
変異型βグルコシダーゼ、バイオマス分解用酵素組成物および糖液の製造方法
【課題】バイオマス存在下で活性が高く、かつ、従来酵素に対し熱安定の高い新規βグルコシダーゼを提供する。
【解決手段】親βグルコシダーゼに対するアミノ酸、Glu39、Asp169、Arg170、Arg220、Tyr227、Glu330の群から選ばれる少なくとも3以上のアミノ酸が、他のアミノ酸への置換および/または欠失してなり、かつセロビオース分解活性を示すことを特徴とするβグルコシダーゼ。
【解決手段】親βグルコシダーゼに対するアミノ酸、Glu39、Asp169、Arg170、Arg220、Tyr227、Glu330の群から選ばれる少なくとも3以上のアミノ酸が、他のアミノ酸への置換および/または欠失してなり、かつセロビオース分解活性を示すことを特徴とするβグルコシダーゼ。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規なβグルコシダーゼ、および、当該βグルコシダーゼを含むバイオマス分解用酵素組成物、およびこれを使用してセルロース由来バイオマスを加水分解することによる糖液の製造方法に関する。
【背景技術】
【0002】
セルロースの糖化には様々な手法があるが、エネルギー使用量が少なく、かつ糖収率の高い酵素糖化法が開発の主流となっている。セルロース分解酵素であるセルラーゼを大別すると、セルロースの結晶領域に作用するセロビオハイドラーゼ、セルロース分子鎖内部から作用して分子量を低減させるエンドグルカナーゼに分けられる。これらセルラーゼは、生成物の一つであるセロビオースによって阻害を受けることが知られている。またβグルコシダーゼは、水溶性オリゴ糖あるいはセロビオースに作用し、そのβ−グリコシド結合を加水分解する反応を触媒する酵素である。特にβグルコシダーゼは、発酵原料として有用なグルコースを十分に得るためには必須の酵素である。また、セロビオハイドラーゼあるいはエンドグルカナーゼは、セルロース分解で生成したセロビオースの蓄積により反応阻害が引き起こされることが知られている。すなわち、βグルコシダーゼは、セルロース分解により生成するセロビオースの蓄積を大幅に低減することができるため、セルロース分解効率を大幅に向上させるといった効果を有する。
【0003】
セルロースは、草本系植物、木本系植物中に多く含まれ、これら植物を総称してセルロース含有バイオマスと呼ぶ。セルロース含有バイオマスは、セルロースに加え、キシラン、アラビナンといったヘミセルロース、およびリグニンを含んでいる。特に、セルロース含有バイオマス中に含まれるリグニンは、芳香族系の高分子化合物であるため、糸状菌由来セルラーゼを使用した酵素糖化において、阻害的に作用することが知られている。リグニンによる糸状菌由来セルラーゼの阻害機構に関して、すべて解明されていないが、リグニンにセルラーゼが吸着し、分解効率が低下することが、要因の一つであると言われている(非特許文献1)。
【0004】
耐熱性酵素は、安定性が高く、高温条件においても長期間活性を保持することから、産業用酵素としての適用が検討されている。こうした耐熱性酵素は、好熱性菌、あるいは超好熱性菌の保有する酵素の中より多くその存在が確認されている。
【0005】
耐熱性のβグルコシダーゼに関しても、数種の好熱性菌あるいは超好熱性菌より同定されており、具体的にはパイロコッカス・フリオサス(Pyrococcus furiosus)、パイロコッカス・ホリコシイ(Pyrococcus horikoshii)、サーモトーガ・マリティマ(Thermotoga maritima)、スルフォロバス・シバタエ(Sulfolobus shibatae)、スルフォロバス・ソルファタリスク(Sulfolobus solfataricus)、クロストリジウム・サーモセラム(Clostridium thermocellum)などの微生物より耐熱性のβグルコシダーゼが同定されている。特にクロストリジウム・サーモセラム(Clostridium thermocellum)由来のβグルコシダーゼは単量体(非特許文献2)、スルフォロバス・ソルファタリスク(Sulfolobus solfataricus)およびパイロコッカス・フリオサス(Pyrococcus furiosus)由来のβグルコシダーゼは4量体を形成する(非特許文献3、非特許文献4)ことが開示されているが、これら構造とその機能との関係は明らかになっていない。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】国際公開第2005/093072号公報
【非特許文献】
【0007】
【非特許文献1】Hetti P.ら,Journal of Biotechnology,107,65−72(2004)
【非特許文献2】Christian P.ら,Trichoderma and Gliocladium:Basic Biology,Taxonomy and Genetics.,vol.1,121−138(1998)
【非特許文献3】Ohba H.ら,Biosci.Biotech.Biochem.,59,1581−1583(1995)
【非特許文献4】Bauer MW.ら,J.Biol.Chem.,vol.271,39,23749−23755(1996)
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の課題は、セルロース含有バイオマスの加水分解における酵素活性を高く保持したβグルコシダーゼを提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、上記の課題を解決すべく、配列番号1に示されるアミノ酸配列を有する従来のβグルコシダーゼ(以下、本明細書において「親βグルコシダーゼ」という。)の特定箇所にアミノ酸変異を導入し、その機能が改良された性質を有する変異体として新規なβグルコシダーゼの取得に成功した。詳細には、本発明者らは、親βグルコシダーゼの三次元構造に着目し、タンパク質結晶構造解析により、その4量体の形成に関わるアミノ酸を特定し、当該アミノ酸に対して選択的に変異を加えることにより、4量体形成が不安定化された新規βグルコシダーゼの取得に成功した。そして本新規βグルコシダーゼは、特に、セルロース含有バイオマスの加水分解において優れた特性を有することを見出した。
【0010】
すなわち、本発明は以下の構成からなる。
[1] 各単量体の間で水素結合またはイオン結合を形成するアミノ酸の位置に、βグルコシダーゼの4量体形成を不安定化させる変異を有し、かつ野生型と同等またはそれ以上のセロビオース分解活性を有することを特徴とする、変異型βグルコシダーゼ。
[2] 配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、39番目のグルタミン酸、169番目のアスパラギン酸、170番目のアルギニン、220番目のアルギニン、227番目のチロシンおよび330番目のグルタミン酸からなる群から選ばれる少なくとも3以上のアミノ酸に相当するアミノ酸の変異を含む、[1]の変異型βグルコシダーゼ。
【0011】
[3] 配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、39番目のグルタミン酸、169番目のアスパラギン酸、170番目のアルギニン、220番目のアルギニン、227番目のチロシンおよび330番目のグルタミン酸からなる群から選ばれる少なくとも3以上のアミノ酸に相当するアミノ酸の、中性側鎖を有するアミノ酸への置換を含む、[1]または[2]の変異型βグルコシダーゼ。
[4] 配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、170番目のアルギニン、220番目のアルギニンおよび227番目のチロシンに相当するアミノ酸の、中性側鎖を有するアミノ酸への置換を含む、[1]〜[3]のいずれかの変異型βグルコシダーゼ。
【0012】
[5] 中性側鎖を有するアミノ酸が、アラニン、フェニルアラニンおよびグリシンからなる群から選択される、[3]または[4]の変異型βグルコシダーゼ。
[6] 配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の170番目のアルギニンに相当するアミノ酸のアラニンへの置換、配列番号1のアミノ酸配列の220番目のアルギニンに相当するアミノ酸のアラニンへの置換および配列番号1のアミノ酸配列の227番目のチロシンに相当するアミノ酸のフェニルアラニンへの置換を含む、[1]〜[5]のいずれかの変異型βグルコシダーゼ。
[7] 配列番号14で示されるアミノ酸配列を含む、[1]〜[6]のいずれかの変異型βグルコシダーゼ。
【0013】
[8] 配列番号1のアミノ酸配列の440番目のロイシン、448番目のアルギニン、449番目のグルタミン酸、459番目のグルタミン酸からなる群から選ばれる1以上のアミノ酸に相当するアミノ酸の変異をさらに含む、[1]〜[7]のいずれかの変異型βグルコシダーゼ。
[9] 配列番号1のアミノ酸配列の448番目のアルギニンに相当するアミノ酸の中性側鎖もしくは酸性側鎖を有するアミノ酸への置換、配列番号1のアミノ酸配列の449番目のグルタミン酸に相当するアミノ酸の中性側鎖もしくは塩基性側鎖を有するアミノ酸への置換および配列番号1のアミノ酸配列の459番目のグルタミン酸に相当するアミノ酸の中性側鎖もしくは塩基性側鎖を有するアミノ酸への置換からなる群から選ばれる1以上の置換を含む、[8]の変異型βグルコシダーゼ。
【0014】
[10] 配列番号35〜38のいずれかで示されるアミノ酸配列を含む、[8]又は[9]の変異型βグルコシダーゼ。
[11] [1]〜[10]のいずれかの変異型βグルコシダーゼをコードするDNA。
[12] 配列番号20、配列番号31〜34のいずれかで示される塩基配列を含む、[11]のDNA。
[13] [11]または[12]のDNAを含む、発現ベクター。
[14] [13]の発現ベクターを用いた形質転換により作製された、形質転換細胞。
[15] [1]〜[10]のいずれかの変異型βグルコシダーゼまたは[14]の形質転換細胞の処理物を含むバイオマス分解用酵素組成物。
[16] セルロース含有バイオマスを[15]のバイオマス分解用酵素組成物を使用して加水分解することを含む、セルロース由来バイオマスより糖液を製造する方法。
【発明の効果】
【0015】
本発明により、セルロース含有バイオマス存在下でのグルコシダーゼ酵素活性が改良された新規変異型βグルコシダーゼを提供することができる。本発明のβグルコシダーゼは、セルロース含有バイオマスの加水分解による糖液の製造に好適に使用することができる。
【図面の簡単な説明】
【0016】
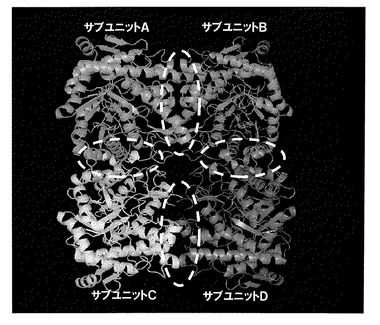
【図1】親βグルコシダーゼの結晶構造解析の結果得られた、親βグルコシダーゼのリボンモデルである。
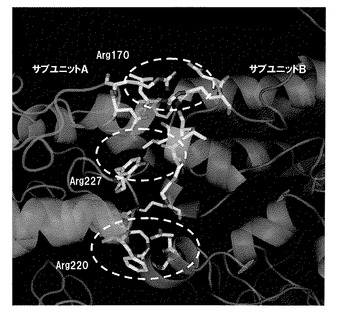
【図2】親βグルコシダーゼのサブユニットA−B間の相互作用部位を拡大したリボンモデルである。
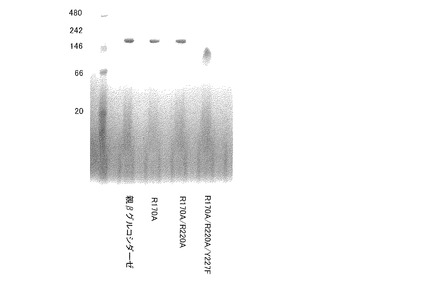
【図3】親βグルコシダーゼおよび各種変異を導入したβグルコシダーゼの、ブルー・ネイティブPAGEによる電気泳動の結果である。
【図4】変異型βグルコシダーゼの結晶構造解析の結果得られた、変異型βグルコシダーゼのリボンモデルである。
【図5】親βグルコシダーゼ、および変異型βグルコシダーゼのゲルろ過による分子量測定の結果である。
【発明を実施するための形態】
【0017】
以下に、本発明を詳細に説明するが、本発明はこれらに限定されない。
(1)βグルコシダーゼ
本発明において「βグルコシダーゼ」とは、糖のβ−グルコシド結合を加水分解する反応を触媒する酵素を意味する。βグルコシダーゼはセロビオースの分解活性が高いことを特徴とする。セロビオース分解活性の測定は、例えば50mM 酢酸−酢酸ナトリウム緩衝液(pH5.0)に溶解したセロビオースの基質溶液に酵素液を添加し、30〜85℃で30分間反応後、必要によりpHを変化させるなどして反応を停止させた後、グルコース定量キットを用いて、反応液中のグルコース濃度を定量することによって行うことができる。βグルコシダーゼとして、EC番号:EC 3.2.1.21として、これに帰属される酵素群が例示されているが、上述したセロビオース分解活性を保有していれば、本発明における「βグルコシダーゼ」に含まれる。
【0018】
(2)親βグルコシダーゼ
本明細書において、親βグルコシダーゼとは、配列番号1で示されるアミノ酸配列を有するβグルコシダーゼであり、セロビオース分解活性を示すものである。本明細書において、「親βグルコシダーゼ」を「野生型」と記載する場合がある。この場合、「親βグルコシダーゼ」および「野生型」なる記載は互換的に使用される。
【0019】
親βグルコシダーゼは、パイロコッカス・フリオサスに由来する酵素である。パイロコッカス・フリオサスは、80−110℃を生育至適温度とする古細菌に分類される微生物で、パイロコッカス・フリオサスは、デンプン、セルロース、マルトース、プルラン、など種々の炭素源を資化することができる微生物である。
【0020】
親βグルコシダーゼは、Kengenら(Eur.J.Biochem.213,305−312,1993年)によって初めて単離精製された酵素である。本文献によると、親βグルコシダーゼは、水溶液中での分子量は約230kDaのホモ4量体で存在することが明示されている。また、SDSなどの界面活性剤によって、この4量体を破壊することができ、各単量体は約58kDaであることが明示されている。さらに、親βグルコシダーゼの至適pHは5.0、等電点は4.4、反応最適温度は102−105℃と記載されている。配列番号1で示される親βグルコシダーゼのアミノ酸配列は、Voorhorstら(J.Bacteriol.177,24,7105−7111,1995年)で同定されたアミノ酸配列に基づくものである。このアミノ酸配列に基づくその分子量は、約54kDa(54.58kDa)と記載されている。
【0021】
(3)本発明の変異型βグルコシダーゼ
本発明の変異型βグルコシダーゼは、水溶液中でβグルコシダーゼ単量体の間で水素結合またはイオン結合を形成するアミノ酸の位置に、βグルコシダーゼの4量体形成を不安定化させる変異を有することを特徴とする。
【0022】
本発明の変異型βグルコシダーゼは、水溶液中で、親βグルコシダーゼが本来形成する4量体と比べて、不安定化された4量体で存在することを特徴とする。親βグルコシダーゼは、一次構造としては約54kDaであるが、水溶液中で安定な4量体を形成し、分子量200kDa以上の分子として溶解していることが知られている(Bauer MW.ら,“Comparison of a β−glucosidase and a β−mannosidase from the hypertherrmophilic archaeon Pyrococcus furiosus”J.Biol.Chem.,vol.271,39,23749−23755(1996))。
【0023】
本発明の変異型βグルコシダーゼが、水溶液中で不安定な4量体を形成していることは、水溶液中での分子量が200kDa未満であることから確認することができる。具体的には、Native−PAGEによる手法でその分子量を確認すると160kDa未満の分子量で検出される。水溶液中4量体が不安定化されていることは、Native−PAGE以外の手法(例えば、ゲル濾過分析、超遠心分析など)を用いて、親βグルコシダーゼと本発明の変異型βグルコシダーゼの水溶液中での挙動を比較することによって、具体的には、水溶液中親βグルコシダーゼの分子量と比較して、本発明のβグルコシダーゼの分子量が小さいことによって、確認することが可能である。
【0024】
下記実施例にて詳述するように、本発明者らは結晶構造解析により、親βグルコシダーゼのアミノ酸配列、すなわち配列番号1で示されるアミノ酸配列において、39番目のグルタミン酸(以下、「Glu39」と記載)、169番目のアスパラギン酸(以下、「Asp169」と記載)、170番目のアルギニン(以下、「Arg170」と記載)、220番目のアルギニン(以下、「Arg220」と記載)、227番目のチロシン(以下、「Try227」と記載)および330番目のグルタミン酸(以下、「Glu330」と記載)が、親βグルコシダーゼの水溶液中における4量体形成に関与していることを明らかにした。すなわち、これらのアミノ酸は、他の親グルコシダーゼの単量体と水溶液中で水素結合またはイオン結合を形成しており、4量体の形成に強く関与しているアミノ酸であることがわかった。本発明における、βグルコシダーゼへのアミノ酸変異の導入の目的は、この4量体の形成に関与するアミノ酸の水素結合またはイオン結合を破壊することにあり、その結果、水溶液中におけるβグルコシダーゼの4量体構造を不安定化させることにある。
【0025】
本発明における、βグルコシダーゼのアミノ酸変異は、βグルコシダーゼの水溶液中における4量体形成に関与しているアミノ酸の置換および/または欠失を含む。
【0026】
好ましくは、本発明の変異型βグルコシダーゼは、配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列を有し、当該配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、Glu39、Asp169、Arg170、Arg220、Tyr227およびGlu330のアミノ酸から選択される少なくとも3以上のアミノ酸に相当するアミノ酸の、中性側鎖を有するアミノ酸への置換を含む。
【0027】
「配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列」は、少なくとも配列番号1のアミノ酸配列で表されるβグルコシダーゼの活性を保持する。
【0028】
ここで「相当するアミノ酸」とは、前記配列同一性を有するアミノ酸配列と配列番号1のアミノ酸配列の立体構造を比較した場合に、前記配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列におけるGlu39、Asp169、Arg170、Arg220、Tyr227およびGlu330と同様の位置に存在し、βグルコシダーゼの水溶液中における4量体形成に関与しているアミノ酸を意味する。「相当するアミノ酸」のアミノ酸種は、配列番号1のアミノ酸配列におけるGlu39、Asp169、Arg170、Arg220、Tyr227およびGlu330とそれぞれ同一である。
【0029】
さらに好ましくは、本発明の変異型βグルコシダーゼは、配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列を有し、当該配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、Arg170、Arg220およびTyr227のアミノ酸に相当するアミノ酸の、中性側鎖を有するアミノ酸への置換を含む。
【0030】
「中性側鎖を有するアミノ酸」としては、グリシン、アスパラギン、グルタミン、セリン、スレオニン、チロシン、システイン、アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファンが挙げられる。好ましくは、「中性側鎖を有するアミノ酸」はアラニン、フェニルアラニンおよびグリシンからなる群から選択される。
【0031】
下記実施例にて詳述するように、本発明者らは結晶構造解析により、配列番号1で示されるアミノ酸配列におけるArg170が、親βグルコシダーゼにおいて、水溶液中4量体を形成する他の全ての親グルコシダーゼ単量体におけるArg170と水素結合またはイオン結合を形成することを明らかとした。すなわち、Arg170に変異を導入することによって、4量体形成に関する水素結合またはイオン結合を効果的に破壊することが可能である。また、下記実施例にて詳述するように、本発明者らは結晶構造解析により、親βグルコシダーゼにおいて、水溶液中、Arg220が、4量体を形成する他の単量体における44番目のグリシン(Gly44)と水素結合を形成していること;また、Tyr227が、4量体を形成する他の単量体における165番目のリシン(Lys165)と水素結合を形成していること、を明らかにした。すなわち、配列番号1で示されるアミノ酸配列において、Arg170に加え、Arg220およびTyr227に変異を導入することによって、4量体形成に関する水素結合またはイオン結合を効果的に破壊することでき、水溶液中でのβグルコシダーゼの4量体構造を不安定化させることが可能である。
【0032】
したがって好ましくは、本発明の変異型βグルコシダーゼは、配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列を有し、当該配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、Arg170に相当するアミノ酸をアラニンへ、Arg220に相当するアミノ酸をアラニンへおよびTyr227に相当するアミノ酸をフェニルアラニンへの置換を含む。
【0033】
特に好ましくは、本発明の変異型βグルコシダーゼは、配列番号14で示されるアミノ酸配列を含む。
【0034】
本発明の変異型βグルコシダーゼは、セロビオース分解活性を保持し得る限り、配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列を有し、当該配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、Glu39、Asp169、Arg170、Arg220、Tyr227およびGlu330に相当するアミノ酸以外の箇所に一または複数のアミノ酸変異を有していても良い。ここで「複数」とは、2〜10個、好ましくは2〜5個、さらに好ましくは2〜3個を意味し、「アミノ酸変異」には、アミノ酸の置換、欠失、挿入および/または付加が含まれる。
【0035】
本発明の変異型βグルコシダーゼは、親βグルコシダーゼと比べて、セルロース含有バイオマス存在下でのβグルコシダーゼ活性が向上している、といった改良された優れた性質を示す。この改良された優れた性質は、親βグルコシダーゼが、水溶液中安定な4量体を形成するのに対して、本発明の変異型βグルコシダーゼでは上記変異導入により、4量体構造が不安定化していることが要因として推定される。
【0036】
本明細書において、本発明の変異型βグルコシダーゼを、「変異体」と記載する場合がある。この場合、「変異型βグルコシダーゼ」および「変異体」なる記載は互換的に使用される。
【0037】
前述した本発明の変異型βグルコシダーゼは、配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列を有し、当該配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、Glu39、Asp169、Arg170、Arg220、Tyr227およびGlu330のアミノ酸から選択される少なくとも3以上のアミノ酸に相当するアミノ酸の、中性側鎖を有するアミノ酸への置換を含み、これらの変異の効果によって4量体化を不安定化させることが可能であるが、これら変異体にさらに変異導入を行ったものでもよい。変異導入の目的としては、さらに活性向上を目的として変異、熱安定性向上を目的とした変異、さらに多量体構造を不安定化することを目的とした変異などを例示することができる。
【0038】
下記実施例にて詳述するように、本発明者らは前述した変異型βグルコシダーゼに関し、さらに結晶構造解析を実施した結果、本発明の変異型βグルコシダーゼは2量体構造を形成していることを明らかにした。この変異型βグルコシダーゼの2量体構造形成に関与するアミノ酸は、配列番号1のアミノ酸配列の440番目のロイシン、448番目のアルギニン、449番目のグルタミン酸、459番目のアルギニンに相当する4つのアミノ酸である。すなわち、これら4つのアミノ酸は、2量体を形成する他方の変異型グルコシダーゼのアミノ酸と水素結合もしくはイオン結合を形成しており、変異型グルコシダーゼの2量体の形成に強く関与する。本発明では、前述した4量体を不安定化する変異に加え、さらに変異型グルコシダーゼが形成する2量体を不安定化する変異をさらに加えてもよい。
【0039】
すなわち、本発明の変異型βグルコシダーゼは、前述した4量体を不安定化する変異に加え、配列番号1のアミノ酸配列の440番目のロイシン、448番目のアルギニン、449番目のグルタミン酸、459番目のグルタミン酸からなる群から選ばれる1以上のアミノ酸に相当するアミノ酸の変異をさらに含む。
【0040】
ここで「相当するアミノ酸」とは、前記配列同一性を有するアミノ酸配列と配列番号1のアミノ酸配列の立体構造を比較した場合に、前記配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の440番目のロイシン、448番目のアルギニン、449番目のグルタミン酸、459番目のグルタミン酸と同様の位置に存在し、変異型βグルコシダーゼの2量体形成に関与しているアミノ酸を意味する。
【0041】
特に好ましくは、前述の変異型βグルコシダーゼのアミノ酸配列において、配列番号1のアミノ酸配列の448番目のアルギニンに相当するアミノ酸の中性側鎖もしくは酸性側鎖を有するアミノ酸への置換、または、449番目のグルタミン酸に相当するアミノ酸の中性側鎖もしくは塩基性側鎖を有するアミノ酸への置換、459番目のアルギニンに相当するアミノ酸の中性側鎖もしくは塩基性側鎖を有するアミノ酸への置換、からなる群から選ばれる1以上のアミノ酸位置における置換を含む。
【0042】
「中性側鎖を有するアミノ酸」としては、グリシン、アスパラギン、グルタミン、セリン、スレオニン、チロシン、システイン、アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファンが挙げられる。好ましくは、「中性側鎖を有するアミノ酸」はアラニン、フェニルアラニンおよびグリシンからなる群から選択される。
【0043】
「酸性側鎖を有するアミノ酸」としては、グルタミン酸、アスパラギン酸、の群から選択される。
【0044】
「塩基性側鎖を有するアミノ酸」としては、アルギニン、リジン、ヒスチジン、の群から選択される。
【0045】
最も好ましくは、前述の変異型βグルコシダーゼのアミノ酸配列において、配列番号1のアミノ酸配列の448番目のアルギニンに相当するアミノ酸のグルタミン酸もしくはグリシン、配列番号1のアミノ酸配列の449番目のグルタミン酸に相当するアミノ酸のアルギニン、配列番号1のアミノ酸配列の459番目のアルギニンに相当するアミノ酸のグリシンへの変異から選ばれる1以上の変異を含む。
【0046】
特に好ましくは、本発明の変異型βグルコシダーゼは、配列番号31〜34で示されるアミノ酸配列を含む。
【0047】
また、こうしたアミノ酸置換による変異に加えて、配列番号1のアミノ酸配列の440番目以降のアミノ酸全部に相当する領域を、あるいは、配列番号1のアミノ酸配列の448番目以降のアミノ酸全部に相当する領域を、あるいは配列番号1のアミノ酸配列の449番目以降のアミノ酸全部に相当する領域を、あるいは配列番号1のアミノ酸配列の459番目以降のアミノ酸全部に相当する領域を、欠失(削除)したものでもよい。これは440番目以降のアミノ酸に相当する配列に関しては、セロビオース分解活性そのものには特段の影響がないためによる。こうした欠失(削除)を行いたい場合は、削除したいアミノ酸の位置に対応する遺伝子においてストップコドンを挿入することで可能になる。
【0048】
(4)本発明の変異型βグルコシダーゼの製造方法
本発明の変異型βグルコシダーゼを製造する方法としては、例えばβグルコシダーゼのアミノ酸配列をコードするDNAを調製し、これを発現ベクターに連結し、発現ベクターを宿主に導入し、異種タンパク質として生産し、単離および精製することで得ることができる。
【0049】
まず上述のDNAの調製方法としては、遺伝子合成により目的のアミノ酸配列をコードするDNAを全合成する方法、配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列をコードするDNAに対して、部位突然変異法により上記所定の位置に変異を導入する方法が例示される。配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列をコードする遺伝子は、このβグルコシダーゼを保有する微生物、特にパイロコッカス・フリオサスより公知の方法に準じてDNAを単離し、PCR等の手法によってDNA増幅させることで取得することができる。
【0050】
上記のようにして調製した変異型βグルコシダーゼをコードするDNAを、制限酵素およびDNAリガーゼを用いて、適当な発現ベクター中のプロモーター下流に連結することにより、当該DNAを含む発現ベクターを製造することができる。発現ベクターとしては、細菌プラスミド、酵母プラスミド、ファージDNA(ラムダファージなど)、レトロウイルス、バキュロウイルス、ワクシニアウイルス、アデノウイルス等のウイルスDNA、SV40の誘導体など、植物細胞用のベクターとしてのアグロバクテリウムなどが挙げられるが、宿主細胞において複製および生存可能ある限り他のいかなるベクターも用いることができる。例えば、宿主が大腸菌である場合、pUC、pET、pBADなどを例示することができる。また、宿主が酵母である場合、pPink−HC、pPink−LC、pPinkα−HC、pPCIZ、pPCIZα、pPCI6、pPCI6α、pFLD1、pFLD1α、pGAPZ、pGAPZα、pPIC9K、pPIC9などが挙げられる。
【0051】
プロモーターとしては、遺伝子の発現に用いる宿主に対応して適切なプロモーターであればいかなるものでもよい。例えば、宿主が大腸菌である場合、lacプロモーター、Trpプロモーター、PLプロモーター、PRプロモーター等が、酵母である場合、AOX1プロモーター、TEF1プロモーター、ADE2プロモーター、CYC1プロモーター、GAL−L1プロモーターなどが挙げられる。
【0052】
本発明において用いる宿主細胞としては、大腸菌、バクテリア細胞、酵母細胞、真菌細胞、昆虫細胞、植物細胞、動物細胞などが好ましい。酵母細胞としては、例えば、ピキア属(Pichia)、サッカロマイセス属(Saccharomyces)、シゾサッカロマイセス属(Schizosaccharomyces)などが挙げられる。真菌細胞としては、アスペルギルス属(Aspergillus)、トリコデルマ属(Trichoderma)などが挙げられる。昆虫細胞としてはSf9など、植物細胞としては双子葉植物など、動物細胞としては、CHO、HeLa、HEK293などが挙げられる。
【0053】
形質転換または、トランスフェクションは、リン酸カルシウム法、電気穿孔法などの公知の方法で行うことができる。本発明のβグルコシダーゼは、上記のように形質転換またはトランスフェクトされた宿主細胞においてプロモーターの制御下にて発現させ、産生物を回収して得ることができる。発現に際しては、宿主細胞を適切な細胞密度まで増殖または成長させた後、温度シフトまたはイソプロピル−1−チオ−β−D−ガラクトシド(IPTG)添加などの化学的誘発手段によってプロモーターを誘発させ、細胞をさらに一定期間培養する。
【0054】
変異型βグルコシダーゼが細胞外に排出される場合には、培地から直接に、また細胞外に存在する場合には、超音波破砕や機械的破砕などの物理的手段もしくは細胞溶解剤などの化学的手段によって、細胞を破壊した後にβグルコシダーゼを精製する。βグルコシダーゼは、組換え細胞の培地から、硫酸アンモニウムもしくはエタノール沈殿、酸抽出、陰イオンもしくは陽イオン交換クロマトグラフィー、逆相高速液体クロマトグラフィー、アフィニティークロマトグラフィー、ゲルろ過クロマトグラフィー、電気泳動などの技術を組み合わせて、部分的にまたは完全に精製することができる。
【0055】
(5)変異型βグルコシダーゼを含むバイオマス分解用酵素組成物
本発明の変異型βグルコシダーゼは、セルロース含有バイオマスの加水分解において、親βグルコシダーゼに対して、高い熱安定性、リグニン耐性を有する。具体的には、親βグルコシダーゼに比べて1.2倍、1.3倍、1.4倍、1.5倍、1.6倍、1.7倍、1.8倍、1.9倍、2倍またはそれ以上の高いセロビオース分解活性を得ることができる。
【0056】
本発明の変異型βグルコシダーゼは、精製されたものであっても、粗精製されたものであっても良い。
【0057】
また、本発明の変異型βグルコシダーゼは、固相に固定化されていても良い。固相としては例えば、ポリアクリルアミドゲル、ポリスチレン樹脂、多孔性ガラス、金属酸化物などが挙げられる(特にこれらに限定されない)。本発明の変異型βグルコシダーゼを固相に固定することによって、連続反復使用が可能となる点において有利である。
【0058】
さらに、上記変異型βグルコシダーゼをコードするDNAを用いて形質転換した細胞の処理物も、粗精製された変異型βグルコシダーゼとして利用することができる。当該「形質転換した細胞の処理物」には、固相に固定化した形質転換細胞、ならびに形質転換細胞の死菌、破砕物、およびそれらを固相に固定化したものなどが含まれる。
【0059】
本発明の変異型βグルコシダーゼは、セルラーゼと混合することで、バイオマス分解用酵素組成物としてセルロース含有バイオマスの加水分解に使用することができる。ここでいうセルラーゼとは、セルロースを分解する活性を有する酵素であれば特に限定されず、一種類以上の混合物であってもよい。このような酵素としては、例えばセルラーゼ、ヘミセルラーゼ、セロビオハイドラーゼ、エンドグルカナーゼ、エキソグルカナーゼ、キシラナーゼ、マンナナーゼなどが挙げられる。
【0060】
上述セルラーゼは、好ましくは糸状菌由来セルラーゼである。糸状菌由来セルラーゼは、少なくともエンドグルカナーゼおよびセロビオハイドラーゼの双方を含んでなる混合物である。さらに効率的なセルロースの糖化を行うためには、エンドグルカナーゼに関して2種以上、および/または、セロビオハイドラーゼに関して2種以上含んでなる糸状菌由来セルラーゼであることが好ましい。前記糸状菌セルラーゼを生産する微生物として、トリコデルマ属(Trichoderma)、アスペルギルス属(Aspergillus)、セルロモナス属(Cellulomonas)、クロストリジウム属(Clostridium)、ストレプトマイセス属(Streptomyces)、フミコラ属(Humicola)、アクレモニウム属(Acremonium)、イルペックス属(Irpex)、ムコール属(Mucor)、タラロマイセス属(Talaromyces)、などの微生物を挙げることができる。これら微生物は、培養液中にセルラーゼを産生するために、その培養液を未精製の糸状菌セルラーゼとしてそのまま使用してもよいし、また培養液を精製し、製剤化したものを糸状菌セルラーゼ混合物として使用してもよい。前記培養液より、糸状菌セルラーゼ混合物を精製し、製剤化したものとして使用する場合、プロテアーゼ阻害剤、分散剤、溶解促進剤、安定化剤など、酵素以外の物質を添加したものを、セルラーゼ製剤として使用してもよい。
【0061】
本発明で使用する糸状菌由来セルラーゼは、トリコデルマ属由来セルラーゼであることが好ましい。トリコデルマ属は、少なくとも2種以上のエンドグルカナーゼ、および、少なくとも2種以上のセロビオハイドラーゼを含んでなるセルラーゼを培養液中に産生し、こうした培養液より調製されたセルラーゼ混合物は本発明に好ましく使用することができる。こうしたトリコデルマ属のうち、より好ましくは、トリコデルマ・リーセイ(Trichoderma reesei)由来のセルラーゼ混合物である。トリコデルマ・リーセイ由来のセルラーゼ混合物としては、トリコデルマ・リーセイQM9414(Trichoderma reesei QM9414)、トリコデルマ・リーセイQM9123(Trichoderma reesei QM9123)、トリコデルマ・リーセイRutC−30(Trichoderma reesei Rut−30)、トリコデルマ・リーセイPC3−7(Trichoderma reesei PC3−7)、トリコデルマ・リーセイCL−847(Trichoderma reesei CL−847)、トリコデルマ・リーセイMCG77(Trichoderma reesei MCG77)、トリコデルマ・リーセイMCG80(Trichoderma reesei MCG80)、トリコデルマ・ビリデQM9123(Trichoderma viride QM9123)に由来するセルラーゼ混合物が挙げられる。また、前記トリコデルマ属に由来し、変異剤あるいは紫外線照射などで変異処理を施し、セルラーゼ生産性が向上した変異株であってもよい。
【0062】
本発明のバイオマス分解用酵素組成物は、広くセルロース含有バイオマスの加水分解に使用することができる。セルロース含有バイオマスとは、少なくともセルロースを含むものであれば限定されない。具体的には、バガス、コーンストーバー、コーンコブ、スイッチグラス、稲藁、麦藁、樹木、木材、廃建材、新聞紙、古紙、パルプなどである。これらセルロース含有バイオマスは、高分子芳香族化合物リグニンやヘミセルロースなどの不純物が含まれているが、前処理として、酸、アルカリ、加圧熱水などを用いて、リグニンやヘミセルロースを部分的に分解したセルロース含有バイオマスを、セルロースとして利用してもよい。
【0063】
本発明におけるセルロース含有バイオマスの酵素処理条件は、処理温度40℃〜60℃、処理pH3〜7、セルロース含有バイオマス固形分濃度0.1〜30%が好ましい。当該範囲に設定することにより、本発明のバイオマス分解用酵素組成物の分解効率を最大限発揮することができる。従来の好熱性菌由来グルコシダーゼは、反応最適温度が100℃付近の酵素もあるが、本発明で使用する好熱性菌由来グルコシダーゼは、40℃〜60℃においても十分高い比活性を示し、セルラーゼ共存下で、セルロース含有バイオマスの分解を効率的に行うことが可能である。この酵素処理は、バッチ式で行っても、連続式で行ってもよい。こうした酵素処理によって得られた加水分解物は、グルコース、キシロースなどの単糖成分を含むため、エタノール、乳酸などの原料糖として使用することが可能である。
【実施例】
【0064】
以下に、実施例を挙げて本発明を具体的に説明する。ただし、本発明はこれらに限定されるものではない。
【0065】
(参考例1)親βグルコシダーゼの大腸菌による組み換え発現調製
親βグルコシダーゼの遺伝子は、配列番号2に示す塩基配列を有する遺伝子を全合成し、pET−11dのNcoIおよびBamHIにLigation High(東洋紡)を使用して連結し、JM109(Takara)に形質転換した。スクリーニングは、アンピシリンを抗生物質として含むLB寒天培地を用いて行った。形質転換されたJM109株より、作製したベクターpET−PfuBGLをミニプレップキット(キアゲン)により単離し、塩基配列解析を行った。pET−PfuBGLは、発現用大腸菌BL21(DE3)pLysS株に形質転換し、BL21−PfuBGL株を作製した。BL21−PfuBGL株を、アンピシリン含有LB培地10mLに植菌し、37℃で一晩振とう培養(前培養)を行った。本培養として、アンピシリン含有LB培地1Lに、前培養で得られた菌体を植菌し、波長600 nmでの吸光度OD600が0.8となるまで振とう培養を行った。その後、最終濃度が0.4mMになるようにイソプロピル−1−チオ−β−D−ガラクトシド(IPTG)を加え、さらに25℃で一晩培養した。培養後、菌体を遠心分離により回収し、50mM トリス−HCl緩衝液(pH8.0)に再懸濁した。この溶液を氷冷しながら、超音波破砕を行い、その上清を無細胞抽出液として遠心分離により回収した。得られた無細胞抽出液を、85℃で15分保温し、該グルコシダーゼ以外の大腸菌に由来するタンパク質を凝集沈殿した。遠心分離により沈殿物を除去し、上清を分画分子量10000の再生セルロース製透析膜(スペクトラム・ラボラトリーズ製)を使用して、50mM 酢酸緩衝液(pH5.0)に透析した。得られたタンパク質溶液を、親βグルコシダーゼとして使用した。調製した親βグルコシダーゼのアミノ酸配列を配列番号1に記載する。
【0066】
(実施例1)親βグルコシダーゼの4量体形成部位の特定
親βグルコシダーゼ(パイロコッカス・フリオサス)のX線結晶構造は既に報告されているが(Thijis K.ら,Biochem.vol.39,No.17(2000))分解能が3.3 Åと低く、またProtein Data Bank(PDB)への登録もされていない。そこで、PfuBGLの三次元構造を決定するため、X線結晶構造解析を試みた。新たな結晶化条件を探索し、リン酸を沈殿剤として結晶化に成功した。大型放射光実験施設SPring−8でX線回折実験を行い、分解能2.5ÅでPfuBGLの構造を決定した。構造決定は分子置換法を用い、モデル分子には配列番号22で示されるアミノ酸配列を有するサーモスファエラ・アグリガンズ(Themosphaera aggregans)由来β−グリコシダーゼ(ThAggBGY、PDB ID:1QVB)を用いた。得られた3次元構造データを構造解析用のソフトウェアCCP4_Contactを用いてサブユニット間の相互作用を分析した。分析の結果得られたリボンモデルを図1に示す。サブユニットA−B間、A−C間で相互作用しており、ホモ4量体を形成していることが確認された(図1)。分析結果より得られたサブユニットA−B間の相互作用に関し、表1にまとめた。
【0067】
【表1】

【0068】
サブユニット間の相互作用のうち、アミノ酸主鎖の窒素原子(N)および酸素原子(O)のみで相互作用しているもの(イタリック表記のアミノ酸)は、どのアミノ酸でも同様な構造を持っているため改変はできない。そのため、サブユニット間の相互作用の改変には側鎖部分が相互作用に関与するGlu39、Asp169、Arg170、Arg220、Tyr227、Glu330への変異導入が有効と考えられた。また、そのうちArg170−Asp169間の結合はイオン結合のため、他の水素結合での相互作用に比べ結合が強く、特に変異導入で大きな効果あることが推定された。また、A−B間の相互作用部位を拡大したリボンモデルを図2に示すが、特に、Arg170、Arg220、Tyr227の残基がインターフェイスエリアが大きく、また結合距離も近いため、変異の導入部位として有力と考えられた。
【0069】
(実施例2)変異体の調製
本発明のβグルコシダーゼ(変異体)は、表2に示すプライマーを使用し、以下の手法で作製した。
【0070】
【表2】
【0071】
配列番号1で示されるアミノ酸配列をコードする遺伝子に対して、配列番号3および4の塩基配列で示されるオリゴヌクレオチドを使用して部位突然変異法を用いてR170A変異体(配列番号16)を作製した。また同様に、配列番号5および6の塩基配列で示されるオリゴヌクレオチドを使用してR220A変異体(配列番号17)、配列番号7および8を使用してY227F変異体(配列番号18)を作製した。また、R170A変異体に対し、配列番号7および8の塩基配列で示されるオリゴヌクレオチドを使用して、R170A/Y227F変異体(配列番号19)を作製した。R170A変異体に対し、配列番号5および6の塩基配列で示されるオリゴヌクレオチドを使用して、R170A/R220A変異体(配列番号21)を作製した。R170A/R220A変異体に対し、配列番号7および8の塩基配列で示されるオリゴヌクレオチドを使用してR170A/R220A/Y227F変異体(配列番号20)を作製した。
【0072】
得られた各遺伝子は、参考例1記載の手順で大腸菌にて発現を実施した。R170A変異体、R220A変異体、Y227F変異体、R170A/Y227F変異体、R170A/R220A変異体、R170A/R220A/Y227F変異体のアミノ酸配列を配列番号9〜14に示す(スタートコドンであるメチオニンは含まない)。前述変異体は、大腸菌にてすべて異種タンパク質として発現可能であることが確認できた。
【0073】
(実施例3)変異体のセロビオース分解活性1
実施例2で得られた変異体(本発明のβグルコシダーゼ)と、参考例1で調整した親βグルコシダーゼのセロビオース分解活性を以下実験にて比較した。基質に10mM セロビオース/50mM 酢酸緩衝液を用い、それぞれ実施例2、参考例1で調製した酵素を終濃度0.23mg/mLで添加し、50℃で酵素反応を30分行った。親βグルコシダーゼにおける50℃の温度条件下で生成したグルコース濃度(g/L)を100%として、各変異体におけるセロビオース分解活性を相対値で表3に示す。
【0074】
【表3】
【0075】
50℃においては、親βグルコシダーゼおよび各変異体で活性に差がないことを確認した。
【0076】
(実施例5)変異体のセロビオース分解活性2
セルロース含有バイオマス存在下での親グルコシダーゼおよび変異体(本発明のβグルコシダーゼ)のセロビオース分解活性を測定した。セルロース含有バイオマスとしては、稲藁を5%希硫酸で150℃10分処理したものを使用した。セルロース含有バイオマス5wt%を懸濁した溶液に、10mM セロビオース/50mM 酢酸緩衝液を添加し反応液とした。この反応液に実施例2、参考例1で調製した酵素を終濃度0.23mg/mLで添加し、50℃で酵素反応を30分行った。親βグルコシダーゼにおける50℃の温度条件下で生成したグルコース濃度(g/L)を100%として、各変異体におけるセロビオース分解活性を相対値で表4に示す。
【0077】
【表4】
【0078】
親グルコシダーゼおよびR170A変異体、R220A変異体、Y227F変異体、R170A/R220A変異体では、セルロース含有バイオマス存在下でのセロビオース分解活性にほとんど差がないことが確認できた。一方でR170A/R220A/Y227F変異体では、親グルコシダーゼあるいはその他の変異体に対して活性が大きく向上することが確認された。
【0079】
(実施例6)ブルー・ネイティブPAGEによる変異体の分子量測定
親βグルコシダーゼ、及び本発明のβグルコシダーゼであるR170A変異体、R170A/R220A変異体、およびR170A/R220A/Y227F変異体が安定な4量体を形成するか否か、ブルー・ネイティブPAGEにより検討を行った。
【0080】
ブルー・ネイティブPAGEは3−12%Native−PAGEビストリスゲル(インビトロジェン)を使用した。親βグルコシダーゼ、R170A変異体、R170A/R220A変異体、およびR170A/R220A/Y227F変異体各1μgを、サンプルバッファー(50mMイミダゾール(pH7.0)、50mM NaCl、5mM 6−アミノヘキサン酸、0.5%クマシーG250、0.5%DDM)に溶解し泳動サンプルとした。分子量マーカーは、NativeMarkプロテインスタンダード(インビトロジェン)を使用した。電気泳動は陽極側に、50mMトリシン、7.5mMイミダゾール(pH7.0)、クマシーG250、陰極側に、25mMイミダゾール(pH7.0)を添加し、150V定電圧で100分電気泳動を行った。バンドの固定化後、G−250の脱色を行った。得られたゲルの写真を図3に示した。
【0081】
親βグルコシダーゼ、R170A変異体およびR170A/R220A変異体に関しては、分子量マーカー146kDaから242kDaの間のほぼ同じ位置にそれぞれのバンドが検出された。親βグルコシダーゼは、4量体形成時の分子量は216kDaであることから、これらは安定な4量体を形成することが確認できた。一方で、本発明のR170A/R220A/Y227F変異体は、分子量マーカー66kDaから146kDaに検出された。すなわち、本発明のR170A/R220A/Y227F変異体は、本来親βグルコシダーゼが形成する4量体形成が不安定化しており、3量体あるいは2量体で水溶液中に存在することが確認された。
【0082】
(実施例7)βグルコシダーゼを含むバイオマス分解用酵素組成物を用いたセルロース含有バイオマスの加水分解
トリコデルマ・リーセイ由来セルラーゼ(セルクラスト、シグマ)、及びグルコシダーゼとして、それぞれ比較例1で調製した親グルコシダーゼ、実施例2で調製した本発明のR170A変異体、R170A/R220A変異体、R170A/R220A/Y227F変異体、とからなるバイオマス分解用酵素組成物を用いた。酵素添加量は、セルラーゼ0.5mg/mL、各種グルコシダーゼ0.005mg/mL(セルラーゼの1/100量)となるよう混合した。
【0083】
セルロース含有バイオマスとしては、稲藁を5%希硫酸で150℃10分処理したものを使用した。加水分解は、50mM 酢酸緩衝液(pH5.0)に5wt%のセルロース含有バイオマスを懸濁したものを基質とした。反応は50℃において28時間まで行い、生成したグルコース濃度の測定を行った。
【0084】
【表5】
【0085】
グルコシダーゼなしに対して、グルコシダーゼを添加することで、グルコース生成量が増大することが確認できた。本発明のR170A/R220A/Y227F変異体では、親βグルコシダーゼ、R170A変異体、A170A/R220A変異体に比べて、反応28時間後におけるグルコース生成量は増えることが確認できた。セルロース含有バイオマスが存在しない時のセロビオース分解活性は、R170A/R220A/Y227F変異体、親βグルコシダーゼ、R170A変異体、A170A/R220A変異体、の間で差はなかった(実施例3)。したがって、本発明の変異体、R170A/R220A/Y227F変異体は、セルロース含有バイオマス存在下での活性が親βグルコシダーゼに対して向上していることが確認できた。
【0086】
(実施例8)R170A/R220A/Y227F変異体の結晶構造解析
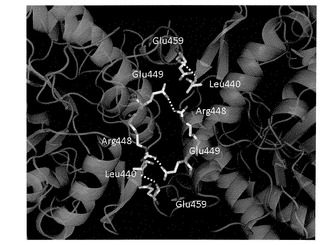
R170A/R220A/Y227F変異体の三次元構造を決定するため、X線結晶構造解析を試みた。新たな結晶化条件を探索し、リン酸を沈殿剤として結晶化に成功した。大型放射光実験施設SPring−8でX線回折実験を行い、分解能2.5オングストロームでPfuBGLの構造を決定した。構造決定は分子置換法を用い、モデル分子には配列番号22で示されるアミノ酸配列を有するサーモスファエラ・アグリガンズ(Themosphaera aggregans)由来β−グリコシダーゼ(ThAggBGY、PDB ID:1QVB)を用いた。得られた3次元構造データを構造解析用のソフトウェアCCP4_Contactを用いてサブユニット間の相互作用を分析した。分析の結果得られたリボンモデルを図4に示す。サブユニットA−C間で相互作用しており、R170A/R220A/Y227F変異体は、ホモ2量体を形成していることが確認された(図4)。分析結果より得られたサブユニットA−C間の相互作用に関し、表6にまとめた。
【0087】
【表6】
【0088】
すなわち、R170A/R220A/Y227F変異体は、Arg448、Leu440、Glu449、Glu459のアミノ酸残基にて水素結合し、2量体を形成していることが判明した。
【0089】
(実施例9)変異体の調製2
本発明の変異型βグルコシダーゼ(R170A/R220A/Y227F変異体)は、実施例8に示すとおり2量体を形成することが判明した。そこで、R170A/R220A/Y227F変異体に対しさらに、Arg448、Glu449、Glu459のいずれか1以上の変異をさらに導入することで、2量体を単量体に改変できるか検討を行った。変異体遺伝子の取得に関しては、表7に示すプライマーを使用し、以下の手法で作製した。
【0090】
【表7】
【0091】
R170A/R220A/Y227F変異体の遺伝子(配列番号20)に対して、配列番号23および24の塩基配列で示されるオリゴヌクレオチド、配列番号25および26の塩基配列で示されるオリゴヌクレオチド、配列番号27および28の塩基配列で示されるオリゴヌクレオチド、配列番号29および30の塩基配列で示されるオリゴヌクレオチド、をそれぞれ使用し、R170A/R220A/Y227F/R448E変異体(配列番号31)、R170A/R220A/Y227F/R448G変異体(配列番号32)、R170A/R220A/Y227F/E449R変異体(配列番号33)、R170A/R220A/Y227F/E459G変異体(配列番号34)遺伝子を作製した。
【0092】
得られた各遺伝子は、参考例1記載の手順で大腸菌にて発現を実施した。R170A/R220A/Y227F/R448E変異体、R170A/R220A/Y227F/R448G変異体、R170A/R220A/Y227F/E449R変異体、R170A/R220A/Y227F/E459G変異体のアミノ酸配列を配列番号35〜38に示す(スタートコドンであるメチオニンは含まない)。前述の変異体は、大腸菌にてすべて異種タンパク質として発現可能であることが確認できた。
【0093】
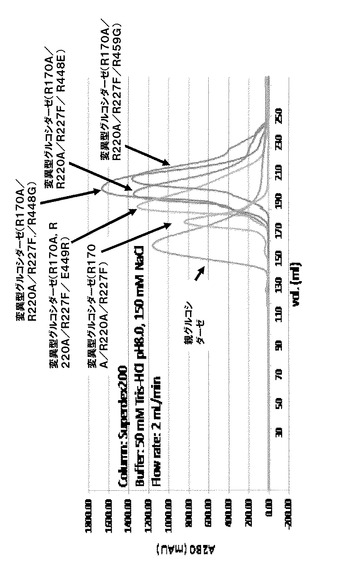
(実施例10)ゲルろ過による分子量の決定
親グルコシダーゼおよび変異体の分子量を決定するために、ゲルろ過による分子量の決定を行った。カラムはHiLood26/60Suuperdex200を使用し、緩衝液は、50mMトリス塩酸塩(pH8)、150mM塩化ナトリウムを使用した。実施例2、参考例1、実施例9で調製した酵素をカラムに添加し、緩衝液2mL/minで分離した。酵素の検出は、吸光度280nmを測定した。分子量は、オブアルブミン(44kDa)、コンアルブミン(75kDa)、アルドラーゼ(158kDa)、フェリチン(440kDa)を分子量マーカーとして使用し、ぞれぞれの酵素成分の溶出時間を元に検量線を作製した。ゲルろ過の結果、測定された分子量(実測分子量、kDa)およびアミノ酸一次構造より推定される分子量(推定分子量、kDa)をそれぞれ表8にまとめた。
【0094】
【表8】
【0095】
親グルコシダーゼに関しては、ほぼ4量体の実測分子量(237kDa)、R170A/R220A/Y227F変異体は、ほぼ2量体の実測分子量(129kDa)を示すことが判明した。一方、R170A/R220A/Y227F変異体に対してさらに変異導入した変異体、R170A/R220A/Y227F/R448E変異体、R170A/R220A/Y227F/E449R変異体、R170A/R220A/ Y227F/R448G変異体、R170A/R220A/Y227F/E459G変異体、に関しては、実測分子量が50kDa〜90kDaの範囲にあり、さらに変異を導入する効果として、完全に単量体化したものと推定された。
【0096】
(実施例11)変異体のセロビオース分解活性3
セルロース含有バイオマス存在下での親グルコシダーゼおよび変異体(本発明のβグルコシダーゼ)のセロビオース分解活性を測定した。セルロース含有バイオマスとしては、稲藁を5%希硫酸で150℃10分処理したものを使用した。セルロース含有バイオマス5wt%を懸濁した溶液に、10mM セロビオース/50mM 酢酸緩衝液を添加し反応液とした。この反応液に実施例9で調製した酵素R170A/R220A/Y227F/R448E変異体、R170A/R220A/Y227F/E449R変異体、R170A/R220A/Y227F/R448G変異体、R170A/R220A/Y227F/E459G変異体、を終濃度0.23mg/mLで添加し、50℃で酵素反応を30分行った。親βグルコシダーゼにおける50℃の温度条件下で生成したグルコース濃度(g/L)を100%として、各変異体におけるセロビオース分解活性を相対値で表9に示す。
【0097】
【表9】
【0098】
R170A/R220A/Y227F/R448E変異体、R170A/R220A/Y227F/E449R変異体、R170A/R220A/ Y227F/R448G変異体、R170A/R220A/Y227F/E459G変異体では、親グルコシダーゼに対して活性が大きく向上することが判明した。また実施例5のR170A/R220A/Y227F変異体(2量体)に対して、実施例9の変異型グルコシダーゼ(単量体)の方が高いことが判明した。
【産業上の利用可能性】
【0099】
本発明におけるβグルコシダーゼは、セルロース含有バイオマス存在下で高いセロビオース分解活性を示すため、バイオマスの加水分解および糖液の製造に使用できる。
【技術分野】
【0001】
本発明は、新規なβグルコシダーゼ、および、当該βグルコシダーゼを含むバイオマス分解用酵素組成物、およびこれを使用してセルロース由来バイオマスを加水分解することによる糖液の製造方法に関する。
【背景技術】
【0002】
セルロースの糖化には様々な手法があるが、エネルギー使用量が少なく、かつ糖収率の高い酵素糖化法が開発の主流となっている。セルロース分解酵素であるセルラーゼを大別すると、セルロースの結晶領域に作用するセロビオハイドラーゼ、セルロース分子鎖内部から作用して分子量を低減させるエンドグルカナーゼに分けられる。これらセルラーゼは、生成物の一つであるセロビオースによって阻害を受けることが知られている。またβグルコシダーゼは、水溶性オリゴ糖あるいはセロビオースに作用し、そのβ−グリコシド結合を加水分解する反応を触媒する酵素である。特にβグルコシダーゼは、発酵原料として有用なグルコースを十分に得るためには必須の酵素である。また、セロビオハイドラーゼあるいはエンドグルカナーゼは、セルロース分解で生成したセロビオースの蓄積により反応阻害が引き起こされることが知られている。すなわち、βグルコシダーゼは、セルロース分解により生成するセロビオースの蓄積を大幅に低減することができるため、セルロース分解効率を大幅に向上させるといった効果を有する。
【0003】
セルロースは、草本系植物、木本系植物中に多く含まれ、これら植物を総称してセルロース含有バイオマスと呼ぶ。セルロース含有バイオマスは、セルロースに加え、キシラン、アラビナンといったヘミセルロース、およびリグニンを含んでいる。特に、セルロース含有バイオマス中に含まれるリグニンは、芳香族系の高分子化合物であるため、糸状菌由来セルラーゼを使用した酵素糖化において、阻害的に作用することが知られている。リグニンによる糸状菌由来セルラーゼの阻害機構に関して、すべて解明されていないが、リグニンにセルラーゼが吸着し、分解効率が低下することが、要因の一つであると言われている(非特許文献1)。
【0004】
耐熱性酵素は、安定性が高く、高温条件においても長期間活性を保持することから、産業用酵素としての適用が検討されている。こうした耐熱性酵素は、好熱性菌、あるいは超好熱性菌の保有する酵素の中より多くその存在が確認されている。
【0005】
耐熱性のβグルコシダーゼに関しても、数種の好熱性菌あるいは超好熱性菌より同定されており、具体的にはパイロコッカス・フリオサス(Pyrococcus furiosus)、パイロコッカス・ホリコシイ(Pyrococcus horikoshii)、サーモトーガ・マリティマ(Thermotoga maritima)、スルフォロバス・シバタエ(Sulfolobus shibatae)、スルフォロバス・ソルファタリスク(Sulfolobus solfataricus)、クロストリジウム・サーモセラム(Clostridium thermocellum)などの微生物より耐熱性のβグルコシダーゼが同定されている。特にクロストリジウム・サーモセラム(Clostridium thermocellum)由来のβグルコシダーゼは単量体(非特許文献2)、スルフォロバス・ソルファタリスク(Sulfolobus solfataricus)およびパイロコッカス・フリオサス(Pyrococcus furiosus)由来のβグルコシダーゼは4量体を形成する(非特許文献3、非特許文献4)ことが開示されているが、これら構造とその機能との関係は明らかになっていない。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】国際公開第2005/093072号公報
【非特許文献】
【0007】
【非特許文献1】Hetti P.ら,Journal of Biotechnology,107,65−72(2004)
【非特許文献2】Christian P.ら,Trichoderma and Gliocladium:Basic Biology,Taxonomy and Genetics.,vol.1,121−138(1998)
【非特許文献3】Ohba H.ら,Biosci.Biotech.Biochem.,59,1581−1583(1995)
【非特許文献4】Bauer MW.ら,J.Biol.Chem.,vol.271,39,23749−23755(1996)
【発明の概要】
【発明が解決しようとする課題】
【0008】
本発明の課題は、セルロース含有バイオマスの加水分解における酵素活性を高く保持したβグルコシダーゼを提供することにある。
【課題を解決するための手段】
【0009】
本発明者らは、上記の課題を解決すべく、配列番号1に示されるアミノ酸配列を有する従来のβグルコシダーゼ(以下、本明細書において「親βグルコシダーゼ」という。)の特定箇所にアミノ酸変異を導入し、その機能が改良された性質を有する変異体として新規なβグルコシダーゼの取得に成功した。詳細には、本発明者らは、親βグルコシダーゼの三次元構造に着目し、タンパク質結晶構造解析により、その4量体の形成に関わるアミノ酸を特定し、当該アミノ酸に対して選択的に変異を加えることにより、4量体形成が不安定化された新規βグルコシダーゼの取得に成功した。そして本新規βグルコシダーゼは、特に、セルロース含有バイオマスの加水分解において優れた特性を有することを見出した。
【0010】
すなわち、本発明は以下の構成からなる。
[1] 各単量体の間で水素結合またはイオン結合を形成するアミノ酸の位置に、βグルコシダーゼの4量体形成を不安定化させる変異を有し、かつ野生型と同等またはそれ以上のセロビオース分解活性を有することを特徴とする、変異型βグルコシダーゼ。
[2] 配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、39番目のグルタミン酸、169番目のアスパラギン酸、170番目のアルギニン、220番目のアルギニン、227番目のチロシンおよび330番目のグルタミン酸からなる群から選ばれる少なくとも3以上のアミノ酸に相当するアミノ酸の変異を含む、[1]の変異型βグルコシダーゼ。
【0011】
[3] 配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、39番目のグルタミン酸、169番目のアスパラギン酸、170番目のアルギニン、220番目のアルギニン、227番目のチロシンおよび330番目のグルタミン酸からなる群から選ばれる少なくとも3以上のアミノ酸に相当するアミノ酸の、中性側鎖を有するアミノ酸への置換を含む、[1]または[2]の変異型βグルコシダーゼ。
[4] 配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、170番目のアルギニン、220番目のアルギニンおよび227番目のチロシンに相当するアミノ酸の、中性側鎖を有するアミノ酸への置換を含む、[1]〜[3]のいずれかの変異型βグルコシダーゼ。
【0012】
[5] 中性側鎖を有するアミノ酸が、アラニン、フェニルアラニンおよびグリシンからなる群から選択される、[3]または[4]の変異型βグルコシダーゼ。
[6] 配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の170番目のアルギニンに相当するアミノ酸のアラニンへの置換、配列番号1のアミノ酸配列の220番目のアルギニンに相当するアミノ酸のアラニンへの置換および配列番号1のアミノ酸配列の227番目のチロシンに相当するアミノ酸のフェニルアラニンへの置換を含む、[1]〜[5]のいずれかの変異型βグルコシダーゼ。
[7] 配列番号14で示されるアミノ酸配列を含む、[1]〜[6]のいずれかの変異型βグルコシダーゼ。
【0013】
[8] 配列番号1のアミノ酸配列の440番目のロイシン、448番目のアルギニン、449番目のグルタミン酸、459番目のグルタミン酸からなる群から選ばれる1以上のアミノ酸に相当するアミノ酸の変異をさらに含む、[1]〜[7]のいずれかの変異型βグルコシダーゼ。
[9] 配列番号1のアミノ酸配列の448番目のアルギニンに相当するアミノ酸の中性側鎖もしくは酸性側鎖を有するアミノ酸への置換、配列番号1のアミノ酸配列の449番目のグルタミン酸に相当するアミノ酸の中性側鎖もしくは塩基性側鎖を有するアミノ酸への置換および配列番号1のアミノ酸配列の459番目のグルタミン酸に相当するアミノ酸の中性側鎖もしくは塩基性側鎖を有するアミノ酸への置換からなる群から選ばれる1以上の置換を含む、[8]の変異型βグルコシダーゼ。
【0014】
[10] 配列番号35〜38のいずれかで示されるアミノ酸配列を含む、[8]又は[9]の変異型βグルコシダーゼ。
[11] [1]〜[10]のいずれかの変異型βグルコシダーゼをコードするDNA。
[12] 配列番号20、配列番号31〜34のいずれかで示される塩基配列を含む、[11]のDNA。
[13] [11]または[12]のDNAを含む、発現ベクター。
[14] [13]の発現ベクターを用いた形質転換により作製された、形質転換細胞。
[15] [1]〜[10]のいずれかの変異型βグルコシダーゼまたは[14]の形質転換細胞の処理物を含むバイオマス分解用酵素組成物。
[16] セルロース含有バイオマスを[15]のバイオマス分解用酵素組成物を使用して加水分解することを含む、セルロース由来バイオマスより糖液を製造する方法。
【発明の効果】
【0015】
本発明により、セルロース含有バイオマス存在下でのグルコシダーゼ酵素活性が改良された新規変異型βグルコシダーゼを提供することができる。本発明のβグルコシダーゼは、セルロース含有バイオマスの加水分解による糖液の製造に好適に使用することができる。
【図面の簡単な説明】
【0016】
【図1】親βグルコシダーゼの結晶構造解析の結果得られた、親βグルコシダーゼのリボンモデルである。
【図2】親βグルコシダーゼのサブユニットA−B間の相互作用部位を拡大したリボンモデルである。
【図3】親βグルコシダーゼおよび各種変異を導入したβグルコシダーゼの、ブルー・ネイティブPAGEによる電気泳動の結果である。
【図4】変異型βグルコシダーゼの結晶構造解析の結果得られた、変異型βグルコシダーゼのリボンモデルである。
【図5】親βグルコシダーゼ、および変異型βグルコシダーゼのゲルろ過による分子量測定の結果である。
【発明を実施するための形態】
【0017】
以下に、本発明を詳細に説明するが、本発明はこれらに限定されない。
(1)βグルコシダーゼ
本発明において「βグルコシダーゼ」とは、糖のβ−グルコシド結合を加水分解する反応を触媒する酵素を意味する。βグルコシダーゼはセロビオースの分解活性が高いことを特徴とする。セロビオース分解活性の測定は、例えば50mM 酢酸−酢酸ナトリウム緩衝液(pH5.0)に溶解したセロビオースの基質溶液に酵素液を添加し、30〜85℃で30分間反応後、必要によりpHを変化させるなどして反応を停止させた後、グルコース定量キットを用いて、反応液中のグルコース濃度を定量することによって行うことができる。βグルコシダーゼとして、EC番号:EC 3.2.1.21として、これに帰属される酵素群が例示されているが、上述したセロビオース分解活性を保有していれば、本発明における「βグルコシダーゼ」に含まれる。
【0018】
(2)親βグルコシダーゼ
本明細書において、親βグルコシダーゼとは、配列番号1で示されるアミノ酸配列を有するβグルコシダーゼであり、セロビオース分解活性を示すものである。本明細書において、「親βグルコシダーゼ」を「野生型」と記載する場合がある。この場合、「親βグルコシダーゼ」および「野生型」なる記載は互換的に使用される。
【0019】
親βグルコシダーゼは、パイロコッカス・フリオサスに由来する酵素である。パイロコッカス・フリオサスは、80−110℃を生育至適温度とする古細菌に分類される微生物で、パイロコッカス・フリオサスは、デンプン、セルロース、マルトース、プルラン、など種々の炭素源を資化することができる微生物である。
【0020】
親βグルコシダーゼは、Kengenら(Eur.J.Biochem.213,305−312,1993年)によって初めて単離精製された酵素である。本文献によると、親βグルコシダーゼは、水溶液中での分子量は約230kDaのホモ4量体で存在することが明示されている。また、SDSなどの界面活性剤によって、この4量体を破壊することができ、各単量体は約58kDaであることが明示されている。さらに、親βグルコシダーゼの至適pHは5.0、等電点は4.4、反応最適温度は102−105℃と記載されている。配列番号1で示される親βグルコシダーゼのアミノ酸配列は、Voorhorstら(J.Bacteriol.177,24,7105−7111,1995年)で同定されたアミノ酸配列に基づくものである。このアミノ酸配列に基づくその分子量は、約54kDa(54.58kDa)と記載されている。
【0021】
(3)本発明の変異型βグルコシダーゼ
本発明の変異型βグルコシダーゼは、水溶液中でβグルコシダーゼ単量体の間で水素結合またはイオン結合を形成するアミノ酸の位置に、βグルコシダーゼの4量体形成を不安定化させる変異を有することを特徴とする。
【0022】
本発明の変異型βグルコシダーゼは、水溶液中で、親βグルコシダーゼが本来形成する4量体と比べて、不安定化された4量体で存在することを特徴とする。親βグルコシダーゼは、一次構造としては約54kDaであるが、水溶液中で安定な4量体を形成し、分子量200kDa以上の分子として溶解していることが知られている(Bauer MW.ら,“Comparison of a β−glucosidase and a β−mannosidase from the hypertherrmophilic archaeon Pyrococcus furiosus”J.Biol.Chem.,vol.271,39,23749−23755(1996))。
【0023】
本発明の変異型βグルコシダーゼが、水溶液中で不安定な4量体を形成していることは、水溶液中での分子量が200kDa未満であることから確認することができる。具体的には、Native−PAGEによる手法でその分子量を確認すると160kDa未満の分子量で検出される。水溶液中4量体が不安定化されていることは、Native−PAGE以外の手法(例えば、ゲル濾過分析、超遠心分析など)を用いて、親βグルコシダーゼと本発明の変異型βグルコシダーゼの水溶液中での挙動を比較することによって、具体的には、水溶液中親βグルコシダーゼの分子量と比較して、本発明のβグルコシダーゼの分子量が小さいことによって、確認することが可能である。
【0024】
下記実施例にて詳述するように、本発明者らは結晶構造解析により、親βグルコシダーゼのアミノ酸配列、すなわち配列番号1で示されるアミノ酸配列において、39番目のグルタミン酸(以下、「Glu39」と記載)、169番目のアスパラギン酸(以下、「Asp169」と記載)、170番目のアルギニン(以下、「Arg170」と記載)、220番目のアルギニン(以下、「Arg220」と記載)、227番目のチロシン(以下、「Try227」と記載)および330番目のグルタミン酸(以下、「Glu330」と記載)が、親βグルコシダーゼの水溶液中における4量体形成に関与していることを明らかにした。すなわち、これらのアミノ酸は、他の親グルコシダーゼの単量体と水溶液中で水素結合またはイオン結合を形成しており、4量体の形成に強く関与しているアミノ酸であることがわかった。本発明における、βグルコシダーゼへのアミノ酸変異の導入の目的は、この4量体の形成に関与するアミノ酸の水素結合またはイオン結合を破壊することにあり、その結果、水溶液中におけるβグルコシダーゼの4量体構造を不安定化させることにある。
【0025】
本発明における、βグルコシダーゼのアミノ酸変異は、βグルコシダーゼの水溶液中における4量体形成に関与しているアミノ酸の置換および/または欠失を含む。
【0026】
好ましくは、本発明の変異型βグルコシダーゼは、配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列を有し、当該配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、Glu39、Asp169、Arg170、Arg220、Tyr227およびGlu330のアミノ酸から選択される少なくとも3以上のアミノ酸に相当するアミノ酸の、中性側鎖を有するアミノ酸への置換を含む。
【0027】
「配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列」は、少なくとも配列番号1のアミノ酸配列で表されるβグルコシダーゼの活性を保持する。
【0028】
ここで「相当するアミノ酸」とは、前記配列同一性を有するアミノ酸配列と配列番号1のアミノ酸配列の立体構造を比較した場合に、前記配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列におけるGlu39、Asp169、Arg170、Arg220、Tyr227およびGlu330と同様の位置に存在し、βグルコシダーゼの水溶液中における4量体形成に関与しているアミノ酸を意味する。「相当するアミノ酸」のアミノ酸種は、配列番号1のアミノ酸配列におけるGlu39、Asp169、Arg170、Arg220、Tyr227およびGlu330とそれぞれ同一である。
【0029】
さらに好ましくは、本発明の変異型βグルコシダーゼは、配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列を有し、当該配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、Arg170、Arg220およびTyr227のアミノ酸に相当するアミノ酸の、中性側鎖を有するアミノ酸への置換を含む。
【0030】
「中性側鎖を有するアミノ酸」としては、グリシン、アスパラギン、グルタミン、セリン、スレオニン、チロシン、システイン、アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファンが挙げられる。好ましくは、「中性側鎖を有するアミノ酸」はアラニン、フェニルアラニンおよびグリシンからなる群から選択される。
【0031】
下記実施例にて詳述するように、本発明者らは結晶構造解析により、配列番号1で示されるアミノ酸配列におけるArg170が、親βグルコシダーゼにおいて、水溶液中4量体を形成する他の全ての親グルコシダーゼ単量体におけるArg170と水素結合またはイオン結合を形成することを明らかとした。すなわち、Arg170に変異を導入することによって、4量体形成に関する水素結合またはイオン結合を効果的に破壊することが可能である。また、下記実施例にて詳述するように、本発明者らは結晶構造解析により、親βグルコシダーゼにおいて、水溶液中、Arg220が、4量体を形成する他の単量体における44番目のグリシン(Gly44)と水素結合を形成していること;また、Tyr227が、4量体を形成する他の単量体における165番目のリシン(Lys165)と水素結合を形成していること、を明らかにした。すなわち、配列番号1で示されるアミノ酸配列において、Arg170に加え、Arg220およびTyr227に変異を導入することによって、4量体形成に関する水素結合またはイオン結合を効果的に破壊することでき、水溶液中でのβグルコシダーゼの4量体構造を不安定化させることが可能である。
【0032】
したがって好ましくは、本発明の変異型βグルコシダーゼは、配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列を有し、当該配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、Arg170に相当するアミノ酸をアラニンへ、Arg220に相当するアミノ酸をアラニンへおよびTyr227に相当するアミノ酸をフェニルアラニンへの置換を含む。
【0033】
特に好ましくは、本発明の変異型βグルコシダーゼは、配列番号14で示されるアミノ酸配列を含む。
【0034】
本発明の変異型βグルコシダーゼは、セロビオース分解活性を保持し得る限り、配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列を有し、当該配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、Glu39、Asp169、Arg170、Arg220、Tyr227およびGlu330に相当するアミノ酸以外の箇所に一または複数のアミノ酸変異を有していても良い。ここで「複数」とは、2〜10個、好ましくは2〜5個、さらに好ましくは2〜3個を意味し、「アミノ酸変異」には、アミノ酸の置換、欠失、挿入および/または付加が含まれる。
【0035】
本発明の変異型βグルコシダーゼは、親βグルコシダーゼと比べて、セルロース含有バイオマス存在下でのβグルコシダーゼ活性が向上している、といった改良された優れた性質を示す。この改良された優れた性質は、親βグルコシダーゼが、水溶液中安定な4量体を形成するのに対して、本発明の変異型βグルコシダーゼでは上記変異導入により、4量体構造が不安定化していることが要因として推定される。
【0036】
本明細書において、本発明の変異型βグルコシダーゼを、「変異体」と記載する場合がある。この場合、「変異型βグルコシダーゼ」および「変異体」なる記載は互換的に使用される。
【0037】
前述した本発明の変異型βグルコシダーゼは、配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列を有し、当該配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、Glu39、Asp169、Arg170、Arg220、Tyr227およびGlu330のアミノ酸から選択される少なくとも3以上のアミノ酸に相当するアミノ酸の、中性側鎖を有するアミノ酸への置換を含み、これらの変異の効果によって4量体化を不安定化させることが可能であるが、これら変異体にさらに変異導入を行ったものでもよい。変異導入の目的としては、さらに活性向上を目的として変異、熱安定性向上を目的とした変異、さらに多量体構造を不安定化することを目的とした変異などを例示することができる。
【0038】
下記実施例にて詳述するように、本発明者らは前述した変異型βグルコシダーゼに関し、さらに結晶構造解析を実施した結果、本発明の変異型βグルコシダーゼは2量体構造を形成していることを明らかにした。この変異型βグルコシダーゼの2量体構造形成に関与するアミノ酸は、配列番号1のアミノ酸配列の440番目のロイシン、448番目のアルギニン、449番目のグルタミン酸、459番目のアルギニンに相当する4つのアミノ酸である。すなわち、これら4つのアミノ酸は、2量体を形成する他方の変異型グルコシダーゼのアミノ酸と水素結合もしくはイオン結合を形成しており、変異型グルコシダーゼの2量体の形成に強く関与する。本発明では、前述した4量体を不安定化する変異に加え、さらに変異型グルコシダーゼが形成する2量体を不安定化する変異をさらに加えてもよい。
【0039】
すなわち、本発明の変異型βグルコシダーゼは、前述した4量体を不安定化する変異に加え、配列番号1のアミノ酸配列の440番目のロイシン、448番目のアルギニン、449番目のグルタミン酸、459番目のグルタミン酸からなる群から選ばれる1以上のアミノ酸に相当するアミノ酸の変異をさらに含む。
【0040】
ここで「相当するアミノ酸」とは、前記配列同一性を有するアミノ酸配列と配列番号1のアミノ酸配列の立体構造を比較した場合に、前記配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の440番目のロイシン、448番目のアルギニン、449番目のグルタミン酸、459番目のグルタミン酸と同様の位置に存在し、変異型βグルコシダーゼの2量体形成に関与しているアミノ酸を意味する。
【0041】
特に好ましくは、前述の変異型βグルコシダーゼのアミノ酸配列において、配列番号1のアミノ酸配列の448番目のアルギニンに相当するアミノ酸の中性側鎖もしくは酸性側鎖を有するアミノ酸への置換、または、449番目のグルタミン酸に相当するアミノ酸の中性側鎖もしくは塩基性側鎖を有するアミノ酸への置換、459番目のアルギニンに相当するアミノ酸の中性側鎖もしくは塩基性側鎖を有するアミノ酸への置換、からなる群から選ばれる1以上のアミノ酸位置における置換を含む。
【0042】
「中性側鎖を有するアミノ酸」としては、グリシン、アスパラギン、グルタミン、セリン、スレオニン、チロシン、システイン、アラニン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファンが挙げられる。好ましくは、「中性側鎖を有するアミノ酸」はアラニン、フェニルアラニンおよびグリシンからなる群から選択される。
【0043】
「酸性側鎖を有するアミノ酸」としては、グルタミン酸、アスパラギン酸、の群から選択される。
【0044】
「塩基性側鎖を有するアミノ酸」としては、アルギニン、リジン、ヒスチジン、の群から選択される。
【0045】
最も好ましくは、前述の変異型βグルコシダーゼのアミノ酸配列において、配列番号1のアミノ酸配列の448番目のアルギニンに相当するアミノ酸のグルタミン酸もしくはグリシン、配列番号1のアミノ酸配列の449番目のグルタミン酸に相当するアミノ酸のアルギニン、配列番号1のアミノ酸配列の459番目のアルギニンに相当するアミノ酸のグリシンへの変異から選ばれる1以上の変異を含む。
【0046】
特に好ましくは、本発明の変異型βグルコシダーゼは、配列番号31〜34で示されるアミノ酸配列を含む。
【0047】
また、こうしたアミノ酸置換による変異に加えて、配列番号1のアミノ酸配列の440番目以降のアミノ酸全部に相当する領域を、あるいは、配列番号1のアミノ酸配列の448番目以降のアミノ酸全部に相当する領域を、あるいは配列番号1のアミノ酸配列の449番目以降のアミノ酸全部に相当する領域を、あるいは配列番号1のアミノ酸配列の459番目以降のアミノ酸全部に相当する領域を、欠失(削除)したものでもよい。これは440番目以降のアミノ酸に相当する配列に関しては、セロビオース分解活性そのものには特段の影響がないためによる。こうした欠失(削除)を行いたい場合は、削除したいアミノ酸の位置に対応する遺伝子においてストップコドンを挿入することで可能になる。
【0048】
(4)本発明の変異型βグルコシダーゼの製造方法
本発明の変異型βグルコシダーゼを製造する方法としては、例えばβグルコシダーゼのアミノ酸配列をコードするDNAを調製し、これを発現ベクターに連結し、発現ベクターを宿主に導入し、異種タンパク質として生産し、単離および精製することで得ることができる。
【0049】
まず上述のDNAの調製方法としては、遺伝子合成により目的のアミノ酸配列をコードするDNAを全合成する方法、配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列をコードするDNAに対して、部位突然変異法により上記所定の位置に変異を導入する方法が例示される。配列番号1のアミノ酸配列と90%、95%、99%、またはそれ以上の配列同一性を有するアミノ酸配列をコードする遺伝子は、このβグルコシダーゼを保有する微生物、特にパイロコッカス・フリオサスより公知の方法に準じてDNAを単離し、PCR等の手法によってDNA増幅させることで取得することができる。
【0050】
上記のようにして調製した変異型βグルコシダーゼをコードするDNAを、制限酵素およびDNAリガーゼを用いて、適当な発現ベクター中のプロモーター下流に連結することにより、当該DNAを含む発現ベクターを製造することができる。発現ベクターとしては、細菌プラスミド、酵母プラスミド、ファージDNA(ラムダファージなど)、レトロウイルス、バキュロウイルス、ワクシニアウイルス、アデノウイルス等のウイルスDNA、SV40の誘導体など、植物細胞用のベクターとしてのアグロバクテリウムなどが挙げられるが、宿主細胞において複製および生存可能ある限り他のいかなるベクターも用いることができる。例えば、宿主が大腸菌である場合、pUC、pET、pBADなどを例示することができる。また、宿主が酵母である場合、pPink−HC、pPink−LC、pPinkα−HC、pPCIZ、pPCIZα、pPCI6、pPCI6α、pFLD1、pFLD1α、pGAPZ、pGAPZα、pPIC9K、pPIC9などが挙げられる。
【0051】
プロモーターとしては、遺伝子の発現に用いる宿主に対応して適切なプロモーターであればいかなるものでもよい。例えば、宿主が大腸菌である場合、lacプロモーター、Trpプロモーター、PLプロモーター、PRプロモーター等が、酵母である場合、AOX1プロモーター、TEF1プロモーター、ADE2プロモーター、CYC1プロモーター、GAL−L1プロモーターなどが挙げられる。
【0052】
本発明において用いる宿主細胞としては、大腸菌、バクテリア細胞、酵母細胞、真菌細胞、昆虫細胞、植物細胞、動物細胞などが好ましい。酵母細胞としては、例えば、ピキア属(Pichia)、サッカロマイセス属(Saccharomyces)、シゾサッカロマイセス属(Schizosaccharomyces)などが挙げられる。真菌細胞としては、アスペルギルス属(Aspergillus)、トリコデルマ属(Trichoderma)などが挙げられる。昆虫細胞としてはSf9など、植物細胞としては双子葉植物など、動物細胞としては、CHO、HeLa、HEK293などが挙げられる。
【0053】
形質転換または、トランスフェクションは、リン酸カルシウム法、電気穿孔法などの公知の方法で行うことができる。本発明のβグルコシダーゼは、上記のように形質転換またはトランスフェクトされた宿主細胞においてプロモーターの制御下にて発現させ、産生物を回収して得ることができる。発現に際しては、宿主細胞を適切な細胞密度まで増殖または成長させた後、温度シフトまたはイソプロピル−1−チオ−β−D−ガラクトシド(IPTG)添加などの化学的誘発手段によってプロモーターを誘発させ、細胞をさらに一定期間培養する。
【0054】
変異型βグルコシダーゼが細胞外に排出される場合には、培地から直接に、また細胞外に存在する場合には、超音波破砕や機械的破砕などの物理的手段もしくは細胞溶解剤などの化学的手段によって、細胞を破壊した後にβグルコシダーゼを精製する。βグルコシダーゼは、組換え細胞の培地から、硫酸アンモニウムもしくはエタノール沈殿、酸抽出、陰イオンもしくは陽イオン交換クロマトグラフィー、逆相高速液体クロマトグラフィー、アフィニティークロマトグラフィー、ゲルろ過クロマトグラフィー、電気泳動などの技術を組み合わせて、部分的にまたは完全に精製することができる。
【0055】
(5)変異型βグルコシダーゼを含むバイオマス分解用酵素組成物
本発明の変異型βグルコシダーゼは、セルロース含有バイオマスの加水分解において、親βグルコシダーゼに対して、高い熱安定性、リグニン耐性を有する。具体的には、親βグルコシダーゼに比べて1.2倍、1.3倍、1.4倍、1.5倍、1.6倍、1.7倍、1.8倍、1.9倍、2倍またはそれ以上の高いセロビオース分解活性を得ることができる。
【0056】
本発明の変異型βグルコシダーゼは、精製されたものであっても、粗精製されたものであっても良い。
【0057】
また、本発明の変異型βグルコシダーゼは、固相に固定化されていても良い。固相としては例えば、ポリアクリルアミドゲル、ポリスチレン樹脂、多孔性ガラス、金属酸化物などが挙げられる(特にこれらに限定されない)。本発明の変異型βグルコシダーゼを固相に固定することによって、連続反復使用が可能となる点において有利である。
【0058】
さらに、上記変異型βグルコシダーゼをコードするDNAを用いて形質転換した細胞の処理物も、粗精製された変異型βグルコシダーゼとして利用することができる。当該「形質転換した細胞の処理物」には、固相に固定化した形質転換細胞、ならびに形質転換細胞の死菌、破砕物、およびそれらを固相に固定化したものなどが含まれる。
【0059】
本発明の変異型βグルコシダーゼは、セルラーゼと混合することで、バイオマス分解用酵素組成物としてセルロース含有バイオマスの加水分解に使用することができる。ここでいうセルラーゼとは、セルロースを分解する活性を有する酵素であれば特に限定されず、一種類以上の混合物であってもよい。このような酵素としては、例えばセルラーゼ、ヘミセルラーゼ、セロビオハイドラーゼ、エンドグルカナーゼ、エキソグルカナーゼ、キシラナーゼ、マンナナーゼなどが挙げられる。
【0060】
上述セルラーゼは、好ましくは糸状菌由来セルラーゼである。糸状菌由来セルラーゼは、少なくともエンドグルカナーゼおよびセロビオハイドラーゼの双方を含んでなる混合物である。さらに効率的なセルロースの糖化を行うためには、エンドグルカナーゼに関して2種以上、および/または、セロビオハイドラーゼに関して2種以上含んでなる糸状菌由来セルラーゼであることが好ましい。前記糸状菌セルラーゼを生産する微生物として、トリコデルマ属(Trichoderma)、アスペルギルス属(Aspergillus)、セルロモナス属(Cellulomonas)、クロストリジウム属(Clostridium)、ストレプトマイセス属(Streptomyces)、フミコラ属(Humicola)、アクレモニウム属(Acremonium)、イルペックス属(Irpex)、ムコール属(Mucor)、タラロマイセス属(Talaromyces)、などの微生物を挙げることができる。これら微生物は、培養液中にセルラーゼを産生するために、その培養液を未精製の糸状菌セルラーゼとしてそのまま使用してもよいし、また培養液を精製し、製剤化したものを糸状菌セルラーゼ混合物として使用してもよい。前記培養液より、糸状菌セルラーゼ混合物を精製し、製剤化したものとして使用する場合、プロテアーゼ阻害剤、分散剤、溶解促進剤、安定化剤など、酵素以外の物質を添加したものを、セルラーゼ製剤として使用してもよい。
【0061】
本発明で使用する糸状菌由来セルラーゼは、トリコデルマ属由来セルラーゼであることが好ましい。トリコデルマ属は、少なくとも2種以上のエンドグルカナーゼ、および、少なくとも2種以上のセロビオハイドラーゼを含んでなるセルラーゼを培養液中に産生し、こうした培養液より調製されたセルラーゼ混合物は本発明に好ましく使用することができる。こうしたトリコデルマ属のうち、より好ましくは、トリコデルマ・リーセイ(Trichoderma reesei)由来のセルラーゼ混合物である。トリコデルマ・リーセイ由来のセルラーゼ混合物としては、トリコデルマ・リーセイQM9414(Trichoderma reesei QM9414)、トリコデルマ・リーセイQM9123(Trichoderma reesei QM9123)、トリコデルマ・リーセイRutC−30(Trichoderma reesei Rut−30)、トリコデルマ・リーセイPC3−7(Trichoderma reesei PC3−7)、トリコデルマ・リーセイCL−847(Trichoderma reesei CL−847)、トリコデルマ・リーセイMCG77(Trichoderma reesei MCG77)、トリコデルマ・リーセイMCG80(Trichoderma reesei MCG80)、トリコデルマ・ビリデQM9123(Trichoderma viride QM9123)に由来するセルラーゼ混合物が挙げられる。また、前記トリコデルマ属に由来し、変異剤あるいは紫外線照射などで変異処理を施し、セルラーゼ生産性が向上した変異株であってもよい。
【0062】
本発明のバイオマス分解用酵素組成物は、広くセルロース含有バイオマスの加水分解に使用することができる。セルロース含有バイオマスとは、少なくともセルロースを含むものであれば限定されない。具体的には、バガス、コーンストーバー、コーンコブ、スイッチグラス、稲藁、麦藁、樹木、木材、廃建材、新聞紙、古紙、パルプなどである。これらセルロース含有バイオマスは、高分子芳香族化合物リグニンやヘミセルロースなどの不純物が含まれているが、前処理として、酸、アルカリ、加圧熱水などを用いて、リグニンやヘミセルロースを部分的に分解したセルロース含有バイオマスを、セルロースとして利用してもよい。
【0063】
本発明におけるセルロース含有バイオマスの酵素処理条件は、処理温度40℃〜60℃、処理pH3〜7、セルロース含有バイオマス固形分濃度0.1〜30%が好ましい。当該範囲に設定することにより、本発明のバイオマス分解用酵素組成物の分解効率を最大限発揮することができる。従来の好熱性菌由来グルコシダーゼは、反応最適温度が100℃付近の酵素もあるが、本発明で使用する好熱性菌由来グルコシダーゼは、40℃〜60℃においても十分高い比活性を示し、セルラーゼ共存下で、セルロース含有バイオマスの分解を効率的に行うことが可能である。この酵素処理は、バッチ式で行っても、連続式で行ってもよい。こうした酵素処理によって得られた加水分解物は、グルコース、キシロースなどの単糖成分を含むため、エタノール、乳酸などの原料糖として使用することが可能である。
【実施例】
【0064】
以下に、実施例を挙げて本発明を具体的に説明する。ただし、本発明はこれらに限定されるものではない。
【0065】
(参考例1)親βグルコシダーゼの大腸菌による組み換え発現調製
親βグルコシダーゼの遺伝子は、配列番号2に示す塩基配列を有する遺伝子を全合成し、pET−11dのNcoIおよびBamHIにLigation High(東洋紡)を使用して連結し、JM109(Takara)に形質転換した。スクリーニングは、アンピシリンを抗生物質として含むLB寒天培地を用いて行った。形質転換されたJM109株より、作製したベクターpET−PfuBGLをミニプレップキット(キアゲン)により単離し、塩基配列解析を行った。pET−PfuBGLは、発現用大腸菌BL21(DE3)pLysS株に形質転換し、BL21−PfuBGL株を作製した。BL21−PfuBGL株を、アンピシリン含有LB培地10mLに植菌し、37℃で一晩振とう培養(前培養)を行った。本培養として、アンピシリン含有LB培地1Lに、前培養で得られた菌体を植菌し、波長600 nmでの吸光度OD600が0.8となるまで振とう培養を行った。その後、最終濃度が0.4mMになるようにイソプロピル−1−チオ−β−D−ガラクトシド(IPTG)を加え、さらに25℃で一晩培養した。培養後、菌体を遠心分離により回収し、50mM トリス−HCl緩衝液(pH8.0)に再懸濁した。この溶液を氷冷しながら、超音波破砕を行い、その上清を無細胞抽出液として遠心分離により回収した。得られた無細胞抽出液を、85℃で15分保温し、該グルコシダーゼ以外の大腸菌に由来するタンパク質を凝集沈殿した。遠心分離により沈殿物を除去し、上清を分画分子量10000の再生セルロース製透析膜(スペクトラム・ラボラトリーズ製)を使用して、50mM 酢酸緩衝液(pH5.0)に透析した。得られたタンパク質溶液を、親βグルコシダーゼとして使用した。調製した親βグルコシダーゼのアミノ酸配列を配列番号1に記載する。
【0066】
(実施例1)親βグルコシダーゼの4量体形成部位の特定
親βグルコシダーゼ(パイロコッカス・フリオサス)のX線結晶構造は既に報告されているが(Thijis K.ら,Biochem.vol.39,No.17(2000))分解能が3.3 Åと低く、またProtein Data Bank(PDB)への登録もされていない。そこで、PfuBGLの三次元構造を決定するため、X線結晶構造解析を試みた。新たな結晶化条件を探索し、リン酸を沈殿剤として結晶化に成功した。大型放射光実験施設SPring−8でX線回折実験を行い、分解能2.5ÅでPfuBGLの構造を決定した。構造決定は分子置換法を用い、モデル分子には配列番号22で示されるアミノ酸配列を有するサーモスファエラ・アグリガンズ(Themosphaera aggregans)由来β−グリコシダーゼ(ThAggBGY、PDB ID:1QVB)を用いた。得られた3次元構造データを構造解析用のソフトウェアCCP4_Contactを用いてサブユニット間の相互作用を分析した。分析の結果得られたリボンモデルを図1に示す。サブユニットA−B間、A−C間で相互作用しており、ホモ4量体を形成していることが確認された(図1)。分析結果より得られたサブユニットA−B間の相互作用に関し、表1にまとめた。
【0067】
【表1】
【0068】
サブユニット間の相互作用のうち、アミノ酸主鎖の窒素原子(N)および酸素原子(O)のみで相互作用しているもの(イタリック表記のアミノ酸)は、どのアミノ酸でも同様な構造を持っているため改変はできない。そのため、サブユニット間の相互作用の改変には側鎖部分が相互作用に関与するGlu39、Asp169、Arg170、Arg220、Tyr227、Glu330への変異導入が有効と考えられた。また、そのうちArg170−Asp169間の結合はイオン結合のため、他の水素結合での相互作用に比べ結合が強く、特に変異導入で大きな効果あることが推定された。また、A−B間の相互作用部位を拡大したリボンモデルを図2に示すが、特に、Arg170、Arg220、Tyr227の残基がインターフェイスエリアが大きく、また結合距離も近いため、変異の導入部位として有力と考えられた。
【0069】
(実施例2)変異体の調製
本発明のβグルコシダーゼ(変異体)は、表2に示すプライマーを使用し、以下の手法で作製した。
【0070】
【表2】
【0071】
配列番号1で示されるアミノ酸配列をコードする遺伝子に対して、配列番号3および4の塩基配列で示されるオリゴヌクレオチドを使用して部位突然変異法を用いてR170A変異体(配列番号16)を作製した。また同様に、配列番号5および6の塩基配列で示されるオリゴヌクレオチドを使用してR220A変異体(配列番号17)、配列番号7および8を使用してY227F変異体(配列番号18)を作製した。また、R170A変異体に対し、配列番号7および8の塩基配列で示されるオリゴヌクレオチドを使用して、R170A/Y227F変異体(配列番号19)を作製した。R170A変異体に対し、配列番号5および6の塩基配列で示されるオリゴヌクレオチドを使用して、R170A/R220A変異体(配列番号21)を作製した。R170A/R220A変異体に対し、配列番号7および8の塩基配列で示されるオリゴヌクレオチドを使用してR170A/R220A/Y227F変異体(配列番号20)を作製した。
【0072】
得られた各遺伝子は、参考例1記載の手順で大腸菌にて発現を実施した。R170A変異体、R220A変異体、Y227F変異体、R170A/Y227F変異体、R170A/R220A変異体、R170A/R220A/Y227F変異体のアミノ酸配列を配列番号9〜14に示す(スタートコドンであるメチオニンは含まない)。前述変異体は、大腸菌にてすべて異種タンパク質として発現可能であることが確認できた。
【0073】
(実施例3)変異体のセロビオース分解活性1
実施例2で得られた変異体(本発明のβグルコシダーゼ)と、参考例1で調整した親βグルコシダーゼのセロビオース分解活性を以下実験にて比較した。基質に10mM セロビオース/50mM 酢酸緩衝液を用い、それぞれ実施例2、参考例1で調製した酵素を終濃度0.23mg/mLで添加し、50℃で酵素反応を30分行った。親βグルコシダーゼにおける50℃の温度条件下で生成したグルコース濃度(g/L)を100%として、各変異体におけるセロビオース分解活性を相対値で表3に示す。
【0074】
【表3】
【0075】
50℃においては、親βグルコシダーゼおよび各変異体で活性に差がないことを確認した。
【0076】
(実施例5)変異体のセロビオース分解活性2
セルロース含有バイオマス存在下での親グルコシダーゼおよび変異体(本発明のβグルコシダーゼ)のセロビオース分解活性を測定した。セルロース含有バイオマスとしては、稲藁を5%希硫酸で150℃10分処理したものを使用した。セルロース含有バイオマス5wt%を懸濁した溶液に、10mM セロビオース/50mM 酢酸緩衝液を添加し反応液とした。この反応液に実施例2、参考例1で調製した酵素を終濃度0.23mg/mLで添加し、50℃で酵素反応を30分行った。親βグルコシダーゼにおける50℃の温度条件下で生成したグルコース濃度(g/L)を100%として、各変異体におけるセロビオース分解活性を相対値で表4に示す。
【0077】
【表4】
【0078】
親グルコシダーゼおよびR170A変異体、R220A変異体、Y227F変異体、R170A/R220A変異体では、セルロース含有バイオマス存在下でのセロビオース分解活性にほとんど差がないことが確認できた。一方でR170A/R220A/Y227F変異体では、親グルコシダーゼあるいはその他の変異体に対して活性が大きく向上することが確認された。
【0079】
(実施例6)ブルー・ネイティブPAGEによる変異体の分子量測定
親βグルコシダーゼ、及び本発明のβグルコシダーゼであるR170A変異体、R170A/R220A変異体、およびR170A/R220A/Y227F変異体が安定な4量体を形成するか否か、ブルー・ネイティブPAGEにより検討を行った。
【0080】
ブルー・ネイティブPAGEは3−12%Native−PAGEビストリスゲル(インビトロジェン)を使用した。親βグルコシダーゼ、R170A変異体、R170A/R220A変異体、およびR170A/R220A/Y227F変異体各1μgを、サンプルバッファー(50mMイミダゾール(pH7.0)、50mM NaCl、5mM 6−アミノヘキサン酸、0.5%クマシーG250、0.5%DDM)に溶解し泳動サンプルとした。分子量マーカーは、NativeMarkプロテインスタンダード(インビトロジェン)を使用した。電気泳動は陽極側に、50mMトリシン、7.5mMイミダゾール(pH7.0)、クマシーG250、陰極側に、25mMイミダゾール(pH7.0)を添加し、150V定電圧で100分電気泳動を行った。バンドの固定化後、G−250の脱色を行った。得られたゲルの写真を図3に示した。
【0081】
親βグルコシダーゼ、R170A変異体およびR170A/R220A変異体に関しては、分子量マーカー146kDaから242kDaの間のほぼ同じ位置にそれぞれのバンドが検出された。親βグルコシダーゼは、4量体形成時の分子量は216kDaであることから、これらは安定な4量体を形成することが確認できた。一方で、本発明のR170A/R220A/Y227F変異体は、分子量マーカー66kDaから146kDaに検出された。すなわち、本発明のR170A/R220A/Y227F変異体は、本来親βグルコシダーゼが形成する4量体形成が不安定化しており、3量体あるいは2量体で水溶液中に存在することが確認された。
【0082】
(実施例7)βグルコシダーゼを含むバイオマス分解用酵素組成物を用いたセルロース含有バイオマスの加水分解
トリコデルマ・リーセイ由来セルラーゼ(セルクラスト、シグマ)、及びグルコシダーゼとして、それぞれ比較例1で調製した親グルコシダーゼ、実施例2で調製した本発明のR170A変異体、R170A/R220A変異体、R170A/R220A/Y227F変異体、とからなるバイオマス分解用酵素組成物を用いた。酵素添加量は、セルラーゼ0.5mg/mL、各種グルコシダーゼ0.005mg/mL(セルラーゼの1/100量)となるよう混合した。
【0083】
セルロース含有バイオマスとしては、稲藁を5%希硫酸で150℃10分処理したものを使用した。加水分解は、50mM 酢酸緩衝液(pH5.0)に5wt%のセルロース含有バイオマスを懸濁したものを基質とした。反応は50℃において28時間まで行い、生成したグルコース濃度の測定を行った。
【0084】
【表5】
【0085】
グルコシダーゼなしに対して、グルコシダーゼを添加することで、グルコース生成量が増大することが確認できた。本発明のR170A/R220A/Y227F変異体では、親βグルコシダーゼ、R170A変異体、A170A/R220A変異体に比べて、反応28時間後におけるグルコース生成量は増えることが確認できた。セルロース含有バイオマスが存在しない時のセロビオース分解活性は、R170A/R220A/Y227F変異体、親βグルコシダーゼ、R170A変異体、A170A/R220A変異体、の間で差はなかった(実施例3)。したがって、本発明の変異体、R170A/R220A/Y227F変異体は、セルロース含有バイオマス存在下での活性が親βグルコシダーゼに対して向上していることが確認できた。
【0086】
(実施例8)R170A/R220A/Y227F変異体の結晶構造解析
R170A/R220A/Y227F変異体の三次元構造を決定するため、X線結晶構造解析を試みた。新たな結晶化条件を探索し、リン酸を沈殿剤として結晶化に成功した。大型放射光実験施設SPring−8でX線回折実験を行い、分解能2.5オングストロームでPfuBGLの構造を決定した。構造決定は分子置換法を用い、モデル分子には配列番号22で示されるアミノ酸配列を有するサーモスファエラ・アグリガンズ(Themosphaera aggregans)由来β−グリコシダーゼ(ThAggBGY、PDB ID:1QVB)を用いた。得られた3次元構造データを構造解析用のソフトウェアCCP4_Contactを用いてサブユニット間の相互作用を分析した。分析の結果得られたリボンモデルを図4に示す。サブユニットA−C間で相互作用しており、R170A/R220A/Y227F変異体は、ホモ2量体を形成していることが確認された(図4)。分析結果より得られたサブユニットA−C間の相互作用に関し、表6にまとめた。
【0087】
【表6】
【0088】
すなわち、R170A/R220A/Y227F変異体は、Arg448、Leu440、Glu449、Glu459のアミノ酸残基にて水素結合し、2量体を形成していることが判明した。
【0089】
(実施例9)変異体の調製2
本発明の変異型βグルコシダーゼ(R170A/R220A/Y227F変異体)は、実施例8に示すとおり2量体を形成することが判明した。そこで、R170A/R220A/Y227F変異体に対しさらに、Arg448、Glu449、Glu459のいずれか1以上の変異をさらに導入することで、2量体を単量体に改変できるか検討を行った。変異体遺伝子の取得に関しては、表7に示すプライマーを使用し、以下の手法で作製した。
【0090】
【表7】
【0091】
R170A/R220A/Y227F変異体の遺伝子(配列番号20)に対して、配列番号23および24の塩基配列で示されるオリゴヌクレオチド、配列番号25および26の塩基配列で示されるオリゴヌクレオチド、配列番号27および28の塩基配列で示されるオリゴヌクレオチド、配列番号29および30の塩基配列で示されるオリゴヌクレオチド、をそれぞれ使用し、R170A/R220A/Y227F/R448E変異体(配列番号31)、R170A/R220A/Y227F/R448G変異体(配列番号32)、R170A/R220A/Y227F/E449R変異体(配列番号33)、R170A/R220A/Y227F/E459G変異体(配列番号34)遺伝子を作製した。
【0092】
得られた各遺伝子は、参考例1記載の手順で大腸菌にて発現を実施した。R170A/R220A/Y227F/R448E変異体、R170A/R220A/Y227F/R448G変異体、R170A/R220A/Y227F/E449R変異体、R170A/R220A/Y227F/E459G変異体のアミノ酸配列を配列番号35〜38に示す(スタートコドンであるメチオニンは含まない)。前述の変異体は、大腸菌にてすべて異種タンパク質として発現可能であることが確認できた。
【0093】
(実施例10)ゲルろ過による分子量の決定
親グルコシダーゼおよび変異体の分子量を決定するために、ゲルろ過による分子量の決定を行った。カラムはHiLood26/60Suuperdex200を使用し、緩衝液は、50mMトリス塩酸塩(pH8)、150mM塩化ナトリウムを使用した。実施例2、参考例1、実施例9で調製した酵素をカラムに添加し、緩衝液2mL/minで分離した。酵素の検出は、吸光度280nmを測定した。分子量は、オブアルブミン(44kDa)、コンアルブミン(75kDa)、アルドラーゼ(158kDa)、フェリチン(440kDa)を分子量マーカーとして使用し、ぞれぞれの酵素成分の溶出時間を元に検量線を作製した。ゲルろ過の結果、測定された分子量(実測分子量、kDa)およびアミノ酸一次構造より推定される分子量(推定分子量、kDa)をそれぞれ表8にまとめた。
【0094】
【表8】
【0095】
親グルコシダーゼに関しては、ほぼ4量体の実測分子量(237kDa)、R170A/R220A/Y227F変異体は、ほぼ2量体の実測分子量(129kDa)を示すことが判明した。一方、R170A/R220A/Y227F変異体に対してさらに変異導入した変異体、R170A/R220A/Y227F/R448E変異体、R170A/R220A/Y227F/E449R変異体、R170A/R220A/ Y227F/R448G変異体、R170A/R220A/Y227F/E459G変異体、に関しては、実測分子量が50kDa〜90kDaの範囲にあり、さらに変異を導入する効果として、完全に単量体化したものと推定された。
【0096】
(実施例11)変異体のセロビオース分解活性3
セルロース含有バイオマス存在下での親グルコシダーゼおよび変異体(本発明のβグルコシダーゼ)のセロビオース分解活性を測定した。セルロース含有バイオマスとしては、稲藁を5%希硫酸で150℃10分処理したものを使用した。セルロース含有バイオマス5wt%を懸濁した溶液に、10mM セロビオース/50mM 酢酸緩衝液を添加し反応液とした。この反応液に実施例9で調製した酵素R170A/R220A/Y227F/R448E変異体、R170A/R220A/Y227F/E449R変異体、R170A/R220A/Y227F/R448G変異体、R170A/R220A/Y227F/E459G変異体、を終濃度0.23mg/mLで添加し、50℃で酵素反応を30分行った。親βグルコシダーゼにおける50℃の温度条件下で生成したグルコース濃度(g/L)を100%として、各変異体におけるセロビオース分解活性を相対値で表9に示す。
【0097】
【表9】
【0098】
R170A/R220A/Y227F/R448E変異体、R170A/R220A/Y227F/E449R変異体、R170A/R220A/ Y227F/R448G変異体、R170A/R220A/Y227F/E459G変異体では、親グルコシダーゼに対して活性が大きく向上することが判明した。また実施例5のR170A/R220A/Y227F変異体(2量体)に対して、実施例9の変異型グルコシダーゼ(単量体)の方が高いことが判明した。
【産業上の利用可能性】
【0099】
本発明におけるβグルコシダーゼは、セルロース含有バイオマス存在下で高いセロビオース分解活性を示すため、バイオマスの加水分解および糖液の製造に使用できる。
【特許請求の範囲】
【請求項1】
各単量体の間で水素結合またはイオン結合を形成するアミノ酸の位置に、βグルコシダーゼの4量体形成を不安定化させる変異を有し、かつ野生型と同等またはそれ以上のセロビオース分解活性を有することを特徴とする、変異型βグルコシダーゼ。
【請求項2】
配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、39番目のグルタミン酸、169番目のアスパラギン酸、170番目のアルギニン、220番目のアルギニン、227番目のチロシンおよび330番目のグルタミン酸からなる群から選ばれる少なくとも3以上のアミノ酸に相当するアミノ酸の変異を含む、請求項1に記載の変異型βグルコシダーゼ。
【請求項3】
配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、39番目のグルタミン酸、169番目のアスパラギン酸、170番目のアルギニン、220番目のアルギニン、227番目のチロシンおよび330番目のグルタミン酸からなる群から選ばれる少なくとも3以上のアミノ酸に相当するアミノ酸の、中性側鎖を有するアミノ酸への置換を含む、請求項1または2に記載の変異型βグルコシダーゼ。
【請求項4】
配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、170番目のアルギニン、220番目のアルギニンおよび227番目のチロシンに相当するアミノ酸の、中性側鎖を有するアミノ酸への置換を含む、請求項1〜3のいずれか1項に記載の変異型βグルコシダーゼ。
【請求項5】
中性側鎖を有するアミノ酸が、アラニン、フェニルアラニンおよびグリシンからなる群から選択される、請求項3または4に記載の変異型βグルコシダーゼ。
【請求項6】
配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の170番目のアルギニンに相当するアミノ酸のアラニンへの置換、配列番号1のアミノ酸配列の220番目のアルギニンに相当するアミノ酸のアラニンへの置換および配列番号1のアミノ酸配列の227番目のチロシンに相当するアミノ酸のフェニルアラニンへの置換を含む、請求項1〜5のいずれか1項に記載の変異型βグルコシダーゼ。
【請求項7】
配列番号14で示されるアミノ酸配列を含む、請求項1〜6のいずれか1項に記載の変異型βグルコシダーゼ。
【請求項8】
配列番号1のアミノ酸配列の440番目のロイシン、448番目のアルギニン、449番目のグルタミン酸、459番目のグルタミン酸からなる群から選ばれる1以上のアミノ酸に相当するアミノ酸の変異をさらに含む、請求項1〜7のいずれか1項に記載の変異型βグルコシダーゼ。
【請求項9】
配列番号1のアミノ酸配列の448番目のアルギニンに相当するアミノ酸の中性側鎖もしくは酸性側鎖を有するアミノ酸への置換、配列番号1のアミノ酸配列の449番目のグルタミン酸に相当するアミノ酸の中性側鎖もしくは塩基性側鎖を有するアミノ酸への置換および配列番号1のアミノ酸配列の459番目のグルタミン酸に相当するアミノ酸の中性側鎖もしくは塩基性側鎖を有するアミノ酸への置換からなる群から選ばれる1以上の置換を含む、請求項8に記載の変異型βグルコシダーゼ。
【請求項10】
配列番号35〜38のいずれかで示されるアミノ酸配列を含む、請求項8又は9に記載の変異型βグルコシダーゼ。
【請求項11】
請求項1〜10のいずれか1項に記載の変異型βグルコシダーゼをコードするDNA。
【請求項12】
配列番号20、配列番号31〜34のいずれかで示される塩基配列を含む、請求項11に記載のDNA。
【請求項13】
請求項11または12に記載のDNAを含む、発現ベクター。
【請求項14】
請求項13に記載の発現ベクターを用いた形質転換により作製された、形質転換細胞。
【請求項15】
請求項1〜10のいずれか1項に記載の変異型βグルコシダーゼまたは請求項14に記載の形質転換細胞の処理物を含むバイオマス分解用酵素組成物。
【請求項16】
セルロース含有バイオマスを請求項15に記載のバイオマス分解用酵素組成物を使用して加水分解することを含む、セルロース由来バイオマスより糖液を製造する方法。
【請求項1】
各単量体の間で水素結合またはイオン結合を形成するアミノ酸の位置に、βグルコシダーゼの4量体形成を不安定化させる変異を有し、かつ野生型と同等またはそれ以上のセロビオース分解活性を有することを特徴とする、変異型βグルコシダーゼ。
【請求項2】
配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、39番目のグルタミン酸、169番目のアスパラギン酸、170番目のアルギニン、220番目のアルギニン、227番目のチロシンおよび330番目のグルタミン酸からなる群から選ばれる少なくとも3以上のアミノ酸に相当するアミノ酸の変異を含む、請求項1に記載の変異型βグルコシダーゼ。
【請求項3】
配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、39番目のグルタミン酸、169番目のアスパラギン酸、170番目のアルギニン、220番目のアルギニン、227番目のチロシンおよび330番目のグルタミン酸からなる群から選ばれる少なくとも3以上のアミノ酸に相当するアミノ酸の、中性側鎖を有するアミノ酸への置換を含む、請求項1または2に記載の変異型βグルコシダーゼ。
【請求項4】
配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の、170番目のアルギニン、220番目のアルギニンおよび227番目のチロシンに相当するアミノ酸の、中性側鎖を有するアミノ酸への置換を含む、請求項1〜3のいずれか1項に記載の変異型βグルコシダーゼ。
【請求項5】
中性側鎖を有するアミノ酸が、アラニン、フェニルアラニンおよびグリシンからなる群から選択される、請求項3または4に記載の変異型βグルコシダーゼ。
【請求項6】
配列番号1のアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を有し、該90%以上の配列同一性を有するアミノ酸配列において、配列番号1のアミノ酸配列の170番目のアルギニンに相当するアミノ酸のアラニンへの置換、配列番号1のアミノ酸配列の220番目のアルギニンに相当するアミノ酸のアラニンへの置換および配列番号1のアミノ酸配列の227番目のチロシンに相当するアミノ酸のフェニルアラニンへの置換を含む、請求項1〜5のいずれか1項に記載の変異型βグルコシダーゼ。
【請求項7】
配列番号14で示されるアミノ酸配列を含む、請求項1〜6のいずれか1項に記載の変異型βグルコシダーゼ。
【請求項8】
配列番号1のアミノ酸配列の440番目のロイシン、448番目のアルギニン、449番目のグルタミン酸、459番目のグルタミン酸からなる群から選ばれる1以上のアミノ酸に相当するアミノ酸の変異をさらに含む、請求項1〜7のいずれか1項に記載の変異型βグルコシダーゼ。
【請求項9】
配列番号1のアミノ酸配列の448番目のアルギニンに相当するアミノ酸の中性側鎖もしくは酸性側鎖を有するアミノ酸への置換、配列番号1のアミノ酸配列の449番目のグルタミン酸に相当するアミノ酸の中性側鎖もしくは塩基性側鎖を有するアミノ酸への置換および配列番号1のアミノ酸配列の459番目のグルタミン酸に相当するアミノ酸の中性側鎖もしくは塩基性側鎖を有するアミノ酸への置換からなる群から選ばれる1以上の置換を含む、請求項8に記載の変異型βグルコシダーゼ。
【請求項10】
配列番号35〜38のいずれかで示されるアミノ酸配列を含む、請求項8又は9に記載の変異型βグルコシダーゼ。
【請求項11】
請求項1〜10のいずれか1項に記載の変異型βグルコシダーゼをコードするDNA。
【請求項12】
配列番号20、配列番号31〜34のいずれかで示される塩基配列を含む、請求項11に記載のDNA。
【請求項13】
請求項11または12に記載のDNAを含む、発現ベクター。
【請求項14】
請求項13に記載の発現ベクターを用いた形質転換により作製された、形質転換細胞。
【請求項15】
請求項1〜10のいずれか1項に記載の変異型βグルコシダーゼまたは請求項14に記載の形質転換細胞の処理物を含むバイオマス分解用酵素組成物。
【請求項16】
セルロース含有バイオマスを請求項15に記載のバイオマス分解用酵素組成物を使用して加水分解することを含む、セルロース由来バイオマスより糖液を製造する方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2012−34690(P2012−34690A)
【公開日】平成24年2月23日(2012.2.23)
【国際特許分類】
【出願番号】特願2011−155792(P2011−155792)
【出願日】平成23年7月14日(2011.7.14)
【出願人】(000003159)東レ株式会社 (7,677)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【公開日】平成24年2月23日(2012.2.23)
【国際特許分類】
【出願日】平成23年7月14日(2011.7.14)
【出願人】(000003159)東レ株式会社 (7,677)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]