
増加した生物学的活性を有する短縮型ケラチノサイト増殖因子(KGF)
【課題】成熟、完全長ヒトKGFと比較して生物活性における増加を示す、成熟、完全長ヒトKGFのフラグメントを提供すること。
【解決手段】単離されたポリペプチドであって、該ポリペプチドは成熟、完全長ヒトKGFのフラグメントであり、ここで該ポリペプチドは、Balb/MK生物活性アッセイにより測定される場合に、成熟、完全長ヒトKGFと比較して生物活性における増加を示し、そして上皮細胞増殖を特異的に刺激し、そして該生物活性における増加は、生物活性における少なくとも約2倍の増加である、単離されたポリペプチド。
【解決手段】単離されたポリペプチドであって、該ポリペプチドは成熟、完全長ヒトKGFのフラグメントであり、ここで該ポリペプチドは、Balb/MK生物活性アッセイにより測定される場合に、成熟、完全長ヒトKGFと比較して生物活性における増加を示し、そして上皮細胞増殖を特異的に刺激し、そして該生物活性における増加は、生物活性における少なくとも約2倍の増加である、単離されたポリペプチド。
【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明は、一般にケラチノサイト増殖因子(「KGF」)に関する。さらに特定的に、本発明は、昆虫細胞系で発現する組換え完全長KGFと比較して、生物学的活性の増加および細胞毒の減少を有する短縮型KGFフラグメントおよびそのアナログに関する。このKGFフラグメントは、本明細書中ではKGFdes1−23と命名され、成熟完全長KGFのN末端の最初の23アミノ酸残基を欠損し、このN末端は、Finch,P.W.ら、Science 245:752−755(1989)に示され、そしてKGFにその上皮細胞特異性を与えると既に考えられているように、N末端からのアミノ酸残基14〜16にグリコシル化部位を含む。
【背景技術】
【0002】
(発明の背景)
KGFは、線維芽細胞成長因子(「FGF」)のファミリーに属し、塩基性FGF(「bFGF」)として示されるものの始原型である。このため、KGFはまた、FGF−6として公知である。他のFGFのように、KGFはヘパリン結合タンパク質であるが、他のFGFと異なり、それは独特な標的細胞特異性を有する。特に、FGFは、一般に、一次または二次中胚葉由来および神経外胚葉由来の種々の細胞型の増殖および分化を刺激し得る。KGFは、Finch,P.W.ら(上記引用文中)で考察されるように、上皮細胞増殖を刺激し得るその能力において他のFGFと同様であるが、内皮細胞または線維芽細胞の増殖を刺激し得ないその能力において他のFGFと同様でない。
【0003】
酸性線維芽細胞成長因子(「aFGF」)および塩基性線維芽細胞成長因子(「bFGF」)を含むFGFは、Gospodarowiczら、Cell.Biol.Rev.25:307−314(1991)、およびBasilicoら、Adv.Cancer
Res.59:115−165(1992)で考察されるように、ヘパリン結合特性を有し、そして腹部および背部の中胚葉の初期胞胚での分化および増殖を誘導する能力を有することが公知である。FGFに対する細胞の応答は、Houら、Science 251:665−668(1991)で考察されるように、細胞表面レセプター(「FGFR」)へのその結合を通じて介在される。このレセプターには、3つの相関型がある。高親和性のFGFRは、Leeら、Science 245:57−60(1989);Dionneら、EMBO 9:2685−2692(1990);Mikiら、Science 251:72−75(1991);Mikiら、Proc.Natl.Acad.Sci.USA 89:246−250(1992);およびDellら、J.Biol.Chem.267:21225−21229(1992)で考察されたように、チロシンキナーゼであり、そしてflgレセプター(「FGFR−1」)、bekレセプター(「FGFR−2」)、およびK Samレセプター(「FGFR−3」)を含む。
【0004】
FGFR−1およびFGFR−2の両者は、中胚葉および神経外胚葉組織で広範囲に発現され、そして両者は、同じ親和性でaFGFおよびbFGFを結合し得る。FGFR−3は、上皮細胞に特異的であるKGFレセプターである。それは、FGFR−2のもう一つの転写物である。Mikiら(上記引用文中)およびDellら(上記引用文中)で考察されたように、aFGFおよびbFGFの両方について高い親和性を示し、そしてKGFについて全く親和性を示さないFGFR−2とは対照的に、FGFR−3は、KGFおよびaFGFをbFGFより約20〜1000倍高い親和性で結合する。
【発明の開示】
【発明が解決しようとする課題】
【0005】
上皮細胞に対するKGFの厳密に制限される活性プロフィールが、例えば、多くのタイプの創傷治癒のための適用ならびに乾癬および基底細胞癌のような表皮の過剰増殖疾患の処置で所望される。現在では、KGFを除いて、高適性マイトジェン因子はこれらの適用のためには存在しない。従って、KGFが、その効力を増加しそしてその細胞毒を減少するように改変されれば、治療用適用のために望ましい。
【0006】
最近、Ronら、J.Biol.Chem.268:2984−2988(1993年2月)により、KGF163がT7原核生物発現系で発現される場合、マイトジェン活性を所有する組換えKGF(「rKGF」)ポリペプチドが得られ得ることが見出された。rKGF分子が、成熟KGF163ポリペプチドのN末端からそれぞれ3、8、27、38、および48アミノ酸残基の欠失により短縮される場合、生じる分子の生物学的活性は変化した。それぞれ、3および8アミノ酸残基の欠失により、生じる分子のマイトジェン活性は、完全長のrKGF(「rKGF163」)と比べて影響し得なかった。しかし、27アミノ酸残基の欠失は、10〜20倍減少したマイトジェン活性を生じた。38および48アミノ酸残基の欠失は、それぞれマイトジェン活性およびヘパリン結合能を完全に失うようになる。従って、Ronらは、rKGF163分子と比べて増加した細胞分裂活性を有するいかなる短縮型KGFフラグメントをも生産し得なかった。
【課題を解決するための手段】
【0007】
(発明の要旨)
本発明の目的の1つは、成熟完全長KGF、KGF163のアミノ酸配列の一部分を含み、そして完全長組換えKGF、rKGF163と比較して少なくとも2倍のマイトジェン活性の増加を有するKGFフラグメントを提供することである。特別な目的は、rKGF163の最初の23個のN末端アミノ酸残基C−N−D−M−T−P−E−Q−M−A−T−N−V−N−C−S−S−P−E−R−H−T−R−を含む配列を欠失しているKGFフラグメントを提供することである。
【0008】
本発明の1つの他の目的は、rKGF163と比較して減少した細胞毒を有するKGFフラグメントを提供することである。さらに他の目的は、上記のKGFフラグメントおよび毒性分子を含む結合体を提供することである。毒性分子は、リシンA、ジフテリアトキシン、およびサポニン(saporin)のうち少なくとも1つであり得る。
【0009】
本発明の他の1つの目的は、上記のようなKGFフラグメント、および薬学的に受容可能なキャリア(例えば、ヒト皮膚への局所適用に適するもの)を含む治療用組成物を提供することである。
【0010】
本発明のさらに他の1つの目的は、上記のKGFフラグメントをコードするヌクレオチド配列からなるDNA分子を提供することである。
【0011】
本発明の他の1つの目的は、上記のKGFフラグメントをコードするDNA分子、およびDNA分子の発現のための調節配列を含む発現ベクターを提供することである。例えば、発現ベクターは、バキュロウイルスであり得る。
【0012】
本発明の他の1つの目的は、上記の発現ベクターで形質転換した宿主細胞を提供することである。例えば、宿主細胞は、細菌細胞、酵母細胞、哺乳動物細胞、または昆虫細胞であり得る。
【0013】
本発明の他の1つの目的は、上記のような形質転換宿主細胞を培養し、そして培養物からKGFフラグメントを単離することにより、KGFフラグメントを生産する方法を提供することである。
【0014】
本発明のさらに他の1つの目的は、上皮細胞成長が所望される領域にKGFフラグメントを適用することにより上皮細胞成長を刺激し、そして細胞を培養し得る方法を提供することである。
【0015】
本発明のさらに他の1つの目的は、処置されるべき創傷の領域に上記の治療組成物を適用して損傷を治癒させることによる、損傷治癒方法を提供することである。
【0016】
本発明のさらに他の1つの目的は、治療すべき領域に上記の結合体を適用することにより表皮の過剰増殖疾患を処置する方法を提供することである。
【0017】
従って、本発明は以下を提供する。
【0018】
1.成熟、完全長ケラチノサイト増殖因子のアミノ酸配列の一部分を含有するケラチノサイト増殖因子フラグメントであって、該一部分が、成熟、組換え、完全長ケラチノサイト増殖因子と比較して少なくとも2倍増加したマイトジェン活性を有し、そして成熟、完全長ケラチノサイト増殖因子の最初の23個のN末端アミノ酸残基を含有する配列を欠く、ケラチノサイト増殖因子フラグメント。
【0019】
2.前記フラグメントが、成熟、組換え、完全長ケラチノサイト増殖因子と比較して7倍増加したマイトジェン活性を有する、項目1に記載のケラチノサイト増殖因子フラグメント。
【0020】
3.前記フラグメントが、成熟、組換え、完全長ケラチノサイト増殖因子と比較して10倍増加したマイトジェン活性を有する、項目1に記載のケラチノサイト増殖因子フラグメント。
【0021】
4.前記フラグメントが、成熟、組換え、完全長ケラチノサイト増殖因子と比較して細胞毒性の減少を示す、項目1に記載のケラチノサイト増殖因子フラグメント。
【0022】
5.(a)成熟、完全長ケラチノサイト増殖因子のアミノ酸配列の一部分を含有するケラチノサイト増殖因子フラグメントであって、該一部分が、成熟、組換え、完全長ケラチノサイト増殖因子と比較して少なくとも2倍増加したマイトジェン活性を有し、そして成熟、完全長ケラチノサイト増殖因子の最初の23個のN末端アミノ酸残基を含有する配列を欠く、ケラチノサイト増殖因子フラグメント、および
(b)毒性分子
を含有する結合体。
【0023】
6.前記毒性分子が、リシンA、ジフテリアトキシンおよびサポニンからなる群から選択れる、項目5に記載の結合体。
【0024】
7.前記フラグメントが、成熟、組換え、完全長ケラチノサイト増殖因子と比較して7倍増加したマイトジェン活性を有する、項目5に記載の結合体。
【0025】
8.前記フラグメントが、成熟、組換え、完全長ケラチノサイト増殖因子と比較して10倍増加したマイトジェン活性を有する、項目5に記載の結合体。
【0026】
9.(a)成熟、完全長ケラチノサイト増殖因子のアミノ酸配列の一部分を含有するケラチノサイト増殖因子フラグメントであって、該一部分が、成熟、組換え、完全長ケラチノサイト増殖因子と比較して少なくとも2倍増加したマイトジェン活性を有するが、成熟、完全長ケラチノサイト増殖因子の最初の23個のN末端アミノ酸残基を含有する配列を欠く、ケラチノサイト増殖因子フラグメント、および
(b)薬剤的に許容し得るキャリアー
を含有する治療組成物。
【0027】
10.項目1に記載のケラチノサイト増殖因子フラグメントをコードするヌクレオチド配列を含有する、DNA分子。
【0028】
11.項目2に記載のケラチノサイト増殖因子フラグメントをコードするヌクレオチド配列を含有する、DNA分子。
【0029】
12.項目3に記載のケラチノサイト増殖因子フラグメントをコードするヌクレオチド配列を含有する、DNA分子。
【0030】
13.項目10に記載のDNA分子および該DNA分子の発現のための調節配列を含有する発現ベクター。
【0031】
14.前記ベクターがバキュロウイルスである項目13に記載の発現ベクター。
【0032】
15.項目13に記載の発現ベクターによって形質転換された宿主細胞。
【0033】
16.前記細胞が、細菌細胞、酵母細胞、哺乳動物細胞および昆虫細胞からなる群から選択される、項目15に記載の宿主細胞。
【0034】
17.項目15に記載の宿主細胞を培養する工程、および該培養物からケラチノサイト増殖因子フラグメントを単離する工程を包含する、ケラチノサイト増殖因子フラグメントを生産する方法。
【0035】
18.項目9に記載の治療組成物を、処置しようとする損傷領域に適用し、そして損傷を治癒し得ることを包含する、損傷治癒の方法。
【0036】
19.項目5に記載の結合体を、処置しようとする領域に適用することを包含する、表皮の過増殖疾患の処置方法。
【0037】
20.前記疾患が、乾癬および基底細胞癌からなる群から選択される疾患である、項目20に記載の処置方法。
【0038】
本発明のさらなる目的、特徴、および利益は、当業者に明らかであり、そしてここに列挙する必要はない。本発明は、当業者に明らかであり、そしてフラグメント、結合体、治療組成物、ベクター、宿主、および使用方法の性質および活性を実質上変化しない上記の変形および改変を含む。
【発明を実施するための最良の形態】
【0039】
(好ましい実施態様の詳細な説明)
本明細書中でKGFフラグメントまたはKGFdes1−23と称される、rKGF163の最初の23個のN末端アミノ酸残基にわたる欠失を有する短縮型非グリコシル化KGFが、rKGF163と比べて上皮細胞上でより大きな生物学的活性および細胞毒の減少を有することを驚くべきことに発見した。一般に、本発明のKGFフラグメントは、上皮細胞の増殖の刺激に対してKGF163の特異性を保持する。
【0040】
従って、本発明の好ましい実施態様は、天然分子がインビボで生産される場合に、通常それに付随する不純物をともなわない新規な短縮型非グリコシル化KGFフラグメント、KGFdes1−23である。このフラグメントは、単一なピークとしてNa Dod SO4/PAGEでその移動度に基づき約18kDaの見かけ上の分子量を有する。Balb/Mk細胞上での精製KGFdes1−23の比活性は、PCT特許出願第WO90/08771号に記載のように、約2.5×107ユニット/ミリグラム(ED5040pg/ml)であり、細胞増殖アッセイで比較する場合、rKGF163タンパク質の比活性、またはaFGFの比活性より約7〜10倍大きく、そしてrKGF163を化学的に規定した培地中でBalb/Mk細胞でDNA合成の開始を試験することによりバイオアッセイする場合より100倍大きい。
【0041】
本発明の他の好ましい実施態様では、KGFdes1−23は、組換えDNA技術により、特に大量生産販売のために生産される。さらに好ましい実施態様では、本発明によるKGFdes1−23またはそのアナログをそれぞれ含む組換えDNA分子および発現ベクターは、当該分野で周知の方法を用いて、標準遺伝子発現技術により実施される。
【0042】
さらなる実施態様では、KGFdes1−23をコードするDNAを含むDNAまたはベクターは、細菌、哺乳動物、酵母、または昆虫細胞発現系で発現され得る。好ましい実施態様では、細菌または酵母細胞の発現系は、KGFdes1−23フラグメントの生産にとって理想である。他の好ましい実施態様では、DNAまたはベクターは、昆虫細胞発現系で発現される。
【0043】
本発明のKGFフラグメントは、レセプター認識部位の同定、およびペプチドアゴニストまたはアンタゴニストの設計に使用され得る。さらに、ケラチノサイトに対するKGFの独特な特異性である、血管内皮細胞または線維芽細胞の増殖を誘導し得ない能力および細胞毒の欠損を考慮して、本発明の他の実施態様では、KGFdes1−23は、好ましくは、特に、皮膚の上皮再形成の促進を所望する場合、創傷治癒適用のために使用される。好ましい実施態様では、KGF des1−23は、角膜の上皮修復で使用される。KGFdes1−23の他の適用は、胃腸管内に見られる表皮細胞に対するその特異性に基づいて想定され得る。
【0044】
皮膚修復について、上皮成長因子(「EGF」)、血小板由来の成長因子(「PDGF」)、および他のFGFのような他の成長因子以外の本明細書におけるKGFフラグメントの選択は、当業者の技術範囲内である。例えば、他の成長因子は、ケラチノサイト増殖を直接的または間接的のいずれかで刺激するのに加えて、線維増殖および脈管形成を誘導する。皮膚修復では、このような特別な活性は瘢痕形成のような所望でない副作用を生成し得る。創傷または手術のいずれかを含む角膜修復では、これらの他の因子の使用は、角膜内に血管侵入を誘導し、所望でない角膜不透明または浮腫を生じる。他方では、KGFは、ケラチノサイトにとって独特な特異性を有し、そして血管内皮細胞または線維芽細胞の増殖を誘導せず、そして従って、これらの特定の創傷治癒適用のために選択した薬剤である。
【0045】
さらに、本発明のKGFフラグメントは、KGFフラグメント/毒素結合体の形態で本発明の他の実施態様で用いられ得る。このような結合体は、上皮から基底層のケラチノサイトの過剰増殖を生じる乾癬のような疾患における疾患のプロセスを減衰させ得るか、または停止し得る。この結合体は、血管内皮細胞および線維芽細胞のような、これらの組織に存在する他の2つの主要な細胞型に影響を及ぼさない。
【0046】
本発明の他の実施態様では、KGFフラグメント/毒素結合体は、腫瘍細胞増殖、例えば、基底細胞癌を根絶するために用いられ得る。
【0047】
本発明の実施は、他に指示がなければ、当該分野の技術範囲内にある分子生物学、微生物学、組換えDNA、および免疫学の従来の技法を使用する。このような技法は文献で十分に説明されている。例えば以下を参照のこと:
【0048】
【表1A】

【0049】
【表1B】
アミノ酸の標準略語は、本明細書中の図1およびその他で用いられる。本明細書中に引用した全ての刊行物、特許、および特許出願は参考として援用される。説明のために、本明細書で用いる用語を下記により特定的に定義する。
【0050】
(定義)
本明細書で用いる用語「ケラチノサイト増殖因子」または「KGF」は、構造的に異なるタンパク質である線維芽細胞成長因子、FGFのファミリーに属するポリペプチドを意味し、このタンパク質は、それらが関連するファミリーの遺伝子にコードされていることを示唆する配列相同性の変化程度を呈示する。KGFは、本明細書の他の所に記載された特性を有し、例えば、FGFR−3に結合し、そして上皮細胞、特にケラチノサイトの成長を刺激し得る。完全長KGFは、163アミノ酸残基からなる。
【0051】
本明細書で用いる本発明のKGFフラグメントは、成熟完全長KGFのアミノ酸配列の一部分からなるポリペプチドである。本明細書で用いるKGFフラグメントのアナログは、KGFフラグメントの特性に実質的に影響しないKGFフラグメントの主要ではない挿入、欠失、または置換を含む。例えば、保存的アミノ酸置換が予測される。このような置換は、例えば、アミノ酸の側鎖に関連するアミノ酸ファミリー内で起こる。アミノ酸のファミリーは、(1)酸性:アスパラギン酸、グルタミン酸;(2)塩基性:リジン、アルギニン、ヒスチジン;(3)非極性:アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン;および(4)非荷電極性:グリシン、アスパラギン、グルタミン、システイン、セリン、トレオニン、チロシンを含む。フェニルアラニン、トリプトファン、およびチロシンは、時折芳香族アミノ酸のファミリーとして共同に分類される。特に、ポリペプチドの活性部位の外側領域でロイシンをイソロイシンまたはバリンで、またはアスパラギン酸をグルタミン酸で、またはトレオニンをセリンで置換する孤立置換、もしくはアミノ酸を構造的な関連アミノ酸で置換する同様の保存的置換からなる保存的アミノ酸置換は、ポリペプチドの特性に大きな影響を有さないことが、一般に認められている。
【0052】
KGFフラグメントの特性は、(i)成熟完全長天然KGF分子と比較して、上皮細胞増殖の刺激を、2〜10倍、好ましくは2倍、最も好ましくは7〜10倍増加するようなその生物学的活性、および(ii)FGFR−3に結合するその能力を含む。
【0053】
本明細書で用いる用語「組換えポリヌクレオチド」は、半合成、または合成起源、もしくはcDNAまたはゲノムDNA(「gDNA」)にコードされたポリヌクレオチドを意味し、それは、(1)天然で会合するポリヌクレオチドの全てまたは一部分と会合しない、(2)天然で連結するポリヌクレオチド以外のポリヌクレオチドに連結する、あるいは(3)天然に存在しない。
【0054】
「レプリコン」は、任意の遺伝要素、例えば、プラスミド、染色体、ウイルス、コスミドなどであり、それは、細胞内でポリヌクレオチド複製の自律単位として挙動する;すなわち、それは、それ自身の制御下で複製し得る。レプリコンは、例えば、選択マーカーを含み得る。
【0055】
「組換えベクター」は、他のポリヌクレオチドセグメントが付加されるレプリコンであり、付加したポリヌクレオチドセグメントの複製および/または発現をもたらす。
【0056】
「調節配列」は、ポリヌクレオチド配列が作動可能に連結されるコーディング配列の発現の調節に必要なポリヌクレオチド配列を意味する。このような調節配列の性質は、コーディング配列が発現されるべき宿主細胞に依存して異なる。原核生物では、このような調節配列は、例えば、一般にプロモーター、リボソーム結合部位、および/または転写終結配列を含む。真核生物では、一般にこのような調節配列は、プロモーターおよび/または転写終結配列を含む。用語「調節配列」はまた、その存在が有益な付加成分、例えば、発現されるべきポリペプチドの分泌をコードする分泌リーダー配列を含み得る。
【0057】
「作動可能に連結された」は、成分が、意図した様式で機能し得るような関係で連結している並置を意味する。例えば、調節配列は、コーディング配列の発現が、調節配列に適合した条件下で達成されるように、それが接続される場合、コーディング配列に「作動可能に連結されて」いる。
【0058】
「コーディング配列」は、適切な調節配列の制御下に置かれた場合、ポリペプチドに翻訳されるポリヌクレオチド配列である。コーディング配列の境界は、一般にその5’末端の翻訳開始コドンおよびその3’末端の翻訳終止コドンにより決定される。コーディング配列は、例えば、cDNA、および組換えポリヌクレオチド配列を含み得る。
【0059】
「PCR」は、Saikiら、Nature 324:163(1986);およびScharfら、Science 233:1076−1078(1986);米国特許第4,683,195号;および米国特許第4,683,202号に記載するポリメラーゼ連鎖反応の技術を意味する。
【0060】
本明細書で用いる用語「ポリペプチド」は、アミノ酸のポリマーを意味し、そして生成物の特定の長さを意味しない。従って、ペプチド、オリゴペプチド、およびタンパク質は、この用語内に包含される。この用語はまた、ポリペプチドの翻訳修飾、例えば、グリコシル化、アセチル化、リン酸化などを包含する。定義内に包含されるものは、置換した結合を有するポリペプチド、および当該分野で公知のその他の改変で、天然に存在するおよび天然に存在しない両方のポリペプチドである。
【0061】
本明細書で用いる「ターミネーター」は、コーディング配列の終止コドンの3’または下流に位置するポリアデニル化および転写終結配列のような調節配列である。
【0062】
「組換え宿主細胞」、「宿主細胞」、「細胞」、「細胞培養物」、および他のこのような用語は、例えば、微生物、昆虫細胞、および哺乳動物細胞を意味し、これは、組換えベクターまたは他の転移DNAを導入するためのレシピエントとして用いられ得るかまたは用いられており、そして形質転換された細胞の子孫を含む。このような子孫は、形態もしくはゲノムDNAまたは全DNAの相補性が元の親と必ずしも同一ではなく、そして天然、偶発的、または意図的な変異によって生成され得る。哺乳動物宿主細胞の例は、チャイニーズハムスター卵巣(「CHO」)およびサル腎臓(「COS」)細胞を含む。
【0063】
本細書で用いる用語「微生物」は、細菌および真菌のような原核生物および真核生物の微生物種を含み、後者は、酵母および糸状菌を含む。
【0064】
本明細書で用いる「形質転換」は、宿主細胞への外因性ポリヌクレオチドの転移を意味し、転移に用いた方法、例えば、感染、直接取り込み、形質導入、F因子交配、マイクロインジェクション、またはエレクトロポレーションにかかわらない。外因性ポリヌクレオチドは、非組み込みベクター、例えば、プラスミドとして維持され得、またはあるいは、宿主ゲノムに組み込まれ得る。
【0065】
ポリペプチドまたはヌクレオチド配列に関する「精製」および「単離」は、示す分子が、同じ種または型の他の生物学的高分子が実質的にない状態で存在することを意味する。本明細書で用いる用語「精製」は、同じ型の生物学的高分子が少なくとも75重量%;好ましくは少なくとも85重量%、さらに好ましくは少なくとも95重量%、最も好ましくは98重量%存在することを意味するが、水、緩衝液、および他の小分子、特に、1000未満の分子量を有する分子もまた存在し得る。
【0066】
「薬学的に受容可能なキャリア」は、抗体、熱などの生成のような所望しない副作用をそれ自身誘導しないヒトへの投与のために、当業者により用いられる任意のキャリアを意味する。適切なキャリアは、典型的には大きく、ゆっくりと代謝される高分子(それはタンパク質、多糖類、ポリ乳酸、ポリグリコール酸、重合アミノ酸、アミノ酸コポリマー、または非活性ウイルス粒子であり得る)である。他のキャリアは、当業者に周知である。好ましくは、キャリアは、チログロブリンを含む。
【0067】
治療用組成物は、典型的には、薬学的に受容可能なキャリア、例えば、水、生理食塩水、グリセロール、エタノールなどを含有し得る。さらに、補助剤、例えば、湿潤剤または乳化剤、pH緩衝化剤などが、このような組成物中に存在し得る。
【0068】
本明細書で用いる「治療有効量」は、所望の結果の生成に効果的な量を意味する。この量は、処置されるべき個体の健康および身体状態、抗体を合成する個々の免疫系の能力、所望の防御程度、製剤、処置する医師の病状の評価、および他の関連因子に依存し得る。この量は、一般検査により決定され得る相対的に広い領域に入り得ることが予期される。
【0069】
KGFが昆虫細胞Spodoptera frugiperda(「SF9」)で、KGF163をコードするcDNAを含む組換えバキュロウイルスAutographa californicaの感染により発現される場合、それらは、KGF163に加えて、単一なグリコシル化部位を含むKGF163N末端ドメインの最初の23アミノ酸残基を欠失するKGFフラグメントを生産する。短縮型の非グリコシル化KGFフラグメントKGFdes1−23またはKGF140は、KGF163として同定された天然の長形態に較べて、標的細胞に対して7〜10倍増加した能力を有する。 KGFdes1−23の標的細胞特異性は、変化しない。さらに、高濃度のKGFフラグメントでは、ケラチノサイトについての毒性効果は、KGF163について観察されたのに較べて全く観察されない。これらの観察は、PCT出願第WO90/08771号で既に提唱されたことに反して、KGFの標的細胞特異性が、そのN末端ドメインの残基にないことを示唆する。さらに、本発明は、KGFのN末端短縮型バージョンが、実際に、治療用適用のためのさらに高い生物学的活性および細胞毒の減少を有する改良KGFバージョンを表すことを示す。
【0070】
大量のKGFフラグメントは、組換えDNA技術により生産され得、組換えDNA技術は、天然供給源からKGFを単離して精製し、そしてそのN末端から最初の23アミノ酸残基を切除するもう一方のプロセスより好ましい。組換え技術により生産されるKGFが、細胞中に通常存在する混入分子の非存在下で、KGFが単離されることを可能にする。実際に、いかなる微量のヒトタンパク質混入物質も完全に含まないKGF組成物は、例えば、細菌細胞、酵母細胞、および昆虫細胞で容易に生産され得る。組換えDNA技術の使用により、部位特異的変異誘発により生産され得る変異型のような、天然で見られないKGFフラグメントもまた、生産され得る。
【0071】
所望の宿主でのKGFフラグメントの発現を可能にする任意のプロモーターが、本発明で用いられ得る。例えば、E.coliでは、lacオペロンの調節プロモーター配列が用いられ得る。他の例は、酵母アルコールデヒドロゲナーゼ(「ADH」)プロモーターであり、それは、ADHプロモーターの活性を調節する上流アクチベーター配列(「UAS」)を有する。さらに、特定のウイルスエンハンサーもまた、本明細書で用いられ得る。このようなエンハンサーは、哺乳動物細胞において増幅するだけでなく、発現を調節する。これらのエンハンサーは、Sassone−CorsiおよびBorelli Trends Genet.2:215(1986);Maniatisら、Science
236:1237(1987)に記載のように、哺乳動物のプロモーター配列に組み込まれ、この哺乳動物プロモーター配列は、ホルモンまたは酵素基質のような誘導物質の存在下でのみ活性化し得る。本発明で用いられ得るプロモーターはまた、バキュロウイルスポリヘドロンハイブリッドプロモーターおよびp10プロモーターを含む。
【0072】
本発明で用いられ得る終結配列の例は、Saccharomyces cerevisiae α因子ターミネーターおよびバキュロウイルスターミネーターである。さらに、特定の宿主細胞で作動可能なウイルスターミネーターもまた、用いられ得る。例えば、SV40ターミネーターは、チャイニーズハムスター卵巣(CHO)細胞で機能する。
【0073】
短縮型タンパク質を設計する場合、いくつかのKGFが、本発明の実施において好ましい。1つは、非グリコシル化KGFであり、例えば、天然KGFの最初の23アミノ酸を切除している非グリコシル化KGFである。好ましい短縮型KGFをコードするcDNAを図1に示す。KGFフラグメントの他の種は、存在するか、または一般的な方法により作製され得る。示された配列において、短縮型タンパク質のための開裂部位が、矢印(a)で示される場所に生じ得、約18,000Daの分子量を有するタンパク質を生じる。図1の配列データーおよび短縮型KGFの示された特徴を利用して、短縮型KGFをコードする他のDNA配列を得ることは、当該分野の技術範囲内にある。例えば、構造遺伝子は、正しいアミノ酸を保持しながら個々のヌクレオチドを変えること、または生物学的活性を損失せずにアミノ酸を改変するようにヌクレオチドを変えることにより操作され得る。ヌクレオチドは、例えば、インビトロでの変異誘発およびプライマー修復を含む公知の技術により置換、挿入、または欠失され得る。構造遺伝子は、その生物学的活性を保持しながら、その3’末端および/またはその5’末端で短縮され得る。
【0074】
KGFフラグメントコーディング配列は、KGFフラグメントをコードするDNA配列を合成すること、もしくは所望の配列の生産のために存在しているまたは天然のKGFコーディング配列を変化することにより構築され得る。天然KGFコーディング配列またはポリペプチドは、天然に生じる。天然KGFのアミノ酸配列を[図1]に示す。合成KGFフラグメントは、Urdeaら、Proc.Natl.Acad.Sci.USA 80:7461(1983)で示唆されるように、KGFの公知のアミノ酸配列に基づき、選択した宿主細胞に好ましいコドンを用いて作製され得る。あるいは、所望のKGFフラグメントコーディング配列は、[図1]に示す核酸配列に基づくプローブを用いて核酸ライブラリーからクローニングされ得る。核酸配列ライブラリーの生産およびプローブ化(probing)のための技術は、例えば、Sambrookら、「Molecular
Cloning:A Laboratory Manual」(New York,Cold Spring Harbor Laboratory,1980)に記載されている。他の組換え技術、例えば、部位特異的変異誘発、PCR、酵素的切断および連結もまた、KGFフラグメントのコーディング配列のアナログおよび誘導体を構築するため用いられ得る。天然KGFポリペプチドコーディング配列は、他のクラスのKGFポリペプチドを作製するために容易に改変され得る。
【0075】
例として、KGFフラグメントのアナログは、KGFフラグメントの活性を損なわない保存的アミノ酸置換により作製される。本発明は、さらにムテイン(mutein)と呼ばれるKGFフラグメントのアナログのサブセットを意図しており、ムテインでは、KGFフラグメントのジスルフィド結合に関与しないシステインが通常は、セリンに置換されている。これらのムテインは、非改変KGFフラグメントより広い温度領域にわたり安定である。本明細書中のムテインはまた、天然KGFフラグメントと比較してアミノ酸欠失を含み得る。ムテインは、少なくとも20%、好ましくは少なくとも50%、および最も好ましくは少なくとも80%の天然KGFフラグメントの活性を保持し得る。ムテインのコーディング配列は、天然KGFフラグメントのコーディング配列のインビトロでの変異誘発により構築され得る。
【0076】
フラグメントは、アミノおよび/またはカルボキシル末端アミノ酸欠失により天然KGF分子と異なる。KGFフラグメントが、N末端グリコシル化部位を含まず、そして天然KGF分子の少なくとも20%のKGF活性を保持するなら、短縮されたアミノ酸の数は重大でない。このようなフラグメントのコーディング配列は、天然KGFコーディング配列またはその変異体由来の所望でないヌクレオチドを開裂することにより容易に構築され得る。
【0077】
本発明の範囲内の発現ベクターは、宿主細胞で作動可能であり、そしてKGFフラグメントまたはアナログのコーディング配列に作動可能に連結されるプロモーターを含む。発現ベクターは、分泌のためのシグナル配列、ターミネーター、選択マーカー、複製起点、および宿主細胞配列と同種(homologous)の配列を任意に含み得る。これらの追加的な要素は、発現を最適化するために含まれ得る。
【0078】
機能的な非天然プロモーターもまた用いられ得、例えば、異なるプロモーターの共通配列に基づいた合成プロモーターであり得る。また、効果的なプロモーターは、異種(heterologous)の発現開始領域と連結した調節領域を含むハイブリッドプロモーターであり得る。ハイブリッドプロモーターの例は、E.coli tac転写活性領域に連結したE.coli lacオペレーター;本明細書に参考として援用される米国特許第4,876,197号および第4,880,734号に記載のような、酵母グリセルアルデヒド−3−ホスフェート−デヒドロゲナーゼ(「GAPDH」)に連結した酵母アルコールデヒドロゲナーゼ(「ADH」)調節配列;およびシミアンウイルスSV40プロモーターに連結したサイトメガロウイルス(「CMV」)エンハンサーである。
【0079】
本発明のKGFフラグメントまたはアナログのコーディング配列はまた、読み取り枠でシグナル配列に結合され得る。シグナル配列は、典型的には、疎水性アミノ酸からなる分泌のためのアミノ酸配列をコードしKGFフラグメントまたはアナログを細胞膜に導く。好ましい実施態様では、プロセッシング部位は、シグナル配列とKGFフラグメントとの間に位置し、インビボまたはインビトロのいずれかでシグナル配列およびKGFフラグメントの間を開裂し得る。適切なシグナル配列は、分泌される内因性宿主細胞タンパク質に対する遺伝子由来であり、例えば、ヨーロッパ特許第12873号および日本特許第62,096,086号に記載の酵母インベルターゼ、米国特許第4,588,684号に記載のA因子遺伝子、およびEP60057に記載のようなインターフェロンシグナル配列である。
【0080】
酵母での発現のために本発明で用いる好ましいクラスの分泌リーダーは、短縮型酵母α因子リーダー配列であり、それは、「プレ」シグナル配列の少なくとも一部分および「プロ」領域を含む。本明細書に参考として援用される米国特許第4,546,083号および第4,870,008号、ならびにヨーロッパ特許第EP324274号に記載のように、本明細書中で使用され得るα因子リーダー配列は、約83アミノ酸残基を有する完全長プレ−プロα因子リーダー、および典型的には、約25〜約50アミノ酸残基を含む短縮型α因子リーダーであり得る。本明細書で用いられ得るα因子リーダーフラグメントを使用する付加的なリーダーは、第二の酵母α因子由来のプロ領域ではなく、第一の酵母シグナル配列のプレ配列で作られるハイブリッドα因子リーダーを含む。(例えば、PCT
WO89/02463を参照のこと。)
(f.発現システム)
KGFフラグメントおよびそのアナログは、当業者に十分に理解されるように、KGFフラグメントまたはアナログをコードするDNA配列がベクターに適切な読み取り枠および方向で作動可能に連結される場合、組換え技術により発現され得る。KGFフラグメントを含む遺伝子構築物を作成する場合、好ましい出発材料は、KGFフラグメントをコードしているcDNAである。KGFフラグメントはまた、所望する場合、開裂され得るハイブリッドタンパク質として生産され得るが、典型的には、短縮型KGF遺伝子は、プロモーターより下流に挿入され得、そして後ろにターミネーター配列が続き得る。一般に、短縮型KGFおよび短縮型KGFポリペプチド誘導体の生産収量を改善する宿主細胞特異的配列が用いられ得、そして適切な制御配列、例えば、エンハンサー配列、ポリアデニル化配列、およびリボソーム結合部位が発現ベクターに付加され得る。一旦、適切なコーディング配列が単離されること、それは、種々の異なる発現系;例えば、哺乳動物細胞、バキュロウイルス、細菌、および酵母で使用され、発現され得る。これらは、以下に考察される。
【0081】
(i.哺乳動物系)
哺乳動物発現系は、当該分野で公知である。哺乳動物プロモーターは、哺乳動物RNAポリメラーゼを結合し、そしてコーディング配列(例えば、構造遺伝子)のmRNAへの下流(3‘)転写を開始し得る任意のDNA配列である。プロモーターは転写開始領域、それは、通常コーディング配列の5’末端に隣接する位置にあり、および転写開始部位の25〜30塩基対(bp)上流に位置するTATAボックスを有し得る。 TATAボックスは、RNA合成を正しい部位で始めるために、RNAポリメラーゼIIに指示すると考えられている。動物細胞のプロモーターはまた、上流のプロモーター要素(典型的には、TATAボックスの100〜200bp上流内に位置する)を含み得る。上流のプロモーター要素は、転写が開始される速度を決定しどちらかの方向に作用し得る[Sambrookら、(1989)「哺乳動物細胞におけるクローン化遺伝子の発現。」Molecular Cloning:A Laboratory Manual,第2版]。
【0082】
哺乳動物のウイルス遺伝子は、しばしば高発現され、そして広い宿主領域を有する;従って、哺乳動物ウイルス遺伝子をコードしている配列は、特に有用なプロモーター配列を提供する。この例として、SV40初期プロモーター、マウス乳ガンウイルスLTRプロモーター、アデノウイルス主要後期プロモーター(AdMLP)、および単純ヘルペスウイルスプロモーターが含まれる。さらに、非ウイルス遺伝子、例えば、ネズミメタロチオネイン遺伝子由来の配列はまた、有用なプロモーター配列を提供する。発現は、構成的か、調節的(誘導性)のいずれかであり得、プロモーターがホルモン応答細胞におけるグルココルチコイドで誘導され得ることに依存する。
【0083】
上記のプロモーター要素と結合したエンハンサー要素(エンハンサー)の存在は、典型的には、発現レベルを増加する。エンハンサーは、同種または異種プロモーターと連結されるとき、正常なRNA開始部位で合成を開始する場合に1000倍まで転写を刺激し得る調節DNA配列である。エンハンサーはまた、それらが転写開始部位から上流または下流に、正常または逆の向きで、もしくはプロモーターから1000ヌクレオチオド以上の距離に位置する場合、活性である[Maniatisら、(1987)Science 236:1237;Albertsら、(1989)Molecular Biology of the Cell,第2版]。ウイルス由来のエンハンサー要素は特に有用である。なぜなら、それらは、典型的には、より広い宿主領域を有するからである。この例として、SV40初期遺伝子エンハンサー[Dijkemaら、(1985)EMBO J.4:761]およびラウス肉腫ウイルスの長末端反復(LTR)由来[Gormanら、(1982b)Proc.Natl.Acad.Sci.79:6777]、およびヒトサイトメガロウイルス[Boshartら、(1985)Cell 41:521]由来のエンハンサー/プロモーターが含まれる。さらに、いくつかのエンハンサーは調節可能であり、そしてホルモンまたは金属イオンのような誘導物質の存在下でのみ活性になる[Sassone−CorsiおよびBorelli(1986)Trends Genet.2:215;Maniatisら、(1987)Science 236:1237]。
【0084】
DNA分子は、哺乳動物細胞において細胞内発現され得る。プロモーター配列は、DNA分子と直接連結し得、その場合、組換えタンパク質のN末端の最初のアミノ酸は常にメチオニンであり、メチオニンは、ATG開始コドンによりコードされている。所望であれば、N末端は、インビトロで臭化シアンとともにインキュベートすることでタンパク質から開裂され得る。
【0085】
あるいは、外来タンパク質はまた、哺乳動物細胞において外来タンパク質の分泌を提供するリーダー配列フラグメントからなる融合タンパク質をコードするキメラDNA分子を作成することにより、細胞から成長培地へ分泌され得る。好ましくは、リーダーフラグメントと、インビボまたはインビトロのいずれかで開裂され得る外来遺伝子との間にコードされるプロセッシング部位がある。リーダー配列フラグメントは、典型的には、細胞からのタンパク質の分泌を指向する疎水性アミノ酸からなるシグナルペプチドをコードする。アデノウイルス三部分(tripartite)リーダーは、哺乳動物細胞において外来タンパク質の分泌を提供するリーダー配列の1例である。
【0086】
典型的には、哺乳動物細胞により認識される転写終結およびポリアデニル化配列は、3’から翻訳終止コドンに位置する調節領域であり、そしてそれ故プロモーター要素とともにコーディング配列に隣接する。成熟mRNAの3’末端は、部位特異的な転写後の開裂およびポリアデニル化により形成される[Birnstielら、(1985)Cell
41:349;ProudfootおよびWhitelaw(1988)「真核生物RNAの終結および3’末端プロセッシング」、Transcription and splicing(B.D.HamesおよびD.M.Glover編);Proudfoot(1989)Trends Biochem.Sci.14:105]。これらの配列は、DNAにコードされるポリペプチドに翻訳され得るmRNAの転写を指向する。転写ターミネーター/ポリアデニル化シグナルの例は、SV40由来のそれらを含む[Sambrookら(1989)「培養哺乳動物細胞におけるクローニングされた遺伝子の発現」、Molecular Cloning:A Laboratory Manual。
【0087】
いくつかの遺伝子は、イントロン(介在配列とも言う)が存在する場合、さらに効果的に発現され得る。しかし、いくつかのcDNAは、スプライシングシグナル(スプライス供与体および受容体部位ともいう)を欠いたベクターから効率的に発現された[GothingおよびSambrook(1981)Nature 293:620を参照のこと]。イントロンは、スプライス供与体および受容体部位を含む、コーディング配列内に介在する非コーディング配列である。それらは、一次転写物のポリアデニル化後「スプライシング」と呼ばれるプロセスにより取り除かれる[Nevins(1983)Annu.Rev.Biochem.52:441;Green(1986)Annu.Rev.Genet.20:671;Padgettら、(1986)Annu.Rev.Biochem.55:1119;KrainerおよびManiatis(1988)「RNAスプライシング」、Transcription and splicing(B.D.HamesおよびD.M.Glover編)。
【0088】
典型的には、プロモーター、ポリアデニル化シグナル、および転写終結配列を含む上記の構成成分は、一緒に発現構築物中に置かれる。エンハンサー、機能的なスプライス供与体および受容体部位を有するイントロン、ならびにリーダー配列はまた、所望する場合、発現構築物中に含まれる。発現構築物は、哺乳動物細胞または細菌のような宿主において安定に維持し得る染色体外要素(例えば、プラスミド)のようなレプリコン中にしばしば維持される。哺乳動物の複製系は、動物ウイルス由来の系を含み、それは、複製にトランスに作用する因子を必要とする。 例えば、SV40[Gluzman(1981)Cell 23:175]のようなパポーバウイルスまたはポリオーマウイルスの複製系を含むプラスミドは、適切なウイルスT抗原の存在下できわめて高いコピー数に複製する。哺乳動物のレプリコンの別の例は、ウシパピローマウイルスおよびエプスタインバーウイルス由来のレプリコンを含む。さらに、レプリコンは、2つの複製系を有し得、従って、例えば、発現のために哺乳動物細胞で、およびクローニングおよび増幅のために原核生物宿主で維持され得る。このような哺乳動物−細菌シャトルベクターの例は、pMT2[Kaufmanら、(1989)Mol.Cell.Biol.9:946およびpHEBO[Shimizuら、(1986)Mol.Cell.Biol.6:1074]を含む。
【0089】
用いた形質転換手順は、形質転換されるべき宿主に依存する。異種ポリヌクレオチドを哺乳動物細胞へ導入するための方法は、当該分野で公知であり、そしてデキストラン仲介トランスフェクション、リン酸カルシウム沈澱、ポリブレン仲介トランスフェクション、プロトプラスト融合、エレクトロポレーション、リポソーム内へのポリヌクレオチドのカプセル化、および核へのDNAの直接マイクロインジェクションを含む。
【0090】
発現のための宿主として利用できる哺乳動物細胞株は当該分野で公知であり、チャイニーズハムスター卵巣(CHO)細胞、HeLa細胞、ベビーハムスター腎臓(BHK)細胞、サル腎臓細胞(COS)、ヒト肝細胞癌細胞(例えば、Hep G2)、および多くのその他の細胞株を含むがこれに限定されない、American Type Cultutre Collection(ATCC)から入手可能な多くの不死化細胞株を含む。
【0091】
(ii.バキュロウイルス系)
KGFをコードしているポリヌクレオチドは、安定な発現ベクター、例えば昆虫細胞発現ベクター中に挿入され得、そしてベクター内で制御要素に作動可能に連結され得る。ベクターの構築は、当該分野で公知の技術を使用して行われる。特に、本発明の目的のため、バキュロウイルス発現ベクターが、Kittsら、BioTechniques 14:810−817(1993)に実質的に従って構築される。
【0092】
簡単に説明すると、バキュロウイルスゲノムの一部(本明細書中では、「バキュロウイルス配列」と呼ばれている)に相同性があり、そしてそれと相同的組換えを行い得るポリヌクレオチド配列を含むトランスファーベクターに、KGF163をコードしている配列を挿入することによって、 KGF発現カセットを最初に構築する。バキュロウイルス配列は、以下により詳しく記載されているように、必須のポリヌクレオチド配列を少なくとも含む。KGFコーディング配列は、バキュロウイルス配列が両端に位置するようにトランスファーベクターに挿入される。
【0093】
KGFコーディング配列を含むトランスファーベクターを、機能的なウイルスの生産に必要な必須のポリヌクレオチド配列を欠いている野生型バキュロウイルスの変異体とともに宿主昆虫細胞中にトランスフェクトする。この点に関して、変異体のバキュロウイルスがKGF発現カセットと組換わるときトランスフェクションに際し機能的なウイルスが宿主細胞中で産生され得る。
【0094】
従って、この様式で産生される機能的なバキュロウイルスは、KGF発現カセットを組み込み、そして組換えKGFおよび組換えKGFdes1−23を産生するために、新たな宿主細胞のトランスフェクションに適切である。あるいは、このプロセスは、KGF163コーディング配列の代わりにKGFdes1−23コーディング配列を用いて実施され得る。
【0095】
一般に、発現系の構成成分は、バキュロウイルスゲノムのフラグメント、および異種遺伝子または発現されるべき遺伝子の挿入に便利な制限部位の両方を含むトランスファーベクター(通常は細菌プラスミド)を含む機能的なバキュロウイルス;トランスファーベクター中のバキュロウイルス特異的フラグメントに相同な配列を有する野生型バキュロウイルス(これは、バキュロウイルスゲノムに異種遺伝子の相同組換えを可能にする);および適切な昆虫宿主細胞および成長培地を含む。
【0096】
短縮型KGF DNA配列をトランスファーベクターに挿入後、ベクターおよび野生型ウイルスゲノムを、ベクターおよびウイルスゲノムが組換えられ得る昆虫宿主細胞にトランスフェクトする。パッケージ化された組換えウイルスを発現し、そして組換えプラークを同定し、そして精製する。バキュロウイルス/昆虫細胞発現系の材料および方法は、前記のInvitorogen,San Diego CAからキット形態(「MaxBac」キット)で市販されている。これらの技術は、一般に、当業者に公知であり、そしてSummersおよびSmith,Texas Agricultural Experiment Station Bulletin No.1555(1987)(本明細書では以後、「SummersおよびSmith」と称する)に十分に記載され、そして参考として援用されている。
【0097】
短縮型KGF DNA配列をバキュロウイルスゲノムに挿入する前に、プロモーター、リーダー(所望の場合)、目的のコーディング配列、および転写終結配列を含む上記の構成成分を、代表的には、中間導入配置構築物(トランスファーベクター)にアセンブルする。この構築物は、単一の遺伝子および作動可能に連結した調節要素;それぞれが作動可能に連結したそれ自身の調節要素のセットを有する複数の遺伝子;または調節要素の同じセットにより調節されているは複数の遺伝子を含み得る。中間導入配置構築物を、しばしば細菌のような宿主で安定に維持し得る染色体外要素(例えば、プラスミド)のようなレプリコン中に維持し得る。レプリコンは複製系を有し、従ってクローニングおよび増幅のために適切な宿主中で、それは維持され得る。
【0098】
現在では、AcNPVに外来遺伝子を導入するために最も一般的に用られるトランスファーベクターはpAc373である。当業者に公知である多くの他のベクターもまた設計された。これらは、例えば、pVL985(これは、ポリヘドリン開始コドンをATGからATTに改変し、そしてATTから32塩基対下流にBamHIクローニング部位を導入する;LuckowおよびSummers,Virology(1989)17:13を参照のこと)を含む。
【0099】
プラスミドはまた、通常ポリヘドリンポリアデニル化シグナル(Millerら(1988)Ann.Rev.Microbiol.,42:177)、ならびにE.coliにおける選択および増殖のための原核生物のアンピシリン耐性(amp)遺伝子および複製起点を含む。
【0100】
バキュロウイルストランスファーベクターは、通常バキュロウイルスプロモーターを含む。バキュロウイルスプロモーターは、バキュロウイルスRNAポリメラーゼを結合し得、そしてコーディング配列(例えば、構造遺伝子)のmRNAへの下流(5’から3’)転写を開始し得る任意のDNA配列である。プロモーターは、通常、コーディング配列の5’末端に隣接して位置する転写開始領域を有する。この転写開始領域は、典型的には、RNAポリメラーゼ結合部位および転写開始部位を含む。バキュロウイルストランスファーベクターはまた、エンハンサーと呼ばれる第二ドメインを有し得、存在する場合は、通常構造遺伝子から遠位にある。発現は、調節され得るか、または構成的であり得る。
【0101】
構造遺伝子は、ウイルス感染サイクルの後期に盛んに転写され、特異的に有用なプロモーター配列を提供する。この例として、ウイルスポリヘドリンタンパク質をコードする遺伝子、Friesenら、(1986)「バキュロウイルス遺伝子発現の調節」、The
Molecular Biology of Baculoviruses(Walter Doerfler編);EPO公開番号第127,839号および第155,476号;およびp10タンパク質をコードする遺伝子、Vlakら、(1988),J.Gen.Virol.69:765由来の配列が含まれる。
【0102】
適切なシグナル配列をコードするDNAは、分泌される昆虫タンパク質または分泌されるバキュロウイルスタンパク質、例えば、バキュロウイルスポリヘドリン遺伝子(Carbonellら、(1988)Gene,73:409)に由来する遺伝子であり得る。あるいは、哺乳動物細胞の翻訳後の修飾(例えば、シグナルペプチド開裂、タンパク質分解開裂、およびリン酸化)のためのシグナルが、昆虫細胞により認識されるようであり、そして分泌および核蓄積に必要なシグナルもまた、無脊椎動物細胞および脊椎動物細胞の間で保存され得るようであるので、昆虫起源でないリーダー、例えば、ヒトα−インターフェロン、Maedaら、(1985)、Nature 315:592;ヒトガストリン放出ペプチド、Lebacq−Verheydenら、(1988)、Molec.Cell.Biol.8:3129;ヒトIL−2、Smithら、(1985)Proc.Nat’l Acad.Sci.USA,82:8404;マウスIL−3、(Miyajimaら、(1987)Gene 58:273;およびヒトグルコセレブロシダーゼ、Martinら、(1988)DNA,7:99をコードする遺伝子由来のリーダーもまた、昆虫で分泌を提供するために使用され得る。
【0103】
組換えポリペプチドまたはポリタンパク質は、細胞内で発現され得、もしくは、適切な調節配列で発現される場合分泌され得る。非融合の外来タンパク質の良好な細胞内発現は、通常、 ATG開始シグナルに先行する適切な翻訳開始シグナルを含む短いリーダー配列を理想的には有する異種遺伝子を必要とする。所望であれば、N末端のメチオニンを、臭化シアンとのインビトロでのインキュベーションにより、成熟タンパク質から開裂し得る。
【0104】
あるいは、天然では分泌されない組換えポリタンパク質またはタンパク質は、昆虫での外来タンパク質の分泌を提供するリーダー配列フラグメントからなる融合タンパク質をコードするキメラDNA分子を作製することにより昆虫細胞から分泌され得る。リーダー配列フラグメントは、典型的には、タンパク質の小胞体中への移動を指向する疎水性アミノ酸からなるシグナルペプチドをコードする。
【0105】
短縮型のKGF DNA配列および/または発現産物前駆体をコードしている遺伝子の挿入後、昆虫細胞宿主は、トランスファーベクターの異種DNAおよび野生型バキュロウイルスのゲノムDNAを用いて同時形質転換され、それは、通常、同時トランスフェクションによる。構築物のプロモーターおよび転写終結配列は、典型的には、2〜5kbセクションのバキュロウイルスゲノムを含む。異種DNAをバキュロウイルスの所望の部位に導入する方法は、当該分野で公知である。(SummersおよびSmith 上記;Juら、(1987);Smithら、Mol.Cell.Biol.(1983)3:2156;ならびにLuckowおよびSummers(1989)参照のこと)。例えば、ポリヘドリン遺伝子のような遺伝子に、相同な二重交差組換えにより挿入され得る;また所望のバキュロウイルス遺伝子中に設計された制限酵素部位にも挿入し得る。Millerら、(1989),Bioessays 4:91。DNA配列が、ポリヘドリン遺伝子の代わりに発現ベクターにクローニングされる場合、その5’および3’の両方にポリヘドリン特異的配列が隣接し、そしてポリヘドリンプロモーターの下流に位置する。
【0106】
新たに形成されたバキュロウイルス発現ベクターは、次いで感染性組換えバキュロウイルスにパッケージ化される。同種の組換えが、低頻度で起こる(約1%と約5%との間);従って、同時トランスフェクション後に産生された多くのウイルスは、なお野生型のままである。それゆえ、組換えウイルスを同定するための方法が必要である。発現系の利点は、組換えウイルスを区別できるようにする視覚化スクリーニングである。ポリヘドリンタンパク質は、天然ウイルスにより産生され、ウイルス感染後、後期に感染細胞の核に非常に高レベルで産生される。蓄積したポリヘドリンタンパク質はまた、包埋粒子を含む封入体を形成する。これらの封入体は、サイズが15μmまでであり、高度に屈折性であり、光学顕微鏡下で容易に視覚化される明るく光った外観を与える。組換えウイルスに感染した細胞は、封入体を欠いている。野生型ウイルスから組換えウイルスを区別するために、トランスフェクション上清を当業者に公知の技術により単層の昆虫細胞上にプラーク化する。すなわち、プラークは、光学顕微鏡下で封入体の存在(野生型ウイルスを示す)または不在(組換えウイルスを示す)についてスクリーニングされる。「Current Protocols in Microbiology」第2巻(Ausubelら編)、16.8(増刊1990年10月);SummersおよびSmith、前述;Millerら(1989)。
【0107】
組換えバキュロウイルス発現ベクターが、いくつかの昆虫細胞に感染させるために開発された。例えば、組換えバキュロウイルスは、とりわけ以下について開発された:
【0108】
【表2】
細胞および細胞培養培地は、バキュロウイルス/発現系におる異種ポリペプチドの指向および融合発現の両方について市販されている;細胞培養技術は、当業者に一般に公知である。例えば、SummersおよびSmith、前述を参照のこと。
【0109】
次いで、改変された昆虫細胞は、適切な栄養培地で成長し得、この培地は、改変された昆虫宿主に存在するプラスミドを安定的に維持し得る。発現産物の遺伝子が、誘導し得る制御下にある場合、宿主は高濃度になるまで成長され得、そして発現を誘導する。あるいは、発現が構成的である場合、産物は連続的に培地中に発現され得、そして栄養培地は連続的に循環されねばならず、その間目的の産物を取り出し、そして枯渇した栄養を増強する。この産物をクロマトグラフィー(例えば、HPLC、アフィニティークロマトグラフィー、イオン交換クロマトグラフィーなど);電気泳動;密度勾配遠心分離;溶媒抽出などのような技術により精製され得る。適切には、必要に応じて、培地に同時に分泌されるか、または昆虫細胞の溶解から生じる任意の昆虫タンパク質を実質的に除去するように、宿主の残骸(例えば、タンパク質、脂質、および多糖)を少なくとも実質的に含まない産物を提供するように、産物をさらに精製する。
【0110】
短縮型KGF発現を得るために、形質転換体由来の組換え宿主細胞を、配列をコードする組換え短縮型KGFの発現を可能にする条件下でインキュベートし得る。これらの条件は、選択される宿主細胞に依存して変化し得る。しかし、条件は、当該分野で公知の事項に基づき、当業者に容易に確認され得る。
【0111】
(iii.細菌系)
細菌発現技術は当該分野で公知である。細菌プロモーターは、細菌RNAポリメラーゼを結合し得、そしてコーディング配列(例えば、構造遺伝子)のmRNAへの下流(3”)転写を開始し得る任意のDNA配列である。プロモーターは、通常コーディング配列の5’末端に隣接して位置する転写開始領域を有する。この転写開始領域は、典型的には、RNAポリメラーゼ結合部位および転写開始部位を含む。細菌プロモーターはまた、オペレーターと呼ばれる第二ドメインを有し得、それは、RNA合成が開始する隣接したRNAポリメラーゼ結合部位とオーバーラップし得る。遺伝子リプレッサータンパク質がオペレーターに結合し得、そしてそれによって特異的な遺伝子の転写を阻害し得るので、オペレーターは、ネガティブに調節された(誘導可能な)転写を可能にする。構成的な発現は、オペレーターのようなネガティブな調節要素の非存在下で生じ得る。さらに、ポジティブな調節は、遺伝子活性化タンパク質結合配列により達成され得、それが存在する場合は、通常、RNAポリメラーゼ結合配列に隣接する(5’側)。遺伝子活性化タンパク質の例は、カタボライトアクチベータータンパク質(CAP)であり、それは、Escherichia coli(E.coli)においてlacオペロンの転写開始を助ける[Raibaudら、(1984)Annu.Rev.Genet.18:173]。従って、調節された発現は、ポジティブまたはネガティブのいずれか、それにより転写促進または減少のいずれであり得る。
【0112】
代謝経路酵素をコードしている配列は、特に有用なプロモーター配列を提供する。この例としては、糖代謝酵素、例えば、ガラクトース、ラクトース(lac)[Changら、(1977)Nature 198:1056]、およびマルトース由来のプロモーター配列を含む。別の例は、トリプトファン(trp)のような生合成酵素由来のプロモーター配列を含む[Goeddelら、(1980)Nuc.Acids Res.8:4057;Yelvertonら、(1981)Nucl.Acids Res.9:731;米国特許第4,738,921号;EPO公開第036776号および第121775号]。g−ラオタマーゼ(g−laotamase)(bla)プロモーター系[Weissmann(1981)「インターフェロンおよび他の間違いのクローニング。」Interferon 3(I.Gresser編)]、バクテリオファージラムダPL[Shimatakeら、(1981)Nature 292:128]およびT5[米国特許第4,689,406号]プロモーター系はまた、有用なプロモーター配列を提供する。
【0113】
さらに、天然にない合成プロモーターがまた、細菌プロモーターとして機能する。例えば、ある細菌またはバクテリオファージプロモーターの転写アクチベーター配列は、他の細菌またはバクテリオファージプロモーターのオペロン配列と結合され得、合成ハイブリッドプロモーターを作製する[米国特許第4,551,433]。例えば、tac−lacプロモーターは、trpプロモーターおよびlacリプレッサーにより調節されるlacオペロン配列の両方を含むハイブリッドtrp−lacプロモーターである[Amannら、(1983)Gene 25:167;de Boerら、(1983)Proc.Natl.Acad.Sci.80:21]。さらに、細菌プロモーターは、細菌RNAポリメラーゼを結合しそして転写を開始する能力を有する非細菌起源の天然に存在するプロモーターを含み得る。非細菌起源の天然に存在するプロモーターはまた、原核生物でいくつかの遺伝子の高レベルの発現を行うために、適合するRNAポリメラーゼと結合され得る。バクテリオファージT7 RNAポリメラーゼ/プロモーター系は、結合プロモーター系の例である[Studierら、(1986)J.Mol.Biol.189:113;Taborら、(1985)Proc Natl.Acad.Sci.82:1074]。さらに、ハイブリッドプロモーターはまた、バクテリオファージプロモーターおよびE.coliオペロン領域(EPO公開第267851号)を含み得る。
【0114】
機能するプロモーター配列に加えて、効率的なリボソーム結合部位はまた、原核生物における外来遺伝子の発現に有用である。E.coliでは、リボソーム結合部位は、Shine−Dalgarno(SD)配列と呼ばれ、そして開始コドン(ATG)および開始コドンの3〜11ヌクレオチド上流に位置する長さが3〜9のヌクレオチドの配列を含む[Shineら、(1975)Nature 254:34]。SD配列は、SD配列とE.coli 16S rRNAの3’末端との間の塩基の対合により、mRNAのリボソームへの結合を促進すると考えられている[Steitzら、(1979)「メッセンジャーRNAにおける遺伝子シグナルおよびヌクレオチド配列」Biological
Regulation and Development:Gene Expression(R.F.Goldberger編)]。弱いリボソーム結合部位を有する真核生物遺伝子および原核生物遺伝子を発現するためには[Sambrookら、(1989)「Escherichia coliにおけるクローニングされた遺伝子の発現」Molecular Cloning:A Laboratory Manual. DNA分子は細胞内に発現され得る。プロモーター配列はまた、DNA分子と直接結合し得、この場合、N末端の最初のアミノ酸は常にメチオニンであり得、メチオニンはATG開始コドンによりコードされる。所望する場合、N末端のメチオニンは、臭化シアンとともにインビトロでインキュベーションすること、もしくは細菌メチオニンN末端ペプチダーゼとともにインビボまたはインビトロのいずれかでインキュベーションすることによりタンパク質から開裂され得る(EPO公開第219237号)。
【0115】
融合タンパク質は、直接発現の代替物を提供する。典型的には、内因性細菌タンパク質のN末端部分または他の安定なタンパク質をコードするDNA配列を、異種コーディング配列の5’末端に融合する。発現の際に、この構築物は、2つのアミノ酸配列の融合を提供する。例えば、バクテリオファージラムダ細胞遺伝子が外来遺伝子の5’末端に連結され得、そしてバクテリア中で発現され得る。生じた融合タンパク質は、好ましくはプロセッシング酵素(Xa因子)が外来遺伝子からバクテリオファージタンパク質を開裂するための部位を保持している[Nagaiら、(1984)Nature 309:810]。融合タンパク質はまた、lacZ[Jiaら、(1987)Gene 60:197];trpE[Allenら、(1987)J.Biotechnol.5:93;Makoffら、(1989)J.Gen.Microbiol.135:11]およびChey[EPO公開第324 647号]遺伝子由来の配列を用いて作成され得る。2つのアミノ酸配列の結合部分でのDNA配列は、開裂可能部位をコードし得るかまたはコードし得ない。他の例は、ユビキチン融合タンパク質である。このような融合タンパク質は、好ましくは、外来タンパク質からユビキチンを開裂するために、プロセッシング酵素(例えばユビキチン特異性プロセッシングプロテアーゼ)のための部位を保有しているユビキチン領域を用いて作成される。この方法により、天然の外来タンパク質を単離し得る[Millerら、(1989)Bio/Technology 7:698]。
【0116】
あるいは、外来タンパク質はまた、細菌中で外来タンパク質の分泌を提供するシグナルペプチド配列のフラグメントからなる融合タンパク質をコードするキメラDNA分子を作成することにより細胞から分泌され得る[米国特許第4,336,336号]。このシグナル配列フラグメントは、典型的には細胞からタンパク質の分泌を指向する疎水性アミノ酸からなるシグナルペプチドをコードする。上記タンパク質は、成長培地中(グラム陽性細菌)、または細胞の内膜と外膜との間に存在する周辺腔(periplasmic space)中(グラム陰性バクテリア)のいずれかに分泌される。 好ましくは、シグナルタンパク質フラグメントと外来遺伝子の間にコードされる、インビボ、またはインビトロで開裂され得るプロセッシング部位が存在する。
【0117】
適当なシグナル配列をコードするDNAは、分泌される細菌タンパク質の遺伝子から由来し得る。例えばE.col外膜タンパク遺伝子(ompA)[Masuiら、(1983):Experimental Manipulation of Gene Expression;Ghrayebら、(1984)EMBOJ.3:2437]およびE.coliアルカリホスファターゼシグナル配列(phoA)[Okaら、(1985)Proc.Natl.Acad.Sci.82:7212]。別の例として、種々のBacilus株由来のα−アミラーゼ遺伝子のシグナル配列を用いて、B.subtilisから異種タンパク質を分泌し得る[Palvaら、(1982)Proc.Natl.Acad.Sci.USA 79:5582;EPO公開 第244042号]。
【0118】
典型的には、細菌により認識される転写終結配列は、翻訳終止コドンの3’側に位置する調節領域であり、従って、プロモーターとともにコーディング配列に隣接する。これらの配列は、DNAによりコードされたポリペプチドに翻訳され得るmRNAの転写を指向する。転写終結配列は、転写の終結を助けるステムループ構造を形成し得る約50ヌクレオチドのDNA配列を頻繁に含む。この例として、E.coliのtrp遺伝子および他の生合成遺伝子のような、強力なプロモーターを有する遺伝子由来の転写終結配列が含まれる。
【0119】
典型的には、プロモーター、シグナル配列(所望であれば)、目的のコーディング配列、および転写終結配列を含む上記の構成成分は、発現構築物中に共に置かれる。発現構築物は、レプリコン、例えば、細菌のような宿主で安定的に維持され得る染色体外要素(例えば、プラスミド)にしばしば維持され得る。レプリコンは、複製系を有し、従って、発現のために、またはクローニングおよび増幅のいずれかのための原核生物宿主で維持され得る。さらに、レプリコンは、高コピー数または低コピー数のいずれかのプラスミドであり得る。高コピー数のプラスミドは、一般に、約5から約200、そして典型的には、約10から約150のコピー数の範囲を有する。高コピー数のプラスミドを含む宿主は、好ましくは、少なくとも約10、およびさらに好ましくは少なくとも約20のプラスミドを含む。高コピー数または低コピー数のベクターのいずれかを、宿主に対するベクターおよび外来タンパク質の影響に依存して選択し得る。
【0120】
あるいは、発現構築物は、組み込みベクターとともに細菌ゲノムに組み込まれ得る。組み込みベクターは、典型的には、ベクターの組み込みを可能にする、細菌染色体に相同な少なくとも1つの配列を含む。組み込みは、ベクター中の相同DNAと細菌染色体との間の組換えから生ずるようである。例えば、種々のBacillus株由来のDNAを用いて構築された組み込みベクターは、Bacillus染色体に組み込まれる(EPO公開第127328号)。組み込みベクターはまた、バクテリオファージまたはトランスポゾン配列を含み得る。
【0121】
典型的には、染色体外および組み込み発現構築物は、選択マーカーを含み得、形質転換された細菌株の選択を可能にする。選択マーカーは、細菌宿主で発現され得、そしてアンピシリン、クロラムフェニコール、エリスロマイシン、カナマイシン(ネオマイシン)、およびテトラサイクリンのような薬剤に対して細菌を耐性にする遺伝子を含み得る[Davisら、(1978)Annu.Rev.Microbiol.32:469]。選択マーカーはまた、生合成遺伝子、例えば、ヒスチジン、トリプトファン、およびロイシン生合成経路の遺伝子を含み得る。
【0122】
あるいは、いくつかの上記の構成成分は、形質転換ベクターに一緒に置かれ得る。形質転換ベクターは、典型的には、上記のように、レプリコン中に維持されるか、または組み込みベクターに発展されるかのいずれかである選択マーカーを含む。
【0123】
発現および形質転換ベクター、染色体外レプリコンまたは組み込みベクターのいずれかは、多くの細菌への形質転換のために開発されている。例えば、発現ベクターは、特に以下の細菌のために開発されている:
【0124】
【表3】
細菌宿主へ外因性DNAを導入する方法は、当該分野で周知であり、そして典型的には、CaCl2または他の試薬(例えば、2価のカチオンおよびDMSO)のいずれかで処理した細菌の形質転換を含む。DNAはまた、エレクトロポレーションにより細菌細胞に導入され得る。形質転換手順は、通常、形質転換されるべき細菌種で変わる。以下の例を参照のこと、
【0125】
【表4】
(iv.酵母発現)
酵母発現系がまた当業者に公知である。酵母プロモーターは、酵母RNAポリメラーゼを結合し得、かつmRNAにコーディング配列(例えば、構造遺伝子)の下流(3’)転写を開始し得る任意のDNA配列である。プロモーターは、通常コーディング配列の5’末端に近接して位置する転写開始領域を有する。この転写開始領域は、典型的には、RNAポリメラーゼ結合部位(「TATA Box」)および転写開始部位を含む。酵母プロモーターはまた、上流アクチベーター配列と呼ばれる第2ドメインを有し、存在する場合、それは、通常、構造遺伝子の遠方にある。UASは、調節(誘導可能)発現を可能にする。構成的な発現はUASの非存在下で起こる。調節発現は、ポジティブまたはネガティブのいずれかであり得、それによって転写の促進または減少のいずれかであり得る。
【0126】
酵母は、活性な代謝経路を有する発酵生物であり、それゆえ代謝経路中の酵素をコードしている配列は、特に有用なプロモーター配列を提供する。この例としては、アルコールデヒドロゲナーゼ(ADH)(EPO公開第284044号)、エノラーゼ、グルコキナーゼ、グルコース−6−ホスフェートイソメラーゼ、グリセルアルデヒド−3−ホスフェートデヒドロゲナーゼ(GAPまたはGAPDH)、ヘキソキナーゼ、ホスホフルクトキナーゼ、3−ホスホグリセレートムターゼ、およびピルビン酸キナーゼ(PyK)(EPO公開第329203号)が含まれる。酸性ホスファターゼをコードしている酵母PHO5遺伝子はまた、有用なプロモーター配列を提供する[Myanoharaら、(1983)Proc.Natl.Acad.Sci.USA 80:1]。
【0127】
さらに、天然には存在しない合成プロモーターがまた酵母プロモーターとして機能する。例えば、一つの酵母プロモーターのUSA配列が、他の酵母プロモーターの転写活性領域と結合され得、合成ハイブリッドプロモーターを生成する。このようなハイブリッドプロモーターの例は、GAP転写活性化領域に連結したADH調節配列を含む(米国特許第4,876,197号および米国特許第4,880,734号)。ハイブリッドプロモーターの他の例は、GAPまたはPyKのような解糖系酵素遺伝子の転写活性化領域と組合せた、ADH2、GAL4、GAL10、またはPHO5遺伝子のいずれかの調節配列からなるプロモーターを含む(EPO公開第164556号)。さらに、酵母プロモーターは、酵母RNAポリメラーゼに結合し、そして転写を開始する能力を有する非酵母起源の天然に存在するプロモーターを含み得る。このようなプロモーターの例は、特に以下を含む
【0128】
【表5】
DNA分子は酵母細胞内で発現され得る。プロモーター配列は、DNA分子と直接連結され得、この場合、組換えタンパク質のN末端の最初のアミノ酸は、常に、ATG開始コドンによりコードされるメチオニンである。所望する場合、N末端のメチオニンを、インビトロで臭化シアンとともにインキュベートすることにより、このタンパク質から開裂し得る。
【0129】
融合タンパク質は、酵母発現系、ならびに哺乳動物、バキュロウイルス、および細菌発現系において代替物を提供する。典型的には、内因性酵母タンパク質または他の安定なタンパク質のN末端部分をコードしているDNA配列を、異種コーディング配列の5’末端に融合する。発現に際して、この構築物は、2つのアミノ酸配列の融合を提供し得る。例えば、酵母またはヒトスーパーオキシドジスムターゼ(SOD)遺伝子は、外来遺伝子の5’末端で結合され得、そして酵母で発現され得た。2つのアミノ酸配列の結合部のDNA配列は、開裂可能部位をコードし得るか、またはコードし得ない。例えば、EPO公開第196056号を参照のこと。他の例は、ユビキチン融合タンパク質である。このような融合タンパク質は、好ましくは、プロセッシング酵素(例えば、ユビキチン特異的プロセッシングプロテアーゼ)が、外来タンパク質からユビキチンを開裂するための部位を保持するユビキチン領域を用いて作成される。従って、この方法により、天然の外来タンパク質を単離し得る(例えば、PCT公開第WO88/024066号参照)。
【0130】
あるいは、外来タンパク質はまた、酵母で外来遺伝子の分泌を提供するリーダー配列フラグメントを含む融合タンパク質をコードするキメラDNA分子を作成することにより細胞から成長培地中に分泌され得る。好ましくは、インビボまたはインビトロのいずれかで開裂され得る、リーダーフラグメントと外来遺伝子との間にコードされるプロセッシング部位がある。リーダー配列フラグメントは、典型的には、細胞からタンパク質の分泌を指向する疎水性アミノ酸を含むシグナルペプチドをコードする。
【0131】
適切なシグナル配列をコードするDNAは、酵母インベルターゼ遺伝子(EPO公開第012873号;日本公開第62,096,086号)およびA因子遺伝子(米国特許第4,588,684号)のような分泌酵母タンパク質の遺伝子由来であり得る。あるいは、酵母における分泌もまた提供する非酵母起源のリーダー(例えば、インターフェロンリーダー)が存在する(EPO公開第060057号)。
【0132】
好ましいクラスの分泌リーダーは、酵母α因子遺伝子のフラグメントを使用しており、それは、「プレ」シグナル配列、および「プロ」領域の両方を含む。使用され得るα因子フラグメントのタイプは、完全長のプレ−プロα因子リーダー(約83アミノ酸残基)および短縮型α因子リーダー(典型的には、約25から約50アミノ酸残基)(米国特許第4,546,083号および米国特許第4,870,008号;EPO公開第324274号)を含む。分泌を提供するα因子リーダーフラグメントを使用する別のリーダーは、第1の酵母のプレ配列を用いるが、第2の酵母α因子由来のプロ領域を用いて作成されるハイブリッドα因子リーダーを含む。例えば、PCT公開第WO89/02463号参照のこと。
【0133】
典型的には、酵母により認識される転写終結配列は、翻訳終止コドンの3’に位置する調節領域であり、そしてそれ故プロモーターとともにコーディング配列に隣接する。これらの配列は、DNAによりコードされるポリペプチドに翻訳され得るmRNAの転写を指向する。転写ターミネーター配列および他の酵母認識終結配列の例は、解糖系酵素についてコードする終結配列である。
【0134】
典型的には、プロモーター、リーダー(所望する場合)、目的のコーディング配列、および転写終結配列を含む上記の構成成分は、発現構築物中に一緒に置かれる。発現構築物は、しばしば、レプリコン、例えば、酵母または細菌のような宿主で安定的に維持され得る染色体外要素(例えば、プラスミド)で維持される。レプリコンは、2つの複製系を有し、従って、例えば、発現するための酵母およびクローニングおよび増幅するための原核生物宿主で維持され得る。このような酵母−細菌シャトルベクターの例は、YEp24[Botsteinら、(1979)Gene 8:17−24];pCl/l[Brakeら、(1984)Proc.Natl.Acad.Sci USA 81:4642−4646];およびYRp17[Stinchcombら、(1982)J.Mol.Biol.158:157]を含む。さらに、レプリコンは、高コピー数または低コピー数のいずれかのプラスミドであり得る。高コピー数のプラスミドは、一般に約5から約200、および典型的には、約10から約150のコピー数の範囲を有する。高コピー数のプラスミドを含む宿主は、好ましくは、少なくとも約10、およびさらに好ましくは少なくとも約20を有する。高コピー数または低コピー数のベクターは、ベクターの影響および宿主での外来タンパク質に依存して選択され得る。例えば、Brakeら、同上を参照のこと。
【0135】
あるいは、発現構築物は、組み込みベクターによって酵母ゲノムに組み込まれ得る。組み込みベクターは、典型的には、ベクターを組み込み得る酵母染色体に相同な少なくとも1つの配列を含み、そして好ましくは、発現構築物に隣接する2つの相同な配列を含む。組み込みは、ベクターおよび酵母染色体における相同なDNAの間での組換えにより生じるようである[Orr−Weaverら、(1983)Methods in Enzymol.101:228−245]。組み込みベクターは、ベクターに包埋するための適切な相同配列を選択することにより酵母の特異的な遺伝子座に指向され得る。 Orr−Weaverら、同上を参照のこと。1またはそれ以上の発現構築物は、生産される組換えタンパク質のレベルができるだけ高くなるように組み込まれる[Rineら、(1983)Proc.Natl.Acad.Sci USA 80:6750]。ベクターに含まれる染色体配列は、完全なベクターの組み込みを生じるベクターの単一セグメントとして、または発現構築物のみからなる安定的な組み込みを生じ得る、染色体の近接したセグメントに相同で、かつベクターで発現構築物の両端に位置する2つのセグメントとしてのいずれかで存在し得る。
【0136】
典型的には、染色体外および組み込み発現構築物は、選択マーカーを含み得、形質転換された酵母株の選択を可能にする。選択マーカーは、酵母宿主で発現され得る生合成遺伝子、例えば、ADE2、HIS4、LEU2、TRP1、およびALG7、ならびにG418耐性遺伝子(ツニカマイシンおよびG418に対する耐性をそれぞれ酵母細胞に供与する)。さらに、適切な選択マーカーはまた、金属のような毒性化合物の存在下で成長する能力を有する酵母を提供し得る。例えば、CUP1の存在は、銅イオンの存在下で酵母を成長させ得る[Buttら、(1987)Microbiol.Rev.51:351]。
【0137】
あるいは、いくつかの上記化合物は、形質転換ベクターに一緒に置かれ得る。形質転換ベクターは、典型的には、上記のように、レプリコンで維持されるか、または組み込みベクターへと展開されるかのいずれかである選択マーカーを含む。
【0138】
発現および形質転換ベクター、染色体外レプリコンまたは組み込みベクターのいずれかは、多くの酵母に形質転換するために発達した。例えば、発現ベクターは、上記のように以下の酵母のために発達した:
【0139】
【表6】
酵母宿主に外因性DNAを導入する方法は、当該分野で周知であり、そして典型的には、スフェロプラストまたはアルカリカチオンで処理したそのままの酵母細胞のいずれかの形質転換を含む。形質転換手順は、通常形質転換されるべき酵母種とともに変化する。以下の例を参照のこと、
【0140】
【表7A】
【0141】
【表7B】
本発明において、選択マーカー、複製起点、および相同宿主細胞の配列が発現ベクターに含まれる。選択マーカーは、潜在的に発現ベクターを含む宿主細胞をスクリーニングするために使用し得る。このようなマーカーは、宿主細胞を、アンピシリン、クロラムフェニコール、エリスロマイシン、ネオマイシンおよびテトラサイクリンのような薬剤に対して耐性にするマーカーを含む。また、マーカーは、宿主細胞の成長に必要なヒスチジン、トリプトファンおよびロイシン経路における生合成遺伝子のような生合成遺伝子を含む。従って、leu(−)宿主細胞を、発現ベクターによる形質転換におけるレシピエントとして使用し、そして例えばロイシンが培地に存在しない場合、leu(+)遺伝子を伴うプラスミドを有する細胞のみが生存する。
【0142】
本発明において、複製起点は発現ベクターに取り込まれ、宿主細胞内で自己複製される。このような複製起点は、細胞内に適切なタンパク質の存在下で、発現ベクターが高いコピー数で再生産され得る複製起点、例えば酵母において有効な2μ配列および自己複製配列;およびCOS−7細胞において有効なウイルスT−抗原の複製起点を含む。
【0143】
本発明の目的のために、発現ベクターは、宿主細胞ゲノムに組み込まれ、または細胞内で自律性を保持し得る。宿主ゲノムへの組み込みのために、本明細書の発現ベクターは、宿主細胞ゲノム内の配列に相同のポリヌクレオチド配列を含み得る。この相同配列は、発現ベクターに連結される必要はない。例えば、発現ベクターは、付随していないジヒドロ葉酸還元酵素遺伝子を介してCHOゲノムに組み込み得る。酵母においては、相同配列は発現カセットに隣接することが好ましい。本発明に特に有用な相同酵母ゲノム配列は、PCT WO90/01800に開示された配列およびGenbank検索番号J01331に記載のHIS4遺伝子配列である。
【0144】
発現ベクターのプロモーター、ターミネーターおよび他の随意エレメントの選択も、選択した宿主細胞に依存し、このことは当業者に既知である。本発明は、選択した宿主細胞には依存しない。簡便性および所望のタンパク質発現のレベルから、至適宿主細胞が指定される。発現のための種々の宿主は当該分野において既知であり、そしてAmerican Type Culture Collection(ATCC)より入手可能である。
【0145】
例えば、KGFフラグメントまたはアナログを発現するための本明細書に適切な細菌宿主として、Campylobacter、Bacillus、Escherichia、Lactobacillus、Pseudomonas、StaphylococcusおよびStreptococcusが挙げられる。以下の属由来の酵母宿主を利用し得る:Candida、Hansenula、Kluyveromyces、Pichia、Saccharomyces、SchizosaccharomycesおよびYarrowia。本明細書において宿主として使用され得る不死化哺乳動物細胞として、CHO細胞、HeLa細胞、ベビーハムスター腎臓(「BHK」)細胞、サル腎臓細胞(「COS」)およびヒト肝細胞癌細胞、例えばHep G2が挙げられる。また多くの昆虫細胞
宿主もKGFフラグメントまたはアナログの発現に適切であって、PCT WO 89/046699;Carbonellら、J.Virol.56:153(1985);Wright Nature 321:718(1986);Smithら、Mol.Cell.Biol.3:2156(1983);および全般的にはFraserら、in vitro Cell.Dev.Biol.25:225(1989)に記載の通り、Aedes aegypti、Autographa californica、Bombyx mori、Drosophila melanogasterおよびSpodoptera frugiperdaが挙げられる。
【0146】
KGFフラグメントまたはアナログを含む発現ベクターは、宿主細胞に挿入される。発現ベクターを宿主細胞に挿入するためには、当該分野で既知の任意の形質転換技術を用い得る。例えば、細菌宿主の形質転換のために、典型的にまずCaCl2または他の試薬(2価カチオンおよびDMSOなど)のいずれかで細菌を処理し、そして外来性DNAを処理した細菌細胞に導入する。DNAは、エレクトロポレーションまたはウイルス感染によっても細菌細胞に導入し得る。本明細書で使用し得る細菌宿主に対する形質転換手順として、Massonら FEMS Microbiol.Lett.60:273(1989);Palvaら Proc.Natl.Acad.Sci.USA 79:5582(1982);欧州特許公開第036,259号および同第063,953号;PCT WO 84/04541、Bacillus)、Millerら Proc.Natl.Acad.Sci.85:856(1988);Wangら J.Bacteriol.172:949(1990)、Campylobacter)、Cohenら Proc.Natl.Acad.Sci.69:2110(1973);Dowerら Nucleic Acids Res.16:6127(1988);Genetic Engineering:Proceedings of the International
Symposium on Genetic Engineering(H.W.BoyerおよびS.Nicosia編)(1978);Mandelら J.Mol.Biol.53:159(1970)のKushner「ColE1由来プラスミドによるEscherichia coli形質転換の改良法(An improved method for transformation of Escherichia coli with ColE1−derived plasmids in;Taketo Biochim.Biophys.Acta 949:318(1988);Escherichia,Chassyら FEMS Microbiol.Lett.44:173(1987)Lactobacillus;Fiedlerら Anal.Biochem 170:38(1988)、Pseudomonas;Augustinら、 FEMS Microbiol.Lett.66:203(1990)、Staphylococcus,Baranyら J.Bacteriol.144:698(1980);Streptococcal Genetics(J.FerrettiおよびR.Curtiss III編)(1987)のHarlander「エレクトロポレーションによるStreptococcus lactisの形質転換(Transformation of Streptococcus lactis by electroporation)」;Perryら Infec.Immun.32:1295(1981);Powellら Appl.Environ.Microbiol.54:655(1988);Somkutiら Proc.4th Evr.Cong.Biotechnology 1:412(1987)、Streptococcus。
【0147】
酵母宿主に対する形質転換法は当該分野で周知であり、典型的にアルカリカチオンで処理したスフェロプラストまたはそのままの酵母細胞のいずれかの形質転換を含む。酵母宿主は、Methods in Enzymology、194巻、1991、「酵母の遺伝学および分子生物学へのガイド(Guide to Yeast Genetics and Molecular Biology)」に記載のエレクトロポレーションによ
っても形質転換し得る。本発明に対して、適用する形質転換手順は、形質転換しようとする酵母種により変化し、Kurtzら Mol.Cell.Biol.6:142(1986);Kunzeら J.Basic Microbiol.25:141(1985);Candida;Gleesonら J.Gen.Microbiol.132:3459(1986);Roggenkampら Mol.Gen.Genet.202:302(1986);Hansenula;Dasら J.Bacteriol.158:1165(1984);De Louvencourtら J.Bacteriol.154:1165(1983);Van den Bergら Bio/Technolgy 8:135(1990);Kluyveromyces;Creggら Mol.Cell.Biol.5:3376(1985);Kunzeら J.Basic Microbiol.25:141(1985);米国特許第4,837,148号および第4,929,555号;Pichia;Hinnenら Proc.Natl.Acad.Sci.USA 75;1929(1978);Itoら J.Bacteriol.153:163(1983)Saccharomyces;BeachおよびNurse
Nature 300:706(1981);Schizosaccharomyces;Davidowら Curr.Genet.10:39(1985);Gaillardinら Curr.Genet.10:49(1985);Yarrowia。
【0148】
異種ポリヌクレオチドを哺乳動物細胞に導入する方法は、当該分野において既知であり、そして例えばウイルス感染、デキストラン介在トランスフェクション、カルシウムリン酸沈殿、ポリブレン介在トランスフェクション、プロトプラスト融合、エレクトロポレーション、ポリヌクレオチドのリポソーム内のカプセル化(encapsulation)、およびDNAの核内へのダイレクトマイクロインジェクションが挙げられる。
【0149】
Smithら、Mol.Cell.Biol.3:2156(1983);およびLucklowおよびSummers、Virology 17:31(1989)に記載の通り、発現ベクターを形成するためにバキュロウイルス属のウイルス内に異種DNAを導入する方法、および昆虫宿主細胞の形質転換の方法も当該分野において既知である。例えば、相同二重交差組換えにより、KGFフラグメントDNAを多角体遺伝子に挿入し得る。Millerら、Bioessays 4:91(1989)に記載の通り、所望のバキュロウイルス遺伝子中に設計された制限酵素部位にも挿入し得る。KGFフラグメントのDNA配列を、発現ベクター内の多角体遺伝子と換えてクローン化した場合、多角体特異配列の両5’および3’に隣接し、そして多角体プロモーターの下流に位置する。
【0150】
本発明の実施態様において、新たに形成されたバキュロウイルス発現ベクターは、次いで感染組換えバキュロウイルス内にパッケージされ得る。バキュロウイルス発現系においては、およそ1%からおよそ5%の間の低い頻度で相同組換えが起こる。従って、通常、トランスフェクション後に生産された大多数のウイルスは、まだ野生型のウイルスである。しかし、組換えウイルスは、既知の方法によって同定し得る。例えば、天然のウイルスは、ウイルス感染の後期の間、感染細胞の核内において、かなり高いレベルで多角体タンパク質を生産する。蓄積した多角体タンパク質は、埋包されたウイルス粒子も含む閉塞体を形成する。これらの閉塞体は、15μmまでのサイズで、屈折性が高いことから明るい光沢の外観で、光学顕微鏡下で容易に可視化される。組換えウイルスが感染した細胞は閉塞体を欠く。組換えウイルスおよび野生型ウイルスはプレートティングすることによって識別され得、トランスフェクション上清またはその希釈液は標準的技術によって昆虫細胞の単層上に存在する。次に「Current Protocols in Microbiology」第2巻(Ausubelら編)の16.8(補遺10,1990)に記載の通り、閉塞体の存在(野生型ウイルスを示す)または非存在(組換えウイルスを示す)について、光学顕微鏡下でプラークをスクリーニングし得る。
【0151】
本明細書において形質転換された宿主細胞が所望のKGFフラグメントを発現しているかどうかを調べるために、当該分野で既知のイムノアッセイおよび活性アッセイを利用し得る。例えば、細胞膜からKGFフラグメントを分離することなく、形質転換された宿主細胞上で免疫蛍光アッセイを実施し得る。本アッセイにおいて、宿主細胞をまず、顕微鏡用スライドまたはマイクロタイターウエルのような固体支持体上に固定する。次に固定化した宿主細胞を抗KGF抗体に暴する。好ましくはアッセイの感度を上昇させるために、固定化した細胞を、抗KGF抗体に結合する標識された2次抗体に暴する。例えば、2次抗体は蛍光性のマーカーで標識し得る。KGFフラグメントを発現する宿主細胞を蛍光的に標識し、顕微鏡下で可視化し得る。
【実施例】
【0152】
(実施例1)
図1に示す通り、cDNAをpAcC13Pst/Not 1発現ベクターにクローニングすることによって、ヒトKGF cDNAをバキュロウイルス発現系に挿入した。Munemitsuら Mol.Cell.Biol.10:5977−5982(1990)に記載の通り、このプラスミドはpAcC12に由来した。
【0153】
Finchら Science 2:752−755(1989)に記載の通り、入手可能なKGFのN末端アミノ酸配列に基づく欠失した18kd KGFについてPCRオリゴヌクレオチドプライマーを設計した。Luckowら Virology 170:31−39(1989)およびQuilliamら Mol.Cell.Biol.10:290102908(1990)に記載の通り、pVL941伝達ベクターの誘導体であるpAcC13発現ベクターへのサブクローニングを容易に行うために、隣接する制限部位(PstIおよびNotI)をプライマーに組み込ませた。この発現ベクターは、pAcC13のPstI/NotIポリリンカー部位にKGFをコードするフラグメントを挿入することによって構築された。KGF cDNAはKGFのプロセシングされた成熟体をコードし、これは163個のアミノ酸残基からなり、14〜16番目の残基に潜在的なNグリコシル化部位を伴った。
【0154】
(コンディション培地の調製)
Spodoptera Frugiperda(Sf9昆虫細胞)に、KGF1−163に対するcDNAを含むバキュロウイルスのAutographa Californicaを感染させた。希釈後、細胞を血清補充物、抗生物質、または殺菌剤非存在下のExcell−400内で48〜72時間培養した。培養液を集め、そして10,000×gで30分間遠心分離して浮遊細胞およびその他の細胞破砕物を除いた。次にコンディション培地を、0.8μフィルター(Millipore)で濾過し、そしてこの培地の約5リットルを、3−kDaの分子量カットオフ値を有するfiltron cassette system(Omega membrane)を用いて200mlに濃縮した。
【0155】
濃縮後、トランスフェクトしたSf9細胞由来のコンディション培地を、10,000×gで20分間の遠心分離によって集め、そして連続ヘパリンセファロース(「HS」)アフィニティークロマトグラフィーによって精製した。本発明の一つの実施態様においては、約1リットルのSf9細胞コンディション培地を、3−kDaのカットオフ値を有するOmega membrane(Filtron)を用いて限外濾過し、200mlの限外濾過濃縮液を生産した。
【0156】
(HSACおよびMono S陽イオン交換クロマトグラフィー)
上清をpH7.2に調整した。30mlの試料を、10mM Tris−HCl pH7.3、 150mM NaClで平衡化したヘパリンセファロース樹脂に添加した。樹脂を、平衡化緩衝液で、吸光度がベースラインに戻るまで大量に洗浄し、そして次に0.
45Mから1Mおよび2Mの上昇NaCl濃度で段階的に溶出させた。細胞増殖アッセイに用いたフラクションからアリコートを分離し、そして最も高い生物活性を有する1M NaClのフラクションをプールした。プールしたフラクションを10mM Tris pH7.2 で5倍に希釈し(最終塩濃度0.2M NaCl)、そして同試料をSuper loopで、FPLCシステムに連結されたMono Sカラム(Pharmacia,Piscataway,NJ)に供した。直線勾配(10mM Tris pH7.3、0.2M NaCl〜10mM Tris pH7.3、1M NaCl)により溶出を行った。分画後、アリコートを生物活性について試験し、活性フラクションをプールした。
【0157】
濃縮液をpH7.3に調整し、そして約30mlのベット容量を含むHSカラムに添加した。カラムを、約2時間4℃で展開させた。カラムを、pH7.3で10mM Tris−HClおよび0.15M NaClを含む150mlの平衡化緩衝液で洗浄した。保持されたタンパク質を0.45M NaClおよび1M NaClで溶出した。溶出中のカラムの流速を、約90ml/時 に調整し、そして3mlのサイズのフラクションを集めた。
【0158】
HSアフィニティークロマトグラフィー工程の生物活性を有する1M NaClフラクションをプールし、そして10mM Tris pH7.3で5倍希釈し、そしてMono S HR 5/5カラム(Pharmacia)に添加した。保持されたタンパク質を、0.2M NaClから1M NaCl勾配を用いて溶出した。活性フラクションのバイオアッセイを、Balb−Mk細胞株を用いて測定した。Mono S 陽イオン交換クロマトグラフィーにより単離された生物活性タンパク質フラクションのSDS PAGE分析を実施した。フラクション37−39およびフラクション41−42をプールし、そして10μlのアリコートをSDSおよび10mM DTTに添加した。これらの試料を加熱して変性させ、そして12%のポリアクリルアミドゲル中で電気泳動を行い、続いて銀染色した。試料が還元状態中において展開された場合でも、または非還元状態中において展開された場合でも、同様の泳動パターンが観察された。25Kdはフラクション37−39に含まれるタンパク質の見かけの分子量で、そして18kdはフラクション41−42に含まれるタンパク質の見かけの分子量であった。
【0159】
HS生物活性フラクションのクロマトグラムは、フラクション31〜47にかけてのBalb/Mk生物活性のプロフィールを示すが、これから、2種類の分子種の存在が明らかであり、一つは0.55M NaClで溶出し、そして他方は0.6M NaClで溶出する。
【0160】
(KGF活性の測定)
得られたフラクション中のKGF活性の存在を、BALB/C−Mk細胞の成長を促進するフラクションの能力により評価した。この点に関して、任意のフラクションの10μlアリコートを1mlのリン酸緩衝生理食塩液(「PBS」)中0.2%ゼラチンに希釈し、そして10μlの希釈フラクションを、各々22mmウエルを含む12−ウエルクラスタープレートに5×103細胞/ウエルで接種されたBALB/C−Mk細胞の成長刺激活性について試験した。全てのKGF活性がカラム内に保持されることが見出され、そして1M NaClで溶出された。
【0161】
HSカラムから1M NaClで溶出されたKGFの生物活性フラクションをさらに、陽イオン交換FPLCカラムクロマトグラフィーによって精製した。これらのフラクションをプールし、10mM Tris、pH7.3で5倍に希釈し、そしてPharmacia製のMono S HR 5/5カラムに直接添加した。
【0162】
Mono S HR 5/5カラムに保持されたタンパク質を、0.2M NaClから1M NaClの勾配で溶出させた。溶出したフラクションの生物活性を、BALB/C−Mk細胞を用いて、上記の通りアッセイした。図3は、フラクション31〜47にわたる活性プロフィールを示す。
【0163】
マイトジェンアッセイにおいて、Bohlenら EMBO 4:1951−1956(1985)、およびGospodarowiczら J.Cell Physiol 127:121−136(1986)、およびBellostaら J.Cell Biol.121:705−713(1993)に記載の通り、ウエル当たり104個のABAE細胞および5×103個のACE細胞を、10%仔牛血清および抗生物質を補充した1mlのDMEM中に密度5×103細胞/ウエルで、12ウエルクラスタープレートに接種した。6時間のインキュベーション後、3重検体のウエルのセットをトリプシン処理し、そしてプレーティング効率を測定するために細胞をカウントした。次に適切な希釈の各試料の10μlアリコートを、0日目、2日目および4日目に3重検体としてシャーレ内のウエルに添加した。培養5日目後、プレートをトリプシン処理し、そしてコールターカウンター(Coulter Electronics,Hialeah,FL)により細胞密度を測定した。
【0164】
0日目および1日おきに、表示した濃度で、KGFまたは塩基性FGFを添加した。培養5日後に、細胞をトリプシン処理し、そしてコールターカウンターを用いて最終的な細胞密度を測定した。
【0165】
(電気泳動(NaDodSO4/PAGE))
Laemmliら Nature 227:680−685(1970)に記載の手順に従い、NaDodSO4によりポリアクリルアミドゲルを調製した。10mM DTT存在下で3分間試料を煮沸し、そして12%ポリアクリルアミドゲル中で電気泳動した。ゲルを固定し、そしてBioRadの試薬およびプロトコールを用い、Merrilら Science 211:1437−1438に考察されている通りに銀染色を行った。適切な分子量マーカーは、Biorad製である。
【0166】
(細胞増殖アッセイ)
カラムフラクションおよび精製試料のマイトジェン活性を、標的細胞としてBalb/Mk細胞を用いることによって測定した。ストック培養物を生育し、そしてGospodarowiczら J.Cell Physiol.142:325−333に記載の通り、10%FCS、50μg/mlゲンタマイシン、0.25μg/mlファンギゾン(fungizone)、および10ng/ml aFGFを補充した低カルシウム改変イーグル培地中に維持した。マイトジェンアッセイ用の細胞を、Gospodarowiczらに記載の通り、10%FCSを補充した1mlの低カルシウムMEM中に5×103〜1×104細胞/ウエルの密度で12ウエルクラスタープレートに接種した。培養5日後に、 Gospodarowiczらに記載の通り、試料の添加および最終細胞密度の測定を行った。
【0167】
最終精製物質のマイトジェン活性を、Gospodarowiczら Proc.Natl.Acad.Sci.73:4120−4124(1976)に記載の通り、成獣ウシ動脈内皮細胞(「ABAE」)および副腎皮質由来毛細血管内皮細胞(「ACE細胞」)上で試験した。ストック培養物を、10%CS、50μg/mlゲンタマイシンを補充したDMEM存在下で維持し、そして0.25μg/mlのファンギゾンを、週1回、1:10の分割比でゼラチン化組織培養シャーレ上を通過させた。
【0168】
当該分野で周知の標準的な方法論を用いて、以下の通り、KGFdcs1−23のNH2末端から1位〜20位の明白なアミノ酸配列を確定した:
S1 Y D M5 E G G D I R V R R L F X R T Q
本発明はまた、KGFdcs1−23をコードするDNAセグメント、およびFGFファミリーの他のメンバーに対してKGF163の非保存性NH2末端特性を欠くKGF由来のより短い他の形状を含む。
【0169】
(タンパク質マイクロシークエンシング)
KGF163およびKGFdcs1−23の2つの名目上100pmolの試料を、エドマン分解、および二フッ化ポリビニリデン(PVDF、Applied Biosystems Biospin)への遠心吸着後のAA分析により分析した。試料をApplied Biosystems 477A 気相タンパク質シークエネーターに添加した。標準的なソフトウエアおよびApplied Biosystems供給の化学試薬を用いて、20回のエドマン分解を行い、そして自動化オンラインHPLCカラム(モデル120A、Applied Biosystems)によりPTHアミノ酸の同定を行った。
【図面の簡単な説明】
【0170】
【図1】図1は、N末端で開始し、そして163アミノ酸残基およびアミノ酸残基14−16に単一のN結合グリコシル化シグナルを含む成熟完全長ヒトKGFポリペプチドのアミノ酸配列を示す。
【図2】図2は、163アミノ酸配列のKGFが挿入されているpAcC13発現ベクターを示す。
【図3】図3は、ヘパリンセファロース(「HS」)カラムで1M NaClを用いて溶出した後の培地で調整されたSF9細胞由来の本発明のKGFフラグメントのさらなる精製を示す。HSカラム由来の生活性画分をプールし、10mM Tris、pH7.3で5倍に希釈し、そしてMono S HR5/5陽イオン交換FPLCにかけた。
【図4】図4は、Balb/Mk細胞株の細胞上で、長形態のKGF(すなわち、rKGF163)および短形態のKGF(すなわちKGFdes1−23)対aFGFの生物学的活性を比較する。
【図5】図5は、大きな血管(A:ABAE細胞)または毛細血管(B:ACE細胞)由来の血管内皮細胞上で、長形態のKGF(すなわち、rKGF163)および短形態のKGF(すなわちKGFdes1−23)対bFGFの生物学的活性を比較する。
【技術分野】
【0001】
(発明の分野)
本発明は、一般にケラチノサイト増殖因子(「KGF」)に関する。さらに特定的に、本発明は、昆虫細胞系で発現する組換え完全長KGFと比較して、生物学的活性の増加および細胞毒の減少を有する短縮型KGFフラグメントおよびそのアナログに関する。このKGFフラグメントは、本明細書中ではKGFdes1−23と命名され、成熟完全長KGFのN末端の最初の23アミノ酸残基を欠損し、このN末端は、Finch,P.W.ら、Science 245:752−755(1989)に示され、そしてKGFにその上皮細胞特異性を与えると既に考えられているように、N末端からのアミノ酸残基14〜16にグリコシル化部位を含む。
【背景技術】
【0002】
(発明の背景)
KGFは、線維芽細胞成長因子(「FGF」)のファミリーに属し、塩基性FGF(「bFGF」)として示されるものの始原型である。このため、KGFはまた、FGF−6として公知である。他のFGFのように、KGFはヘパリン結合タンパク質であるが、他のFGFと異なり、それは独特な標的細胞特異性を有する。特に、FGFは、一般に、一次または二次中胚葉由来および神経外胚葉由来の種々の細胞型の増殖および分化を刺激し得る。KGFは、Finch,P.W.ら(上記引用文中)で考察されるように、上皮細胞増殖を刺激し得るその能力において他のFGFと同様であるが、内皮細胞または線維芽細胞の増殖を刺激し得ないその能力において他のFGFと同様でない。
【0003】
酸性線維芽細胞成長因子(「aFGF」)および塩基性線維芽細胞成長因子(「bFGF」)を含むFGFは、Gospodarowiczら、Cell.Biol.Rev.25:307−314(1991)、およびBasilicoら、Adv.Cancer
Res.59:115−165(1992)で考察されるように、ヘパリン結合特性を有し、そして腹部および背部の中胚葉の初期胞胚での分化および増殖を誘導する能力を有することが公知である。FGFに対する細胞の応答は、Houら、Science 251:665−668(1991)で考察されるように、細胞表面レセプター(「FGFR」)へのその結合を通じて介在される。このレセプターには、3つの相関型がある。高親和性のFGFRは、Leeら、Science 245:57−60(1989);Dionneら、EMBO 9:2685−2692(1990);Mikiら、Science 251:72−75(1991);Mikiら、Proc.Natl.Acad.Sci.USA 89:246−250(1992);およびDellら、J.Biol.Chem.267:21225−21229(1992)で考察されたように、チロシンキナーゼであり、そしてflgレセプター(「FGFR−1」)、bekレセプター(「FGFR−2」)、およびK Samレセプター(「FGFR−3」)を含む。
【0004】
FGFR−1およびFGFR−2の両者は、中胚葉および神経外胚葉組織で広範囲に発現され、そして両者は、同じ親和性でaFGFおよびbFGFを結合し得る。FGFR−3は、上皮細胞に特異的であるKGFレセプターである。それは、FGFR−2のもう一つの転写物である。Mikiら(上記引用文中)およびDellら(上記引用文中)で考察されたように、aFGFおよびbFGFの両方について高い親和性を示し、そしてKGFについて全く親和性を示さないFGFR−2とは対照的に、FGFR−3は、KGFおよびaFGFをbFGFより約20〜1000倍高い親和性で結合する。
【発明の開示】
【発明が解決しようとする課題】
【0005】
上皮細胞に対するKGFの厳密に制限される活性プロフィールが、例えば、多くのタイプの創傷治癒のための適用ならびに乾癬および基底細胞癌のような表皮の過剰増殖疾患の処置で所望される。現在では、KGFを除いて、高適性マイトジェン因子はこれらの適用のためには存在しない。従って、KGFが、その効力を増加しそしてその細胞毒を減少するように改変されれば、治療用適用のために望ましい。
【0006】
最近、Ronら、J.Biol.Chem.268:2984−2988(1993年2月)により、KGF163がT7原核生物発現系で発現される場合、マイトジェン活性を所有する組換えKGF(「rKGF」)ポリペプチドが得られ得ることが見出された。rKGF分子が、成熟KGF163ポリペプチドのN末端からそれぞれ3、8、27、38、および48アミノ酸残基の欠失により短縮される場合、生じる分子の生物学的活性は変化した。それぞれ、3および8アミノ酸残基の欠失により、生じる分子のマイトジェン活性は、完全長のrKGF(「rKGF163」)と比べて影響し得なかった。しかし、27アミノ酸残基の欠失は、10〜20倍減少したマイトジェン活性を生じた。38および48アミノ酸残基の欠失は、それぞれマイトジェン活性およびヘパリン結合能を完全に失うようになる。従って、Ronらは、rKGF163分子と比べて増加した細胞分裂活性を有するいかなる短縮型KGFフラグメントをも生産し得なかった。
【課題を解決するための手段】
【0007】
(発明の要旨)
本発明の目的の1つは、成熟完全長KGF、KGF163のアミノ酸配列の一部分を含み、そして完全長組換えKGF、rKGF163と比較して少なくとも2倍のマイトジェン活性の増加を有するKGFフラグメントを提供することである。特別な目的は、rKGF163の最初の23個のN末端アミノ酸残基C−N−D−M−T−P−E−Q−M−A−T−N−V−N−C−S−S−P−E−R−H−T−R−を含む配列を欠失しているKGFフラグメントを提供することである。
【0008】
本発明の1つの他の目的は、rKGF163と比較して減少した細胞毒を有するKGFフラグメントを提供することである。さらに他の目的は、上記のKGFフラグメントおよび毒性分子を含む結合体を提供することである。毒性分子は、リシンA、ジフテリアトキシン、およびサポニン(saporin)のうち少なくとも1つであり得る。
【0009】
本発明の他の1つの目的は、上記のようなKGFフラグメント、および薬学的に受容可能なキャリア(例えば、ヒト皮膚への局所適用に適するもの)を含む治療用組成物を提供することである。
【0010】
本発明のさらに他の1つの目的は、上記のKGFフラグメントをコードするヌクレオチド配列からなるDNA分子を提供することである。
【0011】
本発明の他の1つの目的は、上記のKGFフラグメントをコードするDNA分子、およびDNA分子の発現のための調節配列を含む発現ベクターを提供することである。例えば、発現ベクターは、バキュロウイルスであり得る。
【0012】
本発明の他の1つの目的は、上記の発現ベクターで形質転換した宿主細胞を提供することである。例えば、宿主細胞は、細菌細胞、酵母細胞、哺乳動物細胞、または昆虫細胞であり得る。
【0013】
本発明の他の1つの目的は、上記のような形質転換宿主細胞を培養し、そして培養物からKGFフラグメントを単離することにより、KGFフラグメントを生産する方法を提供することである。
【0014】
本発明のさらに他の1つの目的は、上皮細胞成長が所望される領域にKGFフラグメントを適用することにより上皮細胞成長を刺激し、そして細胞を培養し得る方法を提供することである。
【0015】
本発明のさらに他の1つの目的は、処置されるべき創傷の領域に上記の治療組成物を適用して損傷を治癒させることによる、損傷治癒方法を提供することである。
【0016】
本発明のさらに他の1つの目的は、治療すべき領域に上記の結合体を適用することにより表皮の過剰増殖疾患を処置する方法を提供することである。
【0017】
従って、本発明は以下を提供する。
【0018】
1.成熟、完全長ケラチノサイト増殖因子のアミノ酸配列の一部分を含有するケラチノサイト増殖因子フラグメントであって、該一部分が、成熟、組換え、完全長ケラチノサイト増殖因子と比較して少なくとも2倍増加したマイトジェン活性を有し、そして成熟、完全長ケラチノサイト増殖因子の最初の23個のN末端アミノ酸残基を含有する配列を欠く、ケラチノサイト増殖因子フラグメント。
【0019】
2.前記フラグメントが、成熟、組換え、完全長ケラチノサイト増殖因子と比較して7倍増加したマイトジェン活性を有する、項目1に記載のケラチノサイト増殖因子フラグメント。
【0020】
3.前記フラグメントが、成熟、組換え、完全長ケラチノサイト増殖因子と比較して10倍増加したマイトジェン活性を有する、項目1に記載のケラチノサイト増殖因子フラグメント。
【0021】
4.前記フラグメントが、成熟、組換え、完全長ケラチノサイト増殖因子と比較して細胞毒性の減少を示す、項目1に記載のケラチノサイト増殖因子フラグメント。
【0022】
5.(a)成熟、完全長ケラチノサイト増殖因子のアミノ酸配列の一部分を含有するケラチノサイト増殖因子フラグメントであって、該一部分が、成熟、組換え、完全長ケラチノサイト増殖因子と比較して少なくとも2倍増加したマイトジェン活性を有し、そして成熟、完全長ケラチノサイト増殖因子の最初の23個のN末端アミノ酸残基を含有する配列を欠く、ケラチノサイト増殖因子フラグメント、および
(b)毒性分子
を含有する結合体。
【0023】
6.前記毒性分子が、リシンA、ジフテリアトキシンおよびサポニンからなる群から選択れる、項目5に記載の結合体。
【0024】
7.前記フラグメントが、成熟、組換え、完全長ケラチノサイト増殖因子と比較して7倍増加したマイトジェン活性を有する、項目5に記載の結合体。
【0025】
8.前記フラグメントが、成熟、組換え、完全長ケラチノサイト増殖因子と比較して10倍増加したマイトジェン活性を有する、項目5に記載の結合体。
【0026】
9.(a)成熟、完全長ケラチノサイト増殖因子のアミノ酸配列の一部分を含有するケラチノサイト増殖因子フラグメントであって、該一部分が、成熟、組換え、完全長ケラチノサイト増殖因子と比較して少なくとも2倍増加したマイトジェン活性を有するが、成熟、完全長ケラチノサイト増殖因子の最初の23個のN末端アミノ酸残基を含有する配列を欠く、ケラチノサイト増殖因子フラグメント、および
(b)薬剤的に許容し得るキャリアー
を含有する治療組成物。
【0027】
10.項目1に記載のケラチノサイト増殖因子フラグメントをコードするヌクレオチド配列を含有する、DNA分子。
【0028】
11.項目2に記載のケラチノサイト増殖因子フラグメントをコードするヌクレオチド配列を含有する、DNA分子。
【0029】
12.項目3に記載のケラチノサイト増殖因子フラグメントをコードするヌクレオチド配列を含有する、DNA分子。
【0030】
13.項目10に記載のDNA分子および該DNA分子の発現のための調節配列を含有する発現ベクター。
【0031】
14.前記ベクターがバキュロウイルスである項目13に記載の発現ベクター。
【0032】
15.項目13に記載の発現ベクターによって形質転換された宿主細胞。
【0033】
16.前記細胞が、細菌細胞、酵母細胞、哺乳動物細胞および昆虫細胞からなる群から選択される、項目15に記載の宿主細胞。
【0034】
17.項目15に記載の宿主細胞を培養する工程、および該培養物からケラチノサイト増殖因子フラグメントを単離する工程を包含する、ケラチノサイト増殖因子フラグメントを生産する方法。
【0035】
18.項目9に記載の治療組成物を、処置しようとする損傷領域に適用し、そして損傷を治癒し得ることを包含する、損傷治癒の方法。
【0036】
19.項目5に記載の結合体を、処置しようとする領域に適用することを包含する、表皮の過増殖疾患の処置方法。
【0037】
20.前記疾患が、乾癬および基底細胞癌からなる群から選択される疾患である、項目20に記載の処置方法。
【0038】
本発明のさらなる目的、特徴、および利益は、当業者に明らかであり、そしてここに列挙する必要はない。本発明は、当業者に明らかであり、そしてフラグメント、結合体、治療組成物、ベクター、宿主、および使用方法の性質および活性を実質上変化しない上記の変形および改変を含む。
【発明を実施するための最良の形態】
【0039】
(好ましい実施態様の詳細な説明)
本明細書中でKGFフラグメントまたはKGFdes1−23と称される、rKGF163の最初の23個のN末端アミノ酸残基にわたる欠失を有する短縮型非グリコシル化KGFが、rKGF163と比べて上皮細胞上でより大きな生物学的活性および細胞毒の減少を有することを驚くべきことに発見した。一般に、本発明のKGFフラグメントは、上皮細胞の増殖の刺激に対してKGF163の特異性を保持する。
【0040】
従って、本発明の好ましい実施態様は、天然分子がインビボで生産される場合に、通常それに付随する不純物をともなわない新規な短縮型非グリコシル化KGFフラグメント、KGFdes1−23である。このフラグメントは、単一なピークとしてNa Dod SO4/PAGEでその移動度に基づき約18kDaの見かけ上の分子量を有する。Balb/Mk細胞上での精製KGFdes1−23の比活性は、PCT特許出願第WO90/08771号に記載のように、約2.5×107ユニット/ミリグラム(ED5040pg/ml)であり、細胞増殖アッセイで比較する場合、rKGF163タンパク質の比活性、またはaFGFの比活性より約7〜10倍大きく、そしてrKGF163を化学的に規定した培地中でBalb/Mk細胞でDNA合成の開始を試験することによりバイオアッセイする場合より100倍大きい。
【0041】
本発明の他の好ましい実施態様では、KGFdes1−23は、組換えDNA技術により、特に大量生産販売のために生産される。さらに好ましい実施態様では、本発明によるKGFdes1−23またはそのアナログをそれぞれ含む組換えDNA分子および発現ベクターは、当該分野で周知の方法を用いて、標準遺伝子発現技術により実施される。
【0042】
さらなる実施態様では、KGFdes1−23をコードするDNAを含むDNAまたはベクターは、細菌、哺乳動物、酵母、または昆虫細胞発現系で発現され得る。好ましい実施態様では、細菌または酵母細胞の発現系は、KGFdes1−23フラグメントの生産にとって理想である。他の好ましい実施態様では、DNAまたはベクターは、昆虫細胞発現系で発現される。
【0043】
本発明のKGFフラグメントは、レセプター認識部位の同定、およびペプチドアゴニストまたはアンタゴニストの設計に使用され得る。さらに、ケラチノサイトに対するKGFの独特な特異性である、血管内皮細胞または線維芽細胞の増殖を誘導し得ない能力および細胞毒の欠損を考慮して、本発明の他の実施態様では、KGFdes1−23は、好ましくは、特に、皮膚の上皮再形成の促進を所望する場合、創傷治癒適用のために使用される。好ましい実施態様では、KGF des1−23は、角膜の上皮修復で使用される。KGFdes1−23の他の適用は、胃腸管内に見られる表皮細胞に対するその特異性に基づいて想定され得る。
【0044】
皮膚修復について、上皮成長因子(「EGF」)、血小板由来の成長因子(「PDGF」)、および他のFGFのような他の成長因子以外の本明細書におけるKGFフラグメントの選択は、当業者の技術範囲内である。例えば、他の成長因子は、ケラチノサイト増殖を直接的または間接的のいずれかで刺激するのに加えて、線維増殖および脈管形成を誘導する。皮膚修復では、このような特別な活性は瘢痕形成のような所望でない副作用を生成し得る。創傷または手術のいずれかを含む角膜修復では、これらの他の因子の使用は、角膜内に血管侵入を誘導し、所望でない角膜不透明または浮腫を生じる。他方では、KGFは、ケラチノサイトにとって独特な特異性を有し、そして血管内皮細胞または線維芽細胞の増殖を誘導せず、そして従って、これらの特定の創傷治癒適用のために選択した薬剤である。
【0045】
さらに、本発明のKGFフラグメントは、KGFフラグメント/毒素結合体の形態で本発明の他の実施態様で用いられ得る。このような結合体は、上皮から基底層のケラチノサイトの過剰増殖を生じる乾癬のような疾患における疾患のプロセスを減衰させ得るか、または停止し得る。この結合体は、血管内皮細胞および線維芽細胞のような、これらの組織に存在する他の2つの主要な細胞型に影響を及ぼさない。
【0046】
本発明の他の実施態様では、KGFフラグメント/毒素結合体は、腫瘍細胞増殖、例えば、基底細胞癌を根絶するために用いられ得る。
【0047】
本発明の実施は、他に指示がなければ、当該分野の技術範囲内にある分子生物学、微生物学、組換えDNA、および免疫学の従来の技法を使用する。このような技法は文献で十分に説明されている。例えば以下を参照のこと:
【0048】
【表1A】
【0049】
【表1B】
アミノ酸の標準略語は、本明細書中の図1およびその他で用いられる。本明細書中に引用した全ての刊行物、特許、および特許出願は参考として援用される。説明のために、本明細書で用いる用語を下記により特定的に定義する。
【0050】
(定義)
本明細書で用いる用語「ケラチノサイト増殖因子」または「KGF」は、構造的に異なるタンパク質である線維芽細胞成長因子、FGFのファミリーに属するポリペプチドを意味し、このタンパク質は、それらが関連するファミリーの遺伝子にコードされていることを示唆する配列相同性の変化程度を呈示する。KGFは、本明細書の他の所に記載された特性を有し、例えば、FGFR−3に結合し、そして上皮細胞、特にケラチノサイトの成長を刺激し得る。完全長KGFは、163アミノ酸残基からなる。
【0051】
本明細書で用いる本発明のKGFフラグメントは、成熟完全長KGFのアミノ酸配列の一部分からなるポリペプチドである。本明細書で用いるKGFフラグメントのアナログは、KGFフラグメントの特性に実質的に影響しないKGFフラグメントの主要ではない挿入、欠失、または置換を含む。例えば、保存的アミノ酸置換が予測される。このような置換は、例えば、アミノ酸の側鎖に関連するアミノ酸ファミリー内で起こる。アミノ酸のファミリーは、(1)酸性:アスパラギン酸、グルタミン酸;(2)塩基性:リジン、アルギニン、ヒスチジン;(3)非極性:アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファン;および(4)非荷電極性:グリシン、アスパラギン、グルタミン、システイン、セリン、トレオニン、チロシンを含む。フェニルアラニン、トリプトファン、およびチロシンは、時折芳香族アミノ酸のファミリーとして共同に分類される。特に、ポリペプチドの活性部位の外側領域でロイシンをイソロイシンまたはバリンで、またはアスパラギン酸をグルタミン酸で、またはトレオニンをセリンで置換する孤立置換、もしくはアミノ酸を構造的な関連アミノ酸で置換する同様の保存的置換からなる保存的アミノ酸置換は、ポリペプチドの特性に大きな影響を有さないことが、一般に認められている。
【0052】
KGFフラグメントの特性は、(i)成熟完全長天然KGF分子と比較して、上皮細胞増殖の刺激を、2〜10倍、好ましくは2倍、最も好ましくは7〜10倍増加するようなその生物学的活性、および(ii)FGFR−3に結合するその能力を含む。
【0053】
本明細書で用いる用語「組換えポリヌクレオチド」は、半合成、または合成起源、もしくはcDNAまたはゲノムDNA(「gDNA」)にコードされたポリヌクレオチドを意味し、それは、(1)天然で会合するポリヌクレオチドの全てまたは一部分と会合しない、(2)天然で連結するポリヌクレオチド以外のポリヌクレオチドに連結する、あるいは(3)天然に存在しない。
【0054】
「レプリコン」は、任意の遺伝要素、例えば、プラスミド、染色体、ウイルス、コスミドなどであり、それは、細胞内でポリヌクレオチド複製の自律単位として挙動する;すなわち、それは、それ自身の制御下で複製し得る。レプリコンは、例えば、選択マーカーを含み得る。
【0055】
「組換えベクター」は、他のポリヌクレオチドセグメントが付加されるレプリコンであり、付加したポリヌクレオチドセグメントの複製および/または発現をもたらす。
【0056】
「調節配列」は、ポリヌクレオチド配列が作動可能に連結されるコーディング配列の発現の調節に必要なポリヌクレオチド配列を意味する。このような調節配列の性質は、コーディング配列が発現されるべき宿主細胞に依存して異なる。原核生物では、このような調節配列は、例えば、一般にプロモーター、リボソーム結合部位、および/または転写終結配列を含む。真核生物では、一般にこのような調節配列は、プロモーターおよび/または転写終結配列を含む。用語「調節配列」はまた、その存在が有益な付加成分、例えば、発現されるべきポリペプチドの分泌をコードする分泌リーダー配列を含み得る。
【0057】
「作動可能に連結された」は、成分が、意図した様式で機能し得るような関係で連結している並置を意味する。例えば、調節配列は、コーディング配列の発現が、調節配列に適合した条件下で達成されるように、それが接続される場合、コーディング配列に「作動可能に連結されて」いる。
【0058】
「コーディング配列」は、適切な調節配列の制御下に置かれた場合、ポリペプチドに翻訳されるポリヌクレオチド配列である。コーディング配列の境界は、一般にその5’末端の翻訳開始コドンおよびその3’末端の翻訳終止コドンにより決定される。コーディング配列は、例えば、cDNA、および組換えポリヌクレオチド配列を含み得る。
【0059】
「PCR」は、Saikiら、Nature 324:163(1986);およびScharfら、Science 233:1076−1078(1986);米国特許第4,683,195号;および米国特許第4,683,202号に記載するポリメラーゼ連鎖反応の技術を意味する。
【0060】
本明細書で用いる用語「ポリペプチド」は、アミノ酸のポリマーを意味し、そして生成物の特定の長さを意味しない。従って、ペプチド、オリゴペプチド、およびタンパク質は、この用語内に包含される。この用語はまた、ポリペプチドの翻訳修飾、例えば、グリコシル化、アセチル化、リン酸化などを包含する。定義内に包含されるものは、置換した結合を有するポリペプチド、および当該分野で公知のその他の改変で、天然に存在するおよび天然に存在しない両方のポリペプチドである。
【0061】
本明細書で用いる「ターミネーター」は、コーディング配列の終止コドンの3’または下流に位置するポリアデニル化および転写終結配列のような調節配列である。
【0062】
「組換え宿主細胞」、「宿主細胞」、「細胞」、「細胞培養物」、および他のこのような用語は、例えば、微生物、昆虫細胞、および哺乳動物細胞を意味し、これは、組換えベクターまたは他の転移DNAを導入するためのレシピエントとして用いられ得るかまたは用いられており、そして形質転換された細胞の子孫を含む。このような子孫は、形態もしくはゲノムDNAまたは全DNAの相補性が元の親と必ずしも同一ではなく、そして天然、偶発的、または意図的な変異によって生成され得る。哺乳動物宿主細胞の例は、チャイニーズハムスター卵巣(「CHO」)およびサル腎臓(「COS」)細胞を含む。
【0063】
本細書で用いる用語「微生物」は、細菌および真菌のような原核生物および真核生物の微生物種を含み、後者は、酵母および糸状菌を含む。
【0064】
本明細書で用いる「形質転換」は、宿主細胞への外因性ポリヌクレオチドの転移を意味し、転移に用いた方法、例えば、感染、直接取り込み、形質導入、F因子交配、マイクロインジェクション、またはエレクトロポレーションにかかわらない。外因性ポリヌクレオチドは、非組み込みベクター、例えば、プラスミドとして維持され得、またはあるいは、宿主ゲノムに組み込まれ得る。
【0065】
ポリペプチドまたはヌクレオチド配列に関する「精製」および「単離」は、示す分子が、同じ種または型の他の生物学的高分子が実質的にない状態で存在することを意味する。本明細書で用いる用語「精製」は、同じ型の生物学的高分子が少なくとも75重量%;好ましくは少なくとも85重量%、さらに好ましくは少なくとも95重量%、最も好ましくは98重量%存在することを意味するが、水、緩衝液、および他の小分子、特に、1000未満の分子量を有する分子もまた存在し得る。
【0066】
「薬学的に受容可能なキャリア」は、抗体、熱などの生成のような所望しない副作用をそれ自身誘導しないヒトへの投与のために、当業者により用いられる任意のキャリアを意味する。適切なキャリアは、典型的には大きく、ゆっくりと代謝される高分子(それはタンパク質、多糖類、ポリ乳酸、ポリグリコール酸、重合アミノ酸、アミノ酸コポリマー、または非活性ウイルス粒子であり得る)である。他のキャリアは、当業者に周知である。好ましくは、キャリアは、チログロブリンを含む。
【0067】
治療用組成物は、典型的には、薬学的に受容可能なキャリア、例えば、水、生理食塩水、グリセロール、エタノールなどを含有し得る。さらに、補助剤、例えば、湿潤剤または乳化剤、pH緩衝化剤などが、このような組成物中に存在し得る。
【0068】
本明細書で用いる「治療有効量」は、所望の結果の生成に効果的な量を意味する。この量は、処置されるべき個体の健康および身体状態、抗体を合成する個々の免疫系の能力、所望の防御程度、製剤、処置する医師の病状の評価、および他の関連因子に依存し得る。この量は、一般検査により決定され得る相対的に広い領域に入り得ることが予期される。
【0069】
KGFが昆虫細胞Spodoptera frugiperda(「SF9」)で、KGF163をコードするcDNAを含む組換えバキュロウイルスAutographa californicaの感染により発現される場合、それらは、KGF163に加えて、単一なグリコシル化部位を含むKGF163N末端ドメインの最初の23アミノ酸残基を欠失するKGFフラグメントを生産する。短縮型の非グリコシル化KGFフラグメントKGFdes1−23またはKGF140は、KGF163として同定された天然の長形態に較べて、標的細胞に対して7〜10倍増加した能力を有する。 KGFdes1−23の標的細胞特異性は、変化しない。さらに、高濃度のKGFフラグメントでは、ケラチノサイトについての毒性効果は、KGF163について観察されたのに較べて全く観察されない。これらの観察は、PCT出願第WO90/08771号で既に提唱されたことに反して、KGFの標的細胞特異性が、そのN末端ドメインの残基にないことを示唆する。さらに、本発明は、KGFのN末端短縮型バージョンが、実際に、治療用適用のためのさらに高い生物学的活性および細胞毒の減少を有する改良KGFバージョンを表すことを示す。
【0070】
大量のKGFフラグメントは、組換えDNA技術により生産され得、組換えDNA技術は、天然供給源からKGFを単離して精製し、そしてそのN末端から最初の23アミノ酸残基を切除するもう一方のプロセスより好ましい。組換え技術により生産されるKGFが、細胞中に通常存在する混入分子の非存在下で、KGFが単離されることを可能にする。実際に、いかなる微量のヒトタンパク質混入物質も完全に含まないKGF組成物は、例えば、細菌細胞、酵母細胞、および昆虫細胞で容易に生産され得る。組換えDNA技術の使用により、部位特異的変異誘発により生産され得る変異型のような、天然で見られないKGFフラグメントもまた、生産され得る。
【0071】
所望の宿主でのKGFフラグメントの発現を可能にする任意のプロモーターが、本発明で用いられ得る。例えば、E.coliでは、lacオペロンの調節プロモーター配列が用いられ得る。他の例は、酵母アルコールデヒドロゲナーゼ(「ADH」)プロモーターであり、それは、ADHプロモーターの活性を調節する上流アクチベーター配列(「UAS」)を有する。さらに、特定のウイルスエンハンサーもまた、本明細書で用いられ得る。このようなエンハンサーは、哺乳動物細胞において増幅するだけでなく、発現を調節する。これらのエンハンサーは、Sassone−CorsiおよびBorelli Trends Genet.2:215(1986);Maniatisら、Science
236:1237(1987)に記載のように、哺乳動物のプロモーター配列に組み込まれ、この哺乳動物プロモーター配列は、ホルモンまたは酵素基質のような誘導物質の存在下でのみ活性化し得る。本発明で用いられ得るプロモーターはまた、バキュロウイルスポリヘドロンハイブリッドプロモーターおよびp10プロモーターを含む。
【0072】
本発明で用いられ得る終結配列の例は、Saccharomyces cerevisiae α因子ターミネーターおよびバキュロウイルスターミネーターである。さらに、特定の宿主細胞で作動可能なウイルスターミネーターもまた、用いられ得る。例えば、SV40ターミネーターは、チャイニーズハムスター卵巣(CHO)細胞で機能する。
【0073】
短縮型タンパク質を設計する場合、いくつかのKGFが、本発明の実施において好ましい。1つは、非グリコシル化KGFであり、例えば、天然KGFの最初の23アミノ酸を切除している非グリコシル化KGFである。好ましい短縮型KGFをコードするcDNAを図1に示す。KGFフラグメントの他の種は、存在するか、または一般的な方法により作製され得る。示された配列において、短縮型タンパク質のための開裂部位が、矢印(a)で示される場所に生じ得、約18,000Daの分子量を有するタンパク質を生じる。図1の配列データーおよび短縮型KGFの示された特徴を利用して、短縮型KGFをコードする他のDNA配列を得ることは、当該分野の技術範囲内にある。例えば、構造遺伝子は、正しいアミノ酸を保持しながら個々のヌクレオチドを変えること、または生物学的活性を損失せずにアミノ酸を改変するようにヌクレオチドを変えることにより操作され得る。ヌクレオチドは、例えば、インビトロでの変異誘発およびプライマー修復を含む公知の技術により置換、挿入、または欠失され得る。構造遺伝子は、その生物学的活性を保持しながら、その3’末端および/またはその5’末端で短縮され得る。
【0074】
KGFフラグメントコーディング配列は、KGFフラグメントをコードするDNA配列を合成すること、もしくは所望の配列の生産のために存在しているまたは天然のKGFコーディング配列を変化することにより構築され得る。天然KGFコーディング配列またはポリペプチドは、天然に生じる。天然KGFのアミノ酸配列を[図1]に示す。合成KGFフラグメントは、Urdeaら、Proc.Natl.Acad.Sci.USA 80:7461(1983)で示唆されるように、KGFの公知のアミノ酸配列に基づき、選択した宿主細胞に好ましいコドンを用いて作製され得る。あるいは、所望のKGFフラグメントコーディング配列は、[図1]に示す核酸配列に基づくプローブを用いて核酸ライブラリーからクローニングされ得る。核酸配列ライブラリーの生産およびプローブ化(probing)のための技術は、例えば、Sambrookら、「Molecular
Cloning:A Laboratory Manual」(New York,Cold Spring Harbor Laboratory,1980)に記載されている。他の組換え技術、例えば、部位特異的変異誘発、PCR、酵素的切断および連結もまた、KGFフラグメントのコーディング配列のアナログおよび誘導体を構築するため用いられ得る。天然KGFポリペプチドコーディング配列は、他のクラスのKGFポリペプチドを作製するために容易に改変され得る。
【0075】
例として、KGFフラグメントのアナログは、KGFフラグメントの活性を損なわない保存的アミノ酸置換により作製される。本発明は、さらにムテイン(mutein)と呼ばれるKGFフラグメントのアナログのサブセットを意図しており、ムテインでは、KGFフラグメントのジスルフィド結合に関与しないシステインが通常は、セリンに置換されている。これらのムテインは、非改変KGFフラグメントより広い温度領域にわたり安定である。本明細書中のムテインはまた、天然KGFフラグメントと比較してアミノ酸欠失を含み得る。ムテインは、少なくとも20%、好ましくは少なくとも50%、および最も好ましくは少なくとも80%の天然KGFフラグメントの活性を保持し得る。ムテインのコーディング配列は、天然KGFフラグメントのコーディング配列のインビトロでの変異誘発により構築され得る。
【0076】
フラグメントは、アミノおよび/またはカルボキシル末端アミノ酸欠失により天然KGF分子と異なる。KGFフラグメントが、N末端グリコシル化部位を含まず、そして天然KGF分子の少なくとも20%のKGF活性を保持するなら、短縮されたアミノ酸の数は重大でない。このようなフラグメントのコーディング配列は、天然KGFコーディング配列またはその変異体由来の所望でないヌクレオチドを開裂することにより容易に構築され得る。
【0077】
本発明の範囲内の発現ベクターは、宿主細胞で作動可能であり、そしてKGFフラグメントまたはアナログのコーディング配列に作動可能に連結されるプロモーターを含む。発現ベクターは、分泌のためのシグナル配列、ターミネーター、選択マーカー、複製起点、および宿主細胞配列と同種(homologous)の配列を任意に含み得る。これらの追加的な要素は、発現を最適化するために含まれ得る。
【0078】
機能的な非天然プロモーターもまた用いられ得、例えば、異なるプロモーターの共通配列に基づいた合成プロモーターであり得る。また、効果的なプロモーターは、異種(heterologous)の発現開始領域と連結した調節領域を含むハイブリッドプロモーターであり得る。ハイブリッドプロモーターの例は、E.coli tac転写活性領域に連結したE.coli lacオペレーター;本明細書に参考として援用される米国特許第4,876,197号および第4,880,734号に記載のような、酵母グリセルアルデヒド−3−ホスフェート−デヒドロゲナーゼ(「GAPDH」)に連結した酵母アルコールデヒドロゲナーゼ(「ADH」)調節配列;およびシミアンウイルスSV40プロモーターに連結したサイトメガロウイルス(「CMV」)エンハンサーである。
【0079】
本発明のKGFフラグメントまたはアナログのコーディング配列はまた、読み取り枠でシグナル配列に結合され得る。シグナル配列は、典型的には、疎水性アミノ酸からなる分泌のためのアミノ酸配列をコードしKGFフラグメントまたはアナログを細胞膜に導く。好ましい実施態様では、プロセッシング部位は、シグナル配列とKGFフラグメントとの間に位置し、インビボまたはインビトロのいずれかでシグナル配列およびKGFフラグメントの間を開裂し得る。適切なシグナル配列は、分泌される内因性宿主細胞タンパク質に対する遺伝子由来であり、例えば、ヨーロッパ特許第12873号および日本特許第62,096,086号に記載の酵母インベルターゼ、米国特許第4,588,684号に記載のA因子遺伝子、およびEP60057に記載のようなインターフェロンシグナル配列である。
【0080】
酵母での発現のために本発明で用いる好ましいクラスの分泌リーダーは、短縮型酵母α因子リーダー配列であり、それは、「プレ」シグナル配列の少なくとも一部分および「プロ」領域を含む。本明細書に参考として援用される米国特許第4,546,083号および第4,870,008号、ならびにヨーロッパ特許第EP324274号に記載のように、本明細書中で使用され得るα因子リーダー配列は、約83アミノ酸残基を有する完全長プレ−プロα因子リーダー、および典型的には、約25〜約50アミノ酸残基を含む短縮型α因子リーダーであり得る。本明細書で用いられ得るα因子リーダーフラグメントを使用する付加的なリーダーは、第二の酵母α因子由来のプロ領域ではなく、第一の酵母シグナル配列のプレ配列で作られるハイブリッドα因子リーダーを含む。(例えば、PCT
WO89/02463を参照のこと。)
(f.発現システム)
KGFフラグメントおよびそのアナログは、当業者に十分に理解されるように、KGFフラグメントまたはアナログをコードするDNA配列がベクターに適切な読み取り枠および方向で作動可能に連結される場合、組換え技術により発現され得る。KGFフラグメントを含む遺伝子構築物を作成する場合、好ましい出発材料は、KGFフラグメントをコードしているcDNAである。KGFフラグメントはまた、所望する場合、開裂され得るハイブリッドタンパク質として生産され得るが、典型的には、短縮型KGF遺伝子は、プロモーターより下流に挿入され得、そして後ろにターミネーター配列が続き得る。一般に、短縮型KGFおよび短縮型KGFポリペプチド誘導体の生産収量を改善する宿主細胞特異的配列が用いられ得、そして適切な制御配列、例えば、エンハンサー配列、ポリアデニル化配列、およびリボソーム結合部位が発現ベクターに付加され得る。一旦、適切なコーディング配列が単離されること、それは、種々の異なる発現系;例えば、哺乳動物細胞、バキュロウイルス、細菌、および酵母で使用され、発現され得る。これらは、以下に考察される。
【0081】
(i.哺乳動物系)
哺乳動物発現系は、当該分野で公知である。哺乳動物プロモーターは、哺乳動物RNAポリメラーゼを結合し、そしてコーディング配列(例えば、構造遺伝子)のmRNAへの下流(3‘)転写を開始し得る任意のDNA配列である。プロモーターは転写開始領域、それは、通常コーディング配列の5’末端に隣接する位置にあり、および転写開始部位の25〜30塩基対(bp)上流に位置するTATAボックスを有し得る。 TATAボックスは、RNA合成を正しい部位で始めるために、RNAポリメラーゼIIに指示すると考えられている。動物細胞のプロモーターはまた、上流のプロモーター要素(典型的には、TATAボックスの100〜200bp上流内に位置する)を含み得る。上流のプロモーター要素は、転写が開始される速度を決定しどちらかの方向に作用し得る[Sambrookら、(1989)「哺乳動物細胞におけるクローン化遺伝子の発現。」Molecular Cloning:A Laboratory Manual,第2版]。
【0082】
哺乳動物のウイルス遺伝子は、しばしば高発現され、そして広い宿主領域を有する;従って、哺乳動物ウイルス遺伝子をコードしている配列は、特に有用なプロモーター配列を提供する。この例として、SV40初期プロモーター、マウス乳ガンウイルスLTRプロモーター、アデノウイルス主要後期プロモーター(AdMLP)、および単純ヘルペスウイルスプロモーターが含まれる。さらに、非ウイルス遺伝子、例えば、ネズミメタロチオネイン遺伝子由来の配列はまた、有用なプロモーター配列を提供する。発現は、構成的か、調節的(誘導性)のいずれかであり得、プロモーターがホルモン応答細胞におけるグルココルチコイドで誘導され得ることに依存する。
【0083】
上記のプロモーター要素と結合したエンハンサー要素(エンハンサー)の存在は、典型的には、発現レベルを増加する。エンハンサーは、同種または異種プロモーターと連結されるとき、正常なRNA開始部位で合成を開始する場合に1000倍まで転写を刺激し得る調節DNA配列である。エンハンサーはまた、それらが転写開始部位から上流または下流に、正常または逆の向きで、もしくはプロモーターから1000ヌクレオチオド以上の距離に位置する場合、活性である[Maniatisら、(1987)Science 236:1237;Albertsら、(1989)Molecular Biology of the Cell,第2版]。ウイルス由来のエンハンサー要素は特に有用である。なぜなら、それらは、典型的には、より広い宿主領域を有するからである。この例として、SV40初期遺伝子エンハンサー[Dijkemaら、(1985)EMBO J.4:761]およびラウス肉腫ウイルスの長末端反復(LTR)由来[Gormanら、(1982b)Proc.Natl.Acad.Sci.79:6777]、およびヒトサイトメガロウイルス[Boshartら、(1985)Cell 41:521]由来のエンハンサー/プロモーターが含まれる。さらに、いくつかのエンハンサーは調節可能であり、そしてホルモンまたは金属イオンのような誘導物質の存在下でのみ活性になる[Sassone−CorsiおよびBorelli(1986)Trends Genet.2:215;Maniatisら、(1987)Science 236:1237]。
【0084】
DNA分子は、哺乳動物細胞において細胞内発現され得る。プロモーター配列は、DNA分子と直接連結し得、その場合、組換えタンパク質のN末端の最初のアミノ酸は常にメチオニンであり、メチオニンは、ATG開始コドンによりコードされている。所望であれば、N末端は、インビトロで臭化シアンとともにインキュベートすることでタンパク質から開裂され得る。
【0085】
あるいは、外来タンパク質はまた、哺乳動物細胞において外来タンパク質の分泌を提供するリーダー配列フラグメントからなる融合タンパク質をコードするキメラDNA分子を作成することにより、細胞から成長培地へ分泌され得る。好ましくは、リーダーフラグメントと、インビボまたはインビトロのいずれかで開裂され得る外来遺伝子との間にコードされるプロセッシング部位がある。リーダー配列フラグメントは、典型的には、細胞からのタンパク質の分泌を指向する疎水性アミノ酸からなるシグナルペプチドをコードする。アデノウイルス三部分(tripartite)リーダーは、哺乳動物細胞において外来タンパク質の分泌を提供するリーダー配列の1例である。
【0086】
典型的には、哺乳動物細胞により認識される転写終結およびポリアデニル化配列は、3’から翻訳終止コドンに位置する調節領域であり、そしてそれ故プロモーター要素とともにコーディング配列に隣接する。成熟mRNAの3’末端は、部位特異的な転写後の開裂およびポリアデニル化により形成される[Birnstielら、(1985)Cell
41:349;ProudfootおよびWhitelaw(1988)「真核生物RNAの終結および3’末端プロセッシング」、Transcription and splicing(B.D.HamesおよびD.M.Glover編);Proudfoot(1989)Trends Biochem.Sci.14:105]。これらの配列は、DNAにコードされるポリペプチドに翻訳され得るmRNAの転写を指向する。転写ターミネーター/ポリアデニル化シグナルの例は、SV40由来のそれらを含む[Sambrookら(1989)「培養哺乳動物細胞におけるクローニングされた遺伝子の発現」、Molecular Cloning:A Laboratory Manual。
【0087】
いくつかの遺伝子は、イントロン(介在配列とも言う)が存在する場合、さらに効果的に発現され得る。しかし、いくつかのcDNAは、スプライシングシグナル(スプライス供与体および受容体部位ともいう)を欠いたベクターから効率的に発現された[GothingおよびSambrook(1981)Nature 293:620を参照のこと]。イントロンは、スプライス供与体および受容体部位を含む、コーディング配列内に介在する非コーディング配列である。それらは、一次転写物のポリアデニル化後「スプライシング」と呼ばれるプロセスにより取り除かれる[Nevins(1983)Annu.Rev.Biochem.52:441;Green(1986)Annu.Rev.Genet.20:671;Padgettら、(1986)Annu.Rev.Biochem.55:1119;KrainerおよびManiatis(1988)「RNAスプライシング」、Transcription and splicing(B.D.HamesおよびD.M.Glover編)。
【0088】
典型的には、プロモーター、ポリアデニル化シグナル、および転写終結配列を含む上記の構成成分は、一緒に発現構築物中に置かれる。エンハンサー、機能的なスプライス供与体および受容体部位を有するイントロン、ならびにリーダー配列はまた、所望する場合、発現構築物中に含まれる。発現構築物は、哺乳動物細胞または細菌のような宿主において安定に維持し得る染色体外要素(例えば、プラスミド)のようなレプリコン中にしばしば維持される。哺乳動物の複製系は、動物ウイルス由来の系を含み、それは、複製にトランスに作用する因子を必要とする。 例えば、SV40[Gluzman(1981)Cell 23:175]のようなパポーバウイルスまたはポリオーマウイルスの複製系を含むプラスミドは、適切なウイルスT抗原の存在下できわめて高いコピー数に複製する。哺乳動物のレプリコンの別の例は、ウシパピローマウイルスおよびエプスタインバーウイルス由来のレプリコンを含む。さらに、レプリコンは、2つの複製系を有し得、従って、例えば、発現のために哺乳動物細胞で、およびクローニングおよび増幅のために原核生物宿主で維持され得る。このような哺乳動物−細菌シャトルベクターの例は、pMT2[Kaufmanら、(1989)Mol.Cell.Biol.9:946およびpHEBO[Shimizuら、(1986)Mol.Cell.Biol.6:1074]を含む。
【0089】
用いた形質転換手順は、形質転換されるべき宿主に依存する。異種ポリヌクレオチドを哺乳動物細胞へ導入するための方法は、当該分野で公知であり、そしてデキストラン仲介トランスフェクション、リン酸カルシウム沈澱、ポリブレン仲介トランスフェクション、プロトプラスト融合、エレクトロポレーション、リポソーム内へのポリヌクレオチドのカプセル化、および核へのDNAの直接マイクロインジェクションを含む。
【0090】
発現のための宿主として利用できる哺乳動物細胞株は当該分野で公知であり、チャイニーズハムスター卵巣(CHO)細胞、HeLa細胞、ベビーハムスター腎臓(BHK)細胞、サル腎臓細胞(COS)、ヒト肝細胞癌細胞(例えば、Hep G2)、および多くのその他の細胞株を含むがこれに限定されない、American Type Cultutre Collection(ATCC)から入手可能な多くの不死化細胞株を含む。
【0091】
(ii.バキュロウイルス系)
KGFをコードしているポリヌクレオチドは、安定な発現ベクター、例えば昆虫細胞発現ベクター中に挿入され得、そしてベクター内で制御要素に作動可能に連結され得る。ベクターの構築は、当該分野で公知の技術を使用して行われる。特に、本発明の目的のため、バキュロウイルス発現ベクターが、Kittsら、BioTechniques 14:810−817(1993)に実質的に従って構築される。
【0092】
簡単に説明すると、バキュロウイルスゲノムの一部(本明細書中では、「バキュロウイルス配列」と呼ばれている)に相同性があり、そしてそれと相同的組換えを行い得るポリヌクレオチド配列を含むトランスファーベクターに、KGF163をコードしている配列を挿入することによって、 KGF発現カセットを最初に構築する。バキュロウイルス配列は、以下により詳しく記載されているように、必須のポリヌクレオチド配列を少なくとも含む。KGFコーディング配列は、バキュロウイルス配列が両端に位置するようにトランスファーベクターに挿入される。
【0093】
KGFコーディング配列を含むトランスファーベクターを、機能的なウイルスの生産に必要な必須のポリヌクレオチド配列を欠いている野生型バキュロウイルスの変異体とともに宿主昆虫細胞中にトランスフェクトする。この点に関して、変異体のバキュロウイルスがKGF発現カセットと組換わるときトランスフェクションに際し機能的なウイルスが宿主細胞中で産生され得る。
【0094】
従って、この様式で産生される機能的なバキュロウイルスは、KGF発現カセットを組み込み、そして組換えKGFおよび組換えKGFdes1−23を産生するために、新たな宿主細胞のトランスフェクションに適切である。あるいは、このプロセスは、KGF163コーディング配列の代わりにKGFdes1−23コーディング配列を用いて実施され得る。
【0095】
一般に、発現系の構成成分は、バキュロウイルスゲノムのフラグメント、および異種遺伝子または発現されるべき遺伝子の挿入に便利な制限部位の両方を含むトランスファーベクター(通常は細菌プラスミド)を含む機能的なバキュロウイルス;トランスファーベクター中のバキュロウイルス特異的フラグメントに相同な配列を有する野生型バキュロウイルス(これは、バキュロウイルスゲノムに異種遺伝子の相同組換えを可能にする);および適切な昆虫宿主細胞および成長培地を含む。
【0096】
短縮型KGF DNA配列をトランスファーベクターに挿入後、ベクターおよび野生型ウイルスゲノムを、ベクターおよびウイルスゲノムが組換えられ得る昆虫宿主細胞にトランスフェクトする。パッケージ化された組換えウイルスを発現し、そして組換えプラークを同定し、そして精製する。バキュロウイルス/昆虫細胞発現系の材料および方法は、前記のInvitorogen,San Diego CAからキット形態(「MaxBac」キット)で市販されている。これらの技術は、一般に、当業者に公知であり、そしてSummersおよびSmith,Texas Agricultural Experiment Station Bulletin No.1555(1987)(本明細書では以後、「SummersおよびSmith」と称する)に十分に記載され、そして参考として援用されている。
【0097】
短縮型KGF DNA配列をバキュロウイルスゲノムに挿入する前に、プロモーター、リーダー(所望の場合)、目的のコーディング配列、および転写終結配列を含む上記の構成成分を、代表的には、中間導入配置構築物(トランスファーベクター)にアセンブルする。この構築物は、単一の遺伝子および作動可能に連結した調節要素;それぞれが作動可能に連結したそれ自身の調節要素のセットを有する複数の遺伝子;または調節要素の同じセットにより調節されているは複数の遺伝子を含み得る。中間導入配置構築物を、しばしば細菌のような宿主で安定に維持し得る染色体外要素(例えば、プラスミド)のようなレプリコン中に維持し得る。レプリコンは複製系を有し、従ってクローニングおよび増幅のために適切な宿主中で、それは維持され得る。
【0098】
現在では、AcNPVに外来遺伝子を導入するために最も一般的に用られるトランスファーベクターはpAc373である。当業者に公知である多くの他のベクターもまた設計された。これらは、例えば、pVL985(これは、ポリヘドリン開始コドンをATGからATTに改変し、そしてATTから32塩基対下流にBamHIクローニング部位を導入する;LuckowおよびSummers,Virology(1989)17:13を参照のこと)を含む。
【0099】
プラスミドはまた、通常ポリヘドリンポリアデニル化シグナル(Millerら(1988)Ann.Rev.Microbiol.,42:177)、ならびにE.coliにおける選択および増殖のための原核生物のアンピシリン耐性(amp)遺伝子および複製起点を含む。
【0100】
バキュロウイルストランスファーベクターは、通常バキュロウイルスプロモーターを含む。バキュロウイルスプロモーターは、バキュロウイルスRNAポリメラーゼを結合し得、そしてコーディング配列(例えば、構造遺伝子)のmRNAへの下流(5’から3’)転写を開始し得る任意のDNA配列である。プロモーターは、通常、コーディング配列の5’末端に隣接して位置する転写開始領域を有する。この転写開始領域は、典型的には、RNAポリメラーゼ結合部位および転写開始部位を含む。バキュロウイルストランスファーベクターはまた、エンハンサーと呼ばれる第二ドメインを有し得、存在する場合は、通常構造遺伝子から遠位にある。発現は、調節され得るか、または構成的であり得る。
【0101】
構造遺伝子は、ウイルス感染サイクルの後期に盛んに転写され、特異的に有用なプロモーター配列を提供する。この例として、ウイルスポリヘドリンタンパク質をコードする遺伝子、Friesenら、(1986)「バキュロウイルス遺伝子発現の調節」、The
Molecular Biology of Baculoviruses(Walter Doerfler編);EPO公開番号第127,839号および第155,476号;およびp10タンパク質をコードする遺伝子、Vlakら、(1988),J.Gen.Virol.69:765由来の配列が含まれる。
【0102】
適切なシグナル配列をコードするDNAは、分泌される昆虫タンパク質または分泌されるバキュロウイルスタンパク質、例えば、バキュロウイルスポリヘドリン遺伝子(Carbonellら、(1988)Gene,73:409)に由来する遺伝子であり得る。あるいは、哺乳動物細胞の翻訳後の修飾(例えば、シグナルペプチド開裂、タンパク質分解開裂、およびリン酸化)のためのシグナルが、昆虫細胞により認識されるようであり、そして分泌および核蓄積に必要なシグナルもまた、無脊椎動物細胞および脊椎動物細胞の間で保存され得るようであるので、昆虫起源でないリーダー、例えば、ヒトα−インターフェロン、Maedaら、(1985)、Nature 315:592;ヒトガストリン放出ペプチド、Lebacq−Verheydenら、(1988)、Molec.Cell.Biol.8:3129;ヒトIL−2、Smithら、(1985)Proc.Nat’l Acad.Sci.USA,82:8404;マウスIL−3、(Miyajimaら、(1987)Gene 58:273;およびヒトグルコセレブロシダーゼ、Martinら、(1988)DNA,7:99をコードする遺伝子由来のリーダーもまた、昆虫で分泌を提供するために使用され得る。
【0103】
組換えポリペプチドまたはポリタンパク質は、細胞内で発現され得、もしくは、適切な調節配列で発現される場合分泌され得る。非融合の外来タンパク質の良好な細胞内発現は、通常、 ATG開始シグナルに先行する適切な翻訳開始シグナルを含む短いリーダー配列を理想的には有する異種遺伝子を必要とする。所望であれば、N末端のメチオニンを、臭化シアンとのインビトロでのインキュベーションにより、成熟タンパク質から開裂し得る。
【0104】
あるいは、天然では分泌されない組換えポリタンパク質またはタンパク質は、昆虫での外来タンパク質の分泌を提供するリーダー配列フラグメントからなる融合タンパク質をコードするキメラDNA分子を作製することにより昆虫細胞から分泌され得る。リーダー配列フラグメントは、典型的には、タンパク質の小胞体中への移動を指向する疎水性アミノ酸からなるシグナルペプチドをコードする。
【0105】
短縮型のKGF DNA配列および/または発現産物前駆体をコードしている遺伝子の挿入後、昆虫細胞宿主は、トランスファーベクターの異種DNAおよび野生型バキュロウイルスのゲノムDNAを用いて同時形質転換され、それは、通常、同時トランスフェクションによる。構築物のプロモーターおよび転写終結配列は、典型的には、2〜5kbセクションのバキュロウイルスゲノムを含む。異種DNAをバキュロウイルスの所望の部位に導入する方法は、当該分野で公知である。(SummersおよびSmith 上記;Juら、(1987);Smithら、Mol.Cell.Biol.(1983)3:2156;ならびにLuckowおよびSummers(1989)参照のこと)。例えば、ポリヘドリン遺伝子のような遺伝子に、相同な二重交差組換えにより挿入され得る;また所望のバキュロウイルス遺伝子中に設計された制限酵素部位にも挿入し得る。Millerら、(1989),Bioessays 4:91。DNA配列が、ポリヘドリン遺伝子の代わりに発現ベクターにクローニングされる場合、その5’および3’の両方にポリヘドリン特異的配列が隣接し、そしてポリヘドリンプロモーターの下流に位置する。
【0106】
新たに形成されたバキュロウイルス発現ベクターは、次いで感染性組換えバキュロウイルスにパッケージ化される。同種の組換えが、低頻度で起こる(約1%と約5%との間);従って、同時トランスフェクション後に産生された多くのウイルスは、なお野生型のままである。それゆえ、組換えウイルスを同定するための方法が必要である。発現系の利点は、組換えウイルスを区別できるようにする視覚化スクリーニングである。ポリヘドリンタンパク質は、天然ウイルスにより産生され、ウイルス感染後、後期に感染細胞の核に非常に高レベルで産生される。蓄積したポリヘドリンタンパク質はまた、包埋粒子を含む封入体を形成する。これらの封入体は、サイズが15μmまでであり、高度に屈折性であり、光学顕微鏡下で容易に視覚化される明るく光った外観を与える。組換えウイルスに感染した細胞は、封入体を欠いている。野生型ウイルスから組換えウイルスを区別するために、トランスフェクション上清を当業者に公知の技術により単層の昆虫細胞上にプラーク化する。すなわち、プラークは、光学顕微鏡下で封入体の存在(野生型ウイルスを示す)または不在(組換えウイルスを示す)についてスクリーニングされる。「Current Protocols in Microbiology」第2巻(Ausubelら編)、16.8(増刊1990年10月);SummersおよびSmith、前述;Millerら(1989)。
【0107】
組換えバキュロウイルス発現ベクターが、いくつかの昆虫細胞に感染させるために開発された。例えば、組換えバキュロウイルスは、とりわけ以下について開発された:
【0108】
【表2】
細胞および細胞培養培地は、バキュロウイルス/発現系におる異種ポリペプチドの指向および融合発現の両方について市販されている;細胞培養技術は、当業者に一般に公知である。例えば、SummersおよびSmith、前述を参照のこと。
【0109】
次いで、改変された昆虫細胞は、適切な栄養培地で成長し得、この培地は、改変された昆虫宿主に存在するプラスミドを安定的に維持し得る。発現産物の遺伝子が、誘導し得る制御下にある場合、宿主は高濃度になるまで成長され得、そして発現を誘導する。あるいは、発現が構成的である場合、産物は連続的に培地中に発現され得、そして栄養培地は連続的に循環されねばならず、その間目的の産物を取り出し、そして枯渇した栄養を増強する。この産物をクロマトグラフィー(例えば、HPLC、アフィニティークロマトグラフィー、イオン交換クロマトグラフィーなど);電気泳動;密度勾配遠心分離;溶媒抽出などのような技術により精製され得る。適切には、必要に応じて、培地に同時に分泌されるか、または昆虫細胞の溶解から生じる任意の昆虫タンパク質を実質的に除去するように、宿主の残骸(例えば、タンパク質、脂質、および多糖)を少なくとも実質的に含まない産物を提供するように、産物をさらに精製する。
【0110】
短縮型KGF発現を得るために、形質転換体由来の組換え宿主細胞を、配列をコードする組換え短縮型KGFの発現を可能にする条件下でインキュベートし得る。これらの条件は、選択される宿主細胞に依存して変化し得る。しかし、条件は、当該分野で公知の事項に基づき、当業者に容易に確認され得る。
【0111】
(iii.細菌系)
細菌発現技術は当該分野で公知である。細菌プロモーターは、細菌RNAポリメラーゼを結合し得、そしてコーディング配列(例えば、構造遺伝子)のmRNAへの下流(3”)転写を開始し得る任意のDNA配列である。プロモーターは、通常コーディング配列の5’末端に隣接して位置する転写開始領域を有する。この転写開始領域は、典型的には、RNAポリメラーゼ結合部位および転写開始部位を含む。細菌プロモーターはまた、オペレーターと呼ばれる第二ドメインを有し得、それは、RNA合成が開始する隣接したRNAポリメラーゼ結合部位とオーバーラップし得る。遺伝子リプレッサータンパク質がオペレーターに結合し得、そしてそれによって特異的な遺伝子の転写を阻害し得るので、オペレーターは、ネガティブに調節された(誘導可能な)転写を可能にする。構成的な発現は、オペレーターのようなネガティブな調節要素の非存在下で生じ得る。さらに、ポジティブな調節は、遺伝子活性化タンパク質結合配列により達成され得、それが存在する場合は、通常、RNAポリメラーゼ結合配列に隣接する(5’側)。遺伝子活性化タンパク質の例は、カタボライトアクチベータータンパク質(CAP)であり、それは、Escherichia coli(E.coli)においてlacオペロンの転写開始を助ける[Raibaudら、(1984)Annu.Rev.Genet.18:173]。従って、調節された発現は、ポジティブまたはネガティブのいずれか、それにより転写促進または減少のいずれであり得る。
【0112】
代謝経路酵素をコードしている配列は、特に有用なプロモーター配列を提供する。この例としては、糖代謝酵素、例えば、ガラクトース、ラクトース(lac)[Changら、(1977)Nature 198:1056]、およびマルトース由来のプロモーター配列を含む。別の例は、トリプトファン(trp)のような生合成酵素由来のプロモーター配列を含む[Goeddelら、(1980)Nuc.Acids Res.8:4057;Yelvertonら、(1981)Nucl.Acids Res.9:731;米国特許第4,738,921号;EPO公開第036776号および第121775号]。g−ラオタマーゼ(g−laotamase)(bla)プロモーター系[Weissmann(1981)「インターフェロンおよび他の間違いのクローニング。」Interferon 3(I.Gresser編)]、バクテリオファージラムダPL[Shimatakeら、(1981)Nature 292:128]およびT5[米国特許第4,689,406号]プロモーター系はまた、有用なプロモーター配列を提供する。
【0113】
さらに、天然にない合成プロモーターがまた、細菌プロモーターとして機能する。例えば、ある細菌またはバクテリオファージプロモーターの転写アクチベーター配列は、他の細菌またはバクテリオファージプロモーターのオペロン配列と結合され得、合成ハイブリッドプロモーターを作製する[米国特許第4,551,433]。例えば、tac−lacプロモーターは、trpプロモーターおよびlacリプレッサーにより調節されるlacオペロン配列の両方を含むハイブリッドtrp−lacプロモーターである[Amannら、(1983)Gene 25:167;de Boerら、(1983)Proc.Natl.Acad.Sci.80:21]。さらに、細菌プロモーターは、細菌RNAポリメラーゼを結合しそして転写を開始する能力を有する非細菌起源の天然に存在するプロモーターを含み得る。非細菌起源の天然に存在するプロモーターはまた、原核生物でいくつかの遺伝子の高レベルの発現を行うために、適合するRNAポリメラーゼと結合され得る。バクテリオファージT7 RNAポリメラーゼ/プロモーター系は、結合プロモーター系の例である[Studierら、(1986)J.Mol.Biol.189:113;Taborら、(1985)Proc Natl.Acad.Sci.82:1074]。さらに、ハイブリッドプロモーターはまた、バクテリオファージプロモーターおよびE.coliオペロン領域(EPO公開第267851号)を含み得る。
【0114】
機能するプロモーター配列に加えて、効率的なリボソーム結合部位はまた、原核生物における外来遺伝子の発現に有用である。E.coliでは、リボソーム結合部位は、Shine−Dalgarno(SD)配列と呼ばれ、そして開始コドン(ATG)および開始コドンの3〜11ヌクレオチド上流に位置する長さが3〜9のヌクレオチドの配列を含む[Shineら、(1975)Nature 254:34]。SD配列は、SD配列とE.coli 16S rRNAの3’末端との間の塩基の対合により、mRNAのリボソームへの結合を促進すると考えられている[Steitzら、(1979)「メッセンジャーRNAにおける遺伝子シグナルおよびヌクレオチド配列」Biological
Regulation and Development:Gene Expression(R.F.Goldberger編)]。弱いリボソーム結合部位を有する真核生物遺伝子および原核生物遺伝子を発現するためには[Sambrookら、(1989)「Escherichia coliにおけるクローニングされた遺伝子の発現」Molecular Cloning:A Laboratory Manual. DNA分子は細胞内に発現され得る。プロモーター配列はまた、DNA分子と直接結合し得、この場合、N末端の最初のアミノ酸は常にメチオニンであり得、メチオニンはATG開始コドンによりコードされる。所望する場合、N末端のメチオニンは、臭化シアンとともにインビトロでインキュベーションすること、もしくは細菌メチオニンN末端ペプチダーゼとともにインビボまたはインビトロのいずれかでインキュベーションすることによりタンパク質から開裂され得る(EPO公開第219237号)。
【0115】
融合タンパク質は、直接発現の代替物を提供する。典型的には、内因性細菌タンパク質のN末端部分または他の安定なタンパク質をコードするDNA配列を、異種コーディング配列の5’末端に融合する。発現の際に、この構築物は、2つのアミノ酸配列の融合を提供する。例えば、バクテリオファージラムダ細胞遺伝子が外来遺伝子の5’末端に連結され得、そしてバクテリア中で発現され得る。生じた融合タンパク質は、好ましくはプロセッシング酵素(Xa因子)が外来遺伝子からバクテリオファージタンパク質を開裂するための部位を保持している[Nagaiら、(1984)Nature 309:810]。融合タンパク質はまた、lacZ[Jiaら、(1987)Gene 60:197];trpE[Allenら、(1987)J.Biotechnol.5:93;Makoffら、(1989)J.Gen.Microbiol.135:11]およびChey[EPO公開第324 647号]遺伝子由来の配列を用いて作成され得る。2つのアミノ酸配列の結合部分でのDNA配列は、開裂可能部位をコードし得るかまたはコードし得ない。他の例は、ユビキチン融合タンパク質である。このような融合タンパク質は、好ましくは、外来タンパク質からユビキチンを開裂するために、プロセッシング酵素(例えばユビキチン特異性プロセッシングプロテアーゼ)のための部位を保有しているユビキチン領域を用いて作成される。この方法により、天然の外来タンパク質を単離し得る[Millerら、(1989)Bio/Technology 7:698]。
【0116】
あるいは、外来タンパク質はまた、細菌中で外来タンパク質の分泌を提供するシグナルペプチド配列のフラグメントからなる融合タンパク質をコードするキメラDNA分子を作成することにより細胞から分泌され得る[米国特許第4,336,336号]。このシグナル配列フラグメントは、典型的には細胞からタンパク質の分泌を指向する疎水性アミノ酸からなるシグナルペプチドをコードする。上記タンパク質は、成長培地中(グラム陽性細菌)、または細胞の内膜と外膜との間に存在する周辺腔(periplasmic space)中(グラム陰性バクテリア)のいずれかに分泌される。 好ましくは、シグナルタンパク質フラグメントと外来遺伝子の間にコードされる、インビボ、またはインビトロで開裂され得るプロセッシング部位が存在する。
【0117】
適当なシグナル配列をコードするDNAは、分泌される細菌タンパク質の遺伝子から由来し得る。例えばE.col外膜タンパク遺伝子(ompA)[Masuiら、(1983):Experimental Manipulation of Gene Expression;Ghrayebら、(1984)EMBOJ.3:2437]およびE.coliアルカリホスファターゼシグナル配列(phoA)[Okaら、(1985)Proc.Natl.Acad.Sci.82:7212]。別の例として、種々のBacilus株由来のα−アミラーゼ遺伝子のシグナル配列を用いて、B.subtilisから異種タンパク質を分泌し得る[Palvaら、(1982)Proc.Natl.Acad.Sci.USA 79:5582;EPO公開 第244042号]。
【0118】
典型的には、細菌により認識される転写終結配列は、翻訳終止コドンの3’側に位置する調節領域であり、従って、プロモーターとともにコーディング配列に隣接する。これらの配列は、DNAによりコードされたポリペプチドに翻訳され得るmRNAの転写を指向する。転写終結配列は、転写の終結を助けるステムループ構造を形成し得る約50ヌクレオチドのDNA配列を頻繁に含む。この例として、E.coliのtrp遺伝子および他の生合成遺伝子のような、強力なプロモーターを有する遺伝子由来の転写終結配列が含まれる。
【0119】
典型的には、プロモーター、シグナル配列(所望であれば)、目的のコーディング配列、および転写終結配列を含む上記の構成成分は、発現構築物中に共に置かれる。発現構築物は、レプリコン、例えば、細菌のような宿主で安定的に維持され得る染色体外要素(例えば、プラスミド)にしばしば維持され得る。レプリコンは、複製系を有し、従って、発現のために、またはクローニングおよび増幅のいずれかのための原核生物宿主で維持され得る。さらに、レプリコンは、高コピー数または低コピー数のいずれかのプラスミドであり得る。高コピー数のプラスミドは、一般に、約5から約200、そして典型的には、約10から約150のコピー数の範囲を有する。高コピー数のプラスミドを含む宿主は、好ましくは、少なくとも約10、およびさらに好ましくは少なくとも約20のプラスミドを含む。高コピー数または低コピー数のベクターのいずれかを、宿主に対するベクターおよび外来タンパク質の影響に依存して選択し得る。
【0120】
あるいは、発現構築物は、組み込みベクターとともに細菌ゲノムに組み込まれ得る。組み込みベクターは、典型的には、ベクターの組み込みを可能にする、細菌染色体に相同な少なくとも1つの配列を含む。組み込みは、ベクター中の相同DNAと細菌染色体との間の組換えから生ずるようである。例えば、種々のBacillus株由来のDNAを用いて構築された組み込みベクターは、Bacillus染色体に組み込まれる(EPO公開第127328号)。組み込みベクターはまた、バクテリオファージまたはトランスポゾン配列を含み得る。
【0121】
典型的には、染色体外および組み込み発現構築物は、選択マーカーを含み得、形質転換された細菌株の選択を可能にする。選択マーカーは、細菌宿主で発現され得、そしてアンピシリン、クロラムフェニコール、エリスロマイシン、カナマイシン(ネオマイシン)、およびテトラサイクリンのような薬剤に対して細菌を耐性にする遺伝子を含み得る[Davisら、(1978)Annu.Rev.Microbiol.32:469]。選択マーカーはまた、生合成遺伝子、例えば、ヒスチジン、トリプトファン、およびロイシン生合成経路の遺伝子を含み得る。
【0122】
あるいは、いくつかの上記の構成成分は、形質転換ベクターに一緒に置かれ得る。形質転換ベクターは、典型的には、上記のように、レプリコン中に維持されるか、または組み込みベクターに発展されるかのいずれかである選択マーカーを含む。
【0123】
発現および形質転換ベクター、染色体外レプリコンまたは組み込みベクターのいずれかは、多くの細菌への形質転換のために開発されている。例えば、発現ベクターは、特に以下の細菌のために開発されている:
【0124】
【表3】
細菌宿主へ外因性DNAを導入する方法は、当該分野で周知であり、そして典型的には、CaCl2または他の試薬(例えば、2価のカチオンおよびDMSO)のいずれかで処理した細菌の形質転換を含む。DNAはまた、エレクトロポレーションにより細菌細胞に導入され得る。形質転換手順は、通常、形質転換されるべき細菌種で変わる。以下の例を参照のこと、
【0125】
【表4】
(iv.酵母発現)
酵母発現系がまた当業者に公知である。酵母プロモーターは、酵母RNAポリメラーゼを結合し得、かつmRNAにコーディング配列(例えば、構造遺伝子)の下流(3’)転写を開始し得る任意のDNA配列である。プロモーターは、通常コーディング配列の5’末端に近接して位置する転写開始領域を有する。この転写開始領域は、典型的には、RNAポリメラーゼ結合部位(「TATA Box」)および転写開始部位を含む。酵母プロモーターはまた、上流アクチベーター配列と呼ばれる第2ドメインを有し、存在する場合、それは、通常、構造遺伝子の遠方にある。UASは、調節(誘導可能)発現を可能にする。構成的な発現はUASの非存在下で起こる。調節発現は、ポジティブまたはネガティブのいずれかであり得、それによって転写の促進または減少のいずれかであり得る。
【0126】
酵母は、活性な代謝経路を有する発酵生物であり、それゆえ代謝経路中の酵素をコードしている配列は、特に有用なプロモーター配列を提供する。この例としては、アルコールデヒドロゲナーゼ(ADH)(EPO公開第284044号)、エノラーゼ、グルコキナーゼ、グルコース−6−ホスフェートイソメラーゼ、グリセルアルデヒド−3−ホスフェートデヒドロゲナーゼ(GAPまたはGAPDH)、ヘキソキナーゼ、ホスホフルクトキナーゼ、3−ホスホグリセレートムターゼ、およびピルビン酸キナーゼ(PyK)(EPO公開第329203号)が含まれる。酸性ホスファターゼをコードしている酵母PHO5遺伝子はまた、有用なプロモーター配列を提供する[Myanoharaら、(1983)Proc.Natl.Acad.Sci.USA 80:1]。
【0127】
さらに、天然には存在しない合成プロモーターがまた酵母プロモーターとして機能する。例えば、一つの酵母プロモーターのUSA配列が、他の酵母プロモーターの転写活性領域と結合され得、合成ハイブリッドプロモーターを生成する。このようなハイブリッドプロモーターの例は、GAP転写活性化領域に連結したADH調節配列を含む(米国特許第4,876,197号および米国特許第4,880,734号)。ハイブリッドプロモーターの他の例は、GAPまたはPyKのような解糖系酵素遺伝子の転写活性化領域と組合せた、ADH2、GAL4、GAL10、またはPHO5遺伝子のいずれかの調節配列からなるプロモーターを含む(EPO公開第164556号)。さらに、酵母プロモーターは、酵母RNAポリメラーゼに結合し、そして転写を開始する能力を有する非酵母起源の天然に存在するプロモーターを含み得る。このようなプロモーターの例は、特に以下を含む
【0128】
【表5】
DNA分子は酵母細胞内で発現され得る。プロモーター配列は、DNA分子と直接連結され得、この場合、組換えタンパク質のN末端の最初のアミノ酸は、常に、ATG開始コドンによりコードされるメチオニンである。所望する場合、N末端のメチオニンを、インビトロで臭化シアンとともにインキュベートすることにより、このタンパク質から開裂し得る。
【0129】
融合タンパク質は、酵母発現系、ならびに哺乳動物、バキュロウイルス、および細菌発現系において代替物を提供する。典型的には、内因性酵母タンパク質または他の安定なタンパク質のN末端部分をコードしているDNA配列を、異種コーディング配列の5’末端に融合する。発現に際して、この構築物は、2つのアミノ酸配列の融合を提供し得る。例えば、酵母またはヒトスーパーオキシドジスムターゼ(SOD)遺伝子は、外来遺伝子の5’末端で結合され得、そして酵母で発現され得た。2つのアミノ酸配列の結合部のDNA配列は、開裂可能部位をコードし得るか、またはコードし得ない。例えば、EPO公開第196056号を参照のこと。他の例は、ユビキチン融合タンパク質である。このような融合タンパク質は、好ましくは、プロセッシング酵素(例えば、ユビキチン特異的プロセッシングプロテアーゼ)が、外来タンパク質からユビキチンを開裂するための部位を保持するユビキチン領域を用いて作成される。従って、この方法により、天然の外来タンパク質を単離し得る(例えば、PCT公開第WO88/024066号参照)。
【0130】
あるいは、外来タンパク質はまた、酵母で外来遺伝子の分泌を提供するリーダー配列フラグメントを含む融合タンパク質をコードするキメラDNA分子を作成することにより細胞から成長培地中に分泌され得る。好ましくは、インビボまたはインビトロのいずれかで開裂され得る、リーダーフラグメントと外来遺伝子との間にコードされるプロセッシング部位がある。リーダー配列フラグメントは、典型的には、細胞からタンパク質の分泌を指向する疎水性アミノ酸を含むシグナルペプチドをコードする。
【0131】
適切なシグナル配列をコードするDNAは、酵母インベルターゼ遺伝子(EPO公開第012873号;日本公開第62,096,086号)およびA因子遺伝子(米国特許第4,588,684号)のような分泌酵母タンパク質の遺伝子由来であり得る。あるいは、酵母における分泌もまた提供する非酵母起源のリーダー(例えば、インターフェロンリーダー)が存在する(EPO公開第060057号)。
【0132】
好ましいクラスの分泌リーダーは、酵母α因子遺伝子のフラグメントを使用しており、それは、「プレ」シグナル配列、および「プロ」領域の両方を含む。使用され得るα因子フラグメントのタイプは、完全長のプレ−プロα因子リーダー(約83アミノ酸残基)および短縮型α因子リーダー(典型的には、約25から約50アミノ酸残基)(米国特許第4,546,083号および米国特許第4,870,008号;EPO公開第324274号)を含む。分泌を提供するα因子リーダーフラグメントを使用する別のリーダーは、第1の酵母のプレ配列を用いるが、第2の酵母α因子由来のプロ領域を用いて作成されるハイブリッドα因子リーダーを含む。例えば、PCT公開第WO89/02463号参照のこと。
【0133】
典型的には、酵母により認識される転写終結配列は、翻訳終止コドンの3’に位置する調節領域であり、そしてそれ故プロモーターとともにコーディング配列に隣接する。これらの配列は、DNAによりコードされるポリペプチドに翻訳され得るmRNAの転写を指向する。転写ターミネーター配列および他の酵母認識終結配列の例は、解糖系酵素についてコードする終結配列である。
【0134】
典型的には、プロモーター、リーダー(所望する場合)、目的のコーディング配列、および転写終結配列を含む上記の構成成分は、発現構築物中に一緒に置かれる。発現構築物は、しばしば、レプリコン、例えば、酵母または細菌のような宿主で安定的に維持され得る染色体外要素(例えば、プラスミド)で維持される。レプリコンは、2つの複製系を有し、従って、例えば、発現するための酵母およびクローニングおよび増幅するための原核生物宿主で維持され得る。このような酵母−細菌シャトルベクターの例は、YEp24[Botsteinら、(1979)Gene 8:17−24];pCl/l[Brakeら、(1984)Proc.Natl.Acad.Sci USA 81:4642−4646];およびYRp17[Stinchcombら、(1982)J.Mol.Biol.158:157]を含む。さらに、レプリコンは、高コピー数または低コピー数のいずれかのプラスミドであり得る。高コピー数のプラスミドは、一般に約5から約200、および典型的には、約10から約150のコピー数の範囲を有する。高コピー数のプラスミドを含む宿主は、好ましくは、少なくとも約10、およびさらに好ましくは少なくとも約20を有する。高コピー数または低コピー数のベクターは、ベクターの影響および宿主での外来タンパク質に依存して選択され得る。例えば、Brakeら、同上を参照のこと。
【0135】
あるいは、発現構築物は、組み込みベクターによって酵母ゲノムに組み込まれ得る。組み込みベクターは、典型的には、ベクターを組み込み得る酵母染色体に相同な少なくとも1つの配列を含み、そして好ましくは、発現構築物に隣接する2つの相同な配列を含む。組み込みは、ベクターおよび酵母染色体における相同なDNAの間での組換えにより生じるようである[Orr−Weaverら、(1983)Methods in Enzymol.101:228−245]。組み込みベクターは、ベクターに包埋するための適切な相同配列を選択することにより酵母の特異的な遺伝子座に指向され得る。 Orr−Weaverら、同上を参照のこと。1またはそれ以上の発現構築物は、生産される組換えタンパク質のレベルができるだけ高くなるように組み込まれる[Rineら、(1983)Proc.Natl.Acad.Sci USA 80:6750]。ベクターに含まれる染色体配列は、完全なベクターの組み込みを生じるベクターの単一セグメントとして、または発現構築物のみからなる安定的な組み込みを生じ得る、染色体の近接したセグメントに相同で、かつベクターで発現構築物の両端に位置する2つのセグメントとしてのいずれかで存在し得る。
【0136】
典型的には、染色体外および組み込み発現構築物は、選択マーカーを含み得、形質転換された酵母株の選択を可能にする。選択マーカーは、酵母宿主で発現され得る生合成遺伝子、例えば、ADE2、HIS4、LEU2、TRP1、およびALG7、ならびにG418耐性遺伝子(ツニカマイシンおよびG418に対する耐性をそれぞれ酵母細胞に供与する)。さらに、適切な選択マーカーはまた、金属のような毒性化合物の存在下で成長する能力を有する酵母を提供し得る。例えば、CUP1の存在は、銅イオンの存在下で酵母を成長させ得る[Buttら、(1987)Microbiol.Rev.51:351]。
【0137】
あるいは、いくつかの上記化合物は、形質転換ベクターに一緒に置かれ得る。形質転換ベクターは、典型的には、上記のように、レプリコンで維持されるか、または組み込みベクターへと展開されるかのいずれかである選択マーカーを含む。
【0138】
発現および形質転換ベクター、染色体外レプリコンまたは組み込みベクターのいずれかは、多くの酵母に形質転換するために発達した。例えば、発現ベクターは、上記のように以下の酵母のために発達した:
【0139】
【表6】
酵母宿主に外因性DNAを導入する方法は、当該分野で周知であり、そして典型的には、スフェロプラストまたはアルカリカチオンで処理したそのままの酵母細胞のいずれかの形質転換を含む。形質転換手順は、通常形質転換されるべき酵母種とともに変化する。以下の例を参照のこと、
【0140】
【表7A】
【0141】
【表7B】
本発明において、選択マーカー、複製起点、および相同宿主細胞の配列が発現ベクターに含まれる。選択マーカーは、潜在的に発現ベクターを含む宿主細胞をスクリーニングするために使用し得る。このようなマーカーは、宿主細胞を、アンピシリン、クロラムフェニコール、エリスロマイシン、ネオマイシンおよびテトラサイクリンのような薬剤に対して耐性にするマーカーを含む。また、マーカーは、宿主細胞の成長に必要なヒスチジン、トリプトファンおよびロイシン経路における生合成遺伝子のような生合成遺伝子を含む。従って、leu(−)宿主細胞を、発現ベクターによる形質転換におけるレシピエントとして使用し、そして例えばロイシンが培地に存在しない場合、leu(+)遺伝子を伴うプラスミドを有する細胞のみが生存する。
【0142】
本発明において、複製起点は発現ベクターに取り込まれ、宿主細胞内で自己複製される。このような複製起点は、細胞内に適切なタンパク質の存在下で、発現ベクターが高いコピー数で再生産され得る複製起点、例えば酵母において有効な2μ配列および自己複製配列;およびCOS−7細胞において有効なウイルスT−抗原の複製起点を含む。
【0143】
本発明の目的のために、発現ベクターは、宿主細胞ゲノムに組み込まれ、または細胞内で自律性を保持し得る。宿主ゲノムへの組み込みのために、本明細書の発現ベクターは、宿主細胞ゲノム内の配列に相同のポリヌクレオチド配列を含み得る。この相同配列は、発現ベクターに連結される必要はない。例えば、発現ベクターは、付随していないジヒドロ葉酸還元酵素遺伝子を介してCHOゲノムに組み込み得る。酵母においては、相同配列は発現カセットに隣接することが好ましい。本発明に特に有用な相同酵母ゲノム配列は、PCT WO90/01800に開示された配列およびGenbank検索番号J01331に記載のHIS4遺伝子配列である。
【0144】
発現ベクターのプロモーター、ターミネーターおよび他の随意エレメントの選択も、選択した宿主細胞に依存し、このことは当業者に既知である。本発明は、選択した宿主細胞には依存しない。簡便性および所望のタンパク質発現のレベルから、至適宿主細胞が指定される。発現のための種々の宿主は当該分野において既知であり、そしてAmerican Type Culture Collection(ATCC)より入手可能である。
【0145】
例えば、KGFフラグメントまたはアナログを発現するための本明細書に適切な細菌宿主として、Campylobacter、Bacillus、Escherichia、Lactobacillus、Pseudomonas、StaphylococcusおよびStreptococcusが挙げられる。以下の属由来の酵母宿主を利用し得る:Candida、Hansenula、Kluyveromyces、Pichia、Saccharomyces、SchizosaccharomycesおよびYarrowia。本明細書において宿主として使用され得る不死化哺乳動物細胞として、CHO細胞、HeLa細胞、ベビーハムスター腎臓(「BHK」)細胞、サル腎臓細胞(「COS」)およびヒト肝細胞癌細胞、例えばHep G2が挙げられる。また多くの昆虫細胞
宿主もKGFフラグメントまたはアナログの発現に適切であって、PCT WO 89/046699;Carbonellら、J.Virol.56:153(1985);Wright Nature 321:718(1986);Smithら、Mol.Cell.Biol.3:2156(1983);および全般的にはFraserら、in vitro Cell.Dev.Biol.25:225(1989)に記載の通り、Aedes aegypti、Autographa californica、Bombyx mori、Drosophila melanogasterおよびSpodoptera frugiperdaが挙げられる。
【0146】
KGFフラグメントまたはアナログを含む発現ベクターは、宿主細胞に挿入される。発現ベクターを宿主細胞に挿入するためには、当該分野で既知の任意の形質転換技術を用い得る。例えば、細菌宿主の形質転換のために、典型的にまずCaCl2または他の試薬(2価カチオンおよびDMSOなど)のいずれかで細菌を処理し、そして外来性DNAを処理した細菌細胞に導入する。DNAは、エレクトロポレーションまたはウイルス感染によっても細菌細胞に導入し得る。本明細書で使用し得る細菌宿主に対する形質転換手順として、Massonら FEMS Microbiol.Lett.60:273(1989);Palvaら Proc.Natl.Acad.Sci.USA 79:5582(1982);欧州特許公開第036,259号および同第063,953号;PCT WO 84/04541、Bacillus)、Millerら Proc.Natl.Acad.Sci.85:856(1988);Wangら J.Bacteriol.172:949(1990)、Campylobacter)、Cohenら Proc.Natl.Acad.Sci.69:2110(1973);Dowerら Nucleic Acids Res.16:6127(1988);Genetic Engineering:Proceedings of the International
Symposium on Genetic Engineering(H.W.BoyerおよびS.Nicosia編)(1978);Mandelら J.Mol.Biol.53:159(1970)のKushner「ColE1由来プラスミドによるEscherichia coli形質転換の改良法(An improved method for transformation of Escherichia coli with ColE1−derived plasmids in;Taketo Biochim.Biophys.Acta 949:318(1988);Escherichia,Chassyら FEMS Microbiol.Lett.44:173(1987)Lactobacillus;Fiedlerら Anal.Biochem 170:38(1988)、Pseudomonas;Augustinら、 FEMS Microbiol.Lett.66:203(1990)、Staphylococcus,Baranyら J.Bacteriol.144:698(1980);Streptococcal Genetics(J.FerrettiおよびR.Curtiss III編)(1987)のHarlander「エレクトロポレーションによるStreptococcus lactisの形質転換(Transformation of Streptococcus lactis by electroporation)」;Perryら Infec.Immun.32:1295(1981);Powellら Appl.Environ.Microbiol.54:655(1988);Somkutiら Proc.4th Evr.Cong.Biotechnology 1:412(1987)、Streptococcus。
【0147】
酵母宿主に対する形質転換法は当該分野で周知であり、典型的にアルカリカチオンで処理したスフェロプラストまたはそのままの酵母細胞のいずれかの形質転換を含む。酵母宿主は、Methods in Enzymology、194巻、1991、「酵母の遺伝学および分子生物学へのガイド(Guide to Yeast Genetics and Molecular Biology)」に記載のエレクトロポレーションによ
っても形質転換し得る。本発明に対して、適用する形質転換手順は、形質転換しようとする酵母種により変化し、Kurtzら Mol.Cell.Biol.6:142(1986);Kunzeら J.Basic Microbiol.25:141(1985);Candida;Gleesonら J.Gen.Microbiol.132:3459(1986);Roggenkampら Mol.Gen.Genet.202:302(1986);Hansenula;Dasら J.Bacteriol.158:1165(1984);De Louvencourtら J.Bacteriol.154:1165(1983);Van den Bergら Bio/Technolgy 8:135(1990);Kluyveromyces;Creggら Mol.Cell.Biol.5:3376(1985);Kunzeら J.Basic Microbiol.25:141(1985);米国特許第4,837,148号および第4,929,555号;Pichia;Hinnenら Proc.Natl.Acad.Sci.USA 75;1929(1978);Itoら J.Bacteriol.153:163(1983)Saccharomyces;BeachおよびNurse
Nature 300:706(1981);Schizosaccharomyces;Davidowら Curr.Genet.10:39(1985);Gaillardinら Curr.Genet.10:49(1985);Yarrowia。
【0148】
異種ポリヌクレオチドを哺乳動物細胞に導入する方法は、当該分野において既知であり、そして例えばウイルス感染、デキストラン介在トランスフェクション、カルシウムリン酸沈殿、ポリブレン介在トランスフェクション、プロトプラスト融合、エレクトロポレーション、ポリヌクレオチドのリポソーム内のカプセル化(encapsulation)、およびDNAの核内へのダイレクトマイクロインジェクションが挙げられる。
【0149】
Smithら、Mol.Cell.Biol.3:2156(1983);およびLucklowおよびSummers、Virology 17:31(1989)に記載の通り、発現ベクターを形成するためにバキュロウイルス属のウイルス内に異種DNAを導入する方法、および昆虫宿主細胞の形質転換の方法も当該分野において既知である。例えば、相同二重交差組換えにより、KGFフラグメントDNAを多角体遺伝子に挿入し得る。Millerら、Bioessays 4:91(1989)に記載の通り、所望のバキュロウイルス遺伝子中に設計された制限酵素部位にも挿入し得る。KGFフラグメントのDNA配列を、発現ベクター内の多角体遺伝子と換えてクローン化した場合、多角体特異配列の両5’および3’に隣接し、そして多角体プロモーターの下流に位置する。
【0150】
本発明の実施態様において、新たに形成されたバキュロウイルス発現ベクターは、次いで感染組換えバキュロウイルス内にパッケージされ得る。バキュロウイルス発現系においては、およそ1%からおよそ5%の間の低い頻度で相同組換えが起こる。従って、通常、トランスフェクション後に生産された大多数のウイルスは、まだ野生型のウイルスである。しかし、組換えウイルスは、既知の方法によって同定し得る。例えば、天然のウイルスは、ウイルス感染の後期の間、感染細胞の核内において、かなり高いレベルで多角体タンパク質を生産する。蓄積した多角体タンパク質は、埋包されたウイルス粒子も含む閉塞体を形成する。これらの閉塞体は、15μmまでのサイズで、屈折性が高いことから明るい光沢の外観で、光学顕微鏡下で容易に可視化される。組換えウイルスが感染した細胞は閉塞体を欠く。組換えウイルスおよび野生型ウイルスはプレートティングすることによって識別され得、トランスフェクション上清またはその希釈液は標準的技術によって昆虫細胞の単層上に存在する。次に「Current Protocols in Microbiology」第2巻(Ausubelら編)の16.8(補遺10,1990)に記載の通り、閉塞体の存在(野生型ウイルスを示す)または非存在(組換えウイルスを示す)について、光学顕微鏡下でプラークをスクリーニングし得る。
【0151】
本明細書において形質転換された宿主細胞が所望のKGFフラグメントを発現しているかどうかを調べるために、当該分野で既知のイムノアッセイおよび活性アッセイを利用し得る。例えば、細胞膜からKGFフラグメントを分離することなく、形質転換された宿主細胞上で免疫蛍光アッセイを実施し得る。本アッセイにおいて、宿主細胞をまず、顕微鏡用スライドまたはマイクロタイターウエルのような固体支持体上に固定する。次に固定化した宿主細胞を抗KGF抗体に暴する。好ましくはアッセイの感度を上昇させるために、固定化した細胞を、抗KGF抗体に結合する標識された2次抗体に暴する。例えば、2次抗体は蛍光性のマーカーで標識し得る。KGFフラグメントを発現する宿主細胞を蛍光的に標識し、顕微鏡下で可視化し得る。
【実施例】
【0152】
(実施例1)
図1に示す通り、cDNAをpAcC13Pst/Not 1発現ベクターにクローニングすることによって、ヒトKGF cDNAをバキュロウイルス発現系に挿入した。Munemitsuら Mol.Cell.Biol.10:5977−5982(1990)に記載の通り、このプラスミドはpAcC12に由来した。
【0153】
Finchら Science 2:752−755(1989)に記載の通り、入手可能なKGFのN末端アミノ酸配列に基づく欠失した18kd KGFについてPCRオリゴヌクレオチドプライマーを設計した。Luckowら Virology 170:31−39(1989)およびQuilliamら Mol.Cell.Biol.10:290102908(1990)に記載の通り、pVL941伝達ベクターの誘導体であるpAcC13発現ベクターへのサブクローニングを容易に行うために、隣接する制限部位(PstIおよびNotI)をプライマーに組み込ませた。この発現ベクターは、pAcC13のPstI/NotIポリリンカー部位にKGFをコードするフラグメントを挿入することによって構築された。KGF cDNAはKGFのプロセシングされた成熟体をコードし、これは163個のアミノ酸残基からなり、14〜16番目の残基に潜在的なNグリコシル化部位を伴った。
【0154】
(コンディション培地の調製)
Spodoptera Frugiperda(Sf9昆虫細胞)に、KGF1−163に対するcDNAを含むバキュロウイルスのAutographa Californicaを感染させた。希釈後、細胞を血清補充物、抗生物質、または殺菌剤非存在下のExcell−400内で48〜72時間培養した。培養液を集め、そして10,000×gで30分間遠心分離して浮遊細胞およびその他の細胞破砕物を除いた。次にコンディション培地を、0.8μフィルター(Millipore)で濾過し、そしてこの培地の約5リットルを、3−kDaの分子量カットオフ値を有するfiltron cassette system(Omega membrane)を用いて200mlに濃縮した。
【0155】
濃縮後、トランスフェクトしたSf9細胞由来のコンディション培地を、10,000×gで20分間の遠心分離によって集め、そして連続ヘパリンセファロース(「HS」)アフィニティークロマトグラフィーによって精製した。本発明の一つの実施態様においては、約1リットルのSf9細胞コンディション培地を、3−kDaのカットオフ値を有するOmega membrane(Filtron)を用いて限外濾過し、200mlの限外濾過濃縮液を生産した。
【0156】
(HSACおよびMono S陽イオン交換クロマトグラフィー)
上清をpH7.2に調整した。30mlの試料を、10mM Tris−HCl pH7.3、 150mM NaClで平衡化したヘパリンセファロース樹脂に添加した。樹脂を、平衡化緩衝液で、吸光度がベースラインに戻るまで大量に洗浄し、そして次に0.
45Mから1Mおよび2Mの上昇NaCl濃度で段階的に溶出させた。細胞増殖アッセイに用いたフラクションからアリコートを分離し、そして最も高い生物活性を有する1M NaClのフラクションをプールした。プールしたフラクションを10mM Tris pH7.2 で5倍に希釈し(最終塩濃度0.2M NaCl)、そして同試料をSuper loopで、FPLCシステムに連結されたMono Sカラム(Pharmacia,Piscataway,NJ)に供した。直線勾配(10mM Tris pH7.3、0.2M NaCl〜10mM Tris pH7.3、1M NaCl)により溶出を行った。分画後、アリコートを生物活性について試験し、活性フラクションをプールした。
【0157】
濃縮液をpH7.3に調整し、そして約30mlのベット容量を含むHSカラムに添加した。カラムを、約2時間4℃で展開させた。カラムを、pH7.3で10mM Tris−HClおよび0.15M NaClを含む150mlの平衡化緩衝液で洗浄した。保持されたタンパク質を0.45M NaClおよび1M NaClで溶出した。溶出中のカラムの流速を、約90ml/時 に調整し、そして3mlのサイズのフラクションを集めた。
【0158】
HSアフィニティークロマトグラフィー工程の生物活性を有する1M NaClフラクションをプールし、そして10mM Tris pH7.3で5倍希釈し、そしてMono S HR 5/5カラム(Pharmacia)に添加した。保持されたタンパク質を、0.2M NaClから1M NaCl勾配を用いて溶出した。活性フラクションのバイオアッセイを、Balb−Mk細胞株を用いて測定した。Mono S 陽イオン交換クロマトグラフィーにより単離された生物活性タンパク質フラクションのSDS PAGE分析を実施した。フラクション37−39およびフラクション41−42をプールし、そして10μlのアリコートをSDSおよび10mM DTTに添加した。これらの試料を加熱して変性させ、そして12%のポリアクリルアミドゲル中で電気泳動を行い、続いて銀染色した。試料が還元状態中において展開された場合でも、または非還元状態中において展開された場合でも、同様の泳動パターンが観察された。25Kdはフラクション37−39に含まれるタンパク質の見かけの分子量で、そして18kdはフラクション41−42に含まれるタンパク質の見かけの分子量であった。
【0159】
HS生物活性フラクションのクロマトグラムは、フラクション31〜47にかけてのBalb/Mk生物活性のプロフィールを示すが、これから、2種類の分子種の存在が明らかであり、一つは0.55M NaClで溶出し、そして他方は0.6M NaClで溶出する。
【0160】
(KGF活性の測定)
得られたフラクション中のKGF活性の存在を、BALB/C−Mk細胞の成長を促進するフラクションの能力により評価した。この点に関して、任意のフラクションの10μlアリコートを1mlのリン酸緩衝生理食塩液(「PBS」)中0.2%ゼラチンに希釈し、そして10μlの希釈フラクションを、各々22mmウエルを含む12−ウエルクラスタープレートに5×103細胞/ウエルで接種されたBALB/C−Mk細胞の成長刺激活性について試験した。全てのKGF活性がカラム内に保持されることが見出され、そして1M NaClで溶出された。
【0161】
HSカラムから1M NaClで溶出されたKGFの生物活性フラクションをさらに、陽イオン交換FPLCカラムクロマトグラフィーによって精製した。これらのフラクションをプールし、10mM Tris、pH7.3で5倍に希釈し、そしてPharmacia製のMono S HR 5/5カラムに直接添加した。
【0162】
Mono S HR 5/5カラムに保持されたタンパク質を、0.2M NaClから1M NaClの勾配で溶出させた。溶出したフラクションの生物活性を、BALB/C−Mk細胞を用いて、上記の通りアッセイした。図3は、フラクション31〜47にわたる活性プロフィールを示す。
【0163】
マイトジェンアッセイにおいて、Bohlenら EMBO 4:1951−1956(1985)、およびGospodarowiczら J.Cell Physiol 127:121−136(1986)、およびBellostaら J.Cell Biol.121:705−713(1993)に記載の通り、ウエル当たり104個のABAE細胞および5×103個のACE細胞を、10%仔牛血清および抗生物質を補充した1mlのDMEM中に密度5×103細胞/ウエルで、12ウエルクラスタープレートに接種した。6時間のインキュベーション後、3重検体のウエルのセットをトリプシン処理し、そしてプレーティング効率を測定するために細胞をカウントした。次に適切な希釈の各試料の10μlアリコートを、0日目、2日目および4日目に3重検体としてシャーレ内のウエルに添加した。培養5日目後、プレートをトリプシン処理し、そしてコールターカウンター(Coulter Electronics,Hialeah,FL)により細胞密度を測定した。
【0164】
0日目および1日おきに、表示した濃度で、KGFまたは塩基性FGFを添加した。培養5日後に、細胞をトリプシン処理し、そしてコールターカウンターを用いて最終的な細胞密度を測定した。
【0165】
(電気泳動(NaDodSO4/PAGE))
Laemmliら Nature 227:680−685(1970)に記載の手順に従い、NaDodSO4によりポリアクリルアミドゲルを調製した。10mM DTT存在下で3分間試料を煮沸し、そして12%ポリアクリルアミドゲル中で電気泳動した。ゲルを固定し、そしてBioRadの試薬およびプロトコールを用い、Merrilら Science 211:1437−1438に考察されている通りに銀染色を行った。適切な分子量マーカーは、Biorad製である。
【0166】
(細胞増殖アッセイ)
カラムフラクションおよび精製試料のマイトジェン活性を、標的細胞としてBalb/Mk細胞を用いることによって測定した。ストック培養物を生育し、そしてGospodarowiczら J.Cell Physiol.142:325−333に記載の通り、10%FCS、50μg/mlゲンタマイシン、0.25μg/mlファンギゾン(fungizone)、および10ng/ml aFGFを補充した低カルシウム改変イーグル培地中に維持した。マイトジェンアッセイ用の細胞を、Gospodarowiczらに記載の通り、10%FCSを補充した1mlの低カルシウムMEM中に5×103〜1×104細胞/ウエルの密度で12ウエルクラスタープレートに接種した。培養5日後に、 Gospodarowiczらに記載の通り、試料の添加および最終細胞密度の測定を行った。
【0167】
最終精製物質のマイトジェン活性を、Gospodarowiczら Proc.Natl.Acad.Sci.73:4120−4124(1976)に記載の通り、成獣ウシ動脈内皮細胞(「ABAE」)および副腎皮質由来毛細血管内皮細胞(「ACE細胞」)上で試験した。ストック培養物を、10%CS、50μg/mlゲンタマイシンを補充したDMEM存在下で維持し、そして0.25μg/mlのファンギゾンを、週1回、1:10の分割比でゼラチン化組織培養シャーレ上を通過させた。
【0168】
当該分野で周知の標準的な方法論を用いて、以下の通り、KGFdcs1−23のNH2末端から1位〜20位の明白なアミノ酸配列を確定した:
S1 Y D M5 E G G D I R V R R L F X R T Q
本発明はまた、KGFdcs1−23をコードするDNAセグメント、およびFGFファミリーの他のメンバーに対してKGF163の非保存性NH2末端特性を欠くKGF由来のより短い他の形状を含む。
【0169】
(タンパク質マイクロシークエンシング)
KGF163およびKGFdcs1−23の2つの名目上100pmolの試料を、エドマン分解、および二フッ化ポリビニリデン(PVDF、Applied Biosystems Biospin)への遠心吸着後のAA分析により分析した。試料をApplied Biosystems 477A 気相タンパク質シークエネーターに添加した。標準的なソフトウエアおよびApplied Biosystems供給の化学試薬を用いて、20回のエドマン分解を行い、そして自動化オンラインHPLCカラム(モデル120A、Applied Biosystems)によりPTHアミノ酸の同定を行った。
【図面の簡単な説明】
【0170】
【図1】図1は、N末端で開始し、そして163アミノ酸残基およびアミノ酸残基14−16に単一のN結合グリコシル化シグナルを含む成熟完全長ヒトKGFポリペプチドのアミノ酸配列を示す。
【図2】図2は、163アミノ酸配列のKGFが挿入されているpAcC13発現ベクターを示す。
【図3】図3は、ヘパリンセファロース(「HS」)カラムで1M NaClを用いて溶出した後の培地で調整されたSF9細胞由来の本発明のKGFフラグメントのさらなる精製を示す。HSカラム由来の生活性画分をプールし、10mM Tris、pH7.3で5倍に希釈し、そしてMono S HR5/5陽イオン交換FPLCにかけた。
【図4】図4は、Balb/Mk細胞株の細胞上で、長形態のKGF(すなわち、rKGF163)および短形態のKGF(すなわちKGFdes1−23)対aFGFの生物学的活性を比較する。
【図5】図5は、大きな血管(A:ABAE細胞)または毛細血管(B:ACE細胞)由来の血管内皮細胞上で、長形態のKGF(すなわち、rKGF163)および短形態のKGF(すなわちKGFdes1−23)対bFGFの生物学的活性を比較する。
【特許請求の範囲】
【請求項1】
以下の式:
【化1】
の配列のアミノ酸24−163の連続するアミノ酸配列からなるポリペプチドと、薬学的に受容可能な賦形剤とを含む、組成物。
【請求項2】
請求項1に記載されたポリペプチドのN末端においてメチオニンを有するポリペプチドを含む、組成物。
【請求項3】
以下の式:
【化2】
の配列のアミノ酸24−163の連続するアミノ酸配列からなるポリペプチド、またはそのN末端にメチオニンを有する化1の配列のアミノ酸24−163の連続するアミノ酸配列からなるポリペプチドを用いる工程を包含する、上皮細胞の増殖を刺激するための医薬の製造方法。
【請求項4】
以下の式:
【化3】
の配列のアミノ酸24−163の連続するアミノ酸配列からなるポリペプチド、またはそのN末端にメチオニンを有する化1の配列のアミノ酸24−163の連続するアミノ酸配列からなるポリペプチドを用いる工程を包含する、創傷治癒のための医薬の製造方法。
【請求項1】
以下の式:
【化1】
の配列のアミノ酸24−163の連続するアミノ酸配列からなるポリペプチドと、薬学的に受容可能な賦形剤とを含む、組成物。
【請求項2】
請求項1に記載されたポリペプチドのN末端においてメチオニンを有するポリペプチドを含む、組成物。
【請求項3】
以下の式:
【化2】
の配列のアミノ酸24−163の連続するアミノ酸配列からなるポリペプチド、またはそのN末端にメチオニンを有する化1の配列のアミノ酸24−163の連続するアミノ酸配列からなるポリペプチドを用いる工程を包含する、上皮細胞の増殖を刺激するための医薬の製造方法。
【請求項4】
以下の式:
【化3】
の配列のアミノ酸24−163の連続するアミノ酸配列からなるポリペプチド、またはそのN末端にメチオニンを有する化1の配列のアミノ酸24−163の連続するアミノ酸配列からなるポリペプチドを用いる工程を包含する、創傷治癒のための医薬の製造方法。
【図1】


【図2】
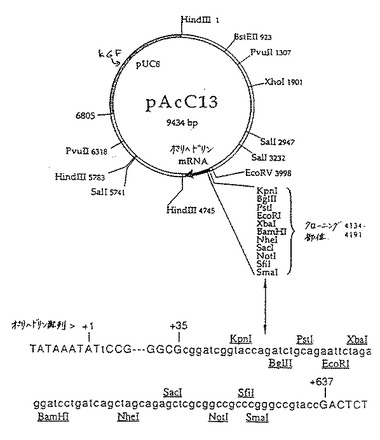
【図3】
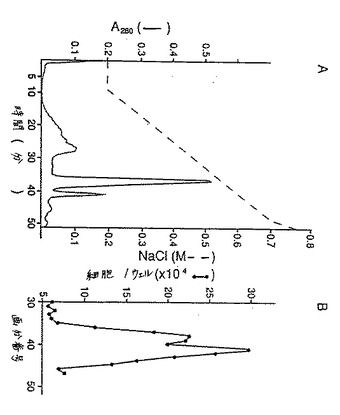
【図4】
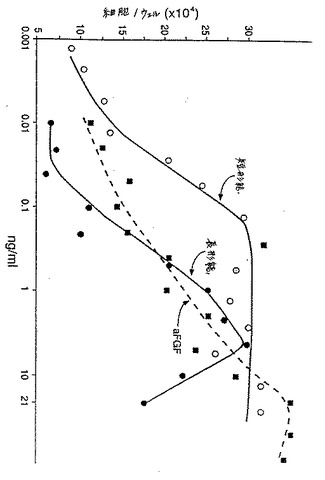
【図5】
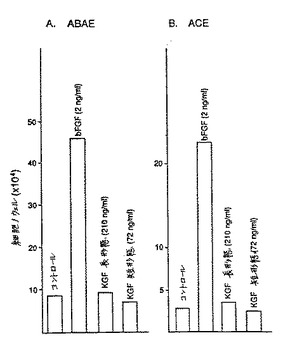
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2009−62396(P2009−62396A)
【公開日】平成21年3月26日(2009.3.26)
【国際特許分類】
【出願番号】特願2008−301630(P2008−301630)
【出願日】平成20年11月26日(2008.11.26)
【分割の表示】特願2005−333438(P2005−333438)の分割
【原出願日】平成6年4月28日(1994.4.28)
【出願人】(591076811)ノバルティス バクシンズ アンド ダイアグノスティックス,インコーポレーテッド (265)
【Fターム(参考)】
【公開日】平成21年3月26日(2009.3.26)
【国際特許分類】
【出願日】平成20年11月26日(2008.11.26)
【分割の表示】特願2005−333438(P2005−333438)の分割
【原出願日】平成6年4月28日(1994.4.28)
【出願人】(591076811)ノバルティス バクシンズ アンド ダイアグノスティックス,インコーポレーテッド (265)
【Fターム(参考)】
[ Back to top ]