
哺乳類様複合N−グリカンを生成する酵母株
哺乳類様複合N−グリカンを含有するかまたは哺乳類グリコシル化経路中の中間体を含有する標的分子を生成するために有用な方法および遺伝子工学的に改変された真菌細胞が、本明細書に記載される。一実施形態において、GlcNAcMan5GlcNAc2 N−グリカンを含むタンパク質を生成できる真菌細胞を生成する方法が提供され、この方法は、a)Man5GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された真菌細胞を提供するステップと、b)該細胞に、GlcNAcトランスフェラーゼIをコードする核酸を導入するステップとを含む。

【発明の詳細な説明】
【技術分野】
【0001】
関連する出願への相互参照
この出願は、2009年11月19日に出願された米国出願第61/262,828号に対する優先権を主張する。この前の出願の開示は、その全体が参考として援用される。
【0002】
技術分野
本発明は、真菌細胞において糖タンパク質を生成するための方法および材料、より具体的には、真菌細胞を遺伝子工学的に改変して、哺乳類様複合N−グリカンを含有するタンパク質または哺乳類グリコシル化経路内の中間体を含有するタンパク質を生成することに関する。
【背景技術】
【0003】
現在開発中のほとんどの生物製剤(例えば組換えタンパク質)を生成するために、高性能の発現系が要求される。これらの生物製剤の多くの生物活性は、それらの翻訳後修飾(例えばリン酸化またはグリコシル化)に依存する。酵母ベースの発現系は、微生物の遺伝子操作および発酵の容易さを、タンパク質を分泌し、修飾する能力と組み合わせる。しかし、酵母細胞で生成される組換え糖タンパク質は、主に不均質な高マンノースおよび過マンノースグリカン構造を示し、これはタンパク質機能、下流のプロセシングおよび後続の治療のための使用にとって、特にグリコシル化が生物学的に重要な役割を演じる場合に有害であり得る。
【発明の概要】
【課題を解決するための手段】
【0004】
本明細書で記載する方法および遺伝子工学的に改変された真菌細胞は、哺乳類様N−グリカンを含有するかまたは哺乳類(例えばヒト)グリコシル化経路内の中間体を含有する標的分子(例えば標的タンパク質)を生成するために用いることができる。このような工学的に改変された細胞から単離された標的分子は、抗体生成、サイトカイン生成を含む生物製剤の用途、およびリソソーム蓄積症のような代謝障害の処置のために用いることができる。
【0005】
一態様では、この文書は、GlcNAcMan5GlcNAc2 N−グリカンを含むタンパク質を生成できる真菌細胞(例えばYarrowia lipolyticaまたはArxula adeninivorans)を生成する方法を特徴とする。方法は、Man5GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された真菌細胞を提供するステップと、細胞に、GlcNAcトランスフェラーゼIをコードする核酸を導入するステップであって、ここで、核酸が、コードされたGlcNAcトランスフェラーゼIを細胞内区画(例えばゴルジ装置)に向かわせる(target)ためのヌクレオチドターゲティング配列を含み、ここで、真菌細胞におけるGlcNAcトランスフェラーゼIの発現が、GlcNAcMan5GlcNAc2 N−グリカンを含むタンパク質を生成するステップとを含む。方法は、細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、細胞は、GlcNAcMan5GlcNAc2 N−グリカンを含むように改変された標的タンパク質を生成するステップをさらに含むことができる。標的タンパク質は、Fc受容体と結合できる。標的タンパク質は、抗体またはその断片であり得る。標的タンパク質は、治療用の糖タンパク質であり得る。標的タンパク質は、インターフェロン−β、GM−CSF、インターフェロンγまたはエリスロポエチンであり得る。
【0006】
Man5GlcNAc2 N−グリカンを含有するタンパク質を生成するように遺伝子工学的に改変された真菌細胞は、OCH1活性を欠損でき、α−1,2−マンノシダーゼをコードする核酸を含み、ここで、α−1,2−マンノシダーゼをコードする核酸は、コードされたα−1,2−マンノシダーゼを小胞体に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含む。ターゲティング配列は、HDEL配列であり得る。
【0007】
方法は、細胞に、マンノシダーゼIIをコードする核酸を導入するステップであって、ここで、マンノシダーゼIIをコードする核酸は、コードされたマンノシダーゼIIをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるマンノシダーゼIIの発現は、GlcNAcMan3GlcNAc2 N−グリカンを含有するタンパク質を生成するステップをさらに含むことができる。
【0008】
方法は、細胞に、ガラクトシルトランスフェラーゼをコードする核酸を導入するステップであって、ここで、ガラクトシルトランスフェラーゼをコードする核酸は、コードされたガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるガラクトシルトランスフェラーゼの発現は、GalGlcNAcMan5GlcNAc2またはGalGlcNAcMan3GlcNAc2 N−グリカンを含有するタンパク質を生成するステップをさらに含むことができる。ガラクトシルトランスフェラーゼは、UDP−Glc−4−エピメラーゼとβ−1,4−ガラクトシルトランスフェラーゼIの触媒ドメインとの融合体であり得る。このような方法は、細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、細胞は、GalGlcNAcMan5GlcNAc2またはGalGlcNAcMan3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質を生成するステップをさらに含むことができる。方法は、GalGlcNAcMan5GlcNAc2またはGalGlcNAcMan3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質を単離するステップを含むことができる。
【0009】
別の態様では、この文書は、GlcNAcMan3GlcNAc2 N−グリカンを含有する標的タンパク質を生成する方法を特徴とする。方法は、GlcNAcトランスフェラーゼIと、α−1,2−マンノシダーゼと、マンノシダーゼIIとをコードする核酸を含むように遺伝子工学的に改変された真菌細胞(例えばYarrowia lipolyticaまたはArxula adeninivorans)を提供するステップであって、ここで、核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるための1つのターゲティング配列をコードする1つのヌクレオチド配列または複数のターゲティング配列をコードする複数のヌクレオチド配列を含み、ここで、真菌細胞が、OCH1活性を欠損しているステップと、細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、細胞が、GlcNAcMan3GlcNAc2 N−グリカンを含有する標的タンパク質を生成するステップとを含む。α−1,2−マンノシダーゼをコードする核酸は、コードされたα−1,2−マンノシダーゼを小胞体に向かわせるための小胞体ターゲティング配列を含むことができる。例えば、ターゲティング配列は、HDEL配列であり得る。GlcNAcトランスフェラーゼIおよびマンノシダーゼIIをコードする核酸は、コードされたGlcNAcトランスフェラーゼIおよびマンノシダーゼIIをゴルジ装置に向かわせるための1つ以上のゴルジターゲティング配列を含むことができる。標的タンパク質は、Fc受容体と結合できる。標的タンパク質は、抗体またはその断片であり得る。標的タンパク質は、治療用の糖タンパク質であり得る。標的タンパク質は、インターフェロン−β、GM−CSF、インターフェロンγまたはエリスロポエチンであり得る。
【0010】
いくつかの実施形態では、方法は、細胞に、ガラクトシルトランスフェラーゼをコードする核酸を導入するステップであって、ここで、ガラクトシルトランスフェラーゼをコードする核酸は、コードされたガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるガラクトシルトランスフェラーゼの発現は、GalGlcNAcMan3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質を生成するステップをさらに含むことができる。GalGlcNAcMan3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質は、真菌細胞から単離できる。
【0011】
この文書は、GlcNAcMan3GlcNAc2 N−グリカンを含有するタンパク質を生成できる真菌細胞(例えばYarrowia lipolyticaまたはArxula adeninivorans)を作製する方法も特徴とする。方法は、Man3GlcNAc2 N−グリカンを含有するタンパク質を生成するように遺伝子工学的に改変された真菌細胞を提供するステップと、細胞に、GlcNAcトランスフェラーゼIをコードする核酸を導入するステップであって、ここで、核酸が、コードされたGlcNAcトランスフェラーゼIを細胞内区画(例えばゴルジ装置)に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるGlcNAcトランスフェラーゼIの発現が、GlcNAcMan3GlcNAc2 N−グリカンを含有するタンパク質を生成するステップとを含む。方法は、細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、細胞は、GlcNAcMan3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質を生成するステップをさらに含むことができる。標的タンパク質は、Fc受容体と結合できる。標的タンパク質は、抗体またはその断片であり得る。標的タンパク質は、治療用の糖タンパク質であり得る。標的タンパク質は、インターフェロン−β、GM−CSF、インターフェロンγまたはエリスロポエチンであり得る。
【0012】
Man3GlcNAc2 N−グリカンを含有するタンパク質を生成するように遺伝子工学的に改変された真菌細胞は、ALG3活性を欠損でき、α−1,2−マンノシダーゼをコードする核酸を含み、ここで、核酸は、コードされたα−1,2−マンノシダーゼを小胞体に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含む。このような真菌細胞は、OCH1活性をさらに欠損でき、かつ/またはα−1,3−グルコシルトランスフェラーゼ(例えばALG6)をコードする核酸をさらに含むことができる。
【0013】
方法は、細胞に、GlcNAcトランスフェラーゼIIをコードする核酸を導入するステップであって、ここで、GlcNAcトランスフェラーゼIIをコードする核酸は、コードされたGlcNAcトランスフェラーゼIIを細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるGlcNAcトランスフェラーゼIIの発現が、GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成するステップをさらに含むことができる。
【0014】
方法は、細胞に、ガラクトシルトランスフェラーゼをコードする核酸を導入するステップであって、ここで、ガラクトシルトランスフェラーゼをコードする核酸は、コードされたガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるガラクトシルトランスフェラーゼの発現は、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成するステップをさらに含むことができる。ガラクトシルトランスフェラーゼは、UDP−Glc−4−エピメラーゼと、β−1,4−ガラクトシルトランスフェラーゼIの触媒ドメインとの融合体であり得る。方法は、細胞に標的タンパク質をコードする核酸を導入するステップであって、ここで、細胞は、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質を生成するステップをさらに含むことができる。
【0015】
方法は、細胞に、グルコシダーゼIIのαおよびβサブユニットをコードする核酸を導入するステップであって、ここで、真菌細胞におけるグルコシダーゼIIのαおよびβサブユニットの発現は、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成するステップをさらに含むことができる。
【0016】
この文書は、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含有する標的タンパク質を生成する方法も特徴とする。方法は、ALG3活性を欠損するように遺伝子工学的に改変され、GlcNAcトランスフェラーゼIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸を含む真菌細胞を提供するステップであって、ここで、GlcNAcトランスフェラーゼIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸が、それぞれのコードされたタンパク質を細胞内区画(例えばゴルジ装置)に向かわせるための1つのターゲティング配列をコードする1つのヌクレオチド配列または複数のターゲティング配列をコードする複数のヌクレオチド配列を含むステップと、細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、細胞が、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含有する標的タンパク質を生成するステップとを含む。真菌細胞は、OCH1活性をさらに欠損でき、かつ/またはALG6のようなα−1,3−グルコシルトランスフェラーゼをコードする核酸をさらに含むことができる。真菌細胞は、グルコシダーゼIIのαおよびβサブユニットをコードする核酸をさらに含むことができ、ここで、真菌細胞におけるグルコシダーゼIIのαおよびβサブユニットの発現は、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含有する標的タンパク質を生成する。
【0017】
別の態様では、この文書は、GlcNAcMan3GlcNAc2 N−グリカンを含有するタンパク質を生成するように遺伝子工学的に改変された単離真菌細胞を特徴とする。真菌細胞は、OCH1活性を欠損でき、α−1,2−マンノシダーゼと、GlcNAcトランスフェラーゼIと、マンノシダーゼIIとをコードする核酸を含み、ここで、α−1,2−マンノシダーゼと、GlcNAcトランスフェラーゼIと、マンノシダーゼIIとをコードする核酸は、それぞれのコードされたタンパク質を細胞内区画に向かわせるための1つのターゲティング配列をコードする1つのヌクレオチド配列または複数のターゲティング配列をコードする複数のヌクレオチド配列を含み、ここで、真菌細胞におけるα−1,2−マンノシダーゼ、GlcNAcトランスフェラーゼIおよびマンノシダーゼIIの発現は、GlcNAcMan3GlcNAc2 N−グリカンを含有するタンパク質を生成する。真菌細胞は、標的タンパク質をコードする核酸をさらに含むことができ、ここで、細胞は、GlcNAcMan3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質を生成する。
【0018】
いくつかの実施形態では、このような真菌細胞は、GlcNAcトランスフェラーゼIIをコードする核酸をさらに含み、ここで、GlcNAcトランスフェラーゼIIをコードする核酸は、コードされたGlcNAcトランスフェラーゼIIを細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるGlcNAcトランスフェラーゼIIの発現は、GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成する。
【0019】
いくつかの実施形態では、このような真菌細胞は、ガラクトシルトランスフェラーゼをコードする核酸をさらに含み、ここで、ガラクトシルトランスフェラーゼをコードする核酸は、コードされたガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるガラクトシルトランスフェラーゼの発現は、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成する。
【0020】
さらに別の態様では、この文書は、GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成するように遺伝子工学的に改変された単離真菌細胞を特徴とする。真菌細胞は、ALG3活性を欠損するように遺伝子工学的に改変され、GlcNAcトランスフェラーゼIとGlcNAcトランスフェラーゼIIとをコードする核酸を含み、ここで、GlcNAcトランスフェラーゼIとGlcNAcトランスフェラーゼIIとをコードする核酸は、それぞれのコードされたタンパク質を細胞内区画に向かわせるための1つのターゲティング配列をコードする1つのヌクレオチド配列または複数のターゲティング配列をコードする複数のヌクレオチド配列を含み、ここで、真菌細胞におけるGlcNAcトランスフェラーゼIおよびGlcNAcトランスフェラーゼIIの発現は、GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成する。遺伝子工学的に改変された真菌細胞は、OCH1活性をさらに欠損でき、かつ/またはα−1,3−グルコシルトランスフェラーゼをコードする核酸をさらに含むことができる。遺伝子工学的に改変された真菌細胞は、標的タンパク質をコードする核酸をさらに含むことができ、ここで、細胞は、GlcNAc2Man3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質を生成する。真菌細胞は、グルコシダーゼIIのαおよびβサブユニットをコードする核酸をさらに含むことができ、ここで、真菌細胞におけるグルコシダーゼIIのαおよびβサブユニットの発現は、GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成する。真菌細胞は、ガラクトシルトランスフェラーゼをコードする核酸をさらに含むことができ、ここで、ガラクトシルトランスフェラーゼをコードする核酸は、コードされたガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるガラクトシルトランスフェラーゼの発現は、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成する。
【0021】
この文書は、相当数のYarrowia lipolytica細胞が、Gal2GlcNac2Man3GlcNAc2 N−グリカンを含有する糖タンパク質を生成するように遺伝子工学的に改変されているYarrowia lipolytica細胞の実質的に純粋な培養物も特徴とする。細胞は、ALG3活性を欠損するように遺伝子工学的に改変され、GlcNAcトランスフェラーゼIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸を含み、ここで、GlcNAcトランスフェラーゼIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸は、それぞれのコードされたタンパク質を細胞内区画に向かわせるための1つのターゲティング配列をコードする1つのヌクレオチド配列または複数のターゲティング配列をコードする複数のヌクレオチド配列を含み、ここで、細胞におけるGlcNAcトランスフェラーゼI、GlcNAcトランスフェラーゼIIおよびガラクトシルトランスフェラーゼの発現は、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成する。遺伝子工学的に改変された真菌細胞は、OCH1活性をさらに欠損でき、かつ/またはα−1,3−グルコシルトランスフェラーゼ(例えばALG6)をコードする核酸をさらに含むことができる。細胞は、グルコシダーゼIIのαおよびβサブユニットをコードする核酸をさらに含むことができ、ここで、真菌細胞におけるグルコシダーゼIIのαおよびβサブユニットの発現は、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含有する標的タンパク質を生成する。
【0022】
別の態様では、この文書は、相当数のYarrowia lipolytica細胞が、Gal2GlcNac2Man3GlcNAc2 N−グリカンを含有する糖タンパク質を生成するように遺伝子工学的に改変されているYarrowia lipolytica細胞の実質的に純粋な培養物であって、ここで、細胞が、OCH1活性を欠損するように遺伝子工学的に改変され、α−1,2−マンノシダーゼと、GlcNAcトランスフェラーゼIと、マンノシダーゼIIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸を含み、ここで、α−1,2−マンノシダーゼと、GlcNAcトランスフェラーゼIと、マンノシダーゼIIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるための1つのターゲティング配列をコードする1つのヌクレオチド配列または複数のターゲティング配列をコードする複数のヌクレオチド配列を含み、ここで、細胞におけるα−1,2−マンノシダーゼ、GlcNAcトランスフェラーゼI、マンノシダーゼII、GlcNAcトランスフェラーゼIIおよびガラクトシルトランスフェラーゼの発現が、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成する培養物を特徴とする。
【0023】
この文書は、糖タンパク質を含み、ここで、糖タンパク質上のN−グリカンの少なくとも50%(例えば少なくとも70%または少なくとも85%)が、GlcNAc2Man3GlcNAc2 N−グリカンである組成物も特徴とする。
【0024】
そうでないと定義しない限り、本明細書で用いる全ての技術用語および科学用語は、本発明が属する当技術分野における当業者により一般的に理解されるのと同じ意味を有する。本明細書で記載するものと同様または等価な方法および材料を、本発明の実施または試験において用いることができるが、例示的な方法および材料は、以下に記載する。本明細書で言及する全ての出版物、特許出願、特許、Genbank(登録商標)受託番号およびその他の参考文献は、それらの全体が参照により組み込まれている。矛盾がある場合には、定義を含む本出願が支配する。材料、方法および実施例は例示のためだけであり、限定することを意図しない。
【0025】
本発明のその他の特徴および利点は、以下の詳細な記載および特許請求の範囲から明らかになる。
【図面の簡単な説明】
【0026】
【図1】図1Aは、Man5GLcNAc2構造およびMan3GlcNAc2構造の図である。図1Bは、プラスミドpYlOCH1 PUT TOPOの模式図である。
【図2】図2は、pold lnuga Yarrowia lipolytica野生型細胞またはΔoch1 pold lnuga Yarrowia lipolytica細胞から得られた分泌タンパク質のN−グリカン分析を示す一連の電気泳動図(electroferogram)である。OCH1不活性化の際の主なN−グリカンは、Man8GlcNAc2になる。分析は、DNAシーケンサー支援蛍光体支援糖質電気泳動(DSΑ−FACE)を用いて行った。「M5」、「M6」、「M8」および「M9」は、ベースのN−アセチルグルコサミン構造にコンジュゲートしたマンノース残基の数である。Y軸は、各N−グリカン構造の量を示す相対蛍光単位を表す。X軸は、キャピラリーを通しての各N−グリカン構造の相対移動度を表す。先頭の電気泳動図は、移動度標準物質として用いるためのデキストランの分析である。
【図3】図3は、プラスミドpYLHUXdL2preManHDELおよびpYLTUXdL2preManHDELの模式図である。
【図4】図4は、ManHDEL(=HDELタグ付きα−1,2−マンノシダーゼ)発現カセット(TEF1またはHp4dのいずれかのプロモーター制御の下で)をG014株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。「Rd」は、図3に示すベクターに存在するゼータ配列による「無作為組み込み」を表す。マンノシダーゼ発現の際の主要なN−グリカンは、Man5GlcNAc2である。これらの株の1つ(G018、表2を参照されたい)からのURA3マーカーのキュアリング(curing)は、N−グリカンプロファイルを変えない。
【図5】図5は、プラスミドJME926 pPTLeu2−ADE2ex−Hp4dManHDEL(Yl)およびOXYP289 pPTAxp1−LEU2ex−Hp4dManHDEL(Yl)についての構築方策の模式図である。ベクターpYLTmAXrGnTIIの構築については、図23を参照されたい。
【図6】図6は、LEU2またはAXP1遺伝子座のいずれかでの標的化組み込み(Tg)により、ManHDEL(=HDELタグ付きα−1,2−マンノシダーゼ)発現カセット(Hp4dプロモーター制御の下で)をG014株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。Man5GlcNAc2が主なN−グリカンになる。
【図7】図7は、Kre2pの100個のN末端アミノ酸とヒトGlcNAcトランスフェラーゼIの触媒ドメインとの間の融合タンパク質のアミノ酸配列(配列番号3)およびYarrowiaコドン最適化ヌクレオチド配列(配列番号4)を示す図である。太字で:融合タンパク質のKre2p部分;通常のフォントで:融合タンパク質のGnT I部分;下線:開始コドンおよび停止コドン。
【図8】図8は、プラスミドpYLTmAXhGnTIおよびpYLHp4mAXhGnTIについての構築方策の模式図である。
【図9】図9は、GnT Iを発現するベクターでの形質転換によりGnT I活性をG036株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。「Rd」は、図8に示すベクターに存在するゼータ配列による「無作為組み込み」を表す。GnT I活性の発現の際の主要なN−グリカンは、GlcNAcMan5GlcNAc2である。α−1,2−マンノシダーゼでのin vitro処理は、プロファイルを著しく変えず、このことは、Man5GlcNAc2以外の高マンノースN−グリカンは少量しか存在しないことを示す。in vitroヘキソサミニダーゼ処理は、GlcNAcMan5GlcNAc2からMan5GlcNAc2へのシフトをもたらす。
【図10−1】図10は、Mnn2pの36個のN末端アミノ酸とDrosophila melanogasterマンノシダーゼIIの触媒ドメインとの間の融合タンパク質のアミノ酸配列(配列番号7)およびYarrowiaコドン最適化ヌクレオチド配列(配列番号8)を示す図である。太字で:融合タンパク質のMnn2p部分;通常のフォントで:融合タンパク質のManII部分;下線:開始コドンおよび停止コドン。
【図10−2】図10は、Mnn2pの36個のN末端アミノ酸とDrosophila melanogasterマンノシダーゼIIの触媒ドメインとの間の融合タンパク質のアミノ酸配列(配列番号7)およびYarrowiaコドン最適化ヌクレオチド配列(配列番号8)を示す図である。太字で:融合タンパク質のMnn2p部分;通常のフォントで:融合タンパク質のManII部分;下線:開始コドンおよび停止コドン。
【図11】図11は、プラスミドpYLTmAXDmManIIおよびpYLTmAXDmManII(LEU2ex)についての構築方策の模式図である。
【図12】図12は、ManII発現ベクターでの形質転換によりManII活性をG040株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。「Rd」は、図11に示すベクターに存在するゼータ配列による「無作為組み込み」を表す。ManII活性の発現の際に、より高い電気泳動移動度を有する新しいピークならびに、Man5GlcNAc2とほぼ同じ位置に泳動される「ショルダー」ピークが出現する。in vitroヘキソサミニダーゼ処理は、これらのピークについて前へのシフトをもたらし(GlcNAcMan5GlcNAc2からMan5GlcNAc2への観察されたシフトの隣りに)、このことは、末端GlcNAcの存在を示し、よって、GlcNAcMan3GlcNAc2およびGlcNAcMan4GlcNAc2のようなピークが同定される。α−1,2−マンノシダーゼでのin vitro処理は、プロファイルを著しく変化させず、このことは、Man5GlcNAc2以外の高マンノースN−グリカンは少量しか存在しないことを示す。
【図13−1】図13は、Mnn2pの46個のN末端アミノ酸とSchizosaccharomyces pombe UDP−Glc−4−エピメラーゼ様タンパク質とヒトβ−1,4−ガラクトシルトランスフェラーゼIの触媒ドメインとの間の融合タンパク質のアミノ酸配列(配列番号9)およびYarrowiaコドン最適化ヌクレオチド配列(配列番号10)である。融合タンパク質のMnn2p部分は配列番号9の1〜46であり、リンカー配列は47〜49および405〜408であり、融合タンパク質のエピメラーゼ配列は50〜404であり、融合タンパク質のManII部分は409〜763である。Mnn2p部分は配列番号10のヌクレオチド1〜138であり、リンカー配列はヌクレオチド139〜147および1213〜1224であり、エピメラーゼ配列はヌクレオチド148〜1212であり、ManII部分は1225〜2289である。開始コドンおよび停止コドンに下線を付す。
【図13−2】図13は、Mnn2pの46個のN末端アミノ酸とSchizosaccharomyces pombe UDP−Glc−4−エピメラーゼ様タンパク質とヒトβ−1,4−ガラクトシルトランスフェラーゼIの触媒ドメインとの間の融合タンパク質のアミノ酸配列(配列番号9)およびYarrowiaコドン最適化ヌクレオチド配列(配列番号10)である。融合タンパク質のMnn2p部分は配列番号9の1〜46であり、リンカー配列は47〜49および405〜408であり、融合タンパク質のエピメラーゼ配列は50〜404であり、融合タンパク質のManII部分は409〜763である。Mnn2p部分は配列番号10のヌクレオチド1〜138であり、リンカー配列はヌクレオチド139〜147および1213〜1224であり、エピメラーゼ配列はヌクレオチド148〜1212であり、ManII部分は1225〜2289である。開始コドンおよび停止コドンに下線を付す。
【図14】図14は、プラスミドpYLTmAXSpGal10hGalTIおよびpYLTmAXSpGal10hGalTI(ADE2ex)についての構築方策の模式図である。
【図15】図15は、Gal10−GalTI活性をG040株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。得られた形質転換体G044を、2つの異なる培地で培養した。「Rd」は、図14に示すベクターに存在するゼータ配列による「無作為組み込み」を表す。Gal10−GalTI活性の発現の際に、Man7GlcNAc2とMan8GlcNAc2との間の位置に泳動される新しいピークが出現する。in vitroガラクトシダーゼ処理は、このピークについての前へのシフトと、GlcNAcMan5GlcNAc2の等しい増加とをもたらす(後者は、ガラクトシダーゼとヘキソサミニダーゼを用いる二重処理によりこのN−グリカンを表すことが確認される)。このことは、末端ガラクトースの存在を示し、よって、GalGlcNAcMan5GlcNAc2としてのG044プロファイルの新しいピークが同定される。α−1,2−マンノシダーゼでのin vitro処理は、Man5GlcNAc2にまでまだ刈り込まれていない(not yet trimmed)多量の高マンノースN−グリカン(特にMan8GlcNAc2)の存在を示す。
【図16】図16は、プラスミドpYLalg3PUT−ALG6の模式図である。
【図17】図17は、pYLalg3PUT−ALG6をG036株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。ALG6の過剰発現は、相当量のグリコシル化ピーク(GlcMan5’GlcNAc2およびGlc2Man5’GlcNAc2)をもたらし、このことは、新生タンパク質に移されたGlc3Man5’GlcNAc2構造が、グルコシダーゼIIによりMan5’GlcNAc2に完全には刈り込まれていないことを示す。増殖培地に依存して、生じるMan5’GlcNAc2は、ER局在化HDELタグ付きT.reeseiα−1,2−マンノシダーゼの作用によりMan3GlcNAc2に部分的(まだいくらかのMan5’GlcNAc2およびMan4’GlcNAc2)、またはほぼ完全に刈り込まれている。Man5’GlcNAc2およびMan4’GlcNAc2のピークは、α−1,2−マンノシダーゼに対するそれらの感受性により、そのままで同定される。キャッピンググルコースにより、GlcMan5’GlcNAc2およびGlc2Man5’GlcNAc2はこの処理に対して非感受性である。ナタマメマンノシダーゼは、遊離のα−1,6結合マンノースを部分的に除去できるが、これは、Man3−5’GlcNAc2をMan1GlcNAc2に変換もする。
【図18】図18は、プラスミドpYLTmAXhGnTI(Hygr ex)についての構築方策の模式図である。
【図19】図19は、GnT I発現ベクターでの形質転換によりGnT I活性をΔalg3−Hp4dALG6株のキュアしていない(G039)かまたはキュアした(G045)いずれかのバージョンに導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。GlcNAcMan3GlcNAc2の産生は、ヘキソサミニダーゼ消化により証明された。新しいピークは、Man3GlcNAc2に完全にシフトして戻った。G048株では、GlcNAcMan3GlcNAc2への変換は完全でなかった。なぜなら、いくらかのMan3GlcNAc2がまだ観察できたからである。この株は、α−1,2−マンノシダーゼ消化により示されるように、いくらかの残りのMan5’GlcNAc2も有する。
【図20】図20は、プラスミドJME925 pPTAde2−URA3ex−Hp4dhGnTIについての構築方策の模式図である。
【図21】図21は、GnT I活性をΔalg3−Hp4dALG6株のキュアしたバージョン(=G045)に導入し、Hp4d駆動型発現構築物をADE2遺伝子座(Tg−ade2)に組み込んだ後のN−グリカンプロファイルを示す一連の電気泳動図である。この培養では、グリコシル化N−グリカンの量は高く、Man4’−5’GlcNAc2からMan3GlcNAc2への変換は、完全でなかった。Man4’GlcNAc2の隣りに泳動される新しいピークが形質転換体G057で観察され、ヘキソサミニダーゼ消化の結果に基づいてGlcNAcMan3GlcNAc2であると示すことができた。この新しいピークは、Man3GlcNAc2に完全にシフトして戻る。
【図22】図22は、Mnn2pの36個のN末端アミノ酸とラットGlcNAcトランスフェラーゼIIの触媒ドメインとの間の融合タンパク質のアミノ酸配列(配列番号17)およびYarrowiaコドン最適化ヌクレオチド配列(配列番号18)である。太字で:融合タンパク質のMnn2p部分;通常のフォントで:融合タンパク質のGnT II部分;下線:開始コドンおよび停止コドン。
【図23】図23は、プラスミドpYLTmAXrGnTIIおよびpYLTmAXrGnTII(ADE2 ex)についての構築方策の模式図である。
【図24】図24は、GnT II活性を、GlcNAcMan3GlcNAc2を合成する株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。得られた株は、G045をGnTIおよびGnT II発現構築物で二重形質転換するか、またはG047をGnTII発現構築物で形質転換することのいずれかにより得た。両方の場合では、GlcNAcMan3GlcNAc2を表すピークはほぼ完全に消失し、約1グルコース単位大きい新しいピークが出現した。ヘキソサミニダーゼ処理は、新しいN−グリカン上での2つの末端GlcNAc残基の存在;約2グルコース単位左へのピークシフトを示し、よって、それは、GlcNAc2Man3GlcNAc2を表す。α−1,2−マンノシダーゼ処理は、大きな違いをもたらさず、このことは、Man4’−5’GlcNAc2が限られた量だけ存在することを示す。
【図25】図25は、グルコシダーゼII活性の発現のためのプラスミドpYLTUXdL2preAnGlcIIおよびpYLeu2ExTEFpreLip2AnGlucIIβの模式図である。
【図26】図26は、プラスミドJME923 pPTura3−LEU2ex−TefL2preAnGlcIIa+b[altl]についての構築方策の模式図である。
【図27】図27は、プラスミドJME923 pPTura3−LEU2ex−Hp4dL2preAnGlcIIa+b[altl]およびZeta−LEU2ex−Hp4dL2preAnGlcIIa+b[alt]についての構築方策の模式図である。
【図28】図28は、グルコシダーゼII活性を、GlcNAcMan3GlcNAc2を合成する株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。得られた株は、gls2αおよびgls2βサブユニットについての二重発現構築物の無作為(G060)または標的化(G061)組み込みのいずれかにより得た。両方の場合では、グルコシル化ピークの低減が観察される。α−1,2−マンノシダーゼ処理は、産生されたMan5’GlcNAc2の全てが、異種HDELタグ付きα−1,2−マンノシダーゼによりMan3GlcNAc2に変換されたのではないことを示す。キャッピンググルコースのために、GlcMan5’GlcNAc2およびGlc2Man5’GlcNAc2は、この処理に非感受性である。ナタマメマンノシダーゼは、残存グルコシル化N−グリカンおよびGlcNAcMan3GlcNAc2の両方における遊離のα−1,6結合マンノースを部分的に除去できる。さらに、この処理は、Man3−5’GlcNAc2をMan1GlcNAc2に変換する。「Rd」は、図27に示すベクターに存在するゼータ配列による「無作為組み込み」を表す。「Tg−ade2」および「Tg−ura3」は、ADE2resp.URA3遺伝子座における標的化組み込みを表す。
【図29】図29は、GlcNAcトランスフェラーゼIIをG061株に導入することにより作製したG070株およびG071株のセクレトーム(secretome)のN−グリカンプロファイルを示す一連の電気泳動図である。N−グリカンを、α−1,2−マンノシダーゼ(全ての末端α−1,2結合マンノース残基を除去する)またはヘキソサミニダーゼ(末端β−1,2結合GlcNAc残基を除去する)のいずれかで処理して、G070およびG071の天然プロファイルにおけるピークを同定した。グルコース含有N−グリカンは、2つの酵素のいずれにも感受性でない。α−1,2−マンノシダーゼ処理は、Man5’GlcNAc2およびMan4GlcNAc2からMan3GlcNAc2への刈り込みをもたらす。ヘキソサミニダーゼ処理は、GlcNAcトランスフェラーゼIおよびIIにより付加されたβ−1,2結合末端GlcNAc残基を除去して、Man3GlcNAc2を産生する。
【図30A】図30Aは、合成プレプロLip2−軽鎖(LC)のヌクレオチド配列(配列番号32)である。
【図30B】図30Bは、合成プレプロLip2−LCのアミノ酸配列(配列番号33)である。
【図31A】図31Aは、合成プレプロLip2−重鎖(HC)のヌクレオチド配列(配列番号34)である。
【図31B】図31Bは、合成プレプロLip2−HCのアミノ酸配列(配列番号35)である。
【図32】図32は、pYLHp4L2preproHerHC/LC(GUT2ex)−ori2で形質転換した、糖鎖工学的に改変したG045株、G057株、G061株およびG071株のSuperT/グリセロール振とうフラスコ培養物のN−グリカンプロファイル分析を示す一連の電気泳動図である。G045株、G057株、G061株およびG071株の記載について、表2を参照されたい。
【図33】図33は、G096流加発酵での異なる時点での機能的ELISAからの結果のグラフである。
【図34A】図34Aは、G096流加発酵における異なる時点でのセクレトームのN−グリカンプロファイル分析を示す一連の電気泳動図である。
【図34B】図34Bは、G096流加発酵における異なる時点でのセクレトームのN−グリカンプロファイル分析を示す一連の電気泳動図である。
【発明を実施するための形態】
【0027】
本明細書で記載する場合、酵母分泌糖タンパク質における哺乳類様複合N−グリカンのin vivo合成は、Man5GlcNAc2基本構造またはMan3GlcNAc2基本構造のいずれかに基づくことができる(図1Aを参照されたい、「Man」はマンノースのことであり、「GlcNAc」はN−グルコサミンのことである)。Man5GlcNAc2基本構造を生成するために、酵母細胞を、α−1,2−マンノシダーゼ活性が細胞内区画で増加し、外鎖伸長(Outer CHain elongation)(OCH1)活性が減少するように工学的に改変できる。Man3GlcNAc2基本構造を生成するために、アスパラギン結合グリコシル化3(ALG3)の活性と、いくつかの実施形態ではOCH1とが減少し、α−1,2−マンノシダーゼの活性と、いくつかの実施形態ではα−1,3−グルコシルトランスフェラーゼの活性とが増加する。このような酵母細胞で生成されたタンパク質のN−グリカンプロファイルは、酵母細胞を、以下の活性の1つ以上を含有するようにさらに工学的に改変することにより変更できる:GlcNAcトランスフェラーゼI(GnT I)活性、マンノシダーゼII活性、GlcNAcトランスフェラーゼII(GnT II)活性、グルコシダーゼII活性およびガラクトシルトランスフェラーゼ(Gal T)活性。例えば、Man5GlcNAc2またはMan3GlcNAc2 N−グリカンを生成する酵母細胞においてGnT Iを発現させることにより、それぞれGlcNAcMan5GlcNAc2またはGlcNAcMan3GlcNAc2 N−グリカンが生成されるように、GlcNAc部分がMan5GlcNAc2またはMan3GlcNAc2 N−グリカンに移動する。GlcNAcMan5GlcNAc2 N−グリカンを生成する細胞では、マンノシダーゼIIを発現させることにより、2つのマンノース残基がGlcNAcMan5GlcNAc2 N−グリカンから除去されて、GlcNAcMan3GlcNAc2 N−グリカンが生成される。GlcNAcMan3GlcNAc2 N−グリカンを生成する細胞では、GnT IIを発現させることにより、別のGlcNAc部分がGlcNAcMan3GlcNAc2 N−グリカンに移動して、GlcNAc2Man3GlcNAc2 N−グリカンが生成される。GlcNAcMan3GlcNAc2またはGlcNAc2Man3GlcNAc2 N−グリカンを生成する細胞でGal Tを発現させることにより、ガラクトースがGlcNAcMan3GlcNAc2またはGlcNAc2Man3GlcNAc2 N−グリカンに移動して、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカンが生成される。いくつかの実施形態では、グルコシダーゼII(例えばαおよびβサブユニットを発現させることにより)を発現させて、Man3GlcNAc2基本構造の生成を増加できる。
【0028】
標的分子
標的分子は、本明細書で用いる場合、遺伝子工学的に改変された細胞(例えばYarrowia lipolytica、Arxula adeninivoransもしくはその他の関連する種二形性酵母細胞(other related species dimorphic yeast cell))のような真菌細胞;植物細胞または動物細胞)においてN−グリコシル化を受ける任意の分子のことをいう。いくつかの実施形態では、標的分子は、Yarrowia lipolyticaまたはArxula adeninivorans(またはその他の関連する種二形性酵母)の分泌経路の1つ以上のステップを介して輸送されて、宿主細胞機構によるそれらのN−グリコシル化をもたらすことができる。標的分子は、内因性または外因性であり得る。
【0029】
適切な標的タンパク質は、病原体タンパク質(例えば破傷風トキソイド;ジフテリアトキソイド;ウイルス表面タンパク質(例えばサイトメガロウイルス(CMV)糖タンパク質B、HおよびgCIII;ヒト免疫不全ウイルス1(HIV−1)エンベロープ糖タンパク質;ラウス肉腫ウイルス(RSV)エンベロープ糖タンパク質;単純ヘルペスウイルス(HSV)エンベロープ糖タンパク質;エプスタインバーウイルス(EBV)エンベロープ糖タンパク質;水痘帯状疱疹ウイルス(VZV)エンベロープ糖タンパク質;ヒトパピローマウイルス(HPV)エンベロープ糖タンパク質;インフルエンザウイルス糖タンパク質;および肝炎ファミリー表面抗原)、リソソームタンパク質(例えばグルコセレブロシダーゼ、セレブロシダーゼまたはガラクトセレブロシダーゼ)、インスリン、グルカゴン、増殖因子、サイトカイン、ケモカイン、Fc受容体と結合するタンパク質、抗体もしくはその断片、またはタンパク質のいずれかと抗体もしくは抗体の断片との融合体(例えばタンパク質−Fc)を含む。増殖因子は、例えば血管内皮増殖因子(VEGF)、インスリン様増殖因子(IGF)、骨形成タンパク質(BMP)、顆粒球コロニー刺激因子(G−CSF)、顆粒球マクロファージコロニー刺激因子(GM−CSF)、神経増殖因子(NGF);ニューロトロフィン、血小板由来増殖因子(PDGF)、エリスロポエチン(EPO)、トロンボポエチン(TPO)、ミオスタチン(GDF−8)、増殖分化因子−9(GDF9)、塩基性線維芽細胞増殖因子(bFGFまたはFGF2)、上皮増殖因子(EGF)、肝細胞増殖因子(HGF)を含む。サイトカインは、例えばインターロイキン(例えばIL−1、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−12、IL−13またはIL−15のようなIL−1〜IL−33)およびインターフェロン(例えばインターフェロンβまたはインターフェロンγ)を含む。ケモカインは、例えばI−309、TCA−3、MCP−1、MIP−1α、MIP−1β、RANTES、C10、MRP−2、MARC、MCP−3、MCP−2、MRP−2、CCF18、MIP−1γ、エオタキシン、MCP−5、MCP−4、NCC−1、Ckβ10、HCC−1、ロイコタクチン−1、LEC、NCC−4、TARC、PARCまたはエオタキシン−2を含む。腫瘍糖タンパク質(例えば腫瘍関連抗原)、例えば癌胎児抗原(CEA)、ヒトムチン、HER−2/neuおよび前立腺特異抗原(PSA)も含まれる[HendersonおよびFinn、Advances in Immunology、62巻、217〜56頁(1996年)]。一実施形態では、標的タンパク質は、抗HER2/neu抗体である。いくつかの実施形態では、標的タンパク質は、リソソーム蓄積症と関連するものであり得、この標的タンパク質は、例えばアルファ−L−イズロニダーゼ、ベータ−D−ガラクトシダーゼ、ベータ−グルコシダーゼ、ベータ−ヘキソサミニダーゼ、ベータ−D−マンノシダーゼ、アルファ−L−フコシダーゼ、アリールスルファターゼB、アリールスルファターゼA、アルファ−N−アセチルガラクトサミニダーゼ、アスパルチルグルコサミニダーゼ、イズロン酸−2−スルファターゼ、アルファ−グルコサミニド−N−アセチルトランスフェラーゼ、ベータ−D−グルクロニダーゼ(glucoronidase)、ヒアルロニダーゼ、アルファ−L−マンノシダーゼ、アルファ−ノイラミニダーゼ、ホスホトランスフェラーゼ、酸性リパーゼ、酸性セラミダーゼ、スフィンゴミエリナーゼ、チオエステラーゼ、カテプシンKおよびリポタンパク質リパーゼを含む。
【0030】
標的タンパク質は、融合タンパク質でもあり得る。融合タンパク質は、例えば(i)本明細書で記載する任意のタンパク質またはその断片と、(ii)抗体またはその断片との融合体を含む。本明細書で使用する場合、用語「抗体断片」とは、(a)抗原結合断片または(b)Fc受容体と相互作用できる抗体のFc部分のことをいう。抗原結合断片は、例えば、Fab、F(ab’)2、Fvおよび単鎖Fv(scFv)断片であり得る。scFv断片は、このscFvが由来する抗体の重鎖および軽鎖の両方の可変領域を含む単一ポリペプチド鎖である。さらに、ダイアボディ[Poljak(1994年)Structure2巻(12号):1121〜1123頁;Hudsonら(1999年)J. Immunol. Methods23巻(1〜2号):177〜189頁]およびイントラボディ[Hustonら(2001年)Hum. Antibodies10巻(3〜4号):127〜142頁;Wheelerら(2003年)Mol. Ther. 8巻(3号):355〜366頁;Stocks(2004年)Drug Discov. Today9巻(22号):960〜966頁]を本発明の方法で用いることができる。
【0031】
標的タンパク質は、ポリマー、キャリア、アジュバント、免疫毒素または検出可能(例えば蛍光、発光もしくは放射活性)部分の1つ以上とつなぐこともできる。例えば、標的タンパク質をポリエチレングリコールとつなぐことができ、これは小さいタンパク質の分子量を増加させ、かつ/または循環滞留時間を増加させるために用いることができる。
【0032】
いくつかの実施形態では、標的分子は、ドリコールであるかまたはドリコールを含むことができる。
【0033】
遺伝子工学的に改変された細胞
本明細書で記載する遺伝子工学的に改変された細胞は、哺乳類様N−グリカンを含有する標的分子、または哺乳類グリコシル化経路内の中間体を含有する標的分子を生成するために用いることができる。例えば、本明細書で記載する場合、1つ以上の酵素をコードする核酸を、細胞が所望のN−グリカン(例えばGlcNAcMan5GlcNAc2、GlcNAcMan3GlcNAc2、GlcNAc2Man3GlcNAc2、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカン)を生成するように真菌細胞に導入できる。つまり、本明細書で記載する実施形態のいずれでも、真菌細胞は、1つの酵素をコードする核酸を含有し得るか、または複数の酵素をコードし得る核酸を含有し得る。このような核酸のそれぞれは、以下で論じるようなターゲティング配列も含有できる。さらに、標的分子をコードする核酸も、標的分子が生成され、所望のN−グリカン(例えばGlcNAcMan5GlcNAc2、GlcNAcMan3GlcNAc2、GlcNAc2Man3GlcNAc2、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカン)を含有するように改変されるように真菌細胞に導入できる。
【0034】
用語「核酸」および「ポリヌクレオチド」は、本明細書で交換可能に用い、cDNA、ゲノムDNA、合成DNAおよび核酸類似体を含有するDNA(またはRNA)を含むRNAならびにDNAの両方のことをいう。核酸は、任意の3次元構造を有することができる。核酸は、2本鎖または1本鎖(すなわちセンス鎖またはアンチセンス鎖)であり得る。核酸の非限定的な例は、遺伝子、遺伝子断片、エキソン、イントロン、メッセンジャーRNA(mRNA)、トランスファーRNA、リボソームRNA、siRNA、マイクロRNA、リボザイム、cDNA、組換えポリヌクレオチド、有枝鎖状ポリヌクレオチド、プラスミド、ベクター、任意の配列の単離DNA、任意の配列の単離RNA、核酸プローブおよびプライマー、ならびに核酸類似体を含む。「ポリペプチド」および「タンパク質」は、本明細書で交換可能に用い、長さまたは翻訳後修飾にかかわらず、アミノ酸の任意のペプチド連結鎖を意味する。
【0035】
「単離核酸」とは、天然で生じるゲノム(例えば酵母ゲノム)中で核酸の一方の側または両側に通常接している核酸を含む、天然で生じるゲノムに存在するその他の核酸分子から分離される核酸のことをいう。用語「単離」は、核酸に関して本明細書で用いる場合、任意の天然で生じない核酸配列も含む。なぜなら、このような天然で生じない配列は天然で見出されず、天然で生じるゲノムにおいて直接隣接する配列を有さないからである。
【0036】
単離核酸は、例えばDNA分子であり得るが、但し、天然で生じるゲノムにおいてそのDNA分子を直接挟む(flanking)ことが通常見出される核酸配列の一方が、除去されているかまたは存在しないことを条件とする。つまり、単離核酸は、限定することなく、他の配列から独立した分離した分子(例えば化学合成核酸、あるいはPCRもしくは制限エンドヌクレアーゼ処理により生成されたcDNAまたはゲノムDNA断片)、ならびにベクター、自己複製プラスミド、ウイルス(例えば任意のパラミクソウイルス、レトロウイルス、レンチウイルス、アデノウイルスまたはヘルペスウイルス)または原核生物もしくは真核生物のゲノムDNAに組み込まれたDNAを含む。さらに、単離核酸は、ハイブリッドまたは融合核酸の一部であるDNA分子のような工学的に改変された核酸を含み得る。例えばcDNAライブラリーもしくはゲノムライブラリーまたはゲノムDNA制限酵素消化物(restriction digenst)を含有するゲルスライス内の数百〜数百万のその他の核酸のうちに存在する1つの核酸は、単離核酸とみなさない。
【0037】
用語「外因性」は、核酸および特定の宿主細胞に関して本明細書で用いる場合、その特定の細胞において天然で見出されるとおりに生じない(そして該細胞から得ることができない)任意の核酸のことをいう。つまり、天然で生じない核酸は、宿主細胞に一旦導入されると、該宿主細胞にとって外因性であるとみなす。天然で生じない核酸は、天然で見出される核酸配列の核酸サブシーケンスまたは断片を含有できるが、この核酸が全体として天然に存在しないことを条件とすることに注意することが重要である。例えば、発現ベクター内のゲノムDNA配列を含有する核酸分子は、天然で生じない核酸であり、よって、宿主細胞に一旦導入されると、該宿主細胞にとって外因性である。なぜなら、この核酸分子は、全体(ゲノムDNA+ベクターDNA)として天然に存在しないからである。つまり、全体として天然に存在しない任意のベクター、自己複製プラスミドまたはウイルス(例えばレトロウイルス、アデノウイルスまたはヘルペスウイルス)は、天然で生じない核酸とみなす。このことは、PCRまたは制限エンドヌクレアーゼ処理により生成されたゲノムDNA断片ならびにcDNAを、これらが天然で見出されない分離した分子として存在するので、天然で生じない核酸とみなすことに従う。このことは、天然で見出されない配置でプロモーター配列およびポリペプチドコード配列(例えばcDNAまたはゲノムDNA)を含有する任意の核酸が、天然で生じない核酸であることにも従う。天然で生じる核酸は、特定の細胞にとって外因性になり得る。例えば、酵母xの細胞から単離された染色体全体は、該染色体が酵母の細胞に一旦導入されると、酵母yの細胞に関して外因性核酸である。
【0038】
遺伝子工学的改変のために適切な細胞は、例えば真菌細胞(例えばYarrowia lipolyticaまたは本明細書で記載する任意のその他の関連する二形性酵母細胞)、植物細胞または動物細胞を含む。細胞は、初代細胞、不死化細胞または形質転換細胞であり得る。細胞は、動物、例えば非ヒト哺乳動物におけるものであり得る。本明細書で特定する遺伝子工学的改変の前のこのような細胞は、例えばAmerican Type Culture Collection(Rockville、MD)のような多様な商業的供給源および研究資源施設から得ることができる。
【0039】
細胞の遺伝子工学的改変は、(i)N−グリコシル化活性を有するタンパク質をコードする内因性遺伝子の欠失;(ii)N−グリコシル化活性を有する(すなわちN−グリコシル化活性を有する変異タンパク質を発現する)タンパク質(例えば内因性または外因性タンパク質)の変異形態をコードする組換え核酸の導入;(iii)N−グリコシル化活性を有するタンパク質の機能的発現に干渉するRNA分子の導入または発現;(iv)N−グリコシル化活性を有する(すなわちN−グリコシル化活性を有するタンパク質を発現する)野生型(例えば内因性または外因性)タンパク質をコードする組換え核酸の導入;または(v)N−グリコシル化活性を有するタンパク質をコードする1つ以上の内因性遺伝子のプロモーター要素またはエンハンサー要素を変更して、そのことによりそれらのコードされるタンパク質の発現を変更することのような遺伝子改変を含み得る。RNA分子は、例えば小型干渉RNA(siRNA)、短いヘアピンRNA(shRNA)、アンチセンスRNAまたはマイクロRNA(miRNA)を含む。項目(ii)は、例えば、内因性遺伝子を、置き換えられるこの内因性遺伝子と比べてより大きいN−グリコシル化活性を有するタンパク質をコードする遺伝子で置き換えることを含む。遺伝子工学的改変は、N−グリコシル化活性を有するタンパク質をコードする内因性遺伝子を、付加(例えば異種配列)、欠失または置換(例えば点変異;保存的または非保存的変異のような変異)を有するタンパク質を生成するように変更することも含む。変異は、特異的に導入できる(例えば部位特異的変異誘発または相同組換え)か、または無作為に導入できる(例えば、細胞は、例えばNewmanおよびFerro−Novick(1987年)J. Cell Biol.105巻(4号):1587頁に記載されるように、化学的に変異誘発させることができる)。
【0040】
本明細書で記載する遺伝子改変は、(i)遺伝子改変された細胞における1種以上のN−グリコシル化活性の増加、(ii)遺伝子改変された細胞における1種以上のN−グリコシル化活性の減少、(iii)遺伝子改変された細胞における1種以上のN−グリコシル化活性の局在化または細胞内分布の変化、あるいは(iv)遺伝子改変された細胞における1種以上のN−グリコシル化活性の比の変化のうちの1つ以上をもたらすことができる。N−グリコシル化活性の量の増加は、N−グリコシル化活性を有する1つ以上のタンパク質の過剰発現、内因性遺伝子のコピー数の増加(例えば遺伝子重複)、または内因性遺伝子によりコードされるタンパク質の発現の増加を刺激する該遺伝子のプロモーターもしくはエンハンサーの変更によることができることが理解される。1種以上のN−グリコシル化活性の減少は、N−グリコシル化を変更する活性を有する1つ以上のタンパク質の変異形態(例えばドミナントネガティブ形態)の過剰発現、N−グリコシル化活性を有する1つ以上のタンパク質の発現を低減する1つ以上の干渉RNA分子の導入または発現、あるいはN−グリコシル化活性を有するタンパク質をコードする1つ以上の内因性遺伝子の欠失によることができる。
【0041】
1つ以上の内因性遺伝子を欠失または破壊する方法は、付随の実施例に記載する。例えば、相同組換えにより遺伝子を破壊するために、「遺伝子置き換え」ベクターを、選択可能マーカー遺伝子を含むような様式で構築できる。選択可能マーカー遺伝子は、5’および3’の両方の端で、相同組換えを媒介するのに十分な長さの遺伝子の部分に作動可能に結合できる。選択可能マーカーは、宿主細胞の栄養要求性を補完するかまたは抗生物質耐性をもたらす任意の数の遺伝子の1つであり得、URA3、LEU2およびHIS3遺伝子を含む。その他の適切な選択可能マーカーは、酵母細胞にクロラムフェニコール耐性を付与するCAT遺伝子、またはβ−ガラクトシダーゼの発現により青色コロニーをもたらすlacZ遺伝子を含む。遺伝子置き換えベクターの直鎖状にしたDNA断片を、次いで、細胞に、当技術分野において周知の方法を用いて導入する(以下を参照されたい)。ゲノムへの直鎖状断片の組み込みおよび遺伝子破壊は、選択マーカーに基づいて決定でき、例えばサザンブロット分析により確認できる。
【0042】
付随の実施例に詳細に示すように、選択におけるその使用の後に、選択可能マーカーを宿主細胞のゲノムから、例えばCre−loxP系により除去できる(以下を参照されたい)。マーカー除去のこのプロセスは、実施例を通して「キュアリング」という。
【0043】
代わりに、遺伝子置き換えベクターは、破壊する遺伝子の部分を含むような様式で構築でき、ここで、この部分は、いずれの内因性遺伝子プロモーター配列がなく、その遺伝子のコード配列をコードしないかまたは該コード配列の不活性断片をコードする。「不活性断片」は、例えば遺伝子の全長コード配列から生成されるタンパク質の活性の約10%未満(例えば約9%未満、約8%未満、約7%未満、約6%未満、約5%未満、約4%未満、約3%未満、約2%未満、約1%未満または0%)を有するタンパク質をコードする遺伝子の断片である。遺伝子のこのような部分は、既知のプロモーター配列は遺伝子配列に作動可能に結合されていないが、停止コドンおよび転写終結配列が遺伝子配列のこの部分と作動可能に結合されているような様式でベクターに挿入される。このベクターは、その後、遺伝子配列のこの部分で直鎖状にして、細胞を形質転換できる。単一の相同組換えにより、この直鎖状にしたベクターを、次いで、その遺伝子の内因性対応物(counterpart)に組み込む。
【0044】
発現ベクターは、自律型または組み込み型であり得る。
【0045】
組換え核酸を、プラスミド、ファージ、トランスポゾン、コスミドまたはウイルス粒子のような発現ベクターの形態で細胞に導入できる。組換え核酸は、染色体外で維持できるか、またはこれを酵母細胞染色体DNAに組み込むことができる。発現ベクターは、選択された条件下で細胞生存性に要求されるタンパク質をコードする選択マーカー遺伝子(例えばウラシル生合成のために必要な酵素をコードするURA3、またはトリプトファン生合成に要求される酵素をコードするTRP1)を含有して、所望の核酸で形質転換されたこれらの細胞の検出および/または選択を許容できる(例えば米国特許第4,704,362号を参照されたい)。発現ベクターは、自己複製配列(ARS)も含むことができる。例えば、米国特許第4,837,148号は、Pichia pastoris中でプラスミドを維持するために適切な手段を提供する自己複製配列について記載している。
【0046】
組み込み型ベクターが、例えば米国特許第4,882,279号に開示されている。組み込み型ベクターは、一般的に、少なくとも第1挿入可能DNA断片と、選択可能マーカー遺伝子と、第2挿入可能DNA断片との連続的に配置された配列を含む。第1および第2の挿入可能DNA断片は、それぞれ、約200(例えば約250、約300、約350、約400、約450、約500または約1000以上)ヌクレオチドの長さであり、形質転換される種のゲノムDNAの部分に相同なヌクレオチド配列を有する。発現のための目的の遺伝子(例えばN−グリコシル化活性を有するタンパク質をコードする遺伝子)を含有するヌクレオチド配列は、第1および第2の挿入可能DNA断片の間で、マーカー遺伝子の前かその後のいずれかにおいて、このベクターに挿入される。組み込み型ベクターを酵母形質転換の前に直鎖状にして、宿主細胞ゲノムへの目的のヌクレオチド配列の組み込みを容易にすることができる。
【0047】
発現ベクターは、それらを酵母で発現させることを可能にする酵母(例えばYarrowia lipolytica、Arxula adeninivoransまたはその他の関連する二形性酵母種)プロモーターの制御下の組換え核酸を特徴とすることができる。適切な酵母プロモーターは、TEF1、HP4D、GAP、POX2、ADC1、TPI1、ADH2、POXおよびGal10プロモーターを含む。例えばMadzakら(2000年)J. Mol. Microbiol. Biotechnol.2巻:207〜216頁;Guarenteら(1982年)Proc. Natl. Acad. Sci. USA79巻(23号):7410頁を参照されたい。さらなる適切なプロモーターは、例えばZhuおよびZhang(1999年)Bioinformatics15巻(7〜8号):608〜611頁および米国特許第6,265,185号に記載されている。発現ベクターが哺乳類細胞のような動物細胞に導入される場合、発現ベクターは、目的の宿主細胞での発現に適切な動物細胞プロモーターの制御下の組換え核酸を特徴とすることができる。哺乳類プロモーターの例は、SV40およびサイトメガロウイルス(CMV)プロモーターを含む。
【0048】
プロモーターは、構成性(constitutive)または誘導性(条件的)であり得る。構成性プロモーターは、その発現が標準的培養条件下で一定であるプロモーターであると理解される。誘導性プロモーターは、1つ以上の誘導指示(induction cue)に応答性であるプロモーターである。例えば、誘導性プロモーターは、化学的に調節され得る(例えばその転写活性が、アルコール、テトラサイクリン、ステロイド、金属またはその他の小分子のような化学的誘導物質の存在または非存在により調節されるプロモーター)か、または物理的に調節され得る(例えばその転写活性が、光または高温もしくは低温のような物理的誘導因子の存在または非存在により調節されるプロモーター)。誘導性プロモーターは、それら自体が化学的もしくは物理的な指示により直接調節される1つ以上の転写因子により間接的に調節されることもできる。
【0049】
細胞の遺伝子工学的改変は、宿主細胞に存在するが、その細胞で通常は発現されないかまたはその細胞で著しいレベルでは発現されない内因性遺伝子(例えばN−グリコシル化活性を有するタンパク質をコードする遺伝子)を活性化することも含む。例えば、内因性遺伝子の調節配列(例えば遺伝子プロモーターまたはエンハンサー)を、作動可能に結合したコード配列が発現の増加を示すように改変できる。相同組換えまたはターゲティングを用いて、遺伝子と通常関連する調節領域を、対応する遺伝子工学的に改変されていない細胞で明らかなレベルよりも高いレベルで遺伝子を発現させるか、または対応する遺伝子工学的に改変されていない細胞で明らかなものとは異なる調節もしくは誘導のパターンを遺伝子が示すようにする調節配列で置き換えるかあるいは不能にすることができる。遺伝子の調節配列(例えばプロモーターまたはエンハンサー)の変更を導入するために適切な方法は、例えば米国特許出願公開第20030147868号に記載されている。
【0050】
その他の遺伝子工学的に作製された改変も、条件的であり得ることが理解される。例えば、遺伝子は、例えばCre−loxP系のような部位特異的DNAリコンビナーゼを用いて条件的に欠失できる(例えばGossenら(2002年)Ann. Rev. Genetics36巻:153〜173頁および米国特許出願公開第20060014264号を参照されたい)。
【0051】
組換え核酸は、本明細書で記載する細胞に、スフェロプラスト技術または全細胞塩化リチウム酵母形質転換法のような様々な方法を用いて導入できる。プラスミドまたは直鎖状核酸ベクターで細胞を形質転換するために有用なその他の方法は、例えば米国特許第4,929,555号;Hinnenら(1978年)Proc. Nat. Acad. Sci. USA75巻:1929頁;Itoら(1983年)J. Bacteriol.153巻:163頁;米国特許第4,879,231号;ならびにSreekrishnaら(1987年)Gene59巻:115頁に記載されている。エレクトロポレーションおよびPEG1000全細胞形質転換手順も、CreggおよびRussel、Methods in Molecular Biology:Pichia Protocols、第3章、Humana Press、Totowa、N.J.、27〜39頁(1998年)に記載されるようにして用いることができる。動物細胞のトランスフェクションは、例えば、ベクターを細胞に、リン酸カルシウム、エレクトロポレーション、熱ショック、リポソーム、またはFUGENE(登録商標)もしくはLIPOFECTAMINE(登録商標)のようなトランスフェクション試薬を用いて、あるいは裸の核酸ベクターを細胞と溶液中で接触させることにより導入することを特徴とすることができる(例えばSambrookら、Molecular Cloning: A Laboratory Manual第2版1、2および3巻、Cold Spring Harbor Laboratory Press: Cold Spring Harbor、New York, USA、1989年11月を参照されたい)。
【0052】
形質転換酵母細胞は、それらに限定されないが、形質転換後の栄養要求性細胞を、要求される生化学的生成物の非存在下で培養すること(細胞の栄養要求性による)、新しい表現型についての選択およびその検出、または形質転換体に含有される耐性遺伝子の非存在下で酵母にとって毒性がある抗生物質の存在下で培養することを含む適当な技術を用いて選択できる。形質転換体は、ゲノムへの発現カセットの組み込みにより選択および/または確認することもでき、これは、例えばサザンブロットまたはPCR分析により評価することができる。
【0053】
目的の標的細胞にベクターを導入する前に、ベクターを、Escherichia coli(E.coli)のような細菌細胞中で増殖(例えば増幅)させることができる。ベクターDNAを、細菌細胞から、細菌環境からのベクターDNAの精製をもたらす当技術分野において公知の方法のいずれかにより単離できる。精製ベクターDNAは、フェノール、クロロホルムおよびエーテルを用いて広く抽出して、プラスミドDNA調製物中にE.coliタンパク質が存在しないことを確実にできる。なぜなら、これらのタンパク質は、哺乳類細胞にとって有毒であり得るからである。
【0054】
遺伝子工学的改変は、本明細書で記載する場合、N−グリコシル化活性を有するタンパク質をコードする任意の数の遺伝子を発現(例えば過剰発現)するか、該遺伝子に改変を導入するか、または該遺伝子を欠失させるために用いることができる。このようなタンパク質は、例えば、OCH1、ALG3、α−1,3−グルコシルトランスフェラーゼ、GnT I、マンノシダーゼII、GnT II、グルコシダーゼIIまたはGal Tを含む。N−グリコシル化活性を有するタンパク質をコードする遺伝子は、このような遺伝子を含有する任意の種に由来し得る。N−グリコシル化活性を有するタンパク質をコードする遺伝子を得ることができる例示的な真菌種は、限定することなく、Pichia anomala、Pichia bovis、Pichia canadensis、Pichia carsonii、Pichia farinose、Pichia fermentans、Pichia fluxuum、Pichia membranaefaciens、Pichia membranaefaciens、Candida valida、Candida albicans、Candida ascalaphidarum、Candida amphixiae、Candida Antarctica、Candida atlantica、Candida atmosphaerica、Candida blattae、Candida carpophila、Candida cerambycidarum、Candida chauliodes、Candida corydalis、Candida dosseyi、Candida dubliniensis、Candida ergatensis、Candida fructus、Candida glabrata、Candida fermentati、Candida guilliermondii、Candida haemulonii、Candida insectamens、Candida insectorum、Candida intermedia、Candida jeffresii、Candida kefyr、Candida krusei、Candida lusitaniae、Candida lyxosophila、Candida maltosa、Candida membranifaciens、Candida milleri、Candida oleophila、Candida oregonensis、Candida parapsilosis、Candida quercitrusa、Candida shehatea、Candida temnochilae、Candida tenuis、Candida tropicalis、Candida tsuchiyae、Candida sinolaborantium、Candida sojae、Candida viswanathii、Candida utilis、Pichia membranaefaciens、Pichia silvestris、Pichia membranaefaciens、Pichia chodati、Pichia membranaefaciens、Pichia menbranaefaciens、Pichia minuscule、Pichia pastoris、Pichia pseudopolymorpha、Pichia quercuum、Pichia robertsii、Pichia saitoi、Pichia silvestrisi、Pichia strasburgensis、Pichia terricola、Pichia vanriji、Pseudozyma Antarctica、Rhodosporidium toruloides、Rhodotorula glutinis、Saccharomyces bayanus、Saccharomyces bayanus、Saccharomyces momdshuricus、Saccharomyces uvarum、Saccharomyces bayanus、Saccharomyces cerevisiae、Saccharomyces bisporus、Saccharomyces chevalieri、Saccharomyces delbrueckii、Saccharomyces exiguous、Saccharomyces fermentati、Saccharomyces fragilis、Saccharomyces marxianus、Saccharomyces mellis、Saccharomyces rosei、Saccharomyces rouxii、Saccharomyces uvarum、Saccharomyces willianus、Saccharomycodes ludwigii、Saccharomycopsis capsularis、Saccharomycopsis fibuligera、Saccharomycopsis fibuligera、Endomyces hordei、Endomycopsis fobuligera. Saturnispora saitoi、Schizosaccharomyces octosporus、Schizosaccharomyces pombe、Schwanniomyces occidentalis、Torulaspora delbrueckii、Torulaspora delbrueckii、Saccharomyces dairensis、Torulaspora delbrueckii、Torulaspora fermentati、Saccharomyces fermentati、Torulaspora delbrueckii、Torulaspora rosei、Saccharomyces rosei、Torulaspora delbrueckii、Saccharomyces rosei、Torulaspora delbrueckii、Saccharomyces delbrueckii、Torulaspora delbrueckii、Saccharomyces delbrueckii、Zygosaccharomyces mongolicus、Dorulaspora globosa、Debaryomyces globosus、Torulopsis globosa、Trichosporon cutaneum、Trigonopsis variabilis、Williopsis californica、Williopsis saturnus、Zygosaccharomyces bisporus、Zygosaccharomyces bisporus、Debaryomyces disporua.、Saccharomyces bisporas、Zygosaccharomyces bisporus、Saccharomyces bisporus、Zygosaccharomyces mellis、Zygosaccharomyces priorianus、Zygosaccharomyces rouxiim、Zygosaccharomyces rouxii、Zygosaccharomyces barkeri、Saccharomyces rouxii、Zygosaccharomyces rouxii、Zygosaccharomyces major、Saccharomyces rousii、Pichia anomala、Pichia bovis、Pichia Canadensis、Pichia carsonii、Pichia farinose、Pichia fermentans、Pichia fluxuum、Pichia membranaefaciens、Pichia pseudopolymorpha、Pichia quercuum、Pichia robertsii、Pseudozyma Antarctica、Rhodosporidium toruloides、Rhodosporidium toruloides、Rhodotorula glutinis、Saccharomyces bayanus、Saccharomyces bayanus、Saccharomyces bisporus、Saccharomyces cerevisiae、Saccharomyces chevalieri、Saccharomyces delbrueckii、Saccharomyces fermentati、Saccharomyces fragilis、Saccharomycodes ludwigii、Schizosaccharomyces pombe、Schwanniomyces occidentalis、Torulaspora delbrueckii、Torulaspora globosa、Trigonopsis variabilis、Williopsis californica、Williopsis saturnus、Zygosaccharomyces bisporus、Zygosaccharomyces mellis、Zygosaccharomyces rouxiiまたは当技術分野で公知であるかもしくは本明細書で記載する任意のその他の真菌(例えば酵母)を含む。例示的な下等真核生物は、それらに限定されないが、Aspergillus caesiellus、Aspergillus candidus、Aspergillus carneus、Aspergillus clavatus、Aspergillus deflectus、Aspergillus flavus、Aspergillus fumigatus、Aspergillus glaucus、Aspergillus nidulans、Aspergillus niger、Aspergillus ochraceus、Aspergillus oryzae、Aspergillus parasiticus、Aspergillus penicilloides、Aspergillus restrictus、Aspergillus sojae、Aspergillus sydowi、Aspergillus tamari、Aspergillus terreus、Aspergillus ustusまたはAspergillus versicolorを含むAspergillusの様々な種も含む。N−グリコシル化活性を有するタンパク質をコードする遺伝子を得ることができる例示的な原生動物の属は、限定することなく、Blastocrithidia、Crithidia、Endotrypanum、Herpetomonas、Leishmania、Leptomonas、Phytomonas、Trypanosoma(例えばT.bruceii、T.gambiense、T.rhodesienseおよびT.cruzi)およびWallaceinaを含む。例えば、GnT Iをコードする遺伝子は、ヒト(Swiss Protein受託番号P26572)、ラット、Arabidopsis、マウスまたはDrosophilaから得ることができ、GntIIをコードする遺伝子は、ヒト、ラット(Swiss Protein受託番号Q09326)、Arabidopsisまたはマウスから得ることができ、Man IIをコードする遺伝子は、ヒト、ラット、Arabidopsis、マウス、Drosophila(Swiss Protein受託番号Q24451)から得ることができ、GalTをコードする遺伝子は、ヒト(Swiss Protein受託番号P15291)、ラット、マウスまたはウシから得ることができる。
【0055】
いくつかの実施形態では、遺伝子工学的に改変された細胞は、OCH1(GenBank受託番号AJ563920)遺伝子またはその遺伝子生成物(mRNAまたはタンパク質)を欠く。いくつかの実施形態では、遺伝子工学的に改変された細胞は、ALG3(Genbank(登録商標)受託番号XM_503488、Genolevures参照:YALI0E03190g)遺伝子またはその遺伝子生成物(mRNAまたはタンパク質)を欠く。いくつかの実施形態では、遺伝子工学的に改変された細胞は、α−1,3−グルコシルトランスフェラーゼ(例えばALG6、Genbank(登録商標)受託番号XM_502922、Genolevures参照:YALI0D17028g)タンパク質を発現(例えば過剰発現)する。いくつかの実施形態では、遺伝子工学的に改変された細胞は、α−1,2−マンノシダーゼ(例えばGenbank受託番号AF212153)タンパク質を発現する。いくつかの実施形態では、遺伝子工学的に改変された細胞は、GlcNAcトランスフェラーゼI(例えばSwiss Prot.受託番号P26572)タンパク質を発現する。いくつかの実施形態では、遺伝子工学的に改変された細胞は、マンノシダーゼIIタンパク質またはその触媒ドメイン(例えばSwiss Prot.受託番号Q24451)を発現する。いくつかの実施形態では、遺伝子工学的に改変された細胞は、ガラクトシルトランスフェラーゼIタンパク質またはその触媒ドメイン(例えばSwiss Prot.受託番号P15291)を発現する。いくつかの実施形態では、遺伝子工学的に改変された細胞は、GlcNAcトランスフェラーゼIIタンパク質またはその触媒ドメイン(例えばSwiss Prot.受託番号Q09326)を発現する。いくつかの実施形態では、遺伝子工学的に改変された細胞は、Yarrowia lipolytica、Trypanosoma bruceiまたはAspergillus nigerのグルコシダーゼIIのようなグルコシダーゼIIのアルファまたはベータサブユニット(またはアルファおよびベータの両方のサブユニット)を発現する。遺伝子工学的に改変された細胞は、これらの改変の任意の組み合わせを有することができる。
【0056】
例えば、いくつかの実施形態では、遺伝子工学的に改変された細胞は、OCH1遺伝子を欠き、α−1,2−マンノシダーゼ、GlcNAcトランスフェラーゼI、マンノシダーゼIIおよびガラクトシルトランスフェラーゼIを発現できる。いくつかの実施形態では、遺伝子工学的に改変された細胞は、ALG3遺伝子を欠き、α−1,2−マンノシダーゼ、GlcNAcトランスフェラーゼI、GlcNAcトランスフェラーゼIおよびガラクトシルトランスフェラーゼIを発現できる。このような遺伝子工学的に改変された細胞は、α−1,3−グルコシルトランスフェラーゼをさらに発現し、かつ/またはグルコシダーゼIIのアルファおよびベータサブユニットを発現し、かつ/またはOCH1遺伝子を欠くことができる。
【0057】
このようなタンパク質の1つ以上は、異種ターゲティング配列を含有する融合タンパク質であり得る。例えば、α−1,2−マンノシダーゼは、HDEL小胞体(ER)保持アミノ酸配列を有することができる(実施例を参照されたい)。N−グリコシル化活性を有する任意のタンパク質を、HDEL配列を含む融合タンパク質中に工学的に作製できることが理解される。その他のタンパク質は、タンパク質をゴルジ装置に向かわせるための異種配列を有することができる。例えば、酵母Kre2p遺伝子によりコードされる最初の100個のN末端アミノ酸、S.cerevisiae Mnn2遺伝子によりコードされる最初の36個のN末端アミノ酸(Swiss Prot.受託番号P38069)またはS.cerevisiae Mnn2p遺伝子によりコードされる最初の46個のN末端アミノ酸を用いて、タンパク質をゴルジに向かわせることができる。そのままで、真菌細胞で発現されるタンパク質をコードする核酸は、コードされたタンパク質を細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含むことができる。例えば、α−1,2−マンノシダーゼをERに向かわせることができ、GnT I、GnTII、マンノシダーゼおよびGal Tをゴルジに向かわせることができる。
【0058】
N−グリコシル化活性を有するタンパク質が、該タンパク質を発現させる細胞とは異なる型(例えば異なる種)のタンパク質である細胞に由来する実施形態では、タンパク質をコードする核酸は、特定の目的の細胞における発現のためにコドン最適化できる。例えば、Trypanosoma bruceiからのN−グリコシル化を有するタンパク質をコードする核酸は、Yarrowia lipolyticaのような酵母細胞での発現のためにコドン最適化できる。このようなコドン最適化は、目的の細胞でのタンパク質の発現を増加させるために有用になり得る。タンパク質をコードする核酸をコドン最適化するための方法は、当技術分野において公知であり、例えばGaoら(Biotechnol. Prog.(2004年)20巻(2号):443〜448頁)、Kotulaら(Nat. Biotechn.(1991年)9巻、1386〜1389頁)およびBennetzenら(J. Biol. Chem.(1982年)257巻(6号):2036〜3031頁)に記載されている。表1は、Yarrowia lipolyticaについてのコドン使用頻度を示す。データは、5,967のコード配列に存在する2,945,919のコドンに由来した。表1の内容は、コドン使用頻度データベース(Codon Usage Database)から得たが、これは、kazusa.or.jp/codon/cgi−bin/showcodon.cgi?species=284591のワールドワイドウェブで見出すことができる。
【0059】
【表1】
いくつかの実施形態では、ヒトタンパク質を細胞に導入でき、N−グリコシル化活性を有する1つ以上の内因性酵母タンパク質を抑制(例えば欠失または変異)できる。真菌グリコシル化経路を「ヒト化」するための技術は、例えばChoiら(2003年)Proc. Natl. Acad. Sci. USA100巻(9号):5022〜5027頁;Verveckenら(2004年)Appl. Environ. Microb.70巻(5号):2639〜2646頁;およびGerngross(2004年)Nature Biotech.22巻(11号):1410〜1414頁に記載されている。
【0060】
遺伝子工学的改変が例えばタンパク質の発現の変化または外因性タンパク質(内因性タンパク質の変異形態を含む)の発現を含む場合、遺伝子工学的に改変された細胞がタンパク質を発現するかを決定するために様々な技術を用いることができる。例えば、タンパク質をコードするmRNAまたはタンパク質自体の存在を、例えばノザンブロットもしくはRT−PCR分析またはウェスタンブロット分析をそれぞれ用いて検出できる。N−グリコシル化活性を有するタンパク質の細胞内局在化は、細胞成分分画(subcellular fractionation)および免疫蛍光を含む様々な技術を用いることにより分析できる。
【0061】
標的分子のグリコシル化を検出する方法は、DNAシーケンサー支援(DSA)、蛍光体支援糖質電気泳動(FACE)または表面増強型レーザー脱離/イオン化飛行時間型質量分析(SELDI−TOF MS)を含む。例えば、分析は、DSA−FACEを利用でき、ここでは、例えば、糖タンパク質を変性させ、その後、例えばメンブレン上に固定化する。糖タンパク質は、次いで、ジチオトレイトール(DTT)またはβ−メルカプトエタノールのような適切な還元剤を用いて還元できる。タンパク質のスルフヒドリル基は、ヨード酢酸のような酸を用いてカルボキシル化できる。次に、N−グリコシダーゼFのような酵素を用いてN−グリカンをタンパク質から切り離すことできる。N−グリカンは、必要に応じて、再構成して、還元的アミノ化により誘導体化できる。誘導体化N−グリカンは、次いで、濃縮できる。N−グリカン分析のために適切な装置は、例えばABI PRISM(登録商標)377 DNAシーケンサー(Applied Biosystems)を含む。データ解析は、例えばGENESCAN(登録商標)3.1ソフトウェア(Applied Biosystems)を用いて行うことができる。必要に応じて、単離マンノタンパク質(mannoprotein)は、1種以上の酵素でさらに処理して、それらのN−グリカン状態を確認できる。N−グリカン分析のさらなる方法は、例えば質量分析(例えばMALDI−TOF−MS)、順相(normal phase)、逆相での高圧液体クロマトグラフィー(HPLC)およびイオン交換クロマトグラフィー(例えばグリカンが標識されていない場合にパルス電流滴定検出を用いて、またグリカンが適当に標識されているならばUV吸収もしくは蛍光を用いて)を含む。Callewaertら(2001年)Glycobiology11巻(4号):275〜281頁およびFreireら(2006年)Bioconjug. Chem.17巻(2号):559〜564頁も参照されたい。
【0062】
遺伝子工学的に改変された細胞の遺伝子改変のいずれかが、誘導指示(例えば化学的または物理的指示)の存在によって誘導性または条件的である場合、遺伝子工学的に改変された細胞は、必要に応じて、核酸の導入前、導入中または導入後に誘導剤の存在下で培養できる。例えば、標的タンパク質をコードする核酸の導入の後に、細胞を、N−グリコシル化活性を有する1つ以上のタンパク質の発現を促進できる化学的誘導剤に曝露できる。複数の誘導指示が、N−グリコシル化活性を有する1つ以上のタンパク質の条件的発現を誘導する場合、細胞は、複数の誘導剤と接触できる。
【0063】
所望のN−グリカンを含むように改変された標的分子は、遺伝子工学的に改変された細胞から単離できる。改変された標的分子を、酵母細胞内で維持し、細胞溶解の際に遊離できるか、または改変標的分子を、細胞からの分子の分泌を誘導するコード配列(外因性核酸に本来備わっているかまたは発現ベクター中に工学的に作製されたかのいずれか)によりもたらされる機構により培養培地中に分泌させることができる。細胞溶解産物または培養培地中の改変標的分子の存在は、その分子の存在を検出するための様々な標準的プロトコールにより確認できる。例えば、変更された標的分子がタンパク質である場合、このようなプロトコールは、それらに限定されないが、変更された標的タンパク質(または標的タンパク質自体)に特異的な抗体を用いる免疫ブロッティングもしくは放射性免疫沈降法、変更された標的タンパク質(または標的タンパク質自体)に特異的なリガンドの結合、または改変された標的タンパク質(または標的タンパク質自体)の特異的酵素活性の試験を含むことができる。
【0064】
いくつかの実施形態では、遺伝子工学的に改変された細胞から単離される標的分子の少なくとも約25%は、所望のN−グリカンを含有する。例えば、遺伝子工学的に改変された細胞から単離される標的分子の少なくとも約27%、少なくとも約30%、少なくとも約35%、少なくとも約40%、少なくとも約45%、少なくとも約50%、少なくとも約55%、少なくとも約60%、少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%または少なくとも約95%または少なくとも約99%が、所望のN−グリカンを含有できる。
【0065】
いくつかの実施形態では、本明細書で記載する方法を用いて生成される標的分子において、糖タンパク質上のN−グリカンの少なくとも50%(例えば少なくとも55、60、65、70、75、80または85%)は、GlcNAc2Man3GlcNAc2 N−グリカンであり得る。GlcNAc2Man3GlcNAc2 N−グリカンのパーセンテージは、DSA−FACE電気泳動図中のピーク面積から見積もることができる。実施例13を参照されたい。
【0066】
いくつかの実施形態では、単離改変標的分子は、凍結、凍結乾燥または固定化して、例えば変更された標的分子が生物活性を保持することを可能にする適当な条件下で貯蔵できる。
【0067】
工学的に改変された細胞の培養
この文書は、本明細書で記載する遺伝子工学的に改変された細胞のいずれかの実質的に純粋な培養物も提供する。本明細書で用いる場合、遺伝子工学的に改変された細胞の「実質的に純粋な培養物」は、この細胞の培養物であって、培養物中の生存細胞の合計数の約40%未満(すなわち約35%;約30%;約25%;約20%;約15%;約10%;約5%;約2%;約1%;約0.5%;約0.25%;約0.1%;約0.01%;約0.001%;約0.0001%またはさらにはそれ未満)が遺伝子工学的に改変された細胞以外の生存細胞、例えば細菌、真菌(酵母を含む)、マイコプラズマまたは原生生物の細胞である培養物である。この状況での用語「約」は、関連するパーセンテージが、特定されたパーセンテージより15%上または下であり得ることを意味する。つまり、例えば、約20%は、17%〜23%であり得る。遺伝子工学的に改変された細胞のこのような培養物は、細胞と、増殖培地、貯蔵培地または輸送培地とを含む。培地は、液体、半固体(例えばゼラチン状培地)、または凍結したものであり得る。培養物は、液体培地中または半固体培地中/上で増殖しているか、あるいは凍結貯蔵培地もしくは輸送培地を含む貯蔵培地または輸送培地中で貯蔵または輸送されている細胞を含む。培養物は、培養容器または貯蔵容器または基材(substrate)(例えば培養皿、フラスコもしくは試験管または貯蔵バイアルもしくは貯蔵試験管)中にある。
【0068】
本明細書で記載する遺伝子工学的に改変された細胞は、例えば、例えばグリセロールもしくはスクロースのような凍結保護物質を含有するバッファー中での凍結細胞懸濁物として、凍結乾燥細胞として貯蔵できる。代わりに、これらは、例えば流動床乾燥もしくは噴霧乾燥または任意のその他の適切な乾燥方法により得られる乾燥細胞調製物として貯蔵できる。
【0069】
変更されたN−グリコシル化分子により処置可能な障害
所望のN−グリカンを含有するように改変された単離標的分子は、代謝障害、がんおよび炎症性障害を含む様々な障害を処置するために用いることができる。
【0070】
(i)代謝障害
代謝障害は、個別のヒト(または動物)細胞内でエネルギーの生成に影響する障害である。ほとんどの代謝障害は遺伝的であるが、いくつかは、食餌、毒素、感染症などの結果として「後天性」であり得る。遺伝性代謝障害は、先天性代謝異常としても公知である。一般的に、遺伝性代謝障害は、遺伝子欠陥により引き起こされ、これにより、細胞の代謝プロセスにおけるいくつかのステップに必要な酵素がなくなるかまたは誤って構築される。代謝障害の最大のクラスは、炭水化物代謝障害、アミノ酸代謝障害、有機酸代謝障害(有機酸性尿症)、脂肪酸酸化およびミトコンドリア代謝障害、ポルフィリン代謝障害、プリンまたはピリミジン代謝障害、ステロイド代謝障害、ミトコンドリア機能障害、ペルオキシソーム機能障害ならびにリソソーム蓄積症(LSD)である。
【0071】
本明細書で記載する1つまた複数のグリコシル化分子(またはその薬学的組成物)の投与により処置できる代謝障害の例は、遺伝性ヘモクロマトーシス、眼皮膚白皮症、プロテインC欠損症、I型遺伝性血管浮腫、先天性スクラーゼ−イソマルターゼ欠損症、クリグラー−ナジャー症候群II型、ラロン症候群、遺伝性ミエロペルオキシダーゼ、原発性甲状腺機能低下症、先天性QT延長症候群、チロキシン(tyroxine)結合グロブリン欠損症、家族性高コレステロール血症、家族性カイロミクロン血症、無ベータリポタンパク質血症、低血漿リポタンパク質Aレベル、肝臓損傷を伴う遺伝性気腫、先天性甲状腺機能低下症、骨形成不全症、遺伝性低フィブリノゲン血症、アルファ−1アンチキモトリプシン欠損症、腎性尿崩症、神経下垂体性尿崩症、アデノシンデアミナーゼ欠損症、ペリツェウスメルツバッヘル病、フォンウィルブランド病IIA型、第V、VIII因子合併欠損症(combined factors V and VIII deficiency)、遅発性脊椎骨端異形成症、コロイデレミア、I細胞病、バッテン病、毛細血管拡張性運動失調、ADPKD−常染色体優性多嚢胞性腎疾患、微絨毛封入体病、結節硬化症、ロウ眼脳腎症候群、筋委縮性側策硬化症、骨髄異形成症候群、不全リンパ球症候群、タンジール病、家族性肝内胆汁うっ滞、X連鎖副腎白質ジストロフィー、スコット症候群、ヘルマンスキー−プドラック症候群1型および2型、ツェルウェガー症候群、近節短縮性点状軟骨異形成症、常染色体劣性原発性高シュウ酸尿症、モーアトラネジャーグ症候群、球脊髄性筋萎縮症(spinal and bullar muscular atrophy)、原発性線毛運動不全(diskenesia)(カルタゲナー症候群)、巨人症および先端巨大症、乳汁漏出症、アジソン病、副腎性男性化、クッシング症候群、ケトアシドーシス、原発性または続発性アルドステロン症、ミラーディーカー症候群、脳回欠損、運動ニューロン疾患、アッシャー症候群、ウィスコット−アルドリッチ症候群、オピッツ(Optiz)症候群、ハンチントン病、遺伝性膵炎、抗リン脂質抗体症候群、オーバーラップ結合組織病、シェーグレン症候群、スティッフマン症候群、ブルガダ症候群、フィンランド型先天性腎炎症候群、デュビン−ジョンソン症候群、X連鎖低リン酸血症(hypophosphosphatemia)、ペンドレッド症候群、新生児持続性高インスリン血症性低血糖症、遺伝性球状赤血球症、無セルロプラスミン血症、乳児性神経セロイドリポフスチン沈着症、偽軟骨形成不全および多発性骨端骨異形成(multiple epiphyseal)、シュタルガルト様黄斑ジストロフィー、X連鎖シャコール−マリー−トゥース病、常染色体優性網膜色素変性、ウォルコット−ラリソン症候群、クッシング病、肢帯筋ジストロフィー、ムコ多糖症IV型、フィンランド型遺伝性家族性アミロイドーシス、アンダーソン病、肉腫、慢性骨髄単球性白血病、心筋症、顔面生殖器異形成、捻転ジストニア(Torsion disease)、ハンチントンおよび脊髄小脳運動失調、遺伝性高ホモシステイン血症(hyperhomosyteinemia)、多発ニューロパチー、下位運動ニューロン疾患、色素性網膜炎、血清反応陰性多発性関節炎、間質性肺線維症、レイノー現象、ウェゲナー(Wegner’s)肉芽腫症、タンパク尿(preoteinuria)、CDG−Ia、CDG−Ib、CDG−Ic、CDG−Id、CDG−Ie、CDG−If、CDG−IIa、CDG−IIb、CDG−IIc、CDG−IId、エーラース−ダンロス症候群、多発性外骨腫症、グリセリ症候群(1型または2型)、あるいはX連鎖非特異性精神遅滞を含み得る。さらに、代謝障害は、それらに限定されないが、ファブリー病、ファーバー病、ゴーシェ病、GM1−ガングリオシド−シス、テイ−サックス病、サンドホフ病、GM2活性化因子病(GM2 activator disease)、クラッベ病、異染性白質ジストロフィー、ニーマン−ピック病(A型、B型およびC型)、ハーラー病、シャイエ病、ハンター病、サンフィリポ病、モルキオ病、マロトー−ラミー病、ヒアルロニダーゼ欠損症、アスパルチルグルコサミン尿症、フコシドーシス、マンノシドーシス、シンドラー病、シアリドーシス1型、ポンぺ病、濃化異骨症(Pycnodysostosis)、セロイドリポフスチン沈着症、コレステロールエステル蓄積症、ウォルマン病、多種スルファターゼ欠損症、ガラクトシアリドーシス、ムコリピド−シス(II型、III型およびIV型)、シスチン症、シアル酸蓄積症、マリネスコ−シェーグレン症候群を伴うカイロミクロン保持病、ヘルマンスキー−プドラック症候群、チェディアック−東症候群、ダノン病または幸福顔貌骨異形成(Geleophysic dysplasia)のようなリソソーム蓄積症も含み得る。
【0072】
代謝障害の症状は多数で多様であり、例えば貧血、疲労、容易に挫傷すること、血液中の血小板低下、肝腫大、脾腫大、骨格脆弱、肺機能障害、感染症(例えば胸部感染症または肺炎)、腎臓機能障害、進行性脳損傷、発作、過剰に粘稠な胎便、咳、喘鳴、過剰な唾液もしくは粘液生成、息切れ、腹痛、腸または消化管(bowel or gut)の閉塞、生殖能力の問題、鼻内のポリープ、ばち指/足指の爪および皮膚、手もしくは足の疼痛、被角血管腫、発汗低下、角膜および水晶体の混濁、白内障、僧帽弁の逸脱および/もしくは逆流、心肥大、温度不耐、歩行困難、嚥下困難、進行性視力喪失、進行性聴力喪失、緊張低下、巨舌症、反射消失、腰痛、睡眠時無呼吸、起坐呼吸、傾眠、脊柱前弯または脊柱側弯の1つ以上を含み得る。欠陥のあるまたは存在しないタンパク質およびその結果としての疾患の表現型(例えば代謝障害の症状の現れ)の性質が多様なので、ある所定の障害は、その特定の障害に特徴的な症状だけを一般的に示す。例えば、ファブリー病の患者は、それらに限定されないが、温度不耐、角膜渦状化、疼痛、皮膚の発疹、悪心または下痢(dirarrhea)のような上記の症状の特定の部分集合を示し得る。ゴーシェ症候群の患者は、巨脾腫、肝硬変、痙攣、緊張亢進、無呼吸、骨粗鬆症または皮膚変色を示し得る。
【0073】
本明細書で記載する1つ以上の分子を投与することに加えて、代謝障害は、適当な栄養素およびビタミン(例えば補助因子療法)、理学療法および疼痛薬物療法により処置することもできる。
【0074】
ある所定の代謝障害の特定の性質に依存して、患者は、これらの症状をいずれの年齢でも示すことができる。多くの場合では、症状は小児期または早期成人期に現れ得る。例えば、ファブリー病の症状は、早い年齢、例えば10または11歳で現れることがある。
【0075】
本明細書で用いる場合、「代謝障害を発生する危険性がある」被験体は、障害を発生する素因、すなわちアルファ−L−イズロニダーゼ、ベータ−D−ガラクトシダーゼ、ベータ−グルコシダーゼ、ベータ−ヘキソサミニダーゼ、ベータ−D−マンノシダーゼ、アルファ−L−フコシダーゼ、アリールスルファターゼB、アリールスルファターゼA、アルファ−N−アセチルガラクトサミニダーゼ、アスパルチルグルコサミニダーゼ、イズロン酸−2−スルファターゼ、アルファ−グルコサミニド−N−アセチルトランスフェラーゼ、ベータ−D−グルクロニダーゼ、ヒアルロニダーゼ、アルファ−L−マンノシダーゼ、アルファ−ノイラミニダーゼ(neurominidase)、ホスホトランスフェラーゼ、酸性リパーゼ、酸性セラミダーゼ、スフィンゴミエリナーゼ(sphinogmyelinase)、チオエステラーゼ、カテプシンKまたはリポタンパク質リパーゼのような酵素の変異の結果として代謝障害を発生する遺伝的素因を有する被験体である。明らかに、「代謝障害を発生する危険性がある」被験体は、目的の種内の被験体全てではない。
【0076】
「障害を有することが疑われる」被験体は、本明細書で記載するもののような障害の1つ以上の症状を有する被験体である。
【0077】
(ii)がん
がんは、細胞の制御されない分裂と、浸潤により隣接組織内で直接成長することまたは転移(がん細胞が血流またはリンパ系を通して輸送される場合)により離れた部位へ植え込まれることのいずれかによりそれらが広がる能力とを特徴とする。がんは、全ての年齢の人が罹患し得るが、危険性は年齢とともに増加する傾向にある。がんの型は、例えば肺がん、乳がん、結腸がん、膵臓がん、腎がん、胃がん、肝臓がん、骨がん、血液のがん、神経組織がん、黒色腫、甲状腺がん、卵巣がん、精巣がん、前立腺がん、子宮頚がん、膣がんまたは膀胱がんを含み得る。
【0078】
本明細書で用いる場合、「がんを発生する危険性がある」被検体は、がんを発生する素因、すなわち腫瘍抑制遺伝子における変異(例えばBRCA1、p53、RBまたはAPCにおける変異)のようながんを発生する遺伝的素因を有するか、またはがんをもたらし得る条件に曝露された被験体である。つまり、被験体は、変異原性または発癌性のレベルの特定の化合物(例えばアクロレイン、ヒ素、ベンゼン、ベンズ{a}アントラセン、ベンゾ{a}ピレン、ポロニウム−210(ラドン)、ウレタンまたは塩化ビニルのような煙草の煙の中の発癌性化合物)に曝露された場合、該被験体は、「がんを発生する危険性がある」ものであり得る。さらに、被験体は、例えば高い線量の紫外線またはX線照射に曝露されたかまたはパピローマウイルス、エプスタイン−バーウイルス、B型肝炎ウイルスまたはヒトT細胞白血病−リンパ腫ウイルスのような腫瘍の原因となる/腫瘍と関連するウイルスに曝露された(例えば感染した)場合、該被験体は、「がんを発生する危険性がある」ことができる。上記のことから、「がんを発生する危険性がある」被験体は、目的の種内の被験体全てではないことが明らかである。
【0079】
「がんを有することが疑われる」被験体は、がんの1つ以上の症状を有する被験体である。がんの症状は当業者に周知であり、限定されることなく、乳房腫瘤、乳頭変化、乳房嚢胞、胸部痛、体重減少、脱力、過剰な疲労、摂食困難、食欲喪失、慢性の咳、息切れの悪化、咳による血液排出(coughing up blood)、血尿、血便、悪心、嘔吐、肝臓転移、肺転移、骨転移、腹部肥満(abdominal fullness)、鼓脹、腹膜腔内の流体、膣出血、便秘、腹部膨満(abdominal distension)、結腸穿孔、急性腹膜炎(感染、発熱、疼痛)、疼痛、吐血(vomiting blood)、重度の発汗、発熱、高い血圧、貧血、下痢、黄疸、めまい、悪寒、筋肉攣縮、結腸転移、肺転移、膀胱転移、肝臓転移、骨転移、腎臓転移および膵臓転移、嚥下困難などを含む。上記のことから、「がんを有することが疑われる」被験体は、目的の種内の被験体全てではないことが明らかである。
【0080】
本明細書で記載する1つ以上の変更されたN−グリコシル化分子の投与に加えて、がんは、化学療法剤、電離放射線、免疫療法剤または温熱療法剤によっても処置できる。化学療法剤は、例えばシスプラチン、カルボプラチン、プロカルバジン、メクロレタミン、シクロホスファミド、カンプトテシン、アドリアマイシン、イホスファミド、メルファラン、クロラムブシル、ブスルファン(bisulfan)、ニトロソウレア(nitrosurea)、ダクチノマイシン、ダウノルビシン、ドキソルビシン、ブレオマイシン、プリカマイシン(plicomycin)、マイトマイシン、エトポシド、ベラパミル(verampil)、ポドフィロトキシン、タモキシフェン、タキソール、トランスプラチナ、5−フルオロウラシル、ビンクリスチン、ビンブラスチンおよびメトトレキセートを含む。
【0081】
(iii)炎症性障害
「炎症性障害」は、本明細書で用いる場合、1つ以上の物質(例えば被験体において天然に生じない物質)が、白血球細胞(例えばB細胞、T細胞、マクロファージ、単球または樹状細胞)の作用により、病的応答、例えば病的免疫応答を不適当に引き起こすプロセスのことをいう。よって、炎症性応答に関与するこのような細胞は、「炎症性細胞」という。不適当に引き起こされる炎症性応答は、外来物質(例えば抗原、ウイルス、細菌、真菌)が被験体内または被験体上に存在しないものであり得る。不適当に引き起こされる応答は、自己成分(例えば自己抗原)が炎症性細胞により標的にされるものであり得る(例えば多発性硬化症のような自己免疫障害)。不適当に引き起こされる応答は、大きさまたは持続期間が不適当である応答、例えばアナフィラキシーでもあり得る。つまり、不適当に標的にされる応答は、微生物感染(例えばウイルス、細菌または真菌)の存在によることができる。炎症性障害の型(例えば自己免疫疾患)は、それらに限定されないが、変形性関節症、関節リウマチ(RA)、脊椎関節症(spondyloarthropathy)、POEMS症候群、クローン病、多中心性キャッスルマン病、全身性エリテマトーデス(SLE)、多発性硬化症(MS)、筋ジストロフィー(MD)、インスリン依存性糖尿病(IDDM)、皮膚筋炎、多発性筋炎、ギランバレー症候群のような炎症性神経障害、ウェゲナー肉芽腫症のような血管炎、結節性多発動脈炎、リウマチ性多発筋痛、側頭動脈炎、シェーグレン症候群、ベーチェット(Bechet’s)病、チャーグ−ストラウス症候群または高安動脈炎を含み得る。鼻炎、副鼻腔炎、じんま疹(urticaria)、じんま疹(hives)、血管浮腫、アトピー性皮膚炎、食物アレルギー(例えばナッツアレルギー)、薬物アレルギー(例えばペニシリン)、昆虫アレルギー(例えばハチ刺傷に対するアレルギー)のような特定の型のアレルギーまたは肥満細胞症も炎症性障害に含まれる。炎症性障害は、潰瘍性大腸炎および喘息も含み得る。
【0082】
「炎症性障害を発生する危険性がある」被験体とは、1種以上の炎症性障害の家族歴(例えば1種以上の炎症性障害に対する遺伝的素因)を有する被験体または1種以上の炎症誘導性状態に曝露された被験体のことをいう。例えば、被験体は、それらに限定されないが、ブドウ球菌エンテロトキシン(SE)、streptococcus pyogenes外毒素(SPE)、staphylococcus aureus毒素ショック症候群毒素(TSST−1)、連鎖球菌分裂促進性外毒素(SME)および連鎖球菌スーパー抗原(SSA)のようなウイルスまたは細菌のスーパー抗原に曝露された可能性がある。上記のことから、「炎症性障害を発生する危険性がある」被験体は、目的の種内の被験体全てではないことが明らかである。
【0083】
「炎症性障害を有することが疑われる」被験体は、炎症性障害の1つ以上の症状を示す被験体である。炎症性障害の症状は当技術分野で周知でありそれらに限定されないが、発赤、腫脹(例えば腫脹関節)、触れると温かい関節、関節痛、こわばり、関節機能の喪失、発熱、悪寒、疲労、エネルギーの喪失、頭痛、食欲喪失、筋肉こわばり、不眠、そう痒、鼻づまり、くしゃみ、咳、めまい、発作または疼痛のような1つ以上の神経性の症状を含む。上記のことから、「炎症性障害を有することが疑われる」被験体は、目的の種内の被験体全てではないことが明らかである。
【0084】
本明細書で記載する1つ以上の分子の投与に加えて、炎症性障害は、非ステロイド性抗炎症薬(NSAID)、疾患修飾性抗リウマチ薬(DMARD)、生体応答修飾物質またはコルチコステロイドによっても処置できる。生体応答修飾物質は、例えば抗TNF剤を含む。抗TNF剤の非限定的な例は、アダリムマブ(adulimumab)、インフリキシマブもしくはエタネルセプトのような可溶性TNF受容体またはTNFに特異的な抗体を含む。
【0085】
変更されたN−グリコシル化分子(またはその薬学的組成物)のいずれかを用いて本明細書で記載する障害のいずれかを処置する(例えばその1つ以上の症状を予防するかまたは改善する)ために適切な方法は、以下の項に示す。
【0086】
薬学的組成物および処置方法
所望のN−グリカンを有するように改変された標的分子は、治療有効量の分子と、1つ以上のアジュバント、賦形剤、キャリアおよび/または希釈剤とを含有する薬学的組成物に組み込むことができる。許容される希釈剤、キャリアおよび賦形剤は、典型的に、受容者の恒常性(例えば電解質バランス)に有害に影響しない。許容されるキャリアは、生体適合性、不活性または生体吸収性の塩、緩衝剤、オリゴ糖もしくは多糖、ポリマー、粘度改善剤、防腐剤などを含む。1つの例示的なキャリアは、生理食塩水である(0.15M NaCl、pH7.0〜7.4)。別の例示的なキャリアは、50mMリン酸ナトリウム、100mM塩化ナトリウムである。薬学的組成物の処方および投与のための技術についてのさらなる詳細は、例えばRemington’s Pharmaceutical Sciences(Maack Publishing Co.、Easton、Pa.)で見出すことができる。補足の活性化合物も組成物に組み込むことができる。
【0087】
N−グリカンを有する分子を含有する薬学的組成物の投与は、全身的または局部的であり得る。薬学的組成物は、それらが非経口および/または非経口でない投与に適切であるように処方できる。具体的な投与様式は、皮下、静脈内、筋肉内、腹腔内、経皮、髄腔内、経口、直腸、頬側、局所、鼻、眼、関節内、動脈内、くも膜下、気管支、リンパ節、膣および子宮内の投与を含む。
【0088】
投与は、薬学的組成物のボーラスの周期的な注射によることができるか、あるいは外的(例えばIVバッグ)もしくは内的(例えば生体浸食性インプラント、バイオ人工臓器(bioartificial organ)または埋め込まれた変更されたN−グリコシル化分子生成細胞のコロニー)であるリザーバからの静脈内または腹腔内投与による不断または連続的であり得る。例えば米国特許第4,407,957号、第5,798,113号および第5,800,828号を参照されたい。薬学的組成物の投与は、ポンプ(例えばAnnals of Pharmacotherapy、27巻:912頁(1993年);Cancer、41巻:1270頁(1993年);Cancer Research、44巻:1698頁(1984年)を参照されたい);マイクロカプセル封入(例えば米国特許第4,352,883号;第4,353,888号;および第5,084,350号を参照されたい);連続放出ポリマーインプラント(例えばSabel、米国特許第4,883,666号を参照されたい);マクロカプセル封入(例えば米国特許第5,284,761号、第5,158,881号、第4,976,859号および第4,968,733号ならびにPCT特許出願公開WO92/19195、WO95/05452を参照されたい);皮下、静脈内、動脈内、筋肉内もしくはその他の適切な部位のいずれかでの注射;またはカプセル、液体、錠剤、丸剤もしくは延長放出処方物での経口投与のような適切な送達手段を用いて達成できる。
【0089】
非経口送達系の例は、エチレン−酢酸ビニルコポリマー粒子、浸透圧ポンプ、埋め込み型注入系、ポンプ送達、カプセル封入細胞送達、リポソーム送達、針送達注射、針なし注射、ネブライザー、エアゾール器(aerosolizer)、エレクトロポレーションおよび経皮パッチを含む。
【0090】
非経口投与のために適切な処方物は、変更されたN−グリコシル化分子の滅菌水性調製物を簡便に含有し、これは、好ましくは、受容者の血液と等張である(例えば生理食塩水溶液)。処方物は、単位用量または複数回用量の形態で提供し得る。
【0091】
経口投与のために適切な処方物は、それぞれが所定量の変更されたN−グリコシル化分子を含有するカプセル剤、カシェ剤、錠剤もしくはロゼンジのような分かれた単位、またはシロップ剤、エリキシル剤、乳剤もしくは飲料(draught)のような水性流体(liquor)または非水性の液体中の懸濁剤で提供し得る。
【0092】
局所投与のために適切なN−グリカンを有する分子は、哺乳動物(例えばヒト患者)に、例えばクリーム、スプレー、フォーム、ゲル、軟膏剤(ointment)、軟膏剤(salve)またはドライラブ(dry rub)として投与できる。ドライラブは、投与部位にて再水和できる。このような分子は、包帯、ガーゼまたはパッチに直接注入(例えば浸して乾燥する)することもでき、これらは、次いで、局所的に施用できる。このような分子は、半液体、ゲル化または完全な液体の状態で、局所投与用の包帯、ガーゼまたはパッチ中に維持することもできる(例えば米国特許第4,307,717号を参照されたい)。
【0093】
治療有効量の薬学的組成物は、それを必要とする被験体に、当業者が確認できる投与量計画(dosage regimen)において投与できる。例えば、組成物は、被験体に、例えば全身的に、1投薬あたり被験体の体重1kgあたり0.01μgから10,000μgまでの投与量で投与できる。別の例では、投与量は、1投薬あたり被験体の体重1kgあたり1μgから100μgまでである。別の例では、投与量は、1投薬あたり被験体の体重1kgあたり1μgから30μgまで、例えば1投薬あたり被験体の体重1kgあたり3μgから10μgまでである。
【0094】
治療効力を最適化するために、N−グリカンを含有する分子は、まず、異なる投薬計画(dosing regimen)で投与できる。単位用量および計画は、例えば哺乳動物の種、その免疫状態、哺乳動物の体重を含む因子に依存する。典型的に、組織におけるこのような分子のレベルは、例えば所定の処置計画の効力を決定するために、臨床試験手順の一部として適当なスクリーニングアッセイを用いて監視できる。
【0095】
分子の投薬の頻度は、医療従事者(例えば医師または看護士)の熟練および臨床的判断の範囲内である。典型的に、投与計画(administration regimen)は、最適投与パラメータを確立し得る臨床試験により確立される。しかし、従事者は、被験体の年齢、健康、体重、性別および医療状態に従ってそのような投与計画を変更してよい。投薬頻度は、処置が予防的または治療的であるかに依存して変更できる。
【0096】
このような分子またはその薬学的組成物の毒性および治療効力は、公知の薬学的手順により、例えば細胞培養または実験動物において決定できる。これらの手順は、例えばLD50(集団の50%に対して致死的な用量)およびED50(集団の50%において治療的に有効な用量)を決定するために用いることができる。毒性作用と治療効果との間の用量の比は、治療指数であり、これは、LD50/ED50の比として表すことができる。高い治療指数を示す薬学的組成物が好ましい。毒性の副作用を示す薬学的組成物を用いることができるが、そのような化合物を罹患組織の部位に向かわせるための送達系を設計して、正常細胞(例えば非標的細胞)への潜在的な損傷を最小限にし、そのことにより副作用を低減するように注意を払うべきである。
【0097】
細胞培養アッセイおよび動物研究から得られたデータは、適当な被験体(例えばヒト患者)において用いるための投与量の範囲を処方する場合に用いることができる。このような薬学的組成物の投与量は、通常、ほとんどまたは全く毒性を有さずにED50を含む循環濃度の範囲内にある。投与量は、採用する投与形態および利用する投与経路に依存してこの範囲内で変更し得る。本明細書で記載するようにして用いる薬学的組成物(例えば被験体における代謝障害を処置するため)について、治療有効用量は、最初に細胞培養アッセイから見積もることができる。用量は、動物モデルにおいて処方して、細胞培養で決定したIC50(すなわち、症状の最大阻害の半分を達成する薬学的組成物の濃度)を含む循環血漿濃度範囲を達成できる。このような情報を用いて、ヒトにおける有用な用量をより正確に決定できる。血漿におけるレベルは、例えば高性能液体クロマトグラフィーにより測定できる。
【0098】
本明細書で定義するように、N−グリカンを含有する分子の「治療有効量」は、処置した被験体において医療的に望ましい結果(例えば代謝障害の1つ以上の症状の改善)を生じることができる分子の量である。治療有効量(すなわち有効投与量)は、被験体または試料重量1キログラムあたりミリグラムまたはマイクログラム量の化合物を含むことができる(例えば1キログラムあたり約1マイクログラムから1キログラムあたり約500ミリグラム、1キログラムあたり約100マイクログラムから1キログラムあたり約5ミリグラムまたは1キログラムあたり約1マイクログラムから1キログラムあたり約50マイクログラム)。
【0099】
被験体は、任意の哺乳動物、例えばヒト(例えばヒト患者)または非ヒト霊長類(例えばチンパンジー、ヒヒまたはサル)、マウス、ラット、ウサギ、モルモット、アレチネズミ、ハムスター、ウマ、ある型の家畜(例えばウシ、ブタ、ヒツジまたはヤギ)、イヌ、ネコまたはクジラであり得る。
【0100】
本明細書で記載する分子またはその薬学的組成物は、被験体に、別の処置、例えば代謝障害(例えばリソソーム蓄積症)のための処置との併用療法として投与できる。例えば併用療法は、被験体(例えばヒト患者)に、代謝障害(例えばリソソーム蓄積症)を有するかまたはその障害を発生する危険性がある(もしくは有することが疑われる)被験体に対して治療利益をもたらす1種以上の追加の薬剤を投与することを含むことができる。つまり、化合物または薬学的組成物と、1種以上の追加の薬剤とは、同時に投与できる。代わりに、分子をまず投与して、1種以上の追加の薬剤を2番目に投与するか、またはその逆が可能である。
【0101】
以前の療法が特に毒性である場合(例えば著しい副作用プロファイルを有する代謝障害の処置)、本明細書で記載する分子の投与を用いて、以前の療法の量を、同じまたは改善された治療利益を与えるのに十分であるが毒性がないレベルまで埋め合わせ(offset)かつ/または少なくすることができることが認識される。
【0102】
本明細書で記載する薬学的組成物のいずれも、投与についての使用説明書と一緒に容器、パックまたはディスペンサーに含めることができる。
【0103】
以下は、本発明の実施の例である。これらは、いずれの様式でも本発明の範囲を限定すると解釈されない。
【実施例】
【0104】
表2は、以下に記載する実験で用いた全ての株のリストを含む。表2では、MH=HDELタグ付きα−1,2−マンノシダーゼ;ζ=ゼータ配列による無作為組み込み;ドッキングΔ=特定の遺伝子座への組み込み;および(H)=ハイグロマイシン耐性。
【0105】
【表2−1】
【0106】
【表2−2】
【0107】
【表2−3】
【0108】
【表2−4】
【0109】
【表2−5】
【0110】
【表2−6】
(実施例1)Yarrowia lipolytica OCH1破壊
糖鎖工学的に改変したタンパク質発現株の作製を、Yarrowia lipolytica pold lnuga株(leu2−、ura3−、gut2−およびade2−の栄養要求性を有する株)で行った。Yarrowia lipolyticaにおいてOCH1(GenBank受託番号AJ563920)遺伝子をノックアウトするための方策は、LIP2遺伝子について記載されたようにして設定した(Fickersら、2003年J Microbiol Methods.55巻(3号):727〜37頁)。OCH1遺伝子について従った遺伝子構築方策は、米国特許出願公開第20090069232−A1号に記載されている。得られたベクターをpYlOCH1 PUT TOPO(図1B)と称した。
【0111】
OCH1 KO断片を、プラスミドから、SpeI/Bst1107I制限消化により単離し、Yarrowia lipolytica pold lnuga株を形質転換した。いくつかのウラシル原栄養性株を得て、PCRによりゲノムDNA(gDNA)上でプライマーYloch1 prom fw(5’−TCGCTATCACGTCTCTAGC−3’、配列番号1)およびYloch1 term rev(5’−ACTCTGTATACTTGTATGTACTGTGAGAC−3’、配列番号2)を用いてスクリーニングして、プラスミドのゲノム組み込みを分析した。正しいサイズの断片(すなわち、2328bp対野生型での1894bp)が、試験したいくつかのクローンについて増幅された。OCH1遺伝子のノックアウトも、増殖培地中に分泌された全糖タンパク質プール(=セクレトーム)のN−グリカン分析により確認した:Man8GlcNAc2構造は、糖プロファイル内の優勢なN−グリカンになった(図2)。このプロファイルは、より高い量のMan9GlcNAc2とともにより高い数のマンノース残基さえ有するいくつかの構造を含有する野生型株のものとは異なる(後者は、Och1p活性の結果として追加のマンノースを含有する可能性が高い)。
【0112】
URA3遺伝子を除去するために、陽性Δoch1クローン(G013と称する、表2を参照されたい)を、Creリコンビナーゼについての発現カセットを含有するエピソームプラスミドpUB4−Cre(Fickersら、2003年、既出)で形質転換した。URA3遺伝子の除去は、プライマーYloch1 prom fwおよびYloch1 term rev(上記を参照されたい)を用いるgDNAのPCRによりスクリーニングした。URA3マーカーが切り出されたクローンでは、2328bpのバンドはもはや増幅されなかった。代わりに、1075bpのPCR断片(excl.URA3)が得られた。陽性クローンは、セクレトームのN−グリカンレベルにおいて確かめられたが、該クローンはキュアしていない株のものと非常によく似たプロファイルを示す(図2)。キュアした株の1つ(G014と称する、表2を参照されたい)を、さらなるN−グリカン工学的改変のために選択した。
【0113】
(実施例2)無作為組み込みまたは標的化/ドッキング組み込みのいずれかによるER保持α−1,2−マンノシダーゼの過剰発現
Δoch1株により発現された糖タンパク質と結合したMan5GlcNAc2の作製を可能にするために、α−1,2−マンノシダーゼを発現させて、Man8GlcNAc2をMan5GlcNAc2(すなわち、ゴルジ型α−1,2−マンノシダーゼ活性)に切断した。このようなマンノシダーゼは、分泌系に向けられるべきである。S.cerevisiaeのプレプロ接合因子と融合し、それをERに局在化させるためにHDEL配列(配列番号21)でタグ付きTrichoderma reeseiのα−1,2−マンノシダーゼ(Genbank受託番号AF212153)は、Pichia pastorisにおいてならびにTrichoderma reeseiおよびAspergillus nigerにおいてin vivoでMan8GlcNAc2をMan5GlcNAc2に刈り込むことができる。HDELタグ付きT.reeseiα−1,2−マンノシダーゼのコドン最適化バージョンがY.lipolytica LIP2プレシグナル配列と融合し、TEFI、Hp4d(Madzakら、2000年、J. Mol. Microbiol. Biotechnol.2巻:207〜216頁)、GAPまたはPOX2プロモーターのいずれかの転写制御下に配置された発現構築物を作製した。これらのプラスミドの構築方策は、米国特許出願公開第20090069232−A1号に記載されている。
【0114】
Hp4d resp.TEFIプロモーターの転写制御下のマンノシダーゼを有するこれらのベクターのうち2つ、pYLHUXdL2preManHDELおよびpYLTUXdL2preManHDEL(図3)を用いて、G014株(実施例1に由来する)を形質転換した。ベクターをNotIで消化して、ゼータ配列によるゲノムへの無作為組み込みを可能にした。URA3原栄養性形質転換体を、N−グリカン分析のために選択した。いくつかの形質転換体は、Man8GlcNAc2からMan5GlcNAc2への明確な変換を示す(図4)。TEF1プロモーター制御下でマンノシダーゼを発現するクローンは、緩慢で塊状の増殖表現型を示したので(これらのクローンのうちの1つをG016と称した)、糖鎖工学的改変のさらなるステップを、遺伝子がHp4d転写制御下にある株のバックグラウンド(strain background)において行った。
【0115】
ManHDELをhp4dプロモーターの制御下で発現する1つの陽性クローン(G018)を選び、これからURA3マーカーを、Cre−リコンビナーゼを発現するプラスミドpRRQ2(Richardら、2001年J. Bacteriol.183巻:3098〜3107頁)の一過性形質転換によりキュアした。いくつかのura3−クローンを手順の後に選択し、セクレトーム上で明確なMan5GlcNAc2プロファイルを示す1つのクローン(G036)を、さらなる工学的改変作業のために用いた(図4)。このクローンのサザン分析により、1つの無作為に組み込まれたマンノシダーゼ発現カセットの存在が明らかになった。このサザン分析は、HindIII消化ゲノムDNAについて、プライマーMan for(5’−GCCTTCCAGACCTCTTGGAACGCCTACCACC−3’、配列番号22)およびMan rev(5’−GCCAGGTGGCCGCCTCGTCGAGAAGAAGATCG−3’、配列番号23)を用いて作製したDIG標識マンノシダーゼ特異的PCR断片を用いて行った。
【0116】
代替の方策では、YarrowiaゲノムのLEU2またはAXP1遺伝子座のいずれかへのHp4d駆動型マンノシダーゼ発現カセットの標的化組み込みを可能にする2つの構築物を作製した。これらのプラスミド、JME926_pPTLeu2−ADE2ex−Hp4dManHDEL(Yl)およびOXYP289_pPTAxp1−LEU2ex−Hp4dManHDEL(Yl)の構築を、図5に記載する。G014株への形質転換の前に、両方の構築物をNotIで消化し、各発現カセットを単離した。選択したADE2原栄養性クローンは、マンノシダーゼ発現カセットをLEU2遺伝子座に潜在的に組み込み、一方、LEU2原栄養体は、カセットをAXP1遺伝子座に潜在的に組み込んだ。形質転換体は、サザン分析により確かめて、ゲノムへの正しいターゲティングを評価した。これは、BamHI消化(LEU2遺伝子座への組み込み)またはHindIII消化(AXP1遺伝子座への組み込み)ゲノムDNAについて、プライマーMan for(5’−GCCTTCCAGACCTCTTGGAACGCCTACCACC−3’、配列番号22)およびMan rev(5’−GCCAGGTGGCCGCCTCGTCGAGAAGAAGATCG−3’、配列番号23)を用いて作製したDIG標識マンノシダーゼ特異的PCR断片を用いて行った。選択したクローンは、分泌された糖タンパク質上に合成されたN−グリカンの性質についても確かめた。ほとんどの場合では、正しく標的にされたHp4d駆動型α−1,2−マンノシダーゼ発現は、優勢にMan5GlcNAc2オリゴ糖の合成をもたらした(図6)。それぞれのターゲティング遺伝子座について、1つのマンノシダーゼ発現クローン(LEU2ドッキングの場合はG046;AXP1ドッキングの場合はG053)を、G046株についてプラスミドpRRQ2およびG053株についてpUB4−Creを用いるCreリコンビナーゼの一過的発現によるキュアリングのために選択した。得られたキュアした株(それぞれG055およびG054)を、サザンブロッティングにより再度確かめ、それらのMan5GlcNAc2プロファイルを、DSΑ−FACEを用いるN−グリカン分析により確認した。
【0117】
(実施例3)GlcNAcトランスフェラーゼIの発現
Yarrowiaコドン最適化配列を、S.cerevisiae Kre2タンパク質(SwissProt 受託番号P27809)の最初の100個のN末端アミノ酸と、それに続くヒトGlcNAcトランスフェラーゼI(SwissProt 受託番号P26572)の触媒ドメインとからなる融合タンパク質(図7、配列番号3および配列番号4)の発現のために作製した。酵母Kre2pの100個のN末端アミノ酸は、触媒GnT Iドメインのためのゴルジ局在化シグナルとして働く。この様式で、GnT I融合タンパク質が、分泌経路において、ER保持HDELタグ付きα−1,2−マンノシダーゼよりも後に局在化されることを確実にして、酵素がタンパク質結合N−グリカンをMan5GlcNAc2からGlcNAcMan5GlcNAc2に変換できるようにする。融合タンパク質の発現のためのコドン最適化合成遺伝子を、TEF1またはHp4dプロモーターのいずれかの転写制御下に配置して、プラスミドpYLTmAXhGnTIおよびpYLHp4mAXhGnTIを得た。構築方策を図8に示す。Kre2−GnT I融合タンパク質の機能的発現は、利用可能なMan5GlcNAc2グリカン上へのβ−1,2結合GlcNAc残基の付加をもたらして、GlcNAcMan5GlcNAc2の合成をもたらすはずである。
【0118】
プラスミドpYLTmAXhGnTIおよびpYLHp4mAXGnTIをNotI消化した後に、その分泌タンパク質上にMan5GlcNAc2 N−グリカンを生成することが公知であるG036株を形質転換した(実施例2を参照されたい)。形質転換体を、ウラシル原栄養性について選択した。これらのクローンのいくつかのセクレトームについてのN−グリコシル化プロファイルの分析は、N−グリカンパターンの明確な変化を示した:Man5GlcNAc2は、著しく低減され、より高い分子量(約1グルコース単位過剰)を有するN−グリカンを表す新しいピークが出現した。単離N−グリカンを、末端β結合GlcNAc残基を除去できる酵素であるナタマメβ−N−アセチルヘキソサミニダーゼで処理すると、新しいN−グリカンがGlcNAcMan5GlcNAc2であることが示された:その新しいピークは消失し、Man5GlcNAc2に完全に変換された(図9)。用いた培養方法に依存して、全N−グリカンプールの約70%がGlcNAcMan5GlcNAc2であることが証明された(利用可能なMan5GlcNAc2のおよそ77%が変換された)。
【0119】
Kre2−GnT I融合タンパク質をTEF1プロモーターの制御下で発現する1つの形質転換体をG040株と命名し、さらなる使用のために選択した。この株のサザンブロットによるゲノム解析は、1つの発現カセットの存在を示した。サザン分析は、BamHI消化ゲノムDNAについて、プライマー5’−GGATGATCACACAATGGCCCTGTTTCTG−3’(配列番号5)および5’−TGCTCTAGACTAGTTCCAAGAGGGGTC−3’(配列番号6)を用いて作製したDIG標識GnT I特異的PCR断片を用いて行った。G040株対1〜3コピー(同じサザンブロットにより確認)のHp4d駆動型Kre2−GnT I発現カセットを有する株のセクレトームについてのグリコシル化プロファイルの分析は、GlcNAc移動能力について著しい違いを示さなかった。
【0120】
(実施例4)マンノシダーゼIIの発現
Yarrowiaコドン最適化配列を、S.cerevisiae Mnn2タンパク質(SwissProt 受託番号P38069)の最初の36個のN末端アミノ酸と、それに続くDrosophila melanogasterマンノシダーゼII(SwissProt 受託番号Q24451)の触媒ドメインとからなる融合タンパク質(図10、配列番号7および配列番号8)の発現のために作製した。酵母Mnn2の36個のN末端アミノ酸は、触媒Man IIドメインのためのゴルジ局在化シグナルとして働く。この様式で、Mnn2−Man II融合タンパク質が、分泌経路において、Kre2−GnT I融合タンパク質と同じまたはそれより後の位置にさえ局在化されることを確実にして、よって、GlcNAcMan5GlcNAc2をGlcNAcMan3GlcNAc2に変換できた。融合タンパク質の発現のためのYarrowiaコドン最適化合成遺伝子を、TEF1プロモーターの転写制御下に配置して、プラスミドpYLTmAXDmManIIおよびpYLTmAXDmManII(LEU2ex)を得た。構築方策を図11に示す。
【0121】
プラスミドpYLTmAXDmManII(LEU2ex)をNotI消化した後に、その分泌タンパク質上にGlcNAcMan5GlcNAc2 N−グリカンを生成することが公知であるG040株を形質転換した(実施例3を参照されたい)。形質転換体を、ロイシン原栄養性について選択した。これらのクローンのいくつかのセクレトームについてのN−グリコシル化プロファイルの分析は、N−グリカンパターンの変化を示した:約2グルコース単位のより低い分子量を有するN−グリカンを表す新しいピークが出現し、これにより、GlcNAcMan3GlcNAc2の形成を示すことができ、よって、部分的マンノシダーゼII活性を示し得た。また、Man5GlcNAc2(すなわちピークのショルダー)とほとんど同じ位置に泳動される別のピークが出現し、これは、潜在的にGlcNAcMan4GlcNAc2を表す。後者の構造は、マンノシダーゼII活性が2つではなく1つのマンノース残基だけを除去した部分的な刈り込み事象の結果である可能性がある。単離N−グリカンをナタマメβ−Ν−アセチルヘキソサミニダーゼで処理すると、グリカンパターンが約1グルコース単位の左側へのシフトをもたらし、よって、末端GlcNAc残基の喪失によるより高い電気泳動移動度をもたらした(図12)。このことにより、機能的マンノシダーゼII活性の発現によるGlcNAcMan5GlcNAc2からのGlcNAcMan4GlcNAc2およびGlcNAcMan3GlcNAc2の作製がさらに確認される。用いた培養方法に依存して、全N−グリカンプールの約15%が、GlcNAcMan3GlcNAc2であることが証明された:利用可能なGlcNAcMan5GlcNAc2のおよそ35%が1または2個のマンノース残基を失い、20%がGlcNAcMan3GlcNAc2に完全に刈り込まれた。
【0122】
(実施例5)ガラクトシルトランスフェラーゼIの発現
末端ガラクトース残基を有するN−グリカンの合成は、分泌経路内の機能的で良好に局在化されたガラクトシルトランスフェラーゼの存在だけでなく、この酵素により用いられるドナー基質であるUDP−Galの利用可能性にも依存する。UDP−GlcとUDP−GlcNAcは、酵母生物のゴルジ装置中で十分に利用可能であると一般的に考えられているが、このことはUDP−Galについてはあまり知られていない。糖鎖工学的改変中の潜在的なUDP−Gal欠乏を克服するために、Pichia pastorisにおいて、Schizosaccharomyces pombe UDP−Glc−4−エピメラーゼ(GAL10様遺伝子SPBC365.14c(SwissProt 受託番号Q9Y7X5)によりコードされる)とヒトβ−1,4−ガラクトシルトランスフェラーゼI(GalT I)(SwissProt 受託番号PI5291)の触媒ドメインとの間の融合タンパク質を酵母ゴルジ装置に向かわせる試みが以前になされた(Bobrowiczら、Glycobiology14巻(9号):757〜766頁、2004年)。分泌経路におけるGal10p−GalT I融合タンパク質の局在化を、好ましくはGlcNAc移動およびマンノシダーゼII活性が分泌のために定められたタンパク質のN−グリカンに対して既に作用した位置にて、S.cerevisiae Mnn2pの最初の46個のN末端アミノ酸をN末端ターゲティングシグナルとして用いることにより遂行した。
【0123】
よって、Yarrowiaコドン最適化配列を、S.cerevisiae Mnn2タンパク質の最初の46個のN末端アミノ酸と、それに続くS.pombe Gal10様タンパク質およびヒトGalT Iの触媒ドメインとからなる融合タンパク質(図13)の発現のために作製した。得られた合成遺伝子を、TEF1プロモーターの転写制御下に配置して、プラスミドpYLTmAXSpGal10hGalTIおよびpYLTmAXSpGal10hGalTI(ADE2ex)を得た。構築方策を図14に示す。
【0124】
プラスミドpYLTmAXSpGal10hGalTI(ADE2ex)をNotI消化した後に、その分泌タンパク質上にGlcNAcMan5GlcNAc2 N−グリカンを生成することが公知であるG040株を形質転換した(実施例3を参照されたい)。形質転換体を、それらのアデニン原栄養性について選択した。これらのクローンのいくつかのセクレトームについてのN−グリコシル化プロファイルの分析は、N−グリカンパターンの変化を示した:Man7GlcNAc2とMan8GlcNAc2との間の位置に泳動される新しいピークが出現する(図15)。N−グリカンを、Streptococcus pneumoniaβ−1,4−ガラクトシダーゼで処理すると、このピークがGalGlcNAcMan5GlcNAc2を表すことが示される。なぜなら、このin vitro消化は、この新しいピークの消失と、GlcNAcMan5GlcNAc2の等しく高い増加とをもたらすからである。
【0125】
この設定を用い、かつ増殖条件に依存して、GlcNAcMan5GlcNAc2の約75%がGalGlcNAcMan5GlcNAc2に変換された。ガラクトシル化構造の全量は、全N−グリカンプールの約25%を占めた。in vitroα−1,2−マンノシダーゼ消化から、しかし、相当量の高マンノースN−グリカンがMan5GlcNAc2に変換されなかったことが明確である(図15)。用いる培養培地に依存して、Man5GlcNAc2からGlcNAcMan5GlcNAc2への変換率も、親のG040株で観察されるものより低い。このことは、このMnn2−Gal10−GalT I融合タンパク質の形質転換体について観察されたより緩慢な増殖速度と関連する可能性が高い。
【0126】
(実施例6)YlALG3のノックアウトおよびYlALG6の同時過剰発現
Man3GlcNAc2プラットフォームの作製を可能にするために、G036株のALG3遺伝子(pold lnuga Δoch1+Hp4d駆動型α−1,2−マンノシダーゼ)は、不活性化する必要がある。このことはER局在化Alg3pα−1,6−マンノシルトランスフェラーゼ活性の喪失をもたらし、脂質結合N−グリカン前駆体構造の組成を変化させる。この構造を新生ポリペプチド鎖のN−グリコシル化部位に移動させることにより、酵母グリコシル化プロファイルを、哺乳類様N−グリカン構造に、マンノシダーゼIIの発現を必要とすることなく変換できる。しかし、この新しい脂質結合構造は、新生ポリペプチドにそれほど効率的に移動されないので、潜在的なタンパク質の低グリコシル化(underglycosylation)をできる限り減らすために、Yarrowia ALG6遺伝子(ER局在化Alg6pα−1,3−グルコシルトランスフェラーゼをコードする)を同時に過剰発現させる必要がある。
【0127】
pYLalg3PUT−ALG6と称するベクター(図16)が以前に構築され、YlALG3のノックアウトおよびそれと同時のYlALG6のHp4d駆動型過剰発現を可能にした。米国特許出願公開第20090069232−A1号を参照されたい。このノックアウト/ノックインカセットを含有するこのベクターのNotI/PacI断片でYarrowia lipolytica G036を形質転換し、形質転換体を、それらのウラシル原栄養性に基づいて選択した。構築物が正しく組み込まれたクローンを、セクレトームについてのN−グリカン分析により直接スクリーニングした。80のスクリーニングしたクローンのうち、2つのクローンが、ERにあるα−1,2−マンノシダーゼを発現する株におけるYlALG3の不活性化に一致し得るN−グリコシル化プロファイルを示した。少量のMan3GlcNAc2グリカンとは別に、まだいくらかのMan4’GlcNAc2およびMan5’GlcNAc2と、相当量のグルコシル化N−グリカン(GlcMan5’GlcNAc2およびGlc2Man5’GlcNAc2)とがあった。後者は、グルコシダーゼIIによる非効率的な刈り込みの結果である(GrinnaおよびRobbins、J. Biol. Chem. 255巻、2255〜2258頁、1980年)。Man4GlcNAc2およびMan5GlcNAc2の構造の性質は、N−グリカンをα−1,2−マンノシダーゼでin vitro処理することにより確認した(図17)。用いた増殖条件に依存して、Man3GlcNAc2のレベルは、全N−グリカンプールの60%まで増加でき、グルコシル化ピークは、α−1,2−マンノシダーゼに対して非感受性であり、ナタマメα−マンノシダーゼ処理(α−1,2−、α−1,3−およびα−1,6結合マンノース残基に対して作用できる特異的でないα−マンノシダーゼ)に対してわずかに感受性であるだけであった。これとは対照的に、後者の酵素は、生じたMan3GlcNAc2をMan1GlcNAc2に変換する(図17)。
【0128】
2つの陽性形質転換体のうちの1つをG039と称し、さらなる糖鎖工学的改変作業のために用いた。この株を、Cre−リコンビナーゼを発現するpRRQ2で一過的に形質転換して、G036をベクターpYLalg3PUT−ALG6で形質転換した際に導入されたURA3マーカーのキュアリングを可能にした。分析は、グリコシル化プロファイルが、キュアリング後に同じままであることを示す。1つのキュアした株を、さらなる使用のために選択して、G045と称した。
【0129】
(実施例7)Man3GlcNAc2生成株でのGlcNAcトランスフェラーゼIの発現
実施例3で行ったことと同様に、GnT I活性の導入を、Kre2−GnT I融合タンパク質の発現により遂行した。GnT Iについてのこのような発現構築物の無作為組み込みは、3つの方法で遂行した:1)キュアしていないG039株(実施例6を参照されたい)を、NotI消化ベクターpYLTmAXhGnTI(Hygr ex)で形質転換し、GnT I発現クローンを、選択プレートに加えられた300μg/mlのハイグロマイシンで生存するそれらの能力に基づいて最初に選択した、2)キュアしたG045株(実施例6を参照されたい)を、NotI消化ベクターpYLTmAXhGnTI(これもまた実施例3を参照されたい)で形質転換し、GnT I発現クローンを、それらのウラシル原栄養性に基づいて最初に選択した、または3)キュアしたG045株(実施例6を参照されたい)を、NotI消化ベクターpYLHp4mAXhGnTIで形質転換し、GnT I発現クローンを、それらのウラシル原栄養性に基づいて最初に選択した。pYLTmAXhGnTI(Hygr ex)についての構築方策を、図18に示す。プラスミドpYLTmAXhGnTI(Hygr ex)およびpYLTmAXhGnTIを用いる場合、GnT Iの発現は、TEF1プロモーターの転写制御下であった;プラスミドpYLHp4mAXhGnTIを用いる場合、GnT I発現は、Hp4dプロモーターの制御下であった。
【0130】
G039をpYLTmAXhGnTI(Hygr ex)で形質転換することにより、期待されたよりも長いインキュベーション期間の後に、培養プレート上に3つのクローンだけが出現した。しかし、これらのクローンのセクレトームのN−グリコシル化プロファイルの分析は、N−グリカンパターンの明確な変化を示した:非形質転換G039株に存在するMan3GlcNAc2は、著しく低減されたかまたはほぼ完全に存在しなかったが、より高い分子量(約1グルコース単位過剰)を有するN−グリカンを表す新しいピークが出現した。単離N−グリカンを、末端β結合GlcNAc残基を除去できる酵素であるナタマメβ−Ν−アセチルヘキソサミニダーゼで処理すると、新しいN−グリカンが実際にGlcNAcMan3GlcNAc2であることが示された。新しいピークは消失し、Man3GlcNAc2に完全に変換された(図19)。評価した形質転換体のうちの1つを、さらなる糖鎖工学的改変作業のために用い、G047と命名した。同様の結果が、キュアしたG045株をpYLTmAXhGnTI(G048)またはpYLHp4mAXhGnTI(G056)で形質転換した場合にも得られた。G056株は、プラスミドpRRQ2を用いるCreリコンビナーゼの一過的発現によるキュアリングのために選択した。得られた株は、G058と称した。
【0131】
用いた培養方法に依存して、G047株の全N−グリカンプールの約70%がGlcNAcMan3GlcNAc2であり、Glc1−2Man5’GlcNAc2がいくらか残存し、Man3GlcNAc2がほとんど存在しないことが証明された(変換率>>90%)(図19)。高い変換率にかかわらず、1コピーのGnT I発現カセットだけがこの株でサザンブロットにより同定できた。サザン分析は、BamHI消化ゲノムDNAについて、プライマー5’−GGATGATCACACAATGGCCCTGTTTCTG−3’(配列番号11)および5’−TGCTCTAGACTAGTTCCAAGAGGGGTC−3’(配列番号12)を用いて作製したDIG標識GnT I特異的PCR断片を用いて行った。
【0132】
代替の方策では、構築物JME925 pPTAde2−URA3ex−Hp4dhGnTIを作製して、YarrowiaゲノムのADE2遺伝子座へのHp4d駆動型GnT I発現カセットの標的化組み込みを可能にした。構築方策を図20に示す。G045株への形質転換の前に、プラスミドをNotI消化し、ターゲティング/発現カセットを単離した。形質転換体を、それらのアデニン原栄養性に基づいて選択した。ADE2遺伝子座への発現カセットの正しい組み込みは、順方向プライマーVerlAde2(5’−CGACGATAGAGCAGGTCTCACTGTTGGGAATGCTG−3’、配列番号13)、逆方向プライマーVer2Ade2(5’−CTACACTGACGAAGTGGACATCCCGGCTTGGACTG−3’、配列番号14)を用いるPCRにより確かめ、サザンブロッティングによりさらに確認した。これは、BamHI/SpeI消化ゲノムDNAについて、プライマー5’−GGATGATCACACAATGGCCCTGTTTCTG−3’(配列番号15)および5’−TGCTCTAGACTAGTTCCAAGAGGGGTC−3’(配列番号16)を用いて作製したDIG標識GnT I特異的PCR断片を用いて行った。セクレトーム上へのGlcNAcMan3GlcNAc2の合成は、N−グリカン分析およびin vitroナタマメβ−Ν−アセチルヘキソサミニダーゼ処理により確認した(図21)。1つのGnT I発現形質転換体(G057と称する)を、プラスミドpRRQ2を用いるCreリコンビナーゼの一過的発現によるキュアリングのために選択した。得られた株は、G059と称した。
【0133】
(実施例8)GlcNAcトランスフェラーゼIIの発現
Yarrowiaコドン最適化配列を、S.cerevisiae Mnn2タンパク質(SwissProt 受託番号P38069)の最初の36個のN末端アミノ酸と、それに続くラットGlcNAcトランスフェラーゼII(GnT II)(SwissProt 受託番号Q09326)の触媒ドメインとからなる融合タンパク質(図22、配列番号17および配列番号18)の発現のために作製した。酵母Mnn2の36個のN末端アミノ酸は、触媒GnT IIドメインのためのゴルジ局在化シグナルとして働く。この様式で、Mnn2−GnT II融合タンパク質が、分泌経路において、Kre2−GnT I(およびMnn2−Man II)融合タンパク質と同じまたはそれより後の位置にさえ局在化されることを確実にして、GlcNAcMan3GlcNAc2をGlcNAc2Man3GlcNAc2に変換できた。融合タンパク質の発現のための合成遺伝子を、TEF1プロモーターの転写制御下に配置して、プラスミドpYLTmAXrGnTIIおよびpYLTmAXrGnTII(ADE2ex)を得た。構築方策を図23に示す。
【0134】
GnT II活性を発現する株を、2つの異なる様式で作製した:1)G045株(実施例6を参照されたい)を、NotI消化pYLTmAXhGnTIおよびNotI消化pYLTmAXrGnTII(ADE2 ex)で同時に形質転換し、形質転換体を、それらのウラシルおよびアデニン原栄養性に基づいて選択した、または2)G047株(実施例7)を、NotI消化pYLTmAXrGnTII(ADE2 ex)で形質転換し、形質転換体を、それらのアデニン原栄養性に基づいて選択した。発現カセットの組み込みを、順方向プライマーTefPromFW5’−GTCCCCGAATTACCTTTCC−3’(配列番号19)および逆方向プライマーLip2TermRV5’−AGGTAGAAGTTGTAAAGAGTG−3’(配列番号20)を用いて確かめた。ナタマメβ−N−アセチルヘキソサミニダーゼでの単離糖のin vitro処理と組み合わせたセクレトームについてのN−グリカン分析は、いくつかの形質転換体が、GlcNAc2Man3GlcNAc2を生成でき、よって機能的GnT II活性を発現できたことを示した(図24)。ある選択された条件では、全N−グリカンプールの約40%が、GlcNAc2Man3GlcNAc2で構成された。基質GlcNAcMan3GlcNAc2からGlcNAc2Man3GlcNAc2への変換率は、90%であった。最終的に選択した株をG050(G045の二重形質転換)およびG051(G047でのGnT II発現)と称した。
【0135】
(実施例9)グルコシダーゼIIアルファおよびベータサブユニット(Gls2αおよびGls2β)の発現
実施例6〜8に記載した実験に基づいて、YlALG3のノックアウトとそれと同時のYlALG6の過剰発現とを含む方策は、1または2つの末端グルコース残基を保持するN−グリカン(Glc1−2Man5’GlcNAc2)の産生をもたらす。これらのグルコース残基の存在は、ER局在化HDELタグ付きα−1,2−マンノシダーゼによるMan3GlcNAc2への変換を妨げる。グルコース残基を除去するために、ER内でのグルコシダーゼII活性を増大させる必要がある。α−1,2−マンノシダーゼ発現がないバックグラウンドでは、Aspergillus nigerグルコシダーゼIIアルファおよびベータサブユニットの過剰発現が、Glc1−2Man5’GlcNAc2からMan5’GlcNAc2への最高の変換をもたらした(米国特許出願公開第20090069232−A1号)。A.niger gls2サブユニットの過剰発現のための構築物を、以下のようにして生成した:1)Yarrowiaコドン最適化cDNAを、成熟(シグナルペプチドを欠く)A.niger gls2αおよびgls2βサブユニットの発現のために作製し、2)cDNAをインフレームでY.lipolytica LIP−2プレ配列にクローニングし、3)得られたLIP2プレgls2αおよびLIP2プレgls2β配列を、構成性TEF1プロモーターの転写制御下にクローニングした。得られたプラスミドを、pYLTUXdL2preAnGlcIIαおよびpYLeu2ExTEFpreLip2AnGlucIIβと称した(図25)。
【0136】
これらのプラスミドに基づいて、A.niger gls2αおよびgls2βサブユニットを、TEF1プロモーター制御(標的化組み込みのためのベクターJME923 pPTura3−LEU2ex−TefL2preAnGlcIIa+b[altl]、図26)またはHp4dプロモーター制御(標的化組み込みのためのベクターJME923 pPTura3−LEU2ex−Hp4dL2preAnGlcIIa+b[altl]および無作為組み込みのためのベクターZeta−LEU2ex−Hp4dL2preAnGlcIIa+b[alt]、図27)のいずれかの下で同時過剰発現するために、新しい構築物を作製した。
【0137】
G057株(実施例7を参照されたい)を、NotI消化プラスミドJME923 pPTura3−LEU2ex−Hp4dL2preAnGlcIIa+b[altl]およびZeta−LEU2ex−Hp4dL2preAnGlcIIa+b[alt]で形質転換し、形質転換体を、それらのロイシン原栄養性に基づいて選択した。いくつかのクローンを、PCRおよびサザン分析によりゲノム学的に分析して、gls2αおよびgls2β発現カセットの組み込みを評価した。gls2αサブユニットについてのPCR分析およびDIGプローブ作製は、プライマーAnGls2α−FW(5’−GCTGGACTCTTCTTCTATCC−3’)(配列番号24)およびAnGls2α−RV(5’−GGTCTCCTTCAGAGACAGG−3’)(配列番号25)を用いて行った。gls2βサブユニットについて、本発明者らは、プライマーAnGls2β−FW(5’−CCAAGTTCTACAAGGACACC−3’)(配列番号26)およびAnGlc2β−RV(5’−CCCTTGACGACCTTAGAGG−3’)(配列番号27)を用いた。二重Hp4dGls2α/β発現カセットの標的化組み込みを確認するためのサザン分析を、gls2αプローブを用いる場合にEco47III消化gDNAについて、およびgls2βプローブを用いる場合にSpeI/SfiI消化gDNAについて行った。選択したクローンの大多数は、URA3遺伝子座への二重発現カセットの正しい組み込みを示した。二重Hp4dGls2α/β発現カセットの無作為組み込みについてのサザン分析は、両方のプローブを用いてPvuI消化gDNAについて確かめた。全ての場合において、1コピーの二重発現カセットだけが組み込まれていた。
【0138】
次に、N−グリカン分析を、二重Hp4dGls2α/β発現カセットが(正しく)組み込まれていることが確認されたいくつかのクローンについて行った。N−グリコシル化を、3日間のファルコン培養の後の全分泌タンパク質について検査した。いくつかのクローンは、グルコシル化糖の著しい低減と、Man3GlcNAc2およびGlcNAcMan3GlcNAc2の増加とを示した。二重発現カセットが一方で無作為に組み込まれたクローン(=G060株)と、他方で標的化された様式で組み込まれたクローン(=G061株)のプロファイルを、図28に示す。2つのより小さいピークは、Man4’GlcNAc2およびMan5’GlcNAc2を表す。なぜなら、これらは、α−1,2−マンノシダーゼおよびナタマメマンノシダーゼでの処理の際にMan3GlcNAc2resp.Man1GlcNAc2にシフトするからである。後者の処理は、残存Glc1−2Man5’GlcNAc2からGlc1−2Man4’GlcNAc2、およびGlcNAcMan3GlcNAc2からGlcNAcMan2GlcNAc2への部分的変換ももたらす。Man4’GlcNAc2およびMan5’GlcNAc2の存在は、しかし、異種的に(heterologously)同時発現されたHDELタグ付きα−1,2−マンノシダーゼによる、Man3GlcNAc2への不完全な変換を示す。同様に、Man3GlcNAc2の存在は、GlcNAcMan3GlcNAc2を得るための組換えヒトGnT IによるGlcNAc残基の不完全な移動を示す。しかし、上記の結果に基づいて(例えば実施例7でのG047培養、図19)、培養条件の違いは、変換率を著しく増加でき、よって、終末の結果を改善できることが明確である。
【0139】
(実施例10)GlcNAcMan3GlcNAc2生成G061株におけるGlcNAcトランスフェラーゼIIの発現
実施例8で記載したようにして、Yarrowiaコドン最適化配列を、S.cerevisiae Mnn2タンパク質(SwissProt 受託番号P38069)の最初の36個のN末端アミノ酸と、それに続くラットGlcNAcトランスフェラーゼII(GnT II)(SwissProt 受託番号Q09326)の触媒ドメインとからなる融合タンパク質(図22、それぞれ配列番号17および配列番号18)の発現のために作製した。酵母Mnn2の36個のN末端アミノ酸は、触媒GnT IIドメインのためのゴルジ局在化シグナルとして働く。この様式で、Mnn2−GnT II融合タンパク質が、分泌経路においてKre2−GnT I融合タンパク質と同じまたはそれより後の位置にさえ局在化されることを確実にして、その結果、GlcNAcMan3GlcNAc2をGlcNAc2Man3GlcNAc2に変換できた。融合タンパク質の発現のための合成遺伝子を、Hp4dプロモーターの転写制御下に配置して、プラスミドpYLHp4mAXrGnTIIを得て、これを、YarrowiaゲノムへのHp4d駆動型GnT II発現カセットの無作為組み込みのために用いた。代替の方策では、構築物OXYP289 pPTAxpl−ADE2ex−Hp4dhGnTIIを作製して、YarrowiaゲノムのAXP1遺伝子座へのHp4d駆動型GnT II発現カセットの標的化組み込みを可能にした。
【0140】
G061株(実施例9を参照されたい)の形質転換の前に、プラスミドをNotI消化し、ターゲティング/発現カセットを単離した。形質転換体を、それらのアデニン原栄養性に基づいて選択した。ADE2遺伝子座への発現カセットの正しい組み込みは、ゲノムDNAをXmnIで消化した後のサザンブロット分析により確認した。GnT IIコード配列に対する特異性を有するDIG標識プローブを、順方向プライマーrGnTII−FW(5’−GACCAGATGCTGCGAAACG−3’)(配列番号28)および逆方向プライマーrGnTII−RV(5’−CTTGACGTCCACCTTGTCG−3’)(配列番号29)を用いて作製した。この方策により、遺伝子がAxp1遺伝子座にうまく組み込まれる場合には、3172bpのバンドが生成される。
【0141】
代替の方策では、Axp1遺伝子座への正しい組み込みは、順方向プライマーAXPVer1b(5’−GCCTGAACGGCACGATGCGATCGTGGCAATCC−3’)(配列番号30)および逆方向プライマーAXPVer2b(5’−CAAGAAGCCTCAGGCTCGGCGAATCTCCATC−3’)(配列番号31)を用いるゲノムDNAについてのPCR反応により検査できる。Axp1遺伝子座へ正しくターゲティングされた場合は、6489bpのPCR断片が予測される。
【0142】
セクレトームについてのN−グリカン分析と、それと組み合わせた単離糖のナタマメβ−Ν−アセチルヘキソサミニダーゼまたはT.reeseiα−1,2−マンノシダーゼでのin vitro処理は、いくつかの形質転換体がGlcNAc2Man3GlcNAc2を生成でき、よって機能的GnT II活性を発現できることを示した(図29)。その分析は、全N−グリカンプールの約25〜30%が、GlcNAc2Man3GlcNAc2からなり、GlcNAcMan3GlcNAc2からGlcNAc2Man3GlcNAc2への変換率が約90%であることを示した。最終的に選択した株をG070(G061へのpYLHp4mAXrGnTIIの組み込み)およびG071(G061へのOXYP289 pPTAxp1−ADE2ex−Hp4dhGnTIIの組み込み)と称した。
【0143】
(実施例11)Yarrowia lipolyticaへの抗HER2重鎖(HC)および軽鎖(LC)の同時Hp4d駆動型発現のためのタンデムプラスミドの構築
抗HER2抗体重鎖および軽鎖についてのアミノ酸配列を、Carterら、Proc Natl Acad Sci USA.、89巻(10号):4285〜4289頁(1992年);およびWardら、Appl Environ Microbiol、70巻(5号):2567〜2576頁(2004年)から得た。関連するアミノ酸配列は、逆翻訳され(reverse translated)、Yarrowia lipoliticaについてコドン最適化され、GenArt、Regensburg Germanyにより合成された。非常に高い(>80%)または非常に低い(<30%)GC含量の領域は、可能であれば回避した。最適化プロセス中に、以下のシス作用性配列モチーフは回避した:内部TATAボックス、カイ部位およびリボソーム進入部位、ATリッチまたはGCリッチ配列ストレッチ、反復配列およびRNA 2次構造ならびに(潜在的)スプライス供与部位および受容部位。異所性タンパク質の分泌を可能にするために、Lip2タンパク質「プレプロ」シグナルのコード配列(ペプチドリンカー「GGG」のコード配列が後に続く)を、コード配列の5’領域に付加した。「GGG」を、正しいKex2プロセシングのための変化を増進するために付加した。図30Aは、合成プレプロLip2−LCのヌクレオチド配列(=750bp)(配列番号32)を含有する。図30Bは、プレプロLip2−LCのアミノ酸配列(=250Aa;MW=27.011Da;pI=8.46)(配列番号33)を含有する。図31Aは、合成プレプロLip2−HCのヌクレオチド配列(=1458bp)(配列番号34)を含有する。図31Bは、プレプロLip2−HCのアミノ酸配列(=486Aa;MW=52.853Da;pI=8.65)(配列番号35)を含有する。プレプロLip2−HCおよび−LCについてのコード配列を、pYLHp4L2preproHerHC/LC(GUT2ex)−ori2と称する同じベクターに導入した。
【0144】
(実施例12)様々な程度の糖鎖工学的改変によるYarrowia lipolytica株での抗HER2抗体HCおよびLCの発現
プラスミドpYLHp4L2preproHerHC&LC(GUT2ex)−ori2をNotIで消化し、HC−/LC−タンデム発現カセットを単離した後に、Yarrowia lipolytica G045株、G057株、G061株およびG071株(表2を参照されたい)を形質転換した。無作為に組み込まれたHC−/LC−発現カセットを含有する形質転換体を、グリセロールを唯一の炭素源として増殖するそれらの能力に基づいて選択した。HCおよびLCの発現解析は、選択した形質転換体を、グリセロールを唯一の炭素源として含有するリッチ培地(SuperT/グリセロール培地:0.5%酵母抽出物;2%麦芽抽出物;1%トリプトン;1.5%グリセロール;200mMリン酸塩緩衝液pH6.8)中で4日間振とうフラスコ培養した後にウェスタンブロッティングにより行った。LC検出は、カッパ遊離軽鎖に対するマウスモノクローナル(4C11)(Abeam)を用いて行い、HC検出は、マウスモノクローナル抗ヒトIgG(γ鎖特異的)(Sigma)を用いて行った。
【0145】
抗HER2抗体生成株のセクレトームのN−グリカンは、いずれのHCおよびLCも発現しない対応する糖鎖工学的に改変した株と同様のプロファイルを示した(図32)。G045、G057、G061およびG071バックグラウンドを有する株でのN−グリカンのパーセンテージは、SuperT/グリセロール培地中での6日間の振とうフラスコ培養の後に決定した。G045バックグラウンドでは、54.6%のN−グリカンがMan3GlcNAc2であった。G057バックグラウンドでは、47.5%のN−グリカンがGlcNAc1Man3GlcNAc2であった。G061バックグラウンドでは、58.9%のN−グリカンがGlcNAc1Man3GlcNAc2であった。G071バックグラウンドでは、37.6%のN−グリカンがGlcNAc2Man3GlcNAc2であった。
【0146】
(実施例13)抗HER2抗体HCおよびLCを発現するGlcNAc2Man3GlcNAc2合成株であるYarrowia G096株の発酵
GlcNAc2Man3GlcNAc2を合成できる株であるYarrowia lipolytica G071のいくつかのpYLHp4L2preproHerHC&LC(GUT2ex)−ori2形質転換体を、HCおよびLC発現レベルについて分析した。これらのクローンのうちの1つであるG096を、さらなる分析のために選択した。
【0147】
発酵は、プロセス制御および管理システム(Lucillus PIMS)を備える14リットル撹拌タンクバイオリアクタ(MAVAG AG)で行った。培地中の相対的酸素分圧、排出ガス中のCO2とO2の濃度、pH値、温度、リアクタ過圧、リアクタ重量、フィード重量および基本重量(base weight)は全てオンラインで監視した。泡の発生は、消泡剤ポリプロピレングリコール(PPG)を加えることにより妨いだ。pHの調整は、25%アンモニア溶液または8.5%リン酸溶液のいずれかの添加により行った。
【0148】
G096の種培養物を28℃にて、リッチ培地を含有する振とうフラスコで増殖させた。種培養物を、無機質培地を含有する発酵槽に接種して、グリセロールを唯一の炭素源として用いて、回分段階(batch phase)を28℃にて無制限増殖で開始した。この段階は、高いバイオマス濃度に迅速に到達するために用いた。この時点以降、プロセスを、0.02の定常増殖速度の指数グリセロール流加(fed batch)(グリセロールを唯一の炭素源およびエネルギー源として用いる;pH6)にシフトした。例として、28℃での流加発酵の結果を以下に記載する。
【0149】
流加段階は、148時間継続した。発酵の異なる時点で試料を採取して、以下のパラメータを追跡した:1)ウェスタンブロットによるLCおよびHCタンパク質主鎖の発現;2)ELISAによる機能的抗HER2抗体の発現;ならびに3)セクレトームのN−グリコシル化プロファイルの進展。全長HC発現レベルは、時点7(39時間)付近で最大に到達し、それ以降ほぼ等しいままである。LC発現は、時点7(39時間)と10(73時間)の間で最大に到達したが、その後の時点で幾分か減少した。いくらかのLC二量体が、時点5(25時間)と9(62時間)の間で生成されたが、これもまたその時点以降で消失した。
【0150】
機能的ELISAを開発して、少なくとも1つの機能的抗原結合ドメインを有する抗HER2抗体の生成を測定した。プレートを、天然HER2抗原の組換えバリアントである組換えヒトErbB2/Fcキメラ(R&D systems)でコーティングした。次いで、異なる時点で採集した培地の希釈物を、コーティングしたプレートに加えた。抗原結合タンパク質の量の評価を、HRPコンジュゲート抗ヒトカッパLC抗体(Sigma)を用いて行った。流加発酵でのErbB2/Fcキメラ結合タンパク質の量の進展(分泌された機能的抗HER2抗体の量の指標)を、図33に示す。データは、抗HER2抗体のレベルの漸増と、生成段階の終末での10〜12mg/Lの最大値とを示す。
【0151】
N−グリカン分析を、流加発酵中のいくつかの時点で採取した試料について行った。結果を図34Aおよび34Bに示す。流加段階の開始時に、相当量のグルコース含有N−グリカンが存在した。時点6以降から(指数的栄養補給の開始後34時間)、グルコシル化N−グリカンのレベルは著しく減少し、採集の時点(時点18、148時間)ではほとんど残っていなかった。このことは、グルコース含有N−グリカンを元来保持するタンパク質が、発酵の終末に薄められことを示した。この時点で、セクレトームから単離されたN−グリカンの約86%が、構造GlcNAc2Man3GlcNAc2を有した。
【0152】
その他の実施形態
本発明を、その詳細な説明と関連付けて記載したが、上記の記載は、例示することを意図し、本発明の範囲を限定することを意図しない(本発明の範囲は、添付の特許請求の範囲により定義される)。その他の態様、利点および改変は、以下の特許請求の範囲内にある。
【技術分野】
【0001】
関連する出願への相互参照
この出願は、2009年11月19日に出願された米国出願第61/262,828号に対する優先権を主張する。この前の出願の開示は、その全体が参考として援用される。
【0002】
技術分野
本発明は、真菌細胞において糖タンパク質を生成するための方法および材料、より具体的には、真菌細胞を遺伝子工学的に改変して、哺乳類様複合N−グリカンを含有するタンパク質または哺乳類グリコシル化経路内の中間体を含有するタンパク質を生成することに関する。
【背景技術】
【0003】
現在開発中のほとんどの生物製剤(例えば組換えタンパク質)を生成するために、高性能の発現系が要求される。これらの生物製剤の多くの生物活性は、それらの翻訳後修飾(例えばリン酸化またはグリコシル化)に依存する。酵母ベースの発現系は、微生物の遺伝子操作および発酵の容易さを、タンパク質を分泌し、修飾する能力と組み合わせる。しかし、酵母細胞で生成される組換え糖タンパク質は、主に不均質な高マンノースおよび過マンノースグリカン構造を示し、これはタンパク質機能、下流のプロセシングおよび後続の治療のための使用にとって、特にグリコシル化が生物学的に重要な役割を演じる場合に有害であり得る。
【発明の概要】
【課題を解決するための手段】
【0004】
本明細書で記載する方法および遺伝子工学的に改変された真菌細胞は、哺乳類様N−グリカンを含有するかまたは哺乳類(例えばヒト)グリコシル化経路内の中間体を含有する標的分子(例えば標的タンパク質)を生成するために用いることができる。このような工学的に改変された細胞から単離された標的分子は、抗体生成、サイトカイン生成を含む生物製剤の用途、およびリソソーム蓄積症のような代謝障害の処置のために用いることができる。
【0005】
一態様では、この文書は、GlcNAcMan5GlcNAc2 N−グリカンを含むタンパク質を生成できる真菌細胞(例えばYarrowia lipolyticaまたはArxula adeninivorans)を生成する方法を特徴とする。方法は、Man5GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された真菌細胞を提供するステップと、細胞に、GlcNAcトランスフェラーゼIをコードする核酸を導入するステップであって、ここで、核酸が、コードされたGlcNAcトランスフェラーゼIを細胞内区画(例えばゴルジ装置)に向かわせる(target)ためのヌクレオチドターゲティング配列を含み、ここで、真菌細胞におけるGlcNAcトランスフェラーゼIの発現が、GlcNAcMan5GlcNAc2 N−グリカンを含むタンパク質を生成するステップとを含む。方法は、細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、細胞は、GlcNAcMan5GlcNAc2 N−グリカンを含むように改変された標的タンパク質を生成するステップをさらに含むことができる。標的タンパク質は、Fc受容体と結合できる。標的タンパク質は、抗体またはその断片であり得る。標的タンパク質は、治療用の糖タンパク質であり得る。標的タンパク質は、インターフェロン−β、GM−CSF、インターフェロンγまたはエリスロポエチンであり得る。
【0006】
Man5GlcNAc2 N−グリカンを含有するタンパク質を生成するように遺伝子工学的に改変された真菌細胞は、OCH1活性を欠損でき、α−1,2−マンノシダーゼをコードする核酸を含み、ここで、α−1,2−マンノシダーゼをコードする核酸は、コードされたα−1,2−マンノシダーゼを小胞体に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含む。ターゲティング配列は、HDEL配列であり得る。
【0007】
方法は、細胞に、マンノシダーゼIIをコードする核酸を導入するステップであって、ここで、マンノシダーゼIIをコードする核酸は、コードされたマンノシダーゼIIをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるマンノシダーゼIIの発現は、GlcNAcMan3GlcNAc2 N−グリカンを含有するタンパク質を生成するステップをさらに含むことができる。
【0008】
方法は、細胞に、ガラクトシルトランスフェラーゼをコードする核酸を導入するステップであって、ここで、ガラクトシルトランスフェラーゼをコードする核酸は、コードされたガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるガラクトシルトランスフェラーゼの発現は、GalGlcNAcMan5GlcNAc2またはGalGlcNAcMan3GlcNAc2 N−グリカンを含有するタンパク質を生成するステップをさらに含むことができる。ガラクトシルトランスフェラーゼは、UDP−Glc−4−エピメラーゼとβ−1,4−ガラクトシルトランスフェラーゼIの触媒ドメインとの融合体であり得る。このような方法は、細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、細胞は、GalGlcNAcMan5GlcNAc2またはGalGlcNAcMan3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質を生成するステップをさらに含むことができる。方法は、GalGlcNAcMan5GlcNAc2またはGalGlcNAcMan3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質を単離するステップを含むことができる。
【0009】
別の態様では、この文書は、GlcNAcMan3GlcNAc2 N−グリカンを含有する標的タンパク質を生成する方法を特徴とする。方法は、GlcNAcトランスフェラーゼIと、α−1,2−マンノシダーゼと、マンノシダーゼIIとをコードする核酸を含むように遺伝子工学的に改変された真菌細胞(例えばYarrowia lipolyticaまたはArxula adeninivorans)を提供するステップであって、ここで、核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるための1つのターゲティング配列をコードする1つのヌクレオチド配列または複数のターゲティング配列をコードする複数のヌクレオチド配列を含み、ここで、真菌細胞が、OCH1活性を欠損しているステップと、細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、細胞が、GlcNAcMan3GlcNAc2 N−グリカンを含有する標的タンパク質を生成するステップとを含む。α−1,2−マンノシダーゼをコードする核酸は、コードされたα−1,2−マンノシダーゼを小胞体に向かわせるための小胞体ターゲティング配列を含むことができる。例えば、ターゲティング配列は、HDEL配列であり得る。GlcNAcトランスフェラーゼIおよびマンノシダーゼIIをコードする核酸は、コードされたGlcNAcトランスフェラーゼIおよびマンノシダーゼIIをゴルジ装置に向かわせるための1つ以上のゴルジターゲティング配列を含むことができる。標的タンパク質は、Fc受容体と結合できる。標的タンパク質は、抗体またはその断片であり得る。標的タンパク質は、治療用の糖タンパク質であり得る。標的タンパク質は、インターフェロン−β、GM−CSF、インターフェロンγまたはエリスロポエチンであり得る。
【0010】
いくつかの実施形態では、方法は、細胞に、ガラクトシルトランスフェラーゼをコードする核酸を導入するステップであって、ここで、ガラクトシルトランスフェラーゼをコードする核酸は、コードされたガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるガラクトシルトランスフェラーゼの発現は、GalGlcNAcMan3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質を生成するステップをさらに含むことができる。GalGlcNAcMan3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質は、真菌細胞から単離できる。
【0011】
この文書は、GlcNAcMan3GlcNAc2 N−グリカンを含有するタンパク質を生成できる真菌細胞(例えばYarrowia lipolyticaまたはArxula adeninivorans)を作製する方法も特徴とする。方法は、Man3GlcNAc2 N−グリカンを含有するタンパク質を生成するように遺伝子工学的に改変された真菌細胞を提供するステップと、細胞に、GlcNAcトランスフェラーゼIをコードする核酸を導入するステップであって、ここで、核酸が、コードされたGlcNAcトランスフェラーゼIを細胞内区画(例えばゴルジ装置)に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるGlcNAcトランスフェラーゼIの発現が、GlcNAcMan3GlcNAc2 N−グリカンを含有するタンパク質を生成するステップとを含む。方法は、細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、細胞は、GlcNAcMan3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質を生成するステップをさらに含むことができる。標的タンパク質は、Fc受容体と結合できる。標的タンパク質は、抗体またはその断片であり得る。標的タンパク質は、治療用の糖タンパク質であり得る。標的タンパク質は、インターフェロン−β、GM−CSF、インターフェロンγまたはエリスロポエチンであり得る。
【0012】
Man3GlcNAc2 N−グリカンを含有するタンパク質を生成するように遺伝子工学的に改変された真菌細胞は、ALG3活性を欠損でき、α−1,2−マンノシダーゼをコードする核酸を含み、ここで、核酸は、コードされたα−1,2−マンノシダーゼを小胞体に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含む。このような真菌細胞は、OCH1活性をさらに欠損でき、かつ/またはα−1,3−グルコシルトランスフェラーゼ(例えばALG6)をコードする核酸をさらに含むことができる。
【0013】
方法は、細胞に、GlcNAcトランスフェラーゼIIをコードする核酸を導入するステップであって、ここで、GlcNAcトランスフェラーゼIIをコードする核酸は、コードされたGlcNAcトランスフェラーゼIIを細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるGlcNAcトランスフェラーゼIIの発現が、GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成するステップをさらに含むことができる。
【0014】
方法は、細胞に、ガラクトシルトランスフェラーゼをコードする核酸を導入するステップであって、ここで、ガラクトシルトランスフェラーゼをコードする核酸は、コードされたガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるガラクトシルトランスフェラーゼの発現は、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成するステップをさらに含むことができる。ガラクトシルトランスフェラーゼは、UDP−Glc−4−エピメラーゼと、β−1,4−ガラクトシルトランスフェラーゼIの触媒ドメインとの融合体であり得る。方法は、細胞に標的タンパク質をコードする核酸を導入するステップであって、ここで、細胞は、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質を生成するステップをさらに含むことができる。
【0015】
方法は、細胞に、グルコシダーゼIIのαおよびβサブユニットをコードする核酸を導入するステップであって、ここで、真菌細胞におけるグルコシダーゼIIのαおよびβサブユニットの発現は、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成するステップをさらに含むことができる。
【0016】
この文書は、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含有する標的タンパク質を生成する方法も特徴とする。方法は、ALG3活性を欠損するように遺伝子工学的に改変され、GlcNAcトランスフェラーゼIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸を含む真菌細胞を提供するステップであって、ここで、GlcNAcトランスフェラーゼIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸が、それぞれのコードされたタンパク質を細胞内区画(例えばゴルジ装置)に向かわせるための1つのターゲティング配列をコードする1つのヌクレオチド配列または複数のターゲティング配列をコードする複数のヌクレオチド配列を含むステップと、細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、細胞が、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含有する標的タンパク質を生成するステップとを含む。真菌細胞は、OCH1活性をさらに欠損でき、かつ/またはALG6のようなα−1,3−グルコシルトランスフェラーゼをコードする核酸をさらに含むことができる。真菌細胞は、グルコシダーゼIIのαおよびβサブユニットをコードする核酸をさらに含むことができ、ここで、真菌細胞におけるグルコシダーゼIIのαおよびβサブユニットの発現は、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含有する標的タンパク質を生成する。
【0017】
別の態様では、この文書は、GlcNAcMan3GlcNAc2 N−グリカンを含有するタンパク質を生成するように遺伝子工学的に改変された単離真菌細胞を特徴とする。真菌細胞は、OCH1活性を欠損でき、α−1,2−マンノシダーゼと、GlcNAcトランスフェラーゼIと、マンノシダーゼIIとをコードする核酸を含み、ここで、α−1,2−マンノシダーゼと、GlcNAcトランスフェラーゼIと、マンノシダーゼIIとをコードする核酸は、それぞれのコードされたタンパク質を細胞内区画に向かわせるための1つのターゲティング配列をコードする1つのヌクレオチド配列または複数のターゲティング配列をコードする複数のヌクレオチド配列を含み、ここで、真菌細胞におけるα−1,2−マンノシダーゼ、GlcNAcトランスフェラーゼIおよびマンノシダーゼIIの発現は、GlcNAcMan3GlcNAc2 N−グリカンを含有するタンパク質を生成する。真菌細胞は、標的タンパク質をコードする核酸をさらに含むことができ、ここで、細胞は、GlcNAcMan3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質を生成する。
【0018】
いくつかの実施形態では、このような真菌細胞は、GlcNAcトランスフェラーゼIIをコードする核酸をさらに含み、ここで、GlcNAcトランスフェラーゼIIをコードする核酸は、コードされたGlcNAcトランスフェラーゼIIを細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるGlcNAcトランスフェラーゼIIの発現は、GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成する。
【0019】
いくつかの実施形態では、このような真菌細胞は、ガラクトシルトランスフェラーゼをコードする核酸をさらに含み、ここで、ガラクトシルトランスフェラーゼをコードする核酸は、コードされたガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるガラクトシルトランスフェラーゼの発現は、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成する。
【0020】
さらに別の態様では、この文書は、GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成するように遺伝子工学的に改変された単離真菌細胞を特徴とする。真菌細胞は、ALG3活性を欠損するように遺伝子工学的に改変され、GlcNAcトランスフェラーゼIとGlcNAcトランスフェラーゼIIとをコードする核酸を含み、ここで、GlcNAcトランスフェラーゼIとGlcNAcトランスフェラーゼIIとをコードする核酸は、それぞれのコードされたタンパク質を細胞内区画に向かわせるための1つのターゲティング配列をコードする1つのヌクレオチド配列または複数のターゲティング配列をコードする複数のヌクレオチド配列を含み、ここで、真菌細胞におけるGlcNAcトランスフェラーゼIおよびGlcNAcトランスフェラーゼIIの発現は、GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成する。遺伝子工学的に改変された真菌細胞は、OCH1活性をさらに欠損でき、かつ/またはα−1,3−グルコシルトランスフェラーゼをコードする核酸をさらに含むことができる。遺伝子工学的に改変された真菌細胞は、標的タンパク質をコードする核酸をさらに含むことができ、ここで、細胞は、GlcNAc2Man3GlcNAc2 N−グリカンを含有するように改変された標的タンパク質を生成する。真菌細胞は、グルコシダーゼIIのαおよびβサブユニットをコードする核酸をさらに含むことができ、ここで、真菌細胞におけるグルコシダーゼIIのαおよびβサブユニットの発現は、GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成する。真菌細胞は、ガラクトシルトランスフェラーゼをコードする核酸をさらに含むことができ、ここで、ガラクトシルトランスフェラーゼをコードする核酸は、コードされたガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、真菌細胞におけるガラクトシルトランスフェラーゼの発現は、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成する。
【0021】
この文書は、相当数のYarrowia lipolytica細胞が、Gal2GlcNac2Man3GlcNAc2 N−グリカンを含有する糖タンパク質を生成するように遺伝子工学的に改変されているYarrowia lipolytica細胞の実質的に純粋な培養物も特徴とする。細胞は、ALG3活性を欠損するように遺伝子工学的に改変され、GlcNAcトランスフェラーゼIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸を含み、ここで、GlcNAcトランスフェラーゼIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸は、それぞれのコードされたタンパク質を細胞内区画に向かわせるための1つのターゲティング配列をコードする1つのヌクレオチド配列または複数のターゲティング配列をコードする複数のヌクレオチド配列を含み、ここで、細胞におけるGlcNAcトランスフェラーゼI、GlcNAcトランスフェラーゼIIおよびガラクトシルトランスフェラーゼの発現は、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含有するタンパク質を生成する。遺伝子工学的に改変された真菌細胞は、OCH1活性をさらに欠損でき、かつ/またはα−1,3−グルコシルトランスフェラーゼ(例えばALG6)をコードする核酸をさらに含むことができる。細胞は、グルコシダーゼIIのαおよびβサブユニットをコードする核酸をさらに含むことができ、ここで、真菌細胞におけるグルコシダーゼIIのαおよびβサブユニットの発現は、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含有する標的タンパク質を生成する。
【0022】
別の態様では、この文書は、相当数のYarrowia lipolytica細胞が、Gal2GlcNac2Man3GlcNAc2 N−グリカンを含有する糖タンパク質を生成するように遺伝子工学的に改変されているYarrowia lipolytica細胞の実質的に純粋な培養物であって、ここで、細胞が、OCH1活性を欠損するように遺伝子工学的に改変され、α−1,2−マンノシダーゼと、GlcNAcトランスフェラーゼIと、マンノシダーゼIIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸を含み、ここで、α−1,2−マンノシダーゼと、GlcNAcトランスフェラーゼIと、マンノシダーゼIIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるための1つのターゲティング配列をコードする1つのヌクレオチド配列または複数のターゲティング配列をコードする複数のヌクレオチド配列を含み、ここで、細胞におけるα−1,2−マンノシダーゼ、GlcNAcトランスフェラーゼI、マンノシダーゼII、GlcNAcトランスフェラーゼIIおよびガラクトシルトランスフェラーゼの発現が、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成する培養物を特徴とする。
【0023】
この文書は、糖タンパク質を含み、ここで、糖タンパク質上のN−グリカンの少なくとも50%(例えば少なくとも70%または少なくとも85%)が、GlcNAc2Man3GlcNAc2 N−グリカンである組成物も特徴とする。
【0024】
そうでないと定義しない限り、本明細書で用いる全ての技術用語および科学用語は、本発明が属する当技術分野における当業者により一般的に理解されるのと同じ意味を有する。本明細書で記載するものと同様または等価な方法および材料を、本発明の実施または試験において用いることができるが、例示的な方法および材料は、以下に記載する。本明細書で言及する全ての出版物、特許出願、特許、Genbank(登録商標)受託番号およびその他の参考文献は、それらの全体が参照により組み込まれている。矛盾がある場合には、定義を含む本出願が支配する。材料、方法および実施例は例示のためだけであり、限定することを意図しない。
【0025】
本発明のその他の特徴および利点は、以下の詳細な記載および特許請求の範囲から明らかになる。
【図面の簡単な説明】
【0026】
【図1】図1Aは、Man5GLcNAc2構造およびMan3GlcNAc2構造の図である。図1Bは、プラスミドpYlOCH1 PUT TOPOの模式図である。
【図2】図2は、pold lnuga Yarrowia lipolytica野生型細胞またはΔoch1 pold lnuga Yarrowia lipolytica細胞から得られた分泌タンパク質のN−グリカン分析を示す一連の電気泳動図(electroferogram)である。OCH1不活性化の際の主なN−グリカンは、Man8GlcNAc2になる。分析は、DNAシーケンサー支援蛍光体支援糖質電気泳動(DSΑ−FACE)を用いて行った。「M5」、「M6」、「M8」および「M9」は、ベースのN−アセチルグルコサミン構造にコンジュゲートしたマンノース残基の数である。Y軸は、各N−グリカン構造の量を示す相対蛍光単位を表す。X軸は、キャピラリーを通しての各N−グリカン構造の相対移動度を表す。先頭の電気泳動図は、移動度標準物質として用いるためのデキストランの分析である。
【図3】図3は、プラスミドpYLHUXdL2preManHDELおよびpYLTUXdL2preManHDELの模式図である。
【図4】図4は、ManHDEL(=HDELタグ付きα−1,2−マンノシダーゼ)発現カセット(TEF1またはHp4dのいずれかのプロモーター制御の下で)をG014株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。「Rd」は、図3に示すベクターに存在するゼータ配列による「無作為組み込み」を表す。マンノシダーゼ発現の際の主要なN−グリカンは、Man5GlcNAc2である。これらの株の1つ(G018、表2を参照されたい)からのURA3マーカーのキュアリング(curing)は、N−グリカンプロファイルを変えない。
【図5】図5は、プラスミドJME926 pPTLeu2−ADE2ex−Hp4dManHDEL(Yl)およびOXYP289 pPTAxp1−LEU2ex−Hp4dManHDEL(Yl)についての構築方策の模式図である。ベクターpYLTmAXrGnTIIの構築については、図23を参照されたい。
【図6】図6は、LEU2またはAXP1遺伝子座のいずれかでの標的化組み込み(Tg)により、ManHDEL(=HDELタグ付きα−1,2−マンノシダーゼ)発現カセット(Hp4dプロモーター制御の下で)をG014株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。Man5GlcNAc2が主なN−グリカンになる。
【図7】図7は、Kre2pの100個のN末端アミノ酸とヒトGlcNAcトランスフェラーゼIの触媒ドメインとの間の融合タンパク質のアミノ酸配列(配列番号3)およびYarrowiaコドン最適化ヌクレオチド配列(配列番号4)を示す図である。太字で:融合タンパク質のKre2p部分;通常のフォントで:融合タンパク質のGnT I部分;下線:開始コドンおよび停止コドン。
【図8】図8は、プラスミドpYLTmAXhGnTIおよびpYLHp4mAXhGnTIについての構築方策の模式図である。
【図9】図9は、GnT Iを発現するベクターでの形質転換によりGnT I活性をG036株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。「Rd」は、図8に示すベクターに存在するゼータ配列による「無作為組み込み」を表す。GnT I活性の発現の際の主要なN−グリカンは、GlcNAcMan5GlcNAc2である。α−1,2−マンノシダーゼでのin vitro処理は、プロファイルを著しく変えず、このことは、Man5GlcNAc2以外の高マンノースN−グリカンは少量しか存在しないことを示す。in vitroヘキソサミニダーゼ処理は、GlcNAcMan5GlcNAc2からMan5GlcNAc2へのシフトをもたらす。
【図10−1】図10は、Mnn2pの36個のN末端アミノ酸とDrosophila melanogasterマンノシダーゼIIの触媒ドメインとの間の融合タンパク質のアミノ酸配列(配列番号7)およびYarrowiaコドン最適化ヌクレオチド配列(配列番号8)を示す図である。太字で:融合タンパク質のMnn2p部分;通常のフォントで:融合タンパク質のManII部分;下線:開始コドンおよび停止コドン。
【図10−2】図10は、Mnn2pの36個のN末端アミノ酸とDrosophila melanogasterマンノシダーゼIIの触媒ドメインとの間の融合タンパク質のアミノ酸配列(配列番号7)およびYarrowiaコドン最適化ヌクレオチド配列(配列番号8)を示す図である。太字で:融合タンパク質のMnn2p部分;通常のフォントで:融合タンパク質のManII部分;下線:開始コドンおよび停止コドン。
【図11】図11は、プラスミドpYLTmAXDmManIIおよびpYLTmAXDmManII(LEU2ex)についての構築方策の模式図である。
【図12】図12は、ManII発現ベクターでの形質転換によりManII活性をG040株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。「Rd」は、図11に示すベクターに存在するゼータ配列による「無作為組み込み」を表す。ManII活性の発現の際に、より高い電気泳動移動度を有する新しいピークならびに、Man5GlcNAc2とほぼ同じ位置に泳動される「ショルダー」ピークが出現する。in vitroヘキソサミニダーゼ処理は、これらのピークについて前へのシフトをもたらし(GlcNAcMan5GlcNAc2からMan5GlcNAc2への観察されたシフトの隣りに)、このことは、末端GlcNAcの存在を示し、よって、GlcNAcMan3GlcNAc2およびGlcNAcMan4GlcNAc2のようなピークが同定される。α−1,2−マンノシダーゼでのin vitro処理は、プロファイルを著しく変化させず、このことは、Man5GlcNAc2以外の高マンノースN−グリカンは少量しか存在しないことを示す。
【図13−1】図13は、Mnn2pの46個のN末端アミノ酸とSchizosaccharomyces pombe UDP−Glc−4−エピメラーゼ様タンパク質とヒトβ−1,4−ガラクトシルトランスフェラーゼIの触媒ドメインとの間の融合タンパク質のアミノ酸配列(配列番号9)およびYarrowiaコドン最適化ヌクレオチド配列(配列番号10)である。融合タンパク質のMnn2p部分は配列番号9の1〜46であり、リンカー配列は47〜49および405〜408であり、融合タンパク質のエピメラーゼ配列は50〜404であり、融合タンパク質のManII部分は409〜763である。Mnn2p部分は配列番号10のヌクレオチド1〜138であり、リンカー配列はヌクレオチド139〜147および1213〜1224であり、エピメラーゼ配列はヌクレオチド148〜1212であり、ManII部分は1225〜2289である。開始コドンおよび停止コドンに下線を付す。
【図13−2】図13は、Mnn2pの46個のN末端アミノ酸とSchizosaccharomyces pombe UDP−Glc−4−エピメラーゼ様タンパク質とヒトβ−1,4−ガラクトシルトランスフェラーゼIの触媒ドメインとの間の融合タンパク質のアミノ酸配列(配列番号9)およびYarrowiaコドン最適化ヌクレオチド配列(配列番号10)である。融合タンパク質のMnn2p部分は配列番号9の1〜46であり、リンカー配列は47〜49および405〜408であり、融合タンパク質のエピメラーゼ配列は50〜404であり、融合タンパク質のManII部分は409〜763である。Mnn2p部分は配列番号10のヌクレオチド1〜138であり、リンカー配列はヌクレオチド139〜147および1213〜1224であり、エピメラーゼ配列はヌクレオチド148〜1212であり、ManII部分は1225〜2289である。開始コドンおよび停止コドンに下線を付す。
【図14】図14は、プラスミドpYLTmAXSpGal10hGalTIおよびpYLTmAXSpGal10hGalTI(ADE2ex)についての構築方策の模式図である。
【図15】図15は、Gal10−GalTI活性をG040株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。得られた形質転換体G044を、2つの異なる培地で培養した。「Rd」は、図14に示すベクターに存在するゼータ配列による「無作為組み込み」を表す。Gal10−GalTI活性の発現の際に、Man7GlcNAc2とMan8GlcNAc2との間の位置に泳動される新しいピークが出現する。in vitroガラクトシダーゼ処理は、このピークについての前へのシフトと、GlcNAcMan5GlcNAc2の等しい増加とをもたらす(後者は、ガラクトシダーゼとヘキソサミニダーゼを用いる二重処理によりこのN−グリカンを表すことが確認される)。このことは、末端ガラクトースの存在を示し、よって、GalGlcNAcMan5GlcNAc2としてのG044プロファイルの新しいピークが同定される。α−1,2−マンノシダーゼでのin vitro処理は、Man5GlcNAc2にまでまだ刈り込まれていない(not yet trimmed)多量の高マンノースN−グリカン(特にMan8GlcNAc2)の存在を示す。
【図16】図16は、プラスミドpYLalg3PUT−ALG6の模式図である。
【図17】図17は、pYLalg3PUT−ALG6をG036株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。ALG6の過剰発現は、相当量のグリコシル化ピーク(GlcMan5’GlcNAc2およびGlc2Man5’GlcNAc2)をもたらし、このことは、新生タンパク質に移されたGlc3Man5’GlcNAc2構造が、グルコシダーゼIIによりMan5’GlcNAc2に完全には刈り込まれていないことを示す。増殖培地に依存して、生じるMan5’GlcNAc2は、ER局在化HDELタグ付きT.reeseiα−1,2−マンノシダーゼの作用によりMan3GlcNAc2に部分的(まだいくらかのMan5’GlcNAc2およびMan4’GlcNAc2)、またはほぼ完全に刈り込まれている。Man5’GlcNAc2およびMan4’GlcNAc2のピークは、α−1,2−マンノシダーゼに対するそれらの感受性により、そのままで同定される。キャッピンググルコースにより、GlcMan5’GlcNAc2およびGlc2Man5’GlcNAc2はこの処理に対して非感受性である。ナタマメマンノシダーゼは、遊離のα−1,6結合マンノースを部分的に除去できるが、これは、Man3−5’GlcNAc2をMan1GlcNAc2に変換もする。
【図18】図18は、プラスミドpYLTmAXhGnTI(Hygr ex)についての構築方策の模式図である。
【図19】図19は、GnT I発現ベクターでの形質転換によりGnT I活性をΔalg3−Hp4dALG6株のキュアしていない(G039)かまたはキュアした(G045)いずれかのバージョンに導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。GlcNAcMan3GlcNAc2の産生は、ヘキソサミニダーゼ消化により証明された。新しいピークは、Man3GlcNAc2に完全にシフトして戻った。G048株では、GlcNAcMan3GlcNAc2への変換は完全でなかった。なぜなら、いくらかのMan3GlcNAc2がまだ観察できたからである。この株は、α−1,2−マンノシダーゼ消化により示されるように、いくらかの残りのMan5’GlcNAc2も有する。
【図20】図20は、プラスミドJME925 pPTAde2−URA3ex−Hp4dhGnTIについての構築方策の模式図である。
【図21】図21は、GnT I活性をΔalg3−Hp4dALG6株のキュアしたバージョン(=G045)に導入し、Hp4d駆動型発現構築物をADE2遺伝子座(Tg−ade2)に組み込んだ後のN−グリカンプロファイルを示す一連の電気泳動図である。この培養では、グリコシル化N−グリカンの量は高く、Man4’−5’GlcNAc2からMan3GlcNAc2への変換は、完全でなかった。Man4’GlcNAc2の隣りに泳動される新しいピークが形質転換体G057で観察され、ヘキソサミニダーゼ消化の結果に基づいてGlcNAcMan3GlcNAc2であると示すことができた。この新しいピークは、Man3GlcNAc2に完全にシフトして戻る。
【図22】図22は、Mnn2pの36個のN末端アミノ酸とラットGlcNAcトランスフェラーゼIIの触媒ドメインとの間の融合タンパク質のアミノ酸配列(配列番号17)およびYarrowiaコドン最適化ヌクレオチド配列(配列番号18)である。太字で:融合タンパク質のMnn2p部分;通常のフォントで:融合タンパク質のGnT II部分;下線:開始コドンおよび停止コドン。
【図23】図23は、プラスミドpYLTmAXrGnTIIおよびpYLTmAXrGnTII(ADE2 ex)についての構築方策の模式図である。
【図24】図24は、GnT II活性を、GlcNAcMan3GlcNAc2を合成する株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。得られた株は、G045をGnTIおよびGnT II発現構築物で二重形質転換するか、またはG047をGnTII発現構築物で形質転換することのいずれかにより得た。両方の場合では、GlcNAcMan3GlcNAc2を表すピークはほぼ完全に消失し、約1グルコース単位大きい新しいピークが出現した。ヘキソサミニダーゼ処理は、新しいN−グリカン上での2つの末端GlcNAc残基の存在;約2グルコース単位左へのピークシフトを示し、よって、それは、GlcNAc2Man3GlcNAc2を表す。α−1,2−マンノシダーゼ処理は、大きな違いをもたらさず、このことは、Man4’−5’GlcNAc2が限られた量だけ存在することを示す。
【図25】図25は、グルコシダーゼII活性の発現のためのプラスミドpYLTUXdL2preAnGlcIIおよびpYLeu2ExTEFpreLip2AnGlucIIβの模式図である。
【図26】図26は、プラスミドJME923 pPTura3−LEU2ex−TefL2preAnGlcIIa+b[altl]についての構築方策の模式図である。
【図27】図27は、プラスミドJME923 pPTura3−LEU2ex−Hp4dL2preAnGlcIIa+b[altl]およびZeta−LEU2ex−Hp4dL2preAnGlcIIa+b[alt]についての構築方策の模式図である。
【図28】図28は、グルコシダーゼII活性を、GlcNAcMan3GlcNAc2を合成する株に導入した後のN−グリカンプロファイルを示す一連の電気泳動図である。得られた株は、gls2αおよびgls2βサブユニットについての二重発現構築物の無作為(G060)または標的化(G061)組み込みのいずれかにより得た。両方の場合では、グルコシル化ピークの低減が観察される。α−1,2−マンノシダーゼ処理は、産生されたMan5’GlcNAc2の全てが、異種HDELタグ付きα−1,2−マンノシダーゼによりMan3GlcNAc2に変換されたのではないことを示す。キャッピンググルコースのために、GlcMan5’GlcNAc2およびGlc2Man5’GlcNAc2は、この処理に非感受性である。ナタマメマンノシダーゼは、残存グルコシル化N−グリカンおよびGlcNAcMan3GlcNAc2の両方における遊離のα−1,6結合マンノースを部分的に除去できる。さらに、この処理は、Man3−5’GlcNAc2をMan1GlcNAc2に変換する。「Rd」は、図27に示すベクターに存在するゼータ配列による「無作為組み込み」を表す。「Tg−ade2」および「Tg−ura3」は、ADE2resp.URA3遺伝子座における標的化組み込みを表す。
【図29】図29は、GlcNAcトランスフェラーゼIIをG061株に導入することにより作製したG070株およびG071株のセクレトーム(secretome)のN−グリカンプロファイルを示す一連の電気泳動図である。N−グリカンを、α−1,2−マンノシダーゼ(全ての末端α−1,2結合マンノース残基を除去する)またはヘキソサミニダーゼ(末端β−1,2結合GlcNAc残基を除去する)のいずれかで処理して、G070およびG071の天然プロファイルにおけるピークを同定した。グルコース含有N−グリカンは、2つの酵素のいずれにも感受性でない。α−1,2−マンノシダーゼ処理は、Man5’GlcNAc2およびMan4GlcNAc2からMan3GlcNAc2への刈り込みをもたらす。ヘキソサミニダーゼ処理は、GlcNAcトランスフェラーゼIおよびIIにより付加されたβ−1,2結合末端GlcNAc残基を除去して、Man3GlcNAc2を産生する。
【図30A】図30Aは、合成プレプロLip2−軽鎖(LC)のヌクレオチド配列(配列番号32)である。
【図30B】図30Bは、合成プレプロLip2−LCのアミノ酸配列(配列番号33)である。
【図31A】図31Aは、合成プレプロLip2−重鎖(HC)のヌクレオチド配列(配列番号34)である。
【図31B】図31Bは、合成プレプロLip2−HCのアミノ酸配列(配列番号35)である。
【図32】図32は、pYLHp4L2preproHerHC/LC(GUT2ex)−ori2で形質転換した、糖鎖工学的に改変したG045株、G057株、G061株およびG071株のSuperT/グリセロール振とうフラスコ培養物のN−グリカンプロファイル分析を示す一連の電気泳動図である。G045株、G057株、G061株およびG071株の記載について、表2を参照されたい。
【図33】図33は、G096流加発酵での異なる時点での機能的ELISAからの結果のグラフである。
【図34A】図34Aは、G096流加発酵における異なる時点でのセクレトームのN−グリカンプロファイル分析を示す一連の電気泳動図である。
【図34B】図34Bは、G096流加発酵における異なる時点でのセクレトームのN−グリカンプロファイル分析を示す一連の電気泳動図である。
【発明を実施するための形態】
【0027】
本明細書で記載する場合、酵母分泌糖タンパク質における哺乳類様複合N−グリカンのin vivo合成は、Man5GlcNAc2基本構造またはMan3GlcNAc2基本構造のいずれかに基づくことができる(図1Aを参照されたい、「Man」はマンノースのことであり、「GlcNAc」はN−グルコサミンのことである)。Man5GlcNAc2基本構造を生成するために、酵母細胞を、α−1,2−マンノシダーゼ活性が細胞内区画で増加し、外鎖伸長(Outer CHain elongation)(OCH1)活性が減少するように工学的に改変できる。Man3GlcNAc2基本構造を生成するために、アスパラギン結合グリコシル化3(ALG3)の活性と、いくつかの実施形態ではOCH1とが減少し、α−1,2−マンノシダーゼの活性と、いくつかの実施形態ではα−1,3−グルコシルトランスフェラーゼの活性とが増加する。このような酵母細胞で生成されたタンパク質のN−グリカンプロファイルは、酵母細胞を、以下の活性の1つ以上を含有するようにさらに工学的に改変することにより変更できる:GlcNAcトランスフェラーゼI(GnT I)活性、マンノシダーゼII活性、GlcNAcトランスフェラーゼII(GnT II)活性、グルコシダーゼII活性およびガラクトシルトランスフェラーゼ(Gal T)活性。例えば、Man5GlcNAc2またはMan3GlcNAc2 N−グリカンを生成する酵母細胞においてGnT Iを発現させることにより、それぞれGlcNAcMan5GlcNAc2またはGlcNAcMan3GlcNAc2 N−グリカンが生成されるように、GlcNAc部分がMan5GlcNAc2またはMan3GlcNAc2 N−グリカンに移動する。GlcNAcMan5GlcNAc2 N−グリカンを生成する細胞では、マンノシダーゼIIを発現させることにより、2つのマンノース残基がGlcNAcMan5GlcNAc2 N−グリカンから除去されて、GlcNAcMan3GlcNAc2 N−グリカンが生成される。GlcNAcMan3GlcNAc2 N−グリカンを生成する細胞では、GnT IIを発現させることにより、別のGlcNAc部分がGlcNAcMan3GlcNAc2 N−グリカンに移動して、GlcNAc2Man3GlcNAc2 N−グリカンが生成される。GlcNAcMan3GlcNAc2またはGlcNAc2Man3GlcNAc2 N−グリカンを生成する細胞でGal Tを発現させることにより、ガラクトースがGlcNAcMan3GlcNAc2またはGlcNAc2Man3GlcNAc2 N−グリカンに移動して、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカンが生成される。いくつかの実施形態では、グルコシダーゼII(例えばαおよびβサブユニットを発現させることにより)を発現させて、Man3GlcNAc2基本構造の生成を増加できる。
【0028】
標的分子
標的分子は、本明細書で用いる場合、遺伝子工学的に改変された細胞(例えばYarrowia lipolytica、Arxula adeninivoransもしくはその他の関連する種二形性酵母細胞(other related species dimorphic yeast cell))のような真菌細胞;植物細胞または動物細胞)においてN−グリコシル化を受ける任意の分子のことをいう。いくつかの実施形態では、標的分子は、Yarrowia lipolyticaまたはArxula adeninivorans(またはその他の関連する種二形性酵母)の分泌経路の1つ以上のステップを介して輸送されて、宿主細胞機構によるそれらのN−グリコシル化をもたらすことができる。標的分子は、内因性または外因性であり得る。
【0029】
適切な標的タンパク質は、病原体タンパク質(例えば破傷風トキソイド;ジフテリアトキソイド;ウイルス表面タンパク質(例えばサイトメガロウイルス(CMV)糖タンパク質B、HおよびgCIII;ヒト免疫不全ウイルス1(HIV−1)エンベロープ糖タンパク質;ラウス肉腫ウイルス(RSV)エンベロープ糖タンパク質;単純ヘルペスウイルス(HSV)エンベロープ糖タンパク質;エプスタインバーウイルス(EBV)エンベロープ糖タンパク質;水痘帯状疱疹ウイルス(VZV)エンベロープ糖タンパク質;ヒトパピローマウイルス(HPV)エンベロープ糖タンパク質;インフルエンザウイルス糖タンパク質;および肝炎ファミリー表面抗原)、リソソームタンパク質(例えばグルコセレブロシダーゼ、セレブロシダーゼまたはガラクトセレブロシダーゼ)、インスリン、グルカゴン、増殖因子、サイトカイン、ケモカイン、Fc受容体と結合するタンパク質、抗体もしくはその断片、またはタンパク質のいずれかと抗体もしくは抗体の断片との融合体(例えばタンパク質−Fc)を含む。増殖因子は、例えば血管内皮増殖因子(VEGF)、インスリン様増殖因子(IGF)、骨形成タンパク質(BMP)、顆粒球コロニー刺激因子(G−CSF)、顆粒球マクロファージコロニー刺激因子(GM−CSF)、神経増殖因子(NGF);ニューロトロフィン、血小板由来増殖因子(PDGF)、エリスロポエチン(EPO)、トロンボポエチン(TPO)、ミオスタチン(GDF−8)、増殖分化因子−9(GDF9)、塩基性線維芽細胞増殖因子(bFGFまたはFGF2)、上皮増殖因子(EGF)、肝細胞増殖因子(HGF)を含む。サイトカインは、例えばインターロイキン(例えばIL−1、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−12、IL−13またはIL−15のようなIL−1〜IL−33)およびインターフェロン(例えばインターフェロンβまたはインターフェロンγ)を含む。ケモカインは、例えばI−309、TCA−3、MCP−1、MIP−1α、MIP−1β、RANTES、C10、MRP−2、MARC、MCP−3、MCP−2、MRP−2、CCF18、MIP−1γ、エオタキシン、MCP−5、MCP−4、NCC−1、Ckβ10、HCC−1、ロイコタクチン−1、LEC、NCC−4、TARC、PARCまたはエオタキシン−2を含む。腫瘍糖タンパク質(例えば腫瘍関連抗原)、例えば癌胎児抗原(CEA)、ヒトムチン、HER−2/neuおよび前立腺特異抗原(PSA)も含まれる[HendersonおよびFinn、Advances in Immunology、62巻、217〜56頁(1996年)]。一実施形態では、標的タンパク質は、抗HER2/neu抗体である。いくつかの実施形態では、標的タンパク質は、リソソーム蓄積症と関連するものであり得、この標的タンパク質は、例えばアルファ−L−イズロニダーゼ、ベータ−D−ガラクトシダーゼ、ベータ−グルコシダーゼ、ベータ−ヘキソサミニダーゼ、ベータ−D−マンノシダーゼ、アルファ−L−フコシダーゼ、アリールスルファターゼB、アリールスルファターゼA、アルファ−N−アセチルガラクトサミニダーゼ、アスパルチルグルコサミニダーゼ、イズロン酸−2−スルファターゼ、アルファ−グルコサミニド−N−アセチルトランスフェラーゼ、ベータ−D−グルクロニダーゼ(glucoronidase)、ヒアルロニダーゼ、アルファ−L−マンノシダーゼ、アルファ−ノイラミニダーゼ、ホスホトランスフェラーゼ、酸性リパーゼ、酸性セラミダーゼ、スフィンゴミエリナーゼ、チオエステラーゼ、カテプシンKおよびリポタンパク質リパーゼを含む。
【0030】
標的タンパク質は、融合タンパク質でもあり得る。融合タンパク質は、例えば(i)本明細書で記載する任意のタンパク質またはその断片と、(ii)抗体またはその断片との融合体を含む。本明細書で使用する場合、用語「抗体断片」とは、(a)抗原結合断片または(b)Fc受容体と相互作用できる抗体のFc部分のことをいう。抗原結合断片は、例えば、Fab、F(ab’)2、Fvおよび単鎖Fv(scFv)断片であり得る。scFv断片は、このscFvが由来する抗体の重鎖および軽鎖の両方の可変領域を含む単一ポリペプチド鎖である。さらに、ダイアボディ[Poljak(1994年)Structure2巻(12号):1121〜1123頁;Hudsonら(1999年)J. Immunol. Methods23巻(1〜2号):177〜189頁]およびイントラボディ[Hustonら(2001年)Hum. Antibodies10巻(3〜4号):127〜142頁;Wheelerら(2003年)Mol. Ther. 8巻(3号):355〜366頁;Stocks(2004年)Drug Discov. Today9巻(22号):960〜966頁]を本発明の方法で用いることができる。
【0031】
標的タンパク質は、ポリマー、キャリア、アジュバント、免疫毒素または検出可能(例えば蛍光、発光もしくは放射活性)部分の1つ以上とつなぐこともできる。例えば、標的タンパク質をポリエチレングリコールとつなぐことができ、これは小さいタンパク質の分子量を増加させ、かつ/または循環滞留時間を増加させるために用いることができる。
【0032】
いくつかの実施形態では、標的分子は、ドリコールであるかまたはドリコールを含むことができる。
【0033】
遺伝子工学的に改変された細胞
本明細書で記載する遺伝子工学的に改変された細胞は、哺乳類様N−グリカンを含有する標的分子、または哺乳類グリコシル化経路内の中間体を含有する標的分子を生成するために用いることができる。例えば、本明細書で記載する場合、1つ以上の酵素をコードする核酸を、細胞が所望のN−グリカン(例えばGlcNAcMan5GlcNAc2、GlcNAcMan3GlcNAc2、GlcNAc2Man3GlcNAc2、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカン)を生成するように真菌細胞に導入できる。つまり、本明細書で記載する実施形態のいずれでも、真菌細胞は、1つの酵素をコードする核酸を含有し得るか、または複数の酵素をコードし得る核酸を含有し得る。このような核酸のそれぞれは、以下で論じるようなターゲティング配列も含有できる。さらに、標的分子をコードする核酸も、標的分子が生成され、所望のN−グリカン(例えばGlcNAcMan5GlcNAc2、GlcNAcMan3GlcNAc2、GlcNAc2Man3GlcNAc2、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカン)を含有するように改変されるように真菌細胞に導入できる。
【0034】
用語「核酸」および「ポリヌクレオチド」は、本明細書で交換可能に用い、cDNA、ゲノムDNA、合成DNAおよび核酸類似体を含有するDNA(またはRNA)を含むRNAならびにDNAの両方のことをいう。核酸は、任意の3次元構造を有することができる。核酸は、2本鎖または1本鎖(すなわちセンス鎖またはアンチセンス鎖)であり得る。核酸の非限定的な例は、遺伝子、遺伝子断片、エキソン、イントロン、メッセンジャーRNA(mRNA)、トランスファーRNA、リボソームRNA、siRNA、マイクロRNA、リボザイム、cDNA、組換えポリヌクレオチド、有枝鎖状ポリヌクレオチド、プラスミド、ベクター、任意の配列の単離DNA、任意の配列の単離RNA、核酸プローブおよびプライマー、ならびに核酸類似体を含む。「ポリペプチド」および「タンパク質」は、本明細書で交換可能に用い、長さまたは翻訳後修飾にかかわらず、アミノ酸の任意のペプチド連結鎖を意味する。
【0035】
「単離核酸」とは、天然で生じるゲノム(例えば酵母ゲノム)中で核酸の一方の側または両側に通常接している核酸を含む、天然で生じるゲノムに存在するその他の核酸分子から分離される核酸のことをいう。用語「単離」は、核酸に関して本明細書で用いる場合、任意の天然で生じない核酸配列も含む。なぜなら、このような天然で生じない配列は天然で見出されず、天然で生じるゲノムにおいて直接隣接する配列を有さないからである。
【0036】
単離核酸は、例えばDNA分子であり得るが、但し、天然で生じるゲノムにおいてそのDNA分子を直接挟む(flanking)ことが通常見出される核酸配列の一方が、除去されているかまたは存在しないことを条件とする。つまり、単離核酸は、限定することなく、他の配列から独立した分離した分子(例えば化学合成核酸、あるいはPCRもしくは制限エンドヌクレアーゼ処理により生成されたcDNAまたはゲノムDNA断片)、ならびにベクター、自己複製プラスミド、ウイルス(例えば任意のパラミクソウイルス、レトロウイルス、レンチウイルス、アデノウイルスまたはヘルペスウイルス)または原核生物もしくは真核生物のゲノムDNAに組み込まれたDNAを含む。さらに、単離核酸は、ハイブリッドまたは融合核酸の一部であるDNA分子のような工学的に改変された核酸を含み得る。例えばcDNAライブラリーもしくはゲノムライブラリーまたはゲノムDNA制限酵素消化物(restriction digenst)を含有するゲルスライス内の数百〜数百万のその他の核酸のうちに存在する1つの核酸は、単離核酸とみなさない。
【0037】
用語「外因性」は、核酸および特定の宿主細胞に関して本明細書で用いる場合、その特定の細胞において天然で見出されるとおりに生じない(そして該細胞から得ることができない)任意の核酸のことをいう。つまり、天然で生じない核酸は、宿主細胞に一旦導入されると、該宿主細胞にとって外因性であるとみなす。天然で生じない核酸は、天然で見出される核酸配列の核酸サブシーケンスまたは断片を含有できるが、この核酸が全体として天然に存在しないことを条件とすることに注意することが重要である。例えば、発現ベクター内のゲノムDNA配列を含有する核酸分子は、天然で生じない核酸であり、よって、宿主細胞に一旦導入されると、該宿主細胞にとって外因性である。なぜなら、この核酸分子は、全体(ゲノムDNA+ベクターDNA)として天然に存在しないからである。つまり、全体として天然に存在しない任意のベクター、自己複製プラスミドまたはウイルス(例えばレトロウイルス、アデノウイルスまたはヘルペスウイルス)は、天然で生じない核酸とみなす。このことは、PCRまたは制限エンドヌクレアーゼ処理により生成されたゲノムDNA断片ならびにcDNAを、これらが天然で見出されない分離した分子として存在するので、天然で生じない核酸とみなすことに従う。このことは、天然で見出されない配置でプロモーター配列およびポリペプチドコード配列(例えばcDNAまたはゲノムDNA)を含有する任意の核酸が、天然で生じない核酸であることにも従う。天然で生じる核酸は、特定の細胞にとって外因性になり得る。例えば、酵母xの細胞から単離された染色体全体は、該染色体が酵母の細胞に一旦導入されると、酵母yの細胞に関して外因性核酸である。
【0038】
遺伝子工学的改変のために適切な細胞は、例えば真菌細胞(例えばYarrowia lipolyticaまたは本明細書で記載する任意のその他の関連する二形性酵母細胞)、植物細胞または動物細胞を含む。細胞は、初代細胞、不死化細胞または形質転換細胞であり得る。細胞は、動物、例えば非ヒト哺乳動物におけるものであり得る。本明細書で特定する遺伝子工学的改変の前のこのような細胞は、例えばAmerican Type Culture Collection(Rockville、MD)のような多様な商業的供給源および研究資源施設から得ることができる。
【0039】
細胞の遺伝子工学的改変は、(i)N−グリコシル化活性を有するタンパク質をコードする内因性遺伝子の欠失;(ii)N−グリコシル化活性を有する(すなわちN−グリコシル化活性を有する変異タンパク質を発現する)タンパク質(例えば内因性または外因性タンパク質)の変異形態をコードする組換え核酸の導入;(iii)N−グリコシル化活性を有するタンパク質の機能的発現に干渉するRNA分子の導入または発現;(iv)N−グリコシル化活性を有する(すなわちN−グリコシル化活性を有するタンパク質を発現する)野生型(例えば内因性または外因性)タンパク質をコードする組換え核酸の導入;または(v)N−グリコシル化活性を有するタンパク質をコードする1つ以上の内因性遺伝子のプロモーター要素またはエンハンサー要素を変更して、そのことによりそれらのコードされるタンパク質の発現を変更することのような遺伝子改変を含み得る。RNA分子は、例えば小型干渉RNA(siRNA)、短いヘアピンRNA(shRNA)、アンチセンスRNAまたはマイクロRNA(miRNA)を含む。項目(ii)は、例えば、内因性遺伝子を、置き換えられるこの内因性遺伝子と比べてより大きいN−グリコシル化活性を有するタンパク質をコードする遺伝子で置き換えることを含む。遺伝子工学的改変は、N−グリコシル化活性を有するタンパク質をコードする内因性遺伝子を、付加(例えば異種配列)、欠失または置換(例えば点変異;保存的または非保存的変異のような変異)を有するタンパク質を生成するように変更することも含む。変異は、特異的に導入できる(例えば部位特異的変異誘発または相同組換え)か、または無作為に導入できる(例えば、細胞は、例えばNewmanおよびFerro−Novick(1987年)J. Cell Biol.105巻(4号):1587頁に記載されるように、化学的に変異誘発させることができる)。
【0040】
本明細書で記載する遺伝子改変は、(i)遺伝子改変された細胞における1種以上のN−グリコシル化活性の増加、(ii)遺伝子改変された細胞における1種以上のN−グリコシル化活性の減少、(iii)遺伝子改変された細胞における1種以上のN−グリコシル化活性の局在化または細胞内分布の変化、あるいは(iv)遺伝子改変された細胞における1種以上のN−グリコシル化活性の比の変化のうちの1つ以上をもたらすことができる。N−グリコシル化活性の量の増加は、N−グリコシル化活性を有する1つ以上のタンパク質の過剰発現、内因性遺伝子のコピー数の増加(例えば遺伝子重複)、または内因性遺伝子によりコードされるタンパク質の発現の増加を刺激する該遺伝子のプロモーターもしくはエンハンサーの変更によることができることが理解される。1種以上のN−グリコシル化活性の減少は、N−グリコシル化を変更する活性を有する1つ以上のタンパク質の変異形態(例えばドミナントネガティブ形態)の過剰発現、N−グリコシル化活性を有する1つ以上のタンパク質の発現を低減する1つ以上の干渉RNA分子の導入または発現、あるいはN−グリコシル化活性を有するタンパク質をコードする1つ以上の内因性遺伝子の欠失によることができる。
【0041】
1つ以上の内因性遺伝子を欠失または破壊する方法は、付随の実施例に記載する。例えば、相同組換えにより遺伝子を破壊するために、「遺伝子置き換え」ベクターを、選択可能マーカー遺伝子を含むような様式で構築できる。選択可能マーカー遺伝子は、5’および3’の両方の端で、相同組換えを媒介するのに十分な長さの遺伝子の部分に作動可能に結合できる。選択可能マーカーは、宿主細胞の栄養要求性を補完するかまたは抗生物質耐性をもたらす任意の数の遺伝子の1つであり得、URA3、LEU2およびHIS3遺伝子を含む。その他の適切な選択可能マーカーは、酵母細胞にクロラムフェニコール耐性を付与するCAT遺伝子、またはβ−ガラクトシダーゼの発現により青色コロニーをもたらすlacZ遺伝子を含む。遺伝子置き換えベクターの直鎖状にしたDNA断片を、次いで、細胞に、当技術分野において周知の方法を用いて導入する(以下を参照されたい)。ゲノムへの直鎖状断片の組み込みおよび遺伝子破壊は、選択マーカーに基づいて決定でき、例えばサザンブロット分析により確認できる。
【0042】
付随の実施例に詳細に示すように、選択におけるその使用の後に、選択可能マーカーを宿主細胞のゲノムから、例えばCre−loxP系により除去できる(以下を参照されたい)。マーカー除去のこのプロセスは、実施例を通して「キュアリング」という。
【0043】
代わりに、遺伝子置き換えベクターは、破壊する遺伝子の部分を含むような様式で構築でき、ここで、この部分は、いずれの内因性遺伝子プロモーター配列がなく、その遺伝子のコード配列をコードしないかまたは該コード配列の不活性断片をコードする。「不活性断片」は、例えば遺伝子の全長コード配列から生成されるタンパク質の活性の約10%未満(例えば約9%未満、約8%未満、約7%未満、約6%未満、約5%未満、約4%未満、約3%未満、約2%未満、約1%未満または0%)を有するタンパク質をコードする遺伝子の断片である。遺伝子のこのような部分は、既知のプロモーター配列は遺伝子配列に作動可能に結合されていないが、停止コドンおよび転写終結配列が遺伝子配列のこの部分と作動可能に結合されているような様式でベクターに挿入される。このベクターは、その後、遺伝子配列のこの部分で直鎖状にして、細胞を形質転換できる。単一の相同組換えにより、この直鎖状にしたベクターを、次いで、その遺伝子の内因性対応物(counterpart)に組み込む。
【0044】
発現ベクターは、自律型または組み込み型であり得る。
【0045】
組換え核酸を、プラスミド、ファージ、トランスポゾン、コスミドまたはウイルス粒子のような発現ベクターの形態で細胞に導入できる。組換え核酸は、染色体外で維持できるか、またはこれを酵母細胞染色体DNAに組み込むことができる。発現ベクターは、選択された条件下で細胞生存性に要求されるタンパク質をコードする選択マーカー遺伝子(例えばウラシル生合成のために必要な酵素をコードするURA3、またはトリプトファン生合成に要求される酵素をコードするTRP1)を含有して、所望の核酸で形質転換されたこれらの細胞の検出および/または選択を許容できる(例えば米国特許第4,704,362号を参照されたい)。発現ベクターは、自己複製配列(ARS)も含むことができる。例えば、米国特許第4,837,148号は、Pichia pastoris中でプラスミドを維持するために適切な手段を提供する自己複製配列について記載している。
【0046】
組み込み型ベクターが、例えば米国特許第4,882,279号に開示されている。組み込み型ベクターは、一般的に、少なくとも第1挿入可能DNA断片と、選択可能マーカー遺伝子と、第2挿入可能DNA断片との連続的に配置された配列を含む。第1および第2の挿入可能DNA断片は、それぞれ、約200(例えば約250、約300、約350、約400、約450、約500または約1000以上)ヌクレオチドの長さであり、形質転換される種のゲノムDNAの部分に相同なヌクレオチド配列を有する。発現のための目的の遺伝子(例えばN−グリコシル化活性を有するタンパク質をコードする遺伝子)を含有するヌクレオチド配列は、第1および第2の挿入可能DNA断片の間で、マーカー遺伝子の前かその後のいずれかにおいて、このベクターに挿入される。組み込み型ベクターを酵母形質転換の前に直鎖状にして、宿主細胞ゲノムへの目的のヌクレオチド配列の組み込みを容易にすることができる。
【0047】
発現ベクターは、それらを酵母で発現させることを可能にする酵母(例えばYarrowia lipolytica、Arxula adeninivoransまたはその他の関連する二形性酵母種)プロモーターの制御下の組換え核酸を特徴とすることができる。適切な酵母プロモーターは、TEF1、HP4D、GAP、POX2、ADC1、TPI1、ADH2、POXおよびGal10プロモーターを含む。例えばMadzakら(2000年)J. Mol. Microbiol. Biotechnol.2巻:207〜216頁;Guarenteら(1982年)Proc. Natl. Acad. Sci. USA79巻(23号):7410頁を参照されたい。さらなる適切なプロモーターは、例えばZhuおよびZhang(1999年)Bioinformatics15巻(7〜8号):608〜611頁および米国特許第6,265,185号に記載されている。発現ベクターが哺乳類細胞のような動物細胞に導入される場合、発現ベクターは、目的の宿主細胞での発現に適切な動物細胞プロモーターの制御下の組換え核酸を特徴とすることができる。哺乳類プロモーターの例は、SV40およびサイトメガロウイルス(CMV)プロモーターを含む。
【0048】
プロモーターは、構成性(constitutive)または誘導性(条件的)であり得る。構成性プロモーターは、その発現が標準的培養条件下で一定であるプロモーターであると理解される。誘導性プロモーターは、1つ以上の誘導指示(induction cue)に応答性であるプロモーターである。例えば、誘導性プロモーターは、化学的に調節され得る(例えばその転写活性が、アルコール、テトラサイクリン、ステロイド、金属またはその他の小分子のような化学的誘導物質の存在または非存在により調節されるプロモーター)か、または物理的に調節され得る(例えばその転写活性が、光または高温もしくは低温のような物理的誘導因子の存在または非存在により調節されるプロモーター)。誘導性プロモーターは、それら自体が化学的もしくは物理的な指示により直接調節される1つ以上の転写因子により間接的に調節されることもできる。
【0049】
細胞の遺伝子工学的改変は、宿主細胞に存在するが、その細胞で通常は発現されないかまたはその細胞で著しいレベルでは発現されない内因性遺伝子(例えばN−グリコシル化活性を有するタンパク質をコードする遺伝子)を活性化することも含む。例えば、内因性遺伝子の調節配列(例えば遺伝子プロモーターまたはエンハンサー)を、作動可能に結合したコード配列が発現の増加を示すように改変できる。相同組換えまたはターゲティングを用いて、遺伝子と通常関連する調節領域を、対応する遺伝子工学的に改変されていない細胞で明らかなレベルよりも高いレベルで遺伝子を発現させるか、または対応する遺伝子工学的に改変されていない細胞で明らかなものとは異なる調節もしくは誘導のパターンを遺伝子が示すようにする調節配列で置き換えるかあるいは不能にすることができる。遺伝子の調節配列(例えばプロモーターまたはエンハンサー)の変更を導入するために適切な方法は、例えば米国特許出願公開第20030147868号に記載されている。
【0050】
その他の遺伝子工学的に作製された改変も、条件的であり得ることが理解される。例えば、遺伝子は、例えばCre−loxP系のような部位特異的DNAリコンビナーゼを用いて条件的に欠失できる(例えばGossenら(2002年)Ann. Rev. Genetics36巻:153〜173頁および米国特許出願公開第20060014264号を参照されたい)。
【0051】
組換え核酸は、本明細書で記載する細胞に、スフェロプラスト技術または全細胞塩化リチウム酵母形質転換法のような様々な方法を用いて導入できる。プラスミドまたは直鎖状核酸ベクターで細胞を形質転換するために有用なその他の方法は、例えば米国特許第4,929,555号;Hinnenら(1978年)Proc. Nat. Acad. Sci. USA75巻:1929頁;Itoら(1983年)J. Bacteriol.153巻:163頁;米国特許第4,879,231号;ならびにSreekrishnaら(1987年)Gene59巻:115頁に記載されている。エレクトロポレーションおよびPEG1000全細胞形質転換手順も、CreggおよびRussel、Methods in Molecular Biology:Pichia Protocols、第3章、Humana Press、Totowa、N.J.、27〜39頁(1998年)に記載されるようにして用いることができる。動物細胞のトランスフェクションは、例えば、ベクターを細胞に、リン酸カルシウム、エレクトロポレーション、熱ショック、リポソーム、またはFUGENE(登録商標)もしくはLIPOFECTAMINE(登録商標)のようなトランスフェクション試薬を用いて、あるいは裸の核酸ベクターを細胞と溶液中で接触させることにより導入することを特徴とすることができる(例えばSambrookら、Molecular Cloning: A Laboratory Manual第2版1、2および3巻、Cold Spring Harbor Laboratory Press: Cold Spring Harbor、New York, USA、1989年11月を参照されたい)。
【0052】
形質転換酵母細胞は、それらに限定されないが、形質転換後の栄養要求性細胞を、要求される生化学的生成物の非存在下で培養すること(細胞の栄養要求性による)、新しい表現型についての選択およびその検出、または形質転換体に含有される耐性遺伝子の非存在下で酵母にとって毒性がある抗生物質の存在下で培養することを含む適当な技術を用いて選択できる。形質転換体は、ゲノムへの発現カセットの組み込みにより選択および/または確認することもでき、これは、例えばサザンブロットまたはPCR分析により評価することができる。
【0053】
目的の標的細胞にベクターを導入する前に、ベクターを、Escherichia coli(E.coli)のような細菌細胞中で増殖(例えば増幅)させることができる。ベクターDNAを、細菌細胞から、細菌環境からのベクターDNAの精製をもたらす当技術分野において公知の方法のいずれかにより単離できる。精製ベクターDNAは、フェノール、クロロホルムおよびエーテルを用いて広く抽出して、プラスミドDNA調製物中にE.coliタンパク質が存在しないことを確実にできる。なぜなら、これらのタンパク質は、哺乳類細胞にとって有毒であり得るからである。
【0054】
遺伝子工学的改変は、本明細書で記載する場合、N−グリコシル化活性を有するタンパク質をコードする任意の数の遺伝子を発現(例えば過剰発現)するか、該遺伝子に改変を導入するか、または該遺伝子を欠失させるために用いることができる。このようなタンパク質は、例えば、OCH1、ALG3、α−1,3−グルコシルトランスフェラーゼ、GnT I、マンノシダーゼII、GnT II、グルコシダーゼIIまたはGal Tを含む。N−グリコシル化活性を有するタンパク質をコードする遺伝子は、このような遺伝子を含有する任意の種に由来し得る。N−グリコシル化活性を有するタンパク質をコードする遺伝子を得ることができる例示的な真菌種は、限定することなく、Pichia anomala、Pichia bovis、Pichia canadensis、Pichia carsonii、Pichia farinose、Pichia fermentans、Pichia fluxuum、Pichia membranaefaciens、Pichia membranaefaciens、Candida valida、Candida albicans、Candida ascalaphidarum、Candida amphixiae、Candida Antarctica、Candida atlantica、Candida atmosphaerica、Candida blattae、Candida carpophila、Candida cerambycidarum、Candida chauliodes、Candida corydalis、Candida dosseyi、Candida dubliniensis、Candida ergatensis、Candida fructus、Candida glabrata、Candida fermentati、Candida guilliermondii、Candida haemulonii、Candida insectamens、Candida insectorum、Candida intermedia、Candida jeffresii、Candida kefyr、Candida krusei、Candida lusitaniae、Candida lyxosophila、Candida maltosa、Candida membranifaciens、Candida milleri、Candida oleophila、Candida oregonensis、Candida parapsilosis、Candida quercitrusa、Candida shehatea、Candida temnochilae、Candida tenuis、Candida tropicalis、Candida tsuchiyae、Candida sinolaborantium、Candida sojae、Candida viswanathii、Candida utilis、Pichia membranaefaciens、Pichia silvestris、Pichia membranaefaciens、Pichia chodati、Pichia membranaefaciens、Pichia menbranaefaciens、Pichia minuscule、Pichia pastoris、Pichia pseudopolymorpha、Pichia quercuum、Pichia robertsii、Pichia saitoi、Pichia silvestrisi、Pichia strasburgensis、Pichia terricola、Pichia vanriji、Pseudozyma Antarctica、Rhodosporidium toruloides、Rhodotorula glutinis、Saccharomyces bayanus、Saccharomyces bayanus、Saccharomyces momdshuricus、Saccharomyces uvarum、Saccharomyces bayanus、Saccharomyces cerevisiae、Saccharomyces bisporus、Saccharomyces chevalieri、Saccharomyces delbrueckii、Saccharomyces exiguous、Saccharomyces fermentati、Saccharomyces fragilis、Saccharomyces marxianus、Saccharomyces mellis、Saccharomyces rosei、Saccharomyces rouxii、Saccharomyces uvarum、Saccharomyces willianus、Saccharomycodes ludwigii、Saccharomycopsis capsularis、Saccharomycopsis fibuligera、Saccharomycopsis fibuligera、Endomyces hordei、Endomycopsis fobuligera. Saturnispora saitoi、Schizosaccharomyces octosporus、Schizosaccharomyces pombe、Schwanniomyces occidentalis、Torulaspora delbrueckii、Torulaspora delbrueckii、Saccharomyces dairensis、Torulaspora delbrueckii、Torulaspora fermentati、Saccharomyces fermentati、Torulaspora delbrueckii、Torulaspora rosei、Saccharomyces rosei、Torulaspora delbrueckii、Saccharomyces rosei、Torulaspora delbrueckii、Saccharomyces delbrueckii、Torulaspora delbrueckii、Saccharomyces delbrueckii、Zygosaccharomyces mongolicus、Dorulaspora globosa、Debaryomyces globosus、Torulopsis globosa、Trichosporon cutaneum、Trigonopsis variabilis、Williopsis californica、Williopsis saturnus、Zygosaccharomyces bisporus、Zygosaccharomyces bisporus、Debaryomyces disporua.、Saccharomyces bisporas、Zygosaccharomyces bisporus、Saccharomyces bisporus、Zygosaccharomyces mellis、Zygosaccharomyces priorianus、Zygosaccharomyces rouxiim、Zygosaccharomyces rouxii、Zygosaccharomyces barkeri、Saccharomyces rouxii、Zygosaccharomyces rouxii、Zygosaccharomyces major、Saccharomyces rousii、Pichia anomala、Pichia bovis、Pichia Canadensis、Pichia carsonii、Pichia farinose、Pichia fermentans、Pichia fluxuum、Pichia membranaefaciens、Pichia pseudopolymorpha、Pichia quercuum、Pichia robertsii、Pseudozyma Antarctica、Rhodosporidium toruloides、Rhodosporidium toruloides、Rhodotorula glutinis、Saccharomyces bayanus、Saccharomyces bayanus、Saccharomyces bisporus、Saccharomyces cerevisiae、Saccharomyces chevalieri、Saccharomyces delbrueckii、Saccharomyces fermentati、Saccharomyces fragilis、Saccharomycodes ludwigii、Schizosaccharomyces pombe、Schwanniomyces occidentalis、Torulaspora delbrueckii、Torulaspora globosa、Trigonopsis variabilis、Williopsis californica、Williopsis saturnus、Zygosaccharomyces bisporus、Zygosaccharomyces mellis、Zygosaccharomyces rouxiiまたは当技術分野で公知であるかもしくは本明細書で記載する任意のその他の真菌(例えば酵母)を含む。例示的な下等真核生物は、それらに限定されないが、Aspergillus caesiellus、Aspergillus candidus、Aspergillus carneus、Aspergillus clavatus、Aspergillus deflectus、Aspergillus flavus、Aspergillus fumigatus、Aspergillus glaucus、Aspergillus nidulans、Aspergillus niger、Aspergillus ochraceus、Aspergillus oryzae、Aspergillus parasiticus、Aspergillus penicilloides、Aspergillus restrictus、Aspergillus sojae、Aspergillus sydowi、Aspergillus tamari、Aspergillus terreus、Aspergillus ustusまたはAspergillus versicolorを含むAspergillusの様々な種も含む。N−グリコシル化活性を有するタンパク質をコードする遺伝子を得ることができる例示的な原生動物の属は、限定することなく、Blastocrithidia、Crithidia、Endotrypanum、Herpetomonas、Leishmania、Leptomonas、Phytomonas、Trypanosoma(例えばT.bruceii、T.gambiense、T.rhodesienseおよびT.cruzi)およびWallaceinaを含む。例えば、GnT Iをコードする遺伝子は、ヒト(Swiss Protein受託番号P26572)、ラット、Arabidopsis、マウスまたはDrosophilaから得ることができ、GntIIをコードする遺伝子は、ヒト、ラット(Swiss Protein受託番号Q09326)、Arabidopsisまたはマウスから得ることができ、Man IIをコードする遺伝子は、ヒト、ラット、Arabidopsis、マウス、Drosophila(Swiss Protein受託番号Q24451)から得ることができ、GalTをコードする遺伝子は、ヒト(Swiss Protein受託番号P15291)、ラット、マウスまたはウシから得ることができる。
【0055】
いくつかの実施形態では、遺伝子工学的に改変された細胞は、OCH1(GenBank受託番号AJ563920)遺伝子またはその遺伝子生成物(mRNAまたはタンパク質)を欠く。いくつかの実施形態では、遺伝子工学的に改変された細胞は、ALG3(Genbank(登録商標)受託番号XM_503488、Genolevures参照:YALI0E03190g)遺伝子またはその遺伝子生成物(mRNAまたはタンパク質)を欠く。いくつかの実施形態では、遺伝子工学的に改変された細胞は、α−1,3−グルコシルトランスフェラーゼ(例えばALG6、Genbank(登録商標)受託番号XM_502922、Genolevures参照:YALI0D17028g)タンパク質を発現(例えば過剰発現)する。いくつかの実施形態では、遺伝子工学的に改変された細胞は、α−1,2−マンノシダーゼ(例えばGenbank受託番号AF212153)タンパク質を発現する。いくつかの実施形態では、遺伝子工学的に改変された細胞は、GlcNAcトランスフェラーゼI(例えばSwiss Prot.受託番号P26572)タンパク質を発現する。いくつかの実施形態では、遺伝子工学的に改変された細胞は、マンノシダーゼIIタンパク質またはその触媒ドメイン(例えばSwiss Prot.受託番号Q24451)を発現する。いくつかの実施形態では、遺伝子工学的に改変された細胞は、ガラクトシルトランスフェラーゼIタンパク質またはその触媒ドメイン(例えばSwiss Prot.受託番号P15291)を発現する。いくつかの実施形態では、遺伝子工学的に改変された細胞は、GlcNAcトランスフェラーゼIIタンパク質またはその触媒ドメイン(例えばSwiss Prot.受託番号Q09326)を発現する。いくつかの実施形態では、遺伝子工学的に改変された細胞は、Yarrowia lipolytica、Trypanosoma bruceiまたはAspergillus nigerのグルコシダーゼIIのようなグルコシダーゼIIのアルファまたはベータサブユニット(またはアルファおよびベータの両方のサブユニット)を発現する。遺伝子工学的に改変された細胞は、これらの改変の任意の組み合わせを有することができる。
【0056】
例えば、いくつかの実施形態では、遺伝子工学的に改変された細胞は、OCH1遺伝子を欠き、α−1,2−マンノシダーゼ、GlcNAcトランスフェラーゼI、マンノシダーゼIIおよびガラクトシルトランスフェラーゼIを発現できる。いくつかの実施形態では、遺伝子工学的に改変された細胞は、ALG3遺伝子を欠き、α−1,2−マンノシダーゼ、GlcNAcトランスフェラーゼI、GlcNAcトランスフェラーゼIおよびガラクトシルトランスフェラーゼIを発現できる。このような遺伝子工学的に改変された細胞は、α−1,3−グルコシルトランスフェラーゼをさらに発現し、かつ/またはグルコシダーゼIIのアルファおよびベータサブユニットを発現し、かつ/またはOCH1遺伝子を欠くことができる。
【0057】
このようなタンパク質の1つ以上は、異種ターゲティング配列を含有する融合タンパク質であり得る。例えば、α−1,2−マンノシダーゼは、HDEL小胞体(ER)保持アミノ酸配列を有することができる(実施例を参照されたい)。N−グリコシル化活性を有する任意のタンパク質を、HDEL配列を含む融合タンパク質中に工学的に作製できることが理解される。その他のタンパク質は、タンパク質をゴルジ装置に向かわせるための異種配列を有することができる。例えば、酵母Kre2p遺伝子によりコードされる最初の100個のN末端アミノ酸、S.cerevisiae Mnn2遺伝子によりコードされる最初の36個のN末端アミノ酸(Swiss Prot.受託番号P38069)またはS.cerevisiae Mnn2p遺伝子によりコードされる最初の46個のN末端アミノ酸を用いて、タンパク質をゴルジに向かわせることができる。そのままで、真菌細胞で発現されるタンパク質をコードする核酸は、コードされたタンパク質を細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含むことができる。例えば、α−1,2−マンノシダーゼをERに向かわせることができ、GnT I、GnTII、マンノシダーゼおよびGal Tをゴルジに向かわせることができる。
【0058】
N−グリコシル化活性を有するタンパク質が、該タンパク質を発現させる細胞とは異なる型(例えば異なる種)のタンパク質である細胞に由来する実施形態では、タンパク質をコードする核酸は、特定の目的の細胞における発現のためにコドン最適化できる。例えば、Trypanosoma bruceiからのN−グリコシル化を有するタンパク質をコードする核酸は、Yarrowia lipolyticaのような酵母細胞での発現のためにコドン最適化できる。このようなコドン最適化は、目的の細胞でのタンパク質の発現を増加させるために有用になり得る。タンパク質をコードする核酸をコドン最適化するための方法は、当技術分野において公知であり、例えばGaoら(Biotechnol. Prog.(2004年)20巻(2号):443〜448頁)、Kotulaら(Nat. Biotechn.(1991年)9巻、1386〜1389頁)およびBennetzenら(J. Biol. Chem.(1982年)257巻(6号):2036〜3031頁)に記載されている。表1は、Yarrowia lipolyticaについてのコドン使用頻度を示す。データは、5,967のコード配列に存在する2,945,919のコドンに由来した。表1の内容は、コドン使用頻度データベース(Codon Usage Database)から得たが、これは、kazusa.or.jp/codon/cgi−bin/showcodon.cgi?species=284591のワールドワイドウェブで見出すことができる。
【0059】
【表1】
いくつかの実施形態では、ヒトタンパク質を細胞に導入でき、N−グリコシル化活性を有する1つ以上の内因性酵母タンパク質を抑制(例えば欠失または変異)できる。真菌グリコシル化経路を「ヒト化」するための技術は、例えばChoiら(2003年)Proc. Natl. Acad. Sci. USA100巻(9号):5022〜5027頁;Verveckenら(2004年)Appl. Environ. Microb.70巻(5号):2639〜2646頁;およびGerngross(2004年)Nature Biotech.22巻(11号):1410〜1414頁に記載されている。
【0060】
遺伝子工学的改変が例えばタンパク質の発現の変化または外因性タンパク質(内因性タンパク質の変異形態を含む)の発現を含む場合、遺伝子工学的に改変された細胞がタンパク質を発現するかを決定するために様々な技術を用いることができる。例えば、タンパク質をコードするmRNAまたはタンパク質自体の存在を、例えばノザンブロットもしくはRT−PCR分析またはウェスタンブロット分析をそれぞれ用いて検出できる。N−グリコシル化活性を有するタンパク質の細胞内局在化は、細胞成分分画(subcellular fractionation)および免疫蛍光を含む様々な技術を用いることにより分析できる。
【0061】
標的分子のグリコシル化を検出する方法は、DNAシーケンサー支援(DSA)、蛍光体支援糖質電気泳動(FACE)または表面増強型レーザー脱離/イオン化飛行時間型質量分析(SELDI−TOF MS)を含む。例えば、分析は、DSA−FACEを利用でき、ここでは、例えば、糖タンパク質を変性させ、その後、例えばメンブレン上に固定化する。糖タンパク質は、次いで、ジチオトレイトール(DTT)またはβ−メルカプトエタノールのような適切な還元剤を用いて還元できる。タンパク質のスルフヒドリル基は、ヨード酢酸のような酸を用いてカルボキシル化できる。次に、N−グリコシダーゼFのような酵素を用いてN−グリカンをタンパク質から切り離すことできる。N−グリカンは、必要に応じて、再構成して、還元的アミノ化により誘導体化できる。誘導体化N−グリカンは、次いで、濃縮できる。N−グリカン分析のために適切な装置は、例えばABI PRISM(登録商標)377 DNAシーケンサー(Applied Biosystems)を含む。データ解析は、例えばGENESCAN(登録商標)3.1ソフトウェア(Applied Biosystems)を用いて行うことができる。必要に応じて、単離マンノタンパク質(mannoprotein)は、1種以上の酵素でさらに処理して、それらのN−グリカン状態を確認できる。N−グリカン分析のさらなる方法は、例えば質量分析(例えばMALDI−TOF−MS)、順相(normal phase)、逆相での高圧液体クロマトグラフィー(HPLC)およびイオン交換クロマトグラフィー(例えばグリカンが標識されていない場合にパルス電流滴定検出を用いて、またグリカンが適当に標識されているならばUV吸収もしくは蛍光を用いて)を含む。Callewaertら(2001年)Glycobiology11巻(4号):275〜281頁およびFreireら(2006年)Bioconjug. Chem.17巻(2号):559〜564頁も参照されたい。
【0062】
遺伝子工学的に改変された細胞の遺伝子改変のいずれかが、誘導指示(例えば化学的または物理的指示)の存在によって誘導性または条件的である場合、遺伝子工学的に改変された細胞は、必要に応じて、核酸の導入前、導入中または導入後に誘導剤の存在下で培養できる。例えば、標的タンパク質をコードする核酸の導入の後に、細胞を、N−グリコシル化活性を有する1つ以上のタンパク質の発現を促進できる化学的誘導剤に曝露できる。複数の誘導指示が、N−グリコシル化活性を有する1つ以上のタンパク質の条件的発現を誘導する場合、細胞は、複数の誘導剤と接触できる。
【0063】
所望のN−グリカンを含むように改変された標的分子は、遺伝子工学的に改変された細胞から単離できる。改変された標的分子を、酵母細胞内で維持し、細胞溶解の際に遊離できるか、または改変標的分子を、細胞からの分子の分泌を誘導するコード配列(外因性核酸に本来備わっているかまたは発現ベクター中に工学的に作製されたかのいずれか)によりもたらされる機構により培養培地中に分泌させることができる。細胞溶解産物または培養培地中の改変標的分子の存在は、その分子の存在を検出するための様々な標準的プロトコールにより確認できる。例えば、変更された標的分子がタンパク質である場合、このようなプロトコールは、それらに限定されないが、変更された標的タンパク質(または標的タンパク質自体)に特異的な抗体を用いる免疫ブロッティングもしくは放射性免疫沈降法、変更された標的タンパク質(または標的タンパク質自体)に特異的なリガンドの結合、または改変された標的タンパク質(または標的タンパク質自体)の特異的酵素活性の試験を含むことができる。
【0064】
いくつかの実施形態では、遺伝子工学的に改変された細胞から単離される標的分子の少なくとも約25%は、所望のN−グリカンを含有する。例えば、遺伝子工学的に改変された細胞から単離される標的分子の少なくとも約27%、少なくとも約30%、少なくとも約35%、少なくとも約40%、少なくとも約45%、少なくとも約50%、少なくとも約55%、少なくとも約60%、少なくとも約65%、少なくとも約70%、少なくとも約75%、少なくとも約80%、少なくとも約85%、少なくとも約90%または少なくとも約95%または少なくとも約99%が、所望のN−グリカンを含有できる。
【0065】
いくつかの実施形態では、本明細書で記載する方法を用いて生成される標的分子において、糖タンパク質上のN−グリカンの少なくとも50%(例えば少なくとも55、60、65、70、75、80または85%)は、GlcNAc2Man3GlcNAc2 N−グリカンであり得る。GlcNAc2Man3GlcNAc2 N−グリカンのパーセンテージは、DSA−FACE電気泳動図中のピーク面積から見積もることができる。実施例13を参照されたい。
【0066】
いくつかの実施形態では、単離改変標的分子は、凍結、凍結乾燥または固定化して、例えば変更された標的分子が生物活性を保持することを可能にする適当な条件下で貯蔵できる。
【0067】
工学的に改変された細胞の培養
この文書は、本明細書で記載する遺伝子工学的に改変された細胞のいずれかの実質的に純粋な培養物も提供する。本明細書で用いる場合、遺伝子工学的に改変された細胞の「実質的に純粋な培養物」は、この細胞の培養物であって、培養物中の生存細胞の合計数の約40%未満(すなわち約35%;約30%;約25%;約20%;約15%;約10%;約5%;約2%;約1%;約0.5%;約0.25%;約0.1%;約0.01%;約0.001%;約0.0001%またはさらにはそれ未満)が遺伝子工学的に改変された細胞以外の生存細胞、例えば細菌、真菌(酵母を含む)、マイコプラズマまたは原生生物の細胞である培養物である。この状況での用語「約」は、関連するパーセンテージが、特定されたパーセンテージより15%上または下であり得ることを意味する。つまり、例えば、約20%は、17%〜23%であり得る。遺伝子工学的に改変された細胞のこのような培養物は、細胞と、増殖培地、貯蔵培地または輸送培地とを含む。培地は、液体、半固体(例えばゼラチン状培地)、または凍結したものであり得る。培養物は、液体培地中または半固体培地中/上で増殖しているか、あるいは凍結貯蔵培地もしくは輸送培地を含む貯蔵培地または輸送培地中で貯蔵または輸送されている細胞を含む。培養物は、培養容器または貯蔵容器または基材(substrate)(例えば培養皿、フラスコもしくは試験管または貯蔵バイアルもしくは貯蔵試験管)中にある。
【0068】
本明細書で記載する遺伝子工学的に改変された細胞は、例えば、例えばグリセロールもしくはスクロースのような凍結保護物質を含有するバッファー中での凍結細胞懸濁物として、凍結乾燥細胞として貯蔵できる。代わりに、これらは、例えば流動床乾燥もしくは噴霧乾燥または任意のその他の適切な乾燥方法により得られる乾燥細胞調製物として貯蔵できる。
【0069】
変更されたN−グリコシル化分子により処置可能な障害
所望のN−グリカンを含有するように改変された単離標的分子は、代謝障害、がんおよび炎症性障害を含む様々な障害を処置するために用いることができる。
【0070】
(i)代謝障害
代謝障害は、個別のヒト(または動物)細胞内でエネルギーの生成に影響する障害である。ほとんどの代謝障害は遺伝的であるが、いくつかは、食餌、毒素、感染症などの結果として「後天性」であり得る。遺伝性代謝障害は、先天性代謝異常としても公知である。一般的に、遺伝性代謝障害は、遺伝子欠陥により引き起こされ、これにより、細胞の代謝プロセスにおけるいくつかのステップに必要な酵素がなくなるかまたは誤って構築される。代謝障害の最大のクラスは、炭水化物代謝障害、アミノ酸代謝障害、有機酸代謝障害(有機酸性尿症)、脂肪酸酸化およびミトコンドリア代謝障害、ポルフィリン代謝障害、プリンまたはピリミジン代謝障害、ステロイド代謝障害、ミトコンドリア機能障害、ペルオキシソーム機能障害ならびにリソソーム蓄積症(LSD)である。
【0071】
本明細書で記載する1つまた複数のグリコシル化分子(またはその薬学的組成物)の投与により処置できる代謝障害の例は、遺伝性ヘモクロマトーシス、眼皮膚白皮症、プロテインC欠損症、I型遺伝性血管浮腫、先天性スクラーゼ−イソマルターゼ欠損症、クリグラー−ナジャー症候群II型、ラロン症候群、遺伝性ミエロペルオキシダーゼ、原発性甲状腺機能低下症、先天性QT延長症候群、チロキシン(tyroxine)結合グロブリン欠損症、家族性高コレステロール血症、家族性カイロミクロン血症、無ベータリポタンパク質血症、低血漿リポタンパク質Aレベル、肝臓損傷を伴う遺伝性気腫、先天性甲状腺機能低下症、骨形成不全症、遺伝性低フィブリノゲン血症、アルファ−1アンチキモトリプシン欠損症、腎性尿崩症、神経下垂体性尿崩症、アデノシンデアミナーゼ欠損症、ペリツェウスメルツバッヘル病、フォンウィルブランド病IIA型、第V、VIII因子合併欠損症(combined factors V and VIII deficiency)、遅発性脊椎骨端異形成症、コロイデレミア、I細胞病、バッテン病、毛細血管拡張性運動失調、ADPKD−常染色体優性多嚢胞性腎疾患、微絨毛封入体病、結節硬化症、ロウ眼脳腎症候群、筋委縮性側策硬化症、骨髄異形成症候群、不全リンパ球症候群、タンジール病、家族性肝内胆汁うっ滞、X連鎖副腎白質ジストロフィー、スコット症候群、ヘルマンスキー−プドラック症候群1型および2型、ツェルウェガー症候群、近節短縮性点状軟骨異形成症、常染色体劣性原発性高シュウ酸尿症、モーアトラネジャーグ症候群、球脊髄性筋萎縮症(spinal and bullar muscular atrophy)、原発性線毛運動不全(diskenesia)(カルタゲナー症候群)、巨人症および先端巨大症、乳汁漏出症、アジソン病、副腎性男性化、クッシング症候群、ケトアシドーシス、原発性または続発性アルドステロン症、ミラーディーカー症候群、脳回欠損、運動ニューロン疾患、アッシャー症候群、ウィスコット−アルドリッチ症候群、オピッツ(Optiz)症候群、ハンチントン病、遺伝性膵炎、抗リン脂質抗体症候群、オーバーラップ結合組織病、シェーグレン症候群、スティッフマン症候群、ブルガダ症候群、フィンランド型先天性腎炎症候群、デュビン−ジョンソン症候群、X連鎖低リン酸血症(hypophosphosphatemia)、ペンドレッド症候群、新生児持続性高インスリン血症性低血糖症、遺伝性球状赤血球症、無セルロプラスミン血症、乳児性神経セロイドリポフスチン沈着症、偽軟骨形成不全および多発性骨端骨異形成(multiple epiphyseal)、シュタルガルト様黄斑ジストロフィー、X連鎖シャコール−マリー−トゥース病、常染色体優性網膜色素変性、ウォルコット−ラリソン症候群、クッシング病、肢帯筋ジストロフィー、ムコ多糖症IV型、フィンランド型遺伝性家族性アミロイドーシス、アンダーソン病、肉腫、慢性骨髄単球性白血病、心筋症、顔面生殖器異形成、捻転ジストニア(Torsion disease)、ハンチントンおよび脊髄小脳運動失調、遺伝性高ホモシステイン血症(hyperhomosyteinemia)、多発ニューロパチー、下位運動ニューロン疾患、色素性網膜炎、血清反応陰性多発性関節炎、間質性肺線維症、レイノー現象、ウェゲナー(Wegner’s)肉芽腫症、タンパク尿(preoteinuria)、CDG−Ia、CDG−Ib、CDG−Ic、CDG−Id、CDG−Ie、CDG−If、CDG−IIa、CDG−IIb、CDG−IIc、CDG−IId、エーラース−ダンロス症候群、多発性外骨腫症、グリセリ症候群(1型または2型)、あるいはX連鎖非特異性精神遅滞を含み得る。さらに、代謝障害は、それらに限定されないが、ファブリー病、ファーバー病、ゴーシェ病、GM1−ガングリオシド−シス、テイ−サックス病、サンドホフ病、GM2活性化因子病(GM2 activator disease)、クラッベ病、異染性白質ジストロフィー、ニーマン−ピック病(A型、B型およびC型)、ハーラー病、シャイエ病、ハンター病、サンフィリポ病、モルキオ病、マロトー−ラミー病、ヒアルロニダーゼ欠損症、アスパルチルグルコサミン尿症、フコシドーシス、マンノシドーシス、シンドラー病、シアリドーシス1型、ポンぺ病、濃化異骨症(Pycnodysostosis)、セロイドリポフスチン沈着症、コレステロールエステル蓄積症、ウォルマン病、多種スルファターゼ欠損症、ガラクトシアリドーシス、ムコリピド−シス(II型、III型およびIV型)、シスチン症、シアル酸蓄積症、マリネスコ−シェーグレン症候群を伴うカイロミクロン保持病、ヘルマンスキー−プドラック症候群、チェディアック−東症候群、ダノン病または幸福顔貌骨異形成(Geleophysic dysplasia)のようなリソソーム蓄積症も含み得る。
【0072】
代謝障害の症状は多数で多様であり、例えば貧血、疲労、容易に挫傷すること、血液中の血小板低下、肝腫大、脾腫大、骨格脆弱、肺機能障害、感染症(例えば胸部感染症または肺炎)、腎臓機能障害、進行性脳損傷、発作、過剰に粘稠な胎便、咳、喘鳴、過剰な唾液もしくは粘液生成、息切れ、腹痛、腸または消化管(bowel or gut)の閉塞、生殖能力の問題、鼻内のポリープ、ばち指/足指の爪および皮膚、手もしくは足の疼痛、被角血管腫、発汗低下、角膜および水晶体の混濁、白内障、僧帽弁の逸脱および/もしくは逆流、心肥大、温度不耐、歩行困難、嚥下困難、進行性視力喪失、進行性聴力喪失、緊張低下、巨舌症、反射消失、腰痛、睡眠時無呼吸、起坐呼吸、傾眠、脊柱前弯または脊柱側弯の1つ以上を含み得る。欠陥のあるまたは存在しないタンパク質およびその結果としての疾患の表現型(例えば代謝障害の症状の現れ)の性質が多様なので、ある所定の障害は、その特定の障害に特徴的な症状だけを一般的に示す。例えば、ファブリー病の患者は、それらに限定されないが、温度不耐、角膜渦状化、疼痛、皮膚の発疹、悪心または下痢(dirarrhea)のような上記の症状の特定の部分集合を示し得る。ゴーシェ症候群の患者は、巨脾腫、肝硬変、痙攣、緊張亢進、無呼吸、骨粗鬆症または皮膚変色を示し得る。
【0073】
本明細書で記載する1つ以上の分子を投与することに加えて、代謝障害は、適当な栄養素およびビタミン(例えば補助因子療法)、理学療法および疼痛薬物療法により処置することもできる。
【0074】
ある所定の代謝障害の特定の性質に依存して、患者は、これらの症状をいずれの年齢でも示すことができる。多くの場合では、症状は小児期または早期成人期に現れ得る。例えば、ファブリー病の症状は、早い年齢、例えば10または11歳で現れることがある。
【0075】
本明細書で用いる場合、「代謝障害を発生する危険性がある」被験体は、障害を発生する素因、すなわちアルファ−L−イズロニダーゼ、ベータ−D−ガラクトシダーゼ、ベータ−グルコシダーゼ、ベータ−ヘキソサミニダーゼ、ベータ−D−マンノシダーゼ、アルファ−L−フコシダーゼ、アリールスルファターゼB、アリールスルファターゼA、アルファ−N−アセチルガラクトサミニダーゼ、アスパルチルグルコサミニダーゼ、イズロン酸−2−スルファターゼ、アルファ−グルコサミニド−N−アセチルトランスフェラーゼ、ベータ−D−グルクロニダーゼ、ヒアルロニダーゼ、アルファ−L−マンノシダーゼ、アルファ−ノイラミニダーゼ(neurominidase)、ホスホトランスフェラーゼ、酸性リパーゼ、酸性セラミダーゼ、スフィンゴミエリナーゼ(sphinogmyelinase)、チオエステラーゼ、カテプシンKまたはリポタンパク質リパーゼのような酵素の変異の結果として代謝障害を発生する遺伝的素因を有する被験体である。明らかに、「代謝障害を発生する危険性がある」被験体は、目的の種内の被験体全てではない。
【0076】
「障害を有することが疑われる」被験体は、本明細書で記載するもののような障害の1つ以上の症状を有する被験体である。
【0077】
(ii)がん
がんは、細胞の制御されない分裂と、浸潤により隣接組織内で直接成長することまたは転移(がん細胞が血流またはリンパ系を通して輸送される場合)により離れた部位へ植え込まれることのいずれかによりそれらが広がる能力とを特徴とする。がんは、全ての年齢の人が罹患し得るが、危険性は年齢とともに増加する傾向にある。がんの型は、例えば肺がん、乳がん、結腸がん、膵臓がん、腎がん、胃がん、肝臓がん、骨がん、血液のがん、神経組織がん、黒色腫、甲状腺がん、卵巣がん、精巣がん、前立腺がん、子宮頚がん、膣がんまたは膀胱がんを含み得る。
【0078】
本明細書で用いる場合、「がんを発生する危険性がある」被検体は、がんを発生する素因、すなわち腫瘍抑制遺伝子における変異(例えばBRCA1、p53、RBまたはAPCにおける変異)のようながんを発生する遺伝的素因を有するか、またはがんをもたらし得る条件に曝露された被験体である。つまり、被験体は、変異原性または発癌性のレベルの特定の化合物(例えばアクロレイン、ヒ素、ベンゼン、ベンズ{a}アントラセン、ベンゾ{a}ピレン、ポロニウム−210(ラドン)、ウレタンまたは塩化ビニルのような煙草の煙の中の発癌性化合物)に曝露された場合、該被験体は、「がんを発生する危険性がある」ものであり得る。さらに、被験体は、例えば高い線量の紫外線またはX線照射に曝露されたかまたはパピローマウイルス、エプスタイン−バーウイルス、B型肝炎ウイルスまたはヒトT細胞白血病−リンパ腫ウイルスのような腫瘍の原因となる/腫瘍と関連するウイルスに曝露された(例えば感染した)場合、該被験体は、「がんを発生する危険性がある」ことができる。上記のことから、「がんを発生する危険性がある」被験体は、目的の種内の被験体全てではないことが明らかである。
【0079】
「がんを有することが疑われる」被験体は、がんの1つ以上の症状を有する被験体である。がんの症状は当業者に周知であり、限定されることなく、乳房腫瘤、乳頭変化、乳房嚢胞、胸部痛、体重減少、脱力、過剰な疲労、摂食困難、食欲喪失、慢性の咳、息切れの悪化、咳による血液排出(coughing up blood)、血尿、血便、悪心、嘔吐、肝臓転移、肺転移、骨転移、腹部肥満(abdominal fullness)、鼓脹、腹膜腔内の流体、膣出血、便秘、腹部膨満(abdominal distension)、結腸穿孔、急性腹膜炎(感染、発熱、疼痛)、疼痛、吐血(vomiting blood)、重度の発汗、発熱、高い血圧、貧血、下痢、黄疸、めまい、悪寒、筋肉攣縮、結腸転移、肺転移、膀胱転移、肝臓転移、骨転移、腎臓転移および膵臓転移、嚥下困難などを含む。上記のことから、「がんを有することが疑われる」被験体は、目的の種内の被験体全てではないことが明らかである。
【0080】
本明細書で記載する1つ以上の変更されたN−グリコシル化分子の投与に加えて、がんは、化学療法剤、電離放射線、免疫療法剤または温熱療法剤によっても処置できる。化学療法剤は、例えばシスプラチン、カルボプラチン、プロカルバジン、メクロレタミン、シクロホスファミド、カンプトテシン、アドリアマイシン、イホスファミド、メルファラン、クロラムブシル、ブスルファン(bisulfan)、ニトロソウレア(nitrosurea)、ダクチノマイシン、ダウノルビシン、ドキソルビシン、ブレオマイシン、プリカマイシン(plicomycin)、マイトマイシン、エトポシド、ベラパミル(verampil)、ポドフィロトキシン、タモキシフェン、タキソール、トランスプラチナ、5−フルオロウラシル、ビンクリスチン、ビンブラスチンおよびメトトレキセートを含む。
【0081】
(iii)炎症性障害
「炎症性障害」は、本明細書で用いる場合、1つ以上の物質(例えば被験体において天然に生じない物質)が、白血球細胞(例えばB細胞、T細胞、マクロファージ、単球または樹状細胞)の作用により、病的応答、例えば病的免疫応答を不適当に引き起こすプロセスのことをいう。よって、炎症性応答に関与するこのような細胞は、「炎症性細胞」という。不適当に引き起こされる炎症性応答は、外来物質(例えば抗原、ウイルス、細菌、真菌)が被験体内または被験体上に存在しないものであり得る。不適当に引き起こされる応答は、自己成分(例えば自己抗原)が炎症性細胞により標的にされるものであり得る(例えば多発性硬化症のような自己免疫障害)。不適当に引き起こされる応答は、大きさまたは持続期間が不適当である応答、例えばアナフィラキシーでもあり得る。つまり、不適当に標的にされる応答は、微生物感染(例えばウイルス、細菌または真菌)の存在によることができる。炎症性障害の型(例えば自己免疫疾患)は、それらに限定されないが、変形性関節症、関節リウマチ(RA)、脊椎関節症(spondyloarthropathy)、POEMS症候群、クローン病、多中心性キャッスルマン病、全身性エリテマトーデス(SLE)、多発性硬化症(MS)、筋ジストロフィー(MD)、インスリン依存性糖尿病(IDDM)、皮膚筋炎、多発性筋炎、ギランバレー症候群のような炎症性神経障害、ウェゲナー肉芽腫症のような血管炎、結節性多発動脈炎、リウマチ性多発筋痛、側頭動脈炎、シェーグレン症候群、ベーチェット(Bechet’s)病、チャーグ−ストラウス症候群または高安動脈炎を含み得る。鼻炎、副鼻腔炎、じんま疹(urticaria)、じんま疹(hives)、血管浮腫、アトピー性皮膚炎、食物アレルギー(例えばナッツアレルギー)、薬物アレルギー(例えばペニシリン)、昆虫アレルギー(例えばハチ刺傷に対するアレルギー)のような特定の型のアレルギーまたは肥満細胞症も炎症性障害に含まれる。炎症性障害は、潰瘍性大腸炎および喘息も含み得る。
【0082】
「炎症性障害を発生する危険性がある」被験体とは、1種以上の炎症性障害の家族歴(例えば1種以上の炎症性障害に対する遺伝的素因)を有する被験体または1種以上の炎症誘導性状態に曝露された被験体のことをいう。例えば、被験体は、それらに限定されないが、ブドウ球菌エンテロトキシン(SE)、streptococcus pyogenes外毒素(SPE)、staphylococcus aureus毒素ショック症候群毒素(TSST−1)、連鎖球菌分裂促進性外毒素(SME)および連鎖球菌スーパー抗原(SSA)のようなウイルスまたは細菌のスーパー抗原に曝露された可能性がある。上記のことから、「炎症性障害を発生する危険性がある」被験体は、目的の種内の被験体全てではないことが明らかである。
【0083】
「炎症性障害を有することが疑われる」被験体は、炎症性障害の1つ以上の症状を示す被験体である。炎症性障害の症状は当技術分野で周知でありそれらに限定されないが、発赤、腫脹(例えば腫脹関節)、触れると温かい関節、関節痛、こわばり、関節機能の喪失、発熱、悪寒、疲労、エネルギーの喪失、頭痛、食欲喪失、筋肉こわばり、不眠、そう痒、鼻づまり、くしゃみ、咳、めまい、発作または疼痛のような1つ以上の神経性の症状を含む。上記のことから、「炎症性障害を有することが疑われる」被験体は、目的の種内の被験体全てではないことが明らかである。
【0084】
本明細書で記載する1つ以上の分子の投与に加えて、炎症性障害は、非ステロイド性抗炎症薬(NSAID)、疾患修飾性抗リウマチ薬(DMARD)、生体応答修飾物質またはコルチコステロイドによっても処置できる。生体応答修飾物質は、例えば抗TNF剤を含む。抗TNF剤の非限定的な例は、アダリムマブ(adulimumab)、インフリキシマブもしくはエタネルセプトのような可溶性TNF受容体またはTNFに特異的な抗体を含む。
【0085】
変更されたN−グリコシル化分子(またはその薬学的組成物)のいずれかを用いて本明細書で記載する障害のいずれかを処置する(例えばその1つ以上の症状を予防するかまたは改善する)ために適切な方法は、以下の項に示す。
【0086】
薬学的組成物および処置方法
所望のN−グリカンを有するように改変された標的分子は、治療有効量の分子と、1つ以上のアジュバント、賦形剤、キャリアおよび/または希釈剤とを含有する薬学的組成物に組み込むことができる。許容される希釈剤、キャリアおよび賦形剤は、典型的に、受容者の恒常性(例えば電解質バランス)に有害に影響しない。許容されるキャリアは、生体適合性、不活性または生体吸収性の塩、緩衝剤、オリゴ糖もしくは多糖、ポリマー、粘度改善剤、防腐剤などを含む。1つの例示的なキャリアは、生理食塩水である(0.15M NaCl、pH7.0〜7.4)。別の例示的なキャリアは、50mMリン酸ナトリウム、100mM塩化ナトリウムである。薬学的組成物の処方および投与のための技術についてのさらなる詳細は、例えばRemington’s Pharmaceutical Sciences(Maack Publishing Co.、Easton、Pa.)で見出すことができる。補足の活性化合物も組成物に組み込むことができる。
【0087】
N−グリカンを有する分子を含有する薬学的組成物の投与は、全身的または局部的であり得る。薬学的組成物は、それらが非経口および/または非経口でない投与に適切であるように処方できる。具体的な投与様式は、皮下、静脈内、筋肉内、腹腔内、経皮、髄腔内、経口、直腸、頬側、局所、鼻、眼、関節内、動脈内、くも膜下、気管支、リンパ節、膣および子宮内の投与を含む。
【0088】
投与は、薬学的組成物のボーラスの周期的な注射によることができるか、あるいは外的(例えばIVバッグ)もしくは内的(例えば生体浸食性インプラント、バイオ人工臓器(bioartificial organ)または埋め込まれた変更されたN−グリコシル化分子生成細胞のコロニー)であるリザーバからの静脈内または腹腔内投与による不断または連続的であり得る。例えば米国特許第4,407,957号、第5,798,113号および第5,800,828号を参照されたい。薬学的組成物の投与は、ポンプ(例えばAnnals of Pharmacotherapy、27巻:912頁(1993年);Cancer、41巻:1270頁(1993年);Cancer Research、44巻:1698頁(1984年)を参照されたい);マイクロカプセル封入(例えば米国特許第4,352,883号;第4,353,888号;および第5,084,350号を参照されたい);連続放出ポリマーインプラント(例えばSabel、米国特許第4,883,666号を参照されたい);マクロカプセル封入(例えば米国特許第5,284,761号、第5,158,881号、第4,976,859号および第4,968,733号ならびにPCT特許出願公開WO92/19195、WO95/05452を参照されたい);皮下、静脈内、動脈内、筋肉内もしくはその他の適切な部位のいずれかでの注射;またはカプセル、液体、錠剤、丸剤もしくは延長放出処方物での経口投与のような適切な送達手段を用いて達成できる。
【0089】
非経口送達系の例は、エチレン−酢酸ビニルコポリマー粒子、浸透圧ポンプ、埋め込み型注入系、ポンプ送達、カプセル封入細胞送達、リポソーム送達、針送達注射、針なし注射、ネブライザー、エアゾール器(aerosolizer)、エレクトロポレーションおよび経皮パッチを含む。
【0090】
非経口投与のために適切な処方物は、変更されたN−グリコシル化分子の滅菌水性調製物を簡便に含有し、これは、好ましくは、受容者の血液と等張である(例えば生理食塩水溶液)。処方物は、単位用量または複数回用量の形態で提供し得る。
【0091】
経口投与のために適切な処方物は、それぞれが所定量の変更されたN−グリコシル化分子を含有するカプセル剤、カシェ剤、錠剤もしくはロゼンジのような分かれた単位、またはシロップ剤、エリキシル剤、乳剤もしくは飲料(draught)のような水性流体(liquor)または非水性の液体中の懸濁剤で提供し得る。
【0092】
局所投与のために適切なN−グリカンを有する分子は、哺乳動物(例えばヒト患者)に、例えばクリーム、スプレー、フォーム、ゲル、軟膏剤(ointment)、軟膏剤(salve)またはドライラブ(dry rub)として投与できる。ドライラブは、投与部位にて再水和できる。このような分子は、包帯、ガーゼまたはパッチに直接注入(例えば浸して乾燥する)することもでき、これらは、次いで、局所的に施用できる。このような分子は、半液体、ゲル化または完全な液体の状態で、局所投与用の包帯、ガーゼまたはパッチ中に維持することもできる(例えば米国特許第4,307,717号を参照されたい)。
【0093】
治療有効量の薬学的組成物は、それを必要とする被験体に、当業者が確認できる投与量計画(dosage regimen)において投与できる。例えば、組成物は、被験体に、例えば全身的に、1投薬あたり被験体の体重1kgあたり0.01μgから10,000μgまでの投与量で投与できる。別の例では、投与量は、1投薬あたり被験体の体重1kgあたり1μgから100μgまでである。別の例では、投与量は、1投薬あたり被験体の体重1kgあたり1μgから30μgまで、例えば1投薬あたり被験体の体重1kgあたり3μgから10μgまでである。
【0094】
治療効力を最適化するために、N−グリカンを含有する分子は、まず、異なる投薬計画(dosing regimen)で投与できる。単位用量および計画は、例えば哺乳動物の種、その免疫状態、哺乳動物の体重を含む因子に依存する。典型的に、組織におけるこのような分子のレベルは、例えば所定の処置計画の効力を決定するために、臨床試験手順の一部として適当なスクリーニングアッセイを用いて監視できる。
【0095】
分子の投薬の頻度は、医療従事者(例えば医師または看護士)の熟練および臨床的判断の範囲内である。典型的に、投与計画(administration regimen)は、最適投与パラメータを確立し得る臨床試験により確立される。しかし、従事者は、被験体の年齢、健康、体重、性別および医療状態に従ってそのような投与計画を変更してよい。投薬頻度は、処置が予防的または治療的であるかに依存して変更できる。
【0096】
このような分子またはその薬学的組成物の毒性および治療効力は、公知の薬学的手順により、例えば細胞培養または実験動物において決定できる。これらの手順は、例えばLD50(集団の50%に対して致死的な用量)およびED50(集団の50%において治療的に有効な用量)を決定するために用いることができる。毒性作用と治療効果との間の用量の比は、治療指数であり、これは、LD50/ED50の比として表すことができる。高い治療指数を示す薬学的組成物が好ましい。毒性の副作用を示す薬学的組成物を用いることができるが、そのような化合物を罹患組織の部位に向かわせるための送達系を設計して、正常細胞(例えば非標的細胞)への潜在的な損傷を最小限にし、そのことにより副作用を低減するように注意を払うべきである。
【0097】
細胞培養アッセイおよび動物研究から得られたデータは、適当な被験体(例えばヒト患者)において用いるための投与量の範囲を処方する場合に用いることができる。このような薬学的組成物の投与量は、通常、ほとんどまたは全く毒性を有さずにED50を含む循環濃度の範囲内にある。投与量は、採用する投与形態および利用する投与経路に依存してこの範囲内で変更し得る。本明細書で記載するようにして用いる薬学的組成物(例えば被験体における代謝障害を処置するため)について、治療有効用量は、最初に細胞培養アッセイから見積もることができる。用量は、動物モデルにおいて処方して、細胞培養で決定したIC50(すなわち、症状の最大阻害の半分を達成する薬学的組成物の濃度)を含む循環血漿濃度範囲を達成できる。このような情報を用いて、ヒトにおける有用な用量をより正確に決定できる。血漿におけるレベルは、例えば高性能液体クロマトグラフィーにより測定できる。
【0098】
本明細書で定義するように、N−グリカンを含有する分子の「治療有効量」は、処置した被験体において医療的に望ましい結果(例えば代謝障害の1つ以上の症状の改善)を生じることができる分子の量である。治療有効量(すなわち有効投与量)は、被験体または試料重量1キログラムあたりミリグラムまたはマイクログラム量の化合物を含むことができる(例えば1キログラムあたり約1マイクログラムから1キログラムあたり約500ミリグラム、1キログラムあたり約100マイクログラムから1キログラムあたり約5ミリグラムまたは1キログラムあたり約1マイクログラムから1キログラムあたり約50マイクログラム)。
【0099】
被験体は、任意の哺乳動物、例えばヒト(例えばヒト患者)または非ヒト霊長類(例えばチンパンジー、ヒヒまたはサル)、マウス、ラット、ウサギ、モルモット、アレチネズミ、ハムスター、ウマ、ある型の家畜(例えばウシ、ブタ、ヒツジまたはヤギ)、イヌ、ネコまたはクジラであり得る。
【0100】
本明細書で記載する分子またはその薬学的組成物は、被験体に、別の処置、例えば代謝障害(例えばリソソーム蓄積症)のための処置との併用療法として投与できる。例えば併用療法は、被験体(例えばヒト患者)に、代謝障害(例えばリソソーム蓄積症)を有するかまたはその障害を発生する危険性がある(もしくは有することが疑われる)被験体に対して治療利益をもたらす1種以上の追加の薬剤を投与することを含むことができる。つまり、化合物または薬学的組成物と、1種以上の追加の薬剤とは、同時に投与できる。代わりに、分子をまず投与して、1種以上の追加の薬剤を2番目に投与するか、またはその逆が可能である。
【0101】
以前の療法が特に毒性である場合(例えば著しい副作用プロファイルを有する代謝障害の処置)、本明細書で記載する分子の投与を用いて、以前の療法の量を、同じまたは改善された治療利益を与えるのに十分であるが毒性がないレベルまで埋め合わせ(offset)かつ/または少なくすることができることが認識される。
【0102】
本明細書で記載する薬学的組成物のいずれも、投与についての使用説明書と一緒に容器、パックまたはディスペンサーに含めることができる。
【0103】
以下は、本発明の実施の例である。これらは、いずれの様式でも本発明の範囲を限定すると解釈されない。
【実施例】
【0104】
表2は、以下に記載する実験で用いた全ての株のリストを含む。表2では、MH=HDELタグ付きα−1,2−マンノシダーゼ;ζ=ゼータ配列による無作為組み込み;ドッキングΔ=特定の遺伝子座への組み込み;および(H)=ハイグロマイシン耐性。
【0105】
【表2−1】
【0106】
【表2−2】
【0107】
【表2−3】
【0108】
【表2−4】
【0109】
【表2−5】
【0110】
【表2−6】
(実施例1)Yarrowia lipolytica OCH1破壊
糖鎖工学的に改変したタンパク質発現株の作製を、Yarrowia lipolytica pold lnuga株(leu2−、ura3−、gut2−およびade2−の栄養要求性を有する株)で行った。Yarrowia lipolyticaにおいてOCH1(GenBank受託番号AJ563920)遺伝子をノックアウトするための方策は、LIP2遺伝子について記載されたようにして設定した(Fickersら、2003年J Microbiol Methods.55巻(3号):727〜37頁)。OCH1遺伝子について従った遺伝子構築方策は、米国特許出願公開第20090069232−A1号に記載されている。得られたベクターをpYlOCH1 PUT TOPO(図1B)と称した。
【0111】
OCH1 KO断片を、プラスミドから、SpeI/Bst1107I制限消化により単離し、Yarrowia lipolytica pold lnuga株を形質転換した。いくつかのウラシル原栄養性株を得て、PCRによりゲノムDNA(gDNA)上でプライマーYloch1 prom fw(5’−TCGCTATCACGTCTCTAGC−3’、配列番号1)およびYloch1 term rev(5’−ACTCTGTATACTTGTATGTACTGTGAGAC−3’、配列番号2)を用いてスクリーニングして、プラスミドのゲノム組み込みを分析した。正しいサイズの断片(すなわち、2328bp対野生型での1894bp)が、試験したいくつかのクローンについて増幅された。OCH1遺伝子のノックアウトも、増殖培地中に分泌された全糖タンパク質プール(=セクレトーム)のN−グリカン分析により確認した:Man8GlcNAc2構造は、糖プロファイル内の優勢なN−グリカンになった(図2)。このプロファイルは、より高い量のMan9GlcNAc2とともにより高い数のマンノース残基さえ有するいくつかの構造を含有する野生型株のものとは異なる(後者は、Och1p活性の結果として追加のマンノースを含有する可能性が高い)。
【0112】
URA3遺伝子を除去するために、陽性Δoch1クローン(G013と称する、表2を参照されたい)を、Creリコンビナーゼについての発現カセットを含有するエピソームプラスミドpUB4−Cre(Fickersら、2003年、既出)で形質転換した。URA3遺伝子の除去は、プライマーYloch1 prom fwおよびYloch1 term rev(上記を参照されたい)を用いるgDNAのPCRによりスクリーニングした。URA3マーカーが切り出されたクローンでは、2328bpのバンドはもはや増幅されなかった。代わりに、1075bpのPCR断片(excl.URA3)が得られた。陽性クローンは、セクレトームのN−グリカンレベルにおいて確かめられたが、該クローンはキュアしていない株のものと非常によく似たプロファイルを示す(図2)。キュアした株の1つ(G014と称する、表2を参照されたい)を、さらなるN−グリカン工学的改変のために選択した。
【0113】
(実施例2)無作為組み込みまたは標的化/ドッキング組み込みのいずれかによるER保持α−1,2−マンノシダーゼの過剰発現
Δoch1株により発現された糖タンパク質と結合したMan5GlcNAc2の作製を可能にするために、α−1,2−マンノシダーゼを発現させて、Man8GlcNAc2をMan5GlcNAc2(すなわち、ゴルジ型α−1,2−マンノシダーゼ活性)に切断した。このようなマンノシダーゼは、分泌系に向けられるべきである。S.cerevisiaeのプレプロ接合因子と融合し、それをERに局在化させるためにHDEL配列(配列番号21)でタグ付きTrichoderma reeseiのα−1,2−マンノシダーゼ(Genbank受託番号AF212153)は、Pichia pastorisにおいてならびにTrichoderma reeseiおよびAspergillus nigerにおいてin vivoでMan8GlcNAc2をMan5GlcNAc2に刈り込むことができる。HDELタグ付きT.reeseiα−1,2−マンノシダーゼのコドン最適化バージョンがY.lipolytica LIP2プレシグナル配列と融合し、TEFI、Hp4d(Madzakら、2000年、J. Mol. Microbiol. Biotechnol.2巻:207〜216頁)、GAPまたはPOX2プロモーターのいずれかの転写制御下に配置された発現構築物を作製した。これらのプラスミドの構築方策は、米国特許出願公開第20090069232−A1号に記載されている。
【0114】
Hp4d resp.TEFIプロモーターの転写制御下のマンノシダーゼを有するこれらのベクターのうち2つ、pYLHUXdL2preManHDELおよびpYLTUXdL2preManHDEL(図3)を用いて、G014株(実施例1に由来する)を形質転換した。ベクターをNotIで消化して、ゼータ配列によるゲノムへの無作為組み込みを可能にした。URA3原栄養性形質転換体を、N−グリカン分析のために選択した。いくつかの形質転換体は、Man8GlcNAc2からMan5GlcNAc2への明確な変換を示す(図4)。TEF1プロモーター制御下でマンノシダーゼを発現するクローンは、緩慢で塊状の増殖表現型を示したので(これらのクローンのうちの1つをG016と称した)、糖鎖工学的改変のさらなるステップを、遺伝子がHp4d転写制御下にある株のバックグラウンド(strain background)において行った。
【0115】
ManHDELをhp4dプロモーターの制御下で発現する1つの陽性クローン(G018)を選び、これからURA3マーカーを、Cre−リコンビナーゼを発現するプラスミドpRRQ2(Richardら、2001年J. Bacteriol.183巻:3098〜3107頁)の一過性形質転換によりキュアした。いくつかのura3−クローンを手順の後に選択し、セクレトーム上で明確なMan5GlcNAc2プロファイルを示す1つのクローン(G036)を、さらなる工学的改変作業のために用いた(図4)。このクローンのサザン分析により、1つの無作為に組み込まれたマンノシダーゼ発現カセットの存在が明らかになった。このサザン分析は、HindIII消化ゲノムDNAについて、プライマーMan for(5’−GCCTTCCAGACCTCTTGGAACGCCTACCACC−3’、配列番号22)およびMan rev(5’−GCCAGGTGGCCGCCTCGTCGAGAAGAAGATCG−3’、配列番号23)を用いて作製したDIG標識マンノシダーゼ特異的PCR断片を用いて行った。
【0116】
代替の方策では、YarrowiaゲノムのLEU2またはAXP1遺伝子座のいずれかへのHp4d駆動型マンノシダーゼ発現カセットの標的化組み込みを可能にする2つの構築物を作製した。これらのプラスミド、JME926_pPTLeu2−ADE2ex−Hp4dManHDEL(Yl)およびOXYP289_pPTAxp1−LEU2ex−Hp4dManHDEL(Yl)の構築を、図5に記載する。G014株への形質転換の前に、両方の構築物をNotIで消化し、各発現カセットを単離した。選択したADE2原栄養性クローンは、マンノシダーゼ発現カセットをLEU2遺伝子座に潜在的に組み込み、一方、LEU2原栄養体は、カセットをAXP1遺伝子座に潜在的に組み込んだ。形質転換体は、サザン分析により確かめて、ゲノムへの正しいターゲティングを評価した。これは、BamHI消化(LEU2遺伝子座への組み込み)またはHindIII消化(AXP1遺伝子座への組み込み)ゲノムDNAについて、プライマーMan for(5’−GCCTTCCAGACCTCTTGGAACGCCTACCACC−3’、配列番号22)およびMan rev(5’−GCCAGGTGGCCGCCTCGTCGAGAAGAAGATCG−3’、配列番号23)を用いて作製したDIG標識マンノシダーゼ特異的PCR断片を用いて行った。選択したクローンは、分泌された糖タンパク質上に合成されたN−グリカンの性質についても確かめた。ほとんどの場合では、正しく標的にされたHp4d駆動型α−1,2−マンノシダーゼ発現は、優勢にMan5GlcNAc2オリゴ糖の合成をもたらした(図6)。それぞれのターゲティング遺伝子座について、1つのマンノシダーゼ発現クローン(LEU2ドッキングの場合はG046;AXP1ドッキングの場合はG053)を、G046株についてプラスミドpRRQ2およびG053株についてpUB4−Creを用いるCreリコンビナーゼの一過的発現によるキュアリングのために選択した。得られたキュアした株(それぞれG055およびG054)を、サザンブロッティングにより再度確かめ、それらのMan5GlcNAc2プロファイルを、DSΑ−FACEを用いるN−グリカン分析により確認した。
【0117】
(実施例3)GlcNAcトランスフェラーゼIの発現
Yarrowiaコドン最適化配列を、S.cerevisiae Kre2タンパク質(SwissProt 受託番号P27809)の最初の100個のN末端アミノ酸と、それに続くヒトGlcNAcトランスフェラーゼI(SwissProt 受託番号P26572)の触媒ドメインとからなる融合タンパク質(図7、配列番号3および配列番号4)の発現のために作製した。酵母Kre2pの100個のN末端アミノ酸は、触媒GnT Iドメインのためのゴルジ局在化シグナルとして働く。この様式で、GnT I融合タンパク質が、分泌経路において、ER保持HDELタグ付きα−1,2−マンノシダーゼよりも後に局在化されることを確実にして、酵素がタンパク質結合N−グリカンをMan5GlcNAc2からGlcNAcMan5GlcNAc2に変換できるようにする。融合タンパク質の発現のためのコドン最適化合成遺伝子を、TEF1またはHp4dプロモーターのいずれかの転写制御下に配置して、プラスミドpYLTmAXhGnTIおよびpYLHp4mAXhGnTIを得た。構築方策を図8に示す。Kre2−GnT I融合タンパク質の機能的発現は、利用可能なMan5GlcNAc2グリカン上へのβ−1,2結合GlcNAc残基の付加をもたらして、GlcNAcMan5GlcNAc2の合成をもたらすはずである。
【0118】
プラスミドpYLTmAXhGnTIおよびpYLHp4mAXGnTIをNotI消化した後に、その分泌タンパク質上にMan5GlcNAc2 N−グリカンを生成することが公知であるG036株を形質転換した(実施例2を参照されたい)。形質転換体を、ウラシル原栄養性について選択した。これらのクローンのいくつかのセクレトームについてのN−グリコシル化プロファイルの分析は、N−グリカンパターンの明確な変化を示した:Man5GlcNAc2は、著しく低減され、より高い分子量(約1グルコース単位過剰)を有するN−グリカンを表す新しいピークが出現した。単離N−グリカンを、末端β結合GlcNAc残基を除去できる酵素であるナタマメβ−N−アセチルヘキソサミニダーゼで処理すると、新しいN−グリカンがGlcNAcMan5GlcNAc2であることが示された:その新しいピークは消失し、Man5GlcNAc2に完全に変換された(図9)。用いた培養方法に依存して、全N−グリカンプールの約70%がGlcNAcMan5GlcNAc2であることが証明された(利用可能なMan5GlcNAc2のおよそ77%が変換された)。
【0119】
Kre2−GnT I融合タンパク質をTEF1プロモーターの制御下で発現する1つの形質転換体をG040株と命名し、さらなる使用のために選択した。この株のサザンブロットによるゲノム解析は、1つの発現カセットの存在を示した。サザン分析は、BamHI消化ゲノムDNAについて、プライマー5’−GGATGATCACACAATGGCCCTGTTTCTG−3’(配列番号5)および5’−TGCTCTAGACTAGTTCCAAGAGGGGTC−3’(配列番号6)を用いて作製したDIG標識GnT I特異的PCR断片を用いて行った。G040株対1〜3コピー(同じサザンブロットにより確認)のHp4d駆動型Kre2−GnT I発現カセットを有する株のセクレトームについてのグリコシル化プロファイルの分析は、GlcNAc移動能力について著しい違いを示さなかった。
【0120】
(実施例4)マンノシダーゼIIの発現
Yarrowiaコドン最適化配列を、S.cerevisiae Mnn2タンパク質(SwissProt 受託番号P38069)の最初の36個のN末端アミノ酸と、それに続くDrosophila melanogasterマンノシダーゼII(SwissProt 受託番号Q24451)の触媒ドメインとからなる融合タンパク質(図10、配列番号7および配列番号8)の発現のために作製した。酵母Mnn2の36個のN末端アミノ酸は、触媒Man IIドメインのためのゴルジ局在化シグナルとして働く。この様式で、Mnn2−Man II融合タンパク質が、分泌経路において、Kre2−GnT I融合タンパク質と同じまたはそれより後の位置にさえ局在化されることを確実にして、よって、GlcNAcMan5GlcNAc2をGlcNAcMan3GlcNAc2に変換できた。融合タンパク質の発現のためのYarrowiaコドン最適化合成遺伝子を、TEF1プロモーターの転写制御下に配置して、プラスミドpYLTmAXDmManIIおよびpYLTmAXDmManII(LEU2ex)を得た。構築方策を図11に示す。
【0121】
プラスミドpYLTmAXDmManII(LEU2ex)をNotI消化した後に、その分泌タンパク質上にGlcNAcMan5GlcNAc2 N−グリカンを生成することが公知であるG040株を形質転換した(実施例3を参照されたい)。形質転換体を、ロイシン原栄養性について選択した。これらのクローンのいくつかのセクレトームについてのN−グリコシル化プロファイルの分析は、N−グリカンパターンの変化を示した:約2グルコース単位のより低い分子量を有するN−グリカンを表す新しいピークが出現し、これにより、GlcNAcMan3GlcNAc2の形成を示すことができ、よって、部分的マンノシダーゼII活性を示し得た。また、Man5GlcNAc2(すなわちピークのショルダー)とほとんど同じ位置に泳動される別のピークが出現し、これは、潜在的にGlcNAcMan4GlcNAc2を表す。後者の構造は、マンノシダーゼII活性が2つではなく1つのマンノース残基だけを除去した部分的な刈り込み事象の結果である可能性がある。単離N−グリカンをナタマメβ−Ν−アセチルヘキソサミニダーゼで処理すると、グリカンパターンが約1グルコース単位の左側へのシフトをもたらし、よって、末端GlcNAc残基の喪失によるより高い電気泳動移動度をもたらした(図12)。このことにより、機能的マンノシダーゼII活性の発現によるGlcNAcMan5GlcNAc2からのGlcNAcMan4GlcNAc2およびGlcNAcMan3GlcNAc2の作製がさらに確認される。用いた培養方法に依存して、全N−グリカンプールの約15%が、GlcNAcMan3GlcNAc2であることが証明された:利用可能なGlcNAcMan5GlcNAc2のおよそ35%が1または2個のマンノース残基を失い、20%がGlcNAcMan3GlcNAc2に完全に刈り込まれた。
【0122】
(実施例5)ガラクトシルトランスフェラーゼIの発現
末端ガラクトース残基を有するN−グリカンの合成は、分泌経路内の機能的で良好に局在化されたガラクトシルトランスフェラーゼの存在だけでなく、この酵素により用いられるドナー基質であるUDP−Galの利用可能性にも依存する。UDP−GlcとUDP−GlcNAcは、酵母生物のゴルジ装置中で十分に利用可能であると一般的に考えられているが、このことはUDP−Galについてはあまり知られていない。糖鎖工学的改変中の潜在的なUDP−Gal欠乏を克服するために、Pichia pastorisにおいて、Schizosaccharomyces pombe UDP−Glc−4−エピメラーゼ(GAL10様遺伝子SPBC365.14c(SwissProt 受託番号Q9Y7X5)によりコードされる)とヒトβ−1,4−ガラクトシルトランスフェラーゼI(GalT I)(SwissProt 受託番号PI5291)の触媒ドメインとの間の融合タンパク質を酵母ゴルジ装置に向かわせる試みが以前になされた(Bobrowiczら、Glycobiology14巻(9号):757〜766頁、2004年)。分泌経路におけるGal10p−GalT I融合タンパク質の局在化を、好ましくはGlcNAc移動およびマンノシダーゼII活性が分泌のために定められたタンパク質のN−グリカンに対して既に作用した位置にて、S.cerevisiae Mnn2pの最初の46個のN末端アミノ酸をN末端ターゲティングシグナルとして用いることにより遂行した。
【0123】
よって、Yarrowiaコドン最適化配列を、S.cerevisiae Mnn2タンパク質の最初の46個のN末端アミノ酸と、それに続くS.pombe Gal10様タンパク質およびヒトGalT Iの触媒ドメインとからなる融合タンパク質(図13)の発現のために作製した。得られた合成遺伝子を、TEF1プロモーターの転写制御下に配置して、プラスミドpYLTmAXSpGal10hGalTIおよびpYLTmAXSpGal10hGalTI(ADE2ex)を得た。構築方策を図14に示す。
【0124】
プラスミドpYLTmAXSpGal10hGalTI(ADE2ex)をNotI消化した後に、その分泌タンパク質上にGlcNAcMan5GlcNAc2 N−グリカンを生成することが公知であるG040株を形質転換した(実施例3を参照されたい)。形質転換体を、それらのアデニン原栄養性について選択した。これらのクローンのいくつかのセクレトームについてのN−グリコシル化プロファイルの分析は、N−グリカンパターンの変化を示した:Man7GlcNAc2とMan8GlcNAc2との間の位置に泳動される新しいピークが出現する(図15)。N−グリカンを、Streptococcus pneumoniaβ−1,4−ガラクトシダーゼで処理すると、このピークがGalGlcNAcMan5GlcNAc2を表すことが示される。なぜなら、このin vitro消化は、この新しいピークの消失と、GlcNAcMan5GlcNAc2の等しく高い増加とをもたらすからである。
【0125】
この設定を用い、かつ増殖条件に依存して、GlcNAcMan5GlcNAc2の約75%がGalGlcNAcMan5GlcNAc2に変換された。ガラクトシル化構造の全量は、全N−グリカンプールの約25%を占めた。in vitroα−1,2−マンノシダーゼ消化から、しかし、相当量の高マンノースN−グリカンがMan5GlcNAc2に変換されなかったことが明確である(図15)。用いる培養培地に依存して、Man5GlcNAc2からGlcNAcMan5GlcNAc2への変換率も、親のG040株で観察されるものより低い。このことは、このMnn2−Gal10−GalT I融合タンパク質の形質転換体について観察されたより緩慢な増殖速度と関連する可能性が高い。
【0126】
(実施例6)YlALG3のノックアウトおよびYlALG6の同時過剰発現
Man3GlcNAc2プラットフォームの作製を可能にするために、G036株のALG3遺伝子(pold lnuga Δoch1+Hp4d駆動型α−1,2−マンノシダーゼ)は、不活性化する必要がある。このことはER局在化Alg3pα−1,6−マンノシルトランスフェラーゼ活性の喪失をもたらし、脂質結合N−グリカン前駆体構造の組成を変化させる。この構造を新生ポリペプチド鎖のN−グリコシル化部位に移動させることにより、酵母グリコシル化プロファイルを、哺乳類様N−グリカン構造に、マンノシダーゼIIの発現を必要とすることなく変換できる。しかし、この新しい脂質結合構造は、新生ポリペプチドにそれほど効率的に移動されないので、潜在的なタンパク質の低グリコシル化(underglycosylation)をできる限り減らすために、Yarrowia ALG6遺伝子(ER局在化Alg6pα−1,3−グルコシルトランスフェラーゼをコードする)を同時に過剰発現させる必要がある。
【0127】
pYLalg3PUT−ALG6と称するベクター(図16)が以前に構築され、YlALG3のノックアウトおよびそれと同時のYlALG6のHp4d駆動型過剰発現を可能にした。米国特許出願公開第20090069232−A1号を参照されたい。このノックアウト/ノックインカセットを含有するこのベクターのNotI/PacI断片でYarrowia lipolytica G036を形質転換し、形質転換体を、それらのウラシル原栄養性に基づいて選択した。構築物が正しく組み込まれたクローンを、セクレトームについてのN−グリカン分析により直接スクリーニングした。80のスクリーニングしたクローンのうち、2つのクローンが、ERにあるα−1,2−マンノシダーゼを発現する株におけるYlALG3の不活性化に一致し得るN−グリコシル化プロファイルを示した。少量のMan3GlcNAc2グリカンとは別に、まだいくらかのMan4’GlcNAc2およびMan5’GlcNAc2と、相当量のグルコシル化N−グリカン(GlcMan5’GlcNAc2およびGlc2Man5’GlcNAc2)とがあった。後者は、グルコシダーゼIIによる非効率的な刈り込みの結果である(GrinnaおよびRobbins、J. Biol. Chem. 255巻、2255〜2258頁、1980年)。Man4GlcNAc2およびMan5GlcNAc2の構造の性質は、N−グリカンをα−1,2−マンノシダーゼでin vitro処理することにより確認した(図17)。用いた増殖条件に依存して、Man3GlcNAc2のレベルは、全N−グリカンプールの60%まで増加でき、グルコシル化ピークは、α−1,2−マンノシダーゼに対して非感受性であり、ナタマメα−マンノシダーゼ処理(α−1,2−、α−1,3−およびα−1,6結合マンノース残基に対して作用できる特異的でないα−マンノシダーゼ)に対してわずかに感受性であるだけであった。これとは対照的に、後者の酵素は、生じたMan3GlcNAc2をMan1GlcNAc2に変換する(図17)。
【0128】
2つの陽性形質転換体のうちの1つをG039と称し、さらなる糖鎖工学的改変作業のために用いた。この株を、Cre−リコンビナーゼを発現するpRRQ2で一過的に形質転換して、G036をベクターpYLalg3PUT−ALG6で形質転換した際に導入されたURA3マーカーのキュアリングを可能にした。分析は、グリコシル化プロファイルが、キュアリング後に同じままであることを示す。1つのキュアした株を、さらなる使用のために選択して、G045と称した。
【0129】
(実施例7)Man3GlcNAc2生成株でのGlcNAcトランスフェラーゼIの発現
実施例3で行ったことと同様に、GnT I活性の導入を、Kre2−GnT I融合タンパク質の発現により遂行した。GnT Iについてのこのような発現構築物の無作為組み込みは、3つの方法で遂行した:1)キュアしていないG039株(実施例6を参照されたい)を、NotI消化ベクターpYLTmAXhGnTI(Hygr ex)で形質転換し、GnT I発現クローンを、選択プレートに加えられた300μg/mlのハイグロマイシンで生存するそれらの能力に基づいて最初に選択した、2)キュアしたG045株(実施例6を参照されたい)を、NotI消化ベクターpYLTmAXhGnTI(これもまた実施例3を参照されたい)で形質転換し、GnT I発現クローンを、それらのウラシル原栄養性に基づいて最初に選択した、または3)キュアしたG045株(実施例6を参照されたい)を、NotI消化ベクターpYLHp4mAXhGnTIで形質転換し、GnT I発現クローンを、それらのウラシル原栄養性に基づいて最初に選択した。pYLTmAXhGnTI(Hygr ex)についての構築方策を、図18に示す。プラスミドpYLTmAXhGnTI(Hygr ex)およびpYLTmAXhGnTIを用いる場合、GnT Iの発現は、TEF1プロモーターの転写制御下であった;プラスミドpYLHp4mAXhGnTIを用いる場合、GnT I発現は、Hp4dプロモーターの制御下であった。
【0130】
G039をpYLTmAXhGnTI(Hygr ex)で形質転換することにより、期待されたよりも長いインキュベーション期間の後に、培養プレート上に3つのクローンだけが出現した。しかし、これらのクローンのセクレトームのN−グリコシル化プロファイルの分析は、N−グリカンパターンの明確な変化を示した:非形質転換G039株に存在するMan3GlcNAc2は、著しく低減されたかまたはほぼ完全に存在しなかったが、より高い分子量(約1グルコース単位過剰)を有するN−グリカンを表す新しいピークが出現した。単離N−グリカンを、末端β結合GlcNAc残基を除去できる酵素であるナタマメβ−Ν−アセチルヘキソサミニダーゼで処理すると、新しいN−グリカンが実際にGlcNAcMan3GlcNAc2であることが示された。新しいピークは消失し、Man3GlcNAc2に完全に変換された(図19)。評価した形質転換体のうちの1つを、さらなる糖鎖工学的改変作業のために用い、G047と命名した。同様の結果が、キュアしたG045株をpYLTmAXhGnTI(G048)またはpYLHp4mAXhGnTI(G056)で形質転換した場合にも得られた。G056株は、プラスミドpRRQ2を用いるCreリコンビナーゼの一過的発現によるキュアリングのために選択した。得られた株は、G058と称した。
【0131】
用いた培養方法に依存して、G047株の全N−グリカンプールの約70%がGlcNAcMan3GlcNAc2であり、Glc1−2Man5’GlcNAc2がいくらか残存し、Man3GlcNAc2がほとんど存在しないことが証明された(変換率>>90%)(図19)。高い変換率にかかわらず、1コピーのGnT I発現カセットだけがこの株でサザンブロットにより同定できた。サザン分析は、BamHI消化ゲノムDNAについて、プライマー5’−GGATGATCACACAATGGCCCTGTTTCTG−3’(配列番号11)および5’−TGCTCTAGACTAGTTCCAAGAGGGGTC−3’(配列番号12)を用いて作製したDIG標識GnT I特異的PCR断片を用いて行った。
【0132】
代替の方策では、構築物JME925 pPTAde2−URA3ex−Hp4dhGnTIを作製して、YarrowiaゲノムのADE2遺伝子座へのHp4d駆動型GnT I発現カセットの標的化組み込みを可能にした。構築方策を図20に示す。G045株への形質転換の前に、プラスミドをNotI消化し、ターゲティング/発現カセットを単離した。形質転換体を、それらのアデニン原栄養性に基づいて選択した。ADE2遺伝子座への発現カセットの正しい組み込みは、順方向プライマーVerlAde2(5’−CGACGATAGAGCAGGTCTCACTGTTGGGAATGCTG−3’、配列番号13)、逆方向プライマーVer2Ade2(5’−CTACACTGACGAAGTGGACATCCCGGCTTGGACTG−3’、配列番号14)を用いるPCRにより確かめ、サザンブロッティングによりさらに確認した。これは、BamHI/SpeI消化ゲノムDNAについて、プライマー5’−GGATGATCACACAATGGCCCTGTTTCTG−3’(配列番号15)および5’−TGCTCTAGACTAGTTCCAAGAGGGGTC−3’(配列番号16)を用いて作製したDIG標識GnT I特異的PCR断片を用いて行った。セクレトーム上へのGlcNAcMan3GlcNAc2の合成は、N−グリカン分析およびin vitroナタマメβ−Ν−アセチルヘキソサミニダーゼ処理により確認した(図21)。1つのGnT I発現形質転換体(G057と称する)を、プラスミドpRRQ2を用いるCreリコンビナーゼの一過的発現によるキュアリングのために選択した。得られた株は、G059と称した。
【0133】
(実施例8)GlcNAcトランスフェラーゼIIの発現
Yarrowiaコドン最適化配列を、S.cerevisiae Mnn2タンパク質(SwissProt 受託番号P38069)の最初の36個のN末端アミノ酸と、それに続くラットGlcNAcトランスフェラーゼII(GnT II)(SwissProt 受託番号Q09326)の触媒ドメインとからなる融合タンパク質(図22、配列番号17および配列番号18)の発現のために作製した。酵母Mnn2の36個のN末端アミノ酸は、触媒GnT IIドメインのためのゴルジ局在化シグナルとして働く。この様式で、Mnn2−GnT II融合タンパク質が、分泌経路において、Kre2−GnT I(およびMnn2−Man II)融合タンパク質と同じまたはそれより後の位置にさえ局在化されることを確実にして、GlcNAcMan3GlcNAc2をGlcNAc2Man3GlcNAc2に変換できた。融合タンパク質の発現のための合成遺伝子を、TEF1プロモーターの転写制御下に配置して、プラスミドpYLTmAXrGnTIIおよびpYLTmAXrGnTII(ADE2ex)を得た。構築方策を図23に示す。
【0134】
GnT II活性を発現する株を、2つの異なる様式で作製した:1)G045株(実施例6を参照されたい)を、NotI消化pYLTmAXhGnTIおよびNotI消化pYLTmAXrGnTII(ADE2 ex)で同時に形質転換し、形質転換体を、それらのウラシルおよびアデニン原栄養性に基づいて選択した、または2)G047株(実施例7)を、NotI消化pYLTmAXrGnTII(ADE2 ex)で形質転換し、形質転換体を、それらのアデニン原栄養性に基づいて選択した。発現カセットの組み込みを、順方向プライマーTefPromFW5’−GTCCCCGAATTACCTTTCC−3’(配列番号19)および逆方向プライマーLip2TermRV5’−AGGTAGAAGTTGTAAAGAGTG−3’(配列番号20)を用いて確かめた。ナタマメβ−N−アセチルヘキソサミニダーゼでの単離糖のin vitro処理と組み合わせたセクレトームについてのN−グリカン分析は、いくつかの形質転換体が、GlcNAc2Man3GlcNAc2を生成でき、よって機能的GnT II活性を発現できたことを示した(図24)。ある選択された条件では、全N−グリカンプールの約40%が、GlcNAc2Man3GlcNAc2で構成された。基質GlcNAcMan3GlcNAc2からGlcNAc2Man3GlcNAc2への変換率は、90%であった。最終的に選択した株をG050(G045の二重形質転換)およびG051(G047でのGnT II発現)と称した。
【0135】
(実施例9)グルコシダーゼIIアルファおよびベータサブユニット(Gls2αおよびGls2β)の発現
実施例6〜8に記載した実験に基づいて、YlALG3のノックアウトとそれと同時のYlALG6の過剰発現とを含む方策は、1または2つの末端グルコース残基を保持するN−グリカン(Glc1−2Man5’GlcNAc2)の産生をもたらす。これらのグルコース残基の存在は、ER局在化HDELタグ付きα−1,2−マンノシダーゼによるMan3GlcNAc2への変換を妨げる。グルコース残基を除去するために、ER内でのグルコシダーゼII活性を増大させる必要がある。α−1,2−マンノシダーゼ発現がないバックグラウンドでは、Aspergillus nigerグルコシダーゼIIアルファおよびベータサブユニットの過剰発現が、Glc1−2Man5’GlcNAc2からMan5’GlcNAc2への最高の変換をもたらした(米国特許出願公開第20090069232−A1号)。A.niger gls2サブユニットの過剰発現のための構築物を、以下のようにして生成した:1)Yarrowiaコドン最適化cDNAを、成熟(シグナルペプチドを欠く)A.niger gls2αおよびgls2βサブユニットの発現のために作製し、2)cDNAをインフレームでY.lipolytica LIP−2プレ配列にクローニングし、3)得られたLIP2プレgls2αおよびLIP2プレgls2β配列を、構成性TEF1プロモーターの転写制御下にクローニングした。得られたプラスミドを、pYLTUXdL2preAnGlcIIαおよびpYLeu2ExTEFpreLip2AnGlucIIβと称した(図25)。
【0136】
これらのプラスミドに基づいて、A.niger gls2αおよびgls2βサブユニットを、TEF1プロモーター制御(標的化組み込みのためのベクターJME923 pPTura3−LEU2ex−TefL2preAnGlcIIa+b[altl]、図26)またはHp4dプロモーター制御(標的化組み込みのためのベクターJME923 pPTura3−LEU2ex−Hp4dL2preAnGlcIIa+b[altl]および無作為組み込みのためのベクターZeta−LEU2ex−Hp4dL2preAnGlcIIa+b[alt]、図27)のいずれかの下で同時過剰発現するために、新しい構築物を作製した。
【0137】
G057株(実施例7を参照されたい)を、NotI消化プラスミドJME923 pPTura3−LEU2ex−Hp4dL2preAnGlcIIa+b[altl]およびZeta−LEU2ex−Hp4dL2preAnGlcIIa+b[alt]で形質転換し、形質転換体を、それらのロイシン原栄養性に基づいて選択した。いくつかのクローンを、PCRおよびサザン分析によりゲノム学的に分析して、gls2αおよびgls2β発現カセットの組み込みを評価した。gls2αサブユニットについてのPCR分析およびDIGプローブ作製は、プライマーAnGls2α−FW(5’−GCTGGACTCTTCTTCTATCC−3’)(配列番号24)およびAnGls2α−RV(5’−GGTCTCCTTCAGAGACAGG−3’)(配列番号25)を用いて行った。gls2βサブユニットについて、本発明者らは、プライマーAnGls2β−FW(5’−CCAAGTTCTACAAGGACACC−3’)(配列番号26)およびAnGlc2β−RV(5’−CCCTTGACGACCTTAGAGG−3’)(配列番号27)を用いた。二重Hp4dGls2α/β発現カセットの標的化組み込みを確認するためのサザン分析を、gls2αプローブを用いる場合にEco47III消化gDNAについて、およびgls2βプローブを用いる場合にSpeI/SfiI消化gDNAについて行った。選択したクローンの大多数は、URA3遺伝子座への二重発現カセットの正しい組み込みを示した。二重Hp4dGls2α/β発現カセットの無作為組み込みについてのサザン分析は、両方のプローブを用いてPvuI消化gDNAについて確かめた。全ての場合において、1コピーの二重発現カセットだけが組み込まれていた。
【0138】
次に、N−グリカン分析を、二重Hp4dGls2α/β発現カセットが(正しく)組み込まれていることが確認されたいくつかのクローンについて行った。N−グリコシル化を、3日間のファルコン培養の後の全分泌タンパク質について検査した。いくつかのクローンは、グルコシル化糖の著しい低減と、Man3GlcNAc2およびGlcNAcMan3GlcNAc2の増加とを示した。二重発現カセットが一方で無作為に組み込まれたクローン(=G060株)と、他方で標的化された様式で組み込まれたクローン(=G061株)のプロファイルを、図28に示す。2つのより小さいピークは、Man4’GlcNAc2およびMan5’GlcNAc2を表す。なぜなら、これらは、α−1,2−マンノシダーゼおよびナタマメマンノシダーゼでの処理の際にMan3GlcNAc2resp.Man1GlcNAc2にシフトするからである。後者の処理は、残存Glc1−2Man5’GlcNAc2からGlc1−2Man4’GlcNAc2、およびGlcNAcMan3GlcNAc2からGlcNAcMan2GlcNAc2への部分的変換ももたらす。Man4’GlcNAc2およびMan5’GlcNAc2の存在は、しかし、異種的に(heterologously)同時発現されたHDELタグ付きα−1,2−マンノシダーゼによる、Man3GlcNAc2への不完全な変換を示す。同様に、Man3GlcNAc2の存在は、GlcNAcMan3GlcNAc2を得るための組換えヒトGnT IによるGlcNAc残基の不完全な移動を示す。しかし、上記の結果に基づいて(例えば実施例7でのG047培養、図19)、培養条件の違いは、変換率を著しく増加でき、よって、終末の結果を改善できることが明確である。
【0139】
(実施例10)GlcNAcMan3GlcNAc2生成G061株におけるGlcNAcトランスフェラーゼIIの発現
実施例8で記載したようにして、Yarrowiaコドン最適化配列を、S.cerevisiae Mnn2タンパク質(SwissProt 受託番号P38069)の最初の36個のN末端アミノ酸と、それに続くラットGlcNAcトランスフェラーゼII(GnT II)(SwissProt 受託番号Q09326)の触媒ドメインとからなる融合タンパク質(図22、それぞれ配列番号17および配列番号18)の発現のために作製した。酵母Mnn2の36個のN末端アミノ酸は、触媒GnT IIドメインのためのゴルジ局在化シグナルとして働く。この様式で、Mnn2−GnT II融合タンパク質が、分泌経路においてKre2−GnT I融合タンパク質と同じまたはそれより後の位置にさえ局在化されることを確実にして、その結果、GlcNAcMan3GlcNAc2をGlcNAc2Man3GlcNAc2に変換できた。融合タンパク質の発現のための合成遺伝子を、Hp4dプロモーターの転写制御下に配置して、プラスミドpYLHp4mAXrGnTIIを得て、これを、YarrowiaゲノムへのHp4d駆動型GnT II発現カセットの無作為組み込みのために用いた。代替の方策では、構築物OXYP289 pPTAxpl−ADE2ex−Hp4dhGnTIIを作製して、YarrowiaゲノムのAXP1遺伝子座へのHp4d駆動型GnT II発現カセットの標的化組み込みを可能にした。
【0140】
G061株(実施例9を参照されたい)の形質転換の前に、プラスミドをNotI消化し、ターゲティング/発現カセットを単離した。形質転換体を、それらのアデニン原栄養性に基づいて選択した。ADE2遺伝子座への発現カセットの正しい組み込みは、ゲノムDNAをXmnIで消化した後のサザンブロット分析により確認した。GnT IIコード配列に対する特異性を有するDIG標識プローブを、順方向プライマーrGnTII−FW(5’−GACCAGATGCTGCGAAACG−3’)(配列番号28)および逆方向プライマーrGnTII−RV(5’−CTTGACGTCCACCTTGTCG−3’)(配列番号29)を用いて作製した。この方策により、遺伝子がAxp1遺伝子座にうまく組み込まれる場合には、3172bpのバンドが生成される。
【0141】
代替の方策では、Axp1遺伝子座への正しい組み込みは、順方向プライマーAXPVer1b(5’−GCCTGAACGGCACGATGCGATCGTGGCAATCC−3’)(配列番号30)および逆方向プライマーAXPVer2b(5’−CAAGAAGCCTCAGGCTCGGCGAATCTCCATC−3’)(配列番号31)を用いるゲノムDNAについてのPCR反応により検査できる。Axp1遺伝子座へ正しくターゲティングされた場合は、6489bpのPCR断片が予測される。
【0142】
セクレトームについてのN−グリカン分析と、それと組み合わせた単離糖のナタマメβ−Ν−アセチルヘキソサミニダーゼまたはT.reeseiα−1,2−マンノシダーゼでのin vitro処理は、いくつかの形質転換体がGlcNAc2Man3GlcNAc2を生成でき、よって機能的GnT II活性を発現できることを示した(図29)。その分析は、全N−グリカンプールの約25〜30%が、GlcNAc2Man3GlcNAc2からなり、GlcNAcMan3GlcNAc2からGlcNAc2Man3GlcNAc2への変換率が約90%であることを示した。最終的に選択した株をG070(G061へのpYLHp4mAXrGnTIIの組み込み)およびG071(G061へのOXYP289 pPTAxp1−ADE2ex−Hp4dhGnTIIの組み込み)と称した。
【0143】
(実施例11)Yarrowia lipolyticaへの抗HER2重鎖(HC)および軽鎖(LC)の同時Hp4d駆動型発現のためのタンデムプラスミドの構築
抗HER2抗体重鎖および軽鎖についてのアミノ酸配列を、Carterら、Proc Natl Acad Sci USA.、89巻(10号):4285〜4289頁(1992年);およびWardら、Appl Environ Microbiol、70巻(5号):2567〜2576頁(2004年)から得た。関連するアミノ酸配列は、逆翻訳され(reverse translated)、Yarrowia lipoliticaについてコドン最適化され、GenArt、Regensburg Germanyにより合成された。非常に高い(>80%)または非常に低い(<30%)GC含量の領域は、可能であれば回避した。最適化プロセス中に、以下のシス作用性配列モチーフは回避した:内部TATAボックス、カイ部位およびリボソーム進入部位、ATリッチまたはGCリッチ配列ストレッチ、反復配列およびRNA 2次構造ならびに(潜在的)スプライス供与部位および受容部位。異所性タンパク質の分泌を可能にするために、Lip2タンパク質「プレプロ」シグナルのコード配列(ペプチドリンカー「GGG」のコード配列が後に続く)を、コード配列の5’領域に付加した。「GGG」を、正しいKex2プロセシングのための変化を増進するために付加した。図30Aは、合成プレプロLip2−LCのヌクレオチド配列(=750bp)(配列番号32)を含有する。図30Bは、プレプロLip2−LCのアミノ酸配列(=250Aa;MW=27.011Da;pI=8.46)(配列番号33)を含有する。図31Aは、合成プレプロLip2−HCのヌクレオチド配列(=1458bp)(配列番号34)を含有する。図31Bは、プレプロLip2−HCのアミノ酸配列(=486Aa;MW=52.853Da;pI=8.65)(配列番号35)を含有する。プレプロLip2−HCおよび−LCについてのコード配列を、pYLHp4L2preproHerHC/LC(GUT2ex)−ori2と称する同じベクターに導入した。
【0144】
(実施例12)様々な程度の糖鎖工学的改変によるYarrowia lipolytica株での抗HER2抗体HCおよびLCの発現
プラスミドpYLHp4L2preproHerHC&LC(GUT2ex)−ori2をNotIで消化し、HC−/LC−タンデム発現カセットを単離した後に、Yarrowia lipolytica G045株、G057株、G061株およびG071株(表2を参照されたい)を形質転換した。無作為に組み込まれたHC−/LC−発現カセットを含有する形質転換体を、グリセロールを唯一の炭素源として増殖するそれらの能力に基づいて選択した。HCおよびLCの発現解析は、選択した形質転換体を、グリセロールを唯一の炭素源として含有するリッチ培地(SuperT/グリセロール培地:0.5%酵母抽出物;2%麦芽抽出物;1%トリプトン;1.5%グリセロール;200mMリン酸塩緩衝液pH6.8)中で4日間振とうフラスコ培養した後にウェスタンブロッティングにより行った。LC検出は、カッパ遊離軽鎖に対するマウスモノクローナル(4C11)(Abeam)を用いて行い、HC検出は、マウスモノクローナル抗ヒトIgG(γ鎖特異的)(Sigma)を用いて行った。
【0145】
抗HER2抗体生成株のセクレトームのN−グリカンは、いずれのHCおよびLCも発現しない対応する糖鎖工学的に改変した株と同様のプロファイルを示した(図32)。G045、G057、G061およびG071バックグラウンドを有する株でのN−グリカンのパーセンテージは、SuperT/グリセロール培地中での6日間の振とうフラスコ培養の後に決定した。G045バックグラウンドでは、54.6%のN−グリカンがMan3GlcNAc2であった。G057バックグラウンドでは、47.5%のN−グリカンがGlcNAc1Man3GlcNAc2であった。G061バックグラウンドでは、58.9%のN−グリカンがGlcNAc1Man3GlcNAc2であった。G071バックグラウンドでは、37.6%のN−グリカンがGlcNAc2Man3GlcNAc2であった。
【0146】
(実施例13)抗HER2抗体HCおよびLCを発現するGlcNAc2Man3GlcNAc2合成株であるYarrowia G096株の発酵
GlcNAc2Man3GlcNAc2を合成できる株であるYarrowia lipolytica G071のいくつかのpYLHp4L2preproHerHC&LC(GUT2ex)−ori2形質転換体を、HCおよびLC発現レベルについて分析した。これらのクローンのうちの1つであるG096を、さらなる分析のために選択した。
【0147】
発酵は、プロセス制御および管理システム(Lucillus PIMS)を備える14リットル撹拌タンクバイオリアクタ(MAVAG AG)で行った。培地中の相対的酸素分圧、排出ガス中のCO2とO2の濃度、pH値、温度、リアクタ過圧、リアクタ重量、フィード重量および基本重量(base weight)は全てオンラインで監視した。泡の発生は、消泡剤ポリプロピレングリコール(PPG)を加えることにより妨いだ。pHの調整は、25%アンモニア溶液または8.5%リン酸溶液のいずれかの添加により行った。
【0148】
G096の種培養物を28℃にて、リッチ培地を含有する振とうフラスコで増殖させた。種培養物を、無機質培地を含有する発酵槽に接種して、グリセロールを唯一の炭素源として用いて、回分段階(batch phase)を28℃にて無制限増殖で開始した。この段階は、高いバイオマス濃度に迅速に到達するために用いた。この時点以降、プロセスを、0.02の定常増殖速度の指数グリセロール流加(fed batch)(グリセロールを唯一の炭素源およびエネルギー源として用いる;pH6)にシフトした。例として、28℃での流加発酵の結果を以下に記載する。
【0149】
流加段階は、148時間継続した。発酵の異なる時点で試料を採取して、以下のパラメータを追跡した:1)ウェスタンブロットによるLCおよびHCタンパク質主鎖の発現;2)ELISAによる機能的抗HER2抗体の発現;ならびに3)セクレトームのN−グリコシル化プロファイルの進展。全長HC発現レベルは、時点7(39時間)付近で最大に到達し、それ以降ほぼ等しいままである。LC発現は、時点7(39時間)と10(73時間)の間で最大に到達したが、その後の時点で幾分か減少した。いくらかのLC二量体が、時点5(25時間)と9(62時間)の間で生成されたが、これもまたその時点以降で消失した。
【0150】
機能的ELISAを開発して、少なくとも1つの機能的抗原結合ドメインを有する抗HER2抗体の生成を測定した。プレートを、天然HER2抗原の組換えバリアントである組換えヒトErbB2/Fcキメラ(R&D systems)でコーティングした。次いで、異なる時点で採集した培地の希釈物を、コーティングしたプレートに加えた。抗原結合タンパク質の量の評価を、HRPコンジュゲート抗ヒトカッパLC抗体(Sigma)を用いて行った。流加発酵でのErbB2/Fcキメラ結合タンパク質の量の進展(分泌された機能的抗HER2抗体の量の指標)を、図33に示す。データは、抗HER2抗体のレベルの漸増と、生成段階の終末での10〜12mg/Lの最大値とを示す。
【0151】
N−グリカン分析を、流加発酵中のいくつかの時点で採取した試料について行った。結果を図34Aおよび34Bに示す。流加段階の開始時に、相当量のグルコース含有N−グリカンが存在した。時点6以降から(指数的栄養補給の開始後34時間)、グルコシル化N−グリカンのレベルは著しく減少し、採集の時点(時点18、148時間)ではほとんど残っていなかった。このことは、グルコース含有N−グリカンを元来保持するタンパク質が、発酵の終末に薄められことを示した。この時点で、セクレトームから単離されたN−グリカンの約86%が、構造GlcNAc2Man3GlcNAc2を有した。
【0152】
その他の実施形態
本発明を、その詳細な説明と関連付けて記載したが、上記の記載は、例示することを意図し、本発明の範囲を限定することを意図しない(本発明の範囲は、添付の特許請求の範囲により定義される)。その他の態様、利点および改変は、以下の特許請求の範囲内にある。
【特許請求の範囲】
【請求項1】
GlcNAcMan5GlcNAc2 N−グリカンを含むタンパク質を生成できる真菌細胞を生成する方法であって、該方法は、
a)Man5GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された真菌細胞を提供するステップと、
b)該細胞に、GlcNAcトランスフェラーゼIをコードする核酸を導入するステップであって、ここで、該核酸が、コードされた該GlcNAcトランスフェラーゼIを細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該GlcNAcトランスフェラーゼIの発現が、GlcNAcMan5GlcNAc2 N−グリカンを含むタンパク質を生成するステップと
を含む方法。
【請求項2】
請求項1に記載の方法であって、該方法は、前記細胞に、標的タンパク質をコードする核酸を導入するステップをさらに含み、ここで、該細胞が、前記GlcNAcMan5GlcNAc2 N−グリカンを含むように改変された該標的タンパク質を生成する、方法。
【請求項3】
前記標的タンパク質が、Fc受容体と結合する、請求項2に記載の方法。
【請求項4】
前記標的タンパク質が、抗体またはその断片である、請求項3に記載の方法。
【請求項5】
前記標的タンパク質が、治療用の糖タンパク質である、請求項2に記載の方法。
【請求項6】
前記標的タンパク質が、インターフェロン−β、GM−CSF、インターフェロンγまたはエリスロポエチンである、請求項2に記載の方法。
【請求項7】
前記細胞内区画が、ゴルジ装置である、請求項1に記載の方法。
【請求項8】
請求項1に記載の方法であって、ここで、Man5GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された前記真菌細胞が、OCH1活性を欠損し、α−1,2−マンノシダーゼをコードする核酸を含み、ここで、該α−1,2−マンノシダーゼをコードする該核酸が、コードされた該α−1,2−マンノシダーゼを小胞体に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含む、方法。
【請求項9】
前記ターゲティング配列が、HDEL配列である、請求項8に記載の方法。
【請求項10】
前記真菌細胞が、Yarrowia lipolyticaまたはArxula adeninivoransである、請求項1に記載の方法。
【請求項11】
請求項1に記載の方法であって、該方法は、前記細胞に、マンノシダーゼIIをコードする核酸を導入するステップであって、ここで、該マンノシダーゼIIをコードする該核酸が、コードされた該マンノシダーゼIIをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該マンノシダーゼIIの発現が、GlcNAcMan3GlcNAc2 N−グリカンを含むタンパク質を生成するステップをさらに含む、方法。
【請求項12】
請求項1または請求項11に記載の方法であって、該方法は、前記細胞に、ガラクトシルトランスフェラーゼをコードする核酸を導入するステップであって、ここで、該ガラクトシルトランスフェラーゼをコードする該核酸が、コードされた該ガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、該真菌細胞における該ガラクトシルトランスフェラーゼの発現が、GalGlcNAcMan5GlcNAc2またはGalGlcNAcMan3GlcNAc2 N−グリカンを含むタンパク質を生成するステップをさらに含む、方法。
【請求項13】
請求項12に記載の方法であって、該方法は、前記細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、該細胞が、前記GalGlcNAcMan5GlcNAc2またはGalGlcNAcMan3GlcNAc2 N−グリカンを含むように改変された該標的タンパク質を生成するステップをさらに含む、方法。
【請求項14】
前記ガラクトシルトランスフェラーゼが、UDP−Glc−4−エピメラーゼと、β−1,4−ガラクトシルトランスフェラーゼIの触媒ドメインとの融合体である、請求項12に記載の方法。
【請求項15】
前記GalGlcNAcMan5GlcNAc2またはGalGlcNAcMan3GlcNAc2 N−グリカンを含むように改変された前記標的タンパク質を単離するステップをさらに含む、請求項13に記載の方法。
【請求項16】
GlcNAcMan3GlcNAc2 N−グリカンを含む標的タンパク質を生成する方法であって、該方法は、
a)GlcNAcトランスフェラーゼIと、α−1,2−マンノシダーゼと、マンノシダーゼIIとをコードする核酸を含むように遺伝子工学的に改変された真菌細胞を提供するステップであって、ここで、該核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞が、OCH1活性を欠損しているステップと、
b)該細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、該細胞が、該GlcNAcMan3GlcNAc2 N−グリカンを含む該標的タンパク質を生成するステップと
を含む方法。
【請求項17】
前記真菌細胞が、Yarrowia lipolyticaまたはArxula adeninivoransである、請求項16に記載の方法。
【請求項18】
前記α−1,2−マンノシダーゼをコードする前記核酸が、コードされた該α−1,2−マンノシダーゼを小胞体に向かわせるための小胞体ターゲティング配列を含む、請求項16に記載の方法。
【請求項19】
前記ターゲティング配列が、HDEL配列である、請求項18に記載の方法。
【請求項20】
前記GlcNAcトランスフェラーゼIおよび前記マンノシダーゼIIをコードする前記核酸が、コードされた該GlcNAcトランスフェラーゼIおよびマンノシダーゼIIをゴルジ装置に向かわせるためのゴルジターゲティング配列をコードするヌクレオチド配列を含む、請求項16に記載の方法。
【請求項21】
前記標的タンパク質が、Fc受容体と結合する、請求項16に記載の方法。
【請求項22】
前記標的タンパク質が、抗体またはその断片である、請求項16に記載の方法。
【請求項23】
前記標的タンパク質が、治療用の糖タンパク質である、請求項16に記載の方法。
【請求項24】
前記標的タンパク質が、インターフェロン−β、GM−CSF、インターフェロンγまたはエリスロポエチンである、請求項16に記載の方法。
【請求項25】
前記細胞に、ガラクトシルトランスフェラーゼをコードする核酸を導入するステップをさらに含む請求項16に記載の方法であって、ここで、該ガラクトシルトランスフェラーゼをコードする該核酸が、コードされた該ガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで該真菌細胞における該ガラクトシルトランスフェラーゼの発現が、GalGlcNAcMan3GlcNAc2 N−グリカンを含むように改変された前記標的タンパク質を生成する、方法。
【請求項26】
前記GalGlcNAcMan3GlcNAc2 N−グリカンを含むように改変された前記標的タンパク質を単離するステップをさらに含む、請求項25に記載の方法。
【請求項27】
GlcNAcMan3GlcNAc2 N−グリカンを含むタンパク質を生成できる真菌細胞を作製する方法であって、該方法は、
a)Man3GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された真菌細胞を提供するステップと、
b)該細胞に、GlcNAcトランスフェラーゼIをコードする核酸を導入するステップであって、ここで、該核酸が、コードされた該GlcNAcトランスフェラーゼIを細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該GlcNAcトランスフェラーゼIの発現が、GlcNAcMan3GlcNAc2 N−グリカンを含むタンパク質を生成するステップと
を含む方法。
【請求項28】
請求項27に記載の方法であって、該方法は、前記細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、該細胞が、前記GlcNAcMan3GlcNAc2 N−グリカンを含むように改変された該標的タンパク質を生成するをさらに含む、方法。
【請求項29】
前記標的タンパク質が、Fc受容体と結合する、請求項27に記載の方法。
【請求項30】
前記標的タンパク質が、抗体またはその断片である、請求項27に記載の方法。
【請求項31】
前記標的タンパク質が、治療用の糖タンパク質である、請求項27に記載の方法。
【請求項32】
前記標的タンパク質が、インターフェロン−β、GM−CSF、インターフェロンγまたはエリスロポエチンである、請求項27に記載の方法。
【請求項33】
前記細胞内区画が、ゴルジ装置である、請求項27に記載の方法。
【請求項34】
請求項27に記載の方法であって、ここで、Man3GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された前記真菌細胞が、ALG3活性を欠損し、α−1,2−マンノシダーゼをコードする核酸を含み、該核酸が、コードされた該α−1,2−マンノシダーゼを小胞体に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含む、方法。
【請求項35】
Man3GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された前記真菌細胞が、OCH1活性をさらに欠損する、請求項34に記載の方法。
【請求項36】
Man3GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された前記真菌細胞が、α−1,3−グルコシルトランスフェラーゼをコードする核酸をさらに含む、請求項34または35に記載の方法。
【請求項37】
前記α−1,3−グルコシルトランスフェラーゼが、ALG6である、請求項36に記載の方法。
【請求項38】
前記真菌細胞が、Yarrowia lipolyticaまたはArxula adeninivoransである、請求項27に記載の方法。
【請求項39】
請求項27に記載の方法であって、該方法は、前記細胞に、GlcNAcトランスフェラーゼIIをコードする核酸を導入するステップであって、ここで、該GlcNAcトランスフェラーゼIIをコードする該核酸が、コードされた該GlcNAcトランスフェラーゼIIを細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該GlcNAcトランスフェラーゼIIの発現が、GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成するステップをさらに含む、方法。
【請求項40】
請求項27または請求項39に記載の方法であって、該方法は、前記細胞に、ガラクトシルトランスフェラーゼをコードする核酸を導入するステップであって、、ここで、該ガラクトシルトランスフェラーゼをコードする該核酸が、コードされた該ガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該ガラクトシルトランスフェラーゼの発現が、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成するステップをさらに含む、方法。
【請求項41】
請求項40に記載の方法であって、該方法は、前記細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、該細胞が、前記GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカンを含むように改変された該標的タンパク質を生成するステップをさらに含む、方法。
【請求項42】
前記ガラクトシルトランスフェラーゼが、UDP−Glc−4−エピメラーゼと、β−1,4−ガラクトシルトランスフェラーゼIの触媒ドメインとの融合体である、請求項40に記載の方法。
【請求項43】
請求項40に記載の方法であって、該方法は、前記細胞に、グルコシダーゼIIのαおよびβサブユニットをコードする核酸を導入するステップであって、ここで、該真菌細胞における該グルコシダーゼIIの該αおよびβサブユニットの発現が、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成するステップをさらに含む、方法。
【請求項44】
Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含む標的タンパク質を生成する方法であって、該方法は、
a)ALG3活性を欠損するように遺伝子工学的に改変され、GlcNAcトランスフェラーゼIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸を含む真菌細胞を提供するステップであって、ここで、該GlcNAcトランスフェラーゼIと、該GlcNAcトランスフェラーゼIIと、該ガラクトシルトランスフェラーゼとをコードする該核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含むステップと、
b)該細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、該細胞が、該Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含む該標的タンパク質を生成するステップと
を含む方法。
【請求項45】
前記真菌細胞が、OCH1活性をさらに欠損する、請求項44に記載の方法。
【請求項46】
前記真菌細胞が、α−1,3−グルコシルトランスフェラーゼをコードする核酸をさらに含む、請求項44または45に記載の方法。
【請求項47】
前記α−1,3−グルコシルトランスフェラーゼをコードする前記核酸が、ALG6である、請求項46に記載の方法。
【請求項48】
請求項44、請求項45または請求項46に記載の方法であって、ここで、前記真菌細胞が、グルコシダーゼIIのαおよびβサブユニットをコードする核酸をさらに含み、ここで、該真菌細胞における該グルコシダーゼIIの該αおよびβサブユニットの発現が、前記Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含む前記標的タンパク質を生成する、方法。
【請求項49】
GlcNAcMan3GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された単離真菌細胞であって、ここで、OCH1活性を欠損し、α−1,2−マンノシダーゼと、GlcNAcトランスフェラーゼIと、マンノシダーゼIIとをコードする核酸を含み、ここで、該α−1,2−マンノシダーゼと、該GlcNAcトランスフェラーゼIと、該マンノシダーゼIIとをコードする該核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該α−1,2−マンノシダーゼ、該GlcNAcトランスフェラーゼIおよび該マンノシダーゼIIの発現が、GlcNAcMan3GlcNAc2 N−グリカンを含むタンパク質を生成する、単離真菌細胞。
【請求項50】
前記遺伝子工学的に改変された真菌細胞が、標的タンパク質をコードする核酸をさらに含み、ここで、該細胞が、前記GlcNAcMan3GlcNAc2 N−グリカンを含むように改変された該標的タンパク質を生成する、請求項49に記載の真菌細胞。
【請求項51】
請求項49または請求項50に記載の真菌細胞であって、ここで、前記遺伝子工学的に改変された真菌細胞が、GlcNAcトランスフェラーゼIIをコードする核酸をさらに含み、ここで、該GlcNAcトランスフェラーゼIIをコードする該核酸が、コードされた該GlcNAcトランスフェラーゼIIを細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該GlcNAcトランスフェラーゼIIの発現が、GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成する、真菌細胞。
【請求項52】
請求項51に記載の真菌細胞であって、該真菌細胞は、ガラクトシルトランスフェラーゼをコードする核酸をさらに含み、ここで、該ガラクトシルトランスフェラーゼをコードする該核酸が、コードされた該ガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該ガラクトシルトランスフェラーゼの発現が、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成する、真菌細胞。
【請求項53】
GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された単離真菌細胞であって、ここで、該真菌細胞は、ALG3活性を欠損するように遺伝子工学的に改変され、GlcNAcトランスフェラーゼIとGlcNAcトランスフェラーゼIIとをコードする核酸を含み、ここで、該GlcNAcトランスフェラーゼIと該GlcNAcトランスフェラーゼIIとをコードする該核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該GlcNAcトランスフェラーゼIおよび該GlcNAcトランスフェラーゼIIの発現が、GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成する、単離真菌細胞。
【請求項54】
前記遺伝子工学的に改変された真菌細胞が、OCH1活性をさらに欠損する、請求項53に記載の真菌細胞。
【請求項55】
前記遺伝子工学的に改変された真菌細胞が、α−1,3−グルコシルトランスフェラーゼをコードする核酸をさらに含む、請求項53または請求項54に記載の真菌細胞。
【請求項56】
請求項53、請求項54または請求項55に記載の真菌細胞であって、ここで、前記遺伝子工学的に改変された真菌細胞が、標的タンパク質をコードする核酸をさらに含み、ここで、該細胞が、前記GlcNAc2Man3GlcNAc2 N−グリカンを含むように改変された該標的タンパク質を生成する、真菌細胞。
【請求項57】
請求項53から56のいずれか一項に記載の真菌細胞であって、該真菌細胞は、グルコシダーゼIIのαおよびβサブユニットをコードする核酸をさらに含み、ここで、該真菌細胞における該グルコシダーゼIIの該αおよびβサブユニットの発現が、前記GlcNAc2Man3GlcNAc2 N−グリカンを含む前記タンパク質を生成する、真菌細胞。
【請求項58】
請求項57に記載の真菌細胞であって、該真菌細胞は、ガラクトシルトランスフェラーゼをコードする核酸をさらに含み、ここで、該ガラクトシルトランスフェラーゼをコードする該核酸が、コードされた該ガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該ガラクトシルトランスフェラーゼの発現が、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成する、真菌細胞。
【請求項59】
相当数のYarrowia lipolytica細胞が、Gal2GlcNac2Man3GlcNAc2 N−グリカンを含む糖タンパク質を生成するように遺伝子工学的に改変されているYarrowia lipolytica細胞の実質的に純粋な培養物であって、ここで、該細胞が、ALG3活性を欠損するように遺伝子工学的に改変され、GlcNAcトランスフェラーゼIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸を含み、ここで、該GlcNAcトランスフェラーゼIと、該GlcNAcトランスフェラーゼIIと、該ガラクトシルトランスフェラーゼとをコードする該核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該細胞における該GlcNAcトランスフェラーゼI、該GlcNAcトランスフェラーゼIIおよび該ガラクトシルトランスフェラーゼの発現が、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成する、実質的に純粋な培養物。
【請求項60】
前記細胞が、OCH1活性をさらに欠損する、請求項49に記載の実質的に純粋な培養物。
【請求項61】
前記細胞が、α−1,3−グルコシルトランスフェラーゼをコードする核酸をさらに含む、請求項59または請求項60に記載の実質的に純粋な培養物。
【請求項62】
前記細胞が、グルコシダーゼIIのαおよびβサブユニットをコードする核酸をさらに含み、ここで、該真菌細胞における該グルコシダーゼIIの該αおよびβサブユニットの発現が、前記Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含む前記標的タンパク質を生成する、請求項59、請求項60または請求項61に記載の実質的に純粋な培養物。
【請求項63】
相当数のYarrowia lipolytica細胞が、Gal2GlcNac2Man3GlcNAc2 N−グリカンを含む糖タンパク質を生成するように遺伝子工学的に改変されているYarrowia lipolytica細胞の実質的に純粋な培養物であって、ここで、該細胞が、OCH1活性を欠損するように遺伝子工学的に改変され、α−1,2−マンノシダーゼと、GlcNAcトランスフェラーゼIと、マンノシダーゼIIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸を含み、ここで、該α−1,2−マンノシダーゼと、該GlcNAcトランスフェラーゼIと、該マンノシダーゼIIと、該GlcNAcトランスフェラーゼIIと、該ガラクトシルトランスフェラーゼとをコードする該核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該細胞における該α−1,2−マンノシダーゼ、該GlcNAcトランスフェラーゼI、該マンノシダーゼII、該GlcNAcトランスフェラーゼIIおよびガラクトシルトランスフェラーゼの発現が、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成する、実質的に純粋な培養物。
【請求項64】
糖タンパク質を含む組成物であって、該糖タンパク質上のN−グリカンの少なくとも50%が、GlcNAc2Man3GlcNAc2 N−グリカンである組成物。
【請求項65】
前記糖タンパク質上のN−グリカンの少なくとも70%が、GlcNAc2Man3GlcNAc2 N−グリカンである、請求項64に記載の組成物。
【請求項66】
前記糖タンパク質上のN−グリカンの少なくとも85%が、GlcNAc2Man3GlcNAc2 N−グリカンである、請求項64に記載の組成物。
【請求項1】
GlcNAcMan5GlcNAc2 N−グリカンを含むタンパク質を生成できる真菌細胞を生成する方法であって、該方法は、
a)Man5GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された真菌細胞を提供するステップと、
b)該細胞に、GlcNAcトランスフェラーゼIをコードする核酸を導入するステップであって、ここで、該核酸が、コードされた該GlcNAcトランスフェラーゼIを細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該GlcNAcトランスフェラーゼIの発現が、GlcNAcMan5GlcNAc2 N−グリカンを含むタンパク質を生成するステップと
を含む方法。
【請求項2】
請求項1に記載の方法であって、該方法は、前記細胞に、標的タンパク質をコードする核酸を導入するステップをさらに含み、ここで、該細胞が、前記GlcNAcMan5GlcNAc2 N−グリカンを含むように改変された該標的タンパク質を生成する、方法。
【請求項3】
前記標的タンパク質が、Fc受容体と結合する、請求項2に記載の方法。
【請求項4】
前記標的タンパク質が、抗体またはその断片である、請求項3に記載の方法。
【請求項5】
前記標的タンパク質が、治療用の糖タンパク質である、請求項2に記載の方法。
【請求項6】
前記標的タンパク質が、インターフェロン−β、GM−CSF、インターフェロンγまたはエリスロポエチンである、請求項2に記載の方法。
【請求項7】
前記細胞内区画が、ゴルジ装置である、請求項1に記載の方法。
【請求項8】
請求項1に記載の方法であって、ここで、Man5GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された前記真菌細胞が、OCH1活性を欠損し、α−1,2−マンノシダーゼをコードする核酸を含み、ここで、該α−1,2−マンノシダーゼをコードする該核酸が、コードされた該α−1,2−マンノシダーゼを小胞体に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含む、方法。
【請求項9】
前記ターゲティング配列が、HDEL配列である、請求項8に記載の方法。
【請求項10】
前記真菌細胞が、Yarrowia lipolyticaまたはArxula adeninivoransである、請求項1に記載の方法。
【請求項11】
請求項1に記載の方法であって、該方法は、前記細胞に、マンノシダーゼIIをコードする核酸を導入するステップであって、ここで、該マンノシダーゼIIをコードする該核酸が、コードされた該マンノシダーゼIIをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該マンノシダーゼIIの発現が、GlcNAcMan3GlcNAc2 N−グリカンを含むタンパク質を生成するステップをさらに含む、方法。
【請求項12】
請求項1または請求項11に記載の方法であって、該方法は、前記細胞に、ガラクトシルトランスフェラーゼをコードする核酸を導入するステップであって、ここで、該ガラクトシルトランスフェラーゼをコードする該核酸が、コードされた該ガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、該真菌細胞における該ガラクトシルトランスフェラーゼの発現が、GalGlcNAcMan5GlcNAc2またはGalGlcNAcMan3GlcNAc2 N−グリカンを含むタンパク質を生成するステップをさらに含む、方法。
【請求項13】
請求項12に記載の方法であって、該方法は、前記細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、該細胞が、前記GalGlcNAcMan5GlcNAc2またはGalGlcNAcMan3GlcNAc2 N−グリカンを含むように改変された該標的タンパク質を生成するステップをさらに含む、方法。
【請求項14】
前記ガラクトシルトランスフェラーゼが、UDP−Glc−4−エピメラーゼと、β−1,4−ガラクトシルトランスフェラーゼIの触媒ドメインとの融合体である、請求項12に記載の方法。
【請求項15】
前記GalGlcNAcMan5GlcNAc2またはGalGlcNAcMan3GlcNAc2 N−グリカンを含むように改変された前記標的タンパク質を単離するステップをさらに含む、請求項13に記載の方法。
【請求項16】
GlcNAcMan3GlcNAc2 N−グリカンを含む標的タンパク質を生成する方法であって、該方法は、
a)GlcNAcトランスフェラーゼIと、α−1,2−マンノシダーゼと、マンノシダーゼIIとをコードする核酸を含むように遺伝子工学的に改変された真菌細胞を提供するステップであって、ここで、該核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞が、OCH1活性を欠損しているステップと、
b)該細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、該細胞が、該GlcNAcMan3GlcNAc2 N−グリカンを含む該標的タンパク質を生成するステップと
を含む方法。
【請求項17】
前記真菌細胞が、Yarrowia lipolyticaまたはArxula adeninivoransである、請求項16に記載の方法。
【請求項18】
前記α−1,2−マンノシダーゼをコードする前記核酸が、コードされた該α−1,2−マンノシダーゼを小胞体に向かわせるための小胞体ターゲティング配列を含む、請求項16に記載の方法。
【請求項19】
前記ターゲティング配列が、HDEL配列である、請求項18に記載の方法。
【請求項20】
前記GlcNAcトランスフェラーゼIおよび前記マンノシダーゼIIをコードする前記核酸が、コードされた該GlcNAcトランスフェラーゼIおよびマンノシダーゼIIをゴルジ装置に向かわせるためのゴルジターゲティング配列をコードするヌクレオチド配列を含む、請求項16に記載の方法。
【請求項21】
前記標的タンパク質が、Fc受容体と結合する、請求項16に記載の方法。
【請求項22】
前記標的タンパク質が、抗体またはその断片である、請求項16に記載の方法。
【請求項23】
前記標的タンパク質が、治療用の糖タンパク質である、請求項16に記載の方法。
【請求項24】
前記標的タンパク質が、インターフェロン−β、GM−CSF、インターフェロンγまたはエリスロポエチンである、請求項16に記載の方法。
【請求項25】
前記細胞に、ガラクトシルトランスフェラーゼをコードする核酸を導入するステップをさらに含む請求項16に記載の方法であって、ここで、該ガラクトシルトランスフェラーゼをコードする該核酸が、コードされた該ガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで該真菌細胞における該ガラクトシルトランスフェラーゼの発現が、GalGlcNAcMan3GlcNAc2 N−グリカンを含むように改変された前記標的タンパク質を生成する、方法。
【請求項26】
前記GalGlcNAcMan3GlcNAc2 N−グリカンを含むように改変された前記標的タンパク質を単離するステップをさらに含む、請求項25に記載の方法。
【請求項27】
GlcNAcMan3GlcNAc2 N−グリカンを含むタンパク質を生成できる真菌細胞を作製する方法であって、該方法は、
a)Man3GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された真菌細胞を提供するステップと、
b)該細胞に、GlcNAcトランスフェラーゼIをコードする核酸を導入するステップであって、ここで、該核酸が、コードされた該GlcNAcトランスフェラーゼIを細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該GlcNAcトランスフェラーゼIの発現が、GlcNAcMan3GlcNAc2 N−グリカンを含むタンパク質を生成するステップと
を含む方法。
【請求項28】
請求項27に記載の方法であって、該方法は、前記細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、該細胞が、前記GlcNAcMan3GlcNAc2 N−グリカンを含むように改変された該標的タンパク質を生成するをさらに含む、方法。
【請求項29】
前記標的タンパク質が、Fc受容体と結合する、請求項27に記載の方法。
【請求項30】
前記標的タンパク質が、抗体またはその断片である、請求項27に記載の方法。
【請求項31】
前記標的タンパク質が、治療用の糖タンパク質である、請求項27に記載の方法。
【請求項32】
前記標的タンパク質が、インターフェロン−β、GM−CSF、インターフェロンγまたはエリスロポエチンである、請求項27に記載の方法。
【請求項33】
前記細胞内区画が、ゴルジ装置である、請求項27に記載の方法。
【請求項34】
請求項27に記載の方法であって、ここで、Man3GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された前記真菌細胞が、ALG3活性を欠損し、α−1,2−マンノシダーゼをコードする核酸を含み、該核酸が、コードされた該α−1,2−マンノシダーゼを小胞体に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含む、方法。
【請求項35】
Man3GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された前記真菌細胞が、OCH1活性をさらに欠損する、請求項34に記載の方法。
【請求項36】
Man3GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された前記真菌細胞が、α−1,3−グルコシルトランスフェラーゼをコードする核酸をさらに含む、請求項34または35に記載の方法。
【請求項37】
前記α−1,3−グルコシルトランスフェラーゼが、ALG6である、請求項36に記載の方法。
【請求項38】
前記真菌細胞が、Yarrowia lipolyticaまたはArxula adeninivoransである、請求項27に記載の方法。
【請求項39】
請求項27に記載の方法であって、該方法は、前記細胞に、GlcNAcトランスフェラーゼIIをコードする核酸を導入するステップであって、ここで、該GlcNAcトランスフェラーゼIIをコードする該核酸が、コードされた該GlcNAcトランスフェラーゼIIを細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該GlcNAcトランスフェラーゼIIの発現が、GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成するステップをさらに含む、方法。
【請求項40】
請求項27または請求項39に記載の方法であって、該方法は、前記細胞に、ガラクトシルトランスフェラーゼをコードする核酸を導入するステップであって、、ここで、該ガラクトシルトランスフェラーゼをコードする該核酸が、コードされた該ガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該ガラクトシルトランスフェラーゼの発現が、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成するステップをさらに含む、方法。
【請求項41】
請求項40に記載の方法であって、該方法は、前記細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、該細胞が、前記GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカンを含むように改変された該標的タンパク質を生成するステップをさらに含む、方法。
【請求項42】
前記ガラクトシルトランスフェラーゼが、UDP−Glc−4−エピメラーゼと、β−1,4−ガラクトシルトランスフェラーゼIの触媒ドメインとの融合体である、請求項40に記載の方法。
【請求項43】
請求項40に記載の方法であって、該方法は、前記細胞に、グルコシダーゼIIのαおよびβサブユニットをコードする核酸を導入するステップであって、ここで、該真菌細胞における該グルコシダーゼIIの該αおよびβサブユニットの発現が、GalGlcNAcMan3GlcNAc2またはGal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成するステップをさらに含む、方法。
【請求項44】
Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含む標的タンパク質を生成する方法であって、該方法は、
a)ALG3活性を欠損するように遺伝子工学的に改変され、GlcNAcトランスフェラーゼIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸を含む真菌細胞を提供するステップであって、ここで、該GlcNAcトランスフェラーゼIと、該GlcNAcトランスフェラーゼIIと、該ガラクトシルトランスフェラーゼとをコードする該核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含むステップと、
b)該細胞に、標的タンパク質をコードする核酸を導入するステップであって、ここで、該細胞が、該Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含む該標的タンパク質を生成するステップと
を含む方法。
【請求項45】
前記真菌細胞が、OCH1活性をさらに欠損する、請求項44に記載の方法。
【請求項46】
前記真菌細胞が、α−1,3−グルコシルトランスフェラーゼをコードする核酸をさらに含む、請求項44または45に記載の方法。
【請求項47】
前記α−1,3−グルコシルトランスフェラーゼをコードする前記核酸が、ALG6である、請求項46に記載の方法。
【請求項48】
請求項44、請求項45または請求項46に記載の方法であって、ここで、前記真菌細胞が、グルコシダーゼIIのαおよびβサブユニットをコードする核酸をさらに含み、ここで、該真菌細胞における該グルコシダーゼIIの該αおよびβサブユニットの発現が、前記Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含む前記標的タンパク質を生成する、方法。
【請求項49】
GlcNAcMan3GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された単離真菌細胞であって、ここで、OCH1活性を欠損し、α−1,2−マンノシダーゼと、GlcNAcトランスフェラーゼIと、マンノシダーゼIIとをコードする核酸を含み、ここで、該α−1,2−マンノシダーゼと、該GlcNAcトランスフェラーゼIと、該マンノシダーゼIIとをコードする該核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該α−1,2−マンノシダーゼ、該GlcNAcトランスフェラーゼIおよび該マンノシダーゼIIの発現が、GlcNAcMan3GlcNAc2 N−グリカンを含むタンパク質を生成する、単離真菌細胞。
【請求項50】
前記遺伝子工学的に改変された真菌細胞が、標的タンパク質をコードする核酸をさらに含み、ここで、該細胞が、前記GlcNAcMan3GlcNAc2 N−グリカンを含むように改変された該標的タンパク質を生成する、請求項49に記載の真菌細胞。
【請求項51】
請求項49または請求項50に記載の真菌細胞であって、ここで、前記遺伝子工学的に改変された真菌細胞が、GlcNAcトランスフェラーゼIIをコードする核酸をさらに含み、ここで、該GlcNAcトランスフェラーゼIIをコードする該核酸が、コードされた該GlcNAcトランスフェラーゼIIを細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該GlcNAcトランスフェラーゼIIの発現が、GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成する、真菌細胞。
【請求項52】
請求項51に記載の真菌細胞であって、該真菌細胞は、ガラクトシルトランスフェラーゼをコードする核酸をさらに含み、ここで、該ガラクトシルトランスフェラーゼをコードする該核酸が、コードされた該ガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該ガラクトシルトランスフェラーゼの発現が、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成する、真菌細胞。
【請求項53】
GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成するように遺伝子工学的に改変された単離真菌細胞であって、ここで、該真菌細胞は、ALG3活性を欠損するように遺伝子工学的に改変され、GlcNAcトランスフェラーゼIとGlcNAcトランスフェラーゼIIとをコードする核酸を含み、ここで、該GlcNAcトランスフェラーゼIと該GlcNAcトランスフェラーゼIIとをコードする該核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該GlcNAcトランスフェラーゼIおよび該GlcNAcトランスフェラーゼIIの発現が、GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成する、単離真菌細胞。
【請求項54】
前記遺伝子工学的に改変された真菌細胞が、OCH1活性をさらに欠損する、請求項53に記載の真菌細胞。
【請求項55】
前記遺伝子工学的に改変された真菌細胞が、α−1,3−グルコシルトランスフェラーゼをコードする核酸をさらに含む、請求項53または請求項54に記載の真菌細胞。
【請求項56】
請求項53、請求項54または請求項55に記載の真菌細胞であって、ここで、前記遺伝子工学的に改変された真菌細胞が、標的タンパク質をコードする核酸をさらに含み、ここで、該細胞が、前記GlcNAc2Man3GlcNAc2 N−グリカンを含むように改変された該標的タンパク質を生成する、真菌細胞。
【請求項57】
請求項53から56のいずれか一項に記載の真菌細胞であって、該真菌細胞は、グルコシダーゼIIのαおよびβサブユニットをコードする核酸をさらに含み、ここで、該真菌細胞における該グルコシダーゼIIの該αおよびβサブユニットの発現が、前記GlcNAc2Man3GlcNAc2 N−グリカンを含む前記タンパク質を生成する、真菌細胞。
【請求項58】
請求項57に記載の真菌細胞であって、該真菌細胞は、ガラクトシルトランスフェラーゼをコードする核酸をさらに含み、ここで、該ガラクトシルトランスフェラーゼをコードする該核酸が、コードされた該ガラクトシルトランスフェラーゼをゴルジ装置に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該真菌細胞における該ガラクトシルトランスフェラーゼの発現が、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成する、真菌細胞。
【請求項59】
相当数のYarrowia lipolytica細胞が、Gal2GlcNac2Man3GlcNAc2 N−グリカンを含む糖タンパク質を生成するように遺伝子工学的に改変されているYarrowia lipolytica細胞の実質的に純粋な培養物であって、ここで、該細胞が、ALG3活性を欠損するように遺伝子工学的に改変され、GlcNAcトランスフェラーゼIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸を含み、ここで、該GlcNAcトランスフェラーゼIと、該GlcNAcトランスフェラーゼIIと、該ガラクトシルトランスフェラーゼとをコードする該核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該細胞における該GlcNAcトランスフェラーゼI、該GlcNAcトランスフェラーゼIIおよび該ガラクトシルトランスフェラーゼの発現が、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成する、実質的に純粋な培養物。
【請求項60】
前記細胞が、OCH1活性をさらに欠損する、請求項49に記載の実質的に純粋な培養物。
【請求項61】
前記細胞が、α−1,3−グルコシルトランスフェラーゼをコードする核酸をさらに含む、請求項59または請求項60に記載の実質的に純粋な培養物。
【請求項62】
前記細胞が、グルコシダーゼIIのαおよびβサブユニットをコードする核酸をさらに含み、ここで、該真菌細胞における該グルコシダーゼIIの該αおよびβサブユニットの発現が、前記Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含む前記標的タンパク質を生成する、請求項59、請求項60または請求項61に記載の実質的に純粋な培養物。
【請求項63】
相当数のYarrowia lipolytica細胞が、Gal2GlcNac2Man3GlcNAc2 N−グリカンを含む糖タンパク質を生成するように遺伝子工学的に改変されているYarrowia lipolytica細胞の実質的に純粋な培養物であって、ここで、該細胞が、OCH1活性を欠損するように遺伝子工学的に改変され、α−1,2−マンノシダーゼと、GlcNAcトランスフェラーゼIと、マンノシダーゼIIと、GlcNAcトランスフェラーゼIIと、ガラクトシルトランスフェラーゼとをコードする核酸を含み、ここで、該α−1,2−マンノシダーゼと、該GlcNAcトランスフェラーゼIと、該マンノシダーゼIIと、該GlcNAcトランスフェラーゼIIと、該ガラクトシルトランスフェラーゼとをコードする該核酸が、それぞれのコードされたタンパク質を細胞内区画に向かわせるためのターゲティング配列をコードするヌクレオチド配列を含み、ここで、該細胞における該α−1,2−マンノシダーゼ、該GlcNAcトランスフェラーゼI、該マンノシダーゼII、該GlcNAcトランスフェラーゼIIおよびガラクトシルトランスフェラーゼの発現が、Gal2GlcNAc2Man3GlcNAc2 N−グリカンを含むタンパク質を生成する、実質的に純粋な培養物。
【請求項64】
糖タンパク質を含む組成物であって、該糖タンパク質上のN−グリカンの少なくとも50%が、GlcNAc2Man3GlcNAc2 N−グリカンである組成物。
【請求項65】
前記糖タンパク質上のN−グリカンの少なくとも70%が、GlcNAc2Man3GlcNAc2 N−グリカンである、請求項64に記載の組成物。
【請求項66】
前記糖タンパク質上のN−グリカンの少なくとも85%が、GlcNAc2Man3GlcNAc2 N−グリカンである、請求項64に記載の組成物。
【図30A】


【図30B】

【図31A】

【図31B】

【図1】
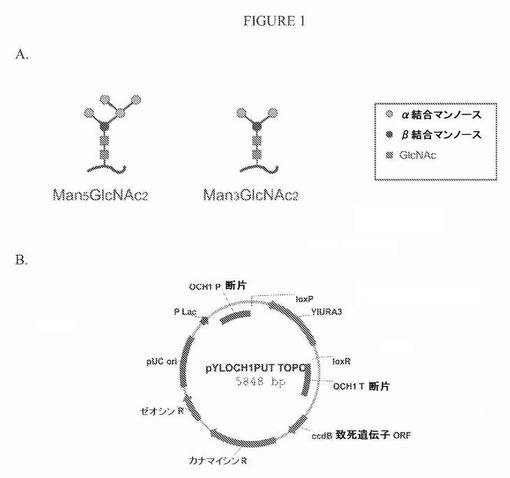
【図2】

【図3】

【図4】

【図5】

【図6】
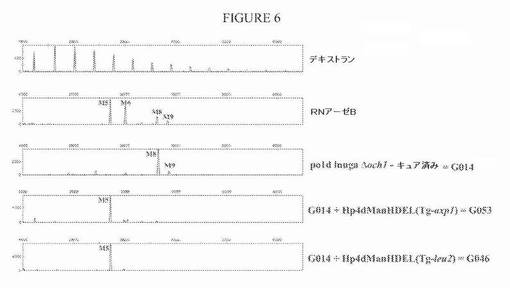
【図7】

【図8】
【図9】
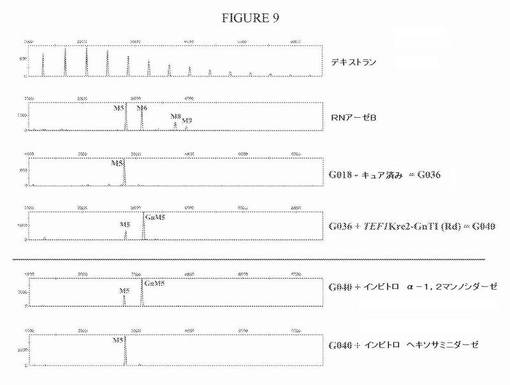
【図10−1】

【図10−2】

【図11】
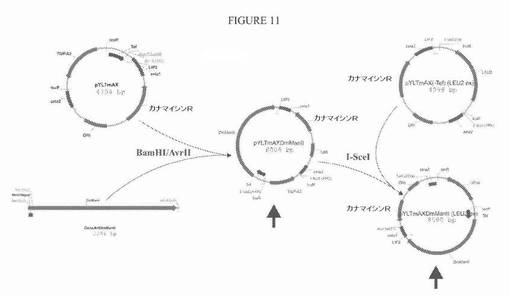
【図12】
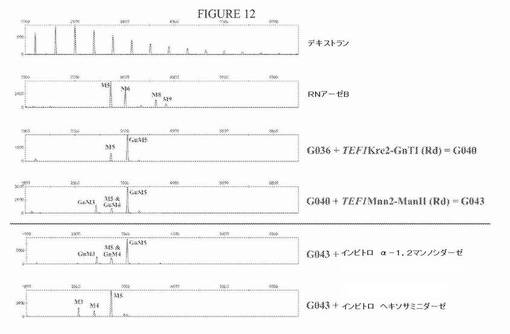
【図13−1】

【図13−2】

【図14】
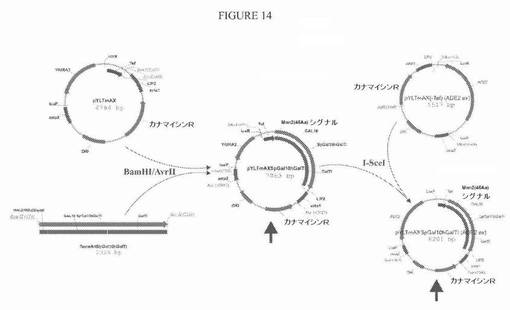
【図15】
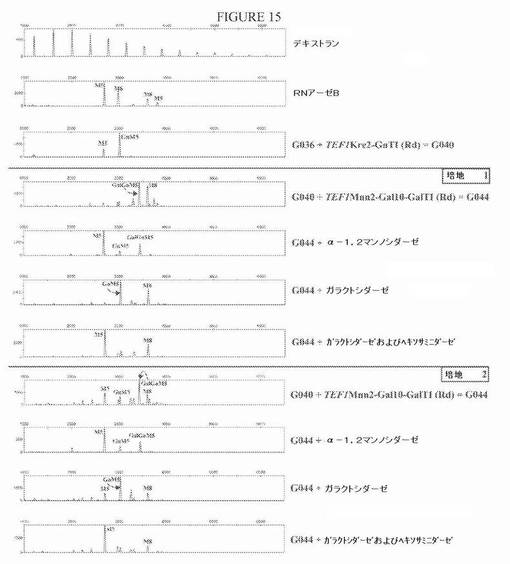
【図16】
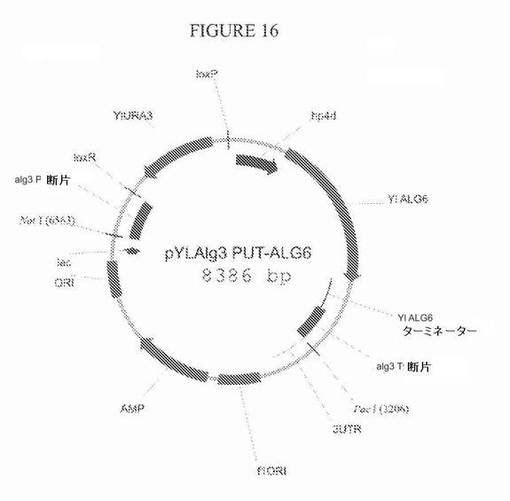
【図17】
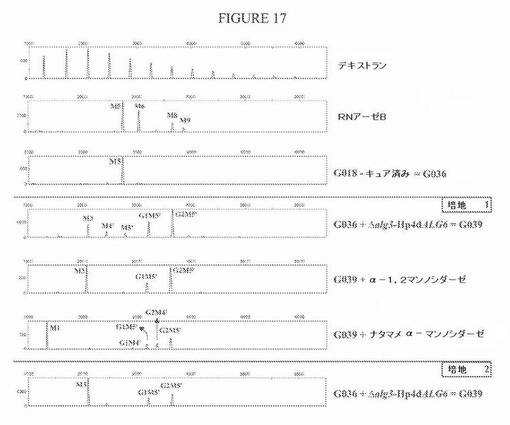
【図18】
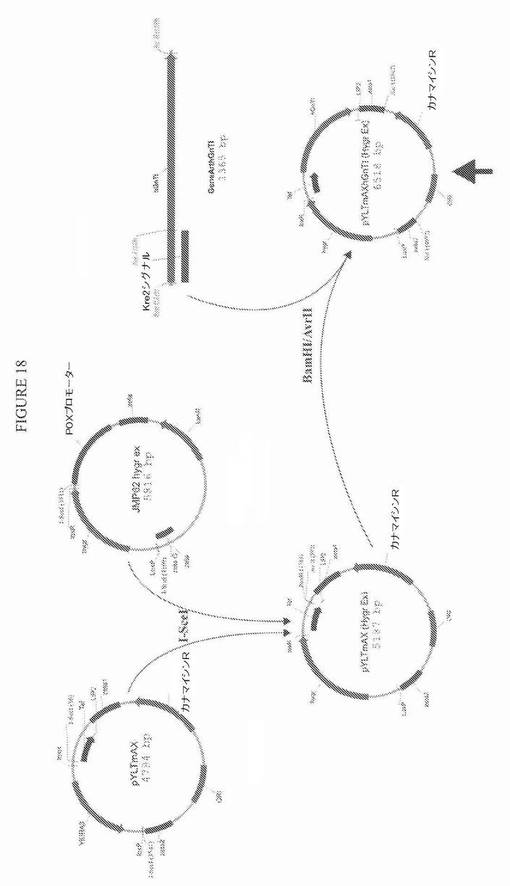
【図19】
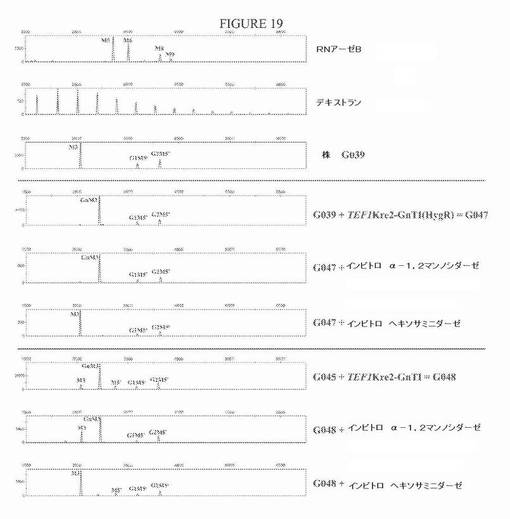
【図20】
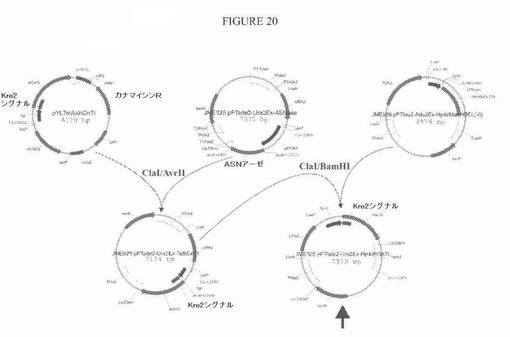
【図21】

【図22】

【図23】
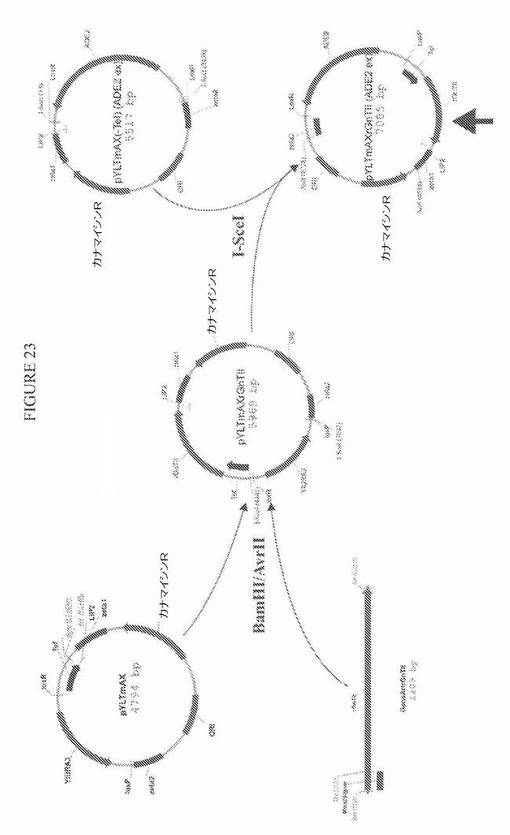
【図24】

【図25】
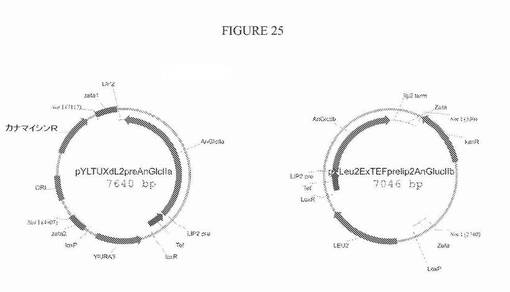
【図26】

【図27】

【図28】
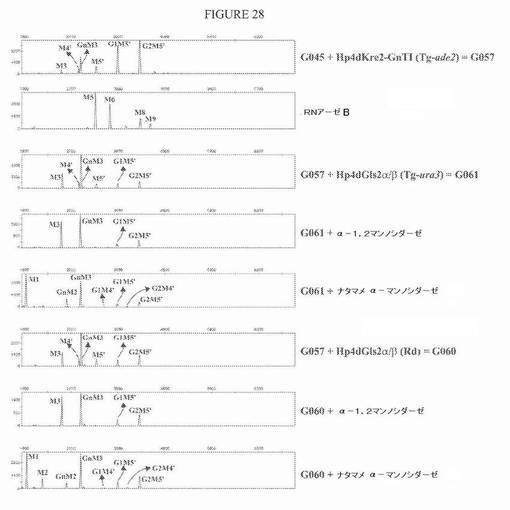
【図29】

【図32】
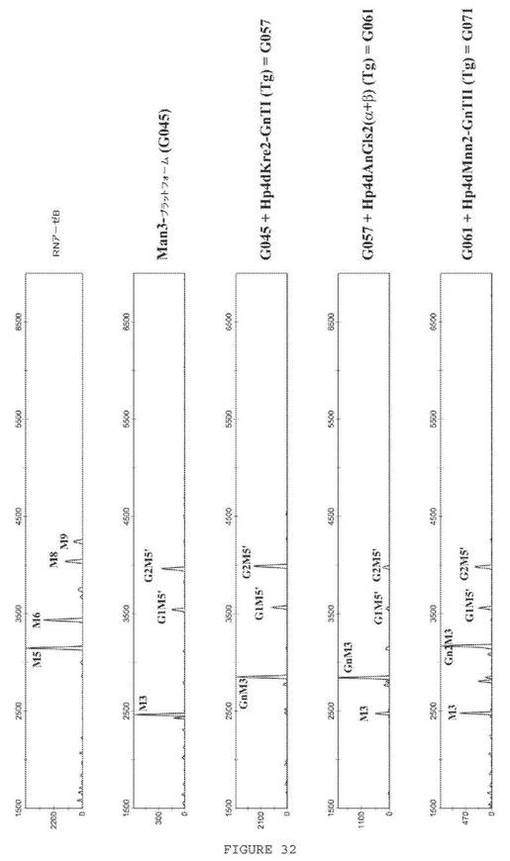
【図33】

【図34A】
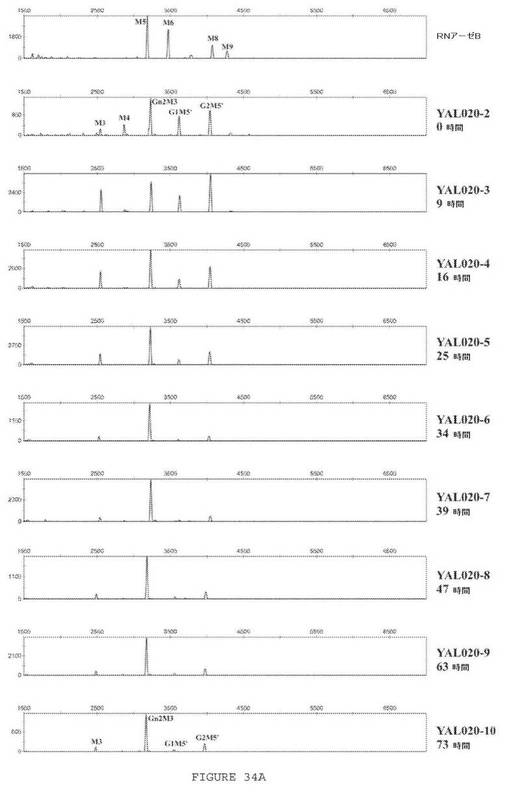
【図34B】

【図30B】
【図31A】
【図31B】
【図1】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10−1】
【図10−2】
【図11】
【図12】
【図13−1】
【図13−2】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図32】
【図33】
【図34A】
【図34B】
【公表番号】特表2013−511272(P2013−511272A)
【公表日】平成25年4月4日(2013.4.4)
【国際特許分類】
【出願番号】特願2012−539436(P2012−539436)
【出願日】平成22年11月19日(2010.11.19)
【国際出願番号】PCT/IB2010/003154
【国際公開番号】WO2011/061629
【国際公開日】平成23年5月26日(2011.5.26)
【出願人】(509275507)オキシレイン ユーケー リミテッド (4)
【Fターム(参考)】
【公表日】平成25年4月4日(2013.4.4)
【国際特許分類】
【出願日】平成22年11月19日(2010.11.19)
【国際出願番号】PCT/IB2010/003154
【国際公開番号】WO2011/061629
【国際公開日】平成23年5月26日(2011.5.26)
【出願人】(509275507)オキシレイン ユーケー リミテッド (4)
【Fターム(参考)】
[ Back to top ]