
多量体構築物
Flt−1のIg様ドメインを有する多量体融合蛋白質は、リンカー部分を包含することにより機能を提供される。融合蛋白質をコードするベクターと融合蛋白質を発現する宿主細胞は、新血管新生に関する病理条件に対して、治験的に新血管新生を防ぐことに使用され得る。そのような条件は、加齢黄斑変性症、がん、乾癬、増殖性の糖尿病網膜症、喘息、骨関節症、および慢性関節リウマチについて効果を包含する。Flt−1のIg様ドメイン、すなわちリンカーおよび多量体化ドメインに利用した多量体化と同じ手法は、細胞外受容体、抗体可変領域、ポリペプチド、サイトカイン、ケモカイン、および成長因子を含む他のポリペプチドでも利用され得る。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、喘息、関節症、癌、および傷の再生などの病理的な新血管新生を治療するために組み換え構築された蛋白質の利用に関連する。
【背景技術】
【0002】
病理性新血管新生は、浮腫性加齢黄斑変性症(AMD)、増殖性糖尿病網膜症、骨関節症、および喘息等の病気を構成する要素の一つである。また、新血管新生は、腫瘍の増殖および転移に重要な役割を果たす。新血管新生は、前駆血管形成因子および抗血管形成因子の厳密な均衡によって制御されている。
【0003】
血管内皮成長因子(VEGF)は、新血管新生に必須であることが知られる。VEGF活性の阻害は、AMD、関節症の動物モデルおよび様々な腫瘍モデルにおいて新血管新生を抑制することが示されている。VEGF活性を阻害する方法は、抗体、受容体融合蛋白質、ペプチド、および小分子を含む。
【0004】
VEGF−R1(Flt−1)およびVEGF−R2(KDR)の蛋白質は、高い結合能でVEGFに結合することが示されている。Flt−1およびKDRの両方は、細胞外領域に7個のIg様ドメインを有する。ドメイン2は、VEGFに結合するために必須であることが示されている。全長、可溶性受容体(ドメイン1−7)、およびドメイン1−3各々ならびにIgGのFcとの融合蛋白質が、効果的にVEGFに結合する。しかしながら、IgGのFcおよびIg様ドメイン2単独との融合は、VEGFに結合できないが、Ig様ドメイン1および2の両方との融合は、結合可能であった。Davis−Smyth,1996。それゆえ、Ig様ドメイン1および3は、ドメイン2を伴うことで効果的なVEGF結合に必要とされるようである。
【発明の開示】
【課題を解決するための手段】
【0005】
(発明の簡潔な要約)
本発明の一の具体例として、融合蛋白質が提供される。融合蛋白質は、式X−Y−Zを有する。Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含む。Yは、5−25個のアミノ酸残基のポリペプチドから必須として成る。Zは、IgG重鎖分子のCH3領域である。
【0006】
本発明の別の具体例は、式X−Y−Zのポリペプチドである。Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含む。Yは、5−25個のアミノ酸残基の空間的な分離を提供するリンカー部分から必須として成る。Zは、IgG重鎖分子のCH3領域である。
【0007】
また、本発明の別の態様としては、式X−Y−Zで示される融合蛋白質である。Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含む。Yは、5−25個のアミノ酸残基のポリペプチドから必須として成る。Zは、抗体分子のFc部位である。
【0008】
式X−Y−Zで示される融合蛋白質もまた提供される。Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドである。Yは、5−25個のアミノ酸残基の空間的分離を提供するリンカー部分から必須として成る。Zは、抗体分子のFc部位である。
【0009】
さらに、本発明の別の態様としては、ポリペプチドXを多量体化する方法である。ポリペプチドXは、ポリペプチドXYZを形成するために、ポリペプチドYを介してポリペプチドZに結合される。Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含む。Yは、5−25個のアミノ酸残基のポリペプチドから必須として成る。Zは、IgG重鎖分子のCH3領域である。形成されるポリペプチドXYZは、多量体化する。
【0010】
さらに本発明の別の具体例としては、ポリペプチドXを多量体化する方法を提供する。ポリペプチドXは、部分Yを介してポリペプチドZに結合されることで、ポリマ−XYZを形成する。Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含む。Yは、5−25個のアミノ酸残基の空間的な分離を提供するリンカー部分から必須として成る。Zは、IgG重鎖分子のCH3領域である。形成されるポリペプチドXYZは、多量体化する。
【0011】
本発明の一の具体例において、核酸分子が提供される。本核酸分子は、VEGF−R1(Flt−1)のIg様ドメイン2、リンカー、および多量体化ドメインを含む融合蛋白質をコードする。融合蛋白質は、配列番号2、8、21、23、および25から成る群から選択される配列を含む。
【0012】
本発明の別の具体例において、融合蛋白質が提供される。本融合蛋白質は、VEGF−R1(Flt−1)のIg様ドメイン2、リンカー、および多量体化ドメインを含む。本融合蛋白質は、配列番号2、8、21、23、および25から成る群から選択される配列を含む。
【0013】
本発明の別の具体例において、in vitroの方法が提供される。本核酸分子は、同定された哺乳類細胞に送達される。この核酸分子は、VEGF−R1(Flt−1)のIg様ドメイン2、リンカー、および多量体化ドメインを含む融合蛋白質をコードする。融合蛋白質は、配列番号2、8、21、23、および25から成る群から選択される配列を含む。融合蛋白質の発現は、プロモーターによって制御される。融合蛋白質を発現する細胞が形成される。
【0014】
さらに、本発明の別の具体例は、融合蛋白質を哺乳類に送達する方法である。融合蛋白質を発現する哺乳類細胞が、哺乳類に送達される。この細胞は、融合蛋白質を発現および分泌し、それによって融合蛋白質を哺乳類動物に供給する。融合蛋白質は、VEGF−R1(Flt−1)のIg様ドメイン2、リンカー、および多量体化ドメインを包含する。融合蛋白質は、配列番号2、8、21、23、および25から成る群から選択される配列を含む。
【0015】
本発明の別の態様は、融合蛋白質を哺乳類動物に供給する方法である。VEGF−R1(Flt−1)のIg様ドメイン2、リンカー、および多量体化ドメインを含む融合蛋白質が、哺乳類動物に送達される。融合蛋白質は、配列番号2、8、21、23、および25から成る群から選択される配列を含む。代替的に、該融合蛋白質をコードする核酸構築物が、哺乳類動物に送達されることも可能であり、そのことにより、融合蛋白質が哺乳類動物によって発現される。
【0016】
これらおよび下記により詳細が記述されている本発明の他の具体例は、血管増殖および炎症に関連する病気を治療する方法および薬剤を当該技術分野に提供するであろう。薬剤は、蛋白質の天然型に関連する上昇した安定性と生物利用を提供するかもしれない。
【0017】
(図の説明)
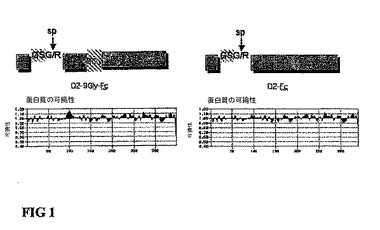
図1。D2−9Gly−Fc構築物における9−Glyリンカーの可撓性領域。KarpusとSchultz(1985年)の方法による予測相対的可撓性は、D2−9Gly−Fc蛋白質中のポリグリシン9mer(9−Gly)のリンカー(アミノ酸94から103)が、9−Glyのリンカーを含まないD2−Fc構築物と比較して、平均(>1)より大きな可撓性を有する領域であることを示す。両方の融合蛋白質は、箱型で囲まれた同一のアミノ酸配列を含む。sp−シグナルペプチド(アミノ酸−24から−1)、Flt−1ドメイン2(アミノ酸1から93)およびIgG1−Fc残基(244アミノ酸)。矢印は、SignalPV2.0programを用いて予測したシグナルペプチダ−ゼ切断領域を表す(Nielsenら,1997)。
【0018】
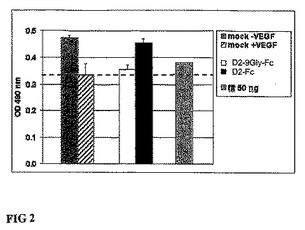
図2。D2−9Gly−Fc対D2−Fcの生物学的活性。293細胞は、飢餓培地(M199+5%FCS)で培養され、CMVプロモーター制御下にあるD2−9Gly−FcおよびD2−Fcの発現カセットを含むプラスミドを移入された。条件培地(CM)は72時間後に回収された。HUVECが、96穴プレート(2E3細胞/穴)の飢餓培地+VEGF(10ng/mL)中に分注され、50ulのCMとVEGF(10ng/mL)が24時間後に添加された。コントロール(+/−VEGF)は、コントロールpEGFP(Clontech;pEGFPは、哺乳類細胞内で、より明るい蛍光およびより高い発現を示すように最適化され、野生型緑色蛍光蛋白質(GFP)の赤色転換変動を有する)に由来するCM中で培養された。陽性対照は、50ngのFlt−1−IgG組み換え体蛋白質(R&D Systems)で処理された。HUVEC数は、CellTiter96(登録商標)AQueous regeant(Promega)を用いて、増殖3日間後に測定された。データは、3種類行われた2回の実験各々のOD490平均値を表す。
【0019】
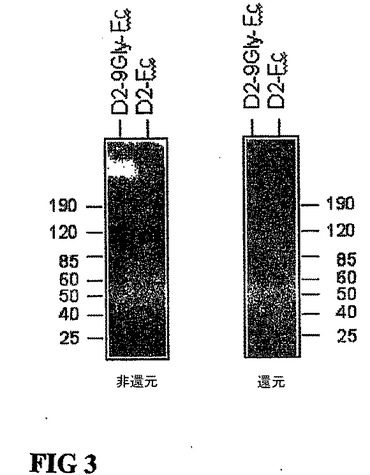
図3。D2−9Gly−FcおよびD2−Fcのウェスタンブロット解析。D2−9Gly−FcおよびD2−Fcの蛋白質両方の大きさは、還元ゲルにおける移動と比較して非還元ゲルにおける移動は2倍になると思われる。D2−9Gly−FcおよびD2−Fcを発現するプラスミドが、293細胞に移入された後、条件培地から蛋白質がロードされ、SDS−電気泳動により分離され、PVDF膜に移行された。ブロットは、ヤギ抗ヒト抗IgG1Fcおよびラビット抗ヤギIgG−HRP抗体を用いて行われた。
【0020】
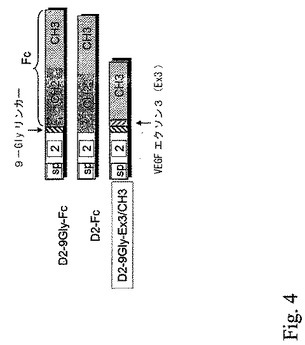
図4。9GlyのリンカーおよびVEGFEx3を含むsFlt−1複合蛋白質。D2−9Gly−Ex3/CH3と過去に構築された蛋白質との構造比較。3個の蛋白質全ては、アミノ酸24個のFlt−1シグナルペプチドおよびアミノ酸93個のFlt−1ドメイン2から成るFlt−1ドメイン2の同一アミノ酸配列を含む。D2−9Gly−Ex3/CH3は、アミノ酸9個の9Glyリンカー、アミノ酸14個のVEGFEx3およびアミノ酸120個のヒトIgG1重鎖FcCH3領域を含む。
【0021】
図5。D2−9Gly−Ex3/CH3対D2−9Gly−Fcの生物学的活性。ドメイン2が9GlyリンカーおよびVEGFのEx3を介してCH3領域に結合されている、蛋白質D2−9Gly−Ex3/CH3は、蛋白質D2−9Gly−FcおよびD2−Fcと比較して効果的にVEGF依存性HUVEC増殖を阻害する。50ngの組み換え体Flt−1−IgG(R&D Systems)をコントロールとして使用した。
【0022】
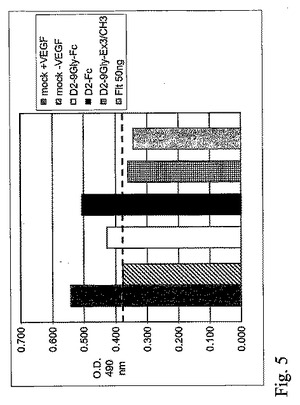
図6。D2−(Gly4Ser)3−Fc蛋白質活性をD2−9Gly−FcおよびD2−9Gly−Ex3/CH3と比較したHUVEC増殖測定。
【0023】
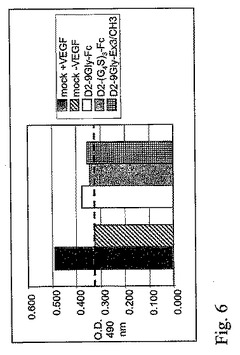
図7。ウェスタンブロット。(1)D2−9Gly−Fc、(2)D2−(G4S)3−Fcおよび(3)EGFP蛋白質を発現するプラスミドを移入された293細胞の条件培地からの蛋白質(非還元と還元)が、SDS−電気泳動によって分離され、PVDF膜に移行された。ブロットは、ヤギ抗ヒトIgG1Fcおよびウサギ抗ヤギIgG−HRP抗体を用いて行われた。
【0024】
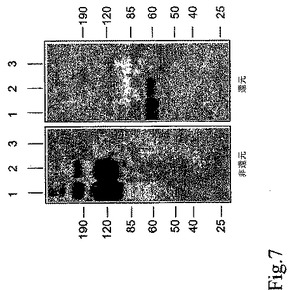
図8。9GlyリンカーもしくはVEGFEx3の有無による蛋白質の組み合わせ。9Glyリンカーおよび/もしくはVEGFEx3、D2−9Gly−CH3、D2−CH3およびD2−Ex3/CH3を有すもしくは有さない、3個の新規蛋白質の構造比較。
【0025】
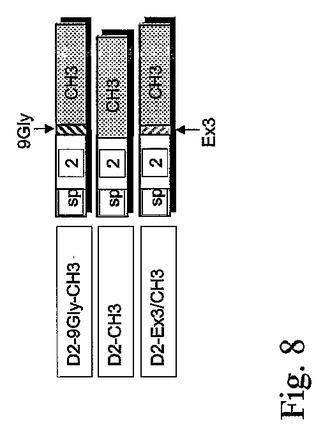
図9。9Gly、Ex3およびCH3の組み合わせを有するFlt−1(D2)構築物におけるHUVEC増殖測定。蛋白質D2−Ex3/CH3、D2−9Gly−CH3およびD2−CH3を含む293細胞(5ul)に由来する条件培地をD2−9Gly−FcおよびD2−9Gly−Ex3/CH3と比較した。
【0026】
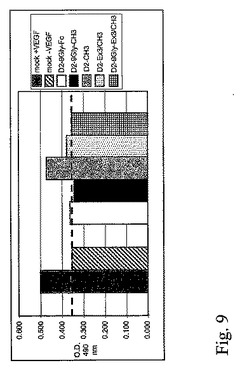
図10。ウェスタンブロット。293細胞に(1)D2−9Gly−Fc、(2)D2−9Gly−CH3(52/26kDa)、および(3)D2−CH3(50/25kDa)を発現するプラスミドが移入された。条件培地(15ulの非還元および/もしくは還元されたCM)中の293細胞に由来する蛋白質は、SDS−電気泳動で分離され、PVDF膜に移行された。ブロットは、抗ヒトVEGF−R1−HRPを用いて行われた(R&D Systems)。
【0027】
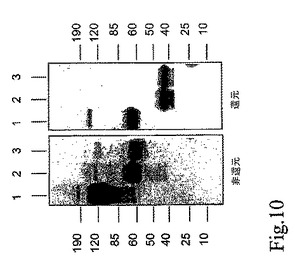
図11。VEGF”in vitro”結合測定。既知濃度のD2−9Gly−FcおよびFlt−1D(1−3)のコントロール可溶性受容体(濃度範囲0.29−150pM)を含む293細胞由来の条件培地が、連続的に希釈され、10pMのVEGFを添加された。非結合VEGFの量がELISAで測定された。D2−9Gly−Fcは、他の構築物すべてより高い結合能でVEGFに結合する。”n”は、(移入および結合測定)非依存的な実験数を表す。
【0028】
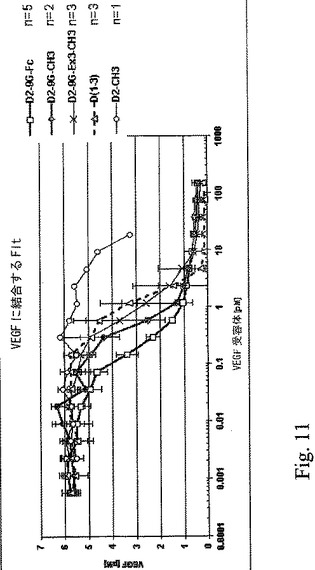
図12。可溶性Flt−1構築物の結合反応速度が、BIAcore機器を用いた表面プラスモン共鳴によって測定された。sFlt−1構築物がセンサーチップ上に固定され、VEGF165が0.2から15nMの濃度範囲で注入された。センサーグラムの値がBIA評価プログラムを用いて求められ、速度定数KaおよびKdが決定され、解離定数(KD)がKd/Ka=KDの割合から計算された。より低いKD値はより高い親和性を意味する。
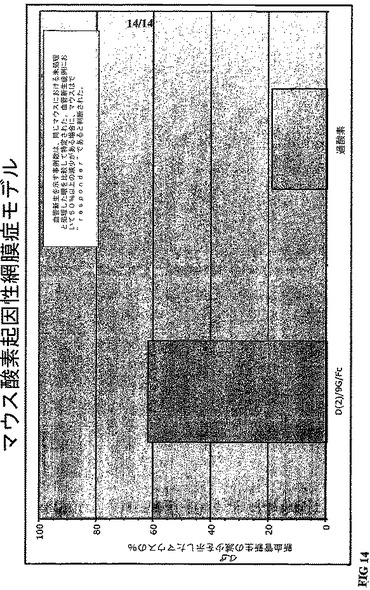
【0029】
図13A−13C。図13Aは、各種リンカーを有するFlt−1構築物の発現レベルを示す。図13Bは、各種リンカーおよびIgG1FcのCH3部分を有するFlt−1構築物の二量体化もしくは多量体化を表す。非還元および還元条件の相違は、蛋白質が多量体化したかである。図13Cは、VEGF存在下のHUVEC増殖測定における条件培地中において、各Flt−1構築物の阻害的な生物活性が存在することを示す。構築物各々は、VEGF非存在下におけるHUVEC増殖レベルに近い阻害活性を示した。
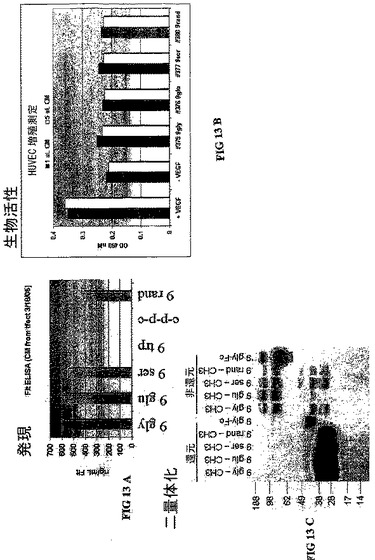
【0030】
図14。網膜新生血管(NV)のマウス酸素誘導型網膜症モデルを用いて、Flt−1構築物の一つがマウスの眼に投与され、新血管新生が特定された。このマウスは高酸素状態に置かれた。血管新生を示す事例数は、同じマウスにおける未処理と処理した眼を比較して特定された。血管新生症例において50%以上の減少がある場合に、マウスは”responder”であると判断された。
【0031】
(発明の詳細な説明)
本発明者の知見は、ドメイン1および3を有さないFlt−1のIg様ドメイン2が、効果的にVEGFに結合し、VEGF依存性内皮細胞の増殖を抑制できることである。ドメイン2は、リンカーを介して共有的に多量体化ドメインに結合され得る。リンカーは、一般的にポリペプチド鎖である。鎖の長さは、6、7、9、11、13、15もしくはそれ以上のアミノ酸残基であり得るが、典型的には5から25残基の間である。長さと側鎖構成物に依存して、リンカーは平均的な可擣性よりも大きな可撓性を有していてもよいが、それは必須ではない。可撓性は、当該技術分野において知られる算法によって計算され得る。多量体化ドメインは、多量体蛋白質の一部分であり、例えば、二量体、三量体、四量体等を形成するサブユニットの連係を増進する。効果的なVEGFの結合および/もしくはVEGF依存性の内皮細胞増殖の抑制に適合可能な組み換え体蛋白質が、配列番号2、8、21、23および25から成る群から選択される。
【0032】
さらに、発明者は、多量体化ドメインおよびリンカーが、様々な他の蛋白質もしくは蛋白質の部位とともに使用され得ることで、多量体化を誘導することを見い出した。このような蛋白質は、多量体化する時に限りリガンドもしくは受容体に結合する蛋白質であるか、または多量体化する時に結合能が上昇する蛋白質であろう。多量体化に適する蛋白質は、細胞外受容体(その一部分を含む)、抗体可変領域、サイトカイン、ケモカイン、および成長因子を包含する。適合する蛋白質は、チロシンキナーゼ受容体およびセリンスレオニンキナーゼ受容体を含む。細胞外受容体の詳細な例としては、EGF受容体、Gタンパク質共役型受容体、FGF受容体、Fc受容体、T細胞受容体等を含む。抗体可変領域の例は、Fab、F(ab’)2、およびScFvを含む。サイトカインの例は、GM−CSF、IL−la、IL−1b、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−10、IL−12、IL−18、IL−21、IL−23、IFN−a、IFN−b、MIP−la、MIP−1b、TGF−b、TNFa、およびTNF−13を含む。ケモカインの例は、BCA−1/BLC、BRAK、ChemokineCC−2、CTACK、CXCL−16、ELC、ENA、ENA−70、ENA−74、ENA−78、Eotaxin、Exodus−2、Fractalkine、GCP−2、GRO、GROalpha(MGSA)、GRO−beta、GRO−gamma、HCC−1、HCC−4、I−309、IP−10、I−TAC、LAG−1、LD78−beta、LEC/NCC−4、LL−37、リンホタクチン(Lymphotactin)、MCP、MCAF(MCP−1)、MCP−2、MCP−3、MCP−4、MDC、MDC、MDC−2、MDC−4、MEC/CCL28、MIG、MIP、MIP−1alpha、MIP−1beta、MIP−1delta、MIP−3/MPIF−1、MIP−3alpha、MIP−3bet、MIP−4(PARC)、MIP−5、NAP−2、PARC、PF−4、RANTES、RANTES−2、SDF−1alpha、SDF−1beta、TARC、およびTECKを含む。成長因子の例は、ヒトアンヒィレギュリン(Amphiregulin)、ヒト血管形成蛋白質(Angiogenesis Protein)、ヒトACE、ヒトアンジオゲニン(Angiogenin)、ヒトアンジオポエチン(Angiopoietin)、ヒトアンジオスタチン(Angiostatin)、ヒトベータセルリン(Betacellulin)、ヒトBMP、ヒトBMP−13/CDMP−2、ヒトBMP−14/CDMP−1、ヒトBMP−2、ヒトBMP−3、ヒトBMP−4、ヒトBMP−5、ヒトBMP−6、ヒトBMP−7、ヒトBMP−8、ヒトBMP−9、ヒトコロニー刺激因子、ヒトflt3−Ligand、ヒトGCSF、ヒトGM−CSF、ヒトM−CSF、ヒト結合組織成長因子(Connective Tissue Growth Factor)、ヒトCripto−1、ヒトCryptic、ヒトECGF、ヒトEGF、ヒトEG−VEGF、ヒトエリスロポエチン(Erythropoietin)、ヒトフェツィン(Fetuin)、ヒトFGF、ヒトFGF−1、ヒトFGF10、ヒトFGF−16、ヒトFGF−17、ヒトFGF−18、ヒトFGF−19、ヒトFGF2、ヒトFGF−20、ヒトFGF−3、ヒトFGF−4、ヒトFGF−5、ヒトFGF−6、ヒトFGF−7/KGF、ヒトFGF−8、ヒトFGF−9、ヒトFGF−酸性、ヒトFGF−塩基性、ヒトGDF−11、ヒトGDF−15、ヒト成長ホルモン遊離因子(Growth Hormone Releasing Factor)、ヒトHB−EGF、ヒトヘレグリン(Heregulin)、ヒトHGF、ヒトIGF、ヒトIGF−I、ヒトIGF−II、ヒトインヒビン(Inhibin)、ヒトKGF、ヒトLCGF、ヒトLIF、ヒト Miscellaneous Growth Factor、ヒトMSP、ヒトミオスチン(Myostatin)、ヒトミオスタチンプロペプチド(Myostatin Propeptide)、ヒト神経成長因子(Nerve Growth Factor)、ヒトオンコスタチン M(Oncostatin M)、ヒトPD−ECGF、ヒトPDGF、ヒトPDGF(AAホモダイマー)、ヒトPDGF(ABホモダイマー)、ヒトPDGF(BBホモダイマー)、ヒトPDGF(CCホモダイマー)、ヒトPIGF、ヒトPIGF、ヒトPIGF−1、ヒトPIGF−2、ヒトSCF、ヒトSMDF、ヒト幹細胞成長因子(Stem Cell Growth Factor)、ヒトSCGF−alpha、ヒトSCGF−beta、ヒトスロンボポエチン(Thrombopoietin)、ヒト形質転換成長因子(Transforming Growth Factor)、ヒトTGF−alpha、ヒトTGF−beta、およびヒトVEGFを含む。
【0033】
Flt−1受容体蛋白質は、7個のIg様ドメインを含む細胞外部分を有する。これらは、ジェンバンク受託番号P17948の残基番号32から123、151から214、230から327,335から421、428から553、556から654、661から747に位置する。配列番号15も参照。残基番号1から26は、シグナル配列を含む。Flt−1蛋白質は、ジェンバンク受託番号NM_002019(配列番号14)で示されるDNA配列によってコードされる。
【0034】
多量体化ドメインは、当該技術分野で知られるように使用され得る。例えば、IgG1もしくはIgG2のラムダ重鎖Fc部分の配列は、CH3単独(アミノ酸371−477)もしくはCH2およびCH3両方のドメイン(アミノ酸247−477)について使用され得る。Ig分子のFc部分は、酵素パパインによって抗体分子全体を切断することで得られる。他の方法を使用することでもこれらの部分が得られる。IgG1のラムダ重鎖の蛋白質配列は、ジェンバンク受託番号Y14737および配列番号10を参照のうえ。例えば、他のIgG型に由来およびIgA、IgM、IgDもしくはIgE抗体に由来するような別のFc領域が使用され得る。VEGFの多量体化領域もまた使用され得る。VEGFをコードするDNA配列は、ジェンバンク受託番号NM003376および配列番号11に記載されている。VEGFのアミノ酸配列は、ジェンバンク受託番号CAC19513および配列番号12に記載されている。VEGF exon3(VEGF Ex3)にコードされるVEGFの多量体化領域(配列番号13)は、VEGF蛋白質のアミノ酸残基75−88(配列番号12)に相当する。多量体化ドメインは、少なくとも5%、10%、20%、30%、40%、50%、60%、70%、80%、85%、90%もしくは95%の単量体融合蛋白質が、多量体に適合する速度で非変性ポリアクリルアミドゲル上を移動することを引き起こす。糖鎖付加は、ゲル中の蛋白質の移動に影響し得る。特定の配列がここで示されるが、対立形質のような変異体もよく使用され得る。典型的なこのような変異体は、開示された配列に少なくとも85%、90%、95%、97%、98%、もしくは99%の同一性を有する。
【0035】
多量体化は、例えば、ここで示されるように還元および非還元ゲルを使用して検証され得る。多量体化はまた、リガンド/受容体に対する蛋白質の上昇した結合能を検出することによって測定され得る。これに関連してBiaCore(登録商標)の表面プラスモン共鳴測定が使用され得る。これらの測定は、センサーチップ表面に近接する水層の屈折率の変化を測定することによって質量の変化を検出する。当該技術分野で知られるいくつかの方法が、多量体化を検出するのに使用され得る。
【0036】
本発明によるリンカー部分は、例えば、5−100個のアミノ酸残基、5−75個のアミノ酸残基、5−50個のアミノ酸残基、5−25個のアミノ酸残基、5−20個のアミノ酸残基、5−15個のアミノ酸残基、5−10個のアミノ酸残基、5−9個のアミノ酸残基を包含され得る。有用なリンカーの例は、gly9(配列番号27)、glu9(配列番号28)、ser9(配列番号29)、gly5cyspro2cys(配列番号30)、(gly4ser)3(配列番号31)、SerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号32)、ProSerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号13)、GlyAspLeuIleTyrArgAsnGlnLys(配列番号26)、およびGly9ProSerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号34)を包含する。使用され得る他のポリペプチドリンカーは、5、7、もしくは30残基を含む異なる長さのポリグリシンを包含する。加えて、Flt−1の他部位、例えば、Flt−1のドメイン3などがリンカーとして使用され得る。配列番号15を参照のうえ。リンカー部分はまた、ポリエチレングリコールのような他の重合体から作製され得る。かかるリンカーは、10から1000個、10から500個、10から250個、10から100個もしくは10から50個のエチレングリコール単量体単位で存在し得る。適合する重合体は、アミノ酸残基が占める適当な範囲の大きさに近い大きさである必要がある。典型的な大きさの重合体は、約10−25オングストロームの空間を提供し得る。
【0037】
本発明による融合蛋白質は、当該技術分野において知られる方法で作製され得る。かかる蛋白質が合成によって、あるいは作製される部分を連結することによって作製され得るが、組み換えによる製法を用いることもできる。融合される遺伝子配列は、組み換えDNAの標準的な方法を利用して作製され得る。融合された遺伝子配列は、融合遺伝子配列を複製するために、ウイルスもしくはプラスミドベクターのようなベクター内に挿入され得る。最終的な受容細胞において機能するプロモーター配列が、融合遺伝子配列の上流に配置され得る。使用されるプロモーターは、構成的、誘導的もしくは抑制的であり得る。それぞれの型の例は、当該技術分野でよく知られている。当該技術分野で公知の方法によってベクターを宿主細胞もしくは哺乳類動物に導入することが可能である。使用に適合するベクターは、アデノウイルス、アデノ随伴ウイルス、レトロウイルス、レンチウイルスおよびプラスミドを含む。仮に、ベクターがウイルスベクターであり、そのベクターがパッケージされる場合、ビリオンが感染細胞で利用され得る。裸のDNAが使用されるなら、特定の宿主細胞に適合する移入もしくは形質転換の手段が使われ得る。重合体、リポゾームもしくはナノ粒子を利用する裸のDNAの製剤形態が、融合遺伝子の送達に利用され得る。本発明による組み換え構築物を形質転換または移入された細胞は、当業者に利便性があるかもしれない。利用され得る典型的な細胞型は、バクテリア、酵母、昆虫および哺乳類細胞を包含する。利便性があることに、哺乳類細胞間では、多くの組織型細胞が、選択可能であるかもしれない。利用され得る典型的な細胞は、線維芽細胞、肝細胞、内皮細胞、幹細胞、造血細胞、上皮細胞、筋細胞、神経細胞およびケラチン生成細胞である。これらの細胞は、in vitroで蛋白質を生成するために利用され得るか、もしくはin vivoでコードされた蛋白質を生成するためにヒトを含む哺乳類動物に送達され得る。この送達方法は、核酸を哺乳類動物に、ウイルスベクターを哺乳類動物に、および融合蛋白質を哺乳類動物に送達することにとって代わるものである。
【0038】
蛋白質もしくは核酸の組成物は、緩衝液のような担体、水溶性もしくは親油性担体、無菌性もしくは菌性、発熱性もしくは非発熱性担体中で存在し得る。非発熱性担体は、注射製剤として有用である。製剤は、例えば、凍結乾燥されたような液状もしくは固形状であり得る。製剤はまた、エアロゾルとして投与され得る。組成物は、一以上の融合蛋白質もしくは一以上の核酸、または融合蛋白質および核酸の両方を包含するであろう。組成物中の融合蛋白質および/もしくは核酸が単一種であり得、その場合には、ホモ多量体蛋白質が形成し、あるいは、組成物中の融合蛋白質および/もしくは核酸が異種であり得、その場合には、ヘテロ多量体蛋白質が形成する。ヘテロ多量体である場合において、典型的には、X部分は融合蛋白質間で異なるが、Z部分は融合蛋白質間で同一である。
【0039】
融合蛋白質は、当該技術分野で知られる方法によって細胞もしくは哺乳類宿主に供給され得る。蛋白質は、細胞もしくは宿主に送達され得る。核酸は、細胞もしくは宿主に投与され得る。形質転換もしくは移入された細胞は、細胞もしくは宿主に投与され得る。後者の場合、同一の遺伝学的背景を有する細胞が、移植の拒絶反応を減らすことに望ましい。
【0040】
哺乳類の宿主動物への送達に適合する細胞は、器官、腫瘍、もしくは細胞系列に由来する哺乳類細胞型を包含する。例えば、ヒト、マウス、ヤギ、ヒツジ、ウシ、ネコ、およびブタの細胞が使用され得る。使用制限がなく、適合する細胞型は、線維芽細胞、幹細胞、ケラチン生成細胞、造血細胞、上皮細胞、筋細胞、神経細胞、および幹細胞を包含する。
【0041】
融合蛋白質もしくは融合蛋白質をコードする核酸の送達手段は、融合蛋白質を発現する細胞の送達、融合蛋白質の送達および融合蛋白質をコードする核酸の送達を包含する。融合蛋白質、細胞、もしくは核酸は、例えば、注射、カテーテル法、もしくは内視鏡によって望ましい器官もしくは腫瘍に直接送達され得る。それらはまた、静脈内、気管支内、腫瘍内、随腔内、筋肉内、眼内、局所、皮下、経皮もしくは経口によって送達され得る。浮腫加齢黄斑変性症、増殖性糖尿病網膜症、慢性関節リウマチ、骨関節症、ブドウ膜炎、喘息および癌を患う患者を効果的に処置することが可能である。これらの処置は、症状および/もしくは病気の重症度および/もしくは病気の進行を改善するであろう。
【0042】
核酸は、望ましいベクターを用いて、哺乳類動物、特にヒトに送達され得る。これらには、アデノウイルスベクター、アデノ随伴ウイルスベクター、レトロウイルスベクター、レンチウイルスベクターおよびプラスミドベクターを包含するウイルス性もしくは非ウイルス性のベクターが含まれる。ウイルスの典型的な型には、HSV(単純ヘルペスウイルス)、アデノウイルス、AAV(アデノ随伴ウイルス)、HIV(ヒト免疫不全ウイルス)、BIV(ウシ免疫不全ウイルス)、およびMLV(マウス白血病ウイルス)を含む。核酸は、ウイルス粒子、リポゾーム、ナノ粒子および混合された重合体を含む十分に効果的な送達レベルを付与する望ましい形式で投与され得る。
【0043】
蛋白質および核酸を組み合わせた処置が使用され得る。例えば、本発明による融合蛋白質が患者に投与され得る。もし、好ましい応答が観察される場合、融合蛋白質をコードする核酸分子が、長期間の効果を調べるために投与され得る。あるいは、蛋白質もしくは核酸が同時、もしくはほぼ同時に投与され得る。別の方法として、リガンドに対する抗体もしくは融合蛋白質が投与され、続けてもしくは同時に、受容体に対する抗体もしくは融合蛋白質が投与され得る。別の選択肢として、一方が抗体をコードし、他方が融合蛋白質をコードする核酸の組み合わせを用いる。本発明のFlt−1構築物(蛋白質もしくは核酸の形のどちらか)との組み合わせによって使用し得る幾つかの抗体は、ベバシマズムおよびラニビズマズであり、両者ともVEGFに結合する。これらは特に、癌および黄斑変性症それぞれを治療するのに有効である。
【0044】
本発明の実施は、別段の表示がない限り、当該技術分野の範囲内である分子生物学(組み換え技術を含む)、微生物学、細胞生物学、生化学および免疫学の一般的技術によって行われる。このような技術は、Molecular Cloning:A Laboratory Manual,Second Edition(Sambrookら,1989);Current Protocols In Molecular Biology(F.M.Ausubelら編,1987);Oligo nucleotide Synthesis(M.J.Gait編,1984);Animal Cell Culture(R.i.Freshney編,1987);Methods In Enzymology(Academic Press,Inc.);Handbook Of Experimental Immunology(D.M.WeiとC.C.Blackwellら編.);Gene Transfer Vectors For Mammalian Cells(J.M.MillerとM.P.Calos編,1987);PCR:The Polymerase Chain Reaction,(Mullisら編,1994);Current Protocols In Immunology(J.E.Coliganら編,1991);Antibodies:A Laboratory Manual(E.HarlowとD.Laneeds.(1988))ならびに、PCR2:A Practical Approach(M.J.MacPherson,B.D.HamesとG.R.Taylor編(1995))のような文献において十分に説明されている。
【0045】
遺伝子送達の担体は、宿主細胞内へ挿入するポリヌクレオチドを運搬することが可能な分子である。遺伝子送達担体の例としては、リポゾーム、リポ蛋白質、ポリペプチド、多糖類、リポ多糖類、人工ウイルス外被、金属粒子およびバクテリアを含む天然型重合体と合成型重合体である生体適合性重合体、バキュロウイルス、アデノウイルスおよびレトロウイルスのようなウイルス、バクテリオファージ、コスミド、プラスミド、真菌ベクターならびに多くの原核生物および真核生物の宿主で発現を示す当該技術分野内で典型的に利用されるその他の組み換え用担体を含み、これらは単一種の蛋白質発現だけではなく、遺伝子治療にも利用され得るかもしれない。
【0046】
遺伝子送達、遺伝子移入およびここで使用するそれらに類似する言葉は、導入に使用される方法に関わりなく、外因性ポリヌクレオチド(時に”トランスジーン”と呼ばれる)を宿主細胞に導入することを意味する。かかる方法は、”裸の”ポリヌクレオチドの送達に使用する技術(ポリヌクレオチドの導入に使用されるエレクトロポレーション、”遺伝子ガン”送達および多くの他技術)だけでなく、ベクター介在遺伝子移入(例えば、ウイルス感染/移入、または多種の他蛋白質由来もしくは脂質由来の遺伝子送達複合体による)のような多くのよく知られている技術を包含する。導入されたポリヌクレオチドは、宿主細胞において安定的にもしくは一時的に維持されるであろう。安定的な維持は、典型的には、導入されるポリヌクレオチドが、宿主細胞に適合可能である複製開始点を含むか、あるいは染色体外レプリコン(例えばプラスミド)のような宿主細胞のレプリコンまたは核もしくはミトコンドリアの染色体に組み入られる必要がある。当該技術分野で周知であり、ここで記述されるように、多くのベクターが哺乳類細胞に遺伝子移入を仲介できることが知られている。
【0047】
外因性ポリヌクレオチドを静脈内、筋肉内、門脈内もしくは別の方法による投与を介して宿主へ送達するために、外因性ポリヌクレオチドが、アデノウイルス、部分的に欠失されたアデノウイルス、完全に欠失されたアデノウイルス、アデノ随伴ウイルス(AAV)、レトロウイルス、レンチウイルス、裸のプラスミド、プラスミド/リポゾーム複合体等のようなベクターに挿入される。本発明で示す方法および組成物で使用され得る発現ベクターは、例えばウイルスベクターを包含する。In vivoとex vivoの両方において、遺伝子治療で最も頻繁に使用される投与方法の一つは、遺伝子送達用ウイルスベクターの使用である。多種類のウイルスが知られ、その多くが遺伝子治療の目的で研究されている。最も一般的に使用されるウイルスベクターは、アデノウイルス、アデノ随伴ウイルス(AAV)およびヒト免疫不全ウイルス(HIV)のようなレンチウイルスを含むレトロウイルスを包含する。
【0048】
アデノウイルスは、非外皮性であり、古典的遺伝学と分子生物学における研究を通してよく特徴づけられた約36kbのゲノムを有する核DNAウイルスである(Hurwitz, M.S., Adenoviruses Virology,3rd edition,Fieldsら編,Raven Press,New York,1996;Hitt, M.M.ら,Adenovirus Vectors,The Development of Human Gene Therapy,Friedman,T.編,Cold Spring Harbor Laboratory Press, New York 1999)。ウイルス遺伝子からウイルス蛋白質が、時間経過によって2つに分かれて生成されることから、ウイルス遺伝子は、早期(E1−E4と表す)および後期(L1−L5と表す)の転写単位に区分けされる。これらの事象の境界は、ウイルス性DNAの複製である。ヒトアデノウイルスは、赤血球の血球凝集、発癌能、DNAおよび蛋白質のアミノ酸組成および相同性、ならびに抗原的関連性を包含する特性に基づいて、多くの血清型(約47個がそれに応じて番号付けられ、A、B、C、D、EおよびFの6グループに区分けされた)に分けられる。
【0049】
組み換え体アデノウイルスベクターは、遺伝子送達の担体としての使用に対していくつかの有利な点を有し、すなわち、それらは、分裂と非分裂細胞の指向性、極小な病原性、ベクターのストックを調製するための高い力価で複製する能力、および大きい挿入物を運搬する能力を包含する(Berkner,K.L.,Curr.Top.Micro.Immunol.158:39−66,1992;Jolly,D.,Cancer Gene Therapy 1:51−64 1994)。擬性アデノウイルスベクター(PAV)および部分的に欠失されたアデノウイルス(”DeAd”と呼ばれる)のように多くのアデノウイルス遺伝子配列欠失を有するアデノウイルスベクターは、核酸の受容細胞への送達に適合した担体として望ましいアデノウイルスの特性を有するように設計されている。
【0050】
特に、擬性アデノウイルスベクター(PAVs)はまた、’弱いアデノウイルス’もしくはミニ−アデノウイルスベクターとして知られ、ベクターの複製およびパッケージに必要である最小のシス作用性ヌクレオチド配列を含み、一以上の導入遺伝子を包含することが可能である、アデノウイルスのゲノムに由来するベクターである(米国特許第5,882,877号には、擬性ベクター(PAV)とPAVを生成する方法を含み、出典明示により、本明細書に一体化させる)。PAVsは、遺伝子送達に適合する担体としてアデノウイルスの望ましい特性を有するように設計されている。アデノウイルスベクターは、ウイルスの増殖に不必要である領域の欠失によって一般的に8kbまでの大きさを挿入することが可能である一方で、最大の運搬能力は、PAVsを包含する多くのウイルスのコード配列を欠失したアデノウイルスベクターを使用することで達成し得る。Gregoryらの米国特許第5,882,877号;Kochanekら,Proc.Natl.Acad.Sci.USA93:5,731−5,736,1996;Parksら,Proc.Natl.Acad.Sci.USA 93:13565−13570,1996;Lieberら,J.Virol.70:8944−8960,1996;Fisherら,Virology 217:11−22,1996;米国特許第5,670,488号;PCT公開番号W096/33,280,1996年10月24日に公開;PCT公開番号W096/40955,1996年12月19日に公開 PCT公開番号W097/25446,1997年7月19日に公開;PCT公開番号 W095/29993,1995年11月9日に公開; PCT公開番号W097/00326,1997年1月3日に公開;Morralら,Hum.Gene Ther.10:2709−2716,1998を参照のうえ。このようなPAVsは、外部からの核酸を約36kbまで受け入れることが可能であり、ベクターの運搬能力は、最適化されているので優位である一方で、ベクターに対する宿主の免疫反応もしくは複製可能なウイルスの生成能力は減少する。PAVベクターは、複製開始点を包含する5’逆方向末端反復(ITR)および3’ITRヌクレオチド配列、ならびにPAVゲノムのパッケージに必要であるシスヌクレオチド配列を含み、例えば、プロモーター、エンハンサー等の適切な調節エレメントを有する一以上の導入遺伝子に適合することが可能である。
【0051】
その他には、部分的に欠失されたアデノウイルスベクターは、ウイルスの複製に必要であるアデノウイルス早期遺伝子の大部分がベクターから欠失し、条件プロモーターの制御下にある生成細胞の染色体内に置かれる部分的欠失(”DeAd”と呼ばれる)ベクターを供給する。生成細胞に配置される欠失可能なアデノウイルス遺伝子は、E1A/E1B、E2、E4(ORF6およびORF6/7のみが細胞内で必要とされる)、pIXおよびpIVa2を包含する。E3もまたベクターから欠失され得るが、ベクターの産生に必須ではないことから、生成細胞から除去することが可能である。主要後期プロモーター(MLP)の制御下にあるアデノウイルス後期遺伝子は、一般的にはベクター内に存在するが、MLPは条件プロモーターに置換され得る。
【0052】
DeAdベクターおよび生成細胞系列の使用に適合可能な条件プロモーターは、以下の性質を有するものを包含する。すなわち、細胞障害性もしくは細胞増殖抑制性のアデノウイルス遺伝子が、細胞を害するレベルで発現しないような、未誘導状態における低い基礎発現、およびベクターの複製と組み立てを支持するウイルス蛋白質が、十分量生成されるような、誘導状態における高いレベルの発現を有するものである。DeAdベクターおよび生成細胞系列における使用に適合する望ましい条件プロモーターは、免疫抑制剤FK506およびラパミシンに基づく二量体化物(dimerizer)遺伝子制御システム、エクジソン遺伝子制御システム、およびテトラサイクリン遺伝子制御システムを包含する。本発明において、Abruzzeseら,Hum.Gene Ther.1999 10:1499−507に記載されているGeneSwitch(登録商標)の技術(Valentis,Inc.,Woodlands,TX)が有用であり、出典明示により本明細書に一体化させる。部分的に欠失されたアデノウイルス発現システムは、さらにWO99/57296で示され、出典明示により本明細書に一体化させる。
【0053】
アデノ随伴ウイルス(AAV)は、ゲノムの大きさが4.6kbである一本鎖のヒトDNAパルボウイルスである。AAVゲノムは2つの主要な遺伝子を含む:rep蛋白質(Rep76、Rep68、Rep52およびRep40)をコードするrep遺伝子ならびにAAVの複製、レスキュー、転写および組込みをコードするcap遺伝子であり、さらに、cap蛋白質はAAVのウイルス粒子を形成する。AAVは、アデノウイルスもしくは他のヘルパーウイルス(例えばヘルペスウイルス)に依存して必須遺伝子産物を供給することにその名前が由来し、それらの産物は、AAVに生産的な感染、すなわち、宿主細胞における自身の再合成を可能にする。ヘルパーウイルスの非存在下では、AAVは、ヘルパーウイルス、一般的にはアデノウイルスに感染している宿主細胞に重複感染することによってレスキューされるまで前駆体ウイルスとして宿主細胞の染色体に組み込まれる(Muzyczka,Curr.Top.Micor.Immunol.158:97−127,1992)。
【0054】
遺伝子移入ベクターとしてのAAVの利点は、いくつかの生物学的な特徴に起因する。AAVゲノムの両末端には、逆位末端配列(ITR)として知られるヌクレオチド配列が存在し、その配列は、ウイルスの複製、レスキュー、パッケージおよび組込みに必要であるシス作動性のヌクレオチド配列を含む。トランス活性におけるrep蛋白質を介するITRの組み込み機能は、ヘルパーウイルス非存在下において、AAVゲノムが感染後に細胞内の染色体に組み込まれることを可能にする。このウイルス独特な特性は、細胞内ゲノムへの遺伝子の利点を含む組み換え体AAVの組み込みを可能にするとともに、遺伝子移入におけるAAVの利用に関連する。それゆえに、遺伝子移入の目的に適した多くの安定な遺伝学的な形質転換が、rAAVベクターの使用によって成し遂げられるかもしれない。さらに、AAVの組み込み部位がよく解明され、その部位はヒト染色体19番に位置する(Kotinら,Proc.Natl.Acad.Sci.87:2211−2215,1990)。この組込み部位の予測可能性は、宿主の遺伝子を活性化もしくは不活性化、またはコード領域を分断してしまうかもしれない細胞内ゲノムへのランダムな挿入事象の危険、rAAvベクターの使用を制限しうる結果を減らす。rAAVベクターのデザインにおけるこの遺伝子の除外は、rAAVベクターとともに観察された変動した組み込みパターンを生じるかもしれない(Ponnazhaganら.,Human Gene Ther.8:275−284,1997)。
【0055】
遺伝子移入のためのAAV利用には、他の有利な点がある。AAV宿主の範囲が広いことである。さらに、AAVは、レトロウイルスと異なり、停止状態および分裂状態の両状態の細胞に感染することが可能である。加えて、AAVは、ヒトの病気に関連せず、レトロウイルス由来の遺伝子移入ベクターで生じる多くの事象を防ぐ。
【0056】
組み換え体rAAVベクターを作製する標準的なアプローチには、一連の細胞内イベントに協調することが必要である:AAVのITR配列に隣接させた導入遺伝子を含むrAAVベクターゲノムの宿主細胞への移入、トランス活性で必要となるAAVのrepおよびcap蛋白質の遺伝子をコードするプラスミドによる宿主細胞への移入、ならびにトランス活性で必要となる非AAVヘルパーの機能を提供するヘルパーウイルスを移入された細胞の感染である(Muzyczka,N.,Curr.Top.Micor.Immunol.158:97−129,1992)。アデノウイルス(もしくは他のヘルパーウイルス)蛋白質は、AAVのrep遺伝子の転写を活性化し、rep蛋白質は、AAVのcap遺伝子の転写を活性化する。cap蛋白質は、rAAVゲノムをrAAVウイルス粒子内にパッケージするのにITR配列を利用する。それゆえ、パッケージの効率は、rAAVベクターゲノムに必要とされるシス活性のパッケージ配列への近接可能性とともに、適当量の構造蛋白質の利用可能性によってある程度決定される。
【0057】
レトロウイルスベクターは、遺伝子送達に用いられる一般的な道具である(Miller,Nature(1992)357:455−460)。再配列されていない単コピー遺伝子を齧歯類、霊長類およびヒト体細胞のような広い範囲内で送達するレトロウイルスベクターの能力は、遺伝子を細胞へ送達するのに最適である。
【0058】
レトロウイルスはRNAウイルスであり、そのゲノムはRNAである。宿主細胞がレトロウイルスに感染すると、ゲノムRNAは、中間体であるDNAに逆転写され、感染細胞の染色体DNAへ非常に効率的に組み込まれる。この組み込まれたDNA中間体は、プロウイルスと呼ばれている。プロウイルスの転写および感染性ウイルスへの構築は、混入するヘルパーウイルスが同時生成することなしに、キャプシド形成を可能にする適当な配列を含む適合するヘルパーウイルスの存在下、もしくは細胞系列で生じる。もしキャプシド形成に必要な配列が適合するベクターとの共移入によって提供されるならば、ヘルパーウイルスは、組み換え体レトロウイルスの生成に必要でない。
【0059】
レトロウイルスのゲノムおよびプロウイルスのDNAは、3個の遺伝子gag、polおよびenvを有し、それらは2個の末端反復(LTR)配列で挟まれている。gap遺伝子は、内部構造(マトリックス、キャプシドおよびヌクレオキャプシド)蛋白質をコードし、pol遺伝子は、RNA依存性DNAポリメラーゼ(逆転写酵素)をコードし、およびenv遺伝子は、ウイルス外皮性糖蛋白質をコードする。5’および3’LTRは、ビリオンRNAの転写およびポリアデニル化を促進する働きがある。LTRは、ウイルスの複製に必要である他のシス活性配列全てを含む。レンチウイルスは、(HIV−1、HIV−2および/もしくはSIV中に)vit、vpr、tat、rev、vpu、nefおよびvpxを含む付加遺伝子を有する。ゲノムの逆転写(tRNAプライマー結合部位)およびウイルスRNAの粒子内への効率的なキャプシド形成(Psi部位)に必要である配列が、5’LTRに隣接する。仮に、キャプシド形成(もしくはレトロウイルスの感染性ビリオンへのパッケージ)に必要な配列がウイルスゲノムから喪失しているなら、ゲノムRNAのキャプシド形成を阻害するシス活性の欠失となる。しかしながら、得られる変異体は、ビリオン蛋白質全ての合成を指示し得る。
【0060】
レンチウイルスは、一般的なレトロウイルス遺伝子gag、polおよびenvに付加して、調節的もしくは構造的な機能を有する遺伝子を包含する複合体レトロウイルスである。より高度な複雑性は、潜伏感染時にレンチウイルスが自身のライフサイクルを調節することを可能にする。典型的なレンチウイルスには、AIDSの病原因子であるヒト免疫不全ウイルス(HIV)がある。in vivoにおいて、HIVは、リンパ球およびマクロファージのような稀にしか分裂しない最終的に分化した細胞に感染し得る。in vitroでは、HIVは、アフィディコリンもしくはガンマ線照射の処理によって細胞周期が停止した状態のHeLa−Cd4もしくはTリンパ球のみならず、初代培養の単球由来マクロファージ(MDM)にも感染し得る。細胞の感染は、標的細胞の核膜孔を介する、活性化したHIV前駆組込み複合体の核移行に依存する。これは、標的細胞の核移入機能を有する複合体における多様で一部重複する分子決定因子の相互作用によって生じる。同定された決定因子は、gapマトリックス(MA)蛋白質における機能的な核局在シグナル(NLS)、核親和性ビリオン関連蛋白質、vpr、およびMA蛋白質におけるC末端のリン酸化チロシン残基を包含する。遺伝子治療に用いられるレトロウイルスは、例えば、米国特許第6,013,516号および米国特許第5,994,136号で示され、出典明示により本明細書に一体化させる。
【0061】
DNAを細胞に送達する他の方法は、送達にウイルスを使用しない方法である。例えば、陽イオン両親媒性化合物が、本発明の核酸を送達するのに使用され得る。生物学的な活性分子の細胞内送達に利用するために設計された化合物は、非極性および極性の両方の環境(例えば、細胞膜、組織液、細胞内構築物、および生物学的活性分子自身内もしくは上)において相互作用する必要があることから、このような化合物は、典型的には、極性および非極性ドメインを含むように設計される。かかるドメイン両方を有する化合物は、両親媒性物質と呼ばれ、このような細胞内送達(in vitroもしくはin vivoのどちらかに応用)を促進する利用が公知されている多くの脂質および合成脂質が、この定義に適合する。このような両親媒性物質の中で特に重要なクラスの一つが、陽イオン両親媒性物質である。一般的に、陽イオン両親媒性物質は、物理学的pH付近において陽に帯電を示し得る極性グループであり、この特性は、例えば、DNAのように負に帯電したポリヌクレオチドを包含する、多種類型の生物学的な活性(医薬上)分子と相互作用するのに重要であることが当該技術分野で理解されている。
【0062】
遺伝子送達用の陽イオン両親媒性物質を含む組成物の利用は、例えば、米国特許第5,049,386号;米国特許第5,279,833号;米国特許第5,650,096号;米国特許第5,747,471号;米国特許第5,767,099号;米国特許第5,910,487号;米国特許第5,719,131号;米国特許第5,840,710号;米国特許第5,783,565号;米国特許第5,925,628号;米国特許第5,912,239号;米国特許第5,942,634号;米国特許第5,948,925号;米国特許第6,022,874号;米国特許第5,994,317号;米国特許第5,861,397号;米国特許第5,952,916号;米国特許第5,948,767号;米国特許第5,939,401号;および米国特許第5,935,936号で示され、出典明示により本明細書に一体化させる。
【0063】
加えて、本発明の核酸は、”裸のDNA”を使用して送達され得る。移入促進蛋白質、ウイルス粒子、リポソーム製剤、荷電脂質およびカルシウムリン酸沈殿剤とは関連せず、プロモーターを結合した望ましいポリペプチドもしくはペプチドをコードする非感染性で非組込み型のDNA配列を送達する方法が、米国特許第5,580,859号;米国特許第5,963,622号;米国特許第5,910,488号に記述され、出典明示により本明細書に一体化させる
【0064】
また、ウイルス性と非ウイルス性の要素を組み合わせる遺伝子移入システムが報告されている。Proc.Natl.Acad.Sci.USA 90:11548;Wuら.(1994) J.Biol.Chem.269:11542;Wagneら.(1992) Proc.Natl.Acad.Sci.USA 89:6099;Yoshimuraら.(1993) J.Biol.Chem.268:2300;Curielら.(1991) Proc.Natl.Acad.Sci.USA 88:8850;Kupferら. (1994) Human Gene Ther. 5:1437およびGottschalkら/.(1994) Gene Ther. 1:185.多くの場合、アデノウイルスが、そのエンドソームの特性を生かして遺伝子送達システムに取り入れられている。報告されたウイルス性と非ウイルス性要素の組み合わせは、一般的には、アデノウイルスの遺伝子送達複合体への共有結合もしくは陽イオン脂質:DNA複合体を伴う非結合アデノウイルスの共内部移行のいずれかに関連する。
【0065】
眼にDNAおよび蛋白質を送達するため、典型的には、局所的な投与が施される。これは、投与する必要のDNA量および全身性副作用を制限するという点で有利である。多くの送達可能な形態が使用され得る。それらは、角膜への点眼を介した遺伝子銃による局所投与、結膜下注射、前房内注射、角膜を介した前房内注射、基質内注射、電磁波を用いた角膜への応用、角膜内注射、網膜下注射、硝子体内注射、および眼球内注射を包含する。代替的には、細胞が、ex vivoで移入もしくは形質導入され、眼内移植によって送達され得る。Auricchio,Mol.Ther.6:490−494,2002;Bennett,Nature Med.2:649−654,1996;Borras,Experimental Eye Research 76:643−652,2003;Chaum,Survey of Ophthalmology 47:449−469,2002;Campochiaro,Expert Opinions in Biological Therapy 2:537−544(2002); Lai,Gene Therapy 9:804 813,2002;Pleyer,Progress in Retinal and Eye Research,22:277−293,2003を参照のうえ。
【0066】
様々な提唱された治療用薬剤および投与の効果が、特定の病気に対応する動物モデルを用いて試験され得る。例えば、未熟児網膜症は、Smith,Investigative Ophthalmology & Visual Science,35:101−111,1994に記載されるようにマウスの酸素誘導型網膜症モデルで試験され得る。マウスにおけるレーザー誘導型脈絡膜新生血管は、加齢黄斑変性症のような病気の際に生じるヒト脈絡膜新生血管(CNV)のモデルとして利用され得る。Tobe,American Journal of Pathology 153:1641−1646,1998。CNVの他のモデルが、霊長類、ラット、ミニブタ、およびウサギにおいて開発された。加齢黄斑変性症のマウスモデルは、遺伝子欠損マウスを用いて開発された。単球走化性蛋白質−1もしくはC−Cケモカイン受容体−2のどちらか一方を欠損させたマウスが加齢黄斑変性症の特性を示す。Ambati,Nature Med.9:1390−1397,2003。
【0067】
本発明は、現時点で発明を実施するのに望ましい様式を包含する特定の例に関して記述されている一方で、当業者は、添付した請求の範囲を説明する本発明の精神および範囲内である上述したシステムおよび技術に対する多くのバリエーションと置き換えが存在することを理解するであろう。
【実施例】
【0068】
実施例1
2個の構築物が作製された。第一に、ポリグリシン9−mer(9Gly)のリンカーを含むD2−9Gly−Fcであり、第二として、9Glyリンカーを除いて同一の配列のD2−Fcである(図1)。
【0069】
我々は、配列解析プログラムMacVector6.5.1(IBI、New Haven、CT)のProtein Analysis Toolboxを用いて、D2−9Gly−FcとD2−Fc蛋白質のアミノ酸配列について解析を行った。D2−9Gly−Fc配列におけるポリグリシン9−merリンカーが、Karpus and Schultz(1985)Naturwiss,72:212−213の可撓性予測法による平均の可撓性より高い領域として同定された。このような領域は、D2−Fc配列中では検出されなかった(図1)。
【0070】
実施例2
我々は、同定された可撓性ポリグリシン9−merリンカーによってIgG1のFc領域に連結させたFlt−1のIg様ドメイン2(D2−9Gly−Fc)を検証した。D2−9Gly−Fc融合蛋白質は、効果的にVEGFに結合し、VEGF依存ヒト臍帯静脈内皮細胞(HUVEC)の増殖を阻害することが可能である。図2参照。対照的に、Flt−1のIgG1様ドメイン2が直接IgG1重鎖(Fc)に結合して、D2−Fcを形成する時には、最少量のVEGF結合のみが観察された。図2参照。IgG1のFcを介した二量体化と可撓性リンカー挿入の両方が、Flt−1ドメイン2のVEGF結合を促進すると考えられる。D2−9Gly−FcおよびD2−Fcにおける二量体型の存在は、ウェスタンブロット解析で確認された。図3参照。
【0071】
実施例3
AAVベクターの硝子体腔内注射(0.0005mLの体積中1x108から1x109個の粒子)が、新生仔(P0)もしくは生後1日後(P1)のC57BL/6系統マウスに施される。網膜新生血管(NV)が、P7の仔とこの仔らを養育中の雌親を5日間過酸素状態におくことによって、C57BL/6系統マウスで誘発される。これらの仔をP12で通常の大気に戻し、P17(NVピーク状態時)で安楽死させる。(Smith LEH、 Weslowski E、 McLellan A、 Kostyk SK、D’Amato R、 Sullivan and D’Amore PA。マウスにおける酸素誘発網膜症。Invest Opth Vis Sci。 1994;35:101−111)眼全体をパラフィンで包埋し、5マイクロン間隔で連続的に横断面を作製する。NVの程度は、100マイクロン毎に選んだ切片において境界膜の内側の内皮細胞核の数を数えることで決定される。
【0072】
抗血管形成薬剤をコードするAAVベクターで処理した動物の同齢集団を、無関連な導入遺伝子をコードするベクターもしくは導入遺伝子をコードしないベクターで処理した同齢集団と比較する。各々の処理した眼における内皮細胞核の平均数が、各々の動物の未処理の眼におけるその数と比較される。
【0073】
実施例4、D2−9Gly−Ex3/CH3の作製
Flt−1のドメイン2がVEGF165の結合に必須であることが示されている。しかし、Flt−1のドメイン2単独ではVEGFのAに結合できないことが示されていた(Davis−Smyth et al., 1996.)。VEGFのAは、二量体として存在する時、酸性残基(成熟蛋白質のアミノ酸63−67)を介してFlt−1に結合し、リガンド誘導型二量体受容体として機能することが可能である(Keytら., 1996)。
【0074】
それゆえ、Flt−1のドメイン2の二量体化は、Flt−1のドメイン2がVEGFのAへ結合する方法として使用されてきた。IgG重鎖の断片との融合は、蛋白質の二量体化に用いられ得る(Davis−Smythら., 1996.)。ここで、我々は、VEGFのA(配列番号12)のアミノ酸75−88(すなわちPSCVPLMRCGGCCN;配列番号13)が、sFlt−1混合蛋白質の生物学的活性を上昇させることを示す。
【0075】
最初に3個の混合蛋白質が設計された。D2−9Gly−Fc、D2−FcおよびD2−9GlyEx3/CH3である(図4)。3個の混合蛋白質全ては、D2−9Gly−Fcのように同一のFlt−1のドメインD2を含む。VEGFへの結合は、ポリグリシン9−mer(9Gly)リンカーを含まないD2−Fcにおいて観察されなかった。3番目の蛋白質であるD2−9Gly−Ex3/CH3は、ポリグリシン9−mer(9Gly)リンカーおよびVEGFの多量体化ドメイン(アミノ酸PSCVPLMRCGGCCN;配列番号13;VEGF Ex3)を包含するが、また、ヒトIgG1重鎖FcのCH3領域(配列番号10のアミノ酸371−477)をも含む。
【0076】
蛋白質D2−Fcは、HUVEC増殖測定(図5)において効果的な阻害活性を示さず、それに伴い、効果的にVEGF165に結合しなかった。しかしながら、3番目の混合蛋白質であるD2−9Gly−Ex3/CH3は、9GlyリンカーおよびVEGF165の二量体化領域(Ex3)の両方を介してCH3領域に融合した、Flt−1のドメイン2を含み、VEGF依存性HUVECの増殖測定法において阻害活性を示した(図5)。これは、この混合蛋白質がVEGF165に効果的に結合することを意味する。
【0077】
実施例5、Flt−1のD2構築物におけるリンカー(Gly4Ser)3の使用
以前に、数個のポリグリシンリンカーの使用が、蛋白質の性状を改変すると報告されている(Mouzら,1996;Qiuら,1998)。次の構築物のために、我々は、別の型のリンカーである15−mer(Gly−Gly−Gly−Gly−Ser)3を試した(Hustonら,1988)。D2−(Gly4Ser)3−Fc蛋白質が作製され、この蛋白質は、Flt−1ドメイン2、(Gly4Ser)3リンカーおよびヒトIgG1重鎖のFc領域を含む。
【0078】
D2−(Gly4Ser)3−Fcは、さらにHUVEC増殖測定法で検査された。D2−(Gly4Ser)3−Fcの生物学的活性は、HUVECの増殖阻害によって測定され、D2−9Gly−FcおよびD2−9Gly−Ex3/CH3の生物学的活性に類似した(図6)。
【0079】
D2−(Gly4Ser)3−Fc構築物は、さらにウェスタンブロットを用いて検査され、D2−9Gly−Fcと比較された(図9)。両方の構築物は、二量体型で多く存在し、単量体型は、還元されたサンプルの分離後に検出された。
【0080】
実施例6、Flt−1(D2)構築物における9GlyもしくはVEGFEx3の役割
可溶性受容体VEGFへの結合に対する9GlyリンカーもしくはVEGF二量体化配列Ex3の役割を調べるために、3個の異なる構築物、D2−9Gly−CH、D2−CH3およびD2−Ex3/CH3が作製された(図8)。3個の構築物全てが作製され、以前の構築物と同様にCMVプロモーターの制御を受けるようにした。これらのVEGF抑制活性はHUVEC増殖測定法で測定された(図9)。
【0081】
IgG1のCH3領域を含む蛋白質のHUVEC増殖測定法は、D2−9Gly−CH3(Ex3無し)および蛋白質D2−Ex3/CH3(9Glyリンカー無し)が、D2−9GlyEx3/CH3と比較して、同様のVEGF抑制能力を有することを示した。しかし、蛋白質D2−CH3は、これら全ての中で最も弱いVEGF阻害剤であると思われた(図9)。
【0082】
移入された293細胞由来条件培地のFlt−1におけるELISAデータでは、D2−9Gly−Ex3/CH3、D2−9Gly−CH3およびD2−Ex3/CH3については、同様のFlt−1レベル(70−90ng/ml)を示し、最小のD2−CH3活性形態については、やや高いレベル(約150ng/ml)を示した。D2−9Gly−CH3およびD2−CH3の構築物のウェスタンブロット(図10)は、非還元条件での二量体型の普及を示している。
【0083】
実施例7、D2−9Gly−Fcは、全ての構築物より一層VEGFに結合する。
VEGF結合測定は、細胞遊離中における可溶性VEGF受容体に対する相対的なVEGF結合能の比較を可能にする。
【0084】
簡潔に説明すると、既知濃度の可溶性受容体(0.29−150pMの濃度範囲)を含む条件培地は、連続的に希釈され、10pMのVEGFを添加された。非結合VEGF量は、ELISAによって測定された。D2−9Gly−Fcは、0.001から0.2pMの受容体濃度において全ての他の構築物より高いVEGF結合能を示す。D2−CH3は、VEGFに対して最も低い結合能を有する(図11)。
【0085】
(参考文献)
Davis−Smythら,EMBO J.15,1996,4919
Huston,J.S.ら(1991)Methods Enzymol.203,46−88
Huston,J.S.ら(1988)Proc.Natl Acad.Sci.Usa,85,5879−5883.
Johnson,S.ら(1991)Methods Enzymol.203,88−98 Karpus,P.A.ら(1985)Naturwiss.,72,212−213.
Keyt,B.A.,ら(1996)J.Biol.Chem.271:5638−5646.Kortt,A.A.ら(1997)Protein Engng,10,423−433.Lee,Y−L.ら(1998)Human Gene Therapy,9,457−465
Mouz N.ら(1996)Proc Nati Acad.Sci.USA,93,9414−9419.
Nielsen,ら(1997)Protein Eng.,10,1
Qiu,H.,ら(1998)J.Biol.Chem.273:11173−11176.
【0086】
【表1】
【表2】
【図面の簡単な説明】
【0087】
【図1】D2−9Gly−Fc構築物における9−Glyリンカーの可撓性領域を示す。
【図2】D2−9Gly−Fc対D2−Fcの生物学的活性を示す。
【図3】D2−9Gly−FcおよびD2−Fcのウェスタンブロット解析を示す。
【図4】D2−9Gly−Ex3/CH3と過去に構築された蛋白質との構造比較を示す。
【図5】D2−9Gly−Ex3/CH3対D2−9Gly−Fcの生物学的活性を示す。
【図6】D2−(Gly4Ser)3−Fc蛋白質活性をD2−9Gly−FcおよびD2−9Gly−Ex3/CH3と比較したHUVEC増殖測定。
【図7】D2−9Gly−Fc、D2−(G4S)3−Fc、およびEGFP蛋白質を用いたウェスタンブロットの結果を示す。
【図8】9Gly−CH3、D2−CH3およびD2−Ex3/CH3を有すもしくは有さない、3個の新規蛋白質の構造比較を示す。
【図9】9Gly、Ex3およびCH3の組み合わせを有するFlt−1(D2)構築物におけるHUVEC増殖測定を示す。
【図10】D2−9Gly−Fc、D2−9Gly−CH3およびD2−CH3を用いたウェスタンブロットの結果を示す。
【図11】VEGF“in vitro”結合測定を示す。
【図12】BIAcore機器を用いた表面プラスモン共鳴によって測定された可溶性Flt−1構築物の結合反応速度を示す。
【図13】各種リンカーを有するFlt−1構築物の発現レベルを示す。
【図14】網膜新生血管(NV)のマウス酸素誘導型網膜症モデルを用いて、Flt−1構築物の一つがマウスの眼に投与され、新血管新生が特定された。
【技術分野】
【0001】
本発明は、喘息、関節症、癌、および傷の再生などの病理的な新血管新生を治療するために組み換え構築された蛋白質の利用に関連する。
【背景技術】
【0002】
病理性新血管新生は、浮腫性加齢黄斑変性症(AMD)、増殖性糖尿病網膜症、骨関節症、および喘息等の病気を構成する要素の一つである。また、新血管新生は、腫瘍の増殖および転移に重要な役割を果たす。新血管新生は、前駆血管形成因子および抗血管形成因子の厳密な均衡によって制御されている。
【0003】
血管内皮成長因子(VEGF)は、新血管新生に必須であることが知られる。VEGF活性の阻害は、AMD、関節症の動物モデルおよび様々な腫瘍モデルにおいて新血管新生を抑制することが示されている。VEGF活性を阻害する方法は、抗体、受容体融合蛋白質、ペプチド、および小分子を含む。
【0004】
VEGF−R1(Flt−1)およびVEGF−R2(KDR)の蛋白質は、高い結合能でVEGFに結合することが示されている。Flt−1およびKDRの両方は、細胞外領域に7個のIg様ドメインを有する。ドメイン2は、VEGFに結合するために必須であることが示されている。全長、可溶性受容体(ドメイン1−7)、およびドメイン1−3各々ならびにIgGのFcとの融合蛋白質が、効果的にVEGFに結合する。しかしながら、IgGのFcおよびIg様ドメイン2単独との融合は、VEGFに結合できないが、Ig様ドメイン1および2の両方との融合は、結合可能であった。Davis−Smyth,1996。それゆえ、Ig様ドメイン1および3は、ドメイン2を伴うことで効果的なVEGF結合に必要とされるようである。
【発明の開示】
【課題を解決するための手段】
【0005】
(発明の簡潔な要約)
本発明の一の具体例として、融合蛋白質が提供される。融合蛋白質は、式X−Y−Zを有する。Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含む。Yは、5−25個のアミノ酸残基のポリペプチドから必須として成る。Zは、IgG重鎖分子のCH3領域である。
【0006】
本発明の別の具体例は、式X−Y−Zのポリペプチドである。Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含む。Yは、5−25個のアミノ酸残基の空間的な分離を提供するリンカー部分から必須として成る。Zは、IgG重鎖分子のCH3領域である。
【0007】
また、本発明の別の態様としては、式X−Y−Zで示される融合蛋白質である。Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含む。Yは、5−25個のアミノ酸残基のポリペプチドから必須として成る。Zは、抗体分子のFc部位である。
【0008】
式X−Y−Zで示される融合蛋白質もまた提供される。Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドである。Yは、5−25個のアミノ酸残基の空間的分離を提供するリンカー部分から必須として成る。Zは、抗体分子のFc部位である。
【0009】
さらに、本発明の別の態様としては、ポリペプチドXを多量体化する方法である。ポリペプチドXは、ポリペプチドXYZを形成するために、ポリペプチドYを介してポリペプチドZに結合される。Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含む。Yは、5−25個のアミノ酸残基のポリペプチドから必須として成る。Zは、IgG重鎖分子のCH3領域である。形成されるポリペプチドXYZは、多量体化する。
【0010】
さらに本発明の別の具体例としては、ポリペプチドXを多量体化する方法を提供する。ポリペプチドXは、部分Yを介してポリペプチドZに結合されることで、ポリマ−XYZを形成する。Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含む。Yは、5−25個のアミノ酸残基の空間的な分離を提供するリンカー部分から必須として成る。Zは、IgG重鎖分子のCH3領域である。形成されるポリペプチドXYZは、多量体化する。
【0011】
本発明の一の具体例において、核酸分子が提供される。本核酸分子は、VEGF−R1(Flt−1)のIg様ドメイン2、リンカー、および多量体化ドメインを含む融合蛋白質をコードする。融合蛋白質は、配列番号2、8、21、23、および25から成る群から選択される配列を含む。
【0012】
本発明の別の具体例において、融合蛋白質が提供される。本融合蛋白質は、VEGF−R1(Flt−1)のIg様ドメイン2、リンカー、および多量体化ドメインを含む。本融合蛋白質は、配列番号2、8、21、23、および25から成る群から選択される配列を含む。
【0013】
本発明の別の具体例において、in vitroの方法が提供される。本核酸分子は、同定された哺乳類細胞に送達される。この核酸分子は、VEGF−R1(Flt−1)のIg様ドメイン2、リンカー、および多量体化ドメインを含む融合蛋白質をコードする。融合蛋白質は、配列番号2、8、21、23、および25から成る群から選択される配列を含む。融合蛋白質の発現は、プロモーターによって制御される。融合蛋白質を発現する細胞が形成される。
【0014】
さらに、本発明の別の具体例は、融合蛋白質を哺乳類に送達する方法である。融合蛋白質を発現する哺乳類細胞が、哺乳類に送達される。この細胞は、融合蛋白質を発現および分泌し、それによって融合蛋白質を哺乳類動物に供給する。融合蛋白質は、VEGF−R1(Flt−1)のIg様ドメイン2、リンカー、および多量体化ドメインを包含する。融合蛋白質は、配列番号2、8、21、23、および25から成る群から選択される配列を含む。
【0015】
本発明の別の態様は、融合蛋白質を哺乳類動物に供給する方法である。VEGF−R1(Flt−1)のIg様ドメイン2、リンカー、および多量体化ドメインを含む融合蛋白質が、哺乳類動物に送達される。融合蛋白質は、配列番号2、8、21、23、および25から成る群から選択される配列を含む。代替的に、該融合蛋白質をコードする核酸構築物が、哺乳類動物に送達されることも可能であり、そのことにより、融合蛋白質が哺乳類動物によって発現される。
【0016】
これらおよび下記により詳細が記述されている本発明の他の具体例は、血管増殖および炎症に関連する病気を治療する方法および薬剤を当該技術分野に提供するであろう。薬剤は、蛋白質の天然型に関連する上昇した安定性と生物利用を提供するかもしれない。
【0017】
(図の説明)
図1。D2−9Gly−Fc構築物における9−Glyリンカーの可撓性領域。KarpusとSchultz(1985年)の方法による予測相対的可撓性は、D2−9Gly−Fc蛋白質中のポリグリシン9mer(9−Gly)のリンカー(アミノ酸94から103)が、9−Glyのリンカーを含まないD2−Fc構築物と比較して、平均(>1)より大きな可撓性を有する領域であることを示す。両方の融合蛋白質は、箱型で囲まれた同一のアミノ酸配列を含む。sp−シグナルペプチド(アミノ酸−24から−1)、Flt−1ドメイン2(アミノ酸1から93)およびIgG1−Fc残基(244アミノ酸)。矢印は、SignalPV2.0programを用いて予測したシグナルペプチダ−ゼ切断領域を表す(Nielsenら,1997)。
【0018】
図2。D2−9Gly−Fc対D2−Fcの生物学的活性。293細胞は、飢餓培地(M199+5%FCS)で培養され、CMVプロモーター制御下にあるD2−9Gly−FcおよびD2−Fcの発現カセットを含むプラスミドを移入された。条件培地(CM)は72時間後に回収された。HUVECが、96穴プレート(2E3細胞/穴)の飢餓培地+VEGF(10ng/mL)中に分注され、50ulのCMとVEGF(10ng/mL)が24時間後に添加された。コントロール(+/−VEGF)は、コントロールpEGFP(Clontech;pEGFPは、哺乳類細胞内で、より明るい蛍光およびより高い発現を示すように最適化され、野生型緑色蛍光蛋白質(GFP)の赤色転換変動を有する)に由来するCM中で培養された。陽性対照は、50ngのFlt−1−IgG組み換え体蛋白質(R&D Systems)で処理された。HUVEC数は、CellTiter96(登録商標)AQueous regeant(Promega)を用いて、増殖3日間後に測定された。データは、3種類行われた2回の実験各々のOD490平均値を表す。
【0019】
図3。D2−9Gly−FcおよびD2−Fcのウェスタンブロット解析。D2−9Gly−FcおよびD2−Fcの蛋白質両方の大きさは、還元ゲルにおける移動と比較して非還元ゲルにおける移動は2倍になると思われる。D2−9Gly−FcおよびD2−Fcを発現するプラスミドが、293細胞に移入された後、条件培地から蛋白質がロードされ、SDS−電気泳動により分離され、PVDF膜に移行された。ブロットは、ヤギ抗ヒト抗IgG1Fcおよびラビット抗ヤギIgG−HRP抗体を用いて行われた。
【0020】
図4。9GlyのリンカーおよびVEGFEx3を含むsFlt−1複合蛋白質。D2−9Gly−Ex3/CH3と過去に構築された蛋白質との構造比較。3個の蛋白質全ては、アミノ酸24個のFlt−1シグナルペプチドおよびアミノ酸93個のFlt−1ドメイン2から成るFlt−1ドメイン2の同一アミノ酸配列を含む。D2−9Gly−Ex3/CH3は、アミノ酸9個の9Glyリンカー、アミノ酸14個のVEGFEx3およびアミノ酸120個のヒトIgG1重鎖FcCH3領域を含む。
【0021】
図5。D2−9Gly−Ex3/CH3対D2−9Gly−Fcの生物学的活性。ドメイン2が9GlyリンカーおよびVEGFのEx3を介してCH3領域に結合されている、蛋白質D2−9Gly−Ex3/CH3は、蛋白質D2−9Gly−FcおよびD2−Fcと比較して効果的にVEGF依存性HUVEC増殖を阻害する。50ngの組み換え体Flt−1−IgG(R&D Systems)をコントロールとして使用した。
【0022】
図6。D2−(Gly4Ser)3−Fc蛋白質活性をD2−9Gly−FcおよびD2−9Gly−Ex3/CH3と比較したHUVEC増殖測定。
【0023】
図7。ウェスタンブロット。(1)D2−9Gly−Fc、(2)D2−(G4S)3−Fcおよび(3)EGFP蛋白質を発現するプラスミドを移入された293細胞の条件培地からの蛋白質(非還元と還元)が、SDS−電気泳動によって分離され、PVDF膜に移行された。ブロットは、ヤギ抗ヒトIgG1Fcおよびウサギ抗ヤギIgG−HRP抗体を用いて行われた。
【0024】
図8。9GlyリンカーもしくはVEGFEx3の有無による蛋白質の組み合わせ。9Glyリンカーおよび/もしくはVEGFEx3、D2−9Gly−CH3、D2−CH3およびD2−Ex3/CH3を有すもしくは有さない、3個の新規蛋白質の構造比較。
【0025】
図9。9Gly、Ex3およびCH3の組み合わせを有するFlt−1(D2)構築物におけるHUVEC増殖測定。蛋白質D2−Ex3/CH3、D2−9Gly−CH3およびD2−CH3を含む293細胞(5ul)に由来する条件培地をD2−9Gly−FcおよびD2−9Gly−Ex3/CH3と比較した。
【0026】
図10。ウェスタンブロット。293細胞に(1)D2−9Gly−Fc、(2)D2−9Gly−CH3(52/26kDa)、および(3)D2−CH3(50/25kDa)を発現するプラスミドが移入された。条件培地(15ulの非還元および/もしくは還元されたCM)中の293細胞に由来する蛋白質は、SDS−電気泳動で分離され、PVDF膜に移行された。ブロットは、抗ヒトVEGF−R1−HRPを用いて行われた(R&D Systems)。
【0027】
図11。VEGF”in vitro”結合測定。既知濃度のD2−9Gly−FcおよびFlt−1D(1−3)のコントロール可溶性受容体(濃度範囲0.29−150pM)を含む293細胞由来の条件培地が、連続的に希釈され、10pMのVEGFを添加された。非結合VEGFの量がELISAで測定された。D2−9Gly−Fcは、他の構築物すべてより高い結合能でVEGFに結合する。”n”は、(移入および結合測定)非依存的な実験数を表す。
【0028】
図12。可溶性Flt−1構築物の結合反応速度が、BIAcore機器を用いた表面プラスモン共鳴によって測定された。sFlt−1構築物がセンサーチップ上に固定され、VEGF165が0.2から15nMの濃度範囲で注入された。センサーグラムの値がBIA評価プログラムを用いて求められ、速度定数KaおよびKdが決定され、解離定数(KD)がKd/Ka=KDの割合から計算された。より低いKD値はより高い親和性を意味する。
【0029】
図13A−13C。図13Aは、各種リンカーを有するFlt−1構築物の発現レベルを示す。図13Bは、各種リンカーおよびIgG1FcのCH3部分を有するFlt−1構築物の二量体化もしくは多量体化を表す。非還元および還元条件の相違は、蛋白質が多量体化したかである。図13Cは、VEGF存在下のHUVEC増殖測定における条件培地中において、各Flt−1構築物の阻害的な生物活性が存在することを示す。構築物各々は、VEGF非存在下におけるHUVEC増殖レベルに近い阻害活性を示した。
【0030】
図14。網膜新生血管(NV)のマウス酸素誘導型網膜症モデルを用いて、Flt−1構築物の一つがマウスの眼に投与され、新血管新生が特定された。このマウスは高酸素状態に置かれた。血管新生を示す事例数は、同じマウスにおける未処理と処理した眼を比較して特定された。血管新生症例において50%以上の減少がある場合に、マウスは”responder”であると判断された。
【0031】
(発明の詳細な説明)
本発明者の知見は、ドメイン1および3を有さないFlt−1のIg様ドメイン2が、効果的にVEGFに結合し、VEGF依存性内皮細胞の増殖を抑制できることである。ドメイン2は、リンカーを介して共有的に多量体化ドメインに結合され得る。リンカーは、一般的にポリペプチド鎖である。鎖の長さは、6、7、9、11、13、15もしくはそれ以上のアミノ酸残基であり得るが、典型的には5から25残基の間である。長さと側鎖構成物に依存して、リンカーは平均的な可擣性よりも大きな可撓性を有していてもよいが、それは必須ではない。可撓性は、当該技術分野において知られる算法によって計算され得る。多量体化ドメインは、多量体蛋白質の一部分であり、例えば、二量体、三量体、四量体等を形成するサブユニットの連係を増進する。効果的なVEGFの結合および/もしくはVEGF依存性の内皮細胞増殖の抑制に適合可能な組み換え体蛋白質が、配列番号2、8、21、23および25から成る群から選択される。
【0032】
さらに、発明者は、多量体化ドメインおよびリンカーが、様々な他の蛋白質もしくは蛋白質の部位とともに使用され得ることで、多量体化を誘導することを見い出した。このような蛋白質は、多量体化する時に限りリガンドもしくは受容体に結合する蛋白質であるか、または多量体化する時に結合能が上昇する蛋白質であろう。多量体化に適する蛋白質は、細胞外受容体(その一部分を含む)、抗体可変領域、サイトカイン、ケモカイン、および成長因子を包含する。適合する蛋白質は、チロシンキナーゼ受容体およびセリンスレオニンキナーゼ受容体を含む。細胞外受容体の詳細な例としては、EGF受容体、Gタンパク質共役型受容体、FGF受容体、Fc受容体、T細胞受容体等を含む。抗体可変領域の例は、Fab、F(ab’)2、およびScFvを含む。サイトカインの例は、GM−CSF、IL−la、IL−1b、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−10、IL−12、IL−18、IL−21、IL−23、IFN−a、IFN−b、MIP−la、MIP−1b、TGF−b、TNFa、およびTNF−13を含む。ケモカインの例は、BCA−1/BLC、BRAK、ChemokineCC−2、CTACK、CXCL−16、ELC、ENA、ENA−70、ENA−74、ENA−78、Eotaxin、Exodus−2、Fractalkine、GCP−2、GRO、GROalpha(MGSA)、GRO−beta、GRO−gamma、HCC−1、HCC−4、I−309、IP−10、I−TAC、LAG−1、LD78−beta、LEC/NCC−4、LL−37、リンホタクチン(Lymphotactin)、MCP、MCAF(MCP−1)、MCP−2、MCP−3、MCP−4、MDC、MDC、MDC−2、MDC−4、MEC/CCL28、MIG、MIP、MIP−1alpha、MIP−1beta、MIP−1delta、MIP−3/MPIF−1、MIP−3alpha、MIP−3bet、MIP−4(PARC)、MIP−5、NAP−2、PARC、PF−4、RANTES、RANTES−2、SDF−1alpha、SDF−1beta、TARC、およびTECKを含む。成長因子の例は、ヒトアンヒィレギュリン(Amphiregulin)、ヒト血管形成蛋白質(Angiogenesis Protein)、ヒトACE、ヒトアンジオゲニン(Angiogenin)、ヒトアンジオポエチン(Angiopoietin)、ヒトアンジオスタチン(Angiostatin)、ヒトベータセルリン(Betacellulin)、ヒトBMP、ヒトBMP−13/CDMP−2、ヒトBMP−14/CDMP−1、ヒトBMP−2、ヒトBMP−3、ヒトBMP−4、ヒトBMP−5、ヒトBMP−6、ヒトBMP−7、ヒトBMP−8、ヒトBMP−9、ヒトコロニー刺激因子、ヒトflt3−Ligand、ヒトGCSF、ヒトGM−CSF、ヒトM−CSF、ヒト結合組織成長因子(Connective Tissue Growth Factor)、ヒトCripto−1、ヒトCryptic、ヒトECGF、ヒトEGF、ヒトEG−VEGF、ヒトエリスロポエチン(Erythropoietin)、ヒトフェツィン(Fetuin)、ヒトFGF、ヒトFGF−1、ヒトFGF10、ヒトFGF−16、ヒトFGF−17、ヒトFGF−18、ヒトFGF−19、ヒトFGF2、ヒトFGF−20、ヒトFGF−3、ヒトFGF−4、ヒトFGF−5、ヒトFGF−6、ヒトFGF−7/KGF、ヒトFGF−8、ヒトFGF−9、ヒトFGF−酸性、ヒトFGF−塩基性、ヒトGDF−11、ヒトGDF−15、ヒト成長ホルモン遊離因子(Growth Hormone Releasing Factor)、ヒトHB−EGF、ヒトヘレグリン(Heregulin)、ヒトHGF、ヒトIGF、ヒトIGF−I、ヒトIGF−II、ヒトインヒビン(Inhibin)、ヒトKGF、ヒトLCGF、ヒトLIF、ヒト Miscellaneous Growth Factor、ヒトMSP、ヒトミオスチン(Myostatin)、ヒトミオスタチンプロペプチド(Myostatin Propeptide)、ヒト神経成長因子(Nerve Growth Factor)、ヒトオンコスタチン M(Oncostatin M)、ヒトPD−ECGF、ヒトPDGF、ヒトPDGF(AAホモダイマー)、ヒトPDGF(ABホモダイマー)、ヒトPDGF(BBホモダイマー)、ヒトPDGF(CCホモダイマー)、ヒトPIGF、ヒトPIGF、ヒトPIGF−1、ヒトPIGF−2、ヒトSCF、ヒトSMDF、ヒト幹細胞成長因子(Stem Cell Growth Factor)、ヒトSCGF−alpha、ヒトSCGF−beta、ヒトスロンボポエチン(Thrombopoietin)、ヒト形質転換成長因子(Transforming Growth Factor)、ヒトTGF−alpha、ヒトTGF−beta、およびヒトVEGFを含む。
【0033】
Flt−1受容体蛋白質は、7個のIg様ドメインを含む細胞外部分を有する。これらは、ジェンバンク受託番号P17948の残基番号32から123、151から214、230から327,335から421、428から553、556から654、661から747に位置する。配列番号15も参照。残基番号1から26は、シグナル配列を含む。Flt−1蛋白質は、ジェンバンク受託番号NM_002019(配列番号14)で示されるDNA配列によってコードされる。
【0034】
多量体化ドメインは、当該技術分野で知られるように使用され得る。例えば、IgG1もしくはIgG2のラムダ重鎖Fc部分の配列は、CH3単独(アミノ酸371−477)もしくはCH2およびCH3両方のドメイン(アミノ酸247−477)について使用され得る。Ig分子のFc部分は、酵素パパインによって抗体分子全体を切断することで得られる。他の方法を使用することでもこれらの部分が得られる。IgG1のラムダ重鎖の蛋白質配列は、ジェンバンク受託番号Y14737および配列番号10を参照のうえ。例えば、他のIgG型に由来およびIgA、IgM、IgDもしくはIgE抗体に由来するような別のFc領域が使用され得る。VEGFの多量体化領域もまた使用され得る。VEGFをコードするDNA配列は、ジェンバンク受託番号NM003376および配列番号11に記載されている。VEGFのアミノ酸配列は、ジェンバンク受託番号CAC19513および配列番号12に記載されている。VEGF exon3(VEGF Ex3)にコードされるVEGFの多量体化領域(配列番号13)は、VEGF蛋白質のアミノ酸残基75−88(配列番号12)に相当する。多量体化ドメインは、少なくとも5%、10%、20%、30%、40%、50%、60%、70%、80%、85%、90%もしくは95%の単量体融合蛋白質が、多量体に適合する速度で非変性ポリアクリルアミドゲル上を移動することを引き起こす。糖鎖付加は、ゲル中の蛋白質の移動に影響し得る。特定の配列がここで示されるが、対立形質のような変異体もよく使用され得る。典型的なこのような変異体は、開示された配列に少なくとも85%、90%、95%、97%、98%、もしくは99%の同一性を有する。
【0035】
多量体化は、例えば、ここで示されるように還元および非還元ゲルを使用して検証され得る。多量体化はまた、リガンド/受容体に対する蛋白質の上昇した結合能を検出することによって測定され得る。これに関連してBiaCore(登録商標)の表面プラスモン共鳴測定が使用され得る。これらの測定は、センサーチップ表面に近接する水層の屈折率の変化を測定することによって質量の変化を検出する。当該技術分野で知られるいくつかの方法が、多量体化を検出するのに使用され得る。
【0036】
本発明によるリンカー部分は、例えば、5−100個のアミノ酸残基、5−75個のアミノ酸残基、5−50個のアミノ酸残基、5−25個のアミノ酸残基、5−20個のアミノ酸残基、5−15個のアミノ酸残基、5−10個のアミノ酸残基、5−9個のアミノ酸残基を包含され得る。有用なリンカーの例は、gly9(配列番号27)、glu9(配列番号28)、ser9(配列番号29)、gly5cyspro2cys(配列番号30)、(gly4ser)3(配列番号31)、SerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号32)、ProSerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号13)、GlyAspLeuIleTyrArgAsnGlnLys(配列番号26)、およびGly9ProSerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号34)を包含する。使用され得る他のポリペプチドリンカーは、5、7、もしくは30残基を含む異なる長さのポリグリシンを包含する。加えて、Flt−1の他部位、例えば、Flt−1のドメイン3などがリンカーとして使用され得る。配列番号15を参照のうえ。リンカー部分はまた、ポリエチレングリコールのような他の重合体から作製され得る。かかるリンカーは、10から1000個、10から500個、10から250個、10から100個もしくは10から50個のエチレングリコール単量体単位で存在し得る。適合する重合体は、アミノ酸残基が占める適当な範囲の大きさに近い大きさである必要がある。典型的な大きさの重合体は、約10−25オングストロームの空間を提供し得る。
【0037】
本発明による融合蛋白質は、当該技術分野において知られる方法で作製され得る。かかる蛋白質が合成によって、あるいは作製される部分を連結することによって作製され得るが、組み換えによる製法を用いることもできる。融合される遺伝子配列は、組み換えDNAの標準的な方法を利用して作製され得る。融合された遺伝子配列は、融合遺伝子配列を複製するために、ウイルスもしくはプラスミドベクターのようなベクター内に挿入され得る。最終的な受容細胞において機能するプロモーター配列が、融合遺伝子配列の上流に配置され得る。使用されるプロモーターは、構成的、誘導的もしくは抑制的であり得る。それぞれの型の例は、当該技術分野でよく知られている。当該技術分野で公知の方法によってベクターを宿主細胞もしくは哺乳類動物に導入することが可能である。使用に適合するベクターは、アデノウイルス、アデノ随伴ウイルス、レトロウイルス、レンチウイルスおよびプラスミドを含む。仮に、ベクターがウイルスベクターであり、そのベクターがパッケージされる場合、ビリオンが感染細胞で利用され得る。裸のDNAが使用されるなら、特定の宿主細胞に適合する移入もしくは形質転換の手段が使われ得る。重合体、リポゾームもしくはナノ粒子を利用する裸のDNAの製剤形態が、融合遺伝子の送達に利用され得る。本発明による組み換え構築物を形質転換または移入された細胞は、当業者に利便性があるかもしれない。利用され得る典型的な細胞型は、バクテリア、酵母、昆虫および哺乳類細胞を包含する。利便性があることに、哺乳類細胞間では、多くの組織型細胞が、選択可能であるかもしれない。利用され得る典型的な細胞は、線維芽細胞、肝細胞、内皮細胞、幹細胞、造血細胞、上皮細胞、筋細胞、神経細胞およびケラチン生成細胞である。これらの細胞は、in vitroで蛋白質を生成するために利用され得るか、もしくはin vivoでコードされた蛋白質を生成するためにヒトを含む哺乳類動物に送達され得る。この送達方法は、核酸を哺乳類動物に、ウイルスベクターを哺乳類動物に、および融合蛋白質を哺乳類動物に送達することにとって代わるものである。
【0038】
蛋白質もしくは核酸の組成物は、緩衝液のような担体、水溶性もしくは親油性担体、無菌性もしくは菌性、発熱性もしくは非発熱性担体中で存在し得る。非発熱性担体は、注射製剤として有用である。製剤は、例えば、凍結乾燥されたような液状もしくは固形状であり得る。製剤はまた、エアロゾルとして投与され得る。組成物は、一以上の融合蛋白質もしくは一以上の核酸、または融合蛋白質および核酸の両方を包含するであろう。組成物中の融合蛋白質および/もしくは核酸が単一種であり得、その場合には、ホモ多量体蛋白質が形成し、あるいは、組成物中の融合蛋白質および/もしくは核酸が異種であり得、その場合には、ヘテロ多量体蛋白質が形成する。ヘテロ多量体である場合において、典型的には、X部分は融合蛋白質間で異なるが、Z部分は融合蛋白質間で同一である。
【0039】
融合蛋白質は、当該技術分野で知られる方法によって細胞もしくは哺乳類宿主に供給され得る。蛋白質は、細胞もしくは宿主に送達され得る。核酸は、細胞もしくは宿主に投与され得る。形質転換もしくは移入された細胞は、細胞もしくは宿主に投与され得る。後者の場合、同一の遺伝学的背景を有する細胞が、移植の拒絶反応を減らすことに望ましい。
【0040】
哺乳類の宿主動物への送達に適合する細胞は、器官、腫瘍、もしくは細胞系列に由来する哺乳類細胞型を包含する。例えば、ヒト、マウス、ヤギ、ヒツジ、ウシ、ネコ、およびブタの細胞が使用され得る。使用制限がなく、適合する細胞型は、線維芽細胞、幹細胞、ケラチン生成細胞、造血細胞、上皮細胞、筋細胞、神経細胞、および幹細胞を包含する。
【0041】
融合蛋白質もしくは融合蛋白質をコードする核酸の送達手段は、融合蛋白質を発現する細胞の送達、融合蛋白質の送達および融合蛋白質をコードする核酸の送達を包含する。融合蛋白質、細胞、もしくは核酸は、例えば、注射、カテーテル法、もしくは内視鏡によって望ましい器官もしくは腫瘍に直接送達され得る。それらはまた、静脈内、気管支内、腫瘍内、随腔内、筋肉内、眼内、局所、皮下、経皮もしくは経口によって送達され得る。浮腫加齢黄斑変性症、増殖性糖尿病網膜症、慢性関節リウマチ、骨関節症、ブドウ膜炎、喘息および癌を患う患者を効果的に処置することが可能である。これらの処置は、症状および/もしくは病気の重症度および/もしくは病気の進行を改善するであろう。
【0042】
核酸は、望ましいベクターを用いて、哺乳類動物、特にヒトに送達され得る。これらには、アデノウイルスベクター、アデノ随伴ウイルスベクター、レトロウイルスベクター、レンチウイルスベクターおよびプラスミドベクターを包含するウイルス性もしくは非ウイルス性のベクターが含まれる。ウイルスの典型的な型には、HSV(単純ヘルペスウイルス)、アデノウイルス、AAV(アデノ随伴ウイルス)、HIV(ヒト免疫不全ウイルス)、BIV(ウシ免疫不全ウイルス)、およびMLV(マウス白血病ウイルス)を含む。核酸は、ウイルス粒子、リポゾーム、ナノ粒子および混合された重合体を含む十分に効果的な送達レベルを付与する望ましい形式で投与され得る。
【0043】
蛋白質および核酸を組み合わせた処置が使用され得る。例えば、本発明による融合蛋白質が患者に投与され得る。もし、好ましい応答が観察される場合、融合蛋白質をコードする核酸分子が、長期間の効果を調べるために投与され得る。あるいは、蛋白質もしくは核酸が同時、もしくはほぼ同時に投与され得る。別の方法として、リガンドに対する抗体もしくは融合蛋白質が投与され、続けてもしくは同時に、受容体に対する抗体もしくは融合蛋白質が投与され得る。別の選択肢として、一方が抗体をコードし、他方が融合蛋白質をコードする核酸の組み合わせを用いる。本発明のFlt−1構築物(蛋白質もしくは核酸の形のどちらか)との組み合わせによって使用し得る幾つかの抗体は、ベバシマズムおよびラニビズマズであり、両者ともVEGFに結合する。これらは特に、癌および黄斑変性症それぞれを治療するのに有効である。
【0044】
本発明の実施は、別段の表示がない限り、当該技術分野の範囲内である分子生物学(組み換え技術を含む)、微生物学、細胞生物学、生化学および免疫学の一般的技術によって行われる。このような技術は、Molecular Cloning:A Laboratory Manual,Second Edition(Sambrookら,1989);Current Protocols In Molecular Biology(F.M.Ausubelら編,1987);Oligo nucleotide Synthesis(M.J.Gait編,1984);Animal Cell Culture(R.i.Freshney編,1987);Methods In Enzymology(Academic Press,Inc.);Handbook Of Experimental Immunology(D.M.WeiとC.C.Blackwellら編.);Gene Transfer Vectors For Mammalian Cells(J.M.MillerとM.P.Calos編,1987);PCR:The Polymerase Chain Reaction,(Mullisら編,1994);Current Protocols In Immunology(J.E.Coliganら編,1991);Antibodies:A Laboratory Manual(E.HarlowとD.Laneeds.(1988))ならびに、PCR2:A Practical Approach(M.J.MacPherson,B.D.HamesとG.R.Taylor編(1995))のような文献において十分に説明されている。
【0045】
遺伝子送達の担体は、宿主細胞内へ挿入するポリヌクレオチドを運搬することが可能な分子である。遺伝子送達担体の例としては、リポゾーム、リポ蛋白質、ポリペプチド、多糖類、リポ多糖類、人工ウイルス外被、金属粒子およびバクテリアを含む天然型重合体と合成型重合体である生体適合性重合体、バキュロウイルス、アデノウイルスおよびレトロウイルスのようなウイルス、バクテリオファージ、コスミド、プラスミド、真菌ベクターならびに多くの原核生物および真核生物の宿主で発現を示す当該技術分野内で典型的に利用されるその他の組み換え用担体を含み、これらは単一種の蛋白質発現だけではなく、遺伝子治療にも利用され得るかもしれない。
【0046】
遺伝子送達、遺伝子移入およびここで使用するそれらに類似する言葉は、導入に使用される方法に関わりなく、外因性ポリヌクレオチド(時に”トランスジーン”と呼ばれる)を宿主細胞に導入することを意味する。かかる方法は、”裸の”ポリヌクレオチドの送達に使用する技術(ポリヌクレオチドの導入に使用されるエレクトロポレーション、”遺伝子ガン”送達および多くの他技術)だけでなく、ベクター介在遺伝子移入(例えば、ウイルス感染/移入、または多種の他蛋白質由来もしくは脂質由来の遺伝子送達複合体による)のような多くのよく知られている技術を包含する。導入されたポリヌクレオチドは、宿主細胞において安定的にもしくは一時的に維持されるであろう。安定的な維持は、典型的には、導入されるポリヌクレオチドが、宿主細胞に適合可能である複製開始点を含むか、あるいは染色体外レプリコン(例えばプラスミド)のような宿主細胞のレプリコンまたは核もしくはミトコンドリアの染色体に組み入られる必要がある。当該技術分野で周知であり、ここで記述されるように、多くのベクターが哺乳類細胞に遺伝子移入を仲介できることが知られている。
【0047】
外因性ポリヌクレオチドを静脈内、筋肉内、門脈内もしくは別の方法による投与を介して宿主へ送達するために、外因性ポリヌクレオチドが、アデノウイルス、部分的に欠失されたアデノウイルス、完全に欠失されたアデノウイルス、アデノ随伴ウイルス(AAV)、レトロウイルス、レンチウイルス、裸のプラスミド、プラスミド/リポゾーム複合体等のようなベクターに挿入される。本発明で示す方法および組成物で使用され得る発現ベクターは、例えばウイルスベクターを包含する。In vivoとex vivoの両方において、遺伝子治療で最も頻繁に使用される投与方法の一つは、遺伝子送達用ウイルスベクターの使用である。多種類のウイルスが知られ、その多くが遺伝子治療の目的で研究されている。最も一般的に使用されるウイルスベクターは、アデノウイルス、アデノ随伴ウイルス(AAV)およびヒト免疫不全ウイルス(HIV)のようなレンチウイルスを含むレトロウイルスを包含する。
【0048】
アデノウイルスは、非外皮性であり、古典的遺伝学と分子生物学における研究を通してよく特徴づけられた約36kbのゲノムを有する核DNAウイルスである(Hurwitz, M.S., Adenoviruses Virology,3rd edition,Fieldsら編,Raven Press,New York,1996;Hitt, M.M.ら,Adenovirus Vectors,The Development of Human Gene Therapy,Friedman,T.編,Cold Spring Harbor Laboratory Press, New York 1999)。ウイルス遺伝子からウイルス蛋白質が、時間経過によって2つに分かれて生成されることから、ウイルス遺伝子は、早期(E1−E4と表す)および後期(L1−L5と表す)の転写単位に区分けされる。これらの事象の境界は、ウイルス性DNAの複製である。ヒトアデノウイルスは、赤血球の血球凝集、発癌能、DNAおよび蛋白質のアミノ酸組成および相同性、ならびに抗原的関連性を包含する特性に基づいて、多くの血清型(約47個がそれに応じて番号付けられ、A、B、C、D、EおよびFの6グループに区分けされた)に分けられる。
【0049】
組み換え体アデノウイルスベクターは、遺伝子送達の担体としての使用に対していくつかの有利な点を有し、すなわち、それらは、分裂と非分裂細胞の指向性、極小な病原性、ベクターのストックを調製するための高い力価で複製する能力、および大きい挿入物を運搬する能力を包含する(Berkner,K.L.,Curr.Top.Micro.Immunol.158:39−66,1992;Jolly,D.,Cancer Gene Therapy 1:51−64 1994)。擬性アデノウイルスベクター(PAV)および部分的に欠失されたアデノウイルス(”DeAd”と呼ばれる)のように多くのアデノウイルス遺伝子配列欠失を有するアデノウイルスベクターは、核酸の受容細胞への送達に適合した担体として望ましいアデノウイルスの特性を有するように設計されている。
【0050】
特に、擬性アデノウイルスベクター(PAVs)はまた、’弱いアデノウイルス’もしくはミニ−アデノウイルスベクターとして知られ、ベクターの複製およびパッケージに必要である最小のシス作用性ヌクレオチド配列を含み、一以上の導入遺伝子を包含することが可能である、アデノウイルスのゲノムに由来するベクターである(米国特許第5,882,877号には、擬性ベクター(PAV)とPAVを生成する方法を含み、出典明示により、本明細書に一体化させる)。PAVsは、遺伝子送達に適合する担体としてアデノウイルスの望ましい特性を有するように設計されている。アデノウイルスベクターは、ウイルスの増殖に不必要である領域の欠失によって一般的に8kbまでの大きさを挿入することが可能である一方で、最大の運搬能力は、PAVsを包含する多くのウイルスのコード配列を欠失したアデノウイルスベクターを使用することで達成し得る。Gregoryらの米国特許第5,882,877号;Kochanekら,Proc.Natl.Acad.Sci.USA93:5,731−5,736,1996;Parksら,Proc.Natl.Acad.Sci.USA 93:13565−13570,1996;Lieberら,J.Virol.70:8944−8960,1996;Fisherら,Virology 217:11−22,1996;米国特許第5,670,488号;PCT公開番号W096/33,280,1996年10月24日に公開;PCT公開番号W096/40955,1996年12月19日に公開 PCT公開番号W097/25446,1997年7月19日に公開;PCT公開番号 W095/29993,1995年11月9日に公開; PCT公開番号W097/00326,1997年1月3日に公開;Morralら,Hum.Gene Ther.10:2709−2716,1998を参照のうえ。このようなPAVsは、外部からの核酸を約36kbまで受け入れることが可能であり、ベクターの運搬能力は、最適化されているので優位である一方で、ベクターに対する宿主の免疫反応もしくは複製可能なウイルスの生成能力は減少する。PAVベクターは、複製開始点を包含する5’逆方向末端反復(ITR)および3’ITRヌクレオチド配列、ならびにPAVゲノムのパッケージに必要であるシスヌクレオチド配列を含み、例えば、プロモーター、エンハンサー等の適切な調節エレメントを有する一以上の導入遺伝子に適合することが可能である。
【0051】
その他には、部分的に欠失されたアデノウイルスベクターは、ウイルスの複製に必要であるアデノウイルス早期遺伝子の大部分がベクターから欠失し、条件プロモーターの制御下にある生成細胞の染色体内に置かれる部分的欠失(”DeAd”と呼ばれる)ベクターを供給する。生成細胞に配置される欠失可能なアデノウイルス遺伝子は、E1A/E1B、E2、E4(ORF6およびORF6/7のみが細胞内で必要とされる)、pIXおよびpIVa2を包含する。E3もまたベクターから欠失され得るが、ベクターの産生に必須ではないことから、生成細胞から除去することが可能である。主要後期プロモーター(MLP)の制御下にあるアデノウイルス後期遺伝子は、一般的にはベクター内に存在するが、MLPは条件プロモーターに置換され得る。
【0052】
DeAdベクターおよび生成細胞系列の使用に適合可能な条件プロモーターは、以下の性質を有するものを包含する。すなわち、細胞障害性もしくは細胞増殖抑制性のアデノウイルス遺伝子が、細胞を害するレベルで発現しないような、未誘導状態における低い基礎発現、およびベクターの複製と組み立てを支持するウイルス蛋白質が、十分量生成されるような、誘導状態における高いレベルの発現を有するものである。DeAdベクターおよび生成細胞系列における使用に適合する望ましい条件プロモーターは、免疫抑制剤FK506およびラパミシンに基づく二量体化物(dimerizer)遺伝子制御システム、エクジソン遺伝子制御システム、およびテトラサイクリン遺伝子制御システムを包含する。本発明において、Abruzzeseら,Hum.Gene Ther.1999 10:1499−507に記載されているGeneSwitch(登録商標)の技術(Valentis,Inc.,Woodlands,TX)が有用であり、出典明示により本明細書に一体化させる。部分的に欠失されたアデノウイルス発現システムは、さらにWO99/57296で示され、出典明示により本明細書に一体化させる。
【0053】
アデノ随伴ウイルス(AAV)は、ゲノムの大きさが4.6kbである一本鎖のヒトDNAパルボウイルスである。AAVゲノムは2つの主要な遺伝子を含む:rep蛋白質(Rep76、Rep68、Rep52およびRep40)をコードするrep遺伝子ならびにAAVの複製、レスキュー、転写および組込みをコードするcap遺伝子であり、さらに、cap蛋白質はAAVのウイルス粒子を形成する。AAVは、アデノウイルスもしくは他のヘルパーウイルス(例えばヘルペスウイルス)に依存して必須遺伝子産物を供給することにその名前が由来し、それらの産物は、AAVに生産的な感染、すなわち、宿主細胞における自身の再合成を可能にする。ヘルパーウイルスの非存在下では、AAVは、ヘルパーウイルス、一般的にはアデノウイルスに感染している宿主細胞に重複感染することによってレスキューされるまで前駆体ウイルスとして宿主細胞の染色体に組み込まれる(Muzyczka,Curr.Top.Micor.Immunol.158:97−127,1992)。
【0054】
遺伝子移入ベクターとしてのAAVの利点は、いくつかの生物学的な特徴に起因する。AAVゲノムの両末端には、逆位末端配列(ITR)として知られるヌクレオチド配列が存在し、その配列は、ウイルスの複製、レスキュー、パッケージおよび組込みに必要であるシス作動性のヌクレオチド配列を含む。トランス活性におけるrep蛋白質を介するITRの組み込み機能は、ヘルパーウイルス非存在下において、AAVゲノムが感染後に細胞内の染色体に組み込まれることを可能にする。このウイルス独特な特性は、細胞内ゲノムへの遺伝子の利点を含む組み換え体AAVの組み込みを可能にするとともに、遺伝子移入におけるAAVの利用に関連する。それゆえに、遺伝子移入の目的に適した多くの安定な遺伝学的な形質転換が、rAAVベクターの使用によって成し遂げられるかもしれない。さらに、AAVの組み込み部位がよく解明され、その部位はヒト染色体19番に位置する(Kotinら,Proc.Natl.Acad.Sci.87:2211−2215,1990)。この組込み部位の予測可能性は、宿主の遺伝子を活性化もしくは不活性化、またはコード領域を分断してしまうかもしれない細胞内ゲノムへのランダムな挿入事象の危険、rAAvベクターの使用を制限しうる結果を減らす。rAAVベクターのデザインにおけるこの遺伝子の除外は、rAAVベクターとともに観察された変動した組み込みパターンを生じるかもしれない(Ponnazhaganら.,Human Gene Ther.8:275−284,1997)。
【0055】
遺伝子移入のためのAAV利用には、他の有利な点がある。AAV宿主の範囲が広いことである。さらに、AAVは、レトロウイルスと異なり、停止状態および分裂状態の両状態の細胞に感染することが可能である。加えて、AAVは、ヒトの病気に関連せず、レトロウイルス由来の遺伝子移入ベクターで生じる多くの事象を防ぐ。
【0056】
組み換え体rAAVベクターを作製する標準的なアプローチには、一連の細胞内イベントに協調することが必要である:AAVのITR配列に隣接させた導入遺伝子を含むrAAVベクターゲノムの宿主細胞への移入、トランス活性で必要となるAAVのrepおよびcap蛋白質の遺伝子をコードするプラスミドによる宿主細胞への移入、ならびにトランス活性で必要となる非AAVヘルパーの機能を提供するヘルパーウイルスを移入された細胞の感染である(Muzyczka,N.,Curr.Top.Micor.Immunol.158:97−129,1992)。アデノウイルス(もしくは他のヘルパーウイルス)蛋白質は、AAVのrep遺伝子の転写を活性化し、rep蛋白質は、AAVのcap遺伝子の転写を活性化する。cap蛋白質は、rAAVゲノムをrAAVウイルス粒子内にパッケージするのにITR配列を利用する。それゆえ、パッケージの効率は、rAAVベクターゲノムに必要とされるシス活性のパッケージ配列への近接可能性とともに、適当量の構造蛋白質の利用可能性によってある程度決定される。
【0057】
レトロウイルスベクターは、遺伝子送達に用いられる一般的な道具である(Miller,Nature(1992)357:455−460)。再配列されていない単コピー遺伝子を齧歯類、霊長類およびヒト体細胞のような広い範囲内で送達するレトロウイルスベクターの能力は、遺伝子を細胞へ送達するのに最適である。
【0058】
レトロウイルスはRNAウイルスであり、そのゲノムはRNAである。宿主細胞がレトロウイルスに感染すると、ゲノムRNAは、中間体であるDNAに逆転写され、感染細胞の染色体DNAへ非常に効率的に組み込まれる。この組み込まれたDNA中間体は、プロウイルスと呼ばれている。プロウイルスの転写および感染性ウイルスへの構築は、混入するヘルパーウイルスが同時生成することなしに、キャプシド形成を可能にする適当な配列を含む適合するヘルパーウイルスの存在下、もしくは細胞系列で生じる。もしキャプシド形成に必要な配列が適合するベクターとの共移入によって提供されるならば、ヘルパーウイルスは、組み換え体レトロウイルスの生成に必要でない。
【0059】
レトロウイルスのゲノムおよびプロウイルスのDNAは、3個の遺伝子gag、polおよびenvを有し、それらは2個の末端反復(LTR)配列で挟まれている。gap遺伝子は、内部構造(マトリックス、キャプシドおよびヌクレオキャプシド)蛋白質をコードし、pol遺伝子は、RNA依存性DNAポリメラーゼ(逆転写酵素)をコードし、およびenv遺伝子は、ウイルス外皮性糖蛋白質をコードする。5’および3’LTRは、ビリオンRNAの転写およびポリアデニル化を促進する働きがある。LTRは、ウイルスの複製に必要である他のシス活性配列全てを含む。レンチウイルスは、(HIV−1、HIV−2および/もしくはSIV中に)vit、vpr、tat、rev、vpu、nefおよびvpxを含む付加遺伝子を有する。ゲノムの逆転写(tRNAプライマー結合部位)およびウイルスRNAの粒子内への効率的なキャプシド形成(Psi部位)に必要である配列が、5’LTRに隣接する。仮に、キャプシド形成(もしくはレトロウイルスの感染性ビリオンへのパッケージ)に必要な配列がウイルスゲノムから喪失しているなら、ゲノムRNAのキャプシド形成を阻害するシス活性の欠失となる。しかしながら、得られる変異体は、ビリオン蛋白質全ての合成を指示し得る。
【0060】
レンチウイルスは、一般的なレトロウイルス遺伝子gag、polおよびenvに付加して、調節的もしくは構造的な機能を有する遺伝子を包含する複合体レトロウイルスである。より高度な複雑性は、潜伏感染時にレンチウイルスが自身のライフサイクルを調節することを可能にする。典型的なレンチウイルスには、AIDSの病原因子であるヒト免疫不全ウイルス(HIV)がある。in vivoにおいて、HIVは、リンパ球およびマクロファージのような稀にしか分裂しない最終的に分化した細胞に感染し得る。in vitroでは、HIVは、アフィディコリンもしくはガンマ線照射の処理によって細胞周期が停止した状態のHeLa−Cd4もしくはTリンパ球のみならず、初代培養の単球由来マクロファージ(MDM)にも感染し得る。細胞の感染は、標的細胞の核膜孔を介する、活性化したHIV前駆組込み複合体の核移行に依存する。これは、標的細胞の核移入機能を有する複合体における多様で一部重複する分子決定因子の相互作用によって生じる。同定された決定因子は、gapマトリックス(MA)蛋白質における機能的な核局在シグナル(NLS)、核親和性ビリオン関連蛋白質、vpr、およびMA蛋白質におけるC末端のリン酸化チロシン残基を包含する。遺伝子治療に用いられるレトロウイルスは、例えば、米国特許第6,013,516号および米国特許第5,994,136号で示され、出典明示により本明細書に一体化させる。
【0061】
DNAを細胞に送達する他の方法は、送達にウイルスを使用しない方法である。例えば、陽イオン両親媒性化合物が、本発明の核酸を送達するのに使用され得る。生物学的な活性分子の細胞内送達に利用するために設計された化合物は、非極性および極性の両方の環境(例えば、細胞膜、組織液、細胞内構築物、および生物学的活性分子自身内もしくは上)において相互作用する必要があることから、このような化合物は、典型的には、極性および非極性ドメインを含むように設計される。かかるドメイン両方を有する化合物は、両親媒性物質と呼ばれ、このような細胞内送達(in vitroもしくはin vivoのどちらかに応用)を促進する利用が公知されている多くの脂質および合成脂質が、この定義に適合する。このような両親媒性物質の中で特に重要なクラスの一つが、陽イオン両親媒性物質である。一般的に、陽イオン両親媒性物質は、物理学的pH付近において陽に帯電を示し得る極性グループであり、この特性は、例えば、DNAのように負に帯電したポリヌクレオチドを包含する、多種類型の生物学的な活性(医薬上)分子と相互作用するのに重要であることが当該技術分野で理解されている。
【0062】
遺伝子送達用の陽イオン両親媒性物質を含む組成物の利用は、例えば、米国特許第5,049,386号;米国特許第5,279,833号;米国特許第5,650,096号;米国特許第5,747,471号;米国特許第5,767,099号;米国特許第5,910,487号;米国特許第5,719,131号;米国特許第5,840,710号;米国特許第5,783,565号;米国特許第5,925,628号;米国特許第5,912,239号;米国特許第5,942,634号;米国特許第5,948,925号;米国特許第6,022,874号;米国特許第5,994,317号;米国特許第5,861,397号;米国特許第5,952,916号;米国特許第5,948,767号;米国特許第5,939,401号;および米国特許第5,935,936号で示され、出典明示により本明細書に一体化させる。
【0063】
加えて、本発明の核酸は、”裸のDNA”を使用して送達され得る。移入促進蛋白質、ウイルス粒子、リポソーム製剤、荷電脂質およびカルシウムリン酸沈殿剤とは関連せず、プロモーターを結合した望ましいポリペプチドもしくはペプチドをコードする非感染性で非組込み型のDNA配列を送達する方法が、米国特許第5,580,859号;米国特許第5,963,622号;米国特許第5,910,488号に記述され、出典明示により本明細書に一体化させる
【0064】
また、ウイルス性と非ウイルス性の要素を組み合わせる遺伝子移入システムが報告されている。Proc.Natl.Acad.Sci.USA 90:11548;Wuら.(1994) J.Biol.Chem.269:11542;Wagneら.(1992) Proc.Natl.Acad.Sci.USA 89:6099;Yoshimuraら.(1993) J.Biol.Chem.268:2300;Curielら.(1991) Proc.Natl.Acad.Sci.USA 88:8850;Kupferら. (1994) Human Gene Ther. 5:1437およびGottschalkら/.(1994) Gene Ther. 1:185.多くの場合、アデノウイルスが、そのエンドソームの特性を生かして遺伝子送達システムに取り入れられている。報告されたウイルス性と非ウイルス性要素の組み合わせは、一般的には、アデノウイルスの遺伝子送達複合体への共有結合もしくは陽イオン脂質:DNA複合体を伴う非結合アデノウイルスの共内部移行のいずれかに関連する。
【0065】
眼にDNAおよび蛋白質を送達するため、典型的には、局所的な投与が施される。これは、投与する必要のDNA量および全身性副作用を制限するという点で有利である。多くの送達可能な形態が使用され得る。それらは、角膜への点眼を介した遺伝子銃による局所投与、結膜下注射、前房内注射、角膜を介した前房内注射、基質内注射、電磁波を用いた角膜への応用、角膜内注射、網膜下注射、硝子体内注射、および眼球内注射を包含する。代替的には、細胞が、ex vivoで移入もしくは形質導入され、眼内移植によって送達され得る。Auricchio,Mol.Ther.6:490−494,2002;Bennett,Nature Med.2:649−654,1996;Borras,Experimental Eye Research 76:643−652,2003;Chaum,Survey of Ophthalmology 47:449−469,2002;Campochiaro,Expert Opinions in Biological Therapy 2:537−544(2002); Lai,Gene Therapy 9:804 813,2002;Pleyer,Progress in Retinal and Eye Research,22:277−293,2003を参照のうえ。
【0066】
様々な提唱された治療用薬剤および投与の効果が、特定の病気に対応する動物モデルを用いて試験され得る。例えば、未熟児網膜症は、Smith,Investigative Ophthalmology & Visual Science,35:101−111,1994に記載されるようにマウスの酸素誘導型網膜症モデルで試験され得る。マウスにおけるレーザー誘導型脈絡膜新生血管は、加齢黄斑変性症のような病気の際に生じるヒト脈絡膜新生血管(CNV)のモデルとして利用され得る。Tobe,American Journal of Pathology 153:1641−1646,1998。CNVの他のモデルが、霊長類、ラット、ミニブタ、およびウサギにおいて開発された。加齢黄斑変性症のマウスモデルは、遺伝子欠損マウスを用いて開発された。単球走化性蛋白質−1もしくはC−Cケモカイン受容体−2のどちらか一方を欠損させたマウスが加齢黄斑変性症の特性を示す。Ambati,Nature Med.9:1390−1397,2003。
【0067】
本発明は、現時点で発明を実施するのに望ましい様式を包含する特定の例に関して記述されている一方で、当業者は、添付した請求の範囲を説明する本発明の精神および範囲内である上述したシステムおよび技術に対する多くのバリエーションと置き換えが存在することを理解するであろう。
【実施例】
【0068】
実施例1
2個の構築物が作製された。第一に、ポリグリシン9−mer(9Gly)のリンカーを含むD2−9Gly−Fcであり、第二として、9Glyリンカーを除いて同一の配列のD2−Fcである(図1)。
【0069】
我々は、配列解析プログラムMacVector6.5.1(IBI、New Haven、CT)のProtein Analysis Toolboxを用いて、D2−9Gly−FcとD2−Fc蛋白質のアミノ酸配列について解析を行った。D2−9Gly−Fc配列におけるポリグリシン9−merリンカーが、Karpus and Schultz(1985)Naturwiss,72:212−213の可撓性予測法による平均の可撓性より高い領域として同定された。このような領域は、D2−Fc配列中では検出されなかった(図1)。
【0070】
実施例2
我々は、同定された可撓性ポリグリシン9−merリンカーによってIgG1のFc領域に連結させたFlt−1のIg様ドメイン2(D2−9Gly−Fc)を検証した。D2−9Gly−Fc融合蛋白質は、効果的にVEGFに結合し、VEGF依存ヒト臍帯静脈内皮細胞(HUVEC)の増殖を阻害することが可能である。図2参照。対照的に、Flt−1のIgG1様ドメイン2が直接IgG1重鎖(Fc)に結合して、D2−Fcを形成する時には、最少量のVEGF結合のみが観察された。図2参照。IgG1のFcを介した二量体化と可撓性リンカー挿入の両方が、Flt−1ドメイン2のVEGF結合を促進すると考えられる。D2−9Gly−FcおよびD2−Fcにおける二量体型の存在は、ウェスタンブロット解析で確認された。図3参照。
【0071】
実施例3
AAVベクターの硝子体腔内注射(0.0005mLの体積中1x108から1x109個の粒子)が、新生仔(P0)もしくは生後1日後(P1)のC57BL/6系統マウスに施される。網膜新生血管(NV)が、P7の仔とこの仔らを養育中の雌親を5日間過酸素状態におくことによって、C57BL/6系統マウスで誘発される。これらの仔をP12で通常の大気に戻し、P17(NVピーク状態時)で安楽死させる。(Smith LEH、 Weslowski E、 McLellan A、 Kostyk SK、D’Amato R、 Sullivan and D’Amore PA。マウスにおける酸素誘発網膜症。Invest Opth Vis Sci。 1994;35:101−111)眼全体をパラフィンで包埋し、5マイクロン間隔で連続的に横断面を作製する。NVの程度は、100マイクロン毎に選んだ切片において境界膜の内側の内皮細胞核の数を数えることで決定される。
【0072】
抗血管形成薬剤をコードするAAVベクターで処理した動物の同齢集団を、無関連な導入遺伝子をコードするベクターもしくは導入遺伝子をコードしないベクターで処理した同齢集団と比較する。各々の処理した眼における内皮細胞核の平均数が、各々の動物の未処理の眼におけるその数と比較される。
【0073】
実施例4、D2−9Gly−Ex3/CH3の作製
Flt−1のドメイン2がVEGF165の結合に必須であることが示されている。しかし、Flt−1のドメイン2単独ではVEGFのAに結合できないことが示されていた(Davis−Smyth et al., 1996.)。VEGFのAは、二量体として存在する時、酸性残基(成熟蛋白質のアミノ酸63−67)を介してFlt−1に結合し、リガンド誘導型二量体受容体として機能することが可能である(Keytら., 1996)。
【0074】
それゆえ、Flt−1のドメイン2の二量体化は、Flt−1のドメイン2がVEGFのAへ結合する方法として使用されてきた。IgG重鎖の断片との融合は、蛋白質の二量体化に用いられ得る(Davis−Smythら., 1996.)。ここで、我々は、VEGFのA(配列番号12)のアミノ酸75−88(すなわちPSCVPLMRCGGCCN;配列番号13)が、sFlt−1混合蛋白質の生物学的活性を上昇させることを示す。
【0075】
最初に3個の混合蛋白質が設計された。D2−9Gly−Fc、D2−FcおよびD2−9GlyEx3/CH3である(図4)。3個の混合蛋白質全ては、D2−9Gly−Fcのように同一のFlt−1のドメインD2を含む。VEGFへの結合は、ポリグリシン9−mer(9Gly)リンカーを含まないD2−Fcにおいて観察されなかった。3番目の蛋白質であるD2−9Gly−Ex3/CH3は、ポリグリシン9−mer(9Gly)リンカーおよびVEGFの多量体化ドメイン(アミノ酸PSCVPLMRCGGCCN;配列番号13;VEGF Ex3)を包含するが、また、ヒトIgG1重鎖FcのCH3領域(配列番号10のアミノ酸371−477)をも含む。
【0076】
蛋白質D2−Fcは、HUVEC増殖測定(図5)において効果的な阻害活性を示さず、それに伴い、効果的にVEGF165に結合しなかった。しかしながら、3番目の混合蛋白質であるD2−9Gly−Ex3/CH3は、9GlyリンカーおよびVEGF165の二量体化領域(Ex3)の両方を介してCH3領域に融合した、Flt−1のドメイン2を含み、VEGF依存性HUVECの増殖測定法において阻害活性を示した(図5)。これは、この混合蛋白質がVEGF165に効果的に結合することを意味する。
【0077】
実施例5、Flt−1のD2構築物におけるリンカー(Gly4Ser)3の使用
以前に、数個のポリグリシンリンカーの使用が、蛋白質の性状を改変すると報告されている(Mouzら,1996;Qiuら,1998)。次の構築物のために、我々は、別の型のリンカーである15−mer(Gly−Gly−Gly−Gly−Ser)3を試した(Hustonら,1988)。D2−(Gly4Ser)3−Fc蛋白質が作製され、この蛋白質は、Flt−1ドメイン2、(Gly4Ser)3リンカーおよびヒトIgG1重鎖のFc領域を含む。
【0078】
D2−(Gly4Ser)3−Fcは、さらにHUVEC増殖測定法で検査された。D2−(Gly4Ser)3−Fcの生物学的活性は、HUVECの増殖阻害によって測定され、D2−9Gly−FcおよびD2−9Gly−Ex3/CH3の生物学的活性に類似した(図6)。
【0079】
D2−(Gly4Ser)3−Fc構築物は、さらにウェスタンブロットを用いて検査され、D2−9Gly−Fcと比較された(図9)。両方の構築物は、二量体型で多く存在し、単量体型は、還元されたサンプルの分離後に検出された。
【0080】
実施例6、Flt−1(D2)構築物における9GlyもしくはVEGFEx3の役割
可溶性受容体VEGFへの結合に対する9GlyリンカーもしくはVEGF二量体化配列Ex3の役割を調べるために、3個の異なる構築物、D2−9Gly−CH、D2−CH3およびD2−Ex3/CH3が作製された(図8)。3個の構築物全てが作製され、以前の構築物と同様にCMVプロモーターの制御を受けるようにした。これらのVEGF抑制活性はHUVEC増殖測定法で測定された(図9)。
【0081】
IgG1のCH3領域を含む蛋白質のHUVEC増殖測定法は、D2−9Gly−CH3(Ex3無し)および蛋白質D2−Ex3/CH3(9Glyリンカー無し)が、D2−9GlyEx3/CH3と比較して、同様のVEGF抑制能力を有することを示した。しかし、蛋白質D2−CH3は、これら全ての中で最も弱いVEGF阻害剤であると思われた(図9)。
【0082】
移入された293細胞由来条件培地のFlt−1におけるELISAデータでは、D2−9Gly−Ex3/CH3、D2−9Gly−CH3およびD2−Ex3/CH3については、同様のFlt−1レベル(70−90ng/ml)を示し、最小のD2−CH3活性形態については、やや高いレベル(約150ng/ml)を示した。D2−9Gly−CH3およびD2−CH3の構築物のウェスタンブロット(図10)は、非還元条件での二量体型の普及を示している。
【0083】
実施例7、D2−9Gly−Fcは、全ての構築物より一層VEGFに結合する。
VEGF結合測定は、細胞遊離中における可溶性VEGF受容体に対する相対的なVEGF結合能の比較を可能にする。
【0084】
簡潔に説明すると、既知濃度の可溶性受容体(0.29−150pMの濃度範囲)を含む条件培地は、連続的に希釈され、10pMのVEGFを添加された。非結合VEGF量は、ELISAによって測定された。D2−9Gly−Fcは、0.001から0.2pMの受容体濃度において全ての他の構築物より高いVEGF結合能を示す。D2−CH3は、VEGFに対して最も低い結合能を有する(図11)。
【0085】
(参考文献)
Davis−Smythら,EMBO J.15,1996,4919
Huston,J.S.ら(1991)Methods Enzymol.203,46−88
Huston,J.S.ら(1988)Proc.Natl Acad.Sci.Usa,85,5879−5883.
Johnson,S.ら(1991)Methods Enzymol.203,88−98 Karpus,P.A.ら(1985)Naturwiss.,72,212−213.
Keyt,B.A.,ら(1996)J.Biol.Chem.271:5638−5646.Kortt,A.A.ら(1997)Protein Engng,10,423−433.Lee,Y−L.ら(1998)Human Gene Therapy,9,457−465
Mouz N.ら(1996)Proc Nati Acad.Sci.USA,93,9414−9419.
Nielsen,ら(1997)Protein Eng.,10,1
Qiu,H.,ら(1998)J.Biol.Chem.273:11173−11176.
【0086】
【表1】
【表2】
【図面の簡単な説明】
【0087】
【図1】D2−9Gly−Fc構築物における9−Glyリンカーの可撓性領域を示す。
【図2】D2−9Gly−Fc対D2−Fcの生物学的活性を示す。
【図3】D2−9Gly−FcおよびD2−Fcのウェスタンブロット解析を示す。
【図4】D2−9Gly−Ex3/CH3と過去に構築された蛋白質との構造比較を示す。
【図5】D2−9Gly−Ex3/CH3対D2−9Gly−Fcの生物学的活性を示す。
【図6】D2−(Gly4Ser)3−Fc蛋白質活性をD2−9Gly−FcおよびD2−9Gly−Ex3/CH3と比較したHUVEC増殖測定。
【図7】D2−9Gly−Fc、D2−(G4S)3−Fc、およびEGFP蛋白質を用いたウェスタンブロットの結果を示す。
【図8】9Gly−CH3、D2−CH3およびD2−Ex3/CH3を有すもしくは有さない、3個の新規蛋白質の構造比較を示す。
【図9】9Gly、Ex3およびCH3の組み合わせを有するFlt−1(D2)構築物におけるHUVEC増殖測定を示す。
【図10】D2−9Gly−Fc、D2−9Gly−CH3およびD2−CH3を用いたウェスタンブロットの結果を示す。
【図11】VEGF“in vitro”結合測定を示す。
【図12】BIAcore機器を用いた表面プラスモン共鳴によって測定された可溶性Flt−1構築物の結合反応速度を示す。
【図13】各種リンカーを有するFlt−1構築物の発現レベルを示す。
【図14】網膜新生血管(NV)のマウス酸素誘導型網膜症モデルを用いて、Flt−1構築物の一つがマウスの眼に投与され、新血管新生が特定された。
【特許請求の範囲】
【請求項1】
式X−Y−Z(式中、Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含み、Yは、5−25個のアミノ酸残基のポリペプチドから必須として成り、Zは、IgG重鎖分子のCH3領域である)で示される、融合蛋白質。
【請求項2】
Xが細胞外受容体を含み、該受容体がチロシンキナ−ゼとセリンスレオニンキナ−ゼ受容体から成る群から選択される、請求項1記載の融合蛋白質。
【請求項3】
Xが、細胞外受容体を含み、該受容体がVEGF受容体である、請求項1記載の融合蛋白質。
【請求項4】
XがVEGF−R1(FLT−1)のIg様ドメイン2である、請求項1記載の融合蛋白質。
【請求項5】
ポリペプチドYが可撓的である、請求項1記載の融合蛋白質。
【請求項6】
ポリペプチドYが、gly9(配列番号27)、glu9(配列番号28)、ser9(配列番号29)、gly5cyspro2cys (配列番号30)、(gly4ser)3(配列番号31)、SerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号32)、ProSerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号13)、Gly−Asp−Leu-Ile−Tyr−Arg−Asn−Gln−Lys(配列番号26)、およびGly9ProSerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号34)から成る群から選択される、請求項1記載の融合蛋白質。
【請求項7】
ZがIgG1のCH3領域である、請求項1記載の融合蛋白質。
【請求項8】
ZがIgG2のCH3領域である、請求項7記載の融合蛋白質。
【請求項9】
さらに一以上の医薬上許容される添加物もしくは担体を含む、請求項1記載の融合蛋白質を含む、組成物。
【請求項10】
液体製剤である、請求項9記載の組成物。
【請求項11】
凍結乾燥製剤である、請求項9記載の組成物。
【請求項12】
該融合蛋白質の多量体を含む組成物であって、請求項1記載の一以上の融合蛋白質を含む組成物。
【請求項13】
ホモ多量体を形成する単一種の融合蛋白質から必須として成る、請求項12記載の組成物。
【請求項14】
組成物中の該融合蛋白質におけるX部分が、異種性である融合蛋白質から必須として成る、請求項12記載の組成物。
【請求項15】
X部分が細胞外受容体であり、該受容体がチロシンキナ−ゼ受容体およびセリンスレオニンキナ−ゼ受容体から成る群から選択される、請求項12記載の組成物。
【請求項16】
X部分が細胞外受容体であり、該受容体がVEGF受容体もしくはその一部である、請求項12記載の組成物。
【請求項17】
X部分がVEGF−R1(FLT−1)のIgG様ドメイン2である、請求項12記載の組成物。
【請求項18】
Yが可撓性を有するポリペプチドである、請求項12記載の組成物。
【請求項19】
Yが、gly9(配列番号27)、glu9(配列番号28)、ser9(配列番号29)、gly5cyspro2cys(配列番号30)、(gly4ser)3(配列番号31)、SerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号32)、ProSerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号13)、Gly−Asp−Leu-Ile−Tyr−ArgAsn−Gln−Lys(配列番号26)、およびGly9ProSerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号34)から成る群から選択されるポリペプチドである、請求項12記載の組成物。
【請求項20】
ZがCH3領域であり、該領域がIgG1のCH3領域である、請求項12記載の組成物。
【請求項21】
さらに一以上の医薬上許容される添加物もしくは担体を含む、請求項12記載の組成物。
【請求項22】
液体製剤である、請求項21記載の組成物。
【請求項23】
凍結乾燥製剤である、請求項21記載の組成物。
【請求項24】
式X−Y−Z(式中、Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含み、Yは、5−25個のアミノ酸残基の空間的分離を与えるリンカー部分から必須として成り、Zは、IgG重鎖分子のCH3領域である)で示されるポリペプチド。
【請求項25】
リンカー部分が、10−100個のエチレングリコール単量体残基のオリゴマーである、請求項24記載のポリペプチド。
【請求項26】
式X−Y−Z、(式中、Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含み、Yは、5−25個のアミノ酸残基のポリペプチドから必須として成り、Zは、抗体分子のFcの一部である)で示される融合蛋白質。
【請求項27】
FcがIgGのFcである、請求項26記載の融合蛋白質。
【請求項28】
IgGのFcがIgG1のFcである、請求項27記載の融合蛋白質。
【請求項29】
さらに一以上の医薬上許容される添加物もしくは担体を含む、請求項26記載の融合蛋白質を含む組成物。
【請求項30】
Zが、IgGのFcである抗体分子のFcの一部である、請求項29記載の組成物。
【請求項31】
Zが、IgG1のFcである抗体分子のFcの一部である、請求項29記載の組成物。
【請求項32】
式X−Y−Z(式中、Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含み、Yは、5−25個のアミノ酸残基の空間的分離を与えるリンカ−部分から必須として成り、Zは、抗体分子のFcの一部である)で示される融合蛋白質。
【請求項33】
FcがIgGのFcである、請求項32記載の融合蛋白質。
【請求項34】
IgGのFcがIgG1のFcである、請求項33記載の融合蛋白質。
【請求項35】
リンカー部分が、10−100個のエチレングリコール単量体残基のオリゴマーである、請求項32記載の融合蛋白質。
【請求項36】
さらに一以上の医薬上許容される添加物または担体を含む、請求項32記載の融合蛋白質を含む組成物。
【請求項37】
Zが、IgGのFcである抗体分子のFcの一部である、請求項36記載の組成物。
【請求項38】
Zが、IgG1のFcである抗体分子のFcの一部である、請求項36記載の組成物。
【請求項39】
ポリペプチドXYZ(Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含み、Yは、5−25個のアミノ酸残基のポリペプチドから必須として成り、Zは、IgG重鎖分子のCH3領域である)を形成するために、ポリペプチドXをポリペプチドYを介してポリペプチドZに結合することを含む、ポリペプチドXを多量体化する、方法。
【請求項40】
結合工程が、X、Y、およびZ各々をコードする核酸分子を単一の翻訳領域として構築することを含み、該ポリペプチドXYZが宿主細胞において核酸構築物から発現される、請求項39記載の方法。
【請求項41】
ポリペプチドXYZ(Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されたポリペプチドを含み、Yは、5−25個のアミノ酸残基の空間的分離を与えるリンカー部分から必須として成り、Zは、IgG重鎖分子のCH3領域である)を形成するために、ポリペプチドXを、部分Yを介してポリペプチドZに結合することを含む、ポリペプチドXを多量体化する、方法。
【請求項42】
リンカー部分が、10−100個のエチレングリコール単量体残基のオリゴマーから必須として成る、請求項41記載の方法。
【請求項43】
請求項1記載の融合蛋白質をコードする核酸分子。
【請求項44】
請求項26記載の融合蛋白質をコードする核酸分子。
【請求項45】
Xが、VEGF−R1(Flt−1)のIg様ドメイン2である、請求項43もしくは44記載の核酸。
【請求項46】
Xが、VEGF−R1(Flt−1)のIg様ドメイン2であり、融合蛋白質が配列番号8、21、および23から成る群から選択される配列を含む、請求項43記載の核酸。
【請求項47】
Xが、VEGF−R1(Flt−1)のIg様ドメイン2であり、融合蛋白質が配列番号2および25から成る群から選択される配列を含む、請求項44記載の核酸。
【請求項48】
Xが、VEGF−R1(Flt−1)のIg様ドメイン2である、請求項1もしくは26記載の融合蛋白質。
【請求項49】
Xが、VEGF−R1(Flt−1)のIg様ドメイン2であり、融合蛋白質が、配列番号8、21、および23から成る群から選択される配列を含む、請求項1記載の融合蛋白質。
【請求項50】
Xが、VEGF−R1(Flt−1)のIg様ドメイン2であり、融合蛋白質が、配列番号2および25から成る群から選択される配列を含む、請求項26記載の融合蛋白質。
【請求項51】
請求項43もしくは44記載の核酸を含む、哺乳類細胞。
【請求項52】
ヒト細胞である、請求項51記載の哺乳類細胞。
【請求項53】
線維芽細胞、肝細胞、内皮細胞、ケラチン生成細胞、造血系細胞、滑膜細胞、上皮細胞、網膜細胞、および幹細胞から成る群から選択される、請求項52記載の哺乳類細胞。
【請求項54】
請求項43もしくは44記載の核酸を、該融合蛋白質を発現する細胞を作製するために同定された哺乳類細胞に送達することを含む、インビトロの方法。
【請求項55】
融合蛋白質が、配列番号2、8、21、23、および25から成る群から選択される配列を含む、請求項54記載の方法。
【請求項56】
さらに該融合蛋白質を発現する細胞を哺乳類動物に送達することを含む、請求項54記載の方法。
【請求項57】
請求項51記載の哺乳類細胞を哺乳類動物に送達することを含む、方法。
【請求項58】
請求項43もしくは44記載の核酸を哺乳類動物に送達し、そのことにより該融合蛋白質が哺乳類動物で発現されることを含む、方法。
【請求項59】
融合蛋白質が、配列番号2、8、21、23、および25から成る群から選択される配列を含む、請求項58記載の方法。
【請求項60】
哺乳類動物が浮腫加齢黄斑変性症もしくは増殖性糖尿病網膜症にかかっている、請求項57記載の方法。
【請求項61】
哺乳類動物が癌にかかっている、請求項58記載の方法。
【請求項62】
哺乳類動物が慢性関節リウマチにかかっている、請求項58記載の方法。
【請求項63】
哺乳類動物が喘息にかかっている、請求項58記載の方法。
【請求項64】
哺乳類動物が骨関節症にかかっている、請求項58記載の方法。
【請求項65】
請求項1もしくは26記載の融合蛋白質を哺乳類動物に送達することを含む、方法。
【請求項66】
哺乳類動物が、浮腫加齢黄斑変性症もしくは増殖性糖尿病網膜症にかかっている、請求項65記載の方法。
【請求項67】
哺乳類動物が癌にかかっている、請求項65記載の方法。
【請求項68】
哺乳類動物が慢性関節リウマチにかかっている、請求項65記載の方法。
【請求項69】
哺乳類動物が喘息にかかっている、請求項65記載の方法。
【請求項70】
哺乳類動物が骨関節症にかかっている、請求項65記載の方法。
【請求項71】
融合蛋白質が、配列番号2、8、21、23、および25から成る群から選択される配列を含む、請求項65記載の方法。
【請求項1】
式X−Y−Z(式中、Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含み、Yは、5−25個のアミノ酸残基のポリペプチドから必須として成り、Zは、IgG重鎖分子のCH3領域である)で示される、融合蛋白質。
【請求項2】
Xが細胞外受容体を含み、該受容体がチロシンキナ−ゼとセリンスレオニンキナ−ゼ受容体から成る群から選択される、請求項1記載の融合蛋白質。
【請求項3】
Xが、細胞外受容体を含み、該受容体がVEGF受容体である、請求項1記載の融合蛋白質。
【請求項4】
XがVEGF−R1(FLT−1)のIg様ドメイン2である、請求項1記載の融合蛋白質。
【請求項5】
ポリペプチドYが可撓的である、請求項1記載の融合蛋白質。
【請求項6】
ポリペプチドYが、gly9(配列番号27)、glu9(配列番号28)、ser9(配列番号29)、gly5cyspro2cys (配列番号30)、(gly4ser)3(配列番号31)、SerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号32)、ProSerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号13)、Gly−Asp−Leu-Ile−Tyr−Arg−Asn−Gln−Lys(配列番号26)、およびGly9ProSerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号34)から成る群から選択される、請求項1記載の融合蛋白質。
【請求項7】
ZがIgG1のCH3領域である、請求項1記載の融合蛋白質。
【請求項8】
ZがIgG2のCH3領域である、請求項7記載の融合蛋白質。
【請求項9】
さらに一以上の医薬上許容される添加物もしくは担体を含む、請求項1記載の融合蛋白質を含む、組成物。
【請求項10】
液体製剤である、請求項9記載の組成物。
【請求項11】
凍結乾燥製剤である、請求項9記載の組成物。
【請求項12】
該融合蛋白質の多量体を含む組成物であって、請求項1記載の一以上の融合蛋白質を含む組成物。
【請求項13】
ホモ多量体を形成する単一種の融合蛋白質から必須として成る、請求項12記載の組成物。
【請求項14】
組成物中の該融合蛋白質におけるX部分が、異種性である融合蛋白質から必須として成る、請求項12記載の組成物。
【請求項15】
X部分が細胞外受容体であり、該受容体がチロシンキナ−ゼ受容体およびセリンスレオニンキナ−ゼ受容体から成る群から選択される、請求項12記載の組成物。
【請求項16】
X部分が細胞外受容体であり、該受容体がVEGF受容体もしくはその一部である、請求項12記載の組成物。
【請求項17】
X部分がVEGF−R1(FLT−1)のIgG様ドメイン2である、請求項12記載の組成物。
【請求項18】
Yが可撓性を有するポリペプチドである、請求項12記載の組成物。
【請求項19】
Yが、gly9(配列番号27)、glu9(配列番号28)、ser9(配列番号29)、gly5cyspro2cys(配列番号30)、(gly4ser)3(配列番号31)、SerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号32)、ProSerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号13)、Gly−Asp−Leu-Ile−Tyr−ArgAsn−Gln−Lys(配列番号26)、およびGly9ProSerCysValProLeuMetArgCysGlyGlyCysCysAsn(配列番号34)から成る群から選択されるポリペプチドである、請求項12記載の組成物。
【請求項20】
ZがCH3領域であり、該領域がIgG1のCH3領域である、請求項12記載の組成物。
【請求項21】
さらに一以上の医薬上許容される添加物もしくは担体を含む、請求項12記載の組成物。
【請求項22】
液体製剤である、請求項21記載の組成物。
【請求項23】
凍結乾燥製剤である、請求項21記載の組成物。
【請求項24】
式X−Y−Z(式中、Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含み、Yは、5−25個のアミノ酸残基の空間的分離を与えるリンカー部分から必須として成り、Zは、IgG重鎖分子のCH3領域である)で示されるポリペプチド。
【請求項25】
リンカー部分が、10−100個のエチレングリコール単量体残基のオリゴマーである、請求項24記載のポリペプチド。
【請求項26】
式X−Y−Z、(式中、Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含み、Yは、5−25個のアミノ酸残基のポリペプチドから必須として成り、Zは、抗体分子のFcの一部である)で示される融合蛋白質。
【請求項27】
FcがIgGのFcである、請求項26記載の融合蛋白質。
【請求項28】
IgGのFcがIgG1のFcである、請求項27記載の融合蛋白質。
【請求項29】
さらに一以上の医薬上許容される添加物もしくは担体を含む、請求項26記載の融合蛋白質を含む組成物。
【請求項30】
Zが、IgGのFcである抗体分子のFcの一部である、請求項29記載の組成物。
【請求項31】
Zが、IgG1のFcである抗体分子のFcの一部である、請求項29記載の組成物。
【請求項32】
式X−Y−Z(式中、Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含み、Yは、5−25個のアミノ酸残基の空間的分離を与えるリンカ−部分から必須として成り、Zは、抗体分子のFcの一部である)で示される融合蛋白質。
【請求項33】
FcがIgGのFcである、請求項32記載の融合蛋白質。
【請求項34】
IgGのFcがIgG1のFcである、請求項33記載の融合蛋白質。
【請求項35】
リンカー部分が、10−100個のエチレングリコール単量体残基のオリゴマーである、請求項32記載の融合蛋白質。
【請求項36】
さらに一以上の医薬上許容される添加物または担体を含む、請求項32記載の融合蛋白質を含む組成物。
【請求項37】
Zが、IgGのFcである抗体分子のFcの一部である、請求項36記載の組成物。
【請求項38】
Zが、IgG1のFcである抗体分子のFcの一部である、請求項36記載の組成物。
【請求項39】
ポリペプチドXYZ(Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されるポリペプチドを含み、Yは、5−25個のアミノ酸残基のポリペプチドから必須として成り、Zは、IgG重鎖分子のCH3領域である)を形成するために、ポリペプチドXをポリペプチドYを介してポリペプチドZに結合することを含む、ポリペプチドXを多量体化する、方法。
【請求項40】
結合工程が、X、Y、およびZ各々をコードする核酸分子を単一の翻訳領域として構築することを含み、該ポリペプチドXYZが宿主細胞において核酸構築物から発現される、請求項39記載の方法。
【請求項41】
ポリペプチドXYZ(Xは、細胞外受容体、抗体可変領域、サイトカイン、ケモカイン、および成長因子から成る群から選択されたポリペプチドを含み、Yは、5−25個のアミノ酸残基の空間的分離を与えるリンカー部分から必須として成り、Zは、IgG重鎖分子のCH3領域である)を形成するために、ポリペプチドXを、部分Yを介してポリペプチドZに結合することを含む、ポリペプチドXを多量体化する、方法。
【請求項42】
リンカー部分が、10−100個のエチレングリコール単量体残基のオリゴマーから必須として成る、請求項41記載の方法。
【請求項43】
請求項1記載の融合蛋白質をコードする核酸分子。
【請求項44】
請求項26記載の融合蛋白質をコードする核酸分子。
【請求項45】
Xが、VEGF−R1(Flt−1)のIg様ドメイン2である、請求項43もしくは44記載の核酸。
【請求項46】
Xが、VEGF−R1(Flt−1)のIg様ドメイン2であり、融合蛋白質が配列番号8、21、および23から成る群から選択される配列を含む、請求項43記載の核酸。
【請求項47】
Xが、VEGF−R1(Flt−1)のIg様ドメイン2であり、融合蛋白質が配列番号2および25から成る群から選択される配列を含む、請求項44記載の核酸。
【請求項48】
Xが、VEGF−R1(Flt−1)のIg様ドメイン2である、請求項1もしくは26記載の融合蛋白質。
【請求項49】
Xが、VEGF−R1(Flt−1)のIg様ドメイン2であり、融合蛋白質が、配列番号8、21、および23から成る群から選択される配列を含む、請求項1記載の融合蛋白質。
【請求項50】
Xが、VEGF−R1(Flt−1)のIg様ドメイン2であり、融合蛋白質が、配列番号2および25から成る群から選択される配列を含む、請求項26記載の融合蛋白質。
【請求項51】
請求項43もしくは44記載の核酸を含む、哺乳類細胞。
【請求項52】
ヒト細胞である、請求項51記載の哺乳類細胞。
【請求項53】
線維芽細胞、肝細胞、内皮細胞、ケラチン生成細胞、造血系細胞、滑膜細胞、上皮細胞、網膜細胞、および幹細胞から成る群から選択される、請求項52記載の哺乳類細胞。
【請求項54】
請求項43もしくは44記載の核酸を、該融合蛋白質を発現する細胞を作製するために同定された哺乳類細胞に送達することを含む、インビトロの方法。
【請求項55】
融合蛋白質が、配列番号2、8、21、23、および25から成る群から選択される配列を含む、請求項54記載の方法。
【請求項56】
さらに該融合蛋白質を発現する細胞を哺乳類動物に送達することを含む、請求項54記載の方法。
【請求項57】
請求項51記載の哺乳類細胞を哺乳類動物に送達することを含む、方法。
【請求項58】
請求項43もしくは44記載の核酸を哺乳類動物に送達し、そのことにより該融合蛋白質が哺乳類動物で発現されることを含む、方法。
【請求項59】
融合蛋白質が、配列番号2、8、21、23、および25から成る群から選択される配列を含む、請求項58記載の方法。
【請求項60】
哺乳類動物が浮腫加齢黄斑変性症もしくは増殖性糖尿病網膜症にかかっている、請求項57記載の方法。
【請求項61】
哺乳類動物が癌にかかっている、請求項58記載の方法。
【請求項62】
哺乳類動物が慢性関節リウマチにかかっている、請求項58記載の方法。
【請求項63】
哺乳類動物が喘息にかかっている、請求項58記載の方法。
【請求項64】
哺乳類動物が骨関節症にかかっている、請求項58記載の方法。
【請求項65】
請求項1もしくは26記載の融合蛋白質を哺乳類動物に送達することを含む、方法。
【請求項66】
哺乳類動物が、浮腫加齢黄斑変性症もしくは増殖性糖尿病網膜症にかかっている、請求項65記載の方法。
【請求項67】
哺乳類動物が癌にかかっている、請求項65記載の方法。
【請求項68】
哺乳類動物が慢性関節リウマチにかかっている、請求項65記載の方法。
【請求項69】
哺乳類動物が喘息にかかっている、請求項65記載の方法。
【請求項70】
哺乳類動物が骨関節症にかかっている、請求項65記載の方法。
【請求項71】
融合蛋白質が、配列番号2、8、21、23、および25から成る群から選択される配列を含む、請求項65記載の方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【公表番号】特表2008−512127(P2008−512127A)
【公表日】平成20年4月24日(2008.4.24)
【国際特許分類】
【出願番号】特願2007−531408(P2007−531408)
【出願日】平成17年9月13日(2005.9.13)
【国際出願番号】PCT/US2005/032320
【国際公開番号】WO2006/031689
【国際公開日】平成18年3月23日(2006.3.23)
【出願人】(593119583)ジェンザイム・コーポレイション (17)
【氏名又は名称原語表記】Genzyme Corporation
【Fターム(参考)】
【公表日】平成20年4月24日(2008.4.24)
【国際特許分類】
【出願日】平成17年9月13日(2005.9.13)
【国際出願番号】PCT/US2005/032320
【国際公開番号】WO2006/031689
【国際公開日】平成18年3月23日(2006.3.23)
【出願人】(593119583)ジェンザイム・コーポレイション (17)
【氏名又は名称原語表記】Genzyme Corporation
【Fターム(参考)】
[ Back to top ]