
多重PCRによるサルモネラ属菌血清型の迅速簡易判別法およびそのプライマーセット
【課題】サルモネラ属菌のうちで主に畜産現場で問題となる血清型(Enteritidis, Infantis, Typhimurium, Choleraesuis, Dublin, Heidelberg, Schwarzengrund)について、迅速でかつ高い感度で簡便に判別できる多重検出法、および、被検試料由来増菌培養液からサルモネラ属菌を単離することなく直接多重PCRを行うことができる簡易判別法の提供。
【解決手段】各血清型に特異的特定配列のプライマーセット、Typhimurium特異的、Infantis特異的、Choleraesuis特異的、Dublin特異的、Heidelberg特異的、及びSchwarzengrund特異的なプライマーセットと共に、サルモネラ属菌特異的なinvA遺伝子増幅用プライマーセットを用いることを特徴とする、多重PCR血清型判別法。
【解決手段】各血清型に特異的特定配列のプライマーセット、Typhimurium特異的、Infantis特異的、Choleraesuis特異的、Dublin特異的、Heidelberg特異的、及びSchwarzengrund特異的なプライマーセットと共に、サルモネラ属菌特異的なinvA遺伝子増幅用プライマーセットを用いることを特徴とする、多重PCR血清型判別法。
【発明の詳細な説明】
【技術分野】
【0001】
家畜生産の現場で問題となるサルモネラ属菌の主要な血清型(Enteritidis, Infantis, Typhimurium, Choleraesuis, Dublin, Heidelberg, Schwarzengrund)についての多重PCRによる簡易的な判別手法に関する。
【背景技術】
【0002】
サルモネラ属菌は、生物学的にはSalmonella enterica及びSalmonella bongoriの2菌種に、S.entericaがさらに6亜種に分類されるが、血清学的には、細胞壁リポ多糖類のO抗原と、鞭毛タンパク質のH抗原の組み合わせで2,500を超える血清型(serovar)が存在している。サルモネラ属菌は、健康なヒトの消化管にはほとんど存在していないが、種々の動物の消化管内に常在菌として存在している場合が多い。これらのサルモネラ属菌は、各血清型によって病原性と宿主域が異なり、強い病原性がある血清型は限られており、ヒトへの感染性が問題となる血清型は少数である。日本家畜伝染病予防法における家畜の監視伝染病としては、S.Gallinarum-Pullorum及びS.Gallinarum-Gallinarumの血清型が引き起こす法定伝染病(ひな白痢、鶏チフス)、S.Abortuequiを原因菌とする届出伝染病の馬パラチフス以外には、Enteritidis、Typhimurium、Choleraesuis及びDublinの血清型が原因で引き起こされる食中毒性サルモネラ症のみが届出伝染病に指定されている。
我が国におけるヒトのサルモネラ属菌由来感染症として、かつては猛威をふるったS.Typhi及びS.Paratyphiが引き起こす腸チフスやパラチフス感染症の発生は上下水道の整備により激減したが、食卓の西洋化に伴いサルモネラ属菌による食中毒の発生件数は増加の一途をたどっている。近年、特に汚染鶏卵を用いた加工食品による大規模食中毒や汚染鶏肉を原因とする食中毒が多発しており、Enteritidisが原因菌である場合が最も多い。Enteritidisは、一般のサルモネラ属菌が食中毒を発症するために105〜106個必要とされるのに対して102個以下でも発症することがあり、2次感染を起こしやすい。上記Typhimurium、Choleraesuis及びDublinの血清型は検出される場合も多く、他に鶏を宿主とするInfantis及びHeidelberg血清型も高頻度で検出される。また、最近ではSchwarzengrund血清型による食中毒も増加傾向にあり、当該血清型は多剤耐性株の割合が高いことが問題となる。
以上のように、食中毒性サルモネラ症の原因菌に関しては、食中毒患者を扱う医療現場では、血清型にあった適切な治療をできるだけ早く施すために、感染サルモネラ属菌の血清型を早急に判別する必要がある。家畜生産現場においても、各農場で分離されたサルモネラ属菌の血清型が監視伝染病に含まれる場合には農場が執るべき対応が異なることから、同様に迅速かつ正確なサルモネラ属菌の血清型の判別が必要である。また、農場にどのようなサルモネラが存在するか把握することにより、外部から通常検出されないサルモネラ属菌が侵入した場合に迅速な対応が可能となる。このように、農場の衛生管理においてサルモネラ属菌の血清型を把握することは非常に重要である。
【0003】
従来、血清型の特定は抗血清との凝集反応を観察する免疫的手法により行われているが、この手法は単離した菌株を用いて抗血清との凝集反応を確認する必要がある。また、サルモネラ属菌においてH抗原は複相性であり、血清型の確定には相誘導を行う必要があるため、確定までには増菌培養の終了後、更に1週間以上の検査期間を要する。さらに、凝集を目視で確認する際、擬陽性による誤判定の可能性があった。
このような背景から、近年PCRによる血清型同定の手法が多数報告されている。血清型を決定するという観点から、O抗原及びH抗原の各タイプ特異的な遺伝子又はその特徴的な一部領域の増幅を行い、増幅産物由来のバンドの出現パターンを解析して判定する手法が報告されている(非特許文献1、2)。これらの手法では、O抗原及びH抗原それぞれの組み合わせで血清型を確定する点では免疫的手法と同様であるため、血清型についての確定的な検査結果を得るには、多重PCRを適用したとしても1株あたり複数回の増幅工程が必要であり、複数の血清型が混入している可能性のある培養溶液の状態では測定できず、被検菌株の単離工程が必須である。
【0004】
抗原関連遺伝子以外の血清型特異的配列を対象として検出を行う手法も報告されているが、既報の手法は単独の血清型を検出対象としたPCR、又は多重PCRを用いるといっても複数の対象遺伝子断片の増幅パターンを解析するものであり、確実に判別できる血清型は2〜3程度であった(非特許文献3−8)。例えばTyphimuriumとHeidelberg(非特許文献3)、CholeraesuisとParatyphi C(非特許文献4)、EnteritidisとTyphimurium(非特許文献5)について、他の血清型との判別が可能な手法が報告されている。Leaderらは多重PCRとキャピラリー電気泳動による分離を併用する判別法を報告しており、16の遺伝子断片の増幅パターンを多重PCRにより解析し、多数の血清型についての判別を可能とした(非特許文献9)。しかし、この手法においても多重PCRは、各血清型に特異的な遺伝子断片を対象としているわけではなく、検出対象の遺伝子断片の増幅パターンで判定を行うため、複数の血清型が混在する場合には誤った判定に繋がる恐れがある。
最近報告された先行特許文献(特許文献1)におけるサルモネラ血清型の迅速判別法は、主要な血清型それぞれに特異的な遺伝子に着目するものであり、分離したサルモネラ属菌についてO抗原を同定した後、各血清型に特異的な3遺伝子全てで増幅が見られたものを当該血清型と判定する手法であるが、O抗原を同定する工程が必須である上に、判定しようとする血清型ごとに複数回のPCRを繰り返す必要がある。同様に複数の血清型のサルモネラ菌株が混入した検体についての判定を行うことができないため、被検菌株の単離が必須である。そのために、解析のために最低でも1週間は必要であった。複数の血清型それぞれに特異的な遺伝子断片の検出を1度の多重PCRによる検出で行おうという試みもある(非特許文献11、12)が、いずれも主要な血清型のEnteritidisとDublin又はEnteritidisとTyphimuriumとが1回の多重PCRでは判別できない不完全なものであった。
したがって、食中毒に関連した病原性の高いサルモネラ菌の主要な血清型である上記Enteritidis、Typhimurium、Choleraesuis、Dublin、Infantis、Heidelberg、Schwarzengrund血清型を1回の多重PCRを利用して検出、判別できる方法であって、迅速、簡便でありながら高感度に判別できる多重検出法が強く望まれていた。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2010−035552号公報
【特許文献2】米国特許第5683883号
【特許文献3】特開2003−199572号公報
【非特許文献】
【0006】
【非特許文献1】Tennant SM et al., PLoS Negl. Trop. Dis. 4: e621 (2010)
【非特許文献2】Hong Y et al., BMC Microbiol. 8: 178 (2008)
【非特許文献3】McCarthy N et al., J Food Prot. 72: 2350-2357 (2009)
【非特許文献4】Woods DF et al., J Clin. Microbiol. 46: 4018-4022 (2008)
【非特許文献5】Trafny EA et al., Lett. Appl. Microbiol. 43: 673-679 (2006)
【非特許文献6】Kardos G et al., Lett. Appl. Microbiol. 45: 421-425 (2007)
【非特許文献7】Kim HJ et al., J Food Prot. 69: 1653-1661 (2006)
【非特許文献8】Wang SJ and Yeh DB, Lett. Appl. Microbiol. 34: 422-427 (2002)
【非特許文献9】Leader BT et al., J Clin. Microbiol. 47: 1290-1299 (2009)
【非特許文献10】Elnifro EM et al., Clin. Microbiol. Rev. 13: 559-570 (2000)
【非特許文献11】de Freitas CG et al., Int J Food Microbiol. 139:15-22(2010)
【非特許文献12】Park SH et al., FEMS Microbiol Lett. 301:137-146(2009)
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明は、サルモネラ属菌のうちでも食中毒の原因菌として主に畜産現場で問題となる血清型(Enteritidis, Infantis, Typhimurium, Choleraesuis, Dublin, Heidelberg, Schwarzengrund)について、1回の多重PCRを利用して検出、判別できる方法であって、より迅速でかつ高い感度で判別できる多重検出法を提供することを目的とする。また、被検試料由来増菌培養液からサルモネラ属菌を単離することなく直接多重PCRを行うことができる簡易判別法を提供することも目的とする。
【課題を解決するための手段】
【0008】
そこで本発明者らは、上記課題を解決するために、多重PCRの利点を最大限に活用するために、対象として選択したEnteritidis、Typhimurium、Choleraesuis、Dublin、Infantis、Heidelberg、Schwarzengrundの7血清型それぞれに特異的な断片を見出して、これら断片を増幅するプライマーセットを設計することができれば、1回のPCR反応によって、これら7血清型を全て検出しそれぞれを判定できる本来の多重PCR法が確立できると発想した。
また、その際にサルモネラ属菌のみに存在する特異的遺伝子も同時に多重PCRで増幅させ、各血清型判別と同時に被検細菌がサルモネラ属菌に属することを確定してしまうことも発想した。
しかし、そのためには、7血清型それぞれに特異的な遺伝子領域を選択するだけではなく、さらにサルモネラ属菌特異的遺伝子を選定して、それぞれの領域を増幅するための計8種類のプライマーセットを設計する必要がある。頭で思い描くことは簡単でも、実際に正確な多重PCRの実現にはプライマー間の干渉が生じないプライマーを設計する必要があり(非特許文献10)、プライマー数が増えるほどその困難性は指数関数的に増大する。また、サルモネラ属菌には非常に多様な血清型が存在するため、血清型特異的な配列を特定し、他の血清型に対する交差反応のないプライマーを作成すること自体も容易なことではない。上記既報の文献(非特許文献3−5、9)でも、1つの血清型に対し複数の増幅産物の出現パターンを検出することで血清型を特定しており、このことは、単一の断片で血清型を特定することが困難であることを示すものでもある。
本発明者らは、このような困難が予想されるにもかかわらず、あえて多重PCR判別法の確立に挑み、鋭意研究の結果、上記7血清型それぞれに特異的で他のサルモネラ属菌血清型に対する交差反応のないプライマーセットを設計すると共に、サルモネラ属菌特異的なinvA遺伝子も同時に検出できるプライマーセットを設計することができた。このことにより、1回の多重PCR工程を行うことで、被検菌株がサルモネラ属菌の上記7血清型のいずれかであるか否かを迅速かつ再現性よく判別することが可能となった。
しかも、この多重PCR判別法は菌株を単離しない増殖培養液の状態でも十分に識別性のある結果を得ることができたことから、増殖培養液に適用する簡易判別法としても用いることができる。
以上の知見を得たことで、本発明を完成することができた。
【0009】
すなわち、本発明は以下に示すとおりである。
〔1〕 下記の(1)〜(8)に示される塩基配列又はそれぞれの相補配列からなるプライマーセットを含むことを特徴とする、サルモネラ属菌血清型の多重PCR判別法用プライマーセット;
(1)invA遺伝子領域を増幅するためのプライマーセットであって、
片方のプライマーが配列番号1に示される塩基配列、又は配列番号1の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号2に示される塩基配列、又は配列番号2の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなる;
(2)Enteritidis特異的なプライマーセットであって、
片方のプライマーが配列番号3に示される塩基配列、又は配列番号3の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号4に示される塩基配列、又は配列番号4の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(3)Typhimurium特異的なプライマーセットであって、
片方のプライマーが配列番号5に示される塩基配列、又は配列番号5の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号6に示される塩基配列、又は配列番号6の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなる;
(4)Infantis特異的なプライマーセットであって、
片方のプライマーが配列番号7に示される塩基配列、又は配列番号7の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号8に示される塩基配列、又は配列番号8の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(5)Choleraesuis特異的なプライマーセットであって、
片方のプライマーが配列番号9に示される塩基配列、又は配列番号9の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号10に示される塩基配列、又は配列番号10の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(6)Dublin特異的なプライマーセットであって、
片方のプライマーが配列番号11に示される塩基配列、又は配列番号11の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号12に示される塩基配列、又は配列番号12の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなる;
(7)Heidelberg特異的なプライマーセットであって、
片方のプライマーが配列番号13に示される塩基配列、又は配列番号13の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号14に示される塩基配列、又は配列番号14の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(8)Schwarzengrund特異的なプライマーセットであって、
片方のプライマーが配列番号15に示される塩基配列、又は配列番号15の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号16に示される塩基配列、又は配列番号16の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである。
〔2〕 被検試料にサルモネラ属菌を増殖させる培養条件下で培養した後、被検菌株を分離して前記〔1〕に記載のプライマーセットを用いて多重PCRを行うことを特徴とする、サルモネラ属菌血清型の多重PCR判別法。
〔3〕 被検試料にサルモネラ属菌を増殖させる培養条件下で培養した後、被検菌株を分離せずに、前記〔1〕に記載のプライマーセットを用いて多重PCRを行うことを特徴とする、サルモネラ属菌血清型の多重PCR判別法。
【発明の効果】
【0010】
本発明の多重PCR判別法は血清型特異的PCR産物の検出により特定の血清型の存否を判定するため、抗血清との凝集反応で見られる擬陽性の可能性が低減される。また、単離したコロニーを用いたコロニーPCRのみならず、増菌培養液より調製したDNAを鋳型としたPCRにより血清型の判別が可能であり、増菌培養後6〜30時間以内に結果が得られるので、特に医療現場での迅速な治療計画や農場での迅速な防疫に有効である。また、複数の血清型について一度の多重PCR実験で判定可能な簡便性は検査コスト低下につながり、農場に存在するサルモネラ属菌の検査を定期的に行うことを容易にする。
【図面の簡単な説明】
【0011】
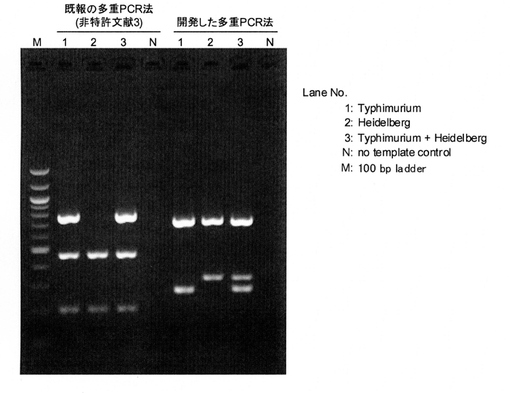
【図1】本発明の多重PCRと従来法との比較
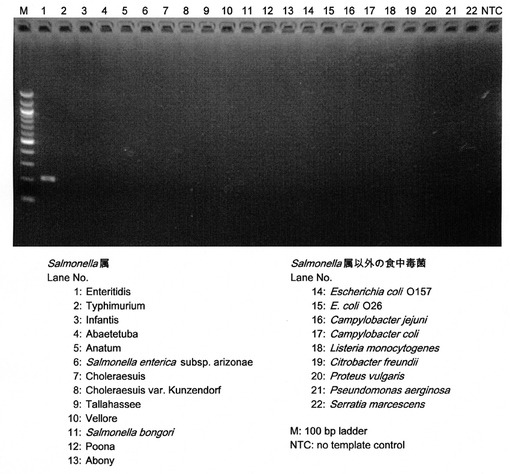
【図2】サルモネラ属菌のEnteritidis血清型特異的プライマーによる増幅
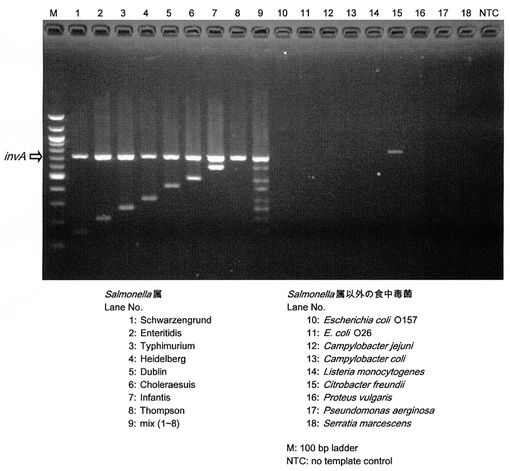
【図3】サルモネラ属菌の各血清型特異的プライマーによる多重PCR
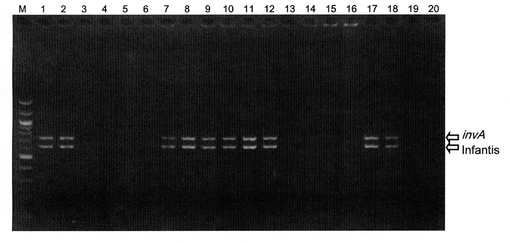
【図4】サルモネラ属菌野生株の各血清型特異的プライマーによる多重PCR
【図5】増殖培養液由来DNAを用いた多重PCRによるサルモネラ属菌の検出と同定
【発明を実施するための形態】
【0012】
1.本発明の「多重PCR血清型判別法」について
本発明において、サルモネラ属菌の「多重PCR血清型判別法」というとき、1回の多重PCR反応で、すなわち、同一のPCR反応チューブ内での、各血清型特異的なプローブセット群に基づくPCR反応(変性工程、アニーリング工程、増幅工程を20サイクル以上繰り返す)のみで、多種類(本発明では7種類)の血清型に属するサルモネラ属菌であることを判別できる方法を指す。多重PCR法を用いる方法であっても従来法のような対象遺伝子の増幅パターンで判別する場合は含まない。
本発明の多重PCR血清型判別法を従来法と比較した図を図1として示す。(なお、図1における比較例としては、増幅パターンで認識する手法を用いている典型例である非特許文献3と比較した。)
【0013】
2.本発明の検出対象となる血清型特異的プライマーセットの構築
(2−1)検出対象の血清型
本発明では、食中毒原因菌として医療現場で注目されている血清型であると共に、特に畜産物、乳製品など食品材料を提供している畜産現場の安全管理においても重要な血清型Enteritidis、Typhimurium、Choleraesuis、Dublin、Infantis、Heidelberg、Schwarzengrundの7血清型を選択した。Enteritidis、Typhimurium、Choleraesuis、Dublinは上述のように届出伝染病の原因菌として指定されており、Infantis、Heidelbergは鶏から高頻度で分離される血清型である。Schwarzengrundは、近年食中毒の発生件数が増加しており、分離株中の多剤耐性株の割合が高いことが報告されている。
【0014】
(2−2)血清型特異的プライマーセットの構築
公的データベースより入手可能であったサルモネラ属菌のゲノム配列について、塩基配列の比較を行った。入手したゲノム配列は以下のとおりである:Choleraesuis str. SC-B67 (accession No.: NC_006905), Dublin str. CT_02021853 (NC_011205), Enteritidis str. P125109 (NC_011294), Gallinarum str. 287/91 (NC_011274), Heidelberg str. SL476 (NC_011083), Paratyphi A str. ATCC 9150 (NC_006511), Paratyphi B str. SPB7 (NC_010102), Paratyphi C strain RKS4594 (NC_012125), Schwarzengrund str. CVM19633 (NC_011094), Typhi str. CT18 (NC_003198), Typhimurium str. LT2 (NC_003197), Typhimurium str. 14028S (CP001363), Typhimurium str. D23580 (FN424405), Typhimurium SL1344 (FQ312003)。
【0015】
ゲノム配列の比較には、公知のソフトウェアであるArtemis Comparison Tool (ACT)を用いた。上記の配列を適宜組み合わせて比較し、各血清型に特異的であると考えられる候補配列を探索した。血清型間での相同性がより低い配列を選抜するため、転写・翻訳の単位に拘らず標的配列を選択した。
選択した部位についてBLAST解析を行い、他属の細菌等を含む既報の配列との相同性が低いことを確認した後、当該部位を検出するプライマーを設計した。Infantisは全ゲノム配列が公開されていなかったため、公開されていた部分配列(accession No.: J03391)を基にプライマーを設計した。
【0016】
また、血清型特異的プライマーを用いて多重PCRを行う際に、同時にサルモネラ属菌存否の判定を行うために、サルモネラ属菌特異的な遺伝子としてinvA遺伝子を選択した。invAはほぼすべてのサルモネラ属菌が保持しており、サルモネラ属菌の簡易的な検出法に応用されている。例えば、先行特許文献にてPCR法やLAMP法での検出に用いるプライマーが報告されている(米国特許5683883、特開2003−199572)。当該invA遺伝子を対象とし、各血清型特異的プライマーとは干渉を起こさずに当該遺伝子の特異的な領域を増幅できるプライマーを設計した。
各プライマー設計にあたっては、多重PCRを行う際にアガロースゲルによる電気泳動で目視による判別が容易となるように各増幅産物の断片長を設定した。具体的には、各増幅産物の断片長がそれぞれに50〜120bp異なるように複数セットずつ設計した。
【0017】
(2−3)設計したプライマーの評価
設計したプライマーは、実施例1の(表1、2)に示すサルモネラ属菌ならびに複数の食中毒菌(Escherichia coli O157, E. coli O26, Campylobacter jejuni, Campylobacter coli, Listeria monocytogenes, Citrobacter freundii, Proteus vulgaris, Pseundomonas aerginosa, Serratia marcescens)のゲノムDNAを鋳型としてPCRを行い、それぞれの血清型に対応して、非特異的な増幅が見られない特異的プライマーが少なくとも1種類以上存在した(同表3、4)。
【0018】
(2−4)本発明の多重プライマー群
本発明の実施例1の(表3、4)に示される各血清型での特異的プライマーから1種類ずつを選択した多重プライマー群の例を同(表5)に示す。
本発明の多重プライマー群はこの1例に限られるものではなく、表3、4に示す他の特異的プライマーセットを組み合わせる場合も含まれる。
また、本発明でプライマーとして用いられるオリゴヌクレオチド断片は、1又は2塩基の範囲内の増減又は変異が導入されても各プライマーにより増幅される対象DNA領域は同一であるから、1又は2塩基の塩基が欠失、置換もしくは付加された配列も許容される。これらプライマーは、例えばビオチンやDIG等の公知の物質により標識してもよい。
【0019】
3.本発明の血清型多重PCR判別法の手順
(3−1)被検試料
被検試料としては、典型的には、家畜生産現場での、家畜の糞便、尿などの排泄物、血液、唾液などの体液、あるいは飼料、飼料添加物などと共に、医療現場での患者からの糞便、尿、血液、組織ホモジェネートなどであるが、畜産加工品、乳製品、鶏卵、卵製品などの飲食品も対象となる。
【0020】
(3−2)サルモネラ属菌の培養方法
これら被検試料を本発明の多重PCRの試料として用いる際に、高濃度のサルモネラ属菌の存在が明らかな場合を除き、まず試料を培地に接種して、サルモネラ属菌に好ましい培養条件下で培養し、サルモネラ属菌を増菌させる。
なお、PCRを進行させるには、プライマーがハイブリダイズできる核酸が数分子から数十分子以上存在すれば十分ではあるので、被検試料から直接核酸抽出してしまうことも可能であるが、再現性の高い結果を得るためには増菌工程を設けることが好ましい。
サルモネラ属菌の増菌培養条件は周知であり、例えば被検試料を緩衝ペプトン水に接種して35℃にて一晩の培養を行った後に、その一部をRV培地に接種して42℃にて一晩の培養を行うことが好ましい。
本発明においては、当該増菌培養液から直ちにゲノムDNAを抽出する下記(3−3)工程に移行することもできるが、各菌株ごとのコロニーを形成させる分離培養に供し、菌株を単離することが好ましい。
その場合の分離培養方法及び分離培養条件は周知であり、例えば、DHL寒天培地に増菌培養液を塗布し、37℃にて一晩の培養を行う。
【0021】
(3−3)被検DNA溶液の調製
分離培養で単離された菌株培養液、又は分離工程を省いた増菌培養液に対して溶菌酵素、界面活性剤、アルカリ等を用いる周知のDNA抽出法を適用する。例えば、培養液を遠心処理して菌体を回収して、細胞溶解液に懸濁し、加熱後遠心分離により回収した上清に対して、フェノール−クロロホルム抽出およびエタノール沈殿によりDNAを回収する。得られたDNAをTE緩衝液に溶解し、PCRに供する。
(3−4)PCR工程
本発明の多重PCRで用いるPCR法では耐熱ポリメラーゼであるTaqDNAポリメラーゼを用い、通常の使用条件の範囲内で増幅させる。具体的には、90〜95℃で熱変成させた後、37〜65℃でアニーリング操作を行い、50〜75℃で重合反応を起こさせ、これを1サイクルとして20から42サイクル行って増幅させる。
(3−5)多重PCRによる増幅産物の検出
本発明の多重PCRによる増幅産物の分子量はいずれも約150〜750bpの範囲内であるため、アガロースゲル電気泳動を用いて検出できる。エチジウムブロマイド等の慣用の染料で染色する。ゲル中のバンドの確認は写真から目視確認してもよいし、ゲル写真を映像データとしてコンピュータに入力し、写真の濃淡を解析することにより確認してもよい。
【実施例】
【0022】
以下、実施例により本発明をより具体的に説明するが、本発明はこれら実施例により何ら限定されるものではない。
なお、本発明で引用した先行文献の記載内容は、本明細書の記載内容として参照されるものとする。
【0023】
[実施例1]各プライマーセットによる増幅の特異性の確認
(1−1)DNA溶液の調製
サルモネラ属菌ならびにサルモネラ属に属しない食中毒菌の単離菌株を増菌培地に接種して培養した。培養液を15,000×gで2分間遠心し、上清を除去して菌体を回収した。回収した菌体を細胞溶解液(PrepMan(登録商標) Ultra Reagent、Applied Biosystems社) 200μlに懸濁し、沸騰水中で10分間加熱した。加熱後の懸濁液を15,000×gで2分間遠心分離し、上清を回収した。上清からフェノール−クロロホルム抽出およびエタノール沈殿によりDNAを回収し、TE緩衝液400μlで溶解した。調製したDNA溶液をPCRに供した。
(1−2)PCR
調製したDNA溶液0.6μlを鋳型として供し、PCRを行った。ExTaq DNAポリメラーゼ(Takara社) 0.375U、1× ExTaq buffer、0.2mM dNTP、ならびに各0.3μMのプライマーを含む15μlの反応液を調製し、DNAサーマルサイクラー(Bio-Rad社製iCycler)を用いて以下の反応を行った。すなわち、94℃にて2分間の熱変性反応を行った後、94℃で30秒間、58℃で30秒間、72℃で40秒間の3ステップからなるサイクリング反応を30回繰り返し、サイクリング反応の終了後に72℃で7分間の伸長反応を行った。
(1−3)検出
PCRによる増幅産物の検出はアガロースゲル電気泳動により行った。アガロースゲルはゲル濃度2%(W/V)とし、100ng/mlの臭化エチジウムを含むものを用いた。TAE溶液の入った泳動槽にゲルを浸し、135Vの定電圧で電気泳動を行った。電気泳動後のゲルにUVランプを照射し、増幅産物の検出ならびにDNA断片長の確認を行った。
(1−4)結果
Enteritidis、Typhimurium、Choleraesuis、Dublin、Infantis、Heidelberg、Schwarzengrundの7血清型について、各血清型に特異的な断片を増幅するプライマーセットをそれぞれ複数セット設計し、増幅の特異性を確認した。サルモネラ属菌標準株44株(表1、2)のゲノムDNAを鋳型として用いた。設計したプライマーセットの中には、標的血清型で増幅産物が検出されないもの、あるいは標的血清型以外のサルモネラ属菌についても増幅産物が検出されるものもあったため、標的血清型のみを特異的に検出するプライマーセットを選抜した。選抜したEnteritidis特異的プライマーセットによるPCRの結果を図2に例示する。選抜したプライマーセットについて、サルモネラ属以外の食中毒菌のゲノムDNAを鋳型としてPCRを行い、増幅産物が検出されないことを確認した(図2、レーン14〜22)。invA検出用プライマーについては、サルモネラ属菌のゲノムDNAを鋳型とした場合のみ増幅産物が検出され、他属の細菌では増幅産物が検出されないことを確認した。
以上の結果から、各血清型について、それぞれ各1セットの血清型特異的なプライマーセットを得ることができた。各血清型の特異的プライマー群及びinvA遺伝子を増幅するプライマーのセットからなる多重PCR判別法用プライマー群を表5に示す。
【0024】
【表1】

【0025】
【表2】
【0026】
【表3】
【0027】
【表4】
【0028】
【表5】
【0029】
[実施例2]多重PCR系の構築
(2−1)多重PCR
実施例1で選抜した各血清型特異的プライマーセット7セット、ならびにinvA遺伝子を増幅するプライマーセットからなる多重PCR判別法用プライマー群(表5)を用いて多重PCRを行った。
鋳型DNA溶液0.6μl、ExTaq DNAポリメラーゼ(Takara社) 0.375U、1×ExTaq buffer、0.2mM dNTP、ならびに各66.7nMのプライマーを含む15μlの反応液を調製し、実施例1と同一の条件で反応を行った。多重PCR用の酵素を用いる場合には、Takara社製Multiplex PCR Assay Kitを用い、鋳型DNA溶液1μlを含む20μlの反応液を調製した。反応条件は、94℃にて1分間の熱変性反応を行った後、94℃で30秒間、58℃で90秒間、72℃で90秒間の3ステップからなるサイクリング反応を30回繰り返し、サイクリング反応の終了後に72℃で10分間の伸長反応を行うこととした。増幅産物は、実施例1と同様のアガロースゲル電気泳動により確認を行った。
(2−2)結果
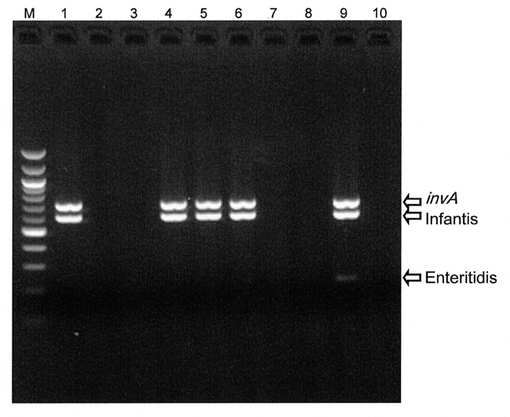
サルモネラ属菌標準株44株、ならびにサルモネラ属以外の食中毒菌のゲノムDNAを鋳型として、多重PCRを行った。汎用的な耐熱性DNAポリメラーゼ(ExTaq)を用いて多重PCRを行ったところ、Schwarzengrund特異的プライマーセットについての感度が低下し、サルモネラ属以外の食中毒菌の一部で非特異的増幅産物が検出される不都合が確認されたので、Multiplex PCR Assay Kitを用いることとした。当該Assay Kitを用いることで上記不都合は解消された。Multiplex PCR Assay Kitを用いて得られた代表的な結果を図3に示す。
サルモネラ属菌標準株のDNAを鋳型としたところ、すべての株においてinvAの増幅産物が検出された(図3、レーン1〜8)。特異的プライマーを構築した7血清型(Enteritidis、Typhimurium、Choleraesuis、Dublin、Infantis、Heidelberg、Schwarzengrund)については、invAの増幅産物に加え、各血清型特異的断片が検出された(図3、レーン1〜7)。それぞれのゲノムDNAを混合して鋳型DNAとして用いたところ、それぞれの血清型に由来する特異的断片が検出され、非特異的産物は検出されなかった(図3、レーン9)。また、サルモネラ属以外の食中毒菌のゲノムDNAを鋳型とした場合には、いずれの増幅産物も検出されなかった(図3、レーン10〜18)。
以上の結果から、構築した多重PCR系は、invAの増幅によるサルモネラ属菌の検出、ならびにEnteritidis、Typhimurium、Choleraesuis、Dublin、Infantis、Heidelberg、Schwarzengrundの7血清型については各血清型特異的な断片の検出による血清型の判別が可能であることが示された。
【0030】
[実施例3]野外単離株を用いた多重PCRの検証
野外検体より単離されたサルモネラ属菌を用い、多重PCRの検証を行った。用いた菌株の血清型ならびに株数を表6に示す。
Enteritidis、Typhimurium、Choleraesuis、Dublin、Infantisについては、従来の手法で決定された血清型と多重PCRの結果が一致していた。I4,5,12:i:−は、検証した2株でいずれもTyphimurium特異的増幅産物を検出した。該血清型はTyphimuriumの単相型の変異株であると言われており、両者は遺伝的に非常に近いと思われる。このため、両者を判別することは非常に困難であることが容易に推測できる。この両者の判別が必要な場合は、さらに他の手法による確認が必要となる。
他の血清型、ならびに血清型が確定されていない株についてはinvAの増幅産物のみを検出した。
以上のように、本手法はTyphimuriumとI4,5,12:i:−の両者がTyphimuriumと判定されるものの、野外単離株の血清型別に有用であることが示された。
【0031】
【表6】
【0032】
[実施例4]コロニーPCRによる野外株の血清型別
(4−1)コロニーPCR
野外から得られた検体から、公定法に準じた作業によりサルモネラ属菌株の分離を行った。すなわち、検体1gあたり9mlのリン酸緩衝ペプトン水を加え、35℃にて24時間の前増菌培養を行った。培養後、0.1mlの前増菌培養液を増菌培養用培地10mlに加え、42℃にて一晩の増菌培養を行った。増菌培養液から1白金耳を寒天培地上に塗布し、一晩培養して菌株を分離した。検出されたコロニーをコロニーPCRに供した。実施例2と同様に調製した15μlの反応液にコロニーの一部を懸濁した後、サーマルサイクラーによって実施例1と同一の条件で反応を行った。増幅産物は実施例1と同様の手法により確認した。
(4−2)結果
野外検体より単離された菌株について、コロニーPCRにより血清型別を行った(図4)。レーン1、2、7、8、9、10、11、12、17、18について、invAならびにInfantis特異的断片が検出され、Salmonella Infantisであることが推測された。抗血清を用いてこれらの菌株の血清型を確定したところ、Infantisであることが確認され、多重PCRの結果と一致した。
また、外見上サルモネラ属菌であることが疑われたコロニーについても同様にコロニーPCRを行ったが、増幅産物は検出されなかった(図4、レーン3〜6、13〜16、19、20)。これらの菌株について16S rDNAの配列を決定したところ、Proteus属の細菌であることが確認された。このため、多重PCRによりサルモネラ属菌特異的に増幅産物が検出されることが示された。
このように、野外検体より単離した菌株について、コロニーPCRによりサルモネラ属菌であることの確認、ならびに簡易的な血清型別が可能であった。
【0033】
[実施例5]増菌培養液由来DNAを用いたサルモネラ血清型迅速判別
(5−1)増菌培養液のDNA溶液の調製
野外検体から実施例4と同様の手法で増菌培養液を調製した。培養液2mlを15,000×gで2分間遠心し、上清を除去して菌体を回収した。回収した菌体を細胞溶解液 (PrepMan(登録商標) Ultra Reagent、Applied Biosystems社) 200μlに懸濁し、沸騰水中で10分間加熱した。加熱後の懸濁液を15,000×gで2分間遠心分離し、上清を回収した。上清からフェノール−クロロホルム抽出およびエタノール沈殿によりDNAを回収し、TE緩衝液400μlに溶解した。得られたDNA溶液を鋳型として多重PCRを行った。
(5−2)結果
実施例4と同一の検体から得られた増菌培養液よりDNAを抽出し、それを鋳型として多重PCRを行った。その結果、5検体にサルモネラ属菌が存在することが確認された(図5、レーン1、4〜6、9)。レーン1、4〜6においてはinvAならびにInfantis特異的断片が検出され、これらの検体中にSalmonella Infantisが存在することが示唆された。実施例3は各検体から得られた単離株(各2株)についてコロニーPCRを行っており、図4の検体1、2(レーン1、2)は図5の検体1(レーン1)から得られた菌株である。このように、検体1、4、5、6については単離菌株を用いたコロニーPCRの結果と一致することが示された。
一方、検体9については、invA、Infantis特異的断片に加えEnteritidis特異的断片が検出された(図5、レーン9)。検体9から得られた分離菌株について、さらに6コロニーを検査したところ、8コロニー中1コロニーがEnteritidisであることが確認された。すなわち、増菌培養液由来DNAを鋳型としたPCRを行うことにより、複数の血清型を検出可能であることが示された。
【産業上の利用可能性】
【0034】
サルモネラ属菌は罹患した宿主の糞便から検出されるため、糞便に存在するサルモネラ属菌の血清型を把握することは疾病の原因特定において重要である。従来法により糞便中に存在するサルモネラ属菌の血清型を同定するには、前述した理由により、採集した糞便検体からサルモネラ属菌を分離する必要がある。一方、本手法は対象とする血清型は限られるものの、ゲノムDNAの抽出が可能であれば様々な検体に応用できるため、糞便の検査にも応用可能である。糞便からより効率的にゲノムDNAを抽出する手法が開発されれば、本手法を用いた検査の感度も向上すると考えられる。また、糞便中にはPCRを阻害する物質が含まれるため、現状では糞便から直接サルモネラ属菌を検出することは不可能であるが、この阻害を回避するPCR酵素やPCR反応緩衝液の開発により、糞便を直接検査する手法の開発に繋がる。
【配列表フリーテキスト】
【0035】
1.invA-F1 AGTGCCGGTTTTATCGTGAC
2.invA-R1 ACCGCCAGACAGTGGTAAAG
・Enteritidis
3.Ent1 TCAAGCATTGATACGGCAAG
4.Ent2 CGCATTCTGCTCAATGAAAA
・Typhimurium
5.Tpm15 GCGTTGTTCCCTTTGTTCTG
6.Tpm16 GACGCTGTTACGAGCCATTT
・Infantis
7.Inf3 GCGGCAGGTTGGTATTATTC
8.Inf4 CTAAACGTTTTCCCGCTGTT
・Choleraesuis
9.Chs1 TCGCACCATGACCAAAAATA
10.Chs2 CACTCCGTCGTTCTGGGTAT
・Dublin
11.Dub11 GCGTGAAGTGGCAACAGTAG
12.Dub12 ATATCGGCCAGTGCAGATTC
・Heidelberg
13.Hdb15 TAACCAAACTCAGCGGCTCT
14.Hdb16 GGGGCGACTGATGCTATAAA
・Schwarzengrund
15.Swg7 GAAGAGAACGCTCGGACAAC
16.Swg8 CAGGCAATTCTCCGTCTCTC
【技術分野】
【0001】
家畜生産の現場で問題となるサルモネラ属菌の主要な血清型(Enteritidis, Infantis, Typhimurium, Choleraesuis, Dublin, Heidelberg, Schwarzengrund)についての多重PCRによる簡易的な判別手法に関する。
【背景技術】
【0002】
サルモネラ属菌は、生物学的にはSalmonella enterica及びSalmonella bongoriの2菌種に、S.entericaがさらに6亜種に分類されるが、血清学的には、細胞壁リポ多糖類のO抗原と、鞭毛タンパク質のH抗原の組み合わせで2,500を超える血清型(serovar)が存在している。サルモネラ属菌は、健康なヒトの消化管にはほとんど存在していないが、種々の動物の消化管内に常在菌として存在している場合が多い。これらのサルモネラ属菌は、各血清型によって病原性と宿主域が異なり、強い病原性がある血清型は限られており、ヒトへの感染性が問題となる血清型は少数である。日本家畜伝染病予防法における家畜の監視伝染病としては、S.Gallinarum-Pullorum及びS.Gallinarum-Gallinarumの血清型が引き起こす法定伝染病(ひな白痢、鶏チフス)、S.Abortuequiを原因菌とする届出伝染病の馬パラチフス以外には、Enteritidis、Typhimurium、Choleraesuis及びDublinの血清型が原因で引き起こされる食中毒性サルモネラ症のみが届出伝染病に指定されている。
我が国におけるヒトのサルモネラ属菌由来感染症として、かつては猛威をふるったS.Typhi及びS.Paratyphiが引き起こす腸チフスやパラチフス感染症の発生は上下水道の整備により激減したが、食卓の西洋化に伴いサルモネラ属菌による食中毒の発生件数は増加の一途をたどっている。近年、特に汚染鶏卵を用いた加工食品による大規模食中毒や汚染鶏肉を原因とする食中毒が多発しており、Enteritidisが原因菌である場合が最も多い。Enteritidisは、一般のサルモネラ属菌が食中毒を発症するために105〜106個必要とされるのに対して102個以下でも発症することがあり、2次感染を起こしやすい。上記Typhimurium、Choleraesuis及びDublinの血清型は検出される場合も多く、他に鶏を宿主とするInfantis及びHeidelberg血清型も高頻度で検出される。また、最近ではSchwarzengrund血清型による食中毒も増加傾向にあり、当該血清型は多剤耐性株の割合が高いことが問題となる。
以上のように、食中毒性サルモネラ症の原因菌に関しては、食中毒患者を扱う医療現場では、血清型にあった適切な治療をできるだけ早く施すために、感染サルモネラ属菌の血清型を早急に判別する必要がある。家畜生産現場においても、各農場で分離されたサルモネラ属菌の血清型が監視伝染病に含まれる場合には農場が執るべき対応が異なることから、同様に迅速かつ正確なサルモネラ属菌の血清型の判別が必要である。また、農場にどのようなサルモネラが存在するか把握することにより、外部から通常検出されないサルモネラ属菌が侵入した場合に迅速な対応が可能となる。このように、農場の衛生管理においてサルモネラ属菌の血清型を把握することは非常に重要である。
【0003】
従来、血清型の特定は抗血清との凝集反応を観察する免疫的手法により行われているが、この手法は単離した菌株を用いて抗血清との凝集反応を確認する必要がある。また、サルモネラ属菌においてH抗原は複相性であり、血清型の確定には相誘導を行う必要があるため、確定までには増菌培養の終了後、更に1週間以上の検査期間を要する。さらに、凝集を目視で確認する際、擬陽性による誤判定の可能性があった。
このような背景から、近年PCRによる血清型同定の手法が多数報告されている。血清型を決定するという観点から、O抗原及びH抗原の各タイプ特異的な遺伝子又はその特徴的な一部領域の増幅を行い、増幅産物由来のバンドの出現パターンを解析して判定する手法が報告されている(非特許文献1、2)。これらの手法では、O抗原及びH抗原それぞれの組み合わせで血清型を確定する点では免疫的手法と同様であるため、血清型についての確定的な検査結果を得るには、多重PCRを適用したとしても1株あたり複数回の増幅工程が必要であり、複数の血清型が混入している可能性のある培養溶液の状態では測定できず、被検菌株の単離工程が必須である。
【0004】
抗原関連遺伝子以外の血清型特異的配列を対象として検出を行う手法も報告されているが、既報の手法は単独の血清型を検出対象としたPCR、又は多重PCRを用いるといっても複数の対象遺伝子断片の増幅パターンを解析するものであり、確実に判別できる血清型は2〜3程度であった(非特許文献3−8)。例えばTyphimuriumとHeidelberg(非特許文献3)、CholeraesuisとParatyphi C(非特許文献4)、EnteritidisとTyphimurium(非特許文献5)について、他の血清型との判別が可能な手法が報告されている。Leaderらは多重PCRとキャピラリー電気泳動による分離を併用する判別法を報告しており、16の遺伝子断片の増幅パターンを多重PCRにより解析し、多数の血清型についての判別を可能とした(非特許文献9)。しかし、この手法においても多重PCRは、各血清型に特異的な遺伝子断片を対象としているわけではなく、検出対象の遺伝子断片の増幅パターンで判定を行うため、複数の血清型が混在する場合には誤った判定に繋がる恐れがある。
最近報告された先行特許文献(特許文献1)におけるサルモネラ血清型の迅速判別法は、主要な血清型それぞれに特異的な遺伝子に着目するものであり、分離したサルモネラ属菌についてO抗原を同定した後、各血清型に特異的な3遺伝子全てで増幅が見られたものを当該血清型と判定する手法であるが、O抗原を同定する工程が必須である上に、判定しようとする血清型ごとに複数回のPCRを繰り返す必要がある。同様に複数の血清型のサルモネラ菌株が混入した検体についての判定を行うことができないため、被検菌株の単離が必須である。そのために、解析のために最低でも1週間は必要であった。複数の血清型それぞれに特異的な遺伝子断片の検出を1度の多重PCRによる検出で行おうという試みもある(非特許文献11、12)が、いずれも主要な血清型のEnteritidisとDublin又はEnteritidisとTyphimuriumとが1回の多重PCRでは判別できない不完全なものであった。
したがって、食中毒に関連した病原性の高いサルモネラ菌の主要な血清型である上記Enteritidis、Typhimurium、Choleraesuis、Dublin、Infantis、Heidelberg、Schwarzengrund血清型を1回の多重PCRを利用して検出、判別できる方法であって、迅速、簡便でありながら高感度に判別できる多重検出法が強く望まれていた。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2010−035552号公報
【特許文献2】米国特許第5683883号
【特許文献3】特開2003−199572号公報
【非特許文献】
【0006】
【非特許文献1】Tennant SM et al., PLoS Negl. Trop. Dis. 4: e621 (2010)
【非特許文献2】Hong Y et al., BMC Microbiol. 8: 178 (2008)
【非特許文献3】McCarthy N et al., J Food Prot. 72: 2350-2357 (2009)
【非特許文献4】Woods DF et al., J Clin. Microbiol. 46: 4018-4022 (2008)
【非特許文献5】Trafny EA et al., Lett. Appl. Microbiol. 43: 673-679 (2006)
【非特許文献6】Kardos G et al., Lett. Appl. Microbiol. 45: 421-425 (2007)
【非特許文献7】Kim HJ et al., J Food Prot. 69: 1653-1661 (2006)
【非特許文献8】Wang SJ and Yeh DB, Lett. Appl. Microbiol. 34: 422-427 (2002)
【非特許文献9】Leader BT et al., J Clin. Microbiol. 47: 1290-1299 (2009)
【非特許文献10】Elnifro EM et al., Clin. Microbiol. Rev. 13: 559-570 (2000)
【非特許文献11】de Freitas CG et al., Int J Food Microbiol. 139:15-22(2010)
【非特許文献12】Park SH et al., FEMS Microbiol Lett. 301:137-146(2009)
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明は、サルモネラ属菌のうちでも食中毒の原因菌として主に畜産現場で問題となる血清型(Enteritidis, Infantis, Typhimurium, Choleraesuis, Dublin, Heidelberg, Schwarzengrund)について、1回の多重PCRを利用して検出、判別できる方法であって、より迅速でかつ高い感度で判別できる多重検出法を提供することを目的とする。また、被検試料由来増菌培養液からサルモネラ属菌を単離することなく直接多重PCRを行うことができる簡易判別法を提供することも目的とする。
【課題を解決するための手段】
【0008】
そこで本発明者らは、上記課題を解決するために、多重PCRの利点を最大限に活用するために、対象として選択したEnteritidis、Typhimurium、Choleraesuis、Dublin、Infantis、Heidelberg、Schwarzengrundの7血清型それぞれに特異的な断片を見出して、これら断片を増幅するプライマーセットを設計することができれば、1回のPCR反応によって、これら7血清型を全て検出しそれぞれを判定できる本来の多重PCR法が確立できると発想した。
また、その際にサルモネラ属菌のみに存在する特異的遺伝子も同時に多重PCRで増幅させ、各血清型判別と同時に被検細菌がサルモネラ属菌に属することを確定してしまうことも発想した。
しかし、そのためには、7血清型それぞれに特異的な遺伝子領域を選択するだけではなく、さらにサルモネラ属菌特異的遺伝子を選定して、それぞれの領域を増幅するための計8種類のプライマーセットを設計する必要がある。頭で思い描くことは簡単でも、実際に正確な多重PCRの実現にはプライマー間の干渉が生じないプライマーを設計する必要があり(非特許文献10)、プライマー数が増えるほどその困難性は指数関数的に増大する。また、サルモネラ属菌には非常に多様な血清型が存在するため、血清型特異的な配列を特定し、他の血清型に対する交差反応のないプライマーを作成すること自体も容易なことではない。上記既報の文献(非特許文献3−5、9)でも、1つの血清型に対し複数の増幅産物の出現パターンを検出することで血清型を特定しており、このことは、単一の断片で血清型を特定することが困難であることを示すものでもある。
本発明者らは、このような困難が予想されるにもかかわらず、あえて多重PCR判別法の確立に挑み、鋭意研究の結果、上記7血清型それぞれに特異的で他のサルモネラ属菌血清型に対する交差反応のないプライマーセットを設計すると共に、サルモネラ属菌特異的なinvA遺伝子も同時に検出できるプライマーセットを設計することができた。このことにより、1回の多重PCR工程を行うことで、被検菌株がサルモネラ属菌の上記7血清型のいずれかであるか否かを迅速かつ再現性よく判別することが可能となった。
しかも、この多重PCR判別法は菌株を単離しない増殖培養液の状態でも十分に識別性のある結果を得ることができたことから、増殖培養液に適用する簡易判別法としても用いることができる。
以上の知見を得たことで、本発明を完成することができた。
【0009】
すなわち、本発明は以下に示すとおりである。
〔1〕 下記の(1)〜(8)に示される塩基配列又はそれぞれの相補配列からなるプライマーセットを含むことを特徴とする、サルモネラ属菌血清型の多重PCR判別法用プライマーセット;
(1)invA遺伝子領域を増幅するためのプライマーセットであって、
片方のプライマーが配列番号1に示される塩基配列、又は配列番号1の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号2に示される塩基配列、又は配列番号2の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなる;
(2)Enteritidis特異的なプライマーセットであって、
片方のプライマーが配列番号3に示される塩基配列、又は配列番号3の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号4に示される塩基配列、又は配列番号4の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(3)Typhimurium特異的なプライマーセットであって、
片方のプライマーが配列番号5に示される塩基配列、又は配列番号5の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号6に示される塩基配列、又は配列番号6の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなる;
(4)Infantis特異的なプライマーセットであって、
片方のプライマーが配列番号7に示される塩基配列、又は配列番号7の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号8に示される塩基配列、又は配列番号8の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(5)Choleraesuis特異的なプライマーセットであって、
片方のプライマーが配列番号9に示される塩基配列、又は配列番号9の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号10に示される塩基配列、又は配列番号10の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(6)Dublin特異的なプライマーセットであって、
片方のプライマーが配列番号11に示される塩基配列、又は配列番号11の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号12に示される塩基配列、又は配列番号12の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなる;
(7)Heidelberg特異的なプライマーセットであって、
片方のプライマーが配列番号13に示される塩基配列、又は配列番号13の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号14に示される塩基配列、又は配列番号14の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(8)Schwarzengrund特異的なプライマーセットであって、
片方のプライマーが配列番号15に示される塩基配列、又は配列番号15の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号16に示される塩基配列、又は配列番号16の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである。
〔2〕 被検試料にサルモネラ属菌を増殖させる培養条件下で培養した後、被検菌株を分離して前記〔1〕に記載のプライマーセットを用いて多重PCRを行うことを特徴とする、サルモネラ属菌血清型の多重PCR判別法。
〔3〕 被検試料にサルモネラ属菌を増殖させる培養条件下で培養した後、被検菌株を分離せずに、前記〔1〕に記載のプライマーセットを用いて多重PCRを行うことを特徴とする、サルモネラ属菌血清型の多重PCR判別法。
【発明の効果】
【0010】
本発明の多重PCR判別法は血清型特異的PCR産物の検出により特定の血清型の存否を判定するため、抗血清との凝集反応で見られる擬陽性の可能性が低減される。また、単離したコロニーを用いたコロニーPCRのみならず、増菌培養液より調製したDNAを鋳型としたPCRにより血清型の判別が可能であり、増菌培養後6〜30時間以内に結果が得られるので、特に医療現場での迅速な治療計画や農場での迅速な防疫に有効である。また、複数の血清型について一度の多重PCR実験で判定可能な簡便性は検査コスト低下につながり、農場に存在するサルモネラ属菌の検査を定期的に行うことを容易にする。
【図面の簡単な説明】
【0011】
【図1】本発明の多重PCRと従来法との比較
【図2】サルモネラ属菌のEnteritidis血清型特異的プライマーによる増幅
【図3】サルモネラ属菌の各血清型特異的プライマーによる多重PCR
【図4】サルモネラ属菌野生株の各血清型特異的プライマーによる多重PCR
【図5】増殖培養液由来DNAを用いた多重PCRによるサルモネラ属菌の検出と同定
【発明を実施するための形態】
【0012】
1.本発明の「多重PCR血清型判別法」について
本発明において、サルモネラ属菌の「多重PCR血清型判別法」というとき、1回の多重PCR反応で、すなわち、同一のPCR反応チューブ内での、各血清型特異的なプローブセット群に基づくPCR反応(変性工程、アニーリング工程、増幅工程を20サイクル以上繰り返す)のみで、多種類(本発明では7種類)の血清型に属するサルモネラ属菌であることを判別できる方法を指す。多重PCR法を用いる方法であっても従来法のような対象遺伝子の増幅パターンで判別する場合は含まない。
本発明の多重PCR血清型判別法を従来法と比較した図を図1として示す。(なお、図1における比較例としては、増幅パターンで認識する手法を用いている典型例である非特許文献3と比較した。)
【0013】
2.本発明の検出対象となる血清型特異的プライマーセットの構築
(2−1)検出対象の血清型
本発明では、食中毒原因菌として医療現場で注目されている血清型であると共に、特に畜産物、乳製品など食品材料を提供している畜産現場の安全管理においても重要な血清型Enteritidis、Typhimurium、Choleraesuis、Dublin、Infantis、Heidelberg、Schwarzengrundの7血清型を選択した。Enteritidis、Typhimurium、Choleraesuis、Dublinは上述のように届出伝染病の原因菌として指定されており、Infantis、Heidelbergは鶏から高頻度で分離される血清型である。Schwarzengrundは、近年食中毒の発生件数が増加しており、分離株中の多剤耐性株の割合が高いことが報告されている。
【0014】
(2−2)血清型特異的プライマーセットの構築
公的データベースより入手可能であったサルモネラ属菌のゲノム配列について、塩基配列の比較を行った。入手したゲノム配列は以下のとおりである:Choleraesuis str. SC-B67 (accession No.: NC_006905), Dublin str. CT_02021853 (NC_011205), Enteritidis str. P125109 (NC_011294), Gallinarum str. 287/91 (NC_011274), Heidelberg str. SL476 (NC_011083), Paratyphi A str. ATCC 9150 (NC_006511), Paratyphi B str. SPB7 (NC_010102), Paratyphi C strain RKS4594 (NC_012125), Schwarzengrund str. CVM19633 (NC_011094), Typhi str. CT18 (NC_003198), Typhimurium str. LT2 (NC_003197), Typhimurium str. 14028S (CP001363), Typhimurium str. D23580 (FN424405), Typhimurium SL1344 (FQ312003)。
【0015】
ゲノム配列の比較には、公知のソフトウェアであるArtemis Comparison Tool (ACT)を用いた。上記の配列を適宜組み合わせて比較し、各血清型に特異的であると考えられる候補配列を探索した。血清型間での相同性がより低い配列を選抜するため、転写・翻訳の単位に拘らず標的配列を選択した。
選択した部位についてBLAST解析を行い、他属の細菌等を含む既報の配列との相同性が低いことを確認した後、当該部位を検出するプライマーを設計した。Infantisは全ゲノム配列が公開されていなかったため、公開されていた部分配列(accession No.: J03391)を基にプライマーを設計した。
【0016】
また、血清型特異的プライマーを用いて多重PCRを行う際に、同時にサルモネラ属菌存否の判定を行うために、サルモネラ属菌特異的な遺伝子としてinvA遺伝子を選択した。invAはほぼすべてのサルモネラ属菌が保持しており、サルモネラ属菌の簡易的な検出法に応用されている。例えば、先行特許文献にてPCR法やLAMP法での検出に用いるプライマーが報告されている(米国特許5683883、特開2003−199572)。当該invA遺伝子を対象とし、各血清型特異的プライマーとは干渉を起こさずに当該遺伝子の特異的な領域を増幅できるプライマーを設計した。
各プライマー設計にあたっては、多重PCRを行う際にアガロースゲルによる電気泳動で目視による判別が容易となるように各増幅産物の断片長を設定した。具体的には、各増幅産物の断片長がそれぞれに50〜120bp異なるように複数セットずつ設計した。
【0017】
(2−3)設計したプライマーの評価
設計したプライマーは、実施例1の(表1、2)に示すサルモネラ属菌ならびに複数の食中毒菌(Escherichia coli O157, E. coli O26, Campylobacter jejuni, Campylobacter coli, Listeria monocytogenes, Citrobacter freundii, Proteus vulgaris, Pseundomonas aerginosa, Serratia marcescens)のゲノムDNAを鋳型としてPCRを行い、それぞれの血清型に対応して、非特異的な増幅が見られない特異的プライマーが少なくとも1種類以上存在した(同表3、4)。
【0018】
(2−4)本発明の多重プライマー群
本発明の実施例1の(表3、4)に示される各血清型での特異的プライマーから1種類ずつを選択した多重プライマー群の例を同(表5)に示す。
本発明の多重プライマー群はこの1例に限られるものではなく、表3、4に示す他の特異的プライマーセットを組み合わせる場合も含まれる。
また、本発明でプライマーとして用いられるオリゴヌクレオチド断片は、1又は2塩基の範囲内の増減又は変異が導入されても各プライマーにより増幅される対象DNA領域は同一であるから、1又は2塩基の塩基が欠失、置換もしくは付加された配列も許容される。これらプライマーは、例えばビオチンやDIG等の公知の物質により標識してもよい。
【0019】
3.本発明の血清型多重PCR判別法の手順
(3−1)被検試料
被検試料としては、典型的には、家畜生産現場での、家畜の糞便、尿などの排泄物、血液、唾液などの体液、あるいは飼料、飼料添加物などと共に、医療現場での患者からの糞便、尿、血液、組織ホモジェネートなどであるが、畜産加工品、乳製品、鶏卵、卵製品などの飲食品も対象となる。
【0020】
(3−2)サルモネラ属菌の培養方法
これら被検試料を本発明の多重PCRの試料として用いる際に、高濃度のサルモネラ属菌の存在が明らかな場合を除き、まず試料を培地に接種して、サルモネラ属菌に好ましい培養条件下で培養し、サルモネラ属菌を増菌させる。
なお、PCRを進行させるには、プライマーがハイブリダイズできる核酸が数分子から数十分子以上存在すれば十分ではあるので、被検試料から直接核酸抽出してしまうことも可能であるが、再現性の高い結果を得るためには増菌工程を設けることが好ましい。
サルモネラ属菌の増菌培養条件は周知であり、例えば被検試料を緩衝ペプトン水に接種して35℃にて一晩の培養を行った後に、その一部をRV培地に接種して42℃にて一晩の培養を行うことが好ましい。
本発明においては、当該増菌培養液から直ちにゲノムDNAを抽出する下記(3−3)工程に移行することもできるが、各菌株ごとのコロニーを形成させる分離培養に供し、菌株を単離することが好ましい。
その場合の分離培養方法及び分離培養条件は周知であり、例えば、DHL寒天培地に増菌培養液を塗布し、37℃にて一晩の培養を行う。
【0021】
(3−3)被検DNA溶液の調製
分離培養で単離された菌株培養液、又は分離工程を省いた増菌培養液に対して溶菌酵素、界面活性剤、アルカリ等を用いる周知のDNA抽出法を適用する。例えば、培養液を遠心処理して菌体を回収して、細胞溶解液に懸濁し、加熱後遠心分離により回収した上清に対して、フェノール−クロロホルム抽出およびエタノール沈殿によりDNAを回収する。得られたDNAをTE緩衝液に溶解し、PCRに供する。
(3−4)PCR工程
本発明の多重PCRで用いるPCR法では耐熱ポリメラーゼであるTaqDNAポリメラーゼを用い、通常の使用条件の範囲内で増幅させる。具体的には、90〜95℃で熱変成させた後、37〜65℃でアニーリング操作を行い、50〜75℃で重合反応を起こさせ、これを1サイクルとして20から42サイクル行って増幅させる。
(3−5)多重PCRによる増幅産物の検出
本発明の多重PCRによる増幅産物の分子量はいずれも約150〜750bpの範囲内であるため、アガロースゲル電気泳動を用いて検出できる。エチジウムブロマイド等の慣用の染料で染色する。ゲル中のバンドの確認は写真から目視確認してもよいし、ゲル写真を映像データとしてコンピュータに入力し、写真の濃淡を解析することにより確認してもよい。
【実施例】
【0022】
以下、実施例により本発明をより具体的に説明するが、本発明はこれら実施例により何ら限定されるものではない。
なお、本発明で引用した先行文献の記載内容は、本明細書の記載内容として参照されるものとする。
【0023】
[実施例1]各プライマーセットによる増幅の特異性の確認
(1−1)DNA溶液の調製
サルモネラ属菌ならびにサルモネラ属に属しない食中毒菌の単離菌株を増菌培地に接種して培養した。培養液を15,000×gで2分間遠心し、上清を除去して菌体を回収した。回収した菌体を細胞溶解液(PrepMan(登録商標) Ultra Reagent、Applied Biosystems社) 200μlに懸濁し、沸騰水中で10分間加熱した。加熱後の懸濁液を15,000×gで2分間遠心分離し、上清を回収した。上清からフェノール−クロロホルム抽出およびエタノール沈殿によりDNAを回収し、TE緩衝液400μlで溶解した。調製したDNA溶液をPCRに供した。
(1−2)PCR
調製したDNA溶液0.6μlを鋳型として供し、PCRを行った。ExTaq DNAポリメラーゼ(Takara社) 0.375U、1× ExTaq buffer、0.2mM dNTP、ならびに各0.3μMのプライマーを含む15μlの反応液を調製し、DNAサーマルサイクラー(Bio-Rad社製iCycler)を用いて以下の反応を行った。すなわち、94℃にて2分間の熱変性反応を行った後、94℃で30秒間、58℃で30秒間、72℃で40秒間の3ステップからなるサイクリング反応を30回繰り返し、サイクリング反応の終了後に72℃で7分間の伸長反応を行った。
(1−3)検出
PCRによる増幅産物の検出はアガロースゲル電気泳動により行った。アガロースゲルはゲル濃度2%(W/V)とし、100ng/mlの臭化エチジウムを含むものを用いた。TAE溶液の入った泳動槽にゲルを浸し、135Vの定電圧で電気泳動を行った。電気泳動後のゲルにUVランプを照射し、増幅産物の検出ならびにDNA断片長の確認を行った。
(1−4)結果
Enteritidis、Typhimurium、Choleraesuis、Dublin、Infantis、Heidelberg、Schwarzengrundの7血清型について、各血清型に特異的な断片を増幅するプライマーセットをそれぞれ複数セット設計し、増幅の特異性を確認した。サルモネラ属菌標準株44株(表1、2)のゲノムDNAを鋳型として用いた。設計したプライマーセットの中には、標的血清型で増幅産物が検出されないもの、あるいは標的血清型以外のサルモネラ属菌についても増幅産物が検出されるものもあったため、標的血清型のみを特異的に検出するプライマーセットを選抜した。選抜したEnteritidis特異的プライマーセットによるPCRの結果を図2に例示する。選抜したプライマーセットについて、サルモネラ属以外の食中毒菌のゲノムDNAを鋳型としてPCRを行い、増幅産物が検出されないことを確認した(図2、レーン14〜22)。invA検出用プライマーについては、サルモネラ属菌のゲノムDNAを鋳型とした場合のみ増幅産物が検出され、他属の細菌では増幅産物が検出されないことを確認した。
以上の結果から、各血清型について、それぞれ各1セットの血清型特異的なプライマーセットを得ることができた。各血清型の特異的プライマー群及びinvA遺伝子を増幅するプライマーのセットからなる多重PCR判別法用プライマー群を表5に示す。
【0024】
【表1】
【0025】
【表2】
【0026】
【表3】
【0027】
【表4】
【0028】
【表5】
【0029】
[実施例2]多重PCR系の構築
(2−1)多重PCR
実施例1で選抜した各血清型特異的プライマーセット7セット、ならびにinvA遺伝子を増幅するプライマーセットからなる多重PCR判別法用プライマー群(表5)を用いて多重PCRを行った。
鋳型DNA溶液0.6μl、ExTaq DNAポリメラーゼ(Takara社) 0.375U、1×ExTaq buffer、0.2mM dNTP、ならびに各66.7nMのプライマーを含む15μlの反応液を調製し、実施例1と同一の条件で反応を行った。多重PCR用の酵素を用いる場合には、Takara社製Multiplex PCR Assay Kitを用い、鋳型DNA溶液1μlを含む20μlの反応液を調製した。反応条件は、94℃にて1分間の熱変性反応を行った後、94℃で30秒間、58℃で90秒間、72℃で90秒間の3ステップからなるサイクリング反応を30回繰り返し、サイクリング反応の終了後に72℃で10分間の伸長反応を行うこととした。増幅産物は、実施例1と同様のアガロースゲル電気泳動により確認を行った。
(2−2)結果
サルモネラ属菌標準株44株、ならびにサルモネラ属以外の食中毒菌のゲノムDNAを鋳型として、多重PCRを行った。汎用的な耐熱性DNAポリメラーゼ(ExTaq)を用いて多重PCRを行ったところ、Schwarzengrund特異的プライマーセットについての感度が低下し、サルモネラ属以外の食中毒菌の一部で非特異的増幅産物が検出される不都合が確認されたので、Multiplex PCR Assay Kitを用いることとした。当該Assay Kitを用いることで上記不都合は解消された。Multiplex PCR Assay Kitを用いて得られた代表的な結果を図3に示す。
サルモネラ属菌標準株のDNAを鋳型としたところ、すべての株においてinvAの増幅産物が検出された(図3、レーン1〜8)。特異的プライマーを構築した7血清型(Enteritidis、Typhimurium、Choleraesuis、Dublin、Infantis、Heidelberg、Schwarzengrund)については、invAの増幅産物に加え、各血清型特異的断片が検出された(図3、レーン1〜7)。それぞれのゲノムDNAを混合して鋳型DNAとして用いたところ、それぞれの血清型に由来する特異的断片が検出され、非特異的産物は検出されなかった(図3、レーン9)。また、サルモネラ属以外の食中毒菌のゲノムDNAを鋳型とした場合には、いずれの増幅産物も検出されなかった(図3、レーン10〜18)。
以上の結果から、構築した多重PCR系は、invAの増幅によるサルモネラ属菌の検出、ならびにEnteritidis、Typhimurium、Choleraesuis、Dublin、Infantis、Heidelberg、Schwarzengrundの7血清型については各血清型特異的な断片の検出による血清型の判別が可能であることが示された。
【0030】
[実施例3]野外単離株を用いた多重PCRの検証
野外検体より単離されたサルモネラ属菌を用い、多重PCRの検証を行った。用いた菌株の血清型ならびに株数を表6に示す。
Enteritidis、Typhimurium、Choleraesuis、Dublin、Infantisについては、従来の手法で決定された血清型と多重PCRの結果が一致していた。I4,5,12:i:−は、検証した2株でいずれもTyphimurium特異的増幅産物を検出した。該血清型はTyphimuriumの単相型の変異株であると言われており、両者は遺伝的に非常に近いと思われる。このため、両者を判別することは非常に困難であることが容易に推測できる。この両者の判別が必要な場合は、さらに他の手法による確認が必要となる。
他の血清型、ならびに血清型が確定されていない株についてはinvAの増幅産物のみを検出した。
以上のように、本手法はTyphimuriumとI4,5,12:i:−の両者がTyphimuriumと判定されるものの、野外単離株の血清型別に有用であることが示された。
【0031】
【表6】
【0032】
[実施例4]コロニーPCRによる野外株の血清型別
(4−1)コロニーPCR
野外から得られた検体から、公定法に準じた作業によりサルモネラ属菌株の分離を行った。すなわち、検体1gあたり9mlのリン酸緩衝ペプトン水を加え、35℃にて24時間の前増菌培養を行った。培養後、0.1mlの前増菌培養液を増菌培養用培地10mlに加え、42℃にて一晩の増菌培養を行った。増菌培養液から1白金耳を寒天培地上に塗布し、一晩培養して菌株を分離した。検出されたコロニーをコロニーPCRに供した。実施例2と同様に調製した15μlの反応液にコロニーの一部を懸濁した後、サーマルサイクラーによって実施例1と同一の条件で反応を行った。増幅産物は実施例1と同様の手法により確認した。
(4−2)結果
野外検体より単離された菌株について、コロニーPCRにより血清型別を行った(図4)。レーン1、2、7、8、9、10、11、12、17、18について、invAならびにInfantis特異的断片が検出され、Salmonella Infantisであることが推測された。抗血清を用いてこれらの菌株の血清型を確定したところ、Infantisであることが確認され、多重PCRの結果と一致した。
また、外見上サルモネラ属菌であることが疑われたコロニーについても同様にコロニーPCRを行ったが、増幅産物は検出されなかった(図4、レーン3〜6、13〜16、19、20)。これらの菌株について16S rDNAの配列を決定したところ、Proteus属の細菌であることが確認された。このため、多重PCRによりサルモネラ属菌特異的に増幅産物が検出されることが示された。
このように、野外検体より単離した菌株について、コロニーPCRによりサルモネラ属菌であることの確認、ならびに簡易的な血清型別が可能であった。
【0033】
[実施例5]増菌培養液由来DNAを用いたサルモネラ血清型迅速判別
(5−1)増菌培養液のDNA溶液の調製
野外検体から実施例4と同様の手法で増菌培養液を調製した。培養液2mlを15,000×gで2分間遠心し、上清を除去して菌体を回収した。回収した菌体を細胞溶解液 (PrepMan(登録商標) Ultra Reagent、Applied Biosystems社) 200μlに懸濁し、沸騰水中で10分間加熱した。加熱後の懸濁液を15,000×gで2分間遠心分離し、上清を回収した。上清からフェノール−クロロホルム抽出およびエタノール沈殿によりDNAを回収し、TE緩衝液400μlに溶解した。得られたDNA溶液を鋳型として多重PCRを行った。
(5−2)結果
実施例4と同一の検体から得られた増菌培養液よりDNAを抽出し、それを鋳型として多重PCRを行った。その結果、5検体にサルモネラ属菌が存在することが確認された(図5、レーン1、4〜6、9)。レーン1、4〜6においてはinvAならびにInfantis特異的断片が検出され、これらの検体中にSalmonella Infantisが存在することが示唆された。実施例3は各検体から得られた単離株(各2株)についてコロニーPCRを行っており、図4の検体1、2(レーン1、2)は図5の検体1(レーン1)から得られた菌株である。このように、検体1、4、5、6については単離菌株を用いたコロニーPCRの結果と一致することが示された。
一方、検体9については、invA、Infantis特異的断片に加えEnteritidis特異的断片が検出された(図5、レーン9)。検体9から得られた分離菌株について、さらに6コロニーを検査したところ、8コロニー中1コロニーがEnteritidisであることが確認された。すなわち、増菌培養液由来DNAを鋳型としたPCRを行うことにより、複数の血清型を検出可能であることが示された。
【産業上の利用可能性】
【0034】
サルモネラ属菌は罹患した宿主の糞便から検出されるため、糞便に存在するサルモネラ属菌の血清型を把握することは疾病の原因特定において重要である。従来法により糞便中に存在するサルモネラ属菌の血清型を同定するには、前述した理由により、採集した糞便検体からサルモネラ属菌を分離する必要がある。一方、本手法は対象とする血清型は限られるものの、ゲノムDNAの抽出が可能であれば様々な検体に応用できるため、糞便の検査にも応用可能である。糞便からより効率的にゲノムDNAを抽出する手法が開発されれば、本手法を用いた検査の感度も向上すると考えられる。また、糞便中にはPCRを阻害する物質が含まれるため、現状では糞便から直接サルモネラ属菌を検出することは不可能であるが、この阻害を回避するPCR酵素やPCR反応緩衝液の開発により、糞便を直接検査する手法の開発に繋がる。
【配列表フリーテキスト】
【0035】
1.invA-F1 AGTGCCGGTTTTATCGTGAC
2.invA-R1 ACCGCCAGACAGTGGTAAAG
・Enteritidis
3.Ent1 TCAAGCATTGATACGGCAAG
4.Ent2 CGCATTCTGCTCAATGAAAA
・Typhimurium
5.Tpm15 GCGTTGTTCCCTTTGTTCTG
6.Tpm16 GACGCTGTTACGAGCCATTT
・Infantis
7.Inf3 GCGGCAGGTTGGTATTATTC
8.Inf4 CTAAACGTTTTCCCGCTGTT
・Choleraesuis
9.Chs1 TCGCACCATGACCAAAAATA
10.Chs2 CACTCCGTCGTTCTGGGTAT
・Dublin
11.Dub11 GCGTGAAGTGGCAACAGTAG
12.Dub12 ATATCGGCCAGTGCAGATTC
・Heidelberg
13.Hdb15 TAACCAAACTCAGCGGCTCT
14.Hdb16 GGGGCGACTGATGCTATAAA
・Schwarzengrund
15.Swg7 GAAGAGAACGCTCGGACAAC
16.Swg8 CAGGCAATTCTCCGTCTCTC
【特許請求の範囲】
【請求項1】
下記の(1)〜(8)に示される塩基配列又はそれぞれの相補配列からなるプライマーセットを含むことを特徴とする、サルモネラ属菌血清型の多重PCR判別法用プライマーセット;
(1)invA遺伝子領域を増幅するためのプライマーセットであって、
片方のプライマーが配列番号1に示される塩基配列、又は配列番号1の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号2に示される塩基配列、又は配列番号2の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなる;
(2)Enteritidis特異的なプライマーセットであって、
片方のプライマーが配列番号3に示される塩基配列、又は配列番号3の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号4に示される塩基配列、又は配列番号4の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(3)Typhimurium特異的なプライマーセットであって、
片方のプライマーが配列番号5に示される塩基配列、又は配列番号5の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号6に示される塩基配列、又は配列番号6の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなる;
(4)Infantis特異的なプライマーセットであって、
片方のプライマーが配列番号7に示される塩基配列、又は配列番号7の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号8に示される塩基配列、又は配列番号8の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(5)Choleraesuis特異的なプライマーセットであって、
片方のプライマーが配列番号9に示される塩基配列、又は配列番号9の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号10に示される塩基配列、又は配列番号10の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(6)Dublin特異的なプライマーセットであって、
片方のプライマーが配列番号11に示される塩基配列、又は配列番号11の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号12に示される塩基配列、又は配列番号12の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなる;
(7)Heidelberg特異的なプライマーセットであって、
片方のプライマーが配列番号13に示される塩基配列、又は配列番号13の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号14に示される塩基配列、又は配列番号14の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(8)Schwarzengrund特異的なプライマーセットであって、
片方のプライマーが配列番号15に示される塩基配列、又は配列番号15の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号16に示される塩基配列、又は配列番号16の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである。
【請求項2】
被検試料にサルモネラ属菌を増殖させる培養条件下で培養した後、被検菌株を分離して請求項1に記載のプライマーセットを用いて多重PCRを行うことを特徴とする、サルモネラ属菌血清型の多重PCR判別法。
【請求項3】
被検試料にサルモネラ属菌を増殖させる培養条件下で培養した後、被検菌株を分離せずに、請求項1に記載のプライマーセットを用いて多重PCRを行うことを特徴とする、サルモネラ属菌血清型の多重PCR判別法。
【請求項1】
下記の(1)〜(8)に示される塩基配列又はそれぞれの相補配列からなるプライマーセットを含むことを特徴とする、サルモネラ属菌血清型の多重PCR判別法用プライマーセット;
(1)invA遺伝子領域を増幅するためのプライマーセットであって、
片方のプライマーが配列番号1に示される塩基配列、又は配列番号1の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号2に示される塩基配列、又は配列番号2の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなる;
(2)Enteritidis特異的なプライマーセットであって、
片方のプライマーが配列番号3に示される塩基配列、又は配列番号3の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号4に示される塩基配列、又は配列番号4の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(3)Typhimurium特異的なプライマーセットであって、
片方のプライマーが配列番号5に示される塩基配列、又は配列番号5の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号6に示される塩基配列、又は配列番号6の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなる;
(4)Infantis特異的なプライマーセットであって、
片方のプライマーが配列番号7に示される塩基配列、又は配列番号7の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号8に示される塩基配列、又は配列番号8の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(5)Choleraesuis特異的なプライマーセットであって、
片方のプライマーが配列番号9に示される塩基配列、又は配列番号9の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号10に示される塩基配列、又は配列番号10の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(6)Dublin特異的なプライマーセットであって、
片方のプライマーが配列番号11に示される塩基配列、又は配列番号11の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号12に示される塩基配列、又は配列番号12の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなる;
(7)Heidelberg特異的なプライマーセットであって、
片方のプライマーが配列番号13に示される塩基配列、又は配列番号13の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号14に示される塩基配列、又は配列番号14の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである;
(8)Schwarzengrund特異的なプライマーセットであって、
片方のプライマーが配列番号15に示される塩基配列、又は配列番号15の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなり、
他方のプライマーが配列番号16に示される塩基配列、又は配列番号16の1もしくは2個の塩基が欠失、置換もしくは付加されている塩基配列からなるプライマーセットである。
【請求項2】
被検試料にサルモネラ属菌を増殖させる培養条件下で培養した後、被検菌株を分離して請求項1に記載のプライマーセットを用いて多重PCRを行うことを特徴とする、サルモネラ属菌血清型の多重PCR判別法。
【請求項3】
被検試料にサルモネラ属菌を増殖させる培養条件下で培養した後、被検菌株を分離せずに、請求項1に記載のプライマーセットを用いて多重PCRを行うことを特徴とする、サルモネラ属菌血清型の多重PCR判別法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2012−170380(P2012−170380A)
【公開日】平成24年9月10日(2012.9.10)
【国際特許分類】
【出願番号】特願2011−34184(P2011−34184)
【出願日】平成23年2月21日(2011.2.21)
【出願人】(000229519)日本ハム株式会社 (57)
【Fターム(参考)】
【公開日】平成24年9月10日(2012.9.10)
【国際特許分類】
【出願日】平成23年2月21日(2011.2.21)
【出願人】(000229519)日本ハム株式会社 (57)
【Fターム(参考)】
[ Back to top ]