
喘息および関連疾患を含むアトピー性アレルギーの治療および診断に有用なIL−9受容体変異体
【課題】アトピー性アレルギーおよび喘息ならびにこれらの関連疾患を診断および/または治療するために有用なIL-9受容体変異体を提供。
【解決手段】ヒト・インターロイキン-9受容体をコードするヌクレオチド配列を有する単離されたDNA分子であって、前記ヌクレオチド配列が1273位にGからAへの核酸変異を含む配列、核酸759-761が欠失された配列、核酸613-617が欠失された配列、核酸435-437に停止コドンを含む配列、核酸613-641が欠失された配列、およびそれらの断片よりなる群から選択される、上記の単離されたDNA分子。
【解決手段】ヒト・インターロイキン-9受容体をコードするヌクレオチド配列を有する単離されたDNA分子であって、前記ヌクレオチド配列が1273位にGからAへの核酸変異を含む配列、核酸759-761が欠失された配列、核酸613-617が欠失された配列、核酸435-437に停止コドンを含む配列、核酸613-641が欠失された配列、およびそれらの断片よりなる群から選択される、上記の単離されたDNA分子。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願の参照)
本出願は、1996年12月2日に出願された米国仮出願第60/032,224号(参照により本明細書に組み入れる)に関連する。また本出願は、米国特許出願第08/697,419号、08/697,360号、08/697,473号、08/697,472号、08/697,471号、08/702,105号、08/702,110号、08/702,168号、および08/697,440号(これらはすべて1996年8月23日に出願された)、そして08/874、503号(これは1997年6月13日に出願された)の記載事項に関連し、これらは参照により本明細書に組み入れる。
【0002】
(発明の分野)
本発明は、IL-9受容体(喘息関連因子2(Asthma Associated Factor 2))(配列番号1)の生物学的変化を記載し、これらの変異体と、喘息、アトピー性アレルギー、および関連疾患への感受性(susceptibility)とを関連付ける。本発明はまた、喘息およびアトピー性アレルギーに対する感受性または耐性の診断のためにこれらのIL-9受容体の配列変異体を利用する方法を教示する。さらに、IL-9活性の調節に依存する喘息用の薬剤の開発において、IL-9受容体変異体を使用する方法が記載される。
【背景技術】
【0003】
炎症は、体の防御システムが外来物質と戦う複雑なプロセスである。外来物質に対する戦いは体の生存のために必要であるが、一部の防御システムは、無害の外来物質に対しても危険なものとして不適当に対応し、その結果、その後の戦いにおいて周りの組織を傷害する。
【0004】
アトピー性アレルギーは、遺伝的背景が、環境からの刺激に対する応答を支配する疾患である。この疾患は、一般に遍在する抗原に応答してIgE抗体を産生するリンパ球の能力が上昇していることが特徴である。これらの抗原による免疫系の活性化はまた、これらの摂取、皮膚を通しての透過、または吸入後に起きるアレルギー性炎症を引き起こす。この免疫活性化が起き、続いて肺の炎症が起きると、この疾患は広く喘息として特徴付けられる。気道での炎症性反応にはある種の細胞が重要であり、これらには、T細胞および抗原提示細胞、IgEを産生するB細胞、および炎症性メディエーターを蓄えIgEに結合する肥満細胞/好塩基球、さらなるメディエーターを放出する好酸球がある。これらの炎症性細胞はアレルギー性炎症の場に蓄積し、これらが放出する有害産物が、この疾患に関連する組織破壊に寄与する。
【0005】
喘息は一般に、気道の炎症性疾患として定義されるが、臨床症状は、間欠的な空気流の遮断から起きる。これは、慢性の障害性の疾患であり、罹患率と重篤度が上昇しているようである1。人口の30〜40%がアトピー性アレルギーに罹っており、児童の15%と成人の5%が喘息に罹っていると推定されている(非特許文献1)1。すなわち、これらの疾患の治療において、医療資源に巨大な負担がかかっている。
【0006】
喘息と関連疾患の診断と治療の両方とも問題がある1。特に炎症を起こした肺の評価は困難なことが多く、炎症の原因が決定できないことが頻繁にある。アトピー性喘息は環境遺伝的疾患であるが、具体的な変異体遺伝子についての知見は最近発見されたばかりである。炎症におけるこれらの遺伝子的変異体の検出方法とその役割、診断および予後は、まだ決定されていない。当該分野で必要なことは、アトピー性疾患に対する感受性/耐性に関与する遺伝子の変異に特異的に関係する、アトピー性喘息の診断を促進する技術の開発である。
【0007】
現代の治療法は、固有の一連の欠点を有する。主要な治療薬であるβ−アゴニストは、症状を低下させ、すなわち肺機能を一過性に改善するが、基礎炎症には作用せず、そのため肺組織は依然として危険な状態にある。さらに、βーアゴニストを使い続けると脱感受性が起き、その効力と安全性が低下する2。基礎炎症を縮小させることができる物質である抗炎症性ステロイドは、免疫抑制から骨喪失にわたる固有の一連の副作用を有する2。
【0008】
従来の治療に起因する問題のために、別の治療方策が評価されている38-39。グリコホリンA37、シクロスポリン38、およびIL-2のノナペプチド断片36はすべて、インターロイキン−2依存性のTリンパ球増殖を阻害する28。しかしこれらは、他の多くの作用を有することが知られている2。例えば、シクロスポリンは、臓器移植後の免疫抑制剤として使用されている。これらの薬剤は、喘息患者の治療においてステロイドの代替となるが36-39、これらはインターロイキン−2依存性Tリンパ球増殖を阻害し、ホメオスタシスに関連して非常に重要と思われる免疫機能を阻害する可能性がある。当該分野で必要なことは、アトピー性喘息の症状ではなくその原因を治療するように具体的に設計された治療薬の開発を促進することである。これらの治療法は、非特異的な治療に関連する毒性を避けるための最も可能性の高い方法である。この治療薬は、免疫機能(例えば、喘息の進展に必要なIL-2介在Tリンパ球活性化)の下流にあり、疾患の偶発性を説明し、アレルギーとの密接な関連を説明する経路を選択的に標的とする。その経路中に生物学的変化がふつうに存在し、その変化を示す個体が、アトピー性喘息の症状以外は免疫無防備状態でも病気でもない場合は、経路は喘息治療の適切な標的であることを、自然は証明している。
【0009】
喘息を含むアトピー性アレルギーの診断および治療に関連する困難さのために、これらの疾患の複雑な病態生理が詳しく研究されている。これらの疾患は、多くの要素からなり多くの型を取るため定義することが困難であるが、喘息患者にはいくつかの共通の特徴が見いだされている。そのような特徴の例には、アレルゲンによる抗原刺激に対する異常な皮膚試験応答、肺の好酸球増加症、気管支過応答性(BHR)、気管支拡張薬可逆性、および空気流遮断がある3-10。喘息に関連する形質のこれらの発現は、定量的または定性的に研究される。
【0010】
多くの場合、IgEレベルの上昇は、BHR、種々の刺激に対する気管支収縮応答の上昇に関係している4,6,8,9。BHRは、気道炎症の存在を反映すると考えられており6,8、喘息の危険因子であると考えられている11-12。BHRは、肺への好酸球浸潤を含む気管支炎症と喘息患者のアレルギー性素因を伴う6,8,13-18。
【0011】
アトピー性喘息に対する遺伝性成分が、多くの研究で記録されている4,10。しかしこれらの疾患は、年令と性、ならびに多くの環境因子(例えば、アレルゲン、ウイルス感染、および汚染物質19-21)に大きく影響されるため、家族研究は解釈することが困難である。さらに、これらの疾患に対する感受性に関連することが知られている生化学的欠陥はないため、突然変異遺伝子およびその異常遺伝子産物は、これらが産生する異常表現型によってのみ認識することができる。
【0012】
IL-9とIL-9受容体(IL-9経路)の機能は、当初認識されていたものより、拡大している。IL-9経路はT細胞増殖の刺激物質として作用するが、このサイトカインはまた、赤血球始原細胞、B細胞、肥満細胞、および胎児胸腺細胞の増殖に介在することが知られている22,23。IL-9経路はIL-3と相乗的に作用して肥満細胞活性化と増殖を引き起こす24。IL-9経路はまた、正常なヒトBリンパ球によるIgE、IgGおよびIgMのIL-4誘導性産生を増強し25、マウスBリンパ球によるIgEおよびIgGのIL-4誘導性放出を増強する26。寄生体感染に対する粘膜の炎症性応答におけるIL-9経路の役割もまた証明されている27,28。
【0013】
しかし、IL-9受容体の配列がアトピー性喘息と気管支過応答性にいかに相関するかは知られていない。IL-9が標的細胞の表面に発現される特異的受容体に結合することは知られている23,29,30。この受容体は実際、2つのタンパク質鎖からなる。1つのタンパク質鎖はIL-9受容体として知られており、IL-9に特異的に結合する。もう1つのタンパク質鎖は、IL-2受容体と共有される23。さらにヒトIL-9受容体をコードするcDNAがクローン化されており、配列が決定されている23,29,30。このcDNAは、マウスIL-9受容体と高い相同性を示す522アミノ酸のタンパク質をコードする。この受容体の細胞外領域は高度に保存されており、マウスとヒトのタンパク質の間で67%の相同性がある。受容体の細胞質領域は、あまり高度に保存されていない。ヒトの細胞質ドメインは、マウス受容体の対応する領域よりはるかに大きい23。
【0014】
IL-9受容体遺伝子は特徴付けもされている30。これは、マウスゲノム中で単一のコピーとして存在し、9個のエキソンと8個のイントロンを含有する30。ヒトのゲノムは、少なくとも4つのIL-9受容体偽遺伝子(pseudogene)を含有する。ヒトのIL-9受容体遺伝子は、性染色体XとYの320 kbの次端部テロメア領域(subtelomeric region)にマッピングされている23。
【0015】
これらの研究にもかかわらず、IL-9受容体遺伝子の変異体は発見されていない。従ってアトピー性アレルギー、喘息、気管支過応答性についての遺伝的情報、およびこれらの疾患の病因におけるIL-9受容体の役割の解明について具体的なニーズがある。この情報は、これらの疾患に関連するこの遺伝子の遺伝的変異体を同定する方法を使用して、アトピー性アレルギーや関連疾患を診断するのに使用することができる。さらに、これらの疾患を治療するための治療薬の開発のために、IL-9受容体変異体を起用する方法についてニーズがある。
【先行技術文献】
【非特許文献】
【0016】
【非特許文献1】Gergen PJ, and Weiss KB: The increasingproblem of asthma in the United States. Am Rev Respir Dis 146: 823-824,1992
【発明の概要】
【課題を解決するための手段】
【0017】
(発明の要約)
本出願人らは、ヒトIL-9受容体(喘息関連因子2またはAAF2としても知られている)の天然の変異体を発見し、これらの変異体を喘息および関連疾患の病態と関連付けた。これらの発見により、アトピー性喘息の診断法および治療薬を発見する方法が開発された。さらに本出願人らは、IL-9受容体は、マウスの多くの抗原誘導性応答(気管支過応答性、気管支洗浄液中の好酸球と細胞数の増加、および血清総IgEの増加を含む)に非常に重要であることを明らかにした。これらの知見は、アトピー性喘息および関連するアレルギー性炎症に典型的である。
【0018】
さらに本出願人らは、GからAへの核酸の変異がcDNA(配列番号2)の1273位に存在し、これはヒトIL-9受容体前駆体タンパク質のコドン344のアルギニンからヒスチジンへの予測されるアミノ酸置換を引き起こす。1個体の両方の対立遺伝子にアルギニン残基が存在する時、これはアトピー性喘息が少ないことに関連している。すなわち本出願人らは、コドン344のアルギニンが1個体で両方のIL-9受容体遺伝子産物中で存在するとき、これにより特徴付けられる非喘息表現型の存在を確認した。さらなる重要な結果として、本出願人らは、コドン344のヒスチジンを特徴とする、喘息性、アトピー性表現型に対する感受性の存在を確認した。すなわち本発明は、そのような配列を有する精製され単離されたDNA分子、ならびにこのDNAによりコードされるタンパク質を包含する。
【0019】
本出願人らはまた、IL-9R前駆体タンパク質の173位のグルタミン残基が欠失しているIL-9Rのスプライス変異体(配列番号3)が存在することを確認した(図5)。さらに本出願人らは、この変異体は、Jak-Stat経路によりシグナルを転写できないこと(図15)、およびIL-9で刺激しても細胞増殖を誘導できないこと(図16)を証明した。従ってこの対立遺伝子を有する個体は、アトピー性喘息および関連疾患に罹りにくいであろう。
【0020】
本出願人らはさらに、イントロン5のチミン残基であるnt-213(エキソン6から213nt上流)が、シトシンヌクレオチドに変換されている、IL-9RゲノムDNAの変異体が存在することを確認した。このような変異はエキソン6の開始点でグルタミン残基を除去するスプライス変異体の頻度を上昇させ得る。
【0021】
さらに本出願人らは、エキソン9の読みとり枠と未熟停止コドンに変異を起こす、エキソン8が欠失しているIL-9Rの変異体(配列番号4)を発見した。このような変異体は、Jak-Stat経路でシグナルを伝達することを妨害し、従ってこの対立遺伝子を有する個体もまた、アトピー性喘息および関連疾患に罹りにくいであろう。
【0022】
IL-9の生物活性は、そのIL-9受容体への結合とその後の特異的細胞での制御シグナル伝搬に起因し、従って、IL-9機能は、IL-9アンタゴニストとIL-9またはその受容体との相互作用により妨害することができる。ダウンレギュレーション、すなわちIL-9により制御される機能の低下は、多くの方法で行われる。受容体へのIL-9の結合を阻止することができるアンタゴニストを投与することは、1つの重要な方法であり、このようなアンタゴニストは、特許請求される本発明の範囲内である。例としては、IL-9受容体の天然に存在する可溶性型のDNA配列によりコードされるポリペプチド産物の投与があり、DNA配列はエキソン2と3からなるポリペプチド(配列番号5)をコードする。他の2つの変異体は、IL-9Rの可溶性型(これは、エキソン2、3および4からなり、ある場合にはエキソン5の異なる読みとり枠からの4つのアミノ酸(配列番号6)(図6)、そして別の場合はエキソン5の異なる読みとり枠の27のアミノ酸(配列番号7)(図7)を含む)を産生する。
【0023】
IL-9受容体経路のアゴニストおよびアンタゴニストの同定方法は、文献に詳細に記載されている受容体−リガンド相互作用を評価することにより特定できる。これらの方法は、化学的スクリーニングと潜在的な治療薬の同定を促進する、高処理量自動アッセイに適合させることができる。アゴニストは、IL-9受容体との特異的な相互作用を同定することにより認識される。100〜1000倍過剰の非標識リガンドを使用しても、標識した推定リガンドに対する結合が存在しないことは、一般に特異的受容体結合の証拠として受け取られている。これらの実験では、多くの標識物および検出スキームを使用することができる。増加する濃度の試験化合物を既知のリガンドおよび受容体に添加しても同様に結合がないことも、アンタゴニストの証拠である。
【0024】
変異体受容体の情報は、受容体結合アッセイの可溶性受容体を作成するのに使用可能な発現ベクターを作製するための手段を提供する。これらの可溶性受容体の突然変異誘発は、リガンドに結合するのにどのアミノ酸が非常に重要であるか、およびどのアミノ酸がアンタゴニストの構造ベースのデザインを補助するかを決定するために使用することができる。
【0025】
ヒトIL-9受容体が欠如した細胞は、変異体受容体を含有する発現ベクターで一過性にまたは安定にトランスフェクションすることができ、IL-9経路活性をアッセイするのに使用することができる。これらの活性は、いずれもIL-9経路が原因であるとされている細胞増殖でもアポトーシスの阻止でもよい。これらの細胞は、前記したように受容体アゴニストおよびアンタゴニストを同定するのに使用することができる。
【0026】
上記方法は、アトピー性喘息や他の関連疾患の治療薬を開発するために、IL-9受容体の変異体型を利用する種々の有効な方法である。
【0027】
IL-9受容体中の単一のヌクレオチド変異体を認識する、アトピー性喘息を診断するために使用できる多くの技術が記載されており、これらはDNA配列決定、制限断片長多型(RFLP)、対立遺伝子特異的オリゴヌクレオチド解析(ASO)、連結連鎖反応(LCR)、化学的切断、および1本鎖コンフォメーション多型解析(SSCP)がある。これらの技術ならびに文献に記載の1つまたはそれ以上の他の技術の使用を、IL-9受容体遺伝子またはmRNA転写物の1つまたはそれ以上の変異を検出するのに使用でき、これらが本発明の範囲内にあることを当業者は容易に理解できるであろう。
【0028】
さらに別の技術(ELISA、免疫沈降、ウェスタン法、および免疫ブロッティング法を含む)を使用して、IL-9受容体のアミノ酸変異体を検出してもよい。すなわち、IL-9受容体の種々の型の構造を特異的に認識するポリクローナル抗体またはモノクローナル抗体もまた、本発明の範囲内にあり、アトピー性喘息や関連疾患に対する感受性または耐性を説明するための有用な診断法である。
【0029】
上記方法は、アトピー性喘息および他の関連疾患を診断するための種々の有効な方法を示す。
【0030】
すなわち本出願人らは、IL-9とその受容体との相互作用を調節することができるアンタゴニストを同定するためにIL-9受容体を使用する方法を提供している。さらに詳しくは、本出願人らは、IL-9経路の発現または機能をダウンレギュレーションするのに充分な量で投与される化合物または物質を同定するために、IL-9受容体の機能をアッセイする方法を提供する。
【0031】
本出願人らは、アトピー性アレルギー、気管支過応答性および喘息におけるIL-9経路の役割を特定したため、アトピー性アレルギー、喘息、および関連疾患に対する感受性と耐性の診断のための方法も提供する。
【0032】
本明細書に取り込まれその一部を構成する添付の図面は、本発明のいくつかの実施態様を例示し、本発明の説明とともに本発明の原理を説明するものである。
【0033】
本発明は、例えば、以下を提供する:
(項目1) ヒト・インターロイキン-9受容体をコードするヌクレオチド配列を有する単離されたDNA分子であって、前記ヌクレオチド配列が1273位にGからAへの核酸変異を含む配列、核酸759-761が欠失された配列(配列番号3)、核酸613-617が欠失された配列(配列番号7)、核酸435-437に停止コドンを含む配列(配列番号5)、核酸613-641が欠失された配列(配列番号6)、およびそれらの断片よりなる群から選択される、上記の単離されたDNA分子。
(項目2) 前記配列が1273位にGからAへの核酸変異を含む項目1に記載の単離されたDNA分子またはその断片。
(項目3) 核酸759-761が欠失された(配列番号3)項目1に記載の単離されたDNA分子またはその断片。
(項目4) 前記配列が核酸435-437に停止コドンを含む(配列番号5)項目1に記載の単離されたDNA分子またはその断片。
(項目5) 核酸613-617が欠失された(配列番号7)項目1に記載の単離されたDNA分子またはその断片。
(項目6) 核酸613-641が欠失された(配列番号6)項目1に記載の単離されたDNA分子またはその断片。
(項目7) 核酸1067-1151が欠失された(配列番号4)項目1に記載の単離されたDNA分子またはその断片。
(項目8) エキソン6から213核酸上流の核酸がTからCに変化した、ヒトIL-9受容体の第5イントロンをコードするヌクレオチド配列を有する単離されたDNA分子。
(項目9) ヒト・インターロイキン-9受容体をコードするヌクレオチド配列を有する単離されたRNA分子であって、前記ヌクレオチド配列が1273位にAからGへの核酸変異を含む配列、核酸759-761が欠失された配列、核酸435-437に停止コドンを含む配列、核酸613-617が欠失された配列、核酸613-641が欠失された配列、核酸1067-1151が欠失された配列、およびそれらの断片よりなる群から選択される、上記の単離されたRNA分子。
(項目10) 前記配列が1273位にAからGへの核酸変異を含む、項目9に記載の単離されたRNA分子またはその断片。
(項目11) 核酸759-761が欠失された、項目9に記載の単離されたRNA分子またはその断片。
(項目12) 前記配列が核酸435-437に停止コドンを含む、項目9に記載の単離されたRNA分子またはその断片。
(項目13) 核酸613-617が欠失された、項目9に記載の単離されたRNA分子またはその断片。
(項目14) 核酸613-641が欠失された、項目9に記載の単離されたRNA分子またはその断片。
(項目15) 核酸1067-1151が欠失された、項目9に記載の単離されたRNA分子またはその断片。
(項目16) ヒト・インターロイキン-9受容体をコードするアミノ酸配列を有する単離されたタンパク質分子であって、前記アミノ酸配列が、344位にヒスチジンを含む配列、グルタミン173が欠失された配列、前記分子がアミノ酸64の後で終結している配列、前記分子が項目4に記載のDNAによりコードされる配列、前記分子が項目5に記載のDNAによりコードされる配列、またはその断片よりなる群から選択される、上記の単離されたタンパク質分子。
(項目17) 前記配列が344位にヒスチジンを含む、項目16に記載の単離されたタンパク質分子またはその断片。
(項目18) グルタミン173が欠失された、項目16に記載の単離されたタンパク質分子またはその断片。
(項目19) 前記分子がアミノ酸64の後で終結している、項目16に記載の単離されたタンパク質分子またはその断片。
(項目20) 前記分子が項目5に記載のDNAによりコードされる、項目16に記載の単離されたタンパク質分子またはその断片。
(項目21) 前記分子が項目6に記載のDNAによりコードされる、項目16に記載の単離されたタンパク質分子またはその断片。
(項目22) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、項目2に記載のDNAの存在または非存在を判定し、前記DNAのみの存在が喘息または関連疾患に対するより高い罹病性を示すものである、上記の方法。
(項目23) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、項目3に記載のDNAの存在または非存在を判定し、前記DNAのみの存在が喘息または関連疾患に対するより高い罹病性を示すものである、上記の方法。
(項目24) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、項目4に記載のDNAの存在または非存在を判定し、前記DNAの存在が喘息または関連疾患に対するより低い罹病性を示すものである、上記の方法。
(項目25) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、項目5に記載のDNAの存在または非存在を判定し、前記DNAの存在が喘息または関連疾患に対するより低い罹病性を示すものである、上記の方法。
(項目26) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、項目6に記載のDNAの存在または非存在を判定し、前記DNAの存在が喘息または関連疾患に対するより低い罹病性を示すものである、上記の方法。
(項目27) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、項目7に記載のDNAの存在または非存在を判定し、前記DNA分子の存在が喘息または関連疾患に対するより低い罹病性を示すものである、上記の方法。
(項目28) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、項目8に記載のDNA分子の存在または非存在を判定し、前記DNAの存在が喘息または関連疾患に対するより低い罹病性を示すものである、上記の方法。
(項目29) IL-9経路のアンタゴニストを同定する方法であって、a)IL-9受容体を発現する細胞を取得し、b)前記細胞を、IL-9と推定上のアンタゴニストの様々な比率の混合物に接触させ、c)Jak-Stat経路のタンパク質のリン酸化の量を測定する、各ステップを含んでなり、リン酸化の低下がIL-9アンタゴニストの存在を示すものである、上記の方法。
(項目30) IL-9受容体がヒトのものである、項目29に記載の方法。
(項目31) IL-9受容体がマウスのものである、項目29に記載の方法。
(項目32) IL-9経路のアンタゴニストを同定する方法であって、a)IL-9受容体を発現する細胞を取得し、b)前記細胞を、デキサメタゾンとIL-9と推定上のアンタゴニストの様々な比率の混合物に接触させ、c)アポトーシスの強度を測定する、各ステップを含んでなり、デキサメタゾン誘発アポトーシスからのIL-9介在レスキューの低下がIL-9アンタゴニストの存在を示すものである、上記の方法。
(項目33) IL-9受容体がヒトのものである、項目32に記載の方法。
(項目34) IL-9受容体がマウスのものである、項目32に記載の方法。
(項目35) ヒトIL-9受容体を発現する細胞がC8166-45、HL60、HL60-clone5、好酸球、好中球、巨核球、およびヒトIL-9RでトランスフェクトされたTS1、KG-1および32D細胞系よりなる群から得られる、項目24または26に記載の方法。
(項目36) マウスIL-9受容体を発現する細胞がTS1、KG-1、32D、好酸球、好中球、および巨核球よりなる群から得られる、項目25または27に記載の方法。
(項目37) ヒトIL-9受容体を発現する細胞がHL60またはHL60-clone5細胞である、項目27または29に記載の方法。
(項目38) ヒトIL-9受容体を発現する細胞がヒトIL-9RでトランスフェクトされたKG-1細胞である、項目27または29に記載の方法。
(項目39) ヒトIL-9受容体を発現する細胞がC8166-45細胞である、項目27または29に記載の方法。
(項目40) ヒトIL-9受容体を発現する細胞が好酸球である、項目27または29に記載の方法。
(項目41) 喘息および喘息関連疾患を軽減させる方法であって、かかる治療を必要とする患者に、有効量の項目18に記載のタンパク質分子を投与することを含んでなる方法。
(項目42) 喘息および喘息関連疾患を軽減させる方法であって、かかる治療を必要とする患者に、有効量の項目19に記載のタンパク質分子を投与することを含んでなる方法。
(項目43) 喘息および喘息関連疾患を軽減させる方法であって、かかる治療を必要とする患者に、有効量の項目20に記載のタンパク質分子を投与することを含んでなる方法。
(項目44) プライマーが配列番号8および配列番号9に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン2を特異的に増幅する方法。
(項目45) プライマーが配列番号10および配列番号11に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン3を特異的に増幅する方法。
(項目46) プライマーが配列番号12および配列番号13に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン4を特異的に増幅する方法。
(項目47) プライマーが配列番号14および配列番号15に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン5を特異的に増幅する方法。
(項目48) プライマーが配列番号16および配列番号17に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン6を特異的に増幅する方法。
(項目49) プライマーが配列番号18および配列番号19に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン7を特異的に増幅する方法。
(項目50) プライマーが配列番号20および配列番号21に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン8を特異的に増幅する方法。
(項目51) プライマーが配列番号22および配列番号23に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン9を特異的に増幅する方法。
(項目52) プライマーが配列番号14および配列番号17に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のイントロン5を特異的に増幅する方法。
(項目53) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、前記DNAによりコードされる非機能性インターロイキン-9受容体をコードするDNAの存在または非存在を判定し、非機能性インターロイキン-9受容体をコードする前記DNAの存在が喘息または関連疾患に対するより低い罹病性を示すものである、上記の方法。
(項目54) 配列番号8、9、10、11、12、13、14、15、16、17、18、19、20、21、22または23よりなる群から選択される真のヒトIL-9受容体を特異的に増幅するためのプライマーを含む組成物。
(項目55) 喘息および喘息関連疾患を軽減させる方法であって、かかる治療を必要とする患者に、有効量の項目21に記載のタンパク質分子を投与することを含んでなる方法。
【図面の簡単な説明】
【0034】
【図1】ヒトIL受容体cDNAの略図。四角は、コード領域を含むエキソン2〜9を示す(スケールは相対的サイズであり、エキソン9の3’非翻訳部分を除いて、破線で示す)。膜貫通領域は、エキソン7によりコードされ、細胞内ドメインはエキソン8と9に、そして細胞外ドメインはエキソン2〜6にコードされる。矢印は、エキソンの部分配列に影響を与える多型または異常スプライスを示す;a)エキソン5の最初の5または29ヌクレオチドの欠失;b)エキソン6の最初の3ヌクレオチドの欠失(コドン173);c)コドン310におけるarg/gly多型;d)コドン344におけるarg/his多型;e)8または9個のセリンからなるコドン410+nの多型;*)エキソン3、4または8の完全な欠失。
【図2−1】コドン344(ヌクレオチド1272〜1274)のArg対立遺伝子とコドン410(ヌクレオチド1470〜1472)で開始する8 Ser/4Asn反復配列とを有する野生型IL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図2−2】コドン344(ヌクレオチド1272〜1274)のArg対立遺伝子とコドン410(ヌクレオチド1470〜1472)で開始する8 Ser/4Asn反復配列とを有する野生型IL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図3−1】コドン344(ヌクレオチド1272〜1274)のHis対立遺伝子とコドン410(ヌクレオチド1470〜1472)で開始する9 Ser/4Asn反復配列とを有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図3−2】コドン344(ヌクレオチド1272〜1274)のHis対立遺伝子とコドン410(ヌクレオチド1470〜1472)で開始する9 Ser/4Asn反復配列とを有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図4】エキソン9でフレームシフトと、11の非野生型アミノ酸および未熟停止コドンの産生とを引き起こす、エキソン8の欠失を有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図5−1】コドン173でグルタミンの欠失を有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図5−2】コドン173でグルタミンの欠失を有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図6】未熟停止コドンと27の非野生型アミノ酸の産生を引き起こす、エキソン5中の代替的なスプライスを有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図7】未熟停止コドンと4つの非野生型アミノ酸の産生を引き起こす、エキソン5中の代替的なスプライスを有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図8】エキソン5の最初のコドンとして停止コドンを産生する、エキソン4の欠失を有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図9】IL-9受容体遺伝子型とアトピー性アレルギーの間の関係を示す表。Arg/Argの個体は、8 Ser/4Asn反復配列を有するArg対立遺伝子についてホモ接合性である。Arg/Hisの個体は、エキソン9中に、8 Ser/4Asn反復配列を有するArg対立遺伝子と、9 Ser/4 Asn反復配列、9 Ser/3 Asn反復配列、および10 Ser/2Asn反復配列を有するHis対立遺伝子についてヘテロ接合性である。His/Hisの個体は、エキソン9中に、9 Ser/4Asn反復配列、9 Ser/3 Asn反復配列、および10 Ser/2 Asn反復配列を有するHis対立遺伝子についてホモ接合性である。Arg/Argの個体は、アトピー性アレルギーから防御される。Arg/HisおよびHis/Hisの個体は、アトピー性アレルギーに対して感受性である(P=0.002)。
【図10】IL-9受容体の発現構築物の地図。
【図11】TK-トランスフェクション細胞系中のC末端抗体プローブを使用する、組換えIL-9受容体タンパク質のウェスタンブロット(左:8 Ser/4Asn反復配列を有するArg対立遺伝子;右:9 Ser/4 Asn反復配列を有するHis対立遺伝子)。
【図12】8 Ser/4Asn反復配列を有するArg344変異体(GR8)と9 Ser/4 Asn反復配列を有するHis344変異体(GH9)の間の移動度の差を示す、TS1細胞中のヒトIL-9受容体変異体の発現。IL-9受容体のこれらの2つの変異体型の翻訳後修飾の差を証明する移動度のシフトがみられる。
【図13】IL-9受容体遺伝子の特異的増幅のためのXY特異的アンプリマー。第9、第10、第16、または第18染色体上の偽遺伝子は、PCRにより増幅されない。(MはマウスDNA、HはヒトDNA、そしてCはハムスターDNAを示す)。
【図14−1】抗ヒトIL-9受容体中和抗体と野生型およびデルタ−Q受容体との免疫反応性。パネルA):COS7細胞を、LXSNベクター単独(AとB)、野生型IL-9R(CとD)、コドン410で開始する9個のSer残基を有する野生型IL-9R(EとF)、Δ-Q 173変異体(GとH)、およびコドン410で開始する9個のSer残基を有するΔ-Q 173(IとJ)で一過性にトランスフェクトし、記載したように(実施例8)MAB290と抗マウスIgGテキサスレッド結合抗体(B、D、F、H、およびJ)で連続的にインキュベートする。DAPI染色(A、C、E、G、およびI)を行って、写真の視野中のすべての細胞を視覚化する。パネルB):A)と同様に行なったが、細胞をまず固定/透過性化し、次にC末端特異的抗体(sc698)でインキュベートし、次に抗ウサギIgGテキサスレッド結合抗体でインキュベートした。横棒=10ミクロン。
【図14−2】抗ヒトIL-9受容体中和抗体と野生型およびデルタ−Q受容体との免疫反応性。パネルA):COS7細胞を、LXSNベクター単独(AとB)、野生型IL-9R(CとD)、コドン410で開始する9個のSer残基を有する野生型IL-9R(EとF)、Δ-Q 173変異体(GとH)、およびコドン410で開始する9個のSer残基を有するΔ-Q 173(IとJ)で一過性にトランスフェクトし、記載したように(実施例8)MAB290と抗マウスIgGテキサスレッド結合抗体(B、D、F、H、およびJ)で連続的にインキュベートする。DAPI染色(A、C、E、G、およびI)を行って、写真の視野中のすべての細胞を視覚化する。パネルB):A)と同様に行なったが、細胞をまず固定/透過性化し、次にC末端特異的抗体(sc698)でインキュベートし、次に抗ウサギIgGテキサスレッド結合抗体でインキュベートした。横棒=10ミクロン。
【図14−3】抗ヒトIL-9受容体中和抗体と野生型およびデルタ−Q受容体との免疫反応性。パネルA):COS7細胞を、LXSNベクター単独(AとB)、野生型IL-9R(CとD)、コドン410で開始する9個のSer残基を有する野生型IL-9R(EとF)、Δ-Q 173変異体(GとH)、およびコドン410で開始する9個のSer残基を有するΔ-Q 173(IとJ)で一過性にトランスフェクトし、記載したように(実施例8)MAB290と抗マウスIgGテキサスレッド結合抗体(B、D、F、H、およびJ)で連続的にインキュベートする。DAPI染色(A、C、E、G、およびI)を行って、写真の視野中のすべての細胞を視覚化する。パネルB):A)と同様に行なったが、細胞をまず固定/透過性化し、次にC末端特異的抗体(sc698)でインキュベートし、次に抗ウサギIgGテキサスレッド結合抗体でインキュベートした。横棒=10ミクロン。
【図14−4】抗ヒトIL-9受容体中和抗体と野生型およびデルタ−Q受容体との免疫反応性。パネルA):COS7細胞を、LXSNベクター単独(AとB)、野生型IL-9R(CとD)、コドン410で開始する9個のSer残基を有する野生型IL-9R(EとF)、Δ-Q 173変異体(GとH)、およびコドン410で開始する9個のSer残基を有するΔ-Q 173(IとJ)で一過性にトランスフェクトし、記載したように(実施例8)MAB290と抗マウスIgGテキサスレッド結合抗体(B、D、F、H、およびJ)で連続的にインキュベートする。DAPI染色(A、C、E、G、およびI)を行って、写真の視野中のすべての細胞を視覚化する。パネルB):A)と同様に行なったが、細胞をまず固定/透過性化し、次にC末端特異的抗体(sc698)でインキュベートし、次に抗ウサギIgGテキサスレッド結合抗体でインキュベートした。横棒=10ミクロン。
【図15−1】ヒトIL-9受容体の異なる変異体のJak、Stat、およびIrsファミリーのメンバーの活性化。GH9、ΔQGR8、またはΔQGH9を発現するTS1細胞を6時間、栄養を欠乏させ、次にサイトカイン無し(−)、マウスIL-9有り(m),またはヒトIL-9有り(h)で5分間処理した。細胞抽出物を、Jak、StatおよびIrsファミリーの異なるメンバーに特異的な種々の抗体で免疫沈降させた。まず免疫ブロットを抗ホスホチロシン抗体と反応させて、チロシン−リン酸化タンパク質のみを検出し、次にストリップし、各タンパク質を免疫沈降させるのに使用したものと同じ抗体で再度釣り上げた。GH9、ΔQGR8、ΔQGH9は、図16に示す通りである。
【図15−2】ヒトIL-9受容体の異なる変異体のJak、Stat、およびIrsファミリーのメンバーの活性化。GH9、ΔQGR8、またはΔQGH9を発現するTS1細胞を6時間、栄養を欠乏させ、次にサイトカイン無し(−)、マウスIL-9有り(m),またはヒトIL-9有り(h)で5分間処理した。細胞抽出物を、Jak、StatおよびIrsファミリーの異なるメンバーに特異的な種々の抗体で免疫沈降させた。まず免疫ブロットを抗ホスホチロシン抗体と反応させて、チロシン−リン酸化タンパク質のみを検出し、次にストリップし、各タンパク質を免疫沈降させるのに使用したものと同じ抗体で再度釣り上げた。GH9、ΔQGR8、ΔQGH9は、図16に示す通りである。
【図16】ヒトIL-9受容体の異なる型を発現するTS1細胞の増殖。細胞をそれぞれ4重測定できるように96ウェルプレート(1000/ウェル)に接種し、サイトカイン無し、マウスIL-9有りまたはヒトIL-9有り(5ng/ml)で処理した。7日後に比色アッセイを行って細胞数を測定し、処理/未処理細胞の比(対照%)を算出して、増殖速度を評価した。LxSN=空ベクターでトランスフェクトした細胞;GR8は野生型IL-9R;GH9は、コドン410で開始する9個のSer残基を有するHis344変異体である;ΔQGR8およびΔQGH9は、それぞれ野生型およびHis344+9-Serバックグランド上のΔQ173変異体である。
【図17】エキソン6から213nt上流の核酸で変異を有するIL-9Rのイントロン5のゲノムDNA配列(ここで、T残基は、矢印で示すようにC残基に変化している)。
【発明を実施するための形態】
【0035】
(発明の詳細な説明)
本出願人らは、IL-9経路と、治療、診断、およびアトピー性喘息と関連疾患を予防または治療するための薬剤の同定方法の開発に使用されるこの経路に影響を与える組成物を解明することにより当該分野のニーズを解決した。
【0036】
喘息は、可逆的な空気流遮断を伴う気道の炎症性障害を含む。アトピー性アレルギーとは、アトピーと関連疾患(喘息、気管支過応答性(BHR)、鼻炎、じん麻疹、腸のアレルギー性炎症性疾患、および種々の型の湿疹を含む)を意味する。アトピーは、環境的アレルゲンに対する過敏症であり、対照と比較して血清総IgEの上昇、またはアレルゲンに対する異常皮膚試験応答として表される。BHRとは、種々の刺激に対する気管支収縮応答の上昇である、気管支過応答性を意味する。
【0037】
アトピー性アレルギーおよび喘息関連疾患を示す個体のDNAを解析することにより、本出願人らは、喘息の発現と相関しうるIL-9受容体(IL-9R)遺伝子中に多型を同定した。IL-9受容体遺伝子(喘息関連因子2またはAAF2としても知られている)は、ヒト骨髄系およびリンパ系の制御を含む多様な機能に関連するサイトカイン受容体である、インターロイキン−9受容体の遺伝子座を意味する。本発明のヒトIL-9受容体遺伝子は、XY染色体の次端部テロメア領域中に見いだされる。
【0038】
本出願人らは、多型とは、通常の配列からの特定DNA配列(「遺伝子座」と呼ばれる)の変化を意味する。一般に、ある遺伝子座を含む2つまたはそれ以上の対立遺伝子が当業者により同定され、少なくとも共通の対立遺伝子が1%またはそれ以上の頻度で存在する時、その遺伝子座を多型であると定義する。
【0039】
IL-9Rは,ヒトゲノムの他の遺伝子座(第9,第10、第16、第18染色体)に、4つの高度に相同性(>90%のヌクレオチド同一性)の非プロセシング偽遺伝子を有するため、標準的プライマーデザインを使用して真のIL-9R(XYq偽常染色体性領域中に位置する、生物学的に機能性のタンパク質をコードする遺伝子)を特異的に増幅することは不可能であった。これらの他の遺伝子の高い同一性のために、標準的プライマーデザインを使用するゲノムPCR増幅は、すべての遺伝子の同時増幅を引き起こし、真の遺伝子の配列解析は不明確なものになる。真のIL-9R構造は、本出願に記載の喘息または癌のような他の疾患(Renauldら、Oncogene, 9:1327-1332, 1994;Grussら、Cancer Res., 52:1026-1031, 1992)に対する素因に関係する可能性があるので、真のIL-9R構造を研究するために、本出願人らは、特異的アンプリマー(amplimer)をデザインした。この特異的プライマーは、4つの偽遺伝子の増幅が無く、IL-9R増幅について信頼できることがわかった。プライマー配列を実施例2に示し、その特異性を図13で証明する。
【0040】
本出願人らはまた、50人のドナーから精製したPBMC(末梢血単核細胞)から抽出したRNAを使用して、IL-9受容体cDNAの全コード領域をRT-PCRにより増幅した。図1は、解析した50人中に見いだされた最も高頻度の変異を示す。エキソン3、4、5、6および8は、完全長cDNAもクローン化できる試料中で、異常スプライシング事象により影響を受けた。一部の転写産物は、エキソン3の完全な欠失を示し、これはフレームシフトを引き起こして、79個の無関係の残基が続いた後に停止コドンを生成する。エキソン4の欠失の場合も,フレームシフトが生じ、エキソン5中の最初のコドンが停止コドンに変換された。一部の他のcDNAでは、エキソン5は最初の5または29ヌクレオチドの部分的欠失を示し、この両方の欠失はフレームシフトを引き起こして、エキソン5内に早い停止コドンを形成した。従ってすべての場合に、推定される末端切断型タンパク質は、細胞外ドメインの大部分ならびに膜貫通および細胞質ドメインの全部を欠失するであろう。もし分泌されるなら、これらの型は可溶性受容体として機能するかも知れない。最後に、コドン173に対応するエキソン6の最初の3個のヌクレオチドは、しばしばスプライシングにより除去され、結果的にこのコドン位置のグルタミンが欠失し、残りのタンパク質配列には他の変異はなかった。このスプライス変異体は、おそらくスプライス変異体の頻度を増加させるゲノムDNAのイントロン5中に見いだされる変異体(配列番号24)に関連する(図17と実施例12)。
【0041】
本出願人らはまた、エキソン9のコード配列に限定される対立遺伝子変異を見いだした。コドン310と410に関係している多型は、すでに開示されている29,30(Kermouni, A.ら、Genomics, 371-382 (1995))。コドン310は,そのコドンの最初のヌクレオチドがアデニンであるかまたはグアニジンであるかに応じて、それぞれアルギニンまたはグリシンをコードする。コドン410(このコドンから「410+n」と呼ぶ)では、それぞれ8個または9個のセリンに翻訳されるであろう8個または9個の一続きのAGCトリヌクレオチド反復配列が開始する。
【0042】
本出願人らは、コドン344に新しい多型を見つけた。ここで2番目のヌクレオチドはアデニンかまたはグアニジンであり、このコドンによりコードされる可能な2つの残基はそれぞれヒスチジンかまたはアルギニンである。さらにコドン344と410+nの間に対応関係が観察され、コドン344のアルギニンが、いつもコドン410+nの8個のセリンとともに存在し、コドン344のヒスチジンは9個のセリンとともに見いだされる。もともとヒト巨核芽球白血病細胞系Mo7eからクローン化されたヒトIL-9受容体cDNAは、コドン410+nに9個のセリンを示し、かつ本出願人らのクローンと異なり、コドン344にアルギニンを示す29。別の巨核芽球白血病細胞系であるUT-7は、同じアルギニン/9−セリン対立遺伝子を有すると報告されている30。本出願人らは、Mo7e細胞系から16のcDNAをクローン化し、6個のクローンはコドン410+nに8個のセリンを有し、コドン344にアルギニンを有することを見いだした。残りの10個のクローンは、公表されている配列を示した。本出願人らはまた、ヒト急性骨髄性白血病細胞系KG-1をゲノタイピングし、これはヒスチジン/9−セリンホモ接合体であることを見つけた。
【0043】
これらのDNA分子と対応するRNAは、例えばSambrookら、Molecular Clonig:A Laboratory Manual,第2版、Cold Spring HarborLaboratory Press (1985)のような当該分野の標準的な方法を使用して単離される。本出願人らは、単離されるとは、DNAに、自然環境中に存在する核酸またはポリペプチドと結合している汚染物質の少なくとも1部が存在しないことを意味する。
【0044】
本発明また、これらのヌクレオチド配列によりコードされるタンパク質を含む。本発明はまた、この分子の断片を含む。本出願人らは、断片とは、全配列の機能を維持する核酸配列の部分であることを意味する。当該分野で公知なように、断片は、欠失、付加、置換および/または修飾から得られる。
【0045】
本発明のIL-9受容体変異体の供給源は、ヒトである。あるいはDNAまたはその断片は、当該分野で公知の方法により合成してもよい。また遺伝子操作技術により、任意の受け入れられている遺伝子操作技術によりDNAを作製して、DNAを発現ビヒクル中にクローニングして、化合物を発現する細胞にビヒクルをトランスフェクトして、化合物を産生することができる。例えば、Sambrookら、Molecular Cloning:A Laboratory Manual,第2版、Cold Spring HarborLaboratory Press (1985)に記載の方法を参照されたい。
【0046】
アトピー性アレルギーおよび喘息様表現型に関連し得る変異体IL-9受容体配列,および喘息様表現型の欠如と関連し得る他の配列の証明は、アトピー性喘息および関連疾患に対する感受性を診断する方法を提供する。ある変異体は、これらの疾患を治療するのに使用できる可溶性受容体を産生することができる。
【0047】
受容体は、分子を認識しこれに結合する可溶性または膜結合成分であり、本発明のIL-9受容体は、IL-9を認識しこれに結合する成分である。IL-9受容体の機能は、IL-9またはIL-9様分子に結合し、特異細胞中でその制御シグナルを伝搬することからなる29,30,34,35。ヒトIL-9は、ヒトIL-9受容体に結合した時、IL-9受容体自身のリン酸化とJak-Stat経路のタンパク質(Jak1、Stat1、Stat3、Stat5、およびIrs2)の活性化を引き起こすことが証明されている(Demoulin, J-B.ら、Molecular and Cellular Biology,p. 4710-4716, Sept.1996)。本出願人らは、IL-9Rまたはその変異体が,これらのタンパク質の活性化においてバイアスを示すかどうかを調べ、この解析をJak3とIrs1に拡大した。Jak3とIrs1を含む経路のすべてのタンパク質は、IL-9R活性化によりリン酸化されることが確認された。また、コドン310、344および410+nで変異を有するIL-9R変異体は、野生型IL-9Rと同じアップレギュレーションを提供することも確認された。従って、本発明の1つの側面は、Jak-Stat経路中の相互作用を阻害するアトピー性喘息の治療用の治療薬である。
【0048】
野生型受容体および他の試験した変異体と異なり、ΔQ173変異体は、Jak-Stat経路中のいかなるタンパク質も活性化することができなかった(図15)。さらにΔQ173変異体は、IL-9刺激により細胞の増殖を援助することができなかった(図16)。従ってΔQ173変異体を発現する個体は、アトピー性喘息や関連疾患に対して感受性である可能性は低い。従って本発明の1つの側面は、アトピー性喘息および関連疾患の治療のためのΔQ173スプライス変異体の発現を増加させる治療薬である。
【0049】
1つの診断的態様は、IL-9受容体遺伝子または転写産物のDNA配列中の変異の認識である。1つの方法は、当業者により理解されるように、充分なハイブリダイゼーション条件下で、本発明のIL-9受容体に相補的な配列を有する核酸分子(プローブとしても知られている)を使用することである。1つの態様において、この核酸分子は、成熟IL-9受容体タンパク質のArg344,またはHis344のコドンに特異的に結合し、別の実施態様では、Arg344とHis344の両方に結合するであろう。さらに別の実施態様では、これはGln173のコドンに結合するであろう。これらの方法は、IL-9受容体の他の変異体を認識するのに使用してもよい。これらの疾患に関連したDNA配列変異を認識する別の方法は、当該分野で公知の複数の方法による直接DNA配列解析である44。別の実施態様は、これらの疾患に関連するIL-9受容体遺伝子のDNA配列変異の検出である40ー44。これらは,ポリメラーゼチェイン反応、制限断片長多型(RFLP)解析、および1本鎖コンフォメーション解析を含む。好適な実施態様において、本出願人らは特に、ASO PCRを使用してHis344対立遺伝子およびArg344対立遺伝子に関連するIL-9受容体中の多型を、遺伝子レベルで認識する方法を具体的に提供する。他の実施態様において、IL-9受容体遺伝子のこれらの対立遺伝子を区別するために、結合チェイン反応を使用することができる。
【0050】
本発明はまた、IL-9およびその受容体のアンタゴニストの同定のための方法を含む。アンタゴニストは、それ自身は薬理活性が欠如しているが、アゴニストの作用を妨害することにより影響を引き起こす化合物である。本発明のアンタゴニストを同定するために、本明細書および引用文献に記載のように,既知のアゴニストとの競合的結合、またはIL-9様機能のダウンレギュレーションについて試験してもよい2,22-35。
【0051】
Tリンパ球の増殖、IgE合成、および肥満細胞からの放出についてのIL-9受容体の既知の役割に基づいてアトピー性アレルギーを治療するのに有用な薬剤についてスクリーニングするのに、具体的な測定法が使用される29,30,33-35。別の測定法は、Mo7e細胞中の複数のタンパク質の急速および一過性のチロシンリン酸化を特異的に誘導するための、ヒトIL-9受容体の能力に関する34。この応答はIL-9受容体の発現と活性化に依存するため、これは、有用な可能性のある化合物の性状解析のための簡便な方法または測定法である。Stat3転写因子のチロシンリン酸化は、IL-9受容体の機能に特異的に関連しているようであり35、この応答は、本発明の範囲内の化合物の性状解析のための簡便な方法または測定法を示す。IL-9受容体の機能を性状解析するためのさらに別の方法は、細胞増殖測定法を用いてヒトIL-9機能を評価するのに使用することができる,ヒト受容体でトランスフェクトされる既知のマウスTS1クローンの使用を含む29。これらの方法は、IL-9受容体のアンタゴニストを同定するために使用することができる。
【0052】
さらなる実施態様において、本発明は、IL-9に結合する可溶性IL-9受容体分子を投与することによる、IL-9発現または機能のダウンレギュレーションを包含する。本出願人らおよびRenauldら29は、可溶性型のIL-9受容体の存在を証明した。この分子は、細胞結合受容体へのIL-9の結合を妨害するのに使用でき、IL-9のアンタゴニストとして作用する。可溶性受容体は、サイトカインまたは他のリガンドに結合してそれらの機能を制御するのに使用されている45。可溶性受容体は、溶液で存在するかまたは膜の外に存在する、膜結合受容体の型である。通常膜に会合している分子のセグメントが存在しないため、可溶性受容体が存在するかも知れない。このセグメントは当該分野では通常、遺伝子の膜貫通ドメイン、またはタンパク質の膜結合セグメントと呼ばれる。すなわち、本発明の1つの実施態様において、可溶性受容体は,膜結合受容体の断片または類似体である。
【0053】
本出願人らは、可溶性受容体として作用することができるタンパク質の産生を引き起こす、ヒトIL-9受容体の3つのスプライス変異体を同定した。1つのスプライス変異体は、エキソン4を欠失させ、これはエキソン5の最初のコドンとして停止コドンとなるフレームシフトを導入した。この変異体は、IL-9/IL-9R相互作用を阻止する抗体と反応性のエピトープを含有する約45残基のペプチドを産生する。他の2つの変異体は、エキソン5に欠失を有し、これはエキソンの初期に未成熟な停止コドンを作成するが、これらの場合エキソン4の欠失はないであろう。これらの変異体は、阻止抗体により認識されるエピトープも含有する約100残基のタンパク質を産生するであろう。
【0054】
可溶性IL-9受容体は、アトピー性喘息および関連疾患を治療するのに有用なアンタゴニストを含む治療薬候補をスクリーニングするのに使用してもよい。例えば、ファージディスプレイを使用するペプチドと1本鎖抗体のスクリーニングは、可溶性受容体を使用して促進することができるであろう。可溶性受容体に結合するファージは単離することができ、この分子は受容体と結合ファージの親和性捕捉により同定することができる。さらに、アトピー性喘息および関連疾患の治療に有用な物質のための化合物のスクリーニングは、可溶性受容体およびアンタゴニストの非存在下で結合するリガンドを取り込むことができる。これらの分子が近傍にあるため、リガンドと受容体相互作用の検出が起きる。アンタゴニストは、これらの相互作用を阻止することにより認識される。
【0055】
さらに本発明は、製薬上許容される担体とともに本発明の化合物を含む医薬組成物を含む。製薬上許容される担体は、無菌の液体、例えば石油、動物、植物または合成起源のものを含む水および油,例えばピーナツ油、大豆油、鉱物油、ゴマ油などがある。医薬組成物を静脈内投与する時は、水が好適な担体である。食塩水溶液ならびにデキストロース水溶液およびグリセロール水溶液もまた、液体担体、特に注射溶液として使用することができる。適当な医薬担体は、Martin, E.W., Remington's Pharmaceutical Sciences(これは特に、参照により本明細書に組み込まれる)に記載されている。
【0056】
本発明の治療法で使用される化合物は、治療される症状、部位特異的治療の必要性、投与される薬剤の量、および同様の事柄などを考慮して、全身性または局所的に投与してもよい。
【0057】
局所投与を使用してもよい。任意の一般的な局所製剤(例えば、液剤、懸濁剤、ゲル剤、軟膏剤、または膏薬など)を使用してもよい。このような局所製剤の調製は、例えばRemington's Pharmaceutical Science、第17版、Mack Publishing Company, Easton,Pa.により例示されるように、医薬製剤の分野で詳しく記載されている。局所投与には、これらの化合物はまた、散剤またはスプレー剤(特にエアゾル型)として投与することができる。好適な実施態様において、本発明の化合物は吸入により投与してもよい。吸入治療法のために、化合物は、用量目盛り付き吸入器、または乾燥粉末吸入器に適した型で投与するのに有用な溶液でもよい。
【0058】
活性成分は、全身性投与に適した医薬組成物で投与される。公知のように、薬剤は、全身投与されるなら、経口投与用に散剤、丸剤、錠剤など、またはシロップ剤もしくはエリキシル剤として調製してもよい。静脈内投与、腹腔内投与または病変内局注投与のためには、化合物は,注射による投与が可能な液剤または懸濁剤として調製される。ある場合には、これらの化合物を坐剤として、または皮下沈降のための長期放出製剤として、または筋肉内注射のための製剤で調製することが有用である。
【0059】
有効な量は、IL-9受容体により調節される機能をダウンレギュレートする量である。具体的な有効量は、症状毎に変化し、ある場合には治療される症状の重篤度および治療に対する患者の感受性により変化する。従って特定の有効量は、常套の実験により、その時間と場所で決定することが最適である。しかし本発明の喘息および関連疾患の治療において、0.001〜5重量%,好ましくは約0.01〜1%を含有する製剤が,通常治療上有効な量であろう。全身投与される時、0.01〜100mg/kg体重/日,しかし好ましくは約0.1〜10mg/kgの量が、多くの場合治療結果に影響を与えるであろう。
【0060】
本出願人らはまた、IL-9受容体により制御される機能をダウンレギュレートする化合物をスクリーニングする方法を提供する。IL-9受容体により発現される機能がダウンレギュレートされたかどうかは、当該分野で標準的な方法を使用して確認してもよい29,30,34,35。具体的な実施態様において、本出願人らは、IL-9に匹敵する機能を有する化合物の同定法を提供する。1つの実施態様において、IL-9受容体の機能は、in vitroで評価される。当業者に公知のように、ヒトIL-9受容体の活性化は、IL-9に応答性のある細胞中の複数のタンパク質の急速かつ一過性のチロシンリン酸化を特異的に誘導する。Stat3転写因子のチロシンリン酸化は、IL-9経路の作用と特異的に関連しているようである。IL-9およびIL-9様分子の機能を特徴付けるための別の方法は、マウスTS1クローンまたはTF1クローン中のIL-9受容体の「安定な発現」に依存し、これは通常ヒト受容体を発現しない。これらのトランスフェクション体は、細胞増殖アッセイを使用して、ヒトIL-9受容体機能を評価するのに使用することができる29。
【0061】
本発明はまた、IL-9受容体変異体を発現する細胞系に基づく飽和性かつ特異的リガンド結合の簡便なスクリーニングアッセイも包含する23,29。IL-9受容体は、広範囲にわたる細胞型(K562、C8166-45、ヒトIL-9受容体でトランスフェクトしたKG-1、B細胞、T細胞、肥満細胞、HL60、HL60-クローン5、ヒトIL-9受容体でトランスフェクトしたTS1、ヒトIL-9受容体でトランスフェクトした32D、好中球、巨核球(UT-7細胞)30、ヒト巨核芽球白血病細胞系Mo7e34、TF129、マクロファージ、好酸球、胎児胸腺細胞、ヒト腎細胞系29330、ならびにマウス32Dおよび胚海馬始原細胞系23,29,30を含む)で発現される。
【0062】
本発明の実施においては、当業者の通常の技術範囲内である、分子生物学,薬学、免疫学、および生化学の通常の用語および技術を使用する。例えば、Sambrookら、Molecular Cloning: A Laboratory Manual,第2版、Cold Spring HarborLaboratory Press、またはAusubelら、Current Protocols in Molecular Biology,John Wiley &Sons, Inc.を参照されたい。
【0063】
なお、我々は、以下の基礎的バックグランド情報を提供する。体の遺伝物質またはDNAは、それぞれが動原体によりつながれた2つのアームを含む46の染色体上に並んでいる。各染色体は、pまたはqと呼ぶセグメントに分かれる。記号pは、動原体から最も近いテロメアまで測定した時に短い方の染色体アームを示す。染色体の長い方のアームは、記号qで示す。染色体上の位置は、染色体の番号(すなわち、染色体5)ならびにpまたはq領域の座標(すなわち、q31-q33)で示される。さらに体には、性染色体XとYがある。減数分裂の間、XおよびY染色体は、偽常染色体領域として知られている領域中でDNA配列情報を交換する。
【0064】
DNA(デオキシリボ核酸)は、ヌクレオチドの2本の相補的な鎖からなり、これらは4つの異なる塩基化合物、アデニン(A)、チミン(T)、シトシン(C)、およびグアニン(G)を含む。一方の鎖のAは他方の鎖のTと結合し、一方の鎖のCは他方の鎖のGと結合して、相補的な「塩基対」を形成し、各対は1本の鎖に1つの塩基を有する。
【0065】
3つのヌクレオチドの連続した群(「コドン」)が、1つのアミノ酸をコードする。例えば、3つのヌクレオチドCAGはアミノ酸グルタミンをコードする。20個の天然に存在するアミノ酸とその1文字コードは以下の通りである:
アラニン Ala A
アルギニン Arg R
アスパラギン Asn N
アスパラギン酸 Asp D
アスパラギンまたは
アスパラギン酸 Asx B
システイン Cys C
グルタミン Gln Q
グルタミン酸 Glu E
グルタミンまたは
グルタミン酸 Glx Z
グリシン Gly G
ヒスチジン His H
イソロイシン Ile I
ロイシン Leu L
リジン Lys K
メチオニン Met M
フェニルアラニン Phe F
プロリン Pro P
セリン Ser S
スレオニン Thr T
トリプトファン Trp W
チロシン Tyr Y
バリン Val V
アミノ酸はタンパク質を構成する。アミノ酸は、親水性(すなわち、水に対する親和性を示す)でもまたは疎水性(すなわち、水を嫌う)でもよい。すなわち、G、A、V、L、I、P、F、Y、W、C、およびMで示されるアミノ酸は疎水性であり、S、Q、K、R、H、D、E、N、およびTで示されるアミノ酸は親水性である。一般にアミノ酸の親水性または疎水性の性質は、ペプチド鎖の折り畳みに影響を与え,従ってタンパク質の3次元構造に影響を与える。
【0066】
DNAは以下のようにタンパク質と関係している:
ゲノムDNA → mRNA → タンパク質
↑
↓
cDNA
ゲノムDNAは、生物の細胞中に存在するすべてのDNA配列を含む。これは、メッセンジャーRNA(「mRNA」)に「転写」される。相補的DNA(「cDNA」)は、mRNAの逆転写により作成される、mRNAの相補的なコピーである。ゲノムDNAと異なり、mRNAとcDNAは両方とも、DNAのタンパク質コード領域またはポリペプチドコード領域(いわゆる「エキソン」)のみを含有する。ゲノムDNAはまた、「イントロン」を含有し、これはタンパク質をコードしない。
【0067】
実際、真核生物遺伝子は、それによりコードされるタンパク質に対して断続的であり、イントロンにより中断されたエキソンからなる。RNAに転写された後、イントロンはスプライシングにより除去されて、成熟メッセンジャーRNA(mRNA)が形成される。エキソンの間のスプライス点は、典型的にはスプライシングプロセスのシグナルとして作用する共通配列により決定される。スプライシングは、一次RNA転写産物からのイントロンの削除と、切断されたイントロンのいずれかの側にある残りのRNAの末端の結合もしくは融合からなる。イントロンの有無、イントロンの組成、および遺伝子当たりのイントロンの数は、同じ種の株の間でも異なり、同じ基本的な機能遺伝子を有する種の間でも異なることがある。ほとんどの場合にイントロンは必須ではなく害はないと考えられているが、この分類は必ずしも絶対的ではない。例えば1つの遺伝子のイントロンは、別のエキソンであることもある。ある場合には、スプライシングの代替的なまたは異なるパターンが、DNAの同じ1つのストレッチから、異なるタンパク質を形成することがある。実際、イントロンの構造的特徴と基礎的スプライシング機構は、異なる種類のイントロンの分類の基礎となる。
【0068】
エキソンについて、これらは、別個のドメインまたはモチーフ、例えばタンパク質の機能性ドメイン、折り畳みドメイン、もしくは構造的エレメント;または短いポリペプチド配列、例えば逆ターン、ループ、グリコシル化シグナルおよび他のシグナル配列、または非構造ポリペプチド、リンカー領域、に対応する。本組合せ法のエキソンモジュールは、天然に存在するエキソン配列または突然変異(例えば、点突然変異、末端切断、融合)を受けている天然に存在するエキソン配列に対応する核酸配列を含有してもよい。
【0069】
DNAの操作にもどると、DNAは、ある既知の部位でDNAを切断する「制限酵素」およびDNAをつなぐDNAリガーゼを使用して、切断、スプライス、および他の操作をすることができる。このような技術は当業者に公知であり、例えばSambrookら、Molecular Cloning:A Laboratory Manual,第2版、Cold Spring HarborLaboratory Press (1985)、またはAusubelら、Current Protocols in Molecular Biology,John Wiley &Sons, Inc. (1994)に記載されている。
【0070】
特定のサイズおよび配列のDNAは、次に「レプリコン」(これは、プラスミド、コスミド,またはウイルスのような、それ自身の制御下で複製ができる遺伝成分である)中に挿入することができる。「組換えベクター」または「発現ベクター」は、DNAセグメントが挿入されるレプリコンであり、DNAの発現(すなわち、DNAによりコードされるタンパク質の産生)を可能にする。発現ベクターは、実験室で作製されるか、他の実験室から入手できるか、または市販品を購入することができる。
【0071】
組換えベクター(当該分野において種々の名前で知られている)は、「形質転換」として一般的に知られているプロセスにより宿主中に導入される。形質転換は、任意の方法(感染、直接摂取、形質導入、F−接合、微量注入法、または電気穿孔法)により、宿主細胞中に外来DNAセグメントを移動することを意味する。
【0072】
単細胞宿主細胞(組換え宿主細胞、細胞および細胞培養物として種々の名前で知られている)は、細菌、酵母、昆虫細胞、植物細胞、哺乳動物細胞、およびヒト細胞を含む。特に好適な実施態様において、宿主細胞は,大腸菌(E. coli )、シュードモナス(Pseudomonas)、バシラス(Bacillus),ストレプトミセス(Streptomyces),酵母、CHO、R1-1、B-W、LH、COS-J、COS-7、BSC1、BSC40、BMT10、およびS69細胞を含む。酵母細胞は特に、サッカロミセス(Saccharomyces),ピキア(Pichia)、カンジダ(Candida)、ハンゼヌラ(Hansenula)、およびトルロプシス(Torulopsis)を含む。
【0073】
当業者は認識できるように、宿主細胞によるDNAセグメントの発現は、適切な制御配列または成分が必要である。制御配列は、使用される宿主細胞により変化するが、例えば原核生物では、プロモーター、リボゾーム結合部位、および/または転写停止部位がある。真核生物では、このような制御配列には、プロモーターおよび/または転写停止部位がある。当業者は認識できるように、これらの制御配列を注意深く選択し配置することにより、ポリペプチドの発現を増強、すなわち標準的なレベルより上昇させることができる。
【0074】
他の実施態様において、使用されるプロモーターには、ヒトサイトメガロウイルス(CMV)プロモーター、テトラサイクリン誘導性プロモーター、サルウイルス(SV40)プロモーター、モロニーマウス白血病ウイルス長い末端反復配列(LTR)プロモーター、糖質コルチコイド誘導性マウス乳癌ウイルス(MMTV)プロモーター、ヘルペスチミジンキナーゼプロモーター、マウスおよびヒトアクチンプロモーター、HTLV1およびHIV IL-9 5'フランキング領域、ヒトおよびマウスIL-9受容体5'フランキング領域、細菌tacプロモーターおよびキイロショウジョウバエ(Drosophila)熱ショックタンパク質足場結合領域(SAR)エンハンサー成分がある。
【0075】
DNAは、ペプチド、オリゴペプチド、およびタンパク質のようなある長さのポリペプチドとして発現される。ポリペプチドはまた、グリコシル化、アセチル化、リン酸化などの翻訳的修飾も含む。
【0076】
本発明に関係のある他の分子生物学的方法は、「連鎖解析」である。連鎖解析は、形質または疾患と相関する染色体または染色体領域を同定するための分析方法である47。染色体は、その上に遺伝子が組み立てられる遺伝の基本的単位である。遺伝子以外に、当業者は、染色体上に「DNAマーカー」を証明している。DNAマーカーは、その同定と配列決定が容易に行なうことができる、DNAの既知の配列である。連鎖解析法は、疾患遺伝子(例えば喘息の感受性に関係する遺伝子)を特定の染色体にマッピングすることに応用されている47,48。
【0077】
本出願人らは、上記および下記のすべての文献を参照により本明細書に組み込む。
【0078】
本明細書の説明および開示された発明の実施から、本発明の他の実施態様は当業者には明らかであろう。明細書および実施例は単に例示のためであり、本発明の範囲は請求の範囲によってのみ示されるものである。
【0079】
背景情報は提供したので、本出願人らは、ここで本発明の好適な面を説明する。
【実施例】
【0080】
(実施例1:IL-9受容体転写産物多型の同定)
Philadelphia, Pennsylvania 地域からの52人の集団を、喘息とアトピーに関してランダムに確認した。総血清IgEを、酵素結合免疫吸着アッセイ(ELISA、Genzyme、Cambridge、Massachusetts)により測定した。
【0081】
ヒトIL-9受容体cDNAの構造型を評価するために、これらの52人の無関係のドナーからPBMCを単離し、PHAとPMAの存在下で培養した(実施例4に記載)。本出願人らの実験室の以前のデータは、分裂促進物質による刺激後6日目に1次培養物中のIL-9受容体メッセージの発現の動力学を証明した。従って本出願人らは、細胞を6日間培養し、この時点で細胞を採取し,実施例5に記載のようにそのRNAとDNAを単離した。
【0082】
RNAを逆転写し、実施例5に記載のように完全長IL-9受容体cDNAに特異的なプライマーを使用してPCRにより増幅した。各個人からの増幅産物をTA PCRクローニングベクターにクローン化し、予測されるインサートを含有する10クローン(消化とゲル電気泳動により測定した)を完全に配列決定し、構造または配列の変化について解析した。
【0083】
上記スクリーニングから7個の主要な変異体を同定した。これらのcDNAは、コドン173の欠失、エキソン4の欠失、エキソン5の2つの別の欠失、エキソン8の欠失、および成熟タンパク質のコドン344でARGからHISへの変異を有する完全長cDNAを示す。これらの遺伝子バックグランドのそれぞれに追加の変異体が存在する。Arg対立遺伝子は、8 Ser/4Asn繰り返しと7 Ser/4 Asn繰り返しに関連し、His対立遺伝子は、9 Ser/4Asn繰り返し、9 Ser/3 Asn繰り返し、および10 Ser/2 Asnに関連する。これらの変異体のすべてを、図1に記載する。
【0084】
変異体を、クローン化cDNAの発現を指令するCMVプロモーターとその後のSV40ポリアデニル化シグナルを含有する真核生物発現ベクターpCEP4(Clontech)中にクローン化した。このベクターはまた、ベクターを含有し、おそらく、クローン化したcDNAをCMVプロモーターの制御下で発現する真核生物細胞の選択に使用される、ヒグロマイシンB耐性遺伝子を含有する。組換えプラスミドを、配列により解析し、正しいcDNAインサートを含有するプラスミドを、真核生物受容体細胞(例えば、実施例3に記載のシリアンハムスター繊維芽細胞TK-ts13、ヒト神経膠芽細胞腫T98G、ヒト骨髄白血病株TF-1、およびマウス骨髄前駆体細胞系32D)にトランスフェクトした。増殖とアポトーシスにおける、IL-9リガンドに対する応答として、機能を生物学的に評価した(実施例7と10)。
【0085】
ARGおよびHIS変異体を含有する完全長IL-9受容体cDNAを行なった実験は、TK-ts13ハムスター繊維芽細胞またはヒトT98G神経膠芽細胞腫細胞である。細胞をトランスフェクトし、48時間後ヒト特異的カルボキシ末端抗体を使用して、ウェスタンブロットとin situ染色により解析した(Santa Cruz)(実施例8)。in situ解析は、ハムスターとヒト系の両方で,両方の型の受容体が発現されるらしいことを証明した(図11と図12)。興味深いことに、ヒトとハムスター系で両方の型のウェスタンブロットは同等レベルで発現され、TS1細胞系中でARGとHIS受容体型との間には移動パターンに差があり(図12)、これは、グリコシル化、リン酸化などの翻訳後修飾の差を示唆する。この生化学的な差は、機能の変化が表現型の変化を引き起こす機構かも知れない。
【0086】
種々の置換の頻度を、一般的集団中の各変異体の普及率の、偏りのない推定値として使用した。アンケートにより遺伝子型を表現型と比較した。アレルギーと喘息の診断は、アンケートを再検討して、医師が行なった。Arg344対立遺伝子についてホモ接合性である個人は、ヘテロ接合性またはホモ接合性His344と比較した時、アレルギーと喘息の証拠を示す可能性が有意に低かった(図9)。
【0087】
(実施例2:IL-9受容体遺伝子のゲノム解析)
アレルギーおよび/または喘息の人のゲノム解析を行うために、X/Y偽常染色体領域上に位置する真のIL-9受容体遺伝子についてPCR特異的プライマーを作成し、かつ染色体9,10、16、および18上に位置する高度に保存されたIL-9受容体偽遺伝子を排除するように,以下の方策を設計した。まず、IL-9受容体遺伝子の2つの公表された偽遺伝子とゲノム配列の間で、配列アライメントを行なった。次に,真の遺伝子と偽遺伝子の間の分岐領域でプライマーを設計し、次にCoreill DNA Repository (Camden,NJ)から入手した単一の染色体特異的ハイブリッドを使用して、PCRにより解析した。プライマーが、XおよびYハイブリッドから正しいサイズの生成物のみを産生するなら、これらをしっかりした増幅について最適化した。いくつかの場合に、分岐領域に向けられたプライマーはXY特異的ではなく、従って本出願人らは、IL-9受容体遺伝子配列に比較して偽遺伝子に対してミスマッチの数を多くするために、特定のプライマー中の追加の塩基の変異を導入した。表1は、XY特異的増幅についてのプライマーの配列と最適アニーリング温度を含む。XY増幅についてこれらのプライマーの特異性を、図13に示す。
表1:IL-9受容体のX/Y特異的アンプリマー
【0088】
【表1】

これらのプライマーは、アトピー性喘息および関連疾患に対する感受性と耐性の診断に使用可能な、これらの遺伝子中のDNA配列の変異を解析するための新規の方法を示す。
【0089】
この技術を使用して、IL-9受容体遺伝子中の配列の変異を検出するために、本出願人らの集団中の人についてDNA配列解析により、各エキソンを調べた44。解析を繰り返して、配列多型をアーチファクトから区別した。受容体変異体とアレルギー性表現型との関連を図9に示す。受容体対立遺伝子の配列を図1〜8に示す。
【0090】
(実施例3:IL-9受容体発現とリガンド結合アッセイ)
市販の技術を使用してサプライヤー(Pierce)の勧めに従って、精製した組換えIL-9および構造または機能がIL-9に似ている可能性のある化合物を、高比活性になるように蛍光標識した。ヒトMo7eとマウス32D細胞を増殖させ、37℃で、それぞれ10%(容量/容量)ウシ胎児血清、50mM 2−メルカプトエタノール、0.55mM L-アルギニン、0.24mM L-アスパラギン、および1.25mM L-グルタミンを補足したダルベッコ改変イーグル培地0.8ml、または10%(容量/容量)ウシ胎児血清、50mM 2−メルカプトエタノール、0.55mM L-アルギニン、0.24mM L-アスパラギン,および1.25mM L-グルタミンを補足したRPMIに再懸濁した。TF1.1(ヒトIL-9受容体を欠失したTF1細胞)、T98G、TK-、およびマウス32D細胞(実施例7と10)を、後述のようにそのまま、またはヒトIL-9受容体構築物でトランスフェクション後に使用した。完全長または末端切断型のIL-9受容体の1つを含有するプラスミドDNAを、pCEP4プラスミド(Clontech)中にクローン化し、Qiagenカラム(Qiagen)を用いて遠心分離により精製した。電気穿孔法の直前に0.4cmのキュベット中の細胞にプラスミドDNA(50μg)を加えた。2重電気パルス(750V/74オーム/40マイクロファラドと100V/74オーム/2100マイクロファラド)後に、直ちに細胞を、IL-9を補足した新鮮な培地に希釈した。
【0091】
ヒグロマイシンB(400μg/ml〜1.6mg/ml)で14日間選択した後に、安定なトランスフェクション細胞が得られた。ヒグロマイシン耐性クローンを、実施例8に記載のようにウェスタンブロットとin situ染色によりIL-9受容体の発現について分析した。
【0092】
蛍光顕微鏡でリアルタイムに、細胞受容体の結合を直接視覚化する。対照細胞(トランスフェクトしてない)と既知のIL-9受容体変異体のそれぞれでトランスフェクトした細胞で、結合とインターナリゼーションを経時的に追跡する。特異的結合を証明するために、過剰の非標識リガンドまたはブロッキング抗体を並行実験で使用する。
【0093】
HAジタグを有するかまたは有さない、アミノ酸44〜270を含む可溶性IL-9受容体をまた、異なる型のヒト標識組換えIL-9とともにインキュベートする。異なる量のFLAGタグ付き(IBI)リガンドを0.5μgの可溶性受容体とともに、PBS中で室温で30分間インキュベートする。1μgの抗HA抗体または抗FLAGモノクローナル抗体(IBI)とともに、EBCバッファー(50mMトリス、pH7.5;0.1M NaCl;0.5% NP40)を加え(300μl),氷上で1時間インキュベートする。各試料に40μlのプロテインAセファロース溶液を加え、4℃で1時間混合した。試料を11,000×Gで1分間遠心分離し、ペレットを500μlのEBCで4回洗浄した。ペレットを26μlの2×SDSバッファーに溶解し、4分間沸騰させ、18%SDSポリアクリルアミドゲルで電気泳動した。ウェスタンブロットを、抗IL-9受容体抗体(Santa Cruz Inc.)またはFLAGタグ付きrhIL-9に対する抗FLAGモノクローナル抗体(IBI)で釣り上げた。治療薬としての候補物質を、リガンド結合の拮抗作用を測定することにより評価する。受容体発現を図11と14に示す。
【0094】
(実施例4:細胞の単離と培養)
製造業者(Pharmacia Biotech,AB Uppsala Sweden)に従ってエンドトキシン試験を行なったFicoll-Paque PLUSを使用して密度勾配遠心分離により、健常ドナーからヒト末梢血単核細胞(PBMC)を単離した。同一遺伝子のヒト血清または熱不活性化FBSを最終濃度10%になるように補足したRPMI-1640(Bethesda ResearchLabs (BRL), Bethesda, MD)7ml中で、PBMC(5×106)、マウス脾臓細胞(5×106)、または5×106個のMo7e細胞を培養した。細胞を、刺激せずに、またはPMA 5μg/ml/PHA 5μg/ml、もしくはPHA 5μg/mlとrhIL2 50ng/ml(R &D Systems, Minneapolis, MN)で刺激して、37℃で24時間培養した。
【0095】
(実施例5:PCR産物のDNAとRNA単離、rtPCR、クローニング、および配列決定)
Nicolaides and Stoeckert46が記載したように、培養したPBMCからマイトジェンで6日間刺激後、細胞質RNAとゲノムDNAを抽出した。各供給源から1μgのRNAを,70℃で>10分間変性し、次に2.5単位のSuperscriptII逆転写酵素(GIBCO, BRL)、1U/lのRNAseインヒビター、2.5mMオリゴd(T)16プライマー、1mMずつのdATP、dCTP、dGTP、dTTP、50mM KCl、10mMトリス塩酸(pH7.0)、25mM MgCl2を使用して、37℃で1時間cDNAに逆転写(V+)した。偽逆転写反応物を陰性対照として使用した。
【0096】
逆転写反応物の20分の1を、6.7mM MgCl2、16.6mM (NH4)2SO4、67mMトリス塩酸(pH8.8)、10mM 2−メルカプトエタノール、6% DMSO、1.25mMずつのdNTP、2.5UのAmplitaq DNAポリメラーゼ、およびヒトcDNA IL-9エキソン2(フォワード5'-GCT GGA CCT TGG AGA GTG-3')(配列番号1)とエキソン9(リバース5'-GTC TCA GAC AAG GGC TCC AG-3')(配列番号2)に相当する300ngずつのオリゴヌクレオチドを、含有するPCR(50μl)中で使用した。反応混合物を以下のPCR条件に付した:95℃で120秒、次に94℃で30秒、58℃で90秒、72℃で90秒を35サイクル。最後に、反応混合物を伸長のために、72℃で15分を1サイクルを行なった。
【0097】
hIL-9受容体cDNAを示すPCR産物を、1.5%アガロースゲルでゲル電気泳動に付し、臭化エチジウム染色を用いて視覚化した。疑似逆転写反応の産物(ここでRNAの代わりにH2Oを使用した)を、すべての実験で陰性対照増幅として使用した。
【0098】
PCR産物を、TAクローニングベクター(Invitrogen, San Diego, CA)中にサブクローニングした。ヒトcDNAの増幅により、1614塩基対の産物が得られた。hIL-9受容体cDNAインサートを含有するプラスミドを、従来法により単離した(Sambrookら、(1989) Molecular Cloning: A Laboratory Manual, 第2版、Cold SpringHarbor Laboratory Press,New York)。増幅後,各インサートを含みかつこれを囲むDNA配列を、PCR誘導性またはクローニング誘導性のエラーの測定のために、自動シーケンサー(ABI 377, Perkin Elmer)で、蛍光ジデオキシターミネーターを使用して、配列決定(Sangerら、1977, Proc.Natl. Acad. Sci. USA 74:5463)により解析した。クローニングおよび/またはポリメラーゼ誘導性の配列エラーの無いhIL-9受容体cDNAインサートを、発現ベクターにサブクローニングした。
【0099】
(実施例6:in vitroでのIL-9受容体構築物のクローニングと発現)
hIL-9受容体を、エピソーム真核細胞発現ベクターpCEP4(Clontech)中にサブクローニングした。標準的方法を使用してCMVプロモーターに対してセンス配向で、pCEP4ポリリンカー中にBamH1-XhoI断片として,インサートをクローン化した(図10)。記載したように、構築物を細胞宿主中で発現させた。
【0100】
(実施例7:Mo7e、32D、TF1.1、TK-ts13、およびT98G)を使用する細胞アッセイ)
変異体IL-9受容体の機能の評価と、アトピー性喘息の治療に有効な化合物のスクリーニングのために、細胞系を使用した。IL-9により誘発される抗アポトーシスまたはベースライン増殖応答に拮抗する能力について、化合物を試験した。ある細胞系でベースライン抗アポトーシスまたは増殖応答がいったん確立されたら、3重の3回反復アッセイでの応答の統計的に有意な低下を、拮抗作用の証拠であると考えた。直接の観察、トリパンブルー染色(当業者に公知の技術)、または酸性ホスファターゼ活性の低下により、細胞傷害性から真の拮抗応答を区別した。他の増殖性物質(例えば、インターロイキン3またはインターロイキン4)に対して活性が証明されるかどうかを評価して、各化合物について拮抗作用の特異性を評価した。
【0101】
適切な培地中で使用するために、組換えIL-9または構造もしくは機能がIL-9に似ている可能性のある化合物を精製し、調製した。推定アゴニストおよびアンタゴニストを、水、食塩水、またはDMSOと水で調製した。細胞はそのまま使用したか、または実施例3に記載のようにIL-9受容体構築物でトランスフェクトした後に使用した。増殖因子を24時間枯渇させた後に、細胞を可変量の精製IL-9または構造もしくは機能がIL-9に似ている可能性のある化合物を加えてまたは加えないでインキュベートした。
【0102】
細胞数の指標として細胞内に存在する酸性ホスファターゼの量を測定する、Abacus Cell Proliferation Kit(Clontech, Palo Alto,CA)を使用して、細胞増殖を評価した。基質p-ニトロフェニルホスフェート(pNPP)を酸性ホスファターゼによりp-ニトロフェノールに変換し、これを酵素濃度の指標として測定した。各ウェルにpNPPを加え、37℃で1時間インキュベートした。次に1Nの水酸化ナトリウムを加えて、酵素反応を停止させ、p-ニトロフェノールの量を、Dynatech 2000プレートリーダー(Dynatech Laboratories, Chantilly, VA)を使用して410nmで定量した。細胞数と吸光度を比較する標準曲線を使用して、アッセイの直線範囲を決定した。吸光度測定値がアッセイの直線範囲内にある時のみ、測定結果を使用した。簡単に説明すると、リガンドを含むかまたは含まない平底マイクロタイタープレート(150または200μlウェル)中の4重の細胞試料を用いて、37℃で72〜96時間アッセイを行なった。存在する細胞の数の尺度として酸性ホスファターゼを使用した。すべての実験は、少なくとも2回繰り返した。
【0103】
アポトーシスの初期マーカーである細胞外ホスファチジルセリンを認識することによりデキサメタゾン誘導性アポトーシスを測定するAnnexin Vキットを使用してサプライヤー(Clontech)が記載するように、アポトーシスを測定した。アポトーシス細胞数は、Annexin V染色細胞のパーセントとして、蛍光顕微鏡によりスコア化した。
【0104】
Mo7e系はヒト巨核芽球細胞系であり、RPMI 1640(GIBCO/BRL, Gaithersburg, MD),20%ウシ胎児血清(Hyclone)および10ng/ml IL-3(R& D Systems,Minneapolis, MN)中で培養した。T98G系は、RPMI 1640(GIBCO/BRL)中で増殖されるヒト神経膠芽細胞腫細胞系である。ハムスター繊維芽細胞TK-ts13系も、マウス骨髄腫前駆体系であるマウス32D細胞系とともに使用し、いずれも10%ウシ胎児血清(Hyclone)を含有するRPMI 1640(GIBCO/BRL)中で培養し、さらに32D細胞系では1ng/mlのIL-3を使用した。TF1.1は,IL-2受容体ガンマサブユニット(ウェスタンブロットとrtPCRにより確認された)を発現することが知られているヒト骨髄白血病系であるが、その先祖(TF1)と比較すると、rtPCR、免疫染色、およびウェスタンブロット分析により,すでにIL-9受容体は持っていない。TF1.1は、RPMI 1640(GIBCO/BRL)と10%ウシ胎児血清(Hyclone)中で培養した。すべての細胞系は、IL-9を含めて複数のサイトカインに応答する。細胞系に栄養を与え,再度72時間毎に2×105細胞/mlで再接種した。
【0105】
細胞を2000rpmで10分間遠心分離し、0.5%ウシ血清アルブミン(GIBCO/BRL, Gaithersburg, MD)とインスリン−トランスフェリン−セレン(ITS)補助因子(GIBCO/BRL, Gaithersburg, MD)を含むRPMI 1640中に再懸濁した。血球計で細胞を数え、1×105細胞/mlに希釈し、96ウエルのマイクロタイタープレートにまいた。各ウェルは、0.15または0.2mlを含有し、細胞により10,000〜50,000細胞/ウェルの最終濃度を与えた。Mo7e細胞を,50ng/mlの幹細胞因子(SCF)(R &D Systems, Minneapolis, MN)単独、50ng/ml SCFプラス50ng/ml IL-3(R& D Systems,Minneapolis, MN)、または50ng/ml SCFプラス50ng/ml IL-9で刺激した。細胞と基本培地のみを含有する対照を含めた。試験化合物(すなわち、組換えIL-9タンパク質、ペプチド、小分子)の連続希釈物を、3重測定で各試験条件に加えた。IL-9受容体でトランスフェクトしなかったTF1.1細胞を、応答と非特異的細胞傷害性の独立した対照として使用した。培養物を5% CO2中で37℃で72〜96時間インキュベートした。
【0106】
(実施例8:トランスフェクトされた細胞系中の外因性IL-9受容体のin situおよびウェスタン分析)
IL-9受容体のin situ染色は以下のように行なった。細胞をカバーグラスの上で24時間増殖させ、次に接着性の細胞を含有するカバーグラスを、カルシウムとマグネシウムを含有するリン酸緩衝化生理食塩水(PBS)(Gibco/BRL)で2回洗浄した。IL-9受容体の細胞内染色のために、細胞を、4%パラホルムアルデヒド/PBSプラス0.1%トリトン-X中で、室温で15分間固定してから、抗ヒトIL-9受容体抗体で処理した。細胞外染色のために、細胞を固定前に抗体で処理した。次に細胞をPBSで2回洗浄し、dH2O中の7.5%BSAにより室温で30分ブロックした。次に、PBSで洗浄した細胞を、1%BSA/PBS中の抗ヒトIL-9受容体(IL-9受容体のカルボキシ末端に対するポリクローナル抗体)の10μg/ml溶液と共に、室温で1時間インキュベートした。PBSで細胞を3回洗浄し、次に1%BSA/PBS中の抗ウサギローダミン結合抗体の10μg/ml溶液中、室温で30分間インキュベートした。次に細胞をPBSで3回洗浄して、1μg/mlのDAPIを使用して室温で1分間カウンター染色した。dH2O中で細胞を3回洗浄し、顕微鏡スライドに固定し、蛍光顕微鏡により分析した。トランスフェクトしたCOS7細胞についての結果を、図14に示す。
【0107】
0.5%溶解バッファー(トリス50mM,NaCl 150mM,NP40)、1mM DTTおよびプロテアーゼインヒビター中の細胞抽出物の直接溶解から得られたタンパク質溶解物について、ウェスタンブロットを行い、5分間沸騰させた。試料を、トリス−グリシンランニングバッファー中の4〜20%トリス−グリシンSDSゲル(Novex)で電気泳動した。次に、Trans Blot II装置(Bio Rad)を使用して、電気ブロットにより、タンパク質をニトロセルロースに移した。移動後、膜を,TBS-T((20mMトリス、137mM NaCl、pH7.6)プラス0.05%ツイーン20)プラス5%ブロット(blotto)中、室温で1時間ブロックした。次に、TBS-T中のIL-9受容体のカルボキシ末端に対するポリクローナル抗体(1μg/ml)を使用して、ブロットを1時間探した。次にTBS-T中でブロットを10分間3回洗浄し、TBS-T中の第2抗ウサギ西洋ワサビペルオキシダーゼ結合抗体(1:10,000)を使用して30分間探した。上記のようにブロットを洗浄し、化学発光基質であるルミノール/エンハンサー溶液(Pierce)と共に室温で5分間インキュベートし、次に1〜60秒間フィルムに露光した。図11と12を参照されたい。
【0108】
(実施例9:真のIL-9Rゲノム増幅法)
IL-9Rは,ヒトゲノムの他の遺伝子座(第9、第10、第16、第18染色体)に4つの高度に相同性(>90%ヌクレオチド同一性)のプロセシングされていない偽遺伝子を有するため、標準的プライマーデザインを使用する真のIL-9R(XYq偽常染色体領域中に位置する生物学的機能性タンパク質をコードする遺伝子)の特異的増幅は不可能であった。これらの他の遺伝子の高度の同一性のために、標準的プライマーデザインを使用するゲノムPCR増幅により、すべての遺伝子が同時増幅され、従って真の遺伝子の配列解析が不明確になった。真のIL-9R構造は、本出願に記載の喘息または癌のような他の疾患(Renauldら、Oncogene, 9:1327-1332, 1994; Grussら、Cancer Res.,52:1026-1031, 1992)に対する素因に関係しうるので、その構造を研究するために、特異的アンプリマーを以下のように設計した:
IL-9R偽遺伝子と真の遺伝子の配列をMac Vectorソフトウェアを使用してアラインメントした。次に、真の遺伝子と偽遺伝子の間で分岐した領域について、各エキソンの周りのイントロン配列を調べた。次にこれらの領域に対してプライマーを設計し、これを使用して、各ヒト染色体を含有するヒト/齧歯類ハイブリッドDNAをPCR増幅した。生成物を3%アガロースゲルで泳動し、4つの偽遺伝子の増幅の無い真のIL-9R増幅について分析した。アニーリング温度とバッファー条件(DMSO含量5〜10%)を変えて、特異的PCR増幅条件も最適化した。まだ偽遺伝子の増幅が存在した場合は、プライマー配列中にヌクレオチド変異を入れて、真の遺伝子と比較して偽遺伝子からのより大きな分岐を生じさせた。プライマー配列を実施例2に示し、その特異性を図13に示す。
【0109】
(実施例10:細胞増殖アッセイとサイトカイン刺激)
種々の型のヒIL-9受容体を発現するTS1細胞の増殖応答を調べるために、細胞をPBSで洗浄し、D-MEM、10%ウシ胎児血清中に再懸濁した。103細胞/ウェルを、96ウェルマイクロプレート中に3回反復で接種し、適宜、組換えヒトIL-9またはマウスIL-9(R &D Systems, Minneapolis, MN)を最終濃度5ng/mlで加えた。酸性ホスファターゼアッセイを使用して7日後に、細胞増殖を評価した。簡単に説明すると、0.1M酢酸ナトリウム(pH5.5),0.1%トリトンX-100および10mM p-ニトロフェニルホスフェート(Sigma 104ホスファターゼ基質)を含有するバッファー50μlを、ウェルに加えた。プレートを室温で1〜0.5時間インキュベートし、10μl/ウェルの1N水酸化ナトリウムで反応を停止させ、Dynatech ModelMR600で410nmで吸光度を読んだ。サイトカイン刺激後のシグナル伝達カスケードのタンパク質のチロシンリン酸化を解析するために、種々の型のヒトIL-9受容体を発現するTS1細胞をPBSで洗浄し、D-MEM,0.5%ウシ血清アルブミンに再懸濁し、37℃で6時間インキュベートした。続いて、20×106細胞をヒトIL-9またはマウスIL-9(100ng/ml)で5分間処理し,直ちに冷PBSで洗浄した。実施例11に記載のように細胞をRIPAバッファーで溶解した。
【0110】
(実施例11:免疫沈降、免疫ブロッティングおよび抗体)
典型的には20〜50×106細胞を、1mlのRIPAバッファー(1%NP40、0.5%デオキシコール酸ナトリウム、0.1%SDS、1mM PMSF、50mMフッ化ナトリウム、1nMオルトバナジン酸ナトリウム、および1×「完全」プロテアーゼインヒビター混合物を含有するPBS、Cat. No. 1697498 Boehringer Mannheim)で溶解し、氷上で45分間インキュベートした。溶解物をEppendorf微量遠心分離機中で20分間遠心分離し、上清を回収し、新しいチューブに移した。免疫沈降のために、1〜5μgの抗体を溶解物に加え、4℃で一晩インキュベートした。20mlのプロテインA+Gアガロース結合ビーズを2時間加え、次にRIPAバッファーを使用して4回洗浄した。ビーズをLaemmliバッファー中に再懸濁し、3分間沸騰してから電気泳動した。タンパク質をImmobilion-P膜(Millipore)に移し、西洋ワサビペルオキシダーゼ結合2次抗体を使用して化学発光検出アッセイ(Pierce)により検出した。マウスおよびヒトIL-9受容体の特異的抗体(sc698),マウスJak1、Irs1、Irs2、Stat1、Stat2、Stat3、Stat4、Stat5、およびホスホチロシン(PY)は、Santa Cruz(Santa Cruz, CA)から購入した。抗Jak3およびモノクローナル抗ヒトIL-9受容体MAB290は、それぞれUpstate BiotechnologyとR & D Systemsから購入した。図15は、ヒトIL-9受容体の異なる変異体による、Jakファミリーのメンバーの活性化を証明する。
【0111】
(実施例12:IL-9受容体ゲノム多型の同定)
既に記載されているように(NicolaidesとStoeckert, Biotechniques 8:154-156, 1990)、志願者ドナーのPBMCからゲノムDNAを単離した。配列番号14と配列番号17のプライマーを使用するPCRにより、ヒトIL-9R遺伝子のイントロン5の配列解析を行ない、分子サイズ約1243塩基対の産物を得た。既に記載されているバッファー(Nicholaidesら、Genomics 30:195-206, 1995)で、94℃で30秒、62℃で1.5分、72℃で1.5分を35サイクル行って増幅した。次に産物を精製し、標準的配列プロトコールを使用して配列決定した。複数の個体のイントロン5からの配列を調べると、エキソン6の配列の-213ヌクレオチド上流にヌクレオチド変化があり、このためチミジン(公表されている配列)がシトシンヌクレオチドに変化していた。この変異の例を図17に示す。
【0112】
本発明を種々の具体的な材料、方法および例を参照して説明し例示したが、本発明は、この目的のために選択された材料の特定の組合せや方法に限定されるものではないことを理解されたい。当業者が理解できるように、このような細部の多くの変更が可能である。
【0113】
【表2】
【0114】
【表3】
【0115】
【表4】
【0116】
【表5】
【0117】
【表6】
【0118】
【表7】
【0119】
【表8】
【0120】
【表9】
【0121】
【表10】
(配列表)
【0122】
【化1】
【0123】
【化2】
【0124】
【化3】
【0125】
【化4】
【0126】
【化5】
【0127】
【化6】
【0128】
【化7】
【0129】
【化8】
【0130】
【化9】
【0131】
【化10】
【0132】
【化11】
【0133】
【化12】
【0134】
【化13】
【0135】
【化14】
【0136】
【化15】
【0137】
【化16】
【0138】
【化17】
【0139】
【化18】
【0140】
【化19】
【0141】
【化20】
【0142】
【化21】
【0143】
【化22】
【0144】
【化23】
【0145】
【化24】
【0146】
【化25】
【0147】
【化26】
【0148】
【化27】
【0149】
【化28】
【技術分野】
【0001】
(関連出願の参照)
本出願は、1996年12月2日に出願された米国仮出願第60/032,224号(参照により本明細書に組み入れる)に関連する。また本出願は、米国特許出願第08/697,419号、08/697,360号、08/697,473号、08/697,472号、08/697,471号、08/702,105号、08/702,110号、08/702,168号、および08/697,440号(これらはすべて1996年8月23日に出願された)、そして08/874、503号(これは1997年6月13日に出願された)の記載事項に関連し、これらは参照により本明細書に組み入れる。
【0002】
(発明の分野)
本発明は、IL-9受容体(喘息関連因子2(Asthma Associated Factor 2))(配列番号1)の生物学的変化を記載し、これらの変異体と、喘息、アトピー性アレルギー、および関連疾患への感受性(susceptibility)とを関連付ける。本発明はまた、喘息およびアトピー性アレルギーに対する感受性または耐性の診断のためにこれらのIL-9受容体の配列変異体を利用する方法を教示する。さらに、IL-9活性の調節に依存する喘息用の薬剤の開発において、IL-9受容体変異体を使用する方法が記載される。
【背景技術】
【0003】
炎症は、体の防御システムが外来物質と戦う複雑なプロセスである。外来物質に対する戦いは体の生存のために必要であるが、一部の防御システムは、無害の外来物質に対しても危険なものとして不適当に対応し、その結果、その後の戦いにおいて周りの組織を傷害する。
【0004】
アトピー性アレルギーは、遺伝的背景が、環境からの刺激に対する応答を支配する疾患である。この疾患は、一般に遍在する抗原に応答してIgE抗体を産生するリンパ球の能力が上昇していることが特徴である。これらの抗原による免疫系の活性化はまた、これらの摂取、皮膚を通しての透過、または吸入後に起きるアレルギー性炎症を引き起こす。この免疫活性化が起き、続いて肺の炎症が起きると、この疾患は広く喘息として特徴付けられる。気道での炎症性反応にはある種の細胞が重要であり、これらには、T細胞および抗原提示細胞、IgEを産生するB細胞、および炎症性メディエーターを蓄えIgEに結合する肥満細胞/好塩基球、さらなるメディエーターを放出する好酸球がある。これらの炎症性細胞はアレルギー性炎症の場に蓄積し、これらが放出する有害産物が、この疾患に関連する組織破壊に寄与する。
【0005】
喘息は一般に、気道の炎症性疾患として定義されるが、臨床症状は、間欠的な空気流の遮断から起きる。これは、慢性の障害性の疾患であり、罹患率と重篤度が上昇しているようである1。人口の30〜40%がアトピー性アレルギーに罹っており、児童の15%と成人の5%が喘息に罹っていると推定されている(非特許文献1)1。すなわち、これらの疾患の治療において、医療資源に巨大な負担がかかっている。
【0006】
喘息と関連疾患の診断と治療の両方とも問題がある1。特に炎症を起こした肺の評価は困難なことが多く、炎症の原因が決定できないことが頻繁にある。アトピー性喘息は環境遺伝的疾患であるが、具体的な変異体遺伝子についての知見は最近発見されたばかりである。炎症におけるこれらの遺伝子的変異体の検出方法とその役割、診断および予後は、まだ決定されていない。当該分野で必要なことは、アトピー性疾患に対する感受性/耐性に関与する遺伝子の変異に特異的に関係する、アトピー性喘息の診断を促進する技術の開発である。
【0007】
現代の治療法は、固有の一連の欠点を有する。主要な治療薬であるβ−アゴニストは、症状を低下させ、すなわち肺機能を一過性に改善するが、基礎炎症には作用せず、そのため肺組織は依然として危険な状態にある。さらに、βーアゴニストを使い続けると脱感受性が起き、その効力と安全性が低下する2。基礎炎症を縮小させることができる物質である抗炎症性ステロイドは、免疫抑制から骨喪失にわたる固有の一連の副作用を有する2。
【0008】
従来の治療に起因する問題のために、別の治療方策が評価されている38-39。グリコホリンA37、シクロスポリン38、およびIL-2のノナペプチド断片36はすべて、インターロイキン−2依存性のTリンパ球増殖を阻害する28。しかしこれらは、他の多くの作用を有することが知られている2。例えば、シクロスポリンは、臓器移植後の免疫抑制剤として使用されている。これらの薬剤は、喘息患者の治療においてステロイドの代替となるが36-39、これらはインターロイキン−2依存性Tリンパ球増殖を阻害し、ホメオスタシスに関連して非常に重要と思われる免疫機能を阻害する可能性がある。当該分野で必要なことは、アトピー性喘息の症状ではなくその原因を治療するように具体的に設計された治療薬の開発を促進することである。これらの治療法は、非特異的な治療に関連する毒性を避けるための最も可能性の高い方法である。この治療薬は、免疫機能(例えば、喘息の進展に必要なIL-2介在Tリンパ球活性化)の下流にあり、疾患の偶発性を説明し、アレルギーとの密接な関連を説明する経路を選択的に標的とする。その経路中に生物学的変化がふつうに存在し、その変化を示す個体が、アトピー性喘息の症状以外は免疫無防備状態でも病気でもない場合は、経路は喘息治療の適切な標的であることを、自然は証明している。
【0009】
喘息を含むアトピー性アレルギーの診断および治療に関連する困難さのために、これらの疾患の複雑な病態生理が詳しく研究されている。これらの疾患は、多くの要素からなり多くの型を取るため定義することが困難であるが、喘息患者にはいくつかの共通の特徴が見いだされている。そのような特徴の例には、アレルゲンによる抗原刺激に対する異常な皮膚試験応答、肺の好酸球増加症、気管支過応答性(BHR)、気管支拡張薬可逆性、および空気流遮断がある3-10。喘息に関連する形質のこれらの発現は、定量的または定性的に研究される。
【0010】
多くの場合、IgEレベルの上昇は、BHR、種々の刺激に対する気管支収縮応答の上昇に関係している4,6,8,9。BHRは、気道炎症の存在を反映すると考えられており6,8、喘息の危険因子であると考えられている11-12。BHRは、肺への好酸球浸潤を含む気管支炎症と喘息患者のアレルギー性素因を伴う6,8,13-18。
【0011】
アトピー性喘息に対する遺伝性成分が、多くの研究で記録されている4,10。しかしこれらの疾患は、年令と性、ならびに多くの環境因子(例えば、アレルゲン、ウイルス感染、および汚染物質19-21)に大きく影響されるため、家族研究は解釈することが困難である。さらに、これらの疾患に対する感受性に関連することが知られている生化学的欠陥はないため、突然変異遺伝子およびその異常遺伝子産物は、これらが産生する異常表現型によってのみ認識することができる。
【0012】
IL-9とIL-9受容体(IL-9経路)の機能は、当初認識されていたものより、拡大している。IL-9経路はT細胞増殖の刺激物質として作用するが、このサイトカインはまた、赤血球始原細胞、B細胞、肥満細胞、および胎児胸腺細胞の増殖に介在することが知られている22,23。IL-9経路はIL-3と相乗的に作用して肥満細胞活性化と増殖を引き起こす24。IL-9経路はまた、正常なヒトBリンパ球によるIgE、IgGおよびIgMのIL-4誘導性産生を増強し25、マウスBリンパ球によるIgEおよびIgGのIL-4誘導性放出を増強する26。寄生体感染に対する粘膜の炎症性応答におけるIL-9経路の役割もまた証明されている27,28。
【0013】
しかし、IL-9受容体の配列がアトピー性喘息と気管支過応答性にいかに相関するかは知られていない。IL-9が標的細胞の表面に発現される特異的受容体に結合することは知られている23,29,30。この受容体は実際、2つのタンパク質鎖からなる。1つのタンパク質鎖はIL-9受容体として知られており、IL-9に特異的に結合する。もう1つのタンパク質鎖は、IL-2受容体と共有される23。さらにヒトIL-9受容体をコードするcDNAがクローン化されており、配列が決定されている23,29,30。このcDNAは、マウスIL-9受容体と高い相同性を示す522アミノ酸のタンパク質をコードする。この受容体の細胞外領域は高度に保存されており、マウスとヒトのタンパク質の間で67%の相同性がある。受容体の細胞質領域は、あまり高度に保存されていない。ヒトの細胞質ドメインは、マウス受容体の対応する領域よりはるかに大きい23。
【0014】
IL-9受容体遺伝子は特徴付けもされている30。これは、マウスゲノム中で単一のコピーとして存在し、9個のエキソンと8個のイントロンを含有する30。ヒトのゲノムは、少なくとも4つのIL-9受容体偽遺伝子(pseudogene)を含有する。ヒトのIL-9受容体遺伝子は、性染色体XとYの320 kbの次端部テロメア領域(subtelomeric region)にマッピングされている23。
【0015】
これらの研究にもかかわらず、IL-9受容体遺伝子の変異体は発見されていない。従ってアトピー性アレルギー、喘息、気管支過応答性についての遺伝的情報、およびこれらの疾患の病因におけるIL-9受容体の役割の解明について具体的なニーズがある。この情報は、これらの疾患に関連するこの遺伝子の遺伝的変異体を同定する方法を使用して、アトピー性アレルギーや関連疾患を診断するのに使用することができる。さらに、これらの疾患を治療するための治療薬の開発のために、IL-9受容体変異体を起用する方法についてニーズがある。
【先行技術文献】
【非特許文献】
【0016】
【非特許文献1】Gergen PJ, and Weiss KB: The increasingproblem of asthma in the United States. Am Rev Respir Dis 146: 823-824,1992
【発明の概要】
【課題を解決するための手段】
【0017】
(発明の要約)
本出願人らは、ヒトIL-9受容体(喘息関連因子2またはAAF2としても知られている)の天然の変異体を発見し、これらの変異体を喘息および関連疾患の病態と関連付けた。これらの発見により、アトピー性喘息の診断法および治療薬を発見する方法が開発された。さらに本出願人らは、IL-9受容体は、マウスの多くの抗原誘導性応答(気管支過応答性、気管支洗浄液中の好酸球と細胞数の増加、および血清総IgEの増加を含む)に非常に重要であることを明らかにした。これらの知見は、アトピー性喘息および関連するアレルギー性炎症に典型的である。
【0018】
さらに本出願人らは、GからAへの核酸の変異がcDNA(配列番号2)の1273位に存在し、これはヒトIL-9受容体前駆体タンパク質のコドン344のアルギニンからヒスチジンへの予測されるアミノ酸置換を引き起こす。1個体の両方の対立遺伝子にアルギニン残基が存在する時、これはアトピー性喘息が少ないことに関連している。すなわち本出願人らは、コドン344のアルギニンが1個体で両方のIL-9受容体遺伝子産物中で存在するとき、これにより特徴付けられる非喘息表現型の存在を確認した。さらなる重要な結果として、本出願人らは、コドン344のヒスチジンを特徴とする、喘息性、アトピー性表現型に対する感受性の存在を確認した。すなわち本発明は、そのような配列を有する精製され単離されたDNA分子、ならびにこのDNAによりコードされるタンパク質を包含する。
【0019】
本出願人らはまた、IL-9R前駆体タンパク質の173位のグルタミン残基が欠失しているIL-9Rのスプライス変異体(配列番号3)が存在することを確認した(図5)。さらに本出願人らは、この変異体は、Jak-Stat経路によりシグナルを転写できないこと(図15)、およびIL-9で刺激しても細胞増殖を誘導できないこと(図16)を証明した。従ってこの対立遺伝子を有する個体は、アトピー性喘息および関連疾患に罹りにくいであろう。
【0020】
本出願人らはさらに、イントロン5のチミン残基であるnt-213(エキソン6から213nt上流)が、シトシンヌクレオチドに変換されている、IL-9RゲノムDNAの変異体が存在することを確認した。このような変異はエキソン6の開始点でグルタミン残基を除去するスプライス変異体の頻度を上昇させ得る。
【0021】
さらに本出願人らは、エキソン9の読みとり枠と未熟停止コドンに変異を起こす、エキソン8が欠失しているIL-9Rの変異体(配列番号4)を発見した。このような変異体は、Jak-Stat経路でシグナルを伝達することを妨害し、従ってこの対立遺伝子を有する個体もまた、アトピー性喘息および関連疾患に罹りにくいであろう。
【0022】
IL-9の生物活性は、そのIL-9受容体への結合とその後の特異的細胞での制御シグナル伝搬に起因し、従って、IL-9機能は、IL-9アンタゴニストとIL-9またはその受容体との相互作用により妨害することができる。ダウンレギュレーション、すなわちIL-9により制御される機能の低下は、多くの方法で行われる。受容体へのIL-9の結合を阻止することができるアンタゴニストを投与することは、1つの重要な方法であり、このようなアンタゴニストは、特許請求される本発明の範囲内である。例としては、IL-9受容体の天然に存在する可溶性型のDNA配列によりコードされるポリペプチド産物の投与があり、DNA配列はエキソン2と3からなるポリペプチド(配列番号5)をコードする。他の2つの変異体は、IL-9Rの可溶性型(これは、エキソン2、3および4からなり、ある場合にはエキソン5の異なる読みとり枠からの4つのアミノ酸(配列番号6)(図6)、そして別の場合はエキソン5の異なる読みとり枠の27のアミノ酸(配列番号7)(図7)を含む)を産生する。
【0023】
IL-9受容体経路のアゴニストおよびアンタゴニストの同定方法は、文献に詳細に記載されている受容体−リガンド相互作用を評価することにより特定できる。これらの方法は、化学的スクリーニングと潜在的な治療薬の同定を促進する、高処理量自動アッセイに適合させることができる。アゴニストは、IL-9受容体との特異的な相互作用を同定することにより認識される。100〜1000倍過剰の非標識リガンドを使用しても、標識した推定リガンドに対する結合が存在しないことは、一般に特異的受容体結合の証拠として受け取られている。これらの実験では、多くの標識物および検出スキームを使用することができる。増加する濃度の試験化合物を既知のリガンドおよび受容体に添加しても同様に結合がないことも、アンタゴニストの証拠である。
【0024】
変異体受容体の情報は、受容体結合アッセイの可溶性受容体を作成するのに使用可能な発現ベクターを作製するための手段を提供する。これらの可溶性受容体の突然変異誘発は、リガンドに結合するのにどのアミノ酸が非常に重要であるか、およびどのアミノ酸がアンタゴニストの構造ベースのデザインを補助するかを決定するために使用することができる。
【0025】
ヒトIL-9受容体が欠如した細胞は、変異体受容体を含有する発現ベクターで一過性にまたは安定にトランスフェクションすることができ、IL-9経路活性をアッセイするのに使用することができる。これらの活性は、いずれもIL-9経路が原因であるとされている細胞増殖でもアポトーシスの阻止でもよい。これらの細胞は、前記したように受容体アゴニストおよびアンタゴニストを同定するのに使用することができる。
【0026】
上記方法は、アトピー性喘息や他の関連疾患の治療薬を開発するために、IL-9受容体の変異体型を利用する種々の有効な方法である。
【0027】
IL-9受容体中の単一のヌクレオチド変異体を認識する、アトピー性喘息を診断するために使用できる多くの技術が記載されており、これらはDNA配列決定、制限断片長多型(RFLP)、対立遺伝子特異的オリゴヌクレオチド解析(ASO)、連結連鎖反応(LCR)、化学的切断、および1本鎖コンフォメーション多型解析(SSCP)がある。これらの技術ならびに文献に記載の1つまたはそれ以上の他の技術の使用を、IL-9受容体遺伝子またはmRNA転写物の1つまたはそれ以上の変異を検出するのに使用でき、これらが本発明の範囲内にあることを当業者は容易に理解できるであろう。
【0028】
さらに別の技術(ELISA、免疫沈降、ウェスタン法、および免疫ブロッティング法を含む)を使用して、IL-9受容体のアミノ酸変異体を検出してもよい。すなわち、IL-9受容体の種々の型の構造を特異的に認識するポリクローナル抗体またはモノクローナル抗体もまた、本発明の範囲内にあり、アトピー性喘息や関連疾患に対する感受性または耐性を説明するための有用な診断法である。
【0029】
上記方法は、アトピー性喘息および他の関連疾患を診断するための種々の有効な方法を示す。
【0030】
すなわち本出願人らは、IL-9とその受容体との相互作用を調節することができるアンタゴニストを同定するためにIL-9受容体を使用する方法を提供している。さらに詳しくは、本出願人らは、IL-9経路の発現または機能をダウンレギュレーションするのに充分な量で投与される化合物または物質を同定するために、IL-9受容体の機能をアッセイする方法を提供する。
【0031】
本出願人らは、アトピー性アレルギー、気管支過応答性および喘息におけるIL-9経路の役割を特定したため、アトピー性アレルギー、喘息、および関連疾患に対する感受性と耐性の診断のための方法も提供する。
【0032】
本明細書に取り込まれその一部を構成する添付の図面は、本発明のいくつかの実施態様を例示し、本発明の説明とともに本発明の原理を説明するものである。
【0033】
本発明は、例えば、以下を提供する:
(項目1) ヒト・インターロイキン-9受容体をコードするヌクレオチド配列を有する単離されたDNA分子であって、前記ヌクレオチド配列が1273位にGからAへの核酸変異を含む配列、核酸759-761が欠失された配列(配列番号3)、核酸613-617が欠失された配列(配列番号7)、核酸435-437に停止コドンを含む配列(配列番号5)、核酸613-641が欠失された配列(配列番号6)、およびそれらの断片よりなる群から選択される、上記の単離されたDNA分子。
(項目2) 前記配列が1273位にGからAへの核酸変異を含む項目1に記載の単離されたDNA分子またはその断片。
(項目3) 核酸759-761が欠失された(配列番号3)項目1に記載の単離されたDNA分子またはその断片。
(項目4) 前記配列が核酸435-437に停止コドンを含む(配列番号5)項目1に記載の単離されたDNA分子またはその断片。
(項目5) 核酸613-617が欠失された(配列番号7)項目1に記載の単離されたDNA分子またはその断片。
(項目6) 核酸613-641が欠失された(配列番号6)項目1に記載の単離されたDNA分子またはその断片。
(項目7) 核酸1067-1151が欠失された(配列番号4)項目1に記載の単離されたDNA分子またはその断片。
(項目8) エキソン6から213核酸上流の核酸がTからCに変化した、ヒトIL-9受容体の第5イントロンをコードするヌクレオチド配列を有する単離されたDNA分子。
(項目9) ヒト・インターロイキン-9受容体をコードするヌクレオチド配列を有する単離されたRNA分子であって、前記ヌクレオチド配列が1273位にAからGへの核酸変異を含む配列、核酸759-761が欠失された配列、核酸435-437に停止コドンを含む配列、核酸613-617が欠失された配列、核酸613-641が欠失された配列、核酸1067-1151が欠失された配列、およびそれらの断片よりなる群から選択される、上記の単離されたRNA分子。
(項目10) 前記配列が1273位にAからGへの核酸変異を含む、項目9に記載の単離されたRNA分子またはその断片。
(項目11) 核酸759-761が欠失された、項目9に記載の単離されたRNA分子またはその断片。
(項目12) 前記配列が核酸435-437に停止コドンを含む、項目9に記載の単離されたRNA分子またはその断片。
(項目13) 核酸613-617が欠失された、項目9に記載の単離されたRNA分子またはその断片。
(項目14) 核酸613-641が欠失された、項目9に記載の単離されたRNA分子またはその断片。
(項目15) 核酸1067-1151が欠失された、項目9に記載の単離されたRNA分子またはその断片。
(項目16) ヒト・インターロイキン-9受容体をコードするアミノ酸配列を有する単離されたタンパク質分子であって、前記アミノ酸配列が、344位にヒスチジンを含む配列、グルタミン173が欠失された配列、前記分子がアミノ酸64の後で終結している配列、前記分子が項目4に記載のDNAによりコードされる配列、前記分子が項目5に記載のDNAによりコードされる配列、またはその断片よりなる群から選択される、上記の単離されたタンパク質分子。
(項目17) 前記配列が344位にヒスチジンを含む、項目16に記載の単離されたタンパク質分子またはその断片。
(項目18) グルタミン173が欠失された、項目16に記載の単離されたタンパク質分子またはその断片。
(項目19) 前記分子がアミノ酸64の後で終結している、項目16に記載の単離されたタンパク質分子またはその断片。
(項目20) 前記分子が項目5に記載のDNAによりコードされる、項目16に記載の単離されたタンパク質分子またはその断片。
(項目21) 前記分子が項目6に記載のDNAによりコードされる、項目16に記載の単離されたタンパク質分子またはその断片。
(項目22) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、項目2に記載のDNAの存在または非存在を判定し、前記DNAのみの存在が喘息または関連疾患に対するより高い罹病性を示すものである、上記の方法。
(項目23) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、項目3に記載のDNAの存在または非存在を判定し、前記DNAのみの存在が喘息または関連疾患に対するより高い罹病性を示すものである、上記の方法。
(項目24) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、項目4に記載のDNAの存在または非存在を判定し、前記DNAの存在が喘息または関連疾患に対するより低い罹病性を示すものである、上記の方法。
(項目25) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、項目5に記載のDNAの存在または非存在を判定し、前記DNAの存在が喘息または関連疾患に対するより低い罹病性を示すものである、上記の方法。
(項目26) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、項目6に記載のDNAの存在または非存在を判定し、前記DNAの存在が喘息または関連疾患に対するより低い罹病性を示すものである、上記の方法。
(項目27) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、項目7に記載のDNAの存在または非存在を判定し、前記DNA分子の存在が喘息または関連疾患に対するより低い罹病性を示すものである、上記の方法。
(項目28) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、項目8に記載のDNA分子の存在または非存在を判定し、前記DNAの存在が喘息または関連疾患に対するより低い罹病性を示すものである、上記の方法。
(項目29) IL-9経路のアンタゴニストを同定する方法であって、a)IL-9受容体を発現する細胞を取得し、b)前記細胞を、IL-9と推定上のアンタゴニストの様々な比率の混合物に接触させ、c)Jak-Stat経路のタンパク質のリン酸化の量を測定する、各ステップを含んでなり、リン酸化の低下がIL-9アンタゴニストの存在を示すものである、上記の方法。
(項目30) IL-9受容体がヒトのものである、項目29に記載の方法。
(項目31) IL-9受容体がマウスのものである、項目29に記載の方法。
(項目32) IL-9経路のアンタゴニストを同定する方法であって、a)IL-9受容体を発現する細胞を取得し、b)前記細胞を、デキサメタゾンとIL-9と推定上のアンタゴニストの様々な比率の混合物に接触させ、c)アポトーシスの強度を測定する、各ステップを含んでなり、デキサメタゾン誘発アポトーシスからのIL-9介在レスキューの低下がIL-9アンタゴニストの存在を示すものである、上記の方法。
(項目33) IL-9受容体がヒトのものである、項目32に記載の方法。
(項目34) IL-9受容体がマウスのものである、項目32に記載の方法。
(項目35) ヒトIL-9受容体を発現する細胞がC8166-45、HL60、HL60-clone5、好酸球、好中球、巨核球、およびヒトIL-9RでトランスフェクトされたTS1、KG-1および32D細胞系よりなる群から得られる、項目24または26に記載の方法。
(項目36) マウスIL-9受容体を発現する細胞がTS1、KG-1、32D、好酸球、好中球、および巨核球よりなる群から得られる、項目25または27に記載の方法。
(項目37) ヒトIL-9受容体を発現する細胞がHL60またはHL60-clone5細胞である、項目27または29に記載の方法。
(項目38) ヒトIL-9受容体を発現する細胞がヒトIL-9RでトランスフェクトされたKG-1細胞である、項目27または29に記載の方法。
(項目39) ヒトIL-9受容体を発現する細胞がC8166-45細胞である、項目27または29に記載の方法。
(項目40) ヒトIL-9受容体を発現する細胞が好酸球である、項目27または29に記載の方法。
(項目41) 喘息および喘息関連疾患を軽減させる方法であって、かかる治療を必要とする患者に、有効量の項目18に記載のタンパク質分子を投与することを含んでなる方法。
(項目42) 喘息および喘息関連疾患を軽減させる方法であって、かかる治療を必要とする患者に、有効量の項目19に記載のタンパク質分子を投与することを含んでなる方法。
(項目43) 喘息および喘息関連疾患を軽減させる方法であって、かかる治療を必要とする患者に、有効量の項目20に記載のタンパク質分子を投与することを含んでなる方法。
(項目44) プライマーが配列番号8および配列番号9に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン2を特異的に増幅する方法。
(項目45) プライマーが配列番号10および配列番号11に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン3を特異的に増幅する方法。
(項目46) プライマーが配列番号12および配列番号13に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン4を特異的に増幅する方法。
(項目47) プライマーが配列番号14および配列番号15に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン5を特異的に増幅する方法。
(項目48) プライマーが配列番号16および配列番号17に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン6を特異的に増幅する方法。
(項目49) プライマーが配列番号18および配列番号19に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン7を特異的に増幅する方法。
(項目50) プライマーが配列番号20および配列番号21に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン8を特異的に増幅する方法。
(項目51) プライマーが配列番号22および配列番号23に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のエキソン9を特異的に増幅する方法。
(項目52) プライマーが配列番号14および配列番号17に示されるものである項目54に記載の組成物を用いるPCRにより、ゲノムDNAから真のヒトIL-9受容体のイントロン5を特異的に増幅する方法。
(項目53) ヒト被験者における喘息または関連疾患に対する罹病性を検出または診断する方法であって、前記DNAによりコードされる非機能性インターロイキン-9受容体をコードするDNAの存在または非存在を判定し、非機能性インターロイキン-9受容体をコードする前記DNAの存在が喘息または関連疾患に対するより低い罹病性を示すものである、上記の方法。
(項目54) 配列番号8、9、10、11、12、13、14、15、16、17、18、19、20、21、22または23よりなる群から選択される真のヒトIL-9受容体を特異的に増幅するためのプライマーを含む組成物。
(項目55) 喘息および喘息関連疾患を軽減させる方法であって、かかる治療を必要とする患者に、有効量の項目21に記載のタンパク質分子を投与することを含んでなる方法。
【図面の簡単な説明】
【0034】
【図1】ヒトIL受容体cDNAの略図。四角は、コード領域を含むエキソン2〜9を示す(スケールは相対的サイズであり、エキソン9の3’非翻訳部分を除いて、破線で示す)。膜貫通領域は、エキソン7によりコードされ、細胞内ドメインはエキソン8と9に、そして細胞外ドメインはエキソン2〜6にコードされる。矢印は、エキソンの部分配列に影響を与える多型または異常スプライスを示す;a)エキソン5の最初の5または29ヌクレオチドの欠失;b)エキソン6の最初の3ヌクレオチドの欠失(コドン173);c)コドン310におけるarg/gly多型;d)コドン344におけるarg/his多型;e)8または9個のセリンからなるコドン410+nの多型;*)エキソン3、4または8の完全な欠失。
【図2−1】コドン344(ヌクレオチド1272〜1274)のArg対立遺伝子とコドン410(ヌクレオチド1470〜1472)で開始する8 Ser/4Asn反復配列とを有する野生型IL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図2−2】コドン344(ヌクレオチド1272〜1274)のArg対立遺伝子とコドン410(ヌクレオチド1470〜1472)で開始する8 Ser/4Asn反復配列とを有する野生型IL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図3−1】コドン344(ヌクレオチド1272〜1274)のHis対立遺伝子とコドン410(ヌクレオチド1470〜1472)で開始する9 Ser/4Asn反復配列とを有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図3−2】コドン344(ヌクレオチド1272〜1274)のHis対立遺伝子とコドン410(ヌクレオチド1470〜1472)で開始する9 Ser/4Asn反復配列とを有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図4】エキソン9でフレームシフトと、11の非野生型アミノ酸および未熟停止コドンの産生とを引き起こす、エキソン8の欠失を有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図5−1】コドン173でグルタミンの欠失を有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図5−2】コドン173でグルタミンの欠失を有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図6】未熟停止コドンと27の非野生型アミノ酸の産生を引き起こす、エキソン5中の代替的なスプライスを有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図7】未熟停止コドンと4つの非野生型アミノ酸の産生を引き起こす、エキソン5中の代替的なスプライスを有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図8】エキソン5の最初のコドンとして停止コドンを産生する、エキソン4の欠失を有するIL-9R前駆体タンパク質の翻訳付きのcDNA配列。
【図9】IL-9受容体遺伝子型とアトピー性アレルギーの間の関係を示す表。Arg/Argの個体は、8 Ser/4Asn反復配列を有するArg対立遺伝子についてホモ接合性である。Arg/Hisの個体は、エキソン9中に、8 Ser/4Asn反復配列を有するArg対立遺伝子と、9 Ser/4 Asn反復配列、9 Ser/3 Asn反復配列、および10 Ser/2Asn反復配列を有するHis対立遺伝子についてヘテロ接合性である。His/Hisの個体は、エキソン9中に、9 Ser/4Asn反復配列、9 Ser/3 Asn反復配列、および10 Ser/2 Asn反復配列を有するHis対立遺伝子についてホモ接合性である。Arg/Argの個体は、アトピー性アレルギーから防御される。Arg/HisおよびHis/Hisの個体は、アトピー性アレルギーに対して感受性である(P=0.002)。
【図10】IL-9受容体の発現構築物の地図。
【図11】TK-トランスフェクション細胞系中のC末端抗体プローブを使用する、組換えIL-9受容体タンパク質のウェスタンブロット(左:8 Ser/4Asn反復配列を有するArg対立遺伝子;右:9 Ser/4 Asn反復配列を有するHis対立遺伝子)。
【図12】8 Ser/4Asn反復配列を有するArg344変異体(GR8)と9 Ser/4 Asn反復配列を有するHis344変異体(GH9)の間の移動度の差を示す、TS1細胞中のヒトIL-9受容体変異体の発現。IL-9受容体のこれらの2つの変異体型の翻訳後修飾の差を証明する移動度のシフトがみられる。
【図13】IL-9受容体遺伝子の特異的増幅のためのXY特異的アンプリマー。第9、第10、第16、または第18染色体上の偽遺伝子は、PCRにより増幅されない。(MはマウスDNA、HはヒトDNA、そしてCはハムスターDNAを示す)。
【図14−1】抗ヒトIL-9受容体中和抗体と野生型およびデルタ−Q受容体との免疫反応性。パネルA):COS7細胞を、LXSNベクター単独(AとB)、野生型IL-9R(CとD)、コドン410で開始する9個のSer残基を有する野生型IL-9R(EとF)、Δ-Q 173変異体(GとH)、およびコドン410で開始する9個のSer残基を有するΔ-Q 173(IとJ)で一過性にトランスフェクトし、記載したように(実施例8)MAB290と抗マウスIgGテキサスレッド結合抗体(B、D、F、H、およびJ)で連続的にインキュベートする。DAPI染色(A、C、E、G、およびI)を行って、写真の視野中のすべての細胞を視覚化する。パネルB):A)と同様に行なったが、細胞をまず固定/透過性化し、次にC末端特異的抗体(sc698)でインキュベートし、次に抗ウサギIgGテキサスレッド結合抗体でインキュベートした。横棒=10ミクロン。
【図14−2】抗ヒトIL-9受容体中和抗体と野生型およびデルタ−Q受容体との免疫反応性。パネルA):COS7細胞を、LXSNベクター単独(AとB)、野生型IL-9R(CとD)、コドン410で開始する9個のSer残基を有する野生型IL-9R(EとF)、Δ-Q 173変異体(GとH)、およびコドン410で開始する9個のSer残基を有するΔ-Q 173(IとJ)で一過性にトランスフェクトし、記載したように(実施例8)MAB290と抗マウスIgGテキサスレッド結合抗体(B、D、F、H、およびJ)で連続的にインキュベートする。DAPI染色(A、C、E、G、およびI)を行って、写真の視野中のすべての細胞を視覚化する。パネルB):A)と同様に行なったが、細胞をまず固定/透過性化し、次にC末端特異的抗体(sc698)でインキュベートし、次に抗ウサギIgGテキサスレッド結合抗体でインキュベートした。横棒=10ミクロン。
【図14−3】抗ヒトIL-9受容体中和抗体と野生型およびデルタ−Q受容体との免疫反応性。パネルA):COS7細胞を、LXSNベクター単独(AとB)、野生型IL-9R(CとD)、コドン410で開始する9個のSer残基を有する野生型IL-9R(EとF)、Δ-Q 173変異体(GとH)、およびコドン410で開始する9個のSer残基を有するΔ-Q 173(IとJ)で一過性にトランスフェクトし、記載したように(実施例8)MAB290と抗マウスIgGテキサスレッド結合抗体(B、D、F、H、およびJ)で連続的にインキュベートする。DAPI染色(A、C、E、G、およびI)を行って、写真の視野中のすべての細胞を視覚化する。パネルB):A)と同様に行なったが、細胞をまず固定/透過性化し、次にC末端特異的抗体(sc698)でインキュベートし、次に抗ウサギIgGテキサスレッド結合抗体でインキュベートした。横棒=10ミクロン。
【図14−4】抗ヒトIL-9受容体中和抗体と野生型およびデルタ−Q受容体との免疫反応性。パネルA):COS7細胞を、LXSNベクター単独(AとB)、野生型IL-9R(CとD)、コドン410で開始する9個のSer残基を有する野生型IL-9R(EとF)、Δ-Q 173変異体(GとH)、およびコドン410で開始する9個のSer残基を有するΔ-Q 173(IとJ)で一過性にトランスフェクトし、記載したように(実施例8)MAB290と抗マウスIgGテキサスレッド結合抗体(B、D、F、H、およびJ)で連続的にインキュベートする。DAPI染色(A、C、E、G、およびI)を行って、写真の視野中のすべての細胞を視覚化する。パネルB):A)と同様に行なったが、細胞をまず固定/透過性化し、次にC末端特異的抗体(sc698)でインキュベートし、次に抗ウサギIgGテキサスレッド結合抗体でインキュベートした。横棒=10ミクロン。
【図15−1】ヒトIL-9受容体の異なる変異体のJak、Stat、およびIrsファミリーのメンバーの活性化。GH9、ΔQGR8、またはΔQGH9を発現するTS1細胞を6時間、栄養を欠乏させ、次にサイトカイン無し(−)、マウスIL-9有り(m),またはヒトIL-9有り(h)で5分間処理した。細胞抽出物を、Jak、StatおよびIrsファミリーの異なるメンバーに特異的な種々の抗体で免疫沈降させた。まず免疫ブロットを抗ホスホチロシン抗体と反応させて、チロシン−リン酸化タンパク質のみを検出し、次にストリップし、各タンパク質を免疫沈降させるのに使用したものと同じ抗体で再度釣り上げた。GH9、ΔQGR8、ΔQGH9は、図16に示す通りである。
【図15−2】ヒトIL-9受容体の異なる変異体のJak、Stat、およびIrsファミリーのメンバーの活性化。GH9、ΔQGR8、またはΔQGH9を発現するTS1細胞を6時間、栄養を欠乏させ、次にサイトカイン無し(−)、マウスIL-9有り(m),またはヒトIL-9有り(h)で5分間処理した。細胞抽出物を、Jak、StatおよびIrsファミリーの異なるメンバーに特異的な種々の抗体で免疫沈降させた。まず免疫ブロットを抗ホスホチロシン抗体と反応させて、チロシン−リン酸化タンパク質のみを検出し、次にストリップし、各タンパク質を免疫沈降させるのに使用したものと同じ抗体で再度釣り上げた。GH9、ΔQGR8、ΔQGH9は、図16に示す通りである。
【図16】ヒトIL-9受容体の異なる型を発現するTS1細胞の増殖。細胞をそれぞれ4重測定できるように96ウェルプレート(1000/ウェル)に接種し、サイトカイン無し、マウスIL-9有りまたはヒトIL-9有り(5ng/ml)で処理した。7日後に比色アッセイを行って細胞数を測定し、処理/未処理細胞の比(対照%)を算出して、増殖速度を評価した。LxSN=空ベクターでトランスフェクトした細胞;GR8は野生型IL-9R;GH9は、コドン410で開始する9個のSer残基を有するHis344変異体である;ΔQGR8およびΔQGH9は、それぞれ野生型およびHis344+9-Serバックグランド上のΔQ173変異体である。
【図17】エキソン6から213nt上流の核酸で変異を有するIL-9Rのイントロン5のゲノムDNA配列(ここで、T残基は、矢印で示すようにC残基に変化している)。
【発明を実施するための形態】
【0035】
(発明の詳細な説明)
本出願人らは、IL-9経路と、治療、診断、およびアトピー性喘息と関連疾患を予防または治療するための薬剤の同定方法の開発に使用されるこの経路に影響を与える組成物を解明することにより当該分野のニーズを解決した。
【0036】
喘息は、可逆的な空気流遮断を伴う気道の炎症性障害を含む。アトピー性アレルギーとは、アトピーと関連疾患(喘息、気管支過応答性(BHR)、鼻炎、じん麻疹、腸のアレルギー性炎症性疾患、および種々の型の湿疹を含む)を意味する。アトピーは、環境的アレルゲンに対する過敏症であり、対照と比較して血清総IgEの上昇、またはアレルゲンに対する異常皮膚試験応答として表される。BHRとは、種々の刺激に対する気管支収縮応答の上昇である、気管支過応答性を意味する。
【0037】
アトピー性アレルギーおよび喘息関連疾患を示す個体のDNAを解析することにより、本出願人らは、喘息の発現と相関しうるIL-9受容体(IL-9R)遺伝子中に多型を同定した。IL-9受容体遺伝子(喘息関連因子2またはAAF2としても知られている)は、ヒト骨髄系およびリンパ系の制御を含む多様な機能に関連するサイトカイン受容体である、インターロイキン−9受容体の遺伝子座を意味する。本発明のヒトIL-9受容体遺伝子は、XY染色体の次端部テロメア領域中に見いだされる。
【0038】
本出願人らは、多型とは、通常の配列からの特定DNA配列(「遺伝子座」と呼ばれる)の変化を意味する。一般に、ある遺伝子座を含む2つまたはそれ以上の対立遺伝子が当業者により同定され、少なくとも共通の対立遺伝子が1%またはそれ以上の頻度で存在する時、その遺伝子座を多型であると定義する。
【0039】
IL-9Rは,ヒトゲノムの他の遺伝子座(第9,第10、第16、第18染色体)に、4つの高度に相同性(>90%のヌクレオチド同一性)の非プロセシング偽遺伝子を有するため、標準的プライマーデザインを使用して真のIL-9R(XYq偽常染色体性領域中に位置する、生物学的に機能性のタンパク質をコードする遺伝子)を特異的に増幅することは不可能であった。これらの他の遺伝子の高い同一性のために、標準的プライマーデザインを使用するゲノムPCR増幅は、すべての遺伝子の同時増幅を引き起こし、真の遺伝子の配列解析は不明確なものになる。真のIL-9R構造は、本出願に記載の喘息または癌のような他の疾患(Renauldら、Oncogene, 9:1327-1332, 1994;Grussら、Cancer Res., 52:1026-1031, 1992)に対する素因に関係する可能性があるので、真のIL-9R構造を研究するために、本出願人らは、特異的アンプリマー(amplimer)をデザインした。この特異的プライマーは、4つの偽遺伝子の増幅が無く、IL-9R増幅について信頼できることがわかった。プライマー配列を実施例2に示し、その特異性を図13で証明する。
【0040】
本出願人らはまた、50人のドナーから精製したPBMC(末梢血単核細胞)から抽出したRNAを使用して、IL-9受容体cDNAの全コード領域をRT-PCRにより増幅した。図1は、解析した50人中に見いだされた最も高頻度の変異を示す。エキソン3、4、5、6および8は、完全長cDNAもクローン化できる試料中で、異常スプライシング事象により影響を受けた。一部の転写産物は、エキソン3の完全な欠失を示し、これはフレームシフトを引き起こして、79個の無関係の残基が続いた後に停止コドンを生成する。エキソン4の欠失の場合も,フレームシフトが生じ、エキソン5中の最初のコドンが停止コドンに変換された。一部の他のcDNAでは、エキソン5は最初の5または29ヌクレオチドの部分的欠失を示し、この両方の欠失はフレームシフトを引き起こして、エキソン5内に早い停止コドンを形成した。従ってすべての場合に、推定される末端切断型タンパク質は、細胞外ドメインの大部分ならびに膜貫通および細胞質ドメインの全部を欠失するであろう。もし分泌されるなら、これらの型は可溶性受容体として機能するかも知れない。最後に、コドン173に対応するエキソン6の最初の3個のヌクレオチドは、しばしばスプライシングにより除去され、結果的にこのコドン位置のグルタミンが欠失し、残りのタンパク質配列には他の変異はなかった。このスプライス変異体は、おそらくスプライス変異体の頻度を増加させるゲノムDNAのイントロン5中に見いだされる変異体(配列番号24)に関連する(図17と実施例12)。
【0041】
本出願人らはまた、エキソン9のコード配列に限定される対立遺伝子変異を見いだした。コドン310と410に関係している多型は、すでに開示されている29,30(Kermouni, A.ら、Genomics, 371-382 (1995))。コドン310は,そのコドンの最初のヌクレオチドがアデニンであるかまたはグアニジンであるかに応じて、それぞれアルギニンまたはグリシンをコードする。コドン410(このコドンから「410+n」と呼ぶ)では、それぞれ8個または9個のセリンに翻訳されるであろう8個または9個の一続きのAGCトリヌクレオチド反復配列が開始する。
【0042】
本出願人らは、コドン344に新しい多型を見つけた。ここで2番目のヌクレオチドはアデニンかまたはグアニジンであり、このコドンによりコードされる可能な2つの残基はそれぞれヒスチジンかまたはアルギニンである。さらにコドン344と410+nの間に対応関係が観察され、コドン344のアルギニンが、いつもコドン410+nの8個のセリンとともに存在し、コドン344のヒスチジンは9個のセリンとともに見いだされる。もともとヒト巨核芽球白血病細胞系Mo7eからクローン化されたヒトIL-9受容体cDNAは、コドン410+nに9個のセリンを示し、かつ本出願人らのクローンと異なり、コドン344にアルギニンを示す29。別の巨核芽球白血病細胞系であるUT-7は、同じアルギニン/9−セリン対立遺伝子を有すると報告されている30。本出願人らは、Mo7e細胞系から16のcDNAをクローン化し、6個のクローンはコドン410+nに8個のセリンを有し、コドン344にアルギニンを有することを見いだした。残りの10個のクローンは、公表されている配列を示した。本出願人らはまた、ヒト急性骨髄性白血病細胞系KG-1をゲノタイピングし、これはヒスチジン/9−セリンホモ接合体であることを見つけた。
【0043】
これらのDNA分子と対応するRNAは、例えばSambrookら、Molecular Clonig:A Laboratory Manual,第2版、Cold Spring HarborLaboratory Press (1985)のような当該分野の標準的な方法を使用して単離される。本出願人らは、単離されるとは、DNAに、自然環境中に存在する核酸またはポリペプチドと結合している汚染物質の少なくとも1部が存在しないことを意味する。
【0044】
本発明また、これらのヌクレオチド配列によりコードされるタンパク質を含む。本発明はまた、この分子の断片を含む。本出願人らは、断片とは、全配列の機能を維持する核酸配列の部分であることを意味する。当該分野で公知なように、断片は、欠失、付加、置換および/または修飾から得られる。
【0045】
本発明のIL-9受容体変異体の供給源は、ヒトである。あるいはDNAまたはその断片は、当該分野で公知の方法により合成してもよい。また遺伝子操作技術により、任意の受け入れられている遺伝子操作技術によりDNAを作製して、DNAを発現ビヒクル中にクローニングして、化合物を発現する細胞にビヒクルをトランスフェクトして、化合物を産生することができる。例えば、Sambrookら、Molecular Cloning:A Laboratory Manual,第2版、Cold Spring HarborLaboratory Press (1985)に記載の方法を参照されたい。
【0046】
アトピー性アレルギーおよび喘息様表現型に関連し得る変異体IL-9受容体配列,および喘息様表現型の欠如と関連し得る他の配列の証明は、アトピー性喘息および関連疾患に対する感受性を診断する方法を提供する。ある変異体は、これらの疾患を治療するのに使用できる可溶性受容体を産生することができる。
【0047】
受容体は、分子を認識しこれに結合する可溶性または膜結合成分であり、本発明のIL-9受容体は、IL-9を認識しこれに結合する成分である。IL-9受容体の機能は、IL-9またはIL-9様分子に結合し、特異細胞中でその制御シグナルを伝搬することからなる29,30,34,35。ヒトIL-9は、ヒトIL-9受容体に結合した時、IL-9受容体自身のリン酸化とJak-Stat経路のタンパク質(Jak1、Stat1、Stat3、Stat5、およびIrs2)の活性化を引き起こすことが証明されている(Demoulin, J-B.ら、Molecular and Cellular Biology,p. 4710-4716, Sept.1996)。本出願人らは、IL-9Rまたはその変異体が,これらのタンパク質の活性化においてバイアスを示すかどうかを調べ、この解析をJak3とIrs1に拡大した。Jak3とIrs1を含む経路のすべてのタンパク質は、IL-9R活性化によりリン酸化されることが確認された。また、コドン310、344および410+nで変異を有するIL-9R変異体は、野生型IL-9Rと同じアップレギュレーションを提供することも確認された。従って、本発明の1つの側面は、Jak-Stat経路中の相互作用を阻害するアトピー性喘息の治療用の治療薬である。
【0048】
野生型受容体および他の試験した変異体と異なり、ΔQ173変異体は、Jak-Stat経路中のいかなるタンパク質も活性化することができなかった(図15)。さらにΔQ173変異体は、IL-9刺激により細胞の増殖を援助することができなかった(図16)。従ってΔQ173変異体を発現する個体は、アトピー性喘息や関連疾患に対して感受性である可能性は低い。従って本発明の1つの側面は、アトピー性喘息および関連疾患の治療のためのΔQ173スプライス変異体の発現を増加させる治療薬である。
【0049】
1つの診断的態様は、IL-9受容体遺伝子または転写産物のDNA配列中の変異の認識である。1つの方法は、当業者により理解されるように、充分なハイブリダイゼーション条件下で、本発明のIL-9受容体に相補的な配列を有する核酸分子(プローブとしても知られている)を使用することである。1つの態様において、この核酸分子は、成熟IL-9受容体タンパク質のArg344,またはHis344のコドンに特異的に結合し、別の実施態様では、Arg344とHis344の両方に結合するであろう。さらに別の実施態様では、これはGln173のコドンに結合するであろう。これらの方法は、IL-9受容体の他の変異体を認識するのに使用してもよい。これらの疾患に関連したDNA配列変異を認識する別の方法は、当該分野で公知の複数の方法による直接DNA配列解析である44。別の実施態様は、これらの疾患に関連するIL-9受容体遺伝子のDNA配列変異の検出である40ー44。これらは,ポリメラーゼチェイン反応、制限断片長多型(RFLP)解析、および1本鎖コンフォメーション解析を含む。好適な実施態様において、本出願人らは特に、ASO PCRを使用してHis344対立遺伝子およびArg344対立遺伝子に関連するIL-9受容体中の多型を、遺伝子レベルで認識する方法を具体的に提供する。他の実施態様において、IL-9受容体遺伝子のこれらの対立遺伝子を区別するために、結合チェイン反応を使用することができる。
【0050】
本発明はまた、IL-9およびその受容体のアンタゴニストの同定のための方法を含む。アンタゴニストは、それ自身は薬理活性が欠如しているが、アゴニストの作用を妨害することにより影響を引き起こす化合物である。本発明のアンタゴニストを同定するために、本明細書および引用文献に記載のように,既知のアゴニストとの競合的結合、またはIL-9様機能のダウンレギュレーションについて試験してもよい2,22-35。
【0051】
Tリンパ球の増殖、IgE合成、および肥満細胞からの放出についてのIL-9受容体の既知の役割に基づいてアトピー性アレルギーを治療するのに有用な薬剤についてスクリーニングするのに、具体的な測定法が使用される29,30,33-35。別の測定法は、Mo7e細胞中の複数のタンパク質の急速および一過性のチロシンリン酸化を特異的に誘導するための、ヒトIL-9受容体の能力に関する34。この応答はIL-9受容体の発現と活性化に依存するため、これは、有用な可能性のある化合物の性状解析のための簡便な方法または測定法である。Stat3転写因子のチロシンリン酸化は、IL-9受容体の機能に特異的に関連しているようであり35、この応答は、本発明の範囲内の化合物の性状解析のための簡便な方法または測定法を示す。IL-9受容体の機能を性状解析するためのさらに別の方法は、細胞増殖測定法を用いてヒトIL-9機能を評価するのに使用することができる,ヒト受容体でトランスフェクトされる既知のマウスTS1クローンの使用を含む29。これらの方法は、IL-9受容体のアンタゴニストを同定するために使用することができる。
【0052】
さらなる実施態様において、本発明は、IL-9に結合する可溶性IL-9受容体分子を投与することによる、IL-9発現または機能のダウンレギュレーションを包含する。本出願人らおよびRenauldら29は、可溶性型のIL-9受容体の存在を証明した。この分子は、細胞結合受容体へのIL-9の結合を妨害するのに使用でき、IL-9のアンタゴニストとして作用する。可溶性受容体は、サイトカインまたは他のリガンドに結合してそれらの機能を制御するのに使用されている45。可溶性受容体は、溶液で存在するかまたは膜の外に存在する、膜結合受容体の型である。通常膜に会合している分子のセグメントが存在しないため、可溶性受容体が存在するかも知れない。このセグメントは当該分野では通常、遺伝子の膜貫通ドメイン、またはタンパク質の膜結合セグメントと呼ばれる。すなわち、本発明の1つの実施態様において、可溶性受容体は,膜結合受容体の断片または類似体である。
【0053】
本出願人らは、可溶性受容体として作用することができるタンパク質の産生を引き起こす、ヒトIL-9受容体の3つのスプライス変異体を同定した。1つのスプライス変異体は、エキソン4を欠失させ、これはエキソン5の最初のコドンとして停止コドンとなるフレームシフトを導入した。この変異体は、IL-9/IL-9R相互作用を阻止する抗体と反応性のエピトープを含有する約45残基のペプチドを産生する。他の2つの変異体は、エキソン5に欠失を有し、これはエキソンの初期に未成熟な停止コドンを作成するが、これらの場合エキソン4の欠失はないであろう。これらの変異体は、阻止抗体により認識されるエピトープも含有する約100残基のタンパク質を産生するであろう。
【0054】
可溶性IL-9受容体は、アトピー性喘息および関連疾患を治療するのに有用なアンタゴニストを含む治療薬候補をスクリーニングするのに使用してもよい。例えば、ファージディスプレイを使用するペプチドと1本鎖抗体のスクリーニングは、可溶性受容体を使用して促進することができるであろう。可溶性受容体に結合するファージは単離することができ、この分子は受容体と結合ファージの親和性捕捉により同定することができる。さらに、アトピー性喘息および関連疾患の治療に有用な物質のための化合物のスクリーニングは、可溶性受容体およびアンタゴニストの非存在下で結合するリガンドを取り込むことができる。これらの分子が近傍にあるため、リガンドと受容体相互作用の検出が起きる。アンタゴニストは、これらの相互作用を阻止することにより認識される。
【0055】
さらに本発明は、製薬上許容される担体とともに本発明の化合物を含む医薬組成物を含む。製薬上許容される担体は、無菌の液体、例えば石油、動物、植物または合成起源のものを含む水および油,例えばピーナツ油、大豆油、鉱物油、ゴマ油などがある。医薬組成物を静脈内投与する時は、水が好適な担体である。食塩水溶液ならびにデキストロース水溶液およびグリセロール水溶液もまた、液体担体、特に注射溶液として使用することができる。適当な医薬担体は、Martin, E.W., Remington's Pharmaceutical Sciences(これは特に、参照により本明細書に組み込まれる)に記載されている。
【0056】
本発明の治療法で使用される化合物は、治療される症状、部位特異的治療の必要性、投与される薬剤の量、および同様の事柄などを考慮して、全身性または局所的に投与してもよい。
【0057】
局所投与を使用してもよい。任意の一般的な局所製剤(例えば、液剤、懸濁剤、ゲル剤、軟膏剤、または膏薬など)を使用してもよい。このような局所製剤の調製は、例えばRemington's Pharmaceutical Science、第17版、Mack Publishing Company, Easton,Pa.により例示されるように、医薬製剤の分野で詳しく記載されている。局所投与には、これらの化合物はまた、散剤またはスプレー剤(特にエアゾル型)として投与することができる。好適な実施態様において、本発明の化合物は吸入により投与してもよい。吸入治療法のために、化合物は、用量目盛り付き吸入器、または乾燥粉末吸入器に適した型で投与するのに有用な溶液でもよい。
【0058】
活性成分は、全身性投与に適した医薬組成物で投与される。公知のように、薬剤は、全身投与されるなら、経口投与用に散剤、丸剤、錠剤など、またはシロップ剤もしくはエリキシル剤として調製してもよい。静脈内投与、腹腔内投与または病変内局注投与のためには、化合物は,注射による投与が可能な液剤または懸濁剤として調製される。ある場合には、これらの化合物を坐剤として、または皮下沈降のための長期放出製剤として、または筋肉内注射のための製剤で調製することが有用である。
【0059】
有効な量は、IL-9受容体により調節される機能をダウンレギュレートする量である。具体的な有効量は、症状毎に変化し、ある場合には治療される症状の重篤度および治療に対する患者の感受性により変化する。従って特定の有効量は、常套の実験により、その時間と場所で決定することが最適である。しかし本発明の喘息および関連疾患の治療において、0.001〜5重量%,好ましくは約0.01〜1%を含有する製剤が,通常治療上有効な量であろう。全身投与される時、0.01〜100mg/kg体重/日,しかし好ましくは約0.1〜10mg/kgの量が、多くの場合治療結果に影響を与えるであろう。
【0060】
本出願人らはまた、IL-9受容体により制御される機能をダウンレギュレートする化合物をスクリーニングする方法を提供する。IL-9受容体により発現される機能がダウンレギュレートされたかどうかは、当該分野で標準的な方法を使用して確認してもよい29,30,34,35。具体的な実施態様において、本出願人らは、IL-9に匹敵する機能を有する化合物の同定法を提供する。1つの実施態様において、IL-9受容体の機能は、in vitroで評価される。当業者に公知のように、ヒトIL-9受容体の活性化は、IL-9に応答性のある細胞中の複数のタンパク質の急速かつ一過性のチロシンリン酸化を特異的に誘導する。Stat3転写因子のチロシンリン酸化は、IL-9経路の作用と特異的に関連しているようである。IL-9およびIL-9様分子の機能を特徴付けるための別の方法は、マウスTS1クローンまたはTF1クローン中のIL-9受容体の「安定な発現」に依存し、これは通常ヒト受容体を発現しない。これらのトランスフェクション体は、細胞増殖アッセイを使用して、ヒトIL-9受容体機能を評価するのに使用することができる29。
【0061】
本発明はまた、IL-9受容体変異体を発現する細胞系に基づく飽和性かつ特異的リガンド結合の簡便なスクリーニングアッセイも包含する23,29。IL-9受容体は、広範囲にわたる細胞型(K562、C8166-45、ヒトIL-9受容体でトランスフェクトしたKG-1、B細胞、T細胞、肥満細胞、HL60、HL60-クローン5、ヒトIL-9受容体でトランスフェクトしたTS1、ヒトIL-9受容体でトランスフェクトした32D、好中球、巨核球(UT-7細胞)30、ヒト巨核芽球白血病細胞系Mo7e34、TF129、マクロファージ、好酸球、胎児胸腺細胞、ヒト腎細胞系29330、ならびにマウス32Dおよび胚海馬始原細胞系23,29,30を含む)で発現される。
【0062】
本発明の実施においては、当業者の通常の技術範囲内である、分子生物学,薬学、免疫学、および生化学の通常の用語および技術を使用する。例えば、Sambrookら、Molecular Cloning: A Laboratory Manual,第2版、Cold Spring HarborLaboratory Press、またはAusubelら、Current Protocols in Molecular Biology,John Wiley &Sons, Inc.を参照されたい。
【0063】
なお、我々は、以下の基礎的バックグランド情報を提供する。体の遺伝物質またはDNAは、それぞれが動原体によりつながれた2つのアームを含む46の染色体上に並んでいる。各染色体は、pまたはqと呼ぶセグメントに分かれる。記号pは、動原体から最も近いテロメアまで測定した時に短い方の染色体アームを示す。染色体の長い方のアームは、記号qで示す。染色体上の位置は、染色体の番号(すなわち、染色体5)ならびにpまたはq領域の座標(すなわち、q31-q33)で示される。さらに体には、性染色体XとYがある。減数分裂の間、XおよびY染色体は、偽常染色体領域として知られている領域中でDNA配列情報を交換する。
【0064】
DNA(デオキシリボ核酸)は、ヌクレオチドの2本の相補的な鎖からなり、これらは4つの異なる塩基化合物、アデニン(A)、チミン(T)、シトシン(C)、およびグアニン(G)を含む。一方の鎖のAは他方の鎖のTと結合し、一方の鎖のCは他方の鎖のGと結合して、相補的な「塩基対」を形成し、各対は1本の鎖に1つの塩基を有する。
【0065】
3つのヌクレオチドの連続した群(「コドン」)が、1つのアミノ酸をコードする。例えば、3つのヌクレオチドCAGはアミノ酸グルタミンをコードする。20個の天然に存在するアミノ酸とその1文字コードは以下の通りである:
アラニン Ala A
アルギニン Arg R
アスパラギン Asn N
アスパラギン酸 Asp D
アスパラギンまたは
アスパラギン酸 Asx B
システイン Cys C
グルタミン Gln Q
グルタミン酸 Glu E
グルタミンまたは
グルタミン酸 Glx Z
グリシン Gly G
ヒスチジン His H
イソロイシン Ile I
ロイシン Leu L
リジン Lys K
メチオニン Met M
フェニルアラニン Phe F
プロリン Pro P
セリン Ser S
スレオニン Thr T
トリプトファン Trp W
チロシン Tyr Y
バリン Val V
アミノ酸はタンパク質を構成する。アミノ酸は、親水性(すなわち、水に対する親和性を示す)でもまたは疎水性(すなわち、水を嫌う)でもよい。すなわち、G、A、V、L、I、P、F、Y、W、C、およびMで示されるアミノ酸は疎水性であり、S、Q、K、R、H、D、E、N、およびTで示されるアミノ酸は親水性である。一般にアミノ酸の親水性または疎水性の性質は、ペプチド鎖の折り畳みに影響を与え,従ってタンパク質の3次元構造に影響を与える。
【0066】
DNAは以下のようにタンパク質と関係している:
ゲノムDNA → mRNA → タンパク質
↑
↓
cDNA
ゲノムDNAは、生物の細胞中に存在するすべてのDNA配列を含む。これは、メッセンジャーRNA(「mRNA」)に「転写」される。相補的DNA(「cDNA」)は、mRNAの逆転写により作成される、mRNAの相補的なコピーである。ゲノムDNAと異なり、mRNAとcDNAは両方とも、DNAのタンパク質コード領域またはポリペプチドコード領域(いわゆる「エキソン」)のみを含有する。ゲノムDNAはまた、「イントロン」を含有し、これはタンパク質をコードしない。
【0067】
実際、真核生物遺伝子は、それによりコードされるタンパク質に対して断続的であり、イントロンにより中断されたエキソンからなる。RNAに転写された後、イントロンはスプライシングにより除去されて、成熟メッセンジャーRNA(mRNA)が形成される。エキソンの間のスプライス点は、典型的にはスプライシングプロセスのシグナルとして作用する共通配列により決定される。スプライシングは、一次RNA転写産物からのイントロンの削除と、切断されたイントロンのいずれかの側にある残りのRNAの末端の結合もしくは融合からなる。イントロンの有無、イントロンの組成、および遺伝子当たりのイントロンの数は、同じ種の株の間でも異なり、同じ基本的な機能遺伝子を有する種の間でも異なることがある。ほとんどの場合にイントロンは必須ではなく害はないと考えられているが、この分類は必ずしも絶対的ではない。例えば1つの遺伝子のイントロンは、別のエキソンであることもある。ある場合には、スプライシングの代替的なまたは異なるパターンが、DNAの同じ1つのストレッチから、異なるタンパク質を形成することがある。実際、イントロンの構造的特徴と基礎的スプライシング機構は、異なる種類のイントロンの分類の基礎となる。
【0068】
エキソンについて、これらは、別個のドメインまたはモチーフ、例えばタンパク質の機能性ドメイン、折り畳みドメイン、もしくは構造的エレメント;または短いポリペプチド配列、例えば逆ターン、ループ、グリコシル化シグナルおよび他のシグナル配列、または非構造ポリペプチド、リンカー領域、に対応する。本組合せ法のエキソンモジュールは、天然に存在するエキソン配列または突然変異(例えば、点突然変異、末端切断、融合)を受けている天然に存在するエキソン配列に対応する核酸配列を含有してもよい。
【0069】
DNAの操作にもどると、DNAは、ある既知の部位でDNAを切断する「制限酵素」およびDNAをつなぐDNAリガーゼを使用して、切断、スプライス、および他の操作をすることができる。このような技術は当業者に公知であり、例えばSambrookら、Molecular Cloning:A Laboratory Manual,第2版、Cold Spring HarborLaboratory Press (1985)、またはAusubelら、Current Protocols in Molecular Biology,John Wiley &Sons, Inc. (1994)に記載されている。
【0070】
特定のサイズおよび配列のDNAは、次に「レプリコン」(これは、プラスミド、コスミド,またはウイルスのような、それ自身の制御下で複製ができる遺伝成分である)中に挿入することができる。「組換えベクター」または「発現ベクター」は、DNAセグメントが挿入されるレプリコンであり、DNAの発現(すなわち、DNAによりコードされるタンパク質の産生)を可能にする。発現ベクターは、実験室で作製されるか、他の実験室から入手できるか、または市販品を購入することができる。
【0071】
組換えベクター(当該分野において種々の名前で知られている)は、「形質転換」として一般的に知られているプロセスにより宿主中に導入される。形質転換は、任意の方法(感染、直接摂取、形質導入、F−接合、微量注入法、または電気穿孔法)により、宿主細胞中に外来DNAセグメントを移動することを意味する。
【0072】
単細胞宿主細胞(組換え宿主細胞、細胞および細胞培養物として種々の名前で知られている)は、細菌、酵母、昆虫細胞、植物細胞、哺乳動物細胞、およびヒト細胞を含む。特に好適な実施態様において、宿主細胞は,大腸菌(E. coli )、シュードモナス(Pseudomonas)、バシラス(Bacillus),ストレプトミセス(Streptomyces),酵母、CHO、R1-1、B-W、LH、COS-J、COS-7、BSC1、BSC40、BMT10、およびS69細胞を含む。酵母細胞は特に、サッカロミセス(Saccharomyces),ピキア(Pichia)、カンジダ(Candida)、ハンゼヌラ(Hansenula)、およびトルロプシス(Torulopsis)を含む。
【0073】
当業者は認識できるように、宿主細胞によるDNAセグメントの発現は、適切な制御配列または成分が必要である。制御配列は、使用される宿主細胞により変化するが、例えば原核生物では、プロモーター、リボゾーム結合部位、および/または転写停止部位がある。真核生物では、このような制御配列には、プロモーターおよび/または転写停止部位がある。当業者は認識できるように、これらの制御配列を注意深く選択し配置することにより、ポリペプチドの発現を増強、すなわち標準的なレベルより上昇させることができる。
【0074】
他の実施態様において、使用されるプロモーターには、ヒトサイトメガロウイルス(CMV)プロモーター、テトラサイクリン誘導性プロモーター、サルウイルス(SV40)プロモーター、モロニーマウス白血病ウイルス長い末端反復配列(LTR)プロモーター、糖質コルチコイド誘導性マウス乳癌ウイルス(MMTV)プロモーター、ヘルペスチミジンキナーゼプロモーター、マウスおよびヒトアクチンプロモーター、HTLV1およびHIV IL-9 5'フランキング領域、ヒトおよびマウスIL-9受容体5'フランキング領域、細菌tacプロモーターおよびキイロショウジョウバエ(Drosophila)熱ショックタンパク質足場結合領域(SAR)エンハンサー成分がある。
【0075】
DNAは、ペプチド、オリゴペプチド、およびタンパク質のようなある長さのポリペプチドとして発現される。ポリペプチドはまた、グリコシル化、アセチル化、リン酸化などの翻訳的修飾も含む。
【0076】
本発明に関係のある他の分子生物学的方法は、「連鎖解析」である。連鎖解析は、形質または疾患と相関する染色体または染色体領域を同定するための分析方法である47。染色体は、その上に遺伝子が組み立てられる遺伝の基本的単位である。遺伝子以外に、当業者は、染色体上に「DNAマーカー」を証明している。DNAマーカーは、その同定と配列決定が容易に行なうことができる、DNAの既知の配列である。連鎖解析法は、疾患遺伝子(例えば喘息の感受性に関係する遺伝子)を特定の染色体にマッピングすることに応用されている47,48。
【0077】
本出願人らは、上記および下記のすべての文献を参照により本明細書に組み込む。
【0078】
本明細書の説明および開示された発明の実施から、本発明の他の実施態様は当業者には明らかであろう。明細書および実施例は単に例示のためであり、本発明の範囲は請求の範囲によってのみ示されるものである。
【0079】
背景情報は提供したので、本出願人らは、ここで本発明の好適な面を説明する。
【実施例】
【0080】
(実施例1:IL-9受容体転写産物多型の同定)
Philadelphia, Pennsylvania 地域からの52人の集団を、喘息とアトピーに関してランダムに確認した。総血清IgEを、酵素結合免疫吸着アッセイ(ELISA、Genzyme、Cambridge、Massachusetts)により測定した。
【0081】
ヒトIL-9受容体cDNAの構造型を評価するために、これらの52人の無関係のドナーからPBMCを単離し、PHAとPMAの存在下で培養した(実施例4に記載)。本出願人らの実験室の以前のデータは、分裂促進物質による刺激後6日目に1次培養物中のIL-9受容体メッセージの発現の動力学を証明した。従って本出願人らは、細胞を6日間培養し、この時点で細胞を採取し,実施例5に記載のようにそのRNAとDNAを単離した。
【0082】
RNAを逆転写し、実施例5に記載のように完全長IL-9受容体cDNAに特異的なプライマーを使用してPCRにより増幅した。各個人からの増幅産物をTA PCRクローニングベクターにクローン化し、予測されるインサートを含有する10クローン(消化とゲル電気泳動により測定した)を完全に配列決定し、構造または配列の変化について解析した。
【0083】
上記スクリーニングから7個の主要な変異体を同定した。これらのcDNAは、コドン173の欠失、エキソン4の欠失、エキソン5の2つの別の欠失、エキソン8の欠失、および成熟タンパク質のコドン344でARGからHISへの変異を有する完全長cDNAを示す。これらの遺伝子バックグランドのそれぞれに追加の変異体が存在する。Arg対立遺伝子は、8 Ser/4Asn繰り返しと7 Ser/4 Asn繰り返しに関連し、His対立遺伝子は、9 Ser/4Asn繰り返し、9 Ser/3 Asn繰り返し、および10 Ser/2 Asnに関連する。これらの変異体のすべてを、図1に記載する。
【0084】
変異体を、クローン化cDNAの発現を指令するCMVプロモーターとその後のSV40ポリアデニル化シグナルを含有する真核生物発現ベクターpCEP4(Clontech)中にクローン化した。このベクターはまた、ベクターを含有し、おそらく、クローン化したcDNAをCMVプロモーターの制御下で発現する真核生物細胞の選択に使用される、ヒグロマイシンB耐性遺伝子を含有する。組換えプラスミドを、配列により解析し、正しいcDNAインサートを含有するプラスミドを、真核生物受容体細胞(例えば、実施例3に記載のシリアンハムスター繊維芽細胞TK-ts13、ヒト神経膠芽細胞腫T98G、ヒト骨髄白血病株TF-1、およびマウス骨髄前駆体細胞系32D)にトランスフェクトした。増殖とアポトーシスにおける、IL-9リガンドに対する応答として、機能を生物学的に評価した(実施例7と10)。
【0085】
ARGおよびHIS変異体を含有する完全長IL-9受容体cDNAを行なった実験は、TK-ts13ハムスター繊維芽細胞またはヒトT98G神経膠芽細胞腫細胞である。細胞をトランスフェクトし、48時間後ヒト特異的カルボキシ末端抗体を使用して、ウェスタンブロットとin situ染色により解析した(Santa Cruz)(実施例8)。in situ解析は、ハムスターとヒト系の両方で,両方の型の受容体が発現されるらしいことを証明した(図11と図12)。興味深いことに、ヒトとハムスター系で両方の型のウェスタンブロットは同等レベルで発現され、TS1細胞系中でARGとHIS受容体型との間には移動パターンに差があり(図12)、これは、グリコシル化、リン酸化などの翻訳後修飾の差を示唆する。この生化学的な差は、機能の変化が表現型の変化を引き起こす機構かも知れない。
【0086】
種々の置換の頻度を、一般的集団中の各変異体の普及率の、偏りのない推定値として使用した。アンケートにより遺伝子型を表現型と比較した。アレルギーと喘息の診断は、アンケートを再検討して、医師が行なった。Arg344対立遺伝子についてホモ接合性である個人は、ヘテロ接合性またはホモ接合性His344と比較した時、アレルギーと喘息の証拠を示す可能性が有意に低かった(図9)。
【0087】
(実施例2:IL-9受容体遺伝子のゲノム解析)
アレルギーおよび/または喘息の人のゲノム解析を行うために、X/Y偽常染色体領域上に位置する真のIL-9受容体遺伝子についてPCR特異的プライマーを作成し、かつ染色体9,10、16、および18上に位置する高度に保存されたIL-9受容体偽遺伝子を排除するように,以下の方策を設計した。まず、IL-9受容体遺伝子の2つの公表された偽遺伝子とゲノム配列の間で、配列アライメントを行なった。次に,真の遺伝子と偽遺伝子の間の分岐領域でプライマーを設計し、次にCoreill DNA Repository (Camden,NJ)から入手した単一の染色体特異的ハイブリッドを使用して、PCRにより解析した。プライマーが、XおよびYハイブリッドから正しいサイズの生成物のみを産生するなら、これらをしっかりした増幅について最適化した。いくつかの場合に、分岐領域に向けられたプライマーはXY特異的ではなく、従って本出願人らは、IL-9受容体遺伝子配列に比較して偽遺伝子に対してミスマッチの数を多くするために、特定のプライマー中の追加の塩基の変異を導入した。表1は、XY特異的増幅についてのプライマーの配列と最適アニーリング温度を含む。XY増幅についてこれらのプライマーの特異性を、図13に示す。
表1:IL-9受容体のX/Y特異的アンプリマー
【0088】
【表1】
これらのプライマーは、アトピー性喘息および関連疾患に対する感受性と耐性の診断に使用可能な、これらの遺伝子中のDNA配列の変異を解析するための新規の方法を示す。
【0089】
この技術を使用して、IL-9受容体遺伝子中の配列の変異を検出するために、本出願人らの集団中の人についてDNA配列解析により、各エキソンを調べた44。解析を繰り返して、配列多型をアーチファクトから区別した。受容体変異体とアレルギー性表現型との関連を図9に示す。受容体対立遺伝子の配列を図1〜8に示す。
【0090】
(実施例3:IL-9受容体発現とリガンド結合アッセイ)
市販の技術を使用してサプライヤー(Pierce)の勧めに従って、精製した組換えIL-9および構造または機能がIL-9に似ている可能性のある化合物を、高比活性になるように蛍光標識した。ヒトMo7eとマウス32D細胞を増殖させ、37℃で、それぞれ10%(容量/容量)ウシ胎児血清、50mM 2−メルカプトエタノール、0.55mM L-アルギニン、0.24mM L-アスパラギン、および1.25mM L-グルタミンを補足したダルベッコ改変イーグル培地0.8ml、または10%(容量/容量)ウシ胎児血清、50mM 2−メルカプトエタノール、0.55mM L-アルギニン、0.24mM L-アスパラギン,および1.25mM L-グルタミンを補足したRPMIに再懸濁した。TF1.1(ヒトIL-9受容体を欠失したTF1細胞)、T98G、TK-、およびマウス32D細胞(実施例7と10)を、後述のようにそのまま、またはヒトIL-9受容体構築物でトランスフェクション後に使用した。完全長または末端切断型のIL-9受容体の1つを含有するプラスミドDNAを、pCEP4プラスミド(Clontech)中にクローン化し、Qiagenカラム(Qiagen)を用いて遠心分離により精製した。電気穿孔法の直前に0.4cmのキュベット中の細胞にプラスミドDNA(50μg)を加えた。2重電気パルス(750V/74オーム/40マイクロファラドと100V/74オーム/2100マイクロファラド)後に、直ちに細胞を、IL-9を補足した新鮮な培地に希釈した。
【0091】
ヒグロマイシンB(400μg/ml〜1.6mg/ml)で14日間選択した後に、安定なトランスフェクション細胞が得られた。ヒグロマイシン耐性クローンを、実施例8に記載のようにウェスタンブロットとin situ染色によりIL-9受容体の発現について分析した。
【0092】
蛍光顕微鏡でリアルタイムに、細胞受容体の結合を直接視覚化する。対照細胞(トランスフェクトしてない)と既知のIL-9受容体変異体のそれぞれでトランスフェクトした細胞で、結合とインターナリゼーションを経時的に追跡する。特異的結合を証明するために、過剰の非標識リガンドまたはブロッキング抗体を並行実験で使用する。
【0093】
HAジタグを有するかまたは有さない、アミノ酸44〜270を含む可溶性IL-9受容体をまた、異なる型のヒト標識組換えIL-9とともにインキュベートする。異なる量のFLAGタグ付き(IBI)リガンドを0.5μgの可溶性受容体とともに、PBS中で室温で30分間インキュベートする。1μgの抗HA抗体または抗FLAGモノクローナル抗体(IBI)とともに、EBCバッファー(50mMトリス、pH7.5;0.1M NaCl;0.5% NP40)を加え(300μl),氷上で1時間インキュベートする。各試料に40μlのプロテインAセファロース溶液を加え、4℃で1時間混合した。試料を11,000×Gで1分間遠心分離し、ペレットを500μlのEBCで4回洗浄した。ペレットを26μlの2×SDSバッファーに溶解し、4分間沸騰させ、18%SDSポリアクリルアミドゲルで電気泳動した。ウェスタンブロットを、抗IL-9受容体抗体(Santa Cruz Inc.)またはFLAGタグ付きrhIL-9に対する抗FLAGモノクローナル抗体(IBI)で釣り上げた。治療薬としての候補物質を、リガンド結合の拮抗作用を測定することにより評価する。受容体発現を図11と14に示す。
【0094】
(実施例4:細胞の単離と培養)
製造業者(Pharmacia Biotech,AB Uppsala Sweden)に従ってエンドトキシン試験を行なったFicoll-Paque PLUSを使用して密度勾配遠心分離により、健常ドナーからヒト末梢血単核細胞(PBMC)を単離した。同一遺伝子のヒト血清または熱不活性化FBSを最終濃度10%になるように補足したRPMI-1640(Bethesda ResearchLabs (BRL), Bethesda, MD)7ml中で、PBMC(5×106)、マウス脾臓細胞(5×106)、または5×106個のMo7e細胞を培養した。細胞を、刺激せずに、またはPMA 5μg/ml/PHA 5μg/ml、もしくはPHA 5μg/mlとrhIL2 50ng/ml(R &D Systems, Minneapolis, MN)で刺激して、37℃で24時間培養した。
【0095】
(実施例5:PCR産物のDNAとRNA単離、rtPCR、クローニング、および配列決定)
Nicolaides and Stoeckert46が記載したように、培養したPBMCからマイトジェンで6日間刺激後、細胞質RNAとゲノムDNAを抽出した。各供給源から1μgのRNAを,70℃で>10分間変性し、次に2.5単位のSuperscriptII逆転写酵素(GIBCO, BRL)、1U/lのRNAseインヒビター、2.5mMオリゴd(T)16プライマー、1mMずつのdATP、dCTP、dGTP、dTTP、50mM KCl、10mMトリス塩酸(pH7.0)、25mM MgCl2を使用して、37℃で1時間cDNAに逆転写(V+)した。偽逆転写反応物を陰性対照として使用した。
【0096】
逆転写反応物の20分の1を、6.7mM MgCl2、16.6mM (NH4)2SO4、67mMトリス塩酸(pH8.8)、10mM 2−メルカプトエタノール、6% DMSO、1.25mMずつのdNTP、2.5UのAmplitaq DNAポリメラーゼ、およびヒトcDNA IL-9エキソン2(フォワード5'-GCT GGA CCT TGG AGA GTG-3')(配列番号1)とエキソン9(リバース5'-GTC TCA GAC AAG GGC TCC AG-3')(配列番号2)に相当する300ngずつのオリゴヌクレオチドを、含有するPCR(50μl)中で使用した。反応混合物を以下のPCR条件に付した:95℃で120秒、次に94℃で30秒、58℃で90秒、72℃で90秒を35サイクル。最後に、反応混合物を伸長のために、72℃で15分を1サイクルを行なった。
【0097】
hIL-9受容体cDNAを示すPCR産物を、1.5%アガロースゲルでゲル電気泳動に付し、臭化エチジウム染色を用いて視覚化した。疑似逆転写反応の産物(ここでRNAの代わりにH2Oを使用した)を、すべての実験で陰性対照増幅として使用した。
【0098】
PCR産物を、TAクローニングベクター(Invitrogen, San Diego, CA)中にサブクローニングした。ヒトcDNAの増幅により、1614塩基対の産物が得られた。hIL-9受容体cDNAインサートを含有するプラスミドを、従来法により単離した(Sambrookら、(1989) Molecular Cloning: A Laboratory Manual, 第2版、Cold SpringHarbor Laboratory Press,New York)。増幅後,各インサートを含みかつこれを囲むDNA配列を、PCR誘導性またはクローニング誘導性のエラーの測定のために、自動シーケンサー(ABI 377, Perkin Elmer)で、蛍光ジデオキシターミネーターを使用して、配列決定(Sangerら、1977, Proc.Natl. Acad. Sci. USA 74:5463)により解析した。クローニングおよび/またはポリメラーゼ誘導性の配列エラーの無いhIL-9受容体cDNAインサートを、発現ベクターにサブクローニングした。
【0099】
(実施例6:in vitroでのIL-9受容体構築物のクローニングと発現)
hIL-9受容体を、エピソーム真核細胞発現ベクターpCEP4(Clontech)中にサブクローニングした。標準的方法を使用してCMVプロモーターに対してセンス配向で、pCEP4ポリリンカー中にBamH1-XhoI断片として,インサートをクローン化した(図10)。記載したように、構築物を細胞宿主中で発現させた。
【0100】
(実施例7:Mo7e、32D、TF1.1、TK-ts13、およびT98G)を使用する細胞アッセイ)
変異体IL-9受容体の機能の評価と、アトピー性喘息の治療に有効な化合物のスクリーニングのために、細胞系を使用した。IL-9により誘発される抗アポトーシスまたはベースライン増殖応答に拮抗する能力について、化合物を試験した。ある細胞系でベースライン抗アポトーシスまたは増殖応答がいったん確立されたら、3重の3回反復アッセイでの応答の統計的に有意な低下を、拮抗作用の証拠であると考えた。直接の観察、トリパンブルー染色(当業者に公知の技術)、または酸性ホスファターゼ活性の低下により、細胞傷害性から真の拮抗応答を区別した。他の増殖性物質(例えば、インターロイキン3またはインターロイキン4)に対して活性が証明されるかどうかを評価して、各化合物について拮抗作用の特異性を評価した。
【0101】
適切な培地中で使用するために、組換えIL-9または構造もしくは機能がIL-9に似ている可能性のある化合物を精製し、調製した。推定アゴニストおよびアンタゴニストを、水、食塩水、またはDMSOと水で調製した。細胞はそのまま使用したか、または実施例3に記載のようにIL-9受容体構築物でトランスフェクトした後に使用した。増殖因子を24時間枯渇させた後に、細胞を可変量の精製IL-9または構造もしくは機能がIL-9に似ている可能性のある化合物を加えてまたは加えないでインキュベートした。
【0102】
細胞数の指標として細胞内に存在する酸性ホスファターゼの量を測定する、Abacus Cell Proliferation Kit(Clontech, Palo Alto,CA)を使用して、細胞増殖を評価した。基質p-ニトロフェニルホスフェート(pNPP)を酸性ホスファターゼによりp-ニトロフェノールに変換し、これを酵素濃度の指標として測定した。各ウェルにpNPPを加え、37℃で1時間インキュベートした。次に1Nの水酸化ナトリウムを加えて、酵素反応を停止させ、p-ニトロフェノールの量を、Dynatech 2000プレートリーダー(Dynatech Laboratories, Chantilly, VA)を使用して410nmで定量した。細胞数と吸光度を比較する標準曲線を使用して、アッセイの直線範囲を決定した。吸光度測定値がアッセイの直線範囲内にある時のみ、測定結果を使用した。簡単に説明すると、リガンドを含むかまたは含まない平底マイクロタイタープレート(150または200μlウェル)中の4重の細胞試料を用いて、37℃で72〜96時間アッセイを行なった。存在する細胞の数の尺度として酸性ホスファターゼを使用した。すべての実験は、少なくとも2回繰り返した。
【0103】
アポトーシスの初期マーカーである細胞外ホスファチジルセリンを認識することによりデキサメタゾン誘導性アポトーシスを測定するAnnexin Vキットを使用してサプライヤー(Clontech)が記載するように、アポトーシスを測定した。アポトーシス細胞数は、Annexin V染色細胞のパーセントとして、蛍光顕微鏡によりスコア化した。
【0104】
Mo7e系はヒト巨核芽球細胞系であり、RPMI 1640(GIBCO/BRL, Gaithersburg, MD),20%ウシ胎児血清(Hyclone)および10ng/ml IL-3(R& D Systems,Minneapolis, MN)中で培養した。T98G系は、RPMI 1640(GIBCO/BRL)中で増殖されるヒト神経膠芽細胞腫細胞系である。ハムスター繊維芽細胞TK-ts13系も、マウス骨髄腫前駆体系であるマウス32D細胞系とともに使用し、いずれも10%ウシ胎児血清(Hyclone)を含有するRPMI 1640(GIBCO/BRL)中で培養し、さらに32D細胞系では1ng/mlのIL-3を使用した。TF1.1は,IL-2受容体ガンマサブユニット(ウェスタンブロットとrtPCRにより確認された)を発現することが知られているヒト骨髄白血病系であるが、その先祖(TF1)と比較すると、rtPCR、免疫染色、およびウェスタンブロット分析により,すでにIL-9受容体は持っていない。TF1.1は、RPMI 1640(GIBCO/BRL)と10%ウシ胎児血清(Hyclone)中で培養した。すべての細胞系は、IL-9を含めて複数のサイトカインに応答する。細胞系に栄養を与え,再度72時間毎に2×105細胞/mlで再接種した。
【0105】
細胞を2000rpmで10分間遠心分離し、0.5%ウシ血清アルブミン(GIBCO/BRL, Gaithersburg, MD)とインスリン−トランスフェリン−セレン(ITS)補助因子(GIBCO/BRL, Gaithersburg, MD)を含むRPMI 1640中に再懸濁した。血球計で細胞を数え、1×105細胞/mlに希釈し、96ウエルのマイクロタイタープレートにまいた。各ウェルは、0.15または0.2mlを含有し、細胞により10,000〜50,000細胞/ウェルの最終濃度を与えた。Mo7e細胞を,50ng/mlの幹細胞因子(SCF)(R &D Systems, Minneapolis, MN)単独、50ng/ml SCFプラス50ng/ml IL-3(R& D Systems,Minneapolis, MN)、または50ng/ml SCFプラス50ng/ml IL-9で刺激した。細胞と基本培地のみを含有する対照を含めた。試験化合物(すなわち、組換えIL-9タンパク質、ペプチド、小分子)の連続希釈物を、3重測定で各試験条件に加えた。IL-9受容体でトランスフェクトしなかったTF1.1細胞を、応答と非特異的細胞傷害性の独立した対照として使用した。培養物を5% CO2中で37℃で72〜96時間インキュベートした。
【0106】
(実施例8:トランスフェクトされた細胞系中の外因性IL-9受容体のin situおよびウェスタン分析)
IL-9受容体のin situ染色は以下のように行なった。細胞をカバーグラスの上で24時間増殖させ、次に接着性の細胞を含有するカバーグラスを、カルシウムとマグネシウムを含有するリン酸緩衝化生理食塩水(PBS)(Gibco/BRL)で2回洗浄した。IL-9受容体の細胞内染色のために、細胞を、4%パラホルムアルデヒド/PBSプラス0.1%トリトン-X中で、室温で15分間固定してから、抗ヒトIL-9受容体抗体で処理した。細胞外染色のために、細胞を固定前に抗体で処理した。次に細胞をPBSで2回洗浄し、dH2O中の7.5%BSAにより室温で30分ブロックした。次に、PBSで洗浄した細胞を、1%BSA/PBS中の抗ヒトIL-9受容体(IL-9受容体のカルボキシ末端に対するポリクローナル抗体)の10μg/ml溶液と共に、室温で1時間インキュベートした。PBSで細胞を3回洗浄し、次に1%BSA/PBS中の抗ウサギローダミン結合抗体の10μg/ml溶液中、室温で30分間インキュベートした。次に細胞をPBSで3回洗浄して、1μg/mlのDAPIを使用して室温で1分間カウンター染色した。dH2O中で細胞を3回洗浄し、顕微鏡スライドに固定し、蛍光顕微鏡により分析した。トランスフェクトしたCOS7細胞についての結果を、図14に示す。
【0107】
0.5%溶解バッファー(トリス50mM,NaCl 150mM,NP40)、1mM DTTおよびプロテアーゼインヒビター中の細胞抽出物の直接溶解から得られたタンパク質溶解物について、ウェスタンブロットを行い、5分間沸騰させた。試料を、トリス−グリシンランニングバッファー中の4〜20%トリス−グリシンSDSゲル(Novex)で電気泳動した。次に、Trans Blot II装置(Bio Rad)を使用して、電気ブロットにより、タンパク質をニトロセルロースに移した。移動後、膜を,TBS-T((20mMトリス、137mM NaCl、pH7.6)プラス0.05%ツイーン20)プラス5%ブロット(blotto)中、室温で1時間ブロックした。次に、TBS-T中のIL-9受容体のカルボキシ末端に対するポリクローナル抗体(1μg/ml)を使用して、ブロットを1時間探した。次にTBS-T中でブロットを10分間3回洗浄し、TBS-T中の第2抗ウサギ西洋ワサビペルオキシダーゼ結合抗体(1:10,000)を使用して30分間探した。上記のようにブロットを洗浄し、化学発光基質であるルミノール/エンハンサー溶液(Pierce)と共に室温で5分間インキュベートし、次に1〜60秒間フィルムに露光した。図11と12を参照されたい。
【0108】
(実施例9:真のIL-9Rゲノム増幅法)
IL-9Rは,ヒトゲノムの他の遺伝子座(第9、第10、第16、第18染色体)に4つの高度に相同性(>90%ヌクレオチド同一性)のプロセシングされていない偽遺伝子を有するため、標準的プライマーデザインを使用する真のIL-9R(XYq偽常染色体領域中に位置する生物学的機能性タンパク質をコードする遺伝子)の特異的増幅は不可能であった。これらの他の遺伝子の高度の同一性のために、標準的プライマーデザインを使用するゲノムPCR増幅により、すべての遺伝子が同時増幅され、従って真の遺伝子の配列解析が不明確になった。真のIL-9R構造は、本出願に記載の喘息または癌のような他の疾患(Renauldら、Oncogene, 9:1327-1332, 1994; Grussら、Cancer Res.,52:1026-1031, 1992)に対する素因に関係しうるので、その構造を研究するために、特異的アンプリマーを以下のように設計した:
IL-9R偽遺伝子と真の遺伝子の配列をMac Vectorソフトウェアを使用してアラインメントした。次に、真の遺伝子と偽遺伝子の間で分岐した領域について、各エキソンの周りのイントロン配列を調べた。次にこれらの領域に対してプライマーを設計し、これを使用して、各ヒト染色体を含有するヒト/齧歯類ハイブリッドDNAをPCR増幅した。生成物を3%アガロースゲルで泳動し、4つの偽遺伝子の増幅の無い真のIL-9R増幅について分析した。アニーリング温度とバッファー条件(DMSO含量5〜10%)を変えて、特異的PCR増幅条件も最適化した。まだ偽遺伝子の増幅が存在した場合は、プライマー配列中にヌクレオチド変異を入れて、真の遺伝子と比較して偽遺伝子からのより大きな分岐を生じさせた。プライマー配列を実施例2に示し、その特異性を図13に示す。
【0109】
(実施例10:細胞増殖アッセイとサイトカイン刺激)
種々の型のヒIL-9受容体を発現するTS1細胞の増殖応答を調べるために、細胞をPBSで洗浄し、D-MEM、10%ウシ胎児血清中に再懸濁した。103細胞/ウェルを、96ウェルマイクロプレート中に3回反復で接種し、適宜、組換えヒトIL-9またはマウスIL-9(R &D Systems, Minneapolis, MN)を最終濃度5ng/mlで加えた。酸性ホスファターゼアッセイを使用して7日後に、細胞増殖を評価した。簡単に説明すると、0.1M酢酸ナトリウム(pH5.5),0.1%トリトンX-100および10mM p-ニトロフェニルホスフェート(Sigma 104ホスファターゼ基質)を含有するバッファー50μlを、ウェルに加えた。プレートを室温で1〜0.5時間インキュベートし、10μl/ウェルの1N水酸化ナトリウムで反応を停止させ、Dynatech ModelMR600で410nmで吸光度を読んだ。サイトカイン刺激後のシグナル伝達カスケードのタンパク質のチロシンリン酸化を解析するために、種々の型のヒトIL-9受容体を発現するTS1細胞をPBSで洗浄し、D-MEM,0.5%ウシ血清アルブミンに再懸濁し、37℃で6時間インキュベートした。続いて、20×106細胞をヒトIL-9またはマウスIL-9(100ng/ml)で5分間処理し,直ちに冷PBSで洗浄した。実施例11に記載のように細胞をRIPAバッファーで溶解した。
【0110】
(実施例11:免疫沈降、免疫ブロッティングおよび抗体)
典型的には20〜50×106細胞を、1mlのRIPAバッファー(1%NP40、0.5%デオキシコール酸ナトリウム、0.1%SDS、1mM PMSF、50mMフッ化ナトリウム、1nMオルトバナジン酸ナトリウム、および1×「完全」プロテアーゼインヒビター混合物を含有するPBS、Cat. No. 1697498 Boehringer Mannheim)で溶解し、氷上で45分間インキュベートした。溶解物をEppendorf微量遠心分離機中で20分間遠心分離し、上清を回収し、新しいチューブに移した。免疫沈降のために、1〜5μgの抗体を溶解物に加え、4℃で一晩インキュベートした。20mlのプロテインA+Gアガロース結合ビーズを2時間加え、次にRIPAバッファーを使用して4回洗浄した。ビーズをLaemmliバッファー中に再懸濁し、3分間沸騰してから電気泳動した。タンパク質をImmobilion-P膜(Millipore)に移し、西洋ワサビペルオキシダーゼ結合2次抗体を使用して化学発光検出アッセイ(Pierce)により検出した。マウスおよびヒトIL-9受容体の特異的抗体(sc698),マウスJak1、Irs1、Irs2、Stat1、Stat2、Stat3、Stat4、Stat5、およびホスホチロシン(PY)は、Santa Cruz(Santa Cruz, CA)から購入した。抗Jak3およびモノクローナル抗ヒトIL-9受容体MAB290は、それぞれUpstate BiotechnologyとR & D Systemsから購入した。図15は、ヒトIL-9受容体の異なる変異体による、Jakファミリーのメンバーの活性化を証明する。
【0111】
(実施例12:IL-9受容体ゲノム多型の同定)
既に記載されているように(NicolaidesとStoeckert, Biotechniques 8:154-156, 1990)、志願者ドナーのPBMCからゲノムDNAを単離した。配列番号14と配列番号17のプライマーを使用するPCRにより、ヒトIL-9R遺伝子のイントロン5の配列解析を行ない、分子サイズ約1243塩基対の産物を得た。既に記載されているバッファー(Nicholaidesら、Genomics 30:195-206, 1995)で、94℃で30秒、62℃で1.5分、72℃で1.5分を35サイクル行って増幅した。次に産物を精製し、標準的配列プロトコールを使用して配列決定した。複数の個体のイントロン5からの配列を調べると、エキソン6の配列の-213ヌクレオチド上流にヌクレオチド変化があり、このためチミジン(公表されている配列)がシトシンヌクレオチドに変化していた。この変異の例を図17に示す。
【0112】
本発明を種々の具体的な材料、方法および例を参照して説明し例示したが、本発明は、この目的のために選択された材料の特定の組合せや方法に限定されるものではないことを理解されたい。当業者が理解できるように、このような細部の多くの変更が可能である。
【0113】
【表2】
【0114】
【表3】
【0115】
【表4】
【0116】
【表5】
【0117】
【表6】
【0118】
【表7】
【0119】
【表8】
【0120】
【表9】
【0121】
【表10】
(配列表)
【0122】
【化1】
【0123】
【化2】
【0124】
【化3】
【0125】
【化4】
【0126】
【化5】
【0127】
【化6】
【0128】
【化7】
【0129】
【化8】
【0130】
【化9】
【0131】
【化10】
【0132】
【化11】
【0133】
【化12】
【0134】
【化13】
【0135】
【化14】
【0136】
【化15】
【0137】
【化16】
【0138】
【化17】
【0139】
【化18】
【0140】
【化19】
【0141】
【化20】
【0142】
【化21】
【0143】
【化22】
【0144】
【化23】
【0145】
【化24】
【0146】
【化25】
【0147】
【化26】
【0148】
【化27】
【0149】
【化28】
【特許請求の範囲】
【請求項1】
明細書に記載の変異ヒト・インターロイキン-9受容体をコードするヌクレオチド配列を有する単離されたDNA分子。
【請求項1】
明細書に記載の変異ヒト・インターロイキン-9受容体をコードするヌクレオチド配列を有する単離されたDNA分子。
【図1】

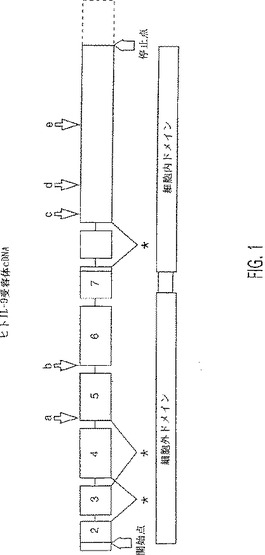
【図2−1】

【図2−2】

【図3−1】

【図3−2】

【図4】

【図5−1】

【図5−2】

【図6】
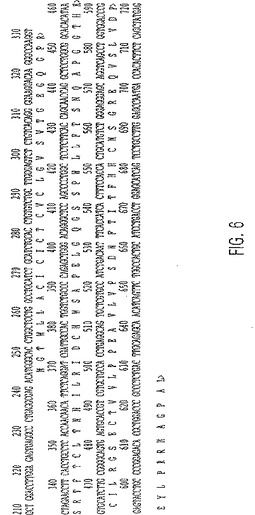
【図7】

【図8】

【図9】
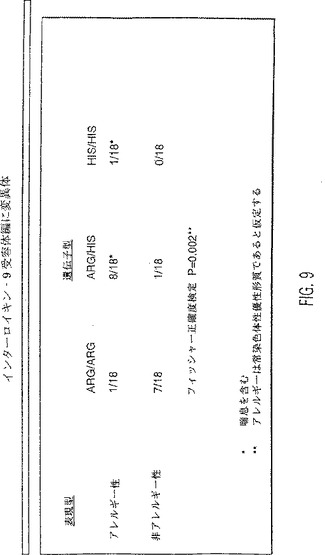
【図10】
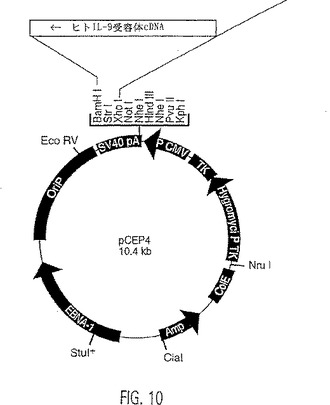
【図11】
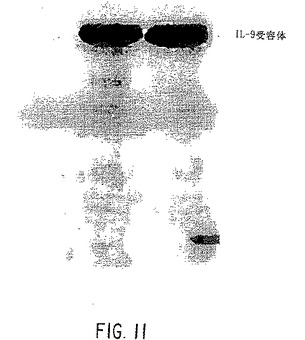
【図12】
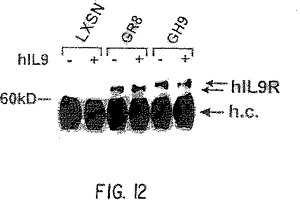
【図13】
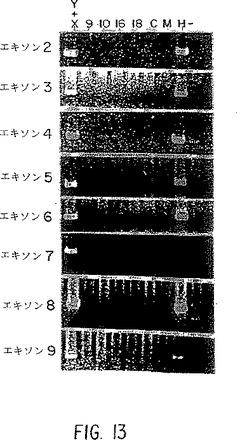
【図14−1】
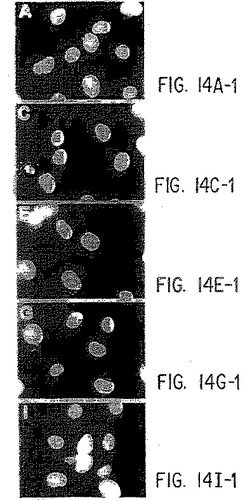
【図14−2】
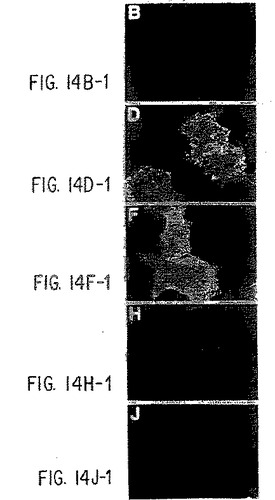
【図14−3】
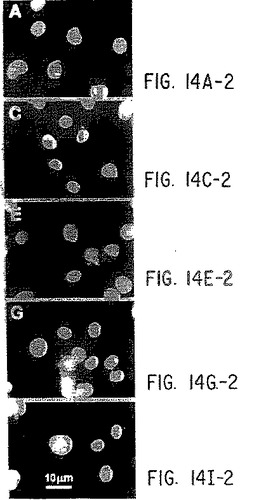
【図14−4】
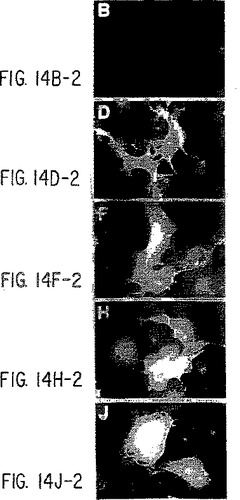
【図15−1】
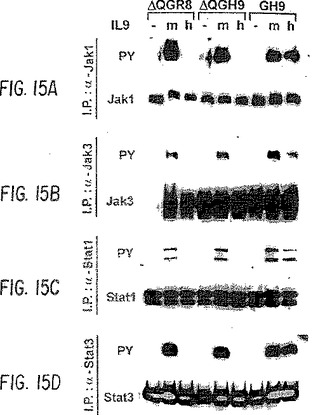
【図15−2】
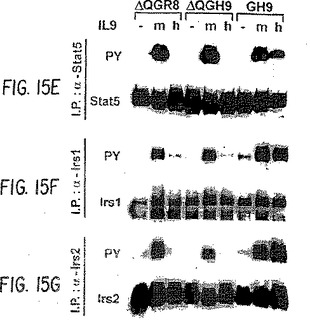
【図16】
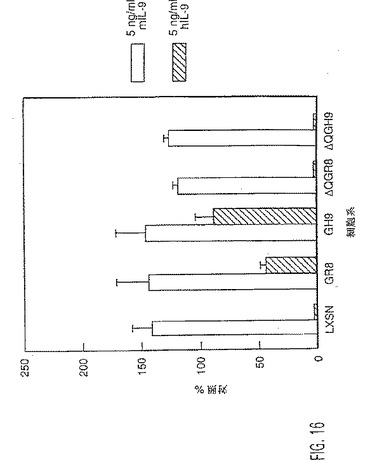
【図17】
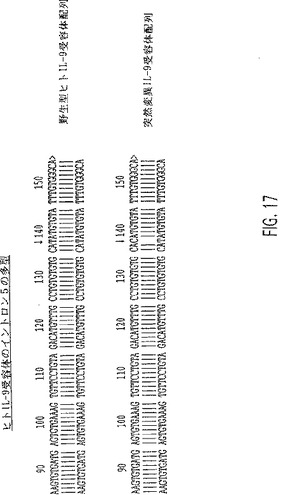
【図2−1】
【図2−2】
【図3−1】
【図3−2】
【図4】
【図5−1】
【図5−2】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14−1】
【図14−2】
【図14−3】
【図14−4】
【図15−1】
【図15−2】
【図16】
【図17】
【公開番号】特開2009−159976(P2009−159976A)
【公開日】平成21年7月23日(2009.7.23)
【国際特許分類】
【出願番号】特願2009−48597(P2009−48597)
【出願日】平成21年3月2日(2009.3.2)
【分割の表示】特願平10−525737の分割
【原出願日】平成9年12月2日(1997.12.2)
【出願人】(501097189)ジェネーラ・コーポレーション (9)
【Fターム(参考)】
【公開日】平成21年7月23日(2009.7.23)
【国際特許分類】
【出願日】平成21年3月2日(2009.3.2)
【分割の表示】特願平10−525737の分割
【原出願日】平成9年12月2日(1997.12.2)
【出願人】(501097189)ジェネーラ・コーポレーション (9)
【Fターム(参考)】
[ Back to top ]