を有する。
【0128】
タグ結合因子は、現在利用可能な任意の種々の方法を使用して固体の基質に固定される。固体の基質は、一般に、タグ結合因子の一部と反応する表面へ化学基を固定する化学試薬に、その基質の全てまたは一部を暴露することによって誘導体化されるかまたは官能基化される。例えば、より長い鎖の部分へ付着するために適している基としては、アミン基、ヒドロキシル基、チオール基、およびカルボキシル基が挙げられる。アミノアルキルシランおよびヒドロキシアルキルシランは、種々の表面(例えば、ガラス表面)を官能基化するために使用され得る。そのような固相のバイオポリマーアレイの構築物は、文献に十分に記載されている。例えば、Merrifield,J.Am.Chem.Soc.85:2149−2154(1963)(例えば、ペプチドの固相合成を記載している);Geysenら、J.Immun.Meth.102:259−274(1987)(ピン(pin)上での固相構成要素の合成を記載している);FrankおよびDoring,Tetrahedron 44:60316040(1988)(セルロースディスク上での種々のペプチド配列の合成を記載している);Fodorら、Science,251:767−777(1991);Sheldonら、Clinical Chemistry 39(4):718−719(1993);およびKozalら、Nature Medicine 2(7):753759(1996)(いずれも、固体の基質へ固定されるバイオポリマーのアレイを記載している)を参照のこと。基質にタグ結合因子を固定するための非化学的アプローチは、他の一般的方法(例えば、加熱、紫外線放射による架橋など)を含む。
【0129】
(T1R3を含むレセプターの免疫学的検出)
核酸ハイブリダイゼーション技術を使用するT1R遺伝子および遺伝子発現の検出に加えて、当業者はまた、本発明のT1R3を含む味レセプターを検出するためにイムノアッセイを使用し得る。そのようなアッセイは、T1R3を含む味レセプターの調節因子についてスクリーニングするため、ならびに治療上の用途および診断上の用途のために有用である。イムノアッセイは、T1R3を含む味レセプターを定性的にまたは定量的に分析するために使用され得る。この適用可能な技術の一般的概要は、HarlowおよびLane,Antibodies:A Laboratory Manual(1988)に見出され得る。
【0130】
(A.抗体の産生)
T1Rタンパク質およびT1R3を含む味レセプターと特異的に反応するポリクローナル抗体およびモノクローナル抗体を産生する方法は、当業者に公知である(例えば、Coligan,Current Protocols in Immunology(1991);HarlowおよびLane(前出);Goding,Monoclonal Antibodies:Principles and Practice(第2版、1986年);ならびに、KohlerおよびMilstein,Nature 256:495−497(1975)を参照のこと)。そのような技術としては、ファージまたは類似のベクター中の組換え型抗体のライブラリー由来の抗体の選択による抗体の調製、ならびに、ウサギまたはマウスを免疫化することによるポリクローナル抗体およびモノクローナル抗体の調製が挙げられる(例えば、Huseら、Science 246:1275−1281(1989);Wardら、Nature 341:544−546(1989)を参照のこと)。
【0131】
T1Rタンパク質またはT1R3を含む味レセプターの一部を含む多数の免疫原が、T1Rタンパク質に特異的に反応する抗体を産生するために使用され得る。例えば、組換え型タンパク質またはその抗原性のフラグメントが、本明細書において記載されるように単離され得る。組換え型タンパク質は、上述のように真核生物細胞または原核生物細胞中に発現され得、そして一般に上述のように精製され得る。組換え型タンパク質は、モノクローナル抗体またはポリクローナル抗体の産生のための好ましい免疫原である。あるいは、本明細書において開示されるような配列に由来し、そしてキャリアタンパク質に接合される合成ペプチドは、免疫原として使用され得る。天然に存在するタンパク質はまた、純粋な形態かまたは純粋ではない形態かのいずれかで使用され得る。次いで、この生成物は、抗体を産生し得る動物に注入される。モノクローナル抗体またはポリクローナル抗体のいずれかが、上記タンパク質を測定するためのイムノアッセイにおけるその後の使用のために産生され得る。
【0132】
ポリクローナル抗体を産生する方法は、当業者に公知である。マウス(例えば、BALB/Cマウス)またはウサギの純系が、標準的なアジュバント(例えば、フロイントアジュバント)および標準的な免疫化プロトコルを使用して上記タンパク質で免疫化される。この動物の免疫原調製物への免疫応答が、テスト採血を採取しそしてβサブユニットへの反応性の力価を決定することによって、モニタリングされる。抗体の免疫原への適切に高い力価が得られる場合、この動物から血液が収集され、そして抗血清が調製される。上記タンパク質に反応する抗体を濃縮するための抗血清のさらなる分画化は、所望される場合に、行われる(HarlowおよびLane(前出)を参照のこと)。
【0133】
モノクローナル抗体は、当業者によく知られた種々の技術によって得られ得る。簡潔にいうと、所望の抗原で免疫化された動物由来の脾臓細胞が、一般に骨髄腫細胞との融合によって不死化される(KohlerおよびMilstein,Eur.J.Immunol.6:511−519(1976)を参照のこと)。不死化の代替的方法としては、エプスタイン−バーウイルス、癌遺伝子、またはレトロウイルスを用いた形質転換、あるいは当該分野において周知の他の方法が挙げられる。1つの不死化細胞から生じたコロニーは、抗原に対する所望の特異性および親和性を有する抗体の産生のためにスクリーニングされ、そしてそのような細胞によって産生についてモノクローナル抗体の収量は、種々の技術(脊椎動物宿主の腹腔内への注射を含む)によって増大され得る。あるいは、当業者は、Huseら、Science 246:1275−1281(1989)によって概説される一般的プロトコルに従ってヒトB細胞由来のDNAライブラリーをスクリーニングすることにより、モノクローナル抗体またはその結合フラグメントをコードするDNA配列を単離し得る。
【0134】
モノクローナル抗体およびポリクローナル血清は、収集されそしてイムノアッセイ(例えば、固体の支持体上に固定された免疫原を用いた固相イムノアッセイ)において免疫原タンパク質に対して力価を測定(titered)される。代表的に、104以上の力価を有するポリクローナル抗血清が、競合結合イムノアッセイを使用して、T1Rではないタンパク質、またはT1R3も含む味レセプターではないタンパク質に対する交差反応性について選択されそしてテストされる。特異的ポリクローナル抗血清およびモノクローナル抗体は、一般的に少なくとも約0.1mM(より一般的には少なくとも約1μM、好ましくは少なくとも約0.1μM以下、そして最も好ましくは0.01μM以下)のKdで結合する。特定のT1R3を含む味レセプターのオルソログ(例えば、ヒトT1R3を含む味レセプター)にのみ特異的な抗体は、1つの種(例えば、非ヒト哺乳動物)から他の交差反応性オルソログを除去することによっても産生され得る。さらに、個々のT1Rタンパク質が、レセプターと個々のT1Rタンパク質との両方へ結合する抗体を除去するために使用され得る。この様式において、特定のレセプターにのみ結合する抗体が得られ得る。
【0135】
一度T1R3を含む味レセプターに対する特異的な抗体が利用可能となった場合、上記タンパク質は、種々のイムノアッセイ方法によって検出され得る。さらに、この抗体は、T1R3を含む味レセプター調節因子として治療用に使用され得る。免疫学的手順およびイムノアッセイ手順の概説については、Basic and Clinical Immunology(StitesおよびTerr(編)、第7版、1991年)を参照のこと。さらに、本発明のイムノアッセイは、Enzyme Immunoassay(Maggio(編)、1980年);およびHarlowおよびLane(前出)中に広範に論評される任意のいくつかの構成において実行され得る。
【0136】
(B.免疫結合アッセイ)
T1R3を含む味レセプターが、任意の多数のよく認識されている免疫結合アッセイ(例えば、米国特許第4,366,241号、同4,376,110号、同4,517,288号、および同4,837,168号を参照のこと)を使用して検出され得、そして定量化され得る。一般のイムノアッセイの概説については、Methods in Cell Biology:Antibodies in Cell Biology,第37巻(Asai(編)、1993年);Basic and Clinical Immunology(StitesおよびTerr(編)、第7版、1991年)もまた参照のこと。免疫結合アッセイ(またはイムノアッセイ)は、代表的に、最適なタンパク質または抗原(この場合、T1R3を含む味レセプターまたはその抗原部分配列)に特異的に結合する抗体を使用する。抗体(例えば、抗T1R3を含む味レセプター)が、任意の多数の当業者に周知の手段のいずれかおよび上述のような手段によって産生され得る。
【0137】
イムノアッセイはまた、多くの場合、上記抗体および抗原によって形成される複合体に特異的に結合しそしてその複合体を標識するために、標識性因子を使用する。この標識性因子は、それ自身、抗体/抗原複合体を含む部分の1つであり得る。従って、この標識性因子は、標識されたT1R3を含む味レセプターであり得るか、または標識された抗T1R3を含む味レセプター抗体であり得る。あるいは、この標識性因子は、抗体/T1R3を含む味レセプター複合体に特異的に結合する第三の部分(例えば、二次抗体)であり得る(二次抗体は、代表的に、一次抗体が由来する種の抗体に特異的である)。免疫グロブリン定常領域(例えば、タンパク質Aまたはタンパク質G)に特異的に結合し得る他のタンパク質もまた、標識性因子として使用され得る。これらのタンパク質は、種々の種に由来する免疫グロブリン定常領域に、強い非免疫原性の反応を示す(例えば、Kronvalら、J.Immunol.111:1401−1406(1973);Akerstromら、J.Immunol.135:2589−2542(1985)を参照のこと)。標識性因子は、別の分子(例えば、ストレプトアビジン)が特異的に結合し得る検出可能部分(例えば、ビオチン)を用いて改変され得る。種々の検出可能部分は、当業者に周知である。
【0138】
このアッセイの全体を通して、インキュベーション工程および/または洗浄工程が、試薬の各組み合わせの後に必要とされ得る。インキュベーション工程は、約5秒から数時間まで(必要に応じて、約5分から約24時間まで)変動し得る。しかし、インキュベーション時間は、このアッセイの構成、抗原、溶液の体積、濃度などに依存する。通常、アッセイは周囲温度で実行されるが、例えば、10℃から40℃の温度の範囲にわたって実行され得る。
【0139】
(非競合アッセイ構成)
サンプル中の、T1R3を含む味レセプターを検出するためのイムノアッセイは、競合アッセイかまたは非競合アッセイのいずれかであり得る。非競合イムノアッセイは、抗原の量が直接測定されるアッセイである。1つの好ましい「サンドイッチ」アッセイにおいて、例えば、抗T1R3を含む味レセプターの抗体は、それらが固定化される固体の基質に直接的に結合され得る。次いで、これらの固定化された抗体は、このテストサンプルに存在するT1R3を含む味レセプターを捕捉する。従って、固定化された、T1R3を含む味レセプターは、次いで、標識性因子(例えば、標識を保有する第二のT1R3を含む味レセプター抗体)によって結合される。あるいは、この第二の抗体は、標識を欠いていてもよいが、それは代わりに、第二の抗体が由来する種の抗体に特異的な、標識された第三の抗体によって結合され得る。第二または第三の抗体は、代表的に、検出可能部分を提供するために、別の分子(例えば、ストレプトアビジン)が特異的に結合する検出可能部分(例えば、ビオチン)によって改変され得る。
【0140】
(競合アッセイ形式)
競合アッセイにおいて、このサンプルに存在するT1R3を含む味レセプターの量は、サンプルに存在する未知のT1R3を含む味レセプターによって抗T1R3を含む味レセプター抗体から置換された(追いやられた)、既知の添加された(外因性)T1R3を含む味レセプターの量を測定することによって間接的に測定される。一競合アッセイにおいて、既知の量のT1R3を含む味レセプターがサンプルに添加され、次いで、このサンプルはT1R3を含む味レセプターに特異的に結合する抗体に接触される。抗体に結合する外因性T1R3を含む味レセプターの量は、このサンプルに存在するT1R3を含む味レセプターの濃度に反比例する。特に好ましい実施形態において、この抗体は、固体の基質上に固定化される。抗体に結合するT1R3を含む味レセプターの量は、T1R3を含む味レセプター/抗体複合体に存在するT1R3を含む味レセプターの量を測定することによってか、または代替的に複合体化されていない残りのタンパク質の量を測定することによってかのいずれかで決定され得る。T1R3を含む味レセプターの量は、標識されたT1R3を含む味レセプター分子を提供することによって検出され得る。
【0141】
ハプテン阻害アッセイは、別の好ましい競合アッセイである。このアッセイにおいて、既知のT1R3を含む味レセプターは、固体の基質上に固定化される。既知の量の抗T1R3を含む味レセプターの抗体が、このサンプルに添加され、次いで、このサンプルは、固定化されたT1R3を含む味レセプターに接触される。既知の固定化されたT1R3を含む味レセプターに結合された抗T1R3を含む味レセプターの抗体の量は、このサンプルに存在するT1R3を含む味レセプターの量に反比例する。ふたたび、固定化された抗体の量は、抗体の固定化された分画かもしくは溶液中に残存する抗体の分画のいずれかを検出することによって検出され得る。検出は、抗体が標識化される直接的なものであるか、または上述のように抗体に特異的に結合する標識化部分のその後の添加による間接的なものであり得る。
【0142】
(交差反応性決定)
競合結合形式におけるイムノアッセイはまた、交差反応性を決定するために使用され得る。例えば、T1R3を含む味レセプターは、固体の支持体に固定化され得る。固定化された抗原への抗血清の結合について競合する、タンパク質(例えば、T1R3を含む味レセプターおよびホモログ)が、アッセイに添加される。この固定化されたタンパク質への抗血清の結合について競合する添加されたタンパク質の能力は、それ自身と競合するT1R3を含む味レセプターの能力と比較される。上記タンパク質のパーセント交差反応性は、標準的な計算を使用して計算される。上に列挙された添加されたタンパク質のそれぞれと10%未満の交差反応性を有する抗血清が、選択されそして集められる。この交差反応する抗体は、添加された考えられたタンパク質(added considered protein)(例えば、遠縁のホモログ)との免疫吸収によって、集められた抗血清から必要に応じて除去される。
【0143】
次いで、上記免疫吸収されそして集められた抗血清を、第二のタンパク質(おそらくT1R3を含む味レセプターの対立遺伝子または多型改変体と考えられる)をその免疫原タンパク質と比較するために、上述のような競合結合イムノアッセイにおいて使用する。この比較を行うために、この2つのタンパク質は、それぞれ、広範な濃度でアッセイされ、そして固定化されたタンパク質へのこの抗血清の結合の50%を阻害するために必要とされる各タンパク質の量が決定される。結合の50%を阻害するために必要とされる第二のタンパク質の量が、結合の50%を阻害するために必要とされるT1R3を含む味レセプターの量の10倍未満の場合には、この第二のタンパク質は、T1R3を含む味レセプター免疫原に対して産生されたポリクローナル抗体に特異的に結合するといわれる。
【0144】
(他のアッセイ形式)
ウェスタンブロット(イムノブロット)分析が、このサンプル中のT1R3を含む味レセプターの存在を検出および定量化するために使用される。この技術は、概して、分子量に基づくゲル電気泳動によるサンプルタンパク質を分離する工程、この分離されたタンパク質を適切な固体の支持体(例えば、ニトロセルロースフィルター、ナイロンフィルター、または誘導化ナイロンフィルター)へ移動する工程、およびT1R3を含む味レセプターに特異的に結合する抗体とともにそのサンプルをインキュベートする工程を包含する。抗T1R3を含む味レセプターの抗体は、固体の支持体上のT1R3を含む味レセプターに特異的に結合する。これらの抗体は、直接的に標識化され得るか、または代替的に、抗T1R3を含む味レセプターの抗体に特異的に結合する、標識化抗体(例えば、標識化ヒツジ抗マウス抗体)を使用して、その後検出され得る。
【0145】
他のアッセイ形式は、特定の分子(例えば、抗体)に結合しそしてカプセル化された試薬またはマーカーを放出するように設計されたリポソームを使用する、リポソームイムノアッセイ(LIA)を含む。次いで、放出された化学物質は、標準的な技術に従って検出される(Monroeら、Amer.Clin.Prod.Rev.5:34−41(1986)を参照のこと)。
【0146】
(非特異的結合の減少)
当業者は、多くの場合、イムノアッセイにおいて非特異的な結合を最小限にすることが望ましいことを理解する。特に、このアッセイが、固体の基質上で固定された抗原または抗体に関連する場合、その基質への非特異的な結合の量を最小限にすることが望ましい。そのような非特異的な結合を減少する手段は、当業者に周知である。代表的に、この技術は、タンパク様の組成物でその基質をコーティングする工程を包含する。特に、タンパク質組成物(例えば、ウシ血清アルブミン(BSA)、無脂粉乳、およびゼラチン)が、広く使用され、粉乳が最も好まれている。
【0147】
(標識)
このアッセイで使用される特定の標識または検出可能なグループは、このアッセイにおいて使用された抗体の特異的な結合を著しく干渉しない限り、本発明の重要な局面ではない。検出可能なグループは、検出可能な物理的もしくは化学的特性を有する任意の物質であり得る。そのような検出可能な標識は、イムノアッセイの分野においてよく開発され、そして概して、そのような方法において有用な任意の標識のほとんどが、本発明に適用され得る。従って、標識は、分光的手段、光化学的手段、生化学的手段、免疫化学的手段、電気的手段、光学的手段、または化学的手段によって検出可能な任意の組成物である。本発明において有用な標識としては、磁気ビーズ(例えば、DYNABEADSTM)、蛍光染料(例えば、フルオレセインイソチオシアネート、テキサスレッド、ローダミンなど)、放射性標識(例えば、3H、125I、35S、14C、または32P)、酵素(例えば、西洋ワサビペルオキシダーゼ、アルカリホスファターゼおよびELISAにおいて一般に使用される他の酵素)、および比色標識(例えば、コロイド金または着色ガラスまたはプラスチックビーズ(例えば、ポリスチレン、ポリプロピレン、ラテックスなど))が挙げられる。
【0148】
その標識は、当該分野に周知の方法によるアッセイの所望の構成要素に直接的にまたは間接的に結合され得る。上に示したように、広範な標識が使用され得、その標識の選択は、必要とされる感度、化合物との接合の容易さ、安定性の必要条件、利用可能な機器、および処理設備に依存する。
【0149】
非放射性標識は、多くの場合、間接的な手段によって付着される。概して、リガンド分子(例えば、ビオチン)は、分子に共有結合性に結合される。次いで、このリガンドは、別の分子(例えば、ストレプトアビジン)に結合し、この分子は、生来に検出可能であるかまたはシグナル系(例えば、検出可能な酵素、蛍光化合物、または化学発光化合物)に共有結合性に結合される。このリガンドおよびこれらの標的は、T1R3を含む味レセプターを認識する抗体、または抗T1R3を含む味レセプターを認識する二次抗体との任意の適切な組み合わせにおいて使用され得る。
【0150】
この分子はまた、シグナルを生成する化合物に直接的に接合され得る(例えば、酵素または発蛍光団との接合によって)。標識として目的の酵素は、主に、ヒドロラーゼ(特に、ホスファターゼ、エステラーゼ、およびグリコシダーゼ)、またはオキシドターゼ(oxidotase)(特に、ペルオキシダーゼ)である。蛍光化合物としては、フルオレセインおよびその誘導体、ローダミンおよびその誘導体、ダンシル、ウンベリフェロンなどが挙げられる。化学発光化合物としては、ルシフェリン、および2,3−ジヒドロフタラジンジオン(dihydrophthalazinedione)(例えば、ルミノール)が挙げられる。使用され得る種々の標識化またはシグナル生成系の概説については、米国特許第4,391,904号を参照のこと。
【0151】
標識を検出する手段は、当業者に周知である。従って、例えば、標識が放射性標識である場合、検出のための手段としては、シンチレーション計数器あるいはオートラジオグラフィー中のような写真フィルムが挙げられる。標識が蛍光標識である場合、それは、適切な光の波長で蛍光色素を励起させ、そして結果として生じる蛍光を検出することによって検出され得る。この蛍光は、写真フィルムによって、電子検出器(例えば、電荷結合デバイス(CCD)または光電子増倍管など)の使用によって、視覚的に検出され得る。同様に、酵素標識は、酵素のための適切な基質を提供し、そして結果として生じた反応産物を検出することによって検出され得る。最終的に、単純な比色標識が、単純に、標識に関連する色を観察することによって、検出され得る。従って、種々のディップスティックアッセイ(dipstick assay)において、接合された金は、多くの場合、ピンク色に見えるが、種々の接合されたビーズは、そのビーズの色に見える。
【0152】
いくつかのアッセイ形式は、標識化構成要素の使用を必要としない。例えば、凝集アッセイ(agglutination assay)は、標的抗体の存在を検出するために使用され得る。この場合において、抗原をコーティングした粒子は、標的抗体を含むサンプルによって凝集される。この形式において、この構成要素は、標識化されることを必要とせず、そして標的抗体の存在は、単純な視覚的検査によって検出される。
【0153】
(薬学的組成物および投与)
薬学的に受容可能なキャリアは、投与される特定の組成物(例えば、核酸、オリゴヌクレオチド、アミノ酸、タンパク質、ペプチド、小有機分子、脂質、炭水化物、単糖類、二糖類、または多糖類、粒子、または変換された細胞)によって、ならびに、その組成物を投与するために使用される特定の方法によって部分的に決定される。従って、本発明の薬学的組成物の多岐にわたる適切な処方物が存在する(例えば、Remington’s Pharmaceutical Sciences,第17版、1989年、を参照のこと)。投与は、任意の都合のよい様式で行われ得る(例えば、注射、経口投与、吸引、経皮的適用、または直腸投与)。
【0154】
経口投与に適している処方物は、(a)溶液(例えば、希釈液(例えば、水、生理食塩水、またはPEG 400)に懸濁している有効量のパッケージ化された核酸);(b)カプセル、小袋(sachet)または錠剤、それぞれが所定の量の活性成分(液体、固体、顆粒またはゼラチンとして)を含んでいる;(c)適切な液体中の懸濁剤;ならびに(d)適切なエマルジョンからなり得る。錠剤形態としては、1つ以上のラクトース、スクロース、マンニトール、ソルビトール、リン酸カルシウム、コーンスターチ、ポテトデンプン、微晶質セルロース、ゼラチン、コロイド二酸化ケイ素、滑石、ステアリン酸マグネシウム、ステアリン酸、および他の賦形剤、着色料、充填剤、結合剤、希釈液、緩衝剤、湿潤剤、保存薬、香料、染料、崩壊剤、および薬学的に適合するキャリアが挙げられ得る。ロゼンジ形態は、香料(例えば、スクロース)中に活性成分を、ならびに不活性塩基中に活性成分を含む香錠(例えば、活性成分に加えて当該分野において公知のキャリアを含んでいる、ゼラチンおよびグリセリンまたはスクロースおよびアカシアエマルジョン、ゲルなど)を含み得る。
【0155】
最適な化合物は、単独でまたは他の適切な構成要素との組み合わせにおいて、吸引を介して投与される、エーロゾル処方物中に作製され得る(すなわち、それらは「噴霧」され得る)。エーロゾル処方物は、加圧された受容可能な推進薬(例えば、ジクロロジフルオロメタン、プロパン、窒素など)中に配置され得る。
【0156】
非経口投与(例えば、関節内(関節(joint)内の)経路、静脈内経路、筋肉経路、皮内経路、腹腔内経路、および皮下経路)に適している処方物は、水溶性および非水溶性の等張無菌注入溶液を含み、この溶液は、抗酸化剤、緩衝液、静菌薬、ならびに意図されるレシピエントの血液とその処方物を等張にする溶質、ならびに水溶性および非水溶性の無菌懸濁液を含み得、この懸濁液は、懸濁剤、溶解剤、シックニング剤(thickening agent)、安定剤、および保存剤を含み得る。本発明の実施において、組成物は、例えば、経口的に、局所的に、腹腔内に、嚢内に(intravesically)、または髄膜内に、静脈内注入によって投与され得る。非経口的投与および静脈内投与が、好ましい投与の方法である。推薦される処方物は、単位用量または複数用量の密封された容器(例えば、アンプルおよびバイアル)中に提供され得る。
【0157】
注入溶液および懸濁液は、既に記載された種類の無菌の粉末、顆粒、および錠剤から調製され得る。エクスビボ治療用の核酸によって変換された細胞もまた、上述のように静脈内にまたは非経口的に投与され得る。
【0158】
患者に投与される用量は、本発明の状況において、長期にわたって患者の有益な治療上の応答をもたらすのに十分であるべきである。この用量は、特定の使用されたベクターの有効性および患者の状態、ならびに処置される患者の体重または表面積によって決定される。用量の大きさはまた、特定のベクターの投与に伴う任意の有害な副作用の存在、性質、および程度、あるいは特定の患者中の変換された細胞型によって決定される。
【0159】
T1R3を含む味レセプターの減少した発現もしくは異常な発現のための状態の処置または予防において投与されるベクターの有効量を決定する際に、医師は、ベクターの循環している血しょうレベル、ベクターの毒性、疾患の進行、および抗ベクター抗体の産生を評価する。概して、ベクター由来の裸の核酸に相当する用量は、代表的な70キログラムの患者について約1μgから100μgまでであり、そしてレトロウイルス粒子を含むベクターの用量は、等量の治療用核酸を産生するために計算される。
【0160】
投与について、本発明の化合物および変換された細胞は、集団のおよび総合的に健康な患者に適用された場合の、インヒビター、ベクター、または変換された細胞型のLD−50、ならびに、種々の濃度のインヒビター、ベクターまたは細胞型の副作用によって決定された頻度で投与され得る。投与は、一回かまたは分割した投与を介して遂行され得る。
【0161】
(細胞トランスフェクションおよび遺伝子治療)
本発明は、インビトロおよびインビボでの細胞のトランスフェクションのためのT1R3を含む味レセプターの核酸を提供する。これらの核酸は、以下に記載されるような標的細胞および生物体のトランスフェクションのための複数の周知のベクターのいずれかへ挿入され得る。この核酸は、そのベクターと標的細胞との相互作用を通じてエクスビボまたはインビボで細胞にトランスフェクトされる。次いで、この核酸は、プロモーターの制御下で、T1Rファミリーの2つのメンバーを同時発現させ、それにより、T1R3を含む味レセプターの欠如、部分的不活性化、または異常な発現の効果を緩和することによって、本発明のT1R3を含む味レセプターを発現する。この組成物は、患者に治療に対する応答を引き起こさせるために十分な量において、患者に投与される。このことを達成するのに適切な量は、「治療有効用量または治療有効量」として規定される。
【0162】
そのような遺伝子治療手順は、複数の状況において、後天的および先天的な遺伝子欠損および他の疾患を矯正するように使用されてきた。ヒトにおいて合成遺伝子を発現する能力は、多くの重要なヒト疾患(他の治療による処置を受けられない多くの疾患を含む)の予防および/または治癒を促進する(遺伝子治療手順の概説については、Anderson,Science 256:808−813(1992);NabelおよびFelgner,TIBTECH 11:211−217(1993);MitaniおよびCaskey,TIBTECH 11:162−166(1993);Mulligan,Science 926−932 (1993);Dillon,TIBTECH 11:167−175(1993);Miller,Nature 357:455−460(1992);Van Brunt,Biotechnology 6(10):1149−1154 (1998);Vigne,Restorative Neurology and Neuroscience 8:35−36(1995);KremerおよびPerricaudet,British Medical Bulletin 51(1):31−44 (1995);Current Topics in Microbiology and Immunology(DoerflerおよびBohm(編)、1995年)中のHaddadaら;およびYuら、Gene Therapy 1:13−26(1994)を参照のこと)。
【0163】
(参考文献)
【0164】
【化001】
【0165】
【化002】
【0166】
【化003】
【0167】
【化004】
【0168】
【化005】
【0169】
【化006】
【実施例】
【0170】
以下の実施例は、説明のために提供されるのであり、本発明の特許請求の範囲を制限するためのものではない。
【0171】
(実施例1)
(結果)
(T1R1 KOマウス、T1R2 KOマウスおよびT1R3 KOマウスの作製)
T1Rレセプターの発現は、舌および口蓋中の味細胞の重複していない3つの主な集団を規定する:T1R1およびT1R3を同時発現する細胞(T1R1+T1R3)、T1R2およびT1R3を同時発現する細胞(T1R2+T1R3)、ならびにT1R3のみを発現する細胞(Nelson,G.ら、Cell,106,381−390(2001))。HEK細胞のT1Rの異種発現の研究は、T1R1およびT1R3は組み合わさって、広範囲にわたるように調節されたL−アミノ酸レセプターを形成し、他方で、T1R2およびT1R3の同時発現は、甘味化合物の全てのクラスに応答する甘味レセプターを生成することを示した(Nelson,G.ら、Cell,106,381−390(2001)、Nelson,G.ら、Nature,416,199−202(2002);Li,X.ら、Proc Natl Acad Sci U S A,99,4692−4696(2002))。T1R3がインビボで甘味レセプターおよびアミノ酸味レセプターの共通の構成要素として機能する場合、このGPCRのノックアウトは、甘味受容能力およびアミノ酸味受容能力を欠くマウスを生成するはずである。対照的に、T1R1またはT1R2のノックアウトは、1つの味様式を選択的に影響すると予測される。
【0172】
インビボでのT1Rの役割を規定するために、本発明者らは、レセプター機能にとって本質的なドメインをコードするエキソンを欠失することによってそれぞれのT1Rを欠くノックアウトマウスを生成した。図1は、KO戦略を示し、そして対応しているホモ接合体KO動物において特異的T1R染色の完全な欠如を示すインサイチュハイブリダイゼーション実験を示す。任意の1つのT1Rの損失が、味細胞の生存率または一体性を影響しないことを確実にするために、本発明者らはまた、コントロールおよびKO動物中の他のT1R、T2R、PLCb2(Rossler,P.ら、Eur J Cell Biol,77,253−261(1998);Zhang,Y.ら、Cell,112,293−301(2003))およびTRPM5(Perez,C.A.ら、Nat Neurosci,5,1169−1176(2002);Zhang,Y.ら、Cell,112,293−301(2003))の発現を比較した。野生型とKO味組織との間のT1R−、T2R、PLCb2およびTRPM5陽性細胞の数においてまたはそれらの分布において、有意差は観察されなかった(図1およびデータは示されていない)。
【0173】
2つの相補的戦略を、遺伝子的に改変されたマウスの味応答をアッセイするために使用した。第一に、本発明者らは、舌の味レセプター細胞を神経支配する主要な神経(鼓索神経)の1つから味覚物質誘導性活動電位を記録した。この生理学的アッセイは、末梢における味覚系の活性をモニタリングし、そして味レセプター細胞の機能の測定を提供する。第二に、本発明者らは、標準的な長期二瓶採取嗜好性アッセイにおける味の選択を測定することによって、または多チャンネル味覚計における即時なめ応答の直接計数によって味に対する行動を調査した(Glendinning,J.I.ら、Chem Senses,27,461−474(2002);Zhang,Y.ら、Cell,112,293−301(2003);実験手順を参照のこと)。この第二の方法は、味覚物質への非常に短い暴露(全体で30分にわたる5秒の事象 対 二瓶嗜好性アッセイについては48時間)により、それゆえ、他の感覚入力の衝撃、ならびにこのアッセイからの摂取後および学習効果を最小限にするという大きな利点を有する。
【0174】
図2は、T1Rのノックアウトは、クエン酸、塩化ナトリウム、および種々の苦味味覚物質に対する生理学的応答または行動学的応答のいずれについても有意な効果を有さないことを示す。これらの結果は、苦味、塩味、および酸味の受容または知覚が、T1Rレセプターと独立した経路を通って作動することを示し、そしてさらに個々の様式が相互に独立して作動する、末梢におけるコーディングモデルを実証する。
【0175】
(T1R1+3は旨味レセプターである)
以前に、Chaudhariらは、代謝生成物産性グルタミン酸レセプター4(mGluR4t)の短縮型改変体を記載し、そしてそれが旨味レセプターとして機能することを示唆した(Chaudhari,N.ら、Nat Neurosci,3,113−119(2000))。本発明者らは,この提唱を多くの理由から満足のいくものではないと見出す。(1)このmGluR4t改変体は、表面標的に必要なmGluR4シグナル配列を欠いている。(2)この推定レセプターはまた、mGluRのグルタメート結合ドメインの結晶構造によって解明されるように、グルタメート認識のために重要なドメインの大きな分画を欠く(Kunishima,N.ら、Nature,407,971−977(2000))。(3)mGluR4t旨味シグナルは、cAMP経路を介して作動すると提唱されてきた(Abaffy,T.ら、Am J Physiol Cell Physiol,284,C1420−1428(2003);Chaudhari,N.ら、Nat Neurosci,3,113−119(2000))。しかし、アミノ酸/旨味は、PLCβ2/TRPM5依存性プロセスである(Zhang,Y.ら、Cell,112,293−301(2003))。(4)旨味は、旨味エンハンサー(IMPおよびGMP)によって強く影響されるが、mGluR4活性は影響されない。(5)最後に、mGluR4 KO動物は、旨味刺激への応答を保持する(Chaudhari,N.およびRoper,S.D.,Ann N Y Acad Sci,855,398−406(1998))。対照的に、最近の証拠は、このT1R1+3アミノ酸レセプターが、哺乳動物旨味(グルタメート)味センサーとして機能し得ることを示唆する:第一に、ヒトおよびげっ歯類T1R1+3レセプターは、げっ歯類とヒトとの間のアミノ酸味の差異を模倣する選択性および感受性の差異を示す(Nelson,G.ら、Nature,416,199−202(2002);Yoshiiら、1986)。第二に、T1R1+3活性は、IMPおよびGMP(2つの最も公知なインビボでの旨味の増強剤)によって依存的に増強される(Nelson,G.ら、Nature,416,199−202(2002);Li,X.ら、Proc Natl Acad Sci U S A,99,4692−4696(2002))。第三に,T1R1+3は、旨味アゴニストL−AspおよびL−AP4の心理物理学的に適切な濃度によって活性化される(Nelson,G.ら、Nature,416,199−202(2002);Li,X.ら、Proc Natl Acad Sci U S A,99,4692−4696(2002))。旨味におけるT1R1+3の役割を厳しく評価するために、本発明者らは、T1R1 KO動物およびT1R3 KO動物を調査した(図1を参照のこと)。
【0176】
そのNa+含有量のため、グルタミン酸一ナトリウム(MSG)は、塩味と旨味の両方を引き出す。それゆえ、本発明者らは、行動学的および電気生理学的研究において、グルタメートの塩味から塩味を単離することを可能にしたいくつかの計画を使用して旨味応答をアッセイした。これらは、MPG(グルタメートのカリウム塩)を使用してナトリウムチャネルブロッカーアミロライドの存在下でMSGをテストする工程、ならびに、旨味エンハンサーIMPの存在下または非存在下で旨味アゴニストAP4およびアスパラギン酸の全てをテストする工程を包含した。図3は、塩効果が最小限にされる場合、T1R3 KOマウスは行動学的誘引性の劇的な損失、およびそれに対応する、全ての旨味味覚物質(グルタメート、アスパルテート、グルタメート+IMP、およびIMPのみを含む)に対する生理学的応答の対応している重大な不足を有することを示す。ごく最近、Damakらは、独立して、T1R3 KO動物を作製したが、複数の旨味レセプターが、彼らのKOマウスの研究において残る顕著なMSG応答として存在するはずであると結論付けた(Damak,S.ら、Science,301,850−853(2003))。特に、KO動物のMSG応答は、IMPと完全に独立であり、IMPの増大は、旨味様式の特徴であることと矛盾する。塩効果は考慮されなかったので、本発明者らは、それらの残存する応答の多くは、旨味よりもむしろ、MSG中のNa+含有量を反映すると疑っている(図3E〜図3FのMSG+IMP対MPG+IMPまたはMSG+IMP+アミロライドに対して応答を比較すること)。
【0177】
T1R1がT1R3と組み合わさって(T1R1+3)哺乳動物旨味レセプターを生成する場合、T1R1のノックアウトもまた、全ての旨味応答を除去するはずである。図3は、まさにこの場合を示す。対照的に、これらの非常に同じ味覚物質は、コントロールとT1R2 KO動物において、正常で、強力な応答を引き起こす。合わせて、これらの結果は、T1R1+3が哺乳動物の旨味レセプターであることを証明する。
【0178】
以前に、本発明者らは、代表的な旨味味覚物質に加えて、マウスT1R1+3レセプターがまた、他のL−アミノ酸によって活性化され、そしてIMPの存在下で広範囲にわたるように調節されたL−アミノ酸センサーとして機能することを示した(Nelson,Gら、Nature,416,199−202(2002))。それゆえ、本発明者らは、IMPの存在下または非存在下におけるL−アミノ酸に対するT1R1 KO動物およびT1R3 KO動物の応答を試験した。実際、アミノ酸味覚物質に対する応答は、T1R1 KO系統およびT1R3 KO系統において重度に欠陥があるが、T1R2 KO系統においては欠陥がなく(図3)、広範なL−アミノ酸およびIMPのための味レセプターとしてのT1R1+3ヘテロマーGPCR複合体を堅固に確立している。興味深いことに、本発明者らがヒトにとって甘く感じる過剰に高濃度のL−アミノ酸(例えば、>300mM Ala、Ser、およびThr)をアッセイした場合、T1R1 KO動物は、わずかな残りの誘引性を保持したが、T1R3 KOマウスでは保持しなかった(図3のパネルDを参照のこと);これらの微量の行動学的応答は、T1R2+3甘味レセプターの活性化を反映する可能性が高い(Nelson,G.ら、Cell,106,381−390(2001);以下を参照のこと)。
【0179】
(T1R2+3およびT1R3は、甘味の受容および知覚のために必要とされる)
T1R2+3は、細胞ベースのアッセイにおいて、天然の糖、人工甘味料、D−アミノ酸および甘味タンパク質を含む種々の化学物質のクラスのためのヘテロマーレセプターとして機能する(Nelson,G.ら、Cell,106,381−390(2001);Li,X.ら、Proc Natl Acad Sci U S A,99,4692−4696(2002))。しかし、複数の研究は、動物が甘味レセプターの別個の型を発現し得ることを示唆している(Schiffman,S.S.ら、Pharmacol Biochem Behav,15,377−388(1981);Ninomiya,Y.ら、J Neurophysiol,81,3087−3091(1999))。インビボでのT1R2+3の役割を規定するために、本発明者らは、機能的T1R2タンパク質およびT1R3タンパク質を欠くノックアウトマウスの甘味応答を調査した。図4および図5は、全てのクラスの甘味味覚物質への応答が、T1R2ノックアウト系統+T1R3ノックアウト系統において劇的に弱められていることを示す。本発明者らは、広範なパネルの糖、人工甘味料およびD−アミノ酸を試験し、そして全ての場合において、応答が重度に障害されていた:行動学的誘引性がほとんど消失し、そして神経応答も大いに減少する。これらの結果は、T1R2+T1R3がインビボでの主要な甘味センサーであることを確証する。
【0180】
特に、非常に高濃度(>300mM)の天然の糖は、T1R2とT1R3との両方の系統において中程度だが検出可能な誘引的応答を誘発したが人工甘味料またはD−アミノ酸では誘発しなかった。従って、さらなる甘味レセプター(すなわち、T1R非依存経路)が存在するか、またはT1R2およびT1R3はまた、それらのヘテロマーのパートナーの存在しない状態で天然の糖に対する低親和性レセプターとしてそれら自身で機能し得る。実際、残存する応答がT1R2またはT1R3に起因する場合、これらのGPCRのダブルノックアウトは、全ての甘味応答を除去するはずである。T1R2およびT1R3遺伝子座は、第4染色体の遠位末端において連鎖されるので(Nelson,G.ら、Cell,106,381−390(2001))、本発明者らは、組み換え型T1R2 KO、T1R3 KOマウスを作製し、次いで、これらを生理学的および行動学的に試験した。図4および図5(赤線)は、T1R2、T1R3ダブルKOマウスが高濃度の糖への全ての応答を失ったことを示す。合わせて、これらの結果は、T1Rのコンビナトリアル集合物のインビボでの重要性を示し、そして全ての甘味受容はT1R2およびT1R3のGPCRを介して作動することを示す。
【0181】
T1R2またはT1R3ホモダイマーレセプターは、野生型マウスの甘味感知において重要な役割を果たすのであろうか?T1R2は、常にT1R3を含む細胞中で発現される(T1R2+3細胞;Nelson,G.ら、Cell,106,381−390(2001))。それゆえ、たとえいくつかのT1R2がこれらの細胞中でT1R3と会合しない場合でも、甘味味覚物質のためのT1R2+3のヘテロマーのレセプターのより高い親和性は、細胞応答を支配しやすい。対照的に、本発明者らは、既に、T1R3がまた、T1R1およびT1R2とは独立して、舌および口蓋上皮の有意な割合の細胞において見出されることを報告した(T1R3単独の細胞;Nelson,G.ら、Cell,106,381−390(2001))。このクラスの細胞は、高濃度の糖を検出し、そしてそれに応答するさらなる手段を動物に提供し得る。T1R3単独で天然の糖のための低い親和性のレセプターとして機能し得ることを示すために、本発明者らは、安定的にT1R3およびT1Rと共役するように操作された最適なGタンパク質キメラを発現するHEK細胞を生成した(実験手順を参照のこと)。図6は、実際、T1R3は単独で非常に高濃度の天然の糖に応答するが、低濃度(<300mM)または人工甘味料には応答しないことを示す。これらの結果は、T1R3が低い親和性の糖レセプターであることを確証し、そしてT1R3単独の細胞はインビボでさらなる甘味センサーとして機能するという仮定を支持する(Nelson,G.ら、Cell,106,381−390(2001))。天然甘味料および人工甘味料を感知するこの部分的な細胞の分離が、なぜ人工甘味料が、高濃度の天然の糖によってもたらされる甘さのレベルを達成しないかを説明することを補助し得る(すなわち、T1R2+3細胞の活性化 対 T1R2+3細胞およびT1R3単独の細胞の活性化)。
【0182】
(T1R2は種特異的甘味嗜好性の範囲を定める)
ヒトは、げっ歯類が味わい得ない複数の天然甘味料および人工甘味料を味わい得る。例えば、モネリン、タウマチン、アスパルテーム、およびネオへスペリジンジヒドロカルコンは、ヒトにはmM以下の濃度で甘いと感じさせるが、他方で、げっ歯類は100倍高い濃度でさえも嗜好性を示さない(Danilova,V.ら、J Neurophysiol,80,2102−2112(1998))。以前に、本発明者らは、げっ歯類T1RおよびヒトT1Rがそれらのアミノ酸配列において30%を超えて類似していないことを報告し、そしてそのような違いは甘味検出における種に特異的な選択性の基礎になっていると仮定した(Nelson,G.ら、Cell,106,381−390(2001);Nelson,G.ら、Nature,416,199−202(2002))。T1R2は排他的に甘味検出に関わるが、他方で、T1R3は甘味認識とアミノ酸認識との両方に関与しているので、本発明者らは、T1R2がインビボでの甘味選択性の特に重要な決定要素であると理由付けした。それゆえ、本発明者らは、T1R2 KOマウス中にヒトT1R2遺伝子を導入することが甘味応答を回復し、そして甘味応答を「ヒト化する」はずであると予測した。
【0183】
本発明者らは、T1R2 KO対立遺伝子であるが、代わりにネイティブ「T1R2細胞」においてヒトT1R2トランスジーンを発現させたホモ接合であるマウスを生成した。T1R2調節配列を含む12kbのゲノムクローンを、hT1R2全長cDNAに融合し、そしてT1R2 KOマウスに導入した。複数の独立した系統を、甘味味覚物質に対するそれらの選択性および感受性についてアッセイした。hTlR2の発現を調査するために、本発明者らは、野生型mT1R2対立遺伝子を保有するトランスジェニック動物で二色蛍光インサイチュハイブリダイゼーション実験を実行した。図7(パネルA〜D)は、ヒトT1R2がT1R2発現細胞中に選択的に発現され、そして甘味機能を効果的に回復することを示す。より重要なことに、ヒトトランスジーンは、ここで、ヒトに甘いと感じさせるが、通常げっ歯類には誘引性ではないいくつかの化合物を検出し、そして応答する能力をこれらのマウスに与える;これらの化合物としては、アスパルテーム、グリチルリジン酸および甘味タンパク質(タウマチンおよびモネリン)が挙げられる。興味深いことに、ヒト化T1R2マウスは、強度の甘味化合物(ネオへスペリジンジヒドロカルコン)になお応答せず、ヒトT1R2およびマウスT1R3 GPCRでトランスフェクトされたHEK細胞も応答しない。しかし、細胞をヒトT1R2およびヒトT1R3でトランスフェクトする場合、この細胞は、ネオへスペリジンジヒドロカルコンに強く応答する。まとめると、これらの実験は、げっ歯類とヒトとの間の甘味選択性および甘味特異性の違いの重要な決定要素がT1Rであることを証明し、そしてさらに、T1R2+3およびT1R2発現細胞をインビボでの甘味の主要なメディエーターとして実証する。最後に、本発明者らは、T1R2とT1R3との両方の多型性が、ヒトの個々の甘味嗜好性の重要な決定要素であることを提案する。
【0184】
(T1R2を発現する細胞は、インビボでの行動学的誘引性をコードする)
味レセプターの活性化は、動物の個々の行動学的応答を誘発する。例えば、T1R2+3レセプターの興奮は、マウスの糖および甘味化合物に対する行動学的誘引性を刺激する。この応答は、これらが発現されるレセプターまたは細胞の特性であろうか?この質問に答える1つの手段は、T1R2+3細胞において味覚系に関連のない新規のレセプターを発現することであり、そしてその選択的刺激が誘引性応答を誘発するか否かを調査することである(Troemel,E.R.ら、Cell 91,161−169(1997))。
【0185】
本発明者らのアプローチは、T1R2+3細胞の内因性シグナル伝達経路へ共役し得るが、非天然のリガンドによって活性化し得るのみであるGPCRの発現を標的にすることであった。レセプター発現の前後にまさにその同じ動物の味応答を調査するために、本発明者らは誘導系を利用した。これを達成するために、本発明者らはトランスジェニックマウスを使用し、このマウスの中で、合成リガンドによってのみ活性化される改変k−オピオイドレセプター(RASSL;Redfern,C.H.ら、Nat Biotechnol,17,165−169(1999))を、Tet−on誘導系の制御の下、T1R2発現細胞を標的にした(実験手順を参照のこと)。
【0186】
図7Eは、非誘導動物、またはドキシサイクリンで処理された野生型コントロールが、完全にk−オピオイドアゴニスト(スピラドリン)に非感受性であることを示す。注目すべきことに、T1R2細胞中のRASSL発現の誘導が、ここで、ナノモルの濃度のスピラドリンに強力に誘引される動物を生成する(図7、赤線)。従って、本発明者らは、T1R2発現細胞の活性化は(それらが発現するレセプターよりも)、マウスにおいて行動学的誘引性を決定することを結論付ける。さらに、これらの結果は、1つの細胞型を活性化することが、特異的味応答を誘発するのに十分であることを明白に示す;それゆえ、活性のコンビナトリアルパターンまたは一時的コーディングを必要とするモデルは、T1R2発現細胞によって媒介される誘引性を説明するために必要とされない。拡張することによって、本発明者らは、上記レセプターの性質に関わらず、ヒトT1R2+3細胞におけるこれらの味シグナル伝達経路の活性が甘味を誘発することを示唆する。
【0187】
複数のレセプターが、哺乳動物の甘味および旨味を媒介するものと提案されている。特に、これらの2つの様式の各々の範囲内でさえも、細胞を透過する味覚物質によって直接的に活性化される細胞内標的を引き起こす数個のGPCR、イオンチャネル、およびモデルが仮定されてきた(Kinnamon,S.C.Neuron,25,507−510(2000);Margolskee,R.F.,J Biol Chem,277,1−4(2002))。本発明者らは、哺乳動物の甘味および旨味のためのレセプターがT1Rであることを証明するために、細胞ベースのアッセイ、遺伝学的アプローチ、生理学的アプローチ、および行動学的アプローチの組み合わせを使用してきた;旨味はT1R1+3のヘテロマーのGPCRによって媒介され、そして甘味は2つのT1Rベースのレセプター(T1R2およびT1R3)(T1R2+3、そして最も可能性の高いものとしては、ホモダイマーのT1R3)によって媒介される。それゆえ、甘味およびアミノ酸味(旨味)(行動学的誘引性を誘発する2つの化学感覚性入力)は、共通のレセプターレパートリーおよび進化の起源を共有する。
【0188】
ヒトT1R1+3レセプターは、他のアミノ酸よりもはるかに効果的にグルタメートおよびアスパラテートによって活性化される(Li,X.ら、Proc Natl Acad Sci U S A,99,4692−4696(2002))。対照的に、マウスT1R1+3レセプターは、細胞ベースのアッセイ(Nelson,G.ら、Nature,416,199−202(2002))とインビボ(本紙)との両方において非常に広範囲のL−アミノ酸を認識する。このT1R1+3レセプターの進化上の役割が、タンパク質の豊富な食料に対する誘引性応答を媒介することであった場合、当業者は、霊長類における2つのアミノ酸のみに対するレセプターの選択性の調整が、実質的にこれらの栄養が豊富な食物を検出する能力を変更したか否かということを問題とし得る。そのようなアミノ酸は、通常、複合体の混合物として見出され、いずれのものも概して適切であり、従って、この味覚物質の選択性の「限定」は食事に影響を顕著に有さないはずである。同じ細胞および同じレセプターが、グルタメート、他のアミノ酸およびIMPを認識すると仮定すると、本発明者らは、げっ歯類において、この旨味様式が、ほとんどのL−アミノ酸を含むように一般化されなければならないこと、そしてげっ歯類における明白なグルタメート味のまさにその概念(Chaudhari,N.ら、Nat Neurosci,3,113−119(2000);Lin,W.ら、J Neurophysiol,89,1434−1439(2003))が再評価される必要があることを示唆する
スプーン一杯の糖または数錠の人工甘味料?本発明者らの日々の経験が、天然の甘味料および人工甘味料は同じ味がしないことを本発明者らに伝えている。この原稿において、本発明者らは、T1R2およびT1R3が、全ての甘味感知の原因であることを示した。それらは、どのように甘味味覚物質の間の知覚された味の違いを明らかにするのであろうか?多くの甘味料は、他の味様式(のためのレセプターを活性化する可能性がある例えば、サッカリンの苦い後味を説明するT2R苦味感知細胞(データは示さず))。それゆえ、1つの甘味分子種の「味」さえも、異なる味様式に調節される細胞の組み合わさった活性を反映し得、そして甘味感知細胞の活性のみを反映し得るわけではない。本発明者らはまた、より高いが、なお生理学的に関連のある濃度の糖(>300mM)において、天然甘味料および人工甘味料が部分的に重複するが、なお別個の甘味レセプター型(T1R2+3およびT1R3のみ)を活性化することを示してきた。
【0189】
本発明者らは、T1Rが2つの主要な誘引性味様式のメディエーターであることを示し、そして、RASSLオピオイドレセプターを発現するマウスがスピラドリン(通常、味がなくそして栄養的に関連のない化合物)に強力に誘引されるようになったことを示し、味わうことが信じることであることを証明した。ここで、苦味、甘味、および旨味のための細胞およびレセプターの発見および機能的特徴付けは、どのように味が末梢においてコードされるかという説得力のある視点を提供する、専用の味レセプター細胞が誘引される行動および回避的な行動を媒介する(例えば、Zhang,Y.ら、Cell,112,293−301(2003)を参照のこと)。
【0190】
(実験手順)
(T1R1、T1R2およびT1R3の遺伝子標的)
T1Rノックアウト動物を作製するために使用される戦略を、図1に示す。T1R1について、7つの予測されたレセプターの膜貫通ドメインをコードするエキソン6を、PGK−neorカセットによって置換した。R1 ES細胞におけるホモログの組み換えを、標的構築物の外でのプローブとの、診断サザンハイブリゼーションによって検出した。2つの標的ESクローンを、C57BL/6胚盤胞中へ注入した。キメラマウスを、C57BL/6マウスと交配させ、そしてホモ接合のノックアウトコロニーを確立する前に、2世代にわたってC57BL/6マウスと後代戻し交雑を行った。
【0191】
T1R2について、類似のアプローチが、エキソン5およびエキソン6を除去した(図1を参照のこと)。キメラ動物を、C57BL/6マウスと交配させ、そして4世代にわたってC57BL/6マウスと後代戻し交雑を行った。T1R3味覚因子(taster)(C57)および非味覚因子(129)対立遺伝子(Nelson,G.ら、Cell,106,381−390(2001))を、EcoRI多型性(T1R3の開始ATGの約12kb上流)に基づき同定した。この研究において使用されたT1R2ノックアウト動物の全てが、T1R3の味覚因子対立遺伝子を保有していた。しかし、非味覚因子T1R3対立遺伝子とホモ接合のT1R2 KOマウスの研究は、定性的に類似の結果を生じた(データは示さず)。T1R3 KOノックアウト動物を作製するために、本発明者らは、エキソン1をエキソン5に置換し、PGK−neorカセットを有するN末端細胞外ドメインをコードした(図1を参照のこと)。キメラマウスを、C57BL/6マウスと交配させ、2世代にわたってC57BL/6マウスと後代戻し交雑を行った。
【0192】
T1Rノックアウトは、正常な生存率、体重、総合的な解剖学的構造および一般的行動を有する。同様に、味レセプター細胞は、全てのノックアウトバックグラウンドにおいて形態学的におよび数的に正常に見える。
【0193】
(インサイチュハイブリダイゼーション)
新鮮凍結切片(16μm/切片)を、シラン化スライドに付着させ、そして既に記載したようにインサイチュハイブリダイゼーションまたは免疫組織化学のために調製した(Hoon,M.A.ら、Cell.,96,541−551(1999))。単一標識インサイチュハイブリダイゼーション(Single label in situ hybridization)を、ジゴキシゲニンで標識されたプローブを使用して実行した;T1R1およびT1R2プローブは、予測された膜貫通ドメインに対してだが、他方で、T1R3およびRASSL(Redfern,C.H.ら、Nat Biotechnol,17,165−169(1999))プローブは完全なコーディング配列を利用した。二重標識蛍光検出は、高いストリンジェンシーにおいて(ハイブリダイゼーション、5×SSC、50%ホルムアミド、65〜72℃;洗浄、0.2×SSC、72℃)、フルオレセイン(全長hT1R2)プローブおよびジゴキシゲニン(全長mT1R2)プローブを使用した。ハイブリダイゼーションを、明白な蛍光基質を用いて検出し(Adler,E.ら、Cell,100,693−702(2000))、そして標識化の特異性をT1R2ノックアウトおよび非トランスジェニックコントロールを使用して確認した。
【0194】
(ヒトT1R2およびRASSLを発現するトランスジェニックマウスの作製)
マウスT1R2の上流の約12kbのゲノムフラグメントを、ヒトT1R2 cDNAに、ならびに、逆テトラサイクリン依存性トランスアクチベーター(rtTA)構築物に融合した(Gossen,M.ら、Curr Opin Biotechnol,5,516−520(1994))。トランスジェニック系統を、FVB/Nマウス由来の接合体の前核注入によって生成した。3つの独立したヒトT1R2トランスジェニック系統は、アスパルテーム(10mM)に対する行動学的誘引性を示した。1つの系統をT1R2ノックアウトバックグラウンド中へ交配し、そして味応答およびトランスジーン発現についてアッセイした。TlR2細胞の外では発現を検出しなかった。TlR2−rtTAトランスジェニック系統をtetO−Ro1/tetO−lacZトランスジェニック動物と交配した(Redfern,C.H.ら、Nat Biotechnol,17,165−169(1999))。ダブルヘテロ接合の子孫を、3日間、ドキシサイクリン処理(6gm/kg)(Bio−Serv)によって誘導し(Gogos,J.A.ら、Cell,103,609−620(2000))、そしてガラクトシダーゼ活性(Zack,D.J.ら、Neuron 6,187−199(1991))および舌および口蓋のRASSL発現について調査した。適切なβガラクトシダーゼ染色およびRASSL発現パターンを示す系統を、行動学的アッセイのために選択した。
【0195】
(行動学的アッセイ)
味覚行動を、マルチチャネル味覚計(Davis MS160−Mouse gustometer;DiLog Instruments,Tallahassee,FL)において即時なめ応答を数えることによって味嗜好性を直接的に測定する短期アッセイを使用してアッセイした。マウスを、既に記載したように訓練し、そして試験した(Zhang,Y.ら、Cell,112,293−301(2003))。個々のマウスを30分間、味覚計に配置し、そして刺激を、その刺激が噴出したマウスのなめる行動により開始される5秒間の試行の間、ランダムな順で提示した。サッカリンナトリウム、グルタメートおよびアスパルテートについて、100μMのアミロライドを全ての溶液(コントロールを含む)に添加し、塩味の効果を最小限にした。データポイントは、適切なコントロール味覚物質のサンプリングに対する、マウスが味覚物質をなめる平均速度(コントロールに対するなめる速さとして規定した比)を示す;なめの抑制を、1からコントロールに対するなめる速さを引いた値、として規定した。ほとんどの場合、上記コントロール味覚物質は、水であったが、アミノ酸+1mM IMP、200mM MSGおよび10mM IMPについては、コントロールは、それぞれ、1mM IMP、200mMグルコン酸ナトリウムおよび10mM CMPであった。
【0196】
標準的な二瓶嗜好性アッセイを、既に記載されたように実行した(Nelson,G.ら、Cell,106,381−390(2001))。TlR2−rtTAおよびtetO−Ro1/tetO−lacZトランスジーンを保有するマウスについて、発現を、行動学的テストの前に3日間およびその間、ドキシサイクリン処理によって誘導した。コントロールは、誘導されていない同じマウス、ならびに、ドキシサイクリンで処理されたT1R2−rtTAトランスジーンのみを保有するマウスをテストする工程を包含した。3つの全てのグループは、スクロースへの正常な応答を示した。
【0197】
本発明者らは、二瓶アッセイにおいてT1R−KO動物が非常に高濃度の天然の糖(>500mM)を含む溶液を同定することを「学習」するように見えることに気付いた;連続した暴露は、検出閾値の減少および嗜好性比の増加をもたらした。マウスは、標準的な二瓶アッセイにおいて、48時間、反復してテスト化合物に暴露されるので、マウスは、味覚物質を水と区別するために、他の感覚の入力(例えば、テクスチャーまたはにおい)を使用し得る。適切に制御されない場合、このことは、味経路を介した行動学的誘引性によるものと容易に誤解され得る。この問題を避けるために、本発明者らは、短期の即時なめ応答アッセイ(上を参照のこと)またはナイーブなノックアウトマウス(すなわち、訓練またはテストのいずれかの間にそのような味覚物質に暴露されないかったマウス)での二瓶アッセイのいずれかを使用した。
【0198】
(神経記録)
舌の刺激および記録手順を、既に記載されているように実行した(Dahl,M.ら、Brain Res,756,22−34(1997);Nelson,G.ら、Nature,416,199−202(2002))。ニューロンのシグナルを、Grass P511 AC増幅器(Astro−Med)を用いて、増幅し(5,000×)、Digidata 1200B A/D変換器(Axon Instruments)でデジタル化し、そして0.5秒の時定数で取り込んだ(r.m.s.電圧)。味刺激を、20秒間隔の4ml/分の一定の流動速度にて提示し、提示の間に人工だ液(Danilova,V.,およびHellekant,G.,BMC Neurosci,4,5.(2003))を用いて2分間リンスすることによって散在させた。全てのデータ分析は、刺激の適用の直後の25秒間にわたって取り込まれた応答を使用した。各実験シリーズは、記録の安定性を確実にするために0.1Mのクエン酸の提示によって分けられる6個の味覚物質の適用からなっていた。0.1Mのクエン酸への平均応答を、各実験シリーズへの応答を標準化するために使用した。
【0199】
神経記録のために使用された味覚物質(最大濃度)は:スクロース、グルコース、マルトース(600mM);サッカリンナトリウム(40mM);AceK(60mM);クエン酸(100mM);NaCl(100mM);NH4Cl(100mM);6−n−プロピル−チオウラシル(10mM)、キニーネ(10mM);シクロヘキシミド(1mM);L−Ser、L−Ala、(30mM、0.5mM IMPが添加されている)、MSGおよびMPG(300mM、0.5mM IMPありまたはなし);D−Ala、D−Phe、およびD−Trp(l00mM)であった。アミロライド(50μM)を、図の説明文中に示されるようにナトリウム応答を減少するために添加した。
【0200】
(T1R異種発現およびカルシウムイメージング)
改変HEK−293細胞(PEAKrapid細胞;Edge BioSystems,MD)を増殖し、T1Rおよび乱交雑Gタンパク質でトランスフェクトし、そして本質的に以前に記載されたように(Nelson,G.ら、Cell,106,381−390(2001))Caイメージングによって味覚物質への機能的応答についてアッセイした。FURA−2負荷およびCa−イメージングにおける小さな違いは、アッセイ緩衝液として0.1% BSA,100μM EGTAおよび200μM CaCl2を含む199(H)Medium(Biosource)を使用する工程、ならびに、FURA−AMエステル切断を可能にする時間を10分まで減少する工程を包含した。このイメージングシステムは,l0×/0.5N.A.蛍光(fluor)対物レンズ(Zeiss)を備えるOlympus IX50顕微鏡、TILLイメージングシステム(TILL Photonics GmbH)、および冷却CCDカメラであった。蛍光イメージの取得および分析は、TILL−Visionソフトウェアを使用した。
【0201】
[Ca2+]iの変化に対するT1R応答の共役を最適化するために、ヒトGα16のC末端残基(Offermanns,S.,およびSimon,M.I.,J Biol Chem,270,15175−15180(1995))を、Gz(Mody,S.M.ら、Mol Pharmacol,57,13−23(2000))、ガストデューシン(McLaughlin,S.K.ら、Nature,357,563−569(1992))またはGαi2に由来する対応する残基で置換した。ガストデューシン(Ggust−25)のC末端の25残基を含むキメラは、一過性トランスフェクションアッセイにおいて、マウスT1R2+3およびT1R1+3の応答を媒介することに特に効果的であると証明し、そしてさらなる研究のために使用された。T1R3およびGgust−25を安定的に発現する細胞系を、ピューロマイシンおよびゼオシン(Invitrogen)選択を使用して確立した。T1R3およびGgust−25を発現する3つの独立の系を、T1R3レセプターの特異性および用量応答を調査するために使用された。スクロースおよびマルトース(>300mM)は、T1R3およびGgust−25に依存する用量依存応答を惹起したが、高濃度の他のいくつかの糖(グルコース、フルクトース、トレハロースおよびガラクトース)を使用する試みは、それらが[Ca2+]iにおける有意なレセプター非依存性の増加を誘発するので、非実用的であると判明した。
【0202】
本明細書において記載される実施例および実施形態は、説明する目的のみのためであり、それを考慮した種々の改変または変化は当業者に示唆され、そして本出願の精神および範囲ならびに添付された特許請求の範囲の範囲内に包含されていることが理解される。本明細書において引用されている全ての刊行物、特許、および特許出願は、あらゆる目的のためにその全体が本明細書において参考として援用されている。
【0203】
(配列表)
(T1R1配列)
(ラットT1R1アミノ酸配列−−配列番号1)
【0204】
【化007】
【0205】
(マウスT1R1アミノ酸配列−−配列番号2)
【0206】
【化008】
【0207】
(ヒトT1R1アミノ酸配列−−配列番号3)
【0208】
【化009】
【0209】
【化010】
【0210】
(ラットT1R1ヌクレオチド配列−−配列番号4)
【0211】
【化011】
【0212】
(マウスT1R1ヌクレオチド配列−−配列番号5)
【0213】
【化012】
【0214】
(ヒトT1R1ヌクレオチド配列−−配列番号6)
【0215】
【化013】
【0216】
【化014】
【0217】
(T1R2配列)
(ラットT1R2アミノ酸配列−−配列番号7)
【0218】
【化015】
【0219】
(マウスT1R2アミノ酸配列−−配列番号8)
【0220】
【化016】
【0221】
(ヒトT1R2アミノ酸配列−−配列番号9)
【0222】
【化017】
【0223】
(ラットT1R2ヌクレオチド配列−−配列番号10)
【0224】
【化018】
【0225】
【化019】
【0226】
(マウスT1R2ヌクレオチド配列−−配列番号11)
【0227】
【化020】
【0228】
【化021】
【0229】
(ヒトT1R2ヌクレオチド配列−−配列番号12)
【0230】
【化022】
【0231】
【化023】
【0232】
(T1R3配列)
(ヒトT1R3ゲノムヌクレオチド配列−−配列番号13)
【0233】
【化024】
【0234】
【化025】
【0235】
(ヒトT1R3 cdsヌクレオチド配列−−配列番号14)
【0236】
【化026】
【0237】
【化027】
【0238】
(ヒトT1R3アミノ酸配列−−配列番号15)
【0239】
【化028】
【0240】
(マウスT1R3 Sac非味覚因子129ゲノムヌクレオチド配列−−配列番号16)
【0241】
【化029】
【0242】
【化030】
【0243】
(マウスT1R3 Sac非味覚因子129 cdsヌクレオチド配列−−配列番号17)
【0244】
【化031】
【0245】
【化032】
【0246】
(マウスT1R3 Sac非味覚因子129アミノ酸配列−−配列番号18)
【0247】
【化033】
【0248】
(マウスT1R3 Sac味覚因子SWR cdsヌクレオチド配列−−配列番号19)
【0249】
【化034】
【0250】
【化035】
【0251】
(マウスT1R3 Sac味覚因子SWRアミノ酸配列−−配列番号20)
【0252】
【化036】
【0253】
(マウスT1R3 Sac味覚因子c57ゲノムヌクレオチド配列−−配列番号21)
【0254】
【化037】
【0255】
【化038】
【0256】
(マウスT1R3 Sac味覚因子C57 cdsヌクレオチド配列−−配列番号22)
【0257】
【化039】
【0258】
(マウスT1R3 Sac味覚因子C57アミノ酸配列−−配列番号23)
【0259】
【化040】
【0260】
【化041】
【0261】
(ラットT1R3 CDSヌクレオチド配列−−配列番号24)
【0262】
【化042】
【0263】
【化043】
【0264】
(ラットT1R3アミノ酸配列−−配列番号25)
【0265】
【化044】
【図面の簡単な説明】
【0266】
【図1A】T1R1、T1R2およびT1R3の標的KO。(a)3つのT1R遺伝子の構造およびノックアウト動物を作製するための戦略を示す模式図。標的構築物は、T1R1およびT1R2の7つの予測された膜貫通へリックスの全て、およびT1R3の細胞外リガンド結合ドメイン全体を除去した。(b)T1Rの野生型動物の味蕾における強い発現、そして対応するノックアウトマウスにおける完全な欠如、を示しているインサイチュハイブリダイゼーション標識。
【図1B】T1R1、T1R2およびT1R3の標的KO。(a)3つのT1R遺伝子の構造およびノックアウト動物を作製するための戦略を示す模式図。標的構築物は、T1R1およびT1R2の7つの予測された膜貫通へリックスの全て、およびT1R3の細胞外リガンド結合ドメイン全体を除去した。(b)T1Rの野生型動物の味蕾における強い発現、そして対応するノックアウトマウスにおける完全な欠如、を示しているインサイチュハイブリダイゼーション標識。
【図2A】T1R変異体は、通常、酸味、塩味および苦味の刺激に応答する。(a)野生型マウス(WT)、T1R1ノックアウトマウス(1−KO)、T1R2ノックアウトマウス(2−KO)およびT1R3ノックアウトマウス(3−KO)は、酸味(100mMクエン酸)、塩味(100mM NaCl)および苦味(10mM PROP)味覚物質に強い神経応答を示す。(b)統合神経応答(例えば、(a)に示されたもの)が、100mMクエン酸によって誘発された応答に対して標準化された;コントロールおよびKO動物は、相互に区別不能である。値は、平均±標準誤差である(n=4)。データは、鼓索神経の応答を示す(詳細については実験手順を参照のこと)。(c)野生型動物およびT1Rノックアウト動物の、味覚嗜好性が、短期接近味試験(brief access taste test)(Zhang,Y.ら、Cell,112,293−301(2003))を使用して水に対して測定された。4つの全ての系統が、酸味、塩味および苦味の刺激に対する正常な応答を示した。値は、平均±標準誤差である(n=7)。類似の結果が、標準的な二瓶嗜好性アッセイ(two bottle preference assay)を使用して得られた(データは示さず)。Cyx、シクロヘキシミド;Den、安息香酸デナトニウム;PROP、6−n−プロピル−チオウラシル;Qui、キニーネ。
【図2BC】T1R変異体は、通常、酸味、塩味および苦味の刺激に応答する。(a)野生型マウス(WT)、T1R1ノックアウトマウス(1−KO)、T1R2ノックアウトマウス(2−KO)およびT1R3ノックアウトマウス(3−KO)は、酸味(100mMクエン酸)、塩味(100mM NaCl)および苦味(10mM PROP)味覚物質に強い神経応答を示す。(b)統合神経応答(例えば、(a)に示されたもの)が、100mMクエン酸によって誘発された応答に対して標準化された;コントロールおよびKO動物は、相互に区別不能である。値は、平均±標準誤差である(n=4)。データは、鼓索神経の応答を示す(詳細については実験手順を参照のこと)。(c)野生型動物およびT1Rノックアウト動物の、味覚嗜好性が、短期接近味試験(Zhang,Y.ら、Cell,112,293−301(2003))を使用して水に対して測定された。4つの全ての系統が、酸味、塩味および苦味の刺激に対する正常な応答を示した。値は、平均±標準誤差である(n=7)。類似の結果が、標準的な二瓶嗜好性アッセイを使用して得られた(データは示さず)。Cyx、シクロヘキシミド;Den、安息香酸デナトニウム;PROP、6−n−プロピル−チオウラシル;Qui、キニーネ。
【図3AD】T1R1+T1R3は、哺乳動物旨味レセプターとして機能する。(a−d)野生型マウス(白丸、点線)、T1R1 KOマウス(青丸およびバー)、T1R2 KOマウス(グレーの丸およびバー)およびT1R3 KOマウス(茶丸およびバー)の味覚嗜好性が、水に対して、短期接近味試験を使用して測定された。T1R2 KOマウスは、野生型コントロールと同等である。対照的に、T1R1およびT1R3ノックアウト動物は、旨味味覚物質(a)MSG+1mM IMP、(b)MSG、(c)IMP、および(d)L−Asp(100mM)、およびAP4(30mM)に対する嗜好性の完全な消失を示す。さらに、両方のノックアウトは、他のアミノ酸応答において顕著な障害を有する。L−Asn(100mM)およびL−Argが、各100mMにて使用された。(e−f)旨味味覚物質およびアミノ酸に対する統合された鼓索神経応答。アミロライドまたはMSGのカリウム塩(MPG)のいずれかを使用することによって、塩効果が回避される場合、T1R1およびT1R3ノックアウトは、(e)旨味アゴニストおよびL−アミノ酸に対する応答を完全に消失されている。対照的に、(f)高濃度の塩が使用される場合(例えば、100mM MSG)、残存応答が検出される。
【図3EF】T1R1+T1R3は、哺乳動物旨味レセプターとして機能する。(a−d)野生型マウス(白丸、点線)、T1R1 KOマウス(青丸およびバー)、T1R2 KOマウス(グレーの丸およびバー)およびT1R3 KOマウス(茶丸およびバー)の味覚嗜好性が、水に対して、短期接近味試験を使用して測定された。T1R2 KOマウスは、野生型コントロールと同等である。対照的に、T1R1およびT1R3ノックアウト動物は、旨味味覚物質(a)MSG+1mM IMP、(b)MSG、(c)IMP、および(d)L−Asp(100mM)、およびAP4(30mM)に対する嗜好性の完全な消失を示す。さらに、両方のノックアウトは、他のアミノ酸応答において顕著な障害を有する。L−Asn(100mM)およびL−Argが、各100mMにて使用された。(e−f)旨味味覚物質およびアミノ酸に対する統合された鼓索神経応答。アミロライドまたはMSGのカリウム塩(MPG)のいずれかを使用することによって、塩効果が回避される場合、T1R1およびT1R3ノックアウトは、(e)旨味アゴニストおよびL−アミノ酸に対する応答を完全に消失されている。対照的に、(f)高濃度の塩が使用される場合(例えば、100mM MSG)、残存応答が検出される。
【図4】T1R2およびT1R3は、甘味の知覚において重要である。(a)野生型マウス(白丸、点線)、T1R1 KOマウス(グレーの丸およびバー)、T1R2 KOマウス(緑の丸およびバー)およびT1R3 KOマウス(茶丸およびバー)の味覚嗜好性が、水に対して、短期接近味試験を使用して測定された。T1R1 KOマウスは、野生型コントロールと同等である。対照的に、T1R2およびT1R3ノックアウト動物は、人工甘味量およびD−アミノ酸に対する嗜好性の完全な消失を示すが、高濃度の天然の糖に対する残存応答を保持する。これらは、マルトース、スクロースおよびグルコースについて、拡大スケールの用量反応として、(b)において強調されている。しかし、T1R2/T1R3ダブルノックアウト動物(赤丸)は、全ての甘味応答の完全な消失を示す。値は、平均±標準誤差である(n=7)。D−AsnおよびD−Pheは、各々100mMであり、そしてD−Trpは、30mMで使用された。
【図5】T1R2およびT1R3は、哺乳動物甘味レセプターをコードする。パネル(a)は、野生型(WT)およびT1Rノックアウト動物(1−KO、2−KO、3−KO)における天然の糖、人工甘味料およびD−アミノ酸への統合された鼓索神経の応答を示す。T1R2およびT1R3ノックアウトは、人工甘味料およびD−アミノ酸に対する応答を完全に消失させているが(赤線)、高濃度の天然の糖への小さな神経応答を示す。しかし、これらは、T1R2/T1R3ダブルノックアウトマウスにおいて完全に消滅した(下部の青線)。パネル(b)は、増大したパネルの味覚物質に対する平均神経応答を示す;野生型、白いバー;T1R2 KO、緑のバー;T1R3KO、茶色のバー;T1R2/T1R3ダブルKO、赤いバー。値は、標準化された鼓索神経応答の平均±標準誤差である(n=4)。
【図6】hT1R3は、高濃度の天然の糖に応答する。乱交雑したGタンパク質Ggust−25(実験手順を参照のこと)およびマウスT1R3 GPCRを同時発現するHEK−293細胞、またはT1R2プラスT1R3を両方同時トランスフェクトされたHEK−293細胞を、種々の甘味化合物を用いて刺激した。上部のパネルは、500mMスクロースによるT1R3発現細胞の刺激の際の、[Ca2+]iの増加(しかし、300mMスクロースでは増加がないこと)を示す。人工甘味料(300mMサッカリン、右のパネル)を用いた場合、またはレセプターもしくはGgust−25のない細胞中において、応答は検出されなかった;スケールは、FURA−2 F340/F380比より決定された[Ca2+]i(nM)を示す。予期されたように、T1R2+T1R3を発現するコントロール細胞(下部のパネル)は、低濃度の天然の甘味料(300mMスクロース)および人工甘味料(30mMサッカリン)に強く応答する。
【図7】T1R2発現細胞の活性は、行動の誘引を誘発する。
【0267】
(a)野生型マウスおよび強力なT1R2プロモーターの制御下でヒトT1R2遺伝子を発現するT1R2 KOマウスが、(b−d)種々のヒト甘味味覚物質への行動の応答について試験された:(b)Ace−K、アセスルファーム−K、(c)アスパルテーム、および(d)MON、モネリン(約10μM);THAU、タウマチン(約5μM);ASP、アスパルテーム(10mM);GA、グリチルリジン酸(500μM);NH、ネオへスペリジンジヒドロカルコン(400μM)。ヒトT1R2味レセプターは、(a)T1R2細胞において選択的に発現され、そして(b)T1R2 KOマウスの甘味応答を効果的に救助する。重要なことに、トランスジーンの存在は(c−d)、トランスジェニック動物の甘味の嗜好性をヒトのようにする。詳細についてはテキストを参照のこと。(e)T1R2細胞中のRASSL(Redfern,C.H.ら、Nat Biotechnol,17,165−169(1999))の発現は、スピラドリンへの特異的な行動の誘引を示す動物を作製する。RASSL発現動物における強い応答を誘発するために必要な濃度の100×においてでさえも、非誘導動物またはコントロールマウスにおいては応答が見られないことに注意すること。値は、平均±標準誤差である(n=7)。
【図8】図8は、hT1R1のヌクレオチド配列(配列番号26)を提供する。
【図9】図9は、hT1R1のアミノ酸配列(配列番号27)を提供する。
【図10】図10は、hT1R2のヌクレオチド配列(配列番号28)を提供する。
【図11】図11は、hT1R2のアミノ酸配列(配列番号29)を提供する。
【図12】図12は、hT1R3のヌクレオチド配列(配列番号30)を提供する。
【図13】図13は、hT1R3のアミノ酸配列(配列番号31)を提供する。
【特許請求の範囲】
【請求項1】
単離されたホモダイマーの味レセプターであって、該レセプターは、2つのT1R3ポリペプチドを含み、ここで、該T1R3ポリペプチドは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる、単離されたホモダイマーの味レセプター。
【請求項2】
前記T1R3ポリペプチドが、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列を含む、請求項1に記載の単離されたレセプター。
【請求項3】
前記T1R3ポリペプチドが、配列番号14、配列番号19、配列番号22、配列番号24、または配列番号30を含むヌクレオチド配列によってコードされる、請求項1に記載の単離されたレセプター。
【請求項4】
前記T1R3ポリペプチドが、非共有結合性に結合される、請求項1に記載の単離されたレセプター。
【請求項5】
前記T1R3ポリペプチドが、共有結合性に結合される、請求項1に記載の単離されたレセプター。
【請求項6】
前記レセプターが、甘味リガンドに結合する、請求項1に記載の単離されたレセプター。
【請求項7】
前記甘味リガンドが、グルコース、フルクトース、ガラクトース、スクロース、マルトース、およびラクトースからなる群より選択される天然に存在する糖である、請求項6に記載の単離されたレセプター。
【請求項8】
前記レセプターが、Gタンパク質共役型レセプター活性を有する、請求項1に記載の単離されたレセプター。
【請求項9】
前記レセプターが、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31に対して生じた抗体に特異的に結合する、請求項1に記載の単離されたレセプター。
【請求項10】
前記T1R3ポリペプチドが、組換え型である、請求項1に記載の単離されたレセプター。
【請求項11】
T1R1またはT1R2を発現しない、請求項10に記載の単離されたレセプターを含む宿主細胞。
【請求項12】
単離されたホモダイマーの味レセプターであって、該レセプターは、2つのT1R3ポリペプチドからなり、ここで、該T1R3ポリペプチドは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列に対して、90%の同一性を有する、ヌクレオチド配列によってコードされる、単離されたホモダイマーの味レセプター。
【請求項13】
単離されたモノマーの味レセプターであって、該レセプターは、1つのT1R3ポリペプチドからなり、ここで、該T1R3ポリペプチドは、ヌクレオチド配列によってコードされ、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズする、単離されたモノマーの味レセプター。
【請求項14】
前記T1R3ポリペプチドが、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列を含む、請求項13に記載の単離されたレセプター。
【請求項15】
前記T1R3ポリペプチドが、配列番号14、配列番号19、配列番号22、配列番号24、または配列番号30を含むヌクレオチド配列によってコードされる、請求項13に記載の単離されたレセプター。
【請求項16】
前記レセプターが、甘味リガンドに結合する、請求項13に記載の単離されたレセプター。
【請求項17】
前記甘味リガンドが、グルコース、フルクトース、ガラクトース、スクロース、マルトース、およびラクトースからなる群より選択される天然に存在する糖である、請求項13に記載の単離されたレセプター。
【請求項18】
前記レセプターが、Gタンパク質共役型レセプター活性を有する、請求項13に記載の単離されたレセプター。
【請求項19】
前記T1R3ポリペプチドが、組換え型である、請求項13に記載の単離されたレセプター。
【請求項20】
T1R1またはT1R2を発現しない、請求項19に記載の単離されたレセプターを含む宿主細胞。
【請求項21】
組換え味レセプターを発現する宿主細胞であって、該レセプターは、T1R3ポリペプチドを含み、ここで、該T1R3ポリペプチドは、ヌクレオチド配列によってコードされ、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズし、ここで、該細胞は、T1R1またはT1R2を発現しない、宿主細胞。
【請求項22】
味細胞において味シグナル変換を調節する化合物を同定する方法であって、該方法は、
(i)2つのT1R3ポリペプチドを含むホモダイマーの味レセプターと該化合物を接触させる工程であって、ここで、該T1R3ポリペプチドは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる、工程;ならびに
(ii)該レセプター上の化合物の機能的効果を決定し、それによって味シグナル変換を調節する化合物を同定する工程
を包含する、方法。
【請求項23】
前記T1R3ポリペプチドが、非共有結合性に結合される、請求項22に記載の方法。
【請求項24】
前記T1R3ポリペプチドが、共有結合性に結合される、請求項22に記載の方法。
【請求項25】
前記レセプターが、組換え型である、請求項22に記載の方法。
【請求項26】
前記レセプターが、Gタンパク質共役型レセプター活性を有する、請求項22に記載の方法。
【請求項27】
前記機能的効果が、インビトロで測定される、請求項22に記載の方法。
【請求項28】
前記機能的効果が、物理的効果である、請求項27に記載の方法。
【請求項29】
前記レセプターが、固相に結合される、請求項27に記載の方法。
【請求項30】
前記機能的効果が、前記レセプターへの化合物の結合を測定することによって決定される、請求項27に記載の方法。
【請求項31】
前記機能的効果が、前記レセプターの細胞外ドメインへの化合物の結合を測定することによって決定される、請求項30に記載の方法。
【請求項32】
前記レセプターが、細胞または細胞膜において発現され、ここで、該細胞は、T1R1またはT1R2を発現しない、請求項22に記載の方法。
【請求項33】
前記機能的効果が、物理的効果である、請求項32に記載の方法。
【請求項34】
前記機能的効果が、前記レセプターへ結合しているリガンドを測定することによって決定される、請求項33に記載の方法。
【請求項35】
前記機能的効果が、前記レセプターの細胞外ドメインへの化合物の結合を測定することによって決定される、請求項34に記載の方法。
【請求項36】
前記機能的効果が、化学的効果または表現型効果である、請求項32に記載の方法。
【請求項37】
前記機能的効果が、細胞内のcAMP、IP3、またはCa2+の変化を測定することによって決定される、請求項36に記載の方法。
【請求項38】
前記細胞が、哺乳動物細胞である、請求項32に記載の方法。
【請求項39】
前記細胞が、ヒト細胞である、請求項38に記載の方法。
【請求項40】
味細胞において味シグナル変換を調節する化合物を同定する方法であって、該方法は、
(i)2つのT1R3ポリペプチドを含むホモダイマーの味レセプターを発現する細胞と、該化合物を接触させる工程であって、ここで、該T1R3ポリペプチドは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされ、ここで、該細胞は、T1R1およびT1R2を発現しない工程;ならびに
(ii)該レセプター上の化合物の機能的効果を決定し、それによって味シグナル変換を調節する化合物を同定する工程
を包含する、方法。
【請求項41】
前記T1R3ポリペプチドが、非共有結合性に結合される、請求項40に記載の方法。
【請求項42】
前記T1R3ポリペプチドが、共有結合性に結合される、請求項40に記載の方法。
【請求項43】
味細胞において味シグナル変換を調節する化合物を同定する方法であって、該方法は、
(i)1つのT1R3ポリペプチドを含むモノマーの味レセプターと、該化合物を接触させる工程であって、ここで、該T1R3ポリペプチドは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる、工程;ならびに
(ii)該レセプター上の化合物の機能的効果を決定し、それによって味シグナル変換を調節する化合物を同定する工程
を包含する、方法。
【請求項44】
味細胞において味シグナル変換を調節する化合物を同定する方法であって、該方法は、
(i)1つのT1R3ポリペプチドを含むモノマーの味レセプターを発現する細胞と、該化合物を接触させる工程であって、ここで、該T1R3ポリペプチドは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされ;ここで、該細胞は、T1R1またはT1R2を発現しない、工程;ならびに
(ii)該レセプター上の化合物の機能的効果を決定し、それによって味シグナル変換を調節する化合物を同定する工程
を包含する、方法。
【請求項45】
味細胞において味シグナル変換を調節する化合物を同定する方法であって、該方法は、
(i)T1R3ポリペプチドを含む味レセプターを発現する細胞と、該化合物を接触させる工程であって、ここで、該T1R3ポリペプチドは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされ;ここで、該細胞は、T1R1およびT1R2を発現しない、工程;ならびに
(ii)該レセプター上の化合物の機能的効果を決定し、それによって味シグナル変換を調節する化合物を同定する工程
を包含する、方法。
【請求項1】
単離されたホモダイマーの味レセプターであって、該レセプターは、2つのT1R3ポリペプチドを含み、ここで、該T1R3ポリペプチドは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる、単離されたホモダイマーの味レセプター。
【請求項2】
前記T1R3ポリペプチドが、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列を含む、請求項1に記載の単離されたレセプター。
【請求項3】
前記T1R3ポリペプチドが、配列番号14、配列番号19、配列番号22、配列番号24、または配列番号30を含むヌクレオチド配列によってコードされる、請求項1に記載の単離されたレセプター。
【請求項4】
前記T1R3ポリペプチドが、非共有結合性に結合される、請求項1に記載の単離されたレセプター。
【請求項5】
前記T1R3ポリペプチドが、共有結合性に結合される、請求項1に記載の単離されたレセプター。
【請求項6】
前記レセプターが、甘味リガンドに結合する、請求項1に記載の単離されたレセプター。
【請求項7】
前記甘味リガンドが、グルコース、フルクトース、ガラクトース、スクロース、マルトース、およびラクトースからなる群より選択される天然に存在する糖である、請求項6に記載の単離されたレセプター。
【請求項8】
前記レセプターが、Gタンパク質共役型レセプター活性を有する、請求項1に記載の単離されたレセプター。
【請求項9】
前記レセプターが、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31に対して生じた抗体に特異的に結合する、請求項1に記載の単離されたレセプター。
【請求項10】
前記T1R3ポリペプチドが、組換え型である、請求項1に記載の単離されたレセプター。
【請求項11】
T1R1またはT1R2を発現しない、請求項10に記載の単離されたレセプターを含む宿主細胞。
【請求項12】
単離されたホモダイマーの味レセプターであって、該レセプターは、2つのT1R3ポリペプチドからなり、ここで、該T1R3ポリペプチドは、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列に対して、90%の同一性を有する、ヌクレオチド配列によってコードされる、単離されたホモダイマーの味レセプター。
【請求項13】
単離されたモノマーの味レセプターであって、該レセプターは、1つのT1R3ポリペプチドからなり、ここで、該T1R3ポリペプチドは、ヌクレオチド配列によってコードされ、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズする、単離されたモノマーの味レセプター。
【請求項14】
前記T1R3ポリペプチドが、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列を含む、請求項13に記載の単離されたレセプター。
【請求項15】
前記T1R3ポリペプチドが、配列番号14、配列番号19、配列番号22、配列番号24、または配列番号30を含むヌクレオチド配列によってコードされる、請求項13に記載の単離されたレセプター。
【請求項16】
前記レセプターが、甘味リガンドに結合する、請求項13に記載の単離されたレセプター。
【請求項17】
前記甘味リガンドが、グルコース、フルクトース、ガラクトース、スクロース、マルトース、およびラクトースからなる群より選択される天然に存在する糖である、請求項13に記載の単離されたレセプター。
【請求項18】
前記レセプターが、Gタンパク質共役型レセプター活性を有する、請求項13に記載の単離されたレセプター。
【請求項19】
前記T1R3ポリペプチドが、組換え型である、請求項13に記載の単離されたレセプター。
【請求項20】
T1R1またはT1R2を発現しない、請求項19に記載の単離されたレセプターを含む宿主細胞。
【請求項21】
組換え味レセプターを発現する宿主細胞であって、該レセプターは、T1R3ポリペプチドを含み、ここで、該T1R3ポリペプチドは、ヌクレオチド配列によってコードされ、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズし、ここで、該細胞は、T1R1またはT1R2を発現しない、宿主細胞。
【請求項22】
味細胞において味シグナル変換を調節する化合物を同定する方法であって、該方法は、
(i)2つのT1R3ポリペプチドを含むホモダイマーの味レセプターと該化合物を接触させる工程であって、ここで、該T1R3ポリペプチドは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる、工程;ならびに
(ii)該レセプター上の化合物の機能的効果を決定し、それによって味シグナル変換を調節する化合物を同定する工程
を包含する、方法。
【請求項23】
前記T1R3ポリペプチドが、非共有結合性に結合される、請求項22に記載の方法。
【請求項24】
前記T1R3ポリペプチドが、共有結合性に結合される、請求項22に記載の方法。
【請求項25】
前記レセプターが、組換え型である、請求項22に記載の方法。
【請求項26】
前記レセプターが、Gタンパク質共役型レセプター活性を有する、請求項22に記載の方法。
【請求項27】
前記機能的効果が、インビトロで測定される、請求項22に記載の方法。
【請求項28】
前記機能的効果が、物理的効果である、請求項27に記載の方法。
【請求項29】
前記レセプターが、固相に結合される、請求項27に記載の方法。
【請求項30】
前記機能的効果が、前記レセプターへの化合物の結合を測定することによって決定される、請求項27に記載の方法。
【請求項31】
前記機能的効果が、前記レセプターの細胞外ドメインへの化合物の結合を測定することによって決定される、請求項30に記載の方法。
【請求項32】
前記レセプターが、細胞または細胞膜において発現され、ここで、該細胞は、T1R1またはT1R2を発現しない、請求項22に記載の方法。
【請求項33】
前記機能的効果が、物理的効果である、請求項32に記載の方法。
【請求項34】
前記機能的効果が、前記レセプターへ結合しているリガンドを測定することによって決定される、請求項33に記載の方法。
【請求項35】
前記機能的効果が、前記レセプターの細胞外ドメインへの化合物の結合を測定することによって決定される、請求項34に記載の方法。
【請求項36】
前記機能的効果が、化学的効果または表現型効果である、請求項32に記載の方法。
【請求項37】
前記機能的効果が、細胞内のcAMP、IP3、またはCa2+の変化を測定することによって決定される、請求項36に記載の方法。
【請求項38】
前記細胞が、哺乳動物細胞である、請求項32に記載の方法。
【請求項39】
前記細胞が、ヒト細胞である、請求項38に記載の方法。
【請求項40】
味細胞において味シグナル変換を調節する化合物を同定する方法であって、該方法は、
(i)2つのT1R3ポリペプチドを含むホモダイマーの味レセプターを発現する細胞と、該化合物を接触させる工程であって、ここで、該T1R3ポリペプチドは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされ、ここで、該細胞は、T1R1およびT1R2を発現しない工程;ならびに
(ii)該レセプター上の化合物の機能的効果を決定し、それによって味シグナル変換を調節する化合物を同定する工程
を包含する、方法。
【請求項41】
前記T1R3ポリペプチドが、非共有結合性に結合される、請求項40に記載の方法。
【請求項42】
前記T1R3ポリペプチドが、共有結合性に結合される、請求項40に記載の方法。
【請求項43】
味細胞において味シグナル変換を調節する化合物を同定する方法であって、該方法は、
(i)1つのT1R3ポリペプチドを含むモノマーの味レセプターと、該化合物を接触させる工程であって、ここで、該T1R3ポリペプチドは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされる、工程;ならびに
(ii)該レセプター上の化合物の機能的効果を決定し、それによって味シグナル変換を調節する化合物を同定する工程
を包含する、方法。
【請求項44】
味細胞において味シグナル変換を調節する化合物を同定する方法であって、該方法は、
(i)1つのT1R3ポリペプチドを含むモノマーの味レセプターを発現する細胞と、該化合物を接触させる工程であって、ここで、該T1R3ポリペプチドは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされ;ここで、該細胞は、T1R1またはT1R2を発現しない、工程;ならびに
(ii)該レセプター上の化合物の機能的効果を決定し、それによって味シグナル変換を調節する化合物を同定する工程
を包含する、方法。
【請求項45】
味細胞において味シグナル変換を調節する化合物を同定する方法であって、該方法は、
(i)T1R3ポリペプチドを含む味レセプターを発現する細胞と、該化合物を接触させる工程であって、ここで、該T1R3ポリペプチドは、高度にストリンジェントなハイブリダイゼーション条件下で、配列番号15、配列番号20、配列番号23、配列番号25、または配列番号31のアミノ酸配列をコードするヌクレオチド配列にハイブリダイズするヌクレオチド配列によってコードされ;ここで、該細胞は、T1R1およびT1R2を発現しない、工程;ならびに
(ii)該レセプター上の化合物の機能的効果を決定し、それによって味シグナル変換を調節する化合物を同定する工程
を包含する、方法。
【図1A】

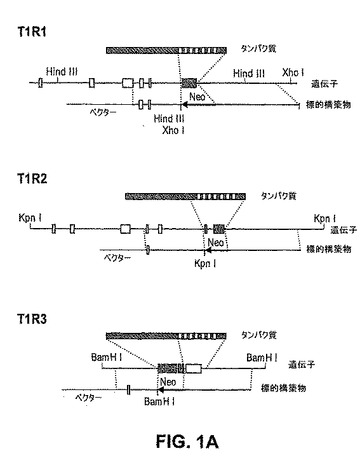
【図1B】
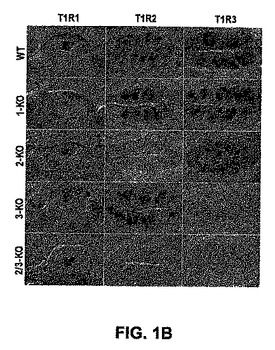
【図2A】
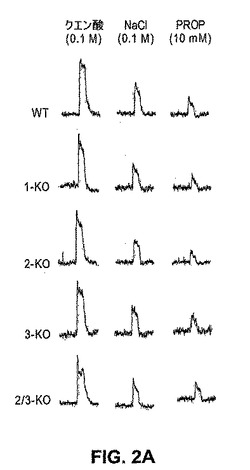
【図2BC】
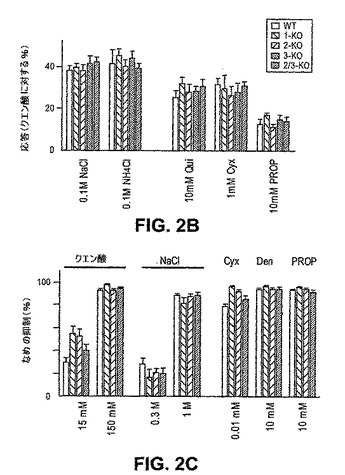
【図3AD】
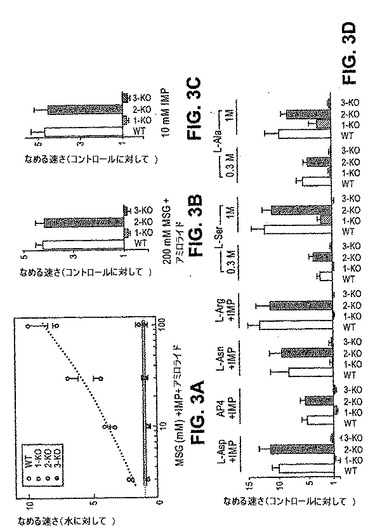
【図3EF】

【図4】
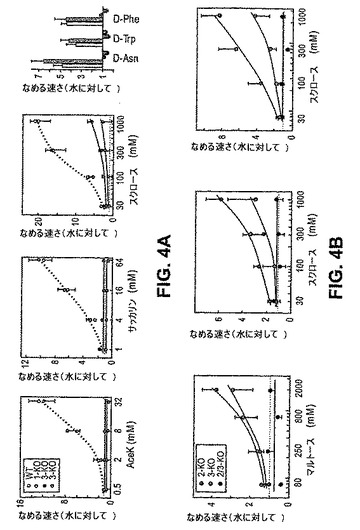
【図5】
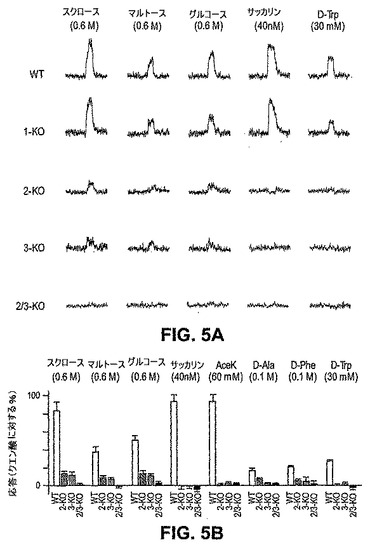
【図6】
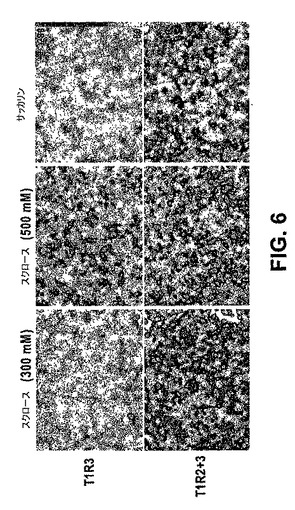
【図7】
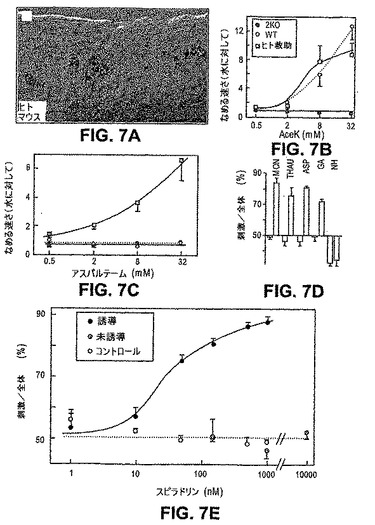
【図8】

【図9】

【図10】

【図11】

【図12】

【図13】

【図1B】
【図2A】
【図2BC】
【図3AD】
【図3EF】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【公表番号】特表2007−507544(P2007−507544A)
【公表日】平成19年3月29日(2007.3.29)
【国際特許分類】
【出願番号】特願2006−534237(P2006−534237)
【出願日】平成16年10月4日(2004.10.4)
【国際出願番号】PCT/US2004/032678
【国際公開番号】WO2005/033125
【国際公開日】平成17年4月14日(2005.4.14)
【出願人】(500025503)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (26)
【出願人】(301008213)ザ ガバメント オブ ザ ユナイテッド ステイツ オブ アメリカ アズ リプレゼンテッド バイ ザ セクレタリー オブ ザ デパートメント オブ ヘルス アンド ヒューマン サービシズ (2)
【Fターム(参考)】
【公表日】平成19年3月29日(2007.3.29)
【国際特許分類】
【出願日】平成16年10月4日(2004.10.4)
【国際出願番号】PCT/US2004/032678
【国際公開番号】WO2005/033125
【国際公開日】平成17年4月14日(2005.4.14)
【出願人】(500025503)ザ リージェンツ オブ ザ ユニバーシティ オブ カリフォルニア (26)
【出願人】(301008213)ザ ガバメント オブ ザ ユナイテッド ステイツ オブ アメリカ アズ リプレゼンテッド バイ ザ セクレタリー オブ ザ デパートメント オブ ヘルス アンド ヒューマン サービシズ (2)
【Fターム(参考)】
[ Back to top ]