
因子H濃縮物の薬剤の形での使用
【課題】新規な因子Hの精製方法の提供。
【解決手段】溶血性尿毒症症候群(HUS)の治療用薬剤の調製のために、ヒト因子Hを使用する。薬剤が、典型的な形のHUSの治療を目的とするものであることが好ましい。また、ヒト因子Hが、細菌、酵母、菌類又は哺乳類の細胞からなる群から選択される細胞において、ヒト因子Hの遺伝子を発現させることによって、遺伝子工学的に調製されることが好ましい。
【解決手段】溶血性尿毒症症候群(HUS)の治療用薬剤の調製のために、ヒト因子Hを使用する。薬剤が、典型的な形のHUSの治療を目的とするものであることが好ましい。また、ヒト因子Hが、細菌、酵母、菌類又は哺乳類の細胞からなる群から選択される細胞において、ヒト因子Hの遺伝子を発現させることによって、遺伝子工学的に調製されることが好ましい。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、凍結の新鮮な血漿から、溶血性尿毒症症候群(Hemolytic Uremic Syndrome(HUS))の治療用の薬剤を作るための因子Hの使用、因子Hの精製方法、並びに当該方法によって得られた因子Hの提供に関する。
【背景技術】
【0002】
溶血性尿毒症症候群(HUS)は、微小血管症性溶血性貧血、血小板減少症及び腎性疾患に関連する症状として知られている。またそれは3歳未満の児童の急性腎不全の主要な原因でもある。
【0003】
HUSには、2つの形式が存在する。HUSの典型的な形式としては、夏期における、血痕下痢の発症後に生じるHUSが挙げられる。典型的にはHUSは感染症の後に発症し、ほとんどのケースでは、腸管病原性大腸菌(特にO−157:H7、ベロ毒素の産生株)による感染症の後に発症する。
【0004】
一方、ある特定の患者では、典型的な形式とは異なる症状を示す場合もある。HUSの非典型的な形式では、発症前の前駆症状が見られず、慢性的に病状が進行し、しばしば慢性腎不全に至る。典型的なHUSはあらゆる年齢層においても発症しうる。その場合、児童におけるHUSの発症率は5%に留まる。その症候群の臨床徴候は、小血管内における血小板立地な微小血塊の形成に起因する。これは特に急性腎疾患を発症させている腎臓の糸球体に影響を及ぼす。HUSは典型的には散発的に発症するが、しばしば家族性である場合もある。これらの両方の症状においても、当該疾患は通常、増悪によって再発するおそれがある。その予後は不良である。更に、腎移移植後においては当該疾患が再発するリスクが高く、ほとんどの場合移植の拒絶反応に至る。
【0005】
HUSは、低補体血症を伴う場合もある。補体は、生物体を感染源から防御する際、また炎症プロセスにおいて不可欠な役割を果たす。それは、多種類の細胞表面マーカー(それらのうちの幾つかは炎症細胞に存在し、その他のものは免疫系細胞に存在する)、並びに宿主細胞を自己免疫から保護する膜における制御タンパク質の両方の血漿タンパク質を含む。補体の血漿タンパク質は合計で約20存在し、酵素として、結合タンパク質として、又はレギュレータ(阻害物質又は活性化物質)として機能する。補体は2つの異なるルート(すなわち従来のルート及び代替のルート、図1参照)により活性化されうる。
【0006】
図1中、酵素的なステップを太い矢印で示す。調節タンパクを枠で示す。膜タンパク質を太字で示し、循環タンパク質をイタリックで示す(そこには因子H(FH)が含まれる)。
【0007】
従来のルートは、外来の粒子と結合する抗体によって活性化される。ゆえにそれは抗体依存性といえる。
【0008】
代替のルートは、微生物の侵入によって活性化される、ゆえにそれは抗体非依存性であり、宿主を細菌感染症から防御する際に極めて重要である。
【0009】
因子Hは、110〜615μg/mLの濃縮で血漿中に存在する155kDaのタンパクである。因子Hは肝臓、マクロファージ、繊維芽細胞、内皮細胞及び血小板において合成される。分泌型のタンパク質は、20の繰り返し単位からなる60アミノ酸からなる。因子Hは補体の代替のルートにおいて中心となるレギュレータである。それは血液中における免疫複合体の形成又は解離速度を制御し、それにより、かかるプロセスにおける平衡状態が調節される。
【0010】
因子Iと共に、因子Hは、遊離型若しくは細胞表面結合型のC3b分子を不活性化する。それにより、抗原抗体複合体からなる免疫複合体(C3b補体の構成成分と複合体を形成する)はもはや、それに続く補体(構成成分C5からC9)のカスケードを活性化することができなくなる。
【0011】
因子Hの機能は、主に以下の3つに分類することができる。
1)第一に、H因子は因子Iの補因子として挙動する。それにより、因子H及び因子IはタンパクC3bの切断を生じさせることにより、補体のC3bタンパクをC3bi(非活性分子)に変化させる。生じる不活性タンパクC3bは補体においてもはやその機能を発揮できなくなり、C3コンベルターゼの形成にもはや関与しなくなる。
2)第二に、H因子は内皮細胞及び血小板への結合機構に関与する。
3)第三に、H因子は補体活性化の代替のルートにおいて、予備成形されたC3コンベルターゼ(C3bBb)の解離に関与する。この最後の活性は、因子Hの分子的整合性に直接依存し、特に因子Hにおける無傷のasn323−asn324結合の存在に依存することを示すものである。
【0012】
すなわち、因子Hの欠損症又は欠如(非典型的なHUSの多くの症例に起因する)によって、補体の活性過剰が生じる(腎生検の間のC3タンパクの堆積、及び血流中に存在するC3タンパク質レベルの減少により、特定の患者において観察される症状)。
【0013】
非典型的なHUSに罹患する特定の患者における低いC3レベルは、疾患の急性期においてのみ観察される。特定の非典型的なHUSの病態における、しばしば低いC3レベルと関連する質的及び量的な因子Hの欠如の果たす役割に関しては、これまで活発な議論なされている(非特許文献1)。
【0014】
H因子欠損症は、低レベルのC3の原因となる代替的な補体ルートを半永久的に活性化させる原因ともなる。
【0015】
非典型的なHUSと、補体(特に因子H)の調節タンパクのコーディング部位との関連を示す証拠が、1番染色体上で発見されている(非特許文献2から4)。
【0016】
また、因子H遺伝子の突然変異が、劣性若しくは優性の常染色体遺伝による、家族性HUSとして同定されている(非特許文献5から8)。
【0017】
非典型的なHUSに罹患する患者の、移植後における再発率は約25%と言われている。再発する場合の予後は悪く、再発に関連して移植片に対する拒絶が通常生じる。
【0018】
(従来技術)HUSにおける補体の役割が公知となる以前から既に、血漿交換の有無を問わない、凍結させた新鮮な血漿を用いた灌流処理が、1970年代から行われている。今日、血漿交換の有無を問わない、凍結させた新鮮な血漿を用いた灌流処理が、HUS治療において一般的に行われている。しかしながら、凍結させた新鮮な血漿の灌流量及び灌流頻度は、未だ経験則に頼っているのが実情である。
【0019】
これらの灌流の適用は、2週間〜1ヵ月に2回、定期的に反復させ、各灌流の適用の際は2〜3時間実施する必要がある。
【0020】
ゆえに、この処理は患者にとって長時間の、反復的な処理を強いるものとなる。
【0021】
また、使用される凍結された新鮮な血漿量は多量であるため、凍結された新鮮な血漿を灌流に適用する際の標準リスクを増加させる。
【0022】
第1に、凍結された新鮮な血漿(FFP)は抗A又は抗Bヘモリシンを含有し、それは同じABO型の患者、あるいは少なくともヘモリシンに対応する抗原A又はBを欠損する患者において保持されなければならない(赤血球の血液適合性とは反対)。これらの要因を考慮に入れなければ、輸血後、受容者側において、ABO式適合性における不適合による赤血球の溶血を生じさせることもありうる。
【0023】
更に、Rhesusシステムの抗原Dに対する同種免疫のいかなるリスクをも回避する目的で、特にリスクの大きい患者(少女、出産期の年齢の女性、多くの輸血を受けている患者)において灌流適用を実施する必要があり、その際、患者及びドナーはこの抗原のレベルで同じ特性を有する必要がある。
【0024】
第2に、FFPのリン酸濃度(特にin viroで弱毒化された血漿(VAP)における)が非常に高い(9〜12mmol/l)ため、FFPによりHUS患者におけるリン酸過剰血症が生じ、また当該HUS患者は腎機能不全に罹患することとなる。VAP中の高いリン酸濃度(高リン酸血症)は、VAPの輸血を受ける患者において生じると考えられ、その原因して主なものとしては、−輸血されるVAPの体積が多量であること、−毎日反復輸血されること、−患者が既に腎機能不全に罹患していること、−患者が既に高リン酸血症に罹患していること、が挙げられる。
【0025】
次に、灌流適用されたFFPの量によっては、タンパクの過剰及び/又はクエン酸の過剰が生じ、それにより循環カルシウムの濃度が減少する。
【0026】
最後に、FFPによってアレルギー、及び感染源の侵入のリスクが生じる。実際、現在行われている検出及び不働化方法は、潜在的に凍結された新鮮な血漿に存在する感染源の検出及び除去を可能にするための充分な感度及び能力を必ずしも有しているわけではない。
【0027】
灌流適用された体積が、利尿によって排出され、通常の動脈圧が維持されるにはあまりに大きい場合、凍結された新鮮な血漿灌流適用と血漿交換との組み合わせは重要である。しかしながらこの組み合わせは、主に血管への介入(中枢ルートが必要)、ロード量の過多、アナフィラキシー反応、凝血塊の問題及びウイルス性疾患の感染などの新たなリスクを生じさせるものである。
【0028】
更に、血漿交換法は幼児に適用するのが困難である。
【0029】
他の治療方法としては、腎臓移植が挙げられる。しかしながら、移植後の再発リスクが非常に高い。
【0030】
更に、aHUS患者(非典型的なHUS)の腎移植後の診断は困難なことが多い。すなわち、再発であるのか、あるいは急性の血管拒絶反応又は移植片のバイオプシーにおける慢性拒絶反応であるのかを識別するのが困難であることが多い。
【0031】
再発の治療方法としては、凍結された新鮮な血漿の灌流適用や、血漿灌流適用の有無にかかわらない血漿交換法などが挙げられるが、その結果を予測することは不可能に近い。これらの予測不可能な結果の原因としては、FFP灌流適用の回数及び量が挙げられ、すなわち、各灌流適用が同種のバッチ由来ではなく、幾つかのドナー由来の血漿の集積物由来であることが主な原因である。
【0032】
因子Hは肝臓で合成されるため、肝臓移植又は肝臓−腎臓の複合移植が提唱されることは一見合理的であるようにも思える。しかしながらかかる移植は、常に内科医及び両親による困難な選択を必要とし、また操作上のリスク及び肝臓移植による拒絶反応のリスクを伴うものである。
【先行技術文献】
【非特許文献】
【0033】
【非特許文献1】Rougier N,Kazatchkine MD,ら、Human complement factor H deficiency associated with hemolytic uremic syndrome,J.Am.Soc.Nephrol.1998、9:2318−2326
【非特許文献2】Norisら、Hypocomplementemia discloses genetic predisposition to hemolytic uremic syndrome and thrombotic thrombocytopenic purpura:role of Factor H abnormalities,J.Am.Soc.Nephrol.1999,10:281−293
【非特許文献3】Warwickerら、Genetic studies into hemolytic uremic syndrome,Kidney Int.,1998、53:836−844
【非特許文献4】Warwickerら、Familial relapsing hemolytic uremic syndrome and complement Factor H deficiency,Nephrol.Dial.Transplant.,1999、14:1229−1233
【非特許文献5】Buddlesら、Complement Factor H gene mutation associated with autosomal recessive atypical hemolytic uremic syndrome,Am.J.Hum.Genet.,2000、66:1721−1722
【非特許文献6】Caprioliら、The molecular basis of familial hemolytic uremic syndrome:mutation analysis of Factor H gene reveals a hot spot in short consensus repeat 20,J.Am.Soc.Nephrol.2001、12:297−307
【非特許文献7】Ohaliら、Hypocomplementemic autosomal recessive hemolytic uremic syndrome with decreased Factor H,Pediatr.Nephrol.1998、12:619−624
【非特許文献8】Yingら、Complement Factor H gene mutation associated with autosomal recessive atypical hemolytic uremic syndrome,Am.J.Hum.Genet.1999、65:1538−1546
【発明の概要】
【発明が解決しようとする課題】
【0034】
従来技術のこれらの欠点を克服するべく、発明者らによる鋭意研究の結果、驚くべきことに、HUSの治療薬の製造に因子Hが利用できることを見出した。
【課題を解決するための手段】
【0035】
因子H、例えば凍結させた新鮮な血漿に由来する因子H濃縮物を用いることにより、注入体積及び注入時間を減少させつつ、HUSに罹患する患者の因子Hの欠損症を、安全かつ安定的に治療することが可能となる。特に、肝臓移植の直後の期間に因子Hを投与することによって、移植された肝臓における低い因子H産生を補填し、それにより移植片における症状の再発及び拒絶反応を防止することが可能となる。
【0036】
本発明はまた、以下のステップを含んでなる因子Hの精製方法の提供に関する。
1)血漿の寒冷型沈降物の上澄を調製するステップと、
2)この上澄をアニオン交換体タイプのゲル/樹脂上のクロマトグラフィに供するステップと、
3)非保持フラクションをヘパリンタイプのグラフト化リガンドを含有するゲル/樹脂上のクロマトグラフィに供するステップと、
4)ステップ(3)のクロマトグラフィの後、非保持フラクションのpHを調整し、ヘパリンタイプのグラフト化リガンドを含有するクロマトグラフ担体ゲル/樹脂へH因子を結合させるステップと、
5)ゲル/樹脂の平衡化用バッファよりもイオン強度の強いバッファでH因子を溶出させるステップと、
6)溶出フラクションを希釈し、更にそれを強酸性のカチオン交換タイプのゲル/樹脂上のクロマトグラフィに供するステップと、
7)ゲル/樹脂の平衡用バッファのよりもイオン強度の強いバッファでH因子を溶出させるステップと、
8)溶出フラクションを希釈し、更にそれを強塩基性のアニオン交換タイプのゲル/樹脂上のクロマトグラフィに供するステップと、
9)ゲル/樹脂を洗浄し、H因子を溶出させるステップと、
10)H因子の濃縮物を調製するステップ。
【図面の簡単な説明】
【0037】
【図1】補体は2つの異なるルート(従来のルート及び代替のルート)。
【図2】因子Hの精製方法のダイアグラム。
【図3】因子HによるC3コンベルターゼの分解。
【発明を実施するための形態】
【0038】
本発明の主要な目的は、溶血性尿毒症症候群(HUS)(特に典型的な形式のHUS又は非典型的な形式のHUS)の治療用薬剤の製造への因子Hの使用方法の提供である。
【0039】
本発明の好ましい実施形態は、溶血性尿毒症症候群の治療用薬剤を調製するための因子Hの使用であり、当該因子Hは、新鮮なヒト血漿から精製されたものであるか、又は当業者に公知の標準的な方法による精製物に由来する血漿のフラクションである。この精製方法は当業者に公知である。例えば、リシン−セファロース、QAE−セファデックス、DEAE−トヨパール、セファクリルS−300及びヒドロキシアパタイトのカラムを用いたクロマトグラフィなどが挙げられる。その詳細は、以下の文献に記載されている:Fearon,J.Immunol.119,1248−1252(1977)、Crossleyら、Biochem.J.,191,173−182(1980)、Nagasawaら、J.Immunol.,125,578−582(1980)、Weilerら、P.N.A.S.,73,3268−3272,(1976)及びWhaleyら、J.Exp.Med.,144,1147−1163(1976)。凍結された新鮮な血漿からの精製により得た因子Hは例えば、因子H濃縮物の形で得られる。
【0040】
本発明の他の実施形態は、溶血性尿毒症症候群の治療用薬剤を調製するための因子Hの使用であり、当該因子Hは、細菌、酵母、菌類又は哺乳類細胞からなる群から選択される細胞内でその遺伝子を発現させる、遺伝子工学的方法によって得られる。
【0041】
本発明の具体的実施形態は、溶血性尿毒症症候群の治療用薬剤を調製するための因子Hの使用であり、上記のように得られた薬剤は、冷凍乾燥させた形態である。
【0042】
本発明の更なる実施形態は、溶血性尿毒症症候群の治療用薬剤を調製するための因子Hの使用であり、上記のように得られた薬剤を、少なくとも1つの感染源を除去するか又は不活性化するための少なくとも1つのプロセスに供することを特徴とする。感染源としては、ウイルス及び従来しない伝染性物質(NCTA)(例えばプリオンタンパク)などが挙げられる。
【0043】
特に、当該薬剤を、ウイルス的に不活性化させるのが好ましい。「ウイルスを不活性化する」とは、当該薬剤を、化学物質(例えば溶剤/洗浄剤)、及び/又は熱(例えば乾熱滅菌又は殺菌処理)、及び/又はナノ濾過などの、当業者に公知の少なくとも1つのウイルス不活性化方法に供することを意味する。これらのいずれかの方法で不活性化できるウイルスとしては、ヒト免疫不全症ウイルス(HIV)、A型肝炎ウイルス(HAV)、B型肝炎ウイルス(HBV)、B19パルボウイルス、細胞拡大ウイルス(CMV)、ブタパルボウイルス、ポリオウイルス、ウシのウイルス性下痢ウイルス(BVDV)などが挙げられる。
【0044】
本発明の別の目的は、冷凍乾燥され、ウイルスを不活性化された、上記の実施形態に係る医薬組成物であり、因子H、並びに薬理学的に許容できる賦形剤及び/又は担体を含んであることを特徴とする。
【0045】
本発明の別の目的は、以下のステップを含んでなる、因子Hの精製方法に関する:
1)血漿の寒冷型沈降物の上澄を調製するステップと、
2)この上澄をアニオン交換体タイプのゲル/樹脂上のクロマトグラフィに供するステップと、
3)非保持フラクションをヘパリンタイプのグラフト化リガンドを含有するゲル/樹脂上のクロマトグラフィに供するステップと、
4)ステップ(3)のクロマトグラフィの後、非保持フラクションのpHを調整し、ヘパリンタイプのグラフト化リガンドを含有するクロマトグラフ担体ゲル/樹脂へH因子を結合させるステップと、
5)ゲル/樹脂の平衡化用バッファよりもイオン強度の強いバッファでH因子を溶出させるステップと、
6)溶出フラクションを希釈し、更にそれを強酸性のカチオン交換タイプのゲル/樹脂上のクロマトグラフィに供するステップと、
7)ゲル/樹脂の平衡用バッファのよりもイオン強度の強いバッファでH因子を溶出させるステップと、
8)溶出フラクションを希釈し、更にそれを強塩基性のアニオン交換タイプのゲル/樹脂上のクロマトグラフィに供するステップと、
9)ゲル/樹脂を洗浄し、H因子を溶出させるステップと、
10)H因子の濃縮物を調製するステップ。
【0046】
本発明の具体的実施形態では、ステップ3における、ヘパリンリガンドがグラフト化されているクロマトグラフ保持体は、セファロースヘパリンゲル/樹脂である。
【0047】
本発明の具体的実施形態では、ステップ4における、ヘパリンリガンドがグラフト化されているクロマトグラフ保持体は、セファロースヘパリンゲル/樹脂である。
【0048】
本発明の具体的実施形態では、ステップ6における、強酸性のカチオン交換タイプのゲル/樹脂上のクロマトグラフィは、SPセファロースタイプのクロマトグラフィである。
【0049】
本発明の具体的実施形態では、ステップ8における、強塩基性のアニオン交換タイプのゲル/樹脂上のクロマトグラフィは、QセファロースFFタイプのクロマトグラフィ又はその均等物である。
【0050】
好適には、ステップ4の非保持フラクションのpHを、pH5.5〜pH6.5の範囲、好ましくはpH6.0となるように調整する。
【0051】
好適には、ステップ8の希釈されたフラクションのpHを、pH6.5〜pH7.5の範囲に調整する。
【0052】
本発明の精製方法は、血漿に由来する因子Hを精製するための、工業的実用化の可能な唯一の方法であると考えられ、当該方法により、精製された因子H濃縮物を、化学物質若しくは合成プロテアーゼに対する阻害剤の非存在下で得ることが可能となり、それにより、これらの阻害剤が最終生産物中に検出されることはなくなる。
【0053】
実際、現在行われているヒト血漿から因子Hを精製する方法は基礎研究において用いられるが、それは沈殿精製技術(例えばPEG、硫酸アンモニウム沈殿)及びプロテアーゼインヒビターを使用して実施されることが多いため、産業的な実施が困難なことが多い。これらのプロテアーゼインヒビターは、因子Hの分子内のアスパラギン323及びアスパラギン324のアミノ酸間の結合を切断する役割を果たすトリプシンタイプのタンパク質(血清及び血漿に存在)の活性を阻害する。したがって、プロテアーゼインヒビターの添加により、この因子のタンパク質分解の減少、及びその安定性の向上に関与する。しかしながら、プロテアーゼインヒビターは毒性化合物であることが多く、治療用途に用いる因子Hの産業的な調製方法へのその使用は不適当である。
【0054】
更に、本発明の方法は、得られる因子Hの濃縮物では、主要な3タイプの活性が保持されるという利点を有し、一方現在行われている技術で得られる因子Hは、かかる活性が保持されない。本発明の方法によって得られた因子Hは、補体の代替ルートにおける中心的な制御活性(HUS、特に非典型的なHUSに罹患する患者に不足すると考えられる活性)を発揮することができる。特に、本発明の方法によって調製される因子Hは、補体の代替ルートにおいても、予備成形されたC3コンベルターゼを分離させる活性を保持しており、HUSの治療の際に、その最大限の機能活性で使用することができる。
【0055】
更なる本発明の方法によって調製される因子H濃縮物は1付近の比活性(AS=0.8〜0.9)を有し、凍結された新鮮な血漿の溶液(AS=0.008)よりも高い効果を有するものの、本願明細書の冒頭で説明したように、治療効果は若干あっても、多くの不利益をもたらすこともある。これらの不利益としては、血漿の投与により、HUSを治療するための不必要な更なるタンパク質(アルブミン、フィブリノーゲンなど)が生体内に導入され、それは一方では、タンパクの過剰ロードに起因する望ましくない反応の誘導や、又は「血清病」として公知のアレルギー反応の誘導の原因ともなる。
【0056】
最後に、血漿に存在する感染性のウイルスの不活性化は、血液製剤中に存在するウイルスの不活性化のために通常用いられる方法よりも通常困難であり、実用的でない。しかしながら、本発明の方法によって調製された因子H濃縮物は、定式化されている試験済みの方法により、ウイルス安全性を担保することができる。
【実施例】
【0057】
<実施例1>:因子Hの精製方法:
因子Hの精製に適用した方法を、図2に図式的に示す。
【0058】
ヒトの凍結された新鮮な血漿を1℃〜6℃の温度で融解させ、寒冷型沈降物を遠心沈降に供し、寒冷型沈降物の不溶性画分と血漿上澄画分とを分離させた。
【0059】
得られた寒冷型沈降物の血漿上澄(因子H濃縮物、約400〜約500mg/Lの濃度で因子Hを含有)を、アニオン交換体タイプの樹脂/ゲル(例えばDEAEセファデックスタイプのゲル/樹脂)を用いたクロマトグラフィに供し、樹脂/ゲル上のこれらの因子を保持させ、血漿上澄からビタミンKに依存する形で因子を分離させた。
【0060】
非保持の血漿上清画分(フラクションA)(因子H濃縮、約400〜約500mg/Lで因子Hを含有)を更に、ヘパリンセファロースFFタイプのゲル/樹脂上のアフィニティークロマトグラフィに供し、樹脂/ゲル中に抗トロンビンIIIを保持させ、抗トロンビンIIIをこのフラクションAから分離させた。
【0061】
この非保持フラクションA(フラクションB)(因子H濃縮物、約300〜約400mg/Lで因子Hを含有)のpHを、pH5.5〜pH6.5の範囲、好ましくはpH6.0に調整した。
【0062】
フラクションB(pHが調整された)を、ヘパリンセファロースFFタイプの第2のゲル/樹脂、又は、ヘパリンタイプのリガンドをグラフト化された他の任意のクロマトグラフ保持体を用いたクロマトグラフィに供した。血漿タンパク分画Bに含まれる大部分のタンパクを更にクロマトグラフィ濾液によって溶出させた。ゲル/樹脂にわずかに吸着したタンパクを、一連の洗浄及び前溶出工程によって除去した。ゲル/樹脂に保持される因子Hは更に、ゲル/樹脂の平衡化に使用するバッファより強いイオン強度を有するバッファを用いて溶出した。
【0063】
因子H(フラクションC)を含有する溶出フラクションを希釈し、更に強酸性のカチオン交換タイプのゲル/樹脂(例えばSPセファロースFfタイプ又はそれと同様のゲル/樹脂)によるクロマトグラフィに供した。ゲル/樹脂にわずかに吸着したタンパクを、一連の洗浄及び前溶出工程によって除去した。ゲル/樹脂に保持される因子Hは更に、ゲル/樹脂の平衡化に使用するバッファより強いイオン強度を有するバッファを用いて溶出した。
【0064】
因子H(フラクションD)を含有する溶出フラクションを更に、溶媒若しくは界面活性剤(例えばポリソルベート80及びTnBP)による処理による、ウイルスの不活性化ステップに供した。かかる処理を行うことにより、ウイルス(特にカプセルを有するタイプのウイルス)を効率的に不活性化することが可能である。
【0065】
更にフラクションDを希釈し、このフラクションのpHを、pH6.5〜pH7.5の範囲となるように調整した。フラクションDを更に、強塩基性のアニオン交換タイプのゲル/樹脂(例えばQセファロースFFタイプ又は当量のゲル/樹脂)を用いたクロマトグラフィに供した。一連の洗浄の後、ゲル/樹脂に保持された因子Hを、ゲル/樹脂の平衡化用のバッファより強いイオン強度を有するバッファを用いて溶出した。
【0066】
上記の界面活性剤タイプの溶剤による処理に用いた、ウイルスの失活のために用いた物質を、クロマトグラフィ工程の間に除去することにより、因子Hの純度が向上した。
【0067】
因子H(フラクションE)を含有する溶出フラクションを更に、約15nmの空隙を有するフィルタを用いたナノ濾過に供し、ウイルスを除去した。このウイルス除去処理により、ウイルス(特に小型のカプセルを有するウイルス)を効率的に除去することができる。得られる溶液(フラクションF)を、最後に濃縮し、限外濾過により調整し、更に0.22μmのフィルタで濾過した。
【0068】
上記の精製方法の収量、及びこの方法により精製された因子Hの比活性を、2つの異なるバッチ毎に測定した。それぞれの結果を表1に示す。比活性(A.S.)を、タンパク質1mg当たりの因子Hタイプの抗原のmgとして表す。
【0069】
【表1】

【0070】
<実施例2>:因子Hの活性の測定方法:
ELISAプレート(96ウェルタイプ)のウェルを、0.2Mの炭酸ナトリウムバッファ中の2.5g/mL(Calbiochem社:ref.341274)の濃度の精製C3bタンパク質溶液で被覆した。その際、溶液100μLをウェルに添加し、プレートを37℃で1時間、及び4℃で一晩静置した。
【0071】
10mMのリン酸ナトリウム緩衝液(pH7.2)、25mMのNaCl、0.1%のTween20の溶液で、300μL/ウェルで3回洗浄した。
【0072】
非特異的な部位を更に、10mMのリン酸ナトリウム緩衝液(pH7.2)、25mMのNaCl、0.05%のTween 20、1%のウシ血清アルブミンを含有する溶液300μL/ウェルで、37℃で1時間静置して飽和させた。次に、上記の洗浄溶液でウェルの洗浄を実施した。
【0073】
以下の成分(20mMのNiCl2のストック溶液(1.5mM最終濃度) 75μL、1mg/mlのB因子(Calbiochem ref.341262) 4μL、1mg/mlのD因子(Calbiochem ref.341273) 3μL、10mMのリン酸ナトリウム緩衝液(pH7.2、25mMのNaCl、4%のウシ血清アルブミンを含有する) 918μL)を含有する100μLの溶液を、各ウェルに添加し、37℃で2時間インキュベートした。
【0074】
10mMのリン酸ナトリウム緩衝液(pH7.2)、25mMのNaCl、0.1%のTween20の溶液で、300μL/ウェルで3回洗浄した。
【0075】
因子H溶液を、20μg/mL、10μg/mL、1μg/mL、0.25μg/mL、0.0625μg/mL、0.015625μg/mL、0.00390625μg/mL及び0.001μg/mLの因子H濃度となるように調製した。各ウェルに100μLの溶液を添加し、37℃で30分間インキュベートした。
【0076】
更に10mMのリン酸ナトリウム、25mMのNaCl、0.1%のTween 20を含有する緩衝溶液(pH7.2)で、300μL/ウェルで3回洗浄した。
【0077】
ヤギ抗ヒトB因子抗体溶液(Calbiochem ref.:341272)を、0.1%のウシ血清アルブミンを含有するPBSバッファ(Sigma社、P−3813)(pH7.4)で1/2,000に希釈し、更に希釈液100μLをウェルに添加し、37℃で1時間インキュベートした。
【0078】
PBS(pH7.2、0.1%のTween20を含有)で、300μL/ウェルで3回洗浄した。次に、0.1%のウシ血清アルブミン含有PBSで1/10,000に希釈した、ペルオキシダーゼ(Calbiochem ref.401515、1mg/mL)標識し抗ヤギウサギ抗体を含有する溶液100μLをウェルに添加し、室温20〜25分間インキュベートした。
【0079】
PBS(pH7.2、0.1%のTween20を含有)で、300μL/ウェルで3回洗浄した。
【0080】
クエン酸ナトリウム溶液中の5mg/10mLのOPDペルオキシダーゼ(Sigma社)基質を最後に100μL/ウェルの量で、更にH2O2を10μLでウェルに添加した。反応混合液を約10分間ウェルと接触させ、更にウェルあたり50μLの4N H2SO4を添加し、反応を停止させた。
【0081】
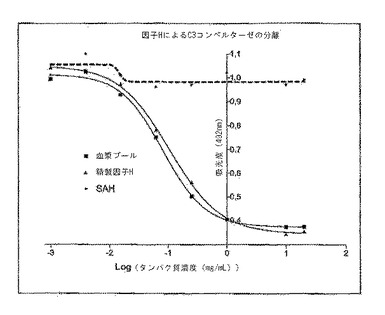
次に、ウェル中の溶液の吸光度を492nmの波長で測定した。その結果を図3に示す。図3に、因子Hの濃度又はタンパク濃度(SAH)に対する吸光度測定値の関係を示すグラフを示す。
【0082】
因子Hの活性を測定するための同様の方法は、McRaeら、The Journal of Immunology、2005、174:6250−6256に記載されている。
【技術分野】
【0001】
本発明は、凍結の新鮮な血漿から、溶血性尿毒症症候群(Hemolytic Uremic Syndrome(HUS))の治療用の薬剤を作るための因子Hの使用、因子Hの精製方法、並びに当該方法によって得られた因子Hの提供に関する。
【背景技術】
【0002】
溶血性尿毒症症候群(HUS)は、微小血管症性溶血性貧血、血小板減少症及び腎性疾患に関連する症状として知られている。またそれは3歳未満の児童の急性腎不全の主要な原因でもある。
【0003】
HUSには、2つの形式が存在する。HUSの典型的な形式としては、夏期における、血痕下痢の発症後に生じるHUSが挙げられる。典型的にはHUSは感染症の後に発症し、ほとんどのケースでは、腸管病原性大腸菌(特にO−157:H7、ベロ毒素の産生株)による感染症の後に発症する。
【0004】
一方、ある特定の患者では、典型的な形式とは異なる症状を示す場合もある。HUSの非典型的な形式では、発症前の前駆症状が見られず、慢性的に病状が進行し、しばしば慢性腎不全に至る。典型的なHUSはあらゆる年齢層においても発症しうる。その場合、児童におけるHUSの発症率は5%に留まる。その症候群の臨床徴候は、小血管内における血小板立地な微小血塊の形成に起因する。これは特に急性腎疾患を発症させている腎臓の糸球体に影響を及ぼす。HUSは典型的には散発的に発症するが、しばしば家族性である場合もある。これらの両方の症状においても、当該疾患は通常、増悪によって再発するおそれがある。その予後は不良である。更に、腎移移植後においては当該疾患が再発するリスクが高く、ほとんどの場合移植の拒絶反応に至る。
【0005】
HUSは、低補体血症を伴う場合もある。補体は、生物体を感染源から防御する際、また炎症プロセスにおいて不可欠な役割を果たす。それは、多種類の細胞表面マーカー(それらのうちの幾つかは炎症細胞に存在し、その他のものは免疫系細胞に存在する)、並びに宿主細胞を自己免疫から保護する膜における制御タンパク質の両方の血漿タンパク質を含む。補体の血漿タンパク質は合計で約20存在し、酵素として、結合タンパク質として、又はレギュレータ(阻害物質又は活性化物質)として機能する。補体は2つの異なるルート(すなわち従来のルート及び代替のルート、図1参照)により活性化されうる。
【0006】
図1中、酵素的なステップを太い矢印で示す。調節タンパクを枠で示す。膜タンパク質を太字で示し、循環タンパク質をイタリックで示す(そこには因子H(FH)が含まれる)。
【0007】
従来のルートは、外来の粒子と結合する抗体によって活性化される。ゆえにそれは抗体依存性といえる。
【0008】
代替のルートは、微生物の侵入によって活性化される、ゆえにそれは抗体非依存性であり、宿主を細菌感染症から防御する際に極めて重要である。
【0009】
因子Hは、110〜615μg/mLの濃縮で血漿中に存在する155kDaのタンパクである。因子Hは肝臓、マクロファージ、繊維芽細胞、内皮細胞及び血小板において合成される。分泌型のタンパク質は、20の繰り返し単位からなる60アミノ酸からなる。因子Hは補体の代替のルートにおいて中心となるレギュレータである。それは血液中における免疫複合体の形成又は解離速度を制御し、それにより、かかるプロセスにおける平衡状態が調節される。
【0010】
因子Iと共に、因子Hは、遊離型若しくは細胞表面結合型のC3b分子を不活性化する。それにより、抗原抗体複合体からなる免疫複合体(C3b補体の構成成分と複合体を形成する)はもはや、それに続く補体(構成成分C5からC9)のカスケードを活性化することができなくなる。
【0011】
因子Hの機能は、主に以下の3つに分類することができる。
1)第一に、H因子は因子Iの補因子として挙動する。それにより、因子H及び因子IはタンパクC3bの切断を生じさせることにより、補体のC3bタンパクをC3bi(非活性分子)に変化させる。生じる不活性タンパクC3bは補体においてもはやその機能を発揮できなくなり、C3コンベルターゼの形成にもはや関与しなくなる。
2)第二に、H因子は内皮細胞及び血小板への結合機構に関与する。
3)第三に、H因子は補体活性化の代替のルートにおいて、予備成形されたC3コンベルターゼ(C3bBb)の解離に関与する。この最後の活性は、因子Hの分子的整合性に直接依存し、特に因子Hにおける無傷のasn323−asn324結合の存在に依存することを示すものである。
【0012】
すなわち、因子Hの欠損症又は欠如(非典型的なHUSの多くの症例に起因する)によって、補体の活性過剰が生じる(腎生検の間のC3タンパクの堆積、及び血流中に存在するC3タンパク質レベルの減少により、特定の患者において観察される症状)。
【0013】
非典型的なHUSに罹患する特定の患者における低いC3レベルは、疾患の急性期においてのみ観察される。特定の非典型的なHUSの病態における、しばしば低いC3レベルと関連する質的及び量的な因子Hの欠如の果たす役割に関しては、これまで活発な議論なされている(非特許文献1)。
【0014】
H因子欠損症は、低レベルのC3の原因となる代替的な補体ルートを半永久的に活性化させる原因ともなる。
【0015】
非典型的なHUSと、補体(特に因子H)の調節タンパクのコーディング部位との関連を示す証拠が、1番染色体上で発見されている(非特許文献2から4)。
【0016】
また、因子H遺伝子の突然変異が、劣性若しくは優性の常染色体遺伝による、家族性HUSとして同定されている(非特許文献5から8)。
【0017】
非典型的なHUSに罹患する患者の、移植後における再発率は約25%と言われている。再発する場合の予後は悪く、再発に関連して移植片に対する拒絶が通常生じる。
【0018】
(従来技術)HUSにおける補体の役割が公知となる以前から既に、血漿交換の有無を問わない、凍結させた新鮮な血漿を用いた灌流処理が、1970年代から行われている。今日、血漿交換の有無を問わない、凍結させた新鮮な血漿を用いた灌流処理が、HUS治療において一般的に行われている。しかしながら、凍結させた新鮮な血漿の灌流量及び灌流頻度は、未だ経験則に頼っているのが実情である。
【0019】
これらの灌流の適用は、2週間〜1ヵ月に2回、定期的に反復させ、各灌流の適用の際は2〜3時間実施する必要がある。
【0020】
ゆえに、この処理は患者にとって長時間の、反復的な処理を強いるものとなる。
【0021】
また、使用される凍結された新鮮な血漿量は多量であるため、凍結された新鮮な血漿を灌流に適用する際の標準リスクを増加させる。
【0022】
第1に、凍結された新鮮な血漿(FFP)は抗A又は抗Bヘモリシンを含有し、それは同じABO型の患者、あるいは少なくともヘモリシンに対応する抗原A又はBを欠損する患者において保持されなければならない(赤血球の血液適合性とは反対)。これらの要因を考慮に入れなければ、輸血後、受容者側において、ABO式適合性における不適合による赤血球の溶血を生じさせることもありうる。
【0023】
更に、Rhesusシステムの抗原Dに対する同種免疫のいかなるリスクをも回避する目的で、特にリスクの大きい患者(少女、出産期の年齢の女性、多くの輸血を受けている患者)において灌流適用を実施する必要があり、その際、患者及びドナーはこの抗原のレベルで同じ特性を有する必要がある。
【0024】
第2に、FFPのリン酸濃度(特にin viroで弱毒化された血漿(VAP)における)が非常に高い(9〜12mmol/l)ため、FFPによりHUS患者におけるリン酸過剰血症が生じ、また当該HUS患者は腎機能不全に罹患することとなる。VAP中の高いリン酸濃度(高リン酸血症)は、VAPの輸血を受ける患者において生じると考えられ、その原因して主なものとしては、−輸血されるVAPの体積が多量であること、−毎日反復輸血されること、−患者が既に腎機能不全に罹患していること、−患者が既に高リン酸血症に罹患していること、が挙げられる。
【0025】
次に、灌流適用されたFFPの量によっては、タンパクの過剰及び/又はクエン酸の過剰が生じ、それにより循環カルシウムの濃度が減少する。
【0026】
最後に、FFPによってアレルギー、及び感染源の侵入のリスクが生じる。実際、現在行われている検出及び不働化方法は、潜在的に凍結された新鮮な血漿に存在する感染源の検出及び除去を可能にするための充分な感度及び能力を必ずしも有しているわけではない。
【0027】
灌流適用された体積が、利尿によって排出され、通常の動脈圧が維持されるにはあまりに大きい場合、凍結された新鮮な血漿灌流適用と血漿交換との組み合わせは重要である。しかしながらこの組み合わせは、主に血管への介入(中枢ルートが必要)、ロード量の過多、アナフィラキシー反応、凝血塊の問題及びウイルス性疾患の感染などの新たなリスクを生じさせるものである。
【0028】
更に、血漿交換法は幼児に適用するのが困難である。
【0029】
他の治療方法としては、腎臓移植が挙げられる。しかしながら、移植後の再発リスクが非常に高い。
【0030】
更に、aHUS患者(非典型的なHUS)の腎移植後の診断は困難なことが多い。すなわち、再発であるのか、あるいは急性の血管拒絶反応又は移植片のバイオプシーにおける慢性拒絶反応であるのかを識別するのが困難であることが多い。
【0031】
再発の治療方法としては、凍結された新鮮な血漿の灌流適用や、血漿灌流適用の有無にかかわらない血漿交換法などが挙げられるが、その結果を予測することは不可能に近い。これらの予測不可能な結果の原因としては、FFP灌流適用の回数及び量が挙げられ、すなわち、各灌流適用が同種のバッチ由来ではなく、幾つかのドナー由来の血漿の集積物由来であることが主な原因である。
【0032】
因子Hは肝臓で合成されるため、肝臓移植又は肝臓−腎臓の複合移植が提唱されることは一見合理的であるようにも思える。しかしながらかかる移植は、常に内科医及び両親による困難な選択を必要とし、また操作上のリスク及び肝臓移植による拒絶反応のリスクを伴うものである。
【先行技術文献】
【非特許文献】
【0033】
【非特許文献1】Rougier N,Kazatchkine MD,ら、Human complement factor H deficiency associated with hemolytic uremic syndrome,J.Am.Soc.Nephrol.1998、9:2318−2326
【非特許文献2】Norisら、Hypocomplementemia discloses genetic predisposition to hemolytic uremic syndrome and thrombotic thrombocytopenic purpura:role of Factor H abnormalities,J.Am.Soc.Nephrol.1999,10:281−293
【非特許文献3】Warwickerら、Genetic studies into hemolytic uremic syndrome,Kidney Int.,1998、53:836−844
【非特許文献4】Warwickerら、Familial relapsing hemolytic uremic syndrome and complement Factor H deficiency,Nephrol.Dial.Transplant.,1999、14:1229−1233
【非特許文献5】Buddlesら、Complement Factor H gene mutation associated with autosomal recessive atypical hemolytic uremic syndrome,Am.J.Hum.Genet.,2000、66:1721−1722
【非特許文献6】Caprioliら、The molecular basis of familial hemolytic uremic syndrome:mutation analysis of Factor H gene reveals a hot spot in short consensus repeat 20,J.Am.Soc.Nephrol.2001、12:297−307
【非特許文献7】Ohaliら、Hypocomplementemic autosomal recessive hemolytic uremic syndrome with decreased Factor H,Pediatr.Nephrol.1998、12:619−624
【非特許文献8】Yingら、Complement Factor H gene mutation associated with autosomal recessive atypical hemolytic uremic syndrome,Am.J.Hum.Genet.1999、65:1538−1546
【発明の概要】
【発明が解決しようとする課題】
【0034】
従来技術のこれらの欠点を克服するべく、発明者らによる鋭意研究の結果、驚くべきことに、HUSの治療薬の製造に因子Hが利用できることを見出した。
【課題を解決するための手段】
【0035】
因子H、例えば凍結させた新鮮な血漿に由来する因子H濃縮物を用いることにより、注入体積及び注入時間を減少させつつ、HUSに罹患する患者の因子Hの欠損症を、安全かつ安定的に治療することが可能となる。特に、肝臓移植の直後の期間に因子Hを投与することによって、移植された肝臓における低い因子H産生を補填し、それにより移植片における症状の再発及び拒絶反応を防止することが可能となる。
【0036】
本発明はまた、以下のステップを含んでなる因子Hの精製方法の提供に関する。
1)血漿の寒冷型沈降物の上澄を調製するステップと、
2)この上澄をアニオン交換体タイプのゲル/樹脂上のクロマトグラフィに供するステップと、
3)非保持フラクションをヘパリンタイプのグラフト化リガンドを含有するゲル/樹脂上のクロマトグラフィに供するステップと、
4)ステップ(3)のクロマトグラフィの後、非保持フラクションのpHを調整し、ヘパリンタイプのグラフト化リガンドを含有するクロマトグラフ担体ゲル/樹脂へH因子を結合させるステップと、
5)ゲル/樹脂の平衡化用バッファよりもイオン強度の強いバッファでH因子を溶出させるステップと、
6)溶出フラクションを希釈し、更にそれを強酸性のカチオン交換タイプのゲル/樹脂上のクロマトグラフィに供するステップと、
7)ゲル/樹脂の平衡用バッファのよりもイオン強度の強いバッファでH因子を溶出させるステップと、
8)溶出フラクションを希釈し、更にそれを強塩基性のアニオン交換タイプのゲル/樹脂上のクロマトグラフィに供するステップと、
9)ゲル/樹脂を洗浄し、H因子を溶出させるステップと、
10)H因子の濃縮物を調製するステップ。
【図面の簡単な説明】
【0037】
【図1】補体は2つの異なるルート(従来のルート及び代替のルート)。
【図2】因子Hの精製方法のダイアグラム。
【図3】因子HによるC3コンベルターゼの分解。
【発明を実施するための形態】
【0038】
本発明の主要な目的は、溶血性尿毒症症候群(HUS)(特に典型的な形式のHUS又は非典型的な形式のHUS)の治療用薬剤の製造への因子Hの使用方法の提供である。
【0039】
本発明の好ましい実施形態は、溶血性尿毒症症候群の治療用薬剤を調製するための因子Hの使用であり、当該因子Hは、新鮮なヒト血漿から精製されたものであるか、又は当業者に公知の標準的な方法による精製物に由来する血漿のフラクションである。この精製方法は当業者に公知である。例えば、リシン−セファロース、QAE−セファデックス、DEAE−トヨパール、セファクリルS−300及びヒドロキシアパタイトのカラムを用いたクロマトグラフィなどが挙げられる。その詳細は、以下の文献に記載されている:Fearon,J.Immunol.119,1248−1252(1977)、Crossleyら、Biochem.J.,191,173−182(1980)、Nagasawaら、J.Immunol.,125,578−582(1980)、Weilerら、P.N.A.S.,73,3268−3272,(1976)及びWhaleyら、J.Exp.Med.,144,1147−1163(1976)。凍結された新鮮な血漿からの精製により得た因子Hは例えば、因子H濃縮物の形で得られる。
【0040】
本発明の他の実施形態は、溶血性尿毒症症候群の治療用薬剤を調製するための因子Hの使用であり、当該因子Hは、細菌、酵母、菌類又は哺乳類細胞からなる群から選択される細胞内でその遺伝子を発現させる、遺伝子工学的方法によって得られる。
【0041】
本発明の具体的実施形態は、溶血性尿毒症症候群の治療用薬剤を調製するための因子Hの使用であり、上記のように得られた薬剤は、冷凍乾燥させた形態である。
【0042】
本発明の更なる実施形態は、溶血性尿毒症症候群の治療用薬剤を調製するための因子Hの使用であり、上記のように得られた薬剤を、少なくとも1つの感染源を除去するか又は不活性化するための少なくとも1つのプロセスに供することを特徴とする。感染源としては、ウイルス及び従来しない伝染性物質(NCTA)(例えばプリオンタンパク)などが挙げられる。
【0043】
特に、当該薬剤を、ウイルス的に不活性化させるのが好ましい。「ウイルスを不活性化する」とは、当該薬剤を、化学物質(例えば溶剤/洗浄剤)、及び/又は熱(例えば乾熱滅菌又は殺菌処理)、及び/又はナノ濾過などの、当業者に公知の少なくとも1つのウイルス不活性化方法に供することを意味する。これらのいずれかの方法で不活性化できるウイルスとしては、ヒト免疫不全症ウイルス(HIV)、A型肝炎ウイルス(HAV)、B型肝炎ウイルス(HBV)、B19パルボウイルス、細胞拡大ウイルス(CMV)、ブタパルボウイルス、ポリオウイルス、ウシのウイルス性下痢ウイルス(BVDV)などが挙げられる。
【0044】
本発明の別の目的は、冷凍乾燥され、ウイルスを不活性化された、上記の実施形態に係る医薬組成物であり、因子H、並びに薬理学的に許容できる賦形剤及び/又は担体を含んであることを特徴とする。
【0045】
本発明の別の目的は、以下のステップを含んでなる、因子Hの精製方法に関する:
1)血漿の寒冷型沈降物の上澄を調製するステップと、
2)この上澄をアニオン交換体タイプのゲル/樹脂上のクロマトグラフィに供するステップと、
3)非保持フラクションをヘパリンタイプのグラフト化リガンドを含有するゲル/樹脂上のクロマトグラフィに供するステップと、
4)ステップ(3)のクロマトグラフィの後、非保持フラクションのpHを調整し、ヘパリンタイプのグラフト化リガンドを含有するクロマトグラフ担体ゲル/樹脂へH因子を結合させるステップと、
5)ゲル/樹脂の平衡化用バッファよりもイオン強度の強いバッファでH因子を溶出させるステップと、
6)溶出フラクションを希釈し、更にそれを強酸性のカチオン交換タイプのゲル/樹脂上のクロマトグラフィに供するステップと、
7)ゲル/樹脂の平衡用バッファのよりもイオン強度の強いバッファでH因子を溶出させるステップと、
8)溶出フラクションを希釈し、更にそれを強塩基性のアニオン交換タイプのゲル/樹脂上のクロマトグラフィに供するステップと、
9)ゲル/樹脂を洗浄し、H因子を溶出させるステップと、
10)H因子の濃縮物を調製するステップ。
【0046】
本発明の具体的実施形態では、ステップ3における、ヘパリンリガンドがグラフト化されているクロマトグラフ保持体は、セファロースヘパリンゲル/樹脂である。
【0047】
本発明の具体的実施形態では、ステップ4における、ヘパリンリガンドがグラフト化されているクロマトグラフ保持体は、セファロースヘパリンゲル/樹脂である。
【0048】
本発明の具体的実施形態では、ステップ6における、強酸性のカチオン交換タイプのゲル/樹脂上のクロマトグラフィは、SPセファロースタイプのクロマトグラフィである。
【0049】
本発明の具体的実施形態では、ステップ8における、強塩基性のアニオン交換タイプのゲル/樹脂上のクロマトグラフィは、QセファロースFFタイプのクロマトグラフィ又はその均等物である。
【0050】
好適には、ステップ4の非保持フラクションのpHを、pH5.5〜pH6.5の範囲、好ましくはpH6.0となるように調整する。
【0051】
好適には、ステップ8の希釈されたフラクションのpHを、pH6.5〜pH7.5の範囲に調整する。
【0052】
本発明の精製方法は、血漿に由来する因子Hを精製するための、工業的実用化の可能な唯一の方法であると考えられ、当該方法により、精製された因子H濃縮物を、化学物質若しくは合成プロテアーゼに対する阻害剤の非存在下で得ることが可能となり、それにより、これらの阻害剤が最終生産物中に検出されることはなくなる。
【0053】
実際、現在行われているヒト血漿から因子Hを精製する方法は基礎研究において用いられるが、それは沈殿精製技術(例えばPEG、硫酸アンモニウム沈殿)及びプロテアーゼインヒビターを使用して実施されることが多いため、産業的な実施が困難なことが多い。これらのプロテアーゼインヒビターは、因子Hの分子内のアスパラギン323及びアスパラギン324のアミノ酸間の結合を切断する役割を果たすトリプシンタイプのタンパク質(血清及び血漿に存在)の活性を阻害する。したがって、プロテアーゼインヒビターの添加により、この因子のタンパク質分解の減少、及びその安定性の向上に関与する。しかしながら、プロテアーゼインヒビターは毒性化合物であることが多く、治療用途に用いる因子Hの産業的な調製方法へのその使用は不適当である。
【0054】
更に、本発明の方法は、得られる因子Hの濃縮物では、主要な3タイプの活性が保持されるという利点を有し、一方現在行われている技術で得られる因子Hは、かかる活性が保持されない。本発明の方法によって得られた因子Hは、補体の代替ルートにおける中心的な制御活性(HUS、特に非典型的なHUSに罹患する患者に不足すると考えられる活性)を発揮することができる。特に、本発明の方法によって調製される因子Hは、補体の代替ルートにおいても、予備成形されたC3コンベルターゼを分離させる活性を保持しており、HUSの治療の際に、その最大限の機能活性で使用することができる。
【0055】
更なる本発明の方法によって調製される因子H濃縮物は1付近の比活性(AS=0.8〜0.9)を有し、凍結された新鮮な血漿の溶液(AS=0.008)よりも高い効果を有するものの、本願明細書の冒頭で説明したように、治療効果は若干あっても、多くの不利益をもたらすこともある。これらの不利益としては、血漿の投与により、HUSを治療するための不必要な更なるタンパク質(アルブミン、フィブリノーゲンなど)が生体内に導入され、それは一方では、タンパクの過剰ロードに起因する望ましくない反応の誘導や、又は「血清病」として公知のアレルギー反応の誘導の原因ともなる。
【0056】
最後に、血漿に存在する感染性のウイルスの不活性化は、血液製剤中に存在するウイルスの不活性化のために通常用いられる方法よりも通常困難であり、実用的でない。しかしながら、本発明の方法によって調製された因子H濃縮物は、定式化されている試験済みの方法により、ウイルス安全性を担保することができる。
【実施例】
【0057】
<実施例1>:因子Hの精製方法:
因子Hの精製に適用した方法を、図2に図式的に示す。
【0058】
ヒトの凍結された新鮮な血漿を1℃〜6℃の温度で融解させ、寒冷型沈降物を遠心沈降に供し、寒冷型沈降物の不溶性画分と血漿上澄画分とを分離させた。
【0059】
得られた寒冷型沈降物の血漿上澄(因子H濃縮物、約400〜約500mg/Lの濃度で因子Hを含有)を、アニオン交換体タイプの樹脂/ゲル(例えばDEAEセファデックスタイプのゲル/樹脂)を用いたクロマトグラフィに供し、樹脂/ゲル上のこれらの因子を保持させ、血漿上澄からビタミンKに依存する形で因子を分離させた。
【0060】
非保持の血漿上清画分(フラクションA)(因子H濃縮、約400〜約500mg/Lで因子Hを含有)を更に、ヘパリンセファロースFFタイプのゲル/樹脂上のアフィニティークロマトグラフィに供し、樹脂/ゲル中に抗トロンビンIIIを保持させ、抗トロンビンIIIをこのフラクションAから分離させた。
【0061】
この非保持フラクションA(フラクションB)(因子H濃縮物、約300〜約400mg/Lで因子Hを含有)のpHを、pH5.5〜pH6.5の範囲、好ましくはpH6.0に調整した。
【0062】
フラクションB(pHが調整された)を、ヘパリンセファロースFFタイプの第2のゲル/樹脂、又は、ヘパリンタイプのリガンドをグラフト化された他の任意のクロマトグラフ保持体を用いたクロマトグラフィに供した。血漿タンパク分画Bに含まれる大部分のタンパクを更にクロマトグラフィ濾液によって溶出させた。ゲル/樹脂にわずかに吸着したタンパクを、一連の洗浄及び前溶出工程によって除去した。ゲル/樹脂に保持される因子Hは更に、ゲル/樹脂の平衡化に使用するバッファより強いイオン強度を有するバッファを用いて溶出した。
【0063】
因子H(フラクションC)を含有する溶出フラクションを希釈し、更に強酸性のカチオン交換タイプのゲル/樹脂(例えばSPセファロースFfタイプ又はそれと同様のゲル/樹脂)によるクロマトグラフィに供した。ゲル/樹脂にわずかに吸着したタンパクを、一連の洗浄及び前溶出工程によって除去した。ゲル/樹脂に保持される因子Hは更に、ゲル/樹脂の平衡化に使用するバッファより強いイオン強度を有するバッファを用いて溶出した。
【0064】
因子H(フラクションD)を含有する溶出フラクションを更に、溶媒若しくは界面活性剤(例えばポリソルベート80及びTnBP)による処理による、ウイルスの不活性化ステップに供した。かかる処理を行うことにより、ウイルス(特にカプセルを有するタイプのウイルス)を効率的に不活性化することが可能である。
【0065】
更にフラクションDを希釈し、このフラクションのpHを、pH6.5〜pH7.5の範囲となるように調整した。フラクションDを更に、強塩基性のアニオン交換タイプのゲル/樹脂(例えばQセファロースFFタイプ又は当量のゲル/樹脂)を用いたクロマトグラフィに供した。一連の洗浄の後、ゲル/樹脂に保持された因子Hを、ゲル/樹脂の平衡化用のバッファより強いイオン強度を有するバッファを用いて溶出した。
【0066】
上記の界面活性剤タイプの溶剤による処理に用いた、ウイルスの失活のために用いた物質を、クロマトグラフィ工程の間に除去することにより、因子Hの純度が向上した。
【0067】
因子H(フラクションE)を含有する溶出フラクションを更に、約15nmの空隙を有するフィルタを用いたナノ濾過に供し、ウイルスを除去した。このウイルス除去処理により、ウイルス(特に小型のカプセルを有するウイルス)を効率的に除去することができる。得られる溶液(フラクションF)を、最後に濃縮し、限外濾過により調整し、更に0.22μmのフィルタで濾過した。
【0068】
上記の精製方法の収量、及びこの方法により精製された因子Hの比活性を、2つの異なるバッチ毎に測定した。それぞれの結果を表1に示す。比活性(A.S.)を、タンパク質1mg当たりの因子Hタイプの抗原のmgとして表す。
【0069】
【表1】
【0070】
<実施例2>:因子Hの活性の測定方法:
ELISAプレート(96ウェルタイプ)のウェルを、0.2Mの炭酸ナトリウムバッファ中の2.5g/mL(Calbiochem社:ref.341274)の濃度の精製C3bタンパク質溶液で被覆した。その際、溶液100μLをウェルに添加し、プレートを37℃で1時間、及び4℃で一晩静置した。
【0071】
10mMのリン酸ナトリウム緩衝液(pH7.2)、25mMのNaCl、0.1%のTween20の溶液で、300μL/ウェルで3回洗浄した。
【0072】
非特異的な部位を更に、10mMのリン酸ナトリウム緩衝液(pH7.2)、25mMのNaCl、0.05%のTween 20、1%のウシ血清アルブミンを含有する溶液300μL/ウェルで、37℃で1時間静置して飽和させた。次に、上記の洗浄溶液でウェルの洗浄を実施した。
【0073】
以下の成分(20mMのNiCl2のストック溶液(1.5mM最終濃度) 75μL、1mg/mlのB因子(Calbiochem ref.341262) 4μL、1mg/mlのD因子(Calbiochem ref.341273) 3μL、10mMのリン酸ナトリウム緩衝液(pH7.2、25mMのNaCl、4%のウシ血清アルブミンを含有する) 918μL)を含有する100μLの溶液を、各ウェルに添加し、37℃で2時間インキュベートした。
【0074】
10mMのリン酸ナトリウム緩衝液(pH7.2)、25mMのNaCl、0.1%のTween20の溶液で、300μL/ウェルで3回洗浄した。
【0075】
因子H溶液を、20μg/mL、10μg/mL、1μg/mL、0.25μg/mL、0.0625μg/mL、0.015625μg/mL、0.00390625μg/mL及び0.001μg/mLの因子H濃度となるように調製した。各ウェルに100μLの溶液を添加し、37℃で30分間インキュベートした。
【0076】
更に10mMのリン酸ナトリウム、25mMのNaCl、0.1%のTween 20を含有する緩衝溶液(pH7.2)で、300μL/ウェルで3回洗浄した。
【0077】
ヤギ抗ヒトB因子抗体溶液(Calbiochem ref.:341272)を、0.1%のウシ血清アルブミンを含有するPBSバッファ(Sigma社、P−3813)(pH7.4)で1/2,000に希釈し、更に希釈液100μLをウェルに添加し、37℃で1時間インキュベートした。
【0078】
PBS(pH7.2、0.1%のTween20を含有)で、300μL/ウェルで3回洗浄した。次に、0.1%のウシ血清アルブミン含有PBSで1/10,000に希釈した、ペルオキシダーゼ(Calbiochem ref.401515、1mg/mL)標識し抗ヤギウサギ抗体を含有する溶液100μLをウェルに添加し、室温20〜25分間インキュベートした。
【0079】
PBS(pH7.2、0.1%のTween20を含有)で、300μL/ウェルで3回洗浄した。
【0080】
クエン酸ナトリウム溶液中の5mg/10mLのOPDペルオキシダーゼ(Sigma社)基質を最後に100μL/ウェルの量で、更にH2O2を10μLでウェルに添加した。反応混合液を約10分間ウェルと接触させ、更にウェルあたり50μLの4N H2SO4を添加し、反応を停止させた。
【0081】
次に、ウェル中の溶液の吸光度を492nmの波長で測定した。その結果を図3に示す。図3に、因子Hの濃度又はタンパク濃度(SAH)に対する吸光度測定値の関係を示すグラフを示す。
【0082】
因子Hの活性を測定するための同様の方法は、McRaeら、The Journal of Immunology、2005、174:6250−6256に記載されている。
【特許請求の範囲】
【請求項1】
溶血性尿毒症症候群(HUS)の治療用薬剤の調製のための、ヒト因子Hの使用。
【請求項2】
前記薬剤が、典型的な形のHUSの治療を目的とするものである、請求項1記載の使用。
【請求項3】
前記薬剤が、非典型的な形のHUSの治療を目的とするものである、請求項1記載の使用。
【請求項4】
前記ヒト因子Hが、細菌、酵母、菌類又は哺乳類の細胞からなる群から選択される細胞において、ヒト因子Hの遺伝子を発現させることによって、遺伝子工学的に調製される、請求項1記載の使用。
【請求項5】
前記薬剤が、冷凍乾燥形態で調製される、請求項1から4のいずれか1項記載の使用。
【請求項6】
前記薬剤が、少なくとも1つの感染源を除去又は不活性化する少なくとも1つの工程に供される、請求項1から5のいずれか1項記載の使用。
【請求項7】
前記薬剤が、ウイルスを不活性化するための少なくとも1つの工程に供される、請求項1から6のいずれか1項記載の使用。
【請求項8】
ヒト因子H、並びに薬理学的に許容できる賦形剤及び/又は担体を含んでなる、ウイルスを不活性され、冷凍乾燥させた医薬組成物。
【請求項1】
溶血性尿毒症症候群(HUS)の治療用薬剤の調製のための、ヒト因子Hの使用。
【請求項2】
前記薬剤が、典型的な形のHUSの治療を目的とするものである、請求項1記載の使用。
【請求項3】
前記薬剤が、非典型的な形のHUSの治療を目的とするものである、請求項1記載の使用。
【請求項4】
前記ヒト因子Hが、細菌、酵母、菌類又は哺乳類の細胞からなる群から選択される細胞において、ヒト因子Hの遺伝子を発現させることによって、遺伝子工学的に調製される、請求項1記載の使用。
【請求項5】
前記薬剤が、冷凍乾燥形態で調製される、請求項1から4のいずれか1項記載の使用。
【請求項6】
前記薬剤が、少なくとも1つの感染源を除去又は不活性化する少なくとも1つの工程に供される、請求項1から5のいずれか1項記載の使用。
【請求項7】
前記薬剤が、ウイルスを不活性化するための少なくとも1つの工程に供される、請求項1から6のいずれか1項記載の使用。
【請求項8】
ヒト因子H、並びに薬理学的に許容できる賦形剤及び/又は担体を含んでなる、ウイルスを不活性され、冷凍乾燥させた医薬組成物。
【図1】


【図2】

【図3】
【図2】
【図3】
【公開番号】特開2012−211189(P2012−211189A)
【公開日】平成24年11月1日(2012.11.1)
【国際特許分類】
【出願番号】特願2012−168827(P2012−168827)
【出願日】平成24年7月30日(2012.7.30)
【分割の表示】特願2008−543870(P2008−543870)の分割
【原出願日】平成18年12月7日(2006.12.7)
【出願人】(508168815)ラボラトワール フランセ デュ フラクションヌメント エ デ バイオテクノロジーズ ソシエテ アノニム (5)
【Fターム(参考)】
【公開日】平成24年11月1日(2012.11.1)
【国際特許分類】
【出願日】平成24年7月30日(2012.7.30)
【分割の表示】特願2008−543870(P2008−543870)の分割
【原出願日】平成18年12月7日(2006.12.7)
【出願人】(508168815)ラボラトワール フランセ デュ フラクションヌメント エ デ バイオテクノロジーズ ソシエテ アノニム (5)
【Fターム(参考)】
[ Back to top ]