
多重特異的脱免疫CD3−結合物質
【課題】ヒトCD3に特異的に結合する第一のドメインと、Igに由来する第二の結合ドメインとを含む細胞障害活性を有するCD3特異的結合構築物を提供する。
【解決手段】CD3特異的結合構築物をコードする核酸配列、該核酸配列を含むベクターおよび宿主細胞、構築物の産生プロセス、ならびに該構築物を含む組成物である。特定の疾患を治療するための薬学的組成物を調製するために該構築物を用いること、特定の疾患を治療する方法、および結合構築物を含むキット。
【解決手段】CD3特異的結合構築物をコードする核酸配列、該核酸配列を含むベクターおよび宿主細胞、構築物の産生プロセス、ならびに該構築物を含む組成物である。特定の疾患を治療するための薬学的組成物を調製するために該構築物を用いること、特定の疾患を治療する方法、および結合構築物を含むキット。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ヒトCD3に特異的に結合する第一のドメインと、Igに由来する第二の結合ドメインとを含む細胞障害活性を有するCD3特異的結合構築物に関する。さらに、本発明のCD3特異的結合構築物をコードする核酸配列を提供する。本発明のさらなる局面は、該核酸配列を含むベクターおよび宿主細胞、本発明の構築物の産生プロセス、ならびに該構築物を含む組成物である。本発明はまた、特定の疾患を治療するための薬学的組成物を調製するために該構築物を用いること、特定の疾患を治療する方法、および本発明の結合構築物を含むキットも提供する。
【背景技術】
【0002】
ヒトCD3は、多分子T細胞複合体の一部としてT細胞上で発現される抗原を指し、これは異なる三つの鎖からなる:CD3-ε、CD3-δ、およびCD3-γ。例えば固定された抗CD3抗体によってT細胞上でCD3が集合すると、T細胞受容体が関与する場合と類似のT細胞活性化が起こるが、これはそのクローンに典型的な特異性とは無関係である;国際公開公報第99/54440号(特許文献1)またはHoffman(1985), J. Immunol. 135:5〜8(非特許文献1)を参照されたい。
【0003】
CD3抗原を特異的に認識する抗体は、先行技術において、例えばTraunecker, EMBO J. 10(1991), 3655〜9(非特許文献2)およびKipriyanov, Int. J. Cancer 77(1998), 763〜772(非特許文献3)において記述されている。最近、CD3に対する抗体が、多様な疾患の治療に提唱されている。これらの抗体または抗体構築物は、欧州特許第1 025 854号(特許文献2)に開示されるように、T細胞枯渇物質またはマイトゲン物質のいずれかとして作用する。ヒトCD3抗原複合体に対して特異的に結合するヒト/齧歯類ハイブリッド抗体は、国際公開公報第00/05268号(特許文献3)に開示され、例えば腎臓、敗血症、および心臓の同種異系移植片の移植後の拒絶事例の治療のための免疫抑制剤として提唱されている。国際公開公報第03/04648号(特許文献4)は、CD3および卵巣癌抗原に対する二重特異性抗体を開示している。さらに、Kufer(1997), Cancer Immunol. Immunother. 45:193〜7(非特許文献4)は、最小の残留癌の治療のためのCD3およびEpCAMに対して特異的な二重特異性抗体に関する。
【0004】
しかし、先行技術のCD3に対する抗体は、ヒト以外の起源に由来している。このために、ヒトにおける治療レジメの一部としてそのような抗CD3抗体を用いる場合には、いくつかの重篤な問題が起こる。
【0005】
そのような問題の一つは、「サイトカイン放出症候群(CRS)」である。CRSは、抗CD3抗体の最初の数回の用量の投与後に認められる臨床症候群であり、CD3に対する多くの抗体がマイトゲン性であるという事実に関連している。インビトロにおいて、CD3に対するマイトゲン抗体は、T細胞増殖およびサイトカイン産生を誘導する。インビボでは、このマイトゲン活性により、抗体の初回注射後1時間以内に、多くのT細胞由来サイトカインを含む大量のサイトカインの放出が起こる。CD3特異的抗体のマイトゲン活性は、単球/マクロファージ依存的であり、これはこれらの細胞によるIL-6およびIL-1βの産生を伴う。
【0006】
CRS症状は、しばしば報告される軽度の「インフルエンザ様」症候群から、報告される回数がより少ない重度の「ショック様」反応(これには、心血管および中枢神経系の症状が含まれることがある)に及ぶ。症状には、とりわけ頭痛、振せん、悪心/嘔吐、下痢、腹痛、倦怠、および筋/関節痛ならびに疼痛、全身の虚弱、心肺事象と共に神経精神学的事象が含まれる。体液過剰負荷の患者および体液過剰負荷ではないように思われる患者において重度の肺浮腫が起こる。特にマウスモノクローナル抗体の治療的治療の障害となるもう一つの重篤な問題は、そのような抗体に対する液性免疫応答の開始であり、それによってヒト抗マウス抗体(「HAMAs」)の産生が起こる(Schroff(1985), Cancer Res. 45:879〜885(非特許文献5), Shawler(1985), J. Immunol. 135:1530〜1535(非特許文献6))。HAMAsは典型的にマウス抗体による治療の2週目のあいだに産生され、マウス抗体を中和し、それによってその意図する標的に対するその結合能を阻害する。HAMA反応は、マウス抗体定常領域(「Fc」)および/またはマウス可変領域(「V」)の性質に依存しうる。
【0007】
先行技術は、非ヒト起源のモノクローナル抗体を改変することによって、HAMAsの産生を減少または阻害する様々なアプローチを含む。
【0008】
そのような抗体の免疫原性を減少させる一つのアプローチは、例えば国際公開公報第91/09968号(特許文献5)および米国特許第6,407,213号(特許文献6)に記述されるようなヒト化である。一般的に、ヒト化は、例えばCDR-移植の場合のように、非ヒト抗体配列を対応するヒト配列に置換することを必要とする。
【0009】
そのような抗体の免疫原性を減少させるもう一つのアプローチは、例えば国際公開公報第00/34317号(特許文献7)、国際公開公報第98/52976号(特許文献8)、国際公開公報第02/079415号(特許文献9)、国際公開公報第02/012899号(特許文献10)、および国際公開公報第02/069232号(特許文献11)に記述されるような、脱免疫である。一般的に、脱免疫は、可能性があるT細胞エピトープ内でのアミノ酸の置換を行うことを必要とする。この方法において、所定の配列が細胞内タンパク質プロセシングの際にT細胞エピトープを生じる可能性は低下する。その上、国際公開公報第92/10755号(特許文献12)は、タンパク質上の抗原決定基を操作するアプローチを記述している。特に、タンパク質をエピトープマッピングして、そのアミノ酸配列を遺伝子操作を通して変化させる。
【0010】
しかし、ヒト化抗体はしばしば、その非ヒト化親抗体と比較してその標的に関する結合親和性の減少を示し、同様にしばしばヒト宿主においてなおいくぶん免疫原性である。
【先行技術文献】
【特許文献】
【0011】
【特許文献1】国際公開公報第99/54440号
【特許文献2】欧州特許第1 025 854号
【特許文献3】国際公開公報第00/05268号
【特許文献4】国際公開公報第03/04648号
【特許文献5】国際公開公報第91/09968号
【特許文献6】米国特許第6,407,213号
【特許文献7】国際公開公報第00/34317号
【特許文献8】国際公開公報第98/52976号
【特許文献9】国際公開公報第02/079415号
【特許文献10】国際公開公報第02/012899号
【特許文献11】国際公開公報第02/069232号
【特許文献12】国際公開公報第92/10755号
【非特許文献】
【0012】
【非特許文献1】Hoffman(1985), J. Immunol. 135:5〜8
【非特許文献2】Traunecker, EMBO J. 10(1991), 3655〜9
【非特許文献3】Kipriyanov, Int. J. Cancer 77(1998), 763〜772
【非特許文献4】Kufer(1997), Cancer Immunol. Immunother. 45:193〜7
【非特許文献5】Schroff(1985), Cancer Res. 45:879〜885
【非特許文献6】Shawler(1985), J. Immunol. 135:1530〜1535
【発明の概要】
【0013】
したがって、本発明の技術的な問題は、T細胞性免疫応答の誘導による、腫瘍様疾患、増殖性障害と共にB細胞関連疾患の治療および/または改善のための手段および方法を提供することであった。上記の手段および方法は、公知の抗体に基づく治療に関して引用された短所を克服しなければならない。
【0014】
この技術的問題に対する解決は、請求の範囲において特徴が述べられている態様を提供することによって達成される。
【0015】
したがって、本発明は、ヒトCD3に対して特異的に結合する第一のドメインとIg由来第二の結合ドメインとを含む細胞障害活性を有するCD3特異的結合構築物に関し、該第一のドメインは脱免疫されて、CDR-H1領域、CDR-H2領域、およびCDR-H3領域を含み、該CDR-H3領域は配列番号:96、108、119、120、121、122、123、124、125、126、または127に記載のアミノ酸配列を含み、ならびに
該第一のドメインはさらに、そのフレームワークH1において配列VKK(Val-Lys-Lys)を含み、フレームワークH1とCDR-H1領域のあいだの移行配列は、配列Ala-Ser-Gly-Tyr-Thr-Phe(ASGYTF;配列番号:233)を含む。
【0016】
意外にも、公知のCDR領域と共にフレームワーク領域およびその対応する移行配列に対して上記で引用した特異的改変を行うと、当初の非脱免疫配列と比較して免疫原性の低下を示すが、その細胞障害活性を保持する、脱免疫されたCD3特異的結合分子が得られることが判明した。必ずしも全ての脱免疫プロトコールによって、明確な細胞障害活性を示す生体活性のある機能的構築物が得られたわけではないことから、この発見は特に意外であった;添付の実施例を参照されたい。さらに、意外にも脱免疫された細胞障害活性を有するCD3結合分子は、生産性の増加を示した。本発明に従って、非脱免疫抗体の特定の配列は、本明細書において先に引用した配列に置換または改変されている。特に、フレームワークH1領域において、当初の配列Leu-Ala-Arg(LAR)は、配列Val-Lys-Lys(VKK)に置換されている。さらに、いくつかの非改変/非脱免疫CD3特異的抗体のフレームワークH1とCDR-H1との移行領域に含まれる配列Thr-Ser-Gly-Tyr-Thr-Phe(TSGYTF)は、本発明に従ってAla-Ser-Gly-Tyr-Thr-Phe(ASGYTF)(配列番号:233)に改変されている(図14を参照されたい)。望ましい本発明のCD3-特異的結合構築物は、それによって第二の結合特異性がIg-由来である少なくとも二つの結合特異性を含むことを特徴とする。さらに、該所望の構築物は、本明細書において先に示された特異的アミノ酸配列を特徴とする。添付の実施例において示されるように、本明細書において提供される構築物は、その改変/脱免疫型においてもなお生物活性を保持している。本実施例はまた、当技術分野において公知の方法(国際公開公報第92/10755号、国際公開公報第00/34317号、国際公開公報第98/52976号、国際公開公報第02/079415号、または国際公開公報第02/012899号)によって決定された脱免疫によって、必ずしも全て生物活性分子が得られたわけではないことを報告する;特に実施例2および5を参照されたい。
【図面の簡単な説明】
【0017】
【図1】非脱免疫抗CD3カセットのDNAおよびアミノ酸配列(配列番号:1および2)。
【図2A−1】A)重鎖VH2(配列番号:70)、VH3(配列番号:72)、VH5(配列番号:74)、およびVH7(配列番号:76)、ならびに軽鎖VL1(配列番号:78)、VL2(配列番号:80)、およびVL3(配列番号:82)のアミノ酸配列。
【図2A−2】2A-1の続きを示す図である。
【図2B−1】B)重鎖VH2(配列番号:69)、VH3(配列番号:71)、VH5(配列番号:73)、およびVH7(配列番号:75)、ならびに軽鎖VL1(配列番号:77)、VL2(配列番号:79)、およびVL3(配列番号:81)のヌクレオチド配列。
【図2B−2】2B-1の続きを示す図である。
【図2B−3】2B-2の続きを示す図である。
【図2C】C)非脱免疫抗CD3の重鎖(それぞれ配列番号:84、90、96)、VH2(それぞれ配列番号:86、94、96)、VH3(それぞれ、配列番号:86、94、96)、VH5(それぞれ、配列番号:88、92、96)、およびVH7(それぞれ、配列番号:88、90、96)ならびに非脱免疫抗CD3の軽鎖(それぞれ、配列番号:98、102、104)、VL1(それぞれ、配列番号:100、102、104)、VL2(それぞれ、配列番号:100、102、104)、およびVL3(それぞれ、配列番号:98、102、104)のCDR1、2、および3のアミノ酸配列。
【図2D−1】D)非脱免疫抗CD3の重鎖(それぞれ配列番号:83、89、95)、VH2(それぞれ配列番号:85、93、95)、VH3(それぞれ、配列番号:85、93、95)、VH5(それぞれ、配列番号:87、91、95)、およびVH7(それぞれ、配列番号:87、89、95)ならびに非脱免疫抗CD3の軽鎖(それぞれ、配列番号:97、101、103)、VL1(それぞれ、配列番号:99、101、103)、VL2(それぞれ、配列番号:99、101、103)、およびVL3(それぞれ、配列番号:97、101、103)のCDR1、2、および3のヌクレオチド配列。
【図2D−2】2D-1の続きを示す図である。
【図3A】A)抗CD3(VH2/VL1)のヌクレオチド配列(配列番号:4)。
【図3B】B)抗CD3(VH2/VL1)のアミノ酸配列(配列番号:5)。
【図3C】C)抗CD3(VH2/VL2)のヌクレオチド配列(配列番号:6)。
【図3D】D)抗CD3(VH2/VL2)のアミノ酸配列(配列番号:7)。
【図3E】E)抗CD3(VH2/VL3)のヌクレオチド配列(配列番号:8)。
【図3F】F)抗CD3(VH2/VL3)のアミノ酸配列(配列番号:9)。
【図4A】A)抗CD3(VH3/VL1)のヌクレオチド配列(配列番号:10)。
【図4B】B)抗CD3(VH3/VL1)のアミノ酸配列(配列番号:11)。
【図4C】C)抗CD3(VH3/VL2)のヌクレオチド配列(配列番号:12)。
【図4D】D)抗CD3(VH3/VL2)のアミノ酸配列(配列番号:13)。
【図4E】E)抗CD3(VH3/VL3)のヌクレオチド配列(配列番号:14)。
【図4F】F)抗CD3(VH3/VL3)のアミノ酸配列(配列番号:15)。
【図5A】A)抗CD3(VH5/VL1)のヌクレオチド配列(配列番号:16)。
【図5B】B)抗CD3(VH5/VL1)のアミノ酸配列(配列番号:17)。
【図5C】C)抗CD3(VH5xVL2)のヌクレオチド配列(配列番号:18)。
【図5D】D)抗CD3(VH5/VL2)のアミノ酸配列(配列番号:19)。
【図5E】E)抗CD3(VH5/VL3)のヌクレオチド配列(配列番号:20)。
【図5F】F)抗CD3(VH5/VL3)のアミノ酸配列(配列番号:21)。
【図6A】A)抗CD3(VH7/VL1)のヌクレオチド配列(配列番号:22)。
【図6B】B)抗CD3(VH7xVL1)のアミノ酸配列(配列番号:23)。
【図6C】C)抗CD3(VH7/VL2)のヌクレオチド配列(配列番号:24)。
【図6D】D)抗CD3(VH7xVL2)のアミノ酸配列(配列番号:25)。
【図6E】E)抗CD3(VH7/VL3)のヌクレオチド配列(配列番号:26)。
【図6F】F)抗CD3(VH7/VL3)のアミノ酸配列(配列番号:27)。
【図7A】異なる脱免疫抗CD3部分との二重特異性抗CD19構築物の結合:抗CD3(VH2/VL1)(配列番号:178)、抗CD3(VH2/VL2)(配列番号:180)、抗CD3(VH2/VL3)(配列番号:182)、抗CD3(VH3/VL1)(配列番号:184)、抗CD3(VH3/VL2)(配列番号:186)、抗CD3(VH3/VL3)(配列番号:188)、抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH5/VK2)(配列番号:192)、抗CD3(VH5/VL3)(配列番号:194)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、抗CD3(VH7/VL3)(配列番号:200)。A)CD3。結合は、CD3濃縮PBMC(A)を用いてFACSに基づくアッセイによって測定した。CD3および二次FITC標識抗マウスIg抗体は、(A)においてネガティブコントロールとして用いた。構築物抗CD19×抗CD3および抗EpCAM(M79)×抗CD3を対照として用いた。MFIは平均蛍光強度を示す。
【図7B】異なる脱免疫抗CD3部分との二重特異性抗CD19構築物の結合:抗CD3(VH2/VL1)(配列番号:178)、抗CD3(VH2/VL2)(配列番号:180)、抗CD3(VH2/VL3)(配列番号:182)、抗CD3(VH3/VL1)(配列番号:184)、抗CD3(VH3/VL2)(配列番号:186)、抗CD3(VH3/VL3)(配列番号:188)、抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH5/VK2)(配列番号:192)、抗CD3(VH5/VL3)(配列番号:194)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、抗CD3(VH7/VL3)(配列番号:200)。B)CD19。結合は、CD19陽性NALM-細胞(B)を用いてFACSに基づくアッセイによって測定した。CD19および二次FITC標識抗マウスIg抗体を(B)においてネガティブコントロールとして用いた。構築物抗CD19×抗CD3および抗EpCAM(M79)×抗CD3を対照として用いた。MFIは平均蛍光強度を示す。
【図8】HCICカラムからの抗CD19×抗CD3タンパク質分画の脱免疫変種の280 nmでの代表的な溶出パターン。700 mlで大きい段差を示す下の線は、20 mM酢酸塩、pH 3.5を含む溶出緩衝液の理論的勾配を示す。280 nmでの高い吸収は、カラムフロースルーにおける非結合タンパク質が原因であった。810.98 mlでの矢印は、溶出した抗CD3分画を示す。
【図9】Ni-キレートHis Trap(登録商標)カラムからの抗CD19×抗CD3タンパク質分画の脱免疫変種の280 nmでの代表的な溶出パターン。85 mlで第一の段差および90 mlで大きい第二の段差を示す下の線は、溶出緩衝液(破線)の理論的勾配を示す。93.16 mlでの矢印は抗CD19×抗CD3構築物を含むタンパク質分画を示す。
【図10】セファデックスS200ゲル濾過カラムからの代表的なタンパク質溶出パターン。分画を0〜130 mlの保持時間から回収した。80.44 mlでのタンパク質ピークは、MW約52 kDに対応し、脱免疫された抗CD19×抗CD3構築物を含む。
【図11】A)抗CD19×抗CD3タンパク質分画の脱免疫変種のSDS-PAGE分析。レーンM:分子量マーカー;レーン1:HCICフロースルー;レーン2:細胞培養上清;レーン3:HCIC溶出液;レーン4:IMACフロースルー;レーン5:IMAC洗浄液;レーン6:IMAC溶出液;レーン7:ゲル濾過溶出液。B)抗CD19×抗CD3タンパク質分画の精製脱免疫変種のウェスタンブロット分析。精製二重特異性タンパク質のウェスタンブロット分析を、His-Tagに対する抗体(PentaHis, Qiagen)およびアルカリホスファターゼ標識ヤギ抗マウスIgを用いて行った。レーンM:分子量マーカー;レーン1:HCICフロースルー;レーン2:細胞培養上清;レーン3:HCIC溶出液;レーン4:IMACフロースルー;レーン5:IMAC洗浄液;レーン6:IMAC溶出液;レーン7:ゲル濾過溶出液。
【図12A】野生型抗CD19×抗CD3構築物と比較した、異なる脱免疫抗CD3部分:抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)を有する精製二重特異性抗CD19構築物の、A)CD3に対する結合。結合は、CD3濃縮PBMCs(A)を用いてFACSに基づくアッセイによって測定した。CD3陽性細胞に対する二次抗体を(A)においてネガティブコントロールとして用いた。構築物抗CD19×抗CD3および抗EpCAM(M79)×抗CD3を対照として用いた。アッセイは、1μg/mlおよび5μg/mlの濃度で行った。MFIは、平均蛍光強度を示す。
【図12B】野生型抗CD19×抗CD3構築物と比較した、異なる脱免疫抗CD3部分:抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)を有する精製二重特異性抗CD19構築物の、B)CD19に対する結合。結合は、CD19陽性NALM細胞(B)を用いてFACSに基づくアッセイによって測定した。CD19陽性細胞に対する二次抗体を(B)においてネガティブコントロールとして用いた。構築物抗CD19×抗CD3および抗EpCAM(M79)×抗CD3を対照として用いた。アッセイは、1μg/mlおよび5μg/mlの濃度で行った。MFIは、平均蛍光強度を示す。
【図13】対照と比較した、異なる脱免疫抗CD3部分抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH5/VL2)(配列番号:192)、抗CD3(VH5/VL3)(配列番号:194)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VK3)(配列番号:200)を有する二重特異性抗CD19構築物の細胞障害アッセイ。
【図14】非脱免疫CD3抗体の可変重鎖領域、VH5(配列番号:74)、VH7(配列番号:76)、VH2(配列番号:70)、およびVH3(配列番号:72)の配列アラインメント。フレームワーク領域(FR1)、相補性決定領域1(CDR1)、フレームワーク領域1(FR1)、相補性決定領域3(CDR3)、およびフレームワーク領域4(FR4)を示す。FR1における配列LARおよびVKK、フレームワーク1とCDR1領域との移行部分での配列ASGYTFおよびASGYTA、FR3での配列LTTDK、ITTDK、およびMTTDT、ならびにFR3での配列MQLS、MELS、およびLQMNを四角で囲んだ。アラインメントは、ベクターNTI AdvanceのAlingn Xプログラム(Informax, Inc., USA)を用いて行った。
【図15A】CD3陽性JurkatおよびEpCAM陽性Kato III細胞における異なる脱免疫抗CD3部分:抗CD3(VH5/VL2)×5-10(配列番号:37)(A)を有する二重特異性抗EpCAM構築物のFACSに基づくアッセイにおける結合分析。右へのシフトは結合を示す。Jurkat細胞において、点線はネガティブコントロールのシフトを示す(二次抗体のみ)、破線は抗EpCAM-抗CD3対照抗体の結合を示し、太い線は対象二重特異性構築物を示す。EpCAM陽性Kato III細胞を用いる結合アッセイにおいて、CD3に対するモノクローナル抗体の代わりにEpCAMに対するモノクローナル抗体を陽性対照として用いた。
【図15B】CD3陽性JurkatおよびEpCAM陽性Kato III細胞における異なる脱免疫抗CD3部分:脱免疫抗CD3(VH5/VL2)×4-7(配列番号:33)(B)を有する二重特異性抗EpCAM構築物のFACSに基づくアッセイにおける結合分析。右へのシフトは結合を示す。Jurkat細胞において、点線はネガティブコントロールのシフトを示す(二次抗体のみ)、破線は抗EpCAM-抗CD3対照抗体の結合を示し、太い線は対象二重特異性構築物を示す。EpCAM陽性Kato III細胞を用いる結合アッセイにおいて、CD3に対するモノクローナル抗体の代わりにEpCAMに対するモノクローナル抗体を陽性対照として用いた。
【図15C】CD3陽性JurkatおよびEpCAM陽性Kato III細胞における異なる脱免疫抗CD3部分:脱免疫抗CD3(VH5/VL2)×3-1(配列番号:31)(C)を有する二重特異性抗EpCAM構築物のFACSに基づくアッセイにおける結合分析。右へのシフトは結合を示す。Jurkat細胞において、点線はネガティブコントロールのシフトを示す(二次抗体のみ)、破線は抗EpCAM-抗CD3対照抗体の結合を示し、太い線は対象二重特異性構築物を示す。EpCAM陽性Kato III細胞を用いる結合アッセイにおいて、CD3に対するモノクローナル抗体の代わりにEpCAMに対するモノクローナル抗体を陽性対照として用いた。
【図15D】CD3陽性JurkatおよびEpCAM陽性Kato III細胞における異なる脱免疫抗CD3部分:脱免疫抗CD3(VH5/VL2)×4-7(VL-VH)(配列番号:35)(D)を有する二重特異性抗EpCAM構築物のFACSに基づくアッセイにおける結合分析。右へのシフトは結合を示す。Jurkat細胞において、点線はネガティブコントロールのシフトを示す(二次抗体のみ)、破線は抗EpCAM-抗CD3対照抗体の結合を示し、太い線は対象二重特異性構築物を示す。EpCAM陽性Kato III細胞を用いる結合アッセイにおいて、CD3に対するモノクローナル抗体の代わりにEpCAMに対するモノクローナル抗体を陽性対照として用いた。
【図15E】CD3陽性JurkatおよびEpCAM陽性Kato III細胞における異なる脱免疫抗CD3部分:脱免疫抗CD3(VH5/VL2)×5-10(VL-VH)(配列番号:39)(E)を有する二重特異性抗EpCAM構築物のFACSに基づくアッセイにおける結合分析。右へのシフトは結合を示す。Jurkat細胞において、点線はネガティブコントロールのシフトを示す(二次抗体のみ)、破線は抗EpCAM-抗CD3対照抗体の結合を示し、太い線は対象二重特異性構築物を示す。EpCAM陽性Kato III細胞を用いる結合アッセイにおいて、CD3に対するモノクローナル抗体の代わりにEpCAMに対するモノクローナル抗体を陽性対照として用いた。
【図16A】CD3陽性JurkatおよびEpCAM陽性Kato細胞における、FACSに基づくアッセイによる異なる脱免疫抗CD3部分:3-1×抗CD3(VH5/VL2)(配列番号:49)(A)を有する二重特異性抗EpCAM構築物の結合分析。右へのシフトは結合を示す。
【図16B】CD3陽性JurkatおよびEpCAM陽性Kato細胞における、FACSに基づくアッセイによる異なる脱免疫抗CD3部分:5-10×抗CD3(VH5/VL2)(配列番号:63)(B)を有する二重特異性抗EpCAM構築物の結合分析。右へのシフトは結合を示す。
【図17】対応する非脱免疫構築物と比較して、N-末端位で脱免疫抗CD3部分(di抗CD3)、抗CD3(VH5/VL2)×3-1(配列番号:31)、抗CD3(VH5/VL2)×5-10(配列番号:37)および抗CD3(VH5/VL2)×4-7(配列番号:33)を有するEpCAM構築物の細胞障害アッセイ。CB15 T細胞クローンおよびCHO-EpCAM細胞をE:T比5:1で用いた。CHO-EpCAM細胞をPKH26色素で染色し、細胞は二重特異性一本鎖抗体をインキュベートした後にFACS分析によって計数した。
【図18】対応する非脱免疫野生型構築物と比較した、C-末端位で脱免疫抗CD3部分、3-1x抗CD3(VH5/VL2)(配列番号:49)および5-10×抗CD3(VH5/VL2)(配列番号:63)を有するEpCAM構築物の細胞障害アッセイ。細胞障害アッセイは図17と同一に行った。
【発明を実施するための形態】
【0018】
本明細書において用いられるように、「細胞障害活性を有するCD3結合構築物」という用語は、T細胞上で発現されるヒトCD3複合体に結合することができ、標的細胞の消失/溶解を誘導することができるCD3特異的構築物に関する。CD3/CD3複合体(例えば、抗体、抗体誘導体、または抗体断片)のCD3特異的結合物質の結合により、当技術分野で公知のようにT細胞の活性化が起こる;国際公開公報第99/54440号を参照されたい。したがって、本発明の構築物は、インビボおよび/またはインビトロで標的細胞を消失/溶解できなければならない。対応する標的細胞は、本発明の構築物の第二のIg-由来結合ドメインによって認識される表面分子を発現する細胞を含む。そのような表面分子の特徴を本明細書において下記に示す。細胞障害性は、当技術分野において公知の方法ならびに本明細書において下記におよび添付の実施例に示す方法によって検出されうる。したがって、そのような方法はとりわけ生理的インビトロアッセイを含む。そのような生理的アッセイは、例えば細胞膜の完全性を失うことによる細胞死をモニターしてもよい(例えば、FACSに基づくヨウ化プロピジウムアッセイ、トリパンブルー流入アッセイ、光度測定酵素放出アッセイ(LDH)、放射線測定51Cr放出アッセイ、蛍光測定ユーロピウム放出およびカルセインAM放出アッセイ)。さらなるアッセイは、例えば、光度測定MTT、XTT、WST-1およびアラマーブルーアッセイによる細胞生存率のモニタリング、放射線測定3H-Thd取り込みアッセイ、細胞分裂活性を測定するクローン原性アッセイ、およびミトコンドリアの膜内外勾配を測定する蛍光測定ローダミン123アッセイを含む。さらに、例えばFACSに基づくホスファチジルセリン曝露アッセイ、ELISAに基づくTUNEL試験、カスパーゼ活性アッセイ(光度測定、蛍光測定、またはELISAに基づく)によって、または細胞形態の変化(収縮、膜の水疱形成)を分析することによってアポトーシスをモニターしてもよい。細胞障害活性は、蛍光に基づく色素の放出のFACSに基づく測定によって分析されることが好ましい。そのようなアッセイにおいて、本発明の細胞障害活性を有する二重特異性CD3結合構築物の第二のドメインに結合する分子を有する蛍光標識細胞(好ましくは、CD19に関してNALM-6細胞、およびEpCAM抗原に関してKato細胞)を、本発明の細胞障害活性を有する二重特異性CD3結合構築物の存在下で、無作為なドナーの単離PBMCsまたは標準化T細胞株と共にインキュベートする。インキュベーション後、蛍光標的細胞から上清への色素の放出を、分光蛍光計によって決定する。本発明の細胞障害活性を有する脱免疫二重特異性CD3結合構築物は、脱免疫されておらず、または標的細胞に対して特異性を有しない類似の構築物の生物活性を測定することによって得られた値と比較することによって特徴が調べられる。
【0019】
「結合する/相互作用する」という用語は、本発明の文脈において使用される場合、少なくとも2つの「抗原相互作用部位」の互いとの結合/相互作用と定義される。「抗原相互作用部位」という用語は、本発明に従って、特異的抗原または特異的抗原の群との特異的相互作用の能力を示すポリペプチドのモチーフと定義される。この結合/相互作用はまた、「特異的認識」と定義されると理解される。「特異的に認識する」という用語は、本発明に従って、抗体分子が、本明細書に定義されるヒト標的分子の各々の少なくとも2つのアミノ酸を特異的に相互作用し、および/またはそれに結合することが可能であることを意味する。抗体は、同じ標的分子上の違ったエピトープを認識、相互作用、および/または、結合することができる。この用語は、抗体分子の特異性、すなわち、本明細書で定義されるヒト標的分子の特異的領域の間を区別するその能力に関する。その特異的抗原との抗原相互作用部位の特異的相互作用は、例えば、抗原のコンホメーションの変化の誘導、抗原のオリゴマー化などに起因して、シグナルの開始を生じ得る。従って、抗原相互作用部位のアミノ酸配列における特異的モチーフおよび抗原は、それらの一次構造、二次構造、または三次構造の結果として、ならびにこの構造の二次的修飾の結果として互いに結合する。
【0020】
「特異的相互作用」という用語は、本発明に従って使用される場合、本発明のCD3特異的結合構築物は、同様の構造の(ポリ)ペプチドと交差反応しないか、または本質的に交差反応しないことを意味する。したがって、本発明の構築物は、ヒトCD3と特異的に結合/相互作用し、かつ、Igに由来する第二の結合ドメインにより、特異な、選択された他の化合物、抗原、細胞表面分子、腫瘍マーカー等と相互作用することができる。Igに由来する第二の結合ドメインに対する分子の特異的な実施例は、下記に示す。
【0021】
研究の際の構築物のパネルの交差反応性は、例えば、従来的な条件下で(例えば、Harlow および Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, 1988 およびUsing Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, 1999を参照されたい)、関心対象の(ポリ)ペプチドに対して、ならびに多数のより多くまたはより少なく(構造的におよび/または機能的に)密接に関連する(ポリ)ペプチドに対して、二重特異性単鎖抗体構築物のこのパネルの結合を評価することによって試験され得る。関心対象の(ポリ)ペプチド/タンパク質に結合するが、例えば心臓組織の細胞といった関心対象の(ポロ)ペプチドと同様の組織によって好ましくは発現する他の(ポリ)ペプチドのいずれかに結合しないか、または本質的に結合しない構築物(つまり、抗体(二重特異性)scFvsなど)のみが、関心対象の(ポリ)ペプチド/タンパク質に特異的であると見なされ、本発明において提供される方法におけるさらなる研究のために選択され、実施例において図示されている。これらの方法は、特に、構造的におよび/または機能的に密接に関連した分子を有する結合研究、ブロッキングおよび競合作用研究を含むことができる。これらの結合研究は、また、FACS解析、表面プラスモン共鳴(SPR(例えばBlAcore(登録商標)により))分析超遠心、等温の滴定熱量測定法、蛍光異方性、蛍光分光法、または、放射性同位元素を使ってラベルされたリガンド結合実験を含む。さらにまた、細胞毒性アッセイ(実施例にて図示したように)および上述したアッセイのような生理学的なアッセイ法が行われても良い。したがって、特異的抗原との抗原相互作用部位の特異的相互作用の例には、そのレセプターについてのリガンドの特異性が含まれてもよい。この定義は、その特異的レセプターを結合する際にシグナルを誘導するリガンドの相互作用を特に含む。対応するリガンドの例には、その特異的サイトカイン-レセプターと相互作用し/結合するサイトカインが含まれる。セレクチンファミリー、インテグリン、およびEGFのような増殖因子のファミリーの抗原のような抗原に対する抗原相互作用部位の結合もまた、この定義に特に含まれる。この定義にまた特に含まれるこの相互作用についての他の例は、抗体の抗原性結合部位との抗原決定基(エピトープ)の相互作用である。
【0022】
「結合する/相互作用する」という用語はまた、直線状エピトープに関連するばかりでなく、ヒト標的分子またはその一部の2つの領域からなる、コンホメーションエピトープ、構造エピトープ、または不連続エピトープに関連し得る。本発明の文脈において、コンホメーションエピトープは、ポリペプチドがネイティブタンパク質にフォールディングするときに分子の表面上で一緒になる、一次配列中で分離した2つ以上の個別のアミノ酸配列によって定義される(Sela, (1969) Science 166, 1365およびLaver, (1990) Cell 61, 553-6)。
【0023】
「不連続エピトープ」という用語は、本発明の文脈において、ポリペプチド鎖の離れた部分からの残基から集められる非線形エピトープを意味する。これらの残基は、ポリペプチド鎖が三次元構造にフォールディングしてコンホメーション/構造エピトープを構成するときに、表面で一緒になる。
【0024】
本発明の構築物はまた、本明細書中に記載されるヒトCD3複合体または本明細書中以下に開示されるようなその部分の2つの領域からなり、および/またはそれを含むコンホメーション/構造エピトープと特異的に結合/相互作用することが想定される。
【0025】
従って、特異性は、当技術分野において公知である方法、および本明細書に開示および記載される方法によって実験的に決定され得る。このような方法には、ウェスタンブロット、ELISA試験、RIA試験、ECL試験、IRMA試験、およびペプチドスキャンが含まれるがこれらに限定されない。
【0026】
「Ig-由来第二の結合ドメイン」という用語は、「免疫グロブリン由来ドメイン」、特に、抗体またはその断片、一本鎖抗体、合成抗体、Fab、F(ab2)'、Fv、もしくはscFv断片等のような抗体断片、またはこれらの任意の化学改変された誘導体を指す。これらの抗体分子は、異なる種に由来してもよく、またはキメラ起源であってもよい。最も好ましくは(本明細書において下記に示すように)、本発明のCD3特異的結合構築物に含まれる該Ig由来第二のドメインは、scFvである。本発明に従って用いられる抗体、抗体構築物、抗体断片、抗体誘導体(全て、Ig-由来)、またはその免疫グロブリン鎖は、当技術分野で公知の通常の技術、例えばアミノ酸枯渇、挿入、置換、付加、および/または組換えおよび/または当技術分野で公知の他の任意の改変を単独または組み合わせて用いることによって、さらに改変することができる。免疫グロブリン鎖のアミノ酸配列の基礎となるDNA配列にそのような改変を導入する方法は当業者に周知である;例えばSambrook(1989)上記引用文、を参照されたい。「Ig-由来ドメイン」という用語は特に、少なくとも一つのCDRを含む(ポリ)ペプチド構築物に関する。引用されたIg-由来ドメインの断片または誘導体は、上記の抗体分子の一部であるおよび/または化学/生化学または分子生物学的方法によって改変された(ポリ)ペプチドを定義する。対応する方法は当技術分野で公知であり、とりわけ実験マニュアルに記述されている(Sambrookら「Molecular Cloning : A Laboratory Manual」、Cold Spring Harbor Laboratory Press, 第二版1989および第三版2001;Gerhardtら「Methods for General and Molecular Bacteriology」、ASM Press, 1994;Lefkovits「Immunology Methods Manual : The Comprehensive Sourcebook of Techniques」、Academic Press, 1997;Golemis「Protein-Protein Interactions : A Molecular Cloning Manual」、Cold Spring Harbor Laboratory Press, 2002を参照されたい)。
【0027】
本明細書において用いられる「脱免疫された」という用語は、本発明のCD3結合構築物の上記で同定された第一のドメインに関し、該第一のドメインは、野生型構築物をヒトにおいて非免疫原性またはより弱い免疫原性にすることによって、当初の野生型構築物と比較して改変されている。本発明に従う野生型構築物は、非ヒト起源の抗体またはその一部(フレームワークおよび/またはCDRsのような)に関する。対応する例は、米国特許第4,361,549号、または国際公開公報第99/54440号に記述されている抗体または抗体断片である。「脱免疫された」という用語はまた、T細胞エピトープの生成傾向の低下を示す構築物に関する。本発明に従って、「T細胞エピトープの生成傾向の低下」という用語は、特異的T細胞活性化に至るT細胞エピトープを除去することに関する。さらに、T細胞エピトープの生成傾向の低下は、T細胞エピトープの生成に寄与するアミノ酸の置換、すなわちT細胞エピトープの形成にとって必須であるアミノ酸の置換を意味する。言い換えれば、T細胞エピトープの生成傾向の低下は、免疫原性の低下または抗原非依存的T細胞増殖の誘導能の低下に関連する。さらに、T細胞エピトープの生成傾向の低下は、脱免疫に関連し、このことは抗原非依存的T細胞増殖を誘導するアミノ酸配列の可能性があるT細胞エピトープの喪失または減少を意味する。本発明に従って、T細胞エピトープの生成傾向が低下しているCD3結合領域は、非脱免疫分子と比較して免疫原性が弱い、または好ましくは非免疫原性であるが、そのCD3結合能をなおも保持している、すなわちCD3に結合する低または非免疫原性抗体構築物である。
【0028】
「T細胞エピトープ」という用語は、細胞内でのペプチド、ポリペプチド、またはタンパク質の分解の際に放出されて、その後T細胞の活性化を誘発するために主要組織適合抗原複合体(MHC)の分子によって提示されうる短いペプチド配列に関する;とりわけ国際公開公報02/066514号を参照されたい。MHCクラスIIによって提示されるペプチドに関して、T細胞のそのような活性化は、次にT細胞の直接刺激によって抗体反応を生じて、該抗体を産生することができる。
【0029】
したがって、ヒトCD3に対して特異的に結合する脱免疫された第一のドメインは、フレームワークH3とH4のあいだに存在する少なくとも上記のCDR-H3を含み、該第一の結合ドメインは、フレームワークH3とH4のあいだに存在する変化していない野生型(wt)-CDR-H3を含む非脱免疫の第一のドメインと比較してT-細胞エピトープの生成傾向の低下を示す。さらに、該脱免疫第一のドメインは、少なくともフレームワークH1とCDR-H1の移行領域において、フレームワークH1とCDR-H1の変化していないwt-H1移行領域を含む非脱免疫第一のドメインと比較してT-細胞エピトープの生成傾向の低下を示す上記の配列モチーフを含む。「T細胞エピトープの生成傾向の低下」および/または「脱免疫」は、当業者に公知の技術によって測定してもよい。好ましくは、タンパク質の脱免疫は、T細胞増殖アッセイによってインビトロで試験してもよい。このアッセイにおいて、世界中でHLA-DR対立遺伝子の>80%を表すドナーからのPBMCを、野生型または脱免疫したペプチドのいずれかに反応した増殖に関してスクリーニングする。理想的に、細胞増殖は、野生型ペプチドを抗原提示細胞にローディングした場合に限って検出される。または、全てのハプロタイプを表すHLA-DR四量体を発現することによって脱免疫を調べてもよい。これらの四量体を、ペプチド結合に関して試験してもよく、増殖アッセイにおいて抗原提示細胞に関するペプチド置換体をローディングしてもよい。脱免疫されたペプチドがHLA-DRハプロタイプにおいて提示されるか否かを調べるために、例えば、PBMC上の蛍光標識ペプチドの結合を測定することができる。さらに、脱免疫は、脱免疫分子に対する抗体が、患者への投与後に形成されたか否かを決定することによって証明することができる。特に好ましい方法は、とりわけ添付の実施例6に示されるT細胞増殖アッセイである。
【0030】
好ましくは、抗体由来分子をフレームワーク領域において脱免疫して、CDR領域のほとんどは、CDR領域の結合親和性が影響を受けないように、T細胞エピトープを誘導する傾向の低下を得るために改変されない。一つのT細胞エピトープの消失でさえも、免疫原性の低下が起こる。好ましくは、分子はVL鎖のCDR2領域において脱免疫され、より好ましくはVH鎖のCDR2領域において、さらにより好ましくはVL鎖のCDR1領域、さらにより好ましくはVH鎖のCDR1領域、より好ましくはVL鎖のフレームワーク領域(FR)、および最も好ましくはVH鎖のフレームワーク領域(FR)において脱免疫される。
【0031】
本明細書において用いられるように「CDR」という用語は、「相補性決定領域」を指し、これは当技術分野で周知である。CDRsは、免疫グロブリンの一部であり、該分子の特異性を決定して、特異的リガンドと接触するT細胞受容体である。CDRsは、分子の最も可変部分であり、これらの分子の多様性に関与している。抗体の三つのCDR領域、CDR1、CDR2、および特にCDRH領域が存在する。
【0032】
より好ましい態様において、T細胞エピトープの生成傾向が低下した該CD3特異的ドメインの該CDR-H1およびCDR-H2領域は、国際公開公報第99/54440号に記述されるように抗体構築物に由来する。さらにより好ましい(および添付の実施例に説明するように)該CDR-H1およびCDR-H2領域と共にCDR-H3領域は、Traunecker(1991), EMBO J. 10, 3655〜3659に記述されるCD3分子に関して特異性を有する抗体/抗体誘導体に由来する。本発明に従って、該CDR-H1、CDR-H2、およびCDR-H3領域は、例えばヒトCD3-ε鎖に関してトランスジェニックであるマウスにおいて、他のTCRサブユニットの状況においてヒトCD3-ε鎖を特異的に認識することができる抗体/抗体誘導体等に由来する。これらのトランスジェニックマウス細胞は、天然またはほぼ天然のコンフォメーションでヒトCD3-εを発現する。
【0033】
本発明に従って、フレーム領域は、抗原に接触する高度可変相補性決定領域(CDRs)のタンパク質骨格構造を提供する免疫グロブリンおよびT細胞受容体のVドメイン(VHまたはVLドメイン)における領域に関する。各Vドメインにおいて、FR1、FR2、FR3、およびFR4と呼ばれる四つのフレームワーク領域が存在する。フレームワーク1は、VドメインのN-末端からCDR1の開始部までの領域を含み、フレームワーク2は、CDR1とCDR2のあいだの領域に関し、フレームワーク3は、CDR2とCDR3のあいだの領域を含み、およびフレームワーク4はCDR3の末端からVドメインのC-末端までの領域を意味する;とりわけJaneway, 「Immunobiology」, Garland Publishing, 2001, 第5版を参照されたい。このように、フレームワーク領域は、VHまたはVLドメインにおけるCDR領域外の全ての領域を含む。さらに、「フレームワークとCDR領域のあいだの移行配列」は、フレームワークとCDR領域とのあいだの直接接合部に関する。特に、「フレームワークとCDR領域のあいだの移行配列」という用語は、CDR領域のN-およびC-末端に直接存在する配列またはCDR領域周囲のアミノ酸を意味する。したがって、フレームワークはまた、異なるCDR領域のあいだの配列を含んでもよい。当業者は、所定の配列からフレームワーク領域、CDRsと共に対応する移行配列を容易に推測することができる;Kabat(1991), 「Sequences of Proteins of Immunological Interest.」、5th edit., NIH Publication no. 91-3242保健社会福祉省、Chothia(1987), J. Mol. Biol. 196, 901〜917およびChothia(1989)Nature 342, 877〜883を参照されたい。
【0034】
本発明の好ましい細胞障害活性CD3特異的結合構築物は、さらに該第一のドメインにおいて、Met-Glu-Leu-Ser(MELS;配列番号:234)を含むフレームワークH3を含む。第一のドメインにおいて配列Ile-Thr-Thr-Asp-Lys(ITTK;配列番号:235)を含むフレームワークH3を含む本発明の構築物は、さらにより好ましい。
【0035】
本発明に従って、ヒトCD3に特異的に結合/相互作用して、T細胞エピトープを産生する傾向が減少した本発明の構築物の第一のドメインは、本明細書において定義されるCDR-H1、CDR-H2、CDR-H3領域を含み、好ましい態様において、先に定義したような、特に配列番号:152もしくは153、156もしくは157、160、もしくは161、および/または164もしくは165のいずれか一つに示されるVH-フレームワーク(フレームワーク1、2、3、4)を含む。したがって、本発明のCD3特異的結合構築物は、ヒトCD3に特異的に結合する第一のドメインを含み、配列番号:152また153に示されるフレームワーク領域、配列番号:156または157に示されるフレームワーク領域2、配列番号:160または161に示されるフレームワーク領域3、および/または配列番号:164または165に示されるフレームワーク領域4を含む。
【0036】
本発明の特に好ましい態様において、細胞障害活性を有する脱免疫CD3特異的結合構築物は、その第一のドメインにおいて(a)配列番号:88に記述されるCDR-H1、および(b)配列番号:90または92に示されるCDR-H2を含む。
【0037】
したがって、改変されたCDR-H1およびCDR-H2領域は、T細胞エピトープをの生成傾向の低下に至り、CD3-ε鎖特異的抗体に由来する。本発明に従って最も好ましくは、該(親)抗体はTCR複合体の状況において提示されるヒトCD3の天然のまたはほぼ天然の構造またはコンフォメーションのエピトープを反映するエピトープに特異的に結合することができなければならない。
【0038】
好ましくは、本発明のCD3特異的結合構築物は、配列番号:74または76に示されるようにVH領域を含む。配列番号:74は、例としての脱免疫可変重鎖領域を示し、同様に配列番号:76も例としての脱免疫可変重鎖領域を示す。
【0039】
好ましくは、本発明のCD3特異的結合構築物は、配列番号:98または100に示されるCDR-L1、配列番号:102に示されるCDR-L2、および/または配列番号:104に示されるCDR-L3を含む。
【0040】
好ましい態様において、本発明のCD3特異的結合構築物は、そのCD3-特異的部分にVL領域を含み、該VL領域は、配列番号:78、配列番号:80、配列番号:82、および配列番号:112からなる群より選択される。配列番号:78において特徴が示されるVL1、配列番号:80において特徴が示されるVL2、および配列番号:82において特徴が示されるVL3は、本発明に従う完全な脱免疫VL領域に関連し、それらは、上記のVH領域との様々な組み合わせにおいて用いてもよい。なおも同様に、非脱免疫VL領域は、本発明に従って上記の脱免疫VH領域と組み合わせてもよいと想像される。本発明の細胞障害活性を有するCD3結合構築物において好ましく用いられる対応する非脱免疫VL-領域を配列番号:112に示す。したがって、本発明のCD3構築物の先に引用した「第一のドメイン」の重鎖部分のみならず、T細胞エピトープの生成傾向が低下するように改変してもよい。同様に、該ドメインは、対応する可変軽鎖部分を含むと想像される。配列番号:78、80、および82は例えば、国際公開公報第99/54440号に開示される構築物のCD3結合部分の脱免疫されたVL1、VL2、およびVL3領域を示す。
【0041】
上記のように、本発明のCD3特異的結合構築物は、最も好ましくは、scFvであるIg由来の第二のドメインを含む。したがって、本発明の最も好ましい態様において、ヒトCD3に対して一つの特異性を有し、さらなる分子/化合物に対するおよび/または相互作用することができる第二のscFvによって媒介されるさらなる特異性を有する、脱免疫された二重特異性一本鎖抗体構築物が提供される。これらのさらなる分子/化合物は、細胞表面分子、腫瘍マーカー、腫瘍抗原等を含んでもよい。そのようなさらなる化合物/分子は本明細書において下記に例示され、特異的構築物も同様に添付の実施例において示され、提供される。
【0042】
「二重特異性一本鎖抗体構築物」という用語は、二つの抗体由来結合ドメイン、好ましくはscFvを含む構築物に関する。該結合ドメインの一つは、ヒトCD3抗原(標的分子1)と特異的に結合/相互作用することができる抗体(またはその一部)、抗体断片、またはその誘導体の可変領域からなる。第二の結合ドメインは、下記のようにもう一つの(ヒト)抗原(標的分子2)と特異的に結合/相互作用することができる抗体、抗体断片、またはその誘導体の可変領域(またはその一部)からなる。したがって、該第二の結合ドメインは、本発明に従って、細胞表面分子および/または腫瘍特異的マーカーに対して特異性を有する抗原-相互作用部位を含む、先に引用したIg由来第二のドメインである。二重特異性構築物、好ましくは該二重特異性一本鎖抗体構築物における該二つのドメイン/領域は、好ましくは一本鎖として互いに共有結合される。この結合は、直接(ドメイン1[T細胞エピトープの生成傾向の低下を含み、およびCDR領域もしくはCDR領域と上記のフレームワーク領域とを含むヒトCD3抗原に対して特異的]−ドメイン2[細胞表面分子および/または腫瘍特異的マーカーに対して特異的]、もしくはドメイン1[細胞表面分子および/または腫瘍特異的マーカーに対して特異的]−ドメイン2[T細胞エピトープの生成傾向の低下を含み、およびCDR領域またはCDR領域と上記のフレームワーク領域とを含む、ヒトCD3抗原に対して特異的]、またはさらなるポリペプチドリンカー配列(ドメイン1−リンカー配列−ドメイン2)のいずれかによって行うことができる。リンカーを用いる事象において、このリンカーは好ましくは、第一および第二のドメインのそれぞれが、互いに独立してその異なる結合特異性を確実に保持することができるために十分な長さおよび配列を有する。先に述べたように、および添付の実施例において示されるように、好ましくは本明細書に定義される少なくとも二つのドメインを含むCD3特異的結合構築物は、「二重特異性一本鎖抗体構築物」であり、最も好ましくは二重特異性一本鎖抗体Fv(scFv)である。該構築物は薬学的組成物の状況において用いられると特に想像される。二重特異性一本鎖分子は当技術分野で公知であり、国際公開公報第99/54440号、Mack, J. Immunol.(1997), 158, 3965〜3970, Mack, PNAS, (1995), 92, 7021〜7025, Kufer, Cancer Immunol. Immunother. (1997), 45, 193〜197, Loffler, Blood, (2000), 95, 6, 2098〜2103, Bruhl, J. Immunol. (2001), 166, 2420〜2426に記述される。本発明の特に好ましい分子フォーマットは、本発明の構築物のCD3特異的結合ドメインが、先に定義された少なくとも一つのVHおよび一つのVL領域を含む、ポリペプチド構築物を提供する。本明細書に記述され、T細胞エピトープの生成傾向が低下したVH領域の他に、該特異的結合構築物は、T細胞エピトープの生成傾向が低下したさらなる領域/ドメインを含んでもよいことに注意されたい。先に述べたように、VL領域および/または対応するフレームワークは、本発明に従って、T細胞エピトープ生成傾向の低下を有するように操作されているアミノ酸の鎖を含んでもよい。scFvフォーマットにおいてリンカードメインによって互いに連結しているVHドメインおよびVLドメインの分子内方向は、引用された二重特異性一本鎖構築物に関して決定的ではない。このように、可能性がある配置(VH-ドメイン−リンカードメイン−VLドメイン;VLドメイン−リンカードメイン−VHドメイン)をいずれも有するscFvは、引用された二重特異性一本鎖構築物の特定の態様である。CD3特異的ドメインは、二重特異性分子のN-末端またはC-末端に存在しうる。各ドメインのVHおよびVL領域は異なる順序で配置することができる(VH−VLまたはVL−VH)。
【0043】
本発明に従って用いられる「一本鎖」という用語は、二重特異性一本鎖構築物の第一および第二のドメインが、好ましくは単一の核酸分子によってコードされる共直線状のアミノ酸配列の形で共有結合していることを意味する。
【0044】
本発明の構築物は、本明細書において定義された第一のドメインおよびIg由来第二のドメインの他に、例えば組換えによって産生された構築物の単離および/または調製のためのさらなるドメインを含んでもよいことに注目されたい。
【0045】
本発明に従って、本発明のCD3構築物のヒトCD3に特異的に結合する上記の第一のドメインのみならず、T細胞エピトープの生成傾向の低下を有してもよいことに注目される。同様に、Ig由来第二のドメインおよび/または結合するリンカー領域は、改変されている、例えばヒト化および/または脱免疫されていると想像される。
【0046】
先に述べたように、脱免疫アプローチは特に、国際公開公報第00/34317号、国際公開公報第98/52976号、国際公開公報第02/079415号、または国際公開公報第02/012899号および添付の実施例に記述されている。これらのアプローチは、可能性があるT細胞エピトープにおいてアミノ酸の置換を行うことを伴う。このように、所定の配列が細胞内タンパク質プロセシングの際にT細胞エピトープを生成する可能性が低下する。さらに、国際公開公報第92/10755号は、タンパク質上の抗原決定基が操作されるアプローチを記述している。特に、タンパク質はエピトープマッピングされて、そのアミノ酸配列は遺伝子操作によって変化している。
【0047】
さらに、「ヒト化アプローチ」は、当技術分野において周知であり、特に抗体分子、例えばIg由来分子に関して記述されている。「ヒト化」という用語は、非ヒト抗体に由来する配列のいくつかの部分を含む非ヒト(例えば、マウス)抗体またはその断片(Fv、Fab、Fab'、F(ab')、scFvs、または抗体の他の抗原結合部分配列)のヒト化型に関する。ヒト化抗体には、ヒト免疫グロブリンの相補性決定領域(CDR)からの残基が、所望の結合特異性、親和性、および結合能を有するマウス、ラット、またはウサギのような非ヒト種のCDRからの残基に置換されているヒト免疫グロブリンが含まれる。一般的に、ヒト化抗体は、CDR領域の全てまたは実質液に全てが非ヒト免疫グロブリンのCDR領域に対応して、およびFR領域の全てまたは実質的に全てがヒト免疫グロブリンコンセンサス配列の領域である、少なくとも一つ、および一般的に二つの可変ドメインの全てまたは実質的に全てを含むであろう。ヒト化抗体は最適には、典型的にヒト免疫グロブリンの免疫グロブリン定常領域(Fc)の少なくとも一部を含むであろう;とりわけJonesら、Nature 321:522〜525(1986)、Presta, Curr. Op. Struct. Biol. 2:593〜596(1992)を参照されたい。非ヒト抗体をヒト化する方法は当技術分野で周知である。一般的にヒト化抗体は、ヒト抗体により厳密に類似するように、非ヒトである起源からそれに導入された一つまたはそれ以上のアミノ酸を有するが、なおも抗体の当初の結合活性を保持している。抗体/抗体分子のヒト化法は、Jonesら、Nature 321:522〜525(1986);Riechmannら、Nature 332:323〜327(1988);およびVerhoeyenら、Science 239:1534〜1536(1988)にさらに詳述されている。ヒト化抗体の特定の例、例えばEpCAMに対する抗体は、当技術分野で公知であり、例えば(LoBuglio, 「Proceedings of the American Society of Clinical Oncology」(抄録)、1997, 1562およびKhor, 「Proceedings of the American Society of Clinical Oncology」(抄録)、1997, 847)を参照されたい。
【0048】
したがって、本発明の状況において、特に、脱免疫され、薬学的組成物において首尾よく用いることができる二重特異性一本鎖抗体構築物が提供される。
【0049】
先に述べたように、上記のCD3特異的結合構築物のIg由来第二のドメインは、細胞表面分子に対する特異性を有する抗原相互作用部位を含んでもよい。
【0050】
本明細書において用いられるように、「細胞表面分子」という用語はまた、細胞の表面上で提示された分子を指す。「細胞表面分子」という用語は、細胞の表面上に提示され、Ig由来結合ドメイン、好ましくは抗体、抗体断片、または誘導体に近づくことができる(インビトロまたはインビボで)ドメインまたはエピトープを含む。先に示したように、最も好ましくは該Ig由来ドメインはscFvである。該細胞表面分子の例は、膜および膜貫通タンパク質、該タンパク質または細胞表面に適合させた分子等である。本発明のさらに好ましい態様に従って、該細胞表面分子は腫瘍特異的マーカーである。本発明の状況において、「腫瘍特異的マーカー」という用語は、腫瘍細胞の表面に提示されるおよび/または存在する分子、または広布に発現されるが、腫瘍細胞の表面上の抗体、抗体断片、または抗体誘導体の結合にとって近づくことができるのみである分子を指す。腫瘍マーカーの例を本明細書において下記に示し、これらはEpCAM、CD19、HER-2、HER-2 neu、HER-3、HER-4、EGFR、PSMA、CEA、MUC-1(ムチン)、MUC2、MUC3、MUC4、MUC5AC、MUC5B、MUC7、Lewis-Y、CD20、CD33、CD30、CD44v6、Wue-1、プラズマ細胞抗原(国際公開公報第01/47953号を参照されたい)、(膜結合型)IgE、黒色腫コンドロイチン硫酸プロテオグリカン(MCSP)、STEAP、メソセリン、前立腺幹細胞抗原(PSCA)、sTn(シアリル化Tn抗原)、FAP(線維芽細胞活性化抗原)、EGFRvIII、Igα、Igβ、MT-MMPs、Cora抗原、EphA2、L6、およびCO-29を含むがこれらに限定されない。
【0051】
本発明のCD3特異的結合構築物のIg由来第二のドメインはまた、EpCAM、CCR5、CD19、HER-2、HER-2 neu、HER-3、HER-4、EGFR、PSMA、CEA、MUC-1(ムチン)、MUC2、MUC3、MUC4、MUC5AC、MUC5B、MUC7、βhCG、Lewis-Y、CD20、CD33、CD30、ガングリオシドGD3、9-O-アセチル-GD3、GM2、Globo H、フコシルGM1、ポリSA、GD2、炭酸脱水酵素IX(MN/CA IX)、CD44v6、Sonic Hedgehog(Shh)、Wue-1、プラズマ細胞抗原(膜結合型)IgE、黒色腫コンドロイチン硫酸プロテオグリカン(MCSP)、CCR8、TNF-α前駆体、STEAP、メソセリン、A33抗原、前立腺幹細胞抗原(PSCA)、Ly-6、デスモグレイン4、E-カドヘリンネオエピトープ、胎児アセチルコリン受容体、CD25、CA19-9マーカー、CA-125マーカー、およびミュラー管阻害物質(MIS)受容体II型、sTn(シアリル化Tn抗原)、FAP(線維芽細胞活性化抗原)、エンドシアリン、EGFRvIII、L6、SAS、CD63、TAG72、TF-抗原、Cora抗原、CD7、CD22、Igα(CD79a)、Igβ(CD79b)、G250、gp100、MT-MMPs、F19-抗原、CO-29およびEphA2からなる群より選択される分子に対して特異性を有する抗原相互作用部位を含んでもよい。
【0052】
本明細書において提供された構築物は、医学的状況において特に有用である。例えば、腫瘍疾患および/またはリンパ腫、特に非ホジキンB-細胞リンパ腫は、ヒトCD3およびCD20(CD3×CD20またはCD20×CD3)に対する本発明の脱免疫(二重特異性)構築物によって治療してもよい。自己免疫疾患は、ヒトCD3およびCD30またはCD19(すなわち、CD3×CD30、CD30×CD3、CD3×CD19またはCD19×CD3)に対する脱免疫(二重特異性)構築物の投与によって治療してもよい。リウマチ性関節炎のみならず他の炎症疾患を、ヒトCD3およびCCR5(CD3×CCR5またはCCR5×CD3)に対する本発明の脱免疫(二重特異性)構築物によって治療してもよい。本明細書において定義し、およびTNF-α前駆体に対する/結合する第二のIg由来ドメインを含む脱免疫CD3特異的結合構築物は、炎症障害の治療または予防において有用となる可能性がある。本明細書において提供され、EpCAM、CD19、HER-2、HER-2 neu、HER-3、HER-4、EGFR、PSMA、CEA、MUC-1(ムチン)、MUC2、MUC3、MUC4、MUC5AC、MUC5B、MUC7、Lewis-Y、CD20、CD33、CD30、CD44v6、Wue-1、プラズマ細胞抗原(国際公開公報第01/47953号を参照されたい)、(膜結合型)IgE、黒色腫コンドロイチン硫酸プロテオグリカン(MCSP)、STEAP、メソセリン、前立腺幹細胞抗原(PSCA)、sTn(シアリル化Tn抗原)、FAP(線維芽細胞活性化抗原)、EGFRvIII、Igα、Igβ、MT-MMPs、Cora抗原、EphA2、L6、およびCO-29に対する/結合する/相互作用する第二のIg由来ドメインを含むCD3構築物は、乳癌、結腸癌、前立腺癌、頭頚部癌、皮膚癌(黒色腫)、尿性器癌、例えば卵巣癌、子宮内膜癌、子宮頚癌および腎臓癌、肺癌、胃癌、小腸癌、肝臓癌、膵臓癌、胆嚢癌、胆管癌、食道癌、唾液腺癌、および甲状腺癌のような腫瘍様疾患、または血液腫瘍、神経膠腫、肉腫、もしくは骨肉腫のような他の腫瘍様疾患の医学的介入において特に有用となる可能性がある。CD3結合構築物の投与はまた、最小の残留疾患、好ましくは初期固形腫瘍、進行固形腫瘍、または転移性固形腫瘍に適応される。
【0053】
同様に添付の実施例において記述されるように、本発明の特に好ましいCD3特異的結合構築物は、T細胞エピトープの生成傾向が低下した先に定義された第一のドメインおよびEpCAMに対して特異性を有する抗原相互作用部位を含む第二のIg由来ドメインを含む。
【0054】
上皮細胞接着分子(17-1A抗原、KSA、EGP40、GA733-2、ks1-4またはesaとも呼ばれるEpCAM)は、特定の上皮および多くのヒト癌において特異的に発現されるアミノ酸314個の40 kDa膜貫通型糖タンパク質である(Balzer, J. Mol. Med. 1999, 77, 699〜712において論評)。EpCAMはマウスモノクローナル抗体17-1A/エドレコロマブ(Goettinger, Int. J. Cancer, 1986;38, 47〜53およびSimon, Proc. Natl. Acad. Sci. USA 1990;87, 2755〜2759)による認識を通して発見され、その後クローニングされた。EpCAMは、内皮細胞を一定方向に非常に秩序だって接着させるために役立つ(Litvinov, J. Cell Biol. 1997, 139, 1337〜1348)。上皮細胞が悪性形質転換すると、急速に増殖する腫瘍細胞により、上皮の高度細胞秩序が放棄される。その結果、EpCAMの表面分布はより制限が少なくなり、分子は腫瘍細胞上でより露出され、腫瘍細胞表面上の抗体、抗体断片、または抗体誘導体の結合にとって近づきやすくなる。その上皮細胞起源のために、ほとんどの癌腫による腫瘍細胞はなおもその表面上にEpCAMを発現する。
【0055】
インビボで、EpCAMの発現は、上皮の増殖の増加に関連し、細胞分化に負に相関する(論評に関してはBalzar, J. Mol. Med., 1999, 77, 699〜712を参照されたい)。EpCAMの発現は本質的に、全ての主要な癌腫について認められる(Balzar, 1999, J. Mol. Med. 77, 699〜712に論評、またはとりわけDe Bree, Nucl. Med. Commun. 1994, 15, 613〜27;Zhang, Clin. Cancer Res. 1998, 4, 295〜302に報告されている)。その広範な発現のために、EpCAMは「汎癌腫」抗原と呼ばれている。多くの場合、腫瘍細胞は、その親上皮または該癌のより攻撃性の低い型よりかなり高い程度にEpCAMを発現することが認められた。例えば、EpCAM発現の増加は、前立腺癌発達の初期事象を表す(Poczatek, J. Urol. 1999, 162, 1462〜1644)。さらに、子宮頚部の扁平上皮癌および腺癌の大多数において、強いEpCAMの発現は、増殖の増加および最終分化に関するマーカーの消失に相関する(Litvinov, Am. J. Pathol, 1996, 148, 865〜75)。乳癌において、腫瘍細胞上でのEpCAMの過剰発現は、生存の予測因子である(Gastl, Lancet, 2000, 356, 1981〜1982)。EpCAMは、頭部、頚部、および肺の扁平上皮癌を有する患者における播種された腫瘍細胞を検出するためのマーカーである(Chaubal, Anticancer Res. 1999, 19, 2237〜2242;Piyathilake, Hum. Pathol. 2000, 31, 482〜487)。表皮、口腔、咽頭蓋、咽頭、喉頭、および食道において認められる正常な扁平上皮は、有意にEpCAMを発現しなかった(Quak, Hybridoma, 1990, 9, 377〜387)。EpCAMは原発、転移、および播種性NSCLC(肺の非小細胞癌細胞(Passlick, Int. J. Cancer, 2000, 87, 548〜552))の大多数において、胃の腺癌および胃食道接合部腺癌(Martin, J. Clin. Pathol. 1999, 52, 701〜4)において、ならびに結腸直腸、膵臓癌および乳癌に由来する細胞株(Szala, Proc. Natl. Acad. Sci. USA 1990, 87, 3542〜6;Packeisen, Hybridoma, 1999, 18, 37〜40)において発現されることが示されている。
【0056】
最も好ましい態様において、EpCAMに対する/結合する第二のIg由来ドメインを含む本発明のCD3特異的結合構築物は、以下からなる群より選択されるアミノ酸配列を含む:
(a)配列番号:31、33、35、37、39、49、55、58、61、63、65、67、237、239、241、243、245、247、249、251、253、255、257、259、261、263、265、267、269、271、273、275、277、279、281、283、285、287、289、291、293、295、297、299、301、303、305、307、309、311、313、315、317、319、321、323、および325のいずれか一つに示されるアミノ酸配列;
(b)配列番号:30、32、34、36、38、48、54、57、60、62、64、66、236、238、240、242、244、246、248、250、252、254、256、258、260、262、264、266、268、270、272、274、276、278、280、282、284、286、288、290、292、294、296、298、300、302、304、306、308、310、312、314、316、318、320、322、および324のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;ならびに
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列。
【0057】
したがって、本発明は、特に好ましい態様において、T細胞エピトープの生成傾向が低下したCD3結合/相互作用部分(「抗CD3」)およびEpCAMに特異的に相互作用/結合する(「抗EpCAM」)さらなる一本鎖部分(Ig由来ドメイン)を含む特異的CD3構築物を提供する。以下の表1A、1B、2A、2B、3A、3B、4A、4B、5A、および5Bは、そのようなCD3およびEpCAM結合構築物の好ましいコンフィギュレーションに関する。
【0058】
EpCAM 3-1、EpCAM 3-5、EpCAM 4-1、EpCAM 4-7およびEpCAM 5-10は、国際公開公報第99/25818号においてファージディスプレイによって単離されたEpCAMに対する特異的一本鎖抗体に関する。
【0059】
表1A、2A、3A、4A、および5Aにおける各構築物は、A-Gと呼ばれる異なる7個のタンパク質モジュールを含む。タンパク質モジュールA-Gは、A-B-C-D-E-F-Gの順でペプチド結合によって一つの連続したポリペプチド鎖において互いに直接共有結合し、N-末端でタンパク質モジュールAとなり、C-末端でタンパク質モジュールGとなる。タンパク質モジュールA、C、EおよびGは、ヒトCD3またはEpCAM抗原に対して特異性を有する抗体のVHまたはVLドメインのいずれかとなりうる抗体可変ドメインを指す。モジュールB、DおよびFはVHおよびVLドメインを接続するリンカーである。
【0060】
タンパク質モジュールAがVH抗体ドメインである場合、タンパク質モジュールCはVLタンパク質ドメインであり、その逆も起こりうる。タンパク質モジュールEがVH抗体ドメインである場合、タンパク質モジュールGはVLタンパク質ドメインであり、その逆もまた同じである。
【0061】
ヒトCD3抗原に対して特異性を有する抗体の脱免疫されたVHドメインは、配列番号:74または76に記載した配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫されたVLドメインは、配列番号:78、80、または82に記載の配列から選択することができる。ヒトEpCAM 3-1、3-5、4-1、4-7、および5-10抗体のVHタンパク質ドメインはそれぞれ、配列番号:137、141、145、149、および133に記載されている。ヒトEpCAM 3-1、3-5、4-1、4-7、および5-10抗体のVLタンパク質ドメインはそれぞれ、配列番号:139、143、147、151、および135に記載されている。
【0062】
タンパク質モジュール対A/CおよびE/Gによって示される抗体可変ドメインの対は、さらなる連結タンパク質モジュールによって結合され、この場合タンパク質モジュールBはモジュール対A/Cを直接結合するために役立ち、タンパク質モジュールFはモジュール対E/Gを直接結合するために役立つ。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体の脱免疫されたVH/VLまたはVL/VHタンパク質ドメインの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:3に記載されるアミノ酸配列を有する。モジュール対A/CまたはE/Gのいずれかが、EpCAM抗原に対して特異性を有する抗体からのVH/VLまたはVL/VHの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:168に記載されるアミノ酸配列を有する。モジュールDはABCおよびEFGモジュール群を結合する。
【0063】
タンパク質モジュールA-B-Cの組み合わせ、およびタンパク質モジュールE-F-Gの組み合わせはそれぞれ、ヒトCD3抗原またはEpCAM抗原のいずれかに対して特異性を有する抗体の一つのscFv断片を構成する。モジュールAおよびCがCD3結合配列を示す場合、タンパク質モジュールA-B-CおよびE-F-Gは、配列番号:176に記載される配列を有するタンパク質モジュールDを通して互いに結合する。一方、モジュールAおよびCがEpCAM結合配列を示す場合、タンパク質モジュールA-B-CおよびE-F-Gのそれぞれの群は、配列番号:174に示される配列を有するタンパク質モジュールDによって互いに結合する。このように、クローニング目的のためにVL鎖の後にさらなるセリンを挿入してもよい。しかし、当業者は、VLドメインを配列番号:176の代わりにその後のVドメインに結合させるために、配列番号:174に示されるリンカーを用いてもよい。タンパク質モジュールDは、タンパク質モジュールCのC-末端をタンパク質モジュールEのN-末端に結合させるように作用する。
【0064】
表1B、2B、3B、4B、および5Bにおける各核酸構築物は、A-Gと呼ばれる異なる7個の核酸モジュールを含む。核酸モジュールA-Gは、A-B-C-D-E-F-Gの順でホスフェートグリコシド結合によって単一の連続ヌクレオチド鎖において互いに直接共有結合しており、それぞれの核酸構築物の5'-末端が核酸モジュールAであって、3'末端が核酸モジュールGである。核酸モジュールA、C、EおよびGは、ヒトCD3またはEpCAM抗原に対して特異性を有する抗体のVHまたはVLドメインのいずれかとなりうる抗体可変ドメインのコード領域を指す。
【0065】
核酸モジュールAがVH抗体ドメインをコードする場合、核酸モジュールCはVLタンパク質ドメインをコードし、その逆も起こりうる。核酸モジュールEがVH抗体ドメインをコードする場合、核酸モジュールGはVLタンパク質ドメインをコードし、その逆も起こりうる。
【0066】
ヒトCD3抗原に対して特異性を有する抗体の脱免疫VHドメインをコードする核酸分子は、配列番号:73または75に記載の配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗原の脱免疫されたVLドメインをコードする核酸分子は、配列番号:77、79、または81記載の配列から選択することができる。ヒトEpCAM 3-1、3-5、4-1、4-7、および5-10抗体のVHタンパク質ドメインをコードする核酸分子はそれぞれ、配列番号:136、140、144、148、および132に記載されている。ヒトEpCAM 3-1、3-5、4-1、4-7、および5-10抗体のVLタンパク質ドメインをコードする核酸分子はそれぞれ、配列番号:138、142、146、150、および134に記載されている。
【0067】
核酸モジュール対A/CおよびE/Gによって示される抗体の可変ドメインをコードする核酸の対は、さらなる連結核酸モジュールによって結合し、この場合核酸モジュールBはモジュール対A/Cを直接結合させるように作用し、核酸モジュールFはモジュール対E/Gを直接結合させるように作用する。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体からの脱免疫されたVH/VLまたはVL/VHタンパク質ドメインの対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:202に記載のヌクレオチド配列を有する。モジュール対A/CまたはE/Gのいずれかが、ヒトEpCAM抗原に対して特異性を有する抗体からのVH/VLまたはVL/VH対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:201に記載のヌクレオチド配列を有する。
【0068】
核酸モジュールA-B-Cの組み合わせと核酸モジュールE-F-Gの組み合わせはそれぞれ、ヒトCD3抗原またはEpCAM抗原に対して特異性を有する抗体の一つのscFv断片を構成する。AおよびCモジュールがCD3結合配列を含む場合、核酸モジュールA-B-CおよびE-F-Gのそれぞれの群は、配列番号:175に記載のヌクレオチド配列を有する核酸モジュールDによって互いに結合する。AおよびCモジュールがEpCAM結合配列を含む場合、核酸モジュールA-B-CおよびE-F-Gのそれぞれの群は、配列番号:173に記載のヌクレオチド配列を有する核酸モジュールDによって互いに結合する。しかし、先に述べたように、セリンをコードするさらなるコドン(配列番号:175における)をクローニング目的のために挿入してもよい。当業者は、VL鎖をコードするヌクレオチド配列を、リンカーの5'末端でセリンをコードするさらなるコドンがなくとも、配列番号:173に示すリンカーによってその後のVドメインに直接結合させてもよい。核酸モジュールDは、核酸モジュールCの3'末端と核酸モジュールEの5'末端とを結合させるように作用する。
【0069】
(表1A)一本鎖抗EpCAM 3-1可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列

【0070】
(表1B)一本鎖抗EpCAM 3-1可変領域を含む脱免疫抗ヒトCD3構築物:ヌクレオチド配列
【0071】
(表2A)一本鎖抗EpCAM 3-5可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列
【0072】
(表2B)一本鎖抗EpCAM 3-5可変領域を含む脱免疫抗ヒトCD3構築物:ヌクレオチド配列
【0073】
(表3A)一本鎖抗EpCAM 4-1可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列
【0074】
(表3B)一本鎖抗EpCAM 4-1可変領域を含む脱免疫抗ヒトCD3構築物:ヌクレオチド配列
【0075】
(表4A)一本鎖抗EpCAM 4-7可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列
【0076】
(表4B)一本鎖抗EpCAM 4-7可変領域を含む脱免疫抗ヒトCD3構築物:ヌクレオチド配列
【0077】
(表5A)一本鎖抗EpCAM 5-10可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列
【0078】
(表5B)一本鎖抗EpCAM 5-10可変領域を含む脱免疫抗ヒトCD3構築物:ヌクレオチド配列
【0079】
最も好ましくは、本発明は、CD3およびEpCAMに対する特異的結合を含み、配列番号:30、31(表1Aおよび1Bの構築物2)、配列番号:48、49(表1A、1Bの構築物5)、配列番号:64、65(表2A、2Bの構築物2)、配列番号:54、55(表2A、2Bの構築物5)、配列番号:66、67(表3A、3Bの構築物2)、配列番号:32、33(表4A、4Bの構築物2)、配列番号:34、35(表4A、4Bの構築物4)、配列番号:60、61(表4A、4Bの構築物5)、配列番号:36、37(表5A、5Bの構築物2)、配列番号:38、39(表5A、5Bの構築物4)、または配列番号:62、63(表5A、5Bの構築物5)を有する二重特異性抗体構築物を提供する。
【0080】
本明細書において上記の構築物に従って、T細胞エピトープ生成傾向が低下して、ヒトCD3に対して特異性を有する少なくとも上記の第一のドメインと、EpCAMに対して特異的である第二のIg由来ドメインとを含む本発明の特に好ましいCD3およびEpCAM結合構築物は、配列番号:31、33、35、37、39、49、55、58、61、63、65、67、237、239、241、243、245、247、249、251、253、255、257、259、261、263、265、267、269、271、273、275、277、279、281、283、285、287、289、291、293、295、297、299、301、303、305、307、309、311、313、315、317、319、321、323、および325に示される。本明細書において定義される該好ましいCD3およびEpCAM結合構築物をコードする対応する核酸分子は、配列番号:30、32、34、36、38、48、54、57、60、62、64、66、236、238、240、242、244、246、248、250、252、254、256、258、260、262、264、266、268、270、272、274、276、278、280、282、284、286、288、290、292、294、296、298、300、302、304、306、308、310、312、314、316、318、320、322、および324を含む。
【0081】
したがって、本発明はまた、以下からなる群より選択される、ヒトCD3に対して特異的に結合してT細胞エピトープの生成傾向が低下している第一のドメインと、EpCAMに対する/結合することができるIg由来第二のドメインとを含む、CD3特異的結合構築物を提供する:
(a)配列番号:31、33、35、37、39、49、55、58、61、63、65、67、237、239、241、243、245、247、249、251、253、255、257、259、261、263、265、267、269、271、273、275、277、279、281、283、285、287、289、291、293、295、297、299、301、303、305、307、309、311、313、315、317、319、321、323、または325のいずれか一つに示されるアミノ酸配列;
(b)配列番号:30、32、34、36、38、48、54、57、60、62、64、66、236、238、240、242、244、246、248、250、252、254、256、258、260、262、264、266、268、270、272、274、276、278、280、282、284、286、288、290、292、294、296、298、300、302、304、306、308、310、312、314、316、318、320、322、または324のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;ならびに
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列。
(d)ストリンジェントなハイブリダイゼーション条件で(b)において定義された核酸配列の相補鎖とハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【0082】
本発明はまた、本明細書において上記の(b)において定義された核酸配列、すなわち配列番号:30、32、34、36、38、48、54、57、60、62、64、66、236、238、240、242、244、246、248、250、252、254、256、258、260、262、264、266、268、270、272、274、276、278、280、282、284、286、288、290、292、294、296、298、300、302、304、306、308、310、312、314、316、318、320、322、または324のいずれか一つに示される核酸配列の相補鎖と、ストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列を含む、ヒトCD3に対して特異的に結合して、T細胞エピトープの生成傾向が低下している第一のドメインと、EpCAMに対する/結合することができるIg由来の第二のドメインとを含むCD3特異的結合構築物を提供する。
【0083】
「ハイブリダイズする」という用語は、本明細書中で使用される場合、本明細書中で定義される脱免疫構築物またはその部分をコードするポリヌクレオチドにハイブリダイズすることが可能であるポリヌクレオチド/核酸配列をいう。従って、このポリヌクレオチドは、それぞれ、RNAまたはDNAの調製物のノーザンブロット分析もしくはサザンブロット分析におけるプローブとして有用であり得、またはそれらのそれぞれのサイズに依存してPCR分析におけるオリゴヌクレオチドプライマーとして使用され得る。好ましくは、このハイブリダイズするポリヌクレオチドは、少なくとも10個、より好ましくは少なくとも15個の長さのヌクレオチドを含むのに対して、プローブとして使用されるための本発明のハイブリダイズするポリヌクレオチドは、少なくとも100個、より好ましくは少なくとも200個の、または最も好ましくは少なくとも500個の長さのヌクレオチドを含む。
【0084】
いかにして核酸分子を用いてハイブリダイゼーション実験を実行するかは周知であり、すなわち、当業者は、本発明に従って彼らがどのハイブリダイゼーション条件を使用しなくてはならないかを知っている。このようなハイブリダイゼーション条件は、Molecular Cloning A Laboratory Manual, Cold Spring Harbor Laboratory (2001) N.Y.などの標準的な教科書において言及されている。本発明に従って好ましいものは、ストリンジェントなハイブリダイゼーション条件下で、本発明のポリヌクレオチドまたはその部分にハイブリダイズすることが可能であるポリヌクレオチドである。
【0085】
「ストリンジェントなハイブリダイゼーション条件」とは、例えば、50%ホルムアミド、5×SSC(750mM NaCl、75mM クエン酸ナトリウム)、50mM リン酸ナトリウム(pH 7.6)、5×デンハルト溶液、10%硫酸デキストラン、および20μg/ml変性剪断サケ精子DNAを含む溶液中での42℃、一晩のインキュベーション、続いて0.1×SSCにおける約65℃でのフィルターの洗浄をいう。よりストリンジェンシーの低いハイブリダイゼーション条件で本発明のポリヌクレオチドにハイブリダイズする核酸分子もまた意図される。ハイブリダイゼーションのストリンジェンシーおよびシグナル検出の変化は、主として、ホルムアミド濃度(より低いホルムアミドのパーセンテージはより低いストリンジェンシーを生じる);塩濃度、または温度の操作を通して達成される。例えば、よりストリンジェンシーの低い条件は、6×SSPE(20×SSPE=3M NaCl;0.2M NaH2PO4;0.02M EDTA, pH 7.4)、0.5% SDS、30%ホルムアミド、100μg/mlサケ精子ブロッキングDNAを含む溶液中での37℃での一晩のインキュベーション;引き続く50℃での1×SSPE、0.1% SDSを用いる洗浄を含む。さらに、なおより低いストリンジェンシーを達成するために、ストリンジェントなハイブリダイゼーション後に実行される洗浄が、より高い塩濃度(例えば、5×SSC)で行われ得る。上記の条件におけるバリエーションが、ハイブリダイゼーション実験におけるバックグラウンドを抑制するために使用される代替的なブロッキング試薬を含めることおよび/またはそれで置換することを通して達成され得ることに注目されたい。代表的なブロッキング試薬には、デンハルト試薬、BLOTTO、ヘパリン、変性サケ精子DNA、および市販の独自製品の処方物が含まれる。特異的ブロッキング試薬を含めることは、適合性に伴う問題に起因して、上記のハイブリダイゼーション条件の修飾を必要とする可能性がある。
【0086】
列挙される核酸分子は、例えば、DNA、cDNA、RNA、または合成的に製造されるDNAもしくはRNA、または単独でもしくは組み合わせてこれらのポリヌクレオチドのいずれかを含む組換え的に製造されるキメラ核酸分子であり得る。
【0087】
本発明において提供された脱免疫CD3およびEpCAM結合構築物は、医学的状況において、例えば腫瘍様疾患、特に乳癌、結腸癌、前立腺癌、頭頚部癌、皮膚癌(黒色腫)、尿性器癌、例えば卵巣癌、子宮内膜癌、子宮頚癌および腎臓癌、肺癌、胃癌、小腸癌、肝臓癌、膵臓癌、胆嚢癌、胆管癌、食道癌、唾液腺癌、および甲状腺癌の予防、治療および/または改善において特に有用である。特に、CD3およびEpCAMに結合する脱免疫構築物は、上皮癌、好ましくは腺癌、または最小の残留疾患、より好ましくは初期固形腫瘍、進行固形腫瘍、または転移性固形腫瘍の治療に用いることができる。
【0088】
本明細書に記述のCD3特異的結合構築物のより特に好ましい態様において、該構築物は、CCR5に対して特異性を有する抗原-相互作用部位を含む第二のIg由来ドメインを含む。
【0089】
ケモカイン受容体CCR5は、前炎症性ケモカインRANTES、MIP1-α、MIP1-β、およびMCP-2に結合する大きいGタンパク質共役7回膜貫通ドメイン受容体ファミリーに属する。ケモカインは、接着分子と共に白血球の浸潤を誘導するように、および組織損傷部位へのその誘導を指示するように作用する。CCR5は、少数のT細胞および単球において発現され、さらに、HIV感染症の過程において初期を支配するM-指向性HIV-1株の主要な共受容体である。
【0090】
ヒト免疫不全ウイルス(HIV)は、細胞表面上の二つの重要な分子であるCD4および共受容体に最初に結合して初めてヒト細胞内に入ることができる。最初に認識される共受容体は、CCR5であり、ウイルスのライフサイクルの後期ではもう一つのケモカイン受容体であるCXCR4はHIV-1の共受容体となる(D'Souza, Nature Med. 2, 1293(1996);Premack, Nature Med. 2, 1174;Fauci, Nature 384, 529(1996))。性的接触によってウイルスのほとんどの伝幡を引き起こすHIV-1株は、M-指向性ウイルスと呼ばれる。これらのHIV-1株(非合胞体誘導(NSI)プライマリウイルスとしても知られる)は、初代培養CD4+ T細胞およびマクロファージにおいて複製することができ、ケモカイン受容体CCR5(および頻度は少ないがCCR3)をその共受容体として用いる。T-指向性ウイルス(時に、合胞体誘導(SI)プライマリウイルスと呼ばれる)はまた、初代培養CD4+ T細胞においても複製できるが、さらにインビトロで確立されたCD4+ T細胞株に感染することができ、それらはケモカイン受容体CXCR4(融合体)を通して行われる。これらのT-指向性株の多くは、CXCR4の他にCCR5を用いることができ、中には、少なくとも特定のインビトロ条件においてCCR5を通してマクロファージに入ることができるものもある(D'Souza, Nature Med. 2, 1293(1996);Premack, Nature Med 2, 1174;Fauci, Nature 384, 529(1996))。
【0091】
他の共受容体がHIV-1の発病に関与しているか否かは解明されていないが、インビトロ試験から、いくつかのT-指向性株に関して他の共受容体の存在が推測されうる。M-指向性HIV-1株は、HIVの性的伝幡の約90%に関係していることから、CCR5は、患者におけるウイルスの主要な共受容体である;CXCR4を用いる(T-指向性)株の伝幡(または全身性の確立)はまれである(D'Souza, Nature Med. 2, 1293(1996);Premack, Nature Med. 2, 1174;Fauci, Nature 384, 529(1996);Paxton, Nature Med. 2, 412(1996);Liu, Cell 86, 367(1996);Samson, Nature 382, 722(1996);Dean, Science 273, 1856(1996);Huang, Nature Med. 2, 1240(1996))。しかし、SIウイルスがインビボで進化すると(またはそれらが伝幡されると)、それらは特にビルレントとなり、より急速な疾患の進行を引き起こす(D'Souza, Nature Med. 2, 1293(1996);Premack, Nature Med 2, 1174;Fauci, Nature 384, 529(1996);Schuitemaker, J. Virol. 66, 1354(1992);Connor, J. Virol. 67, 1772(1993);Richman, J. Infect. Dis. 169, 968(1994);R.I. Connorら、J. Exp. Med. 185, 621(1997);Trkola, Nature 384, 184(1996))。
【0092】
標的細胞上の共受容体分子の数および同一性、ならびにHIV-1株が異なる共受容体を通して細胞に入る能力は、疾患の進行に関する重要な決定因子であるように思われる。これらの要因は、HIV-1感染症の宿主およびウイルス依存的局面の双方に対する主要な影響である。例えば、CCR5におけるホモ接合欠損体(デルタ32)は、インビボおよびインビトロでHIV-1感染症に対する抵抗性と強く相関する。欠損CCR5対立遺伝子に関してヘテロ接合である人は、感染症に対する保護はせいぜい弱く、疾患の進行にごくわずかな遅れを示すに過ぎない(Paxton, Nature Med. 2, 412(1996);Liu, Cell 86, 367(1996);Samson, Nature 382, 722(1996);Dean, Science 273, 1856(1996);Huang, et al., Nature Med. 2, 1240(1996))。しかし、他の要因が活性化CD4+ T細胞上のCCR5発現レベルに影響を及ぼして、それによってインビトロでHIV-1感染症の効率に影響を及ぼしうる(Trkola, Nature 384, 184(1996);Bleul, Proc. Natl. Acad. Sci. USA 94, 1925(1997))。
【0093】
多発性硬化症の場合、CCR5およびCXCR3は、脱髄性脳病変に浸潤するT細胞のみならず、罹患患者の末梢血において主に発現されることが示されている。T細胞の消失は、この自己免疫疾患のT-細胞部分を遮断するであろう。
【0094】
CCR3およびCCR5の高度発現はまた、ホジキン疾患を有する患者に由来するリンパ節のT細胞およびB細胞においても認められた。
【0095】
I型糖尿病は、T細胞媒介自己免疫疾患であると考えられる。膵臓におけるCCR5受容体の発現は、関連する動物モデルにおいてI型糖尿病の進行に関連した(Cameron(2000), J. Immunol. 165, 1102〜1110)。特に、CCR5発現は、インスリン症および自然発生型I型糖尿病の発症に関連した。
【0096】
(ヒト)CCR5に特異的に結合するいくつかの抗体が当技術分野で公知であり、MC-1(Mack(1998), J. Exp. Med. 187, 1215〜1224)またはMC-5(Blanpain(2002)Mol. Biol. Cell. 13:723〜37;Segerer(1999), Kidney Int. 56:52〜64;Kraft(2001)J. Biol. Chem. 14:276:34408〜18)を含む。CCR-5抗体、特にMC-1およびMC-5は、本発明のCD3特異的構築物のIg由来第二のドメイン源として役立つ可能性がある。従って、好ましい態様において、本発明は、第一のドメインがヒトCD3に対する特異性を提供して、T細胞エピトープの生成傾向の低下を有し、該Ig由来第二のドメインが(ヒト)CCR5に対して特異的な抗体に由来する、少なくとも二つのドメインを含む二重特異性構築物に関する。最も好ましくは、そのような構築物は本明細書において定義されるように一本鎖scFvである。
【0097】
MC-1は、ヒトCCR5の第二の細胞外ループの第一の部分に特異的に結合して、添付の実施例に示すようにアカゲザルに由来するCCR5と交叉反応しないことが示された。したがって、本発明のCD3特異的構築物は、例えばCCR5、好ましくはヒトCCR5に対して特異的な抗体のVLおよびVHドメイン(すなわち、Ig-由来第二のドメイン)、ならびにCD3抗原に対して特異的な抗体のVHおよびVLドメインを含むことが好ましい。ヒトCCR5に対して特異的な該抗体は、とりわけMack(1998), J. Exp. Med. 187, 1215〜1224および添付の実施例に記述されているマウス抗ヒトCCR5抗体MC-1である。なおも、MC-5と同様に他の抗CCR5抗体(添付の実施例に特徴が示されているように、およびBlanpain(2002)Mol. Biol. Cell. 13:723〜37;Segerer(1999)Kidney Int. 56:52〜64およびKraft(2001)J. Biol. Chem. 14:276:34408〜18において開示されるように)を、本発明の状況において用いてもよい。
【0098】
本発明の特に好ましい態様において、ヒトCD3に対する/相互作用する脱免疫ドメインと、CCR5に特異的に結合/相互作用する第二のIg-由来ドメインとを含むCD3特異的結合構築物が提供される。そのような構築物を、表6Aおよび6Bに示す。表6Aおよび6BにおけるモジュールA-Gは表1〜5に関して先に述べたように定義することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VHドメインは、配列番号:74または76に記載される配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VLドメインは、配列番号:78、80、または82に記載される配列から選択することができる。ヒトCCR5抗体のVHタンパク質ドメインは、配列番号:129に記載される。ヒトCCR5抗体のVLタンパク質ドメインは、配列番号:131に記載される。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体からの脱免疫VH/VLまたはVL/VHタンパク質ドメインの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:3に記載されるアミノ酸配列を有する。モジュール対A/CまたはE/GのいずれかがEpCAM抗原に対して特異性を有する抗体からのVH/VLまたはVL/VHタンパク質ドメインの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:168に記載のアミノ酸配列を有する。タンパク質モジュールA-B-CおよびE-F-Gのそれぞれの群は、配列番号:174に記載の配列を有するタンパク質モジュールDを通して互いに結合する。しかし、先に述べたように、クローニング目的のためにVLおよびその後のVドメインのあいだにさらなるセリンを導入してもよい(配列番号:176に記載のリンカー)。
【0099】
ヒトCD3抗原に対して特異性を有する抗体の脱免疫VHドメインをコードする核酸分子は、配列番号:73または75に記載の配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VLドメインをコードする核酸分子は、配列番号:77、79、または81に記載の配列から選択することができる。ヒトCCR5抗体のVHタンパク質ドメインをコードする核酸分子は、配列番号:128に記載される。ヒトCCR5抗体のVLタンパク質ドメインをコードする核酸分子は、配列番号:130に記載される。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体からの脱免疫VH/VLまたはVL/VHタンパク質ドメインの対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:202に記載される核酸配列を有する。モジュール対A/CまたはE/GのいずれかがCCR5抗原に対して特異性を有する抗体からのVH/VLまたはVL/VHの対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:201に記載の核酸配列を有する。核酸モジュールA-B-CおよびE-F-Gの群は、配列番号:173に記載の配列を有するタンパク質モジュールDを通して互いに結合する。またはVLとその後のVドメイン(クローニング目的のためにセリン残基をコードするさらなるコドンを含む)とを結合するためにもう一つのリンカー配列番号:175を用いてもよい。
【0100】
(表6A)一本鎖抗CCR5可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列
【0101】
(表6B)一本鎖抗CCR5可変領域を含む脱免疫抗ヒトCD3構築物:核酸配列
構築物 構築物部分の配列番号 脱免疫された抗CD3構築物/特異性
ドメインの配置
【0102】
好ましくは、該構築物は、以下からなる群より選択されるアミノ酸配列を含む:
(a)配列番号:206、208、210、212、214、または216のいずれか一つに示されるアミノ酸配列;
(b)配列番号:205、207、209、211、213、または215のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列;および
(d)(b)において定義された核酸配列の相補鎖とストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【0103】
CCR5およびCD3結合構築物である配列番号:206、208、210は、構築物5を表し、配列番号:212、214、および216は表6の構築物13を表し、異なる三つのVL領域(VL1(配列番号:78)、VL2(配列番号:80)、またはVL3(配列番号:82))を有する。
【0104】
本発明はまた、本明細書において先の(b)において定義した核酸配列、すなわち、配列番号:205、207、209、211、213、または215のいずれか一つに示される核酸配列の相補鎖と、ストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列を含む、ヒトCD3に特異的に結合して、T細胞エピトープの生成傾向が低下した第一のドメインと、CCR5に対する/に結合することができるIg由来第二のドメインとを含むCD3特異的結合構築物を提供する。「ハイブリダイゼーション」および「ストリンジェントな条件」という用語は、本明細書において先に記述されている。対応する定義および態様は、必要な変更を加えて本明細書において適用される。
【0105】
本明細書において提供される脱免疫CD3およびCCR5結合構築物は、ウイルス病、特にHIV感染症およびAIDS、またはリウマチ性関節炎のような自己免疫疾患および/または炎症疾患の医学的介入において特に有用である。
【0106】
もう一つの態様において、本発明は、本発明の構築物のIg由来第二のドメインがCD19に対して特異性を有する抗原相互作用部位を含む、本明細書において先に定義したCD3特異的結合構築物を提供する。
【0107】
CD19は、非常に有用な医学的標的であることが判明した。CD19は、プロB細胞から成熟B細胞へのB細胞系列全体において発現され、細胞表面から消失しない場合、全てのリンパ腫細胞において均一に発現され、幹細胞には存在しない(Haagen, Clin. Exp. Immunol. 90(1992), 368〜75;Uckun, Proc. Natl. Acad. Sci. USA 85(1988), 8603〜7)。CD19に対する抗体とさらなる免疫調節抗体の双方を用いる併用治療が、B細胞の悪性疾患(国際公開公報第02/04021号、US2002006404、US2002028178)および自己免疫疾患(国際公開公報第02/22212号、US2002058029)の治療のために開示されている。国際公開公報第00/67795号は、無痛性の劇症型B-細胞リンパ腫の治療のためと共に急性および慢性型リンパ球性白血病の治療のために、CD19に対する抗体を用いることを開示している。国際公開公報第02/80987号は、B細胞非ホジキンリンパ腫、ホジキンリンパ腫、またはB細胞白血病(例えば、B細胞急性リンパ球性白血病(B-ALL)(例えば、ヘアリーセルリンパ腫)B細胞前駆体急性リンパ球性白血病(プレB-ALL)、B細胞慢性リンパ球性白血病(B-CLL))のような疾患の治療のために抗原CD19に対する抗体に基づいて免疫毒素を治療的に用いることを開示する。
【0108】
本発明の特に好ましい態様において、ヒトCD3に対する/結合する/相互作用する脱免疫ドメインと、CD19に特異的に結合する/相互作用する第二のIg由来ドメインとを含むCD3特異的結合構築物が提供される。そのような構築物を表7Aおよび7Bに示す。表7Aおよび7BにおけるモジュールA-Gは、先の表1〜5に記載されたように定義することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VHドメインは、配列番号:74または76に記載の配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VLドメインは、配列番号:78、80、または82に記載の配列から選択することができる。ヒトCD19抗体のVHタンパク質ドメインは、配列番号:114に記載される。ヒトCCR5抗体のVLタンパク質ドメインは、配列番号:116に記載される。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体からの脱免疫VH/VLまたはVL/VHタンパク質ドメインの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:3に記載されるアミノ酸配列を有する。モジュール対A/CまたはE/GのいずれかがCD19抗原に対して特異性を有する抗体からのVH/VLまたはVL/VHの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:168に記載のアミノ酸配列を有する。タンパク質モジュールA-B-CおよびE-F-Gのそれぞれの群は、配列番号:174に記載の配列を有するタンパク質モジュールDを通して互いに結合する。しかし、先に述べたように、クローニング目的のために(配列番号:176に記載のリンカー)VLとその後のVドメインとのあいだにさらなるセリンを導入してもよい。
【0109】
ヒトCD3抗原に対して特異性を有する抗体の脱免疫VHドメインをコードする核酸分子は、配列番号:73または75に記載の配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VLドメインをコードする核酸分子は、配列番号:77、79、または81に記載の配列から選択することができる。ヒトCD19抗体のVHタンパク質ドメインをコードする核酸分子は、配列番号:113に記載される。ヒトCCR5抗体のVLタンパク質ドメインをコードする核酸分子は、配列番号:115に記載される。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体からの脱免疫VH/VLまたはVL/VHタンパク質ドメインの対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:202に記載されるアミノ酸配列を有する。モジュール対A/CまたはE/GのいずれかがCD19抗原に対して特異性を有する抗体からのVH/VLまたはVL/VHの対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:201に記載の核酸配列を有する。核酸モジュールA-B-CおよびE-F-Gの群は、配列番号:173に記載の配列を有するタンパク質モジュールDを通して互いに結合する。VLとその後のVドメイン(クローニング目的のためにセリン残基をコードするさらなるコドンを含む)とを結合するために,もう一つのリンカー配列番号:175を用いてもよい。
【0110】
(表7A)一本鎖抗CD19可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列
【0111】
(表7B)一本鎖抗CD19可変領域を含む脱免疫抗ヒトCD3構築物:核酸配列
【0112】
より好ましい態様において、本発明は、上記のCD3結合ドメインと、CD19、好ましくはヒトCD19に特異的に結合する/相互作用する第二のIg由来ドメインとを含み、以下からなる群より選択されるアミノ酸配列を含む、脱免疫CD3-特異的結合構築物を提供する:
(a)配列番号:190、192、194、196、198、200、327、329、331、333、335、337、339、341、343、345、347、349、351、353、357、359、361、363、365、367、369、371、373、375、377、379、381、383、385、387、389、391、393、395、397、399、401、403、405、407、または409のいずれか一つに示されるアミノ酸配列;
(b)配列番号:189、191、193、195、197、199、326、328、330、332、334、336、338、340、342、344、346、348、350、352、354、356、358、360、362、364、366、368、370、372、374、376、378、380、382、384、386、388、390、392、394、396、398、400、402、404、406、または408のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;ならびに
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列;
(d)ストリンジェントなハイブリダイゼーション条件で(b)において定義された核酸配列の相補鎖とハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【0113】
本発明に従う好ましいCD19およびCD3結合構築物は、構築物5を表す配列番号:190、192、194であり、および表7の構築物13を表し、異なる三つのVL領域(VL1(配列番号:78)、VL2(配列番号:80)、またはVL3(配列番号:82))を有する配列番号:196、198、および200である。
【0114】
本発明はまた、上記の(b)において定義された核酸配列、すなわち配列番号:189、191、193、195、197、199、326、328、330、332、334、336、338、340、342、344、346、348、350、352、354、356、358、360、362、364、366、368、370、372、374、376、378、380、382、384、386、388、390、392、394、396、398、400、402、404、406、または408のいずれか一つに示される核酸配列の相補鎖とハイブリダイズする核酸配列によってコードされるアミノ酸配列を含む、ヒトCD3に対して特異的に結合し、T細胞エピトープの生成傾向が低下した第一のドメインと、CD19に対する/に結合することができるIg由来第二のドメインとを含むCD3特異的結合構築物を提供する。「ハイブリダイゼーション」および「ストリンジェントな条件」という用語は、本明細書において先に記述されている。対応する定義および態様は、必要な変更を加えて本明細書において適用される。
【0115】
本明細書に開示の脱免疫CD3およびCD19結合構築物は、増殖疾患、腫瘍様疾患、炎症疾患、免疫障害、自己免疫疾患、感染症、ウイルス病、アレルギー反応、寄生虫反応、移植片対宿主病、宿主対移植片病、またはB-細胞悪性疾患、特にB-細胞非ホジキンリンパ腫、ホジキンリンパ腫、またはB細胞白血病(例えば、B細胞急性リンパ球性白血病(B-ALL)(例えば、ヘアリーセルリンパ腫)B細胞前駆体急性リンパ球性白血病(プレB-ALL)、B細胞慢性リンパ球性白血病(B-CLL))の予防、治療、または改善において特に有用である。
【0116】
さらなる態様において、本発明は、ヒトCD3に対して特異的に結合して、T細胞エピトープの生成傾向が低下した第一のドメインと、Ig由来であってCD20に対する特異性を有する抗原相互作用部位を含む第二のドメインとを含む、先に定義されたCD3特異的結合構築物に関する。
【0117】
CD20は、B-リンパ球上に存在する細胞表面タンパク質の一つである。CD20抗原は、B-細胞非ホジキンリンパ腫(NHL)の90%以上におけるリンパ球を含む、正常および悪性プレBおよび成熟Bリンパ球において認められる。抗原は造血幹細胞、活性化Bリンパ球(プラズマ細胞)、および正常組織には存在しない。ほとんどがマウス起源であるいくつかの抗体が記述されている:1F5(Pressら、1987, Blood 69/2, 584〜591)、2B8/C2B8、2H7、1H4(Liuら、1987, J. Immunol. 139, 3521〜3526;Andersonら、1998, 米国特許第5,736,137号;Haismaら、1998, Blood 92, 184〜190;Shanら、1999, J. Immunol. 162, 6589〜6595)。
【0118】
CD20は、担体タンパク質に結合したscFvをコードするDNAのワクチン接種を用いるプラズマ細胞悪性疾患の治療のための免疫治療戦略において(Treonら、2000, Semin Oncol 27(5), 598)記述されており、CD20抗体(IDEC-C2B8)を用いる免疫療法において、非ホジキンB-細胞リンパ腫の治療において有効であることが示されている。CD20抗体は、非ホジキンリンパ腫において有効性および認容性が証明されており、これまで無処置または再発/不応性の無痛性非ホジキンリンパ腫においてそれぞれ、反応率は73%および48%であった(Montserrat, 2003, Semin Oncol. 30(1suppl2), 34〜39)。さらに、CD20抗体は、再発または進行期のB-細胞新生物を治療するために広く用いられており、有効率は約50%である。
【0119】
本発明の特に好ましい態様において、ヒトCD3に対する/と相互作用する脱免疫ドメインと、CD20に特異的に結合する/相互作用する第二のIg由来ドメインとを含むCD3-特異的結合構築物が提供される。そのような構築物を、表8Aおよび8Bに示す。表8Aおよび8BにおけるモジュールA-Gは、表1〜5に関して先に述べた通りに定義することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VHドメインは、配列番号:74または76に記載の配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VLドメインは、配列番号:78、80、または82に記載の配列から選択することができる。ヒトCD20抗体のVHタンパク質ドメインは、配列番号:170に記載される。ヒトCD20抗体のVLタンパク質ドメインは、配列番号:172に記載される。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体からの脱免疫VH/VLまたはVL/VHタンパク質ドメインの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:3に記載されるアミノ酸配列を有する。モジュール対A/CまたはE/GのいずれかがCD20抗原に対して特異性を有する抗体からのVH/VLまたはVL/VHの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:168に記載のアミノ酸配列を有する。タンパク質モジュールA-B-CおよびE-F-Gのそれぞれの群は、配列番号:174に記載の配列を有するタンパク質モジュールDを通して互いに結合する。しかし、先に述べたように、VLとその後のVドメインとのあいだにさらなるセリンをクローニング目的のために(配列番号:176に記載のリンカー)導入してもよい。
【0120】
ヒトCD3抗原に対して特異性を有する抗体の脱免疫VHドメインをコードする核酸分子は、配列番号:73または75に記載の配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VLドメインをコードする核酸分子は、配列番号:77、79、または81に記載の配列から選択することができる。ヒトCD20抗体のVHタンパク質ドメインをコードする核酸分子を、配列番号:169に記載する。ヒトCD20抗体のVLタンパク質ドメインをコードする核酸分子を、配列番号:171に記載する。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体からの脱免疫VH/VLまたはVL/VHタンパク質ドメインの対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:202に記載されるアミノ酸配列を有する。モジュール対A/CまたはE/GのいずれかがCD20抗原に対して特異性を有する抗体からのVH/VLまたはVL/VHの対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:201に記載の核酸配列を有する。核酸モジュールA-B-CおよびE-F-Gの群は、配列番号:173に記載の配列を有するタンパク質モジュールDを通して互いに結合する。VLとその後のVドメインとを結合するためにもう一つのリンカー配列番号:175を用いてもよい(クローニング目的のためにセリン残基をコードするさらなるコドンを含む)。
【0121】
(表8A)一本鎖抗CD20可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列
【0122】
(表8B)一本鎖抗CD20可変領域を含む脱免疫抗ヒトCD3構築物:ヌクレオチド配列
【0123】
より好ましくは、本発明の脱免疫CD3およびCD20結合構築物は、以下からなる群より選択されるアミノ酸配列を含む:
(a)配列番号:218、220、222、224、226、または228のいずれか一つに示されるアミノ酸配列;
(b)配列番号:217、219、221、223、225、または227のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列;および
(d)(b)において定義された核酸配列の相補鎖とストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【0124】
本発明はまた、本明細書において先に定義した核酸配列、すなわち配列番号:217、219、221、223、225、または227のいずれか一つに示される核酸配列の相補鎖とストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列を含む、ヒトCD3に対して特異的に結合し、T細胞エピトープの生成傾向が低下した第一のドメインと、CD20に対する/に結合することができるIg由来第二のドメインとを含むCD3特異的結合構築物を提供する。「ハイブリダイゼーション」および「ストリンジェントな条件」という用語は、本明細書において先に記述されている。対応する定義および態様は、必要に応じて本明細書において適用される。
【0125】
本明細書に記述の脱免疫CD3およびCD20結合構築物は、B-細胞関連障害の治療、予防、および/または改善において、好ましくはリンパ腫の医学的介入において、より好ましくは非ホジキンリンパ腫の治療において用いられると想像される。
【0126】
本発明はまた、本発明のCD3特異的結合分子をコードする核酸配列を提供する。
【0127】
本発明の核酸分子に、調節配列が加えられ得ることは当業者には明らかである。例えば、プロモーター、転写エンハンサー、および/または本発明のポリヌクレオチドの発現の誘導を可能にする配列が利用され得る。適切な誘導系は、例えば、例えばGossen および Bujard (Proc. Natl. Acad. Sci. USA 89 (1992, 5547-5551))およびGossen et al. (Trends Biotech. 12 (1994), 58-62)によって記載されるようなテトラサイクリン調節される遺伝子発現、または、例えばCrook (1989) EMBO J. 8, 513-519によって記載されるようなデキサメタゾン誘導性遺伝子発現系である。
【0128】
また、さらなる目的のために、核酸分子が、例えば、チオエステル結合および/またはヌクレオチドアナログを含み得ることが想定される。この修飾は、細胞中のエンドヌクレアーゼおよび/またはエキソヌクレアーゼに対する核酸分子の安定化のために有用であり得る。この核酸分子は、細胞中でこの核酸分子の転写を可能にするキメラ遺伝子を含む適切なベクターによって転写され得る。この点に関して、このようなポリヌクレオチドが「遺伝子標的化」または「遺伝子治療的」アプローチのために使用され得ることもまた理解される。別の態様において、この核酸分子は標識される。核酸の検出のための方法は当技術分野で周知であり、例えば、サザンブロッティングおよびノーザンブロッティング、PCR、またはプライマー伸長がある。この態様は、遺伝子治療アプローチの間に上記の核酸分子の首尾よい導入を確認するためののスクリーニング方法のために有用であり得る。
【0129】
この核酸分子は、上述の核酸分子のいずれかを単独でまたは組み合わせてのいずれかで含む組換え的に産生されたキメラ核酸分子であり得る。好ましくは、この核酸分子はベクターの一部である。
【0130】
従って、本発明はまた、本発明に記載される核酸分子を含むベクターに関する。
【0131】
多くの適切なベクターが分子生物学における当業者に公知であり、その選択は所望される機能に依存し、これにはプラスミド、コスミド、ウイルス、バクテリオファージ、および遺伝子操作において慣用的に使用される他のベクターが含まれる。種々のプラスミドおよびベクターを構築するために、当業者に周知である方法(例えば、Sambrookら(前掲)およびAusubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.Y. (1989), (1994)を参照されたい)が使用され得る。または、本発明のポリヌクレオチドおよびベクターは、標的細胞への送達のためにリポソームに再構成され得る。以下でさらに詳細に議論されるように、クローニングベクターは、個別のDNAの配列を単離するために使用された。関連する配列は、特定のポリペプチドの発現が必要とされる発現ベクターに移動され得る。代表的なクローニングベクターには、pBluescript SK、pGEM、pUC9、pBR322、およびpGBT9が含まれる。代表的な発現ベクターには、pTRE、pCAL-n-EK、pESP-1、pOP13CATが含まれる。
【0132】
好ましくは、このベクターは、本明細書中に定義された二重特異性単鎖抗体構築物をコードするこの核酸配列に作動可能に連結された調節配列である核酸配列を含む。
【0133】
このような調節配列(制御エレメント)は当業者に公知であり、プロモーター、スプライシングカセット、翻訳開始コドン、ベクターに挿入を導入するための翻訳および挿入部位を含み得る。好ましくは、この核酸分子は、真核生物細胞または原核生物細胞における発現を可能にする、その発現制御配列に作動可能に連結される。
【0134】
このベクターは、本明細書中に定義される二重特異性単鎖抗体構築物をコードする核酸分子を含む発現ベクターであることが想定される。
【0135】
「調節配列」という用語は、それらがライゲーションされるコード配列の発現をもたらすために必要であるDNA配列をいう。このような制御配列の性質は、宿主生物に依存して異なる。原核生物においては、制御配列には、一般的に、プロモーター、リボソーム結合部位、およびターミネーターが含まれる。真核生物においては、一般的に、制御配列は、プロモーター、ターミネーター、およびある場合においては、エンハンサー、トランス活性化因子、または転写因子が含まれる。「制御配列」という用語は、最小限、その存在が発現のために必要である全ての成分を含むことが意図され、さらなる有利な成分もまた含み得る。
【0136】
「作動可能に連結される」という用語は、そのように記載される成分が、それらの意図される様式で機能することを可能にする関連性にある並列をいう。コード配列に「作動可能に連結される」制御配列は、コード配列の発現が制御配列と適合可能な条件下で達成されるようなやり方でライゲーションされる。制御配列がプロモーターである場合、当業者には、二本鎖核酸が好ましく使用されることが明白である。
【0137】
従って、列挙されるベクターは好ましくは発現ベクターである。「発現ベクター」は、選択された宿主を形質転換するために使用され得る構築物であり、選択された宿主中でコード配列の発現を提供する。発現ベクターは、例えば、クローニングベクター、バイナリーベクターまたは組み込みベクターであり得る。発現は、好ましくは翻訳可能なmRNAへの核酸分子の転写を含む。原核生物および/または真核生物細胞における発現を確実にする調節エレメントは当業者に周知である。真核生物細胞の場合、これらは通常、転写の開始を確実にするプロモーターを含み、ならびに任意に、転写の終結および転写の安定化を確実にするポリAシグナルを含む。原核生物宿主細胞において発現を可能にする可能性のある調節エレメントは、例えば、大腸菌(E. coli)におけるPL、lac、trp、またはtacプロモーターを含み、真核生物宿主細胞中での発現を可能にする調節エレメントの例は、酵母におけるAOX1もしくはGAL1プロモーター、CMVプロモーター、SV40プロモーター、RSVプロモーター(ラウス肉腫ウイルス)、CMVエンハンサー、SV40エンハンサー、または哺乳動物および他の動物細胞におけるグロビンイントロンである。
【0138】
転写の開始の原因であるエレメントに加えて、このような調節エレメントはまた、ポリヌクレオチドの下流のSV40-ポリA部位またはtk-ポリA部位などの転写終結シグナルを含み得る。さらに、使用される発現系に依存して、細胞区画にポリペプチドを方向付けるか、または培地にポリペプチドを分泌することが可能であるリーダー配列が、列挙される核酸配列のコード配列に加えられてもよく、これらは当技術分野で周知である;例えば、添付の実施例1も参照されたい。リーダー配列は、適切な段階で、翻訳配列、開始配列、および終結配列と、好ましくは、翻訳されたタンパク質またはその部分の分泌を方向付けることが可能であるリーダー配列と、細胞膜周辺腔または細胞外媒体でアセンブルされる。任意に、異種配列が、所望の性質(例えば、発現された組換え産物の安定化または精製の単純化;前記を参照されたい)を付与するN末端同定ペプチドを含む融合タンパク質をコードし得る。この文脈において、適切な発現ベクターは当技術分野で公知であり、例えば、岡山-バーグcDNA発現ベクターpcDV1(Pharmacia)、pCDM8、pRc/CMV、pcDNA1、pcDNA3(In-vitrogene)、pEF-DHFR、pEF-ADA、またはpEF-neo(Raum et al. Cancer Immunol Immunother (2001) 50(3), 141-150)、またはpSPORT1(GIBCO BRL)である。
【0139】
好ましくは、発現制御配列は、真核生物宿主細胞を形質転換またはトランスフェクトすることが可能であるベクターにおける真核生物プロモーター系であるが、しかし、原核生物宿主のための制御配列もまた使用され得る。一旦、ベクターが適切な宿主に取り込まれると、その宿主は、ヌクレオチド配列の高レベル発現のために適切な条件下で維持され、所望される場合、本発明のポリペプチドの収集および精製が続いて行われ得る;例えば、添付の実施例を参照されたい。
【0140】
使用され得る代替的な発現系は昆虫系である。1つのこのような系において、Autographa californica核多核体病ウイルス(AcNPV)が、Spodoptera frugiperda細胞またはTrichoplusia larvaeにおいて外来性の遺伝子を発現するためのベクターとして使用される。列挙される核酸分子のコード配列は、ポリヘドリン遺伝子などのウイルスの必須でない領域にクローニングされ得、ポリヘドリンプロモーターの制御下に配置され得る。このコード配列の首尾よい挿入はポリヘドリン遺伝子を不活性にし、コートタンパク質の被覆を欠く組換えウイルスを産生する。次いで、組換えウイルスは、本発明のタンパク質が発現されるS. frugiperda細胞またはTrichoplusia larvaeに感染させるために使用される(Smith, j. Virol 46 (1983), 584; Engelhard, Proc. Nat. Acad. Sci. USA 91 (1994), 3224-3227)。
【0141】
さらなる調節エレメントは、転写エンハンサーならびに翻訳エンハンサーを含み得る。有利には、本発明の上記のベクターは、選択可能なおよび/またはスコア付け可能なマーカーを含む。
【0142】
形質転換された細胞、例えば、植物組織および植物細胞の選択のために有用である選択可能なマーカー遺伝子は当業者に周知であり、例えば、dhfr(これはメトレキサートに対する耐性を付与する(Reiss, Plant Physiol. (Life Sci. Adv.)13 (1994), 143-149));npt(これはアミノグリコシドネオマイシン、カナマイシン、およびパロマイシンに対する耐性を付与する(Herrera-Estrella, EMBO J. 2 (1983), 987-995))およびhygro(これはハイグロマイシンに対する耐性を付与する(Marsh, Gene 32 (1984), 481-485))についての選択の基礎として代謝拮抗物質耐性を含む。さらなる選択可能な遺伝子が記載されており、すなわち、以下が記載されている。trpB(これは細胞がトリプトファンの代わりにインドールを利用することを可能にする);hisD(これは細胞がヒスチジンの代わりにヒスチノールを利用することを可能にする)(Hartman, Proc. Natl. Acad. Sci. USA 85 (1988), 8047);マンノース-6-リン酸イソメラーゼ(これは細胞がマンノースを利用することを可能にする)(WO 94/20627)およびODC(オルニチンデカルボキシラーゼ)(これはオルニチンデカルボキシラーゼインヒビター、2-(ジフルオロメチル)-DL-オルニチン、DFMOに対する耐性を付与する)(McConlogue, 1987, Current Communications in Molecular Biology, Cold Spring Harbor Laboratory ed.)またはAspergillus terreusからのデアミナーゼ(これは、ブラスチシジンSに対する耐性を付与する(Tamura, Biosci. Biotechnol. Biochem. 59 (1995), 2336-2338))。
【0143】
有用なスコア付け可能なマーカーもまた当業者に公知であり、市販されている。有利には、このマーカーは、ルシフェラーゼ(Giacomin, Pl. Sci. 116 (1996), 59-72; Scikantha, J. Bact. 178 (1996), 121)グリーン蛍光タンパク質(Gerdes, FEBS Lett. 389 (1996), 44-47)、またはβ-グルクロニダーゼ(Jefferson, EMBO J. 6 (1987), 3901-3907)をコードする遺伝子である。この態様は、列挙されたベクターを含む細胞、組織、および生物体を単純かつ迅速にスクリーニングするために特に有用である。
【0144】
上記のように、列挙された核酸分子は、例えば、精製だけでなく遺伝子治療目的のために、コードされたCD3特異的構築物を細胞中で発現するために単独でまたはベクターの一部として使用され得る。上記の(二重特異性)CD3構築物のいずれか1つをコードするDNA配列を含む核酸分子またはベクターは細胞に導入され、次には関心対象のポリペプチドを産生する。遺伝子治療(これはエクスビボまたはインビボの技術によって細胞に治療遺伝子を導入することに基づく)は、遺伝子移入の最も重要な応用の1つである。インビトロまたはインビボの遺伝子治療のための適切なベクター、方法、または遺伝子送達系は文献に記載されており、当業者に公知である。例えば、以下を参照されたい。列挙された核酸分子およびベクターは、直接的導入のために、またはリポソーム、もしくはウイルスベクター(例えば、アデノウイルス、レトロウイルス)を介する細胞への導入のために設計され得る。好ましくは、この細胞は生殖系列細胞、胚性細胞、または卵細胞、またはそれらから由来する細胞であり、最も好ましくは、幹細胞である。胚性幹細胞の例は、とりわけ、Nagy, Proc. Natl. Acad. Sci. USA 90 (1993), 8424-8428において記載されている幹細胞であり得る。
【0145】
上記に従って、本発明は、本明細書中に定義される二重特異性単鎖抗体構築物のポリペプチド配列をコードする核酸分子を含む、遺伝子操作において慣用的に使用されるベクター(特に、プラスミド、コスミド、ウイルス、およびバクテリオファージ)を誘導するための方法に関する。好ましくは、このベクターは、発現ベクターおよび/または遺伝子移入もしくは標的化ベクターである。レトロウイルス、ワクシニアウイルス、アデノ関連ウイルス、ヘルペスウイルス、またはウシパピローマウイルスなどのウイルスに由来する発現ベクターは、標的とされた細胞集団への列挙されたポリヌクレオチドまたはベクターの送達のために使用され得る。当業者に周知である方法が組換えベクターを構築するために使用され得る;例えば、Sambrook et al. (前掲)、Ausubel(1989、前掲)または他の標準的教科書において記載される技術を参照されたい。または、列挙される核酸分子およびベクターは、標的細胞への送達のためにリポソームに再構成され得る。本発明の核酸分子を含むベクターは、周知の方法(これは細胞宿主の型に依存して変化する)によって宿主細胞に移動され得る。例えば、塩化カルシウムトランスフェクションが原核生物細胞のために利用されるのに対して、リン酸カルシウム処理またはエレクトロポレーションが他の細胞宿主のために使用され得る;Sambrook、前記を参照されたい。
【0146】
列挙されるベクターは、特に、pEF-DHFR、pEF-ADA、またはpEF-neoであり得る。ベクターpEF-DHFR、pEF-ADAおよびpEF-neoは、当該分野において、例えば、Mack et al. (PNAS (1995) 92, 7021-7025)およびRaum et al. (Cancer Immunol Immunother (2001) 50(3), 141-150)ににおいて記載されている。
【0147】
本発明は、ここで定義したようなベクターで形質転換またはトランスフェクトされた宿主をも提供する。この宿主は、少なくとも1つの上記のベクター、または少なくとも1つの上記の核酸分子を、宿主に導入することによって産生され得る。宿主におけるその少なくとも1つのベクターまたは少なくとも1つの核酸分子の存在が、上記の二重特異性単鎖抗体構築物をコードする遺伝子の発現を媒介し得る。
【0148】
宿主に導入される記載された核酸分子またはベクターは、宿主のゲノムに組み込まれ得るか、または染色体外に維持され得るかのいずれかである。
【0149】
宿主は、任意の原型生物または真核生物細胞であり得る。
【0150】
「原核生物」という用語は、本発明のタンパク質の発現のためにDNA分子またはRNA分子で形質転換またはトランスフェクトされ得る全ての細菌を含むことを意味する。原核生物宿主は、グラム陰性細菌ならびにグラム陽性細菌、例えば、大腸菌、S. typhimurium、Serratia marcescensおよびBacillus subtilisなどを含み得る。「真核生物」という用語は、酵母、高等植物、昆虫、および好ましくは哺乳動物の細胞を含むことを意味する。組換え産生手順において利用される宿主に依存して、本発明のポリヌクレオチドによってコードされるタンパク質は、グリコシル化されてもよく、またはグリコシル化されなくてもよい。特に好ましいものは、本発明のポリペプチドのコード配列ならびにそれに遺伝子が融合したN末端FLAG-タグおよび/またはC末端His-タグを含むプラスミドまたはウイルスの使用である。好ましくは、このFLAG-タグの長さは約4〜8アミノ酸であり、最も好ましくは8アミノ酸である。上記のポリヌクレオチドは、当業者に一般的に公知である技術のいずれかを使用して宿主を形質転換またはトランスフェクトするために使用され得る。さらに、融合された、作動可能に連結された遺伝子を調製する方法、ならびに例えば、哺乳動物細胞および細菌中でそれらを発現させる方法は、当技術分野において周知である(Sambrook、前掲)。
【0151】
好ましくは、この宿主は細菌、昆虫、真菌、植物、または動物の細胞である。
【0152】
列挙される宿主は、哺乳動物細胞、より好ましくはヒト細胞またはヒト細胞株であり得ることが特に想定される。
【0153】
特に好ましい宿主細胞には、CHO細胞、COS細胞、ミエローマ細胞株(SP2/0またはNS/0など)が含まれる。実施例で記載されているように、特に好ましい宿主細胞は、COS細胞である。
【0154】
さらなる実施例において、本発明は、該構造物の発現に適した条件下で本発明の細胞、および/または宿主を培養する段階および細胞又は培養液から構造物を単離する段階を含む、上述したCD3特異的構造物の準備工程に関する。
【0155】
形質転換された宿主は、最適な細胞増殖を達成するために、当技術分野において公知である技術に従って、ファーメンター中で増殖され、かつ培養され得る。次いで、本発明のポリペプチドは、増殖培地、細胞溶解物、または細胞膜画分から単離され得る。本発明の、例えば、微生物によって発現されたポリペプチドの単離および精製は、例えば、調製用クロマトグラフィー分離および免疫学的分離(例えば、本発明のポリペプチドのタグに対して指向されるモノクローナル抗体またはポリクローナル抗体の使用を含むもの)などの任意の慣用的な手段によって、または添付の実施例において記載されるようにであり得る。
【0156】
さらに、本発明は、本明細書において定義された(ヒト)CD3特異的結合構築物を含む組成物、または先に開示したプロセスによって産生された(ヒト)CD3特異的結合構築物、本発明の核酸分子、本発明のベクターまたは宿主を提供する。該組成物は、任意で免疫エフェクター細胞の活性化シグナルを提供することができるタンパク質様化合物を含む。最も好ましくは、該組成物は、任意で担体、安定化剤、および/または賦形剤の適した製剤をさらに含む薬学的組成物である。
【0157】
本発明に従って、「薬学的組成物」という用語は、患者、好ましくはヒト患者に投与するための組成物に関する。好ましい態様において、薬学的組成物は、非経口、経皮、腔内、動脈内、髄腔内投与のための、または組織もしくは腫瘍への直接注射による組成物を含む。特に、該薬学的組成物は注入または注射によって患者に投与されると想像される。適した組成物の投与は異なる方法、例えば静脈内、腹腔内、皮下、筋肉内、局所、または皮内投与によって行ってもよい。本発明の薬学的組成物はさらに、薬学的に許容される担体を含んでもよい。適した薬学的担体の例は当技術分野において周知であり、これには、燐酸緩衝生理食塩液、水、油/水乳剤のような乳剤、様々なタイプの湿潤剤、滅菌溶液等が含まれる。そのような担体を含む組成物は、周知の通常の方法によって製剤化することができる。これらの薬学的組成物は、適した用量で被験者に投与することができる。投与レジメは、主治医および臨床的要因によって決定されるであろう。医学の技術分野において周知であるように、任意の患者1人の用量は、患者の体格、体表面積、年齢、投与される特定の化合物、性別、投与期間および経路、全身健康、ならびに同時投与される他の薬物を含む、多くの要因に依存する。一般的に、薬学的組成物の規則的な投与としてのレジメは、1μg〜5 g単位/日の範囲でなければならない。しかし、連続的注入のためのより好ましい用量は、0.01μg〜2 mg、好ましくは0.01μg〜1 mg、より好ましくは0.01μg〜100 μg、さらにより好ましくは0.01μg〜50 μg、および最も好ましくは0.01μg〜10 μg単位/kg体重/時間の範囲であってもよい。特に好ましい用量は、本明細書において下記に引用される。進行は定期的な評価によってモニターすることができる。用量は変化するが、DNAの静脈内投与のための好ましい用量はDNA分子約106〜1012コピーである。本発明の組成物は、局所または全身投与してもよい。投与は一般的に非経口、例えば静脈内投与であろう;DNAはまた、例えば内部または外部標的部位へのバイオリスティック(biolistic)送達、または動脈内の部位にカテーテルによって標的部位に直接投与してもよい。非経口投与のための調製物には、滅菌水溶液、または非水溶液、懸濁液、および乳剤が含まれる。非水性溶媒の例は、プロピレングリコール、ポリエチレングリコール、オリーブ油のような植物油、およびオレイン酸エチルのような注射可能な有機エステルである。水性担体には、生理食塩液および緩衝培地を含む、水、アルコール/水溶液、乳剤、または懸濁液が含まれる。非経口溶媒には、塩化ナトリウム溶液、リンゲルデキストロース、デキストロースおよび塩化ナトリウム、乳酸加リンゲル、または固定油が含まれる。静脈内媒体には、液体および栄養補助剤、電解質補助剤(リンゲルデキストロースに基づくような)等が含まれる。例えば抗菌剤、抗酸化剤、キレート剤、および不活性ガス等のような保存剤および他の添加剤も同様に、存在してもよい。さらに、本発明の薬学的組成物は、例えば好ましくはヒト起源の血清アルブミンまたは免疫グロブリンのような蛋白質様担体を含んでもよい。本発明の薬学的組成物は、二重特異性一本鎖抗体構築物またはこれをコードする核酸分子もしくはベクター(本発明において記述されるように)の他に、薬学的組成物の意図する用途に応じて、さらに生物活性物質を含んでもよいと想像される。そのような物質は、消化管系に作用する薬剤、細胞増殖抑制物質として作用する薬剤、ハイパーウリケミア(hyperurikemia)を予防する薬剤、免疫反応を阻害する薬剤(例えば、コルチコステロイド)、循環系に影響を及ぼす薬剤および/または当技術分野で公知のT-細胞共刺激分子またはサイトカインのような物質であってもよい。
【0158】
本発明の組成物の投与に関して可能性がある適応は、腫瘍様疾患、特に乳癌、結腸癌、前立腺癌、頭頚部癌、皮膚癌(黒色腫)、尿性器癌、例えば卵巣癌、子宮内膜癌、子宮頚癌および腎臓癌、肺癌、胃癌、小腸の癌、肝臓癌、膵臓癌、胆嚢癌、胆管癌、食道癌、唾液腺癌および甲状腺癌のような上皮癌/癌腫、または血液腫瘍、神経膠腫、肉腫、もしくは骨肉腫のような他の腫瘍様疾患である。本発明の組成物の投与は特に、細胞1個の生存によって引き起こされる腫瘍の局所および非局所再発を特徴とする最小残留疾患、好ましくは初期固形腫瘍、進行固形腫瘍、または転移固形腫瘍に適応される。
【0159】
本発明はさらに、他の化合物、例えば免疫エフェクター細胞、細胞増殖、または細胞刺激に関する活性化シグナルを提供することができる分子との同時投与プロトコールを想像する。該分子は、例えばさらにT細胞の一次活性化シグナル(例えば、さらなる共刺激分子:B7ファミリーの分子、Ox40L、4.1 BBL)、またはさらなるサイトカイン:インターロイキン(例えば、IL-2)もしくはNKG-2D関係化合物であってもよい。
【0160】
上記の本発明の組成物は、任意で検出手段および方法をさらに含む診断組成物であってもよい。
【0161】
本明細書において定義されるCD3-特異的構築物はまた、液相において、または固相担体に結合して利用することができるイムノアッセイにおいて用いるために適している。本発明のポリペプチドを利用することができるイムノアッセイの例は、直接または間接的フォーマットのいずれかの競合的および非競合的イムノアッセイである。そのようなイムノアッセイの例は、酵素結合イムノソルベントアッセイ(ELISA)、酵素イムノアッセイ(EIA)、ラジオイムノアッセイ(RIA)、サンドイッチ(免疫測定)アッセイ、およびウェスタンブロットアッセイである。
【0162】
本発明のCD3特異的結合構築物は、異なる多くの担体に結合させて、該ポリペプチドに特異的結合した細胞を単離するために用いることができる。周知の担体の例には、ガラス、ポリスチレン、ポリ塩化ビニル、ポリプロピレン、ポリエチレン、ポリカーボネート、デキストラン、ナイロン、アミロース、天然および改変セルロース、ポリアクリルアミド、アガロース、およびマグネタイトが含まれる。担体の性質は可溶性または不溶性のいずれかとなり得て、例えば本発明の目的に関してビーズとなりうる。
【0163】
当業者に公知の異なる多くの標識および標識法が存在する。本発明において用いることができる標識のタイプの例には、酵素、放射性同位元素、コロイド金属、蛍光化合物、化学発光化合物、および生物発光化合物が含まれる;同様に、本明細書において考察した態様を参照されたい。
【0164】
本発明の最も好ましい態様において、薬学的組成物を調製するために、CD3特異的結合分子、本発明のベクターまたは宿主を用いることが想像される。該薬学的組成物は、増殖疾患、腫瘍様疾患、炎症疾患、免疫障害、自己免疫障害、感染症、ウイルス病、アレルギー反応、寄生虫反応、移植片対宿主病、または宿主対移植片病の予防、治療、または改善において用いてもよい。
【0165】
さらに、本発明に従って、CD19およびCD3結合ドメイン、好ましくは配列番号:190、192、194、196、198、200、327、329、331、333、335、337、339、341、343、345、347、349、351、353、357、359、361、363、365、367、369、371、373、375、377、379、381、383、385、387、389、391、393、395、397、399、401、403、405、407、または409を含む脱免疫構築物は、免疫障害(様々なB細胞悪性疾患)または自己免疫疾患の治療に用いることができ、CCR5およびCD3結合ドメイン、好ましくは配列番号:206、208、210、212、214、または216を含む脱免疫構築物は、腫瘍様疾患、好ましくはリンパ腫、より好ましくは非ホジキンB-細胞リンパ腫の治療に用いることができ、EpCAMおよびCD3結合ドメイン、好ましくは配列番号:31、33、35、37、39、49、55、58、61、63、65、67、237、239、241、243、245、247、249、251、253、255、257、259、261、263、265、267、269、271、273、275、277、279、281、283、285、287、289、291、293、295、297、299、301、303、305、307、309、311、313、315、317、319、321、323、または325を含む脱免疫構築物は、腫瘍様疾患、好ましくは上皮癌の治療に用いることができる。
【0166】
本発明はまた、本発明の(二重特異性)CD3特異的結合分子、本明細書に記述のプロセスによって産生された(二重特異性)CD3特異的結合分子、本発明の核酸分子、ベクターまたは宿主を、そのような予防、治療、または改善を必要とする被験者に投与することを含む、増殖疾患、腫瘍性の疾患、炎症疾患、免疫障害、自己免疫疾患、感染症、ウイルス病、アレルギー反応、寄生虫反応、移植片対宿主病、または宿主対移植片病を予防、治療、または改善する方法に関する。好ましくは該被験者はヒトである。
【0167】
予防、治療、または改善法はまた、さらに、免疫エフェクター細胞の活性化シグナルを提供することができる蛋白質様化合物の投与を含んでもよい。該蛋白質様化合物は、本発明のCD3結合分子、核酸分子、ベクター、または宿主と同時、または非同時に投与してもよい。蛋白質様化合物は、とりわけさらなる共刺激分子:B7ファミリーの分子、Ox40L、4.1 BBLまたはさらなるサイトカイン;インターロイキン(例えば、IL-2)またはNKG-2D結合分子からなる群より選択してもよい。
【0168】
最後に、本発明は、本発明のCD3特異的結合分子、核酸分子、ベクター、または宿主を含むキットを提供する。
【0169】
該キットは、本発明の薬学的組成物を調製するために特に有用であり、とりわけ、注射または注入のために有用な容器からなってもよい。本発明のキットはさらに、任意で医学的または科学的目的を行うために必要な緩衝液、保存液および/または残りの試薬または材料を含むことが都合がよい。さらに、本発明のキットの一部は、バイアルもしくはボトルに個別包装する、または容器もしくは多容器単位と組み合わせて包装することができる。本発明のキットは、とりわけ、本発明の方法を実行するために、例えば研究ツールまたは医学的ツールとして本明細書において言及された多様な適応において都合よく用いることができるであろう。キットの製造は好ましくは、当業者に公知の標準的な技法に従う。
【0170】
これらおよび他の態様は、本発明の記述および実施例によって開示され、含まれる。本発明に従って用いられる抗体、方法、用途、および化合物の任意の一つに関するさらなる文献は、例えばコンピューターを用いて公共のライブラリおよびデータベースから検索してもよい。例えばインターネット上で利用できる公共のデータベース「Medline」を、例えばhttp://www.ncbi.nlm.nih.gov/PubMed/medline.htmlにおいて利用してもよい。http://www.ncbi.nlm.nih.gov/、http://www.infobiogen.fr/、http://www.fmi.ch/biology/research tools.html、http://www.tigr.org/のようなさらなるデータベースおよびアドレスは、当業者に公知であり、同様に、例えばhttp://www.lycos.comを用いて得ることができる。
【実施例】
【0171】
以下の実施例は本発明を説明する。
【0172】
以下の実施例において、多くの一本鎖抗ヒトCD3抗体が、ヒトにおいて免疫原性の低下を示すように操作されている。異なる脱免疫抗ヒトCD3抗体は、互いに結合した異なる4個のVH(VH2(配列番号:69、70)、VH3(配列番号:71、72)、VH5(配列番号:73、74)、およびVH7(配列番号:75、76))、ならびに異なる3個のVL(VL1(配列番号:77、78)、VL2(配列番号:79、80)、およびVL3(配列番号:81、82))領域の組み合わせ12個を含む。上記のVHおよびVL領域のアミノ酸および核酸配列を図3〜6に示す。例として、脱免疫抗CD3一本鎖抗体は、二重特異性産物を形成するために、抗CD19一本鎖抗体、または抗EpCAM一本鎖抗体とを組み合わせた。
【0173】
実施例1.脱免疫抗CD3構築物のクローニングおよび発現
1.1 一本鎖抗体をコードするcDNAの移入
脱免疫されている抗CD3一本鎖抗体をコードするDNAを、本明細書において抗CD3カセットと呼ぶ。この抗CD3カセットは、SGGGGSリンカー(配列番号:176)、抗CD3 VH領域(配列番号:110)、およびアミノ酸14個のGSリンカー(VEGGSGGSGGSGGSGGVDリンカー(配列番号:68))、および抗CD3 VL領域(配列番号:112)からなり、この後にヒスチジン残基6個が続く。上記のDNAをSrf1部位でベクターpPCR-Script-Amp SK(+)(Stratagene)にクローニングした。抗CD3カセットのDNAおよびアミノ酸配列を配列番号:1、配列番号:2および図1に示す。
【0174】
1.2 免疫原性T細胞エピトープの配列のコンピューター分析と脱免疫一本鎖抗体配列のデザイン
国際公開公報第98/52976号に記載の方法によって可能性があるT細胞エピトープを同定するために、抗CD3カセット(配列番号:2)のアミノ酸配列をペプチド糸通しプログラムによって分析した。配列番号:3は、脱免疫リンカー配列および配列番号:68は当初のリンカー配列を示す。
【0175】
1.3 脱免疫一本鎖抗体配列の構築
抗CD3カセットの脱免疫版をオーバーラップPCR組換え法によって構築した。pPCR-S-Amp SK+において、抗CD3カセット(配列番号:1、2)を必要な脱免疫配列への変異誘発のための鋳型として用いた。改変される領域を含む変異原性プライマー対の組を合成した。異なるVH領域4個および異なるVL領域3個を含む、産生された脱免疫配列をNot1からHind III断片としてベクターpPCR-S-Amp SK+にクローニングして、全DNA配列が正確であることを確認した。異なるVH領域4個および異なるVK領域3個を全ての組み合わせ(全体で12)において、PCRまたはVH領域の3'末端に導入した単一のBstEII部位のいずれかを用いて結合した。各組み合わせの全DNA配列が正確であることを確認した。抗CD3構築物の対応する当初の非脱免疫配列(VH:配列番号:110、VL:配列番号:112)を有する異なる脱免疫VH領域(配列番号:70、72、74、および76)およびVL領域(配列番号:78、80、および82)を表9に要約する。
【0176】
(表9)脱免疫VHおよびVL領域の配列番号:
【0177】
1.4 脱免疫一本鎖抗体遺伝子の発現ベクターへの移入
脱免疫抗CD3カセットをBspEIおよびSalIによってpPCR-S-Amp-SK+から切り出して、VLCD19-VHCD19-VHCD3-VLCD3を含む発現ベクターpEFにクローニングした。pEF-DHFRベクターのCD3部分は、BspEI部位からSalI部位への脱免疫抗CD3カセットのそれぞれによって置換して、以下の構築物12個を得た:
pEF抗CD19×抗CD3(VH2/VL1)(配列番号:177、178)
pEF抗CD19×抗CD3(VH2/VL2)(配列番号:179、180)
pEF抗CD19×抗CD3(VH2/VL3)(配列番号:181、182)
pEF抗CD19×抗CD3(VH3/VL1)(配列番号:183、184)
pEF抗CD19×抗CD3(VH3/VL2)(配列番号:185、186)
pEF抗CD19×抗CD3(VH3/VL3)(配列番号:187、188)
pEF抗CD19×抗CD3(VH5/VL1)(配列番号:189、190)
pEF抗CD19×抗CD3(VH5/VL2)(配列番号:191、192)
pEF抗CD19×抗CD3(VH5/VL3)(配列番号:193、194)
pEF抗CD19×抗CD3(VH7/VL1)(配列番号:195、196)
pEF抗CD19×抗CD3(VH7/VL2)(配列番号:197、198)
pEF抗CD19×抗CD3(VH7/VL3)(配列番号:199、200)
【0178】
構築物はさらに、タンパク質を分泌させるためにマウスIgG重鎖リーダーを含む。発現ベクターにおける脱免疫抗CD3カセットのDNA配列を、シークエンシングプライマー(配列番号:28および29)を用いて確認した。BspEI部位からSalI部位までのpEFベクターにおける脱免疫抗CD3カセット12個のDNAおよびアミノ酸配列を、配列番号:177〜200に示す。
【0179】
1.5 抗体構築物の産生
ベクターを大腸菌K12に形質転換した後、異なる発現ベクターのトランスフェクション等級のDNAを調製した。分泌されたタンパク質をCHO-dhfr細胞において産生した。一過性の産生に関して、細胞培養上清をトランスフェクションの2日後に回収して、安定なトランスフェクト細胞を産生するために、細胞をトランスフェクション後2日間、選択培地に入れた。5回継代した後、安定なプールを得た。その後限界希釈によって単一のクローンを同定した。精製プロセスを容易にするために、細胞を無血清培地に適合させた。抗体構築物は上清約1 Lから精製した。産生レベルをELISAにおいて試験した。抗CD19および脱免疫抗CD3構築物を含む異なる構築物のあいだで、分泌された抗体レベルに主要な差を認めなかった。
【0180】
実施例2:結合アッセイ
CD3およびCD19に対する脱免疫構築物の結合効率を分析するために、FACSに基づくアッセイを行った。最初、粗上清をCD3濃縮PBMCまたはCD19陽性NALM-6細胞に対する結合に関して試験した。細胞を非希釈上清と共に8℃で30分インキュベートした。2回の洗浄段階後、細胞を同じ条件で抗His抗体(Qiagen)によって標識した。さらなる洗浄段階の後、構築物の結合をFITC-結合ヒツジ抗マウス抗体(Sigma)によって検出した。細胞をFACS Caliburサイトメーター(B&D)によって分析した。対照として、抗CD19×抗CD3およびGFP-トランスフェクト細胞の上清を含めた。CD3および二次抗体をネガティブコントロールとして用いて、これは平均蛍光強度(MFI)約3.5を示した。抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH5/VL2)(配列番号:192)、抗CD3(VH5/VL3)(配列番号:194)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)を含む抗CD19×抗CD3構築物は、MFI少なくとも90を有し、このように、約25倍強く結合した。非脱免疫抗CD19×抗CD3構築物である陽性対照は、MFI約60に達し、抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH5/VL2)(配列番号:192)、抗CD3(VH5/VL3)(配列番号:194)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)を含む脱免疫構築物が、非常に高い効率でCD3に結合したことを示している。第二の実験において、抗CD19および抗CD3:抗CD3(VH2/VL1)(配列番号:178)、抗CD3(VH2/VL2)(配列番号:180)、抗CD3(VH2/VL3)(配列番号:182)、抗CD3(VH3/VL1)(配列番号:184)、抗CD3(VH3/VL2)(配列番号:186)、および抗CD3(VH3/VL3)(配列番号:188)を含む以下の構築物は、ネガティブコントロールと類似の結合を示した(MFI約6)。
【0181】
FACSに基づく結合アッセイを、CD19に関しても行った。この実験において、CD19および二次抗体はネガティブコントロールであった。本実験において、アッセイした構築物は全てMFI少なくとも80に達したのに対し、ネガティブコントロールのMFIは約3であった。
【0182】
このように、抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH5/VL2)(配列番号:192)、抗CD3(VH5/VL3)(配列番号:194)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)を含む構築物は、非改変抗CD19×抗CD(配列番号:204)3と同程度にCD3およびCD19に結合することが判明した。しかし、構築物抗CD3(VH2/VL1)(配列番号:178)、抗CD3(VH2/VL2)(配列番号:180)、抗CD3(VH2/VL3)(配列番号:182)、抗CD3(VH3/VL1)(配列番号:184)、抗CD3(VH3/VL2)(配列番号:186)、および抗CD3(VH3/VL3)(配列番号:188)は、抗CD3結合能を完全に喪失したが、CD19結合能は完全に保持した(図7)。
【0183】
このように、脱免疫重鎖は、結合の特異性および強度を支配することが証明された。その結果として、VH5およびVH7群を有する抗CD3構築物を精製して細胞障害活性に関して分析した。
【0184】
実施例3.高い結合親和性を示す変種の発現および精製
脱免疫抗CD19×抗CD3タンパク質抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH5/VL2)(配列番号:192)、抗CD3(VH5/VL3)(配列番号:194)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)をチャイニーズハムスター卵巣細胞(CHO)において発現させた。
【0185】
脱免疫抗CD3部分を含む二重特異性一本鎖構築物を精製するために、CHO-CD19細胞をローラーボトルにおいてハイクローンCHO改変DMEM培地(HiQ(登録商標))によって7日間増殖させた後、回収した。細胞を遠心によって除去し、発現されたタンパク質を含む上清を-20℃で保存した。
【0186】
Akta FPLCシステム(登録商標)(Pharmacia)およびUnicornソフトウェア(登録商標)をクロマトグラフィーのために用いた。化学物質は全て研究等級であり、Sigma(Deisenhofen)またはMerck(Darmstadt)から購入した。
【0187】
緩衝液A1(20 mMトリスpH 7.2)によって平衡にしたXK16/60カラム(Pharmacia)にローディングしたMEP Hypercel(登録商標)において、疎水性電荷誘導クロマトグラフィーを行った。細胞培養上清500 mlをカラム(10 ml)に流速3 ml/分で適用した。未結合試料を緩衝液A1によって洗浄し、結合したタンパク質を100%緩衝液B1(20 mM酢酸塩、pH 3.5)によって溶出した。溶出したタンパク質分画をさらなる精製のためにプールした。
【0188】
製造元のプロトコールに従ってNiSO4をローディングしたHisTap(登録商標)カラム(Pharmacia)を用いて、IMACを行った。カラムを緩衝液A2(20 mM NaP、pH 7.5、0.4 M NaCl)によって平衡にして、試料を緩衝液A2によって2:1希釈して、pH 7を得た。試料を流速1 ml/分でカラム(2 ml)に適用して、カラムを緩衝液A2によって洗浄して、未結合試料を除去した。結合したタンパク質を、緩衝液B2(20 mM NaP、pH 7.5、0.4 M NaCl、0.5 Mイミダゾール)の2段階勾配を用いて溶出した。段階1:20%緩衝液B2の10カラム容積;段階2:100%緩衝液B2の10カラム容積。溶出したタンパク質分画をさらなる精製のためにプールした。
【0189】
PBS(Gibco)によって平衡にしたSephadex S200 HiPrep(登録商標)カラム(Pharmacia)においてゲル濾過クロマトグラフィーを行った。溶出したタンパク質試料(流速1 ml/分)に、SDS-PAGEおよび検出のためにウェスタンブロットを行った(図11)。カラムを分子量決定のために予め較正した(分子量マーカーキット、Sigma MW GF-200)。
【0190】
抗CD19×抗CD3タンパク質の脱免疫変種を、疎水性電荷誘導クロマトグラフィー(HCIC)(図8)、固定金属アフィニティクロマトグラフィー(IMAC)(図9)、およびゲル濾過(図10)を含む3段階精製プロセスにおいて単離した。二重特異性構築物は、PBSにおけるゲル濾過によって決定したところ、未変性条件で分子量52 kDaであった。
【0191】
精製二重特異性タンパク質を、成形済みの4〜12%ビストリスゲル(Invitrogen)を用いて還元条件でSDS-PAGEによって分析した。試料の調製および適用は、製造元のプロトコールに従った。分子量はMultiMark(登録商標)タンパク質標準物質(Invitrogen)によって決定した。ゲルをコロイド状クーマシー(Invitrogenのプロトコール)によって染色した。単離されたタンパク質の純度は>95%(図11a)であり、分子の大きさは52 kDaである。
【0192】
さらに、抗CD19×抗CD3タンパク質の脱免疫変種をウェスタンブロットによって特異的に検出した。ウェスタンブロットは、Optitran BA-S83(登録商標)メンブレンおよびInvitrogen Blot Module(登録商標)によって製造元のプロトコールに従って行った。用いた抗体は、Penta His(Quiagen)およびアルカリホスファターゼ(AP)標識ヤギ抗マウスIg(Sigma)であり、染色溶液はBCIP/NBT液(Sigma)であった。主要なシグナルは、SDS-PAGEにおいて52 kDaで主要なバンドに対応することが示された(図11b)。
【0193】
タンパク質アッセイ色素(MicroBCA(登録商標)、Pierce)および標準タンパク質としてIgG(Biorad)を用いてタンパク質濃度を決定した。精製タンパク質変種の最終収率の概要を表10に示し、全ての構築物の生産性が高いことを示し、抗CD3(VH5/VL1)(配列番号:190)に関して924.8 μgの非常に良好な収量を得た。
【0194】
(表10)脱免疫抗CD19抗CD3構築物のタンパク質収量
【0195】
CD19×抗CD3(VH5/VL2)およびCD19×抗CD3(VH7/VL2)構築物の生産性を、対応する非脱免疫構築物と比較した。結果を表11に示す。
【0196】
(表11)対応する非脱免疫構築物と比較した脱免疫二重特異性構築物の収量
【0197】
表11は、脱免疫CD3結合ドメインを含む二重特異性構築物が、対応する非脱免疫構築物よりかなり高い(少なくとも3倍)生産性を有することを明らかに示している。
【0198】
実施例4.抗CD3構築物のFACSに基づく結合アッセイ
抗CD19および抗CD3を含む選択した精製抗体構築物の結合を、実施例2において先に記述するように様々な濃度で検出した。CD3結合アッセイにおいて、ネガティブコントロール二次抗体(抗-His、FITC結合)をCD3陽性細胞と共にインキュベートすると、MFI約2.5を示し、陽性対照である脱免疫抗CD19×抗CD3二重特異性一本鎖抗体は1μg/ml濃度で約70および5μg/ml濃度で約50を示した(図12A)。濃度1μg/mlでは、抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)脱免疫二重特異性抗体はMFI値10〜20を示した;抗CD3(VH5/VL1)(配列番号:190)は最高の値を有した(20)。5μg/mlでは、抗CD3(VH7/VL2)(配列番号:198)はMFI 25に達したが、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL3)(配列番号:200)、および抗CD3(VH5/VL1)(配列番号:190)は、MFI少なくとも40を示し、このように、非脱免疫陽性対照と同じ結合効率を示した。濃度5μg/mlでは、脱免疫抗CD3部分VH5/VL1(配列番号:190)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)を有する強い結合構築物は、非脱免疫抗CD19×抗CD3(配列番号:204)と同様にCD3に結合した。
【0199】
抗体構築物は全て、高い有効性でCD19に結合し、MFIは約200であったが、非脱免疫抗CD19×抗CD3構築物(配列番号:204)は、MFI 80を示した。調べた濃度では異なる構築物に関してCD19結合に関して差を認めなかった(図12B)。
【0200】
実施例5.細胞障害アッセイ
抗CD19×抗CD3は、CD19陽性標的細胞に対するT細胞依存的細胞障害性を媒介する。これを、抗CD19×抗CD3の生物学的有効性の決定に関してインビトロで分析した。
【0201】
この目的に関して、蛍光標識CD19陽性NALM-6標的細胞を、エフェクター細胞としてランダムドナーから単離したPBMCまたはCB15 T細胞(標準化T細胞株)と共に、抗CD19×抗CD3の存在下でインキュベートした。湿潤インキュベーターにおいて37℃で4時間インキュベートした後、標的細胞からの蛍光色素の上清への放出を蛍光分光計において決定する。抗CD19×抗CD3を含まずにインキュベートした標的細胞、およびインキュベーション終了時にサポニンの添加によって完全に溶解した標的細胞をそれぞれ、陰性および陽性対照とした。特定の抗CD19×抗CD3濃度で媒介される特異的細胞障害性を、以下の式によって計算することができる:
。
【0202】
EC50値を特定するために、用量反応を、0.4 pg/ml抗CD19×抗CD3から100 ng/ml抗CD19×抗CD3まで分析した。EC50値は、抗CD19×抗CD3の生物学的有効性を記述するが、絶対値は、エフェクター細胞源に応じて有意に変化するであろう。このように、相対的有効性は、以下の式に基づいて抗CD19×抗CD3参照材料と比較して計算される:
【0203】
抗CD19および脱免疫抗CD3を含む構築物の細胞障害活性を図13に示す。精製された非脱免疫抗CD19×抗CD3を対照として用いた。脱免疫構築物のEC50値は21.9〜81.6 pg/mlの範囲であったが、非脱免疫抗CD19×抗CD3構築物のEC50値は、22.7 pg/mlであった。このように、脱免疫構築物は全て、非脱免疫分子と同等のEC50値であることが判明した。
【0204】
実施例6.T細胞増殖アッセイ
健康なドナー20人をHLA-DRタイピングに基づくT細胞アッセイにおけるスクリーニングのために選択した(表12)。これによって、世界の人口において発現されるDR対立遺伝子の80%より大きい割合に対するT細胞アッセイにおいてペプチドのスクリーニングが可能となる。
【0205】
(表12)脱免疫および非脱免疫抗CD3 scAbから得たペプチドの免疫原性を調べるために用いた健康なドナー20人のHLA DRハプロタイプ
【0206】
6.1 T細胞増殖アッセイ
ペプチドはPepscan(Netherlands)から純度90%より高いものを得た。選択した健康なドナー20人からの末梢血単核球(PBMC)(表12)を用いて1および5μMで1試料あたり3個ずつ個々のペプチドをスクリーニングした。陽性対照ペプチド2個(C32およびC49)およびキーホールリンペットヘモシアニン(KLH)をアッセイに含めた。細胞およびペプチドを7日間インキュベートした後、1μCi/ウェル3H-チミジンによる18時間パルスを用いてT細胞増殖を評価した。これらのデータを以下のような刺激指数として表記した:
刺激指数=試験ペプチドのCPM/無処置対照のCPM
【0207】
T細胞エピトープは、2より大きい刺激指数(SI)を生じるペプチドとして定義される。独立した2回の実験結果は、非脱免疫抗CD3配列におけるMHC結合ペプチド22個中5個が、ヒトT細胞増殖の誘導能(SI>2)を有した。対照的に、対応する脱免疫分子はいずれも、T細胞増殖を誘導しなかった。表13は、独立した2回の実験の平均SI値を示すT細胞増殖アッセイの結果を要約する。
【0208】
データはまた濃度依存的作用を示し、それによって非脱免疫結合分子のそれぞれが、用いた二つの濃度の一つのみ(1μmまたは5μm)においてSI's>2を示した。異なる濃度での反応の差は、個々のペプチドがそれらがT細胞増殖を誘導する最適な濃度を有するであろうという事実によって説明される。この濃度を超えると、増殖は低下しうる(高いペプチド濃度はT細胞増殖に対して阻害作用を有しうる)。これは、場合によっては、増殖がより低い濃度で認められるがより高い濃度では認められないという理由を説明する。経験から、ペプチドがT細胞エピトープを含む場合、T細胞増殖は、用いたペプチド濃度の一つまたは二つにおいて認められるであろう。これらのデータは、脱免疫により抗CD3(VH5/VL2)(配列番号:19)および抗CD3(VH7/VL2)(配列番号:25)からT細胞エピトープが首尾よく除去されたことを証明した。非脱免疫抗CD3配列からのMHC結合ペプチドの約75%がT細胞増殖を誘導できなかったという事実は、これらのペプチドに対するヒト免疫系の寛容によって、またはヒトT細胞レパートリーがこれらの特定のペプチドを認識できないことによって説明することができる。
【0209】
(表13)陽性(SI>2)マウスペプチドおよび対応する脱免疫ペプチドを比較するデータの要約
【0210】
実施例7.非脱免疫抗CD3 VHとの抗CD3(VH5)、抗CD3(VH7)、抗CD3(VH2)、および抗CD3(VH3)の相同性アラインメント
非脱免疫CD3抗体の可変重鎖領域、VH5(配列番号:74)、VH7(配列番号:76)、VH2(配列番号:70)、およびVH3(配列番号:72)を、Vector NTI Advance(Informax, Inc., USA)のAlingnXプログラムを用いて整列させた。用いたClustal Wアルゴリズムは、Nucleic Acid Research, 22(22):4673〜4860, 1994に記述されている。アラインメントを図14に示す。アラインメントから、意外にも良好な結合を示す可変領域VH5およびVH7が、フレームワーク1とCDR1との移行領域で配列ASGYTFを有することが認められうる。さらに、結合を示さないVH領域(VH2(配列番号:70)およびVH3(配列番号:72))が、フレームワーク1とCDR1との移行部で配列ASGYTAを含む。このように、T細胞エピトープの生成傾向およびCD3に対する結合が低下した構築物を得るために、構築物はフレームワーク1とCDR1との移行領域で配列ASGYTFを含まなければならない。意外にも、CD3に結合し、上記の配列ASGYTFを含むT細胞エピトープの生成傾向が低下した重鎖可変領域は良好な結合を示す。
【0211】
実施例8.脱免疫抗CD3および抗EpCAMを含む構築物のクローニング
本発明の脱免疫抗CD3ポリペプチドは、他の標的を有する機能的構築物の一部となりうることを証明するために、脱免疫抗CD3(VH5/VL2)(配列番号:19)および異なる抗EpCAM一本鎖抗体(3-1(配列番号:137、139)、3-5(配列番号:141、143)、4-1(配列番号:145、147)、4-7(配列番号:149、151)、5-10(配列番号:133、135))を含む多くの二重特異性構築物を作製した。
【0212】
8.1 脱免疫抗CD3部分(配列番号:30、31、32、33、34、35、36、37、38、および39)を含むC-末端EpCAM結合物質の同定
8.1.1 抗CD19×抗CD3(VH5/VL2)構築物(配列番号:192)からの脱免疫抗CD3の増幅
N-末端脱免疫抗CD3(VH5/VL2)を、脱免疫(CD19×抗CD3(VH5/VL2)(配列番号:192))を鋳型として、および以下のプライマー(DI CD3 5-2 VH BsrGI
(配列番号:40)、DI CD3 5-2 VL BspEI
(配列番号:41))を用いてPCRによって得た。
【0213】
8.1.2 脱免疫抗CD3×抗EpCAM脱免疫構築物のVHCD3-VLCD3×VHEpCAM-VLEpCAM方向のクローニングおよび発現
脱免疫抗CD3を含む上記のPCR産物を、制限酵素BsrGIおよびBspEIによって切断した後、真核細胞分泌シグナル(リーダーペプチド)のアミノ酸配列を含むbluescript KSベクター(Stratagene, La Jolla, CA)にEcoRI/BsrGI断片としてクローニングした。この構築物をEcoRIおよびBspEIによって切断後、リーダーペプチドと共にそれぞれの抗CD3 scFvを含む得られたDNA断片を、pEF-DHFRベクターにおいてC-末端位で抗EpCAM scFv 3-1、4-7、または5-10を含むEcoRI/BspEI切断プラスミドにクローニングした。シークエンシング(Sequiserve, Vaterstetten)によって二重特異性一本鎖をコードする配列を確認後、プラスミドを真核細胞タンパク質発現のためにDHFR欠損CHO細胞にトランスフェクトした。DHFR欠損CHO細胞における真核細胞タンパク質発現は、Kaufmann, R.J.(1990)Methods Enzymol. 185, 537〜566に記述された通りに行った。
【0214】
8.1.3 脱免疫抗CD3×抗EpCAM脱免疫構築物のVHCD3-VLCD3×VLEpCAM-VHEpCAM方向のクローニングおよび発現
VL-VH方向にアミノ酸15の標準的なリンカーを含む抗EpCAM 4-7(配列番号:168)をPCRによって得た。4-7 VH領域および4-7 VL領域を以下のプライマーによって個々に増幅した(4-7 VL:4-7 VL BspEI 順方向
(配列番号:117)、4-7 VL GS15 逆方向
(配列番号:118)、4-7 VH:4-7 VH GS15 順方向
(配列番号:42)、4-7 VH SalI 逆方向
(配列番号:43))。PCR産物に導入された重なり合う相補的配列を用いて、その後の融合PCRの際に、アミノ酸15個の(G4S1)3(一文字アミノ酸コード)リンカー(標準的なリンカー)(配列番号:168)のコード配列を形成した。この増幅段階は、プライマー対4-7 VL Bsp EI FORおよび4-7 VH SalI REV(配列番号:42および43)を用いて行った。
【0215】
アミノ酸15個の標準的な((G4S1)3)リンカーを含むVL-VH方向の抗EpCAM 5-10を、PCRによって得た。5-10 VH領域および5-10 VL領域を、以下のプライマーによって個々に増幅した(5-10 VL:5-10 VL BspEI 順方向
(配列番号:44)、5-10 VL GS15 逆方向
(配列番号:45)、5-10 VH:5-10 VH GS15 順方向
(配列番号:46)、5-10 VH SalI 逆方向
(配列番号:47))。その後の融合PCRにおいて、PCR産物に導入された重なり合う相補的配列を用いて、アミノ酸15個の(G4S1)3(一文字アミノ酸コード)リンカー(標準的なリンカー)(配列番号:168)のコード配列を形成した。この増幅段階はプライマー対5-10 VL BspEI FORおよび5-10 VH SalI REVによって行った。
【0216】
PCR産物5-10 VL-VHおよび4-7 VL-VHを、抗CD3構築物(VH5/VL2)を含むpEF-DHFRベクターにクローニングした。シークエンシングによって二重特異性一本鎖をコードする配列を確認した後、真核細胞発現のために、プラスミドをDHFR欠損CHO細胞にトランスフェクトした。DHFR欠損CHO細胞における真核細胞タンパク質発現は、Kaufmann R.J.(1990)Methods Enzymol. 185, 537〜566に記述された通りに行った。
【0217】
8.1.4. 脱免疫抗CD3×抗EpCAM構築物のEpCAMおよびCD3に対する結合
N-末端方向で抗CD3部分を有する二重特異性一本鎖抗体のEpCAMおよびCD3に対する結合をFACS分析によって確認した。その目的のために、EpCAM陽性ヒト胃癌細胞株Kato III(ATCC HTB-103)を用いた。抗CD3部分の結合は、Jurkat細胞(ATCC TIB 152)において示された。
【0218】
細胞を供給元の推奨に従って培養して、細胞200000個を10 μg/ml構築物と共に2%FCS(仔ウシ胎児血清)を含むPBS 50μlにおいてインキュベートした。構築物の結合は、2μg/ml抗His抗体(Qiagen, Hilden, FRGから得たPenta-His抗体)によって2%FCSを含むPBSにおいて検出した。第二段階として、ヤギ抗マウスIgGに由来するR-フィコエリスリン結合アフィニティ精製F(ab')2を、2%FCS(Dianova, Hamburg, FRG)を含むPBS 50μlにおいて1:100倍希釈して用いた。試料をFACSscan(BD biosciences, Heidelberg, FRG)において測定した。
【0219】
FACS分析の結果を図15に示す。脱免疫抗CD3部分を含む構築物は全て、EpCAM陽性Kato III細胞上で非脱免疫抗EpCAM(M79)×抗CD3二重特異性一本鎖抗体より強い結合を示した。
【0220】
8.2. 脱免疫抗CD3部分を含むN-末端EpCAM結合物質のクローニング
8.2.1 抗EpCAM×抗CD3構築物のクローニング
8.2.1.1. 脱免疫3-1×抗CD3(VH5/VL2)構築物(配列番号:48、49)のクローニング
脱免疫構築物3-1×抗-CD3(VH5/VL2)(配列番号:48)は、脱免疫構築物抗EpCAM(3-1)×抗CD3に由来した。抗EpCAM抗体3-1のVHおよびVL領域を、配列番号:137および139に示す。プラスミドpEF-DHFR-3-1×抗CD3およびpEF抗CD3(VH5/VL2)(配列番号:192)を、ベクターおよびインサート抗CD3(VH5/VL2)をそれぞれ、単離するためにBspEIおよびSalI(Biolabs)によって消化した。BspEI-SalI消化ベクターを脱燐酸化して、0.7%アガロースゲルにおいて精製し、インサートを1.5%アガロースゲルにおいて精製した。
【0221】
精製断片(BspEI-SalI)を、その後、pEF-DHFRベクターの対応する部位にクローニングした。最終的な3-1×抗CD3(VH5/VL2)構築物(配列番号:48、49)を制限消化物および完全なインサートのDNAシークエンシングによって確認した。
【0222】
非脱免疫3-1×抗CD3構築物のクローニング
3-1×抗CD3(VH5/VL2)構築物のクローニングに関して、対応する非脱免疫構築物を以下のように作製した。
【0223】
非脱免疫3-1×抗CD3分子を構築するために、VH-VL方向のC-末端3-1をPCRによって得た。3-1 VH-VLを二つの部分において含む断片IおよびIIを、プライマー対me 91a(配列番号:53)/me 90(配列番号:52)およびme 83(配列番号:50)/me 84(配列番号:51)をそれぞれ用いてPCRによって増幅した。Expand High Fidelity System of Roche Diagnosticsを用いてホットスタートPCRを行った。20サイクル(94℃/30秒、60℃/1分、72℃/1分)を増幅のために行い、その後72℃で3分を1サイクル行った。
【0224】
PCR断片IおよびIIに1.5%アガロースゲル上での電気泳動を行った。断片を混合して(各1 ng)、完全な3-1を含む断片IIIの増幅のために、プライマー対me 91a(配列番号:53)およびme 84(配列番号:51)によって行う次のPCR反応の鋳型として用いた。PCRは上記の通りに行ったが、アニーリング温度は68℃であった。断片IIIをアガロースゲルにおいて精製して、BsrGIおよびBspEI(Biolabs)によって消化し、精製してその後pEF-DHFR抗EpCAM(M79)×抗CD3構築物の対応する部位にクローニングした。クローニングした領域を制限消化物およびDNAシークエンシングによって確認した。
【0225】
用いたプライマーの配列:
Me 83:
(配列番号:50)Me 84
(配列番号:51)Me 90
(配列番号:52)Me 91a
(配列番号:53)。
【0226】
8.2.1.2 脱免疫3-5×抗CD3(VH5/VL2)構築物(配列番号:54、55)のクローニング
3-5×抗CD3構築物を構築するために、VH-VL方向のC-末端3-5をPCRによって得た。抗EpCAM抗体3-5のVHおよびVL領域を、配列番号:141および143に示す。インサート(3-5)およびベクターをそれぞれ単離するために、プラスミドpEF-DHFR-3-5×抗CD3およびpEF抗CD3(VH5/VL2)(配列番号:192)をEcoRIおよびBspEI(Biolabs)によって消化した。脱燐酸化ベクター(EcoRIおよびBspEI消化)およびインサートをアガロースゲル電気泳動によって精製した。
【0227】
精製された断片(EcoRI-BspEI)をその後、pEF-DHFRベクターの対応する部位にクローニングした。最終的な3-5×抗CD3(VH5/VL2)(配列番号:54)構築物を制限消化物によって確認した。
【0228】
非脱免疫3-5×抗CD3構築物のクローニング
3-5×抗CD3(VH5/VL2)構築物をクローニングするため、対応する非脱免疫構築物を以下のように作製した。
【0229】
3-5を二つの部分で含む断片IおよびIIを、プライマー対me 81(配列番号:56)/me 90(配列番号:52)、およびme 83(配列番号:50)/me 84(配列番号:51)をそれぞれ用いて、3-1×抗CD3に関して記述した条件に従ってPCRによって増幅した。PCR断片IおよびIIを含むアガロースゲル断片を、完全な3-5を含む断片IIIを増幅するために、プライマー対me 81(配列番号:56)およびme 84(配列番号:51)によって再増幅した。PCRは先に記述した通りに実施した。断片IIIをアガロースゲルにおいて精製して、BssHIIおよびBspEI(Biolabs)によって消化して、精製し、その後pEF-DHFRベクターの対応する部位にクローニングした。クローニングした領域を制限消化物およびDNAシークエンシングによって確認した。
Me81プライマー(配列番号:56)の配列:Me 81
【0230】
8.2.1.3 脱免疫4-1×抗CD3(VH5/VL2)構築物(配列番号:57、58)のクローニング
4-1×抗CD3分子を構築するために、VH-VL方向のC-末端4-1をPCRによって得た。抗EpCAM抗体4-1のVHおよびVL領域を配列番号:145および147に示す。プラスミドpEF-DHFR-4-1×抗CD3およびpEF抗CD3(VH5/VL2)(配列番号:192)を、インサート(4-1)およびベクターをそれぞれ単離するために、EcoRIおよびBspEI(Biolabs)によって消化した。脱燐酸化ベクター(EcoRIおよびBspEI消化)およびインサートをアガロースゲル電気泳動によって精製した。
【0231】
精製断片(EcoRI-BspEI)をその後、ベクターの対応する部位にクローニングした。最終的な構築物4-1×抗CD3(VH5/VL2)(配列番号:57)を制限消化物によって確認した。
【0232】
非脱免疫4-1×抗CD3構築物のクローニング
4-1×抗CD3(VH5/VL2)構築物をクローニングするため、対応する非脱免疫構築物を以下のように作製した。
【0233】
4-1を二つの部分で含む断片IおよびIIを、プライマー対me 91a(配列番号:53)/me 90(配列番号:452)およびme 83(配列番号:50)/me 84(配列番号:51)をそれぞれ用いて上記の条件によってPCRによって増幅した。
【0234】
完全な4-1を含む断片IIIを増幅するために、PCR断片IおよびIIを含むアガロースゲル断片を、プライマー対me 92a(配列番号:59)およびme 84(配列番号:51)によって再増幅した。PCRは先に記述した通りに実施したが、アニーリングは68℃で行った。断片IIIをアガロースゲルにおいて精製して、BsrGIおよびBspEI(Biolabs)によって消化して、精製し、その後pEF-DHFR-抗EpCAM(M79)×抗CD3クローニングベクター構築物の対応する部位にクローニングした。クローニングした領域を制限消化物およびDNAシークエンシングによって確認した。
Me92aプライマーの配列(配列番号:59):Me 92a:
【0235】
8.1.2.4 脱免疫4-7×抗CD3(VH5/VL2)構築物(配列番号:60、61)のクローニング
4-7×抗CD3を構築するために、VH-VL方向でのC-末端4-7をPCRによって得た。抗EpCAM抗体4-7のVHおよびVL領域を配列番号:149および151に示す。プラスミドpEF-DHFR-4-7×抗CD3およびpEF-抗CD3VH5/VL2(配列番号:192)を、インサート(4-7)およびベクターをそれぞれ単離するために、EcoRIおよびBspEI(Biolabs)によって消化した。脱燐酸化ベクター(EcoRIおよびBspEI消化)およびインサートをアガロースゲル電気泳動によって精製した。
【0236】
次に、精製断片(EcoRI-BspEI)をpEF-DHFRベクターの対応する部位にクローニングした。最終的な構築物4-7×抗CD3(VH5/VL2)(配列番号:60)を制限消化物によって確認した。
【0237】
非脱免疫構築物4-7×抗CD3のクローニング
4-7×抗CD3(VH5/VL2)構築物をクローニングするために、対応する非脱免疫構築物を以下のように作製した。
【0238】
4-7を二つの部分で含む断片IおよびIIを、プライマー対me 81(配列番号:56)/me 90(配列番号:52)、およびme 83(配列番号:50)/me 84(配列番号:51)をそれぞれ用いて、上記の条件に従ってPCRによって増幅した。PCR断片IおよびIIを含むアガロースゲル断片を、完全な4-7 VHおよびVL鎖を含む断片IIIを増幅するために、プライマー対me 81(配列番号:56)およびme 84(配列番号:51)によって再増幅した。PCRは先に記述した通りに実施した。断片IIIをアガロースゲルにおいて精製して、BssHIIおよびBspEI(Biolabs)によって消化して、精製した後pEF-DHFRベクターの対応する部位にクローニングした。クローニングした領域を制限消化物およびDNAシークエンシングによって確認した。
【0239】
8.1.2.5 脱免疫5-10×抗CD3(VH5/VL2)構築物(配列番号:62、63)のクローニング
5-10×抗CD3分子を構築するために、VH-VL方向でのC-末端5-10をPCRによって得た。抗EpCAM抗体5-10のVHおよびVL領域を配列番号:133および135に示す。プラスミドpEF-DHFR-5-10×抗CD3およびpEF-抗CD3(VH5/VL2)(配列番号:192)を、インサート(5-10)およびベクターをそれぞれ単離するために、EcoRIおよびBspEI(Biolabs)によって消化した。脱燐酸化ベクター(EcoRIおよびBspEI消化)およびインサートをアガロースゲル電気泳動によって精製した。
【0240】
精製断片(EcoRI-BspEI)をその後pEF-DHFRベクターの対応する部位にクローニングした。最終的な構築物5-10×抗CD3(VH5/VL2)(配列番号:62)を制限消化物およびDNAシークエンシングによって確認した。
【0241】
非脱免疫5-10×抗CD3構築物のクローニング
5-10×抗CD3(VH5/VL2)構築物をクローニングするために、対応する非脱免疫構築物を以下のように作製した。
【0242】
5-10を二つの部分で含む断片IおよびIIを、プライマー対me 92a(配列番号:59)/me 90(配列番号:52)、およびme 83(配列番号:50)/me 84(配列番号:51)をそれぞれ用いて、上記の条件に従ってPCRによって増幅した。PCR断片IおよびIIを含むアガロースゲル断片を、完全な5-10を含む断片IIIを増幅するために、プライマー対me 92a(配列番号:59)およびme 84(配列番号:51)によって再増幅した。PCRは先に記述した通りに実施したが、アニーリングは68℃で行った。断片IIIをアガロースゲルにおいて精製して、BsrGIおよびBspEI(Biolabs)によって消化して、精製し、その後pEF-DHFR抗EpCAM(M79)×抗CD3クローニングベクターの対応する部位にクローニングした。クローニングした領域を制限消化物およびDNAシークエンシングによって確認した。
【0243】
8.2.2 N-末端位で抗EpCAMを有する抗EpCAM×脱免疫抗CD3分子の発現
DHFR遺伝子を欠損するCHO細胞を、10%仔ウシ胎児血清(Life Technologies、65℃で30分間熱不活化)およびHT(ヒポキサンチンおよびチミジン;Life Technologies)を添加したαMEM培地(Life Technologies, cat No. 32561)において維持した。Lipofectamine 2000キット(登録商標)(Invitrogen)を用いて製造元によって提供された説明書に従って、細胞にpEF-DHFR-3-1×抗CD3(VH5/VL2)(配列番号:48)およびpEF-DHFR-5-10×抗CD3(VH5/VL2)(配列番号:62)をトランスフェクトした。48時間後、選択培地(熱不活化10%透析仔ウシ胎児血清(Life Technologies)を含むαMEM培地)において選択を行った。3〜4週間後、細胞培養上清を回収して、4℃で300 gで10分間遠心して細胞および細胞の破片を除去した。二重特異性抗体を含む上清を、さらに分析するまで-20℃で保存した。
【0244】
8.2.3 二重特異性抗EpCAM×抗CD3変種の結合アッセイ
Jurkat細胞(CD3結合に関して)およびKato細胞(EpCAM結合に関して)250000個を、二重特異性構築物(それぞれ、pEF-DHFR-3-1×抗CD3(VH5/VL2)(Nr. 50、配列番号:48)およびpEF-DHFR-5-10×抗CD3(VH5/VL2)(Nr.54)(配列番号:62))を含む細胞培養上清(50 μl)と共に独立して4℃で45分間インキュベートした。その後、細胞をFACS緩衝液(1%仔ウシ胎児血清(FCS)および0.05%アジ化ナトリウムを含む燐酸緩衝生理食塩液)において2回洗浄して、マウス抗His抗体(Dianova、DIA 910)と共に4℃で60分間インキュベートした。洗浄段階は上記の通りに行った。
【0245】
細胞を最終的にヤギ抗マウスIg-FITC結合抗体(BD 550003)またはPEに結合させた抗マウスIgG(Sigma, P8547)のいずれかと共に最終的にインキュベートした。洗浄段階後、事象10000個をFACS Calibur(B&D)を用いて分析した。結合アッセイの結果を図16に示す。構築物3-1×抗CD3(VH5/VL2)(配列番号:49)および5-10×抗CD3(配列番号:63)は、Jurkat細胞上でCD3に対して、およびKato細胞上でCD19に対して強い結合を示した。
【0246】
実施例8.3 脱免疫抗CD3部分を有する二重特異性抗EpCAM構築物の精製
脱免疫抗CD3抗原およびEpCAM特異的領域を含む構築物を、固定金属アフィニティクロマトグラフィー(IMAC)およびゲル濾過を含む二段階精製プロセスによって精製した。金属アフィニティクロマトグラフィー(IMAC)およびゲル濾過は実施例3.2に示すように実行した。
【0247】
さらに高分解能の陽イオン交換クロマトグラフィーを、20 mM MES緩衝液pH 5.5によって平衡にしたMiniSカラム(Amersham)において行った。試料を同じ緩衝液によって1:3に希釈してからカラムにローディングした。結合したタンパク質を0〜30%勾配の1 M NaClの平衡緩衝液溶液によって溶出した。溶出したタンパク質分画を生物活性アッセイにおいて試験した。表14は、精製脱免疫EpCAM構築物の収量を示す。構築物は全て、有効に産生することができた。意外にも構築物5-10×抗-CD3(VH5/VL2)(配列番号:63)は、2200 μg/lという極めて良好な収量を有した。
【0248】
(表14)脱免疫EpCAM構築物の収量
【0249】
実施例8.4 脱免疫抗CD3部分による二重特異性抗EpCAM構築物の細胞障害アッセイ
本発明の二重特異性抗体の高い生物活性を確認するために、FACSに基づくアッセイを行った。CHO細胞に上皮細胞接着分子(EpCAM)をトランスフェクトした。CHO-EpCAM細胞と呼ばれるこのトランスフェクションに由来する細胞クローンを実験に用いた。
【0250】
細胞毒性試験のために、CHO-EpCAM(1.5×107)を、PBSで2回洗浄して血清を除き、製造業者の指示書に従ってPKH25色素(Sigma-Aldrich Co.)とともにインキュベートした。細胞染色後にRPMI/10% FCSで2回洗浄した。
【0251】
細胞を計数し、CB15エフェクター細胞と混合した。CD4ポジティブT細胞クローンCB15は、Fickenscher博士(University of Erlangen/Nuernberg, Germany)からご厚意で提供された。細胞を、供給業者によって推奨されるように培養した。得られる細胞懸濁物は、mlあたり400,000標的および2×106エフェクター細胞を含んだ。96ウェル丸底プレート中のウェルあたり、50μlの混合物を使用した。
【0252】
抗体をRPMI/10% FCS中で必要とされる濃度まで希釈し、この溶液の50μlを細胞懸濁物に加えた。標準的な反応は、37℃/5% CO2中で16時間インキュベートした。ヨウ化プロピジウムを1μg/mlの最終濃度で加えた。室温での10分間のインキュベーション後、細胞をFACSによって分析した。PKH26蛍光を、標的細胞のポジティブ同定のために使用した。細胞毒性を、全ての標的細胞にわたるPIポジティブの比として測定した。
【0253】
シグモイド型用量応答曲線は、代表的には、Prismソフトウェア(GraphPad Software Inc., San Diego, USA)によって決定されるように、>0.97のR2値を有した。細胞毒性アッセイの結果は、図17および18に示されている。
【0254】
実施例8.5 CHO細胞におけるEpCAM結合部分と脱免疫CD3結合部分とを含む二重特異性分子の生産性の比較
脱免疫構築物タンパク質の生産性を決定するために、プロテインL ELISAを行った。生産性のデータはバッチ培養から計算した。
【0255】
8.5.1 細胞培養
脱免疫(CHO-DHFR-)および非脱免疫(CHO-DHFR-またはCHO-K1)を産生するCHO細胞株を、HyQ PF CHO LS培地+4 mM L-グルタミンにおいて37℃および5%CO2のCO2インキュベータにおいて培養した。細胞数および生存率はトリパンブルーによって決定した。細胞密度を1〜2×105個mlに設定した。
【0256】
細胞をスピンナフラスコに移して、攪拌培養の条件に調節した。操作パラメータの設定は、CO2インキュベータにおいて通気しながら80 rpm、37℃、および5%CO2であった。培養容積は100〜500 mlの範囲であり、インキュベーション時の細胞密度は1〜2×105個/mlの範囲であった。Tフラスコでのサブ培養に関して、各継代毎に培養物を遠心して、新鮮な予め加温した培地に浮遊させた。細胞密度は1〜2×105個/mlに設定した。
【0257】
生産性のデータの分析に関して(表15)、培地の添加または交換を行わずに細胞を14日(d)まで培養した。細胞数および生存率をトリパンブルー染色によって毎日決定した。上清における産物の濃度をプロテインL ELISAによって分析した。
【0258】
8.5.2 プロテインL ELISA
二重特異性分子の定量的結合は、rプロテインLコーティングマイクロタイタープレートにおいて行った。rプロテインLは、ペプトストレプトコッカス・マグヌス(Peptostreptococcus magnus)によって産生される免疫グロブリン結合プロテインLの組換え型である。これは四つの結合部位を有し、軽鎖(κ)を通して免疫グロブリンに結合する。親抗体のそれぞれ異なる二つの軽鎖からの可変ドメインを含む二重特異性分子は、rプロテインLによって結合される。
【0259】
マイクロタイタープレートをrプロテインLのPBS緩衝液溶液(2μg rプロテインL/ml PBS緩衝液)によって2〜8℃で一晩コーティングした。コーティングの後、残っている吸着部位をブロッキング緩衝液(2%BSAのPBS緩衝液溶液)によってブロックした。次に、プレートを凍結して≦18℃で保存した。使用前、プレートを融解して、洗浄緩衝液(0.05%Tween 20のPBS緩衝液)によって洗浄し、コーティング溶液とブロッキング緩衝液の混合物を除去した。
【0260】
無細胞培養上清の1%BSA+0.01%Tween 20のPBS(希釈緩衝液)による連続希釈液を分析した。二重特異性抗EpCAM(M79)×抗CD3を同等の希釈液において陽性対照として用いた。
【0261】
インキュベーションは2〜8℃で一晩行った。
【0262】
洗浄後、ウサギ抗マウスIgG(希釈緩衝液において1:5,000倍希釈)を加えて、室温で60分間インキュベートした。洗浄後、アルカリホスファターゼ標識ヤギ抗ウサギIgGを加えた(希釈緩衝液において1:1,000倍希釈、室温で60分)。pNPP基質溶液を加えて、3 M NaOHを加えることによって反応を停止させた。ELISAリーダーによって405 nm(参照フィルター492 nm)で吸光度を測定した。
【0263】
(表15)脱免疫抗EpCAM構築物の生産性
【0264】
このように、本発明の5-10×抗CD3(VH5/VL2)構築物は、先行技術の二重特異性脱免疫EpCAMおよびCD3結合抗体よりかなり高い(少なくとも5倍高い)特異的生産性を示した。
【技術分野】
【0001】
本発明は、ヒトCD3に特異的に結合する第一のドメインと、Igに由来する第二の結合ドメインとを含む細胞障害活性を有するCD3特異的結合構築物に関する。さらに、本発明のCD3特異的結合構築物をコードする核酸配列を提供する。本発明のさらなる局面は、該核酸配列を含むベクターおよび宿主細胞、本発明の構築物の産生プロセス、ならびに該構築物を含む組成物である。本発明はまた、特定の疾患を治療するための薬学的組成物を調製するために該構築物を用いること、特定の疾患を治療する方法、および本発明の結合構築物を含むキットも提供する。
【背景技術】
【0002】
ヒトCD3は、多分子T細胞複合体の一部としてT細胞上で発現される抗原を指し、これは異なる三つの鎖からなる:CD3-ε、CD3-δ、およびCD3-γ。例えば固定された抗CD3抗体によってT細胞上でCD3が集合すると、T細胞受容体が関与する場合と類似のT細胞活性化が起こるが、これはそのクローンに典型的な特異性とは無関係である;国際公開公報第99/54440号(特許文献1)またはHoffman(1985), J. Immunol. 135:5〜8(非特許文献1)を参照されたい。
【0003】
CD3抗原を特異的に認識する抗体は、先行技術において、例えばTraunecker, EMBO J. 10(1991), 3655〜9(非特許文献2)およびKipriyanov, Int. J. Cancer 77(1998), 763〜772(非特許文献3)において記述されている。最近、CD3に対する抗体が、多様な疾患の治療に提唱されている。これらの抗体または抗体構築物は、欧州特許第1 025 854号(特許文献2)に開示されるように、T細胞枯渇物質またはマイトゲン物質のいずれかとして作用する。ヒトCD3抗原複合体に対して特異的に結合するヒト/齧歯類ハイブリッド抗体は、国際公開公報第00/05268号(特許文献3)に開示され、例えば腎臓、敗血症、および心臓の同種異系移植片の移植後の拒絶事例の治療のための免疫抑制剤として提唱されている。国際公開公報第03/04648号(特許文献4)は、CD3および卵巣癌抗原に対する二重特異性抗体を開示している。さらに、Kufer(1997), Cancer Immunol. Immunother. 45:193〜7(非特許文献4)は、最小の残留癌の治療のためのCD3およびEpCAMに対して特異的な二重特異性抗体に関する。
【0004】
しかし、先行技術のCD3に対する抗体は、ヒト以外の起源に由来している。このために、ヒトにおける治療レジメの一部としてそのような抗CD3抗体を用いる場合には、いくつかの重篤な問題が起こる。
【0005】
そのような問題の一つは、「サイトカイン放出症候群(CRS)」である。CRSは、抗CD3抗体の最初の数回の用量の投与後に認められる臨床症候群であり、CD3に対する多くの抗体がマイトゲン性であるという事実に関連している。インビトロにおいて、CD3に対するマイトゲン抗体は、T細胞増殖およびサイトカイン産生を誘導する。インビボでは、このマイトゲン活性により、抗体の初回注射後1時間以内に、多くのT細胞由来サイトカインを含む大量のサイトカインの放出が起こる。CD3特異的抗体のマイトゲン活性は、単球/マクロファージ依存的であり、これはこれらの細胞によるIL-6およびIL-1βの産生を伴う。
【0006】
CRS症状は、しばしば報告される軽度の「インフルエンザ様」症候群から、報告される回数がより少ない重度の「ショック様」反応(これには、心血管および中枢神経系の症状が含まれることがある)に及ぶ。症状には、とりわけ頭痛、振せん、悪心/嘔吐、下痢、腹痛、倦怠、および筋/関節痛ならびに疼痛、全身の虚弱、心肺事象と共に神経精神学的事象が含まれる。体液過剰負荷の患者および体液過剰負荷ではないように思われる患者において重度の肺浮腫が起こる。特にマウスモノクローナル抗体の治療的治療の障害となるもう一つの重篤な問題は、そのような抗体に対する液性免疫応答の開始であり、それによってヒト抗マウス抗体(「HAMAs」)の産生が起こる(Schroff(1985), Cancer Res. 45:879〜885(非特許文献5), Shawler(1985), J. Immunol. 135:1530〜1535(非特許文献6))。HAMAsは典型的にマウス抗体による治療の2週目のあいだに産生され、マウス抗体を中和し、それによってその意図する標的に対するその結合能を阻害する。HAMA反応は、マウス抗体定常領域(「Fc」)および/またはマウス可変領域(「V」)の性質に依存しうる。
【0007】
先行技術は、非ヒト起源のモノクローナル抗体を改変することによって、HAMAsの産生を減少または阻害する様々なアプローチを含む。
【0008】
そのような抗体の免疫原性を減少させる一つのアプローチは、例えば国際公開公報第91/09968号(特許文献5)および米国特許第6,407,213号(特許文献6)に記述されるようなヒト化である。一般的に、ヒト化は、例えばCDR-移植の場合のように、非ヒト抗体配列を対応するヒト配列に置換することを必要とする。
【0009】
そのような抗体の免疫原性を減少させるもう一つのアプローチは、例えば国際公開公報第00/34317号(特許文献7)、国際公開公報第98/52976号(特許文献8)、国際公開公報第02/079415号(特許文献9)、国際公開公報第02/012899号(特許文献10)、および国際公開公報第02/069232号(特許文献11)に記述されるような、脱免疫である。一般的に、脱免疫は、可能性があるT細胞エピトープ内でのアミノ酸の置換を行うことを必要とする。この方法において、所定の配列が細胞内タンパク質プロセシングの際にT細胞エピトープを生じる可能性は低下する。その上、国際公開公報第92/10755号(特許文献12)は、タンパク質上の抗原決定基を操作するアプローチを記述している。特に、タンパク質をエピトープマッピングして、そのアミノ酸配列を遺伝子操作を通して変化させる。
【0010】
しかし、ヒト化抗体はしばしば、その非ヒト化親抗体と比較してその標的に関する結合親和性の減少を示し、同様にしばしばヒト宿主においてなおいくぶん免疫原性である。
【先行技術文献】
【特許文献】
【0011】
【特許文献1】国際公開公報第99/54440号
【特許文献2】欧州特許第1 025 854号
【特許文献3】国際公開公報第00/05268号
【特許文献4】国際公開公報第03/04648号
【特許文献5】国際公開公報第91/09968号
【特許文献6】米国特許第6,407,213号
【特許文献7】国際公開公報第00/34317号
【特許文献8】国際公開公報第98/52976号
【特許文献9】国際公開公報第02/079415号
【特許文献10】国際公開公報第02/012899号
【特許文献11】国際公開公報第02/069232号
【特許文献12】国際公開公報第92/10755号
【非特許文献】
【0012】
【非特許文献1】Hoffman(1985), J. Immunol. 135:5〜8
【非特許文献2】Traunecker, EMBO J. 10(1991), 3655〜9
【非特許文献3】Kipriyanov, Int. J. Cancer 77(1998), 763〜772
【非特許文献4】Kufer(1997), Cancer Immunol. Immunother. 45:193〜7
【非特許文献5】Schroff(1985), Cancer Res. 45:879〜885
【非特許文献6】Shawler(1985), J. Immunol. 135:1530〜1535
【発明の概要】
【0013】
したがって、本発明の技術的な問題は、T細胞性免疫応答の誘導による、腫瘍様疾患、増殖性障害と共にB細胞関連疾患の治療および/または改善のための手段および方法を提供することであった。上記の手段および方法は、公知の抗体に基づく治療に関して引用された短所を克服しなければならない。
【0014】
この技術的問題に対する解決は、請求の範囲において特徴が述べられている態様を提供することによって達成される。
【0015】
したがって、本発明は、ヒトCD3に対して特異的に結合する第一のドメインとIg由来第二の結合ドメインとを含む細胞障害活性を有するCD3特異的結合構築物に関し、該第一のドメインは脱免疫されて、CDR-H1領域、CDR-H2領域、およびCDR-H3領域を含み、該CDR-H3領域は配列番号:96、108、119、120、121、122、123、124、125、126、または127に記載のアミノ酸配列を含み、ならびに
該第一のドメインはさらに、そのフレームワークH1において配列VKK(Val-Lys-Lys)を含み、フレームワークH1とCDR-H1領域のあいだの移行配列は、配列Ala-Ser-Gly-Tyr-Thr-Phe(ASGYTF;配列番号:233)を含む。
【0016】
意外にも、公知のCDR領域と共にフレームワーク領域およびその対応する移行配列に対して上記で引用した特異的改変を行うと、当初の非脱免疫配列と比較して免疫原性の低下を示すが、その細胞障害活性を保持する、脱免疫されたCD3特異的結合分子が得られることが判明した。必ずしも全ての脱免疫プロトコールによって、明確な細胞障害活性を示す生体活性のある機能的構築物が得られたわけではないことから、この発見は特に意外であった;添付の実施例を参照されたい。さらに、意外にも脱免疫された細胞障害活性を有するCD3結合分子は、生産性の増加を示した。本発明に従って、非脱免疫抗体の特定の配列は、本明細書において先に引用した配列に置換または改変されている。特に、フレームワークH1領域において、当初の配列Leu-Ala-Arg(LAR)は、配列Val-Lys-Lys(VKK)に置換されている。さらに、いくつかの非改変/非脱免疫CD3特異的抗体のフレームワークH1とCDR-H1との移行領域に含まれる配列Thr-Ser-Gly-Tyr-Thr-Phe(TSGYTF)は、本発明に従ってAla-Ser-Gly-Tyr-Thr-Phe(ASGYTF)(配列番号:233)に改変されている(図14を参照されたい)。望ましい本発明のCD3-特異的結合構築物は、それによって第二の結合特異性がIg-由来である少なくとも二つの結合特異性を含むことを特徴とする。さらに、該所望の構築物は、本明細書において先に示された特異的アミノ酸配列を特徴とする。添付の実施例において示されるように、本明細書において提供される構築物は、その改変/脱免疫型においてもなお生物活性を保持している。本実施例はまた、当技術分野において公知の方法(国際公開公報第92/10755号、国際公開公報第00/34317号、国際公開公報第98/52976号、国際公開公報第02/079415号、または国際公開公報第02/012899号)によって決定された脱免疫によって、必ずしも全て生物活性分子が得られたわけではないことを報告する;特に実施例2および5を参照されたい。
【図面の簡単な説明】
【0017】
【図1】非脱免疫抗CD3カセットのDNAおよびアミノ酸配列(配列番号:1および2)。
【図2A−1】A)重鎖VH2(配列番号:70)、VH3(配列番号:72)、VH5(配列番号:74)、およびVH7(配列番号:76)、ならびに軽鎖VL1(配列番号:78)、VL2(配列番号:80)、およびVL3(配列番号:82)のアミノ酸配列。
【図2A−2】2A-1の続きを示す図である。
【図2B−1】B)重鎖VH2(配列番号:69)、VH3(配列番号:71)、VH5(配列番号:73)、およびVH7(配列番号:75)、ならびに軽鎖VL1(配列番号:77)、VL2(配列番号:79)、およびVL3(配列番号:81)のヌクレオチド配列。
【図2B−2】2B-1の続きを示す図である。
【図2B−3】2B-2の続きを示す図である。
【図2C】C)非脱免疫抗CD3の重鎖(それぞれ配列番号:84、90、96)、VH2(それぞれ配列番号:86、94、96)、VH3(それぞれ、配列番号:86、94、96)、VH5(それぞれ、配列番号:88、92、96)、およびVH7(それぞれ、配列番号:88、90、96)ならびに非脱免疫抗CD3の軽鎖(それぞれ、配列番号:98、102、104)、VL1(それぞれ、配列番号:100、102、104)、VL2(それぞれ、配列番号:100、102、104)、およびVL3(それぞれ、配列番号:98、102、104)のCDR1、2、および3のアミノ酸配列。
【図2D−1】D)非脱免疫抗CD3の重鎖(それぞれ配列番号:83、89、95)、VH2(それぞれ配列番号:85、93、95)、VH3(それぞれ、配列番号:85、93、95)、VH5(それぞれ、配列番号:87、91、95)、およびVH7(それぞれ、配列番号:87、89、95)ならびに非脱免疫抗CD3の軽鎖(それぞれ、配列番号:97、101、103)、VL1(それぞれ、配列番号:99、101、103)、VL2(それぞれ、配列番号:99、101、103)、およびVL3(それぞれ、配列番号:97、101、103)のCDR1、2、および3のヌクレオチド配列。
【図2D−2】2D-1の続きを示す図である。
【図3A】A)抗CD3(VH2/VL1)のヌクレオチド配列(配列番号:4)。
【図3B】B)抗CD3(VH2/VL1)のアミノ酸配列(配列番号:5)。
【図3C】C)抗CD3(VH2/VL2)のヌクレオチド配列(配列番号:6)。
【図3D】D)抗CD3(VH2/VL2)のアミノ酸配列(配列番号:7)。
【図3E】E)抗CD3(VH2/VL3)のヌクレオチド配列(配列番号:8)。
【図3F】F)抗CD3(VH2/VL3)のアミノ酸配列(配列番号:9)。
【図4A】A)抗CD3(VH3/VL1)のヌクレオチド配列(配列番号:10)。
【図4B】B)抗CD3(VH3/VL1)のアミノ酸配列(配列番号:11)。
【図4C】C)抗CD3(VH3/VL2)のヌクレオチド配列(配列番号:12)。
【図4D】D)抗CD3(VH3/VL2)のアミノ酸配列(配列番号:13)。
【図4E】E)抗CD3(VH3/VL3)のヌクレオチド配列(配列番号:14)。
【図4F】F)抗CD3(VH3/VL3)のアミノ酸配列(配列番号:15)。
【図5A】A)抗CD3(VH5/VL1)のヌクレオチド配列(配列番号:16)。
【図5B】B)抗CD3(VH5/VL1)のアミノ酸配列(配列番号:17)。
【図5C】C)抗CD3(VH5xVL2)のヌクレオチド配列(配列番号:18)。
【図5D】D)抗CD3(VH5/VL2)のアミノ酸配列(配列番号:19)。
【図5E】E)抗CD3(VH5/VL3)のヌクレオチド配列(配列番号:20)。
【図5F】F)抗CD3(VH5/VL3)のアミノ酸配列(配列番号:21)。
【図6A】A)抗CD3(VH7/VL1)のヌクレオチド配列(配列番号:22)。
【図6B】B)抗CD3(VH7xVL1)のアミノ酸配列(配列番号:23)。
【図6C】C)抗CD3(VH7/VL2)のヌクレオチド配列(配列番号:24)。
【図6D】D)抗CD3(VH7xVL2)のアミノ酸配列(配列番号:25)。
【図6E】E)抗CD3(VH7/VL3)のヌクレオチド配列(配列番号:26)。
【図6F】F)抗CD3(VH7/VL3)のアミノ酸配列(配列番号:27)。
【図7A】異なる脱免疫抗CD3部分との二重特異性抗CD19構築物の結合:抗CD3(VH2/VL1)(配列番号:178)、抗CD3(VH2/VL2)(配列番号:180)、抗CD3(VH2/VL3)(配列番号:182)、抗CD3(VH3/VL1)(配列番号:184)、抗CD3(VH3/VL2)(配列番号:186)、抗CD3(VH3/VL3)(配列番号:188)、抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH5/VK2)(配列番号:192)、抗CD3(VH5/VL3)(配列番号:194)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、抗CD3(VH7/VL3)(配列番号:200)。A)CD3。結合は、CD3濃縮PBMC(A)を用いてFACSに基づくアッセイによって測定した。CD3および二次FITC標識抗マウスIg抗体は、(A)においてネガティブコントロールとして用いた。構築物抗CD19×抗CD3および抗EpCAM(M79)×抗CD3を対照として用いた。MFIは平均蛍光強度を示す。
【図7B】異なる脱免疫抗CD3部分との二重特異性抗CD19構築物の結合:抗CD3(VH2/VL1)(配列番号:178)、抗CD3(VH2/VL2)(配列番号:180)、抗CD3(VH2/VL3)(配列番号:182)、抗CD3(VH3/VL1)(配列番号:184)、抗CD3(VH3/VL2)(配列番号:186)、抗CD3(VH3/VL3)(配列番号:188)、抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH5/VK2)(配列番号:192)、抗CD3(VH5/VL3)(配列番号:194)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、抗CD3(VH7/VL3)(配列番号:200)。B)CD19。結合は、CD19陽性NALM-細胞(B)を用いてFACSに基づくアッセイによって測定した。CD19および二次FITC標識抗マウスIg抗体を(B)においてネガティブコントロールとして用いた。構築物抗CD19×抗CD3および抗EpCAM(M79)×抗CD3を対照として用いた。MFIは平均蛍光強度を示す。
【図8】HCICカラムからの抗CD19×抗CD3タンパク質分画の脱免疫変種の280 nmでの代表的な溶出パターン。700 mlで大きい段差を示す下の線は、20 mM酢酸塩、pH 3.5を含む溶出緩衝液の理論的勾配を示す。280 nmでの高い吸収は、カラムフロースルーにおける非結合タンパク質が原因であった。810.98 mlでの矢印は、溶出した抗CD3分画を示す。
【図9】Ni-キレートHis Trap(登録商標)カラムからの抗CD19×抗CD3タンパク質分画の脱免疫変種の280 nmでの代表的な溶出パターン。85 mlで第一の段差および90 mlで大きい第二の段差を示す下の線は、溶出緩衝液(破線)の理論的勾配を示す。93.16 mlでの矢印は抗CD19×抗CD3構築物を含むタンパク質分画を示す。
【図10】セファデックスS200ゲル濾過カラムからの代表的なタンパク質溶出パターン。分画を0〜130 mlの保持時間から回収した。80.44 mlでのタンパク質ピークは、MW約52 kDに対応し、脱免疫された抗CD19×抗CD3構築物を含む。
【図11】A)抗CD19×抗CD3タンパク質分画の脱免疫変種のSDS-PAGE分析。レーンM:分子量マーカー;レーン1:HCICフロースルー;レーン2:細胞培養上清;レーン3:HCIC溶出液;レーン4:IMACフロースルー;レーン5:IMAC洗浄液;レーン6:IMAC溶出液;レーン7:ゲル濾過溶出液。B)抗CD19×抗CD3タンパク質分画の精製脱免疫変種のウェスタンブロット分析。精製二重特異性タンパク質のウェスタンブロット分析を、His-Tagに対する抗体(PentaHis, Qiagen)およびアルカリホスファターゼ標識ヤギ抗マウスIgを用いて行った。レーンM:分子量マーカー;レーン1:HCICフロースルー;レーン2:細胞培養上清;レーン3:HCIC溶出液;レーン4:IMACフロースルー;レーン5:IMAC洗浄液;レーン6:IMAC溶出液;レーン7:ゲル濾過溶出液。
【図12A】野生型抗CD19×抗CD3構築物と比較した、異なる脱免疫抗CD3部分:抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)を有する精製二重特異性抗CD19構築物の、A)CD3に対する結合。結合は、CD3濃縮PBMCs(A)を用いてFACSに基づくアッセイによって測定した。CD3陽性細胞に対する二次抗体を(A)においてネガティブコントロールとして用いた。構築物抗CD19×抗CD3および抗EpCAM(M79)×抗CD3を対照として用いた。アッセイは、1μg/mlおよび5μg/mlの濃度で行った。MFIは、平均蛍光強度を示す。
【図12B】野生型抗CD19×抗CD3構築物と比較した、異なる脱免疫抗CD3部分:抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)を有する精製二重特異性抗CD19構築物の、B)CD19に対する結合。結合は、CD19陽性NALM細胞(B)を用いてFACSに基づくアッセイによって測定した。CD19陽性細胞に対する二次抗体を(B)においてネガティブコントロールとして用いた。構築物抗CD19×抗CD3および抗EpCAM(M79)×抗CD3を対照として用いた。アッセイは、1μg/mlおよび5μg/mlの濃度で行った。MFIは、平均蛍光強度を示す。
【図13】対照と比較した、異なる脱免疫抗CD3部分抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH5/VL2)(配列番号:192)、抗CD3(VH5/VL3)(配列番号:194)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VK3)(配列番号:200)を有する二重特異性抗CD19構築物の細胞障害アッセイ。
【図14】非脱免疫CD3抗体の可変重鎖領域、VH5(配列番号:74)、VH7(配列番号:76)、VH2(配列番号:70)、およびVH3(配列番号:72)の配列アラインメント。フレームワーク領域(FR1)、相補性決定領域1(CDR1)、フレームワーク領域1(FR1)、相補性決定領域3(CDR3)、およびフレームワーク領域4(FR4)を示す。FR1における配列LARおよびVKK、フレームワーク1とCDR1領域との移行部分での配列ASGYTFおよびASGYTA、FR3での配列LTTDK、ITTDK、およびMTTDT、ならびにFR3での配列MQLS、MELS、およびLQMNを四角で囲んだ。アラインメントは、ベクターNTI AdvanceのAlingn Xプログラム(Informax, Inc., USA)を用いて行った。
【図15A】CD3陽性JurkatおよびEpCAM陽性Kato III細胞における異なる脱免疫抗CD3部分:抗CD3(VH5/VL2)×5-10(配列番号:37)(A)を有する二重特異性抗EpCAM構築物のFACSに基づくアッセイにおける結合分析。右へのシフトは結合を示す。Jurkat細胞において、点線はネガティブコントロールのシフトを示す(二次抗体のみ)、破線は抗EpCAM-抗CD3対照抗体の結合を示し、太い線は対象二重特異性構築物を示す。EpCAM陽性Kato III細胞を用いる結合アッセイにおいて、CD3に対するモノクローナル抗体の代わりにEpCAMに対するモノクローナル抗体を陽性対照として用いた。
【図15B】CD3陽性JurkatおよびEpCAM陽性Kato III細胞における異なる脱免疫抗CD3部分:脱免疫抗CD3(VH5/VL2)×4-7(配列番号:33)(B)を有する二重特異性抗EpCAM構築物のFACSに基づくアッセイにおける結合分析。右へのシフトは結合を示す。Jurkat細胞において、点線はネガティブコントロールのシフトを示す(二次抗体のみ)、破線は抗EpCAM-抗CD3対照抗体の結合を示し、太い線は対象二重特異性構築物を示す。EpCAM陽性Kato III細胞を用いる結合アッセイにおいて、CD3に対するモノクローナル抗体の代わりにEpCAMに対するモノクローナル抗体を陽性対照として用いた。
【図15C】CD3陽性JurkatおよびEpCAM陽性Kato III細胞における異なる脱免疫抗CD3部分:脱免疫抗CD3(VH5/VL2)×3-1(配列番号:31)(C)を有する二重特異性抗EpCAM構築物のFACSに基づくアッセイにおける結合分析。右へのシフトは結合を示す。Jurkat細胞において、点線はネガティブコントロールのシフトを示す(二次抗体のみ)、破線は抗EpCAM-抗CD3対照抗体の結合を示し、太い線は対象二重特異性構築物を示す。EpCAM陽性Kato III細胞を用いる結合アッセイにおいて、CD3に対するモノクローナル抗体の代わりにEpCAMに対するモノクローナル抗体を陽性対照として用いた。
【図15D】CD3陽性JurkatおよびEpCAM陽性Kato III細胞における異なる脱免疫抗CD3部分:脱免疫抗CD3(VH5/VL2)×4-7(VL-VH)(配列番号:35)(D)を有する二重特異性抗EpCAM構築物のFACSに基づくアッセイにおける結合分析。右へのシフトは結合を示す。Jurkat細胞において、点線はネガティブコントロールのシフトを示す(二次抗体のみ)、破線は抗EpCAM-抗CD3対照抗体の結合を示し、太い線は対象二重特異性構築物を示す。EpCAM陽性Kato III細胞を用いる結合アッセイにおいて、CD3に対するモノクローナル抗体の代わりにEpCAMに対するモノクローナル抗体を陽性対照として用いた。
【図15E】CD3陽性JurkatおよびEpCAM陽性Kato III細胞における異なる脱免疫抗CD3部分:脱免疫抗CD3(VH5/VL2)×5-10(VL-VH)(配列番号:39)(E)を有する二重特異性抗EpCAM構築物のFACSに基づくアッセイにおける結合分析。右へのシフトは結合を示す。Jurkat細胞において、点線はネガティブコントロールのシフトを示す(二次抗体のみ)、破線は抗EpCAM-抗CD3対照抗体の結合を示し、太い線は対象二重特異性構築物を示す。EpCAM陽性Kato III細胞を用いる結合アッセイにおいて、CD3に対するモノクローナル抗体の代わりにEpCAMに対するモノクローナル抗体を陽性対照として用いた。
【図16A】CD3陽性JurkatおよびEpCAM陽性Kato細胞における、FACSに基づくアッセイによる異なる脱免疫抗CD3部分:3-1×抗CD3(VH5/VL2)(配列番号:49)(A)を有する二重特異性抗EpCAM構築物の結合分析。右へのシフトは結合を示す。
【図16B】CD3陽性JurkatおよびEpCAM陽性Kato細胞における、FACSに基づくアッセイによる異なる脱免疫抗CD3部分:5-10×抗CD3(VH5/VL2)(配列番号:63)(B)を有する二重特異性抗EpCAM構築物の結合分析。右へのシフトは結合を示す。
【図17】対応する非脱免疫構築物と比較して、N-末端位で脱免疫抗CD3部分(di抗CD3)、抗CD3(VH5/VL2)×3-1(配列番号:31)、抗CD3(VH5/VL2)×5-10(配列番号:37)および抗CD3(VH5/VL2)×4-7(配列番号:33)を有するEpCAM構築物の細胞障害アッセイ。CB15 T細胞クローンおよびCHO-EpCAM細胞をE:T比5:1で用いた。CHO-EpCAM細胞をPKH26色素で染色し、細胞は二重特異性一本鎖抗体をインキュベートした後にFACS分析によって計数した。
【図18】対応する非脱免疫野生型構築物と比較した、C-末端位で脱免疫抗CD3部分、3-1x抗CD3(VH5/VL2)(配列番号:49)および5-10×抗CD3(VH5/VL2)(配列番号:63)を有するEpCAM構築物の細胞障害アッセイ。細胞障害アッセイは図17と同一に行った。
【発明を実施するための形態】
【0018】
本明細書において用いられるように、「細胞障害活性を有するCD3結合構築物」という用語は、T細胞上で発現されるヒトCD3複合体に結合することができ、標的細胞の消失/溶解を誘導することができるCD3特異的構築物に関する。CD3/CD3複合体(例えば、抗体、抗体誘導体、または抗体断片)のCD3特異的結合物質の結合により、当技術分野で公知のようにT細胞の活性化が起こる;国際公開公報第99/54440号を参照されたい。したがって、本発明の構築物は、インビボおよび/またはインビトロで標的細胞を消失/溶解できなければならない。対応する標的細胞は、本発明の構築物の第二のIg-由来結合ドメインによって認識される表面分子を発現する細胞を含む。そのような表面分子の特徴を本明細書において下記に示す。細胞障害性は、当技術分野において公知の方法ならびに本明細書において下記におよび添付の実施例に示す方法によって検出されうる。したがって、そのような方法はとりわけ生理的インビトロアッセイを含む。そのような生理的アッセイは、例えば細胞膜の完全性を失うことによる細胞死をモニターしてもよい(例えば、FACSに基づくヨウ化プロピジウムアッセイ、トリパンブルー流入アッセイ、光度測定酵素放出アッセイ(LDH)、放射線測定51Cr放出アッセイ、蛍光測定ユーロピウム放出およびカルセインAM放出アッセイ)。さらなるアッセイは、例えば、光度測定MTT、XTT、WST-1およびアラマーブルーアッセイによる細胞生存率のモニタリング、放射線測定3H-Thd取り込みアッセイ、細胞分裂活性を測定するクローン原性アッセイ、およびミトコンドリアの膜内外勾配を測定する蛍光測定ローダミン123アッセイを含む。さらに、例えばFACSに基づくホスファチジルセリン曝露アッセイ、ELISAに基づくTUNEL試験、カスパーゼ活性アッセイ(光度測定、蛍光測定、またはELISAに基づく)によって、または細胞形態の変化(収縮、膜の水疱形成)を分析することによってアポトーシスをモニターしてもよい。細胞障害活性は、蛍光に基づく色素の放出のFACSに基づく測定によって分析されることが好ましい。そのようなアッセイにおいて、本発明の細胞障害活性を有する二重特異性CD3結合構築物の第二のドメインに結合する分子を有する蛍光標識細胞(好ましくは、CD19に関してNALM-6細胞、およびEpCAM抗原に関してKato細胞)を、本発明の細胞障害活性を有する二重特異性CD3結合構築物の存在下で、無作為なドナーの単離PBMCsまたは標準化T細胞株と共にインキュベートする。インキュベーション後、蛍光標的細胞から上清への色素の放出を、分光蛍光計によって決定する。本発明の細胞障害活性を有する脱免疫二重特異性CD3結合構築物は、脱免疫されておらず、または標的細胞に対して特異性を有しない類似の構築物の生物活性を測定することによって得られた値と比較することによって特徴が調べられる。
【0019】
「結合する/相互作用する」という用語は、本発明の文脈において使用される場合、少なくとも2つの「抗原相互作用部位」の互いとの結合/相互作用と定義される。「抗原相互作用部位」という用語は、本発明に従って、特異的抗原または特異的抗原の群との特異的相互作用の能力を示すポリペプチドのモチーフと定義される。この結合/相互作用はまた、「特異的認識」と定義されると理解される。「特異的に認識する」という用語は、本発明に従って、抗体分子が、本明細書に定義されるヒト標的分子の各々の少なくとも2つのアミノ酸を特異的に相互作用し、および/またはそれに結合することが可能であることを意味する。抗体は、同じ標的分子上の違ったエピトープを認識、相互作用、および/または、結合することができる。この用語は、抗体分子の特異性、すなわち、本明細書で定義されるヒト標的分子の特異的領域の間を区別するその能力に関する。その特異的抗原との抗原相互作用部位の特異的相互作用は、例えば、抗原のコンホメーションの変化の誘導、抗原のオリゴマー化などに起因して、シグナルの開始を生じ得る。従って、抗原相互作用部位のアミノ酸配列における特異的モチーフおよび抗原は、それらの一次構造、二次構造、または三次構造の結果として、ならびにこの構造の二次的修飾の結果として互いに結合する。
【0020】
「特異的相互作用」という用語は、本発明に従って使用される場合、本発明のCD3特異的結合構築物は、同様の構造の(ポリ)ペプチドと交差反応しないか、または本質的に交差反応しないことを意味する。したがって、本発明の構築物は、ヒトCD3と特異的に結合/相互作用し、かつ、Igに由来する第二の結合ドメインにより、特異な、選択された他の化合物、抗原、細胞表面分子、腫瘍マーカー等と相互作用することができる。Igに由来する第二の結合ドメインに対する分子の特異的な実施例は、下記に示す。
【0021】
研究の際の構築物のパネルの交差反応性は、例えば、従来的な条件下で(例えば、Harlow および Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, 1988 およびUsing Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, 1999を参照されたい)、関心対象の(ポリ)ペプチドに対して、ならびに多数のより多くまたはより少なく(構造的におよび/または機能的に)密接に関連する(ポリ)ペプチドに対して、二重特異性単鎖抗体構築物のこのパネルの結合を評価することによって試験され得る。関心対象の(ポリ)ペプチド/タンパク質に結合するが、例えば心臓組織の細胞といった関心対象の(ポロ)ペプチドと同様の組織によって好ましくは発現する他の(ポリ)ペプチドのいずれかに結合しないか、または本質的に結合しない構築物(つまり、抗体(二重特異性)scFvsなど)のみが、関心対象の(ポリ)ペプチド/タンパク質に特異的であると見なされ、本発明において提供される方法におけるさらなる研究のために選択され、実施例において図示されている。これらの方法は、特に、構造的におよび/または機能的に密接に関連した分子を有する結合研究、ブロッキングおよび競合作用研究を含むことができる。これらの結合研究は、また、FACS解析、表面プラスモン共鳴(SPR(例えばBlAcore(登録商標)により))分析超遠心、等温の滴定熱量測定法、蛍光異方性、蛍光分光法、または、放射性同位元素を使ってラベルされたリガンド結合実験を含む。さらにまた、細胞毒性アッセイ(実施例にて図示したように)および上述したアッセイのような生理学的なアッセイ法が行われても良い。したがって、特異的抗原との抗原相互作用部位の特異的相互作用の例には、そのレセプターについてのリガンドの特異性が含まれてもよい。この定義は、その特異的レセプターを結合する際にシグナルを誘導するリガンドの相互作用を特に含む。対応するリガンドの例には、その特異的サイトカイン-レセプターと相互作用し/結合するサイトカインが含まれる。セレクチンファミリー、インテグリン、およびEGFのような増殖因子のファミリーの抗原のような抗原に対する抗原相互作用部位の結合もまた、この定義に特に含まれる。この定義にまた特に含まれるこの相互作用についての他の例は、抗体の抗原性結合部位との抗原決定基(エピトープ)の相互作用である。
【0022】
「結合する/相互作用する」という用語はまた、直線状エピトープに関連するばかりでなく、ヒト標的分子またはその一部の2つの領域からなる、コンホメーションエピトープ、構造エピトープ、または不連続エピトープに関連し得る。本発明の文脈において、コンホメーションエピトープは、ポリペプチドがネイティブタンパク質にフォールディングするときに分子の表面上で一緒になる、一次配列中で分離した2つ以上の個別のアミノ酸配列によって定義される(Sela, (1969) Science 166, 1365およびLaver, (1990) Cell 61, 553-6)。
【0023】
「不連続エピトープ」という用語は、本発明の文脈において、ポリペプチド鎖の離れた部分からの残基から集められる非線形エピトープを意味する。これらの残基は、ポリペプチド鎖が三次元構造にフォールディングしてコンホメーション/構造エピトープを構成するときに、表面で一緒になる。
【0024】
本発明の構築物はまた、本明細書中に記載されるヒトCD3複合体または本明細書中以下に開示されるようなその部分の2つの領域からなり、および/またはそれを含むコンホメーション/構造エピトープと特異的に結合/相互作用することが想定される。
【0025】
従って、特異性は、当技術分野において公知である方法、および本明細書に開示および記載される方法によって実験的に決定され得る。このような方法には、ウェスタンブロット、ELISA試験、RIA試験、ECL試験、IRMA試験、およびペプチドスキャンが含まれるがこれらに限定されない。
【0026】
「Ig-由来第二の結合ドメイン」という用語は、「免疫グロブリン由来ドメイン」、特に、抗体またはその断片、一本鎖抗体、合成抗体、Fab、F(ab2)'、Fv、もしくはscFv断片等のような抗体断片、またはこれらの任意の化学改変された誘導体を指す。これらの抗体分子は、異なる種に由来してもよく、またはキメラ起源であってもよい。最も好ましくは(本明細書において下記に示すように)、本発明のCD3特異的結合構築物に含まれる該Ig由来第二のドメインは、scFvである。本発明に従って用いられる抗体、抗体構築物、抗体断片、抗体誘導体(全て、Ig-由来)、またはその免疫グロブリン鎖は、当技術分野で公知の通常の技術、例えばアミノ酸枯渇、挿入、置換、付加、および/または組換えおよび/または当技術分野で公知の他の任意の改変を単独または組み合わせて用いることによって、さらに改変することができる。免疫グロブリン鎖のアミノ酸配列の基礎となるDNA配列にそのような改変を導入する方法は当業者に周知である;例えばSambrook(1989)上記引用文、を参照されたい。「Ig-由来ドメイン」という用語は特に、少なくとも一つのCDRを含む(ポリ)ペプチド構築物に関する。引用されたIg-由来ドメインの断片または誘導体は、上記の抗体分子の一部であるおよび/または化学/生化学または分子生物学的方法によって改変された(ポリ)ペプチドを定義する。対応する方法は当技術分野で公知であり、とりわけ実験マニュアルに記述されている(Sambrookら「Molecular Cloning : A Laboratory Manual」、Cold Spring Harbor Laboratory Press, 第二版1989および第三版2001;Gerhardtら「Methods for General and Molecular Bacteriology」、ASM Press, 1994;Lefkovits「Immunology Methods Manual : The Comprehensive Sourcebook of Techniques」、Academic Press, 1997;Golemis「Protein-Protein Interactions : A Molecular Cloning Manual」、Cold Spring Harbor Laboratory Press, 2002を参照されたい)。
【0027】
本明細書において用いられる「脱免疫された」という用語は、本発明のCD3結合構築物の上記で同定された第一のドメインに関し、該第一のドメインは、野生型構築物をヒトにおいて非免疫原性またはより弱い免疫原性にすることによって、当初の野生型構築物と比較して改変されている。本発明に従う野生型構築物は、非ヒト起源の抗体またはその一部(フレームワークおよび/またはCDRsのような)に関する。対応する例は、米国特許第4,361,549号、または国際公開公報第99/54440号に記述されている抗体または抗体断片である。「脱免疫された」という用語はまた、T細胞エピトープの生成傾向の低下を示す構築物に関する。本発明に従って、「T細胞エピトープの生成傾向の低下」という用語は、特異的T細胞活性化に至るT細胞エピトープを除去することに関する。さらに、T細胞エピトープの生成傾向の低下は、T細胞エピトープの生成に寄与するアミノ酸の置換、すなわちT細胞エピトープの形成にとって必須であるアミノ酸の置換を意味する。言い換えれば、T細胞エピトープの生成傾向の低下は、免疫原性の低下または抗原非依存的T細胞増殖の誘導能の低下に関連する。さらに、T細胞エピトープの生成傾向の低下は、脱免疫に関連し、このことは抗原非依存的T細胞増殖を誘導するアミノ酸配列の可能性があるT細胞エピトープの喪失または減少を意味する。本発明に従って、T細胞エピトープの生成傾向が低下しているCD3結合領域は、非脱免疫分子と比較して免疫原性が弱い、または好ましくは非免疫原性であるが、そのCD3結合能をなおも保持している、すなわちCD3に結合する低または非免疫原性抗体構築物である。
【0028】
「T細胞エピトープ」という用語は、細胞内でのペプチド、ポリペプチド、またはタンパク質の分解の際に放出されて、その後T細胞の活性化を誘発するために主要組織適合抗原複合体(MHC)の分子によって提示されうる短いペプチド配列に関する;とりわけ国際公開公報02/066514号を参照されたい。MHCクラスIIによって提示されるペプチドに関して、T細胞のそのような活性化は、次にT細胞の直接刺激によって抗体反応を生じて、該抗体を産生することができる。
【0029】
したがって、ヒトCD3に対して特異的に結合する脱免疫された第一のドメインは、フレームワークH3とH4のあいだに存在する少なくとも上記のCDR-H3を含み、該第一の結合ドメインは、フレームワークH3とH4のあいだに存在する変化していない野生型(wt)-CDR-H3を含む非脱免疫の第一のドメインと比較してT-細胞エピトープの生成傾向の低下を示す。さらに、該脱免疫第一のドメインは、少なくともフレームワークH1とCDR-H1の移行領域において、フレームワークH1とCDR-H1の変化していないwt-H1移行領域を含む非脱免疫第一のドメインと比較してT-細胞エピトープの生成傾向の低下を示す上記の配列モチーフを含む。「T細胞エピトープの生成傾向の低下」および/または「脱免疫」は、当業者に公知の技術によって測定してもよい。好ましくは、タンパク質の脱免疫は、T細胞増殖アッセイによってインビトロで試験してもよい。このアッセイにおいて、世界中でHLA-DR対立遺伝子の>80%を表すドナーからのPBMCを、野生型または脱免疫したペプチドのいずれかに反応した増殖に関してスクリーニングする。理想的に、細胞増殖は、野生型ペプチドを抗原提示細胞にローディングした場合に限って検出される。または、全てのハプロタイプを表すHLA-DR四量体を発現することによって脱免疫を調べてもよい。これらの四量体を、ペプチド結合に関して試験してもよく、増殖アッセイにおいて抗原提示細胞に関するペプチド置換体をローディングしてもよい。脱免疫されたペプチドがHLA-DRハプロタイプにおいて提示されるか否かを調べるために、例えば、PBMC上の蛍光標識ペプチドの結合を測定することができる。さらに、脱免疫は、脱免疫分子に対する抗体が、患者への投与後に形成されたか否かを決定することによって証明することができる。特に好ましい方法は、とりわけ添付の実施例6に示されるT細胞増殖アッセイである。
【0030】
好ましくは、抗体由来分子をフレームワーク領域において脱免疫して、CDR領域のほとんどは、CDR領域の結合親和性が影響を受けないように、T細胞エピトープを誘導する傾向の低下を得るために改変されない。一つのT細胞エピトープの消失でさえも、免疫原性の低下が起こる。好ましくは、分子はVL鎖のCDR2領域において脱免疫され、より好ましくはVH鎖のCDR2領域において、さらにより好ましくはVL鎖のCDR1領域、さらにより好ましくはVH鎖のCDR1領域、より好ましくはVL鎖のフレームワーク領域(FR)、および最も好ましくはVH鎖のフレームワーク領域(FR)において脱免疫される。
【0031】
本明細書において用いられるように「CDR」という用語は、「相補性決定領域」を指し、これは当技術分野で周知である。CDRsは、免疫グロブリンの一部であり、該分子の特異性を決定して、特異的リガンドと接触するT細胞受容体である。CDRsは、分子の最も可変部分であり、これらの分子の多様性に関与している。抗体の三つのCDR領域、CDR1、CDR2、および特にCDRH領域が存在する。
【0032】
より好ましい態様において、T細胞エピトープの生成傾向が低下した該CD3特異的ドメインの該CDR-H1およびCDR-H2領域は、国際公開公報第99/54440号に記述されるように抗体構築物に由来する。さらにより好ましい(および添付の実施例に説明するように)該CDR-H1およびCDR-H2領域と共にCDR-H3領域は、Traunecker(1991), EMBO J. 10, 3655〜3659に記述されるCD3分子に関して特異性を有する抗体/抗体誘導体に由来する。本発明に従って、該CDR-H1、CDR-H2、およびCDR-H3領域は、例えばヒトCD3-ε鎖に関してトランスジェニックであるマウスにおいて、他のTCRサブユニットの状況においてヒトCD3-ε鎖を特異的に認識することができる抗体/抗体誘導体等に由来する。これらのトランスジェニックマウス細胞は、天然またはほぼ天然のコンフォメーションでヒトCD3-εを発現する。
【0033】
本発明に従って、フレーム領域は、抗原に接触する高度可変相補性決定領域(CDRs)のタンパク質骨格構造を提供する免疫グロブリンおよびT細胞受容体のVドメイン(VHまたはVLドメイン)における領域に関する。各Vドメインにおいて、FR1、FR2、FR3、およびFR4と呼ばれる四つのフレームワーク領域が存在する。フレームワーク1は、VドメインのN-末端からCDR1の開始部までの領域を含み、フレームワーク2は、CDR1とCDR2のあいだの領域に関し、フレームワーク3は、CDR2とCDR3のあいだの領域を含み、およびフレームワーク4はCDR3の末端からVドメインのC-末端までの領域を意味する;とりわけJaneway, 「Immunobiology」, Garland Publishing, 2001, 第5版を参照されたい。このように、フレームワーク領域は、VHまたはVLドメインにおけるCDR領域外の全ての領域を含む。さらに、「フレームワークとCDR領域のあいだの移行配列」は、フレームワークとCDR領域とのあいだの直接接合部に関する。特に、「フレームワークとCDR領域のあいだの移行配列」という用語は、CDR領域のN-およびC-末端に直接存在する配列またはCDR領域周囲のアミノ酸を意味する。したがって、フレームワークはまた、異なるCDR領域のあいだの配列を含んでもよい。当業者は、所定の配列からフレームワーク領域、CDRsと共に対応する移行配列を容易に推測することができる;Kabat(1991), 「Sequences of Proteins of Immunological Interest.」、5th edit., NIH Publication no. 91-3242保健社会福祉省、Chothia(1987), J. Mol. Biol. 196, 901〜917およびChothia(1989)Nature 342, 877〜883を参照されたい。
【0034】
本発明の好ましい細胞障害活性CD3特異的結合構築物は、さらに該第一のドメインにおいて、Met-Glu-Leu-Ser(MELS;配列番号:234)を含むフレームワークH3を含む。第一のドメインにおいて配列Ile-Thr-Thr-Asp-Lys(ITTK;配列番号:235)を含むフレームワークH3を含む本発明の構築物は、さらにより好ましい。
【0035】
本発明に従って、ヒトCD3に特異的に結合/相互作用して、T細胞エピトープを産生する傾向が減少した本発明の構築物の第一のドメインは、本明細書において定義されるCDR-H1、CDR-H2、CDR-H3領域を含み、好ましい態様において、先に定義したような、特に配列番号:152もしくは153、156もしくは157、160、もしくは161、および/または164もしくは165のいずれか一つに示されるVH-フレームワーク(フレームワーク1、2、3、4)を含む。したがって、本発明のCD3特異的結合構築物は、ヒトCD3に特異的に結合する第一のドメインを含み、配列番号:152また153に示されるフレームワーク領域、配列番号:156または157に示されるフレームワーク領域2、配列番号:160または161に示されるフレームワーク領域3、および/または配列番号:164または165に示されるフレームワーク領域4を含む。
【0036】
本発明の特に好ましい態様において、細胞障害活性を有する脱免疫CD3特異的結合構築物は、その第一のドメインにおいて(a)配列番号:88に記述されるCDR-H1、および(b)配列番号:90または92に示されるCDR-H2を含む。
【0037】
したがって、改変されたCDR-H1およびCDR-H2領域は、T細胞エピトープをの生成傾向の低下に至り、CD3-ε鎖特異的抗体に由来する。本発明に従って最も好ましくは、該(親)抗体はTCR複合体の状況において提示されるヒトCD3の天然のまたはほぼ天然の構造またはコンフォメーションのエピトープを反映するエピトープに特異的に結合することができなければならない。
【0038】
好ましくは、本発明のCD3特異的結合構築物は、配列番号:74または76に示されるようにVH領域を含む。配列番号:74は、例としての脱免疫可変重鎖領域を示し、同様に配列番号:76も例としての脱免疫可変重鎖領域を示す。
【0039】
好ましくは、本発明のCD3特異的結合構築物は、配列番号:98または100に示されるCDR-L1、配列番号:102に示されるCDR-L2、および/または配列番号:104に示されるCDR-L3を含む。
【0040】
好ましい態様において、本発明のCD3特異的結合構築物は、そのCD3-特異的部分にVL領域を含み、該VL領域は、配列番号:78、配列番号:80、配列番号:82、および配列番号:112からなる群より選択される。配列番号:78において特徴が示されるVL1、配列番号:80において特徴が示されるVL2、および配列番号:82において特徴が示されるVL3は、本発明に従う完全な脱免疫VL領域に関連し、それらは、上記のVH領域との様々な組み合わせにおいて用いてもよい。なおも同様に、非脱免疫VL領域は、本発明に従って上記の脱免疫VH領域と組み合わせてもよいと想像される。本発明の細胞障害活性を有するCD3結合構築物において好ましく用いられる対応する非脱免疫VL-領域を配列番号:112に示す。したがって、本発明のCD3構築物の先に引用した「第一のドメイン」の重鎖部分のみならず、T細胞エピトープの生成傾向が低下するように改変してもよい。同様に、該ドメインは、対応する可変軽鎖部分を含むと想像される。配列番号:78、80、および82は例えば、国際公開公報第99/54440号に開示される構築物のCD3結合部分の脱免疫されたVL1、VL2、およびVL3領域を示す。
【0041】
上記のように、本発明のCD3特異的結合構築物は、最も好ましくは、scFvであるIg由来の第二のドメインを含む。したがって、本発明の最も好ましい態様において、ヒトCD3に対して一つの特異性を有し、さらなる分子/化合物に対するおよび/または相互作用することができる第二のscFvによって媒介されるさらなる特異性を有する、脱免疫された二重特異性一本鎖抗体構築物が提供される。これらのさらなる分子/化合物は、細胞表面分子、腫瘍マーカー、腫瘍抗原等を含んでもよい。そのようなさらなる化合物/分子は本明細書において下記に例示され、特異的構築物も同様に添付の実施例において示され、提供される。
【0042】
「二重特異性一本鎖抗体構築物」という用語は、二つの抗体由来結合ドメイン、好ましくはscFvを含む構築物に関する。該結合ドメインの一つは、ヒトCD3抗原(標的分子1)と特異的に結合/相互作用することができる抗体(またはその一部)、抗体断片、またはその誘導体の可変領域からなる。第二の結合ドメインは、下記のようにもう一つの(ヒト)抗原(標的分子2)と特異的に結合/相互作用することができる抗体、抗体断片、またはその誘導体の可変領域(またはその一部)からなる。したがって、該第二の結合ドメインは、本発明に従って、細胞表面分子および/または腫瘍特異的マーカーに対して特異性を有する抗原-相互作用部位を含む、先に引用したIg由来第二のドメインである。二重特異性構築物、好ましくは該二重特異性一本鎖抗体構築物における該二つのドメイン/領域は、好ましくは一本鎖として互いに共有結合される。この結合は、直接(ドメイン1[T細胞エピトープの生成傾向の低下を含み、およびCDR領域もしくはCDR領域と上記のフレームワーク領域とを含むヒトCD3抗原に対して特異的]−ドメイン2[細胞表面分子および/または腫瘍特異的マーカーに対して特異的]、もしくはドメイン1[細胞表面分子および/または腫瘍特異的マーカーに対して特異的]−ドメイン2[T細胞エピトープの生成傾向の低下を含み、およびCDR領域またはCDR領域と上記のフレームワーク領域とを含む、ヒトCD3抗原に対して特異的]、またはさらなるポリペプチドリンカー配列(ドメイン1−リンカー配列−ドメイン2)のいずれかによって行うことができる。リンカーを用いる事象において、このリンカーは好ましくは、第一および第二のドメインのそれぞれが、互いに独立してその異なる結合特異性を確実に保持することができるために十分な長さおよび配列を有する。先に述べたように、および添付の実施例において示されるように、好ましくは本明細書に定義される少なくとも二つのドメインを含むCD3特異的結合構築物は、「二重特異性一本鎖抗体構築物」であり、最も好ましくは二重特異性一本鎖抗体Fv(scFv)である。該構築物は薬学的組成物の状況において用いられると特に想像される。二重特異性一本鎖分子は当技術分野で公知であり、国際公開公報第99/54440号、Mack, J. Immunol.(1997), 158, 3965〜3970, Mack, PNAS, (1995), 92, 7021〜7025, Kufer, Cancer Immunol. Immunother. (1997), 45, 193〜197, Loffler, Blood, (2000), 95, 6, 2098〜2103, Bruhl, J. Immunol. (2001), 166, 2420〜2426に記述される。本発明の特に好ましい分子フォーマットは、本発明の構築物のCD3特異的結合ドメインが、先に定義された少なくとも一つのVHおよび一つのVL領域を含む、ポリペプチド構築物を提供する。本明細書に記述され、T細胞エピトープの生成傾向が低下したVH領域の他に、該特異的結合構築物は、T細胞エピトープの生成傾向が低下したさらなる領域/ドメインを含んでもよいことに注意されたい。先に述べたように、VL領域および/または対応するフレームワークは、本発明に従って、T細胞エピトープ生成傾向の低下を有するように操作されているアミノ酸の鎖を含んでもよい。scFvフォーマットにおいてリンカードメインによって互いに連結しているVHドメインおよびVLドメインの分子内方向は、引用された二重特異性一本鎖構築物に関して決定的ではない。このように、可能性がある配置(VH-ドメイン−リンカードメイン−VLドメイン;VLドメイン−リンカードメイン−VHドメイン)をいずれも有するscFvは、引用された二重特異性一本鎖構築物の特定の態様である。CD3特異的ドメインは、二重特異性分子のN-末端またはC-末端に存在しうる。各ドメインのVHおよびVL領域は異なる順序で配置することができる(VH−VLまたはVL−VH)。
【0043】
本発明に従って用いられる「一本鎖」という用語は、二重特異性一本鎖構築物の第一および第二のドメインが、好ましくは単一の核酸分子によってコードされる共直線状のアミノ酸配列の形で共有結合していることを意味する。
【0044】
本発明の構築物は、本明細書において定義された第一のドメインおよびIg由来第二のドメインの他に、例えば組換えによって産生された構築物の単離および/または調製のためのさらなるドメインを含んでもよいことに注目されたい。
【0045】
本発明に従って、本発明のCD3構築物のヒトCD3に特異的に結合する上記の第一のドメインのみならず、T細胞エピトープの生成傾向の低下を有してもよいことに注目される。同様に、Ig由来第二のドメインおよび/または結合するリンカー領域は、改変されている、例えばヒト化および/または脱免疫されていると想像される。
【0046】
先に述べたように、脱免疫アプローチは特に、国際公開公報第00/34317号、国際公開公報第98/52976号、国際公開公報第02/079415号、または国際公開公報第02/012899号および添付の実施例に記述されている。これらのアプローチは、可能性があるT細胞エピトープにおいてアミノ酸の置換を行うことを伴う。このように、所定の配列が細胞内タンパク質プロセシングの際にT細胞エピトープを生成する可能性が低下する。さらに、国際公開公報第92/10755号は、タンパク質上の抗原決定基が操作されるアプローチを記述している。特に、タンパク質はエピトープマッピングされて、そのアミノ酸配列は遺伝子操作によって変化している。
【0047】
さらに、「ヒト化アプローチ」は、当技術分野において周知であり、特に抗体分子、例えばIg由来分子に関して記述されている。「ヒト化」という用語は、非ヒト抗体に由来する配列のいくつかの部分を含む非ヒト(例えば、マウス)抗体またはその断片(Fv、Fab、Fab'、F(ab')、scFvs、または抗体の他の抗原結合部分配列)のヒト化型に関する。ヒト化抗体には、ヒト免疫グロブリンの相補性決定領域(CDR)からの残基が、所望の結合特異性、親和性、および結合能を有するマウス、ラット、またはウサギのような非ヒト種のCDRからの残基に置換されているヒト免疫グロブリンが含まれる。一般的に、ヒト化抗体は、CDR領域の全てまたは実質液に全てが非ヒト免疫グロブリンのCDR領域に対応して、およびFR領域の全てまたは実質的に全てがヒト免疫グロブリンコンセンサス配列の領域である、少なくとも一つ、および一般的に二つの可変ドメインの全てまたは実質的に全てを含むであろう。ヒト化抗体は最適には、典型的にヒト免疫グロブリンの免疫グロブリン定常領域(Fc)の少なくとも一部を含むであろう;とりわけJonesら、Nature 321:522〜525(1986)、Presta, Curr. Op. Struct. Biol. 2:593〜596(1992)を参照されたい。非ヒト抗体をヒト化する方法は当技術分野で周知である。一般的にヒト化抗体は、ヒト抗体により厳密に類似するように、非ヒトである起源からそれに導入された一つまたはそれ以上のアミノ酸を有するが、なおも抗体の当初の結合活性を保持している。抗体/抗体分子のヒト化法は、Jonesら、Nature 321:522〜525(1986);Riechmannら、Nature 332:323〜327(1988);およびVerhoeyenら、Science 239:1534〜1536(1988)にさらに詳述されている。ヒト化抗体の特定の例、例えばEpCAMに対する抗体は、当技術分野で公知であり、例えば(LoBuglio, 「Proceedings of the American Society of Clinical Oncology」(抄録)、1997, 1562およびKhor, 「Proceedings of the American Society of Clinical Oncology」(抄録)、1997, 847)を参照されたい。
【0048】
したがって、本発明の状況において、特に、脱免疫され、薬学的組成物において首尾よく用いることができる二重特異性一本鎖抗体構築物が提供される。
【0049】
先に述べたように、上記のCD3特異的結合構築物のIg由来第二のドメインは、細胞表面分子に対する特異性を有する抗原相互作用部位を含んでもよい。
【0050】
本明細書において用いられるように、「細胞表面分子」という用語はまた、細胞の表面上で提示された分子を指す。「細胞表面分子」という用語は、細胞の表面上に提示され、Ig由来結合ドメイン、好ましくは抗体、抗体断片、または誘導体に近づくことができる(インビトロまたはインビボで)ドメインまたはエピトープを含む。先に示したように、最も好ましくは該Ig由来ドメインはscFvである。該細胞表面分子の例は、膜および膜貫通タンパク質、該タンパク質または細胞表面に適合させた分子等である。本発明のさらに好ましい態様に従って、該細胞表面分子は腫瘍特異的マーカーである。本発明の状況において、「腫瘍特異的マーカー」という用語は、腫瘍細胞の表面に提示されるおよび/または存在する分子、または広布に発現されるが、腫瘍細胞の表面上の抗体、抗体断片、または抗体誘導体の結合にとって近づくことができるのみである分子を指す。腫瘍マーカーの例を本明細書において下記に示し、これらはEpCAM、CD19、HER-2、HER-2 neu、HER-3、HER-4、EGFR、PSMA、CEA、MUC-1(ムチン)、MUC2、MUC3、MUC4、MUC5AC、MUC5B、MUC7、Lewis-Y、CD20、CD33、CD30、CD44v6、Wue-1、プラズマ細胞抗原(国際公開公報第01/47953号を参照されたい)、(膜結合型)IgE、黒色腫コンドロイチン硫酸プロテオグリカン(MCSP)、STEAP、メソセリン、前立腺幹細胞抗原(PSCA)、sTn(シアリル化Tn抗原)、FAP(線維芽細胞活性化抗原)、EGFRvIII、Igα、Igβ、MT-MMPs、Cora抗原、EphA2、L6、およびCO-29を含むがこれらに限定されない。
【0051】
本発明のCD3特異的結合構築物のIg由来第二のドメインはまた、EpCAM、CCR5、CD19、HER-2、HER-2 neu、HER-3、HER-4、EGFR、PSMA、CEA、MUC-1(ムチン)、MUC2、MUC3、MUC4、MUC5AC、MUC5B、MUC7、βhCG、Lewis-Y、CD20、CD33、CD30、ガングリオシドGD3、9-O-アセチル-GD3、GM2、Globo H、フコシルGM1、ポリSA、GD2、炭酸脱水酵素IX(MN/CA IX)、CD44v6、Sonic Hedgehog(Shh)、Wue-1、プラズマ細胞抗原(膜結合型)IgE、黒色腫コンドロイチン硫酸プロテオグリカン(MCSP)、CCR8、TNF-α前駆体、STEAP、メソセリン、A33抗原、前立腺幹細胞抗原(PSCA)、Ly-6、デスモグレイン4、E-カドヘリンネオエピトープ、胎児アセチルコリン受容体、CD25、CA19-9マーカー、CA-125マーカー、およびミュラー管阻害物質(MIS)受容体II型、sTn(シアリル化Tn抗原)、FAP(線維芽細胞活性化抗原)、エンドシアリン、EGFRvIII、L6、SAS、CD63、TAG72、TF-抗原、Cora抗原、CD7、CD22、Igα(CD79a)、Igβ(CD79b)、G250、gp100、MT-MMPs、F19-抗原、CO-29およびEphA2からなる群より選択される分子に対して特異性を有する抗原相互作用部位を含んでもよい。
【0052】
本明細書において提供された構築物は、医学的状況において特に有用である。例えば、腫瘍疾患および/またはリンパ腫、特に非ホジキンB-細胞リンパ腫は、ヒトCD3およびCD20(CD3×CD20またはCD20×CD3)に対する本発明の脱免疫(二重特異性)構築物によって治療してもよい。自己免疫疾患は、ヒトCD3およびCD30またはCD19(すなわち、CD3×CD30、CD30×CD3、CD3×CD19またはCD19×CD3)に対する脱免疫(二重特異性)構築物の投与によって治療してもよい。リウマチ性関節炎のみならず他の炎症疾患を、ヒトCD3およびCCR5(CD3×CCR5またはCCR5×CD3)に対する本発明の脱免疫(二重特異性)構築物によって治療してもよい。本明細書において定義し、およびTNF-α前駆体に対する/結合する第二のIg由来ドメインを含む脱免疫CD3特異的結合構築物は、炎症障害の治療または予防において有用となる可能性がある。本明細書において提供され、EpCAM、CD19、HER-2、HER-2 neu、HER-3、HER-4、EGFR、PSMA、CEA、MUC-1(ムチン)、MUC2、MUC3、MUC4、MUC5AC、MUC5B、MUC7、Lewis-Y、CD20、CD33、CD30、CD44v6、Wue-1、プラズマ細胞抗原(国際公開公報第01/47953号を参照されたい)、(膜結合型)IgE、黒色腫コンドロイチン硫酸プロテオグリカン(MCSP)、STEAP、メソセリン、前立腺幹細胞抗原(PSCA)、sTn(シアリル化Tn抗原)、FAP(線維芽細胞活性化抗原)、EGFRvIII、Igα、Igβ、MT-MMPs、Cora抗原、EphA2、L6、およびCO-29に対する/結合する/相互作用する第二のIg由来ドメインを含むCD3構築物は、乳癌、結腸癌、前立腺癌、頭頚部癌、皮膚癌(黒色腫)、尿性器癌、例えば卵巣癌、子宮内膜癌、子宮頚癌および腎臓癌、肺癌、胃癌、小腸癌、肝臓癌、膵臓癌、胆嚢癌、胆管癌、食道癌、唾液腺癌、および甲状腺癌のような腫瘍様疾患、または血液腫瘍、神経膠腫、肉腫、もしくは骨肉腫のような他の腫瘍様疾患の医学的介入において特に有用となる可能性がある。CD3結合構築物の投与はまた、最小の残留疾患、好ましくは初期固形腫瘍、進行固形腫瘍、または転移性固形腫瘍に適応される。
【0053】
同様に添付の実施例において記述されるように、本発明の特に好ましいCD3特異的結合構築物は、T細胞エピトープの生成傾向が低下した先に定義された第一のドメインおよびEpCAMに対して特異性を有する抗原相互作用部位を含む第二のIg由来ドメインを含む。
【0054】
上皮細胞接着分子(17-1A抗原、KSA、EGP40、GA733-2、ks1-4またはesaとも呼ばれるEpCAM)は、特定の上皮および多くのヒト癌において特異的に発現されるアミノ酸314個の40 kDa膜貫通型糖タンパク質である(Balzer, J. Mol. Med. 1999, 77, 699〜712において論評)。EpCAMはマウスモノクローナル抗体17-1A/エドレコロマブ(Goettinger, Int. J. Cancer, 1986;38, 47〜53およびSimon, Proc. Natl. Acad. Sci. USA 1990;87, 2755〜2759)による認識を通して発見され、その後クローニングされた。EpCAMは、内皮細胞を一定方向に非常に秩序だって接着させるために役立つ(Litvinov, J. Cell Biol. 1997, 139, 1337〜1348)。上皮細胞が悪性形質転換すると、急速に増殖する腫瘍細胞により、上皮の高度細胞秩序が放棄される。その結果、EpCAMの表面分布はより制限が少なくなり、分子は腫瘍細胞上でより露出され、腫瘍細胞表面上の抗体、抗体断片、または抗体誘導体の結合にとって近づきやすくなる。その上皮細胞起源のために、ほとんどの癌腫による腫瘍細胞はなおもその表面上にEpCAMを発現する。
【0055】
インビボで、EpCAMの発現は、上皮の増殖の増加に関連し、細胞分化に負に相関する(論評に関してはBalzar, J. Mol. Med., 1999, 77, 699〜712を参照されたい)。EpCAMの発現は本質的に、全ての主要な癌腫について認められる(Balzar, 1999, J. Mol. Med. 77, 699〜712に論評、またはとりわけDe Bree, Nucl. Med. Commun. 1994, 15, 613〜27;Zhang, Clin. Cancer Res. 1998, 4, 295〜302に報告されている)。その広範な発現のために、EpCAMは「汎癌腫」抗原と呼ばれている。多くの場合、腫瘍細胞は、その親上皮または該癌のより攻撃性の低い型よりかなり高い程度にEpCAMを発現することが認められた。例えば、EpCAM発現の増加は、前立腺癌発達の初期事象を表す(Poczatek, J. Urol. 1999, 162, 1462〜1644)。さらに、子宮頚部の扁平上皮癌および腺癌の大多数において、強いEpCAMの発現は、増殖の増加および最終分化に関するマーカーの消失に相関する(Litvinov, Am. J. Pathol, 1996, 148, 865〜75)。乳癌において、腫瘍細胞上でのEpCAMの過剰発現は、生存の予測因子である(Gastl, Lancet, 2000, 356, 1981〜1982)。EpCAMは、頭部、頚部、および肺の扁平上皮癌を有する患者における播種された腫瘍細胞を検出するためのマーカーである(Chaubal, Anticancer Res. 1999, 19, 2237〜2242;Piyathilake, Hum. Pathol. 2000, 31, 482〜487)。表皮、口腔、咽頭蓋、咽頭、喉頭、および食道において認められる正常な扁平上皮は、有意にEpCAMを発現しなかった(Quak, Hybridoma, 1990, 9, 377〜387)。EpCAMは原発、転移、および播種性NSCLC(肺の非小細胞癌細胞(Passlick, Int. J. Cancer, 2000, 87, 548〜552))の大多数において、胃の腺癌および胃食道接合部腺癌(Martin, J. Clin. Pathol. 1999, 52, 701〜4)において、ならびに結腸直腸、膵臓癌および乳癌に由来する細胞株(Szala, Proc. Natl. Acad. Sci. USA 1990, 87, 3542〜6;Packeisen, Hybridoma, 1999, 18, 37〜40)において発現されることが示されている。
【0056】
最も好ましい態様において、EpCAMに対する/結合する第二のIg由来ドメインを含む本発明のCD3特異的結合構築物は、以下からなる群より選択されるアミノ酸配列を含む:
(a)配列番号:31、33、35、37、39、49、55、58、61、63、65、67、237、239、241、243、245、247、249、251、253、255、257、259、261、263、265、267、269、271、273、275、277、279、281、283、285、287、289、291、293、295、297、299、301、303、305、307、309、311、313、315、317、319、321、323、および325のいずれか一つに示されるアミノ酸配列;
(b)配列番号:30、32、34、36、38、48、54、57、60、62、64、66、236、238、240、242、244、246、248、250、252、254、256、258、260、262、264、266、268、270、272、274、276、278、280、282、284、286、288、290、292、294、296、298、300、302、304、306、308、310、312、314、316、318、320、322、および324のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;ならびに
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列。
【0057】
したがって、本発明は、特に好ましい態様において、T細胞エピトープの生成傾向が低下したCD3結合/相互作用部分(「抗CD3」)およびEpCAMに特異的に相互作用/結合する(「抗EpCAM」)さらなる一本鎖部分(Ig由来ドメイン)を含む特異的CD3構築物を提供する。以下の表1A、1B、2A、2B、3A、3B、4A、4B、5A、および5Bは、そのようなCD3およびEpCAM結合構築物の好ましいコンフィギュレーションに関する。
【0058】
EpCAM 3-1、EpCAM 3-5、EpCAM 4-1、EpCAM 4-7およびEpCAM 5-10は、国際公開公報第99/25818号においてファージディスプレイによって単離されたEpCAMに対する特異的一本鎖抗体に関する。
【0059】
表1A、2A、3A、4A、および5Aにおける各構築物は、A-Gと呼ばれる異なる7個のタンパク質モジュールを含む。タンパク質モジュールA-Gは、A-B-C-D-E-F-Gの順でペプチド結合によって一つの連続したポリペプチド鎖において互いに直接共有結合し、N-末端でタンパク質モジュールAとなり、C-末端でタンパク質モジュールGとなる。タンパク質モジュールA、C、EおよびGは、ヒトCD3またはEpCAM抗原に対して特異性を有する抗体のVHまたはVLドメインのいずれかとなりうる抗体可変ドメインを指す。モジュールB、DおよびFはVHおよびVLドメインを接続するリンカーである。
【0060】
タンパク質モジュールAがVH抗体ドメインである場合、タンパク質モジュールCはVLタンパク質ドメインであり、その逆も起こりうる。タンパク質モジュールEがVH抗体ドメインである場合、タンパク質モジュールGはVLタンパク質ドメインであり、その逆もまた同じである。
【0061】
ヒトCD3抗原に対して特異性を有する抗体の脱免疫されたVHドメインは、配列番号:74または76に記載した配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫されたVLドメインは、配列番号:78、80、または82に記載の配列から選択することができる。ヒトEpCAM 3-1、3-5、4-1、4-7、および5-10抗体のVHタンパク質ドメインはそれぞれ、配列番号:137、141、145、149、および133に記載されている。ヒトEpCAM 3-1、3-5、4-1、4-7、および5-10抗体のVLタンパク質ドメインはそれぞれ、配列番号:139、143、147、151、および135に記載されている。
【0062】
タンパク質モジュール対A/CおよびE/Gによって示される抗体可変ドメインの対は、さらなる連結タンパク質モジュールによって結合され、この場合タンパク質モジュールBはモジュール対A/Cを直接結合するために役立ち、タンパク質モジュールFはモジュール対E/Gを直接結合するために役立つ。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体の脱免疫されたVH/VLまたはVL/VHタンパク質ドメインの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:3に記載されるアミノ酸配列を有する。モジュール対A/CまたはE/Gのいずれかが、EpCAM抗原に対して特異性を有する抗体からのVH/VLまたはVL/VHの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:168に記載されるアミノ酸配列を有する。モジュールDはABCおよびEFGモジュール群を結合する。
【0063】
タンパク質モジュールA-B-Cの組み合わせ、およびタンパク質モジュールE-F-Gの組み合わせはそれぞれ、ヒトCD3抗原またはEpCAM抗原のいずれかに対して特異性を有する抗体の一つのscFv断片を構成する。モジュールAおよびCがCD3結合配列を示す場合、タンパク質モジュールA-B-CおよびE-F-Gは、配列番号:176に記載される配列を有するタンパク質モジュールDを通して互いに結合する。一方、モジュールAおよびCがEpCAM結合配列を示す場合、タンパク質モジュールA-B-CおよびE-F-Gのそれぞれの群は、配列番号:174に示される配列を有するタンパク質モジュールDによって互いに結合する。このように、クローニング目的のためにVL鎖の後にさらなるセリンを挿入してもよい。しかし、当業者は、VLドメインを配列番号:176の代わりにその後のVドメインに結合させるために、配列番号:174に示されるリンカーを用いてもよい。タンパク質モジュールDは、タンパク質モジュールCのC-末端をタンパク質モジュールEのN-末端に結合させるように作用する。
【0064】
表1B、2B、3B、4B、および5Bにおける各核酸構築物は、A-Gと呼ばれる異なる7個の核酸モジュールを含む。核酸モジュールA-Gは、A-B-C-D-E-F-Gの順でホスフェートグリコシド結合によって単一の連続ヌクレオチド鎖において互いに直接共有結合しており、それぞれの核酸構築物の5'-末端が核酸モジュールAであって、3'末端が核酸モジュールGである。核酸モジュールA、C、EおよびGは、ヒトCD3またはEpCAM抗原に対して特異性を有する抗体のVHまたはVLドメインのいずれかとなりうる抗体可変ドメインのコード領域を指す。
【0065】
核酸モジュールAがVH抗体ドメインをコードする場合、核酸モジュールCはVLタンパク質ドメインをコードし、その逆も起こりうる。核酸モジュールEがVH抗体ドメインをコードする場合、核酸モジュールGはVLタンパク質ドメインをコードし、その逆も起こりうる。
【0066】
ヒトCD3抗原に対して特異性を有する抗体の脱免疫VHドメインをコードする核酸分子は、配列番号:73または75に記載の配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗原の脱免疫されたVLドメインをコードする核酸分子は、配列番号:77、79、または81記載の配列から選択することができる。ヒトEpCAM 3-1、3-5、4-1、4-7、および5-10抗体のVHタンパク質ドメインをコードする核酸分子はそれぞれ、配列番号:136、140、144、148、および132に記載されている。ヒトEpCAM 3-1、3-5、4-1、4-7、および5-10抗体のVLタンパク質ドメインをコードする核酸分子はそれぞれ、配列番号:138、142、146、150、および134に記載されている。
【0067】
核酸モジュール対A/CおよびE/Gによって示される抗体の可変ドメインをコードする核酸の対は、さらなる連結核酸モジュールによって結合し、この場合核酸モジュールBはモジュール対A/Cを直接結合させるように作用し、核酸モジュールFはモジュール対E/Gを直接結合させるように作用する。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体からの脱免疫されたVH/VLまたはVL/VHタンパク質ドメインの対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:202に記載のヌクレオチド配列を有する。モジュール対A/CまたはE/Gのいずれかが、ヒトEpCAM抗原に対して特異性を有する抗体からのVH/VLまたはVL/VH対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:201に記載のヌクレオチド配列を有する。
【0068】
核酸モジュールA-B-Cの組み合わせと核酸モジュールE-F-Gの組み合わせはそれぞれ、ヒトCD3抗原またはEpCAM抗原に対して特異性を有する抗体の一つのscFv断片を構成する。AおよびCモジュールがCD3結合配列を含む場合、核酸モジュールA-B-CおよびE-F-Gのそれぞれの群は、配列番号:175に記載のヌクレオチド配列を有する核酸モジュールDによって互いに結合する。AおよびCモジュールがEpCAM結合配列を含む場合、核酸モジュールA-B-CおよびE-F-Gのそれぞれの群は、配列番号:173に記載のヌクレオチド配列を有する核酸モジュールDによって互いに結合する。しかし、先に述べたように、セリンをコードするさらなるコドン(配列番号:175における)をクローニング目的のために挿入してもよい。当業者は、VL鎖をコードするヌクレオチド配列を、リンカーの5'末端でセリンをコードするさらなるコドンがなくとも、配列番号:173に示すリンカーによってその後のVドメインに直接結合させてもよい。核酸モジュールDは、核酸モジュールCの3'末端と核酸モジュールEの5'末端とを結合させるように作用する。
【0069】
(表1A)一本鎖抗EpCAM 3-1可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列
【0070】
(表1B)一本鎖抗EpCAM 3-1可変領域を含む脱免疫抗ヒトCD3構築物:ヌクレオチド配列
【0071】
(表2A)一本鎖抗EpCAM 3-5可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列
【0072】
(表2B)一本鎖抗EpCAM 3-5可変領域を含む脱免疫抗ヒトCD3構築物:ヌクレオチド配列
【0073】
(表3A)一本鎖抗EpCAM 4-1可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列
【0074】
(表3B)一本鎖抗EpCAM 4-1可変領域を含む脱免疫抗ヒトCD3構築物:ヌクレオチド配列
【0075】
(表4A)一本鎖抗EpCAM 4-7可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列
【0076】
(表4B)一本鎖抗EpCAM 4-7可変領域を含む脱免疫抗ヒトCD3構築物:ヌクレオチド配列
【0077】
(表5A)一本鎖抗EpCAM 5-10可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列
【0078】
(表5B)一本鎖抗EpCAM 5-10可変領域を含む脱免疫抗ヒトCD3構築物:ヌクレオチド配列
【0079】
最も好ましくは、本発明は、CD3およびEpCAMに対する特異的結合を含み、配列番号:30、31(表1Aおよび1Bの構築物2)、配列番号:48、49(表1A、1Bの構築物5)、配列番号:64、65(表2A、2Bの構築物2)、配列番号:54、55(表2A、2Bの構築物5)、配列番号:66、67(表3A、3Bの構築物2)、配列番号:32、33(表4A、4Bの構築物2)、配列番号:34、35(表4A、4Bの構築物4)、配列番号:60、61(表4A、4Bの構築物5)、配列番号:36、37(表5A、5Bの構築物2)、配列番号:38、39(表5A、5Bの構築物4)、または配列番号:62、63(表5A、5Bの構築物5)を有する二重特異性抗体構築物を提供する。
【0080】
本明細書において上記の構築物に従って、T細胞エピトープ生成傾向が低下して、ヒトCD3に対して特異性を有する少なくとも上記の第一のドメインと、EpCAMに対して特異的である第二のIg由来ドメインとを含む本発明の特に好ましいCD3およびEpCAM結合構築物は、配列番号:31、33、35、37、39、49、55、58、61、63、65、67、237、239、241、243、245、247、249、251、253、255、257、259、261、263、265、267、269、271、273、275、277、279、281、283、285、287、289、291、293、295、297、299、301、303、305、307、309、311、313、315、317、319、321、323、および325に示される。本明細書において定義される該好ましいCD3およびEpCAM結合構築物をコードする対応する核酸分子は、配列番号:30、32、34、36、38、48、54、57、60、62、64、66、236、238、240、242、244、246、248、250、252、254、256、258、260、262、264、266、268、270、272、274、276、278、280、282、284、286、288、290、292、294、296、298、300、302、304、306、308、310、312、314、316、318、320、322、および324を含む。
【0081】
したがって、本発明はまた、以下からなる群より選択される、ヒトCD3に対して特異的に結合してT細胞エピトープの生成傾向が低下している第一のドメインと、EpCAMに対する/結合することができるIg由来第二のドメインとを含む、CD3特異的結合構築物を提供する:
(a)配列番号:31、33、35、37、39、49、55、58、61、63、65、67、237、239、241、243、245、247、249、251、253、255、257、259、261、263、265、267、269、271、273、275、277、279、281、283、285、287、289、291、293、295、297、299、301、303、305、307、309、311、313、315、317、319、321、323、または325のいずれか一つに示されるアミノ酸配列;
(b)配列番号:30、32、34、36、38、48、54、57、60、62、64、66、236、238、240、242、244、246、248、250、252、254、256、258、260、262、264、266、268、270、272、274、276、278、280、282、284、286、288、290、292、294、296、298、300、302、304、306、308、310、312、314、316、318、320、322、または324のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;ならびに
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列。
(d)ストリンジェントなハイブリダイゼーション条件で(b)において定義された核酸配列の相補鎖とハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【0082】
本発明はまた、本明細書において上記の(b)において定義された核酸配列、すなわち配列番号:30、32、34、36、38、48、54、57、60、62、64、66、236、238、240、242、244、246、248、250、252、254、256、258、260、262、264、266、268、270、272、274、276、278、280、282、284、286、288、290、292、294、296、298、300、302、304、306、308、310、312、314、316、318、320、322、または324のいずれか一つに示される核酸配列の相補鎖と、ストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列を含む、ヒトCD3に対して特異的に結合して、T細胞エピトープの生成傾向が低下している第一のドメインと、EpCAMに対する/結合することができるIg由来の第二のドメインとを含むCD3特異的結合構築物を提供する。
【0083】
「ハイブリダイズする」という用語は、本明細書中で使用される場合、本明細書中で定義される脱免疫構築物またはその部分をコードするポリヌクレオチドにハイブリダイズすることが可能であるポリヌクレオチド/核酸配列をいう。従って、このポリヌクレオチドは、それぞれ、RNAまたはDNAの調製物のノーザンブロット分析もしくはサザンブロット分析におけるプローブとして有用であり得、またはそれらのそれぞれのサイズに依存してPCR分析におけるオリゴヌクレオチドプライマーとして使用され得る。好ましくは、このハイブリダイズするポリヌクレオチドは、少なくとも10個、より好ましくは少なくとも15個の長さのヌクレオチドを含むのに対して、プローブとして使用されるための本発明のハイブリダイズするポリヌクレオチドは、少なくとも100個、より好ましくは少なくとも200個の、または最も好ましくは少なくとも500個の長さのヌクレオチドを含む。
【0084】
いかにして核酸分子を用いてハイブリダイゼーション実験を実行するかは周知であり、すなわち、当業者は、本発明に従って彼らがどのハイブリダイゼーション条件を使用しなくてはならないかを知っている。このようなハイブリダイゼーション条件は、Molecular Cloning A Laboratory Manual, Cold Spring Harbor Laboratory (2001) N.Y.などの標準的な教科書において言及されている。本発明に従って好ましいものは、ストリンジェントなハイブリダイゼーション条件下で、本発明のポリヌクレオチドまたはその部分にハイブリダイズすることが可能であるポリヌクレオチドである。
【0085】
「ストリンジェントなハイブリダイゼーション条件」とは、例えば、50%ホルムアミド、5×SSC(750mM NaCl、75mM クエン酸ナトリウム)、50mM リン酸ナトリウム(pH 7.6)、5×デンハルト溶液、10%硫酸デキストラン、および20μg/ml変性剪断サケ精子DNAを含む溶液中での42℃、一晩のインキュベーション、続いて0.1×SSCにおける約65℃でのフィルターの洗浄をいう。よりストリンジェンシーの低いハイブリダイゼーション条件で本発明のポリヌクレオチドにハイブリダイズする核酸分子もまた意図される。ハイブリダイゼーションのストリンジェンシーおよびシグナル検出の変化は、主として、ホルムアミド濃度(より低いホルムアミドのパーセンテージはより低いストリンジェンシーを生じる);塩濃度、または温度の操作を通して達成される。例えば、よりストリンジェンシーの低い条件は、6×SSPE(20×SSPE=3M NaCl;0.2M NaH2PO4;0.02M EDTA, pH 7.4)、0.5% SDS、30%ホルムアミド、100μg/mlサケ精子ブロッキングDNAを含む溶液中での37℃での一晩のインキュベーション;引き続く50℃での1×SSPE、0.1% SDSを用いる洗浄を含む。さらに、なおより低いストリンジェンシーを達成するために、ストリンジェントなハイブリダイゼーション後に実行される洗浄が、より高い塩濃度(例えば、5×SSC)で行われ得る。上記の条件におけるバリエーションが、ハイブリダイゼーション実験におけるバックグラウンドを抑制するために使用される代替的なブロッキング試薬を含めることおよび/またはそれで置換することを通して達成され得ることに注目されたい。代表的なブロッキング試薬には、デンハルト試薬、BLOTTO、ヘパリン、変性サケ精子DNA、および市販の独自製品の処方物が含まれる。特異的ブロッキング試薬を含めることは、適合性に伴う問題に起因して、上記のハイブリダイゼーション条件の修飾を必要とする可能性がある。
【0086】
列挙される核酸分子は、例えば、DNA、cDNA、RNA、または合成的に製造されるDNAもしくはRNA、または単独でもしくは組み合わせてこれらのポリヌクレオチドのいずれかを含む組換え的に製造されるキメラ核酸分子であり得る。
【0087】
本発明において提供された脱免疫CD3およびEpCAM結合構築物は、医学的状況において、例えば腫瘍様疾患、特に乳癌、結腸癌、前立腺癌、頭頚部癌、皮膚癌(黒色腫)、尿性器癌、例えば卵巣癌、子宮内膜癌、子宮頚癌および腎臓癌、肺癌、胃癌、小腸癌、肝臓癌、膵臓癌、胆嚢癌、胆管癌、食道癌、唾液腺癌、および甲状腺癌の予防、治療および/または改善において特に有用である。特に、CD3およびEpCAMに結合する脱免疫構築物は、上皮癌、好ましくは腺癌、または最小の残留疾患、より好ましくは初期固形腫瘍、進行固形腫瘍、または転移性固形腫瘍の治療に用いることができる。
【0088】
本明細書に記述のCD3特異的結合構築物のより特に好ましい態様において、該構築物は、CCR5に対して特異性を有する抗原-相互作用部位を含む第二のIg由来ドメインを含む。
【0089】
ケモカイン受容体CCR5は、前炎症性ケモカインRANTES、MIP1-α、MIP1-β、およびMCP-2に結合する大きいGタンパク質共役7回膜貫通ドメイン受容体ファミリーに属する。ケモカインは、接着分子と共に白血球の浸潤を誘導するように、および組織損傷部位へのその誘導を指示するように作用する。CCR5は、少数のT細胞および単球において発現され、さらに、HIV感染症の過程において初期を支配するM-指向性HIV-1株の主要な共受容体である。
【0090】
ヒト免疫不全ウイルス(HIV)は、細胞表面上の二つの重要な分子であるCD4および共受容体に最初に結合して初めてヒト細胞内に入ることができる。最初に認識される共受容体は、CCR5であり、ウイルスのライフサイクルの後期ではもう一つのケモカイン受容体であるCXCR4はHIV-1の共受容体となる(D'Souza, Nature Med. 2, 1293(1996);Premack, Nature Med. 2, 1174;Fauci, Nature 384, 529(1996))。性的接触によってウイルスのほとんどの伝幡を引き起こすHIV-1株は、M-指向性ウイルスと呼ばれる。これらのHIV-1株(非合胞体誘導(NSI)プライマリウイルスとしても知られる)は、初代培養CD4+ T細胞およびマクロファージにおいて複製することができ、ケモカイン受容体CCR5(および頻度は少ないがCCR3)をその共受容体として用いる。T-指向性ウイルス(時に、合胞体誘導(SI)プライマリウイルスと呼ばれる)はまた、初代培養CD4+ T細胞においても複製できるが、さらにインビトロで確立されたCD4+ T細胞株に感染することができ、それらはケモカイン受容体CXCR4(融合体)を通して行われる。これらのT-指向性株の多くは、CXCR4の他にCCR5を用いることができ、中には、少なくとも特定のインビトロ条件においてCCR5を通してマクロファージに入ることができるものもある(D'Souza, Nature Med. 2, 1293(1996);Premack, Nature Med 2, 1174;Fauci, Nature 384, 529(1996))。
【0091】
他の共受容体がHIV-1の発病に関与しているか否かは解明されていないが、インビトロ試験から、いくつかのT-指向性株に関して他の共受容体の存在が推測されうる。M-指向性HIV-1株は、HIVの性的伝幡の約90%に関係していることから、CCR5は、患者におけるウイルスの主要な共受容体である;CXCR4を用いる(T-指向性)株の伝幡(または全身性の確立)はまれである(D'Souza, Nature Med. 2, 1293(1996);Premack, Nature Med. 2, 1174;Fauci, Nature 384, 529(1996);Paxton, Nature Med. 2, 412(1996);Liu, Cell 86, 367(1996);Samson, Nature 382, 722(1996);Dean, Science 273, 1856(1996);Huang, Nature Med. 2, 1240(1996))。しかし、SIウイルスがインビボで進化すると(またはそれらが伝幡されると)、それらは特にビルレントとなり、より急速な疾患の進行を引き起こす(D'Souza, Nature Med. 2, 1293(1996);Premack, Nature Med 2, 1174;Fauci, Nature 384, 529(1996);Schuitemaker, J. Virol. 66, 1354(1992);Connor, J. Virol. 67, 1772(1993);Richman, J. Infect. Dis. 169, 968(1994);R.I. Connorら、J. Exp. Med. 185, 621(1997);Trkola, Nature 384, 184(1996))。
【0092】
標的細胞上の共受容体分子の数および同一性、ならびにHIV-1株が異なる共受容体を通して細胞に入る能力は、疾患の進行に関する重要な決定因子であるように思われる。これらの要因は、HIV-1感染症の宿主およびウイルス依存的局面の双方に対する主要な影響である。例えば、CCR5におけるホモ接合欠損体(デルタ32)は、インビボおよびインビトロでHIV-1感染症に対する抵抗性と強く相関する。欠損CCR5対立遺伝子に関してヘテロ接合である人は、感染症に対する保護はせいぜい弱く、疾患の進行にごくわずかな遅れを示すに過ぎない(Paxton, Nature Med. 2, 412(1996);Liu, Cell 86, 367(1996);Samson, Nature 382, 722(1996);Dean, Science 273, 1856(1996);Huang, et al., Nature Med. 2, 1240(1996))。しかし、他の要因が活性化CD4+ T細胞上のCCR5発現レベルに影響を及ぼして、それによってインビトロでHIV-1感染症の効率に影響を及ぼしうる(Trkola, Nature 384, 184(1996);Bleul, Proc. Natl. Acad. Sci. USA 94, 1925(1997))。
【0093】
多発性硬化症の場合、CCR5およびCXCR3は、脱髄性脳病変に浸潤するT細胞のみならず、罹患患者の末梢血において主に発現されることが示されている。T細胞の消失は、この自己免疫疾患のT-細胞部分を遮断するであろう。
【0094】
CCR3およびCCR5の高度発現はまた、ホジキン疾患を有する患者に由来するリンパ節のT細胞およびB細胞においても認められた。
【0095】
I型糖尿病は、T細胞媒介自己免疫疾患であると考えられる。膵臓におけるCCR5受容体の発現は、関連する動物モデルにおいてI型糖尿病の進行に関連した(Cameron(2000), J. Immunol. 165, 1102〜1110)。特に、CCR5発現は、インスリン症および自然発生型I型糖尿病の発症に関連した。
【0096】
(ヒト)CCR5に特異的に結合するいくつかの抗体が当技術分野で公知であり、MC-1(Mack(1998), J. Exp. Med. 187, 1215〜1224)またはMC-5(Blanpain(2002)Mol. Biol. Cell. 13:723〜37;Segerer(1999), Kidney Int. 56:52〜64;Kraft(2001)J. Biol. Chem. 14:276:34408〜18)を含む。CCR-5抗体、特にMC-1およびMC-5は、本発明のCD3特異的構築物のIg由来第二のドメイン源として役立つ可能性がある。従って、好ましい態様において、本発明は、第一のドメインがヒトCD3に対する特異性を提供して、T細胞エピトープの生成傾向の低下を有し、該Ig由来第二のドメインが(ヒト)CCR5に対して特異的な抗体に由来する、少なくとも二つのドメインを含む二重特異性構築物に関する。最も好ましくは、そのような構築物は本明細書において定義されるように一本鎖scFvである。
【0097】
MC-1は、ヒトCCR5の第二の細胞外ループの第一の部分に特異的に結合して、添付の実施例に示すようにアカゲザルに由来するCCR5と交叉反応しないことが示された。したがって、本発明のCD3特異的構築物は、例えばCCR5、好ましくはヒトCCR5に対して特異的な抗体のVLおよびVHドメイン(すなわち、Ig-由来第二のドメイン)、ならびにCD3抗原に対して特異的な抗体のVHおよびVLドメインを含むことが好ましい。ヒトCCR5に対して特異的な該抗体は、とりわけMack(1998), J. Exp. Med. 187, 1215〜1224および添付の実施例に記述されているマウス抗ヒトCCR5抗体MC-1である。なおも、MC-5と同様に他の抗CCR5抗体(添付の実施例に特徴が示されているように、およびBlanpain(2002)Mol. Biol. Cell. 13:723〜37;Segerer(1999)Kidney Int. 56:52〜64およびKraft(2001)J. Biol. Chem. 14:276:34408〜18において開示されるように)を、本発明の状況において用いてもよい。
【0098】
本発明の特に好ましい態様において、ヒトCD3に対する/相互作用する脱免疫ドメインと、CCR5に特異的に結合/相互作用する第二のIg-由来ドメインとを含むCD3特異的結合構築物が提供される。そのような構築物を、表6Aおよび6Bに示す。表6Aおよび6BにおけるモジュールA-Gは表1〜5に関して先に述べたように定義することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VHドメインは、配列番号:74または76に記載される配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VLドメインは、配列番号:78、80、または82に記載される配列から選択することができる。ヒトCCR5抗体のVHタンパク質ドメインは、配列番号:129に記載される。ヒトCCR5抗体のVLタンパク質ドメインは、配列番号:131に記載される。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体からの脱免疫VH/VLまたはVL/VHタンパク質ドメインの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:3に記載されるアミノ酸配列を有する。モジュール対A/CまたはE/GのいずれかがEpCAM抗原に対して特異性を有する抗体からのVH/VLまたはVL/VHタンパク質ドメインの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:168に記載のアミノ酸配列を有する。タンパク質モジュールA-B-CおよびE-F-Gのそれぞれの群は、配列番号:174に記載の配列を有するタンパク質モジュールDを通して互いに結合する。しかし、先に述べたように、クローニング目的のためにVLおよびその後のVドメインのあいだにさらなるセリンを導入してもよい(配列番号:176に記載のリンカー)。
【0099】
ヒトCD3抗原に対して特異性を有する抗体の脱免疫VHドメインをコードする核酸分子は、配列番号:73または75に記載の配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VLドメインをコードする核酸分子は、配列番号:77、79、または81に記載の配列から選択することができる。ヒトCCR5抗体のVHタンパク質ドメインをコードする核酸分子は、配列番号:128に記載される。ヒトCCR5抗体のVLタンパク質ドメインをコードする核酸分子は、配列番号:130に記載される。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体からの脱免疫VH/VLまたはVL/VHタンパク質ドメインの対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:202に記載される核酸配列を有する。モジュール対A/CまたはE/GのいずれかがCCR5抗原に対して特異性を有する抗体からのVH/VLまたはVL/VHの対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:201に記載の核酸配列を有する。核酸モジュールA-B-CおよびE-F-Gの群は、配列番号:173に記載の配列を有するタンパク質モジュールDを通して互いに結合する。またはVLとその後のVドメイン(クローニング目的のためにセリン残基をコードするさらなるコドンを含む)とを結合するためにもう一つのリンカー配列番号:175を用いてもよい。
【0100】
(表6A)一本鎖抗CCR5可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列
【0101】
(表6B)一本鎖抗CCR5可変領域を含む脱免疫抗ヒトCD3構築物:核酸配列
構築物 構築物部分の配列番号 脱免疫された抗CD3構築物/特異性
ドメインの配置
【0102】
好ましくは、該構築物は、以下からなる群より選択されるアミノ酸配列を含む:
(a)配列番号:206、208、210、212、214、または216のいずれか一つに示されるアミノ酸配列;
(b)配列番号:205、207、209、211、213、または215のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列;および
(d)(b)において定義された核酸配列の相補鎖とストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【0103】
CCR5およびCD3結合構築物である配列番号:206、208、210は、構築物5を表し、配列番号:212、214、および216は表6の構築物13を表し、異なる三つのVL領域(VL1(配列番号:78)、VL2(配列番号:80)、またはVL3(配列番号:82))を有する。
【0104】
本発明はまた、本明細書において先の(b)において定義した核酸配列、すなわち、配列番号:205、207、209、211、213、または215のいずれか一つに示される核酸配列の相補鎖と、ストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列を含む、ヒトCD3に特異的に結合して、T細胞エピトープの生成傾向が低下した第一のドメインと、CCR5に対する/に結合することができるIg由来第二のドメインとを含むCD3特異的結合構築物を提供する。「ハイブリダイゼーション」および「ストリンジェントな条件」という用語は、本明細書において先に記述されている。対応する定義および態様は、必要な変更を加えて本明細書において適用される。
【0105】
本明細書において提供される脱免疫CD3およびCCR5結合構築物は、ウイルス病、特にHIV感染症およびAIDS、またはリウマチ性関節炎のような自己免疫疾患および/または炎症疾患の医学的介入において特に有用である。
【0106】
もう一つの態様において、本発明は、本発明の構築物のIg由来第二のドメインがCD19に対して特異性を有する抗原相互作用部位を含む、本明細書において先に定義したCD3特異的結合構築物を提供する。
【0107】
CD19は、非常に有用な医学的標的であることが判明した。CD19は、プロB細胞から成熟B細胞へのB細胞系列全体において発現され、細胞表面から消失しない場合、全てのリンパ腫細胞において均一に発現され、幹細胞には存在しない(Haagen, Clin. Exp. Immunol. 90(1992), 368〜75;Uckun, Proc. Natl. Acad. Sci. USA 85(1988), 8603〜7)。CD19に対する抗体とさらなる免疫調節抗体の双方を用いる併用治療が、B細胞の悪性疾患(国際公開公報第02/04021号、US2002006404、US2002028178)および自己免疫疾患(国際公開公報第02/22212号、US2002058029)の治療のために開示されている。国際公開公報第00/67795号は、無痛性の劇症型B-細胞リンパ腫の治療のためと共に急性および慢性型リンパ球性白血病の治療のために、CD19に対する抗体を用いることを開示している。国際公開公報第02/80987号は、B細胞非ホジキンリンパ腫、ホジキンリンパ腫、またはB細胞白血病(例えば、B細胞急性リンパ球性白血病(B-ALL)(例えば、ヘアリーセルリンパ腫)B細胞前駆体急性リンパ球性白血病(プレB-ALL)、B細胞慢性リンパ球性白血病(B-CLL))のような疾患の治療のために抗原CD19に対する抗体に基づいて免疫毒素を治療的に用いることを開示する。
【0108】
本発明の特に好ましい態様において、ヒトCD3に対する/結合する/相互作用する脱免疫ドメインと、CD19に特異的に結合する/相互作用する第二のIg由来ドメインとを含むCD3特異的結合構築物が提供される。そのような構築物を表7Aおよび7Bに示す。表7Aおよび7BにおけるモジュールA-Gは、先の表1〜5に記載されたように定義することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VHドメインは、配列番号:74または76に記載の配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VLドメインは、配列番号:78、80、または82に記載の配列から選択することができる。ヒトCD19抗体のVHタンパク質ドメインは、配列番号:114に記載される。ヒトCCR5抗体のVLタンパク質ドメインは、配列番号:116に記載される。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体からの脱免疫VH/VLまたはVL/VHタンパク質ドメインの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:3に記載されるアミノ酸配列を有する。モジュール対A/CまたはE/GのいずれかがCD19抗原に対して特異性を有する抗体からのVH/VLまたはVL/VHの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:168に記載のアミノ酸配列を有する。タンパク質モジュールA-B-CおよびE-F-Gのそれぞれの群は、配列番号:174に記載の配列を有するタンパク質モジュールDを通して互いに結合する。しかし、先に述べたように、クローニング目的のために(配列番号:176に記載のリンカー)VLとその後のVドメインとのあいだにさらなるセリンを導入してもよい。
【0109】
ヒトCD3抗原に対して特異性を有する抗体の脱免疫VHドメインをコードする核酸分子は、配列番号:73または75に記載の配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VLドメインをコードする核酸分子は、配列番号:77、79、または81に記載の配列から選択することができる。ヒトCD19抗体のVHタンパク質ドメインをコードする核酸分子は、配列番号:113に記載される。ヒトCCR5抗体のVLタンパク質ドメインをコードする核酸分子は、配列番号:115に記載される。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体からの脱免疫VH/VLまたはVL/VHタンパク質ドメインの対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:202に記載されるアミノ酸配列を有する。モジュール対A/CまたはE/GのいずれかがCD19抗原に対して特異性を有する抗体からのVH/VLまたはVL/VHの対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:201に記載の核酸配列を有する。核酸モジュールA-B-CおよびE-F-Gの群は、配列番号:173に記載の配列を有するタンパク質モジュールDを通して互いに結合する。VLとその後のVドメイン(クローニング目的のためにセリン残基をコードするさらなるコドンを含む)とを結合するために,もう一つのリンカー配列番号:175を用いてもよい。
【0110】
(表7A)一本鎖抗CD19可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列
【0111】
(表7B)一本鎖抗CD19可変領域を含む脱免疫抗ヒトCD3構築物:核酸配列
【0112】
より好ましい態様において、本発明は、上記のCD3結合ドメインと、CD19、好ましくはヒトCD19に特異的に結合する/相互作用する第二のIg由来ドメインとを含み、以下からなる群より選択されるアミノ酸配列を含む、脱免疫CD3-特異的結合構築物を提供する:
(a)配列番号:190、192、194、196、198、200、327、329、331、333、335、337、339、341、343、345、347、349、351、353、357、359、361、363、365、367、369、371、373、375、377、379、381、383、385、387、389、391、393、395、397、399、401、403、405、407、または409のいずれか一つに示されるアミノ酸配列;
(b)配列番号:189、191、193、195、197、199、326、328、330、332、334、336、338、340、342、344、346、348、350、352、354、356、358、360、362、364、366、368、370、372、374、376、378、380、382、384、386、388、390、392、394、396、398、400、402、404、406、または408のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;ならびに
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列;
(d)ストリンジェントなハイブリダイゼーション条件で(b)において定義された核酸配列の相補鎖とハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【0113】
本発明に従う好ましいCD19およびCD3結合構築物は、構築物5を表す配列番号:190、192、194であり、および表7の構築物13を表し、異なる三つのVL領域(VL1(配列番号:78)、VL2(配列番号:80)、またはVL3(配列番号:82))を有する配列番号:196、198、および200である。
【0114】
本発明はまた、上記の(b)において定義された核酸配列、すなわち配列番号:189、191、193、195、197、199、326、328、330、332、334、336、338、340、342、344、346、348、350、352、354、356、358、360、362、364、366、368、370、372、374、376、378、380、382、384、386、388、390、392、394、396、398、400、402、404、406、または408のいずれか一つに示される核酸配列の相補鎖とハイブリダイズする核酸配列によってコードされるアミノ酸配列を含む、ヒトCD3に対して特異的に結合し、T細胞エピトープの生成傾向が低下した第一のドメインと、CD19に対する/に結合することができるIg由来第二のドメインとを含むCD3特異的結合構築物を提供する。「ハイブリダイゼーション」および「ストリンジェントな条件」という用語は、本明細書において先に記述されている。対応する定義および態様は、必要な変更を加えて本明細書において適用される。
【0115】
本明細書に開示の脱免疫CD3およびCD19結合構築物は、増殖疾患、腫瘍様疾患、炎症疾患、免疫障害、自己免疫疾患、感染症、ウイルス病、アレルギー反応、寄生虫反応、移植片対宿主病、宿主対移植片病、またはB-細胞悪性疾患、特にB-細胞非ホジキンリンパ腫、ホジキンリンパ腫、またはB細胞白血病(例えば、B細胞急性リンパ球性白血病(B-ALL)(例えば、ヘアリーセルリンパ腫)B細胞前駆体急性リンパ球性白血病(プレB-ALL)、B細胞慢性リンパ球性白血病(B-CLL))の予防、治療、または改善において特に有用である。
【0116】
さらなる態様において、本発明は、ヒトCD3に対して特異的に結合して、T細胞エピトープの生成傾向が低下した第一のドメインと、Ig由来であってCD20に対する特異性を有する抗原相互作用部位を含む第二のドメインとを含む、先に定義されたCD3特異的結合構築物に関する。
【0117】
CD20は、B-リンパ球上に存在する細胞表面タンパク質の一つである。CD20抗原は、B-細胞非ホジキンリンパ腫(NHL)の90%以上におけるリンパ球を含む、正常および悪性プレBおよび成熟Bリンパ球において認められる。抗原は造血幹細胞、活性化Bリンパ球(プラズマ細胞)、および正常組織には存在しない。ほとんどがマウス起源であるいくつかの抗体が記述されている:1F5(Pressら、1987, Blood 69/2, 584〜591)、2B8/C2B8、2H7、1H4(Liuら、1987, J. Immunol. 139, 3521〜3526;Andersonら、1998, 米国特許第5,736,137号;Haismaら、1998, Blood 92, 184〜190;Shanら、1999, J. Immunol. 162, 6589〜6595)。
【0118】
CD20は、担体タンパク質に結合したscFvをコードするDNAのワクチン接種を用いるプラズマ細胞悪性疾患の治療のための免疫治療戦略において(Treonら、2000, Semin Oncol 27(5), 598)記述されており、CD20抗体(IDEC-C2B8)を用いる免疫療法において、非ホジキンB-細胞リンパ腫の治療において有効であることが示されている。CD20抗体は、非ホジキンリンパ腫において有効性および認容性が証明されており、これまで無処置または再発/不応性の無痛性非ホジキンリンパ腫においてそれぞれ、反応率は73%および48%であった(Montserrat, 2003, Semin Oncol. 30(1suppl2), 34〜39)。さらに、CD20抗体は、再発または進行期のB-細胞新生物を治療するために広く用いられており、有効率は約50%である。
【0119】
本発明の特に好ましい態様において、ヒトCD3に対する/と相互作用する脱免疫ドメインと、CD20に特異的に結合する/相互作用する第二のIg由来ドメインとを含むCD3-特異的結合構築物が提供される。そのような構築物を、表8Aおよび8Bに示す。表8Aおよび8BにおけるモジュールA-Gは、表1〜5に関して先に述べた通りに定義することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VHドメインは、配列番号:74または76に記載の配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VLドメインは、配列番号:78、80、または82に記載の配列から選択することができる。ヒトCD20抗体のVHタンパク質ドメインは、配列番号:170に記載される。ヒトCD20抗体のVLタンパク質ドメインは、配列番号:172に記載される。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体からの脱免疫VH/VLまたはVL/VHタンパク質ドメインの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:3に記載されるアミノ酸配列を有する。モジュール対A/CまたはE/GのいずれかがCD20抗原に対して特異性を有する抗体からのVH/VLまたはVL/VHの対である場合、タンパク質モジュールBまたはFはそれぞれ、配列番号:168に記載のアミノ酸配列を有する。タンパク質モジュールA-B-CおよびE-F-Gのそれぞれの群は、配列番号:174に記載の配列を有するタンパク質モジュールDを通して互いに結合する。しかし、先に述べたように、VLとその後のVドメインとのあいだにさらなるセリンをクローニング目的のために(配列番号:176に記載のリンカー)導入してもよい。
【0120】
ヒトCD3抗原に対して特異性を有する抗体の脱免疫VHドメインをコードする核酸分子は、配列番号:73または75に記載の配列から選択することができる。ヒトCD3抗原に対して特異性を有する抗体の脱免疫VLドメインをコードする核酸分子は、配列番号:77、79、または81に記載の配列から選択することができる。ヒトCD20抗体のVHタンパク質ドメインをコードする核酸分子を、配列番号:169に記載する。ヒトCD20抗体のVLタンパク質ドメインをコードする核酸分子を、配列番号:171に記載する。モジュール対A/CまたはE/Gのいずれかが、ヒトCD3抗原に対して特異性を有する抗体からの脱免疫VH/VLまたはVL/VHタンパク質ドメインの対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:202に記載されるアミノ酸配列を有する。モジュール対A/CまたはE/GのいずれかがCD20抗原に対して特異性を有する抗体からのVH/VLまたはVL/VHの対をコードする核酸を指す場合、核酸モジュールBまたはFはそれぞれ、配列番号:201に記載の核酸配列を有する。核酸モジュールA-B-CおよびE-F-Gの群は、配列番号:173に記載の配列を有するタンパク質モジュールDを通して互いに結合する。VLとその後のVドメインとを結合するためにもう一つのリンカー配列番号:175を用いてもよい(クローニング目的のためにセリン残基をコードするさらなるコドンを含む)。
【0121】
(表8A)一本鎖抗CD20可変領域を含む脱免疫抗ヒトCD3構築物:アミノ酸配列
【0122】
(表8B)一本鎖抗CD20可変領域を含む脱免疫抗ヒトCD3構築物:ヌクレオチド配列
【0123】
より好ましくは、本発明の脱免疫CD3およびCD20結合構築物は、以下からなる群より選択されるアミノ酸配列を含む:
(a)配列番号:218、220、222、224、226、または228のいずれか一つに示されるアミノ酸配列;
(b)配列番号:217、219、221、223、225、または227のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列;および
(d)(b)において定義された核酸配列の相補鎖とストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【0124】
本発明はまた、本明細書において先に定義した核酸配列、すなわち配列番号:217、219、221、223、225、または227のいずれか一つに示される核酸配列の相補鎖とストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列を含む、ヒトCD3に対して特異的に結合し、T細胞エピトープの生成傾向が低下した第一のドメインと、CD20に対する/に結合することができるIg由来第二のドメインとを含むCD3特異的結合構築物を提供する。「ハイブリダイゼーション」および「ストリンジェントな条件」という用語は、本明細書において先に記述されている。対応する定義および態様は、必要に応じて本明細書において適用される。
【0125】
本明細書に記述の脱免疫CD3およびCD20結合構築物は、B-細胞関連障害の治療、予防、および/または改善において、好ましくはリンパ腫の医学的介入において、より好ましくは非ホジキンリンパ腫の治療において用いられると想像される。
【0126】
本発明はまた、本発明のCD3特異的結合分子をコードする核酸配列を提供する。
【0127】
本発明の核酸分子に、調節配列が加えられ得ることは当業者には明らかである。例えば、プロモーター、転写エンハンサー、および/または本発明のポリヌクレオチドの発現の誘導を可能にする配列が利用され得る。適切な誘導系は、例えば、例えばGossen および Bujard (Proc. Natl. Acad. Sci. USA 89 (1992, 5547-5551))およびGossen et al. (Trends Biotech. 12 (1994), 58-62)によって記載されるようなテトラサイクリン調節される遺伝子発現、または、例えばCrook (1989) EMBO J. 8, 513-519によって記載されるようなデキサメタゾン誘導性遺伝子発現系である。
【0128】
また、さらなる目的のために、核酸分子が、例えば、チオエステル結合および/またはヌクレオチドアナログを含み得ることが想定される。この修飾は、細胞中のエンドヌクレアーゼおよび/またはエキソヌクレアーゼに対する核酸分子の安定化のために有用であり得る。この核酸分子は、細胞中でこの核酸分子の転写を可能にするキメラ遺伝子を含む適切なベクターによって転写され得る。この点に関して、このようなポリヌクレオチドが「遺伝子標的化」または「遺伝子治療的」アプローチのために使用され得ることもまた理解される。別の態様において、この核酸分子は標識される。核酸の検出のための方法は当技術分野で周知であり、例えば、サザンブロッティングおよびノーザンブロッティング、PCR、またはプライマー伸長がある。この態様は、遺伝子治療アプローチの間に上記の核酸分子の首尾よい導入を確認するためののスクリーニング方法のために有用であり得る。
【0129】
この核酸分子は、上述の核酸分子のいずれかを単独でまたは組み合わせてのいずれかで含む組換え的に産生されたキメラ核酸分子であり得る。好ましくは、この核酸分子はベクターの一部である。
【0130】
従って、本発明はまた、本発明に記載される核酸分子を含むベクターに関する。
【0131】
多くの適切なベクターが分子生物学における当業者に公知であり、その選択は所望される機能に依存し、これにはプラスミド、コスミド、ウイルス、バクテリオファージ、および遺伝子操作において慣用的に使用される他のベクターが含まれる。種々のプラスミドおよびベクターを構築するために、当業者に周知である方法(例えば、Sambrookら(前掲)およびAusubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.Y. (1989), (1994)を参照されたい)が使用され得る。または、本発明のポリヌクレオチドおよびベクターは、標的細胞への送達のためにリポソームに再構成され得る。以下でさらに詳細に議論されるように、クローニングベクターは、個別のDNAの配列を単離するために使用された。関連する配列は、特定のポリペプチドの発現が必要とされる発現ベクターに移動され得る。代表的なクローニングベクターには、pBluescript SK、pGEM、pUC9、pBR322、およびpGBT9が含まれる。代表的な発現ベクターには、pTRE、pCAL-n-EK、pESP-1、pOP13CATが含まれる。
【0132】
好ましくは、このベクターは、本明細書中に定義された二重特異性単鎖抗体構築物をコードするこの核酸配列に作動可能に連結された調節配列である核酸配列を含む。
【0133】
このような調節配列(制御エレメント)は当業者に公知であり、プロモーター、スプライシングカセット、翻訳開始コドン、ベクターに挿入を導入するための翻訳および挿入部位を含み得る。好ましくは、この核酸分子は、真核生物細胞または原核生物細胞における発現を可能にする、その発現制御配列に作動可能に連結される。
【0134】
このベクターは、本明細書中に定義される二重特異性単鎖抗体構築物をコードする核酸分子を含む発現ベクターであることが想定される。
【0135】
「調節配列」という用語は、それらがライゲーションされるコード配列の発現をもたらすために必要であるDNA配列をいう。このような制御配列の性質は、宿主生物に依存して異なる。原核生物においては、制御配列には、一般的に、プロモーター、リボソーム結合部位、およびターミネーターが含まれる。真核生物においては、一般的に、制御配列は、プロモーター、ターミネーター、およびある場合においては、エンハンサー、トランス活性化因子、または転写因子が含まれる。「制御配列」という用語は、最小限、その存在が発現のために必要である全ての成分を含むことが意図され、さらなる有利な成分もまた含み得る。
【0136】
「作動可能に連結される」という用語は、そのように記載される成分が、それらの意図される様式で機能することを可能にする関連性にある並列をいう。コード配列に「作動可能に連結される」制御配列は、コード配列の発現が制御配列と適合可能な条件下で達成されるようなやり方でライゲーションされる。制御配列がプロモーターである場合、当業者には、二本鎖核酸が好ましく使用されることが明白である。
【0137】
従って、列挙されるベクターは好ましくは発現ベクターである。「発現ベクター」は、選択された宿主を形質転換するために使用され得る構築物であり、選択された宿主中でコード配列の発現を提供する。発現ベクターは、例えば、クローニングベクター、バイナリーベクターまたは組み込みベクターであり得る。発現は、好ましくは翻訳可能なmRNAへの核酸分子の転写を含む。原核生物および/または真核生物細胞における発現を確実にする調節エレメントは当業者に周知である。真核生物細胞の場合、これらは通常、転写の開始を確実にするプロモーターを含み、ならびに任意に、転写の終結および転写の安定化を確実にするポリAシグナルを含む。原核生物宿主細胞において発現を可能にする可能性のある調節エレメントは、例えば、大腸菌(E. coli)におけるPL、lac、trp、またはtacプロモーターを含み、真核生物宿主細胞中での発現を可能にする調節エレメントの例は、酵母におけるAOX1もしくはGAL1プロモーター、CMVプロモーター、SV40プロモーター、RSVプロモーター(ラウス肉腫ウイルス)、CMVエンハンサー、SV40エンハンサー、または哺乳動物および他の動物細胞におけるグロビンイントロンである。
【0138】
転写の開始の原因であるエレメントに加えて、このような調節エレメントはまた、ポリヌクレオチドの下流のSV40-ポリA部位またはtk-ポリA部位などの転写終結シグナルを含み得る。さらに、使用される発現系に依存して、細胞区画にポリペプチドを方向付けるか、または培地にポリペプチドを分泌することが可能であるリーダー配列が、列挙される核酸配列のコード配列に加えられてもよく、これらは当技術分野で周知である;例えば、添付の実施例1も参照されたい。リーダー配列は、適切な段階で、翻訳配列、開始配列、および終結配列と、好ましくは、翻訳されたタンパク質またはその部分の分泌を方向付けることが可能であるリーダー配列と、細胞膜周辺腔または細胞外媒体でアセンブルされる。任意に、異種配列が、所望の性質(例えば、発現された組換え産物の安定化または精製の単純化;前記を参照されたい)を付与するN末端同定ペプチドを含む融合タンパク質をコードし得る。この文脈において、適切な発現ベクターは当技術分野で公知であり、例えば、岡山-バーグcDNA発現ベクターpcDV1(Pharmacia)、pCDM8、pRc/CMV、pcDNA1、pcDNA3(In-vitrogene)、pEF-DHFR、pEF-ADA、またはpEF-neo(Raum et al. Cancer Immunol Immunother (2001) 50(3), 141-150)、またはpSPORT1(GIBCO BRL)である。
【0139】
好ましくは、発現制御配列は、真核生物宿主細胞を形質転換またはトランスフェクトすることが可能であるベクターにおける真核生物プロモーター系であるが、しかし、原核生物宿主のための制御配列もまた使用され得る。一旦、ベクターが適切な宿主に取り込まれると、その宿主は、ヌクレオチド配列の高レベル発現のために適切な条件下で維持され、所望される場合、本発明のポリペプチドの収集および精製が続いて行われ得る;例えば、添付の実施例を参照されたい。
【0140】
使用され得る代替的な発現系は昆虫系である。1つのこのような系において、Autographa californica核多核体病ウイルス(AcNPV)が、Spodoptera frugiperda細胞またはTrichoplusia larvaeにおいて外来性の遺伝子を発現するためのベクターとして使用される。列挙される核酸分子のコード配列は、ポリヘドリン遺伝子などのウイルスの必須でない領域にクローニングされ得、ポリヘドリンプロモーターの制御下に配置され得る。このコード配列の首尾よい挿入はポリヘドリン遺伝子を不活性にし、コートタンパク質の被覆を欠く組換えウイルスを産生する。次いで、組換えウイルスは、本発明のタンパク質が発現されるS. frugiperda細胞またはTrichoplusia larvaeに感染させるために使用される(Smith, j. Virol 46 (1983), 584; Engelhard, Proc. Nat. Acad. Sci. USA 91 (1994), 3224-3227)。
【0141】
さらなる調節エレメントは、転写エンハンサーならびに翻訳エンハンサーを含み得る。有利には、本発明の上記のベクターは、選択可能なおよび/またはスコア付け可能なマーカーを含む。
【0142】
形質転換された細胞、例えば、植物組織および植物細胞の選択のために有用である選択可能なマーカー遺伝子は当業者に周知であり、例えば、dhfr(これはメトレキサートに対する耐性を付与する(Reiss, Plant Physiol. (Life Sci. Adv.)13 (1994), 143-149));npt(これはアミノグリコシドネオマイシン、カナマイシン、およびパロマイシンに対する耐性を付与する(Herrera-Estrella, EMBO J. 2 (1983), 987-995))およびhygro(これはハイグロマイシンに対する耐性を付与する(Marsh, Gene 32 (1984), 481-485))についての選択の基礎として代謝拮抗物質耐性を含む。さらなる選択可能な遺伝子が記載されており、すなわち、以下が記載されている。trpB(これは細胞がトリプトファンの代わりにインドールを利用することを可能にする);hisD(これは細胞がヒスチジンの代わりにヒスチノールを利用することを可能にする)(Hartman, Proc. Natl. Acad. Sci. USA 85 (1988), 8047);マンノース-6-リン酸イソメラーゼ(これは細胞がマンノースを利用することを可能にする)(WO 94/20627)およびODC(オルニチンデカルボキシラーゼ)(これはオルニチンデカルボキシラーゼインヒビター、2-(ジフルオロメチル)-DL-オルニチン、DFMOに対する耐性を付与する)(McConlogue, 1987, Current Communications in Molecular Biology, Cold Spring Harbor Laboratory ed.)またはAspergillus terreusからのデアミナーゼ(これは、ブラスチシジンSに対する耐性を付与する(Tamura, Biosci. Biotechnol. Biochem. 59 (1995), 2336-2338))。
【0143】
有用なスコア付け可能なマーカーもまた当業者に公知であり、市販されている。有利には、このマーカーは、ルシフェラーゼ(Giacomin, Pl. Sci. 116 (1996), 59-72; Scikantha, J. Bact. 178 (1996), 121)グリーン蛍光タンパク質(Gerdes, FEBS Lett. 389 (1996), 44-47)、またはβ-グルクロニダーゼ(Jefferson, EMBO J. 6 (1987), 3901-3907)をコードする遺伝子である。この態様は、列挙されたベクターを含む細胞、組織、および生物体を単純かつ迅速にスクリーニングするために特に有用である。
【0144】
上記のように、列挙された核酸分子は、例えば、精製だけでなく遺伝子治療目的のために、コードされたCD3特異的構築物を細胞中で発現するために単独でまたはベクターの一部として使用され得る。上記の(二重特異性)CD3構築物のいずれか1つをコードするDNA配列を含む核酸分子またはベクターは細胞に導入され、次には関心対象のポリペプチドを産生する。遺伝子治療(これはエクスビボまたはインビボの技術によって細胞に治療遺伝子を導入することに基づく)は、遺伝子移入の最も重要な応用の1つである。インビトロまたはインビボの遺伝子治療のための適切なベクター、方法、または遺伝子送達系は文献に記載されており、当業者に公知である。例えば、以下を参照されたい。列挙された核酸分子およびベクターは、直接的導入のために、またはリポソーム、もしくはウイルスベクター(例えば、アデノウイルス、レトロウイルス)を介する細胞への導入のために設計され得る。好ましくは、この細胞は生殖系列細胞、胚性細胞、または卵細胞、またはそれらから由来する細胞であり、最も好ましくは、幹細胞である。胚性幹細胞の例は、とりわけ、Nagy, Proc. Natl. Acad. Sci. USA 90 (1993), 8424-8428において記載されている幹細胞であり得る。
【0145】
上記に従って、本発明は、本明細書中に定義される二重特異性単鎖抗体構築物のポリペプチド配列をコードする核酸分子を含む、遺伝子操作において慣用的に使用されるベクター(特に、プラスミド、コスミド、ウイルス、およびバクテリオファージ)を誘導するための方法に関する。好ましくは、このベクターは、発現ベクターおよび/または遺伝子移入もしくは標的化ベクターである。レトロウイルス、ワクシニアウイルス、アデノ関連ウイルス、ヘルペスウイルス、またはウシパピローマウイルスなどのウイルスに由来する発現ベクターは、標的とされた細胞集団への列挙されたポリヌクレオチドまたはベクターの送達のために使用され得る。当業者に周知である方法が組換えベクターを構築するために使用され得る;例えば、Sambrook et al. (前掲)、Ausubel(1989、前掲)または他の標準的教科書において記載される技術を参照されたい。または、列挙される核酸分子およびベクターは、標的細胞への送達のためにリポソームに再構成され得る。本発明の核酸分子を含むベクターは、周知の方法(これは細胞宿主の型に依存して変化する)によって宿主細胞に移動され得る。例えば、塩化カルシウムトランスフェクションが原核生物細胞のために利用されるのに対して、リン酸カルシウム処理またはエレクトロポレーションが他の細胞宿主のために使用され得る;Sambrook、前記を参照されたい。
【0146】
列挙されるベクターは、特に、pEF-DHFR、pEF-ADA、またはpEF-neoであり得る。ベクターpEF-DHFR、pEF-ADAおよびpEF-neoは、当該分野において、例えば、Mack et al. (PNAS (1995) 92, 7021-7025)およびRaum et al. (Cancer Immunol Immunother (2001) 50(3), 141-150)ににおいて記載されている。
【0147】
本発明は、ここで定義したようなベクターで形質転換またはトランスフェクトされた宿主をも提供する。この宿主は、少なくとも1つの上記のベクター、または少なくとも1つの上記の核酸分子を、宿主に導入することによって産生され得る。宿主におけるその少なくとも1つのベクターまたは少なくとも1つの核酸分子の存在が、上記の二重特異性単鎖抗体構築物をコードする遺伝子の発現を媒介し得る。
【0148】
宿主に導入される記載された核酸分子またはベクターは、宿主のゲノムに組み込まれ得るか、または染色体外に維持され得るかのいずれかである。
【0149】
宿主は、任意の原型生物または真核生物細胞であり得る。
【0150】
「原核生物」という用語は、本発明のタンパク質の発現のためにDNA分子またはRNA分子で形質転換またはトランスフェクトされ得る全ての細菌を含むことを意味する。原核生物宿主は、グラム陰性細菌ならびにグラム陽性細菌、例えば、大腸菌、S. typhimurium、Serratia marcescensおよびBacillus subtilisなどを含み得る。「真核生物」という用語は、酵母、高等植物、昆虫、および好ましくは哺乳動物の細胞を含むことを意味する。組換え産生手順において利用される宿主に依存して、本発明のポリヌクレオチドによってコードされるタンパク質は、グリコシル化されてもよく、またはグリコシル化されなくてもよい。特に好ましいものは、本発明のポリペプチドのコード配列ならびにそれに遺伝子が融合したN末端FLAG-タグおよび/またはC末端His-タグを含むプラスミドまたはウイルスの使用である。好ましくは、このFLAG-タグの長さは約4〜8アミノ酸であり、最も好ましくは8アミノ酸である。上記のポリヌクレオチドは、当業者に一般的に公知である技術のいずれかを使用して宿主を形質転換またはトランスフェクトするために使用され得る。さらに、融合された、作動可能に連結された遺伝子を調製する方法、ならびに例えば、哺乳動物細胞および細菌中でそれらを発現させる方法は、当技術分野において周知である(Sambrook、前掲)。
【0151】
好ましくは、この宿主は細菌、昆虫、真菌、植物、または動物の細胞である。
【0152】
列挙される宿主は、哺乳動物細胞、より好ましくはヒト細胞またはヒト細胞株であり得ることが特に想定される。
【0153】
特に好ましい宿主細胞には、CHO細胞、COS細胞、ミエローマ細胞株(SP2/0またはNS/0など)が含まれる。実施例で記載されているように、特に好ましい宿主細胞は、COS細胞である。
【0154】
さらなる実施例において、本発明は、該構造物の発現に適した条件下で本発明の細胞、および/または宿主を培養する段階および細胞又は培養液から構造物を単離する段階を含む、上述したCD3特異的構造物の準備工程に関する。
【0155】
形質転換された宿主は、最適な細胞増殖を達成するために、当技術分野において公知である技術に従って、ファーメンター中で増殖され、かつ培養され得る。次いで、本発明のポリペプチドは、増殖培地、細胞溶解物、または細胞膜画分から単離され得る。本発明の、例えば、微生物によって発現されたポリペプチドの単離および精製は、例えば、調製用クロマトグラフィー分離および免疫学的分離(例えば、本発明のポリペプチドのタグに対して指向されるモノクローナル抗体またはポリクローナル抗体の使用を含むもの)などの任意の慣用的な手段によって、または添付の実施例において記載されるようにであり得る。
【0156】
さらに、本発明は、本明細書において定義された(ヒト)CD3特異的結合構築物を含む組成物、または先に開示したプロセスによって産生された(ヒト)CD3特異的結合構築物、本発明の核酸分子、本発明のベクターまたは宿主を提供する。該組成物は、任意で免疫エフェクター細胞の活性化シグナルを提供することができるタンパク質様化合物を含む。最も好ましくは、該組成物は、任意で担体、安定化剤、および/または賦形剤の適した製剤をさらに含む薬学的組成物である。
【0157】
本発明に従って、「薬学的組成物」という用語は、患者、好ましくはヒト患者に投与するための組成物に関する。好ましい態様において、薬学的組成物は、非経口、経皮、腔内、動脈内、髄腔内投与のための、または組織もしくは腫瘍への直接注射による組成物を含む。特に、該薬学的組成物は注入または注射によって患者に投与されると想像される。適した組成物の投与は異なる方法、例えば静脈内、腹腔内、皮下、筋肉内、局所、または皮内投与によって行ってもよい。本発明の薬学的組成物はさらに、薬学的に許容される担体を含んでもよい。適した薬学的担体の例は当技術分野において周知であり、これには、燐酸緩衝生理食塩液、水、油/水乳剤のような乳剤、様々なタイプの湿潤剤、滅菌溶液等が含まれる。そのような担体を含む組成物は、周知の通常の方法によって製剤化することができる。これらの薬学的組成物は、適した用量で被験者に投与することができる。投与レジメは、主治医および臨床的要因によって決定されるであろう。医学の技術分野において周知であるように、任意の患者1人の用量は、患者の体格、体表面積、年齢、投与される特定の化合物、性別、投与期間および経路、全身健康、ならびに同時投与される他の薬物を含む、多くの要因に依存する。一般的に、薬学的組成物の規則的な投与としてのレジメは、1μg〜5 g単位/日の範囲でなければならない。しかし、連続的注入のためのより好ましい用量は、0.01μg〜2 mg、好ましくは0.01μg〜1 mg、より好ましくは0.01μg〜100 μg、さらにより好ましくは0.01μg〜50 μg、および最も好ましくは0.01μg〜10 μg単位/kg体重/時間の範囲であってもよい。特に好ましい用量は、本明細書において下記に引用される。進行は定期的な評価によってモニターすることができる。用量は変化するが、DNAの静脈内投与のための好ましい用量はDNA分子約106〜1012コピーである。本発明の組成物は、局所または全身投与してもよい。投与は一般的に非経口、例えば静脈内投与であろう;DNAはまた、例えば内部または外部標的部位へのバイオリスティック(biolistic)送達、または動脈内の部位にカテーテルによって標的部位に直接投与してもよい。非経口投与のための調製物には、滅菌水溶液、または非水溶液、懸濁液、および乳剤が含まれる。非水性溶媒の例は、プロピレングリコール、ポリエチレングリコール、オリーブ油のような植物油、およびオレイン酸エチルのような注射可能な有機エステルである。水性担体には、生理食塩液および緩衝培地を含む、水、アルコール/水溶液、乳剤、または懸濁液が含まれる。非経口溶媒には、塩化ナトリウム溶液、リンゲルデキストロース、デキストロースおよび塩化ナトリウム、乳酸加リンゲル、または固定油が含まれる。静脈内媒体には、液体および栄養補助剤、電解質補助剤(リンゲルデキストロースに基づくような)等が含まれる。例えば抗菌剤、抗酸化剤、キレート剤、および不活性ガス等のような保存剤および他の添加剤も同様に、存在してもよい。さらに、本発明の薬学的組成物は、例えば好ましくはヒト起源の血清アルブミンまたは免疫グロブリンのような蛋白質様担体を含んでもよい。本発明の薬学的組成物は、二重特異性一本鎖抗体構築物またはこれをコードする核酸分子もしくはベクター(本発明において記述されるように)の他に、薬学的組成物の意図する用途に応じて、さらに生物活性物質を含んでもよいと想像される。そのような物質は、消化管系に作用する薬剤、細胞増殖抑制物質として作用する薬剤、ハイパーウリケミア(hyperurikemia)を予防する薬剤、免疫反応を阻害する薬剤(例えば、コルチコステロイド)、循環系に影響を及ぼす薬剤および/または当技術分野で公知のT-細胞共刺激分子またはサイトカインのような物質であってもよい。
【0158】
本発明の組成物の投与に関して可能性がある適応は、腫瘍様疾患、特に乳癌、結腸癌、前立腺癌、頭頚部癌、皮膚癌(黒色腫)、尿性器癌、例えば卵巣癌、子宮内膜癌、子宮頚癌および腎臓癌、肺癌、胃癌、小腸の癌、肝臓癌、膵臓癌、胆嚢癌、胆管癌、食道癌、唾液腺癌および甲状腺癌のような上皮癌/癌腫、または血液腫瘍、神経膠腫、肉腫、もしくは骨肉腫のような他の腫瘍様疾患である。本発明の組成物の投与は特に、細胞1個の生存によって引き起こされる腫瘍の局所および非局所再発を特徴とする最小残留疾患、好ましくは初期固形腫瘍、進行固形腫瘍、または転移固形腫瘍に適応される。
【0159】
本発明はさらに、他の化合物、例えば免疫エフェクター細胞、細胞増殖、または細胞刺激に関する活性化シグナルを提供することができる分子との同時投与プロトコールを想像する。該分子は、例えばさらにT細胞の一次活性化シグナル(例えば、さらなる共刺激分子:B7ファミリーの分子、Ox40L、4.1 BBL)、またはさらなるサイトカイン:インターロイキン(例えば、IL-2)もしくはNKG-2D関係化合物であってもよい。
【0160】
上記の本発明の組成物は、任意で検出手段および方法をさらに含む診断組成物であってもよい。
【0161】
本明細書において定義されるCD3-特異的構築物はまた、液相において、または固相担体に結合して利用することができるイムノアッセイにおいて用いるために適している。本発明のポリペプチドを利用することができるイムノアッセイの例は、直接または間接的フォーマットのいずれかの競合的および非競合的イムノアッセイである。そのようなイムノアッセイの例は、酵素結合イムノソルベントアッセイ(ELISA)、酵素イムノアッセイ(EIA)、ラジオイムノアッセイ(RIA)、サンドイッチ(免疫測定)アッセイ、およびウェスタンブロットアッセイである。
【0162】
本発明のCD3特異的結合構築物は、異なる多くの担体に結合させて、該ポリペプチドに特異的結合した細胞を単離するために用いることができる。周知の担体の例には、ガラス、ポリスチレン、ポリ塩化ビニル、ポリプロピレン、ポリエチレン、ポリカーボネート、デキストラン、ナイロン、アミロース、天然および改変セルロース、ポリアクリルアミド、アガロース、およびマグネタイトが含まれる。担体の性質は可溶性または不溶性のいずれかとなり得て、例えば本発明の目的に関してビーズとなりうる。
【0163】
当業者に公知の異なる多くの標識および標識法が存在する。本発明において用いることができる標識のタイプの例には、酵素、放射性同位元素、コロイド金属、蛍光化合物、化学発光化合物、および生物発光化合物が含まれる;同様に、本明細書において考察した態様を参照されたい。
【0164】
本発明の最も好ましい態様において、薬学的組成物を調製するために、CD3特異的結合分子、本発明のベクターまたは宿主を用いることが想像される。該薬学的組成物は、増殖疾患、腫瘍様疾患、炎症疾患、免疫障害、自己免疫障害、感染症、ウイルス病、アレルギー反応、寄生虫反応、移植片対宿主病、または宿主対移植片病の予防、治療、または改善において用いてもよい。
【0165】
さらに、本発明に従って、CD19およびCD3結合ドメイン、好ましくは配列番号:190、192、194、196、198、200、327、329、331、333、335、337、339、341、343、345、347、349、351、353、357、359、361、363、365、367、369、371、373、375、377、379、381、383、385、387、389、391、393、395、397、399、401、403、405、407、または409を含む脱免疫構築物は、免疫障害(様々なB細胞悪性疾患)または自己免疫疾患の治療に用いることができ、CCR5およびCD3結合ドメイン、好ましくは配列番号:206、208、210、212、214、または216を含む脱免疫構築物は、腫瘍様疾患、好ましくはリンパ腫、より好ましくは非ホジキンB-細胞リンパ腫の治療に用いることができ、EpCAMおよびCD3結合ドメイン、好ましくは配列番号:31、33、35、37、39、49、55、58、61、63、65、67、237、239、241、243、245、247、249、251、253、255、257、259、261、263、265、267、269、271、273、275、277、279、281、283、285、287、289、291、293、295、297、299、301、303、305、307、309、311、313、315、317、319、321、323、または325を含む脱免疫構築物は、腫瘍様疾患、好ましくは上皮癌の治療に用いることができる。
【0166】
本発明はまた、本発明の(二重特異性)CD3特異的結合分子、本明細書に記述のプロセスによって産生された(二重特異性)CD3特異的結合分子、本発明の核酸分子、ベクターまたは宿主を、そのような予防、治療、または改善を必要とする被験者に投与することを含む、増殖疾患、腫瘍性の疾患、炎症疾患、免疫障害、自己免疫疾患、感染症、ウイルス病、アレルギー反応、寄生虫反応、移植片対宿主病、または宿主対移植片病を予防、治療、または改善する方法に関する。好ましくは該被験者はヒトである。
【0167】
予防、治療、または改善法はまた、さらに、免疫エフェクター細胞の活性化シグナルを提供することができる蛋白質様化合物の投与を含んでもよい。該蛋白質様化合物は、本発明のCD3結合分子、核酸分子、ベクター、または宿主と同時、または非同時に投与してもよい。蛋白質様化合物は、とりわけさらなる共刺激分子:B7ファミリーの分子、Ox40L、4.1 BBLまたはさらなるサイトカイン;インターロイキン(例えば、IL-2)またはNKG-2D結合分子からなる群より選択してもよい。
【0168】
最後に、本発明は、本発明のCD3特異的結合分子、核酸分子、ベクター、または宿主を含むキットを提供する。
【0169】
該キットは、本発明の薬学的組成物を調製するために特に有用であり、とりわけ、注射または注入のために有用な容器からなってもよい。本発明のキットはさらに、任意で医学的または科学的目的を行うために必要な緩衝液、保存液および/または残りの試薬または材料を含むことが都合がよい。さらに、本発明のキットの一部は、バイアルもしくはボトルに個別包装する、または容器もしくは多容器単位と組み合わせて包装することができる。本発明のキットは、とりわけ、本発明の方法を実行するために、例えば研究ツールまたは医学的ツールとして本明細書において言及された多様な適応において都合よく用いることができるであろう。キットの製造は好ましくは、当業者に公知の標準的な技法に従う。
【0170】
これらおよび他の態様は、本発明の記述および実施例によって開示され、含まれる。本発明に従って用いられる抗体、方法、用途、および化合物の任意の一つに関するさらなる文献は、例えばコンピューターを用いて公共のライブラリおよびデータベースから検索してもよい。例えばインターネット上で利用できる公共のデータベース「Medline」を、例えばhttp://www.ncbi.nlm.nih.gov/PubMed/medline.htmlにおいて利用してもよい。http://www.ncbi.nlm.nih.gov/、http://www.infobiogen.fr/、http://www.fmi.ch/biology/research tools.html、http://www.tigr.org/のようなさらなるデータベースおよびアドレスは、当業者に公知であり、同様に、例えばhttp://www.lycos.comを用いて得ることができる。
【実施例】
【0171】
以下の実施例は本発明を説明する。
【0172】
以下の実施例において、多くの一本鎖抗ヒトCD3抗体が、ヒトにおいて免疫原性の低下を示すように操作されている。異なる脱免疫抗ヒトCD3抗体は、互いに結合した異なる4個のVH(VH2(配列番号:69、70)、VH3(配列番号:71、72)、VH5(配列番号:73、74)、およびVH7(配列番号:75、76))、ならびに異なる3個のVL(VL1(配列番号:77、78)、VL2(配列番号:79、80)、およびVL3(配列番号:81、82))領域の組み合わせ12個を含む。上記のVHおよびVL領域のアミノ酸および核酸配列を図3〜6に示す。例として、脱免疫抗CD3一本鎖抗体は、二重特異性産物を形成するために、抗CD19一本鎖抗体、または抗EpCAM一本鎖抗体とを組み合わせた。
【0173】
実施例1.脱免疫抗CD3構築物のクローニングおよび発現
1.1 一本鎖抗体をコードするcDNAの移入
脱免疫されている抗CD3一本鎖抗体をコードするDNAを、本明細書において抗CD3カセットと呼ぶ。この抗CD3カセットは、SGGGGSリンカー(配列番号:176)、抗CD3 VH領域(配列番号:110)、およびアミノ酸14個のGSリンカー(VEGGSGGSGGSGGSGGVDリンカー(配列番号:68))、および抗CD3 VL領域(配列番号:112)からなり、この後にヒスチジン残基6個が続く。上記のDNAをSrf1部位でベクターpPCR-Script-Amp SK(+)(Stratagene)にクローニングした。抗CD3カセットのDNAおよびアミノ酸配列を配列番号:1、配列番号:2および図1に示す。
【0174】
1.2 免疫原性T細胞エピトープの配列のコンピューター分析と脱免疫一本鎖抗体配列のデザイン
国際公開公報第98/52976号に記載の方法によって可能性があるT細胞エピトープを同定するために、抗CD3カセット(配列番号:2)のアミノ酸配列をペプチド糸通しプログラムによって分析した。配列番号:3は、脱免疫リンカー配列および配列番号:68は当初のリンカー配列を示す。
【0175】
1.3 脱免疫一本鎖抗体配列の構築
抗CD3カセットの脱免疫版をオーバーラップPCR組換え法によって構築した。pPCR-S-Amp SK+において、抗CD3カセット(配列番号:1、2)を必要な脱免疫配列への変異誘発のための鋳型として用いた。改変される領域を含む変異原性プライマー対の組を合成した。異なるVH領域4個および異なるVL領域3個を含む、産生された脱免疫配列をNot1からHind III断片としてベクターpPCR-S-Amp SK+にクローニングして、全DNA配列が正確であることを確認した。異なるVH領域4個および異なるVK領域3個を全ての組み合わせ(全体で12)において、PCRまたはVH領域の3'末端に導入した単一のBstEII部位のいずれかを用いて結合した。各組み合わせの全DNA配列が正確であることを確認した。抗CD3構築物の対応する当初の非脱免疫配列(VH:配列番号:110、VL:配列番号:112)を有する異なる脱免疫VH領域(配列番号:70、72、74、および76)およびVL領域(配列番号:78、80、および82)を表9に要約する。
【0176】
(表9)脱免疫VHおよびVL領域の配列番号:
【0177】
1.4 脱免疫一本鎖抗体遺伝子の発現ベクターへの移入
脱免疫抗CD3カセットをBspEIおよびSalIによってpPCR-S-Amp-SK+から切り出して、VLCD19-VHCD19-VHCD3-VLCD3を含む発現ベクターpEFにクローニングした。pEF-DHFRベクターのCD3部分は、BspEI部位からSalI部位への脱免疫抗CD3カセットのそれぞれによって置換して、以下の構築物12個を得た:
pEF抗CD19×抗CD3(VH2/VL1)(配列番号:177、178)
pEF抗CD19×抗CD3(VH2/VL2)(配列番号:179、180)
pEF抗CD19×抗CD3(VH2/VL3)(配列番号:181、182)
pEF抗CD19×抗CD3(VH3/VL1)(配列番号:183、184)
pEF抗CD19×抗CD3(VH3/VL2)(配列番号:185、186)
pEF抗CD19×抗CD3(VH3/VL3)(配列番号:187、188)
pEF抗CD19×抗CD3(VH5/VL1)(配列番号:189、190)
pEF抗CD19×抗CD3(VH5/VL2)(配列番号:191、192)
pEF抗CD19×抗CD3(VH5/VL3)(配列番号:193、194)
pEF抗CD19×抗CD3(VH7/VL1)(配列番号:195、196)
pEF抗CD19×抗CD3(VH7/VL2)(配列番号:197、198)
pEF抗CD19×抗CD3(VH7/VL3)(配列番号:199、200)
【0178】
構築物はさらに、タンパク質を分泌させるためにマウスIgG重鎖リーダーを含む。発現ベクターにおける脱免疫抗CD3カセットのDNA配列を、シークエンシングプライマー(配列番号:28および29)を用いて確認した。BspEI部位からSalI部位までのpEFベクターにおける脱免疫抗CD3カセット12個のDNAおよびアミノ酸配列を、配列番号:177〜200に示す。
【0179】
1.5 抗体構築物の産生
ベクターを大腸菌K12に形質転換した後、異なる発現ベクターのトランスフェクション等級のDNAを調製した。分泌されたタンパク質をCHO-dhfr細胞において産生した。一過性の産生に関して、細胞培養上清をトランスフェクションの2日後に回収して、安定なトランスフェクト細胞を産生するために、細胞をトランスフェクション後2日間、選択培地に入れた。5回継代した後、安定なプールを得た。その後限界希釈によって単一のクローンを同定した。精製プロセスを容易にするために、細胞を無血清培地に適合させた。抗体構築物は上清約1 Lから精製した。産生レベルをELISAにおいて試験した。抗CD19および脱免疫抗CD3構築物を含む異なる構築物のあいだで、分泌された抗体レベルに主要な差を認めなかった。
【0180】
実施例2:結合アッセイ
CD3およびCD19に対する脱免疫構築物の結合効率を分析するために、FACSに基づくアッセイを行った。最初、粗上清をCD3濃縮PBMCまたはCD19陽性NALM-6細胞に対する結合に関して試験した。細胞を非希釈上清と共に8℃で30分インキュベートした。2回の洗浄段階後、細胞を同じ条件で抗His抗体(Qiagen)によって標識した。さらなる洗浄段階の後、構築物の結合をFITC-結合ヒツジ抗マウス抗体(Sigma)によって検出した。細胞をFACS Caliburサイトメーター(B&D)によって分析した。対照として、抗CD19×抗CD3およびGFP-トランスフェクト細胞の上清を含めた。CD3および二次抗体をネガティブコントロールとして用いて、これは平均蛍光強度(MFI)約3.5を示した。抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH5/VL2)(配列番号:192)、抗CD3(VH5/VL3)(配列番号:194)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)を含む抗CD19×抗CD3構築物は、MFI少なくとも90を有し、このように、約25倍強く結合した。非脱免疫抗CD19×抗CD3構築物である陽性対照は、MFI約60に達し、抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH5/VL2)(配列番号:192)、抗CD3(VH5/VL3)(配列番号:194)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)を含む脱免疫構築物が、非常に高い効率でCD3に結合したことを示している。第二の実験において、抗CD19および抗CD3:抗CD3(VH2/VL1)(配列番号:178)、抗CD3(VH2/VL2)(配列番号:180)、抗CD3(VH2/VL3)(配列番号:182)、抗CD3(VH3/VL1)(配列番号:184)、抗CD3(VH3/VL2)(配列番号:186)、および抗CD3(VH3/VL3)(配列番号:188)を含む以下の構築物は、ネガティブコントロールと類似の結合を示した(MFI約6)。
【0181】
FACSに基づく結合アッセイを、CD19に関しても行った。この実験において、CD19および二次抗体はネガティブコントロールであった。本実験において、アッセイした構築物は全てMFI少なくとも80に達したのに対し、ネガティブコントロールのMFIは約3であった。
【0182】
このように、抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH5/VL2)(配列番号:192)、抗CD3(VH5/VL3)(配列番号:194)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)を含む構築物は、非改変抗CD19×抗CD(配列番号:204)3と同程度にCD3およびCD19に結合することが判明した。しかし、構築物抗CD3(VH2/VL1)(配列番号:178)、抗CD3(VH2/VL2)(配列番号:180)、抗CD3(VH2/VL3)(配列番号:182)、抗CD3(VH3/VL1)(配列番号:184)、抗CD3(VH3/VL2)(配列番号:186)、および抗CD3(VH3/VL3)(配列番号:188)は、抗CD3結合能を完全に喪失したが、CD19結合能は完全に保持した(図7)。
【0183】
このように、脱免疫重鎖は、結合の特異性および強度を支配することが証明された。その結果として、VH5およびVH7群を有する抗CD3構築物を精製して細胞障害活性に関して分析した。
【0184】
実施例3.高い結合親和性を示す変種の発現および精製
脱免疫抗CD19×抗CD3タンパク質抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH5/VL2)(配列番号:192)、抗CD3(VH5/VL3)(配列番号:194)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)をチャイニーズハムスター卵巣細胞(CHO)において発現させた。
【0185】
脱免疫抗CD3部分を含む二重特異性一本鎖構築物を精製するために、CHO-CD19細胞をローラーボトルにおいてハイクローンCHO改変DMEM培地(HiQ(登録商標))によって7日間増殖させた後、回収した。細胞を遠心によって除去し、発現されたタンパク質を含む上清を-20℃で保存した。
【0186】
Akta FPLCシステム(登録商標)(Pharmacia)およびUnicornソフトウェア(登録商標)をクロマトグラフィーのために用いた。化学物質は全て研究等級であり、Sigma(Deisenhofen)またはMerck(Darmstadt)から購入した。
【0187】
緩衝液A1(20 mMトリスpH 7.2)によって平衡にしたXK16/60カラム(Pharmacia)にローディングしたMEP Hypercel(登録商標)において、疎水性電荷誘導クロマトグラフィーを行った。細胞培養上清500 mlをカラム(10 ml)に流速3 ml/分で適用した。未結合試料を緩衝液A1によって洗浄し、結合したタンパク質を100%緩衝液B1(20 mM酢酸塩、pH 3.5)によって溶出した。溶出したタンパク質分画をさらなる精製のためにプールした。
【0188】
製造元のプロトコールに従ってNiSO4をローディングしたHisTap(登録商標)カラム(Pharmacia)を用いて、IMACを行った。カラムを緩衝液A2(20 mM NaP、pH 7.5、0.4 M NaCl)によって平衡にして、試料を緩衝液A2によって2:1希釈して、pH 7を得た。試料を流速1 ml/分でカラム(2 ml)に適用して、カラムを緩衝液A2によって洗浄して、未結合試料を除去した。結合したタンパク質を、緩衝液B2(20 mM NaP、pH 7.5、0.4 M NaCl、0.5 Mイミダゾール)の2段階勾配を用いて溶出した。段階1:20%緩衝液B2の10カラム容積;段階2:100%緩衝液B2の10カラム容積。溶出したタンパク質分画をさらなる精製のためにプールした。
【0189】
PBS(Gibco)によって平衡にしたSephadex S200 HiPrep(登録商標)カラム(Pharmacia)においてゲル濾過クロマトグラフィーを行った。溶出したタンパク質試料(流速1 ml/分)に、SDS-PAGEおよび検出のためにウェスタンブロットを行った(図11)。カラムを分子量決定のために予め較正した(分子量マーカーキット、Sigma MW GF-200)。
【0190】
抗CD19×抗CD3タンパク質の脱免疫変種を、疎水性電荷誘導クロマトグラフィー(HCIC)(図8)、固定金属アフィニティクロマトグラフィー(IMAC)(図9)、およびゲル濾過(図10)を含む3段階精製プロセスにおいて単離した。二重特異性構築物は、PBSにおけるゲル濾過によって決定したところ、未変性条件で分子量52 kDaであった。
【0191】
精製二重特異性タンパク質を、成形済みの4〜12%ビストリスゲル(Invitrogen)を用いて還元条件でSDS-PAGEによって分析した。試料の調製および適用は、製造元のプロトコールに従った。分子量はMultiMark(登録商標)タンパク質標準物質(Invitrogen)によって決定した。ゲルをコロイド状クーマシー(Invitrogenのプロトコール)によって染色した。単離されたタンパク質の純度は>95%(図11a)であり、分子の大きさは52 kDaである。
【0192】
さらに、抗CD19×抗CD3タンパク質の脱免疫変種をウェスタンブロットによって特異的に検出した。ウェスタンブロットは、Optitran BA-S83(登録商標)メンブレンおよびInvitrogen Blot Module(登録商標)によって製造元のプロトコールに従って行った。用いた抗体は、Penta His(Quiagen)およびアルカリホスファターゼ(AP)標識ヤギ抗マウスIg(Sigma)であり、染色溶液はBCIP/NBT液(Sigma)であった。主要なシグナルは、SDS-PAGEにおいて52 kDaで主要なバンドに対応することが示された(図11b)。
【0193】
タンパク質アッセイ色素(MicroBCA(登録商標)、Pierce)および標準タンパク質としてIgG(Biorad)を用いてタンパク質濃度を決定した。精製タンパク質変種の最終収率の概要を表10に示し、全ての構築物の生産性が高いことを示し、抗CD3(VH5/VL1)(配列番号:190)に関して924.8 μgの非常に良好な収量を得た。
【0194】
(表10)脱免疫抗CD19抗CD3構築物のタンパク質収量
【0195】
CD19×抗CD3(VH5/VL2)およびCD19×抗CD3(VH7/VL2)構築物の生産性を、対応する非脱免疫構築物と比較した。結果を表11に示す。
【0196】
(表11)対応する非脱免疫構築物と比較した脱免疫二重特異性構築物の収量
【0197】
表11は、脱免疫CD3結合ドメインを含む二重特異性構築物が、対応する非脱免疫構築物よりかなり高い(少なくとも3倍)生産性を有することを明らかに示している。
【0198】
実施例4.抗CD3構築物のFACSに基づく結合アッセイ
抗CD19および抗CD3を含む選択した精製抗体構築物の結合を、実施例2において先に記述するように様々な濃度で検出した。CD3結合アッセイにおいて、ネガティブコントロール二次抗体(抗-His、FITC結合)をCD3陽性細胞と共にインキュベートすると、MFI約2.5を示し、陽性対照である脱免疫抗CD19×抗CD3二重特異性一本鎖抗体は1μg/ml濃度で約70および5μg/ml濃度で約50を示した(図12A)。濃度1μg/mlでは、抗CD3(VH5/VL1)(配列番号:190)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)脱免疫二重特異性抗体はMFI値10〜20を示した;抗CD3(VH5/VL1)(配列番号:190)は最高の値を有した(20)。5μg/mlでは、抗CD3(VH7/VL2)(配列番号:198)はMFI 25に達したが、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL3)(配列番号:200)、および抗CD3(VH5/VL1)(配列番号:190)は、MFI少なくとも40を示し、このように、非脱免疫陽性対照と同じ結合効率を示した。濃度5μg/mlでは、脱免疫抗CD3部分VH5/VL1(配列番号:190)、抗CD3(VH7/VL1)(配列番号:196)、抗CD3(VH7/VL2)(配列番号:198)、および抗CD3(VH7/VL3)(配列番号:200)を有する強い結合構築物は、非脱免疫抗CD19×抗CD3(配列番号:204)と同様にCD3に結合した。
【0199】
抗体構築物は全て、高い有効性でCD19に結合し、MFIは約200であったが、非脱免疫抗CD19×抗CD3構築物(配列番号:204)は、MFI 80を示した。調べた濃度では異なる構築物に関してCD19結合に関して差を認めなかった(図12B)。
【0200】
実施例5.細胞障害アッセイ
抗CD19×抗CD3は、CD19陽性標的細胞に対するT細胞依存的細胞障害性を媒介する。これを、抗CD19×抗CD3の生物学的有効性の決定に関してインビトロで分析した。
【0201】
この目的に関して、蛍光標識CD19陽性NALM-6標的細胞を、エフェクター細胞としてランダムドナーから単離したPBMCまたはCB15 T細胞(標準化T細胞株)と共に、抗CD19×抗CD3の存在下でインキュベートした。湿潤インキュベーターにおいて37℃で4時間インキュベートした後、標的細胞からの蛍光色素の上清への放出を蛍光分光計において決定する。抗CD19×抗CD3を含まずにインキュベートした標的細胞、およびインキュベーション終了時にサポニンの添加によって完全に溶解した標的細胞をそれぞれ、陰性および陽性対照とした。特定の抗CD19×抗CD3濃度で媒介される特異的細胞障害性を、以下の式によって計算することができる:
。
【0202】
EC50値を特定するために、用量反応を、0.4 pg/ml抗CD19×抗CD3から100 ng/ml抗CD19×抗CD3まで分析した。EC50値は、抗CD19×抗CD3の生物学的有効性を記述するが、絶対値は、エフェクター細胞源に応じて有意に変化するであろう。このように、相対的有効性は、以下の式に基づいて抗CD19×抗CD3参照材料と比較して計算される:
【0203】
抗CD19および脱免疫抗CD3を含む構築物の細胞障害活性を図13に示す。精製された非脱免疫抗CD19×抗CD3を対照として用いた。脱免疫構築物のEC50値は21.9〜81.6 pg/mlの範囲であったが、非脱免疫抗CD19×抗CD3構築物のEC50値は、22.7 pg/mlであった。このように、脱免疫構築物は全て、非脱免疫分子と同等のEC50値であることが判明した。
【0204】
実施例6.T細胞増殖アッセイ
健康なドナー20人をHLA-DRタイピングに基づくT細胞アッセイにおけるスクリーニングのために選択した(表12)。これによって、世界の人口において発現されるDR対立遺伝子の80%より大きい割合に対するT細胞アッセイにおいてペプチドのスクリーニングが可能となる。
【0205】
(表12)脱免疫および非脱免疫抗CD3 scAbから得たペプチドの免疫原性を調べるために用いた健康なドナー20人のHLA DRハプロタイプ
【0206】
6.1 T細胞増殖アッセイ
ペプチドはPepscan(Netherlands)から純度90%より高いものを得た。選択した健康なドナー20人からの末梢血単核球(PBMC)(表12)を用いて1および5μMで1試料あたり3個ずつ個々のペプチドをスクリーニングした。陽性対照ペプチド2個(C32およびC49)およびキーホールリンペットヘモシアニン(KLH)をアッセイに含めた。細胞およびペプチドを7日間インキュベートした後、1μCi/ウェル3H-チミジンによる18時間パルスを用いてT細胞増殖を評価した。これらのデータを以下のような刺激指数として表記した:
刺激指数=試験ペプチドのCPM/無処置対照のCPM
【0207】
T細胞エピトープは、2より大きい刺激指数(SI)を生じるペプチドとして定義される。独立した2回の実験結果は、非脱免疫抗CD3配列におけるMHC結合ペプチド22個中5個が、ヒトT細胞増殖の誘導能(SI>2)を有した。対照的に、対応する脱免疫分子はいずれも、T細胞増殖を誘導しなかった。表13は、独立した2回の実験の平均SI値を示すT細胞増殖アッセイの結果を要約する。
【0208】
データはまた濃度依存的作用を示し、それによって非脱免疫結合分子のそれぞれが、用いた二つの濃度の一つのみ(1μmまたは5μm)においてSI's>2を示した。異なる濃度での反応の差は、個々のペプチドがそれらがT細胞増殖を誘導する最適な濃度を有するであろうという事実によって説明される。この濃度を超えると、増殖は低下しうる(高いペプチド濃度はT細胞増殖に対して阻害作用を有しうる)。これは、場合によっては、増殖がより低い濃度で認められるがより高い濃度では認められないという理由を説明する。経験から、ペプチドがT細胞エピトープを含む場合、T細胞増殖は、用いたペプチド濃度の一つまたは二つにおいて認められるであろう。これらのデータは、脱免疫により抗CD3(VH5/VL2)(配列番号:19)および抗CD3(VH7/VL2)(配列番号:25)からT細胞エピトープが首尾よく除去されたことを証明した。非脱免疫抗CD3配列からのMHC結合ペプチドの約75%がT細胞増殖を誘導できなかったという事実は、これらのペプチドに対するヒト免疫系の寛容によって、またはヒトT細胞レパートリーがこれらの特定のペプチドを認識できないことによって説明することができる。
【0209】
(表13)陽性(SI>2)マウスペプチドおよび対応する脱免疫ペプチドを比較するデータの要約
【0210】
実施例7.非脱免疫抗CD3 VHとの抗CD3(VH5)、抗CD3(VH7)、抗CD3(VH2)、および抗CD3(VH3)の相同性アラインメント
非脱免疫CD3抗体の可変重鎖領域、VH5(配列番号:74)、VH7(配列番号:76)、VH2(配列番号:70)、およびVH3(配列番号:72)を、Vector NTI Advance(Informax, Inc., USA)のAlingnXプログラムを用いて整列させた。用いたClustal Wアルゴリズムは、Nucleic Acid Research, 22(22):4673〜4860, 1994に記述されている。アラインメントを図14に示す。アラインメントから、意外にも良好な結合を示す可変領域VH5およびVH7が、フレームワーク1とCDR1との移行領域で配列ASGYTFを有することが認められうる。さらに、結合を示さないVH領域(VH2(配列番号:70)およびVH3(配列番号:72))が、フレームワーク1とCDR1との移行部で配列ASGYTAを含む。このように、T細胞エピトープの生成傾向およびCD3に対する結合が低下した構築物を得るために、構築物はフレームワーク1とCDR1との移行領域で配列ASGYTFを含まなければならない。意外にも、CD3に結合し、上記の配列ASGYTFを含むT細胞エピトープの生成傾向が低下した重鎖可変領域は良好な結合を示す。
【0211】
実施例8.脱免疫抗CD3および抗EpCAMを含む構築物のクローニング
本発明の脱免疫抗CD3ポリペプチドは、他の標的を有する機能的構築物の一部となりうることを証明するために、脱免疫抗CD3(VH5/VL2)(配列番号:19)および異なる抗EpCAM一本鎖抗体(3-1(配列番号:137、139)、3-5(配列番号:141、143)、4-1(配列番号:145、147)、4-7(配列番号:149、151)、5-10(配列番号:133、135))を含む多くの二重特異性構築物を作製した。
【0212】
8.1 脱免疫抗CD3部分(配列番号:30、31、32、33、34、35、36、37、38、および39)を含むC-末端EpCAM結合物質の同定
8.1.1 抗CD19×抗CD3(VH5/VL2)構築物(配列番号:192)からの脱免疫抗CD3の増幅
N-末端脱免疫抗CD3(VH5/VL2)を、脱免疫(CD19×抗CD3(VH5/VL2)(配列番号:192))を鋳型として、および以下のプライマー(DI CD3 5-2 VH BsrGI
(配列番号:40)、DI CD3 5-2 VL BspEI
(配列番号:41))を用いてPCRによって得た。
【0213】
8.1.2 脱免疫抗CD3×抗EpCAM脱免疫構築物のVHCD3-VLCD3×VHEpCAM-VLEpCAM方向のクローニングおよび発現
脱免疫抗CD3を含む上記のPCR産物を、制限酵素BsrGIおよびBspEIによって切断した後、真核細胞分泌シグナル(リーダーペプチド)のアミノ酸配列を含むbluescript KSベクター(Stratagene, La Jolla, CA)にEcoRI/BsrGI断片としてクローニングした。この構築物をEcoRIおよびBspEIによって切断後、リーダーペプチドと共にそれぞれの抗CD3 scFvを含む得られたDNA断片を、pEF-DHFRベクターにおいてC-末端位で抗EpCAM scFv 3-1、4-7、または5-10を含むEcoRI/BspEI切断プラスミドにクローニングした。シークエンシング(Sequiserve, Vaterstetten)によって二重特異性一本鎖をコードする配列を確認後、プラスミドを真核細胞タンパク質発現のためにDHFR欠損CHO細胞にトランスフェクトした。DHFR欠損CHO細胞における真核細胞タンパク質発現は、Kaufmann, R.J.(1990)Methods Enzymol. 185, 537〜566に記述された通りに行った。
【0214】
8.1.3 脱免疫抗CD3×抗EpCAM脱免疫構築物のVHCD3-VLCD3×VLEpCAM-VHEpCAM方向のクローニングおよび発現
VL-VH方向にアミノ酸15の標準的なリンカーを含む抗EpCAM 4-7(配列番号:168)をPCRによって得た。4-7 VH領域および4-7 VL領域を以下のプライマーによって個々に増幅した(4-7 VL:4-7 VL BspEI 順方向
(配列番号:117)、4-7 VL GS15 逆方向
(配列番号:118)、4-7 VH:4-7 VH GS15 順方向
(配列番号:42)、4-7 VH SalI 逆方向
(配列番号:43))。PCR産物に導入された重なり合う相補的配列を用いて、その後の融合PCRの際に、アミノ酸15個の(G4S1)3(一文字アミノ酸コード)リンカー(標準的なリンカー)(配列番号:168)のコード配列を形成した。この増幅段階は、プライマー対4-7 VL Bsp EI FORおよび4-7 VH SalI REV(配列番号:42および43)を用いて行った。
【0215】
アミノ酸15個の標準的な((G4S1)3)リンカーを含むVL-VH方向の抗EpCAM 5-10を、PCRによって得た。5-10 VH領域および5-10 VL領域を、以下のプライマーによって個々に増幅した(5-10 VL:5-10 VL BspEI 順方向
(配列番号:44)、5-10 VL GS15 逆方向
(配列番号:45)、5-10 VH:5-10 VH GS15 順方向
(配列番号:46)、5-10 VH SalI 逆方向
(配列番号:47))。その後の融合PCRにおいて、PCR産物に導入された重なり合う相補的配列を用いて、アミノ酸15個の(G4S1)3(一文字アミノ酸コード)リンカー(標準的なリンカー)(配列番号:168)のコード配列を形成した。この増幅段階はプライマー対5-10 VL BspEI FORおよび5-10 VH SalI REVによって行った。
【0216】
PCR産物5-10 VL-VHおよび4-7 VL-VHを、抗CD3構築物(VH5/VL2)を含むpEF-DHFRベクターにクローニングした。シークエンシングによって二重特異性一本鎖をコードする配列を確認した後、真核細胞発現のために、プラスミドをDHFR欠損CHO細胞にトランスフェクトした。DHFR欠損CHO細胞における真核細胞タンパク質発現は、Kaufmann R.J.(1990)Methods Enzymol. 185, 537〜566に記述された通りに行った。
【0217】
8.1.4. 脱免疫抗CD3×抗EpCAM構築物のEpCAMおよびCD3に対する結合
N-末端方向で抗CD3部分を有する二重特異性一本鎖抗体のEpCAMおよびCD3に対する結合をFACS分析によって確認した。その目的のために、EpCAM陽性ヒト胃癌細胞株Kato III(ATCC HTB-103)を用いた。抗CD3部分の結合は、Jurkat細胞(ATCC TIB 152)において示された。
【0218】
細胞を供給元の推奨に従って培養して、細胞200000個を10 μg/ml構築物と共に2%FCS(仔ウシ胎児血清)を含むPBS 50μlにおいてインキュベートした。構築物の結合は、2μg/ml抗His抗体(Qiagen, Hilden, FRGから得たPenta-His抗体)によって2%FCSを含むPBSにおいて検出した。第二段階として、ヤギ抗マウスIgGに由来するR-フィコエリスリン結合アフィニティ精製F(ab')2を、2%FCS(Dianova, Hamburg, FRG)を含むPBS 50μlにおいて1:100倍希釈して用いた。試料をFACSscan(BD biosciences, Heidelberg, FRG)において測定した。
【0219】
FACS分析の結果を図15に示す。脱免疫抗CD3部分を含む構築物は全て、EpCAM陽性Kato III細胞上で非脱免疫抗EpCAM(M79)×抗CD3二重特異性一本鎖抗体より強い結合を示した。
【0220】
8.2. 脱免疫抗CD3部分を含むN-末端EpCAM結合物質のクローニング
8.2.1 抗EpCAM×抗CD3構築物のクローニング
8.2.1.1. 脱免疫3-1×抗CD3(VH5/VL2)構築物(配列番号:48、49)のクローニング
脱免疫構築物3-1×抗-CD3(VH5/VL2)(配列番号:48)は、脱免疫構築物抗EpCAM(3-1)×抗CD3に由来した。抗EpCAM抗体3-1のVHおよびVL領域を、配列番号:137および139に示す。プラスミドpEF-DHFR-3-1×抗CD3およびpEF抗CD3(VH5/VL2)(配列番号:192)を、ベクターおよびインサート抗CD3(VH5/VL2)をそれぞれ、単離するためにBspEIおよびSalI(Biolabs)によって消化した。BspEI-SalI消化ベクターを脱燐酸化して、0.7%アガロースゲルにおいて精製し、インサートを1.5%アガロースゲルにおいて精製した。
【0221】
精製断片(BspEI-SalI)を、その後、pEF-DHFRベクターの対応する部位にクローニングした。最終的な3-1×抗CD3(VH5/VL2)構築物(配列番号:48、49)を制限消化物および完全なインサートのDNAシークエンシングによって確認した。
【0222】
非脱免疫3-1×抗CD3構築物のクローニング
3-1×抗CD3(VH5/VL2)構築物のクローニングに関して、対応する非脱免疫構築物を以下のように作製した。
【0223】
非脱免疫3-1×抗CD3分子を構築するために、VH-VL方向のC-末端3-1をPCRによって得た。3-1 VH-VLを二つの部分において含む断片IおよびIIを、プライマー対me 91a(配列番号:53)/me 90(配列番号:52)およびme 83(配列番号:50)/me 84(配列番号:51)をそれぞれ用いてPCRによって増幅した。Expand High Fidelity System of Roche Diagnosticsを用いてホットスタートPCRを行った。20サイクル(94℃/30秒、60℃/1分、72℃/1分)を増幅のために行い、その後72℃で3分を1サイクル行った。
【0224】
PCR断片IおよびIIに1.5%アガロースゲル上での電気泳動を行った。断片を混合して(各1 ng)、完全な3-1を含む断片IIIの増幅のために、プライマー対me 91a(配列番号:53)およびme 84(配列番号:51)によって行う次のPCR反応の鋳型として用いた。PCRは上記の通りに行ったが、アニーリング温度は68℃であった。断片IIIをアガロースゲルにおいて精製して、BsrGIおよびBspEI(Biolabs)によって消化し、精製してその後pEF-DHFR抗EpCAM(M79)×抗CD3構築物の対応する部位にクローニングした。クローニングした領域を制限消化物およびDNAシークエンシングによって確認した。
【0225】
用いたプライマーの配列:
Me 83:
(配列番号:50)Me 84
(配列番号:51)Me 90
(配列番号:52)Me 91a
(配列番号:53)。
【0226】
8.2.1.2 脱免疫3-5×抗CD3(VH5/VL2)構築物(配列番号:54、55)のクローニング
3-5×抗CD3構築物を構築するために、VH-VL方向のC-末端3-5をPCRによって得た。抗EpCAM抗体3-5のVHおよびVL領域を、配列番号:141および143に示す。インサート(3-5)およびベクターをそれぞれ単離するために、プラスミドpEF-DHFR-3-5×抗CD3およびpEF抗CD3(VH5/VL2)(配列番号:192)をEcoRIおよびBspEI(Biolabs)によって消化した。脱燐酸化ベクター(EcoRIおよびBspEI消化)およびインサートをアガロースゲル電気泳動によって精製した。
【0227】
精製された断片(EcoRI-BspEI)をその後、pEF-DHFRベクターの対応する部位にクローニングした。最終的な3-5×抗CD3(VH5/VL2)(配列番号:54)構築物を制限消化物によって確認した。
【0228】
非脱免疫3-5×抗CD3構築物のクローニング
3-5×抗CD3(VH5/VL2)構築物をクローニングするため、対応する非脱免疫構築物を以下のように作製した。
【0229】
3-5を二つの部分で含む断片IおよびIIを、プライマー対me 81(配列番号:56)/me 90(配列番号:52)、およびme 83(配列番号:50)/me 84(配列番号:51)をそれぞれ用いて、3-1×抗CD3に関して記述した条件に従ってPCRによって増幅した。PCR断片IおよびIIを含むアガロースゲル断片を、完全な3-5を含む断片IIIを増幅するために、プライマー対me 81(配列番号:56)およびme 84(配列番号:51)によって再増幅した。PCRは先に記述した通りに実施した。断片IIIをアガロースゲルにおいて精製して、BssHIIおよびBspEI(Biolabs)によって消化して、精製し、その後pEF-DHFRベクターの対応する部位にクローニングした。クローニングした領域を制限消化物およびDNAシークエンシングによって確認した。
Me81プライマー(配列番号:56)の配列:Me 81
【0230】
8.2.1.3 脱免疫4-1×抗CD3(VH5/VL2)構築物(配列番号:57、58)のクローニング
4-1×抗CD3分子を構築するために、VH-VL方向のC-末端4-1をPCRによって得た。抗EpCAM抗体4-1のVHおよびVL領域を配列番号:145および147に示す。プラスミドpEF-DHFR-4-1×抗CD3およびpEF抗CD3(VH5/VL2)(配列番号:192)を、インサート(4-1)およびベクターをそれぞれ単離するために、EcoRIおよびBspEI(Biolabs)によって消化した。脱燐酸化ベクター(EcoRIおよびBspEI消化)およびインサートをアガロースゲル電気泳動によって精製した。
【0231】
精製断片(EcoRI-BspEI)をその後、ベクターの対応する部位にクローニングした。最終的な構築物4-1×抗CD3(VH5/VL2)(配列番号:57)を制限消化物によって確認した。
【0232】
非脱免疫4-1×抗CD3構築物のクローニング
4-1×抗CD3(VH5/VL2)構築物をクローニングするため、対応する非脱免疫構築物を以下のように作製した。
【0233】
4-1を二つの部分で含む断片IおよびIIを、プライマー対me 91a(配列番号:53)/me 90(配列番号:452)およびme 83(配列番号:50)/me 84(配列番号:51)をそれぞれ用いて上記の条件によってPCRによって増幅した。
【0234】
完全な4-1を含む断片IIIを増幅するために、PCR断片IおよびIIを含むアガロースゲル断片を、プライマー対me 92a(配列番号:59)およびme 84(配列番号:51)によって再増幅した。PCRは先に記述した通りに実施したが、アニーリングは68℃で行った。断片IIIをアガロースゲルにおいて精製して、BsrGIおよびBspEI(Biolabs)によって消化して、精製し、その後pEF-DHFR-抗EpCAM(M79)×抗CD3クローニングベクター構築物の対応する部位にクローニングした。クローニングした領域を制限消化物およびDNAシークエンシングによって確認した。
Me92aプライマーの配列(配列番号:59):Me 92a:
【0235】
8.1.2.4 脱免疫4-7×抗CD3(VH5/VL2)構築物(配列番号:60、61)のクローニング
4-7×抗CD3を構築するために、VH-VL方向でのC-末端4-7をPCRによって得た。抗EpCAM抗体4-7のVHおよびVL領域を配列番号:149および151に示す。プラスミドpEF-DHFR-4-7×抗CD3およびpEF-抗CD3VH5/VL2(配列番号:192)を、インサート(4-7)およびベクターをそれぞれ単離するために、EcoRIおよびBspEI(Biolabs)によって消化した。脱燐酸化ベクター(EcoRIおよびBspEI消化)およびインサートをアガロースゲル電気泳動によって精製した。
【0236】
次に、精製断片(EcoRI-BspEI)をpEF-DHFRベクターの対応する部位にクローニングした。最終的な構築物4-7×抗CD3(VH5/VL2)(配列番号:60)を制限消化物によって確認した。
【0237】
非脱免疫構築物4-7×抗CD3のクローニング
4-7×抗CD3(VH5/VL2)構築物をクローニングするために、対応する非脱免疫構築物を以下のように作製した。
【0238】
4-7を二つの部分で含む断片IおよびIIを、プライマー対me 81(配列番号:56)/me 90(配列番号:52)、およびme 83(配列番号:50)/me 84(配列番号:51)をそれぞれ用いて、上記の条件に従ってPCRによって増幅した。PCR断片IおよびIIを含むアガロースゲル断片を、完全な4-7 VHおよびVL鎖を含む断片IIIを増幅するために、プライマー対me 81(配列番号:56)およびme 84(配列番号:51)によって再増幅した。PCRは先に記述した通りに実施した。断片IIIをアガロースゲルにおいて精製して、BssHIIおよびBspEI(Biolabs)によって消化して、精製した後pEF-DHFRベクターの対応する部位にクローニングした。クローニングした領域を制限消化物およびDNAシークエンシングによって確認した。
【0239】
8.1.2.5 脱免疫5-10×抗CD3(VH5/VL2)構築物(配列番号:62、63)のクローニング
5-10×抗CD3分子を構築するために、VH-VL方向でのC-末端5-10をPCRによって得た。抗EpCAM抗体5-10のVHおよびVL領域を配列番号:133および135に示す。プラスミドpEF-DHFR-5-10×抗CD3およびpEF-抗CD3(VH5/VL2)(配列番号:192)を、インサート(5-10)およびベクターをそれぞれ単離するために、EcoRIおよびBspEI(Biolabs)によって消化した。脱燐酸化ベクター(EcoRIおよびBspEI消化)およびインサートをアガロースゲル電気泳動によって精製した。
【0240】
精製断片(EcoRI-BspEI)をその後pEF-DHFRベクターの対応する部位にクローニングした。最終的な構築物5-10×抗CD3(VH5/VL2)(配列番号:62)を制限消化物およびDNAシークエンシングによって確認した。
【0241】
非脱免疫5-10×抗CD3構築物のクローニング
5-10×抗CD3(VH5/VL2)構築物をクローニングするために、対応する非脱免疫構築物を以下のように作製した。
【0242】
5-10を二つの部分で含む断片IおよびIIを、プライマー対me 92a(配列番号:59)/me 90(配列番号:52)、およびme 83(配列番号:50)/me 84(配列番号:51)をそれぞれ用いて、上記の条件に従ってPCRによって増幅した。PCR断片IおよびIIを含むアガロースゲル断片を、完全な5-10を含む断片IIIを増幅するために、プライマー対me 92a(配列番号:59)およびme 84(配列番号:51)によって再増幅した。PCRは先に記述した通りに実施したが、アニーリングは68℃で行った。断片IIIをアガロースゲルにおいて精製して、BsrGIおよびBspEI(Biolabs)によって消化して、精製し、その後pEF-DHFR抗EpCAM(M79)×抗CD3クローニングベクターの対応する部位にクローニングした。クローニングした領域を制限消化物およびDNAシークエンシングによって確認した。
【0243】
8.2.2 N-末端位で抗EpCAMを有する抗EpCAM×脱免疫抗CD3分子の発現
DHFR遺伝子を欠損するCHO細胞を、10%仔ウシ胎児血清(Life Technologies、65℃で30分間熱不活化)およびHT(ヒポキサンチンおよびチミジン;Life Technologies)を添加したαMEM培地(Life Technologies, cat No. 32561)において維持した。Lipofectamine 2000キット(登録商標)(Invitrogen)を用いて製造元によって提供された説明書に従って、細胞にpEF-DHFR-3-1×抗CD3(VH5/VL2)(配列番号:48)およびpEF-DHFR-5-10×抗CD3(VH5/VL2)(配列番号:62)をトランスフェクトした。48時間後、選択培地(熱不活化10%透析仔ウシ胎児血清(Life Technologies)を含むαMEM培地)において選択を行った。3〜4週間後、細胞培養上清を回収して、4℃で300 gで10分間遠心して細胞および細胞の破片を除去した。二重特異性抗体を含む上清を、さらに分析するまで-20℃で保存した。
【0244】
8.2.3 二重特異性抗EpCAM×抗CD3変種の結合アッセイ
Jurkat細胞(CD3結合に関して)およびKato細胞(EpCAM結合に関して)250000個を、二重特異性構築物(それぞれ、pEF-DHFR-3-1×抗CD3(VH5/VL2)(Nr. 50、配列番号:48)およびpEF-DHFR-5-10×抗CD3(VH5/VL2)(Nr.54)(配列番号:62))を含む細胞培養上清(50 μl)と共に独立して4℃で45分間インキュベートした。その後、細胞をFACS緩衝液(1%仔ウシ胎児血清(FCS)および0.05%アジ化ナトリウムを含む燐酸緩衝生理食塩液)において2回洗浄して、マウス抗His抗体(Dianova、DIA 910)と共に4℃で60分間インキュベートした。洗浄段階は上記の通りに行った。
【0245】
細胞を最終的にヤギ抗マウスIg-FITC結合抗体(BD 550003)またはPEに結合させた抗マウスIgG(Sigma, P8547)のいずれかと共に最終的にインキュベートした。洗浄段階後、事象10000個をFACS Calibur(B&D)を用いて分析した。結合アッセイの結果を図16に示す。構築物3-1×抗CD3(VH5/VL2)(配列番号:49)および5-10×抗CD3(配列番号:63)は、Jurkat細胞上でCD3に対して、およびKato細胞上でCD19に対して強い結合を示した。
【0246】
実施例8.3 脱免疫抗CD3部分を有する二重特異性抗EpCAM構築物の精製
脱免疫抗CD3抗原およびEpCAM特異的領域を含む構築物を、固定金属アフィニティクロマトグラフィー(IMAC)およびゲル濾過を含む二段階精製プロセスによって精製した。金属アフィニティクロマトグラフィー(IMAC)およびゲル濾過は実施例3.2に示すように実行した。
【0247】
さらに高分解能の陽イオン交換クロマトグラフィーを、20 mM MES緩衝液pH 5.5によって平衡にしたMiniSカラム(Amersham)において行った。試料を同じ緩衝液によって1:3に希釈してからカラムにローディングした。結合したタンパク質を0〜30%勾配の1 M NaClの平衡緩衝液溶液によって溶出した。溶出したタンパク質分画を生物活性アッセイにおいて試験した。表14は、精製脱免疫EpCAM構築物の収量を示す。構築物は全て、有効に産生することができた。意外にも構築物5-10×抗-CD3(VH5/VL2)(配列番号:63)は、2200 μg/lという極めて良好な収量を有した。
【0248】
(表14)脱免疫EpCAM構築物の収量
【0249】
実施例8.4 脱免疫抗CD3部分による二重特異性抗EpCAM構築物の細胞障害アッセイ
本発明の二重特異性抗体の高い生物活性を確認するために、FACSに基づくアッセイを行った。CHO細胞に上皮細胞接着分子(EpCAM)をトランスフェクトした。CHO-EpCAM細胞と呼ばれるこのトランスフェクションに由来する細胞クローンを実験に用いた。
【0250】
細胞毒性試験のために、CHO-EpCAM(1.5×107)を、PBSで2回洗浄して血清を除き、製造業者の指示書に従ってPKH25色素(Sigma-Aldrich Co.)とともにインキュベートした。細胞染色後にRPMI/10% FCSで2回洗浄した。
【0251】
細胞を計数し、CB15エフェクター細胞と混合した。CD4ポジティブT細胞クローンCB15は、Fickenscher博士(University of Erlangen/Nuernberg, Germany)からご厚意で提供された。細胞を、供給業者によって推奨されるように培養した。得られる細胞懸濁物は、mlあたり400,000標的および2×106エフェクター細胞を含んだ。96ウェル丸底プレート中のウェルあたり、50μlの混合物を使用した。
【0252】
抗体をRPMI/10% FCS中で必要とされる濃度まで希釈し、この溶液の50μlを細胞懸濁物に加えた。標準的な反応は、37℃/5% CO2中で16時間インキュベートした。ヨウ化プロピジウムを1μg/mlの最終濃度で加えた。室温での10分間のインキュベーション後、細胞をFACSによって分析した。PKH26蛍光を、標的細胞のポジティブ同定のために使用した。細胞毒性を、全ての標的細胞にわたるPIポジティブの比として測定した。
【0253】
シグモイド型用量応答曲線は、代表的には、Prismソフトウェア(GraphPad Software Inc., San Diego, USA)によって決定されるように、>0.97のR2値を有した。細胞毒性アッセイの結果は、図17および18に示されている。
【0254】
実施例8.5 CHO細胞におけるEpCAM結合部分と脱免疫CD3結合部分とを含む二重特異性分子の生産性の比較
脱免疫構築物タンパク質の生産性を決定するために、プロテインL ELISAを行った。生産性のデータはバッチ培養から計算した。
【0255】
8.5.1 細胞培養
脱免疫(CHO-DHFR-)および非脱免疫(CHO-DHFR-またはCHO-K1)を産生するCHO細胞株を、HyQ PF CHO LS培地+4 mM L-グルタミンにおいて37℃および5%CO2のCO2インキュベータにおいて培養した。細胞数および生存率はトリパンブルーによって決定した。細胞密度を1〜2×105個mlに設定した。
【0256】
細胞をスピンナフラスコに移して、攪拌培養の条件に調節した。操作パラメータの設定は、CO2インキュベータにおいて通気しながら80 rpm、37℃、および5%CO2であった。培養容積は100〜500 mlの範囲であり、インキュベーション時の細胞密度は1〜2×105個/mlの範囲であった。Tフラスコでのサブ培養に関して、各継代毎に培養物を遠心して、新鮮な予め加温した培地に浮遊させた。細胞密度は1〜2×105個/mlに設定した。
【0257】
生産性のデータの分析に関して(表15)、培地の添加または交換を行わずに細胞を14日(d)まで培養した。細胞数および生存率をトリパンブルー染色によって毎日決定した。上清における産物の濃度をプロテインL ELISAによって分析した。
【0258】
8.5.2 プロテインL ELISA
二重特異性分子の定量的結合は、rプロテインLコーティングマイクロタイタープレートにおいて行った。rプロテインLは、ペプトストレプトコッカス・マグヌス(Peptostreptococcus magnus)によって産生される免疫グロブリン結合プロテインLの組換え型である。これは四つの結合部位を有し、軽鎖(κ)を通して免疫グロブリンに結合する。親抗体のそれぞれ異なる二つの軽鎖からの可変ドメインを含む二重特異性分子は、rプロテインLによって結合される。
【0259】
マイクロタイタープレートをrプロテインLのPBS緩衝液溶液(2μg rプロテインL/ml PBS緩衝液)によって2〜8℃で一晩コーティングした。コーティングの後、残っている吸着部位をブロッキング緩衝液(2%BSAのPBS緩衝液溶液)によってブロックした。次に、プレートを凍結して≦18℃で保存した。使用前、プレートを融解して、洗浄緩衝液(0.05%Tween 20のPBS緩衝液)によって洗浄し、コーティング溶液とブロッキング緩衝液の混合物を除去した。
【0260】
無細胞培養上清の1%BSA+0.01%Tween 20のPBS(希釈緩衝液)による連続希釈液を分析した。二重特異性抗EpCAM(M79)×抗CD3を同等の希釈液において陽性対照として用いた。
【0261】
インキュベーションは2〜8℃で一晩行った。
【0262】
洗浄後、ウサギ抗マウスIgG(希釈緩衝液において1:5,000倍希釈)を加えて、室温で60分間インキュベートした。洗浄後、アルカリホスファターゼ標識ヤギ抗ウサギIgGを加えた(希釈緩衝液において1:1,000倍希釈、室温で60分)。pNPP基質溶液を加えて、3 M NaOHを加えることによって反応を停止させた。ELISAリーダーによって405 nm(参照フィルター492 nm)で吸光度を測定した。
【0263】
(表15)脱免疫抗EpCAM構築物の生産性
【0264】
このように、本発明の5-10×抗CD3(VH5/VL2)構築物は、先行技術の二重特異性脱免疫EpCAMおよびCD3結合抗体よりかなり高い(少なくとも5倍高い)特異的生産性を示した。
【特許請求の範囲】
【請求項1】
ヒトCD3に対して特異的に結合する第一のドメインとIg由来第二の結合ドメインとを含む、細胞障害活性CD3特異的結合構築物であって、
該第一のドメインが脱免疫され、CDR-H1領域、CDR-H2領域、およびCDR-H3領域を含み、該CDR-H3領域が配列番号:96、108、119、120、121、122、123、124、125、126、または127に示されるアミノ酸配列を含み、ならびに
該第一のドメインがさらに、そのフレームワークH1において配列VKKを含み、フレームワークH1とCDRH1領域との間の移行配列が配列Ala-Ser-Gly-Tyr-Thr-Phe(ASGYTF;配列番号:233)を含む、CD3特異的結合構築物。
【請求項2】
第一のドメインにおいて、配列Met-Glu-Leu-Ser(MELS;配列番号:234)を含むフレームワークH3をさらに含む、請求項1記載の細胞障害活性CD3特異的結合構築物。
【請求項3】
第一のドメインにおいて、配列Ile-Thr-Thr-Asp-Lys(ITTDK;配列番号:235)を含むフレームワークH3をさらに含む、請求項1または2記載の細胞障害活性CD3特異的結合構築物。
【請求項4】
ヒトCD3に特異的に結合する第一のドメインが、配列番号:152または153に示されるフレームワークH1を含む、請求項1〜3のいずれか一項に記載のCD3特異的結合構築物。
【請求項5】
ヒトCD3に特異的に結合する第一のドメインが、配列番号:156または157に示されるフレームワークH2を含む、請求項1〜4のいずれか一項に記載のCD3特異的結合構築物。
【請求項6】
ヒトCD3に特異的に結合する第一のドメインが、配列番号:160または161に示されるフレームワークH3を含む、請求項1〜5のいずれか一項に記載のCD3特異的結合構築物。
【請求項7】
ヒトCD3に特異的に結合する第一のドメインが、配列番号:164または165に示されるフレームワークH4を含む、請求項1〜6のいずれか一項に記載のCD3特異的結合構築物。
【請求項8】
以下を含む、請求項1〜7のいずれか一項に記載のCD3特異的結合構築物:
(a)配列番号:88に記載のCDR-H1;および
(b)配列番号:90または92に記載のCDR-H2。
【請求項9】
配列番号:74または76に記載のVH領域を含む、請求項1〜8のいずれか一項に記載のCD3特異的結合構築物。
【請求項10】
配列番号:98または100に記載されるCDR-L1を含む、請求項1〜9のいずれか一項に記載のCD3特異的結合構築物。
【請求項11】
配列番号:102に記載のCDR-L2を含む、請求項1〜10のいずれか一項に記載のCD3特異的結合構築物。
【請求項12】
配列番号:104に記載のCDR-L3を含む、請求項1〜11のいずれか一項に記載のCD3特異的結合構築物。
【請求項13】
配列番号:78、配列番号:80、配列番号:82、および配列番号:112からなる群より選択されるVL領域をそのCD3特異的部分に含む、請求項1〜12のいずれか一項に記載のCD3特異的結合構築物。
【請求項14】
Ig由来第二のドメインが、scFvである、請求項1〜13のいずれか一項に記載のCD3特異的結合構築物。
【請求項15】
Ig由来第二のドメインおよび/または連結するリンカー領域がヒト化および/または脱免疫されている、請求項1〜14のいずれか一項に記載のCD3特異的結合構築物。
【請求項16】
Ig由来第二のドメインが、細胞表面分子に対して特異性を有する抗原相互作用部位を含む、請求項1〜15のいずれか一項に記載のCD3特異的結合構築物。
【請求項17】
細胞表面分子が、腫瘍特異的マーカーである、請求項16記載のCD3特異的結合構築物。
【請求項18】
Ig由来第二の結合ドメインが、EpCAM、CCR5、CD19、HER-2、HER-2 neu、HER-3、HER-4、EGFR、PSMA、CEA、MUC-1(ムチン)、MUC2、MUC3、MUC4、MUC5AC、MUC5B、MUC7、βhCG、Lewis-Y、CD20、CD33、CD30、ガングリオシドGD3、9-O-アセチル-GD3、GM2、Globo H、フコシルGM1、ポリSA、GD2、炭酸脱水酵素IX(MN/CA IX)、CD44v6、Sonic Hedgehog(Shh)、Wue-1、プラズマ細胞抗原、(膜結合型)IgE、黒色腫コンドロイチン硫酸プロテオグリカン(MCSP)、CCR8、TNF-α前駆体、STEAP、メソセリン、A33抗原、前立腺幹細胞抗原(PSCA)、Ly-6、デスモグレイン4、E-カドヘリンネオエピトープ、胎児アセチルコリン受容体、CD25、CA19-9マーカー、CA-125マーカー、およびミュラー管阻害物質(MIS)受容体II型、sTn(シアリル化Tn抗原、TAG72)、FAP(線維芽細胞活性化抗原)、エンドシアリン、EGFRvIII、L6、SAS、CD63、TAG72、TF-抗原、Cora抗原、CD7、CD22、Igα、Igβ、G250、gp100、MT-MMPs、F19-抗原、CO-29およびEphA2からなる群より選択される分子に対して特異性を有する抗原相互作用部位を含む、請求項16または17に記載のCD3特異的結合構築物。
【請求項19】
第二のIg由来結合ドメインが、EpCAMに対して特異性を有する抗原相互作用部位を含む、請求項1〜18のいずれか一項に記載のCD3特異的結合構築物。
【請求項20】
以下からなる群より選択されるアミノ酸配列を含む、請求項19記載のCD3特異的結合構築物:
(a)配列番号:31、33、35、37、39、49、55、58、61、63、65、67、237、239、241、243、245、247、249、251、253、255、257、259、261、263、265、267、269、271、273、275、277、279、281、283、285、287、289、291、293、295、297、299、301、303、305、307、309、311、313、315、317、319、321、323、および325のいずれか一つに示されるアミノ酸配列;
(b)配列番号:30、32、34、36、38、48、54、57、60、62、64、66、236、238、240、242、244、246、248、250、252、254、256、258、260、262、264、266、268、270、272、274、276、278、280、282、284、286、288、290、292、294、296、298、300、302、304、306、308、310、312、314、316、318、320、322、および324のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列;ならびに
(d)ストリンジェントなハイブリダイゼーション条件で、(b)において定義される核酸配列の相補鎖とハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【請求項21】
Ig由来第二の結合ドメインが、CCR5に対して特異性を有する抗原相互作用部位を含む、請求項1〜18のいずれか一項に記載のCD3特異的結合構築物。
【請求項22】
以下からなる群より選択されるアミノ酸配列を含む、請求項21記載のCD3特異的結合構築物:
(a)配列番号:206、208、210、212、214、または216のいずれか一つに示されるアミノ酸配列;
(b)配列番号:205、207、209、211、213、または215のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列;および
(d)(b)において定義された核酸配列の相補鎖とストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【請求項23】
Ig由来第二の結合ドメインが、CD19に対して特異性を有する抗原相互作用部位を含む、請求項1〜18のいずれか一項に記載のCD3特異的結合構築物。
【請求項24】
以下からなる群より選択されるアミノ酸配列を含む、請求項23記載のCD3特異的結合構築物:
(a)配列番号:190、192、194、196、198、200、327、329、331、333、335、337、339、341、343、345、347、349、351、353、357、359、361、363、365、367、369、371、373、375、377、379、381、383、385、387、389、391、393、395、397、399、401、403、405、407、または409のいずれか一つに示されるアミノ酸配列;
(b)配列番号:189、191、193、195、197、199、326、328、330、332、334、336、338、340、342、344、346、348、350、352、354、356、358、360、362、364、366、368、370、372、374、376、378、380、382、384、386、388、390、392、394、396、398、400、402、404、406、または408のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列;ならびに
(d)(b)において定義された核酸配列の相補鎖とストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【請求項25】
Ig由来第二の結合ドメインが、CD20に対して特異性を有する抗原相互作用部位を含む、請求項1〜18のいずれか一項に記載のCD3特異的結合構築物。
【請求項26】
以下からなる群より選択されるアミノ酸配列を含む、請求項25記載のCD3特異的結合構築物:
(a)配列番号:218、220、222、224、226、または228のいずれか一つに示されるアミノ酸配列;
(b)配列番号:217、219、221、223、225、または227のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列;および
(d)(b)において定義された核酸配列の相補鎖とストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【請求項27】
請求項1〜26のいずれか一項に記載のCD3特異的結合構築物をコードする核酸配列。
【請求項28】
請求項27記載の核酸配列を含むベクター。
【請求項29】
請求項27記載の核酸配列に機能的に結合した調節配列である核酸配列をさらに含む、請求項28記載のベクター。
【請求項30】
発現ベクターである、請求項28または29記載のベクター。
【請求項31】
請求項28〜30のいずれか一項に記載のベクターによって形質転換またはトランスフェクトした宿主。
【請求項32】
ポリペプチド構築物を発現させて、産生されたポリペプチド構築物を培養物から回収する条件で請求項31記載の宿主を培養する段階を含む、請求項1〜26のいずれか一項に記載のCD3特異的結合構築物を産生するプロセス。
【請求項33】
請求項1〜26のいずれか一項に記載のもしくは請求項32記載のプロセスによって産生されたCD3特異的結合構築物、請求項27記載の核酸分子、請求項28〜30のいずれか一つに記載のベクター、または請求項31記載の宿主、および任意で免疫エフェクター細胞の活性化シグナルを提供することができる蛋白質様化合物、を含む組成物。
【請求項34】
任意で、担体、安定化剤、および/または賦形剤の適した製剤をさらに含む薬学的組成物である、請求項33記載の組成物。
【請求項35】
任意で検出手段および方法をさらに含む診断組成物である、請求項33記載の組成物。
【請求項36】
増殖性疾患、腫瘍性の疾患、炎症疾患、免疫障害、自己免疫疾患、感染症、ウイルス病、アレルギー反応、寄生虫反応、移植片対宿主病、または宿主対移植片病の予防、治療、または改善のための薬学的組成物を調製するために、請求項1〜26のいずれか一項に記載のもしくは請求項32記載のプロセスによって産生されたCD3特異的結合構築物、請求項27記載の核酸分子、請求項28〜30のいずれか一つに記載のベクター、または請求項31記載の宿主の使用。
【請求項37】
請求項1〜26のいずれか一項に記載のもしくは請求項32記載のプロセスによって産生されたCD3特異的結合構築物、請求項27記載の核酸分子、請求項28〜30のいずれか一つに記載のベクター、または請求項31記載の宿主を、そのような予防、治療、または改善を必要とする被験者に投与する段階を含む、増殖性疾患、腫瘍性の疾患、炎症疾患、免疫障害、自己免疫疾患、感染症、ウイルス病、アレルギー反応、寄生虫反応、移植片対宿主病、または宿主対移植片病の予防、治療、または改善のための方法。
【請求項38】
被験者が、ヒトである、請求項37記載の方法。
【請求項39】
免疫エフェクター細胞の活性化シグナルを提供することができる蛋白質様化合物の投与をさらに含む、請求項37または38に記載の方法。
【請求項40】
蛋白質様化合物が、請求項1〜26のいずれか一項に記載のもしくは請求項32記載のプロセスによって産生されたCD3特異的結合構築物、請求項27記載の核酸分子、請求項28〜30のいずれか一つに記載のベクター、または請求項31記載の宿主と同時にまたは非同時に投与される、請求項39記載の方法。
【請求項41】
請求項1〜26のいずれか一項に記載のもしくは請求項32記載のプロセスによって産生されたCD3特異的結合構築物、請求項27記載の核酸分子、請求項28〜30のいずれか一項に記載のベクター、または請求項31記載の宿主を含むキット。
【請求項1】
ヒトCD3に対して特異的に結合する第一のドメインとIg由来第二の結合ドメインとを含む、細胞障害活性CD3特異的結合構築物であって、
該第一のドメインが脱免疫され、CDR-H1領域、CDR-H2領域、およびCDR-H3領域を含み、該CDR-H3領域が配列番号:96、108、119、120、121、122、123、124、125、126、または127に示されるアミノ酸配列を含み、ならびに
該第一のドメインがさらに、そのフレームワークH1において配列VKKを含み、フレームワークH1とCDRH1領域との間の移行配列が配列Ala-Ser-Gly-Tyr-Thr-Phe(ASGYTF;配列番号:233)を含む、CD3特異的結合構築物。
【請求項2】
第一のドメインにおいて、配列Met-Glu-Leu-Ser(MELS;配列番号:234)を含むフレームワークH3をさらに含む、請求項1記載の細胞障害活性CD3特異的結合構築物。
【請求項3】
第一のドメインにおいて、配列Ile-Thr-Thr-Asp-Lys(ITTDK;配列番号:235)を含むフレームワークH3をさらに含む、請求項1または2記載の細胞障害活性CD3特異的結合構築物。
【請求項4】
ヒトCD3に特異的に結合する第一のドメインが、配列番号:152または153に示されるフレームワークH1を含む、請求項1〜3のいずれか一項に記載のCD3特異的結合構築物。
【請求項5】
ヒトCD3に特異的に結合する第一のドメインが、配列番号:156または157に示されるフレームワークH2を含む、請求項1〜4のいずれか一項に記載のCD3特異的結合構築物。
【請求項6】
ヒトCD3に特異的に結合する第一のドメインが、配列番号:160または161に示されるフレームワークH3を含む、請求項1〜5のいずれか一項に記載のCD3特異的結合構築物。
【請求項7】
ヒトCD3に特異的に結合する第一のドメインが、配列番号:164または165に示されるフレームワークH4を含む、請求項1〜6のいずれか一項に記載のCD3特異的結合構築物。
【請求項8】
以下を含む、請求項1〜7のいずれか一項に記載のCD3特異的結合構築物:
(a)配列番号:88に記載のCDR-H1;および
(b)配列番号:90または92に記載のCDR-H2。
【請求項9】
配列番号:74または76に記載のVH領域を含む、請求項1〜8のいずれか一項に記載のCD3特異的結合構築物。
【請求項10】
配列番号:98または100に記載されるCDR-L1を含む、請求項1〜9のいずれか一項に記載のCD3特異的結合構築物。
【請求項11】
配列番号:102に記載のCDR-L2を含む、請求項1〜10のいずれか一項に記載のCD3特異的結合構築物。
【請求項12】
配列番号:104に記載のCDR-L3を含む、請求項1〜11のいずれか一項に記載のCD3特異的結合構築物。
【請求項13】
配列番号:78、配列番号:80、配列番号:82、および配列番号:112からなる群より選択されるVL領域をそのCD3特異的部分に含む、請求項1〜12のいずれか一項に記載のCD3特異的結合構築物。
【請求項14】
Ig由来第二のドメインが、scFvである、請求項1〜13のいずれか一項に記載のCD3特異的結合構築物。
【請求項15】
Ig由来第二のドメインおよび/または連結するリンカー領域がヒト化および/または脱免疫されている、請求項1〜14のいずれか一項に記載のCD3特異的結合構築物。
【請求項16】
Ig由来第二のドメインが、細胞表面分子に対して特異性を有する抗原相互作用部位を含む、請求項1〜15のいずれか一項に記載のCD3特異的結合構築物。
【請求項17】
細胞表面分子が、腫瘍特異的マーカーである、請求項16記載のCD3特異的結合構築物。
【請求項18】
Ig由来第二の結合ドメインが、EpCAM、CCR5、CD19、HER-2、HER-2 neu、HER-3、HER-4、EGFR、PSMA、CEA、MUC-1(ムチン)、MUC2、MUC3、MUC4、MUC5AC、MUC5B、MUC7、βhCG、Lewis-Y、CD20、CD33、CD30、ガングリオシドGD3、9-O-アセチル-GD3、GM2、Globo H、フコシルGM1、ポリSA、GD2、炭酸脱水酵素IX(MN/CA IX)、CD44v6、Sonic Hedgehog(Shh)、Wue-1、プラズマ細胞抗原、(膜結合型)IgE、黒色腫コンドロイチン硫酸プロテオグリカン(MCSP)、CCR8、TNF-α前駆体、STEAP、メソセリン、A33抗原、前立腺幹細胞抗原(PSCA)、Ly-6、デスモグレイン4、E-カドヘリンネオエピトープ、胎児アセチルコリン受容体、CD25、CA19-9マーカー、CA-125マーカー、およびミュラー管阻害物質(MIS)受容体II型、sTn(シアリル化Tn抗原、TAG72)、FAP(線維芽細胞活性化抗原)、エンドシアリン、EGFRvIII、L6、SAS、CD63、TAG72、TF-抗原、Cora抗原、CD7、CD22、Igα、Igβ、G250、gp100、MT-MMPs、F19-抗原、CO-29およびEphA2からなる群より選択される分子に対して特異性を有する抗原相互作用部位を含む、請求項16または17に記載のCD3特異的結合構築物。
【請求項19】
第二のIg由来結合ドメインが、EpCAMに対して特異性を有する抗原相互作用部位を含む、請求項1〜18のいずれか一項に記載のCD3特異的結合構築物。
【請求項20】
以下からなる群より選択されるアミノ酸配列を含む、請求項19記載のCD3特異的結合構築物:
(a)配列番号:31、33、35、37、39、49、55、58、61、63、65、67、237、239、241、243、245、247、249、251、253、255、257、259、261、263、265、267、269、271、273、275、277、279、281、283、285、287、289、291、293、295、297、299、301、303、305、307、309、311、313、315、317、319、321、323、および325のいずれか一つに示されるアミノ酸配列;
(b)配列番号:30、32、34、36、38、48、54、57、60、62、64、66、236、238、240、242、244、246、248、250、252、254、256、258、260、262、264、266、268、270、272、274、276、278、280、282、284、286、288、290、292、294、296、298、300、302、304、306、308、310、312、314、316、318、320、322、および324のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列;ならびに
(d)ストリンジェントなハイブリダイゼーション条件で、(b)において定義される核酸配列の相補鎖とハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【請求項21】
Ig由来第二の結合ドメインが、CCR5に対して特異性を有する抗原相互作用部位を含む、請求項1〜18のいずれか一項に記載のCD3特異的結合構築物。
【請求項22】
以下からなる群より選択されるアミノ酸配列を含む、請求項21記載のCD3特異的結合構築物:
(a)配列番号:206、208、210、212、214、または216のいずれか一つに示されるアミノ酸配列;
(b)配列番号:205、207、209、211、213、または215のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列;および
(d)(b)において定義された核酸配列の相補鎖とストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【請求項23】
Ig由来第二の結合ドメインが、CD19に対して特異性を有する抗原相互作用部位を含む、請求項1〜18のいずれか一項に記載のCD3特異的結合構築物。
【請求項24】
以下からなる群より選択されるアミノ酸配列を含む、請求項23記載のCD3特異的結合構築物:
(a)配列番号:190、192、194、196、198、200、327、329、331、333、335、337、339、341、343、345、347、349、351、353、357、359、361、363、365、367、369、371、373、375、377、379、381、383、385、387、389、391、393、395、397、399、401、403、405、407、または409のいずれか一つに示されるアミノ酸配列;
(b)配列番号:189、191、193、195、197、199、326、328、330、332、334、336、338、340、342、344、346、348、350、352、354、356、358、360、362、364、366、368、370、372、374、376、378、380、382、384、386、388、390、392、394、396、398、400、402、404、406、または408のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列;ならびに
(d)(b)において定義された核酸配列の相補鎖とストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【請求項25】
Ig由来第二の結合ドメインが、CD20に対して特異性を有する抗原相互作用部位を含む、請求項1〜18のいずれか一項に記載のCD3特異的結合構築物。
【請求項26】
以下からなる群より選択されるアミノ酸配列を含む、請求項25記載のCD3特異的結合構築物:
(a)配列番号:218、220、222、224、226、または228のいずれか一つに示されるアミノ酸配列;
(b)配列番号:217、219、221、223、225、または227のいずれか一つに示される核酸配列によってコードされるアミノ酸配列;
(c)(b)のヌクレオチド配列に対する遺伝子コードの結果として縮重している核酸配列によってコードされるアミノ酸配列;および
(d)(b)において定義された核酸配列の相補鎖とストリンジェントなハイブリダイゼーション条件でハイブリダイズする核酸配列によってコードされるアミノ酸配列。
【請求項27】
請求項1〜26のいずれか一項に記載のCD3特異的結合構築物をコードする核酸配列。
【請求項28】
請求項27記載の核酸配列を含むベクター。
【請求項29】
請求項27記載の核酸配列に機能的に結合した調節配列である核酸配列をさらに含む、請求項28記載のベクター。
【請求項30】
発現ベクターである、請求項28または29記載のベクター。
【請求項31】
請求項28〜30のいずれか一項に記載のベクターによって形質転換またはトランスフェクトした宿主。
【請求項32】
ポリペプチド構築物を発現させて、産生されたポリペプチド構築物を培養物から回収する条件で請求項31記載の宿主を培養する段階を含む、請求項1〜26のいずれか一項に記載のCD3特異的結合構築物を産生するプロセス。
【請求項33】
請求項1〜26のいずれか一項に記載のもしくは請求項32記載のプロセスによって産生されたCD3特異的結合構築物、請求項27記載の核酸分子、請求項28〜30のいずれか一つに記載のベクター、または請求項31記載の宿主、および任意で免疫エフェクター細胞の活性化シグナルを提供することができる蛋白質様化合物、を含む組成物。
【請求項34】
任意で、担体、安定化剤、および/または賦形剤の適した製剤をさらに含む薬学的組成物である、請求項33記載の組成物。
【請求項35】
任意で検出手段および方法をさらに含む診断組成物である、請求項33記載の組成物。
【請求項36】
増殖性疾患、腫瘍性の疾患、炎症疾患、免疫障害、自己免疫疾患、感染症、ウイルス病、アレルギー反応、寄生虫反応、移植片対宿主病、または宿主対移植片病の予防、治療、または改善のための薬学的組成物を調製するために、請求項1〜26のいずれか一項に記載のもしくは請求項32記載のプロセスによって産生されたCD3特異的結合構築物、請求項27記載の核酸分子、請求項28〜30のいずれか一つに記載のベクター、または請求項31記載の宿主の使用。
【請求項37】
請求項1〜26のいずれか一項に記載のもしくは請求項32記載のプロセスによって産生されたCD3特異的結合構築物、請求項27記載の核酸分子、請求項28〜30のいずれか一つに記載のベクター、または請求項31記載の宿主を、そのような予防、治療、または改善を必要とする被験者に投与する段階を含む、増殖性疾患、腫瘍性の疾患、炎症疾患、免疫障害、自己免疫疾患、感染症、ウイルス病、アレルギー反応、寄生虫反応、移植片対宿主病、または宿主対移植片病の予防、治療、または改善のための方法。
【請求項38】
被験者が、ヒトである、請求項37記載の方法。
【請求項39】
免疫エフェクター細胞の活性化シグナルを提供することができる蛋白質様化合物の投与をさらに含む、請求項37または38に記載の方法。
【請求項40】
蛋白質様化合物が、請求項1〜26のいずれか一項に記載のもしくは請求項32記載のプロセスによって産生されたCD3特異的結合構築物、請求項27記載の核酸分子、請求項28〜30のいずれか一つに記載のベクター、または請求項31記載の宿主と同時にまたは非同時に投与される、請求項39記載の方法。
【請求項41】
請求項1〜26のいずれか一項に記載のもしくは請求項32記載のプロセスによって産生されたCD3特異的結合構築物、請求項27記載の核酸分子、請求項28〜30のいずれか一項に記載のベクター、または請求項31記載の宿主を含むキット。
【図1】


【図2A−1】

【図2A−2】

【図2B−1】

【図2B−2】

【図2B−3】

【図2C】

【図2D−1】

【図2D−2】

【図3A】

【図3B】

【図3C】

【図3D】

【図3E】

【図3F】

【図4A】

【図4B】

【図4C】
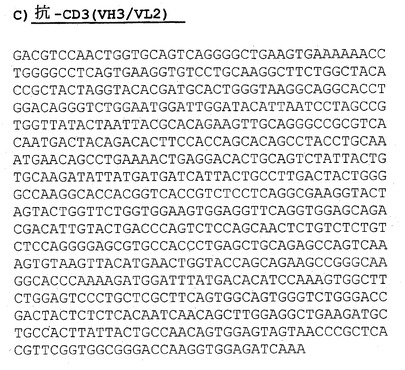
【図4D】

【図4E】

【図4F】

【図5A】

【図5B】

【図5C】

【図5D】

【図5E】

【図5F】

【図6A】

【図6B】

【図6C】

【図6D】

【図6E】

【図6F】

【図7A】
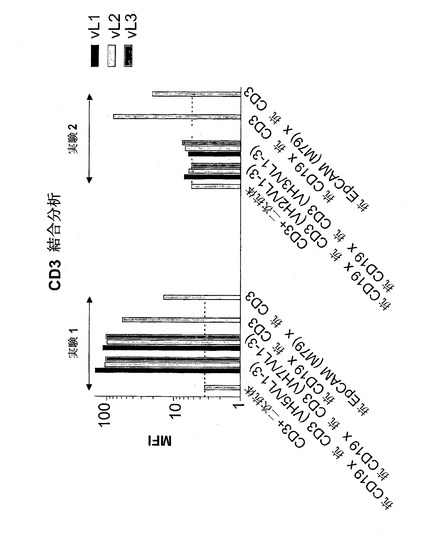
【図7B】
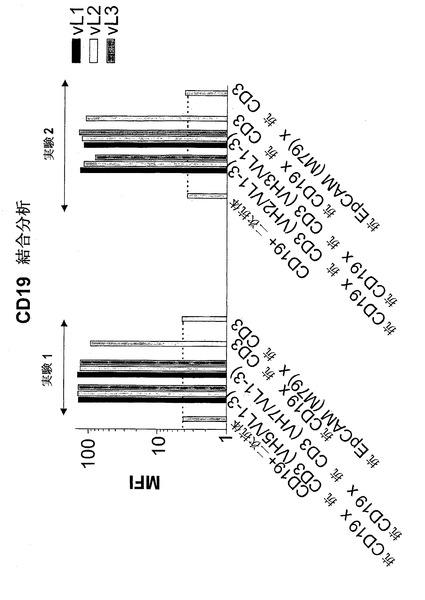
【図8】
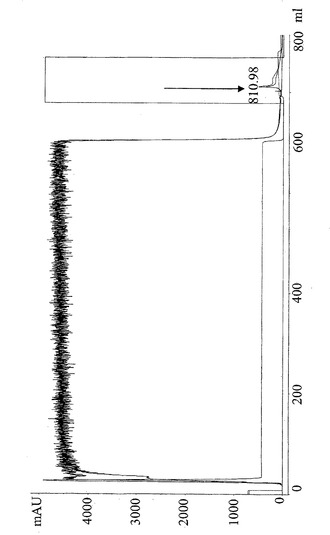
【図9】
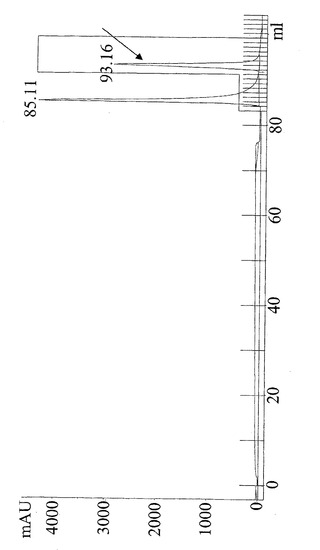
【図10】
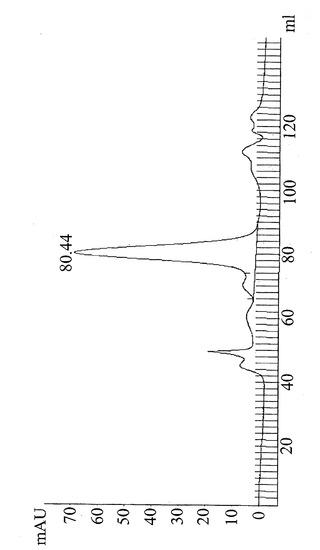
【図11】
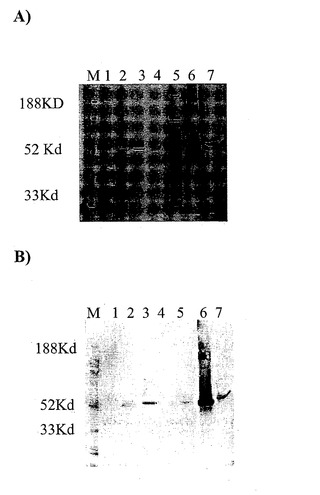
【図12A】
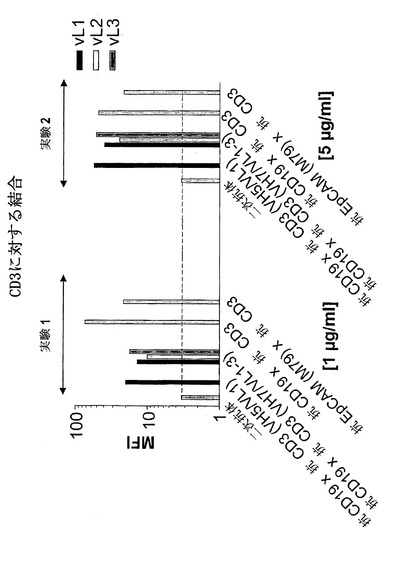
【図12B】
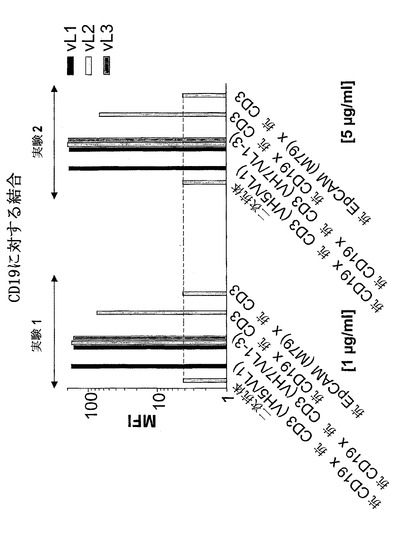
【図13】
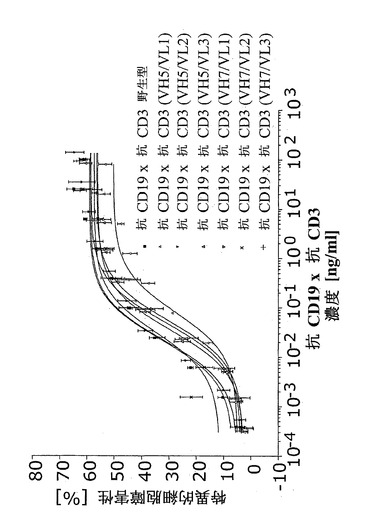
【図14】
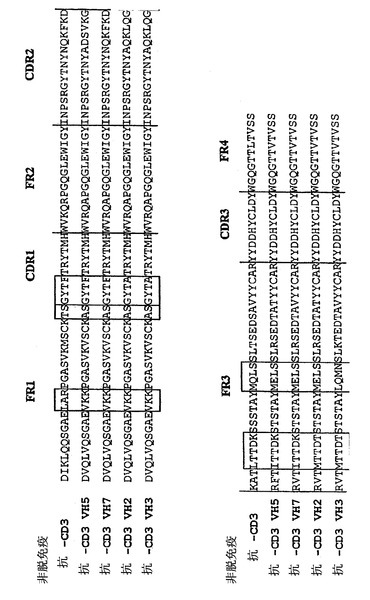
【図15A】
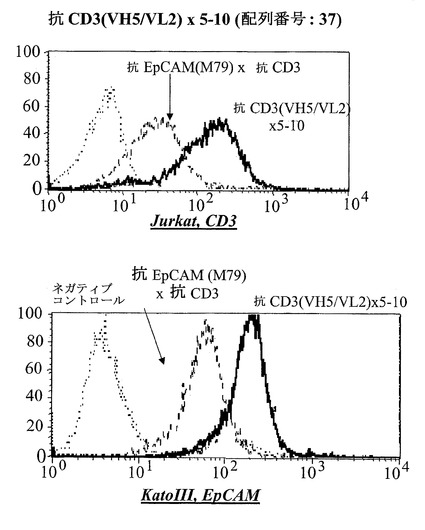
【図15B】
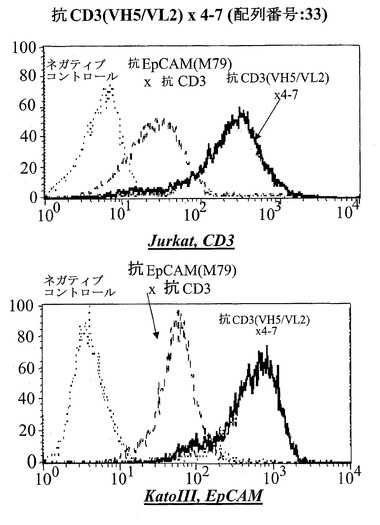
【図15C】
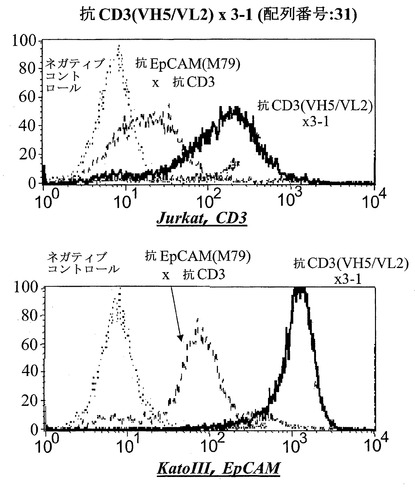
【図15D】
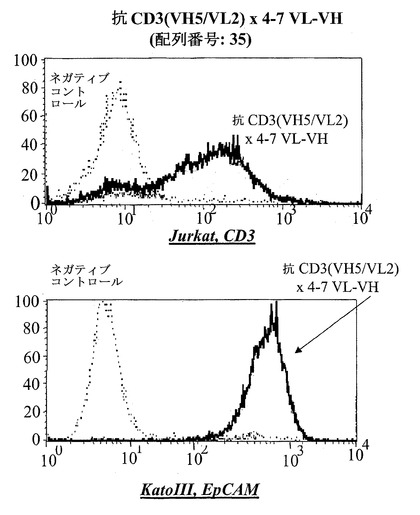
【図15E】
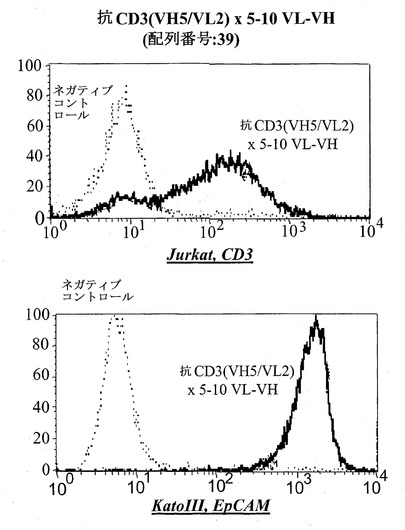
【図16A】
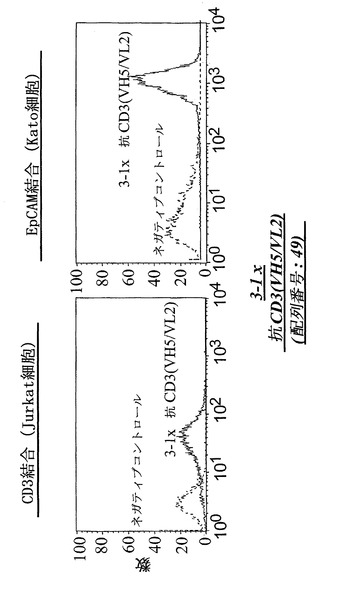
【図16B】
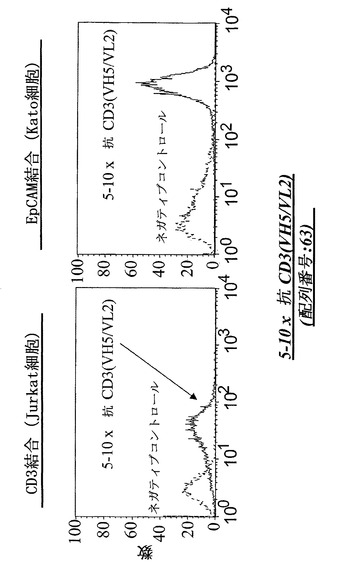
【図17】
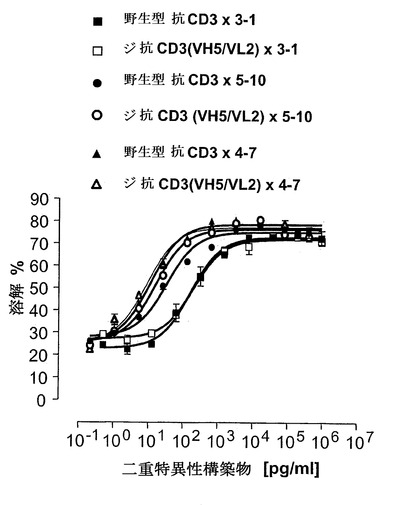
【図18】
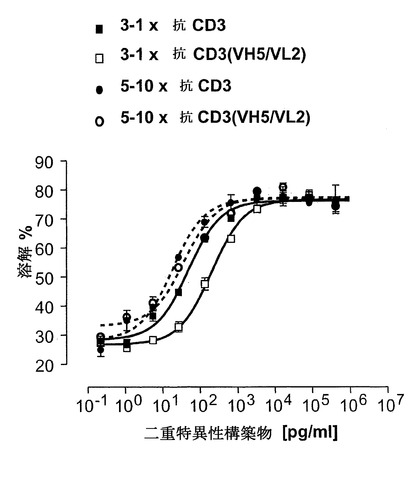
【図2A−1】
【図2A−2】
【図2B−1】
【図2B−2】
【図2B−3】
【図2C】
【図2D−1】
【図2D−2】
【図3A】
【図3B】
【図3C】
【図3D】
【図3E】
【図3F】
【図4A】
【図4B】
【図4C】
【図4D】
【図4E】
【図4F】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図5F】
【図6A】
【図6B】
【図6C】
【図6D】
【図6E】
【図6F】
【図7A】
【図7B】
【図8】
【図9】
【図10】
【図11】
【図12A】
【図12B】
【図13】
【図14】
【図15A】
【図15B】
【図15C】
【図15D】
【図15E】
【図16A】
【図16B】
【図17】
【図18】
【公開番号】特開2012−110340(P2012−110340A)
【公開日】平成24年6月14日(2012.6.14)
【国際特許分類】
【出願番号】特願2012−15443(P2012−15443)
【出願日】平成24年1月27日(2012.1.27)
【分割の表示】特願2006−534709(P2006−534709)の分割
【原出願日】平成16年10月15日(2004.10.15)
【出願人】(505298825)マイクロメット アクツィエン ゲゼルシャフト (14)
【Fターム(参考)】
【公開日】平成24年6月14日(2012.6.14)
【国際特許分類】
【出願日】平成24年1月27日(2012.1.27)
【分割の表示】特願2006−534709(P2006−534709)の分割
【原出願日】平成16年10月15日(2004.10.15)
【出願人】(505298825)マイクロメット アクツィエン ゲゼルシャフト (14)
【Fターム(参考)】
[ Back to top ]