
大腸菌中でプラスミドDNAを製造規模で生産するための流加発酵法及び培地
【課題】均質性が高いプラスミドDNAを高収率で製造する方法を提供すること。
【解決手段】プラスミドを有する大腸菌細胞を最初に前培養で増殖させ、引き続き主培養で発酵させ、得られたプラスミドDNAを回収して精製して、製造規模でプラスミドDNAを生産する方法であって、前記主培養がバッチ期と栄養補給期を含む流加法であり、このとき、a)前記バッチ期の培地及び前記栄養補給期中に添加する培地を化学的に規定し、さらに、b)前記栄養補給期の培地は、i)増殖制限基質を含み、かつii)前記栄養補給期の少なくとも一部分の間、予め規定した指数関数に従う栄養補給速度で添加されることによって、予め規定した値に比増殖速度を維持する、前記方法。
【解決手段】プラスミドを有する大腸菌細胞を最初に前培養で増殖させ、引き続き主培養で発酵させ、得られたプラスミドDNAを回収して精製して、製造規模でプラスミドDNAを生産する方法であって、前記主培養がバッチ期と栄養補給期を含む流加法であり、このとき、a)前記バッチ期の培地及び前記栄養補給期中に添加する培地を化学的に規定し、さらに、b)前記栄養補給期の培地は、i)増殖制限基質を含み、かつii)前記栄養補給期の少なくとも一部分の間、予め規定した指数関数に従う栄養補給速度で添加されることによって、予め規定した値に比増殖速度を維持する、前記方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、プラスミドDNA(pDNA)、特に遺伝子療法及びDNAワクチン接種で使用することを意図したpDNAの生産のための大腸菌の発酵に関する。
【背景技術】
【0002】
最近10年間の遺伝子療法及びDNAワクチン接種の臨床的成功により、pDNAの工業用発酵の要求が生じてきた。
遺伝子療法とは、哺乳動物の細胞内での遺伝子の投与、送達及び発現による病気の治療又は予防である。遺伝子療法の究極的な目標は、遺伝子を添加、補正、又は置換することによって遺伝性及び後天性の両障害を治すことである。基本的に、この目標を達成するための2型の遺伝子療法用ベクター、すなわち不活化ウイルスに基づくウイルスベクターと、プラスミドDNAに基づく非ウイルスベクターがある。本発明は、非ウイルスプラスミドDNAの生産に関する。
抗原をコードするpDNAの筋肉内注射が体液と細胞の両免疫反応を引き出すことが実証されて以来、裸のプラスミドDNAが特に重要になってきた。
プラスミドDNAを製造するための発酵法の所望効率は、発酵ブロスの体積当たり(体積収率)又はバイオマスアリコート(aliquot)当たり(比収率)のpDNAの高い収率によって特徴づけされる。本発明の意味では、収率は体積当たり又は細胞重量当たりのプラスミドDNAの濃度である。高収率を超えると、プラスミドはその無傷の共有結合的に閉じた環状(ccc)又はスーパーコイル型で存在するはずである。本発明の意味では、ccc型の割合を“プラスミド均質性”という。開環状(oc)、線状及び二量体又は多量体型のような他のプラスミド型の濃度は精製プラスミドバルクでは最小限に減らすべきであり、結果として発酵中には望ましくない。
【0003】
治療用プラスミドは3つの基本部分、すなわち真核生物プロモーター、多くはサイトメガロウイルス(CMV)プロモーターの制御下の治療遺伝子(“問題の遺伝子”)、原核生物細胞内での自発繁殖の複製開始点(ori)、及び選択マーカー、通常は抗生物質耐性遺伝子から成る。治療遺伝子は臨床的及び医学的関連性があり、oriと選択マーカーの両者は、プラスミド生産中、特に発酵中に重要な役割を果たす。治療用プラスミドの構築のため、キーとなる因子は、細胞当たり多数のプラスミドコピーを複製する複製開始点の選択である。ほとんどの治療用ベクターはColE1-型oriを有する。pBR322由来のColE1開始点を有するプラスミドは細胞当たり50〜100個のプラスミドというコピー数を達成でき、pUC由来のプラスミドは数百というコピー数を達成することができる。
プラスミドを内部に持つ細胞の形質転換及び選択の際、抗生物質選択マーカー及び抗生物質の使用が必要である。しかし、抗生物質淘汰圧は工業生産時には避けるべきである。
従って、プラスミド損失なしでベクターの安定した繁殖を可能にする発酵法を開発することが望ましい。
【0004】
細菌宿主株の選択は、pDNAの発酵で考慮すべき重要なもう一つの因子である。望ましい宿主表現型として、高い細胞密度に増殖する能力を有するもの、高いプラスミドコピー数を達成する能力を有するもの、最少のプラスミド-フリー細胞を生成する能力を有するもの、該プラスミドの遺伝子変化の可能性が最少のもの、優先的にスーパーコイルのプラスミドを生産するもの、及び通常の精製手順と適合性のものが挙げられる。大部分の大腸菌株を用いてpDNAを繁殖させることができるが、株は得られるpDNAの質と量に影響を及ぼしうる(Schoenfeld et al., 1995)。現在、pDNA製造のために使用する細菌株について理想的であろう遺伝型又は表現型の特徴に関するコンセンサスがない。
【0005】
pDNAの発酵について多くのアプローチが開示されている。提案方法は、発酵に影響を及ぼす細胞と多くの因子に課される制御のレベルについて異なる。低レベル制御は、簡単にプラスミド保有細胞を増殖させることができるが、高レベルの厳重に制御した発酵は、複製を促す特有の手段によってpDNAの高収率を果たす。
実験室規模のpDNA生産では、振とうフラスコ内でのプラスミド保有細胞の培養が最も簡単な方法であるが、通常は低収率となってしまう。振とうフラスコ培養から得られるプラスミド収率は、培養ブロス1L当たり1.5〜7mgの範囲である(O´Kennedy et al., 2003; Reinikainen et al., 1988; O´Kennedy et al., 2000)。振とうフラスコ培養では、不十分な酸素移動及びpH値制御の実現性の欠如などのいくつかの欠点がpDNA収率を制限する。US 6,255,099では、振とうフラスコ培養でさえ、特定の培地組成と緩衝条件で109mg/LまでのpDNA収率を達成できることを示した。
より高量のプラスミドを得るため、制御された発酵槽内で細胞を培養することが示唆されている。すべての栄養素を最初から与え、培養中に栄養素を添加しない簡単な培養法は、“バッチ培養”又は“バッチ発酵”と呼ばれる。制御された発酵槽内でのバッチ法の適用は、体積当たりのpDNA収率の増加をもたらした。プラスミド/宿主の組合せ及び培地によって、このようなバッチ発酵から得られるpDNAの収量は大きく変化しうる。報告されている典型的なプラスミド収率は3.5〜50mg/Lの範囲である(O´Kennedy et al., 2003; WO 96/40905; US 5,487,986; WO 02/064752; Lahijani et al., 1996)。これら培養は炭素源及び窒素源としていわゆる“複合成分”を含有する培地を用いて行われた。これら成分は生物源から得られ;生物源としては、例えば、酵母抽出物、大豆ペプトン又はカゼイン水解物が挙げられる。
【0006】
排他的又は優先的に複合成分から成る培地を“複合培地”と称する。規定部分(規定した炭素源、塩、微量要素、ビタミン)及び複合部分(窒素源)の両部分から構成されている培地を“半規定”培地と称する。 US 5,487,986では、非常に高量の種々の複合成分(全部で50g/L)を用いた。
複合成分を含有する培地は、これら成分が生物材料由来なので、該培地の組成は普通の自然偏差の基礎にあり、培養プロセスの再現性を低くするという欠点を有する。製造業者が生産方法を変更するとき、又は供給業者を変えるとき、同様のことが当てはまる。複合培地成分を使用することのさらなる欠点は、正確な組成についての不確実性(望ましくない物質の存在)、化学量論収率を計算できないこと、無菌化時の望ましくない生成物の形成、不十分な溶解のため難しい取扱い、培地調製中のダスト及び凝集の形成である。発酵中、複合培地はたやすく泡立つ傾向がある。動物起源の複合成分(肉抽出物、カゼイン水解物)は伝播性海綿状脳萎縮の危険のため、pDNA生産には特に望ましくないので、医薬品局によってその使用が制限されている(CBER 1998)。
【0007】
複合培地成分の欠点のため、いかなる複合成分も含有しない培地が開発されている。“規定”又は“合成”培地と呼ばれるこのような培地は、化学的に規定した物質、すなわちグルコース又はグリセロールのような炭素源、塩、ビタミンから、また、ありうる株の栄養要求性に鑑みて、特有のアミノ酸又はチアミンのような他の物質から排他的に構成される。化学的に規定した培地は、その組成が正確に分かるという利点を有する。このことは、良いプロセス分析、発酵モニタリング及び増殖又は産物形成を促す特定物質の特有の添加を可能にする。周知の組成は、増殖の予測及び栄養素を欠いている可能性の同定を容易にする質量バランス計算の樹立を可能にする。複合培地に比し、規定培地による発酵は、高いプロセス整合性とスケールアップ時の改良された成果を示す。規定培地のさらに実務的局面は、良い溶解度、無菌化時に副生物を阻害することの不在、及び培養中の泡形成が少ないことである(Zhang and Greasham, 1999)。
M9(Sambrook and Russel, 2001)のような、pDNA生産に特定して開発したものではない合成培地は低いpDNA収率をもたらしうる(WO 02/064752)。pDNA生産に特定して設計した規定培地によるバッチ発酵では高収率のpDNAが得られた(Wang et al., 2001; WO 02/064752)。後者はpDNA均質性が90%を超えるccc型であることを示した。WO 02/064752及びWangら(2001)によるpDNAの高収率は、ヌクレオシドの生合成基礎単位であるアミノ酸の補充によって、又はヌクレオシドの直接添加によって達成された。
【0008】
バッチ発酵は一般的に簡単かつ短いが、プラスミドDNA収率が制限されることになるという基本的な欠点を有する。これは、バッチ培地内での高い栄養素濃度における基質阻害と塩沈殿が原因である。さらに、バッチ発酵の増殖速度は直接制御できず;それゆえにバッチ発酵は無制限であり、発酵中絶え間なく変化し、1種以上の栄養素が欠失しているとき又は代謝副生物(例えばアセテート)が細胞の増殖を阻害する場合にのみ終止する。
その結果として、pDNA生産でのバイオマス及びプラスミド収率を高めるため、流加発酵(fed-batch fermentation)が開発された。流加発酵はバッチ期後、栄養補給によって培養に1種以上の栄養素を供給する栄養補給期を行う方法である。
ラスミドDNAを生産するための大腸菌の流加発酵では、以下のような種々の戦略が実行されている。
【0009】
一方法は、規定した設定点でプロセスパラメーターを制御するために栄養素を供給することによってフィードバックコントロールアルゴリズムを適用する。従って、フィードバックコントロールは発酵全体にわたって直接細胞活性に関係する。発酵のフィードバックコントロールのために使用されているコントロールパラメーターとしては、pH値、オンライン測定細胞密度又は溶存酸素圧(dissolved oxygen tension)(DOT)が挙げられる。この方法は、供給栄養素によって培養に栄養を過剰に与える危険が少なく、高いバイオマス濃度を得ることができるという利点を有する。
pDNA発酵のため、栄養補給速度によって規定設定点での溶存酸素圧を制御するためのフィードバックアルゴリズムが使用された(WO 99/61633)。
別の方法を適用する場合、より複雑なアルゴリズム、すなわちフィードバック培養法のコントロールパラメーターとしてDOTとpHの両方を使用した(US 5,955,323; Chen et al., 1997)。当該方法では、撹拌速度と濃縮複合培地(グルコース、酵母抽出物)の栄養補給によってDOTを制御し、それによって、同時に水酸化アンモニウムでpHを維持した。
フィードバックアルゴリズムの適用はいくつかの欠点を伴う。1つは、栄養補給速度がDOTのような流動プロセスパラメーターに左右されることである。どんな理由によってもプロセスの刺激はコントロールパラメーターに影響を及ぼしうるので、栄養補給速度に影響を与え、ひいては増殖及びpDNA収率に影響する。例えば、消泡剤を添加しなければならない場合、DOTが変化し(通常減少し)、結果として栄養補給速度が低下する。これは発酵プロセスの再現性を低くする。発酵槽の形状や大きさが異なると異なった酸素移動速度を示すので、プロセスのスケールアップの際にはさらなる困難が生じる。酸素移動速度はDOTにつながるので、大きさを変えた発酵槽のフィードバックコントロール栄養補給速度は異なり、その結果、プロセスは大きさに比例しないだろう。
【0010】
フィードバックコントロールのもう一つの欠点は、比増殖速度を正確に予め規定することも制御することもできないことであり、その結果、産物形成が増殖によって左右されるプロセスでは最適以下のプロセス収率となる。しかし、pDNA発酵では、プラスミドの体積収率及び比収率が比増殖速度に非常に依存することが示された(WO 96/40905; O´Kennedy et al., 2003)。
指数関数に従う栄養補給培地の供給に基づく別の基礎的な栄養補給様式によって、比増殖速度の制御を達成することができる。栄養補給速度は、所望の比増殖速度μに基づいて制御される。規定培地を適用すると、与えられた基質単位Sに基づいて形成されるバイオマスアリコートXを計算することによって、増殖を正確に予測かつ予め規定することができる(バイオマス収量係数YX/Sを考慮して)。
【0011】
WO 96/40905で開示されている発明は、プラスミドDNA生産用の指数関数的流加法を用いて高収率のバイオマスを得るが(1L当たり50gのDCW、乾燥細胞重量)、低いpDNA収率となる(18mg/L;0.36mg/g 乾燥細胞重量)。指数関数的栄養補給の別の例では、30mg/L及び6mg/g DCWというプラスミド収率を達成した(O´Kennedy et al., 2003)。Lahijaniら(1996)により、指数関数的栄養補給と温度制御しうるプラスミド複製の強化を併用することによって、220mg/Lという高いpDNA収率が得られた。これらの指数関数的栄養補給の例では、O´Kennedyら(2003)だけがpDNA均質性について詳述しており、50〜70%のccc型だった。現在、すべての指数関数的流加法がバッチ培地と栄養補給培地の両方で複合成分を使用している。
【発明の概要】
【発明が解決しようとする課題】
【0012】
要するに、治療用プラスミドDNAを製造するための発酵の現在の技術状態は、以下のように特徴づけすることができる:
pDNA生産のために広く適用されているバッチ発酵は、技術的及び経済的な欠点を伴う。バッチ発酵では、複合培地又は半規定培地が使用されることが多く、培養ブロス1L当たり3.5〜68mgの範囲のpDNA収率という結果となる。フィードバックコントロールを適用する流加法は、半規定培地又は複合前培養培地を使用後、主培養で規定培地を用いる。フィードバックアルゴリズムにより、100〜230mg/Lというプラスミド収率が得られる。指数関数的栄養補給発酵は半規定培地を使用する。指数関数的発酵のプラスミド収率は18〜220mg/Lの広い範囲である。一般に、多くのpDNA発酵法は不十分な均質性(すなわち、スーパーコイルプラスミドの割合)に困っている。主培養で規定培地を使用する発酵は例外であり、90%を超えるccc型の割合を達成できる。
【課題を解決するための手段】
【0013】
本発明は、製造規模でプラスミドDNAを生産する方法であって、問題の遺伝子を運ぶプラスミドを有する大腸菌を最初に前培養で増殖させ、引き続き主培養で発酵させる方法に関し、前記主培養はバッチ期と栄養補給期を含む流加法である。バッチ期の培地と栄養補給期中に添加する培地を化学的に規定し、かつ栄養補給期の培地は、該栄養補給期の少なくとも一部分の間、予め規定した指数関数に従う栄養補給速度で添加され、それによって、予め規定した値に比増殖速度を維持する。
本発明の意味では、用語“規定培地”は、化学的に規定した単一成分で排他的に構成される培地を指す。
好ましい態様では、栄養補給培地の添加が指数関数に従う、栄養補給期の前記部分は、該栄養補給期に得られる総乾燥細胞重量の20%を超える量が前記栄養補給期の前記部分中に生成されるような部分である。
好ましい態様では、前培養で用いる培地を含め、すべての培地を化学的に規定する。
本発明の方法では、大腸菌のいずれの株でも使用しうる。本発明で有用な宿主株は、いずれの表現型を有していてもよく;好ましい宿主株は遺伝子relA、endA、及びrecAに突然変異を有する。本発明の最も好ましい態様では、大腸菌株K12 JM108又はその誘導体を使用する。
本発明の方法では、大腸菌内で自発的に複製しうるいずれのプラスミドも使用できる。
好ましいプラスミドは、ColE1-型の複製開始点を有し、最も好ましくは、pUCプラスミド由来のレプリコンである(初めはVieira及びMessing, 1982によって記載された;Yanisch-Perron et al. 1985)。
【0014】
本発明の方法では、バッチ期の培地と栄養補給期の培地を化学的に規定し、好ましくは前培養で使用するものも含め、すべての培地を化学的に規定する。ここで、化学的に規定するとは、これら培地がいかなる複合培地成分も含有しないことを意味する。本発明の方法で使用する培地は、有機炭素源、好ましくはグルコースと、アンモニウム塩のような1種以上の無機窒素源を含む。さらなる成分は、マクロ要素及びミク要素を含有する無機塩である。菌株が特有の栄養素、例えば、チアミンのようなビタミン又はプロリンのようなアミノ酸に対して栄養要求性である場合、これら物質も培地中に存在する。
好ましくは、本発明の培地は、プラスミド保有細胞について淘汰圧を課すために通常使用されるような抗生物質はない。
本発明の好ましい態様では、培地はありうるアミノ酸栄養素要求性と無関係に、イソロイシンを含む。これは、イソロイシンは、菌株がイソロイシンに対して栄養要求性でない場合にも存在してよいことを意味する。菌株がイソロイシン栄養要求性を有する場合、好ましくは前記栄養要求性を補完するために必要な量を超える濃度でイソロイシンが存在する。
増殖速度、ひいては栄養補給速度(これによって増殖速度が維持される)は、細胞の増殖とプラスミド複製を確実にするいずれの値を有していてもよい。
【発明の効果】
【0015】
驚くべきことに、本発明の方法は、600mg/Lを超えるpDNA収率と、45mg/g(乾燥細胞重量)までのpDNA比収率をもたらし、これによって90%を超えるcccというpDNA均質性を達成できることが分かった。
好ましい態様では、低い比増殖速度で操作する発酵では、高コピー数プラスミド(例えばpUC由来)を使用する。
低増殖速度で操作する発酵では、アミノ酸飢餓状態のため、非荷電tRNAが生じてプラスミドコピー数を増やす。relA-ネガティブ大腸菌株を使用すると、代謝の下方制御の点から、細胞はアミノ酸飢餓状態に応答できず、結果としてプラスミド複製が増強される。イソロイシンのさらなる供給はアミノ酸代謝の妨害を持続させ、なおさらにpDNA収率を高める。
驚くべきことに、本発明の方法は、発酵時間全体にわたるpDNAの高い均質性と共に、プラスミドDNAの顕著に高い比収率及び体積収率をもたらすことが分かった。
【図面の簡単な説明】
【0016】
【図1】実施例1の発酵のプラスミドの体積収率と比収率及び増殖の曲線を示す。
【図2】実施例1の発酵の現在の比増殖速度μを示す。
【図3】実施例1の発酵時間にわたる種々のプラスミド型の割合を示す。
【図4】実施例1の発酵終了時のプラスミドの分析クロマトグラムを示す。
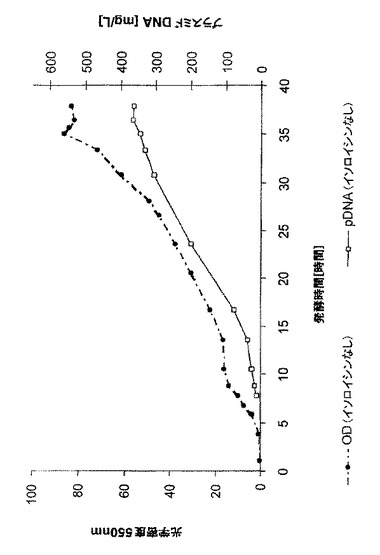
【図5】実施例2のイソロイシンがある場合の光学密度とpDNA体積収率の時間経過を示す。
【図6】実施例2のイソロイシンがない場合の光学密度とpDNA体積収率の時間経過を示す。
【発明を実施するための形態】
【0017】
〔宿主株〕
本発明の方法では、規定培地で培養できるいずれの大腸菌株も使用しうる。好ましくはK-12由来の株を使用する。適切な宿主株の選択は、その宿主の表現型、すなわち該株を特徴づける特異的な突然変異に基づき、及び/又は実験的発酵でいくつかの株を分類学的に比較することに立脚しうる。
本発明の好ましい態様では、菌株はrelA遺伝子に突然変異を有する。この遺伝子はグアノシン四リン酸(ppGpp)の合成に反応性であり、アミノ酸制限時の細胞でのいわゆる“ストリンジェント反応”を引き起こすシグナル分子である。relA+株では、アミノ酸制限がDNAの複製と転写に関与する必須代謝経路の下方制御につながる(Wrobel and Wergrzyn, 1998)。従って、relA遺伝子に突然変異を有する菌株はアミノ酸制限時に増殖し続け、かつプラスミドを複製し続ける。relA遺伝子に突然変異を有することが、化学的に規定した培地で高いプラスミド複製速度で細胞を増殖させるために重要な必須項目であることは分かっている(Hofmann et al., 1989)。
本発明の別の好ましい態様では、relA突然変異に加え、endA(エンドヌクレアーゼA)及びrecA(組換え系)の遺伝子に突然変異を有する宿主株を使用する。これら突然変異はプラスミドの構造の完全性を高めるからである。
好適な宿主株の例は、DH1、DH5、DH5-α、DH10B、JM108、JM109及びHB101である。
本発明の方法では、最も好ましい大腸菌K-12株はJM108(ATCC No. 47107; DSMZ No. 5585;表現型:F-、recA1、endA1、gyrA96、thi-1、hsdR-17、supE44、relA1、λ-及びΔ(lac-proAB);Yanisch-Perron et al., 1985)又はJM108の遺伝子操作で得られるJM108誘導体である。本発明につながる予備試験で行った実験的発酵における宿主スクリーニング中、JM108は、首尾一貫して最高のpDNA収率を示した(体積収率と比収率の両方とも)。
【0018】
〔プラスミド〕
複製開始点と無関係、かつ意図した用途、例えば遺伝子療法ベクターとして又はDNAワクチンとして使用するためのベクターとしての用途と無関係に、大腸菌内で自発的に複製可能ないずれのプラスミドの製造にも本発明の方法を使用できる。例えば、好適なプラスミドはpBR322、pUC18、pUC19、pcDNA3から誘導される(Invitrogen)。
本発明の好ましい態様では、ColE1-型複製開始点を有するプラスミドを用いる。この背後にある論理的根拠は、アミノ酸制限時、アミノ酸で荷電されていない転移RNA(tRNA)が生じることである。これら非荷電tRNAは、プラスミドの複製速度を高めるようにColE1複製開始点と相互作用する。relA-株を使用すると、この作用がより強い(Wrobel and Wergrzyn, 1998)。
本発明の特に好ましい態様では、pUC型のColE1-誘導プラスミド(Vieira and Messing, 1982; Yanisch-Perron et al., 1985)を使用する。このpUCプラスミドは、コピー数を減らす部分Romに突然変異を有するので、普通のColE1-型プラスミドより高いプラスミドコピー数を達成する。rom-プラスミドの複製増強効果は特に、低い比増殖速度で示されたので(Atlung et al., 1999)、本発明では、pUCプラスミドと低い増殖速度培養の組合せが好ましい。
【0019】
〔規定培地〕
主培養のバッチ期及び栄養補給期、好ましくは前培養でも、化学的に規定した培地を使用する。従って、好ましい態様では、全発酵プロセスの間じゅう複合成分を適用しない。
さらに、好ましい態様では、主培養の全体にわたって、プラスミド保有細胞の選択用の抗生物質を使用しない。好ましくは、前培養で用いる培地も抗生物質がない。
前培養及び主培養で用いる培地は同一組成又は異なる組成を有してよい。
前培養、バッチ培地、及び栄養補給培地で有機炭素源を用いる。炭素源は、グルコース、フルクトース、ラクトース、スクロース、アラビノース、グリセロール又は大腸菌によって代謝されうる他のいずれの炭素源でもよく、並びに異なる炭素源の混合物でもよい。
好ましくは、炭素源はグルコースである。
プロセスのそれぞれの時期及び栄養補給培地で用いる炭素源は同一又は異なってよい。
通常、前培養の培地及び主培養のバッチ培地中の炭素源は同一であり、好ましくは、同じ炭素源が栄養補給培地にも含まれる。
バッチ培地では、炭素源の濃度は約1〜約100g/L、好ましくは約10〜約30g/Lの範囲でよい。前培養及び主培養のバッチ培地では炭素源の濃度が同一でよい。
栄養補給期では、栄養補給培地の炭素源は、比増殖速度を制御するための制限基質として働くことが好ましい。本方法の最も簡単な変形では、栄養補給培地は炭素源だけを含有する。
【0020】
栄養補給培地中の炭素源の濃度範囲は約100〜約750g/Lである。グリセロールを用いる場合、グリセロールの濃度は、栄養補給培地中100〜1000g/Lの範囲でよい。好ましくは、炭素源がグルコースであり、約300〜500g/Lの濃度で栄養補給培地中に存在する。
本発明の好ましい態様では、栄養補給培地は増殖制限炭素源と、これとは別に、バッチ培地にさらに存在するいくつか又はすべての培地成分を含有する。
本発明により、前培養及びバッチ培地で初期窒素源として働くように、好ましくはアンモニウム塩又は水酸化アンモニウムを用いる。通常、塩化アンモニウムを使用するが、硫酸アンモニウム、炭酸アンモニウム、リン酸アンモニウム若しくは水酸化アンモニウム又は他のいずれのアンモニウム塩も適切である。アンモニウム濃度が1L当たり約0.1〜約8gのNH4、好ましくはアンモニウム濃度が約0.5〜約2g/L、最も好ましくは約0.6g/Lになるように窒素源の濃度を選択することができる。
【0021】
好ましい態様では、主培養全体にわたって、pH値の制御と窒素の連続供給の両方のためバッチ期と栄養補給期の両期中、水酸化アンモニウムの溶液を別々に供給する。これは最初のバッチ培地にすべてのアンモニウムが与えれらるわけではないという利点を有する。最初のバッチ培地にすべてのアンモニウムが与えれらると培養に毒性効果を及ぼしうる。この好ましい態様では、培地中のアンモニウム濃度は、発酵全体にわたって本質的に一定のままである。水酸化アンモニウムによるpH値の制御は、水酸化ナトリウム又は水酸化カリウムの使用に比べてずっと有益である。水酸化アンモニウムは、浸透ストレスを引き起こして培養を阻害しうる、培地中のナトリウムイオンとカリウムイオンの蓄積と無関係だからである。水酸化アンモニウムの供給は栄養補給と無関係なので、それは増殖速度制限ではなく、かつ栄養補給アルゴリズムの一部でもない。アンモニウム溶液の濃度は約5〜約40%m/mアンモニウムでよい。
【0022】
アミノ酸栄養要求性の宿主株の場合、好ましい態様に従い、宿主株がJM108(その増殖のためにアミノ酸プロリンを必要とする)である場合、培地は(該培地でプロリンの使用は必須である)、1種以上の関連アミノ酸を使用する必要があるかもしれない。さらに、該アミノ酸が栄養補給培地中に存在することも必須である。該アミノ酸が栄養補給溶液中に存在する場合、該アミノ酸は増殖速度を制御するための速度制限因子としても働かず、プラスミドの複製を増強することも意図しない。栄養要求性アミノ酸の濃度範囲は約0.05〜約2g/Lから選択され、好ましくは約0.2g/Lである。
【0023】
本発明の好ましい態様では、培地は、宿主のありうるアミノ酸栄養要求性と無関係に、アミノ酸イソロイシンを含み、イソロイシンはバッチ培養の培地に存在し、好ましくは栄養補給培地にも存在する。任意に、前培養の培地にもイソロイシンが存在してよい。大腸菌K-12株はイソロイシンの生合成に関与する遺伝子の突然変異を有する。この結果、イソロイシンが存在せず、かつ同時にバリンが存在する場合、イソロイシン生合成の抑制となる(Lawther et al., 1981)。従って、バリンはイソロイシン飢餓状態の細胞には毒である。バリンによるイソロイシン代謝との相互作用はバリンの細胞内生成時でさえ見出されたが、この作用はイソロイシンの供給で撤廃された(Andersen et al., 2001)。本発明につながる実験の過程において、バリンの毒性が確認された。さらに、イソロイシンのない規定培地で培養は成長しうるが、イソロイシンの供給がプラスミドDNAの収率を有意に高めることが分かった。従って、規定培地にイソロイシンが存在することは本発明の好ましい態様で必須の特徴である。イソロイシンの濃度範囲は約0.05から約2g/Lの間で選択され、好ましくは約0.1〜約0.3g/L、最も好ましくは約0.2g/Lである。
【0024】
さらに、培地はマクロ要素及びミクロ要素(微量要素)の供給として働く無機塩を含み、かつ任意に、細胞の代謝に正の効果を及ぼす有機物質、例えばクエン酸を含んでよい。
アミノ酸以外、例えば、ビタミンに対する栄養要求性の場合、さらに関連物質が培地に含まれる。宿主株JM108を使用する本発明の態様では、培地にチアミンが存在しなければならない。チアミンは、好ましくは約0.1〜約100mg/Lの濃度、最も好ましくは約1mg/Lの濃度で存在する。
マグネシウムは、通常MgSO4*7H2Oの形態で供給されるが、他のマグネシウム塩も同様に適する。マグネシウム塩の濃度は、通常、約0.1〜約5g/L、好ましくは約0.25g/Lである。
【0025】
リンは、好ましくはKH2PO4及び/又はNa2HPO4*12H2Oの形態で供給されるが、他のリン塩、例えば、K2HPO4、NaH2PO2、Na2HPO4、NH4H2PO4及び/又は(NH4)2HPO4も使用しうる。
リン塩の濃度は、約0.5〜25g/L、好ましくは約2.0〜約15g/L、最も好ましくはそれぞれ約5.0g/LのKH2PO4又は約14.0g/LのNa2HPO4*12H2Oでよい。
カルシウム、鉄、コバルト、マンガン、銅及び亜鉛のような元素は通常以下のように培地中に存在する:約1〜約20の範囲、好ましくは約7.5mg/LのCaCl2*2H2O;約1〜約20の範囲、好ましくは約5.5mg/LのFeSO4*7H2O;約0.5〜約10の範囲、好ましくは約2.5mg/LのCoCl2*6H2O;約0.1〜約5の範囲、好ましくは約2.0mg/LのMnSO4*H2O;約0.05〜約2の範囲、好ましくは約0.25mg/LのCuSO4*5H2O;約0.05〜約2の範囲、好ましくは約0.3mg/LのZnSO4*7H2O。それぞれの元素を含有する他のいずれの塩もそれぞれの濃度範囲で使用できる。
【0026】
上で規定した培地成分及びその濃度は、バッチ培養で使用する培地に関係する。通常、栄養補給培地中の栄養素の濃度は、窒素源(栄養補給期中に別個に添加される)を除き、バッチ培地に比べて約2倍〜約100倍高い。
最小必要条件として、栄養補給培地は増殖制限である量で基質、通常は炭素源を含む。
栄養補給培地は、制限炭素源に加え、バッチ培地に存在するいくつか又はすべての培地成分を含んでよく、或いはこれはあまり好ましくないが、栄養補給培地は、等価な成分、例えば同一元素の異なる塩を含んでもよい。
例として、栄養補給培地の組成は以下のとおり:グルコース 300g/L;MgSO4*7H2O 7.2g/L;L-プロリン 6g/L;L-イソロイシン 6g/L;チアミン塩酸塩 30mg/L、クエン酸 2g/L;KH2PO4 5.4g/L;Na2HPO4*12H2O 14.4g/L;CaCl2*2H2O 220mg/L;FeSO4*7H2O 170mg/L;CoCl2*6H2O 72mg/L、MnSO4*H2O 51mg/L、CuSO4*5H2O 8mg/L及びZnSO4*7H2O 9mg/L。
【0027】
さらなる局面では、本発明は、製造規模で大腸菌内にてプラスミドDNAを生産するための培地に関する。この培地は、以下の成分を含む、化学的に規定した培地である。
a)グルコース、グリセロール、フルクトース、ラクトース、スクロース、アラビノース、又はその混合物から選択される有機炭素源;
b)アンモニウム塩及び水酸化アンモニウムから選択される無機窒素源(該窒素源は、前記培地の成分として存在し、及び/又は発酵中に前記培地に添加される);
c)マクロ要素及びミクロ要素の供給として働く無機イオン;
d)任意に、大腸菌株の栄養要求性を補完する1種以上の物質;及び
e)イソロイシン。
本発明の培地は、本発明の方法のバッチ期及び/又は栄養補給期において、或いは製造規模でプラスミドDNAを生産するための大腸菌を培養するため、すなわち、いずれのバッチ培養又は流加培養でも大腸菌の発酵のための他のいずれの方法での培地としても有用である。
【0028】
さらなる態様では、本発明の培地は、バッチ発酵の開始時に存在し、又は流加発酵のバッチ期の開始時に存在する培地であり、以下の成分を含む:
a)約10〜約30g/Lの濃度のグルコース;
b)アンモニウム濃度が約0.5〜約2g/Lになる濃度のアンモニウム塩又は水酸化アンモニウム;
c)マクロ要素及びミクロ要素の供給として働く無機イオン;
d)任意に、大腸菌株の栄養要求性を補完する1種以上の物質;及び
e)約0.1〜約0.3g/Lの濃度のイソロイシン。
別の態様では、本発明の培地は、流加発酵の栄養補給期中に添加する栄養補給培地であり、以下の成分を含む:
a)約300〜約500g/Lの濃度のグルコース;
b)マクロ要素及びミクロ要素を供給するために働く無機イオン;
c)任意に、大腸菌株の栄養要求性を補完する1種以上の物質;及び
d)約6g/Lの濃度のイソロイシン。
【0029】
〔指数関数的栄養補給手順〕
本発明により、いずれかの所望の予め規定した比増殖速度μで培養を増殖させるため、栄養補給期の少なくとも一部分の間、培地(“栄養補給培地”)の添加を指数関数に従って行う。
この最も一般的な定義では、指数関数は、それぞれ方程式Vt=定数*eμt又はFt=定数*eμtで定義される。この方程式は、関数Vt=定数*eμt+A又はFt=定数*eμt+Aをも包含する。式中、Aは、発酵の規模によって決まる広い範囲で変化しうる値である。好ましくは、A=0である。値Aが0でないと、A=0の関数に比べて上方(A>0)又は下方(A>0)にシフトする関数となる。
【0030】
本発明の好ましい態様では、栄養補給培地を添加する様式を定義する関数は以下のとおりである。

【0031】
この関数中、Vtは、栄養補給開始から計算される時間間隔t[時間]で添加される栄養補給培地の体積[L]である。X0は、栄養補給開始時点のバイオマス乾燥細胞重量の総量[g]である。YX/Sは、バイオマス収量係数(基質1g当たりの乾燥細胞重量のg数)であり、CSは、栄養補給培地中の増殖制限基質(通常、有機炭素源)の濃度[g/L]である。
【0032】
栄養補給様式を定義する関数は、以下のように栄養補給培地の流速で特徴づけすることもできる。
この関数中、Ftは、栄養補給の開始から計算される時点t[時間]で添加される栄養補給培地の流速[L/時間]である。
【0033】
いずれかの所望の予め決定した値で、及び/又は以前の発酵で最適であると決定されたように若しくは実験的発酵の結果に基づいて、比増殖速度μ[時間-1]を選択することができる。バイオマス収量係数は文献から取り出し、又は予備培養実験で決定することができる。光学密度を測定し、又は予備実験の値から取り出し、又はバッチ培地中の初期基質濃度とバイオマス収量係数によって理論値を計算することによって、栄養補給開始時のバイオマスの量X0を決定することができる。
上記方程式に従う、該栄養補給様式の独特な特徴は、栄養補給開始時点と所定時点との間の時間に排他的に依存する。このような栄養補給プロフィールにより、いずれの所望の値でも比増殖速度を予め規定することができる。
種々の方法で、栄養補給を開始する時点を決定することができる。開始時点は、バッチ培地中の炭素源が枯渇したときに、炭素源濃度のオンライン若しくはオフライン測定によって、又は溶存酸素圧の増加(該基質の枯渇を示唆する)によって決定されるように、選定することができる。さらに、特定のバイオマス値(バイオマスのオンライン若しくはオフライン測定によって決定されるような)と相関させるように、又は発酵の開始からの予め規定した時間間隔に応じて、栄養補給の開始時点を選択することができる。
本発明の方法では、指数関数に従った栄養補給培地の供給は種々の方法で達成することができる。本発明の好ましい態様では、栄養補給速度は、発酵槽のプロセスコントロールシステムによって連続的に計算かつ制御され、かつ栄養補給速度は、指数関数に従って連続的に上昇する。それにより、栄養補給培地の質量の流れは、質量流量計(マスフローコントローラー)によって、又は秤で測定される栄養補給容器の重量に応じて培地を供給するポンプによって制御される。
【0034】
本発明の別の態様では、指数関数に従って段階的に栄養補給速度を上昇させることによって、半連続的に栄養補給速度を高める。例えば、所定の時間間隔で、例えば、毎時間栄養補給培地の体積又は質量を再計算することができ、その新しい栄養補給速度を必要な値に調整する。栄養補給速度の上昇は段階的であるが、結果として生じる栄養補給プロフィールは、それでもやはり指数関数的である。
本発明のさらに別の態様では、栄養補給速度を不連続的に上昇させる。これは、規定時点における栄養補給培地の必要量を計算し、その後、(現在のバイオマスに基づいて、選択した時間間隔で予め規定し、又は計算した)増加量の栄養補給培地をパルス様式で添加することによって達成できる。細胞は連続的に栄養補給されないが、それでもなおこの方法は、一定の比増殖速度につながる指数関数的な栄養補給プロフィールをもたらす。
本発明の方法では、増殖とプラスミド複製を確実にするいずれの比増殖速度μも指数関数的栄養補給のために予め規定することができる。最高かつ至適な比増殖速度は、個々の宿主/プラスミドの組合せによって決まり、かつ実験的な流加発酵又は連続(ケモスタット)発酵における各宿主-プラスミド系のケースバイケースの基礎に基づいて決定することができる。大腸菌の発酵に好適な比増殖速度は、約0.03〜約1.5時間-1の範囲である。
本発明の好ましい態様では、約0.05〜約0.15時間-1の低い比増殖速度を使用し、最も好ましくは、増殖速度は約0.1時間-1である。低い比増殖速度の利点は、プラスミドが高いコピー数に複製できるようにし、結果としてバイオマス当たりのプラスミドの比収率が高くなることである。別の利点は、本発明の方法で好ましいrom-ネガティブpUCプラスミドは、低い比増殖速度で複製の増加を示すことである(Atlung et al., 1999)。最も重要なことは、低い比増殖速度での増殖は、アミノ酸で荷電されないtRNAの形成を引き起こす。
このような非荷電tRNAは、高い複製活性につながるように、プラスミドの複製開始点と相互作用する。
【0035】
本発明の一態様では、栄養補給速度は、全栄養補給期の間じゅう、このような指数関数に従う。
別の態様では、栄養補給期は、指数関数期と、栄養補給が非指数関数的な期との組合せを含む。非指数関数的栄養補給様式の例は、線形又はフィードバックコントロール栄養補給様式であり、例えばpH値、オンライン測定した細胞密度又は溶存酸素圧(DOT)によって制御される。
好ましくは、非指数関数的栄養補給は指数関数的栄養補給の後に続く。
指数関数的及び非指数関数的の両栄養補給期を含む本発明の態様では、全体の栄養補給様式は、該栄養補給期で得られる総乾燥細胞重量の約20%より多くが指数関数的栄養補給期中に生成される。例として、指数関数期の比増殖速度が高い場合、すなわち該指数関数が急勾配の場合、最終的に望まれるバイオマス(総乾燥細胞重量)の比率、例えば50%が、栄養補給期が低い比増殖速度で行われるときより早い時点で達成される。
線形栄養補給様式の場合、この様式は線形一定、すなわち栄養補給の流速が経時的に一定であり、或いは線形増加、すなわち流速が一定の勾配で経時的に増加しうる。
この混合栄養補給様式であれば、増殖とプラスミド形成の相互作用がなくなる。指数関数的栄養補給中、短時間で高量のバイオマス(1リットル当たり約30gの乾燥細胞重量)が得られる。線形栄養補給中、プラスミドの蓄積は低い比増殖速度のためである。この栄養補給法により、500〜800mg/Lのプラスミドタイターが得られる。
【0036】
〔発酵の終点〕
製造業者の特有の要求によって、自由に発酵の終了時点を選択できる。製造規模のプラスミド生産のための発酵は一般的に約15〜20時間かかる。発酵中のプラスミド複製は非常に動的であり、結局、発酵時間の中間でpDNAの比収率が非常に上昇することになる。最大ではないが、その体積収率は高い。発酵の目的がプラスミドDNAの可能な最高濃度のバイオマスを得ることである場合、この時点で発酵を終わらせることができる。これは、その後のアルカリリーシス及び精製時に顕著な利点を有する。バイオマス中のpDNA濃度が高いほど、ライセート体積が少なく、かつプロセス時間が短くなる。さらに、プロセスバルク液中のpDNAの濃度は、より高い。
【0037】
最高のpDNA比濃度点を超えて、pDNA複製はまだ起こり、1L当たりのpDNA濃度を高めることとなるが、バイオマス1g当たりの含量は減少する。プロセスの目標が、プラスミドDNAの可能な最高総量を得ることである場合、複製が止むまで発酵を延長してよい。最大ではないが、発酵のこの後期での比収率は、当技術で知られているいずれの方法におけるよりもまだ高い。
この複製挙動は高度のプロセスフレキシビリティーを与え、高い比収率又は高い体積収率の間の選択を可能にし、簡単には発酵時間によって決まる。発酵の後期中、プラスミド均質性の実質的な低減は観察されない。
本発明の方法で得られるプラスミドDNAは、既知の方法によって回収かつ精製される。
プラスミド精製は、典型的には、収集した細胞集団の分解、通常アルカリリーシスによる分解で開始する。その結果、界面活性剤と一緒に強アルカリ性のpH値に細胞を供するので、細胞が溶解してプラスミドが遊離する。その後の酢酸緩衝液による沈殿工程時、タンパク質とゲノムDNAが沈殿し、一方プラスミドDNAは清澄上清中に留まる。その後の精製工程は、主にろ過(限外濾過、ディアフィルトレーション)とクロマトグラフ法を含む。クロマトグラフ法は、例えば、疎水的相互作用、イオン交換又はゲルろ過クロマトグラフィーから選択される。
【0038】
〔実施例1〕
〔プラスミドpRZ-hMCP1を保有する大腸菌JM108の流加発酵〕
20L規模の発酵槽内、プラスミドpRZ-hMCP1を内部にもつ大腸菌K-12株JM108で指数関数的流加発酵を行った(タンク反応器を撹拌した)。このプラスミド(4.9kb)はpUC oriを含有するpcDNA3TM(Invitrogen)の誘導体であり、カナマイシン耐性マーカーである。pRZ-hMCP1の問題の遺伝子は真核生物のCMV(サイトメガロウイルス)プロモーターの転写制御下の単球化学誘引物質タンパク質1である(Furutani et al., 1989)。
前培養のため、300mLの規定培地を含有する整流1000mL振とうフラスコに前記菌株のグリセロールストック(300μL)を接種した。これを回転式振とう機で300rpm及び37℃にて培養した。前培養基は以下のとおり:NH4Cl 2g/L、MgSO4*7H2O 0.24g/L、グルコース 10g/L、L-プロリン 0.2g/L、L-イソロイシン 0.2g/L、チアミン塩酸塩 1mg/L、クエン酸 2g/L、KH2PO4 5.44g/L、Na2HPO4*12H2O 14.38g/L及び微量要素溶液 16.7mL/L。微量要素溶液はHCl(25%) 14.6g/L、CaCl2*2H2O 0.44g/L、FeSO4*7H2O 0.33g/L、CoCl2*6H2O 0.14g/L、MnSO4*H2O 0.10g/L、CuSO4*5H2O 15mg/L及びZnSO4*7H2O 17mg/Lを含有した。
【0039】
前培養がほぼOD=1の光学密度に達したとき、これを発酵槽に移して発酵を始めた。主培養バッチ培地は前培養と同一濃度で同一成分を含有した。発酵槽は、発酵の開始時に7Lのバッチ培地を含有した。
温度を37℃に制御し、0.35バールの背圧で発酵を操作した。1vvm(培地体積及び1分当たり=7L/分当たりの空気の体積)のプロセスエア流量速度で発酵槽を通気した。溶存酸素圧が30%に低下したとき、スターラーの撹拌速度を上げることによって(500〜1000rpm)溶存酸素圧をこの規定点で維持した。撹拌速度の上昇がDOを維持するのに不十分な場合、空気の酸素濃度を純粋な酸素で濃厚にした。水酸化アンモニウムの溶液(25%)で7.0±0.2の設定点にpHを制御した(同時に発酵の間じゅう窒素源として働いた)。必要な場合、25% H2SO4でpHをさらに制御した。
【0040】
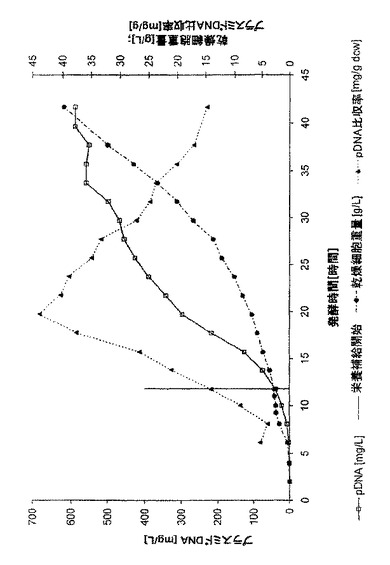
図1に、プラスミドの体積収率と比収率及び増殖の曲線を示す。発酵終了時、590mg/LというpDNAの例外的に高い体積収率が得られた。さらに、発酵時間20時間では、pDNAの比収率が1gの乾燥細胞重量当たり44mgのpDNAという最大値(これも例外的に高い値である)に達した。発酵の終了に向かって、比収率は15mg/g(DCW)に減少した。この比収率及び体積収率の特有の過程は、製造プロセスの必要条件によって、発酵を終わらせる時点を選択できることに関して複数の選択肢という利点を提供する。最高含量のpDNAを有するバイオマスを得なければならない場合、20時間後に発酵を終わらせることができ、3000mg/Lという体積収率となる。プロセスの目標が最高の体積収率である場合、40時間を超えて発酵を延長することができ、約600mg/Lとなるが、比収率は低い。
【0041】
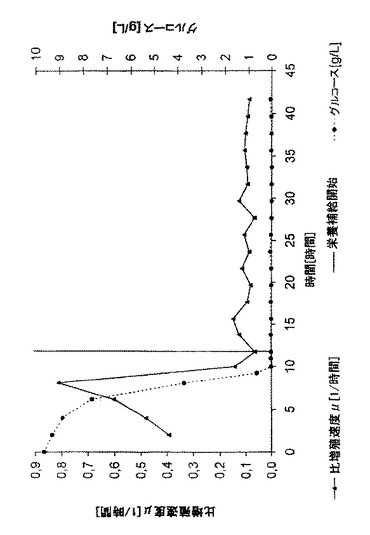
10時間のバッチ培養後、バッチ培地中のグルコースが枯渇した(図2)。これは、急速オフライン測定法で決定した(Yellow Springs Glucose Analyzer, YSI 2700 Select)。
グルコースの枯渇は指数関数的栄養補給期の開始の合図として役立った。栄養補給培地は以下のとおりだった:グルコース 300g/L;MgSO4*7H2O 7.2g/L;L-プロリン 6g/L;L-イソロイシン 6g/L;チアミン塩酸塩 30mg/L;クエン酸 2g/L;KH2PO4 5.4g/L;Na2HPO4*12H2O 14.4g/L;CaCl2*2H2O 220mg/L;FeSO4*7H2O 170mg/L;CoCl2*6H2O 72mg/L、MnSO4*H2O 51mg/L、CuSO4*5H2O 8mg/L及びZnSO4*7H2O 9mg/L。
グルコース枯渇時のバイオマスに基づき(光学密度で推定)、発酵槽のプロセスコントロールシステムによって連続的な指数関数的栄養補給を制御した。
【0042】
0.1時間-1という予め規定した比増殖速度μを得るため栄養補給速度を選択した。質量流量計により栄養補給速度の制御を行った。全発酵の間じゅう、グルコースは限界であり、これは正確な増殖速度制御の必須項目だった。グルコースのオーバーフローの合図であるアセテートは大部分の栄養補給期にわたって有意な量では生じなかった(40時間まで0.5g/L未満)。図1に示されるように、バイオマスは、予め規定した指数関数に正確に従った。発酵時間が40時間を超えると、予測されるバイオマスのコースと実際のバイオマスのコースが逸れたので(図示せず)、発酵を停止した。図2は、現在の比増殖速度μを示す。
この比増殖速度は、得られた乾燥細胞重量から計算され、ほとんどの発酵時間にわたって、予め規定した比増殖速度に従う。
【0043】
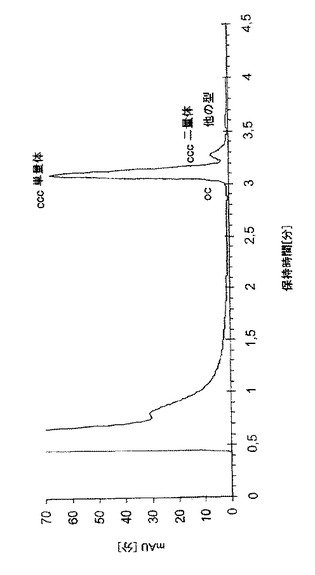
図3は、アニオン交換高速液体クロマトグラフィー(AEX-HPLC)法で分析した、発酵時間にわたる種々のプラスミド型の割合を示す。発酵の主要部の間じゅう、共有結合的に閉じた環状pDNAの割合が90%を超え、42時間の培養時間まで最小89%で維持された。発酵の終了時(42時間)のプラスミドの分析クロマトグラムを図4に示す。
この実施例は、高いpDNA均質性と共に、体積収率と比収率に関して本発明の優れた結果を示す。
【0044】
〔実施例2〕
〔大腸菌JM108の指数関数的流加発酵でプラスミド収率に及ぼすイソロイシンの影響〕 1L規模の選別発酵槽において、実施例1で述べた方法を適用して、増殖とプラスミド生産に及ぼすイソロイシンの効果を示した。実施例1で述べたのと同様に2つの発酵を行った。2つの発酵の相違は、一方の培地はイソロイシンを含むが、他方の培地はイソロイシンを含まないことのみだった。培地の残存組成、並びに培養条件及びμ=0.1時間-1の増殖速度での指数関数的栄養補給の様式は実施例1で述べたのと同一だった。秤及びぜん動ポンプによって栄養補給速度を自動制御した。
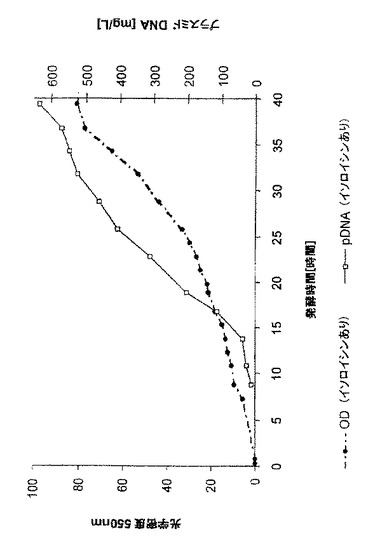
両発酵の光学密度とpDNA体積収率の時間経過を図5(イソロイシンあり)と図6(イソロイシンなし)に示す。両発酵の増殖は、指数関数的栄養補給期中、μ=0.09時間-1という平均比増殖速度でほぼ同一だった。イソロイシンを含有する培地では、pDNA体積収率が633mg/Lであり、イソロイシンがない場合は398mg/Lが得られた。下表1は、発酵パラメーターの概要を与える。両プロセス変形では、pDNA均質性はほぼ90%cccだった。イソロイシンありのpDNAの最大比収率は、イソロイシンがない場合に比べて2倍高かった。
【0045】
表1:
イソロイシンが存在する場合と存在しない場合の指数関数的流加発酵の特性の比較
【0046】
この実施例は、培地中にイソロイシンが存在すると、イソロイシンがない培地から得られるより高い体積収率及び比収率をもたらすことを示す。しかし、イソロイシンがない場合でさえ、発酵のpDNA収率は、既知の方法よりまだ優れる。
【0047】
〔実施例3〕
〔線形栄養補給様式で成功した、指数関数的栄養補給アルゴリズムを用いる、プラスミドpRZ-hMCP1を保有する大腸菌JM108の流加発酵(20Lの発酵槽)〕
この実施例では、実施例1で述べたようにプラスミドpRZ-hMCP1を保有する大腸菌JM108細胞を調製して培養する。実施例1と異なり、栄養補給期を以下の2つの異なる部分に分割する:
(1)比増殖速度μ=0.25時間-1を維持するための栄養補給指数関数に栄養補給速度が従う指数関数的栄養補給期、及び
(2)栄養補給速度が一定値200mL/時間に維持される線形一定栄養補給期。
指数関数的栄養補給から線形栄養補給への切替えを行う時点は、10時間の指数関数的栄養補給後である。線形栄養補給期を選択して10時間とする。このような発酵により、500〜800mgのpDNA/L又は20〜30mgのpDNA/g(DCW)の範囲の体積収率及び比収率が得られる。
【0048】
〔参考文献〕
Andersen, D.C., Swartz, J., Ryll, T., Lin, N., and Snedecor, B. (2001) Metabolic oscillations in an E. coli Fermentation. Biotech. Bioeng. 75:2, 212-218.
Atlung, T., Christensen, B.B., and Hansen, F.G. (1999). Role of the Rom protein in copy number control of plasmid pBR322 at different growth rates in Escherichia coli K-12. Plasmid 41, 110-119.
CBER, Center for Biologics Evaluation and Research, U.S. Food and Drug Administration, Rockville, MD (1998). Guidance for industry - guidance for human somatic cell therapy and gene therapy.
Chen, W., Graham, C., and Ciccarelli, R.B. (1997). Automated fed-batch fermentation with feed-back controls based on dissolved oxygen (DO) and pH for production of DNA vaccines. J. Ind. Microbiol. Biotechnol., 18, 43-48.
Furutani Y., Nomura H., Notake M., Oyamada Y., Fukui T., Yamada M., LarsenC.G., Oppenheim J. J. and Matsushima K. (1989). Cloning and sequencing of the cDNA for human monocyte chemotactic factor (MCAF). Biochem Biophys Res Commun 159(1), 249-55.
Hofmann, K.H., Neubauer, P., Riethdorf, S., and Hecker, M. (1989). Amplification of pBR322 plasmid DNA in Escherichia coli relA strains during batch and fed-batch fermentation. J. Basic Microbiol. 30, 37-41.
Lahijani, R., Hulley, G., Soriano, G., Horn, N.A., and Marquet, M. (1996). High-yield production of pBR322-derived plasmids intended for human gene therapy by employing a temperature-controllable point mutation.
Hum. Gene Ther. 7, 1971-1980.
Lawther, R.P., Calhoun, D.H., Adams, C.W., Hauser, C.A., Gray, J., and Hatfield, G.W. (1981). Molecular basis of valine resistance in Escherichia coli K-12. Proc. Natl. Acad. Sci. USA 78:2, 922-925.
O´Kennedy, R.D., Baldwin, C., and Keshavarz-Moore, E., (2000). Effects of growth medium selection on plasmid DNA production and initial processing steps. J.Biotechnol. 76, 175-183.
O´Kennedy et al (2003). Effects of fermentation strategy on the characteristics of plasmid DNA production. Biotechnol Appl. Biochem. 34, 83-90.
Reinikainen, P., Korpela, K., Nissinen, V., Olkku, J., Soederlund, H., and Markkanen, P. (1988). Escherichia coli plasmid production in Fermenter. Biotech. Bioeng. 33, 386-393.
Sambrook, J., and Russel, D.W. (2001). Molecular Cloning - a Laboratory Manual, 3rd ed., J. Argentine, N. Irwin, K.A. Janssen, S. Curtis, M. Zierler, et. al., eds. (Cold Spring Harbor, NY).
Schoenfeld, T., Mendez, J., Storts, D.R., Portman, E., Patterson, B., Frederiksen,J., and Smith, C. (1995). Effects of bacterial strains carrying the endA1 genotype on DNA quality isolated with Wizard Plasmid Purification System. Promega Notes, 53, 12 - 22.
Ulmer, J.B., Donelly, J.J., and Liu, M.A. (1996). Toward the development of DNA vaccines. Curr. Opin. Biotechnol. 7, 653-658.
Vieira, I. and Messing, I. 81982). The pUC plasmids, an M133mp7-derived system for insertion mutagenesis and sequencing with synthetic universal primers. Gene 19, 259-268.
Wang et al. (2001). Medium design for Plasmid DNA based on stoichimetric model. Proc. Biochem. 36, 1085-1093.
Wrobel, B. and Wegrzyn, G. (1998). Replication regulation of ColE1-like plasmids in amino ascid-starved Escherichia coli. Plasmid 39, 48-62.
Yanisch-Perron, C., Vieira, J., and Messing, J. (1985). Improved M13 phage cloning vectors and host strains: nucleotide sequences of the M13mp18 and pUC19 vectors. Gene 33, 103-119.
Zhang and Greasham (1999). Chemically defined media for commercial fermentations. Appl. Microbiol. Biotechnol. 51, 407-421.
【技術分野】
【0001】
本発明は、プラスミドDNA(pDNA)、特に遺伝子療法及びDNAワクチン接種で使用することを意図したpDNAの生産のための大腸菌の発酵に関する。
【背景技術】
【0002】
最近10年間の遺伝子療法及びDNAワクチン接種の臨床的成功により、pDNAの工業用発酵の要求が生じてきた。
遺伝子療法とは、哺乳動物の細胞内での遺伝子の投与、送達及び発現による病気の治療又は予防である。遺伝子療法の究極的な目標は、遺伝子を添加、補正、又は置換することによって遺伝性及び後天性の両障害を治すことである。基本的に、この目標を達成するための2型の遺伝子療法用ベクター、すなわち不活化ウイルスに基づくウイルスベクターと、プラスミドDNAに基づく非ウイルスベクターがある。本発明は、非ウイルスプラスミドDNAの生産に関する。
抗原をコードするpDNAの筋肉内注射が体液と細胞の両免疫反応を引き出すことが実証されて以来、裸のプラスミドDNAが特に重要になってきた。
プラスミドDNAを製造するための発酵法の所望効率は、発酵ブロスの体積当たり(体積収率)又はバイオマスアリコート(aliquot)当たり(比収率)のpDNAの高い収率によって特徴づけされる。本発明の意味では、収率は体積当たり又は細胞重量当たりのプラスミドDNAの濃度である。高収率を超えると、プラスミドはその無傷の共有結合的に閉じた環状(ccc)又はスーパーコイル型で存在するはずである。本発明の意味では、ccc型の割合を“プラスミド均質性”という。開環状(oc)、線状及び二量体又は多量体型のような他のプラスミド型の濃度は精製プラスミドバルクでは最小限に減らすべきであり、結果として発酵中には望ましくない。
【0003】
治療用プラスミドは3つの基本部分、すなわち真核生物プロモーター、多くはサイトメガロウイルス(CMV)プロモーターの制御下の治療遺伝子(“問題の遺伝子”)、原核生物細胞内での自発繁殖の複製開始点(ori)、及び選択マーカー、通常は抗生物質耐性遺伝子から成る。治療遺伝子は臨床的及び医学的関連性があり、oriと選択マーカーの両者は、プラスミド生産中、特に発酵中に重要な役割を果たす。治療用プラスミドの構築のため、キーとなる因子は、細胞当たり多数のプラスミドコピーを複製する複製開始点の選択である。ほとんどの治療用ベクターはColE1-型oriを有する。pBR322由来のColE1開始点を有するプラスミドは細胞当たり50〜100個のプラスミドというコピー数を達成でき、pUC由来のプラスミドは数百というコピー数を達成することができる。
プラスミドを内部に持つ細胞の形質転換及び選択の際、抗生物質選択マーカー及び抗生物質の使用が必要である。しかし、抗生物質淘汰圧は工業生産時には避けるべきである。
従って、プラスミド損失なしでベクターの安定した繁殖を可能にする発酵法を開発することが望ましい。
【0004】
細菌宿主株の選択は、pDNAの発酵で考慮すべき重要なもう一つの因子である。望ましい宿主表現型として、高い細胞密度に増殖する能力を有するもの、高いプラスミドコピー数を達成する能力を有するもの、最少のプラスミド-フリー細胞を生成する能力を有するもの、該プラスミドの遺伝子変化の可能性が最少のもの、優先的にスーパーコイルのプラスミドを生産するもの、及び通常の精製手順と適合性のものが挙げられる。大部分の大腸菌株を用いてpDNAを繁殖させることができるが、株は得られるpDNAの質と量に影響を及ぼしうる(Schoenfeld et al., 1995)。現在、pDNA製造のために使用する細菌株について理想的であろう遺伝型又は表現型の特徴に関するコンセンサスがない。
【0005】
pDNAの発酵について多くのアプローチが開示されている。提案方法は、発酵に影響を及ぼす細胞と多くの因子に課される制御のレベルについて異なる。低レベル制御は、簡単にプラスミド保有細胞を増殖させることができるが、高レベルの厳重に制御した発酵は、複製を促す特有の手段によってpDNAの高収率を果たす。
実験室規模のpDNA生産では、振とうフラスコ内でのプラスミド保有細胞の培養が最も簡単な方法であるが、通常は低収率となってしまう。振とうフラスコ培養から得られるプラスミド収率は、培養ブロス1L当たり1.5〜7mgの範囲である(O´Kennedy et al., 2003; Reinikainen et al., 1988; O´Kennedy et al., 2000)。振とうフラスコ培養では、不十分な酸素移動及びpH値制御の実現性の欠如などのいくつかの欠点がpDNA収率を制限する。US 6,255,099では、振とうフラスコ培養でさえ、特定の培地組成と緩衝条件で109mg/LまでのpDNA収率を達成できることを示した。
より高量のプラスミドを得るため、制御された発酵槽内で細胞を培養することが示唆されている。すべての栄養素を最初から与え、培養中に栄養素を添加しない簡単な培養法は、“バッチ培養”又は“バッチ発酵”と呼ばれる。制御された発酵槽内でのバッチ法の適用は、体積当たりのpDNA収率の増加をもたらした。プラスミド/宿主の組合せ及び培地によって、このようなバッチ発酵から得られるpDNAの収量は大きく変化しうる。報告されている典型的なプラスミド収率は3.5〜50mg/Lの範囲である(O´Kennedy et al., 2003; WO 96/40905; US 5,487,986; WO 02/064752; Lahijani et al., 1996)。これら培養は炭素源及び窒素源としていわゆる“複合成分”を含有する培地を用いて行われた。これら成分は生物源から得られ;生物源としては、例えば、酵母抽出物、大豆ペプトン又はカゼイン水解物が挙げられる。
【0006】
排他的又は優先的に複合成分から成る培地を“複合培地”と称する。規定部分(規定した炭素源、塩、微量要素、ビタミン)及び複合部分(窒素源)の両部分から構成されている培地を“半規定”培地と称する。 US 5,487,986では、非常に高量の種々の複合成分(全部で50g/L)を用いた。
複合成分を含有する培地は、これら成分が生物材料由来なので、該培地の組成は普通の自然偏差の基礎にあり、培養プロセスの再現性を低くするという欠点を有する。製造業者が生産方法を変更するとき、又は供給業者を変えるとき、同様のことが当てはまる。複合培地成分を使用することのさらなる欠点は、正確な組成についての不確実性(望ましくない物質の存在)、化学量論収率を計算できないこと、無菌化時の望ましくない生成物の形成、不十分な溶解のため難しい取扱い、培地調製中のダスト及び凝集の形成である。発酵中、複合培地はたやすく泡立つ傾向がある。動物起源の複合成分(肉抽出物、カゼイン水解物)は伝播性海綿状脳萎縮の危険のため、pDNA生産には特に望ましくないので、医薬品局によってその使用が制限されている(CBER 1998)。
【0007】
複合培地成分の欠点のため、いかなる複合成分も含有しない培地が開発されている。“規定”又は“合成”培地と呼ばれるこのような培地は、化学的に規定した物質、すなわちグルコース又はグリセロールのような炭素源、塩、ビタミンから、また、ありうる株の栄養要求性に鑑みて、特有のアミノ酸又はチアミンのような他の物質から排他的に構成される。化学的に規定した培地は、その組成が正確に分かるという利点を有する。このことは、良いプロセス分析、発酵モニタリング及び増殖又は産物形成を促す特定物質の特有の添加を可能にする。周知の組成は、増殖の予測及び栄養素を欠いている可能性の同定を容易にする質量バランス計算の樹立を可能にする。複合培地に比し、規定培地による発酵は、高いプロセス整合性とスケールアップ時の改良された成果を示す。規定培地のさらに実務的局面は、良い溶解度、無菌化時に副生物を阻害することの不在、及び培養中の泡形成が少ないことである(Zhang and Greasham, 1999)。
M9(Sambrook and Russel, 2001)のような、pDNA生産に特定して開発したものではない合成培地は低いpDNA収率をもたらしうる(WO 02/064752)。pDNA生産に特定して設計した規定培地によるバッチ発酵では高収率のpDNAが得られた(Wang et al., 2001; WO 02/064752)。後者はpDNA均質性が90%を超えるccc型であることを示した。WO 02/064752及びWangら(2001)によるpDNAの高収率は、ヌクレオシドの生合成基礎単位であるアミノ酸の補充によって、又はヌクレオシドの直接添加によって達成された。
【0008】
バッチ発酵は一般的に簡単かつ短いが、プラスミドDNA収率が制限されることになるという基本的な欠点を有する。これは、バッチ培地内での高い栄養素濃度における基質阻害と塩沈殿が原因である。さらに、バッチ発酵の増殖速度は直接制御できず;それゆえにバッチ発酵は無制限であり、発酵中絶え間なく変化し、1種以上の栄養素が欠失しているとき又は代謝副生物(例えばアセテート)が細胞の増殖を阻害する場合にのみ終止する。
その結果として、pDNA生産でのバイオマス及びプラスミド収率を高めるため、流加発酵(fed-batch fermentation)が開発された。流加発酵はバッチ期後、栄養補給によって培養に1種以上の栄養素を供給する栄養補給期を行う方法である。
ラスミドDNAを生産するための大腸菌の流加発酵では、以下のような種々の戦略が実行されている。
【0009】
一方法は、規定した設定点でプロセスパラメーターを制御するために栄養素を供給することによってフィードバックコントロールアルゴリズムを適用する。従って、フィードバックコントロールは発酵全体にわたって直接細胞活性に関係する。発酵のフィードバックコントロールのために使用されているコントロールパラメーターとしては、pH値、オンライン測定細胞密度又は溶存酸素圧(dissolved oxygen tension)(DOT)が挙げられる。この方法は、供給栄養素によって培養に栄養を過剰に与える危険が少なく、高いバイオマス濃度を得ることができるという利点を有する。
pDNA発酵のため、栄養補給速度によって規定設定点での溶存酸素圧を制御するためのフィードバックアルゴリズムが使用された(WO 99/61633)。
別の方法を適用する場合、より複雑なアルゴリズム、すなわちフィードバック培養法のコントロールパラメーターとしてDOTとpHの両方を使用した(US 5,955,323; Chen et al., 1997)。当該方法では、撹拌速度と濃縮複合培地(グルコース、酵母抽出物)の栄養補給によってDOTを制御し、それによって、同時に水酸化アンモニウムでpHを維持した。
フィードバックアルゴリズムの適用はいくつかの欠点を伴う。1つは、栄養補給速度がDOTのような流動プロセスパラメーターに左右されることである。どんな理由によってもプロセスの刺激はコントロールパラメーターに影響を及ぼしうるので、栄養補給速度に影響を与え、ひいては増殖及びpDNA収率に影響する。例えば、消泡剤を添加しなければならない場合、DOTが変化し(通常減少し)、結果として栄養補給速度が低下する。これは発酵プロセスの再現性を低くする。発酵槽の形状や大きさが異なると異なった酸素移動速度を示すので、プロセスのスケールアップの際にはさらなる困難が生じる。酸素移動速度はDOTにつながるので、大きさを変えた発酵槽のフィードバックコントロール栄養補給速度は異なり、その結果、プロセスは大きさに比例しないだろう。
【0010】
フィードバックコントロールのもう一つの欠点は、比増殖速度を正確に予め規定することも制御することもできないことであり、その結果、産物形成が増殖によって左右されるプロセスでは最適以下のプロセス収率となる。しかし、pDNA発酵では、プラスミドの体積収率及び比収率が比増殖速度に非常に依存することが示された(WO 96/40905; O´Kennedy et al., 2003)。
指数関数に従う栄養補給培地の供給に基づく別の基礎的な栄養補給様式によって、比増殖速度の制御を達成することができる。栄養補給速度は、所望の比増殖速度μに基づいて制御される。規定培地を適用すると、与えられた基質単位Sに基づいて形成されるバイオマスアリコートXを計算することによって、増殖を正確に予測かつ予め規定することができる(バイオマス収量係数YX/Sを考慮して)。
【0011】
WO 96/40905で開示されている発明は、プラスミドDNA生産用の指数関数的流加法を用いて高収率のバイオマスを得るが(1L当たり50gのDCW、乾燥細胞重量)、低いpDNA収率となる(18mg/L;0.36mg/g 乾燥細胞重量)。指数関数的栄養補給の別の例では、30mg/L及び6mg/g DCWというプラスミド収率を達成した(O´Kennedy et al., 2003)。Lahijaniら(1996)により、指数関数的栄養補給と温度制御しうるプラスミド複製の強化を併用することによって、220mg/Lという高いpDNA収率が得られた。これらの指数関数的栄養補給の例では、O´Kennedyら(2003)だけがpDNA均質性について詳述しており、50〜70%のccc型だった。現在、すべての指数関数的流加法がバッチ培地と栄養補給培地の両方で複合成分を使用している。
【発明の概要】
【発明が解決しようとする課題】
【0012】
要するに、治療用プラスミドDNAを製造するための発酵の現在の技術状態は、以下のように特徴づけすることができる:
pDNA生産のために広く適用されているバッチ発酵は、技術的及び経済的な欠点を伴う。バッチ発酵では、複合培地又は半規定培地が使用されることが多く、培養ブロス1L当たり3.5〜68mgの範囲のpDNA収率という結果となる。フィードバックコントロールを適用する流加法は、半規定培地又は複合前培養培地を使用後、主培養で規定培地を用いる。フィードバックアルゴリズムにより、100〜230mg/Lというプラスミド収率が得られる。指数関数的栄養補給発酵は半規定培地を使用する。指数関数的発酵のプラスミド収率は18〜220mg/Lの広い範囲である。一般に、多くのpDNA発酵法は不十分な均質性(すなわち、スーパーコイルプラスミドの割合)に困っている。主培養で規定培地を使用する発酵は例外であり、90%を超えるccc型の割合を達成できる。
【課題を解決するための手段】
【0013】
本発明は、製造規模でプラスミドDNAを生産する方法であって、問題の遺伝子を運ぶプラスミドを有する大腸菌を最初に前培養で増殖させ、引き続き主培養で発酵させる方法に関し、前記主培養はバッチ期と栄養補給期を含む流加法である。バッチ期の培地と栄養補給期中に添加する培地を化学的に規定し、かつ栄養補給期の培地は、該栄養補給期の少なくとも一部分の間、予め規定した指数関数に従う栄養補給速度で添加され、それによって、予め規定した値に比増殖速度を維持する。
本発明の意味では、用語“規定培地”は、化学的に規定した単一成分で排他的に構成される培地を指す。
好ましい態様では、栄養補給培地の添加が指数関数に従う、栄養補給期の前記部分は、該栄養補給期に得られる総乾燥細胞重量の20%を超える量が前記栄養補給期の前記部分中に生成されるような部分である。
好ましい態様では、前培養で用いる培地を含め、すべての培地を化学的に規定する。
本発明の方法では、大腸菌のいずれの株でも使用しうる。本発明で有用な宿主株は、いずれの表現型を有していてもよく;好ましい宿主株は遺伝子relA、endA、及びrecAに突然変異を有する。本発明の最も好ましい態様では、大腸菌株K12 JM108又はその誘導体を使用する。
本発明の方法では、大腸菌内で自発的に複製しうるいずれのプラスミドも使用できる。
好ましいプラスミドは、ColE1-型の複製開始点を有し、最も好ましくは、pUCプラスミド由来のレプリコンである(初めはVieira及びMessing, 1982によって記載された;Yanisch-Perron et al. 1985)。
【0014】
本発明の方法では、バッチ期の培地と栄養補給期の培地を化学的に規定し、好ましくは前培養で使用するものも含め、すべての培地を化学的に規定する。ここで、化学的に規定するとは、これら培地がいかなる複合培地成分も含有しないことを意味する。本発明の方法で使用する培地は、有機炭素源、好ましくはグルコースと、アンモニウム塩のような1種以上の無機窒素源を含む。さらなる成分は、マクロ要素及びミク要素を含有する無機塩である。菌株が特有の栄養素、例えば、チアミンのようなビタミン又はプロリンのようなアミノ酸に対して栄養要求性である場合、これら物質も培地中に存在する。
好ましくは、本発明の培地は、プラスミド保有細胞について淘汰圧を課すために通常使用されるような抗生物質はない。
本発明の好ましい態様では、培地はありうるアミノ酸栄養素要求性と無関係に、イソロイシンを含む。これは、イソロイシンは、菌株がイソロイシンに対して栄養要求性でない場合にも存在してよいことを意味する。菌株がイソロイシン栄養要求性を有する場合、好ましくは前記栄養要求性を補完するために必要な量を超える濃度でイソロイシンが存在する。
増殖速度、ひいては栄養補給速度(これによって増殖速度が維持される)は、細胞の増殖とプラスミド複製を確実にするいずれの値を有していてもよい。
【発明の効果】
【0015】
驚くべきことに、本発明の方法は、600mg/Lを超えるpDNA収率と、45mg/g(乾燥細胞重量)までのpDNA比収率をもたらし、これによって90%を超えるcccというpDNA均質性を達成できることが分かった。
好ましい態様では、低い比増殖速度で操作する発酵では、高コピー数プラスミド(例えばpUC由来)を使用する。
低増殖速度で操作する発酵では、アミノ酸飢餓状態のため、非荷電tRNAが生じてプラスミドコピー数を増やす。relA-ネガティブ大腸菌株を使用すると、代謝の下方制御の点から、細胞はアミノ酸飢餓状態に応答できず、結果としてプラスミド複製が増強される。イソロイシンのさらなる供給はアミノ酸代謝の妨害を持続させ、なおさらにpDNA収率を高める。
驚くべきことに、本発明の方法は、発酵時間全体にわたるpDNAの高い均質性と共に、プラスミドDNAの顕著に高い比収率及び体積収率をもたらすことが分かった。
【図面の簡単な説明】
【0016】
【図1】実施例1の発酵のプラスミドの体積収率と比収率及び増殖の曲線を示す。
【図2】実施例1の発酵の現在の比増殖速度μを示す。
【図3】実施例1の発酵時間にわたる種々のプラスミド型の割合を示す。
【図4】実施例1の発酵終了時のプラスミドの分析クロマトグラムを示す。
【図5】実施例2のイソロイシンがある場合の光学密度とpDNA体積収率の時間経過を示す。
【図6】実施例2のイソロイシンがない場合の光学密度とpDNA体積収率の時間経過を示す。
【発明を実施するための形態】
【0017】
〔宿主株〕
本発明の方法では、規定培地で培養できるいずれの大腸菌株も使用しうる。好ましくはK-12由来の株を使用する。適切な宿主株の選択は、その宿主の表現型、すなわち該株を特徴づける特異的な突然変異に基づき、及び/又は実験的発酵でいくつかの株を分類学的に比較することに立脚しうる。
本発明の好ましい態様では、菌株はrelA遺伝子に突然変異を有する。この遺伝子はグアノシン四リン酸(ppGpp)の合成に反応性であり、アミノ酸制限時の細胞でのいわゆる“ストリンジェント反応”を引き起こすシグナル分子である。relA+株では、アミノ酸制限がDNAの複製と転写に関与する必須代謝経路の下方制御につながる(Wrobel and Wergrzyn, 1998)。従って、relA遺伝子に突然変異を有する菌株はアミノ酸制限時に増殖し続け、かつプラスミドを複製し続ける。relA遺伝子に突然変異を有することが、化学的に規定した培地で高いプラスミド複製速度で細胞を増殖させるために重要な必須項目であることは分かっている(Hofmann et al., 1989)。
本発明の別の好ましい態様では、relA突然変異に加え、endA(エンドヌクレアーゼA)及びrecA(組換え系)の遺伝子に突然変異を有する宿主株を使用する。これら突然変異はプラスミドの構造の完全性を高めるからである。
好適な宿主株の例は、DH1、DH5、DH5-α、DH10B、JM108、JM109及びHB101である。
本発明の方法では、最も好ましい大腸菌K-12株はJM108(ATCC No. 47107; DSMZ No. 5585;表現型:F-、recA1、endA1、gyrA96、thi-1、hsdR-17、supE44、relA1、λ-及びΔ(lac-proAB);Yanisch-Perron et al., 1985)又はJM108の遺伝子操作で得られるJM108誘導体である。本発明につながる予備試験で行った実験的発酵における宿主スクリーニング中、JM108は、首尾一貫して最高のpDNA収率を示した(体積収率と比収率の両方とも)。
【0018】
〔プラスミド〕
複製開始点と無関係、かつ意図した用途、例えば遺伝子療法ベクターとして又はDNAワクチンとして使用するためのベクターとしての用途と無関係に、大腸菌内で自発的に複製可能ないずれのプラスミドの製造にも本発明の方法を使用できる。例えば、好適なプラスミドはpBR322、pUC18、pUC19、pcDNA3から誘導される(Invitrogen)。
本発明の好ましい態様では、ColE1-型複製開始点を有するプラスミドを用いる。この背後にある論理的根拠は、アミノ酸制限時、アミノ酸で荷電されていない転移RNA(tRNA)が生じることである。これら非荷電tRNAは、プラスミドの複製速度を高めるようにColE1複製開始点と相互作用する。relA-株を使用すると、この作用がより強い(Wrobel and Wergrzyn, 1998)。
本発明の特に好ましい態様では、pUC型のColE1-誘導プラスミド(Vieira and Messing, 1982; Yanisch-Perron et al., 1985)を使用する。このpUCプラスミドは、コピー数を減らす部分Romに突然変異を有するので、普通のColE1-型プラスミドより高いプラスミドコピー数を達成する。rom-プラスミドの複製増強効果は特に、低い比増殖速度で示されたので(Atlung et al., 1999)、本発明では、pUCプラスミドと低い増殖速度培養の組合せが好ましい。
【0019】
〔規定培地〕
主培養のバッチ期及び栄養補給期、好ましくは前培養でも、化学的に規定した培地を使用する。従って、好ましい態様では、全発酵プロセスの間じゅう複合成分を適用しない。
さらに、好ましい態様では、主培養の全体にわたって、プラスミド保有細胞の選択用の抗生物質を使用しない。好ましくは、前培養で用いる培地も抗生物質がない。
前培養及び主培養で用いる培地は同一組成又は異なる組成を有してよい。
前培養、バッチ培地、及び栄養補給培地で有機炭素源を用いる。炭素源は、グルコース、フルクトース、ラクトース、スクロース、アラビノース、グリセロール又は大腸菌によって代謝されうる他のいずれの炭素源でもよく、並びに異なる炭素源の混合物でもよい。
好ましくは、炭素源はグルコースである。
プロセスのそれぞれの時期及び栄養補給培地で用いる炭素源は同一又は異なってよい。
通常、前培養の培地及び主培養のバッチ培地中の炭素源は同一であり、好ましくは、同じ炭素源が栄養補給培地にも含まれる。
バッチ培地では、炭素源の濃度は約1〜約100g/L、好ましくは約10〜約30g/Lの範囲でよい。前培養及び主培養のバッチ培地では炭素源の濃度が同一でよい。
栄養補給期では、栄養補給培地の炭素源は、比増殖速度を制御するための制限基質として働くことが好ましい。本方法の最も簡単な変形では、栄養補給培地は炭素源だけを含有する。
【0020】
栄養補給培地中の炭素源の濃度範囲は約100〜約750g/Lである。グリセロールを用いる場合、グリセロールの濃度は、栄養補給培地中100〜1000g/Lの範囲でよい。好ましくは、炭素源がグルコースであり、約300〜500g/Lの濃度で栄養補給培地中に存在する。
本発明の好ましい態様では、栄養補給培地は増殖制限炭素源と、これとは別に、バッチ培地にさらに存在するいくつか又はすべての培地成分を含有する。
本発明により、前培養及びバッチ培地で初期窒素源として働くように、好ましくはアンモニウム塩又は水酸化アンモニウムを用いる。通常、塩化アンモニウムを使用するが、硫酸アンモニウム、炭酸アンモニウム、リン酸アンモニウム若しくは水酸化アンモニウム又は他のいずれのアンモニウム塩も適切である。アンモニウム濃度が1L当たり約0.1〜約8gのNH4、好ましくはアンモニウム濃度が約0.5〜約2g/L、最も好ましくは約0.6g/Lになるように窒素源の濃度を選択することができる。
【0021】
好ましい態様では、主培養全体にわたって、pH値の制御と窒素の連続供給の両方のためバッチ期と栄養補給期の両期中、水酸化アンモニウムの溶液を別々に供給する。これは最初のバッチ培地にすべてのアンモニウムが与えれらるわけではないという利点を有する。最初のバッチ培地にすべてのアンモニウムが与えれらると培養に毒性効果を及ぼしうる。この好ましい態様では、培地中のアンモニウム濃度は、発酵全体にわたって本質的に一定のままである。水酸化アンモニウムによるpH値の制御は、水酸化ナトリウム又は水酸化カリウムの使用に比べてずっと有益である。水酸化アンモニウムは、浸透ストレスを引き起こして培養を阻害しうる、培地中のナトリウムイオンとカリウムイオンの蓄積と無関係だからである。水酸化アンモニウムの供給は栄養補給と無関係なので、それは増殖速度制限ではなく、かつ栄養補給アルゴリズムの一部でもない。アンモニウム溶液の濃度は約5〜約40%m/mアンモニウムでよい。
【0022】
アミノ酸栄養要求性の宿主株の場合、好ましい態様に従い、宿主株がJM108(その増殖のためにアミノ酸プロリンを必要とする)である場合、培地は(該培地でプロリンの使用は必須である)、1種以上の関連アミノ酸を使用する必要があるかもしれない。さらに、該アミノ酸が栄養補給培地中に存在することも必須である。該アミノ酸が栄養補給溶液中に存在する場合、該アミノ酸は増殖速度を制御するための速度制限因子としても働かず、プラスミドの複製を増強することも意図しない。栄養要求性アミノ酸の濃度範囲は約0.05〜約2g/Lから選択され、好ましくは約0.2g/Lである。
【0023】
本発明の好ましい態様では、培地は、宿主のありうるアミノ酸栄養要求性と無関係に、アミノ酸イソロイシンを含み、イソロイシンはバッチ培養の培地に存在し、好ましくは栄養補給培地にも存在する。任意に、前培養の培地にもイソロイシンが存在してよい。大腸菌K-12株はイソロイシンの生合成に関与する遺伝子の突然変異を有する。この結果、イソロイシンが存在せず、かつ同時にバリンが存在する場合、イソロイシン生合成の抑制となる(Lawther et al., 1981)。従って、バリンはイソロイシン飢餓状態の細胞には毒である。バリンによるイソロイシン代謝との相互作用はバリンの細胞内生成時でさえ見出されたが、この作用はイソロイシンの供給で撤廃された(Andersen et al., 2001)。本発明につながる実験の過程において、バリンの毒性が確認された。さらに、イソロイシンのない規定培地で培養は成長しうるが、イソロイシンの供給がプラスミドDNAの収率を有意に高めることが分かった。従って、規定培地にイソロイシンが存在することは本発明の好ましい態様で必須の特徴である。イソロイシンの濃度範囲は約0.05から約2g/Lの間で選択され、好ましくは約0.1〜約0.3g/L、最も好ましくは約0.2g/Lである。
【0024】
さらに、培地はマクロ要素及びミクロ要素(微量要素)の供給として働く無機塩を含み、かつ任意に、細胞の代謝に正の効果を及ぼす有機物質、例えばクエン酸を含んでよい。
アミノ酸以外、例えば、ビタミンに対する栄養要求性の場合、さらに関連物質が培地に含まれる。宿主株JM108を使用する本発明の態様では、培地にチアミンが存在しなければならない。チアミンは、好ましくは約0.1〜約100mg/Lの濃度、最も好ましくは約1mg/Lの濃度で存在する。
マグネシウムは、通常MgSO4*7H2Oの形態で供給されるが、他のマグネシウム塩も同様に適する。マグネシウム塩の濃度は、通常、約0.1〜約5g/L、好ましくは約0.25g/Lである。
【0025】
リンは、好ましくはKH2PO4及び/又はNa2HPO4*12H2Oの形態で供給されるが、他のリン塩、例えば、K2HPO4、NaH2PO2、Na2HPO4、NH4H2PO4及び/又は(NH4)2HPO4も使用しうる。
リン塩の濃度は、約0.5〜25g/L、好ましくは約2.0〜約15g/L、最も好ましくはそれぞれ約5.0g/LのKH2PO4又は約14.0g/LのNa2HPO4*12H2Oでよい。
カルシウム、鉄、コバルト、マンガン、銅及び亜鉛のような元素は通常以下のように培地中に存在する:約1〜約20の範囲、好ましくは約7.5mg/LのCaCl2*2H2O;約1〜約20の範囲、好ましくは約5.5mg/LのFeSO4*7H2O;約0.5〜約10の範囲、好ましくは約2.5mg/LのCoCl2*6H2O;約0.1〜約5の範囲、好ましくは約2.0mg/LのMnSO4*H2O;約0.05〜約2の範囲、好ましくは約0.25mg/LのCuSO4*5H2O;約0.05〜約2の範囲、好ましくは約0.3mg/LのZnSO4*7H2O。それぞれの元素を含有する他のいずれの塩もそれぞれの濃度範囲で使用できる。
【0026】
上で規定した培地成分及びその濃度は、バッチ培養で使用する培地に関係する。通常、栄養補給培地中の栄養素の濃度は、窒素源(栄養補給期中に別個に添加される)を除き、バッチ培地に比べて約2倍〜約100倍高い。
最小必要条件として、栄養補給培地は増殖制限である量で基質、通常は炭素源を含む。
栄養補給培地は、制限炭素源に加え、バッチ培地に存在するいくつか又はすべての培地成分を含んでよく、或いはこれはあまり好ましくないが、栄養補給培地は、等価な成分、例えば同一元素の異なる塩を含んでもよい。
例として、栄養補給培地の組成は以下のとおり:グルコース 300g/L;MgSO4*7H2O 7.2g/L;L-プロリン 6g/L;L-イソロイシン 6g/L;チアミン塩酸塩 30mg/L、クエン酸 2g/L;KH2PO4 5.4g/L;Na2HPO4*12H2O 14.4g/L;CaCl2*2H2O 220mg/L;FeSO4*7H2O 170mg/L;CoCl2*6H2O 72mg/L、MnSO4*H2O 51mg/L、CuSO4*5H2O 8mg/L及びZnSO4*7H2O 9mg/L。
【0027】
さらなる局面では、本発明は、製造規模で大腸菌内にてプラスミドDNAを生産するための培地に関する。この培地は、以下の成分を含む、化学的に規定した培地である。
a)グルコース、グリセロール、フルクトース、ラクトース、スクロース、アラビノース、又はその混合物から選択される有機炭素源;
b)アンモニウム塩及び水酸化アンモニウムから選択される無機窒素源(該窒素源は、前記培地の成分として存在し、及び/又は発酵中に前記培地に添加される);
c)マクロ要素及びミクロ要素の供給として働く無機イオン;
d)任意に、大腸菌株の栄養要求性を補完する1種以上の物質;及び
e)イソロイシン。
本発明の培地は、本発明の方法のバッチ期及び/又は栄養補給期において、或いは製造規模でプラスミドDNAを生産するための大腸菌を培養するため、すなわち、いずれのバッチ培養又は流加培養でも大腸菌の発酵のための他のいずれの方法での培地としても有用である。
【0028】
さらなる態様では、本発明の培地は、バッチ発酵の開始時に存在し、又は流加発酵のバッチ期の開始時に存在する培地であり、以下の成分を含む:
a)約10〜約30g/Lの濃度のグルコース;
b)アンモニウム濃度が約0.5〜約2g/Lになる濃度のアンモニウム塩又は水酸化アンモニウム;
c)マクロ要素及びミクロ要素の供給として働く無機イオン;
d)任意に、大腸菌株の栄養要求性を補完する1種以上の物質;及び
e)約0.1〜約0.3g/Lの濃度のイソロイシン。
別の態様では、本発明の培地は、流加発酵の栄養補給期中に添加する栄養補給培地であり、以下の成分を含む:
a)約300〜約500g/Lの濃度のグルコース;
b)マクロ要素及びミクロ要素を供給するために働く無機イオン;
c)任意に、大腸菌株の栄養要求性を補完する1種以上の物質;及び
d)約6g/Lの濃度のイソロイシン。
【0029】
〔指数関数的栄養補給手順〕
本発明により、いずれかの所望の予め規定した比増殖速度μで培養を増殖させるため、栄養補給期の少なくとも一部分の間、培地(“栄養補給培地”)の添加を指数関数に従って行う。
この最も一般的な定義では、指数関数は、それぞれ方程式Vt=定数*eμt又はFt=定数*eμtで定義される。この方程式は、関数Vt=定数*eμt+A又はFt=定数*eμt+Aをも包含する。式中、Aは、発酵の規模によって決まる広い範囲で変化しうる値である。好ましくは、A=0である。値Aが0でないと、A=0の関数に比べて上方(A>0)又は下方(A>0)にシフトする関数となる。
【0030】
本発明の好ましい態様では、栄養補給培地を添加する様式を定義する関数は以下のとおりである。
【0031】
この関数中、Vtは、栄養補給開始から計算される時間間隔t[時間]で添加される栄養補給培地の体積[L]である。X0は、栄養補給開始時点のバイオマス乾燥細胞重量の総量[g]である。YX/Sは、バイオマス収量係数(基質1g当たりの乾燥細胞重量のg数)であり、CSは、栄養補給培地中の増殖制限基質(通常、有機炭素源)の濃度[g/L]である。
【0032】
栄養補給様式を定義する関数は、以下のように栄養補給培地の流速で特徴づけすることもできる。
この関数中、Ftは、栄養補給の開始から計算される時点t[時間]で添加される栄養補給培地の流速[L/時間]である。
【0033】
いずれかの所望の予め決定した値で、及び/又は以前の発酵で最適であると決定されたように若しくは実験的発酵の結果に基づいて、比増殖速度μ[時間-1]を選択することができる。バイオマス収量係数は文献から取り出し、又は予備培養実験で決定することができる。光学密度を測定し、又は予備実験の値から取り出し、又はバッチ培地中の初期基質濃度とバイオマス収量係数によって理論値を計算することによって、栄養補給開始時のバイオマスの量X0を決定することができる。
上記方程式に従う、該栄養補給様式の独特な特徴は、栄養補給開始時点と所定時点との間の時間に排他的に依存する。このような栄養補給プロフィールにより、いずれの所望の値でも比増殖速度を予め規定することができる。
種々の方法で、栄養補給を開始する時点を決定することができる。開始時点は、バッチ培地中の炭素源が枯渇したときに、炭素源濃度のオンライン若しくはオフライン測定によって、又は溶存酸素圧の増加(該基質の枯渇を示唆する)によって決定されるように、選定することができる。さらに、特定のバイオマス値(バイオマスのオンライン若しくはオフライン測定によって決定されるような)と相関させるように、又は発酵の開始からの予め規定した時間間隔に応じて、栄養補給の開始時点を選択することができる。
本発明の方法では、指数関数に従った栄養補給培地の供給は種々の方法で達成することができる。本発明の好ましい態様では、栄養補給速度は、発酵槽のプロセスコントロールシステムによって連続的に計算かつ制御され、かつ栄養補給速度は、指数関数に従って連続的に上昇する。それにより、栄養補給培地の質量の流れは、質量流量計(マスフローコントローラー)によって、又は秤で測定される栄養補給容器の重量に応じて培地を供給するポンプによって制御される。
【0034】
本発明の別の態様では、指数関数に従って段階的に栄養補給速度を上昇させることによって、半連続的に栄養補給速度を高める。例えば、所定の時間間隔で、例えば、毎時間栄養補給培地の体積又は質量を再計算することができ、その新しい栄養補給速度を必要な値に調整する。栄養補給速度の上昇は段階的であるが、結果として生じる栄養補給プロフィールは、それでもやはり指数関数的である。
本発明のさらに別の態様では、栄養補給速度を不連続的に上昇させる。これは、規定時点における栄養補給培地の必要量を計算し、その後、(現在のバイオマスに基づいて、選択した時間間隔で予め規定し、又は計算した)増加量の栄養補給培地をパルス様式で添加することによって達成できる。細胞は連続的に栄養補給されないが、それでもなおこの方法は、一定の比増殖速度につながる指数関数的な栄養補給プロフィールをもたらす。
本発明の方法では、増殖とプラスミド複製を確実にするいずれの比増殖速度μも指数関数的栄養補給のために予め規定することができる。最高かつ至適な比増殖速度は、個々の宿主/プラスミドの組合せによって決まり、かつ実験的な流加発酵又は連続(ケモスタット)発酵における各宿主-プラスミド系のケースバイケースの基礎に基づいて決定することができる。大腸菌の発酵に好適な比増殖速度は、約0.03〜約1.5時間-1の範囲である。
本発明の好ましい態様では、約0.05〜約0.15時間-1の低い比増殖速度を使用し、最も好ましくは、増殖速度は約0.1時間-1である。低い比増殖速度の利点は、プラスミドが高いコピー数に複製できるようにし、結果としてバイオマス当たりのプラスミドの比収率が高くなることである。別の利点は、本発明の方法で好ましいrom-ネガティブpUCプラスミドは、低い比増殖速度で複製の増加を示すことである(Atlung et al., 1999)。最も重要なことは、低い比増殖速度での増殖は、アミノ酸で荷電されないtRNAの形成を引き起こす。
このような非荷電tRNAは、高い複製活性につながるように、プラスミドの複製開始点と相互作用する。
【0035】
本発明の一態様では、栄養補給速度は、全栄養補給期の間じゅう、このような指数関数に従う。
別の態様では、栄養補給期は、指数関数期と、栄養補給が非指数関数的な期との組合せを含む。非指数関数的栄養補給様式の例は、線形又はフィードバックコントロール栄養補給様式であり、例えばpH値、オンライン測定した細胞密度又は溶存酸素圧(DOT)によって制御される。
好ましくは、非指数関数的栄養補給は指数関数的栄養補給の後に続く。
指数関数的及び非指数関数的の両栄養補給期を含む本発明の態様では、全体の栄養補給様式は、該栄養補給期で得られる総乾燥細胞重量の約20%より多くが指数関数的栄養補給期中に生成される。例として、指数関数期の比増殖速度が高い場合、すなわち該指数関数が急勾配の場合、最終的に望まれるバイオマス(総乾燥細胞重量)の比率、例えば50%が、栄養補給期が低い比増殖速度で行われるときより早い時点で達成される。
線形栄養補給様式の場合、この様式は線形一定、すなわち栄養補給の流速が経時的に一定であり、或いは線形増加、すなわち流速が一定の勾配で経時的に増加しうる。
この混合栄養補給様式であれば、増殖とプラスミド形成の相互作用がなくなる。指数関数的栄養補給中、短時間で高量のバイオマス(1リットル当たり約30gの乾燥細胞重量)が得られる。線形栄養補給中、プラスミドの蓄積は低い比増殖速度のためである。この栄養補給法により、500〜800mg/Lのプラスミドタイターが得られる。
【0036】
〔発酵の終点〕
製造業者の特有の要求によって、自由に発酵の終了時点を選択できる。製造規模のプラスミド生産のための発酵は一般的に約15〜20時間かかる。発酵中のプラスミド複製は非常に動的であり、結局、発酵時間の中間でpDNAの比収率が非常に上昇することになる。最大ではないが、その体積収率は高い。発酵の目的がプラスミドDNAの可能な最高濃度のバイオマスを得ることである場合、この時点で発酵を終わらせることができる。これは、その後のアルカリリーシス及び精製時に顕著な利点を有する。バイオマス中のpDNA濃度が高いほど、ライセート体積が少なく、かつプロセス時間が短くなる。さらに、プロセスバルク液中のpDNAの濃度は、より高い。
【0037】
最高のpDNA比濃度点を超えて、pDNA複製はまだ起こり、1L当たりのpDNA濃度を高めることとなるが、バイオマス1g当たりの含量は減少する。プロセスの目標が、プラスミドDNAの可能な最高総量を得ることである場合、複製が止むまで発酵を延長してよい。最大ではないが、発酵のこの後期での比収率は、当技術で知られているいずれの方法におけるよりもまだ高い。
この複製挙動は高度のプロセスフレキシビリティーを与え、高い比収率又は高い体積収率の間の選択を可能にし、簡単には発酵時間によって決まる。発酵の後期中、プラスミド均質性の実質的な低減は観察されない。
本発明の方法で得られるプラスミドDNAは、既知の方法によって回収かつ精製される。
プラスミド精製は、典型的には、収集した細胞集団の分解、通常アルカリリーシスによる分解で開始する。その結果、界面活性剤と一緒に強アルカリ性のpH値に細胞を供するので、細胞が溶解してプラスミドが遊離する。その後の酢酸緩衝液による沈殿工程時、タンパク質とゲノムDNAが沈殿し、一方プラスミドDNAは清澄上清中に留まる。その後の精製工程は、主にろ過(限外濾過、ディアフィルトレーション)とクロマトグラフ法を含む。クロマトグラフ法は、例えば、疎水的相互作用、イオン交換又はゲルろ過クロマトグラフィーから選択される。
【0038】
〔実施例1〕
〔プラスミドpRZ-hMCP1を保有する大腸菌JM108の流加発酵〕
20L規模の発酵槽内、プラスミドpRZ-hMCP1を内部にもつ大腸菌K-12株JM108で指数関数的流加発酵を行った(タンク反応器を撹拌した)。このプラスミド(4.9kb)はpUC oriを含有するpcDNA3TM(Invitrogen)の誘導体であり、カナマイシン耐性マーカーである。pRZ-hMCP1の問題の遺伝子は真核生物のCMV(サイトメガロウイルス)プロモーターの転写制御下の単球化学誘引物質タンパク質1である(Furutani et al., 1989)。
前培養のため、300mLの規定培地を含有する整流1000mL振とうフラスコに前記菌株のグリセロールストック(300μL)を接種した。これを回転式振とう機で300rpm及び37℃にて培養した。前培養基は以下のとおり:NH4Cl 2g/L、MgSO4*7H2O 0.24g/L、グルコース 10g/L、L-プロリン 0.2g/L、L-イソロイシン 0.2g/L、チアミン塩酸塩 1mg/L、クエン酸 2g/L、KH2PO4 5.44g/L、Na2HPO4*12H2O 14.38g/L及び微量要素溶液 16.7mL/L。微量要素溶液はHCl(25%) 14.6g/L、CaCl2*2H2O 0.44g/L、FeSO4*7H2O 0.33g/L、CoCl2*6H2O 0.14g/L、MnSO4*H2O 0.10g/L、CuSO4*5H2O 15mg/L及びZnSO4*7H2O 17mg/Lを含有した。
【0039】
前培養がほぼOD=1の光学密度に達したとき、これを発酵槽に移して発酵を始めた。主培養バッチ培地は前培養と同一濃度で同一成分を含有した。発酵槽は、発酵の開始時に7Lのバッチ培地を含有した。
温度を37℃に制御し、0.35バールの背圧で発酵を操作した。1vvm(培地体積及び1分当たり=7L/分当たりの空気の体積)のプロセスエア流量速度で発酵槽を通気した。溶存酸素圧が30%に低下したとき、スターラーの撹拌速度を上げることによって(500〜1000rpm)溶存酸素圧をこの規定点で維持した。撹拌速度の上昇がDOを維持するのに不十分な場合、空気の酸素濃度を純粋な酸素で濃厚にした。水酸化アンモニウムの溶液(25%)で7.0±0.2の設定点にpHを制御した(同時に発酵の間じゅう窒素源として働いた)。必要な場合、25% H2SO4でpHをさらに制御した。
【0040】
図1に、プラスミドの体積収率と比収率及び増殖の曲線を示す。発酵終了時、590mg/LというpDNAの例外的に高い体積収率が得られた。さらに、発酵時間20時間では、pDNAの比収率が1gの乾燥細胞重量当たり44mgのpDNAという最大値(これも例外的に高い値である)に達した。発酵の終了に向かって、比収率は15mg/g(DCW)に減少した。この比収率及び体積収率の特有の過程は、製造プロセスの必要条件によって、発酵を終わらせる時点を選択できることに関して複数の選択肢という利点を提供する。最高含量のpDNAを有するバイオマスを得なければならない場合、20時間後に発酵を終わらせることができ、3000mg/Lという体積収率となる。プロセスの目標が最高の体積収率である場合、40時間を超えて発酵を延長することができ、約600mg/Lとなるが、比収率は低い。
【0041】
10時間のバッチ培養後、バッチ培地中のグルコースが枯渇した(図2)。これは、急速オフライン測定法で決定した(Yellow Springs Glucose Analyzer, YSI 2700 Select)。
グルコースの枯渇は指数関数的栄養補給期の開始の合図として役立った。栄養補給培地は以下のとおりだった:グルコース 300g/L;MgSO4*7H2O 7.2g/L;L-プロリン 6g/L;L-イソロイシン 6g/L;チアミン塩酸塩 30mg/L;クエン酸 2g/L;KH2PO4 5.4g/L;Na2HPO4*12H2O 14.4g/L;CaCl2*2H2O 220mg/L;FeSO4*7H2O 170mg/L;CoCl2*6H2O 72mg/L、MnSO4*H2O 51mg/L、CuSO4*5H2O 8mg/L及びZnSO4*7H2O 9mg/L。
グルコース枯渇時のバイオマスに基づき(光学密度で推定)、発酵槽のプロセスコントロールシステムによって連続的な指数関数的栄養補給を制御した。
【0042】
0.1時間-1という予め規定した比増殖速度μを得るため栄養補給速度を選択した。質量流量計により栄養補給速度の制御を行った。全発酵の間じゅう、グルコースは限界であり、これは正確な増殖速度制御の必須項目だった。グルコースのオーバーフローの合図であるアセテートは大部分の栄養補給期にわたって有意な量では生じなかった(40時間まで0.5g/L未満)。図1に示されるように、バイオマスは、予め規定した指数関数に正確に従った。発酵時間が40時間を超えると、予測されるバイオマスのコースと実際のバイオマスのコースが逸れたので(図示せず)、発酵を停止した。図2は、現在の比増殖速度μを示す。
この比増殖速度は、得られた乾燥細胞重量から計算され、ほとんどの発酵時間にわたって、予め規定した比増殖速度に従う。
【0043】
図3は、アニオン交換高速液体クロマトグラフィー(AEX-HPLC)法で分析した、発酵時間にわたる種々のプラスミド型の割合を示す。発酵の主要部の間じゅう、共有結合的に閉じた環状pDNAの割合が90%を超え、42時間の培養時間まで最小89%で維持された。発酵の終了時(42時間)のプラスミドの分析クロマトグラムを図4に示す。
この実施例は、高いpDNA均質性と共に、体積収率と比収率に関して本発明の優れた結果を示す。
【0044】
〔実施例2〕
〔大腸菌JM108の指数関数的流加発酵でプラスミド収率に及ぼすイソロイシンの影響〕 1L規模の選別発酵槽において、実施例1で述べた方法を適用して、増殖とプラスミド生産に及ぼすイソロイシンの効果を示した。実施例1で述べたのと同様に2つの発酵を行った。2つの発酵の相違は、一方の培地はイソロイシンを含むが、他方の培地はイソロイシンを含まないことのみだった。培地の残存組成、並びに培養条件及びμ=0.1時間-1の増殖速度での指数関数的栄養補給の様式は実施例1で述べたのと同一だった。秤及びぜん動ポンプによって栄養補給速度を自動制御した。
両発酵の光学密度とpDNA体積収率の時間経過を図5(イソロイシンあり)と図6(イソロイシンなし)に示す。両発酵の増殖は、指数関数的栄養補給期中、μ=0.09時間-1という平均比増殖速度でほぼ同一だった。イソロイシンを含有する培地では、pDNA体積収率が633mg/Lであり、イソロイシンがない場合は398mg/Lが得られた。下表1は、発酵パラメーターの概要を与える。両プロセス変形では、pDNA均質性はほぼ90%cccだった。イソロイシンありのpDNAの最大比収率は、イソロイシンがない場合に比べて2倍高かった。
【0045】
表1:
イソロイシンが存在する場合と存在しない場合の指数関数的流加発酵の特性の比較
【0046】
この実施例は、培地中にイソロイシンが存在すると、イソロイシンがない培地から得られるより高い体積収率及び比収率をもたらすことを示す。しかし、イソロイシンがない場合でさえ、発酵のpDNA収率は、既知の方法よりまだ優れる。
【0047】
〔実施例3〕
〔線形栄養補給様式で成功した、指数関数的栄養補給アルゴリズムを用いる、プラスミドpRZ-hMCP1を保有する大腸菌JM108の流加発酵(20Lの発酵槽)〕
この実施例では、実施例1で述べたようにプラスミドpRZ-hMCP1を保有する大腸菌JM108細胞を調製して培養する。実施例1と異なり、栄養補給期を以下の2つの異なる部分に分割する:
(1)比増殖速度μ=0.25時間-1を維持するための栄養補給指数関数に栄養補給速度が従う指数関数的栄養補給期、及び
(2)栄養補給速度が一定値200mL/時間に維持される線形一定栄養補給期。
指数関数的栄養補給から線形栄養補給への切替えを行う時点は、10時間の指数関数的栄養補給後である。線形栄養補給期を選択して10時間とする。このような発酵により、500〜800mgのpDNA/L又は20〜30mgのpDNA/g(DCW)の範囲の体積収率及び比収率が得られる。
【0048】
〔参考文献〕
Andersen, D.C., Swartz, J., Ryll, T., Lin, N., and Snedecor, B. (2001) Metabolic oscillations in an E. coli Fermentation. Biotech. Bioeng. 75:2, 212-218.
Atlung, T., Christensen, B.B., and Hansen, F.G. (1999). Role of the Rom protein in copy number control of plasmid pBR322 at different growth rates in Escherichia coli K-12. Plasmid 41, 110-119.
CBER, Center for Biologics Evaluation and Research, U.S. Food and Drug Administration, Rockville, MD (1998). Guidance for industry - guidance for human somatic cell therapy and gene therapy.
Chen, W., Graham, C., and Ciccarelli, R.B. (1997). Automated fed-batch fermentation with feed-back controls based on dissolved oxygen (DO) and pH for production of DNA vaccines. J. Ind. Microbiol. Biotechnol., 18, 43-48.
Furutani Y., Nomura H., Notake M., Oyamada Y., Fukui T., Yamada M., LarsenC.G., Oppenheim J. J. and Matsushima K. (1989). Cloning and sequencing of the cDNA for human monocyte chemotactic factor (MCAF). Biochem Biophys Res Commun 159(1), 249-55.
Hofmann, K.H., Neubauer, P., Riethdorf, S., and Hecker, M. (1989). Amplification of pBR322 plasmid DNA in Escherichia coli relA strains during batch and fed-batch fermentation. J. Basic Microbiol. 30, 37-41.
Lahijani, R., Hulley, G., Soriano, G., Horn, N.A., and Marquet, M. (1996). High-yield production of pBR322-derived plasmids intended for human gene therapy by employing a temperature-controllable point mutation.
Hum. Gene Ther. 7, 1971-1980.
Lawther, R.P., Calhoun, D.H., Adams, C.W., Hauser, C.A., Gray, J., and Hatfield, G.W. (1981). Molecular basis of valine resistance in Escherichia coli K-12. Proc. Natl. Acad. Sci. USA 78:2, 922-925.
O´Kennedy, R.D., Baldwin, C., and Keshavarz-Moore, E., (2000). Effects of growth medium selection on plasmid DNA production and initial processing steps. J.Biotechnol. 76, 175-183.
O´Kennedy et al (2003). Effects of fermentation strategy on the characteristics of plasmid DNA production. Biotechnol Appl. Biochem. 34, 83-90.
Reinikainen, P., Korpela, K., Nissinen, V., Olkku, J., Soederlund, H., and Markkanen, P. (1988). Escherichia coli plasmid production in Fermenter. Biotech. Bioeng. 33, 386-393.
Sambrook, J., and Russel, D.W. (2001). Molecular Cloning - a Laboratory Manual, 3rd ed., J. Argentine, N. Irwin, K.A. Janssen, S. Curtis, M. Zierler, et. al., eds. (Cold Spring Harbor, NY).
Schoenfeld, T., Mendez, J., Storts, D.R., Portman, E., Patterson, B., Frederiksen,J., and Smith, C. (1995). Effects of bacterial strains carrying the endA1 genotype on DNA quality isolated with Wizard Plasmid Purification System. Promega Notes, 53, 12 - 22.
Ulmer, J.B., Donelly, J.J., and Liu, M.A. (1996). Toward the development of DNA vaccines. Curr. Opin. Biotechnol. 7, 653-658.
Vieira, I. and Messing, I. 81982). The pUC plasmids, an M133mp7-derived system for insertion mutagenesis and sequencing with synthetic universal primers. Gene 19, 259-268.
Wang et al. (2001). Medium design for Plasmid DNA based on stoichimetric model. Proc. Biochem. 36, 1085-1093.
Wrobel, B. and Wegrzyn, G. (1998). Replication regulation of ColE1-like plasmids in amino ascid-starved Escherichia coli. Plasmid 39, 48-62.
Yanisch-Perron, C., Vieira, J., and Messing, J. (1985). Improved M13 phage cloning vectors and host strains: nucleotide sequences of the M13mp18 and pUC19 vectors. Gene 33, 103-119.
Zhang and Greasham (1999). Chemically defined media for commercial fermentations. Appl. Microbiol. Biotechnol. 51, 407-421.
【特許請求の範囲】
【請求項1】
問題の遺伝子を運ぶプラスミドを有する大腸菌細胞を最初に前培養で増殖させ、引き続き主培養で発酵させ、得られたプラスミドDNAを回収して精製する、製造規模でプラスミドDNAを生産する方法であって、前記主培養がバッチ期と栄養補給期を含む流加法であり、このとき、
a)前記バッチ期の培地及び前記栄養補給期中に添加する培地を化学的に規定し、さらに、
b)前記栄養補給期の培地は、
i)増殖制限基質を含み、かつ
ii)前記栄養補給期の少なくとも一部分の間、予め規定した指数関数に従う栄養補給速度で添加されることによって、予め規定した値に比増殖速度を維持する、前記方法。
(ここで、前記指数関数は下記式の関数であり、
Ft=定数*eμt
式中、
Ftは、栄養補給培地の流速[L/時間]であり;
μは、比増殖速度[時間-1]であり、かつ
tは、前記栄養補給期の開始からの時間間隔[時間]である。)
【請求項2】
前記栄養補給期の、前記栄養補給速度が前記指数関数に従う前記部分が、前記栄養補給期に得られる総乾燥細胞重量の約20%より多くが前記部分の間に生成されるような部分である、請求項1の方法。
【請求項3】
前記前培養で使用する培地をも化学的に規定する、請求項1の方法。
【請求項4】
relA遺伝子に突然変異がある大腸菌株を使用する、請求項1の方法。
【請求項5】
前記大腸菌株がK-12である、請求項4の方法。
【請求項6】
前記大腸菌K-12株がJM108又はその誘導体である、請求項5の方法。
【請求項7】
前記プラスミドがColE1-型複製開始点を有する、請求項1の方法。
【請求項8】
前記プラスミドがpUCプラスミドである、請求項7の方法。
【請求項9】
前記指数関数が下記式の関数である、請求項1の方法。
(式中、
Ftは、栄養補給培地の流速[L/時間]であり;
X0は、栄養補給期の開始時のバイオマス乾燥細胞重量の総量[g]であり;
YX/Sは、バイオマス収量係数(基質1g当たりの乾燥細胞重量g数)であり;
CSは、前記栄養補給培地中の前記基質の濃度[g/L]であり、かつ
μは、比増殖速度[時間-1]である。)
【請求項10】
前記指数関数に従って前記培地を連続的に添加することによって、前記栄養補給速度を連続的に上昇させる、請求項1の方法。
【請求項11】
前記指数関数に従って段階的に前記培地を添加することによって、前記栄養補給速度を半連続様式で上昇させる、請求項1の方法。
【請求項12】
前記指数関数に従ってパルス式に前記培地を添加することによって、前記栄養補給速度を不連続様式で上昇させる、請求項1の方法。
【請求項13】
前記栄養補給速度の指数関数が、所定時点のバイオマス量の測定に基づく、請求項1の方法。
【請求項14】
前記増殖制限基質が炭素源である、請求項1の方法。
【請求項15】
前記炭素源がグルコースである、請求項14の方法。
【請求項16】
前記増殖速度が約0.03〜約0.2時間-1である、請求項1の方法。
【請求項17】
前記バッチ培養の培地がイソロイシンを含む、請求項1の方法。
【請求項18】
前記栄養補給期の培地もイソロイシンを含む、請求項17の方法。
【請求項19】
前記バッチ培養の培地が、発酵の開始時に、窒素源として1種以上のアンモニウム塩を含む、請求項1の方法。
【請求項20】
前記アンモニウム塩が塩化アンモニウムである、請求項19の方法。
【請求項21】
水酸化アンモニウムを添加することによって、前記主培養のpH値を制御すると同時に、前記水酸化アンモニウムが、前記栄養補給期中、増殖を制限することなく窒素源として働く、請求項1の方法。
【請求項22】
前記培地に抗生物質がない、請求項1の方法。
【請求項23】
前記前培養で使用する培地にも抗生物質がない、請求項22の方法。
【請求項24】
製造規模で大腸菌内にてプラスミドDNAを生産するための培地であって、以下の成分: a)グルコース、グリセロール、フルクトース、ラクトース、スクロース、アラビノース、又はその混合物から選択される有機炭素源;
b)アンモニウム塩及び水酸化アンモニウムから選択される無機窒素源(ここで、前記窒素源は、前記培地の成分として存在し、又は発酵中に前記培地に添加される);
c)マクロ要素及びミクロ要素を供給するために働く無機塩;
d)任意に、前記大腸菌株の栄養要求性を補完する1種以上の物質;及び
e)イソロイシン;
を含有する、化学的に規定した培地。
【請求項25】
バッチ発酵の開始時又は流加発酵のバッチ期の開始時に存在するバッチ培地であり、かつ以下の成分:
a)約10〜約30g/Lの濃度のグルコース;
b)アンモニウム濃度が約0.5〜約2g/Lとなるようなアンモニウム塩又は水酸化アンモニウム;
c)マクロ要素及びミクロ要素の供給として働く無機イオン;
d)任意に、前記大腸菌株の栄養要求性を補完する1種以上の物質;及び
e)約0.1〜約0.3g/Lの濃度のイソロイシン;
を含有する、請求項24の培地。
【請求項26】
流加発酵の栄養補給期中に添加する栄養補給培地であり、かつ以下の成分:
a)約300〜約500g/Lの濃度のグルコース;
b)マクロ要素及びミクロ要素を供給するために働く無機イオン;
c)任意に、前記大腸菌株の栄養要求性を補完する1種以上の物質;及び
d)約6g/Lの濃度のイソロイシン;
を含有する、請求項24の培地。
【請求項1】
問題の遺伝子を運ぶプラスミドを有する大腸菌細胞を最初に前培養で増殖させ、引き続き主培養で発酵させ、得られたプラスミドDNAを回収して精製する、製造規模でプラスミドDNAを生産する方法であって、前記主培養がバッチ期と栄養補給期を含む流加法であり、このとき、
a)前記バッチ期の培地及び前記栄養補給期中に添加する培地を化学的に規定し、さらに、
b)前記栄養補給期の培地は、
i)増殖制限基質を含み、かつ
ii)前記栄養補給期の少なくとも一部分の間、予め規定した指数関数に従う栄養補給速度で添加されることによって、予め規定した値に比増殖速度を維持する、前記方法。
(ここで、前記指数関数は下記式の関数であり、
Ft=定数*eμt
式中、
Ftは、栄養補給培地の流速[L/時間]であり;
μは、比増殖速度[時間-1]であり、かつ
tは、前記栄養補給期の開始からの時間間隔[時間]である。)
【請求項2】
前記栄養補給期の、前記栄養補給速度が前記指数関数に従う前記部分が、前記栄養補給期に得られる総乾燥細胞重量の約20%より多くが前記部分の間に生成されるような部分である、請求項1の方法。
【請求項3】
前記前培養で使用する培地をも化学的に規定する、請求項1の方法。
【請求項4】
relA遺伝子に突然変異がある大腸菌株を使用する、請求項1の方法。
【請求項5】
前記大腸菌株がK-12である、請求項4の方法。
【請求項6】
前記大腸菌K-12株がJM108又はその誘導体である、請求項5の方法。
【請求項7】
前記プラスミドがColE1-型複製開始点を有する、請求項1の方法。
【請求項8】
前記プラスミドがpUCプラスミドである、請求項7の方法。
【請求項9】
前記指数関数が下記式の関数である、請求項1の方法。
(式中、
Ftは、栄養補給培地の流速[L/時間]であり;
X0は、栄養補給期の開始時のバイオマス乾燥細胞重量の総量[g]であり;
YX/Sは、バイオマス収量係数(基質1g当たりの乾燥細胞重量g数)であり;
CSは、前記栄養補給培地中の前記基質の濃度[g/L]であり、かつ
μは、比増殖速度[時間-1]である。)
【請求項10】
前記指数関数に従って前記培地を連続的に添加することによって、前記栄養補給速度を連続的に上昇させる、請求項1の方法。
【請求項11】
前記指数関数に従って段階的に前記培地を添加することによって、前記栄養補給速度を半連続様式で上昇させる、請求項1の方法。
【請求項12】
前記指数関数に従ってパルス式に前記培地を添加することによって、前記栄養補給速度を不連続様式で上昇させる、請求項1の方法。
【請求項13】
前記栄養補給速度の指数関数が、所定時点のバイオマス量の測定に基づく、請求項1の方法。
【請求項14】
前記増殖制限基質が炭素源である、請求項1の方法。
【請求項15】
前記炭素源がグルコースである、請求項14の方法。
【請求項16】
前記増殖速度が約0.03〜約0.2時間-1である、請求項1の方法。
【請求項17】
前記バッチ培養の培地がイソロイシンを含む、請求項1の方法。
【請求項18】
前記栄養補給期の培地もイソロイシンを含む、請求項17の方法。
【請求項19】
前記バッチ培養の培地が、発酵の開始時に、窒素源として1種以上のアンモニウム塩を含む、請求項1の方法。
【請求項20】
前記アンモニウム塩が塩化アンモニウムである、請求項19の方法。
【請求項21】
水酸化アンモニウムを添加することによって、前記主培養のpH値を制御すると同時に、前記水酸化アンモニウムが、前記栄養補給期中、増殖を制限することなく窒素源として働く、請求項1の方法。
【請求項22】
前記培地に抗生物質がない、請求項1の方法。
【請求項23】
前記前培養で使用する培地にも抗生物質がない、請求項22の方法。
【請求項24】
製造規模で大腸菌内にてプラスミドDNAを生産するための培地であって、以下の成分: a)グルコース、グリセロール、フルクトース、ラクトース、スクロース、アラビノース、又はその混合物から選択される有機炭素源;
b)アンモニウム塩及び水酸化アンモニウムから選択される無機窒素源(ここで、前記窒素源は、前記培地の成分として存在し、又は発酵中に前記培地に添加される);
c)マクロ要素及びミクロ要素を供給するために働く無機塩;
d)任意に、前記大腸菌株の栄養要求性を補完する1種以上の物質;及び
e)イソロイシン;
を含有する、化学的に規定した培地。
【請求項25】
バッチ発酵の開始時又は流加発酵のバッチ期の開始時に存在するバッチ培地であり、かつ以下の成分:
a)約10〜約30g/Lの濃度のグルコース;
b)アンモニウム濃度が約0.5〜約2g/Lとなるようなアンモニウム塩又は水酸化アンモニウム;
c)マクロ要素及びミクロ要素の供給として働く無機イオン;
d)任意に、前記大腸菌株の栄養要求性を補完する1種以上の物質;及び
e)約0.1〜約0.3g/Lの濃度のイソロイシン;
を含有する、請求項24の培地。
【請求項26】
流加発酵の栄養補給期中に添加する栄養補給培地であり、かつ以下の成分:
a)約300〜約500g/Lの濃度のグルコース;
b)マクロ要素及びミクロ要素を供給するために働く無機イオン;
c)任意に、前記大腸菌株の栄養要求性を補完する1種以上の物質;及び
d)約6g/Lの濃度のイソロイシン;
を含有する、請求項24の培地。
【図1】

【図2】
【図3】

【図4】
【図5】
【図6】
【図2】
【図3】
【図4】
【図5】
【図6】
【公開番号】特開2011−24596(P2011−24596A)
【公開日】平成23年2月10日(2011.2.10)
【国際特許分類】
【出願番号】特願2010−227565(P2010−227565)
【出願日】平成22年10月7日(2010.10.7)
【分割の表示】特願2007−506711(P2007−506711)の分割
【原出願日】平成17年4月6日(2005.4.6)
【出願人】(504135837)ベーリンガー インゲルハイム エルツェーファウ ゲゼルシャフト ミット ベシュレンクテル ハフツング ウント コンパニー コマンディトゲゼルシャフト (10)
【Fターム(参考)】
【公開日】平成23年2月10日(2011.2.10)
【国際特許分類】
【出願日】平成22年10月7日(2010.10.7)
【分割の表示】特願2007−506711(P2007−506711)の分割
【原出願日】平成17年4月6日(2005.4.6)
【出願人】(504135837)ベーリンガー インゲルハイム エルツェーファウ ゲゼルシャフト ミット ベシュレンクテル ハフツング ウント コンパニー コマンディトゲゼルシャフト (10)
【Fターム(参考)】
[ Back to top ]