
培養培地にプロスタグランジン又はプロスタグランジン類似物を補足することによってインビトロ胚発生を高めるための方法及び組成
哺乳動物胚のインビトロ発生を高める方法が開示されており、胚の完全孵化(すなわち、透明帯から胚の遊離)を促進するのに有効な量において、培養培地にプロスタグランジン、すなわちプロスタグランジン類似物を補足することを有する。培養培地の補足は、好ましくは1又はそれ以上、以下の胚の発生時期の間に行われる、すなわち(a)ゲノムが活性化されて受精卵が桑実胚に発生するとき及び(b)胚の桑実胚から初期胚盤胞への発生の間である。インビトロ受精胚のインビボ着床の可能性及び生児出生の可能性が、その結果高められ、生存に適した妊娠の確立が容易にされる。

【発明の詳細な説明】
【技術分野】
【0001】
連邦政府の支援による研究または開発に関する記載
本発明は、全体的に又は部分的にNational Institute of Child Health and Human Development(NICHD)、契約番号HD01277、から財政的支援を受けて行われるものである。したがって、米国政府が本発明について所定の権利を有する。
【背景技術】
【0002】
発明の分野
本発明は、一般に哺乳動物胚のインビトロ培養のため、及び適した宿主哺乳動物の子宮に培養された胚の着床後における妊娠の達成を高めるための組成及び方法に関する。
【0003】
従来技術の説明
卵管内の環境は、精子の受精の可能性を高め、分裂する胚の発生を促進する。卵管上皮細胞と共培養された胚は、発生、孵化及び着床を向上させたことが示された[Xu、2001年;Xu、2000年]。プロスタサイクリン(PGI2)は、もともと血液及び血管(vascular)のホメオスタシスを維持する点に関与すると考えられている。しかしながら、遺伝子ノックアウトマウスにおける最近の観察結果から示唆されることは、プロスタサイクリンが他の生理的機能を有し得ることである。子宮内膜由来PGI2の重要性が、シクロオキシゲナーゼ(COX)−2ノックアウトマウスにおいて行われた観察によって強調される[Lim、1997年]。COX−2ノックアウトマウスの子宮内膜は、正常には脱落しなかった。結果として、移入された野生型胚(transferred wild type embryo)は、COX−2の欠損した雌マウスには着床できなかった。前述の異常性は、母方に外部からPGI2類似物の投与によってある程度矯正されることができた[Lim、1999年]。
【0004】
着床前胚の発生における卵管由来PGI2の役割が明らかでない理由は、PGI2レセプタ(IP)ノックアウトマウスの一腹の産子数が野生型の産子数と比較されていないからである[Murata、1997年]。しかしながら、ヘテロ接合体IPノックアウトマウスとの交配から生まれる子の遺伝子型の分布がメンデルの法則に従っていなかった。すなわち、ホモ接合体IPノックアウト遺伝子型を有する雄及び雌の子の普及率は、それぞれ37%及び20%であり、予想されるよりも低かった[Murata、1997年]。
【0005】
【特許文献1】米国特許第6100249号公報
【特許文献2】米国特許第5096822号公報
【発明の開示】
【0006】
発明の簡単な概要
最近、我々は、ヒト卵管がたくさんのプロスタサイクリン(PGI2)及びPGE2を合成すること発見した[Huang、2002年]。遺伝子ノックアウトの研究は、子宮内膜由来PGI2が子宮内膜の脱落に必須であることを示唆するが、着床前胚の発生におけるPGI2の効果は報告されていなかった。着床前胚の最適な発生にはPGI2を必要とするという仮説を用いて、マウス胚におけるPGI2の効果が、完全孵化率を基にして検査された。PGI2レセプタ(IP)の発現は、ウエスタンブロット解析及び免疫組織化学によって評価された。PGI2の胚への結合は、ラジオリガンド結合検定法によって確認された。我々の結果は、マウス胚の孵化はアイロプロスト(Iloprost)(ED506.7nM)、すなわち安定化したPGI2合成類似物の補足によって高められることを明らかにしている。8細胞から桑実胚に至る間、又は桑実胚から初期胚盤胞に至る間に、アイロプロストへの暴露が、孵化を促進させるのに重要であった。PGI2に対するこの時期特有の反応が、IPの発生時期特有の発現と同時に起こった。さらなる研究は、高められる胚の孵化を着床及び生児出生の増加と関連づけるために実施され、すなわちアイロプロストを補足された培地中で培養されたマウス胚が、妊娠キャリアに移入された。妊娠嚢及び生存する子の数がコントロール胚のその数と比較された。報告によれば、培養培地の添加物が胎児の体重を増加させるため[DeBaum、2002年;Thompson、1995年;Sinclair、1999年]、子及び胎盤の重量も比較された。我々の結果は、アイロプロストを補足された培養培地中で胚を培養することは、子及び胎盤の重量に影響を与えることなく、マウス胚の着床及び生児出生を高めることを示している。
【0007】
したがって、本発明のある実施例においては、培養培地にプロスタグランジン、例えばPGI2又はPGE2又はその類似物を補足することを含む胚のインビトロ発生を高める方法が提供されている。ある実施例において、プロスタグランジンはPGI2又はPGE2であり、またある実施例では、プロスタグランジン類似物はアイロプロスト又はPGE1である。ある実施例では、方法は、培養培地に胚の完全孵化を促進するのに十分量のプロスタグランジン又は類似物を添加することを含む。好ましくは、「完全孵化」は、胚が透明帯(zona pellucida)から離れることを有する。方法のある実施例において、培養培地の補足は、ゲノムが活性化されて受精卵が桑実胚へ発生する期間である、胚の初期発生時期の間に行われる。胚がヒトである実施例において、初期発生は4乃至8細胞状態で開始する。方法のある実施例において、培養培地の補足は、胚の桑実胚から初期胚盤胞への発生の間に行われる。ある実施例では、胚の初期発生から桑実胚期への発生と、胚の桑実胚から初期胚盤胞への発生の間との両方の間において行われる。方法のある実施例において、胚がマウス起源にある場合、補足する段階は、その2細胞発生の時期に胚を採取後24乃至42時間の間に、培養中の胚をプロスタグランジン又は類似物に暴露することを含む。胚がマウスである方法のある実施例では、桑実胚から初期胚盤胞の間に補足する段階は、その2細胞発生の時期に胚を採取後42乃至72時間の間に、胚を一度にプロスタグランジン又はプロスタグランジン類似物に暴露することを有する。
【0008】
また、本発明のある実施例にしたがって提供されることは、インビトロ受精胚のインビボ着床の可能性を増加させる方法である。「着床の可能性」とは、子宮に着床する胚の能力である。この方法は、胚のインビトロ発生を高めるために、上述の実施例の一つを実行することを含むことにより、培養中の胚の完全な孵化が達せられる、すなわち、別のIVF方法に比べて孵化が高められる。この方法のある実施例にしたがって、完全に又は殆ど孵化された胚が、それから宿主哺乳動物の子宮に導入されて、胚の高められた着床が達せられる。ある実施例では、インビトロで胚の完全孵化は、生存に適した妊娠の確立と相関する。
【0009】
本発明のある実施例において、インビトロで受精された哺乳動物胚の生児出生の可能性を増加させる方法が提供されている。「生児出生の可能性」とは、生児出生を生じる胚の能力である。上述されるように、この方法はプロスタグランジン又はその類似物を有するインビトロ培養によって、胚のインビトロ発生を高めることを有することにより、高められる孵化の可能性、すなわち培養中の胚の完全孵化が達せられる。好ましい実施例において、このような処理は、胚が培養から取り出されてインビボ環境にある後にさえも、完全孵化のための高められる可能性を与えている。この性質は「高められる孵化の可能性」として説明される。孵化される胚、すなわち高められる孵化の可能性を有する胚が、宿主哺乳動物の子宮に移入されて、胚が着床するようになりインビボで成長することにより、生児出生を生じる胚の能力が、孵化の高まりを有せず着床された胚の能力と比較して高められている。
【0010】
本発明のさらにもう一つの実施例においては、インビトロで受精された哺乳動物胚の一群における生児出生率を増加させる方法が、上述の方法に従って胚のインビトロ発生を高めることを有することを提供されていることにより、孵化の高まり、すなわち好ましくは培養中の胚の完全孵化が達せられる。方法は、宿主哺乳動物に胚を導入、又は移入すること、及びそれから胚をインビボで成長できるようにすることを含んでおり、高められる孵化を有する胚の群からの生児出生率は、最初に完全に孵化されていない、すなわち孵化の高まりを受けていない状態で、宿主子宮へ移入される胚の群からの生児出生率よりも非常に高い。
【0011】
なおもさらに、本発明のある実施例に従って提供されていることは、哺乳動物胚のインビトロ発生のための改良された細胞培養培地であり、その改良点は、孵化を高め、また好ましくはインビトロで胚の完全孵化を促進するのに有効な補足量のプロスタグランジン又はその類似物を含むことである。ある実施例において、培地は、ある量のプロスタグランジン又はその類似物を含んでおり、それはインビトロ培養から取り出されて宿主哺乳動物の子宮に移入された後に、胚の完全な孵化の可能性を高めるために有効である。これら及びその他の実施例、本発明の特徴及び利点は、以下の説明及び図を参照することによって明白になるであろう。
【0012】
好ましい実施例の詳細な説明
本開示は、精子及び胚に対するPGI2並びにPGE2の効果の研究及びそれに続く研究の過程で明らかになった。我々の最近の結果は、ヒト[Huang、2002年]及びマウス[Huang、2004年]卵管上皮細胞が、PGI2の合成、すなわちCOX-1又はCOX-2及びPGI2合成酵素に必須の酵素を発現することを示している。たくさんのPGI2又はPGE2は、14C−アラキドン酸がヒト[Huang、2002年]又はマウス卵管[Huang、2004年]から調製されたミクロソームとインキュベーションされたときに産生された。本研究はまた、精子が卵と受精するために卵管末端に移動し、胚発生の最初の72時間は卵管で起こるという事実を考慮した。
【0013】
本研究の一部分として、胚に対するPGI2の影響が、PGI2類似物を伴って培養されたマウス胚の完全な孵化を観察することによって評価された。そのメカニズムをさらに解明するために、IPの発現及び放射ラベルされたアイロプロストによる胚の結合がまた検討された。
【0014】
材料及び方法
試薬の出所及び制度上の承認。
別のものであると記載しない限り、試薬はシグマ社(Sigma Co.)(アメリカリ合衆国、ミズーリ、セント.ルイス)(St.Louis、MO、USA)から購入されたものである。ヒトのサンプルの使用は、ヒューマンサブジェクツの保護のための委員会(the Committee for the Protection of Human Subjects)(Institute Review Boardと同等)によって承認された。実験動物の取扱い及び研究プロトコルは動物福祉委員会によって認定された。
【0015】
マウス胚の採取及び培養。
マウスは、水及び餌へのアクセスが自由で、制御された温度、湿度、及び光周期(12時間明暗周期)条件の下に保った。3週齢のC57B1/6雌マウスはハーラン(Harlan)(アメリカ合衆国、インジアナ、インジアナポリス)(Indianapolis、IN、USA)から購入された。8週齢のC3H雄マウスは最初にハーランから購入され、後にジャクソンラボラトリー(Jackson laboratory)(アメリカ合衆国、メイン、バーハーバー)(Bar Harbor ME、USA)から購入された。雌マウスの過剰排卵は、妊娠雌馬血清ゴナドトロピン(5単位)、及び続く46時間後にヒト絨毛性ゴナドトロピン(hCG、5単位)の腹腔内投与によって行われた。hCGを受けた後、それぞれの雌マウスが一匹の繁殖力のある雄マウスとつがいにされた。48時間後、2細胞胚は卵管から25mMへペス(HEPES)及び1%ウシ血清アルブミン(BSA)(アービン サイエンテイフィツク)(Irvine Scientific)を補足されたα-MEM培地に採取された。
【0016】
胚(群当り17-20個)は、各ウエルに600μLの培地を含む4ウエルデイッシュ(アメリカ合衆国、イリノイス、ネイパービルのナルジ ヌンク インターナショナル ネイパー)(Nalge Nunc International Naper、Naperville、IL、USA)において、37℃で5%CO2の下に培養された。HTF及びα-MEM培地は、96時間連続して用いられて分裂胚の変化する栄養要件を満たした[Gardner、1998年]。HTF培養液(セイジ バイオフアーマ)(SAGE Biopharma)は最初の48時間用い、イーグルの塩類(Earle's salts)及び2mMグルタミンを有するα-MEM培地(アービン サイエンテイフィツク)(Irvine Scientific)は2番目の48時間の間用いた。実験胚は、水中のアイロプロストを受け、コントロール胚は等量の水を受けた。予備実験では、胚の95%以上がKSOM培地(アメリカ合衆国、ニュージャージー、フィリップスバーグのスペシャリテイ−メデイア、セル アンド モレキュラー テクノロジーズ、インク.(Speciality Media、Cell and Molecular Technologies、Inc. Phillipsburg NJ,USA))で培養した胚と同様に、96時間までに胚盤胞になることを示した。96時間培養後、各胚は透明帯(zona pellucida)の有無に対して調べられた。透明帯が完全に無い胚は、完全に孵化したものとして計測された。完全孵化率は、完全に孵化された胚の数を胚の総数で割って求められた。胚の完全孵化を、胚盤胞形成又は胚孵化の代わりに指標として選択したが、この理由は、後者の2マーカーが生存に適した妊娠の確立[Lane、1997年]と相関しないからである。
【0017】
ウエスタンブロット解析(Western blot analysis)。
推定されるマウスIPのアミノ酸配列(417a.a、Genebank_BAA05144)及びヒトIPのアミノ酸配列(386a.a、Genebank_BAA06110)は相同性が高い[Katsuyama、1994年]。予備的な検討では、アフィニテイー精製ポリクローナルペプチド抗体(テキサス大学健康科学センターのケーヘルアン博士(Dr.Ke−He Ruan、University of Texas Health Science Center)から提供)は、マウスIPと交差反応することを確認した。ウエスタンブロット解析は以前に説明したように行われた[Huang、2002年]。簡潔には、60個のマウス胚盤胞由来の全細胞溶解物が、10%アクリルアミドゲル電気泳動(PAGE)によって分離され、ニトロセルロース膜(アメリカ合衆国、ニューハンプシャーのシュレイチャ− アンド シュエル、インク.)(Schleicher & Schuell、Inc.、NH、USA)に転写された。免疫反応タンパクが、抗体とのインキュベーション及び増強された化学発光(アメリカ合衆国、ニュージャージー、ピスカタウエイのアマ−シャム バイオサイエンス)(Amersham Bioscience、Piscataway、NJ、USA)による視覚化によって検出され、またSTORM860レーザースキャナ(アマ−シャム バイオサイエンス)(Amersham Bioscience)を使って検出された。ヒト血小板ミクロソームは、陽性のコントロールとして用いられた。抗体の特異性は、前吸収抗体を用いる平行実験で確かめられた。
【0018】
免疫組織化学、蛍光顕微鏡及び共焦点顕微鏡解析。
マウス胚が4℃で30分間、4%パラホルムアルデヒド中に固定された。リン酸緩衝化生理食塩水(PBS)で3回洗浄後、胚は0.05%のTweenー20、5%の粉末ミルク及び0.1%のTriton X−100含有トリス緩衝化生理食塩水(pH7.4)において、室温で20分間ブロックされた。胚は、2時間ブロッキング緩衝液中でIP抗体(5ng/ml)とインキュベーションされ、それから37℃で30分間、アレクザ(Alexa)488(2.5μg/ml、アメリカ合衆国、アラバマのモレキュラー プローブ(Molecular Probes、AL、USA))と結合したヤギ抗-ラビットIgGとインキュベーションされた。細胞核は、室温で20分間、10μg/mlプロピデウム イオダイド(propdium iodiode)によって対比染色された。胚は、Fluoromount-G(登録商標)(アメリカ合衆国、アラバマ、バーミングハムのサザン バイオテクノロジー アソシエイツ インク.)(Southern Biotechnology Associates Inc.、Birmingham、AL、USA)に埋め込まれた。蛍光顕微鏡解析のために、胚盤胞は封入剤中に置かれてカバースリップが被せられた。共焦点顕微鏡解析のために、およそ50μmのスペーサが、スライドとカバースリップとの間に置かれて胚の3次元形態を保持した。陰性のコントロールには、胚が10ng/mlの非免疫ラビットIgGとインキュベーションされた(すなわち、一次抗体濃度の2倍の濃度)。
【0019】
蛍光顕微鏡解析は、適切なフィルタを備えたツアイス アキシオプラン2(ZeissAxioPlan 2)顕微鏡を用いて行われた。画像は、CCDカメラを用いて捕らえられ、アキシオビジョン プログラム(Vesion 3.0.6)によって処理された。共焦点顕微鏡解析は、オリンパス BX-50顕微鏡に付属されたバイオラッド ラジアンス 2000(BioRad Radiance 2000)共焦点システム(アメリカ合衆国、カルフオル二ア、ハーカルスのバイオラッド ラボラトリーズ)(Bio−Rad Laboratories、Hercules、CA、USA)を使って行われた。その画像は、イメージ プロ プラス プログラム(Image−Pro+ program)(アメリカ合衆国、カルフオル二ア、カールスバッドのメデイア−サイバーネテイックス)(Media−Cybernetics、Carlsbad、CA、USA)を使って処理された。
【0020】
全胚ラジオリガンド結合測定。
3H−アイロプロストの胚盤胞への結合の解析は、わずかに変更修正して以前に説明された [Arbab、2002年] ように行われた。232個が孵化されて孵化する胚盤胞は、結合緩衝液(10mMHEPES、pH7.4中に10mMMnCl2)中で3回洗浄されてから、非標識アイロプロスト(5μM)を有する又は有しない100μlの結合緩衝液に移された。反応は、3H−アイロプロスト(200nM、特異活性11.0Ci/mmol、アマ−シャム バイオサイエンス(Amersham Biosciences))を含む結合緩衝液を加えて、室温で60分間インキュベーションすることによって開始された。その反応は、氷冷洗浄緩衝液(10mMHEPES、pH7.4中に0.01%BSA)に胚盤胞を移すことによって終了された。緩衝液及び胚盤胞は、ガラスフアイバーフィルタ(Whatman GF/C2.4cm)を通してろ過され、フィルタは、2mlの洗浄緩衝液で3回洗浄された。フィルタがオーブン中で乾燥されてから、放射活性が5mlのシンチレーション液で計測されるシンチレーションによって測定された。
【0021】
統計解析
スチュ-デントのt検定又はダネット(Dunnet)検定に従った一元配置分散分析が適所に用いられた。p値<0.05は統計的に有意とみなされた。量反応曲線の作図及びED50値(1.0に設定されるヒル勾配を有して)の計算には、GraphPad Prism(登録商標)Software(アメリカ合衆国、カルフオル二ア、サンデイゴのグラフパッド プリズム ソフトウエア インク.)(GraphPad Prism Software Inc.、San Diego、CA、USA)を備えた。
【0022】
胚移入
マウス胚が採取されて上述のように培養され、その後に本発明者によって公表された[Huangら、2003年]。簡潔には、2細胞胚(C3B6F1)が、hCG投与後42時間、過剰排卵された3週齢のC57Bl/6雌マウスから採取された。胚(群当り14個)は、600μlのHTF培地(アメリカ合衆国、ニュージャージー、ベッドミンスターのセイジ バイオフアーマ)(SAGE Biophama、Bedminster、NJ、USA)を各々含む4ウエルデイッシュ(アメリカ合衆国、イリノイス、ネイパービルのナルジ ヌンク インターナショナル ネイパー)(Nalge Nunc International、Naperville、IL、USA)において、37℃で5%CO2の下に培養された。実験胚は、水中のアイロプロスト(1μM、アメリカ合衆国、ミシガン、アン アーバーのケイメン ケミカル(Caymen Chemical、Ann Arbor、MI、USA))を、またコントロール胚は等量の水を受けた。どちらの場合も水量は培養液の0.1%に満たなかった。
【0023】
胚ドナー(donor)がhCGを受けた翌日に、膣スメアが妊娠可能性キャリア(8週齢のC3B6F1雌マウス)から得られて、発情周期の時期を決定した。発情期のマウスは、精管切除されたICR雄(ハーレン)(Harlan)とつがいにされて翌朝膣栓を調べられた。栓を有する発情期のマウスは0.5日偽妊娠であると指定された。
【0024】
胚移入は、解剖顕微鏡(オリンパスSZ−PT)の下に、2.5日偽妊娠において行われた。外科的麻酔は、ケタミン(ketamine)(200mg/Kg)及びキシラジン(xylazine)(10mg/Kg)(合衆国テキサス州のバーンズ ベトサプライ インク(Burns VetSupply Inc.TX、USA)からの両麻酔薬)の腹腔内投与によって得られた。各子宮角は、2cmの側面切り口からアクセスされた。近位卵管は一対のピンセットで保持されたまま、開口部が30ゲージの皮下針によって、腸間膜の子宮角の遠位端に作り出された。開口部は、135μmの内径を有するトランスフアーピペット(アメリカ合衆国、ニュージャージーのミッドアトランテイックダイアゴノステイックス、インク.)(MidAtiantic Diagnostics、Inc.、NJ、USA)を入らせることができる。0.8μlの移入用培地(25ml HEPES及び1%BSAを加えたα-MEM)中の7個の胚までが、各々の子宮角に移入された。それぞれ移入後、ピペットの内容物が、残留した胚を同定するために、立体写真顕微鏡下で調べられた。側面切り口は、9mmの金属クリップ(アメリカ合衆国、ニュージャージーのクレイ アダムス、パルシッパニー)(Clay Adams、Parsippany、NJ、USA)で閉じられた。一方の子宮角から他方の子宮角への胚の移動によるコントロール胚及び実験胚の混合を避けるために(ドイツのボン大学のアンドレス ツインマー博士との個人情報(Dr.Andreas Zimmer、University of Bon、Germany))、それぞれの妊娠キャリアは、コントロール胚又は実験胚のいずれかを受けた。一定の移入技術を維持するために、胚移入は同じプロトコルに従って、一人の人によって行われる、その人は、判別されている胚が受ける処理には左右されなかった。両群が、研究の過程で(6ヶ月間)得られる経験と等しく利益を得たことを確実にするために、胚は4つのブロックでの処理(コントロール又はアイロプロスト処理)に割り当てられた。
【0025】
着床率の決定
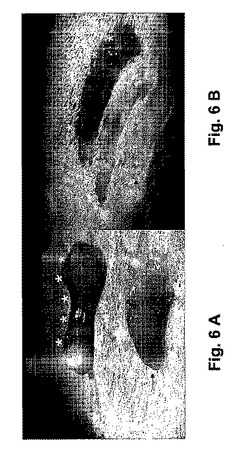
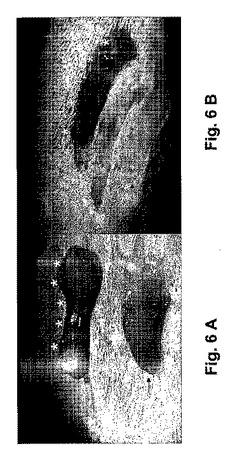
胚移入後72時間目に、着床は、多少変更修正して以前に説明された方法に基いて決定された[Pariaら、1993年]。簡潔には、安楽死前3分間、0.1mlのシカゴブルー(1%)が、妊娠マウスの尾静脈から投与された。そのマウスを殺した後、子宮角が取り出された。子宮角を取り囲む別々のブルーバンド(アスタリスクで示される)が計測された(図6A)。これらのバンドの存在は、着床部位への血管補給の増加を反映している。それから子宮角が開かれ、妊娠嚢が計測された(図6B)。着床率は、移入された胚当りの妊娠嚢の数として表現された。
【0026】
生児出生率と子及び胎盤の重量の決定
予備的な検討は、一腹の大きさが小さいときには、妊娠キャリアが子を共食いするこを示した。子を正確に計測し、体重を量るために、我々は、胚移入後14日目に妊娠キャリアを殺した(妊娠期間の16.5日目、すなわち自然に誕生する2日前)。殺した後、生きている子の数を計測し、個々の子及び胎盤の重量が決定された。全ての子が全体の異常を調べられた。空の妊娠嚢の数がまた、着床率の決定のために計測された。生児出生率は、移入される胚当りの生存する子の数として表現され、着床率は、移入される胚当りの生存する子を有し又は有しない嚢の数として表現された。
【0027】
統計解析
フィッシャーの直接確率検定が、比率を比較するために用いられ、スチュ-ダントのt検定が体重の比較のために用いられた。p値<0.05は統計的に有意であると認められた。GraphPad Instat(登録商標)Software(アメリカ合衆国、カルフオル二ア、サンデイゴのグラフパッド プリズム ソフトウエア インク.)(GraphPad Software Inc.、San Diego、CA、USA)が、統計解析に用いられた。
【0028】
結果
PGI2はマウス胚の完全孵化を高めた。
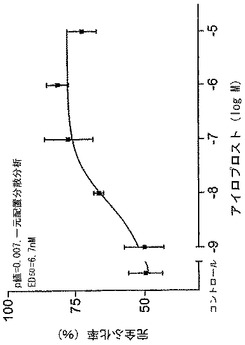
マウス胚の完全孵化が、濃度に依存するかたちでアイロプロストによって高められた。その効果は0.1μM又はそれ以上の濃度(図1) で統計的に有意であった。完全な胚の孵化の最大増加は、1μMで起こっており、実験胚の81±7%(平均値±標準偏差、n=3)が完全に孵化した。対照的に、コントロール胚の49±14%(平均値±標準偏差、n=5)のみが完全に孵化した。量反応曲線からのED50値は6.7nM(図1)であった。飽和濃度依存反応及び6.7nMのED50値は、アイロプロストの効果がレセプタによって仲介されたことを示唆している。
【0029】
高められる孵化への重要なPGI2暴露の継続時間及び胚の発生時期。
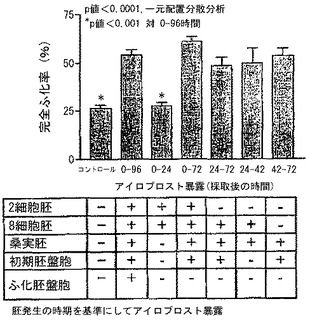
胚が子宮に入る前にいくつかの発生時期を経るので、我々は、アイロプロストに対する発生時期特有の反応を研究した。我々の結果は、96時間培養の間のある期間が、その他の期間よりもより重要であったことを示している。最初の24時間の培養中(この期間の間に、たいていの2細胞の胚は8細胞の胚に発生した)にアイロプロストへの暴露は、孵化を高めなかった(図2)。一方、採取後0−72時間又は24−72時間の間にアイロプロストへの暴露は、0−96時間暴露を行ったものと同じ比率の完全孵化を生じた(図2)。
【0030】
アイロプロストの十分な効果を確保するアイロプロストへの暴露の2つの重要な期間は、採取後24乃至42時間と、42乃至72時間との間にあり、これらはそれぞれ8細胞から桑実胚までと、桑実胚から初期胚盤胞までとの形質転換に対応している(図2の下部の表)。我々のデータは、胚をPGI2に短時間暴露することは、胚が卵管を離れた後でさえも完全孵化のための高められる可能性を与えることを示している。
【0031】
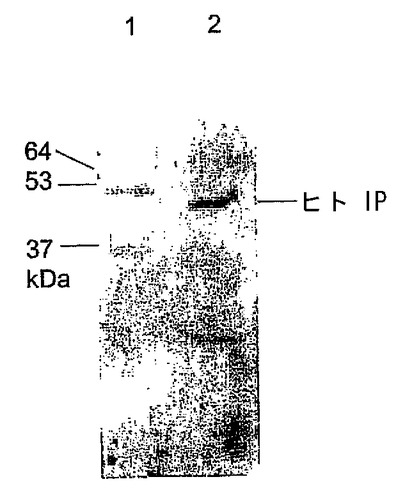
マウス胚はIPを発現する。
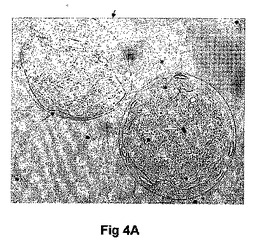
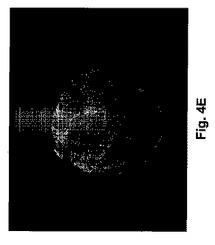
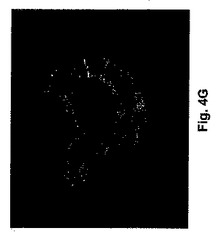
マウス胚の発生時期特有の発現を調査するために、我々は発生の異なる時期において胚盤胞のウエスタンブロット解析及び胚の免疫組織化学解析を行った。ウエスタンブロット解析は期待される分子量のタンパクが、IPに対するアフィニテイ精製抗体によって検出できることを示した(図3)。蛍光顕微鏡は、IP染色が桑実胚及び胚盤胞には存在する(図4A−E)が、未受精卵又は1細胞、2細胞及び8細胞胚(図示せず)には存在しないことを示した。桑実胚及び胚盤胞のIP染色は、同じ細かい網状模様を共有していた。共焦点顕微鏡画像は、IPが栄養外胚葉で好ましく発現されたことを示唆している。したがって、マウス胚によるIPの時期特有発現が、アイロプロストに対する反応と一致した。
【0032】
放射ラベルしたアイロプロストはマウス胚に結合する。
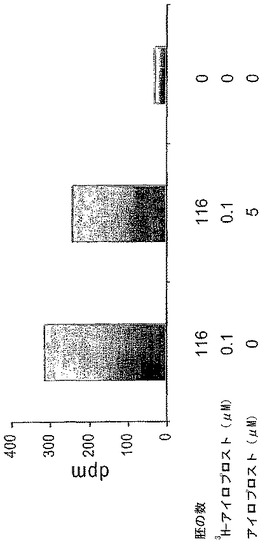
アイロプロストによる胚の完全な孵化の濃度依存性の高まりは、その効果がレセプタ仲介であることを示唆した。我々は、ラジオリガンド結合測定を行って、アイロプロストによるマウス胚の結合をさらに確認した。我々は、0.1μMレベルのリガンドで結合するアイロプロストの量を計ることを選択し、ここで増大された孵化が最初に観察され、およそ3.04fmolの3H−アイロプロストが116胚盤胞に特異的に結合することを見出した(図5)。
【0033】
議論
我々のデータは、PGI2が胚の完全孵化を高めることを最初に示している。これは、ヒト卵管由来の上皮細胞とマウス胚の共培養が、胚細胞数を増加させ、アポトーシスを減少させて、胚の孵化を発生させるという以前の報告と一致している[Piekos、1995年;Xu、2000年;Xu、2001年]。それ故、卵管由来PGI2及び子宮内膜細胞由来PGI2は、同様のパラ分泌方法でこれらの効果を与えている。前者は胚の孵化を促進させ、後者は血小板凝集を妨げる。濃度依存性反応は、レセプタ仲介の事象と一致しており、ED50値6.7nMは報告されているKd、すなわちヒト血小板から可溶化されたIP(8nM)の値と同様であった[Tsai、1989年]。
【0034】
アイロプロストに対する胚の反応は、発生時期特有であり、IPの発現と一致した(図2)。反応性についてのこれらの重要な期間は、受精卵が桑実胚へと発生する期間である、卵管におけるマウス胚の一時滞在と一致した。関連した発生時期が、ヒト及びマウス胚のゲノムの活性化とも一致しており、ゲノムの活性化は、4細胞期及び8細胞期の間と、2細胞期後とにそれぞれ起こる[Tesarik、1986年;Tesarik、1988年;Braude、1988年]。
【0035】
IPがどのようにその効果を発揮するのかという特定の理論に限定されることを望むものではないが、初期発生の間、卵管由来PGI2又はPGE2に暴露された胚が、子宮に達する際に、高められる孵化の可能性を維持することが提案される。したがって、卵管由来PGI2を通じて、胚が卵管中にある間に、着床のために胚自体が積極的に準備している。胚の観点からすれば、卵管由来PGI2は、子宮内膜細胞由来PGI2に相補的であり、受容力を確保するために子宮内膜の脱落を仲介する[Lim、1999年]。
【0036】
特定の理論に限定されることを望むものではないが、PGI2及びPGE2によって仲介される高められた孵化は、胚細胞の数の増加によってもよいことが提案される。様々なメカニズムが、インビトロ及びインビボのマウス胚の孵化に関与している。インビトロの孵化は、胚盤胞の拡張に関与して、透明帯破裂の前に全体的な透明帯の間伐が引き起こるが、インビボの孵化は、子宮の又は栄養外胚葉の分解物質による全体的な透明帯の溶解に関与する[Montag、2000年]。したがって、インビトロで孵化を達するためには十分な量の胚細胞の数が要求される。この点において、われわれの結果は、卵管の上皮細胞と共培養された胚は、より多くの細胞を有し細胞死をより少なくしたという発見と一致しており、孵化を向上させた[Yeung、1992年;Xu、2000年]。その上、PGI2は、栄養外胚葉でトリプシン様プロテアーゼの生産を増加させて透明帯(zona)を溶解することができる[Sawada、1990年;Parona、1986年]。
【0037】
IVF胚(胚当り10−20%)の低着床の可能性[CDC、2001年]は、一つには次善の培養条件に起因すると考えられる[De Vos、2000年]。培養液は、インビトロの胚発生を改良するために変更されてきた[Gardner、1998年]。我々のデータを基に、培地にPGI2及び/又はPGE2類似物、例えばアイロプロスト、PGE1のような物質を補足することで、発生する胚に改良した培養環境を与え、着床の可能性を増加させることができる。
【0038】
要するに、PGI2は、培養されたマウス胚の完全孵化を高めた。マウス胚によるIPの時期特異発現は、PGI2に対する反応と一致した。結局、我々の予備的なデータを基に、PGE2もまた、培養されたマウス胚の完全孵化を高めた。
【0039】
さらなる検討において、我々は、培養培地にPGI2を補足することが、着床の可能性及びマウス胚の生児出生の可能性を高めることを明らかにした。これらの結果は、PGI2は、培養中のマウス胚の完全孵化を高めるという我々の最初の観察を裏付けており [Huang、2003年]、また、卵管由来PGI2が生理的機能を果たすという我々の仮説も裏付けている[Huang、2002年]。
【0040】
表1は、PGI2がマウス胚の着床の可能性を高めることを示す。84個のコントロール胚及び81個のアイロプロスト処理胚が、12匹の妊娠キャリアに移入された。72時間後、よりたくさんの妊娠嚢が、コントロール群よりもアイロプロスト処理群に見出された(76%対42%、相対的危険度1.84、95%信頼区間1.38−2.43、p値<0.0001)。移入時においては、胚の発生時期は同等であった。
【0041】
表2は、PGI2アイロプロストが、マウス胚の生児出生の可能性を高めることを示す。406個のコントロール胚及び415個のアイロプロスト処理胚が、それぞれ30及び31匹の妊娠キャリアに移入された。コントロール及び実験群の生児出生率は、それぞれ28%及び36%であった(p値=0.0017、相対危険度1.28、95%信頼区間:1.044−1.560)。子及び胎盤の重量が比較された。したがって、PGI2は、子又は胎盤のどちらの重量にも影響を与えることなく、マウス胚の生児出生の可能性を増加させた。その上、全体の異常は指摘されなかった。
【0042】
上述したように、およそ70%の実験胚及びコントロール胚が、移入時には桑実胚期にあった(表1)が、移入された実験胚はよりたくさんの妊娠嚢を生じた。これらの結果は、8細胞期と桑実胚期の間にPGI2に暴露することは、マウス胚の完全な孵化を高めるには十分であるという我々の以前の観察を確認するものである[Huang、2003年]。それらはまた、胚栄養因子として作用する卵管由来PGI2と一致しており、この理由は、受精されたマウス卵が、卵管の内部で桑実胚に発生するためである[Snell、1966年]。
【0043】
上述した現象は、PGI2が、結果的に高められる孵化、着床、及び生児出生の増加となる一連の出来事を開始することを示している。この点において、PGI2は血小板活性化因子(PAF)によく似ており、それはPAFに短時間暴露するとマウス[O’Neill、1998年]及びヒト胚[O’Neill、1989年]の着床を増加させた。PGI2によって開始される分子細胞学的事象は、孵化にとっては重要であるに違いない。それらは、胚細胞数の増加[Montag、2000年]、栄養外胚葉でトリプシン様プロテアーゼの産生の増加[Sawada、1990年;Perona、1986年]、栄養外胚葉の高められるナトリウム−カリウム−ATPアーゼ(Na+−K+−ATPase)システムによる高められた胞胚腔の拡張[Biggers、1988年]、又はいまだメカニズムに見出されていないことを含んでいる。
【0044】
コントロール胚の着床率が、移入後72時間又は14日目それぞれを調べたときには同等であった(42%及び46%)。一方、移入後14日目に調べられた実験胚の着床率(59%)は、移入後72時間目に調べられた着床率(76%)よりもかなり小さかった(p=0.038)。よりたくさんの嚢が、着床の位置に群がることによって、実験群に完全に再び吸収されたことはもっともらしい事である。代わって、アイロプロストは、さもなければ着床してなかったいくつかの胚を「救済」し、これら「救済された」胚の一部のみが生存する子へ発生した。この検討と、[Huang、2002年]及びマウス[Huang、2004年]卵管でのたくさんのPGI2生産との結果は、PGI2が、卵管で分泌される胚栄養因子の一つであることを示唆し、したがってPGI2類似物を有する胚の培養培地に補足することは、IVFの効果を改良することを予期させる。
【0045】
IVF培養培地に胚発生又は着床を高める物質を補足することは、たいていの実験室及び非常に注意を払う培地製造業者によって採用されてきた。この懸念事項には、生殖毒物学、奇形形成、及び場合によってはこのような操作の後生的な影響を含んでいる。ヒト血清を補足された培地中で培養された羊胚由来の子羊は、より重みがあり妊娠期間がより長かった[Thompson、1995年;Holm、1996年;Sinclair、1999年]。過剰増殖及び新形成を含む先天性異常症、すなわちベックイス−ワインダーマン(Beckwith−Weindermann)症候群は生殖介助術と関係があった。[DeBaun、2002年;Maher、2003年]。
【0046】
胚及び胎児発生に示したアイロプロスト[Battenfeld、1995年]に関する生殖毒物学の検討は、胚及び胎児発生でウサギ及びサルでは影響されないが、指の変形がラットで観察されることを示した。指の異常は、アイロプロストの先天的奇形形成というよりも、影響を受けた構造から低酸素症を招く子宮胎盤流量の減少によるものらしく、この理由は、アイロプロストは血管拡張剤であり検討された動物が、妊娠期間を通してアイロプロストを受容したからである。ヒドラジン及びジヒドロピリジン類のような異なった化学構造を有する血管拡張剤を用いる場合、同じような問題が引き起こされるであろう。最近の研究では、アイロプロストへの暴露が、着床前の期間の間に行われた。246匹の生存する子において調べると、指の変形又はその他の異常性はなく、2群間の子及び胎盤の重量は同様であった。全てこれらのことを除いて、IVF培地にPGI2類似物を補足することは、実験的プロトコルとみなされるべきである。
【0047】
これらの実例の検討において、我々は、PGI2が胎児又は胎盤のいずれの重量にも影響を与えることなく、マウス胚の着床及び生児出生の可能性を高めること、すなわちプロスタサイクリンは、胎盤又は新生児の重量を増加させないことを明らかにした。これは、着床及び生児出生の高まりとは別の利点であり、プロスタサイクリンが、着床及び生児出生を高める他の代理母とは異なることを示している。着床及び生児出生を高める以前に報告された代理母はまた、新生児の重量を増加させて、ヒト体外受精(IVF)についてこれらの代理母の使用を不適としている。さらに、プロスタサイクリンは、インビボのマウスモデルでは、胎盤及び新生児の重量に影響を与えなかったため、ヒトIVF胚に利用できると考えられている。
【0048】
マウス胚についての前述のインビボ研究で得られた結果は、それらの胚が子宮に移入される前に、プロスタグランジン又はプロスタグランジン類似物を用いてインビトロで同様に処理される場合に、ヒトを含めその他の哺乳動物胚において得られるであろう同様な好ましいインビボの結果を示し、又は予測させるものであるとみなされる。本ヒトIVFは、患者の子宮への8細胞期又は胚盤胞期の胚の移入を含んでいる。この点において、ヒトIVFは、妊娠マウスに対するコントロール、すなわちプロスタサイクリン類似物処理胚への移入を含む我々の実験の計画と同様である。我々のデータは、妊娠キャリアに移入された際に、8細胞期と桑実胚期との間にプロスタサイクリン類似物に暴露された実験胚が、コントロール胚よりも着床及び生児出生率が非常にに高いことを示した(表1及び2、それぞれに示されるように)。
【表1】
【表2】
【0049】
さらに苦心なく、当業者はここまでの説明を用いて、その最大限に本発明を利用できることを確信する。前述の実施例は、例証として解釈されるべきであり、開示の残部をいかように規制するものではない。本発明の好ましい実施例を示され説明したけれども、その変更が、本発明の趣旨及び教示から逸脱することなく当業者によりなされるであろう。ここに説明される実施例は、単なる例示であり限定を意図するものではない。ここに開示された本発明の多くの変形及び変更が、可能であり本発明の範囲のものである。発明の背景における参照例、特にこの出願の優先日後の公開日を有する如何なる参照例の如何なる説明も、本発明に対して従来技術であると認めているものではない。ここに引用した全ての特許、特許出願及び公開の開示は、それらがここでの記載を補う模範的、手法的、その他の詳細を与える限りにおいて、ここに援用される。
【図面の簡単な説明】
【0050】
【図1】プロスタサイクリン(PGI2)及びマウス胚の完全孵化を示したグラフである。2細胞マウス胚が、PGI2類似物(アイロプロスト、1nMから10μMの範囲)で培養された。完全孵化率は96時間後に決定された。3乃至5の個別の実験(各々17−20胚)の結果は、平均±標準偏差として表現される。ED50値はおよそ6.7nMである。
【図2】完全孵化の増大及びプロスタサイクリン(PGI2)暴露の継続時間(胚発生の時期を基にアイロプロスト暴露)を示す棒グラフである。2細胞マウス胚が0.1μMアイロプロスト、すなわちPGI2類似物の存在下で示された期間(2細胞胚の採取後の時間として表現される)培養された。完全孵化率は平均±標準偏差として表現される。胚がアイロプロストに暴露されている間の発生時期が表に記載されている。2期間がアイロプロストの最大限の効果に重要であるようである、すなわち24乃至42時間及び42乃至72時間であり、それぞれ8細胞胚から桑実胚期及び桑実胚期から初期胚盤胞への発生に対応する。いずれの期間の間にアイロプロストに暴露しても、高められる孵化を確実にした。個別の観察の数(18−20胚についてのそれぞれ)は、コントロールで12胚、42−72時間では3胚、その他は4胚であった。コントロール胚の完全孵化率は、図1の孵化率とは異なっており、おそらく雌マウスの個体が変更されたからであろう。
【図3】マウス胚のプロスタサイクリンレセプタ(IP)の発現を示すウエスタンブロットである。60匹のマウス桑実胚(レーン1)から抽出した全細胞溶解物及びヒト血小板のミクロソーム(陽性コントロール、レーン2)のウエスタンブロット解析は、ヒトIPに対する抗体との免疫反応を示した。マウスIPの移動度は、ヒトIPの移動度よりも少なく、予想される分子量のそれぞれ〜50kDa及び〜46kDaと一致した。
【図4A−G】桑実胚及び胚盤胞におけるプロスタサイクリンレセプタ(IP)の発現を示す顕微鏡写真である。2個の桑実胚の位相差(図4A)及びIP染色(図4B)像が示される。透明帯は矢印で示される。図4Cは、胚盤胞における核のプロピジウムイオダイド染色を示す。図4Dにおいて、胚盤胞のIP及びプロピジウムイオダイドが重ね合わせられる。図4Eは胚盤胞のIP染色が網状模様を示すことを示す。IP染色の集合塊が栄養外胚葉のみに見られ、内部細胞塊には染色の増進を示さなかった。図4Fは桑実胚内の内部細胞塊(ICM)の配置を示すスケッチである。図4Gは胚盤胞の共焦点顕微鏡画像であり、栄養外胚葉のみがIP抗体によって染色されることを示す。断面の平面図はパネルF中のラインによって示される。陰性のコントロール、受精卵又はその他の発生時期にある胚(図示せず)にはIP染色はなかった。図中の棒はおよそ20μmである。
【図5】無傷のマウス胚に結合するアイロプロストを示す棒グラフである。116個のマウス胚盤胞が、5μMの非標識アイロプロストの存在又は不在下で、0.1μM3H−アイロプロスト、すなわちPGI2類似物とインキュベーションした。結合した3H−アイロプロスト及び遊離の3H−アイロプロストは膜ろ過によって分離された。
【図6A−B】妊娠マウスの子宮内の妊娠嚢を示す。着床は、着床部位の血管の増加によって決定され、妊娠嚢の存在によって確認された。胚移入後72時間、妊娠マウスは尾静脈から1%シカゴブルー(Chicago Blue)が与えられて3分後に安楽死させられた。図6Aは胚を受け入れた子宮角を取り巻く個々のブルーバンド(アスタリスクによって示される)を示す。図6Bでは、各ブルーバンドは妊娠嚢に対応しており、開放される子宮角ではピンクカラーの組織として現れた。ブルーバンド又は妊娠嚢のどれにも、胚を受け入れない子宮角には見つからなかった(矢印によって印を付けられる)。
【技術分野】
【0001】
連邦政府の支援による研究または開発に関する記載
本発明は、全体的に又は部分的にNational Institute of Child Health and Human Development(NICHD)、契約番号HD01277、から財政的支援を受けて行われるものである。したがって、米国政府が本発明について所定の権利を有する。
【背景技術】
【0002】
発明の分野
本発明は、一般に哺乳動物胚のインビトロ培養のため、及び適した宿主哺乳動物の子宮に培養された胚の着床後における妊娠の達成を高めるための組成及び方法に関する。
【0003】
従来技術の説明
卵管内の環境は、精子の受精の可能性を高め、分裂する胚の発生を促進する。卵管上皮細胞と共培養された胚は、発生、孵化及び着床を向上させたことが示された[Xu、2001年;Xu、2000年]。プロスタサイクリン(PGI2)は、もともと血液及び血管(vascular)のホメオスタシスを維持する点に関与すると考えられている。しかしながら、遺伝子ノックアウトマウスにおける最近の観察結果から示唆されることは、プロスタサイクリンが他の生理的機能を有し得ることである。子宮内膜由来PGI2の重要性が、シクロオキシゲナーゼ(COX)−2ノックアウトマウスにおいて行われた観察によって強調される[Lim、1997年]。COX−2ノックアウトマウスの子宮内膜は、正常には脱落しなかった。結果として、移入された野生型胚(transferred wild type embryo)は、COX−2の欠損した雌マウスには着床できなかった。前述の異常性は、母方に外部からPGI2類似物の投与によってある程度矯正されることができた[Lim、1999年]。
【0004】
着床前胚の発生における卵管由来PGI2の役割が明らかでない理由は、PGI2レセプタ(IP)ノックアウトマウスの一腹の産子数が野生型の産子数と比較されていないからである[Murata、1997年]。しかしながら、ヘテロ接合体IPノックアウトマウスとの交配から生まれる子の遺伝子型の分布がメンデルの法則に従っていなかった。すなわち、ホモ接合体IPノックアウト遺伝子型を有する雄及び雌の子の普及率は、それぞれ37%及び20%であり、予想されるよりも低かった[Murata、1997年]。
【0005】
【特許文献1】米国特許第6100249号公報
【特許文献2】米国特許第5096822号公報
【発明の開示】
【0006】
発明の簡単な概要
最近、我々は、ヒト卵管がたくさんのプロスタサイクリン(PGI2)及びPGE2を合成すること発見した[Huang、2002年]。遺伝子ノックアウトの研究は、子宮内膜由来PGI2が子宮内膜の脱落に必須であることを示唆するが、着床前胚の発生におけるPGI2の効果は報告されていなかった。着床前胚の最適な発生にはPGI2を必要とするという仮説を用いて、マウス胚におけるPGI2の効果が、完全孵化率を基にして検査された。PGI2レセプタ(IP)の発現は、ウエスタンブロット解析及び免疫組織化学によって評価された。PGI2の胚への結合は、ラジオリガンド結合検定法によって確認された。我々の結果は、マウス胚の孵化はアイロプロスト(Iloprost)(ED506.7nM)、すなわち安定化したPGI2合成類似物の補足によって高められることを明らかにしている。8細胞から桑実胚に至る間、又は桑実胚から初期胚盤胞に至る間に、アイロプロストへの暴露が、孵化を促進させるのに重要であった。PGI2に対するこの時期特有の反応が、IPの発生時期特有の発現と同時に起こった。さらなる研究は、高められる胚の孵化を着床及び生児出生の増加と関連づけるために実施され、すなわちアイロプロストを補足された培地中で培養されたマウス胚が、妊娠キャリアに移入された。妊娠嚢及び生存する子の数がコントロール胚のその数と比較された。報告によれば、培養培地の添加物が胎児の体重を増加させるため[DeBaum、2002年;Thompson、1995年;Sinclair、1999年]、子及び胎盤の重量も比較された。我々の結果は、アイロプロストを補足された培養培地中で胚を培養することは、子及び胎盤の重量に影響を与えることなく、マウス胚の着床及び生児出生を高めることを示している。
【0007】
したがって、本発明のある実施例においては、培養培地にプロスタグランジン、例えばPGI2又はPGE2又はその類似物を補足することを含む胚のインビトロ発生を高める方法が提供されている。ある実施例において、プロスタグランジンはPGI2又はPGE2であり、またある実施例では、プロスタグランジン類似物はアイロプロスト又はPGE1である。ある実施例では、方法は、培養培地に胚の完全孵化を促進するのに十分量のプロスタグランジン又は類似物を添加することを含む。好ましくは、「完全孵化」は、胚が透明帯(zona pellucida)から離れることを有する。方法のある実施例において、培養培地の補足は、ゲノムが活性化されて受精卵が桑実胚へ発生する期間である、胚の初期発生時期の間に行われる。胚がヒトである実施例において、初期発生は4乃至8細胞状態で開始する。方法のある実施例において、培養培地の補足は、胚の桑実胚から初期胚盤胞への発生の間に行われる。ある実施例では、胚の初期発生から桑実胚期への発生と、胚の桑実胚から初期胚盤胞への発生の間との両方の間において行われる。方法のある実施例において、胚がマウス起源にある場合、補足する段階は、その2細胞発生の時期に胚を採取後24乃至42時間の間に、培養中の胚をプロスタグランジン又は類似物に暴露することを含む。胚がマウスである方法のある実施例では、桑実胚から初期胚盤胞の間に補足する段階は、その2細胞発生の時期に胚を採取後42乃至72時間の間に、胚を一度にプロスタグランジン又はプロスタグランジン類似物に暴露することを有する。
【0008】
また、本発明のある実施例にしたがって提供されることは、インビトロ受精胚のインビボ着床の可能性を増加させる方法である。「着床の可能性」とは、子宮に着床する胚の能力である。この方法は、胚のインビトロ発生を高めるために、上述の実施例の一つを実行することを含むことにより、培養中の胚の完全な孵化が達せられる、すなわち、別のIVF方法に比べて孵化が高められる。この方法のある実施例にしたがって、完全に又は殆ど孵化された胚が、それから宿主哺乳動物の子宮に導入されて、胚の高められた着床が達せられる。ある実施例では、インビトロで胚の完全孵化は、生存に適した妊娠の確立と相関する。
【0009】
本発明のある実施例において、インビトロで受精された哺乳動物胚の生児出生の可能性を増加させる方法が提供されている。「生児出生の可能性」とは、生児出生を生じる胚の能力である。上述されるように、この方法はプロスタグランジン又はその類似物を有するインビトロ培養によって、胚のインビトロ発生を高めることを有することにより、高められる孵化の可能性、すなわち培養中の胚の完全孵化が達せられる。好ましい実施例において、このような処理は、胚が培養から取り出されてインビボ環境にある後にさえも、完全孵化のための高められる可能性を与えている。この性質は「高められる孵化の可能性」として説明される。孵化される胚、すなわち高められる孵化の可能性を有する胚が、宿主哺乳動物の子宮に移入されて、胚が着床するようになりインビボで成長することにより、生児出生を生じる胚の能力が、孵化の高まりを有せず着床された胚の能力と比較して高められている。
【0010】
本発明のさらにもう一つの実施例においては、インビトロで受精された哺乳動物胚の一群における生児出生率を増加させる方法が、上述の方法に従って胚のインビトロ発生を高めることを有することを提供されていることにより、孵化の高まり、すなわち好ましくは培養中の胚の完全孵化が達せられる。方法は、宿主哺乳動物に胚を導入、又は移入すること、及びそれから胚をインビボで成長できるようにすることを含んでおり、高められる孵化を有する胚の群からの生児出生率は、最初に完全に孵化されていない、すなわち孵化の高まりを受けていない状態で、宿主子宮へ移入される胚の群からの生児出生率よりも非常に高い。
【0011】
なおもさらに、本発明のある実施例に従って提供されていることは、哺乳動物胚のインビトロ発生のための改良された細胞培養培地であり、その改良点は、孵化を高め、また好ましくはインビトロで胚の完全孵化を促進するのに有効な補足量のプロスタグランジン又はその類似物を含むことである。ある実施例において、培地は、ある量のプロスタグランジン又はその類似物を含んでおり、それはインビトロ培養から取り出されて宿主哺乳動物の子宮に移入された後に、胚の完全な孵化の可能性を高めるために有効である。これら及びその他の実施例、本発明の特徴及び利点は、以下の説明及び図を参照することによって明白になるであろう。
【0012】
好ましい実施例の詳細な説明
本開示は、精子及び胚に対するPGI2並びにPGE2の効果の研究及びそれに続く研究の過程で明らかになった。我々の最近の結果は、ヒト[Huang、2002年]及びマウス[Huang、2004年]卵管上皮細胞が、PGI2の合成、すなわちCOX-1又はCOX-2及びPGI2合成酵素に必須の酵素を発現することを示している。たくさんのPGI2又はPGE2は、14C−アラキドン酸がヒト[Huang、2002年]又はマウス卵管[Huang、2004年]から調製されたミクロソームとインキュベーションされたときに産生された。本研究はまた、精子が卵と受精するために卵管末端に移動し、胚発生の最初の72時間は卵管で起こるという事実を考慮した。
【0013】
本研究の一部分として、胚に対するPGI2の影響が、PGI2類似物を伴って培養されたマウス胚の完全な孵化を観察することによって評価された。そのメカニズムをさらに解明するために、IPの発現及び放射ラベルされたアイロプロストによる胚の結合がまた検討された。
【0014】
材料及び方法
試薬の出所及び制度上の承認。
別のものであると記載しない限り、試薬はシグマ社(Sigma Co.)(アメリカリ合衆国、ミズーリ、セント.ルイス)(St.Louis、MO、USA)から購入されたものである。ヒトのサンプルの使用は、ヒューマンサブジェクツの保護のための委員会(the Committee for the Protection of Human Subjects)(Institute Review Boardと同等)によって承認された。実験動物の取扱い及び研究プロトコルは動物福祉委員会によって認定された。
【0015】
マウス胚の採取及び培養。
マウスは、水及び餌へのアクセスが自由で、制御された温度、湿度、及び光周期(12時間明暗周期)条件の下に保った。3週齢のC57B1/6雌マウスはハーラン(Harlan)(アメリカ合衆国、インジアナ、インジアナポリス)(Indianapolis、IN、USA)から購入された。8週齢のC3H雄マウスは最初にハーランから購入され、後にジャクソンラボラトリー(Jackson laboratory)(アメリカ合衆国、メイン、バーハーバー)(Bar Harbor ME、USA)から購入された。雌マウスの過剰排卵は、妊娠雌馬血清ゴナドトロピン(5単位)、及び続く46時間後にヒト絨毛性ゴナドトロピン(hCG、5単位)の腹腔内投与によって行われた。hCGを受けた後、それぞれの雌マウスが一匹の繁殖力のある雄マウスとつがいにされた。48時間後、2細胞胚は卵管から25mMへペス(HEPES)及び1%ウシ血清アルブミン(BSA)(アービン サイエンテイフィツク)(Irvine Scientific)を補足されたα-MEM培地に採取された。
【0016】
胚(群当り17-20個)は、各ウエルに600μLの培地を含む4ウエルデイッシュ(アメリカ合衆国、イリノイス、ネイパービルのナルジ ヌンク インターナショナル ネイパー)(Nalge Nunc International Naper、Naperville、IL、USA)において、37℃で5%CO2の下に培養された。HTF及びα-MEM培地は、96時間連続して用いられて分裂胚の変化する栄養要件を満たした[Gardner、1998年]。HTF培養液(セイジ バイオフアーマ)(SAGE Biopharma)は最初の48時間用い、イーグルの塩類(Earle's salts)及び2mMグルタミンを有するα-MEM培地(アービン サイエンテイフィツク)(Irvine Scientific)は2番目の48時間の間用いた。実験胚は、水中のアイロプロストを受け、コントロール胚は等量の水を受けた。予備実験では、胚の95%以上がKSOM培地(アメリカ合衆国、ニュージャージー、フィリップスバーグのスペシャリテイ−メデイア、セル アンド モレキュラー テクノロジーズ、インク.(Speciality Media、Cell and Molecular Technologies、Inc. Phillipsburg NJ,USA))で培養した胚と同様に、96時間までに胚盤胞になることを示した。96時間培養後、各胚は透明帯(zona pellucida)の有無に対して調べられた。透明帯が完全に無い胚は、完全に孵化したものとして計測された。完全孵化率は、完全に孵化された胚の数を胚の総数で割って求められた。胚の完全孵化を、胚盤胞形成又は胚孵化の代わりに指標として選択したが、この理由は、後者の2マーカーが生存に適した妊娠の確立[Lane、1997年]と相関しないからである。
【0017】
ウエスタンブロット解析(Western blot analysis)。
推定されるマウスIPのアミノ酸配列(417a.a、Genebank_BAA05144)及びヒトIPのアミノ酸配列(386a.a、Genebank_BAA06110)は相同性が高い[Katsuyama、1994年]。予備的な検討では、アフィニテイー精製ポリクローナルペプチド抗体(テキサス大学健康科学センターのケーヘルアン博士(Dr.Ke−He Ruan、University of Texas Health Science Center)から提供)は、マウスIPと交差反応することを確認した。ウエスタンブロット解析は以前に説明したように行われた[Huang、2002年]。簡潔には、60個のマウス胚盤胞由来の全細胞溶解物が、10%アクリルアミドゲル電気泳動(PAGE)によって分離され、ニトロセルロース膜(アメリカ合衆国、ニューハンプシャーのシュレイチャ− アンド シュエル、インク.)(Schleicher & Schuell、Inc.、NH、USA)に転写された。免疫反応タンパクが、抗体とのインキュベーション及び増強された化学発光(アメリカ合衆国、ニュージャージー、ピスカタウエイのアマ−シャム バイオサイエンス)(Amersham Bioscience、Piscataway、NJ、USA)による視覚化によって検出され、またSTORM860レーザースキャナ(アマ−シャム バイオサイエンス)(Amersham Bioscience)を使って検出された。ヒト血小板ミクロソームは、陽性のコントロールとして用いられた。抗体の特異性は、前吸収抗体を用いる平行実験で確かめられた。
【0018】
免疫組織化学、蛍光顕微鏡及び共焦点顕微鏡解析。
マウス胚が4℃で30分間、4%パラホルムアルデヒド中に固定された。リン酸緩衝化生理食塩水(PBS)で3回洗浄後、胚は0.05%のTweenー20、5%の粉末ミルク及び0.1%のTriton X−100含有トリス緩衝化生理食塩水(pH7.4)において、室温で20分間ブロックされた。胚は、2時間ブロッキング緩衝液中でIP抗体(5ng/ml)とインキュベーションされ、それから37℃で30分間、アレクザ(Alexa)488(2.5μg/ml、アメリカ合衆国、アラバマのモレキュラー プローブ(Molecular Probes、AL、USA))と結合したヤギ抗-ラビットIgGとインキュベーションされた。細胞核は、室温で20分間、10μg/mlプロピデウム イオダイド(propdium iodiode)によって対比染色された。胚は、Fluoromount-G(登録商標)(アメリカ合衆国、アラバマ、バーミングハムのサザン バイオテクノロジー アソシエイツ インク.)(Southern Biotechnology Associates Inc.、Birmingham、AL、USA)に埋め込まれた。蛍光顕微鏡解析のために、胚盤胞は封入剤中に置かれてカバースリップが被せられた。共焦点顕微鏡解析のために、およそ50μmのスペーサが、スライドとカバースリップとの間に置かれて胚の3次元形態を保持した。陰性のコントロールには、胚が10ng/mlの非免疫ラビットIgGとインキュベーションされた(すなわち、一次抗体濃度の2倍の濃度)。
【0019】
蛍光顕微鏡解析は、適切なフィルタを備えたツアイス アキシオプラン2(ZeissAxioPlan 2)顕微鏡を用いて行われた。画像は、CCDカメラを用いて捕らえられ、アキシオビジョン プログラム(Vesion 3.0.6)によって処理された。共焦点顕微鏡解析は、オリンパス BX-50顕微鏡に付属されたバイオラッド ラジアンス 2000(BioRad Radiance 2000)共焦点システム(アメリカ合衆国、カルフオル二ア、ハーカルスのバイオラッド ラボラトリーズ)(Bio−Rad Laboratories、Hercules、CA、USA)を使って行われた。その画像は、イメージ プロ プラス プログラム(Image−Pro+ program)(アメリカ合衆国、カルフオル二ア、カールスバッドのメデイア−サイバーネテイックス)(Media−Cybernetics、Carlsbad、CA、USA)を使って処理された。
【0020】
全胚ラジオリガンド結合測定。
3H−アイロプロストの胚盤胞への結合の解析は、わずかに変更修正して以前に説明された [Arbab、2002年] ように行われた。232個が孵化されて孵化する胚盤胞は、結合緩衝液(10mMHEPES、pH7.4中に10mMMnCl2)中で3回洗浄されてから、非標識アイロプロスト(5μM)を有する又は有しない100μlの結合緩衝液に移された。反応は、3H−アイロプロスト(200nM、特異活性11.0Ci/mmol、アマ−シャム バイオサイエンス(Amersham Biosciences))を含む結合緩衝液を加えて、室温で60分間インキュベーションすることによって開始された。その反応は、氷冷洗浄緩衝液(10mMHEPES、pH7.4中に0.01%BSA)に胚盤胞を移すことによって終了された。緩衝液及び胚盤胞は、ガラスフアイバーフィルタ(Whatman GF/C2.4cm)を通してろ過され、フィルタは、2mlの洗浄緩衝液で3回洗浄された。フィルタがオーブン中で乾燥されてから、放射活性が5mlのシンチレーション液で計測されるシンチレーションによって測定された。
【0021】
統計解析
スチュ-デントのt検定又はダネット(Dunnet)検定に従った一元配置分散分析が適所に用いられた。p値<0.05は統計的に有意とみなされた。量反応曲線の作図及びED50値(1.0に設定されるヒル勾配を有して)の計算には、GraphPad Prism(登録商標)Software(アメリカ合衆国、カルフオル二ア、サンデイゴのグラフパッド プリズム ソフトウエア インク.)(GraphPad Prism Software Inc.、San Diego、CA、USA)を備えた。
【0022】
胚移入
マウス胚が採取されて上述のように培養され、その後に本発明者によって公表された[Huangら、2003年]。簡潔には、2細胞胚(C3B6F1)が、hCG投与後42時間、過剰排卵された3週齢のC57Bl/6雌マウスから採取された。胚(群当り14個)は、600μlのHTF培地(アメリカ合衆国、ニュージャージー、ベッドミンスターのセイジ バイオフアーマ)(SAGE Biophama、Bedminster、NJ、USA)を各々含む4ウエルデイッシュ(アメリカ合衆国、イリノイス、ネイパービルのナルジ ヌンク インターナショナル ネイパー)(Nalge Nunc International、Naperville、IL、USA)において、37℃で5%CO2の下に培養された。実験胚は、水中のアイロプロスト(1μM、アメリカ合衆国、ミシガン、アン アーバーのケイメン ケミカル(Caymen Chemical、Ann Arbor、MI、USA))を、またコントロール胚は等量の水を受けた。どちらの場合も水量は培養液の0.1%に満たなかった。
【0023】
胚ドナー(donor)がhCGを受けた翌日に、膣スメアが妊娠可能性キャリア(8週齢のC3B6F1雌マウス)から得られて、発情周期の時期を決定した。発情期のマウスは、精管切除されたICR雄(ハーレン)(Harlan)とつがいにされて翌朝膣栓を調べられた。栓を有する発情期のマウスは0.5日偽妊娠であると指定された。
【0024】
胚移入は、解剖顕微鏡(オリンパスSZ−PT)の下に、2.5日偽妊娠において行われた。外科的麻酔は、ケタミン(ketamine)(200mg/Kg)及びキシラジン(xylazine)(10mg/Kg)(合衆国テキサス州のバーンズ ベトサプライ インク(Burns VetSupply Inc.TX、USA)からの両麻酔薬)の腹腔内投与によって得られた。各子宮角は、2cmの側面切り口からアクセスされた。近位卵管は一対のピンセットで保持されたまま、開口部が30ゲージの皮下針によって、腸間膜の子宮角の遠位端に作り出された。開口部は、135μmの内径を有するトランスフアーピペット(アメリカ合衆国、ニュージャージーのミッドアトランテイックダイアゴノステイックス、インク.)(MidAtiantic Diagnostics、Inc.、NJ、USA)を入らせることができる。0.8μlの移入用培地(25ml HEPES及び1%BSAを加えたα-MEM)中の7個の胚までが、各々の子宮角に移入された。それぞれ移入後、ピペットの内容物が、残留した胚を同定するために、立体写真顕微鏡下で調べられた。側面切り口は、9mmの金属クリップ(アメリカ合衆国、ニュージャージーのクレイ アダムス、パルシッパニー)(Clay Adams、Parsippany、NJ、USA)で閉じられた。一方の子宮角から他方の子宮角への胚の移動によるコントロール胚及び実験胚の混合を避けるために(ドイツのボン大学のアンドレス ツインマー博士との個人情報(Dr.Andreas Zimmer、University of Bon、Germany))、それぞれの妊娠キャリアは、コントロール胚又は実験胚のいずれかを受けた。一定の移入技術を維持するために、胚移入は同じプロトコルに従って、一人の人によって行われる、その人は、判別されている胚が受ける処理には左右されなかった。両群が、研究の過程で(6ヶ月間)得られる経験と等しく利益を得たことを確実にするために、胚は4つのブロックでの処理(コントロール又はアイロプロスト処理)に割り当てられた。
【0025】
着床率の決定
胚移入後72時間目に、着床は、多少変更修正して以前に説明された方法に基いて決定された[Pariaら、1993年]。簡潔には、安楽死前3分間、0.1mlのシカゴブルー(1%)が、妊娠マウスの尾静脈から投与された。そのマウスを殺した後、子宮角が取り出された。子宮角を取り囲む別々のブルーバンド(アスタリスクで示される)が計測された(図6A)。これらのバンドの存在は、着床部位への血管補給の増加を反映している。それから子宮角が開かれ、妊娠嚢が計測された(図6B)。着床率は、移入された胚当りの妊娠嚢の数として表現された。
【0026】
生児出生率と子及び胎盤の重量の決定
予備的な検討は、一腹の大きさが小さいときには、妊娠キャリアが子を共食いするこを示した。子を正確に計測し、体重を量るために、我々は、胚移入後14日目に妊娠キャリアを殺した(妊娠期間の16.5日目、すなわち自然に誕生する2日前)。殺した後、生きている子の数を計測し、個々の子及び胎盤の重量が決定された。全ての子が全体の異常を調べられた。空の妊娠嚢の数がまた、着床率の決定のために計測された。生児出生率は、移入される胚当りの生存する子の数として表現され、着床率は、移入される胚当りの生存する子を有し又は有しない嚢の数として表現された。
【0027】
統計解析
フィッシャーの直接確率検定が、比率を比較するために用いられ、スチュ-ダントのt検定が体重の比較のために用いられた。p値<0.05は統計的に有意であると認められた。GraphPad Instat(登録商標)Software(アメリカ合衆国、カルフオル二ア、サンデイゴのグラフパッド プリズム ソフトウエア インク.)(GraphPad Software Inc.、San Diego、CA、USA)が、統計解析に用いられた。
【0028】
結果
PGI2はマウス胚の完全孵化を高めた。
マウス胚の完全孵化が、濃度に依存するかたちでアイロプロストによって高められた。その効果は0.1μM又はそれ以上の濃度(図1) で統計的に有意であった。完全な胚の孵化の最大増加は、1μMで起こっており、実験胚の81±7%(平均値±標準偏差、n=3)が完全に孵化した。対照的に、コントロール胚の49±14%(平均値±標準偏差、n=5)のみが完全に孵化した。量反応曲線からのED50値は6.7nM(図1)であった。飽和濃度依存反応及び6.7nMのED50値は、アイロプロストの効果がレセプタによって仲介されたことを示唆している。
【0029】
高められる孵化への重要なPGI2暴露の継続時間及び胚の発生時期。
胚が子宮に入る前にいくつかの発生時期を経るので、我々は、アイロプロストに対する発生時期特有の反応を研究した。我々の結果は、96時間培養の間のある期間が、その他の期間よりもより重要であったことを示している。最初の24時間の培養中(この期間の間に、たいていの2細胞の胚は8細胞の胚に発生した)にアイロプロストへの暴露は、孵化を高めなかった(図2)。一方、採取後0−72時間又は24−72時間の間にアイロプロストへの暴露は、0−96時間暴露を行ったものと同じ比率の完全孵化を生じた(図2)。
【0030】
アイロプロストの十分な効果を確保するアイロプロストへの暴露の2つの重要な期間は、採取後24乃至42時間と、42乃至72時間との間にあり、これらはそれぞれ8細胞から桑実胚までと、桑実胚から初期胚盤胞までとの形質転換に対応している(図2の下部の表)。我々のデータは、胚をPGI2に短時間暴露することは、胚が卵管を離れた後でさえも完全孵化のための高められる可能性を与えることを示している。
【0031】
マウス胚はIPを発現する。
マウス胚の発生時期特有の発現を調査するために、我々は発生の異なる時期において胚盤胞のウエスタンブロット解析及び胚の免疫組織化学解析を行った。ウエスタンブロット解析は期待される分子量のタンパクが、IPに対するアフィニテイ精製抗体によって検出できることを示した(図3)。蛍光顕微鏡は、IP染色が桑実胚及び胚盤胞には存在する(図4A−E)が、未受精卵又は1細胞、2細胞及び8細胞胚(図示せず)には存在しないことを示した。桑実胚及び胚盤胞のIP染色は、同じ細かい網状模様を共有していた。共焦点顕微鏡画像は、IPが栄養外胚葉で好ましく発現されたことを示唆している。したがって、マウス胚によるIPの時期特有発現が、アイロプロストに対する反応と一致した。
【0032】
放射ラベルしたアイロプロストはマウス胚に結合する。
アイロプロストによる胚の完全な孵化の濃度依存性の高まりは、その効果がレセプタ仲介であることを示唆した。我々は、ラジオリガンド結合測定を行って、アイロプロストによるマウス胚の結合をさらに確認した。我々は、0.1μMレベルのリガンドで結合するアイロプロストの量を計ることを選択し、ここで増大された孵化が最初に観察され、およそ3.04fmolの3H−アイロプロストが116胚盤胞に特異的に結合することを見出した(図5)。
【0033】
議論
我々のデータは、PGI2が胚の完全孵化を高めることを最初に示している。これは、ヒト卵管由来の上皮細胞とマウス胚の共培養が、胚細胞数を増加させ、アポトーシスを減少させて、胚の孵化を発生させるという以前の報告と一致している[Piekos、1995年;Xu、2000年;Xu、2001年]。それ故、卵管由来PGI2及び子宮内膜細胞由来PGI2は、同様のパラ分泌方法でこれらの効果を与えている。前者は胚の孵化を促進させ、後者は血小板凝集を妨げる。濃度依存性反応は、レセプタ仲介の事象と一致しており、ED50値6.7nMは報告されているKd、すなわちヒト血小板から可溶化されたIP(8nM)の値と同様であった[Tsai、1989年]。
【0034】
アイロプロストに対する胚の反応は、発生時期特有であり、IPの発現と一致した(図2)。反応性についてのこれらの重要な期間は、受精卵が桑実胚へと発生する期間である、卵管におけるマウス胚の一時滞在と一致した。関連した発生時期が、ヒト及びマウス胚のゲノムの活性化とも一致しており、ゲノムの活性化は、4細胞期及び8細胞期の間と、2細胞期後とにそれぞれ起こる[Tesarik、1986年;Tesarik、1988年;Braude、1988年]。
【0035】
IPがどのようにその効果を発揮するのかという特定の理論に限定されることを望むものではないが、初期発生の間、卵管由来PGI2又はPGE2に暴露された胚が、子宮に達する際に、高められる孵化の可能性を維持することが提案される。したがって、卵管由来PGI2を通じて、胚が卵管中にある間に、着床のために胚自体が積極的に準備している。胚の観点からすれば、卵管由来PGI2は、子宮内膜細胞由来PGI2に相補的であり、受容力を確保するために子宮内膜の脱落を仲介する[Lim、1999年]。
【0036】
特定の理論に限定されることを望むものではないが、PGI2及びPGE2によって仲介される高められた孵化は、胚細胞の数の増加によってもよいことが提案される。様々なメカニズムが、インビトロ及びインビボのマウス胚の孵化に関与している。インビトロの孵化は、胚盤胞の拡張に関与して、透明帯破裂の前に全体的な透明帯の間伐が引き起こるが、インビボの孵化は、子宮の又は栄養外胚葉の分解物質による全体的な透明帯の溶解に関与する[Montag、2000年]。したがって、インビトロで孵化を達するためには十分な量の胚細胞の数が要求される。この点において、われわれの結果は、卵管の上皮細胞と共培養された胚は、より多くの細胞を有し細胞死をより少なくしたという発見と一致しており、孵化を向上させた[Yeung、1992年;Xu、2000年]。その上、PGI2は、栄養外胚葉でトリプシン様プロテアーゼの生産を増加させて透明帯(zona)を溶解することができる[Sawada、1990年;Parona、1986年]。
【0037】
IVF胚(胚当り10−20%)の低着床の可能性[CDC、2001年]は、一つには次善の培養条件に起因すると考えられる[De Vos、2000年]。培養液は、インビトロの胚発生を改良するために変更されてきた[Gardner、1998年]。我々のデータを基に、培地にPGI2及び/又はPGE2類似物、例えばアイロプロスト、PGE1のような物質を補足することで、発生する胚に改良した培養環境を与え、着床の可能性を増加させることができる。
【0038】
要するに、PGI2は、培養されたマウス胚の完全孵化を高めた。マウス胚によるIPの時期特異発現は、PGI2に対する反応と一致した。結局、我々の予備的なデータを基に、PGE2もまた、培養されたマウス胚の完全孵化を高めた。
【0039】
さらなる検討において、我々は、培養培地にPGI2を補足することが、着床の可能性及びマウス胚の生児出生の可能性を高めることを明らかにした。これらの結果は、PGI2は、培養中のマウス胚の完全孵化を高めるという我々の最初の観察を裏付けており [Huang、2003年]、また、卵管由来PGI2が生理的機能を果たすという我々の仮説も裏付けている[Huang、2002年]。
【0040】
表1は、PGI2がマウス胚の着床の可能性を高めることを示す。84個のコントロール胚及び81個のアイロプロスト処理胚が、12匹の妊娠キャリアに移入された。72時間後、よりたくさんの妊娠嚢が、コントロール群よりもアイロプロスト処理群に見出された(76%対42%、相対的危険度1.84、95%信頼区間1.38−2.43、p値<0.0001)。移入時においては、胚の発生時期は同等であった。
【0041】
表2は、PGI2アイロプロストが、マウス胚の生児出生の可能性を高めることを示す。406個のコントロール胚及び415個のアイロプロスト処理胚が、それぞれ30及び31匹の妊娠キャリアに移入された。コントロール及び実験群の生児出生率は、それぞれ28%及び36%であった(p値=0.0017、相対危険度1.28、95%信頼区間:1.044−1.560)。子及び胎盤の重量が比較された。したがって、PGI2は、子又は胎盤のどちらの重量にも影響を与えることなく、マウス胚の生児出生の可能性を増加させた。その上、全体の異常は指摘されなかった。
【0042】
上述したように、およそ70%の実験胚及びコントロール胚が、移入時には桑実胚期にあった(表1)が、移入された実験胚はよりたくさんの妊娠嚢を生じた。これらの結果は、8細胞期と桑実胚期の間にPGI2に暴露することは、マウス胚の完全な孵化を高めるには十分であるという我々の以前の観察を確認するものである[Huang、2003年]。それらはまた、胚栄養因子として作用する卵管由来PGI2と一致しており、この理由は、受精されたマウス卵が、卵管の内部で桑実胚に発生するためである[Snell、1966年]。
【0043】
上述した現象は、PGI2が、結果的に高められる孵化、着床、及び生児出生の増加となる一連の出来事を開始することを示している。この点において、PGI2は血小板活性化因子(PAF)によく似ており、それはPAFに短時間暴露するとマウス[O’Neill、1998年]及びヒト胚[O’Neill、1989年]の着床を増加させた。PGI2によって開始される分子細胞学的事象は、孵化にとっては重要であるに違いない。それらは、胚細胞数の増加[Montag、2000年]、栄養外胚葉でトリプシン様プロテアーゼの産生の増加[Sawada、1990年;Perona、1986年]、栄養外胚葉の高められるナトリウム−カリウム−ATPアーゼ(Na+−K+−ATPase)システムによる高められた胞胚腔の拡張[Biggers、1988年]、又はいまだメカニズムに見出されていないことを含んでいる。
【0044】
コントロール胚の着床率が、移入後72時間又は14日目それぞれを調べたときには同等であった(42%及び46%)。一方、移入後14日目に調べられた実験胚の着床率(59%)は、移入後72時間目に調べられた着床率(76%)よりもかなり小さかった(p=0.038)。よりたくさんの嚢が、着床の位置に群がることによって、実験群に完全に再び吸収されたことはもっともらしい事である。代わって、アイロプロストは、さもなければ着床してなかったいくつかの胚を「救済」し、これら「救済された」胚の一部のみが生存する子へ発生した。この検討と、[Huang、2002年]及びマウス[Huang、2004年]卵管でのたくさんのPGI2生産との結果は、PGI2が、卵管で分泌される胚栄養因子の一つであることを示唆し、したがってPGI2類似物を有する胚の培養培地に補足することは、IVFの効果を改良することを予期させる。
【0045】
IVF培養培地に胚発生又は着床を高める物質を補足することは、たいていの実験室及び非常に注意を払う培地製造業者によって採用されてきた。この懸念事項には、生殖毒物学、奇形形成、及び場合によってはこのような操作の後生的な影響を含んでいる。ヒト血清を補足された培地中で培養された羊胚由来の子羊は、より重みがあり妊娠期間がより長かった[Thompson、1995年;Holm、1996年;Sinclair、1999年]。過剰増殖及び新形成を含む先天性異常症、すなわちベックイス−ワインダーマン(Beckwith−Weindermann)症候群は生殖介助術と関係があった。[DeBaun、2002年;Maher、2003年]。
【0046】
胚及び胎児発生に示したアイロプロスト[Battenfeld、1995年]に関する生殖毒物学の検討は、胚及び胎児発生でウサギ及びサルでは影響されないが、指の変形がラットで観察されることを示した。指の異常は、アイロプロストの先天的奇形形成というよりも、影響を受けた構造から低酸素症を招く子宮胎盤流量の減少によるものらしく、この理由は、アイロプロストは血管拡張剤であり検討された動物が、妊娠期間を通してアイロプロストを受容したからである。ヒドラジン及びジヒドロピリジン類のような異なった化学構造を有する血管拡張剤を用いる場合、同じような問題が引き起こされるであろう。最近の研究では、アイロプロストへの暴露が、着床前の期間の間に行われた。246匹の生存する子において調べると、指の変形又はその他の異常性はなく、2群間の子及び胎盤の重量は同様であった。全てこれらのことを除いて、IVF培地にPGI2類似物を補足することは、実験的プロトコルとみなされるべきである。
【0047】
これらの実例の検討において、我々は、PGI2が胎児又は胎盤のいずれの重量にも影響を与えることなく、マウス胚の着床及び生児出生の可能性を高めること、すなわちプロスタサイクリンは、胎盤又は新生児の重量を増加させないことを明らかにした。これは、着床及び生児出生の高まりとは別の利点であり、プロスタサイクリンが、着床及び生児出生を高める他の代理母とは異なることを示している。着床及び生児出生を高める以前に報告された代理母はまた、新生児の重量を増加させて、ヒト体外受精(IVF)についてこれらの代理母の使用を不適としている。さらに、プロスタサイクリンは、インビボのマウスモデルでは、胎盤及び新生児の重量に影響を与えなかったため、ヒトIVF胚に利用できると考えられている。
【0048】
マウス胚についての前述のインビボ研究で得られた結果は、それらの胚が子宮に移入される前に、プロスタグランジン又はプロスタグランジン類似物を用いてインビトロで同様に処理される場合に、ヒトを含めその他の哺乳動物胚において得られるであろう同様な好ましいインビボの結果を示し、又は予測させるものであるとみなされる。本ヒトIVFは、患者の子宮への8細胞期又は胚盤胞期の胚の移入を含んでいる。この点において、ヒトIVFは、妊娠マウスに対するコントロール、すなわちプロスタサイクリン類似物処理胚への移入を含む我々の実験の計画と同様である。我々のデータは、妊娠キャリアに移入された際に、8細胞期と桑実胚期との間にプロスタサイクリン類似物に暴露された実験胚が、コントロール胚よりも着床及び生児出生率が非常にに高いことを示した(表1及び2、それぞれに示されるように)。
【表1】
【表2】
【0049】
さらに苦心なく、当業者はここまでの説明を用いて、その最大限に本発明を利用できることを確信する。前述の実施例は、例証として解釈されるべきであり、開示の残部をいかように規制するものではない。本発明の好ましい実施例を示され説明したけれども、その変更が、本発明の趣旨及び教示から逸脱することなく当業者によりなされるであろう。ここに説明される実施例は、単なる例示であり限定を意図するものではない。ここに開示された本発明の多くの変形及び変更が、可能であり本発明の範囲のものである。発明の背景における参照例、特にこの出願の優先日後の公開日を有する如何なる参照例の如何なる説明も、本発明に対して従来技術であると認めているものではない。ここに引用した全ての特許、特許出願及び公開の開示は、それらがここでの記載を補う模範的、手法的、その他の詳細を与える限りにおいて、ここに援用される。
【図面の簡単な説明】
【0050】
【図1】プロスタサイクリン(PGI2)及びマウス胚の完全孵化を示したグラフである。2細胞マウス胚が、PGI2類似物(アイロプロスト、1nMから10μMの範囲)で培養された。完全孵化率は96時間後に決定された。3乃至5の個別の実験(各々17−20胚)の結果は、平均±標準偏差として表現される。ED50値はおよそ6.7nMである。
【図2】完全孵化の増大及びプロスタサイクリン(PGI2)暴露の継続時間(胚発生の時期を基にアイロプロスト暴露)を示す棒グラフである。2細胞マウス胚が0.1μMアイロプロスト、すなわちPGI2類似物の存在下で示された期間(2細胞胚の採取後の時間として表現される)培養された。完全孵化率は平均±標準偏差として表現される。胚がアイロプロストに暴露されている間の発生時期が表に記載されている。2期間がアイロプロストの最大限の効果に重要であるようである、すなわち24乃至42時間及び42乃至72時間であり、それぞれ8細胞胚から桑実胚期及び桑実胚期から初期胚盤胞への発生に対応する。いずれの期間の間にアイロプロストに暴露しても、高められる孵化を確実にした。個別の観察の数(18−20胚についてのそれぞれ)は、コントロールで12胚、42−72時間では3胚、その他は4胚であった。コントロール胚の完全孵化率は、図1の孵化率とは異なっており、おそらく雌マウスの個体が変更されたからであろう。
【図3】マウス胚のプロスタサイクリンレセプタ(IP)の発現を示すウエスタンブロットである。60匹のマウス桑実胚(レーン1)から抽出した全細胞溶解物及びヒト血小板のミクロソーム(陽性コントロール、レーン2)のウエスタンブロット解析は、ヒトIPに対する抗体との免疫反応を示した。マウスIPの移動度は、ヒトIPの移動度よりも少なく、予想される分子量のそれぞれ〜50kDa及び〜46kDaと一致した。
【図4A−G】桑実胚及び胚盤胞におけるプロスタサイクリンレセプタ(IP)の発現を示す顕微鏡写真である。2個の桑実胚の位相差(図4A)及びIP染色(図4B)像が示される。透明帯は矢印で示される。図4Cは、胚盤胞における核のプロピジウムイオダイド染色を示す。図4Dにおいて、胚盤胞のIP及びプロピジウムイオダイドが重ね合わせられる。図4Eは胚盤胞のIP染色が網状模様を示すことを示す。IP染色の集合塊が栄養外胚葉のみに見られ、内部細胞塊には染色の増進を示さなかった。図4Fは桑実胚内の内部細胞塊(ICM)の配置を示すスケッチである。図4Gは胚盤胞の共焦点顕微鏡画像であり、栄養外胚葉のみがIP抗体によって染色されることを示す。断面の平面図はパネルF中のラインによって示される。陰性のコントロール、受精卵又はその他の発生時期にある胚(図示せず)にはIP染色はなかった。図中の棒はおよそ20μmである。
【図5】無傷のマウス胚に結合するアイロプロストを示す棒グラフである。116個のマウス胚盤胞が、5μMの非標識アイロプロストの存在又は不在下で、0.1μM3H−アイロプロスト、すなわちPGI2類似物とインキュベーションした。結合した3H−アイロプロスト及び遊離の3H−アイロプロストは膜ろ過によって分離された。
【図6A−B】妊娠マウスの子宮内の妊娠嚢を示す。着床は、着床部位の血管の増加によって決定され、妊娠嚢の存在によって確認された。胚移入後72時間、妊娠マウスは尾静脈から1%シカゴブルー(Chicago Blue)が与えられて3分後に安楽死させられた。図6Aは胚を受け入れた子宮角を取り巻く個々のブルーバンド(アスタリスクによって示される)を示す。図6Bでは、各ブルーバンドは妊娠嚢に対応しており、開放される子宮角ではピンクカラーの組織として現れた。ブルーバンド又は妊娠嚢のどれにも、胚を受け入れない子宮角には見つからなかった(矢印によって印を付けられる)。
【特許請求の範囲】
【請求項1】
哺乳動物胚のインビトロ発生を高める方法であって、胚の完全孵化を促進するのに有効な量において、培養培地にプロスタグランジン又はその類似物を補足することを有する、方法。
【請求項2】
前記プロスタグランジンはプロスタサイクリン(PGI2)又はプロスタグランジンE2(PGE2)である請求項1に記載の方法。
【請求項3】
前記プロスタグランジン類似物はアイロプロスト又はプロスタグランジンE1(PGE1)である請求項1に記載の方法。
【請求項4】
培養培地の前記補足は、ゲノムが活性化されて受精卵が桑実胚に発生する期間である胚の初期発生の間に行われる請求項1に記載の方法。
【請求項5】
前記胚はヒトであり、前記初期発生は4乃至8細胞期で開始する請求項1に記載の方法。
【請求項6】
培養培地の前記補足は、胚の桑実胚から初期胚盤胞への発生の間に行われる請求項1に記載の方法。
【請求項7】
培養培地の前記補足は、胚の初期発生から桑実胚期への発生と、胚の桑実胚から初期胚盤胞への発生の間との両方の間において行われる請求項1に記載の方法。
【請求項8】
前記胚はマウスであり、前記補足することは、培養中の胚を、その胚の2細胞発生期に胚の採取後24乃至42時間の間に、プロスタグランジン又は類似物に暴露することを含む請求項1に記載の方法。
【請求項9】
前記胚はマウスであり、前記補足は、胚を、その胚の2細胞発生期に胚の採取後24乃至72時間の間に、前記プロスタグランジン又はその類似物に暴露することを有する請求項1に記載の方法。
【請求項10】
インビトロ受精胚のインビボ着床の可能性を増大させる方法であって、請求項1に記載の方法に従って胚のインビトロ発生を高めることにより、胚の孵化の可能性が高められてインビボ着床の可能性が増大させられることを有する、方法。
【請求項11】
前記胚を完全に孵化させることを有しており、完全孵化は前記胚を透明帯から離すことを有する、請求項9に記載の方法。
【請求項12】
前記胚を宿主哺乳動物の子宮へ着床すること及び前記完全孵化を、生存に適した妊娠の確立と相関させることを有する請求項10に記載の方法。
【請求項13】
インビトロで受精された哺乳動物胚の生児出生の可能性を増加させる方法であって、
請求項1に記載の方法に従って胚のインビトロ発生を高めることにより、高められる孵化、すなわち胚の高められる孵化の可能性が達せられること、
孵化の可能性が高められた胚を宿主哺乳動物の子宮に導入すること、及び
導入された胚を前記子宮内に着床できるようにすること、
とを有し、胚の宿主の子宮内に着床される能力は、このような高まりは有せず同様の宿主の子宮内へ導入される胚の能力と比較して高められる方法。
【請求項14】
インビトロで受精された哺乳動物胚の一群における生児出生率を増加させる方法であって、
請求項1に記載の方法に従って前記胚のインビトロ発生を高めることにより、高められる孵化、すなわち胚の高められる孵化の可能性が達せられること、
高められる孵化の可能性を有する胚を宿主哺乳動物に着床させること、及び
着床された胚をインビボで成長できるようにして、高められる孵化を有する胚、すなわち孵化の可能性が高められた胚からの生児出生率が、このような高まりを有せず着床された胚からの生児出生率よりも非常に高くなるようにすること、
とを有する方法。
【請求項15】
哺乳動物胚のインビトロ初期発生のための改良された細胞培養培地であって、その改良点は、インビトロで前記胚の完全孵化を促進するのに有効な補足量のプロスタグランジン又はその類似物を有する、改良された細胞培養培地。
【請求項16】
プロスタグランジン又はその類似物の前記量は、インビトロ培養から取り出されて宿主哺乳動物の子宮に移入した後の、前記胚の完全孵化の可能性を高めるのに有効である、請求項15に記載の培地。
【請求項1】
哺乳動物胚のインビトロ発生を高める方法であって、胚の完全孵化を促進するのに有効な量において、培養培地にプロスタグランジン又はその類似物を補足することを有する、方法。
【請求項2】
前記プロスタグランジンはプロスタサイクリン(PGI2)又はプロスタグランジンE2(PGE2)である請求項1に記載の方法。
【請求項3】
前記プロスタグランジン類似物はアイロプロスト又はプロスタグランジンE1(PGE1)である請求項1に記載の方法。
【請求項4】
培養培地の前記補足は、ゲノムが活性化されて受精卵が桑実胚に発生する期間である胚の初期発生の間に行われる請求項1に記載の方法。
【請求項5】
前記胚はヒトであり、前記初期発生は4乃至8細胞期で開始する請求項1に記載の方法。
【請求項6】
培養培地の前記補足は、胚の桑実胚から初期胚盤胞への発生の間に行われる請求項1に記載の方法。
【請求項7】
培養培地の前記補足は、胚の初期発生から桑実胚期への発生と、胚の桑実胚から初期胚盤胞への発生の間との両方の間において行われる請求項1に記載の方法。
【請求項8】
前記胚はマウスであり、前記補足することは、培養中の胚を、その胚の2細胞発生期に胚の採取後24乃至42時間の間に、プロスタグランジン又は類似物に暴露することを含む請求項1に記載の方法。
【請求項9】
前記胚はマウスであり、前記補足は、胚を、その胚の2細胞発生期に胚の採取後24乃至72時間の間に、前記プロスタグランジン又はその類似物に暴露することを有する請求項1に記載の方法。
【請求項10】
インビトロ受精胚のインビボ着床の可能性を増大させる方法であって、請求項1に記載の方法に従って胚のインビトロ発生を高めることにより、胚の孵化の可能性が高められてインビボ着床の可能性が増大させられることを有する、方法。
【請求項11】
前記胚を完全に孵化させることを有しており、完全孵化は前記胚を透明帯から離すことを有する、請求項9に記載の方法。
【請求項12】
前記胚を宿主哺乳動物の子宮へ着床すること及び前記完全孵化を、生存に適した妊娠の確立と相関させることを有する請求項10に記載の方法。
【請求項13】
インビトロで受精された哺乳動物胚の生児出生の可能性を増加させる方法であって、
請求項1に記載の方法に従って胚のインビトロ発生を高めることにより、高められる孵化、すなわち胚の高められる孵化の可能性が達せられること、
孵化の可能性が高められた胚を宿主哺乳動物の子宮に導入すること、及び
導入された胚を前記子宮内に着床できるようにすること、
とを有し、胚の宿主の子宮内に着床される能力は、このような高まりは有せず同様の宿主の子宮内へ導入される胚の能力と比較して高められる方法。
【請求項14】
インビトロで受精された哺乳動物胚の一群における生児出生率を増加させる方法であって、
請求項1に記載の方法に従って前記胚のインビトロ発生を高めることにより、高められる孵化、すなわち胚の高められる孵化の可能性が達せられること、
高められる孵化の可能性を有する胚を宿主哺乳動物に着床させること、及び
着床された胚をインビボで成長できるようにして、高められる孵化を有する胚、すなわち孵化の可能性が高められた胚からの生児出生率が、このような高まりを有せず着床された胚からの生児出生率よりも非常に高くなるようにすること、
とを有する方法。
【請求項15】
哺乳動物胚のインビトロ初期発生のための改良された細胞培養培地であって、その改良点は、インビトロで前記胚の完全孵化を促進するのに有効な補足量のプロスタグランジン又はその類似物を有する、改良された細胞培養培地。
【請求項16】
プロスタグランジン又はその類似物の前記量は、インビトロ培養から取り出されて宿主哺乳動物の子宮に移入した後の、前記胚の完全孵化の可能性を高めるのに有効である、請求項15に記載の培地。
【図1】

【図2】
【図3】
【図4A】
【図4B】

【図4C】

【図4D】

【図4E】
【図4F】
【図4G】
【図5】
【図6A】
【図6B】
【図2】
【図3】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【図4F】
【図4G】
【図5】
【図6A】
【図6B】
【公表番号】特表2007−504807(P2007−504807A)
【公表日】平成19年3月8日(2007.3.8)
【国際特許分類】
【出願番号】特願2006−525526(P2006−525526)
【出願日】平成16年9月8日(2004.9.8)
【国際出願番号】PCT/US2004/029167
【国際公開番号】WO2005/026324
【国際公開日】平成17年3月24日(2005.3.24)
【出願人】(505073093)ボード オブ リージェンツ オブ ザ ユニバーシティー オブ テキサス システム (5)
【Fターム(参考)】
【公表日】平成19年3月8日(2007.3.8)
【国際特許分類】
【出願日】平成16年9月8日(2004.9.8)
【国際出願番号】PCT/US2004/029167
【国際公開番号】WO2005/026324
【国際公開日】平成17年3月24日(2005.3.24)
【出願人】(505073093)ボード オブ リージェンツ オブ ザ ユニバーシティー オブ テキサス システム (5)
【Fターム(参考)】
[ Back to top ]