
妊娠関連血漿蛋白−A2(PAPP−A2)
【課題】ヒト胎盤から単離され、配列決定分析により特徴付けられる核酸を提供する。
【解決手段】精製ポリヌクレオチドであって、i)妊娠関連結晶蛋白A2(PAPP−A2)のコード化配列に相当するヌクレオチド1〜5376を含むポリヌクレオチド;およびii)特定のアミノ酸配列を有するポリペプチドをエンコードするポリヌクレオチド;およびiii)ポリヌクレオチド(i)または(ii)によりエンコードされたポリペプチドのフラグメントをエンコードするポリヌクレオチド;およびv)(iii)および(iv)のいずれかで定義されるようなポリヌクレオチドのヌクレオチド配列に縮重されるヌクレオチド配列を含むポリヌクレオチド、およびそのようなポリヌクレオチドの相補鎖、からなる群から選ばれる、精製ポリヌクレオチド。
【解決手段】精製ポリヌクレオチドであって、i)妊娠関連結晶蛋白A2(PAPP−A2)のコード化配列に相当するヌクレオチド1〜5376を含むポリヌクレオチド;およびii)特定のアミノ酸配列を有するポリペプチドをエンコードするポリヌクレオチド;およびiii)ポリヌクレオチド(i)または(ii)によりエンコードされたポリペプチドのフラグメントをエンコードするポリヌクレオチド;およびv)(iii)および(iv)のいずれかで定義されるようなポリヌクレオチドのヌクレオチド配列に縮重されるヌクレオチド配列を含むポリヌクレオチド、およびそのようなポリヌクレオチドの相補鎖、からなる群から選ばれる、精製ポリヌクレオチド。
【発明の詳細な説明】
【技術分野】
【0001】
本発明の属する分野
本発明は、妊娠関連血漿蛋白−A(PAPP−A)と相同性のある新規なポリペプチドに関する。本発明に記載の新規なポリペプチドは、PAPP−A2と示す。本発明は更に、そのようなポリペプチドまたはそのフラグメントをエンコードする核酸配列を含む新規なポリヌクレオチドに関する。
【0002】
本発明は更に、本明細書の以下で定義されるような、新規なポリヌクレオチド(そのフラグメントを含む)を用いる方法、および当該ポリヌクレオチドから生成可能な新規なポリペプチドの使用法に関する。
【0003】
本発明は、組み換えPAPP−A2の発現と精製、およびPAPP−A2に対するポリクローナルおよびモノクローナル抗体の生成、そしてヒトの組織または体液由来の天然のPAPP−A2の精製にも関する。
【0004】
更なる態様において、本発明は、病理的状態に対するマーカーとして、および妊娠している、ならびに妊娠していない個体のPAPP−A2の蛋白分解活性を改変する薬物に対する治療的ターゲットとしてのPAPP−A2の使用に関する。
【背景技術】
【0005】
本発明の背景
妊娠関連血漿蛋白−A(PAPP−A)
PAPP−Aは、1974年に初めて妊婦の血清から胎盤起源と考えられる他の蛋白と一緒に単離された(非特許文献1)。血清中濃度は、妊娠の終了時には約50mg/リットルに達する(非特許文献2;非特許文献3)。PAPP−Aは、高分子量のホモ四量体として最初は特徴付けられた(非特許文献4;非特許文献1;非特許文献5)が、今やPAPP−Aは、主として妊婦の血清および血漿に、好酸球主要塩基性蛋白のプロフォーム(proMBP)との共有結合性のヘテロ四量体の2:2の複合体、PAPP−A/proMBPとして存在することが実証されている(非特許文献6)。妊婦の血清および血漿中において僅か約1%のPAPP−Aが、最近実証されるように(Overgaard et al., 2000, J Biol Chem)ホモ二量体として存在する。PAPP−A/proMBP複合体の存在は、精製されたPAPP−A/proMBPの消化から、ジスルフィド結合によって相互に結合されたPAPP−AおよびproMBPペプチドの単離により一部分は明らかとなった(非特許文献6)。
【0006】
PAPP−A/proMBP複合体のサブユニットは、ジスルフィド結合の還元および変性により不可逆的に分離可能である(非特許文献6)。SDS−PAGEを還元すると、PAPP−Aサブユニットは、見かけの分子量が200kDaであり(非特許文献7)、その1547残基の配列は、クローニングしたcDNAから分かる(非特許文献8)。PAPP−Aは、80残基のpre−pro片(非特許文献9)を含む、pre−pro蛋白(preproPAPP−A)として合成される。PAPP−Aと広範に相同性のある蛋白は、文献上では報告されていないが、PAPP−Aは、482〜492位に伸長した亜鉛結合モチーフ(HEXXHXXGXXH)(非特許文献8に従ってナンバリングしたもの)を含む配列モチーフを含有する。このモチーフおよび構造的に重要なメチオニン残基(PAPP−Aにおいて556位の残基と考えられている)は、厳密には亜鉛ペプチダーゼのスーパーファミリーであるメトジンシン(metzincins):astacins、adamalysins(またはreprolysins)、serralysins、およびmatrixins(マトリックスメタロプロテアーゼまたはMMP’s)内に保持されている(非特許文献10;非特許文献11)。
【0007】
ProMBPサブユニットは、23kDaの計算されたペプチド質量を有する(非特許文献12;非特許文献13)。しかしながらSDS−PAGEにおいて、proMBPは、おそらくその強力で異常なグリコシル化(非特許文献14;非特許文献7)のために、クマシー染色したゲルでは見ることのできない50〜90kDaのスメア(smear)として移動している(非特許文献6)。PAPP−AおよびproMBPは双方とも、インサイツハイブリダイゼーションにより示されるように(非特許文献15)妊娠中に胎盤においてであるが、主に異なる種類の細胞において生成される。RT−PCRによる分析により、PAPP−AおよびproMBPの双方のmRNAが、胎盤中よりは量は少ないけれども、数種の生殖および非生殖組織中に存在することが明らかになった(非特許文献16)。
PAPP−Aの臨床的用途
臨床上、血清PAPP−Aレベルの低下は、ダウン症妊娠の予測値として徐々に利用されてきており(非特許文献17;非特許文献18;非特許文献19;非特許文献20)、血清PAPP−Aレベルは、他の胎児異常においてもまた低下することが示されている(非特許文献21;非特許文献22;非特許文献23)。
【0008】
更に、血管形成後の冠動脈の平滑筋細胞においてはPAPP−A合成が増大し(Bayes−Genis et al., 2000, Arterioscler Thromb Vasc Biol、製本中)、それは、現在臨床的価値の可能性があるために評価されている。妊婦血清におけるproMBPの測定が診断的価値があることもデータは示している(非特許文献24)。
PAPP−Aの蛋白分解活性:IGFBP−4の開裂
ごく最近になって、PAPP−Aの推定のメタロプロテアーゼ活性が、実験で確認されている(非特許文献25)。PAPP−Aは、ヒト線維芽細胞調整培地(HFCM)から一部分精製され、インシュリン様成長因子結合蛋白(IGFBP)−4に対する蛋白分解活性を引き起こすことが示されている。IGFBPは、そのうちの6個が示されているが、IGF−Iおよび−II活性の重要なモジュレータ−である(非特許文献26;非特許文献27)。
【0009】
IGF−Iおよび−IIは、インビボおよびインビトロの双方において、強力な蛋白同化および細胞分裂作用を有する重要なポリペプチドである。IGFBP−4に結合したIGFは、そのレセプターとは相互作用できないが、その結合蛋白が開裂すると、生理活性IGFが遊離される。興味深いことに、PAPP−AによるIGFBP−4の開裂は、厳密にはIGFの存在が必要とされる(非特許文献28;非特許文献25)。PAPP−A分泌は、骨芽細胞および骨髄間質細胞から(非特許文献25)、顆粒膜細胞から(非特許文献29)、および血管平滑筋細胞から(Bayes−Genis et al., 2000, Arterioscler Thromb Vasc Biol, in press)も実証されており、これらはすべて既知のIGF依存性IGFBP−4プロテアーゼ活性を有する。
IGFBP−5
IGFBP−4と同様、IGFBP−5開裂は、多数の組織および調整培地において、同定されていないプロテアーゼにより起こると広範に報告されている(非特許文献30)。
【先行技術文献】
【非特許文献】
【0010】
【非特許文献1】Lin et al., 1974, Am J Obstet Gynecol 118, 223−36
【非特許文献2】Folkersen et al., 1981, Am J Obstet Gynecol 139, 910−4
【非特許文献3】Oxvig et al., 1995, J Biol Chem 270, 13645−51
【非特許文献4】Bischof, 1979, Arch Gynecol 227, 315−26
【非特許文献5】Sinosich, 1990, Electrophoresis 11, 70−8
【非特許文献6】Oxvig et al., 1993, J Biol Chem 268, 12243−6
【非特許文献7】Oxvig et al., 1994, Biochim Biophys Acta 1201, 415−23
【非特許文献8】Kristensen et al., 1994, Biochemistry 33, 1592−8
【非特許文献9】Haaning et al., 1996, Eur J Biochem 237, 159−63
【非特許文献10】Bode et al., 1993, FEBS Lett 331, 134−40
【非特許文献11】Stocker et al., 1995, Protein Sci 4, 823−40
【非特許文献12】Barker et al., 1988, J Exp Med 168, 1493−8
【非特許文献13】McGrogan et al., 1988, J Exp Med 168, 2295−308
【非特許文献14】Oxvig et al., 1994, Biochem Mol Biol Int 33, 329−36
【非特許文献15】Bonno et al., 1994, Lab Invest 71, 560−6
【非特許文献16】Overgaard et al., 1999, Biol Reprod 61, 1083−9
【非特許文献17】Brambati et al., 1993, Br J Obstet Gynaecol 100, 324−6
【非特許文献18】Haddow et al., 1998, N Engl J Med 338, 955−61
【非特許文献19】Wald et al., 1992, Bmj 305, 28
【非特許文献20】Wald et al., 1999, N Engl J Med 341, 461−7
【非特許文献21】Biagiotti et al., 1998, Prenat Diagn 18, 907−13
【非特許文献22】Spencer et al., 2000, Prenat Diagn 20, 411−6
【非特許文献23】Westergaard et al., 1983, Prenat Diagn 3, 225−32
【非特許文献24】Christiansen et al., 1999, Prenat Diagn 19, 905−10
【非特許文献25】Lawrence et al., 1999, Proc Natl Acad Sci U S A 96, 3149−53
【非特許文献26】Fowlkes, 1997, Trends Endocrinol Metab 8, 299−306
【非特許文献27】Rajaram et al., 1997, Endocr Rev 18, 801−31
【非特許文献28】Conover et al., 1993, J Clin Invest 91, 1129−37
【非特許文献29】Conover et al., 1999, J Clin Endocrinol Metab 84, 4742−5
【非特許文献30】Hwa et al., 1999, Endocr Rev 20, 761−87
【発明の概要】
【課題を解決するための手段】
【0011】
本発明の概要
妊娠関連血漿蛋白−A2
本発明に記載の新規な核酸は、ヒト胎盤から単離され、配列決定分析により特徴付けられる。該新規なヌクレオチド配列は、新規なポリペプチド、PAPP−A2をエンコードする。
【0012】
PAPP−A2のアミノ酸配列は、233残基のpre−pro片および1558残基の成熟部分から構成される。PAPP−A2の成熟部分は、PAPP−Aの成熟部分と相同する(およそ45%の同一性)が、そのprepro片は、その2つの蛋白間で全く類似性がない。PAPP−Aと同様、PAPP−A2は、それをメトジンシンスーパーファミリーの推定のメタロプロテアーゼとして分類する保存アミノ酸伸長部を含有する。
【0013】
PAPP−A2は、哺乳類の発現系において発現し、PAPP−A2が活性酵素であることが実証されている。更に、PAPP−A2が、IGFBP−5、インシュリン様成長因子結合蛋白5型を開裂することが実証されている。相対的に、PAPP−AによるIGFBP−4の開裂は、以前から実証されている。
【0014】
完全長の形態のPAPP−A2をエンコードする相補的DNA(cDNA)が、同定され、配列決定され、単離される。該cDNAまたはcDNAの一部分が、組み換え宿主における発現のために発現ベクター中にクローニングされる。該cDNAは、組み換え完全長のPAPP−A2またはPAPP−A2フラグメントを生成するのに有益である。該cDNAおよびそれ由来の組み換えPAPP−A2蛋白は、抗体生成、診断用キット、実験用試薬、およびアッセイのおいて有益となる。
【0015】
該cDNAおよび組み換えPAPP−A2蛋白は、PAPP−A2機能に影響を及ぼす化合物を同定するのにも使用され得る。PAPP−A2アンチセンスオリゴヌクレオチドまたはアンチセンスミメティクスは、臨床上PAPP−A2蛋白の発現を減少させ、それによりPAPP−Aの効果にアンタゴニストとして作用するために有益となり得る。同様に、PAPP−A2コード化配列は、遺伝子治療に対して、ターゲット細胞中にPAPP−A2を導入し、それによりPAPP−A2の効果を促進させるために使用可能である。
【0016】
本発明は更に、細胞中のIGFBP−5蛋白分解活性の減少または削減に対する治療的ターゲットとして使用されるPAPP−A2に関する。
【0017】
診断の目的でのPAPP−A2の使用法を提供することが、更なる本発明の目的である。
【0018】
本発明の他の特徴および利点は、以下に続く図面およびその説明、以下に続く詳細な説明、および請求項から明らかとなるであろう。
【0019】
本発明の好ましい実施形態では、以下の精製ポリヌクレオチドなどが提供される:
(項目1) 精製ポリヌクレオチドであって、
i)受け入れ番号DSM13783下のDSMZに集積された、PAPP−A2のコード化配列に相当する、配列番号1のヌクレオチド1〜5376を含むポリヌクレオチド;および
ii)配列番号2に示されるようなアミノ酸配列を有するポリペプチドをエンコードするポリヌクレオチド;および
iii)ポリヌクレオチド(i)または(ii)によりエンコードされたポリペプチドのフラグメントをエンコードするポリヌクレオチドであって、当該フラグメントが、
a)インシュリン様成長因子結合蛋白5型(IGFBP−5)またはその誘導体、あるいは任意の他の基質に特異的な蛋白分解活性を有し;そして/または
b)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
c)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、ポリヌクレオチド;そして
iv)相補鎖が、(i)、(ii)、および(iii)のいずれかで定義されるようなポリヌクレオチドと、厳密な条件下でハイブリダイズされるポリヌクレオチドであって、当該ポリヌクレオチドが、配列番号2で示されるようなアミノ酸配列を有するポリペプチド、またはそのフラグメントをエンコードし、当該フラグメントが、
a)インシュリン様成長因子結合蛋白5型(IGFBP−5)またはその誘導体、あるいは任意の他の基質に特異的な蛋白分解活性を有し;そして/または
b)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
c)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、ポリヌクレオチド;および
v)(iii)および(iv)のいずれかで定義されるようなポリヌクレオチドのヌクレオチド配列に縮重されるヌクレオチド配列を含むポリヌクレオチド、およびそのようなポリヌクレオチドの相補鎖、からなる群から選ばれる、精製ポリヌクレオチド。
【0020】
(項目2) 項目1に記載で、配列番号1に示されるようなコード化配列を含む、精製ポリヌクレオチド。
【0021】
(項目3) 項目1に記載で、配列番号2に示されるようなアミノ酸配列を有するポリペプチドをエンコードする、ポリヌクレオチド。
【0022】
(項目4) 項目1に記載で、配列番号2に示されるようなアミノ酸配列を有するポリペプチドのフラグメントをエンコードするポリヌクレオチドであって、当該フラグメントが、
a)インシュリン様成長因子結合蛋白5型(IGFBP−5)に特異的な蛋白分解活性を有し;そして/または
b)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
c)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、ポリヌクレオチド。
【0023】
(項目5) 該ポリペプチドの相補鎖が、項目2〜4のいずれかに記載のポリヌクレオチドと、厳密な条件下でハイブリダイズする、項目1に記載のポリヌクレオチド。
【0024】
(項目6) 項目1に記載で、項目3及び4のいずれかに記載のポリヌクレオチドのヌクレオチド配列に縮重されるヌクレオチド配列を含む、ポリヌクレオチド。
【0025】
(項目7) 該ポリヌクレオチドが、項目2〜6のいずれかに記載のポリヌクレオチドの相補鎖を含む、項目1に記載のポリヌクレオチド。
【0026】
(項目8) 3’未翻訳領域、またはそのフラグメント、あるいは配列番号1に相当する、配列番号1の核酸残基5377〜8527を含む更なるポリヌクレオチドに実施可能に結合した、項目1〜7のいずれかに記載のポリヌクレオチド。
【0027】
(項目9) 項目1〜7のいずれかに記載のポリヌクレオチドに実施可能に結合した発現シグナルを含む、発現ベクターの形態の組み換えDNA分子。
【0028】
(項目10) 項目1〜8のいずれかに記載のポリヌクレオチド、又は項目9に記載のベクターを用いてトランスフェクト、あるいは形質転換した、宿主生体。
【0029】
(項目11) 該生体が、哺乳類の生体である、項目10に記載の宿主生体。
【0030】
(項目12) 配列番号2またはそのフラグメントのアミノ酸配列を含むか、あるいはそれらから本質的に構成される単離されたポリペプチドであって、
当該フラグメントが、
i)少なくともインシュリン様成長因子結合蛋白5型(IGFBP−5)に特異的な蛋白分解活性を有し;そして/または
ii)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
iii)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、ポリペプチド。
【0031】
(項目13) 該フラグメントが、任意のプロセシング変異体を含むPAPP−A2の成熟部分に相当する、アミノ酸残基234〜1791を含むか、あるいはそれらから本質的に構成される、項目12に記載のポリペプチド。
【0032】
(項目14) 該フラグメントが、PAPP−A2のprepro部分に相当する、アミノ酸残基1〜233を含むか、あるいはそれらから本質的に構成される、項目12に記載のポリペプチド。
【0033】
(項目15) 該フラグメントが、PAPP−A2のpro部分に相当する、アミノ酸残基23〜233を含むか、あるいはそれらから本質的に構成される、項目12に記載のポリペプチド。
【0034】
(項目16) 該フラグメントが、PAPP−A2のシグナルペプチドまたはリーダー配列に相当する、アミノ酸残基1〜22を含むか、あるいはそれらから本質的に構成される、項目12に記載のポリペプチド。
【0035】
(項目17) 配列番号2のアミノ酸残基234〜1791に相当する、PAPP−A2の成熟部分に実施可能に結合した、項目14〜16のいずれかに記載のポリペプチド。
【0036】
(項目18) 該ポリペプチドが、組み換えポリペプチドである、項目12〜17のいずれかに記載のポリペプチド。
【0037】
(項目19) 該ポリペプチドが、ヒト蛋白、または該ポリペプチドに天然に付随する他の蛋白を含まない、項目12〜18のいずれかに記載のポリペプチド。
【0038】
(項目20) i)項目1〜8のいずれかに記載のポリヌクレオチド、および/またはii)項目9に記載のベクター、および/またはiii)項目10および11のいずれかに記載の宿主生体、および/またはiv)項目12〜19のいずれかに記載のポリペプチドを、生理的に許容される担体と組み合わせて含む、組成物。
【0039】
(項目21) i)項目1〜8のいずれかに記載のポリヌクレオチド、および/またはii)項目9に記載のベクター、および/またはiii)項目10および11のいずれかに記載の宿主生体、および/またはiv)項目12〜19のいずれかに記載のポリペプチドを、薬学的に許容される担体と組み合わせて含む、医薬組成物。
【0040】
(項目22) 項目12に記載のポリペプチドに対して特異性を有する抗体の生成法であって、当該方法が、
i)宿主生体を提供し、
ii)その宿主生体を、項目10に記載のポリペプチドを用いて免疫し、そして
iii)当該抗体を得る、
ことからなるステップを含む、方法。
【0041】
(項目23) 項目12に記載のポリペプチドに対して特異的な結合親和性を有する、抗体。
【0042】
(項目24) 該抗体が、モノクローナル抗体およびポリクローナル抗体からなる群から選ばれる、項目23に記載の抗体。
【0043】
(項目25) 該抗体が、モノクローナルである、項目24に記載の抗体。
【0044】
(項目26) 項目18に記載のポリペプチドの生成法であって、当該方法が、
i)好適な宿主生体を提供し、
ii)ステップi)で提供された宿主生体を、項目1〜8のいずれかに記載のポリヌクレオチド、または項目9に記載のベクターを用いてトランスフェクトまたは形質転換し、 iii)ステップii)で得られた宿主生体を該ポリヌクレオチドまたはベクターによりエンコードされたポリペプチドの発現に好適な条件下で培養し;そして所望により
iv)宿主生体による組み換え発現の結果生じるポリペプチドを、その宿主生体から単離する、
ことからなるステップを含む、方法。
【0045】
(項目27) 該宿主生体が、哺乳類細胞である、項目26に記載の方法。
【0046】
(項目28) アンチセンス法を用いて、細胞中のPAPP−A2の発現を阻害および/または減少させる方法であって、当該方法が、
i)項目7に記載のポリヌクレオチドを提供し、
ii)ステップi)において提供される当該ポリヌクレオチドを用いて、PAPP−A2を発現可能な細胞をトランスフェクトまたは形質転換し、
iii)ステップii)において得られた細胞を、PAPP−A2の発現に関与する当該細胞において、ステップi)で提供されたポリヌクレオチドの相補的なポリヌクレオチドへのハイブリダイゼーションに好適な条件下で培養し、そして
iv)当該細胞においてPAPP−A2の発現を阻害および/または減少させる、
ことからなるステップを含む、方法。
【0047】
(項目29) 該アンチセンスポリヌクレオチドおよび該相補的なポリヌクレオチドが、別々のポリヌクレオチド分子から同時発現される、項目28に記載の方法。
【0048】
(項目30) 個体(individual)から得た生物学的サンプルにおけるPAPP−A2の検出法、あるいはPAPP−A2レベル(level of PAPP−A2)の測定法であって、当該方法が、
i)当該個体から生物学的サンプルを得て、
ii)そのサンプルにおいて、
a)項目12に記載のポリペプチド;および/または
b)PAPP−A2発現に由来するmRNA形態のポリヌクレオチド、および/または
c)好ましくはIGFBP−5、その誘導体の開裂、またはPAPP−A2に対する他の任意の好適な基質を検出することによる、PAPP−A2特異的プロテアーゼ活性、を検出することによりサンプル中のPAPP−A2を検出する、
ことからなるステップを含む、方法。
【0049】
(項目31) 該方法が、ステップii)で検出されたPAPP−A2またはPAPP−A2レベルを、
i)予め決められたPAPP−A2の量および/または濃度;および/または
ii)予め決められたPAPP−A2 mRNAの量および/または濃度;および/または
iii)予め決められたPAPP−A2特異的なプロテアーゼ活性、
からなる群から選ばれる予め決められた値と比較することからなるステップを更に含む、項目30に記載の方法。
【0050】
(項目32) 該予め決められた値が、当該個体の正常な生理学的(physiological)状態を示す、項目31に記載の方法。
【0051】
(項目33) 該生物学的サンプルが、血液、尿、胸腔内液、口腔洗浄液、組織バイオパシー、および卵胞液からなる群から選ばれる、項目30に記載の方法。
【0052】
(項目34) 該PAPP−A2レベルが、PAPP−A2特異的プロテアーゼ活性として測定される、項目30に記載の方法。
【0053】
(項目35) 該PAPP−A2レベルが、PAPP−A2蛋白の量として測定される、項目30に記載の方法。
【0054】
(項目36) 該PAPP−A2レベルが、PAPP−A2のメッセンジャーRNAの量として測定される、項目30に記載の方法。
【0055】
(項目37) 該PAPP−A2蛋白量が、免疫化学的分析により測定される、項目35に記載の方法。
【0056】
(項目38) 該PAPP−A2蛋白量が、少なくとも1つのモノクローナル抗体により検出される、項目37に記載の方法。
【0057】
(項目39) 該PAPP−A2蛋白が、少なくとも1つ更なる成分を含む複合体、好ましくはポリペプチドにおいて検出される、項目30に記載の方法。
【0058】
(項目40) 該PAPP−A2が、PAPP−A2単量体として検出される、項目30に記載の方法。
【0059】
(項目41) 該PAPP−A2が、PAPP−A2二量体として検出される、項目30に記載の方法。
【0060】
(項目42) 個体における病態の診断法であって、当該方法が、
i)項目30〜41のいずれかに記載の方法を実施し、そして
ii)その病態を診断する、
ことからなるステップを含む、方法。
【0061】
(項目43) 該病態が、胎児異常である、項目42に記載の方法。
【0062】
(項目44) 該胎児異常が、トリソミー21、トリソミー18、トリソミー13、および開放型二分脊椎からなる群から選ばれる、項目43に記載の方法。
【0063】
(項目45) 該胎児異常が、子宮外妊娠、開放型二分脊椎、神経管閉鎖障害、腹部壁欠損、Edwards症候群、Pateaus症候群、Turner症候群、モノソミーX、またはKlein−felter’s症候群である、項目43に記載の方法。
【0064】
(項目46) 該病態が、成長促進状態および成長阻害状態からなる群から選ばれる異常な成長状態である、項目43に記載の方法。
【0065】
(項目47) 該病態が、再狭窄、アテローム性心筋梗塞、創傷治癒、線維症、心筋梗塞、骨粗鬆症、リュウマチ性関節炎、多発性骨髄腫、または癌からなる群から選ばれる、項目46に記載の方法。
【0066】
(項目48) 生物学的サンプルにおける項目1に記載のポリヌクレオチドの発現検出法であって、当該方法が:
i)項目1に記載のポリヌクレオチドを推定で含有する生物学的サンプルを提供し、そして
ii)その生物学的サンプルを、i)項目1に記載のポリヌクレオチドに相補的な、およびii)それとハイブリダイズ可能な、鎖を含むポリヌクレオチドと接触させ、そして iii)ハイブリダイゼーションが起こさせ、そして
iv)ステップiii)で得られたハイブリダイゼーション複合体を検出する、
ことからなるステップを含み、
該ハイブリダイゼーション複合体の存在が、項目1に記載のポリヌクレオチド、またはそのフラグメントの生物学的サンプルにおける発現を示している、方法。
【0067】
(項目49) PAPP−A2のプロテアーゼ活性を阻害する薬剤の同定法であって、当該方法が、
i)a)項目12に記載のポリペプチド、およびb)当該ポリペプチドに対して予め決められた基質、およびc)推定の阻害剤をインキュベートし、そして
ii)該基質の蛋白分解が阻害されるかどうかを決定する、
ことからなるステップを含む、方法。
【0068】
(項目50) 該基質が、ポリペプチドを含む、項目49に記載の方法。
【0069】
(項目51) 該基質が、内部でクエンチングする蛍光性ペプチドを含む、項目50に記載の方法。
【0070】
(項目52) 該基質が、IGFBP−5またはそのフラグメントを含むか、あるいはそれから本質的に構成される、項目50に記載の方法。
【0071】
(項目53) 項目49〜52のいずれかの方法のいずれかに従って得られる、阻害剤。
【0072】
(項目54) 病態の処置を必要とする個体においてその病態を処置するための薬物の製造における、項目53に記載の阻害剤の使用。
【0073】
(項目55) PAPP−A2のプロテアーゼ活性を促進する薬剤の同定法に関し、当該方法が、
i)a)項目12に記載のポリペプチド、およびb)当該ポリペプチドに対して予め決められた基質、およびc)推定の促進剤をインキュベートし、そして
ii)該基質の蛋白分解が促進されるかどうかを決定する、
ことからなるステップを含む、方法。
【0074】
(項目56) 該基質が、ポリペプチドを含む、項目53に記載の方法。
【0075】
(項目57) 該基質が、内部でクエンチングする蛍光性ペプチドを含む、項目54に記載の方法。
【0076】
(項目58) 該基質が、IGFBP−5またはそのフラグメントを含むか、あるいはそれから本質的に構成される、項目54に記載の方法。
【0077】
(項目59) 項目54〜57のいずれかの方法のいずれかに従って得られる、促進剤。
【0078】
(項目60) 病態の処置を必要とする個体においてその病態を処置するための薬物の製造における、項目59に記載の促進剤の使用。
【0079】
(項目61) 個体の治療による処置法であって、当該方法が、i)項目21に記載の医薬組成物、および/またはii)項目53に記載の阻害剤、および/または項目59に記載の促進剤を、当該個体に投与することからなるステップを含む、方法。
【0080】
(項目62) PAPP−A2またはPAPP−A2の他の蛋白との複合体の精製法であって、当該方法が、
i)項目12に記載のポリペプチドに対して特異的な結合親和性を有するポリクローナルまたはモノクローナル抗体を提供し、そして
ii)PAPP−A2をアフィニティークロマトグラフィーを用いて精製する、ことからなるステップを含む、方法。
【0081】
(項目63) 個体における該病態の診断法、または当該病態の素因の診断法であって、当該方法が、
a)当該個体からの体液サンプルを提供し;そして
b)当該組体液サンプル中のPAPP−A/proMBP、PAPP−A2/proMBP、PAPP−A/PAPP−A2、PAPP−A/PAPP−A2/proMBP、proMBP/ANG、およびproMBP/ANG/C3dgからなる群から選ばれる複合体のレベルを測定し;そして
c)病態または病態の素因を診断(予め決められた値より上または下の複合体レベルが病態または病態の素因を示す)する、
ことからなるステップを含む、方法。
【0082】
(項目64) 哺乳類の胎児における病態を診断法、または当該病態の素因の診断法であって、当該方法が、
a)当該胎児の母親からの体液サンプルを提供し;そして
b)当該体液サンプル中のPAPP−A/proMBP、PAPP−A2/proMBP、PAPP−A/PAPP−A2、PAPP−A/PAPP−A2/proMBP、proMBP/ANG、およびproMBP/ANG/C3dgからなる群から選ばれる複合体のレベルを測定し;そして
c)病態または病態の素因を診断(予め決められた値より上または下の複合体レベルが病態または病態の素因を示す)する、
ことからなるステップを含む、方法。
【0083】
(項目65) 該病態が、ダウン症、子癇前症、および不安定狭心症および心筋梗塞を含む急性冠症候群からなる群から選ばれる、項目63および64のいずれかに記載の方法。
【0084】
(項目66) 該複合体が、PAPP−A/proMBPであって、該病態が、ダウン症、および不安定狭心症および心筋梗塞を含む急性冠症候群から選ばれる、項目63および64のいずれかに記載の方法。
【0085】
(項目67) 該複合体が、proMBP/ANGであって、該病態が、ダウン症である、項目63および64のいずれかに記載の方法。
【図面の簡単な説明】
【0086】
【図1a】preproPAPP−A2をエンコードするmRNAに相当するcDNAの配列図である。
【図1b】preproPAPP−A2をエンコードするmRNAに相当するcDNAの配列図であり、図1aの続きである。
【図1c】preproPAPP−A2をエンコードするmRNAに相当するcDNAの配列図であり、図1bの続きである。
【図2】アミノ酸配列に翻訳されたときのPAPP−Aと、PAPP−Aの配列伸長部との間の相関関係を示し、またPAPP−Aの中央領域と相同性のあるcDNA配列を得る方法を示す図である。
【図3】preproPAPP−Aと整列したpreproPAPP−A2のアミノ酸配列を示した図である。
【図4】PAPP−A2をウエスタンブロッティングおよびクマシー染色した図である。
【図5】IGFBP−1〜6に対するPAPP−A2活性を示した図である。
【図6】IGFBP−5に対するPAPP−A2の蛋白分解活性を示した図である。
【図7a】PAPP−A2 mRNAコード化領域のcDNA配列を示した図である。
【図7b】PAPP−A2 mRNAコード化領域のcDNA配列を示した図であり、図7aの続きである。
【図8】PAPP−A/proMBP複合体中のPAPP−Aサブユニットのジスルフィド構造、PAPP−Aの推定のドメイン、およびPAPP−Aの遺伝子構造を示す図である。
【発明を実施するための形態】
【0087】
定義
本明細書で使用されるように、PAPP−A2とは、本明細書に定義されるような図1に示されるアミノ酸配列(配列番号2)を有する単離されたPAPP−A2ポリペプチドまたはその変異体をいう。本発明に記載のPAPP−A2またはその変異体は、組み換えDNA技術により生成され得るか、あるいは該PAPP−A2が自然発生し得る。
【0088】
ヌクレオチド配列をエンコードするPAPP−A2とは、本明細書に定義されるような図1に示される配列(配列番号1)を有する単離された核酸をいう。
【0089】
「活性のある」とは、任意の自然発生的なPAPP−A2の生物学的および/または免疫学的活性を保持しているPAPP−A2の形態をいう。
【0090】
「自然発生的なPAPP−A2」とは、遺伝学的に作られたものではないヒト細胞より生成されたPAPP−A2をいい、特にアセチル化、カルボキシル化、グリコシル化、リン酸化反応、脂質付加反応、アシル化、または他のポリペプチドと共有結合または非共有結合性の複合形態を含むがそれに限定されない、ポリペプチドの翻訳後修正により生ずる様々なPAPP−A2が考えられる。
【0091】
「単離されたポリペプチド」とは、ある蛋白質、あるいはその変異体またはフラグメントであり、蛋白化学の分野においては既知の常法により評価されるように特定の調製物のうち90%またはそれ以上の蛋白含量からなる。
【0092】
「誘導体」とは、ユビキチン化、標識化(例えば、放射性ヌクレオチド、様々な酵素等を用いる)、ペグ化(ポリエチレングリコールを用いる誘導体化)といった化学修飾により、またはオルニチンといったアミノ酸の挿入(または化学合成によるアミノ酸の置換)により自然発生的なPAPP−A2から誘導されるポリペプチドをいい、これは通常ヒト蛋白では生じない。
【0093】
「組み換え変異体」とは、組み換えDNA技術により作られた、アミノ酸の挿入、欠失、および置換による自然発生的なPAPP−A2とは異なる任意のポリペプチドをいう。どのアミノが酸残基が、蛋白分解活性または細胞接着性といった目的の活性を失うことなく、置換されたり、付加されたり、欠失されたりし得るかを決定する際の情報は、例えば構造的に類似の蛋白(例えば、他のメトジンシンファミリープテアーゼ)を用いて、既知のジスルフィド構造からなる局部的に相同する蛋白を用いて、または二次構造の予測により、PAPP−A2の一部分の配列を比較することにより見い出し得る。
【0094】
好ましくは、アミノ酸「置換」は、限定ではないが、ロイシンのイソロイシンまたはバリンとの置換、アスパラギン酸のグルタミン酸との置換、あるいはスレオニンのセリンとの置換といった、すなわち1つのアミノ酸を類似の構造的および/または化学的特性を有する別のアミノ酸と置換、すなわち同類アミノ酸置換の結果である。本明細書で使用されるような「置換」なる用語の意味の範囲にある更なる例示および定義は、本明細書の以下の本発明の詳細な説明において提供されている。
【0095】
アミノ酸「挿入」または「欠失」は、典型的には約1個のアミノ酸〜約20個のアミノ酸といった約1個のアミノ酸〜約50個のアミノ酸、例えば約1個のアミノ酸〜約10個のアミノ酸といった約1個のアミノ酸〜約20個のアミノ酸の範囲である。可能な変異体は、組み換えDNA技術を用いて系統的にPAPP−A2分子中にアミノ酸の挿入、欠失、または置換し、その結果生成する組み換え変異体を活性についてアッセイすることにより実験的に決定され得る。
【0096】
所望される場合、「シグナルまたはリーダー配列」は、細胞膜を通るポリペプチド(完全長のPAPP−A2、またはそのPAPP−A2ポリペプチドの一部分)を示し得る。このような配列は、本発明のポリペプチド上に天然に存在し得るか、あるいは組み換えDNA技術により異種の蛋白源から提供され得る。
【0097】
ポリペプチド「フラグメント」、「部分」、または「セグメント」とは、様々な実施態様において、少なくとも約5個のアミノ酸、しばしば少なくとも約7個のアミノ酸、典型的には少なくとも約9〜13個のアミノ酸、例えば少なくとも約17個以上のアミノ酸のアミノ酸残基の伸長部である。それは、長さがインタクトなPAPP−A2までの更に長い残基の伸長部であってもよい。活性となるために、いかなるPAPP−A2ポリペプチドまたはPAPP−A2ポリペプチドフラグメントも、そのままで、あるいはキーホール リムペット ヘモシアニン(keyhole limpet hemocyanin)といったキャリアー蛋白と結合したときに生物学的および/または免疫学的活性を呈するのに十分な長さを有さねばならない。
【0098】
「オリゴヌクレオチド」またはポリヌクレオチド「フラグメント」、「部分」、もしくは「セグメント」は、PAPP−A2ポリペプチドフラグメントの発現に有益な配列をエンコードするPAPP−A2の伸長部である。それはまた、典型的にはmRNAまたはDNA分子の関連する部分を増幅したり明らかにするためのポリメラーゼ鎖反応(PCR)またはハイブリダイゼーション法において使用可能なヌクレオチド残基の伸長部でもあり得る。特に、オリゴヌクレオチドプローブの一方または両方は、いかなる既知のまたは従来技術の分子とも同一性または相補性のないPAPP−A2の一部分と同一または相補的である配列を含むであろう。この目的で、当該オリゴヌクレオチドプローブは、一般的には約10個のヌクレオチド〜50個のヌクレオチド、および好ましくは約15個のヌクレオチド〜約30個ヌクレオチドを含むであろう。
【0099】
本明細書において使用されるように「動物」とは、ヒト、家庭用または農耕用(ネコ、イヌ、ウシ、ヒツジ等)または試験用(マウス、ラット、ラビット等)の種を含むと定義され得る。
【0100】
「組み換え」とはまた、PAPP−A2をエンコードし、組み換えDNA技術を用いて調製されるポリヌクレオチドをいい得る。PAPP−A2をエンコードするDNAは、対立遺伝子または組み換え変異体およびその突然変異体をも含み得る。
【0101】
「核酸プローブ」は、本発明により提供されるPAPP−A2をエンコードするcDNA配列をベースにして調製される。核酸プローブは、約6kbよりも少ないヌクレオチド、通常は約1kbよりも少ないヌクレオチドを有する配列部分を含む。偽陽性部を削除するために適当な試験をした後、これらのプローブをは、PAPP−A2をエンコードするmRNAが細胞または組織中に、または(Walsh et al., 1992, PCR Methods Appl 1, 241−50)に記載されるような細胞または組織から抽出した類似の染色体DNA由来の核酸配列を単離するために存在するかどうかを決定するために使用され得る。プローブは、自然発生的な、まは組み換え1本または2本鎖の核酸から誘導され得るか、あるいは化学的に合成され得る。それらは、ニックトランスレーション法、Klenow フィルイン反応、PCR法、または当業者には周知の他の方法により標識され得る。本発明のプローブ、それらの調製および/または標識化は、(Sambrook et al., 1989);または(Ausubel et al., 1989)に詳しく示されている。
【0102】
別法としては、これらのPAPP−A2または類似のポリペプチドをエンコードする組み換え変異体を合成したり、遺伝子コード中の「重複性」を使用することにより選択したりしてもよい。様々な制限部位を提供するサイレント変化といった様々なコドン置換体が、プラスミドまたはウィルス性ベクター中へのクローニング、あるいは特定の原核または真核系統における発現を最適化するために導入されてもよい。制限されないが活性、鎖間のアフィニティー、あるいはポリペプチドの分解または代謝回転率を含む、ポリペプチドの特性を改変するために突然変異が導入されてもよい。一例としては、より小さい分子量の分子を提供するために、停止コドンをヌクレオチド配列中に挿入してPAPP−A2のサイズを制限することが挙げられる。
【0103】
「発現ベクター」は、遺伝子のクローン化コピーの転写および適当な宿主中におけるmRNAの翻訳に必要なDNA配列として本明細書に定義されている。このようなベクターは、バクテリア、酵母、藍藻類、植物細胞、昆虫細胞、および動物細胞といった様々な宿主において、真核生物遺伝子を発現するために使用可能である。
【0104】
本明細書で使用されるように、「抗体」なる用語は、ポリクローナル抗体およびモノクローナル抗体の両方、ならびに抗原またはハプテンに結合可能なFv、Fab、F(ab)2といったそれらのフラグメントを含む。それは、従来のマウスモノクローナル抗体ならびにヒト抗体、およびヒト以外の抗体のヒト化形態を含み、そしてそれはまた、ファージ抗体ライブラリーから単離した「抗体」をも含む。
【0105】
「リボザイム」とは、RNAの特異的開裂を触媒可能な酵素のRNA分子である。リボザイム作用のメカニズムは、リボザイム分子の相補的なターゲットRNAへの配列特異的なハイブリダイゼーション、次いでヌクレオチド鎖切断型開裂を包含する。本発明の範疇で、PAPP−A2のRNA配列のヌクレオチド鎖切断型開裂を特異的かつ効率的に触媒するハンマーヘッド型リボザイム分子が作られる。いかなる潜在的なRNAターゲット内の特異的なリボザイム開裂部位も、以下の配列、GUA、GUU、およびGUCを含むリボザイム開裂部位に対するターゲット分子をスキャンすることにより最初に同定される。いったん同定されると、開裂部位を含むターゲット遺伝子の領域に相当する15〜20個の間のリボヌクレオチドからなる短いRNA配列が、オリゴヌクレオチド配列を不安定にし得る二次構造といった予測される構造特性について評価され得る。候補ターゲットの適合性もまた、リボヌクレアーゼ蛋白アッセイを用いて、相補的オリゴヌクレオチドとのハイブリダイゼーションのし易さを試験することにより評価され得る。
発明の詳細な説明
PAPP−A2をエンコードするヌクレオチド配列の単離
本発明は1つの態様において、妊娠関連血漿蛋白−A(PAPP−A)と全体的に相同性のある蛋白をエンコードする新規なcDNA配列に関する。この蛋白は、PAPP−A2と示されている。PAPP−A2の完全なヌクレオチド配列は、ヒトの胎盤から単離されたmRNAから得られる(実施例1)。その完全なヌクレオチド配列(配列番号1)および翻訳されたPAPP−A2のアミノ酸配列(配列番号2)の両方が図1に示されている。
【0106】
PAPP−A2のPAPP−Aとの相同性は、図3に示すように2つのアミノ酸配列の整列化により明らかになる。PAPP−A2およびPAPP−Aは、およそ45%のアミノ酸残基を共有する。PAPP−Aの機能(Kristensen et al., 1994, Biochemistry 33, 1592−8;Lawrence et al., 1999, Proc Natl Acad Sci U S A 96, 3149−53;Overgaard et al., 2000, J Biol Chem)に重要と考えられる配列モチーフが、PAPP−A2においても見い出されている。原則的には、PAPP−A2は、733〜743位に伸張された亜鉛結合モチーフ(1文字表記により示されるアミノ酸)を含有する(図2)。このモチーフおよび構造的に重要なメチオニン残基は、厳密的には亜鉛ペプチダーゼのスーパーファミリーである、メトジンシン内に保持されている(Bode et al., 1993, FEBS Lett 331, 134−40;Stocker et al., 1995, Protein Sci 4, 823−40)。
【0107】
PAPP−Aと同様に、PAPP−A2は、prepro蛋白として合成される。PreproPAPP−A2は、1791個のアミノ酸を有する(図1)。PAPP−AおよびPAPP−A2のprepro部分間には、相同性はない。更に、該2つの蛋白のprepro部分は、長さにおいては有意に差はない。PAPP−A2preproペプチドは、233個の残基を有し(図3);PAPP−Apreproペプチドは、80個の残基を有する(Haaning et al., 1996, Eur J Biochem 237, 159−63)。
PAPP−A2をエンコードするヌクレオチド配列の使用
PAPP−A2(またはそのコンプリメント)をエンコードするヌクレオチド配列は、分子生物学の当業者には既知の技術において非常に多くの用途がある。これらの技術としては、ハイブリダイゼーションプローブとしての使用、PCRに対するオリゴマーの構築における使用、PAPP−A2またはそのフラグメントの組み換え生成における使用、およびアンチセンスDNAまたはRNA、それらの化学的類似物(例えばPNAまたはLNA)等の産生における使用が挙げられる。本明細書において開示されるPAPP−A2をエンコードするヌクレオチドの使用は、既知の技術の例示であり、当業者には既知のいかなる技術の使用をも制限する意図はない。更に、本明細書において開示されるヌクレオチド配列は、新しい技術が、例えば、3つの遺伝子コード、特異的塩基対の相互作用が現在既知であるヌクレオチド配列の特性次第であるならば、まだ開発されていない分子生物学的技術において使用され得る。
【0108】
該遺伝子コードの縮重の結果、多数のPAPP−A2をエンコードするヌクレオチド配列が、幾分任意の既知のおよび自然発生的な遺伝子のヌクレオチド配列と最小限の相同性を有して生成され得ることが、当業者により理解されるであろう。本発明は、可能なコドン選択に基づいた組み合わせを選択することにより作成され得るヌクレオチド配列の可能なバリエーションのそれぞれまたはすべてを特に考えている。これらの組み合わせは、自然発生的なPAPP−A2のヌクレオチド配列に適用されるような標準的な3つの遺伝子コードに従って作られ、特に開示されるように、そのバリエーションのすべてが考えら得る。
【0109】
PAPP−A2および/またはその変異体をエンコードするヌクレオチド配列は、好ましくは厳密な条件下で、自然発生的なPAPP−A2のヌクレオチド配列にハイブリダイズ可能であるけれども、PAPP−A2をエンコードするヌクレオチド配列または実質的には異なるコドン使用を有するその誘導体を生成することは有利であり得る。コドンは、特定のコドンが宿主により利用される頻度に従って、特に原核生物または真核生物の発現宿主においてペプチドの発現が起こる速度を上げるよう選択可能である。PAPP−A2をエンコードするヌクレオチド配列および/またはその誘導体を、そのエンコードされたアミノ酸配列を変えないで実質的に変える他の理由としては、自然発生的な配列から生成される転写物より長い半減期といった、より所望される特性を有するRNA転写物の生成が挙げられる。
【0110】
PAPP−A2をエンコードするヌクレオチド配列は、十分に確立された組み換えDNA技術(Sambrook et al., 1989)を用いて、様々な他のヌクレオチド配列に結合され得る。PAPP−A2へ結合するのに有益なヌクレオチド配列としては、例えばプラスミド、コスミド、ラムダ・ファージ誘導体、ファージミド等の当業者には周知の各種クローニングベクターが挙げられる。対象のベクターとしては、発現ベクター、複製ベクター、プローブ産生ベクター、配列決定ベクター等が挙げられる。一般的には、対象のベクターは、少なくとも1つの生体において複製機能の発生源、すなわち簡単な制限エンドヌクレアーゼ感受性部位、および、宿主細胞に対する選択マーカーを含んでいてもよい。
【0111】
本発明の別の態様は、PAPP−A2をエンコードする自然発生的なヌクレオチド配列とハイブリダイズ可能なPAPP−A2特異的核酸ハイブリダイゼーションプローブを提供することである。当該プローブはまた、同様のPAPP−A2をエンコードする配列の検出に使用され得、好ましくは保存領域または活性部位由来の少なくとも50%のヌクレオチドを含有するはずである。本発明のハイブリダイゼーションプローブは、配列番号1のヌクレオチド配列から、またはプロモーター、エンハンサー要素および/またはそれぞれの自然発生的なPAPP−A2を含む可能なイントロンを含む、ゲノム配列から誘導され得る。ハイブリダイゼーションプローブは、32Pまたは35Sといった放射性核種、またはアビジン/ビオチンカップリング系等を介してプローブに結合したアルカリフォスファターゼといった酵素標識を含む、様々なレポーター群により標識され得る。
【0112】
米国特許第4,683,195号および第4,965,188号に記載されるように、PCRは、PAPP−A2をエンコードするヌクレオチド配列に基づいたオリゴヌクレオチドの更なる使用を提供する。PCRにおいて使用される当該プローブは、組み換え起源からなるものであってもよく、化学的に合成されてもよく、あるいはその混合であり、そして診断的用途のための分離したヌクレオチド配列、または非常に関連するゲノム配列の同定のための縮重した可能な配列のプールを含む。
【0113】
PAPP−A2 DNAに対して特異的なハイブリダイゼーションプローブを生成する他の手段としては、PAPP−A2またはPAPP−A2の誘導体をエンコードする核酸配列の、mRNAプローブの生成用ベクター中へのクローニングが挙げられる。そのようなベクターは、当該分野では既知であり、市販で利用可能であり、そしてインビトロでT7またはSP6 RNAポリメラーゼのような適当なRNAポリメラーゼおよび適当に放射活性標識したヌクレオチドの付加によりRNAプローブを合成するのに使用され得る。
【0114】
PAPP−A2をエンコードするDNA配列またはその一部分を完全に合成化学により生成することが可能であり、その後遺伝子をこの特許の出願時に当分野において既知である試薬、ベクター、および細胞を用いて多くの利用可能なDNAベクターのいずれか中に挿入可能である。更に、PAPP−A2配列またはその任意の部分中に変異を導入するのに、合成化学が利用されてもよい。
【0115】
異常レベルのPAPP−A2発現に関係する疾患を検出するためのアッセイにおいて、該ヌクレオチド配列が利用可能である。該ヌクレオチド配列は、当該分野において既知の方法により標識され、ハイブリダイズ条件下で、患者からの体液または組織サンプルに添加可能である。インキュベーション期間後、ヌクレオチドが酵素により標識されるのであれば、そのサンプルを、所望により染料(または現像液を必要とする他の標識)を含有する適合流体で洗浄する。その適合流体を洗い流した後、染料を定量し、スタンダードと比較する。別法としては、PAPP−A2 mRNAのレベルは、固定のプローブを用いるマイクロアレイ法により測定可能である。サンプル中における発現は、(半定量的な)RT−PCRによっても評価可能である。サンプル中における発現は、別法としてハイブリダイゼーションをベースにした方法によっても評価可能である。例えば、インサイツハイブリダイゼーション法は、PAPP−A2 mRNAを検出するのに使用可能である。この方法は、mRNAを合成する細胞を示すという利点を有するが、RT−PCRよりも感受性が低い。
【0116】
アンチセンスRNAとDNA分子、およびPAPP−A2の翻訳を阻害する機能を有するリボザイムを含むオリゴリボヌクレオチド配列が、本発明の範疇に含まれる。アンチセンス法は、当該分野では既知であり、本明細書において適用され得る。本発明のアンチセンスRNAとDNA分子、およびリボザイムが両方とも、RNA分子の合成のために、当該分野で既知の任意の方法により調製され得る。これらは、例えば固相ホスホルアミダイト化学合成といった当該分野では周知のオリゴデオキシリボヌクレオチドを化学的に合成する技術を含む。別法としては、RNA分子は、当該アンチセンスRNA分子をエンコードするDNA配列のインビトロおよびインビボでの転写により産生され得る。そのようなDNA配列は、T7またはSP6ポリメラーゼプロモーターといった好適なRNAポリメラーゼプロモーターを導入する広範なベクター中に導入され得る。別法としては、使用されるプロモーター次第で、構成的にまたは誘導的にアンチセンスRNAを合成するアンチセンスcDNA構築物を、細胞株中に安定に導入可能である。
【0117】
本発明は、他の種から単離した未知のPAPP−A2遺伝子およびそのPAPP−A2遺伝子の対立遺伝子に関し、その中にはPAPP−A2類似遺伝子または相同遺伝子が存在する。本明細書に示されたヒトPAPP−A2クローンの放射活性標識フラグメントを用いて、緩和された厳密条件下においてバクテリオファージcDNAライブラリーが、スクリーニングされ得る。別法として、そのヒトPAPP−A2配列は、PCRプローブとして、またはバクテリオファージcDNAライブラリーをスクリーニングするために使用可能な縮重した、または完全に縮重したオリゴヌクレオチドプローブをデザインするために使用可能である。そのPCR生成物は、サブクローニングされ、増幅された配列がPAPP−A2配列を示すことを保証するために配列決定され得る。そのPCRフラグメントは、増幅フラグメントを放射活性標識し、バクテリオファージcDNAライブラリーをスクリーニングすることにより、完全長のPAPP−A2クローンを単離するために使用され得る。別法として、その標識化フラグメントは、ゲノムライブラリーをスクリーニングするために使用され得る。使用され得るクローニング法の概説については、例えば(Ausubel et al., 1989;Sambrook et al., 1989)が参照される。
組み換えPAPP−A2の発現
生物学的に活性なプロテアーゼを発現するために、その蛋白または機能的等価物に対してコードされるヌクレオチド配列を、適当な発現ベクター、すなわち挿入されたコード化配列の転写および翻訳に必要な要素を含有するベクター中に挿入可能である。例えば、組み換え蛋白は、免疫化して実験試薬としておよび診断キットにおいて抗体を得るために使用可能である。
【0118】
より具体的には、当業者に周知な方法が、PAPP−A2配列および適当な転写/翻訳の制御シグナルを含有する発現ベクターを構築するのに利用可能である。これらの方法としては、インビトロでの組み換えDNA法、合成法、およびインビボでの組み換え/遺伝的組み換えが挙げられる。例えば(Ausubel et al., 1989;Sambrook et al., 1989)に示される方法を参照。
【0119】
更に、PAPP−A2をエンコードする配列のフラグメントを含む発現ベクターが構築されてもよい。特に、これは、免疫用抗原としてのPAPP−A2ポリペプチドの一部分の使用に関連し得る。更に、PAPP−A2またはそのフラグメントのコード化配列が、ベクター中に存在するコード化ヌクレオチド配列を有するフレーム中でクローンされ得、その結果、融合蛋白または「タグ化」PAPP−A2蛋白となる。例えば、当該融合蛋白は、PAPP−A2およびGSTから構成されてもよく、当該タグが、c−mycタグ(検出用)および/またはヒスチジンタグ(精製用)であってもよい。
【0120】
様々な宿主−発現ベクター系が、PAPP−A2コード化配列またはそのフラグメントを発現するために利用され得る。これらは、PAPP−A2コード化配列を含有する組み換えバクテリオファージDNA、プラスミドDNA、またはコスミドDNA発現ベクターを用いて形質転換したバクテリア;PAPP−A2コード化配列を含有する組み換え酵母発現ベクターを用いて形質転換した酵母;PAPP−A2コード化配列を含有する組み換えウィルス発現ベクター(例えばバキュロウィルス)を用いて感染させた昆虫細胞系統;PAPP−A2コード化配列を含有する組み換えウィルス発現ベクター(例えばカリフラワーモザイクウィルス、CaMV;タバコモザイクウィルス、TMV)を用いて感染させた、あるいはPAPP−A2コード化配列を含有する組み換えプラスミド発現ベクター(例えばTiプラスミド)を用いて形質転換した植物細胞系統;または安定に増幅された(CHO/dhfr)、あるいは2つの小さな染色体(例えば、マウスの細胞株)中で不安定に増幅された、いずれかの多数のPAPP−A2 DNAの複製を含有するように作成された細胞株を含む組み換えウィルス発現ベクター(例えば、アデノウィルス、ワクシニアウィルス、ヒト腫瘍細胞)を用いて感染させた動物細胞系統といった微生物体を含むがそれに限定されるわけではない。
【0121】
これらの系統の発現要素は、強度および特性が様々である。利用される宿主/ベクター系統次第で、構成的および誘導的プロモーターを含む、任意の数の好適な転写および翻訳要素が、発現ベクターにおいて利用され得る。例えば、バクテリア系統においてクローニングする場合、バクテリオファージ・ラムダのpL、plac、 ptrp、 ptac(ptrp−lacハイブリッドプロモーター)等といった誘導プロモーターが使用され得;昆虫細胞系統においてクローニングする場合、バキュロウィルス多角体プロモーターといったプロモーターが使用され得;植物細胞系統においてクローニングする場合、植物細胞のゲノムから誘導されるプロモーター(例えば、熱ショックプロモーター;RUBISCOの小さなサブユニットに対するプロモーター;クロロフィルa/b結合蛋白に対するプロモーター)または植物ウィルスから誘導されるプロモーター(例えば、CaMVの35S RNAプロモーター;TMVのコート蛋白プロモーター)が使用され得;哺乳類細胞系統においてクローニングする場合、哺乳類細胞のゲノムから誘導され得プロモーター(例えば、メタロチオネインプロモーター)または哺乳類ウィルスから誘導されるプロモーター(例えば、CMVプロモーター、アデノウィルス後期プロモーター;ワクシンウィルスの7.5Kのプロモーター)使用され得;PAPP−A2 DNAの複数の複製を含有する細胞株を産生する場合、SV40、BPV、およびEBVベースのベクターが適当な選択マーカーと一緒に使用され得る。
【0122】
形質転換、トランスフェクション、感染、プロトプラスト融合、およびエレクトロポレーションを含むがそれに限定されない多数の方法のうちの任意の1つを介して、該発現ベクターを宿主細胞中に導入し得る。その発現ベクターを含有する細胞を、それがPAPP−A2蛋白を生成するかどうかを決定するためにクローンで増殖し、個々に分析する。PAPP−A2を発現する宿主細胞クローンの同定は、抗PAPP−A2抗体との免疫反応性を含むがそれに限定されない数種の手段、および宿主細胞関連PAPP−A2活性の存在によりなされ得る。
【0123】
バクテリア系統において、多数の発現ベクターが、発現されたPAPP−A2の予定の使用される使用次第で有利に選択され得る。例えば、大量のPAPP−A2生成される予定である場合、容易に精製され得る融合蛋白生成物の高レベルの発現を導くベクターが所望となり得る。そのようなベクターとしては、大腸菌発現ベクター、pUR278(Ruther and Muller−Hill, 1983, Embo J 2, 1791−4)が挙げられるがそれに限定されず、当該ベクターにおいてPAPP−A2コード化配列は、ハイブリッドAS−lacZ蛋白を生成するよう、lacZコード化領域を有するフレーム内のベクター中にリゲートされ得る。pGEXベクターはまた、グルタチオンS−トランスフェラーゼ(GST)との融合蛋白として外来のポリペプチドを発現するために使用され得る。一般的に、そのような融合蛋白は可溶性であり、グルタチオン−アガロースビーズへの吸着、次いで遊離型のグルタチオンの存在中での溶出により可溶化細胞から容易に精製可能である。目的のクローン化ポリペプチドがGST部分から遊離可能となるよう、トロンビンまたはXa因子プロテアーゼ開裂部位を含むようにpGEXベクターがデザインされる。酵母において、構成的または誘導的プロモーターを含有する多数のベクターが使用され得る。概説については、(Ausubel et al., 1989;Bitter et al., 1987, Methods Enzymol 153, 516−44;Rosenfeld, 1999, Methods Enzymol 306, 154−69)参照。
【0124】
植物発現ベクターが使用される場合、PAPP−A2コード化配列の発現は、多数のプロモーターのうちのいずれかにより引き起こされ得る。例えば、CaMVの35S RNAおよび19S RNA プロモーターといったウィルスプロモーター(Gmunder and Kohli, 1989, Mol Gen Genet 220, 95−101);あるいは、RUBISCOの小さなサブユニットといった植物プロモーター(Broglie et al., 1984, Science 224, 838−43)が使用され得る。
【0125】
PAPP−A2を発現するために使用可能な代替の発現系は、昆虫系統である。そのような系においては、バキュロウィルスが、外来遺伝子を発現するベクターとして使用される。その後、ウィルスは、昆虫細胞において成育する。PAPP−A2コード化配列が、ウィルスの非必須領域(例えば、多角体遺伝子)中でクローン化されて、バキュロウィルスプロモーターの制御下に置かれてもよい。次いで、これらの組み換えウィルスは、挿入された遺伝子が発現される昆虫細胞を感染するために使用される。例えば(Smith et al., 1983, Mol Cell Biol 3, 2156−65)参照。
【0126】
様々な哺乳類の発現ベクターが、哺乳類細胞中で組み換えPAPP−A2を発現するために使用され得る。組み換えPAPP−A2発現に好適となり得る市販で入手可能な哺乳類の発現ベクターとしては、pMC1neo(Stratagene)、pXT1(Stratagene)、pSG5(Stratagene)、EBO−pSV2−neo(ATTC 37593)、pBPV−1(8−2)(ATCC 37110)、pcDNA3.1およびその誘導体(Stratagene)が挙げられるがそれに限定されない。好適となり得、かつ市販で入手可能である哺乳類種由来の細胞株としては、CV−1、COS−1、COS−7、CHO−K1、3T3、NIH3T3、HeLa、C127I、BS−C−1、MRC−5および293が挙げられるがそれに限定されない。更に、哺乳類の宿主細胞においで、多数のウィルスベースの発現系が利用され得る。発現ベクターとしてアデノウィルスが使用される場合においては、PAPP−A2コード化配列が、アデノウィルス転写/翻訳制御複合体、例えば後期プロモーターおよび3つのリーダー配列にリゲートされ得る。次に、このキメラ遺伝子が、インビトロまたはインビボの組み換えによりアデノウィルスゲノム中に挿入され得る。ウィルスゲノムの非必須領域(例えばE1またはE3領域)への挿入により、生育可能で感染宿主中でPAPP−A2が発現可能な組み換えウィルスとなるであろう。例えば(Logan and Shenk, 1984, Proc Natl Acad Sci U S A 81, 3655−9)参照。別法としては、ワクシニアの7.5Kのプロモーターが使用されてもよい。例えば(Mackett et al., 1982, Proc Natl Acad Sci U S A 79, 7415−9)参照。
【0127】
長期間にわたる高収率の組み換え蛋白生成のために、安定した発現が好ましい。例えば、PAPP−A2を安定に発現する細胞株が作成されてもよい。複製のウィルス源を含有する発現ベクターを用いるよりもむしろ、宿主細胞は、適当な発現制御要素(例えば、プロモーター、エンハンサー、シーケンサー、転写ターミネーター、ポリアデニル化部位等)、および選択マーカーにより制御されたPAPP−A2 DNAを用いて形質転換可能である。外来DNAの導入に続いて、作成された細胞が、栄養強化培地において1〜2日間生育され得、その後、選択培地に交換される。組み換えプラスミド中の選択マーカーは、その選択に耐性を与え、そして細胞が安定にそのプラスミドを染色体に組み入れ、増殖して病巣を形成するのを可能にし、次にクローン化され細胞株中に拡大可能となる。
【0128】
組み換えPAPP−A2の適用のいくつかは、その蛋白を精製されたあるいは一部精製された形態にする必要があり得る。組み換えにより発現したPAPP−A2またはPAPP−A2ポリペプチドのフラグメントは、液体クロマトグラフィーによって単離可能である。当該分野において周知の様々な蛋白精製法としては、例えば(Scopes, 1987)に示されている方法が挙げられる。別法としては、組み換えPAPP−A2融合蛋白、または「タグ化」PAPP−A2がアフィニティークロマトグラフィーにより精製され得る。更に、PAPP−A2対して産生される抗体が、イムノアフィニティークロマトグラフィーによる精製のために使用され得る。
【0129】
PAPP−A2の組み換え変異体が、部位特異的突然変異誘発により生成され得る。PAPP−A2の適用の中には、こういった変異体が、例えば蛋白安定性の増大、または活性の変化のために好ましくなり得るものもある。
PAPP−A2に対する抗体の生成と使用
組み換え蛋白が抗体を産生するのに使用され得る。PAPP−A2に対する単一特異的抗体が、PAPP−A2に対して反応性のある抗体を含有する哺乳類の抗血清から精製可能であり、あるいは標準的な方法を用いてPAPP−A2と反応可能なモノクローナル抗体として調製可能である。
【0130】
本明細書において使用されるように単一特異的抗体は、PAPP−A2に対して同種結合特性を有する単一の抗体種または多数の抗体種として定義される。本明細書において使用されるように同種結合とは、上述に示すように、抗体種のPAPP−A2と関連する抗原といった特異的な抗原またはエピトープへの結合能をいう。PAPP−A2特異的抗体は、マウス、ラット、モルモット、ウサギ、ヤギ、ウマ等といった動物を(ラビットまたはマウスが好ましい)、免疫アジュバント有りあるいは無しのいずれかで、適当なPAPP−A2の濃度を用いて免疫することにより産生される。例えば、PAPP−A2に対して特異的な抗体は、天然および組み換えPAPP−A2の精製用に、実験試薬として、および抗体ベースの診断用キットにおいて使用可能である。
【0131】
PAPP−A2と反応可能なモノクローナル抗体(mAb)は、常法により、例えば同系交配マウスをPAPP−A2で免疫することにより調製可能である。該マウスは、約0.5mlの緩衝液または生理食塩水中において等量の許容可能なアジュバントに組み込まれた約0.1mg〜約10mg、好ましくは約1mgのPAPP−A2を用いて免疫される。フロイントの完全アジュバントが好ましい。該マウスは、0日目に初期免疫を受け、約3〜約30週間休息させる。免疫化マウスは、約0.1〜約10mgのPAPP−A2をリン酸緩衝生理食塩水(PBS)といった緩衝溶液で静注(IV)経路により、1度またはそれ以上追加免疫される。抗体陽性のマウス由来のリンパ節は、当該分野において既知の標準的な方法で免疫化マウスから脾臓を除去することにより得られる。ハイブリドーマ細胞は、安定なハイブリドーマが形成される条件下で、脾リンパ節を適当な融合相手と混合することにより生成される。融合されたハイブリドーマ細胞は、当該分野において既知の方法を用いて、ヒポキサンチン、チミジン、およびアミノプテインを補充したダルベッコ変法イーグル培地(Dulbecco’s Modified Eagles Medium、DMEM)での生育により選択される。約14、18、および21日目で生育陽性のウェルから上清液をを回収し、固相イムノラジオアッセイ(SPIRA)といったイムノアッセイによる抗体生成に対して、抗原としてPAPP−A2を用いてスクリーニングする。mAbのアイソトープを決定するために、オクタロニー(Ouchterlony)沈降アッセイにおいて培養液も試験される。次に抗体陽性ウェルからのハイブリドーマ細胞がクローン化される。詳細については(Peters and Baumgarten, 1992)参照。
【0132】
インビトロでの抗PAPP−A2の生成は、十分な量の特異的mAbを得るために約2%のウシ胎仔血清を含有するDMEMにおけるハイブリドーマの生育により実施される。該mAbは、当該分野において既知の方法により精製される。 腹水またはハイブリドーマ培養液の抗体価は、沈降法、受身凝集反応、酵素免疫測定法(ELISA)(Crowther, 1995)を含むがそれに限定されない様々な血清学的または免疫学的アッセイにより決定される。
【0133】
「モノクローナル抗体」はまた、例えば(Clackson et al., 1991, Nature 352, 624−8;Marks et al., 1991, J Mol Biol 222, 581−97)に示されている方法を用いて、ファージ抗体ライブラリーから単離され得る。同定されたファージ抗体は、バクテリアにおける発現により生成可能である。
【0134】
上述のような方法は、PAPP−A2ポリペプチドフラグメントまたは完全長の発生期のPAPP−A2ポリペプチドに特異的な単一特異的抗体を生成するのに使用され得る。
【0135】
PAPP−A2抗体アフィニティーカラムは、抗体をゲル支持体、たとえばN−ヒドロキシスクシンイミドエステルで予め活性化されているゲル支持体Affigel−10(Biorad)に添加して、該抗体をアガロースゲルビーズ支持体と共有結合を形成させることにより作成することができる。次いで、該抗体は、スペーサーアームによりアミド結合を介して該ゲルに結合される。残りの活性化エステルが、その後1M エタノールアミン HCl(pH8)でクエンチングされる。そのカラムを、水、次いで0.23M グリシン HCl(pH2.6)で洗浄して、いかなる非共役型抗体または外来蛋白をも除去する。次に、そのカラムをリン酸緩衝生理食塩水(pH7.3)中で平衡化し、PAPP−A2またはPAPP−A2フラグメントを含有する細胞培養上清または細胞抽出物をそのカラムにゆっくりと通過させる。次に、そのカラムを洗浄し、蛋白を溶出する。精製されたPAPP−A2蛋白を次に、リン酸緩衝生理食塩水で透析する。
【0136】
ヒト血漿または血清、組織抽出物、あるいはトランスフェクトされていない細胞株由来の培地といった起源(PAPP−A2を内生に分泌する)由来の天然のPAPP−A2も、抗体アフィニティーカラムを使用することにより精製され得る。
【0137】
PAPP−A2に対するポリクローナル抗体またはモノクローナル抗体を用いて、体液または組織、および細胞抽出物中のPAPP−A2抗原の測定に多数のアッセイが実施され得る。診断の目的で、抗体ベースのキットが使用され得る。該アッセイとしては、沈降法、受身凝集反応、酵素免疫測定法(ELISA)、およびラジオイムノアッセイ(RIA)法が挙げられるがそれに限定されない。
【0138】
例えば、そのようなELISAの1つにおいて、サンプル中に存在する抗原が固定化ポリクローナル抗(PAPP−A2)により獲得される場合、サンドイッチアッセイ法が実施可能である。次に、1つまたはそれ以上のモノクローナルPAPP−A2抗体およびペルオキシダーゼ共役抗(マウスIgG)を使用することにより、検出が実施される。別のアッセイにおいては、サンプル中に存在する抗原が、固定化ポリクローナル抗(PAPP−A2)により獲得され、ビオチン化ポリクローナル抗(PAPP−A2)を用いて検出される。更なる実施例および詳細については、(Crowther, 1995)参照。アッセイは、精製されたPAPP−A2を用いて、キャリブレートして一連の希釈により標準曲線を構築することができる。精製された形態での溶液中のPAPP−A2濃度は、アミノ酸分析(Sottrup−Jensen, 1993, Biochem Mol Biol Int 30, 789−94)により正確に測定可能である。
【0139】
PAPP−A2の生物学的活性を阻害するために、ポリクローナル抗体が使用され得る。特異的に、ポリクローナルPAPP−A抗体によるPAPP−AのIGFBP−4蛋白分解活性の阻害(Lawrence et al., 1999, Proc Natl Acad Sci U S A 96, 3149−53)と同様に、PAPP−A2の蛋白分解活性を阻害するために、抗(PAPP−A2)が使用され得る。PAPP−A2の活性に対して、特定のモノクローナル抗体がまた、抑制性となり得る。そのようなモノクローナル抗体は、PAPP−A2の活性部位に非常に近接するエピトープを認識する傾向があるが、その阻害活性は、活性部位の近く以外のエピトープへの結合に基づくものであり得る。抑制性のモノクローナル抗体は、PAPP−A2、PAPP−A2から誘導されたペプチドを有するPAPP−A2フラグメントの免疫により得ることができる。
【0140】
抑制性(モノクローナル)抗体は、PAPP−A2活性を減少させるのに所望となり得る病状において治療的価値を有し得る。
PAPP−A2の活性
PAPP−Aと同様、PAPP−A2は、メトジンシンスーパーファミリーの推定のメタロプロテアーゼ(Stocker et al., 1995, Protein Sci 4, 823−40)として分類される保存アミノ酸分岐鎖を含有する。PAPP−A2がインシュリン様成長因子結合蛋白、(IGFBP)−5の開裂を示すことにより蛋白分解活性を呈する(実施例6.7)ことが、実験で確認されている。
【0141】
一般的に、潜在的な蛋白基質に対するPAPP−A2の蛋白分解活性が、精製または一部分精製されたPAPP−A2を様々な実験条件(例えば、温度、緩衝液組成、イオン強度、およびpH)下でその潜在的な基質とインキュベーションすることにより評価され得る。当該蛋白に対するPAPP−A2の酵素活性は、SDS−PAGE(明確に形成される蛋白分解性フラグメントの分解または遊離が明らかとなる)、または遊離したペプチドの高速液体クロマトグラフィー検出により評価可能である。そのような方法を用いることにより、PAPP−A2の他の基質ターゲットが同定され得る。例えば活性部位の残基が不活性な酵素を獲得するために置換されている様々なPAPP−A2とのインキュベーションが、適切なネガティブコントロールとしての役割を果たす。
【0142】
固相支持体に接着したアミノ酸の可能なすべての組み合わせからなるランダムペプチドライブラリーが、PAPP−A2により開裂可能なペプチドを同定するのに使用され得る。そのようなペプチドの同定は、ペプチドライブラリーを組み換え可溶性PAPP−A2を用いてスクリーニングすることにより達成され得る。酵素の発現および精製法が上述で示され、目的の機能的ドメインによって、組み換えの完全長PAPP−A2またはそのフラグメント、類似物、あるいは誘導体が使用され得る。更なる詳細については、(Meldal, 1998, Methods Mol Biol 87, 65−74;Meldal, 1998, Methods Mol Biol 87, 51−7)参照。あるいは、ペプチド基質が、同定されたPAPP−A2の蛋白基質から誘導されてもよい。
【0143】
別法としては、PAPP−A2により開裂可能なペプチドを同定するのにペプチドライブラリーのファージディスプレイが使用されてもよい(Matthews and Wells, 1993, Science 260, 1113−7)。
【0144】
PAPP−A2基質としての機能するペプチドは、体液または組織、および細胞抽出物におけるPAPP−A2蛋白分解活性の検出用アッセイにおいて機能し得る。蛍光クエンチングに基づいたアッセイにおいて機能するために、基質ペプチドが誘導され得る(Meldal, 1998, Methods Mol Biol 87, 65−74)。そのような、または他の方法に基づいたキットが、PAPP−A2活性の測定が関係する病気において診断目的で使用され得る。
PAPP−A2の活性を改変する試薬の同定
PAPP−A2の蛋白分解活性の検出用アッセイは、上述のように、PAPP−A2の活性を改変する分子の同定法を提供する。そのような分子は、例えば、ペプチド、誘導化ペプチド、ヒドロキサム酸誘導化ペプチド、小さな有機分子、または抗体であってもよい。
【0145】
ペプチドライブラリーのスクリーニングは、PAPP−A2の生物学的活性を改変および/または阻害するよう作用する試薬を見いだすために使用可能である。酵素の発現および精製法は、上述で示されており、目的の機能的ドメインによって、組み換えの完全長PAPP−A2またはそのフラグメント、類似物、あるいは誘導体を発現するために使用され得る。固相支持体に接着したアミノ酸の可能なすべての組み合わせからなるランダムペプチドライブラリーは、PAPP−A2の活性部位または他の部位に結合することにより、PAPP−A2活性を改変および/または阻害可能なペプチドを同定するため使用され得る。例えば、(Meldal, 1998, Methods Mol Biol 87, 75−82)参照。
【0146】
同様に、PAPP−A2の活性に影響を及ぼす低分子量の有機分子を同定するために、コンビナトリアルケミストリーが利用されてもよい。
PAPP−AまたはPAPP−A2の複合体の測定
PAPP−Aは、好酸球主要塩基蛋白(proMBP)のプロフォームとの2:2のジスルフィド結合複合体、PAPP−A/proMBPとして主に妊婦の血清中に存在する。PAPP−A/proMBP複合体に加えて、proMBPは、アンジオテンシン(ANG)との2:2のジスルフィド結合複合体、proMBP/ANGとして循環中に存在し、この複合体のフラクションは、更に補体成分C3dgのフラグメントと複合体化(PROMBP/ANG/C3dg)している(Oxvig, 1995;Christiansen, 2000)。
【0147】
個体の体液中のPAPP−Aおよび/またはPAPP−A2および/またはproMBPを含む複合体のレベルは、病態の素因を示したり、または病態の存在を示したりし得る。従って、本発明は、1つの実施態様において、個体において病態を診断したり、当該病態の素因を診断したりする方法を教示し、その方法は、
a)当該個体から体液サンプルを提供し;そして
b)当該組体液サンプル中のPAPP−A/proMBP、PAPP−A2/proMBP、PAPP−A/PAPP−A2、PAPP−A/PAPP−A2/proMBP、proMBP/ANG、およびproMBP/ANG/C3dgからなる群から選ばれる複合体のレベルを測定し;そして
c)病態または病態の素因を診断(予め決められた値より上または下の複合体レベルが病態または病態の素因を示す)することからなるステップを含む。
【0148】
更に、哺乳類の母親の体液中のPAPP−Aおよび/またはPAPP−A2および/またはproMBPを含む複合体レベルは、当該母親の胎児の病態の素因を示したり、病態の存在を示したりし得る。それゆえ、本発明は、哺乳類の胎児において病態を診断したり、当該病態の素因を診断する方法を提供し、その方法は、
a)当該胎児の母親から体液サンプルを提供し;そして
b)当該体液サンプル中のPAPP−A/proMBP、PAPP−A2/proMBP、PAPP−A/PAPP−A2、PAPP−A/PAPP−A2/proMBP、proMBP/ANG、およびproMBP/ANG/C3dgからなる群から選ばれる複合体のレベルを測定し;そして
c)病態または病態の素因を診断(予め決められた値より上または下の複合体レベルが病態または病態の素因を示す)する
ことからなるステップを含む。
【0149】
特に、本発明の方法に従って、以下に示す複合体のうちの1つまたはそれ以上のレベルが決定され得る:
PAPP−A/proMBP
PAPP−A2およびproMBP(PAPP−A2/proMBP)
PAPP−A2およびPAPP−A(PAPP−A/PAPP−A2)
proMBPを有するPAPP−A/PAPP−A2(PAPP−A/PAPP−A/proMBP)
proMBP/ANG
proMBP/ANG/C3dg
体液サンプル中のPAPP−Aおよび/またはPAPP−A2および/またはproMBPを含む複合体レベルは、当業者には既知の任意の従来法によって決定され得る。例えば、PAPP−A、PAPP−A2、proMBP、ANG、またはC3dgと特異的に相互作用する免疫特異的な試薬といった、測定するのに所望の複合体の1つまたはそれ以上の成分と特異的に相互作用する免疫特異的な試薬の使用を含む方法により、そのレベルは測定可能である。例えば、免疫特異的な試薬は、複合体の個々の成分に特異的な、モノクローナル抗体、ポリクローナル抗体および/またはその抗原結合フラグメントであり得る。
【0150】
そのような方法としては、サンドイッチELISA法が挙げられるがそれに限定されるわけではなく、当該方法においては、複合体の1成分を特異的に認識する免疫特異的な試薬が捕獲抗体として使用され、その複合体が検出抗体として使用されるならば、別の免疫特異的な試薬が別の成分を特異的に認識する。その検出抗体が、直接的または間接的のいずれかで検出可能であるのが好ましく、例えば検出抗体が検出可能なラベルに直接結合してもよく、あるいは検出抗体が、検出可能なラベルと結合した別の試薬と相互作用可能であってもよい。
【0151】
例えば、検出可能なラベルは、蛍光ラベル、色素体、放射活性ラベル、重金属、または酵素であり得る。
【0152】
例えば、体液サンプル中のPAPP−A/proMBP複合体のレベルは、捕獲用のPAPP−A特異的なモノクローナルまたはポリクローナル抗体および検出用のproMBP特異的なモノクローナルまたはポリクローナル抗体を用いて、サンドイッチELISA法により決定され得、あるいは体液サンプル中のproMBP/ANGのレベルは、捕獲用のproMBP特異的なモノクローナルまたはポリクローナル抗体および検出用のANG特異的なモノクローナルまたはポリクローナル抗体を用いてサンドイッチELISA法により決定され得る。
【0153】
病態(clinical condition)は、PAPP−Aおよび/またはPAPP−A2および/またはproMBPを含む複合体のレベルより診断され得る、あるいはPAPP−Aおよび/またはPAPP−A2および/またはproMBPを含む複合体のレベルにより素因が診断され得る任意の病態であり得る。例えば、病態は、ダウン症、子癇前症、および不安定狭心症および心筋梗塞を含む急性冠症候群からなる群から選ばれ得る。
【0154】
体液サンプルは、血清サンプルを含む血液サンプル、尿サンプル、糞サンプル、または羊水サンプルといった任意の有用な体液サンプルであり得る。
【0155】
特に、PAPP−A/proMBPレベルは、病態が、ダウン症、および不安定狭心症および心筋梗塞を含む急性冠症候群からなる群から選ばれる場合に決定され得る。
【0156】
本発明の1つの実施態様において、ダウン症の診断またはダウン症の素因の診断は、PAPP−A/proMBPレベルの決定することを含み、予め決められた値より低いPAPP−A/proMBPレベルは、ダウン症またはダウン症の素因があることを示している。
【0157】
本発明の別に実施態様において、不安定狭心症および心筋梗塞を含む急性冠症候群の診断、または不安定狭心症および心筋梗塞を含む急性冠症候群の素因の診断は、PAPP−A/proMBPレベルを決定することを含み、予め決められた値より高いPAPP−A/proMBPレベルは、不安定狭心症および心筋梗塞を含む急性冠症候群または不安定狭心症および心筋梗塞を含む急性冠症候群の素因があることを示している。
【0158】
更に別の実施態様において、ダウン症の素因を診断する、あるいはダウン症を診断するために、proMBP/ANGレベルが決定され得る。上述の診断法はすべて、1つまたはそれ以上の他の診断法と組み合わせて実施されてもよい。更に、本発明に記載の1以上の異なる診断が実施されてもよく、例えば、1以上の複合体レベルを測定する、あるいは異なる個体サンプルにおいて1つの複合体レベルを測定することが可能である。
天然の蛋白分解性フラグメントを産生するためのPAPP−A2の使用
PAPP−A2は、PAPP−A2により特異的に開裂される蛋白の天然のフラグメントを産生するために使用され得る。IGFBP−5の場合のように(実施例6.7および6.9参照)、そういったフラグメントは、インタクトなIGFBP−5とは異なる生物学的効果を有し得る。精製されたPAPP−A2で開裂後、標準的なクロマトグラフィーによりフラグメントを精製可能である(実施例6.9参照)。
発現用PAPP−A2フラグメントのデザイン
成熟PAPP−Aにおいて見られるシステイン残基がすべて、成熟PAPP−A2においても見られるため(図3参照)、ジスルフィド結合パターンは、共通のシステイン残基のパターンについては、PAPP−A2と同じであると推定することができる。それゆえ、PAPP−Aサブユニットのジスルフィド構造(図8参照)の知見が、すべてのシステイン残基組み合わせが可能であるPAPP−A2フラグメントを合理的にデザインするために利用可能である。PAPP−A2の推定のドメイン境界は、図8に示されるジスルフィド構造に基づいて決めることができる。それらのドメインは、別々にまたは一緒に発現可能である。ドメインが別のPAPP−AサブユニットまたはproMBPに対して鎖間ジスルフィド結合を形成する(図8参照)ことが既知のシステイン残基を含有する場合に、このシステインが突然変異して、例えばセリンまたはアラニン残基となることが必要となり得る。
【0159】
従って、可能な境界領域は、Cys−403とCys−499との間、Cys−828とCys−881との間、Cys−1048とCys−1115との間、Cys−1390とCys−1396との間、Cys−1459とCys−1464との間、Cys−1521とCys−1525との間、Cys−1590とCys−1595との間、Cys−1646とCys−1653との間、およびCys−1729とCys−1773との間(preproPAPP−A2ナンバリングは、図1および3と同様)である。
医薬組成物
IGFBP−5プロテアーゼとしてのPAPP−A2の同定は、治療用ターゲットとしてPAPP−A2を用いることにより、インビボにおいて成長および分化に影響を及ぼす方法を提供する。PAPP−A2の阻害は、バイオアベイラブルなIGF−IおよびIGF−IIの量を減少させると考えられている。例えば、PAPP−A2活性の阻害は、再狭窄、アテローム性動脈硬化、および線維症といった疾患に有益となり得る。アクチベーター、またはPAPP−A2の活性を上昇させる薬剤は、バイオアベイラブルなIGF−IおよびIGF−IIの量を上昇させると考えられている。
【0160】
PAPP−A2活性を変化させる、またはPAPP−A2の細胞表面への接着を変化させる薬剤を、医薬組成物中に組み込むことができる。そのような薬剤は、PAPP−A活性を変化させる、あるいはPAPP−Aの細胞表面への接着を変化させる薬剤と一緒に組み込まれてもよい。PAPP−A2特異的な薬剤とPAPP−A特異的な薬剤との組み合わせは、PAPP−Aに対して示される従来の試薬よりも有効となり得る。PAPP−A2特異的試薬とPAPP−A特異的試薬との組み合わせを薬学的に有効量でそれらを必要とする個体に投与することからなるステップを含む処置法もまた提供されている。
【0161】
一例として、抗PAPP−A2ポリクローナルまたはモノクローナルといった抗体が、薬学的に許容される無毒性の賦形剤または担体との混合により医薬組成物に製剤化することができる。そのような1つまたは複数の組成物は、特に溶液または水性生理緩衝食塩水中に懸濁した形態で経腸投与用に;特に錠剤またはカプセルの形態で経口投与用に;あるいは特に粉末状、点鼻薬、またはエアロゾルの形態で経鼻用に調製されてもよい。他の投与経路用の組成物が、常法を用いて所望されるように調製されてもよい。
【0162】
経腸投与用製剤は、一般的な賦形剤(すなわち、薬学的に許容される担体)として、滅菌水または生理食塩水、ポリエチレングリコールといったポリアルキレングリコール、野菜起源の油、水素化ナフタレン等を含有してもよい。特に、生物適合性、生物分解性の乳酸ポリマー、ラクチド/グリコリドコポリマー、またはポリオキシエチレン−ポリオキシプロピレンコポリマーが、インビボで本発明の化合物の放出を制御する賦形剤の例である。他の好適な経腸的送達システムとしては、エチレン−酢酸ビニルコポリマー粒子、浸透ポンプ、埋込み型注入システム、およびリポソームが挙げられる。吸入投与用製剤は、所望であれば、ラクトースといった賦形剤を含有してもよい。吸入製剤は、例えばポリオキシエチル−9−ラウリルエーテル、グリココール酸、およびデオキシコール酸を含有する水性溶液であってもよく、あるいはそれらが点鼻薬の形態での投与用油状溶液であってもよい。所望であれば、該化合物が経鼻的に適用されるゲルとして製剤化可能である。経腸投与用製剤も、舌下投与用にグリココール酸を含んでいてもよい。
医療用具
本発明は、患者の体内配置(例えば移植)用医療用具も特徴としており、それはPAPP−A2プロテアーゼ活性を阻害または活性化する薬剤を含む。好適な薬剤は、本明細書に記載の方法を用いて容易に同定される。用具は、該薬剤を含んだり、あるいは該試薬をコートすることが可能である。阻害剤の非制限的な例としては、抗−PAPP−A2ポリクローナルまたはモノクローナルといった抗体、あるいは1,10−フェナントロリンといったメタロプロテアーゼ阻害剤が挙げられる。
【0163】
PAPP−A2のIGFBP−5プロテアーゼ活性は、1,10−フェナントロリンにより強く阻害されるが、マトリックスメタロプロテアーゼの組織阻害剤(TIMP’S)によっては阻害されない。他の阻害剤としては、ヒドロキサム酸の誘導体といった小分子が挙げられる。抗PAPP−A2ポリクローナルIgGもまた、HFCMにおいてIGF−依存性−またはIGF−非依存性−IGFBP−5特異的PAPP−A2プロテアーゼ活性を用量依存的に阻害し得る。
【0164】
更に、修飾化ポリペプチドを含むポリペプチド(すなわち、長さまたは翻訳後修飾にかかわらず、任意のアミノ酸)は、阻害剤として機能することができる。本発明に記載の医療用具をコートしたり埋め込むために、PAPP−A2のIGFBP−5プロテアーゼ活性の任意の阻害剤を使用可能である。修飾化ポリペプチドとしては、対応する野生型配列と比較して、アミノ酸配列におけるアミノ酸の置換、欠失、または挿入、ならびに化学修飾が挙げられる。プロテアーゼ抵抗性IGFBP−5が、PAPP−A2のIGFBP−5プロテアーゼ活性の阻害剤そのものではないけれども、それが医療用具をコートしたり埋め込んだりするのに使用される場合、同様の結果が期待される。
【0165】
一例として、PAPP−A2阻害剤を用いた医療用具のコーティングまたは埋め込みは、所望によりPAPP−A阻害剤と組み合わせて、バルーン血管形成後の再狭窄の進行の阻害を助けたり、動脈硬化性プラークサイズの更なる増大を阻害したりすることが可能である。ステント留置による冠動脈血管形成は、冠動脈アテローム性動脈硬化に対する現在の主要な治療法である。冠動脈疾患の血管形成の重要な目的は、急性および慢性の合併症の両方を回避することである。現代の方法は、当面の問題の削減には非常に成功している。残念なことに、ステントされた患者の20〜30%においては、それでもなお再狭窄が起こる。既知の薬理学的治療法(pharmacological intervention)で、再狭窄を回避するのに利用可能なものはない。
【0166】
特定のメカニズムにより結合されないで、ヒトにおける血管形成に応答した新生内膜肥厚に先立って冠動脈平滑筋細胞によるIGFBP−5プロテアーゼ発現が増加すると考えられている。
【0167】
例えば、PAPP−A2活性の促進は、創傷治癒、骨折、骨粗鬆症、または排卵に有益となり得る。骨粗鬆症または骨量減少の他の症状は、骨形成が増大し、骨再吸収が減少することにより利益を受け得る。PAPP−A2活性を促進させる薬剤としては、例えば修飾化IGF、すなわちIGF類似物が可能である。
【0168】
類似物としては、アミノ酸の挿入、欠失、または置換、ならびに化学修飾を含むIGFポリペプチドが挙げられる。アミノ酸の置換としては、同類および非同類アミノ酸置換が挙げられ得る。同類アミノ酸置換は、あるアミノ酸を同じクラスのアミノ酸と置き換え、一方、非同類アミノ酸置換は、あるアミノ酸を異なるクラスのアミノ酸と置き換える。非同類置換の結果、ポリペプチドの疎水性または残りの側鎖のバルクが変化する。更に、非同類置換は、正電荷を減少させ負電荷を導入するなど、ポリペプチドの電荷を本質的に変化させる。非同類置換の例としては、非極性アミノ酸の塩基性アミノ酸への置き換え、または酸性アミノ酸の極性アミノ酸の置き換えが挙げられる。アミノ酸の挿入、欠失、および置換は、当該分野には既知のランダム突然変異誘発、部位特異的突然変異誘発、または他の組み換え法の使用が可能である。
【0169】
例えば、医療用具としては、骨を安定させるために使用される骨プレートまたは骨スクリュー、あるいは体内管腔部の開存性を回復したり維持したりするために典型的には体内で使用されるステントが可能である。例えば、血管は血液の流れを制限する動脈硬化性のプラークにより詰まってくる。典型的にはステントは、体内管腔部内の流動を調節する内部チャンネルを形成する管状構造を有している。ステントの外壁は、体内管腔部の内壁と接合している。病変部内のステントの配置により、体内管腔部の更なる閉塞の回避し、そして継続的な流動を可能にすることを助けることができる。典型的にはステントは、ステントを運ぶカテーテルまたはガイドワイヤーの経皮的挿入により実施される。通常ステントは、膨張可能な構造を有する。所望の部位へ送達する際、ステントはカテーテルに装着されたバルーンにより膨張可能である。別法としては、ステントが、圧縮された状態で鞘または他の制限部内に保持するバイアス構造または弾性構造を有していてもよい。その制限が除かれたときに、ステントは自発的に膨張する。いずれの場合においても、ステント壁は、膨張して体内管腔部の内壁に接合し、そして一般的には所望の位置にステントを固定する。
本発明の記載
第1の態様において、本発明は、
i)受け入れ番号DSM13783下のDSMZに集積された、PAPP−A2のコード化配列に相当する、配列番号1のヌクレオチド1〜5376を含むポリヌクレオチド;および
ii)配列番号2も示されるようなアミノ酸配列を有するポリペプチドをエンコードするポリヌクレオチド;および
iii)ポリヌクレオチド(i)または(ii)によりエンコードされたポリペプチドのフラグメントをエンコードするポリヌクレオチドであって、当該フラグメントが、
a)インシュリン様成長因子結合蛋白5型(IGFBP−5)またはその誘導体、あるいは任意の他の基質に特異的な蛋白分解活性を有し;そして/または
b)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
c)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、ポリヌクレオチド;そして
iv)相補鎖が、(i)、(ii)、および(iii)のいずれかで定義されるようなポリヌクレオチドと、厳密な条件下でハイブリダイズされるポリヌクレオチド
であって、当該ポリヌクレオチドが、配列番号2で示されるようなアミノ酸配列を有するポリペプチド、またはそのフラグメントをエンコードし、当該フラグメントが、
a)インシュリン様成長因子結合蛋白5型(IGFBP−5)またはその誘導体、あるいは任意の他の基質に特異的な蛋白分解活性を有し;そして/または
b)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
c)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、ポリヌクレオチド;および
v)(iii)および(iv)のいずれかで定義されるようなポリヌクレオチドのヌクレオチド配列に縮重されるヌクレオチド配列を含むポリヌクレオチド、
、およびそのようなポリヌクレオチドの相補鎖、からなる群から選ばれる精製ポリヌクレオチドに関する。
【0170】
本明細書において使用されるように、ポリヌクレオチドとは、任意の自然発生的な基本骨格構造を有する任意の自然発生的なポリヌクレオチド、ならびに当該分野においてLNA(ロック型核酸、locked nucleic acid)およびPNA(ペプチド核酸)として既知のヌクレオチドをいう。
【0171】
好ましい実施態様において、精製ポリヌクレオチドは、配列番号1で示されるようなPAPP−A2、ヌクレオチド1〜5376からなるコード化配列、または配列番号2で示されるようなアミノ酸配列をエンコードするヌクレオチド配列を含む。
【0172】
別の好ましい実施態様において、ポリヌクレオチドは、配列番号2で示されるようなアミノ酸配列を有するポリペプチドのフラグメントをエンコードするヌクレオチド配列を含み、当該フラグメントは、
a)インシュリン様成長因子結合蛋白5型(IGFBP−5)またはその誘導体、あるいは任意の他の基質に特異的な蛋白分解活性を有し;そして/または
b)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
c)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する。
【0173】
相補鎖が本発明に記載のポリヌクレオチドと厳密な条件下でハイブリダイズするポリヌクレオチドも提供される。
【0174】
本明細書で使用されるような厳密な条件とは、例えば、Southern E. M., 1975, J. Mol. Biol. 98:503−517により示されるようなサザンブロッティングおよびハイブリダイゼーションに関連して通常適用されるような厳密さをいう。そのような目的のために、プレハイブリダイゼーションおよびハイブリダイゼーションからなるステップを含むことが日常的に実施されている。このようなステップは、通常、“Molecular Cloning/A Laboratory Manual”, Cold Spring HarborにおいてSambrook et al., 1989により示される(これは、参照して本明細書の一部とする)ように、6xSSPE、5% Denhardt’s、0.5% SDS、50% ホルムアミド、100mg/ml 変性サケ精巣DNA(42℃で18時間インキュベーション)を含有する溶液を用いて実施され、次に2xSSCおよび0.5% SDS(室温および37℃)で洗浄し、0.1xSSCおよび0.5% SDS(68℃で30分間インキュベーション)で洗浄する。
【0175】
DNA配列は、様々な方法で使用される。それらは、uHAseのホモログ(例えば、huHAseのホモログ)を同定するためのプローブとして使用され得る。哺乳類のホモログは、相互に実質的な配列類似性、すなわち少なくとも75%、通常少なくとも90%、より一般的には少なくとも95%の配列同一性を有する。配列類似性は、保存モチーフ、コード化領域、フランキング領域等といった比較的長い配列のサブセットであり得る標準配列を基準にして計算される。標準配列は、通常、少なくとも約18ntの長さ、より一般的には少なくとも約30ntの長さとなり、比較される完全長の配列まで延び得る。配列分析に対するアルゴリズムは、当該分野、例えばAltschul et al. 1990 J Mol Biol 215:403−10に記載されるBLASTにおいて既知である。
【0176】
配列類似性を有する核酸が、厳密性の低い条件下、例えば50℃および10倍のSSC(0.9M 生理食塩水/0.09M クエン酸ナトリウム)でハイブリダイゼーションにより検出され、55℃および1倍のSSCで洗浄されるときには結合したままである。配列同一性は、厳密性の高い条件下、例えば50℃またはそれ以上および0.1倍のSSC(9mM 生理食塩水/0.9 Mクエン酸ナトリウム)でハイブリダイゼーションにより検出され得る。プローブ、特にDNA配列の標識化プローブを使用することにより、ホモログまたは関連遺伝子を単離することができる。ホモログ遺伝子の起源は、任意の種、例えば霊長類、特にはヒト;例えばラットおよびマウスといったげっ歯類、イヌ、ネコ、ウシ、オパイン、ウマ、酵母、ショウジョウバエ、シノラブディス(Caenhorabditis)等であり得る。
【0177】
更なる実施態様において、配列番号1にハイブリダイズ可能なポリヌクレオチド、またはそのフラグメントに縮重されるヌクレオチド配列を含むポリヌクレオチドが提供される。
【0178】
縮重は、本明細書において使用されるように、該縮重したポリヌクレオチドから発現されたポリペプチドと関係する活性または機能性に鑑みて定義されており、該ポリヌクレオチドは、i)少なくともインシュリン様成長因子結合蛋白5型(IGFBP−5)特異的なタンパク分解活性を含み、そして/またはii)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体またはその結合フラグメントにより認識され;そして/またはiii)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、のいずれかである。
【0179】
更なる実施態様において、本発明に記載のポリヌクレオチドの相補鎖を含むポリヌクレオチドが提供される。
【0180】
本明細書に記載のポリヌクレオチドは、PAPP−A2の3’未翻訳領域またはそのフラグメントに相当する核酸残基を含む更なるポリヌクレオチドに実施可能に結合され得る。本明細書で使用されるように、その3’未翻訳領域は、配列番号1の5377〜8527の核酸残基を含む。
【0181】
本明細書に記載のポリヌクレオチドに実施可能に結合された発現シグナルを含む発現ベクターの形態で組み換えDNA分子もまた提供される。
【0182】
更なる態様において、本発明に記載のポリヌクレオチドまたは本発明に記載のベクターでトランスフェクトまたは形質転換した宿主生体が提供される。その宿主生体は、例えば哺乳類の細胞株といった哺乳類の生体が好ましい。しかしながら、酵母または真菌類(fungi)といった微生物真核生物もまた使用され得、同様にシラス属(Bacillus)または大腸菌といった微生物原核生物も使用され得る。当業者であれば、特定の細胞における発現に好適なリーダー配列および/またはシグナルペプチドを含む、発現シグナルをいかに選択するかを知っているだろう。当業者であればまた、標準的な分子生物学的方法を使用することにより特定細胞における発現レベルをいかに決定するかを知っているだろう。
【0183】
更なる態様において、本発明は、配列番号2またはそのフラグメントのアミノ酸配列を含むか、あるいはそれらから本質的に構成される単離されたポリペプチドに関し、当該フラグメントは、
a)インシュリン様成長因子結合蛋白5型(IGFBP−5)に特異的な蛋白分解活性を有し;そして/または
b)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
c)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する。
【0184】
本発明の好ましい1つの実施態様において、配列番号2の変異体およびそのフラグメントの変異体もまた提供される。変異体は、予め決められたアミノ酸配列と同一性の程度またはそれらの相同性に基づいて決定され、当該予め決められたアミノ酸配列は、配列番号2であり、またその変異体がフラグメントである場合には、配列番号2のフラグメントである。
【0185】
従って、変異体は、予め決定された配列と、少なくとも75%の配列同一性配列同一性、例えば少なくとも80%の配列同一性、例えば少なくとも85%の配列同一性、例えば少なくとも90%の配列同一性、例えば少なくとも91%の配列同一性、例えば少なくとも92%の配列同一性、例えば少なくとも93%の配列同一性、例えば少なくとも94%の配列同一性、例えば少なくとも95%の配列同一性、例えば少なくとも96%の配列同一性、例えば少なくとも97%の配列同一性、例えば少なくとも98%の配列同一性、例えば少なくとも99%の配列同一性を有するのが好ましい。
【0186】
変異体はまた、本明細書の以下で定義されるような予め決定された数の同類(conservative)アミノ酸置換に基づいて決定される。本明細書で定義されるような同類アミノ酸置換は、1つのアミノ酸(予め決定されたアミノ酸群内)の別のアミノ酸(同一群内)との置換に関し、そのアミノ酸は、同様またはほぼ同様の特性を呈する。
【0187】
本明細書において適用されるような「同類アミノ酸置換」なる用語の意味の範疇で、本明細書の以下で示すアミノ酸群内で、1つのアミノ酸が別のアミノ酸に置換されてもよい:
i)極性の側鎖を有するアミノ酸(Asp、Glu、Lys、Arg、His、Asn、Gln、Ser、Thr、Tyr、およびCys)
ii)非極性の側鎖を有するアミノ酸(Gly、Ala、Val、Leu、Ile、Phe、Trp、Pro、およびMet)
iii)脂肪族の側鎖を有するアミノ酸(Gly、Ala、Val、Leu、Ile)
iv)環状の側鎖を有するアミノ酸(Phe、Tyr、Trp、His、Pro)
v)芳香族側鎖を有するアミノ酸(Phe、Tyr、Trp)
vi)酸性の側鎖を有するアミノ酸(Asp、Glu)
vii)塩基性の側鎖を有するアミノ酸(Lys、Arg、His)
viii)アミド側鎖を有するアミノ酸(Asn、Gln)
ix)ヒドロキシ側鎖を有するアミノ酸(Ser、Thr)
x)硫黄を含有する側鎖を有するアミノ酸(Cys、Met)
xi)中性で弱い疎水性のアミノ酸(Pro、Ala、Gly、Ser、Thr)
xii)親水性で酸性のアミノ酸(Gln、Asn、Glu、Asp)、および
xiii)疎水性アミノ酸(Leu、Ile、Val)。
【0188】
従って、本発明に記載の変異体またはそのフラグメントは、配列が同一の変異体またはそのフラグメント内、あるいは配列が異なる変異体またはそのフラグメント間に、相互に独立して導入された複数の置換体といった、少なくとも1つの置換体を含んでいてもよい。
【0189】
その同一の変異体またはそのフラグメントが、本明細書の上述で定義されるような1つ以上の同類アミノ酸群から1つ以上の同類アミノ酸置換体を含んでいてもよいことが、上述の概要から明らかである。
【0190】
アミノ酸の付加または欠失は、2〜10個のアミノ酸、例えば10〜20個のアミノ酸、例えば20〜30個のアミノ酸、例えば40〜50個のアミノ酸の付加または欠失であり得る。しかしながら、50個以上のアミノ酸の付加または欠失、例えば10〜100個のアミノ酸の付加、例えば100〜150個のアミノ酸の付加、例えば150〜250個のアミノ酸の付加もまた、本発明の範疇に含まれる。
【0191】
いかなる機能的等価物をも含む、本発明に記載のポリペプチドフラグメントは、1つの実施態様において、少なくとも250個の未満のアミノ酸残基、例えば少なくとも240個未満のアミノ酸残基、例えば少なくとも225個未満のアミノ酸残基、例えば少なくとも200個未満のアミノ酸残基、例えば少なくとも180個未満のアミノ酸残基、例えば少なくとも160個未満のアミノ酸残基、例えば少なくとも150個未満のアミノ酸残基、例えば少なくとも140個未満のアミノ酸残基、例えば少なくとも130個未満のアミノ酸残基、例えば少なくとも120個未満のアミノ酸残基、例えば少なくとも110個未満のアミノ酸残基、例えば少なくとも100個未満のアミノ酸残基、例えば少なくとも90個未満のアミノ酸残基、例えば少なくとも85個未満のアミノ酸残基、例えば少なくとも80個未満のアミノ酸残基、例えば少なくとも75個未満のアミノ酸残基、例えば少なくとも70個未満のアミノ酸残基、例えば少なくとも65個未満のアミノ酸残基、例えば少なくとも60個未満のアミノ酸残基、例えば少なくとも55個未満のアミノ酸残基、例えば少なくとも50個未満のアミノ酸残基を含み得る。
【0192】
本発明において使用されるような「機能的等価性」とは、予め決められた配列のフラグメントに相当する機能性を基準にして確立された1つの好ましい実施態様に従う。より具体的には、機能的等価性とは、ポリペプチドフラグメントのIGFBP−5特異的プロテアーゼ活性を示す、および/またはPAPP−A2を認識可能な抗体により認識される、および/またはPAPP−A2に対する親和性を有するレセプターへ結合するのにPAPP−A2と競合する能力として理解されるべきである。
【0193】
PAPP−A2の機能的等価物または変異体は、挿入、欠失、および同類置換を含む置換の数および範囲が増えるにつれて、次第に好ましく予め決められたPAPP−A2配列とは異なるアミノ酸配列を示すと理解される。この相違は、好ましい予め決められた配列とそのフラグメントまたは機能的等価物との間の相同性の減少として測定される。
【0194】
配列番号2のすべてのフラグメントまたは機能的等価物は、本明細書において報告されるようなPAPP−A2の好ましい予め決められた配列に対してそれらが示す相同性の程度にかかわらず、本発明の範疇に含まれる。この理由は、PAPP−A2の領域の幾つかが、その結果生成するフラグメントの結合活性にいかなる有意な影響をも与えることなく最も容易に変異し易い、あるいは完全に欠失可能であるからである。
【0195】
置換により得られる機能的変異体は、天然PAPP−A2活性のある程度の形態または度合を十分に示し得、機能的に類似のアミノ酸側鎖を含む残基が置換されると、相同性は更に低くなる。この点で機能的な類似性とは、疎水性、塩基性、中性または酸性、あるいは立体的なバルクの有無といった側鎖の主要な特性をいう。従って、本発明の1つの実施態様において、同一性の程度は、本発明に記載の好ましい予め決められたフラグメントの変異体または機能的等価物であるフラグメントの原則的な尺度ではない。
【0196】
アミノ酸配列間の相同性は、BLOSUM30、BLOSUM40、BLOSUM45、BLOSUM50、BLOSUM55、BLOSUM60、BLOSUM62、BLOSUM65、BLOSUM70、BLOSUM75、BLOSUM80、BLOSUM85、またはBLOSUM90といった周知のアルゴリズムを用いて計算され得る。
【0197】
配列番号2のフラグメントと少なくともある程度の相同性を共有するフラグメントは、それらが配列番号2の当該フラグメントと、少なくとも約90%相同する、例えば少なくとも92%相同する、例えば少なくとも94%相同する、例えば少なくとも95%相同する、例えば少なくとも96%相同する、例えば少なくとも97%相同する、例えば少なくとも98%相同するときには、本発明の範疇にあると考えられるべきである。本発明の1つの実施態様に従って、相同性の割合を、同一性の割合という。
【0198】
本明細書で使用される意味に従って、機能的等価性を決定するときに考慮に入れられ得る更なる因子は、i)本発明に記載のPAPP−A2フラグメントを検出するた抗血清能、ii)機能的に等価なPAPP−A2フラグメントのバインディングアッセイにおけるPAPP−A2との競合能である。既知のアミノ酸配列内の免疫遺伝学的に活性なアミノ酸の配列を決定する1つの方法は、米国特許5,595,915号においてGeysenにより示されており、これは参照して本明細書に組み込まれている。
【0199】
ペプチドフラグメントの構造および機能相関を決定するために更に好適に適応される方法は、米国特許第6,013,478号に示されており、これは参照して本明細書に組み込まれている。また、アミノ酸配列のレセプター部分への結合のアッセイ法は、当業者には既知である。
【0200】
同類置換は、配列番号2の好ましい予め決められた任意の位置に導入され得、そしてそれは、任意の1以上の位置に非同類置換を導入するのにもまた所望となり得る。
【0201】
PAPP−A2フラグメントの機能的に等価なフラグメントの形成をもたらす非同類置換は、例えば;i)例えば非極性の側鎖(Ala、Leu、Pro、Trp、Val、Ile、Leu、Phe、またはMet)を有する残基が、Gly、Ser、Thr、Cys、Tyr、Asn、またはGlnといった極性の側鎖を有する残基またはAsp、Glu、Arg、またはLysといった帯電したアミノ酸に置換される、あるいは帯電したまたは極性の残基を非極性残基と置換するなど、実質的に極性が異なる;および/またはii)別の残基によるProまたはGlyの置換またはProまたはGlyへの変換といった、ポリペプチド基本骨格配向性への効果が実質的に異なる;および/またはiii)例えばGluまたはAspといった負に帯電した残基のLys、HisまたはArgといった正に帯電した残基への置換(およびその反対)など、実質的に電荷が異なる;および/またはiv)例えばHis、Trp、Phe、またはTyrといった嵩高い残基のマイナーな側鎖を有する残基、例えばAla、GlyまたはSerへの置換(およびその反対)など、実質的に立体的なバルクが異なる。
【0202】
アミノ酸の置換により得られる変異体は、1つの好ましい実施態様において、疎水性および親水性の値、および電荷、サイズ等を含むアミノ酸側鎖置換体との相対的な類似性に基づいて作成され得る。例えば、様々な前述の特性を考慮にいれるアミノ酸置換は、当業者には周知であり、:アルギニンおよびリジン;グルタミン酸およびアスパラギン酸;セリンおよびスレオニン;グルタミンおよびアスパラギン;およびバリン、ロイシンおよびイソロイシンが挙げられる。
【0203】
本明細書に記載の変異体に加えて、立体構造的に類似の変異体は、変異体構造の鍵となる部分を模倣するために形成され得、当該化合物がまた、本発明の変異体と同様に使用されてもよい。これは、当業者には既知のモデリングおよび化学的デザインの技術により達成され得る。そのような立体構造的に類似する構成はすべて、本発明の範疇にあることが理解されよう。
【0204】
更なる実施態様において、本発明は、置換されたアミノ酸の値の+/−2.5以内、例えば+/−2.3以内、例えば+/−2.1以内、例えば+/−2.0以内、例えば+/−1.8以内、例えば+/−1.6以内、例えば+/−1.5以内、例えば+/−1.4以内、例えば+/−1.3以内、例えば+/−1.2以内、例えば+/−1.1以内、例えば+/−1.0以内、例えば+/−0.9以内、例えば+/−0.8以内、例えば+/−0.7以内、例えば+/−0.6以内、例えば+/−0.5以内、例えば+/−0.4以内、例えば+/−0.3以内、例えば+/−0.25以内、例えば+/−0.2以内である親水性または疎水性指標を有する機能的に構成される置換アミノ酸に関する。
【0205】
蛋白に相互作用性の生物学的機能を与える際の親水性および疎水性アミノ酸指標の重要性は、当業者には十分に理解されている(Kyte & Doolit−tle, 1982およびHopp、国特許第4,554,101号、それぞれ参照して本明細書に組み込まれる)。
【0206】
本明細書で使用されるようなアミノ酸疎水性指標値は以下の通りである:イソロイシン(+4.5);バリン(+4.2);ロイシン(+3.8);フェニルアラニン(+2.8);システイン/シスチン(+2.5);メチオニン(+1.9);アラニン(+1.8);グリシン(−0.4);スレオニン(−0.7);セリン(−0.8);トリプトファン(−0.9);チロシン(−1.3);プロリン(−1.6);ヒスチジン(−3.2);グルタミン酸(−3.5);グルタミン(−3.5);アスパラギン酸(−3.5);アスパラギン(−3.5);リジン(−3.9);およびアルギニン(−4.5)(Kyte & Doolittle, 1982)。
【0207】
アミノ酸親水性値を以下の通りである:アルギニン(+3.0);リジン(+3.0);アスパラギン酸(+3.0.+−.1);グルタミン酸(+3.0.+−.1);セリン(+0.3);アスパラギン(+0.2);グルタミン(+0.2);グリシン(0);スレオニン(−0.4);プロリン(−0.5.+−.1);アラニン(−0.5);ヒスチジン(−0.5);システイン(−1.0);メチオニン(−1.3);バリン(−1.5);ロイシン(−1.8);イソロイシン(−1.8);チロシン(−2.3);フェニルアラニン(−2.5);トリプトファン(−3.4)(米国特許第4,554,101号)。
【0208】
本明細書に記載のペプチジル化合物に加えて、立体構造的に類似の化合物が、ペプチド構造の鍵の部分を模倣するために形成され得、そしてその当該化合物はまた、本発明のペプチドと同様に使用され得る。これは、当業者には既知のモデリングおよび化学的デザインの技術により達成され得る。例えば、テトラペプチド構造を模倣するために、例えばジアルギニンペプチド基本骨格のアミノ末端を修飾するようエステル化およびアルキル化が実施されてもよい。そのような立体構造的に類似の構成はすべて、本発明の範疇にあることが理解されよう。
【0209】
N末端をアルキル化およびC末端をエステル化したペプチドもまた、本発明の範疇に包含される。機能的等価物はまた、二量体または無関係な化学的部分を含む、同一または他のPAPP−A2フラグメントおよび/またはPAPP−A2分子を用いて形成される、グリコシル化して共有結合性または凝集性の共役体を含む。そのような機能的等価物は、当該分野において既知の方法により、N末端およびC末端のいずれか一方または両方を含むフラグメントにおいて見られる群への機能性の結合により調製される。
【0210】
機能的等価物が、それゆえカルボキシル末端の脂肪族またはアシルエステルまたはアミド、アルキルアミン、またはカルボキシル側鎖を含む残基に共役した、例えばアスパラギン酸残基でアルキルアミンに共役したフラグメント;水酸基を含む残基のO−アシル誘導体およびアミノ末端のアミノ酸またはアミノ基を含有する残基のN−アシル誘導体、例えばfMet−Leu−Pheまたは免疫原性蛋白を有する共役体を含み得る。アシル基の誘導体は、アルキル部分の群(C3〜C10の直鎖アルキルを含む)から選ばれ、それによりアルカノイル種、および炭素環式またはヘテロ環式化合物を形成し、それによりアロイル種を形成する。反応基は、好ましくは反応側鎖の基を通じて不溶性のマトリクスに架橋結合する蛋白に使用するそれ自身は既知の二官能性化合物である。
【0211】
共有結合性または凝集性の機能的等価物およびその誘導体は、イムノアッセイにおける試薬として、またはアフィニティー精製法に有益である。例えば、本発明に記載のPAPP−A2のフラグメントは、それ自身既知の方法により臭化シアン活性化セファロースに共有結合することにより不可溶化され、抗PAPP−A2抗体または細胞表面レセプターのアッセイまたは精製に使用されるグルタルアルデヒド架橋を用いてまたは用いないでポリオレフィン表面に吸収され得る。フラグメントは、検出可能な基で標識化、例えばクロラミンT法による放射性ヨウ素化される、希土類キレートに共有結合される、または例えば診断アッセイに使用する別の蛍光部分と共役され得る。
【0212】
PAPP−A2の好ましい予め決められたフラグメントの突然変異誘発は、アミノ酸を、通常、約1〜10個程度のアミノ酸残基、好ましくは約1〜5個のアミノ酸残基を挿入する、あるいは約1〜10個の残基、例えば約2〜5個の残基を欠失することにより実施可能である。
【0213】
1つの実施態様において、PAPP−A2のフラグメントは、自動合成により合成される。任意の商業的に利用可能な固相法、例えばMerrifieldの固相合成法が実施され得、その方法において、アミノ酸が、成長するアミノ酸鎖に順次付加される(Merrifield, J. Am. Chem. Soc. 85:2149−2146, 1963参照)。
【0214】
ポリペプチドの自動合成用装置は、Foster City, Calif.のApplied Biosystems, Inc.といった供給業者から市販で入手可能であり、一般的には取扱い説明書に従って操作され得る。固相合成により、本発明に記載の任意のPAPP−A2フラグメントに所望のアミノ酸置換を導入することが可能になる。置換、欠失、相挿入、またはその任意のサブコンビネーションが、組み合わされて機能的等価物の最終配列に到達し得ることが理解されよう。挿入は、例えば疎水性または免疫原性蛋白、または任意のポリペプチドといった担体、または担体と同様の役割をすることが可能な骨格構造を用いたアミノ末端および/またはカルボキシル末端融合を含むと理解されるはずである。
【0215】
本発明によるPAPP−A2のフラグメントのホモ二量体およびヘテロ二量体を含む二量体を含むオリゴマーも提供され、本発明の範疇に該当する。PAPP−A2の機能的等価物および変異体が、他のアミノ酸配列を有する、または天然のPAPP−A2配列を有するホモ二量体またはヘテロ二量体として生成可能である。ヘテロ二量体としては、いかなる生物活性をも有するまたは示す必要がない免疫反応性PAPP−A2フラグメントならびにPAPP−A2フラグメントを含有する二量体である。
【0216】
本発明に記載のPAPP−A2フラグメントは、インビトロおよびインビボの両方で合成され得る。インビトロの合成法は周知であり、PAPP−A2のインビボでの合成法に好適なまたは好適に適応可能な方法は、従来技術でも示されている。インビボで合成される場合、宿主細胞は、PAPP−A2またはそのフラグメントをエンコードするDNAを含有するベクターを用いて形質転換される。ベクターは、複製可能な核酸構築物として定義されている。ベクターは、PAPP−A2の発現を仲介するために使用される。発現ベクターは、複製可能なDNA構築物であり、予め決められたPAPP−A2フラグメントをエンコードする核酸配列、またはインビボで発現可能な任意の機能的等価物が、好適な宿主においてそのフラグメントまたは等価物の発現に影響を与えることが可能な好適なコントロール配列に実施可能に結合される。そのようなコントロール配列は、当該分野においては周知である。
【0217】
多細胞生体由来の細胞培養物は、好ましい宿主細胞である。原則として、脊椎動物培養由来であろうと無脊椎動物培養由来であろうと任意の真核細胞培養物が高等であるほど有効である。例えば有益な宿主細胞株は、VEROおよびHeLa細胞、チャイニーズハムスター卵巣(CHO)細胞株、およびWI38、BHK、COS−7、293およびMDCK細胞株である。好ましい宿主細胞は、内因性のPAPP−A2を合成することが知られている真核細胞である。当該宿主細胞の培養物は、フラグメントの起源として単離および使用され得、あるいは成育状態を促進または阻害することを目的とする治療法、あるいはヒトまたは動物の身体に関して実施される診断方法を含む治療的処置法において使用され得る。
【0218】
特定の実施態様において、本発明は、本発明に記載のポリペプチドフラグメントに関し、PAPP−A2フラグメントは、その任意のプロセシング変異体を含む、PAPP−A2の成熟部分に相当する234〜1791のアミノ酸残基を含むかあるいは本質的に構成される。
【0219】
プロセシング変異体は、シグナルペプチダーゼおよびフリン(furin)を含むがそれに限定されない任意のプロテアーゼにより触媒された、別のプロセシング事象、可能なプロセシング事象の結果生成する変異体である。1つの推定される開裂部位は、233位の後ろに位置し、本明細書の以降において詳細に示されている。別の推定される開裂部位は、RQRRモチーフ(PAPP−A2のアミノ酸配列において196〜199位)の後ろに位置する。プロセシング変異体は、PAPP−A2がヒトまたは動物の組織、血清または体液において発現されるときにインビボでのプロセシングから生じる変異体を含むと理解されるはずである。
【0220】
配列番号2に示される成熟配列(アミノ酸残基234〜1791)から本質的に構成される成熟PAPP−A2アミノ酸配列は、1つの実施態様において、1〜約10個のN末端アミノ酸またはC末端アミノ酸、好ましくは1〜10個のN末端アミノ酸、例えば2〜8個のN末端アミノ酸、例えば3〜6個のN末端アミノ酸の間が欠損しているこの部分の配列を含むことが理解されるはずである。
【0221】
また、本明細書において使用されるように、本質的に構成されるという定義においては、配列番号2に示される成熟配列(アミノ酸残基234〜1791)がそれに加えて更に1〜約10個のN末端アミノ酸またはC末端アミノ酸、好ましくは1〜10個のN末端アミノ酸、例えば2〜8個のN末端アミノ酸、例えば3〜6個のN末端アミノ酸を有することも含まれる。この本質的に構成されるという定義はまた、他の態様において適用されて、PAPP−A2の特定部分と結合して使用されることに制限されない。この定義は、プロセシングされたPAPP−A2がもともと単離されているもの以外の、組織、血清、または体液において別のプロセシングから生じるポリペプチドを含む他のプロセスPAPP−A2ポリペプチドにも適用されるはずである。
【0222】
更に、好ましいフラグメントは、PAPP−A2のprepro部分に相当するアミノ酸残基1〜233、PAPP−A2のpro部分に相当するアミノ酸残基23〜233、PAPP−A2のシグナルペプチドまたはリーダー配列に相当するアミノ酸残基1〜22を含み、あるいはそれらから本質的に構成され、当該配列は、配列番号2のアミノ酸残基234〜1791に相当するPAPP−A2の成熟部分に実施可能に結合される。
【0223】
組み換えPAPP−A2ポリペプチド、またはそのフラグメントも提供され、当該ポリペプチドにはヒト蛋白、あるいは当該ポリペプチドに天然に付随する他の蛋白がないのが好ましい。
【0224】
更なる態様において、i)本発明に記載のポリヌクレオチド、および/またはii)本発明に記載のベクター、および/またはiii)本発明に記載の宿主生体、および/またはiv)本発明に記載のポリペプチドを、生理的に許容される担体と組み合わせて含む組成物が提供される。
【0225】
更に別の態様において、i)本発明に記載のポリヌクレオチド、および/またはii)本発明に記載のベクター、および/またはiii)本発明に記載の宿主生体、および/またはiv)本発明に記載のポリペプチドを、薬学的に許容される担体と組み合わせて含む医薬組成物が提供される。
【0226】
本発明は更に、本発明に記載のPAPP−A2ポリペプチドまたはそのフラグメントに対して特異性を有する抗体の生成法に関し、当該方法は、
i)宿主生体を提供し、
ii)その宿主生体を、請求項10に記載のポリペプチドを用いて免疫し、そして
iii)当該抗体を得る、
ことからなるステップを含む。
【0227】
本発明に記載のPAPP−A2ポリペプチドまたはそのフラグメントに対して特定の結合親和性を有するモノクローナル抗体およびポリクローナル抗体がまた提供される。該抗体は、モノクローナルであるのが好ましい。
【0228】
更なる態様において、本発明に記載のPAPP−A2ポリペプチドの生成法が提供され、当該方法は、
i)好適な宿主生体、好ましくは哺乳類細胞を提供し、
ii)ステップi)で提供された宿主生体を、本発明に記載のポリヌクレオチド、または本発明に記載のベクターを用いてトランスフェクトまたは形質転換し、
iii)ステップii)で得られた宿主生体を該ポリヌクレオチドまたはベクターによりエンコードされたポリペプチドの発現に好適な条件下で培養し;そして所望により
iv)宿主生体による組み換え発現の結果生じるポリペプチドを、その宿主生体から単離する、
ことからなるステップを含む。
【0229】
本発明の更に更なる態様において、アンチセンス法を用いて細胞中のPAPP−A2の発現を阻害および/または減少させる方法が提供され、当該方法は、
i)本発明に記載のアンチセンスポリヌクレオチドを提供し、
ii)ステップi)において提供された当該アンチセンスポリヌクレオチドを用いて、PAPP−A2を発現可能な細胞をトランスフェクトまたは形質転換し、
iii)ステップii)において得られた細胞を、PAPP−A2の発現に関与する当該細胞において、ステップi)で提供されたポリヌクレオチドの相補的なポリヌクレオチドへのハイブリダイゼーションに好適な条件下で培養し、そして
iv)当該細胞においてPAPP−A2の発現を阻害および/または減少させる、
ことからなるステップを含む。
【0230】
該アンチセンスポリヌクレオチドおよび該相補的なポリヌクレオチドは、別々のポリヌクレオチド分子から同時発現されてもよく、あるいはそれらが、同一の分子から発現されてもよい。ハイブリダイゼーションの別法として、該方法は、逆転写PCR法(rtPCT法)の使用を含み得る。
【0231】
本発明の更に別の態様において、個体から得た生物学的サンプルにおけるPAPP−A2の検出法、あるいはPAPP−A2レベルの測定法が提供され、当該方法は、
i)当該個体から生物学的サンプルを得て、
ii)そのサンプルにおいて、
a)PAPP−A2ポリペプチドまたはそのフラグメント、および/または
b)PAPP−A2発現に由来するmRNA形態のポリヌクレオチド、および/または
c)PAPP−A2特異的プロテアーゼ活性、好ましくはIGFBP−5プロテアーゼ活性、またはIGFBP−5の誘導体に対して示される蛋白分解活性を検出することによりサンプル中のPAPP−A2を検出する、
ことからなるステップを含む。
【0232】
該方法は、ステップii)で検出されたPAPP−A2またはPAPP−A2レベルを、
a)PAPP−A2の予め決められた量および/または濃度;および/または
b)PAPP−A2 mRNAの予め決められた量および/または濃度;および/または
c)予め決められたPAPP−A2特異的なプロテアーゼ活性、
からなる群から選ばれる予め決められた値と比較することからなるステップを更に含んでいてもよい。
【0233】
1つの実施態様においてその予め決められた値は、当該個体の正常な生理学的状態を示している。
【0234】
該生物学的サンプルは、血液、尿、胸腔内液、口腔洗浄液、組織バイオパシーおよび卵胞液からなる群から選ばれるのが好ましい。
【0235】
PAPP−A2のレベルが、PAPP−A2蛋白の量として測定される場合、そのPAPP−A2蛋白が免疫化学分析により測定されるのが好ましく、当該方法においてはPAPP−A2蛋白が、少なくとも1つのモノクローナル抗体により検出される。PAPP−A2蛋白はまた、少なくとも1つの更なる成分、好ましくは例えばpro−MBP(pro−主要塩基性蛋白)であるがそれに限定されないポリペプチドを含む複合体において検出され得る。PAPP−A2はまた、PAPP−A2単量体としてあるいはPAPP−A2二量体として検出され得る。
【0236】
本発明の更なる態様は、個体における病態の診断法に関し、当該方法は、 i)PAPP−A2の検出法またはPAPP−A2レベルの測定法を実施し、そして
ii)その病態を診断する
からなるステップを含む。
【0237】
該病態は、例えばトリソミー21、トリソミー18、トリソミー13、および開放型二分脊椎からなる群から選ばれる胎児異常であるがそれに限定されない胎児異常が好ましい。
【0238】
本発明に従って診断可能な更なる胎児異常は、子宮外妊娠、開放型二分脊椎、神経管閉鎖障害、腹部壁欠損、Edwards症候群、Pateaus症候群、Turner症候群、モノソミーX、またはKlein−felter’s症候群である。
【0239】
別の態様において、該病態は、再狭窄、アテローム性心筋梗塞、創傷治癒、線維症、心筋梗塞、骨粗鬆症、リュウマチ性関節炎、多発性骨髄腫、または癌を含むがそれに限定されない成長促進状態および成長阻害状態からなる群から選ばれる異常な成長状態である。
【0240】
本発明の更に更なる態様において、生物学的サンプルにおける本発明に記載のポリヌクレオチドの発現検出法が提供され、当該方法は、
i)本発明に記載のポリヌクレオチドを推定で含有する生物学的サンプルを提供し、そして
ii)その生物学的サンプルを、i)本発明に記載のポリヌクレオチドに相補的な、およびii)それとハイブリダイズ可能な鎖を含むポリヌクレオチドと接触させ、そして
iii)ハイブリダイゼーションが起こさせ、そして
iv)ステップiii)で得られたハイブリダイゼーション複合体を検出する、
ことからなるステップを含み、
当該方法において、ハイブリダイゼーション複合体の存在は、本発明に記載のポリヌクレオチドまたはそのフラグメントの生物学的サンプルにおける発現を示している。
【0241】
本発明の更に更なる態様において、PAPP−A2のプロテアーゼ活性を阻害する薬剤の同定法が提供され、当該方法は、
i)a)本発明に記載のポリペプチドまたはそのフラグメント、およびb)当該ポリペプチドまたはフラグメントに対して予め決められた基質、およびc)推定の阻害剤をインキュベートし、そして
ii)該基質の蛋白分解が阻害されるかどうかを決定する、ことからなるステップを含む。
【0242】
該基質は、内部でクエンチングする蛍光性ペプチドであり得るポリペプチドを含むのが好ましい。1つの好ましい基質は、IGFBP−5またはそのフラグメントを含むか、あるいはそれから本質的に構成される。
【0243】
本発明はまた、PAPP−A2のプロテアーゼ活性を阻害する薬剤を同定する当該方法に従って得られる阻害剤に関する。
【0244】
病態の処置を必要とする個体において、その病態を処置するための薬物の製造における当該提供された阻害剤の使用も提供される。
【0245】
更に更なる態様において、本発明は、PAPP−A2のプロテアーゼ活性を促進可能な薬剤の同定法に関し、当該方法は、
i)a)本発明に記載のポリペプチドまたはフラグメント、およびb)当該ポリペプチドに対して予め決められた基質、およびc)推定の促進剤をインキュベートし、そして
ii)該基質の蛋白分解が促進されるかどうかを決定する、
ことからなるステップを含む。
【0246】
該基質は、内部でクエンチングする蛍光性ペプチドであり得るポリペプチドを含むのが好ましい。IGFBP−5またはそのフラグメントが基質として特に好ましい。
【0247】
PAPP−A2のプロテアーゼ活性を促進可能な薬剤を同定する方法に従って得えられる促進剤も提供され、本発明はまた、病態の処置を必要とする個体においてその病態を処置するための薬物の製造における当該促進剤の使用も提供される。
【0248】
更に別の態様において、個体の治療による処置法が提供され、当該方法は、i)本発明に記載の医薬組成物、および/またはii)本発明に記載の阻害剤、および/または本発明に記載の促進剤を当該個体に投与することからなるステップを含む。
【0249】
更に更なる態様において、PAPP−A2またはPAPP−A2の他の蛋白との複合体の精製法が提供され、当該方法は、
i)本発明に記載のポリペプチド、またはそのフラグメントに対して特異的な結合親和性を有するポリクローナルまたはモノクローナル抗体を提供し、そして
ii)PAPP−A2、またはそのフラグメントをアフィニティークロマトグラフィーにより精製する、
ことからなるステップを含む。
【0250】
本発明がその詳細な説明と組み合わせて示されている一方、前述の説明は、添付の請求の範囲により定義される本発明の範囲を説明し、そして制限しないことが意図されていることが理解される。他の態様、利点、および修飾は、以下に示す請求項の範疇である。
図面の簡単な説明
図1は、preproPAPP−A2をエンコードするmRNAに相当するcDNA配列(5’→3’の方向で)を示す。配列のコード化部分およびその末端のストップコドン(*)だけが示されて、1〜5376がナンバリングされている。preproPAPP−A2の翻訳されたポリペプチド配列も示されている。シグナルペプチド開裂部位は、SignalP V2.0を用いてnt.64−66によりエンコードされたアラニン残基の後となると予測された((Nielsen et al., 1997, Protein Eng 10, 1−6)、http://genome.cbs.dtu.dk/にあるWWW予測サーバー)。preproPAPP−A2のシグナルペプチド(nt.1−66、22残基)が太字で示されている。この図のヌクレオチド配列は、配列番号1のnt.1〜5376を示している。この図の蛋白配列が配列番号2として示されている。
【0251】
図2は、アミノ酸配列に翻訳されたときのPAPP−A(Kristensen et al., 1994, Biochemistry 33, 1592−8)と、PAPP−AのN−末端部(hom−N、受入れ番号 AL031734のコード化部分)およびC−末端部(hom−C、受入れ番号 AL031290のコード化部分)と相同性のある2つのゲノムクローン内に含まれる配列伸長部との間の相関関係を示す図式である。この図はまた、PAPP−Aの中央領域と相同性のあるcDNA配列を得る方法をも示している。Hom−N、hom−C、および中央領域が一緒になって、新規な蛋白、PAPP−A2の完全な配列をエンコードし、これは、PAPP−Aのホモログである。該中央領域は、テンプレートとして特異的に初回刺激を受けて(プライマー、Rt−N−mid)、逆転写したヒト胎盤mRNA、PCR用にプライマー、PR−mid5およびPR−mid3を用いてPCRにより得られた(表1)。完全長のPAPP−A2をエンコードするcDNA構築物を得るために、ゲノムクローンhom−Nおよびhom−Cに相当するcDNAクローンも、テンプレートとして特異的に初回刺激を受けた胎盤mRNA(プライマーは示さず、表1参照)により合成されたcDNAを用いて得た。これは、明細書中に詳細に示されるように、シグナルペプチド伸長部(hom−N中)および停止コドン(hom−Cの3’末端)の同定が必要とされた。プライマーはすべて、表1に示されている。注:ここで示される配列の相対的な位置は、実施される実験に従うものであるが、図は、縮尺に正確には示されていない。
【0252】
図3は、preproPAPP−A2(配列番号2)のアミノ酸配列を、preproPAPP−Aと整列して示している。preproPAPP−A2(PA2)の推定のアミノ酸配列が、CLUSTAL W(Thompson et al., 1994, Nucleic Acids Res 22, 4673−80)を用いてpreproPAPP−A(PA)((Haaning et al., 1996, Eur J Biochem 237, 159−63)、AAC50543)の配列と整列された。PAPP−Aのprepro部分が、PAPP−A2に相当する領域と有意な同一性を示さなかったので、そのアラインメントを、proペプチドの長さの違いを強調するために手動で調節した。矢印は、PAPP−Aについては以前に見い出された成熟蛋白のN末端部(Glu−81)(Kristensen et al., 1994, Biochemistry 33, 1592−8)、そしてPAPP−A2については本発明で見出されたN末端部(Ser−234)を示す。推定のシグナルペプチドは、SignalP V2.0(Nielsen et al., 1997, Protein Eng 10, 1−6)を用いて強く予測されるが、小文字で示している。PAPP−A2のpro部分は、Met−168に相当する候補開始コドンを別に1つ含むが、SignalPを用いてこの残基の後に続くシグナルペプチドは何も予測されなかった。PAPP−Aの配列モチーフ(Kristensen et al., 1994,
Biochemistry 33, 1592−8)もまた、PAPP−A2において見い出され:触媒亜鉛結合モチーフおよび推定のMet−ターンの残基は、両方の配列において下線が引いて、太字にしている。Lin−ノッチモチーフ(LNR1−3)および短いコンセンサスリピート(SCR−1〜5)は四角で囲んでいる。システイン残基は、影付きになっている、成熟PAPP−Aのシステインはすべて、PAPP−A2でも見られる。更に、分泌型のPAPP−A2は4つのシステイン残基(Cys−343、Cys−533、Cys−618、およびCys−1268)を有し、PAPP−Aにはそれに相当するものはない。
【0253】
図4は、ウエスタンブロッティングおよびクマシー染色によるPAPP−A2を示している。トランスフェクトした293T細胞由来の培地は、モノクローナル抗c−mycを用いてウエスタンブロットされる。レーン1、空ベクターでトランスフェクトした細胞;レーン2、c−mycペプチド(pPA2−mH)を用いてC末端をタグ化した野生型PAPP−A2をエンコードするcDNAを用いてトランスフェクトした細胞、非還元型;レーン3、不活性化E734Q変異体(pPA2−KO−mH)を有するPAPP−A2をエンコードするcDNAでトランスフェクトした細胞、非還元型;レーン4、レーン2と同じであるが還元型。可能な自触反応を除去するために、組み換えPAPP−A2を、pPA2−KO−mHでトランスフェクトした細胞の血清フリーの培地からニッケルアフィニティークロマトグラフィーにより精製した(レーン5、還元型)。
【0254】
図5は、IGFBP−1〜6に対するPAPP−A2活性を示す。空ベクター(−)またはPAPP−A2をエンコードするcDNA(pPA2)(+)でトランスフェクトした293T細胞由来の培地を、6つのIGFBP(BP1〜BP6)のそれぞれと一緒にインキュベートし、その活性を放射標識したIGF−IIを用いてリガンドブロッティングにより評価した。IGFBP−5の完全の開裂は、BP5+レーンにおけるシグナルの不在から明らかである。IGBP−3の部分分解もまた、明らかである。
【0255】
図6は、IGFBP−5に対するPAPP−A2の蛋白分解活性を示す。空ベクター(レーン1)、不活性化E734Q変異体を有するPAPP−A2をエンコードするcDNA(pPA2−KO)(レーン2)、または野生型PAPP−A2をエンコードするcDNA(pPA2)(レーン3〜6)でトランスフェクトした293T細胞由来の培地を、C末端をc−mycタグ化したrIGFBP−5と一緒にインキュベートした。蛋白分解活性を、抗c−mycを用いてウエスタンブロッティングにより評価した。「i」は、インタクトなrIGFBP−5を示し;「c」は、検出可能なC末端c−mycタグ化開裂生成物を示す。阻害剤なしの場合において、野生型PAPP−A2は、すべてのrIGFBP−5を分解した(レーン3)。PAPP−A2活性は、10mM フェナントロリン(レーン4)および5mM EDTA(レーン5)により止められるが、100mM 3,4−DCI(レーン6)によっては影響は受けなかった。精製されたrIGFBP−5のクマシー染色したSDS−PAGEが、精製されたPAPP−A2による消化前(レーン7)および後(レーン8)で示されている。抗c−mycを用いた同じ消化のウエスタンブロットも示されている(レーン9)。配列分析により、Ser−142とLys−143の間の部位でPAPP−A2がIGFBP−5を開裂することが明らかになった。
【0256】
図7は、PAPP−A2 mRNAコード化領域のcDNA配列を示しており3’UTRの配列が直接続いている。3’UTRの配列は、実施例6.3で詳細に示されるように得られる。この配列の最初の5376個のヌクレオチド(nt.1〜5376)は、図1および配列番号1(nt.1〜5376)で示されるコード化配列を示す。この配列のヌクレオチド5377〜8527は、配列番号3(nt.5377〜8527)で示されるPAPP−A2 mRNAの3’UTRに相当する。
【0257】
図8は、PAPP−A/proMBP複合体(上の棒状の図)中のPAPP−Aサブユニットのジスルフィド構造を示している。PAPP−A/proMBP複合体をプロテアーゼおよび臭化シアンで分解した後、標準的なHPLCにより、PAPP−A/proMBP複合体起源のシステイン含有ペプチドを単離した。ペプチドは、アミノ酸分析、N末端配列分析により、そして質量分析により同定した(Overgaard, M. T., Oxvig, C.、未公表)。ジスルフィド結合が、細線で示されている。2つのシステイン残基は、proMBPへの鎖間ジスルフィド架橋を形成し、1つがPAPP−Aへの鎖間架橋を形成してそれにより二量体が生じる(図示の通り)。アスタリスクは、パートナーが見い出されていないシステインを示す。成熟PAPP−Aにおいて存在するシステイン残基は、成熟PAPP−A2においても存在している(図3参照)。PAPP−A2ジスルフィドペアは同じであると仮定するのが理にかなっている。それゆえ、この情報は、PAPP−A2の単離されたドメイン(フラグメント)の発現に対して境界領域を決定する際、重要である。PAPP−Aの遺伝子構造も示されている(下の棒状の図)。エクソン/イントロンの境界は、PAPP−A cDNA(AN X68280)のゲノム配列(AN AB020878、AL353141、およびAL137024)との比較に基づいている。中央の棒状の図は、上側および下側の棒状の図の情報に基づいたPAPP−Aの推定のドメインを示している。
【実施例】
【0258】
実施例
6.1.PAPP−A2をエンコードするヌクレオチド配列の同定
本明細書において提供される受入れ番号(AN)は、Gen−Bankまたは他の生物学的配列データベースで示された配列をいう。ANは、特定のANのもとで示される蛋白またはヌクレオチド配列と置き換え可能である。
【0259】
PAPP−Aと相同性のあるDNA配列((Kristensen et al., 1994, Biochemistry 33, 1592−8)、AN CAA48341)について、公式のヌクレオチドデータベースを調査すると、ポリペプチド配列に翻訳された場合、AN AL031734およびAL031290を有する2つのゲノムクローンが明らかとなった。両方とも、ヒト染色体1(1q24)起源である。調査は、デフォルト設定によるhttp://www.ncbi.nlm.nih.gov/BLAST/でtblastnプログラムを用いて、データベースの「nr」コレクションに対して実施された。この実施例において、PAPP−Aは、(Kristensen et al., 1994, Biochemistry 33, 1592−8)に示すように、N末端のGluを残基1としてナンバリングしている。蓄積された配列記録(AN X68280)において、このGluは残基5である。
【0260】
AL031734で報告される配列は、168835の塩基対を含む。全体配列のうち2つの不連続配列伸長部(nt.103432〜103566、および140846〜141919)が一緒に、翻訳されたときにPAPP−Aポリペプチド配列の残基16〜59および59〜413と整列される。AL031290において報告される配列は、121780の塩基対を含む。全体配列のうち4つの不連続配列伸長部(nt.10209〜10358、11752〜11901、20531〜20463、および60536〜60652)が一緒に、翻訳されたときにPAPP−Aポリペプチド配列の残基1313〜1362、1376〜1425、1457〜1479、および1470〜1506と整列される。両方のゲノム配列のコード化領域間の配列伸長部は、非コード化ゲノムDNA(イントロン)または整列しないコード化領域を示す。
【0261】
これらの所見に基づいて、我々は、PAPP−Aと相同性のある新規蛋白、PAPP−A2の存在を仮定した。そのあと、それは、AL031734およびAL031290に報告される2つのゲノム配列により部分的に包含されるPAPP−A2領域の完全なコード化配列を確立した。我々は、これらの連続的配列、hom−Nおよびhom−Cをそれぞれ示した(図2)。しかしまず第1に、我々は、PAPP−Aに対してまた相同性を示し、hom−Nおよびhom−C(図2)の配列に結合するコード化cDNA配列の存在を確立した。使用される重要なプライマーはすべて表1に示されている。1791残基のpreproPAPP−A2をエンコードするcDNA配列全体を、図1に示している。標準的なクローニング法が利用されて、すべてのDNA構築物は、配列決定により分析された。使用される方法は、以下に示される。PAPP−A2という名前は、このDNA配列によりエンコードされた蛋白に対して使用される。
【0262】
Hom−Nとhom−Cとの間の中央領域に相当する連続的なコード化cDNA伸長部のクローニング:該中央領域(図2)を得るために、テンプレートとしてヒト胎盤mRNAおよびプライマー、AL031290から誘導されるRT−N−midを用いて、cDNAを合成した(表1、図2)。このcDNAは、仮定のPAPP−A2の中央領域に相当するcDNAを得るために、PCRにおいてテンプレートとして使用した。PCRプライマーは、PR−mid5およびPR−mid3であった(表1、図2)。得られた中央領域のコード化配列は、図3の残基665〜1572(配列番号1)、全体で908個のアミノ酸に相当する。
【0263】
【表1】

【0264】
PAPP−A2のN末端部に相当する連続的なコード化cDNA伸長部(hom−N)のクローニング:5’方向に更に続くゲノム配列AL031734のマニュアル調査により、PAPP−A残基16〜59に相当する配列伸長部の読み取り枠:Nt.102646〜103566が、メチオニン残基で始まる307残基のポリペプチド配列をエンコードすることが明らかになった。この所見に基づいて、中央領域(上述で詳細に示されるような、RT−N−midで初回刺激を受けた胎盤mRNA)を得るために使用されるcDNAを、hom−Nの連続的なcDNAを得るためのPCRにおいてテンプレートとして使用した。PCRプライマーは:PR−N5およびPR−N3(表1、図2)であった。
【0265】
PAPP−A2のC末端部に相当する連続的なコード化cDNA伸長部(hom−C)のクローニング:AL031290のゲノム配列に適合するヒトEST配列について利用可能なデータベース(デフォルト設定を有するhttp://www.ncbi.nlm.nih.gov/BLAST/でのblastnプログラムを用いて)を調査することにより、伸長部nt.60536〜60652(上述参照)ですでに定義されたAL031290のコード化領域のいくつかとオーバーラップするEST配列が明らかになった。AL031290のnt.62790−62995もまた、胎盤起源のヒトEST配列、AA368081の配列と適合した。ポリペプチド配列に翻訳されたときに、このEST配列は、PAPP−AのC末端部と相同性を示した。更に、停止コドンが、PAPP−Aのアミノ酸1537に相当するコード化配列内に存在した。すなわち、その2つの配列が整列化されるときに、PAPP−A2は、PAPP−AよりもC末端側には存在しない。このことに基づいて、テンプレートとしてヒト胎盤mRNAおよびAL031290起源のプライマー(表1)を用いて、cDNAを合成した。PCRプライマー、PR−C5およびPR−C3(表1、図2)を用いて、hom−Cの連続的cDNAを得るためのPCRにおいてテンプレートとして、このcDNAを使用した。
【0266】
すべてのPCRは、Pfuポリメラーゼ(Stratagene)を用いて実施した。3つのオーバーラップするPAPP−A2 cDNA(hom−N、新規な中央領域、およびhom−C)はすべて、ベクターPCR−BluntII−TOPO(Invitrogen)中でクローン化した。いくつかのクローンを双方向で配列決定した。その構築物は、それぞれp2N、p2Mid、およびp2Cという。PAPP−A2をエンコードする全体的なヌクレオチド配列は、図1(および配列番号1)に示されている。
6.2.PAPP−A2のヌクレオチドおよびアミノ酸配列の分析
成熟PAPP−Aの1547残基のうち、708残基(45.8%)が、preproPAPP−A2と同一である。PAPP−Aのprepro部分とPAPP−A2の残りの(N末端)との間には、十分な同一性はない(図3)。この実施例において、PAPP−Aは、((Haaning et al., 1996, Eur J Biochem 237, 159−63)、AAC50543)に従ってナンバリングしている。
【0267】
PAPP−Aにおいて認識される配列モチーフ(Kristensen et al., 1994, Biochemistry 33, 1592−8)は、PAPP−A2にも存在する:伸長された亜鉛結合連続配列、3つlin−ノッチ反復(LNR1〜3)、および5つの短い連続反復(SCR1〜5)(図3)。更に、PAPP−Aの82個のシステイン残基すべてが、その2つの蛋白の間に保持されて、更に4個のシステインがPAPP−A2ポリペプチド配列中に存在する。
6.3.PAPP−A2 mRNA起源のヒトEST配列の同定
AL031290のゲノム配列と適合するEST配列のクラスターを、PAPP−A2をエンコードする配列の末端部からのおよそ1.2kbで始まり、AL031290のnt.64000〜66000周辺で同定した。PAPP−A2のコード化領域およびこのクラスターに結合するmRNAの存在が、AL031290由来のプライマー(5’−GGAAAGAGCAGAGTTCACCCAT−3’、AL031290のnt.64900−64879)およびPAPP−A2をエンコードする配列(5’−CCGTCTTAGTCCACTGCATCC−3’、AL031290のnt.20499−20519、AF311940のnt.5171−5191)、およびテンプレートとしてオリゴ−dT初回刺激を受けた胎盤cDNAを用いて、PCRにおいて明らかにされた(Overgaard et al., 1999, Biol Reprod 61, 1083−9)。予測されるように、結果の生成物のサイズは、2.2kbであり、更に約3kbの3’UTRを有するPAPP−A2 mRNAの存在が示される。組織間の分配は、表2に示されている。
【0268】
【表2】
【0269】
6.4.組み換えPAPP−A2およびPAPP−A2変異体の哺乳類細胞における発現
以下に示すプラスミド構築物が作成された:
a)pPA2:発現ベクター、pcDNA3.1+においてアミノ酸1−1791をエンコードするpre−pro−PAPP−A2のcDNA配列
b)pPA2−KO:pPA2と同様であるが、PAPP−A2の活性部位のGlu−734がGln残基と置換されたもの(E734Q)
c)pPA2−mH:発現ベクター、pcDNA3.1/Myc−His(−)Aを含有する、アミノ酸1〜1791をエンコードするpre−pro−PAPP−A2のcDNA配列で、後に停止コドンは続くのではなくて、c−mycおよびHisタグが続いているもの
d)pPA2−KO−mH:pPA2−mHと同様であるが、pPA2−KOがE734Q置換されたもの
3つのオーバーラップしているPAPP−A2 cDNAフラグメント(hom−N、中央領域、およびhom−C)を、PAPP−A2をエンコードする一本の連続したcDNA配列の構築に使用した。そのオーバーラップするフラグメントは、ベクター、PCR−BluntII−TOPO(Invitrogen)中にすべて含まれており、上述に詳細に示すように、p2N、p2Mid、およびp2Cという(実施例6.1)。適当な配向のcDNA挿入部を有するp2Nおよびp2Cのクローンを選んだ。
【0270】
pPA2の構築:NotI−BamHIフラグメントをp2Cから切り取り、pBluescriptIISK+(Stratagene)中でクローン化してp2CBlueを得た。NotI−SpeIフラグメントをp2Nから切り取り、SpeI−BclIフラグメントをp2Midから切り取った。それら2つのフラグメントを、1つの反応においてp2CBlueのNotI/BclI部位にリゲートして、PAPP−A2 cDNA全体を含むp2NMidCBlueを得た。pBluescriptIISK+のNotI−ApaIフラグメントを切り取って、哺乳類の発現ベクター、pcDNA3.1+(Invitrogen)のNotI/ApaI部位中にリゲートして、このベクターの修飾型である、pcDNA−NAを得た。その後、完全長のcDNAを、NotIおよびXhoIを用いてp2NMidCBlueから切り取り、pcDNA−NA中でクローン化してpPA2を得た。SpeIとBclIを除いて、使用されるすべての制限部位は、ベクターのマルチクローニング部位中にあり、両方ともp2N、p2Mid、およびp2Cのコード化PAPP−A2配列伸長部の2つのオーバーラップ領域のそれぞれに位置する(それぞれ、図3のnt.2365およびnt.4203)。
【0271】
pPA2−KOの構築:pPA2−KO構築物は、PAPP−A2の活性部位のGlu−734残基がGln残基と置換したpPA2発現構築物の一変異体である。従って、その突然変異体は、E734Qである。pPA2−KO構築物は、テンプレートとしてpPA2を用いて、オーバーラップ伸長PCR法(Ho et al., 1989, Gene 77, 51−9)を用いた部位特異的突然変異誘発により作成した。簡単にいうと、外側のプライマーが、5’−CGCTCAGGGAAGGACAAGGG−3’(5’末端プライマー、配列番号1のnt.976〜995)および5’−CTAGAAGGCACAGTCGAGGC−3’(3’末端プライマーnt.1040〜1021、ベクターpcDNA3.1+の配列)である。オーバーラップする内部プライマーは、5’−TGTCCCACTTGATGGATCATGGTGTCGGTGTGG−3’(配列番号1のnt.2210〜2178、E734Qの結果nt.2200はCではなくてGである)および5’−CCATCAAGTGGGACATGTTCTGGGAC−3’(配列番号1のnt.2196〜2221、E734Qの結果、nt.2200はGではなくてCである)である。その結果生成する突然変異フラグメントを、XbaIおよびXhoIを用いて消化し、pPA2と交換してpPA2−KOを産生した。PCRはすべて、PfuDNAポリメラーゼ(Stratagene)を用いて実施し、全ての構築物を配列分析により確認した。
【0272】
pPA2−mHの構築:2つのプライマー(5’−GAGGGCCTGTGGACCCAGGAG−3’、配列番号1のnt.4906〜4926、および5’−GACGTAAAGCTTCTGATTTTCTTCTGCCTTGG−3’、配列番号1のnt.5373〜5354、HindIII部位、AAGCTT、およびPCR生成物の開裂を促進するnt.GACGTAの前にある)を、PCRにおいて、テンプレートとしてpPA2と一緒に使用して、発現ベクターへのインフレームでリゲーションするためにHindIII部位で置換された停止コドンを有するPAPP−A2のC末端の156残基をエンコードするヌクレオチドフラグメントを産生した。簡単にいうと、PCR生成物をEcoRIとHindIIIを用いて消化し、pcDNA3.1/Myc−His(−)AベクターのEcoRI/HindIII部位中でクローン化してpPA2C−mHを産生した。NotI−XbaIフラグメント(PAPP−A2のN末端部分をエンコードする)、およびXbaI−EcoRIフラグメント(PAPP−A2の残りの中央部分をエンコードする)をpPA2から切り取って、1つの反応においてpPA2C−mHのNotI/EcoRI部位にリゲートした。その結果生成する構築物、pPA2−mHは、PAPP−A2、続いてKLGP残基、mycエピトープ(EQKLISEEDL)、NSAVD残基、および6個のH−残基(アミノ酸は、1文字表記されている)をエンコードした。6個のヒスチジン残基の直後に停止コドンが続いている。
【0273】
pPA2−KO−mHの構築:残基Glu−734をGln残基に置換したpPA2−mH変異体を構築した:pPA2−KOのNotI−KpnIフラグメントを切り取り、pPA2−mHのNotI−KpnI部位と交換してpPA2−KO−mHを産生した。
【0274】
哺乳類細胞における発現:すべての構築物(pPA2、pPA2−KO、pPA2−mH、およびpPA2−KO−mH)ならびに空の発現ベクター(pcDNA3.1+およびpcDNA3.1/Myc−His(−)A)を、組み換えPAPP−A2蛋白の発現のために哺乳類の細胞中に短期間トランスフェクトした。簡単にいうと、ヒトの胎児の腎臓293T細胞(293tsA1609neo)(DuBridge et al., 1987, Mol Cell Biol 7, 379−87)を、10%のウシ胎仔血清、2mM グルタミン、非必須アミノ酸、およびゲンタマイシン(Life Technologies)を補充した高濃度のグルコースDMEM培地中で維持した。細胞を6cmの培養皿上にプレーティングし、QIAprep Spinキット(Qiagen)により調製した10μgのプラスミドDNAを用いたリン酸カルシウム共沈(Pear et al., 1993, Proc Natl Acad Sci U S A 90, 8392−6)により18時間後にトランスフェクトした。更に48時間後、その上清を回収して、血清フリーの培地(293SFMII, Life Technologies)で更に48時間置き換えた。その血清フリーの培地を回収して、遠心分離により除去した。
【0275】
構築物、pPA2−mHおよびpPA2−KO−mHを用いたトランスフェクションの結果生成する組み換え蛋白のウエスタンブロッティングによる分析は、PAPP−A2が、220kDaの蛋白(図2参照)として分泌されることを示した。ジスルフィド結合の還元により、バンド移動には目に見える変化はもたらされなかった。従って、PAPP−Aとは対照的に、PAPP−A2は、単量体として分泌される。
6.5.タグ化PAPP−A2のアフィニティークロマトグラフィーによる精製
金属キレートアフィニティーカラム(2ml、Pharmacia)をニッケルイオンで帯電して、pPA2−KO−mHを用いて短期間トランスフェクトした細胞(実施例6.4参照)由来の血清フリー培地をロードした(50ml)。1M NaClを含有するPBS中での洗浄後、結合蛋白を、10mM EDTAの0.5mlのPBSフラクション中で溶出した。PAPP−A2を含有するフラクションを、SDS−PAGEにより位置決めした(図4、レーン5)。この蛋白は、空ベクターにより同じ方法で処置したトランスフェクトした細胞(偽のトランスフェクタント、mock transfectants)からは見られなかった。
6.6.PAPP−A2のN末端配列分析
構築物、pPA2−KO−mH(実施例6.4および6.5参照)を用いてトランスフェクトした細胞の培地から精製された、C末端側をタグ化したPAPP−A2を還元し、10〜20% SDSゲル上に流して、更にPVDF膜(ProBlott、Biosystemsより提供)上にブロッティングした。4つのレーンのバンドを切り取って、オンラインのHPLCを備えたApplied Biosystems 477Aシーケンサー(Sottrup−Jensen, 1995, Anal Biochem 225, 187−8)で、N末端分析を実施した。PAPP−A2ポリペプチドのSer−234の前で、R(230)VKKの後の開裂の結果得られたおよそ20pmolレベルでN末端配列:Ser−Pro−Pro−Glu−Glu−Ser−Asn(SPPEESN)が見られた。
【0276】
これにより、PAPP−A2がPAPP−Aと同様、prepro蛋白として合成されるという予測が確認される。P1位におけるアルギニン残基の不在は、この開裂の原因となるpro蛋白プロセシング酵素がフリンではなくて、似た別のpro蛋白転換酵素(Nakayama, 1997, Biochem J 327, 625−35)であることを示している。proPAPP−A2の開裂は、典型的にフリン開裂を示すR(196)QRR(Nakayama, 1997, Biochem J 327, 625−35)の後と予想されるかもしれない。我々は、開裂がこの部位で起こり、所見のN末端部が更なるプロセシングの結果によるものであるということを排除することはできない。
6.7.インシュリン様成長因子結合蛋白(IGFBP)−5の開裂
IGFBP−1(HepG2調整培地由来)、rIGFBP−2(GroPep)、rIGFBP−3(Powell博士より贈与)、rIGFBP−4(Austral)、rIGFBP−5(Andress博士より贈与)、およびrIGFBP−6(Austral)に対する活性をアッセイするために、放射標識したIGF−II(Bachem)を用いたリガンドブロッティング(Conover et al., 1993, J Clin Invest 91, 1129−37)を利用した。6つの結合蛋白のうち、IGFBP−5は、完全な開裂を示した(図5)。IGFBP−3は、一部分分解した(図5)。この開裂は、IGFの存在とは無関係であった。実験は、pPA2または空ベクターを用いてトランスフェクトした細胞由来の培地を用いて実施した。
【0277】
更なる分析に対して、組み換えIGFBP−5が哺乳類の細胞中で生成された。簡単にいうと、ヒト胎盤オリゴ−dTで初回刺激したcDNA(Overgaard et al., 1999, Biol Reprod 61, 1083−9)を、テンプレートとして用いて、ヒトIGFBP−5(受け入れ番号 M65062)をエンコードするcDNAを増幅した。XhoI部位(5’−TCCGCTCGAGATGGTGTTGCTCACCGCGGT−3’)およびHindIII部位(5’−CGATAAGCTTCTCAACGTTGCTGCTGTCG−3’)を含有する特異的なプライマーを使用し、その結果のPCR生成物を消化して、pcDNA3.1/Myc−His(−)A(Invitrogen)のXhoI/HindIII部位中でクローン化した。完全長のproIGFBP−5をエンコードした構築物の直後に、残基KLGP、mycエピトープ(EQKLISEEDL)、NSAVD残基、および6個のH−残基(アミノ酸は、一文字表記されている)が続く。その構築物を、配列分析により確認した。トランスフェクション用のプラスミドDNAを、QIAprep Spinキット(Qiagen)により調製した。組み換えIGFBP−5の細胞培養および発現は、上述の実施例6.4.に示される通りに実施した。
【0278】
ウエスタンブロッティングにより開裂分析を実施した(図6)。簡単にいうと、5μLの細胞培養培地中に含まれるような組み換えIGFBP−5を、pPA2、pPA2−KO、または空の発現ベクター(実施例6.4参照)を用いてトランスフェクトされた細胞由来の培養上清(10μL)と一緒にインキュベートした。リン酸緩衝生理食塩水を添加して、50μLの最終容量とした。37℃で12時間インキュベーション後、15μLの反応混合物を、16% SDS−PAGEを還元して分離し、PVDF膜状にブロッティングし、C末端開裂生成物を、ペルオキシダーゼ共役型二次抗体(P260、DAKO)、および化学発光増幅(ECL、Amersham)を使用して、モノクローナル抗c−myc(クローン9E19、ATTC)により検出した。
6.8.PAPP−A2活性の阻害
IGFBP−5に対するPAPP−A2の蛋白分解活性の阻害能について、様々な薬剤を分析した。実験条件は、試験される薬剤を添加することを除いては、本質的には実施例6.7に示される通りであった(図6)。PAPP−A2の蛋白分解活性に影響を与えないと見い出された薬剤としては更に、PMSFおよびアプロチニンが挙げられる。
6.9.IGFBP−5における開裂部位の同定
開裂部位の決定については、精製されたrIGFBP−5(図6、レーン7)を、精製したPAPP−A2で消化し、SDS−PAGE(図6、レーン8)により分析した。ブロットした物質のエドマン分解により、別々の目に見える分解生成物(図6、レーン8)は両方とも、N末端配列、K(144)FVGGA(IGFBP−5は、成熟蛋白のN末端Leuを残基を1としてナンバリングしている)を含んでいることが示された。2つのバンドは両方とも、C末端c−mycタグを含むため(図6、レーン9)、インタクトなC末端開裂フラグメントを示しており;それらは精製されたrIGFBP−5(図6、レーン7)の異質性により、様々にグリコシル化される可能性がある。両方のバンドとも、低レベル(45%)で、IGFBP−5に相当する第2の配列、L(1)GXFVHを含んでいた。Serが存在しないことは、第3のサイクルで予想されるが、Ser−3の炭化水素置換の証拠と考えられた。N末端開裂フラグメント上のO−結合型グリカンが、2つ別々のC末端フラグメントの周りをスメアさせる可能性がある。反応混合物(>100pmol)に関してSDS−PAGE分離をしない配列分析は、等モルで同一の2つのIGFBP−5配列のみを示した。従って、PAPP−A2は、Ser−143とLys−144との間の1つの部位でIGFBP−5を開裂する。
6.10.PAPP−A2がIGFBP−5の蛋白分解を引き起こし得る組織
IGFBP−5に対する蛋白分解活性は、数種の起源、例えば妊婦血清(Claussen et al., 1994, Endocrinology 134, 1964−6)、精漿(Lee et al., 1994, J Clin Endocrinol Metab 79, 1367−72)、平滑筋細胞(Imai et al., 1997, J Clin Invest 100, 2596−605)、顆粒膜細胞(Resnick et al., 1998, Endocrinology 139, 1249−57)、骨肉腫細胞(Conover and Kiefer, 1993, J Clin Endocrinol Metab 76, 1153−9)、およびまたは骨芽細胞(Thrailkill et al., 1995, Endocrinology 136, 3527−33)由来の培養培地、および線維芽細胞(Busby et al., 2000, J Biol Chem)から広範に報告されている。一般的に、IGFBP−5の開裂の原因となるプロテアーゼは、同定されないままである。
【0279】
線維芽細胞および骨芽細胞(Lawrence et al., 1999, Proc Natl Acad Sci U S A 96, 3149−53)と同様、濾胞液(Conover et al., 1999, J Clin Endocrinol Metab 84, 4742−5)、妊婦血清(Overgaard et al., 2000, J Biol Chem)、および血管平滑筋細胞(Bayes−Genis, A., Schwartz, R. S., Ashai, K., Lewis, D. A., Overgaard, M. T., Christiansen, M., Oxvig, C., Holmes, D. R., Jr., and Conover, C. A. Arterioscler. Thromb. Vasc. Biol.、製本中)におけるIGFBP−4プロテアーゼとしてのPAPP−Aの最近の同定は、いくつかの系においてはPAPP−AおよびIGFBP−4を重要な機能ペアとして完全に確立した。PAPP−Aに対して見い出されているような基質以外はいずれも、および他のプロテアーゼはいずれも、生理的にIGFBP−4を開裂することが示されていない。それ故、PAPP−A2とIGFBP−5のペアが、上述のおよび/または他の多数の組織において、類似の役割を果たすものと思われる。興味深いことに、IGFBP−5を平滑筋細胞調整培地と一緒にインキュベーションした結果、Ser−143とLys−144との間が開裂し(Imai et al., 1997, J Clin Invest 100, 2596−605)、これはPAPP−A2により本明細書において見られるのと同一の開裂である。このことは、PAPP−A2をこの組織に対する明らかな候補IGFBP−5プロテアーゼとして直接示唆する。
引用文献
Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z., Miller, W., and Lipman, D. J. (1997). Gapped BLAST and PSI−BLAST: a new generation of protein database search programs. Nucleic Acids Res 25, 3389−402.
Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G., Smith, J. A., and Struhl, K. (1989). Current Protocols in Molecular Biology (New York City, NY: John Wiley & Sons).
Barker, R. L., Gleich, G. J., and Pease, L. R. (1988). Acidic precursor revealed in human eosinophil granule major basic protein cDNA [published erratum appears in J Exp Med 1989 Sep 1;170(3):1057]. J Exp Med 168, 1493−8.
Bayes−Genis, A., Schwartz, R. S., Ashai, K., A., L. D., Overgaard, M. T., Christian−sen, M., Oxvig, C., Holmes Jr, D. R., and Conover, C. A. (2000). Insulin−like growth factor binding protein−4 protease produced by arterial smooth muscle cells in vitro is increased in the coronary artery following angioplasty. Arterioscler Thromb Vasc Biol, in press.
Biagiotti, R., Cariati, E., Brizzi, L., Cappelli, G., and D’Agata, A. (1998). Maternal serum screening for trisomy 18 in the first trimester of pregnancy. Prenat Diagn 18, 907−13.
Bischof, P. (1979). Purification and characterization of pregnancy associated plasma protein A (PAPP−A). Arch Gynecol 227, 315−26.
Bitter, G. A., Egan, K. M., Koski, R. A., Jones, M. O., Elliott, S. G., and Giffin, J. C. (1987). Expression and secretion vectors for yeast. Methods Enzymol 153, 516−44.
Bode, W., Gomis−Ruth, F. X., and Stockler, W. (1993). Astacins, serralysins, snake venom and matrix metalloproteinases exhibit identical zinc−binding environments (HEXXHXXGXXH and Met−turn) and topologies and should be grouped into a common family, the ’metzincins’. FEBS Lett 331, 134−40.
Bonno, M., Oxvig, C., Kephart, G. M., Wagner, J. M., Kristensen, T., Sottrup−Jensen, L., and Gleich, G. J. (1994). Localization of pregnancy−associated plasma protein−A and colocalization of pregnancy−associated plasma protein−A messenger ribonucleic acid and eosinophil granule major basic protein messenger ribonucleic acid in placenta. Lab Invest 71, 560−6.
Brambati, B., Macintosh, M. C., Teisner, B., Maguiness, S., Shrimanker, K., Lanzani, A., Bonacchi, I., Tului, L., Chard, T., and Grudzinskas, J. G. (1993). Low maternal serum levels of pregnancy associated plasma protein A (PAPP−A) in the first trimester in association with abnormal fetal karyotype. Br J Obstet Gynaecol 100, 324−6.
Broglie, R., Coruzzi, G., Fraley, R. T., Rogers, S. G., Horsch, R. B., Niedermeyer, J. G., Fink, C. L., and Chua, N. H. (1984). Light−regulated expression of a pea ribulose−1,5−bisphosphate carboxylase small subunit gene in transformed plant cells. Science 224, 838−43.
Busby, W. H., Jr., Nam, T. J., Moralez, A., Smith, C., Jennings, M., and Clemmons, D. R. (2000). The Complement Component C1s is the Protease That Accounts for Cleavage of Insulin−like Growth Factor Binding Protein−5 in Fibroblast Medium. J Biol Chem.
Christiansen, M., Oxvig, C., Wagner, J. M., Qin, Q. P., Nguyen, T. H., Overgaard, M. T., Larsen, S. O., Sottrup−Jensen, L., Gleich, G. J., and Norgaard−Pedersen, B. (1999). The proform of eosinophil major basic protein: a new maternal serum marker for Down syndrome. Prenat Diagn 19, 905−10.
Christiansen, M., Jaliashvili, I., Overgaard, M.T., Ensinger, C., Oxvig,C. (2000) Quantitation and characterization of pregnancy−associatedcomplexes between angiotensinogen and the proform of eosinophil majorbasic protein in serum and amniotic fluid. Clin. Chem. 46, 1099−1105
Clackson, T., Hoogenboom, H. R., Griffiths, A. D., and Winter, G. (1991). Making antibody fragments using phage display libraries. Nature 352, 624−8.
Claussen, M., Zapf, J., and Braulke, T. (1994). Proteolysis of insulin−like growth factor binding protein−5 by pregnancy serum and amniotic fluid. Endocrinology 134, 1964−6.
Conover, C. A., and Kiefer, M. C. (1993). Regulation and biological effect of endogenous insulin−like growth factor binding protein−5 in human osteoblastic cells. J Clin Endocrinol Metab 76, 1153−9.
Conover, C. A., Kiefer, M. C., and Zapf, J. (1993). Posttranslational regulation of insulin−like growth factor binding protein−4 in normal and transformed human fibroblasts. Insulin−like growth factor dependence and biological studies. J Clin Invest 91, 1129−37.
Conover, C. A., Oxvig, C., Overgaard, M. T., Christiansen, M., and Giudice, L. C. (1999). Evidence that the insulin−like growth factor binding protein−4 protease in human ovarian follicular fluid is pregnancy associated plasma protein−A [In Process Ci−tation]. J Clin Endocrinol Metab 84, 4742−5.
Crowther, J. R. (1995). ELISA. Theory and Practice, Methods in Molecular Biology. vol 42 (Totowa, NJ: Humana Press).
DuBridge, R. B., Tang, P., Hsia, H. C., Leong, P. M., Miller, J. H., and Calos, M. P. (1987). Analysis of mutation in human cells by using an Epstein−Barr virus shuttle system. Mol Cell Biol 7, 379−87.
Folkersen, J., Grudzinskas, J. G., Hindersson, P., Teisner, B., and Westergaard, J. G. (1981). Pregnancy−associated plasma protein A: circulating levels during normal pregnancy. Am J Obstet Gynecol 139, 910−4.
Fowlkes, J. L. (1997). Insulinlike growth factor−binding protein proteolysis. An emerging paradigm in insulin−like growth factor physiology. Trends Endocrinol Metab 8, 299−306.
Gmunder, H., and Kohli, J. (1989). Cauliflower mosaic virus promoters direct efficient expression of a bacterial G418 resistance gene in Schizosaccharomyces pombe. Mol Gen Genet 220, 95−101.
Haddow, J. E., Palomaki, G. E., Knight, G. J., Williams, J., Miller, W. A., and Johnson, A. (1998). Screening of maternal serum for fetal Down’s syndrome in the first trimester. N Engl J Med 338, 955−61.
Ho, S. N., Hunt, H. D., Horton, R. M., Pullen, J. K., and Pease, L. R. (1989). Site−directed mutagenesis by overlap extension using the polymerase chain reaction [see comments]. Gene 77, 51−9.
Hwa, V., Oh, Y., and Rosenfeld, R. G. (1999). The insulin−like growth factor−binding protein (IGFBP) superfamily. Endocr Rev 20, 761−87.
Haaning, J., Oxvig, C., Overgaard, M. T., Ebbesen, P., Kristensen, T., and Sottrup−Jensen, L. (1996). Complete cDNA sequence of the preproform of human pregnancy−associated plasma protein−A. Evidence for expression in the brain and induction by cAMP. Eur J Biochem 237, 159−63.
Imai, Y., Busby, W. H., Jr., Smith, C. E., Clarke, J. B., Garmong, A. J., Horwitz, G. D., Rees, C., and Clemmons, D. R. (1997). Protease−resistant form of insulin−like growth factor−binding protein 5 is an inhibitor of insulin−like growth factor−I actions on porcine smooth muscle cells in culture. J Clin Invest 100, 2596−605.
Kristensen, T., Oxvig, C., Sand, O., Moller, N. P., and Sottrup−Jensen, L. (1994). Amino acid sequence of human pregnancy−associated plasma protein−A derived from cloned cDNA. Biochemistry 33, 1592−8.
Lawrence, J. B., Oxvig, C., Overgaard, M. T., Sottrup−Jensen, L., Gleich, G. J., Hays, L. G., Yates, J. R., 3rd, and Conover, C. A. (1999). The insulin−like growth factor (IGF)−dependent IGF binding protein−4 protease secreted by human fibroblasts is pregnancy−associated plasma protein−A. Proc Natl Acad Sci U S A 96, 3149−53.
Lee, K. O., Oh, Y., Giudice, L. C., Cohen, P., Peehl, D. M., and Rosenfeld, R. G. (1994). Identification of insulin−like growth factor−binding protein−3 (IGFBP−3) fragments and IGFBP−5 proteolytic activity in human seminal plasma: a comparison of normal and vasectomized patients. J Clin Endocrinol Metab 79, 1367−72.
Lin, T. M., Galbert, S. P., Kiefer, D., Spellacy, W. N., and Gall, S. (1974). Characterization of four human pregnancy−associated plasma proteins. Am J Obstet Gynecol 118, 223−36.
Logan, J., and Shenk, T. (1984). Adenovirus tripartite leader sequence enhances translation of mRNAs late after infection. Proc Natl Acad Sci U S A 81, 3655−9.
Mackett, M., Smith, G. L., and Moss, B. (1982). Vaccinia virus: a selectable eukaryotic cloning and expression vector. Proc Natl Acad Sci U S A 79, 7415−9.
Marks, J. D., Hoogenboom, H. R., Bonnert, T. P., McCafferty, J., Griffiths, A. D., and Winter, G. (1991). By−passing immunization. Human antibodies from V−gene libraries displayed on phage. J Mol Biol 222, 581−97.
Matthews, D. J., and Wells, J. A. (1993). Substrate phage: selection of protease substrates by monovalent phage display. Science 260, 1113−7.
McGrogan, M., Simonsen, C., Scott, R., Griffith, J., Ellis, N., Kennedy, J., Campanelli, D., Nathan, C., and Gabay, J. (1988). Isolation of a complementary DNA clone encoding a precursor to human eosinophil major basic protein. J Exp Med 168, 2295−308.
Meldal, M. (1998). Intramolecular fluorescence−quenched substrate libraries. Methods Mol Biol 87, 65−74.
Meldal, M. (1998). Introduction to combinatorial solid−phase assays for enzyme activity and inhibition. Methods Mol Biol 87, 51−7.
Meldal, M. (1998). The solid−phase enzyme inhibitor library assay. Methods Mol Biol 87, 75−82.
Nakayama, K. (1997). Furin: a mammalian subtilisin/Kex2p−like endoprotease involved in processing of a wide variety of precursor proteins. Biochem J 327, 625−35.
Nielsen, H., Engelbrecht, J., Brunak, S., and von Heijne, G. (1997). Identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavage sites. Protein Eng 10, 1−6.
Overgaard, M. T., Haaning, J., Boldt, H. B., Olsen, I. M., Laursen, L., Christiansen, M., Gleich, G. J., Sottrup−Jensen, L., Conover, C. A., and Oxvig, C. (2000). Expression of Recombinant Human Pregnancy−Associated Plasma Protein−A and Identification of the Proform of Eosinophil Major Basic Protein as its Physiological Inhibitor. J Biol Chem.
Overgaard, M. T., Oxvig, C., Christiansen, M., Lawrence, J. B., Conover, C. A., Gleich, G. J., Sottrup−Jensen, L., and Haaning, J. (1999). Messenger ribonucleic acid levels of pregnancy−associated plasma protein−A and the proform of eosinophil major basic protein: expression in human reproductive and nonreproductive tissues. Biol Reprod 61, 1083−9.
Oxvig, C., Haaning, J., Hojrup, P., and Sottrup−Jensen, L. (1994). Location and nature of carbohydrate groups in proform of human major basic protein isolated from pregnancy serum. Biochem Mol Biol Int 33, 329−36.
Oxvig, C., Haaning, J., Kristensen, L., Wagner, J. M., Rubin, I., Stigbrand, T., Gleich, G. J., and Sottrup−Jensen, L. (1995). Identification of angiotensinogen and complement C3dg as novel proteins binding the proform of eosinophil major basic protein in human pregnancy serum and plasma. J Biol Chem 270, 13645−51.
Oxvig, C., Sand, O., Kristensen, T., Gleich, G. J., and Sottrup−Jensen, L. (1993). Circulating human pregnancy−associated plasma protein−A is disulfide−bridged to the proform of eosinophil major basic protein. J Biol Chem 268, 12243−6.
Oxvig, C., Sand, O., Kristensen, T., Kristensen, L., and Sottrup−Jensen, L. (1994). Isolation and characterization of circulating complex between human pregnancy−associated plasma protein−A and proform of eosinophil major basic protein. Biochim Biophys Acta 1201, 415−23.
Pear, W. S., Nolan, G. P., Scott, M. L., and Baltimore, D. (1993). Production of high−titer helper−free retroviruses by transient transfection. Proc Natl Acad Sci U S A 90, 8392−6.
Peters, J. H., and Baumgarten, H. (1992). Monoclonal Antibodies (Offersheim, Germany: Springer−Verlag).
Rajaram, S., Baylink, D. J., and Mohan, S. (1997). Insulin−like growth factor−binding proteins in serum and other biological fluids: regulation and functions. Endocr Rev 18, 801−31.
Resnick, C. E., Fielder, P. J., Rosenfeld, R. G., and Adashi, E. Y. (1998). Characterization and hormonal regulation of a rat ovarian insulin−like growth factor binding protein−5 endopeptidase: an FSH−inducible granulosa cell−derived metalloprotease. Endocrinology 139, 1249−57.
Rosenfeld, S. A. (1999). Use of Pichia pastoris for expression of recombinant proteins. Methods Enzymol 306, 154−69.
Ruther, U., and Muller−Hill, B. (1983). Easy identification of cDNA clones. Embo J 2, 1791−4.
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). Molecular Cloning. A Laboratory Manual (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press).
Scopes, R. K. (1987). Protein Purification. Principles and Practice. (Harrisonburg, VA: Springer−Verlag).
Sinosich, M. J. (1990). Molecular characterization of pregnancy−associated plasma protein−A by electrophoresis. Electrophoresis 11, 70−8.
Smith, G. E., Summers, M. D., and Fraser, M. J. (1983). Production of human beta interferon in insect cells infected with a baculovirus expression vector. Mol Cell Biol 3, 2156−65.
Sottrup−Jensen, L. (1993). Determination of halfcystine in proteins as cysteine from reducing hydrolyzates. Biochem Mol Biol Int 30, 789−94.
Sottrup−Jensen, L. (1995). A low−pH reverse−phase high−performance liquid chromatography system for analysis of the phenylthiohydantoins of S−carboxymethylcysteine and S−carboxyamidomethylcysteine. Anal Biochem 225, 187−8.
Spencer, K., Ong, C., Skentou, H., A, W. L., and K, H. N. (2000). Screening for tri−somy 13 by fetal nuchal translucency and maternal serum free beta−hCG and PAPP−A at 10−14 weeks of gestation. Prenat Diagn 20, 411−6.
Stocker, W., Grams, F., Baumann, U., Reinemer, P., Gomis−Ruth, F. X., McKay, D. B., and Bode, W. (1995). The metzincins−−topological and sequential relations between the astacins, adamalysins, serralysins, and matrixins (collagenases) define a superfamily of zinc−peptidases. Protein Sci 4, 823−40.
Thompson, J. D., Higgins, D. G., and Gibson, T. J. (1994). CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position−specific gap penalties and weight matrix choice. Nucleic Acids Res 22, 4673−80.
Thrailkill, K. M., Quarles, L. D., Nagase, H., Suzuki, K., Serra, D. M., and Fowlkes, J. L. (1995). Characterization of insulin−like growth factor−binding protein 5−degrading proteases produced throughout murine osteoblast differentiation. Endocrinology 136, 3527−33.
Wald, N., Stone, R., Cuckle, H. S., Grudzinskas, J. G., Barkai, G., Brambati, B., Teisner, B., and Fuhrmann, W. (1992). First trimester concentrations of pregnancy associated plasma protein A and placental protein 14 in Down’s syndrome [see comments]. Bmj 305, 28.
Wald, N. J., Watt, H. C., and Hackshaw, A. K. (1999). Integrated screening for Down’s syndrome on the basis of tests performed during the first and second trimesters [see comments]. N Engl J Med 341, 461−7.
Walsh, P. S., Erlich, H. A., and Higuchi, R. (1992). Preferential PCR amplification of alleles: mechanisms and solutions. PCR Methods Appl 1, 241−50.
Westergaard, J. G., Chemnitz, J., Teisner, B., Poulsen, H. K., Ipsen, L., Beck, B., and Grudzinskas, J. G. (1983). Pregnancy−associated plasma protein A: a possible marker in the classification and prenatal diagnosis of Cornelia de Lange syndrome. Prenat Diagn 3, 225−32.
【0280】
【表3】
【受託番号】
【0281】
DSM13783
【技術分野】
【0001】
本発明の属する分野
本発明は、妊娠関連血漿蛋白−A(PAPP−A)と相同性のある新規なポリペプチドに関する。本発明に記載の新規なポリペプチドは、PAPP−A2と示す。本発明は更に、そのようなポリペプチドまたはそのフラグメントをエンコードする核酸配列を含む新規なポリヌクレオチドに関する。
【0002】
本発明は更に、本明細書の以下で定義されるような、新規なポリヌクレオチド(そのフラグメントを含む)を用いる方法、および当該ポリヌクレオチドから生成可能な新規なポリペプチドの使用法に関する。
【0003】
本発明は、組み換えPAPP−A2の発現と精製、およびPAPP−A2に対するポリクローナルおよびモノクローナル抗体の生成、そしてヒトの組織または体液由来の天然のPAPP−A2の精製にも関する。
【0004】
更なる態様において、本発明は、病理的状態に対するマーカーとして、および妊娠している、ならびに妊娠していない個体のPAPP−A2の蛋白分解活性を改変する薬物に対する治療的ターゲットとしてのPAPP−A2の使用に関する。
【背景技術】
【0005】
本発明の背景
妊娠関連血漿蛋白−A(PAPP−A)
PAPP−Aは、1974年に初めて妊婦の血清から胎盤起源と考えられる他の蛋白と一緒に単離された(非特許文献1)。血清中濃度は、妊娠の終了時には約50mg/リットルに達する(非特許文献2;非特許文献3)。PAPP−Aは、高分子量のホモ四量体として最初は特徴付けられた(非特許文献4;非特許文献1;非特許文献5)が、今やPAPP−Aは、主として妊婦の血清および血漿に、好酸球主要塩基性蛋白のプロフォーム(proMBP)との共有結合性のヘテロ四量体の2:2の複合体、PAPP−A/proMBPとして存在することが実証されている(非特許文献6)。妊婦の血清および血漿中において僅か約1%のPAPP−Aが、最近実証されるように(Overgaard et al., 2000, J Biol Chem)ホモ二量体として存在する。PAPP−A/proMBP複合体の存在は、精製されたPAPP−A/proMBPの消化から、ジスルフィド結合によって相互に結合されたPAPP−AおよびproMBPペプチドの単離により一部分は明らかとなった(非特許文献6)。
【0006】
PAPP−A/proMBP複合体のサブユニットは、ジスルフィド結合の還元および変性により不可逆的に分離可能である(非特許文献6)。SDS−PAGEを還元すると、PAPP−Aサブユニットは、見かけの分子量が200kDaであり(非特許文献7)、その1547残基の配列は、クローニングしたcDNAから分かる(非特許文献8)。PAPP−Aは、80残基のpre−pro片(非特許文献9)を含む、pre−pro蛋白(preproPAPP−A)として合成される。PAPP−Aと広範に相同性のある蛋白は、文献上では報告されていないが、PAPP−Aは、482〜492位に伸長した亜鉛結合モチーフ(HEXXHXXGXXH)(非特許文献8に従ってナンバリングしたもの)を含む配列モチーフを含有する。このモチーフおよび構造的に重要なメチオニン残基(PAPP−Aにおいて556位の残基と考えられている)は、厳密には亜鉛ペプチダーゼのスーパーファミリーであるメトジンシン(metzincins):astacins、adamalysins(またはreprolysins)、serralysins、およびmatrixins(マトリックスメタロプロテアーゼまたはMMP’s)内に保持されている(非特許文献10;非特許文献11)。
【0007】
ProMBPサブユニットは、23kDaの計算されたペプチド質量を有する(非特許文献12;非特許文献13)。しかしながらSDS−PAGEにおいて、proMBPは、おそらくその強力で異常なグリコシル化(非特許文献14;非特許文献7)のために、クマシー染色したゲルでは見ることのできない50〜90kDaのスメア(smear)として移動している(非特許文献6)。PAPP−AおよびproMBPは双方とも、インサイツハイブリダイゼーションにより示されるように(非特許文献15)妊娠中に胎盤においてであるが、主に異なる種類の細胞において生成される。RT−PCRによる分析により、PAPP−AおよびproMBPの双方のmRNAが、胎盤中よりは量は少ないけれども、数種の生殖および非生殖組織中に存在することが明らかになった(非特許文献16)。
PAPP−Aの臨床的用途
臨床上、血清PAPP−Aレベルの低下は、ダウン症妊娠の予測値として徐々に利用されてきており(非特許文献17;非特許文献18;非特許文献19;非特許文献20)、血清PAPP−Aレベルは、他の胎児異常においてもまた低下することが示されている(非特許文献21;非特許文献22;非特許文献23)。
【0008】
更に、血管形成後の冠動脈の平滑筋細胞においてはPAPP−A合成が増大し(Bayes−Genis et al., 2000, Arterioscler Thromb Vasc Biol、製本中)、それは、現在臨床的価値の可能性があるために評価されている。妊婦血清におけるproMBPの測定が診断的価値があることもデータは示している(非特許文献24)。
PAPP−Aの蛋白分解活性:IGFBP−4の開裂
ごく最近になって、PAPP−Aの推定のメタロプロテアーゼ活性が、実験で確認されている(非特許文献25)。PAPP−Aは、ヒト線維芽細胞調整培地(HFCM)から一部分精製され、インシュリン様成長因子結合蛋白(IGFBP)−4に対する蛋白分解活性を引き起こすことが示されている。IGFBPは、そのうちの6個が示されているが、IGF−Iおよび−II活性の重要なモジュレータ−である(非特許文献26;非特許文献27)。
【0009】
IGF−Iおよび−IIは、インビボおよびインビトロの双方において、強力な蛋白同化および細胞分裂作用を有する重要なポリペプチドである。IGFBP−4に結合したIGFは、そのレセプターとは相互作用できないが、その結合蛋白が開裂すると、生理活性IGFが遊離される。興味深いことに、PAPP−AによるIGFBP−4の開裂は、厳密にはIGFの存在が必要とされる(非特許文献28;非特許文献25)。PAPP−A分泌は、骨芽細胞および骨髄間質細胞から(非特許文献25)、顆粒膜細胞から(非特許文献29)、および血管平滑筋細胞から(Bayes−Genis et al., 2000, Arterioscler Thromb Vasc Biol, in press)も実証されており、これらはすべて既知のIGF依存性IGFBP−4プロテアーゼ活性を有する。
IGFBP−5
IGFBP−4と同様、IGFBP−5開裂は、多数の組織および調整培地において、同定されていないプロテアーゼにより起こると広範に報告されている(非特許文献30)。
【先行技術文献】
【非特許文献】
【0010】
【非特許文献1】Lin et al., 1974, Am J Obstet Gynecol 118, 223−36
【非特許文献2】Folkersen et al., 1981, Am J Obstet Gynecol 139, 910−4
【非特許文献3】Oxvig et al., 1995, J Biol Chem 270, 13645−51
【非特許文献4】Bischof, 1979, Arch Gynecol 227, 315−26
【非特許文献5】Sinosich, 1990, Electrophoresis 11, 70−8
【非特許文献6】Oxvig et al., 1993, J Biol Chem 268, 12243−6
【非特許文献7】Oxvig et al., 1994, Biochim Biophys Acta 1201, 415−23
【非特許文献8】Kristensen et al., 1994, Biochemistry 33, 1592−8
【非特許文献9】Haaning et al., 1996, Eur J Biochem 237, 159−63
【非特許文献10】Bode et al., 1993, FEBS Lett 331, 134−40
【非特許文献11】Stocker et al., 1995, Protein Sci 4, 823−40
【非特許文献12】Barker et al., 1988, J Exp Med 168, 1493−8
【非特許文献13】McGrogan et al., 1988, J Exp Med 168, 2295−308
【非特許文献14】Oxvig et al., 1994, Biochem Mol Biol Int 33, 329−36
【非特許文献15】Bonno et al., 1994, Lab Invest 71, 560−6
【非特許文献16】Overgaard et al., 1999, Biol Reprod 61, 1083−9
【非特許文献17】Brambati et al., 1993, Br J Obstet Gynaecol 100, 324−6
【非特許文献18】Haddow et al., 1998, N Engl J Med 338, 955−61
【非特許文献19】Wald et al., 1992, Bmj 305, 28
【非特許文献20】Wald et al., 1999, N Engl J Med 341, 461−7
【非特許文献21】Biagiotti et al., 1998, Prenat Diagn 18, 907−13
【非特許文献22】Spencer et al., 2000, Prenat Diagn 20, 411−6
【非特許文献23】Westergaard et al., 1983, Prenat Diagn 3, 225−32
【非特許文献24】Christiansen et al., 1999, Prenat Diagn 19, 905−10
【非特許文献25】Lawrence et al., 1999, Proc Natl Acad Sci U S A 96, 3149−53
【非特許文献26】Fowlkes, 1997, Trends Endocrinol Metab 8, 299−306
【非特許文献27】Rajaram et al., 1997, Endocr Rev 18, 801−31
【非特許文献28】Conover et al., 1993, J Clin Invest 91, 1129−37
【非特許文献29】Conover et al., 1999, J Clin Endocrinol Metab 84, 4742−5
【非特許文献30】Hwa et al., 1999, Endocr Rev 20, 761−87
【発明の概要】
【課題を解決するための手段】
【0011】
本発明の概要
妊娠関連血漿蛋白−A2
本発明に記載の新規な核酸は、ヒト胎盤から単離され、配列決定分析により特徴付けられる。該新規なヌクレオチド配列は、新規なポリペプチド、PAPP−A2をエンコードする。
【0012】
PAPP−A2のアミノ酸配列は、233残基のpre−pro片および1558残基の成熟部分から構成される。PAPP−A2の成熟部分は、PAPP−Aの成熟部分と相同する(およそ45%の同一性)が、そのprepro片は、その2つの蛋白間で全く類似性がない。PAPP−Aと同様、PAPP−A2は、それをメトジンシンスーパーファミリーの推定のメタロプロテアーゼとして分類する保存アミノ酸伸長部を含有する。
【0013】
PAPP−A2は、哺乳類の発現系において発現し、PAPP−A2が活性酵素であることが実証されている。更に、PAPP−A2が、IGFBP−5、インシュリン様成長因子結合蛋白5型を開裂することが実証されている。相対的に、PAPP−AによるIGFBP−4の開裂は、以前から実証されている。
【0014】
完全長の形態のPAPP−A2をエンコードする相補的DNA(cDNA)が、同定され、配列決定され、単離される。該cDNAまたはcDNAの一部分が、組み換え宿主における発現のために発現ベクター中にクローニングされる。該cDNAは、組み換え完全長のPAPP−A2またはPAPP−A2フラグメントを生成するのに有益である。該cDNAおよびそれ由来の組み換えPAPP−A2蛋白は、抗体生成、診断用キット、実験用試薬、およびアッセイのおいて有益となる。
【0015】
該cDNAおよび組み換えPAPP−A2蛋白は、PAPP−A2機能に影響を及ぼす化合物を同定するのにも使用され得る。PAPP−A2アンチセンスオリゴヌクレオチドまたはアンチセンスミメティクスは、臨床上PAPP−A2蛋白の発現を減少させ、それによりPAPP−Aの効果にアンタゴニストとして作用するために有益となり得る。同様に、PAPP−A2コード化配列は、遺伝子治療に対して、ターゲット細胞中にPAPP−A2を導入し、それによりPAPP−A2の効果を促進させるために使用可能である。
【0016】
本発明は更に、細胞中のIGFBP−5蛋白分解活性の減少または削減に対する治療的ターゲットとして使用されるPAPP−A2に関する。
【0017】
診断の目的でのPAPP−A2の使用法を提供することが、更なる本発明の目的である。
【0018】
本発明の他の特徴および利点は、以下に続く図面およびその説明、以下に続く詳細な説明、および請求項から明らかとなるであろう。
【0019】
本発明の好ましい実施形態では、以下の精製ポリヌクレオチドなどが提供される:
(項目1) 精製ポリヌクレオチドであって、
i)受け入れ番号DSM13783下のDSMZに集積された、PAPP−A2のコード化配列に相当する、配列番号1のヌクレオチド1〜5376を含むポリヌクレオチド;および
ii)配列番号2に示されるようなアミノ酸配列を有するポリペプチドをエンコードするポリヌクレオチド;および
iii)ポリヌクレオチド(i)または(ii)によりエンコードされたポリペプチドのフラグメントをエンコードするポリヌクレオチドであって、当該フラグメントが、
a)インシュリン様成長因子結合蛋白5型(IGFBP−5)またはその誘導体、あるいは任意の他の基質に特異的な蛋白分解活性を有し;そして/または
b)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
c)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、ポリヌクレオチド;そして
iv)相補鎖が、(i)、(ii)、および(iii)のいずれかで定義されるようなポリヌクレオチドと、厳密な条件下でハイブリダイズされるポリヌクレオチドであって、当該ポリヌクレオチドが、配列番号2で示されるようなアミノ酸配列を有するポリペプチド、またはそのフラグメントをエンコードし、当該フラグメントが、
a)インシュリン様成長因子結合蛋白5型(IGFBP−5)またはその誘導体、あるいは任意の他の基質に特異的な蛋白分解活性を有し;そして/または
b)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
c)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、ポリヌクレオチド;および
v)(iii)および(iv)のいずれかで定義されるようなポリヌクレオチドのヌクレオチド配列に縮重されるヌクレオチド配列を含むポリヌクレオチド、およびそのようなポリヌクレオチドの相補鎖、からなる群から選ばれる、精製ポリヌクレオチド。
【0020】
(項目2) 項目1に記載で、配列番号1に示されるようなコード化配列を含む、精製ポリヌクレオチド。
【0021】
(項目3) 項目1に記載で、配列番号2に示されるようなアミノ酸配列を有するポリペプチドをエンコードする、ポリヌクレオチド。
【0022】
(項目4) 項目1に記載で、配列番号2に示されるようなアミノ酸配列を有するポリペプチドのフラグメントをエンコードするポリヌクレオチドであって、当該フラグメントが、
a)インシュリン様成長因子結合蛋白5型(IGFBP−5)に特異的な蛋白分解活性を有し;そして/または
b)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
c)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、ポリヌクレオチド。
【0023】
(項目5) 該ポリペプチドの相補鎖が、項目2〜4のいずれかに記載のポリヌクレオチドと、厳密な条件下でハイブリダイズする、項目1に記載のポリヌクレオチド。
【0024】
(項目6) 項目1に記載で、項目3及び4のいずれかに記載のポリヌクレオチドのヌクレオチド配列に縮重されるヌクレオチド配列を含む、ポリヌクレオチド。
【0025】
(項目7) 該ポリヌクレオチドが、項目2〜6のいずれかに記載のポリヌクレオチドの相補鎖を含む、項目1に記載のポリヌクレオチド。
【0026】
(項目8) 3’未翻訳領域、またはそのフラグメント、あるいは配列番号1に相当する、配列番号1の核酸残基5377〜8527を含む更なるポリヌクレオチドに実施可能に結合した、項目1〜7のいずれかに記載のポリヌクレオチド。
【0027】
(項目9) 項目1〜7のいずれかに記載のポリヌクレオチドに実施可能に結合した発現シグナルを含む、発現ベクターの形態の組み換えDNA分子。
【0028】
(項目10) 項目1〜8のいずれかに記載のポリヌクレオチド、又は項目9に記載のベクターを用いてトランスフェクト、あるいは形質転換した、宿主生体。
【0029】
(項目11) 該生体が、哺乳類の生体である、項目10に記載の宿主生体。
【0030】
(項目12) 配列番号2またはそのフラグメントのアミノ酸配列を含むか、あるいはそれらから本質的に構成される単離されたポリペプチドであって、
当該フラグメントが、
i)少なくともインシュリン様成長因子結合蛋白5型(IGFBP−5)に特異的な蛋白分解活性を有し;そして/または
ii)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
iii)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、ポリペプチド。
【0031】
(項目13) 該フラグメントが、任意のプロセシング変異体を含むPAPP−A2の成熟部分に相当する、アミノ酸残基234〜1791を含むか、あるいはそれらから本質的に構成される、項目12に記載のポリペプチド。
【0032】
(項目14) 該フラグメントが、PAPP−A2のprepro部分に相当する、アミノ酸残基1〜233を含むか、あるいはそれらから本質的に構成される、項目12に記載のポリペプチド。
【0033】
(項目15) 該フラグメントが、PAPP−A2のpro部分に相当する、アミノ酸残基23〜233を含むか、あるいはそれらから本質的に構成される、項目12に記載のポリペプチド。
【0034】
(項目16) 該フラグメントが、PAPP−A2のシグナルペプチドまたはリーダー配列に相当する、アミノ酸残基1〜22を含むか、あるいはそれらから本質的に構成される、項目12に記載のポリペプチド。
【0035】
(項目17) 配列番号2のアミノ酸残基234〜1791に相当する、PAPP−A2の成熟部分に実施可能に結合した、項目14〜16のいずれかに記載のポリペプチド。
【0036】
(項目18) 該ポリペプチドが、組み換えポリペプチドである、項目12〜17のいずれかに記載のポリペプチド。
【0037】
(項目19) 該ポリペプチドが、ヒト蛋白、または該ポリペプチドに天然に付随する他の蛋白を含まない、項目12〜18のいずれかに記載のポリペプチド。
【0038】
(項目20) i)項目1〜8のいずれかに記載のポリヌクレオチド、および/またはii)項目9に記載のベクター、および/またはiii)項目10および11のいずれかに記載の宿主生体、および/またはiv)項目12〜19のいずれかに記載のポリペプチドを、生理的に許容される担体と組み合わせて含む、組成物。
【0039】
(項目21) i)項目1〜8のいずれかに記載のポリヌクレオチド、および/またはii)項目9に記載のベクター、および/またはiii)項目10および11のいずれかに記載の宿主生体、および/またはiv)項目12〜19のいずれかに記載のポリペプチドを、薬学的に許容される担体と組み合わせて含む、医薬組成物。
【0040】
(項目22) 項目12に記載のポリペプチドに対して特異性を有する抗体の生成法であって、当該方法が、
i)宿主生体を提供し、
ii)その宿主生体を、項目10に記載のポリペプチドを用いて免疫し、そして
iii)当該抗体を得る、
ことからなるステップを含む、方法。
【0041】
(項目23) 項目12に記載のポリペプチドに対して特異的な結合親和性を有する、抗体。
【0042】
(項目24) 該抗体が、モノクローナル抗体およびポリクローナル抗体からなる群から選ばれる、項目23に記載の抗体。
【0043】
(項目25) 該抗体が、モノクローナルである、項目24に記載の抗体。
【0044】
(項目26) 項目18に記載のポリペプチドの生成法であって、当該方法が、
i)好適な宿主生体を提供し、
ii)ステップi)で提供された宿主生体を、項目1〜8のいずれかに記載のポリヌクレオチド、または項目9に記載のベクターを用いてトランスフェクトまたは形質転換し、 iii)ステップii)で得られた宿主生体を該ポリヌクレオチドまたはベクターによりエンコードされたポリペプチドの発現に好適な条件下で培養し;そして所望により
iv)宿主生体による組み換え発現の結果生じるポリペプチドを、その宿主生体から単離する、
ことからなるステップを含む、方法。
【0045】
(項目27) 該宿主生体が、哺乳類細胞である、項目26に記載の方法。
【0046】
(項目28) アンチセンス法を用いて、細胞中のPAPP−A2の発現を阻害および/または減少させる方法であって、当該方法が、
i)項目7に記載のポリヌクレオチドを提供し、
ii)ステップi)において提供される当該ポリヌクレオチドを用いて、PAPP−A2を発現可能な細胞をトランスフェクトまたは形質転換し、
iii)ステップii)において得られた細胞を、PAPP−A2の発現に関与する当該細胞において、ステップi)で提供されたポリヌクレオチドの相補的なポリヌクレオチドへのハイブリダイゼーションに好適な条件下で培養し、そして
iv)当該細胞においてPAPP−A2の発現を阻害および/または減少させる、
ことからなるステップを含む、方法。
【0047】
(項目29) 該アンチセンスポリヌクレオチドおよび該相補的なポリヌクレオチドが、別々のポリヌクレオチド分子から同時発現される、項目28に記載の方法。
【0048】
(項目30) 個体(individual)から得た生物学的サンプルにおけるPAPP−A2の検出法、あるいはPAPP−A2レベル(level of PAPP−A2)の測定法であって、当該方法が、
i)当該個体から生物学的サンプルを得て、
ii)そのサンプルにおいて、
a)項目12に記載のポリペプチド;および/または
b)PAPP−A2発現に由来するmRNA形態のポリヌクレオチド、および/または
c)好ましくはIGFBP−5、その誘導体の開裂、またはPAPP−A2に対する他の任意の好適な基質を検出することによる、PAPP−A2特異的プロテアーゼ活性、を検出することによりサンプル中のPAPP−A2を検出する、
ことからなるステップを含む、方法。
【0049】
(項目31) 該方法が、ステップii)で検出されたPAPP−A2またはPAPP−A2レベルを、
i)予め決められたPAPP−A2の量および/または濃度;および/または
ii)予め決められたPAPP−A2 mRNAの量および/または濃度;および/または
iii)予め決められたPAPP−A2特異的なプロテアーゼ活性、
からなる群から選ばれる予め決められた値と比較することからなるステップを更に含む、項目30に記載の方法。
【0050】
(項目32) 該予め決められた値が、当該個体の正常な生理学的(physiological)状態を示す、項目31に記載の方法。
【0051】
(項目33) 該生物学的サンプルが、血液、尿、胸腔内液、口腔洗浄液、組織バイオパシー、および卵胞液からなる群から選ばれる、項目30に記載の方法。
【0052】
(項目34) 該PAPP−A2レベルが、PAPP−A2特異的プロテアーゼ活性として測定される、項目30に記載の方法。
【0053】
(項目35) 該PAPP−A2レベルが、PAPP−A2蛋白の量として測定される、項目30に記載の方法。
【0054】
(項目36) 該PAPP−A2レベルが、PAPP−A2のメッセンジャーRNAの量として測定される、項目30に記載の方法。
【0055】
(項目37) 該PAPP−A2蛋白量が、免疫化学的分析により測定される、項目35に記載の方法。
【0056】
(項目38) 該PAPP−A2蛋白量が、少なくとも1つのモノクローナル抗体により検出される、項目37に記載の方法。
【0057】
(項目39) 該PAPP−A2蛋白が、少なくとも1つ更なる成分を含む複合体、好ましくはポリペプチドにおいて検出される、項目30に記載の方法。
【0058】
(項目40) 該PAPP−A2が、PAPP−A2単量体として検出される、項目30に記載の方法。
【0059】
(項目41) 該PAPP−A2が、PAPP−A2二量体として検出される、項目30に記載の方法。
【0060】
(項目42) 個体における病態の診断法であって、当該方法が、
i)項目30〜41のいずれかに記載の方法を実施し、そして
ii)その病態を診断する、
ことからなるステップを含む、方法。
【0061】
(項目43) 該病態が、胎児異常である、項目42に記載の方法。
【0062】
(項目44) 該胎児異常が、トリソミー21、トリソミー18、トリソミー13、および開放型二分脊椎からなる群から選ばれる、項目43に記載の方法。
【0063】
(項目45) 該胎児異常が、子宮外妊娠、開放型二分脊椎、神経管閉鎖障害、腹部壁欠損、Edwards症候群、Pateaus症候群、Turner症候群、モノソミーX、またはKlein−felter’s症候群である、項目43に記載の方法。
【0064】
(項目46) 該病態が、成長促進状態および成長阻害状態からなる群から選ばれる異常な成長状態である、項目43に記載の方法。
【0065】
(項目47) 該病態が、再狭窄、アテローム性心筋梗塞、創傷治癒、線維症、心筋梗塞、骨粗鬆症、リュウマチ性関節炎、多発性骨髄腫、または癌からなる群から選ばれる、項目46に記載の方法。
【0066】
(項目48) 生物学的サンプルにおける項目1に記載のポリヌクレオチドの発現検出法であって、当該方法が:
i)項目1に記載のポリヌクレオチドを推定で含有する生物学的サンプルを提供し、そして
ii)その生物学的サンプルを、i)項目1に記載のポリヌクレオチドに相補的な、およびii)それとハイブリダイズ可能な、鎖を含むポリヌクレオチドと接触させ、そして iii)ハイブリダイゼーションが起こさせ、そして
iv)ステップiii)で得られたハイブリダイゼーション複合体を検出する、
ことからなるステップを含み、
該ハイブリダイゼーション複合体の存在が、項目1に記載のポリヌクレオチド、またはそのフラグメントの生物学的サンプルにおける発現を示している、方法。
【0067】
(項目49) PAPP−A2のプロテアーゼ活性を阻害する薬剤の同定法であって、当該方法が、
i)a)項目12に記載のポリペプチド、およびb)当該ポリペプチドに対して予め決められた基質、およびc)推定の阻害剤をインキュベートし、そして
ii)該基質の蛋白分解が阻害されるかどうかを決定する、
ことからなるステップを含む、方法。
【0068】
(項目50) 該基質が、ポリペプチドを含む、項目49に記載の方法。
【0069】
(項目51) 該基質が、内部でクエンチングする蛍光性ペプチドを含む、項目50に記載の方法。
【0070】
(項目52) 該基質が、IGFBP−5またはそのフラグメントを含むか、あるいはそれから本質的に構成される、項目50に記載の方法。
【0071】
(項目53) 項目49〜52のいずれかの方法のいずれかに従って得られる、阻害剤。
【0072】
(項目54) 病態の処置を必要とする個体においてその病態を処置するための薬物の製造における、項目53に記載の阻害剤の使用。
【0073】
(項目55) PAPP−A2のプロテアーゼ活性を促進する薬剤の同定法に関し、当該方法が、
i)a)項目12に記載のポリペプチド、およびb)当該ポリペプチドに対して予め決められた基質、およびc)推定の促進剤をインキュベートし、そして
ii)該基質の蛋白分解が促進されるかどうかを決定する、
ことからなるステップを含む、方法。
【0074】
(項目56) 該基質が、ポリペプチドを含む、項目53に記載の方法。
【0075】
(項目57) 該基質が、内部でクエンチングする蛍光性ペプチドを含む、項目54に記載の方法。
【0076】
(項目58) 該基質が、IGFBP−5またはそのフラグメントを含むか、あるいはそれから本質的に構成される、項目54に記載の方法。
【0077】
(項目59) 項目54〜57のいずれかの方法のいずれかに従って得られる、促進剤。
【0078】
(項目60) 病態の処置を必要とする個体においてその病態を処置するための薬物の製造における、項目59に記載の促進剤の使用。
【0079】
(項目61) 個体の治療による処置法であって、当該方法が、i)項目21に記載の医薬組成物、および/またはii)項目53に記載の阻害剤、および/または項目59に記載の促進剤を、当該個体に投与することからなるステップを含む、方法。
【0080】
(項目62) PAPP−A2またはPAPP−A2の他の蛋白との複合体の精製法であって、当該方法が、
i)項目12に記載のポリペプチドに対して特異的な結合親和性を有するポリクローナルまたはモノクローナル抗体を提供し、そして
ii)PAPP−A2をアフィニティークロマトグラフィーを用いて精製する、ことからなるステップを含む、方法。
【0081】
(項目63) 個体における該病態の診断法、または当該病態の素因の診断法であって、当該方法が、
a)当該個体からの体液サンプルを提供し;そして
b)当該組体液サンプル中のPAPP−A/proMBP、PAPP−A2/proMBP、PAPP−A/PAPP−A2、PAPP−A/PAPP−A2/proMBP、proMBP/ANG、およびproMBP/ANG/C3dgからなる群から選ばれる複合体のレベルを測定し;そして
c)病態または病態の素因を診断(予め決められた値より上または下の複合体レベルが病態または病態の素因を示す)する、
ことからなるステップを含む、方法。
【0082】
(項目64) 哺乳類の胎児における病態を診断法、または当該病態の素因の診断法であって、当該方法が、
a)当該胎児の母親からの体液サンプルを提供し;そして
b)当該体液サンプル中のPAPP−A/proMBP、PAPP−A2/proMBP、PAPP−A/PAPP−A2、PAPP−A/PAPP−A2/proMBP、proMBP/ANG、およびproMBP/ANG/C3dgからなる群から選ばれる複合体のレベルを測定し;そして
c)病態または病態の素因を診断(予め決められた値より上または下の複合体レベルが病態または病態の素因を示す)する、
ことからなるステップを含む、方法。
【0083】
(項目65) 該病態が、ダウン症、子癇前症、および不安定狭心症および心筋梗塞を含む急性冠症候群からなる群から選ばれる、項目63および64のいずれかに記載の方法。
【0084】
(項目66) 該複合体が、PAPP−A/proMBPであって、該病態が、ダウン症、および不安定狭心症および心筋梗塞を含む急性冠症候群から選ばれる、項目63および64のいずれかに記載の方法。
【0085】
(項目67) 該複合体が、proMBP/ANGであって、該病態が、ダウン症である、項目63および64のいずれかに記載の方法。
【図面の簡単な説明】
【0086】
【図1a】preproPAPP−A2をエンコードするmRNAに相当するcDNAの配列図である。
【図1b】preproPAPP−A2をエンコードするmRNAに相当するcDNAの配列図であり、図1aの続きである。
【図1c】preproPAPP−A2をエンコードするmRNAに相当するcDNAの配列図であり、図1bの続きである。
【図2】アミノ酸配列に翻訳されたときのPAPP−Aと、PAPP−Aの配列伸長部との間の相関関係を示し、またPAPP−Aの中央領域と相同性のあるcDNA配列を得る方法を示す図である。
【図3】preproPAPP−Aと整列したpreproPAPP−A2のアミノ酸配列を示した図である。
【図4】PAPP−A2をウエスタンブロッティングおよびクマシー染色した図である。
【図5】IGFBP−1〜6に対するPAPP−A2活性を示した図である。
【図6】IGFBP−5に対するPAPP−A2の蛋白分解活性を示した図である。
【図7a】PAPP−A2 mRNAコード化領域のcDNA配列を示した図である。
【図7b】PAPP−A2 mRNAコード化領域のcDNA配列を示した図であり、図7aの続きである。
【図8】PAPP−A/proMBP複合体中のPAPP−Aサブユニットのジスルフィド構造、PAPP−Aの推定のドメイン、およびPAPP−Aの遺伝子構造を示す図である。
【発明を実施するための形態】
【0087】
定義
本明細書で使用されるように、PAPP−A2とは、本明細書に定義されるような図1に示されるアミノ酸配列(配列番号2)を有する単離されたPAPP−A2ポリペプチドまたはその変異体をいう。本発明に記載のPAPP−A2またはその変異体は、組み換えDNA技術により生成され得るか、あるいは該PAPP−A2が自然発生し得る。
【0088】
ヌクレオチド配列をエンコードするPAPP−A2とは、本明細書に定義されるような図1に示される配列(配列番号1)を有する単離された核酸をいう。
【0089】
「活性のある」とは、任意の自然発生的なPAPP−A2の生物学的および/または免疫学的活性を保持しているPAPP−A2の形態をいう。
【0090】
「自然発生的なPAPP−A2」とは、遺伝学的に作られたものではないヒト細胞より生成されたPAPP−A2をいい、特にアセチル化、カルボキシル化、グリコシル化、リン酸化反応、脂質付加反応、アシル化、または他のポリペプチドと共有結合または非共有結合性の複合形態を含むがそれに限定されない、ポリペプチドの翻訳後修正により生ずる様々なPAPP−A2が考えられる。
【0091】
「単離されたポリペプチド」とは、ある蛋白質、あるいはその変異体またはフラグメントであり、蛋白化学の分野においては既知の常法により評価されるように特定の調製物のうち90%またはそれ以上の蛋白含量からなる。
【0092】
「誘導体」とは、ユビキチン化、標識化(例えば、放射性ヌクレオチド、様々な酵素等を用いる)、ペグ化(ポリエチレングリコールを用いる誘導体化)といった化学修飾により、またはオルニチンといったアミノ酸の挿入(または化学合成によるアミノ酸の置換)により自然発生的なPAPP−A2から誘導されるポリペプチドをいい、これは通常ヒト蛋白では生じない。
【0093】
「組み換え変異体」とは、組み換えDNA技術により作られた、アミノ酸の挿入、欠失、および置換による自然発生的なPAPP−A2とは異なる任意のポリペプチドをいう。どのアミノが酸残基が、蛋白分解活性または細胞接着性といった目的の活性を失うことなく、置換されたり、付加されたり、欠失されたりし得るかを決定する際の情報は、例えば構造的に類似の蛋白(例えば、他のメトジンシンファミリープテアーゼ)を用いて、既知のジスルフィド構造からなる局部的に相同する蛋白を用いて、または二次構造の予測により、PAPP−A2の一部分の配列を比較することにより見い出し得る。
【0094】
好ましくは、アミノ酸「置換」は、限定ではないが、ロイシンのイソロイシンまたはバリンとの置換、アスパラギン酸のグルタミン酸との置換、あるいはスレオニンのセリンとの置換といった、すなわち1つのアミノ酸を類似の構造的および/または化学的特性を有する別のアミノ酸と置換、すなわち同類アミノ酸置換の結果である。本明細書で使用されるような「置換」なる用語の意味の範囲にある更なる例示および定義は、本明細書の以下の本発明の詳細な説明において提供されている。
【0095】
アミノ酸「挿入」または「欠失」は、典型的には約1個のアミノ酸〜約20個のアミノ酸といった約1個のアミノ酸〜約50個のアミノ酸、例えば約1個のアミノ酸〜約10個のアミノ酸といった約1個のアミノ酸〜約20個のアミノ酸の範囲である。可能な変異体は、組み換えDNA技術を用いて系統的にPAPP−A2分子中にアミノ酸の挿入、欠失、または置換し、その結果生成する組み換え変異体を活性についてアッセイすることにより実験的に決定され得る。
【0096】
所望される場合、「シグナルまたはリーダー配列」は、細胞膜を通るポリペプチド(完全長のPAPP−A2、またはそのPAPP−A2ポリペプチドの一部分)を示し得る。このような配列は、本発明のポリペプチド上に天然に存在し得るか、あるいは組み換えDNA技術により異種の蛋白源から提供され得る。
【0097】
ポリペプチド「フラグメント」、「部分」、または「セグメント」とは、様々な実施態様において、少なくとも約5個のアミノ酸、しばしば少なくとも約7個のアミノ酸、典型的には少なくとも約9〜13個のアミノ酸、例えば少なくとも約17個以上のアミノ酸のアミノ酸残基の伸長部である。それは、長さがインタクトなPAPP−A2までの更に長い残基の伸長部であってもよい。活性となるために、いかなるPAPP−A2ポリペプチドまたはPAPP−A2ポリペプチドフラグメントも、そのままで、あるいはキーホール リムペット ヘモシアニン(keyhole limpet hemocyanin)といったキャリアー蛋白と結合したときに生物学的および/または免疫学的活性を呈するのに十分な長さを有さねばならない。
【0098】
「オリゴヌクレオチド」またはポリヌクレオチド「フラグメント」、「部分」、もしくは「セグメント」は、PAPP−A2ポリペプチドフラグメントの発現に有益な配列をエンコードするPAPP−A2の伸長部である。それはまた、典型的にはmRNAまたはDNA分子の関連する部分を増幅したり明らかにするためのポリメラーゼ鎖反応(PCR)またはハイブリダイゼーション法において使用可能なヌクレオチド残基の伸長部でもあり得る。特に、オリゴヌクレオチドプローブの一方または両方は、いかなる既知のまたは従来技術の分子とも同一性または相補性のないPAPP−A2の一部分と同一または相補的である配列を含むであろう。この目的で、当該オリゴヌクレオチドプローブは、一般的には約10個のヌクレオチド〜50個のヌクレオチド、および好ましくは約15個のヌクレオチド〜約30個ヌクレオチドを含むであろう。
【0099】
本明細書において使用されるように「動物」とは、ヒト、家庭用または農耕用(ネコ、イヌ、ウシ、ヒツジ等)または試験用(マウス、ラット、ラビット等)の種を含むと定義され得る。
【0100】
「組み換え」とはまた、PAPP−A2をエンコードし、組み換えDNA技術を用いて調製されるポリヌクレオチドをいい得る。PAPP−A2をエンコードするDNAは、対立遺伝子または組み換え変異体およびその突然変異体をも含み得る。
【0101】
「核酸プローブ」は、本発明により提供されるPAPP−A2をエンコードするcDNA配列をベースにして調製される。核酸プローブは、約6kbよりも少ないヌクレオチド、通常は約1kbよりも少ないヌクレオチドを有する配列部分を含む。偽陽性部を削除するために適当な試験をした後、これらのプローブをは、PAPP−A2をエンコードするmRNAが細胞または組織中に、または(Walsh et al., 1992, PCR Methods Appl 1, 241−50)に記載されるような細胞または組織から抽出した類似の染色体DNA由来の核酸配列を単離するために存在するかどうかを決定するために使用され得る。プローブは、自然発生的な、まは組み換え1本または2本鎖の核酸から誘導され得るか、あるいは化学的に合成され得る。それらは、ニックトランスレーション法、Klenow フィルイン反応、PCR法、または当業者には周知の他の方法により標識され得る。本発明のプローブ、それらの調製および/または標識化は、(Sambrook et al., 1989);または(Ausubel et al., 1989)に詳しく示されている。
【0102】
別法としては、これらのPAPP−A2または類似のポリペプチドをエンコードする組み換え変異体を合成したり、遺伝子コード中の「重複性」を使用することにより選択したりしてもよい。様々な制限部位を提供するサイレント変化といった様々なコドン置換体が、プラスミドまたはウィルス性ベクター中へのクローニング、あるいは特定の原核または真核系統における発現を最適化するために導入されてもよい。制限されないが活性、鎖間のアフィニティー、あるいはポリペプチドの分解または代謝回転率を含む、ポリペプチドの特性を改変するために突然変異が導入されてもよい。一例としては、より小さい分子量の分子を提供するために、停止コドンをヌクレオチド配列中に挿入してPAPP−A2のサイズを制限することが挙げられる。
【0103】
「発現ベクター」は、遺伝子のクローン化コピーの転写および適当な宿主中におけるmRNAの翻訳に必要なDNA配列として本明細書に定義されている。このようなベクターは、バクテリア、酵母、藍藻類、植物細胞、昆虫細胞、および動物細胞といった様々な宿主において、真核生物遺伝子を発現するために使用可能である。
【0104】
本明細書で使用されるように、「抗体」なる用語は、ポリクローナル抗体およびモノクローナル抗体の両方、ならびに抗原またはハプテンに結合可能なFv、Fab、F(ab)2といったそれらのフラグメントを含む。それは、従来のマウスモノクローナル抗体ならびにヒト抗体、およびヒト以外の抗体のヒト化形態を含み、そしてそれはまた、ファージ抗体ライブラリーから単離した「抗体」をも含む。
【0105】
「リボザイム」とは、RNAの特異的開裂を触媒可能な酵素のRNA分子である。リボザイム作用のメカニズムは、リボザイム分子の相補的なターゲットRNAへの配列特異的なハイブリダイゼーション、次いでヌクレオチド鎖切断型開裂を包含する。本発明の範疇で、PAPP−A2のRNA配列のヌクレオチド鎖切断型開裂を特異的かつ効率的に触媒するハンマーヘッド型リボザイム分子が作られる。いかなる潜在的なRNAターゲット内の特異的なリボザイム開裂部位も、以下の配列、GUA、GUU、およびGUCを含むリボザイム開裂部位に対するターゲット分子をスキャンすることにより最初に同定される。いったん同定されると、開裂部位を含むターゲット遺伝子の領域に相当する15〜20個の間のリボヌクレオチドからなる短いRNA配列が、オリゴヌクレオチド配列を不安定にし得る二次構造といった予測される構造特性について評価され得る。候補ターゲットの適合性もまた、リボヌクレアーゼ蛋白アッセイを用いて、相補的オリゴヌクレオチドとのハイブリダイゼーションのし易さを試験することにより評価され得る。
発明の詳細な説明
PAPP−A2をエンコードするヌクレオチド配列の単離
本発明は1つの態様において、妊娠関連血漿蛋白−A(PAPP−A)と全体的に相同性のある蛋白をエンコードする新規なcDNA配列に関する。この蛋白は、PAPP−A2と示されている。PAPP−A2の完全なヌクレオチド配列は、ヒトの胎盤から単離されたmRNAから得られる(実施例1)。その完全なヌクレオチド配列(配列番号1)および翻訳されたPAPP−A2のアミノ酸配列(配列番号2)の両方が図1に示されている。
【0106】
PAPP−A2のPAPP−Aとの相同性は、図3に示すように2つのアミノ酸配列の整列化により明らかになる。PAPP−A2およびPAPP−Aは、およそ45%のアミノ酸残基を共有する。PAPP−Aの機能(Kristensen et al., 1994, Biochemistry 33, 1592−8;Lawrence et al., 1999, Proc Natl Acad Sci U S A 96, 3149−53;Overgaard et al., 2000, J Biol Chem)に重要と考えられる配列モチーフが、PAPP−A2においても見い出されている。原則的には、PAPP−A2は、733〜743位に伸張された亜鉛結合モチーフ(1文字表記により示されるアミノ酸)を含有する(図2)。このモチーフおよび構造的に重要なメチオニン残基は、厳密的には亜鉛ペプチダーゼのスーパーファミリーである、メトジンシン内に保持されている(Bode et al., 1993, FEBS Lett 331, 134−40;Stocker et al., 1995, Protein Sci 4, 823−40)。
【0107】
PAPP−Aと同様に、PAPP−A2は、prepro蛋白として合成される。PreproPAPP−A2は、1791個のアミノ酸を有する(図1)。PAPP−AおよびPAPP−A2のprepro部分間には、相同性はない。更に、該2つの蛋白のprepro部分は、長さにおいては有意に差はない。PAPP−A2preproペプチドは、233個の残基を有し(図3);PAPP−Apreproペプチドは、80個の残基を有する(Haaning et al., 1996, Eur J Biochem 237, 159−63)。
PAPP−A2をエンコードするヌクレオチド配列の使用
PAPP−A2(またはそのコンプリメント)をエンコードするヌクレオチド配列は、分子生物学の当業者には既知の技術において非常に多くの用途がある。これらの技術としては、ハイブリダイゼーションプローブとしての使用、PCRに対するオリゴマーの構築における使用、PAPP−A2またはそのフラグメントの組み換え生成における使用、およびアンチセンスDNAまたはRNA、それらの化学的類似物(例えばPNAまたはLNA)等の産生における使用が挙げられる。本明細書において開示されるPAPP−A2をエンコードするヌクレオチドの使用は、既知の技術の例示であり、当業者には既知のいかなる技術の使用をも制限する意図はない。更に、本明細書において開示されるヌクレオチド配列は、新しい技術が、例えば、3つの遺伝子コード、特異的塩基対の相互作用が現在既知であるヌクレオチド配列の特性次第であるならば、まだ開発されていない分子生物学的技術において使用され得る。
【0108】
該遺伝子コードの縮重の結果、多数のPAPP−A2をエンコードするヌクレオチド配列が、幾分任意の既知のおよび自然発生的な遺伝子のヌクレオチド配列と最小限の相同性を有して生成され得ることが、当業者により理解されるであろう。本発明は、可能なコドン選択に基づいた組み合わせを選択することにより作成され得るヌクレオチド配列の可能なバリエーションのそれぞれまたはすべてを特に考えている。これらの組み合わせは、自然発生的なPAPP−A2のヌクレオチド配列に適用されるような標準的な3つの遺伝子コードに従って作られ、特に開示されるように、そのバリエーションのすべてが考えら得る。
【0109】
PAPP−A2および/またはその変異体をエンコードするヌクレオチド配列は、好ましくは厳密な条件下で、自然発生的なPAPP−A2のヌクレオチド配列にハイブリダイズ可能であるけれども、PAPP−A2をエンコードするヌクレオチド配列または実質的には異なるコドン使用を有するその誘導体を生成することは有利であり得る。コドンは、特定のコドンが宿主により利用される頻度に従って、特に原核生物または真核生物の発現宿主においてペプチドの発現が起こる速度を上げるよう選択可能である。PAPP−A2をエンコードするヌクレオチド配列および/またはその誘導体を、そのエンコードされたアミノ酸配列を変えないで実質的に変える他の理由としては、自然発生的な配列から生成される転写物より長い半減期といった、より所望される特性を有するRNA転写物の生成が挙げられる。
【0110】
PAPP−A2をエンコードするヌクレオチド配列は、十分に確立された組み換えDNA技術(Sambrook et al., 1989)を用いて、様々な他のヌクレオチド配列に結合され得る。PAPP−A2へ結合するのに有益なヌクレオチド配列としては、例えばプラスミド、コスミド、ラムダ・ファージ誘導体、ファージミド等の当業者には周知の各種クローニングベクターが挙げられる。対象のベクターとしては、発現ベクター、複製ベクター、プローブ産生ベクター、配列決定ベクター等が挙げられる。一般的には、対象のベクターは、少なくとも1つの生体において複製機能の発生源、すなわち簡単な制限エンドヌクレアーゼ感受性部位、および、宿主細胞に対する選択マーカーを含んでいてもよい。
【0111】
本発明の別の態様は、PAPP−A2をエンコードする自然発生的なヌクレオチド配列とハイブリダイズ可能なPAPP−A2特異的核酸ハイブリダイゼーションプローブを提供することである。当該プローブはまた、同様のPAPP−A2をエンコードする配列の検出に使用され得、好ましくは保存領域または活性部位由来の少なくとも50%のヌクレオチドを含有するはずである。本発明のハイブリダイゼーションプローブは、配列番号1のヌクレオチド配列から、またはプロモーター、エンハンサー要素および/またはそれぞれの自然発生的なPAPP−A2を含む可能なイントロンを含む、ゲノム配列から誘導され得る。ハイブリダイゼーションプローブは、32Pまたは35Sといった放射性核種、またはアビジン/ビオチンカップリング系等を介してプローブに結合したアルカリフォスファターゼといった酵素標識を含む、様々なレポーター群により標識され得る。
【0112】
米国特許第4,683,195号および第4,965,188号に記載されるように、PCRは、PAPP−A2をエンコードするヌクレオチド配列に基づいたオリゴヌクレオチドの更なる使用を提供する。PCRにおいて使用される当該プローブは、組み換え起源からなるものであってもよく、化学的に合成されてもよく、あるいはその混合であり、そして診断的用途のための分離したヌクレオチド配列、または非常に関連するゲノム配列の同定のための縮重した可能な配列のプールを含む。
【0113】
PAPP−A2 DNAに対して特異的なハイブリダイゼーションプローブを生成する他の手段としては、PAPP−A2またはPAPP−A2の誘導体をエンコードする核酸配列の、mRNAプローブの生成用ベクター中へのクローニングが挙げられる。そのようなベクターは、当該分野では既知であり、市販で利用可能であり、そしてインビトロでT7またはSP6 RNAポリメラーゼのような適当なRNAポリメラーゼおよび適当に放射活性標識したヌクレオチドの付加によりRNAプローブを合成するのに使用され得る。
【0114】
PAPP−A2をエンコードするDNA配列またはその一部分を完全に合成化学により生成することが可能であり、その後遺伝子をこの特許の出願時に当分野において既知である試薬、ベクター、および細胞を用いて多くの利用可能なDNAベクターのいずれか中に挿入可能である。更に、PAPP−A2配列またはその任意の部分中に変異を導入するのに、合成化学が利用されてもよい。
【0115】
異常レベルのPAPP−A2発現に関係する疾患を検出するためのアッセイにおいて、該ヌクレオチド配列が利用可能である。該ヌクレオチド配列は、当該分野において既知の方法により標識され、ハイブリダイズ条件下で、患者からの体液または組織サンプルに添加可能である。インキュベーション期間後、ヌクレオチドが酵素により標識されるのであれば、そのサンプルを、所望により染料(または現像液を必要とする他の標識)を含有する適合流体で洗浄する。その適合流体を洗い流した後、染料を定量し、スタンダードと比較する。別法としては、PAPP−A2 mRNAのレベルは、固定のプローブを用いるマイクロアレイ法により測定可能である。サンプル中における発現は、(半定量的な)RT−PCRによっても評価可能である。サンプル中における発現は、別法としてハイブリダイゼーションをベースにした方法によっても評価可能である。例えば、インサイツハイブリダイゼーション法は、PAPP−A2 mRNAを検出するのに使用可能である。この方法は、mRNAを合成する細胞を示すという利点を有するが、RT−PCRよりも感受性が低い。
【0116】
アンチセンスRNAとDNA分子、およびPAPP−A2の翻訳を阻害する機能を有するリボザイムを含むオリゴリボヌクレオチド配列が、本発明の範疇に含まれる。アンチセンス法は、当該分野では既知であり、本明細書において適用され得る。本発明のアンチセンスRNAとDNA分子、およびリボザイムが両方とも、RNA分子の合成のために、当該分野で既知の任意の方法により調製され得る。これらは、例えば固相ホスホルアミダイト化学合成といった当該分野では周知のオリゴデオキシリボヌクレオチドを化学的に合成する技術を含む。別法としては、RNA分子は、当該アンチセンスRNA分子をエンコードするDNA配列のインビトロおよびインビボでの転写により産生され得る。そのようなDNA配列は、T7またはSP6ポリメラーゼプロモーターといった好適なRNAポリメラーゼプロモーターを導入する広範なベクター中に導入され得る。別法としては、使用されるプロモーター次第で、構成的にまたは誘導的にアンチセンスRNAを合成するアンチセンスcDNA構築物を、細胞株中に安定に導入可能である。
【0117】
本発明は、他の種から単離した未知のPAPP−A2遺伝子およびそのPAPP−A2遺伝子の対立遺伝子に関し、その中にはPAPP−A2類似遺伝子または相同遺伝子が存在する。本明細書に示されたヒトPAPP−A2クローンの放射活性標識フラグメントを用いて、緩和された厳密条件下においてバクテリオファージcDNAライブラリーが、スクリーニングされ得る。別法として、そのヒトPAPP−A2配列は、PCRプローブとして、またはバクテリオファージcDNAライブラリーをスクリーニングするために使用可能な縮重した、または完全に縮重したオリゴヌクレオチドプローブをデザインするために使用可能である。そのPCR生成物は、サブクローニングされ、増幅された配列がPAPP−A2配列を示すことを保証するために配列決定され得る。そのPCRフラグメントは、増幅フラグメントを放射活性標識し、バクテリオファージcDNAライブラリーをスクリーニングすることにより、完全長のPAPP−A2クローンを単離するために使用され得る。別法として、その標識化フラグメントは、ゲノムライブラリーをスクリーニングするために使用され得る。使用され得るクローニング法の概説については、例えば(Ausubel et al., 1989;Sambrook et al., 1989)が参照される。
組み換えPAPP−A2の発現
生物学的に活性なプロテアーゼを発現するために、その蛋白または機能的等価物に対してコードされるヌクレオチド配列を、適当な発現ベクター、すなわち挿入されたコード化配列の転写および翻訳に必要な要素を含有するベクター中に挿入可能である。例えば、組み換え蛋白は、免疫化して実験試薬としておよび診断キットにおいて抗体を得るために使用可能である。
【0118】
より具体的には、当業者に周知な方法が、PAPP−A2配列および適当な転写/翻訳の制御シグナルを含有する発現ベクターを構築するのに利用可能である。これらの方法としては、インビトロでの組み換えDNA法、合成法、およびインビボでの組み換え/遺伝的組み換えが挙げられる。例えば(Ausubel et al., 1989;Sambrook et al., 1989)に示される方法を参照。
【0119】
更に、PAPP−A2をエンコードする配列のフラグメントを含む発現ベクターが構築されてもよい。特に、これは、免疫用抗原としてのPAPP−A2ポリペプチドの一部分の使用に関連し得る。更に、PAPP−A2またはそのフラグメントのコード化配列が、ベクター中に存在するコード化ヌクレオチド配列を有するフレーム中でクローンされ得、その結果、融合蛋白または「タグ化」PAPP−A2蛋白となる。例えば、当該融合蛋白は、PAPP−A2およびGSTから構成されてもよく、当該タグが、c−mycタグ(検出用)および/またはヒスチジンタグ(精製用)であってもよい。
【0120】
様々な宿主−発現ベクター系が、PAPP−A2コード化配列またはそのフラグメントを発現するために利用され得る。これらは、PAPP−A2コード化配列を含有する組み換えバクテリオファージDNA、プラスミドDNA、またはコスミドDNA発現ベクターを用いて形質転換したバクテリア;PAPP−A2コード化配列を含有する組み換え酵母発現ベクターを用いて形質転換した酵母;PAPP−A2コード化配列を含有する組み換えウィルス発現ベクター(例えばバキュロウィルス)を用いて感染させた昆虫細胞系統;PAPP−A2コード化配列を含有する組み換えウィルス発現ベクター(例えばカリフラワーモザイクウィルス、CaMV;タバコモザイクウィルス、TMV)を用いて感染させた、あるいはPAPP−A2コード化配列を含有する組み換えプラスミド発現ベクター(例えばTiプラスミド)を用いて形質転換した植物細胞系統;または安定に増幅された(CHO/dhfr)、あるいは2つの小さな染色体(例えば、マウスの細胞株)中で不安定に増幅された、いずれかの多数のPAPP−A2 DNAの複製を含有するように作成された細胞株を含む組み換えウィルス発現ベクター(例えば、アデノウィルス、ワクシニアウィルス、ヒト腫瘍細胞)を用いて感染させた動物細胞系統といった微生物体を含むがそれに限定されるわけではない。
【0121】
これらの系統の発現要素は、強度および特性が様々である。利用される宿主/ベクター系統次第で、構成的および誘導的プロモーターを含む、任意の数の好適な転写および翻訳要素が、発現ベクターにおいて利用され得る。例えば、バクテリア系統においてクローニングする場合、バクテリオファージ・ラムダのpL、plac、 ptrp、 ptac(ptrp−lacハイブリッドプロモーター)等といった誘導プロモーターが使用され得;昆虫細胞系統においてクローニングする場合、バキュロウィルス多角体プロモーターといったプロモーターが使用され得;植物細胞系統においてクローニングする場合、植物細胞のゲノムから誘導されるプロモーター(例えば、熱ショックプロモーター;RUBISCOの小さなサブユニットに対するプロモーター;クロロフィルa/b結合蛋白に対するプロモーター)または植物ウィルスから誘導されるプロモーター(例えば、CaMVの35S RNAプロモーター;TMVのコート蛋白プロモーター)が使用され得;哺乳類細胞系統においてクローニングする場合、哺乳類細胞のゲノムから誘導され得プロモーター(例えば、メタロチオネインプロモーター)または哺乳類ウィルスから誘導されるプロモーター(例えば、CMVプロモーター、アデノウィルス後期プロモーター;ワクシンウィルスの7.5Kのプロモーター)使用され得;PAPP−A2 DNAの複数の複製を含有する細胞株を産生する場合、SV40、BPV、およびEBVベースのベクターが適当な選択マーカーと一緒に使用され得る。
【0122】
形質転換、トランスフェクション、感染、プロトプラスト融合、およびエレクトロポレーションを含むがそれに限定されない多数の方法のうちの任意の1つを介して、該発現ベクターを宿主細胞中に導入し得る。その発現ベクターを含有する細胞を、それがPAPP−A2蛋白を生成するかどうかを決定するためにクローンで増殖し、個々に分析する。PAPP−A2を発現する宿主細胞クローンの同定は、抗PAPP−A2抗体との免疫反応性を含むがそれに限定されない数種の手段、および宿主細胞関連PAPP−A2活性の存在によりなされ得る。
【0123】
バクテリア系統において、多数の発現ベクターが、発現されたPAPP−A2の予定の使用される使用次第で有利に選択され得る。例えば、大量のPAPP−A2生成される予定である場合、容易に精製され得る融合蛋白生成物の高レベルの発現を導くベクターが所望となり得る。そのようなベクターとしては、大腸菌発現ベクター、pUR278(Ruther and Muller−Hill, 1983, Embo J 2, 1791−4)が挙げられるがそれに限定されず、当該ベクターにおいてPAPP−A2コード化配列は、ハイブリッドAS−lacZ蛋白を生成するよう、lacZコード化領域を有するフレーム内のベクター中にリゲートされ得る。pGEXベクターはまた、グルタチオンS−トランスフェラーゼ(GST)との融合蛋白として外来のポリペプチドを発現するために使用され得る。一般的に、そのような融合蛋白は可溶性であり、グルタチオン−アガロースビーズへの吸着、次いで遊離型のグルタチオンの存在中での溶出により可溶化細胞から容易に精製可能である。目的のクローン化ポリペプチドがGST部分から遊離可能となるよう、トロンビンまたはXa因子プロテアーゼ開裂部位を含むようにpGEXベクターがデザインされる。酵母において、構成的または誘導的プロモーターを含有する多数のベクターが使用され得る。概説については、(Ausubel et al., 1989;Bitter et al., 1987, Methods Enzymol 153, 516−44;Rosenfeld, 1999, Methods Enzymol 306, 154−69)参照。
【0124】
植物発現ベクターが使用される場合、PAPP−A2コード化配列の発現は、多数のプロモーターのうちのいずれかにより引き起こされ得る。例えば、CaMVの35S RNAおよび19S RNA プロモーターといったウィルスプロモーター(Gmunder and Kohli, 1989, Mol Gen Genet 220, 95−101);あるいは、RUBISCOの小さなサブユニットといった植物プロモーター(Broglie et al., 1984, Science 224, 838−43)が使用され得る。
【0125】
PAPP−A2を発現するために使用可能な代替の発現系は、昆虫系統である。そのような系においては、バキュロウィルスが、外来遺伝子を発現するベクターとして使用される。その後、ウィルスは、昆虫細胞において成育する。PAPP−A2コード化配列が、ウィルスの非必須領域(例えば、多角体遺伝子)中でクローン化されて、バキュロウィルスプロモーターの制御下に置かれてもよい。次いで、これらの組み換えウィルスは、挿入された遺伝子が発現される昆虫細胞を感染するために使用される。例えば(Smith et al., 1983, Mol Cell Biol 3, 2156−65)参照。
【0126】
様々な哺乳類の発現ベクターが、哺乳類細胞中で組み換えPAPP−A2を発現するために使用され得る。組み換えPAPP−A2発現に好適となり得る市販で入手可能な哺乳類の発現ベクターとしては、pMC1neo(Stratagene)、pXT1(Stratagene)、pSG5(Stratagene)、EBO−pSV2−neo(ATTC 37593)、pBPV−1(8−2)(ATCC 37110)、pcDNA3.1およびその誘導体(Stratagene)が挙げられるがそれに限定されない。好適となり得、かつ市販で入手可能である哺乳類種由来の細胞株としては、CV−1、COS−1、COS−7、CHO−K1、3T3、NIH3T3、HeLa、C127I、BS−C−1、MRC−5および293が挙げられるがそれに限定されない。更に、哺乳類の宿主細胞においで、多数のウィルスベースの発現系が利用され得る。発現ベクターとしてアデノウィルスが使用される場合においては、PAPP−A2コード化配列が、アデノウィルス転写/翻訳制御複合体、例えば後期プロモーターおよび3つのリーダー配列にリゲートされ得る。次に、このキメラ遺伝子が、インビトロまたはインビボの組み換えによりアデノウィルスゲノム中に挿入され得る。ウィルスゲノムの非必須領域(例えばE1またはE3領域)への挿入により、生育可能で感染宿主中でPAPP−A2が発現可能な組み換えウィルスとなるであろう。例えば(Logan and Shenk, 1984, Proc Natl Acad Sci U S A 81, 3655−9)参照。別法としては、ワクシニアの7.5Kのプロモーターが使用されてもよい。例えば(Mackett et al., 1982, Proc Natl Acad Sci U S A 79, 7415−9)参照。
【0127】
長期間にわたる高収率の組み換え蛋白生成のために、安定した発現が好ましい。例えば、PAPP−A2を安定に発現する細胞株が作成されてもよい。複製のウィルス源を含有する発現ベクターを用いるよりもむしろ、宿主細胞は、適当な発現制御要素(例えば、プロモーター、エンハンサー、シーケンサー、転写ターミネーター、ポリアデニル化部位等)、および選択マーカーにより制御されたPAPP−A2 DNAを用いて形質転換可能である。外来DNAの導入に続いて、作成された細胞が、栄養強化培地において1〜2日間生育され得、その後、選択培地に交換される。組み換えプラスミド中の選択マーカーは、その選択に耐性を与え、そして細胞が安定にそのプラスミドを染色体に組み入れ、増殖して病巣を形成するのを可能にし、次にクローン化され細胞株中に拡大可能となる。
【0128】
組み換えPAPP−A2の適用のいくつかは、その蛋白を精製されたあるいは一部精製された形態にする必要があり得る。組み換えにより発現したPAPP−A2またはPAPP−A2ポリペプチドのフラグメントは、液体クロマトグラフィーによって単離可能である。当該分野において周知の様々な蛋白精製法としては、例えば(Scopes, 1987)に示されている方法が挙げられる。別法としては、組み換えPAPP−A2融合蛋白、または「タグ化」PAPP−A2がアフィニティークロマトグラフィーにより精製され得る。更に、PAPP−A2対して産生される抗体が、イムノアフィニティークロマトグラフィーによる精製のために使用され得る。
【0129】
PAPP−A2の組み換え変異体が、部位特異的突然変異誘発により生成され得る。PAPP−A2の適用の中には、こういった変異体が、例えば蛋白安定性の増大、または活性の変化のために好ましくなり得るものもある。
PAPP−A2に対する抗体の生成と使用
組み換え蛋白が抗体を産生するのに使用され得る。PAPP−A2に対する単一特異的抗体が、PAPP−A2に対して反応性のある抗体を含有する哺乳類の抗血清から精製可能であり、あるいは標準的な方法を用いてPAPP−A2と反応可能なモノクローナル抗体として調製可能である。
【0130】
本明細書において使用されるように単一特異的抗体は、PAPP−A2に対して同種結合特性を有する単一の抗体種または多数の抗体種として定義される。本明細書において使用されるように同種結合とは、上述に示すように、抗体種のPAPP−A2と関連する抗原といった特異的な抗原またはエピトープへの結合能をいう。PAPP−A2特異的抗体は、マウス、ラット、モルモット、ウサギ、ヤギ、ウマ等といった動物を(ラビットまたはマウスが好ましい)、免疫アジュバント有りあるいは無しのいずれかで、適当なPAPP−A2の濃度を用いて免疫することにより産生される。例えば、PAPP−A2に対して特異的な抗体は、天然および組み換えPAPP−A2の精製用に、実験試薬として、および抗体ベースの診断用キットにおいて使用可能である。
【0131】
PAPP−A2と反応可能なモノクローナル抗体(mAb)は、常法により、例えば同系交配マウスをPAPP−A2で免疫することにより調製可能である。該マウスは、約0.5mlの緩衝液または生理食塩水中において等量の許容可能なアジュバントに組み込まれた約0.1mg〜約10mg、好ましくは約1mgのPAPP−A2を用いて免疫される。フロイントの完全アジュバントが好ましい。該マウスは、0日目に初期免疫を受け、約3〜約30週間休息させる。免疫化マウスは、約0.1〜約10mgのPAPP−A2をリン酸緩衝生理食塩水(PBS)といった緩衝溶液で静注(IV)経路により、1度またはそれ以上追加免疫される。抗体陽性のマウス由来のリンパ節は、当該分野において既知の標準的な方法で免疫化マウスから脾臓を除去することにより得られる。ハイブリドーマ細胞は、安定なハイブリドーマが形成される条件下で、脾リンパ節を適当な融合相手と混合することにより生成される。融合されたハイブリドーマ細胞は、当該分野において既知の方法を用いて、ヒポキサンチン、チミジン、およびアミノプテインを補充したダルベッコ変法イーグル培地(Dulbecco’s Modified Eagles Medium、DMEM)での生育により選択される。約14、18、および21日目で生育陽性のウェルから上清液をを回収し、固相イムノラジオアッセイ(SPIRA)といったイムノアッセイによる抗体生成に対して、抗原としてPAPP−A2を用いてスクリーニングする。mAbのアイソトープを決定するために、オクタロニー(Ouchterlony)沈降アッセイにおいて培養液も試験される。次に抗体陽性ウェルからのハイブリドーマ細胞がクローン化される。詳細については(Peters and Baumgarten, 1992)参照。
【0132】
インビトロでの抗PAPP−A2の生成は、十分な量の特異的mAbを得るために約2%のウシ胎仔血清を含有するDMEMにおけるハイブリドーマの生育により実施される。該mAbは、当該分野において既知の方法により精製される。 腹水またはハイブリドーマ培養液の抗体価は、沈降法、受身凝集反応、酵素免疫測定法(ELISA)(Crowther, 1995)を含むがそれに限定されない様々な血清学的または免疫学的アッセイにより決定される。
【0133】
「モノクローナル抗体」はまた、例えば(Clackson et al., 1991, Nature 352, 624−8;Marks et al., 1991, J Mol Biol 222, 581−97)に示されている方法を用いて、ファージ抗体ライブラリーから単離され得る。同定されたファージ抗体は、バクテリアにおける発現により生成可能である。
【0134】
上述のような方法は、PAPP−A2ポリペプチドフラグメントまたは完全長の発生期のPAPP−A2ポリペプチドに特異的な単一特異的抗体を生成するのに使用され得る。
【0135】
PAPP−A2抗体アフィニティーカラムは、抗体をゲル支持体、たとえばN−ヒドロキシスクシンイミドエステルで予め活性化されているゲル支持体Affigel−10(Biorad)に添加して、該抗体をアガロースゲルビーズ支持体と共有結合を形成させることにより作成することができる。次いで、該抗体は、スペーサーアームによりアミド結合を介して該ゲルに結合される。残りの活性化エステルが、その後1M エタノールアミン HCl(pH8)でクエンチングされる。そのカラムを、水、次いで0.23M グリシン HCl(pH2.6)で洗浄して、いかなる非共役型抗体または外来蛋白をも除去する。次に、そのカラムをリン酸緩衝生理食塩水(pH7.3)中で平衡化し、PAPP−A2またはPAPP−A2フラグメントを含有する細胞培養上清または細胞抽出物をそのカラムにゆっくりと通過させる。次に、そのカラムを洗浄し、蛋白を溶出する。精製されたPAPP−A2蛋白を次に、リン酸緩衝生理食塩水で透析する。
【0136】
ヒト血漿または血清、組織抽出物、あるいはトランスフェクトされていない細胞株由来の培地といった起源(PAPP−A2を内生に分泌する)由来の天然のPAPP−A2も、抗体アフィニティーカラムを使用することにより精製され得る。
【0137】
PAPP−A2に対するポリクローナル抗体またはモノクローナル抗体を用いて、体液または組織、および細胞抽出物中のPAPP−A2抗原の測定に多数のアッセイが実施され得る。診断の目的で、抗体ベースのキットが使用され得る。該アッセイとしては、沈降法、受身凝集反応、酵素免疫測定法(ELISA)、およびラジオイムノアッセイ(RIA)法が挙げられるがそれに限定されない。
【0138】
例えば、そのようなELISAの1つにおいて、サンプル中に存在する抗原が固定化ポリクローナル抗(PAPP−A2)により獲得される場合、サンドイッチアッセイ法が実施可能である。次に、1つまたはそれ以上のモノクローナルPAPP−A2抗体およびペルオキシダーゼ共役抗(マウスIgG)を使用することにより、検出が実施される。別のアッセイにおいては、サンプル中に存在する抗原が、固定化ポリクローナル抗(PAPP−A2)により獲得され、ビオチン化ポリクローナル抗(PAPP−A2)を用いて検出される。更なる実施例および詳細については、(Crowther, 1995)参照。アッセイは、精製されたPAPP−A2を用いて、キャリブレートして一連の希釈により標準曲線を構築することができる。精製された形態での溶液中のPAPP−A2濃度は、アミノ酸分析(Sottrup−Jensen, 1993, Biochem Mol Biol Int 30, 789−94)により正確に測定可能である。
【0139】
PAPP−A2の生物学的活性を阻害するために、ポリクローナル抗体が使用され得る。特異的に、ポリクローナルPAPP−A抗体によるPAPP−AのIGFBP−4蛋白分解活性の阻害(Lawrence et al., 1999, Proc Natl Acad Sci U S A 96, 3149−53)と同様に、PAPP−A2の蛋白分解活性を阻害するために、抗(PAPP−A2)が使用され得る。PAPP−A2の活性に対して、特定のモノクローナル抗体がまた、抑制性となり得る。そのようなモノクローナル抗体は、PAPP−A2の活性部位に非常に近接するエピトープを認識する傾向があるが、その阻害活性は、活性部位の近く以外のエピトープへの結合に基づくものであり得る。抑制性のモノクローナル抗体は、PAPP−A2、PAPP−A2から誘導されたペプチドを有するPAPP−A2フラグメントの免疫により得ることができる。
【0140】
抑制性(モノクローナル)抗体は、PAPP−A2活性を減少させるのに所望となり得る病状において治療的価値を有し得る。
PAPP−A2の活性
PAPP−Aと同様、PAPP−A2は、メトジンシンスーパーファミリーの推定のメタロプロテアーゼ(Stocker et al., 1995, Protein Sci 4, 823−40)として分類される保存アミノ酸分岐鎖を含有する。PAPP−A2がインシュリン様成長因子結合蛋白、(IGFBP)−5の開裂を示すことにより蛋白分解活性を呈する(実施例6.7)ことが、実験で確認されている。
【0141】
一般的に、潜在的な蛋白基質に対するPAPP−A2の蛋白分解活性が、精製または一部分精製されたPAPP−A2を様々な実験条件(例えば、温度、緩衝液組成、イオン強度、およびpH)下でその潜在的な基質とインキュベーションすることにより評価され得る。当該蛋白に対するPAPP−A2の酵素活性は、SDS−PAGE(明確に形成される蛋白分解性フラグメントの分解または遊離が明らかとなる)、または遊離したペプチドの高速液体クロマトグラフィー検出により評価可能である。そのような方法を用いることにより、PAPP−A2の他の基質ターゲットが同定され得る。例えば活性部位の残基が不活性な酵素を獲得するために置換されている様々なPAPP−A2とのインキュベーションが、適切なネガティブコントロールとしての役割を果たす。
【0142】
固相支持体に接着したアミノ酸の可能なすべての組み合わせからなるランダムペプチドライブラリーが、PAPP−A2により開裂可能なペプチドを同定するのに使用され得る。そのようなペプチドの同定は、ペプチドライブラリーを組み換え可溶性PAPP−A2を用いてスクリーニングすることにより達成され得る。酵素の発現および精製法が上述で示され、目的の機能的ドメインによって、組み換えの完全長PAPP−A2またはそのフラグメント、類似物、あるいは誘導体が使用され得る。更なる詳細については、(Meldal, 1998, Methods Mol Biol 87, 65−74;Meldal, 1998, Methods Mol Biol 87, 51−7)参照。あるいは、ペプチド基質が、同定されたPAPP−A2の蛋白基質から誘導されてもよい。
【0143】
別法としては、PAPP−A2により開裂可能なペプチドを同定するのにペプチドライブラリーのファージディスプレイが使用されてもよい(Matthews and Wells, 1993, Science 260, 1113−7)。
【0144】
PAPP−A2基質としての機能するペプチドは、体液または組織、および細胞抽出物におけるPAPP−A2蛋白分解活性の検出用アッセイにおいて機能し得る。蛍光クエンチングに基づいたアッセイにおいて機能するために、基質ペプチドが誘導され得る(Meldal, 1998, Methods Mol Biol 87, 65−74)。そのような、または他の方法に基づいたキットが、PAPP−A2活性の測定が関係する病気において診断目的で使用され得る。
PAPP−A2の活性を改変する試薬の同定
PAPP−A2の蛋白分解活性の検出用アッセイは、上述のように、PAPP−A2の活性を改変する分子の同定法を提供する。そのような分子は、例えば、ペプチド、誘導化ペプチド、ヒドロキサム酸誘導化ペプチド、小さな有機分子、または抗体であってもよい。
【0145】
ペプチドライブラリーのスクリーニングは、PAPP−A2の生物学的活性を改変および/または阻害するよう作用する試薬を見いだすために使用可能である。酵素の発現および精製法は、上述で示されており、目的の機能的ドメインによって、組み換えの完全長PAPP−A2またはそのフラグメント、類似物、あるいは誘導体を発現するために使用され得る。固相支持体に接着したアミノ酸の可能なすべての組み合わせからなるランダムペプチドライブラリーは、PAPP−A2の活性部位または他の部位に結合することにより、PAPP−A2活性を改変および/または阻害可能なペプチドを同定するため使用され得る。例えば、(Meldal, 1998, Methods Mol Biol 87, 75−82)参照。
【0146】
同様に、PAPP−A2の活性に影響を及ぼす低分子量の有機分子を同定するために、コンビナトリアルケミストリーが利用されてもよい。
PAPP−AまたはPAPP−A2の複合体の測定
PAPP−Aは、好酸球主要塩基蛋白(proMBP)のプロフォームとの2:2のジスルフィド結合複合体、PAPP−A/proMBPとして主に妊婦の血清中に存在する。PAPP−A/proMBP複合体に加えて、proMBPは、アンジオテンシン(ANG)との2:2のジスルフィド結合複合体、proMBP/ANGとして循環中に存在し、この複合体のフラクションは、更に補体成分C3dgのフラグメントと複合体化(PROMBP/ANG/C3dg)している(Oxvig, 1995;Christiansen, 2000)。
【0147】
個体の体液中のPAPP−Aおよび/またはPAPP−A2および/またはproMBPを含む複合体のレベルは、病態の素因を示したり、または病態の存在を示したりし得る。従って、本発明は、1つの実施態様において、個体において病態を診断したり、当該病態の素因を診断したりする方法を教示し、その方法は、
a)当該個体から体液サンプルを提供し;そして
b)当該組体液サンプル中のPAPP−A/proMBP、PAPP−A2/proMBP、PAPP−A/PAPP−A2、PAPP−A/PAPP−A2/proMBP、proMBP/ANG、およびproMBP/ANG/C3dgからなる群から選ばれる複合体のレベルを測定し;そして
c)病態または病態の素因を診断(予め決められた値より上または下の複合体レベルが病態または病態の素因を示す)することからなるステップを含む。
【0148】
更に、哺乳類の母親の体液中のPAPP−Aおよび/またはPAPP−A2および/またはproMBPを含む複合体レベルは、当該母親の胎児の病態の素因を示したり、病態の存在を示したりし得る。それゆえ、本発明は、哺乳類の胎児において病態を診断したり、当該病態の素因を診断する方法を提供し、その方法は、
a)当該胎児の母親から体液サンプルを提供し;そして
b)当該体液サンプル中のPAPP−A/proMBP、PAPP−A2/proMBP、PAPP−A/PAPP−A2、PAPP−A/PAPP−A2/proMBP、proMBP/ANG、およびproMBP/ANG/C3dgからなる群から選ばれる複合体のレベルを測定し;そして
c)病態または病態の素因を診断(予め決められた値より上または下の複合体レベルが病態または病態の素因を示す)する
ことからなるステップを含む。
【0149】
特に、本発明の方法に従って、以下に示す複合体のうちの1つまたはそれ以上のレベルが決定され得る:
PAPP−A/proMBP
PAPP−A2およびproMBP(PAPP−A2/proMBP)
PAPP−A2およびPAPP−A(PAPP−A/PAPP−A2)
proMBPを有するPAPP−A/PAPP−A2(PAPP−A/PAPP−A/proMBP)
proMBP/ANG
proMBP/ANG/C3dg
体液サンプル中のPAPP−Aおよび/またはPAPP−A2および/またはproMBPを含む複合体レベルは、当業者には既知の任意の従来法によって決定され得る。例えば、PAPP−A、PAPP−A2、proMBP、ANG、またはC3dgと特異的に相互作用する免疫特異的な試薬といった、測定するのに所望の複合体の1つまたはそれ以上の成分と特異的に相互作用する免疫特異的な試薬の使用を含む方法により、そのレベルは測定可能である。例えば、免疫特異的な試薬は、複合体の個々の成分に特異的な、モノクローナル抗体、ポリクローナル抗体および/またはその抗原結合フラグメントであり得る。
【0150】
そのような方法としては、サンドイッチELISA法が挙げられるがそれに限定されるわけではなく、当該方法においては、複合体の1成分を特異的に認識する免疫特異的な試薬が捕獲抗体として使用され、その複合体が検出抗体として使用されるならば、別の免疫特異的な試薬が別の成分を特異的に認識する。その検出抗体が、直接的または間接的のいずれかで検出可能であるのが好ましく、例えば検出抗体が検出可能なラベルに直接結合してもよく、あるいは検出抗体が、検出可能なラベルと結合した別の試薬と相互作用可能であってもよい。
【0151】
例えば、検出可能なラベルは、蛍光ラベル、色素体、放射活性ラベル、重金属、または酵素であり得る。
【0152】
例えば、体液サンプル中のPAPP−A/proMBP複合体のレベルは、捕獲用のPAPP−A特異的なモノクローナルまたはポリクローナル抗体および検出用のproMBP特異的なモノクローナルまたはポリクローナル抗体を用いて、サンドイッチELISA法により決定され得、あるいは体液サンプル中のproMBP/ANGのレベルは、捕獲用のproMBP特異的なモノクローナルまたはポリクローナル抗体および検出用のANG特異的なモノクローナルまたはポリクローナル抗体を用いてサンドイッチELISA法により決定され得る。
【0153】
病態(clinical condition)は、PAPP−Aおよび/またはPAPP−A2および/またはproMBPを含む複合体のレベルより診断され得る、あるいはPAPP−Aおよび/またはPAPP−A2および/またはproMBPを含む複合体のレベルにより素因が診断され得る任意の病態であり得る。例えば、病態は、ダウン症、子癇前症、および不安定狭心症および心筋梗塞を含む急性冠症候群からなる群から選ばれ得る。
【0154】
体液サンプルは、血清サンプルを含む血液サンプル、尿サンプル、糞サンプル、または羊水サンプルといった任意の有用な体液サンプルであり得る。
【0155】
特に、PAPP−A/proMBPレベルは、病態が、ダウン症、および不安定狭心症および心筋梗塞を含む急性冠症候群からなる群から選ばれる場合に決定され得る。
【0156】
本発明の1つの実施態様において、ダウン症の診断またはダウン症の素因の診断は、PAPP−A/proMBPレベルの決定することを含み、予め決められた値より低いPAPP−A/proMBPレベルは、ダウン症またはダウン症の素因があることを示している。
【0157】
本発明の別に実施態様において、不安定狭心症および心筋梗塞を含む急性冠症候群の診断、または不安定狭心症および心筋梗塞を含む急性冠症候群の素因の診断は、PAPP−A/proMBPレベルを決定することを含み、予め決められた値より高いPAPP−A/proMBPレベルは、不安定狭心症および心筋梗塞を含む急性冠症候群または不安定狭心症および心筋梗塞を含む急性冠症候群の素因があることを示している。
【0158】
更に別の実施態様において、ダウン症の素因を診断する、あるいはダウン症を診断するために、proMBP/ANGレベルが決定され得る。上述の診断法はすべて、1つまたはそれ以上の他の診断法と組み合わせて実施されてもよい。更に、本発明に記載の1以上の異なる診断が実施されてもよく、例えば、1以上の複合体レベルを測定する、あるいは異なる個体サンプルにおいて1つの複合体レベルを測定することが可能である。
天然の蛋白分解性フラグメントを産生するためのPAPP−A2の使用
PAPP−A2は、PAPP−A2により特異的に開裂される蛋白の天然のフラグメントを産生するために使用され得る。IGFBP−5の場合のように(実施例6.7および6.9参照)、そういったフラグメントは、インタクトなIGFBP−5とは異なる生物学的効果を有し得る。精製されたPAPP−A2で開裂後、標準的なクロマトグラフィーによりフラグメントを精製可能である(実施例6.9参照)。
発現用PAPP−A2フラグメントのデザイン
成熟PAPP−Aにおいて見られるシステイン残基がすべて、成熟PAPP−A2においても見られるため(図3参照)、ジスルフィド結合パターンは、共通のシステイン残基のパターンについては、PAPP−A2と同じであると推定することができる。それゆえ、PAPP−Aサブユニットのジスルフィド構造(図8参照)の知見が、すべてのシステイン残基組み合わせが可能であるPAPP−A2フラグメントを合理的にデザインするために利用可能である。PAPP−A2の推定のドメイン境界は、図8に示されるジスルフィド構造に基づいて決めることができる。それらのドメインは、別々にまたは一緒に発現可能である。ドメインが別のPAPP−AサブユニットまたはproMBPに対して鎖間ジスルフィド結合を形成する(図8参照)ことが既知のシステイン残基を含有する場合に、このシステインが突然変異して、例えばセリンまたはアラニン残基となることが必要となり得る。
【0159】
従って、可能な境界領域は、Cys−403とCys−499との間、Cys−828とCys−881との間、Cys−1048とCys−1115との間、Cys−1390とCys−1396との間、Cys−1459とCys−1464との間、Cys−1521とCys−1525との間、Cys−1590とCys−1595との間、Cys−1646とCys−1653との間、およびCys−1729とCys−1773との間(preproPAPP−A2ナンバリングは、図1および3と同様)である。
医薬組成物
IGFBP−5プロテアーゼとしてのPAPP−A2の同定は、治療用ターゲットとしてPAPP−A2を用いることにより、インビボにおいて成長および分化に影響を及ぼす方法を提供する。PAPP−A2の阻害は、バイオアベイラブルなIGF−IおよびIGF−IIの量を減少させると考えられている。例えば、PAPP−A2活性の阻害は、再狭窄、アテローム性動脈硬化、および線維症といった疾患に有益となり得る。アクチベーター、またはPAPP−A2の活性を上昇させる薬剤は、バイオアベイラブルなIGF−IおよびIGF−IIの量を上昇させると考えられている。
【0160】
PAPP−A2活性を変化させる、またはPAPP−A2の細胞表面への接着を変化させる薬剤を、医薬組成物中に組み込むことができる。そのような薬剤は、PAPP−A活性を変化させる、あるいはPAPP−Aの細胞表面への接着を変化させる薬剤と一緒に組み込まれてもよい。PAPP−A2特異的な薬剤とPAPP−A特異的な薬剤との組み合わせは、PAPP−Aに対して示される従来の試薬よりも有効となり得る。PAPP−A2特異的試薬とPAPP−A特異的試薬との組み合わせを薬学的に有効量でそれらを必要とする個体に投与することからなるステップを含む処置法もまた提供されている。
【0161】
一例として、抗PAPP−A2ポリクローナルまたはモノクローナルといった抗体が、薬学的に許容される無毒性の賦形剤または担体との混合により医薬組成物に製剤化することができる。そのような1つまたは複数の組成物は、特に溶液または水性生理緩衝食塩水中に懸濁した形態で経腸投与用に;特に錠剤またはカプセルの形態で経口投与用に;あるいは特に粉末状、点鼻薬、またはエアロゾルの形態で経鼻用に調製されてもよい。他の投与経路用の組成物が、常法を用いて所望されるように調製されてもよい。
【0162】
経腸投与用製剤は、一般的な賦形剤(すなわち、薬学的に許容される担体)として、滅菌水または生理食塩水、ポリエチレングリコールといったポリアルキレングリコール、野菜起源の油、水素化ナフタレン等を含有してもよい。特に、生物適合性、生物分解性の乳酸ポリマー、ラクチド/グリコリドコポリマー、またはポリオキシエチレン−ポリオキシプロピレンコポリマーが、インビボで本発明の化合物の放出を制御する賦形剤の例である。他の好適な経腸的送達システムとしては、エチレン−酢酸ビニルコポリマー粒子、浸透ポンプ、埋込み型注入システム、およびリポソームが挙げられる。吸入投与用製剤は、所望であれば、ラクトースといった賦形剤を含有してもよい。吸入製剤は、例えばポリオキシエチル−9−ラウリルエーテル、グリココール酸、およびデオキシコール酸を含有する水性溶液であってもよく、あるいはそれらが点鼻薬の形態での投与用油状溶液であってもよい。所望であれば、該化合物が経鼻的に適用されるゲルとして製剤化可能である。経腸投与用製剤も、舌下投与用にグリココール酸を含んでいてもよい。
医療用具
本発明は、患者の体内配置(例えば移植)用医療用具も特徴としており、それはPAPP−A2プロテアーゼ活性を阻害または活性化する薬剤を含む。好適な薬剤は、本明細書に記載の方法を用いて容易に同定される。用具は、該薬剤を含んだり、あるいは該試薬をコートすることが可能である。阻害剤の非制限的な例としては、抗−PAPP−A2ポリクローナルまたはモノクローナルといった抗体、あるいは1,10−フェナントロリンといったメタロプロテアーゼ阻害剤が挙げられる。
【0163】
PAPP−A2のIGFBP−5プロテアーゼ活性は、1,10−フェナントロリンにより強く阻害されるが、マトリックスメタロプロテアーゼの組織阻害剤(TIMP’S)によっては阻害されない。他の阻害剤としては、ヒドロキサム酸の誘導体といった小分子が挙げられる。抗PAPP−A2ポリクローナルIgGもまた、HFCMにおいてIGF−依存性−またはIGF−非依存性−IGFBP−5特異的PAPP−A2プロテアーゼ活性を用量依存的に阻害し得る。
【0164】
更に、修飾化ポリペプチドを含むポリペプチド(すなわち、長さまたは翻訳後修飾にかかわらず、任意のアミノ酸)は、阻害剤として機能することができる。本発明に記載の医療用具をコートしたり埋め込むために、PAPP−A2のIGFBP−5プロテアーゼ活性の任意の阻害剤を使用可能である。修飾化ポリペプチドとしては、対応する野生型配列と比較して、アミノ酸配列におけるアミノ酸の置換、欠失、または挿入、ならびに化学修飾が挙げられる。プロテアーゼ抵抗性IGFBP−5が、PAPP−A2のIGFBP−5プロテアーゼ活性の阻害剤そのものではないけれども、それが医療用具をコートしたり埋め込んだりするのに使用される場合、同様の結果が期待される。
【0165】
一例として、PAPP−A2阻害剤を用いた医療用具のコーティングまたは埋め込みは、所望によりPAPP−A阻害剤と組み合わせて、バルーン血管形成後の再狭窄の進行の阻害を助けたり、動脈硬化性プラークサイズの更なる増大を阻害したりすることが可能である。ステント留置による冠動脈血管形成は、冠動脈アテローム性動脈硬化に対する現在の主要な治療法である。冠動脈疾患の血管形成の重要な目的は、急性および慢性の合併症の両方を回避することである。現代の方法は、当面の問題の削減には非常に成功している。残念なことに、ステントされた患者の20〜30%においては、それでもなお再狭窄が起こる。既知の薬理学的治療法(pharmacological intervention)で、再狭窄を回避するのに利用可能なものはない。
【0166】
特定のメカニズムにより結合されないで、ヒトにおける血管形成に応答した新生内膜肥厚に先立って冠動脈平滑筋細胞によるIGFBP−5プロテアーゼ発現が増加すると考えられている。
【0167】
例えば、PAPP−A2活性の促進は、創傷治癒、骨折、骨粗鬆症、または排卵に有益となり得る。骨粗鬆症または骨量減少の他の症状は、骨形成が増大し、骨再吸収が減少することにより利益を受け得る。PAPP−A2活性を促進させる薬剤としては、例えば修飾化IGF、すなわちIGF類似物が可能である。
【0168】
類似物としては、アミノ酸の挿入、欠失、または置換、ならびに化学修飾を含むIGFポリペプチドが挙げられる。アミノ酸の置換としては、同類および非同類アミノ酸置換が挙げられ得る。同類アミノ酸置換は、あるアミノ酸を同じクラスのアミノ酸と置き換え、一方、非同類アミノ酸置換は、あるアミノ酸を異なるクラスのアミノ酸と置き換える。非同類置換の結果、ポリペプチドの疎水性または残りの側鎖のバルクが変化する。更に、非同類置換は、正電荷を減少させ負電荷を導入するなど、ポリペプチドの電荷を本質的に変化させる。非同類置換の例としては、非極性アミノ酸の塩基性アミノ酸への置き換え、または酸性アミノ酸の極性アミノ酸の置き換えが挙げられる。アミノ酸の挿入、欠失、および置換は、当該分野には既知のランダム突然変異誘発、部位特異的突然変異誘発、または他の組み換え法の使用が可能である。
【0169】
例えば、医療用具としては、骨を安定させるために使用される骨プレートまたは骨スクリュー、あるいは体内管腔部の開存性を回復したり維持したりするために典型的には体内で使用されるステントが可能である。例えば、血管は血液の流れを制限する動脈硬化性のプラークにより詰まってくる。典型的にはステントは、体内管腔部内の流動を調節する内部チャンネルを形成する管状構造を有している。ステントの外壁は、体内管腔部の内壁と接合している。病変部内のステントの配置により、体内管腔部の更なる閉塞の回避し、そして継続的な流動を可能にすることを助けることができる。典型的にはステントは、ステントを運ぶカテーテルまたはガイドワイヤーの経皮的挿入により実施される。通常ステントは、膨張可能な構造を有する。所望の部位へ送達する際、ステントはカテーテルに装着されたバルーンにより膨張可能である。別法としては、ステントが、圧縮された状態で鞘または他の制限部内に保持するバイアス構造または弾性構造を有していてもよい。その制限が除かれたときに、ステントは自発的に膨張する。いずれの場合においても、ステント壁は、膨張して体内管腔部の内壁に接合し、そして一般的には所望の位置にステントを固定する。
本発明の記載
第1の態様において、本発明は、
i)受け入れ番号DSM13783下のDSMZに集積された、PAPP−A2のコード化配列に相当する、配列番号1のヌクレオチド1〜5376を含むポリヌクレオチド;および
ii)配列番号2も示されるようなアミノ酸配列を有するポリペプチドをエンコードするポリヌクレオチド;および
iii)ポリヌクレオチド(i)または(ii)によりエンコードされたポリペプチドのフラグメントをエンコードするポリヌクレオチドであって、当該フラグメントが、
a)インシュリン様成長因子結合蛋白5型(IGFBP−5)またはその誘導体、あるいは任意の他の基質に特異的な蛋白分解活性を有し;そして/または
b)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
c)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、ポリヌクレオチド;そして
iv)相補鎖が、(i)、(ii)、および(iii)のいずれかで定義されるようなポリヌクレオチドと、厳密な条件下でハイブリダイズされるポリヌクレオチド
であって、当該ポリヌクレオチドが、配列番号2で示されるようなアミノ酸配列を有するポリペプチド、またはそのフラグメントをエンコードし、当該フラグメントが、
a)インシュリン様成長因子結合蛋白5型(IGFBP−5)またはその誘導体、あるいは任意の他の基質に特異的な蛋白分解活性を有し;そして/または
b)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
c)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、ポリヌクレオチド;および
v)(iii)および(iv)のいずれかで定義されるようなポリヌクレオチドのヌクレオチド配列に縮重されるヌクレオチド配列を含むポリヌクレオチド、
、およびそのようなポリヌクレオチドの相補鎖、からなる群から選ばれる精製ポリヌクレオチドに関する。
【0170】
本明細書において使用されるように、ポリヌクレオチドとは、任意の自然発生的な基本骨格構造を有する任意の自然発生的なポリヌクレオチド、ならびに当該分野においてLNA(ロック型核酸、locked nucleic acid)およびPNA(ペプチド核酸)として既知のヌクレオチドをいう。
【0171】
好ましい実施態様において、精製ポリヌクレオチドは、配列番号1で示されるようなPAPP−A2、ヌクレオチド1〜5376からなるコード化配列、または配列番号2で示されるようなアミノ酸配列をエンコードするヌクレオチド配列を含む。
【0172】
別の好ましい実施態様において、ポリヌクレオチドは、配列番号2で示されるようなアミノ酸配列を有するポリペプチドのフラグメントをエンコードするヌクレオチド配列を含み、当該フラグメントは、
a)インシュリン様成長因子結合蛋白5型(IGFBP−5)またはその誘導体、あるいは任意の他の基質に特異的な蛋白分解活性を有し;そして/または
b)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
c)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する。
【0173】
相補鎖が本発明に記載のポリヌクレオチドと厳密な条件下でハイブリダイズするポリヌクレオチドも提供される。
【0174】
本明細書で使用されるような厳密な条件とは、例えば、Southern E. M., 1975, J. Mol. Biol. 98:503−517により示されるようなサザンブロッティングおよびハイブリダイゼーションに関連して通常適用されるような厳密さをいう。そのような目的のために、プレハイブリダイゼーションおよびハイブリダイゼーションからなるステップを含むことが日常的に実施されている。このようなステップは、通常、“Molecular Cloning/A Laboratory Manual”, Cold Spring HarborにおいてSambrook et al., 1989により示される(これは、参照して本明細書の一部とする)ように、6xSSPE、5% Denhardt’s、0.5% SDS、50% ホルムアミド、100mg/ml 変性サケ精巣DNA(42℃で18時間インキュベーション)を含有する溶液を用いて実施され、次に2xSSCおよび0.5% SDS(室温および37℃)で洗浄し、0.1xSSCおよび0.5% SDS(68℃で30分間インキュベーション)で洗浄する。
【0175】
DNA配列は、様々な方法で使用される。それらは、uHAseのホモログ(例えば、huHAseのホモログ)を同定するためのプローブとして使用され得る。哺乳類のホモログは、相互に実質的な配列類似性、すなわち少なくとも75%、通常少なくとも90%、より一般的には少なくとも95%の配列同一性を有する。配列類似性は、保存モチーフ、コード化領域、フランキング領域等といった比較的長い配列のサブセットであり得る標準配列を基準にして計算される。標準配列は、通常、少なくとも約18ntの長さ、より一般的には少なくとも約30ntの長さとなり、比較される完全長の配列まで延び得る。配列分析に対するアルゴリズムは、当該分野、例えばAltschul et al. 1990 J Mol Biol 215:403−10に記載されるBLASTにおいて既知である。
【0176】
配列類似性を有する核酸が、厳密性の低い条件下、例えば50℃および10倍のSSC(0.9M 生理食塩水/0.09M クエン酸ナトリウム)でハイブリダイゼーションにより検出され、55℃および1倍のSSCで洗浄されるときには結合したままである。配列同一性は、厳密性の高い条件下、例えば50℃またはそれ以上および0.1倍のSSC(9mM 生理食塩水/0.9 Mクエン酸ナトリウム)でハイブリダイゼーションにより検出され得る。プローブ、特にDNA配列の標識化プローブを使用することにより、ホモログまたは関連遺伝子を単離することができる。ホモログ遺伝子の起源は、任意の種、例えば霊長類、特にはヒト;例えばラットおよびマウスといったげっ歯類、イヌ、ネコ、ウシ、オパイン、ウマ、酵母、ショウジョウバエ、シノラブディス(Caenhorabditis)等であり得る。
【0177】
更なる実施態様において、配列番号1にハイブリダイズ可能なポリヌクレオチド、またはそのフラグメントに縮重されるヌクレオチド配列を含むポリヌクレオチドが提供される。
【0178】
縮重は、本明細書において使用されるように、該縮重したポリヌクレオチドから発現されたポリペプチドと関係する活性または機能性に鑑みて定義されており、該ポリヌクレオチドは、i)少なくともインシュリン様成長因子結合蛋白5型(IGFBP−5)特異的なタンパク分解活性を含み、そして/またはii)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体またはその結合フラグメントにより認識され;そして/またはiii)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、のいずれかである。
【0179】
更なる実施態様において、本発明に記載のポリヌクレオチドの相補鎖を含むポリヌクレオチドが提供される。
【0180】
本明細書に記載のポリヌクレオチドは、PAPP−A2の3’未翻訳領域またはそのフラグメントに相当する核酸残基を含む更なるポリヌクレオチドに実施可能に結合され得る。本明細書で使用されるように、その3’未翻訳領域は、配列番号1の5377〜8527の核酸残基を含む。
【0181】
本明細書に記載のポリヌクレオチドに実施可能に結合された発現シグナルを含む発現ベクターの形態で組み換えDNA分子もまた提供される。
【0182】
更なる態様において、本発明に記載のポリヌクレオチドまたは本発明に記載のベクターでトランスフェクトまたは形質転換した宿主生体が提供される。その宿主生体は、例えば哺乳類の細胞株といった哺乳類の生体が好ましい。しかしながら、酵母または真菌類(fungi)といった微生物真核生物もまた使用され得、同様にシラス属(Bacillus)または大腸菌といった微生物原核生物も使用され得る。当業者であれば、特定の細胞における発現に好適なリーダー配列および/またはシグナルペプチドを含む、発現シグナルをいかに選択するかを知っているだろう。当業者であればまた、標準的な分子生物学的方法を使用することにより特定細胞における発現レベルをいかに決定するかを知っているだろう。
【0183】
更なる態様において、本発明は、配列番号2またはそのフラグメントのアミノ酸配列を含むか、あるいはそれらから本質的に構成される単離されたポリペプチドに関し、当該フラグメントは、
a)インシュリン様成長因子結合蛋白5型(IGFBP−5)に特異的な蛋白分解活性を有し;そして/または
b)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
c)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する。
【0184】
本発明の好ましい1つの実施態様において、配列番号2の変異体およびそのフラグメントの変異体もまた提供される。変異体は、予め決められたアミノ酸配列と同一性の程度またはそれらの相同性に基づいて決定され、当該予め決められたアミノ酸配列は、配列番号2であり、またその変異体がフラグメントである場合には、配列番号2のフラグメントである。
【0185】
従って、変異体は、予め決定された配列と、少なくとも75%の配列同一性配列同一性、例えば少なくとも80%の配列同一性、例えば少なくとも85%の配列同一性、例えば少なくとも90%の配列同一性、例えば少なくとも91%の配列同一性、例えば少なくとも92%の配列同一性、例えば少なくとも93%の配列同一性、例えば少なくとも94%の配列同一性、例えば少なくとも95%の配列同一性、例えば少なくとも96%の配列同一性、例えば少なくとも97%の配列同一性、例えば少なくとも98%の配列同一性、例えば少なくとも99%の配列同一性を有するのが好ましい。
【0186】
変異体はまた、本明細書の以下で定義されるような予め決定された数の同類(conservative)アミノ酸置換に基づいて決定される。本明細書で定義されるような同類アミノ酸置換は、1つのアミノ酸(予め決定されたアミノ酸群内)の別のアミノ酸(同一群内)との置換に関し、そのアミノ酸は、同様またはほぼ同様の特性を呈する。
【0187】
本明細書において適用されるような「同類アミノ酸置換」なる用語の意味の範疇で、本明細書の以下で示すアミノ酸群内で、1つのアミノ酸が別のアミノ酸に置換されてもよい:
i)極性の側鎖を有するアミノ酸(Asp、Glu、Lys、Arg、His、Asn、Gln、Ser、Thr、Tyr、およびCys)
ii)非極性の側鎖を有するアミノ酸(Gly、Ala、Val、Leu、Ile、Phe、Trp、Pro、およびMet)
iii)脂肪族の側鎖を有するアミノ酸(Gly、Ala、Val、Leu、Ile)
iv)環状の側鎖を有するアミノ酸(Phe、Tyr、Trp、His、Pro)
v)芳香族側鎖を有するアミノ酸(Phe、Tyr、Trp)
vi)酸性の側鎖を有するアミノ酸(Asp、Glu)
vii)塩基性の側鎖を有するアミノ酸(Lys、Arg、His)
viii)アミド側鎖を有するアミノ酸(Asn、Gln)
ix)ヒドロキシ側鎖を有するアミノ酸(Ser、Thr)
x)硫黄を含有する側鎖を有するアミノ酸(Cys、Met)
xi)中性で弱い疎水性のアミノ酸(Pro、Ala、Gly、Ser、Thr)
xii)親水性で酸性のアミノ酸(Gln、Asn、Glu、Asp)、および
xiii)疎水性アミノ酸(Leu、Ile、Val)。
【0188】
従って、本発明に記載の変異体またはそのフラグメントは、配列が同一の変異体またはそのフラグメント内、あるいは配列が異なる変異体またはそのフラグメント間に、相互に独立して導入された複数の置換体といった、少なくとも1つの置換体を含んでいてもよい。
【0189】
その同一の変異体またはそのフラグメントが、本明細書の上述で定義されるような1つ以上の同類アミノ酸群から1つ以上の同類アミノ酸置換体を含んでいてもよいことが、上述の概要から明らかである。
【0190】
アミノ酸の付加または欠失は、2〜10個のアミノ酸、例えば10〜20個のアミノ酸、例えば20〜30個のアミノ酸、例えば40〜50個のアミノ酸の付加または欠失であり得る。しかしながら、50個以上のアミノ酸の付加または欠失、例えば10〜100個のアミノ酸の付加、例えば100〜150個のアミノ酸の付加、例えば150〜250個のアミノ酸の付加もまた、本発明の範疇に含まれる。
【0191】
いかなる機能的等価物をも含む、本発明に記載のポリペプチドフラグメントは、1つの実施態様において、少なくとも250個の未満のアミノ酸残基、例えば少なくとも240個未満のアミノ酸残基、例えば少なくとも225個未満のアミノ酸残基、例えば少なくとも200個未満のアミノ酸残基、例えば少なくとも180個未満のアミノ酸残基、例えば少なくとも160個未満のアミノ酸残基、例えば少なくとも150個未満のアミノ酸残基、例えば少なくとも140個未満のアミノ酸残基、例えば少なくとも130個未満のアミノ酸残基、例えば少なくとも120個未満のアミノ酸残基、例えば少なくとも110個未満のアミノ酸残基、例えば少なくとも100個未満のアミノ酸残基、例えば少なくとも90個未満のアミノ酸残基、例えば少なくとも85個未満のアミノ酸残基、例えば少なくとも80個未満のアミノ酸残基、例えば少なくとも75個未満のアミノ酸残基、例えば少なくとも70個未満のアミノ酸残基、例えば少なくとも65個未満のアミノ酸残基、例えば少なくとも60個未満のアミノ酸残基、例えば少なくとも55個未満のアミノ酸残基、例えば少なくとも50個未満のアミノ酸残基を含み得る。
【0192】
本発明において使用されるような「機能的等価性」とは、予め決められた配列のフラグメントに相当する機能性を基準にして確立された1つの好ましい実施態様に従う。より具体的には、機能的等価性とは、ポリペプチドフラグメントのIGFBP−5特異的プロテアーゼ活性を示す、および/またはPAPP−A2を認識可能な抗体により認識される、および/またはPAPP−A2に対する親和性を有するレセプターへ結合するのにPAPP−A2と競合する能力として理解されるべきである。
【0193】
PAPP−A2の機能的等価物または変異体は、挿入、欠失、および同類置換を含む置換の数および範囲が増えるにつれて、次第に好ましく予め決められたPAPP−A2配列とは異なるアミノ酸配列を示すと理解される。この相違は、好ましい予め決められた配列とそのフラグメントまたは機能的等価物との間の相同性の減少として測定される。
【0194】
配列番号2のすべてのフラグメントまたは機能的等価物は、本明細書において報告されるようなPAPP−A2の好ましい予め決められた配列に対してそれらが示す相同性の程度にかかわらず、本発明の範疇に含まれる。この理由は、PAPP−A2の領域の幾つかが、その結果生成するフラグメントの結合活性にいかなる有意な影響をも与えることなく最も容易に変異し易い、あるいは完全に欠失可能であるからである。
【0195】
置換により得られる機能的変異体は、天然PAPP−A2活性のある程度の形態または度合を十分に示し得、機能的に類似のアミノ酸側鎖を含む残基が置換されると、相同性は更に低くなる。この点で機能的な類似性とは、疎水性、塩基性、中性または酸性、あるいは立体的なバルクの有無といった側鎖の主要な特性をいう。従って、本発明の1つの実施態様において、同一性の程度は、本発明に記載の好ましい予め決められたフラグメントの変異体または機能的等価物であるフラグメントの原則的な尺度ではない。
【0196】
アミノ酸配列間の相同性は、BLOSUM30、BLOSUM40、BLOSUM45、BLOSUM50、BLOSUM55、BLOSUM60、BLOSUM62、BLOSUM65、BLOSUM70、BLOSUM75、BLOSUM80、BLOSUM85、またはBLOSUM90といった周知のアルゴリズムを用いて計算され得る。
【0197】
配列番号2のフラグメントと少なくともある程度の相同性を共有するフラグメントは、それらが配列番号2の当該フラグメントと、少なくとも約90%相同する、例えば少なくとも92%相同する、例えば少なくとも94%相同する、例えば少なくとも95%相同する、例えば少なくとも96%相同する、例えば少なくとも97%相同する、例えば少なくとも98%相同するときには、本発明の範疇にあると考えられるべきである。本発明の1つの実施態様に従って、相同性の割合を、同一性の割合という。
【0198】
本明細書で使用される意味に従って、機能的等価性を決定するときに考慮に入れられ得る更なる因子は、i)本発明に記載のPAPP−A2フラグメントを検出するた抗血清能、ii)機能的に等価なPAPP−A2フラグメントのバインディングアッセイにおけるPAPP−A2との競合能である。既知のアミノ酸配列内の免疫遺伝学的に活性なアミノ酸の配列を決定する1つの方法は、米国特許5,595,915号においてGeysenにより示されており、これは参照して本明細書に組み込まれている。
【0199】
ペプチドフラグメントの構造および機能相関を決定するために更に好適に適応される方法は、米国特許第6,013,478号に示されており、これは参照して本明細書に組み込まれている。また、アミノ酸配列のレセプター部分への結合のアッセイ法は、当業者には既知である。
【0200】
同類置換は、配列番号2の好ましい予め決められた任意の位置に導入され得、そしてそれは、任意の1以上の位置に非同類置換を導入するのにもまた所望となり得る。
【0201】
PAPP−A2フラグメントの機能的に等価なフラグメントの形成をもたらす非同類置換は、例えば;i)例えば非極性の側鎖(Ala、Leu、Pro、Trp、Val、Ile、Leu、Phe、またはMet)を有する残基が、Gly、Ser、Thr、Cys、Tyr、Asn、またはGlnといった極性の側鎖を有する残基またはAsp、Glu、Arg、またはLysといった帯電したアミノ酸に置換される、あるいは帯電したまたは極性の残基を非極性残基と置換するなど、実質的に極性が異なる;および/またはii)別の残基によるProまたはGlyの置換またはProまたはGlyへの変換といった、ポリペプチド基本骨格配向性への効果が実質的に異なる;および/またはiii)例えばGluまたはAspといった負に帯電した残基のLys、HisまたはArgといった正に帯電した残基への置換(およびその反対)など、実質的に電荷が異なる;および/またはiv)例えばHis、Trp、Phe、またはTyrといった嵩高い残基のマイナーな側鎖を有する残基、例えばAla、GlyまたはSerへの置換(およびその反対)など、実質的に立体的なバルクが異なる。
【0202】
アミノ酸の置換により得られる変異体は、1つの好ましい実施態様において、疎水性および親水性の値、および電荷、サイズ等を含むアミノ酸側鎖置換体との相対的な類似性に基づいて作成され得る。例えば、様々な前述の特性を考慮にいれるアミノ酸置換は、当業者には周知であり、:アルギニンおよびリジン;グルタミン酸およびアスパラギン酸;セリンおよびスレオニン;グルタミンおよびアスパラギン;およびバリン、ロイシンおよびイソロイシンが挙げられる。
【0203】
本明細書に記載の変異体に加えて、立体構造的に類似の変異体は、変異体構造の鍵となる部分を模倣するために形成され得、当該化合物がまた、本発明の変異体と同様に使用されてもよい。これは、当業者には既知のモデリングおよび化学的デザインの技術により達成され得る。そのような立体構造的に類似する構成はすべて、本発明の範疇にあることが理解されよう。
【0204】
更なる実施態様において、本発明は、置換されたアミノ酸の値の+/−2.5以内、例えば+/−2.3以内、例えば+/−2.1以内、例えば+/−2.0以内、例えば+/−1.8以内、例えば+/−1.6以内、例えば+/−1.5以内、例えば+/−1.4以内、例えば+/−1.3以内、例えば+/−1.2以内、例えば+/−1.1以内、例えば+/−1.0以内、例えば+/−0.9以内、例えば+/−0.8以内、例えば+/−0.7以内、例えば+/−0.6以内、例えば+/−0.5以内、例えば+/−0.4以内、例えば+/−0.3以内、例えば+/−0.25以内、例えば+/−0.2以内である親水性または疎水性指標を有する機能的に構成される置換アミノ酸に関する。
【0205】
蛋白に相互作用性の生物学的機能を与える際の親水性および疎水性アミノ酸指標の重要性は、当業者には十分に理解されている(Kyte & Doolit−tle, 1982およびHopp、国特許第4,554,101号、それぞれ参照して本明細書に組み込まれる)。
【0206】
本明細書で使用されるようなアミノ酸疎水性指標値は以下の通りである:イソロイシン(+4.5);バリン(+4.2);ロイシン(+3.8);フェニルアラニン(+2.8);システイン/シスチン(+2.5);メチオニン(+1.9);アラニン(+1.8);グリシン(−0.4);スレオニン(−0.7);セリン(−0.8);トリプトファン(−0.9);チロシン(−1.3);プロリン(−1.6);ヒスチジン(−3.2);グルタミン酸(−3.5);グルタミン(−3.5);アスパラギン酸(−3.5);アスパラギン(−3.5);リジン(−3.9);およびアルギニン(−4.5)(Kyte & Doolittle, 1982)。
【0207】
アミノ酸親水性値を以下の通りである:アルギニン(+3.0);リジン(+3.0);アスパラギン酸(+3.0.+−.1);グルタミン酸(+3.0.+−.1);セリン(+0.3);アスパラギン(+0.2);グルタミン(+0.2);グリシン(0);スレオニン(−0.4);プロリン(−0.5.+−.1);アラニン(−0.5);ヒスチジン(−0.5);システイン(−1.0);メチオニン(−1.3);バリン(−1.5);ロイシン(−1.8);イソロイシン(−1.8);チロシン(−2.3);フェニルアラニン(−2.5);トリプトファン(−3.4)(米国特許第4,554,101号)。
【0208】
本明細書に記載のペプチジル化合物に加えて、立体構造的に類似の化合物が、ペプチド構造の鍵の部分を模倣するために形成され得、そしてその当該化合物はまた、本発明のペプチドと同様に使用され得る。これは、当業者には既知のモデリングおよび化学的デザインの技術により達成され得る。例えば、テトラペプチド構造を模倣するために、例えばジアルギニンペプチド基本骨格のアミノ末端を修飾するようエステル化およびアルキル化が実施されてもよい。そのような立体構造的に類似の構成はすべて、本発明の範疇にあることが理解されよう。
【0209】
N末端をアルキル化およびC末端をエステル化したペプチドもまた、本発明の範疇に包含される。機能的等価物はまた、二量体または無関係な化学的部分を含む、同一または他のPAPP−A2フラグメントおよび/またはPAPP−A2分子を用いて形成される、グリコシル化して共有結合性または凝集性の共役体を含む。そのような機能的等価物は、当該分野において既知の方法により、N末端およびC末端のいずれか一方または両方を含むフラグメントにおいて見られる群への機能性の結合により調製される。
【0210】
機能的等価物が、それゆえカルボキシル末端の脂肪族またはアシルエステルまたはアミド、アルキルアミン、またはカルボキシル側鎖を含む残基に共役した、例えばアスパラギン酸残基でアルキルアミンに共役したフラグメント;水酸基を含む残基のO−アシル誘導体およびアミノ末端のアミノ酸またはアミノ基を含有する残基のN−アシル誘導体、例えばfMet−Leu−Pheまたは免疫原性蛋白を有する共役体を含み得る。アシル基の誘導体は、アルキル部分の群(C3〜C10の直鎖アルキルを含む)から選ばれ、それによりアルカノイル種、および炭素環式またはヘテロ環式化合物を形成し、それによりアロイル種を形成する。反応基は、好ましくは反応側鎖の基を通じて不溶性のマトリクスに架橋結合する蛋白に使用するそれ自身は既知の二官能性化合物である。
【0211】
共有結合性または凝集性の機能的等価物およびその誘導体は、イムノアッセイにおける試薬として、またはアフィニティー精製法に有益である。例えば、本発明に記載のPAPP−A2のフラグメントは、それ自身既知の方法により臭化シアン活性化セファロースに共有結合することにより不可溶化され、抗PAPP−A2抗体または細胞表面レセプターのアッセイまたは精製に使用されるグルタルアルデヒド架橋を用いてまたは用いないでポリオレフィン表面に吸収され得る。フラグメントは、検出可能な基で標識化、例えばクロラミンT法による放射性ヨウ素化される、希土類キレートに共有結合される、または例えば診断アッセイに使用する別の蛍光部分と共役され得る。
【0212】
PAPP−A2の好ましい予め決められたフラグメントの突然変異誘発は、アミノ酸を、通常、約1〜10個程度のアミノ酸残基、好ましくは約1〜5個のアミノ酸残基を挿入する、あるいは約1〜10個の残基、例えば約2〜5個の残基を欠失することにより実施可能である。
【0213】
1つの実施態様において、PAPP−A2のフラグメントは、自動合成により合成される。任意の商業的に利用可能な固相法、例えばMerrifieldの固相合成法が実施され得、その方法において、アミノ酸が、成長するアミノ酸鎖に順次付加される(Merrifield, J. Am. Chem. Soc. 85:2149−2146, 1963参照)。
【0214】
ポリペプチドの自動合成用装置は、Foster City, Calif.のApplied Biosystems, Inc.といった供給業者から市販で入手可能であり、一般的には取扱い説明書に従って操作され得る。固相合成により、本発明に記載の任意のPAPP−A2フラグメントに所望のアミノ酸置換を導入することが可能になる。置換、欠失、相挿入、またはその任意のサブコンビネーションが、組み合わされて機能的等価物の最終配列に到達し得ることが理解されよう。挿入は、例えば疎水性または免疫原性蛋白、または任意のポリペプチドといった担体、または担体と同様の役割をすることが可能な骨格構造を用いたアミノ末端および/またはカルボキシル末端融合を含むと理解されるはずである。
【0215】
本発明によるPAPP−A2のフラグメントのホモ二量体およびヘテロ二量体を含む二量体を含むオリゴマーも提供され、本発明の範疇に該当する。PAPP−A2の機能的等価物および変異体が、他のアミノ酸配列を有する、または天然のPAPP−A2配列を有するホモ二量体またはヘテロ二量体として生成可能である。ヘテロ二量体としては、いかなる生物活性をも有するまたは示す必要がない免疫反応性PAPP−A2フラグメントならびにPAPP−A2フラグメントを含有する二量体である。
【0216】
本発明に記載のPAPP−A2フラグメントは、インビトロおよびインビボの両方で合成され得る。インビトロの合成法は周知であり、PAPP−A2のインビボでの合成法に好適なまたは好適に適応可能な方法は、従来技術でも示されている。インビボで合成される場合、宿主細胞は、PAPP−A2またはそのフラグメントをエンコードするDNAを含有するベクターを用いて形質転換される。ベクターは、複製可能な核酸構築物として定義されている。ベクターは、PAPP−A2の発現を仲介するために使用される。発現ベクターは、複製可能なDNA構築物であり、予め決められたPAPP−A2フラグメントをエンコードする核酸配列、またはインビボで発現可能な任意の機能的等価物が、好適な宿主においてそのフラグメントまたは等価物の発現に影響を与えることが可能な好適なコントロール配列に実施可能に結合される。そのようなコントロール配列は、当該分野においては周知である。
【0217】
多細胞生体由来の細胞培養物は、好ましい宿主細胞である。原則として、脊椎動物培養由来であろうと無脊椎動物培養由来であろうと任意の真核細胞培養物が高等であるほど有効である。例えば有益な宿主細胞株は、VEROおよびHeLa細胞、チャイニーズハムスター卵巣(CHO)細胞株、およびWI38、BHK、COS−7、293およびMDCK細胞株である。好ましい宿主細胞は、内因性のPAPP−A2を合成することが知られている真核細胞である。当該宿主細胞の培養物は、フラグメントの起源として単離および使用され得、あるいは成育状態を促進または阻害することを目的とする治療法、あるいはヒトまたは動物の身体に関して実施される診断方法を含む治療的処置法において使用され得る。
【0218】
特定の実施態様において、本発明は、本発明に記載のポリペプチドフラグメントに関し、PAPP−A2フラグメントは、その任意のプロセシング変異体を含む、PAPP−A2の成熟部分に相当する234〜1791のアミノ酸残基を含むかあるいは本質的に構成される。
【0219】
プロセシング変異体は、シグナルペプチダーゼおよびフリン(furin)を含むがそれに限定されない任意のプロテアーゼにより触媒された、別のプロセシング事象、可能なプロセシング事象の結果生成する変異体である。1つの推定される開裂部位は、233位の後ろに位置し、本明細書の以降において詳細に示されている。別の推定される開裂部位は、RQRRモチーフ(PAPP−A2のアミノ酸配列において196〜199位)の後ろに位置する。プロセシング変異体は、PAPP−A2がヒトまたは動物の組織、血清または体液において発現されるときにインビボでのプロセシングから生じる変異体を含むと理解されるはずである。
【0220】
配列番号2に示される成熟配列(アミノ酸残基234〜1791)から本質的に構成される成熟PAPP−A2アミノ酸配列は、1つの実施態様において、1〜約10個のN末端アミノ酸またはC末端アミノ酸、好ましくは1〜10個のN末端アミノ酸、例えば2〜8個のN末端アミノ酸、例えば3〜6個のN末端アミノ酸の間が欠損しているこの部分の配列を含むことが理解されるはずである。
【0221】
また、本明細書において使用されるように、本質的に構成されるという定義においては、配列番号2に示される成熟配列(アミノ酸残基234〜1791)がそれに加えて更に1〜約10個のN末端アミノ酸またはC末端アミノ酸、好ましくは1〜10個のN末端アミノ酸、例えば2〜8個のN末端アミノ酸、例えば3〜6個のN末端アミノ酸を有することも含まれる。この本質的に構成されるという定義はまた、他の態様において適用されて、PAPP−A2の特定部分と結合して使用されることに制限されない。この定義は、プロセシングされたPAPP−A2がもともと単離されているもの以外の、組織、血清、または体液において別のプロセシングから生じるポリペプチドを含む他のプロセスPAPP−A2ポリペプチドにも適用されるはずである。
【0222】
更に、好ましいフラグメントは、PAPP−A2のprepro部分に相当するアミノ酸残基1〜233、PAPP−A2のpro部分に相当するアミノ酸残基23〜233、PAPP−A2のシグナルペプチドまたはリーダー配列に相当するアミノ酸残基1〜22を含み、あるいはそれらから本質的に構成され、当該配列は、配列番号2のアミノ酸残基234〜1791に相当するPAPP−A2の成熟部分に実施可能に結合される。
【0223】
組み換えPAPP−A2ポリペプチド、またはそのフラグメントも提供され、当該ポリペプチドにはヒト蛋白、あるいは当該ポリペプチドに天然に付随する他の蛋白がないのが好ましい。
【0224】
更なる態様において、i)本発明に記載のポリヌクレオチド、および/またはii)本発明に記載のベクター、および/またはiii)本発明に記載の宿主生体、および/またはiv)本発明に記載のポリペプチドを、生理的に許容される担体と組み合わせて含む組成物が提供される。
【0225】
更に別の態様において、i)本発明に記載のポリヌクレオチド、および/またはii)本発明に記載のベクター、および/またはiii)本発明に記載の宿主生体、および/またはiv)本発明に記載のポリペプチドを、薬学的に許容される担体と組み合わせて含む医薬組成物が提供される。
【0226】
本発明は更に、本発明に記載のPAPP−A2ポリペプチドまたはそのフラグメントに対して特異性を有する抗体の生成法に関し、当該方法は、
i)宿主生体を提供し、
ii)その宿主生体を、請求項10に記載のポリペプチドを用いて免疫し、そして
iii)当該抗体を得る、
ことからなるステップを含む。
【0227】
本発明に記載のPAPP−A2ポリペプチドまたはそのフラグメントに対して特定の結合親和性を有するモノクローナル抗体およびポリクローナル抗体がまた提供される。該抗体は、モノクローナルであるのが好ましい。
【0228】
更なる態様において、本発明に記載のPAPP−A2ポリペプチドの生成法が提供され、当該方法は、
i)好適な宿主生体、好ましくは哺乳類細胞を提供し、
ii)ステップi)で提供された宿主生体を、本発明に記載のポリヌクレオチド、または本発明に記載のベクターを用いてトランスフェクトまたは形質転換し、
iii)ステップii)で得られた宿主生体を該ポリヌクレオチドまたはベクターによりエンコードされたポリペプチドの発現に好適な条件下で培養し;そして所望により
iv)宿主生体による組み換え発現の結果生じるポリペプチドを、その宿主生体から単離する、
ことからなるステップを含む。
【0229】
本発明の更に更なる態様において、アンチセンス法を用いて細胞中のPAPP−A2の発現を阻害および/または減少させる方法が提供され、当該方法は、
i)本発明に記載のアンチセンスポリヌクレオチドを提供し、
ii)ステップi)において提供された当該アンチセンスポリヌクレオチドを用いて、PAPP−A2を発現可能な細胞をトランスフェクトまたは形質転換し、
iii)ステップii)において得られた細胞を、PAPP−A2の発現に関与する当該細胞において、ステップi)で提供されたポリヌクレオチドの相補的なポリヌクレオチドへのハイブリダイゼーションに好適な条件下で培養し、そして
iv)当該細胞においてPAPP−A2の発現を阻害および/または減少させる、
ことからなるステップを含む。
【0230】
該アンチセンスポリヌクレオチドおよび該相補的なポリヌクレオチドは、別々のポリヌクレオチド分子から同時発現されてもよく、あるいはそれらが、同一の分子から発現されてもよい。ハイブリダイゼーションの別法として、該方法は、逆転写PCR法(rtPCT法)の使用を含み得る。
【0231】
本発明の更に別の態様において、個体から得た生物学的サンプルにおけるPAPP−A2の検出法、あるいはPAPP−A2レベルの測定法が提供され、当該方法は、
i)当該個体から生物学的サンプルを得て、
ii)そのサンプルにおいて、
a)PAPP−A2ポリペプチドまたはそのフラグメント、および/または
b)PAPP−A2発現に由来するmRNA形態のポリヌクレオチド、および/または
c)PAPP−A2特異的プロテアーゼ活性、好ましくはIGFBP−5プロテアーゼ活性、またはIGFBP−5の誘導体に対して示される蛋白分解活性を検出することによりサンプル中のPAPP−A2を検出する、
ことからなるステップを含む。
【0232】
該方法は、ステップii)で検出されたPAPP−A2またはPAPP−A2レベルを、
a)PAPP−A2の予め決められた量および/または濃度;および/または
b)PAPP−A2 mRNAの予め決められた量および/または濃度;および/または
c)予め決められたPAPP−A2特異的なプロテアーゼ活性、
からなる群から選ばれる予め決められた値と比較することからなるステップを更に含んでいてもよい。
【0233】
1つの実施態様においてその予め決められた値は、当該個体の正常な生理学的状態を示している。
【0234】
該生物学的サンプルは、血液、尿、胸腔内液、口腔洗浄液、組織バイオパシーおよび卵胞液からなる群から選ばれるのが好ましい。
【0235】
PAPP−A2のレベルが、PAPP−A2蛋白の量として測定される場合、そのPAPP−A2蛋白が免疫化学分析により測定されるのが好ましく、当該方法においてはPAPP−A2蛋白が、少なくとも1つのモノクローナル抗体により検出される。PAPP−A2蛋白はまた、少なくとも1つの更なる成分、好ましくは例えばpro−MBP(pro−主要塩基性蛋白)であるがそれに限定されないポリペプチドを含む複合体において検出され得る。PAPP−A2はまた、PAPP−A2単量体としてあるいはPAPP−A2二量体として検出され得る。
【0236】
本発明の更なる態様は、個体における病態の診断法に関し、当該方法は、 i)PAPP−A2の検出法またはPAPP−A2レベルの測定法を実施し、そして
ii)その病態を診断する
からなるステップを含む。
【0237】
該病態は、例えばトリソミー21、トリソミー18、トリソミー13、および開放型二分脊椎からなる群から選ばれる胎児異常であるがそれに限定されない胎児異常が好ましい。
【0238】
本発明に従って診断可能な更なる胎児異常は、子宮外妊娠、開放型二分脊椎、神経管閉鎖障害、腹部壁欠損、Edwards症候群、Pateaus症候群、Turner症候群、モノソミーX、またはKlein−felter’s症候群である。
【0239】
別の態様において、該病態は、再狭窄、アテローム性心筋梗塞、創傷治癒、線維症、心筋梗塞、骨粗鬆症、リュウマチ性関節炎、多発性骨髄腫、または癌を含むがそれに限定されない成長促進状態および成長阻害状態からなる群から選ばれる異常な成長状態である。
【0240】
本発明の更に更なる態様において、生物学的サンプルにおける本発明に記載のポリヌクレオチドの発現検出法が提供され、当該方法は、
i)本発明に記載のポリヌクレオチドを推定で含有する生物学的サンプルを提供し、そして
ii)その生物学的サンプルを、i)本発明に記載のポリヌクレオチドに相補的な、およびii)それとハイブリダイズ可能な鎖を含むポリヌクレオチドと接触させ、そして
iii)ハイブリダイゼーションが起こさせ、そして
iv)ステップiii)で得られたハイブリダイゼーション複合体を検出する、
ことからなるステップを含み、
当該方法において、ハイブリダイゼーション複合体の存在は、本発明に記載のポリヌクレオチドまたはそのフラグメントの生物学的サンプルにおける発現を示している。
【0241】
本発明の更に更なる態様において、PAPP−A2のプロテアーゼ活性を阻害する薬剤の同定法が提供され、当該方法は、
i)a)本発明に記載のポリペプチドまたはそのフラグメント、およびb)当該ポリペプチドまたはフラグメントに対して予め決められた基質、およびc)推定の阻害剤をインキュベートし、そして
ii)該基質の蛋白分解が阻害されるかどうかを決定する、ことからなるステップを含む。
【0242】
該基質は、内部でクエンチングする蛍光性ペプチドであり得るポリペプチドを含むのが好ましい。1つの好ましい基質は、IGFBP−5またはそのフラグメントを含むか、あるいはそれから本質的に構成される。
【0243】
本発明はまた、PAPP−A2のプロテアーゼ活性を阻害する薬剤を同定する当該方法に従って得られる阻害剤に関する。
【0244】
病態の処置を必要とする個体において、その病態を処置するための薬物の製造における当該提供された阻害剤の使用も提供される。
【0245】
更に更なる態様において、本発明は、PAPP−A2のプロテアーゼ活性を促進可能な薬剤の同定法に関し、当該方法は、
i)a)本発明に記載のポリペプチドまたはフラグメント、およびb)当該ポリペプチドに対して予め決められた基質、およびc)推定の促進剤をインキュベートし、そして
ii)該基質の蛋白分解が促進されるかどうかを決定する、
ことからなるステップを含む。
【0246】
該基質は、内部でクエンチングする蛍光性ペプチドであり得るポリペプチドを含むのが好ましい。IGFBP−5またはそのフラグメントが基質として特に好ましい。
【0247】
PAPP−A2のプロテアーゼ活性を促進可能な薬剤を同定する方法に従って得えられる促進剤も提供され、本発明はまた、病態の処置を必要とする個体においてその病態を処置するための薬物の製造における当該促進剤の使用も提供される。
【0248】
更に別の態様において、個体の治療による処置法が提供され、当該方法は、i)本発明に記載の医薬組成物、および/またはii)本発明に記載の阻害剤、および/または本発明に記載の促進剤を当該個体に投与することからなるステップを含む。
【0249】
更に更なる態様において、PAPP−A2またはPAPP−A2の他の蛋白との複合体の精製法が提供され、当該方法は、
i)本発明に記載のポリペプチド、またはそのフラグメントに対して特異的な結合親和性を有するポリクローナルまたはモノクローナル抗体を提供し、そして
ii)PAPP−A2、またはそのフラグメントをアフィニティークロマトグラフィーにより精製する、
ことからなるステップを含む。
【0250】
本発明がその詳細な説明と組み合わせて示されている一方、前述の説明は、添付の請求の範囲により定義される本発明の範囲を説明し、そして制限しないことが意図されていることが理解される。他の態様、利点、および修飾は、以下に示す請求項の範疇である。
図面の簡単な説明
図1は、preproPAPP−A2をエンコードするmRNAに相当するcDNA配列(5’→3’の方向で)を示す。配列のコード化部分およびその末端のストップコドン(*)だけが示されて、1〜5376がナンバリングされている。preproPAPP−A2の翻訳されたポリペプチド配列も示されている。シグナルペプチド開裂部位は、SignalP V2.0を用いてnt.64−66によりエンコードされたアラニン残基の後となると予測された((Nielsen et al., 1997, Protein Eng 10, 1−6)、http://genome.cbs.dtu.dk/にあるWWW予測サーバー)。preproPAPP−A2のシグナルペプチド(nt.1−66、22残基)が太字で示されている。この図のヌクレオチド配列は、配列番号1のnt.1〜5376を示している。この図の蛋白配列が配列番号2として示されている。
【0251】
図2は、アミノ酸配列に翻訳されたときのPAPP−A(Kristensen et al., 1994, Biochemistry 33, 1592−8)と、PAPP−AのN−末端部(hom−N、受入れ番号 AL031734のコード化部分)およびC−末端部(hom−C、受入れ番号 AL031290のコード化部分)と相同性のある2つのゲノムクローン内に含まれる配列伸長部との間の相関関係を示す図式である。この図はまた、PAPP−Aの中央領域と相同性のあるcDNA配列を得る方法をも示している。Hom−N、hom−C、および中央領域が一緒になって、新規な蛋白、PAPP−A2の完全な配列をエンコードし、これは、PAPP−Aのホモログである。該中央領域は、テンプレートとして特異的に初回刺激を受けて(プライマー、Rt−N−mid)、逆転写したヒト胎盤mRNA、PCR用にプライマー、PR−mid5およびPR−mid3を用いてPCRにより得られた(表1)。完全長のPAPP−A2をエンコードするcDNA構築物を得るために、ゲノムクローンhom−Nおよびhom−Cに相当するcDNAクローンも、テンプレートとして特異的に初回刺激を受けた胎盤mRNA(プライマーは示さず、表1参照)により合成されたcDNAを用いて得た。これは、明細書中に詳細に示されるように、シグナルペプチド伸長部(hom−N中)および停止コドン(hom−Cの3’末端)の同定が必要とされた。プライマーはすべて、表1に示されている。注:ここで示される配列の相対的な位置は、実施される実験に従うものであるが、図は、縮尺に正確には示されていない。
【0252】
図3は、preproPAPP−A2(配列番号2)のアミノ酸配列を、preproPAPP−Aと整列して示している。preproPAPP−A2(PA2)の推定のアミノ酸配列が、CLUSTAL W(Thompson et al., 1994, Nucleic Acids Res 22, 4673−80)を用いてpreproPAPP−A(PA)((Haaning et al., 1996, Eur J Biochem 237, 159−63)、AAC50543)の配列と整列された。PAPP−Aのprepro部分が、PAPP−A2に相当する領域と有意な同一性を示さなかったので、そのアラインメントを、proペプチドの長さの違いを強調するために手動で調節した。矢印は、PAPP−Aについては以前に見い出された成熟蛋白のN末端部(Glu−81)(Kristensen et al., 1994, Biochemistry 33, 1592−8)、そしてPAPP−A2については本発明で見出されたN末端部(Ser−234)を示す。推定のシグナルペプチドは、SignalP V2.0(Nielsen et al., 1997, Protein Eng 10, 1−6)を用いて強く予測されるが、小文字で示している。PAPP−A2のpro部分は、Met−168に相当する候補開始コドンを別に1つ含むが、SignalPを用いてこの残基の後に続くシグナルペプチドは何も予測されなかった。PAPP−Aの配列モチーフ(Kristensen et al., 1994,
Biochemistry 33, 1592−8)もまた、PAPP−A2において見い出され:触媒亜鉛結合モチーフおよび推定のMet−ターンの残基は、両方の配列において下線が引いて、太字にしている。Lin−ノッチモチーフ(LNR1−3)および短いコンセンサスリピート(SCR−1〜5)は四角で囲んでいる。システイン残基は、影付きになっている、成熟PAPP−Aのシステインはすべて、PAPP−A2でも見られる。更に、分泌型のPAPP−A2は4つのシステイン残基(Cys−343、Cys−533、Cys−618、およびCys−1268)を有し、PAPP−Aにはそれに相当するものはない。
【0253】
図4は、ウエスタンブロッティングおよびクマシー染色によるPAPP−A2を示している。トランスフェクトした293T細胞由来の培地は、モノクローナル抗c−mycを用いてウエスタンブロットされる。レーン1、空ベクターでトランスフェクトした細胞;レーン2、c−mycペプチド(pPA2−mH)を用いてC末端をタグ化した野生型PAPP−A2をエンコードするcDNAを用いてトランスフェクトした細胞、非還元型;レーン3、不活性化E734Q変異体(pPA2−KO−mH)を有するPAPP−A2をエンコードするcDNAでトランスフェクトした細胞、非還元型;レーン4、レーン2と同じであるが還元型。可能な自触反応を除去するために、組み換えPAPP−A2を、pPA2−KO−mHでトランスフェクトした細胞の血清フリーの培地からニッケルアフィニティークロマトグラフィーにより精製した(レーン5、還元型)。
【0254】
図5は、IGFBP−1〜6に対するPAPP−A2活性を示す。空ベクター(−)またはPAPP−A2をエンコードするcDNA(pPA2)(+)でトランスフェクトした293T細胞由来の培地を、6つのIGFBP(BP1〜BP6)のそれぞれと一緒にインキュベートし、その活性を放射標識したIGF−IIを用いてリガンドブロッティングにより評価した。IGFBP−5の完全の開裂は、BP5+レーンにおけるシグナルの不在から明らかである。IGBP−3の部分分解もまた、明らかである。
【0255】
図6は、IGFBP−5に対するPAPP−A2の蛋白分解活性を示す。空ベクター(レーン1)、不活性化E734Q変異体を有するPAPP−A2をエンコードするcDNA(pPA2−KO)(レーン2)、または野生型PAPP−A2をエンコードするcDNA(pPA2)(レーン3〜6)でトランスフェクトした293T細胞由来の培地を、C末端をc−mycタグ化したrIGFBP−5と一緒にインキュベートした。蛋白分解活性を、抗c−mycを用いてウエスタンブロッティングにより評価した。「i」は、インタクトなrIGFBP−5を示し;「c」は、検出可能なC末端c−mycタグ化開裂生成物を示す。阻害剤なしの場合において、野生型PAPP−A2は、すべてのrIGFBP−5を分解した(レーン3)。PAPP−A2活性は、10mM フェナントロリン(レーン4)および5mM EDTA(レーン5)により止められるが、100mM 3,4−DCI(レーン6)によっては影響は受けなかった。精製されたrIGFBP−5のクマシー染色したSDS−PAGEが、精製されたPAPP−A2による消化前(レーン7)および後(レーン8)で示されている。抗c−mycを用いた同じ消化のウエスタンブロットも示されている(レーン9)。配列分析により、Ser−142とLys−143の間の部位でPAPP−A2がIGFBP−5を開裂することが明らかになった。
【0256】
図7は、PAPP−A2 mRNAコード化領域のcDNA配列を示しており3’UTRの配列が直接続いている。3’UTRの配列は、実施例6.3で詳細に示されるように得られる。この配列の最初の5376個のヌクレオチド(nt.1〜5376)は、図1および配列番号1(nt.1〜5376)で示されるコード化配列を示す。この配列のヌクレオチド5377〜8527は、配列番号3(nt.5377〜8527)で示されるPAPP−A2 mRNAの3’UTRに相当する。
【0257】
図8は、PAPP−A/proMBP複合体(上の棒状の図)中のPAPP−Aサブユニットのジスルフィド構造を示している。PAPP−A/proMBP複合体をプロテアーゼおよび臭化シアンで分解した後、標準的なHPLCにより、PAPP−A/proMBP複合体起源のシステイン含有ペプチドを単離した。ペプチドは、アミノ酸分析、N末端配列分析により、そして質量分析により同定した(Overgaard, M. T., Oxvig, C.、未公表)。ジスルフィド結合が、細線で示されている。2つのシステイン残基は、proMBPへの鎖間ジスルフィド架橋を形成し、1つがPAPP−Aへの鎖間架橋を形成してそれにより二量体が生じる(図示の通り)。アスタリスクは、パートナーが見い出されていないシステインを示す。成熟PAPP−Aにおいて存在するシステイン残基は、成熟PAPP−A2においても存在している(図3参照)。PAPP−A2ジスルフィドペアは同じであると仮定するのが理にかなっている。それゆえ、この情報は、PAPP−A2の単離されたドメイン(フラグメント)の発現に対して境界領域を決定する際、重要である。PAPP−Aの遺伝子構造も示されている(下の棒状の図)。エクソン/イントロンの境界は、PAPP−A cDNA(AN X68280)のゲノム配列(AN AB020878、AL353141、およびAL137024)との比較に基づいている。中央の棒状の図は、上側および下側の棒状の図の情報に基づいたPAPP−Aの推定のドメインを示している。
【実施例】
【0258】
実施例
6.1.PAPP−A2をエンコードするヌクレオチド配列の同定
本明細書において提供される受入れ番号(AN)は、Gen−Bankまたは他の生物学的配列データベースで示された配列をいう。ANは、特定のANのもとで示される蛋白またはヌクレオチド配列と置き換え可能である。
【0259】
PAPP−Aと相同性のあるDNA配列((Kristensen et al., 1994, Biochemistry 33, 1592−8)、AN CAA48341)について、公式のヌクレオチドデータベースを調査すると、ポリペプチド配列に翻訳された場合、AN AL031734およびAL031290を有する2つのゲノムクローンが明らかとなった。両方とも、ヒト染色体1(1q24)起源である。調査は、デフォルト設定によるhttp://www.ncbi.nlm.nih.gov/BLAST/でtblastnプログラムを用いて、データベースの「nr」コレクションに対して実施された。この実施例において、PAPP−Aは、(Kristensen et al., 1994, Biochemistry 33, 1592−8)に示すように、N末端のGluを残基1としてナンバリングしている。蓄積された配列記録(AN X68280)において、このGluは残基5である。
【0260】
AL031734で報告される配列は、168835の塩基対を含む。全体配列のうち2つの不連続配列伸長部(nt.103432〜103566、および140846〜141919)が一緒に、翻訳されたときにPAPP−Aポリペプチド配列の残基16〜59および59〜413と整列される。AL031290において報告される配列は、121780の塩基対を含む。全体配列のうち4つの不連続配列伸長部(nt.10209〜10358、11752〜11901、20531〜20463、および60536〜60652)が一緒に、翻訳されたときにPAPP−Aポリペプチド配列の残基1313〜1362、1376〜1425、1457〜1479、および1470〜1506と整列される。両方のゲノム配列のコード化領域間の配列伸長部は、非コード化ゲノムDNA(イントロン)または整列しないコード化領域を示す。
【0261】
これらの所見に基づいて、我々は、PAPP−Aと相同性のある新規蛋白、PAPP−A2の存在を仮定した。そのあと、それは、AL031734およびAL031290に報告される2つのゲノム配列により部分的に包含されるPAPP−A2領域の完全なコード化配列を確立した。我々は、これらの連続的配列、hom−Nおよびhom−Cをそれぞれ示した(図2)。しかしまず第1に、我々は、PAPP−Aに対してまた相同性を示し、hom−Nおよびhom−C(図2)の配列に結合するコード化cDNA配列の存在を確立した。使用される重要なプライマーはすべて表1に示されている。1791残基のpreproPAPP−A2をエンコードするcDNA配列全体を、図1に示している。標準的なクローニング法が利用されて、すべてのDNA構築物は、配列決定により分析された。使用される方法は、以下に示される。PAPP−A2という名前は、このDNA配列によりエンコードされた蛋白に対して使用される。
【0262】
Hom−Nとhom−Cとの間の中央領域に相当する連続的なコード化cDNA伸長部のクローニング:該中央領域(図2)を得るために、テンプレートとしてヒト胎盤mRNAおよびプライマー、AL031290から誘導されるRT−N−midを用いて、cDNAを合成した(表1、図2)。このcDNAは、仮定のPAPP−A2の中央領域に相当するcDNAを得るために、PCRにおいてテンプレートとして使用した。PCRプライマーは、PR−mid5およびPR−mid3であった(表1、図2)。得られた中央領域のコード化配列は、図3の残基665〜1572(配列番号1)、全体で908個のアミノ酸に相当する。
【0263】
【表1】
【0264】
PAPP−A2のN末端部に相当する連続的なコード化cDNA伸長部(hom−N)のクローニング:5’方向に更に続くゲノム配列AL031734のマニュアル調査により、PAPP−A残基16〜59に相当する配列伸長部の読み取り枠:Nt.102646〜103566が、メチオニン残基で始まる307残基のポリペプチド配列をエンコードすることが明らかになった。この所見に基づいて、中央領域(上述で詳細に示されるような、RT−N−midで初回刺激を受けた胎盤mRNA)を得るために使用されるcDNAを、hom−Nの連続的なcDNAを得るためのPCRにおいてテンプレートとして使用した。PCRプライマーは:PR−N5およびPR−N3(表1、図2)であった。
【0265】
PAPP−A2のC末端部に相当する連続的なコード化cDNA伸長部(hom−C)のクローニング:AL031290のゲノム配列に適合するヒトEST配列について利用可能なデータベース(デフォルト設定を有するhttp://www.ncbi.nlm.nih.gov/BLAST/でのblastnプログラムを用いて)を調査することにより、伸長部nt.60536〜60652(上述参照)ですでに定義されたAL031290のコード化領域のいくつかとオーバーラップするEST配列が明らかになった。AL031290のnt.62790−62995もまた、胎盤起源のヒトEST配列、AA368081の配列と適合した。ポリペプチド配列に翻訳されたときに、このEST配列は、PAPP−AのC末端部と相同性を示した。更に、停止コドンが、PAPP−Aのアミノ酸1537に相当するコード化配列内に存在した。すなわち、その2つの配列が整列化されるときに、PAPP−A2は、PAPP−AよりもC末端側には存在しない。このことに基づいて、テンプレートとしてヒト胎盤mRNAおよびAL031290起源のプライマー(表1)を用いて、cDNAを合成した。PCRプライマー、PR−C5およびPR−C3(表1、図2)を用いて、hom−Cの連続的cDNAを得るためのPCRにおいてテンプレートとして、このcDNAを使用した。
【0266】
すべてのPCRは、Pfuポリメラーゼ(Stratagene)を用いて実施した。3つのオーバーラップするPAPP−A2 cDNA(hom−N、新規な中央領域、およびhom−C)はすべて、ベクターPCR−BluntII−TOPO(Invitrogen)中でクローン化した。いくつかのクローンを双方向で配列決定した。その構築物は、それぞれp2N、p2Mid、およびp2Cという。PAPP−A2をエンコードする全体的なヌクレオチド配列は、図1(および配列番号1)に示されている。
6.2.PAPP−A2のヌクレオチドおよびアミノ酸配列の分析
成熟PAPP−Aの1547残基のうち、708残基(45.8%)が、preproPAPP−A2と同一である。PAPP−Aのprepro部分とPAPP−A2の残りの(N末端)との間には、十分な同一性はない(図3)。この実施例において、PAPP−Aは、((Haaning et al., 1996, Eur J Biochem 237, 159−63)、AAC50543)に従ってナンバリングしている。
【0267】
PAPP−Aにおいて認識される配列モチーフ(Kristensen et al., 1994, Biochemistry 33, 1592−8)は、PAPP−A2にも存在する:伸長された亜鉛結合連続配列、3つlin−ノッチ反復(LNR1〜3)、および5つの短い連続反復(SCR1〜5)(図3)。更に、PAPP−Aの82個のシステイン残基すべてが、その2つの蛋白の間に保持されて、更に4個のシステインがPAPP−A2ポリペプチド配列中に存在する。
6.3.PAPP−A2 mRNA起源のヒトEST配列の同定
AL031290のゲノム配列と適合するEST配列のクラスターを、PAPP−A2をエンコードする配列の末端部からのおよそ1.2kbで始まり、AL031290のnt.64000〜66000周辺で同定した。PAPP−A2のコード化領域およびこのクラスターに結合するmRNAの存在が、AL031290由来のプライマー(5’−GGAAAGAGCAGAGTTCACCCAT−3’、AL031290のnt.64900−64879)およびPAPP−A2をエンコードする配列(5’−CCGTCTTAGTCCACTGCATCC−3’、AL031290のnt.20499−20519、AF311940のnt.5171−5191)、およびテンプレートとしてオリゴ−dT初回刺激を受けた胎盤cDNAを用いて、PCRにおいて明らかにされた(Overgaard et al., 1999, Biol Reprod 61, 1083−9)。予測されるように、結果の生成物のサイズは、2.2kbであり、更に約3kbの3’UTRを有するPAPP−A2 mRNAの存在が示される。組織間の分配は、表2に示されている。
【0268】
【表2】
【0269】
6.4.組み換えPAPP−A2およびPAPP−A2変異体の哺乳類細胞における発現
以下に示すプラスミド構築物が作成された:
a)pPA2:発現ベクター、pcDNA3.1+においてアミノ酸1−1791をエンコードするpre−pro−PAPP−A2のcDNA配列
b)pPA2−KO:pPA2と同様であるが、PAPP−A2の活性部位のGlu−734がGln残基と置換されたもの(E734Q)
c)pPA2−mH:発現ベクター、pcDNA3.1/Myc−His(−)Aを含有する、アミノ酸1〜1791をエンコードするpre−pro−PAPP−A2のcDNA配列で、後に停止コドンは続くのではなくて、c−mycおよびHisタグが続いているもの
d)pPA2−KO−mH:pPA2−mHと同様であるが、pPA2−KOがE734Q置換されたもの
3つのオーバーラップしているPAPP−A2 cDNAフラグメント(hom−N、中央領域、およびhom−C)を、PAPP−A2をエンコードする一本の連続したcDNA配列の構築に使用した。そのオーバーラップするフラグメントは、ベクター、PCR−BluntII−TOPO(Invitrogen)中にすべて含まれており、上述に詳細に示すように、p2N、p2Mid、およびp2Cという(実施例6.1)。適当な配向のcDNA挿入部を有するp2Nおよびp2Cのクローンを選んだ。
【0270】
pPA2の構築:NotI−BamHIフラグメントをp2Cから切り取り、pBluescriptIISK+(Stratagene)中でクローン化してp2CBlueを得た。NotI−SpeIフラグメントをp2Nから切り取り、SpeI−BclIフラグメントをp2Midから切り取った。それら2つのフラグメントを、1つの反応においてp2CBlueのNotI/BclI部位にリゲートして、PAPP−A2 cDNA全体を含むp2NMidCBlueを得た。pBluescriptIISK+のNotI−ApaIフラグメントを切り取って、哺乳類の発現ベクター、pcDNA3.1+(Invitrogen)のNotI/ApaI部位中にリゲートして、このベクターの修飾型である、pcDNA−NAを得た。その後、完全長のcDNAを、NotIおよびXhoIを用いてp2NMidCBlueから切り取り、pcDNA−NA中でクローン化してpPA2を得た。SpeIとBclIを除いて、使用されるすべての制限部位は、ベクターのマルチクローニング部位中にあり、両方ともp2N、p2Mid、およびp2Cのコード化PAPP−A2配列伸長部の2つのオーバーラップ領域のそれぞれに位置する(それぞれ、図3のnt.2365およびnt.4203)。
【0271】
pPA2−KOの構築:pPA2−KO構築物は、PAPP−A2の活性部位のGlu−734残基がGln残基と置換したpPA2発現構築物の一変異体である。従って、その突然変異体は、E734Qである。pPA2−KO構築物は、テンプレートとしてpPA2を用いて、オーバーラップ伸長PCR法(Ho et al., 1989, Gene 77, 51−9)を用いた部位特異的突然変異誘発により作成した。簡単にいうと、外側のプライマーが、5’−CGCTCAGGGAAGGACAAGGG−3’(5’末端プライマー、配列番号1のnt.976〜995)および5’−CTAGAAGGCACAGTCGAGGC−3’(3’末端プライマーnt.1040〜1021、ベクターpcDNA3.1+の配列)である。オーバーラップする内部プライマーは、5’−TGTCCCACTTGATGGATCATGGTGTCGGTGTGG−3’(配列番号1のnt.2210〜2178、E734Qの結果nt.2200はCではなくてGである)および5’−CCATCAAGTGGGACATGTTCTGGGAC−3’(配列番号1のnt.2196〜2221、E734Qの結果、nt.2200はGではなくてCである)である。その結果生成する突然変異フラグメントを、XbaIおよびXhoIを用いて消化し、pPA2と交換してpPA2−KOを産生した。PCRはすべて、PfuDNAポリメラーゼ(Stratagene)を用いて実施し、全ての構築物を配列分析により確認した。
【0272】
pPA2−mHの構築:2つのプライマー(5’−GAGGGCCTGTGGACCCAGGAG−3’、配列番号1のnt.4906〜4926、および5’−GACGTAAAGCTTCTGATTTTCTTCTGCCTTGG−3’、配列番号1のnt.5373〜5354、HindIII部位、AAGCTT、およびPCR生成物の開裂を促進するnt.GACGTAの前にある)を、PCRにおいて、テンプレートとしてpPA2と一緒に使用して、発現ベクターへのインフレームでリゲーションするためにHindIII部位で置換された停止コドンを有するPAPP−A2のC末端の156残基をエンコードするヌクレオチドフラグメントを産生した。簡単にいうと、PCR生成物をEcoRIとHindIIIを用いて消化し、pcDNA3.1/Myc−His(−)AベクターのEcoRI/HindIII部位中でクローン化してpPA2C−mHを産生した。NotI−XbaIフラグメント(PAPP−A2のN末端部分をエンコードする)、およびXbaI−EcoRIフラグメント(PAPP−A2の残りの中央部分をエンコードする)をpPA2から切り取って、1つの反応においてpPA2C−mHのNotI/EcoRI部位にリゲートした。その結果生成する構築物、pPA2−mHは、PAPP−A2、続いてKLGP残基、mycエピトープ(EQKLISEEDL)、NSAVD残基、および6個のH−残基(アミノ酸は、1文字表記されている)をエンコードした。6個のヒスチジン残基の直後に停止コドンが続いている。
【0273】
pPA2−KO−mHの構築:残基Glu−734をGln残基に置換したpPA2−mH変異体を構築した:pPA2−KOのNotI−KpnIフラグメントを切り取り、pPA2−mHのNotI−KpnI部位と交換してpPA2−KO−mHを産生した。
【0274】
哺乳類細胞における発現:すべての構築物(pPA2、pPA2−KO、pPA2−mH、およびpPA2−KO−mH)ならびに空の発現ベクター(pcDNA3.1+およびpcDNA3.1/Myc−His(−)A)を、組み換えPAPP−A2蛋白の発現のために哺乳類の細胞中に短期間トランスフェクトした。簡単にいうと、ヒトの胎児の腎臓293T細胞(293tsA1609neo)(DuBridge et al., 1987, Mol Cell Biol 7, 379−87)を、10%のウシ胎仔血清、2mM グルタミン、非必須アミノ酸、およびゲンタマイシン(Life Technologies)を補充した高濃度のグルコースDMEM培地中で維持した。細胞を6cmの培養皿上にプレーティングし、QIAprep Spinキット(Qiagen)により調製した10μgのプラスミドDNAを用いたリン酸カルシウム共沈(Pear et al., 1993, Proc Natl Acad Sci U S A 90, 8392−6)により18時間後にトランスフェクトした。更に48時間後、その上清を回収して、血清フリーの培地(293SFMII, Life Technologies)で更に48時間置き換えた。その血清フリーの培地を回収して、遠心分離により除去した。
【0275】
構築物、pPA2−mHおよびpPA2−KO−mHを用いたトランスフェクションの結果生成する組み換え蛋白のウエスタンブロッティングによる分析は、PAPP−A2が、220kDaの蛋白(図2参照)として分泌されることを示した。ジスルフィド結合の還元により、バンド移動には目に見える変化はもたらされなかった。従って、PAPP−Aとは対照的に、PAPP−A2は、単量体として分泌される。
6.5.タグ化PAPP−A2のアフィニティークロマトグラフィーによる精製
金属キレートアフィニティーカラム(2ml、Pharmacia)をニッケルイオンで帯電して、pPA2−KO−mHを用いて短期間トランスフェクトした細胞(実施例6.4参照)由来の血清フリー培地をロードした(50ml)。1M NaClを含有するPBS中での洗浄後、結合蛋白を、10mM EDTAの0.5mlのPBSフラクション中で溶出した。PAPP−A2を含有するフラクションを、SDS−PAGEにより位置決めした(図4、レーン5)。この蛋白は、空ベクターにより同じ方法で処置したトランスフェクトした細胞(偽のトランスフェクタント、mock transfectants)からは見られなかった。
6.6.PAPP−A2のN末端配列分析
構築物、pPA2−KO−mH(実施例6.4および6.5参照)を用いてトランスフェクトした細胞の培地から精製された、C末端側をタグ化したPAPP−A2を還元し、10〜20% SDSゲル上に流して、更にPVDF膜(ProBlott、Biosystemsより提供)上にブロッティングした。4つのレーンのバンドを切り取って、オンラインのHPLCを備えたApplied Biosystems 477Aシーケンサー(Sottrup−Jensen, 1995, Anal Biochem 225, 187−8)で、N末端分析を実施した。PAPP−A2ポリペプチドのSer−234の前で、R(230)VKKの後の開裂の結果得られたおよそ20pmolレベルでN末端配列:Ser−Pro−Pro−Glu−Glu−Ser−Asn(SPPEESN)が見られた。
【0276】
これにより、PAPP−A2がPAPP−Aと同様、prepro蛋白として合成されるという予測が確認される。P1位におけるアルギニン残基の不在は、この開裂の原因となるpro蛋白プロセシング酵素がフリンではなくて、似た別のpro蛋白転換酵素(Nakayama, 1997, Biochem J 327, 625−35)であることを示している。proPAPP−A2の開裂は、典型的にフリン開裂を示すR(196)QRR(Nakayama, 1997, Biochem J 327, 625−35)の後と予想されるかもしれない。我々は、開裂がこの部位で起こり、所見のN末端部が更なるプロセシングの結果によるものであるということを排除することはできない。
6.7.インシュリン様成長因子結合蛋白(IGFBP)−5の開裂
IGFBP−1(HepG2調整培地由来)、rIGFBP−2(GroPep)、rIGFBP−3(Powell博士より贈与)、rIGFBP−4(Austral)、rIGFBP−5(Andress博士より贈与)、およびrIGFBP−6(Austral)に対する活性をアッセイするために、放射標識したIGF−II(Bachem)を用いたリガンドブロッティング(Conover et al., 1993, J Clin Invest 91, 1129−37)を利用した。6つの結合蛋白のうち、IGFBP−5は、完全な開裂を示した(図5)。IGFBP−3は、一部分分解した(図5)。この開裂は、IGFの存在とは無関係であった。実験は、pPA2または空ベクターを用いてトランスフェクトした細胞由来の培地を用いて実施した。
【0277】
更なる分析に対して、組み換えIGFBP−5が哺乳類の細胞中で生成された。簡単にいうと、ヒト胎盤オリゴ−dTで初回刺激したcDNA(Overgaard et al., 1999, Biol Reprod 61, 1083−9)を、テンプレートとして用いて、ヒトIGFBP−5(受け入れ番号 M65062)をエンコードするcDNAを増幅した。XhoI部位(5’−TCCGCTCGAGATGGTGTTGCTCACCGCGGT−3’)およびHindIII部位(5’−CGATAAGCTTCTCAACGTTGCTGCTGTCG−3’)を含有する特異的なプライマーを使用し、その結果のPCR生成物を消化して、pcDNA3.1/Myc−His(−)A(Invitrogen)のXhoI/HindIII部位中でクローン化した。完全長のproIGFBP−5をエンコードした構築物の直後に、残基KLGP、mycエピトープ(EQKLISEEDL)、NSAVD残基、および6個のH−残基(アミノ酸は、一文字表記されている)が続く。その構築物を、配列分析により確認した。トランスフェクション用のプラスミドDNAを、QIAprep Spinキット(Qiagen)により調製した。組み換えIGFBP−5の細胞培養および発現は、上述の実施例6.4.に示される通りに実施した。
【0278】
ウエスタンブロッティングにより開裂分析を実施した(図6)。簡単にいうと、5μLの細胞培養培地中に含まれるような組み換えIGFBP−5を、pPA2、pPA2−KO、または空の発現ベクター(実施例6.4参照)を用いてトランスフェクトされた細胞由来の培養上清(10μL)と一緒にインキュベートした。リン酸緩衝生理食塩水を添加して、50μLの最終容量とした。37℃で12時間インキュベーション後、15μLの反応混合物を、16% SDS−PAGEを還元して分離し、PVDF膜状にブロッティングし、C末端開裂生成物を、ペルオキシダーゼ共役型二次抗体(P260、DAKO)、および化学発光増幅(ECL、Amersham)を使用して、モノクローナル抗c−myc(クローン9E19、ATTC)により検出した。
6.8.PAPP−A2活性の阻害
IGFBP−5に対するPAPP−A2の蛋白分解活性の阻害能について、様々な薬剤を分析した。実験条件は、試験される薬剤を添加することを除いては、本質的には実施例6.7に示される通りであった(図6)。PAPP−A2の蛋白分解活性に影響を与えないと見い出された薬剤としては更に、PMSFおよびアプロチニンが挙げられる。
6.9.IGFBP−5における開裂部位の同定
開裂部位の決定については、精製されたrIGFBP−5(図6、レーン7)を、精製したPAPP−A2で消化し、SDS−PAGE(図6、レーン8)により分析した。ブロットした物質のエドマン分解により、別々の目に見える分解生成物(図6、レーン8)は両方とも、N末端配列、K(144)FVGGA(IGFBP−5は、成熟蛋白のN末端Leuを残基を1としてナンバリングしている)を含んでいることが示された。2つのバンドは両方とも、C末端c−mycタグを含むため(図6、レーン9)、インタクトなC末端開裂フラグメントを示しており;それらは精製されたrIGFBP−5(図6、レーン7)の異質性により、様々にグリコシル化される可能性がある。両方のバンドとも、低レベル(45%)で、IGFBP−5に相当する第2の配列、L(1)GXFVHを含んでいた。Serが存在しないことは、第3のサイクルで予想されるが、Ser−3の炭化水素置換の証拠と考えられた。N末端開裂フラグメント上のO−結合型グリカンが、2つ別々のC末端フラグメントの周りをスメアさせる可能性がある。反応混合物(>100pmol)に関してSDS−PAGE分離をしない配列分析は、等モルで同一の2つのIGFBP−5配列のみを示した。従って、PAPP−A2は、Ser−143とLys−144との間の1つの部位でIGFBP−5を開裂する。
6.10.PAPP−A2がIGFBP−5の蛋白分解を引き起こし得る組織
IGFBP−5に対する蛋白分解活性は、数種の起源、例えば妊婦血清(Claussen et al., 1994, Endocrinology 134, 1964−6)、精漿(Lee et al., 1994, J Clin Endocrinol Metab 79, 1367−72)、平滑筋細胞(Imai et al., 1997, J Clin Invest 100, 2596−605)、顆粒膜細胞(Resnick et al., 1998, Endocrinology 139, 1249−57)、骨肉腫細胞(Conover and Kiefer, 1993, J Clin Endocrinol Metab 76, 1153−9)、およびまたは骨芽細胞(Thrailkill et al., 1995, Endocrinology 136, 3527−33)由来の培養培地、および線維芽細胞(Busby et al., 2000, J Biol Chem)から広範に報告されている。一般的に、IGFBP−5の開裂の原因となるプロテアーゼは、同定されないままである。
【0279】
線維芽細胞および骨芽細胞(Lawrence et al., 1999, Proc Natl Acad Sci U S A 96, 3149−53)と同様、濾胞液(Conover et al., 1999, J Clin Endocrinol Metab 84, 4742−5)、妊婦血清(Overgaard et al., 2000, J Biol Chem)、および血管平滑筋細胞(Bayes−Genis, A., Schwartz, R. S., Ashai, K., Lewis, D. A., Overgaard, M. T., Christiansen, M., Oxvig, C., Holmes, D. R., Jr., and Conover, C. A. Arterioscler. Thromb. Vasc. Biol.、製本中)におけるIGFBP−4プロテアーゼとしてのPAPP−Aの最近の同定は、いくつかの系においてはPAPP−AおよびIGFBP−4を重要な機能ペアとして完全に確立した。PAPP−Aに対して見い出されているような基質以外はいずれも、および他のプロテアーゼはいずれも、生理的にIGFBP−4を開裂することが示されていない。それ故、PAPP−A2とIGFBP−5のペアが、上述のおよび/または他の多数の組織において、類似の役割を果たすものと思われる。興味深いことに、IGFBP−5を平滑筋細胞調整培地と一緒にインキュベーションした結果、Ser−143とLys−144との間が開裂し(Imai et al., 1997, J Clin Invest 100, 2596−605)、これはPAPP−A2により本明細書において見られるのと同一の開裂である。このことは、PAPP−A2をこの組織に対する明らかな候補IGFBP−5プロテアーゼとして直接示唆する。
引用文献
Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z., Miller, W., and Lipman, D. J. (1997). Gapped BLAST and PSI−BLAST: a new generation of protein database search programs. Nucleic Acids Res 25, 3389−402.
Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G., Smith, J. A., and Struhl, K. (1989). Current Protocols in Molecular Biology (New York City, NY: John Wiley & Sons).
Barker, R. L., Gleich, G. J., and Pease, L. R. (1988). Acidic precursor revealed in human eosinophil granule major basic protein cDNA [published erratum appears in J Exp Med 1989 Sep 1;170(3):1057]. J Exp Med 168, 1493−8.
Bayes−Genis, A., Schwartz, R. S., Ashai, K., A., L. D., Overgaard, M. T., Christian−sen, M., Oxvig, C., Holmes Jr, D. R., and Conover, C. A. (2000). Insulin−like growth factor binding protein−4 protease produced by arterial smooth muscle cells in vitro is increased in the coronary artery following angioplasty. Arterioscler Thromb Vasc Biol, in press.
Biagiotti, R., Cariati, E., Brizzi, L., Cappelli, G., and D’Agata, A. (1998). Maternal serum screening for trisomy 18 in the first trimester of pregnancy. Prenat Diagn 18, 907−13.
Bischof, P. (1979). Purification and characterization of pregnancy associated plasma protein A (PAPP−A). Arch Gynecol 227, 315−26.
Bitter, G. A., Egan, K. M., Koski, R. A., Jones, M. O., Elliott, S. G., and Giffin, J. C. (1987). Expression and secretion vectors for yeast. Methods Enzymol 153, 516−44.
Bode, W., Gomis−Ruth, F. X., and Stockler, W. (1993). Astacins, serralysins, snake venom and matrix metalloproteinases exhibit identical zinc−binding environments (HEXXHXXGXXH and Met−turn) and topologies and should be grouped into a common family, the ’metzincins’. FEBS Lett 331, 134−40.
Bonno, M., Oxvig, C., Kephart, G. M., Wagner, J. M., Kristensen, T., Sottrup−Jensen, L., and Gleich, G. J. (1994). Localization of pregnancy−associated plasma protein−A and colocalization of pregnancy−associated plasma protein−A messenger ribonucleic acid and eosinophil granule major basic protein messenger ribonucleic acid in placenta. Lab Invest 71, 560−6.
Brambati, B., Macintosh, M. C., Teisner, B., Maguiness, S., Shrimanker, K., Lanzani, A., Bonacchi, I., Tului, L., Chard, T., and Grudzinskas, J. G. (1993). Low maternal serum levels of pregnancy associated plasma protein A (PAPP−A) in the first trimester in association with abnormal fetal karyotype. Br J Obstet Gynaecol 100, 324−6.
Broglie, R., Coruzzi, G., Fraley, R. T., Rogers, S. G., Horsch, R. B., Niedermeyer, J. G., Fink, C. L., and Chua, N. H. (1984). Light−regulated expression of a pea ribulose−1,5−bisphosphate carboxylase small subunit gene in transformed plant cells. Science 224, 838−43.
Busby, W. H., Jr., Nam, T. J., Moralez, A., Smith, C., Jennings, M., and Clemmons, D. R. (2000). The Complement Component C1s is the Protease That Accounts for Cleavage of Insulin−like Growth Factor Binding Protein−5 in Fibroblast Medium. J Biol Chem.
Christiansen, M., Oxvig, C., Wagner, J. M., Qin, Q. P., Nguyen, T. H., Overgaard, M. T., Larsen, S. O., Sottrup−Jensen, L., Gleich, G. J., and Norgaard−Pedersen, B. (1999). The proform of eosinophil major basic protein: a new maternal serum marker for Down syndrome. Prenat Diagn 19, 905−10.
Christiansen, M., Jaliashvili, I., Overgaard, M.T., Ensinger, C., Oxvig,C. (2000) Quantitation and characterization of pregnancy−associatedcomplexes between angiotensinogen and the proform of eosinophil majorbasic protein in serum and amniotic fluid. Clin. Chem. 46, 1099−1105
Clackson, T., Hoogenboom, H. R., Griffiths, A. D., and Winter, G. (1991). Making antibody fragments using phage display libraries. Nature 352, 624−8.
Claussen, M., Zapf, J., and Braulke, T. (1994). Proteolysis of insulin−like growth factor binding protein−5 by pregnancy serum and amniotic fluid. Endocrinology 134, 1964−6.
Conover, C. A., and Kiefer, M. C. (1993). Regulation and biological effect of endogenous insulin−like growth factor binding protein−5 in human osteoblastic cells. J Clin Endocrinol Metab 76, 1153−9.
Conover, C. A., Kiefer, M. C., and Zapf, J. (1993). Posttranslational regulation of insulin−like growth factor binding protein−4 in normal and transformed human fibroblasts. Insulin−like growth factor dependence and biological studies. J Clin Invest 91, 1129−37.
Conover, C. A., Oxvig, C., Overgaard, M. T., Christiansen, M., and Giudice, L. C. (1999). Evidence that the insulin−like growth factor binding protein−4 protease in human ovarian follicular fluid is pregnancy associated plasma protein−A [In Process Ci−tation]. J Clin Endocrinol Metab 84, 4742−5.
Crowther, J. R. (1995). ELISA. Theory and Practice, Methods in Molecular Biology. vol 42 (Totowa, NJ: Humana Press).
DuBridge, R. B., Tang, P., Hsia, H. C., Leong, P. M., Miller, J. H., and Calos, M. P. (1987). Analysis of mutation in human cells by using an Epstein−Barr virus shuttle system. Mol Cell Biol 7, 379−87.
Folkersen, J., Grudzinskas, J. G., Hindersson, P., Teisner, B., and Westergaard, J. G. (1981). Pregnancy−associated plasma protein A: circulating levels during normal pregnancy. Am J Obstet Gynecol 139, 910−4.
Fowlkes, J. L. (1997). Insulinlike growth factor−binding protein proteolysis. An emerging paradigm in insulin−like growth factor physiology. Trends Endocrinol Metab 8, 299−306.
Gmunder, H., and Kohli, J. (1989). Cauliflower mosaic virus promoters direct efficient expression of a bacterial G418 resistance gene in Schizosaccharomyces pombe. Mol Gen Genet 220, 95−101.
Haddow, J. E., Palomaki, G. E., Knight, G. J., Williams, J., Miller, W. A., and Johnson, A. (1998). Screening of maternal serum for fetal Down’s syndrome in the first trimester. N Engl J Med 338, 955−61.
Ho, S. N., Hunt, H. D., Horton, R. M., Pullen, J. K., and Pease, L. R. (1989). Site−directed mutagenesis by overlap extension using the polymerase chain reaction [see comments]. Gene 77, 51−9.
Hwa, V., Oh, Y., and Rosenfeld, R. G. (1999). The insulin−like growth factor−binding protein (IGFBP) superfamily. Endocr Rev 20, 761−87.
Haaning, J., Oxvig, C., Overgaard, M. T., Ebbesen, P., Kristensen, T., and Sottrup−Jensen, L. (1996). Complete cDNA sequence of the preproform of human pregnancy−associated plasma protein−A. Evidence for expression in the brain and induction by cAMP. Eur J Biochem 237, 159−63.
Imai, Y., Busby, W. H., Jr., Smith, C. E., Clarke, J. B., Garmong, A. J., Horwitz, G. D., Rees, C., and Clemmons, D. R. (1997). Protease−resistant form of insulin−like growth factor−binding protein 5 is an inhibitor of insulin−like growth factor−I actions on porcine smooth muscle cells in culture. J Clin Invest 100, 2596−605.
Kristensen, T., Oxvig, C., Sand, O., Moller, N. P., and Sottrup−Jensen, L. (1994). Amino acid sequence of human pregnancy−associated plasma protein−A derived from cloned cDNA. Biochemistry 33, 1592−8.
Lawrence, J. B., Oxvig, C., Overgaard, M. T., Sottrup−Jensen, L., Gleich, G. J., Hays, L. G., Yates, J. R., 3rd, and Conover, C. A. (1999). The insulin−like growth factor (IGF)−dependent IGF binding protein−4 protease secreted by human fibroblasts is pregnancy−associated plasma protein−A. Proc Natl Acad Sci U S A 96, 3149−53.
Lee, K. O., Oh, Y., Giudice, L. C., Cohen, P., Peehl, D. M., and Rosenfeld, R. G. (1994). Identification of insulin−like growth factor−binding protein−3 (IGFBP−3) fragments and IGFBP−5 proteolytic activity in human seminal plasma: a comparison of normal and vasectomized patients. J Clin Endocrinol Metab 79, 1367−72.
Lin, T. M., Galbert, S. P., Kiefer, D., Spellacy, W. N., and Gall, S. (1974). Characterization of four human pregnancy−associated plasma proteins. Am J Obstet Gynecol 118, 223−36.
Logan, J., and Shenk, T. (1984). Adenovirus tripartite leader sequence enhances translation of mRNAs late after infection. Proc Natl Acad Sci U S A 81, 3655−9.
Mackett, M., Smith, G. L., and Moss, B. (1982). Vaccinia virus: a selectable eukaryotic cloning and expression vector. Proc Natl Acad Sci U S A 79, 7415−9.
Marks, J. D., Hoogenboom, H. R., Bonnert, T. P., McCafferty, J., Griffiths, A. D., and Winter, G. (1991). By−passing immunization. Human antibodies from V−gene libraries displayed on phage. J Mol Biol 222, 581−97.
Matthews, D. J., and Wells, J. A. (1993). Substrate phage: selection of protease substrates by monovalent phage display. Science 260, 1113−7.
McGrogan, M., Simonsen, C., Scott, R., Griffith, J., Ellis, N., Kennedy, J., Campanelli, D., Nathan, C., and Gabay, J. (1988). Isolation of a complementary DNA clone encoding a precursor to human eosinophil major basic protein. J Exp Med 168, 2295−308.
Meldal, M. (1998). Intramolecular fluorescence−quenched substrate libraries. Methods Mol Biol 87, 65−74.
Meldal, M. (1998). Introduction to combinatorial solid−phase assays for enzyme activity and inhibition. Methods Mol Biol 87, 51−7.
Meldal, M. (1998). The solid−phase enzyme inhibitor library assay. Methods Mol Biol 87, 75−82.
Nakayama, K. (1997). Furin: a mammalian subtilisin/Kex2p−like endoprotease involved in processing of a wide variety of precursor proteins. Biochem J 327, 625−35.
Nielsen, H., Engelbrecht, J., Brunak, S., and von Heijne, G. (1997). Identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavage sites. Protein Eng 10, 1−6.
Overgaard, M. T., Haaning, J., Boldt, H. B., Olsen, I. M., Laursen, L., Christiansen, M., Gleich, G. J., Sottrup−Jensen, L., Conover, C. A., and Oxvig, C. (2000). Expression of Recombinant Human Pregnancy−Associated Plasma Protein−A and Identification of the Proform of Eosinophil Major Basic Protein as its Physiological Inhibitor. J Biol Chem.
Overgaard, M. T., Oxvig, C., Christiansen, M., Lawrence, J. B., Conover, C. A., Gleich, G. J., Sottrup−Jensen, L., and Haaning, J. (1999). Messenger ribonucleic acid levels of pregnancy−associated plasma protein−A and the proform of eosinophil major basic protein: expression in human reproductive and nonreproductive tissues. Biol Reprod 61, 1083−9.
Oxvig, C., Haaning, J., Hojrup, P., and Sottrup−Jensen, L. (1994). Location and nature of carbohydrate groups in proform of human major basic protein isolated from pregnancy serum. Biochem Mol Biol Int 33, 329−36.
Oxvig, C., Haaning, J., Kristensen, L., Wagner, J. M., Rubin, I., Stigbrand, T., Gleich, G. J., and Sottrup−Jensen, L. (1995). Identification of angiotensinogen and complement C3dg as novel proteins binding the proform of eosinophil major basic protein in human pregnancy serum and plasma. J Biol Chem 270, 13645−51.
Oxvig, C., Sand, O., Kristensen, T., Gleich, G. J., and Sottrup−Jensen, L. (1993). Circulating human pregnancy−associated plasma protein−A is disulfide−bridged to the proform of eosinophil major basic protein. J Biol Chem 268, 12243−6.
Oxvig, C., Sand, O., Kristensen, T., Kristensen, L., and Sottrup−Jensen, L. (1994). Isolation and characterization of circulating complex between human pregnancy−associated plasma protein−A and proform of eosinophil major basic protein. Biochim Biophys Acta 1201, 415−23.
Pear, W. S., Nolan, G. P., Scott, M. L., and Baltimore, D. (1993). Production of high−titer helper−free retroviruses by transient transfection. Proc Natl Acad Sci U S A 90, 8392−6.
Peters, J. H., and Baumgarten, H. (1992). Monoclonal Antibodies (Offersheim, Germany: Springer−Verlag).
Rajaram, S., Baylink, D. J., and Mohan, S. (1997). Insulin−like growth factor−binding proteins in serum and other biological fluids: regulation and functions. Endocr Rev 18, 801−31.
Resnick, C. E., Fielder, P. J., Rosenfeld, R. G., and Adashi, E. Y. (1998). Characterization and hormonal regulation of a rat ovarian insulin−like growth factor binding protein−5 endopeptidase: an FSH−inducible granulosa cell−derived metalloprotease. Endocrinology 139, 1249−57.
Rosenfeld, S. A. (1999). Use of Pichia pastoris for expression of recombinant proteins. Methods Enzymol 306, 154−69.
Ruther, U., and Muller−Hill, B. (1983). Easy identification of cDNA clones. Embo J 2, 1791−4.
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). Molecular Cloning. A Laboratory Manual (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press).
Scopes, R. K. (1987). Protein Purification. Principles and Practice. (Harrisonburg, VA: Springer−Verlag).
Sinosich, M. J. (1990). Molecular characterization of pregnancy−associated plasma protein−A by electrophoresis. Electrophoresis 11, 70−8.
Smith, G. E., Summers, M. D., and Fraser, M. J. (1983). Production of human beta interferon in insect cells infected with a baculovirus expression vector. Mol Cell Biol 3, 2156−65.
Sottrup−Jensen, L. (1993). Determination of halfcystine in proteins as cysteine from reducing hydrolyzates. Biochem Mol Biol Int 30, 789−94.
Sottrup−Jensen, L. (1995). A low−pH reverse−phase high−performance liquid chromatography system for analysis of the phenylthiohydantoins of S−carboxymethylcysteine and S−carboxyamidomethylcysteine. Anal Biochem 225, 187−8.
Spencer, K., Ong, C., Skentou, H., A, W. L., and K, H. N. (2000). Screening for tri−somy 13 by fetal nuchal translucency and maternal serum free beta−hCG and PAPP−A at 10−14 weeks of gestation. Prenat Diagn 20, 411−6.
Stocker, W., Grams, F., Baumann, U., Reinemer, P., Gomis−Ruth, F. X., McKay, D. B., and Bode, W. (1995). The metzincins−−topological and sequential relations between the astacins, adamalysins, serralysins, and matrixins (collagenases) define a superfamily of zinc−peptidases. Protein Sci 4, 823−40.
Thompson, J. D., Higgins, D. G., and Gibson, T. J. (1994). CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position−specific gap penalties and weight matrix choice. Nucleic Acids Res 22, 4673−80.
Thrailkill, K. M., Quarles, L. D., Nagase, H., Suzuki, K., Serra, D. M., and Fowlkes, J. L. (1995). Characterization of insulin−like growth factor−binding protein 5−degrading proteases produced throughout murine osteoblast differentiation. Endocrinology 136, 3527−33.
Wald, N., Stone, R., Cuckle, H. S., Grudzinskas, J. G., Barkai, G., Brambati, B., Teisner, B., and Fuhrmann, W. (1992). First trimester concentrations of pregnancy associated plasma protein A and placental protein 14 in Down’s syndrome [see comments]. Bmj 305, 28.
Wald, N. J., Watt, H. C., and Hackshaw, A. K. (1999). Integrated screening for Down’s syndrome on the basis of tests performed during the first and second trimesters [see comments]. N Engl J Med 341, 461−7.
Walsh, P. S., Erlich, H. A., and Higuchi, R. (1992). Preferential PCR amplification of alleles: mechanisms and solutions. PCR Methods Appl 1, 241−50.
Westergaard, J. G., Chemnitz, J., Teisner, B., Poulsen, H. K., Ipsen, L., Beck, B., and Grudzinskas, J. G. (1983). Pregnancy−associated plasma protein A: a possible marker in the classification and prenatal diagnosis of Cornelia de Lange syndrome. Prenat Diagn 3, 225−32.
【0280】
【表3】
【受託番号】
【0281】
DSM13783
【特許請求の範囲】
【請求項1】
配列番号2またはそのフラグメントのアミノ酸配列を含むか、あるいはそれらから本質的に構成される単離されたポリペプチドに対して特異的な結合親和性を有する抗体であって、当該フラグメントが、
i)少なくともインシュリン様成長因子結合蛋白5型(IGFBP−5)に特異的な蛋白分解活性を有し;そして/または
ii)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
iii)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、抗体。
【請求項2】
該抗体が、モノクローナル抗体およびポリクローナル抗体からなる群から選ばれる、請求項1に記載の抗体。
【請求項3】
該抗体が、モノクローナルである、請求項2に記載の抗体。
【請求項4】
個体から得た生物学的サンプルにおけるPAPP−A2の検出法、あるいはPAPP−A2レベルの測定法であって、当該方法が、
i)当該個体から生物学的サンプルを提供して、
ii)
a)配列番号2またはそのフラグメントのアミノ酸配列を含むか、あるいはそれらから本質的に構成される単離されたポリペプチドであって、当該フラグメントが、
少なくともインシュリン様成長因子結合蛋白5型(IGFBP−5)に特異的な蛋白分解活性を有し;そして/または
配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、ポリペプチド;および/または
b)PAPP−A2発現に由来するmRNA形態のポリヌクレオチド、および/または
c)好ましくはIGFBP−5、その誘導体、またはPAPP−A2に対する他の任意の好適な基質の開裂を検出することによる、PAPP−A2特異的プロテアーゼ活性、を検出することによりサンプル中のPAPP−A2を検出する、
ことからなるステップを含む、方法。
【請求項5】
該方法が、ステップii)で検出されたPAPP−A2またはPAPP−A2レベルを、
i)予め決められたPAPP−A2の量および/または濃度;および/または
ii)予め決められたPAPP−A2 mRNAの量および/または濃度;および/または
iii)予め決められたPAPP−A2特異的なプロテアーゼ活性、
からなる群から選ばれる予め決められた値と比較することからなる更なるステップを含む、請求項4に記載の方法。
【請求項6】
該予め決められた値が、当該個体の正常な生理学的状態を示す、請求項51に記載の方法。
【請求項7】
該生物学的サンプルが、血液、尿、胸腔内液、口腔洗浄液、組織バイオパシー、および卵胞液からなる群から選ばれる、請求項4に記載の方法。
【請求項8】
該PAPP−A2レベルが、PAPP−A2特異的プロテアーゼ活性として測定される、請求項4に記載の方法。
【請求項9】
該PAPP−A2レベルが、PAPP−A2蛋白の量として測定される、請求項4に記載の方法。
【請求項10】
該PAPP−A2レベルが、PAPP−A2のメッセンジャーRNAの量として測定される、請求項4に記載の方法。
【請求項11】
該PAPP−A2蛋白量が、免疫化学的分析により測定される、請求項9に記載の方法。
【請求項12】
該PAPP−A2蛋白量が、少なくとも1つのモノクローナル抗体により検出される、請求項11に記載の方法。
【請求項13】
該PAPP−A2蛋白が、少なくとも1つ更なる成分、好ましくはポリペプチドを含む複合体において検出される、請求項4に記載の方法。
【請求項14】
該PAPP−A2が、PAPP−A2単量体として検出される、請求項4に記載の方法。
【請求項15】
該PAPP−A2が、PAPP−A2二量体として検出される、請求項4に記載の方法。
【請求項1】
配列番号2またはそのフラグメントのアミノ酸配列を含むか、あるいはそれらから本質的に構成される単離されたポリペプチドに対して特異的な結合親和性を有する抗体であって、当該フラグメントが、
i)少なくともインシュリン様成長因子結合蛋白5型(IGFBP−5)に特異的な蛋白分解活性を有し;そして/または
ii)配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
iii)当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、抗体。
【請求項2】
該抗体が、モノクローナル抗体およびポリクローナル抗体からなる群から選ばれる、請求項1に記載の抗体。
【請求項3】
該抗体が、モノクローナルである、請求項2に記載の抗体。
【請求項4】
個体から得た生物学的サンプルにおけるPAPP−A2の検出法、あるいはPAPP−A2レベルの測定法であって、当該方法が、
i)当該個体から生物学的サンプルを提供して、
ii)
a)配列番号2またはそのフラグメントのアミノ酸配列を含むか、あるいはそれらから本質的に構成される単離されたポリペプチドであって、当該フラグメントが、
少なくともインシュリン様成長因子結合蛋白5型(IGFBP−5)に特異的な蛋白分解活性を有し;そして/または
配列番号2に示されるようなアミノ酸配列を有するポリペプチドを認識可能な抗体、またはその結合フラグメントにより認識され;そして/または
当該ポリペプチドに対して親和性を有する細胞表面レセプターに結合するのに、配列番号2に示されるようなアミノ酸配列を有するポリペプチドと競合する、ポリペプチド;および/または
b)PAPP−A2発現に由来するmRNA形態のポリヌクレオチド、および/または
c)好ましくはIGFBP−5、その誘導体、またはPAPP−A2に対する他の任意の好適な基質の開裂を検出することによる、PAPP−A2特異的プロテアーゼ活性、を検出することによりサンプル中のPAPP−A2を検出する、
ことからなるステップを含む、方法。
【請求項5】
該方法が、ステップii)で検出されたPAPP−A2またはPAPP−A2レベルを、
i)予め決められたPAPP−A2の量および/または濃度;および/または
ii)予め決められたPAPP−A2 mRNAの量および/または濃度;および/または
iii)予め決められたPAPP−A2特異的なプロテアーゼ活性、
からなる群から選ばれる予め決められた値と比較することからなる更なるステップを含む、請求項4に記載の方法。
【請求項6】
該予め決められた値が、当該個体の正常な生理学的状態を示す、請求項51に記載の方法。
【請求項7】
該生物学的サンプルが、血液、尿、胸腔内液、口腔洗浄液、組織バイオパシー、および卵胞液からなる群から選ばれる、請求項4に記載の方法。
【請求項8】
該PAPP−A2レベルが、PAPP−A2特異的プロテアーゼ活性として測定される、請求項4に記載の方法。
【請求項9】
該PAPP−A2レベルが、PAPP−A2蛋白の量として測定される、請求項4に記載の方法。
【請求項10】
該PAPP−A2レベルが、PAPP−A2のメッセンジャーRNAの量として測定される、請求項4に記載の方法。
【請求項11】
該PAPP−A2蛋白量が、免疫化学的分析により測定される、請求項9に記載の方法。
【請求項12】
該PAPP−A2蛋白量が、少なくとも1つのモノクローナル抗体により検出される、請求項11に記載の方法。
【請求項13】
該PAPP−A2蛋白が、少なくとも1つ更なる成分、好ましくはポリペプチドを含む複合体において検出される、請求項4に記載の方法。
【請求項14】
該PAPP−A2が、PAPP−A2単量体として検出される、請求項4に記載の方法。
【請求項15】
該PAPP−A2が、PAPP−A2二量体として検出される、請求項4に記載の方法。
【図1a】


【図1b】

【図1c】

【図2】

【図3】

【図4】

【図5】

【図6】

【図7a】

【図7b】

【図8】
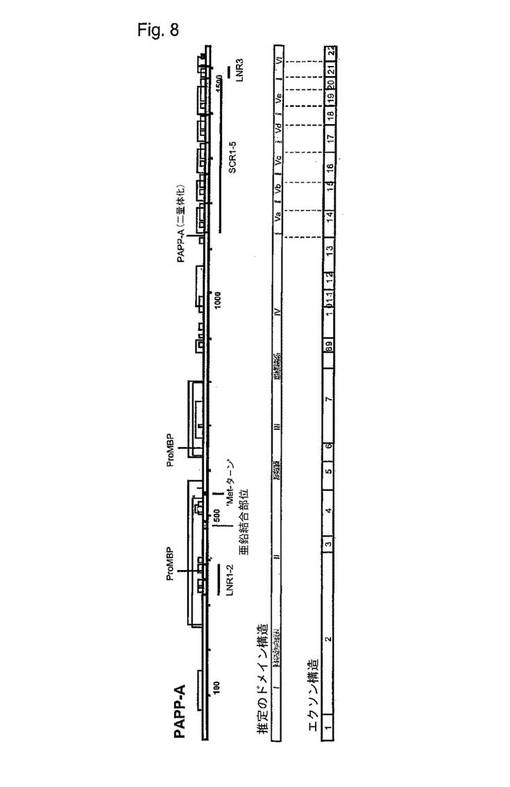
【図1b】
【図1c】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7a】
【図7b】
【図8】
【公開番号】特開2012−229235(P2012−229235A)
【公開日】平成24年11月22日(2012.11.22)
【国際特許分類】
【出願番号】特願2012−147230(P2012−147230)
【出願日】平成24年6月29日(2012.6.29)
【分割の表示】特願2009−291514(P2009−291514)の分割
【原出願日】平成13年10月19日(2001.10.19)
【出願人】(503148155)
【氏名又は名称原語表記】Como Biotech ApS
【Fターム(参考)】
【公開日】平成24年11月22日(2012.11.22)
【国際特許分類】
【出願日】平成24年6月29日(2012.6.29)
【分割の表示】特願2009−291514(P2009−291514)の分割
【原出願日】平成13年10月19日(2001.10.19)
【出願人】(503148155)
【氏名又は名称原語表記】Como Biotech ApS
【Fターム(参考)】
[ Back to top ]