
大腸菌シグナル配列を用いて抗体断片を分泌する発現ベクター及び抗体断片の大量生産方法
【課題】 本発明の目的は、大腸菌シグナル配列を用いて抗体断片を抗原−結合親和力を有する水溶性異型接合体形態で過剰発現させ得る組換え発現ベクター及び前記発現ベクターにより形質転換された微生物を提供することである。
【解決手段】 大腸菌耐熱性エンテロトキシンシグナル配列変異体または大腸菌外膜タンパク質A(OmpA)シグナル配列に融合された抗体断片を水溶性異型接合体タンパク質の形態に発現・分泌させ得る組換え発現ベクターは、前記発現ベクターにより形質転換された微生物を培養し、前記形質転換体から培地内に分泌された抗体断片を回収することによって抗体断片の大量生産に有効である。

【解決手段】 大腸菌耐熱性エンテロトキシンシグナル配列変異体または大腸菌外膜タンパク質A(OmpA)シグナル配列に融合された抗体断片を水溶性異型接合体タンパク質の形態に発現・分泌させ得る組換え発現ベクターは、前記発現ベクターにより形質転換された微生物を培養し、前記形質転換体から培地内に分泌された抗体断片を回収することによって抗体断片の大量生産に有効である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、水溶性異型接合体(heterozygote)の形態で大腸菌シグナル配列に融合された抗体断片を分泌的に発現する組換え発現ベクター;前記発現ベクターにより形質転換された微生物;及び前記形質転換された微生物を用いて抗体断片を大量生産する方法に関する。
【背景技術】
【0002】
本発明において、「抗体断片」は、抗体の抗原−結合領域またはその可変領域を含む、抗体の一部を意味するものである。抗体断片の例としては、Fab’、Fab’、F(ab’)2及びscFv断片などがある。抗体をパパイン(papain)で切断すると、Fab’断片と呼ばれる二つの同一の抗原−結合断片と残りのFc断片とが生成する。抗体をペプシン(pepsin)で処理すると、二つの抗原−結合部位を有し、抗原と依然として架橋結合を形成し得るF(ab’)2断片が生成する。Fab断片は、さらに軽鎖の定常ドメイン及び重鎖の第1定常ドメイン(CH1)を含む。Fab’断片は、Fab断片と異なって、抗体ヒンジ(hinge)領域から一つ以上のシステインを含む重鎖CH1ドメインのカルボキシ末端にいくつかの残基が付加されている。
【0003】
抗原結合能力の増加、抗原性の変化、または二重特異的抗体の製造などで抗体の固有性質を調制するために、遺伝子組換え方法を用いた抗体と抗体断片の生産が研究されている。その結果、大腸菌(E.coli)発現系を用いて抗体と抗体断片を生産する方法が開発された。大腸菌の発現系を用いた生産方法は既存の動物細胞を用いた方法に比べて次のようなメリットを提供する:1)発現ベクターの製作が容易であるため、発現可否を容易に検証することができる;2)大腸菌は、成長速度が速いため、低費用で抗体の大量生産が可能であるので、これは研究用試料確保の面で有利である;3)比較的に単純な発酵方法を適用することができ、他の宿主細胞を用いる場合よりも一層容易に商業化できる。
【0004】
抗体−コード遺伝子を大腸菌内に導入して発現させた事例は多くの文献で報告されている(Cabilly et al.,Proc.Natl.Acad.Sci.USA81: 3273-3277,1984,Boss et al.Nucleic Acids Res.12: 3791-3806,1984)。これらの文献は、抗体分子が多様な収率で細胞質内で発現され、また、大腸菌が免疫グロブリンの軽鎖を分泌的に発現させること (Zemel-Dreasen et al., Gene 315-322, 1984)、または抗体断片遺伝子をアルカリ・ホスファターゼ(alkaline phosphatase)あるいはベータラクタマーゼ(β−lactamase)シグナル配列に融合された抗体断片をペリプラズム空間(periplasmic space)に分泌させるための宿主細胞として使用可能であることを開示している(Pluckthun et al., Cold Spring Harbor Symposiua on Quantitative Biology Volume LII、105-112、1987)。国際出願公開第WO92/01059号は、癌細胞の特定部位を認識する抗体であるFab’−コード遺伝子を、大腸菌外膜タンパク質A(OmpA)シグナル配列に融合された融合タンパク質の形態で発現されることを教示しており;米国特許第5,648,237号は、大腸菌のエンテロトキシンシグナル配列に融合されたFab’−コード遺伝子がPhoAプロモーターの調節下で発現されることを開示している。
【0005】
発現された抗体断片は野生型抗体と類似した抗原−結合能を示すが、それは野生型抗体よりも小さいため、局所浸透力が高く(Blumenthal et al., Ad. DrugDel. Rev.4: 279, 1990)、またFc領域を含んでいないため、人体に何等の副作用も引き起こさない。よって、抗体断片は診断試薬の製造や、治療用抗体の開発に利用可能であり、大腸菌からの大量生産は経済的な面で非常に有利である。野生型抗体から抗体断片を製造する場合、野生型抗体はタンパク質分解酵素で切断して精製しなければならないが、これは煩わしくて経済的にも不利である。
【0006】
しかしながら、抗体断片を大腸菌で生産する場合には様々な問題点がある。まず、すべての抗体断片が意図した治療あるいは診断的目的を満たす程度の量で発現されず、場合によっては培養過程を何度も繰り返しなければならない。また、抗体断片に機能的活性を付与するためには、重鎖と軽鎖が単一細胞内で共に発現され、これらの間に形成されたジスルフィド結合によって異型接合体の形態を維持しなければならないが、この際抗体断片は、タンパク質が大腸菌で過剰発現される場合に頻繁に見られる封入体の形態ではなく、高水溶性の形態で発現されることが求められる。このような問題点を解決するため、シグナル配列(signal sequence)を用いて発現された抗体断片をペリプラズム空間に分泌させる方法に関する研究が活発に行われてきた。しかし、シグナル配列に融合されたすべての抗体断片が大腸菌のペリプラズム空間に分泌的に過剰発現されるのではなく、発現させようとする抗体の遺伝子配列によって発現された抗体の量において大きく異なる(Kelly et,al.Biochemistry 31: 5434-5441,1992; Humphreys et,al.Protein Expression and Purification 26: 309-320,2002)。発現ベクター内の構造遺伝子の相違または重鎖遺伝子と軽鎖遺伝子との距離などによって抗体断片の発現パターンが変動することが報告されている(国際特許公開第WO01/94585号)。このような事実から、標的抗体断片の発現のためには、標的抗体遺伝子と発現に必要な宿主の構造遺伝子との相互作用、及びシグナル配列を用いる場合における標的抗体遺伝子とシグナル配列との相互作用における適切なバランスが要求される。
【0007】
ここに、本発明者らは、大腸菌で抗体断片を抗原結合を有する水溶性異型接合体の形態で過剰発現させるために鋭意研究に努めた結果、大腸菌耐熱性エンテロトキシンシグナル配列変異体または大腸菌外膜タンパク質A(OmpA)シグナル配列を用いて、目的抗体断片である腫瘍壊死因子−アルファ(TNF−α)のヒト抗体Fab’を分泌的に発現する組換え発現ベクターを製造することによって、目的とする抗原−結合親和力を有する水溶性異型接合体の形態で抗体断片を大腸菌内で発現する方法を開発することで本発明に至った。
【発明の概要】
【0008】
したがって、本発明の目的は、大腸菌シグナル配列を用いて抗体断片を抗原−結合親和力を有する水溶性異型接合体形態で過剰発現させ得る組換え発現ベクター及び前記発現ベクターにより形質転換された微生物を提供することである。
本発明の他の目的は、前記形質転換された微生物を用いて抗体断片を大量に生産する方法を提供することである。
【0009】
前記目的を達成するために、本発明は、第1大腸菌シグナル配列に融合された抗体断片の軽鎖をコードする遺伝子と、第2大腸菌シグナル配列に融合された抗体断片の重鎖をコードする遺伝子とを含む組換え発現ベクターであって、前記組換え発現ベクターから発現された抗体断片が培養培地内に分泌されることを特徴とする組換え発現ベクターを提供する。
【0010】
また、本発明は、前記組換え発現ベクターを製造する段階;前記発現ベクターで微生物を形質転換させる段階;前記微生物を培地で培養する段階;及び前記微生物から培地内に分泌された抗体断片を回収する段階を含む、抗体断片の生産方法を提供する。
本発明の前記および他の目的および特徴は、下記の図面を伴う本発明の詳細な説明によって明白になる。
【発明の詳細な説明】
【0011】
本発明は、第1大腸菌シグナル配列に融合された抗体断片の軽鎖をコードする遺伝子及び第2大腸菌シグナル配列に融合された抗体断片の重鎖をコードする遺伝子を含み、発現ベクターから発現された抗体断片を培養培地内に分泌することを特徴とする組換え発現ベクター;及び前記発現ベクターにより形質転換された微生物を提供する。
【0012】
野生型抗体と機能的に類似した抗体断片を分泌的に発現できる前記発現ベクターは、抗体断片の軽鎖及び重鎖をコードする遺伝子をそれぞれ大腸菌耐熱性エンテロトキシンシグナル配列変異体または大腸菌外膜タンパク質A(OmpA)シグナル配列に融合させて製造される。
【0013】
本発明で抗体断片は、例えばFab’、Fab’、F(ab’)2またはscFvであり、好ましくはFab’である。Fab断片は軽鎖の定常ドメイン及び重鎖の第1定常ドメイン(CH1)を含む。Fab断片は、抗体のヒンジ(hinge)領域から一つ以上のシステインを含む重鎖CH1ドメインのカルボキシ末端にいくつかの残基が付加されている点がFab断片と異なる。また、F(ab’)2断片は二つのFab’分子がジスルフィド結合によって連結されて製造され、scFv断片は軽鎖と重鎖の可変領域だけがペプチドリンカー(peptide linker)によって連結されて単一ポリペプチド鎖の形態で存在する。
【0014】
本発明に使用可能な抗体断片は、抗原に選択的に結合できるすべての抗体を含み、この場合、抗原はT−細胞、内皮細胞あるいは癌細胞マーカーのような細胞表面に結合している抗原であるか、あるいは、細胞と結合していない水溶性抗原である。細胞表面に結合されている抗原としては、β1インテグリン(integrins)、P−セレクチン(P−selectin)、E−セレクチン、CD2、CD3、CD4、CD5、CD7、CD8、CD11a、CD11b、CD18、CD19、CD20、CD23、CD25、CD33、CD38、CD40、CD45、CD69、MHCクラスI、II、VEGF、及びその他の受容体が含まれるが、これらに限定されない。水溶性抗原としては、IL−1、2、3、4、5、6、8及びIL−12のようなインターロイキン類;サイトメガロウイルス抗原のようなウイルス抗原;IgEのような免疫グロブリン;INF−α、β及びγのようなインターフェロン;及び腫瘍壊死因子−α及びβ、血小板由来成長因子−α及びβのような細胞の成長及び分化に影響を及ぼす因子が含まれるが、これらに限定されない。好ましい実施例で、本発明は、抗−腫瘍壊死因子−アルファ(TNF−α)の抗体断片の軽鎖及び重鎖をコードする遺伝子を用いる。TNF−αは、単球細胞や大食細胞(macrophage)のような免疫関連細胞によって生成されて過剰発現時に炎症を引き起こすサイトカイン類として知られているIL−1、IL−6及びIL−8の生成を誘導し、軟骨組織を破壊するタンパク質分解酵素の分泌を促進して関節炎を起こす主な媒介体として知られている。関節炎治療剤としての抗−腫瘍壊死因子−アルファ抗体のFab’は、単クローン抗体から得られ、好ましくは、ヒト化抗体、より好ましくは、完全なヒト抗体から得られる。
【0015】
抗−腫瘍壊死因子−アルファのいくつかのキメラ抗体あるいはヒト化抗体の相補性決定領域(complementarity determining regions,CDRs)をコードする遺伝子は既に報告されており、完全なヒト抗体の塩基配列も決まっている(国際特許第WO97/2913号)。このような情報を参考に、ヌクレオチド合成器を用いて抗体断片をコードする遺伝子を人工的に合成し、一連のアニーリング(annealing)とリゲーション(ligation)の過程、またはリゲーション連鎖反応(LCR,ligation-chain reaction)とポリメラーゼ連鎖反応(PCR,polymerase chain reaction)を通じて、Fabの重鎖と軽鎖をコードする上で合成した遺伝子をクローニングできる。
【0016】
好ましい実施例で、本発明は配列番号:1〜6に示される塩基配列を有するヒト抗−腫瘍壊死因子−アルファFab’の軽鎖の可変領域をコードする遺伝子断片と、配列番号:7〜12に示される塩基配列を有する重鎖の可変領域をコードする遺伝子断片とをヌクレオチド合成器で合成した後、これらの断片をPCRによって連結させて軽鎖及び重鎖の可変領域を製造する。また、軽鎖の定常領域と、IgG1のCH1及びヒンジ領域を含む重鎖の定常領域とを含む遺伝子断片は、人の血液から抽出したRNA全体を鋳型とし、ヒト抗−腫瘍壊死因子−アルファFab’の軽鎖および重鎖の定常領域に対して特異的なプライマー対を用いたRT−PCRによって製造される。このように製造された軽鎖及び重鎖の定常領域をコードする遺伝子断片のそれぞれを適当な発現ベクターにクローニングして、軽鎖の定常領域をコードする遺伝子を含むプラスミドpBLC及び重鎖の定常領域をコードする遺伝子を含むpBHCを製造する(図1及び2参照)。塩基配列分析の結果、前記プラスミドpBLCに挿入された重鎖の定常領域をコードする遺伝子は、配列番号:32に示す塩基配列を有し、プラスミドpBHCに挿入された重鎖の定常領域をコードする遺伝子は、配列番号:33に示す塩基配列を有することが確認された。
【0017】
発現させようとする抗体断片は、重鎖と軽鎖で構成された二つの異なる種類のタンパク質間の異型接合体として存在する。よって、抗原−結合親和力を有する活性型の異型接合体としての抗体断片を得るためには、重鎖と軽鎖が同等の水準で発現されなければならず、発現ベクターから発現された重鎖と軽鎖との間の正確な位置でジスルフィド結合が形成されなければならない。また、抗体断片が大腸菌内で発現される時、細胞質内に発現されたタンパク質は封入体の形態で存在し得る。この場合、重鎖と軽鎖のジスルフィド結合が正確な位置で生成されないこともあり、水溶性状態で発現されても、精製段階で細胞を完全に破壊できないという問題が起こり得る。このような問題点を解決するために、本発明は、大腸菌シグナル配列を用いて、ベクターから発現された抗体断片を大腸菌のペリプラズム空間あるいは培養培地に分泌させる組換え発現ベクターを提供する。
【0018】
発現されたタンパク質が培養培地内に存在すると、発現されたタンパク質が細菌細胞内に存在する際に要求される細胞を破壊しなければならないという煩わしい過程を行わなくても良い。また、発現されたタンパク質がペリプラズム空間に存在するため、回収した細胞の浸透圧衝撃のような過程を省略できる。
【0019】
さらに、酸化環境は異型接合体を形成するようにするジスルフィド結合を容易にするため、発現されたタンパク質が細胞の細胞質内の非−酸化環境に存在するよりもペリプラズム空間あるいは培養培地内に存在するのが有利である。
【0020】
本発明の組換え発現ベクターに使用可能な大腸菌シグナル配列は、大腸菌耐熱性エンテロトキシンシグナル配列、OmpAシグナル配列、ベータラクタマーゼシグナル配列、GeneIIIシグナル配列、PelBシグナル配列またはこれらの変異体などであり、好ましくは大腸菌耐熱性エンテロトキシンシグナル配列変異体またはOmpAシグナル配列であるが、これらに限定されない。
【0021】
本発明の好ましい実施例で、発現ベクターから発現された活性型の抗体断片Fab’を培養培地内に分泌させるために用いられた大腸菌耐熱性エンテロトキシンシグナル配列変異体は、大腸菌で様々な異種タンパク質を分泌・発現させるために本発明者らが開発したものである。前記大腸菌耐熱性エンテロトキシンシグナル配列変異体は、配列番号:17に示される塩基配列を有し、最適効率で大腸菌内で様々な異種タンパク質を発現・分泌できることが報告されている(韓国特許第316347号)。本発明で、発現ベクターから発現された活性型の抗体断片Fab’を培養培地内に分泌させ得る組換え発現ベクターは、抗体断片の重鎖及び軽鎖をコードする遺伝子をそれぞれ大腸菌シグナル配列、例えば、大腸菌耐熱性エンテロトキシンシグナル配列変異体または外膜タンパク質Aシグナル配列に融合させることによって製造される。
【0022】
抗体断片の重鎖及び軽鎖をコードする遺伝子を一つの発現ベクター内で発現させるために、本発明は、一つのプロモーターの下に二つのタンパク質を適切な間隔をおいて配置して同時に発現させるダイ−シストロニック体系(di−cistronic system)を用いる発現ベクターと、独立された各プロモーターに遺伝子を連結して二つのタンパク質を独立的に発現できる二重−プロモーター体系(dual−promoter system)を用いる発現ベクターとの二つの種類の発現ベクターを提供する。
【0023】
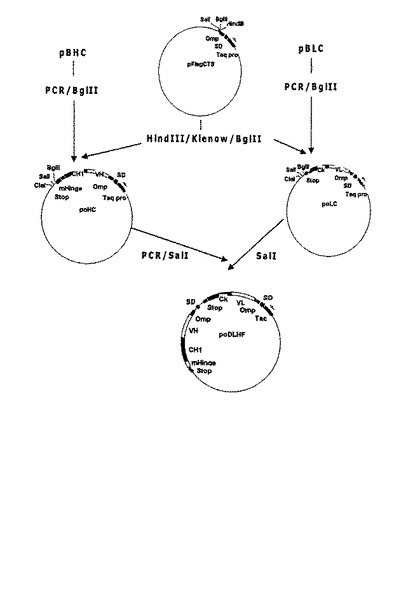
まず、ヒト成長ホルモン(hGH)をコードする遺伝子断片を、大腸菌耐熱性エンテロトキシンシグナル配列及びhGHを含む発現ベクターpT14S1SH−4T20V22Q(韓国特許第316347号)から除去する。プラスミドpBLCから得た軽鎖をコードする遺伝子断片と、プラスミドpBHCから得た重鎖をコードする遺伝子断片とを、前記発現ベクターpT14S1SH−4T20V22Qに挿入し、耐熱性エンテロトキシンシグナル配列変異体に融合された重鎖をコードする遺伝子断片を含む発現ベクターpmsHC、及び耐熱性エンテロトキシンシグナル配列変異体に融合された軽鎖をコードする遺伝子断片を含む発現ベクターpmsLCを製作する。重鎖と軽鎖をコードする遺伝子のそれぞれが大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合され、単一プロモーターが重鎖及び軽鎖をコードする遺伝子の発現を調節するダイ−シストロニック発現ベクターを製作するために、プラスミドpmsHCからPCRによって得た重鎖をコードする遺伝子断片をプラスミドpmsLCに挿入して発現ベクターpsDLHF_B及びpsDLHF_Bpを製造する(図3及び4参照)。
【0024】
また、本発明は前記のような方法によって、軽鎖及び重鎖をコードする遺伝子のそれぞれが大腸菌OmpAシグナル配列(配列番号:24)に融合され、単一Tacプロモーターが軽鎖及び重鎖をコードする遺伝子の発現を調節する発現ベクターpoDLHFを製作する(図5参照)。
【0025】
前記で製作された発現ベクターpsDLHF_B、psDLHF_Bp及びpoDLHFはシグナル配列に融合された抗−腫瘍壊死因子−アルファの重鎖及び軽鎖のそれぞれが単一プロモーターの調節下で発現・分泌されるダイ−シストロニック発現ベクターである。前記発現ベクターのそれぞれを大腸菌BL21(DE3)に形質転換させて製造した大腸菌形質転換体BL21(DE3)/psDLHF_B(HM10920)、BL21(DE3)/psDLHF_BP(HM10921)及びBL21/poDLHF(HM10922)はそれぞれ、特許手続きの目的のための微生物寄託の国際的認識に関するブダペスト条約に従って、寄託番号KCCM−10509、KCCM−10510及びKCCM−10511として韓国微生物保存センター(KCCM)に2003年10月2日付で寄託された。
【0026】
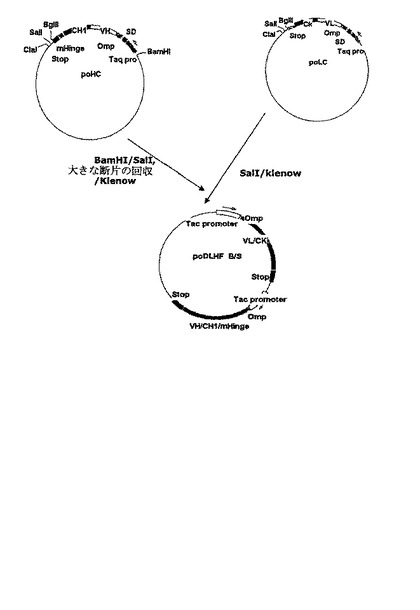
上述したように、単一プロモーターの調節下で異なる二つのタンパク質を分泌的に発現させる場合、プロモーターに近いタンパク質の発現効率はプロモーターに遠いタンパク質の発現効率と異なり得る(Humphreys et, al. Protein Expression and Purification 26: 309-320, 2002)。本発明は、抗−腫瘍壊死因子−アルファFab’の重鎖と軽鎖の発現及び分泌が独立したプロモーターによって個別的に調節可能な二重−プロモーターシステムを用いる発現ベクターを提供する。好ましい実施例で、本発明は、抗−腫瘍壊死因子−アルファFab’の鎖及び軽鎖をコードする遺伝子をそれぞれOmpAシグナル配列に融合して、Tacプロモーターが軽鎖及び重鎖をコードする遺伝子の発現及び分泌を調節する発現ベクターpoDLHF_B/Sを製作する(図6参照)。前記発現ベクターpoDLHF_B/Sにより大腸菌BL21を形質転換させて製造した大腸菌形質転換体BL21/poDLHF_B/S(HM10923)は、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約に基づき、寄託番号KCCM−10512として韓国微生物保存センター(KCCM)に2003年10月2日付に寄託された。
【0027】
抗体断片のような異型接合体の形成は前記異型接合体の二つの断片の一方が過剰発現された時に抑制されるので、二つのタンパク質の断片は互いに同様の水準で発現されなければならない。ここに、本発明では、重鎖と軽鎖をコードする遺伝子をそれぞれ互いに異なるシグナル配列に融合させて抗体断片を分泌的に発現させ得る組換え発現ベクターを提供する。
【0028】
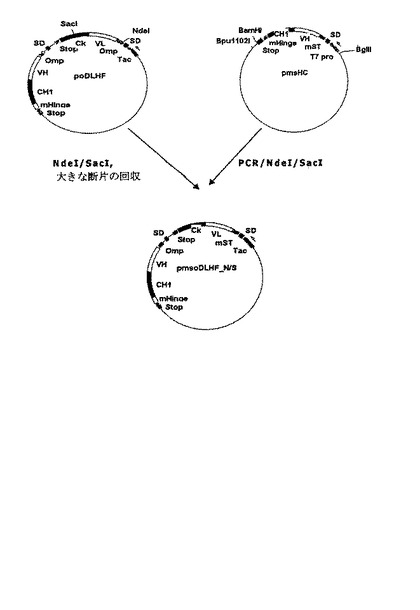
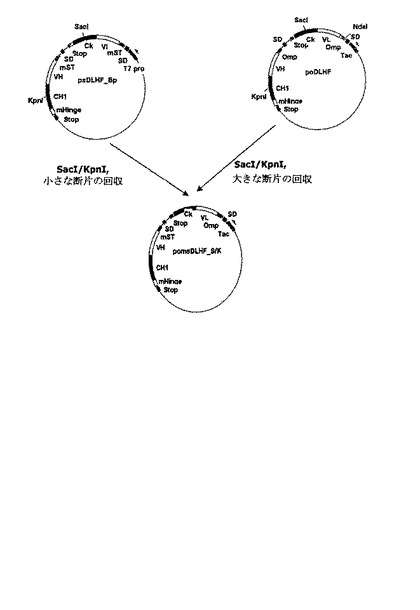
好ましい実施例で、本発明は、抗−腫瘍壊死因子−アルファFab’の軽鎖をコードする遺伝子は耐熱性エンテロトキシンシグナル配列変異体に融合され、その重鎖をコードする遺伝子はOmpAシグナル配列に融合され、単一T7プロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節する発現ベクターpmsoDLHF_N/Sを製作する(図7参照)。また、本発明で、発現ベクターpmsoDLHF_S/Kは、抗−腫瘍壊死因子−アルファFab’の重鎖をコードする遺伝子は耐熱性エンテロトキシンシグナル配列変異体に融合され、その軽鎖をコードする遺伝子はOmpAシグナル配列に融合され、単一Tacプロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節するように製作される(図8参照)。前記発現ベクターpmsoDLHF_N/S及びpmsoDLHF_S/Kをそれぞれ大腸菌BL21に形質転換させて製造した大腸菌形質転換体BL21/pmsoDLHF_N/S(HM10924)及びBL21/pmsoDLHF_S/K(HM10925)のそれぞれは、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約に基づき、寄託番号KCCM−10513及びKCCM−10516として韓国微生物保存センター(KCCM)に2003年10月2日付で寄託された。
【0029】
本発明は、前記のように製作された組換え発現ベクターを適当な宿主細胞に導入して製造された組換え微生物を提供する。本発明で使用可能な宿主細胞としては、大腸菌W3110、RV308、BL21、BL21(DE3)などを含み、好ましくは大腸菌BL21であるが、これらに限定されない。前記発現ベクターにより形質転換された組換え微生物を発酵槽を用いて高濃度で培養(OD600=120〜150)する場合、抗−腫瘍壊死因子−アルファの抗体断片は培養培地1L当たり100〜500mg以上の高収率で培養培地内に分泌的に生産されることが確認された(表1参照)。このように大量生産された抗−腫瘍壊死因子−アルファの抗体断片はタンパク質−Gアフィニティーカラムによって部分的に精製できる。精製された抗−腫瘍壊死因子−アルファの抗体断片を還元条件下及び非−還元条件下で行ったSDS−PAGEで分析した結果、抗体断片が水溶性異型接合体形態で形成されたことを確認した(図9参照)。また、結合抗原として抗−腫瘍壊死因子−アルファを用いたELISAは、抗−腫瘍壊死因子−アルファの抗体断片が腫瘍壊死因子−アルファ抗原に対して優れた結合親和力を有することを示した(図10参照)。
【0030】
本発明は、第1大腸菌シグナル配列に融合された抗体断片の軽鎖をコードする遺伝子及び第2大腸菌シグナル配列に融合された抗体断片の重鎖をコードする遺伝子を含む発現ベクターを製造する段階;前記発現ベクターで微生物を形質転換させる段階;前記微生物を培地で培養する段階;及び 前記微生物から培地内に分泌された抗体断片を回収する段階を含む、抗体断片の生産方法を提供する。
【0031】
本発明の組換え発現ベクターで使用可能な大腸菌シグナル配列としては、大腸菌耐熱性エンテロトキシンシグナル配列、OmpAシグナル配列、ベータラクタマーゼシグナル配列、GeneIIIシグナル配列、PelBシグナル配列またはこれらの変異体を含み、好ましくは配列番号:17に示される塩基配列を有する大腸菌耐熱性エンテロトキシンシグナル配列変異体または配列番号:23に示される塩基配列を有する大腸菌外膜タンパク質Aシグナル配列を用いるが、これらに限定されない。
【0032】
前記組換え発現ベクターにより形質転換された微生物を適当な培地で予備培養した後、この予備培養液を発酵槽に接種して30〜35℃範囲の温度で空気と接触させながら本−培養を実施する。600nmでOD値が80に達すると、誘導物質(inducer)を発酵槽に添加して標的抗体断片の発現を誘導する。本発明で使用可能な誘導物質としては、IPTGが好ましいが、これに限定されない。600nmでOD値が120〜140に逹するまで、40〜45時間更に培養する。前記培養液を遠心分離して上澄液を分離した後、これをアフィニティーカラムクロマトグラフィーで抗体断片を精製する。前記形質転換された微生物は、大腸菌BL21(DE3)/psDLHF_B(HM10920)(KCCM-10509)、BL21(DE3)/psDLHF_BP(HM10921)(KCCM-10510)、BL21/poDLHF(HM10922)(KCCM-10511)、BL21/poDLHF_B/S(HM10923)(KCCM-10512)、BL21/pmsoDLHF_N/S(HM10924)(KCCM-10513)、好ましくはBL21/pmsoDLHF_S/K(HM10925)(KCCM-10516)であるが、これらに限定されない。
【0033】
本発明によって発現された抗体断片は宿主細胞のペリプラズム空間または培養培地内に分泌されるため、抗体断片は浸透圧ショック処理のような工程を経なとも回収できる。
【0034】
本発明の方法によって発現され得る目標抗体断片は、好ましくはFab’、Fab’F(ab’)2またはscFvなどであるが、抗−腫瘍壊死因子−アルファの抗体断片に限定されるものではなく、よって、診断または治療を目的とする任意のすべての抗体、例えば、抗癌効果がある抗体断片または過剰−発現された場合、人体に致命的なダメージを与える様々なサイトカインに対する抗体断片の生産に適用できる。
【0035】
前述したように、大腸菌耐熱性エンテロトキシンシグナル配列変異体または大腸菌外膜タンパク質Aシグナル配列を含む本発明の組換え発現ベクターは、抗体断片を抗原−結合親和力を有する水溶性異型接合体の形態で発現させ得る。前記発現ベクターにより形質転換された微生物は抗体断片を宿主細胞のペリプラズム空間または培養培地に分泌させることが可能であるため、発現された抗体断片は大量生産に適する方法で容易に回収できる。従って、本発明による方法は診断試薬または治療用抗体の製造に有効に利用可能である。
【0036】
下記実施例は本発明をさらに詳細に説明するためのものであり、本発明の範囲を制限しない。
【0037】
実施例1:抗−腫瘍壊死因子−アルファFab’をコードする遺伝子のクローニング
国際特許第WO97/29131号に記載された塩基配列を参考にして、ヒト抗−腫瘍壊死因子−アルファFab’の重鎖と軽鎖の可変領域をヌクレオチド合成器を用いて合成した。軽鎖の可変領域をクローニングするための遺伝子断片は配列番号:1〜6の塩基配列を有し、重鎖の可変領域をクローニングするための遺伝子断片は配列番号:7〜12の塩基配列を有した。この後のクローニング過程を容易にするために、前記重鎖及び軽鎖をコードする遺伝子のそれぞれの一番目である遺伝子断片(即ち、配列番号:1及び7)にそれぞれHindIII及びPuvI制限酵素認識部位を挿入した。前記合成した重鎖及び軽鎖の遺伝子断片のそれぞれを一つのチューブに混ぜた後、これらを一つの遺伝子配列で連結させるために通常的なPCRを行って、軽鎖及び重鎖の可変領域をコードする遺伝子を製造した。前記PCR反応で、遺伝子断片のそれぞれは鋳型のみならずプライマーとして作用可能であり、従って、これらは互いにアニーリングされ、約320mer程度の断片を形成した。前記PCR反応混合物は、それぞれの遺伝子断片100pmole、5ulの2mMdNTP混合液、2.5単位のpfuDNAポリメラーゼ(Stratagen社)及び5ulのpfu反応緩衝溶液(100mM Tris-HCl(pH8.3), 15mM MgCl2, 500mM KCl)を含み、蒸留水を加えて最終体積が50ulになるようにした。前記PCRはDNAサーマルサイクラー(DNA thermocycler, Perkin-Elmer社)を用いて94℃で5分間初期変性させた後、94℃で60秒、60℃で60秒及び72℃で60秒の反応を30回行った。
【0038】
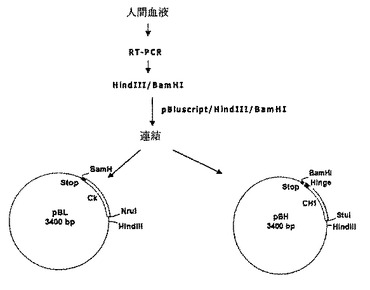
ヒト項−腫瘍壊死因子−アルファFab’の軽鎖の定常領域とIgG1のCH1とヒンジ領域とを含んだ重鎖の定常領域をクローニングするため,ヒト血液の血球細胞からRNAを抽出した後、次のようにRT−PCRを行った。すべてのRNAは、QiampRNA血液キット(Qiagen社)を用いて約6mlの血液から抽出されて、RT−PCRの鋳型として用いられた。前記重鎖及び軽鎖をコードする遺伝子のそれぞれは、重鎖増幅のための配列番号:13及び14に示されるプライマー対と、軽鎖増幅のための配列番号:15及び16に示されるプライマー対を用いたOne−StepRT−PCRキット(Qiagen社)を用いて増幅された。この際、この後のクローニング過程を容易にするために、配列番号:26の重鎖5’−プライマーにはHindIII及びStuI制限酵素認識部位を挿入し、配列番号:27の重鎖3’−プライマーには終止コドンを含むBamHI制限酵素認識部位を挿入し;配列番号:28の軽鎖5’−プライマーにはHindIII及びNruI制限酵素認識部位を挿入し、配列番号:29の軽鎖3’−プライマーには終止コドンを含むBamHI制限酵素認識部位を挿入した。これから増幅された重鎖及び軽鎖の定常領域をコードするPCR産物をそれぞれHindIIIとBamHIで切断した後、同一制限酵素で処理されたプラスミドpBluscriptSK(-)(Stratagen社)に挿入してプラスミドpBH及びpBLを製造した(図1)。DNA塩基配列の分析によって、プラスミドpBH及びpBLにそれぞれの重鎖及び軽鎖の定常領域が正確に挿入されたことが確認された。
【0039】
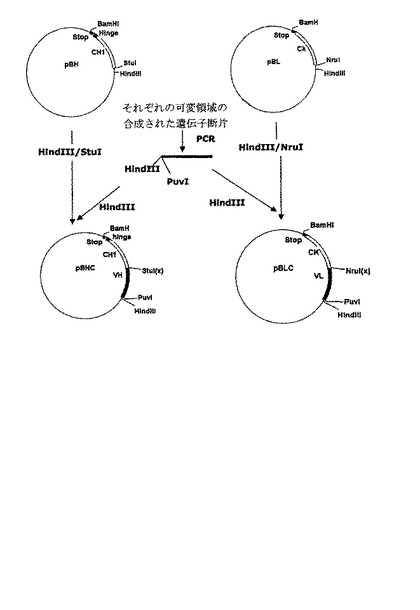
前記重鎖及び軽鎖の可変領域をこれらに相応する定常領域に連結するため、前記でPCRによって合成された重鎖及び軽鎖の可変領域それぞれをHindIII制限酵素で切断した。酵素処理された重鎖の可変領域は制限酵素HindIII/StuIで予め処理されたプラスミドpBHに挿入され、酵素処理された軽鎖の可変領域は、制限酵素HindIII/NruIで予め処理されたプラスミドpBLに挿入されて、重鎖及び軽鎖の可変領域のそれぞれがこれらに相応する定常領域に連結されたプラスミドpBHC及びpBLCを製作した(図2)。前記プラスミドpBHC及びpBLCにそれぞれ挿入された重鎖と軽鎖はそれぞれ配列番号:32及び33に示された塩基配列を有する。
【0040】
実施例2:抗−腫瘍壊死因子−アルファFab’遺伝子発現ベクターの製作
<2−1> 大腸菌耐熱性エンテロトキシンシグナル配列変異体及び抗−腫瘍壊死因子−アルファFab’遺伝子を含む発現ベクターの製作
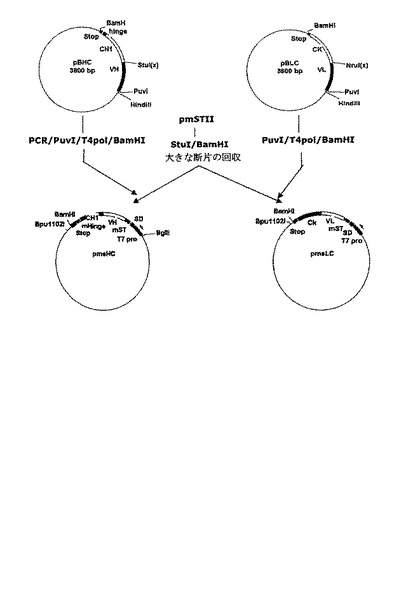
大腸菌耐熱性エンテロトキシンシグナル配列変異体を用いた発現ベクターを製作するため、ヒト成長ホルモン(hGH)を発現する発現ベクターpT14S1SH-4T20V22Q(韓国特許第316347号)を用いた。前記発現ベクターは配列番号:17に示される塩基配列を有する耐熱性エンテロトキシンシグナル配列変異体を含む。この発現ベクターからhGHを除去し前記実施例1で製造された重鎖及び軽鎖をコードする遺伝子断片を挿入するために、配列番号:18及び19のプライマー対を用いた部位特異的突然変異誘発法(site-directed mutagenesis)によってプラスミドpT14S1SH-4T20V22Qの耐熱性エンテロトキシンシグナル配列変異体とhGH遺伝子の融合部位にStuI制限酵素認識部位を挿入した。塩基配列分析法によってStuI制限酵素認識部位が成功的に標的位置に生成されたことが確認した。前記プラスミドpT14S1SH-4T20V22Q内のStuI制限酵素認識部位を有するプラスミドをpmSTIIと命名した。
【0041】
前記プラスミドpmSTIIをStuI/BamHI制限酵素で処理した後、アガロースゲル電気泳動を行って、hGH部位が除去され大腸菌耐熱性エンテロトキシンシグナル配列変異体を含む大きい断片(4.7kb)をゲルから回収した。前記実施例1で製造された軽鎖をコードする遺伝子断片を有するプラスミドpBLCをPuvI制限酵素で処理した後、これらの両末端をT4DNAポリメラーゼを用いて平滑末端にした。これから得たプラスミドをBamHI制限酵素で処理して約720bpの軽鎖をコードする遺伝子断片を回収した。また、重鎖をコードする遺伝子断片を含むプラスミドpBHCを鋳型として配列番号:1及び20のプライマー対を用いてPCRを行った。これから増幅されたPCR産物を、前記軽鎖をコードする遺伝子断片の製造に用いられた同一酵素で処理して約700bpの重鎖をコードする遺伝子断片を回収した。配列番号:20のプライマー対は重鎖ヒンジ領域の一部を含む変形されたヒンジ配列であるEPKSCDKTHTCAA−終止コドンが重鎖をコードする遺伝子断片に挿入されてBamHI制限酵素認識部位を含むように考案されているため、この後の過程でFab’発現ベクターの製作に有用に利用できる。このように用意された重鎖及び軽鎖をコードする遺伝子断片のそれぞれをプラスミドpmSTII断片に挿入し、耐熱性エンテロトキシンシグナル配列変異体に融合された軽鎖をコードする遺伝子断片を含む発現ベクターpmsLCのみならず、耐熱性エンテロトキシンシグナル配列変異体に融合された重鎖をコードする遺伝子断片を含む発現ベクターpmsHCを製作した(図3)。
【0042】
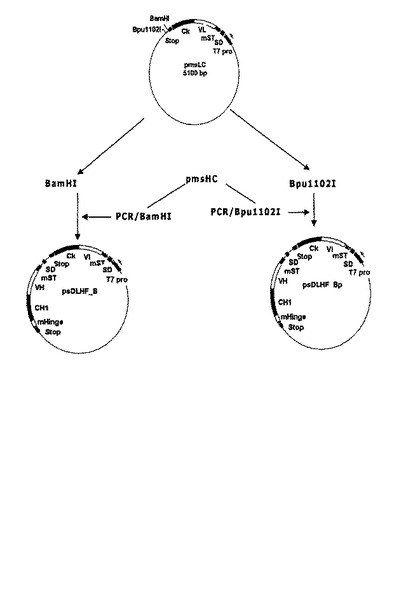
重鎖と軽鎖が一つのプロモーターによって調節されて発現・分泌されるダイ−シストロニック(di−cistronic)発現ベクターを製作するために、耐熱性エンテロトキシンシグナル配列変異体に融合された重鎖をコードする遺伝子断片を含むプラスミドpmsHCを鋳型とし、配列番号:21及び20のプライマー対と配列番号:22及び23のプライマー対を用いてPCRを行った。配列番号:21の5’−プライマーは重鎖をコードする遺伝子断片の発現に必要であるシャインダルガノ(Shine-Dalgarno,SD)配列からPCR反応が開始するように考案されており、増幅された産物を容易にクローニングするためにBamHI制限酵素認識部位を含んでいる。また、配列番号:22及び23のプライマー対は、クローニング部位を異ならせるために、BamHIではなくBpuI制限酵素認識部位を含むことを除けば、配列番号:21及び20のプライマー対と同一の塩基配列を有している。各プライマー対を用いて増幅された重鎖をコードする遺伝子断片(一方はBamHI制限酵素認識部位を含み、他方はBpuI制限酵素認識部位を含む)をBamHIとBpuI制限酵素でそれぞれ処理した。
【0043】
耐熱性エンテロトキシンシグナル配列変異体に融合された軽鎖をコードする遺伝子断片を含むプラスミドpmsLCでBamHIとBpuI制限酵素認識部位はすべて軽鎖遺伝子の終止コドンの下流に位置し、BpuIの制限酵素認識部位はBamHIより約60ヌクレオチド以上下流に存在するようにした。前記プラスミドpmsLCをBamHIとBpuI制限酵素で処理した後、その部位にBamHIまたはBpuI制限酵素で処理された重鎖をコードする遺伝子断片を挿入した。プラスミドpmsLCのBamHI制限酵素部位内に挿入された重鎖遺伝子を含むFab’発現ベクターをpsDLHF_Bpと命名した(図4)。これらの発現ベクターの場合、重鎖及び軽鎖をコードする遺伝子はそれぞれ、大腸菌エンテロトキシンシグナル配列に融合され、単一T7プロモーターがこれらの重鎖及び軽鎖をコードする遺伝子の発現を調節する。前記発現ベクターそれぞれを大腸菌BL21(DE3)に形質転換させて大腸菌形質転換体BL21(DE3)/ psDLHF_B (HM10920) 及び BL21(DE3)/psDLHF_BP(HM10921)を製造し、これらを特許手続上の微生物の寄託の国際的承認に関するブダペスト条約に基づき、寄託番号KCCM−10509及びKCCM−10510として韓国微細物保存センター(KCCM)に2003年10月2日付で寄託した。
【0044】
<2−2> 大腸菌外膜タンパク質A(OmpA)シグナル配列及び抗−腫瘍壊死因子−アルファFab’を含む発現ベクター製作
大腸菌OmpAシグナル配列(配列番号:24)を用いて抗−腫瘍壊死因子−アルファの重鎖及び軽鎖をコードする遺伝子を分泌的に発現する発現ベクターを次のように製作した。
【0045】
まず、TacプロモーターとOmpAシグナル配列とを含むプラスミドpFlag.CTS(Eastman Chemical Company)をHindIIIで処理し、前記プラスミドのHindIII制限酵素認識部位をクレノウ DNA ポリメラーゼ(Klenow DNA polymerase, NEB, New Egland Biolab USA)で処理して平滑末端にした後、BglII制限酵素で切断して5’−末端は平滑末端を、3’−末端は付着端を有するベクター断片を製作した。また、OmpAシグナル配列と、重鎖及び軽鎖をコードする遺伝子断片とを連結するため、重鎖をコードする遺伝子断片を含むプラスミドpBHCを鋳型とし、配列番号:25及び26のプライマー対を用いたPCRを行って重鎖の可変領域と定常領域、そして変形されたヒンジ配列と終止コドンを含むPCR産物を増幅した。この際、3’−末端に相応する配列番号:26のプライマー対はBglII制限酵素認識部位を挿入するように考案された。このように増幅された重鎖をコードする遺伝子断片を制限酵素BglIIで切断した後、この断片を前記でBglIIによって予め切断されたプラスミドpFlag.CTSに挿入してOmpAシグナル配列に融合された重鎖をコードする遺伝子断片を含むプラスミドpoHCを製作した。前記記載の同一方法に従って、軽鎖をコードする遺伝子断片を含むプラスミドpBLCを鋳型とし、配列番号:27及び28のプライマー対を用いたPCRを行った後、このように増幅されたPCR産物を同一制限酵素処理過程を経ってプラスミドpFlag.CTSに挿入してOmpAシグナル配列に融合された軽鎖をコードする遺伝子断片を含むプラスミドpoLCを製作した。
【0046】
それぞれOmpAシグナル配列に融合された重鎖及び軽鎖をコードする遺伝子が単一Tacプロモーターによって調節されて、発現分泌されるようにダイ−シストロニック発現ベクターを製造するため、鋳型としてプラスミドpoHCを用い、配列番号:29及び30のプライマー対を用いてPCRを行った。これから増幅されたPCR産物は、両末端にSalI制限酵素認識部位を有し、プロモーター領域を除いた重鎖の発現に必要であるシャインダルガノ配列とOmpAシグナル配列とを含む。このPCR産物を制限酵素SalIで切断してから回収した遺伝子断片を同一制限酵素で予め処理されたプラスミドpoLCに挿入してFab’遺伝子を発現する発現ベクターpoDLHFを製作した(図5)。前記発現ベクターpoDLHFで大腸菌BL21を形質転換させて大腸菌形質転換体BL21/poDLHF(HM10922)を製造し、これを韓国微細物保存センター(KCCM)に寄託番号KCCM−10511として2003年10月2日付で寄託した。
【0047】
<2−3> 二重プロモーター(dual−promoter)によって調節される発現ベクターの製作
抗−腫瘍壊死因子−アルファFab’の重鎖と軽鎖をそれぞれ独立したプロモーターによって個別的に発現・分泌させ得る二重−プロモーターシステムを有する発現ベクターを次のように製作した。OmpAシグナル配列に融合された抗−腫瘍壊死因子−アルファFab’の 重鎖をコードする遺伝子断片を含む実施例<2−2>のプラスミドpoHCをBamHIとSalIで処理した。前記酵素−処理されたプラスミドのアガロースゲル電気泳動を行い、抗−腫瘍壊死因子−アルファFab’の重鎖をコードする遺伝子断片、Tacプロモーター及びOmpAシグナル配列を含む、より小さな断片(1.2kb)をゲルから回収した。前記断片の両末端をクレノウ DNA ポリメラーゼで処理して平滑末端にした。また、抗−腫瘍壊死因子−アルファFab’の軽鎖をコードする遺伝子断片を含む実施例<2−2>のプラスミドpoLCをSalIで処理した後、両末端を前記と同一の方法によって平滑末端に作った。それから、重鎖をコードする遺伝子断片をここに挿入してOmpAシグナル配列に融合された軽鎖及び重鎖をコードする遺伝子のそれぞれが独立的にTacプロモーターの調節によって発現分泌される発現ベクターpoDLHF_B/Sを製作した(図6)。前記発現ベクターpoDLHF_B/Sで大腸菌BL21を形質転換させて大腸菌形質転換体BL21/poDLHF_B/S(HM10923)を製造し、これを韓国微生物保存センター(KCCM)に寄託番号KCCM−10512として2003年10月2日付で寄託した。
【0048】
<2−4> 各鎖の発現及び分泌が互いに異なるシグナル配列によって調節される発現ベクターの製作
<2−4−1>
抗−腫瘍壊死因子−アルファFab’α軽鎖をコードする遺伝子は耐熱性エンテロトキシンシグナル配列変異体に融合され、重鎖をコードする遺伝子はOmpAシグナル配列に融合されて、これらの発現及び分泌が単一プロモーターによって調節される発現ベクターを次のように製作した。
【0049】
単一Tacプロモーターの調節によってOmpAシグナル配列に融合された重鎖及び軽鎖をコードする遺伝子がそれぞれ発現される実施例<2−2>の発現ベクターpoDLHFをNdeIとSacIで処理した後、このベクターのアガロースゲル電気泳動を行って、比較的に大きな遺伝子断片(6.6kb)をゲルから回収した。NdeIの認識部位はOmpAシグナル配列の一番目のアミノ酸であるメチオニンに位置し、SacIの認識部位は軽鎖の定常領域に位置した。従って、前記発現ベクターをNdeI及びSacIで処理すると、OmpAシグナル配列は含まないが、軽鎖の可変領域の全部と軽鎖の定常領域の一部を含む遺伝子断片が回収できる。
【0050】
重鎖をコードする遺伝子断片が耐熱性エンテロトキシンシグナル配列変異体に融合されたプラスミドであるpmsLCを鋳型とし、配列番号:31及び20のプライマー対を用いてPCRを行った。配列番号:31の5’−プライマーはNdeI認識部位を含み、耐熱性エンテロトキシンシグナル配列の一番目のアミノ酸に結合して、配列番号:20の3’−プライマーはSacI認識部位を含む。これから増幅されたPCR産物をNdeIとSacIで処理した後、欠失した軽鎖領域を有するプラスミドpoDLHFに挿入して発現ベクターpmsoDLHF_N/Sを製作した(図7)。前記発現ベクター pmsoDLHF_N/Sを大腸菌BL21に形質組転換させて大腸菌形質転換体BL21/pmsoDLHF_N/S(HM10924)を製造し、これを韓国微細物保存センターに寄託番号KCCM−10513として2003年10月2日付で寄託した。
【0051】
<2−4−2>
前記実施例<2−4−1>で製造された発現ベクターとは異なって、抗−腫瘍壊死因子−アルファFab’の重鎖をコードする遺伝子は耐熱性エンテロトキシンシグナル配列変異体に融合され、軽鎖をコードする遺伝子はOmpAシグナル配列に融合されて、単一プロモーターによってこれらの発現及び分泌が調節される発現ベクターを次のように製作した。
【0052】
実施例<2−1>の発現ベクターpsDLHF_BpをSacIとKpnIで切断した後、アガロースゲル電気泳動を行って小さな遺伝子断片(0.6kb)をゲルから回収した。SacIの認識部位は軽鎖の定常領域に存在し、KpnIの認識部位は重鎖の定常領域に存在するため、回収された断片は耐熱性エンテロトキシンシグナル配列変異体に融合された軽鎖遺伝子断片を含んでいた。重鎖及び軽鎖をコードする遺伝子のそれぞれがOmpAシグナル配列に融合され、単一Tacプロモーターによって発現されるように製作された発現ベクターpoDLHFを同一制限酵素で処理した後、アガロースゲル電気泳動を行った。次いで、サイズが大きい遺伝子断片(7.4kb)をゲルから回収し、これを上で回収した軽鎖をコードする遺伝子断片に連結して発現ベクターpmsoDLHF_S/Kを製作した(図8)。前記発現ベクターpmsoDLHF_S/Kを大腸菌BL21に形質組転換させて大腸菌形質転換体BL21/pmsoDLHF_S/K(HM10925)を製造し、これを韓国微生物保存センターに寄託番号KCCM−10516として2003年10月2日付で寄託した。
【0053】
実施例3:抗−腫瘍壊死因子−アルファFab’遺伝子の発現
前記実施例2で製造された大腸菌形質転換体HM10920〜HM10925をそれぞれ5L体積の発酵槽(Marubishi)に接種して発酵させた後、大腸菌形質転換による抗−腫瘍壊死因子−アルファFab’の発現有無を次のように調査した。
【0054】
LB培地100mlに前記形質転換体それぞれを一晩中振盪予備培養してから、発酵槽に接種して培養を進めた。発酵槽の温度は35℃あるいは30℃に維持し、嫌気状態で進むのを防止するために空気を20vvmで投入し、培地を500rpmで撹拌した。微生物の成長のために補助エネルギー源としてグルコース(glucose)と酵母エキス(yeast extract)を微生物の発酵状態によって供給した。600nmでOD値が80に達した時、誘導物質(inducer)であるIPTGを発酵槽に添加した。600nmでOD値が120〜140に達するまで、前記発酵を40〜45時間行った。
【0055】
前記醗酵培養液を遠心分離(20,000g、30分)して上澄液のみを取って、それぞれの形質転換体から発現された抗−腫瘍壊死因子−アルファFab’の量を測定するために前記上澄液をELISAで分析した。ヒト抗体CH1に対する単クローン抗体(MAB1304,Chemicon international)を10mMカーボネイト緩衝溶液(pH 9.6)に1ug/mlの濃度で溶解させた後、96−ウェルプレートをウェル当たり200ngの量で前記抗体溶液で4℃で一晩中コーティングした後、 PBS−T溶液(137mM NaCl, 2mM KCl, 10 mM Na2HPO4, 2 mM KH2PO4 0.05% トゥイーン20)で3回洗浄した。ウシ血清アルブミンを1%の濃度でPBS−T溶液に溶解させたブロッキング溶液を用意した。前記ブロッキング溶液250ulを各ウェルに点滴した後、前記ウェルプレートを常温で1時間放置しておいて、同一のPBS−T溶液で3回洗浄した。標準液と試料液を、適当な濃度にPBS−T溶液で希釈した後、抗体がコーティングされたウェルに添加した。前記ウェルプレートを常温で1時間放置させて反応させてから、再度PBS−T溶液で3回洗浄した。標準液は、ヒト免疫グロブリンG(IB−グロブリンA、緑十字 PBM)をタンパク質分解酵素であるパパインで切断し、一連のカラムを用いてFabを精製し、前記FabをPBS−T溶液に1ng/mlの濃度で溶解させ、二倍ずつ希釈して製造した。ヒト軽鎖の定常領域に対する多クローン抗体にパラクサイドが共有結合された、コンジュゲート試料(A7164、Sigma社)は、ブロッキング溶液で1000:1に希釈して用いた。希釈された200ulの試料を各ウェルに点滴した後、前記ウェルプレートを1時間常温で反応させた。反応が完了した後、各ウェルをPBS−T溶液で3回洗浄した。発色溶液AとB(カラー−A−安定化パラクサイド溶液及びカラーB−安定化クロモゲン溶液、DY 999、R&D Systems社)を同量で混合した後、各ウェルに前記発色溶液混合物を200ulずつ添加して前記ウェルプレートを30分間放置した。次いで、2M硫酸を各ウェルに50ulずつ添加して反応を停止させた。450nmの標準液と試料液の吸光度を測定するために、前記ウェルプレートをマイクロプレートリーダー(Molecular Device社)で分析した。その結果を下記表1に示した。
【0056】
【表1】
【0057】
実施例4;タンパク質発現の確認
<4−1>SDS−PAGE
前記実施例3で製造された大腸菌形質転換体それぞれから発現された抗−腫瘍壊死因子−アルファFab’を検出するために,発現されたタンパク質を、Fabに親和力を有するタンパク質−Gカラムを用いて、次の方法で部分的に精製した後、SDS−PAGE電気泳動を行った。
【0058】
ハイトラップタンパク質G HPカラム(Hitrap Protein G HP、Amersharm Bioscience社)に10カラム体積の平衡緩衝溶液(20mM リン酸ナトリウム、pH7.0)を1分当たり1mlの流速で流し、充分に平衡化させた後、前記実施例3から得た醗酵培養上澄液を前記カラムにローディングした。試料のローディングが完了した後、前記カラムを10カラム体積の平衡緩衝溶液で洗浄してから、カラムに吸着された抗−腫瘍壊死因子−アルファFab’を5カラム体積の溶出液(0.1M グリシン HClpH2.7)で溶出した。
【0059】
このように溶出された抗−腫瘍壊死因子−アルファFab’は、二つの異なるタンパク質、即ち、ジスルフィド結合によって連結された重鎖と軽鎖の結合によって形成された異型接合体であるため、還元条件下でのSDS−PAGEと非−還元条件下でのSDS−PAGEとにおける移動様式が異なるようになる。図9は、前記と同様に精製された抗−腫瘍壊死因子−アルファFab’を15%クライテリオンゲル(criteriongel、Bio-Rad社)を用いて非−還元状態のSDS−PAGEゲルと還元状態のSDS−PAGEゲルに展開させてその移動様式を比べたものである。図9で、レーン2及び3は還元条件下でSDS−PAGEゲルに展開したものであり、レーン4及び5は非−還元条件下でSDS−PAGEゲルに展開したものである。レーン1は、予め染色された低い範囲標準タンパク質マーカー(prestained low-range standard marker, Bio-Rad社)であり;レーン2及び4は、実施例3で用いられたFab標準タンパク質であり;レーン3及び5は、大腸菌形質転換体HM10922(KCCM−10511)を醗酵させ生産された精製試料である。
【0060】
図9に示すように,還元条件下でのSDS−PAGEにおいては重鎖と軽鎖との結合が還元的に切断され、重鎖及び軽鎖は略同一大きさの個別的な単量体として移動する(重鎖の分子量[約24kDa]は軽鎖の分子量[約23kDa]と類似しており,これらはゲル上で一つの分子のように見える)。一方、重鎖及び軽鎖は、非−還元条件下でのSDS−PAGEにおいてはジスルフィド結合で連結された異型接合体として存在し、その結果、約47kDaの移動距離を示した。
【0061】
<4−2> N−末端配列分析
前記実施例<4−1>のSDS−PAGEによって決定された約47kDaの分子量を有する異型接合体のアミノ酸配列を分析するため、基礎科学支援研究所ソウル支店に前記タンパク質のN−末端配列分析を依頼し、分析する試料は次のように用意した。
【0062】
PVDF膜(BioRad社)をメタノールに約2−3秒間漬けて活性化させてから、ブロッキング溶液(170mM グリシン,25mM Tris・HCl(pH8),20% メタノール)に充分に濡らした。実施例<4−1>の非−還元条件下で展開されたSDS−PAGEゲルを前記と同様に用意されたPVDF膜にブロッティングキット(HoeferSemi-Dry Transfer unit, Amersham)を用いて1時間程度ブロッティングを行った。PVDF膜へ移動されたタンパク質をクマシーブルーR−250(Amnesco社)で3−4秒間染色してから、脱色液(水:酢酸:メタノール=5:1:4)で洗浄した。洗浄された膜からタンパク質を含む部位をはさみで切り取って、N−末端配列分析に用いた。
【0063】
抗−腫瘍壊死因子−アルファFab’の軽鎖のアミノ酸配列はAsp-Ile-Gln-Met-Thr-Gln-Ser(配列番号:34)であり、重鎖のアミノ酸配列はGlu-Val-Gln-Leu-Glu-Val-Asp-Ser(配列番号:35)である。アミノ酸配列分析の結果、本発明の大腸菌形質転換体から分泌的に発現された約47kDaの分子量を有する異型接合体のN−末端は、Asp/Glu-Ile/Val-Gln-Met/Leu-Thr/Val-Gln/Asp-Ser(配列番号:36)であった。前記結果は、本発明によって生産された抗−腫瘍壊死因子−アルファFab’の軽鎖及び重鎖が正常な異型接合体形態で存在することを明確にするものである。
【0064】
<4−3>活性酵素免疫測定(ELISA)
前記実施例3で、大腸菌形質転換体から発現された抗−腫瘍壊死因子−アルファFab’が腫瘍壊死因子−アルファ抗原と結合するか否かを確認するため、次のように、活性酵素免疫測定分析法(ELISA)を行った。
【0065】
実施例<4−1>から得た抗−腫瘍壊死因子−アルファFab’の濃度を280nmでの吸収係数(extinction coefficient=1.43, Humphreys et,al. Protein Expression and Purification 26: 309-320, 2002)を決めることにより測定し、その後、PBS−T緩衝溶液に10ng/mlの濃度で溶解させた。また、腫瘍壊死因子−アルファに結合する標準抗体として、腫瘍壊死因子−アルファと結合し関節炎治療剤としての商業的効能が立証されたヒューミラ(humira,Abbott社)を同一緩衝溶液を用いて同一濃度で製造した。実施例3に記述と同一の方法でウェルプレートを腫瘍壊死因子−アルファにコーティングした後、動物細胞から生産された標準抗体と本発明の大腸菌形質転換体から生産されたFab’とが同一の濃度で同一の結合親和力を現わすかどうかを調査した。この際、標準抗体(対照群)はヒト免疫グロブリンGをタンパク質分解酵素であるパパインで切断した後、一連のカラムで精製して用意したFabを用いた。
【0066】
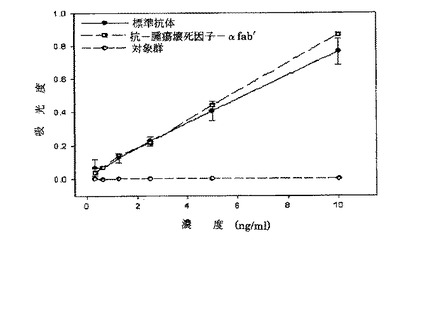
図10に示したように、対照群として用いられたFab標準タンパク質は、抗−腫瘍壊死因子−アルファに対して結合親和力を全く示さないのに対して,本発明によって大腸菌形質転換体から生産された抗−腫瘍壊死因子−アルファFab’希釈倍数に比例して腫瘍壊死因子−アルファと結合し、標準抗体と同一又はより高い吸光度を現わした。
【0067】
本発明を前記具体的な実施例と関連して記述しましたが、添付された特許請求の範囲によって定義された本発明の範囲内で当分野の熟練者が本発明を多様に変形及び変化させ得ることを理解しなければならない。
【図面の簡単な説明】
【0068】
【図1】図1は、重鎖と軽鎖の定常領域のそれぞれをプラスミドpBluescript SK(−)に挿入して、プラスミドpBH及びpBLの製作過程を示す図である。
【図2】図2は、重鎖と軽鎖の可変領域をそれぞれプラスミドpBH及びpBLに挿入してプラスミドpBHC及びpBLCの製作過程を示す図である。
【図3】図3は、T7プロモーターと大腸菌耐熱性エンテロトキシンシグナル配列に融合された抗体の軽鎖をコードする遺伝子とを含む発現ベクターpmsLC、及びT7プロモーターと大腸菌耐熱性エンテロトキシンシグナル配列に融合された抗体の重鎖をコードする遺伝子とを含む発現ベクターpmsHCのそれぞれの製作過程を示す図である。
【図4】図4は、単一T7プロモーターの調節によって軽鎖と重鎖を個別的に発現・分泌するダイ−シストロニック発現ベクターpsDLHF_B及びpsDLHF−Bpの製作過程を示す図である。
【図5】図5は、Tacプロモーターと大腸菌外膜タンパク質A(OmpA)シグナル配列に融合された抗体の重鎖及び軽鎖をコードする遺伝子とを含む発現ベクターpoDLHFの製作過程を示す図である。
【図6】図6は、個別のTacプロモーターの調節によって重鎖及び軽鎖を発現して分泌する発現ベクターpoDLHF_B/Sの製作過程を示す図である。
【図7】図7は、抗体の軽鎖をコードする遺伝子が大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合され、抗体の重鎖をコードする遺伝子が大腸菌外膜タンパク質A(OmpA)シグナル配列に融合されており、これらの発現及び分泌が単一のTacプロモーターによって調節される発現ベクターpmsoDLHF_N/Sの製作過程を示す図である。
【図8】図8は、抗体の軽鎖をコードする遺伝子が大腸菌外膜タンパク質A(OmpA)シグナル配列変異体に融合されており、抗体の重鎖をコードする遺伝子が大腸菌耐熱性エンテロトキシンシグナル配列に融合され、これらの発現及び分泌が単一のTacプロモーターによって調節される、発現ベクターpmsoDLHF_S/Kの製作過程を示す図である。
【図9】図9は、還元条件下及び非還元条件下で行ったSDS−PAGE分析によって得られた大腸菌形質転換体から発現された抗−腫瘍壊死因子−αFab’の発現パターンを比較した結果を示す図である。
【図10】図10は、大腸菌形質転換体から発現された抗−腫瘍壊死因子−αFab’が腫瘍壊死因子−αと結合するか否かを調べるためのELISAの結果を示す図である。
【技術分野】
【0001】
本発明は、水溶性異型接合体(heterozygote)の形態で大腸菌シグナル配列に融合された抗体断片を分泌的に発現する組換え発現ベクター;前記発現ベクターにより形質転換された微生物;及び前記形質転換された微生物を用いて抗体断片を大量生産する方法に関する。
【背景技術】
【0002】
本発明において、「抗体断片」は、抗体の抗原−結合領域またはその可変領域を含む、抗体の一部を意味するものである。抗体断片の例としては、Fab’、Fab’、F(ab’)2及びscFv断片などがある。抗体をパパイン(papain)で切断すると、Fab’断片と呼ばれる二つの同一の抗原−結合断片と残りのFc断片とが生成する。抗体をペプシン(pepsin)で処理すると、二つの抗原−結合部位を有し、抗原と依然として架橋結合を形成し得るF(ab’)2断片が生成する。Fab断片は、さらに軽鎖の定常ドメイン及び重鎖の第1定常ドメイン(CH1)を含む。Fab’断片は、Fab断片と異なって、抗体ヒンジ(hinge)領域から一つ以上のシステインを含む重鎖CH1ドメインのカルボキシ末端にいくつかの残基が付加されている。
【0003】
抗原結合能力の増加、抗原性の変化、または二重特異的抗体の製造などで抗体の固有性質を調制するために、遺伝子組換え方法を用いた抗体と抗体断片の生産が研究されている。その結果、大腸菌(E.coli)発現系を用いて抗体と抗体断片を生産する方法が開発された。大腸菌の発現系を用いた生産方法は既存の動物細胞を用いた方法に比べて次のようなメリットを提供する:1)発現ベクターの製作が容易であるため、発現可否を容易に検証することができる;2)大腸菌は、成長速度が速いため、低費用で抗体の大量生産が可能であるので、これは研究用試料確保の面で有利である;3)比較的に単純な発酵方法を適用することができ、他の宿主細胞を用いる場合よりも一層容易に商業化できる。
【0004】
抗体−コード遺伝子を大腸菌内に導入して発現させた事例は多くの文献で報告されている(Cabilly et al.,Proc.Natl.Acad.Sci.USA81: 3273-3277,1984,Boss et al.Nucleic Acids Res.12: 3791-3806,1984)。これらの文献は、抗体分子が多様な収率で細胞質内で発現され、また、大腸菌が免疫グロブリンの軽鎖を分泌的に発現させること (Zemel-Dreasen et al., Gene 315-322, 1984)、または抗体断片遺伝子をアルカリ・ホスファターゼ(alkaline phosphatase)あるいはベータラクタマーゼ(β−lactamase)シグナル配列に融合された抗体断片をペリプラズム空間(periplasmic space)に分泌させるための宿主細胞として使用可能であることを開示している(Pluckthun et al., Cold Spring Harbor Symposiua on Quantitative Biology Volume LII、105-112、1987)。国際出願公開第WO92/01059号は、癌細胞の特定部位を認識する抗体であるFab’−コード遺伝子を、大腸菌外膜タンパク質A(OmpA)シグナル配列に融合された融合タンパク質の形態で発現されることを教示しており;米国特許第5,648,237号は、大腸菌のエンテロトキシンシグナル配列に融合されたFab’−コード遺伝子がPhoAプロモーターの調節下で発現されることを開示している。
【0005】
発現された抗体断片は野生型抗体と類似した抗原−結合能を示すが、それは野生型抗体よりも小さいため、局所浸透力が高く(Blumenthal et al., Ad. DrugDel. Rev.4: 279, 1990)、またFc領域を含んでいないため、人体に何等の副作用も引き起こさない。よって、抗体断片は診断試薬の製造や、治療用抗体の開発に利用可能であり、大腸菌からの大量生産は経済的な面で非常に有利である。野生型抗体から抗体断片を製造する場合、野生型抗体はタンパク質分解酵素で切断して精製しなければならないが、これは煩わしくて経済的にも不利である。
【0006】
しかしながら、抗体断片を大腸菌で生産する場合には様々な問題点がある。まず、すべての抗体断片が意図した治療あるいは診断的目的を満たす程度の量で発現されず、場合によっては培養過程を何度も繰り返しなければならない。また、抗体断片に機能的活性を付与するためには、重鎖と軽鎖が単一細胞内で共に発現され、これらの間に形成されたジスルフィド結合によって異型接合体の形態を維持しなければならないが、この際抗体断片は、タンパク質が大腸菌で過剰発現される場合に頻繁に見られる封入体の形態ではなく、高水溶性の形態で発現されることが求められる。このような問題点を解決するため、シグナル配列(signal sequence)を用いて発現された抗体断片をペリプラズム空間に分泌させる方法に関する研究が活発に行われてきた。しかし、シグナル配列に融合されたすべての抗体断片が大腸菌のペリプラズム空間に分泌的に過剰発現されるのではなく、発現させようとする抗体の遺伝子配列によって発現された抗体の量において大きく異なる(Kelly et,al.Biochemistry 31: 5434-5441,1992; Humphreys et,al.Protein Expression and Purification 26: 309-320,2002)。発現ベクター内の構造遺伝子の相違または重鎖遺伝子と軽鎖遺伝子との距離などによって抗体断片の発現パターンが変動することが報告されている(国際特許公開第WO01/94585号)。このような事実から、標的抗体断片の発現のためには、標的抗体遺伝子と発現に必要な宿主の構造遺伝子との相互作用、及びシグナル配列を用いる場合における標的抗体遺伝子とシグナル配列との相互作用における適切なバランスが要求される。
【0007】
ここに、本発明者らは、大腸菌で抗体断片を抗原結合を有する水溶性異型接合体の形態で過剰発現させるために鋭意研究に努めた結果、大腸菌耐熱性エンテロトキシンシグナル配列変異体または大腸菌外膜タンパク質A(OmpA)シグナル配列を用いて、目的抗体断片である腫瘍壊死因子−アルファ(TNF−α)のヒト抗体Fab’を分泌的に発現する組換え発現ベクターを製造することによって、目的とする抗原−結合親和力を有する水溶性異型接合体の形態で抗体断片を大腸菌内で発現する方法を開発することで本発明に至った。
【発明の概要】
【0008】
したがって、本発明の目的は、大腸菌シグナル配列を用いて抗体断片を抗原−結合親和力を有する水溶性異型接合体形態で過剰発現させ得る組換え発現ベクター及び前記発現ベクターにより形質転換された微生物を提供することである。
本発明の他の目的は、前記形質転換された微生物を用いて抗体断片を大量に生産する方法を提供することである。
【0009】
前記目的を達成するために、本発明は、第1大腸菌シグナル配列に融合された抗体断片の軽鎖をコードする遺伝子と、第2大腸菌シグナル配列に融合された抗体断片の重鎖をコードする遺伝子とを含む組換え発現ベクターであって、前記組換え発現ベクターから発現された抗体断片が培養培地内に分泌されることを特徴とする組換え発現ベクターを提供する。
【0010】
また、本発明は、前記組換え発現ベクターを製造する段階;前記発現ベクターで微生物を形質転換させる段階;前記微生物を培地で培養する段階;及び前記微生物から培地内に分泌された抗体断片を回収する段階を含む、抗体断片の生産方法を提供する。
本発明の前記および他の目的および特徴は、下記の図面を伴う本発明の詳細な説明によって明白になる。
【発明の詳細な説明】
【0011】
本発明は、第1大腸菌シグナル配列に融合された抗体断片の軽鎖をコードする遺伝子及び第2大腸菌シグナル配列に融合された抗体断片の重鎖をコードする遺伝子を含み、発現ベクターから発現された抗体断片を培養培地内に分泌することを特徴とする組換え発現ベクター;及び前記発現ベクターにより形質転換された微生物を提供する。
【0012】
野生型抗体と機能的に類似した抗体断片を分泌的に発現できる前記発現ベクターは、抗体断片の軽鎖及び重鎖をコードする遺伝子をそれぞれ大腸菌耐熱性エンテロトキシンシグナル配列変異体または大腸菌外膜タンパク質A(OmpA)シグナル配列に融合させて製造される。
【0013】
本発明で抗体断片は、例えばFab’、Fab’、F(ab’)2またはscFvであり、好ましくはFab’である。Fab断片は軽鎖の定常ドメイン及び重鎖の第1定常ドメイン(CH1)を含む。Fab断片は、抗体のヒンジ(hinge)領域から一つ以上のシステインを含む重鎖CH1ドメインのカルボキシ末端にいくつかの残基が付加されている点がFab断片と異なる。また、F(ab’)2断片は二つのFab’分子がジスルフィド結合によって連結されて製造され、scFv断片は軽鎖と重鎖の可変領域だけがペプチドリンカー(peptide linker)によって連結されて単一ポリペプチド鎖の形態で存在する。
【0014】
本発明に使用可能な抗体断片は、抗原に選択的に結合できるすべての抗体を含み、この場合、抗原はT−細胞、内皮細胞あるいは癌細胞マーカーのような細胞表面に結合している抗原であるか、あるいは、細胞と結合していない水溶性抗原である。細胞表面に結合されている抗原としては、β1インテグリン(integrins)、P−セレクチン(P−selectin)、E−セレクチン、CD2、CD3、CD4、CD5、CD7、CD8、CD11a、CD11b、CD18、CD19、CD20、CD23、CD25、CD33、CD38、CD40、CD45、CD69、MHCクラスI、II、VEGF、及びその他の受容体が含まれるが、これらに限定されない。水溶性抗原としては、IL−1、2、3、4、5、6、8及びIL−12のようなインターロイキン類;サイトメガロウイルス抗原のようなウイルス抗原;IgEのような免疫グロブリン;INF−α、β及びγのようなインターフェロン;及び腫瘍壊死因子−α及びβ、血小板由来成長因子−α及びβのような細胞の成長及び分化に影響を及ぼす因子が含まれるが、これらに限定されない。好ましい実施例で、本発明は、抗−腫瘍壊死因子−アルファ(TNF−α)の抗体断片の軽鎖及び重鎖をコードする遺伝子を用いる。TNF−αは、単球細胞や大食細胞(macrophage)のような免疫関連細胞によって生成されて過剰発現時に炎症を引き起こすサイトカイン類として知られているIL−1、IL−6及びIL−8の生成を誘導し、軟骨組織を破壊するタンパク質分解酵素の分泌を促進して関節炎を起こす主な媒介体として知られている。関節炎治療剤としての抗−腫瘍壊死因子−アルファ抗体のFab’は、単クローン抗体から得られ、好ましくは、ヒト化抗体、より好ましくは、完全なヒト抗体から得られる。
【0015】
抗−腫瘍壊死因子−アルファのいくつかのキメラ抗体あるいはヒト化抗体の相補性決定領域(complementarity determining regions,CDRs)をコードする遺伝子は既に報告されており、完全なヒト抗体の塩基配列も決まっている(国際特許第WO97/2913号)。このような情報を参考に、ヌクレオチド合成器を用いて抗体断片をコードする遺伝子を人工的に合成し、一連のアニーリング(annealing)とリゲーション(ligation)の過程、またはリゲーション連鎖反応(LCR,ligation-chain reaction)とポリメラーゼ連鎖反応(PCR,polymerase chain reaction)を通じて、Fabの重鎖と軽鎖をコードする上で合成した遺伝子をクローニングできる。
【0016】
好ましい実施例で、本発明は配列番号:1〜6に示される塩基配列を有するヒト抗−腫瘍壊死因子−アルファFab’の軽鎖の可変領域をコードする遺伝子断片と、配列番号:7〜12に示される塩基配列を有する重鎖の可変領域をコードする遺伝子断片とをヌクレオチド合成器で合成した後、これらの断片をPCRによって連結させて軽鎖及び重鎖の可変領域を製造する。また、軽鎖の定常領域と、IgG1のCH1及びヒンジ領域を含む重鎖の定常領域とを含む遺伝子断片は、人の血液から抽出したRNA全体を鋳型とし、ヒト抗−腫瘍壊死因子−アルファFab’の軽鎖および重鎖の定常領域に対して特異的なプライマー対を用いたRT−PCRによって製造される。このように製造された軽鎖及び重鎖の定常領域をコードする遺伝子断片のそれぞれを適当な発現ベクターにクローニングして、軽鎖の定常領域をコードする遺伝子を含むプラスミドpBLC及び重鎖の定常領域をコードする遺伝子を含むpBHCを製造する(図1及び2参照)。塩基配列分析の結果、前記プラスミドpBLCに挿入された重鎖の定常領域をコードする遺伝子は、配列番号:32に示す塩基配列を有し、プラスミドpBHCに挿入された重鎖の定常領域をコードする遺伝子は、配列番号:33に示す塩基配列を有することが確認された。
【0017】
発現させようとする抗体断片は、重鎖と軽鎖で構成された二つの異なる種類のタンパク質間の異型接合体として存在する。よって、抗原−結合親和力を有する活性型の異型接合体としての抗体断片を得るためには、重鎖と軽鎖が同等の水準で発現されなければならず、発現ベクターから発現された重鎖と軽鎖との間の正確な位置でジスルフィド結合が形成されなければならない。また、抗体断片が大腸菌内で発現される時、細胞質内に発現されたタンパク質は封入体の形態で存在し得る。この場合、重鎖と軽鎖のジスルフィド結合が正確な位置で生成されないこともあり、水溶性状態で発現されても、精製段階で細胞を完全に破壊できないという問題が起こり得る。このような問題点を解決するために、本発明は、大腸菌シグナル配列を用いて、ベクターから発現された抗体断片を大腸菌のペリプラズム空間あるいは培養培地に分泌させる組換え発現ベクターを提供する。
【0018】
発現されたタンパク質が培養培地内に存在すると、発現されたタンパク質が細菌細胞内に存在する際に要求される細胞を破壊しなければならないという煩わしい過程を行わなくても良い。また、発現されたタンパク質がペリプラズム空間に存在するため、回収した細胞の浸透圧衝撃のような過程を省略できる。
【0019】
さらに、酸化環境は異型接合体を形成するようにするジスルフィド結合を容易にするため、発現されたタンパク質が細胞の細胞質内の非−酸化環境に存在するよりもペリプラズム空間あるいは培養培地内に存在するのが有利である。
【0020】
本発明の組換え発現ベクターに使用可能な大腸菌シグナル配列は、大腸菌耐熱性エンテロトキシンシグナル配列、OmpAシグナル配列、ベータラクタマーゼシグナル配列、GeneIIIシグナル配列、PelBシグナル配列またはこれらの変異体などであり、好ましくは大腸菌耐熱性エンテロトキシンシグナル配列変異体またはOmpAシグナル配列であるが、これらに限定されない。
【0021】
本発明の好ましい実施例で、発現ベクターから発現された活性型の抗体断片Fab’を培養培地内に分泌させるために用いられた大腸菌耐熱性エンテロトキシンシグナル配列変異体は、大腸菌で様々な異種タンパク質を分泌・発現させるために本発明者らが開発したものである。前記大腸菌耐熱性エンテロトキシンシグナル配列変異体は、配列番号:17に示される塩基配列を有し、最適効率で大腸菌内で様々な異種タンパク質を発現・分泌できることが報告されている(韓国特許第316347号)。本発明で、発現ベクターから発現された活性型の抗体断片Fab’を培養培地内に分泌させ得る組換え発現ベクターは、抗体断片の重鎖及び軽鎖をコードする遺伝子をそれぞれ大腸菌シグナル配列、例えば、大腸菌耐熱性エンテロトキシンシグナル配列変異体または外膜タンパク質Aシグナル配列に融合させることによって製造される。
【0022】
抗体断片の重鎖及び軽鎖をコードする遺伝子を一つの発現ベクター内で発現させるために、本発明は、一つのプロモーターの下に二つのタンパク質を適切な間隔をおいて配置して同時に発現させるダイ−シストロニック体系(di−cistronic system)を用いる発現ベクターと、独立された各プロモーターに遺伝子を連結して二つのタンパク質を独立的に発現できる二重−プロモーター体系(dual−promoter system)を用いる発現ベクターとの二つの種類の発現ベクターを提供する。
【0023】
まず、ヒト成長ホルモン(hGH)をコードする遺伝子断片を、大腸菌耐熱性エンテロトキシンシグナル配列及びhGHを含む発現ベクターpT14S1SH−4T20V22Q(韓国特許第316347号)から除去する。プラスミドpBLCから得た軽鎖をコードする遺伝子断片と、プラスミドpBHCから得た重鎖をコードする遺伝子断片とを、前記発現ベクターpT14S1SH−4T20V22Qに挿入し、耐熱性エンテロトキシンシグナル配列変異体に融合された重鎖をコードする遺伝子断片を含む発現ベクターpmsHC、及び耐熱性エンテロトキシンシグナル配列変異体に融合された軽鎖をコードする遺伝子断片を含む発現ベクターpmsLCを製作する。重鎖と軽鎖をコードする遺伝子のそれぞれが大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合され、単一プロモーターが重鎖及び軽鎖をコードする遺伝子の発現を調節するダイ−シストロニック発現ベクターを製作するために、プラスミドpmsHCからPCRによって得た重鎖をコードする遺伝子断片をプラスミドpmsLCに挿入して発現ベクターpsDLHF_B及びpsDLHF_Bpを製造する(図3及び4参照)。
【0024】
また、本発明は前記のような方法によって、軽鎖及び重鎖をコードする遺伝子のそれぞれが大腸菌OmpAシグナル配列(配列番号:24)に融合され、単一Tacプロモーターが軽鎖及び重鎖をコードする遺伝子の発現を調節する発現ベクターpoDLHFを製作する(図5参照)。
【0025】
前記で製作された発現ベクターpsDLHF_B、psDLHF_Bp及びpoDLHFはシグナル配列に融合された抗−腫瘍壊死因子−アルファの重鎖及び軽鎖のそれぞれが単一プロモーターの調節下で発現・分泌されるダイ−シストロニック発現ベクターである。前記発現ベクターのそれぞれを大腸菌BL21(DE3)に形質転換させて製造した大腸菌形質転換体BL21(DE3)/psDLHF_B(HM10920)、BL21(DE3)/psDLHF_BP(HM10921)及びBL21/poDLHF(HM10922)はそれぞれ、特許手続きの目的のための微生物寄託の国際的認識に関するブダペスト条約に従って、寄託番号KCCM−10509、KCCM−10510及びKCCM−10511として韓国微生物保存センター(KCCM)に2003年10月2日付で寄託された。
【0026】
上述したように、単一プロモーターの調節下で異なる二つのタンパク質を分泌的に発現させる場合、プロモーターに近いタンパク質の発現効率はプロモーターに遠いタンパク質の発現効率と異なり得る(Humphreys et, al. Protein Expression and Purification 26: 309-320, 2002)。本発明は、抗−腫瘍壊死因子−アルファFab’の重鎖と軽鎖の発現及び分泌が独立したプロモーターによって個別的に調節可能な二重−プロモーターシステムを用いる発現ベクターを提供する。好ましい実施例で、本発明は、抗−腫瘍壊死因子−アルファFab’の鎖及び軽鎖をコードする遺伝子をそれぞれOmpAシグナル配列に融合して、Tacプロモーターが軽鎖及び重鎖をコードする遺伝子の発現及び分泌を調節する発現ベクターpoDLHF_B/Sを製作する(図6参照)。前記発現ベクターpoDLHF_B/Sにより大腸菌BL21を形質転換させて製造した大腸菌形質転換体BL21/poDLHF_B/S(HM10923)は、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約に基づき、寄託番号KCCM−10512として韓国微生物保存センター(KCCM)に2003年10月2日付に寄託された。
【0027】
抗体断片のような異型接合体の形成は前記異型接合体の二つの断片の一方が過剰発現された時に抑制されるので、二つのタンパク質の断片は互いに同様の水準で発現されなければならない。ここに、本発明では、重鎖と軽鎖をコードする遺伝子をそれぞれ互いに異なるシグナル配列に融合させて抗体断片を分泌的に発現させ得る組換え発現ベクターを提供する。
【0028】
好ましい実施例で、本発明は、抗−腫瘍壊死因子−アルファFab’の軽鎖をコードする遺伝子は耐熱性エンテロトキシンシグナル配列変異体に融合され、その重鎖をコードする遺伝子はOmpAシグナル配列に融合され、単一T7プロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節する発現ベクターpmsoDLHF_N/Sを製作する(図7参照)。また、本発明で、発現ベクターpmsoDLHF_S/Kは、抗−腫瘍壊死因子−アルファFab’の重鎖をコードする遺伝子は耐熱性エンテロトキシンシグナル配列変異体に融合され、その軽鎖をコードする遺伝子はOmpAシグナル配列に融合され、単一Tacプロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節するように製作される(図8参照)。前記発現ベクターpmsoDLHF_N/S及びpmsoDLHF_S/Kをそれぞれ大腸菌BL21に形質転換させて製造した大腸菌形質転換体BL21/pmsoDLHF_N/S(HM10924)及びBL21/pmsoDLHF_S/K(HM10925)のそれぞれは、特許手続上の微生物の寄託の国際的承認に関するブダペスト条約に基づき、寄託番号KCCM−10513及びKCCM−10516として韓国微生物保存センター(KCCM)に2003年10月2日付で寄託された。
【0029】
本発明は、前記のように製作された組換え発現ベクターを適当な宿主細胞に導入して製造された組換え微生物を提供する。本発明で使用可能な宿主細胞としては、大腸菌W3110、RV308、BL21、BL21(DE3)などを含み、好ましくは大腸菌BL21であるが、これらに限定されない。前記発現ベクターにより形質転換された組換え微生物を発酵槽を用いて高濃度で培養(OD600=120〜150)する場合、抗−腫瘍壊死因子−アルファの抗体断片は培養培地1L当たり100〜500mg以上の高収率で培養培地内に分泌的に生産されることが確認された(表1参照)。このように大量生産された抗−腫瘍壊死因子−アルファの抗体断片はタンパク質−Gアフィニティーカラムによって部分的に精製できる。精製された抗−腫瘍壊死因子−アルファの抗体断片を還元条件下及び非−還元条件下で行ったSDS−PAGEで分析した結果、抗体断片が水溶性異型接合体形態で形成されたことを確認した(図9参照)。また、結合抗原として抗−腫瘍壊死因子−アルファを用いたELISAは、抗−腫瘍壊死因子−アルファの抗体断片が腫瘍壊死因子−アルファ抗原に対して優れた結合親和力を有することを示した(図10参照)。
【0030】
本発明は、第1大腸菌シグナル配列に融合された抗体断片の軽鎖をコードする遺伝子及び第2大腸菌シグナル配列に融合された抗体断片の重鎖をコードする遺伝子を含む発現ベクターを製造する段階;前記発現ベクターで微生物を形質転換させる段階;前記微生物を培地で培養する段階;及び 前記微生物から培地内に分泌された抗体断片を回収する段階を含む、抗体断片の生産方法を提供する。
【0031】
本発明の組換え発現ベクターで使用可能な大腸菌シグナル配列としては、大腸菌耐熱性エンテロトキシンシグナル配列、OmpAシグナル配列、ベータラクタマーゼシグナル配列、GeneIIIシグナル配列、PelBシグナル配列またはこれらの変異体を含み、好ましくは配列番号:17に示される塩基配列を有する大腸菌耐熱性エンテロトキシンシグナル配列変異体または配列番号:23に示される塩基配列を有する大腸菌外膜タンパク質Aシグナル配列を用いるが、これらに限定されない。
【0032】
前記組換え発現ベクターにより形質転換された微生物を適当な培地で予備培養した後、この予備培養液を発酵槽に接種して30〜35℃範囲の温度で空気と接触させながら本−培養を実施する。600nmでOD値が80に達すると、誘導物質(inducer)を発酵槽に添加して標的抗体断片の発現を誘導する。本発明で使用可能な誘導物質としては、IPTGが好ましいが、これに限定されない。600nmでOD値が120〜140に逹するまで、40〜45時間更に培養する。前記培養液を遠心分離して上澄液を分離した後、これをアフィニティーカラムクロマトグラフィーで抗体断片を精製する。前記形質転換された微生物は、大腸菌BL21(DE3)/psDLHF_B(HM10920)(KCCM-10509)、BL21(DE3)/psDLHF_BP(HM10921)(KCCM-10510)、BL21/poDLHF(HM10922)(KCCM-10511)、BL21/poDLHF_B/S(HM10923)(KCCM-10512)、BL21/pmsoDLHF_N/S(HM10924)(KCCM-10513)、好ましくはBL21/pmsoDLHF_S/K(HM10925)(KCCM-10516)であるが、これらに限定されない。
【0033】
本発明によって発現された抗体断片は宿主細胞のペリプラズム空間または培養培地内に分泌されるため、抗体断片は浸透圧ショック処理のような工程を経なとも回収できる。
【0034】
本発明の方法によって発現され得る目標抗体断片は、好ましくはFab’、Fab’F(ab’)2またはscFvなどであるが、抗−腫瘍壊死因子−アルファの抗体断片に限定されるものではなく、よって、診断または治療を目的とする任意のすべての抗体、例えば、抗癌効果がある抗体断片または過剰−発現された場合、人体に致命的なダメージを与える様々なサイトカインに対する抗体断片の生産に適用できる。
【0035】
前述したように、大腸菌耐熱性エンテロトキシンシグナル配列変異体または大腸菌外膜タンパク質Aシグナル配列を含む本発明の組換え発現ベクターは、抗体断片を抗原−結合親和力を有する水溶性異型接合体の形態で発現させ得る。前記発現ベクターにより形質転換された微生物は抗体断片を宿主細胞のペリプラズム空間または培養培地に分泌させることが可能であるため、発現された抗体断片は大量生産に適する方法で容易に回収できる。従って、本発明による方法は診断試薬または治療用抗体の製造に有効に利用可能である。
【0036】
下記実施例は本発明をさらに詳細に説明するためのものであり、本発明の範囲を制限しない。
【0037】
実施例1:抗−腫瘍壊死因子−アルファFab’をコードする遺伝子のクローニング
国際特許第WO97/29131号に記載された塩基配列を参考にして、ヒト抗−腫瘍壊死因子−アルファFab’の重鎖と軽鎖の可変領域をヌクレオチド合成器を用いて合成した。軽鎖の可変領域をクローニングするための遺伝子断片は配列番号:1〜6の塩基配列を有し、重鎖の可変領域をクローニングするための遺伝子断片は配列番号:7〜12の塩基配列を有した。この後のクローニング過程を容易にするために、前記重鎖及び軽鎖をコードする遺伝子のそれぞれの一番目である遺伝子断片(即ち、配列番号:1及び7)にそれぞれHindIII及びPuvI制限酵素認識部位を挿入した。前記合成した重鎖及び軽鎖の遺伝子断片のそれぞれを一つのチューブに混ぜた後、これらを一つの遺伝子配列で連結させるために通常的なPCRを行って、軽鎖及び重鎖の可変領域をコードする遺伝子を製造した。前記PCR反応で、遺伝子断片のそれぞれは鋳型のみならずプライマーとして作用可能であり、従って、これらは互いにアニーリングされ、約320mer程度の断片を形成した。前記PCR反応混合物は、それぞれの遺伝子断片100pmole、5ulの2mMdNTP混合液、2.5単位のpfuDNAポリメラーゼ(Stratagen社)及び5ulのpfu反応緩衝溶液(100mM Tris-HCl(pH8.3), 15mM MgCl2, 500mM KCl)を含み、蒸留水を加えて最終体積が50ulになるようにした。前記PCRはDNAサーマルサイクラー(DNA thermocycler, Perkin-Elmer社)を用いて94℃で5分間初期変性させた後、94℃で60秒、60℃で60秒及び72℃で60秒の反応を30回行った。
【0038】
ヒト項−腫瘍壊死因子−アルファFab’の軽鎖の定常領域とIgG1のCH1とヒンジ領域とを含んだ重鎖の定常領域をクローニングするため,ヒト血液の血球細胞からRNAを抽出した後、次のようにRT−PCRを行った。すべてのRNAは、QiampRNA血液キット(Qiagen社)を用いて約6mlの血液から抽出されて、RT−PCRの鋳型として用いられた。前記重鎖及び軽鎖をコードする遺伝子のそれぞれは、重鎖増幅のための配列番号:13及び14に示されるプライマー対と、軽鎖増幅のための配列番号:15及び16に示されるプライマー対を用いたOne−StepRT−PCRキット(Qiagen社)を用いて増幅された。この際、この後のクローニング過程を容易にするために、配列番号:26の重鎖5’−プライマーにはHindIII及びStuI制限酵素認識部位を挿入し、配列番号:27の重鎖3’−プライマーには終止コドンを含むBamHI制限酵素認識部位を挿入し;配列番号:28の軽鎖5’−プライマーにはHindIII及びNruI制限酵素認識部位を挿入し、配列番号:29の軽鎖3’−プライマーには終止コドンを含むBamHI制限酵素認識部位を挿入した。これから増幅された重鎖及び軽鎖の定常領域をコードするPCR産物をそれぞれHindIIIとBamHIで切断した後、同一制限酵素で処理されたプラスミドpBluscriptSK(-)(Stratagen社)に挿入してプラスミドpBH及びpBLを製造した(図1)。DNA塩基配列の分析によって、プラスミドpBH及びpBLにそれぞれの重鎖及び軽鎖の定常領域が正確に挿入されたことが確認された。
【0039】
前記重鎖及び軽鎖の可変領域をこれらに相応する定常領域に連結するため、前記でPCRによって合成された重鎖及び軽鎖の可変領域それぞれをHindIII制限酵素で切断した。酵素処理された重鎖の可変領域は制限酵素HindIII/StuIで予め処理されたプラスミドpBHに挿入され、酵素処理された軽鎖の可変領域は、制限酵素HindIII/NruIで予め処理されたプラスミドpBLに挿入されて、重鎖及び軽鎖の可変領域のそれぞれがこれらに相応する定常領域に連結されたプラスミドpBHC及びpBLCを製作した(図2)。前記プラスミドpBHC及びpBLCにそれぞれ挿入された重鎖と軽鎖はそれぞれ配列番号:32及び33に示された塩基配列を有する。
【0040】
実施例2:抗−腫瘍壊死因子−アルファFab’遺伝子発現ベクターの製作
<2−1> 大腸菌耐熱性エンテロトキシンシグナル配列変異体及び抗−腫瘍壊死因子−アルファFab’遺伝子を含む発現ベクターの製作
大腸菌耐熱性エンテロトキシンシグナル配列変異体を用いた発現ベクターを製作するため、ヒト成長ホルモン(hGH)を発現する発現ベクターpT14S1SH-4T20V22Q(韓国特許第316347号)を用いた。前記発現ベクターは配列番号:17に示される塩基配列を有する耐熱性エンテロトキシンシグナル配列変異体を含む。この発現ベクターからhGHを除去し前記実施例1で製造された重鎖及び軽鎖をコードする遺伝子断片を挿入するために、配列番号:18及び19のプライマー対を用いた部位特異的突然変異誘発法(site-directed mutagenesis)によってプラスミドpT14S1SH-4T20V22Qの耐熱性エンテロトキシンシグナル配列変異体とhGH遺伝子の融合部位にStuI制限酵素認識部位を挿入した。塩基配列分析法によってStuI制限酵素認識部位が成功的に標的位置に生成されたことが確認した。前記プラスミドpT14S1SH-4T20V22Q内のStuI制限酵素認識部位を有するプラスミドをpmSTIIと命名した。
【0041】
前記プラスミドpmSTIIをStuI/BamHI制限酵素で処理した後、アガロースゲル電気泳動を行って、hGH部位が除去され大腸菌耐熱性エンテロトキシンシグナル配列変異体を含む大きい断片(4.7kb)をゲルから回収した。前記実施例1で製造された軽鎖をコードする遺伝子断片を有するプラスミドpBLCをPuvI制限酵素で処理した後、これらの両末端をT4DNAポリメラーゼを用いて平滑末端にした。これから得たプラスミドをBamHI制限酵素で処理して約720bpの軽鎖をコードする遺伝子断片を回収した。また、重鎖をコードする遺伝子断片を含むプラスミドpBHCを鋳型として配列番号:1及び20のプライマー対を用いてPCRを行った。これから増幅されたPCR産物を、前記軽鎖をコードする遺伝子断片の製造に用いられた同一酵素で処理して約700bpの重鎖をコードする遺伝子断片を回収した。配列番号:20のプライマー対は重鎖ヒンジ領域の一部を含む変形されたヒンジ配列であるEPKSCDKTHTCAA−終止コドンが重鎖をコードする遺伝子断片に挿入されてBamHI制限酵素認識部位を含むように考案されているため、この後の過程でFab’発現ベクターの製作に有用に利用できる。このように用意された重鎖及び軽鎖をコードする遺伝子断片のそれぞれをプラスミドpmSTII断片に挿入し、耐熱性エンテロトキシンシグナル配列変異体に融合された軽鎖をコードする遺伝子断片を含む発現ベクターpmsLCのみならず、耐熱性エンテロトキシンシグナル配列変異体に融合された重鎖をコードする遺伝子断片を含む発現ベクターpmsHCを製作した(図3)。
【0042】
重鎖と軽鎖が一つのプロモーターによって調節されて発現・分泌されるダイ−シストロニック(di−cistronic)発現ベクターを製作するために、耐熱性エンテロトキシンシグナル配列変異体に融合された重鎖をコードする遺伝子断片を含むプラスミドpmsHCを鋳型とし、配列番号:21及び20のプライマー対と配列番号:22及び23のプライマー対を用いてPCRを行った。配列番号:21の5’−プライマーは重鎖をコードする遺伝子断片の発現に必要であるシャインダルガノ(Shine-Dalgarno,SD)配列からPCR反応が開始するように考案されており、増幅された産物を容易にクローニングするためにBamHI制限酵素認識部位を含んでいる。また、配列番号:22及び23のプライマー対は、クローニング部位を異ならせるために、BamHIではなくBpuI制限酵素認識部位を含むことを除けば、配列番号:21及び20のプライマー対と同一の塩基配列を有している。各プライマー対を用いて増幅された重鎖をコードする遺伝子断片(一方はBamHI制限酵素認識部位を含み、他方はBpuI制限酵素認識部位を含む)をBamHIとBpuI制限酵素でそれぞれ処理した。
【0043】
耐熱性エンテロトキシンシグナル配列変異体に融合された軽鎖をコードする遺伝子断片を含むプラスミドpmsLCでBamHIとBpuI制限酵素認識部位はすべて軽鎖遺伝子の終止コドンの下流に位置し、BpuIの制限酵素認識部位はBamHIより約60ヌクレオチド以上下流に存在するようにした。前記プラスミドpmsLCをBamHIとBpuI制限酵素で処理した後、その部位にBamHIまたはBpuI制限酵素で処理された重鎖をコードする遺伝子断片を挿入した。プラスミドpmsLCのBamHI制限酵素部位内に挿入された重鎖遺伝子を含むFab’発現ベクターをpsDLHF_Bpと命名した(図4)。これらの発現ベクターの場合、重鎖及び軽鎖をコードする遺伝子はそれぞれ、大腸菌エンテロトキシンシグナル配列に融合され、単一T7プロモーターがこれらの重鎖及び軽鎖をコードする遺伝子の発現を調節する。前記発現ベクターそれぞれを大腸菌BL21(DE3)に形質転換させて大腸菌形質転換体BL21(DE3)/ psDLHF_B (HM10920) 及び BL21(DE3)/psDLHF_BP(HM10921)を製造し、これらを特許手続上の微生物の寄託の国際的承認に関するブダペスト条約に基づき、寄託番号KCCM−10509及びKCCM−10510として韓国微細物保存センター(KCCM)に2003年10月2日付で寄託した。
【0044】
<2−2> 大腸菌外膜タンパク質A(OmpA)シグナル配列及び抗−腫瘍壊死因子−アルファFab’を含む発現ベクター製作
大腸菌OmpAシグナル配列(配列番号:24)を用いて抗−腫瘍壊死因子−アルファの重鎖及び軽鎖をコードする遺伝子を分泌的に発現する発現ベクターを次のように製作した。
【0045】
まず、TacプロモーターとOmpAシグナル配列とを含むプラスミドpFlag.CTS(Eastman Chemical Company)をHindIIIで処理し、前記プラスミドのHindIII制限酵素認識部位をクレノウ DNA ポリメラーゼ(Klenow DNA polymerase, NEB, New Egland Biolab USA)で処理して平滑末端にした後、BglII制限酵素で切断して5’−末端は平滑末端を、3’−末端は付着端を有するベクター断片を製作した。また、OmpAシグナル配列と、重鎖及び軽鎖をコードする遺伝子断片とを連結するため、重鎖をコードする遺伝子断片を含むプラスミドpBHCを鋳型とし、配列番号:25及び26のプライマー対を用いたPCRを行って重鎖の可変領域と定常領域、そして変形されたヒンジ配列と終止コドンを含むPCR産物を増幅した。この際、3’−末端に相応する配列番号:26のプライマー対はBglII制限酵素認識部位を挿入するように考案された。このように増幅された重鎖をコードする遺伝子断片を制限酵素BglIIで切断した後、この断片を前記でBglIIによって予め切断されたプラスミドpFlag.CTSに挿入してOmpAシグナル配列に融合された重鎖をコードする遺伝子断片を含むプラスミドpoHCを製作した。前記記載の同一方法に従って、軽鎖をコードする遺伝子断片を含むプラスミドpBLCを鋳型とし、配列番号:27及び28のプライマー対を用いたPCRを行った後、このように増幅されたPCR産物を同一制限酵素処理過程を経ってプラスミドpFlag.CTSに挿入してOmpAシグナル配列に融合された軽鎖をコードする遺伝子断片を含むプラスミドpoLCを製作した。
【0046】
それぞれOmpAシグナル配列に融合された重鎖及び軽鎖をコードする遺伝子が単一Tacプロモーターによって調節されて、発現分泌されるようにダイ−シストロニック発現ベクターを製造するため、鋳型としてプラスミドpoHCを用い、配列番号:29及び30のプライマー対を用いてPCRを行った。これから増幅されたPCR産物は、両末端にSalI制限酵素認識部位を有し、プロモーター領域を除いた重鎖の発現に必要であるシャインダルガノ配列とOmpAシグナル配列とを含む。このPCR産物を制限酵素SalIで切断してから回収した遺伝子断片を同一制限酵素で予め処理されたプラスミドpoLCに挿入してFab’遺伝子を発現する発現ベクターpoDLHFを製作した(図5)。前記発現ベクターpoDLHFで大腸菌BL21を形質転換させて大腸菌形質転換体BL21/poDLHF(HM10922)を製造し、これを韓国微細物保存センター(KCCM)に寄託番号KCCM−10511として2003年10月2日付で寄託した。
【0047】
<2−3> 二重プロモーター(dual−promoter)によって調節される発現ベクターの製作
抗−腫瘍壊死因子−アルファFab’の重鎖と軽鎖をそれぞれ独立したプロモーターによって個別的に発現・分泌させ得る二重−プロモーターシステムを有する発現ベクターを次のように製作した。OmpAシグナル配列に融合された抗−腫瘍壊死因子−アルファFab’の 重鎖をコードする遺伝子断片を含む実施例<2−2>のプラスミドpoHCをBamHIとSalIで処理した。前記酵素−処理されたプラスミドのアガロースゲル電気泳動を行い、抗−腫瘍壊死因子−アルファFab’の重鎖をコードする遺伝子断片、Tacプロモーター及びOmpAシグナル配列を含む、より小さな断片(1.2kb)をゲルから回収した。前記断片の両末端をクレノウ DNA ポリメラーゼで処理して平滑末端にした。また、抗−腫瘍壊死因子−アルファFab’の軽鎖をコードする遺伝子断片を含む実施例<2−2>のプラスミドpoLCをSalIで処理した後、両末端を前記と同一の方法によって平滑末端に作った。それから、重鎖をコードする遺伝子断片をここに挿入してOmpAシグナル配列に融合された軽鎖及び重鎖をコードする遺伝子のそれぞれが独立的にTacプロモーターの調節によって発現分泌される発現ベクターpoDLHF_B/Sを製作した(図6)。前記発現ベクターpoDLHF_B/Sで大腸菌BL21を形質転換させて大腸菌形質転換体BL21/poDLHF_B/S(HM10923)を製造し、これを韓国微生物保存センター(KCCM)に寄託番号KCCM−10512として2003年10月2日付で寄託した。
【0048】
<2−4> 各鎖の発現及び分泌が互いに異なるシグナル配列によって調節される発現ベクターの製作
<2−4−1>
抗−腫瘍壊死因子−アルファFab’α軽鎖をコードする遺伝子は耐熱性エンテロトキシンシグナル配列変異体に融合され、重鎖をコードする遺伝子はOmpAシグナル配列に融合されて、これらの発現及び分泌が単一プロモーターによって調節される発現ベクターを次のように製作した。
【0049】
単一Tacプロモーターの調節によってOmpAシグナル配列に融合された重鎖及び軽鎖をコードする遺伝子がそれぞれ発現される実施例<2−2>の発現ベクターpoDLHFをNdeIとSacIで処理した後、このベクターのアガロースゲル電気泳動を行って、比較的に大きな遺伝子断片(6.6kb)をゲルから回収した。NdeIの認識部位はOmpAシグナル配列の一番目のアミノ酸であるメチオニンに位置し、SacIの認識部位は軽鎖の定常領域に位置した。従って、前記発現ベクターをNdeI及びSacIで処理すると、OmpAシグナル配列は含まないが、軽鎖の可変領域の全部と軽鎖の定常領域の一部を含む遺伝子断片が回収できる。
【0050】
重鎖をコードする遺伝子断片が耐熱性エンテロトキシンシグナル配列変異体に融合されたプラスミドであるpmsLCを鋳型とし、配列番号:31及び20のプライマー対を用いてPCRを行った。配列番号:31の5’−プライマーはNdeI認識部位を含み、耐熱性エンテロトキシンシグナル配列の一番目のアミノ酸に結合して、配列番号:20の3’−プライマーはSacI認識部位を含む。これから増幅されたPCR産物をNdeIとSacIで処理した後、欠失した軽鎖領域を有するプラスミドpoDLHFに挿入して発現ベクターpmsoDLHF_N/Sを製作した(図7)。前記発現ベクター pmsoDLHF_N/Sを大腸菌BL21に形質組転換させて大腸菌形質転換体BL21/pmsoDLHF_N/S(HM10924)を製造し、これを韓国微細物保存センターに寄託番号KCCM−10513として2003年10月2日付で寄託した。
【0051】
<2−4−2>
前記実施例<2−4−1>で製造された発現ベクターとは異なって、抗−腫瘍壊死因子−アルファFab’の重鎖をコードする遺伝子は耐熱性エンテロトキシンシグナル配列変異体に融合され、軽鎖をコードする遺伝子はOmpAシグナル配列に融合されて、単一プロモーターによってこれらの発現及び分泌が調節される発現ベクターを次のように製作した。
【0052】
実施例<2−1>の発現ベクターpsDLHF_BpをSacIとKpnIで切断した後、アガロースゲル電気泳動を行って小さな遺伝子断片(0.6kb)をゲルから回収した。SacIの認識部位は軽鎖の定常領域に存在し、KpnIの認識部位は重鎖の定常領域に存在するため、回収された断片は耐熱性エンテロトキシンシグナル配列変異体に融合された軽鎖遺伝子断片を含んでいた。重鎖及び軽鎖をコードする遺伝子のそれぞれがOmpAシグナル配列に融合され、単一Tacプロモーターによって発現されるように製作された発現ベクターpoDLHFを同一制限酵素で処理した後、アガロースゲル電気泳動を行った。次いで、サイズが大きい遺伝子断片(7.4kb)をゲルから回収し、これを上で回収した軽鎖をコードする遺伝子断片に連結して発現ベクターpmsoDLHF_S/Kを製作した(図8)。前記発現ベクターpmsoDLHF_S/Kを大腸菌BL21に形質組転換させて大腸菌形質転換体BL21/pmsoDLHF_S/K(HM10925)を製造し、これを韓国微生物保存センターに寄託番号KCCM−10516として2003年10月2日付で寄託した。
【0053】
実施例3:抗−腫瘍壊死因子−アルファFab’遺伝子の発現
前記実施例2で製造された大腸菌形質転換体HM10920〜HM10925をそれぞれ5L体積の発酵槽(Marubishi)に接種して発酵させた後、大腸菌形質転換による抗−腫瘍壊死因子−アルファFab’の発現有無を次のように調査した。
【0054】
LB培地100mlに前記形質転換体それぞれを一晩中振盪予備培養してから、発酵槽に接種して培養を進めた。発酵槽の温度は35℃あるいは30℃に維持し、嫌気状態で進むのを防止するために空気を20vvmで投入し、培地を500rpmで撹拌した。微生物の成長のために補助エネルギー源としてグルコース(glucose)と酵母エキス(yeast extract)を微生物の発酵状態によって供給した。600nmでOD値が80に達した時、誘導物質(inducer)であるIPTGを発酵槽に添加した。600nmでOD値が120〜140に達するまで、前記発酵を40〜45時間行った。
【0055】
前記醗酵培養液を遠心分離(20,000g、30分)して上澄液のみを取って、それぞれの形質転換体から発現された抗−腫瘍壊死因子−アルファFab’の量を測定するために前記上澄液をELISAで分析した。ヒト抗体CH1に対する単クローン抗体(MAB1304,Chemicon international)を10mMカーボネイト緩衝溶液(pH 9.6)に1ug/mlの濃度で溶解させた後、96−ウェルプレートをウェル当たり200ngの量で前記抗体溶液で4℃で一晩中コーティングした後、 PBS−T溶液(137mM NaCl, 2mM KCl, 10 mM Na2HPO4, 2 mM KH2PO4 0.05% トゥイーン20)で3回洗浄した。ウシ血清アルブミンを1%の濃度でPBS−T溶液に溶解させたブロッキング溶液を用意した。前記ブロッキング溶液250ulを各ウェルに点滴した後、前記ウェルプレートを常温で1時間放置しておいて、同一のPBS−T溶液で3回洗浄した。標準液と試料液を、適当な濃度にPBS−T溶液で希釈した後、抗体がコーティングされたウェルに添加した。前記ウェルプレートを常温で1時間放置させて反応させてから、再度PBS−T溶液で3回洗浄した。標準液は、ヒト免疫グロブリンG(IB−グロブリンA、緑十字 PBM)をタンパク質分解酵素であるパパインで切断し、一連のカラムを用いてFabを精製し、前記FabをPBS−T溶液に1ng/mlの濃度で溶解させ、二倍ずつ希釈して製造した。ヒト軽鎖の定常領域に対する多クローン抗体にパラクサイドが共有結合された、コンジュゲート試料(A7164、Sigma社)は、ブロッキング溶液で1000:1に希釈して用いた。希釈された200ulの試料を各ウェルに点滴した後、前記ウェルプレートを1時間常温で反応させた。反応が完了した後、各ウェルをPBS−T溶液で3回洗浄した。発色溶液AとB(カラー−A−安定化パラクサイド溶液及びカラーB−安定化クロモゲン溶液、DY 999、R&D Systems社)を同量で混合した後、各ウェルに前記発色溶液混合物を200ulずつ添加して前記ウェルプレートを30分間放置した。次いで、2M硫酸を各ウェルに50ulずつ添加して反応を停止させた。450nmの標準液と試料液の吸光度を測定するために、前記ウェルプレートをマイクロプレートリーダー(Molecular Device社)で分析した。その結果を下記表1に示した。
【0056】
【表1】
【0057】
実施例4;タンパク質発現の確認
<4−1>SDS−PAGE
前記実施例3で製造された大腸菌形質転換体それぞれから発現された抗−腫瘍壊死因子−アルファFab’を検出するために,発現されたタンパク質を、Fabに親和力を有するタンパク質−Gカラムを用いて、次の方法で部分的に精製した後、SDS−PAGE電気泳動を行った。
【0058】
ハイトラップタンパク質G HPカラム(Hitrap Protein G HP、Amersharm Bioscience社)に10カラム体積の平衡緩衝溶液(20mM リン酸ナトリウム、pH7.0)を1分当たり1mlの流速で流し、充分に平衡化させた後、前記実施例3から得た醗酵培養上澄液を前記カラムにローディングした。試料のローディングが完了した後、前記カラムを10カラム体積の平衡緩衝溶液で洗浄してから、カラムに吸着された抗−腫瘍壊死因子−アルファFab’を5カラム体積の溶出液(0.1M グリシン HClpH2.7)で溶出した。
【0059】
このように溶出された抗−腫瘍壊死因子−アルファFab’は、二つの異なるタンパク質、即ち、ジスルフィド結合によって連結された重鎖と軽鎖の結合によって形成された異型接合体であるため、還元条件下でのSDS−PAGEと非−還元条件下でのSDS−PAGEとにおける移動様式が異なるようになる。図9は、前記と同様に精製された抗−腫瘍壊死因子−アルファFab’を15%クライテリオンゲル(criteriongel、Bio-Rad社)を用いて非−還元状態のSDS−PAGEゲルと還元状態のSDS−PAGEゲルに展開させてその移動様式を比べたものである。図9で、レーン2及び3は還元条件下でSDS−PAGEゲルに展開したものであり、レーン4及び5は非−還元条件下でSDS−PAGEゲルに展開したものである。レーン1は、予め染色された低い範囲標準タンパク質マーカー(prestained low-range standard marker, Bio-Rad社)であり;レーン2及び4は、実施例3で用いられたFab標準タンパク質であり;レーン3及び5は、大腸菌形質転換体HM10922(KCCM−10511)を醗酵させ生産された精製試料である。
【0060】
図9に示すように,還元条件下でのSDS−PAGEにおいては重鎖と軽鎖との結合が還元的に切断され、重鎖及び軽鎖は略同一大きさの個別的な単量体として移動する(重鎖の分子量[約24kDa]は軽鎖の分子量[約23kDa]と類似しており,これらはゲル上で一つの分子のように見える)。一方、重鎖及び軽鎖は、非−還元条件下でのSDS−PAGEにおいてはジスルフィド結合で連結された異型接合体として存在し、その結果、約47kDaの移動距離を示した。
【0061】
<4−2> N−末端配列分析
前記実施例<4−1>のSDS−PAGEによって決定された約47kDaの分子量を有する異型接合体のアミノ酸配列を分析するため、基礎科学支援研究所ソウル支店に前記タンパク質のN−末端配列分析を依頼し、分析する試料は次のように用意した。
【0062】
PVDF膜(BioRad社)をメタノールに約2−3秒間漬けて活性化させてから、ブロッキング溶液(170mM グリシン,25mM Tris・HCl(pH8),20% メタノール)に充分に濡らした。実施例<4−1>の非−還元条件下で展開されたSDS−PAGEゲルを前記と同様に用意されたPVDF膜にブロッティングキット(HoeferSemi-Dry Transfer unit, Amersham)を用いて1時間程度ブロッティングを行った。PVDF膜へ移動されたタンパク質をクマシーブルーR−250(Amnesco社)で3−4秒間染色してから、脱色液(水:酢酸:メタノール=5:1:4)で洗浄した。洗浄された膜からタンパク質を含む部位をはさみで切り取って、N−末端配列分析に用いた。
【0063】
抗−腫瘍壊死因子−アルファFab’の軽鎖のアミノ酸配列はAsp-Ile-Gln-Met-Thr-Gln-Ser(配列番号:34)であり、重鎖のアミノ酸配列はGlu-Val-Gln-Leu-Glu-Val-Asp-Ser(配列番号:35)である。アミノ酸配列分析の結果、本発明の大腸菌形質転換体から分泌的に発現された約47kDaの分子量を有する異型接合体のN−末端は、Asp/Glu-Ile/Val-Gln-Met/Leu-Thr/Val-Gln/Asp-Ser(配列番号:36)であった。前記結果は、本発明によって生産された抗−腫瘍壊死因子−アルファFab’の軽鎖及び重鎖が正常な異型接合体形態で存在することを明確にするものである。
【0064】
<4−3>活性酵素免疫測定(ELISA)
前記実施例3で、大腸菌形質転換体から発現された抗−腫瘍壊死因子−アルファFab’が腫瘍壊死因子−アルファ抗原と結合するか否かを確認するため、次のように、活性酵素免疫測定分析法(ELISA)を行った。
【0065】
実施例<4−1>から得た抗−腫瘍壊死因子−アルファFab’の濃度を280nmでの吸収係数(extinction coefficient=1.43, Humphreys et,al. Protein Expression and Purification 26: 309-320, 2002)を決めることにより測定し、その後、PBS−T緩衝溶液に10ng/mlの濃度で溶解させた。また、腫瘍壊死因子−アルファに結合する標準抗体として、腫瘍壊死因子−アルファと結合し関節炎治療剤としての商業的効能が立証されたヒューミラ(humira,Abbott社)を同一緩衝溶液を用いて同一濃度で製造した。実施例3に記述と同一の方法でウェルプレートを腫瘍壊死因子−アルファにコーティングした後、動物細胞から生産された標準抗体と本発明の大腸菌形質転換体から生産されたFab’とが同一の濃度で同一の結合親和力を現わすかどうかを調査した。この際、標準抗体(対照群)はヒト免疫グロブリンGをタンパク質分解酵素であるパパインで切断した後、一連のカラムで精製して用意したFabを用いた。
【0066】
図10に示したように、対照群として用いられたFab標準タンパク質は、抗−腫瘍壊死因子−アルファに対して結合親和力を全く示さないのに対して,本発明によって大腸菌形質転換体から生産された抗−腫瘍壊死因子−アルファFab’希釈倍数に比例して腫瘍壊死因子−アルファと結合し、標準抗体と同一又はより高い吸光度を現わした。
【0067】
本発明を前記具体的な実施例と関連して記述しましたが、添付された特許請求の範囲によって定義された本発明の範囲内で当分野の熟練者が本発明を多様に変形及び変化させ得ることを理解しなければならない。
【図面の簡単な説明】
【0068】
【図1】図1は、重鎖と軽鎖の定常領域のそれぞれをプラスミドpBluescript SK(−)に挿入して、プラスミドpBH及びpBLの製作過程を示す図である。
【図2】図2は、重鎖と軽鎖の可変領域をそれぞれプラスミドpBH及びpBLに挿入してプラスミドpBHC及びpBLCの製作過程を示す図である。
【図3】図3は、T7プロモーターと大腸菌耐熱性エンテロトキシンシグナル配列に融合された抗体の軽鎖をコードする遺伝子とを含む発現ベクターpmsLC、及びT7プロモーターと大腸菌耐熱性エンテロトキシンシグナル配列に融合された抗体の重鎖をコードする遺伝子とを含む発現ベクターpmsHCのそれぞれの製作過程を示す図である。
【図4】図4は、単一T7プロモーターの調節によって軽鎖と重鎖を個別的に発現・分泌するダイ−シストロニック発現ベクターpsDLHF_B及びpsDLHF−Bpの製作過程を示す図である。
【図5】図5は、Tacプロモーターと大腸菌外膜タンパク質A(OmpA)シグナル配列に融合された抗体の重鎖及び軽鎖をコードする遺伝子とを含む発現ベクターpoDLHFの製作過程を示す図である。
【図6】図6は、個別のTacプロモーターの調節によって重鎖及び軽鎖を発現して分泌する発現ベクターpoDLHF_B/Sの製作過程を示す図である。
【図7】図7は、抗体の軽鎖をコードする遺伝子が大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合され、抗体の重鎖をコードする遺伝子が大腸菌外膜タンパク質A(OmpA)シグナル配列に融合されており、これらの発現及び分泌が単一のTacプロモーターによって調節される発現ベクターpmsoDLHF_N/Sの製作過程を示す図である。
【図8】図8は、抗体の軽鎖をコードする遺伝子が大腸菌外膜タンパク質A(OmpA)シグナル配列変異体に融合されており、抗体の重鎖をコードする遺伝子が大腸菌耐熱性エンテロトキシンシグナル配列に融合され、これらの発現及び分泌が単一のTacプロモーターによって調節される、発現ベクターpmsoDLHF_S/Kの製作過程を示す図である。
【図9】図9は、還元条件下及び非還元条件下で行ったSDS−PAGE分析によって得られた大腸菌形質転換体から発現された抗−腫瘍壊死因子−αFab’の発現パターンを比較した結果を示す図である。
【図10】図10は、大腸菌形質転換体から発現された抗−腫瘍壊死因子−αFab’が腫瘍壊死因子−αと結合するか否かを調べるためのELISAの結果を示す図である。
【特許請求の範囲】
【請求項1】
1)第1大腸菌シグナル配列に融合された抗体断片の軽鎖をコードする遺伝子と第2大腸菌シグナル配列に融合された抗体断片の重鎖をコードする遺伝子とを含む発現ベクターを製作する段階;
2)前記発現ベクターにより微生物を形質転換する段階;
3)前記形質転換された微生物を培地で培養する段階;及び
4)前記培地内に分泌された抗体断片を回収する段階を含む抗体断片を生産する方法。
【請求項2】
前記抗体断片が、キメラ抗体、ヒト化抗体またはヒト抗体から得られることを特徴とする請求項1に記載の方法。
【請求項3】
前記抗体断片が、Fab’、Fab’、F(ab’)2及びscFvからなる群から選ばれることを特徴とする請求項1に記載の方法。
【請求項4】
前記第1及び第2大腸菌シグナル配列のそれぞれが、大腸菌耐熱性エンテロトキシンシグナル配列、外膜タンパク質Aシグナル配列、βラクタマーゼシグナル配列、GeneIIIシグナル配列、PelBシグナル配列及びこれらの変異体からなる群から選ばれることを特徴とする請求項1に記載方法。
【請求項5】
前記第1及び第2大腸菌シグナル配列のそれぞれが、配列番号:17に示される塩基配列を有する大腸菌耐熱性エンテロトキシンシグナル配列変異体、または配列番号:23に示される塩基配列を有する大腸菌外膜タンパク質Aシグナル配列であることを特徴とする請求項4に記載の方法。
【請求項6】
前記軽鎖及び重鎖をコードする遺伝子のそれぞれが大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合され、単一プロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節することを特徴とする請求項5に記載の方法。
【請求項7】
前記軽鎖及び重鎖をコードする遺伝子のそれぞれが大腸菌外膜タンパク質Aシグナル配列に融合され、前記単一プロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節することを特徴とする請求項5に記載の方法。
【請求項8】
前記軽鎖をコードする遺伝子は大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合され、前記重鎖をコードする遺伝子は大腸菌外膜タンパク質Aシグナル配列に融合され、前記単一プロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節することを特徴とする請求項5に記載の方法。
【請求項9】
前記軽鎖をコードする遺伝子は大腸菌外膜タンパク質Aシグナル配列に融合され、前記重鎖をコードする遺伝子は大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合されて、前記単一プロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節することを特徴とする請求項5に記載の方法。
【請求項10】
前記軽鎖及び重鎖をコードする遺伝子のそれぞれが、大腸菌外膜タンパク質Aシグナル配列に融合され、2つのプロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節することを特徴とする請求項5に記載の方法。
【請求項11】
前記プロモーターがT7またはTacプロモーターであることを特徴とする請求項6ないし請求項10のいずれかに記載の方法。
【請求項12】
前記抗体断片が抗−腫瘍壊死因子−アルファ(anti-tumor necrosis factor-alpha, anti-TNF-α)であることを特徴とする請求項1に記載の方法。
【請求項13】
前記発現ベクターがpsDLHF_B、psDLHF_Bp、poDLHF、pmsoDLHF_B/S、pmsoDLHF_N/S及びpmsoDLHF_S/Kからなる群から選ばれることを特徴とする請求項1に記載の方法。
【請求項14】
前記微生物が大腸菌であることを特徴とする請求項1に記載の方法。
【請求項15】
前記発現ベクターにより形質転換された微生物が大腸菌BL21(DE3)/psDLHF_B(HM10920)(KCCM-10509)、BL21(DE3)/psDLHF_BP(HM10921)(KCCM-10510)、BL21/poDLHF(HM10922)(KCCM-10511)、BL21/poDLHF_B/S(HM10923)(KCCM-10512)、BL21/pmsoDLHF_N/S(HM10924)(KCCM-10513)及びBL21/pmsoDLHF_S/K(HM10925)(KCCM-10516)からなる群から選ばれることを特徴とする請求項14に記載の方法。
【請求項16】
抗体断片の軽鎖をコードする遺伝子は第1大腸菌シグナル配列に融合され、抗体断片の重鎖をコードする遺伝子は第2大腸菌シグナル配列に融合され、発現ベクターから発現された抗体断片が培養培地内に分泌される発現ベクター。
【請求項17】
前記抗体断片が、キメラ抗体、ヒト化抗体またはヒト抗体であることを特徴とする請求項16に記載の発現ベクター。
【請求項18】
前記抗体断片が、Fab’、Fab’、F(ab’)2及びscFvからなる群から選ばれることを特徴とする請求項16に記載の発現ベクター。
【請求項19】
前記第1及び第2大腸菌シグナル配列のそれぞれが、大腸菌耐熱性エンテロトキシンシグナル配列、外膜タンパク質Aシグナル配列、ベータラクタマーゼシグナル配列、GeneIIIシグナル配列、PelBシグナル配列及びこれらの変異体からなる群から選ばれることを特徴とする請求項16に記載の発現ベクター。
【請求項20】
前記第1及び第2大腸菌シグナル配列のそれぞれが、配列番号:17に示される塩基配列を有する大腸菌耐熱性エンテロトキシンシグナル配列変異体、または配列番号:23に示される塩基配列を有する大腸菌外膜タンパク質Aシグナル配列であることを特徴とする請求項19に記載の発現ベクター。
【請求項21】
前記軽鎖及び重鎖をコードする遺伝子のそれぞれが大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合され、単一プロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節することを特徴とする請求項20に記載の発現ベクター。
【請求項22】
前記抗体断片が抗−腫瘍壊死因子−アルファの断片であることを特徴とする請求項21に記載の発現ベクター。
【請求項23】
前記単一プロモーターがT7プロモーターであることを特徴とする請求項22に記載の発現ベクター。
【請求項24】
psDLHF_BまたはpsDLHF_Bpであることを特徴とする請求項23に記載の発現ベクター。
【請求項25】
前記軽鎖及び重鎖をコードする遺伝子のそれぞれが大腸菌外膜タンパク質Aシグナル配列に融合され、単一プロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節することを特徴とする請求項20に記載の発現ベクター。
【請求項26】
前記抗体断片が抗−腫瘍壊死因子−アルファの断片であることを特徴とする請求項25に記載の発現ベクター。
【請求項27】
前記単一プロモーターがTacプロモーターであることを特徴とする請求項26に記載の発現ベクター。
【請求項28】
poDLHFであることを特徴とする請求項27に記載の発現ベクター。
【請求項29】
前記軽鎖及び重鎖をコードする遺伝子のそれぞれが大腸菌外膜タンパク質Aシグナル配列に融合され、二つのプロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を独立的に調節することを特徴とする請求項20に記載の発現ベクター。
【請求項30】
前記抗体断片が抗−腫瘍壊死因子−アルファの断片であることを特徴とする請求項29に記載の発現ベクター。
【請求項31】
前記プロモーターがTacプロモーターであることを特徴とする請求項30に記載の発現ベクター。
【請求項32】
poDLHF_B/Sであることを特徴とする請求項31に記載の発現ベクター。
【請求項33】
前記軽鎖をコードする遺伝子は大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合され、前記重鎖をコードする遺伝子は大腸菌外膜タンパク質Aシグナル配列に融合され、単一プロモーターが前記軽鎖及び重鎖遺伝子の発現を調節することを特徴とする請求項20に記載の発現ベクター。
【請求項34】
前記抗体断片が抗−腫瘍壊死因子−アルファの断片であることを特徴とする請求項33に記載の発現ベクター。
【請求項35】
前記単一プロモーターがTacプロモーターであることを特徴とする請求項34に記載の発現ベクター。
【請求項36】
pmsoDLHF_N/Sであることを特徴とする請求項35に記載の発現ベクター。
【請求項37】
前記軽鎖をコードする遺伝子は大腸菌外膜タンパク質Aシグナル配列に融合され、前記重鎖をコードする遺伝子は大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合され、単一プロモーターが前記軽鎖及び重鎖遺伝子の発現を調節することを特徴とする請求項20に記載の発現ベクター。
【請求項38】
前記抗体断片が抗−腫瘍壊死因子−アルファの断片であることを特徴とする請求項37に記載の発現ベクター。
【請求項39】
前記単一プロモーターがTacプロモーターであることを特徴とする請求項38に記載の発現ベクター。
【請求項40】
pmsoDLHF_S/Kであることを特徴とする請求項39に記載の発現ベクター。
【請求項41】
請求項16に記載の発現ベクターにより形質転換された微生物。
【請求項42】
大腸菌であることを特徴とする請求項41記載の微生物。
【請求項43】
大腸菌BL21(DE3)/psDLHF_B(HM10920)(KCCM-10509)、BL21(DE3)/psDLHF_BP(HM10921)(KCCM-10510)、BL21/poDLHF(HM10922)(KCCM-10511)、BL21/poDLHF_B/S(HM10923)(KCCM-10512)、BL21/pmsoDLHF_N/S(HM10924)(KCCM-10513)、及びBL21/pmsoDLHF_S/K(HM10925)(KCCM-10516)からなる群から選ばれることを特徴とする請求項42記載の微生物。
【請求項1】
1)第1大腸菌シグナル配列に融合された抗体断片の軽鎖をコードする遺伝子と第2大腸菌シグナル配列に融合された抗体断片の重鎖をコードする遺伝子とを含む発現ベクターを製作する段階;
2)前記発現ベクターにより微生物を形質転換する段階;
3)前記形質転換された微生物を培地で培養する段階;及び
4)前記培地内に分泌された抗体断片を回収する段階を含む抗体断片を生産する方法。
【請求項2】
前記抗体断片が、キメラ抗体、ヒト化抗体またはヒト抗体から得られることを特徴とする請求項1に記載の方法。
【請求項3】
前記抗体断片が、Fab’、Fab’、F(ab’)2及びscFvからなる群から選ばれることを特徴とする請求項1に記載の方法。
【請求項4】
前記第1及び第2大腸菌シグナル配列のそれぞれが、大腸菌耐熱性エンテロトキシンシグナル配列、外膜タンパク質Aシグナル配列、βラクタマーゼシグナル配列、GeneIIIシグナル配列、PelBシグナル配列及びこれらの変異体からなる群から選ばれることを特徴とする請求項1に記載方法。
【請求項5】
前記第1及び第2大腸菌シグナル配列のそれぞれが、配列番号:17に示される塩基配列を有する大腸菌耐熱性エンテロトキシンシグナル配列変異体、または配列番号:23に示される塩基配列を有する大腸菌外膜タンパク質Aシグナル配列であることを特徴とする請求項4に記載の方法。
【請求項6】
前記軽鎖及び重鎖をコードする遺伝子のそれぞれが大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合され、単一プロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節することを特徴とする請求項5に記載の方法。
【請求項7】
前記軽鎖及び重鎖をコードする遺伝子のそれぞれが大腸菌外膜タンパク質Aシグナル配列に融合され、前記単一プロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節することを特徴とする請求項5に記載の方法。
【請求項8】
前記軽鎖をコードする遺伝子は大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合され、前記重鎖をコードする遺伝子は大腸菌外膜タンパク質Aシグナル配列に融合され、前記単一プロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節することを特徴とする請求項5に記載の方法。
【請求項9】
前記軽鎖をコードする遺伝子は大腸菌外膜タンパク質Aシグナル配列に融合され、前記重鎖をコードする遺伝子は大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合されて、前記単一プロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節することを特徴とする請求項5に記載の方法。
【請求項10】
前記軽鎖及び重鎖をコードする遺伝子のそれぞれが、大腸菌外膜タンパク質Aシグナル配列に融合され、2つのプロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節することを特徴とする請求項5に記載の方法。
【請求項11】
前記プロモーターがT7またはTacプロモーターであることを特徴とする請求項6ないし請求項10のいずれかに記載の方法。
【請求項12】
前記抗体断片が抗−腫瘍壊死因子−アルファ(anti-tumor necrosis factor-alpha, anti-TNF-α)であることを特徴とする請求項1に記載の方法。
【請求項13】
前記発現ベクターがpsDLHF_B、psDLHF_Bp、poDLHF、pmsoDLHF_B/S、pmsoDLHF_N/S及びpmsoDLHF_S/Kからなる群から選ばれることを特徴とする請求項1に記載の方法。
【請求項14】
前記微生物が大腸菌であることを特徴とする請求項1に記載の方法。
【請求項15】
前記発現ベクターにより形質転換された微生物が大腸菌BL21(DE3)/psDLHF_B(HM10920)(KCCM-10509)、BL21(DE3)/psDLHF_BP(HM10921)(KCCM-10510)、BL21/poDLHF(HM10922)(KCCM-10511)、BL21/poDLHF_B/S(HM10923)(KCCM-10512)、BL21/pmsoDLHF_N/S(HM10924)(KCCM-10513)及びBL21/pmsoDLHF_S/K(HM10925)(KCCM-10516)からなる群から選ばれることを特徴とする請求項14に記載の方法。
【請求項16】
抗体断片の軽鎖をコードする遺伝子は第1大腸菌シグナル配列に融合され、抗体断片の重鎖をコードする遺伝子は第2大腸菌シグナル配列に融合され、発現ベクターから発現された抗体断片が培養培地内に分泌される発現ベクター。
【請求項17】
前記抗体断片が、キメラ抗体、ヒト化抗体またはヒト抗体であることを特徴とする請求項16に記載の発現ベクター。
【請求項18】
前記抗体断片が、Fab’、Fab’、F(ab’)2及びscFvからなる群から選ばれることを特徴とする請求項16に記載の発現ベクター。
【請求項19】
前記第1及び第2大腸菌シグナル配列のそれぞれが、大腸菌耐熱性エンテロトキシンシグナル配列、外膜タンパク質Aシグナル配列、ベータラクタマーゼシグナル配列、GeneIIIシグナル配列、PelBシグナル配列及びこれらの変異体からなる群から選ばれることを特徴とする請求項16に記載の発現ベクター。
【請求項20】
前記第1及び第2大腸菌シグナル配列のそれぞれが、配列番号:17に示される塩基配列を有する大腸菌耐熱性エンテロトキシンシグナル配列変異体、または配列番号:23に示される塩基配列を有する大腸菌外膜タンパク質Aシグナル配列であることを特徴とする請求項19に記載の発現ベクター。
【請求項21】
前記軽鎖及び重鎖をコードする遺伝子のそれぞれが大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合され、単一プロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節することを特徴とする請求項20に記載の発現ベクター。
【請求項22】
前記抗体断片が抗−腫瘍壊死因子−アルファの断片であることを特徴とする請求項21に記載の発現ベクター。
【請求項23】
前記単一プロモーターがT7プロモーターであることを特徴とする請求項22に記載の発現ベクター。
【請求項24】
psDLHF_BまたはpsDLHF_Bpであることを特徴とする請求項23に記載の発現ベクター。
【請求項25】
前記軽鎖及び重鎖をコードする遺伝子のそれぞれが大腸菌外膜タンパク質Aシグナル配列に融合され、単一プロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を調節することを特徴とする請求項20に記載の発現ベクター。
【請求項26】
前記抗体断片が抗−腫瘍壊死因子−アルファの断片であることを特徴とする請求項25に記載の発現ベクター。
【請求項27】
前記単一プロモーターがTacプロモーターであることを特徴とする請求項26に記載の発現ベクター。
【請求項28】
poDLHFであることを特徴とする請求項27に記載の発現ベクター。
【請求項29】
前記軽鎖及び重鎖をコードする遺伝子のそれぞれが大腸菌外膜タンパク質Aシグナル配列に融合され、二つのプロモーターが前記軽鎖及び重鎖をコードする遺伝子の発現を独立的に調節することを特徴とする請求項20に記載の発現ベクター。
【請求項30】
前記抗体断片が抗−腫瘍壊死因子−アルファの断片であることを特徴とする請求項29に記載の発現ベクター。
【請求項31】
前記プロモーターがTacプロモーターであることを特徴とする請求項30に記載の発現ベクター。
【請求項32】
poDLHF_B/Sであることを特徴とする請求項31に記載の発現ベクター。
【請求項33】
前記軽鎖をコードする遺伝子は大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合され、前記重鎖をコードする遺伝子は大腸菌外膜タンパク質Aシグナル配列に融合され、単一プロモーターが前記軽鎖及び重鎖遺伝子の発現を調節することを特徴とする請求項20に記載の発現ベクター。
【請求項34】
前記抗体断片が抗−腫瘍壊死因子−アルファの断片であることを特徴とする請求項33に記載の発現ベクター。
【請求項35】
前記単一プロモーターがTacプロモーターであることを特徴とする請求項34に記載の発現ベクター。
【請求項36】
pmsoDLHF_N/Sであることを特徴とする請求項35に記載の発現ベクター。
【請求項37】
前記軽鎖をコードする遺伝子は大腸菌外膜タンパク質Aシグナル配列に融合され、前記重鎖をコードする遺伝子は大腸菌耐熱性エンテロトキシンシグナル配列変異体に融合され、単一プロモーターが前記軽鎖及び重鎖遺伝子の発現を調節することを特徴とする請求項20に記載の発現ベクター。
【請求項38】
前記抗体断片が抗−腫瘍壊死因子−アルファの断片であることを特徴とする請求項37に記載の発現ベクター。
【請求項39】
前記単一プロモーターがTacプロモーターであることを特徴とする請求項38に記載の発現ベクター。
【請求項40】
pmsoDLHF_S/Kであることを特徴とする請求項39に記載の発現ベクター。
【請求項41】
請求項16に記載の発現ベクターにより形質転換された微生物。
【請求項42】
大腸菌であることを特徴とする請求項41記載の微生物。
【請求項43】
大腸菌BL21(DE3)/psDLHF_B(HM10920)(KCCM-10509)、BL21(DE3)/psDLHF_BP(HM10921)(KCCM-10510)、BL21/poDLHF(HM10922)(KCCM-10511)、BL21/poDLHF_B/S(HM10923)(KCCM-10512)、BL21/pmsoDLHF_N/S(HM10924)(KCCM-10513)、及びBL21/pmsoDLHF_S/K(HM10925)(KCCM-10516)からなる群から選ばれることを特徴とする請求項42記載の微生物。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
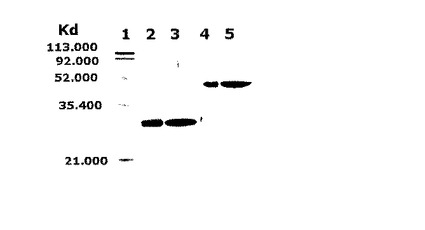
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公表番号】特表2007−508032(P2007−508032A)
【公表日】平成19年4月5日(2007.4.5)
【国際特許分類】
【出願番号】特願2006−535264(P2006−535264)
【出願日】平成16年10月14日(2004.10.14)
【国際出願番号】PCT/KR2004/002625
【国際公開番号】WO2005/038031
【国際公開日】平成17年4月28日(2005.4.28)
【出願人】(599139534)ハンミ ファーム. シーオー., エルティーディー. (56)
【Fターム(参考)】
【公表日】平成19年4月5日(2007.4.5)
【国際特許分類】
【出願日】平成16年10月14日(2004.10.14)
【国際出願番号】PCT/KR2004/002625
【国際公開番号】WO2005/038031
【国際公開日】平成17年4月28日(2005.4.28)
【出願人】(599139534)ハンミ ファーム. シーオー., エルティーディー. (56)
【Fターム(参考)】
[ Back to top ]