
変異グルコース脱水素酵素
【課題】CyGDH(QY)よりも、グルコースに対する基質特異性の向上したCyGDHを提供する。
【解決手段】特定の配列と少なくとも80%の同一性を有するアミノ酸配列を有し、かつ、グルコース脱水素酵素活性を有する変異グルコース脱水素酵素であって、前記アミノ酸配列の326位、365位および472位に相当するアミノ酸残基が、それぞれグルタミン、チロシンおよびチロシンで置換されており、グルコースに対する基質特異性が向上し、二糖類に対する反応性が低下したことを特徴とする、変異グルコース脱水素酵素。
【解決手段】特定の配列と少なくとも80%の同一性を有するアミノ酸配列を有し、かつ、グルコース脱水素酵素活性を有する変異グルコース脱水素酵素であって、前記アミノ酸配列の326位、365位および472位に相当するアミノ酸残基が、それぞれグルタミン、チロシンおよびチロシンで置換されており、グルコースに対する基質特異性が向上し、二糖類に対する反応性が低下したことを特徴とする、変異グルコース脱水素酵素。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は基質特異性の向上した変異グルコース脱水素酵素に関する。詳細には、本発明は、シトクロムCを含むグルコース脱水素酵素(以下CyGDHとも称する)を構成するαサブユニットのアミノ酸残基に変異導入した変異型αサブユニットを含むグルコース脱水素酵素およびその遺伝子に関する。本発明のグルコース脱水素酵素は、グルコースセンサ、及びグルコースアッセイキット等に好適に使用することができ、生化学分野、臨床分野等で有用である。
【背景技術】
【0002】
現在、野生型のCyGDHや、ピロロキノリンキノンを補酵素とするPQQGDHが、自己血糖測定センサに用いられている。野生型のCyGDHやPQQGDHは、グルコースのみならずマルトースにも反応することから、患者の血中マルトース濃度が高い場合においては、正確な血糖値の測定ができないという欠点があった。特に日本や英国においては、輸液中のエネルギー物質としてマルトースが用いられており、実際に、腹膜透析等により輸液を投与している低血糖患者が、PQQGDH型の血糖センサで高血糖と誤認されたことによる事故も発生している。
野生型CyGDHは、グルコースに対してマルトースの反応性が高く、グルコース濃度50mg/dL時において、マルトース濃度が100mg/dL時の血糖値計測結果は、170%高値を示す。
【0003】
このような状況に鑑み、CyGDHの変異体酵素(αサブユニットのアミノ酸残基の326位と365位に変異導入し、それぞれグルタミン(Q)、チロシン(Y)に置換した変異体グルコース脱水素酵素(以下CyGDH(QY)または単にQY、QYAとも称する)が発明されており(特許文献1)、グルコース濃度50mg/dL時において、マルトース濃度が100mg/dL時の血糖値計測結果が、36%の高値まで抑えることが出来ている。しかしながら、マルトース影響回避効果が完全なものとは言えなかった。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】国際公開第2006/137283号
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明は、CyGDH(QY)よりも、グルコースに対する基質特異性の向上したCyGDHを提供することを課題とする。
【課題を解決するための手段】
【0006】
本発明者は、上記課題を解決するために鋭意検討を行った結果、CyGDHを構成するαサブユニットのアミノ酸残基のうち、326位および365位に相当する部位をそれぞれグルタミンおよびチロシンで置換し、さらに472位に相当する部位をチロシンで置換することにより、基質特異性がCyGDH(QY)と比較してさらに向上すること、そして、GDHのホモログにおいても326位、365位および472位に相当する部位をそれぞれグルタミン、チロシンおよびチロシンで置換することが同様の効果をもたらすことを見出し、本発明を完成するに至った。
【0007】
すなわち、本発明は以下のとおりである。
(1)配列番号3と少なくとも80%の同一性を有するアミノ酸配列を有し、かつ、グルコース脱水素酵素活性を有する変異グルコース脱水素酵素であって、
前記アミノ酸配列の326位、365位および472位に相当するアミノ酸残基が、それぞれグルタミン、チロシンおよびチロシンで置換されており、
グルコースに対する基質特異性が向上し、二糖類に対する反応性が低下したことを特徴とする、変異グルコース脱水素酵素。
(2)配列番号3と少なくとも90%の同一性を有するアミノ酸配列を有する、(1)に記載の変異グルコース脱水素酵素。
(3)326位、365位および472位に相当する位置以外は配列番号3に示すアミノ酸配列を有する、(1)または(2)に記載の変異グルコース脱水素酵素。
(4)326位、365位および472位に相当する位置以外は配列番号7に示すアミノ酸配列を有する、(1)または(2)に記載の変異グルコース脱水素酵素。
(5)326位、365位および472位に相当する位置以外は配列番号8に示すアミノ酸配列を有する、(1)または(2)に記載の変異グルコース脱水素酵素。
(6)326位、365位および472位に相当する位置以外は配列番号9に示すアミノ酸配列を有する、(1)または(2)に記載の変異グルコース脱水素酵素。
(7)326位、365位および472位に相当する位置以外は配列番号10に示すアミノ酸配列を有する、(1)または(2)に記載の変異グルコース脱水素酵素。
(8)前記アミノ酸配列において326位および365位に相当するアミノ酸残基がそれぞれグルタミンおよびチロシンで置換されているが472位は置換されていないグルコース脱水素酵素と比較して、グルコースに対する基質特異性が向上し、二糖類に対する反応性が低下したことを特徴とする、(1)〜(7)のいずれか1項に記載の変異グルコース脱水素酵素。
(9)二糖類がマルトースであることを特徴とする(1)〜(8)のいずれか1項に記載の変異グルコース脱水素酵素。
(10)(1)〜(9)のいずれか1項に記載の変異グルコース脱水素酵素と、電子伝達サブユニットとを少なくとも含む変異グルコース脱水素酵素複合体。
(11)電子伝達サブユニットがシトクロムCであることを特徴とする(10)に記載のグルコース脱水素酵素複合体。
(12)(1)〜(9)のいずれか1項に記載の変異グルコース脱水素酵素をコードするDNA。
(13)(12)に記載のDNAを保持し、(1)〜(9)のいずれか1項に記載の変異グルコース脱水素酵素、あるいは(10)または(11)に記載の変異グルコース脱水素酵素複合体を産生する微生物。
(14)(1)〜(9)のいずれか1項に記載の変異グルコース脱水素酵素、(10)または(11)に記載の変異グルコース脱水素酵素複合体、あるいは(13)に記載の微生物を含む、グルコースアッセイキット。
(15)(1)〜(9)のいずれか1項に記載の変異グルコース脱水素酵素、(10)または(11)に記載の変異グルコース脱水素酵素複合体、あるいは(13)に記載の微生物を含む、グルコースセンサ。
【図面の簡単な説明】
【0008】
【図1】グルコースセンサの構造を示す図。
【図2】グルコースセンサの各試薬部を示す図。
【図3】比色式センサでの血糖測定における、グルコース濃度50mg/dLでの、マルトース濃度(100mg/dL、200mg/dL、300mg/dL)の血糖値に対する影響を示す図。
【図4】電極式センサでの血糖測定における、グルコース濃度50mg/dLでの、マルトース濃度(100mg/dL、200mg/dL、300mg/dL)の血糖値に対する影響を示す図。
【図5】電極式グルコースセンサの構造を示す図。
【発明を実施するための形態】
【0009】
以下、本発明を詳細に説明する。
本発明の変異GDHは、野生型GDHのαサブユニットに特定の変異を導入することにより、製造することができる。野生型GDHとしては、ブルクホリデリア・セパシアが産生するGDHが挙げられる。ブルクホリデリア・セパシアのGDHとしては、ブルクホリデリア・セパシアKS1株、JCM2800株又はJCM2801株が産生するGDHが挙げられる。KS1株は、平成12年9月25日に独立行政法人産業技術総合研究所特許生物寄託センター(〒305−8566 日本国茨城県つくば市東1丁目1番地1 中央第6)に、受託番号第FERM BP−7306として寄託されている。
【0010】
KS1株のGDH αサブユニット遺伝子、及びβサブユニット遺伝子の一部を含む染色体DNA断片の塩基配列を配列番号1に示す(米国特許出願公開第2004/0023330号)。この塩基配列には3つのオープンリーディングフレーム(ORF)が存在し、5’末端側から2番目及び3番目のORFは、それぞれαサブユニット(配列番号3)、及びβサブユニット(配列番号4)をコードしている。また、1番目のORFはγサブユニット(配列番号2)をコードしていると推定される。また、配列番号5に、βサブユニット遺伝子全長を含む断片の塩基配列を示す。さらに、βサブユニットのアミノ酸配列を配列番号6に示す(欧州特許出願公開第1498484号)。配列番号6において、アミノ酸番号1〜22はシグナルペプチドであると推定される。
【0011】
また、これらの他に、ブルクホリデリア・セパシアKS1株のGDHのホモログである、ブルクホルデリア・セノセパシアJ2315株のputative oxidoreductase(配列番号7)、ブルクホルデリア・タイランデンシスTXDOH株のhypothetical protein BthaT#07876(配列番号8)、ラルストニア・ピッケティ12D株のFAD dependent oxidoreductase(配列番号9)、ラルストニア・ソラナセアラムIPO1609株のtransmembrane dehydrogenase(配列番号10)およびブルクホルデリア・フィトフィルマンスPsJN株のglucose-methanol-choline oxidoreductase(配列番号11)のそれぞれのαサブユニットも、ブルクホリデリア・セパシアKS1株のGDHと同様に用いることができる。
なお、配列番号7〜11に示したアミノ酸配列については、いずれもアメリカの国立生物工学情報センター(National Center for Biotechnology Information, NCBI)のデータベースに登録されている。それぞれの登録番号は、配列番号7がYP_002234347、配列番号8がZP_02370914、配列番号9がYP_002980762、配列番号10がYP_002260434、配列番号11がYP_001890482である。
また、ブルクホルデリア・セノセパシアJ2315株は、LMG 16656、ATCC BAA-245、CCM 4899、CCUG 48434、NCTC 13227として寄託されている。ブルクホルデリア・フィトフィルマンスPsJN株は、LMG 22487、CCUG 49060として寄託されている。
【0012】
また、ブルクホリデリア・セパシアKS1株と同属である、ブルクホリデリア・セパシア株、例えばJCM2800、JCM2801、JCM5506、JCM5507、IFO14595由来のそれぞれのGDHのαサブユニット(配列番号12〜16)も、ブルクホリデリア・セパシアKS1株のGDHと同様に用いることができる。なお、JCM2800、JCM2801、JCM5506、およびJCM5507は、独立行政法人理化学研究所微生物系統保存施設(Japan Collection of Microorganisms, JCM)に保存されている。IFO14595は、財団法人発酵研究所(IFO)に保存されている。
【0013】
本発明の変異GDHは、αサブユニットのみであってもよいし、αサブユニットとβサブユニットとの複合体、αサブユニットとγサブユニットとの複合体、又はαサブユニット、βサブユニット及びγサブユニットからなる複合体であってもよい。本明細書において、βサブユニットを含むGDH複合体をCyGDHと称し、βサブユニットを含まないGDH複合体をGDHと称する。本発明の変異GDHは、いずれの場合もαサブユニットに特定の変異(326位、365位及び472位における変異、またはこれらに対応する位置における変異)が導入されたものであるが、この特定の変異の他に、保存的な変異を有していてもよい。また、他のサブユニットは野生型であっても、保存的な変異を有するものであってもよい。尚、「保存的変異」とは、GDH活性に実質的に影響しない変異をいう。
【0014】
本発明の変異型αサブユニットは、好ましくは、前記特定の変異以外は、配列番号3、7〜11に示すアミノ酸配列を有する。また、変異型αサブユニットは、GDH活性を有する限り、前述したような保存的変異を有していてもよい。すなわち、配列番号3、7〜11のアミノ酸配列において、前記特定の変異以外に、1又は複数のアミノ酸残基が置換、欠失、挿入、又は付加されたアミノ酸配列を有するタンパク質であってもよい。尚、配列番号3には、配列番号1の塩基配列によってコードされ得るアミノ酸配列を示してあるが、N末端のメチオニン残基は、翻訳後に脱落している可能性がある。前記「1又は複数」とは、好ましくは1〜10個、より好ましくは1〜5個、特に好ましくは1〜3個である。なお、本発明の変異型αサブユニットは、配列番号3に示すアミノ酸配列に対して、少なくとも80%、好ましくは85%、より好ましくは90%のアミノ酸同一性を有する。
【0015】
また、βサブユニットは、典型的には配列番号6のアミノ酸配列を有する。しかし、CyGDHのβサブユニットとして機能し得る限り、配列番号6のアミノ酸番号23〜425からなるアミノ酸配列において、1又は複数のアミノ酸残基が置換、欠失、挿入、又は付加されたアミノ酸配列を有するタンパク質であってもよい。また、CyGDHのβサブユニットとして機能し得る限り、KS1株以外のβサブユニットおよび同βサブユニットのアミノ酸配列において、1又は複数のアミノ酸残基が置換、欠失、挿入、又は付加されたアミノ酸配列を有するタンパク質であってもよい。前記「1又は複数」とは、好ましくは1〜20個、より好ましくは1〜10個、特に好ましくは1〜5個である。
尚、CyGDHのβサブユニットとして機能するとは、αサブユニットとともに複合体を形成したときに同複合体のGDH活性を損なわずに電子伝達サブユニット、すなわち、チトクロームCとして機能することをいう。
【0016】
野生型αサブユニット遺伝子として具体的には、配列番号1の塩基番号764〜2380からなる塩基配列を含むDNAが挙げられる。また、αサブユニット遺伝子は、配列番号1の塩基配列の塩基番号764〜2380からなる塩基配列を有するDNA、又はこの配列から調製され得るプローブとストリンジェントな条件下でハイブリダイズし、かつ、GDH活性を有するタンパク質をコードするDNAであってもよい。
【0017】
また、βサブユニット遺伝子として具体的には、配列番号5の塩基番号187〜1398からなる塩基配列を含むDNAが挙げられる。またβサブユニット遺伝子は、配列番号5の塩基番号187〜1398からなる塩基配列を有するDNA、又はこの配列から調製され得るプローブとストリンジェントな条件下でハイブリダイズし、かつ、βサブユニットとして機能し得るタンパク質をコードするDNAであってもよい。
【0018】
前記ストリンジェントな条件としては、好ましくは80%、より好ましくは90%以上、特に好ましくは95%以上の相同性を有するDNA同士がハイブリダイズする条件、具体的には、0.1×SSC、0.1%SDS、60℃が挙げられる。
【0019】
αサブユニット遺伝子及びβサブユニット遺伝子は、例えば、ブルクホルデリア・セパシアKS1株の染色体DNAを鋳型とするPCRによって、取得することができる。PCR用プライマーは、前記の塩基配列に基づいて化学合成することによって調製することができる。また、前記配列に基づいて作製したオリゴヌクレオチドをプローブとするハイブリダイゼーションによって、ブルクホルデリア・セパシアKS1株の染色体DNAから取得することもできる。なお、KS1株以外のブルクホルデリア・セノセパシアJ2315株、ブルクホルデリア・タイランデンシスTXDOH株、ラルストニア・ピッケティ12D株、ラルストニア・ソラナセアラムIPO1609株およびブルクホルデリア・フィトフィルマンスPsJN株も用いることができる。
【0020】
本発明の変異GDHは、上記のような野生型GDH又は保存的変異を有するGDHが特定の変異を有することによって、グルコースに対する基質特異性が向上したものである。「グルコースに対する基質特異性が向上した」とは、グルコースに対する反応性を実質的に維持したまま、他の単糖類、二糖類又はオリゴ糖等の糖類、例えばマルトース、ガラクトース、キシロース等に対する反応性が低下したこと、あるいは、グルコースに対する反応性が他の糖類に対する反応性に比べて向上したことを含む。例えば、グルコースに対する反応性が低下しても、他の糖類に対する反応性がそれ以上に低下すれば、グルコースに対する基質特異性は向上する。また、他の糖類に対する反応性が上昇しても、グルコースに対する基質特異性がそれ以上に上昇すれば、グルコースに対する基質特異性は向上する。具体的には例えば、野生型酵素に対する変異型酵素の基質特異性(グルコースに対する比活性と、他の糖類、例えばマルトース、に対する比活性との比)の向上(下記式で表される)が、10%以上、好ましくは20%以上、より好ましくは40%以上であれば、グルコースに対する基質特異性は向上している。例えば、基質特異性が、野生型酵素では60%、変異型GDHでは40%の場合、グルコース以外の糖類に対する反応性は、33%低下している。
基質特異性=(グルコース以外の糖類に対する比活性/グルコースに対する比活性)×100)
基質特異性の向上=(A−B)×100/A
A:野生型酵素の基質特異性
B:変異型酵素の基質特異性
また、変異型GDHは、マルトースに対する反応性(比活性)が、グルコースに対する反応性(比活性)の1%以下、好ましくは0.5%以下である。
【0021】
本発明の変異型GDHは、好ましくは、配列番号3に示すアミノ酸配列の326位および365位に相当するアミノ酸残基がそれぞれグルタミンおよびチロシンで置換されているが472位は置換されていないグルコース脱水素酵素と比較して、グルコースに対する基質特異性が向上し、二糖類に対する反応性が低下している。
【0022】
本発明における変異とは、具体的には以下のとおりである。
(1)配列番号3に示すアミノ酸配列の326位に相当するセリン残基のグルタミンへの置換。
(2)配列番号3に示すアミノ酸配列の365位に相当するセリン残基のチロシンへの置換。
(3)配列番号3に示すアミノ酸配列の472位に相当するアラニン残基のチロシンへの置換。
【0023】
上記アミノ酸置換変異の位置は、配列番号3、すなわちブルクホリデリア・セパシアKS1株の野生型GDH αサブユニットのアミノ酸配列における位置であるが、配列番号3のアミノ酸配列において、前記特定の変異以外に、1又は複数のアミノ酸残基が置換、欠失、挿入、又は付加されたアミノ酸配列を有するGDH αサブユニットのホモログ又はバリアントにおいては、配列番号3のアミノ酸配列とのアラインメントにおいて、前記アミノ酸置換の位置に相当する位置を示す。例えば、1〜364位の領域において1個のアミノ酸残基の欠失を有するGDH αサブユニットの保存的バリアントにおいては、365位とは、前記バリアントの364位を示す。
【0024】
配列番号7においては、配列番号3の326位、365位および472位に相当する残基はそれぞれ326位、365位および472位で同じである。
【0025】
配列番号8においては、配列番号3の326位、365位および472位に相当する残基はそれぞれ324位、363位および470位である。
【0026】
配列番号9においては、配列番号3の326位、365位および472位に相当する残基はそれぞれ327位、366位および473位である。
【0027】
配列番号10においては、配列番号3の326位、365位および472位に相当する残基はそれぞれ327位、366位および473位である。
【0028】
配列番号11においては、配列番号3の326位、365位および472位に相当する残基はそれぞれ322位、361位および466位である。
【0029】
なお、配列番号3に示すアミノ酸配列は、配列番号7〜11に対して、それぞれ、96%、93%、82%、82%、63%のアミノ酸配列同一性を有する。
以下の表1において、配列番号3,7〜11のアミノ酸配列アライメントを示す。
【0030】
【表1−1】

【0031】
【表1−2】
【0032】
本発明者は、変異体グルコース脱水素酵素(CyGDH(QY))と比較して、グルコースに対する基質特異性を増加すべく、遺伝的アルゴリズムを用いて、最適な326位、365位および472位の組み合わせについて検討した。その結果、基質特異性を向上することができる組み合わせを発見した。
【0033】
本発明の変異型GDHの好ましい変異の態様を以下に示す。
(数字はアミノ酸配列における位置を、アミノ酸残基は前記位置における置換後のアミノ酸残基を示し、「+」は同時に2つのアミノ酸置換を有することを示す。)
326Gln+365Tyr+472Tyr
【0034】
所望の変異を有するGDH αサブユニットは、GDH αサブユニットをコードするDNA(αサブユニット遺伝子)に、部位特異的変異法によって、所望のアミノ酸変異に対応するヌクレオチド変異を導入し、得られる変異DNAを適当な発現系を用いて発現させることによって、取得することができる。また、変異GDH αサブユニットをコードするDNAを、βサブユニットをコードするDNA(βサブユニット遺伝子)、又はβサブユニット遺伝子とγサブユニットをコードするDNA(γサブユニット遺伝子)とともに発現させることによって、変異CyGDH複合体を取得することができる。尚、GDH
αサブユニットをコードするDNAへの変異の導入は、γサブユニット、αサブユニット及びβサブユニットをこの順にコードするポリシストロニックなDNA断片を用いてもよい。
【0035】
変異が導入されたGDH αサブユニット又はCyGDH複合体の糖類に対する基質特異性は、実施例に記載された方法によって各種糖類に対する反応性を調べ、野生型GDH
αサブユニット又は野生型CyGDH複合体の反応性と比較することよって、決定することができる。
【0036】
γサブユニット、αサブユニット及びβサブユニットをこの順にコードするポリシストロニックなDNA断片は、例えば、ブルクホルデリア・セパシアKS1株の染色体DNAを鋳型とし、配列番号19、20の塩基配列を有するオリゴヌクレオチドをプライマーとするPCRによって取得することができる(後記実施例参照)。
【0037】
GDHの各サブユニットの遺伝子の取得、変異の導入、遺伝子の発現等に用いるベクターとしては、例えばエシェリヒア属細菌で機能するベクター、具体的にはpTrc99A、pBR322、pUC18、pUC118、pUC19、pUC119、pACYC184、pBBR122等が挙げられる。遺伝子の発現に用いるプロモーターとしては、例えばlac、trp、tac、trc、PL、tet、PhoA等が挙げられる。また、プロモーターを含む発現ベクターの適当な部位に、αサブユニット遺伝子又は他のサブユニット遺伝子を挿入することによって、これらの遺伝子のベクターへの挿入とプロモーターの連結とを同じ工程で行うことができる。このような発現ベクターとしては、pTrc99A、pBluescript、pKK223−3等が挙げられる。
【0038】
また、αサブユニット遺伝子又は他のサブユニット遺伝子は、発現可能な形態で宿主微生物の染色体DNA中に組み込まれてもよい。
【0039】
組換えベクターで微生物を形質転換するには、例えばカルシウム処理によるコンピテントセル法、プロトプラスト法又はエレクトロポレーション法等が挙げられる。
【0040】
宿主微生物としては、バチルス・サブチリス等のバチルス属細菌、サッカロマイセス・セレビシエ等の酵母、アスペルギルス・ニガー等の糸状菌が挙げられるが、これらに限られず、異種タンパク質生産に適した宿主微生物であれば用いることができる。
【0041】
本発明の変異αサブユニットもしくは変異CyGDH複合体又はこれらを発現する微生物は、グルコースセンサの酵素電極、又はグルコースのアッセイキットの構成要素として用いることができる。ブルクホルデリア・セパシアの野生型GDHを用いたグルコースセンサ及びグルコースアッセイキットは、米国特許公開第2004/0023330A1に記載されている。本発明の変異GDHも、同様にして使用することができる。
【実施例】
【0042】
次に、実施例を挙げて本発明をさらに具体的に説明するが、本発明はこれら実施例に何ら限定されるものではない。
【0043】
〔実施例1〕ブルクホルデリア・セパシアのGDHまたはCyGDHを発現するプラスミド
ブルクホルデリア・セパシアのGDHを発現するプラスミドとして、GDHのαサブユニット及びγサブユニットを発現するプラスミド、並びに、CyGDHを発現するプラスミドとして、αサブユニット、βサブユニット及びγサブユニットを発現するプラスミドを用意した。
【0044】
<1>GDHのαサブユニット及びγサブユニットを発現するプラスミド
αサブユニット及びγサブユニットを発現するプラスミドとしては、WO02/036779(EP1331272A1、US2004023330A1、CN1484703Aに対応)に記載のプラスミドpTrc99A/γ+αを使用した。同プラスミドは、ブルクホルデリア・セパシアKS1株(FERM BP−7306)染色体DNAから単離された、GDH γサブユニット構造遺伝子とαサブユニット構造遺伝子を連続して含むDNA断片が、ベクターpTrc99Aのクローニング部位であるNcoI/HindIIIに挿入されてなるプラスミドである。本プラスミド中のGDHγα遺伝子は、trcプロモーターによって制御される。pTrc99A/γ+αは、アンピシリン耐性遺伝子を保持している。
前記プラスミドpTrc99A/γ+αを鋳型として、以下の配列を有するオリゴヌクレオチドをプライマーとするPCRにより、GDHのαサブユニットC末端に6個のヒスチジン残基が付加されたDNA断片を含むプラスミド全体を増幅した。
【0045】
〔フォワードプライマー〕
5’−ACCACCACTGATAAGGAGGTCTGACCGTGCGGAAATCTAC−3’(配列番号17)
〔リバースプライマー〕
5’−AGCCTGTGCGACTTCTTCCTTCAGCGATCGGTGGTGGTGG−3’(配列番号18)
【0046】
増幅した断片の両端を平滑末端化した後、5’末端をリン酸化し、ライゲーションによって環状化した。得られた組換えベクターでエシェリヒア・コリDH5αを形質転換し、アンピシリン50μg/mLを含むLB寒天培地で生じるコロニーを採取した。得られた形質転換体を液体のLB培地で培養してプラスミドを抽出し、その挿入DNA断片を解析したところ、約2.1kbの挿入断片が確認された。本プラスミド中のGDHの各構造遺伝子は、trcプロモーターによって制御される。本プラスミドは、アンピシリン耐性遺伝子を保持している。
【0047】
<2>CyGDHのαサブユニット、βサブユニット及びγサブユニットを発現するプラスミド
CyGDHのαサブユニット、βサブユニット及びγサブユニットを発現するプラスミドは、以下のようにして調製した。
【0048】
(1)ブルクホルデリア・セパシア KS1株からの染色体DNAの調製
ブルクホルデリア・セパシア KS1株より染色体遺伝子を常法に従って調製した。すなわち、同菌株をTL液体倍地(ポリペプトン 10g、酵母抽出液 1g、NaCl 5g、KH2PO4 2g、グルコース 5g;1L、pH 7.2)を用いて、34℃で一晩振盪した。増殖した菌体を遠心分離により回収した。この菌体を10mM NaCl、20mM Tris−HCl(pH8.0)、1mM EDTA、0.5% SDS、100μg/mlのプロテイナーゼKを含む溶液に懸濁し、50℃で6時間処理した。ここに等量のフェノール−クロロホルムを加えて室温で10分間撹拌した後、遠心分離により上清を回収した。これに終濃度0.3Mになるように酢酸ナトリウムを加え、2倍量のエタノールを重層して中間層に染色体DNAを析出させた。これをガラス棒を用いてすくいとり、70%エタノールで洗浄した後、適当量のTEバッファーに溶解させ、染色体DNA溶液とした。
【0049】
(2)CyGDHのγサブユニット、αサブユニット及びβサブユニットをコードするDNA断片の調製
前記染色体DNAを鋳型として、以下の配列を有するオリゴヌクレオチドをプライマーとするPCRにより、CyGDHのγサブユニット、αサブユニット及びβサブユニットをコードするDNA断片を増幅した。
【0050】
〔フォワードプライマー〕
5’−CATGCCATGGCACACAACGACAACAC−3’(配列番号19)〔リバースプライマー〕
5’−GTCGACGATCTTCTTCCAGCCGAACATCAC−3’(配列番号20)
【0051】
増幅した断片のC末端側を平滑末端化した後、N末端側をNcoIで消化し、同様に処理したpTrc99Aにライゲーションした。得られた組換えベクターでエシェリヒア・コリDH5αを形質転換し、アンピシリン50μg/mLを含むLB寒天培地で生じるコロニーを採取した。得られた形質転換体を液体のLB培地で培養してプラスミドを抽出し、その挿入DNA断片を解析したところ、約3.8kbの挿入断片が確認された。本プラスミドをpTrc99Aγαβと命名した。本プラスミド中のCyGDHの各構造遺伝子は、trcプロモーターによって制御される。pTrc99Aγαβは、アンピシリン耐性遺伝子及びカナマイシン耐性遺伝子を保持している。
【0052】
〔実施例2〕CyGDH αサブユニット遺伝子への変異導入による基質相互作用部位の探索
(1)326位、365位、および472位への変異導入
実施例1で得られたpTrc99Aγαβに含まれるGDH αサブユニット遺伝子に、同遺伝子がコードするαサブユニットの326位のセリン残基、365位のセリン残基、および472位のアラニン残基を他のアミノ酸残基に置換されるように変異を導入した。
具体的には、市販の部位特異的変異導入キット(Stratagene社、QuikChangeII Site−Directed Mutagenesis Kit)を用いて、実施例1に記載のプラスミドpTrc99A/γ+α及びpTrc99Aγαβに含まれるGDH αサブユニット遺伝子の326位のセリンのコドン(TCG)、365位のセリンのコドン(TCG)、および472位のアラニンのコドン(GCG)を他のアミノ酸のコドンに置換した。
【0053】
前記アミノ酸残基置換に用いたフォワードプライマーおよびリバースプライマーの配列を以下の表2に示す。また、3変異作製のために用いたリバースプライマーを表3に表す。
尚、変異を表す表記において、数字はアミノ酸配列における位置を、数字の前のアルファベットはアミノ酸置換前のアミノ酸残基を、数字の後のアルファベットはアミノ酸置換後のアミノ酸残基を示す。例えば、R53Fは、53位のアルギニンからフェニルアラニンへの置換を示す。
【0054】
PCR反応は、以下の反応組成で、95℃、30秒の後、95℃ 30秒、55℃ 1分、68℃ 8分を15サイクル繰り返し、68℃ 30分の反応を行った後、4℃で保持した。
【0055】
〔反応液組成〕
鋳型DNA(5ng/μl) 2μl
(3変異導入pTrc99A/γ+α及びpTrc99Aγαβ)
10×反応緩衝液 5μl
フォワードプライマー(100ng/μl) 1.25μl
リバースプライマー(100ng/μl) 1.25μl
dNTP 1μl
蒸留水 38.5μl
DNAポリメラーゼ 1μl
合計 50μl
【0056】
PCR反応の後、反応液にDpnIを0.5μl添加し、37℃で1時間インキュベートし、鋳型プラスミドを分解させた。
得られた反応液で、エシェリヒア・コリDH5α(supE44, ΔlacU169(φ80lacZΔM15), hsdR17, recAi, endA1, gyrA96, thi−1, relA1)のコンピンテントセルを形質転換した。アンピシリン(50μg/ml)およびカナマイシン(30μg/ml)を含むLB寒天培地(バクトリプトン1%、酵母エキス0.5%、塩化ナトリウム1%、寒天1.5%)上に生育してきたコロニー数個からプラスミドDNAを調製し、配列解析を行って、GDH αサブユニット遺伝子に目的の変異が導入されたことを確認した。
【0057】
【表2−1】
【表2−2】
【0058】
【表3−1】
【表3−2】
【0059】
〔実施例3〕変異GDHの基質特異性の解析
実施例2で得られた変異GDH発現プラスミドを用いて、変異GDHを製造し、基質特異性を検討した。
【0060】
(1)培養
変異導入したエシェリヒア・コリDH5α株を、各々2mlのLB培地(アンピシリン50μg/ml及びカナマイシン30μg/ml含有)で、L字管を用いて37℃で一晩振とう培養した。それらの培養液を、150mlのLB培地(アンピシリン50μg/mlおよびカナマイシン30μg/ml含有)を含む500mlの坂口フラスコに植菌し、37℃で振とう培養した。培養開始から3時間後にIPTG(イソプロピル−β−D−チオガラクトピラノシド)を終濃度0.1mMになるように添加し、さらに2時間培養した。
【0061】
(2)粗酵素サンプルの調製
前記で培養した培養液から菌体を集め、洗浄後、湿菌体0.3mgあたり1mlの0.2% Triton X100を含む10mMリン酸カリウムバッファー(PPB)(pH7.0)で菌体を懸濁し、超音波破砕した。この懸濁液遠心分離(10000r.p.m、10分、4℃)して残渣を除去した後、上清を超遠心分離(50,000r.p.m.、60分、4℃)し、得られた上清(水溶性画分)を、粗酵素サンプルとした。また、このサンプルを、通常の疎水性クロマトグラフィー(カラム名:オクチルセファロース(アマシャム・バイオサイエンス製)およびイオン交換クロマトグラフィー(Q−セファロース(アマシャム・バイオサイエンス製)により精製して、精製酵素サンプルを得た。目的とする酵素画分の決定は、GDH活性を指標として行った。
【0062】
なお、Histagを有するタンパク質の精製は、以下のようにして行った。
培養した培養液から菌体を集め、洗浄後、湿菌体0.3mgあたり1mlの0.5M塩化ナトリウム、20mMイミダゾールを含む20mMリン酸ナトリウムバッファー(pH7.5)で菌体を懸濁し、超音波破砕した。この懸濁液遠心分離(10000r.p.m、10分、4℃)して残渣を除去した後、上清を超遠心分離(50,000r.p.m.、60分、4℃)し、得られた上清(水溶性画分)を、粗酵素サンプルとした。このサンプルを、0.5M塩化ナトリウム、20mMイミダゾールを含む20mMリン酸ナトリウムバッファー(pH7.5)で平衡化したHistrap FFカラム(アマシャム・バイオサイエンス製)に添加し、0.5M塩化ナトリウム、60mMイミダゾールを含む20mMリン酸ナトリウムバッファー(pH7.5)で洗浄した後、0.5M塩化ナトリウム、150mMイミダゾールを含む20mMリン酸ナトリウムバッファー(pH7.5)で溶出させ、精製酵素サンプルを得た。目的とする酵素画分の決定は、GDH活性を指標として行った。
【0063】
(3)GDH活性の測定
前記粗酵素サンプル8μlに、活性測定用試薬(12μlの600mM メチルフェナジンメトサルフェート(PMS)、120μlの6 mM 2,6−ジクロロフェノールインドフェノール(DCIP)に、0.2(w/v)% Triton X−100 10mM PPBを加え、全量480μlとした溶液)を8μl添加した。これをアルミブロック恒温槽を用いて、各反応温度で1分間プレインキュベートした後、素早く各濃度の基質(グルコース又はマルトース)、または蒸留水を8μl加えて攪拌し、分光光度計を用いてDCIP由来の吸収波長である600 nmの吸光度を測定した。各試薬の終濃度はDCIP:0.06mM、PMS、0.6mM。基質の終濃度は10mMまたは5mMである。
結果を、表4に示す。尚、野生型GDHの反応比は、10mMまたは5mMの基質濃度において、それぞれ23.32%および18.88%であった。
【0064】
【表4】
【0065】
表中、QYAは、326位および365位に相当するアミノ酸残基がそれぞれグルタミンおよびチロシンで置換されているが、472位は置換されていない(すなわちアラニン残基のままである)変異グルコース脱水素酵素(すなわち、GDH(QY))を示す。
【0066】
この結果、326Glu+365Arg+472Tyr、326Gln+365Tyr+472Arg、326Lys+365Tyr+472Thr、326Ile+365Phe+472Lys、326Arg+365Arg+472Tyr、326Lys+365Tyr+472Arg、326Lys+365Tyr+472Ser、326Gln+365Tyr+472Tyrが、グルコースに対する反応性を維持しつつ、マルトースとの反応性が低下する大変効果の高い変異であった。
すなわち、326位、365位、および472位の3箇所の変異の組み合わせのうちいくつかの組み合わせにおいて、変異GDH(QY)と同等またはそれ以上の相乗効果が確認された。
【0067】
〔実施例4〕精製酵素の調製(αサブユニットおよびγサブユニットの複合体)
実施例3で基質特異性の改善が認められた8つの変異GDHについて精製を実施した。方法は実施例3に記載の通りに行なった。各精製酵素のグルコースおよびマルトースに対する比活性(U/mg)、反応比(マルトースに対する比活性/グルコースに対する比活性)、グルコースに対するKm値およびVmaxを表5に示す。
その結果、QYAのマルトースとの反応比が、10mMおよび5mMにおいて、それぞれ1%台であるのに対し、ERYおよびKYTでは、反応比が1%以下まで低下し、QYYでは、マルトースとの反応比が0.3%台まで低下していた。
【0068】
【表5】
【0069】
〔実施例5〕精製酵素の調製(αサブユニット、βサブユニット、およびγサブユニットの複合体)
実施例3で基質特異性の改善が認められた変異CyGDH(QYY)について精製を実施した。方法は実施例3に記載の通りに行なった。各精製酵素のグルコースに対する比活性(U/mg)を表6に示す。
その結果、αサブユニット、βサブユニット、およびγサブユニットの複合体の形態においては、QYY(326Gln+365Tyr+472Tyr)は、QYA(326Gln+365Tyr)と比較して、マルトースとの反応性が約1/2〜1/3に低下していた。
【0070】
【表6】
【0071】
〔実施例6〕変異酵素(QYX変異)の基質特異性の解析
実施例2と同様にして、pTrc99A/γ+αに含まれるGDH αサブユニット遺伝子に、同遺伝子がコードするαサブユニットの326位のセリン残基および365位のセリン残基をそれぞれグルタミンおよびチロシンに置換されるように、そして472位のアラニン残基を他のアミノ酸残基に置換されるように変異を導入した。得られた変異酵素発現プラスミドを用いて、実施例3と同様にして、変異酵素を製造し、基質特異性を検討した。結果を以下の表7に示す。
【0072】
【表7】
【0073】
その結果、QYD、QYG、QYK、QYL、QYP、QYQ、QYR、QYT、QYV、およびQYYが、グルコースに対する反応性を維持しつつ、マルトースとの反応性が低下する変異であることが認められた。
【0074】
[実施例7]精製酵素の調製(αサブユニットおよびγサブユニットの複合体)
実施例6で基質特異性の改善が認められた10種類の変異GDH(QYD、QYG、QYK、QYL、QYP、QYQ、QYR、QYT、QYV、およびQYY)について精製を実施した。方法は実施例3に記載の通りに行なった。各精製酵素のグルコースおよびマルトースに対する比活性(U/mg)、反応比(マルトースに対する比活性/グルコースに対する比活性)、グルコースに対するKm値およびVmaxを表8に示す。
その結果、いずれの変異GDHについてもマルトースとの反応比は低いものの、QYYを除いて酵素活性が低下しており、グルコース濃度の測定には適していないことが示唆される。すなわち、QYXで示される10種類の変異GDHのうちで、QYYが最もグルコース濃度の測定に適していることが示唆される。
【0075】
【表8−1】
【表8−2】
【0076】
〔実施例8〕変異CyGDHを用いた血糖測定用比色式センサの作製
最も比活性が優れていたQYYに関して、実施例5で得られた精製変異酵素を用いてグルコースセンサを作製した。
【0077】
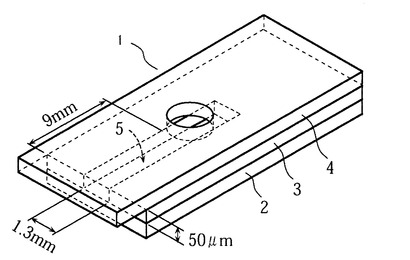
図1に示す基本構成を有するグルコースセンサを作製した。すなわち、前記グルコースセンサは、透明基板2に対してスペーサー3を介して透明カバー4(材質PET)を積層した形態を有し、各要素2〜4によってキャピラリ5が規定されている。キャピラリ5の寸法は、1.3mm×9mm×50μmである(図1)。透明基板2および透明カバー4は、厚みが250μmであるPETにより形成し、スペーサー3は黒色の両面テープにより構成した。
【0078】
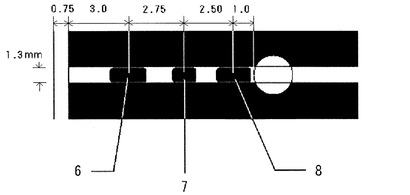
グルコースセンサは、図2に示す第1試薬部〜第3試薬部を有し、それぞれ成分及び塗布量を表9に示す。表中、「Ru」は、ヘキサルテニウムアンミン錯体(Ru(NH3)6Cl3)を、ACESはN−(2−acetamido)−2−aminoethanesulfonic acidを、WST−4は2−Benzothiazolyl−3−(4−carboxy−2−methoxyphenyl)−5−[4−(2−sulfoethylcarbamoyl)phenyl]−2H−tetrazoliumを、SWNはLithium Magnesium Sodium Silicateを、各々示す。
【0079】
【表9】
【0080】
上記グルコースセンサのキャピラリに測定試料を供給し、その時点から5秒後のエンドポイントの吸光度をプロットした。各回の吸光度の測定においては、第3試薬部に対して、キャピラリの高さ方向に沿って光を照射し、そのときにグルコースセンサを透過した光を受光した。光の照射は、発光ダイオードを用いて630nmの光を照射することにより行なった。透過光は、フォトダイオードにおいて受光した。
【0081】
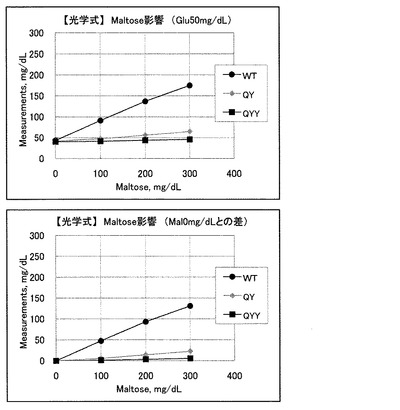
マルトース影響に関してであるが、グルコース50mg/dlにマルトースを添加した場合、野生型ではマルトース濃度に依存して吸光度が大きく増加し、マルトースと強く反応していることが示唆される。一方、変異CyGDH(QY)を用いたセンサでは、マルトース濃度に応じた吸光度の増加が抑えられており、マルトースの影響が少なくなっていることがわかる。変異CyGDH(QYY)を用いたセンサでは、さらに、マルトース濃度に応じた吸光度の増加が抑えられており、マルトースの影響が少なくなっていることがわかる。これらのデータを見かけの血糖上昇値に換算した結果を図3に示す。
野生型酵素を用いたセンサでは、低血糖(50mg/dlグルコース)がマルトースの混入によって正常域以上の(174.8mg/dlグルコース)に見かけ上表示されてしまう(変化率は300%)。また、改変CyGDH(QY)を用いたセンサであっても、マルトースの混入によって64.8mg/dlグルコースに見かけ上表示されてしまう(変化率は54%)。一方、改変CyGDH(QYY)を用いたセンサでは、マルトースが最大300mg/dl混入している場合でも、見かけの血糖値が最大でも46.1mg/dlまでしか上昇せず、影響は大幅に押さえられていると言える(変化率は14%)。結論としてはQYY変異体が、グルコースに対する反応性(直線性)とマルトース影響の観点から最適であることが示唆される。
【0082】
以上の結果から明らかなように、変異CyGDH(QYY)を用いたグルコースセンサは、変異CyGDH(QY)を用いたグルコースセンサよりも、マルトースとの反応性が大きく低下している。この変異CyGDH(QYY)を用いたグルコースセンサを用いれば、病院等で投与される血中マルトース濃度の上限値である200mg/dlにおいては正誤差は無いと言え、上限値以上の300mg/dlにおいても低血糖(50mg/dl以下)が正常値や高血糖と判定されること無く、安全な治療が行なえる。
【0083】
〔実施例9〕変異CyGDHを用いた電極式血糖センサの作製
最も比活性が優れていたQYYに関して、実施例5で得られた精製変異酵素を用いて電極式グルコースセンサを作製した。
【0084】
(作製手順)
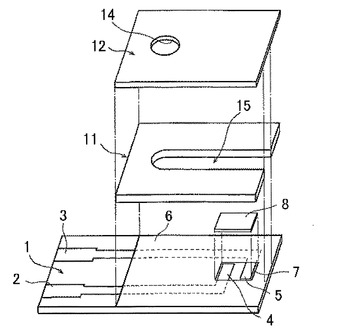
電極式グルコースセンサの作製方法を、図5を参照して以下に示す。絶縁基板1として、PET製基板(長さ28mm、幅7mm、厚み250μm)を準備し、その一方の表面に、カーボンインクのスクリーン印刷により、リード部2および3をそれぞれ有する作用極4および対極5からなるカーボン電極系を形成した。つぎに、絶縁性ペーストを調製し、これを前記電極上にスクリーン印刷して絶縁層6とし、前記絶縁層を形成しない部分を検出部およびリード部とした。次に、合成スメクタイトである商品名「ルーセンタイトSWN」(コープケミカル社製)0.6gを精製水100mLに懸濁し、約8〜24時間攪拌した。この合成スメクタイト懸濁液10mLに、10%(w/v)CHAPS(同仁化学研究所製)水溶液0.1mL、1.0M ACES緩衝液(pH7.4:同仁化学研究所製)5.0mLおよび精製水4.0mLをこの順序で添加し、さらにメディエータとして[Ru(NH3)6]Cl3(アルドリッチ社製)1.0gを混合した。この混合液1.0μLを、検出部に分注した。そして、これを、30℃、相対湿度10%の条件下で10分間乾燥させて、試薬層7を形成した。さらに、前記試薬層7上に酵素試薬層8を形成した。これは、2700U/mLの本願発明のグルコース脱水素酵素水溶液1.0μLを、検出部の試薬層の上に分注して、30℃、相対湿度10%の条件下で10分間乾燥させて形成した。最後に、スリット15を有するスペーサー11を絶縁層上に配置し、さらに、前記スペーサー上に空気孔14となる貫通孔を有するカバー12を配置してバイオセンサを作製した。
【0085】
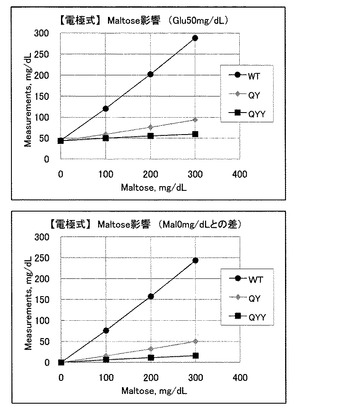
マルトース影響に関してであるが、グルコース50mg/dlにマルトースを添加した場合、野生型ではマルトース濃度に依存して電流値が大きく増加し、マルトースと強く反応していることが示唆される。一方、変異CyGDH(QY)を用いたセンサでは、マルトース濃度に応じた電流値の増加が抑えられており、マルトースの影響が少なくなっていることがわかる。変異CyGDH(QYY)を用いたセンサでは、さらに、マルトース濃度に応じた電流値の増加が抑えられており、マルトースの影響が少なくなっていることがわかる。これらのデータを見かけの血糖上昇値に換算した結果を図4に示す。
野生型酵素を用いたセンサでは、低血糖(50mg/dlグルコース)がマルトースの混入によって正常域以上の(288.2mg/dlグルコース)に見かけ上表示されてしまう(変化率は549%)。また、改変CyGDH(QY)を用いたセンサであっても、マルトースの混入によって93.9mg/dlグルコースに見かけ上表示されてしまう(変化率は114%)。一方、改変CyGDH(QYY)を用いたセンサでは、マルトースが最大300mg/dl混入している場合でも、見かけの血糖値が最大でも60.0mg/dlまでしか上昇せず、影響は大幅に押さえられていると言える(変化率は37%)。結論としてはQYY変異体が、グルコースに対する反応性(直線性)とマルトース影響の観点から最適であることが示唆される。
【0086】
以上の結果から明らかなように、変異CyGDH(QYY)を用いたグルコースセンサは、変異CyGDH(QY)を用いたグルコースセンサよりも、マルトースとの反応性が大きく低下している。この変異CyGDH(QYY)を用いたグルコースセンサを用いれば、病院等で投与される血中マルトース濃度の上限値である200mg/dlにおいては正誤差は無いと言え、上限値以上の300mg/dlにおいても低血糖(50mg/dl以下)が正常値や高血糖と判定されること無く、安全な治療が行なえる。
【0087】
〔実施例10〕GDHホモログへの変異導入
実施例1と同様にして、ブルクホルデリア・セノセパシアJ2315株のputative oxidoreductase、ブルクホルデリア・タイランデンシスTXDOH株のhypothetical protein BthaT#07876、ラルストニア・ピッケティ12D株のFAD dependent oxidoreductase、ラルストニア・ソラナセアラムIPO1609株のtransmembrane dehydrogenaseおよびブルクホルデリア・フィトフィルマンスPsJN株のglucose-methanol-choline oxidoreductaseの各αサブユニットおよびγサブユニットを発現するプラスミドを用意した。
【0088】
これらの発現プラスミドはγサブユニット構造遺伝子とαサブユニット構造遺伝子を連続して含むDNA断片が、ベクターpET30c(+)(Novagen社)のクローニング部位であるNdeI/HindIIIに挿入されてなるプラスミドである。本プラスミド中のγα遺伝子は、T7プロモーターによって制御される。本プラスミドは、カナマイシン耐性遺伝子を保持している。
【0089】
上記プラスミドに含まれるαサブユニット遺伝子に対し、ブルクホリデリア・セパシアKS1株の野生型GDH αサブユニットにおける326位、365位、および472位に相当する残基がそれぞれグルタミン、チロシン、チロシンに置換されるように、市販の部位特異的変異導入キット(Stratagene社、QuikChangeII Site−Directed Mutagenesis Kit)を用いて変異を導入した。
【0090】
前記アミノ酸残基置換に用いたフォワードプライマーの配列を以下に示す。リバースプライマーの配列は、フォワードプライマーの完全相補鎖とした。
尚、変異を表す表記において、数字はアミノ酸配列における位置を、数字の前のアルファベットはアミノ酸置換前のアミノ酸残基を、数字の後のアルファベットはアミノ酸置換後のアミノ酸残基を示す。例えば、R53Fは、53位のアルギニンからフェニルアラニンへの置換を示す。
【0091】
PCR反応は、以下の反応組成で、95℃、30秒の後、95℃ 30秒、55℃ 1分、68℃ 8分を15サイクル繰り返し、68℃ 30分の反応を行った後、4℃で保持した。
【0092】
〔反応液組成〕
鋳型DNA(5ng/μl) 2μl
(3変異導入pTrc99A/γ+α及びpTrc99Aγαβ)
10×反応緩衝液 5μl
フォワードプライマー(100ng/μl) 1.25μl
リバースプライマー(100ng/μl) 1.25μl
dNTP 1μl
蒸留水 38.5μl
DNAポリメラーゼ 1μl
合計 50μl
【0093】
PCR反応の後、反応液にDpnIを0.5μl添加し、37℃で1時間インキュベートし、鋳型プラスミドを分解させた。
得られた反応液で、エシェリヒア・コリDH5α(supE44, ΔlacU169(φ80lacZΔM15), hsdR17, recAi, endA1, gyrA96, thi−1, relA1)のコンピンテントセルを形質転換した。アンピシリン(50μg/ml)およびカナマイシン(30μg/ml)を含むLB寒天培地(バクトリプトン1%、酵母エキス0.5%、塩化ナトリウム1%、寒天1.5%)上に生育してきたコロニー数個からプラスミドDNAを調製し、配列解析を行って、各αサブユニット遺伝子に目的の変異が導入されたことを確認した。
【0094】
【表10−1】
【表10−2】
【0095】
〔実施例11〕変異酵素の基質特異性の解析
実施例10で得られた変異酵素発現プラスミドを用いて、変異酵素を製造し、基質特異性を検討した。
【0096】
(1)培養
変異酵素発現プラスミドでエシェリヒア・コリBL21(DE3)(F−, dcm, ompT, hsdS(rB− mB−), gal, λ(DE3))のコンピンテントセルを形質転換した。得られた形質転換体を各々3mlのLB培地(カナマイシン25μg/ml含有)で、L字管を用いて37℃で一晩振とう培養した。それらの培養液を、100mlのLB培地(0.5%グリセロール、0.05%グルコース、0.2%ラクトース、100mM PO4、 25mM SO4、50mM NH4、100mM Na、50mM K、1mM MgSO4およびカナマイシン25μg/ml含有)を含む500mlの坂口フラスコに植菌し、20℃で24時間振とう培養した。
【0097】
(2)粗酵素サンプルの調製
前記で培養した培養液から菌体を集め、洗浄後、湿菌体0.1gあたり1mlの10mMリン酸カリウムバッファー(PPB)(pH7.0)で菌体を懸濁し、超音波破砕した。この懸濁液を遠心分離(10000r.p.m、10分、4℃)して残渣を除去した後、上清を超遠心分離(50,000r.p.m.、60分、4℃)し、得られた上清(水溶性画分)を、粗酵素サンプルとした。
【0098】
(3)GDH活性の測定
前記粗酵素サンプル10μlに、活性測定用試薬(94μlの600mM メチルフェナジンメトサルフェート(PMS)、9.4μlの6mM 2,6−ジクロロフェノールインドフェノール(DCIP)に、10mM PPBを加え、全量8mlとした溶液)を170μl添加した。これに各濃度の基質(グルコース又はマルトース)、または蒸留水を20μl加えて攪拌し、分光光度計を用いてDCIP由来の吸収波長である600nmの吸光度を測定した。各試薬の終濃度はDCIP:0.06mM、PMS:0.6mMである。
結果を表11に示す。いずれの変異酵素も、変異導入前の野生型酵素に比べてマルトースへの反応性が低下しており、グルコースに対する基質特異性が向上していることが確認された。
【0099】
【表11】
【0100】
以上の結果より、ブルクホルデリア・セパシア以外のGDHホモログにおいても、QYY変異が有効であることが示唆される。
【産業上の利用可能性】
【0101】
本発明の変異GDHは、グルコースに対する基質特異性が向上しており、グルコースセンサ等を用いたグルコースの測定に好適に使用することができる。
【技術分野】
【0001】
本発明は基質特異性の向上した変異グルコース脱水素酵素に関する。詳細には、本発明は、シトクロムCを含むグルコース脱水素酵素(以下CyGDHとも称する)を構成するαサブユニットのアミノ酸残基に変異導入した変異型αサブユニットを含むグルコース脱水素酵素およびその遺伝子に関する。本発明のグルコース脱水素酵素は、グルコースセンサ、及びグルコースアッセイキット等に好適に使用することができ、生化学分野、臨床分野等で有用である。
【背景技術】
【0002】
現在、野生型のCyGDHや、ピロロキノリンキノンを補酵素とするPQQGDHが、自己血糖測定センサに用いられている。野生型のCyGDHやPQQGDHは、グルコースのみならずマルトースにも反応することから、患者の血中マルトース濃度が高い場合においては、正確な血糖値の測定ができないという欠点があった。特に日本や英国においては、輸液中のエネルギー物質としてマルトースが用いられており、実際に、腹膜透析等により輸液を投与している低血糖患者が、PQQGDH型の血糖センサで高血糖と誤認されたことによる事故も発生している。
野生型CyGDHは、グルコースに対してマルトースの反応性が高く、グルコース濃度50mg/dL時において、マルトース濃度が100mg/dL時の血糖値計測結果は、170%高値を示す。
【0003】
このような状況に鑑み、CyGDHの変異体酵素(αサブユニットのアミノ酸残基の326位と365位に変異導入し、それぞれグルタミン(Q)、チロシン(Y)に置換した変異体グルコース脱水素酵素(以下CyGDH(QY)または単にQY、QYAとも称する)が発明されており(特許文献1)、グルコース濃度50mg/dL時において、マルトース濃度が100mg/dL時の血糖値計測結果が、36%の高値まで抑えることが出来ている。しかしながら、マルトース影響回避効果が完全なものとは言えなかった。
【先行技術文献】
【特許文献】
【0004】
【特許文献1】国際公開第2006/137283号
【発明の概要】
【発明が解決しようとする課題】
【0005】
本発明は、CyGDH(QY)よりも、グルコースに対する基質特異性の向上したCyGDHを提供することを課題とする。
【課題を解決するための手段】
【0006】
本発明者は、上記課題を解決するために鋭意検討を行った結果、CyGDHを構成するαサブユニットのアミノ酸残基のうち、326位および365位に相当する部位をそれぞれグルタミンおよびチロシンで置換し、さらに472位に相当する部位をチロシンで置換することにより、基質特異性がCyGDH(QY)と比較してさらに向上すること、そして、GDHのホモログにおいても326位、365位および472位に相当する部位をそれぞれグルタミン、チロシンおよびチロシンで置換することが同様の効果をもたらすことを見出し、本発明を完成するに至った。
【0007】
すなわち、本発明は以下のとおりである。
(1)配列番号3と少なくとも80%の同一性を有するアミノ酸配列を有し、かつ、グルコース脱水素酵素活性を有する変異グルコース脱水素酵素であって、
前記アミノ酸配列の326位、365位および472位に相当するアミノ酸残基が、それぞれグルタミン、チロシンおよびチロシンで置換されており、
グルコースに対する基質特異性が向上し、二糖類に対する反応性が低下したことを特徴とする、変異グルコース脱水素酵素。
(2)配列番号3と少なくとも90%の同一性を有するアミノ酸配列を有する、(1)に記載の変異グルコース脱水素酵素。
(3)326位、365位および472位に相当する位置以外は配列番号3に示すアミノ酸配列を有する、(1)または(2)に記載の変異グルコース脱水素酵素。
(4)326位、365位および472位に相当する位置以外は配列番号7に示すアミノ酸配列を有する、(1)または(2)に記載の変異グルコース脱水素酵素。
(5)326位、365位および472位に相当する位置以外は配列番号8に示すアミノ酸配列を有する、(1)または(2)に記載の変異グルコース脱水素酵素。
(6)326位、365位および472位に相当する位置以外は配列番号9に示すアミノ酸配列を有する、(1)または(2)に記載の変異グルコース脱水素酵素。
(7)326位、365位および472位に相当する位置以外は配列番号10に示すアミノ酸配列を有する、(1)または(2)に記載の変異グルコース脱水素酵素。
(8)前記アミノ酸配列において326位および365位に相当するアミノ酸残基がそれぞれグルタミンおよびチロシンで置換されているが472位は置換されていないグルコース脱水素酵素と比較して、グルコースに対する基質特異性が向上し、二糖類に対する反応性が低下したことを特徴とする、(1)〜(7)のいずれか1項に記載の変異グルコース脱水素酵素。
(9)二糖類がマルトースであることを特徴とする(1)〜(8)のいずれか1項に記載の変異グルコース脱水素酵素。
(10)(1)〜(9)のいずれか1項に記載の変異グルコース脱水素酵素と、電子伝達サブユニットとを少なくとも含む変異グルコース脱水素酵素複合体。
(11)電子伝達サブユニットがシトクロムCであることを特徴とする(10)に記載のグルコース脱水素酵素複合体。
(12)(1)〜(9)のいずれか1項に記載の変異グルコース脱水素酵素をコードするDNA。
(13)(12)に記載のDNAを保持し、(1)〜(9)のいずれか1項に記載の変異グルコース脱水素酵素、あるいは(10)または(11)に記載の変異グルコース脱水素酵素複合体を産生する微生物。
(14)(1)〜(9)のいずれか1項に記載の変異グルコース脱水素酵素、(10)または(11)に記載の変異グルコース脱水素酵素複合体、あるいは(13)に記載の微生物を含む、グルコースアッセイキット。
(15)(1)〜(9)のいずれか1項に記載の変異グルコース脱水素酵素、(10)または(11)に記載の変異グルコース脱水素酵素複合体、あるいは(13)に記載の微生物を含む、グルコースセンサ。
【図面の簡単な説明】
【0008】
【図1】グルコースセンサの構造を示す図。
【図2】グルコースセンサの各試薬部を示す図。
【図3】比色式センサでの血糖測定における、グルコース濃度50mg/dLでの、マルトース濃度(100mg/dL、200mg/dL、300mg/dL)の血糖値に対する影響を示す図。
【図4】電極式センサでの血糖測定における、グルコース濃度50mg/dLでの、マルトース濃度(100mg/dL、200mg/dL、300mg/dL)の血糖値に対する影響を示す図。
【図5】電極式グルコースセンサの構造を示す図。
【発明を実施するための形態】
【0009】
以下、本発明を詳細に説明する。
本発明の変異GDHは、野生型GDHのαサブユニットに特定の変異を導入することにより、製造することができる。野生型GDHとしては、ブルクホリデリア・セパシアが産生するGDHが挙げられる。ブルクホリデリア・セパシアのGDHとしては、ブルクホリデリア・セパシアKS1株、JCM2800株又はJCM2801株が産生するGDHが挙げられる。KS1株は、平成12年9月25日に独立行政法人産業技術総合研究所特許生物寄託センター(〒305−8566 日本国茨城県つくば市東1丁目1番地1 中央第6)に、受託番号第FERM BP−7306として寄託されている。
【0010】
KS1株のGDH αサブユニット遺伝子、及びβサブユニット遺伝子の一部を含む染色体DNA断片の塩基配列を配列番号1に示す(米国特許出願公開第2004/0023330号)。この塩基配列には3つのオープンリーディングフレーム(ORF)が存在し、5’末端側から2番目及び3番目のORFは、それぞれαサブユニット(配列番号3)、及びβサブユニット(配列番号4)をコードしている。また、1番目のORFはγサブユニット(配列番号2)をコードしていると推定される。また、配列番号5に、βサブユニット遺伝子全長を含む断片の塩基配列を示す。さらに、βサブユニットのアミノ酸配列を配列番号6に示す(欧州特許出願公開第1498484号)。配列番号6において、アミノ酸番号1〜22はシグナルペプチドであると推定される。
【0011】
また、これらの他に、ブルクホリデリア・セパシアKS1株のGDHのホモログである、ブルクホルデリア・セノセパシアJ2315株のputative oxidoreductase(配列番号7)、ブルクホルデリア・タイランデンシスTXDOH株のhypothetical protein BthaT#07876(配列番号8)、ラルストニア・ピッケティ12D株のFAD dependent oxidoreductase(配列番号9)、ラルストニア・ソラナセアラムIPO1609株のtransmembrane dehydrogenase(配列番号10)およびブルクホルデリア・フィトフィルマンスPsJN株のglucose-methanol-choline oxidoreductase(配列番号11)のそれぞれのαサブユニットも、ブルクホリデリア・セパシアKS1株のGDHと同様に用いることができる。
なお、配列番号7〜11に示したアミノ酸配列については、いずれもアメリカの国立生物工学情報センター(National Center for Biotechnology Information, NCBI)のデータベースに登録されている。それぞれの登録番号は、配列番号7がYP_002234347、配列番号8がZP_02370914、配列番号9がYP_002980762、配列番号10がYP_002260434、配列番号11がYP_001890482である。
また、ブルクホルデリア・セノセパシアJ2315株は、LMG 16656、ATCC BAA-245、CCM 4899、CCUG 48434、NCTC 13227として寄託されている。ブルクホルデリア・フィトフィルマンスPsJN株は、LMG 22487、CCUG 49060として寄託されている。
【0012】
また、ブルクホリデリア・セパシアKS1株と同属である、ブルクホリデリア・セパシア株、例えばJCM2800、JCM2801、JCM5506、JCM5507、IFO14595由来のそれぞれのGDHのαサブユニット(配列番号12〜16)も、ブルクホリデリア・セパシアKS1株のGDHと同様に用いることができる。なお、JCM2800、JCM2801、JCM5506、およびJCM5507は、独立行政法人理化学研究所微生物系統保存施設(Japan Collection of Microorganisms, JCM)に保存されている。IFO14595は、財団法人発酵研究所(IFO)に保存されている。
【0013】
本発明の変異GDHは、αサブユニットのみであってもよいし、αサブユニットとβサブユニットとの複合体、αサブユニットとγサブユニットとの複合体、又はαサブユニット、βサブユニット及びγサブユニットからなる複合体であってもよい。本明細書において、βサブユニットを含むGDH複合体をCyGDHと称し、βサブユニットを含まないGDH複合体をGDHと称する。本発明の変異GDHは、いずれの場合もαサブユニットに特定の変異(326位、365位及び472位における変異、またはこれらに対応する位置における変異)が導入されたものであるが、この特定の変異の他に、保存的な変異を有していてもよい。また、他のサブユニットは野生型であっても、保存的な変異を有するものであってもよい。尚、「保存的変異」とは、GDH活性に実質的に影響しない変異をいう。
【0014】
本発明の変異型αサブユニットは、好ましくは、前記特定の変異以外は、配列番号3、7〜11に示すアミノ酸配列を有する。また、変異型αサブユニットは、GDH活性を有する限り、前述したような保存的変異を有していてもよい。すなわち、配列番号3、7〜11のアミノ酸配列において、前記特定の変異以外に、1又は複数のアミノ酸残基が置換、欠失、挿入、又は付加されたアミノ酸配列を有するタンパク質であってもよい。尚、配列番号3には、配列番号1の塩基配列によってコードされ得るアミノ酸配列を示してあるが、N末端のメチオニン残基は、翻訳後に脱落している可能性がある。前記「1又は複数」とは、好ましくは1〜10個、より好ましくは1〜5個、特に好ましくは1〜3個である。なお、本発明の変異型αサブユニットは、配列番号3に示すアミノ酸配列に対して、少なくとも80%、好ましくは85%、より好ましくは90%のアミノ酸同一性を有する。
【0015】
また、βサブユニットは、典型的には配列番号6のアミノ酸配列を有する。しかし、CyGDHのβサブユニットとして機能し得る限り、配列番号6のアミノ酸番号23〜425からなるアミノ酸配列において、1又は複数のアミノ酸残基が置換、欠失、挿入、又は付加されたアミノ酸配列を有するタンパク質であってもよい。また、CyGDHのβサブユニットとして機能し得る限り、KS1株以外のβサブユニットおよび同βサブユニットのアミノ酸配列において、1又は複数のアミノ酸残基が置換、欠失、挿入、又は付加されたアミノ酸配列を有するタンパク質であってもよい。前記「1又は複数」とは、好ましくは1〜20個、より好ましくは1〜10個、特に好ましくは1〜5個である。
尚、CyGDHのβサブユニットとして機能するとは、αサブユニットとともに複合体を形成したときに同複合体のGDH活性を損なわずに電子伝達サブユニット、すなわち、チトクロームCとして機能することをいう。
【0016】
野生型αサブユニット遺伝子として具体的には、配列番号1の塩基番号764〜2380からなる塩基配列を含むDNAが挙げられる。また、αサブユニット遺伝子は、配列番号1の塩基配列の塩基番号764〜2380からなる塩基配列を有するDNA、又はこの配列から調製され得るプローブとストリンジェントな条件下でハイブリダイズし、かつ、GDH活性を有するタンパク質をコードするDNAであってもよい。
【0017】
また、βサブユニット遺伝子として具体的には、配列番号5の塩基番号187〜1398からなる塩基配列を含むDNAが挙げられる。またβサブユニット遺伝子は、配列番号5の塩基番号187〜1398からなる塩基配列を有するDNA、又はこの配列から調製され得るプローブとストリンジェントな条件下でハイブリダイズし、かつ、βサブユニットとして機能し得るタンパク質をコードするDNAであってもよい。
【0018】
前記ストリンジェントな条件としては、好ましくは80%、より好ましくは90%以上、特に好ましくは95%以上の相同性を有するDNA同士がハイブリダイズする条件、具体的には、0.1×SSC、0.1%SDS、60℃が挙げられる。
【0019】
αサブユニット遺伝子及びβサブユニット遺伝子は、例えば、ブルクホルデリア・セパシアKS1株の染色体DNAを鋳型とするPCRによって、取得することができる。PCR用プライマーは、前記の塩基配列に基づいて化学合成することによって調製することができる。また、前記配列に基づいて作製したオリゴヌクレオチドをプローブとするハイブリダイゼーションによって、ブルクホルデリア・セパシアKS1株の染色体DNAから取得することもできる。なお、KS1株以外のブルクホルデリア・セノセパシアJ2315株、ブルクホルデリア・タイランデンシスTXDOH株、ラルストニア・ピッケティ12D株、ラルストニア・ソラナセアラムIPO1609株およびブルクホルデリア・フィトフィルマンスPsJN株も用いることができる。
【0020】
本発明の変異GDHは、上記のような野生型GDH又は保存的変異を有するGDHが特定の変異を有することによって、グルコースに対する基質特異性が向上したものである。「グルコースに対する基質特異性が向上した」とは、グルコースに対する反応性を実質的に維持したまま、他の単糖類、二糖類又はオリゴ糖等の糖類、例えばマルトース、ガラクトース、キシロース等に対する反応性が低下したこと、あるいは、グルコースに対する反応性が他の糖類に対する反応性に比べて向上したことを含む。例えば、グルコースに対する反応性が低下しても、他の糖類に対する反応性がそれ以上に低下すれば、グルコースに対する基質特異性は向上する。また、他の糖類に対する反応性が上昇しても、グルコースに対する基質特異性がそれ以上に上昇すれば、グルコースに対する基質特異性は向上する。具体的には例えば、野生型酵素に対する変異型酵素の基質特異性(グルコースに対する比活性と、他の糖類、例えばマルトース、に対する比活性との比)の向上(下記式で表される)が、10%以上、好ましくは20%以上、より好ましくは40%以上であれば、グルコースに対する基質特異性は向上している。例えば、基質特異性が、野生型酵素では60%、変異型GDHでは40%の場合、グルコース以外の糖類に対する反応性は、33%低下している。
基質特異性=(グルコース以外の糖類に対する比活性/グルコースに対する比活性)×100)
基質特異性の向上=(A−B)×100/A
A:野生型酵素の基質特異性
B:変異型酵素の基質特異性
また、変異型GDHは、マルトースに対する反応性(比活性)が、グルコースに対する反応性(比活性)の1%以下、好ましくは0.5%以下である。
【0021】
本発明の変異型GDHは、好ましくは、配列番号3に示すアミノ酸配列の326位および365位に相当するアミノ酸残基がそれぞれグルタミンおよびチロシンで置換されているが472位は置換されていないグルコース脱水素酵素と比較して、グルコースに対する基質特異性が向上し、二糖類に対する反応性が低下している。
【0022】
本発明における変異とは、具体的には以下のとおりである。
(1)配列番号3に示すアミノ酸配列の326位に相当するセリン残基のグルタミンへの置換。
(2)配列番号3に示すアミノ酸配列の365位に相当するセリン残基のチロシンへの置換。
(3)配列番号3に示すアミノ酸配列の472位に相当するアラニン残基のチロシンへの置換。
【0023】
上記アミノ酸置換変異の位置は、配列番号3、すなわちブルクホリデリア・セパシアKS1株の野生型GDH αサブユニットのアミノ酸配列における位置であるが、配列番号3のアミノ酸配列において、前記特定の変異以外に、1又は複数のアミノ酸残基が置換、欠失、挿入、又は付加されたアミノ酸配列を有するGDH αサブユニットのホモログ又はバリアントにおいては、配列番号3のアミノ酸配列とのアラインメントにおいて、前記アミノ酸置換の位置に相当する位置を示す。例えば、1〜364位の領域において1個のアミノ酸残基の欠失を有するGDH αサブユニットの保存的バリアントにおいては、365位とは、前記バリアントの364位を示す。
【0024】
配列番号7においては、配列番号3の326位、365位および472位に相当する残基はそれぞれ326位、365位および472位で同じである。
【0025】
配列番号8においては、配列番号3の326位、365位および472位に相当する残基はそれぞれ324位、363位および470位である。
【0026】
配列番号9においては、配列番号3の326位、365位および472位に相当する残基はそれぞれ327位、366位および473位である。
【0027】
配列番号10においては、配列番号3の326位、365位および472位に相当する残基はそれぞれ327位、366位および473位である。
【0028】
配列番号11においては、配列番号3の326位、365位および472位に相当する残基はそれぞれ322位、361位および466位である。
【0029】
なお、配列番号3に示すアミノ酸配列は、配列番号7〜11に対して、それぞれ、96%、93%、82%、82%、63%のアミノ酸配列同一性を有する。
以下の表1において、配列番号3,7〜11のアミノ酸配列アライメントを示す。
【0030】
【表1−1】
【0031】
【表1−2】
【0032】
本発明者は、変異体グルコース脱水素酵素(CyGDH(QY))と比較して、グルコースに対する基質特異性を増加すべく、遺伝的アルゴリズムを用いて、最適な326位、365位および472位の組み合わせについて検討した。その結果、基質特異性を向上することができる組み合わせを発見した。
【0033】
本発明の変異型GDHの好ましい変異の態様を以下に示す。
(数字はアミノ酸配列における位置を、アミノ酸残基は前記位置における置換後のアミノ酸残基を示し、「+」は同時に2つのアミノ酸置換を有することを示す。)
326Gln+365Tyr+472Tyr
【0034】
所望の変異を有するGDH αサブユニットは、GDH αサブユニットをコードするDNA(αサブユニット遺伝子)に、部位特異的変異法によって、所望のアミノ酸変異に対応するヌクレオチド変異を導入し、得られる変異DNAを適当な発現系を用いて発現させることによって、取得することができる。また、変異GDH αサブユニットをコードするDNAを、βサブユニットをコードするDNA(βサブユニット遺伝子)、又はβサブユニット遺伝子とγサブユニットをコードするDNA(γサブユニット遺伝子)とともに発現させることによって、変異CyGDH複合体を取得することができる。尚、GDH
αサブユニットをコードするDNAへの変異の導入は、γサブユニット、αサブユニット及びβサブユニットをこの順にコードするポリシストロニックなDNA断片を用いてもよい。
【0035】
変異が導入されたGDH αサブユニット又はCyGDH複合体の糖類に対する基質特異性は、実施例に記載された方法によって各種糖類に対する反応性を調べ、野生型GDH
αサブユニット又は野生型CyGDH複合体の反応性と比較することよって、決定することができる。
【0036】
γサブユニット、αサブユニット及びβサブユニットをこの順にコードするポリシストロニックなDNA断片は、例えば、ブルクホルデリア・セパシアKS1株の染色体DNAを鋳型とし、配列番号19、20の塩基配列を有するオリゴヌクレオチドをプライマーとするPCRによって取得することができる(後記実施例参照)。
【0037】
GDHの各サブユニットの遺伝子の取得、変異の導入、遺伝子の発現等に用いるベクターとしては、例えばエシェリヒア属細菌で機能するベクター、具体的にはpTrc99A、pBR322、pUC18、pUC118、pUC19、pUC119、pACYC184、pBBR122等が挙げられる。遺伝子の発現に用いるプロモーターとしては、例えばlac、trp、tac、trc、PL、tet、PhoA等が挙げられる。また、プロモーターを含む発現ベクターの適当な部位に、αサブユニット遺伝子又は他のサブユニット遺伝子を挿入することによって、これらの遺伝子のベクターへの挿入とプロモーターの連結とを同じ工程で行うことができる。このような発現ベクターとしては、pTrc99A、pBluescript、pKK223−3等が挙げられる。
【0038】
また、αサブユニット遺伝子又は他のサブユニット遺伝子は、発現可能な形態で宿主微生物の染色体DNA中に組み込まれてもよい。
【0039】
組換えベクターで微生物を形質転換するには、例えばカルシウム処理によるコンピテントセル法、プロトプラスト法又はエレクトロポレーション法等が挙げられる。
【0040】
宿主微生物としては、バチルス・サブチリス等のバチルス属細菌、サッカロマイセス・セレビシエ等の酵母、アスペルギルス・ニガー等の糸状菌が挙げられるが、これらに限られず、異種タンパク質生産に適した宿主微生物であれば用いることができる。
【0041】
本発明の変異αサブユニットもしくは変異CyGDH複合体又はこれらを発現する微生物は、グルコースセンサの酵素電極、又はグルコースのアッセイキットの構成要素として用いることができる。ブルクホルデリア・セパシアの野生型GDHを用いたグルコースセンサ及びグルコースアッセイキットは、米国特許公開第2004/0023330A1に記載されている。本発明の変異GDHも、同様にして使用することができる。
【実施例】
【0042】
次に、実施例を挙げて本発明をさらに具体的に説明するが、本発明はこれら実施例に何ら限定されるものではない。
【0043】
〔実施例1〕ブルクホルデリア・セパシアのGDHまたはCyGDHを発現するプラスミド
ブルクホルデリア・セパシアのGDHを発現するプラスミドとして、GDHのαサブユニット及びγサブユニットを発現するプラスミド、並びに、CyGDHを発現するプラスミドとして、αサブユニット、βサブユニット及びγサブユニットを発現するプラスミドを用意した。
【0044】
<1>GDHのαサブユニット及びγサブユニットを発現するプラスミド
αサブユニット及びγサブユニットを発現するプラスミドとしては、WO02/036779(EP1331272A1、US2004023330A1、CN1484703Aに対応)に記載のプラスミドpTrc99A/γ+αを使用した。同プラスミドは、ブルクホルデリア・セパシアKS1株(FERM BP−7306)染色体DNAから単離された、GDH γサブユニット構造遺伝子とαサブユニット構造遺伝子を連続して含むDNA断片が、ベクターpTrc99Aのクローニング部位であるNcoI/HindIIIに挿入されてなるプラスミドである。本プラスミド中のGDHγα遺伝子は、trcプロモーターによって制御される。pTrc99A/γ+αは、アンピシリン耐性遺伝子を保持している。
前記プラスミドpTrc99A/γ+αを鋳型として、以下の配列を有するオリゴヌクレオチドをプライマーとするPCRにより、GDHのαサブユニットC末端に6個のヒスチジン残基が付加されたDNA断片を含むプラスミド全体を増幅した。
【0045】
〔フォワードプライマー〕
5’−ACCACCACTGATAAGGAGGTCTGACCGTGCGGAAATCTAC−3’(配列番号17)
〔リバースプライマー〕
5’−AGCCTGTGCGACTTCTTCCTTCAGCGATCGGTGGTGGTGG−3’(配列番号18)
【0046】
増幅した断片の両端を平滑末端化した後、5’末端をリン酸化し、ライゲーションによって環状化した。得られた組換えベクターでエシェリヒア・コリDH5αを形質転換し、アンピシリン50μg/mLを含むLB寒天培地で生じるコロニーを採取した。得られた形質転換体を液体のLB培地で培養してプラスミドを抽出し、その挿入DNA断片を解析したところ、約2.1kbの挿入断片が確認された。本プラスミド中のGDHの各構造遺伝子は、trcプロモーターによって制御される。本プラスミドは、アンピシリン耐性遺伝子を保持している。
【0047】
<2>CyGDHのαサブユニット、βサブユニット及びγサブユニットを発現するプラスミド
CyGDHのαサブユニット、βサブユニット及びγサブユニットを発現するプラスミドは、以下のようにして調製した。
【0048】
(1)ブルクホルデリア・セパシア KS1株からの染色体DNAの調製
ブルクホルデリア・セパシア KS1株より染色体遺伝子を常法に従って調製した。すなわち、同菌株をTL液体倍地(ポリペプトン 10g、酵母抽出液 1g、NaCl 5g、KH2PO4 2g、グルコース 5g;1L、pH 7.2)を用いて、34℃で一晩振盪した。増殖した菌体を遠心分離により回収した。この菌体を10mM NaCl、20mM Tris−HCl(pH8.0)、1mM EDTA、0.5% SDS、100μg/mlのプロテイナーゼKを含む溶液に懸濁し、50℃で6時間処理した。ここに等量のフェノール−クロロホルムを加えて室温で10分間撹拌した後、遠心分離により上清を回収した。これに終濃度0.3Mになるように酢酸ナトリウムを加え、2倍量のエタノールを重層して中間層に染色体DNAを析出させた。これをガラス棒を用いてすくいとり、70%エタノールで洗浄した後、適当量のTEバッファーに溶解させ、染色体DNA溶液とした。
【0049】
(2)CyGDHのγサブユニット、αサブユニット及びβサブユニットをコードするDNA断片の調製
前記染色体DNAを鋳型として、以下の配列を有するオリゴヌクレオチドをプライマーとするPCRにより、CyGDHのγサブユニット、αサブユニット及びβサブユニットをコードするDNA断片を増幅した。
【0050】
〔フォワードプライマー〕
5’−CATGCCATGGCACACAACGACAACAC−3’(配列番号19)〔リバースプライマー〕
5’−GTCGACGATCTTCTTCCAGCCGAACATCAC−3’(配列番号20)
【0051】
増幅した断片のC末端側を平滑末端化した後、N末端側をNcoIで消化し、同様に処理したpTrc99Aにライゲーションした。得られた組換えベクターでエシェリヒア・コリDH5αを形質転換し、アンピシリン50μg/mLを含むLB寒天培地で生じるコロニーを採取した。得られた形質転換体を液体のLB培地で培養してプラスミドを抽出し、その挿入DNA断片を解析したところ、約3.8kbの挿入断片が確認された。本プラスミドをpTrc99Aγαβと命名した。本プラスミド中のCyGDHの各構造遺伝子は、trcプロモーターによって制御される。pTrc99Aγαβは、アンピシリン耐性遺伝子及びカナマイシン耐性遺伝子を保持している。
【0052】
〔実施例2〕CyGDH αサブユニット遺伝子への変異導入による基質相互作用部位の探索
(1)326位、365位、および472位への変異導入
実施例1で得られたpTrc99Aγαβに含まれるGDH αサブユニット遺伝子に、同遺伝子がコードするαサブユニットの326位のセリン残基、365位のセリン残基、および472位のアラニン残基を他のアミノ酸残基に置換されるように変異を導入した。
具体的には、市販の部位特異的変異導入キット(Stratagene社、QuikChangeII Site−Directed Mutagenesis Kit)を用いて、実施例1に記載のプラスミドpTrc99A/γ+α及びpTrc99Aγαβに含まれるGDH αサブユニット遺伝子の326位のセリンのコドン(TCG)、365位のセリンのコドン(TCG)、および472位のアラニンのコドン(GCG)を他のアミノ酸のコドンに置換した。
【0053】
前記アミノ酸残基置換に用いたフォワードプライマーおよびリバースプライマーの配列を以下の表2に示す。また、3変異作製のために用いたリバースプライマーを表3に表す。
尚、変異を表す表記において、数字はアミノ酸配列における位置を、数字の前のアルファベットはアミノ酸置換前のアミノ酸残基を、数字の後のアルファベットはアミノ酸置換後のアミノ酸残基を示す。例えば、R53Fは、53位のアルギニンからフェニルアラニンへの置換を示す。
【0054】
PCR反応は、以下の反応組成で、95℃、30秒の後、95℃ 30秒、55℃ 1分、68℃ 8分を15サイクル繰り返し、68℃ 30分の反応を行った後、4℃で保持した。
【0055】
〔反応液組成〕
鋳型DNA(5ng/μl) 2μl
(3変異導入pTrc99A/γ+α及びpTrc99Aγαβ)
10×反応緩衝液 5μl
フォワードプライマー(100ng/μl) 1.25μl
リバースプライマー(100ng/μl) 1.25μl
dNTP 1μl
蒸留水 38.5μl
DNAポリメラーゼ 1μl
合計 50μl
【0056】
PCR反応の後、反応液にDpnIを0.5μl添加し、37℃で1時間インキュベートし、鋳型プラスミドを分解させた。
得られた反応液で、エシェリヒア・コリDH5α(supE44, ΔlacU169(φ80lacZΔM15), hsdR17, recAi, endA1, gyrA96, thi−1, relA1)のコンピンテントセルを形質転換した。アンピシリン(50μg/ml)およびカナマイシン(30μg/ml)を含むLB寒天培地(バクトリプトン1%、酵母エキス0.5%、塩化ナトリウム1%、寒天1.5%)上に生育してきたコロニー数個からプラスミドDNAを調製し、配列解析を行って、GDH αサブユニット遺伝子に目的の変異が導入されたことを確認した。
【0057】
【表2−1】
【表2−2】
【0058】
【表3−1】
【表3−2】
【0059】
〔実施例3〕変異GDHの基質特異性の解析
実施例2で得られた変異GDH発現プラスミドを用いて、変異GDHを製造し、基質特異性を検討した。
【0060】
(1)培養
変異導入したエシェリヒア・コリDH5α株を、各々2mlのLB培地(アンピシリン50μg/ml及びカナマイシン30μg/ml含有)で、L字管を用いて37℃で一晩振とう培養した。それらの培養液を、150mlのLB培地(アンピシリン50μg/mlおよびカナマイシン30μg/ml含有)を含む500mlの坂口フラスコに植菌し、37℃で振とう培養した。培養開始から3時間後にIPTG(イソプロピル−β−D−チオガラクトピラノシド)を終濃度0.1mMになるように添加し、さらに2時間培養した。
【0061】
(2)粗酵素サンプルの調製
前記で培養した培養液から菌体を集め、洗浄後、湿菌体0.3mgあたり1mlの0.2% Triton X100を含む10mMリン酸カリウムバッファー(PPB)(pH7.0)で菌体を懸濁し、超音波破砕した。この懸濁液遠心分離(10000r.p.m、10分、4℃)して残渣を除去した後、上清を超遠心分離(50,000r.p.m.、60分、4℃)し、得られた上清(水溶性画分)を、粗酵素サンプルとした。また、このサンプルを、通常の疎水性クロマトグラフィー(カラム名:オクチルセファロース(アマシャム・バイオサイエンス製)およびイオン交換クロマトグラフィー(Q−セファロース(アマシャム・バイオサイエンス製)により精製して、精製酵素サンプルを得た。目的とする酵素画分の決定は、GDH活性を指標として行った。
【0062】
なお、Histagを有するタンパク質の精製は、以下のようにして行った。
培養した培養液から菌体を集め、洗浄後、湿菌体0.3mgあたり1mlの0.5M塩化ナトリウム、20mMイミダゾールを含む20mMリン酸ナトリウムバッファー(pH7.5)で菌体を懸濁し、超音波破砕した。この懸濁液遠心分離(10000r.p.m、10分、4℃)して残渣を除去した後、上清を超遠心分離(50,000r.p.m.、60分、4℃)し、得られた上清(水溶性画分)を、粗酵素サンプルとした。このサンプルを、0.5M塩化ナトリウム、20mMイミダゾールを含む20mMリン酸ナトリウムバッファー(pH7.5)で平衡化したHistrap FFカラム(アマシャム・バイオサイエンス製)に添加し、0.5M塩化ナトリウム、60mMイミダゾールを含む20mMリン酸ナトリウムバッファー(pH7.5)で洗浄した後、0.5M塩化ナトリウム、150mMイミダゾールを含む20mMリン酸ナトリウムバッファー(pH7.5)で溶出させ、精製酵素サンプルを得た。目的とする酵素画分の決定は、GDH活性を指標として行った。
【0063】
(3)GDH活性の測定
前記粗酵素サンプル8μlに、活性測定用試薬(12μlの600mM メチルフェナジンメトサルフェート(PMS)、120μlの6 mM 2,6−ジクロロフェノールインドフェノール(DCIP)に、0.2(w/v)% Triton X−100 10mM PPBを加え、全量480μlとした溶液)を8μl添加した。これをアルミブロック恒温槽を用いて、各反応温度で1分間プレインキュベートした後、素早く各濃度の基質(グルコース又はマルトース)、または蒸留水を8μl加えて攪拌し、分光光度計を用いてDCIP由来の吸収波長である600 nmの吸光度を測定した。各試薬の終濃度はDCIP:0.06mM、PMS、0.6mM。基質の終濃度は10mMまたは5mMである。
結果を、表4に示す。尚、野生型GDHの反応比は、10mMまたは5mMの基質濃度において、それぞれ23.32%および18.88%であった。
【0064】
【表4】
【0065】
表中、QYAは、326位および365位に相当するアミノ酸残基がそれぞれグルタミンおよびチロシンで置換されているが、472位は置換されていない(すなわちアラニン残基のままである)変異グルコース脱水素酵素(すなわち、GDH(QY))を示す。
【0066】
この結果、326Glu+365Arg+472Tyr、326Gln+365Tyr+472Arg、326Lys+365Tyr+472Thr、326Ile+365Phe+472Lys、326Arg+365Arg+472Tyr、326Lys+365Tyr+472Arg、326Lys+365Tyr+472Ser、326Gln+365Tyr+472Tyrが、グルコースに対する反応性を維持しつつ、マルトースとの反応性が低下する大変効果の高い変異であった。
すなわち、326位、365位、および472位の3箇所の変異の組み合わせのうちいくつかの組み合わせにおいて、変異GDH(QY)と同等またはそれ以上の相乗効果が確認された。
【0067】
〔実施例4〕精製酵素の調製(αサブユニットおよびγサブユニットの複合体)
実施例3で基質特異性の改善が認められた8つの変異GDHについて精製を実施した。方法は実施例3に記載の通りに行なった。各精製酵素のグルコースおよびマルトースに対する比活性(U/mg)、反応比(マルトースに対する比活性/グルコースに対する比活性)、グルコースに対するKm値およびVmaxを表5に示す。
その結果、QYAのマルトースとの反応比が、10mMおよび5mMにおいて、それぞれ1%台であるのに対し、ERYおよびKYTでは、反応比が1%以下まで低下し、QYYでは、マルトースとの反応比が0.3%台まで低下していた。
【0068】
【表5】
【0069】
〔実施例5〕精製酵素の調製(αサブユニット、βサブユニット、およびγサブユニットの複合体)
実施例3で基質特異性の改善が認められた変異CyGDH(QYY)について精製を実施した。方法は実施例3に記載の通りに行なった。各精製酵素のグルコースに対する比活性(U/mg)を表6に示す。
その結果、αサブユニット、βサブユニット、およびγサブユニットの複合体の形態においては、QYY(326Gln+365Tyr+472Tyr)は、QYA(326Gln+365Tyr)と比較して、マルトースとの反応性が約1/2〜1/3に低下していた。
【0070】
【表6】
【0071】
〔実施例6〕変異酵素(QYX変異)の基質特異性の解析
実施例2と同様にして、pTrc99A/γ+αに含まれるGDH αサブユニット遺伝子に、同遺伝子がコードするαサブユニットの326位のセリン残基および365位のセリン残基をそれぞれグルタミンおよびチロシンに置換されるように、そして472位のアラニン残基を他のアミノ酸残基に置換されるように変異を導入した。得られた変異酵素発現プラスミドを用いて、実施例3と同様にして、変異酵素を製造し、基質特異性を検討した。結果を以下の表7に示す。
【0072】
【表7】
【0073】
その結果、QYD、QYG、QYK、QYL、QYP、QYQ、QYR、QYT、QYV、およびQYYが、グルコースに対する反応性を維持しつつ、マルトースとの反応性が低下する変異であることが認められた。
【0074】
[実施例7]精製酵素の調製(αサブユニットおよびγサブユニットの複合体)
実施例6で基質特異性の改善が認められた10種類の変異GDH(QYD、QYG、QYK、QYL、QYP、QYQ、QYR、QYT、QYV、およびQYY)について精製を実施した。方法は実施例3に記載の通りに行なった。各精製酵素のグルコースおよびマルトースに対する比活性(U/mg)、反応比(マルトースに対する比活性/グルコースに対する比活性)、グルコースに対するKm値およびVmaxを表8に示す。
その結果、いずれの変異GDHについてもマルトースとの反応比は低いものの、QYYを除いて酵素活性が低下しており、グルコース濃度の測定には適していないことが示唆される。すなわち、QYXで示される10種類の変異GDHのうちで、QYYが最もグルコース濃度の測定に適していることが示唆される。
【0075】
【表8−1】
【表8−2】
【0076】
〔実施例8〕変異CyGDHを用いた血糖測定用比色式センサの作製
最も比活性が優れていたQYYに関して、実施例5で得られた精製変異酵素を用いてグルコースセンサを作製した。
【0077】
図1に示す基本構成を有するグルコースセンサを作製した。すなわち、前記グルコースセンサは、透明基板2に対してスペーサー3を介して透明カバー4(材質PET)を積層した形態を有し、各要素2〜4によってキャピラリ5が規定されている。キャピラリ5の寸法は、1.3mm×9mm×50μmである(図1)。透明基板2および透明カバー4は、厚みが250μmであるPETにより形成し、スペーサー3は黒色の両面テープにより構成した。
【0078】
グルコースセンサは、図2に示す第1試薬部〜第3試薬部を有し、それぞれ成分及び塗布量を表9に示す。表中、「Ru」は、ヘキサルテニウムアンミン錯体(Ru(NH3)6Cl3)を、ACESはN−(2−acetamido)−2−aminoethanesulfonic acidを、WST−4は2−Benzothiazolyl−3−(4−carboxy−2−methoxyphenyl)−5−[4−(2−sulfoethylcarbamoyl)phenyl]−2H−tetrazoliumを、SWNはLithium Magnesium Sodium Silicateを、各々示す。
【0079】
【表9】
【0080】
上記グルコースセンサのキャピラリに測定試料を供給し、その時点から5秒後のエンドポイントの吸光度をプロットした。各回の吸光度の測定においては、第3試薬部に対して、キャピラリの高さ方向に沿って光を照射し、そのときにグルコースセンサを透過した光を受光した。光の照射は、発光ダイオードを用いて630nmの光を照射することにより行なった。透過光は、フォトダイオードにおいて受光した。
【0081】
マルトース影響に関してであるが、グルコース50mg/dlにマルトースを添加した場合、野生型ではマルトース濃度に依存して吸光度が大きく増加し、マルトースと強く反応していることが示唆される。一方、変異CyGDH(QY)を用いたセンサでは、マルトース濃度に応じた吸光度の増加が抑えられており、マルトースの影響が少なくなっていることがわかる。変異CyGDH(QYY)を用いたセンサでは、さらに、マルトース濃度に応じた吸光度の増加が抑えられており、マルトースの影響が少なくなっていることがわかる。これらのデータを見かけの血糖上昇値に換算した結果を図3に示す。
野生型酵素を用いたセンサでは、低血糖(50mg/dlグルコース)がマルトースの混入によって正常域以上の(174.8mg/dlグルコース)に見かけ上表示されてしまう(変化率は300%)。また、改変CyGDH(QY)を用いたセンサであっても、マルトースの混入によって64.8mg/dlグルコースに見かけ上表示されてしまう(変化率は54%)。一方、改変CyGDH(QYY)を用いたセンサでは、マルトースが最大300mg/dl混入している場合でも、見かけの血糖値が最大でも46.1mg/dlまでしか上昇せず、影響は大幅に押さえられていると言える(変化率は14%)。結論としてはQYY変異体が、グルコースに対する反応性(直線性)とマルトース影響の観点から最適であることが示唆される。
【0082】
以上の結果から明らかなように、変異CyGDH(QYY)を用いたグルコースセンサは、変異CyGDH(QY)を用いたグルコースセンサよりも、マルトースとの反応性が大きく低下している。この変異CyGDH(QYY)を用いたグルコースセンサを用いれば、病院等で投与される血中マルトース濃度の上限値である200mg/dlにおいては正誤差は無いと言え、上限値以上の300mg/dlにおいても低血糖(50mg/dl以下)が正常値や高血糖と判定されること無く、安全な治療が行なえる。
【0083】
〔実施例9〕変異CyGDHを用いた電極式血糖センサの作製
最も比活性が優れていたQYYに関して、実施例5で得られた精製変異酵素を用いて電極式グルコースセンサを作製した。
【0084】
(作製手順)
電極式グルコースセンサの作製方法を、図5を参照して以下に示す。絶縁基板1として、PET製基板(長さ28mm、幅7mm、厚み250μm)を準備し、その一方の表面に、カーボンインクのスクリーン印刷により、リード部2および3をそれぞれ有する作用極4および対極5からなるカーボン電極系を形成した。つぎに、絶縁性ペーストを調製し、これを前記電極上にスクリーン印刷して絶縁層6とし、前記絶縁層を形成しない部分を検出部およびリード部とした。次に、合成スメクタイトである商品名「ルーセンタイトSWN」(コープケミカル社製)0.6gを精製水100mLに懸濁し、約8〜24時間攪拌した。この合成スメクタイト懸濁液10mLに、10%(w/v)CHAPS(同仁化学研究所製)水溶液0.1mL、1.0M ACES緩衝液(pH7.4:同仁化学研究所製)5.0mLおよび精製水4.0mLをこの順序で添加し、さらにメディエータとして[Ru(NH3)6]Cl3(アルドリッチ社製)1.0gを混合した。この混合液1.0μLを、検出部に分注した。そして、これを、30℃、相対湿度10%の条件下で10分間乾燥させて、試薬層7を形成した。さらに、前記試薬層7上に酵素試薬層8を形成した。これは、2700U/mLの本願発明のグルコース脱水素酵素水溶液1.0μLを、検出部の試薬層の上に分注して、30℃、相対湿度10%の条件下で10分間乾燥させて形成した。最後に、スリット15を有するスペーサー11を絶縁層上に配置し、さらに、前記スペーサー上に空気孔14となる貫通孔を有するカバー12を配置してバイオセンサを作製した。
【0085】
マルトース影響に関してであるが、グルコース50mg/dlにマルトースを添加した場合、野生型ではマルトース濃度に依存して電流値が大きく増加し、マルトースと強く反応していることが示唆される。一方、変異CyGDH(QY)を用いたセンサでは、マルトース濃度に応じた電流値の増加が抑えられており、マルトースの影響が少なくなっていることがわかる。変異CyGDH(QYY)を用いたセンサでは、さらに、マルトース濃度に応じた電流値の増加が抑えられており、マルトースの影響が少なくなっていることがわかる。これらのデータを見かけの血糖上昇値に換算した結果を図4に示す。
野生型酵素を用いたセンサでは、低血糖(50mg/dlグルコース)がマルトースの混入によって正常域以上の(288.2mg/dlグルコース)に見かけ上表示されてしまう(変化率は549%)。また、改変CyGDH(QY)を用いたセンサであっても、マルトースの混入によって93.9mg/dlグルコースに見かけ上表示されてしまう(変化率は114%)。一方、改変CyGDH(QYY)を用いたセンサでは、マルトースが最大300mg/dl混入している場合でも、見かけの血糖値が最大でも60.0mg/dlまでしか上昇せず、影響は大幅に押さえられていると言える(変化率は37%)。結論としてはQYY変異体が、グルコースに対する反応性(直線性)とマルトース影響の観点から最適であることが示唆される。
【0086】
以上の結果から明らかなように、変異CyGDH(QYY)を用いたグルコースセンサは、変異CyGDH(QY)を用いたグルコースセンサよりも、マルトースとの反応性が大きく低下している。この変異CyGDH(QYY)を用いたグルコースセンサを用いれば、病院等で投与される血中マルトース濃度の上限値である200mg/dlにおいては正誤差は無いと言え、上限値以上の300mg/dlにおいても低血糖(50mg/dl以下)が正常値や高血糖と判定されること無く、安全な治療が行なえる。
【0087】
〔実施例10〕GDHホモログへの変異導入
実施例1と同様にして、ブルクホルデリア・セノセパシアJ2315株のputative oxidoreductase、ブルクホルデリア・タイランデンシスTXDOH株のhypothetical protein BthaT#07876、ラルストニア・ピッケティ12D株のFAD dependent oxidoreductase、ラルストニア・ソラナセアラムIPO1609株のtransmembrane dehydrogenaseおよびブルクホルデリア・フィトフィルマンスPsJN株のglucose-methanol-choline oxidoreductaseの各αサブユニットおよびγサブユニットを発現するプラスミドを用意した。
【0088】
これらの発現プラスミドはγサブユニット構造遺伝子とαサブユニット構造遺伝子を連続して含むDNA断片が、ベクターpET30c(+)(Novagen社)のクローニング部位であるNdeI/HindIIIに挿入されてなるプラスミドである。本プラスミド中のγα遺伝子は、T7プロモーターによって制御される。本プラスミドは、カナマイシン耐性遺伝子を保持している。
【0089】
上記プラスミドに含まれるαサブユニット遺伝子に対し、ブルクホリデリア・セパシアKS1株の野生型GDH αサブユニットにおける326位、365位、および472位に相当する残基がそれぞれグルタミン、チロシン、チロシンに置換されるように、市販の部位特異的変異導入キット(Stratagene社、QuikChangeII Site−Directed Mutagenesis Kit)を用いて変異を導入した。
【0090】
前記アミノ酸残基置換に用いたフォワードプライマーの配列を以下に示す。リバースプライマーの配列は、フォワードプライマーの完全相補鎖とした。
尚、変異を表す表記において、数字はアミノ酸配列における位置を、数字の前のアルファベットはアミノ酸置換前のアミノ酸残基を、数字の後のアルファベットはアミノ酸置換後のアミノ酸残基を示す。例えば、R53Fは、53位のアルギニンからフェニルアラニンへの置換を示す。
【0091】
PCR反応は、以下の反応組成で、95℃、30秒の後、95℃ 30秒、55℃ 1分、68℃ 8分を15サイクル繰り返し、68℃ 30分の反応を行った後、4℃で保持した。
【0092】
〔反応液組成〕
鋳型DNA(5ng/μl) 2μl
(3変異導入pTrc99A/γ+α及びpTrc99Aγαβ)
10×反応緩衝液 5μl
フォワードプライマー(100ng/μl) 1.25μl
リバースプライマー(100ng/μl) 1.25μl
dNTP 1μl
蒸留水 38.5μl
DNAポリメラーゼ 1μl
合計 50μl
【0093】
PCR反応の後、反応液にDpnIを0.5μl添加し、37℃で1時間インキュベートし、鋳型プラスミドを分解させた。
得られた反応液で、エシェリヒア・コリDH5α(supE44, ΔlacU169(φ80lacZΔM15), hsdR17, recAi, endA1, gyrA96, thi−1, relA1)のコンピンテントセルを形質転換した。アンピシリン(50μg/ml)およびカナマイシン(30μg/ml)を含むLB寒天培地(バクトリプトン1%、酵母エキス0.5%、塩化ナトリウム1%、寒天1.5%)上に生育してきたコロニー数個からプラスミドDNAを調製し、配列解析を行って、各αサブユニット遺伝子に目的の変異が導入されたことを確認した。
【0094】
【表10−1】
【表10−2】
【0095】
〔実施例11〕変異酵素の基質特異性の解析
実施例10で得られた変異酵素発現プラスミドを用いて、変異酵素を製造し、基質特異性を検討した。
【0096】
(1)培養
変異酵素発現プラスミドでエシェリヒア・コリBL21(DE3)(F−, dcm, ompT, hsdS(rB− mB−), gal, λ(DE3))のコンピンテントセルを形質転換した。得られた形質転換体を各々3mlのLB培地(カナマイシン25μg/ml含有)で、L字管を用いて37℃で一晩振とう培養した。それらの培養液を、100mlのLB培地(0.5%グリセロール、0.05%グルコース、0.2%ラクトース、100mM PO4、 25mM SO4、50mM NH4、100mM Na、50mM K、1mM MgSO4およびカナマイシン25μg/ml含有)を含む500mlの坂口フラスコに植菌し、20℃で24時間振とう培養した。
【0097】
(2)粗酵素サンプルの調製
前記で培養した培養液から菌体を集め、洗浄後、湿菌体0.1gあたり1mlの10mMリン酸カリウムバッファー(PPB)(pH7.0)で菌体を懸濁し、超音波破砕した。この懸濁液を遠心分離(10000r.p.m、10分、4℃)して残渣を除去した後、上清を超遠心分離(50,000r.p.m.、60分、4℃)し、得られた上清(水溶性画分)を、粗酵素サンプルとした。
【0098】
(3)GDH活性の測定
前記粗酵素サンプル10μlに、活性測定用試薬(94μlの600mM メチルフェナジンメトサルフェート(PMS)、9.4μlの6mM 2,6−ジクロロフェノールインドフェノール(DCIP)に、10mM PPBを加え、全量8mlとした溶液)を170μl添加した。これに各濃度の基質(グルコース又はマルトース)、または蒸留水を20μl加えて攪拌し、分光光度計を用いてDCIP由来の吸収波長である600nmの吸光度を測定した。各試薬の終濃度はDCIP:0.06mM、PMS:0.6mMである。
結果を表11に示す。いずれの変異酵素も、変異導入前の野生型酵素に比べてマルトースへの反応性が低下しており、グルコースに対する基質特異性が向上していることが確認された。
【0099】
【表11】
【0100】
以上の結果より、ブルクホルデリア・セパシア以外のGDHホモログにおいても、QYY変異が有効であることが示唆される。
【産業上の利用可能性】
【0101】
本発明の変異GDHは、グルコースに対する基質特異性が向上しており、グルコースセンサ等を用いたグルコースの測定に好適に使用することができる。
【特許請求の範囲】
【請求項1】
配列番号3と少なくとも80%の同一性を有するアミノ酸配列を有し、かつ、グルコース脱水素酵素活性を有する変異グルコース脱水素酵素であって、
前記アミノ酸配列の326位、365位および472位に相当するアミノ酸残基が、それぞれグルタミン、チロシンおよびチロシンで置換されており、
グルコースに対する基質特異性が向上し、二糖類に対する反応性が低下したことを特徴とする、変異グルコース脱水素酵素。
【請求項2】
配列番号3と少なくとも90%の同一性を有するアミノ酸配列を有する、請求項1に記載の変異グルコース脱水素酵素。
【請求項3】
326位、365位および472位に相当する位置以外は配列番号3に示すアミノ酸配列を有する、請求項1または2に記載の変異グルコース脱水素酵素。
【請求項4】
326位、365位および472位に相当する位置以外は配列番号7に示すアミノ酸配列を有する、請求項1または2に記載の変異グルコース脱水素酵素。
【請求項5】
326位、365位および472位に相当する位置以外は配列番号8に示すアミノ酸配列を有する、請求項1または2に記載の変異グルコース脱水素酵素。
【請求項6】
326位、365位および472位に相当する位置以外は配列番号9に示すアミノ酸配列を有する、請求項1または2に記載の変異グルコース脱水素酵素。
【請求項7】
326位、365位および472位に相当する位置以外は配列番号10に示すアミノ酸配列を有する、請求項1または2に記載の変異グルコース脱水素酵素。
【請求項8】
前記アミノ酸配列において326位および365位に相当するアミノ酸残基がそれぞれグルタミンおよびチロシンで置換されているが472位は置換されていないグルコース脱水素酵素と比較して、グルコースに対する基質特異性が向上し、二糖類に対する反応性が低下したことを特徴とする、請求項1〜7のいずれか1項に記載の変異グルコース脱水素酵素。
【請求項9】
二糖類がマルトースであることを特徴とする請求項1〜8のいずれか1項に記載の変異グルコース脱水素酵素。
【請求項10】
請求項1〜9のいずれか1項に記載の変異グルコース脱水素酵素と、電子伝達サブユニットとを少なくとも含む変異グルコース脱水素酵素複合体。
【請求項11】
電子伝達サブユニットがシトクロムCであることを特徴とする請求項10に記載のグルコース脱水素酵素複合体。
【請求項12】
請求項1〜9のいずれか1項に記載の変異グルコース脱水素酵素をコードするDNA。
【請求項13】
請求項12に記載のDNAを保持し、請求項1〜9のいずれか1項に記載の変異グルコース脱水素酵素、あるいは請求項10または11に記載の変異グルコース脱水素酵素複合体を産生する微生物。
【請求項14】
請求項1〜9のいずれか1項に記載の変異グルコース脱水素酵素、請求項10または11に記載の変異グルコース脱水素酵素複合体、あるいは請求項13に記載の微生物を含む、グルコースアッセイキット。
【請求項15】
請求項1〜9のいずれか1項に記載の変異グルコース脱水素酵素、請求項10または11に記載の変異グルコース脱水素酵素複合体、あるいは請求項13に記載の微生物を含む、グルコースセンサ。
【請求項1】
配列番号3と少なくとも80%の同一性を有するアミノ酸配列を有し、かつ、グルコース脱水素酵素活性を有する変異グルコース脱水素酵素であって、
前記アミノ酸配列の326位、365位および472位に相当するアミノ酸残基が、それぞれグルタミン、チロシンおよびチロシンで置換されており、
グルコースに対する基質特異性が向上し、二糖類に対する反応性が低下したことを特徴とする、変異グルコース脱水素酵素。
【請求項2】
配列番号3と少なくとも90%の同一性を有するアミノ酸配列を有する、請求項1に記載の変異グルコース脱水素酵素。
【請求項3】
326位、365位および472位に相当する位置以外は配列番号3に示すアミノ酸配列を有する、請求項1または2に記載の変異グルコース脱水素酵素。
【請求項4】
326位、365位および472位に相当する位置以外は配列番号7に示すアミノ酸配列を有する、請求項1または2に記載の変異グルコース脱水素酵素。
【請求項5】
326位、365位および472位に相当する位置以外は配列番号8に示すアミノ酸配列を有する、請求項1または2に記載の変異グルコース脱水素酵素。
【請求項6】
326位、365位および472位に相当する位置以外は配列番号9に示すアミノ酸配列を有する、請求項1または2に記載の変異グルコース脱水素酵素。
【請求項7】
326位、365位および472位に相当する位置以外は配列番号10に示すアミノ酸配列を有する、請求項1または2に記載の変異グルコース脱水素酵素。
【請求項8】
前記アミノ酸配列において326位および365位に相当するアミノ酸残基がそれぞれグルタミンおよびチロシンで置換されているが472位は置換されていないグルコース脱水素酵素と比較して、グルコースに対する基質特異性が向上し、二糖類に対する反応性が低下したことを特徴とする、請求項1〜7のいずれか1項に記載の変異グルコース脱水素酵素。
【請求項9】
二糖類がマルトースであることを特徴とする請求項1〜8のいずれか1項に記載の変異グルコース脱水素酵素。
【請求項10】
請求項1〜9のいずれか1項に記載の変異グルコース脱水素酵素と、電子伝達サブユニットとを少なくとも含む変異グルコース脱水素酵素複合体。
【請求項11】
電子伝達サブユニットがシトクロムCであることを特徴とする請求項10に記載のグルコース脱水素酵素複合体。
【請求項12】
請求項1〜9のいずれか1項に記載の変異グルコース脱水素酵素をコードするDNA。
【請求項13】
請求項12に記載のDNAを保持し、請求項1〜9のいずれか1項に記載の変異グルコース脱水素酵素、あるいは請求項10または11に記載の変異グルコース脱水素酵素複合体を産生する微生物。
【請求項14】
請求項1〜9のいずれか1項に記載の変異グルコース脱水素酵素、請求項10または11に記載の変異グルコース脱水素酵素複合体、あるいは請求項13に記載の微生物を含む、グルコースアッセイキット。
【請求項15】
請求項1〜9のいずれか1項に記載の変異グルコース脱水素酵素、請求項10または11に記載の変異グルコース脱水素酵素複合体、あるいは請求項13に記載の微生物を含む、グルコースセンサ。
【図1】

【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2012−90563(P2012−90563A)
【公開日】平成24年5月17日(2012.5.17)
【国際特許分類】
【出願番号】特願2010−240426(P2010−240426)
【出願日】平成22年10月27日(2010.10.27)
【出願人】(000141897)アークレイ株式会社 (288)
【出願人】(510285805)合同会社バイオエンジニアリング研究所 (2)
【Fターム(参考)】
【公開日】平成24年5月17日(2012.5.17)
【国際特許分類】
【出願日】平成22年10月27日(2010.10.27)
【出願人】(000141897)アークレイ株式会社 (288)
【出願人】(510285805)合同会社バイオエンジニアリング研究所 (2)
【Fターム(参考)】
[ Back to top ]