
増殖連動型のコハク酸塩生成のための方法と生物
本発明は、酵素の活性が低下したときに、増殖連動型のコハク酸塩の生成と関連する酵素をコード化する1つ以上の遺伝子破壊で構成される非自然発生的な微生物を提供し、それにより1つ以上の遺伝子破壊は、非自然発生的な微生物に安定的な増殖連動型のコハク酸塩の生成を与える。また、コハク酸塩生成を微生物の増殖と強制的に連動させる代謝改変の組であって、

からなる遺伝子の組またはそのオーソログから選択される1つ以上の遺伝子破壊からなる代謝改変の組で構成される非自然発生的な微生物であって、安定的な増殖連動型のコハク酸塩の生成を示す微生物も提供される。
からなる遺伝子の組またはそのオーソログから選択される1つ以上の遺伝子破壊からなる代謝改変の組で構成される非自然発生的な微生物であって、安定的な増殖連動型のコハク酸塩の生成を示す微生物も提供される。
【発明の詳細な説明】
【背景技術】
【0001】
発明の背景
本発明は、概してコンピュータを利用した生物の設計に関し、特に増殖連動型のコハク酸塩の生成のための選択された遺伝子型を有する生物に関する。
【0002】
コハク酸塩は、食品、製薬、洗剤、および高分子工業における汎用化学物質の前駆体としての用途から、膨大な商業的利益のある化合物である。発酵により生成されたコハク酸塩は、1,4ブタンジオールおよび関連生成物、テトラヒドロフラン、γ−ブチロラクトン、n−メチルピロリジノン(NMP)および2−ピロリジノンを含む工業製品を年間2.7×108 kg以上供給することが潜在的に可能である(非特許文献1)。コハク酸の基本的な化学的性質は、石油化学的に生成されたマレイン酸/無水物の性質と類似しており、従ってその生成コストのみが、コハク酸が発展性のある数々の市場に急速に展開するのを阻んでいる。生物学的なコハク酸塩の生成は、糖発酵中に温室効果ガスCO2がコハク酸塩に固定されなければならないグリーンプロセスでもある。これらの理由から、1,4−二塩基酸(コハク酸塩、フマル酸塩、およびリンゴ酸塩)は、評価された300の候補の中から、エネルギー省により、バイオプロセスの開発の最優先事項として特定された12の化合物に含まれる。
【0003】
コハク酸塩などの中心的代謝化合物は、しばしば基礎代謝中に構成的に生成されるため、代謝工学のよい初期標的となる。先天的に炭水化物からコハク酸塩を生成できる生物の例としては、アナエロビオスピルリム・サクシニシプロデュセンス(Anaerobiospirillum succiniciproducens)(非特許文献2、非特許文献3、非特許文献4)、アクチノバチルス・スクシノゲネス(Actinobacillus succinogenes)(非特許文献5、非特許文献6)、および最近配列が決定されたウシ管腔内細菌のマンヘミア・サクシニシプロデュセンス(Mannheimia succiniciproducens)(非特許文献7、非特許文献8、非特許文献9)が含まれる。さらに報告のいくつかは、さまざまな代謝工学的戦略によって、より高いコハク酸塩の収率を有する大腸菌(Escherichia coli)株の構成を達成したと主張している。これらの取り組みは、コハク酸塩への炭素流束のチャネリングの増加、および補助因子NADHの可用性に焦点を当ててきた。例えば、PEPカルボキシラーゼ(ppc)の過剰発現(非特許文献10)、およびRhizobium etliピルビン酸塩カルボキシラーゼ(pyc)の発現(非特許文献11)は、TCA回路のコハク酸塩部分への流束を増加させることにより、大腸菌(E.coli)でのコハク酸塩の収率それぞれ0.30 g/gおよび0.17 g/gをもたらした。さらに、乳酸塩脱水素酵素(ldh)およびピルビン酸塩ギ酸塩リアーゼ(pfl)が欠損した大腸菌(E.coli)突然変異体(NZN111株)は、大腸菌(E.coli)(非特許文献12、および非特許文献13)、あるいはブタ回虫(Ascaris suum)(非特許文献14)のリンゴ酸酵素の過剰発現との組み合わせで、コハク酸塩収率の向上を達成した。のちにホスホトランスフェラーゼ系のptsG遺伝子にマップされた、NZN111におけるさらなる染色体偶発突然変異は(非特許文献15)、1 mol/molグルコース(0.66 g/g)の嫌気的なコハク酸塩の収率を有するAFP111株をもたらした(非特許文献16)。インゲン根粒菌(R.etli)ピルビン酸塩カルボキシラーゼの存在下または不在下でのNZN111およびAFP111株のさまざまな特性は、嫌気および二相条件(嫌気的生成が後続する好気性増殖)のもとで調査されてきた(非特許文献17)。非特許文献18では、約0.96 g/gの収率となった。その他の取り組みでは、好気的に0.91 mol/mol(0.60 g/g)を達成できる大腸菌(E.coli)のコハク酸塩生成菌株が得られた(Linら、Metab En
g、(2005年)。また近刊では、嫌気的に1.6 mol/mol(1.0 g/g)となっている(非特許文献19)。
【0004】
しかしながら、より高いコハク酸塩の収率を生み出す一部の細菌の菌株の開発を称している上記の取り組みおよび報告にもかかわらず、採用された方法には、商業目的での適用性を妨げるいくつかの欠点がある。以下にさらに詳しく記載されるように、上記の方法で生成される菌株は、未変化あるいは野生型の親菌株に有利にはたらく選択圧により、一般的に商業用の発酵プロセスでは不安定である。
【非特許文献1】Zeikusら、Appl Microbiol Biotechnol、51:545−552(1999年)
【非特許文献2】Samuelovら、Appl Environ Microbiol、65:2260−63(1999年)
【非特許文献3】Leeら、Appl Microbiol Biotechnol、54:23−27(2000年)
【非特許文献4】Leeら、Biotechnol Lett、25:111−14(2003年)
【非特許文献5】Guettlerら、Int J Syst Bacteriol、49:207−16(1999年)
【非特許文献6】Urbanceら、Appl Microbiol Biotechnol、65:664−70(2004年)
【非特許文献7】Leeら、Bioprocess Biosyst Eng、26:63−7(2003年)
【非特許文献8】Hongら、Nat Biotechnol、22:1275−81(2004年)
【非特許文献9】Leeら、Appl Microbiol Biotechnol、58:663−8(2002年)
【非特許文献10】Millardら、Appl Environ Microbiol、62:1808−10(1996年)
【非特許文献11】Gokarnら、Biotechnol Lett、20:795−798(1998年)
【非特許文献12】Stolsら、Appl Environ Microbiol、63:2695−701(1997年)
【非特許文献13】Hongら、Biotechnol Bioeng、74:89−95(2001年)
【非特許文献14】Stolsら、Appl Biochem Biotechnol、63−65:153−8(1997年)
【非特許文献15】Chatterjeeら、Appl Environ Microbiol、67:148−54(2001年)
【非特許文献16】Donnellyら、Appl Biochem Biotechnol、70−72:187−98(1998年)
【非特許文献17】Vemuriら、Appl Environ Microbiol、68:1715−27、18(2002年)
【非特許文献18】Vemuriら、J Ind Microbiol Biotechnol、28:325−32(2002年)
【非特許文献19】Sanchezら、Metab Eng、7:229−39(2005年)
【発明の開示】
【発明が解決しようとする課題】
【0005】
従って、所望の生成物の生合成を最適培養条件に強制的に結び付ける、商業的に有益な
特徴を有する微生物に対する必要性が存在する。本発明は、この必要性を満たし、かつ関連する利益をも提供する。
【課題を解決するための手段】
【0006】
(発明の要旨)
本発明は、酵素の活性が低下したときに、増殖連動型のコハク酸塩の生成と関連する酵素をコード化する1つ以上の遺伝子破壊で構成される非自然発生的な微生物を提供し、それにより1つ以上の遺伝子破壊は、非自然発生的な微生物に安定的な増殖連動型のコハク酸塩の生成を与える。また、コハク酸生成を微生物の増殖と強制的に連動させる代謝改変の組であって、(a)adhE、ldhA、(b)adhE、ldhA、ackA−pta、(c)pfl、ldhA、(d)pfl、ldhA、adhE、(e)ackA−pta、pykF、atpF、sdhA、(f)ackA−pta、pykF、ptsG、または(g)ackA−pta、pykF、ptsG、adhE、ldhAからなる遺伝子の組あるいはそのオーソログから選択される1つ以上の遺伝子破壊を備える代謝改変の組で構成される非自然発生的な生物であって、安定的な増殖連動型のコハク酸塩の生成を示す微生物も提供される。さらに、pykA、atpH、sdhB、またはdhaKLMから選択される少なくとも1つの遺伝子破壊をさらに含む、代謝改変(e)ackA−pta、pykF、atpF、sdhAをコード化する遺伝子を有する非自然発生的な微生物、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊をさらに含む、代謝改変(f)ackA−pta、pykF、ptsGをコード化する遺伝子を有する非自然発生的な微生物、あるいはpykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊をさらに含む、代謝改変(g)ackA−pta、pykF、ptsG、adhE、ldhAをコード化する遺伝子を有する非自然発生的な微生物も提供される。破壊は、完全な遺伝子破壊とすることができ、また非自然発生的な生物は、さまざまな原核または真核微生物を含むことができる。安定的な増殖連動型のコハク酸塩の生成を有する非自然発生的な微生物を生成する方法も提供される。この方法は、(a)指数増殖中にコハク酸塩の生成を要する代謝改変の組をコンピュータで同定するステップと、(b)コハク酸塩の生成を要する代謝改変の組を含有するように微生物を遺伝子改変するステップとで構成される。
【発明を実施するための最良の形態】
【0007】
(発明の詳細な説明)
本発明は、増殖連動型のコハク酸塩の生成を有する細胞や生物の設計と生成を対象としている。1つの実施例では、本発明は、コハク酸塩の最適生成のための代謝設計を同定する、大腸菌(Escherichia coli)代謝のコンピュータを利用した化学量論モデルを基に、最適化によるアプローチを利用する。二段階プログラミングフレームワークOptKnockは、反復アルゴリズムに適用され、遺伝子破壊の複数の組を予測するが、これは集合的に増殖連動型のコハク酸塩の生成をもたらす。ここに記載される結果は、戦略的に配置された遺伝子欠損または遺伝子の機能破壊の組み合わせが、大腸菌(Escherichia coli)およびその他の細胞または生物のコハク酸塩の生成能力を有意に向上させることを示す。コンピュータを利用した設計のための増殖連動型のコハク酸塩の生成は、設計された代謝遺伝子型を有する菌株の構成により確認される。これらの代謝的に変更された細胞または生物は、適応進化を受け、増殖連動型のコハク酸塩の生成をさらに増大することもできる。
【0008】
さらなる実施例では、本発明は、より促進されたコハク酸塩の生成特性を有する代謝的に変更された微生物菌株を開発するための、統合された計算と工学のプラットフォームを対象としている。プラットフォームの計算要素によって同定された菌株は、コハク酸塩の生成促進をもたらす予測された代謝改変を遺伝子操作することにより、実際の生成に適用される。コハク酸塩の生成は、微生物の最適増殖に共役され、発酵時のこの生成物の収率
を最適化する。またさらなる実施例では、増殖連動型のコハク酸塩の生成を示す菌株は、さらに適応進化を受け、生成物の生合成をさらに増大する。適応進化後の増殖連動型のコハク酸塩の生成のレベルは、システムの計算要素により推測することもできるが、この特定の実施例では、コハク酸塩のレベルの上昇は、進化のあとにのみ実現される。
【0009】
ここで使用される「非自然発生的な」という用語は、本発明の微生物に関連して使用される場合、微生物が言及されている種の野生型菌株では通常見られない遺伝子変更を少なくとも1つ有することを意味する。遺伝子変更は、遺伝子欠損または遺伝物質のその他なんらかの機能破壊としうる。
【0010】
ここで使用される「微生物」という用語は、微視的な大きさの原核または真核の細胞または生物を意味する。この用語は、全ての種の細菌および酵母または菌類などの真核生物を含むものとする。この用語は、生化学物質の生成用に培養できる任意の種の細胞培養物も含む。
【0011】
ここで使用される「増殖連動型の」という用語は、生化学物質の生成に関連して使用される場合、言及されている生化学物質の生合成が、微生物の増殖相中に強制的に生成される生成物であることを意味する。
【0012】
ここで使用される「代謝改変」という用語は、その自然発生状態から変更された生化学反応を指す。代謝改変は例えば、反応に参加している酵素をコード化する1つ以上の遺伝子の機能破壊による生化学反応活性の排除を含むことができる。典型的な代謝改変の組は表1に示されている。かかる代謝改変により特定される個々の反応、およびその対応する遺伝子相補体は、表2で大腸菌(E.coli)について例示されている。これらの反応で使用される反応物質および生成物は、表3に例示されている。
【0013】
ここで使用される「コハク酸塩」という用語は、クレブス回路およびさまざまな発酵プロセスにおいて形成されるジカルボン酸HOOCCH2CH2COOHを意味する。ここで使用される「コハク酸塩」という用語は、「コハク酸」という用語と同義である。化学的には、コハク酸塩は、コハク酸の塩またはエステルに該当する。従って、コハク酸塩およびコハク酸は、溶液のpHによって2つの形状のうちいずれかで存在することができる同じ化合物を指す。
【0014】
ここで使用される「遺伝子破壊」という用語、あるいはその文法的に対応する語は、コード化された遺伝子産物を不活性にさせる遺伝子変更を意味する。遺伝子変更は、例えば全遺伝子の欠損、転写または翻訳に必要な調整配列の欠損、切断遺伝子産物をもたらす遺伝子の一部の欠損とすることができ、またはコード化された遺伝子産物を不活性化するさまざまな突然変異戦略のいずれかによるものとしうる。特に有用な遺伝子破壊の1つの方法は完全な遺伝子欠損であるが、これは本発明の非自然発生的な微生物における遺伝的復帰の発生を低下あるいは排除するためである。
【0015】
ここで使用される「安定的な」という用語は、生化学的生成物の増殖連動型の生成に関連して使用される場合、増殖と生化学合成との間の連動を失わずに、5世代を超えて培養できる微生物に言及している。一般的に、安定的な増殖連動型の生化学的生成は10世代を超え、具体的には、安定的な増殖連動型の生化学的生成は約25世代を超え、さらに具体的には、安定的な増殖連動型の生化学的生成は50世代を超え、無限を含む。安定的な増殖連動型の生化学物質の生成は、例えば、代謝改変内のそれぞれの反応を触媒する酵素をコード化する遺伝子の欠損によって達成することができる。増殖連動型の生化学物質の生成の安定性は、複数の欠損によって強化することができ、それぞれの破壊された活性に対する複数の補償的な復帰の可能性を有意に減少させる。
【0016】
当業者は、ここに例示される代謝改変は、大腸菌(E.coli)遺伝子およびその対応する代謝反応に関連して記載されることを理解するであろう。しかし、多種多様な生物の完全なゲノム配列決定と、ゲノミクス分野での高いスキルとを前提にすると、当業者はここで提供される教示と指針を、原則としてその他全ての生物に容易に適用できるであろう。例えば、ここで例示される大腸菌(E.coli)代謝変調は、同じまたは類似の遺伝子破壊を他の種に組み込むことにより、他の種に容易に適用できる。かかる破壊は、例えば、全般的には種のホモログの遺伝子変更、また具体的にはオーソログ、パラログ、または非オーソロガス遺伝子置換体を含むことができる。
【0017】
オーソログは、直系の関係を持つ1つまたは複数の遺伝子であり、異なる生物において実質的には同様または同一の機能に関与する。例えば、マウスエポキシド加水分解酵素およびヒトエポキシド加水分解酵素は、エポキシドの加水分解の生物学的機能に対するオーソログであると考えられる。遺伝子は、例えばそれらが相同であること、あるいは共通祖先からの進化による関係を持つことを示すのに十分なだけの配列類似性を共有する場合、直系の関係を持つ。遺伝子は、一次配列類似性が同定不能である限りでは、共通祖先から進化したことを示すのに十分なだけの三次元構造を共有するが、必ずしも配列類似性を共有しない場合にもオーソログであると考えられる。オーソロガスである遺伝子は、約25%〜100%のアミノ酸配列同一性を有するタンパク質をコード化することができる。25%未満のアミノ酸類似性を共有するタンパク質をコード化する遺伝子は、その三次元構造も類似性を示している場合、直系で生じたと考えられる。組織プラスミノゲン活性化因子とエラスターゼを含む、セリンプロテアーゼ系の酵素は、共通祖先からの直系で生じたと考えられている。
【0018】
オーソログは、例えば進化を通じて、構造または全体的活性が分岐した遺伝子、またはそのコード化された遺伝子産物を含む。例えば、1つの種が2つの機能を示す遺伝子産物をコード化し、かかる機能が第2の種において異なる遺伝子に分かれている場合、この3つの遺伝子およびその対応する産物はオーソログであると考えられる。生化学的生成物の増殖連動型の生成については、当業者は、破壊される代謝活性を有するオーソロガス遺伝子が、非自然発生的な微生物の構成のために選定されることを理解するであろう。分離可能な活性を示すオーソログの例は、異なる活性が、2つ以上の種の間あるいは単一の種の中で異なる遺伝子産物に分かれているものがある。具体例としては、2種類のセリンプロテアーゼ活性であるエラスターゼタンパク質分解とプラスミノーゲンタンパク質分解の、プラスミノーゲン活性化因子およびエラスターゼとしての別個の分子への分離がある。第2の例としては、マイコプラズマ5’−3’エクソヌクレアーゼとショウジョウバエDNAポリメラーゼIII活性の分離がある。第1の種のDNAポリメラーゼは、第2の種のエクソヌクレアーゼまたはポリメラーゼの双方またはいずれかのオーソログであると考えられ、逆もまた同様である。
【0019】
対照的に、パラログは、例えば進化を後続する複製によって関連付けられるホモログであり、類似または共通であるが同一でない機能を有する。パラログは、例えば同種または異種に起源あるいは由来する。例えば、ミクロソームエポキシド加水分解酵素(エポキシド加水分解酵素I)と可溶性エポキシド加水分解酵素(エポキシド加水分解酵素II)は、共通祖先から共進化し、同種内で異なる反応を触媒し、異なる機能を有する異なる2つの酵素を表わすため、パラログであると考えられる。パラログは、お互いに著しい配列類似性を有する、同種を起源とするタンパク質であり、これらが相同である、あるいは共通祖先からの共進化による関係を持つことを示している。パラロガスなタンパク質ファミリーの群には、HipAホモログ、ルシフェラーゼ遺伝子、ペプチダーゼなどが含まれる。
【0020】
非オーソロガス遺伝子置換体は、異種における言及されている遺伝子機能を代理できる
、1つの種からの非オーソロガス遺伝子である。代理には例えば、異種における言及されている機能と比較して実質的には同様のまたは類似の機能を、起源種内で行うことができることが含まれる。一般的に、非オーソロガス遺伝子置換体は、言及されている機能をコード化する既知の遺伝子と構造的に関連するとして同定可能であるが、構造的にはそれほど関連していないが、機能的に類似している遺伝子とその対応する遺伝子産物であっても、ここで使用されるこの用語の意味に含まれる。機能的類似性は、例えば、非オーソロガス遺伝子の活性部位または結合領域に、代理されようとする機能をコード化する遺伝子と比較して、少なくともいくつかの構造的類似性を必要とする。従って非オーソロガス遺伝子は、例えばパラログまたは非関連遺伝子を含む。
【0021】
従って、増殖連動型の生化学物質の生成を有する本発明の非自然発生的な微生物を同定および構成する際、当業者は、ここで提供される教示と指針を特定の種に適用することにより、代謝改変の同定はオーソログの同定と破壊を含むべきであると理解するであろう。パラログおよび非オーソロガス遺伝子置換体またはそのいずれかが、類似あるいは実質的に類似する代謝反応を触媒する酵素をコード化する言及される微生物に存在する範囲で、当業者は、これらの進化的に関連する遺伝子を排除して、酵素活性におけるいかなる機能的冗長性も設計された代謝改変を妨げないよう確保することもできる。
【0022】
オーソログ、パラログ、および非オーソロガス遺伝子置換体は、当業者には周知の方法により判定できる。例えば、2つのポリペプチドの核酸またはアミノ酸配列を検査することで、比較される配列間の配列同一性および類似性が明らかになる。当業者は、かかる類似性に基づいて、その類似性が、タンパク質が共通祖先からの進化によって関連付けられることを示すのに十分なほど高いかどうか判定することができる。Align、BLAST、Clustal Wなど、当業者には周知のアルゴリズムは、粗配列類似性または同一性を比較および判定し、また重量あるいはスコアを指定できる配列間の差異の存在または有意性も判定する。かかるアルゴリズムは当技術分野でも知られており、ヌクレオチド配列類似性または同一性を判定するために、同様に適用することができる。近縁性を判定するための十分な類似性のパラメータは、統計的類似性、またはランダムポリペプチドで類似適合が見られる可能性を計算するための周知の方法、および判定された適合の有意性に基づいて計算される。2つ以上の配列のコンピュータ比較は、必要に応じて、当業者によって視覚的に最適化することもできる。関連する遺伝子産物またはタンパク質は、例えば25%〜100%の配列同一性など、高い類似性を有することが予想できる。関連していないタンパク質は、十分な大きさのデータベースが読み取られた場合、偶発すると予想されるのものと本質的に同様の同一性を有する(約5%)。5%〜24%の間の配列は、比較される配列が関連していると判断するのに十分な相同関係を示しても、示さなくてもよい。かかる適合の有意性を判定するためのさらなる統計的解析は、データセットのサイズを考慮して、これらの配列の関連性を判定するために実施することができる。
【0023】
BLASTアルゴリズムを用いて2つ以上の配列の近縁性を判定するための典型的なパラメータは、例えば以下に記載されるものとできる。つまり、BLASTPバージョン2.0.8(1999年1月5日)およびパラメータ(Matrix:0 BLOSUM62;gap open:11;gap extension:1;x_dropoff:50;expect:10.0;wordsize:3;filter:on)を用いてアミノ酸配列アラインメントを行うことができる。核酸配列アラインメントは、BLASTNバージョン2.0.6(1998年9月16日)およびパラメータ(Match:1;mismatch:−2;gap open:5;gap extension:2;x_dropoff:50;expect:10.0;wordsize:11;filter:off)を用いて行うことができる。当業者は、例えば比較の厳密性を上げるあるいは下げるために、上記のパラメータにどのような変更を加えることができるか理解し、2つ以上の配列の近縁性を判定するであろう。
【0024】
本発明は、安定的な増殖連動型のコハク酸塩の生成を有する非自然発生的な微生物を生成する方法を提供する。方法は、(a)指数増殖中にコハク酸塩の生成を要する代謝改変の組をコンピュータで同定するステップと、(b)コハク酸塩の生成を要する代謝改変の組を含有するように、微生物を遺伝子改変するステップとで構成される。
【0025】
バイオプロセスに対して考慮すべき重要な点は、バッチ式あるいは連続式の発酵方式のいずれを使用するかである。生成される生成物の量に影響するこの2方式間の1つの差異は、バッチ方式には、指数増殖相に加え、前処理、誘導期、定常期が存在することである。対照的に、連続式プロセスは、絶えず指数増殖の状態を維持し、正しく操作すれば、一度に何ヶ月もの間稼働することができる。増殖関連および混合増殖関連の生成物の形成では、連続式プロセスは、前処理、誘導期、定常期の排除により、ずっと高い生成性(希釈率×細胞質量)を提供する。例えば、以下の妥当な想定では、
【0026】
【数1】
【0027】
連続式プロセスにより向上した生成性は、8倍と推定されている(Shulerら、Prentice Hall,Inc.:ニュージャージー州アッパーサドルリバー、245−247)。
【0028】
生成性における圧倒的な優位性にもかかわらず、多くの理由から、連続式プロセスよりもバッチ式プロセスの方がかなり多く実施されている。まず、非増殖関連の生成物形成(ペニシリンなど)では、バッチシステムの生成性は、連続式プロセスの生成性を有意に超えるが、これは後者が非常に低い希釈率で稼働しなければならないためである。次に、生成菌株は一般的に、生化学物質またはタンパク質生成能力を向上させるために、その遺伝物質に変更を行なっている。これらの特異化された菌株はその親相補体よりも成長が遅い可能性が高いので、ケモスタット(連続モードで操作される発酵槽)などを使用する連続式プロセスは、最も急速に増殖する細胞に高い淘汰圧が課される。所望の過剰生成表現型をもたらす組み換えDNAを含む、あるいは点突然変異を有する組み換えDNAは、元の生成性が劣る親菌株への復帰突然変異を起こしやすい。単一の遺伝子欠損を有する菌株が、野生型の増殖表現型を復帰させる可能性が高い、補償的な突然変異を発現することもある。より成長の速い細胞は通常、より生成性の高い細胞と制限栄養物質に関して競合しないので、大幅に生成性を低下させる。一方バッチ式プロセスは、各サイクルの終了時に細胞を再利用することによらずに利用できる世代の数を制限し、従ってその野生型の表現型に復帰する生成菌株の確率を減少させる。最後に、連続式プロセスは、機器の不具合や異生物の混入などの技術的障害により、長期の操作がより困難である。またかかる不具合の結果は、連続式プロセスでは回分培養よりもはるかに重大である。
【0029】
特殊化学製品やタンパク質の少量の生成では、連続式プロセスの生成性向上は、菌株の安定性と信頼性に関連するリスクを上回ることはまれである。しかし、コハク酸塩などの大量の増殖関連生成物の場合は、連続式プロセスの生成性の向上は、バッチ式プロセスに比較して、著しい経済的利益をもたらす可能性がある。連続式バイオプロセス操作に関連する技術的障害は常に存在するが、菌株の安定性問題は、代謝経路を変更して、負の選択圧を減少または回避し、指数増殖相での対象生成物の生成を支持する代謝工学的戦略によって克服できる。
【0030】
増殖連動型の生成物の生成を支持する代謝変調を同定し、設計するための1つの計算方法は、OptKnock計算フレームワークである(Burgardら、Biotechnol Bioeng、84:647−57(2003年))。OptKnockは、対象生成物を過剰生成する遺伝的に安定した微生物をもたらす遺伝子欠損戦略を示唆する、代謝のモデル化とシミュレーションのプログラムである。具体的には、このフレームワークは、微生物の完全な代謝および生化学ネットワークまたはそのいずれかを検討して、所望の生化学物質を細胞増殖の強制的な副生成物にさせる遺伝子操作を示唆する。戦略的に配置された遺伝子欠損またはその他の機能的遺伝子破壊によって、生化学的生成を細胞増殖に連動させることにより、バイオリアクタに長時間入れた後、操作された菌株に課された増殖淘汰圧は、強制的な増殖連動型の生化学的生成の結果として、実績の向上をもたらす。最後に、遺伝子欠損が構成された場合、設計された菌株がその野生型の状態に復帰する可能性はごくわずかであるが、これはOptKnockによって選択された遺伝子がゲノムから完全に除外されるためである。
【0031】
つまり、OptKnockという用語は、ここでは細胞代謝をモデル化するための計算方法およびシステムを指して使用される。OptKnockプログラムは、流束均衡解析(FBA)モデルに特定の制約を組み込むモデルと方法のフレームワークに関する。これらの制約には、例えば定性的反応速度情報、定性的規制情報、およびDNAマイクロアレイ実験データ、またはそのいずれかが含まれる。OptKnockは、例えば、流束均衡モデルによって得られた流束境界を狭め、続いて遺伝子付加または欠損の存在下での代謝ネットワークの性能限界を探ることにより、さまざまな代謝問題の解も計算する。OptKnock計算フレームワークは、代謝ネットワークの性能限界の効果的なクエリを可能にするモデル形式の構築を可能にし、もたらされた混合整数線形計画問題を解決する方法を提供する。ここでOptKnockと称される代謝のモデル化とシミュレーションの方法は、例えば米国特許出願第10/043,440号(2002年1月10日出願)、および国際特許第PCT/US02/00660号(2002年1月10日出願)に記載されている。
【0032】
生成物の増殖連動型の生成を支持する代謝変調を同定および設計するための別の計算方法は、SimPheny(登録商標)と称される代謝のモデル化およびシミュレーションシステムである。この計算方法とシステムは、例えば米国特許出願第10/173,547号(2002年6月14日出願)、および国際特許出願第PCT/US03/18838号(2003年6月13日出願)に記載されている。
【0033】
SimPheny(登録商標)は、コンピュータでネットワークモデルを作り出し、生物系の化学反応を通して質量、エネルギー、または電荷の流束をシミュレートして、その生物系における化学反応のあらゆる機能性を含む解空間を規定し、それによりその生物系に許容される活性の範囲を判定するために使用できる計算システムである。このアプローチは、含まれる反応の既知の化学量論、および反応による最大流束に関連する反応熱力学および容量制約などの制約によって、解空間が規定されるため、制約に基づくモデル化と称される。これらの制約によって規定される空間は、生物系またはその生化学成分の表現型の能力および性質を判定するために問い合わせることができる。凸解析、線形計画法、および例えばSchillingら、J.Theor.Biol.203:229−248(2000年);Schillingら、Biotech.Bioeng.71:286−306(2000年)、およびSchillingら、Biotech.Prog.15:288−295(1999年)などに記載される極限経路(extreme pathways)の計算は、かかる表現型の能力を判定するために用いることができる。
【0034】
上述のように、本発明に適用可能な計算プログラムで使用される制約に基づく1つの方
法は、流束均衡解析である。定常状態条件における流束均衡に基づく流束均衡解析は、例えばVarmaおよびPalsson、Biotech.Bioeng.12:994−998(1994年)に記載されているように行うことができる。流束均衡法は、例えば、FellおよびSmall、J.Biochem.138:781−786(1986年)に記載されている脂肪細胞代謝(MajewskiおよびDomach、Biotech.Bioeng.35:732−738(1990年))に記載されているATP極大化条件下での大腸菌(E.coli)からの酢酸塩分泌、あるいはVanrolleghemら、Biotech.Prog.12:434−448(1996年)に記載されている酵母によるエタノール分泌のシステム特性をシミュレートまたは予測するために、反応ネットワークに適用されてきた。さらにこの方法を用いて、多様な単一炭素源における大腸菌(E.coli)の増殖、またEdwardsおよびPalsson、Proc.Natl.Acad.Sci.97:5528−5533(2000年);EdwardsおよびPalsson、J.Bio.Chem.274:17410−17416(1999年);Edwardsら、Nature Biotech.19:125−130(2001年)に記載されるようにH.インフルエンザ菌(H.influenzae)の代謝を予測またはシミュレートすることもできる。
【0035】
解空間が規定されると、さまざまな条件下での可能な解を判定するために解析することができる。生物系には適応性があり、異なる多くの方式で同じ結果に辿り着くことができるため、この計算法は生物学的現実に合致する。生物系は、全生命系が直面しなければならない基本的制約によって制限されてきた進化機構を通して設計されている。従って、制約に基づくモデル化戦略は、これらの一般的現実を包含する。さらに、制約の強化によりネットワークモデルにさらなる制限を持続的に課す能力は、解空間のサイズの減少につながり、それにより生理学的機能または表現型の推測精度を向上させる。
【0036】
ここで提供される教示および指針を前提に、当業者は、代謝のモデル化およびシミュレーションのためのさまざまな計算フレームワークを利用し、生化学的生成物を設計して増殖連動型の生成を実施することができるであろう。かかる代謝のモデル化およびシミュレーションの方法には、例えばSimPheny(登録商標)およびOptKnockとして上記に例示される計算システムが含まれる。本発明の説明を簡単にする目的で、方法と菌株は、モデル化とシミュレーションのためのOptKnock計算フレームワークに関連して記載される。当業者は、OptKnockを使用しての代謝変調の同定、設計、および実施を、当技術分野で周知のその他のかかる代謝のモデル化およびシミュレーションの計算フレームワークおよび方法にどう適用するかを理解するであろう。
【0037】
増殖を生化学的生成物の生成に強制的に連動させる細胞または生物の能力は、コンピュータによるモデルを使用して計算された典型的な代謝ネットワークの生化学的生成の限界に照らして説明することができる。これらの限界は、制限基質の摂取速度をその実験的に測定された値に修正し、それぞれの到達可能な増殖レベルで生化学的生成の最大および最小速度を計算することによって得られる。図1に示されるように、目的の生化学物質の生成は、一般的に、細胞内の資源に関してバイオマス形成と直接競合する。これらの状況もとで、強化された生化学的生成速度は必然的に最大増殖速度には及ばなくなる。OptKnockなど上記の代謝のモデル化およびシミュレーションプログラムにより示唆される遺伝子破壊は、図1に示されるように、野生型菌株の代謝挙動の変化を強いる許容解の境界を制限するよう設計されている。所定の菌株の実際の解の境界は、基質の摂取速度(1つまたは複数)が増加あるいは減少すると拡大または縮小するが、それぞれの実験値は、その計算された解の境界内に位置する。これらのような座標は、設計された菌株がその性能限界にどの程度近づいているかの正確な予測を可能とし、改善の余地がどの程度であるかも示される。
【0038】
OptKnock数学的フレームワークは、図1で説明されるように、増殖連動型の生化学的生成をもたらす遺伝子欠損を特定するために、ここで例示されている。この手順は、支配する物理化学的制約を連続的に課すことによって、細胞系が示すことができる可能な表現型の範囲を狭める、制約に基づく代謝のモデル化を基礎としている(Priceら、Nat Rev Microbiol、2:886−97(2004年))。上述のように、制約に基づくモデルおよびシミュレーションは当技術分野で周知であり、一般的に、ネットワークの化学量論を前提に、特定細胞目標の最適化を引き合いに出して、考えられる流束分布を示唆する。
【0039】
つまり、代謝物の組N={1,…,N}と代謝反応の組M={1,…,M}で構成される定常状態の代謝ネットワークの総反応流束として定量化された細胞目標の極大化は、数学的に以下のように表現される。
【0040】
【数2】
【0041】
式中、Sijは反応jにおける代謝物iの化学量論係数であり、vjは反応jの流束であり、v基質取り込みは制限基質(1つまたは複数)の想定あるいは測定された摂取速度(1つまたは複数)を表わし、vatp維持は非増殖関連のATP維持要求である。ベクトルvは内向きおよび外向き流束の双方を含む。この研究では、細胞目標は、しばしばバイオマス形成に必要とされる比率での生合成前駆体の放出となるよう想定される(Neidhardt、F.C.ら、第2版、1996年、ワシントンD.C.:ASM Press.2v.(xx、2822、lxxvi))。流束は一般的に、バイオマス形成が、生成されたバイオマス g/gDW時間または1/時間として表現されるように、1gDW時間(乾燥重量グラム×時間)当たりで報告される。
【0042】
遺伝子欠損のモデル化、従って反応の排除は、まず制約に基づくアプローチへのフレームワークに2値変数の組み込みを行う(Burgardら、Biotechnol Bioeng、74:364−375(2001年);Burgardら、Biotechnol Prog、17:791−797(2001年))。これらの2値変数、
【0043】
【数3】
【0044】
は、反応jが活性の場合は1の値を、不活性の場合は0の値を想定する。以下の制約、
【0045】
【数4】
【0046】
によって、反応流束vjが、変数yj=0の場合のみゼロに設定されることが確保される。
あるいは、yj=1のとき、vjは下限vjminと上限vjmaxとの間の任意の値を想定できる。ここでvjminおよびvjmaxは、上記のネットワーク制約の対象となるあらゆる反応流束をそれぞれ最小化、最大化することにより同定される(Mahadevanら、Metab Eng、5:264−76(2003年))。
【0047】
最適な遺伝子/反応破壊は、得られたネットワークの最適増殖解が関心化学物質を過剰生成するように、活性反応(yj=1)の組を選定する二段階最適化問題を解決することによって同定される。概略的に、この二段階最適化問題は図2に説明されている。数学的には、この二段階最適化問題は以下の二段階混合整数最適化問題として表現される。
【0048】
【数5】
【0049】
式中、v化学物質は、例えばコハク酸塩またはその他の生化学的生成物など所望の対象生成物の生成であり、Kは許容される遺伝子破壊の数である。K=0の設定は全ネットワー
クの最大バイオマス解を返し、K=1の設定は得られたネットワークがその最大バイオマス収率を前提として最大過剰生成を伴うように、単一遺伝子/反応破壊(yj=0)を同定する。最後の制約によって、得られたネットワークが最小バイオマス収率を満たすことが確保される。Burgardら、Biotechnol Bioeng、84:647−57(2003年)は、モデル形成および解法についてのより詳細な説明を提供している。何百もの2値変数を含む問題は、GAMS(Brookeら、GAMS Development Corporation(1998年))を通してアクセスできるCPLEX8.0(GAMS:The Solver Manuals。2003年:GAMS
Development Corporation)のモデル化環境をIBM RS6000−270ワークステーション上で用いて、数分から数時間程度で解決できる。OptKnockフレームワークは、生化学物質過剰生成のための有望な遺伝子欠損戦略を同定することが既に可能で(Burgardら、Biotechnol Bioeng、84:647−57(2003年);Pharkyaら、Biotechnol Bioeng、84:887−899(2003年))、代謝および規制モデル化のフレームワークのさらなる改善を必然的に網羅する体系的なフレームワークを確立する。
【0050】
前述の二段階OptKnock問題のいかなる解も、1組の破壊される代謝反応を提供する。その組内の各反応の排除、あるいは代謝改変により、生物の増殖相において強制的な生成物としてコハク酸塩をもたらすことができる。反応が既知であるため、二段階OptKnock問題の解は、反応の組内の各反応を触媒する1つ以上の酵素をコード化する1つまたは複数の関連遺伝子も提供する。反応の組および各反応に参加する酵素をコード化する対応する遺伝子の同定は、通常は自動プロセスであり、反応と、酵素とコード化遺伝子との間の関係を有する反応データベースとの相互関係によって達成される。
【0051】
同定されると、増殖連動型のコハク酸塩の生成を達成するために破壊される反応の組は、対象細胞または生物において、その組内の各代謝反応をコード化する少なくとも1つの遺伝子の機能破壊によって実施される。前述のように、反応の組の機能破壊を達成するための1つの特に有用な手段は、コード化遺伝子それぞれの欠損によるものである。しかしいくつかの例では、例えば突然変異、プロモータや調整因子のcis結合部位などの調整領域の欠損、任意の数の位置でのコード化配列の切断など、その他の遺伝子異常によって反応を破壊することが有益となり得る。遺伝子の組の完全な欠損に満たないこととなるこれら後者の異常は、例えば、コハク酸塩共役の迅速な評価が望まれる場合、または遺伝子の復帰が起こる可能性が低い場合に有用となる。
【0052】
増殖連動型のコハク酸塩またはその他の生化学生成物の生成をもたらし得るさらなる反応の組の破壊または代謝改変をもたらす、上記の二段階OptKnock問題のさらなる生成的な解を同定するために、整数切捨てと称される最適化方法が実施できる。この方法は、各反復に整数切捨てと称されるさらなる制約を組み込み、上記に例示されるOptKnock問題を反復して解決することにより進行する。整数切捨て制約は、解法が、生成物生合成を増殖に強制的に共役する、以前の任意の反復において同定されたのと全く同じ反応の組を選定するのを効果的に防止する。例えば、以前に同定された増殖連動型の代謝改変が、破壊される反応1、2、および3を特定する場合、y1+y2+y331という制約は同じ反応が後の解で同時に検討されるのを防止する。整数切捨て方法は、当技術分野で周知であり、例えば、Burgardら、Biotechnol Prog、17:791−797(2001年)に記載されている。代謝のモデル化およびシミュレーションのためのOptKnock計算フレームワークとの組み合わせで、その使用に関連してここに記載されている全ての方法と同様に、反復計算解析における冗長性を減少させる整数切捨て方法も、例えばSimPhenyを含む当技術分野で周知の他の計算フレームワークとともに適用することができる。
【0053】
上記の形式の制約は、以前に同定された組を含む、より大きい反応の組の同定を防ぐ。例えば、さらなる反復で上記の整数切捨て最適化方法を使用すると、破壊される反応1、2、および3を特定した4反応の組は、これらの反応が以前に同定されているため、同定することが防がれる。生成物の増殖連動型の生成につながる全ての可能な反応の組を確実に同定するために、整数切捨て方法の修正が行われた。
【0054】
つまり、修正された整数切捨て法は、反復「ゼロ」で始まり、これは野生型ネットワークの最適増殖で目的の生化学物質の最大生成を計算する。この計算は、K=0でのOptKnock解に相当する。次に単一の遺伝子破壊が検討され、2つのパラメータの組、objstoreiterおよびystoreiter,jが導入されて、各反復iterで目標機能(vchemical)および反応オン−オフ情報(yj)がそれぞれ保存される。次に、各反復で、OptKnock公式に以下の制約が引き続き付与される。
【0055】
【数6】
【0056】
上記の方程式では、εとMはそれぞれ小さい数および大きい数である。一般的には約0.01に設定でき、Mは約1000に設定できる。しかし、これらの数値よりも小さい数や大きい数も使用できる。Mは、制約が以前に同定された遺伝子破壊戦略のみを拘束することができることを確実にし、一方εは、以前に同定された戦略への遺伝子破壊の追加が、最適増殖での生化学的生成において少なくともεの増加をもたらさなければならないことを確実にする。このアプローチは単一の欠損戦略が野生型菌株を改良するのに失敗したときは、常に2つの欠損へと移行する。2つの欠損戦略が野生型菌株を改良しない場合は、次に3つの欠損が検討され、以下同様である。最適増殖での所望の生化学的生成として表わされる最終結果は、少なくとも1つの遺伝子破壊の分だけお互いに異なる別個の欠損戦略の順位付けられたリストである。この最適化手順、および破壊された場合に生化学的生成物の増殖連動型の生成をもたらす多種多様な反応の組の同定は、下記の例にさらに詳しく例示される。例では、増殖連動型のコハク酸塩の生成がさらに例示される。しかし、ここで提供される教示および指針を前提に、当業者は、ここで例示される方法および代謝工学設計が、細胞または微生物増殖の任意の生化学的生成物への強制的共役に、同等に適用可能であることを理解するであろう。
【0057】
上記に例示され下記の例でさらに説明される方法を使用して、本発明の方法は、対象生化学的生成物の生成を、同定された遺伝子変更を有するように遺伝子操作された細胞または生物の増殖に強制的に共役することを可能にする。この関連で、コハク酸塩の生成を微生物の増殖に強制的に共役する代謝変調が同定されている。同定された代謝変調によって構成された微生物菌株は、指数増殖相において高いレベルのコハク酸塩を生成する。これらの菌株は、連続式発酵プロセスでの商業用のコハク酸塩の生成において、前述の負の選択圧を受けることなく有利に使用できる。
【0058】
従って、本発明の方法は、OptKnockまたはSimPhenyから選択されるコンピュータを利用した方法によって同定される代謝改変の組を提供する。この代謝改変の組は、例えば遺伝子欠損による破壊などの1つ以上の代謝反応の機能破壊を含むことができる。代謝改変は、表1に挙げられる代謝改変の組から選択できる。
【0059】
安定的な増殖連動型のコハク酸塩の生成を有する非自然発生的な微生物を生成する方法も提供される。この方法は、(a)指数増殖中にコハク酸塩の生成を要する代謝改変の組をコンピュータで同定するステップと、(b)コハク酸塩の生成を要する代謝改変の組を含有するように微生物を遺伝子改変し、遺伝子改変された微生物を培養するステップとで構成される。培養は、コハク酸塩の生成を要する条件下で、遺伝子改変された微生物を適応進化させるステップを含むことができる。本発明の方法は、細菌、酵母、および菌、またその他さまざまな細胞と微生物に適用可能である。細菌は、例えば、大腸菌(E.coli)、アナエロビオスピルリム・サクシニシプロデュセンス(A.succiniciproducens)、アクチノバチルス・スクシノゲネス(A.succinogenes)、マンヘミア・サクシニシプロデュセンス(M.succiniciproducens)、およびインゲン根粒菌(R.etli)を含むことができる。
【0060】
本発明の方法によって生成された微生物がさらに提供される。さらに本発明は、増殖連動型のコハク酸塩の生成に関連する酵素をコード化する1つ以上の遺伝子破壊を備え、安定的な増殖連動型のコハク酸塩の生成を示す非自然発生的な微生物を提供する。本発明の非自然発生的な微生物は、コハク酸塩生成を微生物の増殖と強制的に連動させる酵素をコード化する遺伝子において、遺伝子破壊が酵素の活性を低下させたときに発生する1つ以上の遺伝子破壊を含み、それにより1つ以上の遺伝子破壊は、非自然発生的な微生物に安定的な増殖連動型のコハク酸塩の生成を与える。
【0061】
非自然発生的な微生物は、表1に挙げられる代謝改変に含まれる1つ以上の遺伝子破壊を有することができる。1つ以上の遺伝子破壊は、欠損であることができる。本発明の非自然発生的な微生物は、表1に挙げられる代謝改変を有する微生物の群から選択できる。本発明の非自然発生的な微生物は、細菌、酵母、菌類、または発酵プロセスに適用可能なその他さまざまな任意の微生物を含む。細菌の例には、大腸菌(E.coli)、アナエロビオスピルリム・サクシニシプロデュセンス(A.succiniciproducens)、アクチノバチルス・スクシノゲネス(A.succinogenes)、マンヘミア・サクシニシプロデュセンス(M.succiniciproducens)、インゲン根粒菌(R.etli)、枯草菌(Bacillus subtilis)、コリネバクテリウム・グルタミクム(Corynebacterium glutamicum)、グルコノバクテル・オキシダンス(Gluconobacter oxydans)、ザイモモナス・モビリス(Zymomonas mobilis)、ラクトコッカス・ラクティス(Lactococcus lactis)、ラクトバチルス・プランタルム(Lactobacillus plantarum)、ストレプトマイセス・コエリコロル(Streptomyces coelicolor)、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、シュードモナス・フルオレッセンス(Pseudomonas fluorescens)、およびシュードモナス・プチダ(Pseudomonas putida)から選択される種が含まれる。酵母の例には、サッカロマイセス・ケレウィシエ(Saccharomyces
cerevisiae)、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)、クリベロマイセスラクティス(Kluyveromyces lactis)、クリベロマイセス・マルキシアヌス(Kluyveromyces marxianus)、アスペルギルス・テレウス(Aspergillus terreus)、クロコウジカビ(Aspergillus niger)、およびピキア・パストリス(Pichia pastoris)から選択される種が含まれる。
【0062】
増殖連動型のコハク酸塩の生成を有する微生物は、ここで大腸菌(E.coli)の遺伝的背景に関連して例示される。しかし395の微生物ゲノムと、多種の酵母、菌類、植物、および哺乳類ゲノムを含む550種を超える完全なゲノム配列が利用可能であり、(
中でも半数以上はNCBIなどの公共データベースで入手可能である)、例えばオーソログ、パラログ、および非オーソロガス遺伝子置換体を含む代替種の1つ以上の遺伝子のホモログ、および生物間での遺伝子変更の置き換えの同定は日常的で、当技術分野で周知である。従って、大腸菌(E.coli)など特定の生物に関連してここで記載されている増殖連動型のコハク酸塩の生成を可能にする代謝変調は、原核および真核生物などを含む他の微生物にも容易に適用できる。ここで提供される教示および指針を前提に、当業者は1つの生物において例示される代謝変調が他の生物にも同等に適用できることを理解するであろう。
【0063】
例えばコハク酸塩の生成は、大腸菌(E.coli)において、ここでADHErと称される反応を触媒する酵素をコード化している1つ以上の遺伝子、およびLDH_Dと称される反応を触媒する酵素をコード化している1つ以上の遺伝子の欠損または機能除去により、指数増殖と連動させることができる。表2に示されるように、adhE反応を触媒する酵素をコード化する大腸菌(E.coli)遺伝子は、adhEまたはb1241である。また、LDH_D反応を触媒する酵素をコード化する2つの大腸菌(E.coli)遺伝子も表2に示されている。これら2つのLDH_D関連遺伝子は、b2133およびb1380である。b1380の一般名はldhAである。b2133遺伝子はldhAのオーソログである。増殖連動型のコハク酸塩の生成を示す代謝的に操作された大腸菌(E.coli)を生成するために、ADHErおよびLDH_D反応それぞれを触媒する少なくとも1つの酵素をコード化する遺伝子は、機能的に破壊されていないければならない。かかる破壊は、例えばb1241遺伝子(adhE)、およびb1380遺伝子(ldhA)、そのオーソログb2133、またはb1241とb2133の両方のいずれかを削除することによって行うことができる。大腸菌(E.coli)以外の細胞または生物における増殖連動型のコハク酸塩の生成では、関心種におけるADHErおよびLDH_Dに相当する反応をコード化する遺伝子は、機能的に破壊することができる。類似代謝経路を有する生物の場合、かかる破壊は、例えばb1241と相同の種、およびb2133またはb1380遺伝子のいずれかを削除することによって達成できる。前述のように、かかる相同体はオーソログおよび非オーソロガス遺伝子置換体またはそのいずれかを含むことができる。関心種に代替代謝経路が存在する場合などいくつかの場合には、機能破壊は例えば、言及される反応に代わる類似するが非同一の代謝反応を触媒するパラログの欠損によって達成できる。異なる生物間の代謝ネットワークの間には一部の差異があるため、当業者は、異なる生物間で破壊される実際の遺伝子は異なってよいことを理解するであろう。しかしここで提供される教示と指針を前提に、当業者は、本発明の方法が、生物間の同種の代謝変調を同定し、コハク酸塩の生合成の増殖への共役を促進する関心種の生物を構成するために、全ての微生物に適用できることも理解するであろう。
【0064】
本発明は、本文書において、一般に代謝反応、反応物質、またはその生成物に関連して、あるいは特に前述の代謝反応、反応物質、または生成物に関連する1つ以上の遺伝子に関連して説明される。本文書において特に明記しない限り、当業者は反応への言及は、その反応の反応物質および生成物への言及ともなることを理解するであろう。同様に、本文書において特に明記しない限り、反応物質または生成物への言及は、その反応にも言及し、これらの代謝要素のいずれかに対するその言及は、遺伝子または言及される反応、反応物質、または生成物の触媒となる酵素をコード化する1つまたは複数の遺伝子にも言及する。同様に、周知の代謝生化学、酵素学、およびゲノミクスの分野を前提に、ここでの遺伝子への言及は、対応するコード化された酵素とそれが触媒する反応、およびその反応の反応物質と生成物への言及ともなる。前述されさらに後述されるように、例となる反応、反応の学名、反応物質、生成物、補助因子、および増殖連動型のコハク酸塩の生成に関与する反応を触媒する酵素をコード化する遺伝子は、表1、2、および3に記載されている。
【0065】
本発明は、増殖連動型のコハク酸塩の生成を有する微生物を提供する。コハク酸塩の生成は、細胞の代謝経路を遺伝的に変更することによって、微生物の指数増殖相と強制的に関連付けられる。遺伝子変更は、コハク酸塩を増殖相における強制的な生成物とする。指数増殖における高いレベルのコハク酸塩の生合成につながる代謝変調または変換の組は、表1に例示されている。組内の各変更は、機能的に破壊されるべき必要な代謝反応に対応している。各組内の全ての反応の機能破壊は、増殖相中に、操作された菌株によるコハク酸塩の強制的な生成をもたらす。言及される表1の変更に対応する反応、および大腸菌(E.coli)においてそれらを潜在的にコード化する1つまたは複数の遺伝子は、表2に記載されている。表3は、表2の反応で言及される反応物質、補助因子、および生成物の完全な生化学名を提供している。
【0066】
例えば、表1に例示される各菌株では、増殖共役するコハク酸塩の生成のために発生させることができる代謝変調は、各行に示されている。これらの変更は、1〜6、あるいはそれ以上の反応の機能破壊を含む。具体的には、非自然発生的な代謝の遺伝子型を有する187の菌株が表1に例示されている。これらそれぞれの非自然発生的な変更は、適切な培養条件下で、微生物の指数増殖相中に、野生型菌株よりも向上したレベルのコハク酸塩の生成をもたらす。適切な培養条件には、例えば特定の炭素源または反応物質の可用性や、適応進化など、下記の例でさらに例示されるものが含まれる。
【0067】
ここで提供される教示と指針を前提に、当業者は、酵素反応を破壊するために、反応に関与する1つ以上の酵素の触媒活性を破壊することが必要であることを理解するであろう。破壊は、例えばコード化遺伝子の欠損、またはコード化遺伝子配列のちの1つ以上への遺伝子変更の組み込みを含むさまざまな手段によって行うことができる。破壊の対象となるコード化遺伝子は、触媒活性に関与する酵素をコード化する遺伝子のうちの1つ、いくつか、あるいは全てとできる。例えば、単一の酵素が対象となる触媒活性に関与する場合、破壊は、コード化遺伝子産物の触媒活性を減少または破壊する遺伝子変更によって行うことができる。同様に、単一の酵素が、異種結合を含む多重結合の場合、破壊は、コード化遺伝子産物の1つまたは全てのサブユニットの機能を減少または破壊する遺伝子変更によって行うことができる。活性の破壊は、活性複合体を形成するための1つ以上のサブユニットの結合活性の損失、または多量体の触媒サブユニットの破壊、あるいはその両方によって達成できる。多量体タンパク質の会合と活性のその他の機能を対象として、本発明の代謝反応を破壊することもできる。かかるその他の機能は、当業者には周知である。さらに、単一のポリペプチドまたは多量体の機能のいくつかまたは全ては、本発明の反応または代謝改変に関与する1つ以上の酵素の触媒活性を減少させるか、または無効にするために、本発明に従って破壊することができる。同様に、本発明の反応または代謝改変に関与するいくつかまたは全ては、対象とされる反応が破壊されるとすれば、破壊することができる。
【0068】
ここで提供される教示と指針を前提に、当業者は、共通遺伝子によって、また類似あるいは実質的に同様の活性を示すその遺伝子の1つ以上のオーソログによってコード化される反応を減少または排除することによって、酵素反応を破壊できることも理解するであろう。共通遺伝子および全てのオーソログの両方を減少させることで、対象とされる反応の完全な停止をもたらすことができる。しかし、共通遺伝子または1つ以上のオーソログのいずれかの破壊は、増殖のコハク酸塩の生合成への共役を推進するのに十分なだけの、対象とされる反応の触媒活性の減少をもたらすことができる。多様な代謝改変の対象となる触媒活性をコード化する共通遺伝子、およびそのオーソログの双方がここに例示されている。当業者は、対象とされる代謝反応の酵素をコード化する遺伝子のいくつかまたは全ての破壊は、本発明の方法において実施でき、また増殖連動型のコハク酸塩の生成を達成するために本発明の非自然発生的な微生物に組み込むことができることを理解するであろう。
【0069】
従って本発明は、コハク酸塩生成を前記微生物の増殖と強制的に連動させる代謝改変の組を有する非自然発生的な微生物をさらに提供する。この代謝改変の組は、
【0070】
【化3】
【0071】
のいずれかで構成される反応の組から選択される各反応を触媒する酵素をコード化する1つ以上の遺伝子破壊を含み、前記微生物は安定的な増殖連動型のコハク酸塩の生成を示す。特定の反応を触媒する役割を果たす酵素をコード化する遺伝子の一般名は、括弧内に記されている。代謝改変(e)PTAr、PYK、ATPS4r、SUCD1i、(f)PTAr、PYK、GLCpts、または(g)PTAr、PYK、GLCpts、ADHEr、LDH_Dを有する非自然発生的な微生物は、反応DHAPT(dha)を触媒する酵素をコード化する少なくとも1つの遺伝子破壊をさらに含むことができる。pykAおよびpykFは、潜在的にPYK反応を実行することができる個々の酵素をコード化する遺伝子であることに留意されたい。従って、pykAおよびpykFの少なくとも1つ、あるいは両方を除去して、PYKがコハク酸塩の生成が細胞増殖に連動しなくなることを阻止する必要がある。あるいは、反応PFL、ATPS4r、SUCD1i、およびDHAPTは、複数の遺伝子にコード化されるタンパク質複合体によって実行される。pfl、atp、sdh、またはdha遺伝子クラスタから、1つまたは遺伝子の組み合わせをそれぞれ削除することは、従って、ATPS4r、SUCD1i、またはDHAPT反応を破壊するのに十分である。
【0072】
つまり、上記に例示される遺伝子、およびそれらの遺伝子の多量体内のそれらの同種サブユニット、それらのオーソログ、およびそれらの遺伝子産物に触媒される反応との関連に関して、ADHrは1つの遺伝子b1241(adhE)にコード化される酵素によって触媒される。LDH_Dは、オーソログb2133を有する1つの遺伝子b1380(ldhA)の産物によってコード化される。PFL活性は、4つの遺伝子b0902、0903、b3952、およびb3951(集合的にpflとして表わす)にコード化される酵素サブユニットを必要とする。b3114は、b0903のオーソログである。PTArは、オーソログb2458を有する1つの遺伝子b2297(ackA−pta)の産物にコード化される。PYKは、2つの異なるオーソロガス遺伝子b1854(pykA)およびb1676(pykF)の産物によってコード化される。これら双方は、大腸菌(E.coli)において活性であることが示されている。SUCD1i活性は、4つの遺伝子b0721、b0722、b0723、およびb0724(集合的にsdhとして表わす)にコード化される酵素サブユニットを必要とする。ATPS4rは、6つの遺伝子b3731〜b3739(集合的にatpとして表わす)にコード化されるマルチサブユニット酵素によって触媒される。DHAPT活性は、5つの遺伝子b1198、b1199、b1200、b2415、およびb2416(集合的にdhaとして表わす)にコード化される酵素サブユニットを必要とする。GLCpts活性は、9つの遺伝子b2
415、b2416、b2417、b1817、b1818、b1819、b1101、b0679、およびb1621(集合的にptsGとして表わす)にコード化される酵素サブユニットを必要とする。反応ATPS4r、SUCD1i、PFL、およびDHAPTは、複数の遺伝子にコード化されるタンパク質複合体によって実行されるので、atp、sdh、pfl、またはdha遺伝子クラスタから1つまたは遺伝子の組み合わせをそれぞれ削除することは、従って、反応を破壊するのに十分である。その他の事例では、文献の情報に基づき、大腸菌(E.coli)において一次反応活性を担う遺伝子が選定された。
【0073】
従って本発明は、コハク酸塩生成を微生物の増殖と強制的に連動させる代謝改変の組を有する非自然発生的な微生物も提供するが、この代謝改変の組は、(a)adhE、ldhA;(b)adhE、ldhA、ackA−pta;(c)pfl、ldhA;(d)pfl、ldhA、adhE;(e)ackA−pta、pykF、atpF、sdhA;(f)ackA−pta、pykF、ptsG、または(g)ackA−pta、pykF、ptsG、adhE、ldhA、のいずれかを含む遺伝子の組またはそのオーソログから選択される1つ以上の遺伝子破壊で構成され、この微生物は安定的な増殖連動型のコハク酸塩の生成を示す。さらに、pykA、atpH、sdhB、またはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成され、代謝改変(e)ackA−pta、pykF、atpF、sdhAをコード化する遺伝子を有する非自然発生的な微生物、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成される代謝改変(f)ackA−pta、pykF、ptsGをコード化する遺伝子を有する非自然発生的な微生物、あるいはpykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成される代謝改変(g)ackA−pta、pykF、ptsG、adhE、ldhAをコード化する遺伝子を有する非自然発生的な微生物も提供される。
【0074】
本発明の非自然発生的な微生物は、増殖連動型のコハク酸塩の生成に使用できる。本発明の増殖連動型のコハク酸塩の生成株を用いて、商業的な量を含み基本的に任意の量を合成することができる。本発明の微生物はコハク酸塩の増殖に強制的に連動させるため、連続式または近連続式の増殖プロセスが、生合成によるコハク酸塩の生成に特に有用である。かかる連続式や近連続式の増殖プロセスは、上述のとおりであり、また下記の例で示されている。連続式や近連続式の微生物増殖プロセスは、当技術分野で周知でもある。つまり、連続式や近連続式の増殖プロセスは、指数増殖または対数増殖において微生物を維持することを伴う。手順には、EvolugatorTMエボリューションマシン(Evolugate LLC、フロリダ州ゲーンズビル)、発酵槽などの装置の使用が含まれる。さらに、微好気的条件下での振盪フラスコ発酵および増殖も利用できる。ここで提供される教示と指針を前提に、当業者は、増殖連動型のコハク酸塩の生成を行う微生物は、当技術分野で周知である多種多様なプロセスや装置を使用して、多種多様な条件下で、多種多様な設定で使用できることを理解するであろう。
【0075】
一般的に、連続式および/または近連続式のコハク酸塩生成は、本発明の非自然発生的な増殖連動型のコハク酸塩の生成を行う生物を十分な栄養物と培地で培養して、指数増殖相において増殖を維持またはほぼ維持することを含む。かかる条件下での連続培養は、例えば1、2、3、4、5、6、または7日、あるいはそれ以上を含むことができる。さらに連続培養は、1、2、3、4、または5週、あるいはそれ以上の週から数ヶ月までを含むことができる。連続や近連続培養条件は、これら例示される期間の間の全ての時間間隔も含み得ることを理解すべきである。
【0076】
コハク酸塩は、上記に例示されている連続や近連続培養の期間中の任意の時点で採取または単離できる。下記の例で示されるように、微生物が連続や近連続の増殖相に長く維持
されるほど、比例的に大きい量のコハク酸塩が生成できる。
【0077】
従って本発明は、微生物の増殖に共役されたコハク酸塩の生成方法を提供する。この方法は、(a)指数増殖相のもと、十分な量の栄養物と培地の中で、
【0078】
【化4】
【0079】
のいずれかからなる遺伝子の組またはそのオーソログから選択される1つ以上の遺伝子破壊で構成されるコハク酸塩生成を微生物の増殖と強制的に連動させる代謝改変の組で構成される非自然発生的な微生物を培養するステップであり、微生物は安定的な増殖連動型のコハク酸塩の生成を示すステップと、(b)非自然発生的な微生物から生成されたコハク酸塩を単離するステップとで構成される。代謝改変(5)ackA−pta、pykF、atpF、sdhAをコード化する遺伝子は、pykA、atpH、sdhBまたはdhaKLMから選択される少なくとも1つの遺伝子破壊をさらに備えることができる。代謝改変(6)ackA−pta、pykF、ptsGをコード化する遺伝子は、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊をさらに備えることができる。代謝改変(7)ackA−pta、pykF、ptsG、adhE、ldhAをコード化する遺伝子は、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊をさらに備えることができる。
【0080】
本発明のさまざまな実施例の作用に実質的に影響しない修正も、ここに提供される本発明の定義の範囲に含まれることは当然である。従って、以下の例は本発明を説明することを目的とし、制限することを目的としていない。
【実施例】
【0081】
実施例I
増殖連動型のコハク酸塩の生成を有する微生物
本実施例では、前述の方法によって同定した代謝工学的戦略について述べる。全体では、百以上の戦略が同定されている。増殖連動型のコハク酸塩の生成を引き起こす同定された代謝変調の詳細な一覧は、以下の表1〜3(本実施例の後)に見ることができ、OptKnockにより同定された反応欠損の組み合わせ(表1)、表1において言及される各反応と一致する化学量論および遺伝子(表2)、および表2の略語と一致する代謝物名の一覧(表3)を記述する。特にここで述べる方法を実証するために有益な設計は、(1)直感的設計(低リスク)、(2)非直感的好気性設計(中リスク)、(3)非直感的嫌気性設計(より高リスク)の3項目に分類される。戦略によって、10 mmol/gDW/hrの基底グルコース摂取速度に対する、好気性または嫌気性条件のいずれかを検討する。各設計に対する解の境界は、実現可能な増殖速度ごとに、別々にコハク酸塩生成を極大化および極小化することで、各設計に対する解の境界が与えられる。解の境界の右端部は、最大バイオマス収率と同義の「最適増殖」解に一致する。システムの直線性のため、グルコース収率が任意の基礎とは異なる場合、その結果は比例的に変わる。増殖連動型の
コハク酸塩の生成を持つと同定された菌株の作成および培養の手順も、さらに以下で述べる。
【0082】
直感的設計
OptKnock技術により同定された4つの直感的菌株設計に対する生化学的生成を、図3で述べる。境界は豊富な二酸化炭素の存在下での嫌気条件を想定する。これらの菌株(すなわち、pfl、ldh、adhEの名が付けられた菌株以外の全て)のうち3つは、その他で作成されているため、それらの菌株の評価に関連するリスクは最小とみなした。しかし菌株を特異的に作成するその3つ全てに、増殖共役コハク酸塩の生成は見られず、コハク酸塩の安定的な増殖共役の生成も見られなかった。さらに、菌株を特異的に作成するその3つ全てが、本明細書で述べるような、さらに増殖連動型のコハク酸塩生成を増大するような適応進化にはさらされていなかった。したがって本発明のadhE、ldh;adhE、ldh、pta、およびpfl、ldhの菌株は、以前に作成された菌株とは別個である。
【0083】
示唆した設計の簡単な説明を次に述べる。示唆した各突然変異体の最大増殖シミュレーションに一致する交換流束を表4に示す。
【0084】
adhE、ldhA(青)−この特定の突然変異体は、SanchezらのMetab
Eng、7:229−39(2005年)およびSanchezらのBiotechnol Prog、21:358−65(2005年)にて言及され、SBS110MGと呼ばれている。異種性のpycは、その最終生成菌株内で発現した。著者はpycなしでコントロールプラスミドを持つ二重突然変異体に対しわずか0.2 mol/molのコハク酸塩収率を報告した。Pycプラスミドがあると、収率は1.3 mol/molにまで増加した。0.9 mol/molの収率は進化後と予想される。
【0085】
adhE、ldhA、pta(赤)−これらの欠損は、SanchezらのMetab
Eng、7:229−39(2005年)に記載の最も嫌気性の強い生成菌株に存在する。実際の生成菌株もpycプラスミドを内部に持ち、別の遺伝子破壊iclRというグリオシキル酸短絡回路を抑圧する調節遺伝子を含む。シミュレーションで、グリオシキル酸短絡回路の重要性は最小であることが明らかとなる。pycの過剰発現が、おそらく1.6 mol/molという高い収率が発表された理由であろう。コンピュータを利用した解析に基づくと、トリプル欠損変異体の収率は適応進化後には0.9 mol/molと予測され、過去にSanchezらのMetab Eng、7:229−39(2005年)で観察された1.6 mol/molから著しく減少する。
【0086】
pfl、ldhA(灰色)−pfl、ldhA菌株はDonnellyら、Appl Biochem Biotechnol,70−72:187−98(1998年)および最初のOptKnock論文であるBurgardら、Biotechnol Bioeng,84:647−57(2003年)にて言及された。NZN111と呼ばれる菌株は、グルコースを発酵させることができなかったが、最終的にはグルコースの発酵を許すptsG内で自然突然変異が起こった。Hongら、Biotechnol Bioeng、 74:89−95(2001年)のHongおよびLeeは、リンゴ酸酵素をNZN111内で過剰発現し、30〜40%のコハク酸塩収率を達成した。終点でのコハク酸塩生成は小さいはずであるため、NZN111またはその他全てのpfl、ldhでの進化実験は特に有益である。
【0087】
pfl、ldhA、adhE(緑)−この突然変異体は、最初のOptKnock論文のBurgardら、Biotechnol Bioeng、 84:647−57(2003年)にて示唆された。この菌株は評価の終点で主要生成物としてコハク酸塩および
酢酸塩を伴い、〜1.2 mol/molのグルコース収率があるはずである。この菌株は進化の終点で最高のコハク酸塩収率を持つはずであるため、興味深い。ギ酸塩の生成は等量を減少させる原因で、それゆえコハク酸塩形成に有利と広くみなされているため、pfl欠損は半直感的である。ギ酸塩脱水素酵素はシミュレーションでは嫌気的に機能しないため、化学量論モデルではこの点を議論する。SBS110MG(ldhA、adhE突然変異体)を含むpycからfdhF(ギ酸塩脱水素酵素)を除去すると、コハク酸塩収率は1.2〜0.9 mol/molに減少した(Sanchezら、Biotechnol Prog,21:358−65(2005年))。
表4:嫌気条件および基底グルコース摂取率10 mmol/(gDW・hr)を想定した最適増殖用交換流束。正の値は生成を示し、負の値は消費を示す。基底グルコース摂取率(すなわち、10)で入力値を割ることで、全化学物質のモル収率を得ることができる。
【0088】
【表4】
【0089】
非直感的好気性設計
この菌株設計には、好気性条件下での増殖連動型のコハク酸塩の生成のために6つの欠損(すなわち、ackA−pta、pykA、pykF、atpFH、sdhAB、dhaKLM)を必要とする。この欠損によって不可能となる反応には、PTAr、PYK、ATPS4r、SUCD1iおよびDHAPTが含まれる。菌株設計に対する生化学的生成の制限を図4に示し、最適増殖シミュレーション結果を図5で提供する。欠損によって理論上の最大コハク酸塩収率における減少が引き起こされることに留意する。にもかかわらず、この設計によって好気性の生化学的生成に対するatpFHの欠損のさらなる特徴付けが許容されるため、この設計は特に有益である。
【0090】
dhaKLM欠損の背後にある論理的根拠には、シミュレーションの場合を含め、F6PA(f6p≪dha+g3p)およびDHAPT(dha+pep(R)dhap+pyr)の反応によって大きな流束を獲得することである。これらの反応によって、PEPをピルビン酸塩に変換する手段が用意されるが、コハク酸塩形成には所望され得ない。PEPはかなりの高エネルギー化合物であるため、この経路はエネルギー性の観点から現実的であるように思われる。加えて、両反応の原因である酵素が特徴付けられている(Schurmannら、J Biol Chem,276:11055−61(2001年)、
Gutknechtら、Embo J,20:2480−6(2001年))。これらの酵素はおそらく完全な解糖流束を持ち得ないであろうが、最も保守的な方法は、これらの流束がdhaKLMを除去することで機能しなくなることを保証することである。実際には、この欠損は必要としてもよく、不必要としてもよい。シミュレーションでも、ptsGはdhaKLMに代わるものとして取り除くことができることが明らかになったが、グルコース摂取のためのへキソキナーゼへの完全な依存により、ptsGおよびatpFH両方の欠損を内部に持つ菌株は、増殖が困難になることもある。
表5:基底グルコース摂取率10 mmol/(gDW・hr)を想定した最適増殖シミュレーション用交換流束。正の値は生成を示し、負の値は消費を示す。基底グルコース摂取率(すなわち、10)で入力値を割ることで、全化学物質のモル収率を得ることができる。
【0091】
【表5】
【0092】
非直感的嫌気性設計
この菌株は、嫌気性条件下での増殖連動型コハク酸塩形成のため、5つの欠損(すなわち、ackA−pta、pykA、pykF、ptsG、dhaKLM)を必要とする。欠損によって不可能とされた反応にはPTAr、PYK、GLCpts、およびDHAPTを含む。また、DHAPT欠損(すなわち、dhaKLM)は、実際には任意とできる。欠損組は結果として、ピルビン酸塩の非定型の原因への強制依存のような(例えば、エントナー−ドウドロフ経路、セリンデアミナーゼ、リンゴ酸塩 酵素など)著しい代謝変更を引き起こすと予想される。ptsG、pykA、pykFの突然変異体は、グルコース最小培地で増殖されるとき、生存不可能となりうるため、この設計では幾つかの弱点が示される(Ponceら、J Bacteriol,177:5719−22 (1995年))。シミュレーションはこの発見と矛盾し、グルコースに関する好気性増殖では高
いバイオマス収率が可能であることを示す。この結果は、pykA、pykF、ptsGの突然変異体に対し予測される増殖への障害が起こった場合、欠損に適応をする菌株を強化するため進化戦略が適用できることを示す。
【0093】
嫌気的には、グルコース摂取率が10 mmol/gDW/hrと想定した場合、この菌株が標準の7.6 mmol/gDW/hrの非増殖関連のATP維持必要量に合致する点では不十分であることが、シミュレーションで明らかになる。図5では計算解の境界上のATP維持必要量を緩和する影響を検討する。ここでは、このペンタ−突然変異体が嫌気的に増殖できる場合(例えば、細胞に維持エネルギー必要量を減らすよう強制する増殖選択圧力を適用する)、最適増殖では極度に高いコハク酸塩収率が伴わなくてはならないということが示される。したがって、この設計にはそのリスクを凌ぐ巨大な潜在的上昇傾向がある。加えて、無益サイクルの導入を利用し、図5で描写した解の境界の最も左方の上部で表示した理論上のコハク酸塩収率へ近づけるよう、この菌株の性能を促進することができる。エネルギー維持必要量および嫌気条件なしを想定する最大増殖シミュレーションの交換流束を、表6に示す。
表6:嫌気条件および基底グルコース摂取率10 mmol/(gDW・hr)および非増殖関連維持必要量ゼロを想定した最適嫌気性増殖シミュレーション用交換流束。正の値は生成を示し、負の値は消費を示す。基底グルコース摂取率(すなわち、10)で入力値を割ることで、全化学物質のモル収率を得ることができる。
【0094】
【表6】
【0095】
新しく作成された菌株が好気性環境下で生存可能だが、嫌気条件下では十分には生存可能ではないある特定の例では、通気率の減少にかかわる進化戦略が適用できる。コハク酸
塩生成限界に関し酸素摂取率が徐々に減少する影響を、図6に描写する。通気性の減少によって、理論上の最大コハク酸塩収率(上方左)にますます近い最適増殖点が導かれることに留意する。図7で示されるように、細胞増殖に対しより緊密に共役するコハク酸塩形成は、adhEおよびldhのさらなる欠損で達成することができる。これらの遺伝子はそれぞれADHErおよびLDH_D反応を触媒する。また好気性増殖相(黒線)、次に酸素制限生成相(色付き線)に関する二相発酵戦略が適用できることも、嫌気および好気条件下で、adhE、dhaKLM、ldhを持つ、または持たない菌株の生存率は、さらに示している。
【0096】
増殖連動型のコハク酸塩菌株の生成およびバリデーション
本発明の方法のコンピュータ予測を確認するために、菌株を形成、進化、および試験する。1週間から6週間、一般的には約3週間の増殖条件づけ相を、より複雑な設計に割り当てる。時間がより短い期間には、それに対応してより簡単な設計を条件づけることができる。
【0097】
菌株形成:大腸菌K−12 MG1655は欠損を導入する1つの基準野生型菌株として使用する。例遺伝子破壊を、例えば一つ一つ幾つかの欠損が蓄積されたままの受容菌株へ組み込む。形成された突然変異体が元の野生型に戻る可能性を避けるために、欠損方法論で除去を目的とした遺伝子を完全に取り除く。1つの研究では、4つの予測した非直感的設計のうち2つが形成され、増殖連動型のコハク酸塩の生成を検定する。ここでは、多くても3つの欠損(1)pfl、ldhAおよび(2)pfl、ldhA、adhEを必要とする2つの菌株を形成することができる。別の研究では、4つの共通遺伝子破壊ackA−pta、pykA、pykF、dhaKLMを形成する。さらに別の研究では、atpFHおよびsdhABの追加欠損を行い、その一方で代替の研究ではptsGの追加欠損を行う。非直感的設計の確証試験のため、多くて7つの欠損を取り込む。全体では、すべての望ましい非直感的菌株および研究のために、多くて10個の欠損を行うことができる。
【0098】
適応進化:適応進化手順は、定常期が達成される前の新鮮な培地への回分培養の連続継代による遷延性の指数増殖における細胞の維持に関係する。細胞は希釈し新鮮な培地に移される(すなわち、2 g/L炭素現にM9最小培地)前に、短時間で中定常増殖(A600=0.5)に達することができる。この過程を繰り返し、約500世代までにする。培養試料を取り出し、液体窒素で凍らせ、最適培養密度を進化の過程の間中毎日記録する。表7に各進化に必要な条件を要約する。過去にDonnellyら、Appl Biochem Biotechnol 70−72:187−98(1998年)、Vemuriら、Appl Environ Microbiol 68:1715−27(2002年)で目撃された進化的パターンにおける相違のため、進化は3通り(したがって、全18進化)で行い、それにより潜在的に他より上位の生成の品質を持つ1つの菌株となる可能性がある。適応進化段階には2ヶ月またはそれ以上を要する。また例えば菌株設計によっては適応進化段階が2ヶ月未満のこともある。
表7:各進化に対する望ましい条件。進化は3通りで実行する。
【0099】
【表7】
【0100】
バリデーション:増殖速度(GR)、基質摂取率(SUR)および酸素摂取率(OUR)(好気性の場合)は、進化の過程の間中、10日ごとに標本抽出を行う。前培養を一晩増殖させ、指数増殖中に測定を行う新鮮な回分培養用の種菌として使用する。分光光度計(A600およびA420)を使用して最適密度を測定し、GR、HPLCで時間をかけ炭素源をモニターしSURを判定し、およびポーラログラフの溶解した酸素プローブを使用して呼吸計内で溶解した酸素欠損を測定してOURを判定する。コハク酸塩および副生成物生成をHPLCまたは酵素試験で数量化する。測定は10日の間隔で3通りの進化菌株ごとに行う。試験は進化と同時に進めることができる。
【0101】
菌株安定性の評価:OptKnock法の背後にある増殖共役の生化学的生成概念の結果として、細胞遺伝学的に安定した過剰産生株が発生する。最高のコハク酸塩生成特性および少なくともモデル予測との満足な一致を有する菌株を、1ヶ月ケモスタット内で増殖し、その長期安定性を確認する。M9最小培地を使用し、1.3−Lベンチトップ発酵槽(New Brunswick Scientific、ニュージャージー州、エジソン)内で、約600 mLの稼働範囲で、ケモスタット培養を行う。炭素源濃度を調節し、A600=1.0に一致するするよう細胞密度を達成し、希釈率を最大培養率の約80%に設定し洗い流しの可能性を避ける。無菌空気を好気性増殖に使用し、攪拌速度を使い溶解酸素を飽和の>95%に維持する。OptKnockが嫌気性増殖環境を提案する場合は、窒素/CO2の混合物と共に反応器を連続的に散布し、DOレベルを欠損レベルより確実に下に保つ。CO2も炭酸水素ナトリウム(NaHCO3)の形で培地内に用意する。上記に記載したように各週ごとに代謝挙動を検定する。
【0102】
結論
本明細書に記載するのは、有用な遺伝子欠損目標を産生するためのOptKnock方法論の応用である。コハク酸塩生成および大腸菌(E.Coli)増殖間の結合を確立するため、多様な欠損戦略が列挙されてきた。本方法論は幅広い種類の細胞および大腸菌以外の微生物に適用が可能であり、代謝モデル化およびOptKnock以外のシミュレーションシステムに利用することもできる。2つの相対的には直感的な好気性設計(以下3および4)、1つの非直感的好気性設計(5)、および1つの非直感的嫌気性設計(6)の作成およびバリデーションも記載する。その手順には、各生体触媒の生成特性を実証し強化する適応進化の性能の詳細な評価も含まれる。具体的には、増殖連動型のコハク酸塩の生成として同定された7つの菌種の設計を選択し、それを実行することでコハク酸塩の生合成の結合を導くことができる。
実験のバリデーション用に選択された欠損戦略:
【0103】
【化5】
【0104】
ここに記載された分析でも、進化的方法によって示唆された菌株全てのコハク酸塩生成特性を向上できることが示唆されている。本明細書に記載された分析結果も、一覧の代謝改変の完全な一組未満を取り込んだとき、上記に記載した特定の菌株は、まだ増殖連動型のコハク酸塩生成を示すことを示唆している。したがって、以下の一覧では各設計のために必要な欠損の一最小組に特化する。
上記設計のために必要な欠損の最小組:
【0105】
【化6】
【0106】
sdhおよびatpはコハク酸塩脱水素酵素(SUCD1i)およびATP合成酵素(ATPS4r)活性をそれぞれ大腸菌から排除することができる欠損全てを表す。本明細書に記載された組み合わされた計算および工学プラットフォームは、通常化学量論モデル生物全てに適用可能であり、本明細書で提供された教示および助言によって、当業者は増殖連動型のどんな生化学的生成物をも生成に導く代謝変調のため、何組もの遺伝子操作を設計および実行することができるだろう。
【0107】
実施例II
増殖連動型のコハク酸塩の生成を有する微生物
本実施例では、増殖連動型のコハク酸塩の生成のため、実施例Iに記載のコンピュータを利用した設計した菌株2つの作成および性能について記載する。
【0108】
以下に記載するのは、先述の菌株設計2つを含む生物を作成および特徴付けるのに使用する方法である。つまり、adhE、pflA、およびldh 内に欠損を含むAB3と名づけられた菌株、一方はackA−pta、dhaKLM、ptsG、pykAおよびpykF内に欠損を含むAB4と名づけられた第二の菌株で、両方共に先に記述されている。
【0109】
菌株特徴付け前進化
第一の設定にはpfl、ldh、およびadhEの同時除去が関与する。提唱されるpfl、ldh、adhEの三重突然変異体の理論上の生成制限は、可能な増殖速度ごとにコハク酸塩収率を別々に極大化および最小化することで得られ、図3で野生型菌株の理論上の生成制限と比較する。緑と黒の直線で包囲された領域は、理論上は大腸菌に達成可能なコハク酸塩およびバイオマス生成率を表示する。生成制限の最も右側の部分は「最適増殖」解に一致し、最大バイオマス収率と同義である。システムの直線性のため、グルコー
ス接種率が任意の基礎と異なる場合は、その結果は比例的に変わる。実施例1に先だって記載したように、pf、ldh、adhEの三重突然変異体は完全な嫌気姓発酵が可能であり、その発酵は最適増殖で1.2 mol/molのコハク酸塩モル濃度収率に導かれると予測される。コンピュータで予測した最大値である25%以上の増殖速度で、ノンゼロコハク酸塩収率が不可欠となり、コハク酸塩の生成はこの菌株で緊密に細胞増殖に結合していると予測されることにも留意する。最後に、理論上の最大コハク酸塩収率は細胞破壊からの影響は受けず、したがってこの設計戦略はこの菌株のさらなる代謝工学に負の影響は与えない。
【0110】
第二のOptKnock由来の設計は、嫌気条件下での増殖連動型のコハク酸塩形成のために、5つの欠損(すなわち、ackA−pta、pykA、pykF、ptsG、dhaKLM)を必要とする。先に実施例に記載したように、欠損組は結果として、ピルビン酸塩の非定型の原因への強制依存のような(例、エントナー−ドウドロフ経路、セリンデアミナーゼ、リンゴ酸酵素など)著しい代謝変更を引き起こすと予想される。
【0111】
つまり、欠損を導入する基準野生型菌株として大腸菌K−12MG1655を使用し、菌株を形成する。遺伝子破壊を一つ一つ、幾つかの欠損が蓄積された受容菌株へ組み込む。Datsenkoら、Proc. Nat. Acad. Sci. USA、 6640−6645(2000年)による、DatsenkoおよびWannerのλ Redリコンビナーゼシステムを通して相同組換えを使用しインフレームの欠損を取り込み、菌株を形成した。各欠損後に薬剤抵抗性マーカーは残らず、目標の菌株に多重突然変異が蓄積される。加えて、目的とした遺伝子を完全に取り除き、形成された突然変異体が元の野生型に戻る可能性を避ける。第一の菌株はpfl、ldh、およびadhE内に欠損を持って形成され、一方第二の菌株はackA−pta、pykA、pykF、dhaKLM、およびptsG内に欠損を持つ。図8aで、これらの菌株を形成するために使用された戦略の概要を述べる。
【0112】
進化の前後両方で全ての菌株で振盪フラスコ発酵を行い、菌株の性能を数量化した。最初窒素で培地を散布し、それから隔壁およびクリンプキャップでフラスコを密封し、嫌気条件を得た。増殖が嫌気的に観察されなかった菌株には、通気を限定するための隔壁を通る小さな穴を作り、微好気的条件を適用した。pH7.0(6.78 g/L Na2HPO4、3.0 g/L KH2PO4、0.5 g/L NaCl、1.0 g/L NH4Cl、1 mM MgSO4、0.1 mM CaCl2)の、グルコース2 g/LおよびNaHCO3 20mMで補われたM9最小培地を使用し、実験全てを行った。前培養を一晩増殖させ、指数増殖中に測定を行う新鮮な回分培養用の種菌として使用した。分光光度計(600 nm)を使用して光学密度を測定し増殖速度を判定し、時間をかけて炭素源欠損をモニターしグルコース摂取率を判定した。Linら、Biotechnol. Bioeng.、 775−779(2005年)に従い、グルコースおよびエタノールにた屈折率検出器を、有機酸にはUV検出器を使用し、HPX−087カラム(BioRad)つきのHPLC(島津)でコハク酸塩、副生成物、および残渣グルコースを数量化した。各菌株に対し三通りの培養を行い、報告した測定はその3つの平均である。
【0113】
適応進化手順の実施に関する定量的結論を出すため、進化の前に野生型および操作された菌株の性能を特徴付けた。表8でこの分析からの結果を見ることができる。中間AB1およびAB2(図8a参照)を含む、本研究で形成した多重菌に対し株振盪フラスコ特性を行ったことに留意する。
図8:操作された菌株の増殖速度、加えて消費されたグルコース、形成されたバイオマス、および生成された発酵生成物の量。全データは3つ以上の培養を平均したものであり、信頼区間95%を示唆する。N.D.は測定なし。
【0114】
【表8】
【0115】
操作された菌株は、野生型および変質した発酵特性より遅い増殖速度を示した。AB1およびAB3は厳しい嫌気条件下では、あまり増殖できなかったため、代わりに上記に記載したのと同様に微好気的に増殖した。この観察は、同じ遺伝子型AB1に関する文献報告、NZN111、Stolsら、Appl Environ Microbiol、63:2695−701(1997年)、Hongら、Biotechnol Bioeng、74:89−95(2001年)、Stolsら、Appl Biochem Biotechnol、63−65:153−8(1997年)に一致する。これらの菌株は、この比較的大量の代謝性複製生物によって示唆されるように、ピルピン酸にボトルネックを有するようである。
【0116】
AdhE欠損によってAB3内で消費されるグルコースの量は著しく減少するが、エタノール生成も排除され、その結果より高いコハク酸塩収率となった。突然変異体は成長率および最終バイオマス濃度にかなりの影響を与えたが、菌株AB2およびAB4に振盪フラスコ特徴づけを行うと、完全な嫌気性増殖の間の野生型MG1655と比較し、コハク酸塩収率が約4倍および10倍の増加を示した(図8b)。
【0117】
適応進化菌株
新しく形成された突然変異体の増殖で観察されたのは、以下モデル化分析により先験的に予測したものあり、そのためそれからEvolugatorTM マシーンを使用し菌株を進化させた。つまり、野生型および両方の操作した菌株で適応進化を行った。適応進化手順は、EvolugatorTM進化機械を使用し、Evolugate、LLC(フロリダ州、ゲーンズビル)によって実行された。定常期が達成される前に、新鮮な倍地への回分培養の連続経路による遷延性指数増殖中、この装置で細胞を維持する。最適密度測定および液体取扱いを機械化することで、Evolugatorは大量の培養液量を使用し高速で連続導入を行うことができ、したがって細胞適応度の進化のため、ケモスタットの効率に近づく。しかし、単一の容器内に細胞を維持するケモスタットとは対照的に、Evolugatorは細区画された管のスプール領域内を1つの「反応器」から次へと移動することで機能し、したがって壁増殖に対する選択は排除される。培養の試料を採取し、液体窒素で冷凍し、進化の過程の間中毎日最適培養密度を記録した。
【0118】
AB3の嫌気性増殖は研究室では無視できたが、進化過程の最初の2、3日の間に徐々に酸素付加を減少させ、その条件下での増殖能力を発展させた。大腸菌を嫌気的に進化させることができると実証する対照実験として、野生型MG1655を15日の期間Evolugatorにかけた。図9aに見られるように、日ごとに必要な欠損の数によって推定すると、2.2 hr−1から0.9 hr−1へと約2倍、時間が減少した。さらに
その増殖発展を特徴付けるため、本来の菌株および進化させた(EVG09)菌株の3通りの培養を行った。野生型菌株の培養率は0.351±0.002 hr−1、EVG09の培養率は0.521±0.049 hr−1であった。野生型菌株で観察された満足いく増殖発展を与え、それからAB3菌株を進化させると、図9bに見られるように同じような2倍の時間の減少という結果となった。注目すべき2倍の時間減少が両方の菌株に対し観察された。
【0119】
さらなる菌株の特徴づけ
上記に記載した進化前の元の菌株に行ったように、進化させたAB3を振盪フラスコ発酵で特徴づけた。完全な嫌気性増殖はこの菌株に対しては有意ではなく、EvolugatorTM増殖環境は完全には嫌気性ではないことが示唆された。しかし、EvolugatorTMから取られた試料の生成物特性から、かなりの量の発酵生成物が菌株AB3によって生成されたことが明らかとなり、その進化は完全な好気性からはほど遠く、おそらく微好気性だということが示された(図10)。実行の最後でピルビン酸塩の量はゼロへと下がったため、EvolugatorTM内での時間によって進化前のAB3菌株内に存在したかなりのピルビン酸塩のボトルネックが軽減されていたことを、これらの結果は示している。エタノール、乳酸塩、またはコハク酸塩の生成とは異なり、NADは再生されないため、ピルビン酸塩の分泌は生物に対しほとんど利点はなかい。加えて、酢酸塩の生成とは異なり、エネルギーは再生されない。
【0120】
工業発酵は完全な嫌気条件下ではほとんど実行されない。それゆえ、進化後のAB3菌株を微好気性増殖下でさらに特徴付けた。図11にこれらの増殖条件下での生成物特性を示し、進化前の親株と比較したとき、より高いコハク酸塩形成へ転換が明らかとなる。図12は、発酵生成物におけるコハク酸塩の質量パーセンテージが4倍以上増加し、ピルビン酸塩および酢酸塩においてはそれぞれ約50%および40%の減少を伴うことを示している。コハク酸塩の最終モル収率は1.1 mol/molグルコースで、進化後のコハク酸塩菌株のモデルによって予測された1.2 mol/molグルコース収率に非常に類似していた(上記記述および図3の突然変異菌株の最右境界、参照)。Evolugatorから直接取り出した試料とは対照的に、進化前AB3菌株よりは少ないものの、まだ相当なピルビン酸塩の蓄積があった。微好気性振盪フラスコ内にはおそらくより多くのO2が存在するということを、この結果は示している。特に発展させた嫌気性条件下で、さらに進化の時間を取ることで、例えばピルビン酸塩のボトルネックを一斉に排除することができる。生物を細胞増殖に対し最適化した後、コンピュータ予測はコハク酸塩2 moleから酢酸塩1 moleの発酵特性を示し、さらに菌株特徴づけデータは予測され菌株の性能に到達できることを実証している。
【0121】
結論
本実施例では、増殖連動型のコハク酸塩の生成のためにOptKnock計算枠組みによりコンピュータ設計された、実施例Iに記載した2つの大腸菌株の作成および性能の特徴づけを示す。AB3と名づけられた第一の菌株はadhE、pflA、およびldhA内に欠損を含み、一方第二の菌株AB4はackA−pta、dhaKLM、ptsG、pykA,、およびpykF内に欠損を含む。これらの菌株は、形成中に創造された親菌株AB1およびAB2と同様、欠損が導入された野生型大腸菌株MG1655以上のコハク酸塩収率の増加を示した。実際、AB4菌株はコハク酸塩収率において野生型菌株以上の約10倍の増加を示した。
【0122】
増殖速度を増加させるためEvolugatorTM技術を使用し、野生型大腸菌株MG1655およびMG1655も適応進化を受けた。野生型菌株に対しては、嫌気条件下で進化手順によって増殖速度を約50%向上したが、予想されたように、最終生成物特性に重大な影響はなかった。菌株AB3に対しては、進化段階により増殖速度およびコハク
酸塩収率の両方を増加させ、一方副生成物ピルビン酸塩の分泌はかなり減少した。本研究ではさらにOptKnockコンピュータ法で、結果として大腸菌内のコハク酸塩生成を増加させる遺伝子欠損の組み合わせが同定できるということを実証している。そのうえ、OptKnock設計菌株の性能をコンピュータで予測された過剰生成表現型へ向上させるため適応進化を利用できることを、本発明ではさらに実証している。
【0123】
【表1−1】
【0124】
【表1−2】
【0125】
【表1−3】
【0126】
【表1−4】
【0127】
【表1−5】
【0128】
【表2−1】
【0129】
【表2−2】
【0130】
【表3−1】
【0131】
【表3−2】
【0132】
本明細書を通じ、様々な出版物を括弧内で参考文献として挙げた。本発明に関連する最新技術をより完全に記載するために、これらの出版物全体の開示を参照することで、それにより本明細書内に含まれる。
【0133】
本発明は開示された実施例を参照し記載されているが、上記の詳細で具体的な例示および研究が本発明を例証だけするものであることが、当業者には容易に理解される。当然のことながら、本発明の精神から逸脱することなく様々な変更が行われうる。それゆえ、本発明は以下の請求項によってのみ限定される。
【図面の簡単な説明】
【0134】
【図1】図1は、生化学的生成および細胞増殖間のトレードオフの推定される描写を示す。点AおよびBはそれぞれ、野生型および突然変異菌株の最大バイオマス解を表す。ここで留意すべきは、突然変異菌株は増殖連動型の生成を示すことである。
【図2】図2は、OptKnockの二段階最適化構造を示す。内側の課題によって、ある特定の細胞目標の最適化に基づく流束の配分を行う。それから内側の課題の最適化を可能とする重要な反応への接近を制限することで、外側の問題によって生物工学的目標(例えば、化合物過剰生成)を極大化する。
【図3】図3は、野生型と比較した、4つの突然変異菌株の増殖速度境界に対するコハク酸塩を示す。10 mmol/(gDW・hr)の基底グルコース摂取速度の摂取速度に従い、完全な嫌気条件を想定する。全ての菌株が外側境界に続き、どの境界も野生型菌株に一致するが、adhE、ldh、pta(第一分岐、赤)、pfl ldh、adhE(第二分岐、緑)、adhE、ldh(第三分岐、青)、そしてpfl、ldh(第四分岐、灰色)の順で、左から下方へと分岐する。
【図4】図4は、野生型と比較した、OptKnock由来の突然変異菌株(ackA−pta、atpFH、pykA、pykF、dhaKLM、sdhAB)の増殖速度境界に対するコハク酸塩を示す。10 mmol/(gDW・hr)の基底グルコース摂取速度を想定する。菌株は、ackA−pta,、atpFH、pykA、pykF、dhaKLM、sdhAB(嫌気)(第一直線、緑)、ackA−pta、atpFH、pykA、pykF、dhaKLM、sdhAB(嫌気)(第二直線、赤)、野生型(嫌気)(第三直線、青)、および野生型(嫌気)(第四直線、黒)の順で左から始まる直線に一致する。
【図5】図5は、OptKnock由来の突然変異菌株(ackA−pta、pykA、pykF、ptsG、dhaKLM)の増殖速度境界に対するコハク酸塩を示し、様々な非増殖関連のATP維持必要量に対する野生型と比較した。10 mmol/(gDW・hr)の基底グルコース摂取速度を想定する。菌株は、ackA−pta、pykA、pykF、ptsG、dhaKLM(ATPM=6)(y軸と外側境界との交差、緑)、ackA−pta、pykA、pykF、ptsG、dhaKLM(ATPM=3)(第二直線、赤)、ackA−pta、pykA、pykF、ptsG、dhaKLM(ATPM=0)(第三直線、青)、および野生型(外側境界、黒)の順で、左から始まる直線に一致する。
【図6】図6は、様々な酸素化率での、OptKnock由来の突然変異菌株(ackA−pta、pykA、pykF、ptsG、dhaKLM)の増殖速度境界に対するコハク酸塩を示す。典型的な7.6 mmol/(gDW・hr)の維持エネルギー必要量を課す。10 mmol/(gDW・hr)の基底グルコース摂取速度を想定する。菌株は、ackA−pta、pykA、pykF、ptsG、dhaKLM(O2<2)(第一直線、緑)、ackA−pta、pykA、pykF、ptsG、dhaKLM(O2<5)(第二直線、赤)、ackA−pta、pykA、pykF、ptsG、dhaKLM(O2<10)(第三直線、青)、およびpta、ptsG、f6pa、pyk(O2無制限)(外側境界、黒)の順で、左から始まる直線に一致する。
【図7】図7は、様々な酸素化率での、OptKnock由来の突然変異菌株(ackA−pta、pykA、pykF、ptsG、dhaKLM、ldh、adhE)の増殖速度境界に対するコハク酸塩を示す。典型的な7.6 mmol/(gDW・hr)の維持エネルギー必要量を課す。10 mmol/(gDW・hr)の基底グルコース摂取速度を想定する。菌株は、ackA−pta、pykA、pykF、ptsG、dhaKLM、adhE、ldh(O2<2)(第一直線、緑)、ackA−pta、pykA、pykF、ptsG、dhaKLM、adhE、ldh(O2<5)(第二直線、赤)、ackA−pta、pykA、pykF、ptsG、dhaKLM、adhE、ldh(O2<10)(第三直線、青)、およびackA−pta、pykA、pykF、ptsG、dhaKLM、adhE、ldh(O2無制限)(外側境界、黒)の順で、左から始まる直線に一致する。
【図8】図8は、OptKnock設計のコハク酸塩菌株の進化前を示す。(a)は菌株作成戦略である。AB1およびAB2は中間的な菌株で、各菌株の系譜における最終的な遺伝子破壊の効果を評価するよう特徴付けられてもいる。(b)は3つの独立培養を平均した全実験にわたるコハク酸塩の重量収率である。
【図9】図9は、希釈の頻度から判定した、菌株MG1655およびAB3のおおよその倍加時間を示し、進化の過程全体を通してプロットした。
【図10】図10は、菌株AB3の進化中にEvolugatorから得た試料の生成物特性を示す。ダイヤ(青)がコハク酸塩、三角(赤)がギ酸塩、丸(緑)が酢酸塩である。
【図11】図11は、大腸菌株を微好気性培養した後に存在したコハク酸塩およびほかの発酵生成物の質量(a)およびモル濃度(b)の収率を示し、代謝された全グルコースに関連する。左は野生型MG1655、中央が進化前のAB3、右が進化後のAB3である。
【図12】図12は、進化前(左のバー)および進化後(右のバー)AB3菌株の微好気性培養が見られた後、存在する発酵生成物の質量パーセントを示す。
【背景技術】
【0001】
発明の背景
本発明は、概してコンピュータを利用した生物の設計に関し、特に増殖連動型のコハク酸塩の生成のための選択された遺伝子型を有する生物に関する。
【0002】
コハク酸塩は、食品、製薬、洗剤、および高分子工業における汎用化学物質の前駆体としての用途から、膨大な商業的利益のある化合物である。発酵により生成されたコハク酸塩は、1,4ブタンジオールおよび関連生成物、テトラヒドロフラン、γ−ブチロラクトン、n−メチルピロリジノン(NMP)および2−ピロリジノンを含む工業製品を年間2.7×108 kg以上供給することが潜在的に可能である(非特許文献1)。コハク酸の基本的な化学的性質は、石油化学的に生成されたマレイン酸/無水物の性質と類似しており、従ってその生成コストのみが、コハク酸が発展性のある数々の市場に急速に展開するのを阻んでいる。生物学的なコハク酸塩の生成は、糖発酵中に温室効果ガスCO2がコハク酸塩に固定されなければならないグリーンプロセスでもある。これらの理由から、1,4−二塩基酸(コハク酸塩、フマル酸塩、およびリンゴ酸塩)は、評価された300の候補の中から、エネルギー省により、バイオプロセスの開発の最優先事項として特定された12の化合物に含まれる。
【0003】
コハク酸塩などの中心的代謝化合物は、しばしば基礎代謝中に構成的に生成されるため、代謝工学のよい初期標的となる。先天的に炭水化物からコハク酸塩を生成できる生物の例としては、アナエロビオスピルリム・サクシニシプロデュセンス(Anaerobiospirillum succiniciproducens)(非特許文献2、非特許文献3、非特許文献4)、アクチノバチルス・スクシノゲネス(Actinobacillus succinogenes)(非特許文献5、非特許文献6)、および最近配列が決定されたウシ管腔内細菌のマンヘミア・サクシニシプロデュセンス(Mannheimia succiniciproducens)(非特許文献7、非特許文献8、非特許文献9)が含まれる。さらに報告のいくつかは、さまざまな代謝工学的戦略によって、より高いコハク酸塩の収率を有する大腸菌(Escherichia coli)株の構成を達成したと主張している。これらの取り組みは、コハク酸塩への炭素流束のチャネリングの増加、および補助因子NADHの可用性に焦点を当ててきた。例えば、PEPカルボキシラーゼ(ppc)の過剰発現(非特許文献10)、およびRhizobium etliピルビン酸塩カルボキシラーゼ(pyc)の発現(非特許文献11)は、TCA回路のコハク酸塩部分への流束を増加させることにより、大腸菌(E.coli)でのコハク酸塩の収率それぞれ0.30 g/gおよび0.17 g/gをもたらした。さらに、乳酸塩脱水素酵素(ldh)およびピルビン酸塩ギ酸塩リアーゼ(pfl)が欠損した大腸菌(E.coli)突然変異体(NZN111株)は、大腸菌(E.coli)(非特許文献12、および非特許文献13)、あるいはブタ回虫(Ascaris suum)(非特許文献14)のリンゴ酸酵素の過剰発現との組み合わせで、コハク酸塩収率の向上を達成した。のちにホスホトランスフェラーゼ系のptsG遺伝子にマップされた、NZN111におけるさらなる染色体偶発突然変異は(非特許文献15)、1 mol/molグルコース(0.66 g/g)の嫌気的なコハク酸塩の収率を有するAFP111株をもたらした(非特許文献16)。インゲン根粒菌(R.etli)ピルビン酸塩カルボキシラーゼの存在下または不在下でのNZN111およびAFP111株のさまざまな特性は、嫌気および二相条件(嫌気的生成が後続する好気性増殖)のもとで調査されてきた(非特許文献17)。非特許文献18では、約0.96 g/gの収率となった。その他の取り組みでは、好気的に0.91 mol/mol(0.60 g/g)を達成できる大腸菌(E.coli)のコハク酸塩生成菌株が得られた(Linら、Metab En
g、(2005年)。また近刊では、嫌気的に1.6 mol/mol(1.0 g/g)となっている(非特許文献19)。
【0004】
しかしながら、より高いコハク酸塩の収率を生み出す一部の細菌の菌株の開発を称している上記の取り組みおよび報告にもかかわらず、採用された方法には、商業目的での適用性を妨げるいくつかの欠点がある。以下にさらに詳しく記載されるように、上記の方法で生成される菌株は、未変化あるいは野生型の親菌株に有利にはたらく選択圧により、一般的に商業用の発酵プロセスでは不安定である。
【非特許文献1】Zeikusら、Appl Microbiol Biotechnol、51:545−552(1999年)
【非特許文献2】Samuelovら、Appl Environ Microbiol、65:2260−63(1999年)
【非特許文献3】Leeら、Appl Microbiol Biotechnol、54:23−27(2000年)
【非特許文献4】Leeら、Biotechnol Lett、25:111−14(2003年)
【非特許文献5】Guettlerら、Int J Syst Bacteriol、49:207−16(1999年)
【非特許文献6】Urbanceら、Appl Microbiol Biotechnol、65:664−70(2004年)
【非特許文献7】Leeら、Bioprocess Biosyst Eng、26:63−7(2003年)
【非特許文献8】Hongら、Nat Biotechnol、22:1275−81(2004年)
【非特許文献9】Leeら、Appl Microbiol Biotechnol、58:663−8(2002年)
【非特許文献10】Millardら、Appl Environ Microbiol、62:1808−10(1996年)
【非特許文献11】Gokarnら、Biotechnol Lett、20:795−798(1998年)
【非特許文献12】Stolsら、Appl Environ Microbiol、63:2695−701(1997年)
【非特許文献13】Hongら、Biotechnol Bioeng、74:89−95(2001年)
【非特許文献14】Stolsら、Appl Biochem Biotechnol、63−65:153−8(1997年)
【非特許文献15】Chatterjeeら、Appl Environ Microbiol、67:148−54(2001年)
【非特許文献16】Donnellyら、Appl Biochem Biotechnol、70−72:187−98(1998年)
【非特許文献17】Vemuriら、Appl Environ Microbiol、68:1715−27、18(2002年)
【非特許文献18】Vemuriら、J Ind Microbiol Biotechnol、28:325−32(2002年)
【非特許文献19】Sanchezら、Metab Eng、7:229−39(2005年)
【発明の開示】
【発明が解決しようとする課題】
【0005】
従って、所望の生成物の生合成を最適培養条件に強制的に結び付ける、商業的に有益な
特徴を有する微生物に対する必要性が存在する。本発明は、この必要性を満たし、かつ関連する利益をも提供する。
【課題を解決するための手段】
【0006】
(発明の要旨)
本発明は、酵素の活性が低下したときに、増殖連動型のコハク酸塩の生成と関連する酵素をコード化する1つ以上の遺伝子破壊で構成される非自然発生的な微生物を提供し、それにより1つ以上の遺伝子破壊は、非自然発生的な微生物に安定的な増殖連動型のコハク酸塩の生成を与える。また、コハク酸生成を微生物の増殖と強制的に連動させる代謝改変の組であって、(a)adhE、ldhA、(b)adhE、ldhA、ackA−pta、(c)pfl、ldhA、(d)pfl、ldhA、adhE、(e)ackA−pta、pykF、atpF、sdhA、(f)ackA−pta、pykF、ptsG、または(g)ackA−pta、pykF、ptsG、adhE、ldhAからなる遺伝子の組あるいはそのオーソログから選択される1つ以上の遺伝子破壊を備える代謝改変の組で構成される非自然発生的な生物であって、安定的な増殖連動型のコハク酸塩の生成を示す微生物も提供される。さらに、pykA、atpH、sdhB、またはdhaKLMから選択される少なくとも1つの遺伝子破壊をさらに含む、代謝改変(e)ackA−pta、pykF、atpF、sdhAをコード化する遺伝子を有する非自然発生的な微生物、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊をさらに含む、代謝改変(f)ackA−pta、pykF、ptsGをコード化する遺伝子を有する非自然発生的な微生物、あるいはpykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊をさらに含む、代謝改変(g)ackA−pta、pykF、ptsG、adhE、ldhAをコード化する遺伝子を有する非自然発生的な微生物も提供される。破壊は、完全な遺伝子破壊とすることができ、また非自然発生的な生物は、さまざまな原核または真核微生物を含むことができる。安定的な増殖連動型のコハク酸塩の生成を有する非自然発生的な微生物を生成する方法も提供される。この方法は、(a)指数増殖中にコハク酸塩の生成を要する代謝改変の組をコンピュータで同定するステップと、(b)コハク酸塩の生成を要する代謝改変の組を含有するように微生物を遺伝子改変するステップとで構成される。
【発明を実施するための最良の形態】
【0007】
(発明の詳細な説明)
本発明は、増殖連動型のコハク酸塩の生成を有する細胞や生物の設計と生成を対象としている。1つの実施例では、本発明は、コハク酸塩の最適生成のための代謝設計を同定する、大腸菌(Escherichia coli)代謝のコンピュータを利用した化学量論モデルを基に、最適化によるアプローチを利用する。二段階プログラミングフレームワークOptKnockは、反復アルゴリズムに適用され、遺伝子破壊の複数の組を予測するが、これは集合的に増殖連動型のコハク酸塩の生成をもたらす。ここに記載される結果は、戦略的に配置された遺伝子欠損または遺伝子の機能破壊の組み合わせが、大腸菌(Escherichia coli)およびその他の細胞または生物のコハク酸塩の生成能力を有意に向上させることを示す。コンピュータを利用した設計のための増殖連動型のコハク酸塩の生成は、設計された代謝遺伝子型を有する菌株の構成により確認される。これらの代謝的に変更された細胞または生物は、適応進化を受け、増殖連動型のコハク酸塩の生成をさらに増大することもできる。
【0008】
さらなる実施例では、本発明は、より促進されたコハク酸塩の生成特性を有する代謝的に変更された微生物菌株を開発するための、統合された計算と工学のプラットフォームを対象としている。プラットフォームの計算要素によって同定された菌株は、コハク酸塩の生成促進をもたらす予測された代謝改変を遺伝子操作することにより、実際の生成に適用される。コハク酸塩の生成は、微生物の最適増殖に共役され、発酵時のこの生成物の収率
を最適化する。またさらなる実施例では、増殖連動型のコハク酸塩の生成を示す菌株は、さらに適応進化を受け、生成物の生合成をさらに増大する。適応進化後の増殖連動型のコハク酸塩の生成のレベルは、システムの計算要素により推測することもできるが、この特定の実施例では、コハク酸塩のレベルの上昇は、進化のあとにのみ実現される。
【0009】
ここで使用される「非自然発生的な」という用語は、本発明の微生物に関連して使用される場合、微生物が言及されている種の野生型菌株では通常見られない遺伝子変更を少なくとも1つ有することを意味する。遺伝子変更は、遺伝子欠損または遺伝物質のその他なんらかの機能破壊としうる。
【0010】
ここで使用される「微生物」という用語は、微視的な大きさの原核または真核の細胞または生物を意味する。この用語は、全ての種の細菌および酵母または菌類などの真核生物を含むものとする。この用語は、生化学物質の生成用に培養できる任意の種の細胞培養物も含む。
【0011】
ここで使用される「増殖連動型の」という用語は、生化学物質の生成に関連して使用される場合、言及されている生化学物質の生合成が、微生物の増殖相中に強制的に生成される生成物であることを意味する。
【0012】
ここで使用される「代謝改変」という用語は、その自然発生状態から変更された生化学反応を指す。代謝改変は例えば、反応に参加している酵素をコード化する1つ以上の遺伝子の機能破壊による生化学反応活性の排除を含むことができる。典型的な代謝改変の組は表1に示されている。かかる代謝改変により特定される個々の反応、およびその対応する遺伝子相補体は、表2で大腸菌(E.coli)について例示されている。これらの反応で使用される反応物質および生成物は、表3に例示されている。
【0013】
ここで使用される「コハク酸塩」という用語は、クレブス回路およびさまざまな発酵プロセスにおいて形成されるジカルボン酸HOOCCH2CH2COOHを意味する。ここで使用される「コハク酸塩」という用語は、「コハク酸」という用語と同義である。化学的には、コハク酸塩は、コハク酸の塩またはエステルに該当する。従って、コハク酸塩およびコハク酸は、溶液のpHによって2つの形状のうちいずれかで存在することができる同じ化合物を指す。
【0014】
ここで使用される「遺伝子破壊」という用語、あるいはその文法的に対応する語は、コード化された遺伝子産物を不活性にさせる遺伝子変更を意味する。遺伝子変更は、例えば全遺伝子の欠損、転写または翻訳に必要な調整配列の欠損、切断遺伝子産物をもたらす遺伝子の一部の欠損とすることができ、またはコード化された遺伝子産物を不活性化するさまざまな突然変異戦略のいずれかによるものとしうる。特に有用な遺伝子破壊の1つの方法は完全な遺伝子欠損であるが、これは本発明の非自然発生的な微生物における遺伝的復帰の発生を低下あるいは排除するためである。
【0015】
ここで使用される「安定的な」という用語は、生化学的生成物の増殖連動型の生成に関連して使用される場合、増殖と生化学合成との間の連動を失わずに、5世代を超えて培養できる微生物に言及している。一般的に、安定的な増殖連動型の生化学的生成は10世代を超え、具体的には、安定的な増殖連動型の生化学的生成は約25世代を超え、さらに具体的には、安定的な増殖連動型の生化学的生成は50世代を超え、無限を含む。安定的な増殖連動型の生化学物質の生成は、例えば、代謝改変内のそれぞれの反応を触媒する酵素をコード化する遺伝子の欠損によって達成することができる。増殖連動型の生化学物質の生成の安定性は、複数の欠損によって強化することができ、それぞれの破壊された活性に対する複数の補償的な復帰の可能性を有意に減少させる。
【0016】
当業者は、ここに例示される代謝改変は、大腸菌(E.coli)遺伝子およびその対応する代謝反応に関連して記載されることを理解するであろう。しかし、多種多様な生物の完全なゲノム配列決定と、ゲノミクス分野での高いスキルとを前提にすると、当業者はここで提供される教示と指針を、原則としてその他全ての生物に容易に適用できるであろう。例えば、ここで例示される大腸菌(E.coli)代謝変調は、同じまたは類似の遺伝子破壊を他の種に組み込むことにより、他の種に容易に適用できる。かかる破壊は、例えば、全般的には種のホモログの遺伝子変更、また具体的にはオーソログ、パラログ、または非オーソロガス遺伝子置換体を含むことができる。
【0017】
オーソログは、直系の関係を持つ1つまたは複数の遺伝子であり、異なる生物において実質的には同様または同一の機能に関与する。例えば、マウスエポキシド加水分解酵素およびヒトエポキシド加水分解酵素は、エポキシドの加水分解の生物学的機能に対するオーソログであると考えられる。遺伝子は、例えばそれらが相同であること、あるいは共通祖先からの進化による関係を持つことを示すのに十分なだけの配列類似性を共有する場合、直系の関係を持つ。遺伝子は、一次配列類似性が同定不能である限りでは、共通祖先から進化したことを示すのに十分なだけの三次元構造を共有するが、必ずしも配列類似性を共有しない場合にもオーソログであると考えられる。オーソロガスである遺伝子は、約25%〜100%のアミノ酸配列同一性を有するタンパク質をコード化することができる。25%未満のアミノ酸類似性を共有するタンパク質をコード化する遺伝子は、その三次元構造も類似性を示している場合、直系で生じたと考えられる。組織プラスミノゲン活性化因子とエラスターゼを含む、セリンプロテアーゼ系の酵素は、共通祖先からの直系で生じたと考えられている。
【0018】
オーソログは、例えば進化を通じて、構造または全体的活性が分岐した遺伝子、またはそのコード化された遺伝子産物を含む。例えば、1つの種が2つの機能を示す遺伝子産物をコード化し、かかる機能が第2の種において異なる遺伝子に分かれている場合、この3つの遺伝子およびその対応する産物はオーソログであると考えられる。生化学的生成物の増殖連動型の生成については、当業者は、破壊される代謝活性を有するオーソロガス遺伝子が、非自然発生的な微生物の構成のために選定されることを理解するであろう。分離可能な活性を示すオーソログの例は、異なる活性が、2つ以上の種の間あるいは単一の種の中で異なる遺伝子産物に分かれているものがある。具体例としては、2種類のセリンプロテアーゼ活性であるエラスターゼタンパク質分解とプラスミノーゲンタンパク質分解の、プラスミノーゲン活性化因子およびエラスターゼとしての別個の分子への分離がある。第2の例としては、マイコプラズマ5’−3’エクソヌクレアーゼとショウジョウバエDNAポリメラーゼIII活性の分離がある。第1の種のDNAポリメラーゼは、第2の種のエクソヌクレアーゼまたはポリメラーゼの双方またはいずれかのオーソログであると考えられ、逆もまた同様である。
【0019】
対照的に、パラログは、例えば進化を後続する複製によって関連付けられるホモログであり、類似または共通であるが同一でない機能を有する。パラログは、例えば同種または異種に起源あるいは由来する。例えば、ミクロソームエポキシド加水分解酵素(エポキシド加水分解酵素I)と可溶性エポキシド加水分解酵素(エポキシド加水分解酵素II)は、共通祖先から共進化し、同種内で異なる反応を触媒し、異なる機能を有する異なる2つの酵素を表わすため、パラログであると考えられる。パラログは、お互いに著しい配列類似性を有する、同種を起源とするタンパク質であり、これらが相同である、あるいは共通祖先からの共進化による関係を持つことを示している。パラロガスなタンパク質ファミリーの群には、HipAホモログ、ルシフェラーゼ遺伝子、ペプチダーゼなどが含まれる。
【0020】
非オーソロガス遺伝子置換体は、異種における言及されている遺伝子機能を代理できる
、1つの種からの非オーソロガス遺伝子である。代理には例えば、異種における言及されている機能と比較して実質的には同様のまたは類似の機能を、起源種内で行うことができることが含まれる。一般的に、非オーソロガス遺伝子置換体は、言及されている機能をコード化する既知の遺伝子と構造的に関連するとして同定可能であるが、構造的にはそれほど関連していないが、機能的に類似している遺伝子とその対応する遺伝子産物であっても、ここで使用されるこの用語の意味に含まれる。機能的類似性は、例えば、非オーソロガス遺伝子の活性部位または結合領域に、代理されようとする機能をコード化する遺伝子と比較して、少なくともいくつかの構造的類似性を必要とする。従って非オーソロガス遺伝子は、例えばパラログまたは非関連遺伝子を含む。
【0021】
従って、増殖連動型の生化学物質の生成を有する本発明の非自然発生的な微生物を同定および構成する際、当業者は、ここで提供される教示と指針を特定の種に適用することにより、代謝改変の同定はオーソログの同定と破壊を含むべきであると理解するであろう。パラログおよび非オーソロガス遺伝子置換体またはそのいずれかが、類似あるいは実質的に類似する代謝反応を触媒する酵素をコード化する言及される微生物に存在する範囲で、当業者は、これらの進化的に関連する遺伝子を排除して、酵素活性におけるいかなる機能的冗長性も設計された代謝改変を妨げないよう確保することもできる。
【0022】
オーソログ、パラログ、および非オーソロガス遺伝子置換体は、当業者には周知の方法により判定できる。例えば、2つのポリペプチドの核酸またはアミノ酸配列を検査することで、比較される配列間の配列同一性および類似性が明らかになる。当業者は、かかる類似性に基づいて、その類似性が、タンパク質が共通祖先からの進化によって関連付けられることを示すのに十分なほど高いかどうか判定することができる。Align、BLAST、Clustal Wなど、当業者には周知のアルゴリズムは、粗配列類似性または同一性を比較および判定し、また重量あるいはスコアを指定できる配列間の差異の存在または有意性も判定する。かかるアルゴリズムは当技術分野でも知られており、ヌクレオチド配列類似性または同一性を判定するために、同様に適用することができる。近縁性を判定するための十分な類似性のパラメータは、統計的類似性、またはランダムポリペプチドで類似適合が見られる可能性を計算するための周知の方法、および判定された適合の有意性に基づいて計算される。2つ以上の配列のコンピュータ比較は、必要に応じて、当業者によって視覚的に最適化することもできる。関連する遺伝子産物またはタンパク質は、例えば25%〜100%の配列同一性など、高い類似性を有することが予想できる。関連していないタンパク質は、十分な大きさのデータベースが読み取られた場合、偶発すると予想されるのものと本質的に同様の同一性を有する(約5%)。5%〜24%の間の配列は、比較される配列が関連していると判断するのに十分な相同関係を示しても、示さなくてもよい。かかる適合の有意性を判定するためのさらなる統計的解析は、データセットのサイズを考慮して、これらの配列の関連性を判定するために実施することができる。
【0023】
BLASTアルゴリズムを用いて2つ以上の配列の近縁性を判定するための典型的なパラメータは、例えば以下に記載されるものとできる。つまり、BLASTPバージョン2.0.8(1999年1月5日)およびパラメータ(Matrix:0 BLOSUM62;gap open:11;gap extension:1;x_dropoff:50;expect:10.0;wordsize:3;filter:on)を用いてアミノ酸配列アラインメントを行うことができる。核酸配列アラインメントは、BLASTNバージョン2.0.6(1998年9月16日)およびパラメータ(Match:1;mismatch:−2;gap open:5;gap extension:2;x_dropoff:50;expect:10.0;wordsize:11;filter:off)を用いて行うことができる。当業者は、例えば比較の厳密性を上げるあるいは下げるために、上記のパラメータにどのような変更を加えることができるか理解し、2つ以上の配列の近縁性を判定するであろう。
【0024】
本発明は、安定的な増殖連動型のコハク酸塩の生成を有する非自然発生的な微生物を生成する方法を提供する。方法は、(a)指数増殖中にコハク酸塩の生成を要する代謝改変の組をコンピュータで同定するステップと、(b)コハク酸塩の生成を要する代謝改変の組を含有するように、微生物を遺伝子改変するステップとで構成される。
【0025】
バイオプロセスに対して考慮すべき重要な点は、バッチ式あるいは連続式の発酵方式のいずれを使用するかである。生成される生成物の量に影響するこの2方式間の1つの差異は、バッチ方式には、指数増殖相に加え、前処理、誘導期、定常期が存在することである。対照的に、連続式プロセスは、絶えず指数増殖の状態を維持し、正しく操作すれば、一度に何ヶ月もの間稼働することができる。増殖関連および混合増殖関連の生成物の形成では、連続式プロセスは、前処理、誘導期、定常期の排除により、ずっと高い生成性(希釈率×細胞質量)を提供する。例えば、以下の妥当な想定では、
【0026】
【数1】
【0027】
連続式プロセスにより向上した生成性は、8倍と推定されている(Shulerら、Prentice Hall,Inc.:ニュージャージー州アッパーサドルリバー、245−247)。
【0028】
生成性における圧倒的な優位性にもかかわらず、多くの理由から、連続式プロセスよりもバッチ式プロセスの方がかなり多く実施されている。まず、非増殖関連の生成物形成(ペニシリンなど)では、バッチシステムの生成性は、連続式プロセスの生成性を有意に超えるが、これは後者が非常に低い希釈率で稼働しなければならないためである。次に、生成菌株は一般的に、生化学物質またはタンパク質生成能力を向上させるために、その遺伝物質に変更を行なっている。これらの特異化された菌株はその親相補体よりも成長が遅い可能性が高いので、ケモスタット(連続モードで操作される発酵槽)などを使用する連続式プロセスは、最も急速に増殖する細胞に高い淘汰圧が課される。所望の過剰生成表現型をもたらす組み換えDNAを含む、あるいは点突然変異を有する組み換えDNAは、元の生成性が劣る親菌株への復帰突然変異を起こしやすい。単一の遺伝子欠損を有する菌株が、野生型の増殖表現型を復帰させる可能性が高い、補償的な突然変異を発現することもある。より成長の速い細胞は通常、より生成性の高い細胞と制限栄養物質に関して競合しないので、大幅に生成性を低下させる。一方バッチ式プロセスは、各サイクルの終了時に細胞を再利用することによらずに利用できる世代の数を制限し、従ってその野生型の表現型に復帰する生成菌株の確率を減少させる。最後に、連続式プロセスは、機器の不具合や異生物の混入などの技術的障害により、長期の操作がより困難である。またかかる不具合の結果は、連続式プロセスでは回分培養よりもはるかに重大である。
【0029】
特殊化学製品やタンパク質の少量の生成では、連続式プロセスの生成性向上は、菌株の安定性と信頼性に関連するリスクを上回ることはまれである。しかし、コハク酸塩などの大量の増殖関連生成物の場合は、連続式プロセスの生成性の向上は、バッチ式プロセスに比較して、著しい経済的利益をもたらす可能性がある。連続式バイオプロセス操作に関連する技術的障害は常に存在するが、菌株の安定性問題は、代謝経路を変更して、負の選択圧を減少または回避し、指数増殖相での対象生成物の生成を支持する代謝工学的戦略によって克服できる。
【0030】
増殖連動型の生成物の生成を支持する代謝変調を同定し、設計するための1つの計算方法は、OptKnock計算フレームワークである(Burgardら、Biotechnol Bioeng、84:647−57(2003年))。OptKnockは、対象生成物を過剰生成する遺伝的に安定した微生物をもたらす遺伝子欠損戦略を示唆する、代謝のモデル化とシミュレーションのプログラムである。具体的には、このフレームワークは、微生物の完全な代謝および生化学ネットワークまたはそのいずれかを検討して、所望の生化学物質を細胞増殖の強制的な副生成物にさせる遺伝子操作を示唆する。戦略的に配置された遺伝子欠損またはその他の機能的遺伝子破壊によって、生化学的生成を細胞増殖に連動させることにより、バイオリアクタに長時間入れた後、操作された菌株に課された増殖淘汰圧は、強制的な増殖連動型の生化学的生成の結果として、実績の向上をもたらす。最後に、遺伝子欠損が構成された場合、設計された菌株がその野生型の状態に復帰する可能性はごくわずかであるが、これはOptKnockによって選択された遺伝子がゲノムから完全に除外されるためである。
【0031】
つまり、OptKnockという用語は、ここでは細胞代謝をモデル化するための計算方法およびシステムを指して使用される。OptKnockプログラムは、流束均衡解析(FBA)モデルに特定の制約を組み込むモデルと方法のフレームワークに関する。これらの制約には、例えば定性的反応速度情報、定性的規制情報、およびDNAマイクロアレイ実験データ、またはそのいずれかが含まれる。OptKnockは、例えば、流束均衡モデルによって得られた流束境界を狭め、続いて遺伝子付加または欠損の存在下での代謝ネットワークの性能限界を探ることにより、さまざまな代謝問題の解も計算する。OptKnock計算フレームワークは、代謝ネットワークの性能限界の効果的なクエリを可能にするモデル形式の構築を可能にし、もたらされた混合整数線形計画問題を解決する方法を提供する。ここでOptKnockと称される代謝のモデル化とシミュレーションの方法は、例えば米国特許出願第10/043,440号(2002年1月10日出願)、および国際特許第PCT/US02/00660号(2002年1月10日出願)に記載されている。
【0032】
生成物の増殖連動型の生成を支持する代謝変調を同定および設計するための別の計算方法は、SimPheny(登録商標)と称される代謝のモデル化およびシミュレーションシステムである。この計算方法とシステムは、例えば米国特許出願第10/173,547号(2002年6月14日出願)、および国際特許出願第PCT/US03/18838号(2003年6月13日出願)に記載されている。
【0033】
SimPheny(登録商標)は、コンピュータでネットワークモデルを作り出し、生物系の化学反応を通して質量、エネルギー、または電荷の流束をシミュレートして、その生物系における化学反応のあらゆる機能性を含む解空間を規定し、それによりその生物系に許容される活性の範囲を判定するために使用できる計算システムである。このアプローチは、含まれる反応の既知の化学量論、および反応による最大流束に関連する反応熱力学および容量制約などの制約によって、解空間が規定されるため、制約に基づくモデル化と称される。これらの制約によって規定される空間は、生物系またはその生化学成分の表現型の能力および性質を判定するために問い合わせることができる。凸解析、線形計画法、および例えばSchillingら、J.Theor.Biol.203:229−248(2000年);Schillingら、Biotech.Bioeng.71:286−306(2000年)、およびSchillingら、Biotech.Prog.15:288−295(1999年)などに記載される極限経路(extreme pathways)の計算は、かかる表現型の能力を判定するために用いることができる。
【0034】
上述のように、本発明に適用可能な計算プログラムで使用される制約に基づく1つの方
法は、流束均衡解析である。定常状態条件における流束均衡に基づく流束均衡解析は、例えばVarmaおよびPalsson、Biotech.Bioeng.12:994−998(1994年)に記載されているように行うことができる。流束均衡法は、例えば、FellおよびSmall、J.Biochem.138:781−786(1986年)に記載されている脂肪細胞代謝(MajewskiおよびDomach、Biotech.Bioeng.35:732−738(1990年))に記載されているATP極大化条件下での大腸菌(E.coli)からの酢酸塩分泌、あるいはVanrolleghemら、Biotech.Prog.12:434−448(1996年)に記載されている酵母によるエタノール分泌のシステム特性をシミュレートまたは予測するために、反応ネットワークに適用されてきた。さらにこの方法を用いて、多様な単一炭素源における大腸菌(E.coli)の増殖、またEdwardsおよびPalsson、Proc.Natl.Acad.Sci.97:5528−5533(2000年);EdwardsおよびPalsson、J.Bio.Chem.274:17410−17416(1999年);Edwardsら、Nature Biotech.19:125−130(2001年)に記載されるようにH.インフルエンザ菌(H.influenzae)の代謝を予測またはシミュレートすることもできる。
【0035】
解空間が規定されると、さまざまな条件下での可能な解を判定するために解析することができる。生物系には適応性があり、異なる多くの方式で同じ結果に辿り着くことができるため、この計算法は生物学的現実に合致する。生物系は、全生命系が直面しなければならない基本的制約によって制限されてきた進化機構を通して設計されている。従って、制約に基づくモデル化戦略は、これらの一般的現実を包含する。さらに、制約の強化によりネットワークモデルにさらなる制限を持続的に課す能力は、解空間のサイズの減少につながり、それにより生理学的機能または表現型の推測精度を向上させる。
【0036】
ここで提供される教示および指針を前提に、当業者は、代謝のモデル化およびシミュレーションのためのさまざまな計算フレームワークを利用し、生化学的生成物を設計して増殖連動型の生成を実施することができるであろう。かかる代謝のモデル化およびシミュレーションの方法には、例えばSimPheny(登録商標)およびOptKnockとして上記に例示される計算システムが含まれる。本発明の説明を簡単にする目的で、方法と菌株は、モデル化とシミュレーションのためのOptKnock計算フレームワークに関連して記載される。当業者は、OptKnockを使用しての代謝変調の同定、設計、および実施を、当技術分野で周知のその他のかかる代謝のモデル化およびシミュレーションの計算フレームワークおよび方法にどう適用するかを理解するであろう。
【0037】
増殖を生化学的生成物の生成に強制的に連動させる細胞または生物の能力は、コンピュータによるモデルを使用して計算された典型的な代謝ネットワークの生化学的生成の限界に照らして説明することができる。これらの限界は、制限基質の摂取速度をその実験的に測定された値に修正し、それぞれの到達可能な増殖レベルで生化学的生成の最大および最小速度を計算することによって得られる。図1に示されるように、目的の生化学物質の生成は、一般的に、細胞内の資源に関してバイオマス形成と直接競合する。これらの状況もとで、強化された生化学的生成速度は必然的に最大増殖速度には及ばなくなる。OptKnockなど上記の代謝のモデル化およびシミュレーションプログラムにより示唆される遺伝子破壊は、図1に示されるように、野生型菌株の代謝挙動の変化を強いる許容解の境界を制限するよう設計されている。所定の菌株の実際の解の境界は、基質の摂取速度(1つまたは複数)が増加あるいは減少すると拡大または縮小するが、それぞれの実験値は、その計算された解の境界内に位置する。これらのような座標は、設計された菌株がその性能限界にどの程度近づいているかの正確な予測を可能とし、改善の余地がどの程度であるかも示される。
【0038】
OptKnock数学的フレームワークは、図1で説明されるように、増殖連動型の生化学的生成をもたらす遺伝子欠損を特定するために、ここで例示されている。この手順は、支配する物理化学的制約を連続的に課すことによって、細胞系が示すことができる可能な表現型の範囲を狭める、制約に基づく代謝のモデル化を基礎としている(Priceら、Nat Rev Microbiol、2:886−97(2004年))。上述のように、制約に基づくモデルおよびシミュレーションは当技術分野で周知であり、一般的に、ネットワークの化学量論を前提に、特定細胞目標の最適化を引き合いに出して、考えられる流束分布を示唆する。
【0039】
つまり、代謝物の組N={1,…,N}と代謝反応の組M={1,…,M}で構成される定常状態の代謝ネットワークの総反応流束として定量化された細胞目標の極大化は、数学的に以下のように表現される。
【0040】
【数2】
【0041】
式中、Sijは反応jにおける代謝物iの化学量論係数であり、vjは反応jの流束であり、v基質取り込みは制限基質(1つまたは複数)の想定あるいは測定された摂取速度(1つまたは複数)を表わし、vatp維持は非増殖関連のATP維持要求である。ベクトルvは内向きおよび外向き流束の双方を含む。この研究では、細胞目標は、しばしばバイオマス形成に必要とされる比率での生合成前駆体の放出となるよう想定される(Neidhardt、F.C.ら、第2版、1996年、ワシントンD.C.:ASM Press.2v.(xx、2822、lxxvi))。流束は一般的に、バイオマス形成が、生成されたバイオマス g/gDW時間または1/時間として表現されるように、1gDW時間(乾燥重量グラム×時間)当たりで報告される。
【0042】
遺伝子欠損のモデル化、従って反応の排除は、まず制約に基づくアプローチへのフレームワークに2値変数の組み込みを行う(Burgardら、Biotechnol Bioeng、74:364−375(2001年);Burgardら、Biotechnol Prog、17:791−797(2001年))。これらの2値変数、
【0043】
【数3】
【0044】
は、反応jが活性の場合は1の値を、不活性の場合は0の値を想定する。以下の制約、
【0045】
【数4】
【0046】
によって、反応流束vjが、変数yj=0の場合のみゼロに設定されることが確保される。
あるいは、yj=1のとき、vjは下限vjminと上限vjmaxとの間の任意の値を想定できる。ここでvjminおよびvjmaxは、上記のネットワーク制約の対象となるあらゆる反応流束をそれぞれ最小化、最大化することにより同定される(Mahadevanら、Metab Eng、5:264−76(2003年))。
【0047】
最適な遺伝子/反応破壊は、得られたネットワークの最適増殖解が関心化学物質を過剰生成するように、活性反応(yj=1)の組を選定する二段階最適化問題を解決することによって同定される。概略的に、この二段階最適化問題は図2に説明されている。数学的には、この二段階最適化問題は以下の二段階混合整数最適化問題として表現される。
【0048】
【数5】
【0049】
式中、v化学物質は、例えばコハク酸塩またはその他の生化学的生成物など所望の対象生成物の生成であり、Kは許容される遺伝子破壊の数である。K=0の設定は全ネットワー
クの最大バイオマス解を返し、K=1の設定は得られたネットワークがその最大バイオマス収率を前提として最大過剰生成を伴うように、単一遺伝子/反応破壊(yj=0)を同定する。最後の制約によって、得られたネットワークが最小バイオマス収率を満たすことが確保される。Burgardら、Biotechnol Bioeng、84:647−57(2003年)は、モデル形成および解法についてのより詳細な説明を提供している。何百もの2値変数を含む問題は、GAMS(Brookeら、GAMS Development Corporation(1998年))を通してアクセスできるCPLEX8.0(GAMS:The Solver Manuals。2003年:GAMS
Development Corporation)のモデル化環境をIBM RS6000−270ワークステーション上で用いて、数分から数時間程度で解決できる。OptKnockフレームワークは、生化学物質過剰生成のための有望な遺伝子欠損戦略を同定することが既に可能で(Burgardら、Biotechnol Bioeng、84:647−57(2003年);Pharkyaら、Biotechnol Bioeng、84:887−899(2003年))、代謝および規制モデル化のフレームワークのさらなる改善を必然的に網羅する体系的なフレームワークを確立する。
【0050】
前述の二段階OptKnock問題のいかなる解も、1組の破壊される代謝反応を提供する。その組内の各反応の排除、あるいは代謝改変により、生物の増殖相において強制的な生成物としてコハク酸塩をもたらすことができる。反応が既知であるため、二段階OptKnock問題の解は、反応の組内の各反応を触媒する1つ以上の酵素をコード化する1つまたは複数の関連遺伝子も提供する。反応の組および各反応に参加する酵素をコード化する対応する遺伝子の同定は、通常は自動プロセスであり、反応と、酵素とコード化遺伝子との間の関係を有する反応データベースとの相互関係によって達成される。
【0051】
同定されると、増殖連動型のコハク酸塩の生成を達成するために破壊される反応の組は、対象細胞または生物において、その組内の各代謝反応をコード化する少なくとも1つの遺伝子の機能破壊によって実施される。前述のように、反応の組の機能破壊を達成するための1つの特に有用な手段は、コード化遺伝子それぞれの欠損によるものである。しかしいくつかの例では、例えば突然変異、プロモータや調整因子のcis結合部位などの調整領域の欠損、任意の数の位置でのコード化配列の切断など、その他の遺伝子異常によって反応を破壊することが有益となり得る。遺伝子の組の完全な欠損に満たないこととなるこれら後者の異常は、例えば、コハク酸塩共役の迅速な評価が望まれる場合、または遺伝子の復帰が起こる可能性が低い場合に有用となる。
【0052】
増殖連動型のコハク酸塩またはその他の生化学生成物の生成をもたらし得るさらなる反応の組の破壊または代謝改変をもたらす、上記の二段階OptKnock問題のさらなる生成的な解を同定するために、整数切捨てと称される最適化方法が実施できる。この方法は、各反復に整数切捨てと称されるさらなる制約を組み込み、上記に例示されるOptKnock問題を反復して解決することにより進行する。整数切捨て制約は、解法が、生成物生合成を増殖に強制的に共役する、以前の任意の反復において同定されたのと全く同じ反応の組を選定するのを効果的に防止する。例えば、以前に同定された増殖連動型の代謝改変が、破壊される反応1、2、および3を特定する場合、y1+y2+y331という制約は同じ反応が後の解で同時に検討されるのを防止する。整数切捨て方法は、当技術分野で周知であり、例えば、Burgardら、Biotechnol Prog、17:791−797(2001年)に記載されている。代謝のモデル化およびシミュレーションのためのOptKnock計算フレームワークとの組み合わせで、その使用に関連してここに記載されている全ての方法と同様に、反復計算解析における冗長性を減少させる整数切捨て方法も、例えばSimPhenyを含む当技術分野で周知の他の計算フレームワークとともに適用することができる。
【0053】
上記の形式の制約は、以前に同定された組を含む、より大きい反応の組の同定を防ぐ。例えば、さらなる反復で上記の整数切捨て最適化方法を使用すると、破壊される反応1、2、および3を特定した4反応の組は、これらの反応が以前に同定されているため、同定することが防がれる。生成物の増殖連動型の生成につながる全ての可能な反応の組を確実に同定するために、整数切捨て方法の修正が行われた。
【0054】
つまり、修正された整数切捨て法は、反復「ゼロ」で始まり、これは野生型ネットワークの最適増殖で目的の生化学物質の最大生成を計算する。この計算は、K=0でのOptKnock解に相当する。次に単一の遺伝子破壊が検討され、2つのパラメータの組、objstoreiterおよびystoreiter,jが導入されて、各反復iterで目標機能(vchemical)および反応オン−オフ情報(yj)がそれぞれ保存される。次に、各反復で、OptKnock公式に以下の制約が引き続き付与される。
【0055】
【数6】
【0056】
上記の方程式では、εとMはそれぞれ小さい数および大きい数である。一般的には約0.01に設定でき、Mは約1000に設定できる。しかし、これらの数値よりも小さい数や大きい数も使用できる。Mは、制約が以前に同定された遺伝子破壊戦略のみを拘束することができることを確実にし、一方εは、以前に同定された戦略への遺伝子破壊の追加が、最適増殖での生化学的生成において少なくともεの増加をもたらさなければならないことを確実にする。このアプローチは単一の欠損戦略が野生型菌株を改良するのに失敗したときは、常に2つの欠損へと移行する。2つの欠損戦略が野生型菌株を改良しない場合は、次に3つの欠損が検討され、以下同様である。最適増殖での所望の生化学的生成として表わされる最終結果は、少なくとも1つの遺伝子破壊の分だけお互いに異なる別個の欠損戦略の順位付けられたリストである。この最適化手順、および破壊された場合に生化学的生成物の増殖連動型の生成をもたらす多種多様な反応の組の同定は、下記の例にさらに詳しく例示される。例では、増殖連動型のコハク酸塩の生成がさらに例示される。しかし、ここで提供される教示および指針を前提に、当業者は、ここで例示される方法および代謝工学設計が、細胞または微生物増殖の任意の生化学的生成物への強制的共役に、同等に適用可能であることを理解するであろう。
【0057】
上記に例示され下記の例でさらに説明される方法を使用して、本発明の方法は、対象生化学的生成物の生成を、同定された遺伝子変更を有するように遺伝子操作された細胞または生物の増殖に強制的に共役することを可能にする。この関連で、コハク酸塩の生成を微生物の増殖に強制的に共役する代謝変調が同定されている。同定された代謝変調によって構成された微生物菌株は、指数増殖相において高いレベルのコハク酸塩を生成する。これらの菌株は、連続式発酵プロセスでの商業用のコハク酸塩の生成において、前述の負の選択圧を受けることなく有利に使用できる。
【0058】
従って、本発明の方法は、OptKnockまたはSimPhenyから選択されるコンピュータを利用した方法によって同定される代謝改変の組を提供する。この代謝改変の組は、例えば遺伝子欠損による破壊などの1つ以上の代謝反応の機能破壊を含むことができる。代謝改変は、表1に挙げられる代謝改変の組から選択できる。
【0059】
安定的な増殖連動型のコハク酸塩の生成を有する非自然発生的な微生物を生成する方法も提供される。この方法は、(a)指数増殖中にコハク酸塩の生成を要する代謝改変の組をコンピュータで同定するステップと、(b)コハク酸塩の生成を要する代謝改変の組を含有するように微生物を遺伝子改変し、遺伝子改変された微生物を培養するステップとで構成される。培養は、コハク酸塩の生成を要する条件下で、遺伝子改変された微生物を適応進化させるステップを含むことができる。本発明の方法は、細菌、酵母、および菌、またその他さまざまな細胞と微生物に適用可能である。細菌は、例えば、大腸菌(E.coli)、アナエロビオスピルリム・サクシニシプロデュセンス(A.succiniciproducens)、アクチノバチルス・スクシノゲネス(A.succinogenes)、マンヘミア・サクシニシプロデュセンス(M.succiniciproducens)、およびインゲン根粒菌(R.etli)を含むことができる。
【0060】
本発明の方法によって生成された微生物がさらに提供される。さらに本発明は、増殖連動型のコハク酸塩の生成に関連する酵素をコード化する1つ以上の遺伝子破壊を備え、安定的な増殖連動型のコハク酸塩の生成を示す非自然発生的な微生物を提供する。本発明の非自然発生的な微生物は、コハク酸塩生成を微生物の増殖と強制的に連動させる酵素をコード化する遺伝子において、遺伝子破壊が酵素の活性を低下させたときに発生する1つ以上の遺伝子破壊を含み、それにより1つ以上の遺伝子破壊は、非自然発生的な微生物に安定的な増殖連動型のコハク酸塩の生成を与える。
【0061】
非自然発生的な微生物は、表1に挙げられる代謝改変に含まれる1つ以上の遺伝子破壊を有することができる。1つ以上の遺伝子破壊は、欠損であることができる。本発明の非自然発生的な微生物は、表1に挙げられる代謝改変を有する微生物の群から選択できる。本発明の非自然発生的な微生物は、細菌、酵母、菌類、または発酵プロセスに適用可能なその他さまざまな任意の微生物を含む。細菌の例には、大腸菌(E.coli)、アナエロビオスピルリム・サクシニシプロデュセンス(A.succiniciproducens)、アクチノバチルス・スクシノゲネス(A.succinogenes)、マンヘミア・サクシニシプロデュセンス(M.succiniciproducens)、インゲン根粒菌(R.etli)、枯草菌(Bacillus subtilis)、コリネバクテリウム・グルタミクム(Corynebacterium glutamicum)、グルコノバクテル・オキシダンス(Gluconobacter oxydans)、ザイモモナス・モビリス(Zymomonas mobilis)、ラクトコッカス・ラクティス(Lactococcus lactis)、ラクトバチルス・プランタルム(Lactobacillus plantarum)、ストレプトマイセス・コエリコロル(Streptomyces coelicolor)、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、シュードモナス・フルオレッセンス(Pseudomonas fluorescens)、およびシュードモナス・プチダ(Pseudomonas putida)から選択される種が含まれる。酵母の例には、サッカロマイセス・ケレウィシエ(Saccharomyces
cerevisiae)、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)、クリベロマイセスラクティス(Kluyveromyces lactis)、クリベロマイセス・マルキシアヌス(Kluyveromyces marxianus)、アスペルギルス・テレウス(Aspergillus terreus)、クロコウジカビ(Aspergillus niger)、およびピキア・パストリス(Pichia pastoris)から選択される種が含まれる。
【0062】
増殖連動型のコハク酸塩の生成を有する微生物は、ここで大腸菌(E.coli)の遺伝的背景に関連して例示される。しかし395の微生物ゲノムと、多種の酵母、菌類、植物、および哺乳類ゲノムを含む550種を超える完全なゲノム配列が利用可能であり、(
中でも半数以上はNCBIなどの公共データベースで入手可能である)、例えばオーソログ、パラログ、および非オーソロガス遺伝子置換体を含む代替種の1つ以上の遺伝子のホモログ、および生物間での遺伝子変更の置き換えの同定は日常的で、当技術分野で周知である。従って、大腸菌(E.coli)など特定の生物に関連してここで記載されている増殖連動型のコハク酸塩の生成を可能にする代謝変調は、原核および真核生物などを含む他の微生物にも容易に適用できる。ここで提供される教示および指針を前提に、当業者は1つの生物において例示される代謝変調が他の生物にも同等に適用できることを理解するであろう。
【0063】
例えばコハク酸塩の生成は、大腸菌(E.coli)において、ここでADHErと称される反応を触媒する酵素をコード化している1つ以上の遺伝子、およびLDH_Dと称される反応を触媒する酵素をコード化している1つ以上の遺伝子の欠損または機能除去により、指数増殖と連動させることができる。表2に示されるように、adhE反応を触媒する酵素をコード化する大腸菌(E.coli)遺伝子は、adhEまたはb1241である。また、LDH_D反応を触媒する酵素をコード化する2つの大腸菌(E.coli)遺伝子も表2に示されている。これら2つのLDH_D関連遺伝子は、b2133およびb1380である。b1380の一般名はldhAである。b2133遺伝子はldhAのオーソログである。増殖連動型のコハク酸塩の生成を示す代謝的に操作された大腸菌(E.coli)を生成するために、ADHErおよびLDH_D反応それぞれを触媒する少なくとも1つの酵素をコード化する遺伝子は、機能的に破壊されていないければならない。かかる破壊は、例えばb1241遺伝子(adhE)、およびb1380遺伝子(ldhA)、そのオーソログb2133、またはb1241とb2133の両方のいずれかを削除することによって行うことができる。大腸菌(E.coli)以外の細胞または生物における増殖連動型のコハク酸塩の生成では、関心種におけるADHErおよびLDH_Dに相当する反応をコード化する遺伝子は、機能的に破壊することができる。類似代謝経路を有する生物の場合、かかる破壊は、例えばb1241と相同の種、およびb2133またはb1380遺伝子のいずれかを削除することによって達成できる。前述のように、かかる相同体はオーソログおよび非オーソロガス遺伝子置換体またはそのいずれかを含むことができる。関心種に代替代謝経路が存在する場合などいくつかの場合には、機能破壊は例えば、言及される反応に代わる類似するが非同一の代謝反応を触媒するパラログの欠損によって達成できる。異なる生物間の代謝ネットワークの間には一部の差異があるため、当業者は、異なる生物間で破壊される実際の遺伝子は異なってよいことを理解するであろう。しかしここで提供される教示と指針を前提に、当業者は、本発明の方法が、生物間の同種の代謝変調を同定し、コハク酸塩の生合成の増殖への共役を促進する関心種の生物を構成するために、全ての微生物に適用できることも理解するであろう。
【0064】
本発明は、本文書において、一般に代謝反応、反応物質、またはその生成物に関連して、あるいは特に前述の代謝反応、反応物質、または生成物に関連する1つ以上の遺伝子に関連して説明される。本文書において特に明記しない限り、当業者は反応への言及は、その反応の反応物質および生成物への言及ともなることを理解するであろう。同様に、本文書において特に明記しない限り、反応物質または生成物への言及は、その反応にも言及し、これらの代謝要素のいずれかに対するその言及は、遺伝子または言及される反応、反応物質、または生成物の触媒となる酵素をコード化する1つまたは複数の遺伝子にも言及する。同様に、周知の代謝生化学、酵素学、およびゲノミクスの分野を前提に、ここでの遺伝子への言及は、対応するコード化された酵素とそれが触媒する反応、およびその反応の反応物質と生成物への言及ともなる。前述されさらに後述されるように、例となる反応、反応の学名、反応物質、生成物、補助因子、および増殖連動型のコハク酸塩の生成に関与する反応を触媒する酵素をコード化する遺伝子は、表1、2、および3に記載されている。
【0065】
本発明は、増殖連動型のコハク酸塩の生成を有する微生物を提供する。コハク酸塩の生成は、細胞の代謝経路を遺伝的に変更することによって、微生物の指数増殖相と強制的に関連付けられる。遺伝子変更は、コハク酸塩を増殖相における強制的な生成物とする。指数増殖における高いレベルのコハク酸塩の生合成につながる代謝変調または変換の組は、表1に例示されている。組内の各変更は、機能的に破壊されるべき必要な代謝反応に対応している。各組内の全ての反応の機能破壊は、増殖相中に、操作された菌株によるコハク酸塩の強制的な生成をもたらす。言及される表1の変更に対応する反応、および大腸菌(E.coli)においてそれらを潜在的にコード化する1つまたは複数の遺伝子は、表2に記載されている。表3は、表2の反応で言及される反応物質、補助因子、および生成物の完全な生化学名を提供している。
【0066】
例えば、表1に例示される各菌株では、増殖共役するコハク酸塩の生成のために発生させることができる代謝変調は、各行に示されている。これらの変更は、1〜6、あるいはそれ以上の反応の機能破壊を含む。具体的には、非自然発生的な代謝の遺伝子型を有する187の菌株が表1に例示されている。これらそれぞれの非自然発生的な変更は、適切な培養条件下で、微生物の指数増殖相中に、野生型菌株よりも向上したレベルのコハク酸塩の生成をもたらす。適切な培養条件には、例えば特定の炭素源または反応物質の可用性や、適応進化など、下記の例でさらに例示されるものが含まれる。
【0067】
ここで提供される教示と指針を前提に、当業者は、酵素反応を破壊するために、反応に関与する1つ以上の酵素の触媒活性を破壊することが必要であることを理解するであろう。破壊は、例えばコード化遺伝子の欠損、またはコード化遺伝子配列のちの1つ以上への遺伝子変更の組み込みを含むさまざまな手段によって行うことができる。破壊の対象となるコード化遺伝子は、触媒活性に関与する酵素をコード化する遺伝子のうちの1つ、いくつか、あるいは全てとできる。例えば、単一の酵素が対象となる触媒活性に関与する場合、破壊は、コード化遺伝子産物の触媒活性を減少または破壊する遺伝子変更によって行うことができる。同様に、単一の酵素が、異種結合を含む多重結合の場合、破壊は、コード化遺伝子産物の1つまたは全てのサブユニットの機能を減少または破壊する遺伝子変更によって行うことができる。活性の破壊は、活性複合体を形成するための1つ以上のサブユニットの結合活性の損失、または多量体の触媒サブユニットの破壊、あるいはその両方によって達成できる。多量体タンパク質の会合と活性のその他の機能を対象として、本発明の代謝反応を破壊することもできる。かかるその他の機能は、当業者には周知である。さらに、単一のポリペプチドまたは多量体の機能のいくつかまたは全ては、本発明の反応または代謝改変に関与する1つ以上の酵素の触媒活性を減少させるか、または無効にするために、本発明に従って破壊することができる。同様に、本発明の反応または代謝改変に関与するいくつかまたは全ては、対象とされる反応が破壊されるとすれば、破壊することができる。
【0068】
ここで提供される教示と指針を前提に、当業者は、共通遺伝子によって、また類似あるいは実質的に同様の活性を示すその遺伝子の1つ以上のオーソログによってコード化される反応を減少または排除することによって、酵素反応を破壊できることも理解するであろう。共通遺伝子および全てのオーソログの両方を減少させることで、対象とされる反応の完全な停止をもたらすことができる。しかし、共通遺伝子または1つ以上のオーソログのいずれかの破壊は、増殖のコハク酸塩の生合成への共役を推進するのに十分なだけの、対象とされる反応の触媒活性の減少をもたらすことができる。多様な代謝改変の対象となる触媒活性をコード化する共通遺伝子、およびそのオーソログの双方がここに例示されている。当業者は、対象とされる代謝反応の酵素をコード化する遺伝子のいくつかまたは全ての破壊は、本発明の方法において実施でき、また増殖連動型のコハク酸塩の生成を達成するために本発明の非自然発生的な微生物に組み込むことができることを理解するであろう。
【0069】
従って本発明は、コハク酸塩生成を前記微生物の増殖と強制的に連動させる代謝改変の組を有する非自然発生的な微生物をさらに提供する。この代謝改変の組は、
【0070】
【化3】
【0071】
のいずれかで構成される反応の組から選択される各反応を触媒する酵素をコード化する1つ以上の遺伝子破壊を含み、前記微生物は安定的な増殖連動型のコハク酸塩の生成を示す。特定の反応を触媒する役割を果たす酵素をコード化する遺伝子の一般名は、括弧内に記されている。代謝改変(e)PTAr、PYK、ATPS4r、SUCD1i、(f)PTAr、PYK、GLCpts、または(g)PTAr、PYK、GLCpts、ADHEr、LDH_Dを有する非自然発生的な微生物は、反応DHAPT(dha)を触媒する酵素をコード化する少なくとも1つの遺伝子破壊をさらに含むことができる。pykAおよびpykFは、潜在的にPYK反応を実行することができる個々の酵素をコード化する遺伝子であることに留意されたい。従って、pykAおよびpykFの少なくとも1つ、あるいは両方を除去して、PYKがコハク酸塩の生成が細胞増殖に連動しなくなることを阻止する必要がある。あるいは、反応PFL、ATPS4r、SUCD1i、およびDHAPTは、複数の遺伝子にコード化されるタンパク質複合体によって実行される。pfl、atp、sdh、またはdha遺伝子クラスタから、1つまたは遺伝子の組み合わせをそれぞれ削除することは、従って、ATPS4r、SUCD1i、またはDHAPT反応を破壊するのに十分である。
【0072】
つまり、上記に例示される遺伝子、およびそれらの遺伝子の多量体内のそれらの同種サブユニット、それらのオーソログ、およびそれらの遺伝子産物に触媒される反応との関連に関して、ADHrは1つの遺伝子b1241(adhE)にコード化される酵素によって触媒される。LDH_Dは、オーソログb2133を有する1つの遺伝子b1380(ldhA)の産物によってコード化される。PFL活性は、4つの遺伝子b0902、0903、b3952、およびb3951(集合的にpflとして表わす)にコード化される酵素サブユニットを必要とする。b3114は、b0903のオーソログである。PTArは、オーソログb2458を有する1つの遺伝子b2297(ackA−pta)の産物にコード化される。PYKは、2つの異なるオーソロガス遺伝子b1854(pykA)およびb1676(pykF)の産物によってコード化される。これら双方は、大腸菌(E.coli)において活性であることが示されている。SUCD1i活性は、4つの遺伝子b0721、b0722、b0723、およびb0724(集合的にsdhとして表わす)にコード化される酵素サブユニットを必要とする。ATPS4rは、6つの遺伝子b3731〜b3739(集合的にatpとして表わす)にコード化されるマルチサブユニット酵素によって触媒される。DHAPT活性は、5つの遺伝子b1198、b1199、b1200、b2415、およびb2416(集合的にdhaとして表わす)にコード化される酵素サブユニットを必要とする。GLCpts活性は、9つの遺伝子b2
415、b2416、b2417、b1817、b1818、b1819、b1101、b0679、およびb1621(集合的にptsGとして表わす)にコード化される酵素サブユニットを必要とする。反応ATPS4r、SUCD1i、PFL、およびDHAPTは、複数の遺伝子にコード化されるタンパク質複合体によって実行されるので、atp、sdh、pfl、またはdha遺伝子クラスタから1つまたは遺伝子の組み合わせをそれぞれ削除することは、従って、反応を破壊するのに十分である。その他の事例では、文献の情報に基づき、大腸菌(E.coli)において一次反応活性を担う遺伝子が選定された。
【0073】
従って本発明は、コハク酸塩生成を微生物の増殖と強制的に連動させる代謝改変の組を有する非自然発生的な微生物も提供するが、この代謝改変の組は、(a)adhE、ldhA;(b)adhE、ldhA、ackA−pta;(c)pfl、ldhA;(d)pfl、ldhA、adhE;(e)ackA−pta、pykF、atpF、sdhA;(f)ackA−pta、pykF、ptsG、または(g)ackA−pta、pykF、ptsG、adhE、ldhA、のいずれかを含む遺伝子の組またはそのオーソログから選択される1つ以上の遺伝子破壊で構成され、この微生物は安定的な増殖連動型のコハク酸塩の生成を示す。さらに、pykA、atpH、sdhB、またはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成され、代謝改変(e)ackA−pta、pykF、atpF、sdhAをコード化する遺伝子を有する非自然発生的な微生物、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成される代謝改変(f)ackA−pta、pykF、ptsGをコード化する遺伝子を有する非自然発生的な微生物、あるいはpykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成される代謝改変(g)ackA−pta、pykF、ptsG、adhE、ldhAをコード化する遺伝子を有する非自然発生的な微生物も提供される。
【0074】
本発明の非自然発生的な微生物は、増殖連動型のコハク酸塩の生成に使用できる。本発明の増殖連動型のコハク酸塩の生成株を用いて、商業的な量を含み基本的に任意の量を合成することができる。本発明の微生物はコハク酸塩の増殖に強制的に連動させるため、連続式または近連続式の増殖プロセスが、生合成によるコハク酸塩の生成に特に有用である。かかる連続式や近連続式の増殖プロセスは、上述のとおりであり、また下記の例で示されている。連続式や近連続式の微生物増殖プロセスは、当技術分野で周知でもある。つまり、連続式や近連続式の増殖プロセスは、指数増殖または対数増殖において微生物を維持することを伴う。手順には、EvolugatorTMエボリューションマシン(Evolugate LLC、フロリダ州ゲーンズビル)、発酵槽などの装置の使用が含まれる。さらに、微好気的条件下での振盪フラスコ発酵および増殖も利用できる。ここで提供される教示と指針を前提に、当業者は、増殖連動型のコハク酸塩の生成を行う微生物は、当技術分野で周知である多種多様なプロセスや装置を使用して、多種多様な条件下で、多種多様な設定で使用できることを理解するであろう。
【0075】
一般的に、連続式および/または近連続式のコハク酸塩生成は、本発明の非自然発生的な増殖連動型のコハク酸塩の生成を行う生物を十分な栄養物と培地で培養して、指数増殖相において増殖を維持またはほぼ維持することを含む。かかる条件下での連続培養は、例えば1、2、3、4、5、6、または7日、あるいはそれ以上を含むことができる。さらに連続培養は、1、2、3、4、または5週、あるいはそれ以上の週から数ヶ月までを含むことができる。連続や近連続培養条件は、これら例示される期間の間の全ての時間間隔も含み得ることを理解すべきである。
【0076】
コハク酸塩は、上記に例示されている連続や近連続培養の期間中の任意の時点で採取または単離できる。下記の例で示されるように、微生物が連続や近連続の増殖相に長く維持
されるほど、比例的に大きい量のコハク酸塩が生成できる。
【0077】
従って本発明は、微生物の増殖に共役されたコハク酸塩の生成方法を提供する。この方法は、(a)指数増殖相のもと、十分な量の栄養物と培地の中で、
【0078】
【化4】
【0079】
のいずれかからなる遺伝子の組またはそのオーソログから選択される1つ以上の遺伝子破壊で構成されるコハク酸塩生成を微生物の増殖と強制的に連動させる代謝改変の組で構成される非自然発生的な微生物を培養するステップであり、微生物は安定的な増殖連動型のコハク酸塩の生成を示すステップと、(b)非自然発生的な微生物から生成されたコハク酸塩を単離するステップとで構成される。代謝改変(5)ackA−pta、pykF、atpF、sdhAをコード化する遺伝子は、pykA、atpH、sdhBまたはdhaKLMから選択される少なくとも1つの遺伝子破壊をさらに備えることができる。代謝改変(6)ackA−pta、pykF、ptsGをコード化する遺伝子は、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊をさらに備えることができる。代謝改変(7)ackA−pta、pykF、ptsG、adhE、ldhAをコード化する遺伝子は、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊をさらに備えることができる。
【0080】
本発明のさまざまな実施例の作用に実質的に影響しない修正も、ここに提供される本発明の定義の範囲に含まれることは当然である。従って、以下の例は本発明を説明することを目的とし、制限することを目的としていない。
【実施例】
【0081】
実施例I
増殖連動型のコハク酸塩の生成を有する微生物
本実施例では、前述の方法によって同定した代謝工学的戦略について述べる。全体では、百以上の戦略が同定されている。増殖連動型のコハク酸塩の生成を引き起こす同定された代謝変調の詳細な一覧は、以下の表1〜3(本実施例の後)に見ることができ、OptKnockにより同定された反応欠損の組み合わせ(表1)、表1において言及される各反応と一致する化学量論および遺伝子(表2)、および表2の略語と一致する代謝物名の一覧(表3)を記述する。特にここで述べる方法を実証するために有益な設計は、(1)直感的設計(低リスク)、(2)非直感的好気性設計(中リスク)、(3)非直感的嫌気性設計(より高リスク)の3項目に分類される。戦略によって、10 mmol/gDW/hrの基底グルコース摂取速度に対する、好気性または嫌気性条件のいずれかを検討する。各設計に対する解の境界は、実現可能な増殖速度ごとに、別々にコハク酸塩生成を極大化および極小化することで、各設計に対する解の境界が与えられる。解の境界の右端部は、最大バイオマス収率と同義の「最適増殖」解に一致する。システムの直線性のため、グルコース収率が任意の基礎とは異なる場合、その結果は比例的に変わる。増殖連動型の
コハク酸塩の生成を持つと同定された菌株の作成および培養の手順も、さらに以下で述べる。
【0082】
直感的設計
OptKnock技術により同定された4つの直感的菌株設計に対する生化学的生成を、図3で述べる。境界は豊富な二酸化炭素の存在下での嫌気条件を想定する。これらの菌株(すなわち、pfl、ldh、adhEの名が付けられた菌株以外の全て)のうち3つは、その他で作成されているため、それらの菌株の評価に関連するリスクは最小とみなした。しかし菌株を特異的に作成するその3つ全てに、増殖共役コハク酸塩の生成は見られず、コハク酸塩の安定的な増殖共役の生成も見られなかった。さらに、菌株を特異的に作成するその3つ全てが、本明細書で述べるような、さらに増殖連動型のコハク酸塩生成を増大するような適応進化にはさらされていなかった。したがって本発明のadhE、ldh;adhE、ldh、pta、およびpfl、ldhの菌株は、以前に作成された菌株とは別個である。
【0083】
示唆した設計の簡単な説明を次に述べる。示唆した各突然変異体の最大増殖シミュレーションに一致する交換流束を表4に示す。
【0084】
adhE、ldhA(青)−この特定の突然変異体は、SanchezらのMetab
Eng、7:229−39(2005年)およびSanchezらのBiotechnol Prog、21:358−65(2005年)にて言及され、SBS110MGと呼ばれている。異種性のpycは、その最終生成菌株内で発現した。著者はpycなしでコントロールプラスミドを持つ二重突然変異体に対しわずか0.2 mol/molのコハク酸塩収率を報告した。Pycプラスミドがあると、収率は1.3 mol/molにまで増加した。0.9 mol/molの収率は進化後と予想される。
【0085】
adhE、ldhA、pta(赤)−これらの欠損は、SanchezらのMetab
Eng、7:229−39(2005年)に記載の最も嫌気性の強い生成菌株に存在する。実際の生成菌株もpycプラスミドを内部に持ち、別の遺伝子破壊iclRというグリオシキル酸短絡回路を抑圧する調節遺伝子を含む。シミュレーションで、グリオシキル酸短絡回路の重要性は最小であることが明らかとなる。pycの過剰発現が、おそらく1.6 mol/molという高い収率が発表された理由であろう。コンピュータを利用した解析に基づくと、トリプル欠損変異体の収率は適応進化後には0.9 mol/molと予測され、過去にSanchezらのMetab Eng、7:229−39(2005年)で観察された1.6 mol/molから著しく減少する。
【0086】
pfl、ldhA(灰色)−pfl、ldhA菌株はDonnellyら、Appl Biochem Biotechnol,70−72:187−98(1998年)および最初のOptKnock論文であるBurgardら、Biotechnol Bioeng,84:647−57(2003年)にて言及された。NZN111と呼ばれる菌株は、グルコースを発酵させることができなかったが、最終的にはグルコースの発酵を許すptsG内で自然突然変異が起こった。Hongら、Biotechnol Bioeng、 74:89−95(2001年)のHongおよびLeeは、リンゴ酸酵素をNZN111内で過剰発現し、30〜40%のコハク酸塩収率を達成した。終点でのコハク酸塩生成は小さいはずであるため、NZN111またはその他全てのpfl、ldhでの進化実験は特に有益である。
【0087】
pfl、ldhA、adhE(緑)−この突然変異体は、最初のOptKnock論文のBurgardら、Biotechnol Bioeng、 84:647−57(2003年)にて示唆された。この菌株は評価の終点で主要生成物としてコハク酸塩および
酢酸塩を伴い、〜1.2 mol/molのグルコース収率があるはずである。この菌株は進化の終点で最高のコハク酸塩収率を持つはずであるため、興味深い。ギ酸塩の生成は等量を減少させる原因で、それゆえコハク酸塩形成に有利と広くみなされているため、pfl欠損は半直感的である。ギ酸塩脱水素酵素はシミュレーションでは嫌気的に機能しないため、化学量論モデルではこの点を議論する。SBS110MG(ldhA、adhE突然変異体)を含むpycからfdhF(ギ酸塩脱水素酵素)を除去すると、コハク酸塩収率は1.2〜0.9 mol/molに減少した(Sanchezら、Biotechnol Prog,21:358−65(2005年))。
表4:嫌気条件および基底グルコース摂取率10 mmol/(gDW・hr)を想定した最適増殖用交換流束。正の値は生成を示し、負の値は消費を示す。基底グルコース摂取率(すなわち、10)で入力値を割ることで、全化学物質のモル収率を得ることができる。
【0088】
【表4】
【0089】
非直感的好気性設計
この菌株設計には、好気性条件下での増殖連動型のコハク酸塩の生成のために6つの欠損(すなわち、ackA−pta、pykA、pykF、atpFH、sdhAB、dhaKLM)を必要とする。この欠損によって不可能となる反応には、PTAr、PYK、ATPS4r、SUCD1iおよびDHAPTが含まれる。菌株設計に対する生化学的生成の制限を図4に示し、最適増殖シミュレーション結果を図5で提供する。欠損によって理論上の最大コハク酸塩収率における減少が引き起こされることに留意する。にもかかわらず、この設計によって好気性の生化学的生成に対するatpFHの欠損のさらなる特徴付けが許容されるため、この設計は特に有益である。
【0090】
dhaKLM欠損の背後にある論理的根拠には、シミュレーションの場合を含め、F6PA(f6p≪dha+g3p)およびDHAPT(dha+pep(R)dhap+pyr)の反応によって大きな流束を獲得することである。これらの反応によって、PEPをピルビン酸塩に変換する手段が用意されるが、コハク酸塩形成には所望され得ない。PEPはかなりの高エネルギー化合物であるため、この経路はエネルギー性の観点から現実的であるように思われる。加えて、両反応の原因である酵素が特徴付けられている(Schurmannら、J Biol Chem,276:11055−61(2001年)、
Gutknechtら、Embo J,20:2480−6(2001年))。これらの酵素はおそらく完全な解糖流束を持ち得ないであろうが、最も保守的な方法は、これらの流束がdhaKLMを除去することで機能しなくなることを保証することである。実際には、この欠損は必要としてもよく、不必要としてもよい。シミュレーションでも、ptsGはdhaKLMに代わるものとして取り除くことができることが明らかになったが、グルコース摂取のためのへキソキナーゼへの完全な依存により、ptsGおよびatpFH両方の欠損を内部に持つ菌株は、増殖が困難になることもある。
表5:基底グルコース摂取率10 mmol/(gDW・hr)を想定した最適増殖シミュレーション用交換流束。正の値は生成を示し、負の値は消費を示す。基底グルコース摂取率(すなわち、10)で入力値を割ることで、全化学物質のモル収率を得ることができる。
【0091】
【表5】
【0092】
非直感的嫌気性設計
この菌株は、嫌気性条件下での増殖連動型コハク酸塩形成のため、5つの欠損(すなわち、ackA−pta、pykA、pykF、ptsG、dhaKLM)を必要とする。欠損によって不可能とされた反応にはPTAr、PYK、GLCpts、およびDHAPTを含む。また、DHAPT欠損(すなわち、dhaKLM)は、実際には任意とできる。欠損組は結果として、ピルビン酸塩の非定型の原因への強制依存のような(例えば、エントナー−ドウドロフ経路、セリンデアミナーゼ、リンゴ酸塩 酵素など)著しい代謝変更を引き起こすと予想される。ptsG、pykA、pykFの突然変異体は、グルコース最小培地で増殖されるとき、生存不可能となりうるため、この設計では幾つかの弱点が示される(Ponceら、J Bacteriol,177:5719−22 (1995年))。シミュレーションはこの発見と矛盾し、グルコースに関する好気性増殖では高
いバイオマス収率が可能であることを示す。この結果は、pykA、pykF、ptsGの突然変異体に対し予測される増殖への障害が起こった場合、欠損に適応をする菌株を強化するため進化戦略が適用できることを示す。
【0093】
嫌気的には、グルコース摂取率が10 mmol/gDW/hrと想定した場合、この菌株が標準の7.6 mmol/gDW/hrの非増殖関連のATP維持必要量に合致する点では不十分であることが、シミュレーションで明らかになる。図5では計算解の境界上のATP維持必要量を緩和する影響を検討する。ここでは、このペンタ−突然変異体が嫌気的に増殖できる場合(例えば、細胞に維持エネルギー必要量を減らすよう強制する増殖選択圧力を適用する)、最適増殖では極度に高いコハク酸塩収率が伴わなくてはならないということが示される。したがって、この設計にはそのリスクを凌ぐ巨大な潜在的上昇傾向がある。加えて、無益サイクルの導入を利用し、図5で描写した解の境界の最も左方の上部で表示した理論上のコハク酸塩収率へ近づけるよう、この菌株の性能を促進することができる。エネルギー維持必要量および嫌気条件なしを想定する最大増殖シミュレーションの交換流束を、表6に示す。
表6:嫌気条件および基底グルコース摂取率10 mmol/(gDW・hr)および非増殖関連維持必要量ゼロを想定した最適嫌気性増殖シミュレーション用交換流束。正の値は生成を示し、負の値は消費を示す。基底グルコース摂取率(すなわち、10)で入力値を割ることで、全化学物質のモル収率を得ることができる。
【0094】
【表6】
【0095】
新しく作成された菌株が好気性環境下で生存可能だが、嫌気条件下では十分には生存可能ではないある特定の例では、通気率の減少にかかわる進化戦略が適用できる。コハク酸
塩生成限界に関し酸素摂取率が徐々に減少する影響を、図6に描写する。通気性の減少によって、理論上の最大コハク酸塩収率(上方左)にますます近い最適増殖点が導かれることに留意する。図7で示されるように、細胞増殖に対しより緊密に共役するコハク酸塩形成は、adhEおよびldhのさらなる欠損で達成することができる。これらの遺伝子はそれぞれADHErおよびLDH_D反応を触媒する。また好気性増殖相(黒線)、次に酸素制限生成相(色付き線)に関する二相発酵戦略が適用できることも、嫌気および好気条件下で、adhE、dhaKLM、ldhを持つ、または持たない菌株の生存率は、さらに示している。
【0096】
増殖連動型のコハク酸塩菌株の生成およびバリデーション
本発明の方法のコンピュータ予測を確認するために、菌株を形成、進化、および試験する。1週間から6週間、一般的には約3週間の増殖条件づけ相を、より複雑な設計に割り当てる。時間がより短い期間には、それに対応してより簡単な設計を条件づけることができる。
【0097】
菌株形成:大腸菌K−12 MG1655は欠損を導入する1つの基準野生型菌株として使用する。例遺伝子破壊を、例えば一つ一つ幾つかの欠損が蓄積されたままの受容菌株へ組み込む。形成された突然変異体が元の野生型に戻る可能性を避けるために、欠損方法論で除去を目的とした遺伝子を完全に取り除く。1つの研究では、4つの予測した非直感的設計のうち2つが形成され、増殖連動型のコハク酸塩の生成を検定する。ここでは、多くても3つの欠損(1)pfl、ldhAおよび(2)pfl、ldhA、adhEを必要とする2つの菌株を形成することができる。別の研究では、4つの共通遺伝子破壊ackA−pta、pykA、pykF、dhaKLMを形成する。さらに別の研究では、atpFHおよびsdhABの追加欠損を行い、その一方で代替の研究ではptsGの追加欠損を行う。非直感的設計の確証試験のため、多くて7つの欠損を取り込む。全体では、すべての望ましい非直感的菌株および研究のために、多くて10個の欠損を行うことができる。
【0098】
適応進化:適応進化手順は、定常期が達成される前の新鮮な培地への回分培養の連続継代による遷延性の指数増殖における細胞の維持に関係する。細胞は希釈し新鮮な培地に移される(すなわち、2 g/L炭素現にM9最小培地)前に、短時間で中定常増殖(A600=0.5)に達することができる。この過程を繰り返し、約500世代までにする。培養試料を取り出し、液体窒素で凍らせ、最適培養密度を進化の過程の間中毎日記録する。表7に各進化に必要な条件を要約する。過去にDonnellyら、Appl Biochem Biotechnol 70−72:187−98(1998年)、Vemuriら、Appl Environ Microbiol 68:1715−27(2002年)で目撃された進化的パターンにおける相違のため、進化は3通り(したがって、全18進化)で行い、それにより潜在的に他より上位の生成の品質を持つ1つの菌株となる可能性がある。適応進化段階には2ヶ月またはそれ以上を要する。また例えば菌株設計によっては適応進化段階が2ヶ月未満のこともある。
表7:各進化に対する望ましい条件。進化は3通りで実行する。
【0099】
【表7】
【0100】
バリデーション:増殖速度(GR)、基質摂取率(SUR)および酸素摂取率(OUR)(好気性の場合)は、進化の過程の間中、10日ごとに標本抽出を行う。前培養を一晩増殖させ、指数増殖中に測定を行う新鮮な回分培養用の種菌として使用する。分光光度計(A600およびA420)を使用して最適密度を測定し、GR、HPLCで時間をかけ炭素源をモニターしSURを判定し、およびポーラログラフの溶解した酸素プローブを使用して呼吸計内で溶解した酸素欠損を測定してOURを判定する。コハク酸塩および副生成物生成をHPLCまたは酵素試験で数量化する。測定は10日の間隔で3通りの進化菌株ごとに行う。試験は進化と同時に進めることができる。
【0101】
菌株安定性の評価:OptKnock法の背後にある増殖共役の生化学的生成概念の結果として、細胞遺伝学的に安定した過剰産生株が発生する。最高のコハク酸塩生成特性および少なくともモデル予測との満足な一致を有する菌株を、1ヶ月ケモスタット内で増殖し、その長期安定性を確認する。M9最小培地を使用し、1.3−Lベンチトップ発酵槽(New Brunswick Scientific、ニュージャージー州、エジソン)内で、約600 mLの稼働範囲で、ケモスタット培養を行う。炭素源濃度を調節し、A600=1.0に一致するするよう細胞密度を達成し、希釈率を最大培養率の約80%に設定し洗い流しの可能性を避ける。無菌空気を好気性増殖に使用し、攪拌速度を使い溶解酸素を飽和の>95%に維持する。OptKnockが嫌気性増殖環境を提案する場合は、窒素/CO2の混合物と共に反応器を連続的に散布し、DOレベルを欠損レベルより確実に下に保つ。CO2も炭酸水素ナトリウム(NaHCO3)の形で培地内に用意する。上記に記載したように各週ごとに代謝挙動を検定する。
【0102】
結論
本明細書に記載するのは、有用な遺伝子欠損目標を産生するためのOptKnock方法論の応用である。コハク酸塩生成および大腸菌(E.Coli)増殖間の結合を確立するため、多様な欠損戦略が列挙されてきた。本方法論は幅広い種類の細胞および大腸菌以外の微生物に適用が可能であり、代謝モデル化およびOptKnock以外のシミュレーションシステムに利用することもできる。2つの相対的には直感的な好気性設計(以下3および4)、1つの非直感的好気性設計(5)、および1つの非直感的嫌気性設計(6)の作成およびバリデーションも記載する。その手順には、各生体触媒の生成特性を実証し強化する適応進化の性能の詳細な評価も含まれる。具体的には、増殖連動型のコハク酸塩の生成として同定された7つの菌種の設計を選択し、それを実行することでコハク酸塩の生合成の結合を導くことができる。
実験のバリデーション用に選択された欠損戦略:
【0103】
【化5】
【0104】
ここに記載された分析でも、進化的方法によって示唆された菌株全てのコハク酸塩生成特性を向上できることが示唆されている。本明細書に記載された分析結果も、一覧の代謝改変の完全な一組未満を取り込んだとき、上記に記載した特定の菌株は、まだ増殖連動型のコハク酸塩生成を示すことを示唆している。したがって、以下の一覧では各設計のために必要な欠損の一最小組に特化する。
上記設計のために必要な欠損の最小組:
【0105】
【化6】
【0106】
sdhおよびatpはコハク酸塩脱水素酵素(SUCD1i)およびATP合成酵素(ATPS4r)活性をそれぞれ大腸菌から排除することができる欠損全てを表す。本明細書に記載された組み合わされた計算および工学プラットフォームは、通常化学量論モデル生物全てに適用可能であり、本明細書で提供された教示および助言によって、当業者は増殖連動型のどんな生化学的生成物をも生成に導く代謝変調のため、何組もの遺伝子操作を設計および実行することができるだろう。
【0107】
実施例II
増殖連動型のコハク酸塩の生成を有する微生物
本実施例では、増殖連動型のコハク酸塩の生成のため、実施例Iに記載のコンピュータを利用した設計した菌株2つの作成および性能について記載する。
【0108】
以下に記載するのは、先述の菌株設計2つを含む生物を作成および特徴付けるのに使用する方法である。つまり、adhE、pflA、およびldh 内に欠損を含むAB3と名づけられた菌株、一方はackA−pta、dhaKLM、ptsG、pykAおよびpykF内に欠損を含むAB4と名づけられた第二の菌株で、両方共に先に記述されている。
【0109】
菌株特徴付け前進化
第一の設定にはpfl、ldh、およびadhEの同時除去が関与する。提唱されるpfl、ldh、adhEの三重突然変異体の理論上の生成制限は、可能な増殖速度ごとにコハク酸塩収率を別々に極大化および最小化することで得られ、図3で野生型菌株の理論上の生成制限と比較する。緑と黒の直線で包囲された領域は、理論上は大腸菌に達成可能なコハク酸塩およびバイオマス生成率を表示する。生成制限の最も右側の部分は「最適増殖」解に一致し、最大バイオマス収率と同義である。システムの直線性のため、グルコー
ス接種率が任意の基礎と異なる場合は、その結果は比例的に変わる。実施例1に先だって記載したように、pf、ldh、adhEの三重突然変異体は完全な嫌気姓発酵が可能であり、その発酵は最適増殖で1.2 mol/molのコハク酸塩モル濃度収率に導かれると予測される。コンピュータで予測した最大値である25%以上の増殖速度で、ノンゼロコハク酸塩収率が不可欠となり、コハク酸塩の生成はこの菌株で緊密に細胞増殖に結合していると予測されることにも留意する。最後に、理論上の最大コハク酸塩収率は細胞破壊からの影響は受けず、したがってこの設計戦略はこの菌株のさらなる代謝工学に負の影響は与えない。
【0110】
第二のOptKnock由来の設計は、嫌気条件下での増殖連動型のコハク酸塩形成のために、5つの欠損(すなわち、ackA−pta、pykA、pykF、ptsG、dhaKLM)を必要とする。先に実施例に記載したように、欠損組は結果として、ピルビン酸塩の非定型の原因への強制依存のような(例、エントナー−ドウドロフ経路、セリンデアミナーゼ、リンゴ酸酵素など)著しい代謝変更を引き起こすと予想される。
【0111】
つまり、欠損を導入する基準野生型菌株として大腸菌K−12MG1655を使用し、菌株を形成する。遺伝子破壊を一つ一つ、幾つかの欠損が蓄積された受容菌株へ組み込む。Datsenkoら、Proc. Nat. Acad. Sci. USA、 6640−6645(2000年)による、DatsenkoおよびWannerのλ Redリコンビナーゼシステムを通して相同組換えを使用しインフレームの欠損を取り込み、菌株を形成した。各欠損後に薬剤抵抗性マーカーは残らず、目標の菌株に多重突然変異が蓄積される。加えて、目的とした遺伝子を完全に取り除き、形成された突然変異体が元の野生型に戻る可能性を避ける。第一の菌株はpfl、ldh、およびadhE内に欠損を持って形成され、一方第二の菌株はackA−pta、pykA、pykF、dhaKLM、およびptsG内に欠損を持つ。図8aで、これらの菌株を形成するために使用された戦略の概要を述べる。
【0112】
進化の前後両方で全ての菌株で振盪フラスコ発酵を行い、菌株の性能を数量化した。最初窒素で培地を散布し、それから隔壁およびクリンプキャップでフラスコを密封し、嫌気条件を得た。増殖が嫌気的に観察されなかった菌株には、通気を限定するための隔壁を通る小さな穴を作り、微好気的条件を適用した。pH7.0(6.78 g/L Na2HPO4、3.0 g/L KH2PO4、0.5 g/L NaCl、1.0 g/L NH4Cl、1 mM MgSO4、0.1 mM CaCl2)の、グルコース2 g/LおよびNaHCO3 20mMで補われたM9最小培地を使用し、実験全てを行った。前培養を一晩増殖させ、指数増殖中に測定を行う新鮮な回分培養用の種菌として使用した。分光光度計(600 nm)を使用して光学密度を測定し増殖速度を判定し、時間をかけて炭素源欠損をモニターしグルコース摂取率を判定した。Linら、Biotechnol. Bioeng.、 775−779(2005年)に従い、グルコースおよびエタノールにた屈折率検出器を、有機酸にはUV検出器を使用し、HPX−087カラム(BioRad)つきのHPLC(島津)でコハク酸塩、副生成物、および残渣グルコースを数量化した。各菌株に対し三通りの培養を行い、報告した測定はその3つの平均である。
【0113】
適応進化手順の実施に関する定量的結論を出すため、進化の前に野生型および操作された菌株の性能を特徴付けた。表8でこの分析からの結果を見ることができる。中間AB1およびAB2(図8a参照)を含む、本研究で形成した多重菌に対し株振盪フラスコ特性を行ったことに留意する。
図8:操作された菌株の増殖速度、加えて消費されたグルコース、形成されたバイオマス、および生成された発酵生成物の量。全データは3つ以上の培養を平均したものであり、信頼区間95%を示唆する。N.D.は測定なし。
【0114】
【表8】
【0115】
操作された菌株は、野生型および変質した発酵特性より遅い増殖速度を示した。AB1およびAB3は厳しい嫌気条件下では、あまり増殖できなかったため、代わりに上記に記載したのと同様に微好気的に増殖した。この観察は、同じ遺伝子型AB1に関する文献報告、NZN111、Stolsら、Appl Environ Microbiol、63:2695−701(1997年)、Hongら、Biotechnol Bioeng、74:89−95(2001年)、Stolsら、Appl Biochem Biotechnol、63−65:153−8(1997年)に一致する。これらの菌株は、この比較的大量の代謝性複製生物によって示唆されるように、ピルピン酸にボトルネックを有するようである。
【0116】
AdhE欠損によってAB3内で消費されるグルコースの量は著しく減少するが、エタノール生成も排除され、その結果より高いコハク酸塩収率となった。突然変異体は成長率および最終バイオマス濃度にかなりの影響を与えたが、菌株AB2およびAB4に振盪フラスコ特徴づけを行うと、完全な嫌気性増殖の間の野生型MG1655と比較し、コハク酸塩収率が約4倍および10倍の増加を示した(図8b)。
【0117】
適応進化菌株
新しく形成された突然変異体の増殖で観察されたのは、以下モデル化分析により先験的に予測したものあり、そのためそれからEvolugatorTM マシーンを使用し菌株を進化させた。つまり、野生型および両方の操作した菌株で適応進化を行った。適応進化手順は、EvolugatorTM進化機械を使用し、Evolugate、LLC(フロリダ州、ゲーンズビル)によって実行された。定常期が達成される前に、新鮮な倍地への回分培養の連続経路による遷延性指数増殖中、この装置で細胞を維持する。最適密度測定および液体取扱いを機械化することで、Evolugatorは大量の培養液量を使用し高速で連続導入を行うことができ、したがって細胞適応度の進化のため、ケモスタットの効率に近づく。しかし、単一の容器内に細胞を維持するケモスタットとは対照的に、Evolugatorは細区画された管のスプール領域内を1つの「反応器」から次へと移動することで機能し、したがって壁増殖に対する選択は排除される。培養の試料を採取し、液体窒素で冷凍し、進化の過程の間中毎日最適培養密度を記録した。
【0118】
AB3の嫌気性増殖は研究室では無視できたが、進化過程の最初の2、3日の間に徐々に酸素付加を減少させ、その条件下での増殖能力を発展させた。大腸菌を嫌気的に進化させることができると実証する対照実験として、野生型MG1655を15日の期間Evolugatorにかけた。図9aに見られるように、日ごとに必要な欠損の数によって推定すると、2.2 hr−1から0.9 hr−1へと約2倍、時間が減少した。さらに
その増殖発展を特徴付けるため、本来の菌株および進化させた(EVG09)菌株の3通りの培養を行った。野生型菌株の培養率は0.351±0.002 hr−1、EVG09の培養率は0.521±0.049 hr−1であった。野生型菌株で観察された満足いく増殖発展を与え、それからAB3菌株を進化させると、図9bに見られるように同じような2倍の時間の減少という結果となった。注目すべき2倍の時間減少が両方の菌株に対し観察された。
【0119】
さらなる菌株の特徴づけ
上記に記載した進化前の元の菌株に行ったように、進化させたAB3を振盪フラスコ発酵で特徴づけた。完全な嫌気性増殖はこの菌株に対しては有意ではなく、EvolugatorTM増殖環境は完全には嫌気性ではないことが示唆された。しかし、EvolugatorTMから取られた試料の生成物特性から、かなりの量の発酵生成物が菌株AB3によって生成されたことが明らかとなり、その進化は完全な好気性からはほど遠く、おそらく微好気性だということが示された(図10)。実行の最後でピルビン酸塩の量はゼロへと下がったため、EvolugatorTM内での時間によって進化前のAB3菌株内に存在したかなりのピルビン酸塩のボトルネックが軽減されていたことを、これらの結果は示している。エタノール、乳酸塩、またはコハク酸塩の生成とは異なり、NADは再生されないため、ピルビン酸塩の分泌は生物に対しほとんど利点はなかい。加えて、酢酸塩の生成とは異なり、エネルギーは再生されない。
【0120】
工業発酵は完全な嫌気条件下ではほとんど実行されない。それゆえ、進化後のAB3菌株を微好気性増殖下でさらに特徴付けた。図11にこれらの増殖条件下での生成物特性を示し、進化前の親株と比較したとき、より高いコハク酸塩形成へ転換が明らかとなる。図12は、発酵生成物におけるコハク酸塩の質量パーセンテージが4倍以上増加し、ピルビン酸塩および酢酸塩においてはそれぞれ約50%および40%の減少を伴うことを示している。コハク酸塩の最終モル収率は1.1 mol/molグルコースで、進化後のコハク酸塩菌株のモデルによって予測された1.2 mol/molグルコース収率に非常に類似していた(上記記述および図3の突然変異菌株の最右境界、参照)。Evolugatorから直接取り出した試料とは対照的に、進化前AB3菌株よりは少ないものの、まだ相当なピルビン酸塩の蓄積があった。微好気性振盪フラスコ内にはおそらくより多くのO2が存在するということを、この結果は示している。特に発展させた嫌気性条件下で、さらに進化の時間を取ることで、例えばピルビン酸塩のボトルネックを一斉に排除することができる。生物を細胞増殖に対し最適化した後、コンピュータ予測はコハク酸塩2 moleから酢酸塩1 moleの発酵特性を示し、さらに菌株特徴づけデータは予測され菌株の性能に到達できることを実証している。
【0121】
結論
本実施例では、増殖連動型のコハク酸塩の生成のためにOptKnock計算枠組みによりコンピュータ設計された、実施例Iに記載した2つの大腸菌株の作成および性能の特徴づけを示す。AB3と名づけられた第一の菌株はadhE、pflA、およびldhA内に欠損を含み、一方第二の菌株AB4はackA−pta、dhaKLM、ptsG、pykA,、およびpykF内に欠損を含む。これらの菌株は、形成中に創造された親菌株AB1およびAB2と同様、欠損が導入された野生型大腸菌株MG1655以上のコハク酸塩収率の増加を示した。実際、AB4菌株はコハク酸塩収率において野生型菌株以上の約10倍の増加を示した。
【0122】
増殖速度を増加させるためEvolugatorTM技術を使用し、野生型大腸菌株MG1655およびMG1655も適応進化を受けた。野生型菌株に対しては、嫌気条件下で進化手順によって増殖速度を約50%向上したが、予想されたように、最終生成物特性に重大な影響はなかった。菌株AB3に対しては、進化段階により増殖速度およびコハク
酸塩収率の両方を増加させ、一方副生成物ピルビン酸塩の分泌はかなり減少した。本研究ではさらにOptKnockコンピュータ法で、結果として大腸菌内のコハク酸塩生成を増加させる遺伝子欠損の組み合わせが同定できるということを実証している。そのうえ、OptKnock設計菌株の性能をコンピュータで予測された過剰生成表現型へ向上させるため適応進化を利用できることを、本発明ではさらに実証している。
【0123】
【表1−1】
【0124】
【表1−2】
【0125】
【表1−3】
【0126】
【表1−4】
【0127】
【表1−5】
【0128】
【表2−1】
【0129】
【表2−2】
【0130】
【表3−1】
【0131】
【表3−2】
【0132】
本明細書を通じ、様々な出版物を括弧内で参考文献として挙げた。本発明に関連する最新技術をより完全に記載するために、これらの出版物全体の開示を参照することで、それにより本明細書内に含まれる。
【0133】
本発明は開示された実施例を参照し記載されているが、上記の詳細で具体的な例示および研究が本発明を例証だけするものであることが、当業者には容易に理解される。当然のことながら、本発明の精神から逸脱することなく様々な変更が行われうる。それゆえ、本発明は以下の請求項によってのみ限定される。
【図面の簡単な説明】
【0134】
【図1】図1は、生化学的生成および細胞増殖間のトレードオフの推定される描写を示す。点AおよびBはそれぞれ、野生型および突然変異菌株の最大バイオマス解を表す。ここで留意すべきは、突然変異菌株は増殖連動型の生成を示すことである。
【図2】図2は、OptKnockの二段階最適化構造を示す。内側の課題によって、ある特定の細胞目標の最適化に基づく流束の配分を行う。それから内側の課題の最適化を可能とする重要な反応への接近を制限することで、外側の問題によって生物工学的目標(例えば、化合物過剰生成)を極大化する。
【図3】図3は、野生型と比較した、4つの突然変異菌株の増殖速度境界に対するコハク酸塩を示す。10 mmol/(gDW・hr)の基底グルコース摂取速度の摂取速度に従い、完全な嫌気条件を想定する。全ての菌株が外側境界に続き、どの境界も野生型菌株に一致するが、adhE、ldh、pta(第一分岐、赤)、pfl ldh、adhE(第二分岐、緑)、adhE、ldh(第三分岐、青)、そしてpfl、ldh(第四分岐、灰色)の順で、左から下方へと分岐する。
【図4】図4は、野生型と比較した、OptKnock由来の突然変異菌株(ackA−pta、atpFH、pykA、pykF、dhaKLM、sdhAB)の増殖速度境界に対するコハク酸塩を示す。10 mmol/(gDW・hr)の基底グルコース摂取速度を想定する。菌株は、ackA−pta,、atpFH、pykA、pykF、dhaKLM、sdhAB(嫌気)(第一直線、緑)、ackA−pta、atpFH、pykA、pykF、dhaKLM、sdhAB(嫌気)(第二直線、赤)、野生型(嫌気)(第三直線、青)、および野生型(嫌気)(第四直線、黒)の順で左から始まる直線に一致する。
【図5】図5は、OptKnock由来の突然変異菌株(ackA−pta、pykA、pykF、ptsG、dhaKLM)の増殖速度境界に対するコハク酸塩を示し、様々な非増殖関連のATP維持必要量に対する野生型と比較した。10 mmol/(gDW・hr)の基底グルコース摂取速度を想定する。菌株は、ackA−pta、pykA、pykF、ptsG、dhaKLM(ATPM=6)(y軸と外側境界との交差、緑)、ackA−pta、pykA、pykF、ptsG、dhaKLM(ATPM=3)(第二直線、赤)、ackA−pta、pykA、pykF、ptsG、dhaKLM(ATPM=0)(第三直線、青)、および野生型(外側境界、黒)の順で、左から始まる直線に一致する。
【図6】図6は、様々な酸素化率での、OptKnock由来の突然変異菌株(ackA−pta、pykA、pykF、ptsG、dhaKLM)の増殖速度境界に対するコハク酸塩を示す。典型的な7.6 mmol/(gDW・hr)の維持エネルギー必要量を課す。10 mmol/(gDW・hr)の基底グルコース摂取速度を想定する。菌株は、ackA−pta、pykA、pykF、ptsG、dhaKLM(O2<2)(第一直線、緑)、ackA−pta、pykA、pykF、ptsG、dhaKLM(O2<5)(第二直線、赤)、ackA−pta、pykA、pykF、ptsG、dhaKLM(O2<10)(第三直線、青)、およびpta、ptsG、f6pa、pyk(O2無制限)(外側境界、黒)の順で、左から始まる直線に一致する。
【図7】図7は、様々な酸素化率での、OptKnock由来の突然変異菌株(ackA−pta、pykA、pykF、ptsG、dhaKLM、ldh、adhE)の増殖速度境界に対するコハク酸塩を示す。典型的な7.6 mmol/(gDW・hr)の維持エネルギー必要量を課す。10 mmol/(gDW・hr)の基底グルコース摂取速度を想定する。菌株は、ackA−pta、pykA、pykF、ptsG、dhaKLM、adhE、ldh(O2<2)(第一直線、緑)、ackA−pta、pykA、pykF、ptsG、dhaKLM、adhE、ldh(O2<5)(第二直線、赤)、ackA−pta、pykA、pykF、ptsG、dhaKLM、adhE、ldh(O2<10)(第三直線、青)、およびackA−pta、pykA、pykF、ptsG、dhaKLM、adhE、ldh(O2無制限)(外側境界、黒)の順で、左から始まる直線に一致する。
【図8】図8は、OptKnock設計のコハク酸塩菌株の進化前を示す。(a)は菌株作成戦略である。AB1およびAB2は中間的な菌株で、各菌株の系譜における最終的な遺伝子破壊の効果を評価するよう特徴付けられてもいる。(b)は3つの独立培養を平均した全実験にわたるコハク酸塩の重量収率である。
【図9】図9は、希釈の頻度から判定した、菌株MG1655およびAB3のおおよその倍加時間を示し、進化の過程全体を通してプロットした。
【図10】図10は、菌株AB3の進化中にEvolugatorから得た試料の生成物特性を示す。ダイヤ(青)がコハク酸塩、三角(赤)がギ酸塩、丸(緑)が酢酸塩である。
【図11】図11は、大腸菌株を微好気性培養した後に存在したコハク酸塩およびほかの発酵生成物の質量(a)およびモル濃度(b)の収率を示し、代謝された全グルコースに関連する。左は野生型MG1655、中央が進化前のAB3、右が進化後のAB3である。
【図12】図12は、進化前(左のバー)および進化後(右のバー)AB3菌株の微好気性培養が見られた後、存在する発酵生成物の質量パーセントを示す。
【特許請求の範囲】
【請求項1】
1つ以上の遺伝子破壊で構成される非自然発生的な微生物であって、前記1つ以上の遺伝子破壊は、前記コハク酸塩生成を微生物の増殖と強制的に連動させる酵素をコード化する遺伝子において、前記遺伝子破壊が前記酵素の活性を低下させたときに発生し、それにより前記1つ以上の遺伝子破壊は、前記非自然発生的な微生物に安定的な増殖連動型のコハク酸塩の生成を与える。
【請求項2】
前記1つ以上の遺伝子破壊が、表1に挙げられる代謝改変で構成される請求項1の非自然発生的な微生物。
【請求項3】
前記1つ以上の遺伝子破壊が、前記1つ以上の遺伝子の欠損で構成される請求項1の非自然発生的な微生物。
【請求項4】
前記非自然発生的な微生物が、表1に挙げられる代謝改変を有する微生物の群から選択される、請求項1の非自然発生的な微生物。
【請求項5】
前記微生物が、細菌、酵母、または菌で構成される請求項1の非自然発生的な微生物。
【請求項6】
前記細菌は、大腸菌(E.coli)、アナエロビオスピルリム・サクシニシプロデュセンス(A.succiniciproducens)、アクチノバチルス・スクシノゲネス(A.succinogenes)、マンヘミア・サクシニシプロデュセンス(M.succiniciproducens)、インゲン根粒菌(R.etli)、枯草菌(Bacillus subtilis)、コリネバクテリウム・グルタミクム(Corynebacterium glutamicum)、グルコノバクテル・オキシダンス(Gluconobacter oxydans)、ザイモモナス・モビリス(Zymomonas mobilis)、ラクトコッカス・ラクティス(Lactococcus lactis)、ラクトバチルス・プランタルム(Lactobacillus plantarum)、ストレプトマイセス・コエリコロル(Streptomyces coelicolor)、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、シュードモナス・フルオレッセンス(Pseudomonas fluorescens)、およびシュードモナス・プチダ(Pseudomonas putida)から選択される種で構成される請求項5の非自然発生的な微生物。
【請求項7】
前記酵母は、サッカロマイセス・ケレウィシエ(Saccharomyces cerevisiae)、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)、クリベロマイセスラクティス(Kluyveromyces lactis)、クリベロマイセス・マルキシアヌス(Kluyveromyces marxianus)、アスペルギルス・テレウス(Aspergillus terreus)、クロコウジカビ(Aspergillus niger)、およびピキア・パストリス(Pichia pastoris)から選択される種で構成される請求項5の非自然発生的な微生物。
【請求項8】
前記コハク酸塩生成を微生物の増殖と強制的に連動させる代謝改変の組で構成される非自然発生的な微生物であって、前記代謝改変の組は、
【化1】
のいずれかからなる遺伝子の組またはそのオーソログから選択される1つ以上の遺伝子破壊で構成され、前記微生物は安定的な増殖連動型のコハク酸塩の生成を示す。
【請求項9】
前記代謝改変(e)ackA−pta、pykF、atpF、sdhAをコード化している前記遺伝子は、pykA、atpH、sdhB、またはdhaKLMから選択される少なくとも1つの遺伝子破壊をさらに備える、請求項8の非自然発生的な微生物。
【請求項10】
前記代謝改変(f)ackA−pta、pykF、ptsGをコード化している前記遺伝子が、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成される請求項8の非自然発生的な微生物。
【請求項11】
前記代謝改変(g)ackA−pta、pykF、ptsG、adhE、ldhAをコード化している前記遺伝子が、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成される請求項8の非自然発生的な微生物。
【請求項12】
前記1つ以上の遺伝子破壊が、欠損で構成される請求項8の非自然発生的な微生物。
【請求項13】
前記微生物が、細菌、酵母、または菌で構成される請求項8の非自然発生的な微生物。
【請求項14】
前記細菌が、大腸菌(E.coli)、アナエロビオスピルリム・サクシニシプロデュセンス(A.succiniciproducens)、アクチノバチルス・スクシノゲネス(A.succinogenes)、マンヘミア・サクシニシプロデュセンス(M.succiniciproducens)、インゲン根粒菌(R.etli)、枯草菌(Bacillus subtilis)、コリネバクテリウム・グルタミクム(Corynebacterium glutamicum)、グルコノバクテル・オキシダンス(Gluconobacter oxydans)、ザイモモナス・モビリス(Zymomonas mobilis)、ラクトコッカス・ラクティス(Lactococcus lactis)、ラクトバチルス・プランタルム(Lactobacillus plantarum)、ストレプトマイセス・コエリコロル(Streptomyces coelicolor)、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、シュードモナス・フルオレッセンス(Pseudomonas fluorescens)、およびシュードモナス・プチダ(Pseudomonas putida)から選択される種で構成される請求項13の非自然発生的な微生物。
【請求項15】
前記酵母が、サッカロマイセス・ケレウィシエ(Saccharomyces cerevisiae)、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)、クリベロマイセスラクティス(Kluyveromyces lactis)、クリベロマイセス・マルキシアヌス(Kluyveromyces marxianus)、アスペルギルス・テレウス(Aspergillus terreus)、クロコウジカビ(Aspergillus niger)、およびピキア・パストリス(Pichia pastoris)から選択される種で構成される請求項13の非自然発生的な微生物。
【請求項16】
(a) 指数増殖中にコハク酸塩の生成を要する代謝改変の組をコンピュータで同定するステップと、
(b) 前記コハク酸塩の生成を要する代謝改変の組を含有するように、微生物を遺伝子改変するステップ
で構成される、安定的な増殖連動型のコハク酸塩の生成を有する非自然発生的な微生物を生成する方法。
【請求項17】
前記代謝改変の組は、OptKnockまたはSimPhenyから選択されるコンピュータを利用した方法により同定される、請求項16の方法。
【請求項18】
前記代謝改変の組は、1つ以上の代謝反応の機能破壊で構成される請求項16の方法。
【請求項19】
前記代謝改変は、遺伝子欠損により破壊されている、請求項18の方法。
【請求項20】
前記非自然発生的な微生物は、表1に挙げられる代謝改変の組から選択される代謝改変を有する微生物で構成される請求項18の方法。
【請求項21】
前記遺伝子改変された微生物を培養するステップをさらに含む、請求項16の方法。
【請求項22】
コハク酸塩の生成を要する条件下で、前記遺伝子改変された微生物を適応進化させるステップをさらに含む、請求項21の方法。
【請求項23】
前記非自然発生的な微生物は、細菌、酵母、または菌で構成される請求項16の方法。
【請求項24】
前記細菌が、大腸菌(E.coli)、アナエロビオスピルリム・サクシニシプロデュセンス(A.succiniciproducens)、アクチノバチルス・スクシノゲネス(A.succinogenes)、マンヘミア・サクシニシプロデュセンス(M.succiniciproducens)、インゲン根粒菌(R.etli)、枯草菌(Bacillus subtilis)、コリネバクテリウム・グルタミクム(Corynebacterium glutamicum)、グルコノバクテル・オキシダンス(Gluconobacter oxydans)、ザイモモナス・モビリス(Zymomonas mobilis)、ラクトコッカス・ラクティス(Lactococcus lactis)、ラクトバチルス・プランタルム(Lactobacillus plantarum)、ストレプトマイセス・コエリコロル(Streptomyces coelicolor)、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、シュードモナス・フルオレッセンス(Pseudomonas fluorescens)、およびシュードモナス・プチダ(Pseudomonas putida)から選択される種で構成される請求項23の方法。
【請求項25】
前記酵母が、サッカロマイセス・ケレウィシエ(Saccharomyces cerevisiae)、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)、クリベロマイセスラクティス(Kluyveromyces lactis)、クリベロマイセス・マルキシアヌス(Kluyveromyces marxianus)、アスペルギルス・テレウス(Aspergillus terreus)、クロコウジカビ(Aspergillus niger)、およびピキア・パストリス(Pichia pastoris)から選択される種で構成される請求項23の非自然発生的な微生物。
【請求項26】
請求項16または22の方法により生成される微生物。
【請求項27】
(a) 前記コハク酸塩生成を微生物の増殖と強制的に連動させる代謝改変の組であって、
【化2】
のいずれかからなる遺伝子の組またはそのオーソログから選択される1つ以上の遺伝子破壊を備える前記代謝改変の組で構成される非自然発生的な微生物を、指数増殖相のもとで十分な量の栄養物と培地の中で培養するステップであり、前記微生物は安定的な増殖連動型のコハク酸塩の生成を示すステップと、
(b) 前記非自然発生的な微生物から生成されたコハク酸塩を単離するステップ
で構成される微生物の増殖に共役するコハク酸塩を生成する方法。
【請求項28】
前記代謝改変(5)ackA−pta、pykF、atpF、sdhAをコード化する前記遺伝子が、pykA、atpH、sdhB、またはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成される請求項27の方法。
【請求項29】
前記代謝改変(6)ackA−pta、pykF、ptsGをコード化する前記遺伝子が、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成される請求項27の方法。
【請求項30】
前記代謝改変(7)ackA−pta、pykF、ptsG、adhE、ldhAをコード化する前記遺伝子が、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成される請求項27の方法。
【請求項1】
1つ以上の遺伝子破壊で構成される非自然発生的な微生物であって、前記1つ以上の遺伝子破壊は、前記コハク酸塩生成を微生物の増殖と強制的に連動させる酵素をコード化する遺伝子において、前記遺伝子破壊が前記酵素の活性を低下させたときに発生し、それにより前記1つ以上の遺伝子破壊は、前記非自然発生的な微生物に安定的な増殖連動型のコハク酸塩の生成を与える。
【請求項2】
前記1つ以上の遺伝子破壊が、表1に挙げられる代謝改変で構成される請求項1の非自然発生的な微生物。
【請求項3】
前記1つ以上の遺伝子破壊が、前記1つ以上の遺伝子の欠損で構成される請求項1の非自然発生的な微生物。
【請求項4】
前記非自然発生的な微生物が、表1に挙げられる代謝改変を有する微生物の群から選択される、請求項1の非自然発生的な微生物。
【請求項5】
前記微生物が、細菌、酵母、または菌で構成される請求項1の非自然発生的な微生物。
【請求項6】
前記細菌は、大腸菌(E.coli)、アナエロビオスピルリム・サクシニシプロデュセンス(A.succiniciproducens)、アクチノバチルス・スクシノゲネス(A.succinogenes)、マンヘミア・サクシニシプロデュセンス(M.succiniciproducens)、インゲン根粒菌(R.etli)、枯草菌(Bacillus subtilis)、コリネバクテリウム・グルタミクム(Corynebacterium glutamicum)、グルコノバクテル・オキシダンス(Gluconobacter oxydans)、ザイモモナス・モビリス(Zymomonas mobilis)、ラクトコッカス・ラクティス(Lactococcus lactis)、ラクトバチルス・プランタルム(Lactobacillus plantarum)、ストレプトマイセス・コエリコロル(Streptomyces coelicolor)、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、シュードモナス・フルオレッセンス(Pseudomonas fluorescens)、およびシュードモナス・プチダ(Pseudomonas putida)から選択される種で構成される請求項5の非自然発生的な微生物。
【請求項7】
前記酵母は、サッカロマイセス・ケレウィシエ(Saccharomyces cerevisiae)、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)、クリベロマイセスラクティス(Kluyveromyces lactis)、クリベロマイセス・マルキシアヌス(Kluyveromyces marxianus)、アスペルギルス・テレウス(Aspergillus terreus)、クロコウジカビ(Aspergillus niger)、およびピキア・パストリス(Pichia pastoris)から選択される種で構成される請求項5の非自然発生的な微生物。
【請求項8】
前記コハク酸塩生成を微生物の増殖と強制的に連動させる代謝改変の組で構成される非自然発生的な微生物であって、前記代謝改変の組は、
【化1】
のいずれかからなる遺伝子の組またはそのオーソログから選択される1つ以上の遺伝子破壊で構成され、前記微生物は安定的な増殖連動型のコハク酸塩の生成を示す。
【請求項9】
前記代謝改変(e)ackA−pta、pykF、atpF、sdhAをコード化している前記遺伝子は、pykA、atpH、sdhB、またはdhaKLMから選択される少なくとも1つの遺伝子破壊をさらに備える、請求項8の非自然発生的な微生物。
【請求項10】
前記代謝改変(f)ackA−pta、pykF、ptsGをコード化している前記遺伝子が、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成される請求項8の非自然発生的な微生物。
【請求項11】
前記代謝改変(g)ackA−pta、pykF、ptsG、adhE、ldhAをコード化している前記遺伝子が、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成される請求項8の非自然発生的な微生物。
【請求項12】
前記1つ以上の遺伝子破壊が、欠損で構成される請求項8の非自然発生的な微生物。
【請求項13】
前記微生物が、細菌、酵母、または菌で構成される請求項8の非自然発生的な微生物。
【請求項14】
前記細菌が、大腸菌(E.coli)、アナエロビオスピルリム・サクシニシプロデュセンス(A.succiniciproducens)、アクチノバチルス・スクシノゲネス(A.succinogenes)、マンヘミア・サクシニシプロデュセンス(M.succiniciproducens)、インゲン根粒菌(R.etli)、枯草菌(Bacillus subtilis)、コリネバクテリウム・グルタミクム(Corynebacterium glutamicum)、グルコノバクテル・オキシダンス(Gluconobacter oxydans)、ザイモモナス・モビリス(Zymomonas mobilis)、ラクトコッカス・ラクティス(Lactococcus lactis)、ラクトバチルス・プランタルム(Lactobacillus plantarum)、ストレプトマイセス・コエリコロル(Streptomyces coelicolor)、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、シュードモナス・フルオレッセンス(Pseudomonas fluorescens)、およびシュードモナス・プチダ(Pseudomonas putida)から選択される種で構成される請求項13の非自然発生的な微生物。
【請求項15】
前記酵母が、サッカロマイセス・ケレウィシエ(Saccharomyces cerevisiae)、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)、クリベロマイセスラクティス(Kluyveromyces lactis)、クリベロマイセス・マルキシアヌス(Kluyveromyces marxianus)、アスペルギルス・テレウス(Aspergillus terreus)、クロコウジカビ(Aspergillus niger)、およびピキア・パストリス(Pichia pastoris)から選択される種で構成される請求項13の非自然発生的な微生物。
【請求項16】
(a) 指数増殖中にコハク酸塩の生成を要する代謝改変の組をコンピュータで同定するステップと、
(b) 前記コハク酸塩の生成を要する代謝改変の組を含有するように、微生物を遺伝子改変するステップ
で構成される、安定的な増殖連動型のコハク酸塩の生成を有する非自然発生的な微生物を生成する方法。
【請求項17】
前記代謝改変の組は、OptKnockまたはSimPhenyから選択されるコンピュータを利用した方法により同定される、請求項16の方法。
【請求項18】
前記代謝改変の組は、1つ以上の代謝反応の機能破壊で構成される請求項16の方法。
【請求項19】
前記代謝改変は、遺伝子欠損により破壊されている、請求項18の方法。
【請求項20】
前記非自然発生的な微生物は、表1に挙げられる代謝改変の組から選択される代謝改変を有する微生物で構成される請求項18の方法。
【請求項21】
前記遺伝子改変された微生物を培養するステップをさらに含む、請求項16の方法。
【請求項22】
コハク酸塩の生成を要する条件下で、前記遺伝子改変された微生物を適応進化させるステップをさらに含む、請求項21の方法。
【請求項23】
前記非自然発生的な微生物は、細菌、酵母、または菌で構成される請求項16の方法。
【請求項24】
前記細菌が、大腸菌(E.coli)、アナエロビオスピルリム・サクシニシプロデュセンス(A.succiniciproducens)、アクチノバチルス・スクシノゲネス(A.succinogenes)、マンヘミア・サクシニシプロデュセンス(M.succiniciproducens)、インゲン根粒菌(R.etli)、枯草菌(Bacillus subtilis)、コリネバクテリウム・グルタミクム(Corynebacterium glutamicum)、グルコノバクテル・オキシダンス(Gluconobacter oxydans)、ザイモモナス・モビリス(Zymomonas mobilis)、ラクトコッカス・ラクティス(Lactococcus lactis)、ラクトバチルス・プランタルム(Lactobacillus plantarum)、ストレプトマイセス・コエリコロル(Streptomyces coelicolor)、クロストリジウム・アセトブチリカム(Clostridium acetobutylicum)、シュードモナス・フルオレッセンス(Pseudomonas fluorescens)、およびシュードモナス・プチダ(Pseudomonas putida)から選択される種で構成される請求項23の方法。
【請求項25】
前記酵母が、サッカロマイセス・ケレウィシエ(Saccharomyces cerevisiae)、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)、クリベロマイセスラクティス(Kluyveromyces lactis)、クリベロマイセス・マルキシアヌス(Kluyveromyces marxianus)、アスペルギルス・テレウス(Aspergillus terreus)、クロコウジカビ(Aspergillus niger)、およびピキア・パストリス(Pichia pastoris)から選択される種で構成される請求項23の非自然発生的な微生物。
【請求項26】
請求項16または22の方法により生成される微生物。
【請求項27】
(a) 前記コハク酸塩生成を微生物の増殖と強制的に連動させる代謝改変の組であって、
【化2】
のいずれかからなる遺伝子の組またはそのオーソログから選択される1つ以上の遺伝子破壊を備える前記代謝改変の組で構成される非自然発生的な微生物を、指数増殖相のもとで十分な量の栄養物と培地の中で培養するステップであり、前記微生物は安定的な増殖連動型のコハク酸塩の生成を示すステップと、
(b) 前記非自然発生的な微生物から生成されたコハク酸塩を単離するステップ
で構成される微生物の増殖に共役するコハク酸塩を生成する方法。
【請求項28】
前記代謝改変(5)ackA−pta、pykF、atpF、sdhAをコード化する前記遺伝子が、pykA、atpH、sdhB、またはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成される請求項27の方法。
【請求項29】
前記代謝改変(6)ackA−pta、pykF、ptsGをコード化する前記遺伝子が、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成される請求項27の方法。
【請求項30】
前記代謝改変(7)ackA−pta、pykF、ptsG、adhE、ldhAをコード化する前記遺伝子が、pykAまたはdhaKLMから選択される少なくとも1つの遺伝子破壊でさらに構成される請求項27の方法。
【図1】

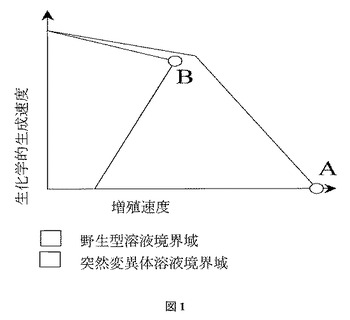
【図2】
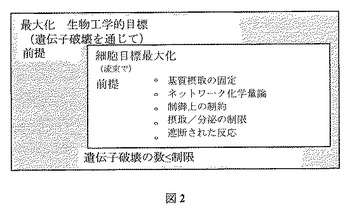
【図3】
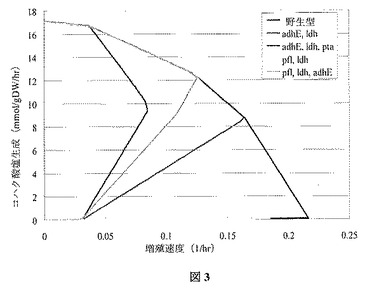
【図4】
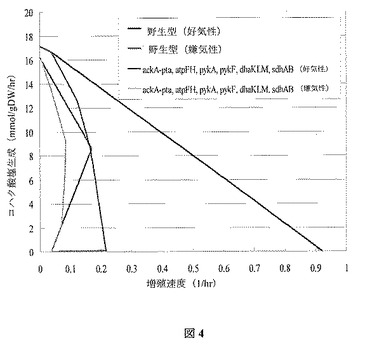
【図5】
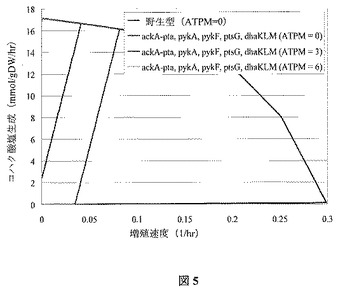
【図6】
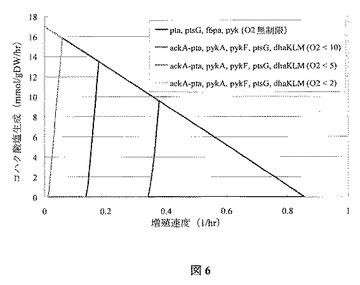
【図7】
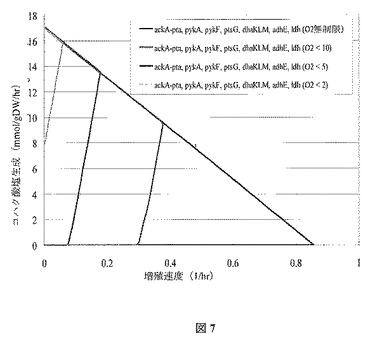
【図8】
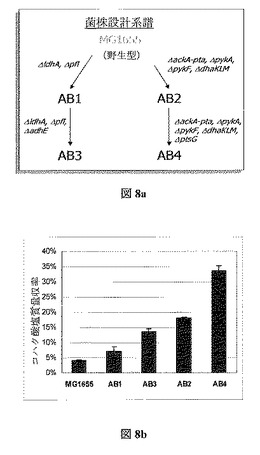
【図9】
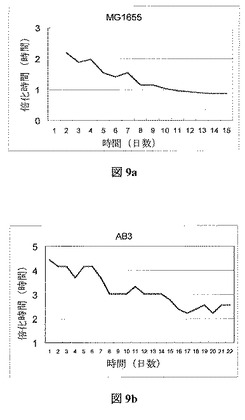
【図10】
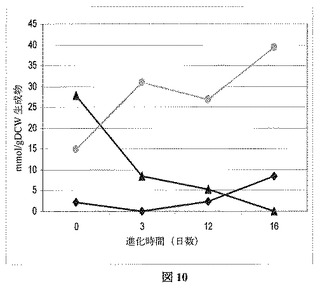
【図11】
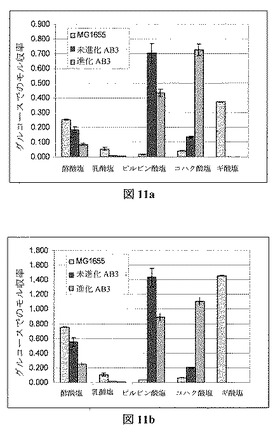
【図12】
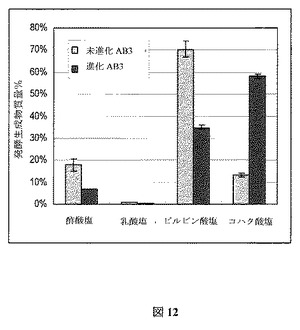
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公表番号】特表2009−507493(P2009−507493A)
【公表日】平成21年2月26日(2009.2.26)
【国際特許分類】
【出願番号】特願2008−530032(P2008−530032)
【出願日】平成18年9月8日(2006.9.8)
【国際出願番号】PCT/US2006/035569
【国際公開番号】WO2007/030830
【国際公開日】平成19年3月15日(2007.3.15)
【出願人】(504356052)ジェノマティカ・インコーポレイテッド (8)
【氏名又は名称原語表記】Genomatica, Inc.
【Fターム(参考)】
【公表日】平成21年2月26日(2009.2.26)
【国際特許分類】
【出願日】平成18年9月8日(2006.9.8)
【国際出願番号】PCT/US2006/035569
【国際公開番号】WO2007/030830
【国際公開日】平成19年3月15日(2007.3.15)
【出願人】(504356052)ジェノマティカ・インコーポレイテッド (8)
【氏名又は名称原語表記】Genomatica, Inc.
【Fターム(参考)】
[ Back to top ]