
天然の免疫グロブリン形式を有する容易に単離される二重特異性抗体
単離が容易である二重特異性抗体の形式が提供され、その形式は、CH3ドメイン内を識別的に改変されている免疫グロブリン重鎖可変ドメインを含み、ここで、その識別的改変は、そのCH3改変に関して免疫原性でないかまたは実質的に免疫原性でなく、それらの改変の少なくとも1つは、プロテインAなどの親和性試薬に対する二重特異性抗体の親和性に違いをもたらし、その二重特異性抗体は、プロテインAに対するその親和性に基づいて、破壊された細胞、培地、または抗体の混合物から単離可能である。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、重鎖のヘテロ二量体を有する、抗原結合タンパク質または抗体、すなわち、親和性試薬に対する免疫グロブリン重鎖の親和性と改変免疫グロブリン重鎖または変異免疫グロブリン重鎖の親和性との違いに基づいて抗原結合タンパク質の単離を可能にする、少なくとも1つのアミノ酸が異なる2つの免疫グロブリン重鎖を有する、抗原結合タンパク質または抗体に関する。本発明はまた、プロテインAに対するIgG領域の結合性の違いによる迅速な単離を可能にする、プロテインAに対して異なる親和性を有するIgG CH2およびCH3領域を有する抗原結合タンパク質(二重特異性抗体を含む)にも関する。
【背景技術】
【0002】
背景
抗体は、標的抗原に対する独特の結合特異性ならびに抗原とは無関係な機序を介して免疫系と相互作用する能力を保有する多機能性分子である。癌に対して現在使用されている多くの生物学的治療薬は、標的にされている癌細胞上で代表的に過剰発現されている抗原に対するモノクローナル抗体である。そのような抗体は、腫瘍細胞に結合すると、抗体依存性細胞傷害(ADCC)または補体依存性細胞傷害(CDC)の引き金を引くことがある。残念なことに、癌性細胞は、これらの正常な免疫応答を抑制する機序を発達させることが多い。
【0003】
近年、2以上の抗原結合特異性を有する抗体様の治療薬、例えば、二重特異性抗体を開発する試みが進行中である。癌治療の場合、多重特異的形式によって、例えば、その分子が腫瘍細胞抗原を標的にするという1つの特異性、通常免疫系が利用できない反応の引き金を引くというその他の特異性を使用することが可能になり得る。二重特異性抗体は、2成分のヘテロ二量体レセプター系(この2成分のヘテロ二量体レセプター系は、それらの天然のリガンドが、そのレセプター系に結合して、その両方の成分と1つになるとき、そのリガンドによって正常に活性化される)に対する代用リガンドとしての使用法も見出し得る。
【0004】
複数の結合特異性を有する分子によって与えられる治療の機会に応える数多くの形式が当該分野で開発されている。理想的には、そのような分子は、作製しやすくかつ精製しやすく、好ましいインビボ特性、例えば、意図される目的にとって適切な薬物動態、最小の免疫原性、および望まれる場合、従来の抗体のエフェクター機能を有する、よく機能するタンパク質であるべきである。
【0005】
二重特異性抗体を生成する(単一細胞内で2つの異なる抗体を発現させる)最も単刀直入な方法は、複数の種を生じることである。なぜなら、それぞれの重鎖が、ホモ二量体とヘテロ二量体の両方を形成するが、ヘテロ二量体だけが望まれるからである。また、軽鎖および重鎖は、不適切に対形成することがある。種々の方法でこれらの問題に対処するように試みられた形式のいくつかの例を以下に記載する。
【0006】
二重特異性T細胞Engager(BiTE)分子(例えば、Wolf,E.ら(2005)Drug Discovery Today 10:1237−1244(非特許文献1))を参照のこと)に対して使用される1つの形式は、一本鎖可変フラグメント(scFv)モジュールに基づく。scFvは、通常適切に折り畳まれ得る可撓性リンカーを介して融合された抗体の軽鎖および重鎖の可変領域からなり、それらの領域は、同族抗原に結合することができる。BiTEでは、特異性の異なる2つのscFvが一本鎖にタンデムにつながれている(図1Aを参照のこと)。この配置は、2コピーの同じ重鎖可変領域を有する分子の生成を妨げる。さらに、それぞれの軽鎖および重鎖の正しい対形成を確実にするようにリンカー配置が設計される。
【0007】
BiTE形式は、いくつかの欠点を有する。第1に、scFv分子は、凝集する傾向があるとよく知られている。また、scFvリンカーの免疫原性は、通説では低いが、BiTEに対する抗体が産生される可能性を除外することはできない。また、BiTE形式ではFc部が存在しないので、その血清半減期は非常に短くなり、高頻度の反復投与またはポンプによる持続注入の面倒な問題が伴う。最後に、Fcが存在しないことにより、Fc媒介性のエフェクター機能が存在しないことも当然伴う(そのエフェクター機能が存在しないことは、いくつかの状況では有益であり得るが)。
【0008】
第2の形式(図1B)は、マウスとラットのモノクローナル抗体のハイブリッドであり、従来のプロテインAアフィニティークロマトグラフィの変法に依存するものである(例えば、Lindhofer,H.ら(1995)J.Immunol.155:219−225(非特許文献2))を参照のこと)。この形式では、マウスIgG2a抗体およびラットIgG2b抗体を、同じ細胞内で同時に産生させる(例えば、2つのハイブリドーマのクアドローマ融合物として、または操作されたCHO細胞内で)。各抗体の軽鎖は、それらの同族の種の重鎖と優先的に会合するので、3つの異なる種の抗体:2つの親抗体、およびFc部によって会合している、その2つの抗体の各々の1つの重鎖/軽鎖対を含むその2つの抗体のヘテロ二量体だけが組み立てられ得る。所望のヘテロ二量体は、そのプロテインAへの結合特性が親抗体の結合特性と異なる(すなわち、ラットIgG2bは、プロテインAに結合せず、一方、マウスIgG2aは、プロテインAに結合する)ので、この混合物から容易に精製され得る。それゆえ、マウス−ラットヘテロ二量体は、プロテインAに結合するが、マウスIgG2aホモ二量体よりも高いpHで溶出することから、これにより、二重特異性ヘテロ二量体を選択的に精製することが可能になる。BiTE形式と同様に、このハイブリッド形式は、2つの一価抗原結合部位を有する。
【0009】
マウス/ラットハイブリッドの欠点は、それが非ヒトであるので、有害な副作用(例えば、「HAMA」または「HARA」反応)を有し得、そして/またはその治療薬を中和し得る免疫応答を患者に誘発する可能性があるという点である。
【0010】
「ノブ・イントゥ・ホール(knobs−into−holes)」と呼ばれる第3の形式(図1C)は、二重特異性抗体の作製にとって有用である可能性があると従来技術において考察されている(米国特許第7,183,076号(特許文献1))。このストラテジーでは、一方が「ノブ」を突出させ、他方が相補的な「ホール」となるように、2つの抗体のFc部を操作する。それらの重鎖は、同じ細胞において産生されると、操作された「ノブ」と操作された「ホール」との会合によって、ホモ二量体ではなくヘテロ二量体を優先的に形成すると言われている。異なる特異性を有するが同一の軽鎖を採用する抗体を選択することによって、正しい軽鎖−重鎖の対形成の問題が対処される。
【0011】
この形式の欠点は、「ノブ・イントゥ・ホール」ストラテジーが、かなりの量の望ましくないホモ二量体を産生し得るがゆえに、さらなる精製工程が必要となるという点である。この難点は、汚染している種の特性の多くが所望の種とほぼ同一であるという事実によって深刻になっている。「ノブ」および「ホール」を生成する変異によって外来配列が導入されるので、操作された形状は、潜在的に免疫原性でもあり得る。
【0012】
依然として、二重特異性抗体の形式、特に、上で述べた欠点の一部または全部を最小にする、治療に役立つ用途のための二重特異性抗体の形式が必要とされている。
【先行技術文献】
【特許文献】
【0013】
【特許文献1】米国特許第7,183,076号明細書
【非特許文献】
【0014】
【非特許文献1】Wolf,E.ら(2005)Drug Discovery Today 10:1237−1244
【非特許文献2】Lindhofer,H.ら(1995)J.Immunol.155:219−225
【発明の概要】
【課題を解決するための手段】
【0015】
概要
本発明は、二重特異性抗原結合タンパク質において少なくとも1つのアミノ酸が異なる2つの免疫グロブリンCH3重鎖定常ドメイン配列を使用することに少なくとも部分的に基づく。少なくとも1つのアミノ酸が異なることにより、CH3ドメイン配列が親和性物質に結合する能力に違いがもたらされるので、そのタンパク質を単離する能力が改善される。
【0016】
1つの局面において、第1および第2のポリペプチドを含む抗原結合タンパク質が提供され、その第1のポリペプチドは、N末端からC末端に向かって、第1の抗原に選択的に結合する第1の抗原結合領域、それに続く、IgG1(配列番号1)、IgG2(配列番号3)IgG4(配列番号5)およびそれらの組み合わせから選択されるヒトIgGの第1のCH3領域を含む定常領域を含み;そして第2のポリペプチドは、N末端からC末端に向かって、第2の抗原に選択的に結合する第2の抗原結合領域、それに続くIgG1、IgG2、IgG4およびそれらの組み合わせから選択されるヒトIgGの第2のCH3領域を含む定常領域を含み、ここで、その第2のCH3領域は、プロテインAへの第2のCH3ドメインの結合を減少させるかまたは排除する改変を含む。
【0017】
1つの実施形態において、第2のCH3領域は、95R改変(IMGTエキソンナンバリングにおいて;EUナンバリングでは435R)を含む。別の実施形態において、第2のCH3領域は、96F改変(IMGT;EUでは436F)をさらに含む。特定の実施形態において、第2のCH3領域は、配列番号2、配列番号4および配列番号6から選択される。
【0018】
1つの実施形態において、第2のCH3領域は、改変されたヒトIgG1(配列番号2)に由来し、D16E、L18M、N44S、K52N、V57MおよびV82I(IMGT;EUではD356E、L358M、N384S、K392N、V397MおよびV422I)からなる群から選択される改変をさらに含む。
【0019】
1つの実施形態において、第2のCH3領域は、改変されたヒトIgG2(配列番号4)に由来し、N44S、K52NおよびV82I(IMGT;EUではN384S、K392NおよびV422I)からなる群から選択される改変をさらに含む。
【0020】
1つの実施形態において、第2のCH3領域は、改変されたヒトIgG4(配列番号6)に由来し、Q15R、N44S、K52N、V57M、R69K、E79QおよびV82I(IMGT;EUではQ355R、N384S、K392N、V397M、R409K、E419QおよびV422I)からなる群から選択される改変をさらに含む。
【0021】
1つの実施形態において、CH3ドメインは、ヒトIgG1、ヒトIgG2、ヒトIgG3およびヒトIgG4のうちの2つ以上の配列を含むキメラドメインである。
【0022】
1つの実施形態において、CH3ドメインは、ヒトIgG1、ヒトIgG2またはヒトIgG4に由来し、本抗原結合タンパク質は、CH1ドメインおよびCH2ドメインをさらに含み、ここで、そのCH1ドメインおよびCH2ドメインは、独立して、ヒトIgG1 CH1もしくはCH2ドメイン、ヒトIgG2 CH1もしくはCH2ドメイン、またはキメラヒト/ヒトIgG1/IgG2もしくはキメラヒト/ヒトIgG1/IgG3もしくはキメラヒト/ヒトIgG2/IgG3ドメインもしくはキメラヒト/ヒトIgG1/IgG4もしくはキメラIgG3/IgG4もしくはキメラIgG2/IgG4ドメインからなる群から選択される。特定の実施形態において、そのキメラIgG1/IgG2、IgG1/IgG3、IgG2/IgG3、IgG1/IgG4、IgG3/IgG4およびIgG2/IgG4ドメインは、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない。
【0023】
1つの実施形態において、本抗原結合タンパク質は、免疫グロブリン軽鎖をさらに含む。1つの実施形態において、その免疫グロブリン軽鎖は、ヒトラムダ軽鎖およびヒトカッパー軽鎖から選択される。
【0024】
1つの実施形態において、第1および第2の抗原結合領域の各々は、少なくとも1つのCDRを含み、別の実施形態では、少なくとも2つのCDRを含み、別の実施形態では、各々は、3つのCDRを含む。特定の実施形態において、そのCDRは、免疫グロブリン重鎖に由来する。別の特定の実施形態において、その重鎖は、ヒト重鎖である。
【0025】
1つの実施形態において、第1の抗原結合領域は、第1の免疫グロブリン重鎖可変ドメインを含み、第2の抗原結合領域は、第2の免疫グロブリン重鎖可変ドメインを含む。
【0026】
1つの実施形態において、第1および第2の免疫グロブリン重鎖可変ドメインは、独立して、マウス、ラット、ハムスター、ウサギ、サル、類人猿およびヒトのドメインから選択される。
【0027】
1つの実施形態において、第1および第2の免疫グロブリン重鎖可変ドメインは、独立して、ヒトCDR、マウスCDR、ラットCDR、ウサギCDR、サルCDR、類人猿CDRおよびヒト化CDRを含む。1つの実施形態において、そのCDRは、ヒトのCDRであり、体細胞変異している。
【0028】
1つの実施形態において、第1および第2の免疫グロブリン重鎖可変ドメインは、ヒトフレームワーク領域(FR)を含む。1つの実施形態において、そのヒトFRは、体細胞変異したヒトFRである。
【0029】
1つの実施形態において、第1および/または第2の抗原結合領域は、抗体可変領域を含むファージライブラリーを目的の抗原に対する反応性についてスクリーニングすることによって得られる。別の実施形態において、第1および/または第2の抗原結合領域は、非ヒト動物(例えば、マウス、ラット、ウサギ、サルまたは類人猿)を目的の抗原で免疫し、目的の抗原に特異的な可変領域をコードする抗体可変領域の核酸配列を同定することによって得られる。特定の実施形態において、その非ヒト動物は、1つ以上のヒト免疫グロブリン可変領域の遺伝子を含む。別の特定の実施形態において、1つ以上のヒト免疫グロブリン可変領域の遺伝子は、染色体外に、内在性の免疫グロブリン遺伝子座における置換として、または非ヒト動物のゲノムにランダムにインテグレートされたトランスジーンとして、非ヒト動物に存在する。1つの実施形態において、第1および/または第2の抗原結合領域は、ハイブリドーマまたはクアドローマから得られ、別の実施形態では、免疫された非ヒト動物の免疫細胞を、細胞選別を用いてスクリーニングすることによって得られる。
【0030】
1つの実施形態において、本抗原結合タンパク質は、二重特異性抗体である。1つの実施形態において、その二重特異性抗体は、二重特異性の完全ヒト抗体であり、各エピトープに対して独立してマイクロモル濃度、ナノモル濃度またはピコモル濃度の範囲の親和性を有する。
【0031】
1つの実施形態において、本抗原結合タンパク質は、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない。特定の実施形態において、本抗原結合タンパク質は、非天然のヒトT細胞エピトープを有しない。1つの実施形態において、CH3領域の改変は、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない。特定の実施形態において、CH3領域の改変は、非天然のヒトT細胞エピトープをもたらさない。
【0032】
1つの実施形態において、本抗原結合タンパク質は、重鎖を含み、ここで、その重鎖は、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない。1つの実施形態において、その重鎖は、非天然のT細胞エピトープを含まないアミノ酸配列を有する。1つの実施形態において、その重鎖は、タンパク質分解によってヒトにおいて免疫原性である約9アミノ酸のアミノ酸配列が形成され得ないアミノ酸配列を含む。特定の実施形態において、上記ヒトは、本抗原結合タンパク質で処置されたヒトである。1つの実施形態において、その重鎖は、タンパク質分解によってヒトにおいて免疫原性である約13〜約17アミノ酸のアミノ酸配列が形成され得ないアミノ酸配列を含む。特定の実施形態において、上記ヒトは、本抗原結合タンパク質で処置されているヒトである。
【0033】
1つの局面において、本明細書中に記載されるようなCH2改変および/またはCH3改変を含む二重特異性結合タンパク質が提供され、ここで、その二重特異性結合タンパク質は、B細胞上の抗原を特異的に認識する第1の結合部分、およびT細胞上の抗原を特異的に認識する第2の結合部分を含む。
【0034】
1つの実施形態において、結合タンパク質は、二重特異性抗体である。特定の実施形態において、その二重特異性抗体は、ヒトIgG1重鎖およびヒトIgG1ΔAdp重鎖を含む。1つの実施形態において、第1の結合部分は、CD20を特異的に認識するヒト重鎖可変ドメインである。1つの実施形態において、第2の結合部分は、CD3を特異的に認識するヒト重鎖可変ドメインである。1つの実施形態において、上記二重特異性抗体は、Raji死滅アッセイにおいて約2.8〜3.2×10−12Mまたは約2.8〜3.0×10−12MのEC50を示し、バイスタンダー細胞死滅アッセイにおいて約1〜10%または1〜5%を越えないバイスタンダー死滅を示し、ここで、そのバイスタンダー細胞は、CD20エピトープを含まない。特定の実施形態において、バイスタンダー細胞は、293細胞である。別の特定の実施形態において、上記アッセイにおけるバイスタンダー細胞死滅は、約10−8M〜約10−14Mという二重特異性抗体の濃度にわたって測定される。
【0035】
1つの局面において、二重特異性抗体を作製するための方法が提供され、その方法は:第1のエピトープを認識する第1の可変ドメインを含む第1の免疫グロブリン重鎖をコードする核酸配列を得る工程(ここで、その第1の免疫グロブリン重鎖は、IgG1、IgG2もしくはIgG4のアイソタイプ定常ドメインまたはそのキメラアイソタイプ定常ドメインを含む);第2のエピトープを認識する第2の可変ドメインを含む第2の免疫グロブリン重鎖をコードする第2の核酸配列を得る工程(ここで、その第2の免疫グロブリン重鎖は、プロテインAへの結合を無くすかまたは減少させる改変をそのCH3ドメインに含む、IgG1、IgG2もしくはIgG4のアイソタイプ定常ドメインまたはそのキメラアイソタイプ定常ドメインを含む);第1および第2の免疫グロブリン重鎖と対形成する免疫グロブリン軽鎖をコードする第3の核酸配列を得る工程;その第1、第2および第3の核酸配列を哺乳動物細胞に導入する工程;その細胞に免疫グロブリンを発現させる工程、およびプロテインAを用いてその免疫グロブリンを単離する工程を包含する。
【0036】
1つの実施形態において、上記細胞は、CHO、COS、293、HeLaおよびウイルス核酸配列を発現する網膜細胞(例えば、PERC.6(商標)細胞)から選択される。
【0037】
1つの局面において、抗原に結合する第1の特異性およびレセプターを活性化させる第2の特異性を備える二重特異性抗原結合タンパク質が提供され、ここで、その二重特異性抗原結合タンパク質は、プロテインAに結合する決定基を含む第1のIgG1、IgG2またはIgG4 CH3ドメインを含む第1のポリペプチド、およびそのプロテインAに結合する決定基を欠く第2のIgG1、IgG3またはIgG4 CH3ドメインを含む第2のポリペプチドを含む。
【0038】
1つの実施形態において、上記レセプターを活性化する第2の特異性のおかげで、上記レセプターはモル濃度、ミリモル濃度、マイクロモル濃度、ナノモル濃度またはピコモル濃度の範囲であるKDで結合する。
【0039】
1つの実施形態において、上記第2の特異性のおかげで、Gタンパク質共役レセプター、チロシンキナーゼレセプター、インテグリンレセプターおよびtoll様レセプターから選択されるレセプターに結合する。
【0040】
1つの実施形態において、上記第2の特異性によって、上記レセプターと接触し、そのレセプターまたはそれと物理的に会合するサブユニットもしくはタンパク質が、セリン、トレオニンまたはチロシンのリン酸化を起こすか;ヌクレオチド(例えば、cAMP、cADPまたはcGMP)の環化(cyclicization)を引き起こすか;ホスファチジルイノシトールまたはその誘導体(例えば、IP3またはPIP3)の産生を引き起こすか;脂質セカンドメッセンジャー(例えば、ジアシルグリセロール(diacylclycerol)、セラミド、リゾホスファチジン酸、エイコサノイド)の産生を引き起こすか;脱リン酸化(例えば、ホスファターゼ活性)を引き起こすか;脂質のリン酸化を引き起こしてセカンドメッセンジャーを形成するか;セカンドメッセンジャーの加水分解を引き起こすか;タンパク質分解を引き起こすか;レドックスシグナル伝達を引き起こすか;細胞小器官(例えば、核)へのタンパク質のトランスロケーションを引き起こすか;レセプターが(それ自体と)凝集して、ホモ多量体を形成するかまたは(他のレセプターと)ヘテロ多量体を形成するか;あるいは膜貫通型チャネルの開口または閉鎖を引き起こす。
【0041】
1つの局面において、二重特異性抗体を作製するための方法が提供され、その方法は:クアドローマから目的の二重特異性抗体を単離する工程を包含し、ここで、その目的の二重特異性抗体は、IgG1、IgG2またはIgG4のアイソタイプである第1の重鎖、プロテインAへの結合を無くすかまたは減少させる改変をそのCH3ドメインに含む定常ドメインを有するIgG1、IgG2またはIgG4のアイソタイプである第2の重鎖を含み、ここで、その目的の二重特異性抗体は、プロテインAを用いて単離される。
【0042】
1つの局面において、二重特異性抗体を作製するための方法が提供され、その方法は、破壊された細胞または抗体の混合物から、識別的に改変された(differentially modified)IgG1、IgG2またはIgG4のCH3ドメインを有する二重特異性抗体を単離する工程を包含し、ここで、その識別的に改変されたCH3ドメインは、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でなく、その改変は、プロテインAに対するモノマーの親和性が互いに異なるヘテロ二量体の重鎖を有する二重特異性抗体をもたらし、その二重特異性抗体は、プロテインAを用いて、破壊された細胞または上記混合物から単離される。
【0043】
1つの実施形態において、本二重特異性抗体は、プロテインA親和性支持体を用いて単離され、ここで、その二重特異性抗体は、約3.9〜約4.4、約4.0〜約4.3、約4.1〜約4.2のpH、またはpH約4.2で溶出する。1つの実施形態において、本二重特異性抗体は、約4、4.1、4.2、4.3、4.4または4.5のpHで溶出する。
【0044】
1つの実施形態において、本二重特異性抗体は、プロテインA親和性支持体およびpH勾配またはpH段階(pH step)を用いて単離され、ここで、そのpH勾配またはpH段階は、イオン調節物質(ionic modifier)を含む。特定の実施形態において、イオン調節物質は、約0.5〜約1.0モル濃度の濃度で存在する。特定の実施形態において、イオン調節物質は、塩である。1つの実施形態において、イオン調節物質は、酢酸のベリリウム塩、リチウム塩、ナトリウム塩およびカリウム塩;重炭酸ナトリウムおよび重炭酸カリウム;炭酸リチウム、炭酸ナトリウム、炭酸カリウムおよび炭酸セシウム;塩化リチウム、塩化ナトリウム、塩化カリウム、塩化セシウムおよび塩化マグネシウム;フッ化ナトリウムおよびフッ化カリウム;硝酸ナトリウム、硝酸カリウムおよび硝酸カルシウム;リン酸ナトリウムおよびリン酸カリウム;ならびに硫酸カルシウムおよび硫酸マグネシウムからなる群から選択される。特定の実施形態において、イオン調節物質は、アルカリ金属またはアルカリ土類金属のハロゲン化物塩である。特定の実施形態において、イオン調節物質は、塩化ナトリウムである。
【0045】
1つの局面において、Fcを含む結合タンパク質(ここで、そのFcは、ヘテロ二量体のFcを形成するように、本明細書中に記載されるように改変された第1のCH3ドメインおよび改変されていない第2のCH3を含む)、ここで、その識別的改変は、識別的改変を有しない対応する結合タンパク質よりも0.5、0.6、0.7、0.8、0.9、1.0、1.2、1.3または1.4高いpH単位で、プロテインA親和性材料から溶出する結合タンパク質をもたらす。
【0046】
1つの実施形態において、識別的に改変された結合タンパク質は、約4.2のpHで溶出するのに対し、改変されていない結合タンパク質は、約3のpHで溶出する。1つの実施形態において、識別的に改変された結合タンパク質は、約4.5のpHで溶出するのに対し、改変されていない結合タンパク質は、約3.5のpHで溶出する。1つの実施形態において、識別的に改変された結合タンパク質は、約4のpHで溶出するのに対し、改変されていない結合タンパク質は、約2.8〜3.5、2.8〜3.2または2.8〜3のpHで溶出する。1つの実施形態において、識別的に改変された結合タンパク質は、約4.2のpHで溶出するのに対し、改変されていない結合タンパク質は、約2.8のpHで溶出する。1つの実施形態において、識別的に改変された結合タンパク質は、約4.4のpHで溶出するのに対し、改変されていない結合タンパク質は、約3.6のpHで溶出する。これらの実施形態において、「改変されていない」とは、両方のCH3ドメインにおいて、435(EUナンバリング)に改変を有しないこと、または435および436(EUナンバリング)に改変を有しないことを指す。
【0047】
本明細書中に記載される任意の実施形態および局面は、別段示されないかまたは文脈から明らかでない限り、互いと同時に使用され得る。次の記載を検討することにより、他の実施形態が当業者に明らかになるだろう。
【図面の簡単な説明】
【0048】
【図1】図1は、3つの二重特異性抗体形式:(A)二重特異性T細胞Engager(BiTE);(B)マウス−ラットハイブリッド;および(C)通常の軽鎖を有するノブ・イントゥ・ホールを図示している。
【図2】図2は、FcΔAdp改変を図示している:(A)ヒトIgG1(配列番号1)およびIgG3(配列番号3)のFc領域のアラインメント(FcΔAdp改変を四角で示している);(B)FcΔAdp二重特異性抗体の模式的表示。
【図2B】図2は、FcΔAdp改変を図示している:(A)ヒトIgG1(配列番号1)およびIgG3(配列番号3)のFc領域のアラインメント(FcΔAdp改変を四角で示している);(B)FcΔAdp二重特異性抗体の模式的表示。
【図3】図3は、ΔAdpジペプチド改変有りおよび無しのIgG1、IgG2およびIgG4のヒトCH3ドメイン(IMGTエキソンナンバリングおよびEUナンバリングを使用)、ならびにIgG3のアラインメントを図示している。
【図4】図4は、段階勾配を利用した溶出プロファイルを示している、二重特異性抗体を単離するためのプロテインAカラムのトレースを示している。
【図5】図5は、図4に示されるクロマトグラフ分離から溶出されたカラム画分のIL4−RaおよびIL−6Ra BIACORE(商標)結合プロファイルを示している。画分中の抗体を、固定化された抗Fc抗体で捕捉し、次いで、捕捉された抗体への結合について、可溶性IL−4RaまたはIL−6Raをアッセイした。
【図6】図6は、FcΔAdp二重特異性抗体(IL−6RΔ/IL−4R)、FcΔAdpホモ二量体(IL−6RΔ/IL−6RΔ)、野生型CH3配列を有するIgG1抗体(IL−4R/IL−4R)およびコントロール単一特異的抗体の薬物動態学的プロファイルを示している。
【図7】図7は、Raji細胞死滅アッセイにおけるCD20×CD3ΔAdp二重特異性抗体の有効性を図示している。
【図8】図8は、CD20×CD3ΔAdp二重特異性抗体を用いたバイスタンダー(bystander)細胞(293)死滅アッセイを図示している。
【図9】図9は、種々のmFcヘテロ二量体を使用した発現実験についての結果を示している。パネルA:ホモ二量体のmIgG2aおよびホモ二量体のIgG2aPTTTKからのヘテロ二量体のmIgG2a/mIgG2aPTTTKのpH分離のウエスタンブロット;パネルB:ホモ二量体のmIgG2aおよびホモ二量体のIgG2aTTTKからのヘテロ二量体のmIgG2a/mIgG2aTTTKのpH分離のウエスタンブロット;IP=投入物;FT=フロースルー;W2=2回目の洗浄物(1×PBS pH7.2);E1=1回目の溶出物(20mMクエン酸Na、1M NaCl pH5.5);E2=2回目の溶出物(20mMクエン酸Na、1M NaCl;57%pH5.5+43%pH2.6);E3=3回目の溶出物(20mMクエン酸Na、1M NaCl pH2.6)。
【図10】図10は、4:1というIFNAR1構築物:IFNAR2構築物の比を用いたときの、アイソタイプの混合物(例えば、mIgG2aおよびmIgG1)のヘテロ二量体の形態に対する変異体IgG2aのヘテロ二量体の優先的な形態を図示している。レーン1:IFNAR1−IgG2a:IFNAR2−IgG1;レーン2:INFAR1−IgG2a:IFNAR2−IgG2aTTT;レーン3:IFNAR1−IgG2a:IFNAR2−IgG2aTTTK;レーン4:IFNAR1−IgG2a:IFNAR2−IgG2aPTTTK;レーン5:IFNAR1−IgG2a:IFNAR2−IgG2aRF。
【発明を実施するための形態】
【0049】
詳細な説明
特定の方法および記載される実験条件は変動し得るので、本発明は、そのような方法および条件に限定されない。本発明の範囲は、請求項によって定義されるので、本明細書中で使用される用語は、特定の実施形態だけを記載する目的であって、限定する意図はないことも理解される。
【0050】
別段定義されない限り、本明細書中で使用される専門用語および科学用語のすべては、本発明が属する分野の当業者が通常理解している意味と同じ意味を有する。本明細書中に記載される方法および材料と類似または等価な任意の方法および材料を、本発明の実施または試験において使用することができるが、ここでは特定の方法および材料が記載される。言及されるすべての刊行物が、本明細書に参照によって援用される。
【0051】
用語「抗体」は、本明細書中で使用されるとき、ジスルフィド結合によって相互に接続された4つのポリペプチド鎖、すなわち2つの重(H)鎖および2つの軽(L)鎖を含む免疫グロブリン分子を含む。各重鎖は、重鎖可変領域(本明細書中でHCVRまたはVHと省略される)および重鎖定常領域を含む。重鎖定常領域は、3つのドメインCH1、CH2およびCH3を含む。各軽鎖は、軽鎖可変領域(本明細書中でLCVRまたはVLと省略される)および軽鎖定常領域を含む。軽鎖定常領域は、1つのドメインCLを含む。VHおよびVL領域は、フレームワーク領域(FR)と呼ばれるより保存された領域とともに点在する、相補性決定領域(CDR)と呼ばれる超可変性の領域にさらに細分され得る。VHおよびVLの各々は、アミノ末端からカルボキシ末端に向かって以下の順序:FR1、CDR1、FR2、CDR2、FR3、CDR3、FR4で配置される3つのCDRおよび4つのFRから構成される(重鎖CDRは、HCDR1、HCDR2およびHCDR3と省略され得る;軽鎖CDRは、LCDR1、LCDR2およびLCDR3と省略され得る)。用語「高親和性」抗体とは、表面プラズモン共鳴、例えば、BIACORE(商標)または溶液親和性(solution−affinity)ELISAによって測定されたとき、その標的に対して少なくとも10−9M、少なくとも10−10M;少なくとも10−11M;または少なくとも10−12Mの結合親和性を有する抗体のことを指す。
【0052】
句「抗原結合タンパク質」は、少なくとも1つのCDRを有し、かつ抗原を選択的に認識することができる、すなわち、少なくともマイクロモル濃度の範囲のKDで抗原に結合することができるタンパク質を含む。治療用の抗原結合タンパク質(例えば、治療用抗体)は、ナノモル濃度またはピコモル濃度の範囲のKDを必要とすることが多い。「抗原結合タンパク質」はまた、本明細書中に記載されるような第1および第2のCH3ドメイン、ならびに第1のタンパク質認識ドメインまたはリガンド認識ドメイン、および第2のタンパク質認識ドメインまたはリガンド認識ドメインを含むタンパク質を含み、ここで、その第1のタンパク質認識ドメインまたはリガンド認識ドメインおよび第2のタンパク質認識ドメインまたはリガンド認識ドメインの各々は、独立して、同じタンパク質もしくはリガンドを認識するか、または一緒に同じタンパク質もしくはリガンドを認識するか、あるいは各々独立して、異なるタンパク質またはリガンドを認識する。そのようなタンパク質の1つの例は、融合タンパク質(ヘテロ−またはホモ−)二量体を含むイムノアドヘシンであり、ここで、その二量体のポリペプチドは、レセプター構成要素またはリガンド構成要素を含む融合ポリペプチドであり、そのリガンド構成要素は、レセプターに結合するアミノ酸配列を含む。
【0053】
句「二重特異性抗体」は、2つ以上のエピトープに選択的に結合することができる抗体を含む。二重特異性抗体は、一般に、2つの異なる重鎖を含み、各重鎖は、異なるエピトープ(2つの異なる分子(例えば、抗原)上のエピトープまたは同じ分子(例えば、同じ抗原)上のエピトープ)に特異的に結合する。二重特異性抗体が、2つの異なるエピトープ(第1のエピトープおよび第2のエピトープ)に選択的に結合することができる場合、第1のエピトープに対する第1の重鎖の親和性は、一般に、第2のエピトープに対する第1の重鎖の親和性よりも少なくとも1桁から2桁または3桁または4桁低く、逆もまた同じである。二重特異性抗体によって認識されるエピトープは、同じ標的上または異なる標的上(例えば、同じまたは異なるタンパク質上)に存在し得る。二重特異性抗体は、例えば、同じ抗原の異なるエピトープを認識する重鎖を組み合わせることによって作製され得る。例えば、同じ抗原の異なるエピトープを認識する重鎖可変配列をコードする核酸配列が、種々の重鎖定常領域をコードする核酸配列に融合され得、そのような配列が、免疫グロブリン軽鎖を発現する細胞において発現され得る。代表的な二重特異性抗体は、2つの重鎖(各々が、3つの重鎖CDR、それに続いて(N末端からC末端に向かって)CH1ドメイン、ヒンジ、CH2ドメインおよびCH3ドメインを有する)、および抗原結合特異性を与えないが各重鎖と会合し得る免疫グロブリン軽鎖、または各重鎖と会合し得、かつ重鎖抗原結合領域によって結合される1つ以上のエピトープに結合し得る免疫グロブリン軽鎖、または各重鎖と会合し得、かつ一方または両方のエピトープへの一方または両方の重鎖の(or)結合を可能にする免疫グロブリン軽鎖を有する。
【0054】
用語「細胞」は、組換え核酸配列を発現するのに適した任意の細胞を含む。細胞には、原核生物および真核生物の細胞(単細胞または多細胞)、細菌細胞(例えば、大腸菌、Bacillus spp.、Streptomyces spp.などの系統)、マイコバクテリア細胞、真菌細胞、酵母細胞(例えば、S.cerevisiae、S.pombe、P.pastoris、P.methanolicaなど)、植物細胞、昆虫細胞(例えば、SF−9、SF−21、バキュロウイルス感染昆虫細胞、Trichoplusia niなど)、非ヒト動物細胞、ヒト細胞または細胞融合物(例えば、ハイブリドーマまたはクアドローマ)が含まれる。いくつかの実施形態において、細胞は、ヒト、サル、類人猿、ハムスター、ラットまたはマウス細胞である。いくつかの実施形態において、細胞は、真核生物細胞であり、以下の細胞:CHO(例えば、CHO K1、DXB−11CHO、Veggie−CHO)、COS(例えば、COS−7)、網膜細胞、Vero、CV1、腎臓(例えば、HEK293、293EBNA、MSR293、MDCK、HaK、BHK)、HeLa、HepG2、WI38、MRC5、Colo205、HB8065、HL−60(例えば、BHK21)、Jurkat、Daudi、A431(表皮)、CV−1、U937、3T3、L細胞、C127細胞、SP2/0、NS−0、MMT060562、セルトリ細胞、BRL3A細胞、HT1080細胞、ミエローマ細胞、腫瘍細胞、および上述の細胞に由来する細胞株から選択される。いくつかの実施形態において、細胞は、1つ以上のウイルス遺伝子を含む(例えば、ウイルス遺伝子を発現する網膜細胞(例えば、PER.C6(商標)細胞))。
【0055】
句「相補性決定領域」または用語「CDR」は、通常(すなわち、野生型動物において)免疫グロブリン分子(例えば、抗体またはT細胞レセプター)の軽鎖または重鎖の可変領域内の2つのフレームワーク領域の間に見られる、生物の免疫グロブリン遺伝子の核酸配列によってコードされるアミノ酸配列を含む。CDRは、例えば、生殖細胞系列の配列または再配列された配列もしくは再配列されていない配列によって、例えば、ナイーブB細胞もしくは成熟B細胞またはナイーブT細胞もしくは成熟T細胞によって、コードされ得る。いくつかの状況において(例えば、CDR3の場合)、CDRは、連続していない(例えば、再配列されていない核酸配列では連続していない)が、例えば、それらの配列のスプライシングまたは接続(例えば、重鎖CDR3を形成するV−D−J組換え)の結果としてB細胞の核酸配列では連続している、2つ以上の配列(例えば、生殖細胞系列の配列)によってコードされ得る。
【0056】
句「重鎖」または「免疫グロブリン重鎖」は、任意の生物由来の免疫グロブリン重鎖定常領域配列を含み、別段明記されない限り、重鎖可変ドメインを含む。重鎖可変ドメインは、別段明記されない限り、3つの重鎖CDRおよび4つのFR領域を含む。重鎖のフラグメントとしては、CDR、CDRおよびFR、ならびにそれらの組み合わせが挙げられる。代表的な重鎖は、可変ドメインの後、(N末端からC末端に向かって)CH1ドメイン、ヒンジ、CH2ドメインおよびCH3ドメインを有する。重鎖の機能的なフラグメントには、抗原を特異的に認識することができ(例えば、マイクロモル濃度、ナノモル濃度またはピコモル濃度の範囲のKDで抗原を認識することができ)、発現して細胞から分泌されることができ、かつ、少なくとも1つのCDRを含む、フラグメントが含まれる。
【0057】
句「Fc含有タンパク質」は、抗体、二重特異性抗体、イムノアドヘシン、ならびに免疫グロブリンCH2およびCH3領域の少なくとも機能的な一部を含む他の結合タンパク質を含む。「機能的な一部」とは、Fcレセプター(例えば、FcγR;またはFcRn、すなわち、新生児Fcレセプター)に結合し得、そして/または補体の活性化に関与し得る、CH2およびCH3領域のことを指す。そのCH2およびCH3領域が、いずれのFcレセプターにも結合できなくし、また、補体を活性化できなくする、欠失、置換および/もしくは挿入または他の改変を含む場合、そのCH2およびCH3領域は、機能的でない。
【0058】
Fc含有タンパク質は、改変が結合タンパク質の1つ以上のエフェクター機能に影響する場合(例えば、FcγR結合性、FcRn結合性ひいては半減期、および/またはCDC活性に影響する改変)を含む改変を免疫グロブリンドメインに含み得る。そのような改変としては、免疫グロブリン定常領域のEUナンバリングに準拠した以下の改変およびそれらの組み合わせ:238、239、248、249、250、252、254、255、256、258、265、267、268、269、270、272、276、278、280、283、285、286、289、290、292、293、294、295、296、297、298、301、303、305、307、308、309、311、312、315、318、320、322、324、326、327、328、329、330、331、332、333、334、335、337、338、339、340、342、344、356、358、359、360、361、362、373、375、376、378、380、382、383、384、386、388、389、398、414、416、419、428、430、433、434、435、437、438および439が挙げられるが、これらに限定されない。
【0059】
例えば、限定するわけではないが、結合タンパク質は、Fc含有タンパク質であり、長い血清半減期(列挙される改変を有しない同じFc含有タンパク質と比べて)を示し、250位(例えば、EまたはQ);250位および428位(例えば、LまたはF);252位(例えば、L/Y/F/WまたはT)、254位(例えば、SまたはT)および256位(例えば、S/R/Q/E/DまたはT)に改変を有するか;または428位および/もしくは433位(例えば、L/R/SI/P/QまたはK)および/もしくは434位(例えば、H/FまたはY)に改変を有するか;または250位および/もしくは428位に改変を有するか;または307位もしくは308位(例えば、308F、V308F)および434位に改変を有する。別の例では、改変は、428L(例えば、M428L)および434S(例えば、N434S)改変;428L、259I(例えば、V259I)および308F(例えば、V308F)改変;433K(例えば、H433K)および434(例えば、434Y)改変;252、254および256(例えば、252Y、254Tおよび256E)改変;250Qおよび428L改変(例えば、T250QおよびM428L);307および/または308改変(例えば、308Fまたは308P)を含み得る。
【0060】
句「イオン調節物質」は、タンパク質間の非特異的な(すなわち、非親和性の)イオン性相互作用の影響を減少させるかまたはその影響を乱す部分を含む。「イオン調節物質」としては、例えば、塩、酢酸、重炭酸、炭酸、ハロゲン(例えば、塩化物またはフッ化物)、硝酸、リン酸または硫酸とI族およびII族金属とのイオン性の組み合わせ物が挙げられる。「イオン調節物質」の非限定的な例示的リストとしては、酢酸のベリリウム塩、リチウム塩、ナトリウム塩およびカリウム塩;重炭酸ナトリウムおよび重炭酸カリウム;炭酸リチウム、炭酸ナトリウム、炭酸カリウムおよび炭酸セシウム;塩化リチウム、塩化ナトリウム、塩化カリウム、塩化セシウムおよび塩化マグネシウム;フッ化ナトリウムおよびフッ化カリウム;硝酸ナトリウム、硝酸カリウムおよび硝酸カルシウム;リン酸ナトリウムおよびリン酸カリウム;ならびに硫酸カルシウムおよび硫酸マグネシウムが挙げられる。「イオン調節物質」は、pH勾配もしくはpH段階に加えられたとき、または「イオン調節物質」でのプロテインA支持体の平衡化のとき、イオン性の相互作用に影響する部分を含み、pH段階またはpH勾配を適用することにより、ホモ二量体のIgGの溶出とヘテロ二量体のIgGの溶出との間(例えば、野生型ヒトIgGの溶出と、本明細書中に記載されるようなそのCH3ドメインの1つ以上の改変を有すること以外は同じIgGの溶出との間)のpH単位の間隔が広がる。「イオン調節物質」の適当な濃度は、最大のpHの間隔が所与のpH段階またはpH勾配に達するまで「イオン調節物質」の濃度を上げながら、同じカラム、pH段階またはpH勾配を使用して、その濃度によって決定され得る。
【0061】
句「軽鎖」は、任意の生物由来の免疫グロブリン軽鎖定常領域配列を含み、別段明記されない限り、ヒトカッパー軽鎖およびヒトラムダ軽鎖を含む。軽鎖可変(VL)ドメインは、別段明記されない限り、代表的には、3つの軽鎖CDRおよび4つのフレームワーク(FR)領域を含む。一般に、全長軽鎖は、アミノ末端からカルボキシル末端に向かって、FR1−CDR1−FR2−CDR2−FR3−CDR3−FR4を含むVLドメイン、および軽鎖定常ドメインを含む。本発明と共に使用され得る軽鎖は、例えば、本抗原結合タンパク質によって選択的に結合される第1または第2の抗原に選択的に結合しない軽鎖を含む。適当な軽鎖には、既存の抗体ライブラリー(ウェットライブラリーまたはインシリコ)において最もよく使用される軽鎖についてスクリーニングすることによって特定され得る軽鎖が含まれ、ここで、その軽鎖は、本抗原結合タンパク質の抗原結合ドメインの親和性および/または選択性を実質的に干渉しない。適当な軽鎖には、本抗原結合タンパク質の抗原結合領域によって結合される一方または両方のエピトープに結合し得る軽鎖が含まれる。
【0062】
句「マイクロモル濃度範囲」は、1〜999マイクロモル濃度を意味することを意図している;句「ナノモル濃度範囲」は、1〜999ナノモル濃度を意味することを意図している;句「ピコモル濃度範囲」は、1〜999ピコモル濃度を意味することを意図している。
【0063】
句「体細胞変異した」は、クラススイッチを起こしたB細胞由来の核酸配列に対する言及を含み、ここで、クラススイッチしたB細胞における免疫グロブリン可変領域(例えば、重鎖可変ドメインまたは重鎖CDR配列もしくは重鎖FR配列を含む領域)の核酸配列は、クラススイッチ前のB細胞における核酸配列と同一ではない(例えば、クラススイッチを起こしていないB細胞とクラススイッチを起こしたB細胞との間でCDR核酸配列またはフレームワーク核酸配列が異なる)。「体細胞変異した」は、親和性成熟していないB細胞における対応する配列(すなわち、生殖細胞系列の細胞のゲノムにおける配列)と同一でない親和性成熟したB細胞由来の核酸配列に対する言及を含む。句「体細胞変異した」は、B細胞を目的の抗原に曝露した後のB細胞由来の核酸配列に対する言及も含み、ここで、その核酸配列は、そのB細胞をその目的の抗原に曝露する前の対応する核酸配列と異なる。句「体細胞変異した」とは、抗原チャレンジに応答したヒト免疫グロブリン可変領域の核酸配列を有する動物、例えばマウスにおいて産生され、そのような動物において本質的に有効な選択プロセスから生じる、抗体由来の配列のことを指す。
【0064】
句「可変ドメイン」は、(別段示されない限り)以下のアミノ酸領域を順にN末端からC末端に向かって含む免疫グロブリン軽鎖または重鎖(所望のとおり改変されたもの)のアミノ酸配列を含む:FR1、CDR1、FR2、CDR2、FR3、CDR3、FR4。「可変ドメイン」は、二重のベータシート構造を有する基準のドメイン(VHまたはVL)に折り畳むことができるアミノ酸配列を含み、ここで、そのベータシートは、第1のベータシートの残基と第2のベータシートの残基との間のジスルフィド結合によって接続されている。
【0065】
改変されたIgG CH3領域を有する二重特異性抗体
本発明者らは、通常の軽鎖ストラテジーを、ヒト抗体の構成要素とともに使用され得る選択的なプロテインA精製スキームの実行と組み合わせる新規形式を開発した。
【0066】
ヒトIgG3は、プロテインAに結合しないので、マウス−ラットハイブリッドに対して使用される精製ストラテジーと類似の精製ストラテジーにおいて他の3つのヒトIgGサブクラスのうちのいずれかと一緒に使用することができるという可能性があることは、以前から有名だった(Lindhofer,H.ら(1995)J.Immunol.155:219−225))。しかしながら、4つすべてのヒトIgGサブクラスの配列が、高度に相同であるにもかかわらず、IgG1、IgG2およびIgG4のFc部が、どれだけ容易にIgG3とヘテロ二量体を形成するかは知られていない;ある特定の状況下では(例えば、クアドローマからの単離では)、ホモ二量体の優先的な形成だけこそが所望のヘテロ二量体の総収量に対して悪影響を及ぼし得る。IgG3のヒンジ領域と他のサブクラスのヒンジ領域との差異を補うために追加の改変が必要となることもある。エフェクター機能に対する潜在的な影響があるので、いくつかの状況では、完全なIgG3 Fcの存在を必要としないことが好ましい場合もある。
【0067】
それゆえ、本発明者らは、プロテインA結合の偶然にも単純な決定基を活用する「最小」形式を考案した。IgG3がプロテインAに結合できないことは単一のアミノ酸残基Arg435(EUナンバリング;IMGTではArg95)(他のIgGサブクラスでは、対応する位置はヒスチジン残基で占められている)によって判定されることが報告されている(Jendeberg,L.ら(1997)J.Immunological Meth.201:25−34))。それゆえ、IgG3の代わりに、His435がArgに変異したIgG1配列を使用することが可能である。よって、IgG1における単一の点変異が、新しい精製スキームに適用できる異なる結合親和性をもたらすのに十分であるはずである。この改変は、プロテインAに結合することができないことを表すIgG1ΔAと呼ばれる(そして同様に、IgG2ΔAおよびIgG4ΔA、またはより一般的にはFcΔA)。
【0068】
しかしながら、明記される点変異が、その変異によって新規ペプチド配列を導入することから、それは、潜在的に免疫原性であり得る。この点変異は、理論上、MHCクラスII分子上にのせられて、T細胞に提示され、その結果として、免疫応答を誘発し得る。この不測の困難を回避するために、ジペプチド変異H435R/Y436F(EUナンバリング;IMGTではH95R/Y96F)が使用され得る。その結果生じるこの変更の近傍における配列は、IgG3の配列と同一となり(図2Aを参照のこと)、ゆえに、T細胞への提示に利用可能な非天然の短いペプチドが存在しないだろうことから、免疫学的に「隠されている」と予想され得る。この二重変異体がなおもプロテインAに結合しないことが報告されている(Jendeberg,L.ら(1997)J.Immunological Meth.201:25−34)。最後に、そのジペプチド変異は、Fc二量体の界面を形成するいずれの残基も含まないので、ヘテロ二量体の形成が干渉される可能性は低い。このジペプチド変異は、「IgG1ΔAdp」(そして同様にIgG2ΔAdp、IgG4ΔAdpおよびFcΔAdp)と命名される。IgG1、IgG2およびIgG4におけるジペプチド改変の配置は、IMGTエキソンナンバリングおよびEUナンバリングを示しつつ、野生型ヒトIgG CH3ドメイン配列、ならびにhIgG3と共に示されている、図3のIgG1ΔAdp、IgG2ΔAdpおよびIgG4ΔAdpと表されている配列に示されている。
【0069】
FcΔAdp改変は、Fc二量体の界面を形成すると考えられているいずれの残基も含まないので、そのFcΔAdp改変がヘテロ二量体の形成を干渉する可能性は低い。FcΔAdpは、そのように最小であるので、同様に、他の操作されたFc形態に組み込まれる可能性もあり得る。IgG2ΔAdpおよびIgG4ΔAdpは、後者の各々に関連するエフェクター機能(またはその欠損)が望まれる状況において有益であり得る。
【0070】
要約すれば、上に記載された二重特異性抗体形式は、同じ軽鎖を使用する特異性の異なる2つの抗体を含み、ここで、それらのうちの1つのFc領域は、FcΔAdp形式に改変されている(図2Bを参照のこと)。その配置は、天然のヒト抗体の配置であり、ゆえに、凝集する傾向が低いこと、インビボ安定性、最小の免疫原性、抗体の体内分布特性に類似した体内分布特性、良好な薬物動態、および必要に応じてエフェクター機能をはじめとした、その好ましい特性を共有するはずである。そのような二重特異性抗体を単離するための、実行することが比較的迅速かつ単純な方法が提供される。
【0071】
改変されたマウスIgG CH領域を有する二重特異性結合タンパク質
本発明者らは、CH3ドメインにおける1つ以上のアミノ酸に関してヘテロ二量体である免疫グロブリン重鎖(またはその機能的なCH2およびCH3含有フラグメント)を含む結合タンパク質を容易に単離するための方法を考案した。マウスIgG CHドメインの改変を慎重に選択し、特定の分離技術を適用することにより、それらの改変を含まないホモ二量体およびヘテロ二量体から2つの識別的に改変されたマウスCH領域を含む結合タンパク質を容易に単離することができるようになる。
【0072】
247にプロリン、252、254および256にトレオニン、ならびに258にリジンを含むマウスIgG1は、プロテインAと弱くしか結合しない。しかしながら、マウスIgG2aおよびIgG2bは、それらの位置に異なる残基を含み(IgG2bの256位および258位を除いて)、マウスIgG2aおよび2bは、プロテインAに十分に結合する。重鎖がヘテロ二量体である抗体を作製するための方法において2つのマウスIgGのCH領域を識別的に改変することにより、そのような抗体に識別的なプロテインA結合特性が付与され得る。このように、IgG1のホモ二量体(プロテインAには、たとえ結合するとしてもかなり弱くしか結合しない)であるか、またはマウスIgG2aのホモ二量体であるか、マウスIgG2bのホモ二量体であるか、またはIgG2a/IgG2bのヘテロ二量体であるかに関係なく、任意のマウスIgGホモ二量体から、改変されたヘテロ二量体を容易に(ready)分離することができる識別的なプロテインA単離スキームが考案される。例えば、2つの異なる重鎖可変ドメインを有するが、同じアイソタイプ、例えば、IgG2aの二重特異性抗体は、その重鎖配列を使用する適当な発現系において発現され得、ここで、IgG2a CH領域のうちのただ1つが、プロテインA結合決定基を減少させるかまたは排除するように改変される。このように、IgG2a CH領域のうちの1つだけが、プロテインAに対する実質的な親和性を示し、改変されていないIgG2aと改変されたIgG2aとの二量体から形成される任意の抗体は、改変されたヘテロ二量体から容易に単離され得る。
【0073】
様々な実施形態において、Fc二量体の単一のCH領域が、改変されたCH領域を含むのに対し、Fc二量体の他方のCHは、それらを欠いている、抗体。そのマウスIgG CH領域は:252T、254Tおよび256T;252T、254T、256Tおよび258K;247P、252T、254T、256Tおよび258K;435Rおよび436F;252T、254T、256T、435Rおよび436F;252T、254T、256T、258K、435Rおよび436F;24tP、252T、254T、256T、258K、435Rおよび436F;ならびに435Rからなる群から選択される特定の位置(EUナンバリング)に特定のアミノ酸を含むように改変される。特定の実施形態において、M252T、S254T、S256T;M252T、S254T、S256T、I258K;I247P、M252T、S254T、S256T、I258K;H435R、H436F;M252T、S254T、S256T、H435R、H436F;M252T、S254T、S256T、I258K、H435R、H436F;I247P、M252T、S254T、S256T、I258K、H435R、H436F;およびH435Rからなる群から選択される特定の基の改変が行われる。
【0074】
ヘテロ二量体のマウスIgGに基づく結合タンパク質は、種々の用途のために使用され得る。例えば、それらは、マウス定常ドメインを有する二重特異性抗体を単離する方法を可能にし、ここで、その改変は、1つ以上のマウスFcレセプターへのその抗体の結合を干渉しないか、または実質的に干渉せず、その抗体は、例えば、ADCCまたはCDCに関与し得、また、同じまたは異なる標的上の2つ以上のエピトープに結合し得る。
【0075】
1つの局面において、第1のマウスIgG CH領域および第2のマウスIgG CH領域を含む結合タンパク質を単離するための方法、ここで、その第1のIgG CH領域は、第1のマウスIgG CH領域のプロテインA結合親和性を減少させるかまたは排除するが第2のマウスIgG CH領域のプロテインA結合親和性を減少させないかまたは排除しないように改変され(第2のIgG CH領域は改変されない)、その結合タンパク質は、第1のエピトープに結合する第1の結合部分および第2のエピトープに結合する第2の結合部分を含む。
【0076】
1つの実施形態において、改変は、Fcレセプターへの結合タンパク質の結合親和性を変更しないかまたは実質的に変更しない。1つの実施形態において、本結合タンパク質は、本結合タンパク質のFcレセプターへの親和性を増加させるかまたは減少させる改変を含む。
【0077】
1つの実施形態において、改変は、その改変を有しない対応する結合タンパク質と比べて、天然のマウスFcγRレセプターおよび/または天然のマウスFcRnを含むマウスにおける結合タンパク質の血清半減期を変更しないかまたは実質的に変更しない。
【0078】
1つの実施形態において、改変は、その改変を有しない対応する結合タンパク質と比べて、天然の高親和性および低親和性のマウスFcγRレセプターならびに/またはFcRnレセプターの置き換えを含むマウスにおける本結合タンパク質の血清半減期を変更しないかまたは実質的に変更しない。
【0079】
1つの実施形態において、第1および第2のエピトープは、異なるものであり、異なる細胞上または異なるタンパク質上に存在する。1つの実施形態において、第1のエピトープおよび第2のエピトープは、異なるものであり、同じ細胞上または同じタンパク質上に存在する。
【0080】
1つの実施形態において、Fcレセプターは、高親和性Fcレセプター、低親和性FcレセプターおよびFcRnから選択される。特定の実施形態において、Fcレセプターは、マウスFcRn、マウスFcγR、マウスFcγRIIB、マウスFcγRIII、マウスFcγRIVおよびそれらの組み合わせのうちの1つ以上から選択される。特定の実施形態において、Fcレセプターは、ヒトFcRn、ヒトFcγR、ヒトFcγRIIB、ヒトFcγRIIC、ヒトFcγRIIIB、ヒトFcγRIIIA、ヒトFcγRIIAおよびそれらの組み合わせのうちの1つ以上から(form)選択される。
【0081】
免疫原性
本発明の多くの実施形態の利点の1つは、プロテインAに対する結合性の違いに基づいて容易に単離可能であり、かつヒトにおいて免疫原性でないかまたは実質的に免疫原性でない、二重特異性抗体をもたらす改変を使用することができることである。この特性のおかげで、そのような実施形態は、ヒトへの治療に使用するための二重特異性抗体を作製する際、および例えば、免疫原性でないかまたは実質的に免疫原性でないイムノアドヘシンを作製する際(ヒト結合部分、すなわち、ヒトレセプター構成要素および/またはヒトリガンドを使用するとき)に、特に有用になる。この特性は、IgG1、IgG2およびIgG3のH95R/Y96F(IMGTナンバリング)改変を伴うCH3ドメイン、ならびに異なるIgGアイソタイプの野生型配列を反映して改変された位置をもたらすさらなる改変を含むCH3ドメインを有する二重特異性抗体に関連する。よって、特定のIgGアイソタイプに関連する改変は、天然には見られないが、改変された配列は、異なるIgGアイソタイプの野生型配列と局所的に同一であり、その改変は、免疫原性であるかまたは実質的に免疫原性であると予想されない。配列が、任意の天然の配列と局所的に同一でなかったとしても、その改変は、免疫原性でない可能性もあり;そのような改変は、等しく有用であり得る。ゆえに、最小の点変異H95R(IMGTナンバリング)は、免疫原性でない場合、本発明の適当な実施形態であり得る。
【0082】
よって、重鎖定常ドメインに関してヒトにおいて免疫原性でないかまたは実質的に免疫原性でないが、親和性試薬(例えば、プロテインA)に対する重鎖定常ドメインの親和性に違いをもたらす改変を含む重鎖定常ドメインの1つ以上の識別的改変を有する、二重特異性抗体が提供される。その改変は、本明細書中に開示される改変を含む。特定の実施形態において、CH3ドメインに関してヒトにおいて免疫原性でないかまたは実質的に免疫原性でないが、重鎖が識別的に改変された二重特異性抗体は、以下の改変のうちの1つを含む(または、別の実施形態では、以下の改変のうちの1つから本質的になる)CH3ドメインを含むヒトIgG1、IgG2またはIgG4である:H95RまたはH95RおよびY96F(IMGTナンバリング)。
【0083】
本二重特異性抗体は、ヒトのIgG1、IgG2およびIgG4のアイソフォームに対する許容度が任意の著しい程度にまで壊されていないヒトに対して免疫原性でないかまたは実質的に免疫原性でないと予想される。
【0084】
具体的には、MHCクラスII分子の結合溝(binding groove)は、T細胞レセプターの可変ループによって認識される主要な決定基を含む9merを収容するものであり、ペプチドの長さが9merに届かない任意の天然の配列である場合、免疫応答が誘発される可能性は低いとみられるので、FcΔAdp改変は、免疫学的に「隠されている」と予想される。しかしながら、9merより長いペプチド(通常、約13〜17mer)は、MHCクラスIIによって結合され、突出したセグメントが潜在的に結合に影響し得る可能性がある。ゆえに、より長い非天然の配列を排除する追加の改変(FcΔAdp改変に対して追加の改変)が、免疫原性に対する可能性をさらに低下させ得る。1つの特定の例は、最小の非天然のペプチドの長さを、IgG1ΔAdpでは14残基から39残基に伸ばし、および同じように定義されるIgG2ΔAdpでは14残基から43残基に伸ばす、V422I(EU;IMGTナンバリングではV82I)の改変である。別の例は、IgG4ΔAdpにおいて10残基から14残基に上記長さを伸ばすL445P(EU;IMGTナンバリングではL105P)の改変である。
【0085】
薬物動態
プロテインAに対する結合部位は、免疫グロブリンに長い血清半減期を与えるのに関与すると考えられている新生児FcレセプターFcRNに対する結合部位と重複する。ゆえに、ヒトIgG3が、他のIgGサブクラス(約21日)よりも短い血清半減期を有する(約7日)ことを考えると、プロテインA結合部位の近傍の改変は、ここで提案される形式がIgG1、2および4よりも短い血清半減期を有し得る可能性を高める。His435に影響するいくつかのFc変異体は、FcRNに結合しないことおよびマウスにおいてより短い半減期を有することが示されている。しかしながら、薬物動態学的解析から、IgG1ΔA/IgG1ヘテロ二量体の血清半減期が、IgG1ホモ二量体の血清半減期と大きく異ならないことが示されている(実施例2を参照のこと)。よって、IgG1ΔAdp変異は、IgG1の長い半減期をなおも保ちつつ、プロテインA結合性をなくすという利点を有する。
【0086】
したがって、1つの実施形態において、本明細書中に記載されるようなCH3ドメインの改変を含む二重特異性抗原結合タンパク質が提供され、ここで、その抗原結合タンパク質は、CH3ドメインに改変を有しない同じ二重特異性抗原結合タンパク質と等価な薬物動態学的プロファイルを示す。1つの実施形態において、IgG1ΔA/IgG1ヘテロ二量体のFcを含む二重特異性抗体が提供され、ここで、その二重特異性抗体は、その他の点は同一であるがIgG3 CH3ドメインを含むかまたはその他の点は同一であるが少なくとも1つのIgG3重鎖を含む二重特異性抗体よりも、約1.5倍、約2倍、約2.5倍または約3倍長い血清半減期を有する。1つの実施形態において、IgG1ΔA/IgG1ヘテロ二量体のFcを含む二重特異性抗体が提供され、ここで、その二重特異性抗体は、IgG1ΔA改変を有しない二重特異性抗体(すなわち、IgG1ホモ二量体二重特異性抗体)とほぼ同じ血清半減期を示す。
【0087】
免疫グロブリン重鎖
所望の特徴(例えば、所望の特異性、所望の親和性、所望の機能性、例えば、遮断、非遮断、阻害、活性化など)を備えた二重特異性抗体を作製するために使用することができる免疫グロブリン重鎖可変領域は、当該分野で公知の任意の方法を用いて作製され得る。次いで、所望の重鎖は、本明細書中に記載される所望の重鎖定常領域を有する構築物内に可変領域を含む核酸配列をクローニングすることによって構築され得る。
【0088】
1つの実施形態において、第1の重鎖は、第1の抗原で免疫された第1の動物の成熟B細胞のゲノムに由来する核酸によってコードされる可変領域を含み、その第1の重鎖は、第1の抗原を特異的に認識する。特定の実施形態において、第2の重鎖は、第2の抗原で免疫された第2の動物の成熟B細胞のゲノムに由来する核酸によってコードされる可変領域を含み、その第2の重鎖は、第2の抗原を特異的に認識する。
【0089】
1つの実施形態において、第1の動物および/または第2の動物は、再配列されていないヒト免疫グロブリン重鎖可変領域を含む遺伝子改変動物である。1つの実施形態において、第1の動物および/または第2の動物は、再配列されていないヒト免疫グロブリン重鎖可変領域およびヒト免疫グロブリン定常領域を含む遺伝子改変動物である。1つの実施形態において、第1の動物および/または第2の動物は、再配列されていないヒト免疫グロブリン重鎖可変領域を含む遺伝子改変マウスである。
【0090】
免疫グロブリン重鎖可変領域配列は、当該分野で公知の他の任意の方法、例えば、ファージディスプレイによって得ることができ、それによって得られた配列を使用することにより、任意の適当な重鎖、例えば、本明細書中に記載されるようにCH3ドメインが改変された重鎖をコードする核酸に連結され、発現構築物内に置かれ、そして例えば、適当な軽鎖の存在下において、重鎖を作製することができる細胞に運ばれる、核酸構築物が作製され得る。
【0091】
免疫グロブリン軽鎖
2つの異なるエピトープ(または2つの異なる抗原)を認識する2つの重鎖を含む二重特異性抗体は、同じ軽鎖(すなわち、同一の可変ドメインおよび定常ドメインを有する軽鎖)と対形成し得る場合、より容易に単離される。重鎖可変ドメインの選択性および/またはその標的抗原との親和性を干渉せずにまたは実質的に干渉せずに、特異性が異なる2つの重鎖と対形成し得る軽鎖を作製するための種々の方法が当該分野で公知である。
【0092】
あるアプローチにおいて、軽鎖は、すべての軽鎖可変ドメインに対する使用統計を調査し、ヒト抗体において最も頻繁に使用される軽鎖を特定し、その軽鎖を特異性が異なる2つの重鎖と対形成させることによって選択され得る。
【0093】
別のアプローチでは、軽鎖は、ファージディスプレイライブラリー(例えば、ヒト軽鎖可変領域配列を含むファージディスプレイライブラリー、例えば、ヒトScFvライブラリー)内の軽鎖配列を観察し、そのライブラリーから最もよく使用される軽鎖可変領域を選択することによって選択され得る。
【0094】
別のアプローチでは、軽鎖は、プローブとして両方の重鎖の重鎖可変配列を用いて軽鎖可変配列のファージディスプレイライブラリーをアッセイすることによって選択され得る。両方の重鎖可変配列と会合する軽鎖は、それらの重鎖に対する軽鎖として選択され、両方のエピトープに対する結合および/または活性化を可能にする。
【0095】
別のアプローチでは、軽鎖は、公知の軽鎖と所望の重鎖とを組み合わせ、得られた二重特異性抗体を、結合特異性、親和性および/または活性化能力についてアッセイすることによって選択され得る。
【0096】
軽鎖を選択するための任意のアプローチにおいて困難に直面する(例えば、軽鎖が、一方もしくは両方の重鎖とその抗原との結合を干渉するか、または軽鎖が、一方もしくは両方の重鎖と十分に会合しない)程度まで、その軽鎖は、重鎖の同族の軽鎖と整列することができ、両方の重鎖の同族の軽鎖と共通の配列特徴とよりマッチするように軽鎖において改変が行われる。免疫原性の変更を最小にする必要がある場合、免疫原性の可能性を評価する(すなわち、インシリコならびにウェットアッセイ)ための当該分野で公知のパラメータおよび方法に基づいて、タンパク分解性プロセシングがT細胞エピトープを生成する可能性が低くなるように、改変は、好ましくは公知のヒト軽鎖配列に存在する配列をもたらす。
【0097】
抗体および結合タンパク質
上記組成物および方法は、ヒト二重特異性抗体、すなわち、ヒト定常ドメインおよびヒト可変ドメインを含む二重特異性抗体を作製する際に特に有用である。いくつかの実施形態において、ヒト抗体には、ヒト生殖細胞系列の免疫グロブリン配列に由来する重鎖可変ドメインおよび重鎖定常ドメイン、いくつかの実施形態では、体細胞変異したヒト免疫グロブリン配列(例えば、ヒト免疫グロブリン遺伝子配列を含む動物において産生されるヒト免疫グロブリン配列)に由来する重鎖可変ドメインおよび重鎖定常ドメインを有する抗体が含まれる。いくつかの実施形態において、ヒト可変領域および/またはヒト定常領域は、ヒト生殖細胞系列の免疫グロブリン配列によってコードされないアミノ酸残基、または例えばCDR、特にCDR3における、組換えおよび/もしくはスプライシングの結果としてコードされるアミノ酸残基を含み得る。ヒト抗体は、マウスなどの別の哺乳動物種の生殖細胞系列に由来するCDR配列がヒトフレームワーク配列に移植された抗体を含むと意図されない。それらの抗体は、ヒト化抗体またはキメラ抗体と呼ばれる。ヒト抗体には、例えば、ランダム突然変異誘発または部位特異的突然変異誘発によってインビトロにおいて導入された変異を含む抗体が含まれるが、その変異は、好ましくは、ヒトにおいて免疫原性でない。
【0098】
上記方法および組成物は、キメラ抗体、好ましくは、ヒトにおいて免疫原性でないかまたは低免疫原性のキメラ抗体を作製するために使用され得る。キメラ抗体は、重鎖可変領域またはフレームワークもしくはCDRあるいは重鎖定常領域または重鎖定常ドメインのうちの1つが、異なる種(例えば、ヒトおよびマウス、またはヒトおよび霊長類)に由来する抗体である。いくつかの実施形態において、キメラ抗体には、非ヒト起源(例えば、マウス)の重鎖可変領域およびヒト起源の重鎖定常領域を有する抗体が含まれる。いくつかの実施形態において、キメラ抗体には、ヒト起源の重鎖可変領域および非ヒト(例えば、マウス)起源の重鎖定常領域を有する抗体が含まれる。様々な実施形態において、マウス起源の領域は、体細胞超変異を有するかまたは有しない、マウス免疫グロブリン生殖細胞系列の配列と同一であるかまたは実質的に同一である。キメラ抗体には、ヒト免疫グロブリン生殖細胞系列の配列と同一であるかまたは実質的に同一である軽鎖定常領域、および非ヒト(例えば、マウス)重鎖またはキメラヒト/非ヒト重鎖を有する抗体も含まれる。キメラ抗体には、非ヒト(例えば、マウス)免疫グロブリン生殖細胞系列の配列と同一であるかまたは実質的に同一である軽鎖定常ドメイン、およびヒトまたはキメラ非ヒト/ヒト重鎖を有する抗体が含まれる。
【0099】
いくつかの実施形態において、上記組成物および方法は、親和性成熟抗体を作製するためのものである。いくつかの実施形態において、親和性成熟抗体は、変更を有しない実質的に同一の抗体と比べて、その標的抗原に対してより高い親和性(例えば、ナノモル濃度またはピコモル濃度の範囲のKD)をもたらす1つ以上の変更を1つ以上のCDRに含む。親和性成熟抗体は、当該分野で公知の任意の適当な方法によって、例えば、CDRおよび/またはフレームワーク領域のランダム突然変異誘発もしくは部位特異的突然変異誘発、その後の親和性スクリーニング、VHドメインシャフリングなどによって、作製され得る。
【0100】
いくつかの実施形態において、抗体は、中和抗体である。中和抗体には、抗原の生物学的活性を中和すること、阻害することまたは妨害することができる抗体が含まれる。中和抗体には、抗原に結合したときに、インビボおよびインビトロにおいて、その抗原の天然の標的に対してその抗原が作用することができる能力を妨害するかまたは減少させる抗体が含まれる。中和抗体の例としては、タンパク質リガンドが生物学的レセプターに結合するのを妨害する、そのレセプターのリガンドに対する抗体、または生物学的レセプターがそのリガンドに結合するのを妨害する、そのレセプターに対する抗体が挙げられ、ここで、その抗体の非存在下でのリガンドの結合によって、そのレセプターは細胞の内部に変化を起こす。抗体が中和抗体であるか否かの判定は、一般的に、抗原の生物学的活性に対する抗体の影響を測定する機能的アッセイの実施を必要とする。
【0101】
本発明の方法および組成物は、抗体および他の結合タンパク質に対する種々の用途でも有用である。いくつかの有用な用途の短い説明をここに提供する。
【0102】
細胞上の抗原、例えば、CD20を標的にし、また、T細胞上の抗原、例えば、CD3を標的にする、腫瘍抗原およびT細胞抗原に対する結合特異性を備える二重特異性結合タンパク質が作製され得る。このように、その二重特異性抗体は、患者内の目的の細胞を標的にし(例えば、リンパ腫患者におけるB細胞、CD20結合を介して)、かつその患者のT細胞を標的にする。その二重特異性抗体は、様々な実施形態において、CD3に結合したときにT細胞を活性化し、ゆえにT細胞活性化を特定の選択された腫瘍細胞に結合するように設計される。
【0103】
2つの結合部分を含み、その各々が同じ細胞の表面上の結合パートナーに対するものである(すなわち、各々が異なる標的に対するものである)二重特異性結合タンパク質も作製され得る。この設計は、同じ細胞の表面上に両方の標的を発現する特定の細胞または細胞型を標的にすることに特にふさわしい。標的は、他の細胞上に個別に現れ得るが、これらの結合タンパク質の結合部分は、各結合部分が、比較的低い親和性(例えば、低マイクロモル濃度または高ナノモル濃度、例えば、100ナノモル濃度を越えるKD、例えば、500、600、700、800ナノモル濃度)でその標的に結合するように選択される。このように、標的への長期にわたる結合は、2つの標的が同じ細胞上で近接して存在する状況においてのみ好ましい。
【0104】
同じ標的に結合する2つの結合部分(各々、同じ標的の異なるエピトープに結合する)を含む二重特異性結合タンパク質が作製され得る。この設計は、特に、結合タンパク質で標的を遮断するのを成功させる確率を最大にするのにふさわしい。複数の細胞外ループ、例えば、膜貫通チャネルまたは細胞表面レセプターの複数の細胞外ループが、同じ二重特異性結合分子によって標的にされ得る。
【0105】
クラスター化して免疫シグナル伝達の負の制御因子を活性化することにより免疫抑制をもたらす2つの結合部分を含む二重特異性結合タンパク質が作製され得る。シスでの抑制(Repression in cis)は、標的が同じ細胞上に存在する場合に達成され得る;トランスでの抑制(repression in trans)は、標的が異なる細胞上に存在する場合に達成され得る。シスでの抑制は、例えば、FelD1に対する免疫応答をダウンレギュレートするために、抗IgGRIIb結合部分および抗FelD1結合部分を有する二重特異性結合タンパク質を用いて達成され得る(IgGRIIbは、FelD1の存在下でのみクラスター化する)。トランスでの抑制は、例えば、抗BTLA結合部分、および目的の組織特異的抗原に特異的に結合する結合部分を有する二重特異性結合タンパク質を用いて達成され得(阻害性BTLA分子のクラスター化は、選択された標的組織だけで生じる)、これは、自己免疫疾患に対処できる可能性がある。
【0106】
多成分性のレセプターを活性化する二重特異性結合タンパク質が作製され得る。この設計では、レセプターの2成分に対する2つの結合部分が、そのレセプターに結合して、そのレセプターを架橋し、そのレセプターからのシグナル伝達を活性化する。これは、例えば、IFNAR1に結合する結合部分およびIFNAR2に結合する結合部分を有する二重特異性結合タンパク質を用いて行われ得、ここで、結合によって、そのレセプターが架橋される。そのような二重特異性結合タンパク質は、インターフェロン処置に対する代替物を提供し得る。
【0107】
半透性障壁、例えば、血液脳関門を越えて結合部分を輸送する二重特異性結合タンパク質が作製され得る。この設計では、一方の結合部分は、特定の選択的な障壁を通過し得る標的に結合する;他方の結合部分は、治療活性を有する分子を標的にし、ここで、その治療活性を有する標的分子は、通常はその障壁を通過することができない。この種の二重特異性結合タンパク質は、治療薬が他の方法では到達しなかった組織にその治療薬を持って行くために有用である。いくつかの例としては、腸または肺に治療薬を輸送するためにpIGRレセプターを標的にすること、または血液脳関門を越えて治療薬を輸送するためにトランスフェリンレセプターを標的にすることが挙げられる。
【0108】
特定の細胞または細胞型に結合部分を輸送する二重特異性結合タンパク質が作製され得る。この設計では、一方の結合部分が、細胞内に容易に内部移行される細胞表面タンパク質(例えば、レセプター)を標的にする。他方の結合部分は、細胞内タンパク質を標的にし、ここで、その細胞内タンパク質の結合は、治療効果をもたらす。
【0109】
二重特異性結合タンパク質は、食作用性免疫細胞の表面レセプターおよび感染性病原体(例えば、酵母または細菌)の表面分子に結合することにより、その感染性病原体が食作用性免疫細胞の近くに来て、その病原体のファゴサイトーシスを促進する。そのような設計の例は、CD64またはCD89分子および病原体を標的にする二重特異性抗体(antibodiy)であり得る。
【0110】
二重特異性結合タンパク質は、1つの結合部分として抗体可変領域および第2の結合部分として非Ig部分を有する。その抗体可変領域は、標的化を達成するのに対し、非Ig部分は、Fcに連結されたエフェクターまたはトキシンである。このように、リガンド(例えば、エフェクターまたはトキシン)は、抗体可変領域に結合した標的に送達される。
【0111】
二重特異性結合タンパク質は、任意の2つのタンパク質部分が、Fcの状況において互いに近くに置かれ得るように、2つの部分(各々が、Ig領域(例えば、CH2およびCH3領域を含むIg配列)に結合している)を有する。この設計の例としては、捕捉、例えば、ホモ二量体またはヘテロ二量体の捕捉分子が挙げられる。
【0112】
核酸
モノクローナル抗体をコードする核酸配列は、当該分野で公知の任意の適当な方法によって得ることができる。モノクローナル抗体(およびそれらの核酸配列)を得るための適当な方法の例としては、例えば、ハイブリドーマ法(例えば、Kohlerら(1975)Nature 256:495−497を参照のこと)またはファージ抗体ライブラリー(例えば、Clacksonら(1991)Nature 352:624−628を参照のこと)による方法が挙げられる。
【0113】
様々な実施形態において、免疫グロブリン重鎖可変ドメインは、遺伝子改変動物またはトランスジェニック動物の核酸配列から得られる。いくつかの実施形態において、それらの領域は、ヒト免疫グロブリンのミニ遺伝子座(minilocus)を含む動物から得られる。いくつかの実施形態において、それらの領域は、免疫グロブリン配列をコードする1つ以上の核酸を含む1つ以上の染色体外核酸を含むマウスから得られる。様々な実施形態において、上記動物は、再配列されていない1つ以上のヒト免疫グロブリン核酸配列を有し得る。いくつかの実施形態において、上記動物は、ヒト軽鎖可変領域核酸配列を含み、いくつかの実施形態において、ヒト重鎖可変配列を含み、いくつかの実施形態において、重鎖可変配列と軽鎖可変配列の両方を含み、そしていくつかの実施形態において、ヒト定常領域配列をさらに含む。特定の実施形態において、上記核酸配列は、内在性のマウス重鎖可変遺伝子セグメントおよび軽鎖可変遺伝子セグメントがヒト重鎖可変遺伝子セグメントおよび軽鎖可変遺伝子セグメントで置き換えられているマウスから得られる。
【0114】
いくつかの実施形態において、上記核酸配列は、そのような動物のナイーブB細胞またはナイーブT細胞から得られる。他の実施形態において、上記核酸配列は、目的の抗原で免疫された動物のB細胞またはT細胞から得られる。
【0115】
様々な実施形態において、上記核酸配列は、プライマー(例えば、1つ以上のFR、連結配列または定常配列を含む縮重プライマーのセットが挙げられる)でその核酸配列を増幅することによって、細胞から得られる。
【0116】
様々な実施形態において、免疫グロブリン重鎖可変ドメインは、目的の抗原で免疫された動物の核酸から得られる。例えば、非ヒトトランスジェニック動物または非ヒト遺伝子改変動物を、目的の抗原で免疫することにより(例えば、その動物を、その抗原またはその抗原を有する細胞またはその抗原の発現可能な形態をコードする核酸に曝露することにより)、その動物に免疫応答を起こさせ、その動物から免疫細胞(例えば、B細胞)を単離し、必要に応じて、それらの細胞を不死化し、そしてそれらの細胞をスクリーニングすることにより、その抗原との反応性を特定し、ならびに/または抗体の状況で置かれたときその抗原を認識することができる免疫グロブリン可変領域をコードする核酸配列を同定し、そして/もしくは単離する。いくつかの実施形態において、上記細胞は、B細胞である。いくつかの実施形態において、免疫された動物のB細胞を用いることによりハイブリドーマを作製し、その抗原のエピトープを特異的に認識する抗体を発現するB細胞を特定し、そのエピトープを認識する可変領域のアミノ酸配列をコードする核酸配列を特定し、そして/または単離する。
【0117】
いくつかの実施形態において、上記核酸は、ヒト、非ヒト霊長類(例えば、チンパンジーなどの類人猿)、サル(例えば、カニクイザル(cynomologous)またはアカゲザル)、げっ歯類(例えば、マウス、ラット、ハムスター)、ロバ、ヤギ、ヒツジなどから得られる。
【0118】
いくつかの実施形態において、重鎖は、ヒト細胞から得られる配列を含む。例えば、ヒト胎児細胞が、インビトロで抗原に曝露され、適当な宿主動物(例えば、SCIDマウス)に入れられる。
【0119】
いくつかの実施形態において、上記核酸は、ベクターを用いて細胞に導入される。ベクターとしては、例えば、プラスミド、コスミド、レトロウイルス、アデノウイルス、アデノ随伴ウイルス、植物ウイルス、YAC、BAC、EBV由来エピソームが挙げられる。
【0120】
いくつかの実施形態において、上記核酸は、発現ベクターまたは発現構築物に存在する。いくつかの実施形態において、その発現ベクターまたは発現構築物は、目的の核酸配列が適当な細胞において適当な条件下で発現することができるように、その目的の核酸配列に作動可能に連結されたプロモーターを含むベクターである。発現ベクターまたは発現構築物は、リーダー配列、エンハンサー、転写または翻訳を増強するプロモーターエレメント、転写停止配列、スプライシング配列、転写増強イントロン、IRESエレメント、マーカー遺伝子、選択配列、リコンビナーゼ認識部位、ホモロジーアーム(homology arm)、ウイルス配列、オペレーター(例えば、原核生物のオペレーター)などを含み得る。いくつかの実施形態において、発現ベクターは、誘導可能な発現を可能にするエレメント、例えば、真核生物のプロモーターに作動可能に連結された原核生物のオペレーターを含む。いくつかの実施形態において、発現は、発現誘導因子を加えたときに誘導される。他の実施形態において、発現は、発現阻害剤を除去したときに誘導される。いくつかの実施形態において、発現は、温度変化によって誘導される。
【0121】
いくつかの実施形態において、1つ以上の重鎖ヌクレオチド配列は、同じベクター上に存在する。いくつかの実施形態において、本明細書中の重鎖核酸配列および軽鎖核酸配列は、同じベクター上に存在する。1つの実施形態において、2つの重鎖核酸配列および1つの軽鎖核酸配列は、同じベクター上に存在する。
【0122】
いくつかの実施形態において、核酸は、ウイルス由来の1つ以上の核酸を含む細胞内で発現される。特定の実施形態において、そのウイルスは、アデノウイルス、アデノ随伴ウイルス、SV−40、エプスタイン・バーウイルス、レトロウイルス、レンチウイルス、バキュロウイルス、コロナウイルス、単純ヘルペスウイルス、ポリオウイルス、セムリキ森林ウイルス、シンドビスウイルスおよびワクシニアウイルスから選択される。
【0123】
宿主細胞は、目的の核酸を発現するように形質転換され得る細胞である。様々な実施形態において、形質転換は、細胞が外来性核酸(例えば、天然ではその細胞内に見られない核酸、または天然でその細胞内に見られる核酸配列に対応する1つ以上の追加のコピーの核酸)を含むように細胞の核酸含有量を変化させることを含む。細胞の核酸含有量は、当該分野で公知の任意の適当な方法によって、例えば、その核酸を細胞のゲノムにインテグレートすることによって、またはその核酸を染色体外もしくはゲノム外の形態で細胞内に入れることによって、変更され得る。いくつかの実施形態において、細胞の核酸含有量は、その細胞が目的の核酸を一過性に発現するように変更され得るか、または、核酸含有量は、その細胞が目的の核酸を安定的に発現するように変更され得る。いくつかの実施形態において、その細胞の遺伝子含有量の変更は、細胞が分裂するときに受け継がれる。
【0124】
二重特異性抗原結合タンパク質の単離
適当な改変のセットが、本明細書中の情報に基づいて選択されたら、当該分野で公知の方法を用いて二重特異性抗原結合タンパク質を単離する試みを行った。公開された方法を単に適用することによって、すべての場合において満足な分離が提供されるわけではなかった。
【0125】
Lindhoferらは、プロテインAに結合する1つの重鎖(マウスIgG)およびプロテインAに結合しない1つの重鎖(ラットIgG)を有するヘテロ二量体の重鎖を有する二重特異性抗体を作製し、そのラット/マウスヘテロ二量体二重特異性抗体を溶出するために中性からpH5.8までのpH段階勾配、次いで、マウス/マウスホモ二量体を溶出するために5.8から3.5までのpH段階を用いて、そのヘテロ二量体をマウス/マウスおよびラット/ラット二量体のクアドローマ混合物から分離することに成功した。(Lindhoferら(1995)Preferential species−restricted heavy/light chain pairing in rat/mouse quadromas:Implications for a single−step purification of bispecific antibodies.J.Immunol.155(1):219−225を参照のこと)。
【0126】
上記のLindhoferのアプローチは、IgG1 CH3ドメインのうちの一方がH435R/Y436Fジペプチド改変を含むという事実を別にすれば同一である2つのIgG1重鎖を有するIgG1ヘテロ二量体からIgG1ホモ二量体を分離することに適用したときは、失敗した。本発明者らは、直線のpH勾配では、そのジペプチド改変されたIgG1がpH約3.9で溶出されるのに対し、IgG1ホモ二量体は、pH約3.7で溶出されることを見出した。このpHの差異では、Lindhofer法を用いて上記ホモ二量体からヘテロ二量体を十分に分離するには不十分であると考えられた。この差異は、予測可能様式では再現性が無かった。
【0127】
特定のpH段階またはpH勾配を維持するために必要とされる緩衝液強度によって与えられる比較的実質的なイオン強度を使用するクロマトグラフィの実行において、クロマトグラフィ上の挙動の変動が観察された。しかし、有機修飾剤(organic modifier)(1−プロパノール)を加えることによって、十分な分離は達成されなかった。いくぶん驚いたことに、いくつかのクロマトグラフィの実行については、代わりに、0.5モル濃度〜1.0モル濃度のイオン調節物質(例えば、NaCl)を加えることによって、ホモ二量体のIgG1およびヘテロ二量体のIgG1の分離が急激かつ予想外に改善された。イオン調節物質の添加によって、溶出に対するpH範囲が広がり(イオン調節物質を用いたときは1.2pH単位だったが、イオン調節物質を用いないときは0.2pH単位だった)、pH段階勾配によって2種の分離に成功することができた。しかしながら、他の実行では、たった約150mMのNaCl濃度を用いただけで、十分な分離が達成された(実施例4を参照のこと)。十分な分離が達成され得るのを確実にするために、1つの実施形態において、約0.5〜約1.0モル濃度のイオン調節物質の存在下で二重特異性抗原結合タンパク質の単離が行われる。
【0128】
したがって、1つの実施形態において、本明細書中に記載されるような改変を含む1つの鎖を有するヘテロ二量体のIgGを含む二重特異性抗原結合タンパク質を分離するための方法は、イオン調節物質の存在下においてpH勾配を使用する工程を包含する。1つの実施形態において、そのイオン調節物質は、プロテインA支持体からのIgGホモ二量体の溶出と、本明細書中に記載されるような(すなわち、CH3改変を有する)IgGヘテロ二量体の溶出との間のpHの差を最大にするのに十分な濃度で存在する。特定の実施形態において、イオン調節物質は、約0.5〜約1.0モル濃度の濃度で存在する。別の特定の実施形態において、イオン調節物質は、約0.15〜約0.5モル濃度の濃度で存在する。
【0129】
1つの実施形態において、イオン調節物質は、塩である。1つの実施形態において、イオン調節物質は、アルカリ金属またはアルカリ土類金属およびハロゲンの塩である。特定の実施形態において、その塩は、アルカリ金属またはアルカリ土類金属の塩化物塩、例えば、NaCl、KCl、LiCl、CaCl2、MgCl2である。特定の実施形態において、塩は、約0.5、0.6、0.7、0.8、0.9または1.0のモル濃度で存在する。
【0130】
1つの実施形態において、pH勾配は、pH約3.9〜pH約4.5、別の実施形態では、pH約4.0〜pH約4.4、別の実施形態では、pH約4.1〜pH約4.3である。特定の実施形態において、勾配は、直線勾配である。
【0131】
1つの実施形態において、pH勾配は、段階勾配である。1つの実施形態において、その方法は、pH約3.9、pH約4.0、pH約4.1、pH約4.2、pH約4.3またはpH約4.4の段階を、平衡化されたプロテインAカラム(例えば、PBSまたは別の適当な緩衝液もしくは液体で平衡化されたカラム)に適用する工程を包含する。特定の実施形態において、その段階は、pH約4.2である。
【0132】
1つの実施形態において、ヘテロ二量体のIgG CH3ドメインを含む二重特異性抗体は、非ヘテロ二量体のIgGを実質的に含まない1つ以上の画分にプロテインA支持体から溶出する。特定の実施形態において、溶出された二重特異性抗体画分は、非ヘテロ二量体の抗体である全タンパク質の約1重量%未満、0.5重量%未満または0.1重量%未満を構成する。
【実施例】
【0133】
以下の実施例は、本発明の方法および組成物を作製する方法および使用する方法を当業者に対して説明するために提案されるものであって、本発明者らが本発明者らの発明と考える範囲を限定する意図はない。使用される数値(例えば、量、温度など)に関する精度を保証する努力がなされているが、いくらかの実験誤差および偏差が釈明されるだろう。別段示されない限り、部は、重量部であり、分子量は、平均分子量であり、温度は、摂氏温度であり、圧力は、大気圧または大気圧付近である。
【0134】
実施例1:二重特異性IL−4Ra/IL−6Ra抗原結合タンパク質
ヒトIgG1アイソタイプの2つの公知抗体(1つは、IL−4Raに対する抗体で、1つは、IL−6Raに対する抗体)が、4つのアミノ酸だけが異なる軽鎖を有することが見出された。共発現実験から、抗IL−4Ra抗体の軽鎖が、IL−6Ra由来の軽鎖で置き換えられ得、IL−4Raに対する高親和性結合をなおも維持し、ゆえに抗IL−4Ra重鎖および抗IL−6Ra重鎖ならびに同じ軽鎖を用いて二重特異性抗体を作製することが可能になることが明らかになった。したがって、IL−6Ra抗体の重鎖をFcΔAdp型に改変した(すなわち、IMGTエキソンナンバリングではH95R/Y96FのCH3ジペプチド改変)。
【0135】
次いで、抗IL−6Ra軽鎖をCHO細胞において抗IL4Ra/Fcおよび抗IL6Ra/FcΔAdp重鎖と共発現させ、これらの細胞からのコンディションドメディウムをプロテインAクロマトグラフィに供した。ホモ二量体とヘテロ二量体との混合物を含む細胞上清をプロテインAカラムに充填した後、それぞれpH6.0、pH4.2およびpH3.0で3相となるように2つの緩衝液(A:100mMクエン酸Na、150mM NaCl,pH6.0およびB:100mMクエン酸Na、150mM NaCl,pH3.0;図4を参照のこと)の組み合わせを変化させることによって作製されたpH段階勾配を用いて溶出を行った。図4において、IL−4Rは、抗IL−4Raを表し、IL−6RΔは、抗IL−6Ra(IgG1ΔAdp)を表す。示されているカラム画分を、IL−6RaおよびIL−4Raタンパク質への結合についてアッセイした(図5を参照のこと)。段階溶出を行ったところ、pH4.2で溶出する1つのピークおよびpH3.0における第2のピークが生じた(図4)。BIACORE(商標)解析から、予想のとおり、フロースルー材料は、可溶性IL−6Raに結合することができたが、IL−4Raには結合できなかったことが示された(図5)。pH4.2のピークに対応する画分は、ほぼ等量のIL−6RaおよびIL−4Raに結合することができ、これは、ヘテロ二量体と矛盾しない。pH3.0で溶出するピークは、IL−4Raにだけ結合することができ、IL−6Raには結合できず、このことから、予想される抗IL−4Raホモ二量体に対応する。このことから、ヘテロ二量体の二重特異性抗体が、単純なpH段階勾配を伴うプロテインAクロマトグラフィを使用して効率的に単離され得ることが立証された。
【0136】
実施例2:FcΔAdpタンパク質の薬物動態
FcΔAdp改変が、ヘテロ二量体のFc/FcΔAdpを含む分子の薬物動態に影響したか否かを試験するために、上に記載された抗IL−4Ra/抗IL−6Raの精製されたヘテロ二量体種をマウスに注射し、28日間にわたって血清中のヒト免疫グロブリン濃度を測定した(図6;表1)。そのヘテロ二量体の血清半減期は、約10日であり、これは、野生型の血清半減期と同様だった。このことから、FcΔAdp改変は、血清半減期に検出可能な影響を及ぼさないことが立証された。
【0137】
【表1】

実施例3:二重特異性CD20/CD3抗原結合タンパク質
公知の抗ヒトCD20抗体の重鎖は、公知の(活性化する)抗ヒトCD3抗体の軽鎖と共発現したときも、依然としてCD20に結合することができることが見出された。次いで、その抗CD3軽鎖を抗CD20/Fc重鎖、抗CD3/FcΔAdp重鎖または両方の重鎖と共発現させた。次いで、得られたホモ二量体およびヘテロ二量体の混合集団をバイオアッセイに使用して、CD20を発現している標的細胞を殺滅する能力を測定した(図7)。簡潔には、2×107個のヒトPBMC細胞を、6×107個のCD3×CD28ビーズ(Invitrogen)で72時間活性化した。次いで、30単位のIL−2(R&D Systems)を加え、その細胞をさらに24時間インキュベートした。次いで、その細胞を0.5×106/mLの濃度に分け、さらに30UのIL−2を加えた。次いで、その細胞をさらに48時間インキュベートし、バイオアッセイに使用した。バイオアッセイの当日、CD20を発現している2×106個/mLの標的細胞(Raji)を、8μMのカルセイン−AM(Invitrogen)で30分間標識した。洗浄した標的細胞を、活性化されたhPBMC細胞に、示されている量の抗体含有上清を含む総体積200マイクロリットルにおいて標的:エフェクター細胞(1ウェルあたり合計220,000個の細胞)1:10の比で加えた。細胞を2時間インキュベートし、上清を回収し、蛍光を定量化した。特異的な蛍光と最大蛍光との比を計算することによって、細胞傷害性を測定した。CD20抗体単独(抗CD3軽鎖を用いた)と抗CD3抗体の両方が、標的細胞の殺滅を誘発できなかった;それら2つの試薬を混合したときでさえ、効果がなかった。しかしながら、3つすべての構成要素を共発現させると、有意な殺滅が観察されたことから、その効果はヘテロ二量体の二重特異性種に起因したことが示唆される。一過性にトランスフェクトされたCHO細胞の上清中の二重特異性抗体の推定量に基づいて、この効果に対するEC50が、約15pMであると推定された。
【0138】
実施例4:精製された二重特異性CD20/CD3抗原結合タンパク質を用いた細胞殺滅特異性
実施例3に記載されたようなトランスフェクションからのCHO細胞上清を、溶出のために段階勾配を利用するプロテインAアフィニティークロマトグラフィに供した。その段階勾配は、それぞれpH5.2、pH4.2およびpH2.8で3相となるように2つの緩衝液(A:20mMクエン酸Na、1M NaCl,pH5.2;B:20mMクエン酸Na、1M NaCl,pH2.7)の組み合わせを変化させることによって作製された。次いで、pH4.2で溶出するピークからのタンパク質を、実施例3に記載されたような細胞死滅アッセイにおいて使用した。3pMというEC50で標的細胞の殺滅が観察された(図8)。観察された殺滅の標的特異性を調べる追加の細胞傷害性アッセイを行った。この実験では、CD20を発現する標識された標的細胞(Raji)またはCD20を有しない標識された標的細胞(293)を、活性化されたヒトPBMCとともにインキュベートした。各標的細胞型をそのアッセイに単独でまたは他のタイプの未標識の標的細胞と組み合わせて加えた。すべての場合において、CD20を発現している標的細胞は、3pMというEC50で特異的に殺滅されたが、CD20陰性細胞株は、殺滅されなかった。
【0139】
実施例5:識別的に改変されたhIgG2 FcのプロテインA分離
識別的に改変されたヘテロ二量体のヒトIgG2 Fc/ΔAdpFcおよび改変されていないヒトホモ二量体のIgG2 Fc/Fcを、まず、プロテインAカラム(rProteinA FF,GE)に通す結合・洗浄(bind−and−wash)プロセスによって濃縮した。hIgG2 Fc/FcからhIgG2 Fc/ΔAdpFcをさらに分離するために、以下のとおり、SMART(商標)システム(GE)を使用して段階勾配溶出を行った。この溶媒系は、溶媒A(PBS,1×)、溶媒B(20mMクエン酸ナトリウムおよび1M NaCl,pH5.5)および溶媒C(20mMクエン酸ナトリウムおよび1M NaCl,pH2.5)からなるものだった。溶出を、最初の20分間の100%Aによるアイソクラティック溶出から開始し、続いて、20分後に素早く100%Bに切り換えた。次いで、次の10分間にわたって、33.5%Cおよび66.5%Bに向かって直線勾配を開始した;第1のピーク(Fc/ΔAdpFc)が完全に溶出するまで、Cの濃度を20分間33.5%に維持した。続いて、33.5%Cから100%Cへの直線勾配を30分間行った。流速を250マイクロリットル/分に維持し、UV検出器によって280nmでクロマトグラムを検出した。hIgG2 Fc/ΔAdpFcは、pH4.5で溶出し、一方、hIgG2は、pH3.5で溶出した。
【0140】
実施例6:識別的に改変されたhIgG4 FcのプロテインA分離
識別的に改変されたヘテロ二量体のヒトIgG4(Fc/ΔAdpFc)および改変されていないホモ二量体のIgG4(Fc/Fc)をまず、プロテインAカラム(rProteinA FF,GE)に通す結合・洗浄プロセスによって濃縮した。hIgG4 Fc/FcからhIgG4 Fc/ΔAdpFcをさらに分離するために、以下のとおり、SMART(商標)システム(GE)を使用して段階勾配溶出を行った。この溶媒系は、溶媒A(PBS,1×)、溶媒B(20mMクエン酸ナトリウムおよび1M NaCl,pH5.1)および溶媒C(20mMクエン酸ナトリウムおよび1M NaCl,pH2.8)からなるものだった。溶出を、最初の20分間の100%Aによるアイソクラティック溶出から開始し、続いて、20分後に素早く100%Bに切り換えた。次いで、次の10分間にわたって、50%Cおよび50%Bに向かって直線勾配を開始した;第1のピーク(Fc/ΔAdpFc)が完全に溶出するまで、Cの濃度を20分間50%に維持した。続いて、50%Cから100%Cへの直線勾配を30分間行った。流速を250マイクロリットル/分に維持し、UV検出器によって280nmでクロマトグラムを検出した。hIgG4 Fc/ΔdpFcは、pH約4で溶出し、一方、ホモ二量体は、pH約4からpH2.8までの勾配中に溶出した。
【0141】
実施例7;識別的に改変されたhIgG1 CD3×CD20のプロテインA分離
識別的に改変されたヘテロ二量体の抗hCD3×CD20 IgG1(Fc/ΔAdpFc)および改変されていないホモ二量体の抗hCD20を、以下のとおり、1mLのrProtein AFF(GE Biosciences)カラムにおいて分離した。この溶媒系は、緩衝液A1(PBS 1×)、緩衝液A2(20mMクエン酸ナトリウムおよび1M NaCl pH5.1)、緩衝液B(20mMクエン酸ナトリウムおよび1M NaCl pH2.8)だった。混合サンプルを結合させ、PBSおよび緩衝液A2で洗浄した。段階を用いてpH4.2に達したところ、それにより、二重特異性CD3*×CD20 IgG1(Fc/ΔAdpFc)が溶出し、次いで、pH4.2からpH2.8までの直線勾配により、ホモ二量体の抗hCD20 IgG1が溶出した。
【0142】
実施例8:改変されたCH3のFcレセプターに対する結合親和性
ΔAdp改変(H435RおよびY436F,EUナンバリング)を有するヒトIgG1アイソタイプ二重特異性抗体の種々のヒトFcレセプターに対する結合親和性を、Biacore(商標)定常状態平衡結合アッセイにおいて試験した。
【0143】
簡潔には、アミンに結合した抗ペンタ−his mAb(Qiagen)を有するカルボキシメチル化デキストラン(CM5)チップを使用して、ヒトFcレセプターの様々な構築物を捕捉した。以下のhisタグ化Fcレセプター外部ドメインを、抗ペンタ−hisでコーティングされた種々のCM5チップの表面に結合させた:FcγRI、FcγRIIA(R131多形)、FcγRIIBおよびFcγRIIIB(各々、R&D Systemsから入手した);およびRcγRIIA(H131多形)、FcγRIIIA(V176多形)およびRcγRIIIA(F176多形)(各々、Regeneronにて製造されたもの)。高親和性レセプターFcγRI外部ドメインについては3つの濃度(25nM、50nMおよび100nM)で、および低親和性FcγRレセプター外部ドメインについては5マイクロモル濃度〜39ナノモル濃度で抗体を上記表面の上に通過させ、会合速度定数および解離速度定数(kaおよびkd)の値を測定し、それを用いてそれらの抗体に対する平衡解離定数(KD)を算出した。pH7.2のHBS−T緩衝液を使用して、室温にて結合研究を行った。コントロール抗体(hmAb)、抗CD20および抗CD3Δdp改変ならびにCD20×CD3Δdp二重特異性抗体に対するKDを測定した。抗CD3Δdp抗体に対するKD値は、改変されていないhIgG1アイソタイプ抗体と比べて、試験されたいずれのFcレセプターに対する結合についても有意差を明らかにしなかった(表2)。
【0144】
【表2】
実施例9:hFcRnマウスにおける二重特異性hIgG1ΔAdpの薬物動態
二重特異性抗hCD3/hCD20 IgG1ΔAdp抗体およびその抗体に関するコントロール(抗hCD3 IgGおよび抗hCD3 IgGΔAdpホモ二量体)の薬物動態学的クリアランス速度を、野生型(WT)マウスおよびhFcRn遺伝子によるマウスFcRnの置き換えについてホモ接合性のマウス(hFcRnマウス)において測定した。野生型マウスおよびhFcRnマウスは、C57BL6(75%)および129Sv(25%)を含むバックグラウンドの交雑系統由来のものだった。IgG1アイソタイプマッチコントロール抗体を投与されるWTマウスの1つのコホートの場合(このコホートは3匹のマウスを含んだ)を除いて、コホートは、WTマウスまたはhFcRnマウスの各々を4匹含んだ。1mg/kgのアイソタイプマッチ(hIgG1)コントロール、抗hCD3×CD20IgG1ΔAdp二重特異性、抗hCD3IgG1または抗hCD3IgG1ΔAdpホモ二量体をマウスに投与した。すべての被験物質を皮下に投与した。血液(Bleeds)を0時間、6時間、1日、2日、3日、4日、7日、10日、14日、21日および30日の時点で回収した。
【0145】
ヒト抗体の血清レベルをサンドイッチELISAによって測定した。簡潔には、ヤギポリクローナル抗ヒトIgG(Fc特異的)抗体(Jackson ImmunoResearch)を、1マイクログラム/mLの濃度で96ウェルプレートにコーティングし、4℃で一晩インキュベートした。そのプレートをBSAでブロッキングした後、6点段階希釈物中の血清サンプルおよび12点段階希釈物中のそれぞれの抗体の対照基準を、そのプレートに加え、室温で1時間インキュベートした。洗浄して未結合の抗体を除去した後、捕捉されたヒト抗体を、西洋ワサビペルオキシダーゼ(HRP)で結合体化された同じヤギポリクローナル抗ヒトIgG(Fc特異的)抗体(Jackson ImmunoResearch)を用いて検出し、製造者の推奨に従って、標準的な比色テトラメチルベンジジン(TMB)基質によって発色させた。450nmにおける吸光度をプレートリーダーにて記録し、そのサンプルプレートにおいて作成された基準検量線を用いて、血清サンプル中のhIgGの濃度を算出した。
【0146】
試験された30日間にわたる4つのIgG1抗体の血清半減期に有意差は観察されなかった。特に、ΔAdp改変を有するIgG1抗体と野生型IgG1抗体との間に有意差は観察されなかった。野生型(mFcRn)マウスまたはヒト化FcRn(hFcRn)を有するマウスを用いたとき、それらの抗体の間に差は観察されなかった。予想の通り、hFcRnマウスは、野生型マウスよりもわずかに速いクリアランスを示した。結果を表3に示す。
【0147】
【表3】
実施例10:低塩緩衝液における大量単離
二重特異性CD3×CD20ΔAdp抗体を、本発明に従って大量培養物から単離した。簡潔には、二重特異性抗hCD3×CD20ΔAdp(CD3重鎖に改変)抗体を発現しているCHO−K1細胞株を11リットルのバイオリアクターにおいて培養した。二重特異性抗体を産出する細胞は、約8.25×106細胞/mLの密度まで成長し、約250〜350mg抗体/Lが得られた。対照的に、コントロール抗hCD3抗体は、約100〜150mg/Lで得られた。
【0148】
10mMリン酸ナトリウム、0.5M NaCl,pH7.2で平衡化されたMabSelect SuRe(商標)樹脂(GE)(床の高さ20cm,内径1cm)において抗体を単離し、浄化された細胞培養物を19g/Lで充填し、そのカラムを3カラム体積の10mMリン酸ナトリウム、0.5M NaCl,pH7.2で洗浄した後、2カラム体積の20mMリン酸ナトリウム,pH7.2(NaClなし)で洗浄した。40mM酢酸塩,pH3.0で抗体を溶出した。
【0149】
単一特異的な抗CD30抗体は、pH3.6で溶出し、一方、二重特異性抗hCD3×CD20DAdpは、pH4.4で溶出した。
【0150】
実施例11:中性付近pHによるマウスヘテロ二量体の選択的プロテインA溶出
野生型または変異体(TTTKまたはPTTK)mIgG2a Fcと融合された、ヒトIFNAR1(hIFNAR1)およびヒトIFNAR2(hIFNAR2)細胞外ドメインに対する発現構築物でCHO−K1細胞を一過性にトランスフェクトした。等量の2つの発現プラスミドで細胞をトランスフェクトすることによって、hIFNAR1−mFcとhIFNAR2−mFcとの比を1:1に保った。トランスフェクションの4日後に培養液を回収し、それを、0.2mLのNAb Protein A Plus(商標)スピンカラム(Thermo Scientific/Pierce)を使用するプロテインA精製に供した。簡潔には、カラムを1×PBS,pH7.2で平衡化した。1mlのCHO−K1培養液を、室温で10分間、プロテインA樹脂とともにインキュベートした。次いで、そのカラムを1×PBS,pH7.2で3回洗浄した。結合したタンパク質を、1M NaClを含む20mMクエン酸ナトリウム緩衝液を用いて溶出した。pHを下げながら0.4mLの溶出緩衝液を使用して、溶出を3回行った。異なる画分中のタンパク質を、ウエスタンブロット解析によって検出した。
【0151】
結果から、pH勾配溶出を用いたとき、野生型mIgG2aのホモ二量体から野生型およびstar変異体mIgG2aのヘテロ二量体を分離することが可能であることが示される(図9の両方のゲルにおける画分E1を参照のこと)。
【0152】
実施例12;アイソタイプヘテロ二量体に対するmIgG2a変異体の優先的なヘテロ二量体形成
C末端のマウスFc(mIgG2aまたはmIgG1)を有するヒトI型インターフェロンレセプター(hIFNAR1およびhIFNAR2)の細胞外ドメインの哺乳動物発現用のDNAプラスミドを構築した。部位特異的突然変異誘発を用いて、mIgG2a配列内に変異を導入した。これらの変異体は、TTT=M252T、S254T、S256T;TTTK=M252T、S254T、S256T、I258K;PTTTK=I247P、M252T、S254T、S256T、I258K;RF=H435R、H436Fである。CHO−K1細胞を、上記発現構築物で一過性にトランスフェクトした。hIFNAR2−mFc(mIgG1または変異体mIgG2a)よりも4倍多いhIFNAR1−mFc発現プラスミド(hIFNAR1−mIgG2a)で細胞をトランスフェクトすることによって、IFNAR1−mFcとIFNAR2−mFcとの比を4:1に保った。トランスフェクションの4日後に培養液を回収し、mFcタンパク質をウエスタンブロット解析によって検出した。
【0153】
結果から、mIgG2aとmIgG1とのヘテロ二量体の形成が、野生型mIgG2aとmIgG2a変異体とのヘテロ二量体の形成よりもかなり効率的でないことが示される(図10のレーン1とレーン2〜5とを比較のこと)。この実験では、4:1の比のIFNAR1構築物:IFNAR2構築物を使用することにより、過剰な野生型IgG2a構築物を維持した。
【技術分野】
【0001】
発明の分野
本発明は、重鎖のヘテロ二量体を有する、抗原結合タンパク質または抗体、すなわち、親和性試薬に対する免疫グロブリン重鎖の親和性と改変免疫グロブリン重鎖または変異免疫グロブリン重鎖の親和性との違いに基づいて抗原結合タンパク質の単離を可能にする、少なくとも1つのアミノ酸が異なる2つの免疫グロブリン重鎖を有する、抗原結合タンパク質または抗体に関する。本発明はまた、プロテインAに対するIgG領域の結合性の違いによる迅速な単離を可能にする、プロテインAに対して異なる親和性を有するIgG CH2およびCH3領域を有する抗原結合タンパク質(二重特異性抗体を含む)にも関する。
【背景技術】
【0002】
背景
抗体は、標的抗原に対する独特の結合特異性ならびに抗原とは無関係な機序を介して免疫系と相互作用する能力を保有する多機能性分子である。癌に対して現在使用されている多くの生物学的治療薬は、標的にされている癌細胞上で代表的に過剰発現されている抗原に対するモノクローナル抗体である。そのような抗体は、腫瘍細胞に結合すると、抗体依存性細胞傷害(ADCC)または補体依存性細胞傷害(CDC)の引き金を引くことがある。残念なことに、癌性細胞は、これらの正常な免疫応答を抑制する機序を発達させることが多い。
【0003】
近年、2以上の抗原結合特異性を有する抗体様の治療薬、例えば、二重特異性抗体を開発する試みが進行中である。癌治療の場合、多重特異的形式によって、例えば、その分子が腫瘍細胞抗原を標的にするという1つの特異性、通常免疫系が利用できない反応の引き金を引くというその他の特異性を使用することが可能になり得る。二重特異性抗体は、2成分のヘテロ二量体レセプター系(この2成分のヘテロ二量体レセプター系は、それらの天然のリガンドが、そのレセプター系に結合して、その両方の成分と1つになるとき、そのリガンドによって正常に活性化される)に対する代用リガンドとしての使用法も見出し得る。
【0004】
複数の結合特異性を有する分子によって与えられる治療の機会に応える数多くの形式が当該分野で開発されている。理想的には、そのような分子は、作製しやすくかつ精製しやすく、好ましいインビボ特性、例えば、意図される目的にとって適切な薬物動態、最小の免疫原性、および望まれる場合、従来の抗体のエフェクター機能を有する、よく機能するタンパク質であるべきである。
【0005】
二重特異性抗体を生成する(単一細胞内で2つの異なる抗体を発現させる)最も単刀直入な方法は、複数の種を生じることである。なぜなら、それぞれの重鎖が、ホモ二量体とヘテロ二量体の両方を形成するが、ヘテロ二量体だけが望まれるからである。また、軽鎖および重鎖は、不適切に対形成することがある。種々の方法でこれらの問題に対処するように試みられた形式のいくつかの例を以下に記載する。
【0006】
二重特異性T細胞Engager(BiTE)分子(例えば、Wolf,E.ら(2005)Drug Discovery Today 10:1237−1244(非特許文献1))を参照のこと)に対して使用される1つの形式は、一本鎖可変フラグメント(scFv)モジュールに基づく。scFvは、通常適切に折り畳まれ得る可撓性リンカーを介して融合された抗体の軽鎖および重鎖の可変領域からなり、それらの領域は、同族抗原に結合することができる。BiTEでは、特異性の異なる2つのscFvが一本鎖にタンデムにつながれている(図1Aを参照のこと)。この配置は、2コピーの同じ重鎖可変領域を有する分子の生成を妨げる。さらに、それぞれの軽鎖および重鎖の正しい対形成を確実にするようにリンカー配置が設計される。
【0007】
BiTE形式は、いくつかの欠点を有する。第1に、scFv分子は、凝集する傾向があるとよく知られている。また、scFvリンカーの免疫原性は、通説では低いが、BiTEに対する抗体が産生される可能性を除外することはできない。また、BiTE形式ではFc部が存在しないので、その血清半減期は非常に短くなり、高頻度の反復投与またはポンプによる持続注入の面倒な問題が伴う。最後に、Fcが存在しないことにより、Fc媒介性のエフェクター機能が存在しないことも当然伴う(そのエフェクター機能が存在しないことは、いくつかの状況では有益であり得るが)。
【0008】
第2の形式(図1B)は、マウスとラットのモノクローナル抗体のハイブリッドであり、従来のプロテインAアフィニティークロマトグラフィの変法に依存するものである(例えば、Lindhofer,H.ら(1995)J.Immunol.155:219−225(非特許文献2))を参照のこと)。この形式では、マウスIgG2a抗体およびラットIgG2b抗体を、同じ細胞内で同時に産生させる(例えば、2つのハイブリドーマのクアドローマ融合物として、または操作されたCHO細胞内で)。各抗体の軽鎖は、それらの同族の種の重鎖と優先的に会合するので、3つの異なる種の抗体:2つの親抗体、およびFc部によって会合している、その2つの抗体の各々の1つの重鎖/軽鎖対を含むその2つの抗体のヘテロ二量体だけが組み立てられ得る。所望のヘテロ二量体は、そのプロテインAへの結合特性が親抗体の結合特性と異なる(すなわち、ラットIgG2bは、プロテインAに結合せず、一方、マウスIgG2aは、プロテインAに結合する)ので、この混合物から容易に精製され得る。それゆえ、マウス−ラットヘテロ二量体は、プロテインAに結合するが、マウスIgG2aホモ二量体よりも高いpHで溶出することから、これにより、二重特異性ヘテロ二量体を選択的に精製することが可能になる。BiTE形式と同様に、このハイブリッド形式は、2つの一価抗原結合部位を有する。
【0009】
マウス/ラットハイブリッドの欠点は、それが非ヒトであるので、有害な副作用(例えば、「HAMA」または「HARA」反応)を有し得、そして/またはその治療薬を中和し得る免疫応答を患者に誘発する可能性があるという点である。
【0010】
「ノブ・イントゥ・ホール(knobs−into−holes)」と呼ばれる第3の形式(図1C)は、二重特異性抗体の作製にとって有用である可能性があると従来技術において考察されている(米国特許第7,183,076号(特許文献1))。このストラテジーでは、一方が「ノブ」を突出させ、他方が相補的な「ホール」となるように、2つの抗体のFc部を操作する。それらの重鎖は、同じ細胞において産生されると、操作された「ノブ」と操作された「ホール」との会合によって、ホモ二量体ではなくヘテロ二量体を優先的に形成すると言われている。異なる特異性を有するが同一の軽鎖を採用する抗体を選択することによって、正しい軽鎖−重鎖の対形成の問題が対処される。
【0011】
この形式の欠点は、「ノブ・イントゥ・ホール」ストラテジーが、かなりの量の望ましくないホモ二量体を産生し得るがゆえに、さらなる精製工程が必要となるという点である。この難点は、汚染している種の特性の多くが所望の種とほぼ同一であるという事実によって深刻になっている。「ノブ」および「ホール」を生成する変異によって外来配列が導入されるので、操作された形状は、潜在的に免疫原性でもあり得る。
【0012】
依然として、二重特異性抗体の形式、特に、上で述べた欠点の一部または全部を最小にする、治療に役立つ用途のための二重特異性抗体の形式が必要とされている。
【先行技術文献】
【特許文献】
【0013】
【特許文献1】米国特許第7,183,076号明細書
【非特許文献】
【0014】
【非特許文献1】Wolf,E.ら(2005)Drug Discovery Today 10:1237−1244
【非特許文献2】Lindhofer,H.ら(1995)J.Immunol.155:219−225
【発明の概要】
【課題を解決するための手段】
【0015】
概要
本発明は、二重特異性抗原結合タンパク質において少なくとも1つのアミノ酸が異なる2つの免疫グロブリンCH3重鎖定常ドメイン配列を使用することに少なくとも部分的に基づく。少なくとも1つのアミノ酸が異なることにより、CH3ドメイン配列が親和性物質に結合する能力に違いがもたらされるので、そのタンパク質を単離する能力が改善される。
【0016】
1つの局面において、第1および第2のポリペプチドを含む抗原結合タンパク質が提供され、その第1のポリペプチドは、N末端からC末端に向かって、第1の抗原に選択的に結合する第1の抗原結合領域、それに続く、IgG1(配列番号1)、IgG2(配列番号3)IgG4(配列番号5)およびそれらの組み合わせから選択されるヒトIgGの第1のCH3領域を含む定常領域を含み;そして第2のポリペプチドは、N末端からC末端に向かって、第2の抗原に選択的に結合する第2の抗原結合領域、それに続くIgG1、IgG2、IgG4およびそれらの組み合わせから選択されるヒトIgGの第2のCH3領域を含む定常領域を含み、ここで、その第2のCH3領域は、プロテインAへの第2のCH3ドメインの結合を減少させるかまたは排除する改変を含む。
【0017】
1つの実施形態において、第2のCH3領域は、95R改変(IMGTエキソンナンバリングにおいて;EUナンバリングでは435R)を含む。別の実施形態において、第2のCH3領域は、96F改変(IMGT;EUでは436F)をさらに含む。特定の実施形態において、第2のCH3領域は、配列番号2、配列番号4および配列番号6から選択される。
【0018】
1つの実施形態において、第2のCH3領域は、改変されたヒトIgG1(配列番号2)に由来し、D16E、L18M、N44S、K52N、V57MおよびV82I(IMGT;EUではD356E、L358M、N384S、K392N、V397MおよびV422I)からなる群から選択される改変をさらに含む。
【0019】
1つの実施形態において、第2のCH3領域は、改変されたヒトIgG2(配列番号4)に由来し、N44S、K52NおよびV82I(IMGT;EUではN384S、K392NおよびV422I)からなる群から選択される改変をさらに含む。
【0020】
1つの実施形態において、第2のCH3領域は、改変されたヒトIgG4(配列番号6)に由来し、Q15R、N44S、K52N、V57M、R69K、E79QおよびV82I(IMGT;EUではQ355R、N384S、K392N、V397M、R409K、E419QおよびV422I)からなる群から選択される改変をさらに含む。
【0021】
1つの実施形態において、CH3ドメインは、ヒトIgG1、ヒトIgG2、ヒトIgG3およびヒトIgG4のうちの2つ以上の配列を含むキメラドメインである。
【0022】
1つの実施形態において、CH3ドメインは、ヒトIgG1、ヒトIgG2またはヒトIgG4に由来し、本抗原結合タンパク質は、CH1ドメインおよびCH2ドメインをさらに含み、ここで、そのCH1ドメインおよびCH2ドメインは、独立して、ヒトIgG1 CH1もしくはCH2ドメイン、ヒトIgG2 CH1もしくはCH2ドメイン、またはキメラヒト/ヒトIgG1/IgG2もしくはキメラヒト/ヒトIgG1/IgG3もしくはキメラヒト/ヒトIgG2/IgG3ドメインもしくはキメラヒト/ヒトIgG1/IgG4もしくはキメラIgG3/IgG4もしくはキメラIgG2/IgG4ドメインからなる群から選択される。特定の実施形態において、そのキメラIgG1/IgG2、IgG1/IgG3、IgG2/IgG3、IgG1/IgG4、IgG3/IgG4およびIgG2/IgG4ドメインは、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない。
【0023】
1つの実施形態において、本抗原結合タンパク質は、免疫グロブリン軽鎖をさらに含む。1つの実施形態において、その免疫グロブリン軽鎖は、ヒトラムダ軽鎖およびヒトカッパー軽鎖から選択される。
【0024】
1つの実施形態において、第1および第2の抗原結合領域の各々は、少なくとも1つのCDRを含み、別の実施形態では、少なくとも2つのCDRを含み、別の実施形態では、各々は、3つのCDRを含む。特定の実施形態において、そのCDRは、免疫グロブリン重鎖に由来する。別の特定の実施形態において、その重鎖は、ヒト重鎖である。
【0025】
1つの実施形態において、第1の抗原結合領域は、第1の免疫グロブリン重鎖可変ドメインを含み、第2の抗原結合領域は、第2の免疫グロブリン重鎖可変ドメインを含む。
【0026】
1つの実施形態において、第1および第2の免疫グロブリン重鎖可変ドメインは、独立して、マウス、ラット、ハムスター、ウサギ、サル、類人猿およびヒトのドメインから選択される。
【0027】
1つの実施形態において、第1および第2の免疫グロブリン重鎖可変ドメインは、独立して、ヒトCDR、マウスCDR、ラットCDR、ウサギCDR、サルCDR、類人猿CDRおよびヒト化CDRを含む。1つの実施形態において、そのCDRは、ヒトのCDRであり、体細胞変異している。
【0028】
1つの実施形態において、第1および第2の免疫グロブリン重鎖可変ドメインは、ヒトフレームワーク領域(FR)を含む。1つの実施形態において、そのヒトFRは、体細胞変異したヒトFRである。
【0029】
1つの実施形態において、第1および/または第2の抗原結合領域は、抗体可変領域を含むファージライブラリーを目的の抗原に対する反応性についてスクリーニングすることによって得られる。別の実施形態において、第1および/または第2の抗原結合領域は、非ヒト動物(例えば、マウス、ラット、ウサギ、サルまたは類人猿)を目的の抗原で免疫し、目的の抗原に特異的な可変領域をコードする抗体可変領域の核酸配列を同定することによって得られる。特定の実施形態において、その非ヒト動物は、1つ以上のヒト免疫グロブリン可変領域の遺伝子を含む。別の特定の実施形態において、1つ以上のヒト免疫グロブリン可変領域の遺伝子は、染色体外に、内在性の免疫グロブリン遺伝子座における置換として、または非ヒト動物のゲノムにランダムにインテグレートされたトランスジーンとして、非ヒト動物に存在する。1つの実施形態において、第1および/または第2の抗原結合領域は、ハイブリドーマまたはクアドローマから得られ、別の実施形態では、免疫された非ヒト動物の免疫細胞を、細胞選別を用いてスクリーニングすることによって得られる。
【0030】
1つの実施形態において、本抗原結合タンパク質は、二重特異性抗体である。1つの実施形態において、その二重特異性抗体は、二重特異性の完全ヒト抗体であり、各エピトープに対して独立してマイクロモル濃度、ナノモル濃度またはピコモル濃度の範囲の親和性を有する。
【0031】
1つの実施形態において、本抗原結合タンパク質は、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない。特定の実施形態において、本抗原結合タンパク質は、非天然のヒトT細胞エピトープを有しない。1つの実施形態において、CH3領域の改変は、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない。特定の実施形態において、CH3領域の改変は、非天然のヒトT細胞エピトープをもたらさない。
【0032】
1つの実施形態において、本抗原結合タンパク質は、重鎖を含み、ここで、その重鎖は、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない。1つの実施形態において、その重鎖は、非天然のT細胞エピトープを含まないアミノ酸配列を有する。1つの実施形態において、その重鎖は、タンパク質分解によってヒトにおいて免疫原性である約9アミノ酸のアミノ酸配列が形成され得ないアミノ酸配列を含む。特定の実施形態において、上記ヒトは、本抗原結合タンパク質で処置されたヒトである。1つの実施形態において、その重鎖は、タンパク質分解によってヒトにおいて免疫原性である約13〜約17アミノ酸のアミノ酸配列が形成され得ないアミノ酸配列を含む。特定の実施形態において、上記ヒトは、本抗原結合タンパク質で処置されているヒトである。
【0033】
1つの局面において、本明細書中に記載されるようなCH2改変および/またはCH3改変を含む二重特異性結合タンパク質が提供され、ここで、その二重特異性結合タンパク質は、B細胞上の抗原を特異的に認識する第1の結合部分、およびT細胞上の抗原を特異的に認識する第2の結合部分を含む。
【0034】
1つの実施形態において、結合タンパク質は、二重特異性抗体である。特定の実施形態において、その二重特異性抗体は、ヒトIgG1重鎖およびヒトIgG1ΔAdp重鎖を含む。1つの実施形態において、第1の結合部分は、CD20を特異的に認識するヒト重鎖可変ドメインである。1つの実施形態において、第2の結合部分は、CD3を特異的に認識するヒト重鎖可変ドメインである。1つの実施形態において、上記二重特異性抗体は、Raji死滅アッセイにおいて約2.8〜3.2×10−12Mまたは約2.8〜3.0×10−12MのEC50を示し、バイスタンダー細胞死滅アッセイにおいて約1〜10%または1〜5%を越えないバイスタンダー死滅を示し、ここで、そのバイスタンダー細胞は、CD20エピトープを含まない。特定の実施形態において、バイスタンダー細胞は、293細胞である。別の特定の実施形態において、上記アッセイにおけるバイスタンダー細胞死滅は、約10−8M〜約10−14Mという二重特異性抗体の濃度にわたって測定される。
【0035】
1つの局面において、二重特異性抗体を作製するための方法が提供され、その方法は:第1のエピトープを認識する第1の可変ドメインを含む第1の免疫グロブリン重鎖をコードする核酸配列を得る工程(ここで、その第1の免疫グロブリン重鎖は、IgG1、IgG2もしくはIgG4のアイソタイプ定常ドメインまたはそのキメラアイソタイプ定常ドメインを含む);第2のエピトープを認識する第2の可変ドメインを含む第2の免疫グロブリン重鎖をコードする第2の核酸配列を得る工程(ここで、その第2の免疫グロブリン重鎖は、プロテインAへの結合を無くすかまたは減少させる改変をそのCH3ドメインに含む、IgG1、IgG2もしくはIgG4のアイソタイプ定常ドメインまたはそのキメラアイソタイプ定常ドメインを含む);第1および第2の免疫グロブリン重鎖と対形成する免疫グロブリン軽鎖をコードする第3の核酸配列を得る工程;その第1、第2および第3の核酸配列を哺乳動物細胞に導入する工程;その細胞に免疫グロブリンを発現させる工程、およびプロテインAを用いてその免疫グロブリンを単離する工程を包含する。
【0036】
1つの実施形態において、上記細胞は、CHO、COS、293、HeLaおよびウイルス核酸配列を発現する網膜細胞(例えば、PERC.6(商標)細胞)から選択される。
【0037】
1つの局面において、抗原に結合する第1の特異性およびレセプターを活性化させる第2の特異性を備える二重特異性抗原結合タンパク質が提供され、ここで、その二重特異性抗原結合タンパク質は、プロテインAに結合する決定基を含む第1のIgG1、IgG2またはIgG4 CH3ドメインを含む第1のポリペプチド、およびそのプロテインAに結合する決定基を欠く第2のIgG1、IgG3またはIgG4 CH3ドメインを含む第2のポリペプチドを含む。
【0038】
1つの実施形態において、上記レセプターを活性化する第2の特異性のおかげで、上記レセプターはモル濃度、ミリモル濃度、マイクロモル濃度、ナノモル濃度またはピコモル濃度の範囲であるKDで結合する。
【0039】
1つの実施形態において、上記第2の特異性のおかげで、Gタンパク質共役レセプター、チロシンキナーゼレセプター、インテグリンレセプターおよびtoll様レセプターから選択されるレセプターに結合する。
【0040】
1つの実施形態において、上記第2の特異性によって、上記レセプターと接触し、そのレセプターまたはそれと物理的に会合するサブユニットもしくはタンパク質が、セリン、トレオニンまたはチロシンのリン酸化を起こすか;ヌクレオチド(例えば、cAMP、cADPまたはcGMP)の環化(cyclicization)を引き起こすか;ホスファチジルイノシトールまたはその誘導体(例えば、IP3またはPIP3)の産生を引き起こすか;脂質セカンドメッセンジャー(例えば、ジアシルグリセロール(diacylclycerol)、セラミド、リゾホスファチジン酸、エイコサノイド)の産生を引き起こすか;脱リン酸化(例えば、ホスファターゼ活性)を引き起こすか;脂質のリン酸化を引き起こしてセカンドメッセンジャーを形成するか;セカンドメッセンジャーの加水分解を引き起こすか;タンパク質分解を引き起こすか;レドックスシグナル伝達を引き起こすか;細胞小器官(例えば、核)へのタンパク質のトランスロケーションを引き起こすか;レセプターが(それ自体と)凝集して、ホモ多量体を形成するかまたは(他のレセプターと)ヘテロ多量体を形成するか;あるいは膜貫通型チャネルの開口または閉鎖を引き起こす。
【0041】
1つの局面において、二重特異性抗体を作製するための方法が提供され、その方法は:クアドローマから目的の二重特異性抗体を単離する工程を包含し、ここで、その目的の二重特異性抗体は、IgG1、IgG2またはIgG4のアイソタイプである第1の重鎖、プロテインAへの結合を無くすかまたは減少させる改変をそのCH3ドメインに含む定常ドメインを有するIgG1、IgG2またはIgG4のアイソタイプである第2の重鎖を含み、ここで、その目的の二重特異性抗体は、プロテインAを用いて単離される。
【0042】
1つの局面において、二重特異性抗体を作製するための方法が提供され、その方法は、破壊された細胞または抗体の混合物から、識別的に改変された(differentially modified)IgG1、IgG2またはIgG4のCH3ドメインを有する二重特異性抗体を単離する工程を包含し、ここで、その識別的に改変されたCH3ドメインは、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でなく、その改変は、プロテインAに対するモノマーの親和性が互いに異なるヘテロ二量体の重鎖を有する二重特異性抗体をもたらし、その二重特異性抗体は、プロテインAを用いて、破壊された細胞または上記混合物から単離される。
【0043】
1つの実施形態において、本二重特異性抗体は、プロテインA親和性支持体を用いて単離され、ここで、その二重特異性抗体は、約3.9〜約4.4、約4.0〜約4.3、約4.1〜約4.2のpH、またはpH約4.2で溶出する。1つの実施形態において、本二重特異性抗体は、約4、4.1、4.2、4.3、4.4または4.5のpHで溶出する。
【0044】
1つの実施形態において、本二重特異性抗体は、プロテインA親和性支持体およびpH勾配またはpH段階(pH step)を用いて単離され、ここで、そのpH勾配またはpH段階は、イオン調節物質(ionic modifier)を含む。特定の実施形態において、イオン調節物質は、約0.5〜約1.0モル濃度の濃度で存在する。特定の実施形態において、イオン調節物質は、塩である。1つの実施形態において、イオン調節物質は、酢酸のベリリウム塩、リチウム塩、ナトリウム塩およびカリウム塩;重炭酸ナトリウムおよび重炭酸カリウム;炭酸リチウム、炭酸ナトリウム、炭酸カリウムおよび炭酸セシウム;塩化リチウム、塩化ナトリウム、塩化カリウム、塩化セシウムおよび塩化マグネシウム;フッ化ナトリウムおよびフッ化カリウム;硝酸ナトリウム、硝酸カリウムおよび硝酸カルシウム;リン酸ナトリウムおよびリン酸カリウム;ならびに硫酸カルシウムおよび硫酸マグネシウムからなる群から選択される。特定の実施形態において、イオン調節物質は、アルカリ金属またはアルカリ土類金属のハロゲン化物塩である。特定の実施形態において、イオン調節物質は、塩化ナトリウムである。
【0045】
1つの局面において、Fcを含む結合タンパク質(ここで、そのFcは、ヘテロ二量体のFcを形成するように、本明細書中に記載されるように改変された第1のCH3ドメインおよび改変されていない第2のCH3を含む)、ここで、その識別的改変は、識別的改変を有しない対応する結合タンパク質よりも0.5、0.6、0.7、0.8、0.9、1.0、1.2、1.3または1.4高いpH単位で、プロテインA親和性材料から溶出する結合タンパク質をもたらす。
【0046】
1つの実施形態において、識別的に改変された結合タンパク質は、約4.2のpHで溶出するのに対し、改変されていない結合タンパク質は、約3のpHで溶出する。1つの実施形態において、識別的に改変された結合タンパク質は、約4.5のpHで溶出するのに対し、改変されていない結合タンパク質は、約3.5のpHで溶出する。1つの実施形態において、識別的に改変された結合タンパク質は、約4のpHで溶出するのに対し、改変されていない結合タンパク質は、約2.8〜3.5、2.8〜3.2または2.8〜3のpHで溶出する。1つの実施形態において、識別的に改変された結合タンパク質は、約4.2のpHで溶出するのに対し、改変されていない結合タンパク質は、約2.8のpHで溶出する。1つの実施形態において、識別的に改変された結合タンパク質は、約4.4のpHで溶出するのに対し、改変されていない結合タンパク質は、約3.6のpHで溶出する。これらの実施形態において、「改変されていない」とは、両方のCH3ドメインにおいて、435(EUナンバリング)に改変を有しないこと、または435および436(EUナンバリング)に改変を有しないことを指す。
【0047】
本明細書中に記載される任意の実施形態および局面は、別段示されないかまたは文脈から明らかでない限り、互いと同時に使用され得る。次の記載を検討することにより、他の実施形態が当業者に明らかになるだろう。
【図面の簡単な説明】
【0048】
【図1】図1は、3つの二重特異性抗体形式:(A)二重特異性T細胞Engager(BiTE);(B)マウス−ラットハイブリッド;および(C)通常の軽鎖を有するノブ・イントゥ・ホールを図示している。
【図2】図2は、FcΔAdp改変を図示している:(A)ヒトIgG1(配列番号1)およびIgG3(配列番号3)のFc領域のアラインメント(FcΔAdp改変を四角で示している);(B)FcΔAdp二重特異性抗体の模式的表示。
【図2B】図2は、FcΔAdp改変を図示している:(A)ヒトIgG1(配列番号1)およびIgG3(配列番号3)のFc領域のアラインメント(FcΔAdp改変を四角で示している);(B)FcΔAdp二重特異性抗体の模式的表示。
【図3】図3は、ΔAdpジペプチド改変有りおよび無しのIgG1、IgG2およびIgG4のヒトCH3ドメイン(IMGTエキソンナンバリングおよびEUナンバリングを使用)、ならびにIgG3のアラインメントを図示している。
【図4】図4は、段階勾配を利用した溶出プロファイルを示している、二重特異性抗体を単離するためのプロテインAカラムのトレースを示している。
【図5】図5は、図4に示されるクロマトグラフ分離から溶出されたカラム画分のIL4−RaおよびIL−6Ra BIACORE(商標)結合プロファイルを示している。画分中の抗体を、固定化された抗Fc抗体で捕捉し、次いで、捕捉された抗体への結合について、可溶性IL−4RaまたはIL−6Raをアッセイした。
【図6】図6は、FcΔAdp二重特異性抗体(IL−6RΔ/IL−4R)、FcΔAdpホモ二量体(IL−6RΔ/IL−6RΔ)、野生型CH3配列を有するIgG1抗体(IL−4R/IL−4R)およびコントロール単一特異的抗体の薬物動態学的プロファイルを示している。
【図7】図7は、Raji細胞死滅アッセイにおけるCD20×CD3ΔAdp二重特異性抗体の有効性を図示している。
【図8】図8は、CD20×CD3ΔAdp二重特異性抗体を用いたバイスタンダー(bystander)細胞(293)死滅アッセイを図示している。
【図9】図9は、種々のmFcヘテロ二量体を使用した発現実験についての結果を示している。パネルA:ホモ二量体のmIgG2aおよびホモ二量体のIgG2aPTTTKからのヘテロ二量体のmIgG2a/mIgG2aPTTTKのpH分離のウエスタンブロット;パネルB:ホモ二量体のmIgG2aおよびホモ二量体のIgG2aTTTKからのヘテロ二量体のmIgG2a/mIgG2aTTTKのpH分離のウエスタンブロット;IP=投入物;FT=フロースルー;W2=2回目の洗浄物(1×PBS pH7.2);E1=1回目の溶出物(20mMクエン酸Na、1M NaCl pH5.5);E2=2回目の溶出物(20mMクエン酸Na、1M NaCl;57%pH5.5+43%pH2.6);E3=3回目の溶出物(20mMクエン酸Na、1M NaCl pH2.6)。
【図10】図10は、4:1というIFNAR1構築物:IFNAR2構築物の比を用いたときの、アイソタイプの混合物(例えば、mIgG2aおよびmIgG1)のヘテロ二量体の形態に対する変異体IgG2aのヘテロ二量体の優先的な形態を図示している。レーン1:IFNAR1−IgG2a:IFNAR2−IgG1;レーン2:INFAR1−IgG2a:IFNAR2−IgG2aTTT;レーン3:IFNAR1−IgG2a:IFNAR2−IgG2aTTTK;レーン4:IFNAR1−IgG2a:IFNAR2−IgG2aPTTTK;レーン5:IFNAR1−IgG2a:IFNAR2−IgG2aRF。
【発明を実施するための形態】
【0049】
詳細な説明
特定の方法および記載される実験条件は変動し得るので、本発明は、そのような方法および条件に限定されない。本発明の範囲は、請求項によって定義されるので、本明細書中で使用される用語は、特定の実施形態だけを記載する目的であって、限定する意図はないことも理解される。
【0050】
別段定義されない限り、本明細書中で使用される専門用語および科学用語のすべては、本発明が属する分野の当業者が通常理解している意味と同じ意味を有する。本明細書中に記載される方法および材料と類似または等価な任意の方法および材料を、本発明の実施または試験において使用することができるが、ここでは特定の方法および材料が記載される。言及されるすべての刊行物が、本明細書に参照によって援用される。
【0051】
用語「抗体」は、本明細書中で使用されるとき、ジスルフィド結合によって相互に接続された4つのポリペプチド鎖、すなわち2つの重(H)鎖および2つの軽(L)鎖を含む免疫グロブリン分子を含む。各重鎖は、重鎖可変領域(本明細書中でHCVRまたはVHと省略される)および重鎖定常領域を含む。重鎖定常領域は、3つのドメインCH1、CH2およびCH3を含む。各軽鎖は、軽鎖可変領域(本明細書中でLCVRまたはVLと省略される)および軽鎖定常領域を含む。軽鎖定常領域は、1つのドメインCLを含む。VHおよびVL領域は、フレームワーク領域(FR)と呼ばれるより保存された領域とともに点在する、相補性決定領域(CDR)と呼ばれる超可変性の領域にさらに細分され得る。VHおよびVLの各々は、アミノ末端からカルボキシ末端に向かって以下の順序:FR1、CDR1、FR2、CDR2、FR3、CDR3、FR4で配置される3つのCDRおよび4つのFRから構成される(重鎖CDRは、HCDR1、HCDR2およびHCDR3と省略され得る;軽鎖CDRは、LCDR1、LCDR2およびLCDR3と省略され得る)。用語「高親和性」抗体とは、表面プラズモン共鳴、例えば、BIACORE(商標)または溶液親和性(solution−affinity)ELISAによって測定されたとき、その標的に対して少なくとも10−9M、少なくとも10−10M;少なくとも10−11M;または少なくとも10−12Mの結合親和性を有する抗体のことを指す。
【0052】
句「抗原結合タンパク質」は、少なくとも1つのCDRを有し、かつ抗原を選択的に認識することができる、すなわち、少なくともマイクロモル濃度の範囲のKDで抗原に結合することができるタンパク質を含む。治療用の抗原結合タンパク質(例えば、治療用抗体)は、ナノモル濃度またはピコモル濃度の範囲のKDを必要とすることが多い。「抗原結合タンパク質」はまた、本明細書中に記載されるような第1および第2のCH3ドメイン、ならびに第1のタンパク質認識ドメインまたはリガンド認識ドメイン、および第2のタンパク質認識ドメインまたはリガンド認識ドメインを含むタンパク質を含み、ここで、その第1のタンパク質認識ドメインまたはリガンド認識ドメインおよび第2のタンパク質認識ドメインまたはリガンド認識ドメインの各々は、独立して、同じタンパク質もしくはリガンドを認識するか、または一緒に同じタンパク質もしくはリガンドを認識するか、あるいは各々独立して、異なるタンパク質またはリガンドを認識する。そのようなタンパク質の1つの例は、融合タンパク質(ヘテロ−またはホモ−)二量体を含むイムノアドヘシンであり、ここで、その二量体のポリペプチドは、レセプター構成要素またはリガンド構成要素を含む融合ポリペプチドであり、そのリガンド構成要素は、レセプターに結合するアミノ酸配列を含む。
【0053】
句「二重特異性抗体」は、2つ以上のエピトープに選択的に結合することができる抗体を含む。二重特異性抗体は、一般に、2つの異なる重鎖を含み、各重鎖は、異なるエピトープ(2つの異なる分子(例えば、抗原)上のエピトープまたは同じ分子(例えば、同じ抗原)上のエピトープ)に特異的に結合する。二重特異性抗体が、2つの異なるエピトープ(第1のエピトープおよび第2のエピトープ)に選択的に結合することができる場合、第1のエピトープに対する第1の重鎖の親和性は、一般に、第2のエピトープに対する第1の重鎖の親和性よりも少なくとも1桁から2桁または3桁または4桁低く、逆もまた同じである。二重特異性抗体によって認識されるエピトープは、同じ標的上または異なる標的上(例えば、同じまたは異なるタンパク質上)に存在し得る。二重特異性抗体は、例えば、同じ抗原の異なるエピトープを認識する重鎖を組み合わせることによって作製され得る。例えば、同じ抗原の異なるエピトープを認識する重鎖可変配列をコードする核酸配列が、種々の重鎖定常領域をコードする核酸配列に融合され得、そのような配列が、免疫グロブリン軽鎖を発現する細胞において発現され得る。代表的な二重特異性抗体は、2つの重鎖(各々が、3つの重鎖CDR、それに続いて(N末端からC末端に向かって)CH1ドメイン、ヒンジ、CH2ドメインおよびCH3ドメインを有する)、および抗原結合特異性を与えないが各重鎖と会合し得る免疫グロブリン軽鎖、または各重鎖と会合し得、かつ重鎖抗原結合領域によって結合される1つ以上のエピトープに結合し得る免疫グロブリン軽鎖、または各重鎖と会合し得、かつ一方または両方のエピトープへの一方または両方の重鎖の(or)結合を可能にする免疫グロブリン軽鎖を有する。
【0054】
用語「細胞」は、組換え核酸配列を発現するのに適した任意の細胞を含む。細胞には、原核生物および真核生物の細胞(単細胞または多細胞)、細菌細胞(例えば、大腸菌、Bacillus spp.、Streptomyces spp.などの系統)、マイコバクテリア細胞、真菌細胞、酵母細胞(例えば、S.cerevisiae、S.pombe、P.pastoris、P.methanolicaなど)、植物細胞、昆虫細胞(例えば、SF−9、SF−21、バキュロウイルス感染昆虫細胞、Trichoplusia niなど)、非ヒト動物細胞、ヒト細胞または細胞融合物(例えば、ハイブリドーマまたはクアドローマ)が含まれる。いくつかの実施形態において、細胞は、ヒト、サル、類人猿、ハムスター、ラットまたはマウス細胞である。いくつかの実施形態において、細胞は、真核生物細胞であり、以下の細胞:CHO(例えば、CHO K1、DXB−11CHO、Veggie−CHO)、COS(例えば、COS−7)、網膜細胞、Vero、CV1、腎臓(例えば、HEK293、293EBNA、MSR293、MDCK、HaK、BHK)、HeLa、HepG2、WI38、MRC5、Colo205、HB8065、HL−60(例えば、BHK21)、Jurkat、Daudi、A431(表皮)、CV−1、U937、3T3、L細胞、C127細胞、SP2/0、NS−0、MMT060562、セルトリ細胞、BRL3A細胞、HT1080細胞、ミエローマ細胞、腫瘍細胞、および上述の細胞に由来する細胞株から選択される。いくつかの実施形態において、細胞は、1つ以上のウイルス遺伝子を含む(例えば、ウイルス遺伝子を発現する網膜細胞(例えば、PER.C6(商標)細胞))。
【0055】
句「相補性決定領域」または用語「CDR」は、通常(すなわち、野生型動物において)免疫グロブリン分子(例えば、抗体またはT細胞レセプター)の軽鎖または重鎖の可変領域内の2つのフレームワーク領域の間に見られる、生物の免疫グロブリン遺伝子の核酸配列によってコードされるアミノ酸配列を含む。CDRは、例えば、生殖細胞系列の配列または再配列された配列もしくは再配列されていない配列によって、例えば、ナイーブB細胞もしくは成熟B細胞またはナイーブT細胞もしくは成熟T細胞によって、コードされ得る。いくつかの状況において(例えば、CDR3の場合)、CDRは、連続していない(例えば、再配列されていない核酸配列では連続していない)が、例えば、それらの配列のスプライシングまたは接続(例えば、重鎖CDR3を形成するV−D−J組換え)の結果としてB細胞の核酸配列では連続している、2つ以上の配列(例えば、生殖細胞系列の配列)によってコードされ得る。
【0056】
句「重鎖」または「免疫グロブリン重鎖」は、任意の生物由来の免疫グロブリン重鎖定常領域配列を含み、別段明記されない限り、重鎖可変ドメインを含む。重鎖可変ドメインは、別段明記されない限り、3つの重鎖CDRおよび4つのFR領域を含む。重鎖のフラグメントとしては、CDR、CDRおよびFR、ならびにそれらの組み合わせが挙げられる。代表的な重鎖は、可変ドメインの後、(N末端からC末端に向かって)CH1ドメイン、ヒンジ、CH2ドメインおよびCH3ドメインを有する。重鎖の機能的なフラグメントには、抗原を特異的に認識することができ(例えば、マイクロモル濃度、ナノモル濃度またはピコモル濃度の範囲のKDで抗原を認識することができ)、発現して細胞から分泌されることができ、かつ、少なくとも1つのCDRを含む、フラグメントが含まれる。
【0057】
句「Fc含有タンパク質」は、抗体、二重特異性抗体、イムノアドヘシン、ならびに免疫グロブリンCH2およびCH3領域の少なくとも機能的な一部を含む他の結合タンパク質を含む。「機能的な一部」とは、Fcレセプター(例えば、FcγR;またはFcRn、すなわち、新生児Fcレセプター)に結合し得、そして/または補体の活性化に関与し得る、CH2およびCH3領域のことを指す。そのCH2およびCH3領域が、いずれのFcレセプターにも結合できなくし、また、補体を活性化できなくする、欠失、置換および/もしくは挿入または他の改変を含む場合、そのCH2およびCH3領域は、機能的でない。
【0058】
Fc含有タンパク質は、改変が結合タンパク質の1つ以上のエフェクター機能に影響する場合(例えば、FcγR結合性、FcRn結合性ひいては半減期、および/またはCDC活性に影響する改変)を含む改変を免疫グロブリンドメインに含み得る。そのような改変としては、免疫グロブリン定常領域のEUナンバリングに準拠した以下の改変およびそれらの組み合わせ:238、239、248、249、250、252、254、255、256、258、265、267、268、269、270、272、276、278、280、283、285、286、289、290、292、293、294、295、296、297、298、301、303、305、307、308、309、311、312、315、318、320、322、324、326、327、328、329、330、331、332、333、334、335、337、338、339、340、342、344、356、358、359、360、361、362、373、375、376、378、380、382、383、384、386、388、389、398、414、416、419、428、430、433、434、435、437、438および439が挙げられるが、これらに限定されない。
【0059】
例えば、限定するわけではないが、結合タンパク質は、Fc含有タンパク質であり、長い血清半減期(列挙される改変を有しない同じFc含有タンパク質と比べて)を示し、250位(例えば、EまたはQ);250位および428位(例えば、LまたはF);252位(例えば、L/Y/F/WまたはT)、254位(例えば、SまたはT)および256位(例えば、S/R/Q/E/DまたはT)に改変を有するか;または428位および/もしくは433位(例えば、L/R/SI/P/QまたはK)および/もしくは434位(例えば、H/FまたはY)に改変を有するか;または250位および/もしくは428位に改変を有するか;または307位もしくは308位(例えば、308F、V308F)および434位に改変を有する。別の例では、改変は、428L(例えば、M428L)および434S(例えば、N434S)改変;428L、259I(例えば、V259I)および308F(例えば、V308F)改変;433K(例えば、H433K)および434(例えば、434Y)改変;252、254および256(例えば、252Y、254Tおよび256E)改変;250Qおよび428L改変(例えば、T250QおよびM428L);307および/または308改変(例えば、308Fまたは308P)を含み得る。
【0060】
句「イオン調節物質」は、タンパク質間の非特異的な(すなわち、非親和性の)イオン性相互作用の影響を減少させるかまたはその影響を乱す部分を含む。「イオン調節物質」としては、例えば、塩、酢酸、重炭酸、炭酸、ハロゲン(例えば、塩化物またはフッ化物)、硝酸、リン酸または硫酸とI族およびII族金属とのイオン性の組み合わせ物が挙げられる。「イオン調節物質」の非限定的な例示的リストとしては、酢酸のベリリウム塩、リチウム塩、ナトリウム塩およびカリウム塩;重炭酸ナトリウムおよび重炭酸カリウム;炭酸リチウム、炭酸ナトリウム、炭酸カリウムおよび炭酸セシウム;塩化リチウム、塩化ナトリウム、塩化カリウム、塩化セシウムおよび塩化マグネシウム;フッ化ナトリウムおよびフッ化カリウム;硝酸ナトリウム、硝酸カリウムおよび硝酸カルシウム;リン酸ナトリウムおよびリン酸カリウム;ならびに硫酸カルシウムおよび硫酸マグネシウムが挙げられる。「イオン調節物質」は、pH勾配もしくはpH段階に加えられたとき、または「イオン調節物質」でのプロテインA支持体の平衡化のとき、イオン性の相互作用に影響する部分を含み、pH段階またはpH勾配を適用することにより、ホモ二量体のIgGの溶出とヘテロ二量体のIgGの溶出との間(例えば、野生型ヒトIgGの溶出と、本明細書中に記載されるようなそのCH3ドメインの1つ以上の改変を有すること以外は同じIgGの溶出との間)のpH単位の間隔が広がる。「イオン調節物質」の適当な濃度は、最大のpHの間隔が所与のpH段階またはpH勾配に達するまで「イオン調節物質」の濃度を上げながら、同じカラム、pH段階またはpH勾配を使用して、その濃度によって決定され得る。
【0061】
句「軽鎖」は、任意の生物由来の免疫グロブリン軽鎖定常領域配列を含み、別段明記されない限り、ヒトカッパー軽鎖およびヒトラムダ軽鎖を含む。軽鎖可変(VL)ドメインは、別段明記されない限り、代表的には、3つの軽鎖CDRおよび4つのフレームワーク(FR)領域を含む。一般に、全長軽鎖は、アミノ末端からカルボキシル末端に向かって、FR1−CDR1−FR2−CDR2−FR3−CDR3−FR4を含むVLドメイン、および軽鎖定常ドメインを含む。本発明と共に使用され得る軽鎖は、例えば、本抗原結合タンパク質によって選択的に結合される第1または第2の抗原に選択的に結合しない軽鎖を含む。適当な軽鎖には、既存の抗体ライブラリー(ウェットライブラリーまたはインシリコ)において最もよく使用される軽鎖についてスクリーニングすることによって特定され得る軽鎖が含まれ、ここで、その軽鎖は、本抗原結合タンパク質の抗原結合ドメインの親和性および/または選択性を実質的に干渉しない。適当な軽鎖には、本抗原結合タンパク質の抗原結合領域によって結合される一方または両方のエピトープに結合し得る軽鎖が含まれる。
【0062】
句「マイクロモル濃度範囲」は、1〜999マイクロモル濃度を意味することを意図している;句「ナノモル濃度範囲」は、1〜999ナノモル濃度を意味することを意図している;句「ピコモル濃度範囲」は、1〜999ピコモル濃度を意味することを意図している。
【0063】
句「体細胞変異した」は、クラススイッチを起こしたB細胞由来の核酸配列に対する言及を含み、ここで、クラススイッチしたB細胞における免疫グロブリン可変領域(例えば、重鎖可変ドメインまたは重鎖CDR配列もしくは重鎖FR配列を含む領域)の核酸配列は、クラススイッチ前のB細胞における核酸配列と同一ではない(例えば、クラススイッチを起こしていないB細胞とクラススイッチを起こしたB細胞との間でCDR核酸配列またはフレームワーク核酸配列が異なる)。「体細胞変異した」は、親和性成熟していないB細胞における対応する配列(すなわち、生殖細胞系列の細胞のゲノムにおける配列)と同一でない親和性成熟したB細胞由来の核酸配列に対する言及を含む。句「体細胞変異した」は、B細胞を目的の抗原に曝露した後のB細胞由来の核酸配列に対する言及も含み、ここで、その核酸配列は、そのB細胞をその目的の抗原に曝露する前の対応する核酸配列と異なる。句「体細胞変異した」とは、抗原チャレンジに応答したヒト免疫グロブリン可変領域の核酸配列を有する動物、例えばマウスにおいて産生され、そのような動物において本質的に有効な選択プロセスから生じる、抗体由来の配列のことを指す。
【0064】
句「可変ドメイン」は、(別段示されない限り)以下のアミノ酸領域を順にN末端からC末端に向かって含む免疫グロブリン軽鎖または重鎖(所望のとおり改変されたもの)のアミノ酸配列を含む:FR1、CDR1、FR2、CDR2、FR3、CDR3、FR4。「可変ドメイン」は、二重のベータシート構造を有する基準のドメイン(VHまたはVL)に折り畳むことができるアミノ酸配列を含み、ここで、そのベータシートは、第1のベータシートの残基と第2のベータシートの残基との間のジスルフィド結合によって接続されている。
【0065】
改変されたIgG CH3領域を有する二重特異性抗体
本発明者らは、通常の軽鎖ストラテジーを、ヒト抗体の構成要素とともに使用され得る選択的なプロテインA精製スキームの実行と組み合わせる新規形式を開発した。
【0066】
ヒトIgG3は、プロテインAに結合しないので、マウス−ラットハイブリッドに対して使用される精製ストラテジーと類似の精製ストラテジーにおいて他の3つのヒトIgGサブクラスのうちのいずれかと一緒に使用することができるという可能性があることは、以前から有名だった(Lindhofer,H.ら(1995)J.Immunol.155:219−225))。しかしながら、4つすべてのヒトIgGサブクラスの配列が、高度に相同であるにもかかわらず、IgG1、IgG2およびIgG4のFc部が、どれだけ容易にIgG3とヘテロ二量体を形成するかは知られていない;ある特定の状況下では(例えば、クアドローマからの単離では)、ホモ二量体の優先的な形成だけこそが所望のヘテロ二量体の総収量に対して悪影響を及ぼし得る。IgG3のヒンジ領域と他のサブクラスのヒンジ領域との差異を補うために追加の改変が必要となることもある。エフェクター機能に対する潜在的な影響があるので、いくつかの状況では、完全なIgG3 Fcの存在を必要としないことが好ましい場合もある。
【0067】
それゆえ、本発明者らは、プロテインA結合の偶然にも単純な決定基を活用する「最小」形式を考案した。IgG3がプロテインAに結合できないことは単一のアミノ酸残基Arg435(EUナンバリング;IMGTではArg95)(他のIgGサブクラスでは、対応する位置はヒスチジン残基で占められている)によって判定されることが報告されている(Jendeberg,L.ら(1997)J.Immunological Meth.201:25−34))。それゆえ、IgG3の代わりに、His435がArgに変異したIgG1配列を使用することが可能である。よって、IgG1における単一の点変異が、新しい精製スキームに適用できる異なる結合親和性をもたらすのに十分であるはずである。この改変は、プロテインAに結合することができないことを表すIgG1ΔAと呼ばれる(そして同様に、IgG2ΔAおよびIgG4ΔA、またはより一般的にはFcΔA)。
【0068】
しかしながら、明記される点変異が、その変異によって新規ペプチド配列を導入することから、それは、潜在的に免疫原性であり得る。この点変異は、理論上、MHCクラスII分子上にのせられて、T細胞に提示され、その結果として、免疫応答を誘発し得る。この不測の困難を回避するために、ジペプチド変異H435R/Y436F(EUナンバリング;IMGTではH95R/Y96F)が使用され得る。その結果生じるこの変更の近傍における配列は、IgG3の配列と同一となり(図2Aを参照のこと)、ゆえに、T細胞への提示に利用可能な非天然の短いペプチドが存在しないだろうことから、免疫学的に「隠されている」と予想され得る。この二重変異体がなおもプロテインAに結合しないことが報告されている(Jendeberg,L.ら(1997)J.Immunological Meth.201:25−34)。最後に、そのジペプチド変異は、Fc二量体の界面を形成するいずれの残基も含まないので、ヘテロ二量体の形成が干渉される可能性は低い。このジペプチド変異は、「IgG1ΔAdp」(そして同様にIgG2ΔAdp、IgG4ΔAdpおよびFcΔAdp)と命名される。IgG1、IgG2およびIgG4におけるジペプチド改変の配置は、IMGTエキソンナンバリングおよびEUナンバリングを示しつつ、野生型ヒトIgG CH3ドメイン配列、ならびにhIgG3と共に示されている、図3のIgG1ΔAdp、IgG2ΔAdpおよびIgG4ΔAdpと表されている配列に示されている。
【0069】
FcΔAdp改変は、Fc二量体の界面を形成すると考えられているいずれの残基も含まないので、そのFcΔAdp改変がヘテロ二量体の形成を干渉する可能性は低い。FcΔAdpは、そのように最小であるので、同様に、他の操作されたFc形態に組み込まれる可能性もあり得る。IgG2ΔAdpおよびIgG4ΔAdpは、後者の各々に関連するエフェクター機能(またはその欠損)が望まれる状況において有益であり得る。
【0070】
要約すれば、上に記載された二重特異性抗体形式は、同じ軽鎖を使用する特異性の異なる2つの抗体を含み、ここで、それらのうちの1つのFc領域は、FcΔAdp形式に改変されている(図2Bを参照のこと)。その配置は、天然のヒト抗体の配置であり、ゆえに、凝集する傾向が低いこと、インビボ安定性、最小の免疫原性、抗体の体内分布特性に類似した体内分布特性、良好な薬物動態、および必要に応じてエフェクター機能をはじめとした、その好ましい特性を共有するはずである。そのような二重特異性抗体を単離するための、実行することが比較的迅速かつ単純な方法が提供される。
【0071】
改変されたマウスIgG CH領域を有する二重特異性結合タンパク質
本発明者らは、CH3ドメインにおける1つ以上のアミノ酸に関してヘテロ二量体である免疫グロブリン重鎖(またはその機能的なCH2およびCH3含有フラグメント)を含む結合タンパク質を容易に単離するための方法を考案した。マウスIgG CHドメインの改変を慎重に選択し、特定の分離技術を適用することにより、それらの改変を含まないホモ二量体およびヘテロ二量体から2つの識別的に改変されたマウスCH領域を含む結合タンパク質を容易に単離することができるようになる。
【0072】
247にプロリン、252、254および256にトレオニン、ならびに258にリジンを含むマウスIgG1は、プロテインAと弱くしか結合しない。しかしながら、マウスIgG2aおよびIgG2bは、それらの位置に異なる残基を含み(IgG2bの256位および258位を除いて)、マウスIgG2aおよび2bは、プロテインAに十分に結合する。重鎖がヘテロ二量体である抗体を作製するための方法において2つのマウスIgGのCH領域を識別的に改変することにより、そのような抗体に識別的なプロテインA結合特性が付与され得る。このように、IgG1のホモ二量体(プロテインAには、たとえ結合するとしてもかなり弱くしか結合しない)であるか、またはマウスIgG2aのホモ二量体であるか、マウスIgG2bのホモ二量体であるか、またはIgG2a/IgG2bのヘテロ二量体であるかに関係なく、任意のマウスIgGホモ二量体から、改変されたヘテロ二量体を容易に(ready)分離することができる識別的なプロテインA単離スキームが考案される。例えば、2つの異なる重鎖可変ドメインを有するが、同じアイソタイプ、例えば、IgG2aの二重特異性抗体は、その重鎖配列を使用する適当な発現系において発現され得、ここで、IgG2a CH領域のうちのただ1つが、プロテインA結合決定基を減少させるかまたは排除するように改変される。このように、IgG2a CH領域のうちの1つだけが、プロテインAに対する実質的な親和性を示し、改変されていないIgG2aと改変されたIgG2aとの二量体から形成される任意の抗体は、改変されたヘテロ二量体から容易に単離され得る。
【0073】
様々な実施形態において、Fc二量体の単一のCH領域が、改変されたCH領域を含むのに対し、Fc二量体の他方のCHは、それらを欠いている、抗体。そのマウスIgG CH領域は:252T、254Tおよび256T;252T、254T、256Tおよび258K;247P、252T、254T、256Tおよび258K;435Rおよび436F;252T、254T、256T、435Rおよび436F;252T、254T、256T、258K、435Rおよび436F;24tP、252T、254T、256T、258K、435Rおよび436F;ならびに435Rからなる群から選択される特定の位置(EUナンバリング)に特定のアミノ酸を含むように改変される。特定の実施形態において、M252T、S254T、S256T;M252T、S254T、S256T、I258K;I247P、M252T、S254T、S256T、I258K;H435R、H436F;M252T、S254T、S256T、H435R、H436F;M252T、S254T、S256T、I258K、H435R、H436F;I247P、M252T、S254T、S256T、I258K、H435R、H436F;およびH435Rからなる群から選択される特定の基の改変が行われる。
【0074】
ヘテロ二量体のマウスIgGに基づく結合タンパク質は、種々の用途のために使用され得る。例えば、それらは、マウス定常ドメインを有する二重特異性抗体を単離する方法を可能にし、ここで、その改変は、1つ以上のマウスFcレセプターへのその抗体の結合を干渉しないか、または実質的に干渉せず、その抗体は、例えば、ADCCまたはCDCに関与し得、また、同じまたは異なる標的上の2つ以上のエピトープに結合し得る。
【0075】
1つの局面において、第1のマウスIgG CH領域および第2のマウスIgG CH領域を含む結合タンパク質を単離するための方法、ここで、その第1のIgG CH領域は、第1のマウスIgG CH領域のプロテインA結合親和性を減少させるかまたは排除するが第2のマウスIgG CH領域のプロテインA結合親和性を減少させないかまたは排除しないように改変され(第2のIgG CH領域は改変されない)、その結合タンパク質は、第1のエピトープに結合する第1の結合部分および第2のエピトープに結合する第2の結合部分を含む。
【0076】
1つの実施形態において、改変は、Fcレセプターへの結合タンパク質の結合親和性を変更しないかまたは実質的に変更しない。1つの実施形態において、本結合タンパク質は、本結合タンパク質のFcレセプターへの親和性を増加させるかまたは減少させる改変を含む。
【0077】
1つの実施形態において、改変は、その改変を有しない対応する結合タンパク質と比べて、天然のマウスFcγRレセプターおよび/または天然のマウスFcRnを含むマウスにおける結合タンパク質の血清半減期を変更しないかまたは実質的に変更しない。
【0078】
1つの実施形態において、改変は、その改変を有しない対応する結合タンパク質と比べて、天然の高親和性および低親和性のマウスFcγRレセプターならびに/またはFcRnレセプターの置き換えを含むマウスにおける本結合タンパク質の血清半減期を変更しないかまたは実質的に変更しない。
【0079】
1つの実施形態において、第1および第2のエピトープは、異なるものであり、異なる細胞上または異なるタンパク質上に存在する。1つの実施形態において、第1のエピトープおよび第2のエピトープは、異なるものであり、同じ細胞上または同じタンパク質上に存在する。
【0080】
1つの実施形態において、Fcレセプターは、高親和性Fcレセプター、低親和性FcレセプターおよびFcRnから選択される。特定の実施形態において、Fcレセプターは、マウスFcRn、マウスFcγR、マウスFcγRIIB、マウスFcγRIII、マウスFcγRIVおよびそれらの組み合わせのうちの1つ以上から選択される。特定の実施形態において、Fcレセプターは、ヒトFcRn、ヒトFcγR、ヒトFcγRIIB、ヒトFcγRIIC、ヒトFcγRIIIB、ヒトFcγRIIIA、ヒトFcγRIIAおよびそれらの組み合わせのうちの1つ以上から(form)選択される。
【0081】
免疫原性
本発明の多くの実施形態の利点の1つは、プロテインAに対する結合性の違いに基づいて容易に単離可能であり、かつヒトにおいて免疫原性でないかまたは実質的に免疫原性でない、二重特異性抗体をもたらす改変を使用することができることである。この特性のおかげで、そのような実施形態は、ヒトへの治療に使用するための二重特異性抗体を作製する際、および例えば、免疫原性でないかまたは実質的に免疫原性でないイムノアドヘシンを作製する際(ヒト結合部分、すなわち、ヒトレセプター構成要素および/またはヒトリガンドを使用するとき)に、特に有用になる。この特性は、IgG1、IgG2およびIgG3のH95R/Y96F(IMGTナンバリング)改変を伴うCH3ドメイン、ならびに異なるIgGアイソタイプの野生型配列を反映して改変された位置をもたらすさらなる改変を含むCH3ドメインを有する二重特異性抗体に関連する。よって、特定のIgGアイソタイプに関連する改変は、天然には見られないが、改変された配列は、異なるIgGアイソタイプの野生型配列と局所的に同一であり、その改変は、免疫原性であるかまたは実質的に免疫原性であると予想されない。配列が、任意の天然の配列と局所的に同一でなかったとしても、その改変は、免疫原性でない可能性もあり;そのような改変は、等しく有用であり得る。ゆえに、最小の点変異H95R(IMGTナンバリング)は、免疫原性でない場合、本発明の適当な実施形態であり得る。
【0082】
よって、重鎖定常ドメインに関してヒトにおいて免疫原性でないかまたは実質的に免疫原性でないが、親和性試薬(例えば、プロテインA)に対する重鎖定常ドメインの親和性に違いをもたらす改変を含む重鎖定常ドメインの1つ以上の識別的改変を有する、二重特異性抗体が提供される。その改変は、本明細書中に開示される改変を含む。特定の実施形態において、CH3ドメインに関してヒトにおいて免疫原性でないかまたは実質的に免疫原性でないが、重鎖が識別的に改変された二重特異性抗体は、以下の改変のうちの1つを含む(または、別の実施形態では、以下の改変のうちの1つから本質的になる)CH3ドメインを含むヒトIgG1、IgG2またはIgG4である:H95RまたはH95RおよびY96F(IMGTナンバリング)。
【0083】
本二重特異性抗体は、ヒトのIgG1、IgG2およびIgG4のアイソフォームに対する許容度が任意の著しい程度にまで壊されていないヒトに対して免疫原性でないかまたは実質的に免疫原性でないと予想される。
【0084】
具体的には、MHCクラスII分子の結合溝(binding groove)は、T細胞レセプターの可変ループによって認識される主要な決定基を含む9merを収容するものであり、ペプチドの長さが9merに届かない任意の天然の配列である場合、免疫応答が誘発される可能性は低いとみられるので、FcΔAdp改変は、免疫学的に「隠されている」と予想される。しかしながら、9merより長いペプチド(通常、約13〜17mer)は、MHCクラスIIによって結合され、突出したセグメントが潜在的に結合に影響し得る可能性がある。ゆえに、より長い非天然の配列を排除する追加の改変(FcΔAdp改変に対して追加の改変)が、免疫原性に対する可能性をさらに低下させ得る。1つの特定の例は、最小の非天然のペプチドの長さを、IgG1ΔAdpでは14残基から39残基に伸ばし、および同じように定義されるIgG2ΔAdpでは14残基から43残基に伸ばす、V422I(EU;IMGTナンバリングではV82I)の改変である。別の例は、IgG4ΔAdpにおいて10残基から14残基に上記長さを伸ばすL445P(EU;IMGTナンバリングではL105P)の改変である。
【0085】
薬物動態
プロテインAに対する結合部位は、免疫グロブリンに長い血清半減期を与えるのに関与すると考えられている新生児FcレセプターFcRNに対する結合部位と重複する。ゆえに、ヒトIgG3が、他のIgGサブクラス(約21日)よりも短い血清半減期を有する(約7日)ことを考えると、プロテインA結合部位の近傍の改変は、ここで提案される形式がIgG1、2および4よりも短い血清半減期を有し得る可能性を高める。His435に影響するいくつかのFc変異体は、FcRNに結合しないことおよびマウスにおいてより短い半減期を有することが示されている。しかしながら、薬物動態学的解析から、IgG1ΔA/IgG1ヘテロ二量体の血清半減期が、IgG1ホモ二量体の血清半減期と大きく異ならないことが示されている(実施例2を参照のこと)。よって、IgG1ΔAdp変異は、IgG1の長い半減期をなおも保ちつつ、プロテインA結合性をなくすという利点を有する。
【0086】
したがって、1つの実施形態において、本明細書中に記載されるようなCH3ドメインの改変を含む二重特異性抗原結合タンパク質が提供され、ここで、その抗原結合タンパク質は、CH3ドメインに改変を有しない同じ二重特異性抗原結合タンパク質と等価な薬物動態学的プロファイルを示す。1つの実施形態において、IgG1ΔA/IgG1ヘテロ二量体のFcを含む二重特異性抗体が提供され、ここで、その二重特異性抗体は、その他の点は同一であるがIgG3 CH3ドメインを含むかまたはその他の点は同一であるが少なくとも1つのIgG3重鎖を含む二重特異性抗体よりも、約1.5倍、約2倍、約2.5倍または約3倍長い血清半減期を有する。1つの実施形態において、IgG1ΔA/IgG1ヘテロ二量体のFcを含む二重特異性抗体が提供され、ここで、その二重特異性抗体は、IgG1ΔA改変を有しない二重特異性抗体(すなわち、IgG1ホモ二量体二重特異性抗体)とほぼ同じ血清半減期を示す。
【0087】
免疫グロブリン重鎖
所望の特徴(例えば、所望の特異性、所望の親和性、所望の機能性、例えば、遮断、非遮断、阻害、活性化など)を備えた二重特異性抗体を作製するために使用することができる免疫グロブリン重鎖可変領域は、当該分野で公知の任意の方法を用いて作製され得る。次いで、所望の重鎖は、本明細書中に記載される所望の重鎖定常領域を有する構築物内に可変領域を含む核酸配列をクローニングすることによって構築され得る。
【0088】
1つの実施形態において、第1の重鎖は、第1の抗原で免疫された第1の動物の成熟B細胞のゲノムに由来する核酸によってコードされる可変領域を含み、その第1の重鎖は、第1の抗原を特異的に認識する。特定の実施形態において、第2の重鎖は、第2の抗原で免疫された第2の動物の成熟B細胞のゲノムに由来する核酸によってコードされる可変領域を含み、その第2の重鎖は、第2の抗原を特異的に認識する。
【0089】
1つの実施形態において、第1の動物および/または第2の動物は、再配列されていないヒト免疫グロブリン重鎖可変領域を含む遺伝子改変動物である。1つの実施形態において、第1の動物および/または第2の動物は、再配列されていないヒト免疫グロブリン重鎖可変領域およびヒト免疫グロブリン定常領域を含む遺伝子改変動物である。1つの実施形態において、第1の動物および/または第2の動物は、再配列されていないヒト免疫グロブリン重鎖可変領域を含む遺伝子改変マウスである。
【0090】
免疫グロブリン重鎖可変領域配列は、当該分野で公知の他の任意の方法、例えば、ファージディスプレイによって得ることができ、それによって得られた配列を使用することにより、任意の適当な重鎖、例えば、本明細書中に記載されるようにCH3ドメインが改変された重鎖をコードする核酸に連結され、発現構築物内に置かれ、そして例えば、適当な軽鎖の存在下において、重鎖を作製することができる細胞に運ばれる、核酸構築物が作製され得る。
【0091】
免疫グロブリン軽鎖
2つの異なるエピトープ(または2つの異なる抗原)を認識する2つの重鎖を含む二重特異性抗体は、同じ軽鎖(すなわち、同一の可変ドメインおよび定常ドメインを有する軽鎖)と対形成し得る場合、より容易に単離される。重鎖可変ドメインの選択性および/またはその標的抗原との親和性を干渉せずにまたは実質的に干渉せずに、特異性が異なる2つの重鎖と対形成し得る軽鎖を作製するための種々の方法が当該分野で公知である。
【0092】
あるアプローチにおいて、軽鎖は、すべての軽鎖可変ドメインに対する使用統計を調査し、ヒト抗体において最も頻繁に使用される軽鎖を特定し、その軽鎖を特異性が異なる2つの重鎖と対形成させることによって選択され得る。
【0093】
別のアプローチでは、軽鎖は、ファージディスプレイライブラリー(例えば、ヒト軽鎖可変領域配列を含むファージディスプレイライブラリー、例えば、ヒトScFvライブラリー)内の軽鎖配列を観察し、そのライブラリーから最もよく使用される軽鎖可変領域を選択することによって選択され得る。
【0094】
別のアプローチでは、軽鎖は、プローブとして両方の重鎖の重鎖可変配列を用いて軽鎖可変配列のファージディスプレイライブラリーをアッセイすることによって選択され得る。両方の重鎖可変配列と会合する軽鎖は、それらの重鎖に対する軽鎖として選択され、両方のエピトープに対する結合および/または活性化を可能にする。
【0095】
別のアプローチでは、軽鎖は、公知の軽鎖と所望の重鎖とを組み合わせ、得られた二重特異性抗体を、結合特異性、親和性および/または活性化能力についてアッセイすることによって選択され得る。
【0096】
軽鎖を選択するための任意のアプローチにおいて困難に直面する(例えば、軽鎖が、一方もしくは両方の重鎖とその抗原との結合を干渉するか、または軽鎖が、一方もしくは両方の重鎖と十分に会合しない)程度まで、その軽鎖は、重鎖の同族の軽鎖と整列することができ、両方の重鎖の同族の軽鎖と共通の配列特徴とよりマッチするように軽鎖において改変が行われる。免疫原性の変更を最小にする必要がある場合、免疫原性の可能性を評価する(すなわち、インシリコならびにウェットアッセイ)ための当該分野で公知のパラメータおよび方法に基づいて、タンパク分解性プロセシングがT細胞エピトープを生成する可能性が低くなるように、改変は、好ましくは公知のヒト軽鎖配列に存在する配列をもたらす。
【0097】
抗体および結合タンパク質
上記組成物および方法は、ヒト二重特異性抗体、すなわち、ヒト定常ドメインおよびヒト可変ドメインを含む二重特異性抗体を作製する際に特に有用である。いくつかの実施形態において、ヒト抗体には、ヒト生殖細胞系列の免疫グロブリン配列に由来する重鎖可変ドメインおよび重鎖定常ドメイン、いくつかの実施形態では、体細胞変異したヒト免疫グロブリン配列(例えば、ヒト免疫グロブリン遺伝子配列を含む動物において産生されるヒト免疫グロブリン配列)に由来する重鎖可変ドメインおよび重鎖定常ドメインを有する抗体が含まれる。いくつかの実施形態において、ヒト可変領域および/またはヒト定常領域は、ヒト生殖細胞系列の免疫グロブリン配列によってコードされないアミノ酸残基、または例えばCDR、特にCDR3における、組換えおよび/もしくはスプライシングの結果としてコードされるアミノ酸残基を含み得る。ヒト抗体は、マウスなどの別の哺乳動物種の生殖細胞系列に由来するCDR配列がヒトフレームワーク配列に移植された抗体を含むと意図されない。それらの抗体は、ヒト化抗体またはキメラ抗体と呼ばれる。ヒト抗体には、例えば、ランダム突然変異誘発または部位特異的突然変異誘発によってインビトロにおいて導入された変異を含む抗体が含まれるが、その変異は、好ましくは、ヒトにおいて免疫原性でない。
【0098】
上記方法および組成物は、キメラ抗体、好ましくは、ヒトにおいて免疫原性でないかまたは低免疫原性のキメラ抗体を作製するために使用され得る。キメラ抗体は、重鎖可変領域またはフレームワークもしくはCDRあるいは重鎖定常領域または重鎖定常ドメインのうちの1つが、異なる種(例えば、ヒトおよびマウス、またはヒトおよび霊長類)に由来する抗体である。いくつかの実施形態において、キメラ抗体には、非ヒト起源(例えば、マウス)の重鎖可変領域およびヒト起源の重鎖定常領域を有する抗体が含まれる。いくつかの実施形態において、キメラ抗体には、ヒト起源の重鎖可変領域および非ヒト(例えば、マウス)起源の重鎖定常領域を有する抗体が含まれる。様々な実施形態において、マウス起源の領域は、体細胞超変異を有するかまたは有しない、マウス免疫グロブリン生殖細胞系列の配列と同一であるかまたは実質的に同一である。キメラ抗体には、ヒト免疫グロブリン生殖細胞系列の配列と同一であるかまたは実質的に同一である軽鎖定常領域、および非ヒト(例えば、マウス)重鎖またはキメラヒト/非ヒト重鎖を有する抗体も含まれる。キメラ抗体には、非ヒト(例えば、マウス)免疫グロブリン生殖細胞系列の配列と同一であるかまたは実質的に同一である軽鎖定常ドメイン、およびヒトまたはキメラ非ヒト/ヒト重鎖を有する抗体が含まれる。
【0099】
いくつかの実施形態において、上記組成物および方法は、親和性成熟抗体を作製するためのものである。いくつかの実施形態において、親和性成熟抗体は、変更を有しない実質的に同一の抗体と比べて、その標的抗原に対してより高い親和性(例えば、ナノモル濃度またはピコモル濃度の範囲のKD)をもたらす1つ以上の変更を1つ以上のCDRに含む。親和性成熟抗体は、当該分野で公知の任意の適当な方法によって、例えば、CDRおよび/またはフレームワーク領域のランダム突然変異誘発もしくは部位特異的突然変異誘発、その後の親和性スクリーニング、VHドメインシャフリングなどによって、作製され得る。
【0100】
いくつかの実施形態において、抗体は、中和抗体である。中和抗体には、抗原の生物学的活性を中和すること、阻害することまたは妨害することができる抗体が含まれる。中和抗体には、抗原に結合したときに、インビボおよびインビトロにおいて、その抗原の天然の標的に対してその抗原が作用することができる能力を妨害するかまたは減少させる抗体が含まれる。中和抗体の例としては、タンパク質リガンドが生物学的レセプターに結合するのを妨害する、そのレセプターのリガンドに対する抗体、または生物学的レセプターがそのリガンドに結合するのを妨害する、そのレセプターに対する抗体が挙げられ、ここで、その抗体の非存在下でのリガンドの結合によって、そのレセプターは細胞の内部に変化を起こす。抗体が中和抗体であるか否かの判定は、一般的に、抗原の生物学的活性に対する抗体の影響を測定する機能的アッセイの実施を必要とする。
【0101】
本発明の方法および組成物は、抗体および他の結合タンパク質に対する種々の用途でも有用である。いくつかの有用な用途の短い説明をここに提供する。
【0102】
細胞上の抗原、例えば、CD20を標的にし、また、T細胞上の抗原、例えば、CD3を標的にする、腫瘍抗原およびT細胞抗原に対する結合特異性を備える二重特異性結合タンパク質が作製され得る。このように、その二重特異性抗体は、患者内の目的の細胞を標的にし(例えば、リンパ腫患者におけるB細胞、CD20結合を介して)、かつその患者のT細胞を標的にする。その二重特異性抗体は、様々な実施形態において、CD3に結合したときにT細胞を活性化し、ゆえにT細胞活性化を特定の選択された腫瘍細胞に結合するように設計される。
【0103】
2つの結合部分を含み、その各々が同じ細胞の表面上の結合パートナーに対するものである(すなわち、各々が異なる標的に対するものである)二重特異性結合タンパク質も作製され得る。この設計は、同じ細胞の表面上に両方の標的を発現する特定の細胞または細胞型を標的にすることに特にふさわしい。標的は、他の細胞上に個別に現れ得るが、これらの結合タンパク質の結合部分は、各結合部分が、比較的低い親和性(例えば、低マイクロモル濃度または高ナノモル濃度、例えば、100ナノモル濃度を越えるKD、例えば、500、600、700、800ナノモル濃度)でその標的に結合するように選択される。このように、標的への長期にわたる結合は、2つの標的が同じ細胞上で近接して存在する状況においてのみ好ましい。
【0104】
同じ標的に結合する2つの結合部分(各々、同じ標的の異なるエピトープに結合する)を含む二重特異性結合タンパク質が作製され得る。この設計は、特に、結合タンパク質で標的を遮断するのを成功させる確率を最大にするのにふさわしい。複数の細胞外ループ、例えば、膜貫通チャネルまたは細胞表面レセプターの複数の細胞外ループが、同じ二重特異性結合分子によって標的にされ得る。
【0105】
クラスター化して免疫シグナル伝達の負の制御因子を活性化することにより免疫抑制をもたらす2つの結合部分を含む二重特異性結合タンパク質が作製され得る。シスでの抑制(Repression in cis)は、標的が同じ細胞上に存在する場合に達成され得る;トランスでの抑制(repression in trans)は、標的が異なる細胞上に存在する場合に達成され得る。シスでの抑制は、例えば、FelD1に対する免疫応答をダウンレギュレートするために、抗IgGRIIb結合部分および抗FelD1結合部分を有する二重特異性結合タンパク質を用いて達成され得る(IgGRIIbは、FelD1の存在下でのみクラスター化する)。トランスでの抑制は、例えば、抗BTLA結合部分、および目的の組織特異的抗原に特異的に結合する結合部分を有する二重特異性結合タンパク質を用いて達成され得(阻害性BTLA分子のクラスター化は、選択された標的組織だけで生じる)、これは、自己免疫疾患に対処できる可能性がある。
【0106】
多成分性のレセプターを活性化する二重特異性結合タンパク質が作製され得る。この設計では、レセプターの2成分に対する2つの結合部分が、そのレセプターに結合して、そのレセプターを架橋し、そのレセプターからのシグナル伝達を活性化する。これは、例えば、IFNAR1に結合する結合部分およびIFNAR2に結合する結合部分を有する二重特異性結合タンパク質を用いて行われ得、ここで、結合によって、そのレセプターが架橋される。そのような二重特異性結合タンパク質は、インターフェロン処置に対する代替物を提供し得る。
【0107】
半透性障壁、例えば、血液脳関門を越えて結合部分を輸送する二重特異性結合タンパク質が作製され得る。この設計では、一方の結合部分は、特定の選択的な障壁を通過し得る標的に結合する;他方の結合部分は、治療活性を有する分子を標的にし、ここで、その治療活性を有する標的分子は、通常はその障壁を通過することができない。この種の二重特異性結合タンパク質は、治療薬が他の方法では到達しなかった組織にその治療薬を持って行くために有用である。いくつかの例としては、腸または肺に治療薬を輸送するためにpIGRレセプターを標的にすること、または血液脳関門を越えて治療薬を輸送するためにトランスフェリンレセプターを標的にすることが挙げられる。
【0108】
特定の細胞または細胞型に結合部分を輸送する二重特異性結合タンパク質が作製され得る。この設計では、一方の結合部分が、細胞内に容易に内部移行される細胞表面タンパク質(例えば、レセプター)を標的にする。他方の結合部分は、細胞内タンパク質を標的にし、ここで、その細胞内タンパク質の結合は、治療効果をもたらす。
【0109】
二重特異性結合タンパク質は、食作用性免疫細胞の表面レセプターおよび感染性病原体(例えば、酵母または細菌)の表面分子に結合することにより、その感染性病原体が食作用性免疫細胞の近くに来て、その病原体のファゴサイトーシスを促進する。そのような設計の例は、CD64またはCD89分子および病原体を標的にする二重特異性抗体(antibodiy)であり得る。
【0110】
二重特異性結合タンパク質は、1つの結合部分として抗体可変領域および第2の結合部分として非Ig部分を有する。その抗体可変領域は、標的化を達成するのに対し、非Ig部分は、Fcに連結されたエフェクターまたはトキシンである。このように、リガンド(例えば、エフェクターまたはトキシン)は、抗体可変領域に結合した標的に送達される。
【0111】
二重特異性結合タンパク質は、任意の2つのタンパク質部分が、Fcの状況において互いに近くに置かれ得るように、2つの部分(各々が、Ig領域(例えば、CH2およびCH3領域を含むIg配列)に結合している)を有する。この設計の例としては、捕捉、例えば、ホモ二量体またはヘテロ二量体の捕捉分子が挙げられる。
【0112】
核酸
モノクローナル抗体をコードする核酸配列は、当該分野で公知の任意の適当な方法によって得ることができる。モノクローナル抗体(およびそれらの核酸配列)を得るための適当な方法の例としては、例えば、ハイブリドーマ法(例えば、Kohlerら(1975)Nature 256:495−497を参照のこと)またはファージ抗体ライブラリー(例えば、Clacksonら(1991)Nature 352:624−628を参照のこと)による方法が挙げられる。
【0113】
様々な実施形態において、免疫グロブリン重鎖可変ドメインは、遺伝子改変動物またはトランスジェニック動物の核酸配列から得られる。いくつかの実施形態において、それらの領域は、ヒト免疫グロブリンのミニ遺伝子座(minilocus)を含む動物から得られる。いくつかの実施形態において、それらの領域は、免疫グロブリン配列をコードする1つ以上の核酸を含む1つ以上の染色体外核酸を含むマウスから得られる。様々な実施形態において、上記動物は、再配列されていない1つ以上のヒト免疫グロブリン核酸配列を有し得る。いくつかの実施形態において、上記動物は、ヒト軽鎖可変領域核酸配列を含み、いくつかの実施形態において、ヒト重鎖可変配列を含み、いくつかの実施形態において、重鎖可変配列と軽鎖可変配列の両方を含み、そしていくつかの実施形態において、ヒト定常領域配列をさらに含む。特定の実施形態において、上記核酸配列は、内在性のマウス重鎖可変遺伝子セグメントおよび軽鎖可変遺伝子セグメントがヒト重鎖可変遺伝子セグメントおよび軽鎖可変遺伝子セグメントで置き換えられているマウスから得られる。
【0114】
いくつかの実施形態において、上記核酸配列は、そのような動物のナイーブB細胞またはナイーブT細胞から得られる。他の実施形態において、上記核酸配列は、目的の抗原で免疫された動物のB細胞またはT細胞から得られる。
【0115】
様々な実施形態において、上記核酸配列は、プライマー(例えば、1つ以上のFR、連結配列または定常配列を含む縮重プライマーのセットが挙げられる)でその核酸配列を増幅することによって、細胞から得られる。
【0116】
様々な実施形態において、免疫グロブリン重鎖可変ドメインは、目的の抗原で免疫された動物の核酸から得られる。例えば、非ヒトトランスジェニック動物または非ヒト遺伝子改変動物を、目的の抗原で免疫することにより(例えば、その動物を、その抗原またはその抗原を有する細胞またはその抗原の発現可能な形態をコードする核酸に曝露することにより)、その動物に免疫応答を起こさせ、その動物から免疫細胞(例えば、B細胞)を単離し、必要に応じて、それらの細胞を不死化し、そしてそれらの細胞をスクリーニングすることにより、その抗原との反応性を特定し、ならびに/または抗体の状況で置かれたときその抗原を認識することができる免疫グロブリン可変領域をコードする核酸配列を同定し、そして/もしくは単離する。いくつかの実施形態において、上記細胞は、B細胞である。いくつかの実施形態において、免疫された動物のB細胞を用いることによりハイブリドーマを作製し、その抗原のエピトープを特異的に認識する抗体を発現するB細胞を特定し、そのエピトープを認識する可変領域のアミノ酸配列をコードする核酸配列を特定し、そして/または単離する。
【0117】
いくつかの実施形態において、上記核酸は、ヒト、非ヒト霊長類(例えば、チンパンジーなどの類人猿)、サル(例えば、カニクイザル(cynomologous)またはアカゲザル)、げっ歯類(例えば、マウス、ラット、ハムスター)、ロバ、ヤギ、ヒツジなどから得られる。
【0118】
いくつかの実施形態において、重鎖は、ヒト細胞から得られる配列を含む。例えば、ヒト胎児細胞が、インビトロで抗原に曝露され、適当な宿主動物(例えば、SCIDマウス)に入れられる。
【0119】
いくつかの実施形態において、上記核酸は、ベクターを用いて細胞に導入される。ベクターとしては、例えば、プラスミド、コスミド、レトロウイルス、アデノウイルス、アデノ随伴ウイルス、植物ウイルス、YAC、BAC、EBV由来エピソームが挙げられる。
【0120】
いくつかの実施形態において、上記核酸は、発現ベクターまたは発現構築物に存在する。いくつかの実施形態において、その発現ベクターまたは発現構築物は、目的の核酸配列が適当な細胞において適当な条件下で発現することができるように、その目的の核酸配列に作動可能に連結されたプロモーターを含むベクターである。発現ベクターまたは発現構築物は、リーダー配列、エンハンサー、転写または翻訳を増強するプロモーターエレメント、転写停止配列、スプライシング配列、転写増強イントロン、IRESエレメント、マーカー遺伝子、選択配列、リコンビナーゼ認識部位、ホモロジーアーム(homology arm)、ウイルス配列、オペレーター(例えば、原核生物のオペレーター)などを含み得る。いくつかの実施形態において、発現ベクターは、誘導可能な発現を可能にするエレメント、例えば、真核生物のプロモーターに作動可能に連結された原核生物のオペレーターを含む。いくつかの実施形態において、発現は、発現誘導因子を加えたときに誘導される。他の実施形態において、発現は、発現阻害剤を除去したときに誘導される。いくつかの実施形態において、発現は、温度変化によって誘導される。
【0121】
いくつかの実施形態において、1つ以上の重鎖ヌクレオチド配列は、同じベクター上に存在する。いくつかの実施形態において、本明細書中の重鎖核酸配列および軽鎖核酸配列は、同じベクター上に存在する。1つの実施形態において、2つの重鎖核酸配列および1つの軽鎖核酸配列は、同じベクター上に存在する。
【0122】
いくつかの実施形態において、核酸は、ウイルス由来の1つ以上の核酸を含む細胞内で発現される。特定の実施形態において、そのウイルスは、アデノウイルス、アデノ随伴ウイルス、SV−40、エプスタイン・バーウイルス、レトロウイルス、レンチウイルス、バキュロウイルス、コロナウイルス、単純ヘルペスウイルス、ポリオウイルス、セムリキ森林ウイルス、シンドビスウイルスおよびワクシニアウイルスから選択される。
【0123】
宿主細胞は、目的の核酸を発現するように形質転換され得る細胞である。様々な実施形態において、形質転換は、細胞が外来性核酸(例えば、天然ではその細胞内に見られない核酸、または天然でその細胞内に見られる核酸配列に対応する1つ以上の追加のコピーの核酸)を含むように細胞の核酸含有量を変化させることを含む。細胞の核酸含有量は、当該分野で公知の任意の適当な方法によって、例えば、その核酸を細胞のゲノムにインテグレートすることによって、またはその核酸を染色体外もしくはゲノム外の形態で細胞内に入れることによって、変更され得る。いくつかの実施形態において、細胞の核酸含有量は、その細胞が目的の核酸を一過性に発現するように変更され得るか、または、核酸含有量は、その細胞が目的の核酸を安定的に発現するように変更され得る。いくつかの実施形態において、その細胞の遺伝子含有量の変更は、細胞が分裂するときに受け継がれる。
【0124】
二重特異性抗原結合タンパク質の単離
適当な改変のセットが、本明細書中の情報に基づいて選択されたら、当該分野で公知の方法を用いて二重特異性抗原結合タンパク質を単離する試みを行った。公開された方法を単に適用することによって、すべての場合において満足な分離が提供されるわけではなかった。
【0125】
Lindhoferらは、プロテインAに結合する1つの重鎖(マウスIgG)およびプロテインAに結合しない1つの重鎖(ラットIgG)を有するヘテロ二量体の重鎖を有する二重特異性抗体を作製し、そのラット/マウスヘテロ二量体二重特異性抗体を溶出するために中性からpH5.8までのpH段階勾配、次いで、マウス/マウスホモ二量体を溶出するために5.8から3.5までのpH段階を用いて、そのヘテロ二量体をマウス/マウスおよびラット/ラット二量体のクアドローマ混合物から分離することに成功した。(Lindhoferら(1995)Preferential species−restricted heavy/light chain pairing in rat/mouse quadromas:Implications for a single−step purification of bispecific antibodies.J.Immunol.155(1):219−225を参照のこと)。
【0126】
上記のLindhoferのアプローチは、IgG1 CH3ドメインのうちの一方がH435R/Y436Fジペプチド改変を含むという事実を別にすれば同一である2つのIgG1重鎖を有するIgG1ヘテロ二量体からIgG1ホモ二量体を分離することに適用したときは、失敗した。本発明者らは、直線のpH勾配では、そのジペプチド改変されたIgG1がpH約3.9で溶出されるのに対し、IgG1ホモ二量体は、pH約3.7で溶出されることを見出した。このpHの差異では、Lindhofer法を用いて上記ホモ二量体からヘテロ二量体を十分に分離するには不十分であると考えられた。この差異は、予測可能様式では再現性が無かった。
【0127】
特定のpH段階またはpH勾配を維持するために必要とされる緩衝液強度によって与えられる比較的実質的なイオン強度を使用するクロマトグラフィの実行において、クロマトグラフィ上の挙動の変動が観察された。しかし、有機修飾剤(organic modifier)(1−プロパノール)を加えることによって、十分な分離は達成されなかった。いくぶん驚いたことに、いくつかのクロマトグラフィの実行については、代わりに、0.5モル濃度〜1.0モル濃度のイオン調節物質(例えば、NaCl)を加えることによって、ホモ二量体のIgG1およびヘテロ二量体のIgG1の分離が急激かつ予想外に改善された。イオン調節物質の添加によって、溶出に対するpH範囲が広がり(イオン調節物質を用いたときは1.2pH単位だったが、イオン調節物質を用いないときは0.2pH単位だった)、pH段階勾配によって2種の分離に成功することができた。しかしながら、他の実行では、たった約150mMのNaCl濃度を用いただけで、十分な分離が達成された(実施例4を参照のこと)。十分な分離が達成され得るのを確実にするために、1つの実施形態において、約0.5〜約1.0モル濃度のイオン調節物質の存在下で二重特異性抗原結合タンパク質の単離が行われる。
【0128】
したがって、1つの実施形態において、本明細書中に記載されるような改変を含む1つの鎖を有するヘテロ二量体のIgGを含む二重特異性抗原結合タンパク質を分離するための方法は、イオン調節物質の存在下においてpH勾配を使用する工程を包含する。1つの実施形態において、そのイオン調節物質は、プロテインA支持体からのIgGホモ二量体の溶出と、本明細書中に記載されるような(すなわち、CH3改変を有する)IgGヘテロ二量体の溶出との間のpHの差を最大にするのに十分な濃度で存在する。特定の実施形態において、イオン調節物質は、約0.5〜約1.0モル濃度の濃度で存在する。別の特定の実施形態において、イオン調節物質は、約0.15〜約0.5モル濃度の濃度で存在する。
【0129】
1つの実施形態において、イオン調節物質は、塩である。1つの実施形態において、イオン調節物質は、アルカリ金属またはアルカリ土類金属およびハロゲンの塩である。特定の実施形態において、その塩は、アルカリ金属またはアルカリ土類金属の塩化物塩、例えば、NaCl、KCl、LiCl、CaCl2、MgCl2である。特定の実施形態において、塩は、約0.5、0.6、0.7、0.8、0.9または1.0のモル濃度で存在する。
【0130】
1つの実施形態において、pH勾配は、pH約3.9〜pH約4.5、別の実施形態では、pH約4.0〜pH約4.4、別の実施形態では、pH約4.1〜pH約4.3である。特定の実施形態において、勾配は、直線勾配である。
【0131】
1つの実施形態において、pH勾配は、段階勾配である。1つの実施形態において、その方法は、pH約3.9、pH約4.0、pH約4.1、pH約4.2、pH約4.3またはpH約4.4の段階を、平衡化されたプロテインAカラム(例えば、PBSまたは別の適当な緩衝液もしくは液体で平衡化されたカラム)に適用する工程を包含する。特定の実施形態において、その段階は、pH約4.2である。
【0132】
1つの実施形態において、ヘテロ二量体のIgG CH3ドメインを含む二重特異性抗体は、非ヘテロ二量体のIgGを実質的に含まない1つ以上の画分にプロテインA支持体から溶出する。特定の実施形態において、溶出された二重特異性抗体画分は、非ヘテロ二量体の抗体である全タンパク質の約1重量%未満、0.5重量%未満または0.1重量%未満を構成する。
【実施例】
【0133】
以下の実施例は、本発明の方法および組成物を作製する方法および使用する方法を当業者に対して説明するために提案されるものであって、本発明者らが本発明者らの発明と考える範囲を限定する意図はない。使用される数値(例えば、量、温度など)に関する精度を保証する努力がなされているが、いくらかの実験誤差および偏差が釈明されるだろう。別段示されない限り、部は、重量部であり、分子量は、平均分子量であり、温度は、摂氏温度であり、圧力は、大気圧または大気圧付近である。
【0134】
実施例1:二重特異性IL−4Ra/IL−6Ra抗原結合タンパク質
ヒトIgG1アイソタイプの2つの公知抗体(1つは、IL−4Raに対する抗体で、1つは、IL−6Raに対する抗体)が、4つのアミノ酸だけが異なる軽鎖を有することが見出された。共発現実験から、抗IL−4Ra抗体の軽鎖が、IL−6Ra由来の軽鎖で置き換えられ得、IL−4Raに対する高親和性結合をなおも維持し、ゆえに抗IL−4Ra重鎖および抗IL−6Ra重鎖ならびに同じ軽鎖を用いて二重特異性抗体を作製することが可能になることが明らかになった。したがって、IL−6Ra抗体の重鎖をFcΔAdp型に改変した(すなわち、IMGTエキソンナンバリングではH95R/Y96FのCH3ジペプチド改変)。
【0135】
次いで、抗IL−6Ra軽鎖をCHO細胞において抗IL4Ra/Fcおよび抗IL6Ra/FcΔAdp重鎖と共発現させ、これらの細胞からのコンディションドメディウムをプロテインAクロマトグラフィに供した。ホモ二量体とヘテロ二量体との混合物を含む細胞上清をプロテインAカラムに充填した後、それぞれpH6.0、pH4.2およびpH3.0で3相となるように2つの緩衝液(A:100mMクエン酸Na、150mM NaCl,pH6.0およびB:100mMクエン酸Na、150mM NaCl,pH3.0;図4を参照のこと)の組み合わせを変化させることによって作製されたpH段階勾配を用いて溶出を行った。図4において、IL−4Rは、抗IL−4Raを表し、IL−6RΔは、抗IL−6Ra(IgG1ΔAdp)を表す。示されているカラム画分を、IL−6RaおよびIL−4Raタンパク質への結合についてアッセイした(図5を参照のこと)。段階溶出を行ったところ、pH4.2で溶出する1つのピークおよびpH3.0における第2のピークが生じた(図4)。BIACORE(商標)解析から、予想のとおり、フロースルー材料は、可溶性IL−6Raに結合することができたが、IL−4Raには結合できなかったことが示された(図5)。pH4.2のピークに対応する画分は、ほぼ等量のIL−6RaおよびIL−4Raに結合することができ、これは、ヘテロ二量体と矛盾しない。pH3.0で溶出するピークは、IL−4Raにだけ結合することができ、IL−6Raには結合できず、このことから、予想される抗IL−4Raホモ二量体に対応する。このことから、ヘテロ二量体の二重特異性抗体が、単純なpH段階勾配を伴うプロテインAクロマトグラフィを使用して効率的に単離され得ることが立証された。
【0136】
実施例2:FcΔAdpタンパク質の薬物動態
FcΔAdp改変が、ヘテロ二量体のFc/FcΔAdpを含む分子の薬物動態に影響したか否かを試験するために、上に記載された抗IL−4Ra/抗IL−6Raの精製されたヘテロ二量体種をマウスに注射し、28日間にわたって血清中のヒト免疫グロブリン濃度を測定した(図6;表1)。そのヘテロ二量体の血清半減期は、約10日であり、これは、野生型の血清半減期と同様だった。このことから、FcΔAdp改変は、血清半減期に検出可能な影響を及ぼさないことが立証された。
【0137】
【表1】
実施例3:二重特異性CD20/CD3抗原結合タンパク質
公知の抗ヒトCD20抗体の重鎖は、公知の(活性化する)抗ヒトCD3抗体の軽鎖と共発現したときも、依然としてCD20に結合することができることが見出された。次いで、その抗CD3軽鎖を抗CD20/Fc重鎖、抗CD3/FcΔAdp重鎖または両方の重鎖と共発現させた。次いで、得られたホモ二量体およびヘテロ二量体の混合集団をバイオアッセイに使用して、CD20を発現している標的細胞を殺滅する能力を測定した(図7)。簡潔には、2×107個のヒトPBMC細胞を、6×107個のCD3×CD28ビーズ(Invitrogen)で72時間活性化した。次いで、30単位のIL−2(R&D Systems)を加え、その細胞をさらに24時間インキュベートした。次いで、その細胞を0.5×106/mLの濃度に分け、さらに30UのIL−2を加えた。次いで、その細胞をさらに48時間インキュベートし、バイオアッセイに使用した。バイオアッセイの当日、CD20を発現している2×106個/mLの標的細胞(Raji)を、8μMのカルセイン−AM(Invitrogen)で30分間標識した。洗浄した標的細胞を、活性化されたhPBMC細胞に、示されている量の抗体含有上清を含む総体積200マイクロリットルにおいて標的:エフェクター細胞(1ウェルあたり合計220,000個の細胞)1:10の比で加えた。細胞を2時間インキュベートし、上清を回収し、蛍光を定量化した。特異的な蛍光と最大蛍光との比を計算することによって、細胞傷害性を測定した。CD20抗体単独(抗CD3軽鎖を用いた)と抗CD3抗体の両方が、標的細胞の殺滅を誘発できなかった;それら2つの試薬を混合したときでさえ、効果がなかった。しかしながら、3つすべての構成要素を共発現させると、有意な殺滅が観察されたことから、その効果はヘテロ二量体の二重特異性種に起因したことが示唆される。一過性にトランスフェクトされたCHO細胞の上清中の二重特異性抗体の推定量に基づいて、この効果に対するEC50が、約15pMであると推定された。
【0138】
実施例4:精製された二重特異性CD20/CD3抗原結合タンパク質を用いた細胞殺滅特異性
実施例3に記載されたようなトランスフェクションからのCHO細胞上清を、溶出のために段階勾配を利用するプロテインAアフィニティークロマトグラフィに供した。その段階勾配は、それぞれpH5.2、pH4.2およびpH2.8で3相となるように2つの緩衝液(A:20mMクエン酸Na、1M NaCl,pH5.2;B:20mMクエン酸Na、1M NaCl,pH2.7)の組み合わせを変化させることによって作製された。次いで、pH4.2で溶出するピークからのタンパク質を、実施例3に記載されたような細胞死滅アッセイにおいて使用した。3pMというEC50で標的細胞の殺滅が観察された(図8)。観察された殺滅の標的特異性を調べる追加の細胞傷害性アッセイを行った。この実験では、CD20を発現する標識された標的細胞(Raji)またはCD20を有しない標識された標的細胞(293)を、活性化されたヒトPBMCとともにインキュベートした。各標的細胞型をそのアッセイに単独でまたは他のタイプの未標識の標的細胞と組み合わせて加えた。すべての場合において、CD20を発現している標的細胞は、3pMというEC50で特異的に殺滅されたが、CD20陰性細胞株は、殺滅されなかった。
【0139】
実施例5:識別的に改変されたhIgG2 FcのプロテインA分離
識別的に改変されたヘテロ二量体のヒトIgG2 Fc/ΔAdpFcおよび改変されていないヒトホモ二量体のIgG2 Fc/Fcを、まず、プロテインAカラム(rProteinA FF,GE)に通す結合・洗浄(bind−and−wash)プロセスによって濃縮した。hIgG2 Fc/FcからhIgG2 Fc/ΔAdpFcをさらに分離するために、以下のとおり、SMART(商標)システム(GE)を使用して段階勾配溶出を行った。この溶媒系は、溶媒A(PBS,1×)、溶媒B(20mMクエン酸ナトリウムおよび1M NaCl,pH5.5)および溶媒C(20mMクエン酸ナトリウムおよび1M NaCl,pH2.5)からなるものだった。溶出を、最初の20分間の100%Aによるアイソクラティック溶出から開始し、続いて、20分後に素早く100%Bに切り換えた。次いで、次の10分間にわたって、33.5%Cおよび66.5%Bに向かって直線勾配を開始した;第1のピーク(Fc/ΔAdpFc)が完全に溶出するまで、Cの濃度を20分間33.5%に維持した。続いて、33.5%Cから100%Cへの直線勾配を30分間行った。流速を250マイクロリットル/分に維持し、UV検出器によって280nmでクロマトグラムを検出した。hIgG2 Fc/ΔAdpFcは、pH4.5で溶出し、一方、hIgG2は、pH3.5で溶出した。
【0140】
実施例6:識別的に改変されたhIgG4 FcのプロテインA分離
識別的に改変されたヘテロ二量体のヒトIgG4(Fc/ΔAdpFc)および改変されていないホモ二量体のIgG4(Fc/Fc)をまず、プロテインAカラム(rProteinA FF,GE)に通す結合・洗浄プロセスによって濃縮した。hIgG4 Fc/FcからhIgG4 Fc/ΔAdpFcをさらに分離するために、以下のとおり、SMART(商標)システム(GE)を使用して段階勾配溶出を行った。この溶媒系は、溶媒A(PBS,1×)、溶媒B(20mMクエン酸ナトリウムおよび1M NaCl,pH5.1)および溶媒C(20mMクエン酸ナトリウムおよび1M NaCl,pH2.8)からなるものだった。溶出を、最初の20分間の100%Aによるアイソクラティック溶出から開始し、続いて、20分後に素早く100%Bに切り換えた。次いで、次の10分間にわたって、50%Cおよび50%Bに向かって直線勾配を開始した;第1のピーク(Fc/ΔAdpFc)が完全に溶出するまで、Cの濃度を20分間50%に維持した。続いて、50%Cから100%Cへの直線勾配を30分間行った。流速を250マイクロリットル/分に維持し、UV検出器によって280nmでクロマトグラムを検出した。hIgG4 Fc/ΔdpFcは、pH約4で溶出し、一方、ホモ二量体は、pH約4からpH2.8までの勾配中に溶出した。
【0141】
実施例7;識別的に改変されたhIgG1 CD3×CD20のプロテインA分離
識別的に改変されたヘテロ二量体の抗hCD3×CD20 IgG1(Fc/ΔAdpFc)および改変されていないホモ二量体の抗hCD20を、以下のとおり、1mLのrProtein AFF(GE Biosciences)カラムにおいて分離した。この溶媒系は、緩衝液A1(PBS 1×)、緩衝液A2(20mMクエン酸ナトリウムおよび1M NaCl pH5.1)、緩衝液B(20mMクエン酸ナトリウムおよび1M NaCl pH2.8)だった。混合サンプルを結合させ、PBSおよび緩衝液A2で洗浄した。段階を用いてpH4.2に達したところ、それにより、二重特異性CD3*×CD20 IgG1(Fc/ΔAdpFc)が溶出し、次いで、pH4.2からpH2.8までの直線勾配により、ホモ二量体の抗hCD20 IgG1が溶出した。
【0142】
実施例8:改変されたCH3のFcレセプターに対する結合親和性
ΔAdp改変(H435RおよびY436F,EUナンバリング)を有するヒトIgG1アイソタイプ二重特異性抗体の種々のヒトFcレセプターに対する結合親和性を、Biacore(商標)定常状態平衡結合アッセイにおいて試験した。
【0143】
簡潔には、アミンに結合した抗ペンタ−his mAb(Qiagen)を有するカルボキシメチル化デキストラン(CM5)チップを使用して、ヒトFcレセプターの様々な構築物を捕捉した。以下のhisタグ化Fcレセプター外部ドメインを、抗ペンタ−hisでコーティングされた種々のCM5チップの表面に結合させた:FcγRI、FcγRIIA(R131多形)、FcγRIIBおよびFcγRIIIB(各々、R&D Systemsから入手した);およびRcγRIIA(H131多形)、FcγRIIIA(V176多形)およびRcγRIIIA(F176多形)(各々、Regeneronにて製造されたもの)。高親和性レセプターFcγRI外部ドメインについては3つの濃度(25nM、50nMおよび100nM)で、および低親和性FcγRレセプター外部ドメインについては5マイクロモル濃度〜39ナノモル濃度で抗体を上記表面の上に通過させ、会合速度定数および解離速度定数(kaおよびkd)の値を測定し、それを用いてそれらの抗体に対する平衡解離定数(KD)を算出した。pH7.2のHBS−T緩衝液を使用して、室温にて結合研究を行った。コントロール抗体(hmAb)、抗CD20および抗CD3Δdp改変ならびにCD20×CD3Δdp二重特異性抗体に対するKDを測定した。抗CD3Δdp抗体に対するKD値は、改変されていないhIgG1アイソタイプ抗体と比べて、試験されたいずれのFcレセプターに対する結合についても有意差を明らかにしなかった(表2)。
【0144】
【表2】
実施例9:hFcRnマウスにおける二重特異性hIgG1ΔAdpの薬物動態
二重特異性抗hCD3/hCD20 IgG1ΔAdp抗体およびその抗体に関するコントロール(抗hCD3 IgGおよび抗hCD3 IgGΔAdpホモ二量体)の薬物動態学的クリアランス速度を、野生型(WT)マウスおよびhFcRn遺伝子によるマウスFcRnの置き換えについてホモ接合性のマウス(hFcRnマウス)において測定した。野生型マウスおよびhFcRnマウスは、C57BL6(75%)および129Sv(25%)を含むバックグラウンドの交雑系統由来のものだった。IgG1アイソタイプマッチコントロール抗体を投与されるWTマウスの1つのコホートの場合(このコホートは3匹のマウスを含んだ)を除いて、コホートは、WTマウスまたはhFcRnマウスの各々を4匹含んだ。1mg/kgのアイソタイプマッチ(hIgG1)コントロール、抗hCD3×CD20IgG1ΔAdp二重特異性、抗hCD3IgG1または抗hCD3IgG1ΔAdpホモ二量体をマウスに投与した。すべての被験物質を皮下に投与した。血液(Bleeds)を0時間、6時間、1日、2日、3日、4日、7日、10日、14日、21日および30日の時点で回収した。
【0145】
ヒト抗体の血清レベルをサンドイッチELISAによって測定した。簡潔には、ヤギポリクローナル抗ヒトIgG(Fc特異的)抗体(Jackson ImmunoResearch)を、1マイクログラム/mLの濃度で96ウェルプレートにコーティングし、4℃で一晩インキュベートした。そのプレートをBSAでブロッキングした後、6点段階希釈物中の血清サンプルおよび12点段階希釈物中のそれぞれの抗体の対照基準を、そのプレートに加え、室温で1時間インキュベートした。洗浄して未結合の抗体を除去した後、捕捉されたヒト抗体を、西洋ワサビペルオキシダーゼ(HRP)で結合体化された同じヤギポリクローナル抗ヒトIgG(Fc特異的)抗体(Jackson ImmunoResearch)を用いて検出し、製造者の推奨に従って、標準的な比色テトラメチルベンジジン(TMB)基質によって発色させた。450nmにおける吸光度をプレートリーダーにて記録し、そのサンプルプレートにおいて作成された基準検量線を用いて、血清サンプル中のhIgGの濃度を算出した。
【0146】
試験された30日間にわたる4つのIgG1抗体の血清半減期に有意差は観察されなかった。特に、ΔAdp改変を有するIgG1抗体と野生型IgG1抗体との間に有意差は観察されなかった。野生型(mFcRn)マウスまたはヒト化FcRn(hFcRn)を有するマウスを用いたとき、それらの抗体の間に差は観察されなかった。予想の通り、hFcRnマウスは、野生型マウスよりもわずかに速いクリアランスを示した。結果を表3に示す。
【0147】
【表3】
実施例10:低塩緩衝液における大量単離
二重特異性CD3×CD20ΔAdp抗体を、本発明に従って大量培養物から単離した。簡潔には、二重特異性抗hCD3×CD20ΔAdp(CD3重鎖に改変)抗体を発現しているCHO−K1細胞株を11リットルのバイオリアクターにおいて培養した。二重特異性抗体を産出する細胞は、約8.25×106細胞/mLの密度まで成長し、約250〜350mg抗体/Lが得られた。対照的に、コントロール抗hCD3抗体は、約100〜150mg/Lで得られた。
【0148】
10mMリン酸ナトリウム、0.5M NaCl,pH7.2で平衡化されたMabSelect SuRe(商標)樹脂(GE)(床の高さ20cm,内径1cm)において抗体を単離し、浄化された細胞培養物を19g/Lで充填し、そのカラムを3カラム体積の10mMリン酸ナトリウム、0.5M NaCl,pH7.2で洗浄した後、2カラム体積の20mMリン酸ナトリウム,pH7.2(NaClなし)で洗浄した。40mM酢酸塩,pH3.0で抗体を溶出した。
【0149】
単一特異的な抗CD30抗体は、pH3.6で溶出し、一方、二重特異性抗hCD3×CD20DAdpは、pH4.4で溶出した。
【0150】
実施例11:中性付近pHによるマウスヘテロ二量体の選択的プロテインA溶出
野生型または変異体(TTTKまたはPTTK)mIgG2a Fcと融合された、ヒトIFNAR1(hIFNAR1)およびヒトIFNAR2(hIFNAR2)細胞外ドメインに対する発現構築物でCHO−K1細胞を一過性にトランスフェクトした。等量の2つの発現プラスミドで細胞をトランスフェクトすることによって、hIFNAR1−mFcとhIFNAR2−mFcとの比を1:1に保った。トランスフェクションの4日後に培養液を回収し、それを、0.2mLのNAb Protein A Plus(商標)スピンカラム(Thermo Scientific/Pierce)を使用するプロテインA精製に供した。簡潔には、カラムを1×PBS,pH7.2で平衡化した。1mlのCHO−K1培養液を、室温で10分間、プロテインA樹脂とともにインキュベートした。次いで、そのカラムを1×PBS,pH7.2で3回洗浄した。結合したタンパク質を、1M NaClを含む20mMクエン酸ナトリウム緩衝液を用いて溶出した。pHを下げながら0.4mLの溶出緩衝液を使用して、溶出を3回行った。異なる画分中のタンパク質を、ウエスタンブロット解析によって検出した。
【0151】
結果から、pH勾配溶出を用いたとき、野生型mIgG2aのホモ二量体から野生型およびstar変異体mIgG2aのヘテロ二量体を分離することが可能であることが示される(図9の両方のゲルにおける画分E1を参照のこと)。
【0152】
実施例12;アイソタイプヘテロ二量体に対するmIgG2a変異体の優先的なヘテロ二量体形成
C末端のマウスFc(mIgG2aまたはmIgG1)を有するヒトI型インターフェロンレセプター(hIFNAR1およびhIFNAR2)の細胞外ドメインの哺乳動物発現用のDNAプラスミドを構築した。部位特異的突然変異誘発を用いて、mIgG2a配列内に変異を導入した。これらの変異体は、TTT=M252T、S254T、S256T;TTTK=M252T、S254T、S256T、I258K;PTTTK=I247P、M252T、S254T、S256T、I258K;RF=H435R、H436Fである。CHO−K1細胞を、上記発現構築物で一過性にトランスフェクトした。hIFNAR2−mFc(mIgG1または変異体mIgG2a)よりも4倍多いhIFNAR1−mFc発現プラスミド(hIFNAR1−mIgG2a)で細胞をトランスフェクトすることによって、IFNAR1−mFcとIFNAR2−mFcとの比を4:1に保った。トランスフェクションの4日後に培養液を回収し、mFcタンパク質をウエスタンブロット解析によって検出した。
【0153】
結果から、mIgG2aとmIgG1とのヘテロ二量体の形成が、野生型mIgG2aとmIgG2a変異体とのヘテロ二量体の形成よりもかなり効率的でないことが示される(図10のレーン1とレーン2〜5とを比較のこと)。この実験では、4:1の比のIFNAR1構築物:IFNAR2構築物を使用することにより、過剰な野生型IgG2a構築物を維持した。
【特許請求の範囲】
【請求項1】
a.N末端からC末端に向かって、第1のエピトープに選択的に結合する第1のエピトープ結合領域、IgG1、IgG2およびIgG4から選択されるヒトIgGの第1のCH3領域を含む免疫グロブリン定常領域を含む、第1のポリペプチド;ならびに
b.N末端からC末端に向かって、第2のエピトープに選択的に結合する第2のエピトープ結合領域、IgG1、IgG2およびIgG4から選択されるヒトIgGの第2のCH3領域を含む免疫グロブリン定常領域を含む、第2のポリペプチド;
を含むヘテロ二量体の二重特異性抗原結合タンパク質であって、ここで、前記第2のCH3領域は、プロテインAへの該第2のCH3ドメインの結合を減少させるかまたは排除する改変を含む、ヘテロ二量体の二重特異性抗原結合タンパク質。
【請求項2】
前記第1のポリペプチドおよび前記第2のポリペプチドが、ヒトIgG重鎖である、請求項1に記載の二重特異性タンパク質。
【請求項3】
免疫グロブリン軽鎖をさらに含む、請求項1に記載の二重特異性タンパク質。
【請求項4】
前記免疫グロブリン軽鎖が、ヒト免疫グロブリン軽鎖である、請求項3に記載の二重特異性タンパク質。
【請求項5】
前記第1のポリペプチドおよび前記第2のポリペプチドが各々、ヒトIgG1重鎖である、請求項1に記載の二重特異性タンパク質。
【請求項6】
前記改変が、IMGTエキソンナンバリングシステムにおける(a)95Rならびに(b)95Rおよび96F、またはEUナンバリングシステムにおける(a’)435Rならびに(b’)435Rおよび436Fからなる群から選択される、請求項1に記載の二重特異性タンパク質。
【請求項7】
IMGTエキソンナンバリングシステムにおける16E、18M、44S、52N、57Mおよび82I、またはEUナンバリングシステムにおける356E、358M、384S、392N、397Mおよび422Iからなる群から選択される1から5つの改変をさらに含む、請求項6に記載の二重特異性タンパク質。
【請求項8】
前記二重特異性抗体のCH3ドメインが、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない、請求項6に記載の二重特異性タンパク質。
【請求項9】
前記二重特異性抗体のCH3ドメインが、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない、請求項7に記載の二重特異性タンパク質。
【請求項10】
二重特異性抗体を作製するための方法であって、前記方法は:
a.第1のエピトープを認識する第1の可変ドメインを含む第1の免疫グロブリン重鎖をコードする核酸配列を得る工程であって、ここで、該第1の免疫グロブリン重鎖は、IgG1、IgG2またはIgG4のアイソタイプ定常ドメインを含む、工程;
b.第2のエピトープを認識する第2の可変ドメインを含む第2の免疫グロブリン重鎖をコードする第2の核酸配列を得る工程であって、ここで、該第2の免疫グロブリン重鎖は、プロテインAへの結合を無くすかまたは減少させる改変をそのCH3ドメインに含むIgG1、IgG2またはIgG4のアイソタイプ定常ドメインを含む、工程;
c.該第1の免疫グロブリン重鎖および該第2の免疫グロブリン重鎖と対形成する免疫グロブリン軽鎖をコードする第3の核酸配列を得る工程;
d.該第1の核酸配列、該第2の核酸配列および該第3の核酸配列を哺乳動物細胞に導入する工程;
e.該細胞が二重特異性抗体を発現するのを可能にする工程;ならびに
f.該二重特異性抗体がプロテインAに結合する能力に基づいて該二重特異性抗体を単離する工程
を包含する、方法。
【請求項11】
前記改変が、IMGTエキソンナンバリングシステムにおける(a)95Rならびに(b)95Rおよび96F、またはEUナンバリングシステムにおける(a’)435Rならびに(b’)435Rおよび436Fからなる群から選択される、請求項10に記載の方法。
【請求項12】
IMGTエキソンナンバリングシステムにおける16E、18M、44S、52N、57Mおよび82I、またはEUナンバリングシステムにおける356E、358M、384S、392N、397Mおよび422Iからなる群から選択される1から5つの改変をさらに含む、請求項11に記載の方法。
【請求項13】
前記二重特異性抗体の前記CH3ドメインが、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない、請求項11に記載の方法。
【請求項14】
前記二重特異性抗体の前記CH3ドメインが、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない、請求項12に記載の方法。
【請求項15】
前記二重特異性抗体が、プロテインAを備える固体支持体で単離される、請求項10に記載の方法。
【請求項16】
前記固体支持体が、プロテインAアフィニティーカラムを含み、前記二重特異性抗体が、pH勾配を使用して単離される、請求項15に記載の方法。
【請求項17】
前記pH勾配が、pH3とpH5との間の1またはそれより高いpH段階を含む段階勾配である、請求項16に記載の方法。
【請求項18】
二重特異性抗体を単離するための方法であって、
破壊された細胞または抗体の混合物から、識別的に改変されたIgG1、IgG2またはIgG4のCH3ドメインを有する二重特異性抗体を単離する工程を包含し、ここで、該識別的に改変されたCH3ドメインは、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でなく、該改変は、プロテインAに対するモノマーの親和性が互いに異なるヘテロ二量体の重鎖定常領域を有する二重特異性抗体をもたらし、該二重特異性抗体は、プロテインAに対するその親和性に基づいて、該破壊された細胞または該混合物から単離される、方法。
【請求項19】
前記ヘテロ二量体の重鎖定常領域の一方のモノマーが、ヒトIgG1であり、該ヘテロ二量体の重鎖定常領域の他方のモノマーが、IMGTエキソンナンバリングシステムにおける(a)H95Rならびに(b)H95RおよびY96F、またはEUナンバリングシステムにおける(a’)H435Rならびに(b’)H435RおよびY436Fからなる群から選択される改変を含む改変されたヒトIgG1である、請求項18に記載の方法。
【請求項20】
前記改変されたヒトIgG1が、IMGTエキソンナンバリングシステムにおけるD16E、L18M、N44S、K52N、V57MおよびV82I、またはEUナンバリングシステムにおけるD356E、L358M、N384S、K392N、V397MおよびV422Iからなる群から選択される改変をさらに含む、請求項19に記載の方法。
【請求項1】
a.N末端からC末端に向かって、第1のエピトープに選択的に結合する第1のエピトープ結合領域、IgG1、IgG2およびIgG4から選択されるヒトIgGの第1のCH3領域を含む免疫グロブリン定常領域を含む、第1のポリペプチド;ならびに
b.N末端からC末端に向かって、第2のエピトープに選択的に結合する第2のエピトープ結合領域、IgG1、IgG2およびIgG4から選択されるヒトIgGの第2のCH3領域を含む免疫グロブリン定常領域を含む、第2のポリペプチド;
を含むヘテロ二量体の二重特異性抗原結合タンパク質であって、ここで、前記第2のCH3領域は、プロテインAへの該第2のCH3ドメインの結合を減少させるかまたは排除する改変を含む、ヘテロ二量体の二重特異性抗原結合タンパク質。
【請求項2】
前記第1のポリペプチドおよび前記第2のポリペプチドが、ヒトIgG重鎖である、請求項1に記載の二重特異性タンパク質。
【請求項3】
免疫グロブリン軽鎖をさらに含む、請求項1に記載の二重特異性タンパク質。
【請求項4】
前記免疫グロブリン軽鎖が、ヒト免疫グロブリン軽鎖である、請求項3に記載の二重特異性タンパク質。
【請求項5】
前記第1のポリペプチドおよび前記第2のポリペプチドが各々、ヒトIgG1重鎖である、請求項1に記載の二重特異性タンパク質。
【請求項6】
前記改変が、IMGTエキソンナンバリングシステムにおける(a)95Rならびに(b)95Rおよび96F、またはEUナンバリングシステムにおける(a’)435Rならびに(b’)435Rおよび436Fからなる群から選択される、請求項1に記載の二重特異性タンパク質。
【請求項7】
IMGTエキソンナンバリングシステムにおける16E、18M、44S、52N、57Mおよび82I、またはEUナンバリングシステムにおける356E、358M、384S、392N、397Mおよび422Iからなる群から選択される1から5つの改変をさらに含む、請求項6に記載の二重特異性タンパク質。
【請求項8】
前記二重特異性抗体のCH3ドメインが、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない、請求項6に記載の二重特異性タンパク質。
【請求項9】
前記二重特異性抗体のCH3ドメインが、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない、請求項7に記載の二重特異性タンパク質。
【請求項10】
二重特異性抗体を作製するための方法であって、前記方法は:
a.第1のエピトープを認識する第1の可変ドメインを含む第1の免疫グロブリン重鎖をコードする核酸配列を得る工程であって、ここで、該第1の免疫グロブリン重鎖は、IgG1、IgG2またはIgG4のアイソタイプ定常ドメインを含む、工程;
b.第2のエピトープを認識する第2の可変ドメインを含む第2の免疫グロブリン重鎖をコードする第2の核酸配列を得る工程であって、ここで、該第2の免疫グロブリン重鎖は、プロテインAへの結合を無くすかまたは減少させる改変をそのCH3ドメインに含むIgG1、IgG2またはIgG4のアイソタイプ定常ドメインを含む、工程;
c.該第1の免疫グロブリン重鎖および該第2の免疫グロブリン重鎖と対形成する免疫グロブリン軽鎖をコードする第3の核酸配列を得る工程;
d.該第1の核酸配列、該第2の核酸配列および該第3の核酸配列を哺乳動物細胞に導入する工程;
e.該細胞が二重特異性抗体を発現するのを可能にする工程;ならびに
f.該二重特異性抗体がプロテインAに結合する能力に基づいて該二重特異性抗体を単離する工程
を包含する、方法。
【請求項11】
前記改変が、IMGTエキソンナンバリングシステムにおける(a)95Rならびに(b)95Rおよび96F、またはEUナンバリングシステムにおける(a’)435Rならびに(b’)435Rおよび436Fからなる群から選択される、請求項10に記載の方法。
【請求項12】
IMGTエキソンナンバリングシステムにおける16E、18M、44S、52N、57Mおよび82I、またはEUナンバリングシステムにおける356E、358M、384S、392N、397Mおよび422Iからなる群から選択される1から5つの改変をさらに含む、請求項11に記載の方法。
【請求項13】
前記二重特異性抗体の前記CH3ドメインが、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない、請求項11に記載の方法。
【請求項14】
前記二重特異性抗体の前記CH3ドメインが、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でない、請求項12に記載の方法。
【請求項15】
前記二重特異性抗体が、プロテインAを備える固体支持体で単離される、請求項10に記載の方法。
【請求項16】
前記固体支持体が、プロテインAアフィニティーカラムを含み、前記二重特異性抗体が、pH勾配を使用して単離される、請求項15に記載の方法。
【請求項17】
前記pH勾配が、pH3とpH5との間の1またはそれより高いpH段階を含む段階勾配である、請求項16に記載の方法。
【請求項18】
二重特異性抗体を単離するための方法であって、
破壊された細胞または抗体の混合物から、識別的に改変されたIgG1、IgG2またはIgG4のCH3ドメインを有する二重特異性抗体を単離する工程を包含し、ここで、該識別的に改変されたCH3ドメインは、ヒトにおいて免疫原性でないかまたは実質的に免疫原性でなく、該改変は、プロテインAに対するモノマーの親和性が互いに異なるヘテロ二量体の重鎖定常領域を有する二重特異性抗体をもたらし、該二重特異性抗体は、プロテインAに対するその親和性に基づいて、該破壊された細胞または該混合物から単離される、方法。
【請求項19】
前記ヘテロ二量体の重鎖定常領域の一方のモノマーが、ヒトIgG1であり、該ヘテロ二量体の重鎖定常領域の他方のモノマーが、IMGTエキソンナンバリングシステムにおける(a)H95Rならびに(b)H95RおよびY96F、またはEUナンバリングシステムにおける(a’)H435Rならびに(b’)H435RおよびY436Fからなる群から選択される改変を含む改変されたヒトIgG1である、請求項18に記載の方法。
【請求項20】
前記改変されたヒトIgG1が、IMGTエキソンナンバリングシステムにおけるD16E、L18M、N44S、K52N、V57MおよびV82I、またはEUナンバリングシステムにおけるD356E、L358M、N384S、K392N、V397MおよびV422Iからなる群から選択される改変をさらに含む、請求項19に記載の方法。
【図1】

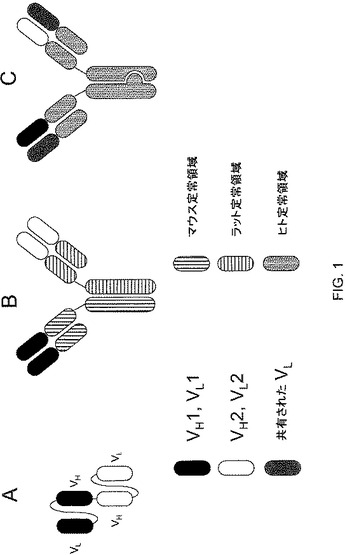
【図2】
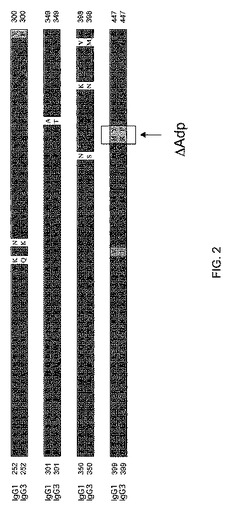
【図2B】
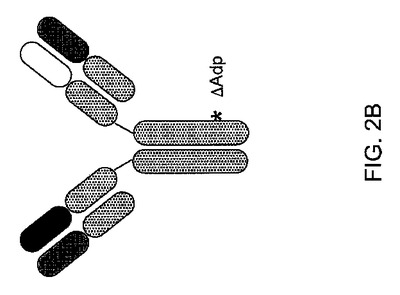
【図3】

【図4】
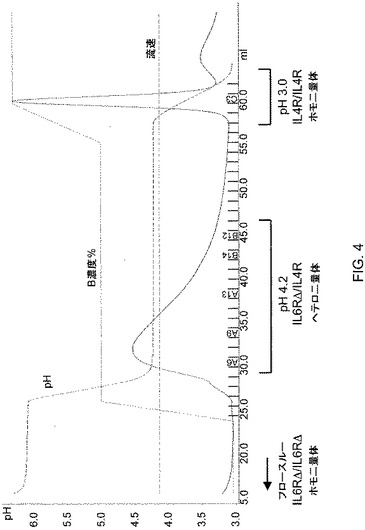
【図5】
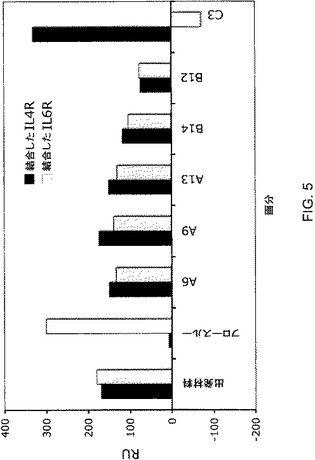
【図6】
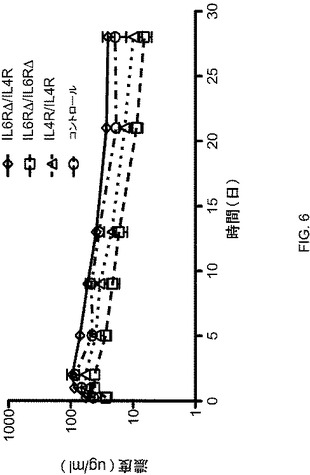
【図7】
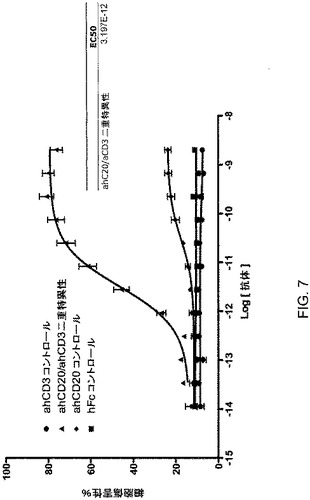
【図8】
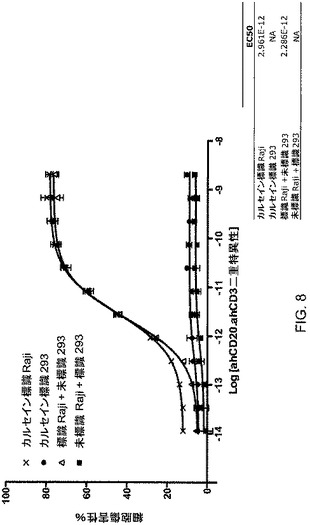
【図9】
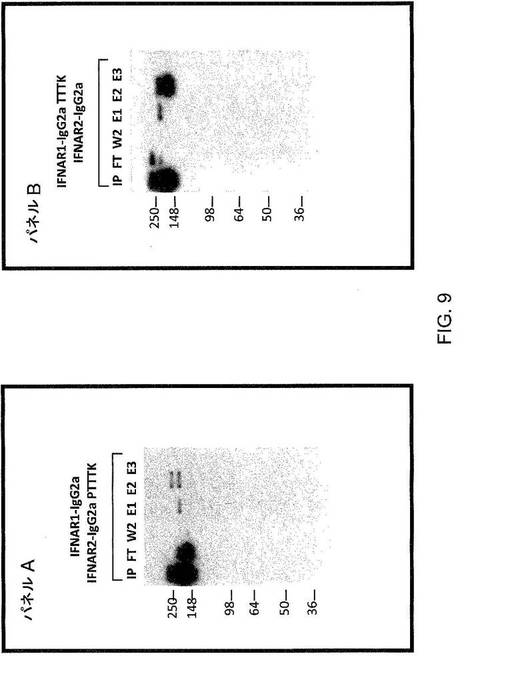
【図10】
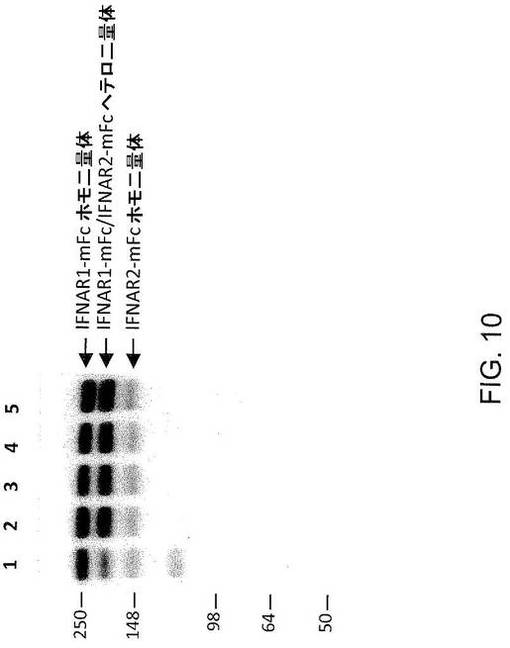
【図2】
【図2B】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公表番号】特表2012−531439(P2012−531439A)
【公表日】平成24年12月10日(2012.12.10)
【国際特許分類】
【出願番号】特願2012−517780(P2012−517780)
【出願日】平成22年6月25日(2010.6.25)
【国際出願番号】PCT/US2010/040028
【国際公開番号】WO2010/151792
【国際公開日】平成22年12月29日(2010.12.29)
【出願人】(597160510)リジェネロン・ファーマシューティカルズ・インコーポレイテッド (50)
【氏名又は名称原語表記】REGENERON PHARMACEUTICALS, INC.
【Fターム(参考)】
【公表日】平成24年12月10日(2012.12.10)
【国際特許分類】
【出願日】平成22年6月25日(2010.6.25)
【国際出願番号】PCT/US2010/040028
【国際公開番号】WO2010/151792
【国際公開日】平成22年12月29日(2010.12.29)
【出願人】(597160510)リジェネロン・ファーマシューティカルズ・インコーポレイテッド (50)
【氏名又は名称原語表記】REGENERON PHARMACEUTICALS, INC.
【Fターム(参考)】
[ Back to top ]