
多様性の生成方法
【課題】特定の核酸領域の指向性構成的高頻度突然変異を起こすことができる抗体産生細胞系の製造方法の開発。
【解決手段】a)クローン細胞集団をV遺伝子の多様性についてスクリーニングする工程;b)V遺伝子の多様性を示す1種以上の細胞を単離し、選別された細胞のV遺伝子と他の遺伝子における突然変異の蓄積速度を比較する工程;c)V遺伝子の突然変異率が他の遺伝子の突然変異率を上回っている細胞を選別する工程を含む上記方法。
【解決手段】a)クローン細胞集団をV遺伝子の多様性についてスクリーニングする工程;b)V遺伝子の多様性を示す1種以上の細胞を単離し、選別された細胞のV遺伝子と他の遺伝子における突然変異の蓄積速度を比較する工程;c)V遺伝子の突然変異率が他の遺伝子の突然変異率を上回っている細胞を選別する工程を含む上記方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、抗体産生細胞の生来の体細胞高頻度突然変異能を利用することによる遺伝子または遺伝子産物における多様性の生成方法、ならびに所定の遺伝子産物において多様性を生成できる細胞系に関する。
【背景技術】
【0002】
遺伝子産物において多様性を生成させるための多くのin vitro法は、非常に多数の突然変異体を作製し、次にそれらを効果的な選別技術を用いて選別することに基づいている。例えば、ファージ提示法は、提示されたタンパク質の選別を可能にする媒介手段が提供された場合に非常にうまくいっている(Smith, 1985; Bassら, 1990;McCaffertyら, 1990;概説としては、ClacksonおよびWells, 1994を参照されたい)。同様に、特定のペプチドリガンドが、lacリプレッサーLaclのC末端に連結されたペプチドの大きなライブラリーを用いる親和性選別法により、受容体への結合について選別されている(Cullら, 1992)。大腸菌で発現させた場合、リプレッサータンパク質は、そのリガンドを、コード化プラスミド上のlacオペレーター配列への結合によって該プラスミドに物理的に連結させる。更に、完全にin vitroのポリソーム提示系についても報告されており(Mattheakisら, 1994)、その場合、新生ペプチドはリボソームを介してそれらをコードするRNAに物理的に結合される。
【0003】
in vivoでは、抗体特異性の一次レパートリーは、免疫グロブリンV、DおよびJ遺伝子セグメントの結合を含むDNA再構成のプロセスにより作製される。マウスおよびヒトでは、抗原と出会った後、その抗原により起動されたB細胞内で再構成されたV遺伝子が、今度は体細胞高頻度突然変異による第2段階の多様化を起こす。この高頻度突然変異によって第二のレパートリーが生成され、そのレパートリーから良好な結合特異性が選別され、それによって液性免疫応答の親和性成熟が可能になる。
【0004】
現在までの人為的な選別系は、ひとえに、第一次の突然変異および選別に基づくものであり、これは、概念としては、所望の活性を有する遺伝子産物の選別がなされる遺伝子産物突然変異体の「固定された」レパートリーが得られるという点で、自然の抗体産生で起こるV-D-J再構成の第1段階と似ている。
【0005】
in vitroでのRNAの選別および進化(EllingtonおよびSzostak, 1990)は、SELEX(指数的濃化(enrichment)によるリガンドの体系的進化)(TuerkおよびGold, 1990)とも呼ばれることがあり、これは結合活性と化学的活性の両者についての選別を可能にするが、但し、核酸についてしかできない。選別を結合について行う場合、核酸のプールを固定化された基質と共にインキュベートする。未結合のものを洗い流し、次に結合しているものを遊離させ、増幅し、このプロセス全体を反復段階として繰り返して、より良好な結合配列を濃化する。この方法は、触媒作用を持つRNAおよびDNAの単離にも適用できる(GreenおよびSzostak, 1992;概説については、ChapmanおよびSzostak, 1994;Joyce, 1994;Goldら, 1995;Moore, 1995を参照されたい)。したがって、SELEXでは、所望の活性を繰り返し工程により向上させることができるが、その範囲は核酸の調製に限られている。
【0006】
しかし、人為的な選別系は、自然の免疫系の場合とは違って、手軽な形態の「親和性成熟」や繰り返し工程によるレパートリーの作製・開発にはあまり適さない。この理由の1つは、突然変異をそれらが必要とされる分子の領域にターゲティングすることは難しいので、それに続いて突然変異と選別を繰り返しても、活性が向上した分子を十分な効率で単離することはできないからである。
【0007】
自然の抗体産生において親和性成熟の際に起こる体細胞高頻度突然変異プロセスについての知見の多くは、in vivoでの高頻度突然変異の際に起こった突然変異の分析から導き出されたものである(概説については、NeubergerおよびMilstein, 1995;WeillおよびReynaud, 1996;Parham, 1998を参照されたい)。これらの突然変異の大部分は、段階的に導入された一ヌクレオチド置換である。それらは、(特徴的なホットスポットと共にではあるが)再構成されたVドメイン全体に散らばっており、それら置換は、塩基のトランジションに偏りを示す。それらの突然変異の殆どは、(B細胞の分化・増殖の他の段階の間ではなく)胚中心でのB細胞増殖の間に蓄積し、その場合、高頻度突然変異期の間のV遺伝子へのヌクレオチド置換の組み込み率は10-4〜10-3/bp/世代であると見積もられる(McKeanら, 1984; Berek & Milstein, 1988)。
【0008】
リンパ球様細胞系によって高頻度突然変異を研究するための扱い易い系が得られる可能性について、何年も前から検討されてきた(CoffinoおよびScharff, 1971;Adetugboら, 1977;Bruggemannら, 1982)。明らかに、重要なことは、研究対象の細胞系におけるV遺伝子の突然変異率が、実行可能なアッセイを提供するのに十分なほど高いものであり、しかも、突然変異が局在性の抗体高頻度突然変異機構によって真に生じたものであって、多くの腫瘍と特徴的に関連しているような全体的な突然変異率の増大を反映しているのではないと確信できるくらい十分高いことである。マウスのプレB細胞系および形質細胞腫細胞系におけるVH内の停止コドンの復帰変異(reversion)をモニタリングすることによって、突然変異が広く研究されている(Wablら, 1985;Chuiら, 1995;Zhuら, 1995;Greenら, 1998により概説されている)。発現されたV遺伝子の直接配列決定という別の戦略を行ったところ、幾つかの濾胞性リンパ腫、バーキットリンパ腫およびホジキンリンパ腫におけるVH遺伝子の多様化が、最初のトランスフォーメーション事象の後で継続し得ることが示された(BahlerおよびLevy, 1992;Jainら, 1994;Chapmanら, 1995および1996;Braeuningerら, 1997)。また、直接配列決定からは、クローン化濾胞性リンパ腫系では突然変異の存在率(prevalence)が低いことも明らかになっており、これは、VHの多様化がin vitroでも継続可能であることを立証している(Wuら, 1995)。上記で引用した細胞系での構成的突然変異についてのどの報告からも、観察された突然変異が、自然の抗体の多様化で見られるようなV遺伝子に集中している指向性高頻度突然変異(directed hypermutation)の結果であって、異なる系統に由来する多くの腫瘍細胞系において記載されているような突然変異に対する全体的な感受性とは異なる、ということの証拠は得られていない。
【0009】
近年、Denepouxら(1997)が、抗免疫グロブリン抗体および活性化T細胞の存在下で細胞を培養することによって、細胞系で高頻度突然変異を誘導した。しかし、観察された高頻度突然変異は誘発型であって構成的ではないことが述べられている。
【発明の概要】
【0010】
本発明の第1の態様において、標的核酸領域の指向性構成的高頻度突然変異を起こすことができるリンパ球様細胞系の製造方法であって、細胞集団を進行中の標的配列の多様化についてスクリーニングし、標的核酸の突然変異率が他の核酸の突然変異率よりも100倍以上上回っている細胞を選別することを含む上記方法が提供される。
【0011】
本明細書中で用いられる「指向性構成的高頻度突然変異(directed constitutive hypermutation)」とは、本明細書で報告する実験で初めて見出だされたものであり、特定の細胞系の、内在性DNAまたは導入遺伝子DNAの1箇所以上の特定の部位の核酸配列の改変を構成的様式で(すなわち、外的な刺激を与える必要なしに)引き起こす能力をいう。指向性構成的高頻度突然変異を起こすことができる細胞では、内在性DNAまたは導入遺伝子DNAの特定の部位以外の配列は、バックグラウンド突然変異率を上回る突然変異率とはならない。
【0012】
「標的核酸領域」とは、指向性構成的高頻度突然変異が起こる、本発明による細胞中の核酸の配列または領域である。標的核酸は、遺伝子産物をコードする1つ以上の転写ユニットを含むことができ、その転写ユニットはその細胞に対して同種であっても異種であってもよい。典型的な標的核酸領域は、免疫グロブリン産生細胞に見られる免疫グロブリンV遺伝子である。これらの遺伝子は、後記で更に詳細に説明されている高頻度突然変異収集エレメント(hypermutation-recruiting elements)の影響下にあり、そのエレメントによって高頻度突然変異が問題の遺伝子座で誘導される。その他の標的核酸配列は、例えば、遺伝子座内の高頻度突然変異収集エレメントを含むV遺伝子転写ユニットを、別の所望の転写ユニットで置換することにより、あるいは高頻度突然変異収集エレメントを含む人工遺伝子を構築することにより、構築できる。
【0013】
「高頻度突然変異」とは、細胞中の核酸の、バックグランドよりも高い率での突然変異をいう。好ましくは、高頻度突然変異とは、10-5〜10-3/bp/世代の突然変異率をいう。これは、バックグランド突然変異率(10-9〜10-10の突然変異/bp/世代のオーダー:Drakeら, 1988)や、PCRで見られる自然発生的突然変異率を大幅に上回る。Pfuポリメラーゼを用いて増幅を30サイクル行うと、その産物には1 bp当たり0.05×10-3未満の突然変異が起こるが、これは本発明の場合に観察される突然変異の1/100未満である(Lundbergら, 1991)。
【0014】
高頻度突然変異は、天然における免疫グロブリン可変鎖(V)遺伝子生成の一部である。したがって、本発明によれば、細胞系は、好ましくは、少なくとも1つの免疫グロブリンV遺伝子を生成できる免疫グロブリン産生細胞系である。V遺伝子は、可変軽鎖(VL)遺伝子であっても可変重鎖(VH)遺伝子であってもよく、免疫グロブリン分子全体の一部として生成されてもよい。それは、抗体、T細胞受容体、または免疫グロブリンスーパーファミリーの他のメンバーに由来するV遺伝子でありうる。免疫グロブリンスーパーファミリーのメンバーは、免疫系における広範な役割(例えば、抗体、T細胞受容体分子など)、細胞接着への関与(例えばICAM分子)、ならびに細胞内シグナル伝達(例えば、PDGF受容体などの受容体分子)を含むin vivoでの細胞性および非細胞性相互作用の多くの局面に関与する。したがって、本発明による好ましい細胞系は、B細胞から誘導されるものである。本発明によれば、V領域遺伝子に高頻度突然変異を起こす能力は保持しているが他の遺伝子には高頻度突然変異を生じない抗体産生B細胞由来の細胞系が単離できることが確認された。
【0015】
1つの好ましい実施形態において、本発明による細胞は、in vivoで高頻度突然変異が起こる細胞から誘導されたものか、あるいは該細胞に関連するものである。in vivoで高頻度突然変異を起こす細胞は、例えば、B細胞などの免疫グロブリン発現細胞である。リンパ腫細胞は、Ig発現細胞の腫瘍であり、これは、本発明による構成的に高頻度突然変異を起こす細胞系を単離するための特に優れた候補である。
【0016】
本明細書中で用いられる「進行中の標的配列の多様化についてのスクリーニング」とは、試験される細胞系の標的核酸領域における高頻度突然変異の存在の判定をいう。これは、直接配列決定法、またはMutSアッセイ(Jollyら, 1997)のような間接法、または免疫グロブリン欠損変異体の生成のモニタリングなどの種々のやり方で実施できる。この手順に従って選別された細胞は、標的配列の多様化を示す細胞である。
【0017】
本発明の方法による選別に供される細胞集団は、種々の細胞型および/もしくは種々の標的配列を含む、ポリクローナル細胞集団または(単)クローン細胞集団でありうる。
【0018】
クローン細胞集団とは、単一のクローンから誘導される細胞の集団であって、細胞がそこで起こっている突然変異以外は同一であるようになっているものである。クローン細胞集団の使用では、好ましくは、V遺伝子の高頻度突然変異を誘導する目的では他の細胞型(例えば活性化T細胞)との共培養は行わない。
【0019】
本発明による細胞は、高頻度突然変異を引き起こすために誘導工程の使用には依存しない。
【0020】
好ましくは、本発明においてスクリーニングされるクローン細胞集団は、B細胞から誘導される。有利には、それは、バーキットリンパ腫細胞系、濾胞性リンパ腫細胞系またはびまん性大細胞リンパ腫細胞系などのリンパ腫細胞系である。
【0021】
好ましくは、本発明による方法は、更に、標的配列の多様化を示す1種以上の細胞を単離する工程、および、単離した細胞において、標的配列内の突然変異の蓄積速度を非標的配列内のものと比較する工程を含む。
【0022】
本発明の特徴は、高頻度突然変異が、特定の(標的)核酸領域だけで誘導され、通常はこれらの領域以外では見られないことである。したがって、特異性は、本発明の方法の一部として、標的配列以外の配列の突然変異率をアッセイすることによってアッセイされる。C領域遺伝子(天然では高頻度突然変異に暴露されない)は、そうした技法に有利に用いることができるが、特異的な高頻度突然変異を受けない他の核酸領域もまた使用可能である。高頻度突然変異は配列依存的ではないので、比較のために選択された核酸領域の実際の配列そのものは重要でない。しかし、それは、後記で説明するように、高頻度突然変異を誘導する配列の制御を受けていてはならない。便利なことに、バックグラウンド突然変異は、例えばHPRT遺伝子座における彷徨解析(fluctuation analysis)により評価できる[LuriaおよびDelbreck, (1943);CapizziおよびJameson, (1973)を参照されたい]。
【0023】
標的領域の突然変異が非標的領域の突然変異を上回っている細胞が、本発明による特定の核酸領域の指向性構成的高頻度突然変異を起こすことができる細胞である。V領域遺伝子の突然変異が他の遺伝子の突然変異を上回っている係数は様々であるが、一般には少なくとも102、有利には103、好ましくは104以上のオーダーである。
【0024】
全体的な突然変異率および多様性は、例えば、突然変異誘発原の投与、または配列改変遺伝子(例えばターミナルデオキシヌクレオチジルトランスフェラーゼ(TdT))の発現によって増大し得る。しかし、高頻度突然変異とバックグラウンドとの差は、そうした方法では増大しないと予想される。
【0025】
本発明による好ましい細胞は、そこで見られる体細胞高頻度突然変異率を上げるために、遺伝子の欠失、変換もしくは挿入などの遺伝子操作に供することができる。例えば、本発明による細胞は、RAD51パラログ体の1コピー以上を欠失していてもよい。
【0026】
その細胞は、適切な脊椎動物細胞であればよく、例えば哺乳動物の細胞および鳥類の細胞が挙げられる。
【0027】
本発明の第2の態様において、所望の活性を有する遺伝子産物の製造方法であって、以下の工程a)〜c):
a) 本発明の第1の態様による細胞の集団において、高頻度突然変異を誘導する配列に機能し得る形で連結された該遺伝子産物をコードする核酸を発現させる工程、
b) 該細胞集団中で、該所望の活性を有する突然変異した遺伝子産物を発現する細胞または細胞群を同定する工程、
c) 工程(b)で同定された細胞または細胞群から1種以上のクローン細胞集団を樹立し、そのクローン細胞集団から、該所望の活性が向上している遺伝子産物を発現する細胞または細胞群を選別する工程、
を含む上記方法が提供される。
【0028】
上記のパートa)による細胞の集団は、V領域遺伝子の構成的高頻度突然変異を起こすことができ本発明の第1の態様による方法によって同定される細胞を含むクローン細胞集団またはポリクローナル細胞集団である。したがって、遺伝子産物は、後記で更に詳細に説明するように、内在性の免疫グロブリンポリペプチド、操作された内在性の遺伝子により発現される遺伝子産物、または体細胞高頻度突然変異を誘導する調節配列に機能し得る形で連結された異種転写ユニットにより発現される遺伝子産物、とすることができる。
【0029】
本発明の細胞内で発現され高頻度突然変異を起こす核酸は、内在性V領域のような内在性の領域、または本発明の細胞系に挿入された異種の領域であってよい。これは、例えば、高頻度突然変異を誘導する内在性の調節配列を保持したまま、内在性V領域が異種転写ユニット(例えば異種V領域)によって置換されている形態;または、高頻度突然変異を誘導するための細胞自身の調節配列の制御下で、該細胞へ異種転写ユニットが挿入されている形態を取ることができ、その場合、その転写ユニットはV領域遺伝子または任意の他の所望の遺伝子産物をコードしていればよい。本発明による核酸は、後記で更に詳細に説明されている。
【0030】
上記の工程b)において、細胞は、所望の遺伝子産物活性についてスクリーニングされる。これは、例えば免疫グロブリンの場合、結合活性とすることができる。また、適切なアッセイ法を用いて、酵素活性などの他の活性を評価することも可能である。遺伝子産物が細胞表面に提示される場合、所望の活性をもたらす細胞は、細胞表面での活性を(例えば蛍光発光によって)検出することにより、あるいは該細胞を表面の遺伝子産物を介して支持体に固定化することにより単離できる。活性が増殖培地中に分泌される場合、あるいはそれぞれ個々の細胞についてではなく細胞培地全体についてしか評価できない場合は、その所望の活性を有する遺伝子産物を分泌する細胞の同定確率を上げるために、工程a)から複数のクローン集団を樹立することが有利である。用いられる選別系は、細胞の増殖能・突然変異能に影響を及ぼさないことが有利である。
【0031】
好ましくは、この段階(および工程c)において、より良好な、向上した、またはより望ましい活性を有する遺伝子産物を発現する細胞を選別する。そのような活性は、例えば、所与のリガンドに対するより高度な親和性結合、またはより有効な酵素活性である。したがって、この方法によれば、所望の活性の定性的および/または定量的評価に基づいて、細胞を選別できる。
【0032】
本発明の第3の態様において、所望の活性を有する遺伝子産物の製造における、特定の核酸領域の指向性構成的高頻度突然変異を起こすことができる細胞の使用が提供される。
【0033】
本発明による使用において、所望の活性を有する遺伝子産物をコードする核酸は、細胞内での高頻度突然変異を誘導する調節配列に機能し得る形で連結されている。したがって、その細胞を連続的に作製することにより、その核酸配列の突然変異体が得られ、それを本発明の方法によりスクリーニングして、有利な特性を有する突然変異体を単離する。
【0034】
更に別の態様において、本発明は、本発明による指向性構成的高頻度突然変異を起こすことができる細胞に関する。好ましくは、この細胞は、遺伝子操作されたニワトリDT40細胞である。上記で記載したように、1つ以上のDNA修復遺伝子が操作できる。好ましくは、1つ以上のRad51遺伝子が操作されている。これらの遺伝子はダウンレギュレートまたは欠失していることが有利である。好ましくは、その遺伝子はRad51b遺伝子またはRad51c遺伝子である。
【0035】
1つの非常に好ましい実施形態において、本発明は、Δxrcc2 DT40およびΔxrcc3 DT40からなる群から選択される細胞を提供する。
【0036】
図面の説明
図1。バーキット系におけるVHの多様性
(A) 4種の散発性バーキットリンパ腫系の再構成されたVH遺伝子における配列多様性を円グラフとして示したもの。各細胞系について配列決定を行ったM13クローンの数を円の中心に示す。円弧の様々な大きさは、0個、1個、2個等(図示のとおり)の突然変異によりコンセンサスと区別される配列の割合を示す。
【0037】
(B) 初期Ramos培養物において同定されたVH突然変異の推定される系統的関係(dynastic relationship)。それぞれの丸(突然変異の程度に比例した濃淡を付けた)は、丸の中に示す数の蓄積された突然変異を有する別個の配列を示す。
【0038】
(C) 再構成されたVλ遺伝子における突然変異の存在率。Ramosにおいて、2種のVλの再構成が同定される。多様性および生殖細胞系の起源の割り当ては、図1Aと同様にして示してある。
【0039】
(D) 初期Ramos培養物のVH領域およびCμ領域における突然変異の存在率の比較。円グラフは、図1Aと同様にして示してある。
【0040】
図2。Ramosにおける構成的なVH多様化
(A) MutSアッセイにより評価した多様化。直接クローニングおよび配列決定により推定される各集団における突然変異の存在率を示す。
【0041】
(B) 3種の独立したRamosクローンの子孫から推定される系統的関係。
【0042】
図3。RamosのVHに沿った未選択のヌクレオチド置換の分布。
【0043】
図4。Ramosにおける高頻度突然変異により、様々な復帰可能なIgM欠損変異体が生じる。
【0044】
(A) IgM欠損変異体の単離を示すスキーム。
【0045】
(B) 複数のナンセンス突然変異がVHの不活性化に関与し得ることを示す表。これら2種の集団において停止がみとめられるそれぞれのVHコドンの位置を示す。
【0046】
(C) IgM欠損変異体の復帰変異率の表。
【0047】
(D) IgM欠損誘導体における停止コドン周辺の配列。
【0048】
図5。TdTを発現するRamosトランスフェクト体におけるIgM欠損変異体
(A) 3種のpSV-pβG/TdTおよび3種のRamosの対照トランスフェクト体におけるTdTの発現のウエスタンブロット分析。
【0049】
(B) IgM欠損変異体を生じる独立した突然変異事象を示す円グラフ。
【0050】
図6。一ヌクレオチド置換以外のVHにおける突然変異をまとめた配列表。
【0051】
図7。抗イディオタイプ(抗-Id1)結合特異性が失われているRamos細胞のVH遺伝子から単離した配列の比較。出発集団のコンセンサスとは異なるヌクレオチド置換は太字で示す。推定アミノ酸変化も太字で示す。
【0052】
図8。5ラウンドにわたる反復的な選別による、新規な結合特異性を有する免疫グロブリンを製造するためのRamos細胞の濃化を示す棒グラフ。
【0053】
図9。ビーズ:細胞の比率を増大させることによる新規な特異性(ストレプトアビジン)を結合するRamos細胞の回収の向上を示す棒グラフ。
【0054】
図10。標的抗原の濃度を増大させることによる、新規な結合特異性を有するRamos細胞の回収の増大を示すグラフ。
【0055】
図11。ストレプトアビジン結合Ramos細胞から得られたVH配列。出発集団のVH配列と比較した場合に見られるヌクレオチドの変化および推定アミノ酸の変化を太字で示す。
【0056】
図12。対照の培地および未選択のRamos細胞の上清に対する、ストレプトアビジン結合に対する選別プロセスの第4、第6および第7ラウンドにて選別された細胞の上清中のIgMの量。
【0057】
図13。図12の上清から得られたIgMのストレプトアビジン結合。
【0058】
図14。表面プラズモン共鳴により分析した、ストレプトアビジン結合に対する選別の第4および第6ラウンドから得られた上清のストレプトアビジン結合。
【0059】
図15。第4および第6ラウンドで選別された細胞のストレプトアビジン-FITCへの結合のFACS分析。
【0060】
図16。第6ラウンドで選別されたIgMのVHおよびVLの配列。
【0061】
図17。ストレプトアビジンに対して選別された親和性成熟Ramos細胞のFACS分析。
【0062】
図18。親和性成熟Ramos細胞のELISA。
【0063】
図19。野生型および修復欠損DT40におけるsIgM欠損変異体
(A) 野生型および修復欠損細胞におけるsIgMの異質性(heterogeneity)のフローサイトメトリー分析。
【0064】
(B) sIgM欠損変異体の生成頻度の彷徨解析。
【0065】
図20。DT40のsIgM変異体からクローニングしたVλ配列の分析。
【0066】
図21。1ヶ月間クローン性増殖させた後の、ソーティングされていないDT40集団のIg配列の分析。
【0067】
図22。DNA-PK、Ku70およびRad51Bが欠損しているDT40細胞のsIgM欠損変異体の分析。
【0068】
図23。ラット免疫グロブリンのイディオタイプに特異的なDT40由来の一連のIgM。細胞をビオチン化ラットS7 IgG2aと、モノクローナル抗体(Ab)/FITC-ストレプトアビジンとの凝集物を用いる6ラウンドの選別に供してDT-Ab3を得て、次に、これを更に3ラウンドの直接PE-Abコンジュゲートを用いるソーティングに供してDTAb6を得る。(a) ビオチン化Ab/FITC-Strep(ストレプトアビジン)凝集物およびPE-Abの、DT-Ab3細胞およびDT-Ab6細胞への結合の分析。(b) S7 Abで被覆したプレートを用いるELISAによりモニタリングした、DTAb3、5および6細胞の上清中のIgMのS7 Abへの結合、ならびに抗ニワトリIgMを用いた検出。上清は、総IgM力価について調整した。種々のラット・ハイブリドーマおよび各種のイソ型のキメラmAb(G. ButcherおよびM Bruggemannから恵贈されたもの(図示せず))へのDT-Ab6 IgMによる結合は検出されなかった。(c) DT-Ab6 IgMの、プレート上に被覆されたS7ラットIgG Abへの結合は、S7 Ab自身による競合を受けるが、正常なラット血清によって競合を受けることはほとんどない。S7(0.5mg/mlのストック)およびラット血清の競合物は、図示の希釈率で用いた。(d) DT-Ab6 IgMは、S7ハイブリドーマ細胞を染色するのに使用できる。染色は、固定した透過化細胞のフローサイトメトリーにより、FITC-コンジュゲート抗ニワトリIgMを用いて検出することにより検出した。(e) 元の0.2mg/mlのストック(PE-S7 Ab;Pharamacia)の各種の希釈液を用いたPE-AbコンジュゲートによるDT-Ab3細胞およびDT-Ab6細胞の染色の比較。(f) 親和性の測定。DT-Ab6細胞(106個)を、各種容量の0.2ピコモルのPE-コンジュゲートS7 Abと共にインキュベートして、図示のPE-Ab濃度を得た。洗浄した後で、細胞の平均蛍光強度(MFI)を測定した。(g) DT-Ab6と親集団とのIgVH/VL配列の比較。
【0069】
図24。DT40由来の一連のプロテインA特異的IgM。(a) 108個の細胞を106個のビーズと共にインキュベートした後で回収される細胞の数を示す、誘導化した磁性ビーズに結合しているDT40変異体の選別。DT-P1およびP2は、ビオチン化プロテインAで被覆したストレプトアビジン-ビーズで選別され、一方、DT-P3は、DT-P2から、プロテインAで直接被覆したトシル化磁性ビーズで選別された。(b) ビオチン化プロテインAとFITC-ストレプトアビジンとの凝集物による、FITCと直接コンジュゲートさせたプロテインAによる、およびFITC-抗IgMによる、各種のDT-P細胞のサブクローンの染色の比較。(c) 検出にニワトリIgMに特異的なマウスIgM mAb(Southern Biotechnology)を用いたELISAによりモニタリングした、親DT40およびDT-P14バルク集団の上清中のIgMの、プロテインAで被覆したプレートへの結合。対照として、総IgM力価も比較した。(d) DT-P4、-P6およびP7のクローンの上清から得られたIgM(親DT40からのものではない)は、プロテインA-セファロース(Sepharose)上に保持されているニワトリIgMを検出するためのウエスタンブロットを用いてプロテインA上で精製することができる。(e) [35S]プロテインAのDTP細胞への結合。親DT40(DT)またはDT-P細胞(0.1ml中に5×105個)を、[35S]プロテインA(48,000の結合可能なcpm;200〜2000Ci/mmol)と共に、氷上で1時間インキュベートした。PBSで1回洗浄した後、結合したcpmを測定した。(f) 親和性の測定。DT-P14細胞(106個)を各種の容量の0.7ピコモルの[35S]プロテインA(7×1010の結合可能なcpm/μmole)と共にインキュベートして、図示したプロテインA濃度を得た。洗浄した後、結合したcpmを測定した。(g) FITCプロテインAによるDT-P4細胞およびDT-P14細胞の染色は、1mg/mlの未標識ウサギIgGの存在下(P4+、P14+)で染色することにより増強される。(h) DT-Pサブクローン由来のIgV遺伝子の配列。
【0070】
図25。DT40の抗原選別により、特異性への成熟の可能性を有する多重反応性のIgMを得ることができる。DT-H10細胞は、親Xrcc2-欠損DT40のプールから、ヒト血清アルブミン(HAS)が結合しているカルボキシル化磁性ビーズを用いて、連続した10ラウンドの選別により得た。DT-T10細胞およびDT-O6細胞は、FITC-ストレプトアビジンとビオチン化サイログロブリン(DT-T細胞)もしくはオボアルブミン(DT-O細胞)との凝集物で染色した細胞のフローサイトメトリーによる選別を連続したラウンドで行うことにより得た。各ラウンドでは、集団の最も明るい2%を採取した。(a) 図示の試薬(Cy-strep:サイクローム(cychrome)-ストレプトアビジン)で染色したDT-H6細胞およびDT-T6細胞のフローサイトメトリー分析。(b) DT-H6細胞およびDT-T6細胞の抗原結合は、sIgM欠損変異体について濃化してあるDT-H6およびDT-T6の亜集団の二次元フローサイトメトリー分析で確認されるように、表面IgMにより仲介される。(c) 図示の抗原に特異的なニワトリIgMをモニタリングした、DTH6およびDT-T6の培養上清のELISA。(d) DT-O6細胞(オボアルブミン凝集物を用いて選別)もまたTg(ならびに、数種の他の被験抗原(図示せず))に結合するが、FITC-Ovabright/サイクローム-ストレプトアビジンdull細胞について連続ソーティングを行ったところ、Ovaに対する特異性がより高い集団が得られる。
【0071】
図26。天然に存在する構成的に高頻度突然変異を起こすBL細胞系の分析。
【図面の簡単な説明】
【0072】
【図1−1】バーキット系におけるVHの多様性を示す。
【図1−2】バーキット系におけるVHの多様性を示す。
【図1−3】バーキット系におけるVHの多様性を示す。
【図2】Ramosにおける構成的なVH多様化を示す。
【図3】RamosのVHに沿った未選択のヌクレオチド置換の分布を示す。
【図4】Ramosにおける高頻度突然変異により、様々な復帰可能なIgM欠損変異体が生じることを示す。
【図5】TdTを発現するRamosトランスフェクト体におけるIgM欠損変異体を示す。
【図6】一ヌクレオチド置換以外のVHにおける突然変異をまとめた配列表を示す。
【図7】抗イディオタイプ(抗-Id1)結合特異性が失われているRamos細胞のVH遺伝子から単離した配列の比較を示す。
【図8】5ラウンドにわたる反復的な選別による、新規な結合特異性を有する免疫グロブリンを製造するためのRamos細胞の濃化を示す棒グラフを示す。
【図9】ビーズ:細胞の比率を増大させることによる新規な特異性(ストレプトアビジン)を結合するRamos細胞の回収の向上を示す棒グラフを示す。
【図10】標的抗原の濃度を増大させることによる、新規な結合特異性を有するRamos細胞の回収の増大を示すグラフを示す。
【図11−1】ストレプトアビジン結合Ramos細胞から得られたVH配列を示す。
【図11−2】ストレプトアビジン結合Ramos細胞から得られたVH配列を示す。
【図12】対照の培地および未選択のRamos細胞の上清に対する、ストレプトアビジン結合に基づく選別プロセスの第4、第6および第7ラウンドにて選別された細胞の上清中のIgMの量を示す。
【図13】図12の上清からのIgMのストレプトアビジン結合を示す。
【図14】表面プラズモン共鳴により分析した、ストレプトアビジン結合に対する選別の第4および第6ラウンドから得た上清のストレプトアビジン結合を示す。
【図15】第4および第6ウランドで選別された細胞のストレプトアビジン-FITCへの結合のFACS分析を示す。
【図16−1】第6ラウンドで選別されたIgMのVHおよびVLの配列を示す。
【図16−2】第6ラウンドで選別されたIgMのVHおよびVLの配列を示す。
【図17】ストレプトアビジンに対する選別された親和性成熟Ramos細胞のFACS分析を示す。
【図18】親和性成熟Ramos細胞のELISAを示す。
【図19】野生型および修復欠損DT40におけるsIgM欠損変異体を示す。
【図20】DT40のsIgM変異体からクローニングしたVλ配列の分析を示す。
【図21−1】1ヶ月間クローン性増殖させた後の、ソーティングされていないDT40集団のIg配列の分析を示す。
【図21−2】1ヶ月間クローン性増殖させた後の、ソーティングされていないDT40集団のIg配列の分析を示す。
【図22】DNA-PK、Ku70およびRad51Bが欠損しているDT40細胞のsIgM欠損変異体の分析を示す。
【図23】ラット免疫グロブリンのイディオタイプに特異的なDT40由来の一連のIgMを示す。
【図24−1】DT40由来の一連のプロテインA特異的IgMを示す。
【図24−2】DT40由来の一連のプロテインA特異的IgMを示す。
【図25−1】DT40の抗原選別により、特異性への成熟の可能性を有する多重反応性のIgMを得ることができることを示す。
【図25−2】DT40の抗原選別により、特異性への成熟の可能性を有する多重反応性のIgMを得ることができることを示す。
【図26−1】天然に存在する構成的に高頻度突然変異を起こすBL細胞系の分析を示す。
【図26−2】天然に存在する構成的に高頻度突然変異を起こすBL細胞系の分析を示す。
【発明を実施するための形態】
【0073】
発明の詳細な説明
本発明は、選択された核酸領域を構成的に高頻度突然変異させる細胞系を、初めて利用できるようにした。これにより、自然の抗体産生における親和性成熟によく似た技法によって、突然変異した遺伝子産物を産生する系の設計が可能になる。Ramosバーキット系は、in vitro培養物中で、その再構成された免疫グロブリンV遺伝子を構成的に多様化する。この高頻度突然変異には、活性化T細胞や外来的に添加されるサイトカインによる刺激も、更にはB細胞抗原受容体の維持も必要としない。
【0074】
その突然変異率(0.2〜1×10-4/bp/世代の範囲)は、未選択の突然変異の大きなデータベースを容易に蓄積させるのに十分高いものであり、したがって、Ramosにおける高頻度突然変異が、in vivoでの免疫グロブリンV遺伝子の高頻度突然変異と古典的に関連づけられている特徴(突然変異をV遺伝子へ選択的にターゲッティングすること;一ヌクレオチド置換の段階的な蓄積;トランジションへの偏り;特徴的な突然変異のホットスポット)の多くを示すことは明らかである。データベースにおける未選択の突然変異の大部分は一ヌクレオチド置換であるが、欠失および重複(時には隣接するヌクレオチド置換が共に存在する)が検出可能である。そのような欠失および重複もまた、in vivoでの高頻度突然変異の結果として生じることが提唱されている(Wilsonら, 1998;Goosensら, 1998;Wu & Kaartinen, 1995)。
【0075】
選択された核酸領域で構成的に高頻度突然変異を起こす細胞の単離は、B細胞などの抗体産生細胞から誘導される細胞系でV遺伝子の突然変異をモニタリングすることに基づく。本発明で用いられる選別方法は、多くの方法で構成できる。
【0076】
高頻度突然変異細胞の選別
高頻度突然変異細胞は、標的配列の配列決定、発現欠損突然変異体についての選別、細菌性MutSタンパク質を用いるアッセイ、および遺伝子産物の活性の変化についての選別などの種々の技法により、細胞集団から選別できる。
【0077】
標的核酸の高頻度突然変異の特徴の一つは、このプロセスにより、停止コドンが、高頻度突然変異が起こらない場合に見られるよりもずっと高い頻度でその標的配列に導入されることである。これによって、細胞からの遺伝子産物の産生が欠損する。この欠損を、核酸配列が高頻度突然変異を起こしている細胞の同定に利用できる。
【0078】
本発明の1つの好ましい実施形態において、標的核酸は免疫グロブリンをコードする。免疫グロブリンの欠損は、免疫グロブリンを培地に分泌する細胞、および免疫グロブリンが細胞表面に提示されている細胞の両者について検出できる。免疫グロブリンが細胞表面に存在するのであれば、それが存在しないことは、例えばFACS分析、免疫蛍光顕微鏡法、または支持体へのリガンドの固定により、個々の細胞について同定すればよい。1つの好ましい実施形態では、細胞を抗原被覆磁性ビーズと混合して、沈降したら、細胞懸濁液から、目的とする特異性を有する免疫グロブリンが表面上に提示されている全ての細胞を除去すればよい。
【0079】
この技法は、抗体やT細胞受容体などを含むいずれの免疫グロブリン分子にも適用できる。免疫グロブリン分子の選別は、本発明に従ってアッセイすることが望まれるクローン細胞集団の性質に応じて決まってくる。
【0080】
あるいはまた、本発明による細胞は、V遺伝子などの標的核酸の配列決定、および配列比較による突然変異の検出によって選別することも可能である。この方法は、スループット(処理量)を増大させるために自動化することができる。
【0081】
更なる実施形態では、V遺伝子を高頻度突然変異させる細胞を、クローン細胞集団中で産生された免疫グロブリンにおける抗原結合活性の変化を評価することによって検出できる。例えば、特定の単位量の細胞培地または抽出物により結合される抗原の量を評価して、特定の結合活性を保持している細胞により産生された免疫グロブリンの割合を求めることができる。V遺伝子が突然変異すると、結合活性は変化し、特定の抗原に結合している産生された免疫グロブリンの割合は減少する。
【0082】
あるいはまた、細胞を、新規な結合親和性を生み出す能力についても同様にして評価でき、これは、例えば、初めは結合していない抗原または抗原の混合物にそれらを暴露し、高頻度突然変異の結果として結合親和性が生み出されるか否かを観察することにより行うことができる。
【0083】
更に別の実施形態では、細菌性MutSアッセイを用いて、標的核酸における配列の変異を検出することができる。MutSタンパク質は、核酸ハイブリッド中のミスマッチに結合する。親の核酸と突然変異した可能性のある子孫との間でヘテロ二本鎖を形成させることによりミスマッチ形成の程度、ひいては核酸の突然変異の程度が評価できる。
【0084】
標的核酸が免疫グロブリン以外の遺伝子産物をコードする場合、選別は、結合以外の機能の欠損または変化についてスクリーニングすることにより行ってもよい。例えば、酵素活性の欠損または変化についてスクリーニングしてもよい。
【0085】
配列の高頻度突然変異をターゲッティングする細胞を、他の核酸領域における突然変異について評価する。アッセイするのに都合の良い領域は、免疫グロブリン遺伝子の定常(C)領域である。C領域は、本発明による指向性高頻度突然変異を起こさない。C領域の評価は、好ましくは配列決定および比較により行われる。何故ならば、これが、突然変異が存在しないことを判定するための最も確実な方法だからである。しかし、他の技法を用いることも可能であり、例えば、高頻度突然変異事象により破壊される可能性のあるC領域の活性(例えば補体結合)の保持についてのモニタリングが挙げられる。
【0086】
細胞の遺伝子操作
本発明による高頻度突然変異細胞は、IgのV領域における高頻度突然変異率を増大するように遺伝子操作されている細胞から選別できる。突然変異率のモジュレーションを担う遺伝子は、一般に、細胞における核酸修復手順に含まれる。本発明に従って操作される遺伝子は、アップレギュレート、ダウンレギュレートまたは欠失されていてもよい。
【0087】
アップレギュレーションまたはダウンレギュレーションとは、問題の遺伝子によりコードされる遺伝子産物の活性が、少なくとも10%、好ましくは25%、更に好ましくは40%、50%、60%、70%、80%、90%、95%、99%、またはそれ以上増大または低下していることをいう。勿論、アップレギュレーションは、200%や500%などの100%を超える活性の増大を示してもよい。100%ダウンレギュレーションされている遺伝子は、機能的に欠失しており、これを本明細書中では「欠失している」ともいう。
【0088】
本発明により操作される好ましい遺伝子としては、Rad51遺伝子、特にxrcc2、xrcc3およびRad51b遺伝子の類似体および/またはパラログ体が含まれる。
【0089】
Rad51の類似体および/またはパラログ体は、ダウンレギュレートされていることが有利であり、好ましくは欠失している。1つ以上のRad51パラログ体のダウンレギュレーションまたは欠失により、本発明による高頻度突然変異率の増大が起こる。好ましくは、2つ以上のRad51遺伝子(その類似体および/またはパラログ体を含む)がダウンレギュレートされているか欠失している。
【0090】
1つの非常に好ましい実施形態では、ニワトリDT40細胞系などの鳥類の細胞系が、xrcc2および/またはxrcc3の欠失によって改変されている。Δxrcc2 DT40ならびにΔxrcc3-DT40は、本発明に従って単離される構成的に高頻度突然変異を起こす細胞系である。
【0091】
内在性遺伝子産物の改変
免疫グロブリンV領域遺伝子などの内在性遺伝子に構成的に高頻度突然変異を起こす細胞系が得られたことから、本発明は、新規な特性を有する遺伝子産物を産生させるための、構成的高頻度突然変異による内在性遺伝子産物の改変(adaptation)を提供する。例えば、本発明は、新規な結合特異性または改変された結合親和性を有する免疫グロブリンの産生を提供する。
【0092】
実際に、高頻度突然変異のプロセスを用いて、免疫グロブリン分子において向上した、または新規な結合特異性を生じさせる。したがって、目的とする抗原に結合できる免疫グロブリンを産生する本発明による細胞を選別し、次に、これらの細胞を増殖させて更なる突然変異体を産生させることにより、目的とする抗原への結合が向上している免疫グロブリンを発現する細胞を単離できる。
【0093】
目的の特異性を有する突然変異体を単離するために、種々の選別手順が適用できる。これらとしては、蛍光活性化セルソーティング(FACS)、磁性粒子を用いる細胞分離、抗原クロマトグラフィー法、および他の細胞分離法(例えばポリスチレンビーズの使用)が挙げられる。
【0094】
磁性による捕捉を用いる細胞の分離は、対象とする抗原を磁性粒子またはビーズにコンジュゲートさせることにより達成できる。例えば、その抗原を、Miltenyi Biotec GmbHにより供給されるような超常磁性鉄-デキストラン粒子またはビーズにコンジュゲートさせる。次に、これらのコンジュゲート粒子またはビーズを、多様な表面免疫グロブリンを発現しうる細胞集団と混合する。特定の細胞がその抗原と結合できる免疫グロブリンを発現する場合、それは、この相互作用により磁性ビーズと複合体化するだろう。次に、磁性粒子を固定している懸濁液に磁場を印加し、共有結合により結合している抗原を介してそれらに結合している細胞を保持する。次に、ビーズと結合しない未結合細胞を洗い流し、対象とする抗原に結合する能力に基づいて純粋に単離された細胞の集団を残存させる。試薬およびキットは、そのような一段階単離を行うための様々な供給元から入手可能であり、例えばDynalビーズ(Dynal AS; http://www.dynal.no)、MACS-磁性セルソーティング(Miltenyi Biotec GmbH; http://www.miltenyibiotec.com)、CliniMACS(AmCell; http://www.amcell.com)、ならびにBiomag, Amerlex-Mビーズなどが挙げられる。
【0095】
蛍光活性化セルソーティング(FACS)は、異なる表面分子(例えば表面に提示される免疫グロブリン)に基づく細胞の単離に使用できる。ソーティング(分別)しようとするサンプル中または集団中の細胞を、細胞表面分子に結合する特定の蛍光試薬で染色する。これらの試薬は、フルオレセイン、テキサスレッド、マラカイトグリーン、緑色蛍光タンパク質(GFP)または当業者に公知の任意の他の蛍光団などの蛍光マーカーに(直接または間接的に)結合している対象とする抗原である。次に、細胞集団をFACS装置の振動フローチャンバーに導入する。このチャンバーを通過する細胞の流れを、PBS(リン酸緩衝化生理食塩水)のような緩衝液のシース(sheath)に入れる。この流れにレーザー光を照射し、各細胞を蛍光(蛍光標識抗原の結合を示す)について測定する。細胞流の振動により、その細胞流は液滴になり、それは小さな電荷を持つ。これらの液滴に対して、コンピューター制御下で電気的偏向板により方向を誘導して、蛍光標識抗原に対する親和性に相応して異なる細胞集団を回収することができる。このようにして、対象とする抗原に対して異なる親和性を示す細胞集団を、その抗原に結合しない細胞から容易に分離できる。FACSで用いるFACS装置および試薬は、Becton-Dickinsonなどの世界規模の供給元や、Arizona Research Laboratories (http://www.arl.arizona.edu/facs/)などのサービス会社から広く入手できる。
【0096】
特定の抗原に対する細胞表面タンパク質の親和性に応じて細胞の集団を分離するのに使用可能な別の方法は、アフィニティクロマトグラフィーである。この方法では、適切な樹脂(例えばCL-600セファロース、Pharmacia Inc.)を適当な抗原に共有結合させる。この樹脂をカラムに詰め、混合細胞集団をそのカラムに通す。適当時間(例えば20分間)にわたりインキュベートした後、未結合細胞を(例えば)PBS緩衝液を用いて洗い流す。これにより、対象とする抗原に結合した免疫グロブリンを発現する細胞のサブセットだけが残存し、次に、これらの細胞を、(例えば)対象とする抗原を過剰量で用いて、または該抗原を該樹脂から酵素的もしくは化学的に切断することにより、カラムから溶出させる。これは、X因子、トロンビンまたは当業者に公知の他の特定のプロテアーゼなどの特定のプロテアーゼを用いて、予め抗原−樹脂複合体に組み込まれている適当な切断部位により該抗原をカラムから切り離すことによって行うことができる。あるいはまた、トリプシンなどの非特異的プロテアーゼを用いて、該抗原を樹脂から除去して、その対象とする抗原に対して親和性を示す細胞集団を遊離させることも可能である。
【0097】
異種転写ユニットの挿入
任意の所与の抗原に結合できる抗体変異体を迅速に選別する可能性を最大化するため、または非免疫グロブリン遺伝子についての高頻度突然変異系を開発するために、多くの技法を用いて、本発明による細胞を、それらの高頻度突然変異能が引き出されるように遺伝子操作することができる。
【0098】
第1の実施形態では、導入遺伝子を本発明による細胞にトランスフェクトして、その導入遺伝子が指向性高頻度突然変異事象の標的となるようする。
【0099】
本明細書中で用いられる「導入遺伝子」とは、例えばトランスフェクションまたはトランスダクションにより細胞に挿入される核酸分子である。例えば、「導入遺伝子」は、上記で述べたような異種転写ユニットを含むものであってもよく、それは細胞のゲノムの所望の位置に挿入できる。
【0100】
導入遺伝子を細胞に送達するのに用いられるプラスミドは、慣用の構造のものであり、プロモーターの制御下に目的の遺伝子産物をコードするコード配列を含む。本発明による細胞におけるベクターからの遺伝子の転写は、ポリオーマウイルス、アデノウイルス、鶏痘ウイルス、ウシ乳頭腫ウイルス、トリ肉腫ウイルス、サイトメガロウイルス(CMV)、レトロウイルスおよびサルウイルス40(SV40)などのウイルスのゲノム由来、アクチンプロモーターまたは例えばリボソームタンパク質プロモーターである非常に強力なプロモーターなどの異種哺乳動物プロモーター由来、ならびに通常は異種コード配列に結合しているプロモーター由来のプロモーターによって制御できるが、但し、そうしたプロモーターは本発明の宿主系と適合するものである。
【0101】
本発明による細胞による異種コード配列の転写は、ベクターにエンハンサー配列を挿入することによって増大させることができる。エンハンサーは、方向および位置には比較的依存しない。哺乳動物遺伝子(例えば、エラスターゼおよびグロビン)に由来する多くのエンハンサー配列が知られている。しかし、典型的には、真核細胞ウイルス由来のエンハンサーが用いられる。例としては、複製起点の後ろ側のSV40エンハンサー(bp 100-270)およびCMV初期プロモーターエンハンサーが挙げられる。エンハンサーは、ベクター中のコード配列に対して5’側または3’側の位置でスプライシングされ得るが、好ましくは、プロモーターの5’側の部位に位置する。
【0102】
有利には、真核性発現ベクターは、遺伝子座調節領域(LCR)を含み得る。LCRは、宿主細胞のクロマチンに組み込まれた導入遺伝子の高レベルの組込み部位に依存しない発現を誘導でき、これは、遺伝子治療用途のために設計されたベクターにおける、またはトランスジェニック動物における、ベクターの染色体組込みが起こっている永続的にトランスフェクトされた真核細胞系において異種コード配列を発現させようとする場合には特に重要である。
【0103】
真核性発現ベクターは、転写の終結およびmRNAの安定化のために必要な配列も含んでいる。そのような配列は、真核性またはウイルス性のDNAまたはcDNAの5’側および3’側非翻訳領域から一般に入手可能である。これらの領域は、mRNAの非翻訳部分においてポリアデニル化断片として転写されるヌクレオチドセグメントを含む。
【0104】
発現ベクターとしては、そのようなDNAを発現させることができる調節配列(例えばプロモーター領域)と機能し得る形で連結された目的の遺伝子産物をコードするコード配列を発現できる任意のベクターが挙げられる。したがって、発現ベクターとは、適当な宿主に導入されるとクローン化DNAの発現が起こる組換えDNAまたはRNA構築物をいい、例えば、プラスミド、ファージ、組換えウイルスまたは他のベクターが挙げられる。適当な発現ベクターは当業者には周知であり、真核細胞および/もしくは原核細胞において複製可能なもの、ならびにエピゾームを保持するもの、または宿主細胞のゲノムに組み込まれるものが挙げられる。例えば、異種コード配列をコードするDNAを、哺乳動物細胞におけるcDNAの発現に適するベクター、例えばpEVRF(Matthiasら, 1989)などのCMVエンハンサーをベースとするベクターに挿入できる。
【0105】
本発明によるベクターの構築には、慣用のライゲーション法が用いられる。単離したプラスミドまたはDNA断片を切断し、あつらえ、所望の形態に再連結して、必要とするプラスミドを作製する。所望により、構築したプラスミド中の配列が正しいことを確認するための分析を公知の方法で行う。発現ベクターを構築し、in vitroで転写産物を調製し、DNAを宿主細胞に導入し、遺伝子産物の発現および機能を評価するための分析を行うための好適な方法は、当業者には周知である。遺伝子の存在、増幅および/または発現は、例えば、適当に標識したプローブ(本発明で提供される配列に基づいたものでありうる)を用いて、慣用のサザンブロッティング、mRNAの転写を定量するためのノーザンブロッティング、ドットブロッティング(DNAもしくはRNAの分析)またはin situハイブリダイゼーションにより、直接サンプル中で測定できる。当業者であれば、必要に応じてこれらの方法にどのように変更を加えればよいか容易に判るであろう。
【0106】
第1の実施形態の1つの別形態では、本発明による導入遺伝子は、高頻度突然変異を誘導する配列も含む。そうした配列は特性決定されており、Klixら, (1998)およびSharpeら, (1991)(参照により本明細書に組み入れられる)で述べられている配列が挙げられる。つまり、遺伝子産物を発現し、且つその遺伝子産物をコードする転写ユニットに高頻度突然変異を誘導することができる遺伝子座全体が細胞に導入される。したがって、高頻度突然変異を誘導する転写ユニットおよび配列が、その細胞に対して外来性である。しかし、外来性ではあっても、高頻度突然変異を誘導する配列そのものは、細胞において天然に見られる高頻度突然変異を誘導する配列と類似または同一であってもよい。
【0107】
第2の実施形態では、内在性V遺伝子またはそのセグメントを、相同組換えにより、例えばLox/Cre系もしくは類似の技法を使用する遺伝子ターゲティングにより、または自然発生的に内在性V遺伝子が欠失している高頻度突然変異を起こす細胞系への挿入によって、異種V遺伝子と置換する。あるいはまた、高頻度突然変異には再構成されたV遺伝子近傍での二本鎖の破断が伴う、という知見を利用して、V領域遺伝子を置換することも可能である。
【0108】
以下、次の実施例において本発明を更に詳細に説明するが、それらは単に説明を目的とする。
【実施例】
【0109】
実施例1:高頻度突然変異を起こす細胞の選別
in vitroで高頻度突然変異を起こす細胞についてスクリーニングするために、数種のヒト・バーキットリンパ腫においてクローン性増殖の際に蓄積する多様性の程度を評価する。バーキット系BL2、BL41およびBL70はG. Lenoir(IARC, Lyon, フランス)から恵贈されたもの、Ramos(Kleinら, 1975)はD. Fearon(Cambridge, UK)から恵贈されたものである。それらの再構成されたVH遺伝子を、複数のVHファミリーのプライマーをJHコンセンサスオリゴヌクレオチドと共に用いて、ゲノムDNAからPCR増幅する。再構成されたVHセグメントの増幅は、Pfuポリメラーゼ、ならびに主要なヒトVHファミリーの各々について設計された14のプライマー(Tomlinson, 1997)のうちの1つ、および6つのヒトJHセグメント全てにアニーリングするコンセンサスJHバックプライマー(JOL48、5’-GCGGTACCTGAGGAGACGGTGACC-3’、 C. Jollyからの恵贈)を用いて達成される。RamosのVHのゲノムDNAからの増幅は、オリゴヌクレオチドRVHFOR(5’-CCCCAAGCTTCCCAGGTGCAGCTACAGCAG)およびJOL48を用いて行う。発現されたVH-CμcDNAの増幅は、RVHFORおよびCμ2BACK(5'-CCCCGGTACCAGATGAGCTTGGACTTGCGG)を用いて行う。ゲノムのCμ1/2領域は、Cμ2BACKおよびCμ1FOR(5'-CCCCAAGCTTCGGGAGTGCATCCGCCCCAACCCTT)を用いて増幅する。Ramosの機能性のCμ対立遺伝子は、Cμ2の8位のヌクレオチドとしてCを含んでおり、これに対して非機能性の対立遺伝子上ではTである。再構成されたVλは、5’-CCCCAAGCTTCCCAGTCTGCCCTGACTCAGおよび5’-CCCCTCTAGACCACCTAGGACGGTC-AGCTTを用いて増幅する。PCR産物は、QIAquick(Qiagen)スピンカラムを用いて精製し、M13中にクローニングした後でABI377シーケンサーを用いて配列決定する。突然変異は、GAP4アライメントプログラム(Bonfieldら, 1995)を用いて求められる。
【0110】
クローン化PCR産物の配列決定を行ったところ、Ramos細胞系でかなりの多様性(VHでの存在率は2.8×10-3の突然変異/bp)が示されるが、有意な異質性(heterogeneity)はBL41ならびにBL2でも見られる。図1Aを参照されたい。4種の散発性バーキットリンパ腫系の再構成されたVH遺伝子の配列多様性は、円グラフに示すとおりである。各細胞系における再構成されたVH遺伝子を、PCR増幅し、M13中にクローニングする。各細胞系について、最も多数のM13クローンに共通する配列としてコンセンサスをとり、さらにヒト免疫グロブリン配列のVBASEデータベース(Tomlinson, 1997)を用いて最も近くマッチングすることに基づいて生殖細胞系の対応物(各グラフの上部に示す)を割り当てる。ここで用いたRamosのVHコンセンサス配列は、Chapmanら(1996)により決定された配列とは3つの位置で、Ratech(1992)により決定された配列とは5つの位置で、それと最も近縁の生殖細胞系の対応物VH4(DP-63)とは6つの位置で異なっている。
【0111】
RamosにおけるVHの多様性の解析は、9回の別個のPCR増幅から得られる産物の配列決定により拡張される。これによって、集団中の突然変異したクローン同士の、個々のヌクレオチド置換の独立した反復の推定数が最小になるような有望な系統的(dynastic)関係の推定が可能になる(図1B)。9回の別個のPCR増幅から得られた315個のM13VHクローンを配列決定する。その系統は、同定された配列だけを含む(推定上の中間型は含まない)。個々の突然変異は、フォーマット「C230」に従って設計され、ここで、230とはRamosのVHのヌクレオチド位置(図3に示す番号付け)であり、「C」とは、その位置の新規な塩基を示す。同じヌクレオチド置換が独立して起こる回数を最小化することを、基準として系統を推定するのに用いた。分岐の大部分は、別々のPCR増幅によりもたらされる独立したメンバーを含む。まれな欠失および重複は、それぞれ接頭語「x」および「d」で示す。矢印は、2つの突然変異を強調しており(264位での置換により、停止コドンおよび184位での重複が生じる)、系統樹内でのその位置は、突然変異が、機能性重鎖の発現が欠損した後も蓄積し続けることができることを示している。
【0112】
PCRのアーチファクト(artefact)は、突然変異のデータベースに殆ど影響を及ぼさない。ヌクレオチド置換の存在率が、対照のPCR増幅で見られる存在率(0.05×10-3/bp)よりも遥かに高いことだけでなく、別個の増幅において同一の突然変異を生じたクローン(ならびに系統的に関連するもの)が見られる。多くの場合、ある系統内の世代同士は、一ヌクレオチド置換だけ異なっており、このことから、突然変異の各ラウンドにおいて少数の置換しか導入されていないことが示される。
【0113】
Vλ再構成の解析から、Ramosが、Vλ2.2-16の、読み枠が合っている(in-frame)再構成(Chapmanら, 1996により記載されている)およびVλ2.2-25の読み枠が合わない(out-of-frame)再構成を含んでいることが明らかになる。双方の再構成されたVλには突然変異による多様性が存在するが、非機能性の対立遺伝子では、それより高度の多様性が蓄積している(図1C)。
【0114】
抗体の高頻度突然変異の古典的な特徴といえば、突然変異の多くはV領域に蓄積し、C領域には殆ど蓄積しないことである。これは、RamosのIgH遺伝子座に蓄積している突然変異でも明らかである(図1D)。VH、Cμ1およびCμ2の最初の87ヌクレオチドに及ぶcDNAインサートを含むM13クローンは、初期Ramos培養物からPCRにより作製される。円グラフ(図1Aに示すようなもの)には、Cμ1の始点から伸びるCμの380ヌクレオチドのストレッチと比較した場合の、VHの341ヌクレオチドのストレッチ内で同定された突然変異の程度を示す。
【0115】
Ramosにより産生されたIgM免疫グロブリンは、細胞表面上および培地中(分泌形態)の双方に存在する。培地を分析したところ、Ramosが免疫グロブリン分子を非常に高濃度(1μg/ml程度)で分泌することが判明する。したがって、Ramosは、製造のために発現細胞系または細菌に免疫グロブリン遺伝子を再クローニングする必要がなくなるレベルまで免疫グロブリンを分泌することができる。
【0116】
実施例2:RamosにおけるVHの多様化は構成的である
V遺伝子の多様化が進行中であるか否かを調べるために、細胞をクローングし、in vitroでの培養期間の後で、VHの多様性をMutSベースのアッセイを用いて評価する。RamosのVHを、上記で記載したように、5’末端にビオチン化塩基を含むオリゴヌクレオチドを用いてPCR増幅し、精製する。変性/再生(99℃で3分;75℃で90分)の後、突然変異の程度を、先に記載(Jollyら, 1997)されているような細菌性ミスマッチ修復タンパク質MutS(濾紙に結合させてある)へのミスマッチヘテロ二本鎖物質の結合をECLによる検出によりモニタリングすることによって、評価する。
【0117】
その結果から、VHの多様化が、まさに進行中であることが示される(図2Aを参照)。限界希釈クローニングした後で1ヶ月間または3ヶ月間培養したRamos細胞からDNAを抽出する。再構成されたVHを、ビオチン化オリゴヌクレオチドを用いてPCR増幅した後、変性/再生させる。次にミスマッチヘテロ二本鎖を、先に記載(Jollyら, 1997)されているような固定化MutSへの結合により検出する。再生されたDNAのアリコートをメンブレンに直接結合させて、マッチしたDNAの負荷を確認する(全DNA対照)。比較のために、細菌性プラスミド鋳型ならびに初期Ramos培養物から増幅したRamosのVHについて行うアッセイを含める。
【0118】
4週間(Rc1)および6週間(Rc13および14)にわたり増殖させたRamos培養物からVH遺伝子をPCR増幅する(図2B)。各クローンの突然変異率を示すが、これは、クローニングの4週間後または6週間後の独立したVHの突然変異の存在率を、一世代時間が24時間であることに基づいて推定した細胞分裂の回数で除算することにより算出される。配列から、段階的な突然変異の蓄積が明らかになり、その際の突然変異率は約0.24×10-4の突然変異/bp/世代である。
【0119】
RamosにおけるVHの突然変異率を他の細胞系におけるVHの突然変異率と直接比較することは容易ではない。何故ならば、単一の前駆細胞からクローン性増殖させた後に得られるVH全体に取り込まれた未選択の突然変異により決定されるような、他の系における突然変異率に関する情報は殆ど無いからである。しかし、50個のBL2前駆細胞を2週間にわたり増殖させた後の突然変異の存在率が、突然変異誘発の条件下で測定されている(2.7×10-3の突然変異/bp; Denepouxら, 1997)。通常の培養条件下でRamosを用いて行った同様の実験から、突然変異の存在率が2.3×10-3の突然変異/bpであることが判明している。サイトカイン、ヘルパーT細胞などを与えることによる突然変異率を増大させるための様々な試みは、うまく行かないことが判っている。したがって、BL2細胞の特異的誘発により達成可能な突然異率は、RamosにおけるVHの構成的な突然変異率とほぼ同じであると思われる。
【0120】
実施例3:RamosにおけるVHの突然変異の性質の検討
初期Ramos培養物において検出したもの(141の異なる配列からのもの)を、特定の選択を行わない種々の実験で培養した4種のサブクローン(更に別の135の異なる配列からのもの)において検出したものと組み合わせた、突然変異事象のデータベースを作成する。このデータベースは、個々の配列セットを系統的関係(図1Bの説明文に詳細に説明されている)に組み立てた後で作成して、個々の突然変異細胞のクローン性増殖によって特定の突然変異事象が複数回カウントされてしまわないことを確実にする。ここで、この340の異なった、そしておそらくは未選択の突然変異事象(200は初期Ramos培養物によりもたらされたものであり、140は増殖させたサブクローンによりもたらされたもの)の複合的なデータベースの解析を説明する。初期集団およびサブクローン集団を別々に解析しても、同じ結論が得られる。
【0121】
突然変異の圧倒的大部分(340のうち333)は、一ヌクレオチド置換である。少数の欠失(4)および重複(3)が見られるが、鋳型のない挿入は見られない。これらの事象については、後記で更に詳細に述べる。隣接位置にヌクレオチド置換を示す配列は5つしかない。しかし、これら5つの場合のうち3つにおいて、その系統では、隣接する置換が逐次に取り込まれていることが判明している。このように、隣接位置でヌクレオチド置換が同時に作られることは稀な事象である。
【0122】
VHに沿った突然変異の分布は高度に非ランダムである(図3を参照)。独立して起こっている塩基置換を、各ヌクレオチド位置に示す。CDR1および2の位置を示す。ヌクレオチドの位置は、配列決定用プライマーの3’末端から番号付けし、+1位のヌクレオチドは第7コドンの最初の塩基に相当する。コドンはKabatに従って番号付けしている。イタリック体で示す突然変異(15位、193位、195位および237位のヌクレオチド位置)は、突然変異したサブクローンで起こっている置換であり、その位置の配列は示したコンセンサスに復帰している。
【0123】
主要なホットスポットはSer82aコドンのGおよびCのヌクレオチドにあり、これは他のVH遺伝子における主要な内因性の突然変異ホットスポットとして先に同定されており(Wagnerら, 1995;Jollyら, 1996)、RGYWのコンセンサスと一致している(RogozinおよびKolchanov, 1992;Betzら, 1993)。多くのVH遺伝子における主要な内因性の突然変異ホットスポットはSer31にあるが、このコドンは、その位置にGlyを有するRamosのコンセンサスVH(またはその生殖細胞系のVH)には存在しない。個々のヌクレオチド置換は、トランジションに著しい偏りを示す(無作為に予想される33%ではなく、51%)。また、GおよびCをターゲティングすることについて著しい選択性を示し、これはターゲティングされたヌクレオチドの82%を占める(表1)。
【表1】

【0124】
実施例4:IgM欠損による高頻度突然変異を起こす細胞の選別
Ramnos変異体の分析から、不活性化されたVHを有しているに違いない幾つかの突然変異が明らかになり(図1Bを参照)、このことから、細胞がIgM発現を欠損しても依然として生存可能である可能性があることが示唆される。これが正しいならば、Igの発現の欠損は、構成的に高頻度突然変異を起こすB細胞系を選別するための容易な尺度となるだろう。
【0125】
Ramos培養物の分析から、それが8%の表面IgM−細胞を含んでいることが判明する。そのようなIgM欠損変異体は、以下のようにして、in vitro培養の際に作製される。出発物質のRamos培養物を、pSV2neoプラスミドでトランスフェクトし、96ウェルプレートに希釈し、選択培地で増殖するクローンを増殖させる。最初のトランスフェクションから6ヵ月後に増殖させたクローンについてフローサイトメトリーを行ったところ、IgM欠損変異体の存在が明らかになり、ここで示す2種のクローン集団(Rc13および14)の16%および18%を占めている(図4A)。ソーティングを1ラウンド行うことにより濃化して、87%(Rc13)および76%(Rc14)の表面IgMネガティブ細胞を含む亜集団を得る。これらの亜集団における再構成されたVH遺伝子をPCR増幅した後で配列決定を行うことにより、クローン化したVHセグメントの75%(Rc13)および67%(Rc14)が、分析した341ヌクレオチドのVHストレッチ内にナンセンス(stop)、欠失(del)または重複(dup)の突然変異を含んでいたことが示される。クローンの残りは野生型(wt)と名づけるが、可能性のあるVH不活性化ミスセンス突然変異を区別する試みは行わない。Rc13集団において同定された4つの欠失および3つの重複は全て異なっているが、欠失を有すると判定された7つのRc14の配列のうち、異なる突然変異はたった4つである。欠失および重複の性質を図6に示す。各事象は、文字の後ろに数字を付けて表示する。文字は、突然変異の起源を示す(A、BおよびCはクローン化TdT-対照トランスフェクト体であり、D、EおよびFはTdT+トランスフェクト体であり、Uは初期の未選択Ramos培養物において同定された事象を示す)。数字は、配列文字列(string)の最初のヌクレオチド位置を示す。欠失しているヌクレオチドはラインの上に明記し、付加されたヌクレオチド(重複または鋳型のない挿入)はラインの下に明記する。一ヌクレオチド置換は丸で囲み、新たな塩基が明記してある。VH起源の重複したセグメントには下線が引いてある。鋳型のない欠失は太字にしてある。幾つかの欠失または重複では、その事象は起源が不明の1個のヌクレオチドと隣接している。そのような隣接した変化は、(鋳型のない挿入ではなく)ヌクレオチド置換により十分に起こり得るものであり、したがって、これらの事象は別個にグループ化される。欠失/重複の一端または他端への一塩基置換(丸で囲む)の割り当ては、任意である場合が多い。
【0126】
IgM−細胞は、ソーティングを1ラウンド行って濃化した後、PCR増幅およびそれらのVHセグメントのクローニングを行う。配列から、かなりの範囲のVH不活性化突然変異(停止コドンおよびフレームシフト)が明らかになるが(図4)、様々な不活性化突然変異は、クローン性増殖のわずか6週間後にソーティングしたIgM欠損変異体においても示されている(図5を参照)。図5Aで、3種のpSV-pβG/TdTおよび3種のRamosの対照トランスフェクト体におけるTdTの発現を、核タンパク質抽出物のウエスタンブロッティング分析により比較する。Nalm6(TdT-ポジティブなヒト・プレB細胞リンパ腫)およびHMy2(TdT-ネガティブな成熟ヒトBリンパ腫)を対照とした。
【0127】
図5Bでは、IgM欠損変異体を生じる独立した突然変異事象を示す円グラフを示す。クローニング後に6週間培養した上記の3種のTdT+および3種のTdT−対照トランスフェクト体をソーティグすることによりIgM−変異体(集団の1〜5%を占める)が得られる。ソーティングした亜集団におけるVH領域をPCR増幅し、配列決定する。円グラフは、VH不活性化を生じる突然変異のタイプ、ならびに別々にプールされたTdT+およびTdT−のIgM−亜集団から得られたデータを示している。略語は図4Aのとおりであるが、但し、「ins」は明らかに鋳型のない(非鋳型)ヌクレオチド挿入を含むクローンを示す。複数のヌクレオチド非鋳型挿入と共に欠失または重複を含むクローンだけが、このグラフの「ins」の区分に含まれる。明らかに異なる突然変異事象だけを算出する。それにより、TdT+ IgM欠損亜集団において同定された77の異なるVH不活性化突然変異のうち、30の異なる停止コドン突然変異が同定される。1つのRamosトランスフェクト体から誘導されるIgM欠損集団内で同じ停止コドンが独立して作られている場合は、それに下線を引いてある。
【0128】
停止コドンは様々な位置に作られるが(図4B)、ランダムには位置していない。図4Bに、Rc13およびRc14のIgM欠損集団で見られる停止コドンの性質をまとめてある。少なくとも8種の独立した突然変異事象により、Rc13のデータベースの27の非機能性VH配列のうち20を占めるナンセンス突然変異が生じる。最低で10の独立した突然変異事象から、Rc14のデータベースの22の非機能性VH配列のうち15を占めるナンセンス突然変異が生じる。各停止コドンの後ろのカッコ内の数字は、当該停止コドンを保有するデータベース内の配列の数を示し、その後ろに、更に別の突然変異に基づいて区別される異なったこれらの配列の数を示す。4種の他のクローン集団から選別されるIgM欠損変異体における停止コドンの分析から、VH内で更に5つの位置で停止コドンが作られることが判明する。6つの独立した実験で得られたデータにおいて、停止コドンの創出は、可能性のある39の部位のうちの16に限られる。これらの選択された部位でのDNA配列は、(コード鎖または非コード鎖のいずれにおいても)RGYWコンセンサスの方に偏っている。
【0129】
驚くことではないが、欠失および挿入は、未選択のRamos培養物における突然変異のほんの一部にすぎないが(上記を参照)、それらは、VH不活性化突然変異に注目した場合、非常に重大な意義がある。IgM欠損変異体の大部分がVH自体における停止コドン/フレームシフト突然変異により説明できることは、注目に値する。このことは、Ramosにおける高頻度突然変異が主に免疫グロブリンのVドメインを標的としており、確かにCドメインや、それが突然変異すると表面IgM−表現型が生じる他の遺伝子(例えばIgα/Igβストレッチ)は全く標的としていない、という提唱を更に支持している。また、RamosのVHが、その生産的に再構成されたVλよりもしばしば高頻度突然変異の標的とされる可能性も十分にあり、この結論は、初期培養物中の突然変異パターンにより支持される(図1C)。
【0130】
Ig欠損変異体の検出による細胞の選別は、それらの変異体が復帰変異する(すなわちそれらの内在性のIg発現能を再獲得する)能力を有する場合に特に有用である。より初期に樹立した系統(図1B)からは、IgM欠損細胞が生じ得ることだけでなくそれらに更なる突然変異が起こる可能性があることも示唆される。このことを確認するために、Rc13からソーティングされたIgM欠損変異体を限界希釈によりクローニングする。クローニングから3週間後、IgM−サブクローン中のIgM+復帰変異体の存在を、5×104個の細胞の細胞質免疫蛍光分析によりスクリーニングする。それらの存在率を示す(図4C)。次に、これらのIgM+復帰変異体を、1ラウンドのソーティングで濃化し、クローン性IgM−変異体のVH配列をそのIgM+復帰変異体の子孫のVH配列と比較する。
【0131】
10の増殖させたクローン集団の細胞質免疫蛍光から、IgM+復帰変異体が様々な存在率(0.005%〜1.2%;図4C)で存在することが明らかになり、これによって彷徨解析により1×10-4の突然変異/bp/世代の突然変異率であることが算出できる。これは、未選択の突然変異の直接分析により算出された率(0.25×10-4の突然変異/bp/世代;上記を参照)よりも多少高い。これは、おそらく、異なるIgM欠損クローンが、破壊している突然変異の性質に応じて異なる率で復帰していることをある程度反映していると思われる。事実、Rc13のIgM欠損誘導体における停止コドン周辺の配列からは、TAG32が、内在性の突然変異ホットスポット(Betzら, 1993)の大部分を占めるRGYWコンセンサス(R=プリン、Y=ピリミジン、W=AまたはT;RogozinおよびKolchanov, 1992)と十分に一致するが、TAA33およびTGA36では一致しないことが明らかになる。
【0132】
実施例5:新規なIg結合活性の選別
新規な結合親和性が生じることを実証するために設計された実験では、後記に記載するRamos細胞系の多くのメンバーが、Ramosの表面IgMに対して特異的に誘導された抗イディオタイプ抗体(抗Id1および抗Id2)に結合する膜IgM分子を発現することに注目する。しかし、少数の細胞は表面IgMを保持するが、抗イディオタイプ抗体に結合できない。これは、表面IgM分子における結合親和性が、抗体には結合しないように変化したことによる。表面IgMを発現する細胞は、抗体に結合できず、本発明による1ラウンドのセルソーティングで選別できる。
【0133】
これは、表面IgMを保持しているにもかかわらず抗Id2への結合能を欠損しているμ-ポジティブ/id-ネガティブクローンをELISAにより単離することにより実証される。これらのクローンを配列決定したところ、6つの独立したクローンにおいて、保存されたVH残基であるK70が次のようにN、MまたはRへと突然変異していることが示される。
【0134】
クローン 突然変異
2 K70N AAG-AAC
S77N AGC-AAC
4 K70M AAG-ATG
9 S59R AGT-AGG
K70N AAG-AAC
10 K70N AAG-AAC
12 K70N AAG-AAC
13 K70R AAG-AGG
軽鎖では突然変異は見られなかった。したがって、産生されたIg分子が親クローンと比較して一塩基対の変異を有する突然変異体をRamos細胞系から選別できることは明らかである。
【0135】
抗Id1を用いて、Igμ定常領域の発現は維持しているが抗イディオタイプ抗体への結合を欠損している同様の細胞集団を単離する。これらの細胞は、ソーティングサイトメトリーにより濃化し、VHの配列を決定する(図7)。このことにより、出発集団のコンセンサス配列と比較した場合に、6つの突然変異が判明する。これらの突然変異のうち2つは、CDR3近傍でアミノ酸配列の変化をもたらす(95にてR→T、98にてP→H)。したがって、免疫グロブリン分子におけるより微細な変化の選別は、結合の欠損についてアッセイすることによって選択可能である。
【0136】
更に別の実施形態では、本発明による高頻度突然変異を起こす細胞を洗浄し、PBS/BSAに再懸濁し(0.25ml中に108個の細胞)、10%(v/v)の抗原被覆磁性ビーズを含む同容量のPBS/BSAと混合する。この実験では、ストレプトアビジン被覆磁性ビーズ(Dynal)を用いる。ローラー上で4℃にて30分間混合した後、ビーズをPBS/BSAで3回洗浄する。各回ごとに、ビーズを磁石で沈降させ、未結合の細胞を除去する。次に、残った細胞を96ウェルプレートに接種し、108個まで増殖させた後、更にもう1ラウンドの選別に供する。細胞の増殖(構成的に進行中の高頻度突然変異を伴う)および選別を複数ラウンド行う。複数ラウンドの選別を行った後、ビーズに結合した細胞の割合(最初はバックグラウンドレベルである0.02%またはそれに近いレベル)が増大し始める。
【0137】
4ラウンド後、ストレプトアビジン結合細胞の濃化が見られる。これを第5ラウンドでも繰り返す(図8)。回収率のパーセンテージが低いことは、ビーズが細胞で飽和していることを反映している。何故ならば、細胞:ビーズ比を大過剰から1:2に変更することにより、第5ラウンドのストレプトアビジン結合細胞から約20%の回収が可能になるからである(図9)。このことから、4ラウンドの反復的な選別により、高頻度突然変異を起こすRamos細胞系から新規な結合特異性をうまく選別できることが実証される。
【0138】
ストレプトアビジン結合細胞由来の重鎖および軽鎖のヌクレオチド配列決定により、出発集団のコンセンサス配列と比較した場合に、VHのCDR3で1つのアミノ酸の変化、そしてVLで4つの変化(FR1に1つ、CDR1に2つ、CDR2に1つ)があることが推定される(図11)。
【0139】
ストレプトアビジンの結合が表面免疫グロブリンの発現に依存することを確実にするために、ストレプトアビジン結合細胞の免疫グロブリンネガティブ変異体をソーティングサイトメトリーにより濃化する。これにより、過剰のビーズを用いた場合でもストレプトアビジン結合細胞の回収率は著しく低下する。ソーティングされたネガティブ細胞からDynal-ストレプトアビジンビーズにより回収された細胞は、事実、Igμポジティブであり、これがおそらくソーティングされた免疫グロブリンネガティブ細胞集団に混入しているIgμストレプトアビジン結合細胞の効率的な回収を説明すると思われる。
【0140】
予備のデータから、回収の効率は、ビーズ上のストレプトアビジンの濃度が低下するに従って低下することが示唆される(図9)。これは、ある濃度範囲のストレプトアビジンと共にインキュベートしたビーズを用いてストレプトアビジン結合細胞の回収率をアッセイすることにより確認される(図10)。結合している集団から回収可能な細胞のパーセンテージは、ビーズと細胞との割合により求められる。この実験では、ビーズ:細胞の比率は1:1未満とする。
【0141】
更なる一連の実験では、更に2ラウンドの選別を完了させ、つまり、全体で7ラウンドとする。これは、ビーズに結合するストレプトアビジンの濃度を、第5ラウンドでの50μg/mlを第7ラウンドでは10μg/mlへと下げることにより達成される。IgMの分泌レベルは、第4〜7ラウンドで選別された集団と同等であるが(図12)、ELISAにより評価した場合のストレプトアビジン結合は、第6および第7ラウンドでは第4ラウンドと比較して明らかに大きく増大している(図13)。
【0142】
これは、ストレプトアビジンで被覆されたBiaCoreチップでの表面プラズモン共鳴により結合を評価することによって確認される(図14)。第7ラウンドから得た上清を、Aの時点で注入してチップに流し、Bの時点で中止する。Cの時点で、抗ヒトIgMを注入して、ストレプトアビジンに結合している物質がIgMであることを実証する。勾配A-Bは会合定数を表わし、勾配B-Cは解離定数を表わす。BiaCoreをトレースすることにより、第6ラウンドの上清が、第4ラウンドの集団から単離されたものや未選択のRamos細胞よりも優れた結合特性を示すことが明らかになる。
【0143】
選別プロセスの第6ラウンドから得られる抗体もまた、第4ラウンドと比較して向上した結合を示す。図15に示すように、第6ラウンドでの選別から得られた細胞のストレプトアビジン-FITC凝集物への結合(蛍光団をビオチン化タンパク質と共にプレインキュベートすることにより形成される)は、FACSにより可視化できる。第4ラウンドの集団、未選択のRamos細胞またはIgMネガティブRamosへの結合は見られず、このことは、ストレプトアビジン結合の成熟を示している。
【0144】
凝集していないストレプトアビジン-FITCを使用すると、同様の結果は得られず、第6ラウンドの細胞の大部分が結合しない。これは、ELISAによるデータと一致して、ストレプトアビジンへの結合が、一連の抗原に結合する抗体の親和力によるものであって、一価の親和性によるものではないことを示唆している。親和性がより高い結合体は、凝集していないストレプトアビジン-FITCへの結合についてソーティングすることにより単離できる。
【0145】
第4ラウンドの細胞の場合を上回る、第6ラウンドの細胞で見られる結合の増大に関与する突然変異を突き止めるために、軽鎖および重鎖抗体遺伝子をPCRにより増幅し、次に配列決定する。第4ラウンドの細胞と比較して、重鎖遺伝子の変化は見られず、突然変異R103Sは保存されている。軽鎖でも、V23FおよびG24Cの突然変異は保存されているが、46位に更に別の突然変異が存在している。野生型のRamosはこの位置にアスパラギン酸を有するが、第6ラウンドの細胞はアラニンを有する。この位置での変化は、抗原結合に影響を及ぼすと予測される。何故ならば、この領域の残基は軽鎖のCDR2に関与するからである(図16)。D46Aの突然変異はおそらく、第6ラウンドの細胞で見られるストレプトアビジンへの結合の観察された増大に関与すると思われる。
【0146】
実施例6:Ramosのストレプトアビジン結合体のin vitro成熟
Ram B→Ram C(FITC-ポリ-ストレプトアビジンを用いた選別)
約5×107個のRam B細胞(ストレプトアビジン被覆ミクロビーズに結合させるためにRamos細胞系から誘導したもの)をPBSで洗浄し、ポリ-ストレプトアビジン-FITCを含む1mlのPBS/BSA溶液中で氷上で30分間インキュベートする(ポリ-ストレプトアビジン-FITCは、ストレプトアビジン FITC(20μg/mlのタンパク質含量)をビオチン化タンパク質(10μg/ml)に添加し、氷上で数分間インキュベートした後、細胞を添加することにより作製される)。
【0147】
次に、細胞を氷冷PBSで軽く洗浄し、遠心沈降させ、500μlのPBSに再懸濁する。
【0148】
最も蛍光性が高い1%の細胞をMoFloセルソーターでソーティングし、この細胞集団を組織培養培地に戻し、約5×107細胞まで増殖させ、この手順を繰り返す。
【0149】
ポリ-ストレプトアビジン-FITCを用いて4ラウンドにわたりソーティングした後、細胞はストレプトアビジン-FITCに弱く結合する。このRamos細胞集団からの発現された免疫グロブリンV領域の配列からは、重鎖V領域の第3フレームワーク中のアミノ酸番号82aがセリンからアルギニンへと変化していることが判明する。この細胞集団をRam Cと呼ぶ。
【0150】
Ram C→Ram D(FITC-ストレプトアビジンを用いた選別)
次の数ラウンドのセルソーティングは、上記で記載されているようにして行うが、ここではストレプトアビジン-FITC(20μg/mlのタンパク質含量)を用いる。
【0151】
ストレプトアビジン-FITCを用いてソーティングを3ラウンド行った後、ソーティングされた細胞集団(Ram Dと呼ぶ)は、FACSによりアッセイしたところ、ストレプトアビジン FITCに、より強く結合している。発現されたV遺伝子の配列から、更なるアミノ酸の変化が明らかになる。第3フレームワークにおいて、元々はセリンであった65位のアミノ酸がアルギニンへと変化している。
【0152】
Ram D→Ram E(FITC-ストレプトアビジンおよび未標識ストレプトアビジン競合を用いた選別)
次のソーティングは、ストレプトアビジン-FITCを用いて、上記で記載されているのと同様にして行う。しかし、細胞を氷上で30分間染色した後、細胞を氷冷PBSで一回洗浄し、次に0.5mg/mlのストレプトアビジンに再懸濁し、氷上で20分間インキュベートする。これは、既に結合しているストレプトアビジン-FITCに対して競合させて、強く結合しているストレプトアビジン-FITCだけが残るようにするためである。次に、細胞を氷冷PBSで1回洗浄し、500μlのPBSに再懸濁した後で、上記で記載したようにして最も蛍光性が高い1%の集団をソーティングする。
【0153】
このソーティングプロトコールを更に2回繰り返した後、Ramos細胞集団(Ram E)は、ストレプトアビジン-FITCに極めて強く結合すると思われる。これらの細胞は、発現された重鎖V遺伝子の第1フレームワーク中で別のアミノ酸の変化を獲得している。すなわち、10位のアミノ酸がグリシンからアルギニンへと変化している。更に、18番目の残基がロイシンからメチオニンへと変化している。
【0154】
Ramos細胞におけるストレプトアビジン成熟の結果を図17に示す。
【0155】
ELISAによる比較
各種のRamos細胞集団の上清を用いて行ったELISAアッセイにより、Ramos細胞から発現され分泌されたIgM抗体が、in vitroで成熟して、ストレプトアビジンに対する強い親和性を獲得していることが確認される。結果を図18に示す。
【0156】
実施例7:高頻度突然変異誘導配列を含む導入遺伝子の構築
Ig遺伝子の遺伝子座の特定のエレメントが、in vivoでの高頻度突然変異事象の誘導に必要であることが知られている。例えば、イントロンエンハンサーおよびマトリックス結合領域であるEi/MARは、極めて重要な役割を担っていることが実証されている(Betzら1994)。更に、3’側エンハンサーであるE3’が重要であることが知られている(Goyenecheaら, 1997)。しかし、本発明者らは、これらのエレメントは必要ではあるが、導入遺伝子内での高頻度突然変異を誘導するには不十分であることを示した。
【0157】
これに対して、Ei/MARおよびE3'を、更なるJκ-CκイントロンDNAおよびCκと共に提供すれば、高頻度突然変異性を十分にもたらすことができる。βG-Cκ導入遺伝子は、PCRで作製した0.96KbのKpnI-SpeI β-グロビン断片(これはβ-グロビン転写開始部位を基準にして-104から+863まで及ぶものであり、その末端に人工的なKpnIおよびSpeI制限部位を有している)をLκΔ[3’Fl](Betzら, 1994)の部分断片(これはMaxら(1081)の配列のヌクレオチド2314からEi/MAR、CκおよびE3’まで及ぶものであり、3’Fl欠失を含んでいる)に結合することにより構築される。
【0158】
高頻度突然変異は、Pfuポリメラーゼを用いてPCR増幅した導入遺伝子のセグメントを配列決定することにより評価される。増幅された領域は、転写開始部位のすぐ上流からJκ5の300ヌクレオチド下流までに及ぶ。
【0159】
このキメラ導入遺伝子は、正常なIgκ導入遺伝子で見られるものと同様の頻度で蓄積するヌクレオチド置換による突然変異の良好な標的になる。この導入遺伝子は、これまでに記載されているものの中で最も小さく、高頻度突然変異を効率的に収集するものであり、その結果から、Cκを含みそれに隣接する領域のどこかに位置する複数の配列が組み合わさって、β-グロビン/Igκキメラの5’末端へ高頻度突然変異を収集することが示される。
【0160】
したがって、高頻度突然変異の収集は、高頻度突然変異ドメインの3’末端に向かって位置する配列によってのみ誘導され得る。しかし、プロモーターの近傍にある正常なIg遺伝子内の突然変異ドメインの5’側境界は、転写開始部位の100〜200ヌクレオチド程度下流である。開始部位に対する突然変異ドメインの5’側境界のこの配置は、β-グロビン遺伝子がプロモーターおよび突然変異ドメインの大部分の双方をもたらす場合は、βG-Cκ導入遺伝子でもそのまま存在する。これらの結果は、他の導入遺伝子を用いて得られた知見と一致しており、それが突然変異ドメインの5’側境界を規定するプロモーターそれ自体の位置であることを示している。
【0161】
κ調節エレメントの(全てではないとしても)幾つかが突然変異の収集に関与する様式についての最も簡単な説明は、それらが、高頻度突然変異開始因子を転写開始複合体に組み込むことにより作動することを提唱することである。転写調節エレメントとしてのエンハンサーについての古典的な研究を用いて類推すれば、Igκエンハンサーは、位置や方向に依存しない様式で高頻度突然変異の調節因子(regulators)として働き得る。事実、βG-Cκ導入遺伝子を用いて得られたデータ、ならびにE3’がよりCκの近くへ移動した、という先の結果(Betzら, 1994)から、E3’の高頻度突然変異増強活性が、突然変異ドメインに対するその位置に対しても方向に対しても特に感受性ではないことが判明する。
【0162】
Ei/MARは、通常、突然変異ドメインの3’末端に向かって位置している。Ei/MARの欠失は、突然変異ターゲティングの効率を著しく低下させるが、それがプロモーターの上流の位置(したがって、転写される領域の外側)に復活すると、突然変異の部分的な救済が起こるが、突然変異ドメインの5’側境界の位置には見かけ上は影響を及ぼさない。これらの結果の独立した確認が、トランスジェニックマウスにおいて、第2の導入遺伝子tk-neo::Cκを用いて得られた。ここでは、neo転写ユニット(HSVtkプロモーターの制御下にある)が、胚性幹細胞においてジーンターゲティングすることによりCκエキソンに組み込まれる(Zouら, 1995)。このマウスでは、Vκ-Jκ結合の後、Igκ Ei/MARは両側を転写ドメイン(上流側はV遺伝子、下流側はtk::neo)で挟まれる。neo挿入についてホモ接合型であるマウス由来のソーティングした胚中心B細胞からtk-neo遺伝子をPCR増幅する。
【0163】
tk-neo::Cκマウスにおけるtk-neoインサートについて、増幅された領域は、(プラスミドpMCNeo(GenbankアクセッションU43611)中で番号付けした場合の)残基607から1417の及び、629位から1329位までのヌクレオチド配列を決定する。tk-neo::Cκマウスにおける内在性VJκの再構成物の突然変異頻度は、Meyerら, 1996に記載されているのと同様の戦略を用いて決定する。内在性VJκ5の再構成物は、Vκ FR3コンセンサスフォワードプライマー(GGACTGCAGTCAGGTTCAGTGGCAGTGGG)と、Jκクラスターの下流から後方にプライミングするオリゴヌクレオチドLκFOR(Gonzalez-FernandezおよびMilstein, (1993) PNAS (USA) 90:9862-9866)を用いて増幅する。
【0164】
tk-neoの突然変異レベルは低く、同じ細胞集団において再構成されたVκ遺伝子の3’側隣接領域よりも間違いなく低い効率で突然変異の標的となるが、neo遺伝子インサート内の突然変異ドメインは、(正常なV遺伝子の場合と同様に)Ei/MARがプロモーターの上流にあるにもかかわらず、転写開始部位の100ヌクレオチド以上下流から始まると思われる。
【0165】
従って、構成的高頻度突然変異を起こす細胞系において高頻度突然変異を誘導できる導入遺伝子は、Ei/MAR、E3'、および本明細書中で定義されるようなJκの下流に見られる調節エレメントを用いて構築できる。更に、導入遺伝子は、内在性V遺伝子の置換または内在性V遺伝子への挿入(tk-neo ::Cκマウスの場合と同様)により、あるいは、目的のコード配列をJκイントロンと連結させること(βG-Cκ導入遺伝子の場合と同様)により構築できる。
【0166】
実施例8:構成的高頻度突然変異を起こす細胞系の選別
上記で記載したようなごく一部のV遺伝子の変換事象により、非機能性Ig遺伝子が、最も高頻度にはフレームシフト突然変異の導入により作製され得る。したがって、ニワトリ・ファブリキウス嚢リンパ腫細胞系DT40におけるsIgM欠損変異体の作製を用いて、IgV遺伝子の変換活性の初期指標を得ることができる。親DT40系と比較して、Rad54が欠損している突然変異体は、かなり低下した割合でsIgM欠損変異体を示す(図19)。複数のクローンについて彷徨解析を行うと、ΔRAD54系は、親DT40系の場合のほぼ1/10の頻度でsIgM欠損変異体を生じ、一方、ΔRAD52系は、野生型細胞とほぼ同様の頻度でsIgM欠損変異体を生じることが明らかになる(図19)。これらの知見は、ΔRAD54細胞およびΔRAD52-DT40細胞における遺伝子変換に関するこれまでの知見(Bezzubovaら, 1997;Yamaguchi-Iwaiら, 1998)と一致している。
【0167】
この分析を、Xrcc2およびXrcc3が欠損しているDT40細胞に適用する。これらのRad51パラログ体は、DNA損傷修復の組換え依存的経路において役割を果たしていることが提唱されている(Liuら, 1998; Johnsonら, 1999;Brennemanら, 2000;Takataら, 2001)。ΔXRCC2系およびΔXRCC3系は、sIgM欠損変異体の存在量の低下を引き起こすのではなく、親の系よりもずっと高い欠損変異体の蓄積を示す(図19)。ΔXRCC2-DT40の場合、ヒトβ-グロビンプロモーターの制御下でのヒトXrcc2 cDNAのトランスフェクションにより、sIgM欠損変異体の生じる頻度が野生型の値近くまで復帰する。図19に、野生型および修復欠損のDT40細胞によるsIgM欠損変異体の生成を示す。単一のsIgM+の正常(WT)または修復欠損(ΔRAD54、ΔRAD52、ΔXRCC2、ΔXRCC3)DT40細胞を1ヶ月間クローン性増殖させたものから誘導した培養物におけるsIgM発現の異質性のフローサイトメトリー分析をパネル(a)に示す。3種の代表的なsIgM+前駆体クローンから誘導した培養物の分析を、修復欠損DT40の各タイプについて示す。各分析におけるsIgM−細胞のパーセンテージを、sIgM+のピークの中心の1/8として設定した蛍光ゲートと共に示す。パネル(b)は、sIgM欠損変異体の生成頻度の彷徨解析を示す。sIgM欠損変異体の存在量は、1ヶ月間クローン性増殖させた後のsIgM+単一細胞から誘導した複数の並行した培養物において測定する。各データセットの上にパーセンテージ中央値を注記し、破線で示してある。分析した[pβG-hXRCC2]ΔXRCC2トランスフェクト体は、彷徨解析において6.4%および10.2%のsIgM−細胞を含むsIgM+ DT40-ΔXRCC2サブクローンにpβG-hXRCC2をトランスフェクトすることにより作製する。複数の独立したsIgM+クローン(異なるが、同様の祖先Vλ配列を有する)について全分析を行ったところ、各修復欠損系では、1ヵ月後にsIgM欠損変異体が生じる頻度の平均中央値は、WT(0.4%)、ΔRAD(0.07%)、ΔRAD52(0.4%)、ΔXRCC2(6%)およびΔXCRCC3(2%)である。
【0168】
Xrcc2およびXrcc3の双方の欠損は、染色体の不安定性に関係するが(Liuら, 1998);Cuiら, 1999;Deansら, 2000;Griffinら, 2000)、sIgM欠損変異体の頻度の増大は、Ig遺伝子座内での全体的な再構成または欠失を反映している可能性がある。しかし、ΔXRCC3-DT40の24種のsIgM−サブクローンのサザンブロット分析からは、再構成されたVλを含む6kbのSalI-BamHI断片の欠失や改変は明らかになっていない。
【0169】
したがって、V遺伝子内のより局在化された突然変異がsIgM発現の欠損を説明できるか否かを確認するために、1ヵ月間増殖させた後で野生型DT40、ΔXRCC2-DT40およびΔXRCC3-DT40サブクローンからソーティングしたsIgM−細胞の集団における再構成されたVλセグメントをクローニングし、配列決定する。
【0170】
細胞培養、トランスフェクションおよび分析
DT40サブクローンCL18およびその突然変異体を、7%のウシ胎仔血清、3%のニワトリ血清(Life Technologies)、50μMの2-メルカプトエタノール、ペニシリンおよびストレプトマイシンを補充したRPMI 1640中で37℃にて10%CO2中で増殖させる。細胞密度は、培養物を毎日分割することにより0.2〜1.0×10-6個/mlに維持した。ターゲティングされた遺伝子破壊を保有するDT40誘導体の作製については、別の文献に記載されている(Bezzubovaら, 1997;Yamaguchi-Iwaiら, 1998;Takataら, 1998, 2000, 2001)。β-グロビンプロモーターの制御下にあるXRCC2のオープンリーディングフレーム(HeLaのcDNAからクローニングされたもの)を含んでいるpSV2-neoベースのプラスミドを保有するΔXRCC2-DT40のトランスフェクト体を、エレクトロポレーションにより作製する。
【0171】
CL18は、DT40のsIgM−サブクローンであり、本明細書で記載されているDNA修復突然変異体の親クローンである。複数のsIgM+サブクローンを、FITC-コンジュゲートヤギ抗-ニワトリIgM(Bethyl Laboratories)で染色した後、野生型および修復欠損突然変異体の両者からMo-Flo(Cytomation)ソーターを用いて得る。この研究で用いたsIgM+ DT40-CL18由来の修復欠損細胞の全てにより発現される初期のVλ配列に違いは殆どない。何故ならば、ほぼ全てのsIgM+誘導体が、ψV8ドナー(これはフレームシフトしたCL18のCDR1と最も近縁である)を用いた遺伝子変換によって元のCL18のVλフレームシフトを復元しているからである。
【0172】
突然変異の分析
ゲノムDNAを、5000個の細胞同等物から、Pfu Turbo(Stratagene)ポリメラーゼおよびホットスタートタッチダウンPCR[95℃で1分、68〜60℃(サイクル毎に1℃ずつ下げる)で1分、72℃で1分30秒を8サイクル;94℃で30秒、60℃で1分、72℃で1分30秒を22サイクル]を用いてPCR増幅する。再構成されたVλはCVLF6(5’-CAGGAGCTCGCGGGGCCGTCACTGATTGCCG;リーダーVλイントロン内でプライミングする)およびCVLR3(5’-GCGCAAGCTTCCCCAGCCTGCCGCCAAGTCCAAG;Jλの3’側から逆方向にプライミングする)を用いて;再構成されていないVλ1はCVLF6およびCVLURR1(5’-GGAATTCTCAGTGGGAGCAGGAGCAG)を用いて;再構成されたVH遺伝子はCVH1F1(5’-CGGGAGCTCCGTCAGCGCTCTCTGTCC)およびCJH1R1(5’-GGGGTACCCGGAGGAGACGATGACTTCGG)を用いて;Cλ領域はCJCIR1F(5’-GCAGTTCAAGAATTCCTCGCTGG;Jλ-Cλイントロンの内部からプライミングする)およびCCMUCLAR(5’-GGAGCCATCGATCACCCAATCCAC;Cλの内部から逆方向にプライミングする)を用いて、増幅する。QIAquickスピンカラム(Qiagen)で精製した後、PCR産物を適当な制限酵素で切断し、pBluescriptSKにクローニングし、T3もしくはT7プライマーおよびABI377シーケンサー(Applied Biosystems)を用いて配列決定する。GAP4との配列アライメント(Bonfieldら, 1995)により、各クローンのコンセンサス配列からの変化が同定できる。
【0173】
全ての配列の変化は、3つのカテゴリー(遺伝子変換、点突然変異、または不明確であるカテゴリー)のうちの1つに割り当てられる。この識別は、遺伝子変換の供与体として働き得るVλの偽遺伝子の公表されている配列に基づく。そのような供与体の配列のデータベースはReynaudら(1987)から取るが、DT40において発現されたIgλに対応する(Kimら, 1990)Igλ G4対立遺伝子に関して修正(McCormackら, 1993)を加えている(この研究において同定された配列/遺伝子変換は、このψVλ配列のデータベースの有効性を支持するものであった)。それぞれの突然変異について、Vλ偽遺伝子のデータベースを可能性のある供与体について検索する。9bp以上の文字列を含む偽遺伝子の供与体が見つからない場合には、鋳型のない点突然変異として分類する。そのような文字列が同定され、同じ供与体により説明可能な更に別の突然変異が存在する場合は、これら全ての突然変異は、1つの遺伝子変換事象に割り当てられる。更に別の突然変異が存在しない場合、単離された突然変異は何らかの変換機構により生じたか、あるいは鋳型がなかった可能性があり、したがって、不明確であるとして分類される。
【0174】
複数の野生型DT40クローンからソーティングされたsIgM−亜集団からクローニングしたVλ配列については、図20に示すように、67%が突然変異を含んでおり、多く(73%)の場合、これらの突然変異によって、Vλは明らかに非機能性になっている。おそらく、残存するsIgM−細胞の大部分が、VH、またはVλの配列決定領域の外側に、不活性化突然変異を保有している。図20に、sIgM欠損変異体からクローニングしたVλ配列の分析を示す。パネル(a)は、1ヶ月間クローン性増殖させた後の正常DT40細胞またはXrcc2欠損DT40細胞の親sIgM+クローンからソーティングされたsIgM欠損細胞から得たVλ配列の比較である。横線はそれぞれ、上記で記載されているようにして点突然変異(棒付の丸)、遺伝子変換域(線上の横棒)、または点突然変異もしくは遺伝子変換の結果であると思われる一ヌクレオチド置換(不明確、縦棒)として分類された突然変異を含む再構成されたVλ1/Jλ(427bp)を表わす。線にかかっている四角の白枠は欠失を表わし、三角は重複を表わす。円グラフをパネル(b)に示すが、これには1ヶ月間クローン性増殖させた後の野生型、ΔXRCC2またはΔXRCC3 DT40 sIgM+クローンから誘導したソーティングされたsIgM欠損集団の中で、異なる数の点突然変異(PM)、遺伝子変換(GC)または起源が不明確な突然変異(Amb)を保有するVλ配列の割合が示されている。それらの円弧の大きさは、円の周囲に示す数の突然変異を保有する配列の数に比例する。分析したVλ配列の総数は、各円の中心に示し、データは、野生型DT40の4種のサブクローン、ΔXRCC2-DT40の2種のサブクローンおよびΔXRCC3-DT40の3種のサブクローンの分析から集計したものである。欠失、重複および挿入は、この分析から除外されている。野生型細胞では、更に6つの欠失、1つの重複および1つの挿入が存在する。ΔXRCC2-DT40では他の事象は起こっておらず、ΔXRCC3-DT40のデータベースでは1bpの欠失および1bpの挿入がそれぞれ一例ずつある。
【0175】
同定された不活性化突然変異を含む、野生型、ΔXRCC2(ΔX2)およびΔXRCC3(ΔX3) DT40細胞におけるVλ遺伝子の不活性化の原因[ミスセンス突然変異(黒);遺伝子変換に伴うフレームシフト(白);欠失、挿入もしくは重複に伴うフレームシフト(灰色)]を、全配列に対するパーセンテージとして表してパネル(b)に示す。それぞれの不活性化突然変異に伴う更に別の突然変異事象を(d)に示す。データは、それぞれの不活性化突然変異に伴う更に別の突然変異の平均数と、パネル(c)で示す更なる突然変異のタイプとで表わす。したがって、ΔXRCC2-DT40は、指標となる不活性化突然変異に加えて、平均で1.2個の追加の点突然変異を有するが、野生型DT40ではわずか0.07である。
【0176】
上記で詳細に述べたように、突然変異は、上流のVλ偽遺伝子を鋳型とする遺伝子変換に起因し得るもの、鋳型のない点突然変異に起因し得るもの、または不明確であるカテゴリーに入れられるもの、として分類できる。不活性化突然変異の多く(67%)は遺伝子変換によるものであるが、一部(15%)は鋳型のない点突然変異により生じる停止コドンであり、このことから、本明細書中および他の文献(Buersteddeら, (1985);Kimら, 1990)に見られるDT40細胞での低い点突然変異の頻度はPCRのアーチファクトではなく、低頻度の点突然変異は、実際に野生型DT40での遺伝子変換に伴って生じることが明らかになったことが示される。
【0177】
著しく異なる突然変異パターンは、ΔXRCC2-DT40由来のsIgM欠損変異体のVλ配列で見られる。ほぼ全ての配列が点突然変異を有しており、典型的には、配列当たり複数の点突然変異が含まれる。また、点突然変異への実質的なシフトは、sIgM− ΔXRCC3-DT40細胞由来の配列でも見られる。したがって、野生型DT40におけるVλ不活性化突然変異は、おそらく読み枠が合わない遺伝子変換域を反映していると思われ、ΔXRCC2/3では、それはミスセンス突然変異であると思われる(図20c)。更に、ΔXRCC2-DT40またはΔXRCC3-DT40のソーティングされたsIgM欠損変異体から得られた非機能性Vλ配列の多く(ΔXRCC2-DT40では53%、ΔXRCC3-DT40では64%)は、Vλ不活性化突然変異に加えて、更なる点突然変異を含んでいるが、そのような同乗は、親DT40系由来の非機能性Vλ配列ではごく稀にしか見られない(7%、図20d)。
【0178】
これらの知見は全て、ΔXRCC2/3-DT40細胞におけるsIgM欠損変異体の高い存在率が、これらの細胞における自然発生的なIgV遺伝子の高頻度突然変異の非常に高い頻度をそのまま反映していることを示唆している。図21に、1ヶ月クローン性増殖した後のソーティングされていないDT40集団からクローニングしたIg配列の分析を示す。代表的な野生型およびΔXRCC2 DT40クローンから得たVλ配列を、図20と同様の記号と共にパネル(a)に示す。パネル(b)には、図示されている異なる数の各種タイプの突然変異を含むVλ配列の割合を示す円グラフを示す。データは、独立したクローン:野生型(2種のクローン)、ΔXRCC2(4種のクローン)およびΔXRCC3(2種のクローン)の分析からプールしたものである。図示されている突然変異に加えて、1方のΔXRCC2-DT40配列は、リーダーイントロン内に、供与体である偽遺伝子からの明らかに鋳型のない2bpの挿入を含み、1方のΔXRCC3-DT40配列も、リーダーイントロン内に一塩基対の欠失を含んでいた。
【0179】
ΔXRCC2-DT40の他の遺伝子座にある突然変異をパネル(c)に示す。円グラフには、再構成された重鎖の再構成されたVH(CDR1からJHの末端まで及ぶ272bp)、除外された対立遺伝子上の再構成されていないVλ1(458bp)、およびCλの近傍(Jλ-CλイントロンからCλの最初の132bpまで及ぶ425bp)内に突然変異を含むΔXRCC2-DT40細胞を1ヶ月増殖させたものから誘導される配列の割合を示す。既知のVH偽遺伝子配列の分析(Reynaudら, 1989)からは、再構成されたVHで見られる突然変異のすべてが遺伝子変異によるというわけではないことが示されており、そのことから、それらは点突然変異によるものであるがこの割り当てが完全に決定的であると見なすことはできないことが強く示唆される。これらのデータセットにおける突然変異の存在率は、VHでは1.6×10-3の突然変異/bp、再構成されていないVλ1では0.03×10-3、Cλでは0.13×10-3であり、これと比較して、ΔXRCC2-DT40内の再構成されたVλ1における点突然変異では2.0×10-3、野生型DT40内の再構成されたVλ1における点突然変異では0.13×10-3、およびバックグランドPCRエラーでは0.04×10-3である。
【0180】
Vλ1に入っている点突然変異の分布をパネル(d)に示す。ΔXRCC2-DT40のコンセンサスは大文字で示し、第1の塩基は、リーダーイントロンの76番目の塩基対に相当する。ΔXRCC3-DT40のコンセンサスに見られるバリエーションは、下にイタリック体の文字で示す。突然変異はコンセンサスの上に小文字で示し、ΔXRCC2-DT40に由来するものは黒で、ΔXRCC3-DT40に由来するものはミッドグレイ色で示す。点突然変異および不明確であるカテゴリーに分類される全ての突然変異を含めた。クローン性増殖については、先に記載(Takataら, 1998)されているようにして較正を行っており、したがって、それぞれの小文字は独立した突然変異事象を表わす。27の突然変異(そのため、158の元のデータベースから除去されている)の大部分は、7つの主要なホットスポットの1つに存在する。クローン性についての較正により何らかの歪みが生じる場合、ホットスポットの優先性(dominance)が過少に見積もられることになる。7つの主要なホットスポット(5以上の突然変異が蓄積していることによって同定される)のうち、5つは、黒枠で示される2本の鎖の一方にあるAGYコンセンサス配列と一致する。パネル(e)に示されるヌクレオチド置換の存在率(131の独立した事象からなるデータベースに対するパーセンテージとして表わされる)は、未選択のΔXRCC2-DT40およびΔXRCC3-DT40に由来する配列内の点突然変異から推定される。ΔXRCC2/ΔXRCC3のデータベースをそれぞれ分析した場合、同様の存在率パターンが示される。
【0181】
野生型細胞およびΔXRCC2/3-DT40細胞における自然発生的なVλの突然変異頻度は、サブクローニングの後で1ヶ月間増殖させた全ての(ソーティングされていない)DT40集団から再構成されたVλセグメントをPCR増幅することによって分析される。その結果から、実際に、ΔXRCC2細胞およびΔXRCC3細胞では、親DT40と比べて突然変異の自然発生的蓄積がより高レベルで起こっていることが明らかになる(図21a、b)。ΔXRCC2-DT40細胞では、突然変異は、Vλに約0.4×10-4/bp/世代の速度(分裂時間を約12時間とする)で蓄積し、この値は、構成的に突然変異を起こすヒト・バーキットリンパ腫系Ramosで見られるものとほぼ同じである。
【0182】
ヒトおよびマウスにおける胚中心B細胞での体細胞高頻度突然変異は、主に、再構成された免疫グロブリンのVHセグメントおよびVLセグメントを標的とする。同様の状況が、ΔXRCC2-DT40細胞での点突然変異にも当てはまる。したがって、生産的に再構成されたVH1遺伝子でも有意なレベルで見かけの点突然変異が見られる(図3c)。しかし、これは、全体的な突然変異誘発表現型を反映してはいない。何故ならば、突然変異の蓄積は、Cλでは、再構成されたVλの場合よりもずっと少なく、また除外された対立遺伝子上の再構成されていないVλでも少ないからであり、この場合、見かけの突然変異率はPCR増幅自体が原因となってバックグラウンドレベルを超えない(図21c)。
【0183】
ΔXRCC2-DT40細胞内のVλドメインでの突然変異の分布は、非常に非ランダムである。この突然変異は主に一ヌクレオチド置換であり、2本のDNA鎖の一方におけるAGY(Y=ピリミジン)コンセンサスに一致するホットスポットで選択的な蓄積を示す(図21d)。また、それらは圧倒的(96%)にG/Cで起こっている。ΔXRCC2-DT40細胞におけるこのG/Cに偏ったホットスポット集中型の高頻度突然変異は、ヌクレオチドのトランジションへの偏りは多少低いものの、培養したヒト・バーキットリンパ腫細胞において記載されているV遺伝子の高頻度突然変異のパターンならびにカエル、サメおよびMsh欠損マウスで生じるそのパターン(Radaら, 1998;Diazら, 2001)と非常に類似している。ヒトおよび正常なマウスにおいてin vivoで起こるIgV遺伝子の高頻度突然変異は、先に述べられているように、A/Tを標的とする機構と協働して作用するこのホットスポット集中型のG/Cに偏った成分により達成されると思われる(図21e)。
【0184】
したがって、DT40ニワトリ・ファブリキウス嚢リンパ腫系は、通常は遺伝子変換により低頻度のIgV多様化を示すが、Xrcc2またはXrcc3を欠失させることにより、高頻度の構成的なIgV遺伝子の体細胞突然変異(ヒトB細胞リンパ腫モデルで起こるものと本質的に類似している)を誘導することができる。このことは、IgV遺伝子の変換および高頻度突然変異が、一般的なDNA損傷を解決するための別の方法となり得るという以前の考え(Maizelsら, 1995;Weillら, 1996)を強く支持する。近年のデータから、損傷の始まりは二本鎖の破断であることがよくあるということが示唆され(Sale & Neuberger, 1998;Papavasilouら, 2000;Brossら, 2000)、したがって、Xrcc2およびXrcc3の両者が、DNA破壊の修復の組換え依存的経路に関与していることは重要と思われる(Liuら, 1998;Johnsonら, 1999;Pierceら, 1999;Brennermanら, 2000;Takataら, 2001)。事実、DT40細胞におけるIgV遺伝子の高頻度突然変異の同様の誘導は、その産物が破断の組換え依存的修復に関与する別の遺伝子(RAD51B)(Takataら, 2000)を欠失させることによって達成されるが、非相同的末端結合に関与するKu70およびDNA-PKCSの遺伝子を欠失させた場合には達成されない。図22に、DNA-PK、Ku70およびRad51Bが欠損しているDT40細胞におけるsIgM欠損変異体の分析を示す。1ヶ月間クローン性増殖した後のsIgM欠損変異体の生成頻度の彷徨解析をパネル(a)に示す。比較のために、野生型およびΔXRCC2のDT40を用いて得られた中央値を入れてある。図示されている各種のタイプの突然変異を含む2種のsIgM+ Rad51B欠損DT40クローンから誘導したsIgM欠損変異体から増幅したVλ配列の割合を示す円グラフをパネル(b)に示す。さらに、1つの配列は9bpの欠失を含み、1つは4bpの重複を含み、1つは一塩基対の挿入を含んでいた。
【0185】
しかし、この結果は、Xrcc2の不在下で、通常は遺伝子変換により解消される損傷が、代わりに体細胞高頻度突然変異をもたらすプロセスによって解消されることを単に示唆しているわけではない。第1に、ΔXRCC2-DT40細胞は、レベルは多少低いものの、IgV遺伝子の変換を行う能力を保持している(図21b)。第2に、ΔXRCC2-DT40細胞における高頻度突然変異の頻度は、親のDT40系における遺伝子変換の頻度よりもほぼ一桁高い。したがって、正常なDT40細胞では、IgV遺伝子の損傷のほんの一部が、上流の偽遺伝子由来の鋳型のある修復に供されるにすぎず、そのため、遺伝子変換事象が見られることになると思われる。本発明者らは、損傷の大部分が、姉妹染色分体上に位置する同一のV遺伝子を鋳型として用いる組換え修復に供され、したがって、それらは「隠されて」いると考える。これは、高頻度突然変異を起こす哺乳動物B細胞における検出可能なIgV遺伝子の破壊がG2/S期に限定されることを見出したPapavasiliouおよびSchatz(2000)の知見と一致する。本発明者らは、Xrcc2、Xrcc3またはRad51Bが存在しない場合は、この「隠された」姉妹染色分体依存的組換え修復が逆用され、その結果として高頻度突然変異が起こることを提唱する。この高頻度突然変異が、Xrcc2/3が存在しない場合に姉妹染色分体依存的組換え修復が間違いを起こしやすくなることを反映しているのかどうか、それとも、それがそのような修復の阻害を反映しており、それにより別の鋳型のない破壊解消機構を示しているのかどうか、ということは、検討を要する問題である。この問題は、高頻度突然変異の機構を理解する上で重要であるばかりでなく、Rad51パラログ体の生理学的機能の見通しを与えてくれる。
【0186】
実施例9:Δxrcc2 DT40 IgMの親和性成熟
数ヶ月間増殖させたXrcc2欠損DT40細胞の集団を用いて、これらの細胞におけるユニークなVHDJH/VLJL再構成に対する高頻度突然変異の作用が、被験抗原のかなりの割合に対して抗体の成熟系統を進化させるのに十分な機能的多様性をもたらすことができるか否かを判定した。選別には2つの方法を用いた。第1の方法では、細胞を、FITC-ストレプトアビジンを各種のビオチン化抗原(カゼイン、インスリン、オボアルブミン(Ova)、サイログロブリン(Tg)およびラット・モノクローナル抗体(Ab))と混合することにより形成される可溶性凝集物と共にインキュベートした。次に、結合している変異体をフローサイトメトリーにより濃化した。あるいはまた、変異体は、S. アウレウス(S. aureus)のプロテインAまたはヒト血清アルブミン(HAS)で被覆した磁性ビーズを用いて選別した。試験した抗原はいずれも、検出可能な親DT40またはその分泌されたIgMに対する結合を示さなかった。選別を6ラウンド連続して行った後で、本質的に3つのタイプの結果が得られた。すなわち、インスリンおよびカゼインの場合、結合している変異体が濃化されたという証拠はなかった。ラット・モノクローナル抗体およびプロテインAの場合、特異的な結合体が得られた。HAS、OvaおよびTgの場合、結合体は得られたが、これらは様々な程度で多重反応性(polyreactivity)を示した。したがって、選別をビオチン化ラットIgG mAb S7とFITC-ストレプトアビジンとの凝集物を用いて行った場合(図23)、特異的な結合は、既に第3ラウンドまでに示された。それに続く濃化は、フィコエリトリン(PE)に直接コンジュゲートさせたラットIgG S7 mAbを用いて行った。第6ラウンドで得られた細胞(DT-Ab6)は、種々の他の試薬で染色されないことにより判定される通り、ラットIgG S7 mAbに特異的であった。培養上清のELISAから、DT-Ab6細胞によるS7 mAbの結合がDT-Ab6 IgM自身によりもたらされることが実証された(図23b)。このDT-Ab6 IgMは、実際に抗-S7イディオタイプである。それは、精製されたS7 mAbを認識する(そして、S7ハイブリドーマ透過化細胞を染色するのに使用できる)が、この相互作用は、他のラット免疫グロブリンによる競合を受けない(図23c、d)。DT-Ab6細胞は、高希釈率のPES7 mAbコンジュゲートで染色できた。このことから、高親和性の相互作用が示唆される。分子PE-S7 mAb:DT-Ab6細胞を固定比率で、しかし容量を変えて染色を行うことにより、5.8 nMの範囲の親和性が推定された(図23f)。
【0187】
配列分析から、DT-Ab6 VH/VLが、親DT40の配列と比較して、合計19のアミノ酸置換を含むことが判明した(図23g)。
【0188】
ビオチン化プロテインAで被覆したストレプトアビジンビーズを用いて行ったビーズ選別では、結合している変異体は第2ラウンドまでに現われた(DT-P2;図24a)。しかし、DT-P2集団中の細胞の多くは、ストレプトアビジンもプロテインAも提示していない種々の他のタイプのビーズにも結合した、という点に問題があった。プロテインAを直接コンジュゲートさせたトシル化ビーズを用いて更に濃化したところ、FITCコンジュゲートプロテインA凝集物で染色可能な細胞(DT-P4)の集団が得られた(図24b)。フローサイトメトリーの使用によるこの凝集物への結合に対する一連の選別により、凝集していないFITC-プロテインAで僅かに染色可能な集団(DT-P9)が得られた。次に、FITC-プロテインAを用いて更にソーティングしたところ、もっと明るく染色される子孫が得られた(図24b)。DT-P14細胞により(ならびにそのいくつかの前駆細胞により)分泌されたIgMは、ELISAおよび免疫沈降法の両者により判定したところ、プロテインAに良好に結合した(図24c、d)。放射標識したプロテインAを用いた直接結合アッセイから、DT-P9とDT-P14の間で親和性の実質的な増大が起こったことが示され(図24e)、これはフローサイトメトリー分析(図24b)と一致する。この増大は、完全に、VHのCDR2に隣接したAla→Val置換によるものであり(図24h)、これにより、プロテインAに対する見掛けの親和性が0.32nMであるIgMが生じた(図24f)。興味深いことに、DT-PのIgMとプロテインAとの相互作用は、十分に特性付けが行われているプロテインAとウサギIgGのFc部分との相互作用とは、DT-P/プロテインAの相互作用がIgM分子のC部分ではなくV部分とのものである(突然変異はCμではなくVHで見られる)という事実、ならびに、高濃度のウサギIgGはFITC-プロテインAによるDT-P4/P9細胞の染色を阻害しないので、おそらくプロテインA上の異なる部位がDT-P IgMおよびウサギIgGとの相互作用に用いられるという事実において異なる(図24g)。実際に、染色の増強が見られ、おそらくこれは、ウサギIgGによるプロテインAの凝集によるものと考えられる。
【0189】
DT-P14のVH/VLの配列から、親DT40配列と比較して、17のアミノ酸置換が明らかになっている。これらの突然変異のうち5つは、ラット・イディオタイプ特異的変異体DT-Ab6と共通する。しかし、この5つの突然変異のうち少なくとも4つは、(Xrcc2欠損DT40細胞のプールにおける共通の突然変異した前駆体からの派生していることを反映しているのではなく)その2つの系統において独立して起こったはずである。何故ならば、DT-P14とDT-Ab6とに共通するこれら4つの突然変異体は、DT-P4では見られないからである。置換を繰り返すこの傾向は、DT40における突然変異の殆どが、ホットスポット集中型のGCに偏った第1段階の突然変異(本発明者らがA/Tに偏った第2段階によるものとした突然変異の範囲の一部を欠いている;SaleおよびNeuberger, 1998;Radaら, 1998)に限定されていることをある程度反映していると思われるが、それらが、抗体構造を成熟させやすくすることによって利点を付与している可能性もある。Xrcc2欠損DT40細胞は(体細胞高頻度突然変異よりもずっと低い頻度ではあるが)IgV遺伝子の変換を行う能力を保持しているが、DT-P14細胞およびDT-Ab6細胞におけるV遺伝子の配列(図24h)を生殖細胞系のVセグメントの配列(Reynaudら, 1987, 1989)と比較したところ、その変化は、(全部でないとしても)大部分が遺伝子変換ではなく点突然変異によるものであることが判明したことは注目に値する。HSA誘導体化カルボキシル化磁性ビーズを用いた選別を6ラウンド行った後で得られた細胞(DT-H6)は、FITC-HASだけでなく、FITC-Tg、FITC-Ovaおよびサイクローム-コンジュゲートストレプトアビジンによっても染色された(DT-H6細胞集団がこれらの他の抗原に暴露されたことはない、という事実にもかかわらず)。HASを用いて更に選別したところ、多重特異性染色(polyspecific staining)の明るさが単純に増大した(図25a)。FITC-ストレプトアビジンとビオチン化Tgもしくはビオチン化Ovaとの複合体を用いたフローサイトメトリーによる濃化に供した細胞集団では、類似しているが異なる多重特異性パターンが示された(図25a、d)。
【0190】
亜集団を分析したところ、この見掛けの多重特異性が細胞の異質性を反映していないことが示された。sIgM欠損変異体には抗原結合活性が欠けているので、多重特異的染色は表面IgM自体によって仲介された。更に、抗原結合は、培養物上清のELISAにより容易に検出される(図25b、c)。
【0191】
多重反応性抗体はヒトおよびマウスにおいて(血清中およびハイブリドーマ間で)十分に報告されており、それらが単一特異性(monospecificity)が進化するための良好な開始点を構成するか否かという問題が提起されてきている(CasaliおよびSchettino, 1996;BouvetおよびDighiero, 1998)。in vitroでの選別系でも同じ問題が出てきている(自己免疫を回避するという制約はないが)。したがって、Ovaに対する特異性を増大させる方向にDT-O6集団を進化させることが可能か否かを、フローサイトメトリーによるFITC-Ovabright/サイクローム-ストレプトアビジンdull細胞の濃化によって試験した。ソーティングを複数ラウンド行った後、FITC-Ovaに対してより高い特異性を示す細胞を得た(図25d)。多重特異的結合集団が親DT40集団よりも良好な特異性進化の開始点を提供するか否かを確認することは興味深い。
【0192】
ここで示す結果から、高頻度突然変異を起こす細胞系が、in vitroでの抗体の誘導および反復的な成熟の双方に使用可能であることがはっきりと実証される。DT40の遺伝学的な扱いやすさを考慮すれば、この用途をトランスフェクトしたIgV遺伝子にまで拡大し、そうすることにより既存の抗体の親和性を成熟させることが可能である。更に、高頻度突然変異機構は、再構成されたIgVセグメントの代わりに配置された異種の遺伝子を標的とすることができるので(Yelamosら, 1995)、この戦略を、他のリガンド/受容体ペアの成熟にまで拡張できる公算は十分にあると思われる。抗体のde novo選別については、単一のVH/VL再構成に体細胞高頻度突然変異を作用させることにより、試験した7種の抗体のうち2種に対して高親和性の結合物を選別して成熟させること、ならびにそれ以外のもののうち3種に対する初期の低親和性結合の兆候(成熟可能ではないが)を得ることが可能なレパートリーが得られたことは注目すべきである。これは、抗原親和性が非常に低くても、選別を開始するには十分であるという事実を反映している。かくして、これらの結合部位は成熟可能である。明らかに、この方法を、高親和性のヒト・モノクローナル抗体のin vitro産生が可能になるように外挿したい場合は、DT40の遺伝学的易操作性を利用して、2つ以上のVH/VL再構成を含む一次レパートリーを作製することが有利であろう。ここで得られた結果から、この再構成の一次レパートリーはin vivoで用いられる一次レパートリーよりも数桁少ないことが示される。
【0193】
実施例10:天然に存在する、構成的に高頻度突然変異を起こすEBVポジティブBL細胞系の単離
天然に存在するEBV+BL細胞系を調べたところ、それら(例えば、Akata, BL74, Chep, Daudi, Raji,およびWan)の多くの中に、明らかに同定可能なsIgM欠損変異体の集団は存在しないことが判明した。しかし、これらEBV+細胞系のうち2つ(ELI-BLおよびBL16)において明確なsIgM−/low集団が確認され、このことから、本来備わっている高頻度突然変異能であることが示唆される。Ramos、EHRB、ELI-BLおよびBL16のsIgM発現プロファイルを図26aに示す。sIgM−/low細胞集団は枠で囲み、その中の細胞のパーセンテージを示す。各ドットが1個の細胞を示す。BL16における相当大きなsIgM−/low集団の一部は、それほど強くは染色にポジティブではない細胞によるものであり、これもまた、彷徨解析を妨害したことに注意されたい。ELI-BLは2型のEBVを含み、胚中心B細胞に類似しており、EBNA1および非コードEBERおよびBam A RNAのみから構成される潜伏遺伝子レパートリーを発現する(Roweら,1987)。BL16もまた2型ウイルスを含んでいるが、ELI-BLとは違って、よりLCLに類似していると思われ、完全に潜伏している遺伝子のレパートリーを発現する(Rooneyら, 1984;Roweら, 1987)。
【0194】
ELI-BLおよびBL16の培養物中に明確なsIgM−/low集団が見られるが、これらの変異体が真の高頻度突然変異に起因しているのか否かを検討することは重要である。これは、彷徨解析により検討した。簡単に説明すると、(サブ)クローンを24ウェルまたは48ウェルのプレートに移し、新鮮な培地と共に3〜8週間維持し、細胞(1〜2.5×105個)をPBS/3%FBS中で2回洗浄し、適切な抗体もしくは抗体の組合せ(後記)で染色し(氷上で30分間)、再度洗浄した後、少なくとも104個の細胞をフローサイトメトリー(FACSCalibur、Becton Dickinson)により分析することにより分析した。使用した抗体は、R-フィコエリトリン-コンジュゲートしたヤギ抗-ヒトIgM(μ鎖特異的;Sigma)、フルオレセインイソチオシアネート(FITC)-コンジュゲートしたマウス・モノクローナル抗-Ramosイディオタイプ[ZL16/1(Zhangら,1995);M. CraggおよびM. J. Glennie(Tenovus Research Laboratory, Southampton)から恵贈]、およびFITC-コンジュゲートしたヤギ抗-マウスIgM(Southern Biotechnology Associates, Inc.)とした。データを得て、CellQuestソフトウェア(Becton Dickinson)を用いて解析した。
【0195】
特に指示がない限り、彷徨解析で比較した細胞は並行して誘導し、培養し、分析した。同様にして誘導した多くの(サブ)クローンにおけるsIgM欠損変異体のパーセンテージ中央値(平均値ではない)を細胞の体細胞高頻度突然変異能の指標として用いて、早期突然変異事象の影響を最低限に抑える。ELI-BLサブクローンの彷徨解析を行ったところ、sIgM−/low変異体が、実際に、in vitro培養の際に実際に高頻度で生成されていることが判明し(図26b;それぞれの×印は、単一のサブクローンを1ヶ月間増殖させた後のsIgM−/lowウィンドウに含まれる細胞のパーセンテージを表わす;パーセンテージ中央値を示す)、BL16サブクローンの場合、VH配列の分析から、この不安定さが体細胞高頻度突然変異を反映していることが確認された(図26c)。塩基置換突然変異は、大文字のトリプレットで示されている338bpのコンセンサスDNA配列の上に小文字で示す。相補性決定領域および部分PCRプライマー配列には、それぞれ下線をひき、太字にしてある。対応するアミノ酸配列は、大文字1文字によって示す。このコンセンサス配列は、GenBankのエントリー配列gi.2253343とは2つの位置で異なっている[TCA(Ser20)−TCTおよびAGC(Ser55)−ACC(Thr)]。
【0196】
複数の塩基置換突然変異を有する数種の配列を含むVH配列の相当な多様性および全体として高いVH突然変異の頻度から、BL16において高頻度突然変異が進行中であることが示された。更に、比較的少数のVH配列しかサンプリングしなかったにもかかわらず、1つの系統的関係が推定できた[Gly54での第1の突然変異(GGT−GAT);Val92での第2の突然変異(GTG−ATG)]。最後に、Ramosと同様に、BL16のVHの塩基置換突然変異の多く(24/33、すなわち73%)がGまたはCのヌクレオチドで起こり、相補性決定領域内(図26cの下線部)に集中していた。したがって、2種の天然のEBV+ BL細胞系(一方は限定的な潜伏遺伝子レパートリーを発現し、他方は全ての組合せを発現する)では、進行中の高頻度突然変異の幾つかの特徴が区別できる。したがって、体細胞高頻度突然変異が、EBVの存在下でも衰えることなく進行可能であることは明らかであった。
【0197】
参考文献
Adetugbo, K., Milstein, C. and Secher, D.S. (1977). Molecular analysis of spontaneous somatic mutants. Nature 265, 299-304.
Bahler, D.W. and Levy, R. (1992). Clonal evolution of a follicular lymphoma: Evidence for antigen selection. Proc. Natl. Acad. Sci. USA 89, 6770-6774.
Bass, S., R. Greene, and J.A. Wells. (1990). Hormone Phage: An Enrichment Method for Variant Proteins With Altered Binding Properties. Proteins. 8, 309-314.
Berek, C. and Milstein, C. (1988). The dynamic nature of the antibody repertoire. Immunol. Rev. 105, 5-26.
Betz, A. G., Milstein, C., Gonzalez-Fernandez, A., Pannell, R., Larson, T. and Neuberger, M. S., Cell 1994. 77: 239-248.
Betz, A.G., Neuberger, M.S. and Milstein, C. (1993). Discriminating intrinsic and antigen-selected mutational hotspots in immunoglobulin V genes. Immunol. Today 14, 405-411.
Bezzubova, O., Silbergleit, A., Yamaguchi-Iwai, Y., Takeda, S. & Buerstedde, J. M. Reduced X-ray resistance and homologous recombination frequencies in a RAD54-/- mutant of the chicken DT40 cell line. Cell 89, 185-193. (1997).
Bonfield, J.K., Smith, K.F. and Staden, R. (1995). A new DNA-sequence assembly program. Nucleic Acids Res. 23, 4992-99.
Bouvet J.P. & Dighiero, G. From natural polyreactive autoantibodies to a la carte monoreactive antibodies to infectious agents: is it a small world after all? Infect. Immun. 66, 1-4 (1998).
Braeuninger, A., Kuppers, R., Strickler, J.G., Wacker, H.-H., Rajewsky, K. and Hansmann, M.-L. (1997). Hodgkin and Reed-Sternberg cells in lymphocyte predominant disease represent clonal populations of germinal center-derived tumor B cells. Proc. Natl. Acad. Sci. USA 94, 9337-9342.
Brenneman, M. A., Weiss, A. E., Nickoloff, J. A. & Chen, D. J. XRCC3 is required for efficient repair of chromosome breaks by homologous recombination. Mutat Res 459, 89-97. (2000).
Bross, L., Fukita, Y., McBlane, F., Demolliere, C., Rajewsky, K. & Jacobs, H. DNA double-strand breaks in immunoglobulin genes undergoing somatic hypermutation. Immunity 13, 589-597. (2000)
Bruggemann, M., Radbruch, A. and Rajewsky, K. (1982). Immunoglobulin V region variants in hybridoma cells. I. Isolation of a variant with altered idiotypic and antigen binding specificity. EMBO J. 1, 629-634.
Buerstedde, J. M., Reynaud, C.A., Humphries, E.H., Olson, W., Ewert, D.L. & Weill, J.C. Light chain gene conversion continues at high rate in an ALV-induced cell line. Embo J 9, 921-927. (1990)
Capizzi and Jameson, (1973) Mutat. Res. 17:147-8
Casali P. & Schettino, E.W. Structure and function of natural antibodies. Curr. Top. Microbiol. Immunol. 10, 167-179 (1996).
Chapman, C.J., Mockridge, C.I., Rowe, M., Rickinson, A.B. and Stevenson, F.K. (1995). Analysis of VH genes used by neoplastic B cells in endemic Burkitt’s lymphoma shows somatic hypermutation and intraclonal heterogeneity. Blood 85, 2176-2181.
Chapman, K.B. and Szostak, J.W. (1994) Curr. op. Struct. Biol., 4, 618-622.
Chui, Y.-L., Lozano, F., Jarvis, J.M., Pannell, R. and Milstein, C. (1995). A reporter gene to analyse the hypermutation of immunoglobulin genes. J. Mol. Biol. 249, 555-563.
Clackson, T. and Wells, J.A. (1994) Trends Biotechnol, 12, 173-84.
Coffino, P. and Scharff, M.D. (1971). Rate of somatic mutation in immunoglobulin production by mouse myeloma cells. Proc. Natl. Acad. Sci. USA 68, 219-223.
Cui, X., Brenneman, M., Meyne, J., Oshimura, M., Goodwin, E.H. & Chen D.J. The XRCC2 and XRCC3 repair genes are required for chromosome stability in mammalian cells. Mutat Res 434, 75-88. (1999).
Cull, M.G., Miller, J.F. and Schatz, P.J. (1992) Proc Natl Acad Sci U S A, 89, 1865-9.
Deans, B., Griffin, C. S., Maconochie, M. & Thacker, J. Xrcc2 is required for genetic stability, embryonic neurogenesis and viability in mice. Embo J 19, 6675-6685. (2000).
Denepoux, S., Razanajaona, D., Blanchard, D., Meffre, G., Capra, J.D., Banchereau, J and Lebecque, S. (1997). Induction of somatic mutation in a human B cell line in vitro. Immunity 6, 35-46.
Diaz, M., Flajnik, M. F. & Klinman, N. Evolution and the molecular basis of somatic hypermutation of antigen receptor genes. Philos Trans R Soc Lond B Biol Sci 356, 67-72. (2001)
Drake, J.W. (1998) Genetics 148:1667-1686.
Ellington, A.D. and Szostak, J.W. (1990) Nature, 346, 81822.
Gold, L., Polisky, B., Uhlenbeck, 0. and Yarus, M. (1995) Annu Rev Biochem, 64, 763-97.
Goossens, T., Klein, U. and Kuppers, R. (1998). Frequent occurrence of deletions and duplications during somatic hypermutation: Implications for oncogenic translocations and heavy chain disease. Proc. Natl. Acad. Sci. USA 95, 2463-2468
Goyenechea, B., Klix, N., Yelamos, J., Williams, G. T., Riddell, A., Neuberger, M. S. and Milstein, C., EMBO J. 1997. 16, 3987-3994
Green, N.S., Lin, M.M. and Scharff, M.D. (1998). Immunoglobulin hypermutation in cultured cells. Immunol. Rev. 162, 77-87.
Green, R. and Szostak, J.W. (1992) Science, 258, 1910-5.
Griffin, C. S., Simpson, P. J., Wilson, C. R. & Thacker, J. Mammalian recombination-repair genes XRCC2 and XRCC3 promote correct chromosome segregation. Nat Cell Biol 2, 757-761. (2000)
Jain, R., Roncella, S., Hashimoto, S., Carbone, A., Franco di Celle, P., Foa, R., Ferrarini, M and Chiorazzi, N. (1994). A potential role for antigen selection in the clonal evolution of Burkitt’s lymphoma. J. Immunol. 153, 45-52.
Johnson, R. D., Liu, N. & Jasin, M. Mammalian XRCC2 promotes the repair of DNA double-strand breaks by homologous recombination. Nature 401, 397-399. (1999).
Jolly, C.J., Klix, N. and Neuberger, M.S. (1997). Rapid methods for the analysis of immunoglobulin gene hypermutation: application to transgenic and gene targeted mice. Nucleic Acids Res. 25, 1913-1919.
Jolly, C.J., Wagner, S.D., Rada, C.A., Klix, N., Milstein, C. and Neuberger, M.S. (1996). The targeting of somatic hypermutation. Semin. Immunol. 8, 159-168.
Joyce, G.F. (1994) Curr. op. Structural Biol., 4, 331-336.
Kim, S., Humphries, E. H., Tjoelker, L., Carlson, L. & Thompson, C. B. Ongoing diversification of the rearranged immunoglobulin light-chain gene in a bursal lymphoma cell line. Mol Cell Biol 10, 3224-3231. (1990)
Klein, G., Giovanella, B., Westman, A., Stehlin, J. and Mumford, D. (1975). An EBV negative cell line established from a American Burkitt lymphoma; receptor characteristics, EBV infectability and permanent conversion into EBV-positive sublines by in vitro infection. Intervirology 5, 319-334.
Klix et al., (1998) Eur. J. Immunol. 28:317-326.
Liu, N., Lamerdin, J.E., Tebbs, R.S., Schild, D., Tucker, J.D., Shen, M.R., Brookman, K.W., Siciliano, M.J., Walter, C.A., Fan, W., Narayana, L.S., Zhou, Z.Q., Adamson, A.W., Sorenson, K.J., Chen, D.J., Jones, N.J. & Thompson, L.H. XRCC2 and XRCC3, new human Rad51-family members, promote chromosome stability and protect against DNA cross-links and other damages. Mol Cell 1, 783-793. (1998).
Lundberg, K.S., et al., (1991) Gene 108:1-6.
Luria and Delbreck., (1943) Genetics 28:491-511
Maizels, N. Somatic hypermutation: how many mechanisms diversify V region sequences? Cell 83, 9-12. (1995).
Mattheakis, L.C., Bhatt, R.R. and Dower, W.J. (1994) Proc Natl Acad Sci U S A, 91, 9022-6.
Matthias, et al., (1989) NAR 17, 6418.
Max, E. E., Maizel, J. V. and Leder, P., J. Biol. Chem. 1981. 256: 5116-5120.
McCafferty, J., Griffiths, A.D., Winter, G. and Chiswell, D.J. (1990) Nature, 348, 552-4.
McCormack, W. T., Hurley, E. A. & Thompson, C. B. Germ line maintenance of the pseudogene donor pool for somatic immunoglobulin gene conversion in chickens. Mol Cell Biol 13, 821-830. (1993)
McKean, D., Huppi, K., Bell, M., Staudt, L., Gerhard, W. and Weigert, M. (1984). Generation of antibody diversity in the immune response of BALB/c mice to influenza virus haemagglutinin. Proc. Natl. Acad. Sci. USA 81, 3180-3184.
Moore, M.J. (1995) Nature, 374, 766-7.
Neuberger, M.S. and Milstein, C. (1995). Somatic hypermutation. Curr. Opin. Immunol. 7, 248-254.
Papavasiliou, F. N. & Schatz, D. G. Cell-cycle-regulated DNA double-stranded breaks in somatic hypermutation of immunoglobulin genes. Nature 408, 216-221. (2000).
Parham, P. (ed). (1998). Somatic hypermutation of immunoglobulin genes. Immunological Reviews, Vol. 162 (Copenhagen, Denmark: Munksgaard).
Pierce, A. J., Johnson, R. D., Thompson, L. H. & Jasin, M. XRCC3 promotes omology-directed repair of DNA damage in mammalian cells. Genes Dev 13, 2633-2638. (1999).
Rada, C., Ehrenstein, M. R., Neuberger, M. S. & Milstein, C. Hot spot focusing of somatic hypermutation in MSH2-deficient mice suggests two stages of mutational targeting. Immunity 9, 135-141. (1998).
Ratech, H. (1992). Rapid cloning of rearranged immunoglobulin heavy chain genes from human B-cell lines using anchored polymerase chain reaction. Biochem. Biophys. Res. Commun. 182, 1260-1263
Reynaud, C. A., Anquez, V., Grimal, H. & Weill, J. C. A hyperconversion mechanism generates the chicken light chain preimmune repertoire. Cell 48, 379-388. (1987).
Reynaud, C.A., Dahan, A., Anquez, V. & Weill, J.C. Somatic hyperconversion diversifies the single VH gene of the chicken with a high incidence in the D region. Cell 59, 171-183 (1989)
Rogozin, I.B. and Kolchanov, N.A. (1992). Somatic hypermutation of immunoglobulin genes. II. Influence of neighbouring base sequences on mutagenesis. Biochem. Biophys. Acta 1171, 11-18.
Rooney, C. M., A. B. Rickinson, D. J. Moss, G. M. Lenoir, and M. A. Epstein. 1984. Paired Epstein-Barr virus-carrying lymphoma and lymphoblastoid cell lines from Burkitt's lymphoma patients: comparative sensitivity to non-specific and to allo-specific cytotoxic responses in vitro. Int J Cancer. 34:339-48
Rowe, M., D. T. Rowe, C. D. Gregory, L. S. Young, P. J. Farrell, H. Rupani, and A. B. Rickinson. 1987. Differences in B cell growth phenotype reflect novel patterns of Epstein-Barr virus latent gene expression in Burkitt's lymphoma cells. Embo J. 6:2743-51
Sale, J. E. & Neuberger, M. S. TdT-accessible breaks are scattered over the immunoglobulin V domain in a constitutively hypermutating B cell line. Immunity 9, 859-869. (1998).
Sharpe et al., (1991) EMBO J. 10:2139-2145.
Smith, G.P. (1985) Science, 228, 1315-7.
Takata, M., Sasaki, M.S., Sonoda, E., Fukushima, T., Morrison, C., Albala, J.S., Swagemakers, S.M., Kanaar, R., Thompson, L.H. & Takeda, S. The Rad51 paralog Rad51B promotes homologous recombinational repair. Mol Cell Biol 20, 6476-6482. (2000).
Takata, M., Sasaki, M.S., Sonoda, E., Morrison, C., Hashimoto, M., Utsumi, H., Yamaguchi-Iwai, Y., Shinohara, A. & Takeda, S. Homologous recombination and non-homologous end-joining pathways of DNA double-strand break repair have overlapping roles in the maintenance of chromosomal integrity in vertebrate cells. Embo J 17, 5497-5508. (1998)
Takata, M., Sasaki, M.S., Tachiiri, S., Fukushima, T., Sonoda, E., Schild, D., Thompson, L.H. & Takeda, S. Chromosome Instability and Defective Recombinational Repair in Knockout Mutants of the Five Rad51 Paralogs. Mol Cell Biol 21, 2858-2866. (2001).
Tomlinson, I.M. (1997). V Base database of human antibody genes. Medical Research Council, Centre for Protein Engineering, UK. http://www.mrc-cpe.cam.ac.uk/
Tuerk, C. and Gold, L. (1990) Science, 249, 505-10.
Wabl, M., Burrows, P.D., von Gabain, A. and Steinberg, C.M. (1985). Hypermutation at the immunoglobulin heavy chain locus in a pre-B-cell line. Proc. Natl. Acad. Sci. USA 82, 479-482.
Wagner, S.D., Milstein, C. and Neuberger, M.S. (1995). Codon bias targets mutation. Nature 376, 732.
Weill, J. C. & Reynaud, C. A. Rearrangement/hypermutation/gene conversion: when, where and why? Immunol Today 17, 92-97. (1996)
Wilson, P.C., de Boutellier, O., Liu, Y-J., Potter, K., Banchereau, J., Capra, J.D. and Pascual, V. (1998). Somatic hypermutation introduces insertions and deletions into immunoglobulin V genes. J. Exp. Med. 187, 59-70.
Wu, H., and Kaartinen, M., (1995) Scand. J. Immunol. 42:52-59.
Wu, H., Pelkonen, E., Knuutila, S and Kaartinen M. (1995). A human follicular lymphoma B cell line hypermutates its functional immunoglobulin genes in vitro. Eur. J. Immunol. 25, 3263-3269.
Yamaguchi-Iwai, Y., Sonoda, E., Buerstedde, J.M., Bezzubova, O., Morrison, C., Takata, M., Shinohara, A. & Takeda, S. Homologous recombination, but not DNA repair, is reduced in vertebrate cells deficient in RAD52. Mol Cell Biol 18, 6430-6435. (1998).
Yelamos, J., Klix, N., Goyenechea, B., Lozano, F., Chui, Y.L., Gonzalez Fernandez, A., Pannell, R., Neuberger, M.S. & Milstein, C (1995). Targeting of non-Ig sequences in place of the V segment by somatic hypermutation. Nature 376, 225-229.
Zhang, L., R. R. French, H. T. Chan, T. L. O'Keefe, M. S. Cragg, M. J. Power, and M. J. Glennie. 1995. The development of anti-CD79 monoclonal antibodies for treatment of B-cell neoplastic disease. Ther Immunol. 2:191-202
Zhu, M., Rabinowitz, J.L., Green, N.S., Kobrin, B.J. and Scharff, M.D. (1995) A well-differentiated B cell line is permissive for somatic mutation of a transfected immunoglobulin heavy-chain gene. Proc. Natl. Acad. Sci. USA 92, 2810-2814.
Zou, X., Xian, J., Popov, A. V., Rosewell, I. R., Muller, M. and Bruggemann, M., Eur. J. Immunol. 1995. 25: 2154-62.
【技術分野】
【0001】
本発明は、抗体産生細胞の生来の体細胞高頻度突然変異能を利用することによる遺伝子または遺伝子産物における多様性の生成方法、ならびに所定の遺伝子産物において多様性を生成できる細胞系に関する。
【背景技術】
【0002】
遺伝子産物において多様性を生成させるための多くのin vitro法は、非常に多数の突然変異体を作製し、次にそれらを効果的な選別技術を用いて選別することに基づいている。例えば、ファージ提示法は、提示されたタンパク質の選別を可能にする媒介手段が提供された場合に非常にうまくいっている(Smith, 1985; Bassら, 1990;McCaffertyら, 1990;概説としては、ClacksonおよびWells, 1994を参照されたい)。同様に、特定のペプチドリガンドが、lacリプレッサーLaclのC末端に連結されたペプチドの大きなライブラリーを用いる親和性選別法により、受容体への結合について選別されている(Cullら, 1992)。大腸菌で発現させた場合、リプレッサータンパク質は、そのリガンドを、コード化プラスミド上のlacオペレーター配列への結合によって該プラスミドに物理的に連結させる。更に、完全にin vitroのポリソーム提示系についても報告されており(Mattheakisら, 1994)、その場合、新生ペプチドはリボソームを介してそれらをコードするRNAに物理的に結合される。
【0003】
in vivoでは、抗体特異性の一次レパートリーは、免疫グロブリンV、DおよびJ遺伝子セグメントの結合を含むDNA再構成のプロセスにより作製される。マウスおよびヒトでは、抗原と出会った後、その抗原により起動されたB細胞内で再構成されたV遺伝子が、今度は体細胞高頻度突然変異による第2段階の多様化を起こす。この高頻度突然変異によって第二のレパートリーが生成され、そのレパートリーから良好な結合特異性が選別され、それによって液性免疫応答の親和性成熟が可能になる。
【0004】
現在までの人為的な選別系は、ひとえに、第一次の突然変異および選別に基づくものであり、これは、概念としては、所望の活性を有する遺伝子産物の選別がなされる遺伝子産物突然変異体の「固定された」レパートリーが得られるという点で、自然の抗体産生で起こるV-D-J再構成の第1段階と似ている。
【0005】
in vitroでのRNAの選別および進化(EllingtonおよびSzostak, 1990)は、SELEX(指数的濃化(enrichment)によるリガンドの体系的進化)(TuerkおよびGold, 1990)とも呼ばれることがあり、これは結合活性と化学的活性の両者についての選別を可能にするが、但し、核酸についてしかできない。選別を結合について行う場合、核酸のプールを固定化された基質と共にインキュベートする。未結合のものを洗い流し、次に結合しているものを遊離させ、増幅し、このプロセス全体を反復段階として繰り返して、より良好な結合配列を濃化する。この方法は、触媒作用を持つRNAおよびDNAの単離にも適用できる(GreenおよびSzostak, 1992;概説については、ChapmanおよびSzostak, 1994;Joyce, 1994;Goldら, 1995;Moore, 1995を参照されたい)。したがって、SELEXでは、所望の活性を繰り返し工程により向上させることができるが、その範囲は核酸の調製に限られている。
【0006】
しかし、人為的な選別系は、自然の免疫系の場合とは違って、手軽な形態の「親和性成熟」や繰り返し工程によるレパートリーの作製・開発にはあまり適さない。この理由の1つは、突然変異をそれらが必要とされる分子の領域にターゲティングすることは難しいので、それに続いて突然変異と選別を繰り返しても、活性が向上した分子を十分な効率で単離することはできないからである。
【0007】
自然の抗体産生において親和性成熟の際に起こる体細胞高頻度突然変異プロセスについての知見の多くは、in vivoでの高頻度突然変異の際に起こった突然変異の分析から導き出されたものである(概説については、NeubergerおよびMilstein, 1995;WeillおよびReynaud, 1996;Parham, 1998を参照されたい)。これらの突然変異の大部分は、段階的に導入された一ヌクレオチド置換である。それらは、(特徴的なホットスポットと共にではあるが)再構成されたVドメイン全体に散らばっており、それら置換は、塩基のトランジションに偏りを示す。それらの突然変異の殆どは、(B細胞の分化・増殖の他の段階の間ではなく)胚中心でのB細胞増殖の間に蓄積し、その場合、高頻度突然変異期の間のV遺伝子へのヌクレオチド置換の組み込み率は10-4〜10-3/bp/世代であると見積もられる(McKeanら, 1984; Berek & Milstein, 1988)。
【0008】
リンパ球様細胞系によって高頻度突然変異を研究するための扱い易い系が得られる可能性について、何年も前から検討されてきた(CoffinoおよびScharff, 1971;Adetugboら, 1977;Bruggemannら, 1982)。明らかに、重要なことは、研究対象の細胞系におけるV遺伝子の突然変異率が、実行可能なアッセイを提供するのに十分なほど高いものであり、しかも、突然変異が局在性の抗体高頻度突然変異機構によって真に生じたものであって、多くの腫瘍と特徴的に関連しているような全体的な突然変異率の増大を反映しているのではないと確信できるくらい十分高いことである。マウスのプレB細胞系および形質細胞腫細胞系におけるVH内の停止コドンの復帰変異(reversion)をモニタリングすることによって、突然変異が広く研究されている(Wablら, 1985;Chuiら, 1995;Zhuら, 1995;Greenら, 1998により概説されている)。発現されたV遺伝子の直接配列決定という別の戦略を行ったところ、幾つかの濾胞性リンパ腫、バーキットリンパ腫およびホジキンリンパ腫におけるVH遺伝子の多様化が、最初のトランスフォーメーション事象の後で継続し得ることが示された(BahlerおよびLevy, 1992;Jainら, 1994;Chapmanら, 1995および1996;Braeuningerら, 1997)。また、直接配列決定からは、クローン化濾胞性リンパ腫系では突然変異の存在率(prevalence)が低いことも明らかになっており、これは、VHの多様化がin vitroでも継続可能であることを立証している(Wuら, 1995)。上記で引用した細胞系での構成的突然変異についてのどの報告からも、観察された突然変異が、自然の抗体の多様化で見られるようなV遺伝子に集中している指向性高頻度突然変異(directed hypermutation)の結果であって、異なる系統に由来する多くの腫瘍細胞系において記載されているような突然変異に対する全体的な感受性とは異なる、ということの証拠は得られていない。
【0009】
近年、Denepouxら(1997)が、抗免疫グロブリン抗体および活性化T細胞の存在下で細胞を培養することによって、細胞系で高頻度突然変異を誘導した。しかし、観察された高頻度突然変異は誘発型であって構成的ではないことが述べられている。
【発明の概要】
【0010】
本発明の第1の態様において、標的核酸領域の指向性構成的高頻度突然変異を起こすことができるリンパ球様細胞系の製造方法であって、細胞集団を進行中の標的配列の多様化についてスクリーニングし、標的核酸の突然変異率が他の核酸の突然変異率よりも100倍以上上回っている細胞を選別することを含む上記方法が提供される。
【0011】
本明細書中で用いられる「指向性構成的高頻度突然変異(directed constitutive hypermutation)」とは、本明細書で報告する実験で初めて見出だされたものであり、特定の細胞系の、内在性DNAまたは導入遺伝子DNAの1箇所以上の特定の部位の核酸配列の改変を構成的様式で(すなわち、外的な刺激を与える必要なしに)引き起こす能力をいう。指向性構成的高頻度突然変異を起こすことができる細胞では、内在性DNAまたは導入遺伝子DNAの特定の部位以外の配列は、バックグラウンド突然変異率を上回る突然変異率とはならない。
【0012】
「標的核酸領域」とは、指向性構成的高頻度突然変異が起こる、本発明による細胞中の核酸の配列または領域である。標的核酸は、遺伝子産物をコードする1つ以上の転写ユニットを含むことができ、その転写ユニットはその細胞に対して同種であっても異種であってもよい。典型的な標的核酸領域は、免疫グロブリン産生細胞に見られる免疫グロブリンV遺伝子である。これらの遺伝子は、後記で更に詳細に説明されている高頻度突然変異収集エレメント(hypermutation-recruiting elements)の影響下にあり、そのエレメントによって高頻度突然変異が問題の遺伝子座で誘導される。その他の標的核酸配列は、例えば、遺伝子座内の高頻度突然変異収集エレメントを含むV遺伝子転写ユニットを、別の所望の転写ユニットで置換することにより、あるいは高頻度突然変異収集エレメントを含む人工遺伝子を構築することにより、構築できる。
【0013】
「高頻度突然変異」とは、細胞中の核酸の、バックグランドよりも高い率での突然変異をいう。好ましくは、高頻度突然変異とは、10-5〜10-3/bp/世代の突然変異率をいう。これは、バックグランド突然変異率(10-9〜10-10の突然変異/bp/世代のオーダー:Drakeら, 1988)や、PCRで見られる自然発生的突然変異率を大幅に上回る。Pfuポリメラーゼを用いて増幅を30サイクル行うと、その産物には1 bp当たり0.05×10-3未満の突然変異が起こるが、これは本発明の場合に観察される突然変異の1/100未満である(Lundbergら, 1991)。
【0014】
高頻度突然変異は、天然における免疫グロブリン可変鎖(V)遺伝子生成の一部である。したがって、本発明によれば、細胞系は、好ましくは、少なくとも1つの免疫グロブリンV遺伝子を生成できる免疫グロブリン産生細胞系である。V遺伝子は、可変軽鎖(VL)遺伝子であっても可変重鎖(VH)遺伝子であってもよく、免疫グロブリン分子全体の一部として生成されてもよい。それは、抗体、T細胞受容体、または免疫グロブリンスーパーファミリーの他のメンバーに由来するV遺伝子でありうる。免疫グロブリンスーパーファミリーのメンバーは、免疫系における広範な役割(例えば、抗体、T細胞受容体分子など)、細胞接着への関与(例えばICAM分子)、ならびに細胞内シグナル伝達(例えば、PDGF受容体などの受容体分子)を含むin vivoでの細胞性および非細胞性相互作用の多くの局面に関与する。したがって、本発明による好ましい細胞系は、B細胞から誘導されるものである。本発明によれば、V領域遺伝子に高頻度突然変異を起こす能力は保持しているが他の遺伝子には高頻度突然変異を生じない抗体産生B細胞由来の細胞系が単離できることが確認された。
【0015】
1つの好ましい実施形態において、本発明による細胞は、in vivoで高頻度突然変異が起こる細胞から誘導されたものか、あるいは該細胞に関連するものである。in vivoで高頻度突然変異を起こす細胞は、例えば、B細胞などの免疫グロブリン発現細胞である。リンパ腫細胞は、Ig発現細胞の腫瘍であり、これは、本発明による構成的に高頻度突然変異を起こす細胞系を単離するための特に優れた候補である。
【0016】
本明細書中で用いられる「進行中の標的配列の多様化についてのスクリーニング」とは、試験される細胞系の標的核酸領域における高頻度突然変異の存在の判定をいう。これは、直接配列決定法、またはMutSアッセイ(Jollyら, 1997)のような間接法、または免疫グロブリン欠損変異体の生成のモニタリングなどの種々のやり方で実施できる。この手順に従って選別された細胞は、標的配列の多様化を示す細胞である。
【0017】
本発明の方法による選別に供される細胞集団は、種々の細胞型および/もしくは種々の標的配列を含む、ポリクローナル細胞集団または(単)クローン細胞集団でありうる。
【0018】
クローン細胞集団とは、単一のクローンから誘導される細胞の集団であって、細胞がそこで起こっている突然変異以外は同一であるようになっているものである。クローン細胞集団の使用では、好ましくは、V遺伝子の高頻度突然変異を誘導する目的では他の細胞型(例えば活性化T細胞)との共培養は行わない。
【0019】
本発明による細胞は、高頻度突然変異を引き起こすために誘導工程の使用には依存しない。
【0020】
好ましくは、本発明においてスクリーニングされるクローン細胞集団は、B細胞から誘導される。有利には、それは、バーキットリンパ腫細胞系、濾胞性リンパ腫細胞系またはびまん性大細胞リンパ腫細胞系などのリンパ腫細胞系である。
【0021】
好ましくは、本発明による方法は、更に、標的配列の多様化を示す1種以上の細胞を単離する工程、および、単離した細胞において、標的配列内の突然変異の蓄積速度を非標的配列内のものと比較する工程を含む。
【0022】
本発明の特徴は、高頻度突然変異が、特定の(標的)核酸領域だけで誘導され、通常はこれらの領域以外では見られないことである。したがって、特異性は、本発明の方法の一部として、標的配列以外の配列の突然変異率をアッセイすることによってアッセイされる。C領域遺伝子(天然では高頻度突然変異に暴露されない)は、そうした技法に有利に用いることができるが、特異的な高頻度突然変異を受けない他の核酸領域もまた使用可能である。高頻度突然変異は配列依存的ではないので、比較のために選択された核酸領域の実際の配列そのものは重要でない。しかし、それは、後記で説明するように、高頻度突然変異を誘導する配列の制御を受けていてはならない。便利なことに、バックグラウンド突然変異は、例えばHPRT遺伝子座における彷徨解析(fluctuation analysis)により評価できる[LuriaおよびDelbreck, (1943);CapizziおよびJameson, (1973)を参照されたい]。
【0023】
標的領域の突然変異が非標的領域の突然変異を上回っている細胞が、本発明による特定の核酸領域の指向性構成的高頻度突然変異を起こすことができる細胞である。V領域遺伝子の突然変異が他の遺伝子の突然変異を上回っている係数は様々であるが、一般には少なくとも102、有利には103、好ましくは104以上のオーダーである。
【0024】
全体的な突然変異率および多様性は、例えば、突然変異誘発原の投与、または配列改変遺伝子(例えばターミナルデオキシヌクレオチジルトランスフェラーゼ(TdT))の発現によって増大し得る。しかし、高頻度突然変異とバックグラウンドとの差は、そうした方法では増大しないと予想される。
【0025】
本発明による好ましい細胞は、そこで見られる体細胞高頻度突然変異率を上げるために、遺伝子の欠失、変換もしくは挿入などの遺伝子操作に供することができる。例えば、本発明による細胞は、RAD51パラログ体の1コピー以上を欠失していてもよい。
【0026】
その細胞は、適切な脊椎動物細胞であればよく、例えば哺乳動物の細胞および鳥類の細胞が挙げられる。
【0027】
本発明の第2の態様において、所望の活性を有する遺伝子産物の製造方法であって、以下の工程a)〜c):
a) 本発明の第1の態様による細胞の集団において、高頻度突然変異を誘導する配列に機能し得る形で連結された該遺伝子産物をコードする核酸を発現させる工程、
b) 該細胞集団中で、該所望の活性を有する突然変異した遺伝子産物を発現する細胞または細胞群を同定する工程、
c) 工程(b)で同定された細胞または細胞群から1種以上のクローン細胞集団を樹立し、そのクローン細胞集団から、該所望の活性が向上している遺伝子産物を発現する細胞または細胞群を選別する工程、
を含む上記方法が提供される。
【0028】
上記のパートa)による細胞の集団は、V領域遺伝子の構成的高頻度突然変異を起こすことができ本発明の第1の態様による方法によって同定される細胞を含むクローン細胞集団またはポリクローナル細胞集団である。したがって、遺伝子産物は、後記で更に詳細に説明するように、内在性の免疫グロブリンポリペプチド、操作された内在性の遺伝子により発現される遺伝子産物、または体細胞高頻度突然変異を誘導する調節配列に機能し得る形で連結された異種転写ユニットにより発現される遺伝子産物、とすることができる。
【0029】
本発明の細胞内で発現され高頻度突然変異を起こす核酸は、内在性V領域のような内在性の領域、または本発明の細胞系に挿入された異種の領域であってよい。これは、例えば、高頻度突然変異を誘導する内在性の調節配列を保持したまま、内在性V領域が異種転写ユニット(例えば異種V領域)によって置換されている形態;または、高頻度突然変異を誘導するための細胞自身の調節配列の制御下で、該細胞へ異種転写ユニットが挿入されている形態を取ることができ、その場合、その転写ユニットはV領域遺伝子または任意の他の所望の遺伝子産物をコードしていればよい。本発明による核酸は、後記で更に詳細に説明されている。
【0030】
上記の工程b)において、細胞は、所望の遺伝子産物活性についてスクリーニングされる。これは、例えば免疫グロブリンの場合、結合活性とすることができる。また、適切なアッセイ法を用いて、酵素活性などの他の活性を評価することも可能である。遺伝子産物が細胞表面に提示される場合、所望の活性をもたらす細胞は、細胞表面での活性を(例えば蛍光発光によって)検出することにより、あるいは該細胞を表面の遺伝子産物を介して支持体に固定化することにより単離できる。活性が増殖培地中に分泌される場合、あるいはそれぞれ個々の細胞についてではなく細胞培地全体についてしか評価できない場合は、その所望の活性を有する遺伝子産物を分泌する細胞の同定確率を上げるために、工程a)から複数のクローン集団を樹立することが有利である。用いられる選別系は、細胞の増殖能・突然変異能に影響を及ぼさないことが有利である。
【0031】
好ましくは、この段階(および工程c)において、より良好な、向上した、またはより望ましい活性を有する遺伝子産物を発現する細胞を選別する。そのような活性は、例えば、所与のリガンドに対するより高度な親和性結合、またはより有効な酵素活性である。したがって、この方法によれば、所望の活性の定性的および/または定量的評価に基づいて、細胞を選別できる。
【0032】
本発明の第3の態様において、所望の活性を有する遺伝子産物の製造における、特定の核酸領域の指向性構成的高頻度突然変異を起こすことができる細胞の使用が提供される。
【0033】
本発明による使用において、所望の活性を有する遺伝子産物をコードする核酸は、細胞内での高頻度突然変異を誘導する調節配列に機能し得る形で連結されている。したがって、その細胞を連続的に作製することにより、その核酸配列の突然変異体が得られ、それを本発明の方法によりスクリーニングして、有利な特性を有する突然変異体を単離する。
【0034】
更に別の態様において、本発明は、本発明による指向性構成的高頻度突然変異を起こすことができる細胞に関する。好ましくは、この細胞は、遺伝子操作されたニワトリDT40細胞である。上記で記載したように、1つ以上のDNA修復遺伝子が操作できる。好ましくは、1つ以上のRad51遺伝子が操作されている。これらの遺伝子はダウンレギュレートまたは欠失していることが有利である。好ましくは、その遺伝子はRad51b遺伝子またはRad51c遺伝子である。
【0035】
1つの非常に好ましい実施形態において、本発明は、Δxrcc2 DT40およびΔxrcc3 DT40からなる群から選択される細胞を提供する。
【0036】
図面の説明
図1。バーキット系におけるVHの多様性
(A) 4種の散発性バーキットリンパ腫系の再構成されたVH遺伝子における配列多様性を円グラフとして示したもの。各細胞系について配列決定を行ったM13クローンの数を円の中心に示す。円弧の様々な大きさは、0個、1個、2個等(図示のとおり)の突然変異によりコンセンサスと区別される配列の割合を示す。
【0037】
(B) 初期Ramos培養物において同定されたVH突然変異の推定される系統的関係(dynastic relationship)。それぞれの丸(突然変異の程度に比例した濃淡を付けた)は、丸の中に示す数の蓄積された突然変異を有する別個の配列を示す。
【0038】
(C) 再構成されたVλ遺伝子における突然変異の存在率。Ramosにおいて、2種のVλの再構成が同定される。多様性および生殖細胞系の起源の割り当ては、図1Aと同様にして示してある。
【0039】
(D) 初期Ramos培養物のVH領域およびCμ領域における突然変異の存在率の比較。円グラフは、図1Aと同様にして示してある。
【0040】
図2。Ramosにおける構成的なVH多様化
(A) MutSアッセイにより評価した多様化。直接クローニングおよび配列決定により推定される各集団における突然変異の存在率を示す。
【0041】
(B) 3種の独立したRamosクローンの子孫から推定される系統的関係。
【0042】
図3。RamosのVHに沿った未選択のヌクレオチド置換の分布。
【0043】
図4。Ramosにおける高頻度突然変異により、様々な復帰可能なIgM欠損変異体が生じる。
【0044】
(A) IgM欠損変異体の単離を示すスキーム。
【0045】
(B) 複数のナンセンス突然変異がVHの不活性化に関与し得ることを示す表。これら2種の集団において停止がみとめられるそれぞれのVHコドンの位置を示す。
【0046】
(C) IgM欠損変異体の復帰変異率の表。
【0047】
(D) IgM欠損誘導体における停止コドン周辺の配列。
【0048】
図5。TdTを発現するRamosトランスフェクト体におけるIgM欠損変異体
(A) 3種のpSV-pβG/TdTおよび3種のRamosの対照トランスフェクト体におけるTdTの発現のウエスタンブロット分析。
【0049】
(B) IgM欠損変異体を生じる独立した突然変異事象を示す円グラフ。
【0050】
図6。一ヌクレオチド置換以外のVHにおける突然変異をまとめた配列表。
【0051】
図7。抗イディオタイプ(抗-Id1)結合特異性が失われているRamos細胞のVH遺伝子から単離した配列の比較。出発集団のコンセンサスとは異なるヌクレオチド置換は太字で示す。推定アミノ酸変化も太字で示す。
【0052】
図8。5ラウンドにわたる反復的な選別による、新規な結合特異性を有する免疫グロブリンを製造するためのRamos細胞の濃化を示す棒グラフ。
【0053】
図9。ビーズ:細胞の比率を増大させることによる新規な特異性(ストレプトアビジン)を結合するRamos細胞の回収の向上を示す棒グラフ。
【0054】
図10。標的抗原の濃度を増大させることによる、新規な結合特異性を有するRamos細胞の回収の増大を示すグラフ。
【0055】
図11。ストレプトアビジン結合Ramos細胞から得られたVH配列。出発集団のVH配列と比較した場合に見られるヌクレオチドの変化および推定アミノ酸の変化を太字で示す。
【0056】
図12。対照の培地および未選択のRamos細胞の上清に対する、ストレプトアビジン結合に対する選別プロセスの第4、第6および第7ラウンドにて選別された細胞の上清中のIgMの量。
【0057】
図13。図12の上清から得られたIgMのストレプトアビジン結合。
【0058】
図14。表面プラズモン共鳴により分析した、ストレプトアビジン結合に対する選別の第4および第6ラウンドから得られた上清のストレプトアビジン結合。
【0059】
図15。第4および第6ラウンドで選別された細胞のストレプトアビジン-FITCへの結合のFACS分析。
【0060】
図16。第6ラウンドで選別されたIgMのVHおよびVLの配列。
【0061】
図17。ストレプトアビジンに対して選別された親和性成熟Ramos細胞のFACS分析。
【0062】
図18。親和性成熟Ramos細胞のELISA。
【0063】
図19。野生型および修復欠損DT40におけるsIgM欠損変異体
(A) 野生型および修復欠損細胞におけるsIgMの異質性(heterogeneity)のフローサイトメトリー分析。
【0064】
(B) sIgM欠損変異体の生成頻度の彷徨解析。
【0065】
図20。DT40のsIgM変異体からクローニングしたVλ配列の分析。
【0066】
図21。1ヶ月間クローン性増殖させた後の、ソーティングされていないDT40集団のIg配列の分析。
【0067】
図22。DNA-PK、Ku70およびRad51Bが欠損しているDT40細胞のsIgM欠損変異体の分析。
【0068】
図23。ラット免疫グロブリンのイディオタイプに特異的なDT40由来の一連のIgM。細胞をビオチン化ラットS7 IgG2aと、モノクローナル抗体(Ab)/FITC-ストレプトアビジンとの凝集物を用いる6ラウンドの選別に供してDT-Ab3を得て、次に、これを更に3ラウンドの直接PE-Abコンジュゲートを用いるソーティングに供してDTAb6を得る。(a) ビオチン化Ab/FITC-Strep(ストレプトアビジン)凝集物およびPE-Abの、DT-Ab3細胞およびDT-Ab6細胞への結合の分析。(b) S7 Abで被覆したプレートを用いるELISAによりモニタリングした、DTAb3、5および6細胞の上清中のIgMのS7 Abへの結合、ならびに抗ニワトリIgMを用いた検出。上清は、総IgM力価について調整した。種々のラット・ハイブリドーマおよび各種のイソ型のキメラmAb(G. ButcherおよびM Bruggemannから恵贈されたもの(図示せず))へのDT-Ab6 IgMによる結合は検出されなかった。(c) DT-Ab6 IgMの、プレート上に被覆されたS7ラットIgG Abへの結合は、S7 Ab自身による競合を受けるが、正常なラット血清によって競合を受けることはほとんどない。S7(0.5mg/mlのストック)およびラット血清の競合物は、図示の希釈率で用いた。(d) DT-Ab6 IgMは、S7ハイブリドーマ細胞を染色するのに使用できる。染色は、固定した透過化細胞のフローサイトメトリーにより、FITC-コンジュゲート抗ニワトリIgMを用いて検出することにより検出した。(e) 元の0.2mg/mlのストック(PE-S7 Ab;Pharamacia)の各種の希釈液を用いたPE-AbコンジュゲートによるDT-Ab3細胞およびDT-Ab6細胞の染色の比較。(f) 親和性の測定。DT-Ab6細胞(106個)を、各種容量の0.2ピコモルのPE-コンジュゲートS7 Abと共にインキュベートして、図示のPE-Ab濃度を得た。洗浄した後で、細胞の平均蛍光強度(MFI)を測定した。(g) DT-Ab6と親集団とのIgVH/VL配列の比較。
【0069】
図24。DT40由来の一連のプロテインA特異的IgM。(a) 108個の細胞を106個のビーズと共にインキュベートした後で回収される細胞の数を示す、誘導化した磁性ビーズに結合しているDT40変異体の選別。DT-P1およびP2は、ビオチン化プロテインAで被覆したストレプトアビジン-ビーズで選別され、一方、DT-P3は、DT-P2から、プロテインAで直接被覆したトシル化磁性ビーズで選別された。(b) ビオチン化プロテインAとFITC-ストレプトアビジンとの凝集物による、FITCと直接コンジュゲートさせたプロテインAによる、およびFITC-抗IgMによる、各種のDT-P細胞のサブクローンの染色の比較。(c) 検出にニワトリIgMに特異的なマウスIgM mAb(Southern Biotechnology)を用いたELISAによりモニタリングした、親DT40およびDT-P14バルク集団の上清中のIgMの、プロテインAで被覆したプレートへの結合。対照として、総IgM力価も比較した。(d) DT-P4、-P6およびP7のクローンの上清から得られたIgM(親DT40からのものではない)は、プロテインA-セファロース(Sepharose)上に保持されているニワトリIgMを検出するためのウエスタンブロットを用いてプロテインA上で精製することができる。(e) [35S]プロテインAのDTP細胞への結合。親DT40(DT)またはDT-P細胞(0.1ml中に5×105個)を、[35S]プロテインA(48,000の結合可能なcpm;200〜2000Ci/mmol)と共に、氷上で1時間インキュベートした。PBSで1回洗浄した後、結合したcpmを測定した。(f) 親和性の測定。DT-P14細胞(106個)を各種の容量の0.7ピコモルの[35S]プロテインA(7×1010の結合可能なcpm/μmole)と共にインキュベートして、図示したプロテインA濃度を得た。洗浄した後、結合したcpmを測定した。(g) FITCプロテインAによるDT-P4細胞およびDT-P14細胞の染色は、1mg/mlの未標識ウサギIgGの存在下(P4+、P14+)で染色することにより増強される。(h) DT-Pサブクローン由来のIgV遺伝子の配列。
【0070】
図25。DT40の抗原選別により、特異性への成熟の可能性を有する多重反応性のIgMを得ることができる。DT-H10細胞は、親Xrcc2-欠損DT40のプールから、ヒト血清アルブミン(HAS)が結合しているカルボキシル化磁性ビーズを用いて、連続した10ラウンドの選別により得た。DT-T10細胞およびDT-O6細胞は、FITC-ストレプトアビジンとビオチン化サイログロブリン(DT-T細胞)もしくはオボアルブミン(DT-O細胞)との凝集物で染色した細胞のフローサイトメトリーによる選別を連続したラウンドで行うことにより得た。各ラウンドでは、集団の最も明るい2%を採取した。(a) 図示の試薬(Cy-strep:サイクローム(cychrome)-ストレプトアビジン)で染色したDT-H6細胞およびDT-T6細胞のフローサイトメトリー分析。(b) DT-H6細胞およびDT-T6細胞の抗原結合は、sIgM欠損変異体について濃化してあるDT-H6およびDT-T6の亜集団の二次元フローサイトメトリー分析で確認されるように、表面IgMにより仲介される。(c) 図示の抗原に特異的なニワトリIgMをモニタリングした、DTH6およびDT-T6の培養上清のELISA。(d) DT-O6細胞(オボアルブミン凝集物を用いて選別)もまたTg(ならびに、数種の他の被験抗原(図示せず))に結合するが、FITC-Ovabright/サイクローム-ストレプトアビジンdull細胞について連続ソーティングを行ったところ、Ovaに対する特異性がより高い集団が得られる。
【0071】
図26。天然に存在する構成的に高頻度突然変異を起こすBL細胞系の分析。
【図面の簡単な説明】
【0072】
【図1−1】バーキット系におけるVHの多様性を示す。
【図1−2】バーキット系におけるVHの多様性を示す。
【図1−3】バーキット系におけるVHの多様性を示す。
【図2】Ramosにおける構成的なVH多様化を示す。
【図3】RamosのVHに沿った未選択のヌクレオチド置換の分布を示す。
【図4】Ramosにおける高頻度突然変異により、様々な復帰可能なIgM欠損変異体が生じることを示す。
【図5】TdTを発現するRamosトランスフェクト体におけるIgM欠損変異体を示す。
【図6】一ヌクレオチド置換以外のVHにおける突然変異をまとめた配列表を示す。
【図7】抗イディオタイプ(抗-Id1)結合特異性が失われているRamos細胞のVH遺伝子から単離した配列の比較を示す。
【図8】5ラウンドにわたる反復的な選別による、新規な結合特異性を有する免疫グロブリンを製造するためのRamos細胞の濃化を示す棒グラフを示す。
【図9】ビーズ:細胞の比率を増大させることによる新規な特異性(ストレプトアビジン)を結合するRamos細胞の回収の向上を示す棒グラフを示す。
【図10】標的抗原の濃度を増大させることによる、新規な結合特異性を有するRamos細胞の回収の増大を示すグラフを示す。
【図11−1】ストレプトアビジン結合Ramos細胞から得られたVH配列を示す。
【図11−2】ストレプトアビジン結合Ramos細胞から得られたVH配列を示す。
【図12】対照の培地および未選択のRamos細胞の上清に対する、ストレプトアビジン結合に基づく選別プロセスの第4、第6および第7ラウンドにて選別された細胞の上清中のIgMの量を示す。
【図13】図12の上清からのIgMのストレプトアビジン結合を示す。
【図14】表面プラズモン共鳴により分析した、ストレプトアビジン結合に対する選別の第4および第6ラウンドから得た上清のストレプトアビジン結合を示す。
【図15】第4および第6ウランドで選別された細胞のストレプトアビジン-FITCへの結合のFACS分析を示す。
【図16−1】第6ラウンドで選別されたIgMのVHおよびVLの配列を示す。
【図16−2】第6ラウンドで選別されたIgMのVHおよびVLの配列を示す。
【図17】ストレプトアビジンに対する選別された親和性成熟Ramos細胞のFACS分析を示す。
【図18】親和性成熟Ramos細胞のELISAを示す。
【図19】野生型および修復欠損DT40におけるsIgM欠損変異体を示す。
【図20】DT40のsIgM変異体からクローニングしたVλ配列の分析を示す。
【図21−1】1ヶ月間クローン性増殖させた後の、ソーティングされていないDT40集団のIg配列の分析を示す。
【図21−2】1ヶ月間クローン性増殖させた後の、ソーティングされていないDT40集団のIg配列の分析を示す。
【図22】DNA-PK、Ku70およびRad51Bが欠損しているDT40細胞のsIgM欠損変異体の分析を示す。
【図23】ラット免疫グロブリンのイディオタイプに特異的なDT40由来の一連のIgMを示す。
【図24−1】DT40由来の一連のプロテインA特異的IgMを示す。
【図24−2】DT40由来の一連のプロテインA特異的IgMを示す。
【図25−1】DT40の抗原選別により、特異性への成熟の可能性を有する多重反応性のIgMを得ることができることを示す。
【図25−2】DT40の抗原選別により、特異性への成熟の可能性を有する多重反応性のIgMを得ることができることを示す。
【図26−1】天然に存在する構成的に高頻度突然変異を起こすBL細胞系の分析を示す。
【図26−2】天然に存在する構成的に高頻度突然変異を起こすBL細胞系の分析を示す。
【発明を実施するための形態】
【0073】
発明の詳細な説明
本発明は、選択された核酸領域を構成的に高頻度突然変異させる細胞系を、初めて利用できるようにした。これにより、自然の抗体産生における親和性成熟によく似た技法によって、突然変異した遺伝子産物を産生する系の設計が可能になる。Ramosバーキット系は、in vitro培養物中で、その再構成された免疫グロブリンV遺伝子を構成的に多様化する。この高頻度突然変異には、活性化T細胞や外来的に添加されるサイトカインによる刺激も、更にはB細胞抗原受容体の維持も必要としない。
【0074】
その突然変異率(0.2〜1×10-4/bp/世代の範囲)は、未選択の突然変異の大きなデータベースを容易に蓄積させるのに十分高いものであり、したがって、Ramosにおける高頻度突然変異が、in vivoでの免疫グロブリンV遺伝子の高頻度突然変異と古典的に関連づけられている特徴(突然変異をV遺伝子へ選択的にターゲッティングすること;一ヌクレオチド置換の段階的な蓄積;トランジションへの偏り;特徴的な突然変異のホットスポット)の多くを示すことは明らかである。データベースにおける未選択の突然変異の大部分は一ヌクレオチド置換であるが、欠失および重複(時には隣接するヌクレオチド置換が共に存在する)が検出可能である。そのような欠失および重複もまた、in vivoでの高頻度突然変異の結果として生じることが提唱されている(Wilsonら, 1998;Goosensら, 1998;Wu & Kaartinen, 1995)。
【0075】
選択された核酸領域で構成的に高頻度突然変異を起こす細胞の単離は、B細胞などの抗体産生細胞から誘導される細胞系でV遺伝子の突然変異をモニタリングすることに基づく。本発明で用いられる選別方法は、多くの方法で構成できる。
【0076】
高頻度突然変異細胞の選別
高頻度突然変異細胞は、標的配列の配列決定、発現欠損突然変異体についての選別、細菌性MutSタンパク質を用いるアッセイ、および遺伝子産物の活性の変化についての選別などの種々の技法により、細胞集団から選別できる。
【0077】
標的核酸の高頻度突然変異の特徴の一つは、このプロセスにより、停止コドンが、高頻度突然変異が起こらない場合に見られるよりもずっと高い頻度でその標的配列に導入されることである。これによって、細胞からの遺伝子産物の産生が欠損する。この欠損を、核酸配列が高頻度突然変異を起こしている細胞の同定に利用できる。
【0078】
本発明の1つの好ましい実施形態において、標的核酸は免疫グロブリンをコードする。免疫グロブリンの欠損は、免疫グロブリンを培地に分泌する細胞、および免疫グロブリンが細胞表面に提示されている細胞の両者について検出できる。免疫グロブリンが細胞表面に存在するのであれば、それが存在しないことは、例えばFACS分析、免疫蛍光顕微鏡法、または支持体へのリガンドの固定により、個々の細胞について同定すればよい。1つの好ましい実施形態では、細胞を抗原被覆磁性ビーズと混合して、沈降したら、細胞懸濁液から、目的とする特異性を有する免疫グロブリンが表面上に提示されている全ての細胞を除去すればよい。
【0079】
この技法は、抗体やT細胞受容体などを含むいずれの免疫グロブリン分子にも適用できる。免疫グロブリン分子の選別は、本発明に従ってアッセイすることが望まれるクローン細胞集団の性質に応じて決まってくる。
【0080】
あるいはまた、本発明による細胞は、V遺伝子などの標的核酸の配列決定、および配列比較による突然変異の検出によって選別することも可能である。この方法は、スループット(処理量)を増大させるために自動化することができる。
【0081】
更なる実施形態では、V遺伝子を高頻度突然変異させる細胞を、クローン細胞集団中で産生された免疫グロブリンにおける抗原結合活性の変化を評価することによって検出できる。例えば、特定の単位量の細胞培地または抽出物により結合される抗原の量を評価して、特定の結合活性を保持している細胞により産生された免疫グロブリンの割合を求めることができる。V遺伝子が突然変異すると、結合活性は変化し、特定の抗原に結合している産生された免疫グロブリンの割合は減少する。
【0082】
あるいはまた、細胞を、新規な結合親和性を生み出す能力についても同様にして評価でき、これは、例えば、初めは結合していない抗原または抗原の混合物にそれらを暴露し、高頻度突然変異の結果として結合親和性が生み出されるか否かを観察することにより行うことができる。
【0083】
更に別の実施形態では、細菌性MutSアッセイを用いて、標的核酸における配列の変異を検出することができる。MutSタンパク質は、核酸ハイブリッド中のミスマッチに結合する。親の核酸と突然変異した可能性のある子孫との間でヘテロ二本鎖を形成させることによりミスマッチ形成の程度、ひいては核酸の突然変異の程度が評価できる。
【0084】
標的核酸が免疫グロブリン以外の遺伝子産物をコードする場合、選別は、結合以外の機能の欠損または変化についてスクリーニングすることにより行ってもよい。例えば、酵素活性の欠損または変化についてスクリーニングしてもよい。
【0085】
配列の高頻度突然変異をターゲッティングする細胞を、他の核酸領域における突然変異について評価する。アッセイするのに都合の良い領域は、免疫グロブリン遺伝子の定常(C)領域である。C領域は、本発明による指向性高頻度突然変異を起こさない。C領域の評価は、好ましくは配列決定および比較により行われる。何故ならば、これが、突然変異が存在しないことを判定するための最も確実な方法だからである。しかし、他の技法を用いることも可能であり、例えば、高頻度突然変異事象により破壊される可能性のあるC領域の活性(例えば補体結合)の保持についてのモニタリングが挙げられる。
【0086】
細胞の遺伝子操作
本発明による高頻度突然変異細胞は、IgのV領域における高頻度突然変異率を増大するように遺伝子操作されている細胞から選別できる。突然変異率のモジュレーションを担う遺伝子は、一般に、細胞における核酸修復手順に含まれる。本発明に従って操作される遺伝子は、アップレギュレート、ダウンレギュレートまたは欠失されていてもよい。
【0087】
アップレギュレーションまたはダウンレギュレーションとは、問題の遺伝子によりコードされる遺伝子産物の活性が、少なくとも10%、好ましくは25%、更に好ましくは40%、50%、60%、70%、80%、90%、95%、99%、またはそれ以上増大または低下していることをいう。勿論、アップレギュレーションは、200%や500%などの100%を超える活性の増大を示してもよい。100%ダウンレギュレーションされている遺伝子は、機能的に欠失しており、これを本明細書中では「欠失している」ともいう。
【0088】
本発明により操作される好ましい遺伝子としては、Rad51遺伝子、特にxrcc2、xrcc3およびRad51b遺伝子の類似体および/またはパラログ体が含まれる。
【0089】
Rad51の類似体および/またはパラログ体は、ダウンレギュレートされていることが有利であり、好ましくは欠失している。1つ以上のRad51パラログ体のダウンレギュレーションまたは欠失により、本発明による高頻度突然変異率の増大が起こる。好ましくは、2つ以上のRad51遺伝子(その類似体および/またはパラログ体を含む)がダウンレギュレートされているか欠失している。
【0090】
1つの非常に好ましい実施形態では、ニワトリDT40細胞系などの鳥類の細胞系が、xrcc2および/またはxrcc3の欠失によって改変されている。Δxrcc2 DT40ならびにΔxrcc3-DT40は、本発明に従って単離される構成的に高頻度突然変異を起こす細胞系である。
【0091】
内在性遺伝子産物の改変
免疫グロブリンV領域遺伝子などの内在性遺伝子に構成的に高頻度突然変異を起こす細胞系が得られたことから、本発明は、新規な特性を有する遺伝子産物を産生させるための、構成的高頻度突然変異による内在性遺伝子産物の改変(adaptation)を提供する。例えば、本発明は、新規な結合特異性または改変された結合親和性を有する免疫グロブリンの産生を提供する。
【0092】
実際に、高頻度突然変異のプロセスを用いて、免疫グロブリン分子において向上した、または新規な結合特異性を生じさせる。したがって、目的とする抗原に結合できる免疫グロブリンを産生する本発明による細胞を選別し、次に、これらの細胞を増殖させて更なる突然変異体を産生させることにより、目的とする抗原への結合が向上している免疫グロブリンを発現する細胞を単離できる。
【0093】
目的の特異性を有する突然変異体を単離するために、種々の選別手順が適用できる。これらとしては、蛍光活性化セルソーティング(FACS)、磁性粒子を用いる細胞分離、抗原クロマトグラフィー法、および他の細胞分離法(例えばポリスチレンビーズの使用)が挙げられる。
【0094】
磁性による捕捉を用いる細胞の分離は、対象とする抗原を磁性粒子またはビーズにコンジュゲートさせることにより達成できる。例えば、その抗原を、Miltenyi Biotec GmbHにより供給されるような超常磁性鉄-デキストラン粒子またはビーズにコンジュゲートさせる。次に、これらのコンジュゲート粒子またはビーズを、多様な表面免疫グロブリンを発現しうる細胞集団と混合する。特定の細胞がその抗原と結合できる免疫グロブリンを発現する場合、それは、この相互作用により磁性ビーズと複合体化するだろう。次に、磁性粒子を固定している懸濁液に磁場を印加し、共有結合により結合している抗原を介してそれらに結合している細胞を保持する。次に、ビーズと結合しない未結合細胞を洗い流し、対象とする抗原に結合する能力に基づいて純粋に単離された細胞の集団を残存させる。試薬およびキットは、そのような一段階単離を行うための様々な供給元から入手可能であり、例えばDynalビーズ(Dynal AS; http://www.dynal.no)、MACS-磁性セルソーティング(Miltenyi Biotec GmbH; http://www.miltenyibiotec.com)、CliniMACS(AmCell; http://www.amcell.com)、ならびにBiomag, Amerlex-Mビーズなどが挙げられる。
【0095】
蛍光活性化セルソーティング(FACS)は、異なる表面分子(例えば表面に提示される免疫グロブリン)に基づく細胞の単離に使用できる。ソーティング(分別)しようとするサンプル中または集団中の細胞を、細胞表面分子に結合する特定の蛍光試薬で染色する。これらの試薬は、フルオレセイン、テキサスレッド、マラカイトグリーン、緑色蛍光タンパク質(GFP)または当業者に公知の任意の他の蛍光団などの蛍光マーカーに(直接または間接的に)結合している対象とする抗原である。次に、細胞集団をFACS装置の振動フローチャンバーに導入する。このチャンバーを通過する細胞の流れを、PBS(リン酸緩衝化生理食塩水)のような緩衝液のシース(sheath)に入れる。この流れにレーザー光を照射し、各細胞を蛍光(蛍光標識抗原の結合を示す)について測定する。細胞流の振動により、その細胞流は液滴になり、それは小さな電荷を持つ。これらの液滴に対して、コンピューター制御下で電気的偏向板により方向を誘導して、蛍光標識抗原に対する親和性に相応して異なる細胞集団を回収することができる。このようにして、対象とする抗原に対して異なる親和性を示す細胞集団を、その抗原に結合しない細胞から容易に分離できる。FACSで用いるFACS装置および試薬は、Becton-Dickinsonなどの世界規模の供給元や、Arizona Research Laboratories (http://www.arl.arizona.edu/facs/)などのサービス会社から広く入手できる。
【0096】
特定の抗原に対する細胞表面タンパク質の親和性に応じて細胞の集団を分離するのに使用可能な別の方法は、アフィニティクロマトグラフィーである。この方法では、適切な樹脂(例えばCL-600セファロース、Pharmacia Inc.)を適当な抗原に共有結合させる。この樹脂をカラムに詰め、混合細胞集団をそのカラムに通す。適当時間(例えば20分間)にわたりインキュベートした後、未結合細胞を(例えば)PBS緩衝液を用いて洗い流す。これにより、対象とする抗原に結合した免疫グロブリンを発現する細胞のサブセットだけが残存し、次に、これらの細胞を、(例えば)対象とする抗原を過剰量で用いて、または該抗原を該樹脂から酵素的もしくは化学的に切断することにより、カラムから溶出させる。これは、X因子、トロンビンまたは当業者に公知の他の特定のプロテアーゼなどの特定のプロテアーゼを用いて、予め抗原−樹脂複合体に組み込まれている適当な切断部位により該抗原をカラムから切り離すことによって行うことができる。あるいはまた、トリプシンなどの非特異的プロテアーゼを用いて、該抗原を樹脂から除去して、その対象とする抗原に対して親和性を示す細胞集団を遊離させることも可能である。
【0097】
異種転写ユニットの挿入
任意の所与の抗原に結合できる抗体変異体を迅速に選別する可能性を最大化するため、または非免疫グロブリン遺伝子についての高頻度突然変異系を開発するために、多くの技法を用いて、本発明による細胞を、それらの高頻度突然変異能が引き出されるように遺伝子操作することができる。
【0098】
第1の実施形態では、導入遺伝子を本発明による細胞にトランスフェクトして、その導入遺伝子が指向性高頻度突然変異事象の標的となるようする。
【0099】
本明細書中で用いられる「導入遺伝子」とは、例えばトランスフェクションまたはトランスダクションにより細胞に挿入される核酸分子である。例えば、「導入遺伝子」は、上記で述べたような異種転写ユニットを含むものであってもよく、それは細胞のゲノムの所望の位置に挿入できる。
【0100】
導入遺伝子を細胞に送達するのに用いられるプラスミドは、慣用の構造のものであり、プロモーターの制御下に目的の遺伝子産物をコードするコード配列を含む。本発明による細胞におけるベクターからの遺伝子の転写は、ポリオーマウイルス、アデノウイルス、鶏痘ウイルス、ウシ乳頭腫ウイルス、トリ肉腫ウイルス、サイトメガロウイルス(CMV)、レトロウイルスおよびサルウイルス40(SV40)などのウイルスのゲノム由来、アクチンプロモーターまたは例えばリボソームタンパク質プロモーターである非常に強力なプロモーターなどの異種哺乳動物プロモーター由来、ならびに通常は異種コード配列に結合しているプロモーター由来のプロモーターによって制御できるが、但し、そうしたプロモーターは本発明の宿主系と適合するものである。
【0101】
本発明による細胞による異種コード配列の転写は、ベクターにエンハンサー配列を挿入することによって増大させることができる。エンハンサーは、方向および位置には比較的依存しない。哺乳動物遺伝子(例えば、エラスターゼおよびグロビン)に由来する多くのエンハンサー配列が知られている。しかし、典型的には、真核細胞ウイルス由来のエンハンサーが用いられる。例としては、複製起点の後ろ側のSV40エンハンサー(bp 100-270)およびCMV初期プロモーターエンハンサーが挙げられる。エンハンサーは、ベクター中のコード配列に対して5’側または3’側の位置でスプライシングされ得るが、好ましくは、プロモーターの5’側の部位に位置する。
【0102】
有利には、真核性発現ベクターは、遺伝子座調節領域(LCR)を含み得る。LCRは、宿主細胞のクロマチンに組み込まれた導入遺伝子の高レベルの組込み部位に依存しない発現を誘導でき、これは、遺伝子治療用途のために設計されたベクターにおける、またはトランスジェニック動物における、ベクターの染色体組込みが起こっている永続的にトランスフェクトされた真核細胞系において異種コード配列を発現させようとする場合には特に重要である。
【0103】
真核性発現ベクターは、転写の終結およびmRNAの安定化のために必要な配列も含んでいる。そのような配列は、真核性またはウイルス性のDNAまたはcDNAの5’側および3’側非翻訳領域から一般に入手可能である。これらの領域は、mRNAの非翻訳部分においてポリアデニル化断片として転写されるヌクレオチドセグメントを含む。
【0104】
発現ベクターとしては、そのようなDNAを発現させることができる調節配列(例えばプロモーター領域)と機能し得る形で連結された目的の遺伝子産物をコードするコード配列を発現できる任意のベクターが挙げられる。したがって、発現ベクターとは、適当な宿主に導入されるとクローン化DNAの発現が起こる組換えDNAまたはRNA構築物をいい、例えば、プラスミド、ファージ、組換えウイルスまたは他のベクターが挙げられる。適当な発現ベクターは当業者には周知であり、真核細胞および/もしくは原核細胞において複製可能なもの、ならびにエピゾームを保持するもの、または宿主細胞のゲノムに組み込まれるものが挙げられる。例えば、異種コード配列をコードするDNAを、哺乳動物細胞におけるcDNAの発現に適するベクター、例えばpEVRF(Matthiasら, 1989)などのCMVエンハンサーをベースとするベクターに挿入できる。
【0105】
本発明によるベクターの構築には、慣用のライゲーション法が用いられる。単離したプラスミドまたはDNA断片を切断し、あつらえ、所望の形態に再連結して、必要とするプラスミドを作製する。所望により、構築したプラスミド中の配列が正しいことを確認するための分析を公知の方法で行う。発現ベクターを構築し、in vitroで転写産物を調製し、DNAを宿主細胞に導入し、遺伝子産物の発現および機能を評価するための分析を行うための好適な方法は、当業者には周知である。遺伝子の存在、増幅および/または発現は、例えば、適当に標識したプローブ(本発明で提供される配列に基づいたものでありうる)を用いて、慣用のサザンブロッティング、mRNAの転写を定量するためのノーザンブロッティング、ドットブロッティング(DNAもしくはRNAの分析)またはin situハイブリダイゼーションにより、直接サンプル中で測定できる。当業者であれば、必要に応じてこれらの方法にどのように変更を加えればよいか容易に判るであろう。
【0106】
第1の実施形態の1つの別形態では、本発明による導入遺伝子は、高頻度突然変異を誘導する配列も含む。そうした配列は特性決定されており、Klixら, (1998)およびSharpeら, (1991)(参照により本明細書に組み入れられる)で述べられている配列が挙げられる。つまり、遺伝子産物を発現し、且つその遺伝子産物をコードする転写ユニットに高頻度突然変異を誘導することができる遺伝子座全体が細胞に導入される。したがって、高頻度突然変異を誘導する転写ユニットおよび配列が、その細胞に対して外来性である。しかし、外来性ではあっても、高頻度突然変異を誘導する配列そのものは、細胞において天然に見られる高頻度突然変異を誘導する配列と類似または同一であってもよい。
【0107】
第2の実施形態では、内在性V遺伝子またはそのセグメントを、相同組換えにより、例えばLox/Cre系もしくは類似の技法を使用する遺伝子ターゲティングにより、または自然発生的に内在性V遺伝子が欠失している高頻度突然変異を起こす細胞系への挿入によって、異種V遺伝子と置換する。あるいはまた、高頻度突然変異には再構成されたV遺伝子近傍での二本鎖の破断が伴う、という知見を利用して、V領域遺伝子を置換することも可能である。
【0108】
以下、次の実施例において本発明を更に詳細に説明するが、それらは単に説明を目的とする。
【実施例】
【0109】
実施例1:高頻度突然変異を起こす細胞の選別
in vitroで高頻度突然変異を起こす細胞についてスクリーニングするために、数種のヒト・バーキットリンパ腫においてクローン性増殖の際に蓄積する多様性の程度を評価する。バーキット系BL2、BL41およびBL70はG. Lenoir(IARC, Lyon, フランス)から恵贈されたもの、Ramos(Kleinら, 1975)はD. Fearon(Cambridge, UK)から恵贈されたものである。それらの再構成されたVH遺伝子を、複数のVHファミリーのプライマーをJHコンセンサスオリゴヌクレオチドと共に用いて、ゲノムDNAからPCR増幅する。再構成されたVHセグメントの増幅は、Pfuポリメラーゼ、ならびに主要なヒトVHファミリーの各々について設計された14のプライマー(Tomlinson, 1997)のうちの1つ、および6つのヒトJHセグメント全てにアニーリングするコンセンサスJHバックプライマー(JOL48、5’-GCGGTACCTGAGGAGACGGTGACC-3’、 C. Jollyからの恵贈)を用いて達成される。RamosのVHのゲノムDNAからの増幅は、オリゴヌクレオチドRVHFOR(5’-CCCCAAGCTTCCCAGGTGCAGCTACAGCAG)およびJOL48を用いて行う。発現されたVH-CμcDNAの増幅は、RVHFORおよびCμ2BACK(5'-CCCCGGTACCAGATGAGCTTGGACTTGCGG)を用いて行う。ゲノムのCμ1/2領域は、Cμ2BACKおよびCμ1FOR(5'-CCCCAAGCTTCGGGAGTGCATCCGCCCCAACCCTT)を用いて増幅する。Ramosの機能性のCμ対立遺伝子は、Cμ2の8位のヌクレオチドとしてCを含んでおり、これに対して非機能性の対立遺伝子上ではTである。再構成されたVλは、5’-CCCCAAGCTTCCCAGTCTGCCCTGACTCAGおよび5’-CCCCTCTAGACCACCTAGGACGGTC-AGCTTを用いて増幅する。PCR産物は、QIAquick(Qiagen)スピンカラムを用いて精製し、M13中にクローニングした後でABI377シーケンサーを用いて配列決定する。突然変異は、GAP4アライメントプログラム(Bonfieldら, 1995)を用いて求められる。
【0110】
クローン化PCR産物の配列決定を行ったところ、Ramos細胞系でかなりの多様性(VHでの存在率は2.8×10-3の突然変異/bp)が示されるが、有意な異質性(heterogeneity)はBL41ならびにBL2でも見られる。図1Aを参照されたい。4種の散発性バーキットリンパ腫系の再構成されたVH遺伝子の配列多様性は、円グラフに示すとおりである。各細胞系における再構成されたVH遺伝子を、PCR増幅し、M13中にクローニングする。各細胞系について、最も多数のM13クローンに共通する配列としてコンセンサスをとり、さらにヒト免疫グロブリン配列のVBASEデータベース(Tomlinson, 1997)を用いて最も近くマッチングすることに基づいて生殖細胞系の対応物(各グラフの上部に示す)を割り当てる。ここで用いたRamosのVHコンセンサス配列は、Chapmanら(1996)により決定された配列とは3つの位置で、Ratech(1992)により決定された配列とは5つの位置で、それと最も近縁の生殖細胞系の対応物VH4(DP-63)とは6つの位置で異なっている。
【0111】
RamosにおけるVHの多様性の解析は、9回の別個のPCR増幅から得られる産物の配列決定により拡張される。これによって、集団中の突然変異したクローン同士の、個々のヌクレオチド置換の独立した反復の推定数が最小になるような有望な系統的(dynastic)関係の推定が可能になる(図1B)。9回の別個のPCR増幅から得られた315個のM13VHクローンを配列決定する。その系統は、同定された配列だけを含む(推定上の中間型は含まない)。個々の突然変異は、フォーマット「C230」に従って設計され、ここで、230とはRamosのVHのヌクレオチド位置(図3に示す番号付け)であり、「C」とは、その位置の新規な塩基を示す。同じヌクレオチド置換が独立して起こる回数を最小化することを、基準として系統を推定するのに用いた。分岐の大部分は、別々のPCR増幅によりもたらされる独立したメンバーを含む。まれな欠失および重複は、それぞれ接頭語「x」および「d」で示す。矢印は、2つの突然変異を強調しており(264位での置換により、停止コドンおよび184位での重複が生じる)、系統樹内でのその位置は、突然変異が、機能性重鎖の発現が欠損した後も蓄積し続けることができることを示している。
【0112】
PCRのアーチファクト(artefact)は、突然変異のデータベースに殆ど影響を及ぼさない。ヌクレオチド置換の存在率が、対照のPCR増幅で見られる存在率(0.05×10-3/bp)よりも遥かに高いことだけでなく、別個の増幅において同一の突然変異を生じたクローン(ならびに系統的に関連するもの)が見られる。多くの場合、ある系統内の世代同士は、一ヌクレオチド置換だけ異なっており、このことから、突然変異の各ラウンドにおいて少数の置換しか導入されていないことが示される。
【0113】
Vλ再構成の解析から、Ramosが、Vλ2.2-16の、読み枠が合っている(in-frame)再構成(Chapmanら, 1996により記載されている)およびVλ2.2-25の読み枠が合わない(out-of-frame)再構成を含んでいることが明らかになる。双方の再構成されたVλには突然変異による多様性が存在するが、非機能性の対立遺伝子では、それより高度の多様性が蓄積している(図1C)。
【0114】
抗体の高頻度突然変異の古典的な特徴といえば、突然変異の多くはV領域に蓄積し、C領域には殆ど蓄積しないことである。これは、RamosのIgH遺伝子座に蓄積している突然変異でも明らかである(図1D)。VH、Cμ1およびCμ2の最初の87ヌクレオチドに及ぶcDNAインサートを含むM13クローンは、初期Ramos培養物からPCRにより作製される。円グラフ(図1Aに示すようなもの)には、Cμ1の始点から伸びるCμの380ヌクレオチドのストレッチと比較した場合の、VHの341ヌクレオチドのストレッチ内で同定された突然変異の程度を示す。
【0115】
Ramosにより産生されたIgM免疫グロブリンは、細胞表面上および培地中(分泌形態)の双方に存在する。培地を分析したところ、Ramosが免疫グロブリン分子を非常に高濃度(1μg/ml程度)で分泌することが判明する。したがって、Ramosは、製造のために発現細胞系または細菌に免疫グロブリン遺伝子を再クローニングする必要がなくなるレベルまで免疫グロブリンを分泌することができる。
【0116】
実施例2:RamosにおけるVHの多様化は構成的である
V遺伝子の多様化が進行中であるか否かを調べるために、細胞をクローングし、in vitroでの培養期間の後で、VHの多様性をMutSベースのアッセイを用いて評価する。RamosのVHを、上記で記載したように、5’末端にビオチン化塩基を含むオリゴヌクレオチドを用いてPCR増幅し、精製する。変性/再生(99℃で3分;75℃で90分)の後、突然変異の程度を、先に記載(Jollyら, 1997)されているような細菌性ミスマッチ修復タンパク質MutS(濾紙に結合させてある)へのミスマッチヘテロ二本鎖物質の結合をECLによる検出によりモニタリングすることによって、評価する。
【0117】
その結果から、VHの多様化が、まさに進行中であることが示される(図2Aを参照)。限界希釈クローニングした後で1ヶ月間または3ヶ月間培養したRamos細胞からDNAを抽出する。再構成されたVHを、ビオチン化オリゴヌクレオチドを用いてPCR増幅した後、変性/再生させる。次にミスマッチヘテロ二本鎖を、先に記載(Jollyら, 1997)されているような固定化MutSへの結合により検出する。再生されたDNAのアリコートをメンブレンに直接結合させて、マッチしたDNAの負荷を確認する(全DNA対照)。比較のために、細菌性プラスミド鋳型ならびに初期Ramos培養物から増幅したRamosのVHについて行うアッセイを含める。
【0118】
4週間(Rc1)および6週間(Rc13および14)にわたり増殖させたRamos培養物からVH遺伝子をPCR増幅する(図2B)。各クローンの突然変異率を示すが、これは、クローニングの4週間後または6週間後の独立したVHの突然変異の存在率を、一世代時間が24時間であることに基づいて推定した細胞分裂の回数で除算することにより算出される。配列から、段階的な突然変異の蓄積が明らかになり、その際の突然変異率は約0.24×10-4の突然変異/bp/世代である。
【0119】
RamosにおけるVHの突然変異率を他の細胞系におけるVHの突然変異率と直接比較することは容易ではない。何故ならば、単一の前駆細胞からクローン性増殖させた後に得られるVH全体に取り込まれた未選択の突然変異により決定されるような、他の系における突然変異率に関する情報は殆ど無いからである。しかし、50個のBL2前駆細胞を2週間にわたり増殖させた後の突然変異の存在率が、突然変異誘発の条件下で測定されている(2.7×10-3の突然変異/bp; Denepouxら, 1997)。通常の培養条件下でRamosを用いて行った同様の実験から、突然変異の存在率が2.3×10-3の突然変異/bpであることが判明している。サイトカイン、ヘルパーT細胞などを与えることによる突然変異率を増大させるための様々な試みは、うまく行かないことが判っている。したがって、BL2細胞の特異的誘発により達成可能な突然異率は、RamosにおけるVHの構成的な突然変異率とほぼ同じであると思われる。
【0120】
実施例3:RamosにおけるVHの突然変異の性質の検討
初期Ramos培養物において検出したもの(141の異なる配列からのもの)を、特定の選択を行わない種々の実験で培養した4種のサブクローン(更に別の135の異なる配列からのもの)において検出したものと組み合わせた、突然変異事象のデータベースを作成する。このデータベースは、個々の配列セットを系統的関係(図1Bの説明文に詳細に説明されている)に組み立てた後で作成して、個々の突然変異細胞のクローン性増殖によって特定の突然変異事象が複数回カウントされてしまわないことを確実にする。ここで、この340の異なった、そしておそらくは未選択の突然変異事象(200は初期Ramos培養物によりもたらされたものであり、140は増殖させたサブクローンによりもたらされたもの)の複合的なデータベースの解析を説明する。初期集団およびサブクローン集団を別々に解析しても、同じ結論が得られる。
【0121】
突然変異の圧倒的大部分(340のうち333)は、一ヌクレオチド置換である。少数の欠失(4)および重複(3)が見られるが、鋳型のない挿入は見られない。これらの事象については、後記で更に詳細に述べる。隣接位置にヌクレオチド置換を示す配列は5つしかない。しかし、これら5つの場合のうち3つにおいて、その系統では、隣接する置換が逐次に取り込まれていることが判明している。このように、隣接位置でヌクレオチド置換が同時に作られることは稀な事象である。
【0122】
VHに沿った突然変異の分布は高度に非ランダムである(図3を参照)。独立して起こっている塩基置換を、各ヌクレオチド位置に示す。CDR1および2の位置を示す。ヌクレオチドの位置は、配列決定用プライマーの3’末端から番号付けし、+1位のヌクレオチドは第7コドンの最初の塩基に相当する。コドンはKabatに従って番号付けしている。イタリック体で示す突然変異(15位、193位、195位および237位のヌクレオチド位置)は、突然変異したサブクローンで起こっている置換であり、その位置の配列は示したコンセンサスに復帰している。
【0123】
主要なホットスポットはSer82aコドンのGおよびCのヌクレオチドにあり、これは他のVH遺伝子における主要な内因性の突然変異ホットスポットとして先に同定されており(Wagnerら, 1995;Jollyら, 1996)、RGYWのコンセンサスと一致している(RogozinおよびKolchanov, 1992;Betzら, 1993)。多くのVH遺伝子における主要な内因性の突然変異ホットスポットはSer31にあるが、このコドンは、その位置にGlyを有するRamosのコンセンサスVH(またはその生殖細胞系のVH)には存在しない。個々のヌクレオチド置換は、トランジションに著しい偏りを示す(無作為に予想される33%ではなく、51%)。また、GおよびCをターゲティングすることについて著しい選択性を示し、これはターゲティングされたヌクレオチドの82%を占める(表1)。
【表1】
【0124】
実施例4:IgM欠損による高頻度突然変異を起こす細胞の選別
Ramnos変異体の分析から、不活性化されたVHを有しているに違いない幾つかの突然変異が明らかになり(図1Bを参照)、このことから、細胞がIgM発現を欠損しても依然として生存可能である可能性があることが示唆される。これが正しいならば、Igの発現の欠損は、構成的に高頻度突然変異を起こすB細胞系を選別するための容易な尺度となるだろう。
【0125】
Ramos培養物の分析から、それが8%の表面IgM−細胞を含んでいることが判明する。そのようなIgM欠損変異体は、以下のようにして、in vitro培養の際に作製される。出発物質のRamos培養物を、pSV2neoプラスミドでトランスフェクトし、96ウェルプレートに希釈し、選択培地で増殖するクローンを増殖させる。最初のトランスフェクションから6ヵ月後に増殖させたクローンについてフローサイトメトリーを行ったところ、IgM欠損変異体の存在が明らかになり、ここで示す2種のクローン集団(Rc13および14)の16%および18%を占めている(図4A)。ソーティングを1ラウンド行うことにより濃化して、87%(Rc13)および76%(Rc14)の表面IgMネガティブ細胞を含む亜集団を得る。これらの亜集団における再構成されたVH遺伝子をPCR増幅した後で配列決定を行うことにより、クローン化したVHセグメントの75%(Rc13)および67%(Rc14)が、分析した341ヌクレオチドのVHストレッチ内にナンセンス(stop)、欠失(del)または重複(dup)の突然変異を含んでいたことが示される。クローンの残りは野生型(wt)と名づけるが、可能性のあるVH不活性化ミスセンス突然変異を区別する試みは行わない。Rc13集団において同定された4つの欠失および3つの重複は全て異なっているが、欠失を有すると判定された7つのRc14の配列のうち、異なる突然変異はたった4つである。欠失および重複の性質を図6に示す。各事象は、文字の後ろに数字を付けて表示する。文字は、突然変異の起源を示す(A、BおよびCはクローン化TdT-対照トランスフェクト体であり、D、EおよびFはTdT+トランスフェクト体であり、Uは初期の未選択Ramos培養物において同定された事象を示す)。数字は、配列文字列(string)の最初のヌクレオチド位置を示す。欠失しているヌクレオチドはラインの上に明記し、付加されたヌクレオチド(重複または鋳型のない挿入)はラインの下に明記する。一ヌクレオチド置換は丸で囲み、新たな塩基が明記してある。VH起源の重複したセグメントには下線が引いてある。鋳型のない欠失は太字にしてある。幾つかの欠失または重複では、その事象は起源が不明の1個のヌクレオチドと隣接している。そのような隣接した変化は、(鋳型のない挿入ではなく)ヌクレオチド置換により十分に起こり得るものであり、したがって、これらの事象は別個にグループ化される。欠失/重複の一端または他端への一塩基置換(丸で囲む)の割り当ては、任意である場合が多い。
【0126】
IgM−細胞は、ソーティングを1ラウンド行って濃化した後、PCR増幅およびそれらのVHセグメントのクローニングを行う。配列から、かなりの範囲のVH不活性化突然変異(停止コドンおよびフレームシフト)が明らかになるが(図4)、様々な不活性化突然変異は、クローン性増殖のわずか6週間後にソーティングしたIgM欠損変異体においても示されている(図5を参照)。図5Aで、3種のpSV-pβG/TdTおよび3種のRamosの対照トランスフェクト体におけるTdTの発現を、核タンパク質抽出物のウエスタンブロッティング分析により比較する。Nalm6(TdT-ポジティブなヒト・プレB細胞リンパ腫)およびHMy2(TdT-ネガティブな成熟ヒトBリンパ腫)を対照とした。
【0127】
図5Bでは、IgM欠損変異体を生じる独立した突然変異事象を示す円グラフを示す。クローニング後に6週間培養した上記の3種のTdT+および3種のTdT−対照トランスフェクト体をソーティグすることによりIgM−変異体(集団の1〜5%を占める)が得られる。ソーティングした亜集団におけるVH領域をPCR増幅し、配列決定する。円グラフは、VH不活性化を生じる突然変異のタイプ、ならびに別々にプールされたTdT+およびTdT−のIgM−亜集団から得られたデータを示している。略語は図4Aのとおりであるが、但し、「ins」は明らかに鋳型のない(非鋳型)ヌクレオチド挿入を含むクローンを示す。複数のヌクレオチド非鋳型挿入と共に欠失または重複を含むクローンだけが、このグラフの「ins」の区分に含まれる。明らかに異なる突然変異事象だけを算出する。それにより、TdT+ IgM欠損亜集団において同定された77の異なるVH不活性化突然変異のうち、30の異なる停止コドン突然変異が同定される。1つのRamosトランスフェクト体から誘導されるIgM欠損集団内で同じ停止コドンが独立して作られている場合は、それに下線を引いてある。
【0128】
停止コドンは様々な位置に作られるが(図4B)、ランダムには位置していない。図4Bに、Rc13およびRc14のIgM欠損集団で見られる停止コドンの性質をまとめてある。少なくとも8種の独立した突然変異事象により、Rc13のデータベースの27の非機能性VH配列のうち20を占めるナンセンス突然変異が生じる。最低で10の独立した突然変異事象から、Rc14のデータベースの22の非機能性VH配列のうち15を占めるナンセンス突然変異が生じる。各停止コドンの後ろのカッコ内の数字は、当該停止コドンを保有するデータベース内の配列の数を示し、その後ろに、更に別の突然変異に基づいて区別される異なったこれらの配列の数を示す。4種の他のクローン集団から選別されるIgM欠損変異体における停止コドンの分析から、VH内で更に5つの位置で停止コドンが作られることが判明する。6つの独立した実験で得られたデータにおいて、停止コドンの創出は、可能性のある39の部位のうちの16に限られる。これらの選択された部位でのDNA配列は、(コード鎖または非コード鎖のいずれにおいても)RGYWコンセンサスの方に偏っている。
【0129】
驚くことではないが、欠失および挿入は、未選択のRamos培養物における突然変異のほんの一部にすぎないが(上記を参照)、それらは、VH不活性化突然変異に注目した場合、非常に重大な意義がある。IgM欠損変異体の大部分がVH自体における停止コドン/フレームシフト突然変異により説明できることは、注目に値する。このことは、Ramosにおける高頻度突然変異が主に免疫グロブリンのVドメインを標的としており、確かにCドメインや、それが突然変異すると表面IgM−表現型が生じる他の遺伝子(例えばIgα/Igβストレッチ)は全く標的としていない、という提唱を更に支持している。また、RamosのVHが、その生産的に再構成されたVλよりもしばしば高頻度突然変異の標的とされる可能性も十分にあり、この結論は、初期培養物中の突然変異パターンにより支持される(図1C)。
【0130】
Ig欠損変異体の検出による細胞の選別は、それらの変異体が復帰変異する(すなわちそれらの内在性のIg発現能を再獲得する)能力を有する場合に特に有用である。より初期に樹立した系統(図1B)からは、IgM欠損細胞が生じ得ることだけでなくそれらに更なる突然変異が起こる可能性があることも示唆される。このことを確認するために、Rc13からソーティングされたIgM欠損変異体を限界希釈によりクローニングする。クローニングから3週間後、IgM−サブクローン中のIgM+復帰変異体の存在を、5×104個の細胞の細胞質免疫蛍光分析によりスクリーニングする。それらの存在率を示す(図4C)。次に、これらのIgM+復帰変異体を、1ラウンドのソーティングで濃化し、クローン性IgM−変異体のVH配列をそのIgM+復帰変異体の子孫のVH配列と比較する。
【0131】
10の増殖させたクローン集団の細胞質免疫蛍光から、IgM+復帰変異体が様々な存在率(0.005%〜1.2%;図4C)で存在することが明らかになり、これによって彷徨解析により1×10-4の突然変異/bp/世代の突然変異率であることが算出できる。これは、未選択の突然変異の直接分析により算出された率(0.25×10-4の突然変異/bp/世代;上記を参照)よりも多少高い。これは、おそらく、異なるIgM欠損クローンが、破壊している突然変異の性質に応じて異なる率で復帰していることをある程度反映していると思われる。事実、Rc13のIgM欠損誘導体における停止コドン周辺の配列からは、TAG32が、内在性の突然変異ホットスポット(Betzら, 1993)の大部分を占めるRGYWコンセンサス(R=プリン、Y=ピリミジン、W=AまたはT;RogozinおよびKolchanov, 1992)と十分に一致するが、TAA33およびTGA36では一致しないことが明らかになる。
【0132】
実施例5:新規なIg結合活性の選別
新規な結合親和性が生じることを実証するために設計された実験では、後記に記載するRamos細胞系の多くのメンバーが、Ramosの表面IgMに対して特異的に誘導された抗イディオタイプ抗体(抗Id1および抗Id2)に結合する膜IgM分子を発現することに注目する。しかし、少数の細胞は表面IgMを保持するが、抗イディオタイプ抗体に結合できない。これは、表面IgM分子における結合親和性が、抗体には結合しないように変化したことによる。表面IgMを発現する細胞は、抗体に結合できず、本発明による1ラウンドのセルソーティングで選別できる。
【0133】
これは、表面IgMを保持しているにもかかわらず抗Id2への結合能を欠損しているμ-ポジティブ/id-ネガティブクローンをELISAにより単離することにより実証される。これらのクローンを配列決定したところ、6つの独立したクローンにおいて、保存されたVH残基であるK70が次のようにN、MまたはRへと突然変異していることが示される。
【0134】
クローン 突然変異
2 K70N AAG-AAC
S77N AGC-AAC
4 K70M AAG-ATG
9 S59R AGT-AGG
K70N AAG-AAC
10 K70N AAG-AAC
12 K70N AAG-AAC
13 K70R AAG-AGG
軽鎖では突然変異は見られなかった。したがって、産生されたIg分子が親クローンと比較して一塩基対の変異を有する突然変異体をRamos細胞系から選別できることは明らかである。
【0135】
抗Id1を用いて、Igμ定常領域の発現は維持しているが抗イディオタイプ抗体への結合を欠損している同様の細胞集団を単離する。これらの細胞は、ソーティングサイトメトリーにより濃化し、VHの配列を決定する(図7)。このことにより、出発集団のコンセンサス配列と比較した場合に、6つの突然変異が判明する。これらの突然変異のうち2つは、CDR3近傍でアミノ酸配列の変化をもたらす(95にてR→T、98にてP→H)。したがって、免疫グロブリン分子におけるより微細な変化の選別は、結合の欠損についてアッセイすることによって選択可能である。
【0136】
更に別の実施形態では、本発明による高頻度突然変異を起こす細胞を洗浄し、PBS/BSAに再懸濁し(0.25ml中に108個の細胞)、10%(v/v)の抗原被覆磁性ビーズを含む同容量のPBS/BSAと混合する。この実験では、ストレプトアビジン被覆磁性ビーズ(Dynal)を用いる。ローラー上で4℃にて30分間混合した後、ビーズをPBS/BSAで3回洗浄する。各回ごとに、ビーズを磁石で沈降させ、未結合の細胞を除去する。次に、残った細胞を96ウェルプレートに接種し、108個まで増殖させた後、更にもう1ラウンドの選別に供する。細胞の増殖(構成的に進行中の高頻度突然変異を伴う)および選別を複数ラウンド行う。複数ラウンドの選別を行った後、ビーズに結合した細胞の割合(最初はバックグラウンドレベルである0.02%またはそれに近いレベル)が増大し始める。
【0137】
4ラウンド後、ストレプトアビジン結合細胞の濃化が見られる。これを第5ラウンドでも繰り返す(図8)。回収率のパーセンテージが低いことは、ビーズが細胞で飽和していることを反映している。何故ならば、細胞:ビーズ比を大過剰から1:2に変更することにより、第5ラウンドのストレプトアビジン結合細胞から約20%の回収が可能になるからである(図9)。このことから、4ラウンドの反復的な選別により、高頻度突然変異を起こすRamos細胞系から新規な結合特異性をうまく選別できることが実証される。
【0138】
ストレプトアビジン結合細胞由来の重鎖および軽鎖のヌクレオチド配列決定により、出発集団のコンセンサス配列と比較した場合に、VHのCDR3で1つのアミノ酸の変化、そしてVLで4つの変化(FR1に1つ、CDR1に2つ、CDR2に1つ)があることが推定される(図11)。
【0139】
ストレプトアビジンの結合が表面免疫グロブリンの発現に依存することを確実にするために、ストレプトアビジン結合細胞の免疫グロブリンネガティブ変異体をソーティングサイトメトリーにより濃化する。これにより、過剰のビーズを用いた場合でもストレプトアビジン結合細胞の回収率は著しく低下する。ソーティングされたネガティブ細胞からDynal-ストレプトアビジンビーズにより回収された細胞は、事実、Igμポジティブであり、これがおそらくソーティングされた免疫グロブリンネガティブ細胞集団に混入しているIgμストレプトアビジン結合細胞の効率的な回収を説明すると思われる。
【0140】
予備のデータから、回収の効率は、ビーズ上のストレプトアビジンの濃度が低下するに従って低下することが示唆される(図9)。これは、ある濃度範囲のストレプトアビジンと共にインキュベートしたビーズを用いてストレプトアビジン結合細胞の回収率をアッセイすることにより確認される(図10)。結合している集団から回収可能な細胞のパーセンテージは、ビーズと細胞との割合により求められる。この実験では、ビーズ:細胞の比率は1:1未満とする。
【0141】
更なる一連の実験では、更に2ラウンドの選別を完了させ、つまり、全体で7ラウンドとする。これは、ビーズに結合するストレプトアビジンの濃度を、第5ラウンドでの50μg/mlを第7ラウンドでは10μg/mlへと下げることにより達成される。IgMの分泌レベルは、第4〜7ラウンドで選別された集団と同等であるが(図12)、ELISAにより評価した場合のストレプトアビジン結合は、第6および第7ラウンドでは第4ラウンドと比較して明らかに大きく増大している(図13)。
【0142】
これは、ストレプトアビジンで被覆されたBiaCoreチップでの表面プラズモン共鳴により結合を評価することによって確認される(図14)。第7ラウンドから得た上清を、Aの時点で注入してチップに流し、Bの時点で中止する。Cの時点で、抗ヒトIgMを注入して、ストレプトアビジンに結合している物質がIgMであることを実証する。勾配A-Bは会合定数を表わし、勾配B-Cは解離定数を表わす。BiaCoreをトレースすることにより、第6ラウンドの上清が、第4ラウンドの集団から単離されたものや未選択のRamos細胞よりも優れた結合特性を示すことが明らかになる。
【0143】
選別プロセスの第6ラウンドから得られる抗体もまた、第4ラウンドと比較して向上した結合を示す。図15に示すように、第6ラウンドでの選別から得られた細胞のストレプトアビジン-FITC凝集物への結合(蛍光団をビオチン化タンパク質と共にプレインキュベートすることにより形成される)は、FACSにより可視化できる。第4ラウンドの集団、未選択のRamos細胞またはIgMネガティブRamosへの結合は見られず、このことは、ストレプトアビジン結合の成熟を示している。
【0144】
凝集していないストレプトアビジン-FITCを使用すると、同様の結果は得られず、第6ラウンドの細胞の大部分が結合しない。これは、ELISAによるデータと一致して、ストレプトアビジンへの結合が、一連の抗原に結合する抗体の親和力によるものであって、一価の親和性によるものではないことを示唆している。親和性がより高い結合体は、凝集していないストレプトアビジン-FITCへの結合についてソーティングすることにより単離できる。
【0145】
第4ラウンドの細胞の場合を上回る、第6ラウンドの細胞で見られる結合の増大に関与する突然変異を突き止めるために、軽鎖および重鎖抗体遺伝子をPCRにより増幅し、次に配列決定する。第4ラウンドの細胞と比較して、重鎖遺伝子の変化は見られず、突然変異R103Sは保存されている。軽鎖でも、V23FおよびG24Cの突然変異は保存されているが、46位に更に別の突然変異が存在している。野生型のRamosはこの位置にアスパラギン酸を有するが、第6ラウンドの細胞はアラニンを有する。この位置での変化は、抗原結合に影響を及ぼすと予測される。何故ならば、この領域の残基は軽鎖のCDR2に関与するからである(図16)。D46Aの突然変異はおそらく、第6ラウンドの細胞で見られるストレプトアビジンへの結合の観察された増大に関与すると思われる。
【0146】
実施例6:Ramosのストレプトアビジン結合体のin vitro成熟
Ram B→Ram C(FITC-ポリ-ストレプトアビジンを用いた選別)
約5×107個のRam B細胞(ストレプトアビジン被覆ミクロビーズに結合させるためにRamos細胞系から誘導したもの)をPBSで洗浄し、ポリ-ストレプトアビジン-FITCを含む1mlのPBS/BSA溶液中で氷上で30分間インキュベートする(ポリ-ストレプトアビジン-FITCは、ストレプトアビジン FITC(20μg/mlのタンパク質含量)をビオチン化タンパク質(10μg/ml)に添加し、氷上で数分間インキュベートした後、細胞を添加することにより作製される)。
【0147】
次に、細胞を氷冷PBSで軽く洗浄し、遠心沈降させ、500μlのPBSに再懸濁する。
【0148】
最も蛍光性が高い1%の細胞をMoFloセルソーターでソーティングし、この細胞集団を組織培養培地に戻し、約5×107細胞まで増殖させ、この手順を繰り返す。
【0149】
ポリ-ストレプトアビジン-FITCを用いて4ラウンドにわたりソーティングした後、細胞はストレプトアビジン-FITCに弱く結合する。このRamos細胞集団からの発現された免疫グロブリンV領域の配列からは、重鎖V領域の第3フレームワーク中のアミノ酸番号82aがセリンからアルギニンへと変化していることが判明する。この細胞集団をRam Cと呼ぶ。
【0150】
Ram C→Ram D(FITC-ストレプトアビジンを用いた選別)
次の数ラウンドのセルソーティングは、上記で記載されているようにして行うが、ここではストレプトアビジン-FITC(20μg/mlのタンパク質含量)を用いる。
【0151】
ストレプトアビジン-FITCを用いてソーティングを3ラウンド行った後、ソーティングされた細胞集団(Ram Dと呼ぶ)は、FACSによりアッセイしたところ、ストレプトアビジン FITCに、より強く結合している。発現されたV遺伝子の配列から、更なるアミノ酸の変化が明らかになる。第3フレームワークにおいて、元々はセリンであった65位のアミノ酸がアルギニンへと変化している。
【0152】
Ram D→Ram E(FITC-ストレプトアビジンおよび未標識ストレプトアビジン競合を用いた選別)
次のソーティングは、ストレプトアビジン-FITCを用いて、上記で記載されているのと同様にして行う。しかし、細胞を氷上で30分間染色した後、細胞を氷冷PBSで一回洗浄し、次に0.5mg/mlのストレプトアビジンに再懸濁し、氷上で20分間インキュベートする。これは、既に結合しているストレプトアビジン-FITCに対して競合させて、強く結合しているストレプトアビジン-FITCだけが残るようにするためである。次に、細胞を氷冷PBSで1回洗浄し、500μlのPBSに再懸濁した後で、上記で記載したようにして最も蛍光性が高い1%の集団をソーティングする。
【0153】
このソーティングプロトコールを更に2回繰り返した後、Ramos細胞集団(Ram E)は、ストレプトアビジン-FITCに極めて強く結合すると思われる。これらの細胞は、発現された重鎖V遺伝子の第1フレームワーク中で別のアミノ酸の変化を獲得している。すなわち、10位のアミノ酸がグリシンからアルギニンへと変化している。更に、18番目の残基がロイシンからメチオニンへと変化している。
【0154】
Ramos細胞におけるストレプトアビジン成熟の結果を図17に示す。
【0155】
ELISAによる比較
各種のRamos細胞集団の上清を用いて行ったELISAアッセイにより、Ramos細胞から発現され分泌されたIgM抗体が、in vitroで成熟して、ストレプトアビジンに対する強い親和性を獲得していることが確認される。結果を図18に示す。
【0156】
実施例7:高頻度突然変異誘導配列を含む導入遺伝子の構築
Ig遺伝子の遺伝子座の特定のエレメントが、in vivoでの高頻度突然変異事象の誘導に必要であることが知られている。例えば、イントロンエンハンサーおよびマトリックス結合領域であるEi/MARは、極めて重要な役割を担っていることが実証されている(Betzら1994)。更に、3’側エンハンサーであるE3’が重要であることが知られている(Goyenecheaら, 1997)。しかし、本発明者らは、これらのエレメントは必要ではあるが、導入遺伝子内での高頻度突然変異を誘導するには不十分であることを示した。
【0157】
これに対して、Ei/MARおよびE3'を、更なるJκ-CκイントロンDNAおよびCκと共に提供すれば、高頻度突然変異性を十分にもたらすことができる。βG-Cκ導入遺伝子は、PCRで作製した0.96KbのKpnI-SpeI β-グロビン断片(これはβ-グロビン転写開始部位を基準にして-104から+863まで及ぶものであり、その末端に人工的なKpnIおよびSpeI制限部位を有している)をLκΔ[3’Fl](Betzら, 1994)の部分断片(これはMaxら(1081)の配列のヌクレオチド2314からEi/MAR、CκおよびE3’まで及ぶものであり、3’Fl欠失を含んでいる)に結合することにより構築される。
【0158】
高頻度突然変異は、Pfuポリメラーゼを用いてPCR増幅した導入遺伝子のセグメントを配列決定することにより評価される。増幅された領域は、転写開始部位のすぐ上流からJκ5の300ヌクレオチド下流までに及ぶ。
【0159】
このキメラ導入遺伝子は、正常なIgκ導入遺伝子で見られるものと同様の頻度で蓄積するヌクレオチド置換による突然変異の良好な標的になる。この導入遺伝子は、これまでに記載されているものの中で最も小さく、高頻度突然変異を効率的に収集するものであり、その結果から、Cκを含みそれに隣接する領域のどこかに位置する複数の配列が組み合わさって、β-グロビン/Igκキメラの5’末端へ高頻度突然変異を収集することが示される。
【0160】
したがって、高頻度突然変異の収集は、高頻度突然変異ドメインの3’末端に向かって位置する配列によってのみ誘導され得る。しかし、プロモーターの近傍にある正常なIg遺伝子内の突然変異ドメインの5’側境界は、転写開始部位の100〜200ヌクレオチド程度下流である。開始部位に対する突然変異ドメインの5’側境界のこの配置は、β-グロビン遺伝子がプロモーターおよび突然変異ドメインの大部分の双方をもたらす場合は、βG-Cκ導入遺伝子でもそのまま存在する。これらの結果は、他の導入遺伝子を用いて得られた知見と一致しており、それが突然変異ドメインの5’側境界を規定するプロモーターそれ自体の位置であることを示している。
【0161】
κ調節エレメントの(全てではないとしても)幾つかが突然変異の収集に関与する様式についての最も簡単な説明は、それらが、高頻度突然変異開始因子を転写開始複合体に組み込むことにより作動することを提唱することである。転写調節エレメントとしてのエンハンサーについての古典的な研究を用いて類推すれば、Igκエンハンサーは、位置や方向に依存しない様式で高頻度突然変異の調節因子(regulators)として働き得る。事実、βG-Cκ導入遺伝子を用いて得られたデータ、ならびにE3’がよりCκの近くへ移動した、という先の結果(Betzら, 1994)から、E3’の高頻度突然変異増強活性が、突然変異ドメインに対するその位置に対しても方向に対しても特に感受性ではないことが判明する。
【0162】
Ei/MARは、通常、突然変異ドメインの3’末端に向かって位置している。Ei/MARの欠失は、突然変異ターゲティングの効率を著しく低下させるが、それがプロモーターの上流の位置(したがって、転写される領域の外側)に復活すると、突然変異の部分的な救済が起こるが、突然変異ドメインの5’側境界の位置には見かけ上は影響を及ぼさない。これらの結果の独立した確認が、トランスジェニックマウスにおいて、第2の導入遺伝子tk-neo::Cκを用いて得られた。ここでは、neo転写ユニット(HSVtkプロモーターの制御下にある)が、胚性幹細胞においてジーンターゲティングすることによりCκエキソンに組み込まれる(Zouら, 1995)。このマウスでは、Vκ-Jκ結合の後、Igκ Ei/MARは両側を転写ドメイン(上流側はV遺伝子、下流側はtk::neo)で挟まれる。neo挿入についてホモ接合型であるマウス由来のソーティングした胚中心B細胞からtk-neo遺伝子をPCR増幅する。
【0163】
tk-neo::Cκマウスにおけるtk-neoインサートについて、増幅された領域は、(プラスミドpMCNeo(GenbankアクセッションU43611)中で番号付けした場合の)残基607から1417の及び、629位から1329位までのヌクレオチド配列を決定する。tk-neo::Cκマウスにおける内在性VJκの再構成物の突然変異頻度は、Meyerら, 1996に記載されているのと同様の戦略を用いて決定する。内在性VJκ5の再構成物は、Vκ FR3コンセンサスフォワードプライマー(GGACTGCAGTCAGGTTCAGTGGCAGTGGG)と、Jκクラスターの下流から後方にプライミングするオリゴヌクレオチドLκFOR(Gonzalez-FernandezおよびMilstein, (1993) PNAS (USA) 90:9862-9866)を用いて増幅する。
【0164】
tk-neoの突然変異レベルは低く、同じ細胞集団において再構成されたVκ遺伝子の3’側隣接領域よりも間違いなく低い効率で突然変異の標的となるが、neo遺伝子インサート内の突然変異ドメインは、(正常なV遺伝子の場合と同様に)Ei/MARがプロモーターの上流にあるにもかかわらず、転写開始部位の100ヌクレオチド以上下流から始まると思われる。
【0165】
従って、構成的高頻度突然変異を起こす細胞系において高頻度突然変異を誘導できる導入遺伝子は、Ei/MAR、E3'、および本明細書中で定義されるようなJκの下流に見られる調節エレメントを用いて構築できる。更に、導入遺伝子は、内在性V遺伝子の置換または内在性V遺伝子への挿入(tk-neo ::Cκマウスの場合と同様)により、あるいは、目的のコード配列をJκイントロンと連結させること(βG-Cκ導入遺伝子の場合と同様)により構築できる。
【0166】
実施例8:構成的高頻度突然変異を起こす細胞系の選別
上記で記載したようなごく一部のV遺伝子の変換事象により、非機能性Ig遺伝子が、最も高頻度にはフレームシフト突然変異の導入により作製され得る。したがって、ニワトリ・ファブリキウス嚢リンパ腫細胞系DT40におけるsIgM欠損変異体の作製を用いて、IgV遺伝子の変換活性の初期指標を得ることができる。親DT40系と比較して、Rad54が欠損している突然変異体は、かなり低下した割合でsIgM欠損変異体を示す(図19)。複数のクローンについて彷徨解析を行うと、ΔRAD54系は、親DT40系の場合のほぼ1/10の頻度でsIgM欠損変異体を生じ、一方、ΔRAD52系は、野生型細胞とほぼ同様の頻度でsIgM欠損変異体を生じることが明らかになる(図19)。これらの知見は、ΔRAD54細胞およびΔRAD52-DT40細胞における遺伝子変換に関するこれまでの知見(Bezzubovaら, 1997;Yamaguchi-Iwaiら, 1998)と一致している。
【0167】
この分析を、Xrcc2およびXrcc3が欠損しているDT40細胞に適用する。これらのRad51パラログ体は、DNA損傷修復の組換え依存的経路において役割を果たしていることが提唱されている(Liuら, 1998; Johnsonら, 1999;Brennemanら, 2000;Takataら, 2001)。ΔXRCC2系およびΔXRCC3系は、sIgM欠損変異体の存在量の低下を引き起こすのではなく、親の系よりもずっと高い欠損変異体の蓄積を示す(図19)。ΔXRCC2-DT40の場合、ヒトβ-グロビンプロモーターの制御下でのヒトXrcc2 cDNAのトランスフェクションにより、sIgM欠損変異体の生じる頻度が野生型の値近くまで復帰する。図19に、野生型および修復欠損のDT40細胞によるsIgM欠損変異体の生成を示す。単一のsIgM+の正常(WT)または修復欠損(ΔRAD54、ΔRAD52、ΔXRCC2、ΔXRCC3)DT40細胞を1ヶ月間クローン性増殖させたものから誘導した培養物におけるsIgM発現の異質性のフローサイトメトリー分析をパネル(a)に示す。3種の代表的なsIgM+前駆体クローンから誘導した培養物の分析を、修復欠損DT40の各タイプについて示す。各分析におけるsIgM−細胞のパーセンテージを、sIgM+のピークの中心の1/8として設定した蛍光ゲートと共に示す。パネル(b)は、sIgM欠損変異体の生成頻度の彷徨解析を示す。sIgM欠損変異体の存在量は、1ヶ月間クローン性増殖させた後のsIgM+単一細胞から誘導した複数の並行した培養物において測定する。各データセットの上にパーセンテージ中央値を注記し、破線で示してある。分析した[pβG-hXRCC2]ΔXRCC2トランスフェクト体は、彷徨解析において6.4%および10.2%のsIgM−細胞を含むsIgM+ DT40-ΔXRCC2サブクローンにpβG-hXRCC2をトランスフェクトすることにより作製する。複数の独立したsIgM+クローン(異なるが、同様の祖先Vλ配列を有する)について全分析を行ったところ、各修復欠損系では、1ヵ月後にsIgM欠損変異体が生じる頻度の平均中央値は、WT(0.4%)、ΔRAD(0.07%)、ΔRAD52(0.4%)、ΔXRCC2(6%)およびΔXCRCC3(2%)である。
【0168】
Xrcc2およびXrcc3の双方の欠損は、染色体の不安定性に関係するが(Liuら, 1998);Cuiら, 1999;Deansら, 2000;Griffinら, 2000)、sIgM欠損変異体の頻度の増大は、Ig遺伝子座内での全体的な再構成または欠失を反映している可能性がある。しかし、ΔXRCC3-DT40の24種のsIgM−サブクローンのサザンブロット分析からは、再構成されたVλを含む6kbのSalI-BamHI断片の欠失や改変は明らかになっていない。
【0169】
したがって、V遺伝子内のより局在化された突然変異がsIgM発現の欠損を説明できるか否かを確認するために、1ヵ月間増殖させた後で野生型DT40、ΔXRCC2-DT40およびΔXRCC3-DT40サブクローンからソーティングしたsIgM−細胞の集団における再構成されたVλセグメントをクローニングし、配列決定する。
【0170】
細胞培養、トランスフェクションおよび分析
DT40サブクローンCL18およびその突然変異体を、7%のウシ胎仔血清、3%のニワトリ血清(Life Technologies)、50μMの2-メルカプトエタノール、ペニシリンおよびストレプトマイシンを補充したRPMI 1640中で37℃にて10%CO2中で増殖させる。細胞密度は、培養物を毎日分割することにより0.2〜1.0×10-6個/mlに維持した。ターゲティングされた遺伝子破壊を保有するDT40誘導体の作製については、別の文献に記載されている(Bezzubovaら, 1997;Yamaguchi-Iwaiら, 1998;Takataら, 1998, 2000, 2001)。β-グロビンプロモーターの制御下にあるXRCC2のオープンリーディングフレーム(HeLaのcDNAからクローニングされたもの)を含んでいるpSV2-neoベースのプラスミドを保有するΔXRCC2-DT40のトランスフェクト体を、エレクトロポレーションにより作製する。
【0171】
CL18は、DT40のsIgM−サブクローンであり、本明細書で記載されているDNA修復突然変異体の親クローンである。複数のsIgM+サブクローンを、FITC-コンジュゲートヤギ抗-ニワトリIgM(Bethyl Laboratories)で染色した後、野生型および修復欠損突然変異体の両者からMo-Flo(Cytomation)ソーターを用いて得る。この研究で用いたsIgM+ DT40-CL18由来の修復欠損細胞の全てにより発現される初期のVλ配列に違いは殆どない。何故ならば、ほぼ全てのsIgM+誘導体が、ψV8ドナー(これはフレームシフトしたCL18のCDR1と最も近縁である)を用いた遺伝子変換によって元のCL18のVλフレームシフトを復元しているからである。
【0172】
突然変異の分析
ゲノムDNAを、5000個の細胞同等物から、Pfu Turbo(Stratagene)ポリメラーゼおよびホットスタートタッチダウンPCR[95℃で1分、68〜60℃(サイクル毎に1℃ずつ下げる)で1分、72℃で1分30秒を8サイクル;94℃で30秒、60℃で1分、72℃で1分30秒を22サイクル]を用いてPCR増幅する。再構成されたVλはCVLF6(5’-CAGGAGCTCGCGGGGCCGTCACTGATTGCCG;リーダーVλイントロン内でプライミングする)およびCVLR3(5’-GCGCAAGCTTCCCCAGCCTGCCGCCAAGTCCAAG;Jλの3’側から逆方向にプライミングする)を用いて;再構成されていないVλ1はCVLF6およびCVLURR1(5’-GGAATTCTCAGTGGGAGCAGGAGCAG)を用いて;再構成されたVH遺伝子はCVH1F1(5’-CGGGAGCTCCGTCAGCGCTCTCTGTCC)およびCJH1R1(5’-GGGGTACCCGGAGGAGACGATGACTTCGG)を用いて;Cλ領域はCJCIR1F(5’-GCAGTTCAAGAATTCCTCGCTGG;Jλ-Cλイントロンの内部からプライミングする)およびCCMUCLAR(5’-GGAGCCATCGATCACCCAATCCAC;Cλの内部から逆方向にプライミングする)を用いて、増幅する。QIAquickスピンカラム(Qiagen)で精製した後、PCR産物を適当な制限酵素で切断し、pBluescriptSKにクローニングし、T3もしくはT7プライマーおよびABI377シーケンサー(Applied Biosystems)を用いて配列決定する。GAP4との配列アライメント(Bonfieldら, 1995)により、各クローンのコンセンサス配列からの変化が同定できる。
【0173】
全ての配列の変化は、3つのカテゴリー(遺伝子変換、点突然変異、または不明確であるカテゴリー)のうちの1つに割り当てられる。この識別は、遺伝子変換の供与体として働き得るVλの偽遺伝子の公表されている配列に基づく。そのような供与体の配列のデータベースはReynaudら(1987)から取るが、DT40において発現されたIgλに対応する(Kimら, 1990)Igλ G4対立遺伝子に関して修正(McCormackら, 1993)を加えている(この研究において同定された配列/遺伝子変換は、このψVλ配列のデータベースの有効性を支持するものであった)。それぞれの突然変異について、Vλ偽遺伝子のデータベースを可能性のある供与体について検索する。9bp以上の文字列を含む偽遺伝子の供与体が見つからない場合には、鋳型のない点突然変異として分類する。そのような文字列が同定され、同じ供与体により説明可能な更に別の突然変異が存在する場合は、これら全ての突然変異は、1つの遺伝子変換事象に割り当てられる。更に別の突然変異が存在しない場合、単離された突然変異は何らかの変換機構により生じたか、あるいは鋳型がなかった可能性があり、したがって、不明確であるとして分類される。
【0174】
複数の野生型DT40クローンからソーティングされたsIgM−亜集団からクローニングしたVλ配列については、図20に示すように、67%が突然変異を含んでおり、多く(73%)の場合、これらの突然変異によって、Vλは明らかに非機能性になっている。おそらく、残存するsIgM−細胞の大部分が、VH、またはVλの配列決定領域の外側に、不活性化突然変異を保有している。図20に、sIgM欠損変異体からクローニングしたVλ配列の分析を示す。パネル(a)は、1ヶ月間クローン性増殖させた後の正常DT40細胞またはXrcc2欠損DT40細胞の親sIgM+クローンからソーティングされたsIgM欠損細胞から得たVλ配列の比較である。横線はそれぞれ、上記で記載されているようにして点突然変異(棒付の丸)、遺伝子変換域(線上の横棒)、または点突然変異もしくは遺伝子変換の結果であると思われる一ヌクレオチド置換(不明確、縦棒)として分類された突然変異を含む再構成されたVλ1/Jλ(427bp)を表わす。線にかかっている四角の白枠は欠失を表わし、三角は重複を表わす。円グラフをパネル(b)に示すが、これには1ヶ月間クローン性増殖させた後の野生型、ΔXRCC2またはΔXRCC3 DT40 sIgM+クローンから誘導したソーティングされたsIgM欠損集団の中で、異なる数の点突然変異(PM)、遺伝子変換(GC)または起源が不明確な突然変異(Amb)を保有するVλ配列の割合が示されている。それらの円弧の大きさは、円の周囲に示す数の突然変異を保有する配列の数に比例する。分析したVλ配列の総数は、各円の中心に示し、データは、野生型DT40の4種のサブクローン、ΔXRCC2-DT40の2種のサブクローンおよびΔXRCC3-DT40の3種のサブクローンの分析から集計したものである。欠失、重複および挿入は、この分析から除外されている。野生型細胞では、更に6つの欠失、1つの重複および1つの挿入が存在する。ΔXRCC2-DT40では他の事象は起こっておらず、ΔXRCC3-DT40のデータベースでは1bpの欠失および1bpの挿入がそれぞれ一例ずつある。
【0175】
同定された不活性化突然変異を含む、野生型、ΔXRCC2(ΔX2)およびΔXRCC3(ΔX3) DT40細胞におけるVλ遺伝子の不活性化の原因[ミスセンス突然変異(黒);遺伝子変換に伴うフレームシフト(白);欠失、挿入もしくは重複に伴うフレームシフト(灰色)]を、全配列に対するパーセンテージとして表してパネル(b)に示す。それぞれの不活性化突然変異に伴う更に別の突然変異事象を(d)に示す。データは、それぞれの不活性化突然変異に伴う更に別の突然変異の平均数と、パネル(c)で示す更なる突然変異のタイプとで表わす。したがって、ΔXRCC2-DT40は、指標となる不活性化突然変異に加えて、平均で1.2個の追加の点突然変異を有するが、野生型DT40ではわずか0.07である。
【0176】
上記で詳細に述べたように、突然変異は、上流のVλ偽遺伝子を鋳型とする遺伝子変換に起因し得るもの、鋳型のない点突然変異に起因し得るもの、または不明確であるカテゴリーに入れられるもの、として分類できる。不活性化突然変異の多く(67%)は遺伝子変換によるものであるが、一部(15%)は鋳型のない点突然変異により生じる停止コドンであり、このことから、本明細書中および他の文献(Buersteddeら, (1985);Kimら, 1990)に見られるDT40細胞での低い点突然変異の頻度はPCRのアーチファクトではなく、低頻度の点突然変異は、実際に野生型DT40での遺伝子変換に伴って生じることが明らかになったことが示される。
【0177】
著しく異なる突然変異パターンは、ΔXRCC2-DT40由来のsIgM欠損変異体のVλ配列で見られる。ほぼ全ての配列が点突然変異を有しており、典型的には、配列当たり複数の点突然変異が含まれる。また、点突然変異への実質的なシフトは、sIgM− ΔXRCC3-DT40細胞由来の配列でも見られる。したがって、野生型DT40におけるVλ不活性化突然変異は、おそらく読み枠が合わない遺伝子変換域を反映していると思われ、ΔXRCC2/3では、それはミスセンス突然変異であると思われる(図20c)。更に、ΔXRCC2-DT40またはΔXRCC3-DT40のソーティングされたsIgM欠損変異体から得られた非機能性Vλ配列の多く(ΔXRCC2-DT40では53%、ΔXRCC3-DT40では64%)は、Vλ不活性化突然変異に加えて、更なる点突然変異を含んでいるが、そのような同乗は、親DT40系由来の非機能性Vλ配列ではごく稀にしか見られない(7%、図20d)。
【0178】
これらの知見は全て、ΔXRCC2/3-DT40細胞におけるsIgM欠損変異体の高い存在率が、これらの細胞における自然発生的なIgV遺伝子の高頻度突然変異の非常に高い頻度をそのまま反映していることを示唆している。図21に、1ヶ月クローン性増殖した後のソーティングされていないDT40集団からクローニングしたIg配列の分析を示す。代表的な野生型およびΔXRCC2 DT40クローンから得たVλ配列を、図20と同様の記号と共にパネル(a)に示す。パネル(b)には、図示されている異なる数の各種タイプの突然変異を含むVλ配列の割合を示す円グラフを示す。データは、独立したクローン:野生型(2種のクローン)、ΔXRCC2(4種のクローン)およびΔXRCC3(2種のクローン)の分析からプールしたものである。図示されている突然変異に加えて、1方のΔXRCC2-DT40配列は、リーダーイントロン内に、供与体である偽遺伝子からの明らかに鋳型のない2bpの挿入を含み、1方のΔXRCC3-DT40配列も、リーダーイントロン内に一塩基対の欠失を含んでいた。
【0179】
ΔXRCC2-DT40の他の遺伝子座にある突然変異をパネル(c)に示す。円グラフには、再構成された重鎖の再構成されたVH(CDR1からJHの末端まで及ぶ272bp)、除外された対立遺伝子上の再構成されていないVλ1(458bp)、およびCλの近傍(Jλ-CλイントロンからCλの最初の132bpまで及ぶ425bp)内に突然変異を含むΔXRCC2-DT40細胞を1ヶ月増殖させたものから誘導される配列の割合を示す。既知のVH偽遺伝子配列の分析(Reynaudら, 1989)からは、再構成されたVHで見られる突然変異のすべてが遺伝子変異によるというわけではないことが示されており、そのことから、それらは点突然変異によるものであるがこの割り当てが完全に決定的であると見なすことはできないことが強く示唆される。これらのデータセットにおける突然変異の存在率は、VHでは1.6×10-3の突然変異/bp、再構成されていないVλ1では0.03×10-3、Cλでは0.13×10-3であり、これと比較して、ΔXRCC2-DT40内の再構成されたVλ1における点突然変異では2.0×10-3、野生型DT40内の再構成されたVλ1における点突然変異では0.13×10-3、およびバックグランドPCRエラーでは0.04×10-3である。
【0180】
Vλ1に入っている点突然変異の分布をパネル(d)に示す。ΔXRCC2-DT40のコンセンサスは大文字で示し、第1の塩基は、リーダーイントロンの76番目の塩基対に相当する。ΔXRCC3-DT40のコンセンサスに見られるバリエーションは、下にイタリック体の文字で示す。突然変異はコンセンサスの上に小文字で示し、ΔXRCC2-DT40に由来するものは黒で、ΔXRCC3-DT40に由来するものはミッドグレイ色で示す。点突然変異および不明確であるカテゴリーに分類される全ての突然変異を含めた。クローン性増殖については、先に記載(Takataら, 1998)されているようにして較正を行っており、したがって、それぞれの小文字は独立した突然変異事象を表わす。27の突然変異(そのため、158の元のデータベースから除去されている)の大部分は、7つの主要なホットスポットの1つに存在する。クローン性についての較正により何らかの歪みが生じる場合、ホットスポットの優先性(dominance)が過少に見積もられることになる。7つの主要なホットスポット(5以上の突然変異が蓄積していることによって同定される)のうち、5つは、黒枠で示される2本の鎖の一方にあるAGYコンセンサス配列と一致する。パネル(e)に示されるヌクレオチド置換の存在率(131の独立した事象からなるデータベースに対するパーセンテージとして表わされる)は、未選択のΔXRCC2-DT40およびΔXRCC3-DT40に由来する配列内の点突然変異から推定される。ΔXRCC2/ΔXRCC3のデータベースをそれぞれ分析した場合、同様の存在率パターンが示される。
【0181】
野生型細胞およびΔXRCC2/3-DT40細胞における自然発生的なVλの突然変異頻度は、サブクローニングの後で1ヶ月間増殖させた全ての(ソーティングされていない)DT40集団から再構成されたVλセグメントをPCR増幅することによって分析される。その結果から、実際に、ΔXRCC2細胞およびΔXRCC3細胞では、親DT40と比べて突然変異の自然発生的蓄積がより高レベルで起こっていることが明らかになる(図21a、b)。ΔXRCC2-DT40細胞では、突然変異は、Vλに約0.4×10-4/bp/世代の速度(分裂時間を約12時間とする)で蓄積し、この値は、構成的に突然変異を起こすヒト・バーキットリンパ腫系Ramosで見られるものとほぼ同じである。
【0182】
ヒトおよびマウスにおける胚中心B細胞での体細胞高頻度突然変異は、主に、再構成された免疫グロブリンのVHセグメントおよびVLセグメントを標的とする。同様の状況が、ΔXRCC2-DT40細胞での点突然変異にも当てはまる。したがって、生産的に再構成されたVH1遺伝子でも有意なレベルで見かけの点突然変異が見られる(図3c)。しかし、これは、全体的な突然変異誘発表現型を反映してはいない。何故ならば、突然変異の蓄積は、Cλでは、再構成されたVλの場合よりもずっと少なく、また除外された対立遺伝子上の再構成されていないVλでも少ないからであり、この場合、見かけの突然変異率はPCR増幅自体が原因となってバックグラウンドレベルを超えない(図21c)。
【0183】
ΔXRCC2-DT40細胞内のVλドメインでの突然変異の分布は、非常に非ランダムである。この突然変異は主に一ヌクレオチド置換であり、2本のDNA鎖の一方におけるAGY(Y=ピリミジン)コンセンサスに一致するホットスポットで選択的な蓄積を示す(図21d)。また、それらは圧倒的(96%)にG/Cで起こっている。ΔXRCC2-DT40細胞におけるこのG/Cに偏ったホットスポット集中型の高頻度突然変異は、ヌクレオチドのトランジションへの偏りは多少低いものの、培養したヒト・バーキットリンパ腫細胞において記載されているV遺伝子の高頻度突然変異のパターンならびにカエル、サメおよびMsh欠損マウスで生じるそのパターン(Radaら, 1998;Diazら, 2001)と非常に類似している。ヒトおよび正常なマウスにおいてin vivoで起こるIgV遺伝子の高頻度突然変異は、先に述べられているように、A/Tを標的とする機構と協働して作用するこのホットスポット集中型のG/Cに偏った成分により達成されると思われる(図21e)。
【0184】
したがって、DT40ニワトリ・ファブリキウス嚢リンパ腫系は、通常は遺伝子変換により低頻度のIgV多様化を示すが、Xrcc2またはXrcc3を欠失させることにより、高頻度の構成的なIgV遺伝子の体細胞突然変異(ヒトB細胞リンパ腫モデルで起こるものと本質的に類似している)を誘導することができる。このことは、IgV遺伝子の変換および高頻度突然変異が、一般的なDNA損傷を解決するための別の方法となり得るという以前の考え(Maizelsら, 1995;Weillら, 1996)を強く支持する。近年のデータから、損傷の始まりは二本鎖の破断であることがよくあるということが示唆され(Sale & Neuberger, 1998;Papavasilouら, 2000;Brossら, 2000)、したがって、Xrcc2およびXrcc3の両者が、DNA破壊の修復の組換え依存的経路に関与していることは重要と思われる(Liuら, 1998;Johnsonら, 1999;Pierceら, 1999;Brennermanら, 2000;Takataら, 2001)。事実、DT40細胞におけるIgV遺伝子の高頻度突然変異の同様の誘導は、その産物が破断の組換え依存的修復に関与する別の遺伝子(RAD51B)(Takataら, 2000)を欠失させることによって達成されるが、非相同的末端結合に関与するKu70およびDNA-PKCSの遺伝子を欠失させた場合には達成されない。図22に、DNA-PK、Ku70およびRad51Bが欠損しているDT40細胞におけるsIgM欠損変異体の分析を示す。1ヶ月間クローン性増殖した後のsIgM欠損変異体の生成頻度の彷徨解析をパネル(a)に示す。比較のために、野生型およびΔXRCC2のDT40を用いて得られた中央値を入れてある。図示されている各種のタイプの突然変異を含む2種のsIgM+ Rad51B欠損DT40クローンから誘導したsIgM欠損変異体から増幅したVλ配列の割合を示す円グラフをパネル(b)に示す。さらに、1つの配列は9bpの欠失を含み、1つは4bpの重複を含み、1つは一塩基対の挿入を含んでいた。
【0185】
しかし、この結果は、Xrcc2の不在下で、通常は遺伝子変換により解消される損傷が、代わりに体細胞高頻度突然変異をもたらすプロセスによって解消されることを単に示唆しているわけではない。第1に、ΔXRCC2-DT40細胞は、レベルは多少低いものの、IgV遺伝子の変換を行う能力を保持している(図21b)。第2に、ΔXRCC2-DT40細胞における高頻度突然変異の頻度は、親のDT40系における遺伝子変換の頻度よりもほぼ一桁高い。したがって、正常なDT40細胞では、IgV遺伝子の損傷のほんの一部が、上流の偽遺伝子由来の鋳型のある修復に供されるにすぎず、そのため、遺伝子変換事象が見られることになると思われる。本発明者らは、損傷の大部分が、姉妹染色分体上に位置する同一のV遺伝子を鋳型として用いる組換え修復に供され、したがって、それらは「隠されて」いると考える。これは、高頻度突然変異を起こす哺乳動物B細胞における検出可能なIgV遺伝子の破壊がG2/S期に限定されることを見出したPapavasiliouおよびSchatz(2000)の知見と一致する。本発明者らは、Xrcc2、Xrcc3またはRad51Bが存在しない場合は、この「隠された」姉妹染色分体依存的組換え修復が逆用され、その結果として高頻度突然変異が起こることを提唱する。この高頻度突然変異が、Xrcc2/3が存在しない場合に姉妹染色分体依存的組換え修復が間違いを起こしやすくなることを反映しているのかどうか、それとも、それがそのような修復の阻害を反映しており、それにより別の鋳型のない破壊解消機構を示しているのかどうか、ということは、検討を要する問題である。この問題は、高頻度突然変異の機構を理解する上で重要であるばかりでなく、Rad51パラログ体の生理学的機能の見通しを与えてくれる。
【0186】
実施例9:Δxrcc2 DT40 IgMの親和性成熟
数ヶ月間増殖させたXrcc2欠損DT40細胞の集団を用いて、これらの細胞におけるユニークなVHDJH/VLJL再構成に対する高頻度突然変異の作用が、被験抗原のかなりの割合に対して抗体の成熟系統を進化させるのに十分な機能的多様性をもたらすことができるか否かを判定した。選別には2つの方法を用いた。第1の方法では、細胞を、FITC-ストレプトアビジンを各種のビオチン化抗原(カゼイン、インスリン、オボアルブミン(Ova)、サイログロブリン(Tg)およびラット・モノクローナル抗体(Ab))と混合することにより形成される可溶性凝集物と共にインキュベートした。次に、結合している変異体をフローサイトメトリーにより濃化した。あるいはまた、変異体は、S. アウレウス(S. aureus)のプロテインAまたはヒト血清アルブミン(HAS)で被覆した磁性ビーズを用いて選別した。試験した抗原はいずれも、検出可能な親DT40またはその分泌されたIgMに対する結合を示さなかった。選別を6ラウンド連続して行った後で、本質的に3つのタイプの結果が得られた。すなわち、インスリンおよびカゼインの場合、結合している変異体が濃化されたという証拠はなかった。ラット・モノクローナル抗体およびプロテインAの場合、特異的な結合体が得られた。HAS、OvaおよびTgの場合、結合体は得られたが、これらは様々な程度で多重反応性(polyreactivity)を示した。したがって、選別をビオチン化ラットIgG mAb S7とFITC-ストレプトアビジンとの凝集物を用いて行った場合(図23)、特異的な結合は、既に第3ラウンドまでに示された。それに続く濃化は、フィコエリトリン(PE)に直接コンジュゲートさせたラットIgG S7 mAbを用いて行った。第6ラウンドで得られた細胞(DT-Ab6)は、種々の他の試薬で染色されないことにより判定される通り、ラットIgG S7 mAbに特異的であった。培養上清のELISAから、DT-Ab6細胞によるS7 mAbの結合がDT-Ab6 IgM自身によりもたらされることが実証された(図23b)。このDT-Ab6 IgMは、実際に抗-S7イディオタイプである。それは、精製されたS7 mAbを認識する(そして、S7ハイブリドーマ透過化細胞を染色するのに使用できる)が、この相互作用は、他のラット免疫グロブリンによる競合を受けない(図23c、d)。DT-Ab6細胞は、高希釈率のPES7 mAbコンジュゲートで染色できた。このことから、高親和性の相互作用が示唆される。分子PE-S7 mAb:DT-Ab6細胞を固定比率で、しかし容量を変えて染色を行うことにより、5.8 nMの範囲の親和性が推定された(図23f)。
【0187】
配列分析から、DT-Ab6 VH/VLが、親DT40の配列と比較して、合計19のアミノ酸置換を含むことが判明した(図23g)。
【0188】
ビオチン化プロテインAで被覆したストレプトアビジンビーズを用いて行ったビーズ選別では、結合している変異体は第2ラウンドまでに現われた(DT-P2;図24a)。しかし、DT-P2集団中の細胞の多くは、ストレプトアビジンもプロテインAも提示していない種々の他のタイプのビーズにも結合した、という点に問題があった。プロテインAを直接コンジュゲートさせたトシル化ビーズを用いて更に濃化したところ、FITCコンジュゲートプロテインA凝集物で染色可能な細胞(DT-P4)の集団が得られた(図24b)。フローサイトメトリーの使用によるこの凝集物への結合に対する一連の選別により、凝集していないFITC-プロテインAで僅かに染色可能な集団(DT-P9)が得られた。次に、FITC-プロテインAを用いて更にソーティングしたところ、もっと明るく染色される子孫が得られた(図24b)。DT-P14細胞により(ならびにそのいくつかの前駆細胞により)分泌されたIgMは、ELISAおよび免疫沈降法の両者により判定したところ、プロテインAに良好に結合した(図24c、d)。放射標識したプロテインAを用いた直接結合アッセイから、DT-P9とDT-P14の間で親和性の実質的な増大が起こったことが示され(図24e)、これはフローサイトメトリー分析(図24b)と一致する。この増大は、完全に、VHのCDR2に隣接したAla→Val置換によるものであり(図24h)、これにより、プロテインAに対する見掛けの親和性が0.32nMであるIgMが生じた(図24f)。興味深いことに、DT-PのIgMとプロテインAとの相互作用は、十分に特性付けが行われているプロテインAとウサギIgGのFc部分との相互作用とは、DT-P/プロテインAの相互作用がIgM分子のC部分ではなくV部分とのものである(突然変異はCμではなくVHで見られる)という事実、ならびに、高濃度のウサギIgGはFITC-プロテインAによるDT-P4/P9細胞の染色を阻害しないので、おそらくプロテインA上の異なる部位がDT-P IgMおよびウサギIgGとの相互作用に用いられるという事実において異なる(図24g)。実際に、染色の増強が見られ、おそらくこれは、ウサギIgGによるプロテインAの凝集によるものと考えられる。
【0189】
DT-P14のVH/VLの配列から、親DT40配列と比較して、17のアミノ酸置換が明らかになっている。これらの突然変異のうち5つは、ラット・イディオタイプ特異的変異体DT-Ab6と共通する。しかし、この5つの突然変異のうち少なくとも4つは、(Xrcc2欠損DT40細胞のプールにおける共通の突然変異した前駆体からの派生していることを反映しているのではなく)その2つの系統において独立して起こったはずである。何故ならば、DT-P14とDT-Ab6とに共通するこれら4つの突然変異体は、DT-P4では見られないからである。置換を繰り返すこの傾向は、DT40における突然変異の殆どが、ホットスポット集中型のGCに偏った第1段階の突然変異(本発明者らがA/Tに偏った第2段階によるものとした突然変異の範囲の一部を欠いている;SaleおよびNeuberger, 1998;Radaら, 1998)に限定されていることをある程度反映していると思われるが、それらが、抗体構造を成熟させやすくすることによって利点を付与している可能性もある。Xrcc2欠損DT40細胞は(体細胞高頻度突然変異よりもずっと低い頻度ではあるが)IgV遺伝子の変換を行う能力を保持しているが、DT-P14細胞およびDT-Ab6細胞におけるV遺伝子の配列(図24h)を生殖細胞系のVセグメントの配列(Reynaudら, 1987, 1989)と比較したところ、その変化は、(全部でないとしても)大部分が遺伝子変換ではなく点突然変異によるものであることが判明したことは注目に値する。HSA誘導体化カルボキシル化磁性ビーズを用いた選別を6ラウンド行った後で得られた細胞(DT-H6)は、FITC-HASだけでなく、FITC-Tg、FITC-Ovaおよびサイクローム-コンジュゲートストレプトアビジンによっても染色された(DT-H6細胞集団がこれらの他の抗原に暴露されたことはない、という事実にもかかわらず)。HASを用いて更に選別したところ、多重特異性染色(polyspecific staining)の明るさが単純に増大した(図25a)。FITC-ストレプトアビジンとビオチン化Tgもしくはビオチン化Ovaとの複合体を用いたフローサイトメトリーによる濃化に供した細胞集団では、類似しているが異なる多重特異性パターンが示された(図25a、d)。
【0190】
亜集団を分析したところ、この見掛けの多重特異性が細胞の異質性を反映していないことが示された。sIgM欠損変異体には抗原結合活性が欠けているので、多重特異的染色は表面IgM自体によって仲介された。更に、抗原結合は、培養物上清のELISAにより容易に検出される(図25b、c)。
【0191】
多重反応性抗体はヒトおよびマウスにおいて(血清中およびハイブリドーマ間で)十分に報告されており、それらが単一特異性(monospecificity)が進化するための良好な開始点を構成するか否かという問題が提起されてきている(CasaliおよびSchettino, 1996;BouvetおよびDighiero, 1998)。in vitroでの選別系でも同じ問題が出てきている(自己免疫を回避するという制約はないが)。したがって、Ovaに対する特異性を増大させる方向にDT-O6集団を進化させることが可能か否かを、フローサイトメトリーによるFITC-Ovabright/サイクローム-ストレプトアビジンdull細胞の濃化によって試験した。ソーティングを複数ラウンド行った後、FITC-Ovaに対してより高い特異性を示す細胞を得た(図25d)。多重特異的結合集団が親DT40集団よりも良好な特異性進化の開始点を提供するか否かを確認することは興味深い。
【0192】
ここで示す結果から、高頻度突然変異を起こす細胞系が、in vitroでの抗体の誘導および反復的な成熟の双方に使用可能であることがはっきりと実証される。DT40の遺伝学的な扱いやすさを考慮すれば、この用途をトランスフェクトしたIgV遺伝子にまで拡大し、そうすることにより既存の抗体の親和性を成熟させることが可能である。更に、高頻度突然変異機構は、再構成されたIgVセグメントの代わりに配置された異種の遺伝子を標的とすることができるので(Yelamosら, 1995)、この戦略を、他のリガンド/受容体ペアの成熟にまで拡張できる公算は十分にあると思われる。抗体のde novo選別については、単一のVH/VL再構成に体細胞高頻度突然変異を作用させることにより、試験した7種の抗体のうち2種に対して高親和性の結合物を選別して成熟させること、ならびにそれ以外のもののうち3種に対する初期の低親和性結合の兆候(成熟可能ではないが)を得ることが可能なレパートリーが得られたことは注目すべきである。これは、抗原親和性が非常に低くても、選別を開始するには十分であるという事実を反映している。かくして、これらの結合部位は成熟可能である。明らかに、この方法を、高親和性のヒト・モノクローナル抗体のin vitro産生が可能になるように外挿したい場合は、DT40の遺伝学的易操作性を利用して、2つ以上のVH/VL再構成を含む一次レパートリーを作製することが有利であろう。ここで得られた結果から、この再構成の一次レパートリーはin vivoで用いられる一次レパートリーよりも数桁少ないことが示される。
【0193】
実施例10:天然に存在する、構成的に高頻度突然変異を起こすEBVポジティブBL細胞系の単離
天然に存在するEBV+BL細胞系を調べたところ、それら(例えば、Akata, BL74, Chep, Daudi, Raji,およびWan)の多くの中に、明らかに同定可能なsIgM欠損変異体の集団は存在しないことが判明した。しかし、これらEBV+細胞系のうち2つ(ELI-BLおよびBL16)において明確なsIgM−/low集団が確認され、このことから、本来備わっている高頻度突然変異能であることが示唆される。Ramos、EHRB、ELI-BLおよびBL16のsIgM発現プロファイルを図26aに示す。sIgM−/low細胞集団は枠で囲み、その中の細胞のパーセンテージを示す。各ドットが1個の細胞を示す。BL16における相当大きなsIgM−/low集団の一部は、それほど強くは染色にポジティブではない細胞によるものであり、これもまた、彷徨解析を妨害したことに注意されたい。ELI-BLは2型のEBVを含み、胚中心B細胞に類似しており、EBNA1および非コードEBERおよびBam A RNAのみから構成される潜伏遺伝子レパートリーを発現する(Roweら,1987)。BL16もまた2型ウイルスを含んでいるが、ELI-BLとは違って、よりLCLに類似していると思われ、完全に潜伏している遺伝子のレパートリーを発現する(Rooneyら, 1984;Roweら, 1987)。
【0194】
ELI-BLおよびBL16の培養物中に明確なsIgM−/low集団が見られるが、これらの変異体が真の高頻度突然変異に起因しているのか否かを検討することは重要である。これは、彷徨解析により検討した。簡単に説明すると、(サブ)クローンを24ウェルまたは48ウェルのプレートに移し、新鮮な培地と共に3〜8週間維持し、細胞(1〜2.5×105個)をPBS/3%FBS中で2回洗浄し、適切な抗体もしくは抗体の組合せ(後記)で染色し(氷上で30分間)、再度洗浄した後、少なくとも104個の細胞をフローサイトメトリー(FACSCalibur、Becton Dickinson)により分析することにより分析した。使用した抗体は、R-フィコエリトリン-コンジュゲートしたヤギ抗-ヒトIgM(μ鎖特異的;Sigma)、フルオレセインイソチオシアネート(FITC)-コンジュゲートしたマウス・モノクローナル抗-Ramosイディオタイプ[ZL16/1(Zhangら,1995);M. CraggおよびM. J. Glennie(Tenovus Research Laboratory, Southampton)から恵贈]、およびFITC-コンジュゲートしたヤギ抗-マウスIgM(Southern Biotechnology Associates, Inc.)とした。データを得て、CellQuestソフトウェア(Becton Dickinson)を用いて解析した。
【0195】
特に指示がない限り、彷徨解析で比較した細胞は並行して誘導し、培養し、分析した。同様にして誘導した多くの(サブ)クローンにおけるsIgM欠損変異体のパーセンテージ中央値(平均値ではない)を細胞の体細胞高頻度突然変異能の指標として用いて、早期突然変異事象の影響を最低限に抑える。ELI-BLサブクローンの彷徨解析を行ったところ、sIgM−/low変異体が、実際に、in vitro培養の際に実際に高頻度で生成されていることが判明し(図26b;それぞれの×印は、単一のサブクローンを1ヶ月間増殖させた後のsIgM−/lowウィンドウに含まれる細胞のパーセンテージを表わす;パーセンテージ中央値を示す)、BL16サブクローンの場合、VH配列の分析から、この不安定さが体細胞高頻度突然変異を反映していることが確認された(図26c)。塩基置換突然変異は、大文字のトリプレットで示されている338bpのコンセンサスDNA配列の上に小文字で示す。相補性決定領域および部分PCRプライマー配列には、それぞれ下線をひき、太字にしてある。対応するアミノ酸配列は、大文字1文字によって示す。このコンセンサス配列は、GenBankのエントリー配列gi.2253343とは2つの位置で異なっている[TCA(Ser20)−TCTおよびAGC(Ser55)−ACC(Thr)]。
【0196】
複数の塩基置換突然変異を有する数種の配列を含むVH配列の相当な多様性および全体として高いVH突然変異の頻度から、BL16において高頻度突然変異が進行中であることが示された。更に、比較的少数のVH配列しかサンプリングしなかったにもかかわらず、1つの系統的関係が推定できた[Gly54での第1の突然変異(GGT−GAT);Val92での第2の突然変異(GTG−ATG)]。最後に、Ramosと同様に、BL16のVHの塩基置換突然変異の多く(24/33、すなわち73%)がGまたはCのヌクレオチドで起こり、相補性決定領域内(図26cの下線部)に集中していた。したがって、2種の天然のEBV+ BL細胞系(一方は限定的な潜伏遺伝子レパートリーを発現し、他方は全ての組合せを発現する)では、進行中の高頻度突然変異の幾つかの特徴が区別できる。したがって、体細胞高頻度突然変異が、EBVの存在下でも衰えることなく進行可能であることは明らかであった。
【0197】
参考文献
Adetugbo, K., Milstein, C. and Secher, D.S. (1977). Molecular analysis of spontaneous somatic mutants. Nature 265, 299-304.
Bahler, D.W. and Levy, R. (1992). Clonal evolution of a follicular lymphoma: Evidence for antigen selection. Proc. Natl. Acad. Sci. USA 89, 6770-6774.
Bass, S., R. Greene, and J.A. Wells. (1990). Hormone Phage: An Enrichment Method for Variant Proteins With Altered Binding Properties. Proteins. 8, 309-314.
Berek, C. and Milstein, C. (1988). The dynamic nature of the antibody repertoire. Immunol. Rev. 105, 5-26.
Betz, A. G., Milstein, C., Gonzalez-Fernandez, A., Pannell, R., Larson, T. and Neuberger, M. S., Cell 1994. 77: 239-248.
Betz, A.G., Neuberger, M.S. and Milstein, C. (1993). Discriminating intrinsic and antigen-selected mutational hotspots in immunoglobulin V genes. Immunol. Today 14, 405-411.
Bezzubova, O., Silbergleit, A., Yamaguchi-Iwai, Y., Takeda, S. & Buerstedde, J. M. Reduced X-ray resistance and homologous recombination frequencies in a RAD54-/- mutant of the chicken DT40 cell line. Cell 89, 185-193. (1997).
Bonfield, J.K., Smith, K.F. and Staden, R. (1995). A new DNA-sequence assembly program. Nucleic Acids Res. 23, 4992-99.
Bouvet J.P. & Dighiero, G. From natural polyreactive autoantibodies to a la carte monoreactive antibodies to infectious agents: is it a small world after all? Infect. Immun. 66, 1-4 (1998).
Braeuninger, A., Kuppers, R., Strickler, J.G., Wacker, H.-H., Rajewsky, K. and Hansmann, M.-L. (1997). Hodgkin and Reed-Sternberg cells in lymphocyte predominant disease represent clonal populations of germinal center-derived tumor B cells. Proc. Natl. Acad. Sci. USA 94, 9337-9342.
Brenneman, M. A., Weiss, A. E., Nickoloff, J. A. & Chen, D. J. XRCC3 is required for efficient repair of chromosome breaks by homologous recombination. Mutat Res 459, 89-97. (2000).
Bross, L., Fukita, Y., McBlane, F., Demolliere, C., Rajewsky, K. & Jacobs, H. DNA double-strand breaks in immunoglobulin genes undergoing somatic hypermutation. Immunity 13, 589-597. (2000)
Bruggemann, M., Radbruch, A. and Rajewsky, K. (1982). Immunoglobulin V region variants in hybridoma cells. I. Isolation of a variant with altered idiotypic and antigen binding specificity. EMBO J. 1, 629-634.
Buerstedde, J. M., Reynaud, C.A., Humphries, E.H., Olson, W., Ewert, D.L. & Weill, J.C. Light chain gene conversion continues at high rate in an ALV-induced cell line. Embo J 9, 921-927. (1990)
Capizzi and Jameson, (1973) Mutat. Res. 17:147-8
Casali P. & Schettino, E.W. Structure and function of natural antibodies. Curr. Top. Microbiol. Immunol. 10, 167-179 (1996).
Chapman, C.J., Mockridge, C.I., Rowe, M., Rickinson, A.B. and Stevenson, F.K. (1995). Analysis of VH genes used by neoplastic B cells in endemic Burkitt’s lymphoma shows somatic hypermutation and intraclonal heterogeneity. Blood 85, 2176-2181.
Chapman, K.B. and Szostak, J.W. (1994) Curr. op. Struct. Biol., 4, 618-622.
Chui, Y.-L., Lozano, F., Jarvis, J.M., Pannell, R. and Milstein, C. (1995). A reporter gene to analyse the hypermutation of immunoglobulin genes. J. Mol. Biol. 249, 555-563.
Clackson, T. and Wells, J.A. (1994) Trends Biotechnol, 12, 173-84.
Coffino, P. and Scharff, M.D. (1971). Rate of somatic mutation in immunoglobulin production by mouse myeloma cells. Proc. Natl. Acad. Sci. USA 68, 219-223.
Cui, X., Brenneman, M., Meyne, J., Oshimura, M., Goodwin, E.H. & Chen D.J. The XRCC2 and XRCC3 repair genes are required for chromosome stability in mammalian cells. Mutat Res 434, 75-88. (1999).
Cull, M.G., Miller, J.F. and Schatz, P.J. (1992) Proc Natl Acad Sci U S A, 89, 1865-9.
Deans, B., Griffin, C. S., Maconochie, M. & Thacker, J. Xrcc2 is required for genetic stability, embryonic neurogenesis and viability in mice. Embo J 19, 6675-6685. (2000).
Denepoux, S., Razanajaona, D., Blanchard, D., Meffre, G., Capra, J.D., Banchereau, J and Lebecque, S. (1997). Induction of somatic mutation in a human B cell line in vitro. Immunity 6, 35-46.
Diaz, M., Flajnik, M. F. & Klinman, N. Evolution and the molecular basis of somatic hypermutation of antigen receptor genes. Philos Trans R Soc Lond B Biol Sci 356, 67-72. (2001)
Drake, J.W. (1998) Genetics 148:1667-1686.
Ellington, A.D. and Szostak, J.W. (1990) Nature, 346, 81822.
Gold, L., Polisky, B., Uhlenbeck, 0. and Yarus, M. (1995) Annu Rev Biochem, 64, 763-97.
Goossens, T., Klein, U. and Kuppers, R. (1998). Frequent occurrence of deletions and duplications during somatic hypermutation: Implications for oncogenic translocations and heavy chain disease. Proc. Natl. Acad. Sci. USA 95, 2463-2468
Goyenechea, B., Klix, N., Yelamos, J., Williams, G. T., Riddell, A., Neuberger, M. S. and Milstein, C., EMBO J. 1997. 16, 3987-3994
Green, N.S., Lin, M.M. and Scharff, M.D. (1998). Immunoglobulin hypermutation in cultured cells. Immunol. Rev. 162, 77-87.
Green, R. and Szostak, J.W. (1992) Science, 258, 1910-5.
Griffin, C. S., Simpson, P. J., Wilson, C. R. & Thacker, J. Mammalian recombination-repair genes XRCC2 and XRCC3 promote correct chromosome segregation. Nat Cell Biol 2, 757-761. (2000)
Jain, R., Roncella, S., Hashimoto, S., Carbone, A., Franco di Celle, P., Foa, R., Ferrarini, M and Chiorazzi, N. (1994). A potential role for antigen selection in the clonal evolution of Burkitt’s lymphoma. J. Immunol. 153, 45-52.
Johnson, R. D., Liu, N. & Jasin, M. Mammalian XRCC2 promotes the repair of DNA double-strand breaks by homologous recombination. Nature 401, 397-399. (1999).
Jolly, C.J., Klix, N. and Neuberger, M.S. (1997). Rapid methods for the analysis of immunoglobulin gene hypermutation: application to transgenic and gene targeted mice. Nucleic Acids Res. 25, 1913-1919.
Jolly, C.J., Wagner, S.D., Rada, C.A., Klix, N., Milstein, C. and Neuberger, M.S. (1996). The targeting of somatic hypermutation. Semin. Immunol. 8, 159-168.
Joyce, G.F. (1994) Curr. op. Structural Biol., 4, 331-336.
Kim, S., Humphries, E. H., Tjoelker, L., Carlson, L. & Thompson, C. B. Ongoing diversification of the rearranged immunoglobulin light-chain gene in a bursal lymphoma cell line. Mol Cell Biol 10, 3224-3231. (1990)
Klein, G., Giovanella, B., Westman, A., Stehlin, J. and Mumford, D. (1975). An EBV negative cell line established from a American Burkitt lymphoma; receptor characteristics, EBV infectability and permanent conversion into EBV-positive sublines by in vitro infection. Intervirology 5, 319-334.
Klix et al., (1998) Eur. J. Immunol. 28:317-326.
Liu, N., Lamerdin, J.E., Tebbs, R.S., Schild, D., Tucker, J.D., Shen, M.R., Brookman, K.W., Siciliano, M.J., Walter, C.A., Fan, W., Narayana, L.S., Zhou, Z.Q., Adamson, A.W., Sorenson, K.J., Chen, D.J., Jones, N.J. & Thompson, L.H. XRCC2 and XRCC3, new human Rad51-family members, promote chromosome stability and protect against DNA cross-links and other damages. Mol Cell 1, 783-793. (1998).
Lundberg, K.S., et al., (1991) Gene 108:1-6.
Luria and Delbreck., (1943) Genetics 28:491-511
Maizels, N. Somatic hypermutation: how many mechanisms diversify V region sequences? Cell 83, 9-12. (1995).
Mattheakis, L.C., Bhatt, R.R. and Dower, W.J. (1994) Proc Natl Acad Sci U S A, 91, 9022-6.
Matthias, et al., (1989) NAR 17, 6418.
Max, E. E., Maizel, J. V. and Leder, P., J. Biol. Chem. 1981. 256: 5116-5120.
McCafferty, J., Griffiths, A.D., Winter, G. and Chiswell, D.J. (1990) Nature, 348, 552-4.
McCormack, W. T., Hurley, E. A. & Thompson, C. B. Germ line maintenance of the pseudogene donor pool for somatic immunoglobulin gene conversion in chickens. Mol Cell Biol 13, 821-830. (1993)
McKean, D., Huppi, K., Bell, M., Staudt, L., Gerhard, W. and Weigert, M. (1984). Generation of antibody diversity in the immune response of BALB/c mice to influenza virus haemagglutinin. Proc. Natl. Acad. Sci. USA 81, 3180-3184.
Moore, M.J. (1995) Nature, 374, 766-7.
Neuberger, M.S. and Milstein, C. (1995). Somatic hypermutation. Curr. Opin. Immunol. 7, 248-254.
Papavasiliou, F. N. & Schatz, D. G. Cell-cycle-regulated DNA double-stranded breaks in somatic hypermutation of immunoglobulin genes. Nature 408, 216-221. (2000).
Parham, P. (ed). (1998). Somatic hypermutation of immunoglobulin genes. Immunological Reviews, Vol. 162 (Copenhagen, Denmark: Munksgaard).
Pierce, A. J., Johnson, R. D., Thompson, L. H. & Jasin, M. XRCC3 promotes omology-directed repair of DNA damage in mammalian cells. Genes Dev 13, 2633-2638. (1999).
Rada, C., Ehrenstein, M. R., Neuberger, M. S. & Milstein, C. Hot spot focusing of somatic hypermutation in MSH2-deficient mice suggests two stages of mutational targeting. Immunity 9, 135-141. (1998).
Ratech, H. (1992). Rapid cloning of rearranged immunoglobulin heavy chain genes from human B-cell lines using anchored polymerase chain reaction. Biochem. Biophys. Res. Commun. 182, 1260-1263
Reynaud, C. A., Anquez, V., Grimal, H. & Weill, J. C. A hyperconversion mechanism generates the chicken light chain preimmune repertoire. Cell 48, 379-388. (1987).
Reynaud, C.A., Dahan, A., Anquez, V. & Weill, J.C. Somatic hyperconversion diversifies the single VH gene of the chicken with a high incidence in the D region. Cell 59, 171-183 (1989)
Rogozin, I.B. and Kolchanov, N.A. (1992). Somatic hypermutation of immunoglobulin genes. II. Influence of neighbouring base sequences on mutagenesis. Biochem. Biophys. Acta 1171, 11-18.
Rooney, C. M., A. B. Rickinson, D. J. Moss, G. M. Lenoir, and M. A. Epstein. 1984. Paired Epstein-Barr virus-carrying lymphoma and lymphoblastoid cell lines from Burkitt's lymphoma patients: comparative sensitivity to non-specific and to allo-specific cytotoxic responses in vitro. Int J Cancer. 34:339-48
Rowe, M., D. T. Rowe, C. D. Gregory, L. S. Young, P. J. Farrell, H. Rupani, and A. B. Rickinson. 1987. Differences in B cell growth phenotype reflect novel patterns of Epstein-Barr virus latent gene expression in Burkitt's lymphoma cells. Embo J. 6:2743-51
Sale, J. E. & Neuberger, M. S. TdT-accessible breaks are scattered over the immunoglobulin V domain in a constitutively hypermutating B cell line. Immunity 9, 859-869. (1998).
Sharpe et al., (1991) EMBO J. 10:2139-2145.
Smith, G.P. (1985) Science, 228, 1315-7.
Takata, M., Sasaki, M.S., Sonoda, E., Fukushima, T., Morrison, C., Albala, J.S., Swagemakers, S.M., Kanaar, R., Thompson, L.H. & Takeda, S. The Rad51 paralog Rad51B promotes homologous recombinational repair. Mol Cell Biol 20, 6476-6482. (2000).
Takata, M., Sasaki, M.S., Sonoda, E., Morrison, C., Hashimoto, M., Utsumi, H., Yamaguchi-Iwai, Y., Shinohara, A. & Takeda, S. Homologous recombination and non-homologous end-joining pathways of DNA double-strand break repair have overlapping roles in the maintenance of chromosomal integrity in vertebrate cells. Embo J 17, 5497-5508. (1998)
Takata, M., Sasaki, M.S., Tachiiri, S., Fukushima, T., Sonoda, E., Schild, D., Thompson, L.H. & Takeda, S. Chromosome Instability and Defective Recombinational Repair in Knockout Mutants of the Five Rad51 Paralogs. Mol Cell Biol 21, 2858-2866. (2001).
Tomlinson, I.M. (1997). V Base database of human antibody genes. Medical Research Council, Centre for Protein Engineering, UK. http://www.mrc-cpe.cam.ac.uk/
Tuerk, C. and Gold, L. (1990) Science, 249, 505-10.
Wabl, M., Burrows, P.D., von Gabain, A. and Steinberg, C.M. (1985). Hypermutation at the immunoglobulin heavy chain locus in a pre-B-cell line. Proc. Natl. Acad. Sci. USA 82, 479-482.
Wagner, S.D., Milstein, C. and Neuberger, M.S. (1995). Codon bias targets mutation. Nature 376, 732.
Weill, J. C. & Reynaud, C. A. Rearrangement/hypermutation/gene conversion: when, where and why? Immunol Today 17, 92-97. (1996)
Wilson, P.C., de Boutellier, O., Liu, Y-J., Potter, K., Banchereau, J., Capra, J.D. and Pascual, V. (1998). Somatic hypermutation introduces insertions and deletions into immunoglobulin V genes. J. Exp. Med. 187, 59-70.
Wu, H., and Kaartinen, M., (1995) Scand. J. Immunol. 42:52-59.
Wu, H., Pelkonen, E., Knuutila, S and Kaartinen M. (1995). A human follicular lymphoma B cell line hypermutates its functional immunoglobulin genes in vitro. Eur. J. Immunol. 25, 3263-3269.
Yamaguchi-Iwai, Y., Sonoda, E., Buerstedde, J.M., Bezzubova, O., Morrison, C., Takata, M., Shinohara, A. & Takeda, S. Homologous recombination, but not DNA repair, is reduced in vertebrate cells deficient in RAD52. Mol Cell Biol 18, 6430-6435. (1998).
Yelamos, J., Klix, N., Goyenechea, B., Lozano, F., Chui, Y.L., Gonzalez Fernandez, A., Pannell, R., Neuberger, M.S. & Milstein, C (1995). Targeting of non-Ig sequences in place of the V segment by somatic hypermutation. Nature 376, 225-229.
Zhang, L., R. R. French, H. T. Chan, T. L. O'Keefe, M. S. Cragg, M. J. Power, and M. J. Glennie. 1995. The development of anti-CD79 monoclonal antibodies for treatment of B-cell neoplastic disease. Ther Immunol. 2:191-202
Zhu, M., Rabinowitz, J.L., Green, N.S., Kobrin, B.J. and Scharff, M.D. (1995) A well-differentiated B cell line is permissive for somatic mutation of a transfected immunoglobulin heavy-chain gene. Proc. Natl. Acad. Sci. USA 92, 2810-2814.
Zou, X., Xian, J., Popov, A. V., Rosewell, I. R., Muller, M. and Bruggemann, M., Eur. J. Immunol. 1995. 25: 2154-62.
【特許請求の範囲】
【請求項1】
標的核酸領域の指向性構成的高頻度突然変異を起こすことができる細胞系の製造方法であって、細胞集団を進行中の標的配列の多様化についてスクリーニングし、標的核酸の突然変異率が他の核酸の突然変異率よりも100倍以上上回っている細胞を選別することを含む上記方法。
【請求項2】
細胞系がリンパ球様細胞系である、請求項1に記載の方法。
【請求項3】
細胞系が免疫グロブリン発現細胞に由来する、請求項2に記載の方法。
【請求項4】
細胞系が、標的核酸領域を、該領域の突然変異体を含む細胞の選別を容易にするような様式で発現する、請求項1〜3のいずれか一項に記載の方法。
【請求項5】
細胞系が、標的核酸領域によりコードされる遺伝子産物を細胞表面で発現する、請求項4に記載の方法。
【請求項6】
細胞系が、in vivoで高頻度突然変異を起こす細胞型に由来または関連する、請求項1〜5のいずれか一項に記載の方法。
【請求項7】
細胞系が、バーキットリンパ腫、濾胞性リンパ腫またはびまん性大細胞リンパ腫の細胞系である、請求項6に記載の方法。
【請求項8】
標的配列の多様化を示す1種以上の細胞を単離する工程と、その単離された細胞において、標的配列における突然変異の蓄積速度を非標的配列における突然変異の蓄積速度と比較する工程とを更に含む、請求項1〜7のいずれか一項に記載の方法。
【請求項9】
標的配列が免疫グロブリンのV遺伝子配列である、請求項1〜8のいずれか一項に記載の方法。
【請求項10】
細胞が、発現される免疫グロブリンの欠損を評価することによってスクリーニングされる、請求項9に記載の方法。
【請求項11】
細胞が、標的配列の直接配列決定により突然変異率を評価することによってスクリーニングされる、請求項1〜9のいずれか一項に記載の方法。
【請求項12】
細胞が免疫蛍光法によりスクリーニングされる、請求項1〜9のいずれか一項に記載の方法。
【請求項13】
細胞における突然変異率が、突然変異誘発原の投与、または配列改変性の遺伝子産物の発現によりモジュレートされる、請求項1〜12のいずれか一項に記載の方法。
【請求項14】
細胞における突然変異率が、遺伝子操作によりモジュレートされる、請求項1〜13のいずれか一項に記載の方法。
【請求項15】
DNA修復に関与する1種以上の遺伝子が操作されている、請求項14に記載の方法。
【請求項16】
前記1種以上の遺伝子がRad51の類似体および/またはパラログ体である、請求項15に記載の方法。
【請求項17】
前記遺伝子が、Rad51b、Rad51c、ならびにそれらの類似体および/またはパラログ体からなる群から選択される、請求項15に記載の方法。
【請求項18】
所望の活性を有する遺伝子産物の製造方法であって、以下のa)〜c)の工程:
a) 請求項1に記載の細胞の集団において、高頻度突然変異を誘導する配列に機能し得る形で連結された該遺伝子産物をコードする核酸を発現させる工程、
b) 該細胞の集団内で、該所望の活性を有する突然変異した遺伝子産物を発現する細胞または細胞群を同定する工程、
c) 工程b)で同定された細胞または細胞群から、1種以上のクローン細胞集団を樹立し、そして、そのクローン集団から、該所望の活性が向上している遺伝子産物を発現する細胞または細胞群を選別する工程、
を含む上記方法。
【請求項19】
前記細胞または細胞群が、内在性V遺伝子座に構成的高頻度突然変異を誘導する、請求項18に記載の方法。
【請求項20】
高頻度突然変異を誘導する調節配列が、J遺伝子クラスターの下流に存在する配列から選択される、請求項18または19に記載の方法。
【請求項21】
調節配列が、Klixら, (1998) Eur J. Immunol. 28:317-326に従って定義されるエレメントEi/MAR、Cκ+隣接領域およびE3'を含む、請求項20に記載の方法。
【請求項22】
高頻度突然変異を誘導する調節配列に機能し得る形で連結された核酸領域が、前記細胞または細胞群に挿入された外来性の配列である、請求項18〜21のいずれか一項に記載の方法。
【請求項23】
外来性の配列が、前記細胞または細胞群と同種であり高頻度突然変異を誘導する調節配列に、機能し得る形で連結された異種コード配列を含む、請求項22に記載の方法。
【請求項24】
内在性V領域コード配列が、異種コード配列によって置換されている、請求項23に記載の方法。
【請求項25】
遺伝子産物が免疫グロブリンである、請求項18〜24のいずれか一項に記載の方法。
【請求項26】
遺伝子産物がDNA結合タンパク質である、請求項18〜25のいずれか一項に記載の方法。
【請求項27】
所望の活性が結合活性である、請求項18〜26のいずれか一項に記載の方法。
【請求項28】
遺伝子産物が酵素である、請求項18〜27のいずれか一項に記載の方法。
【請求項29】
工程b)およびc)を反復的に繰り返す、請求項18〜28のいずれか一項に記載の方法。
【請求項30】
所望の活性を有する遺伝子産物の調製における、特定の核酸領域の指向性構成的高頻度突然変異を起こすことができる細胞の使用。
【請求項31】
指向性構成的高頻度突然変異を起こすことができる、遺伝子操作されたニワトリ・ファブリキウス嚢リンパ腫細胞系である細胞。
【請求項32】
指向性構成的高頻度突然変異を起こすことができる、遺伝子操作されたニワトリDT40細胞である細胞。
【請求項33】
Δxrcc2 DT40およびΔxrcc3 DT40からなる群から選択される、請求項32に記載の細胞。
【請求項1】
標的核酸領域の指向性構成的高頻度突然変異を起こすことができる細胞系の製造方法であって、細胞集団を進行中の標的配列の多様化についてスクリーニングし、標的核酸の突然変異率が他の核酸の突然変異率よりも100倍以上上回っている細胞を選別することを含む上記方法。
【請求項2】
細胞系がリンパ球様細胞系である、請求項1に記載の方法。
【請求項3】
細胞系が免疫グロブリン発現細胞に由来する、請求項2に記載の方法。
【請求項4】
細胞系が、標的核酸領域を、該領域の突然変異体を含む細胞の選別を容易にするような様式で発現する、請求項1〜3のいずれか一項に記載の方法。
【請求項5】
細胞系が、標的核酸領域によりコードされる遺伝子産物を細胞表面で発現する、請求項4に記載の方法。
【請求項6】
細胞系が、in vivoで高頻度突然変異を起こす細胞型に由来または関連する、請求項1〜5のいずれか一項に記載の方法。
【請求項7】
細胞系が、バーキットリンパ腫、濾胞性リンパ腫またはびまん性大細胞リンパ腫の細胞系である、請求項6に記載の方法。
【請求項8】
標的配列の多様化を示す1種以上の細胞を単離する工程と、その単離された細胞において、標的配列における突然変異の蓄積速度を非標的配列における突然変異の蓄積速度と比較する工程とを更に含む、請求項1〜7のいずれか一項に記載の方法。
【請求項9】
標的配列が免疫グロブリンのV遺伝子配列である、請求項1〜8のいずれか一項に記載の方法。
【請求項10】
細胞が、発現される免疫グロブリンの欠損を評価することによってスクリーニングされる、請求項9に記載の方法。
【請求項11】
細胞が、標的配列の直接配列決定により突然変異率を評価することによってスクリーニングされる、請求項1〜9のいずれか一項に記載の方法。
【請求項12】
細胞が免疫蛍光法によりスクリーニングされる、請求項1〜9のいずれか一項に記載の方法。
【請求項13】
細胞における突然変異率が、突然変異誘発原の投与、または配列改変性の遺伝子産物の発現によりモジュレートされる、請求項1〜12のいずれか一項に記載の方法。
【請求項14】
細胞における突然変異率が、遺伝子操作によりモジュレートされる、請求項1〜13のいずれか一項に記載の方法。
【請求項15】
DNA修復に関与する1種以上の遺伝子が操作されている、請求項14に記載の方法。
【請求項16】
前記1種以上の遺伝子がRad51の類似体および/またはパラログ体である、請求項15に記載の方法。
【請求項17】
前記遺伝子が、Rad51b、Rad51c、ならびにそれらの類似体および/またはパラログ体からなる群から選択される、請求項15に記載の方法。
【請求項18】
所望の活性を有する遺伝子産物の製造方法であって、以下のa)〜c)の工程:
a) 請求項1に記載の細胞の集団において、高頻度突然変異を誘導する配列に機能し得る形で連結された該遺伝子産物をコードする核酸を発現させる工程、
b) 該細胞の集団内で、該所望の活性を有する突然変異した遺伝子産物を発現する細胞または細胞群を同定する工程、
c) 工程b)で同定された細胞または細胞群から、1種以上のクローン細胞集団を樹立し、そして、そのクローン集団から、該所望の活性が向上している遺伝子産物を発現する細胞または細胞群を選別する工程、
を含む上記方法。
【請求項19】
前記細胞または細胞群が、内在性V遺伝子座に構成的高頻度突然変異を誘導する、請求項18に記載の方法。
【請求項20】
高頻度突然変異を誘導する調節配列が、J遺伝子クラスターの下流に存在する配列から選択される、請求項18または19に記載の方法。
【請求項21】
調節配列が、Klixら, (1998) Eur J. Immunol. 28:317-326に従って定義されるエレメントEi/MAR、Cκ+隣接領域およびE3'を含む、請求項20に記載の方法。
【請求項22】
高頻度突然変異を誘導する調節配列に機能し得る形で連結された核酸領域が、前記細胞または細胞群に挿入された外来性の配列である、請求項18〜21のいずれか一項に記載の方法。
【請求項23】
外来性の配列が、前記細胞または細胞群と同種であり高頻度突然変異を誘導する調節配列に、機能し得る形で連結された異種コード配列を含む、請求項22に記載の方法。
【請求項24】
内在性V領域コード配列が、異種コード配列によって置換されている、請求項23に記載の方法。
【請求項25】
遺伝子産物が免疫グロブリンである、請求項18〜24のいずれか一項に記載の方法。
【請求項26】
遺伝子産物がDNA結合タンパク質である、請求項18〜25のいずれか一項に記載の方法。
【請求項27】
所望の活性が結合活性である、請求項18〜26のいずれか一項に記載の方法。
【請求項28】
遺伝子産物が酵素である、請求項18〜27のいずれか一項に記載の方法。
【請求項29】
工程b)およびc)を反復的に繰り返す、請求項18〜28のいずれか一項に記載の方法。
【請求項30】
所望の活性を有する遺伝子産物の調製における、特定の核酸領域の指向性構成的高頻度突然変異を起こすことができる細胞の使用。
【請求項31】
指向性構成的高頻度突然変異を起こすことができる、遺伝子操作されたニワトリ・ファブリキウス嚢リンパ腫細胞系である細胞。
【請求項32】
指向性構成的高頻度突然変異を起こすことができる、遺伝子操作されたニワトリDT40細胞である細胞。
【請求項33】
Δxrcc2 DT40およびΔxrcc3 DT40からなる群から選択される、請求項32に記載の細胞。
【図1−1】

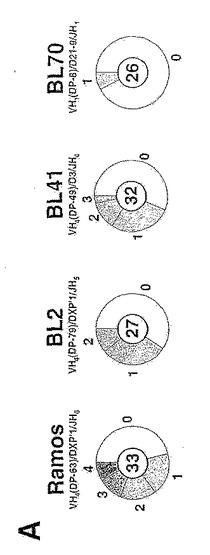
【図1−2】
【図1−3】
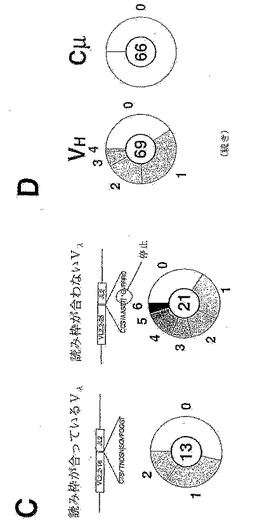
【図2】
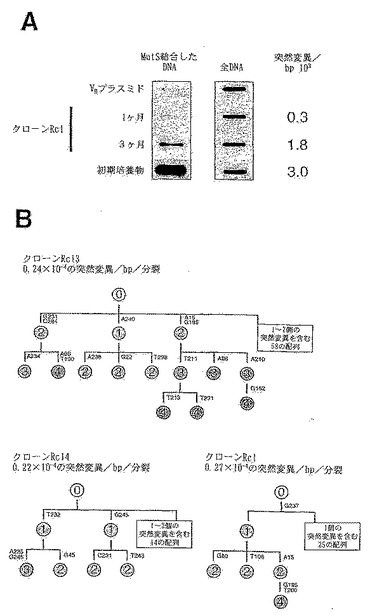
【図3】

【図4】
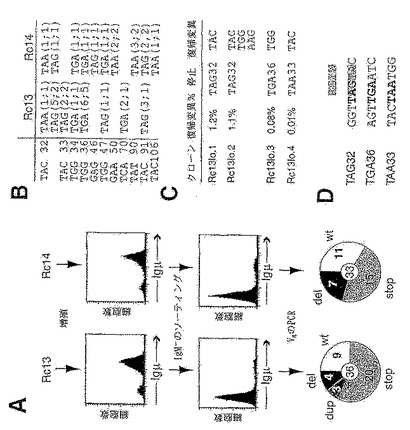
【図5】

【図6】
【図7】

【図8】
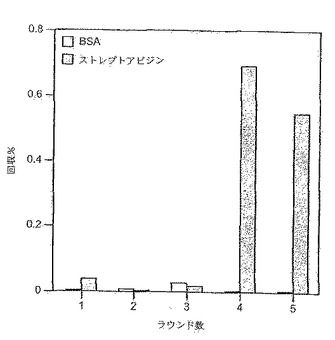
【図9】
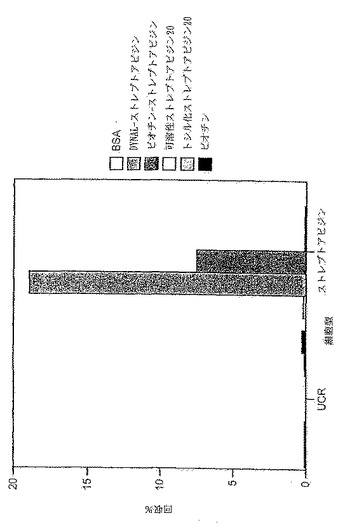
【図10】
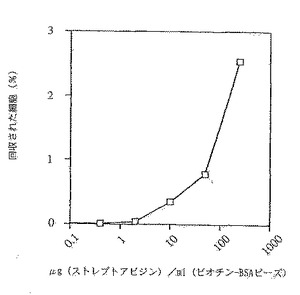
【図11−1】

【図11−2】

【図12】
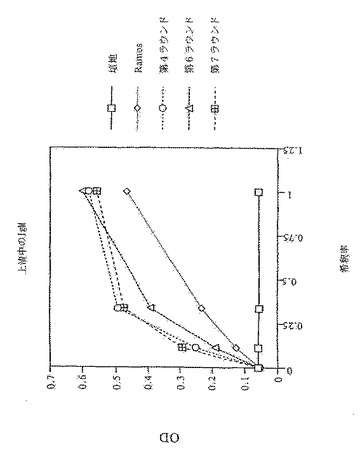
【図13】
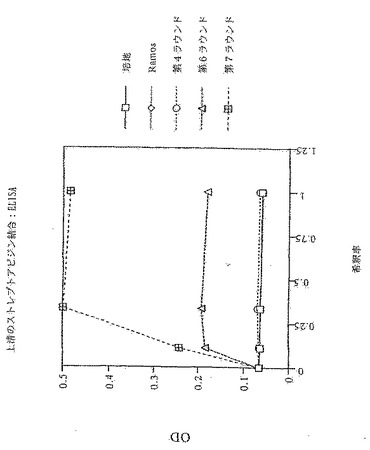
【図14】
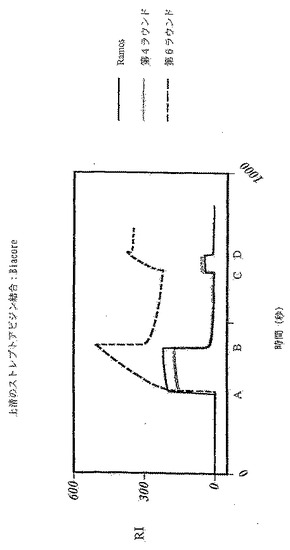
【図15】
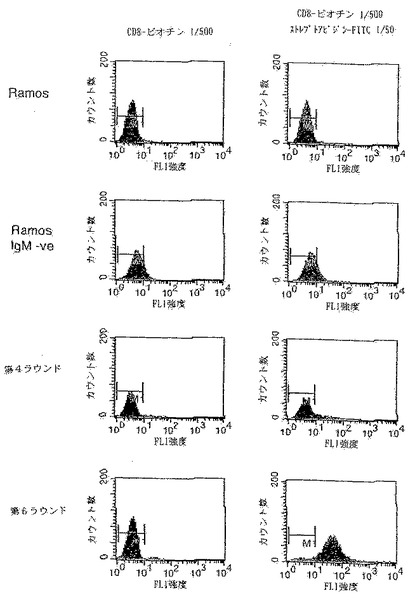
【図16−1】

【図16−2】

【図17】
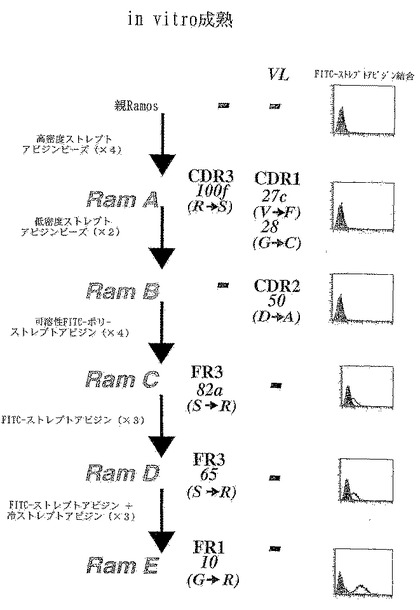
【図18】
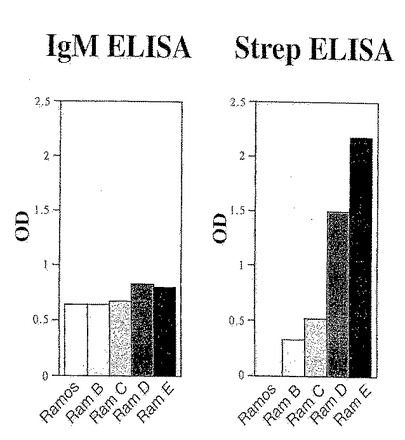
【図19】
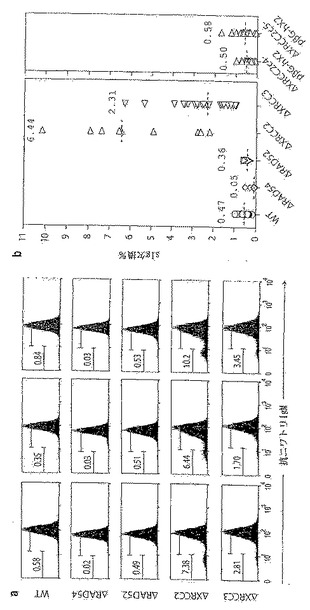
【図20】
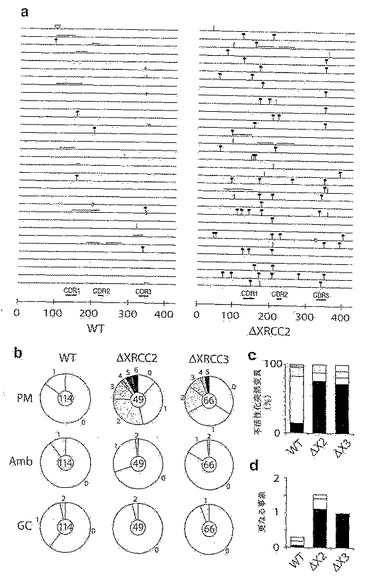
【図21−1】
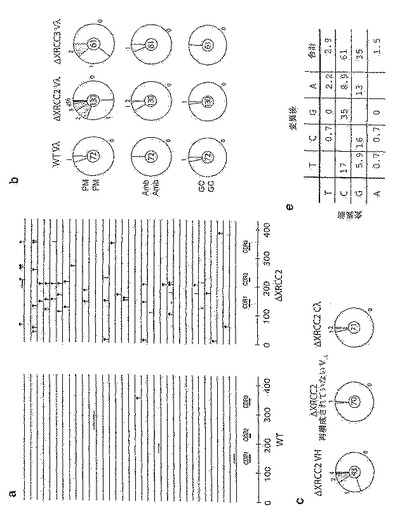
【図21−2】
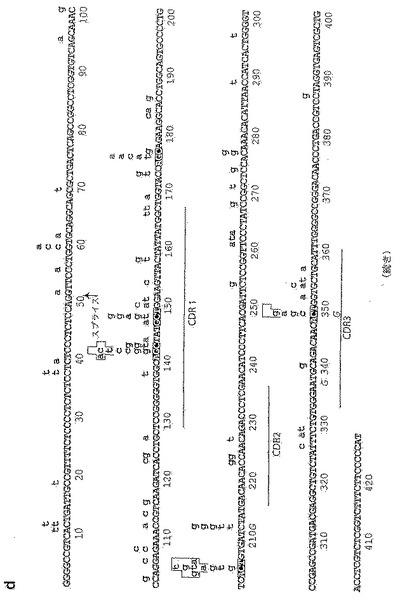
【図22】
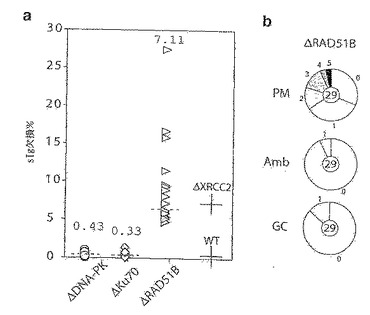
【図23】
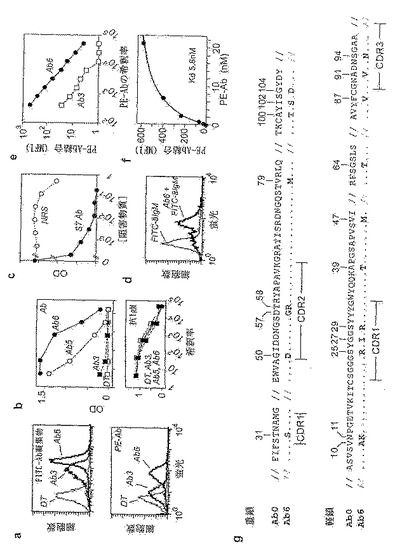
【図24−1】
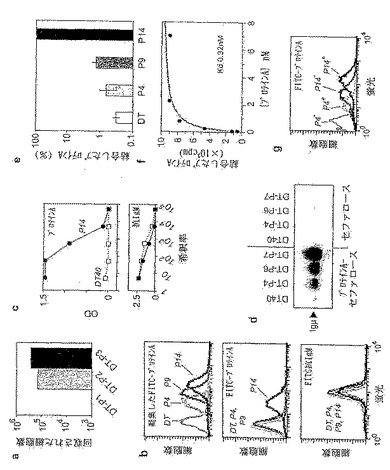
【図24−2】
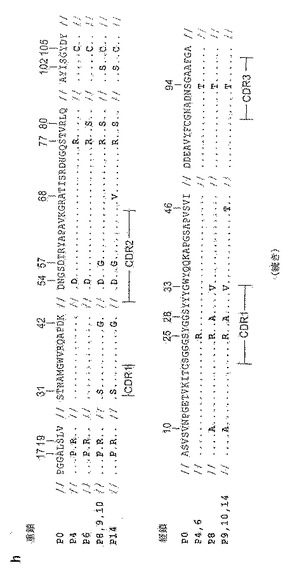
【図25−1】
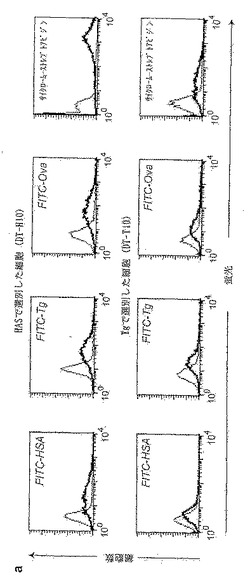
【図25−2】

【図26−1】
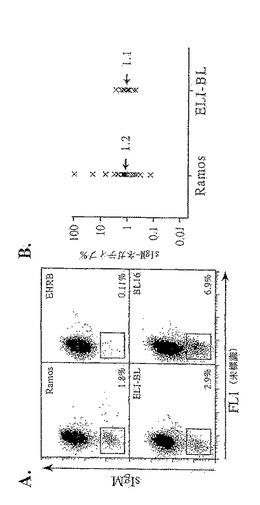
【図26−2】

【図1−2】
【図1−3】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11−1】
【図11−2】
【図12】
【図13】
【図14】
【図15】
【図16−1】
【図16−2】
【図17】
【図18】
【図19】
【図20】
【図21−1】
【図21−2】
【図22】
【図23】
【図24−1】
【図24−2】
【図25−1】
【図25−2】
【図26−1】
【図26−2】
【公開番号】特開2012−165756(P2012−165756A)
【公開日】平成24年9月6日(2012.9.6)
【国際特許分類】
【出願番号】特願2012−106705(P2012−106705)
【出願日】平成24年5月8日(2012.5.8)
【分割の表示】特願2009−232144(P2009−232144)の分割
【原出願日】平成14年6月11日(2002.6.11)
【出願人】(597166578)メディカル リサーチ カウンシル (60)
【Fターム(参考)】
【公開日】平成24年9月6日(2012.9.6)
【国際特許分類】
【出願日】平成24年5月8日(2012.5.8)
【分割の表示】特願2009−232144(P2009−232144)の分割
【原出願日】平成14年6月11日(2002.6.11)
【出願人】(597166578)メディカル リサーチ カウンシル (60)
【Fターム(参考)】
[ Back to top ]