
変異型キシリトールデヒドロゲナーゼ酵素、これを産生する微生物、該酵素または微生物を用いたキシリトールをキシルロースに変換する方法
【課題】 野生型キシリトールデヒドロゲナーゼ(XDH)の補酵素要求性をニコチンアミドアデニンジヌクレオチドリン酸(NADP)型に改良するとともに耐熱性を向上させ、キシリトールからキシルロースへの変換効率を高めた変異型XDHを提供すること。
【解決手段】 野生型XDHの補酵素要求性をNADP要求性に変えるために、そのアミノ酸配列の207番目のアスパラギン酸をアラニンに、208番目のイソロイシンをアルギニンに、209番目のフェニルアラニンをスレオニンもしくはチロシンに、211番目のアスパラギンをアルギニンに置換し、耐熱性を向上させるために、96番目のセリンをシステインに、99番目のセリンをシステインに、102番目のチロシンをシステインに置換して、構造安定化亜鉛結合部位を導入する。
【解決手段】 野生型XDHの補酵素要求性をNADP要求性に変えるために、そのアミノ酸配列の207番目のアスパラギン酸をアラニンに、208番目のイソロイシンをアルギニンに、209番目のフェニルアラニンをスレオニンもしくはチロシンに、211番目のアスパラギンをアルギニンに置換し、耐熱性を向上させるために、96番目のセリンをシステインに、99番目のセリンをシステインに、102番目のチロシンをシステインに置換して、構造安定化亜鉛結合部位を導入する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、キシリトールを高効率でキシルロースへ変換することができる変異型キシリトールデヒドロゲナーゼ(Xylitol dehydrogenase; 以下XDH)酵素、これをコードする遺伝子、該遺伝子を含むベクター、該ベクターを含む微生物、及び該酵素または該微生物を用いてキシリトールを高効率でキシルロースに変換する方法を提供するものである。さらに詳細には本発明は、木質などに含まれるセルロース、ヘミセルロースのうち、これまで利用する手段が少なかったヘミセルロース中に多量に含まれているキシロースを、バイオマス液体燃料として注目されているエタノールへと変換する酵素系の中で、これまでキシリトールからキシルロースへの変換効率が低かったキシリトールデヒドロゲナーゼ(XDH)酵素のアミノ酸配列を変異させ、補酵素要求性をニコチンアミドアデニンジヌクレオチド(NAD+)からニコチンアミドアデニンジヌクレオチドリン酸(NADP+)要求性へと改良し、および/または、構造安定化亜鉛結合部位を導入して耐熱性を改良したXDHを提供すること、これを用いてキシリトールからキシルロースへの高効率変換を可能とし、これによりキシロースを高効率でエタノールへと変換する方法を提供するものである。
【背景技術】
【0002】
地球上のバイオマスの大部分を占める木質系バイオマスは、リグノセルロースから構成されており、リグノセルロースはセルロース(約45%)とヘミセルロース(約30%)、リグニン(約25%)に分類される。このうち、セルロースやリグニンは短時間で超臨界水を用いたグルコースへの変換などにより、近年、盛んにそのエネルギー利用が研究されている。ヘミセルロースは酸加水分解あるいは酵素分解によって容易にキシロースなどの単糖類に分解することが出来る。また、ヘミセルロースの大部分(非特許文献1参照)を占め、容易に調達できるキシロースを液体燃料へと効率よく変換させることは、エネルギー問題の点からも重要な課題と考えられている。
これまで、酵母などの多くの微生物を用いてセルロースなどのヘキソースをエタノールへと嫌気的に効率よく変換させることには成功しているが、キシロースなどのペントースは高い効率で変換できないことが知られている。
【0003】
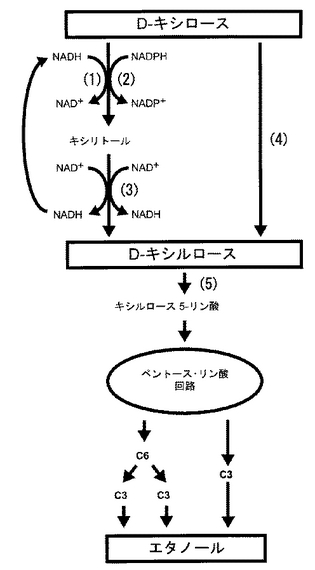
キシロースをエタノールへと変換する経路として、図1に示したように、主に二つの経路が知られている。一つは、キシロースからキシルロースへとキシロースイソメラーゼ(Xylose isomerase; 以下XI)により補酵素に依存せずに一段階で変換する方法である。もう一つは、キシロースリダクターゼ(Xylose reductase; 以下XR)によりキシロースをキシリトールへと変換させた後、キシリトールをキシリトールデヒドロゲナーゼ(XDH)によりキシルロースへと変換する方法である。この時、変換には補酵素を必要とする。両方法とも、キシロースをキシルロースへと変換する事が出来れば、ペントース・リン酸回路を経由してエタノールへと容易に変換することが出来る。
【0004】
数多くのバクテリアや菌類で、キシロースからキシルロースへとXIにより一段階で変換する経路を通って、キシロースをエタノールへと変換できることが知られている。例えば、Streptomyces sp.(ストレプトミセス)やActinoplanes sp.(アクチノプラネス)のようなバクテリアは、XIによりキシロースをキシルロースへと変換した後、ペントース・リン酸回路を経てエタノールへと変換できることが知られている(非特許文献2参照)。しかし、その効率は非常に低い(特許文献1参照)。これは、有機酸が副生成物として生成することが原因ではないかと考えられている(特許文献2参照)。これらのことから、キシロースからキシルロースへとXIにより一段階で変換する方法は工業的に利用されるには至っていない。
【0005】
二段階でキシロースからキシルロースへと変換する微生物として、Pichia stipitis(ピチア スチピチス)、Candida shehatae(カンジダ シェハタエ)、Pachysolen tannophilus(パチソレン タンフィルス)等が知られている(非特許文献3,4参照)。これらの微生物を用いることで、キシロースをエタノールへと変換することにある程度の成功をしている。しかし、これらの微生物もしくは酵素を用いた方法は、純粋なキシロースを用いた場合の結果であり、キシロース以外の化学物質の存在下で、キシロースをキシルロースへと変換させた場合、その収率は大きく低下することが知られている(非特許文献5,6参照)。通常、木質などからキシロースを生産する際、種々の化学物質を使用する。これらの物質を取り除くことは不可能ではないが、この操作により、大幅なコスト増となる。
微生物を用いてキシロースをキシリトールへと変換させることは、これまでに多くの成功例がある(非特許文献7,8参照)。従って、キシリトールからキシルロースを効率よく変換できる酵素系もしくはその酵素系を有する微生物を用いれば、高効率でキシロースをエタノールへと変換できる可能性がある。
【0006】
遺伝子工学的手法により酵母の遺伝子を変異させ、キシロースをエタノールへと高効率で変換させる検討も行われてきた。例えば、C. S. Gongら(特許文献3参照)は、紫外線照射により遺伝子を変異させた酵母を用いて、キシロースを代謝させることに成功している。しかし、どの遺伝子を変異させたかなどの遺伝子工学的な情報は何ら開示されていない。
キシロースをエタノールに変換させるために、キシロースをキシルロースへと補酵素に依存せずに直接変換できるXI遺伝子を大腸菌や酵母に導入した例も報告されている(非特許文献9参照)。しかし、大腸菌や酵母に、これらの遺伝子を導入し、発現させた場合の変換効率は非常に低かった。
【0007】
酵母、Saccharomyces cerevisiae(サッカロミセス セレビシエ)は、グルコースを高効率でエタノールに変換できることが知られているが、出発原料としてキシロースを用いた時には、エタノールに変換することは出来ない。これは、S. cerevisiaeがキシロースをキシルロースに代謝することが出来ないことが原因であり、もしこれを代謝することが出来れば、この微生物が持つ高いエタノール生産能力、高いエタノール耐性能力を用い、非常に効率良くキシロースをエタノールに変換できるのではないかと考えられ、キシロースをキシルロースへと変換する際の酵素系の改良が種々検討されてきた。しかし、S. cerevisiaeのXR(非特許文献10,11参照)およびP. stipitisのXDH(非特許文献12参照)の遺伝子を変異させ、その遺伝子を酵母に導入し、嫌気性条件下でキシロースからエタノールを得ることに成功したことが報告されているが(非特許文献13−15参照)(特許文献4,5参照)、その変換効率は低く、工業的生産にまでは至っていない。
【0008】
タンパク質、特に工業的に有用な酵素の安定化は実用化に向けた至上命題の1つである。無機触媒に比べて生体高分子である酵素は一般に熱に弱く、その失活の多くは不可逆である。安定性を上昇させ耐久寿命を延ばすことで頻繁な酵素の添加を防ぐことができ、大幅な経費節減を実現できる。安定化した酵素は、種々の分野に応用されている。例えば、洗剤などに添加するタンパク質分解酵素、固定化酵素、アフィニティークロマトグラフィーなどである。安定化を行う方法としては、アミノ酸配列を無作為に変異させた後、高い安定性を持つ酵素をスクリーニングする方法、耐熱性を有する酵素のアミノ酸配列、構造などを参考に酵素のアミノ酸配列を改変させる方法、固定支持体に酵素を結合させる方法、高分子架橋型ゲルで酵素を包み込む方法等、種々の方法が考案されている。
【0009】
アミノ酸配列を変異させることで酵素の改変を行う方法は、大きく2つに分類できる。
第一の方法は、無作為にアミノ酸を置換させるランダム変異(Random mutagenesis)である。遺伝子シャフリング(gene-shuffling)やエラープローン(error-prone)PCRを用いて膨大な酵素変異体プールを構築し、その中から目的の性質に改変された変異体をスクリーニングする方法である。この方法の最大の利点は多くの変異体集団が効率的に得られる点にあり、ターゲット酵素の活性部位等の情報が全く無い場合にも適応できる可能性さえある。一方で、適切なスクリーニング系が必須なのが欠点である。例えば、安定化した中温性酵素変異体を得る場合であれば、そのホモログ酵素をコードする遺伝子の欠損した好熱菌株に構築した変異体プールを導入し、高温下での栄養要求性の復帰等でセレクション(選択)する。
【0010】
第二の方法は、変異させる部位を最初から特定する部位特異的変異(Site-directed mutagenesis)である。この方法では、ターゲット酵素そのものか少なくともそのホモログの高次構造や反応機構に関する情報に基づいて、変異させるべき部位を決定する。ランダム変異に比べて変異体作製の効率は大幅に下がり変異体の性質が予想通りに改変される保証もないが、上記のようなセレクション系が無い場合や他の類似酵素で成功例がある場合はランダム変異よりもむしろ近道なケースもある。
部位特異的変異による酵素の熱安定性の向上の研究は、主に異なる温度で生育する生物から単離されたホモログ酵素のアミノ酸配列の比較から始まった。好熱性生物由来の酵素には、特定のアミノ酸残基の含有率の偏りや全体の電荷の分布、またループの挿入や欠損などに一定の特徴が見られる。また最近では好熱性、中温性、好冷性酵素の結晶構造の比較から、水素結合や疎水性コアなどの分子内相互作用や分子表面の面積や電荷分布の詳細な比較が可能になり、こうした情報に基づいて部位特異的変異体が設計されるようになっている。しかしながら、現在のところではそうした改変の統一原理というものはなく、ターゲット酵素ごとに安定性向上に使われる戦略は異なっている。
【0011】
部位特異的変異の“決め打ち”として最も成功を収めた例としては、バクテリオファージ・T4リゾチームへのジスルフィド結合の導入を挙げることができる。構造情報から注意深く選ばれた部位に最終的に3本のジスルフィド結合を導入し、野生型酵素に比べて23℃もTm値(含まれるα-へリックス構造の半分が崩壊する温度)の上昇した変異体を得ることに成功した(非特許文献16)。このような共有結合の導入は、ランダム変異ではまず起こり得ないであろうし、その期待できる効果は少数の水素結合の追加に比べてずっと大きいだろう。これに準ずるような変異導入の戦略は、安定性の向上した部位特異的変異体の効率的な構築に応用可能である。
【0012】
アルコール脱水素酵素(ADH)は、最も大きなタンパク質ファミリーの1つである。この酵素が触媒する化学反応はいたって簡単である。基質のアルコールは、そのヒドロキシル基が結合している炭素原子の水素が酸化型補酵素NAD(P)+へ転移し(hydryl-transfer)、ヒドロキシル基からプロトンが除去されアルデヒドに酸化される。この酵素は亜鉛原子を活性中心に持ち(触媒亜鉛(catalytic zinc))、これは3つの高度に保存されたアミノ酸残基と1つの水分子によって配位している。この3つの残基はどれか1つが欠けてもその酵素活性に致命的な影響がある。一方、この触媒亜鉛(catalytic zinc)に加えてもう1つの亜鉛分子を持つADHが数多く知られている。この亜鉛には4つの保存されたシステイン残基が配位しており、構造亜鉛(structural zinc)(この明細書においては「構造安定化亜鉛」と称することもある。)と呼ばれる。構造亜鉛(structural zinc)の有無と、ADHの由来となる生物種やその高次構造の間には明瞭な関係はない。触媒亜鉛(catalytic zinc)に比べて構造亜鉛(structural zinc)の役割はよく分かっていない。実際、構造亜鉛(structural zinc)の有無によらずその結合しているループの構造は高度に保存されている。しかしながら、2つのADHファミリー酵素で4つのシステイン残基のどれかひとつを変異させても、触媒亜鉛同様その酵素活性に致命的な影響があることが報告されており、何らかの高次構造維持に役立っていると考えられてきた。
【0013】
【特許文献1】米国特許第4,368,268号明細書
【特許文献2】米国特許第1,857,429号明細書
【特許文献3】米国特許第4,511,656号明細書
【特許文献4】米国特許第6,582,944号明細書
【特許文献5】米国特許第5,866,382号明細書
【特許文献6】欧州特許第0329822号明細書
【0014】
【非特許文献1】M. R. Ladischら (1983) Enzyme Microb. Technol., 5: 82-102
【非特許文献2】K. Skoogら (1988) Enzyme Microb. Technol., 10: 66-78
【非特許文献3】P. J. Sliningerら (1987) Enzyme Microb. Technol., 9: 5-15
【非特許文献4】B. A. Priorら (1989) Proc. Biochem., 2: 21-32
【非特許文献5】T. Lindenら (1989) Enzyme Microb. Technol., 11: 583-589
【非特許文献6】T. Lindenら (1989) Biotechnol. Techniques, 3: 189-192
【非特許文献7】H. Onishiら (1966) Agr. Biol. Chem., 30: 1139-1144
【非特許文献8】M. F. S. Barbosaら (1988) J. Ind. Microbiol., 3: 241-251
【非特許文献9】A. V. Sarthyら (1987) Appl. Environ. Microbiol., 53: 1996-2000
【非特許文献10】Takamaら (1991) Appl. Biochem. Biotechnol., 327: 27-28
【非特許文献11】Hallbornら (1991) Bio Technol., 9: 1090
【非特許文献12】Koetterら (1990) Curr. Gen et., 18: 493-500
【非特許文献13】Y. S. Jinら (2000) J. Microbiol. Biotechnol., 10:564-567
【非特許文献14】Koetterら (1990) Curr. Gen et., 18: 493-500
【非特許文献15】Hallbornら (1991) Bio Technol., 9: 1090
【非特許文献16】Matsumuraら (1989) Nature, 342: 291
【発明の開示】
【発明が解決しようとする課題】
【0015】
木質などのバイオマス資源のうち、30%以上を占めるヘミセルロースを加水分解することによって容易に得ることが出来るキシロースを、高効率でエタノールへと変換し、エネルギー問題を解決するための1つの手段を提供することは重要な課題である。そのためには、セルロースなどのヘキソースを高効率でエタノールへと変換でき、エタノール耐性の高い酵母に、キシロースなどのペントースを原料として高効率でエタノールへと変換できる能力を与えるのが最も理想的である。これまでに、キシロースをキシリトールへと高効率で変換できる酵母は開発されているが、キシリトールをキシルロースへと高効率で変換するXDHおよび酵母は開発されていない。
【0016】
本発明の課題は、野生型のXDHを参考に高効率でキシリトールをキシルロースへと変換するXDHを開発することである。XRは、前述のように、高効率でキシロースをキシリトールに変換することが出来るが、この際に補酵素として、NADP+を使用する。しかし、XDHは補酵素としてNAD+を使用する。この様に両酵素の補酵素に対する要求性が異なっていることが、補酵素供給の量的バランスを崩し、その結果、キシリトールからキシルロースへの変換が効率よく進行せず、最終的にキシロースからエタノールへの変換効率が低くなると推定される。これらのことを考慮すると、XDHの補酵素要求性を変えて、XRと同じNADP+要求性にすれば、高効率でキシリトールをキシロースに変換でき、この部分が効率よく進行すれば、キシロースをエタノールに高い効率で変換できると考えられる。
従って、本発明の第1の目的は、XDH遺伝子を遺伝子工学的に変異させて、補酵素NADP+要求性の変異型XDHを提供すること、及び該変異型XDHをコードする遺伝子を提供することである。
【0017】
また、酵素を工業的規模で使用する場合には、その酵素の安定性を向上させることは、非常に重要な課題と考えられる。これは、酵素を微生物中に組み込んで使用する場合でも、酵素を固定支持担体に結合させて使用する固定化酵素として使用する場合でも同様である。酵素は、化学物質、例えば、本発明の場合、キシロース、キシリトールやエタノール等や熱に安定である方が好ましい。特に、熱に対する安定性は、系の反応温度が高いほど一般的に反応速度が高く、単位時間あたりの収量が多くなるので重要である。また、微生物に酵素を組み込んだ場合、一般に代謝の進行に伴って培養槽の温度が高くなり、温度上昇に伴って酵素が失活するので、この点からも高い耐熱能力を酵素に持たせることは非常に重要である。
従って本発明の第2の目的は、高い耐熱性を持つ変異型XDHを提供すること、及び該変異型XDHをコードする遺伝子を提供することである。
本発明の他の目的は、キシリトールを高効率でキシルロースへ変換することができる変異型XDHをコードする遺伝子を含むベクター、該ベクターを含む微生物、及び該酵素または該微生物を用いてキシリトールをキシルロースに高効率で変換する方法を提供することである。
本発明のさらに他の目的は、該変異型XDHを含むキシロースをエタノールに変換するための酵素系、該酵素系を産生する微生物、該酵素系または該微生物を使用することを特徴とするキシロースをエタノールに変換する方法を提供することである。
【課題を解決するための手段】
【0018】
1.野生型キシリトールデヒドロゲナーゼの少なくとも1つのアミノ酸を他のアミノ酸に置換し、補酵素としてニコチンアミドアデニンジヌクレオチドリン酸が使用可能となるように変異させた変異型キシリトールデヒドロゲナーゼ。
2.野生型キシリトールデヒドロゲナーゼが、Pichia stipitis(ピチア スチピチス)由来のものである上記1記載の変異型キシリトールデヒドロゲナーゼ。
3.野生型キシリトールデヒドロゲナーゼの207番目から211番目に対応するアミノ酸の少なくとも1つを他のアミノ酸に置換した上記2記載の変異型キシリトールデヒドロゲナーゼ。
4.野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、および209番目のフェニルアラニンをセリンもしくはスレオニンに置換した上記3記載の変異型キシリトールデヒドロゲナーゼ。
5.野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、および209番目のフェニルアラニンをセリン、および211番目のアスパラギンをアルギニンに置換した上記3または4記載の変異型キシリトールデヒドロゲナーゼ。
6.野生型キシリトールデヒドロゲナーゼの少なくとも1つのアミノ酸を他のアミノ酸に置換し、構造安定化亜鉛結合部位が導入されたことを特徴とする変異型キシリトールデヒドロゲナーゼ。
7.他のアミノ酸がシステインである上記6記載の変異型キシリトールデヒドロゲナーゼ。
8.野生型キシリトールデヒドロゲナーゼの96番目のセリン、99番目のセリン、および102番目のチロシンの少なくとも1つをシステインに置換した上記7記載の変異型キシリトールデヒドロゲナーゼ。
9.野生型キシリトールデヒドロゲナーゼの96番目のセリン、99番目のセリン、および102番目のチロシンのすべてをシステインに置換した上記7記載の変異型キシリトールデヒドロゲナーゼ。
10.野生型キシリトールデヒドロゲナーゼの少なくとも2つのアミノ酸を他のアミノ酸に置換し、補酵素としてニコチンアミドアデニンジヌクレオチドリン酸が使用可能となるように変異させ、かつ構造安定化亜鉛結合部位が導入されたことを特徴とする変異型キシリトールデヒドロゲナーゼ。
11.野生型キシリトールデヒドロゲナーゼが、Pichia stipitis(ピチア スチピチス)由来のものである上記10記載の変異型キシリトールデヒドロゲナーゼ。
12.野生型キシリトールデヒドロゲナーゼの207番目から211番目に対応するアミノ酸の少なくとも1つを他のアミノ酸に置換し、かつ96番目のセリン、99番目のセリン、および102番目のチロシンの少なくとも1つをシステインに置換した上記11記載の変異型キシリトールデヒドロゲナーゼ。
13.野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、209番目のフェニルアラニンをセリンもしくはスレオニン、96番目のセリンをシステイン、99番目のセリンをシステイン、および102番目のチロシンをシステインにそれぞれ置換した上記12記載の変異型キシリトールデヒドロゲナーゼ。
14.野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、209番目のフェニルアラニンをセリン、211番目のアスパラギンをアルギニン、96番目のセリンをシステイン、99番目のセリンをシステイン、および102番目のチロシンをシステインにそれぞれ置換した上記12記載の変異型キシリトールデヒドロゲナーゼ。
15.上記1〜14のいずれか1項記載の変異型キシリトールデヒドロゲナーゼをコードする遺伝子。
16.上記15記載の遺伝子を含むベクター。
17.上記16記載のベクターにより形質転換された微生物。
18.微生物が酵母または大腸菌である上記17記載の微生物。
19.上記1〜14のいずれか1項記載の変異型キシリトールデヒドロゲナーゼを使用することを特徴とするキシリトールをキシルロースに変換する方法。
20.上記17または18記載の微生物を使用することを特徴とするキシリトールをキシルロースに変換する方法。
21.上記1〜14のいずれか1項記載の変異型キシリトールデヒドロゲナーゼを含むキシロースをエタノールに変換するための酵素系。
22.上記21記載の酵素系を産生する微生物。
23.微生物が酵母または大腸菌である上記22記載の微生物。
24.上記22または23記載の微生物を使用することを特徴とするキシロースをエタノールに変換する方法。
【発明の効果】
【0019】
本発明によって改良された変異型XDHは、高効率で、キシリトールをキシルロースへと変換できる。従って、この酵素をコードする遺伝子が組み込まれ、該酵素を産生する酵母を用いることで、キシロースからエタノールへと効率よく変換でき、バイオマスとしてこれまで利用されることが少なかったヘミセルロースを次世代の液体エネルギーとして期待されているエタノールへと高効率で変換することが出来る。
【発明を実施するための最良の形態】
【0020】
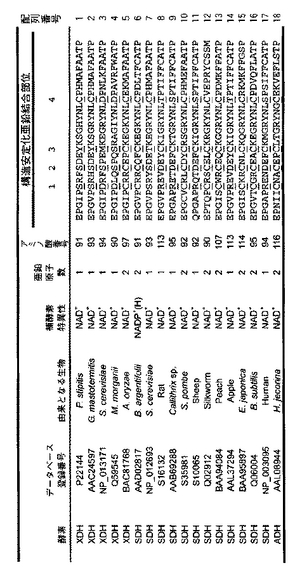
種々の微生物由来のXDHおよびソルビトールデヒドロゲナーゼ(sorbitol dehydrogenase; 以下SDH)のアミノ酸配列および補酵素依存性を比較したところ(図2B)、昆虫Bemisia argentifolii(ベミシア アルゲンチフォリ)由来のSDHのみが補酵素としてNADP+を使用していた。そのアミノ酸配列を比較したところ、種々特徴的なアミノ酸配列が見られた。最も特徴的なのが、この昆虫が持つSDHの199番目のアミノ酸に対応する部分で、NAD+を補酵素とするXDHあるいはSDHの場合にはアスパラギン酸が使用されているが、NADP+を補酵素とするSDHの場合には、アラニンが用いられていることである。そこで、Pichia stipitisのXDHのこの部位のホモロジー対応するアミノ酸、すなわち、アスパラギン酸をアラニンに、イソロイシンをアルギニンに変えることによって、P. stipitisのXDHがNADP+を補酵素として使用できるのではないかと考えた。
構造亜鉛(structural zinc)を含まないADHの場合、4つのシステイン残基は他の残基に置換されているか、特定の置換される数や置換後の残基に特定のパターンはない。この残基を再びシステインに置き換えた時、野生型酵素に新たな亜鉛原子を導入でき、それに伴う安定性の上昇と付加的な活性の上昇が期待できる。
【0021】
XDHは一般に補酵素としてNAD+を要求し、キシリトールをキシルロースに変換する反応を触媒する。P. stipitisのXDH (PsXDH)の亜鉛含有量は報告されていないが、触媒亜鉛及び構造亜鉛(catalyticおよび structural zinc)を含むB. argentifolii由来のSDHと亜鉛結合部位を比較すると(図2A)、触媒亜鉛(catalytic zinc)の結合部位は完全に保存されていることから少なくとも触媒亜鉛(catalytic zinc)は含まれていると思われる。一方、PsXDHの構造亜鉛(structural zinc)結合部位では4つのシステインのうち1つのみ(Cys110)が保存されており、他の3つはそれぞれSer96、Ser99、Tyr102に置換されていた。そこで、これらの残基を全てシステインに置換することを試みた。
本酵素の野生型は完全なNAD+依存性であるが、Pichia stipitisのキシロース還元酵素(キシロースをキシリトールに変換する反応を触媒)はNADPH依存性である。この両酵素の異なる補酵素依存性が、酵母を用いたキシロースからのエタノール生産の効率を低下させているとの知見に基づいて、本発明者はNADP+に補酵素依存性が逆転したXDH変異体を作製した。野生型(WT)に加えて、これらの変異体、特にARSとARSdRにも構造亜鉛(structural zinc)の導入を試みた。
【0022】
本発明で使用する変異の対象となるXDHは、どの様な生物から得られたものを用いてもかまわない。例えば、同じ酵母であるGalactocandida mastotermitis(ガラクトカンジダ マストテルミシチス)由来のものを用いることが出来る。この中で特に好ましいのは、Pichia stipitisである。それは、S. cerevisiaeに導入されるXRとXDHとしてはP. stipitis由来のものが最もよく用いられており、その発酵条件等が詳細に検討されているからである。また、現時点におけるエタノール生産の最高収率はこの系によって達成されているからである。
【0023】
遺伝子増幅法としては、PCR(Polymerase chain reaction)法の他に、リプレゼンテーショナル・ディフェレンス・アナリシス(RDA)法(N. Lisitsynら (1993) Science, 259: 946)、その改良法(N. G. Gurskayaら (1996) Anal. Biochem., 240: 90)、Nucleic Acid Sequence-Based Amplification (NASBA)法(特許文献6参照)等を使用することが出来、遺伝子を増幅することが出来れば特に制限はない。
遺伝子に変異を導入する方法としては、特に限定しないが、合成DNAプライマーを用いたPCR法などを使用することが出来る。
XDHの補酵素要求性はNADP+が使用可能になればよく、特に制限はしないが、補酵素要求性は、NAD+のkcat/Kmに対して、NADP+のkcat/Kmが好ましくは10倍から100倍であれば良く、300倍から1000倍であることがさらに好ましい。
【0024】
NADP+を補酵素として使用することが出来れば、そのアミノ酸配列もしくは塩基配列は、特に制限しないが、XDHのアミノ酸配列の207番目から211番目に対応するアミノ酸の少なくとも1つを他のアミノ酸、例えば、アラニン、アルギニン、セリンまたはスレオニンに置換させたものが好ましい。XDHのアミノ酸配列で207番目のアスパラギン酸をアラニンに置換させたもの、208番目のイソロイシンをアルギニンに置換させたもの、アミノ酸配列の209番目のフェニルアラニンをセリンもしくはスレオニンに置換させたもの、およびアミノ酸配列の211番目のアスパラギンをアルギニンに置換させたものが特に好ましい。
XDH遺伝子を組み込む生物は特に限定しないが、生産されるエタノールに対する耐性などの理由から、酵母、特にSaccharomyces属酵母を使用することが好ましい。
本発明で使用する変異の対象となるADHは、構造亜鉛(structural zinc)を含まないものであればどの様な生物から得られたものを用いてもかまわない。この中で特に好ましいのは、酵母Pichia stipitis由来のキシリトール脱水素酵素(XDH)である。その理由として、本酵素が酵母を用いたキシロースからエタノール生産における必須酵素であり工業的有用性が高いことが挙げられる。
【0025】
構造安定化亜鉛結合部位の酵素への導入は、酵素の構造、特に、耐熱性を向上させるものであれば、酵素のアミノ酸配列のどの位置に導入しても良いし、システインを少なくとも1つ、好ましくは3つ以上導入することが良い。好ましくは、XDHのアミノ酸配列の96番目のセリン、99番目のセリン及び102番目のチロシンの少なくとも1つをシステインに置換したXDHである。最も好ましいものは、96番目のセリンをシステイン、99番目のセリンをシステイン、及び102番目のチロシンをシステインに置換したXDHである。また、構造を安定化、特に熱に対する安定性を向上させるものであればどの様なものをXDHに導入しても良い。例えば、構造安定化亜鉛結合部位以外にループ領域の可塑性を減少させるためのプロリン残基の導入などがある。
【0026】
本発明で言うところの、固定支持体上に結合した酵素とは、一般的に固定化酵素もしくはアフィニティクロマトグラフィーとしての使用のことであり、例えば、千畑一郎編集、「固定化酵素」、講談社サイエンティフィク;福井三郎ら編集、「酵素工学」、東京化学同人;山崎誠ら編集、「アフィニティクロマトグラフィー」、講談社サイエンティフィク、等に記されている固定化酵素、アフィニティクロマトグラフィーのことを言う。
【実施例】
【0027】
次に、実施例により本発明を説明するが、本発明の範囲はこれら実施例に限定されるものではない。
実施例1 種々の微生物由来のXDHおよびSDHのアミノ酸配列および補酵素依存性の配列依存性
図2Aおよび図2Bに種々の微生物由来のXDHおよびSDHのアミノ酸配列および補酵素依存性の配列依存性の表を示した。
(1)構造安定化亜鉛結合部位について
図2Aにおいて、構造安定化亜鉛結合部位と予測されるサイト1−4のアミノ酸残基に下線を付した。サイト1−4で示される構造安定化亜鉛結合部位は、下に示した6つのパターンに分類できる。
S-S-Y-C (P. stipitis由来XDH)
D-S-M-D (G. mastotermitis由来XDHの精製酵素の解析から、含まれる亜鉛イオンは1個のみ(触媒亜鉛)であることが既知)
C-C-C-C (B. argentifolii由来SDHの結晶構造解析により構造安定化亜鉛があることが既知)
S-S-T-C
R-D-C-T
R-D-C-S (human(ヒト)由来SDHの結晶構造解析により構造安定化亜鉛がないことが既知)
【0028】
これらの比較から、S-S-Y-C型のP. stipitis由来XDHにはR-D-C-S型のhuman由来SDHと同様に構造安定化亜鉛は存在しないことが推測される。また同時に、C-C-C-C型からの推定、およびこれまでに報告された、他の構造安定化亜鉛を含む酵素の上記4つのシステインの部位特異的変異体の解析の結果、どのシステインが1つでも欠けても酵素活性が著しく減少することが報告されていることを考慮すると、構造安定化亜鉛の結合には4つのシステイン残基が必須であることが示唆される。すなわち、P. stipitis由来XDHの擬似結合部位にB. argentifolii由来SDHと同様に4つのシステイン残基、すなわち、96番目のセリンをシステイン、99番目のセリンをシステイン、102番目のチロシンをシステインに置換することで、3つのシステイン残基を新たに導入することで、新たな亜鉛イオンを導入できる可能性がある。これにより、野生型に比べて構造の安定化が期待できる。
これまでに、構造化亜鉛を導入する試みは全くなされておらず新たな試みである。
【0029】
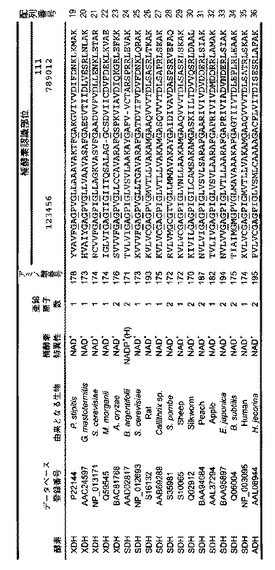
(2)補酵素認識部位について
図2Bの配列の比較から下線を付したサイト1−12に特徴的なアミノ酸配列を見出した。これらのサイトの配列から以下のことを考察した。
サイト1-6: NAD+、NADP+依存性酵素によらず共通
サイト7: NAD+依存性酵素→アスパラギン酸、NADP+依存性酵素→アラニン
サイト8: NAD+依存性酵素→疎水性残基、NADP+依存性酵素→塩基性残基(アルギニン)
サイト9: NAD+、NADP+依存性酵素によらずセリンか疎水性残基
サイト10: 統一性なし
サイト11: NAD+依存性酵素→統一性なし、 NADP+依存性酵素→塩基性残基(アルギニン)
サイト12: NAD+、NADP+依存性酵素によらず塩基性残基(リジンかアルギニン)
以上の比較より、新規NADP+依存性XDHの創生のために導入する変異部位のターゲットはサイト7、8、11が適している。さらにサイト9も候補に加えたが、これはサイト7、8の近傍ということで多重変異にしたときの効果を期待した。
【0030】
実施例2 野生型PsXDH遺伝子の作製
GeneBankに登録されているPichia stipitis XDH遺伝子(登録番号X55392)を参考にして、下記の2つのプライマーを設計した。
ATGACTGCTAACCCTTCCTTGGTGTTG (27量体)(配列番号37)
TTACTCAGGGCCGTCAATGAGACACTTG (28量体)(配列番号38)
PCRは、KOD-Plus DNAポリメラーゼ(東洋紡株式会社)を用いて行われた。10pmolの各プライマーと100ngのP. stipitisゲノムDNAを使い、変性を94℃で15秒間、アニーリングを50℃で30秒間、伸長反応を68℃で1.5分間の条件でPsXDH遺伝子を増幅した。得られたDNA断片を、プラスミドpBluescript SK(-)のSmaI制限酵素切断部位に導入し、これをpBS-XYL2と名付けた。
次に、PsXDH遺伝子の5'末端にBamHI切断部位、3'末端にPstI切断部位を導入するために、2つのプライマー、
CATACggatccgACTGCTAACCCTTCCTTGG (31量体)(配列番号39)
CTTGGctgcagTTACTCAGGGCCGTCAATGAGACAC (36量体)(配列番号40)
を準備し、pBS-XYL2プラスミドを用いてPCRを行った。ここで、上記プライマーの小文字の部位は、それぞれBamHIとPstIの切断部位を示す。増幅したDNA断片は、BamHIとPstIで切断した後、同様に処理したプラスミドpQE81-L (Qiagen)に導入した。このプラスミドは、アミノ酸末端に6個のヒスチジンタグを付けてタンパク質を発現させることができる。
【0031】
実施例3 変異型PsXDH遺伝子の作製
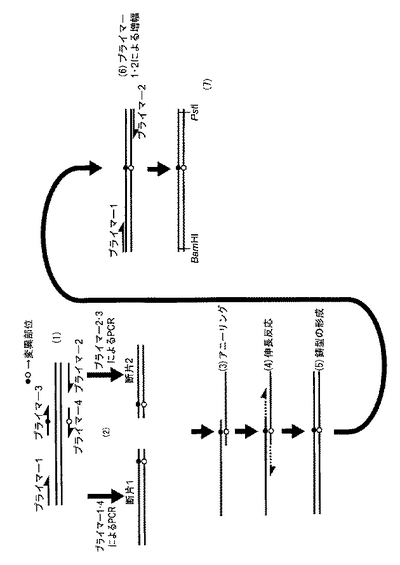
野生型PsXDH遺伝子の作製と同様の方法でプラスミドを作製した。この時、部位特異的にDNA塩基配列の変異を導入するために、two-round PCR法を用いた。使用したDNAの塩基配列を図3A,Bに示した。以下、図4を用いてtwo-round PCR法の原理を概略する。
(1)まず、置換させたいアミノ酸残基に対応する塩基(●、○で示す)を変異させた2本のプライマー、センス鎖(プライマー3)とアンチセンス鎖(プライマー4)、を用意する。プライマー1、2は、野生型PsXDH遺伝子を発現ベクターに組み込むときに用いたもので、5'と3'末端にそれぞれBamHIとPstIの制限酵素サイトを含む。
(2)「1st PCR」
野生型XDH遺伝子を鋳型として、プライマー1と4、プライマー2と3を用いてそれぞれ独立にPCRを行う。結果として、XDH遺伝子の一部が増幅される(断片1、2)。断片を電気泳動で精製することで、鋳型を完全に除くことが重要である。
「2nd PCR」
2つのDNA断片1、2、およびプライマー1、2を混合してPCRを行う。
(3)2つのDNA断片はプライマー3、4の領域をどちらも含んでいるので、断片1のセンス鎖と断片2のアンチセンス鎖はこの部分でアニーリングする。
(4)それぞれの鎖がプライマーかつ鋳型として伸長反応が起こる。ちなみに、(3)では断片1のアンチセンス鎖と断片2のセンス鎖がアニールしたものも生じるが、DNAポリメラーゼの特性により伸長反応は起きない。
(5)最終的に変異の導入されたXDH遺伝子が生じる。
(6)これは、共存するプライマー1、2によりさらに増幅される。
(7)最終的に両末端に制限酵素切断部位を持つ変異XDH遺伝子が作られる。
【0032】
実施例4 ヒスチジンタグのついたPsXDHの発現と精製
野生型PsXDHもしくは変異型PsXDH遺伝子を含むプラスミドを導入した大腸菌DH5αを、50mg/lのアンピシリンを含むスーパーブロス(Super broth) (1リットル当たり12gのペプトン、24gのイースト・エクストラクト、5mlのグリセロール、3.81gのKH2PO4、12.5gのK2HPO4、pH7.0)を用いて、37℃で600nmの吸光度0.6まで培養した。その後、PsXDHを発現誘導するために、最終濃度1mMになるようにイソプロピル−β−D−チオガラクトピラノシド (IPTG)を添加しさらに6時間培養を続けた。培養終了後に遠心分離で集菌し、培養液1リットル当たり20mlのバッファーA (2mMのMgCl2、0.3MのNaCl、10mMのキシリトール、10mMの2-メルカプトエタノール、10mMのイミダゾールを含むpH8.0の50mM リン酸ナトリウム) に懸濁した。この懸濁液にニワトリ卵白リゾチームを最終濃度2mg/mlになるように添加し、4℃で2時間緩やかに振盪させ溶菌させた。さらに超音波処理により破砕した後、細胞溶解物を4℃で2時間、105000×gで遠心し、その上澄みを同様の条件でさらに6時間遠心した。
【0033】
細胞抽出液からのPsXDHの精製は、AKTA purifier system (Amersham Biosciences)のクロマトグラフィーシステムで4℃で行った。バッファーAで平衡化したNi-NTA Superflow column (Qiagen)にアプライした。バッファーAで洗浄した後、バッファーB (10% (v/v) のグリセロールを含み、10mMのイミダゾールの代わりに50mMのイミダゾールを含むバッファーA) で洗浄した。PsXDHは、バッファーC (50mMのイミダゾールの代わりに250mMのイミダゾールを含むバッファーB)で溶出させた。PsXDHを含む分画をCentriplus YM-30 (Millipore)で濃縮し、バッファーD (2mMのMgCl2、10mMのキシリトール、1mMのジチオスレイトール (DTT)、50% (v/v)のグリセロールを含むpH8.0の50mM リン酸ナトリウム)に透析した。精製した酵素は、使うまで−35℃で保存した。精製度の確認は、12% 濃度のアクリルアミドゲルを用いたSDS-ゲル電気泳動によって行った。図5Aに精製したPsXDH (各10μg)の電気泳動を示した。泳動した全てのタンパク質は、XDHの推定分子量である40,000ダルトン付近に唯一のバンドが確認された。これは、全てのタンパク質が十分に精製されたことを示している。
【0034】
実施例5 酵素活性の測定
PsXDHの活性測定は、反応によって生成されるNAD(P)Hの特異的な340nmの吸収度の増加を35℃でモニターすることによって行った。適量のPsXDHを、50mM MgCl2、300mMのキシリトールを含む50mM Tris-HClバッファー (900μl) に加えた。酵素反応は、100μlの20mM NAD(P)+を加えることで開始した。この酵素の1ユニットは、1μmolのNAD(P)Hを1分間に生成するために必要な量と定義した。Kmおよびkcatは、種々の濃度の基質、補酵素を用いたLineweaver-Burkプロットから算出した。タンパク濃度は、ウシ血清アルブミンを標準物質としてLowry法によって決定した。
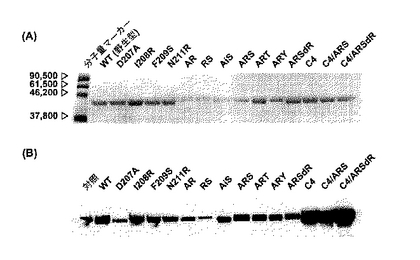
図6Aに酵素比活性の結果を示した。WT、すなわち野生型PsXDHの比活性は、P. stipitisから直接精製したXDH同様 (Rizziら (1989) J. Ferment. Bioeng., 67: 20-24参照) NAD+に極めて高い特異性を示した。NADP+ / NAD+の値は、0.001であった。一方、変異体ARS、ART、ARSdRは逆にNADP+に高い活性を示しNADP+ / NAD+の値はそれぞれ7.3、6.8、9.9であった。さらに、WTのNAD+に対する比活性に比べてARS、ART、ARSdRのNADP+に対する比活性は1.7-2.3倍上昇した。
図6Bに野生型および変異体酵素の代謝効率(kcat/Km)を示した。比活性と同様に、ARS、ART、ARSdRのkcat/KmNADは185、1190、83であり、WT (2760)に比べて十分低かった。一方、ARS、ART、ARSdRのkcat/KmNADPは2830、3100、2700であり、WT (0.6)に比べて4500-5200倍上昇した。これらの変異体のkcat/KmNADPは、WTのkcat/KmNADと同等かそれ以上であった。
最終的に、ARS、ART、ARSdRのNADP+のNAD+に対する代謝効率の比([kcat/KmNADP] / [kcat/KmNAD])は、WTに比べて劇的に上昇した(12000-150000倍)。
【0035】
実施例6 変異型PsXDH遺伝子C4の作製
PsXDH遺伝子の擬似構造亜鉛(structural zinc)結合部位に3つのシステイン残基を導入するために、以下の2つのプライマーを設計した。
GGTATTCCATGTAGATTCTGTGACGAATGTAAGAGCGG (38量体)(配列番号51)
CCGCTCTTACATTCGTCACAGAATCTACATGGAATACC (38量体)(配列番号62)
ここで、上記プライマーの下線を付した部位は変異導入箇所を示す。
さらに、実施例2でPsXDH遺伝子の5'末端にBamHI切断部位、3'末端にPstI切断部位を導入するため作製した2本のプライマー(配列番号39および配列番号40)と、実施例2で作製した野生型PsXDH遺伝子が組み込まれたプラスミドpQE81-Lを用いて、実施例3に従い変異体C4遺伝子を作製した (図3A,B)。この遺伝子はWT遺伝子同様、プラスミドpQE81-Lに導入された。
【0036】
実施例7 変異型PsXDH遺伝子C4/ARSとC4/ARSdRの作製(図3A,B)
ARSとARSdR変異体にC4と同様の変異を導入するために、以下のようにXDH遺伝子内に存在する制限酵素部位を用いたDNA断片交換を行った。
C4とARS、ARSdRの変異部位の間にはKpnIの切断部位が1箇所ある。これらの遺伝子はBamHIとPstIの制限酵素サイトでプラスミドpQE81-Lに導入されている。
C4、ARS、ARSdRを組み込んだプラスミドpQE81-LをそれぞれBamHIとKpnIで切断する。
ARS、ARSdRの上流半分に相当するDNA断片を、C4の上流半分を除去したプラスミドpQE81-Lに導入し、C4/ARSとC4/ARSdR遺伝子を含むプラスミドpQE81-Lを構築する。
C4、C4/ARS、C4/ARSdR変異型XDHは大腸菌DH5αに形質転換され、実施例4に従い精製した。SDS-ゲル電気泳動の結果は、これらのタンパク質はWT同様に精製されたことを示す(図5A)。
【0037】
実施例8 C4、C4/ARS、C4/ARSdR変異型XDHの酵素活性の測定
実施例4に従いその酵素活性を測定した。C4は、WTと比較してNAD+、NADP+の両方の比活性(図6A)、代謝効率(図6B)に大きな変化を及ぼさなかった。すなわち、NADP+のNAD+に対する代謝効率の比([kcat/KmNADP] / [kcat/KmNAD])は、WTとほぼ同じであり十分にNAD+依存性であった。
一方、C4/ARS、C4/ARSdRのNADP+に対するkcatは、ARS、ARSdRに比べてさらにそれぞれ5倍、4倍上昇し、そのkcat/KmNADPも約2倍増加した。一方で、kcat/KmNADはARS、ARSdR同様低い値を維持した。結局、C4/ARS、C4/ARSdRの([kcat/KmNADP] / [kcat/KmNAD])は、WTに比べて710000倍、510000倍という劇的な上昇となり、これはARS、ARSdRよりさらにNADP+に対する特異性が増したことを示している。
【0038】
実施例9 WTおよび変異型PsXDHの円二色性による熱安定性の測定
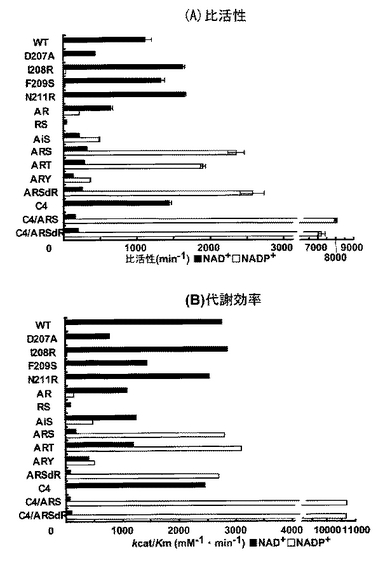
2mMのMgCl2、10mMのキシリトール、1mMのジチオスレイトール (DTT)、50% (v/v)のグリセロールを含むpH8.0の50mM リン酸ナトリウムバッファー中で−35℃で保存されていた精製酵素は、2mMのMgCl2と1mMのジチオスレイトール (DTT)を含むpH8.0の50mM リン酸ナトリウムバッファーに対して透析された。その後、同じバッファーで1mg/mlのタンパク質濃度に合わせた。温度変化(温度範囲; 10-70℃、温度上昇速度; 1℃/min)に伴う二次構造(特にα-へリックス)の崩壊は、固定波長(220nm)における円二色性(CD)スペクトルによって2mm石英セルを用いて追跡した。熱安定性は、全体のα-ヘリックスの半分が崩壊する温度(Tm)で比較した(図7)。
C4のTmは、WTに比べて4.4℃上昇した。同様に、ARSからC4/ARS、ARSdRからC4/ARSdRへの変異もまたTmの上昇をもたらし、特に前者の場合7.7℃も増加した。これらの結果は、C4の変異の導入は明らかにPsXDHの熱安定性を増加させることを示している。
【0039】
実施例10 熱変性に伴う酵素活性の消失の測定
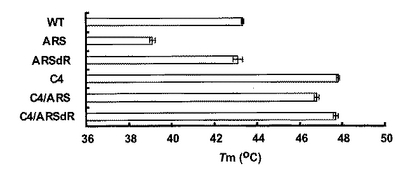
保存されていた精製酵素は、2mMのMgCl2と1mMのジチオスレイトール (DTT)を含むpH8.0の50mM リン酸ナトリウムバッファー中で透析した。その後、同じバッファーで活性測定可能な適当なタンパク質濃度に合わせた。酵素溶液は、いろいろな温度で10分間インキュベート後すみやかに氷上に戻し10分間放置した。その後、WTとC4は2mM NAD+、C4/ARSとC4/ARSdRは2mM NADP+を補酵素として、35℃で残存活性を測定した。測定値は、インキュベートしていない酵素の活性を100%とした相対値で表した(図8)。
例えば、45℃でのインキュベートによってWT(−○−)、ARS(−△−)、ARSdR(−□−)は完全に失活する。一方、同じ処理後でもC4(−●−)、C4/ARS(−▲−)、C4/ARSdR(−■−)は50-70%の活性を維持していた。CDスペクトル解析同様、この結果もまたC4の変異がPsXDHの安定性を上昇させることを示している。
【0040】
実施例11 WTおよび変異型PsXDHの亜鉛原子の定量
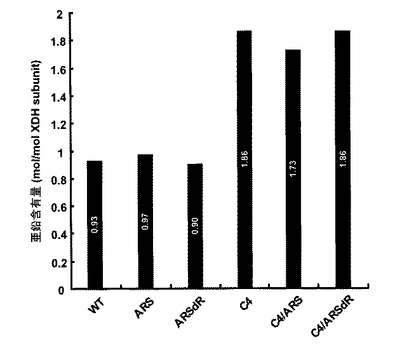
保存していた精製酵素は、微量の混入金属イオンを除くために、50mM リン酸ナトリウムバッファー(pH 8.0)で平衡化したHiLoad 16/60 Superdex 200 pg カラム(Amersham Biosciences)でゲル濾過した。その後、同じバッファーで約0.5mg/mlのタンパク質濃度に合わせた。酵素中の亜鉛の定量は、原子吸光スペクトル(波長; 約214.1nm)で行った。標準亜鉛溶液で検量線を作成し、それに基づいて各酵素溶液の測定値から亜鉛濃度を算出した。これを、各酵素溶液のタンパク質濃度から求めたサブユニット濃度で割ることで、1サブユニットあたりの亜鉛原子の数を算出した(図9)。
WTの亜鉛含有量は、0.93 (mol of zinc / mol of subunit)でありその値は1.0に極めて近かった。同様にARS、ARSdRではそれぞれ0.97、0.90であり、これらの結果はWTには1つのみの亜鉛原子(おそらく触媒亜鉛(catalytic zinc)に相当)が含まれており、またARS、ARSdRの変異ではこれに変化を及ぼさないことを示している。一方、C4、C4/ARS、C4/ARSdRの亜鉛含有量はそれぞれ1.86、1.73、1.86 (mol of zinc / mol of subunit)であり2.0に近かった。これは、C4の変異によってPsXDHが新たな亜鉛原子を獲得したことを強く示唆した。
【0041】
実施例12 WTおよび変異型PsXDHの大腸菌内での発現量の比較
実施例3に従い準備した細胞溶解液を各10μgを、0.1μgの精製されたWT(対照)と共に12% 濃度のアクリルアミドゲルを用いたSDS-ゲル電気泳動によって分離した。ゲル内のタンパク質をニトロセルロース膜(Hybond-C; Amersham Biosciences)にエレクトロブロッティングした後、WTおよび変異型PsXDHのヒスチジンタグに対するモノクローナル抗体(RGS His HRP antibody; Qiagen)と交差反応させた。本抗体は西洋ワサビ由来ペルオキシダーゼとのフュージョン抗体であり、ECL-ウエスタンブロッティング定量システム(Amersham Biosciences) による蛍光発色で同定した(図5B)。
大腸菌の中でのC4の発現量は、WTと比べて約3倍となった。一方、ARS、ARSdRの発現量はWTの約1/2だった。この低い発現量はC4変異を導入することで増加に転じ、C4変異体と同程度の高発現になった。これは、in vitroにおける安定性とよく相関していた。
【0042】
WT対C4.
この結果は、システインを他の残基に置換することで、構造亜鉛(structural zinc)を除去しようと試みた結果と大きく異なっていた。この場合、4つのシステインのどれか1つの欠損でもその変異体の酵素活性は野生型に比べて全く無くなる(Jelokov痰 1994) Eur. J. Biochem., 225: 1015-1019)か4%以下に落ち込む(Wangら(1999) Biosic. Biotech. Biochem., 63: 2216-2218)。これは、構造亜鉛(structural zinc)が高次構造維持とそれに伴う活性の維持の両方に関与していることを示唆する。一方、WTにもう1つの 亜鉛原子を加えてもその活性には何ら変化が無い。C4の中の付加された亜鉛原子は、安定性のみに寄与する「真の」構造亜鉛(structural zinc)と言うことができる。
【0043】
ARS対C4/ARSおよびC4/ARSdR
構造亜鉛(structural zinc)の導入は、C4の熱安定性の上昇という性質をそのまま維持しつつARS、ARSdRのNADP+依存性の効果をさらに増強した。加えて、このin vitroでの安定化はin vivoにおける変異体酵素の高発現を導いているように思われる。この傾向は、これらの変異体を酵母内で発現させたときも保持されると思われ、細胞内でのタンパク質の寿命を延ばし、より効率的なエタノール生産に役立つことが期待される。
【図面の簡単な説明】
【0044】
【図1】キシロースのエタノールへの異性化工程を示す。 (1)〜(5)は酵素名を示す。(1)NADH依存性キシロースリダクターゼ (XR)、(2)NADPH依存性キシロースリダクターゼ、(3)NAD+依存性キシリトールデヒドロゲナーゼ (XDH)、(4)キシロースイソメラーゼ (XI)、(5)キシルロキナーゼ (XK)
【図2A】XDH/SDHタンパク質ファミリーの構造安定化亜鉛結合部位の多重アミノ酸配列アライメントを示す。
【図2B】XDH/SDHタンパク質ファミリーの補酵素認識部位の多重アミノ酸配列アライメントを示す。
【図3A】P. stipitis XDH及びその変異体の構造安定化亜鉛結合部位及び補酵素認識部位のアミノ酸配列、及びXDH変異体をコードする遺伝子を作成するためのPCRプライマー(センス鎖)を示す。「・」は、P. stipitis XDHと同じアミノ酸残基である。下線は、変異を導入した部分である。
【図3B】P. stipitis XDH及びその変異体の構造安定化亜鉛結合部位及び補酵素認識部位のアミノ酸配列、及びXDH変異体をコードする遺伝子を作成するためのPCRプライマー(アンチセンス鎖)を示す。「・」は、P. stipitis XDHと同じアミノ酸残基である。下線は、変異を導入した部分である。
【図4】XDH遺伝子への部位特異的変異の導入に使用した、"2 round-PCR法"と呼ばれる方法を説明する図面である。
【図5】図5Aは、野生型および構築したP. stipitis XDH変異体の精製度を確認するためのSDS-ポリアクリルアミドゲル電気泳動(12%ゲル使用)パターンを示す。全てのタンパク質(10μg)は、分子量マーカーと共に泳動した。図5Bは、抗ヒスチジンタグ抗体を用いた細胞抽出液中のXDHの定量。全ての細胞抽出液は(10μg)、コントロールである精製PsXDH WT(0.1μg)と共に解析された。
【図6】図6Aは、野生型および変異型XDHの比活性を示す。図6Bは、野生型および変異型XDHの代謝効率を示す。
【図7】CDによる野生型および変異型XDHの熱安定性を示す。
【図8】野生型および変異型XDHの熱失活曲線を示す。WT(−○−)、ARS(−△−)、ARSdR(−□−)、 C4(−●−)、C4/ARS(−▲−)、C4/ARSdR(−■−)。
【図9】野生型および変異型XDHの亜鉛含有量を示す。
【技術分野】
【0001】
本発明は、キシリトールを高効率でキシルロースへ変換することができる変異型キシリトールデヒドロゲナーゼ(Xylitol dehydrogenase; 以下XDH)酵素、これをコードする遺伝子、該遺伝子を含むベクター、該ベクターを含む微生物、及び該酵素または該微生物を用いてキシリトールを高効率でキシルロースに変換する方法を提供するものである。さらに詳細には本発明は、木質などに含まれるセルロース、ヘミセルロースのうち、これまで利用する手段が少なかったヘミセルロース中に多量に含まれているキシロースを、バイオマス液体燃料として注目されているエタノールへと変換する酵素系の中で、これまでキシリトールからキシルロースへの変換効率が低かったキシリトールデヒドロゲナーゼ(XDH)酵素のアミノ酸配列を変異させ、補酵素要求性をニコチンアミドアデニンジヌクレオチド(NAD+)からニコチンアミドアデニンジヌクレオチドリン酸(NADP+)要求性へと改良し、および/または、構造安定化亜鉛結合部位を導入して耐熱性を改良したXDHを提供すること、これを用いてキシリトールからキシルロースへの高効率変換を可能とし、これによりキシロースを高効率でエタノールへと変換する方法を提供するものである。
【背景技術】
【0002】
地球上のバイオマスの大部分を占める木質系バイオマスは、リグノセルロースから構成されており、リグノセルロースはセルロース(約45%)とヘミセルロース(約30%)、リグニン(約25%)に分類される。このうち、セルロースやリグニンは短時間で超臨界水を用いたグルコースへの変換などにより、近年、盛んにそのエネルギー利用が研究されている。ヘミセルロースは酸加水分解あるいは酵素分解によって容易にキシロースなどの単糖類に分解することが出来る。また、ヘミセルロースの大部分(非特許文献1参照)を占め、容易に調達できるキシロースを液体燃料へと効率よく変換させることは、エネルギー問題の点からも重要な課題と考えられている。
これまで、酵母などの多くの微生物を用いてセルロースなどのヘキソースをエタノールへと嫌気的に効率よく変換させることには成功しているが、キシロースなどのペントースは高い効率で変換できないことが知られている。
【0003】
キシロースをエタノールへと変換する経路として、図1に示したように、主に二つの経路が知られている。一つは、キシロースからキシルロースへとキシロースイソメラーゼ(Xylose isomerase; 以下XI)により補酵素に依存せずに一段階で変換する方法である。もう一つは、キシロースリダクターゼ(Xylose reductase; 以下XR)によりキシロースをキシリトールへと変換させた後、キシリトールをキシリトールデヒドロゲナーゼ(XDH)によりキシルロースへと変換する方法である。この時、変換には補酵素を必要とする。両方法とも、キシロースをキシルロースへと変換する事が出来れば、ペントース・リン酸回路を経由してエタノールへと容易に変換することが出来る。
【0004】
数多くのバクテリアや菌類で、キシロースからキシルロースへとXIにより一段階で変換する経路を通って、キシロースをエタノールへと変換できることが知られている。例えば、Streptomyces sp.(ストレプトミセス)やActinoplanes sp.(アクチノプラネス)のようなバクテリアは、XIによりキシロースをキシルロースへと変換した後、ペントース・リン酸回路を経てエタノールへと変換できることが知られている(非特許文献2参照)。しかし、その効率は非常に低い(特許文献1参照)。これは、有機酸が副生成物として生成することが原因ではないかと考えられている(特許文献2参照)。これらのことから、キシロースからキシルロースへとXIにより一段階で変換する方法は工業的に利用されるには至っていない。
【0005】
二段階でキシロースからキシルロースへと変換する微生物として、Pichia stipitis(ピチア スチピチス)、Candida shehatae(カンジダ シェハタエ)、Pachysolen tannophilus(パチソレン タンフィルス)等が知られている(非特許文献3,4参照)。これらの微生物を用いることで、キシロースをエタノールへと変換することにある程度の成功をしている。しかし、これらの微生物もしくは酵素を用いた方法は、純粋なキシロースを用いた場合の結果であり、キシロース以外の化学物質の存在下で、キシロースをキシルロースへと変換させた場合、その収率は大きく低下することが知られている(非特許文献5,6参照)。通常、木質などからキシロースを生産する際、種々の化学物質を使用する。これらの物質を取り除くことは不可能ではないが、この操作により、大幅なコスト増となる。
微生物を用いてキシロースをキシリトールへと変換させることは、これまでに多くの成功例がある(非特許文献7,8参照)。従って、キシリトールからキシルロースを効率よく変換できる酵素系もしくはその酵素系を有する微生物を用いれば、高効率でキシロースをエタノールへと変換できる可能性がある。
【0006】
遺伝子工学的手法により酵母の遺伝子を変異させ、キシロースをエタノールへと高効率で変換させる検討も行われてきた。例えば、C. S. Gongら(特許文献3参照)は、紫外線照射により遺伝子を変異させた酵母を用いて、キシロースを代謝させることに成功している。しかし、どの遺伝子を変異させたかなどの遺伝子工学的な情報は何ら開示されていない。
キシロースをエタノールに変換させるために、キシロースをキシルロースへと補酵素に依存せずに直接変換できるXI遺伝子を大腸菌や酵母に導入した例も報告されている(非特許文献9参照)。しかし、大腸菌や酵母に、これらの遺伝子を導入し、発現させた場合の変換効率は非常に低かった。
【0007】
酵母、Saccharomyces cerevisiae(サッカロミセス セレビシエ)は、グルコースを高効率でエタノールに変換できることが知られているが、出発原料としてキシロースを用いた時には、エタノールに変換することは出来ない。これは、S. cerevisiaeがキシロースをキシルロースに代謝することが出来ないことが原因であり、もしこれを代謝することが出来れば、この微生物が持つ高いエタノール生産能力、高いエタノール耐性能力を用い、非常に効率良くキシロースをエタノールに変換できるのではないかと考えられ、キシロースをキシルロースへと変換する際の酵素系の改良が種々検討されてきた。しかし、S. cerevisiaeのXR(非特許文献10,11参照)およびP. stipitisのXDH(非特許文献12参照)の遺伝子を変異させ、その遺伝子を酵母に導入し、嫌気性条件下でキシロースからエタノールを得ることに成功したことが報告されているが(非特許文献13−15参照)(特許文献4,5参照)、その変換効率は低く、工業的生産にまでは至っていない。
【0008】
タンパク質、特に工業的に有用な酵素の安定化は実用化に向けた至上命題の1つである。無機触媒に比べて生体高分子である酵素は一般に熱に弱く、その失活の多くは不可逆である。安定性を上昇させ耐久寿命を延ばすことで頻繁な酵素の添加を防ぐことができ、大幅な経費節減を実現できる。安定化した酵素は、種々の分野に応用されている。例えば、洗剤などに添加するタンパク質分解酵素、固定化酵素、アフィニティークロマトグラフィーなどである。安定化を行う方法としては、アミノ酸配列を無作為に変異させた後、高い安定性を持つ酵素をスクリーニングする方法、耐熱性を有する酵素のアミノ酸配列、構造などを参考に酵素のアミノ酸配列を改変させる方法、固定支持体に酵素を結合させる方法、高分子架橋型ゲルで酵素を包み込む方法等、種々の方法が考案されている。
【0009】
アミノ酸配列を変異させることで酵素の改変を行う方法は、大きく2つに分類できる。
第一の方法は、無作為にアミノ酸を置換させるランダム変異(Random mutagenesis)である。遺伝子シャフリング(gene-shuffling)やエラープローン(error-prone)PCRを用いて膨大な酵素変異体プールを構築し、その中から目的の性質に改変された変異体をスクリーニングする方法である。この方法の最大の利点は多くの変異体集団が効率的に得られる点にあり、ターゲット酵素の活性部位等の情報が全く無い場合にも適応できる可能性さえある。一方で、適切なスクリーニング系が必須なのが欠点である。例えば、安定化した中温性酵素変異体を得る場合であれば、そのホモログ酵素をコードする遺伝子の欠損した好熱菌株に構築した変異体プールを導入し、高温下での栄養要求性の復帰等でセレクション(選択)する。
【0010】
第二の方法は、変異させる部位を最初から特定する部位特異的変異(Site-directed mutagenesis)である。この方法では、ターゲット酵素そのものか少なくともそのホモログの高次構造や反応機構に関する情報に基づいて、変異させるべき部位を決定する。ランダム変異に比べて変異体作製の効率は大幅に下がり変異体の性質が予想通りに改変される保証もないが、上記のようなセレクション系が無い場合や他の類似酵素で成功例がある場合はランダム変異よりもむしろ近道なケースもある。
部位特異的変異による酵素の熱安定性の向上の研究は、主に異なる温度で生育する生物から単離されたホモログ酵素のアミノ酸配列の比較から始まった。好熱性生物由来の酵素には、特定のアミノ酸残基の含有率の偏りや全体の電荷の分布、またループの挿入や欠損などに一定の特徴が見られる。また最近では好熱性、中温性、好冷性酵素の結晶構造の比較から、水素結合や疎水性コアなどの分子内相互作用や分子表面の面積や電荷分布の詳細な比較が可能になり、こうした情報に基づいて部位特異的変異体が設計されるようになっている。しかしながら、現在のところではそうした改変の統一原理というものはなく、ターゲット酵素ごとに安定性向上に使われる戦略は異なっている。
【0011】
部位特異的変異の“決め打ち”として最も成功を収めた例としては、バクテリオファージ・T4リゾチームへのジスルフィド結合の導入を挙げることができる。構造情報から注意深く選ばれた部位に最終的に3本のジスルフィド結合を導入し、野生型酵素に比べて23℃もTm値(含まれるα-へリックス構造の半分が崩壊する温度)の上昇した変異体を得ることに成功した(非特許文献16)。このような共有結合の導入は、ランダム変異ではまず起こり得ないであろうし、その期待できる効果は少数の水素結合の追加に比べてずっと大きいだろう。これに準ずるような変異導入の戦略は、安定性の向上した部位特異的変異体の効率的な構築に応用可能である。
【0012】
アルコール脱水素酵素(ADH)は、最も大きなタンパク質ファミリーの1つである。この酵素が触媒する化学反応はいたって簡単である。基質のアルコールは、そのヒドロキシル基が結合している炭素原子の水素が酸化型補酵素NAD(P)+へ転移し(hydryl-transfer)、ヒドロキシル基からプロトンが除去されアルデヒドに酸化される。この酵素は亜鉛原子を活性中心に持ち(触媒亜鉛(catalytic zinc))、これは3つの高度に保存されたアミノ酸残基と1つの水分子によって配位している。この3つの残基はどれか1つが欠けてもその酵素活性に致命的な影響がある。一方、この触媒亜鉛(catalytic zinc)に加えてもう1つの亜鉛分子を持つADHが数多く知られている。この亜鉛には4つの保存されたシステイン残基が配位しており、構造亜鉛(structural zinc)(この明細書においては「構造安定化亜鉛」と称することもある。)と呼ばれる。構造亜鉛(structural zinc)の有無と、ADHの由来となる生物種やその高次構造の間には明瞭な関係はない。触媒亜鉛(catalytic zinc)に比べて構造亜鉛(structural zinc)の役割はよく分かっていない。実際、構造亜鉛(structural zinc)の有無によらずその結合しているループの構造は高度に保存されている。しかしながら、2つのADHファミリー酵素で4つのシステイン残基のどれかひとつを変異させても、触媒亜鉛同様その酵素活性に致命的な影響があることが報告されており、何らかの高次構造維持に役立っていると考えられてきた。
【0013】
【特許文献1】米国特許第4,368,268号明細書
【特許文献2】米国特許第1,857,429号明細書
【特許文献3】米国特許第4,511,656号明細書
【特許文献4】米国特許第6,582,944号明細書
【特許文献5】米国特許第5,866,382号明細書
【特許文献6】欧州特許第0329822号明細書
【0014】
【非特許文献1】M. R. Ladischら (1983) Enzyme Microb. Technol., 5: 82-102
【非特許文献2】K. Skoogら (1988) Enzyme Microb. Technol., 10: 66-78
【非特許文献3】P. J. Sliningerら (1987) Enzyme Microb. Technol., 9: 5-15
【非特許文献4】B. A. Priorら (1989) Proc. Biochem., 2: 21-32
【非特許文献5】T. Lindenら (1989) Enzyme Microb. Technol., 11: 583-589
【非特許文献6】T. Lindenら (1989) Biotechnol. Techniques, 3: 189-192
【非特許文献7】H. Onishiら (1966) Agr. Biol. Chem., 30: 1139-1144
【非特許文献8】M. F. S. Barbosaら (1988) J. Ind. Microbiol., 3: 241-251
【非特許文献9】A. V. Sarthyら (1987) Appl. Environ. Microbiol., 53: 1996-2000
【非特許文献10】Takamaら (1991) Appl. Biochem. Biotechnol., 327: 27-28
【非特許文献11】Hallbornら (1991) Bio Technol., 9: 1090
【非特許文献12】Koetterら (1990) Curr. Gen et., 18: 493-500
【非特許文献13】Y. S. Jinら (2000) J. Microbiol. Biotechnol., 10:564-567
【非特許文献14】Koetterら (1990) Curr. Gen et., 18: 493-500
【非特許文献15】Hallbornら (1991) Bio Technol., 9: 1090
【非特許文献16】Matsumuraら (1989) Nature, 342: 291
【発明の開示】
【発明が解決しようとする課題】
【0015】
木質などのバイオマス資源のうち、30%以上を占めるヘミセルロースを加水分解することによって容易に得ることが出来るキシロースを、高効率でエタノールへと変換し、エネルギー問題を解決するための1つの手段を提供することは重要な課題である。そのためには、セルロースなどのヘキソースを高効率でエタノールへと変換でき、エタノール耐性の高い酵母に、キシロースなどのペントースを原料として高効率でエタノールへと変換できる能力を与えるのが最も理想的である。これまでに、キシロースをキシリトールへと高効率で変換できる酵母は開発されているが、キシリトールをキシルロースへと高効率で変換するXDHおよび酵母は開発されていない。
【0016】
本発明の課題は、野生型のXDHを参考に高効率でキシリトールをキシルロースへと変換するXDHを開発することである。XRは、前述のように、高効率でキシロースをキシリトールに変換することが出来るが、この際に補酵素として、NADP+を使用する。しかし、XDHは補酵素としてNAD+を使用する。この様に両酵素の補酵素に対する要求性が異なっていることが、補酵素供給の量的バランスを崩し、その結果、キシリトールからキシルロースへの変換が効率よく進行せず、最終的にキシロースからエタノールへの変換効率が低くなると推定される。これらのことを考慮すると、XDHの補酵素要求性を変えて、XRと同じNADP+要求性にすれば、高効率でキシリトールをキシロースに変換でき、この部分が効率よく進行すれば、キシロースをエタノールに高い効率で変換できると考えられる。
従って、本発明の第1の目的は、XDH遺伝子を遺伝子工学的に変異させて、補酵素NADP+要求性の変異型XDHを提供すること、及び該変異型XDHをコードする遺伝子を提供することである。
【0017】
また、酵素を工業的規模で使用する場合には、その酵素の安定性を向上させることは、非常に重要な課題と考えられる。これは、酵素を微生物中に組み込んで使用する場合でも、酵素を固定支持担体に結合させて使用する固定化酵素として使用する場合でも同様である。酵素は、化学物質、例えば、本発明の場合、キシロース、キシリトールやエタノール等や熱に安定である方が好ましい。特に、熱に対する安定性は、系の反応温度が高いほど一般的に反応速度が高く、単位時間あたりの収量が多くなるので重要である。また、微生物に酵素を組み込んだ場合、一般に代謝の進行に伴って培養槽の温度が高くなり、温度上昇に伴って酵素が失活するので、この点からも高い耐熱能力を酵素に持たせることは非常に重要である。
従って本発明の第2の目的は、高い耐熱性を持つ変異型XDHを提供すること、及び該変異型XDHをコードする遺伝子を提供することである。
本発明の他の目的は、キシリトールを高効率でキシルロースへ変換することができる変異型XDHをコードする遺伝子を含むベクター、該ベクターを含む微生物、及び該酵素または該微生物を用いてキシリトールをキシルロースに高効率で変換する方法を提供することである。
本発明のさらに他の目的は、該変異型XDHを含むキシロースをエタノールに変換するための酵素系、該酵素系を産生する微生物、該酵素系または該微生物を使用することを特徴とするキシロースをエタノールに変換する方法を提供することである。
【課題を解決するための手段】
【0018】
1.野生型キシリトールデヒドロゲナーゼの少なくとも1つのアミノ酸を他のアミノ酸に置換し、補酵素としてニコチンアミドアデニンジヌクレオチドリン酸が使用可能となるように変異させた変異型キシリトールデヒドロゲナーゼ。
2.野生型キシリトールデヒドロゲナーゼが、Pichia stipitis(ピチア スチピチス)由来のものである上記1記載の変異型キシリトールデヒドロゲナーゼ。
3.野生型キシリトールデヒドロゲナーゼの207番目から211番目に対応するアミノ酸の少なくとも1つを他のアミノ酸に置換した上記2記載の変異型キシリトールデヒドロゲナーゼ。
4.野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、および209番目のフェニルアラニンをセリンもしくはスレオニンに置換した上記3記載の変異型キシリトールデヒドロゲナーゼ。
5.野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、および209番目のフェニルアラニンをセリン、および211番目のアスパラギンをアルギニンに置換した上記3または4記載の変異型キシリトールデヒドロゲナーゼ。
6.野生型キシリトールデヒドロゲナーゼの少なくとも1つのアミノ酸を他のアミノ酸に置換し、構造安定化亜鉛結合部位が導入されたことを特徴とする変異型キシリトールデヒドロゲナーゼ。
7.他のアミノ酸がシステインである上記6記載の変異型キシリトールデヒドロゲナーゼ。
8.野生型キシリトールデヒドロゲナーゼの96番目のセリン、99番目のセリン、および102番目のチロシンの少なくとも1つをシステインに置換した上記7記載の変異型キシリトールデヒドロゲナーゼ。
9.野生型キシリトールデヒドロゲナーゼの96番目のセリン、99番目のセリン、および102番目のチロシンのすべてをシステインに置換した上記7記載の変異型キシリトールデヒドロゲナーゼ。
10.野生型キシリトールデヒドロゲナーゼの少なくとも2つのアミノ酸を他のアミノ酸に置換し、補酵素としてニコチンアミドアデニンジヌクレオチドリン酸が使用可能となるように変異させ、かつ構造安定化亜鉛結合部位が導入されたことを特徴とする変異型キシリトールデヒドロゲナーゼ。
11.野生型キシリトールデヒドロゲナーゼが、Pichia stipitis(ピチア スチピチス)由来のものである上記10記載の変異型キシリトールデヒドロゲナーゼ。
12.野生型キシリトールデヒドロゲナーゼの207番目から211番目に対応するアミノ酸の少なくとも1つを他のアミノ酸に置換し、かつ96番目のセリン、99番目のセリン、および102番目のチロシンの少なくとも1つをシステインに置換した上記11記載の変異型キシリトールデヒドロゲナーゼ。
13.野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、209番目のフェニルアラニンをセリンもしくはスレオニン、96番目のセリンをシステイン、99番目のセリンをシステイン、および102番目のチロシンをシステインにそれぞれ置換した上記12記載の変異型キシリトールデヒドロゲナーゼ。
14.野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、209番目のフェニルアラニンをセリン、211番目のアスパラギンをアルギニン、96番目のセリンをシステイン、99番目のセリンをシステイン、および102番目のチロシンをシステインにそれぞれ置換した上記12記載の変異型キシリトールデヒドロゲナーゼ。
15.上記1〜14のいずれか1項記載の変異型キシリトールデヒドロゲナーゼをコードする遺伝子。
16.上記15記載の遺伝子を含むベクター。
17.上記16記載のベクターにより形質転換された微生物。
18.微生物が酵母または大腸菌である上記17記載の微生物。
19.上記1〜14のいずれか1項記載の変異型キシリトールデヒドロゲナーゼを使用することを特徴とするキシリトールをキシルロースに変換する方法。
20.上記17または18記載の微生物を使用することを特徴とするキシリトールをキシルロースに変換する方法。
21.上記1〜14のいずれか1項記載の変異型キシリトールデヒドロゲナーゼを含むキシロースをエタノールに変換するための酵素系。
22.上記21記載の酵素系を産生する微生物。
23.微生物が酵母または大腸菌である上記22記載の微生物。
24.上記22または23記載の微生物を使用することを特徴とするキシロースをエタノールに変換する方法。
【発明の効果】
【0019】
本発明によって改良された変異型XDHは、高効率で、キシリトールをキシルロースへと変換できる。従って、この酵素をコードする遺伝子が組み込まれ、該酵素を産生する酵母を用いることで、キシロースからエタノールへと効率よく変換でき、バイオマスとしてこれまで利用されることが少なかったヘミセルロースを次世代の液体エネルギーとして期待されているエタノールへと高効率で変換することが出来る。
【発明を実施するための最良の形態】
【0020】
種々の微生物由来のXDHおよびソルビトールデヒドロゲナーゼ(sorbitol dehydrogenase; 以下SDH)のアミノ酸配列および補酵素依存性を比較したところ(図2B)、昆虫Bemisia argentifolii(ベミシア アルゲンチフォリ)由来のSDHのみが補酵素としてNADP+を使用していた。そのアミノ酸配列を比較したところ、種々特徴的なアミノ酸配列が見られた。最も特徴的なのが、この昆虫が持つSDHの199番目のアミノ酸に対応する部分で、NAD+を補酵素とするXDHあるいはSDHの場合にはアスパラギン酸が使用されているが、NADP+を補酵素とするSDHの場合には、アラニンが用いられていることである。そこで、Pichia stipitisのXDHのこの部位のホモロジー対応するアミノ酸、すなわち、アスパラギン酸をアラニンに、イソロイシンをアルギニンに変えることによって、P. stipitisのXDHがNADP+を補酵素として使用できるのではないかと考えた。
構造亜鉛(structural zinc)を含まないADHの場合、4つのシステイン残基は他の残基に置換されているか、特定の置換される数や置換後の残基に特定のパターンはない。この残基を再びシステインに置き換えた時、野生型酵素に新たな亜鉛原子を導入でき、それに伴う安定性の上昇と付加的な活性の上昇が期待できる。
【0021】
XDHは一般に補酵素としてNAD+を要求し、キシリトールをキシルロースに変換する反応を触媒する。P. stipitisのXDH (PsXDH)の亜鉛含有量は報告されていないが、触媒亜鉛及び構造亜鉛(catalyticおよび structural zinc)を含むB. argentifolii由来のSDHと亜鉛結合部位を比較すると(図2A)、触媒亜鉛(catalytic zinc)の結合部位は完全に保存されていることから少なくとも触媒亜鉛(catalytic zinc)は含まれていると思われる。一方、PsXDHの構造亜鉛(structural zinc)結合部位では4つのシステインのうち1つのみ(Cys110)が保存されており、他の3つはそれぞれSer96、Ser99、Tyr102に置換されていた。そこで、これらの残基を全てシステインに置換することを試みた。
本酵素の野生型は完全なNAD+依存性であるが、Pichia stipitisのキシロース還元酵素(キシロースをキシリトールに変換する反応を触媒)はNADPH依存性である。この両酵素の異なる補酵素依存性が、酵母を用いたキシロースからのエタノール生産の効率を低下させているとの知見に基づいて、本発明者はNADP+に補酵素依存性が逆転したXDH変異体を作製した。野生型(WT)に加えて、これらの変異体、特にARSとARSdRにも構造亜鉛(structural zinc)の導入を試みた。
【0022】
本発明で使用する変異の対象となるXDHは、どの様な生物から得られたものを用いてもかまわない。例えば、同じ酵母であるGalactocandida mastotermitis(ガラクトカンジダ マストテルミシチス)由来のものを用いることが出来る。この中で特に好ましいのは、Pichia stipitisである。それは、S. cerevisiaeに導入されるXRとXDHとしてはP. stipitis由来のものが最もよく用いられており、その発酵条件等が詳細に検討されているからである。また、現時点におけるエタノール生産の最高収率はこの系によって達成されているからである。
【0023】
遺伝子増幅法としては、PCR(Polymerase chain reaction)法の他に、リプレゼンテーショナル・ディフェレンス・アナリシス(RDA)法(N. Lisitsynら (1993) Science, 259: 946)、その改良法(N. G. Gurskayaら (1996) Anal. Biochem., 240: 90)、Nucleic Acid Sequence-Based Amplification (NASBA)法(特許文献6参照)等を使用することが出来、遺伝子を増幅することが出来れば特に制限はない。
遺伝子に変異を導入する方法としては、特に限定しないが、合成DNAプライマーを用いたPCR法などを使用することが出来る。
XDHの補酵素要求性はNADP+が使用可能になればよく、特に制限はしないが、補酵素要求性は、NAD+のkcat/Kmに対して、NADP+のkcat/Kmが好ましくは10倍から100倍であれば良く、300倍から1000倍であることがさらに好ましい。
【0024】
NADP+を補酵素として使用することが出来れば、そのアミノ酸配列もしくは塩基配列は、特に制限しないが、XDHのアミノ酸配列の207番目から211番目に対応するアミノ酸の少なくとも1つを他のアミノ酸、例えば、アラニン、アルギニン、セリンまたはスレオニンに置換させたものが好ましい。XDHのアミノ酸配列で207番目のアスパラギン酸をアラニンに置換させたもの、208番目のイソロイシンをアルギニンに置換させたもの、アミノ酸配列の209番目のフェニルアラニンをセリンもしくはスレオニンに置換させたもの、およびアミノ酸配列の211番目のアスパラギンをアルギニンに置換させたものが特に好ましい。
XDH遺伝子を組み込む生物は特に限定しないが、生産されるエタノールに対する耐性などの理由から、酵母、特にSaccharomyces属酵母を使用することが好ましい。
本発明で使用する変異の対象となるADHは、構造亜鉛(structural zinc)を含まないものであればどの様な生物から得られたものを用いてもかまわない。この中で特に好ましいのは、酵母Pichia stipitis由来のキシリトール脱水素酵素(XDH)である。その理由として、本酵素が酵母を用いたキシロースからエタノール生産における必須酵素であり工業的有用性が高いことが挙げられる。
【0025】
構造安定化亜鉛結合部位の酵素への導入は、酵素の構造、特に、耐熱性を向上させるものであれば、酵素のアミノ酸配列のどの位置に導入しても良いし、システインを少なくとも1つ、好ましくは3つ以上導入することが良い。好ましくは、XDHのアミノ酸配列の96番目のセリン、99番目のセリン及び102番目のチロシンの少なくとも1つをシステインに置換したXDHである。最も好ましいものは、96番目のセリンをシステイン、99番目のセリンをシステイン、及び102番目のチロシンをシステインに置換したXDHである。また、構造を安定化、特に熱に対する安定性を向上させるものであればどの様なものをXDHに導入しても良い。例えば、構造安定化亜鉛結合部位以外にループ領域の可塑性を減少させるためのプロリン残基の導入などがある。
【0026】
本発明で言うところの、固定支持体上に結合した酵素とは、一般的に固定化酵素もしくはアフィニティクロマトグラフィーとしての使用のことであり、例えば、千畑一郎編集、「固定化酵素」、講談社サイエンティフィク;福井三郎ら編集、「酵素工学」、東京化学同人;山崎誠ら編集、「アフィニティクロマトグラフィー」、講談社サイエンティフィク、等に記されている固定化酵素、アフィニティクロマトグラフィーのことを言う。
【実施例】
【0027】
次に、実施例により本発明を説明するが、本発明の範囲はこれら実施例に限定されるものではない。
実施例1 種々の微生物由来のXDHおよびSDHのアミノ酸配列および補酵素依存性の配列依存性
図2Aおよび図2Bに種々の微生物由来のXDHおよびSDHのアミノ酸配列および補酵素依存性の配列依存性の表を示した。
(1)構造安定化亜鉛結合部位について
図2Aにおいて、構造安定化亜鉛結合部位と予測されるサイト1−4のアミノ酸残基に下線を付した。サイト1−4で示される構造安定化亜鉛結合部位は、下に示した6つのパターンに分類できる。
S-S-Y-C (P. stipitis由来XDH)
D-S-M-D (G. mastotermitis由来XDHの精製酵素の解析から、含まれる亜鉛イオンは1個のみ(触媒亜鉛)であることが既知)
C-C-C-C (B. argentifolii由来SDHの結晶構造解析により構造安定化亜鉛があることが既知)
S-S-T-C
R-D-C-T
R-D-C-S (human(ヒト)由来SDHの結晶構造解析により構造安定化亜鉛がないことが既知)
【0028】
これらの比較から、S-S-Y-C型のP. stipitis由来XDHにはR-D-C-S型のhuman由来SDHと同様に構造安定化亜鉛は存在しないことが推測される。また同時に、C-C-C-C型からの推定、およびこれまでに報告された、他の構造安定化亜鉛を含む酵素の上記4つのシステインの部位特異的変異体の解析の結果、どのシステインが1つでも欠けても酵素活性が著しく減少することが報告されていることを考慮すると、構造安定化亜鉛の結合には4つのシステイン残基が必須であることが示唆される。すなわち、P. stipitis由来XDHの擬似結合部位にB. argentifolii由来SDHと同様に4つのシステイン残基、すなわち、96番目のセリンをシステイン、99番目のセリンをシステイン、102番目のチロシンをシステインに置換することで、3つのシステイン残基を新たに導入することで、新たな亜鉛イオンを導入できる可能性がある。これにより、野生型に比べて構造の安定化が期待できる。
これまでに、構造化亜鉛を導入する試みは全くなされておらず新たな試みである。
【0029】
(2)補酵素認識部位について
図2Bの配列の比較から下線を付したサイト1−12に特徴的なアミノ酸配列を見出した。これらのサイトの配列から以下のことを考察した。
サイト1-6: NAD+、NADP+依存性酵素によらず共通
サイト7: NAD+依存性酵素→アスパラギン酸、NADP+依存性酵素→アラニン
サイト8: NAD+依存性酵素→疎水性残基、NADP+依存性酵素→塩基性残基(アルギニン)
サイト9: NAD+、NADP+依存性酵素によらずセリンか疎水性残基
サイト10: 統一性なし
サイト11: NAD+依存性酵素→統一性なし、 NADP+依存性酵素→塩基性残基(アルギニン)
サイト12: NAD+、NADP+依存性酵素によらず塩基性残基(リジンかアルギニン)
以上の比較より、新規NADP+依存性XDHの創生のために導入する変異部位のターゲットはサイト7、8、11が適している。さらにサイト9も候補に加えたが、これはサイト7、8の近傍ということで多重変異にしたときの効果を期待した。
【0030】
実施例2 野生型PsXDH遺伝子の作製
GeneBankに登録されているPichia stipitis XDH遺伝子(登録番号X55392)を参考にして、下記の2つのプライマーを設計した。
ATGACTGCTAACCCTTCCTTGGTGTTG (27量体)(配列番号37)
TTACTCAGGGCCGTCAATGAGACACTTG (28量体)(配列番号38)
PCRは、KOD-Plus DNAポリメラーゼ(東洋紡株式会社)を用いて行われた。10pmolの各プライマーと100ngのP. stipitisゲノムDNAを使い、変性を94℃で15秒間、アニーリングを50℃で30秒間、伸長反応を68℃で1.5分間の条件でPsXDH遺伝子を増幅した。得られたDNA断片を、プラスミドpBluescript SK(-)のSmaI制限酵素切断部位に導入し、これをpBS-XYL2と名付けた。
次に、PsXDH遺伝子の5'末端にBamHI切断部位、3'末端にPstI切断部位を導入するために、2つのプライマー、
CATACggatccgACTGCTAACCCTTCCTTGG (31量体)(配列番号39)
CTTGGctgcagTTACTCAGGGCCGTCAATGAGACAC (36量体)(配列番号40)
を準備し、pBS-XYL2プラスミドを用いてPCRを行った。ここで、上記プライマーの小文字の部位は、それぞれBamHIとPstIの切断部位を示す。増幅したDNA断片は、BamHIとPstIで切断した後、同様に処理したプラスミドpQE81-L (Qiagen)に導入した。このプラスミドは、アミノ酸末端に6個のヒスチジンタグを付けてタンパク質を発現させることができる。
【0031】
実施例3 変異型PsXDH遺伝子の作製
野生型PsXDH遺伝子の作製と同様の方法でプラスミドを作製した。この時、部位特異的にDNA塩基配列の変異を導入するために、two-round PCR法を用いた。使用したDNAの塩基配列を図3A,Bに示した。以下、図4を用いてtwo-round PCR法の原理を概略する。
(1)まず、置換させたいアミノ酸残基に対応する塩基(●、○で示す)を変異させた2本のプライマー、センス鎖(プライマー3)とアンチセンス鎖(プライマー4)、を用意する。プライマー1、2は、野生型PsXDH遺伝子を発現ベクターに組み込むときに用いたもので、5'と3'末端にそれぞれBamHIとPstIの制限酵素サイトを含む。
(2)「1st PCR」
野生型XDH遺伝子を鋳型として、プライマー1と4、プライマー2と3を用いてそれぞれ独立にPCRを行う。結果として、XDH遺伝子の一部が増幅される(断片1、2)。断片を電気泳動で精製することで、鋳型を完全に除くことが重要である。
「2nd PCR」
2つのDNA断片1、2、およびプライマー1、2を混合してPCRを行う。
(3)2つのDNA断片はプライマー3、4の領域をどちらも含んでいるので、断片1のセンス鎖と断片2のアンチセンス鎖はこの部分でアニーリングする。
(4)それぞれの鎖がプライマーかつ鋳型として伸長反応が起こる。ちなみに、(3)では断片1のアンチセンス鎖と断片2のセンス鎖がアニールしたものも生じるが、DNAポリメラーゼの特性により伸長反応は起きない。
(5)最終的に変異の導入されたXDH遺伝子が生じる。
(6)これは、共存するプライマー1、2によりさらに増幅される。
(7)最終的に両末端に制限酵素切断部位を持つ変異XDH遺伝子が作られる。
【0032】
実施例4 ヒスチジンタグのついたPsXDHの発現と精製
野生型PsXDHもしくは変異型PsXDH遺伝子を含むプラスミドを導入した大腸菌DH5αを、50mg/lのアンピシリンを含むスーパーブロス(Super broth) (1リットル当たり12gのペプトン、24gのイースト・エクストラクト、5mlのグリセロール、3.81gのKH2PO4、12.5gのK2HPO4、pH7.0)を用いて、37℃で600nmの吸光度0.6まで培養した。その後、PsXDHを発現誘導するために、最終濃度1mMになるようにイソプロピル−β−D−チオガラクトピラノシド (IPTG)を添加しさらに6時間培養を続けた。培養終了後に遠心分離で集菌し、培養液1リットル当たり20mlのバッファーA (2mMのMgCl2、0.3MのNaCl、10mMのキシリトール、10mMの2-メルカプトエタノール、10mMのイミダゾールを含むpH8.0の50mM リン酸ナトリウム) に懸濁した。この懸濁液にニワトリ卵白リゾチームを最終濃度2mg/mlになるように添加し、4℃で2時間緩やかに振盪させ溶菌させた。さらに超音波処理により破砕した後、細胞溶解物を4℃で2時間、105000×gで遠心し、その上澄みを同様の条件でさらに6時間遠心した。
【0033】
細胞抽出液からのPsXDHの精製は、AKTA purifier system (Amersham Biosciences)のクロマトグラフィーシステムで4℃で行った。バッファーAで平衡化したNi-NTA Superflow column (Qiagen)にアプライした。バッファーAで洗浄した後、バッファーB (10% (v/v) のグリセロールを含み、10mMのイミダゾールの代わりに50mMのイミダゾールを含むバッファーA) で洗浄した。PsXDHは、バッファーC (50mMのイミダゾールの代わりに250mMのイミダゾールを含むバッファーB)で溶出させた。PsXDHを含む分画をCentriplus YM-30 (Millipore)で濃縮し、バッファーD (2mMのMgCl2、10mMのキシリトール、1mMのジチオスレイトール (DTT)、50% (v/v)のグリセロールを含むpH8.0の50mM リン酸ナトリウム)に透析した。精製した酵素は、使うまで−35℃で保存した。精製度の確認は、12% 濃度のアクリルアミドゲルを用いたSDS-ゲル電気泳動によって行った。図5Aに精製したPsXDH (各10μg)の電気泳動を示した。泳動した全てのタンパク質は、XDHの推定分子量である40,000ダルトン付近に唯一のバンドが確認された。これは、全てのタンパク質が十分に精製されたことを示している。
【0034】
実施例5 酵素活性の測定
PsXDHの活性測定は、反応によって生成されるNAD(P)Hの特異的な340nmの吸収度の増加を35℃でモニターすることによって行った。適量のPsXDHを、50mM MgCl2、300mMのキシリトールを含む50mM Tris-HClバッファー (900μl) に加えた。酵素反応は、100μlの20mM NAD(P)+を加えることで開始した。この酵素の1ユニットは、1μmolのNAD(P)Hを1分間に生成するために必要な量と定義した。Kmおよびkcatは、種々の濃度の基質、補酵素を用いたLineweaver-Burkプロットから算出した。タンパク濃度は、ウシ血清アルブミンを標準物質としてLowry法によって決定した。
図6Aに酵素比活性の結果を示した。WT、すなわち野生型PsXDHの比活性は、P. stipitisから直接精製したXDH同様 (Rizziら (1989) J. Ferment. Bioeng., 67: 20-24参照) NAD+に極めて高い特異性を示した。NADP+ / NAD+の値は、0.001であった。一方、変異体ARS、ART、ARSdRは逆にNADP+に高い活性を示しNADP+ / NAD+の値はそれぞれ7.3、6.8、9.9であった。さらに、WTのNAD+に対する比活性に比べてARS、ART、ARSdRのNADP+に対する比活性は1.7-2.3倍上昇した。
図6Bに野生型および変異体酵素の代謝効率(kcat/Km)を示した。比活性と同様に、ARS、ART、ARSdRのkcat/KmNADは185、1190、83であり、WT (2760)に比べて十分低かった。一方、ARS、ART、ARSdRのkcat/KmNADPは2830、3100、2700であり、WT (0.6)に比べて4500-5200倍上昇した。これらの変異体のkcat/KmNADPは、WTのkcat/KmNADと同等かそれ以上であった。
最終的に、ARS、ART、ARSdRのNADP+のNAD+に対する代謝効率の比([kcat/KmNADP] / [kcat/KmNAD])は、WTに比べて劇的に上昇した(12000-150000倍)。
【0035】
実施例6 変異型PsXDH遺伝子C4の作製
PsXDH遺伝子の擬似構造亜鉛(structural zinc)結合部位に3つのシステイン残基を導入するために、以下の2つのプライマーを設計した。
GGTATTCCATGTAGATTCTGTGACGAATGTAAGAGCGG (38量体)(配列番号51)
CCGCTCTTACATTCGTCACAGAATCTACATGGAATACC (38量体)(配列番号62)
ここで、上記プライマーの下線を付した部位は変異導入箇所を示す。
さらに、実施例2でPsXDH遺伝子の5'末端にBamHI切断部位、3'末端にPstI切断部位を導入するため作製した2本のプライマー(配列番号39および配列番号40)と、実施例2で作製した野生型PsXDH遺伝子が組み込まれたプラスミドpQE81-Lを用いて、実施例3に従い変異体C4遺伝子を作製した (図3A,B)。この遺伝子はWT遺伝子同様、プラスミドpQE81-Lに導入された。
【0036】
実施例7 変異型PsXDH遺伝子C4/ARSとC4/ARSdRの作製(図3A,B)
ARSとARSdR変異体にC4と同様の変異を導入するために、以下のようにXDH遺伝子内に存在する制限酵素部位を用いたDNA断片交換を行った。
C4とARS、ARSdRの変異部位の間にはKpnIの切断部位が1箇所ある。これらの遺伝子はBamHIとPstIの制限酵素サイトでプラスミドpQE81-Lに導入されている。
C4、ARS、ARSdRを組み込んだプラスミドpQE81-LをそれぞれBamHIとKpnIで切断する。
ARS、ARSdRの上流半分に相当するDNA断片を、C4の上流半分を除去したプラスミドpQE81-Lに導入し、C4/ARSとC4/ARSdR遺伝子を含むプラスミドpQE81-Lを構築する。
C4、C4/ARS、C4/ARSdR変異型XDHは大腸菌DH5αに形質転換され、実施例4に従い精製した。SDS-ゲル電気泳動の結果は、これらのタンパク質はWT同様に精製されたことを示す(図5A)。
【0037】
実施例8 C4、C4/ARS、C4/ARSdR変異型XDHの酵素活性の測定
実施例4に従いその酵素活性を測定した。C4は、WTと比較してNAD+、NADP+の両方の比活性(図6A)、代謝効率(図6B)に大きな変化を及ぼさなかった。すなわち、NADP+のNAD+に対する代謝効率の比([kcat/KmNADP] / [kcat/KmNAD])は、WTとほぼ同じであり十分にNAD+依存性であった。
一方、C4/ARS、C4/ARSdRのNADP+に対するkcatは、ARS、ARSdRに比べてさらにそれぞれ5倍、4倍上昇し、そのkcat/KmNADPも約2倍増加した。一方で、kcat/KmNADはARS、ARSdR同様低い値を維持した。結局、C4/ARS、C4/ARSdRの([kcat/KmNADP] / [kcat/KmNAD])は、WTに比べて710000倍、510000倍という劇的な上昇となり、これはARS、ARSdRよりさらにNADP+に対する特異性が増したことを示している。
【0038】
実施例9 WTおよび変異型PsXDHの円二色性による熱安定性の測定
2mMのMgCl2、10mMのキシリトール、1mMのジチオスレイトール (DTT)、50% (v/v)のグリセロールを含むpH8.0の50mM リン酸ナトリウムバッファー中で−35℃で保存されていた精製酵素は、2mMのMgCl2と1mMのジチオスレイトール (DTT)を含むpH8.0の50mM リン酸ナトリウムバッファーに対して透析された。その後、同じバッファーで1mg/mlのタンパク質濃度に合わせた。温度変化(温度範囲; 10-70℃、温度上昇速度; 1℃/min)に伴う二次構造(特にα-へリックス)の崩壊は、固定波長(220nm)における円二色性(CD)スペクトルによって2mm石英セルを用いて追跡した。熱安定性は、全体のα-ヘリックスの半分が崩壊する温度(Tm)で比較した(図7)。
C4のTmは、WTに比べて4.4℃上昇した。同様に、ARSからC4/ARS、ARSdRからC4/ARSdRへの変異もまたTmの上昇をもたらし、特に前者の場合7.7℃も増加した。これらの結果は、C4の変異の導入は明らかにPsXDHの熱安定性を増加させることを示している。
【0039】
実施例10 熱変性に伴う酵素活性の消失の測定
保存されていた精製酵素は、2mMのMgCl2と1mMのジチオスレイトール (DTT)を含むpH8.0の50mM リン酸ナトリウムバッファー中で透析した。その後、同じバッファーで活性測定可能な適当なタンパク質濃度に合わせた。酵素溶液は、いろいろな温度で10分間インキュベート後すみやかに氷上に戻し10分間放置した。その後、WTとC4は2mM NAD+、C4/ARSとC4/ARSdRは2mM NADP+を補酵素として、35℃で残存活性を測定した。測定値は、インキュベートしていない酵素の活性を100%とした相対値で表した(図8)。
例えば、45℃でのインキュベートによってWT(−○−)、ARS(−△−)、ARSdR(−□−)は完全に失活する。一方、同じ処理後でもC4(−●−)、C4/ARS(−▲−)、C4/ARSdR(−■−)は50-70%の活性を維持していた。CDスペクトル解析同様、この結果もまたC4の変異がPsXDHの安定性を上昇させることを示している。
【0040】
実施例11 WTおよび変異型PsXDHの亜鉛原子の定量
保存していた精製酵素は、微量の混入金属イオンを除くために、50mM リン酸ナトリウムバッファー(pH 8.0)で平衡化したHiLoad 16/60 Superdex 200 pg カラム(Amersham Biosciences)でゲル濾過した。その後、同じバッファーで約0.5mg/mlのタンパク質濃度に合わせた。酵素中の亜鉛の定量は、原子吸光スペクトル(波長; 約214.1nm)で行った。標準亜鉛溶液で検量線を作成し、それに基づいて各酵素溶液の測定値から亜鉛濃度を算出した。これを、各酵素溶液のタンパク質濃度から求めたサブユニット濃度で割ることで、1サブユニットあたりの亜鉛原子の数を算出した(図9)。
WTの亜鉛含有量は、0.93 (mol of zinc / mol of subunit)でありその値は1.0に極めて近かった。同様にARS、ARSdRではそれぞれ0.97、0.90であり、これらの結果はWTには1つのみの亜鉛原子(おそらく触媒亜鉛(catalytic zinc)に相当)が含まれており、またARS、ARSdRの変異ではこれに変化を及ぼさないことを示している。一方、C4、C4/ARS、C4/ARSdRの亜鉛含有量はそれぞれ1.86、1.73、1.86 (mol of zinc / mol of subunit)であり2.0に近かった。これは、C4の変異によってPsXDHが新たな亜鉛原子を獲得したことを強く示唆した。
【0041】
実施例12 WTおよび変異型PsXDHの大腸菌内での発現量の比較
実施例3に従い準備した細胞溶解液を各10μgを、0.1μgの精製されたWT(対照)と共に12% 濃度のアクリルアミドゲルを用いたSDS-ゲル電気泳動によって分離した。ゲル内のタンパク質をニトロセルロース膜(Hybond-C; Amersham Biosciences)にエレクトロブロッティングした後、WTおよび変異型PsXDHのヒスチジンタグに対するモノクローナル抗体(RGS His HRP antibody; Qiagen)と交差反応させた。本抗体は西洋ワサビ由来ペルオキシダーゼとのフュージョン抗体であり、ECL-ウエスタンブロッティング定量システム(Amersham Biosciences) による蛍光発色で同定した(図5B)。
大腸菌の中でのC4の発現量は、WTと比べて約3倍となった。一方、ARS、ARSdRの発現量はWTの約1/2だった。この低い発現量はC4変異を導入することで増加に転じ、C4変異体と同程度の高発現になった。これは、in vitroにおける安定性とよく相関していた。
【0042】
WT対C4.
この結果は、システインを他の残基に置換することで、構造亜鉛(structural zinc)を除去しようと試みた結果と大きく異なっていた。この場合、4つのシステインのどれか1つの欠損でもその変異体の酵素活性は野生型に比べて全く無くなる(Jelokov痰 1994) Eur. J. Biochem., 225: 1015-1019)か4%以下に落ち込む(Wangら(1999) Biosic. Biotech. Biochem., 63: 2216-2218)。これは、構造亜鉛(structural zinc)が高次構造維持とそれに伴う活性の維持の両方に関与していることを示唆する。一方、WTにもう1つの 亜鉛原子を加えてもその活性には何ら変化が無い。C4の中の付加された亜鉛原子は、安定性のみに寄与する「真の」構造亜鉛(structural zinc)と言うことができる。
【0043】
ARS対C4/ARSおよびC4/ARSdR
構造亜鉛(structural zinc)の導入は、C4の熱安定性の上昇という性質をそのまま維持しつつARS、ARSdRのNADP+依存性の効果をさらに増強した。加えて、このin vitroでの安定化はin vivoにおける変異体酵素の高発現を導いているように思われる。この傾向は、これらの変異体を酵母内で発現させたときも保持されると思われ、細胞内でのタンパク質の寿命を延ばし、より効率的なエタノール生産に役立つことが期待される。
【図面の簡単な説明】
【0044】
【図1】キシロースのエタノールへの異性化工程を示す。 (1)〜(5)は酵素名を示す。(1)NADH依存性キシロースリダクターゼ (XR)、(2)NADPH依存性キシロースリダクターゼ、(3)NAD+依存性キシリトールデヒドロゲナーゼ (XDH)、(4)キシロースイソメラーゼ (XI)、(5)キシルロキナーゼ (XK)
【図2A】XDH/SDHタンパク質ファミリーの構造安定化亜鉛結合部位の多重アミノ酸配列アライメントを示す。
【図2B】XDH/SDHタンパク質ファミリーの補酵素認識部位の多重アミノ酸配列アライメントを示す。
【図3A】P. stipitis XDH及びその変異体の構造安定化亜鉛結合部位及び補酵素認識部位のアミノ酸配列、及びXDH変異体をコードする遺伝子を作成するためのPCRプライマー(センス鎖)を示す。「・」は、P. stipitis XDHと同じアミノ酸残基である。下線は、変異を導入した部分である。
【図3B】P. stipitis XDH及びその変異体の構造安定化亜鉛結合部位及び補酵素認識部位のアミノ酸配列、及びXDH変異体をコードする遺伝子を作成するためのPCRプライマー(アンチセンス鎖)を示す。「・」は、P. stipitis XDHと同じアミノ酸残基である。下線は、変異を導入した部分である。
【図4】XDH遺伝子への部位特異的変異の導入に使用した、"2 round-PCR法"と呼ばれる方法を説明する図面である。
【図5】図5Aは、野生型および構築したP. stipitis XDH変異体の精製度を確認するためのSDS-ポリアクリルアミドゲル電気泳動(12%ゲル使用)パターンを示す。全てのタンパク質(10μg)は、分子量マーカーと共に泳動した。図5Bは、抗ヒスチジンタグ抗体を用いた細胞抽出液中のXDHの定量。全ての細胞抽出液は(10μg)、コントロールである精製PsXDH WT(0.1μg)と共に解析された。
【図6】図6Aは、野生型および変異型XDHの比活性を示す。図6Bは、野生型および変異型XDHの代謝効率を示す。
【図7】CDによる野生型および変異型XDHの熱安定性を示す。
【図8】野生型および変異型XDHの熱失活曲線を示す。WT(−○−)、ARS(−△−)、ARSdR(−□−)、 C4(−●−)、C4/ARS(−▲−)、C4/ARSdR(−■−)。
【図9】野生型および変異型XDHの亜鉛含有量を示す。
【特許請求の範囲】
【請求項1】
野生型キシリトールデヒドロゲナーゼの少なくとも1つのアミノ酸を他のアミノ酸に置換し、補酵素としてニコチンアミドアデニンジヌクレオチドリン酸が使用可能となるように変異させた変異型キシリトールデヒドロゲナーゼ。
【請求項2】
野生型キシリトールデヒドロゲナーゼが、Pichia stipitis(ピチア スチピチス)由来のものである請求項1記載の変異型キシリトールデヒドロゲナーゼ。
【請求項3】
野生型キシリトールデヒドロゲナーゼの207番目から211番目に対応するアミノ酸の少なくとも1つを他のアミノ酸に置換した請求項2記載の変異型キシリトールデヒドロゲナーゼ。
【請求項4】
野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、および209番目のフェニルアラニンをセリンもしくはスレオニンに置換した請求項3記載の変異型キシリトールデヒドロゲナーゼ。
【請求項5】
野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、および209番目のフェニルアラニンをセリン、および211番目のアスパラギンをアルギニンに置換した請求項3または4記載の変異型キシリトールデヒドロゲナーゼ。
【請求項6】
野生型キシリトールデヒドロゲナーゼの少なくとも1つのアミノ酸を他のアミノ酸に置換し、構造安定化亜鉛結合部位が導入されたことを特徴とする変異型キシリトールデヒドロゲナーゼ。
【請求項7】
他のアミノ酸がシステインである請求項6記載の変異型キシリトールデヒドロゲナーゼ。
【請求項8】
野生型キシリトールデヒドロゲナーゼの96番目のセリン、99番目のセリン、および102番目のチロシンの少なくとも1つをシステインに置換した請求項7記載の変異型キシリトールデヒドロゲナーゼ。
【請求項9】
野生型キシリトールデヒドロゲナーゼの96番目のセリン、99番目のセリン、および102番目のチロシンのすべてをシステインに置換した請求項7記載の変異型キシリトールデヒドロゲナーゼ。
【請求項10】
野生型キシリトールデヒドロゲナーゼの少なくとも2つのアミノ酸を他のアミノ酸に置換し、補酵素としてニコチンアミドアデニンジヌクレオチドリン酸が使用可能となるように変異させ、かつ構造安定化亜鉛結合部位が導入されたことを特徴とする変異型キシリトールデヒドロゲナーゼ。
【請求項11】
野生型キシリトールデヒドロゲナーゼが、Pichia stipitis(ピチア スチピチス)由来のものである請求項10記載の変異型キシリトールデヒドロゲナーゼ。
【請求項12】
野生型キシリトールデヒドロゲナーゼの207番目から211番目に対応するアミノ酸の少なくとも1つを他のアミノ酸に置換し、かつ96番目のセリン、99番目のセリン、および102番目のチロシンの少なくとも1つをシステインに置換した請求項11記載の変異型キシリトールデヒドロゲナーゼ。
【請求項13】
野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、209番目のフェニルアラニンをセリンもしくはスレオニン、96番目のセリンをシステイン、99番目のセリンをシステイン、および102番目のチロシンをシステインにそれぞれ置換した請求項12記載の変異型キシリトールデヒドロゲナーゼ。
【請求項14】
野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、209番目のフェニルアラニンをセリン、211番目のアスパラギンをアルギニン、96番目のセリンをシステイン、99番目のセリンをシステイン、および102番目のチロシンをシステインにそれぞれ置換した請求項12記載の変異型キシリトールデヒドロゲナーゼ。
【請求項15】
請求項1〜14のいずれか1項記載の変異型キシリトールデヒドロゲナーゼをコードする遺伝子。
【請求項16】
請求項15記載の遺伝子を含むベクター。
【請求項17】
請求項16記載のベクターにより形質転換された微生物。
【請求項18】
微生物が酵母または大腸菌である請求項17記載の微生物。
【請求項19】
請求項1〜14のいずれか1項記載の変異型キシリトールデヒドロゲナーゼを使用することを特徴とするキシリトールをキシルロースに変換する方法。
【請求項20】
請求項17または18記載の微生物を使用することを特徴とするキシリトールをキシルロースに変換する方法。
【請求項21】
請求項1〜14のいずれか1項記載の変異型キシリトールデヒドロゲナーゼを含むキシロースをエタノールに変換するための酵素系。
【請求項22】
請求項21記載の酵素系を産生する微生物。
【請求項23】
微生物が酵母または大腸菌である請求項22記載の微生物。
【請求項24】
請求項21記載の酵素系、または請求項22もしくは23記載の微生物を使用することを特徴とするキシロースをエタノールに変換する方法。
【請求項1】
野生型キシリトールデヒドロゲナーゼの少なくとも1つのアミノ酸を他のアミノ酸に置換し、補酵素としてニコチンアミドアデニンジヌクレオチドリン酸が使用可能となるように変異させた変異型キシリトールデヒドロゲナーゼ。
【請求項2】
野生型キシリトールデヒドロゲナーゼが、Pichia stipitis(ピチア スチピチス)由来のものである請求項1記載の変異型キシリトールデヒドロゲナーゼ。
【請求項3】
野生型キシリトールデヒドロゲナーゼの207番目から211番目に対応するアミノ酸の少なくとも1つを他のアミノ酸に置換した請求項2記載の変異型キシリトールデヒドロゲナーゼ。
【請求項4】
野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、および209番目のフェニルアラニンをセリンもしくはスレオニンに置換した請求項3記載の変異型キシリトールデヒドロゲナーゼ。
【請求項5】
野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、および209番目のフェニルアラニンをセリン、および211番目のアスパラギンをアルギニンに置換した請求項3または4記載の変異型キシリトールデヒドロゲナーゼ。
【請求項6】
野生型キシリトールデヒドロゲナーゼの少なくとも1つのアミノ酸を他のアミノ酸に置換し、構造安定化亜鉛結合部位が導入されたことを特徴とする変異型キシリトールデヒドロゲナーゼ。
【請求項7】
他のアミノ酸がシステインである請求項6記載の変異型キシリトールデヒドロゲナーゼ。
【請求項8】
野生型キシリトールデヒドロゲナーゼの96番目のセリン、99番目のセリン、および102番目のチロシンの少なくとも1つをシステインに置換した請求項7記載の変異型キシリトールデヒドロゲナーゼ。
【請求項9】
野生型キシリトールデヒドロゲナーゼの96番目のセリン、99番目のセリン、および102番目のチロシンのすべてをシステインに置換した請求項7記載の変異型キシリトールデヒドロゲナーゼ。
【請求項10】
野生型キシリトールデヒドロゲナーゼの少なくとも2つのアミノ酸を他のアミノ酸に置換し、補酵素としてニコチンアミドアデニンジヌクレオチドリン酸が使用可能となるように変異させ、かつ構造安定化亜鉛結合部位が導入されたことを特徴とする変異型キシリトールデヒドロゲナーゼ。
【請求項11】
野生型キシリトールデヒドロゲナーゼが、Pichia stipitis(ピチア スチピチス)由来のものである請求項10記載の変異型キシリトールデヒドロゲナーゼ。
【請求項12】
野生型キシリトールデヒドロゲナーゼの207番目から211番目に対応するアミノ酸の少なくとも1つを他のアミノ酸に置換し、かつ96番目のセリン、99番目のセリン、および102番目のチロシンの少なくとも1つをシステインに置換した請求項11記載の変異型キシリトールデヒドロゲナーゼ。
【請求項13】
野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、209番目のフェニルアラニンをセリンもしくはスレオニン、96番目のセリンをシステイン、99番目のセリンをシステイン、および102番目のチロシンをシステインにそれぞれ置換した請求項12記載の変異型キシリトールデヒドロゲナーゼ。
【請求項14】
野生型キシリトールデヒドロゲナーゼの207番目のアスパラギン酸をアラニン、208番目のイソロイシンをアルギニン、209番目のフェニルアラニンをセリン、211番目のアスパラギンをアルギニン、96番目のセリンをシステイン、99番目のセリンをシステイン、および102番目のチロシンをシステインにそれぞれ置換した請求項12記載の変異型キシリトールデヒドロゲナーゼ。
【請求項15】
請求項1〜14のいずれか1項記載の変異型キシリトールデヒドロゲナーゼをコードする遺伝子。
【請求項16】
請求項15記載の遺伝子を含むベクター。
【請求項17】
請求項16記載のベクターにより形質転換された微生物。
【請求項18】
微生物が酵母または大腸菌である請求項17記載の微生物。
【請求項19】
請求項1〜14のいずれか1項記載の変異型キシリトールデヒドロゲナーゼを使用することを特徴とするキシリトールをキシルロースに変換する方法。
【請求項20】
請求項17または18記載の微生物を使用することを特徴とするキシリトールをキシルロースに変換する方法。
【請求項21】
請求項1〜14のいずれか1項記載の変異型キシリトールデヒドロゲナーゼを含むキシロースをエタノールに変換するための酵素系。
【請求項22】
請求項21記載の酵素系を産生する微生物。
【請求項23】
微生物が酵母または大腸菌である請求項22記載の微生物。
【請求項24】
請求項21記載の酵素系、または請求項22もしくは23記載の微生物を使用することを特徴とするキシロースをエタノールに変換する方法。
【図1】


【図2A】
【図2B】
【図3A】

【図3B】

【図4】
【図5】
【図6】
【図7】
【図8】

【図9】
【図2A】
【図2B】
【図3A】
【図3B】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公開番号】特開2006−6213(P2006−6213A)
【公開日】平成18年1月12日(2006.1.12)
【国際特許分類】
【出願番号】特願2004−188417(P2004−188417)
【出願日】平成16年6月25日(2004.6.25)
【出願人】(591159251)信和化工株式会社 (10)
【出願人】(504132272)国立大学法人京都大学 (1,269)
【Fターム(参考)】
【公開日】平成18年1月12日(2006.1.12)
【国際特許分類】
【出願日】平成16年6月25日(2004.6.25)
【出願人】(591159251)信和化工株式会社 (10)
【出願人】(504132272)国立大学法人京都大学 (1,269)
【Fターム(参考)】
[ Back to top ]