
変異型ルシフェラーゼ
【課題】ルシフェラーゼ酵素が、対応する野生型ルシフェラーゼと比較して異なる波長の光を発することができ、および/または対応する野生型ルシフェラーゼと比較して熱安定性が向上している組換えタンパク質を提供する。
【解決手段】野生型ルシフェラーゼと少なくとも60%の類似性を有する組換えタンパク質であって、この酵素の配列中、Photinus pyralisのルシフェラーゼ中の残基357に対応するアミノ酸残基が対応する野生型ルシフェラーゼと比較して変異させられている。変異型ルシフェラーゼは、発せられる光の大きな(50nm)波長のシフトを生み出すことができ、優れた熱安定性を有し、その結果生、カラーシフトは、補酵素Aを加えることによって逆転させることができる。これらの性質によって、この変異体はさまざまなアッセイにおいて非常に有用である。
【解決手段】野生型ルシフェラーゼと少なくとも60%の類似性を有する組換えタンパク質であって、この酵素の配列中、Photinus pyralisのルシフェラーゼ中の残基357に対応するアミノ酸残基が対応する野生型ルシフェラーゼと比較して変異させられている。変異型ルシフェラーゼは、発せられる光の大きな(50nm)波長のシフトを生み出すことができ、優れた熱安定性を有し、その結果生、カラーシフトは、補酵素Aを加えることによって逆転させることができる。これらの性質によって、この変異体はさまざまなアッセイにおいて非常に有用である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、新規なタンパク質、詳細には対応する野生型酵素と比較して特徴的な性質を示す変異型ルシフェラーゼ酵素、これらのタンパク質をエンコードするDNA、アッセイにおけるこれらの酵素の使用、およびこれらを含む試験用キットに関する。
【背景技術】
【0002】
ホタルのルシフェラーゼは、ATP、Mg2+、および分子状酸素の存在下でルシフェリンの酸化を触媒し、その結果光を生み出す。この反応は、約0.88の量的収率を有する。この発光性が、ATPレベルが測定される、広範囲の照度アッセイにおける使用につながる。このようなアッセイの例には、EP−B−680515およびWO96/02665の記載に基づくアッセイがあるが、他の多くのアッセイも研究室において日常的に使用されている。
【0003】
昆虫の身体、特にホタルまたはツチボタルなどの甲虫から、ルシフェラーゼを直接得ることができる。ルシフェラーゼがそこから得られている特定の種には、日本のGENJIまたはKEIKEホタル、Luciola cruciataおよびLuciola lateralis、東ヨーロッパのホタルLuciola mingrelica、北アメリカのホタルPhotinus pyralis、およびツチボタルLampyris noctilucaがある。
【発明の概要】
【発明が解決しようとする課題】
【0004】
しかしながら、これらの酵素をエンコードしている遺伝子の多くはクローン化および配列決定されており、組換えDNA技術を使用して生成させることもできる。酵素をエンコードしている組換えDNA配列を使用して、大腸菌などの微生物を形質転換し、したがってこれが所望の酵素産物を発現する。
【0005】
研究室におけるアッセイにおいて使用されるとき、これらの酵素によって発せられる光の色は非常に似通っている。波長が変えられれば、特定の検出器によってより容易な読み取りのために、またはたとえば同じサンプル中の異なるエベントを監視するために多数のレポーターが必要なシステムにおいて使用するために、役に立つであろう。レポーター分子を区別する1つの方法は、特定の波長で光を発するルシフェラーゼ分子を使用することである。異なる種の甲虫またはツチボタルに由来するルシフェラーゼを含むレポーター分子を使用することによって、このことを達成することができる。しかしながら、組換えDNA技術を使用して変異型ルシフェラーゼを生成することが代替的な戦略であり、その結果シグナルの波長のバリエーションを生み出す。このような変異体の例は、WO95/18853において提供されている。
【0006】
さらに、野生および組換え型のルシフェラーゼの熱安定性は、それらが約30℃を超える温度、特に35℃を超える温度にさらされると、非常に急速に活性を失うようなものである。酵素が高い周囲温度で使用または保存されると、またはたとえば反応速度を増大させるために高温の反応条件下でアッセイが行なわれると、この不安定性は問題を引き起す。
【0007】
熱安定性が増大した変異型ルシフェラーゼは、EP−A−524448およびWO/95/25798から知られている。これらの第1のものは、日本のホタルのルシフェラーゼ中の位置217において変異(詳細にはスレオニン残基がイソロイシン残基で置き換えられている)を有する、変異型ルシフェラーゼを記載している。後者のものは、Photinus pyralis、Luciola mingrelica、Luciola cruciataまたはLuciola lateralisからのルシフェラーゼと60%を超える類似性を有する変異型ルシフェラーゼを記載しているが、Photinus pyralisの残基354またはLuciola種の356に対応するアミノ酸残基がグルタメート以外に変異しており、詳細にはグルタメート、アスパルテート、プロリンまたはグリシン以外に変異している。
【0008】
同時係属の英国特許出願No.9823468.5およびそれに由来する国際特許出願は、このような変異体をさらに記載している。この場合、ルシフェラーゼ活性があり、Photinus pyralis、Luciola mingrelica、Luciola cruciataまたはLuciola lateralis酵素などのルシフェラーゼからの野生型ルシフェラーゼと少なくとも60%の類似性を有するタンパク質が記載されているが、これらのタンパク質はタンパク質中のさまざまな位置に、特に以下のものを含めた変異を含む。
【0009】
(a)Photinus pyralisのルシフェラーゼ中の残基214、Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基216に対応するアミノ酸残基、または (b)Photinus pyralisのルシフェラーゼ中の残基232、Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基234に対応するアミノ酸残基、または (c)Photinus pyralisのルシフェラーゼ中の残基295、Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基297に対応するアミノ酸残基。
【課題を解決するための手段】
【0010】
本出願人は、ルシフェラーゼタンパク質中の異なる位置でアミノ酸を変異させる(または導入する)ことによって、発せられる光の波長の大きなシフトを得ることができることと、および/または酵素の熱安定性が改善されることを発見した。さらに、グローキネティクスが妨げられているインビボでのアッセイにおいて、またはCoAまたは他の「グローキネティクス誘導」化合物が存在しないインビトロアッセイにおいて、発せられるプロトンの流れが改善され酵素がより適合し得る。
【0011】
本発明は、ルシフェラーゼ活性、および野生型ルシフェラーゼと少なくとも60%の類似性を有する組換えタンパク質を提供する。この酵素の配列中において、Photinus pyralisのルシフェラーゼ中の残基357に対応するアミノ酸残基が対応する野生型ルシフェラーゼと比較して変異しており、その結果そのルシフェラーゼ酵素は、対応する野生型ルシフェラーゼと比較して異なる波長の光を発することができ、および/または対応する野生型ルシフェラーゼと比較して熱安定性が向上している。
【図面の簡単な説明】
【0012】
【図1】残存活性%対本発明によるいくつかの変異型酵素の45℃でのインキュベーションの時間を示す対数グラフである。
【図2(a)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素は組換え野生型Photinus pyralisのルシフェラーゼである。
【図2(b)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素はD357K変異型である。
【図2(c)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素はD357N変異型である。
【図2(d)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素はD357W変異型である。
【図2(e)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素はD357I変異型である。
【図2(f)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素はD357F変異型である。
【図2(g)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素はD357Y変異型である。
【図2(h)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素は二重変異型E354I+D357Yである。
【図3】残存活性%対3つの変異型酵素E354I、D357Yおよび二重変異型(DM)E354I/D357Yの時間を示すグラフである。
【図4(a)】組換え野生型酵素の発光スペクトルを示す。
【図4(b)】二重変異型(DM)E354I/D357Yの発光スペクトルを示す。
【図5】組換え野生型(◆)r−wtおよびD357K変異型酵素(■)の光量子発光の速度減衰を示すグラフである。
【図6】分子のモデル図を示し、ルシフェラーゼ酵素中の考えられるCoA結合ポケットを示す。
【図7a】LB上で成長させた変異体P. pyralisのルシフェラーゼD357Yを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【図7b】最小培地および酢酸ナトリウム上で成長させた変異体P. pyralisのルシフェラーゼD357Yを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【図7c】最小培地およびグルコース上で成長させた変異体P. pyralisのルシフェラーゼD357Yを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【図8(a)】LB上で成長させた変異体P. pyralisのルシフェラーゼE354K/D357Mを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【図8(b)】最小培地および酢酸ナトリウム上で成長させた変異体P. pyralisのルシフェラーゼE354K/D357Mを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【図8(c)】最小培地およびグルコース上で成長させた変異体P. pyralisのルシフェラーゼE354K/D357Mを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【図9】変異体P. pyralisのルシフェラーゼD357Yによって発せられた光のスペクトル分布に対するCoAの影響を示すグラフである。
【図10】変異体P. pyralisのルシフェラーゼD357Yによって発せられた光のスペクトル分布に対するCoAの影響を示すグラフである。
【図11(a)】変異体P. pyralisのルシフェラーゼE354I/D357Yによって発せられた光のスペクトル分布に対するCoAの影響を示すグラフである。
【図11(b)】変異体P. pyralisのルシフェラーゼE354I/D357Yによって発せられた光のスペクトル分布に対するCoAの影響および正規化したデータを示すグラフである。
【図12】合成ルシフェラーゼ遺伝子の構築において使用された制限部位の修飾を示す。
【図13】ルシフェラーゼ遺伝子の合成において使用する構築体を示す。
【図14】合成ルシフェラーゼ遺伝子のcDNA配列(配列番号1)(リボソーム結合部位の一部を形成し、コードしていないヌクレオチド1〜8含む)、および上向き矢印で示されたメチオニン残基で始まるエンコードされているアミノ酸配列(配列番号2)を示す。
【図15】合成遺伝子によってエンコードされている変異体を含む、変異体の50℃における熱安定性を示す。
【発明を実施するための形態】
【0013】
本発明の組換え体の基本部分を形成することができる野生型ルシフェラーゼ配列にはPhotinus pyralis、Luciola mingrelica、Luciola cruciataまたはLuciola lateralis、Hotaria paroula、Pyrophorus plagiophthalamus Lampyris noctiluca、Pyrocoelia nayako、Photinus pennsylvanicaまたはPhrixothrix(railroad−worms Biochem.38(1999)8271-8279を参照のこと)がある。
【0014】
基質D−ルシフェリン(4,5−ジヒドロ−2−〔6−ヒドロキシ−2−ベンゾチアゾリル〕−4−チアゾールカルボン酸)を使用して、発光を生み出すことができる種からの生物発光酵素は、本発明の変異型酵素の基本部分を形成することができる。
【0015】
本発明の組換え体の基本部分を形成することができる特定の野生型ルシフェラーゼ配列にはPhotinus pyralis、Luciola mingrelica、Luciola cruciataまたはLuciola lateralis、Hotaria paroula、Pyrophorus plagiophthalamus Lampyris noctiluca、Pyrocoelia nayakoおよびPhotinus pennsylvanicaがある。
【0016】
詳細には、このルシフェラーゼはPhotinus pyralis、Luciola mingrelica、Luciola cruciataまたはLuciola lateralisの酵素から得られる酵素である。Luciola mingrelica、Luciola cruciataまたはLuciola lateralisの酵素では、適切なアミノ酸残基は配列中の位置359に存在する。
【0017】
さまざまなルシフェラーゼすべての配列によって、これらは、これらの間で高い類似性を有して高度に保存されていることが示されている。このことは、配列を調べることによって、酵素配列間の対応する領域を容易に決定することができ、最も類似な領域を検出できることを意味するが、必要ならば、さまざまな配列間の対応する領域または個々のアミノ酸を決定するために、市販のソフトウェア(たとえばUniversity of Wisconsin Genetics Computer Groupからの「Bestfit」;Devereux他(1984)Nucleic Acid Research 12:387−395を参照のこと)を使用することができる。あるいはまたはさらに、この酵素の配列およびナンバリングを示し、本出願に関してこのナンバリングシステムを使用する、L.Ye他のBiochim.Biophys Acta 1339(1997)39−52を参照することによって対応する酸を決定することができる。
【0018】
Photinus pyralisのルシフェラーゼ中の残基357に対応するアミノ酸残基の可能な変化に関して、大部分の野生型配列はこの位置に酸残基(アスパラギン酸またはグルタミン酸)を有する。この例外はPhotinus pennsylvanicaのある形のルシフェラーゼであり、これにおいては、対応する残基(355)は非極性残基バリンであり、またはPhrixothrixのある形のルシフェラーゼでは、この場合対応する位置はPvGRまたはPhRE中のV354であり、L354ロイシンである。したがって一般に、この位置で置換アミノ酸として使用されるアミノ酸は、アスパラギン酸、グルタミン酸、バリンまたはロイシン以外である。
【0019】
したがって、多くの場合、酸性アミノ酸残基は、リジンまたはアルギニンなどの塩基性アミノ酸、ロイシン、バリンまたはイソロイシンなどの非極性アミノ酸、チロシン、アスパラギン、グルタミン、フェニルアラニン、セリン、トリプトファンまたはスレオニンなどの帯電していない極性アミノ酸を含めた酸性ではない残基で置き換えられる。特に、チロシン、アスパラギン、セリンまたはスレオニンなどの帯電していない極性アミノ酸でこれを置き換えることができる。この位置での置換に関して特に好ましいアミノ酸残基は、チロシン、フェニルアラニンまたはトリプトファンであり、チロシンが最も好ましい。一般的に言うと、この位置での芳香族残基によって最大のシフトが生じ、熱安定性を保つこともできる。
【0020】
野生型配列が酸性ではないアミノ酸残基をこの位置に含む場合、これらを酸性ではない異なる残基に適切に変異させる。
【0021】
このようにして酵素を変異させることによって、ルシフェラーゼによって発せられる光の波長が、ある場合はスペクトルの赤端に向かって50nmまでシフトすることが分かった。このように、D357Y変異型Photinus pyralisルシフェラーゼは、562nmの波長で発光する野生型酵素と比較して、約612nmの波長で発光する。
【0022】
50nmという波長のシフトは、この強度のシフトはスペクトル的に容易に決定することができるから、アッセイ用途において使用するためのかなりの能力を有する。たとえばWO95/18853に記載されているように、遺伝子発現の研究では異なる色のルシフェラーゼをレポーター分子として使用することができ、これによって2つ以上の遺伝子を同時に監視することが可能になる。標識としてルシフェラーゼを用いて、多数の検体試験を行うこともできる。
【0023】
この場合の光が深い赤色であるという事実は、アッセイ法において特に有用である。光の短い波長を吸収することができる色素または他の化合物を含むATPに関する溶液を分析するとき、赤色の変異体が有用であろう。たとえば、赤色の溶液は赤色の光を吸収しないであろう。しばしばこのような分析の対象となる赤色の溶液の例には、赤色のpH指標物を含む可能性がある血液サンプルまたは真核細胞培養培地の溶液がある。
【0024】
ルシフェラーゼなどの比色剤の混合物を使用するとき、特にサンプル中の他の試薬が緑のシグナルを生成する場合には、深い赤色のシグナルを生成させる能力が有用であろう。光電極スペクトル分析において使用する光電子増倍管をセットして、1つのサンプル中で生み出される1つまたは2つのピークを検出することができる。言い換えれば、同じサンプル中で、赤色からの光量子の流れと緑色のエミッターを区別することが可能である。
【0025】
さらに、補因子補酵素A(CoA)の存在によって、波長のシフトが影響を受け得ることが分かった。この特長によって、補因子についてのアッセイにおいてこの酵素を使用することができる可能性が生じる。
【0026】
以下に記載するように、発光される光のインビトロでのスペクトルに対する補因子補酵素Aの影響を調べた。補酵素Aの濃度が増大すると、スペクトルの分布が変化し、CoAの最大濃度において、610nmの明確なピークを有する590〜630nmの領域の波長がスペクトルの主となる。
【0027】
したがって、本発明の他の態様によれば、CoAのサンプル中の存在を判定するためのアッセイが提供される。このアッセイは、CoAを含む疑いのあるサンプルに前述のルシフェラーゼと共にルシフェラーゼ/ルシフェリン反応を引き起こすために必要とされる他の試薬を加えること、サンプルから発光される光の波長を測定すること、およびこれをCoAの存在または不在と関連付けることを含む。
【0028】
このようなアッセイは、細胞の、たとえば微生物または真核細胞の成長または活性の状態の検出において有用であり得る。
【0029】
たとえば、大腸菌細胞中のCoAの濃度は比較的高く、代謝状況により大幅に変化する。発光の波長はCoAの濃度に応じて変化するので、本発明の変異型酵素を使用して、生物の代謝状況、特にインビボでのCoAの濃度を監視することができる。このようなアッセイは、CoAが抗生物質(たとえばストレプトミセス)の生成における重要な主要代謝産物である状況において、特に有用であり得る。細胞のCoA濃度は、また脂肪酸の生合成の重要な指標であり、また細胞の飢餓状況により変化する。発ガンおよび糖尿病などのいくつかの代謝障害は、脂肪酸の代謝産物の異常、したがって異常なCoAレベルを示す。このような状態の診断において、本発明のアッセイを使用することができる。たとえば、アッセイ中で使用する本発明のルシフェラーゼから発せられる光の波長を測定することによって、患者からの血液サンプルなどの細胞サンプルおよびCoAレベルを測定することができる。この結果を健康な細胞のサンプルから得られた結果と比較して、波長が変化して、変化したCoAレベルが存在するかどうか測定することができる。これは、患者の疾患状態の指標であり得る。アッセイの前に、知られている溶解剤を使用して細胞を適切に溶解させる。
【0030】
位置357のアミノ酸残基は補酵素Aの結合部位と重大な関連があると信じられている。ルシフェラーゼ酵素の表面が1オングストローム(Å)の分解能まで輪郭が示されて(Tripos Ltd.のSYBL protein nodelling softwareを使用して)、小さな極性ポケットが見い出された。このポケットは残基H310、E354およびD357によって内側を覆われているようであり、8から10Åであると測定される。分子の上部から見ると、このポケットは残基H310、E354、D357およびI232によって内側を覆われている大きなポケットの一部分であるようである。残基H310およびE354は裂け目を越えてブリッジを形成しているようであり、2つの小さなポケットの外観を与えている(図8を参照のこと)。
【0031】
理論によって縛られることなく、この酵素が溶液中にあるとき、このブリッジ残基は切り離れるほどに充分に柔軟であり、CoA結合を可能にするより大きなポケット(深さ12Åまで、幅8Åまで)を提供するようにみえる。このことは、エネルギー計算と一致する。
【0032】
本発明のホタルのルシフェラーゼの変異体を発現する大腸菌細胞を異なる炭素源上で成長させると、インビボでの発せられる光のスペクトルの変化が観察された。富化培地(LB)から、唯一の炭素源としてアセテートまたはグルコースを有する限定最小培地への変換により、発せられる光は長い波長にシフトし、より短い波長からの貢献が減少した。このことは、発せられる光の波長を調節する、アッセイ目的のための他の手段を提供しよう。
【0033】
このタンパク質中の357位置の変異は、向上した熱安定性を生じることが分かった。
【0034】
このタンパク質のルシフェラーゼ活性が著しくは損ねられなければ、このタンパク質は配列中に他の変異を含んでもよい。この変異は酵素の性質を適切に向上させるか、またはある程度酵素を所望の目的に、より良く適合させる。このことは、変異が熱安定性および/またはカラーシフトの性質、および/またはこの酵素のATPに対するKmが向上させることを、意味しよう。カラーシフトを生じさせる変異の例は、WO95/18853に記載されている。Km値に影響を与える変異は、たとえばWO96/223776および国際特許出願No.PCT/GB98/01026に記載されている。
【0035】
一般に、諸性質の変化の点において、変異の影響は追加的なものであることが分かった。
【0036】
本発明の変異型ルシフェラーゼは、野生型ルシフェラーゼと比較して熱安定性を増大させる他の特定の変異を含んでもよい。詳細には、(a)Photinus pyralisのルシフェラーゼのアミノ酸354(Luciolaのルシフェラーゼ中の356)に対応するアミノ酸残基が変異している、(b)Photinus pyralisのルシフェラーゼ中の位置215または(Luciolaのルシフェラーゼ中の217)に対応するアミノ酸残基が、異なる疎水性アミノ酸である、(c)Photinus pyralisのルシフェラーゼ中の残基214またはLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基216に対応するアミノ酸残基、(d)Photinus pyralisのルシフェラーゼ中の残基232またはLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基234に対応するアミノ酸残基、(e)Photinus pyralisのルシフェラーゼ中の残基295またはLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基297に対応するアミノ酸残基、(f)Photinus pyralisのルシフェラーゼ中のアミノ酸14、またはLuciola mingrelicaの残基16、またはLuciola cruciataまたはLuciola lateralisの17に対応するアミノ酸残基、(g)Photinus pyralisのルシフェラーゼ中のアミノ酸35、またはLuciola mingrelicaの残基37、またはLuciola cruciataまたはLuciola lateralisの残基38に対応するアミノ酸残基、(h)Photinus pyralisのルシフェラーゼのアミノ酸残基105、またはLuciola mingrelicaの残基106、Luciola cruciataまたはLuciola lateralisの107、またはLuciola lateralis遺伝子の108に対応するアミノ酸残基、(i)Photinus pyralisのルシフェラーゼ中のアミノ酸残基234またはLuciola mingrelica、Luciola cruciataまたはLuciola lateralisの残基236に対応するアミノ酸残基、(j)Photinus pyralisのルシフェラーゼ中のアミノ酸残基420またはLuciola mingrelica、Luciola cruciataまたはLuciola lateralisの残基422に対応するアミノ酸残基、(k)Photinus pyralisのルシフェラーゼ中のアミノ酸残基310またはLuciola mingrelica、Luciola cruciataまたはLuciola lateralisの残基312に対応するアミノ酸残基、の少なくとも1つが対応する野生型配列中にあるアミノ酸と異なり、このルシフェラーゼ酵素は、野生型ルシフェラーゼのこの位置での対応アミノ酸を有する酵素と比較して、増大した熱安定性を有する。
【0037】
このように、本発明のタンパク質の好ましい例は、2つ以上のアミノ酸が、たとえば100個までのアミノ酸残基が、好ましくは40個を超えないアミノ酸が、より好ましくは30個までのアミノ酸が、適切な野生型酵素中の対応する位置のアミノ酸と異なる変異した野生型ルシフェラーゼである。
【0038】
したがって、好ましい一実施形態では、本発明のタンパク質はPhotinus pyralisのルシフェラーゼを含み、前述の357位置の変異に加えて、以下のものを少なくとも1つ含む。
【0039】
(a)Photinus pyralisのルシフェラーゼのアミノ酸354に対応するアミノ酸残基がグルタメート以外である、(b)Photinus pyralisのルシフェラーゼ中の位置215に対応するアミノ酸残基がアラニン以外の疎水性アミノ酸である、(c)Photinus pyralisのルシフェラーゼ中の残基214に対応するアミノ酸残基がスレオニン以外である、(d)Photinus pyralisのルシフェラーゼ中の残基232に対応するアミノ酸残基がイソロイシン以外である、(e)Photinus pyralisのルシフェラーゼ中の残基295に対応するアミノ酸残基がフェニルアラニン以外である、(f)Photinus pyralisのルシフェラーゼのアミノ酸14に対応するアミノ酸残基がフェニルアラニン以外である、(g)Photinus pyralisのルシフェラーゼのアミノ酸35に対応するアミノ酸残基がロイシン以外である、(h)Photinus pyralisのルシフェラーゼのアミノ酸残基105に対応するアミノ酸残基がアラニン以外である、(i)Photinus pyralisのルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基がアスパラギン酸以外である、(j)Photinus pyralisのルシフェラーゼのアミノ酸残基420に対応するアミノ酸残基がセリン以外である、(k)Photinus pyralisのルシフェラーゼのアミノ酸残基310に対応するアミノ酸残基がヒスチジン以外である。
【0040】
あるいは、本発明のタンパク質は、Luciola mingrelica、Luciola cruciataまたはLuciola lateralis酵素のタンパク質のルシフェラーゼ配列を含み、前述の359位置の変異に加えて、以下のものを少なくとも1つ含む。
【0041】
(a)Photinus pyralisのルシフェラーゼのアミノ酸356に対応するアミノ酸残基がグルタメート以外である、(b)Photinus pyralisのルシフェラーゼ中の位置215に対応するアミノ酸残基がアラニンまたはスレオニン以外の疎水性アミノ酸である、(c)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基216に対応するアミノ酸残基がグリシン(Luciola mingrelicaベースの配列の場合)またはアパラギン(Luciola cruciataまたはLuciola lateralisベースの配列の場合)以外である、(d)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基234に対応するアミノ酸残基がセリン以外である、(e)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基297に対応するアミノ酸残基がロイシン以外である、(f)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのアミノ酸16に対応するアミノ酸残基がフェニルアラニン以外である、(g)Luciola mingrelicaの残基37、またはLuciola cruciataまたはLuciola lateralisの38に対応するアミノ酸残基がリジン以外である、(h)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのアミノ酸残基106に対応するアミノ酸残基がグリシン以外である、(i)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのアミノ酸残基236に対応するアミノ酸残基がグリシン以外である、(j)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisの残基422に対応するアミノ酸残基がスレオニン以外である、(k)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのアミノ酸残基312に対応するアミノ酸残基がスレオニン(Luciola mingrelicaベースの配列の場合)またはバリン(Luciola cruciataまたはLuciola lateralisベースの配列の場合)以外である。
【0042】
いずれの場合も、熱安定性を向上させる特定の置換アミノ酸を、以下に例示する一般的な方法によって決定することができる。それぞれの場合において、異なる置換が熱安定性を向上させ得よう。当業者によって理解されるように、置換は、元のタンパク質または適切に変異したタンパク質をエンコードしているDNAの部位特異的変異によって行い得る。この場合本発明は、熱安定性を伴う位置の特定を伴っている。
【0043】
しかしながら、一般に、野生型のアミノ酸と異なる性質のアミノ酸に置換することを考慮することが望ましいであろう。したがって、親水性アミノ酸残基を疎水性アミノ酸残基で置換することができるのが好ましい場合もある(逆も然り)。同様に、酸性アミノ酸残基を塩基性残基で置換することができる。
【0044】
たとえば、このタンパク質は、ルシフェラーゼ活性、およびPhotinus pyralis、Luciola mingrelica、Luciola cruciataまたはLuciola lateralis酵素からのルシフェラーゼと少なくとも60%の類似性を有するタンパク質を含んでよい。この酵素の配列中では、以下の少くとも一つが変異され、(a)Photinus pyralisのルシフェラーゼの残基214、Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基216に対応するアミノ酸残基が変異され、およびPhotinus pyralisのルシフェラーゼの場合はスレオニン以外である、または(b)Photinus pyralisのルシフェラーゼの残基232、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基234に対応するアミノ酸残基が変異され、Photinus pyralisのルシフェラーゼの場合はイソロイシン以外である、または(c)Photinus pyralisのルシフェラーゼの残基295、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基297に対応するアミノ酸残基が変異され、たとえばPhotinus pyralisのルシフェラーゼの場合はフェニルアラニン以外である。このルシフェラーゼ酵素は、野生型ルシフェラーゼと比較して熱安定性が増大している。
【0045】
さまざまなルシフェラーゼのすべての配列は、これらはそれらの間で高い類似性を有し、高度に保存されていることを示している。これは、この配列間の対応する領域は、この配列を調べることによって容易に決定することができ、最も類似の、領域を検出できることを意味する。しかし、必要ならば、市販のソフトウェア(たとえばUniversity of Wisconsin Genetics Computer Groupからの「Bestfit」;Devereux他(1984)Nucleic Acid Research 12:387-395を参照のこと)を使用して、さまざまな配列間の対応する領域または特定のアミノ酸決定することができる。あるいはまたはさらに、L.Ye他のBiochim.Biophys Acta 1339(1997)39-52を参照することによって対応する酸を決定することができる。
【0046】
Photinus pyralisのルシフェラーゼ中の残基214に対応するアミノ酸残基の可能な変化について、極性アミノ酸スレオニンが、アラニン、グリシン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファンまたはシステインなどの非極性アミノ酸で適切に置き換えられる。Photinus pyralis中の残基214に対応する、スレオニン残基に関する特に好ましい置換はアラニンである。より好ましい置換はシステインである。しかしながら、この位置のアスパラギンなどの異なる極性残基が、この位置にスレオニンを有する対応する酵素の熱安定性を向上させる可能性もある。野生型ルシフェラーゼ酵素中のこの位置に現れる他のアミノ酸にはグリシン(Luciola mingrelica、Hotaria paroula)、アスパラギン(Pyrophorus plagiophthalamus、GR、YG、YEおよびOR、Luciola cruciata、Luciola lateralis、Lampyris noctiluca、Pyrocelia nayako Photinus pennsylvanica LY、KW、J19)およびセリン(Phrixothix)がある。有利には、アラニンおよびシステインなどの非極性または異なる非極性側鎖でこれらを置換することができる。
【0047】
Photinus pyralisのルシフェラーゼ中の残基232に対応するアミノ酸残基の可能な変化に関しては、非極性アミノ酸イソロイシンが、アラニン、グリシン、バリン、ロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファンまたはシステインなどの異なる非極性アミノ酸で適切に置き換えられる。野生型ルシフェラーゼ酵素中のこの位置に現れる他のアミノ酸には、セリンおよびアスパラギンがある。適切には、これらの極性残基が前述のような非極性残基で置換される。Photinus pyralisのルシフェラーゼ中の残基232、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisの基の部分の残基234に対応する残基に関する特に好ましい置換はアラニンである。
【0048】
Photinus pyralisのルシフェラーゼ中の残基295、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基297に対応するアミノ酸残基の変化が、タンパク質の熱安定性に影響を及ぼそう(これはPhrixothixのルシフェラーゼ中の位置292に対応する。)。一般に、この位置のアミノ酸は、非極性アミノ酸フェニルアラニンまたはロイシンである。これらは、異なる非極性アミノ酸に、適切に変えられる。たとえばPhotinus pyralisでは、非極性アミノ酸フェニルアラニンが、アラニン、ロイシン、グリシン、バリン、イソロイシン、プロリン、メチオニン、トリプトファンまたはシステインなどの異なる非極性アミノ酸で適切に置き換えられる。Photinus pyralisのルシフェラーゼ中の残基214に対応するフェニルアラニン残基に関する特に好ましい置換はロイシンである。
【0049】
Photinus pyralisのルシフェラーゼのアミノ酸14、またはLuciolaのルシフェラーゼのアミノ酸16(Phrixothrixのルシフェラーゼの13)に対応するアミノ酸残基における変異も可能である。このアミノ酸残基(通常はフェニルアラニンであるが、ロイシン、セリン、アルギニンまたはある場合はチロシンであってもよい)を異なるアミノ酸、特にアラニン、バリン、ロイシン、イソロイシン、プロリン、メチオニンまたはトリプトファン、好ましくはアラニンなどの異なる非極性アミノ酸に適切に変化させる。
【0050】
Photinus pyralisのルシフェラーゼのアミノ酸35、またはLuciola mingrelicaのルシフェラーゼのアミノ酸残基37(他のLuciola spp.中の38)に対応するアミノ酸残基における変異も効果的であろう。このアミノ酸は野生型酵素の間で変化し、ロイシン(Photinus pyralis)だけでなくリジン、ヒスチジン、グリシン、アラニン、グルタミンおよびアスパラギン酸をこの位置に含み得る。適切には、この位置のアミノ残基は、アラニン、バリン、フェニルアラニン、イソロイシン、プロリン、メチオニンまたはトリプトファンなどの非極性アミノ酸残基または異なる非極性アミノ酸で置換される。この位置の好ましいアミノ酸はアラニンであり、これは野生型酵素と異なる。
【0051】
Photinus pyralisの配列の位置14に対応するアミノ酸における変異、および/またはPhotinus pyralisのルシフェラーゼのアミノ酸35に対応するアミノ酸残基における変異は、酵素中の唯一の変異ではないことが好ましい。
【0052】
これらは、前述の変異の他のもの、特にPhotinus pyralisのルシフェラーゼの位置214、395または232に対応する位置での変異を伴うことが適切である。
【0053】
Photinus pyralisのルシフェラーゼ中の残基105、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基106(Phrixothix中の102)に対応するアミノ酸残基の変化が、タンパク質の熱安定性に影響を及ぼそう。一般に、この位置のアミノ酸は、非極性アミノ酸アラニンまたはグリシン、またはPhrixothix中のセリンである。これらは、異なる非極性アミノ酸に、適切に変化させる。たとえばPhotinus pyralisでは、非極性アミノ酸アラニンが、フェニルアラニン、ロイシン、グリシン、バリン、イソロイシン、プロリン、メチオニン、トリプトファンなどの異なる非極性アミノ酸で適切に置き換えられる。Photinus pyralisのルシフェラーゼ中の残基105に対応するアラニン残基に関する特に好ましい置換はバリンである。
【0054】
Photinus pyralisのルシフェラーゼ中の残基234、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基236(Phrixothix中の231)に対応するアミノ酸残基の変化が、タンパク質の熱安定性に影響を及ぼそう。一般に、この位置のアミノ酸は、アスパラギン酸またはグリシンであり、グルタミンまたはスレオニンである場合もある。これらは、非極性または適切な異なる非極性アミノ酸に、適切に変化させる。たとえば、Photinus pyralisでは、アミノ酸残基はアスパラギン酸であり、アラニン、ロイシン、グリシン、バリン、イソロイシン、プロリン、メチオニンまたはトリプトファンなどの非極性アミノ酸で適切に置き換えられる。Photinus pyralisのルシフェラーゼ中の残基234に対応するフェニルアラニン残基に関する特に好ましい置換はグリシンである。グリシンなどの非極性アミノ酸残基がこの位置に存在する場合(たとえばLuciolaのルシフェラーゼ中で)、異なる非極性アミノ酸でこれを置換することができよう。
【0055】
Photinus pyralisのルシフェラーゼ中の残基420、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基422(Phrixothixグリーン中の417およびPhrixothixレッド中の418)に対応するアミノ酸残基の変化は、タンパク質の熱安定性に影響を及ぼそう。一般に、この位置のアミノ酸は、帯電していない極性アミノ酸セリンまたはスレオニンまたはグリシンである。これらは、異なる帯電していない極性アミノ酸に、適切に変化させる。たとえば、Photinus pyralisではセリンを、アスパラギン、グルタミン、スレオニンまたはチロシン、特にスレオニンで適切に置き換えることができよう。
【0056】
Photinus pyralisのルシフェラーゼ中の残基310、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基312に対応するアミノ酸残基の変化は、タンパク質の熱安定性に影響を及ぼそう。この位置のアミノ酸残基は、知られているルシフェラーゼタンパク質の間で変化し、Photinus pyralis、Pyrocelia nayako、Lampyris noctilucaおよびある形のPhotinus pennsylvanicaのルシフェラーゼではヒスチジン、Luciola mingrelica、Hotaria paroulaおよびPhrixothix(この場合それはアミノ酸307である)のルシフェラーゼではスレオニン、Luciola cruciataおよびLuciola lateralisではバリン、およびある種のPyrophorus plagiophthalamusのルシフェラーゼではアスパラギンである。したがって、一般に、この位置のアミノ酸は、酵素の熱安定性を増大させる異なるアミノ酸残基に変化させ得る親水性アミノ酸である。Photinus pyralisのルシフェラーゼ中の残基310に対応するヒスチジン残基に関する特に好ましい置換はアルギニンである。
【0057】
他の変異がこの酵素中に存在してもよい。たとえば好ましい実施形態では、タンパク質は、グルタメートから変化した、Photinus pyralisのルシフェラーゼのアミノ酸354(Luciolaのルシフェラーゼ中の356)に対応するこの位置でのアミノ酸残基を有し、特にグリシン、プロリンまたはアスパラギン酸以外のアミノ酸へ変化したこの位置でのアミノ酸も有する。適切にはこの位置のアミノ酸はトリプトファン、バリン、ロイシン、イソロイシンおよびアスパラギンであり、最も好ましくはリジンまたはアルギニンである。この変異はWO95/25798中に記載されている。この位置の疎水性残基は、この酵素の波長シフトを向上させることが分かっており、さらに、位置354における大きな疎水性(VまたはI)、極性(N)または正に帯電している(KまたはR)アミノ酸の存在は熱安定性を向上させる。
【0058】
他の実施形態では、EP−A−052448に記載されているように、このタンパク質は、Luciolaルシフェラーゼ中のアミノ酸217(Photinus pyralis中の215)に対応する位置において、疎水性アミノ酸に、特にイソロイシン、ロイシンまたはバリンに変化したアミノ酸を有している。
【0059】
本発明のタンパク質は、野生型および組換えルシフェラーゼ酵素の両方を含む。これらは、野生型酵素中に存在する少なくとも60%のアミノ酸が本発明のタンパク質中に存在するという意味で、Photinus pyralis、Luciola mingrelica、Luciola cruciataまたはLuciola lateralisの酵素の配列などの野生型配列と少なくとも60%の類似性を有する。このようなタンパク質は、前述の野生型酵素とより大きな類似性、詳細には少なくとも70%、より好ましくは少なくとも80%および最も好ましくは少なくとも90%を有し得る。類似なタンパク質はこのタイプであり、対立遺伝子変異体、他の昆虫種からのタンパク質、および組換えによって生成された酵素を含む。野生型酵素をエンコードしている配列とストリンジエントハイブリダイゼーション条件下でハイブリダイズする核酸によってエンコードされている点で、これらのタンパク質は容易に同定することができる。このような条件は当業者によってよく理解されていよう。たとえばSambrook他(1989)Molecular Cloning、Cold Spring Harbor Laboratory Pressの中で例示されている。一般的に、低ストリンジェンシー条件はほぼ周囲温度から約65℃での3×SCC、高ストリンジェンシー条件は約65℃での0.1×SCCとして定義される。SSCは0.15M NaCl、0.015Mクエン酸三ナトリウムのバッファーの名前である。3×SCCはSSCなどの3倍強い。
【0060】
詳細には、LipmanおよびPearsonによって記載されている多重アラインメント法(Lipman、D.J.and Pearson、W.R.(1985)Rapid and Sensitive Protein Similarity Searches、Science、vol227、pp1435-1441)を使用して、特定の配列と本発明の配列との類似性を評価することができる。「最適化した」パーセンテージスコアは、Lipman-Pearsonアルゴリズムについて以下のパラメータを用いて計算すべきである。ktup=1、ギャップペナルティ=4およびギャップペナルティレングス=12。類似性を評価するための配列は「試験配列」として使用すべきであり、これは、Photinus pyralisの配列またはYe他に記録されている任意の他の配列などの比較用の塩基配列を、最初にこのアルゴリズムに入れるべきであることを意味する。
【0061】
本発明のタンパク質の特定の例は、前述の1つまたは複数の変異を有する野生型ルシフェラーゼ配列である。
【0062】
さらに本発明は、前述のルシフェラーゼをエンコードしている核酸を提供する。適切には、この核酸は当分野でよく知られる野生型配列をベースとする。アミノ酸配列中の所望の変異を行うための適切な変異は、遺伝コードの知識に基づいて容易に明らかであろう。
【0063】
本発明の好ましい実施形態では、核酸は合成遺伝子である。適切には、合成遺伝子を工作して、高度に発現された遺伝子中ではほとんど見られないコドンを大腸菌などの一般的な発現宿主から取り除き、これと同時に甲虫のルシフェラーゼをコードしている遺伝子中ではほとんど見られないコドンの導入を避ける。この手法は、この新しい遺伝子は大腸菌および昆虫の発現系の両方に最適なコドン使用を有することを確実にする。
【0064】
たとえば、可能なかぎり、アミノ酸arg、leu、ile、glyおよびproのコドンを、CGTまたはCGC(arg)、CTG、CTTまたはCTC(leu)、ATCまたはATT(ile)、GGTまたはGGC(gly)、およびCCG、CCAまたはCCT(pro)に変化させ、したがって希なコドンが取り除かれる。以下(配列番号1)および図14に例示する合成遺伝子の場合、これによって62個の新しく一般的なコドン(全体の11%)を生み出す合計139のサイレント変異を生じた。図14に示す最初の8個のヌクレオチドは、リボゾーム結合部位の一部を形成し、したがってコードしていない。コード配列は、上向き矢印によって示されるメチオニン残基で始まる。このコード配列および非常に類似した配列、たとえば少なくとも90%の類似性、好ましくは95%の類似性を有する配列は、本発明の好ましい態様を形成する。
【0065】
合成アセンブリを生成するときに使用することができる他の有用な性質は、新しい独特な制限部位の導入である。これらの部位は遺伝子の変異誘発、特にコンビナトリアルカセット変異をより簡単でより効率的にする。詳細には、酵素中にサブドメインBをコードするcDNA内に、独特な制限部位を作製することが望ましいであろう。さらに、遺伝子の3'端部に独特な制限部位を作製して、ペルオキソーム標的配列の簡単な融合および/または除去を可能にすることが有利であろう。
【0066】
以後に示す例では、9つの新しい独特な制限部位を工作し(大部分遺伝子の中央の3番目)、独特なHind III部位を遺伝子の3'端部に生成させ、簡単なC末端融合を可能にした(図12)。
【0067】
最後に、合成遺伝子の使用は、遺伝子産物の熱安定性を増大させるための、またはそうでない場合は産物の性質を望むように改変するための、変異の導入を可能にする。たとえば、以後に示す実施例では、サイレントではない3つの変異を工作して、熱安定アミノ酸の変化T214C、E354KおよびD357Fをポリペプチドに導入した。
【0068】
本発明の核酸を、プロモーター、エンハンサー、ターミネーターなどの調節要素の調節下で、プラスミドなどの発現ベクターに適切に導入する。次いでこれらのベクターは、宿主細胞、たとえば植物または動物細胞などの原核または真核細胞(詳細には大腸菌などの原核細胞である)を形質転換させるために、細胞が所望のルシフェラーゼ酵素を発現するように使用することができる。当分野でよく知られる条件を使用して、このようにして形質転換した細胞を培養することによりルシフェラーゼ酵素が生成されよう。次いでこれを培養培地から分離することができる。細胞が植物細胞または動物細胞である場合、前記細胞から植物または動物を繁殖させることができる。したがって植物からタンパク質を抽出することができ、トランスジェニック動物の場合は、ミルクからタンパク質を回収することができる。ベクター、形質転換した細胞、トランスジェニック植物および動物、およびこれらの細胞を培養することによって酵素を生成させる方法は、いずれも本発明の他の態様を形成する。
【0069】
以下に記載するように、ランダム突然変異誘発によって、Photinus pyralisD357Y突然変異型ルシフェラーゼを作製した。D357Yの1点での変異は発せられる光の波長の大きなカラーシフトを生み出し、また野生型ルシフェラーゼより熱安定性が高いことが分かった。さらなる調査によって、この位置でのある範囲の置換によって優れた熱安定性、および/または大きなカラーシフトが生み出さることが明らかになっている。
【0070】
本発明の範囲内に入るPhotinus pyralisの変異型酵素の特定の例には以下のもの、または、他の種のルシフェラーゼであれば、これらの任意の均等物がある。
【0071】
【表1】

【0072】
上記変異体を作製するための変異は、部位特異的変異法、(PCR)またはコンビナトリアルカセット変異によって、プラスミドpET23上のルシフェラーゼ遺伝子に導入した。適切な変異を行うための、PCR反応に加えられたオリゴヌクレオチドは以下に示す。
【0073】
354および215位置での点変異の影響は追加的なものであることが以前に報告している。本発明は、3つ以上のこのような変異を組み合わせる可能性を提供して、大きなカラーシフトを有する変異型酵素中における高い熱安定性を提供する。
【0074】
本発明のルシフェラーゼタンパク質は、シグナル手段としてルシフェラーゼ/ルシフェリン反応を使用するいかなる生物発光アッセイにおいても、有利に使用できよう。文献中で知られている多くのこのようなアッセイが存在する。したがって、このようなアッセイを行う目的で作製されたキット中このタンパク質が含まれよう。場合によってはルシフェリンおよび特定のアッセイを行うために必要とされる任意の他の試薬も含まれる。
【0075】
ここで本発明を、例示として添付の図面を参照することによって詳細に記載する。
【0076】
図1は、残存活性%対本発明によるいくつかの変異型酵素の45℃でのインキュベーションの時間を示す対数グラフである。
【0077】
図2は、D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。この場合、使用した酵素は(a)組換え野生型Photinus pyralisのルシフェラーゼ、(b)D357K変異型、(c)D357N変異型、(d)D357W変異型、(e)D357I変異型、(f)D357F変異型、(g)D357Y変異型および(h)二重変異型E354I+D357Yである。
【0078】
図3は、残存活性%対3つの変異型酵素E354I、D357Yおよび二重変異型(DM)E354I/D357Yの時間を示すグラフである。
【0079】
図4は、(a)組換え野生型酵素および(b)二重変異型(DM)E354I/D357Yの発光スペクトルを示す。
【0080】
図5は、組換え野生型(◆)r−wtおよびD357K変異型酵素(■)の光量子発光の速度減衰を示すグラフである。
【0081】
図6は分子モデル図を示し、ルシフェラーゼ酵素中の考えられるCoA結合ポケットを示す。
【0082】
図7は、(a)LB上で成長させた、(b)最小培地および酢酸ナトリウム上で成長させた、(c)最小培地およびグルコース上で成長させた、変異体P.pyralisルシフェラーゼD357Yを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【0083】
図8は、(a)LB上で成長させた、(b)最小培地および酢酸ナトリウム上で成長させた、(c)最小培地およびグルコース上で成長させた、変異体P. pyralisのルシフェラーゼE354K/D357Mを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【0084】
図9は、変異体P. pyralisルシフェラーゼD357Yによって発せられた光のスペクトル分布に対するCoAの影響を示すグラフである。
【0085】
図10は、変異体P. pyralisルシフェラーゼD357Yによって発せられた光のスペクトル分布に対するCoAの影響の正規化したデータを示すグラフである。
【0086】
図11は、変異体P. pyralisルシフェラーゼE354I/D357Yによって発せられた光のスペクトル分布に対するCoAの影響(図11a)および正規化したデータ(図11b)を示すグラフである。
【0087】
図12は、合成ルシフェラーゼ遺伝子の構築において使用された制限部位の修飾を示す。
【0088】
図13は、ルシフェラーゼ遺伝子の合成において使用された構築体を示す。
【0089】
図14は、合成ルシフェラーゼ遺伝子のcDNA配列(配列番号1)(リボソーム結合部位の一部を形成し、コードしていないヌクレオチド1〜8を含む)、および上向き矢印で示されたメチオニン残基で始まるエンコードされているアミノ酸配列(配列番号2)を示す。
【0090】
図15は、合成遺伝子によってエンコードされている変異体を含む、変異体の50℃における熱安定性を示す。
【0091】
実施例1
変異型ルシフェラーゼの同定および特徴付け エラープロンPCR〔M.Fromant他、Anal.Biochem.(1995)224、347-353〕を使用して作製した、ホタル(Photinus pyralis)ルシフェラーゼの2つのライブラリーを調製した。完全長のluc遺伝子のエラープロンPCR産物からなる1つのライブラリーを、T7発現系pET23aにクローン化した(Novagen Inc.、Madison、WI、U.S.A)。199〜35個のアミノ酸をカバーする、luc遺伝子の短い断片のエラープロンPCR産物からなる第2のライブラリーを、ベクターpBSK(+)中にクローン化した(Stratagene、La Jolla、CA、U.S.A)。
【0092】
pET23aライブラリーを、大腸菌菌株BL21(DE3)中で発現させた(大腸菌B F dcm ompt hsdS(rB−mB−)galλ(DE3))。
【0093】
pBSK(+)ライブラリーを、HB101 大腸菌細胞(supE44 aral4 galK2 lacY1 Δ(gpt−proA)62 rpsL20(Strr)xyl−5 mtl−1 recAl3 Δ(mrcC−mrr)HsdS−(r−m−))中で発現させた。pET23aおよびpBSK(+)は両方共にβ−ラクタマーゼの遺伝子を保有し、このプラスミドを有する大腸菌細胞にアンピシリン耐性を与える。
【0094】
BIORAD E.coli Pulserを使用するエレクトロポレーションによって、作製したライブラリーを用いて、大腸菌株細胞を形質転換させ、50μg/mlの濃度のアンピシリンを含むLB寒天上で37℃で一晩生育させた。細胞をナイロン膜(Osmonics、Minnetonka、Minnesota、U.S.A)に移し、ルシフェリン溶液(クエン酸ナトリウムバッファー100mM、pH5.0中に500μM D−ルシフェリン、カリウム塩、)をスプレーした。AlphImager(商標)1200DocumentationおよびAnalysis System(Flowgen、Lichfield、Staffordshire、UK)を使用して、コロニーを観察した。これによって特定の時間で発せられる生物発光が取り込まれて、コロニーによって発せられる光の像を生み出す。発光の明るさを、ルシフェラーゼの熱安定性の指標として記録した。
【0095】
次いで、熱安定性についてコロニーを選別した。発せられる光の明るさに基づいてコロニーを選択し、さらなる特徴付けのために単離した。いくつかの選別では、選別の前に大腸菌コロニーを42℃で2時間インキュベートし、その結果熱安定性の変異体を選択することができた。最初の選別から単離したコロニーをナイロン膜の上に載せ、アンピシリンを含むLB培地上で一晩生育させた。パッチをルシフェリン溶液でスプレーし、AlphImager(商標)中で観察した。この第2の選別は、ルシフェラーゼ活性のインビトロでの分析用のクローンを陽性に同定するのに役立った。可能性ある熱安定性酵素を発現する大腸菌のクローンを、ルシフェラーゼ活性および熱安定性についてインビトロでアッセイした。
【0096】
室温で、Promega Luciferase Assay System(Promega Corporation、Madison、WI、U.S.A)を使用して、ルシフェラーゼ活性についてインビトロでのアッセイを行った。
【0097】
粗細胞抽出物10μlをPromega Luciferase Assay Cocktail(1:2に希釈)に加えることによって、ルシフェラーゼ反応を開始させた。Biotrace M3 照度計を使用して、生じた生物発光を測定した。
【0098】
Promega technical bulletein no.101に記載されているように、粗細胞抽出物を調製した。細胞培養物溶解試薬(25mM トリスリン酸塩 pH7.8、2mM ジチオスレイトール(DTT)、2mM 1,2−ジアミノシクロヘキサン−N,N,N',N'−テトラ酢酸、10%グリセロール、1%トリトン X−100、1.25mg/ml hen リソザイム)中で10分間かけて室温で、一晩培養した大腸菌のアリコートを溶解させた。次いで粗溶解物をアッセイ前に氷上に保存した。
【0099】
時間依存不活性化実験において、酵素の諸性質をさらに試験した。粗細胞抽出物のアリコート50μlを含むエッペンドルフチューブを、水浴中において所与の温度でインキュベートした。設定したいくつかのタイムポイントで、チューブを取り除きアッセイ前に氷上で冷却した。残存ルシフェラーゼ活性を、元の活性のパーセンテージとして表した。
【0100】
残存活性%対インキュベーション時間の対数グラフをプロットし、t1/2値を計算するために使用した。T1/2とは、酵素の、所与の温度でインキュベーション後の、その元の活性の50%を失うのにかかる時間のことである。残存活性%に対する時間の対数グラフから、37℃での粗抽出物中のT1/2値(活性が元の活性の50%に低下する時間)を決定した(示さず)。
【0101】
上に述べたように決定した最も熱安定性のあるルシフェラーゼを発現する大腸菌クローンからのプラスミドDNAを配列決定して、酵素の熱安定性を担う変異を決定した。
【0102】
QIAGEN QIAprep Spin Miniprep Kit(QIAGEN Ltd、Crawley、W.Sussex、UK)、次にミクロ遠心分離を使用するためのプロトコル(QIAprep Miniprep Handbook04/98)を使用して、プラスミドDNAを作製した。
【0103】
DNAの配列決定はすべて、ABI PRISM(商標)377 DNA Sequencerおよびジデオキシ鎖停止法〔F.Sanger他、Proc.Natl.Acad.Sci.U.S.A 74、(1977)5463−5467〕に基づくABI PRISM(商標)BigDye(商標)Terminator Cycle Sequencer Ready Reaction Kit(Perkin Elmer Applied Biosystems)を使用して、Babraham Technix、Cambridge、UKによって行った。
【0104】
この作業の結果、新規な変異体D357Yを特定した。
【0105】
ルシフェラーゼの結晶構造〔E.Conti他、Structure、4(1996)287−298〕は、位置357はタンパク質の表面に適合しており位置354に近く、熱安定性およびスペクトルの諸性質の両方に影響を及ぼす可能性があることを示す。これは、この領域は酵素の熱安定性の点で重要である可能性があることを示す。
【0106】
D357Yは非常に熱安定性のある変異体であって、最も熱安定性のあるルシフェラーゼであり、ただ1つのアミノ酸が変化している。
【0107】
実施例2
他の357変異体を作製するための部位特異的変異 357位置における異なる変異体を評価するために、Stratagene QuikChange(商標)Site−Directed Mutagenesis Kit(Stratagene、La Jolla、CA、U.S.A)を使用して、部位特異的変異を行った。プラスミドpPW601a J54(PJW、MoD Report、3/96)を、すべての部位特異的変異において使用した。変異誘発反応のすべての産物を、大腸菌菌株XL1−Blue〔e14−(mcrA−)Δ(mcrCB−hsdSMR−mrr)171 endA1 supE44 thi−1 gyrA96 relAl lac recB recJ sbcC umuC::Tn5(Kanr)uvrC〔F'proAB laclqzΔm15 Tn 10 (Tetr)Amy Camr〕に形質転換した。Sigma−Genosys Ltd.、Cambridge、UKによってオリゴヌクレオチドプライマーを合成し、インテリジェントドーピングシステム〔A.R Arkin他、Bio−technology、(1992)10、297−300、W、.Hung他、Anal.Biochem.(1992)10、454−457〕を使用して設計し、それぞれのアミノ酸置換に関する個々のプライマーを使用せずに、これを使用して縮重オリゴヌクレオチドプライマーを設計し、可能性ある変異体の群を作成した。
【0108】
このようにして、アミノ酸置換ルシフェラーゼ変異体のライブラリーを作成した。
【0109】
以下のオリゴヌクレオチド(およびその相補的なパートナー)を使用した。
【0110】
【化1】
【0111】
変異体のライブラリーを、前述のように熱安定性に関して選別した。以下の等式〔S.Climie他、J.Biol.Chem.265(1990)18776−18779〕を使用して、選別したコロニーの数を計算した。
【0112】
N=〔1n(1−p)〕/〔1n((n−1)/n〕
Nが選別されるコロニーの数の場合、nは標的位置における可能性あるコドンの数であり、Pは混合物中のそれぞれのコドンが、少なくとも1度の選別用にサンプル採取される確率である。この計算はP=0.95に基づいていた。部位特異的変異から得た変異体をルシフェラーゼ活性に関してアッセイし、時間依存熱不活性化実験において特徴付けした。
【0113】
このようにして望ましいとした変異体を、アンピシリンを含みA260 ?0.5の400mlのLB培地中で生育させた。次いでイソプロピルβ−チオガラクトシダーゼ(IPTG)を加え最終濃度1mMにすることによって、ルシフェラーゼ発現を誘導した。細胞を30℃で撹拌しながら3時間インキュベートし、次いで遠心分離によって採取した。生じた細胞のペレットを、Maxi−Scale Bacterial Protein Extractionに関するB−PER(商標)のプロトコルに従って、B−PER(商標)Protein Extraction Reagent、(Pierce Chemical Company、Rochford、U.S.A)10ml、DTT1mM中に再懸濁させて、粗抽出物を作成した。再構築Sigma Protease Inhibitor Cocktail、500μl(Product No.P8465、Sigma、Saint Louis、Missouri、U.S.A)をB−PER(商標)溶液に加えて、内生プロテアーゼを阻害した。次いで細胞の溶解物を、30000gで30分間遠心分離した。
【0114】
粗抽出物の上澄みを、硫酸アンモニウムを用いて分画した。30%と55%の間の飽和状態で沈殿した分画は、ルシフェラーゼ活性を含んでいた。この物質を0.5ml Tris HCl pH8.0、1mM DTTに再懸濁させ、熱不活性化およびスペクトルの実験用に使用した。
【0115】
変換D357L、T、V、W、R、I、S、K、NおよびFを導入した。粗抽出物のインビトロでの熱不活性化実験において、これらの変異体を特徴付けた。
【0116】
部分精製した抽出物を、熱不活性化バッファー:飽和した硫酸アンモニウム10%、1mMジチオスレイトールおよび0.2%BSAを含む50mM リン酸カリウムバッファー pH7.8中11に1の比率で希釈した。
【0117】
タンパク質溶液のアリコート110μlを、アッセイ前に設定した時間だけ40℃または45℃でインキュベートし、氷上で冷却した。次いでPromega Luciferase Assay Reagent(1:2に希釈)を使用して、実施例1に記載するようにルシフェラーゼ活性を測定した。
【0118】
その結果を表2および3、および図1中に示す。未精製抽出物のT1/2値を40℃(表2)および45℃(表3)において決定した。
【0119】
【表2】
【0120】
【表3】
【0121】
すべての置換が、組換え野生型と比較して、向上した熱安定性を示した。
【0122】
実施例3
発せられる光の波長の変化 位置357におけるアミノ酸置換が、インビトロで酵素によって発せられる光のスペクトルに影響を及ぼすことも観察した。実施例2に記載の大腸菌細胞培養物のアリコート(250μl)を37℃で一晩生育させ、ミクロ遠心分離においてスピンダウンさせ、上澄みを取り除いた。D−ルシフェリン150μlを含むクエン酸バッファー(pH5.0)中で、異なる変異型ルシフェラーゼを発現する細胞をインキュベートし、SPECTRAmax(登録商標)Microplate Spectrofluorometer、(Molecular Devices Crop.California、U.S.A)を使用して発光スペクトルを測定することによって、インビトロでの反応から発せられる光を分析した。変異体D357Y、FおよびIに関して、スペクトルのピークおよび波長の分布の大きな変化を観察した(図2(a)〜(g))。これらの結果を以下の表4に要約する。
【0123】
さらに、インビボでの変異体の発光を、暗室で肉眼によって評価した。D357変異体は、その発光スペクトルにおいてさまざまな色を示した。特にD357Y、FおよびIは、発せられる光のより長い波長への大幅なシフトを示した。
【0124】
ある場合(たとえばD357F)、光の発光の色の変化はλmaxのシフトによるだけでなく、可視光線の異なる波長からのスペクトルへの貢献の違いによるようである。
【0125】
【表4】
【0126】
ある種の357変異体のインビボでの発光のλmaxの比較のために、組換え野生型(r−wt)酵素を使用した。D357Y、FおよびIは、その波長最大値の大幅なシフトを示した。
【0127】
実施例4
CoAの存在または不在下での酵素の諸性質 実施例1に記載したようにして、硫酸アンモニウム沈殿によってD357Yを部分的に精製した。この部分的に精製したD357Y酵素(5μl)をPromega Luciferase Assay Reagent150μlと混合させた。他のアリコートをCoAは存在しない(25mM トリストリシン pH7.8、5.0mM MgSO4、0.1mM EDTA、2mM DTT、470μM D−ルシフェリン、530μM ATP)同等のアッセイ用バッファーと混合させた。2つの反応物の発光スペクトルを測定した。これを図9および10に示す。
【0128】
これらのスペクトルは、CoAの存在または不在下での生物発光の顕著な違い、およびλmaxの大幅なシフトを示す。CoAのルシフェラーゼ反応のキネテイクスに対する影響を、RLUスケール(RLU−相対的光単位)の違いによって見ることもできる。
【0129】
この発光の違いによって、CoAの存在を検出するためのアッセイにおいてこの酵素を使用する可能性が生じる。
【0130】
実施例5
二重変異体の作製および諸性質 実施例2に記載するようにして、部位特異的変異を使用して、熱安定性および発光の色に対するあらゆる累積的影響を調べるために、E354I+D357Yの二重変異体を工作した。
【0131】
部分精製した二重変異体E354I+D357Yを、熱不活性化バッファー:飽和した硫酸アンモニウム10%、1mMジチオスレイトールおよび0.2%BSAを含む50mM リン酸カリウムバッファー pH7.8中11に1の比率で希釈した。
【0132】
タンパク質溶液のアリコート110μlを、アッセイ前に設定した時間だけ40℃または45℃でインキュベートし、氷上で冷却した。次いでPromega Luciferase Assay Reagent(1:2に希釈)を使用して、前述のようにルシフェラーゼ活性を測定した。
【0133】
二重変異体は、単一の変異体E354IおよびD357Yそれぞれと比較して、熱安定性の顕著な増大を示した(図3を参照)。部分精製した二重変異体の熱不活性化実験により、この変異体の熱安定性の増大を確認し、45℃での不活性化において7.7分のt1/2値を与えた。
【0134】
E354IおよびD357Yの個々の変異体より、この二重変異体は、より深い赤色の発光を示し、発光の色の添加性を示すことが示された。
【0135】
実施例3に記載のアッセイ用バッファーを使用して、組換え野生型および二重変異体E354I+D357Yの粗抽出物の発光スペクトルも測定した。
【0136】
インビボで測定した発光スペクトルは、611nmのλmaxを与えた。しかしながら、このスペクトルは、波長の赤い領域からの発光に大いに貢献し、肉眼によって視覚化するとより深い赤色の外観となる。未精製抽出物の発光スペクトルは、rWTと比較して、スペクトル形状の明確な変化および44nmの波長シフトを示した(図4を参照)。
【0137】
インビボでの二重変異体の発光スペクトルは、発光(613nm)のピーク波長の帯幅をシャープにし、および540〜560nmの範囲の光の波長の貢献の低下させる両方を示す。
【0138】
これらの変異の劇的な効果によって、この酵素のこの領域の生物発光の色への重要性が示される。
【0139】
実施例6
改善された光量子の流れ 変異体D357Kを発現している大腸菌細胞のインビボでの生物発光は、この位置の他の変異体と比較して、非常に鮮やかであることを観察した。時間における光量子の発光の速度を測定することができる光度計を使用して、この酵素のフラッシュキネテイクスを分析した。組換え野生型酵素または変異体D357を含む大腸菌無細胞抽出物のアリコートを、グローキネティクスを助長するいかなる試薬(たとえば補酵素A)も含まないルシフェラーゼアッセイ用カクテルに加えた。時間(15秒)における光量子の発光の減衰の速度を両方の酵素について測定し、変異体D357Kは大幅に遅いことを観察した(図4)。言い換えれば、変異型酵素は、反応の少なくとも最初の15秒間において、組換え野生型酵素よりも低い程度で阻害される反応キネテイクスを有する。
【0140】
実施例7
位置E354およびD357でのコンビナトリアルカセット変異 ステップ1 カセット変異用のプラスミドpPW601aJ54の工作 2対の合成オリゴヌクレオチド(以下参照)を使用して、2つの新しい独特な制限部位をプラスミドpPW601a/J54内のluc遺伝子中に導入した。合計6個のサイレント変異を、63塩基対隔ててこの遺伝子中のSpeIおよびKpnI制限部位に導入した。これらの新しい部位を含むプラスミドを、pPW601aJ54SpeI/KpnIと呼んだ。これらの制限部位の存在および近接性は、コンビナトリアルカセット変異を使用して、ホタルルシフェラーゼの主要配列中のアミノ酸の位置E354およびD357での、ランダムな置換の影響を探索することを可能にする。
【0141】
【化2】
【0142】
太線で強調したヌクレオチドはエンドヌクレアーゼ認識部位を形成し、および上側のケースのこれらはこの部位を作製するために必要なポイント変異の位置を形成する。
【0143】
ステップ2 カセットの設計およびライブラリーの構築 1対の合成オリゴヌクレオチドを合成した。このオリゴヌクレオチドを次いでアニールし、新しい制限部位で消化されたプラスミドpPW601aJ54SpeI/KpnIに直接連結することができる二本鎖カセットが作製された。このカセットは、天然に存在する20個のアミノ酸の考えられるすべての組み合わせを、この一次配列中の位置E354およびD357において導入できるように設計された。
【0144】
【化3】
ループライブラリーオリゴヌクレオチドのそれぞれ2μgを、50mM Tris−HCl pH7.4、25mM NaClを含むバッファー中に混合し、3分間100℃に加熱した。次いでこの溶液をヒーティングブロック中で<50℃までゆっくりと冷却して、相補的な配列をアニールさせた。次いでアニールしたオリゴヌクレオチドをプラスミドpPW601aJ54SpeI/KpnIに連結させ、SpeIおよびKpnIを用いてこれを消化した。次いでこの連結反応のアリコートを使用して、エレクトロポレーションを使用する大腸菌HB101細胞を形質転換した。エレクトロポレーションの後、形質転換した細胞をアンピシリン50μg/mlを含むLB寒天プレート上に載せ、37℃で一晩生育させた。翌日、869個のコロニーをプレートからランダムに採取し、これを使用してアンピシリンを含むLB1mlを96スクエアウエルのプレート(Beckman)に接種した。このプレートをカバーし、振とうしながら37℃で細胞を一晩生育させた。
【0145】
ステップ3 ランダムに選択したクローンのインビボでの選別 翌朝、定常期の一晩培養したもののアリコート50μlを、2つの透明プラスチック製丸底96ウエルのミクロタイタープレート(Dynex)に移した。1つのプレートをカバーし、加熱したブロック上で8分間インキュベートし(ブロック表面温度45℃)、一方もう1つのブロックは37℃に保った。次いで0.5mMのD−ルシフェリンを含む100mMのクエン酸ナトリウムバッファー(pH5.0)50μlをウエルに加え、次いでこのプレートをvideo camera imager capture system(Alpha Imager)に移すことによって、インビボでの両方のプレートからの細胞中のシフェラーゼ活性を室温で検出し記録した。加熱および調節された培養物によって発せられた光を1〜2分かけて積分し、その像を熱性紙フィルム上に記録した。
【0146】
フィルム上で記録した像の明るさの判定によって、最大の生物発光を示した79個の培養物を、選別の第2ラウンド用に選択した。今回は培養物を、アッセイの前にヒーティングブロック上で16分間インキュベートした。インビボでの熱安定性選別から選択した55個のクローンの内、インビボでのスペクトル分析用に25個を選択した。これらのクローンをLB中において37℃で一晩生育させ、翌朝一晩培養したもの200μlを翌朝遠心分離し、大腸菌細胞のペレットを0.5mMのD−ルシフェリンを含む100mMのクエン酸ナトリウムバッファー(pH5.0)150μl中に再懸濁させた。次いで再懸濁させた細胞を白いプラスチック製のミクロタイタープレート中に置き、変異型ルシフェラーゼのそれぞれによって発せられるインビボでの生物発光のスペクトルを、Molecular Devices Spectramax、96ウエルプレートの蛍光光度計を使用して分析した。それらの結果を以下の表1に要約する。
【0147】
ステップ4 変異の同定 インビボでの選別によって選択された25個のクローンからプラスミドDNAを調製し、遺伝子特異的配列決定プライマーを使用して配列決定した。一次配列中の位置354および357においてアミノ酸の変化を生じた変異を同定した。ある変異体は、位置I351にアミノ酸の置換をもたらした(表5)。
【0148】
【表5】
【0149】
いくつかの変異型ルシフェラーゼを熱安定性のインビボアッセイから選択した。これらのルシフェラーゼの大部分は、また発せられる光のインビボスペクトルにおいて大きな変化を示し、多くは光の長い波長(>580nm)からのより大きな貢献を示した。いくつかのスペクトルは、610〜614nmというただ1つのピーク周辺で非常に狭い帯幅を示す。
【0150】
E354およびD357のそれぞれ疎水性および芳香族アミノ酸での置き換え(たとえばE354V、D357Y)は、インビボでのスペクトルの大きく変化をもたらし、612nm周辺で、狭い帯幅の単位ピークを示す。
【0151】
実施例8
熱安定性に関するインビボ選別 選択したクローンの無細胞抽出物を溶解によって調製し、それぞれの抽出物からのルシフェラーゼの熱安定性を、熱不活性化実験において測定した。それぞれの抽出物50μlをエッペンドルフチューブ中に置き、4、9および16分間、45℃に加熱した水浴中でインキュベートした。適切なタイムポイントにおいてアリコートを取り、残存ルシフェラーゼ活性を測定した。表6は、すべての変異型酵素および組換え野生型について残存活性%対時間を示す。
【0152】
これらの結果は、最も熱安定性であるルシフェラーゼは、位置357に芳香アミノ酸(Y、FまたはW)、および位置354に大きな疎水性(VまたはI)、極性(N)または正に帯電している(KまたはR)アミノ酸を有するルシフェラーゼであることを示す。
【0153】
【表6】
【0154】
実施例9
発せられる光のインビボでのスペクトルに対する成長条件の影響 変異型ルシフェラーゼE354K+D357M(前述の7のもの)を発現している大腸菌BL21(DE3)細胞から発せられる、光のスペクトルに対する異なる炭素源の影響を調べた。
【0155】
細胞の培養物50mlをLB培地上で対数期中期まで生育させ、次いで遠心分離によって採取した。細胞ペレットを1mlの滅菌蒸留水中に再懸濁させ、次いでこの懸濁液のアリコート100ulを使用して、新鮮なLB5ml、M9最小培地+2mM酢酸ナトリウムまたはM9最小培地+2mMグルコースを25mlの滅菌チューブ中に接種した。この培養物を振とうしながら37℃で成長させ続け、90分(D357Y)および120分(酵素7)後、細胞のアリコート200ulを取り遠心分離し、0.5mMのD−ルシフェリンを含む100mMのクエン酸ナトリウムバッファー(pH5.0)150ul中に再懸濁させた。次いで再懸濁させた細胞をミクロタイタープレート中に置き、変異型ルシフェラーゼのそれぞれによって発せられる生物発光のインビボスペクトルを、Molecular Devices Spectramax、96ウエルプレートの蛍光光度計を使用して分析した。それらの結果を図7および8に示す。
【0156】
これらの結果は、富化培地(LB)(図7a、8a)からの、唯一の炭素源としてアセテート(図7b、8b)またはグルコース(図7c、8c)のいずれかを有する限定最小培地への変化は、発せられる光がより長い波長にシフトと、より短い波長からの貢献の低下をもたらすことを示す。
【0157】
実施例10
組換え野生型および変異型ルシフェラーゼの精製およびスペクトルの特徴付け 組換え野生型Photinus pyralis酵素および変異型ルシフェラーゼD357YおよびE354I+D357Yを、生物発光反応のスペクトルに対する補因子補酵素Aの影響を分析するために、均一になるまで精製した。3つのルシフェラーゼすべてを、嫌気性真菌Piromyces equiiからの143アミノ酸炭水化物結合モジュール(CBM)への融合体として、精製した。このCBMは、酸膨張性セルロースおよび可溶性炭水化物ガラクトマナンおよびグルコマナンに選択的に結合し、簡単な単一ステップのアフィニティ精製スキームのベースを形成することが示されている。
【0158】
CBMに融合したルシフェラーゼは、粗無細胞抽出物中でセルロースに結合することができ、これを洗浄し、次いで可溶性多糖を使用して選択的に溶離させた。このようにして精製した融合タンパク質をアッセイにおいて使用して、異なる量の補酵素Aを含む反応中で発せられた光の波長を測定した。異なる量の補酵素Aを含むアッセイ試薬100μl(25mM トリストリシン pH7.8、5.0mM MgSO4、0.1mM EDTA、530μM ATPおよび470μM D−ルシフェリン)に、酵素(5μl)を加えた。図9〜11は、精製したルシフェラーゼD357およびE354I+D357Yによって発せられた光のスペクトルに対する、補酵素Aの増大した濃度の影響を示す。
【0159】
真菌CBMのC末端に融合しホタルのルシフェラーゼを発現している大腸菌細胞によって発せられる、生物発光のスペクトルのインビボでのアッセイは、元のルシフェラーゼを発現している細胞といかなる顕著な違いも示さなかった。同様に、精製組換えルシフェラーゼの市販品(Promega)によって発せられる、生物発光のスペクトルのインビトロアッセイは、融合タンパク質によって発せられたスペクトルと同一であった。
【0160】
したがって、観察した違いはCoAの濃度に伴われたものである。補酵素Aの濃度が増大すると、スペクトルの分布が変化し、CoAの最大濃度では、スペクトルは、610nmでの明確なピークを有する590〜630nmの領域の波長が主となる。スペクトルのシフトは二重変異体に関して最も顕著であり、波長610nmという唯一のピーク周辺で帯幅の大幅な狭まりが存在する(図11)。
【0161】
実施例12
214C/354K/357Fを有するように変異させた、合成Photinus pyralisルシフェラーゼの生産 前述の合成戦略を使用して、オリゴヌクレオチド対から合成luc遺伝子を設計し組み立てた。遺伝子配列を工作して、アミノ酸214C、354Kおよび357Fを有するルシフェラーゼを作製した。
【0162】
重複合成オリゴヌクレオチド29対をSigma−Genosys Ltdによって合成し、PAGEによって精製し、約550bpの3つのアセンブリに連結させた(IDRIS1、2および3、図13)。次いでそれぞれのアセンブリをベクターpBSK(+)別々に連結させ、生じた構築体を使用して大腸菌XL1−Blue細胞を形質転換させた。組み立てた挿入体を含むクローンからプラスミドDNAを作製し、配列決定してORFの適合度を確認した。アセンブリ中のn−1個のオリゴヌクレオチドの存在(オリゴ合成の副産物)によって、構築プロセスが複雑になった。DNAの配列決定によってIDRIS2の1つの正確なアセンブリを同定し、PCRを使用して、構築体の5'端に1つの塩基対欠失体を含むIDRIS3の1アセンブリを正しくした。IDRIS1を含むプラスミドの、IDRIS2および3との混合物を連結させることによって、完全なORFのアセンブリを得た。
【0163】
次いで連結したDNAを使用して大腸菌XL1−Blue細胞を形質転換させ、インビボアッセイを使用して活性酵素を発現しているクローンを選択した。数個のクローンを選択し、配列決定して図14に示す配列を有する合成luc遺伝子の存在および適合度を確認した。完全なORFをIDRIS(FA)と呼んだ。
【0164】
合成遺伝子をベクターpBSK(+)内のポリリンカー中のBamH IとSal I部位の間に組立てた。この位置において、この遺伝子はアルファペプチドとはフレームが同じでなく、lacプロモーターからは相当な距離がある。しかしながら、充分なルシフェラーゼが生成されて、酵素の予備的特徴付けが可能である。Promega溶解法を使用して一晩培養したものから、IDRIS(FA)を発現している大腸菌XL1−Blue細胞の粗無細胞抽出物を作製した。
【0165】
次いで抽出物中の酵素の熱安定性を50℃で20分間試験し、熱安定性の変異体E354I+D357Yと比較した。新しいコドンを最適化させた三重変異体は、変異体E354I+D357Yよりも大幅に熱安定性であった(図15)。
(配列表)
【技術分野】
【0001】
本発明は、新規なタンパク質、詳細には対応する野生型酵素と比較して特徴的な性質を示す変異型ルシフェラーゼ酵素、これらのタンパク質をエンコードするDNA、アッセイにおけるこれらの酵素の使用、およびこれらを含む試験用キットに関する。
【背景技術】
【0002】
ホタルのルシフェラーゼは、ATP、Mg2+、および分子状酸素の存在下でルシフェリンの酸化を触媒し、その結果光を生み出す。この反応は、約0.88の量的収率を有する。この発光性が、ATPレベルが測定される、広範囲の照度アッセイにおける使用につながる。このようなアッセイの例には、EP−B−680515およびWO96/02665の記載に基づくアッセイがあるが、他の多くのアッセイも研究室において日常的に使用されている。
【0003】
昆虫の身体、特にホタルまたはツチボタルなどの甲虫から、ルシフェラーゼを直接得ることができる。ルシフェラーゼがそこから得られている特定の種には、日本のGENJIまたはKEIKEホタル、Luciola cruciataおよびLuciola lateralis、東ヨーロッパのホタルLuciola mingrelica、北アメリカのホタルPhotinus pyralis、およびツチボタルLampyris noctilucaがある。
【発明の概要】
【発明が解決しようとする課題】
【0004】
しかしながら、これらの酵素をエンコードしている遺伝子の多くはクローン化および配列決定されており、組換えDNA技術を使用して生成させることもできる。酵素をエンコードしている組換えDNA配列を使用して、大腸菌などの微生物を形質転換し、したがってこれが所望の酵素産物を発現する。
【0005】
研究室におけるアッセイにおいて使用されるとき、これらの酵素によって発せられる光の色は非常に似通っている。波長が変えられれば、特定の検出器によってより容易な読み取りのために、またはたとえば同じサンプル中の異なるエベントを監視するために多数のレポーターが必要なシステムにおいて使用するために、役に立つであろう。レポーター分子を区別する1つの方法は、特定の波長で光を発するルシフェラーゼ分子を使用することである。異なる種の甲虫またはツチボタルに由来するルシフェラーゼを含むレポーター分子を使用することによって、このことを達成することができる。しかしながら、組換えDNA技術を使用して変異型ルシフェラーゼを生成することが代替的な戦略であり、その結果シグナルの波長のバリエーションを生み出す。このような変異体の例は、WO95/18853において提供されている。
【0006】
さらに、野生および組換え型のルシフェラーゼの熱安定性は、それらが約30℃を超える温度、特に35℃を超える温度にさらされると、非常に急速に活性を失うようなものである。酵素が高い周囲温度で使用または保存されると、またはたとえば反応速度を増大させるために高温の反応条件下でアッセイが行なわれると、この不安定性は問題を引き起す。
【0007】
熱安定性が増大した変異型ルシフェラーゼは、EP−A−524448およびWO/95/25798から知られている。これらの第1のものは、日本のホタルのルシフェラーゼ中の位置217において変異(詳細にはスレオニン残基がイソロイシン残基で置き換えられている)を有する、変異型ルシフェラーゼを記載している。後者のものは、Photinus pyralis、Luciola mingrelica、Luciola cruciataまたはLuciola lateralisからのルシフェラーゼと60%を超える類似性を有する変異型ルシフェラーゼを記載しているが、Photinus pyralisの残基354またはLuciola種の356に対応するアミノ酸残基がグルタメート以外に変異しており、詳細にはグルタメート、アスパルテート、プロリンまたはグリシン以外に変異している。
【0008】
同時係属の英国特許出願No.9823468.5およびそれに由来する国際特許出願は、このような変異体をさらに記載している。この場合、ルシフェラーゼ活性があり、Photinus pyralis、Luciola mingrelica、Luciola cruciataまたはLuciola lateralis酵素などのルシフェラーゼからの野生型ルシフェラーゼと少なくとも60%の類似性を有するタンパク質が記載されているが、これらのタンパク質はタンパク質中のさまざまな位置に、特に以下のものを含めた変異を含む。
【0009】
(a)Photinus pyralisのルシフェラーゼ中の残基214、Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基216に対応するアミノ酸残基、または (b)Photinus pyralisのルシフェラーゼ中の残基232、Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基234に対応するアミノ酸残基、または (c)Photinus pyralisのルシフェラーゼ中の残基295、Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基297に対応するアミノ酸残基。
【課題を解決するための手段】
【0010】
本出願人は、ルシフェラーゼタンパク質中の異なる位置でアミノ酸を変異させる(または導入する)ことによって、発せられる光の波長の大きなシフトを得ることができることと、および/または酵素の熱安定性が改善されることを発見した。さらに、グローキネティクスが妨げられているインビボでのアッセイにおいて、またはCoAまたは他の「グローキネティクス誘導」化合物が存在しないインビトロアッセイにおいて、発せられるプロトンの流れが改善され酵素がより適合し得る。
【0011】
本発明は、ルシフェラーゼ活性、および野生型ルシフェラーゼと少なくとも60%の類似性を有する組換えタンパク質を提供する。この酵素の配列中において、Photinus pyralisのルシフェラーゼ中の残基357に対応するアミノ酸残基が対応する野生型ルシフェラーゼと比較して変異しており、その結果そのルシフェラーゼ酵素は、対応する野生型ルシフェラーゼと比較して異なる波長の光を発することができ、および/または対応する野生型ルシフェラーゼと比較して熱安定性が向上している。
【図面の簡単な説明】
【0012】
【図1】残存活性%対本発明によるいくつかの変異型酵素の45℃でのインキュベーションの時間を示す対数グラフである。
【図2(a)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素は組換え野生型Photinus pyralisのルシフェラーゼである。
【図2(b)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素はD357K変異型である。
【図2(c)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素はD357N変異型である。
【図2(d)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素はD357W変異型である。
【図2(e)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素はD357I変異型である。
【図2(f)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素はD357F変異型である。
【図2(g)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素はD357Y変異型である。
【図2(h)】D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。使用した酵素は二重変異型E354I+D357Yである。
【図3】残存活性%対3つの変異型酵素E354I、D357Yおよび二重変異型(DM)E354I/D357Yの時間を示すグラフである。
【図4(a)】組換え野生型酵素の発光スペクトルを示す。
【図4(b)】二重変異型(DM)E354I/D357Yの発光スペクトルを示す。
【図5】組換え野生型(◆)r−wtおよびD357K変異型酵素(■)の光量子発光の速度減衰を示すグラフである。
【図6】分子のモデル図を示し、ルシフェラーゼ酵素中の考えられるCoA結合ポケットを示す。
【図7a】LB上で成長させた変異体P. pyralisのルシフェラーゼD357Yを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【図7b】最小培地および酢酸ナトリウム上で成長させた変異体P. pyralisのルシフェラーゼD357Yを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【図7c】最小培地およびグルコース上で成長させた変異体P. pyralisのルシフェラーゼD357Yを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【図8(a)】LB上で成長させた変異体P. pyralisのルシフェラーゼE354K/D357Mを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【図8(b)】最小培地および酢酸ナトリウム上で成長させた変異体P. pyralisのルシフェラーゼE354K/D357Mを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【図8(c)】最小培地およびグルコース上で成長させた変異体P. pyralisのルシフェラーゼE354K/D357Mを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【図9】変異体P. pyralisのルシフェラーゼD357Yによって発せられた光のスペクトル分布に対するCoAの影響を示すグラフである。
【図10】変異体P. pyralisのルシフェラーゼD357Yによって発せられた光のスペクトル分布に対するCoAの影響を示すグラフである。
【図11(a)】変異体P. pyralisのルシフェラーゼE354I/D357Yによって発せられた光のスペクトル分布に対するCoAの影響を示すグラフである。
【図11(b)】変異体P. pyralisのルシフェラーゼE354I/D357Yによって発せられた光のスペクトル分布に対するCoAの影響および正規化したデータを示すグラフである。
【図12】合成ルシフェラーゼ遺伝子の構築において使用された制限部位の修飾を示す。
【図13】ルシフェラーゼ遺伝子の合成において使用する構築体を示す。
【図14】合成ルシフェラーゼ遺伝子のcDNA配列(配列番号1)(リボソーム結合部位の一部を形成し、コードしていないヌクレオチド1〜8含む)、および上向き矢印で示されたメチオニン残基で始まるエンコードされているアミノ酸配列(配列番号2)を示す。
【図15】合成遺伝子によってエンコードされている変異体を含む、変異体の50℃における熱安定性を示す。
【発明を実施するための形態】
【0013】
本発明の組換え体の基本部分を形成することができる野生型ルシフェラーゼ配列にはPhotinus pyralis、Luciola mingrelica、Luciola cruciataまたはLuciola lateralis、Hotaria paroula、Pyrophorus plagiophthalamus Lampyris noctiluca、Pyrocoelia nayako、Photinus pennsylvanicaまたはPhrixothrix(railroad−worms Biochem.38(1999)8271-8279を参照のこと)がある。
【0014】
基質D−ルシフェリン(4,5−ジヒドロ−2−〔6−ヒドロキシ−2−ベンゾチアゾリル〕−4−チアゾールカルボン酸)を使用して、発光を生み出すことができる種からの生物発光酵素は、本発明の変異型酵素の基本部分を形成することができる。
【0015】
本発明の組換え体の基本部分を形成することができる特定の野生型ルシフェラーゼ配列にはPhotinus pyralis、Luciola mingrelica、Luciola cruciataまたはLuciola lateralis、Hotaria paroula、Pyrophorus plagiophthalamus Lampyris noctiluca、Pyrocoelia nayakoおよびPhotinus pennsylvanicaがある。
【0016】
詳細には、このルシフェラーゼはPhotinus pyralis、Luciola mingrelica、Luciola cruciataまたはLuciola lateralisの酵素から得られる酵素である。Luciola mingrelica、Luciola cruciataまたはLuciola lateralisの酵素では、適切なアミノ酸残基は配列中の位置359に存在する。
【0017】
さまざまなルシフェラーゼすべての配列によって、これらは、これらの間で高い類似性を有して高度に保存されていることが示されている。このことは、配列を調べることによって、酵素配列間の対応する領域を容易に決定することができ、最も類似な領域を検出できることを意味するが、必要ならば、さまざまな配列間の対応する領域または個々のアミノ酸を決定するために、市販のソフトウェア(たとえばUniversity of Wisconsin Genetics Computer Groupからの「Bestfit」;Devereux他(1984)Nucleic Acid Research 12:387−395を参照のこと)を使用することができる。あるいはまたはさらに、この酵素の配列およびナンバリングを示し、本出願に関してこのナンバリングシステムを使用する、L.Ye他のBiochim.Biophys Acta 1339(1997)39−52を参照することによって対応する酸を決定することができる。
【0018】
Photinus pyralisのルシフェラーゼ中の残基357に対応するアミノ酸残基の可能な変化に関して、大部分の野生型配列はこの位置に酸残基(アスパラギン酸またはグルタミン酸)を有する。この例外はPhotinus pennsylvanicaのある形のルシフェラーゼであり、これにおいては、対応する残基(355)は非極性残基バリンであり、またはPhrixothrixのある形のルシフェラーゼでは、この場合対応する位置はPvGRまたはPhRE中のV354であり、L354ロイシンである。したがって一般に、この位置で置換アミノ酸として使用されるアミノ酸は、アスパラギン酸、グルタミン酸、バリンまたはロイシン以外である。
【0019】
したがって、多くの場合、酸性アミノ酸残基は、リジンまたはアルギニンなどの塩基性アミノ酸、ロイシン、バリンまたはイソロイシンなどの非極性アミノ酸、チロシン、アスパラギン、グルタミン、フェニルアラニン、セリン、トリプトファンまたはスレオニンなどの帯電していない極性アミノ酸を含めた酸性ではない残基で置き換えられる。特に、チロシン、アスパラギン、セリンまたはスレオニンなどの帯電していない極性アミノ酸でこれを置き換えることができる。この位置での置換に関して特に好ましいアミノ酸残基は、チロシン、フェニルアラニンまたはトリプトファンであり、チロシンが最も好ましい。一般的に言うと、この位置での芳香族残基によって最大のシフトが生じ、熱安定性を保つこともできる。
【0020】
野生型配列が酸性ではないアミノ酸残基をこの位置に含む場合、これらを酸性ではない異なる残基に適切に変異させる。
【0021】
このようにして酵素を変異させることによって、ルシフェラーゼによって発せられる光の波長が、ある場合はスペクトルの赤端に向かって50nmまでシフトすることが分かった。このように、D357Y変異型Photinus pyralisルシフェラーゼは、562nmの波長で発光する野生型酵素と比較して、約612nmの波長で発光する。
【0022】
50nmという波長のシフトは、この強度のシフトはスペクトル的に容易に決定することができるから、アッセイ用途において使用するためのかなりの能力を有する。たとえばWO95/18853に記載されているように、遺伝子発現の研究では異なる色のルシフェラーゼをレポーター分子として使用することができ、これによって2つ以上の遺伝子を同時に監視することが可能になる。標識としてルシフェラーゼを用いて、多数の検体試験を行うこともできる。
【0023】
この場合の光が深い赤色であるという事実は、アッセイ法において特に有用である。光の短い波長を吸収することができる色素または他の化合物を含むATPに関する溶液を分析するとき、赤色の変異体が有用であろう。たとえば、赤色の溶液は赤色の光を吸収しないであろう。しばしばこのような分析の対象となる赤色の溶液の例には、赤色のpH指標物を含む可能性がある血液サンプルまたは真核細胞培養培地の溶液がある。
【0024】
ルシフェラーゼなどの比色剤の混合物を使用するとき、特にサンプル中の他の試薬が緑のシグナルを生成する場合には、深い赤色のシグナルを生成させる能力が有用であろう。光電極スペクトル分析において使用する光電子増倍管をセットして、1つのサンプル中で生み出される1つまたは2つのピークを検出することができる。言い換えれば、同じサンプル中で、赤色からの光量子の流れと緑色のエミッターを区別することが可能である。
【0025】
さらに、補因子補酵素A(CoA)の存在によって、波長のシフトが影響を受け得ることが分かった。この特長によって、補因子についてのアッセイにおいてこの酵素を使用することができる可能性が生じる。
【0026】
以下に記載するように、発光される光のインビトロでのスペクトルに対する補因子補酵素Aの影響を調べた。補酵素Aの濃度が増大すると、スペクトルの分布が変化し、CoAの最大濃度において、610nmの明確なピークを有する590〜630nmの領域の波長がスペクトルの主となる。
【0027】
したがって、本発明の他の態様によれば、CoAのサンプル中の存在を判定するためのアッセイが提供される。このアッセイは、CoAを含む疑いのあるサンプルに前述のルシフェラーゼと共にルシフェラーゼ/ルシフェリン反応を引き起こすために必要とされる他の試薬を加えること、サンプルから発光される光の波長を測定すること、およびこれをCoAの存在または不在と関連付けることを含む。
【0028】
このようなアッセイは、細胞の、たとえば微生物または真核細胞の成長または活性の状態の検出において有用であり得る。
【0029】
たとえば、大腸菌細胞中のCoAの濃度は比較的高く、代謝状況により大幅に変化する。発光の波長はCoAの濃度に応じて変化するので、本発明の変異型酵素を使用して、生物の代謝状況、特にインビボでのCoAの濃度を監視することができる。このようなアッセイは、CoAが抗生物質(たとえばストレプトミセス)の生成における重要な主要代謝産物である状況において、特に有用であり得る。細胞のCoA濃度は、また脂肪酸の生合成の重要な指標であり、また細胞の飢餓状況により変化する。発ガンおよび糖尿病などのいくつかの代謝障害は、脂肪酸の代謝産物の異常、したがって異常なCoAレベルを示す。このような状態の診断において、本発明のアッセイを使用することができる。たとえば、アッセイ中で使用する本発明のルシフェラーゼから発せられる光の波長を測定することによって、患者からの血液サンプルなどの細胞サンプルおよびCoAレベルを測定することができる。この結果を健康な細胞のサンプルから得られた結果と比較して、波長が変化して、変化したCoAレベルが存在するかどうか測定することができる。これは、患者の疾患状態の指標であり得る。アッセイの前に、知られている溶解剤を使用して細胞を適切に溶解させる。
【0030】
位置357のアミノ酸残基は補酵素Aの結合部位と重大な関連があると信じられている。ルシフェラーゼ酵素の表面が1オングストローム(Å)の分解能まで輪郭が示されて(Tripos Ltd.のSYBL protein nodelling softwareを使用して)、小さな極性ポケットが見い出された。このポケットは残基H310、E354およびD357によって内側を覆われているようであり、8から10Åであると測定される。分子の上部から見ると、このポケットは残基H310、E354、D357およびI232によって内側を覆われている大きなポケットの一部分であるようである。残基H310およびE354は裂け目を越えてブリッジを形成しているようであり、2つの小さなポケットの外観を与えている(図8を参照のこと)。
【0031】
理論によって縛られることなく、この酵素が溶液中にあるとき、このブリッジ残基は切り離れるほどに充分に柔軟であり、CoA結合を可能にするより大きなポケット(深さ12Åまで、幅8Åまで)を提供するようにみえる。このことは、エネルギー計算と一致する。
【0032】
本発明のホタルのルシフェラーゼの変異体を発現する大腸菌細胞を異なる炭素源上で成長させると、インビボでの発せられる光のスペクトルの変化が観察された。富化培地(LB)から、唯一の炭素源としてアセテートまたはグルコースを有する限定最小培地への変換により、発せられる光は長い波長にシフトし、より短い波長からの貢献が減少した。このことは、発せられる光の波長を調節する、アッセイ目的のための他の手段を提供しよう。
【0033】
このタンパク質中の357位置の変異は、向上した熱安定性を生じることが分かった。
【0034】
このタンパク質のルシフェラーゼ活性が著しくは損ねられなければ、このタンパク質は配列中に他の変異を含んでもよい。この変異は酵素の性質を適切に向上させるか、またはある程度酵素を所望の目的に、より良く適合させる。このことは、変異が熱安定性および/またはカラーシフトの性質、および/またはこの酵素のATPに対するKmが向上させることを、意味しよう。カラーシフトを生じさせる変異の例は、WO95/18853に記載されている。Km値に影響を与える変異は、たとえばWO96/223776および国際特許出願No.PCT/GB98/01026に記載されている。
【0035】
一般に、諸性質の変化の点において、変異の影響は追加的なものであることが分かった。
【0036】
本発明の変異型ルシフェラーゼは、野生型ルシフェラーゼと比較して熱安定性を増大させる他の特定の変異を含んでもよい。詳細には、(a)Photinus pyralisのルシフェラーゼのアミノ酸354(Luciolaのルシフェラーゼ中の356)に対応するアミノ酸残基が変異している、(b)Photinus pyralisのルシフェラーゼ中の位置215または(Luciolaのルシフェラーゼ中の217)に対応するアミノ酸残基が、異なる疎水性アミノ酸である、(c)Photinus pyralisのルシフェラーゼ中の残基214またはLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基216に対応するアミノ酸残基、(d)Photinus pyralisのルシフェラーゼ中の残基232またはLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基234に対応するアミノ酸残基、(e)Photinus pyralisのルシフェラーゼ中の残基295またはLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基297に対応するアミノ酸残基、(f)Photinus pyralisのルシフェラーゼ中のアミノ酸14、またはLuciola mingrelicaの残基16、またはLuciola cruciataまたはLuciola lateralisの17に対応するアミノ酸残基、(g)Photinus pyralisのルシフェラーゼ中のアミノ酸35、またはLuciola mingrelicaの残基37、またはLuciola cruciataまたはLuciola lateralisの残基38に対応するアミノ酸残基、(h)Photinus pyralisのルシフェラーゼのアミノ酸残基105、またはLuciola mingrelicaの残基106、Luciola cruciataまたはLuciola lateralisの107、またはLuciola lateralis遺伝子の108に対応するアミノ酸残基、(i)Photinus pyralisのルシフェラーゼ中のアミノ酸残基234またはLuciola mingrelica、Luciola cruciataまたはLuciola lateralisの残基236に対応するアミノ酸残基、(j)Photinus pyralisのルシフェラーゼ中のアミノ酸残基420またはLuciola mingrelica、Luciola cruciataまたはLuciola lateralisの残基422に対応するアミノ酸残基、(k)Photinus pyralisのルシフェラーゼ中のアミノ酸残基310またはLuciola mingrelica、Luciola cruciataまたはLuciola lateralisの残基312に対応するアミノ酸残基、の少なくとも1つが対応する野生型配列中にあるアミノ酸と異なり、このルシフェラーゼ酵素は、野生型ルシフェラーゼのこの位置での対応アミノ酸を有する酵素と比較して、増大した熱安定性を有する。
【0037】
このように、本発明のタンパク質の好ましい例は、2つ以上のアミノ酸が、たとえば100個までのアミノ酸残基が、好ましくは40個を超えないアミノ酸が、より好ましくは30個までのアミノ酸が、適切な野生型酵素中の対応する位置のアミノ酸と異なる変異した野生型ルシフェラーゼである。
【0038】
したがって、好ましい一実施形態では、本発明のタンパク質はPhotinus pyralisのルシフェラーゼを含み、前述の357位置の変異に加えて、以下のものを少なくとも1つ含む。
【0039】
(a)Photinus pyralisのルシフェラーゼのアミノ酸354に対応するアミノ酸残基がグルタメート以外である、(b)Photinus pyralisのルシフェラーゼ中の位置215に対応するアミノ酸残基がアラニン以外の疎水性アミノ酸である、(c)Photinus pyralisのルシフェラーゼ中の残基214に対応するアミノ酸残基がスレオニン以外である、(d)Photinus pyralisのルシフェラーゼ中の残基232に対応するアミノ酸残基がイソロイシン以外である、(e)Photinus pyralisのルシフェラーゼ中の残基295に対応するアミノ酸残基がフェニルアラニン以外である、(f)Photinus pyralisのルシフェラーゼのアミノ酸14に対応するアミノ酸残基がフェニルアラニン以外である、(g)Photinus pyralisのルシフェラーゼのアミノ酸35に対応するアミノ酸残基がロイシン以外である、(h)Photinus pyralisのルシフェラーゼのアミノ酸残基105に対応するアミノ酸残基がアラニン以外である、(i)Photinus pyralisのルシフェラーゼのアミノ酸残基234に対応するアミノ酸残基がアスパラギン酸以外である、(j)Photinus pyralisのルシフェラーゼのアミノ酸残基420に対応するアミノ酸残基がセリン以外である、(k)Photinus pyralisのルシフェラーゼのアミノ酸残基310に対応するアミノ酸残基がヒスチジン以外である。
【0040】
あるいは、本発明のタンパク質は、Luciola mingrelica、Luciola cruciataまたはLuciola lateralis酵素のタンパク質のルシフェラーゼ配列を含み、前述の359位置の変異に加えて、以下のものを少なくとも1つ含む。
【0041】
(a)Photinus pyralisのルシフェラーゼのアミノ酸356に対応するアミノ酸残基がグルタメート以外である、(b)Photinus pyralisのルシフェラーゼ中の位置215に対応するアミノ酸残基がアラニンまたはスレオニン以外の疎水性アミノ酸である、(c)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基216に対応するアミノ酸残基がグリシン(Luciola mingrelicaベースの配列の場合)またはアパラギン(Luciola cruciataまたはLuciola lateralisベースの配列の場合)以外である、(d)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基234に対応するアミノ酸残基がセリン以外である、(e)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基297に対応するアミノ酸残基がロイシン以外である、(f)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのアミノ酸16に対応するアミノ酸残基がフェニルアラニン以外である、(g)Luciola mingrelicaの残基37、またはLuciola cruciataまたはLuciola lateralisの38に対応するアミノ酸残基がリジン以外である、(h)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのアミノ酸残基106に対応するアミノ酸残基がグリシン以外である、(i)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのアミノ酸残基236に対応するアミノ酸残基がグリシン以外である、(j)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisの残基422に対応するアミノ酸残基がスレオニン以外である、(k)Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのアミノ酸残基312に対応するアミノ酸残基がスレオニン(Luciola mingrelicaベースの配列の場合)またはバリン(Luciola cruciataまたはLuciola lateralisベースの配列の場合)以外である。
【0042】
いずれの場合も、熱安定性を向上させる特定の置換アミノ酸を、以下に例示する一般的な方法によって決定することができる。それぞれの場合において、異なる置換が熱安定性を向上させ得よう。当業者によって理解されるように、置換は、元のタンパク質または適切に変異したタンパク質をエンコードしているDNAの部位特異的変異によって行い得る。この場合本発明は、熱安定性を伴う位置の特定を伴っている。
【0043】
しかしながら、一般に、野生型のアミノ酸と異なる性質のアミノ酸に置換することを考慮することが望ましいであろう。したがって、親水性アミノ酸残基を疎水性アミノ酸残基で置換することができるのが好ましい場合もある(逆も然り)。同様に、酸性アミノ酸残基を塩基性残基で置換することができる。
【0044】
たとえば、このタンパク質は、ルシフェラーゼ活性、およびPhotinus pyralis、Luciola mingrelica、Luciola cruciataまたはLuciola lateralis酵素からのルシフェラーゼと少なくとも60%の類似性を有するタンパク質を含んでよい。この酵素の配列中では、以下の少くとも一つが変異され、(a)Photinus pyralisのルシフェラーゼの残基214、Luciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基216に対応するアミノ酸残基が変異され、およびPhotinus pyralisのルシフェラーゼの場合はスレオニン以外である、または(b)Photinus pyralisのルシフェラーゼの残基232、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基234に対応するアミノ酸残基が変異され、Photinus pyralisのルシフェラーゼの場合はイソロイシン以外である、または(c)Photinus pyralisのルシフェラーゼの残基295、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼの残基297に対応するアミノ酸残基が変異され、たとえばPhotinus pyralisのルシフェラーゼの場合はフェニルアラニン以外である。このルシフェラーゼ酵素は、野生型ルシフェラーゼと比較して熱安定性が増大している。
【0045】
さまざまなルシフェラーゼのすべての配列は、これらはそれらの間で高い類似性を有し、高度に保存されていることを示している。これは、この配列間の対応する領域は、この配列を調べることによって容易に決定することができ、最も類似の、領域を検出できることを意味する。しかし、必要ならば、市販のソフトウェア(たとえばUniversity of Wisconsin Genetics Computer Groupからの「Bestfit」;Devereux他(1984)Nucleic Acid Research 12:387-395を参照のこと)を使用して、さまざまな配列間の対応する領域または特定のアミノ酸決定することができる。あるいはまたはさらに、L.Ye他のBiochim.Biophys Acta 1339(1997)39-52を参照することによって対応する酸を決定することができる。
【0046】
Photinus pyralisのルシフェラーゼ中の残基214に対応するアミノ酸残基の可能な変化について、極性アミノ酸スレオニンが、アラニン、グリシン、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファンまたはシステインなどの非極性アミノ酸で適切に置き換えられる。Photinus pyralis中の残基214に対応する、スレオニン残基に関する特に好ましい置換はアラニンである。より好ましい置換はシステインである。しかしながら、この位置のアスパラギンなどの異なる極性残基が、この位置にスレオニンを有する対応する酵素の熱安定性を向上させる可能性もある。野生型ルシフェラーゼ酵素中のこの位置に現れる他のアミノ酸にはグリシン(Luciola mingrelica、Hotaria paroula)、アスパラギン(Pyrophorus plagiophthalamus、GR、YG、YEおよびOR、Luciola cruciata、Luciola lateralis、Lampyris noctiluca、Pyrocelia nayako Photinus pennsylvanica LY、KW、J19)およびセリン(Phrixothix)がある。有利には、アラニンおよびシステインなどの非極性または異なる非極性側鎖でこれらを置換することができる。
【0047】
Photinus pyralisのルシフェラーゼ中の残基232に対応するアミノ酸残基の可能な変化に関しては、非極性アミノ酸イソロイシンが、アラニン、グリシン、バリン、ロイシン、プロリン、フェニルアラニン、メチオニン、トリプトファンまたはシステインなどの異なる非極性アミノ酸で適切に置き換えられる。野生型ルシフェラーゼ酵素中のこの位置に現れる他のアミノ酸には、セリンおよびアスパラギンがある。適切には、これらの極性残基が前述のような非極性残基で置換される。Photinus pyralisのルシフェラーゼ中の残基232、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisの基の部分の残基234に対応する残基に関する特に好ましい置換はアラニンである。
【0048】
Photinus pyralisのルシフェラーゼ中の残基295、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基297に対応するアミノ酸残基の変化が、タンパク質の熱安定性に影響を及ぼそう(これはPhrixothixのルシフェラーゼ中の位置292に対応する。)。一般に、この位置のアミノ酸は、非極性アミノ酸フェニルアラニンまたはロイシンである。これらは、異なる非極性アミノ酸に、適切に変えられる。たとえばPhotinus pyralisでは、非極性アミノ酸フェニルアラニンが、アラニン、ロイシン、グリシン、バリン、イソロイシン、プロリン、メチオニン、トリプトファンまたはシステインなどの異なる非極性アミノ酸で適切に置き換えられる。Photinus pyralisのルシフェラーゼ中の残基214に対応するフェニルアラニン残基に関する特に好ましい置換はロイシンである。
【0049】
Photinus pyralisのルシフェラーゼのアミノ酸14、またはLuciolaのルシフェラーゼのアミノ酸16(Phrixothrixのルシフェラーゼの13)に対応するアミノ酸残基における変異も可能である。このアミノ酸残基(通常はフェニルアラニンであるが、ロイシン、セリン、アルギニンまたはある場合はチロシンであってもよい)を異なるアミノ酸、特にアラニン、バリン、ロイシン、イソロイシン、プロリン、メチオニンまたはトリプトファン、好ましくはアラニンなどの異なる非極性アミノ酸に適切に変化させる。
【0050】
Photinus pyralisのルシフェラーゼのアミノ酸35、またはLuciola mingrelicaのルシフェラーゼのアミノ酸残基37(他のLuciola spp.中の38)に対応するアミノ酸残基における変異も効果的であろう。このアミノ酸は野生型酵素の間で変化し、ロイシン(Photinus pyralis)だけでなくリジン、ヒスチジン、グリシン、アラニン、グルタミンおよびアスパラギン酸をこの位置に含み得る。適切には、この位置のアミノ残基は、アラニン、バリン、フェニルアラニン、イソロイシン、プロリン、メチオニンまたはトリプトファンなどの非極性アミノ酸残基または異なる非極性アミノ酸で置換される。この位置の好ましいアミノ酸はアラニンであり、これは野生型酵素と異なる。
【0051】
Photinus pyralisの配列の位置14に対応するアミノ酸における変異、および/またはPhotinus pyralisのルシフェラーゼのアミノ酸35に対応するアミノ酸残基における変異は、酵素中の唯一の変異ではないことが好ましい。
【0052】
これらは、前述の変異の他のもの、特にPhotinus pyralisのルシフェラーゼの位置214、395または232に対応する位置での変異を伴うことが適切である。
【0053】
Photinus pyralisのルシフェラーゼ中の残基105、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基106(Phrixothix中の102)に対応するアミノ酸残基の変化が、タンパク質の熱安定性に影響を及ぼそう。一般に、この位置のアミノ酸は、非極性アミノ酸アラニンまたはグリシン、またはPhrixothix中のセリンである。これらは、異なる非極性アミノ酸に、適切に変化させる。たとえばPhotinus pyralisでは、非極性アミノ酸アラニンが、フェニルアラニン、ロイシン、グリシン、バリン、イソロイシン、プロリン、メチオニン、トリプトファンなどの異なる非極性アミノ酸で適切に置き換えられる。Photinus pyralisのルシフェラーゼ中の残基105に対応するアラニン残基に関する特に好ましい置換はバリンである。
【0054】
Photinus pyralisのルシフェラーゼ中の残基234、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基236(Phrixothix中の231)に対応するアミノ酸残基の変化が、タンパク質の熱安定性に影響を及ぼそう。一般に、この位置のアミノ酸は、アスパラギン酸またはグリシンであり、グルタミンまたはスレオニンである場合もある。これらは、非極性または適切な異なる非極性アミノ酸に、適切に変化させる。たとえば、Photinus pyralisでは、アミノ酸残基はアスパラギン酸であり、アラニン、ロイシン、グリシン、バリン、イソロイシン、プロリン、メチオニンまたはトリプトファンなどの非極性アミノ酸で適切に置き換えられる。Photinus pyralisのルシフェラーゼ中の残基234に対応するフェニルアラニン残基に関する特に好ましい置換はグリシンである。グリシンなどの非極性アミノ酸残基がこの位置に存在する場合(たとえばLuciolaのルシフェラーゼ中で)、異なる非極性アミノ酸でこれを置換することができよう。
【0055】
Photinus pyralisのルシフェラーゼ中の残基420、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基422(Phrixothixグリーン中の417およびPhrixothixレッド中の418)に対応するアミノ酸残基の変化は、タンパク質の熱安定性に影響を及ぼそう。一般に、この位置のアミノ酸は、帯電していない極性アミノ酸セリンまたはスレオニンまたはグリシンである。これらは、異なる帯電していない極性アミノ酸に、適切に変化させる。たとえば、Photinus pyralisではセリンを、アスパラギン、グルタミン、スレオニンまたはチロシン、特にスレオニンで適切に置き換えることができよう。
【0056】
Photinus pyralisのルシフェラーゼ中の残基310、およびLuciola mingrelica、Luciola cruciataまたはLuciola lateralisのルシフェラーゼ中の残基312に対応するアミノ酸残基の変化は、タンパク質の熱安定性に影響を及ぼそう。この位置のアミノ酸残基は、知られているルシフェラーゼタンパク質の間で変化し、Photinus pyralis、Pyrocelia nayako、Lampyris noctilucaおよびある形のPhotinus pennsylvanicaのルシフェラーゼではヒスチジン、Luciola mingrelica、Hotaria paroulaおよびPhrixothix(この場合それはアミノ酸307である)のルシフェラーゼではスレオニン、Luciola cruciataおよびLuciola lateralisではバリン、およびある種のPyrophorus plagiophthalamusのルシフェラーゼではアスパラギンである。したがって、一般に、この位置のアミノ酸は、酵素の熱安定性を増大させる異なるアミノ酸残基に変化させ得る親水性アミノ酸である。Photinus pyralisのルシフェラーゼ中の残基310に対応するヒスチジン残基に関する特に好ましい置換はアルギニンである。
【0057】
他の変異がこの酵素中に存在してもよい。たとえば好ましい実施形態では、タンパク質は、グルタメートから変化した、Photinus pyralisのルシフェラーゼのアミノ酸354(Luciolaのルシフェラーゼ中の356)に対応するこの位置でのアミノ酸残基を有し、特にグリシン、プロリンまたはアスパラギン酸以外のアミノ酸へ変化したこの位置でのアミノ酸も有する。適切にはこの位置のアミノ酸はトリプトファン、バリン、ロイシン、イソロイシンおよびアスパラギンであり、最も好ましくはリジンまたはアルギニンである。この変異はWO95/25798中に記載されている。この位置の疎水性残基は、この酵素の波長シフトを向上させることが分かっており、さらに、位置354における大きな疎水性(VまたはI)、極性(N)または正に帯電している(KまたはR)アミノ酸の存在は熱安定性を向上させる。
【0058】
他の実施形態では、EP−A−052448に記載されているように、このタンパク質は、Luciolaルシフェラーゼ中のアミノ酸217(Photinus pyralis中の215)に対応する位置において、疎水性アミノ酸に、特にイソロイシン、ロイシンまたはバリンに変化したアミノ酸を有している。
【0059】
本発明のタンパク質は、野生型および組換えルシフェラーゼ酵素の両方を含む。これらは、野生型酵素中に存在する少なくとも60%のアミノ酸が本発明のタンパク質中に存在するという意味で、Photinus pyralis、Luciola mingrelica、Luciola cruciataまたはLuciola lateralisの酵素の配列などの野生型配列と少なくとも60%の類似性を有する。このようなタンパク質は、前述の野生型酵素とより大きな類似性、詳細には少なくとも70%、より好ましくは少なくとも80%および最も好ましくは少なくとも90%を有し得る。類似なタンパク質はこのタイプであり、対立遺伝子変異体、他の昆虫種からのタンパク質、および組換えによって生成された酵素を含む。野生型酵素をエンコードしている配列とストリンジエントハイブリダイゼーション条件下でハイブリダイズする核酸によってエンコードされている点で、これらのタンパク質は容易に同定することができる。このような条件は当業者によってよく理解されていよう。たとえばSambrook他(1989)Molecular Cloning、Cold Spring Harbor Laboratory Pressの中で例示されている。一般的に、低ストリンジェンシー条件はほぼ周囲温度から約65℃での3×SCC、高ストリンジェンシー条件は約65℃での0.1×SCCとして定義される。SSCは0.15M NaCl、0.015Mクエン酸三ナトリウムのバッファーの名前である。3×SCCはSSCなどの3倍強い。
【0060】
詳細には、LipmanおよびPearsonによって記載されている多重アラインメント法(Lipman、D.J.and Pearson、W.R.(1985)Rapid and Sensitive Protein Similarity Searches、Science、vol227、pp1435-1441)を使用して、特定の配列と本発明の配列との類似性を評価することができる。「最適化した」パーセンテージスコアは、Lipman-Pearsonアルゴリズムについて以下のパラメータを用いて計算すべきである。ktup=1、ギャップペナルティ=4およびギャップペナルティレングス=12。類似性を評価するための配列は「試験配列」として使用すべきであり、これは、Photinus pyralisの配列またはYe他に記録されている任意の他の配列などの比較用の塩基配列を、最初にこのアルゴリズムに入れるべきであることを意味する。
【0061】
本発明のタンパク質の特定の例は、前述の1つまたは複数の変異を有する野生型ルシフェラーゼ配列である。
【0062】
さらに本発明は、前述のルシフェラーゼをエンコードしている核酸を提供する。適切には、この核酸は当分野でよく知られる野生型配列をベースとする。アミノ酸配列中の所望の変異を行うための適切な変異は、遺伝コードの知識に基づいて容易に明らかであろう。
【0063】
本発明の好ましい実施形態では、核酸は合成遺伝子である。適切には、合成遺伝子を工作して、高度に発現された遺伝子中ではほとんど見られないコドンを大腸菌などの一般的な発現宿主から取り除き、これと同時に甲虫のルシフェラーゼをコードしている遺伝子中ではほとんど見られないコドンの導入を避ける。この手法は、この新しい遺伝子は大腸菌および昆虫の発現系の両方に最適なコドン使用を有することを確実にする。
【0064】
たとえば、可能なかぎり、アミノ酸arg、leu、ile、glyおよびproのコドンを、CGTまたはCGC(arg)、CTG、CTTまたはCTC(leu)、ATCまたはATT(ile)、GGTまたはGGC(gly)、およびCCG、CCAまたはCCT(pro)に変化させ、したがって希なコドンが取り除かれる。以下(配列番号1)および図14に例示する合成遺伝子の場合、これによって62個の新しく一般的なコドン(全体の11%)を生み出す合計139のサイレント変異を生じた。図14に示す最初の8個のヌクレオチドは、リボゾーム結合部位の一部を形成し、したがってコードしていない。コード配列は、上向き矢印によって示されるメチオニン残基で始まる。このコード配列および非常に類似した配列、たとえば少なくとも90%の類似性、好ましくは95%の類似性を有する配列は、本発明の好ましい態様を形成する。
【0065】
合成アセンブリを生成するときに使用することができる他の有用な性質は、新しい独特な制限部位の導入である。これらの部位は遺伝子の変異誘発、特にコンビナトリアルカセット変異をより簡単でより効率的にする。詳細には、酵素中にサブドメインBをコードするcDNA内に、独特な制限部位を作製することが望ましいであろう。さらに、遺伝子の3'端部に独特な制限部位を作製して、ペルオキソーム標的配列の簡単な融合および/または除去を可能にすることが有利であろう。
【0066】
以後に示す例では、9つの新しい独特な制限部位を工作し(大部分遺伝子の中央の3番目)、独特なHind III部位を遺伝子の3'端部に生成させ、簡単なC末端融合を可能にした(図12)。
【0067】
最後に、合成遺伝子の使用は、遺伝子産物の熱安定性を増大させるための、またはそうでない場合は産物の性質を望むように改変するための、変異の導入を可能にする。たとえば、以後に示す実施例では、サイレントではない3つの変異を工作して、熱安定アミノ酸の変化T214C、E354KおよびD357Fをポリペプチドに導入した。
【0068】
本発明の核酸を、プロモーター、エンハンサー、ターミネーターなどの調節要素の調節下で、プラスミドなどの発現ベクターに適切に導入する。次いでこれらのベクターは、宿主細胞、たとえば植物または動物細胞などの原核または真核細胞(詳細には大腸菌などの原核細胞である)を形質転換させるために、細胞が所望のルシフェラーゼ酵素を発現するように使用することができる。当分野でよく知られる条件を使用して、このようにして形質転換した細胞を培養することによりルシフェラーゼ酵素が生成されよう。次いでこれを培養培地から分離することができる。細胞が植物細胞または動物細胞である場合、前記細胞から植物または動物を繁殖させることができる。したがって植物からタンパク質を抽出することができ、トランスジェニック動物の場合は、ミルクからタンパク質を回収することができる。ベクター、形質転換した細胞、トランスジェニック植物および動物、およびこれらの細胞を培養することによって酵素を生成させる方法は、いずれも本発明の他の態様を形成する。
【0069】
以下に記載するように、ランダム突然変異誘発によって、Photinus pyralisD357Y突然変異型ルシフェラーゼを作製した。D357Yの1点での変異は発せられる光の波長の大きなカラーシフトを生み出し、また野生型ルシフェラーゼより熱安定性が高いことが分かった。さらなる調査によって、この位置でのある範囲の置換によって優れた熱安定性、および/または大きなカラーシフトが生み出さることが明らかになっている。
【0070】
本発明の範囲内に入るPhotinus pyralisの変異型酵素の特定の例には以下のもの、または、他の種のルシフェラーゼであれば、これらの任意の均等物がある。
【0071】
【表1】
【0072】
上記変異体を作製するための変異は、部位特異的変異法、(PCR)またはコンビナトリアルカセット変異によって、プラスミドpET23上のルシフェラーゼ遺伝子に導入した。適切な変異を行うための、PCR反応に加えられたオリゴヌクレオチドは以下に示す。
【0073】
354および215位置での点変異の影響は追加的なものであることが以前に報告している。本発明は、3つ以上のこのような変異を組み合わせる可能性を提供して、大きなカラーシフトを有する変異型酵素中における高い熱安定性を提供する。
【0074】
本発明のルシフェラーゼタンパク質は、シグナル手段としてルシフェラーゼ/ルシフェリン反応を使用するいかなる生物発光アッセイにおいても、有利に使用できよう。文献中で知られている多くのこのようなアッセイが存在する。したがって、このようなアッセイを行う目的で作製されたキット中このタンパク質が含まれよう。場合によってはルシフェリンおよび特定のアッセイを行うために必要とされる任意の他の試薬も含まれる。
【0075】
ここで本発明を、例示として添付の図面を参照することによって詳細に記載する。
【0076】
図1は、残存活性%対本発明によるいくつかの変異型酵素の45℃でのインキュベーションの時間を示す対数グラフである。
【0077】
図2は、D−ルシフェリンを有するクエン酸バッファー中で、ルシフェラーゼ酵素を発現する大腸菌細胞をインキュベートすることによって得られたスペクトルのピークを示す。この場合、使用した酵素は(a)組換え野生型Photinus pyralisのルシフェラーゼ、(b)D357K変異型、(c)D357N変異型、(d)D357W変異型、(e)D357I変異型、(f)D357F変異型、(g)D357Y変異型および(h)二重変異型E354I+D357Yである。
【0078】
図3は、残存活性%対3つの変異型酵素E354I、D357Yおよび二重変異型(DM)E354I/D357Yの時間を示すグラフである。
【0079】
図4は、(a)組換え野生型酵素および(b)二重変異型(DM)E354I/D357Yの発光スペクトルを示す。
【0080】
図5は、組換え野生型(◆)r−wtおよびD357K変異型酵素(■)の光量子発光の速度減衰を示すグラフである。
【0081】
図6は分子モデル図を示し、ルシフェラーゼ酵素中の考えられるCoA結合ポケットを示す。
【0082】
図7は、(a)LB上で成長させた、(b)最小培地および酢酸ナトリウム上で成長させた、(c)最小培地およびグルコース上で成長させた、変異体P.pyralisルシフェラーゼD357Yを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【0083】
図8は、(a)LB上で成長させた、(b)最小培地および酢酸ナトリウム上で成長させた、(c)最小培地およびグルコース上で成長させた、変異体P. pyralisのルシフェラーゼE354K/D357Mを発現する大腸菌細胞によって発せられたインビボでの生物発光スペクトルを示す。
【0084】
図9は、変異体P. pyralisルシフェラーゼD357Yによって発せられた光のスペクトル分布に対するCoAの影響を示すグラフである。
【0085】
図10は、変異体P. pyralisルシフェラーゼD357Yによって発せられた光のスペクトル分布に対するCoAの影響の正規化したデータを示すグラフである。
【0086】
図11は、変異体P. pyralisルシフェラーゼE354I/D357Yによって発せられた光のスペクトル分布に対するCoAの影響(図11a)および正規化したデータ(図11b)を示すグラフである。
【0087】
図12は、合成ルシフェラーゼ遺伝子の構築において使用された制限部位の修飾を示す。
【0088】
図13は、ルシフェラーゼ遺伝子の合成において使用された構築体を示す。
【0089】
図14は、合成ルシフェラーゼ遺伝子のcDNA配列(配列番号1)(リボソーム結合部位の一部を形成し、コードしていないヌクレオチド1〜8を含む)、および上向き矢印で示されたメチオニン残基で始まるエンコードされているアミノ酸配列(配列番号2)を示す。
【0090】
図15は、合成遺伝子によってエンコードされている変異体を含む、変異体の50℃における熱安定性を示す。
【0091】
実施例1
変異型ルシフェラーゼの同定および特徴付け エラープロンPCR〔M.Fromant他、Anal.Biochem.(1995)224、347-353〕を使用して作製した、ホタル(Photinus pyralis)ルシフェラーゼの2つのライブラリーを調製した。完全長のluc遺伝子のエラープロンPCR産物からなる1つのライブラリーを、T7発現系pET23aにクローン化した(Novagen Inc.、Madison、WI、U.S.A)。199〜35個のアミノ酸をカバーする、luc遺伝子の短い断片のエラープロンPCR産物からなる第2のライブラリーを、ベクターpBSK(+)中にクローン化した(Stratagene、La Jolla、CA、U.S.A)。
【0092】
pET23aライブラリーを、大腸菌菌株BL21(DE3)中で発現させた(大腸菌B F dcm ompt hsdS(rB−mB−)galλ(DE3))。
【0093】
pBSK(+)ライブラリーを、HB101 大腸菌細胞(supE44 aral4 galK2 lacY1 Δ(gpt−proA)62 rpsL20(Strr)xyl−5 mtl−1 recAl3 Δ(mrcC−mrr)HsdS−(r−m−))中で発現させた。pET23aおよびpBSK(+)は両方共にβ−ラクタマーゼの遺伝子を保有し、このプラスミドを有する大腸菌細胞にアンピシリン耐性を与える。
【0094】
BIORAD E.coli Pulserを使用するエレクトロポレーションによって、作製したライブラリーを用いて、大腸菌株細胞を形質転換させ、50μg/mlの濃度のアンピシリンを含むLB寒天上で37℃で一晩生育させた。細胞をナイロン膜(Osmonics、Minnetonka、Minnesota、U.S.A)に移し、ルシフェリン溶液(クエン酸ナトリウムバッファー100mM、pH5.0中に500μM D−ルシフェリン、カリウム塩、)をスプレーした。AlphImager(商標)1200DocumentationおよびAnalysis System(Flowgen、Lichfield、Staffordshire、UK)を使用して、コロニーを観察した。これによって特定の時間で発せられる生物発光が取り込まれて、コロニーによって発せられる光の像を生み出す。発光の明るさを、ルシフェラーゼの熱安定性の指標として記録した。
【0095】
次いで、熱安定性についてコロニーを選別した。発せられる光の明るさに基づいてコロニーを選択し、さらなる特徴付けのために単離した。いくつかの選別では、選別の前に大腸菌コロニーを42℃で2時間インキュベートし、その結果熱安定性の変異体を選択することができた。最初の選別から単離したコロニーをナイロン膜の上に載せ、アンピシリンを含むLB培地上で一晩生育させた。パッチをルシフェリン溶液でスプレーし、AlphImager(商標)中で観察した。この第2の選別は、ルシフェラーゼ活性のインビトロでの分析用のクローンを陽性に同定するのに役立った。可能性ある熱安定性酵素を発現する大腸菌のクローンを、ルシフェラーゼ活性および熱安定性についてインビトロでアッセイした。
【0096】
室温で、Promega Luciferase Assay System(Promega Corporation、Madison、WI、U.S.A)を使用して、ルシフェラーゼ活性についてインビトロでのアッセイを行った。
【0097】
粗細胞抽出物10μlをPromega Luciferase Assay Cocktail(1:2に希釈)に加えることによって、ルシフェラーゼ反応を開始させた。Biotrace M3 照度計を使用して、生じた生物発光を測定した。
【0098】
Promega technical bulletein no.101に記載されているように、粗細胞抽出物を調製した。細胞培養物溶解試薬(25mM トリスリン酸塩 pH7.8、2mM ジチオスレイトール(DTT)、2mM 1,2−ジアミノシクロヘキサン−N,N,N',N'−テトラ酢酸、10%グリセロール、1%トリトン X−100、1.25mg/ml hen リソザイム)中で10分間かけて室温で、一晩培養した大腸菌のアリコートを溶解させた。次いで粗溶解物をアッセイ前に氷上に保存した。
【0099】
時間依存不活性化実験において、酵素の諸性質をさらに試験した。粗細胞抽出物のアリコート50μlを含むエッペンドルフチューブを、水浴中において所与の温度でインキュベートした。設定したいくつかのタイムポイントで、チューブを取り除きアッセイ前に氷上で冷却した。残存ルシフェラーゼ活性を、元の活性のパーセンテージとして表した。
【0100】
残存活性%対インキュベーション時間の対数グラフをプロットし、t1/2値を計算するために使用した。T1/2とは、酵素の、所与の温度でインキュベーション後の、その元の活性の50%を失うのにかかる時間のことである。残存活性%に対する時間の対数グラフから、37℃での粗抽出物中のT1/2値(活性が元の活性の50%に低下する時間)を決定した(示さず)。
【0101】
上に述べたように決定した最も熱安定性のあるルシフェラーゼを発現する大腸菌クローンからのプラスミドDNAを配列決定して、酵素の熱安定性を担う変異を決定した。
【0102】
QIAGEN QIAprep Spin Miniprep Kit(QIAGEN Ltd、Crawley、W.Sussex、UK)、次にミクロ遠心分離を使用するためのプロトコル(QIAprep Miniprep Handbook04/98)を使用して、プラスミドDNAを作製した。
【0103】
DNAの配列決定はすべて、ABI PRISM(商標)377 DNA Sequencerおよびジデオキシ鎖停止法〔F.Sanger他、Proc.Natl.Acad.Sci.U.S.A 74、(1977)5463−5467〕に基づくABI PRISM(商標)BigDye(商標)Terminator Cycle Sequencer Ready Reaction Kit(Perkin Elmer Applied Biosystems)を使用して、Babraham Technix、Cambridge、UKによって行った。
【0104】
この作業の結果、新規な変異体D357Yを特定した。
【0105】
ルシフェラーゼの結晶構造〔E.Conti他、Structure、4(1996)287−298〕は、位置357はタンパク質の表面に適合しており位置354に近く、熱安定性およびスペクトルの諸性質の両方に影響を及ぼす可能性があることを示す。これは、この領域は酵素の熱安定性の点で重要である可能性があることを示す。
【0106】
D357Yは非常に熱安定性のある変異体であって、最も熱安定性のあるルシフェラーゼであり、ただ1つのアミノ酸が変化している。
【0107】
実施例2
他の357変異体を作製するための部位特異的変異 357位置における異なる変異体を評価するために、Stratagene QuikChange(商標)Site−Directed Mutagenesis Kit(Stratagene、La Jolla、CA、U.S.A)を使用して、部位特異的変異を行った。プラスミドpPW601a J54(PJW、MoD Report、3/96)を、すべての部位特異的変異において使用した。変異誘発反応のすべての産物を、大腸菌菌株XL1−Blue〔e14−(mcrA−)Δ(mcrCB−hsdSMR−mrr)171 endA1 supE44 thi−1 gyrA96 relAl lac recB recJ sbcC umuC::Tn5(Kanr)uvrC〔F'proAB laclqzΔm15 Tn 10 (Tetr)Amy Camr〕に形質転換した。Sigma−Genosys Ltd.、Cambridge、UKによってオリゴヌクレオチドプライマーを合成し、インテリジェントドーピングシステム〔A.R Arkin他、Bio−technology、(1992)10、297−300、W、.Hung他、Anal.Biochem.(1992)10、454−457〕を使用して設計し、それぞれのアミノ酸置換に関する個々のプライマーを使用せずに、これを使用して縮重オリゴヌクレオチドプライマーを設計し、可能性ある変異体の群を作成した。
【0108】
このようにして、アミノ酸置換ルシフェラーゼ変異体のライブラリーを作成した。
【0109】
以下のオリゴヌクレオチド(およびその相補的なパートナー)を使用した。
【0110】
【化1】
【0111】
変異体のライブラリーを、前述のように熱安定性に関して選別した。以下の等式〔S.Climie他、J.Biol.Chem.265(1990)18776−18779〕を使用して、選別したコロニーの数を計算した。
【0112】
N=〔1n(1−p)〕/〔1n((n−1)/n〕
Nが選別されるコロニーの数の場合、nは標的位置における可能性あるコドンの数であり、Pは混合物中のそれぞれのコドンが、少なくとも1度の選別用にサンプル採取される確率である。この計算はP=0.95に基づいていた。部位特異的変異から得た変異体をルシフェラーゼ活性に関してアッセイし、時間依存熱不活性化実験において特徴付けした。
【0113】
このようにして望ましいとした変異体を、アンピシリンを含みA260 ?0.5の400mlのLB培地中で生育させた。次いでイソプロピルβ−チオガラクトシダーゼ(IPTG)を加え最終濃度1mMにすることによって、ルシフェラーゼ発現を誘導した。細胞を30℃で撹拌しながら3時間インキュベートし、次いで遠心分離によって採取した。生じた細胞のペレットを、Maxi−Scale Bacterial Protein Extractionに関するB−PER(商標)のプロトコルに従って、B−PER(商標)Protein Extraction Reagent、(Pierce Chemical Company、Rochford、U.S.A)10ml、DTT1mM中に再懸濁させて、粗抽出物を作成した。再構築Sigma Protease Inhibitor Cocktail、500μl(Product No.P8465、Sigma、Saint Louis、Missouri、U.S.A)をB−PER(商標)溶液に加えて、内生プロテアーゼを阻害した。次いで細胞の溶解物を、30000gで30分間遠心分離した。
【0114】
粗抽出物の上澄みを、硫酸アンモニウムを用いて分画した。30%と55%の間の飽和状態で沈殿した分画は、ルシフェラーゼ活性を含んでいた。この物質を0.5ml Tris HCl pH8.0、1mM DTTに再懸濁させ、熱不活性化およびスペクトルの実験用に使用した。
【0115】
変換D357L、T、V、W、R、I、S、K、NおよびFを導入した。粗抽出物のインビトロでの熱不活性化実験において、これらの変異体を特徴付けた。
【0116】
部分精製した抽出物を、熱不活性化バッファー:飽和した硫酸アンモニウム10%、1mMジチオスレイトールおよび0.2%BSAを含む50mM リン酸カリウムバッファー pH7.8中11に1の比率で希釈した。
【0117】
タンパク質溶液のアリコート110μlを、アッセイ前に設定した時間だけ40℃または45℃でインキュベートし、氷上で冷却した。次いでPromega Luciferase Assay Reagent(1:2に希釈)を使用して、実施例1に記載するようにルシフェラーゼ活性を測定した。
【0118】
その結果を表2および3、および図1中に示す。未精製抽出物のT1/2値を40℃(表2)および45℃(表3)において決定した。
【0119】
【表2】
【0120】
【表3】
【0121】
すべての置換が、組換え野生型と比較して、向上した熱安定性を示した。
【0122】
実施例3
発せられる光の波長の変化 位置357におけるアミノ酸置換が、インビトロで酵素によって発せられる光のスペクトルに影響を及ぼすことも観察した。実施例2に記載の大腸菌細胞培養物のアリコート(250μl)を37℃で一晩生育させ、ミクロ遠心分離においてスピンダウンさせ、上澄みを取り除いた。D−ルシフェリン150μlを含むクエン酸バッファー(pH5.0)中で、異なる変異型ルシフェラーゼを発現する細胞をインキュベートし、SPECTRAmax(登録商標)Microplate Spectrofluorometer、(Molecular Devices Crop.California、U.S.A)を使用して発光スペクトルを測定することによって、インビトロでの反応から発せられる光を分析した。変異体D357Y、FおよびIに関して、スペクトルのピークおよび波長の分布の大きな変化を観察した(図2(a)〜(g))。これらの結果を以下の表4に要約する。
【0123】
さらに、インビボでの変異体の発光を、暗室で肉眼によって評価した。D357変異体は、その発光スペクトルにおいてさまざまな色を示した。特にD357Y、FおよびIは、発せられる光のより長い波長への大幅なシフトを示した。
【0124】
ある場合(たとえばD357F)、光の発光の色の変化はλmaxのシフトによるだけでなく、可視光線の異なる波長からのスペクトルへの貢献の違いによるようである。
【0125】
【表4】
【0126】
ある種の357変異体のインビボでの発光のλmaxの比較のために、組換え野生型(r−wt)酵素を使用した。D357Y、FおよびIは、その波長最大値の大幅なシフトを示した。
【0127】
実施例4
CoAの存在または不在下での酵素の諸性質 実施例1に記載したようにして、硫酸アンモニウム沈殿によってD357Yを部分的に精製した。この部分的に精製したD357Y酵素(5μl)をPromega Luciferase Assay Reagent150μlと混合させた。他のアリコートをCoAは存在しない(25mM トリストリシン pH7.8、5.0mM MgSO4、0.1mM EDTA、2mM DTT、470μM D−ルシフェリン、530μM ATP)同等のアッセイ用バッファーと混合させた。2つの反応物の発光スペクトルを測定した。これを図9および10に示す。
【0128】
これらのスペクトルは、CoAの存在または不在下での生物発光の顕著な違い、およびλmaxの大幅なシフトを示す。CoAのルシフェラーゼ反応のキネテイクスに対する影響を、RLUスケール(RLU−相対的光単位)の違いによって見ることもできる。
【0129】
この発光の違いによって、CoAの存在を検出するためのアッセイにおいてこの酵素を使用する可能性が生じる。
【0130】
実施例5
二重変異体の作製および諸性質 実施例2に記載するようにして、部位特異的変異を使用して、熱安定性および発光の色に対するあらゆる累積的影響を調べるために、E354I+D357Yの二重変異体を工作した。
【0131】
部分精製した二重変異体E354I+D357Yを、熱不活性化バッファー:飽和した硫酸アンモニウム10%、1mMジチオスレイトールおよび0.2%BSAを含む50mM リン酸カリウムバッファー pH7.8中11に1の比率で希釈した。
【0132】
タンパク質溶液のアリコート110μlを、アッセイ前に設定した時間だけ40℃または45℃でインキュベートし、氷上で冷却した。次いでPromega Luciferase Assay Reagent(1:2に希釈)を使用して、前述のようにルシフェラーゼ活性を測定した。
【0133】
二重変異体は、単一の変異体E354IおよびD357Yそれぞれと比較して、熱安定性の顕著な増大を示した(図3を参照)。部分精製した二重変異体の熱不活性化実験により、この変異体の熱安定性の増大を確認し、45℃での不活性化において7.7分のt1/2値を与えた。
【0134】
E354IおよびD357Yの個々の変異体より、この二重変異体は、より深い赤色の発光を示し、発光の色の添加性を示すことが示された。
【0135】
実施例3に記載のアッセイ用バッファーを使用して、組換え野生型および二重変異体E354I+D357Yの粗抽出物の発光スペクトルも測定した。
【0136】
インビボで測定した発光スペクトルは、611nmのλmaxを与えた。しかしながら、このスペクトルは、波長の赤い領域からの発光に大いに貢献し、肉眼によって視覚化するとより深い赤色の外観となる。未精製抽出物の発光スペクトルは、rWTと比較して、スペクトル形状の明確な変化および44nmの波長シフトを示した(図4を参照)。
【0137】
インビボでの二重変異体の発光スペクトルは、発光(613nm)のピーク波長の帯幅をシャープにし、および540〜560nmの範囲の光の波長の貢献の低下させる両方を示す。
【0138】
これらの変異の劇的な効果によって、この酵素のこの領域の生物発光の色への重要性が示される。
【0139】
実施例6
改善された光量子の流れ 変異体D357Kを発現している大腸菌細胞のインビボでの生物発光は、この位置の他の変異体と比較して、非常に鮮やかであることを観察した。時間における光量子の発光の速度を測定することができる光度計を使用して、この酵素のフラッシュキネテイクスを分析した。組換え野生型酵素または変異体D357を含む大腸菌無細胞抽出物のアリコートを、グローキネティクスを助長するいかなる試薬(たとえば補酵素A)も含まないルシフェラーゼアッセイ用カクテルに加えた。時間(15秒)における光量子の発光の減衰の速度を両方の酵素について測定し、変異体D357Kは大幅に遅いことを観察した(図4)。言い換えれば、変異型酵素は、反応の少なくとも最初の15秒間において、組換え野生型酵素よりも低い程度で阻害される反応キネテイクスを有する。
【0140】
実施例7
位置E354およびD357でのコンビナトリアルカセット変異 ステップ1 カセット変異用のプラスミドpPW601aJ54の工作 2対の合成オリゴヌクレオチド(以下参照)を使用して、2つの新しい独特な制限部位をプラスミドpPW601a/J54内のluc遺伝子中に導入した。合計6個のサイレント変異を、63塩基対隔ててこの遺伝子中のSpeIおよびKpnI制限部位に導入した。これらの新しい部位を含むプラスミドを、pPW601aJ54SpeI/KpnIと呼んだ。これらの制限部位の存在および近接性は、コンビナトリアルカセット変異を使用して、ホタルルシフェラーゼの主要配列中のアミノ酸の位置E354およびD357での、ランダムな置換の影響を探索することを可能にする。
【0141】
【化2】
【0142】
太線で強調したヌクレオチドはエンドヌクレアーゼ認識部位を形成し、および上側のケースのこれらはこの部位を作製するために必要なポイント変異の位置を形成する。
【0143】
ステップ2 カセットの設計およびライブラリーの構築 1対の合成オリゴヌクレオチドを合成した。このオリゴヌクレオチドを次いでアニールし、新しい制限部位で消化されたプラスミドpPW601aJ54SpeI/KpnIに直接連結することができる二本鎖カセットが作製された。このカセットは、天然に存在する20個のアミノ酸の考えられるすべての組み合わせを、この一次配列中の位置E354およびD357において導入できるように設計された。
【0144】
【化3】
ループライブラリーオリゴヌクレオチドのそれぞれ2μgを、50mM Tris−HCl pH7.4、25mM NaClを含むバッファー中に混合し、3分間100℃に加熱した。次いでこの溶液をヒーティングブロック中で<50℃までゆっくりと冷却して、相補的な配列をアニールさせた。次いでアニールしたオリゴヌクレオチドをプラスミドpPW601aJ54SpeI/KpnIに連結させ、SpeIおよびKpnIを用いてこれを消化した。次いでこの連結反応のアリコートを使用して、エレクトロポレーションを使用する大腸菌HB101細胞を形質転換した。エレクトロポレーションの後、形質転換した細胞をアンピシリン50μg/mlを含むLB寒天プレート上に載せ、37℃で一晩生育させた。翌日、869個のコロニーをプレートからランダムに採取し、これを使用してアンピシリンを含むLB1mlを96スクエアウエルのプレート(Beckman)に接種した。このプレートをカバーし、振とうしながら37℃で細胞を一晩生育させた。
【0145】
ステップ3 ランダムに選択したクローンのインビボでの選別 翌朝、定常期の一晩培養したもののアリコート50μlを、2つの透明プラスチック製丸底96ウエルのミクロタイタープレート(Dynex)に移した。1つのプレートをカバーし、加熱したブロック上で8分間インキュベートし(ブロック表面温度45℃)、一方もう1つのブロックは37℃に保った。次いで0.5mMのD−ルシフェリンを含む100mMのクエン酸ナトリウムバッファー(pH5.0)50μlをウエルに加え、次いでこのプレートをvideo camera imager capture system(Alpha Imager)に移すことによって、インビボでの両方のプレートからの細胞中のシフェラーゼ活性を室温で検出し記録した。加熱および調節された培養物によって発せられた光を1〜2分かけて積分し、その像を熱性紙フィルム上に記録した。
【0146】
フィルム上で記録した像の明るさの判定によって、最大の生物発光を示した79個の培養物を、選別の第2ラウンド用に選択した。今回は培養物を、アッセイの前にヒーティングブロック上で16分間インキュベートした。インビボでの熱安定性選別から選択した55個のクローンの内、インビボでのスペクトル分析用に25個を選択した。これらのクローンをLB中において37℃で一晩生育させ、翌朝一晩培養したもの200μlを翌朝遠心分離し、大腸菌細胞のペレットを0.5mMのD−ルシフェリンを含む100mMのクエン酸ナトリウムバッファー(pH5.0)150μl中に再懸濁させた。次いで再懸濁させた細胞を白いプラスチック製のミクロタイタープレート中に置き、変異型ルシフェラーゼのそれぞれによって発せられるインビボでの生物発光のスペクトルを、Molecular Devices Spectramax、96ウエルプレートの蛍光光度計を使用して分析した。それらの結果を以下の表1に要約する。
【0147】
ステップ4 変異の同定 インビボでの選別によって選択された25個のクローンからプラスミドDNAを調製し、遺伝子特異的配列決定プライマーを使用して配列決定した。一次配列中の位置354および357においてアミノ酸の変化を生じた変異を同定した。ある変異体は、位置I351にアミノ酸の置換をもたらした(表5)。
【0148】
【表5】
【0149】
いくつかの変異型ルシフェラーゼを熱安定性のインビボアッセイから選択した。これらのルシフェラーゼの大部分は、また発せられる光のインビボスペクトルにおいて大きな変化を示し、多くは光の長い波長(>580nm)からのより大きな貢献を示した。いくつかのスペクトルは、610〜614nmというただ1つのピーク周辺で非常に狭い帯幅を示す。
【0150】
E354およびD357のそれぞれ疎水性および芳香族アミノ酸での置き換え(たとえばE354V、D357Y)は、インビボでのスペクトルの大きく変化をもたらし、612nm周辺で、狭い帯幅の単位ピークを示す。
【0151】
実施例8
熱安定性に関するインビボ選別 選択したクローンの無細胞抽出物を溶解によって調製し、それぞれの抽出物からのルシフェラーゼの熱安定性を、熱不活性化実験において測定した。それぞれの抽出物50μlをエッペンドルフチューブ中に置き、4、9および16分間、45℃に加熱した水浴中でインキュベートした。適切なタイムポイントにおいてアリコートを取り、残存ルシフェラーゼ活性を測定した。表6は、すべての変異型酵素および組換え野生型について残存活性%対時間を示す。
【0152】
これらの結果は、最も熱安定性であるルシフェラーゼは、位置357に芳香アミノ酸(Y、FまたはW)、および位置354に大きな疎水性(VまたはI)、極性(N)または正に帯電している(KまたはR)アミノ酸を有するルシフェラーゼであることを示す。
【0153】
【表6】
【0154】
実施例9
発せられる光のインビボでのスペクトルに対する成長条件の影響 変異型ルシフェラーゼE354K+D357M(前述の7のもの)を発現している大腸菌BL21(DE3)細胞から発せられる、光のスペクトルに対する異なる炭素源の影響を調べた。
【0155】
細胞の培養物50mlをLB培地上で対数期中期まで生育させ、次いで遠心分離によって採取した。細胞ペレットを1mlの滅菌蒸留水中に再懸濁させ、次いでこの懸濁液のアリコート100ulを使用して、新鮮なLB5ml、M9最小培地+2mM酢酸ナトリウムまたはM9最小培地+2mMグルコースを25mlの滅菌チューブ中に接種した。この培養物を振とうしながら37℃で成長させ続け、90分(D357Y)および120分(酵素7)後、細胞のアリコート200ulを取り遠心分離し、0.5mMのD−ルシフェリンを含む100mMのクエン酸ナトリウムバッファー(pH5.0)150ul中に再懸濁させた。次いで再懸濁させた細胞をミクロタイタープレート中に置き、変異型ルシフェラーゼのそれぞれによって発せられる生物発光のインビボスペクトルを、Molecular Devices Spectramax、96ウエルプレートの蛍光光度計を使用して分析した。それらの結果を図7および8に示す。
【0156】
これらの結果は、富化培地(LB)(図7a、8a)からの、唯一の炭素源としてアセテート(図7b、8b)またはグルコース(図7c、8c)のいずれかを有する限定最小培地への変化は、発せられる光がより長い波長にシフトと、より短い波長からの貢献の低下をもたらすことを示す。
【0157】
実施例10
組換え野生型および変異型ルシフェラーゼの精製およびスペクトルの特徴付け 組換え野生型Photinus pyralis酵素および変異型ルシフェラーゼD357YおよびE354I+D357Yを、生物発光反応のスペクトルに対する補因子補酵素Aの影響を分析するために、均一になるまで精製した。3つのルシフェラーゼすべてを、嫌気性真菌Piromyces equiiからの143アミノ酸炭水化物結合モジュール(CBM)への融合体として、精製した。このCBMは、酸膨張性セルロースおよび可溶性炭水化物ガラクトマナンおよびグルコマナンに選択的に結合し、簡単な単一ステップのアフィニティ精製スキームのベースを形成することが示されている。
【0158】
CBMに融合したルシフェラーゼは、粗無細胞抽出物中でセルロースに結合することができ、これを洗浄し、次いで可溶性多糖を使用して選択的に溶離させた。このようにして精製した融合タンパク質をアッセイにおいて使用して、異なる量の補酵素Aを含む反応中で発せられた光の波長を測定した。異なる量の補酵素Aを含むアッセイ試薬100μl(25mM トリストリシン pH7.8、5.0mM MgSO4、0.1mM EDTA、530μM ATPおよび470μM D−ルシフェリン)に、酵素(5μl)を加えた。図9〜11は、精製したルシフェラーゼD357およびE354I+D357Yによって発せられた光のスペクトルに対する、補酵素Aの増大した濃度の影響を示す。
【0159】
真菌CBMのC末端に融合しホタルのルシフェラーゼを発現している大腸菌細胞によって発せられる、生物発光のスペクトルのインビボでのアッセイは、元のルシフェラーゼを発現している細胞といかなる顕著な違いも示さなかった。同様に、精製組換えルシフェラーゼの市販品(Promega)によって発せられる、生物発光のスペクトルのインビトロアッセイは、融合タンパク質によって発せられたスペクトルと同一であった。
【0160】
したがって、観察した違いはCoAの濃度に伴われたものである。補酵素Aの濃度が増大すると、スペクトルの分布が変化し、CoAの最大濃度では、スペクトルは、610nmでの明確なピークを有する590〜630nmの領域の波長が主となる。スペクトルのシフトは二重変異体に関して最も顕著であり、波長610nmという唯一のピーク周辺で帯幅の大幅な狭まりが存在する(図11)。
【0161】
実施例12
214C/354K/357Fを有するように変異させた、合成Photinus pyralisルシフェラーゼの生産 前述の合成戦略を使用して、オリゴヌクレオチド対から合成luc遺伝子を設計し組み立てた。遺伝子配列を工作して、アミノ酸214C、354Kおよび357Fを有するルシフェラーゼを作製した。
【0162】
重複合成オリゴヌクレオチド29対をSigma−Genosys Ltdによって合成し、PAGEによって精製し、約550bpの3つのアセンブリに連結させた(IDRIS1、2および3、図13)。次いでそれぞれのアセンブリをベクターpBSK(+)別々に連結させ、生じた構築体を使用して大腸菌XL1−Blue細胞を形質転換させた。組み立てた挿入体を含むクローンからプラスミドDNAを作製し、配列決定してORFの適合度を確認した。アセンブリ中のn−1個のオリゴヌクレオチドの存在(オリゴ合成の副産物)によって、構築プロセスが複雑になった。DNAの配列決定によってIDRIS2の1つの正確なアセンブリを同定し、PCRを使用して、構築体の5'端に1つの塩基対欠失体を含むIDRIS3の1アセンブリを正しくした。IDRIS1を含むプラスミドの、IDRIS2および3との混合物を連結させることによって、完全なORFのアセンブリを得た。
【0163】
次いで連結したDNAを使用して大腸菌XL1−Blue細胞を形質転換させ、インビボアッセイを使用して活性酵素を発現しているクローンを選択した。数個のクローンを選択し、配列決定して図14に示す配列を有する合成luc遺伝子の存在および適合度を確認した。完全なORFをIDRIS(FA)と呼んだ。
【0164】
合成遺伝子をベクターpBSK(+)内のポリリンカー中のBamH IとSal I部位の間に組立てた。この位置において、この遺伝子はアルファペプチドとはフレームが同じでなく、lacプロモーターからは相当な距離がある。しかしながら、充分なルシフェラーゼが生成されて、酵素の予備的特徴付けが可能である。Promega溶解法を使用して一晩培養したものから、IDRIS(FA)を発現している大腸菌XL1−Blue細胞の粗無細胞抽出物を作製した。
【0165】
次いで抽出物中の酵素の熱安定性を50℃で20分間試験し、熱安定性の変異体E354I+D357Yと比較した。新しいコドンを最適化させた三重変異体は、変異体E354I+D357Yよりも大幅に熱安定性であった(図15)。
(配列表)
【特許請求の範囲】
【請求項1】
ルシフェラーゼ活性を有し、野生型ルシフェラーゼと少なくとも60% の類似性を有する組換えタンパク質であって、該酵素の配列においてPhotinus pyralis ルシフェラーゼ中の残基357に対応するアミノ酸残基が、対応する野生型ルシフェラーゼと比較して変異しており、その結果このルシフェラーゼ酵素が、対応する野生型ルシフェラーゼと比較して異なる波長の光を発することができ、および/または対応する野生型ルシフェラーゼと比較して熱安定性が向上している、前記組換えタンパク質。
【請求項2】
前記野生型ルシフェラーゼ配列が、Photinus pyralis、Luciola mingrel ica 、Luciola cruciata またはLuciola lateralis、Hotaria parvula、Pyrophorus plag iophthalamus、Lampyris noctiluca、Pyrocoelia miyako、Phrixothrix またはPhotinus pennsylvanicaからのルシフェラーゼのものである、請求項1に記載の組換えタンパク質。
【請求項3】
前記野生型ルシフェラーゼ配列が、Photinus pyralis、Luciola mingrel ica、Luciola cruciata またはLuciola lateralis の酵素から得られる酵素の配列である、請求項2に記載の組換えタンパク質。
【請求項4】
前記野生型ルシフェラーゼ配列が、Photinus pyralis、Luciola mingreli ca、Luciola cruciata またはLuciola lateralis、Hotaria parvula、Pyrophorus plagio phthalamus、Lampyris noctilucaまたはPyrocoelia miyako からのルシフェラーゼのものであり、Photinus pyralis ルシフェラーゼ中の残基357に対応するアミノ酸残基がアスパラギン酸またはグルタミン酸以外である、請求項1または請求項2に記載の組換えタンパク質。
【請求項5】
前記野生型ルシフェラーゼ配列が、Photuris pennsylvanica からのルシフェラーゼのものであり、Photinus pyralisルシフェラーゼ中の残基357に対応するアミノ酸残基がバリン以外である、請求項1または請求項2に記載の組換えタンパク質。
【請求項6】
Photinus pyralis ルシフェラーゼ中の残基357に対応するアミノ酸残基がアスパラギン酸、グルタミン酸またはバリン以外である、請求項1から5のいずれか一項に記載の組換えタンパク質。
【請求項7】
前記野生型ルシフェラーゼ配列が、Phrixothrix からのルシフェラーゼであり、Photinus pyralis ルシフェラーゼ中の残基357に対応するアミノ酸残基がロイシン以外である、請求項1または請求項2に記載の組換えタンパク質。
【請求項8】
Photinus pyralis ルシフェラーゼ中の残基357に対応するアミノ酸残基が、チロシン、フェニルアラニン、トリプトファン、アスパラギン、グルタミン、セリンまたはスレオニンである、請求項1〜7のいずれか1 項に記載の組換えタンパク質。
【請求項9】
Photinus pyralis ルシフェラーゼ中の残基357に対応するアミノ酸残基がチロシンである、請求項8に記載の組換えタンパク質。
【請求項10】
タンパク質がPhotinus pyralis、Luciola mingrelica、Luciola crucia ta またはLuciola lateralis からのルシフェラーゼと少なくとも80%の類似性を有する、請求項1〜9のいずれか一項に記載のタンパク質。
【請求項11】
野生型ルシフェラーゼと比較して、少なくとも1 つの以下の変異を有するタンパク質であって、
(a) Photinus pyralis ルシフェラーゼのアミノ酸354(Luciola ルシフェラーゼ中の356)に対応するアミノ酸残基が変異している;
(b) Photinus pyralis ルシフェラーゼ中の215位(Luciola ルシフェラーゼ中の217位)に対応するアミノ酸残基が、異なる疎水性アミノ酸である; あるいは
(c) Photinus pyralis ルシフェラーゼ中の残基214、またはLuciola mingrelica 、Luciola cruciata もしくはLuciola lateralis のルシフェラーゼの残基216に対応するアミノ酸残基;
(d) Photinus pyralis ルシフェラーゼ中の残基232、またはLuciola mingreli ca、 Luciola cruciata もしくはLuciola lateralis のルシフェラーゼの残基234に対応するアミノ酸残基;
(e) Photinus pyralis ルシフェラーゼ中の残基295、またはLuciola mingreli ca 、Luciola cruciata もしくはLuciola lateralis のルシフェラーゼの残基297に対応するアミノ酸残基;
(f) Photinus pyralis ルシフェラーゼ中のアミノ酸14、またはLuciola mingrelica の残基16、またはLuciola cruciata もしくはLuciola lateralis の残基16に対応するアミノ酸残基;
(g) Photinus pyralis ルシフェラーゼ中のアミノ酸35、またはLuciola mingre lica の残基37、またはLuciola cruciata もしくはLuciola lateralis の残基38に対応するアミノ酸残基;
(h) Photinus pyralis ルシフェラーゼのアミノ酸残基105、またはLuciola mi ngrelica の残基106、Luciola cruciata の残基107もしくはLuciola lateralis 遺伝子の残基108に対応するアミノ酸残基;
(i) Photinus pyralis ルシフェラーゼのアミノ酸残基234、またはLuciola mi ngrelica 、Luciola cruciata もしくはLuciola lateralis の残基236に対応するアミノ酸残基;
(j) Photinus pyralis ルシフェラーゼのアミノ酸残基420、またはLuciola mi ngrelica 、Luciola cruciata もしくはLuciola lateralis の残基422に対応するアミノ酸残基;
(k) Photinus pyralis ルシフェラーゼのアミノ酸残基310 、またはLuciola mingrelica、Luciola cruciata もしくはLuciola lateralis の残基312に対応するアミノ酸残基、
が対応する野生型配列中に見出されるアミノ酸とは異なっており、ルシフェラーゼ酵素が、対応する野生型ルシフェラーゼのこの位置でのアミノ酸を有する酵素と比較して増大した熱安定性を有することを特徴とする、請求項1 〜 1 0 のいずれか一項に記載のタンパク質。
【請求項12】
Photinus pyralis ルシフェラーゼのアミノ酸354(Luciola ルシフェラーゼ中の356)に対応するアミノ酸残基が変異している、請求項1〜11のいずれか一項に記載のタンパク質。
【請求項13】
Photinus pyralis ルシフェラーゼ中の残基214、またはLuciola min grelica、Luciola cruciata もしくはLuciola lateralis のルシフェラーゼの残基216 に対応するアミノ酸残基が、異なる疎水性アミノ酸に変異している、請求項12に記載のタンパク質。
【請求項14】
請求項1〜13のいずれか一項に記載のルシフェラーゼをコードしている核酸。
【請求項15】
合成遺伝子を含む請求項14に記載の核酸。
【請求項16】
特定の発現宿主用にコドンの使用を最適化し、および/または独特の制限部位を導入した、請求項14に記載の核酸。
【請求項17】
配列番号1のヌクレオチド9〜1661、またはそれと少なくとも90%の類似性を有する配列を含む、請求項14または請求項15に記載の核酸。
【請求項18】
請求項14〜17のいずれか一項に記載の核酸を含むベクター。
【請求項19】
請求項18に記載のベクターで形質転換された細胞。
【請求項20】
請求項1〜13のいずれか一項に記載のタンパク質を生成するための方法であって、請求項19に記載の細胞の培養を含む、前記方法。
【請求項21】
生物発光アッセイにおける、請求項1〜13のいずれか一項に記載のタンパク質の使用。
【請求項22】
請求項1〜13のいずれか一項に記載のタンパク質を含むキット。
【請求項23】
ルシフェリンをさらに含む請求項22に記載のキット。
【請求項24】
サンプル中のCoAの存在を判定するためのアッセイであって、CoAを含む疑いのあるサンプルに、前述の請求項1〜11のいずれか一項に記載のルシフェラーゼと共にルシフェラーゼ/ルシフェリン反応を引き起こすのに必要とされる他の試薬を加えること、サンプルから発せられる光の波長を測定すること、及びこれをCoAの存在または不在と関連付けること、を包含する、前記アッセイ。
【請求項25】
糖尿病の診断に使用するための請求項24に記載のアッセイ。
【請求項1】
ルシフェラーゼ活性を有し、野生型ルシフェラーゼと少なくとも60% の類似性を有する組換えタンパク質であって、該酵素の配列においてPhotinus pyralis ルシフェラーゼ中の残基357に対応するアミノ酸残基が、対応する野生型ルシフェラーゼと比較して変異しており、その結果このルシフェラーゼ酵素が、対応する野生型ルシフェラーゼと比較して異なる波長の光を発することができ、および/または対応する野生型ルシフェラーゼと比較して熱安定性が向上している、前記組換えタンパク質。
【請求項2】
前記野生型ルシフェラーゼ配列が、Photinus pyralis、Luciola mingrel ica 、Luciola cruciata またはLuciola lateralis、Hotaria parvula、Pyrophorus plag iophthalamus、Lampyris noctiluca、Pyrocoelia miyako、Phrixothrix またはPhotinus pennsylvanicaからのルシフェラーゼのものである、請求項1に記載の組換えタンパク質。
【請求項3】
前記野生型ルシフェラーゼ配列が、Photinus pyralis、Luciola mingrel ica、Luciola cruciata またはLuciola lateralis の酵素から得られる酵素の配列である、請求項2に記載の組換えタンパク質。
【請求項4】
前記野生型ルシフェラーゼ配列が、Photinus pyralis、Luciola mingreli ca、Luciola cruciata またはLuciola lateralis、Hotaria parvula、Pyrophorus plagio phthalamus、Lampyris noctilucaまたはPyrocoelia miyako からのルシフェラーゼのものであり、Photinus pyralis ルシフェラーゼ中の残基357に対応するアミノ酸残基がアスパラギン酸またはグルタミン酸以外である、請求項1または請求項2に記載の組換えタンパク質。
【請求項5】
前記野生型ルシフェラーゼ配列が、Photuris pennsylvanica からのルシフェラーゼのものであり、Photinus pyralisルシフェラーゼ中の残基357に対応するアミノ酸残基がバリン以外である、請求項1または請求項2に記載の組換えタンパク質。
【請求項6】
Photinus pyralis ルシフェラーゼ中の残基357に対応するアミノ酸残基がアスパラギン酸、グルタミン酸またはバリン以外である、請求項1から5のいずれか一項に記載の組換えタンパク質。
【請求項7】
前記野生型ルシフェラーゼ配列が、Phrixothrix からのルシフェラーゼであり、Photinus pyralis ルシフェラーゼ中の残基357に対応するアミノ酸残基がロイシン以外である、請求項1または請求項2に記載の組換えタンパク質。
【請求項8】
Photinus pyralis ルシフェラーゼ中の残基357に対応するアミノ酸残基が、チロシン、フェニルアラニン、トリプトファン、アスパラギン、グルタミン、セリンまたはスレオニンである、請求項1〜7のいずれか1 項に記載の組換えタンパク質。
【請求項9】
Photinus pyralis ルシフェラーゼ中の残基357に対応するアミノ酸残基がチロシンである、請求項8に記載の組換えタンパク質。
【請求項10】
タンパク質がPhotinus pyralis、Luciola mingrelica、Luciola crucia ta またはLuciola lateralis からのルシフェラーゼと少なくとも80%の類似性を有する、請求項1〜9のいずれか一項に記載のタンパク質。
【請求項11】
野生型ルシフェラーゼと比較して、少なくとも1 つの以下の変異を有するタンパク質であって、
(a) Photinus pyralis ルシフェラーゼのアミノ酸354(Luciola ルシフェラーゼ中の356)に対応するアミノ酸残基が変異している;
(b) Photinus pyralis ルシフェラーゼ中の215位(Luciola ルシフェラーゼ中の217位)に対応するアミノ酸残基が、異なる疎水性アミノ酸である; あるいは
(c) Photinus pyralis ルシフェラーゼ中の残基214、またはLuciola mingrelica 、Luciola cruciata もしくはLuciola lateralis のルシフェラーゼの残基216に対応するアミノ酸残基;
(d) Photinus pyralis ルシフェラーゼ中の残基232、またはLuciola mingreli ca、 Luciola cruciata もしくはLuciola lateralis のルシフェラーゼの残基234に対応するアミノ酸残基;
(e) Photinus pyralis ルシフェラーゼ中の残基295、またはLuciola mingreli ca 、Luciola cruciata もしくはLuciola lateralis のルシフェラーゼの残基297に対応するアミノ酸残基;
(f) Photinus pyralis ルシフェラーゼ中のアミノ酸14、またはLuciola mingrelica の残基16、またはLuciola cruciata もしくはLuciola lateralis の残基16に対応するアミノ酸残基;
(g) Photinus pyralis ルシフェラーゼ中のアミノ酸35、またはLuciola mingre lica の残基37、またはLuciola cruciata もしくはLuciola lateralis の残基38に対応するアミノ酸残基;
(h) Photinus pyralis ルシフェラーゼのアミノ酸残基105、またはLuciola mi ngrelica の残基106、Luciola cruciata の残基107もしくはLuciola lateralis 遺伝子の残基108に対応するアミノ酸残基;
(i) Photinus pyralis ルシフェラーゼのアミノ酸残基234、またはLuciola mi ngrelica 、Luciola cruciata もしくはLuciola lateralis の残基236に対応するアミノ酸残基;
(j) Photinus pyralis ルシフェラーゼのアミノ酸残基420、またはLuciola mi ngrelica 、Luciola cruciata もしくはLuciola lateralis の残基422に対応するアミノ酸残基;
(k) Photinus pyralis ルシフェラーゼのアミノ酸残基310 、またはLuciola mingrelica、Luciola cruciata もしくはLuciola lateralis の残基312に対応するアミノ酸残基、
が対応する野生型配列中に見出されるアミノ酸とは異なっており、ルシフェラーゼ酵素が、対応する野生型ルシフェラーゼのこの位置でのアミノ酸を有する酵素と比較して増大した熱安定性を有することを特徴とする、請求項1 〜 1 0 のいずれか一項に記載のタンパク質。
【請求項12】
Photinus pyralis ルシフェラーゼのアミノ酸354(Luciola ルシフェラーゼ中の356)に対応するアミノ酸残基が変異している、請求項1〜11のいずれか一項に記載のタンパク質。
【請求項13】
Photinus pyralis ルシフェラーゼ中の残基214、またはLuciola min grelica、Luciola cruciata もしくはLuciola lateralis のルシフェラーゼの残基216 に対応するアミノ酸残基が、異なる疎水性アミノ酸に変異している、請求項12に記載のタンパク質。
【請求項14】
請求項1〜13のいずれか一項に記載のルシフェラーゼをコードしている核酸。
【請求項15】
合成遺伝子を含む請求項14に記載の核酸。
【請求項16】
特定の発現宿主用にコドンの使用を最適化し、および/または独特の制限部位を導入した、請求項14に記載の核酸。
【請求項17】
配列番号1のヌクレオチド9〜1661、またはそれと少なくとも90%の類似性を有する配列を含む、請求項14または請求項15に記載の核酸。
【請求項18】
請求項14〜17のいずれか一項に記載の核酸を含むベクター。
【請求項19】
請求項18に記載のベクターで形質転換された細胞。
【請求項20】
請求項1〜13のいずれか一項に記載のタンパク質を生成するための方法であって、請求項19に記載の細胞の培養を含む、前記方法。
【請求項21】
生物発光アッセイにおける、請求項1〜13のいずれか一項に記載のタンパク質の使用。
【請求項22】
請求項1〜13のいずれか一項に記載のタンパク質を含むキット。
【請求項23】
ルシフェリンをさらに含む請求項22に記載のキット。
【請求項24】
サンプル中のCoAの存在を判定するためのアッセイであって、CoAを含む疑いのあるサンプルに、前述の請求項1〜11のいずれか一項に記載のルシフェラーゼと共にルシフェラーゼ/ルシフェリン反応を引き起こすのに必要とされる他の試薬を加えること、サンプルから発せられる光の波長を測定すること、及びこれをCoAの存在または不在と関連付けること、を包含する、前記アッセイ。
【請求項25】
糖尿病の診断に使用するための請求項24に記載のアッセイ。
【図1】

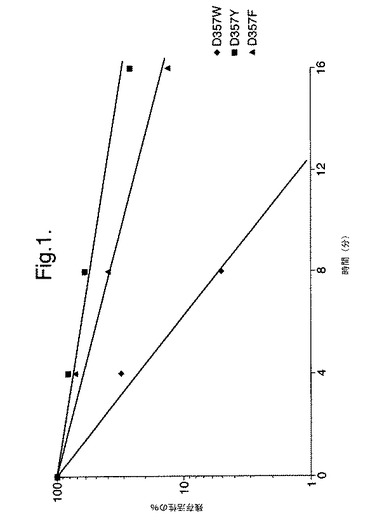
【図2(a)】
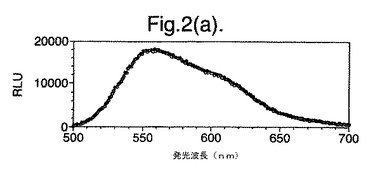
【図2(b)】
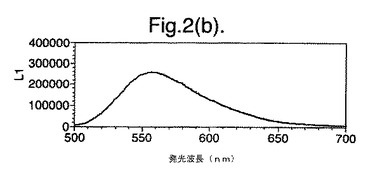
【図2(c)】
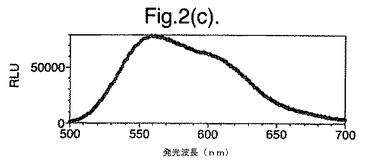
【図2(d)】
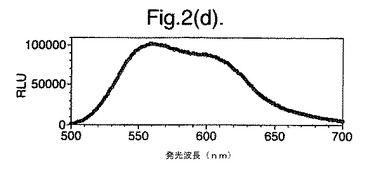
【図2(e)】
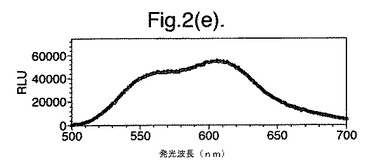
【図2(f)】
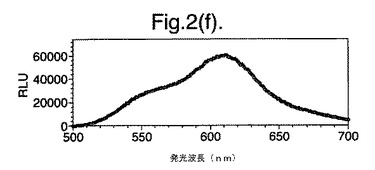
【図2(g)】
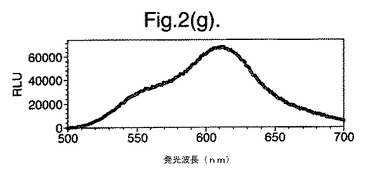
【図2(h)】
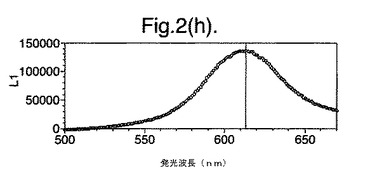
【図3】
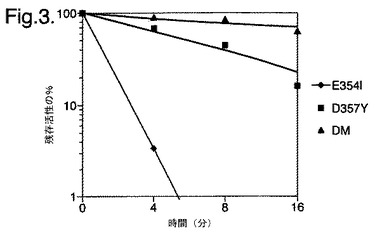
【図4(a)】
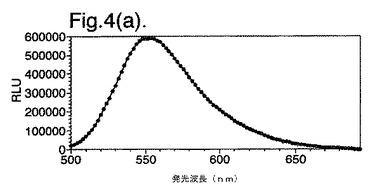
【図4(b)】
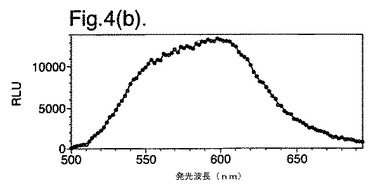
【図5】
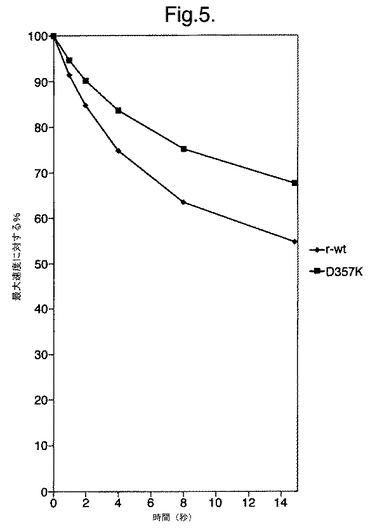
【図6】
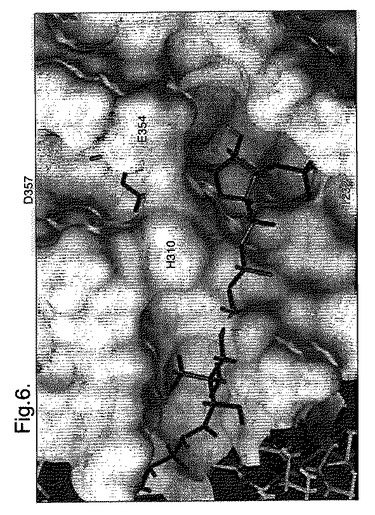
【図7a】
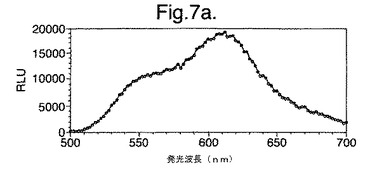
【図7b】
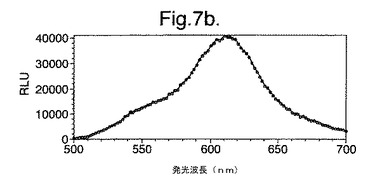
【図7c】
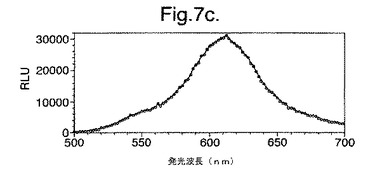
【図8(a)】
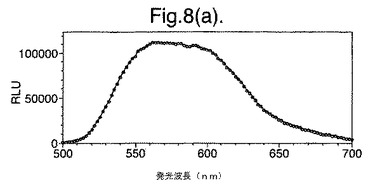
【図8(b)】
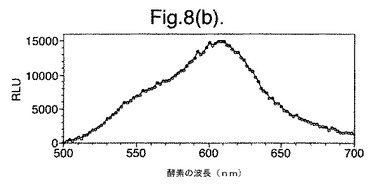
【図8(c)】
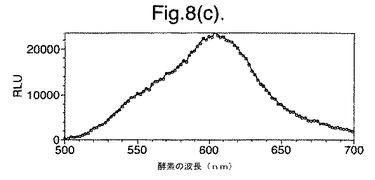
【図9】
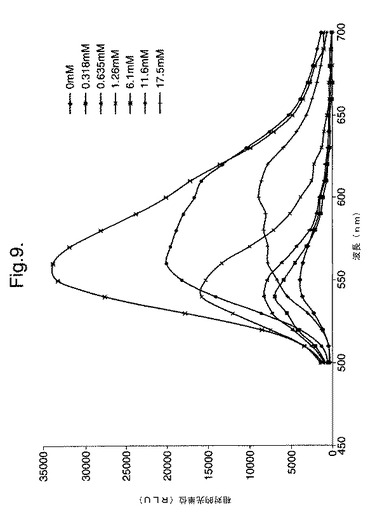
【図10】
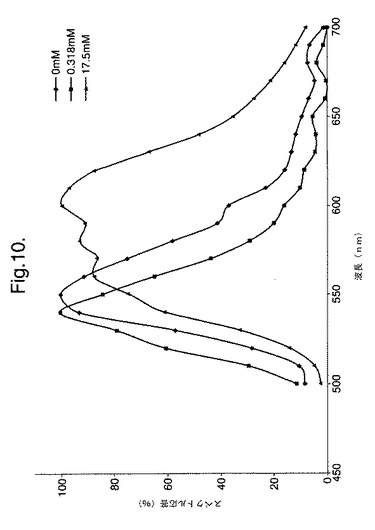
【図11(a)】
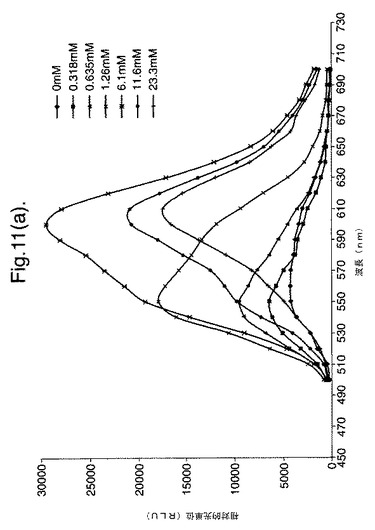
【図11(b)】
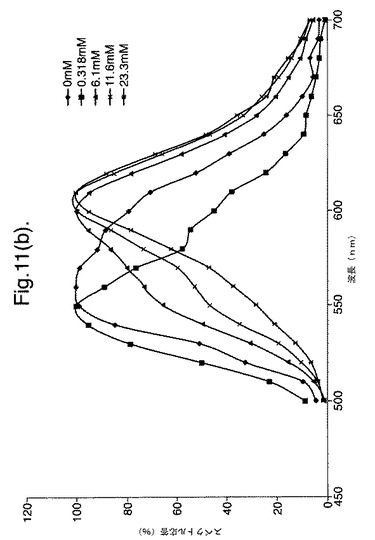
【図12】
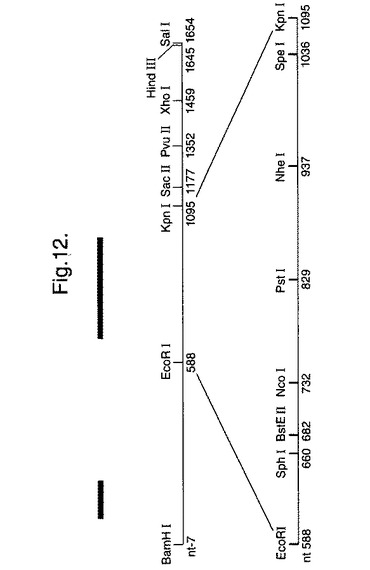
【図13】
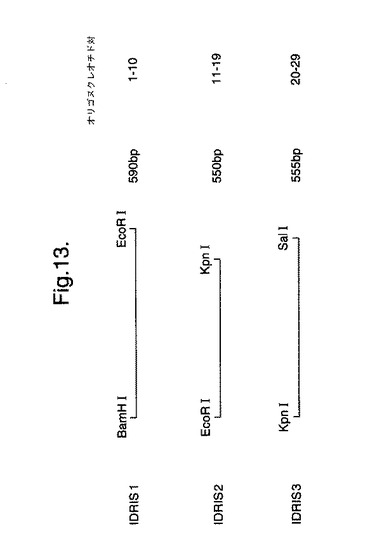
【図14−1】
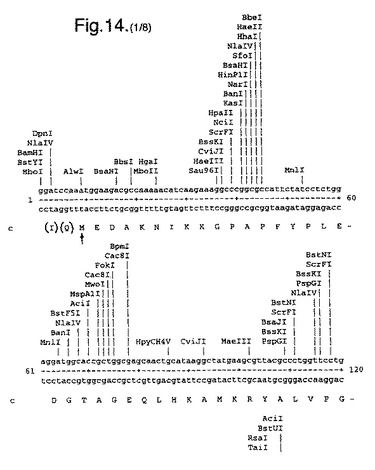
【図14−2】

【図14−3】

【図14−4】
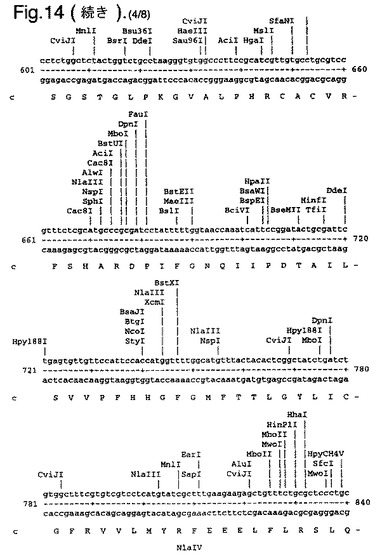
【図14−5】
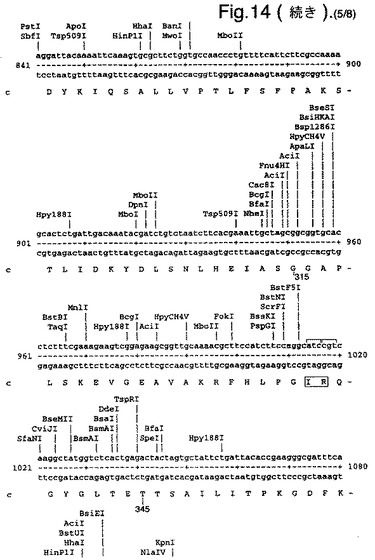
【図14−6】
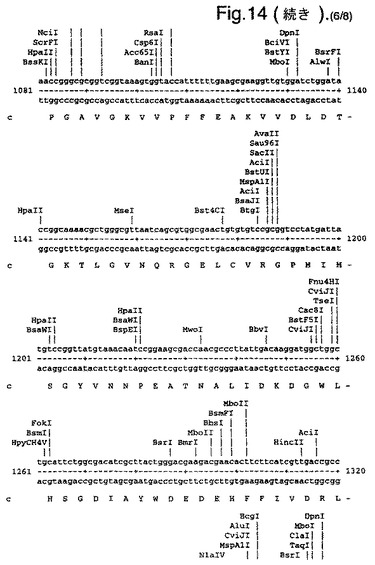
【図14−7】

【図14−8】
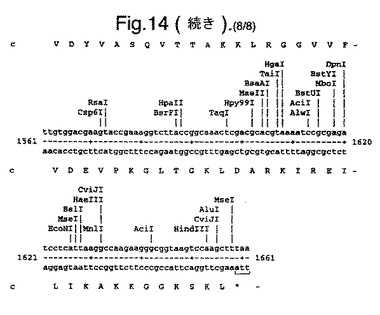
【図15】
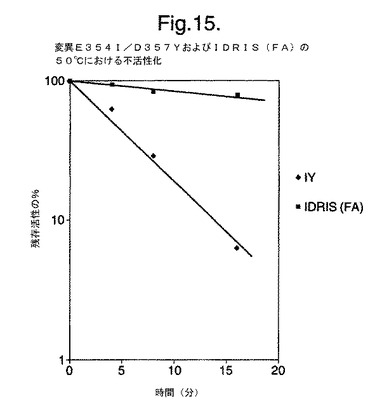
【図2(a)】
【図2(b)】
【図2(c)】
【図2(d)】
【図2(e)】
【図2(f)】
【図2(g)】
【図2(h)】
【図3】
【図4(a)】
【図4(b)】
【図5】
【図6】
【図7a】
【図7b】
【図7c】
【図8(a)】
【図8(b)】
【図8(c)】
【図9】
【図10】
【図11(a)】
【図11(b)】
【図12】
【図13】
【図14−1】
【図14−2】
【図14−3】
【図14−4】
【図14−5】
【図14−6】
【図14−7】
【図14−8】
【図15】
【公開番号】特開2011−83289(P2011−83289A)
【公開日】平成23年4月28日(2011.4.28)
【国際特許分類】
【出願番号】特願2011−791(P2011−791)
【出願日】平成23年1月5日(2011.1.5)
【分割の表示】特願2001−533163(P2001−533163)の分割
【原出願日】平成12年10月26日(2000.10.26)
【出願人】(505005049)スリーエム イノベイティブ プロパティズ カンパニー (2,080)
【Fターム(参考)】
【公開日】平成23年4月28日(2011.4.28)
【国際特許分類】
【出願日】平成23年1月5日(2011.1.5)
【分割の表示】特願2001−533163(P2001−533163)の分割
【原出願日】平成12年10月26日(2000.10.26)
【出願人】(505005049)スリーエム イノベイティブ プロパティズ カンパニー (2,080)
【Fターム(参考)】
[ Back to top ]