
多価組換え抗原のコンビナトリアルベースの生産のための方法および組成物
本発明は、糸状菌ヘテロカリオンを用いて、多価組換えワクチンを速やかに生産するための方法および組成物を提供する。本発明は、病原性生物由来の抗原の変異体をコードする組換えDNA分子が導入された二つまたはそれ以上の親株の組み合わせより産生された糸状菌ヘテロカリオンの使用に依拠する。結果的に生じるワクチンが多価である。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願
本出願は、全体として参照により本明細書によって組み入れられる、2004年10月15日に提出された米国仮出願第60/619,364号と関連する。
【0002】
技術分野
開示される発明は、分子生物学の分野、および病原性生物に対する多価ワクチンの生産に関する。本発明の一つの態様は、多価ワクチンの集団を生産するのに使用され得るヘテロカリオン性(heterokaryotic)糸状菌において抗原をコードするヌクレオチド配列の集団を提供する方法および組成物を、具体的に提供する。
【背景技術】
【0003】
背景技術
ワクチンは、現今、様々な方法によって生産される。典型的には、インフルエンザワクチンは、受精した鶏卵を用いて生産される。米国においては、the Centers for Disease Controlが、特定のインフルエンザの季節において、攻撃する可能性が最も高いウイルスを代表すると考えられる3種類のウイルス株を選択するであろう。選択されたウイルスの試料は、所望の抗原性の特徴を所有する種ウイルスストックとして、製造業者に提供される。種ウイルスは受精した鶏卵中に注入される。これらの卵は、インフルエンザウイルスが増殖する間インキュベーションされる。適当な期間の後、卵が開けられて、卵白が採取される。この試料がウイルスを含む。ウイルスは卵材料より精製されて、不活性化される。個々のウイルスストックは、その後、典型的には三価のワクチンである、共通インフルエンザワクチンを創出するために混合される。
【0004】
ワクチンバッチ全体に欠陥を生じさせ得る様々な問題が起こり得る。例えば、2004年には、滅菌に伴う問題が、カイロン(Chiron)のワクチン生産施設の認証撤回をもたらす。この状況は、伝統的なワクチン生産法がいかに信頼できない可能性があるかを例証する。更に、現今のインフルエンザワクチン生産法は、各年何億個という鶏卵を使用する。保管、取り扱い、および加工の段階は、時間を消費し、かつ大きな労働力を要する。その上、長い生産時間を考慮すると、インフルエンザの季節の間にインフルエンザウイルスの新たな株が優勢になった場合、現今の卵に基づく生産法は、新しいワクチンが生産されるのに数ヶ月要するであろう。
【0005】
これらの限界を考慮して、インフルエンザワクチンのような抗原性材料を生産する、より柔軟な、かつ有効な方法が非常に必要とされる。
【0006】
タンパク質の組換え真菌発現
真菌における異種遺伝子のクローニングおよび発現は、様々な有用タンパク質を生産するのに使用されてきた。例えば:Lambowitz、米国特許第4,486,533号は、ミトコンドリアプラスミドDNAによる糸状菌のためのDNAベクターの自律複製、ならびに、ニューロスポラ属(Neurospora)への異種遺伝子の導入および発現を開示する;Yeltonら、米国特許第4,816,405号は、大量の所望の異種タンパク質を生産し、かつ分泌するために、糸状子嚢菌の重要な株の修飾を可能にする道具および系を開示する;Buxtonら、米国特許第4,885,249号は、宿主アスペルギルス・ニガー(Aspergillus niger)細胞へ組み込まれ得る選択マーカーを含むDNAベクターによるA.ニガーの形質転換を開示する;および、McKnightら、米国特許第4,935,349号は、アスペルギルス属(Aspergillus)および他の糸状菌において、異種遺伝子の発現を指示し得るプロモーターを含む、アスペルギルス属において高等真核生物遺伝子を発現するための方法を開示する。同様の技術は、ニューロスポラ・クラッサ(Neurospora crassa)("N. クラッサ")においてアミノ酸輸送に関与するmtr遺伝子をクローン化し、かつ、クローン化DNAの、インビボにおいてこの遺伝子に隣接するゲノムマーカーへの強い関連を立証するために使用されてきた。Stuart, W. D. et al., Genome (1988)30:198-203; Koo, K. and Stuart, W. D. Genome (1991)34:644-651。
【0007】
糸状菌は、真核生物タンパク質を生産するのに使用するための良い候補者となる多くの特徴を所有する。糸状菌は、複雑なタンパク質を分泌し得る;ジスルフィド結合形成を含む3次元タンパク質を正しくフォールディングする;翻訳後に、タンパク質をタンパク質分解的に切り詰める;かつ、N-結合型およびO-結合型グリコシル化反応を用いてタンパク質をグリコシル化する。これらの能力は、この生物群を分泌組換えタンパク質の生産のための魅力的な宿主としてきた(MacKenzie, D. A. et al., J Gen Microbial (1993) 139:2295-2307; Peberdy, J.F., Trends in BioTechnology (1994) 12:50-57)。
【0008】
ニューロスポラ・クラッサは、組換え同種および異種タンパク質生産のための宿主細胞として使用されてきた(Carattoli, A., et al., Proc Nat Acad Sci USA, (1995) 92:6612-6616; Yamashita, R. A. et al., Fungal Genetics Newsletter (1995 Suppl.) 42A; Kato, E. et al., Fungal Genetics Newsletter (1995 Suppl.) 42A; Buczynski, S. et al. Fungal Genetics Newsletter (1995 Suppl.) 42A, Nakano, E. T. et al. Fungal Genetics Newsletter (1995 Suppl.) 40:54 0)。加えて、ニューロスポラ・クラッサは、ヘテロカリオン(heterokaryon)の手段により、組換えヘテロダイマーおよびマルチマー結合タンパク質を発現するための宿主細胞として使用されてきた(米国特許第5,643,745号、1997年7月Stuart435/69.1)。
【発明の開示】
【0009】
発明の概要
本発明は、多価ワクチンを生産するヘテロカリオン糸状菌を提供する。
【0010】
本発明の個々のヘテロカリオンは、第一および第二の、および三価ワクチンの場合には第三の親真菌株、ならびに、より高次レベルのワクチン価の場合は、付加される抗原の各セットに対して一つの付加的な親株を融合することによって産生され、各親株は、多価ワクチンの抗原の自然に見出される変異体、多価ワクチンの抗原の合理的に設計された変異体、または、多価ワクチンの抗原のランダムに産生された変異体をコードする発現ユニットと同様に、ヘテロカリオン性状態を維持するために必要なマーカーを含む。このように、自然の変異体に加えて、化学的、物理的、もしくは部位特異的突然変異誘発、または他の技術を通して産生された変異体が、生産され得る。
【0011】
本発明のヘテロカリオンは、定義された抗原の所望の多価ワクチンを選択的に生産するために有用である。
【0012】
上記に基づいて、本発明は、多価ワクチンの変異体を生産するヘテロカリオンを提供する。
【0013】
発明の開示
「ヘテロカリオン」(またはヘテロカリオン性細胞)は、二つ(またはそれ以上)の糸状菌親株の融合より形成される細胞であり、こうして各ヘテロカリオン細胞は二つ(またはそれ以上)の遺伝的に異なる核を含む。ヘテロカリオンは、すべてのヘテロカリオン適合性アリルについて一般に同型接合である(tol遺伝子が存在する場合、交配型アリルを除いて)親株由来の核を含む。少なくとも10個の染色体座(het-c、het-d、het-e、het-i、het-5、het-6、het-7、het-8、het-9、およびhet-10)が、ヘテロカリオン不適合性について同定されてきており、かつ、それ以上が存在することが推測されている。Peris et al., "Chromosomal Loci of Neurospora crassa", Microbiological Reviews (1982) 46:426-570, at 478。
【0014】
本発明は、ヘテロカリオン性糸状菌を用いて多価ワクチンの集団を生産するための方法および組成物を提供することによって、米国特許第5,643,745号、第5,683,899号、および第6,268,140号において開示された研究を進歩させる。そのような方法および組成物は、抗ウイルスワクチン、抗細菌ワクチン、および抗真菌ワクチンのような多価ワクチンの発見および生産において有用である。
【0015】
好ましい態様の記述
本発明は、多価ワクチンの集団を産生するための新規の方法および組成物を提供する。多価ワクチンは、定義された抗原の、順序づけられた、またはランダムな組み合わせより産生される。多価ワクチンを取得するために、多価ワクチンの定義された抗原をコードする第一、第二、および時には第三またはそれ以上のDNA分子の集団を、第一、第二の真菌の、および時には第三またはそれ以上の宿主親に導入する段階、第一、第二、および時には第三またはそれ以上の宿主真菌親を用いてヘテロカリオン性真菌株を形成する段階、および、その後、適切である場合は、所望の性質を有する多価ワクチンの生産のために、結果として生じたヘテロカリオンをDNAをコードするサブユニットが発現する条件の下で培養する段階、および、結果として生じたヘテロカリオンを更にスクリーニングする段階を含む方法において、ヘテロカリオンが取得される。各要素、すなわち、真菌親、DNA分子、および融合方法は、以下に詳細に記述される。
【0016】
糸状菌の性質およびヘテロカリオン形成のためのバックグラウンド要求性
真菌は、糸状多核鎖をもたらす単一の単核細胞、酵母細胞、多様な胞子を伴う子実体、および/または有性的に分化された細胞として生育し得る。それらはまた、多核形態においても存在し得る。カビとしての真菌の成長形態の主な要素は、直径約2μm〜10μmの分岐した管状構造である菌糸である。菌糸は、先端での伸長(頂端生長)および側面の分岐を生ずることによって、増殖する。このように、コロニーが増殖するとき、菌糸は多量の絡み合う繊維を形成する。
【0017】
いくつかの菌糸は、栄養を吸収するために、真菌が増殖している培養培地中に貫通し、一方、培地の表面より上に突き出した菌糸は、「気中菌糸体」を構成する。大抵のコロニーは、不規則な、乾燥した糸状マットとして、液体または固体培地の表面に増殖する。大抵の種においては、菌糸は、「隔壁」と呼ばれる横断壁によって分割される。しかしながら、これらの隔壁は、細かい中心細孔を有する。こうして、隔壁を有する菌糸でさえ、連続的な多量の細胞質に埋め込まれた複数の核を有し、かつ、事実上、移動可能な細胞質において核の多様性を含む。
【0018】
「糸状菌」という用語は、多量の分岐し、連結した繊維を通して菌糸体を形成し得、かつ、横断壁によって遮断されるが、横断壁における穿孔のために区画間の細胞質の輸送を許容し得る真菌のことを指す。これらの真菌の多くは、有性的に繁殖した際に、嚢の中に減数分裂した胞子を形成する。メカニズムは完全に理解されていないが、適切な刺激で、生殖は無性的に起こり得る。この生殖の様式において、「分生子」として公知である胞子は、繊維に沿った様々な場所で形成される出芽した突出物の先端で、外面的に生ずる。
【0019】
本発明のヘテロカリオンパネルを産生するのに使用される糸状菌は、一般に、藻菌類(Phycomycetes)、子嚢菌綱(Ascomycetes)、担子菌綱(Basidiomycetes)、および不完全菌類(Deuteromycetes)である。藻菌類は、いくつかの隔壁を有する糸状菌と同様に、隔壁を有しないすべての糸状菌を含む。それらの無性胞子は様々な種類があり、分化した柄の末端に形成される嚢の中に含まれる胞子嚢胞子を含む。異なる種は、異なる有性サイクルを有する。
【0020】
子嚢菌綱は、子嚢胞子として公知である有性胞子を含む嚢様構造である子嚢によって、他の真菌より識別される。子嚢胞子は、交配、雄および雌の核の融合、2回の減数分裂、および通常1回の最終的な有糸分裂の最終産物である。担子菌綱は、分化した構造の表面上に形成する有性胞子によって識別される。不完全菌類は、有性相が未だ観察されていないため、「不完全な真菌」としてしばしば称される。それらの菌糸は隔壁を有し、かつ、分生子の形態は、子嚢菌綱のものに類似している。
【0021】
好ましい糸状菌は、子嚢菌綱の群のものであり、より好ましくは、ニューロスポラ属、アスペルギルス属、フザリウム属(Fusarium)、トリコデルマ属(Tricoderma)、クリソスポリウム属(Chrysosporium)、およびペニシリウム属(Penicillium)由来である。ニューロスポラ属由来の特に有用な種は、N.インターメディア(N. intermedia)、N.クラッサ、N.シトプーラ(N. sitopula)、およびN.テトラスポラ(N. tetraspora)を含み、そのうち最も好ましい種はN.クラッサである。アスペルギルス属の有用な種は、A.ニデュランス(A. nidulans)、A.ニガー、A.テレウス(A.terreus)、およびA.フメガテュス(A.fumegatus)を含む。
【0022】
糸状菌の栄養成長は、細胞分裂(有糸分裂)に伴う核分裂を含む。この型の細胞分裂は、無性生殖、すなわち、配偶子の関与無し、および分生子経由の核融合無しの、新たなクローンの形成からなる。例えば、ニューロスポラ属の種は、核に、それぞれがシングルコピーを有する7本の異なる染色体を含み、すなわち、栄養体は一倍体である。この一倍体状態が、菌糸体増殖の間、および分生子の形成を通した無性生殖の間、典型的に維持される。
【0023】
有性生殖もまた起こり得、その時に異なる交配型の二つの一倍体細胞(菌糸または分生子)が融合して、二つの別個の核を含むジカリオン性細胞を形成する。こうして二つの一倍体核は同一の細胞質に共存し、しばらくの間は、同調して多少分裂する。しかしながら、細胞が子嚢胞子の形成を開始する場合は、二つの異なる一倍体核は、相同な染色体対を含む二倍体核を形成するように実際に融合し得る。この二倍体細胞が、その後減数分裂を始める。
【0024】
「ヘテロカリオン」(またはヘテロカリオン性細胞)は、二つの(またはそれ以上の)遺伝的に異なる核を伴う細胞である。本発明のヘテロカリオンは、すべてのヘテロカリオン適合性アリルについて同型接合である(tol遺伝子が存在する場合、交配型アリルを除いて)細胞由来の核を含まなければいけない。例えば、ニューロスポラ属において、少なくとも10個の染色体座(het-c、het-d、het-e、het-i、het-5、het-6、het-7、het-8、het-9、およびhet-10)が、ヘテロカリオン不適合性について同定されてきており、かつ、それ以上が存在することが推測されている。Perkins et al., "Chromosomal Loci of Neurospora crassa", Microbiological Reviews (1982) 46:426-570, at 478。
【0025】
二つの株が一つまた複数のhet座で異なるアリルを有する場合、それらは安定なヘテロカリオンを形成することができない。異なった菌糸の融合後、または細胞質もしくは抽出物の異なった株へのマイクロインジェクション後に、原形質殺傷(protoplasmic killing)が起きる。hetの一つまたは複数のアリルに対して重複(duplication)(部分的二倍体)が異型接合である場合、増殖が阻害されて、高度に異常となる。いくつかのヘテロカリオン適合性座(特にhet-c、-d、-e、および-i)は、ヘテロカリオン試験によって最初に定義された。het-5からhet-10の座は、het座での差異が自然の集団において共通しているため、重複を使用して検出された。同文献。
【0026】
いくらかの緩慢なヘテロカリオン性増殖が起きる可能性があるが、交配型アリル「A」および「a」はまた、N.クラッサにおいてhet遺伝子として作用する。マイクロインジェクション実験は、殺傷反応におけるタンパク質の関与を示している。このように、反対の交配型はまた、ヘテロカリオン性の無胞子菌糸の増殖と関連づけられる複雑な事象にとって一般に重要である。同文献436および478。しかしながら、tol遺伝子が存在する場合、反対の交配型アリルAおよびaと関連づけられる栄養(ヘテロカリオン)不適合性は、有性適合性が影響を及ぼされること無しに抑制される。このように、(tol; A+a;a)ヘテロカリオンは、他のhet座が同一のアリル(またはコンアレリック(conallelic))であれば完全に適合性であり、かつ安定であり得、tol遺伝子が存在するときA/a重複は正常に増殖する。
【0027】
適合性座についてコンアレリックである二つの異なる株由来の菌糸が提供される場合、それらは、同一の培地で増殖させられるとき、特に以下に記述されるように融合が強いられるとき、融合する可能性がある。結果として生じた培養物は、その後、共通の菌糸体マットの共有された細胞質において循環する双方の株由来の核を含むであろう。
【0028】
定義された抗原の混合集団をコードする発現ユニットの構築
本発明を記述するにあたって、以下の用語法が、下記に述べられた定義に従って使用されるであろう:
【0029】
本発明は、糸状菌ヘテロカリオンにおける「異種多価ワクチン」の生産を含む。本文脈において、「異種」とは、タンパク質が通常はその真菌によって生産されないことを意味する。「多価」とは、最終的なワクチン産物が、少なくとも二つの抗原または抗原変異体から構成されることを意味する。産物は、完全に異なる抗原を含むヘテロ多価ワクチンであってもよいし、または、単一のサブユニットの変異体から構成されるホモ多価でもあり得る。多価ワクチンの例は、細胞表面由来の組換え抗原、ウイルスコートタンパク質、特定の病原性タンパク質抗原などの混合物を含むが、それらに限定されない。
【0030】
「抗原をコードするヌクレオチド配列」は、適切な制御配列に作動可能に連結されたとき、転写産物がポリペプチドに翻訳される配列の部分である。コード配列の境界は、5'(アミノ)末端で開始コドン、および3'(カルボキシ)末端で翻訳終止コドンによって決定される。このコード配列は、例えば、原核生物遺伝子、真核生物mRNAからのcDNA、(真菌のような)真核生物DNAからのゲノムDNA配列由来であり得、または、合成DNAを含んでもよい。ポリアデニル化シグナルおよび転写終結配列は、通常コード配列の3'に位置するであろう。
【0031】
コード配列は、適切な宿主細胞において制御配列がコード配列の発現に影響を及ぼすとき、制御配列に「作動可能に連結」される。
【0032】
「発現ユニット」は、適切な宿主生物体において適切な条件の下で、作動可能に連結された配列の転写および翻訳を指示する「制御配列または領域」に作動可能に連結されたコード配列を含むDNA分子である。
【0033】
細胞は、外因性DNAが宿主細胞膜内に導入されているとき、外因性DNAによって「形質転換」されている。細菌のような原核生物については、外因性DNAはプラスミドのようなエピソーム要素上に維持されてもよい。糸状菌は核を有する(真核生物である)ため、大抵の安定に形質転換された真菌宿主細胞は、染色体に組み込まれた外因性DNAを含み、そのために染色体複製を通して娘細胞によって承継される。
【0034】
「組換え宿主」は、組換え技術によって調製されたDNA配列で形質転換されている、形質転換される、または形質転換されるであろう細胞のことを指し、かつ、元来形質転換された細胞ならびにそれらの培養物および子孫を含む。
【0035】
種々の方法が、1)多価ワクチンの抗原サブユニットの自然に見出される変異体、2)多価ワクチンの抗原のランダムに産生された、もしくは選択された変異体、または、3)多価ワクチンの抗原の合理的に設計された、もしくは選択された変異体、をコードするDNA分子の集団を産生するために使用され得る。以下において、インフルエンザワクチンが、例証的な例として使用される。当業者は、下記に概観される方法、または、当技術分野において公知である同等の方法を、サブユニットをコードするDNA分子の集団を産生するために容易に使用し得る。
【0036】
自然の不均一性を有する抗原の自然に見出される抗原変異体をコードするDNA分子の集団は、標準的なcDNA産生/クローニング技術を用いて生産され得る。一般に、mRNAおよびウイルスゲノムRNAの集団は、最初に病原体より、または、例えばインフルエンザワクチンの場合は、直接ウイルス自体のゲノムRNAより単離される。単離されたRNA分子の集団は、その後、RTPCRのような当技術分野で公知のクローニング方法において、cDNA分子の産生のための鋳型として使用される。このように生産されたcDNA分子の集団は、下記に記述されるように、適当な発現ユニットに挿入され得る。
【0037】
または、タンパク質配列が公知である抗原については、人工cDNA配列が、当技術分野に公知である方法を用いて、糸状菌宿主株によって自身の内因性タンパク質を生産するために高頻度に使用されるコドンを組み込んで、産生され得る。
【0038】
加えて、部位特異的またはランダムな突然変異誘発が、特定のサブユニットの自然に見出される変異体を生産するために、多価ワクチンの抗原をコードする単離された、または人工のcDNA分子について行われ得る。ランダムまたは部位特異的なミスマッチPCRプライミング、リンカースキャニング突然変異誘発、または化学的および物理的突然変異誘発のような手順が、抗原の合理的に設計された、またはランダムに産生された変異体をコードするDNA分子の集団を産生するために容易に使用され得る。例えば、ランダムに産生された、または合理的に設計されたPCRプライマーは、抗原コード配列において、ランダムな、または標的とされた不均一性を産生するために使用され得る。
【0039】
本明細書において使用されるとき、タンパク質フォールディングまたは特定の標的残基もしくは領域の選択のような選択基準が、変異体を産生する、または変異体をコードするDNA分子を選択する際に使用される場合、変異体が合理的に設計されると言われる。変異体をコードするDNA分子を産生する、または選択するときに選択基準が使用されない場合、変異体がランダムに産生されると言われる。
【0040】
多価ワクチンサブユニットにおいて不均一性を産生するための好ましい標的部位は、免疫原性エピトープおよび周囲のアミノ酸配列である。インフルエンザ抗原遺伝子の場合、この型の変異の創出は、各抗原の公知である天然の可変領域に集中するであろう。例として、公知の変異のライブラリーを生産するために、可変領域における各アミノ酸を一つずつ変化させ得るであろう。
【0041】
多価ワクチンの抗原をコードする発現ユニットの構築
多価ワクチンの抗原をコードするヌクレオチド分子を含む発現ユニットは、周知の技術を用いて構築される。一般に、発現ユニットは、サブユニットをコードする配列を、最終的な糸状菌宿主においてサブユニットをコードする配列の発現を方向付ける制御配列との作動可能な連結に置くことによって、産生される。
【0042】
構成的または誘導的いずれかの様式において、作動可能に連結されたタンパク質をコードする配列の発現を方向付けるために、様々な制御要素が、現在当技術分野において公知である。制御配列の選択は、使用される真菌株、真菌を培養するために使用される条件、所望のタンパク質発現レベル、および必要とされる発現の性質(例えば、誘導的対構成的)に基づくであろう。当業者は、本ヘテロカリオンパネルに使用される発現ユニットを産生するために、当技術分野において公知である制御配列を容易に利用し得る。
【0043】
タンパク質をコードする配列の転写および翻訳を方向付ける配列に加えて、本発明の発現ユニットは、抗原の細胞外への搬出を方向付ける発現制御要素であるシグナル配列を更に制御してもよい。糸状菌において公知である分泌シグナルの総説は、Dalbey R. E., et al., TIBS 17:474-478(1992)によって提供される。当業者は、分泌シグナルを含む発現ユニットを、容易に産生し得る。
【0044】
本発明の発現ユニットのもう一つの形態は、宿主細胞もしくは異種膜アンカー配列への融合を経て細胞膜へ、または、宿主細胞もしくは異種細胞表面分子への融合によって細胞表面へ、抗原を方向付ける融合タンパク質を含んでもよい。
【0045】
一つの応用において、組換えユニットが、発現ユニットの代わりに産生される。そのような使用において、サブユニットをコードする配列、または抗原をコードする配列の断片は、宿主真菌株における組込み部位に相同である配列を含むDNA領域によって隣接される。その後、相同な配列は、組換えユニットと宿主染色体との間での相同組換えを刺激し、かつ方向付けるように使用される。組換えユニットが使用されるとき、宿主株は、好ましくは、標的組換えのために使用される配列が続く発現制御要素を含む発現ユニットで、最初に形質転換される。例えば、インフルエンザ抗原が宿主真菌に導入され得、その後、相同組換えユニットが、宿主染色体の標的領域内に異種性を導入するために使用され得る。
【0046】
最終的な真菌細胞を形質転換し得る中間ベクターを生産するために、中間宿主が時々使用される。その時、所望の糸状菌宿主を形質転換するために使用され得る所望のDNA量を取得するために、中間細菌形質転換体が増殖され得る。中間ベクターとして役立ち得る共通に入手可能な細菌ベクターの例は、例えば、pBR322、pUC8、およびpUC9を含む。追加的な有用な中間ベクターは、pHY201、pKBY2、pTZ18R、pX182、およびpCVN2.9、pN807、pN846を含む。
【0047】
もう一つの態様において、関心対象の抗原または複数の抗原は、EASのような真菌ハイドロフォビン(hydrophobin)との融合タンパク質として発現される。真菌ハイドロフォビンは、典型的には大量に発現され、かつ分泌される。そして、真菌の表面は、ハイドロフォビンでコートされる。これらのタンパク質はまた、溶液中で凝集することが公知である。この凝集の特徴により、抗原性粒子を形成する凝集組換えタンパク質の調製が可能である。これらのタンパク質は、精製され、抗原として使用され得る。多数の抗原が同一の培養において発現されるとき、多価抗原性粒子が生産される。
【0048】
本発明のこの記述および開示は、本発明の精神および範囲内であるすべての態様に及ぶことを意図すると理解されるであろう。例えば、分子の抗原性に実質的に影響を及ぼすこと無く、オープンリーディングフレームのアミノ酸配列内でアミノ酸配列を挿入、欠失、または置換することは、当技術分野の知識内であり、かつ、自然に見出されるサブユニットへの欠失、付加、または置換で産生され得るような多価抗原は、本発明に含まれる。
【0049】
親株の性質
本発明のヘテロカリオンを作製することにおいて使用される各親真菌株は、多価ワクチンの抗原をコードするDNA分子の集団のメンバーを含んでいるであろうため、一つの真菌親は、所望の多価ワクチンの第一の抗原または抗原群をコードする、第一のDNA分子の集団のメンバーを含むように修飾された核を有するであろうし、かつ、それぞれの次の真菌親は、所望の多価ワクチンの異なる抗原または抗原群をコードする、異なるDNA分子の集団のメンバーを含むように修飾された核を有するであろう。例えば、二価のインフルエンザワクチンを生産するヘテロカリオンを生産するために、一つの真菌親は、インフルエンザA型H1 N1からのcDNA由来の抗原群を生産するであろうし、一方、他方の真菌親は、インフルエンザA型H3 N2からのcDNA由来の抗原群を生産するであろう。三価のインフルエンザワクチンを生産するためには、一つの真菌親は、インフルエンザA型H1 N1からのcDNA由来の抗原群を生産するであろうし、第二の真菌親は、インフルエンザA型H3 N2からのcDNA由来の抗原群を生産するであろうし、かつ、第三の真菌親は、インフルエンザB型H NからのcDNA由来の抗原群を生産するであろう。ヌクレオチドおよびタンパク質配列は、公的に入手可能である。例えば、HA遺伝子についての例示的な配列は、H3N2(AY738729)、インフルエンザAウイルス(A/Leningrad/54/1(H1N1))ノイラミニダーゼ遺伝子(M38309)、インフルエンザAウイルス(A/Swine/Ontario/42729A/01(H3N3))ノイラミニダーゼ(NA)遺伝子(AY619975)、インフルエンザAウイルス(A/Swine/Ontario/K01477/01(H3N3))ノイラミニダーゼ(NA)遺伝子(AY619966)、インフルエンザAウイルス(A/Swine/Saskatchewan/18789/02(H1N1))ノイラミニダーゼ(NA)遺伝子(AY619960)、インフルエンザAウイルス(A/Puerto Rico/8/34/Mount Sinai(H1N1))セグメント6(NC004523)、インフルエンザAウイルス(A/mallard/Alberta/211/98(H1N1))ノイラミニダーゼ(NA)遺伝子(AY633214)、インフルエンザAウイルス(A/mallard/Alberta/99/91(H1N1))ノイラミニダーゼ(NA)遺伝子(AY207541)、およびインフルエンザAウイルス(A/duck/Miyagil/9/77(H1N1)ノイラミニダーゼ(NA)遺伝子(AY207534)を含む。
【0050】
各親株の核は、上記に記述されたように、抗原または抗原群をコードするDNA分子を含むように修飾されるのに加えて、ヘテロカリオンを形成するのに使用される条件の下での生存のために、真菌を、第二の核、および/または、第三の核および/または付加的な核の存在に依存するようにする特徴をもたらすゲノムを含まなければならない。このように、各親の核は、第二の核、および/または、第三の核および/または付加的な核も存在しなければ、それが含まれる真菌が培養条件下で生存することができないようにするであろう特徴を付与する。例えば、特定の栄養を要求する親が、この要求性を有さない親とともに、その栄養を欠損する培地上で培養されてもよい。菌糸融合が起こる場合は、第二の親の核が、この栄養の非存在下で生存する能力を付与する。そしてまた、第二の親も、第一の親によって要求されない異なる栄養を要求してもよい。その時、双方の核を含む真菌のみが、双方の栄養を欠損するときに生存し得る。
【0051】
要求される栄養は、真菌株細胞が、増殖のために必要とする、または、存在しないと、真菌株が増殖したり、もしくは生存したりする能力を深刻に損なう任意の物質であり得る。有用な栄養要求性および関連性のある変異株の例は、以下を含む:
(1)ヒスチジン(his-1〜his-7変異株)、プロリン(aga変異株)、アルギニン(arg-11変異株)、シトルリン(arg-11変異株)、アスパラギン(asn変異株)、コリン(chol-1およびchol-2変異株)、システイン(cys-1変異株)、グルタミン(gln-1変異株)、ロイシン(leu-1〜leu-4)、リジン(lys-2、-4、および-5)、メチオチン(mac変異株、ならびにmet-6、-9、および-10変異株)、およびスレオニン(thr-2および-3変異株)のようなアミノ酸;
(2)p-アミノ安息香酸、チロシン、トリプトファン、およびフェニルアラニンの混合物(aro-6、aro-7、およびaro-8を除くすべてのaro株によって要求される)、トリプトファンおよびフェニルアラニンの混合物(aro-6変異株に要求される)、イソロイシンおよびバリンの混合物(ilv-1、-2、および-3に要求される)、および、フェニルアラニンおよびチロシンの混合物(pt変異株に要求される)のような、芳香族アミノ酸の混合物;
(3)パントテン酸(pan-1変異株)およびチアミン(thi-2およびthi-4変異株)のようなビタミン;
(4)アデニン(ad-2〜ad-4、およびad-8変異株)、ヒポキサンチン(ad-2およびad-3変異株)、イノシン、および、グアニンまたはグアノシン(gua-1または-2変異株)のようなプリン塩基;
(5)ウラシル(pyr-1〜pyr-6)のようなピリミジン塩基;
(6)飽和脂肪酸(cel変異株)、または、9位もしくは11位いずれかでシス構造において二重結合を有するC16もしくはC18脂肪酸、9位でのトランス配置における二重結合を伴う脂肪酸、および、メチレン架橋により分断される多数のシス二重結合を伴う脂肪酸(ufa-1および-2)のような不飽和脂肪酸;
(7)カリウム(trk)のような、生理学的に重要なイオン;
(8)イノシトール(acu変異株およびinl変異株)およびグリセロールのような糖アルコール;および
(9)酢酸塩(ace変異株)、I-ケトグルタル酸塩、コハク酸塩、リンゴ酸塩、ギ酸塩、またはホルムアルデヒド(for変異株)、p-アミノ安息香酸(pab-1、-2、および-3変異株)、およびスルホンアミド(35℃でのsfo変異株)のような、他の有機物。
【0052】
栄養要求性に基づく一つの具体的な例は、酵素オルニチントランスカルバミラーゼをコードするArg B+遺伝子である。この酵素は、野生型のA.ニガーに存在する。この酵素を欠損する変異株(Arg B-株)は、紫外線照射処理のような通常の非特異的技術によって調製され得、続いてアルギニンを含む培地上で増殖する能力とカップルする最少培地上での非増殖能力に基づいてスクリーニングされる。このゲノムを含む真菌は、Arg B+核も含む場合に最少培地上で増殖するであろう。
【0053】
様々な細胞毒性剤の任意の一つへの耐性を付与する遺伝子もまた、ヘテロカリオン形成を強いるために有用である。例えば、代替的な態様において、親の一つは、栄養についての要求性、ならびに、有毒な化学物質、抗生物質もしくはウイルス、または、他の親が感受性である予め決定された温度範囲のような過酷な環境条件によって誘導される毒効果への耐性を有し得る。
【0054】
毒効果を発揮し得る有毒な化学物質の具体的な例は、アクリフラビン(一般にacrによって耐性が付与され、acr-4およびacr-6による耐性に要求されるshg遺伝子の存在を伴う);3-アミノ-1,2,4-トリアゾール(acr-2、atr-1、cpc、leu-1、またはleu-2によって耐性が付与される);マラカイトグリーンのような色素(acr-3によって耐性が付与される);カフェイン(caf-1によって耐性が付与される);プリン類似体(8-アザアデニンおよび2,6-ジアミノプリンへの耐性はaza-1によって付与される;8-アザアデニンおよび8-アザグアニンへの耐性はaza-2によって付与される;8-アザグアニンおよび6-メルカプトプリンへの耐性はaza-3によって付与される;6-メチルプリンへの耐性はmep(3)およびmep(10)によって付与される);シアン化物(増殖の最初の24時間においてcni-1によって非感受性が付与される);テトラゾリウム(cya-6およびcya-7によって耐性が付与される);シクロヘキシミド(cyh-1、-2、および-3によって耐性が付与される);クロム酸塩(cys-13によって耐性が付与される);2-デオキシ-D-グルコース(dgrによって耐性が付与される);エデイン(edeine)(edr-1、および-2によって耐性が付与される);エチオニン(eth-1によって、p-フルオロフェニルアラニンの存在下ではnapによって、および、エチオニンがD型である場合はoxDによって耐性が付与される);5-フルオロデオキシウリジン、5-フルオロウラシル、および5-フルオロウリジン(3種すべてへの耐性はfdu-2によって付与される;5-フルオロウラシルへの耐性はアンモニアを含まない最少培地においてuc-5によって付与される;5-フルオロデオキシウリジンおよび5-フルオロウリジンへの耐性はud-1によって付与される)、ならびにフルオロフェニルアラニン(ある条件の下でfpr-1〜fpr-6によって耐性が付与される)のようなフルオロ化合物;8-アザアデニン(mtsによって耐性が付与される);メチルメタンスルホン酸塩(upr-1に非感受性またはわずかに感受性);塩化デカリニウム、臭化セチルトリメチルアンモニウム、および塩化ベンザルコニウム(sur-1によって耐性が付与される)のような表面活性剤;およびバナジン酸塩(vanによって耐性が付与される)のような金属イオン、を含む。
【0055】
典型的に毒効果を発揮する抗生物質の例は、ベノミル(メチル-1-ブチルカルバモルベンズイミダゾール-2-イルカルバメート)(Bmlによって耐性が付与される);アンチマイシンA(増殖の最初の24時間において、cni-1によって非感受性が付与される);ナイスタチン(erg-1および-3によって耐性が付与される)のようなポリエン抗生物質;および、オリゴマイエン(oliによって耐性が付与される)を含む。
【0056】
また、高いまたは低い温度、酸素の欠乏(anによって耐性が付与される)、恒常的な光(lis-1、-2、および-3によって耐性が付与される)または光の欠如、UV照射、電離放射線、および、高いまたは低い浸透圧のような、様々な環境条件における極端な状態への耐性を付与する遺伝子も、有用である。特に好ましい態様において、毒効果への耐性は、ハイグロマイシンのような抗生物質への耐性である。
【0057】
本発明において一般に有用な株は、綿栓をしたテストローブにおいて、株の表現型に依存して添加される、例えば、ヒスチジン、アルギニン、および/またはイノシトールのような栄養補助剤を伴う1X Vogel's Minimal Medium(N培地)上で、増殖させられ得る。典型的な株は、例えば、the Fungal Genetics Stock Center ("FGSC")およびD. D. Perkins、Stanford Universityより取得されてもよい。有用であると信用されるもう一つのN.クラッサ株は、M246-89601-2A(Dr. Mary Case, University of Georgia, Athensより取得される)である。この株は、安定なqa-2変異(M246)、arom-9変異(M6-11)、およびinos(io601)変異を含む、野生型74Aの誘導体である。qa-2、arom-9の二重変異株は、生合成および異化デヒドロクイナーゼ(dehydroquinase)活性の双方を欠損し、かつ、例えば、約80μg/mlの濃度のフェニルアラニンのような芳香族アミノ酸の栄養補助剤無しの最少培地上では、増殖することができない。
【0058】
A.ニガーの有用株(ATCC 46951)はまた、フザリウム属、ゲラシノスポラ(Gelasinospora)、およびソルダリア・フィミコラ(Sordaria fimicola)の株と同様に、the Fungal Genetics Stock Centerより入手可能であるし、または、UV光で突然変異を起こさせて、定義された最少培地において増殖のためにオルニチンまたはアルギニンを要求する分離菌を形成させることによって、調製され得る。オルニチンカルバモイルトランスフェラーゼを欠損するこの株は、arg B(350(-)52)と呼ばれてきた。A.ニガーまたはA.ニデュランスを増殖させるための培地は、Cove, Biochim Biophys Acta (1966) 113:51-56によって記述される。
【0059】
標準的な手順が、株の維持および分生子の調製のために一般に使用される(Davis and de Serres, Methods Enzymol (1971) 17A: 79-141)。菌糸体は、Lambowitz et al. J Cell Biol (1979) 82:17-31に記述されるように、液体培養において約14時間(25℃)で典型的に増殖する。宿主株は、例えば、ヒスチジン;アルギニン;phe、tyr、および/またはtrp(各約80μg/ml);p-アミノ安息香酸(約2μg/ml);およびイノシトール(約0.2mg/ml)のような、適切な栄養が添加されたVogel'sまたはFriesいずれかの最少培地において一般に増殖し得る。
【0060】
所望の特徴を伴う多くの真菌株は、公的に入手可能である。しかしながら、容易に入手可能でない場合は、当業者は、所望の特徴を提供する所望の変異株または操作された核のいずれかを選抜するために、当技術分野において周知である選択技術を使用し得る。例証的な親の組み合わせが、下記の表に示される。
【0061】
【表1】

【0062】
表に見られるように、様々な相補的特徴/特性の組み合わせが、様々な融合条件に適合するように選択され得る。一般に、栄養要求性は変異体株によって明示され、一方、ある物質に抵抗する能力は、耐性遺伝子のための発現系を伴う核の修飾によってより便利に付与されてもよい。または、栄養要求性は、形質転換ベクターを用いる相同組換えのような組換え技術を用いて影響を及ぼされ得、かつ、耐性は、毒条件が存在する条件の下で変異によって付与され得る。
【0063】
本発明の一つの態様において、宿主細胞は、形質転換のためにスフェロプラストに変換される。スフェロプラストが使用されるとき、好ましい方法またはそれらの調製は、細胞壁の酵素的消化、例えば、キチナーゼ/グルタマーゼ混合物を用いることによる。酵素的消化に適する酵素の選択は、当技術分野内である。有用な酵素は、複雑な多糖類を消化し得るものであり、広く様々な真菌種の真菌スフェロプラストを調製するのに効果的であるとして公知のものの中に見出される。適した酵素の具体的例は、Novozyme234(不純な酵素の混合物)およびβ-グルクロウイダーゼ(Beta-glucurouidase)を含む。その他の適した方法が、スフェロプラストを形成するのに使用されてもよい。しかしながら、ベクターの使用による細胞壁貫通に適した方法が同定される場合、真菌宿主の細胞全体が、スフェロプラストと共に、またはその代わりに使用されてもよい。
【0064】
多価ワクチンの特定のサブユニットをコードするDNAについての発現ユニットを含むように真菌宿主株の核を修飾するために、本発明の実践は、別の方法で指示されていない限り、当技術分野内である分子生物学、微生物学、および組換えDNA技術を使用する。そのような技術は、文献において十分に説明されている。例えば、Maniatis et al., Molecular Cloning: A Laboratory Manual (1982); D. N. Gover et al. DNA Cloning: A Practical Approach (1985) Volumes I and II; Oligonucleotide Synthesis (M. J. Gait ed. 1984); Nuclei Acid Hybridization (Hames et al. eds. 1985); Transcription and Translation (Hames et al. eds. 1984); Animal Cell Culture (R.I. Freshhey ed. 1986); Immobilized Cells and Enzymes (IRL Press 1986); B. Perbat, A Practical Guide to Molecular Cloning (1984)を参照されたい。
【0065】
N.クラッサの形質転換のための一般的手順
いったん多価抗原をコードするDNA分子の集団が発現ユニット中に配置されると、DNA分子は、Smartの"Heterologous dimeric proteins produced in heterokaryons"によって記述されるように、糸状菌の親宿主株を形質転換するために使用される。ニューロスポラ・クラッサの株は、the Fungal Genetics Stock Centerより公的に入手可能であるが、独立して調製された株もまた使用され得る。Stadler et al. Genetics (1966) 54:677-685およびHaas et al. Genetics (1952) 37:217-26によって例証されるように、変異株が新たに単離されてもよい。有用な株はまた、Stanford UniversityのD.D.Perkinsより取得され得る。株は、典型的には、綿栓された試験管において、1X Vogel's Minimal Medium("N 培地")上で、株の表現型に依存して添加される適切な栄養補助剤と共に増殖される。
【0066】
スフェロプラストは、形質転換のための被験体として使用される。分生子のスフェロプラストを形成するために、真菌は、綿栓をされた4個〜5個の125mlエルレンマイヤーフラスコにおいて、適切な栄養補助剤を伴う25mlの固体N培地上に接種される。その培養物が、室温で5〜7日間増殖させられる。
【0067】
分生子は、各フラスコに10mlのN培地を添加し、綿栓を戻し、かつフラスコを旋回することによって採取される。数分間、固体が、静置するようにされる。分生子の混合物が、エルレンマイヤーフラスコの口に掛かったオートクレーブされたチーズクロスバッグに注がれて、一つまたは複数のゴムバンドで締められる。濾過物が回収され、分生子の濃度が、鎖は一つとして数えられて、血球計算盤勘定によって決定される。
【0068】
2x109の量の分生子が、1.5%スクロースおよび適切な栄養補助剤を含む150mlの液体N培地に添加される。分生子は、75%以上が発芽して、かつ発芽管が分生子の直径の1〜4倍の長さになるまで、綿栓されたフラスコにおいて、5〜6時間、室温で振盪(150〜200rpm)しながら発芽させられる。細胞は、約1500〜2000rpmで10分間遠心分離することによって採取される。細胞ペレットは、水で3回すすがれる。
【0069】
その後、ペレットは10mlの1.0Mソルビトールに再懸濁されて、形成されるときのスフェロプラストの「破裂」を防ぐように、等浸透圧条件下で、酵素を用いた丈夫な分生子細胞壁の酵素的除去によって、スフェロプラストが調製される。プロトコルは、Vollmer and Yanofsky, Proc Natl Acad Sci USA (1986) 83:4869-73の方法より適応される。
【0070】
具体的には、滅菌された250mlエルレンマイヤーフラスコにおいて、分生子の懸濁液が、一般に、商品名Novozyme 234でInterspexにより販売されている50mgの固体酵素に添加される。混合物は、細胞壁を消化するために約1時間(4±10分)、30℃で振盪(100rpm)される。スフェロプラスト形成の過程は、混合物のわずかなアリコートをカバーガラスの下、顕微鏡で検査することによってモニターされる。スフェロプラストは、水がカバーグラスの一端に加えられたときに浸透圧的に溶解するため、検出され得る。スフェロプラスト形成の後期には、たびたび過程がモニターされるべきである。
【0071】
スフェロプラスト混合物は、滅菌された15mlコニカル遠心チューブに注がれて、卓上遠心機のスイングバケットで、500rpmで(10分間)遠心分離することによって、スフェロプラストが回収される。生じたペレットが、10の1.0Mソルビトールで2回、その後、以下のSTC溶液で1回すすがれる:91g ソルビトール;50mM Tris. Cl;50mM CaCl2;pHを8.0に調整するのに十分なNaOH;および500mlにするのに適量。
【0072】
最終的なスフェロプラストペレットは、16.0ml STC、200μl DMSO、および4mlの以下のPTC溶液の混合物に懸濁される:200gのSigmaにより商品名"4000"として販売されるポリエチレングリコール;50mM Tris.Cl;50mM CaCl2;pHを8.0に調整するのに十分なNaOH;および50mlにするのに適量。
【0073】
結果として生じたスフェロプラストの懸濁液は、直接使用され得、または1.0mlのアリコートで-80℃で凍結保存され得る。
【0074】
滅菌された15mlスクリューキャップチューブにおいて、2.0μlの50mM スペルミジン溶液、ベノミル耐性(通常約1.0mg/mlの濃度)のような選択可能なマーカーに加えて所望の多価ワクチンの抗原のための発現系を含むような、トランスフェクションされる5.0μlのプラスミドDNA、および5.0μlの5mg/mlヘパリン溶液が、チューブを軽くはじくことによって混合される。スペルミジン溶液は、12.73mgのスペルミジンを1.0mlのTEに溶解し、pHを8.0に合わせることによって調製され、-20℃で保存され得る。ヘパリン溶液は、50mgのヘパリンナトリウム塩を10mlのSTCに溶解することによって調製され、凍結アリコートとして保存され得る。
【0075】
チューブの内容物は、卓上遠心機で短く回転され(パルスされ)、その後アイスバス上に置かれる。約50〜100μlの融解されたスフェロプラストが、チューブに添加される。混合物は、その後氷上で約30分インキュベーションされるが、約20分の氷上でのインキュベーション期間が好結果である。約1mlのPTCが添加され、チューブを軽くはじくことによってよく混合される。混合物は、約20分間室温でさらにインキュベーションされる。
【0076】
再生「上層」寒天は、以下を混合することによって調製される:20ml 50x Vogel's Minimal Medium;825mlの水;182g ソルビトール;および28g 寒天。上層寒天はオートクレーブされ、100mlの10x FIGS溶液(5g/lフルクトース、2g/l イノシトール、2g/l グルコース、および200ソルボースを含む)が添加される。15mlの上層寒天は、50℃〜55℃でインキュベーションされ、スフェロプラストおよびプラスミドDNAを含むチューブに注がれる。軽くはじくこと、および2〜3回チューブを逆さにすることによって内容物が素早く混合され、その後、プレーティングされた「下層」寒天の層の上に均等に注がれる。
【0077】
「下層」寒天は、1xN培地に任意の要求される栄養補助剤を混合し;オートクレーブし;かつ、10x FIGSおよびベノミル(ベノミル耐性がマーカーとして使用される場合)を最終濃度がそれぞれ1xおよび0.5μg/mlになるように添加することによって、調製される。25mlの容量の「下層」寒天が、ペトリプレートに注がれて、固められる。
【0078】
上層寒天が下層寒天の上に注がれた後、炎をあてることによって泡が除去される。プレートは、上層寒天が凝固するまで(約5分)、直立位置で保持される。上層寒天が時期尚早に固まる傾向がある場合、下層寒天プレートは予め温められ得る。いったん上層寒天が凝固したら、プレートは、逆向きで、30℃でインキュベーションされる。
【0079】
N.クラッサ形質転換体の選択のために、このように、宿主は適切な培地(形質転換された細胞のみが利用し得る組成物を有する、または、形質転換された細胞のみが耐性である抗生物質を含む)上で培養され、約34℃でインキュベーションされる。好結果な形質転換の徴候は、プレーティング後約24〜36時間で見られ得る。安定な形質転換体は、一般に、増殖の3日後に記録される。形質転換株を検出するためのインキュベーション期間は、宿主株および表現型マーカーに依存して変化するであろう。
【0080】
選択された形質転換体は、適切なELISA、コロニーブロット免疫学的検定、制限酵素解析、フィルターハイブリダイゼーション、ネステッド・デリーション・サブクローニングなどのような標準的な方法によって、所望の抗原サブユニットの発現についてスクリーニングされ得る。
【0081】
本発明において、上記に記述された組換え技術が、所望の組換え抗原または抗原群を発現する真菌宿主株を生産するために使用され、各宿主株は、指定された条件下で増殖に否定的に影響を及ぼすが、一つまたは複数の他の核によって付与される特性によって修正され得る一つまたは複数の特徴を有する。
【0082】
結果として生じた宿主株は、本発明のヘテロカリオンを形成するために使用される親である。
【0083】
または、エレクトロポレーション過程が、ニューロスポラ・クラッサのような糸状菌の新たに採取された分生子を形質転換するために使用され得る(Van, D. C. Fungal Genetics Newsletter No. 42A (Supplement)(1995))。一般に、分生子は7〜28日間の培養より採取される。細胞は、1M ソルビトール溶液中で洗浄されて、最終濃度2.5x109細胞/mlで懸濁される。約5μgの線状化DNAが分生子懸濁液のアリコートに添加され、この一部がエレクトロポレーションキュベット、例えば間隙が0.2cmのエレクトロポレーションキュベットの底に入れられる。InVitrogen Electroporator IIのようなエレクトロポレーターは、約7.25kb/cmの電圧勾配で、約71μFおよび約200オームの設定でセットされる。エレクトロポレーションに続いて、本質的に上記に記述されたように、細胞は上層寒天ありまたは無しで、適切な培地上にプレーティングされる。
【0084】
形質転換に続いて、各分子変異体由来の安定な生産株が、各個々の事例において使用される特定の宿主細胞および発現ユニットのための選択培地上で培養を拡大することによって、確立される。
【0085】
ヘテロカリオンの生産
真菌宿主株のすべてが、すべてのヘテロカリオン適合性アリルに関してコンアレリックであるように選択されているため(上記で説明されたように、tol遺伝子が存在する場合は交配アリルを除いて)、いずれの宿主株も単独では生存し得ない条件の下で宿主株を一緒に培養した際、真菌が融合して本発明のヘテロカリオン性真菌が形成される。菌糸の融合によって、宿主真菌の異なる一倍体核が、共通の細胞質において共存することになる。任意の理論に束縛されることを望まないが、出願人らは、膜融合が、各細胞中の膜内粒子の凝集に起因し、タンパク質を含まない領域間で可能な細胞の接触を起こすと信じる。その後、接触領域における脂質の再編成が、完全な融合に導く。
【0086】
各親は、多価ワクチンの異なる抗原を生産する核を含むため、結果として生じたヘテロカリオンは、複数の抗原を含む完全な多価ワクチンを生産し得る。
【0087】
このように生じたヘテロカリオンは、ほぼ同一の割合で分裂する核を伴って安定である。
【0088】
定義された多価ワクチンを生産するヘテロカリオンの産生
本発明の組成物および方法は、病原性生物由来の免疫原性抗原を発現するヘテロカリオンを使用する。上記に記述されたように、該ヘテロカリオンは、二つまたはそれ以上の宿主株より産生され、各ヘテロカリオンは異なる多価ワクチンを生産する。
【0089】
一つの例は、インフルエンザ抗原変異体を生産するヘテロカリオンである。そのような株を産生するために、各個々の親株の分生子懸濁液が、栄養の補助剤無しの固体培地基質の上で、または、耐性遺伝子を伴う宿主細胞の場合には、細胞毒性剤を含む培地において、一緒に混合される。予め決定された親株の組み合わせより作製されたヘテロカリオンが形成されてもよいし、または、ヘテロカリオンのライブラリーが、マイクロタイタープレートもしくは他の便利なフォーマットを用いたマトリックスにおいて形成されてもよい。入手可能な多くの組み合わせのうちのいくつかが以下の表、表2に例証されるが、例のために示されるものであって、限定的であることを全く意味しない。
【0090】
(表2)多価ワクチンの生産のためのヘテロカリオンの産生
親宿主細胞が以下のインフルエンザ株由来の遺伝子を含む
AH1N1 もしくは AH3N2 もしくは BHN
抗原性変異体を伴い a,b,c,... d,e,f,... g,h,i,...
単一のワクチン生産株は使用者の選択に基づいて産生され得る
以下を含むヘテロカリオン: 株c + 株f + 株h
またはワクチン生産株のパネルが次のように産生され得る
以下を含むヘテロカリオン1: 株a + 株d + 株g
以下を含むヘテロカリオン2: 株a + 株e + 株h
以下を含むヘテロカリオン3: 株b + 株f + 株i
その他、望ましい場合は、すべての可能な組み合わせで
ここにおいて:‘A’=インフルエンザA型;‘B’=インフルエンザB型;‘H’=血球凝集素;‘N’=ノイラミニダーゼ;“変異体”=インフルエンザ型および株のHおよびN抗原の抗原性クラスの異なる組み合わせ。
【0091】
典型的な最少培地は、以下を含む:リットルあたり、5.0g デキストロース、50.0mlの塩溶液(下記)、1.0ml 微量要素(下記)、および、培地が固体形態となる場合は12.5g 寒天(pH6.5に調整)。塩溶液は、以下を含む:120.0g NaNO3、10.4g KCl、10.4g MgSO4、および30.4g KH2PO4。
【0092】
微量要素溶液は、以下を含む:1.1g (NH4)6Mo7O24.4H20、11.0g H3BO3、1.6g CoCl2 6H2O、1.6g CuSO4、50.0g Na2EDTA、5.0g FeSO4.7H2O、5.0g MnCl24H2O、および22.0g ZnSO47H2O (pH 6.5)。
【0093】
このように、ヘテロカリオン性状態においてヘテロカリオン性糸状菌を維持するために、外部の強制が維持される。最少培地においてヘテロカリオン性真菌細胞を増殖させることは、株が一緒に存続するように「強いる」。交配型が反対の場合は、tol遺伝子の存在が、安定な(A+a)ヘテロカリオンを維持するために使用され得る。
【0094】
多価ワクチンは、抗原の生産に有利な条件下で本発明のヘテロカリオンを培養することによって生産される。多価ワクチン抗原は、適合した標準的な技術に従って、もちろん、抗原の構造を保存するのに必要なように、培養より回収され、かつ精製されてもよい。
【0095】
好ましくは、ヘテロカリオン性糸状菌は、所望の多価ワクチンを直接最少増殖培地中に分泌するように宿主が培養されることを許容する発現ユニットを所持し、そのために多価ワクチンは細胞を含まない培地より直接精製され得る。もしくは、ヘテロカリオン性糸状菌は、細胞表面に抗原を方向付ける発現ユニットを所持する。細胞内に生産された多価抗原は、細胞溶解物より単離され得る。例えば、分子サイズ排除、イオン交換クロマトグラフィー、HPLC、アフィニティークロマトグラフィー、疎水的相互作用クロマトグラフィーなどのような、公知の過程に従う有用な精製方法は、当技術分野内である。
【0096】
抗原
開示された発明は、病原体、特に抗原性の性質を変化させる能力を示す病原体に対する、ワクチンのような抗原性組成物の調製法に向けられる。真核生物、ウイルス、細菌、および真菌抗原はすべて、記述された発明での使用として企図される。本発明についてのもう一つの使用は、いくらかの異なる抗原に対して同時に抗原性組成物を調製することである。本発明の好ましい態様は、多価インフルエンザワクチンの調製法に関する。インフルエンザウイルスは、絶えず突然変異し、かつ、新たな株を産生する。したがって、本発明が任意のインフルエンザ株、または、更に言えば任意の病原体株に適用可能であることから、開示された発明における使用について個々の遺伝子を列挙することは、不必要に限定的であろう。
【0097】
インフルエンザウイルスには、A、B、およびCの3種類の型が存在する。AおよびB型ウイルスは、ほぼ毎年の冬、流行病を引き起こし、一方C型ウイルスは、軽い呼吸器疾患を引き起こすのみで、臨床的に重要と考えられていない。インフルエンザA型ウイルスは、ウイルス表面上の2種類のタンパク質、血球凝集素(HA)およびノイラミニダーゼ(NA)に基づいて、亜類型に分類される。ヒトに感染する現今のインフルエンザAウイルスの亜類型は、A(H1N1)およびA(H3N2)である。同様にヒトに感染することが公知であるトリインフルエンザウイルス(H5N1)に関しては、目下多くの議論が存在する。
【0098】
インフルエンザウイルスは、誰もが知っているように、おびただしい抗原性変動を受ける。ウイルスの抗原性変動をモニターするために世界中で莫大な量の努力がささげられ、それによって、現われる任意の新たな変異体株が同定され得、かつ、その後、所定のインフルエンザの季節に人々に感染する可能性を有する株に最も厳密に合う最新のワクチンを生産するために使用され得る。関心対象のインフルエンザ株の同定について、開示された発明の方法が、その後ワクチンを調製するために使用され得る多価抗原性材料を産生するために使用され得る。
【0099】
もう一つの態様は、ヒトにおいてマラリアを引き起こす4種類の種を含む原生動物の属であるプラスモディウム属(Plasmodium)に対する多価ワクチンを調製するために方向付けられる。4種類の例は、三日熱マラリア原虫(Plasmodium vivax)および熱帯熱マラリア原虫(Plasmodium falciparum)を含む。多価ワクチンが企図される病原体の一般的一覧は、ヒトパピローマウイルス16型、18型、および31型、ヒト免疫不全ウイルス(HIV)、ヘルペス水痘ウイルス、麻疹ウイルス、エプスタイン・バーウイルス、RS(respiratory syncytial)ウイルス、パラインフルエンザ3型、単純ヘルペス1型ウイルス、および単純ヘルペス2型ウイルスを含む。開示された発明に対する標的として適する抗原は、宿主において免疫応答を誘発し得るこれらの生物由来の任意のタンパク質を含む。
【0100】
分泌抗原についてのアッセイ
ヘテロカリオン宿主は、固体最少培地上で保存され得、かつ、また、多価抗原の発現のために有利な条件下で、液体最少培地上で培養され得る。2〜7日の増殖に続いて、その後液体培地が滅菌条件下で収集され得、かつ、ELISA、PAGE、キャピラリー電気泳動、および分光光度法を含むが、それらに限定されない標準的な解析方法によって、特異的な所望の各抗原の存在について試験され得る。
【0101】
多価ワクチンの望ましい変異体を生産する培養の同定の際に、固体培地上で保存された細胞は、標準的な方法によってより大きな発酵培養に拡大され得る。使用される特定の真菌宿主について最適な条件がいかなるものであっても、その下で培養される場合、この拡大された宿主培養は、更なる研究評価および来たるべき組換えワクチンとしての使用に十分な量の、所望の産物を生産するであろう。
【0102】
細胞表面に方向付けられた抗原についてのアッセイ
ヘテロカリオンは、真菌胞子の表面上にウイルス抗原をディスプレイする融合タンパク質を発現するように構築され得る。通常真菌細胞表面に方向付けられる天然のタンパク質、例えば、ニューロスポラ属のMTRタンパク質のようなパーミアーゼタンパク質と同様に、ニューロスポラ属のEASタンパク質のようなハイドロフォビンタンパク質。これらのタンパク質は、ウイルス抗原との細胞表面に方向付けられた融合パートナーとして有用である。融合タンパク質は、当業者に周知の手段によって構築され、かつ発現される。細胞表面上に抗原を発現する個々の株は、標準的な解析方法によってアッセイされ得る。これらの方法は、ウイルス抗原に特異的な抗体による直接的または間接的細胞標識、続く様々な手段による特異的結合の検出を含むが、それらに限定されない。そのような手段は、ELISA、可視または蛍光分光光度法、およびフローサイトメトリーを含むが、それらに限定されない。抗原を発現する個々の株は、その後、融合されて、個々に発現された表面抗原の同時発現について再試験されてもよく、こうして、直接または精製後いずれかに組換えワクチンに有用な、免疫系に関する多価標的を結果的に生じる。
【0103】
非分泌抗原についてのアッセイ
抗原が分泌されない場合、各液体培養における細胞集団が除去され、標準的な方法によって粉砕され、かつ細胞上清および残骸が、望ましい性質を伴う多価ワクチンについてアッセイされ得る。いったん所望の変異体を生産する株が標準的な方法によって同定されると、固体培地上に保存された株が、接種し、かつ拡大培養を作製するために使用され得る。再び、特定のヘテロカリオンのための最適条件下で増殖されると、この拡大された宿主培養は、更なる評価および使用に十分な量、所望の産物を生産するであろう。
【0104】
以下の実施例は、本発明を例証するために提供されるが、限定はしない。
【0105】
実施例1
合成HA遺伝子構築物
合成遺伝子(A/New Caledonial/20/1999/H1N1のHA0、A/Vietnam/1194/2004/H5N1のHA、およびA/Vietnam/1194/2004/H5N1のM1)は、以下の方法によって設計された:
【0106】
各遺伝子について、アミノ酸配列が公的NCBIデータベースより取得された。HAおよびHA0遺伝子において、天然のリーダー配列が真菌シグナル配列に置換された。この配列を使用して、真菌発現のための遺伝子が逆翻訳されて、かつ、ニューロスポラ・クラッサのコドン優先度を用いてコドンが最適化された。配列は、新生mRNAにおいて二次構造を減少させるために計算された、低い自由エネルギーの形態に変更された。結果として生じた遺伝子は、イントロンスプライシングの供与および受容部位について、一連の検索をされた。そのような部位を除去するための配列の変更の後、配列はまた、転写終結部位について確認され、かつ、任意の偶然の部位が除去された。
【0107】
その後、最適化された配列が配列決定された。結果として生じたDNAは、大腸菌(E. coli)にサブクローン化され、その後、合成における誤りについて確認するために配列決定された。配列確定の後、DNAは発現ベクター(HAおよびHA0遺伝子についてはpHDKXL1、M1遺伝子についてはpALGAM)にサブクローン化され、ニューロスポラ属に形質転換された。HAおよびHA0遺伝子は、ニューロスポラ属のHis-3座での組込みのために標的とされ、かつ、M1遺伝子は、ニューロスポラ属のAm遺伝子での組込みのために標的とされた。
【0108】
実施例2
N.クラッサにおけるHA0の発現
タンパク質生産を促進するために付着された真菌シグナル配列を伴う合成血球凝集素0(HA0)遺伝子をコードする発現ベクターは、実施例1において論じられたように産生された。
【0109】
形質転換体は、Histidine-3での変異を有する宿主株を用いて、ヒスチジン原栄養性(pHDKXL1/HAまたはHA0形質転換体)、またはハイグロマイシンB耐性(pALGAM/M1形質転換体)いずれかについて選択された。選択培地上に繰り返しストリーキングすることによる形質転換された株の純化の後、形質転換体は、ELISAにより遺伝子の発現についてスクリーニングされた。振盪フラスコ由来の培地において、分泌されたインフルエンザ抗原が検出された。125mlエルレンマイヤーに25mlの最少Vogel塩に加えて0.5%酵母エキスを含むフラスコが、約100万個分生子胞子/mlで接種された。試料は、26℃、200rpmで振盪しながら、または同一の温度での静置培養いずれかで、増殖させられた。試料は、増殖の、2、3、4、5、および6日後に採取された。ELISAは、BIODESIGNより購入された、インフルエンザタンパク質に対する抗体を用いて検出された。標準として使用された対照抗原は、Protein Sciences Corporationより購入された。ELISA陽性の試料が、標準的な方法および試薬を用いるウェスタンブロットによって再スクリーニングされた。
【0110】
様々な増殖条件が試験された。具体的には、振盪および静置培養が、具体的に試験された。典型的には、酵母は2〜6日間培養されて、解析のために試料が採取された。
【0111】
試料はSDS-PAGEによって分画されて、イメージングのためにニトロセルロースにブロットされた。血球凝集素は、ヤギポリクローナル抗HA(H1N1)抗体、続いて、抗ヤギIg-アルカリホスファターゼ抱合体を用いて検出された。結合は、比色検出によって測定された。
【0112】
N.クラッサにおける合成HA0の発現のウエスタンブロット検出が図7に示される。図8は、2種類の異なるHA0クローン由来のクマシーブルー染色SDS-PAGEゲルを示す。図9は、HA05の静置培養発現のウエスタンブロットを示す。このデータは、真菌においてインフルエンザタンパク質を組換え的に発現する本系の能力を示す。
【0113】
実施例3
真菌ヘテロカリオンにおけるインフルエンザ抗原の多価発現
発現ベクターは、実施例1において論じられた方法に従って調製される。DNAは、エレクトロポレーションによってニューロスポラ属に導入されて、形質転換体は実施例2の方法に従って選択される。発現ベクターがHA0、HA、およびM1マトリックスタンパク質をコードするため、インフルエンザウイルスタンパク質の多価混合物が培養より生産される。
【図面の簡単な説明】
【0114】
【図1】合成血球凝集素0(HA0)遺伝子(A/New Caledonial/20/1999/H1N1)のヌクレオチド配列(SEQ ID NO:1)および真菌発現のための概念的な翻訳を示す。開始コドンに下線が引かれている。
【図2】合成血球凝集素0(HA0)遺伝子によりコードされるアミノ酸配列(SEQ ID NO:2)を示す。真菌のシグナル配列が太字で示される。
【図3】合成血球凝集素(HA)遺伝子(A/Vietnam/1194/2004/H5N1)のヌクレオチド配列(SEQ ID NO:3)を示す。開始コドンに下線が引かれている。
【図4】合成血球凝集素(HA)遺伝子によってコードされるアミノ酸配列(SEQ ID NO:4)を示す。真菌のシグナル配列が太字で示される。
【図5】合成M1マトリックスタンパク質(A/Vietnam/1194/2004/H5N1)のヌクレオチド配列(SEQ ID NO:5)を示す。開始コドンに下線が引かれている。
【図6】M1遺伝子によってコードされるアミノ酸配列(SEQ ID NO:6)を示す。
【図7】N.クラッサにおける合成HA0の発現のウエスタンブロット検出を示す。株HAO5およびHAO8の2日振盪フラスコ培養由来の培地が、形質転換されていない対照(C)、ならびに50および200ngの対照HA(A/New Caldonia 20/99, Protein Sciences)と同時にSDS PAGEによって分画されて、ニトロセルロースにブロットされた。血球凝集素が、ヤギポリクローナル抗HA(H1N1)抗体(BioDesign)、続いて抗ヤギIg‐アルカリホスファターゼ抱合体および比色検出を用いて検出された。
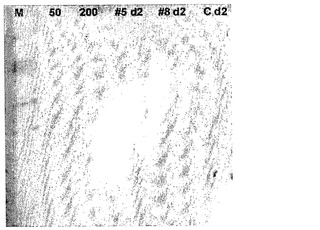
【図8】HAO5およびHAO8の、2日振盪フラスコ培養由来の培地の、クマシーブリリアントブルー染色ゲルを示す。比較的小さな分泌タンパク質が、これらの条件の下で振盪フラスコ中に2日間で生産される。
【図9】HAO5の静置培養発現を示す。HAO5の6日静置培養由来の培地のウエスタンブロット。HAタンパク質は、上記の図7のように検出された。この実験において、検出された主なバンドは、対照HAタンパク質(C)の72kDAに対して、〜57kDa(アミノ酸組成に基づいて予想されるサイズ)である。
【技術分野】
【0001】
関連出願
本出願は、全体として参照により本明細書によって組み入れられる、2004年10月15日に提出された米国仮出願第60/619,364号と関連する。
【0002】
技術分野
開示される発明は、分子生物学の分野、および病原性生物に対する多価ワクチンの生産に関する。本発明の一つの態様は、多価ワクチンの集団を生産するのに使用され得るヘテロカリオン性(heterokaryotic)糸状菌において抗原をコードするヌクレオチド配列の集団を提供する方法および組成物を、具体的に提供する。
【背景技術】
【0003】
背景技術
ワクチンは、現今、様々な方法によって生産される。典型的には、インフルエンザワクチンは、受精した鶏卵を用いて生産される。米国においては、the Centers for Disease Controlが、特定のインフルエンザの季節において、攻撃する可能性が最も高いウイルスを代表すると考えられる3種類のウイルス株を選択するであろう。選択されたウイルスの試料は、所望の抗原性の特徴を所有する種ウイルスストックとして、製造業者に提供される。種ウイルスは受精した鶏卵中に注入される。これらの卵は、インフルエンザウイルスが増殖する間インキュベーションされる。適当な期間の後、卵が開けられて、卵白が採取される。この試料がウイルスを含む。ウイルスは卵材料より精製されて、不活性化される。個々のウイルスストックは、その後、典型的には三価のワクチンである、共通インフルエンザワクチンを創出するために混合される。
【0004】
ワクチンバッチ全体に欠陥を生じさせ得る様々な問題が起こり得る。例えば、2004年には、滅菌に伴う問題が、カイロン(Chiron)のワクチン生産施設の認証撤回をもたらす。この状況は、伝統的なワクチン生産法がいかに信頼できない可能性があるかを例証する。更に、現今のインフルエンザワクチン生産法は、各年何億個という鶏卵を使用する。保管、取り扱い、および加工の段階は、時間を消費し、かつ大きな労働力を要する。その上、長い生産時間を考慮すると、インフルエンザの季節の間にインフルエンザウイルスの新たな株が優勢になった場合、現今の卵に基づく生産法は、新しいワクチンが生産されるのに数ヶ月要するであろう。
【0005】
これらの限界を考慮して、インフルエンザワクチンのような抗原性材料を生産する、より柔軟な、かつ有効な方法が非常に必要とされる。
【0006】
タンパク質の組換え真菌発現
真菌における異種遺伝子のクローニングおよび発現は、様々な有用タンパク質を生産するのに使用されてきた。例えば:Lambowitz、米国特許第4,486,533号は、ミトコンドリアプラスミドDNAによる糸状菌のためのDNAベクターの自律複製、ならびに、ニューロスポラ属(Neurospora)への異種遺伝子の導入および発現を開示する;Yeltonら、米国特許第4,816,405号は、大量の所望の異種タンパク質を生産し、かつ分泌するために、糸状子嚢菌の重要な株の修飾を可能にする道具および系を開示する;Buxtonら、米国特許第4,885,249号は、宿主アスペルギルス・ニガー(Aspergillus niger)細胞へ組み込まれ得る選択マーカーを含むDNAベクターによるA.ニガーの形質転換を開示する;および、McKnightら、米国特許第4,935,349号は、アスペルギルス属(Aspergillus)および他の糸状菌において、異種遺伝子の発現を指示し得るプロモーターを含む、アスペルギルス属において高等真核生物遺伝子を発現するための方法を開示する。同様の技術は、ニューロスポラ・クラッサ(Neurospora crassa)("N. クラッサ")においてアミノ酸輸送に関与するmtr遺伝子をクローン化し、かつ、クローン化DNAの、インビボにおいてこの遺伝子に隣接するゲノムマーカーへの強い関連を立証するために使用されてきた。Stuart, W. D. et al., Genome (1988)30:198-203; Koo, K. and Stuart, W. D. Genome (1991)34:644-651。
【0007】
糸状菌は、真核生物タンパク質を生産するのに使用するための良い候補者となる多くの特徴を所有する。糸状菌は、複雑なタンパク質を分泌し得る;ジスルフィド結合形成を含む3次元タンパク質を正しくフォールディングする;翻訳後に、タンパク質をタンパク質分解的に切り詰める;かつ、N-結合型およびO-結合型グリコシル化反応を用いてタンパク質をグリコシル化する。これらの能力は、この生物群を分泌組換えタンパク質の生産のための魅力的な宿主としてきた(MacKenzie, D. A. et al., J Gen Microbial (1993) 139:2295-2307; Peberdy, J.F., Trends in BioTechnology (1994) 12:50-57)。
【0008】
ニューロスポラ・クラッサは、組換え同種および異種タンパク質生産のための宿主細胞として使用されてきた(Carattoli, A., et al., Proc Nat Acad Sci USA, (1995) 92:6612-6616; Yamashita, R. A. et al., Fungal Genetics Newsletter (1995 Suppl.) 42A; Kato, E. et al., Fungal Genetics Newsletter (1995 Suppl.) 42A; Buczynski, S. et al. Fungal Genetics Newsletter (1995 Suppl.) 42A, Nakano, E. T. et al. Fungal Genetics Newsletter (1995 Suppl.) 40:54 0)。加えて、ニューロスポラ・クラッサは、ヘテロカリオン(heterokaryon)の手段により、組換えヘテロダイマーおよびマルチマー結合タンパク質を発現するための宿主細胞として使用されてきた(米国特許第5,643,745号、1997年7月Stuart435/69.1)。
【発明の開示】
【0009】
発明の概要
本発明は、多価ワクチンを生産するヘテロカリオン糸状菌を提供する。
【0010】
本発明の個々のヘテロカリオンは、第一および第二の、および三価ワクチンの場合には第三の親真菌株、ならびに、より高次レベルのワクチン価の場合は、付加される抗原の各セットに対して一つの付加的な親株を融合することによって産生され、各親株は、多価ワクチンの抗原の自然に見出される変異体、多価ワクチンの抗原の合理的に設計された変異体、または、多価ワクチンの抗原のランダムに産生された変異体をコードする発現ユニットと同様に、ヘテロカリオン性状態を維持するために必要なマーカーを含む。このように、自然の変異体に加えて、化学的、物理的、もしくは部位特異的突然変異誘発、または他の技術を通して産生された変異体が、生産され得る。
【0011】
本発明のヘテロカリオンは、定義された抗原の所望の多価ワクチンを選択的に生産するために有用である。
【0012】
上記に基づいて、本発明は、多価ワクチンの変異体を生産するヘテロカリオンを提供する。
【0013】
発明の開示
「ヘテロカリオン」(またはヘテロカリオン性細胞)は、二つ(またはそれ以上)の糸状菌親株の融合より形成される細胞であり、こうして各ヘテロカリオン細胞は二つ(またはそれ以上)の遺伝的に異なる核を含む。ヘテロカリオンは、すべてのヘテロカリオン適合性アリルについて一般に同型接合である(tol遺伝子が存在する場合、交配型アリルを除いて)親株由来の核を含む。少なくとも10個の染色体座(het-c、het-d、het-e、het-i、het-5、het-6、het-7、het-8、het-9、およびhet-10)が、ヘテロカリオン不適合性について同定されてきており、かつ、それ以上が存在することが推測されている。Peris et al., "Chromosomal Loci of Neurospora crassa", Microbiological Reviews (1982) 46:426-570, at 478。
【0014】
本発明は、ヘテロカリオン性糸状菌を用いて多価ワクチンの集団を生産するための方法および組成物を提供することによって、米国特許第5,643,745号、第5,683,899号、および第6,268,140号において開示された研究を進歩させる。そのような方法および組成物は、抗ウイルスワクチン、抗細菌ワクチン、および抗真菌ワクチンのような多価ワクチンの発見および生産において有用である。
【0015】
好ましい態様の記述
本発明は、多価ワクチンの集団を産生するための新規の方法および組成物を提供する。多価ワクチンは、定義された抗原の、順序づけられた、またはランダムな組み合わせより産生される。多価ワクチンを取得するために、多価ワクチンの定義された抗原をコードする第一、第二、および時には第三またはそれ以上のDNA分子の集団を、第一、第二の真菌の、および時には第三またはそれ以上の宿主親に導入する段階、第一、第二、および時には第三またはそれ以上の宿主真菌親を用いてヘテロカリオン性真菌株を形成する段階、および、その後、適切である場合は、所望の性質を有する多価ワクチンの生産のために、結果として生じたヘテロカリオンをDNAをコードするサブユニットが発現する条件の下で培養する段階、および、結果として生じたヘテロカリオンを更にスクリーニングする段階を含む方法において、ヘテロカリオンが取得される。各要素、すなわち、真菌親、DNA分子、および融合方法は、以下に詳細に記述される。
【0016】
糸状菌の性質およびヘテロカリオン形成のためのバックグラウンド要求性
真菌は、糸状多核鎖をもたらす単一の単核細胞、酵母細胞、多様な胞子を伴う子実体、および/または有性的に分化された細胞として生育し得る。それらはまた、多核形態においても存在し得る。カビとしての真菌の成長形態の主な要素は、直径約2μm〜10μmの分岐した管状構造である菌糸である。菌糸は、先端での伸長(頂端生長)および側面の分岐を生ずることによって、増殖する。このように、コロニーが増殖するとき、菌糸は多量の絡み合う繊維を形成する。
【0017】
いくつかの菌糸は、栄養を吸収するために、真菌が増殖している培養培地中に貫通し、一方、培地の表面より上に突き出した菌糸は、「気中菌糸体」を構成する。大抵のコロニーは、不規則な、乾燥した糸状マットとして、液体または固体培地の表面に増殖する。大抵の種においては、菌糸は、「隔壁」と呼ばれる横断壁によって分割される。しかしながら、これらの隔壁は、細かい中心細孔を有する。こうして、隔壁を有する菌糸でさえ、連続的な多量の細胞質に埋め込まれた複数の核を有し、かつ、事実上、移動可能な細胞質において核の多様性を含む。
【0018】
「糸状菌」という用語は、多量の分岐し、連結した繊維を通して菌糸体を形成し得、かつ、横断壁によって遮断されるが、横断壁における穿孔のために区画間の細胞質の輸送を許容し得る真菌のことを指す。これらの真菌の多くは、有性的に繁殖した際に、嚢の中に減数分裂した胞子を形成する。メカニズムは完全に理解されていないが、適切な刺激で、生殖は無性的に起こり得る。この生殖の様式において、「分生子」として公知である胞子は、繊維に沿った様々な場所で形成される出芽した突出物の先端で、外面的に生ずる。
【0019】
本発明のヘテロカリオンパネルを産生するのに使用される糸状菌は、一般に、藻菌類(Phycomycetes)、子嚢菌綱(Ascomycetes)、担子菌綱(Basidiomycetes)、および不完全菌類(Deuteromycetes)である。藻菌類は、いくつかの隔壁を有する糸状菌と同様に、隔壁を有しないすべての糸状菌を含む。それらの無性胞子は様々な種類があり、分化した柄の末端に形成される嚢の中に含まれる胞子嚢胞子を含む。異なる種は、異なる有性サイクルを有する。
【0020】
子嚢菌綱は、子嚢胞子として公知である有性胞子を含む嚢様構造である子嚢によって、他の真菌より識別される。子嚢胞子は、交配、雄および雌の核の融合、2回の減数分裂、および通常1回の最終的な有糸分裂の最終産物である。担子菌綱は、分化した構造の表面上に形成する有性胞子によって識別される。不完全菌類は、有性相が未だ観察されていないため、「不完全な真菌」としてしばしば称される。それらの菌糸は隔壁を有し、かつ、分生子の形態は、子嚢菌綱のものに類似している。
【0021】
好ましい糸状菌は、子嚢菌綱の群のものであり、より好ましくは、ニューロスポラ属、アスペルギルス属、フザリウム属(Fusarium)、トリコデルマ属(Tricoderma)、クリソスポリウム属(Chrysosporium)、およびペニシリウム属(Penicillium)由来である。ニューロスポラ属由来の特に有用な種は、N.インターメディア(N. intermedia)、N.クラッサ、N.シトプーラ(N. sitopula)、およびN.テトラスポラ(N. tetraspora)を含み、そのうち最も好ましい種はN.クラッサである。アスペルギルス属の有用な種は、A.ニデュランス(A. nidulans)、A.ニガー、A.テレウス(A.terreus)、およびA.フメガテュス(A.fumegatus)を含む。
【0022】
糸状菌の栄養成長は、細胞分裂(有糸分裂)に伴う核分裂を含む。この型の細胞分裂は、無性生殖、すなわち、配偶子の関与無し、および分生子経由の核融合無しの、新たなクローンの形成からなる。例えば、ニューロスポラ属の種は、核に、それぞれがシングルコピーを有する7本の異なる染色体を含み、すなわち、栄養体は一倍体である。この一倍体状態が、菌糸体増殖の間、および分生子の形成を通した無性生殖の間、典型的に維持される。
【0023】
有性生殖もまた起こり得、その時に異なる交配型の二つの一倍体細胞(菌糸または分生子)が融合して、二つの別個の核を含むジカリオン性細胞を形成する。こうして二つの一倍体核は同一の細胞質に共存し、しばらくの間は、同調して多少分裂する。しかしながら、細胞が子嚢胞子の形成を開始する場合は、二つの異なる一倍体核は、相同な染色体対を含む二倍体核を形成するように実際に融合し得る。この二倍体細胞が、その後減数分裂を始める。
【0024】
「ヘテロカリオン」(またはヘテロカリオン性細胞)は、二つの(またはそれ以上の)遺伝的に異なる核を伴う細胞である。本発明のヘテロカリオンは、すべてのヘテロカリオン適合性アリルについて同型接合である(tol遺伝子が存在する場合、交配型アリルを除いて)細胞由来の核を含まなければいけない。例えば、ニューロスポラ属において、少なくとも10個の染色体座(het-c、het-d、het-e、het-i、het-5、het-6、het-7、het-8、het-9、およびhet-10)が、ヘテロカリオン不適合性について同定されてきており、かつ、それ以上が存在することが推測されている。Perkins et al., "Chromosomal Loci of Neurospora crassa", Microbiological Reviews (1982) 46:426-570, at 478。
【0025】
二つの株が一つまた複数のhet座で異なるアリルを有する場合、それらは安定なヘテロカリオンを形成することができない。異なった菌糸の融合後、または細胞質もしくは抽出物の異なった株へのマイクロインジェクション後に、原形質殺傷(protoplasmic killing)が起きる。hetの一つまたは複数のアリルに対して重複(duplication)(部分的二倍体)が異型接合である場合、増殖が阻害されて、高度に異常となる。いくつかのヘテロカリオン適合性座(特にhet-c、-d、-e、および-i)は、ヘテロカリオン試験によって最初に定義された。het-5からhet-10の座は、het座での差異が自然の集団において共通しているため、重複を使用して検出された。同文献。
【0026】
いくらかの緩慢なヘテロカリオン性増殖が起きる可能性があるが、交配型アリル「A」および「a」はまた、N.クラッサにおいてhet遺伝子として作用する。マイクロインジェクション実験は、殺傷反応におけるタンパク質の関与を示している。このように、反対の交配型はまた、ヘテロカリオン性の無胞子菌糸の増殖と関連づけられる複雑な事象にとって一般に重要である。同文献436および478。しかしながら、tol遺伝子が存在する場合、反対の交配型アリルAおよびaと関連づけられる栄養(ヘテロカリオン)不適合性は、有性適合性が影響を及ぼされること無しに抑制される。このように、(tol; A+a;a)ヘテロカリオンは、他のhet座が同一のアリル(またはコンアレリック(conallelic))であれば完全に適合性であり、かつ安定であり得、tol遺伝子が存在するときA/a重複は正常に増殖する。
【0027】
適合性座についてコンアレリックである二つの異なる株由来の菌糸が提供される場合、それらは、同一の培地で増殖させられるとき、特に以下に記述されるように融合が強いられるとき、融合する可能性がある。結果として生じた培養物は、その後、共通の菌糸体マットの共有された細胞質において循環する双方の株由来の核を含むであろう。
【0028】
定義された抗原の混合集団をコードする発現ユニットの構築
本発明を記述するにあたって、以下の用語法が、下記に述べられた定義に従って使用されるであろう:
【0029】
本発明は、糸状菌ヘテロカリオンにおける「異種多価ワクチン」の生産を含む。本文脈において、「異種」とは、タンパク質が通常はその真菌によって生産されないことを意味する。「多価」とは、最終的なワクチン産物が、少なくとも二つの抗原または抗原変異体から構成されることを意味する。産物は、完全に異なる抗原を含むヘテロ多価ワクチンであってもよいし、または、単一のサブユニットの変異体から構成されるホモ多価でもあり得る。多価ワクチンの例は、細胞表面由来の組換え抗原、ウイルスコートタンパク質、特定の病原性タンパク質抗原などの混合物を含むが、それらに限定されない。
【0030】
「抗原をコードするヌクレオチド配列」は、適切な制御配列に作動可能に連結されたとき、転写産物がポリペプチドに翻訳される配列の部分である。コード配列の境界は、5'(アミノ)末端で開始コドン、および3'(カルボキシ)末端で翻訳終止コドンによって決定される。このコード配列は、例えば、原核生物遺伝子、真核生物mRNAからのcDNA、(真菌のような)真核生物DNAからのゲノムDNA配列由来であり得、または、合成DNAを含んでもよい。ポリアデニル化シグナルおよび転写終結配列は、通常コード配列の3'に位置するであろう。
【0031】
コード配列は、適切な宿主細胞において制御配列がコード配列の発現に影響を及ぼすとき、制御配列に「作動可能に連結」される。
【0032】
「発現ユニット」は、適切な宿主生物体において適切な条件の下で、作動可能に連結された配列の転写および翻訳を指示する「制御配列または領域」に作動可能に連結されたコード配列を含むDNA分子である。
【0033】
細胞は、外因性DNAが宿主細胞膜内に導入されているとき、外因性DNAによって「形質転換」されている。細菌のような原核生物については、外因性DNAはプラスミドのようなエピソーム要素上に維持されてもよい。糸状菌は核を有する(真核生物である)ため、大抵の安定に形質転換された真菌宿主細胞は、染色体に組み込まれた外因性DNAを含み、そのために染色体複製を通して娘細胞によって承継される。
【0034】
「組換え宿主」は、組換え技術によって調製されたDNA配列で形質転換されている、形質転換される、または形質転換されるであろう細胞のことを指し、かつ、元来形質転換された細胞ならびにそれらの培養物および子孫を含む。
【0035】
種々の方法が、1)多価ワクチンの抗原サブユニットの自然に見出される変異体、2)多価ワクチンの抗原のランダムに産生された、もしくは選択された変異体、または、3)多価ワクチンの抗原の合理的に設計された、もしくは選択された変異体、をコードするDNA分子の集団を産生するために使用され得る。以下において、インフルエンザワクチンが、例証的な例として使用される。当業者は、下記に概観される方法、または、当技術分野において公知である同等の方法を、サブユニットをコードするDNA分子の集団を産生するために容易に使用し得る。
【0036】
自然の不均一性を有する抗原の自然に見出される抗原変異体をコードするDNA分子の集団は、標準的なcDNA産生/クローニング技術を用いて生産され得る。一般に、mRNAおよびウイルスゲノムRNAの集団は、最初に病原体より、または、例えばインフルエンザワクチンの場合は、直接ウイルス自体のゲノムRNAより単離される。単離されたRNA分子の集団は、その後、RTPCRのような当技術分野で公知のクローニング方法において、cDNA分子の産生のための鋳型として使用される。このように生産されたcDNA分子の集団は、下記に記述されるように、適当な発現ユニットに挿入され得る。
【0037】
または、タンパク質配列が公知である抗原については、人工cDNA配列が、当技術分野に公知である方法を用いて、糸状菌宿主株によって自身の内因性タンパク質を生産するために高頻度に使用されるコドンを組み込んで、産生され得る。
【0038】
加えて、部位特異的またはランダムな突然変異誘発が、特定のサブユニットの自然に見出される変異体を生産するために、多価ワクチンの抗原をコードする単離された、または人工のcDNA分子について行われ得る。ランダムまたは部位特異的なミスマッチPCRプライミング、リンカースキャニング突然変異誘発、または化学的および物理的突然変異誘発のような手順が、抗原の合理的に設計された、またはランダムに産生された変異体をコードするDNA分子の集団を産生するために容易に使用され得る。例えば、ランダムに産生された、または合理的に設計されたPCRプライマーは、抗原コード配列において、ランダムな、または標的とされた不均一性を産生するために使用され得る。
【0039】
本明細書において使用されるとき、タンパク質フォールディングまたは特定の標的残基もしくは領域の選択のような選択基準が、変異体を産生する、または変異体をコードするDNA分子を選択する際に使用される場合、変異体が合理的に設計されると言われる。変異体をコードするDNA分子を産生する、または選択するときに選択基準が使用されない場合、変異体がランダムに産生されると言われる。
【0040】
多価ワクチンサブユニットにおいて不均一性を産生するための好ましい標的部位は、免疫原性エピトープおよび周囲のアミノ酸配列である。インフルエンザ抗原遺伝子の場合、この型の変異の創出は、各抗原の公知である天然の可変領域に集中するであろう。例として、公知の変異のライブラリーを生産するために、可変領域における各アミノ酸を一つずつ変化させ得るであろう。
【0041】
多価ワクチンの抗原をコードする発現ユニットの構築
多価ワクチンの抗原をコードするヌクレオチド分子を含む発現ユニットは、周知の技術を用いて構築される。一般に、発現ユニットは、サブユニットをコードする配列を、最終的な糸状菌宿主においてサブユニットをコードする配列の発現を方向付ける制御配列との作動可能な連結に置くことによって、産生される。
【0042】
構成的または誘導的いずれかの様式において、作動可能に連結されたタンパク質をコードする配列の発現を方向付けるために、様々な制御要素が、現在当技術分野において公知である。制御配列の選択は、使用される真菌株、真菌を培養するために使用される条件、所望のタンパク質発現レベル、および必要とされる発現の性質(例えば、誘導的対構成的)に基づくであろう。当業者は、本ヘテロカリオンパネルに使用される発現ユニットを産生するために、当技術分野において公知である制御配列を容易に利用し得る。
【0043】
タンパク質をコードする配列の転写および翻訳を方向付ける配列に加えて、本発明の発現ユニットは、抗原の細胞外への搬出を方向付ける発現制御要素であるシグナル配列を更に制御してもよい。糸状菌において公知である分泌シグナルの総説は、Dalbey R. E., et al., TIBS 17:474-478(1992)によって提供される。当業者は、分泌シグナルを含む発現ユニットを、容易に産生し得る。
【0044】
本発明の発現ユニットのもう一つの形態は、宿主細胞もしくは異種膜アンカー配列への融合を経て細胞膜へ、または、宿主細胞もしくは異種細胞表面分子への融合によって細胞表面へ、抗原を方向付ける融合タンパク質を含んでもよい。
【0045】
一つの応用において、組換えユニットが、発現ユニットの代わりに産生される。そのような使用において、サブユニットをコードする配列、または抗原をコードする配列の断片は、宿主真菌株における組込み部位に相同である配列を含むDNA領域によって隣接される。その後、相同な配列は、組換えユニットと宿主染色体との間での相同組換えを刺激し、かつ方向付けるように使用される。組換えユニットが使用されるとき、宿主株は、好ましくは、標的組換えのために使用される配列が続く発現制御要素を含む発現ユニットで、最初に形質転換される。例えば、インフルエンザ抗原が宿主真菌に導入され得、その後、相同組換えユニットが、宿主染色体の標的領域内に異種性を導入するために使用され得る。
【0046】
最終的な真菌細胞を形質転換し得る中間ベクターを生産するために、中間宿主が時々使用される。その時、所望の糸状菌宿主を形質転換するために使用され得る所望のDNA量を取得するために、中間細菌形質転換体が増殖され得る。中間ベクターとして役立ち得る共通に入手可能な細菌ベクターの例は、例えば、pBR322、pUC8、およびpUC9を含む。追加的な有用な中間ベクターは、pHY201、pKBY2、pTZ18R、pX182、およびpCVN2.9、pN807、pN846を含む。
【0047】
もう一つの態様において、関心対象の抗原または複数の抗原は、EASのような真菌ハイドロフォビン(hydrophobin)との融合タンパク質として発現される。真菌ハイドロフォビンは、典型的には大量に発現され、かつ分泌される。そして、真菌の表面は、ハイドロフォビンでコートされる。これらのタンパク質はまた、溶液中で凝集することが公知である。この凝集の特徴により、抗原性粒子を形成する凝集組換えタンパク質の調製が可能である。これらのタンパク質は、精製され、抗原として使用され得る。多数の抗原が同一の培養において発現されるとき、多価抗原性粒子が生産される。
【0048】
本発明のこの記述および開示は、本発明の精神および範囲内であるすべての態様に及ぶことを意図すると理解されるであろう。例えば、分子の抗原性に実質的に影響を及ぼすこと無く、オープンリーディングフレームのアミノ酸配列内でアミノ酸配列を挿入、欠失、または置換することは、当技術分野の知識内であり、かつ、自然に見出されるサブユニットへの欠失、付加、または置換で産生され得るような多価抗原は、本発明に含まれる。
【0049】
親株の性質
本発明のヘテロカリオンを作製することにおいて使用される各親真菌株は、多価ワクチンの抗原をコードするDNA分子の集団のメンバーを含んでいるであろうため、一つの真菌親は、所望の多価ワクチンの第一の抗原または抗原群をコードする、第一のDNA分子の集団のメンバーを含むように修飾された核を有するであろうし、かつ、それぞれの次の真菌親は、所望の多価ワクチンの異なる抗原または抗原群をコードする、異なるDNA分子の集団のメンバーを含むように修飾された核を有するであろう。例えば、二価のインフルエンザワクチンを生産するヘテロカリオンを生産するために、一つの真菌親は、インフルエンザA型H1 N1からのcDNA由来の抗原群を生産するであろうし、一方、他方の真菌親は、インフルエンザA型H3 N2からのcDNA由来の抗原群を生産するであろう。三価のインフルエンザワクチンを生産するためには、一つの真菌親は、インフルエンザA型H1 N1からのcDNA由来の抗原群を生産するであろうし、第二の真菌親は、インフルエンザA型H3 N2からのcDNA由来の抗原群を生産するであろうし、かつ、第三の真菌親は、インフルエンザB型H NからのcDNA由来の抗原群を生産するであろう。ヌクレオチドおよびタンパク質配列は、公的に入手可能である。例えば、HA遺伝子についての例示的な配列は、H3N2(AY738729)、インフルエンザAウイルス(A/Leningrad/54/1(H1N1))ノイラミニダーゼ遺伝子(M38309)、インフルエンザAウイルス(A/Swine/Ontario/42729A/01(H3N3))ノイラミニダーゼ(NA)遺伝子(AY619975)、インフルエンザAウイルス(A/Swine/Ontario/K01477/01(H3N3))ノイラミニダーゼ(NA)遺伝子(AY619966)、インフルエンザAウイルス(A/Swine/Saskatchewan/18789/02(H1N1))ノイラミニダーゼ(NA)遺伝子(AY619960)、インフルエンザAウイルス(A/Puerto Rico/8/34/Mount Sinai(H1N1))セグメント6(NC004523)、インフルエンザAウイルス(A/mallard/Alberta/211/98(H1N1))ノイラミニダーゼ(NA)遺伝子(AY633214)、インフルエンザAウイルス(A/mallard/Alberta/99/91(H1N1))ノイラミニダーゼ(NA)遺伝子(AY207541)、およびインフルエンザAウイルス(A/duck/Miyagil/9/77(H1N1)ノイラミニダーゼ(NA)遺伝子(AY207534)を含む。
【0050】
各親株の核は、上記に記述されたように、抗原または抗原群をコードするDNA分子を含むように修飾されるのに加えて、ヘテロカリオンを形成するのに使用される条件の下での生存のために、真菌を、第二の核、および/または、第三の核および/または付加的な核の存在に依存するようにする特徴をもたらすゲノムを含まなければならない。このように、各親の核は、第二の核、および/または、第三の核および/または付加的な核も存在しなければ、それが含まれる真菌が培養条件下で生存することができないようにするであろう特徴を付与する。例えば、特定の栄養を要求する親が、この要求性を有さない親とともに、その栄養を欠損する培地上で培養されてもよい。菌糸融合が起こる場合は、第二の親の核が、この栄養の非存在下で生存する能力を付与する。そしてまた、第二の親も、第一の親によって要求されない異なる栄養を要求してもよい。その時、双方の核を含む真菌のみが、双方の栄養を欠損するときに生存し得る。
【0051】
要求される栄養は、真菌株細胞が、増殖のために必要とする、または、存在しないと、真菌株が増殖したり、もしくは生存したりする能力を深刻に損なう任意の物質であり得る。有用な栄養要求性および関連性のある変異株の例は、以下を含む:
(1)ヒスチジン(his-1〜his-7変異株)、プロリン(aga変異株)、アルギニン(arg-11変異株)、シトルリン(arg-11変異株)、アスパラギン(asn変異株)、コリン(chol-1およびchol-2変異株)、システイン(cys-1変異株)、グルタミン(gln-1変異株)、ロイシン(leu-1〜leu-4)、リジン(lys-2、-4、および-5)、メチオチン(mac変異株、ならびにmet-6、-9、および-10変異株)、およびスレオニン(thr-2および-3変異株)のようなアミノ酸;
(2)p-アミノ安息香酸、チロシン、トリプトファン、およびフェニルアラニンの混合物(aro-6、aro-7、およびaro-8を除くすべてのaro株によって要求される)、トリプトファンおよびフェニルアラニンの混合物(aro-6変異株に要求される)、イソロイシンおよびバリンの混合物(ilv-1、-2、および-3に要求される)、および、フェニルアラニンおよびチロシンの混合物(pt変異株に要求される)のような、芳香族アミノ酸の混合物;
(3)パントテン酸(pan-1変異株)およびチアミン(thi-2およびthi-4変異株)のようなビタミン;
(4)アデニン(ad-2〜ad-4、およびad-8変異株)、ヒポキサンチン(ad-2およびad-3変異株)、イノシン、および、グアニンまたはグアノシン(gua-1または-2変異株)のようなプリン塩基;
(5)ウラシル(pyr-1〜pyr-6)のようなピリミジン塩基;
(6)飽和脂肪酸(cel変異株)、または、9位もしくは11位いずれかでシス構造において二重結合を有するC16もしくはC18脂肪酸、9位でのトランス配置における二重結合を伴う脂肪酸、および、メチレン架橋により分断される多数のシス二重結合を伴う脂肪酸(ufa-1および-2)のような不飽和脂肪酸;
(7)カリウム(trk)のような、生理学的に重要なイオン;
(8)イノシトール(acu変異株およびinl変異株)およびグリセロールのような糖アルコール;および
(9)酢酸塩(ace変異株)、I-ケトグルタル酸塩、コハク酸塩、リンゴ酸塩、ギ酸塩、またはホルムアルデヒド(for変異株)、p-アミノ安息香酸(pab-1、-2、および-3変異株)、およびスルホンアミド(35℃でのsfo変異株)のような、他の有機物。
【0052】
栄養要求性に基づく一つの具体的な例は、酵素オルニチントランスカルバミラーゼをコードするArg B+遺伝子である。この酵素は、野生型のA.ニガーに存在する。この酵素を欠損する変異株(Arg B-株)は、紫外線照射処理のような通常の非特異的技術によって調製され得、続いてアルギニンを含む培地上で増殖する能力とカップルする最少培地上での非増殖能力に基づいてスクリーニングされる。このゲノムを含む真菌は、Arg B+核も含む場合に最少培地上で増殖するであろう。
【0053】
様々な細胞毒性剤の任意の一つへの耐性を付与する遺伝子もまた、ヘテロカリオン形成を強いるために有用である。例えば、代替的な態様において、親の一つは、栄養についての要求性、ならびに、有毒な化学物質、抗生物質もしくはウイルス、または、他の親が感受性である予め決定された温度範囲のような過酷な環境条件によって誘導される毒効果への耐性を有し得る。
【0054】
毒効果を発揮し得る有毒な化学物質の具体的な例は、アクリフラビン(一般にacrによって耐性が付与され、acr-4およびacr-6による耐性に要求されるshg遺伝子の存在を伴う);3-アミノ-1,2,4-トリアゾール(acr-2、atr-1、cpc、leu-1、またはleu-2によって耐性が付与される);マラカイトグリーンのような色素(acr-3によって耐性が付与される);カフェイン(caf-1によって耐性が付与される);プリン類似体(8-アザアデニンおよび2,6-ジアミノプリンへの耐性はaza-1によって付与される;8-アザアデニンおよび8-アザグアニンへの耐性はaza-2によって付与される;8-アザグアニンおよび6-メルカプトプリンへの耐性はaza-3によって付与される;6-メチルプリンへの耐性はmep(3)およびmep(10)によって付与される);シアン化物(増殖の最初の24時間においてcni-1によって非感受性が付与される);テトラゾリウム(cya-6およびcya-7によって耐性が付与される);シクロヘキシミド(cyh-1、-2、および-3によって耐性が付与される);クロム酸塩(cys-13によって耐性が付与される);2-デオキシ-D-グルコース(dgrによって耐性が付与される);エデイン(edeine)(edr-1、および-2によって耐性が付与される);エチオニン(eth-1によって、p-フルオロフェニルアラニンの存在下ではnapによって、および、エチオニンがD型である場合はoxDによって耐性が付与される);5-フルオロデオキシウリジン、5-フルオロウラシル、および5-フルオロウリジン(3種すべてへの耐性はfdu-2によって付与される;5-フルオロウラシルへの耐性はアンモニアを含まない最少培地においてuc-5によって付与される;5-フルオロデオキシウリジンおよび5-フルオロウリジンへの耐性はud-1によって付与される)、ならびにフルオロフェニルアラニン(ある条件の下でfpr-1〜fpr-6によって耐性が付与される)のようなフルオロ化合物;8-アザアデニン(mtsによって耐性が付与される);メチルメタンスルホン酸塩(upr-1に非感受性またはわずかに感受性);塩化デカリニウム、臭化セチルトリメチルアンモニウム、および塩化ベンザルコニウム(sur-1によって耐性が付与される)のような表面活性剤;およびバナジン酸塩(vanによって耐性が付与される)のような金属イオン、を含む。
【0055】
典型的に毒効果を発揮する抗生物質の例は、ベノミル(メチル-1-ブチルカルバモルベンズイミダゾール-2-イルカルバメート)(Bmlによって耐性が付与される);アンチマイシンA(増殖の最初の24時間において、cni-1によって非感受性が付与される);ナイスタチン(erg-1および-3によって耐性が付与される)のようなポリエン抗生物質;および、オリゴマイエン(oliによって耐性が付与される)を含む。
【0056】
また、高いまたは低い温度、酸素の欠乏(anによって耐性が付与される)、恒常的な光(lis-1、-2、および-3によって耐性が付与される)または光の欠如、UV照射、電離放射線、および、高いまたは低い浸透圧のような、様々な環境条件における極端な状態への耐性を付与する遺伝子も、有用である。特に好ましい態様において、毒効果への耐性は、ハイグロマイシンのような抗生物質への耐性である。
【0057】
本発明において一般に有用な株は、綿栓をしたテストローブにおいて、株の表現型に依存して添加される、例えば、ヒスチジン、アルギニン、および/またはイノシトールのような栄養補助剤を伴う1X Vogel's Minimal Medium(N培地)上で、増殖させられ得る。典型的な株は、例えば、the Fungal Genetics Stock Center ("FGSC")およびD. D. Perkins、Stanford Universityより取得されてもよい。有用であると信用されるもう一つのN.クラッサ株は、M246-89601-2A(Dr. Mary Case, University of Georgia, Athensより取得される)である。この株は、安定なqa-2変異(M246)、arom-9変異(M6-11)、およびinos(io601)変異を含む、野生型74Aの誘導体である。qa-2、arom-9の二重変異株は、生合成および異化デヒドロクイナーゼ(dehydroquinase)活性の双方を欠損し、かつ、例えば、約80μg/mlの濃度のフェニルアラニンのような芳香族アミノ酸の栄養補助剤無しの最少培地上では、増殖することができない。
【0058】
A.ニガーの有用株(ATCC 46951)はまた、フザリウム属、ゲラシノスポラ(Gelasinospora)、およびソルダリア・フィミコラ(Sordaria fimicola)の株と同様に、the Fungal Genetics Stock Centerより入手可能であるし、または、UV光で突然変異を起こさせて、定義された最少培地において増殖のためにオルニチンまたはアルギニンを要求する分離菌を形成させることによって、調製され得る。オルニチンカルバモイルトランスフェラーゼを欠損するこの株は、arg B(350(-)52)と呼ばれてきた。A.ニガーまたはA.ニデュランスを増殖させるための培地は、Cove, Biochim Biophys Acta (1966) 113:51-56によって記述される。
【0059】
標準的な手順が、株の維持および分生子の調製のために一般に使用される(Davis and de Serres, Methods Enzymol (1971) 17A: 79-141)。菌糸体は、Lambowitz et al. J Cell Biol (1979) 82:17-31に記述されるように、液体培養において約14時間(25℃)で典型的に増殖する。宿主株は、例えば、ヒスチジン;アルギニン;phe、tyr、および/またはtrp(各約80μg/ml);p-アミノ安息香酸(約2μg/ml);およびイノシトール(約0.2mg/ml)のような、適切な栄養が添加されたVogel'sまたはFriesいずれかの最少培地において一般に増殖し得る。
【0060】
所望の特徴を伴う多くの真菌株は、公的に入手可能である。しかしながら、容易に入手可能でない場合は、当業者は、所望の特徴を提供する所望の変異株または操作された核のいずれかを選抜するために、当技術分野において周知である選択技術を使用し得る。例証的な親の組み合わせが、下記の表に示される。
【0061】
【表1】
【0062】
表に見られるように、様々な相補的特徴/特性の組み合わせが、様々な融合条件に適合するように選択され得る。一般に、栄養要求性は変異体株によって明示され、一方、ある物質に抵抗する能力は、耐性遺伝子のための発現系を伴う核の修飾によってより便利に付与されてもよい。または、栄養要求性は、形質転換ベクターを用いる相同組換えのような組換え技術を用いて影響を及ぼされ得、かつ、耐性は、毒条件が存在する条件の下で変異によって付与され得る。
【0063】
本発明の一つの態様において、宿主細胞は、形質転換のためにスフェロプラストに変換される。スフェロプラストが使用されるとき、好ましい方法またはそれらの調製は、細胞壁の酵素的消化、例えば、キチナーゼ/グルタマーゼ混合物を用いることによる。酵素的消化に適する酵素の選択は、当技術分野内である。有用な酵素は、複雑な多糖類を消化し得るものであり、広く様々な真菌種の真菌スフェロプラストを調製するのに効果的であるとして公知のものの中に見出される。適した酵素の具体的例は、Novozyme234(不純な酵素の混合物)およびβ-グルクロウイダーゼ(Beta-glucurouidase)を含む。その他の適した方法が、スフェロプラストを形成するのに使用されてもよい。しかしながら、ベクターの使用による細胞壁貫通に適した方法が同定される場合、真菌宿主の細胞全体が、スフェロプラストと共に、またはその代わりに使用されてもよい。
【0064】
多価ワクチンの特定のサブユニットをコードするDNAについての発現ユニットを含むように真菌宿主株の核を修飾するために、本発明の実践は、別の方法で指示されていない限り、当技術分野内である分子生物学、微生物学、および組換えDNA技術を使用する。そのような技術は、文献において十分に説明されている。例えば、Maniatis et al., Molecular Cloning: A Laboratory Manual (1982); D. N. Gover et al. DNA Cloning: A Practical Approach (1985) Volumes I and II; Oligonucleotide Synthesis (M. J. Gait ed. 1984); Nuclei Acid Hybridization (Hames et al. eds. 1985); Transcription and Translation (Hames et al. eds. 1984); Animal Cell Culture (R.I. Freshhey ed. 1986); Immobilized Cells and Enzymes (IRL Press 1986); B. Perbat, A Practical Guide to Molecular Cloning (1984)を参照されたい。
【0065】
N.クラッサの形質転換のための一般的手順
いったん多価抗原をコードするDNA分子の集団が発現ユニット中に配置されると、DNA分子は、Smartの"Heterologous dimeric proteins produced in heterokaryons"によって記述されるように、糸状菌の親宿主株を形質転換するために使用される。ニューロスポラ・クラッサの株は、the Fungal Genetics Stock Centerより公的に入手可能であるが、独立して調製された株もまた使用され得る。Stadler et al. Genetics (1966) 54:677-685およびHaas et al. Genetics (1952) 37:217-26によって例証されるように、変異株が新たに単離されてもよい。有用な株はまた、Stanford UniversityのD.D.Perkinsより取得され得る。株は、典型的には、綿栓された試験管において、1X Vogel's Minimal Medium("N 培地")上で、株の表現型に依存して添加される適切な栄養補助剤と共に増殖される。
【0066】
スフェロプラストは、形質転換のための被験体として使用される。分生子のスフェロプラストを形成するために、真菌は、綿栓をされた4個〜5個の125mlエルレンマイヤーフラスコにおいて、適切な栄養補助剤を伴う25mlの固体N培地上に接種される。その培養物が、室温で5〜7日間増殖させられる。
【0067】
分生子は、各フラスコに10mlのN培地を添加し、綿栓を戻し、かつフラスコを旋回することによって採取される。数分間、固体が、静置するようにされる。分生子の混合物が、エルレンマイヤーフラスコの口に掛かったオートクレーブされたチーズクロスバッグに注がれて、一つまたは複数のゴムバンドで締められる。濾過物が回収され、分生子の濃度が、鎖は一つとして数えられて、血球計算盤勘定によって決定される。
【0068】
2x109の量の分生子が、1.5%スクロースおよび適切な栄養補助剤を含む150mlの液体N培地に添加される。分生子は、75%以上が発芽して、かつ発芽管が分生子の直径の1〜4倍の長さになるまで、綿栓されたフラスコにおいて、5〜6時間、室温で振盪(150〜200rpm)しながら発芽させられる。細胞は、約1500〜2000rpmで10分間遠心分離することによって採取される。細胞ペレットは、水で3回すすがれる。
【0069】
その後、ペレットは10mlの1.0Mソルビトールに再懸濁されて、形成されるときのスフェロプラストの「破裂」を防ぐように、等浸透圧条件下で、酵素を用いた丈夫な分生子細胞壁の酵素的除去によって、スフェロプラストが調製される。プロトコルは、Vollmer and Yanofsky, Proc Natl Acad Sci USA (1986) 83:4869-73の方法より適応される。
【0070】
具体的には、滅菌された250mlエルレンマイヤーフラスコにおいて、分生子の懸濁液が、一般に、商品名Novozyme 234でInterspexにより販売されている50mgの固体酵素に添加される。混合物は、細胞壁を消化するために約1時間(4±10分)、30℃で振盪(100rpm)される。スフェロプラスト形成の過程は、混合物のわずかなアリコートをカバーガラスの下、顕微鏡で検査することによってモニターされる。スフェロプラストは、水がカバーグラスの一端に加えられたときに浸透圧的に溶解するため、検出され得る。スフェロプラスト形成の後期には、たびたび過程がモニターされるべきである。
【0071】
スフェロプラスト混合物は、滅菌された15mlコニカル遠心チューブに注がれて、卓上遠心機のスイングバケットで、500rpmで(10分間)遠心分離することによって、スフェロプラストが回収される。生じたペレットが、10の1.0Mソルビトールで2回、その後、以下のSTC溶液で1回すすがれる:91g ソルビトール;50mM Tris. Cl;50mM CaCl2;pHを8.0に調整するのに十分なNaOH;および500mlにするのに適量。
【0072】
最終的なスフェロプラストペレットは、16.0ml STC、200μl DMSO、および4mlの以下のPTC溶液の混合物に懸濁される:200gのSigmaにより商品名"4000"として販売されるポリエチレングリコール;50mM Tris.Cl;50mM CaCl2;pHを8.0に調整するのに十分なNaOH;および50mlにするのに適量。
【0073】
結果として生じたスフェロプラストの懸濁液は、直接使用され得、または1.0mlのアリコートで-80℃で凍結保存され得る。
【0074】
滅菌された15mlスクリューキャップチューブにおいて、2.0μlの50mM スペルミジン溶液、ベノミル耐性(通常約1.0mg/mlの濃度)のような選択可能なマーカーに加えて所望の多価ワクチンの抗原のための発現系を含むような、トランスフェクションされる5.0μlのプラスミドDNA、および5.0μlの5mg/mlヘパリン溶液が、チューブを軽くはじくことによって混合される。スペルミジン溶液は、12.73mgのスペルミジンを1.0mlのTEに溶解し、pHを8.0に合わせることによって調製され、-20℃で保存され得る。ヘパリン溶液は、50mgのヘパリンナトリウム塩を10mlのSTCに溶解することによって調製され、凍結アリコートとして保存され得る。
【0075】
チューブの内容物は、卓上遠心機で短く回転され(パルスされ)、その後アイスバス上に置かれる。約50〜100μlの融解されたスフェロプラストが、チューブに添加される。混合物は、その後氷上で約30分インキュベーションされるが、約20分の氷上でのインキュベーション期間が好結果である。約1mlのPTCが添加され、チューブを軽くはじくことによってよく混合される。混合物は、約20分間室温でさらにインキュベーションされる。
【0076】
再生「上層」寒天は、以下を混合することによって調製される:20ml 50x Vogel's Minimal Medium;825mlの水;182g ソルビトール;および28g 寒天。上層寒天はオートクレーブされ、100mlの10x FIGS溶液(5g/lフルクトース、2g/l イノシトール、2g/l グルコース、および200ソルボースを含む)が添加される。15mlの上層寒天は、50℃〜55℃でインキュベーションされ、スフェロプラストおよびプラスミドDNAを含むチューブに注がれる。軽くはじくこと、および2〜3回チューブを逆さにすることによって内容物が素早く混合され、その後、プレーティングされた「下層」寒天の層の上に均等に注がれる。
【0077】
「下層」寒天は、1xN培地に任意の要求される栄養補助剤を混合し;オートクレーブし;かつ、10x FIGSおよびベノミル(ベノミル耐性がマーカーとして使用される場合)を最終濃度がそれぞれ1xおよび0.5μg/mlになるように添加することによって、調製される。25mlの容量の「下層」寒天が、ペトリプレートに注がれて、固められる。
【0078】
上層寒天が下層寒天の上に注がれた後、炎をあてることによって泡が除去される。プレートは、上層寒天が凝固するまで(約5分)、直立位置で保持される。上層寒天が時期尚早に固まる傾向がある場合、下層寒天プレートは予め温められ得る。いったん上層寒天が凝固したら、プレートは、逆向きで、30℃でインキュベーションされる。
【0079】
N.クラッサ形質転換体の選択のために、このように、宿主は適切な培地(形質転換された細胞のみが利用し得る組成物を有する、または、形質転換された細胞のみが耐性である抗生物質を含む)上で培養され、約34℃でインキュベーションされる。好結果な形質転換の徴候は、プレーティング後約24〜36時間で見られ得る。安定な形質転換体は、一般に、増殖の3日後に記録される。形質転換株を検出するためのインキュベーション期間は、宿主株および表現型マーカーに依存して変化するであろう。
【0080】
選択された形質転換体は、適切なELISA、コロニーブロット免疫学的検定、制限酵素解析、フィルターハイブリダイゼーション、ネステッド・デリーション・サブクローニングなどのような標準的な方法によって、所望の抗原サブユニットの発現についてスクリーニングされ得る。
【0081】
本発明において、上記に記述された組換え技術が、所望の組換え抗原または抗原群を発現する真菌宿主株を生産するために使用され、各宿主株は、指定された条件下で増殖に否定的に影響を及ぼすが、一つまたは複数の他の核によって付与される特性によって修正され得る一つまたは複数の特徴を有する。
【0082】
結果として生じた宿主株は、本発明のヘテロカリオンを形成するために使用される親である。
【0083】
または、エレクトロポレーション過程が、ニューロスポラ・クラッサのような糸状菌の新たに採取された分生子を形質転換するために使用され得る(Van, D. C. Fungal Genetics Newsletter No. 42A (Supplement)(1995))。一般に、分生子は7〜28日間の培養より採取される。細胞は、1M ソルビトール溶液中で洗浄されて、最終濃度2.5x109細胞/mlで懸濁される。約5μgの線状化DNAが分生子懸濁液のアリコートに添加され、この一部がエレクトロポレーションキュベット、例えば間隙が0.2cmのエレクトロポレーションキュベットの底に入れられる。InVitrogen Electroporator IIのようなエレクトロポレーターは、約7.25kb/cmの電圧勾配で、約71μFおよび約200オームの設定でセットされる。エレクトロポレーションに続いて、本質的に上記に記述されたように、細胞は上層寒天ありまたは無しで、適切な培地上にプレーティングされる。
【0084】
形質転換に続いて、各分子変異体由来の安定な生産株が、各個々の事例において使用される特定の宿主細胞および発現ユニットのための選択培地上で培養を拡大することによって、確立される。
【0085】
ヘテロカリオンの生産
真菌宿主株のすべてが、すべてのヘテロカリオン適合性アリルに関してコンアレリックであるように選択されているため(上記で説明されたように、tol遺伝子が存在する場合は交配アリルを除いて)、いずれの宿主株も単独では生存し得ない条件の下で宿主株を一緒に培養した際、真菌が融合して本発明のヘテロカリオン性真菌が形成される。菌糸の融合によって、宿主真菌の異なる一倍体核が、共通の細胞質において共存することになる。任意の理論に束縛されることを望まないが、出願人らは、膜融合が、各細胞中の膜内粒子の凝集に起因し、タンパク質を含まない領域間で可能な細胞の接触を起こすと信じる。その後、接触領域における脂質の再編成が、完全な融合に導く。
【0086】
各親は、多価ワクチンの異なる抗原を生産する核を含むため、結果として生じたヘテロカリオンは、複数の抗原を含む完全な多価ワクチンを生産し得る。
【0087】
このように生じたヘテロカリオンは、ほぼ同一の割合で分裂する核を伴って安定である。
【0088】
定義された多価ワクチンを生産するヘテロカリオンの産生
本発明の組成物および方法は、病原性生物由来の免疫原性抗原を発現するヘテロカリオンを使用する。上記に記述されたように、該ヘテロカリオンは、二つまたはそれ以上の宿主株より産生され、各ヘテロカリオンは異なる多価ワクチンを生産する。
【0089】
一つの例は、インフルエンザ抗原変異体を生産するヘテロカリオンである。そのような株を産生するために、各個々の親株の分生子懸濁液が、栄養の補助剤無しの固体培地基質の上で、または、耐性遺伝子を伴う宿主細胞の場合には、細胞毒性剤を含む培地において、一緒に混合される。予め決定された親株の組み合わせより作製されたヘテロカリオンが形成されてもよいし、または、ヘテロカリオンのライブラリーが、マイクロタイタープレートもしくは他の便利なフォーマットを用いたマトリックスにおいて形成されてもよい。入手可能な多くの組み合わせのうちのいくつかが以下の表、表2に例証されるが、例のために示されるものであって、限定的であることを全く意味しない。
【0090】
(表2)多価ワクチンの生産のためのヘテロカリオンの産生
親宿主細胞が以下のインフルエンザ株由来の遺伝子を含む
AH1N1 もしくは AH3N2 もしくは BHN
抗原性変異体を伴い a,b,c,... d,e,f,... g,h,i,...
単一のワクチン生産株は使用者の選択に基づいて産生され得る
以下を含むヘテロカリオン: 株c + 株f + 株h
またはワクチン生産株のパネルが次のように産生され得る
以下を含むヘテロカリオン1: 株a + 株d + 株g
以下を含むヘテロカリオン2: 株a + 株e + 株h
以下を含むヘテロカリオン3: 株b + 株f + 株i
その他、望ましい場合は、すべての可能な組み合わせで
ここにおいて:‘A’=インフルエンザA型;‘B’=インフルエンザB型;‘H’=血球凝集素;‘N’=ノイラミニダーゼ;“変異体”=インフルエンザ型および株のHおよびN抗原の抗原性クラスの異なる組み合わせ。
【0091】
典型的な最少培地は、以下を含む:リットルあたり、5.0g デキストロース、50.0mlの塩溶液(下記)、1.0ml 微量要素(下記)、および、培地が固体形態となる場合は12.5g 寒天(pH6.5に調整)。塩溶液は、以下を含む:120.0g NaNO3、10.4g KCl、10.4g MgSO4、および30.4g KH2PO4。
【0092】
微量要素溶液は、以下を含む:1.1g (NH4)6Mo7O24.4H20、11.0g H3BO3、1.6g CoCl2 6H2O、1.6g CuSO4、50.0g Na2EDTA、5.0g FeSO4.7H2O、5.0g MnCl24H2O、および22.0g ZnSO47H2O (pH 6.5)。
【0093】
このように、ヘテロカリオン性状態においてヘテロカリオン性糸状菌を維持するために、外部の強制が維持される。最少培地においてヘテロカリオン性真菌細胞を増殖させることは、株が一緒に存続するように「強いる」。交配型が反対の場合は、tol遺伝子の存在が、安定な(A+a)ヘテロカリオンを維持するために使用され得る。
【0094】
多価ワクチンは、抗原の生産に有利な条件下で本発明のヘテロカリオンを培養することによって生産される。多価ワクチン抗原は、適合した標準的な技術に従って、もちろん、抗原の構造を保存するのに必要なように、培養より回収され、かつ精製されてもよい。
【0095】
好ましくは、ヘテロカリオン性糸状菌は、所望の多価ワクチンを直接最少増殖培地中に分泌するように宿主が培養されることを許容する発現ユニットを所持し、そのために多価ワクチンは細胞を含まない培地より直接精製され得る。もしくは、ヘテロカリオン性糸状菌は、細胞表面に抗原を方向付ける発現ユニットを所持する。細胞内に生産された多価抗原は、細胞溶解物より単離され得る。例えば、分子サイズ排除、イオン交換クロマトグラフィー、HPLC、アフィニティークロマトグラフィー、疎水的相互作用クロマトグラフィーなどのような、公知の過程に従う有用な精製方法は、当技術分野内である。
【0096】
抗原
開示された発明は、病原体、特に抗原性の性質を変化させる能力を示す病原体に対する、ワクチンのような抗原性組成物の調製法に向けられる。真核生物、ウイルス、細菌、および真菌抗原はすべて、記述された発明での使用として企図される。本発明についてのもう一つの使用は、いくらかの異なる抗原に対して同時に抗原性組成物を調製することである。本発明の好ましい態様は、多価インフルエンザワクチンの調製法に関する。インフルエンザウイルスは、絶えず突然変異し、かつ、新たな株を産生する。したがって、本発明が任意のインフルエンザ株、または、更に言えば任意の病原体株に適用可能であることから、開示された発明における使用について個々の遺伝子を列挙することは、不必要に限定的であろう。
【0097】
インフルエンザウイルスには、A、B、およびCの3種類の型が存在する。AおよびB型ウイルスは、ほぼ毎年の冬、流行病を引き起こし、一方C型ウイルスは、軽い呼吸器疾患を引き起こすのみで、臨床的に重要と考えられていない。インフルエンザA型ウイルスは、ウイルス表面上の2種類のタンパク質、血球凝集素(HA)およびノイラミニダーゼ(NA)に基づいて、亜類型に分類される。ヒトに感染する現今のインフルエンザAウイルスの亜類型は、A(H1N1)およびA(H3N2)である。同様にヒトに感染することが公知であるトリインフルエンザウイルス(H5N1)に関しては、目下多くの議論が存在する。
【0098】
インフルエンザウイルスは、誰もが知っているように、おびただしい抗原性変動を受ける。ウイルスの抗原性変動をモニターするために世界中で莫大な量の努力がささげられ、それによって、現われる任意の新たな変異体株が同定され得、かつ、その後、所定のインフルエンザの季節に人々に感染する可能性を有する株に最も厳密に合う最新のワクチンを生産するために使用され得る。関心対象のインフルエンザ株の同定について、開示された発明の方法が、その後ワクチンを調製するために使用され得る多価抗原性材料を産生するために使用され得る。
【0099】
もう一つの態様は、ヒトにおいてマラリアを引き起こす4種類の種を含む原生動物の属であるプラスモディウム属(Plasmodium)に対する多価ワクチンを調製するために方向付けられる。4種類の例は、三日熱マラリア原虫(Plasmodium vivax)および熱帯熱マラリア原虫(Plasmodium falciparum)を含む。多価ワクチンが企図される病原体の一般的一覧は、ヒトパピローマウイルス16型、18型、および31型、ヒト免疫不全ウイルス(HIV)、ヘルペス水痘ウイルス、麻疹ウイルス、エプスタイン・バーウイルス、RS(respiratory syncytial)ウイルス、パラインフルエンザ3型、単純ヘルペス1型ウイルス、および単純ヘルペス2型ウイルスを含む。開示された発明に対する標的として適する抗原は、宿主において免疫応答を誘発し得るこれらの生物由来の任意のタンパク質を含む。
【0100】
分泌抗原についてのアッセイ
ヘテロカリオン宿主は、固体最少培地上で保存され得、かつ、また、多価抗原の発現のために有利な条件下で、液体最少培地上で培養され得る。2〜7日の増殖に続いて、その後液体培地が滅菌条件下で収集され得、かつ、ELISA、PAGE、キャピラリー電気泳動、および分光光度法を含むが、それらに限定されない標準的な解析方法によって、特異的な所望の各抗原の存在について試験され得る。
【0101】
多価ワクチンの望ましい変異体を生産する培養の同定の際に、固体培地上で保存された細胞は、標準的な方法によってより大きな発酵培養に拡大され得る。使用される特定の真菌宿主について最適な条件がいかなるものであっても、その下で培養される場合、この拡大された宿主培養は、更なる研究評価および来たるべき組換えワクチンとしての使用に十分な量の、所望の産物を生産するであろう。
【0102】
細胞表面に方向付けられた抗原についてのアッセイ
ヘテロカリオンは、真菌胞子の表面上にウイルス抗原をディスプレイする融合タンパク質を発現するように構築され得る。通常真菌細胞表面に方向付けられる天然のタンパク質、例えば、ニューロスポラ属のMTRタンパク質のようなパーミアーゼタンパク質と同様に、ニューロスポラ属のEASタンパク質のようなハイドロフォビンタンパク質。これらのタンパク質は、ウイルス抗原との細胞表面に方向付けられた融合パートナーとして有用である。融合タンパク質は、当業者に周知の手段によって構築され、かつ発現される。細胞表面上に抗原を発現する個々の株は、標準的な解析方法によってアッセイされ得る。これらの方法は、ウイルス抗原に特異的な抗体による直接的または間接的細胞標識、続く様々な手段による特異的結合の検出を含むが、それらに限定されない。そのような手段は、ELISA、可視または蛍光分光光度法、およびフローサイトメトリーを含むが、それらに限定されない。抗原を発現する個々の株は、その後、融合されて、個々に発現された表面抗原の同時発現について再試験されてもよく、こうして、直接または精製後いずれかに組換えワクチンに有用な、免疫系に関する多価標的を結果的に生じる。
【0103】
非分泌抗原についてのアッセイ
抗原が分泌されない場合、各液体培養における細胞集団が除去され、標準的な方法によって粉砕され、かつ細胞上清および残骸が、望ましい性質を伴う多価ワクチンについてアッセイされ得る。いったん所望の変異体を生産する株が標準的な方法によって同定されると、固体培地上に保存された株が、接種し、かつ拡大培養を作製するために使用され得る。再び、特定のヘテロカリオンのための最適条件下で増殖されると、この拡大された宿主培養は、更なる評価および使用に十分な量、所望の産物を生産するであろう。
【0104】
以下の実施例は、本発明を例証するために提供されるが、限定はしない。
【0105】
実施例1
合成HA遺伝子構築物
合成遺伝子(A/New Caledonial/20/1999/H1N1のHA0、A/Vietnam/1194/2004/H5N1のHA、およびA/Vietnam/1194/2004/H5N1のM1)は、以下の方法によって設計された:
【0106】
各遺伝子について、アミノ酸配列が公的NCBIデータベースより取得された。HAおよびHA0遺伝子において、天然のリーダー配列が真菌シグナル配列に置換された。この配列を使用して、真菌発現のための遺伝子が逆翻訳されて、かつ、ニューロスポラ・クラッサのコドン優先度を用いてコドンが最適化された。配列は、新生mRNAにおいて二次構造を減少させるために計算された、低い自由エネルギーの形態に変更された。結果として生じた遺伝子は、イントロンスプライシングの供与および受容部位について、一連の検索をされた。そのような部位を除去するための配列の変更の後、配列はまた、転写終結部位について確認され、かつ、任意の偶然の部位が除去された。
【0107】
その後、最適化された配列が配列決定された。結果として生じたDNAは、大腸菌(E. coli)にサブクローン化され、その後、合成における誤りについて確認するために配列決定された。配列確定の後、DNAは発現ベクター(HAおよびHA0遺伝子についてはpHDKXL1、M1遺伝子についてはpALGAM)にサブクローン化され、ニューロスポラ属に形質転換された。HAおよびHA0遺伝子は、ニューロスポラ属のHis-3座での組込みのために標的とされ、かつ、M1遺伝子は、ニューロスポラ属のAm遺伝子での組込みのために標的とされた。
【0108】
実施例2
N.クラッサにおけるHA0の発現
タンパク質生産を促進するために付着された真菌シグナル配列を伴う合成血球凝集素0(HA0)遺伝子をコードする発現ベクターは、実施例1において論じられたように産生された。
【0109】
形質転換体は、Histidine-3での変異を有する宿主株を用いて、ヒスチジン原栄養性(pHDKXL1/HAまたはHA0形質転換体)、またはハイグロマイシンB耐性(pALGAM/M1形質転換体)いずれかについて選択された。選択培地上に繰り返しストリーキングすることによる形質転換された株の純化の後、形質転換体は、ELISAにより遺伝子の発現についてスクリーニングされた。振盪フラスコ由来の培地において、分泌されたインフルエンザ抗原が検出された。125mlエルレンマイヤーに25mlの最少Vogel塩に加えて0.5%酵母エキスを含むフラスコが、約100万個分生子胞子/mlで接種された。試料は、26℃、200rpmで振盪しながら、または同一の温度での静置培養いずれかで、増殖させられた。試料は、増殖の、2、3、4、5、および6日後に採取された。ELISAは、BIODESIGNより購入された、インフルエンザタンパク質に対する抗体を用いて検出された。標準として使用された対照抗原は、Protein Sciences Corporationより購入された。ELISA陽性の試料が、標準的な方法および試薬を用いるウェスタンブロットによって再スクリーニングされた。
【0110】
様々な増殖条件が試験された。具体的には、振盪および静置培養が、具体的に試験された。典型的には、酵母は2〜6日間培養されて、解析のために試料が採取された。
【0111】
試料はSDS-PAGEによって分画されて、イメージングのためにニトロセルロースにブロットされた。血球凝集素は、ヤギポリクローナル抗HA(H1N1)抗体、続いて、抗ヤギIg-アルカリホスファターゼ抱合体を用いて検出された。結合は、比色検出によって測定された。
【0112】
N.クラッサにおける合成HA0の発現のウエスタンブロット検出が図7に示される。図8は、2種類の異なるHA0クローン由来のクマシーブルー染色SDS-PAGEゲルを示す。図9は、HA05の静置培養発現のウエスタンブロットを示す。このデータは、真菌においてインフルエンザタンパク質を組換え的に発現する本系の能力を示す。
【0113】
実施例3
真菌ヘテロカリオンにおけるインフルエンザ抗原の多価発現
発現ベクターは、実施例1において論じられた方法に従って調製される。DNAは、エレクトロポレーションによってニューロスポラ属に導入されて、形質転換体は実施例2の方法に従って選択される。発現ベクターがHA0、HA、およびM1マトリックスタンパク質をコードするため、インフルエンザウイルスタンパク質の多価混合物が培養より生産される。
【図面の簡単な説明】
【0114】
【図1】合成血球凝集素0(HA0)遺伝子(A/New Caledonial/20/1999/H1N1)のヌクレオチド配列(SEQ ID NO:1)および真菌発現のための概念的な翻訳を示す。開始コドンに下線が引かれている。
【図2】合成血球凝集素0(HA0)遺伝子によりコードされるアミノ酸配列(SEQ ID NO:2)を示す。真菌のシグナル配列が太字で示される。
【図3】合成血球凝集素(HA)遺伝子(A/Vietnam/1194/2004/H5N1)のヌクレオチド配列(SEQ ID NO:3)を示す。開始コドンに下線が引かれている。
【図4】合成血球凝集素(HA)遺伝子によってコードされるアミノ酸配列(SEQ ID NO:4)を示す。真菌のシグナル配列が太字で示される。
【図5】合成M1マトリックスタンパク質(A/Vietnam/1194/2004/H5N1)のヌクレオチド配列(SEQ ID NO:5)を示す。開始コドンに下線が引かれている。
【図6】M1遺伝子によってコードされるアミノ酸配列(SEQ ID NO:6)を示す。
【図7】N.クラッサにおける合成HA0の発現のウエスタンブロット検出を示す。株HAO5およびHAO8の2日振盪フラスコ培養由来の培地が、形質転換されていない対照(C)、ならびに50および200ngの対照HA(A/New Caldonia 20/99, Protein Sciences)と同時にSDS PAGEによって分画されて、ニトロセルロースにブロットされた。血球凝集素が、ヤギポリクローナル抗HA(H1N1)抗体(BioDesign)、続いて抗ヤギIg‐アルカリホスファターゼ抱合体および比色検出を用いて検出された。
【図8】HAO5およびHAO8の、2日振盪フラスコ培養由来の培地の、クマシーブリリアントブルー染色ゲルを示す。比較的小さな分泌タンパク質が、これらの条件の下で振盪フラスコ中に2日間で生産される。
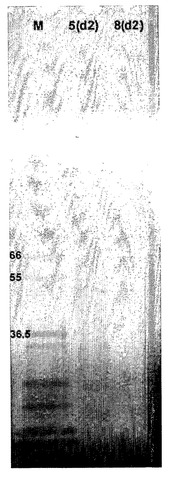
【図9】HAO5の静置培養発現を示す。HAO5の6日静置培養由来の培地のウエスタンブロット。HAタンパク質は、上記の図7のように検出された。この実験において、検出された主なバンドは、対照HAタンパク質(C)の72kDAに対して、〜57kDa(アミノ酸組成に基づいて予想されるサイズ)である。
【特許請求の範囲】
【請求項1】
二つまたはそれ以上の親真菌株を融合させることによって形成され、生存のためにすべての親真菌の核の存在を要求し、該親真菌株それぞれが病原性生物由来の抗原の変異体をコードする外因的に供給された核酸分子を含み、かつ該親真菌株がすべてのヘテロカリオン適合性アリルについて同型接合である、抗原の多価組換え変異体を生産する、糸状菌ヘテロカリオン。
【請求項2】
多価抗原が、可溶性タンパク質として培養培地中に分泌される、請求項1記載のヘテロカリオン。
【請求項3】
多価抗原が、粒子に凝集する、請求項2記載のヘテロカリオン。
【請求項4】
多価抗原が、真菌ヘテロカリオンの表面上にディスプレイされる、請求項1記載のヘテロカリオン。
【請求項5】
多価抗原が、真菌ヘテロカリオンの細胞質内に保持される、請求項1記載のヘテロカリオン。
【請求項6】
病原性生物が、ウイルスである、請求項1記載のヘテロカリオン。
【請求項7】
ウイルスが、インフルエンザウイルス、ヒトパピローマウイルス16型、ヒトパピローマウイルス18型、ヒトパピローマウイルス31型、ヘルペス水痘ウイルス、麻疹ウイルス、エプスタイン・バーウイルス、RSウイルス、パラインフルエンザ3型、単純ヘルペス1型ウイルス、および単純ヘルペス2型ウイルスより構成される群のうち一つである、請求項1記載のヘテロカリオン。
【請求項8】
ウイルス抗原が、インフルエンザA型およびインフルエンザB型由来である、請求項1記載のヘテロカリオン。
【請求項9】
抗原が、インフルエンザA型およびB型の血球凝集素およびノイラミニダーゼの変異体を含む、請求項4記載のヘテロカリオン。
【請求項10】
インフルエンザA型およびB型の血球凝集素およびノイラミニダーゼの各変異体が、自然に見出される変異体である、請求項1記載のヘテロカリオン。
【請求項11】
インフルエンザA型およびインフルエンザB型の血球凝集素およびノイラミニダーゼの各変異体が、自然に見出されるサブユニット変異体でない、請求項1記載のヘテロカリオン。
【請求項12】
病原性生物が、細菌である、請求項1記載のヘテロカリオン。
【請求項13】
病原性生物が、真菌である、請求項1記載のヘテロカリオン。
【請求項14】
病原性生物が、ウイルス、細菌、および真菌生物の任意およびすべての組み合わせの混合物を含む、請求項1記載のヘテロカリオン。
【請求項15】
外因的に供給された核酸分子が多価ワクチンを構成するように発現される条件下で、請求項1記載のヘテロカリオンを培養する段階を含む、多価ワクチンを生産する方法。
【請求項1】
二つまたはそれ以上の親真菌株を融合させることによって形成され、生存のためにすべての親真菌の核の存在を要求し、該親真菌株それぞれが病原性生物由来の抗原の変異体をコードする外因的に供給された核酸分子を含み、かつ該親真菌株がすべてのヘテロカリオン適合性アリルについて同型接合である、抗原の多価組換え変異体を生産する、糸状菌ヘテロカリオン。
【請求項2】
多価抗原が、可溶性タンパク質として培養培地中に分泌される、請求項1記載のヘテロカリオン。
【請求項3】
多価抗原が、粒子に凝集する、請求項2記載のヘテロカリオン。
【請求項4】
多価抗原が、真菌ヘテロカリオンの表面上にディスプレイされる、請求項1記載のヘテロカリオン。
【請求項5】
多価抗原が、真菌ヘテロカリオンの細胞質内に保持される、請求項1記載のヘテロカリオン。
【請求項6】
病原性生物が、ウイルスである、請求項1記載のヘテロカリオン。
【請求項7】
ウイルスが、インフルエンザウイルス、ヒトパピローマウイルス16型、ヒトパピローマウイルス18型、ヒトパピローマウイルス31型、ヘルペス水痘ウイルス、麻疹ウイルス、エプスタイン・バーウイルス、RSウイルス、パラインフルエンザ3型、単純ヘルペス1型ウイルス、および単純ヘルペス2型ウイルスより構成される群のうち一つである、請求項1記載のヘテロカリオン。
【請求項8】
ウイルス抗原が、インフルエンザA型およびインフルエンザB型由来である、請求項1記載のヘテロカリオン。
【請求項9】
抗原が、インフルエンザA型およびB型の血球凝集素およびノイラミニダーゼの変異体を含む、請求項4記載のヘテロカリオン。
【請求項10】
インフルエンザA型およびB型の血球凝集素およびノイラミニダーゼの各変異体が、自然に見出される変異体である、請求項1記載のヘテロカリオン。
【請求項11】
インフルエンザA型およびインフルエンザB型の血球凝集素およびノイラミニダーゼの各変異体が、自然に見出されるサブユニット変異体でない、請求項1記載のヘテロカリオン。
【請求項12】
病原性生物が、細菌である、請求項1記載のヘテロカリオン。
【請求項13】
病原性生物が、真菌である、請求項1記載のヘテロカリオン。
【請求項14】
病原性生物が、ウイルス、細菌、および真菌生物の任意およびすべての組み合わせの混合物を含む、請求項1記載のヘテロカリオン。
【請求項15】
外因的に供給された核酸分子が多価ワクチンを構成するように発現される条件下で、請求項1記載のヘテロカリオンを培養する段階を含む、多価ワクチンを生産する方法。
【図1】


【図2】

【図3】

【図4】

【図5】

【図6】

【図7】
【図8】
【図9】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公表番号】特表2008−516609(P2008−516609A)
【公表日】平成20年5月22日(2008.5.22)
【国際特許分類】
【出願番号】特願2007−536999(P2007−536999)
【出願日】平成17年10月17日(2005.10.17)
【国際出願番号】PCT/US2005/037249
【国際公開番号】WO2006/044796
【国際公開日】平成18年4月27日(2006.4.27)
【出願人】(507124911)ニュージェネシス コーポレイション (2)
【Fターム(参考)】
【公表日】平成20年5月22日(2008.5.22)
【国際特許分類】
【出願日】平成17年10月17日(2005.10.17)
【国際出願番号】PCT/US2005/037249
【国際公開番号】WO2006/044796
【国際公開日】平成18年4月27日(2006.4.27)
【出願人】(507124911)ニュージェネシス コーポレイション (2)
【Fターム(参考)】
[ Back to top ]